
グリコアルカロイド生合成酵素活性を有するタンパク質とそれをコードする遺伝子
【課題】バレイショ等のナス科植物(Solanaceae)のグリコアルカロイド生合成酵素のDNAの提供。
【解決手段】バレイショ等のナス科植物のグリコアルカロイド生合成酵素酵素活性を有するタンパク質、及び該タンパク質をコードする遺伝子を用いて新規の生物を作成・検定する方法。
【解決手段】バレイショ等のナス科植物のグリコアルカロイド生合成酵素酵素活性を有するタンパク質、及び該タンパク質をコードする遺伝子を用いて新規の生物を作成・検定する方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、バレイショ等ナス科植物における特徴的なグリコアルカロイド化合物を生産する製造方法、グリコアルカロイド生合成酵素、当該グリコアルカロイド生合成酵素をコードするDNA、および当該DNAを用いた新たなバレイショ等ナス科植物の育種、選抜方法、ならびにグリコアルカロイドを生産しないバレイショ等ナス科植物に関する。
【背景技術】
【0002】
グリコアルカロイドはステロイダルアルカロイドとも呼ばれる一群の植物由来の化合物である。構造的には炭素鎖が27のイソプレノイドにN原子を含むものであり、ナス属植物の422種が含まれることが報告されている(非特許文献1の7.8章)。ナス属以外のナス科植物ではユリ科でもグリコアルカロイドを含むものが知られている。その中でも重要なものとしてナス科ナス属のバレイショ(Solanum tuberosum)のチャコニンおよびソラニン、ならびにトマト(Solanum lycopersicum)のトマチンである。
【0003】
バレイショは、トモロコシ、イネ、コムギに次いで世界第四番目の生産量を示す作物であるが、塊茎から出る芽や地上部植物は有毒なチャコニンおよびソラニンを含んでいることは周知の事実である。チャコニンやソラニンにより腹痛、めまい、軽い意識障害等の中毒症状を引き起こす。塊茎も、傷害を受けることや太陽光に曝されることでチャコニンおよびソラニンを容易に蓄積するため、塊茎の管理を誤ると中毒事故を起こす危険がある。
【0004】
これらの中毒事故はしばしば起きており、最近では2009年7月16日に日本国奈良市の小学校でグリコアルカロイドの中毒事件が発生している(Asahi.com報道)。バレイショの塊茎は暗所で保存すること等により20mg/100g以下のグリコアルカロイドになるように管理されているため通常は安全な食品である。しかしながら、上記のような中毒事故の危険性を考慮すると、バイレイショにおけるグリコアルカロイドを低減させることはバレイショの育種、生産、貯蔵、輸送、販売、購買などあらゆるバレイショを扱っている関係者の関心事である。しかし現在まで達成することはできてはいない。その理由としてはグリコアルカロイドのない野生種のバレイショがないこと、グリコアルカロイド生合成経路が未確定であり(非特許文献1の図7.24A,B、非特許文献2)、生合成経路に関与する遺伝子の同定が進まなかったことにある。
【0005】
グリコアルカロイドはコリンエステラーゼ阻害活性や膜破壊効果等の毒性を持つが、それ以外に、抗癌性活性、肝臓保護効果、鎮痙効果、免疫系促進効果、抗カビ性効果、抗原虫性効果、殺貝剤活性などの薬用作用が知られている(非特許文献1)。トマトではグリコアルカロイドの代謝産物であるesculeoside Aが抗動脈硬化作用を示すことも報告されている(非特許文献3)。しかし生合成経路が不明であることから、代謝産物を抑制することや効率よく生産する研究・開発はほとんど進んでこなかった。
【0006】
近年、アグリコン以降の糖転移過程で遺伝子の報告が幾つか報告されている(非特許文献4−6)。しかし、非特許文献4ではアグリコンであるソラニジンからのγソラニンへの経路であるUDP-ガラクトシルトランスフェラーゼの遺伝子を報告し、その抑制株を報告しているが、チャコニンは全く抑制できていない(非特許文献4の図2)。非特許文献4ではソラニジンからのγチャコニンへの経路であるUDP-グルコシルトランスフェラーゼの遺伝子を報告し、その抑制株を報告しているが、チャコニンおよびソラニンいずれも殆ど抑制できていない(非特許文献5の図5)。非特許文献6ではβチャコニンからαチャコニン、βソラニンからαソラニンへの経路であるラムノシルトランスフェラーゼの遺伝子を報告しているが、α体は減少しているがβ体やγ体は増加している。このように糖転移過程を抑制してもグリコアルカロイド分子種を変化させることはできるが、グリコアルカロイドの総量を調節することは極めて難しいことがわかる。
【0007】
植物ステロールや植物ホルモンの生合成遺伝子を過剰発現することでグリコアルカロイドの低下を試みた報告がある(非特許文献7)。しかしグリコアルカロイド量は多くても半分程度にしか減少できていない(非特許文献7の図5)。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Eich, Soloanaceae and Convolvulaceae: Secondary Metabolite (2008), Springer
【非特許文献2】Ginzbergら、 Poteto Research (2009) 52: 1-15
【非特許文献3】藤原ら、 2008年農芸化学会大会要旨 2B07p22
【非特許文献4】McCueら、 Plant Sci.(2005) 168: 267-273
【非特許文献5】McCueら、Phytochemistry (2006) 67: 1590-1597
【非特許文献6】McCueら、Phytochemistry (1998) 68: 327-334
【非特許文献7】Arnqvistら、 Plant Physiol. (2003) 131: 1792-1799
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、バレイショ等ナス科植物における特徴的なグリコアルカロイド化合物を生産する製造方法、グリコアルカロイド生合成酵素、当該グリコアルカロイド生合成酵素をコードするDNA、および当該DNAを用いた新たなバレイショ等ナス科植物の育種、選抜方法、ならびにグリコアルカロイドを産生しないバレイショ等ナス科植物の提供を課題とする。
【課題を解決するための手段】
【0010】
本発明者は、上記課題を解決すべく鋭意研究を重ねた。本発明者は、まずアグリコンを生成する以前の段階に着目した。この生合成経路の候補遺伝子をin silicoで探しだし、その一部分を用いてRNAiを起こす形で発現させることで内在性の候補遺伝子の発現を抑制させた。その結果、形質転換体の中で顕著にグリコアルカロイド含量が低下しているバレイショを得ることに成功するとともにグリコアルカロイド生合成酵素遺伝子を同定することができた。また、本遺伝子の発現が抑制された植物を選抜することでグリコアルカロイドのないバレイショ等ナス科植物を得ることを示した。さらに本遺伝子を発現させることで新規のグリコアルカロイド化合物の生産が可能になり、同遺伝子のゲノム配列を各種のバレイショ等ナス科植物で比較することにより多型を解析することが可能となり新規に育種されたバレイショ等ナス科植物を系呈することが可能になることを示し、本発明を完成させるに至った。同様にしてトマトにおいても内在性の遺伝子を抑制することでグリコアルカロイド含量の低減されたトマトを作出することができた。
【0011】
即ち、本発明は以下の発明を包含する。
[1] 以下の(a)または(b)のタンパク質:
(a) 配列番号1に示すアミノ酸配列からなるタンパク質;および
(b) 配列番号1に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質。
[2] 以下の(c)〜(f)のいずれかのDNAからなる遺伝子;
(c) 配列番号2に示す塩基配列からなるDNA;
(d) 配列番号2に示す塩基配列からなるDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;
(e) 配列番号2に示す塩基配列と80%以上の配列同一性を有する塩基配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;および
(f) 配列番号2に示す塩基配列の縮重異性体からなるDNA。
【0012】
[3] 以下の(g)又は(h)のタンパク質:
(g) 配列番号3に示すアミノ酸配列からなるタンパク質;および
(h) 配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質。
[4] 以下の(i)〜(l)のいずれかのDNAからなる遺伝子:
(i) 配列番号4に示す塩基配列からなるDNA;
(j) 配列番号4に示す塩基配列からなるDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;
(k) 配列番号4に示す塩基配列と80%以上の相同性を有する塩基配列からなり、かつ、
グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;および
(l) 配列番号4に示す塩基配列の縮重異性体からなるDNA。
[5] [2]または[4]の遺伝子を含有する組換えベクター。
[6] [5]の組換えベクターを導入した形質転換体。
[7] 植物体である[6]の形質転換体。
【0013】
[8] (i) ゲノムDNAまたはRNAである核酸を植物から単離する工程、
(ii) (i)の核酸がRNAである場合に逆転写しcDNAを合成する工程、
(iii) (i)または(ii)の工程で得られたDNAから配列番号2、配列番号4または配列番号5に示す塩基配列を含有する遺伝子断片を増幅する工程、ならびに
(iv) DNA中に突然変異および/または多型の存在を決定する工程、
とを含む、植物におけるグリコアルカロイド生合成酵素をコードする遺伝子の突然変異および/または多型の存在を検出する方法。
[9] 植物がバレイショ等のナス科植物である[8]の方法。
[10] [8]または[9]の方法によってグリコアルカロイド生合成酵素をコードする遺伝子の突然変異および/または多型を検出し、突然変異および/または多型を有する植物体を選抜する方法。
[11] [10]の方法により選抜された、グリコアルカロイド生合成酵素をコードする遺伝子に突然変異および/または多型を有する植物体。
[12] バレイショ等のナス科植物である[11]の植物体。
[13] グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはコードするグリコアルカロイド生合成酵素の活性が、既存品種に対して変化している植物を選抜する、[8]または[9]の植物体を選抜する方法。
[14] [13]の方法によって選抜された、グリコアルカロイド生合成酵素をコードする遺伝子の発現能が既存品種に対して変化しているか、またはグリコアルカロイド生合成酵素の活性が既存品種に対して変化している植物体。
[15] バレイショ等のナス科植物である[14]の植物体。
【発明の効果】
【0014】
本発明によれば、バレイショ等ナス科植物における特徴的なグリコアルカロイド化合物生合成活性を有するタンパク質とそれをコードする遺伝子の活性発現の調整を行うことができ、即ち当該遺伝子の活性が調整された植物を作成する方法、グリコアルカロイドを生産しないバレイショ等ナス科植物が提供される。本発明によりグリコアルカロイド化合物の含有に特徴があるバレイショ等ナス科植物の育種が可能になる。本発明の酵素により、様々な有用な生理活性を示すグリコアルカロイド化合物を大量かつ安価に生産できる。
【図面の簡単な説明】
【0015】
【図1−1】バレイショとトマトの生合成遺伝子Eの相同性をDNA解析ソフトGENETYX(ゼネティックス社)で解析した結果を示す。全般で非常に高い相同性が認められる。
【図1−2】バレイショとトマトの生合成遺伝子Eの相同性をDNA解析ソフトGENETYX(ゼネティックス社)で解析した結果を示す(図1−1の続き)。
【図1−3】バレイショとトマトの生合成遺伝子Eの相同性をDNA解析ソフトGENETYX(ゼネティックス社)で解析した結果を示す(図1−2の続き)。
【図2】候補E遺伝子抑制用ベクターの構造を示す。図2には、導入する遺伝子部分のT-DNAのライトボーダー(RB)、レフトボーダー(LB)の内部の構造、制限酵素部位を示す。
【図3】A)α−ソラニンおよびB)α−チャコニンのプロトン付加親イオンピーク(M+H)+を示すMSスペクトルの図である。
【図4】A)α-ソラニン、B)α-チャコニンおよびC)ブラシノライドの検量線(LC-MS定量分析系)を示す図である。
【図5】標品(α-ソラニン、α-チャコニンおよびブラシノライド)のLC-MSクロマトグラムを示す図である。
【図6】茎由来の試料におけるα-ソラニン、α-チャコニンおよびブラシノライドのLC-MSクロマトグラムを示す図である。
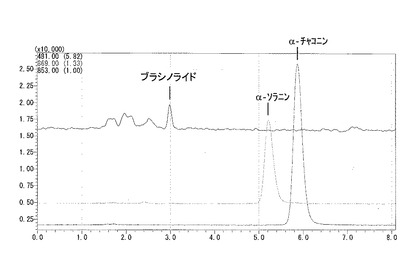
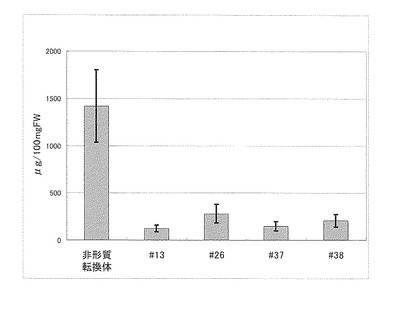
【図7】バレイショ形質転換体のin vitro茎のグリコアルカロイド含量を示す。エラーバーは標準偏差を示す。
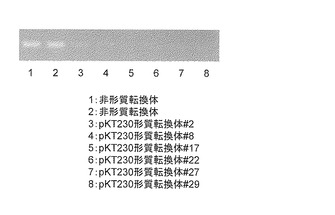
【図8】バレイショ形質転換体のin vitro茎から抽出したmRNAに対するRT-PCRの結果を示す。
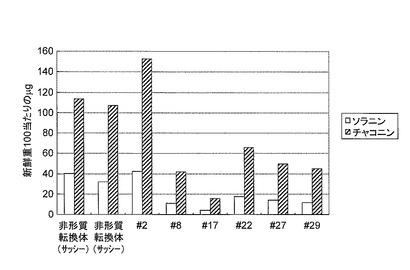
【図9】バレイショ形質転換体の塊茎の表皮のグリコアルカロイド含量を示す。エラーバーは標準偏差を示す。
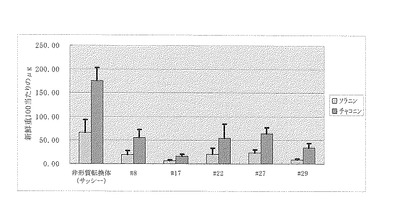
【図10】トマト形質転換体の若い葉のグリコアルカロイド含量を示す。エラーバーは標準偏差を示す。
【発明を実施するための形態】
【0016】
以下、本発明を詳細に説明する。
1.新規のグリコアルカロイド生合成酵素
本発明のタンパク質・酵素は、バレイショ等ナス科植物(Solanaceae)に含まれるグリコアルカロイド生合成酵素である。バレイショ等ナス科には、バレイショ(Solanum tuberosum)、トマト(Solanum lycopersicum)、ナス(Solanum melongena)、トウガラシ(Capsium annum)等が含まれる。また、本発明の酵素は、膜結合型のチトクロームP450モノオキシダーゼである。本発明の酵素により得られるグリコアルカロイドは、バレイショ等のナス科植物に合成されるグリコアルカロイドが含まれ、例えばバレイショのチャコニン及びソラニン等のグリコアルカロイド、トマトのトマチン等のグリコアルカロイドが挙げられる。
【0017】
本発明のグリコアルカロイド生合成酵素の基質となる好ましいステロイド化合物としては、コレステロール類が挙げられる。コレステロール類としては、コレステロール、シトステロール、カンペステロール、スティグマステロール、ブラシカステロールなどが挙げられる。本発明のグリコアルカロイド生合成酵素はこれらに水酸基を転移する水酸化酵素である。
【0018】
本発明の酵素の全長アミノ酸配列は、配列番号1または3に示される。さらに、本発明のタンパク質は、配列番号1に示されるアミノ酸配列または配列番号3に示されるアミノ酸配列と実質的に同一のアミノ酸配列を有し、グリコアルカロイド生合成酵素活性を有するタンパク質を包含する。ここで、実質的に同一のアミノ酸配列としては、当該アミノ酸配列に対して1または数個(1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個、さらに好ましくは1〜3個、さらに好ましくは1個もしくは2個)のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列、または当該アミノ酸配列と、BLAST(Basic Local Alignment Search Tool at the National Center for Biological Information(米国国立生物学情報センターの基本ローカルアラインメント検索ツール))等(例えば、デフォルトすなわち初期設定のパラメータ)を用いて計算したときに、少なくとも85%以上、好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%以上の配列同一性を有しているアミノ酸配列が挙げられる。
【0019】
本発明のグリコアルカロイド生合成酵素は、植物体から単離された天然のグリコアルカロイド生合成酵素および遺伝子工学の手法により製造されたリコンビナントのグリコアルカロイド生合成酵素を含む。
【0020】
2.グリコアルカロイド生合成酵素をコードする遺伝子
本発明の遺伝子は、ステロイド化合物に水酸基を結合する活性を持つグリコアルカロイド生合成酵素をコードする遺伝子であり、上記のグリコアルカロイド生合成酵素活性を有するタンパク質をコードする遺伝子である。
【0021】
本発明の遺伝子のDNAの塩基配列は、配列番号2または4に示される。さらに、配列番号2または4に示される塩基配列に相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズするDNA、配列番号2または4に示される塩基配列と、BLAST(Basic Local Alignment Search Tool at the National Center for Biological Information(米国国立生物学情報センターの基本ローカルアラインメント検索ツール))等(例えば、デフォルトすなわち初期設定のパラメータを用いて)を用いて計算したときに、少なくとも85%以上、好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%以上の配列同一性を有しているDNA、または前記DNAによりコードされるタンパク質のアミノ酸配列に対して1または数個(1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個、さらに好ましくは1〜3個、さらに好ましくは1個もしくは2個)のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするDNAであって、グリコアルカロイド生合成酵素活性を有するタンパク質を有するタンパクをコードするDNAを包含する。ここで、「ストリンジェントな条件」とは、例えば、「1XSSC、0.1% SDS、37℃」程度の条件であり、より厳しい条件としては「0.5XSSC、0.1% SDS、42℃」程度の条件であり、さらに厳しい条件としては「0.2XSSC、0.1% SDS、65℃」程度の条件である。さらに、本発明の遺伝子は、配列番号2または4に示す塩基配列の縮重異性体からなるDNAを包含する。
【0022】
3.組換えベクター
本発明のベクターは、上記配列番号2または配列番号4のDNAが挿入された組換えベクターである。ベクターとしては公知の酵母用、植物細胞用、昆虫細胞用等のものを広く使用できる。公知の酵母用ベクターとしてはpDR196、pYES-DEST 52、Yip5、Yrp17、Yep24などが挙げられ、公知の植物細胞用ベクターとしては、pGWB vector、pBiEl2-GUS、pIG121-Hm、pBI121、pBiHyg-HSE、pB119、pBI101、pGV3850、pABH-Hm1などが挙げられ、公知の昆虫細胞用ベクターとしては、pBM030、pBM034、pBK283などが挙げられる。本発明において使用されるベクターには、プロモーター、ターミネーター、エンハンサー等の遺伝子の発現や抑制に関する構成要素が組込まれ、必要に応じて、選択マーカー(例えば、薬物耐性遺伝子、抗生物質耐性遺伝子、レポーター遺伝子)を含有する。遺伝子の発現や抑制に関する構成要素は、その性質に応じて、それぞれが機能し得る形で組換えベクターに組み込まれることが好ましい。そのような操作は、当業者であれば適切に行うことができる。
【0023】
4.形質転換体
本発明の形質転換体は、本発明の組換えベクターを保持する形質転換体である。形質転換体は、酵素をコードする遺伝子を挿入した組換えベクターを、目的遺伝子が発現し得るように宿主中に導入することにより得ることができる。宿主は、ベクターに適したものを使用すればよい。例えば、酵母、植物細胞、昆虫細胞(Sf9など)、植物ウイルスなどが挙げられる。好ましくは、酵母、植物細胞または植物ウイルスなどが挙げられる。組換えベクターの導入方法は、微生物にDNAを導入する方法であれば特に限定されるものではない。例えばカルシウムイオンを用いる方法[Cohen, S.N.et al.:Proc. Natl. Acad. Sci., USA, 69:2110(1972)]、エレクトロポレーション法、トリペアレンタルメイティング(tri-parental mating)法等が挙げられる。また、形質転換植物体を作製する方法として、ウイルス、アグロバクテリウムのTiプラスミド、Riプラスミド等をベクターとして用いる方法が挙げられる。宿主植物としては、イネ、ムギ、トウモロコシ等の単子葉植物、ダイズ、ナタネ、トマト、バレイショ等の双子葉植物が挙げられる。形質転換植物体は、本発明の遺伝子で形質転換した植物細胞を再生させることにより得ることができる。植物細胞からの植物体の再生は公知の方法により行うことができる。
【0024】
5.グリコアルカロイド生合成酵素の製造とグリコアルカロイド化合物の生産方法
本発明のグリコアルカロイド生合成酵素は膜結合型のチトクロームP450モノオキシダーゼであり、通常の植物体から回収することができる[Collu ら, 2001, FEBS Lett. 508:215-220など]。さらには、例えば、本発明の遺伝子で形質転換した酵母等の微生物や昆虫細胞発現系を用いた大量生産により製造することができ、昆虫細胞の例としては、Morikawaら[2006, Plant Cell 18:1008-1022]のものが挙げられる。
【0025】
これらの系を使って、高い活性を持ったタンパク質として発現できるため、形質転換酵母や昆虫細胞培養液に前記グリコアルカロイド生合成酵素の基質を添加することにより、グリコアルカロイド化合物を生産することができる。例えば、形質転換酵母の培養液にコレステロール類を基質として投与することにより、水酸化されたコレステロール類を効率的に大量に生産することが可能である。酵母がサイトゾルにDMAPPを生合成する経路(メバロン酸経路)を有していることや、大腸菌にメバロン酸経路を導入することで前駆体や基質を生産することが可能にしたことが報告されている[原田と三沢2009 Aug 12. Epub Appl Microbiol Biotechnol.]。この方法を利用することで他の遺伝子と膜結合型のチトクロームP450モノオキシダーゼを同時に発現しグリコアルカロイドを生産することが可能となる。これらの膜結合型のチトクロームP450モノオキシダーゼを発現し代謝物を得た例としては大腸菌ではChangら[2007 Nat. Chem. Biol. 3:274-277]の報告、酵母では関ら[2008 PNAS 105:14204-14209]の報告がある。このような方法を組み合わせることでグリコアルカロイド化合物を生産することが可能となる。
【0026】
6.遺伝子変異、多型個体、遺伝子発現変異の選抜
本発明は、植物におけるグリコアルカロイド生合成酵素遺伝子突然変異、一塩基多型(SNP)等の多型、遺伝子発現変異の存在を検出するための方法を提供する。変異個体は放射線によるもの、化学処理によるもの、UV照射によるもの、自然突然変異によるものであっても構わない。
【0027】
この方法には、ゲノムDNAやRNAを変異個体や様々な品種や育成個体の植物から単離し、後者は逆転写しcDNAを合成する工程と、DNA増幅技術の使用によりDNAからグリコアルカロイド生合成酵素遺伝子を含有する遺伝子断片を増幅する工程と、このDNA中に突然変異の存在を決定する工程が含まれる。DNAやRNAを抽出する方法には市販のキット(例えばDNeasyやRNeasy(キアゲン社)など)が使用できる。cDNAを合成する方法も市販キット(例えばスーパースクリプト ファーストストランド システム(インビトロジェン社)など)を使うことができる。DNA増幅技術の使用により遺伝子断片を増幅する方法としては、いわゆるPCR法やLAMP法などの技術を用いることができる。これらは継続的なポリメラーゼ反応により特異的なDNA配列の増幅(つまり、コピー数を増やすこと)を達成するためにポリメラーゼを使用することを基にした、一群の技術を意味する。この反応は、クローニングの代わりに使用することができるが、必要であるのは、核酸配列に関する情報のみである。DNAの増幅を行うために、増幅しようとするDNAの配列に相補的なプライマーを設計する。次にそのプライマーを自動DNA合成により作成する。DNA増幅方法は、当技術分野で周知であり、本明細書中で与えられる教示及び指示に基づき、当業者であれば容易に行うことができる。いくつかのPCR法(ならびに関連技術)は、例えば、米国特許第4,683,195号、同第4,683,202号、同第4,800,159号、同第4,965,188号、およびInnisら編、PCR Protocols:A guide to method and applicationsで述べられている。
【0028】
DNA中に突然変異や多型の存在を決定する工程では塩基配列の決定(アプライドバイオシステムズ社)やミスマッチペアの片側を切断する酵素を用いて突然変異体を検出するTILLING法(Till et al., 2003, Genome Res 13:524-530)など変異遺伝子と正常遺伝子の相同性を利用し検出する方法を用いればよい。これらは該技術から得られた配列データを遺伝子部分に関する配列番号2、配列番号4または配列番号5に表される塩基配列と比較することで行うことができる。
【0029】
mRNA量の違いを決定する工程では上記cDNAに対し、配列番号2または配列番号4に表される塩基配列に基づいて作製したプライマーを利用してリアルタイムPCR法(ロシュ・ダイアグノスティックス社ライトサイクラーなど)等の定量的PCRを採用すればよい。その後、例えば、品種「サッシー」から得られたcDNAの量と比較することでmRNA量の違いを決定することができる。
【0030】
特に好ましい実施形態において、上記で定義したグリコアルカロイド生合成酵素遺伝子の変異の存在の決定方法を、ナス科植物(Solanaceae)のバレイショ(Solanum tuberosum)から得られた材料に適用する。
【0031】
上記の突然変異および/または多型を決定する方法により、グリコアルカロイド生合成酵素をコードする遺伝子の突然変異や多型を塩基レベルで同定することができ、さらにグリコアルカロイド生合成酵素をコードする遺伝子に突然変異および/または多型を有する植物体を選抜することができる。本発明はこのようにして得られたグリコアルカロイド生合成酵素をコードする遺伝子に突然変異や多型を有する植物体を包含する。
【0032】
また、突然変異や多型の決定やmRNA量の違いの決定、さらに方法について後述するグリコアルカロイド含量の分析により、グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはグリコアルカロイド生合成酵素の活性が変化している植物を選抜することが可能になる。
【0033】
ここで、グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはグリコアルカロイド生合成酵素の活性の変化とは、人為的突然変異等の突然変異による遺伝子の発現能またはグリコアルカロイド生合成酵素の活性の改変および多型による遺伝子の発現能またはグリコアルカロイド生合成酵素の活性が異なっていることを含む。
【0034】
ある植物のグリコアルカロイド生合成酵素活性の突然変異による改変は、その植物の種に含まれる既存品種に対する改変をいい、既存品種には野生型も含まれるが、自然状態で出現した野生種であっても、すでに産業上利用されている品種でなければ既存品種には含めない。既存の品種は、グリコアルカロイド生合成酵素活性が改変された植物が得られたときに存在するすべての品種をいい、交配、遺伝子操作等の人為的操作により作出された品種を含む。また、活性の改変において、すべての既存品種に対して、活性が変化している必要はなく、特定の既存品種に対して改変されていれば、「グリコアルカロイド生合成酵素の活性が改変された植物」に含まれる。「グリコアルカロイド生合成酵素の活性が改変された植物」は、人為的操作を受けず自然状態で突然変異により活性が改変された植物も含み、本発明の方法により、自然状態で活性が変化した植物を選抜することができ、新たな品種として確立することもできる。また、ある既存品種に変異誘発処理を行い、グリコアルカロイド生合成酵素の活性が改変された植物を作出した場合、比較対象は変異誘発処理を行った品種と同じ既存品種でもよいし、それ以外の他の既存品種でもよい。また、自然界からの選抜あるいは変異誘発処理により作出された、グリコアルカロイド生合成酵素をコードする遺伝子に突然変異や多型を有する植物を交配することにより、グリコアルカロイド生合成酵素をコードする遺伝子の変異が固定されグリコアルカロイド生合成酵素遺伝子の発現能またはグリコアルカロイド生合成酵素活性が改変された植物新品種として得ることもできる。
【0035】
例えば、植物がバレイショ(Solanum tuberosum)の場合、既存品種として、「シンシア」、「サッシー」、「シェリー」、「男爵」、「メークイーン」、「さやか(農林登録番号:農林36号)」等がある。ここで、グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはグリコアルカロイド生合成酵素の活性が既存品種に対して改変された植物とは、既存品種に対してグリコアルカロイド生合成酵素をコードする遺伝子の発現能が増強した植物および低下した植物を含み、さらに、グリコアルカロイド生合成酵素の活性が既存品種に対して上昇した植物および低下した植物を含む。本発明は、このようなグリコアルカロイド生合成酵素をコードする遺伝子の発現能またはグリコアルカロイド生合成酵素の活性が既存品種に対して改変された植物体も包含する。
【0036】
特に有毒物質であるグリコアルカロイド生合成酵素の活性が低下した植物が好ましい。このような植物体は、グリコアルカロイド生合成酵素の合成量が低いか、又は合成できず、植物体中のグリコアルカロイド生合成酵素の含量が低いか、又はグリコアルカロイド合成酵素が存在せず、あるいはグリコアルカロイド合成酵素の活性が低いか又は喪失している。その結果、植物体内のグリコアルカロイド含量も低いか、あるいはグリコアルカロイドが存在しない。例えば、バレイショの場合はチャコニン及びソラニン等のグリコアルカロイドが合成されず、バレイショの塊茎内においてチャコニン及びソラニン等のグリコアルカロイドの合成量及び存在量が低い。また、トマトの場合は、トマチン等のグリコアルカロイドが合成されず、トマトの実内においてトマチン等のグリコアルカロイドの合成量及び存在量が低い。
【0037】
バレイショの場合、グリコアルカロイド合成酵素の活性が低いか、喪失した植物体は、塊茎内でチャコニン及びソラニン等のグリコアルカロイドが合成されないか、又は上記の既存品種に比べて塊茎内で合成されるチャコニン及びソラニン等のグリコアルカロイドが少なく、塊茎内に存在するチャコニン及びソラニン等のグリコアルカロイドの量も低い。
【0038】
7.グリコアルカロイドの分析および精製
グリコアルカロイド含量の分析方法および精製方法については、Matsudaら(Phytochem. Anal. 15:121-124, 2004)やKozukueら(J. Agric. Food Chem. 52: 2079-2083, 2004)等液体クロマトグラフィーを用いた方法が知られている。しかし、サンプルの前処理が煩雑であったり、検出限界が十分に高いものではない、強酸を用いるためカラムや装置への負荷が大きいといった課題がある。よって本発明では、GA類を効率的に精製し、且つ高い精度で分析できるアルカリ耐性の逆相クロマトグラフィー用カラムを用いた液体クロマトグラフィーを用いた方法(該方法は、日本国特許出願 特願2009-170317として出願済み)を用いることが出来る。実施例5に本方法のジャガイモでの適用例を示す。
【0039】
本方法に用いるカラムとしては、アルカリ耐性に優れたカラムであれば利用することができる。アルカリ耐性に優れたカラムとしては、例えばエチレン架橋カラムを用いることができる。好ましくは、XBridge(商標)ブランド(Waters社)のカラムを用いる。特に好ましくはWaters XBridgeTM Shield RP18(Waters社)およびWaters XBridgeTM C18である。本方法において、XBridgeTMShield RP18カラムは1サンプルあたりの所要時間が短く、一方、Waters XBridgeTMC18カラムは耐久性が高いといった有利点をそれぞれ有する。
【0040】
液体クロマトグラフィーに用いる移動相は、アルカリ性緩衝液を用いることができる。好ましくは、揮発性のアルカリ性緩衝液を用いる。液体クロマトグラフィーにより精製した試料を質量分析に供する場合に、移動相に揮発性のアルカリ性緩衝液を用いると試料中に当該アルカリ性緩衝液が残留しないために都合が良い。揮発性のアルカリ性緩衝液としては例えば、トリエチルアミン、炭酸水素アンモニウムなどを用いることができるが、好ましくは緩衝効果の高い炭酸水素アンモニウムを用いる。
【0041】
移動相に用いられる炭酸水素アンモニウムの濃度は、5〜20mM、好ましくは5〜15mM、さらに好ましくは10mMである。炭酸水素アンモニウムのpHは、好ましくはpH 9.0〜11.0、さらに好ましくはpH 10.0にすることが可能である。移動相のpHを10.0とすることによって、炭酸水素アンモニウムの緩衝能がより高まる。
【0042】
GA類は、移動相にアルカリ性緩衝液と有機溶媒を用いて、アイソクラティック法により溶出しても良いし、グラジェント法により溶出しても良いが、操作が簡便なアイソクラティック法により溶出するのが好ましい。
【0043】
移動相に用いられる有機溶媒としては、例えば、メタノール、エタノール、テトラヒドロフラン(THF)、アセトニトリル(MeCN)等を用いることができるが、これらに限定されない。好ましくは、MeCNを用いる。
【0044】
アイソクラティック法においては、アルカリ性緩衝液と有機溶媒、好ましくは炭酸水素アンモニウム水とMeCNとを、30〜70:70〜30、好ましくは40〜60:60〜40の割合で、所望のGA類に応じて適宜用いる。例えば、所望のGA類がα−ソラニンまたはα−チャコニンである場合には40:60の割合で、所望のGA類がα−トマチンである場合には60:40の割合でアルカリ性緩衝液と有機溶媒、好ましくは炭酸水素アンモニウム水とMeCNとを用いる。
【0045】
液体クロマトグラフィーは市販のHPLC装置を用いて行うことができ、カラムの平衡化や流速はカラムサイズや試料の容量によって適宜設定することができる。
【0046】
液体クロマトグラフィーを行い、得られた画分は、後述の質量分析やUVまたは多波長検出器等を用いて分析することができる。
【0047】
植物由来の試料は、液体クロマトグラフィーに付す前に、予め以下の前処理を行い粗精製しておくことが好ましい。
【0048】
植物由来の試料は、GA類に加え、様々な高分子夾雑物(デンプン、タンパク質、セルロース等)を含有する。したがって、GA類を効率的に精製かつ高精度に分析するために、試料中に含まれる高分子夾雑物を除去し、GA類を粗精製および洗浄する必要がある。
【0049】
高分子夾雑物を除去する方法としては、当業者にとって一般的な方法、例えば、アルコール沈殿法を用いることができる。アルコールは、エタノールまたはメタノールを用いることができるが、メタノールが好ましい。この際、GA類を塩として効率よく抽出するためにアルコールには酸を添加する。用いることができる酸としては、酢酸、塩酸、ギ酸等が挙げられるがこれらに限定されない。好ましくはギ酸を添加する。アルコールへの酸の添加量は、所望のGA類が破壊されない範囲で適宜設定することができ、ギ酸であれば0.1〜2%(v/v)、好ましくは0.1%(v/v)となるようにアルコールに添加する。ギ酸以外の酸を用いる際には、上記添加したギ酸と同等の規定度になるまで添加することができる。
【0050】
なお従来の試料調製方法(Matsudaら, Phytochem. Anal. 15: 121-124, 2004を参照)では、試料を長時間ホモジネートし、さらに試料中に大量に含まれるデンプン等の高分子夾雑物を除去するために遠心分離を複数回行い、さらにその後、ろ過に供することを含む、長時間かつ煩雑な前処理を要していた。一方、本方法における調製方法では、上記のように短時間で破砕した植物片よりアルコール沈殿でデンプン等の高分子夾雑物を除去できるため、短時間かつ容易に試料調製が可能である。
【0051】
アルコール沈殿後、GA類を含む上清を0.1〜2%(v/v)のギ酸、酢酸等の酸、好ましくは0.1%(v/v)ギ酸で希釈し、上記の条件で液体クロマトグラフィーに付す。
【0052】
液体クロマトグラフィーにより精製した画分は、さらに質量分析に付すことができる。この場合、液体クロマトグラフィーと質量分析を連結した手法であるLC-MS法により行えばよい。
【0053】
質量分析は、単収束扇形磁場型質量分析法、二重収束扇形磁場型質量分析法、四重極型質量分析法、四重極イオントラップ型質量分析法、飛行時間型質量分析法、イオンサイクロトロン型質量分析法(フーリエ変換型質量分析法)などを用いて行うことができる。
【0054】
質量分析において試料をイオン化させる方法としては、EI(electron ionization:電子イオン化)法、CI(chemical ionization:化学イオン化)法、DEI(desorption electron ionization:脱離電子イオン化)法、DCI(desorption chemical ionization:脱離化学イオン化)法、FAB(fast atom bombardment:高速原子衝撃)法、FRIT-FAB(FRIT-fast atom bombardment:フリット高速原子衝撃)法、ESI(electrospray ionization:エレクトロスプレーイオン化)法、MALDI(matrix-assisted laser desorption ionization:マトリックス支援レーザー脱離イオン化)法を用いることができる。
【0055】
質量分析における諸条件については、実施例に具体的に記載するが、当業者であれば分析対象となるGA類に応じて、適宜条件を設定することが可能である。
【0056】
LC-MS法を用いてGA類の分析標品を分析し、当業者にとって一般的な方法に従って検量線を作成することが可能である。ジャガイモ由来の試料において、特にα−ソラニンおよびα−チャコニンの分析系では、内部標準物質としてβ−D−グルコサミンペンタアセテートを利用することが可能であるが、α−ソラニンおよびα−チャコニンと同様にステロイド骨格を有するブラシノライドを用いることが好ましい。一方、トマト由来の試料において、特にα−トマチンの分析系では水溶性アミンを用いることが好ましい。内部標準物質として利用し得る水溶性アミンとして、セリンメチルエステルおよびアラニンメチルエステルが挙げられるが、アラニンメチルエステルはカラム保持が強いために特に好ましい。したがって、ジャガイモ由来の試料においてはブラシノライドを、またトマト由来の試料においてはアラニンメチルエステルを利用することによって、定量分析の信頼性を顕著に高めることができる。
【0057】
本方法においては、HPLCに汎用されるサイズのカラムを用いることができるため、本条件はこのままUVまたは多波長検出器による分析にも使用することができる。
【実施例】
【0058】
以下、本発明を、実施例を示してより詳しく説明するが、本発明はこれらに限定されるものではない。
(実施例1)グリコアルカロイド生合成候補遺伝子Eの全長配列の取得
バレイショ(Solanum tuberosum)の品種「サッシー」の萌芽からmRNAの抽出をRNeasy(キアゲン社)で行った。全cDNAの合成はスーパースクリプト ファーストストランド システム(インビトロジェン社)を用いて行った。グリコアルカロイドのアグリコンはコレステロールからできるといわれているが確証はない(非特許文献1)。しかし、近縁の化合物から作られると仮定しても幾つかの水酸化の過程が必要になる。水酸化の過程には少なくともチトクロームP450型モノオキシゲナーゼ、ジオキシゲナーゼ、NADPH-flavin リダクターゼの3種の可能性が考えられる。この中からP450型を標的に考え、バレイショの発現する遺伝子は公開されている情報のDFCI Potato Gene Index (http://compbio.dfci.harvard.edu/tgi/plant.html) Release 11.0から萌芽で多くのESTクローンが単離されている遺伝子TC155233に注目した。
【0059】
この配列を元にプライマー[U890: GAGGCTAAGAAAAAGAGAGAGAGA (配列番号6)、U889:CGTTCTACAAAAACATCCAATTT (配列番号7)]を用いてPCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃3分)を30回、72℃10分)を行った。増幅産物をTOPOTAクローニングキットシークエンシング用(インビトロジェン社)を用いてクローニングした。さらにABI310(アプライドバイオシステムズ社)を用いて塩基配列を決定した。ORFを含む部分を配列番号2に、cDNA配列からコードされる酵素のアミノ酸配列を配列番号1に示す。
【0060】
なお、トマトの相同遺伝子は、ナス科ゲノムネットワーク(http://solgenomics.net/index.pl)の、SGN-U583521に相当する。ORFを含む部分を配列番号4に、cDNA配列からコードされる酵素のアミノ酸配列を配列番号3に示す。これらの遺伝子の塩基配列を比較したところ相同性は95%であった。このトマトの相同遺伝子のゲノム配列は同じくナス科ゲノムネットワークのSL1.00sc03540としてゲノム構造が掲載され7つのイントロンを含むことが報告されている。しかし、同ホームページには、なんら機能に関する報告はない(図1)。
【0061】
(実施例2)グリコアルカロイド生合成候補遺伝子Eのゲノム遺伝子の単離
ゲノムDNAをRNeasy(キアゲン社)で「サッシー」から抽出した。実施例1と同じプライマー並びに(U904: TGATAAGGAAATCCTGGGAGA(配列番号8)、U901: AGAGAAGCCATGAAGGATGG(配列番号9))を用いて、さらに第2イントロンは酵素をPrimeSTAR HS DNA Polymerase(タカラバイオ社)とプライマー(U898: GAAATACGCTACTACGGAAGAACC(配列番号10)とU899: CGTCATTTGCCTAATCTCATC(配列番号11))を用いてPCRを行い、全長ゲノムDNAの塩基配列を決定した(配列番号5)。イントロンは7箇所あることが明らかになった。
【0062】
(実施例3)グリコアルカロイド生合成候補遺伝子Eの抑制形質転換体を作成するためのベクター構築
遺伝子を形質転換によって抑制する方法としては、強力なプロモーターで駆動する構成を持つ逆方向の相補鎖遺伝子断片の発現(植物で一般的にRNAi法と呼ばれる)で行った[Chuangと Meyerowitz Proc Natl Acad Sci U S A., 97, 4985-90 (2000)、WesleyらPlant J., 27, 581-90 (2001)]。実施例1で取得した全長cDNAに対し、プライマー[U675: GAGCTCTAGAGGTTTGGGACAGGAGGAAT (配列番号12)、U676: GGATCCATATGCAAGCCTGTGCATCTTAT (配列番号13)]を用いてPCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃30秒)を30回、72℃10分)を行い、遺伝子断片を取得した。バイナリーベクターpKT11(特開2001-161373号公報)を基本として、カルフラワーモザイクウイルスの35S RNAプロモーター、当該遺伝子断片を順方向、シロイヌナズナのフィトエンデサチュラーゼ遺伝子(AT4g14210)の第3イントロン、当該遺伝子断片を逆方向、ノパリン合成酵素遺伝子のターミネーターの順に連結を行い、植物形質転換用ベクターpKT230を作成した(図2)。
【0063】
(実施例4)バレイショ形質転換植物体の作出
実施例3で作製したベクターをエレクトロポレーション法(GelvinとSchilperoor編, Plant Molecular Biology Manual, C2, 1-32 (1994), Kluwer Academic Publishers)により、アグロバクテリウム・ツメファシエンスGV3110株に導入した。ベクターを含むアグロバクテリウム・ツメファシエンスGV3110株を、50ppmのカナマイシンを含むYEB液体培地[5g/lビ−フエキス、1g/l酵母エキス、5g/lペプトン、5g/lスクロ−ス、2mM硫酸マグネシウム(pH7.2)]にて28℃、12時間振とう培養した。培養液1.5 mlを10,000rpm、3分間遠心して集菌後、カナマイシンを除くために1mlのLB培地で洗浄した。更に10,000rpm、3分間遠心して集菌後、1.5 mlの3%蔗糖を含むMS培地[Murashige & Skoog, Physiol. Plant., 15, 473-497 (1962)]に再懸濁し、感染用菌液とした。
【0064】
バレイショの形質転換は[門馬(1990)植物組織培養7:57-63]に従い実施した。バレイショ品種「サッシー」(キリンアグリバイオ社)から得られたマイクロチューバーを2〜3mmにスライスし、アグロバクテリウム感染用の材料とした。これを上記のアグロバクテリウムの菌液に浸した後、滅菌済みの濾紙上に置いて過剰のアグロバクテリウムを除いた。シャーレ内のMS培地(Zeatin 1ppm, IAA 0.1ppm, アセトシリンゴン100μM、及び寒天0.8%を含む)上に置き、培養は3日間25℃、16時間照明(光量子束密度32μE/m2s)/8時間無照明の条件下で行った。ついで、アセトシリンゴンの代わりにカルベニシリン250ppmを含んだ培地で1週間培養した。その後、さらにカナマイシン50 ppmを含む培地上に移し、2週間ごとに継代した。この間に不定芽が形成し、シュートを生じた。伸張したシュートをカルベニシン250 ppm及びカナマイシン100 ppmを含み、植物生長調節物質を含まないMS培地に置床した。発根したシュートをカナマイシン耐性の生長した植物体の中から外来遺伝子としてカナマイシン耐性遺伝子を含有する個体を、PCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃1分)を30回、72℃10分)を行うことで検出し、該再分化植物体が形質転換植物体であることを確認した。ここで、カナマイシン耐性遺伝子の配列を特異的に増幅するプライマーとして、TAAAGCACGAGGAAGCGGT(配列番号14)、及びGCACAACAGACAATCGGCT(配列番号15)を用いた。以上から、ベクターpKT230が導入されたバレイショの形質転換植物体30系統を取得した。
【0065】
(実施例5)形質転換植物体のグリコアルカロイド含量と候補遺伝子Eの発現解析
実施例4で得られた30個体のin vitro茎を継代後一ヶ月伸張させ、その部分2-4本をまとめて約100mgにしアルカリ耐性の逆相クロマトグラフィー用カラムを用いた液体クロマトグラフィーを用いた以下の方法(該方法は、日本国特許出願 特願2009-170317として出願済み)によりグリコアルカロイド含量を測定した。
【0066】
バレイショに含まれるGA類(α-ソラニン、α-チャコニン)の分析
1. 試料の調製
実施例4で得られた30個体のin vitro茎を継代後一ヶ月伸張させ、その部分を2-4本をまとめて約100mgにし0.1%ギ酸 in 80%MeOH aq. 990μLおよび内部標準としてブラシノライド(ブラシノ)10μg/10μLを添加し、ミキサーミルで破砕した(1/25 sec, 5 min, 4℃)。得られた破砕物を遠心分離(10,000 rpm, 5 min)に供しアルコール沈殿を行った後、上清25μLを分取し、0.1%ギ酸水で500μLにフィルアップした。これを試料として、以下の条件でLC-MSに用いた。LC-MS装置は、LCMS-2010EV(島津製作所)を用いた。
【0067】
2. LC-MS条件
(i)LC条件
LC系には、アルカリ耐性に優れたエチレン架橋カラム(XBridgeTM Shield RP18-5(φ2.1×150 mm, Waters社))を採用した。移動相には、移動相A:10 mM炭酸水素アンモニウム水(pH 10)および移動相B:MeCNを、上記試料溶媒についてA:B=40:60の割合でアイソクラティック条件で用いた。その他の条件は以下のものを用いた:
流速:0.2 mL/min
カラムオーブン:40℃
【0068】
(ii)MS条件
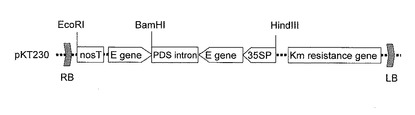
まず各成分のMSスペクトルをスキャンモードで確認し(図3参照)、その結果として検出法:SIMモード
m/z:481(ブラシノライド)、869(α−ソラニン)、853(α−チャコニン)
を用いた。
その他MS条件は以下のものを用いた。
MS検出:ポジティブイオンモード
イオン化法:ESI
イベント時間:1 sec
検出器電圧:1.5 kV
分析時間:8 min
【0069】
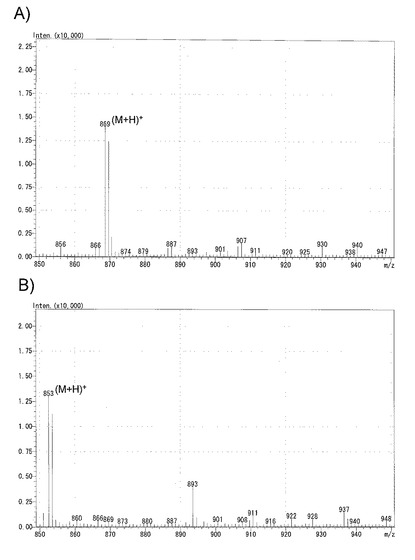
3. 標品α-ソラニン、α-チャコニン、ブラシノライドを使っての検量線作成
α−ソラニン(和光純薬)2mgとα−チャコニン(シグマアルドリッチ)2mgをそれぞれ1mLの0.1%(v/v)ギ酸水に溶解した(それぞれ2μg/μL溶液)。2種類の溶液を等容で混和し、α−ソラニンとα−チャコニンが1μg/μL(=1000ng/μL)の溶液を調製した。これを0.1%(v/v)ギ酸水で段階的に10倍希釈後、LC-MSに供して検量線を作製した。また、両物質の検出限界値を求めた。
【0070】
ブラシノライド(ブラシノ)については1mgを1mLのMeOH溶液に溶解した(1μg/μg)。これを50%(v/v)含水MeOHで段階的に10倍希釈後、LC-MSに供して検量線を作製した。
【0071】
α−ソラニン、α−チャコニンおよびブラシノライドのそれぞれについて作成した検量線を図4に示す。α−ソラニンとα−チャコニンについては図4に示したように、0.05〜50ngの範囲で良好な直線性が得られ、信頼係数は0.99以上であった。両物質ともに100ngを超えるとシグナルがサチュレートして、直線性が失われた。また両物質の検出限界は0.02ng(2μLインジェクト)であった。
【0072】
一方、ブラシノライドは2〜200ngの範囲で良好な直線性が得られた(図4を参照)。500ng以上ではやはりシグナルがサチュレートした。
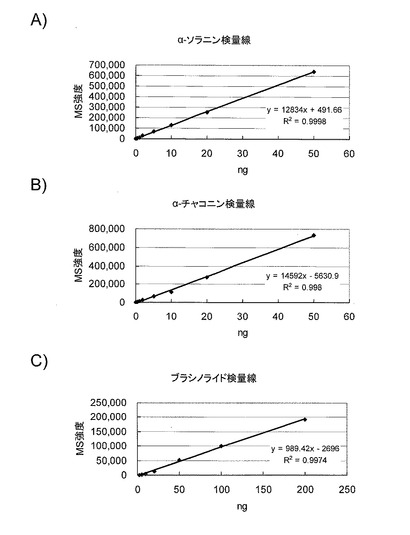
【0073】
標品α-ソラニン、α-チャコニン、ブラシノライドを用いての典型的クロマトグラムを図5に示した。
【0074】
4. ブラシノライドを内部標準としたバレイショ中のGA類のLC-MS分析
上記1.にて調製した各試料の10μLまたは20μLを上記条件を用いたLC-MS系にインジェクトした。
【0075】
内部標準ブラシノライドの回収率は50〜110%であった。ブラシノライドの定量値で補
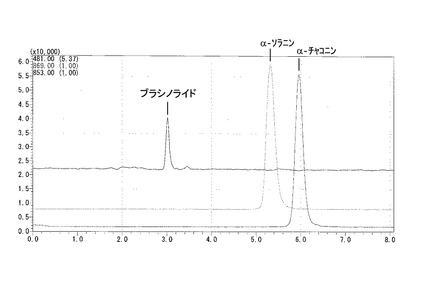
正を行い、各試料に含まれるα-ソラニンおよびα-チャコニンの量について上記検量線を用いて定量し、試料100 mg(FW)あたりに含まれる量として算出した。
当該分析における典型的クロマトグラムを図6に示す。
【0076】
30個体のうち5系統(#8, #17, #22, #27, #29)においてはグリコアルカロイドの蓄積が再現性よく低かったことから、低くなかった1系統(#2)と遺伝子を導入していない対照の個体2つを同じくin vitro茎を液体窒素で粉砕し、半分をグリコアルカロイド含量の測定、半分をmRNAの抽出をRNeasy(キアゲン社)で行い、全cDNAの合成はスーパースクリプト ファーストストランド システム(インビトロジェン社)を用いて行った。これらの個体はグリコアルカロイドの蓄積が非形質転換体(2個体)と比較して極めて低く(図7)、さらにプライマー [U887: TAAGGGACTCAAGGCTCGAA (配列番号16)、U886: TTCCTCTTTGGCTTTCTCCA (配列番号17)]を用いたRT-PCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃3分)を25回、72℃5分)の結果、mRNAの発現はいずれの個体も極めて少ないか観察できなかった(図8)。このことから、候補遺伝子Eの遺伝子の発現を抑制することによってグリコアルカロイドの蓄積が極端に減少することが明らかとなり、候補遺伝子Eはグリコアルカロイド生合成酵素をコードする遺伝子であることが明らかとなった。非形質転換体とともに、これら5系統のin vitro植物を増殖し、各3個体を市販されている野菜用の培養土に馴化しバイオハザード温室で定法に従い栽培し塊茎を収穫した。この5系統の各個体(#8, #17, #22, #27, #29)は非形質転換体と同等の生育を示し、同等の塊茎を収穫することができた(表1)。
【0077】
【表1】

【0078】
さらに収穫した塊茎各3つの中央部表皮を約1mmで剥離し同様にグリコアルカロイド含量を解析した。その結果、驚くべきことに、塊茎でのグリコアルカロイドは極めて低く、同様な方法で測定した、グリコアルカロイドの低い品種として知られている「さやか」と比較しても、それ以下であることが確認できた(図9)。
【0079】
(実施例6)トマト形質転換植物体の作出
トマトの形質転換は[Sunら (2006) Plant Cell Physiol. 47:426-431.]に従い実施した。(実施例3)で作製したベクターpKT230を含むアグロバクテリウム・ツメファシエンスAGL0株を培養し感染用菌液とした。トマト(Solanum lycopersicum)実験系統「マイクロトム」の無菌播種植物体の子葉を5mm以下の切片を、上記のアグロバクテリウム懸濁液に浸し、10分間感染した後、滅菌済みの濾紙上に葉を置いて過剰のアグロバクテリウムを除いた。シャーレ内の共存MS培地(ゼアチン1.5mg/l、アセトシリンゴン40μM及びゲルライト0.3%を含む)[Murashige & Skoog, Physiol. Plant., 15, 473-497 (1962)]上に葉を置き、シャーレを暗所で3日間25℃で培養した。切片は選択MS培地1(ゼアチン1.5mg/l、カナマイシン100mg/l、オーグメンチン375mg/l及びゲルライト0.3%を含む)で25℃、16時間照明(光量子束密度32μE/m2s)/8時間無照明の条件下で2週間ごとに継代した。この間に不定芽が形成し、シュートを生じた。さらにシュートを伸張させるため、選択MS培地2(ゼアチン1.0mg/l、カナマイシン100mg/l、オーグメンチン375mg/l及びゲルライト0.3%を含む)に移植し、伸張したシュートは選択1/2濃度MS培地(カナマイシン100mg/l、オーグメンチン375mg/l及びゲルライト0.3%を含む)で発根させた。シュートをカナマイシン耐性の生長した植物体の中から外来遺伝子としてカナマイシン耐性遺伝子を含有する個体を、PCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃1分)を30回、72℃10分)を行うことで検出し、該再分化植物体が形質転換植物体であることを確認した。ここで、カナマイシン耐性遺伝子の配列を特異的に増幅するプライマーとして、TAAAGCACGAGGAAGCGGT(配列番号18)、及びGCACAACAGACAATCGGCT(配列番号19)を用いた。以上から、ベクターpKT230が導入されたトマトの形質転換植物体13系統を取得した。得られた13個体を温室に馴化し約1ヶ月栽培し、新しく展開した若い葉の3枚から各約100mg秤量し、バレイショと同様にアルカリ耐性の逆相クロマトグラフィー用カラムを用いた液体クロマトグラフィーを用いた実施例5の方法によりグリコアルカロイド含量を測定した。ただし、分析条件は、移動相には、移動相A:10 mM炭酸水素アンモニウム水(pH 10)および移動相B:MeCNを、上記試料溶媒についてA:B=60:40の割合でアイソクラティック条件を用いた。13系統のうち4系統は対照の1/5である新鮮重100mgあたり280μg以下と顕著にトマチン含量が低かった(図10)。
【0080】
(実施例7)グリコアルカロイド生合成候補遺伝子E変異植物のスクリーニング
バレイショの品種「サッシー」を量子ビーム照射(NIRS-HIMAC照射装置、アルゴンイオンビーム500MeV/核子を0.1から3Gyまたは、ネオンイオンビーム400Mev/核子を0.2から3Gy、または炭素イオンビーム290MeV/核子を0.5Gyから5Gy)で変異処理を行ったin vitroの植物体(キリンアグリバイオ(株)岡村主研分譲)の10個体からそれぞれ葉を採取しDNeasyでゲノムDNAを採取した。これをプライマー[U890: GAGGCTAAGAAAAAGAGAGAGAGA (配列番号6)、U889:CGTTCTACAAAAACATCCAATTT (配列番号7)U904: TGATAAGGAAATCCTGGGAGA(配列番号8)、U901: AGAGAAGCCATGAAGGATGG(配列番号9))、さらに第2イントロンは酵素をPrimeSTAR HS DNA Polymerase(タカラバイオ社)とプライマー(U898: GAAATACGCTACTACGGAAGAACC(配列番号10)とU899: CGTCATTTGCCTAATCTCATC(配列番号11)]を用いて構造遺伝子を、PCRを行い、遺伝子領域を取得した。さらにTOPOTAクローニングキットシークエンシング用を用いてクローニングした。さらにABI310を用いて塩基配列を決定した。その結果、今回分譲を受けた10株の中に変異遺伝子を持つ系統は存在しないことがわかった。しかし、十分な変異処理を施した植物に対し、この操作を繰り返すことで変異遺伝子を持つ植物を獲得することは可能である。
【産業上の利用可能性】
【0081】
本発明のグリコアルカロイド生合成酵素及びその遺伝子を用いる生物作成・検定方法は、植物等の生物を用いた、グリコアルカロイド化合物の生産の開発、バレイショ等ナス科植物品種の選抜に有用である。
【配列表フリーテキスト】
【0082】
配列番号6〜19 プライマー
【技術分野】
【0001】
本発明は、バレイショ等ナス科植物における特徴的なグリコアルカロイド化合物を生産する製造方法、グリコアルカロイド生合成酵素、当該グリコアルカロイド生合成酵素をコードするDNA、および当該DNAを用いた新たなバレイショ等ナス科植物の育種、選抜方法、ならびにグリコアルカロイドを生産しないバレイショ等ナス科植物に関する。
【背景技術】
【0002】
グリコアルカロイドはステロイダルアルカロイドとも呼ばれる一群の植物由来の化合物である。構造的には炭素鎖が27のイソプレノイドにN原子を含むものであり、ナス属植物の422種が含まれることが報告されている(非特許文献1の7.8章)。ナス属以外のナス科植物ではユリ科でもグリコアルカロイドを含むものが知られている。その中でも重要なものとしてナス科ナス属のバレイショ(Solanum tuberosum)のチャコニンおよびソラニン、ならびにトマト(Solanum lycopersicum)のトマチンである。
【0003】
バレイショは、トモロコシ、イネ、コムギに次いで世界第四番目の生産量を示す作物であるが、塊茎から出る芽や地上部植物は有毒なチャコニンおよびソラニンを含んでいることは周知の事実である。チャコニンやソラニンにより腹痛、めまい、軽い意識障害等の中毒症状を引き起こす。塊茎も、傷害を受けることや太陽光に曝されることでチャコニンおよびソラニンを容易に蓄積するため、塊茎の管理を誤ると中毒事故を起こす危険がある。
【0004】
これらの中毒事故はしばしば起きており、最近では2009年7月16日に日本国奈良市の小学校でグリコアルカロイドの中毒事件が発生している(Asahi.com報道)。バレイショの塊茎は暗所で保存すること等により20mg/100g以下のグリコアルカロイドになるように管理されているため通常は安全な食品である。しかしながら、上記のような中毒事故の危険性を考慮すると、バイレイショにおけるグリコアルカロイドを低減させることはバレイショの育種、生産、貯蔵、輸送、販売、購買などあらゆるバレイショを扱っている関係者の関心事である。しかし現在まで達成することはできてはいない。その理由としてはグリコアルカロイドのない野生種のバレイショがないこと、グリコアルカロイド生合成経路が未確定であり(非特許文献1の図7.24A,B、非特許文献2)、生合成経路に関与する遺伝子の同定が進まなかったことにある。
【0005】
グリコアルカロイドはコリンエステラーゼ阻害活性や膜破壊効果等の毒性を持つが、それ以外に、抗癌性活性、肝臓保護効果、鎮痙効果、免疫系促進効果、抗カビ性効果、抗原虫性効果、殺貝剤活性などの薬用作用が知られている(非特許文献1)。トマトではグリコアルカロイドの代謝産物であるesculeoside Aが抗動脈硬化作用を示すことも報告されている(非特許文献3)。しかし生合成経路が不明であることから、代謝産物を抑制することや効率よく生産する研究・開発はほとんど進んでこなかった。
【0006】
近年、アグリコン以降の糖転移過程で遺伝子の報告が幾つか報告されている(非特許文献4−6)。しかし、非特許文献4ではアグリコンであるソラニジンからのγソラニンへの経路であるUDP-ガラクトシルトランスフェラーゼの遺伝子を報告し、その抑制株を報告しているが、チャコニンは全く抑制できていない(非特許文献4の図2)。非特許文献4ではソラニジンからのγチャコニンへの経路であるUDP-グルコシルトランスフェラーゼの遺伝子を報告し、その抑制株を報告しているが、チャコニンおよびソラニンいずれも殆ど抑制できていない(非特許文献5の図5)。非特許文献6ではβチャコニンからαチャコニン、βソラニンからαソラニンへの経路であるラムノシルトランスフェラーゼの遺伝子を報告しているが、α体は減少しているがβ体やγ体は増加している。このように糖転移過程を抑制してもグリコアルカロイド分子種を変化させることはできるが、グリコアルカロイドの総量を調節することは極めて難しいことがわかる。
【0007】
植物ステロールや植物ホルモンの生合成遺伝子を過剰発現することでグリコアルカロイドの低下を試みた報告がある(非特許文献7)。しかしグリコアルカロイド量は多くても半分程度にしか減少できていない(非特許文献7の図5)。
【先行技術文献】
【非特許文献】
【0008】
【非特許文献1】Eich, Soloanaceae and Convolvulaceae: Secondary Metabolite (2008), Springer
【非特許文献2】Ginzbergら、 Poteto Research (2009) 52: 1-15
【非特許文献3】藤原ら、 2008年農芸化学会大会要旨 2B07p22
【非特許文献4】McCueら、 Plant Sci.(2005) 168: 267-273
【非特許文献5】McCueら、Phytochemistry (2006) 67: 1590-1597
【非特許文献6】McCueら、Phytochemistry (1998) 68: 327-334
【非特許文献7】Arnqvistら、 Plant Physiol. (2003) 131: 1792-1799
【発明の概要】
【発明が解決しようとする課題】
【0009】
本発明は、バレイショ等ナス科植物における特徴的なグリコアルカロイド化合物を生産する製造方法、グリコアルカロイド生合成酵素、当該グリコアルカロイド生合成酵素をコードするDNA、および当該DNAを用いた新たなバレイショ等ナス科植物の育種、選抜方法、ならびにグリコアルカロイドを産生しないバレイショ等ナス科植物の提供を課題とする。
【課題を解決するための手段】
【0010】
本発明者は、上記課題を解決すべく鋭意研究を重ねた。本発明者は、まずアグリコンを生成する以前の段階に着目した。この生合成経路の候補遺伝子をin silicoで探しだし、その一部分を用いてRNAiを起こす形で発現させることで内在性の候補遺伝子の発現を抑制させた。その結果、形質転換体の中で顕著にグリコアルカロイド含量が低下しているバレイショを得ることに成功するとともにグリコアルカロイド生合成酵素遺伝子を同定することができた。また、本遺伝子の発現が抑制された植物を選抜することでグリコアルカロイドのないバレイショ等ナス科植物を得ることを示した。さらに本遺伝子を発現させることで新規のグリコアルカロイド化合物の生産が可能になり、同遺伝子のゲノム配列を各種のバレイショ等ナス科植物で比較することにより多型を解析することが可能となり新規に育種されたバレイショ等ナス科植物を系呈することが可能になることを示し、本発明を完成させるに至った。同様にしてトマトにおいても内在性の遺伝子を抑制することでグリコアルカロイド含量の低減されたトマトを作出することができた。
【0011】
即ち、本発明は以下の発明を包含する。
[1] 以下の(a)または(b)のタンパク質:
(a) 配列番号1に示すアミノ酸配列からなるタンパク質;および
(b) 配列番号1に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質。
[2] 以下の(c)〜(f)のいずれかのDNAからなる遺伝子;
(c) 配列番号2に示す塩基配列からなるDNA;
(d) 配列番号2に示す塩基配列からなるDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;
(e) 配列番号2に示す塩基配列と80%以上の配列同一性を有する塩基配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;および
(f) 配列番号2に示す塩基配列の縮重異性体からなるDNA。
【0012】
[3] 以下の(g)又は(h)のタンパク質:
(g) 配列番号3に示すアミノ酸配列からなるタンパク質;および
(h) 配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質。
[4] 以下の(i)〜(l)のいずれかのDNAからなる遺伝子:
(i) 配列番号4に示す塩基配列からなるDNA;
(j) 配列番号4に示す塩基配列からなるDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;
(k) 配列番号4に示す塩基配列と80%以上の相同性を有する塩基配列からなり、かつ、
グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;および
(l) 配列番号4に示す塩基配列の縮重異性体からなるDNA。
[5] [2]または[4]の遺伝子を含有する組換えベクター。
[6] [5]の組換えベクターを導入した形質転換体。
[7] 植物体である[6]の形質転換体。
【0013】
[8] (i) ゲノムDNAまたはRNAである核酸を植物から単離する工程、
(ii) (i)の核酸がRNAである場合に逆転写しcDNAを合成する工程、
(iii) (i)または(ii)の工程で得られたDNAから配列番号2、配列番号4または配列番号5に示す塩基配列を含有する遺伝子断片を増幅する工程、ならびに
(iv) DNA中に突然変異および/または多型の存在を決定する工程、
とを含む、植物におけるグリコアルカロイド生合成酵素をコードする遺伝子の突然変異および/または多型の存在を検出する方法。
[9] 植物がバレイショ等のナス科植物である[8]の方法。
[10] [8]または[9]の方法によってグリコアルカロイド生合成酵素をコードする遺伝子の突然変異および/または多型を検出し、突然変異および/または多型を有する植物体を選抜する方法。
[11] [10]の方法により選抜された、グリコアルカロイド生合成酵素をコードする遺伝子に突然変異および/または多型を有する植物体。
[12] バレイショ等のナス科植物である[11]の植物体。
[13] グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはコードするグリコアルカロイド生合成酵素の活性が、既存品種に対して変化している植物を選抜する、[8]または[9]の植物体を選抜する方法。
[14] [13]の方法によって選抜された、グリコアルカロイド生合成酵素をコードする遺伝子の発現能が既存品種に対して変化しているか、またはグリコアルカロイド生合成酵素の活性が既存品種に対して変化している植物体。
[15] バレイショ等のナス科植物である[14]の植物体。
【発明の効果】
【0014】
本発明によれば、バレイショ等ナス科植物における特徴的なグリコアルカロイド化合物生合成活性を有するタンパク質とそれをコードする遺伝子の活性発現の調整を行うことができ、即ち当該遺伝子の活性が調整された植物を作成する方法、グリコアルカロイドを生産しないバレイショ等ナス科植物が提供される。本発明によりグリコアルカロイド化合物の含有に特徴があるバレイショ等ナス科植物の育種が可能になる。本発明の酵素により、様々な有用な生理活性を示すグリコアルカロイド化合物を大量かつ安価に生産できる。
【図面の簡単な説明】
【0015】
【図1−1】バレイショとトマトの生合成遺伝子Eの相同性をDNA解析ソフトGENETYX(ゼネティックス社)で解析した結果を示す。全般で非常に高い相同性が認められる。
【図1−2】バレイショとトマトの生合成遺伝子Eの相同性をDNA解析ソフトGENETYX(ゼネティックス社)で解析した結果を示す(図1−1の続き)。
【図1−3】バレイショとトマトの生合成遺伝子Eの相同性をDNA解析ソフトGENETYX(ゼネティックス社)で解析した結果を示す(図1−2の続き)。
【図2】候補E遺伝子抑制用ベクターの構造を示す。図2には、導入する遺伝子部分のT-DNAのライトボーダー(RB)、レフトボーダー(LB)の内部の構造、制限酵素部位を示す。
【図3】A)α−ソラニンおよびB)α−チャコニンのプロトン付加親イオンピーク(M+H)+を示すMSスペクトルの図である。
【図4】A)α-ソラニン、B)α-チャコニンおよびC)ブラシノライドの検量線(LC-MS定量分析系)を示す図である。
【図5】標品(α-ソラニン、α-チャコニンおよびブラシノライド)のLC-MSクロマトグラムを示す図である。
【図6】茎由来の試料におけるα-ソラニン、α-チャコニンおよびブラシノライドのLC-MSクロマトグラムを示す図である。
【図7】バレイショ形質転換体のin vitro茎のグリコアルカロイド含量を示す。エラーバーは標準偏差を示す。
【図8】バレイショ形質転換体のin vitro茎から抽出したmRNAに対するRT-PCRの結果を示す。
【図9】バレイショ形質転換体の塊茎の表皮のグリコアルカロイド含量を示す。エラーバーは標準偏差を示す。
【図10】トマト形質転換体の若い葉のグリコアルカロイド含量を示す。エラーバーは標準偏差を示す。
【発明を実施するための形態】
【0016】
以下、本発明を詳細に説明する。
1.新規のグリコアルカロイド生合成酵素
本発明のタンパク質・酵素は、バレイショ等ナス科植物(Solanaceae)に含まれるグリコアルカロイド生合成酵素である。バレイショ等ナス科には、バレイショ(Solanum tuberosum)、トマト(Solanum lycopersicum)、ナス(Solanum melongena)、トウガラシ(Capsium annum)等が含まれる。また、本発明の酵素は、膜結合型のチトクロームP450モノオキシダーゼである。本発明の酵素により得られるグリコアルカロイドは、バレイショ等のナス科植物に合成されるグリコアルカロイドが含まれ、例えばバレイショのチャコニン及びソラニン等のグリコアルカロイド、トマトのトマチン等のグリコアルカロイドが挙げられる。
【0017】
本発明のグリコアルカロイド生合成酵素の基質となる好ましいステロイド化合物としては、コレステロール類が挙げられる。コレステロール類としては、コレステロール、シトステロール、カンペステロール、スティグマステロール、ブラシカステロールなどが挙げられる。本発明のグリコアルカロイド生合成酵素はこれらに水酸基を転移する水酸化酵素である。
【0018】
本発明の酵素の全長アミノ酸配列は、配列番号1または3に示される。さらに、本発明のタンパク質は、配列番号1に示されるアミノ酸配列または配列番号3に示されるアミノ酸配列と実質的に同一のアミノ酸配列を有し、グリコアルカロイド生合成酵素活性を有するタンパク質を包含する。ここで、実質的に同一のアミノ酸配列としては、当該アミノ酸配列に対して1または数個(1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個、さらに好ましくは1〜3個、さらに好ましくは1個もしくは2個)のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列、または当該アミノ酸配列と、BLAST(Basic Local Alignment Search Tool at the National Center for Biological Information(米国国立生物学情報センターの基本ローカルアラインメント検索ツール))等(例えば、デフォルトすなわち初期設定のパラメータ)を用いて計算したときに、少なくとも85%以上、好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%以上の配列同一性を有しているアミノ酸配列が挙げられる。
【0019】
本発明のグリコアルカロイド生合成酵素は、植物体から単離された天然のグリコアルカロイド生合成酵素および遺伝子工学の手法により製造されたリコンビナントのグリコアルカロイド生合成酵素を含む。
【0020】
2.グリコアルカロイド生合成酵素をコードする遺伝子
本発明の遺伝子は、ステロイド化合物に水酸基を結合する活性を持つグリコアルカロイド生合成酵素をコードする遺伝子であり、上記のグリコアルカロイド生合成酵素活性を有するタンパク質をコードする遺伝子である。
【0021】
本発明の遺伝子のDNAの塩基配列は、配列番号2または4に示される。さらに、配列番号2または4に示される塩基配列に相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズするDNA、配列番号2または4に示される塩基配列と、BLAST(Basic Local Alignment Search Tool at the National Center for Biological Information(米国国立生物学情報センターの基本ローカルアラインメント検索ツール))等(例えば、デフォルトすなわち初期設定のパラメータを用いて)を用いて計算したときに、少なくとも85%以上、好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%以上の配列同一性を有しているDNA、または前記DNAによりコードされるタンパク質のアミノ酸配列に対して1または数個(1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個、さらに好ましくは1〜3個、さらに好ましくは1個もしくは2個)のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列からなるタンパク質をコードするDNAであって、グリコアルカロイド生合成酵素活性を有するタンパク質を有するタンパクをコードするDNAを包含する。ここで、「ストリンジェントな条件」とは、例えば、「1XSSC、0.1% SDS、37℃」程度の条件であり、より厳しい条件としては「0.5XSSC、0.1% SDS、42℃」程度の条件であり、さらに厳しい条件としては「0.2XSSC、0.1% SDS、65℃」程度の条件である。さらに、本発明の遺伝子は、配列番号2または4に示す塩基配列の縮重異性体からなるDNAを包含する。
【0022】
3.組換えベクター
本発明のベクターは、上記配列番号2または配列番号4のDNAが挿入された組換えベクターである。ベクターとしては公知の酵母用、植物細胞用、昆虫細胞用等のものを広く使用できる。公知の酵母用ベクターとしてはpDR196、pYES-DEST 52、Yip5、Yrp17、Yep24などが挙げられ、公知の植物細胞用ベクターとしては、pGWB vector、pBiEl2-GUS、pIG121-Hm、pBI121、pBiHyg-HSE、pB119、pBI101、pGV3850、pABH-Hm1などが挙げられ、公知の昆虫細胞用ベクターとしては、pBM030、pBM034、pBK283などが挙げられる。本発明において使用されるベクターには、プロモーター、ターミネーター、エンハンサー等の遺伝子の発現や抑制に関する構成要素が組込まれ、必要に応じて、選択マーカー(例えば、薬物耐性遺伝子、抗生物質耐性遺伝子、レポーター遺伝子)を含有する。遺伝子の発現や抑制に関する構成要素は、その性質に応じて、それぞれが機能し得る形で組換えベクターに組み込まれることが好ましい。そのような操作は、当業者であれば適切に行うことができる。
【0023】
4.形質転換体
本発明の形質転換体は、本発明の組換えベクターを保持する形質転換体である。形質転換体は、酵素をコードする遺伝子を挿入した組換えベクターを、目的遺伝子が発現し得るように宿主中に導入することにより得ることができる。宿主は、ベクターに適したものを使用すればよい。例えば、酵母、植物細胞、昆虫細胞(Sf9など)、植物ウイルスなどが挙げられる。好ましくは、酵母、植物細胞または植物ウイルスなどが挙げられる。組換えベクターの導入方法は、微生物にDNAを導入する方法であれば特に限定されるものではない。例えばカルシウムイオンを用いる方法[Cohen, S.N.et al.:Proc. Natl. Acad. Sci., USA, 69:2110(1972)]、エレクトロポレーション法、トリペアレンタルメイティング(tri-parental mating)法等が挙げられる。また、形質転換植物体を作製する方法として、ウイルス、アグロバクテリウムのTiプラスミド、Riプラスミド等をベクターとして用いる方法が挙げられる。宿主植物としては、イネ、ムギ、トウモロコシ等の単子葉植物、ダイズ、ナタネ、トマト、バレイショ等の双子葉植物が挙げられる。形質転換植物体は、本発明の遺伝子で形質転換した植物細胞を再生させることにより得ることができる。植物細胞からの植物体の再生は公知の方法により行うことができる。
【0024】
5.グリコアルカロイド生合成酵素の製造とグリコアルカロイド化合物の生産方法
本発明のグリコアルカロイド生合成酵素は膜結合型のチトクロームP450モノオキシダーゼであり、通常の植物体から回収することができる[Collu ら, 2001, FEBS Lett. 508:215-220など]。さらには、例えば、本発明の遺伝子で形質転換した酵母等の微生物や昆虫細胞発現系を用いた大量生産により製造することができ、昆虫細胞の例としては、Morikawaら[2006, Plant Cell 18:1008-1022]のものが挙げられる。
【0025】
これらの系を使って、高い活性を持ったタンパク質として発現できるため、形質転換酵母や昆虫細胞培養液に前記グリコアルカロイド生合成酵素の基質を添加することにより、グリコアルカロイド化合物を生産することができる。例えば、形質転換酵母の培養液にコレステロール類を基質として投与することにより、水酸化されたコレステロール類を効率的に大量に生産することが可能である。酵母がサイトゾルにDMAPPを生合成する経路(メバロン酸経路)を有していることや、大腸菌にメバロン酸経路を導入することで前駆体や基質を生産することが可能にしたことが報告されている[原田と三沢2009 Aug 12. Epub Appl Microbiol Biotechnol.]。この方法を利用することで他の遺伝子と膜結合型のチトクロームP450モノオキシダーゼを同時に発現しグリコアルカロイドを生産することが可能となる。これらの膜結合型のチトクロームP450モノオキシダーゼを発現し代謝物を得た例としては大腸菌ではChangら[2007 Nat. Chem. Biol. 3:274-277]の報告、酵母では関ら[2008 PNAS 105:14204-14209]の報告がある。このような方法を組み合わせることでグリコアルカロイド化合物を生産することが可能となる。
【0026】
6.遺伝子変異、多型個体、遺伝子発現変異の選抜
本発明は、植物におけるグリコアルカロイド生合成酵素遺伝子突然変異、一塩基多型(SNP)等の多型、遺伝子発現変異の存在を検出するための方法を提供する。変異個体は放射線によるもの、化学処理によるもの、UV照射によるもの、自然突然変異によるものであっても構わない。
【0027】
この方法には、ゲノムDNAやRNAを変異個体や様々な品種や育成個体の植物から単離し、後者は逆転写しcDNAを合成する工程と、DNA増幅技術の使用によりDNAからグリコアルカロイド生合成酵素遺伝子を含有する遺伝子断片を増幅する工程と、このDNA中に突然変異の存在を決定する工程が含まれる。DNAやRNAを抽出する方法には市販のキット(例えばDNeasyやRNeasy(キアゲン社)など)が使用できる。cDNAを合成する方法も市販キット(例えばスーパースクリプト ファーストストランド システム(インビトロジェン社)など)を使うことができる。DNA増幅技術の使用により遺伝子断片を増幅する方法としては、いわゆるPCR法やLAMP法などの技術を用いることができる。これらは継続的なポリメラーゼ反応により特異的なDNA配列の増幅(つまり、コピー数を増やすこと)を達成するためにポリメラーゼを使用することを基にした、一群の技術を意味する。この反応は、クローニングの代わりに使用することができるが、必要であるのは、核酸配列に関する情報のみである。DNAの増幅を行うために、増幅しようとするDNAの配列に相補的なプライマーを設計する。次にそのプライマーを自動DNA合成により作成する。DNA増幅方法は、当技術分野で周知であり、本明細書中で与えられる教示及び指示に基づき、当業者であれば容易に行うことができる。いくつかのPCR法(ならびに関連技術)は、例えば、米国特許第4,683,195号、同第4,683,202号、同第4,800,159号、同第4,965,188号、およびInnisら編、PCR Protocols:A guide to method and applicationsで述べられている。
【0028】
DNA中に突然変異や多型の存在を決定する工程では塩基配列の決定(アプライドバイオシステムズ社)やミスマッチペアの片側を切断する酵素を用いて突然変異体を検出するTILLING法(Till et al., 2003, Genome Res 13:524-530)など変異遺伝子と正常遺伝子の相同性を利用し検出する方法を用いればよい。これらは該技術から得られた配列データを遺伝子部分に関する配列番号2、配列番号4または配列番号5に表される塩基配列と比較することで行うことができる。
【0029】
mRNA量の違いを決定する工程では上記cDNAに対し、配列番号2または配列番号4に表される塩基配列に基づいて作製したプライマーを利用してリアルタイムPCR法(ロシュ・ダイアグノスティックス社ライトサイクラーなど)等の定量的PCRを採用すればよい。その後、例えば、品種「サッシー」から得られたcDNAの量と比較することでmRNA量の違いを決定することができる。
【0030】
特に好ましい実施形態において、上記で定義したグリコアルカロイド生合成酵素遺伝子の変異の存在の決定方法を、ナス科植物(Solanaceae)のバレイショ(Solanum tuberosum)から得られた材料に適用する。
【0031】
上記の突然変異および/または多型を決定する方法により、グリコアルカロイド生合成酵素をコードする遺伝子の突然変異や多型を塩基レベルで同定することができ、さらにグリコアルカロイド生合成酵素をコードする遺伝子に突然変異および/または多型を有する植物体を選抜することができる。本発明はこのようにして得られたグリコアルカロイド生合成酵素をコードする遺伝子に突然変異や多型を有する植物体を包含する。
【0032】
また、突然変異や多型の決定やmRNA量の違いの決定、さらに方法について後述するグリコアルカロイド含量の分析により、グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはグリコアルカロイド生合成酵素の活性が変化している植物を選抜することが可能になる。
【0033】
ここで、グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはグリコアルカロイド生合成酵素の活性の変化とは、人為的突然変異等の突然変異による遺伝子の発現能またはグリコアルカロイド生合成酵素の活性の改変および多型による遺伝子の発現能またはグリコアルカロイド生合成酵素の活性が異なっていることを含む。
【0034】
ある植物のグリコアルカロイド生合成酵素活性の突然変異による改変は、その植物の種に含まれる既存品種に対する改変をいい、既存品種には野生型も含まれるが、自然状態で出現した野生種であっても、すでに産業上利用されている品種でなければ既存品種には含めない。既存の品種は、グリコアルカロイド生合成酵素活性が改変された植物が得られたときに存在するすべての品種をいい、交配、遺伝子操作等の人為的操作により作出された品種を含む。また、活性の改変において、すべての既存品種に対して、活性が変化している必要はなく、特定の既存品種に対して改変されていれば、「グリコアルカロイド生合成酵素の活性が改変された植物」に含まれる。「グリコアルカロイド生合成酵素の活性が改変された植物」は、人為的操作を受けず自然状態で突然変異により活性が改変された植物も含み、本発明の方法により、自然状態で活性が変化した植物を選抜することができ、新たな品種として確立することもできる。また、ある既存品種に変異誘発処理を行い、グリコアルカロイド生合成酵素の活性が改変された植物を作出した場合、比較対象は変異誘発処理を行った品種と同じ既存品種でもよいし、それ以外の他の既存品種でもよい。また、自然界からの選抜あるいは変異誘発処理により作出された、グリコアルカロイド生合成酵素をコードする遺伝子に突然変異や多型を有する植物を交配することにより、グリコアルカロイド生合成酵素をコードする遺伝子の変異が固定されグリコアルカロイド生合成酵素遺伝子の発現能またはグリコアルカロイド生合成酵素活性が改変された植物新品種として得ることもできる。
【0035】
例えば、植物がバレイショ(Solanum tuberosum)の場合、既存品種として、「シンシア」、「サッシー」、「シェリー」、「男爵」、「メークイーン」、「さやか(農林登録番号:農林36号)」等がある。ここで、グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはグリコアルカロイド生合成酵素の活性が既存品種に対して改変された植物とは、既存品種に対してグリコアルカロイド生合成酵素をコードする遺伝子の発現能が増強した植物および低下した植物を含み、さらに、グリコアルカロイド生合成酵素の活性が既存品種に対して上昇した植物および低下した植物を含む。本発明は、このようなグリコアルカロイド生合成酵素をコードする遺伝子の発現能またはグリコアルカロイド生合成酵素の活性が既存品種に対して改変された植物体も包含する。
【0036】
特に有毒物質であるグリコアルカロイド生合成酵素の活性が低下した植物が好ましい。このような植物体は、グリコアルカロイド生合成酵素の合成量が低いか、又は合成できず、植物体中のグリコアルカロイド生合成酵素の含量が低いか、又はグリコアルカロイド合成酵素が存在せず、あるいはグリコアルカロイド合成酵素の活性が低いか又は喪失している。その結果、植物体内のグリコアルカロイド含量も低いか、あるいはグリコアルカロイドが存在しない。例えば、バレイショの場合はチャコニン及びソラニン等のグリコアルカロイドが合成されず、バレイショの塊茎内においてチャコニン及びソラニン等のグリコアルカロイドの合成量及び存在量が低い。また、トマトの場合は、トマチン等のグリコアルカロイドが合成されず、トマトの実内においてトマチン等のグリコアルカロイドの合成量及び存在量が低い。
【0037】
バレイショの場合、グリコアルカロイド合成酵素の活性が低いか、喪失した植物体は、塊茎内でチャコニン及びソラニン等のグリコアルカロイドが合成されないか、又は上記の既存品種に比べて塊茎内で合成されるチャコニン及びソラニン等のグリコアルカロイドが少なく、塊茎内に存在するチャコニン及びソラニン等のグリコアルカロイドの量も低い。
【0038】
7.グリコアルカロイドの分析および精製
グリコアルカロイド含量の分析方法および精製方法については、Matsudaら(Phytochem. Anal. 15:121-124, 2004)やKozukueら(J. Agric. Food Chem. 52: 2079-2083, 2004)等液体クロマトグラフィーを用いた方法が知られている。しかし、サンプルの前処理が煩雑であったり、検出限界が十分に高いものではない、強酸を用いるためカラムや装置への負荷が大きいといった課題がある。よって本発明では、GA類を効率的に精製し、且つ高い精度で分析できるアルカリ耐性の逆相クロマトグラフィー用カラムを用いた液体クロマトグラフィーを用いた方法(該方法は、日本国特許出願 特願2009-170317として出願済み)を用いることが出来る。実施例5に本方法のジャガイモでの適用例を示す。
【0039】
本方法に用いるカラムとしては、アルカリ耐性に優れたカラムであれば利用することができる。アルカリ耐性に優れたカラムとしては、例えばエチレン架橋カラムを用いることができる。好ましくは、XBridge(商標)ブランド(Waters社)のカラムを用いる。特に好ましくはWaters XBridgeTM Shield RP18(Waters社)およびWaters XBridgeTM C18である。本方法において、XBridgeTMShield RP18カラムは1サンプルあたりの所要時間が短く、一方、Waters XBridgeTMC18カラムは耐久性が高いといった有利点をそれぞれ有する。
【0040】
液体クロマトグラフィーに用いる移動相は、アルカリ性緩衝液を用いることができる。好ましくは、揮発性のアルカリ性緩衝液を用いる。液体クロマトグラフィーにより精製した試料を質量分析に供する場合に、移動相に揮発性のアルカリ性緩衝液を用いると試料中に当該アルカリ性緩衝液が残留しないために都合が良い。揮発性のアルカリ性緩衝液としては例えば、トリエチルアミン、炭酸水素アンモニウムなどを用いることができるが、好ましくは緩衝効果の高い炭酸水素アンモニウムを用いる。
【0041】
移動相に用いられる炭酸水素アンモニウムの濃度は、5〜20mM、好ましくは5〜15mM、さらに好ましくは10mMである。炭酸水素アンモニウムのpHは、好ましくはpH 9.0〜11.0、さらに好ましくはpH 10.0にすることが可能である。移動相のpHを10.0とすることによって、炭酸水素アンモニウムの緩衝能がより高まる。
【0042】
GA類は、移動相にアルカリ性緩衝液と有機溶媒を用いて、アイソクラティック法により溶出しても良いし、グラジェント法により溶出しても良いが、操作が簡便なアイソクラティック法により溶出するのが好ましい。
【0043】
移動相に用いられる有機溶媒としては、例えば、メタノール、エタノール、テトラヒドロフラン(THF)、アセトニトリル(MeCN)等を用いることができるが、これらに限定されない。好ましくは、MeCNを用いる。
【0044】
アイソクラティック法においては、アルカリ性緩衝液と有機溶媒、好ましくは炭酸水素アンモニウム水とMeCNとを、30〜70:70〜30、好ましくは40〜60:60〜40の割合で、所望のGA類に応じて適宜用いる。例えば、所望のGA類がα−ソラニンまたはα−チャコニンである場合には40:60の割合で、所望のGA類がα−トマチンである場合には60:40の割合でアルカリ性緩衝液と有機溶媒、好ましくは炭酸水素アンモニウム水とMeCNとを用いる。
【0045】
液体クロマトグラフィーは市販のHPLC装置を用いて行うことができ、カラムの平衡化や流速はカラムサイズや試料の容量によって適宜設定することができる。
【0046】
液体クロマトグラフィーを行い、得られた画分は、後述の質量分析やUVまたは多波長検出器等を用いて分析することができる。
【0047】
植物由来の試料は、液体クロマトグラフィーに付す前に、予め以下の前処理を行い粗精製しておくことが好ましい。
【0048】
植物由来の試料は、GA類に加え、様々な高分子夾雑物(デンプン、タンパク質、セルロース等)を含有する。したがって、GA類を効率的に精製かつ高精度に分析するために、試料中に含まれる高分子夾雑物を除去し、GA類を粗精製および洗浄する必要がある。
【0049】
高分子夾雑物を除去する方法としては、当業者にとって一般的な方法、例えば、アルコール沈殿法を用いることができる。アルコールは、エタノールまたはメタノールを用いることができるが、メタノールが好ましい。この際、GA類を塩として効率よく抽出するためにアルコールには酸を添加する。用いることができる酸としては、酢酸、塩酸、ギ酸等が挙げられるがこれらに限定されない。好ましくはギ酸を添加する。アルコールへの酸の添加量は、所望のGA類が破壊されない範囲で適宜設定することができ、ギ酸であれば0.1〜2%(v/v)、好ましくは0.1%(v/v)となるようにアルコールに添加する。ギ酸以外の酸を用いる際には、上記添加したギ酸と同等の規定度になるまで添加することができる。
【0050】
なお従来の試料調製方法(Matsudaら, Phytochem. Anal. 15: 121-124, 2004を参照)では、試料を長時間ホモジネートし、さらに試料中に大量に含まれるデンプン等の高分子夾雑物を除去するために遠心分離を複数回行い、さらにその後、ろ過に供することを含む、長時間かつ煩雑な前処理を要していた。一方、本方法における調製方法では、上記のように短時間で破砕した植物片よりアルコール沈殿でデンプン等の高分子夾雑物を除去できるため、短時間かつ容易に試料調製が可能である。
【0051】
アルコール沈殿後、GA類を含む上清を0.1〜2%(v/v)のギ酸、酢酸等の酸、好ましくは0.1%(v/v)ギ酸で希釈し、上記の条件で液体クロマトグラフィーに付す。
【0052】
液体クロマトグラフィーにより精製した画分は、さらに質量分析に付すことができる。この場合、液体クロマトグラフィーと質量分析を連結した手法であるLC-MS法により行えばよい。
【0053】
質量分析は、単収束扇形磁場型質量分析法、二重収束扇形磁場型質量分析法、四重極型質量分析法、四重極イオントラップ型質量分析法、飛行時間型質量分析法、イオンサイクロトロン型質量分析法(フーリエ変換型質量分析法)などを用いて行うことができる。
【0054】
質量分析において試料をイオン化させる方法としては、EI(electron ionization:電子イオン化)法、CI(chemical ionization:化学イオン化)法、DEI(desorption electron ionization:脱離電子イオン化)法、DCI(desorption chemical ionization:脱離化学イオン化)法、FAB(fast atom bombardment:高速原子衝撃)法、FRIT-FAB(FRIT-fast atom bombardment:フリット高速原子衝撃)法、ESI(electrospray ionization:エレクトロスプレーイオン化)法、MALDI(matrix-assisted laser desorption ionization:マトリックス支援レーザー脱離イオン化)法を用いることができる。
【0055】
質量分析における諸条件については、実施例に具体的に記載するが、当業者であれば分析対象となるGA類に応じて、適宜条件を設定することが可能である。
【0056】
LC-MS法を用いてGA類の分析標品を分析し、当業者にとって一般的な方法に従って検量線を作成することが可能である。ジャガイモ由来の試料において、特にα−ソラニンおよびα−チャコニンの分析系では、内部標準物質としてβ−D−グルコサミンペンタアセテートを利用することが可能であるが、α−ソラニンおよびα−チャコニンと同様にステロイド骨格を有するブラシノライドを用いることが好ましい。一方、トマト由来の試料において、特にα−トマチンの分析系では水溶性アミンを用いることが好ましい。内部標準物質として利用し得る水溶性アミンとして、セリンメチルエステルおよびアラニンメチルエステルが挙げられるが、アラニンメチルエステルはカラム保持が強いために特に好ましい。したがって、ジャガイモ由来の試料においてはブラシノライドを、またトマト由来の試料においてはアラニンメチルエステルを利用することによって、定量分析の信頼性を顕著に高めることができる。
【0057】
本方法においては、HPLCに汎用されるサイズのカラムを用いることができるため、本条件はこのままUVまたは多波長検出器による分析にも使用することができる。
【実施例】
【0058】
以下、本発明を、実施例を示してより詳しく説明するが、本発明はこれらに限定されるものではない。
(実施例1)グリコアルカロイド生合成候補遺伝子Eの全長配列の取得
バレイショ(Solanum tuberosum)の品種「サッシー」の萌芽からmRNAの抽出をRNeasy(キアゲン社)で行った。全cDNAの合成はスーパースクリプト ファーストストランド システム(インビトロジェン社)を用いて行った。グリコアルカロイドのアグリコンはコレステロールからできるといわれているが確証はない(非特許文献1)。しかし、近縁の化合物から作られると仮定しても幾つかの水酸化の過程が必要になる。水酸化の過程には少なくともチトクロームP450型モノオキシゲナーゼ、ジオキシゲナーゼ、NADPH-flavin リダクターゼの3種の可能性が考えられる。この中からP450型を標的に考え、バレイショの発現する遺伝子は公開されている情報のDFCI Potato Gene Index (http://compbio.dfci.harvard.edu/tgi/plant.html) Release 11.0から萌芽で多くのESTクローンが単離されている遺伝子TC155233に注目した。
【0059】
この配列を元にプライマー[U890: GAGGCTAAGAAAAAGAGAGAGAGA (配列番号6)、U889:CGTTCTACAAAAACATCCAATTT (配列番号7)]を用いてPCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃3分)を30回、72℃10分)を行った。増幅産物をTOPOTAクローニングキットシークエンシング用(インビトロジェン社)を用いてクローニングした。さらにABI310(アプライドバイオシステムズ社)を用いて塩基配列を決定した。ORFを含む部分を配列番号2に、cDNA配列からコードされる酵素のアミノ酸配列を配列番号1に示す。
【0060】
なお、トマトの相同遺伝子は、ナス科ゲノムネットワーク(http://solgenomics.net/index.pl)の、SGN-U583521に相当する。ORFを含む部分を配列番号4に、cDNA配列からコードされる酵素のアミノ酸配列を配列番号3に示す。これらの遺伝子の塩基配列を比較したところ相同性は95%であった。このトマトの相同遺伝子のゲノム配列は同じくナス科ゲノムネットワークのSL1.00sc03540としてゲノム構造が掲載され7つのイントロンを含むことが報告されている。しかし、同ホームページには、なんら機能に関する報告はない(図1)。
【0061】
(実施例2)グリコアルカロイド生合成候補遺伝子Eのゲノム遺伝子の単離
ゲノムDNAをRNeasy(キアゲン社)で「サッシー」から抽出した。実施例1と同じプライマー並びに(U904: TGATAAGGAAATCCTGGGAGA(配列番号8)、U901: AGAGAAGCCATGAAGGATGG(配列番号9))を用いて、さらに第2イントロンは酵素をPrimeSTAR HS DNA Polymerase(タカラバイオ社)とプライマー(U898: GAAATACGCTACTACGGAAGAACC(配列番号10)とU899: CGTCATTTGCCTAATCTCATC(配列番号11))を用いてPCRを行い、全長ゲノムDNAの塩基配列を決定した(配列番号5)。イントロンは7箇所あることが明らかになった。
【0062】
(実施例3)グリコアルカロイド生合成候補遺伝子Eの抑制形質転換体を作成するためのベクター構築
遺伝子を形質転換によって抑制する方法としては、強力なプロモーターで駆動する構成を持つ逆方向の相補鎖遺伝子断片の発現(植物で一般的にRNAi法と呼ばれる)で行った[Chuangと Meyerowitz Proc Natl Acad Sci U S A., 97, 4985-90 (2000)、WesleyらPlant J., 27, 581-90 (2001)]。実施例1で取得した全長cDNAに対し、プライマー[U675: GAGCTCTAGAGGTTTGGGACAGGAGGAAT (配列番号12)、U676: GGATCCATATGCAAGCCTGTGCATCTTAT (配列番号13)]を用いてPCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃30秒)を30回、72℃10分)を行い、遺伝子断片を取得した。バイナリーベクターpKT11(特開2001-161373号公報)を基本として、カルフラワーモザイクウイルスの35S RNAプロモーター、当該遺伝子断片を順方向、シロイヌナズナのフィトエンデサチュラーゼ遺伝子(AT4g14210)の第3イントロン、当該遺伝子断片を逆方向、ノパリン合成酵素遺伝子のターミネーターの順に連結を行い、植物形質転換用ベクターpKT230を作成した(図2)。
【0063】
(実施例4)バレイショ形質転換植物体の作出
実施例3で作製したベクターをエレクトロポレーション法(GelvinとSchilperoor編, Plant Molecular Biology Manual, C2, 1-32 (1994), Kluwer Academic Publishers)により、アグロバクテリウム・ツメファシエンスGV3110株に導入した。ベクターを含むアグロバクテリウム・ツメファシエンスGV3110株を、50ppmのカナマイシンを含むYEB液体培地[5g/lビ−フエキス、1g/l酵母エキス、5g/lペプトン、5g/lスクロ−ス、2mM硫酸マグネシウム(pH7.2)]にて28℃、12時間振とう培養した。培養液1.5 mlを10,000rpm、3分間遠心して集菌後、カナマイシンを除くために1mlのLB培地で洗浄した。更に10,000rpm、3分間遠心して集菌後、1.5 mlの3%蔗糖を含むMS培地[Murashige & Skoog, Physiol. Plant., 15, 473-497 (1962)]に再懸濁し、感染用菌液とした。
【0064】
バレイショの形質転換は[門馬(1990)植物組織培養7:57-63]に従い実施した。バレイショ品種「サッシー」(キリンアグリバイオ社)から得られたマイクロチューバーを2〜3mmにスライスし、アグロバクテリウム感染用の材料とした。これを上記のアグロバクテリウムの菌液に浸した後、滅菌済みの濾紙上に置いて過剰のアグロバクテリウムを除いた。シャーレ内のMS培地(Zeatin 1ppm, IAA 0.1ppm, アセトシリンゴン100μM、及び寒天0.8%を含む)上に置き、培養は3日間25℃、16時間照明(光量子束密度32μE/m2s)/8時間無照明の条件下で行った。ついで、アセトシリンゴンの代わりにカルベニシリン250ppmを含んだ培地で1週間培養した。その後、さらにカナマイシン50 ppmを含む培地上に移し、2週間ごとに継代した。この間に不定芽が形成し、シュートを生じた。伸張したシュートをカルベニシン250 ppm及びカナマイシン100 ppmを含み、植物生長調節物質を含まないMS培地に置床した。発根したシュートをカナマイシン耐性の生長した植物体の中から外来遺伝子としてカナマイシン耐性遺伝子を含有する個体を、PCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃1分)を30回、72℃10分)を行うことで検出し、該再分化植物体が形質転換植物体であることを確認した。ここで、カナマイシン耐性遺伝子の配列を特異的に増幅するプライマーとして、TAAAGCACGAGGAAGCGGT(配列番号14)、及びGCACAACAGACAATCGGCT(配列番号15)を用いた。以上から、ベクターpKT230が導入されたバレイショの形質転換植物体30系統を取得した。
【0065】
(実施例5)形質転換植物体のグリコアルカロイド含量と候補遺伝子Eの発現解析
実施例4で得られた30個体のin vitro茎を継代後一ヶ月伸張させ、その部分2-4本をまとめて約100mgにしアルカリ耐性の逆相クロマトグラフィー用カラムを用いた液体クロマトグラフィーを用いた以下の方法(該方法は、日本国特許出願 特願2009-170317として出願済み)によりグリコアルカロイド含量を測定した。
【0066】
バレイショに含まれるGA類(α-ソラニン、α-チャコニン)の分析
1. 試料の調製
実施例4で得られた30個体のin vitro茎を継代後一ヶ月伸張させ、その部分を2-4本をまとめて約100mgにし0.1%ギ酸 in 80%MeOH aq. 990μLおよび内部標準としてブラシノライド(ブラシノ)10μg/10μLを添加し、ミキサーミルで破砕した(1/25 sec, 5 min, 4℃)。得られた破砕物を遠心分離(10,000 rpm, 5 min)に供しアルコール沈殿を行った後、上清25μLを分取し、0.1%ギ酸水で500μLにフィルアップした。これを試料として、以下の条件でLC-MSに用いた。LC-MS装置は、LCMS-2010EV(島津製作所)を用いた。
【0067】
2. LC-MS条件
(i)LC条件
LC系には、アルカリ耐性に優れたエチレン架橋カラム(XBridgeTM Shield RP18-5(φ2.1×150 mm, Waters社))を採用した。移動相には、移動相A:10 mM炭酸水素アンモニウム水(pH 10)および移動相B:MeCNを、上記試料溶媒についてA:B=40:60の割合でアイソクラティック条件で用いた。その他の条件は以下のものを用いた:
流速:0.2 mL/min
カラムオーブン:40℃
【0068】
(ii)MS条件
まず各成分のMSスペクトルをスキャンモードで確認し(図3参照)、その結果として検出法:SIMモード
m/z:481(ブラシノライド)、869(α−ソラニン)、853(α−チャコニン)
を用いた。
その他MS条件は以下のものを用いた。
MS検出:ポジティブイオンモード
イオン化法:ESI
イベント時間:1 sec
検出器電圧:1.5 kV
分析時間:8 min
【0069】
3. 標品α-ソラニン、α-チャコニン、ブラシノライドを使っての検量線作成
α−ソラニン(和光純薬)2mgとα−チャコニン(シグマアルドリッチ)2mgをそれぞれ1mLの0.1%(v/v)ギ酸水に溶解した(それぞれ2μg/μL溶液)。2種類の溶液を等容で混和し、α−ソラニンとα−チャコニンが1μg/μL(=1000ng/μL)の溶液を調製した。これを0.1%(v/v)ギ酸水で段階的に10倍希釈後、LC-MSに供して検量線を作製した。また、両物質の検出限界値を求めた。
【0070】
ブラシノライド(ブラシノ)については1mgを1mLのMeOH溶液に溶解した(1μg/μg)。これを50%(v/v)含水MeOHで段階的に10倍希釈後、LC-MSに供して検量線を作製した。
【0071】
α−ソラニン、α−チャコニンおよびブラシノライドのそれぞれについて作成した検量線を図4に示す。α−ソラニンとα−チャコニンについては図4に示したように、0.05〜50ngの範囲で良好な直線性が得られ、信頼係数は0.99以上であった。両物質ともに100ngを超えるとシグナルがサチュレートして、直線性が失われた。また両物質の検出限界は0.02ng(2μLインジェクト)であった。
【0072】
一方、ブラシノライドは2〜200ngの範囲で良好な直線性が得られた(図4を参照)。500ng以上ではやはりシグナルがサチュレートした。
【0073】
標品α-ソラニン、α-チャコニン、ブラシノライドを用いての典型的クロマトグラムを図5に示した。
【0074】
4. ブラシノライドを内部標準としたバレイショ中のGA類のLC-MS分析
上記1.にて調製した各試料の10μLまたは20μLを上記条件を用いたLC-MS系にインジェクトした。
【0075】
内部標準ブラシノライドの回収率は50〜110%であった。ブラシノライドの定量値で補
正を行い、各試料に含まれるα-ソラニンおよびα-チャコニンの量について上記検量線を用いて定量し、試料100 mg(FW)あたりに含まれる量として算出した。
当該分析における典型的クロマトグラムを図6に示す。
【0076】
30個体のうち5系統(#8, #17, #22, #27, #29)においてはグリコアルカロイドの蓄積が再現性よく低かったことから、低くなかった1系統(#2)と遺伝子を導入していない対照の個体2つを同じくin vitro茎を液体窒素で粉砕し、半分をグリコアルカロイド含量の測定、半分をmRNAの抽出をRNeasy(キアゲン社)で行い、全cDNAの合成はスーパースクリプト ファーストストランド システム(インビトロジェン社)を用いて行った。これらの個体はグリコアルカロイドの蓄積が非形質転換体(2個体)と比較して極めて低く(図7)、さらにプライマー [U887: TAAGGGACTCAAGGCTCGAA (配列番号16)、U886: TTCCTCTTTGGCTTTCTCCA (配列番号17)]を用いたRT-PCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃3分)を25回、72℃5分)の結果、mRNAの発現はいずれの個体も極めて少ないか観察できなかった(図8)。このことから、候補遺伝子Eの遺伝子の発現を抑制することによってグリコアルカロイドの蓄積が極端に減少することが明らかとなり、候補遺伝子Eはグリコアルカロイド生合成酵素をコードする遺伝子であることが明らかとなった。非形質転換体とともに、これら5系統のin vitro植物を増殖し、各3個体を市販されている野菜用の培養土に馴化しバイオハザード温室で定法に従い栽培し塊茎を収穫した。この5系統の各個体(#8, #17, #22, #27, #29)は非形質転換体と同等の生育を示し、同等の塊茎を収穫することができた(表1)。
【0077】
【表1】
【0078】
さらに収穫した塊茎各3つの中央部表皮を約1mmで剥離し同様にグリコアルカロイド含量を解析した。その結果、驚くべきことに、塊茎でのグリコアルカロイドは極めて低く、同様な方法で測定した、グリコアルカロイドの低い品種として知られている「さやか」と比較しても、それ以下であることが確認できた(図9)。
【0079】
(実施例6)トマト形質転換植物体の作出
トマトの形質転換は[Sunら (2006) Plant Cell Physiol. 47:426-431.]に従い実施した。(実施例3)で作製したベクターpKT230を含むアグロバクテリウム・ツメファシエンスAGL0株を培養し感染用菌液とした。トマト(Solanum lycopersicum)実験系統「マイクロトム」の無菌播種植物体の子葉を5mm以下の切片を、上記のアグロバクテリウム懸濁液に浸し、10分間感染した後、滅菌済みの濾紙上に葉を置いて過剰のアグロバクテリウムを除いた。シャーレ内の共存MS培地(ゼアチン1.5mg/l、アセトシリンゴン40μM及びゲルライト0.3%を含む)[Murashige & Skoog, Physiol. Plant., 15, 473-497 (1962)]上に葉を置き、シャーレを暗所で3日間25℃で培養した。切片は選択MS培地1(ゼアチン1.5mg/l、カナマイシン100mg/l、オーグメンチン375mg/l及びゲルライト0.3%を含む)で25℃、16時間照明(光量子束密度32μE/m2s)/8時間無照明の条件下で2週間ごとに継代した。この間に不定芽が形成し、シュートを生じた。さらにシュートを伸張させるため、選択MS培地2(ゼアチン1.0mg/l、カナマイシン100mg/l、オーグメンチン375mg/l及びゲルライト0.3%を含む)に移植し、伸張したシュートは選択1/2濃度MS培地(カナマイシン100mg/l、オーグメンチン375mg/l及びゲルライト0.3%を含む)で発根させた。シュートをカナマイシン耐性の生長した植物体の中から外来遺伝子としてカナマイシン耐性遺伝子を含有する個体を、PCR(条件:95℃5分、(95℃30秒、55℃30秒、72℃1分)を30回、72℃10分)を行うことで検出し、該再分化植物体が形質転換植物体であることを確認した。ここで、カナマイシン耐性遺伝子の配列を特異的に増幅するプライマーとして、TAAAGCACGAGGAAGCGGT(配列番号18)、及びGCACAACAGACAATCGGCT(配列番号19)を用いた。以上から、ベクターpKT230が導入されたトマトの形質転換植物体13系統を取得した。得られた13個体を温室に馴化し約1ヶ月栽培し、新しく展開した若い葉の3枚から各約100mg秤量し、バレイショと同様にアルカリ耐性の逆相クロマトグラフィー用カラムを用いた液体クロマトグラフィーを用いた実施例5の方法によりグリコアルカロイド含量を測定した。ただし、分析条件は、移動相には、移動相A:10 mM炭酸水素アンモニウム水(pH 10)および移動相B:MeCNを、上記試料溶媒についてA:B=60:40の割合でアイソクラティック条件を用いた。13系統のうち4系統は対照の1/5である新鮮重100mgあたり280μg以下と顕著にトマチン含量が低かった(図10)。
【0080】
(実施例7)グリコアルカロイド生合成候補遺伝子E変異植物のスクリーニング
バレイショの品種「サッシー」を量子ビーム照射(NIRS-HIMAC照射装置、アルゴンイオンビーム500MeV/核子を0.1から3Gyまたは、ネオンイオンビーム400Mev/核子を0.2から3Gy、または炭素イオンビーム290MeV/核子を0.5Gyから5Gy)で変異処理を行ったin vitroの植物体(キリンアグリバイオ(株)岡村主研分譲)の10個体からそれぞれ葉を採取しDNeasyでゲノムDNAを採取した。これをプライマー[U890: GAGGCTAAGAAAAAGAGAGAGAGA (配列番号6)、U889:CGTTCTACAAAAACATCCAATTT (配列番号7)U904: TGATAAGGAAATCCTGGGAGA(配列番号8)、U901: AGAGAAGCCATGAAGGATGG(配列番号9))、さらに第2イントロンは酵素をPrimeSTAR HS DNA Polymerase(タカラバイオ社)とプライマー(U898: GAAATACGCTACTACGGAAGAACC(配列番号10)とU899: CGTCATTTGCCTAATCTCATC(配列番号11)]を用いて構造遺伝子を、PCRを行い、遺伝子領域を取得した。さらにTOPOTAクローニングキットシークエンシング用を用いてクローニングした。さらにABI310を用いて塩基配列を決定した。その結果、今回分譲を受けた10株の中に変異遺伝子を持つ系統は存在しないことがわかった。しかし、十分な変異処理を施した植物に対し、この操作を繰り返すことで変異遺伝子を持つ植物を獲得することは可能である。
【産業上の利用可能性】
【0081】
本発明のグリコアルカロイド生合成酵素及びその遺伝子を用いる生物作成・検定方法は、植物等の生物を用いた、グリコアルカロイド化合物の生産の開発、バレイショ等ナス科植物品種の選抜に有用である。
【配列表フリーテキスト】
【0082】
配列番号6〜19 プライマー
【特許請求の範囲】
【請求項1】
以下の(a)または(b)のタンパク質:
(a) 配列番号1に示すアミノ酸配列からなるタンパク質;および
(b) 配列番号1に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質。
【請求項2】
以下の(c)〜(f)のいずれかのDNAからなる遺伝子;
(c) 配列番号2に示す塩基配列からなるDNA;
(d) 配列番号2に示す塩基配列からなるDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;
(e) 配列番号2に示す塩基配列と80%以上の配列同一性を有する塩基配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;および
(f) 配列番号2に示す塩基配列の縮重異性体からなるDNA。
【請求項3】
以下の(g)又は(h)のタンパク質:
(g) 配列番号3に示すアミノ酸配列からなるタンパク質;および
(h) 配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質。
【請求項4】
以下の(i)〜(l)のいずれかのDNAからなる遺伝子:
(i) 配列番号4に示す塩基配列からなるDNA;
(j) 配列番号4に示す塩基配列からなるDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;
(k) 配列番号4に示す塩基配列と80%以上の相同性を有する塩基配列からなり、かつ、
グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;および
(l) 配列番号4に示す塩基配列の縮重異性体からなるDNA。
【請求項5】
請求項2または4に記載の遺伝子を含有する組換えベクター。
【請求項6】
請求項5に記載の組換えベクターを導入した形質転換体。
【請求項7】
植物体である請求項6記載の形質転換体。
【請求項8】
(i) ゲノムDNAまたはRNAである核酸を植物から単離する工程、
(ii) (i)の核酸がRNAである場合に逆転写しcDNAを合成する工程、
(iii) (i)または(ii)の工程で得られたDNAから配列番号2、配列番号4または配列番号5に示す塩基配列を含有する遺伝子断片を増幅する工程、ならびに
(iv) DNA中に突然変異および/または多型の存在を決定する工程、
とを含む、植物におけるグリコアルカロイド生合成酵素をコードする遺伝子の突然変異および/または多型の存在を検出する方法。
【請求項9】
植物がナス科植物である請求項8に記載の方法。
【請求項10】
請求項8または9に記載の方法によってグリコアルカロイド生合成酵素をコードする遺伝子の突然変異および/または多型を検出し、突然変異および/または多型を有する植物体を選抜する方法。
【請求項11】
請求項10に記載の方法により選抜された、グリコアルカロイド生合成酵素をコードする遺伝子に突然変異および/または多型を有する植物体。
【請求項12】
ナス科植物である請求項11記載の植物体。
【請求項13】
グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはコードするグリコアルカロイド生合成酵素の活性が、既存品種に対して変化している植物を選抜する、請求項8または9に記載の植物体を選抜する方法。
【請求項14】
請求項13に記載の方法によって選抜された、グリコアルカロイド生合成酵素をコードする遺伝子の発現能が既存品種に対して変化しているか、またはグリコアルカロイド生合成酵素の活性が既存品種に対して変化している植物体。
【請求項15】
ナス科植物である請求項14記載の植物体。
【請求項1】
以下の(a)または(b)のタンパク質:
(a) 配列番号1に示すアミノ酸配列からなるタンパク質;および
(b) 配列番号1に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質。
【請求項2】
以下の(c)〜(f)のいずれかのDNAからなる遺伝子;
(c) 配列番号2に示す塩基配列からなるDNA;
(d) 配列番号2に示す塩基配列からなるDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;
(e) 配列番号2に示す塩基配列と80%以上の配列同一性を有する塩基配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;および
(f) 配列番号2に示す塩基配列の縮重異性体からなるDNA。
【請求項3】
以下の(g)又は(h)のタンパク質:
(g) 配列番号3に示すアミノ酸配列からなるタンパク質;および
(h) 配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入、又は付加されたアミノ酸配列からなり、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質。
【請求項4】
以下の(i)〜(l)のいずれかのDNAからなる遺伝子:
(i) 配列番号4に示す塩基配列からなるDNA;
(j) 配列番号4に示す塩基配列からなるDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;
(k) 配列番号4に示す塩基配列と80%以上の相同性を有する塩基配列からなり、かつ、
グリコアルカロイド生合成酵素活性を有するタンパク質をコードするDNA;および
(l) 配列番号4に示す塩基配列の縮重異性体からなるDNA。
【請求項5】
請求項2または4に記載の遺伝子を含有する組換えベクター。
【請求項6】
請求項5に記載の組換えベクターを導入した形質転換体。
【請求項7】
植物体である請求項6記載の形質転換体。
【請求項8】
(i) ゲノムDNAまたはRNAである核酸を植物から単離する工程、
(ii) (i)の核酸がRNAである場合に逆転写しcDNAを合成する工程、
(iii) (i)または(ii)の工程で得られたDNAから配列番号2、配列番号4または配列番号5に示す塩基配列を含有する遺伝子断片を増幅する工程、ならびに
(iv) DNA中に突然変異および/または多型の存在を決定する工程、
とを含む、植物におけるグリコアルカロイド生合成酵素をコードする遺伝子の突然変異および/または多型の存在を検出する方法。
【請求項9】
植物がナス科植物である請求項8に記載の方法。
【請求項10】
請求項8または9に記載の方法によってグリコアルカロイド生合成酵素をコードする遺伝子の突然変異および/または多型を検出し、突然変異および/または多型を有する植物体を選抜する方法。
【請求項11】
請求項10に記載の方法により選抜された、グリコアルカロイド生合成酵素をコードする遺伝子に突然変異および/または多型を有する植物体。
【請求項12】
ナス科植物である請求項11記載の植物体。
【請求項13】
グリコアルカロイド生合成酵素をコードする遺伝子の発現能またはコードするグリコアルカロイド生合成酵素の活性が、既存品種に対して変化している植物を選抜する、請求項8または9に記載の植物体を選抜する方法。
【請求項14】
請求項13に記載の方法によって選抜された、グリコアルカロイド生合成酵素をコードする遺伝子の発現能が既存品種に対して変化しているか、またはグリコアルカロイド生合成酵素の活性が既存品種に対して変化している植物体。
【請求項15】
ナス科植物である請求項14記載の植物体。
【図1−1】


【図1−2】

【図1−3】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図1−2】
【図1−3】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2012−50374(P2012−50374A)
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【出願番号】特願2010−194590(P2010−194590)
【出願日】平成22年8月31日(2010.8.31)
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【出願日】平成22年8月31日(2010.8.31)
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
[ Back to top ]