
グルタミニル、及びグルタミン酸シクラーゼのエフェクターの使用
【課題】なし
【解決手段】本発明は、哺乳動物グルタミニルシクラーゼ(QC、EC 2.3.2.5)の新規の
生理学的基質、QCの新しいエフェクター、そのようなエフェクターのスクリーニング方
法、及びQC活性の調整によって治療可能な病態の治療のための、そのようなエフェクタ
ー、及びそのようなエフェクターを含む医薬組成物を提供する。さらに、好ましい組成物
は、QC、及びDP IV活性の調整によって治療可能な病態の治療、又は緩和のために、DP
IV、又はDP IV様酵素のインヒビターを含む。
【解決手段】本発明は、哺乳動物グルタミニルシクラーゼ(QC、EC 2.3.2.5)の新規の
生理学的基質、QCの新しいエフェクター、そのようなエフェクターのスクリーニング方
法、及びQC活性の調整によって治療可能な病態の治療のための、そのようなエフェクタ
ー、及びそのようなエフェクターを含む医薬組成物を提供する。さらに、好ましい組成物
は、QC、及びDP IV活性の調整によって治療可能な病態の治療、又は緩和のために、DP
IV、又はDP IV様酵素のインヒビターを含む。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、アンモニアの遊離下における、N末端グルタミン残基のピログルタミン酸(
5-オクソ-プロリン、pGlu*)への分子内環化反応、及び水の遊離下における、N末端グル
タミン酸残基のピログルタミン酸への分子内環化反応を触媒する、グルタミニルシクラー
ゼ(QC、EC 2.3.2.5)に関するものである。
本発明は、哺乳動物QCを金属酵素として同定し、哺乳動物における新規の生理学的基
質、及びQC活性の調整によって治療可能な病態の治療のための、QCのエフェクターの
使用、及びQCのエフェクターを含む医薬組成物を提供する。さらに、金属相互作用が、
QCインヒビターの開発に有用なアプローチであることが示される。
好ましい実施態様において、本発明は、QC、及び/又はDP IV活性の調整によって治
療可能な病態の治療、又は緩和のために、DP IV、又はDP IV様酵素のインヒビターを組合
せるQC活性のエフェクターの使用を提供する。
また、QC活性のエフェクターの同定、及び選定のためのスクリーニング方法を提供す
る。
【背景技術】
【0002】
(背景)
グルタミニルシクラーゼ(QC、EC 2.3.2.5)は、アンモニアの遊離を伴う、N末端グ
ルタミン残基のピログルタミン酸(pGlu*)への分子内環化反応を触媒する。QCは、196
3年にMesserが熱帯植物パパイヤ(Carica papaya)のラテックスからはじめて精製された
(Messer, Mの論文、1963 Nature 4874, 1299)。24年後、類似の酵素活性が、動物脳
下垂体において発見された(Busby, W. H. J.らの論文、1987 J Biol Chem 262, 8532-85
36;Fischer, W. H. 及びSpiess, J.の論文1987 Proc Natl Acad Sci U S A 84, 3628-36
32)。該哺乳動物QCにおいて、QCによる pGluへのGln の転換は、TRH 及びGnRHの前
駆体に見ることができる(Busby, W. H. J.らの論文1987 J Biol Chem 262, 8532-8536;
Fischer, W. H. 及びSpiess, J.の論文、1987 Proc Natl Acad Sci U S A 84, 3628-3632
)。その上、QCの初期局在実験によって、ウシ脳下垂体における触媒の推定的産物との
同一場所での局在が明らかになり、さらに、ペプチドホルモン合成における示唆された機
能が裏付けられた(Bockers, T. M.らの論文、1995 J Neuroendocrinol 7, 445-453)。
これに対し、植物QCの生理学的機能は、明らかでない。パパイヤ由来の該酵素の場合、
病原微生物に対する植物防御における役割が示唆された(El Moussaoui, A.らの論文、20
01 Cell Mol Life Sci 58, 556-570)。近年、配列比較によって、他の植物からの推定上
QCが同定された(Dahl, S. W.らの論文、2000 Protein Expr Purif 20, 27-36)。これ
らの酵素の生理学的機能は、依然として明らかとなっていない。
【0003】
植物、及び動物由来の公知のQCは、基質N末端のL-グルタミンに高い特異性を示し、
その速度論的挙動は、ミカエリス−メンテン式に従うことが見い出された(Pohl, T.らの
論文、1991 Proc Natl Acad Sci U S A 88, 10059-10063;Consalvo, A. P.らの論文、19
88 Anal Biochem 175, 131-138;Gololobov, M. Y.らの論文、1996 Biol Chem Hoppe Sey
ler 377, 395-398)。しかしながら、パパイヤ由来QC、及び保存性の高い哺乳動物由来
のQCの一次構造の比較では、配列の相同性がないことが明らかになった(Dahl, S. W.
らの論文、2000 Protein Expr Purif 20, 27-36)。該植物QCが、新しい酵素ファミリ
ーに属していると思われるのに対し(Dahl, S. W.らの論文、2000 Protein Expr Purif 2
0, 27-36)、哺乳動物QCは細菌アミノペプチダーゼと顕著な配列相同性をもつことが見
い出され(Bateman, R. C.らの論文、2001 Biochemistry 40, 11246-11250)、植物、及
び動物由来のQCは異なる進化上の起源をもつという結論が導かれた。
【0004】
欧州特許出願公開第02 011 349.4号には、昆虫グルタミニルシクラーゼをコードしてい
るポリヌクレオチド、それによってコードされるポリペプチドが開示されている。さらに
、この出願は、該発明のポリヌクレオチドを含む発現ベクターを含む宿主細胞を提供する
。昆虫QCを含む精製ポリペプチド、及び宿主細胞は、グルタミニルシクラーゼ活性を低
下させる薬剤のスクリーニング方法において有用である。そのような薬剤は、殺虫剤と同
様に有用であると記載されている。
【0005】
アルツハイマー病(AD)は、神経細胞変性、活性化アストロサイト、及び活性化マイク
ログリアが密接に関連する細胞外アミロイド斑の異常蓄積が特徴である(Terry, R. D.、
及びKatzman, R.の論文、1983 Ann Neurol 14, 497-506;Glenner, G. G.、及びWong, C.
W.の論文、1984 Biochem Biophys Res Comm 120, 885-890;Intagaki, S.らの論文、198
9 J Neuroimmunol 24, 173-182;Funato, H.らの論文、1998 Am J Pathol 152, 983-992
;Selkoe, D. J.の論文、2001 Physiol Rev 81, 741-766)。アミロイド-β(Aβ)ペプ
チドは老人斑の主要構成成分であり、ADの発病、及び進行、遺伝学的研究が支持する仮説
に直接関与していると考えられている(Glenner, G. G.、及びWong, C. W.の論文、1984
Biochem Biophys Res Comm 120, 885-890;Borchelt, D. R.らの論文、1996 Neuron 17,
1005-1013;Lemere, C. A.らの論文、1996 Nat Med 2, 1146-1150;Mann, D. M.、及びIw
atsubo, T.の論文、1996 Neurodegeneration 5, 115-120;Citron, M.らの論文、1997 Na
t Med 3, 67-72;Selkoe, D. J.の論文、2001 Physiol Rev 81, 741-766)。Aβは、β-
アミロイド前駆体タンパク質(APP)のタンパク質から(Kang, J.らの論文、1987 Nature
325, 733-736;Selkoe, D. J. 1998 Trends Cell Biol 8, 447-453)、AβのN末端でベ
ータセクレターゼ、及びC末端でのガンマセクレターゼにより配列切断されるタンパク質
分解プロセスによって生成される(Haass, C.、及びSelkoe, D. J.の論文 1993 Cell 75,
1039-1042;Simons, M.らの論文、1996 J Neurosci 16 899-908)。N末端がL-Aspで始
まる主要なAβペプチド(Aβ1-42/40)に加えて、N末端切断型の多異種混合が老人斑に
生じる。そのような短縮型ペプチドは、フルレングス型に比べてインビトロでより神経毒
性が高く、より急速に凝集することが報告された(Pike, C. J.らの論文、1995 J Biol C
hem 270 23895-23898)。N末端切断型ペプチドは、家族性アルツハイマー病(FAD)患者
において過剰生成されること(Saido, T. C.らの論文、1995 Neuron 14, 457-466;Russo
, C.らの論文、2000 Nature 405, 531-532)、ダウン症の脳で早期に出現し、加齢ととも
に増加することが知られている(Russo, C.らの論文、1997 FEBS Lett 409, 411-416, Ru
sso, C.らの論文、2001 Neurobiol Dis 8, 173-180;Tekirian, T. L.らの論文、1998 J
Neuropathol Exp Neurol 57, 76-94)。最後に、それらの量は該疾患の進行性重症度を反
映する(Russo, C.らの論文、1997 FEBS Lett 409, 411-416)。さらに、付加的翻訳後プ
ロセスは、1及び7位のアスパラギン酸の異性化、又はラセミ化によって、及び残基3及
び11位のグルタミン酸の環化によって、N末端を改質し得る。3位にピログルタミン酸
を含む異性体[pGlu3]Aβ3-40/42は、老人斑内のN末端切断種の顕著な型で、−全Aβ量
の約50%−を占め(Mori, H.らの論文、1992 J Biol Chem 267, 17082-17086;Saido,
T. C.らの論文、1995 Neuron 14, 457-466;Russo, C.らの論文、1997 FEBS Lett 409, 4
11-416;Tekirian, T. L.らの論文、1998 J Neuropathol Exp Neurol 57, 76-94;Geddes
, J. W.らの論文、1999 Neurobiol Aging 20, 75-79;Harigaya, Y.らの論文、2000 Bioc
hem Biophys Res Commun 276, 422-427)、かつ、それはアミロイド傷害以前にも存在す
る(Lalowski, M.らの論文、1996 J Biol Chem 271, 33623-33631)。AβN3(pE)ペプチ
ドの蓄積は、凝集を増進しほとんどのアミノペプチダーゼに対する耐性を付与する、構造
上の改質によるものと思われる(Saido, T. C.らの論文、1995 Neuron 14, 457-466 ;Te
kirian, T. L.らの論文、1999 J Neurochem 73, 1584-1589)。この証拠は、アルツハイ
マー病の発症機序におけるAβN3(pE)ペプチドのきわめて重要な役割の手掛かりを提供
する。しかしながら、それらの神経毒性、及び凝集特性は比較的ほとんど知られていない
(He, W.、及びBarrow, C. J.の論文、1999 Biochemistry 38, 10871-10877;Tekirian,
T. L.らの論文、1999 J Neurochem 73, 1584-1589)。さらに、活性化グリアは厳密に老
人斑に関連しており、アミロイド沈着の凝集に積極的に寄与しうるが、該イソ型のグリア
細胞への作用、及び該ペプチドへのグリア反応は全く知られていない。近年の研究では、
Aβ1-42、Aβ1-40、[pGlu3]Aβ3-42、及び[pGlu3]Aβ3-40ペプチドの毒性、凝集特性、及
び異化を神経細胞培養、及びグリア細胞培養で調べ、ピログルタミン酸改質がAβ-ペプチ
ドの毒性を高め、さらに培養アストロサイトによる該ペプチドの分解を阻害することが示
された。Shirotaniらは、インビトロでシンドビスウィルスにより感染した初代皮質神経
細胞における[pGlu3]Aβ-ペプチドの産生を調べた。彼らは、アミノ酸置換、及びアミノ
酸欠失により、[pGlu3]Aβの潜在前駆体をコードする相補的DNAを作成した。本来の前駆
体のグルタミン酸の代わりにN末端グルタミン残基ではじまる1種の人工前駆体では、グ
ルタミニルシクラーゼによるピログルタミン酸への自然転換、又は酵素的転換が示唆され
た。[pGlu3]Aβの本来の前駆体における3位のN末端グルタミン酸の環化機構は、インビ
ボでは確定されなかった(Shirotani, K.、Tsubuki, S.、Lee, H. J.、Maruyama, K.、及
びSaido, T. C.の論文、(2002)Neurosci Lett 327, 25-28)。
【0006】
ジペプチジルペプチダーゼIV(DP IV)は、腎臓、肝臓、及び腸を含む多様な人体組織
で見出される、プロリンの後(より少ない程度にアラニンの後、セリンの後、又はグリシ
ンの後)を開裂するセリンプロテアーゼであり、かつペプチド鎖からN末端のジペプチド
を開裂する。最近、DP IVが神経ペプチド代謝、T細胞活性化、癌細胞の内皮への接着、
及びHIVのリンパ球様細胞への侵入に重要な役割を果たすことが示された。それについて
は、国際公開第02/34242号、国際公開第02/34243号、国際公開第03/002595号、及び国
際公開第03/002596号を参照されたい。
【0007】
DP IVインヒビターは、グルコース耐性障害、及び糖尿病の治療に有用であり得ること
が知られている(国際特許出願、公開番号、国際公開第99/61431号、Pederson, R. A.ら
の論文(1998 Diabetes 47, 1253-1258)、及びPauly, R. P.らの論文(1999 Metabolism
48, 385-389))。特に、国際公開第99/61431号では、アミノ酸残基、及びチアゾリジ
ン、又はピロリジン群、及びその塩、特にL-threo-イソロイシルチアゾリン、L-allo-イ
ソロイシルチアゾリン、L-threo-イソロイシルピロリジン、L-allo-イソロイシルチアゾ
リン、L-allo-イソロイシルピロリジン、及びその塩を含むDP IVインヒビターを開示する
。
【0008】
さらなる低分子量ジペプチジルペプチダーゼIVインヒビターの例には、テトラヒドロイ
ソキノリン-3-カルボキサミド誘導体、N-置換2-シアノピロール、及びピロリジン、N-(N
'-置換グリシル)2-シアノピロリジン、N-(置換グリシル)-チアゾリジン、N-(置換グ
リシル)-4-シアノチアゾリジン、アミノ-アシル-ボロノ-プロピル-インヒビター、シク
ロプロピル-融合ピロリジン、及び六員環化合物のような薬剤がある。ジペプチジルペプ
チダーゼIVインヒビターは、下記の文献に記載されている:米国特許第6,380,398号、米
国特許第6,011,155号、米国特許第6,107,317号、米国特許第6,110,949号、米国特許第6,1
24,305号、米国特許第6,172,081号、国際公開第 95/15309号、国際公開第99/61431号、
国際公開第99/67278号、国際公開第99/67279号、ドイツ特許第198 34 591号公報、国際
公開第97/40832号、ドイツ特許第196 16 486 C 2号公報、国際公開第98/19998号、国際
公開第00/07617号、国際公開第99/38501号、国際公開第99/46272号、国際公開第99/3
8501号、国際公開第01/68603号、国際公開第01/40180号、国際公開第01/8133号、国際
公開第01/81304号、国際公開第01/55105、国際公開第02/02560号、及び国際公開第02
/14271号、国際公開第02/04610号、国際公開第02/051836号、国際公開第02/068420号
、国際公開第02/076450号、国際公開第02/083128号、国際公開第02/38541号、国際公
開第03/000180号、国際公開第03/000181号、国際公開第03/000250号、国際公開第03/
002530号、国際公開第03/002531号、国際公開第03/002553号、国際公開第03/002593号
、国際公開第03/004496号、国際公開第03/024942号、及び国際公開第03/024965号であ
る。これらの文献に教示されている内容は、その全体、特にインヒビター、その定義、使
用、及び産生に関して、引用により本明細書に取り込まれるものとする。
【発明の概要】
【発明が解決しようとする課題】
【0009】
(本発明の要旨)
本発明は、哺乳動物QCの新規生理学的基質、Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu
11]Aβ11-40/42、[Gln11]Aβ11-40/42、[Gln1]ガストリン、[Gln1]ニューロテンシン、
[Gln1]FPP、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 18、[G
ln1]フラクタルカイン、[Gln1]オレキシンA、[Gln3]グルカゴン3-29、及び[Gln5]サブス
タンスP5-11、及びQC活性の調節によって治療可能な病態の治療のための、QCエフェ
クターの使用、及びQCエフェクターを含む医薬組成物を提供する。
【0010】
阻害研究により、ヒトQCは、金属依存性トランスフェラーゼであることが示された。
QCのアポ酵素は、亜鉛イオンにより最も効果的に活性化され得て、かつ、亜鉛依存性ア
ミノペプチダーゼの金属結合モチーフは、ヒトQCにも存在する。金属と結合する活性部
位と相互作用する化合物は、有力なインヒビターである。
【0011】
予想外に、組み換えQCは、脳抽出物由来QCの活性と同様に、N末端グルタミニルと
同様グルタミン酸環化反応の両方を触媒することが示された。最も特筆すべきことは、シ
クラーゼ触媒のGlu1-転換がpH 6.0付近で活発なのに対し、pGlu-誘導体へのGln1-転換は
至適pH 8.0付近で起こる。pGlu-Aβに関連するペプチドの産生は、組み換えヒトQC、及
びブタ脳下垂体抽出物由来QCの活性阻害によって抑制できるので、該酵素QCは、アル
ツハイマー病治療に用いる薬剤開発の標的である。
【0012】
QC活性のエフェクターを哺乳動物に投与することによって、アルツハイマー病、ダウ
ン症、ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は伴わない、潰瘍性疾患及び胃
癌、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、悪
性転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性
免疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネル
ギー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害か
ら選択される病態を、予防、又は緩和、又は治療することが可能である。
【0013】
さらに、哺乳動物へのQC活性のエフェクターの投与により、胃腸管細胞増殖、好まし
くは、胃粘膜細胞、上皮細胞の増殖、急性の酸分泌、及び酸産生壁細胞、及びヒスタミン
分泌クロム親和性様細胞分化を刺激促進することが可能である。
さらに、哺乳動物へのQC活性のエフェクターの投与により、骨髄性前駆細胞増殖を抑
制することが可能である。
さらに、QCインヒビターの投与は、男性生殖能力の抑制をもたらすことができる。
【0014】
好ましい実施態様において、本発明は、QC、及び/又はDP IV活性を調整することに
よって治療可能な病態の治療、又は緩和のために、DP IV、又はDP IV様酵素のインヒビタ
ーと組合せる、QC活性のエフェクターの使用を提供する。
本発明は、任意に慣用の担体、及び/又は賦形剤と共に少なくとも1種のQCのエフェ
クターを含むか;又は任意に慣用の担体、及び/又は賦形剤と共に少なくとも1種のQC
のエフェクターを、少なくとも1種のDP IVインヒビターと共に含む、非経口、経腸、又
は経口投与に用いる医薬組成物を提供する。
また、QCエフェクターの同定、及び選定のためのスクリーニング方法を提供する。
【0015】
(ペプチド配列)
本明細書中で言及し、かつ使用したペプチドは、下記の配列を有する。
Aβ1-42:
Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-
Ala-Glu-Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val-
Ile-Ala
Aβ1-40:
Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-
Ala-Glu-Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val
Aβ3-42:
Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-Glu-
Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val-Ile-Ala
Aβ3-40:
Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-Glu-
Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val
Aβ1-11a:
Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-NH2
Aβ3-11a:
Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-NH2
【0016】
Aβ1-21a:
Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-
Ala-NH2
Aβ3-21a:
Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-NH2
Gln3-Aβ3-40:
Gln-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-Glu-
Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val
Gln3-Aβ3-21a:
Gln-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-NH2
Gln3-Aβ1-11a:
Asp-Ala-Gln-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-NH2
Gln3-Aβ3-11a:
Gln-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-NH2
【課題を解決するための手段】
【0017】
(発明の詳細な説明)
本発明は、a)インビボQC活性の調整によって治療可能な、哺乳動物の疾患の治療、
及び/又は、b)QC活性の調整によって引き起こされる、pGlu含有ペプチドの活性に基
づく生理的プロセスの調整のための、グルタミニルシクラーゼ(QC)のエフェクターを
提供する。
さらに、本発明は哺乳動物におけるグルタミニルシクラーゼ(QC、EC 2.3.2.5)、及
び/又はQC様酵素の阻害に用いる化合物を提供し、かつQC活性に関わる病態の治療の
ための、QC活性のインヒビターの使用を提供する。
【0018】
また、本発明はアルツハイマー病、及びダウン症の新規な治療方法を提供する。アルツ
ハイマー病、及びダウン症の脳に沈着したアミロイドβ-ペプチドのN末端は、ピログル
タミン酸を有する。pGluの産生は、該疾患の発症、及び進行における重要な事象である。
その理由は、改質されたアミロイドβ-ペプチドは、β-アミロイドの凝集、及び毒性を増
大し、疾患の発病、及び進行を悪化させ得るからである(Russo, C.らの論文、2002 J Ne
urochem 82, 1480-1489)。
【0019】
これに対し、本来のAβ-ペプチド(3-40/42)においては、グルタミン酸がN末端アミ
ノ酸として存在する。今日までに公知のGluのpGluへの酵素転換は存在しない。さらに、G
luペプチドのpGluペプチドへの自然環化反応は、今までのところ観察されていない。した
がって、本発明の一態様は、アルツハイマー病、及びダウン症におけるQCの役割を決定
することであった。この一態様においては、3位のアミノ酸としてグルタミン酸の代わり
にグルタミンを含む、Aβ3-11、及びAβ1-11の合成、これらの改質アミロイドβ-ペプチ
ドのQC、DP IV、及びDP IV様酵素、及びアミノペプチダーゼに対する基質特性の決定、
及び、QCインヒビターを使用して、該アミロイドβ-誘導体ペプチド1-11、及び3-11の
N末端グルタミン残基からpGluの産生を抑制し、解決することに向けられている。該結果
を、実施例8に示す。適用された方法は、実施例3に記載されている。
【0020】
今日まで、該疾患の進行におけるQCの関与が全く示唆されていない。その理由は、グ
ルタミン酸がAβ(3-40/42、及び11-40/42)におけるN末端アミノ酸だからである。し
かし、QCはペプチドのN末端でpGluを産生できる唯一公知の酵素である。本発明の他の
形態は下記の知見、及び発見に関与している:
a)副反応として、QCはごく低速度でグルタミン酸のピログルタミン酸への環化反応を
触媒する、
b)APP、又はその後に形成されるアミロイドβ-ペプチドのグルタミン酸は、未知の酵素
活性により翻訳後にグルタミンに転換され、かつ第二工程として、QCはグルタミンのピ
ログルタミン酸への環化反応を、アミロイドβ-ペプチドN末端のプロセス後、触媒する
、
c)グルタミン酸は化学触媒、又は自己触媒により、翻訳後にグルタミンに転換され、か
つ続いて、該アミロイドβ-ペプチドN末端のプロセス後、QCは、グルタミンのピログ
ルタミン酸への環化反応を触媒する、
d)アミロイドβ-タンパク質をコードするAPP遺伝子において、3位のGluの代わりにGln
をもたらす突然変異がある。翻訳、及び該N末端のプロセシング後、QCはグルタミンの
ピログルタミン酸への該環化反応を触媒する、
e)グルタミンは、未知の酵素活性の異常により、APPの未完成のペプチド鎖に取り込ま
れ、かつ続いて、該アミロイドβ-ペプチドN末端のプロセシング後、QCは、N末端グ
ルタミンのピログルタミン酸への環化反応を触媒する。
【0021】
QCは上記全5事例における重要な工程、すなわちアミロイドβ-ペプチドの凝集を促
進するピログルタミン酸の産生に関与している。このように、QCの阻害は、環化反応が
起こる機構に関わらず、アルツハイマー病、及びダウン症の発症と進行を起こす、斑形成
型Aβ3-40/41/43、又はAβ11-40/41/43の沈着の予防を可能にする。
グルタミン酸は、アミロイドβ-ペプチドの3、11、及び22位に見い出される。そ
の中で、22位のグルタミン酸(E)からグルタミン(Q)への突然変異(アミロイド前駆
体タンパク質APP 693、スイスプロットP05067に相当)は、いわゆるオランダ型脳動脈ア
ミロイドーシス突然変異として記載されている。
3、11、及び22位にピログルタミン酸を有するβ-アミロイドペプチドは、A β1-4
0/42/43に比べて、細胞毒性、及び疎水性がより高いことが記載されている(Saido T.C
.の論文、2000 Medical Hypotheses 54(3):427-429)。
【0022】
多種のN末端変異は、異なる部位でのβ-セクレターゼ酵素、β-部位アミロイド前駆体
タンパク質-切断酵素(BACE)(Huse J.T.らの論文、2002 J. Biol. Chem. 277(18):1
6278-16284)、及び/又はアミノペプチダーゼのプロセシングによって、産生が可能であ
る。全ての場合において、先に記載したa)〜e)の経路によって環化反応が起こり得る
。
【0023】
今までのところ、経路a)に相当する、未知のグルタミルシクラーゼ(EC)による、
Glu1ペプチドのpGluペプチドへの酵素的転換を支持する実験的証拠はない(Garden, R. W
.、Moroz, T. P.、Gleeson, J. M.、Floyd, P. D.、Li, L. J.、Rubakhin, S. S.、及びS
weedler, J. V.の論文(1999)J Neurochem 72, 676-681;Hosoda R.らの論文(1998)J
Neuropathol Exp Neurol. 57, 1089-1095)。今日までに、弱アルカリpH条件下で、N末
端側がプロトン化され、かつ負電荷のGlu1γ-カルボン酸部分をもつGlu1ペプチドを、環
化できるような酵素活性は同定されていない。
【0024】
Gln1基質に対するQC活性は、pH 7.0以下で劇的に低下する。これに対し、Glu1転換は
酸性反応条件で起こり得る(Iwatsubo, T.、Saido, T. C.、Mann, D. M.、Lee, V. M.、
及びTrojanowski, J. Q.の論文、(1996)Am J Pathol 149, 1823-1830;Russo, C.、Sai
do, T. C.、DeBusk, L. M.、Tabaton, M.、Gambetti, P.、及びTeller, J. K.の論文、(
1977)FEBS Lett 409, 411-416;Russo, C.、Salis, S.、Dolcini, V.、Venezia, V.、So
ng, X. H.、Teller, J. K.、及びSchettini, G.の論文(2001)Neurobiol Dis 8, 173-1
80;Tekirian, T. L.、Saido, T. C.、Markesbery, W. R.、Russell, M. J.、Wekstein,
D. R.、Patel, E.、及びGeddes, J. W.の論文(1998)J Neuropathol Exp Neurol. 57, 7
6-94;Russo, C.、Violani, E.、Salis, S.、Venezia, V.、Dolcini, V.、Damonte, G.、
Benatti, U.、DArrigo, C.、Patrone, E.、Carlo, P.、及びSchettini, G.の論文(2002
)J Neurochem 82, 1480-1489;Hosoda, R.、Saido, T. C.、Otvos, L., Jr.、Arai, T.
、Mann, D. M.、Lee, V. M.、Trojanowski, J. Q.、及びIwatsubo, T.の論文(1998)J N
europathol Exp Neurol. 57, 1089-1095;Garden, R. W.、Moroz, T. P.、Gleeson, J. M
.、Floyd, P. D.、Li, L. J.、Rubakhin, S. S.、及びSweedler, J. V.の論文(1999)J
Neurochem 72, 676-681)。
【0025】
本発明に従って、QCが弱酸性条件下でアミロイド-β誘導ペプチドを認識し、かつ代
謝回転できるか否かを調べた。したがって、該酵素の潜在的基質として、[Gln3]Aβ1-11a
、Aβ3-11a、[Gln3]Aβ3-11a、Aβ3-21a、[Gln3]Aβ3-21a、及び[Gln3]Aβ3-40 を合成し
、かつ調べた。これらの配列は、翻訳後のGluアミド化が原因で起こり得る、本来のN末
端、及びC末端切断型[Glu3]Aβペプチド、及び[Gln3]Aβペプチドを模倣するように選択
された。
【0026】
本発明において、パパイヤ、及びヒトQCがグルタミニル、及びグルタミン酸環化反応
の両方を触媒することを示した。明らかに、QCの主要生理学的機能は、ホルモン分泌プ
ロセスに先立つ、又はその間の、グルタミン環化反応による内分泌細胞内のホルモン成熟
を完成させることである。そのような分泌小胞は、酸性pHであることが知られている。従
って、pH 5.0〜7.0の狭いpH範囲内での該酵素の副活性は、Glu-Aβペプチドをも転換する
、新規に発見されたグルタミルシクラーゼの活性であり得る。しかしながら、Gln環化に
比べてGlu環化はかなり遅く起こるので、該グルタミルシクラーゼが有意義な生理的役割
を果たすかどうかは、疑問の余地がある。
【0027】
しかしながら、神経変性疾患の病理においては、該グルタミル環化反応は有意義である
。該酵素反応のpH依存性を研究し、我々はプロトン化されていないN末端がGln1ペプチド
の環化反応に不可欠であり、かつ従って、該基質のpKa値は、QC触媒のpKa値と同一であ
ることを見いだした(図17参照)。このように、QCは、プロトン化されていないα-
アミノ部分の、アミド化により親電子的に活性化されたγ-カルボニル炭素への分子内求
核攻撃を安定化させる(スキーム1)。
【0028】
N末端グルタミン含有ペプチドに存在する一価の電荷に対して、Glu含有ペプチドにお
けるN末端Glu残基は、中性付近pHで、大部分は二価に荷電される。グルタミン酸は、γ-
カルボキシル、及びα-アミノ部位に対し、各々、約4.2、及び7.5のpKa値を示した。すな
わち、中性、及びそれ以上のpHでは、該α-アミノ窒素は、部分的に、又は全部プロトン
化されず、かつ求核的であるが、γ-カルボキシル基はプロトン化されず、かつそれによ
り親電子的カルボニル活性を発揮しない。したがって、分子内環化反応は不可能である。
【0029】
しかしながら、その各々のpKa値の間、pH約5.2〜6.5の範囲において、全N末端Glu含有
ペプチドの約1〜10%(-NH2)、又は10〜1%(-COOH)の濃度で、2官能基は両方非イオ
ン化型で存在する。その結果として、弱酸性pH範囲間で、電荷していない両基を持つN末
端Glu含有ペプチド種が存在し、かつ従って、QCがpGluペプチドへの分子内環化反応の
中間体を安定化させることが可能である。すなわち、もし、γ-カルボキシル基がプロト
ン化していると、カルボニル炭素は親電子的で、プロトン化されていないα-アミノ基に
よる親核攻撃が可能になる。このpHにおいて、水酸基イオンは脱離基として機能する(ス
キーム3)。これらの仮説は、QCが触媒するGlu-βNAの転換のために得られたpH依存性
のデータによって確認された(実施例11参照)。QCによるGln-βNAのグルタミン転換に
対し、該触媒の至適pHはpH 6.0付近の酸性範囲、すなわち、基質分子種がプロトン化され
たγ-カルボキシル基、及びプロトン化されていないα-アミノ基を同時に十分に持って存
在するようなpH範囲に移動する。さらに、速度論的に決定したpKa値7.55±0.02は、滴定
により決定されたGlu-βNAのα-アミノ基のpKa値に極めてよく一致している(7.57±0.05
)。
【0030】
生理学的に、pH 6.0で、QCが触媒するグルタミン酸環化反応の二次速度定数(又は特
異性定数、kcat/KM)は、グルタミン環化反応の該定数より、8,000倍程度遅くなり得る
(図17)。しかしながら、Glu-βNA、及びGln-βNAの両モデル基質の非酵素的転換がご
くわずかであることは、本発明において観察されたごくわずかなpGluペプチド産生と一致
する。それゆえ、QCによるpGluの産生において、酵素対非酵素の速度定数の比から、少
なくとも108の加速を推定することが可能である(該酵素触媒の該二次速度定数を、各非
酵素環化反応の一次速度定数と比較すると、Gln転換、及びGlu転換における触媒有効係数
は、各々109〜1010 M-1である)。これらのデータからの結論は、pGlu産生をもたらす酵
素経路は、インビボでのみあり得ると思われる。
【0031】
QCは脳内に非常に豊富に存在しており、かつ最近見い出された、30μM(Gln-)TRH様
ペプチドを成熟するための高代謝回転(Prokai, L.、Prokai-Tatrai, K.、Ouyang, X.、K
im, H. S.、Wu, W. M.、Zharikova, A.、及びBodor, N.の論文、(1999)J Med Chem 42,
4563-4571)であるから、同様の反応条件下での適切なグルタミン酸基質には、約100時
間の環化反応半減期を予測し得る。さらに、分泌経路における脳QC/ECの局在化、お
よび位置を考慮すると、実際のインビボ酵素、及び基質濃度、及び反応条件は、正常細胞
の酵素的環化反応において、なおさら好都合であり得る。かつ、N末端GluがGlnに転換さ
れているなら、QCにより調節される、かなり急速なpGluの産生を予想し得る。インビト
ロにおいて、両反応はQC/EC活性のインヒビターを適用することによって抑制された
(図9、10、及び15)。
【0032】
要約すると、本発明は、脳内にかなり豊富に存在するヒトQCが、Glu-Aβ、及びGln-A
β前駆体から、アルツハイマー病で見い出される斑沈着の50%以上を構成する、アミロ
イド生成性pGlu-Aβ-ペプチドを産生する有望な触媒であることを示した。これらの知見
は、QC/ECを老人斑形成に関わる一因子と同定し、かつ従って、アルツハイマー病の
治療における新規な薬剤標的と同定した。
【0033】
本発明の第二の実施態様において、アミロイドβ-誘導ペプチドが、ジルペプチダーゼI
V(DP IV)、又はDP IV様酵素、好ましくはジペプチジルペプチダーゼII(DP II)の基質
であることを見い出された。DP IV、DP II、又はDP IV様酵素は、改質したアミロイドβ-
ペプチド(1-11)のN末端からジペプチドを放出し、グルタミンをN末端アミノ酸残基に
持つアミロイドβ-ペプチド(3-11)を産生する。その結果は、実施例8に示した。
【0034】
DP II、DP IV、又は他のDP IV様酵素の切断に先立ち、アスパラギン酸(アミロイドβ-
ペプチドの1番めの残基)、及びアラニン(アミロイドβ-ペプチドの2番めの残基)間
のペプチド結合は、文献に記載されているように、異性化されイソアスパルチル残基を生
じ得る(Kuo, Y.-M.、Emmerling, M. R.、Woods, A. S.、Cotter, R. J.、及びRoher, A.
E.の論文、(1997)BBRC 237, 188-191;Shimizu, T.、Watanabe, A.、Ogawara, M.、Mo
ri, H.、及びShirasawa, T.の論文、(2000)Arch. Biochem. Biophys. 381, 225-234)
。
これらのイソアスパラチル残基は、該アミロイドβ-ペプチドにアミノペプチダーゼ分
解に対する耐性を与え、かつその結果として、核化老人斑は多量のisoAsp1アミロイドベ
ータペプチドを含んでおり、それはN末端における低下した代謝回転を示唆する。
【0035】
しかしながら、本発明において初めて、N末端ジペプチドH-isoAsp1-Ala2-OHが、特に
酸性条件下でジペプチダーゼにより放出されることを示した。さらにまた、異性化はβ-
セクレターゼによる切断に先行することが可能であり、かつ該異性化はタンパク質分解を
促進し、isoAsp1アミロイドβ-ペプチドのN末端イソアスパルチル結合の遊離を可能にし
、次いでDP II、DPIV、又は DP IV様酵素により代謝回転され得ることが示された(Moman
d, J.、及びClarke, S.(1987)Biochemistry 26, 7798-7805;Kuo, Y.-M.、Emmerling,
M. R.、Woods, A. S.、Cotter, R. J.、Roher, A. E.の論文、(1997)BBRC 237, 188-19
1)。従って、イソアスパルチル産生の阻害は、β-セクレターゼによる切断の減少を可能
にし、かつ順に、アミロイドβ-ペプチドの産生の減少を可能にし得る。その上、isoAsp1
アミロイドβ-ペプチドのDP II、DPIV、又は DP IV様酵素による代謝回転の阻害は、[Glu
3]Aβの露出を防げ、QC/EC触媒による[pGlu3]Aβ産生を抑制し得る。
本発明の第三の実施態様において、DP IV活性のインヒビター、及びQCのインヒビタ
ーとの組み合わせを、アルツハイマー病、及びダウン症の治療のために使用することがで
きる。
【0036】
DP IV、及び/又は DP IV様酵素、及びQCを組合せた効果は、以下に示すとおりであ
る:
a)DP IV、及び/又は DP IV様酵素はAβ1-40/42を切断し、H-Asp-Ala-OHを含むジペプ
チド、及びAβ3-40/42が放出される、
b)副反応として、QCは低速度でグルタミン酸のピログルタミン酸への環化反応を触媒
する、
c)グルタミン酸は、未知の酵素活性により翻訳後にグルタミンに転換され、かつ続いて
、アミロイドβ-ペプチドN末端のプロセス後、QCはグルタミンのピログルタミン酸へ
の環化反応を触媒する、
d)グルタミン酸は化学触媒、又は自己触媒により、翻訳後にグルタミンに転換され、か
つ第二工程として、アミロイドβ-ペプチドN末端のプロセシング後、QCはグルタミン
のピログルタミン酸への環化反応を触媒する、
e)アミロイドβ-タンパク質をコードするAPP遺伝子において、Aβの3位のGluの代わり
にGlnをもたらす突然変異がある。翻訳、及びN末端のプロセシング後、QCはグルタミ
ンのピログルタミン酸への環化反応を触媒する、
f)グルタミンは、未知の酵素活性の異常により未完成のペプチド鎖に取り込まれ、かつ
続いて、アミロイドβ-ペプチドN末端のプロセス後、QCはグルタミンのピログルタミ
ン酸への環化反応を触媒する。
【0037】
また、QC活性への該N末端Glnの露出は、異なるペプチダーゼ活性によって、引き起
こし得る。アミノペプチダーゼは、Aβ1-40/41/43のN末端から、Asp、及びAlaを順次
取り去ることが可能で、従って、環化反応され易い3番目のアミノ酸が露出される。DP I
、DP II、DP IV、DP 8、DP 9、及びDP 10のようなジペプチジルペプチダーゼは、一工程
でAsp-Alaを除去する。従って、アミノペプチダーゼ、又はジペプチジルペプチダーゼの
活性の阻害は、Aβ3-40/41/43の産生を防止するのに有用である。
【0038】
DP IV、及び/又はDP IV様酵素のインヒビター、及びQCの活性低下エフェクターとの
組合せ効果を、下記の経路で示す:
a)DP IV、及び/又はDP IV様酵素は、Aβ1-40/42のAβ3-40/42への転換を阻害する。
b)それにより、グルタミン酸のN末端露出を妨げ、酵素による、又は化学触媒によるグ
ルタミンへの転換はなく、続いてピログルタミン酸産生することは不可能である。
c)さらに、QCのインヒビターは、残基が改質されたAβ3-40/42分子、及びAPP遺伝子
の突然変異により産生されたAβ3-40/42分子からのピログルタミン酸形成を阻害する。
本発明の範囲内で、DP IV、又はDP IV様酵素、及びQCとの類似の組合せ効果は、さら
にグルカゴン、CCケモカイン、及びサブスタンスPのような、ペプチドホルモンで示さ
れた。
【0039】
グルカゴンは、すい臓のランゲルハンス島アルファ細胞から放出される、29アミノ酸ポ
リペプチドであり、肝臓でのグリコーゲン分解、及びグルコース新生を刺激促進すること
により、血糖値を維持するよう作用する。その重要性にかかわらず、体内でのグルカゴン
排除をつかさどる機構に関しては見解の一致を見ないままである。Pospisilikらは、高感
度の質量分光学的技術を用い、グルカゴンの酵素代謝を測定し、分子産物を同定した。グ
ルカゴンを精製されたブタジペプチジルペプチダーゼIV(DP IV)とインキュベートする
ことにより、グルカゴン3-29、及びグルカゴン5-29が生じた。ヒト血清では、グルカゴン
3-29への分解後速やかにグルカゴンのN末端環化反応が起こり、さらなる加水分解を抑制
した。グルカゴンのバイオアッセイにおいて、精製DP IV、又は、正常ラット血清とイン
キュベートの後、高血糖活性の有意な低下が示されたのに対し、DP IV欠損ラット血清と
の同様のインキュベートでは、グルカゴン生物活性の低下は示されなかった。質量分析、
およびバイオアッセイによってモニターすると、分解は、DP IV特異的インヒビター、イ
ソロイシルチアゾリジンにより阻害された。これらの結果により、DP IVがグルカゴンの
分解、及び不活性化に関係する主要酵素と同定した。これらの知見は、ヒト血漿中のグル
カゴンレベルの決定に、重要な影響をもたらす(Pospisilikらの論文、Regul Pept 2001
Jan 12;96(3):133-41)。
【0040】
当初、ヒト単球走化性タンパク質(MCP)-2は、MCP-1、及びMCP-3と共に生成されるケ
モカインとして、刺激促進された骨肉腫細胞から精製されてきた。Von Coillieら(Van C
oillie, E.らの論文、1998 Biochemistry 37, 12672-12680)は、ヒト精巣cDNAライブラ
リーから5'末端伸張型MCP-2 cDNAをクローニングした。それは76残基のMCP-2タンパク
質をコードしたが、報告されている骨髄由来MCP-2 cDNA配列とはコドン46で異なり、Gl
nの代わりにLysをコードした。一塩基変異多型(SNP)により生じた、該MCP-2Lys46変異
体をMCP-2Gln46と生物学的に比較した。該コード領域をバクテリア発現ベクターpHEN1に
サブクローンし、かつ大腸菌(Escherichia coli)の形質転換後、該2種のMCP-2変異体を
ペリプラズムから回収した。エドマン分解によって、N末端がpGluの代わりにGln残基で
あることが明らかになった。rMCP-2Gln46、及びrMCP-2Lys46、及びそのN末端が環化した
該対応型を、単球細胞上でカルシウム流動、及びアッセイで、分析した。rMCP-2Gln46、
及びrMCP-2Lys46 異性体間で、生物学活性には有意な差はなかった。しかしながら、両MC
P-2において、N末端ピログルタミン酸が遊走性に不可欠であるが、カルシウム流動には
不可欠でないことが示された。セリンプロテアーゼCD26/ジペプチジルペプチダーゼIV(
CD26/DPP IV)による、rMCP-2Lys46のN末端切断型は、アミノ末端Gln-Proジペプチドの
放出をもたらすが、アミノ末端pGluを含む人工MCP-2は、影響を受けなかった。CD26/DPP
IVで切断されたrMCP-2Lys46(3-76)は、遊走性、及びシグナル伝達アッセイの両方でほ
ぼ完全に不活性であった。これらの観察は、MCP-2のNH2末端pGluが遊走活性に不可欠であ
ること、また、それが該タンパク質をCD26/DPP IVによる分解から保護することを示唆し
た(van Coillie, E.らの論文、Biochemistry 1998 37, 12672-80)。
【0041】
本発明の範囲内で、LC/MS分析により、グルカゴン3-29(Pospisilikらの論文、2001)
、及びMCP-2異性体(van Coillieらの論文、1998)のN末端ピログルタミン酸残基の産生
がQCにより触媒されることが決定された。
さらに、LC/MS実験により、サブスタンスPからLys-Pro、及びArg-Proの2種のジペプ
チドのN末端DP IV触媒による除去後、残存[Gln5]サブスタンスP5-11は、QCにより[pGl
u5]サブスタンスP5-11へ転換されることを証明した。
【0042】
DP IVインヒビターは、国際公開第99/61431号において開示されている。特に、アミノ
酸残基、及びチアゾリジン、又はピロリジン群、及びその塩を含むDP IVインヒビターが
開示されており、特にL-threo-イソロイシルチアゾリン、L-allo-イソロイシルチアゾリ
ン、L-threo-イソロイシルピロリジン、L-allo-イソロイシルチアゾリン、L-allo-イソロ
イシルピロリジン、及びその塩である。
【0043】
低分子量ジペプチジルペプチダーゼIVインヒビターのさらなる例には、テトラヒドロイ
ソキノリン-3-カルボキサミド誘導体、N-置換 2-シアノピロール、及びピロリジン、N-(
N'-置換グリシル)2-シアノピロリジン、N-(置換グリシル)-チアゾリジン、N-(置換グ
リシル)-4-シアノチアゾリジン、アミノ-アシル-ボロノ-プロピル-インヒビター、シク
ロプロピル-融合ピロリジン、及び六員環化合物のような薬剤がある。ジペプチジルペプ
チダーゼIVインヒビターは、下記の文献に記載されている:米国特許第6,380,398号、米
国特許第6,011,155号、米国特許第6,107,317号、米国特許第6,110,949号、米国特許第6,1
24,305号、米国特許第6,172,081号、国際公開第95/15309号、国際公開第99/61431号、
国際公開第99/67278号、国際公開第99/67279号、ドイツ特許第198 34 591号公報、国際
公開第97/40832号、ドイツ特許第196 16 486 C 2号公報、国際公開第98/19998号、国際
公開第00/07617号、国際公開第99/38501号、国際公開第99/46272号、国際公開第99/3
8501号、国際公開第01/68603号、国際公開第01/40180号、国際公開第01/8133号、国際
公開第01/81304号、国際公開第01/55105、国際公開第02/02560号、及び国際公開第02
/14271号、国際公開第02/04610号、国際公開第02/051836号、国際公開第02/068420号
、国際公開第02/076450号、国際公開第02/083128号、国際公開第02/38541号、国際公
開第03/000180号、国際公開第03/000181号、国際公開第03/000250号、国際公開第03/
002530号、国際公開第03/002531号、国際公開第03/002553号、国際公開第03/002593号
、国際公開第03/004496号、国際公開第03/024942号、及び国際公開第03/024965号であ
る。これらの文献に教示されている内容は、その全体、特にインヒビター、その定義、使
用、及び産生に関して、引用により本明細書に取り込まれるものとする。
【0044】
QCのエフェクターと組合せる使用が好ましいのは、Hughesらの論文(1999 Biochemis
try 38 11597-11603)により開示されたNVP-DPP728A(1-[[[2-[{5-シアノピリジン-2-イ
ル}アミノ]エチル]アミノ]アセチル]-2-シアノ-(S)-ピロリジン)(Novartis社)、Hug
hesら(アメリカ糖尿病協会会議、2002年、要旨番号272)により開示されたLAF-237(1-[
(3-ヒドロキシ-アダマンチル-1-アミノ)-アセチル]-プロリジン-2(S)-カルボニトリ
ル)(Novartis社)、Yamadaらの論文(1998 Bioorg Med Chem Lett 8, 1537-1540)に
開示されたTSL-225(トリプトフィル-1,2,3,4-テトラヒドロイソキノリン-3-カルボン酸
)、Asworthらの論文(1996 Bioorg Med Chem Lett 6, 1163-1166、及び2745-2748)に
開示された2-シアノピロリジド、及び4-シアノピロリジド、Sudreらの論文(2002 Diabet
es 51, 1461-1469)によって開示されたFE-999011(Ferring社)、及び国際公開第01/34
594号で開示された化合物(Guilford)のようなDP IVインヒビターがあり、上記の文献で
設定した投与量を採用する。
【0045】
QCのエフェクターと組合せる使用がより好ましいDP IVインヒビターは、ジペプチド
化合物であり、そのアミノ酸は、例えば、ロイシン、バリン、グルタミン、グルタミン酸
、プロリン、イソロイシン、アスパラギン、及びアスパラギン酸のような天然アミノ酸、
から選択されるのが好ましい。本発明で使用されるジペプチド様化合物は、10μMの(ジ
ペプチド化合物の)濃度で、血漿ジペプチジルペプチダーゼIV、又はDP IVアナログ酵素
活性の少なくとも10%、特に少なくとも40%活性の低下を示す。また、インビボでは、多
くの場合少なくとも60%、又は少なくとも70%の活性の低下が所望される。また、好まし
い化合物では、最高20%、又は30%の活性の低下が示され得る。
【0046】
好ましいジペプチド化合物は、N-バリルプロリル、O-ベンゾイルヒドロキシルアミン、
アラニルピロリジン、L-allo-イソロイシルチアゾリジン、L-threo-イソロイシルピロリ
ジンのようなイソロイシルチアゾリジン、及びその塩、特にフマル酸塩、及び、L-allo-
イソロイシルピロリジン、及びその塩である。特に好ましい化合物は、グルタミニルピロ
リジン、及びグルタミニルチアゾリジン、H-Asn-ピロリジン、H-Asn-チアゾリジン、H-As
p-ピロリジン、H-Asp-チアゾリジン、H-Asp(NHOH)-ピロリジン、H-Asp(NHOH)-チアゾ
リジン、H-Glu-ピロリジン、H-Glu-チアゾリジン、H-Glu(NHOH)-ピロリジン、H-Glu(N
HOH)-チアゾリジン、H-His-ピロリジン、H-His-チアゾリジン、H-Pro-ピロリジン、H-Pr
o-チアゾリジン、H-Ile-アジジジン、H-Ile-ピロリジン、H-L-allo-Ile-チアゾリジン、H
-Val-ピロリジン、及びH-Val-チアゾリジン、及び医薬として許容し得るその塩である。
これらの化合物は、国際公開第99/61431号、及び欧州特許出願公開第1 304 327号に記載
されている。
【0047】
さらに本発明は、ジペプチジルペプチダーゼIV触媒の競合的調整に有用な基質様ペプチ
ド化合物と組合せるQCのエフェクターの使用を提供する。好ましいペプチド化合物は、
2-アミノオクタン酸-Pro-Ile、Abu-Pro-Ile、Aib-Pro-Ile、Aze-Pro-Ile、Cha-Pro-Ile、
Ile-Hyp-Ile、Ile-Pro-allo-Ile、Ile-Pro-t-ブチル-Gly、Ile-Pro-Val、Nle-Pro-Ile、N
va-Pro-Ile、Orn-Pro-Ile、Phe-Pro-Ile、Phg-Pro-Ile、Pip-Pro-Ile、Ser(Bzl)-Pro-I
le、Ser(P)-Pro-Ile、Ser-Pro-Ile、t-ブチル-Gly-Pro-D-Val、t-ブチル-Gly-Pro-Gly
、t-ブチル-Gly-Pro-Ile、t-ブチル-Gly-Pro-Ile-アミド、t-ブチル-Gly-Pro-t-ブチル-G
ly、t-ブチル-Gly-Pro-Val、Thr-Pro-Ile、Tic-Pro-Ile、Trp-Pro-Ile、Tyr(P)-Pro-Il
e、Tyr-Pro-allo-Ile、Val-Pro-allo-Ile、Val-Pro-t-ブチル-Gly、Val-Pro-Val、及びそ
の医薬として許容し得るその塩である。そのうち、t-ブチル-Glyは、下記式のように定義
し、かつ、Ser(Bzl)、及びSer(P)は、それぞれベンジル-セリン、及びホスホリル-セ
リンと定義する。Tyr(P)は、ホスホリル-チロシンと定義する。これらの化合物は、国
際公開第03/002593号に開示されている。
【0048】
【化1】

【0049】
本発明に従ってQCのエフェクターと組合せて使用でき、さらに好ましいDP IVインヒビ
ターは、ペプチジルケトンである。例を挙げると、下記化合物、及びその医薬として許容
し得るその塩である:
2-メチルカルボニル-1-N-[(L)-アラニル-(L)-バリニル]-(2S)-ピロリジン臭化水素
酸塩、2-メチル)カルボニル-1-N-[(L)-バリニル-(L)-プロリル-(L)-バリニル]-(
2S)-ピロリジン臭化水素酸塩、
-[(アセチル-オキシ-メチル)カルボニル]-1-N-[(L)-アラニル-(L)-バリニル]-(2S
)-ピロリジン臭化水素酸塩、
2-[ベンゾイル-オキシ-メチル)カルボニル]-1-N-[{(L)-アラニル}-(L)-バリニル]-
(2S)-ピロリジン臭化水素酸塩、
2-{[(2,6-ジクロロベンジル)チオメチル]カルボニル}-1-N-[{(L)-アラニル}-(L)-
バリニル]-(2S)-ピロリジン、
2-[ベンゾイ-ルオキシ-メチル)カルボニル]-1-N-[グリシル-(L)-バリニル]-(2S)-ピ
ロリジン臭化水素酸塩、
2-[([1,3]-チアゾールチアゾール-2-イル)カルボニル]-1-N-[{(L)-アラニル}-(L)-
バリニル]-(2S)-ピロリジントリフルオロ酢酸塩、
2-[(ベンゾチアゾールチアゾール-2-イル)カルボニル]-1-N-[N-{(L)-アラニル}-(L
)-バリニル]-(2S)-ピロリジントリフルオロ酢酸塩、
2-[(-ベンゾチアゾールチアゾール-2-イル)カルボニル]-1-N-[{(L)-アラニル}-グリ
シル]-(2S)-ピロリジントリフルオロ酢酸塩、及び
2-[(ピリジン-2-イル)カルボニル]-1-N-[N-{(L)-アラニル}-(L)-バリニル]-(2S)
-ピロリジントリフルオロ酢酸塩である。これらの化合物は、国際公開第03/033524号に
開示されている。
【0050】
さらに、本発明に従って、置換アミノケトンをQCエフェクターと組合せて使用するこ
とが可能である。好ましい置換ケトンを挙げると、
1-シクロペンチル-3-メチル-1-オクソ-2-ペンタンアミウムクロライド、
1-シクロペンチル-3-メチル-1-オクソ-2-ブタンアミウムクロライド、
1-シクロペンチル-3,3-ジメチル-1-オクソ-2-ブタンアミウムクロライド、
1-シクロヘキシル-3,3-ジメチル-1-オクソ-2-ブタンアミウムクロライド、
3-(シクロペンチルカルボニル)-1,2,3,4-テトラヒドロイソキノリニウムクロライド、
N-(2-シクロペンチル-2-オクソエチル)シクロヘキサンアミウムクロライド、及び
その医薬として許容し得るその塩がある。
【0051】
当初、希少なプロリン特異的プロテアーゼグループのうちで、DP IVは、唯一膜結合性
酵素で、ポリペプチド鎖のアミノ末端でその端から2番目残基のプロリンに特異性がある
と考えられていた。しかしながら、DP IVと構造的に相同性はないが、同様の酵素活性を
もつ、他の分子が同定された。今までに同定されたDP IV様酵素は、例えば、Sedo、及びM
alik(Sedo、及びMalikの論文、2001, Biochim Biophys Acta, 36506, 1-10)、及びAbbo
tt、及びGorrell(Abbott, C.A.、及びGorrell, M.Dの著書、エクトペプチダーゼ(Langn
e、及びAnsorge編集、Kluwer Academic/Plenum Publishers, New York中、171-195ペー
ジ))による総説に記載されている繊維芽活性化タンパク質α(fibroblast activation
protein α)、ジペプチジルペプチダーゼIV β、ジペプチジルアミノペプチダーゼ様タ
ンパク質、N-アセチル化α結合型酸性ジペプチダーゼ、休止細胞プロリンジペプチダーゼ
(quiescent cell proline dipeptidase)、ジペプチジルペプチダーゼII、アトラクチン
、及びジペプチジルペプチダーゼIV 関連タンパク質(DPP 8)、DPL1(DPX、DP6)、DPL2
、及びDPP 9 がある。最近、ジペプチジルペプチダーゼ10(DPP 10)のクローニング、及
び特性が報告された(Qi, S.Y.らの論文、Biochemical Journal Immediate Publication
において、2003年3月28日に原稿番号BJ20021914として発表)。
【0052】
本明細書中で使われる用語、エフェクターは、酵素に結合し、インビボ、及び/又はイ
ンビトロでその活性を上昇、又は低下させる分子として定義する。酵素によっては、その
触媒活性に影響する低分子の結合部位を有し;刺激促進分子はアクチベーターと呼ばれる
。酵素は1種以上のアクチベーター、又はインヒビターを認識するのための複数の結合部
位を有すことすらあり得る。酵素は様々な分子の濃度を感知し、かつその情報を使用し、
それ自身の活性を変えることが可能である。
【0053】
エフェクターが酵素活性を調整可能なのは、酵素が活性型と不活性型の両構造を呈する
からであり:アクチベーターは、正のエフェクターであり、インヒビターは負のエフェク
ターである。エフェクターは、酵素の活性部位においてだけでなく、調節部位、又はアロ
ステリック部位においても作用し、この用語は、調節部位が触媒部位から異なる酵素の要
素であることを強調し、触媒部位における基質、及びインヒビター間の競合による調節形
態と区別するために用いられる(Darnell, J.、Lodish, H.、及びBaltimore, D. の著書
、分子生物学第2版(1990, Molecular Cell Biology 2nd Edition, Scientific American
Books, New York)63ページ)。
【0054】
本発明のペプチドにおいて、各アミノ酸残基は、以下の慣用一覧表に従い、アミノ酸の
慣用名に相当する一文字、又は三文字表示で表す。
アミノ酸 一文字表記 三文字表記
アラニン A Ala
アルギニン R Arg
アスパラギン N Asn
アスパラギン酸 D Asp
システイン C Cys
グルタミン Q Gln
グルタミン酸 E Glu
グリシン G Gly
ヒスチジン H His
イソロイシン I Ile
ロイシン L Leu
リシン K Lys
メチオニン M Met
フェニルアラニン F Phe
プロリン P Pro
セリン S Ser
トレオニン T Thr
トリプトファン W Trp
チロシン Y Tyr
バリン V Val
【0055】
本明細書中で使用する用語“QC”とは、グルタミニルシクラーゼ(QC)、及びQC
様酵素を含む。さらに、QC、及びQC様酵素は同一の、又は同様の酵素活性を有し、Q
C活性と定義する。この点において、根本的に、QC様酵素は、分子構造上QCから異な
ることが可能である。
【0056】
本明細書中で使用する用語“QC活性”とは、アンモニア遊離下における、N末端グル
タミン残基のピログルタミン酸(pGlu*)への、又はN末端L-ホモグルタミン、又はL-β
ホモグルタミンの環状ピロ-ホモグルタミン誘導体への分子内環化反応と定義する。それ
については、スキーム1、及び2を参照されたい。
【0057】
スキーム1:QCによるグルタミンの環化反応
【化2】
【0058】
スキーム2:QCによるL-ホモグルタミンの環化反応
【化3】
【0059】
本明細書中で使用する用語“EC”とは、QC、及びQC様酵素のグルタミン酸シクラ
ーゼ(EC)ような副活性を含み、さらにEC活性として定義する。
本明細書中で使用する用語“EC活性”とは、QCによるN末端グルタミン酸残基の、
ピログルタミン酸(pGlu*)への分子間環化と定義する。それについては、スキーム3を
参照されたい。
本明細書中で使用する用語“金属依存性酵素”とは、その触媒機能を果たすために結合
した金属イオンとの結合を必要とする、かつ/又は、触媒活性の構造を形成するために結
合した金属イオンを必要とする酵素と定義する。
【0060】
酵素に結合して、かつその活性を上昇させたり、又は低下させる分子は、“エフェクタ
ー”と呼ばれる。エフェクターが酵素活性を調整可能なのは、酵素が活性型と不活性型の
両構造を呈するからであり:アクチベーターは、正のエフェクターであり;インヒビター
は負のエフェクターである。エフェクターは、調節部位、又はアロステリック部位(ギリ
シャ語の“もう一つの形”由来)において結合し、この用語は、調節部位が触媒部位から
異なる酵素の要素であることを強調し、触媒部位における基質、及びインヒビター間の競
合による調節形態と区別するために用いられる。本発明の個々の実施態様によって、アク
チベーター、又はインヒビターのどちらかが好まれる。
【0061】
スキーム3:QCによる、荷電していないグルタミルペプチドのN末端環化反応(EC)
【化4】
【0062】
本発明のもう一つの態様は、QCの新規の生理学的基質の同定である。これらは、実施
例5に記載した哺乳動物ペプチドでの環化反応実験を行い、同定した。それに先立ち、ヒ
トQC、及びパパイヤQCを実施例1に記載されるように精製した。該適用方法は実施例
2に記載し、かつ使用した該ペプチド合成は実施例6に概要を示す。該研究の結果は、表
1に示す。
【0063】
【表1】
【0064】
すべての分析は、実施例4に示されるように、ヒト、又は植物QCのいずれかの最適活
性、及び安定性の範囲で行った。N末端にグルタミン残基をもち、かつそれゆえQC酵素
の基質である、生理活性ペプチドのアミノ酸配列は表2に記載する:
【0065】
【表2】
【0066】
第四の実施態様において、[Gln1]ガストリン(長さ17、及び34アミノ酸)、[Gln1]ニュ
ーロテンシン、及び[Gln1]FPPペプチドを、QCの新規生理的基質として同定した。ガス
トリン、ニューロテンシン、及びFPPは、N末端にpGlu残基を含有する。すべてのペプチ
ドで、N末端pGlu残基が、QCによりN末端のグルタミンから産生されることを示した。
結果として、N末端グルタミン残基のpGluへの転換により、その生物学機能の点で活性化
される。
【0067】
経上皮変換細胞、特にガストリン(G)細胞は、胃への食物到来とともに胃酸分泌を調
節する。最近の研究により、複数の活性産物がガストリン前駆体より産生され、かつガス
トリン生合成には、複数のコントロール箇所があることが示された。生合成前駆体、及び
中間体(プロガストリン、及びGly-ガストリン)は、推定成長因子であり;その産物、ア
ミド化されたガストリンは、上皮細胞の増殖、酸産生壁細胞、及びヒスタミン分泌クロム
親和性様(ECL)細胞の分化、及びECL細胞でのヒスタミン合成、及び貯蔵に関わる遺伝子
の発現を、急性の酸分泌刺激促進と同様に調節する。また、ガストリンは、上皮成長因子
(EGF)ファミリーメンバーの産生を刺激促進し、順に壁細胞機能を阻害し、表面上皮細
胞の増殖を促進する。血漿ガストリン濃度は、ピロリ菌を保有し、十二指腸潰瘍、及び胃
癌のリスクが高いことが公知の個体で上昇する(Dockray, G.J.の論文、1999 J Physiol
15, 315-324)。
【0068】
管腔G細胞から放出されるペプチドホルモンガストリンは、CCK-2レセプターを介して、
酸分泌粘膜中のECL細胞由来のヒスタミン合成、及び放出を刺激促進することが知られて
いる。動員されたヒスタミンは、壁細胞にあるH(2)レセプターに結合することにより、
酸分泌を引き起こす。最近の研究により、ガストリンは、完全にアミド化された、及び、
プロセスが少ない型(プロガストリン、及びグリシン延長型ガストリン)の両方で、胃腸
管の成長因子であることが示唆された。アミド化されたガストリンの主要な栄養作用は、
胃の酸分泌粘膜に対してであり、胃幹細胞、及びECL細胞の細胞増殖の促進を起こし、壁
、及びECL細胞塊増大を生じることが確立している。一方、プロセスが少ないガストリン
(例えば、グリシン延長ガストリン)の主要栄養標的は、結腸粘膜であるように思われる
(Koh, T.J.、及びChen, D.の論文、2000 Regul Pept 93, 37-44)。
【0069】
第五の実施態様において、本発明は、胃腸管細胞増殖、特に、胃粘膜細胞及び上皮細胞
の増殖、酸産生壁細胞、及びヒスタミン分泌クロム親和性様(ECL)細胞の分化、及びECL
細胞内のヒスタミン生合成、及び貯蔵に関連した遺伝子の発現を刺激し、並びに、活性型
[pGlu1]-ガストリンの濃度を維持、又は上昇させることによる、哺乳動物における急性
の酸分泌をするために、QCの活性上昇エフェクターの使用を提供する。
第六の実施態様において、本発明は、不活性型[Gln1]のガストリンの活性型[pGlu1]ガ
ストリンへの転換率を低下させることによる、哺乳動物におけるピロリ菌を伴う、又は伴
なわない十二指腸潰瘍、及び胃癌の治療のために、QCの活性低下エフェクターの使用を
提供する。
【0070】
ニューロテンシン(NT)は、統合失調症の病態生理学に関わる、特に神経伝達物質シ
ステムを調整する神経ペプチドで、今までに、該疾患では調節障害が起こっていることが
示された。脳脊髄液(CSF)中のNT濃度を測定する臨床研究では、脳脊髄液(CSF)中の
NT濃度が低下している統合失調症患者の一部は、効果的な抗精神病剤治療で、回復する
ことを示した。また、抗精神病剤の作用機構におけるNTシステムの関与に一致した多数
の証拠がある。中枢系に投与されたNTの行動学的、及び生化学効果は、抗精神病剤を全
身に投与した該効果に著しく類似し、かつ抗精神病剤は、NT神経伝達を上昇させる。こ
の知見の連鎖から、NTが内在性の抗精神病剤として機能するという仮説が導かれた。さ
らに、典型、及び非典型抗精神病剤は、黒質線条体系、及び中脳辺縁系のドーパミン経路
の終末領域のNT神経伝達を異なった形で改質し、かつ該効果で、副作用障害、及び有効
性が予想可能である。
【0071】
第七の実施態様において、本発明は、抗精神病剤の調剤、及び/又は哺乳動物における
統合失調症治療のために、QCの活性上昇エフェクターの使用を提供する。該QCのエフ
ェクターは、活性型[pGlu1]ニューロテンシン濃度を維持、又は上昇させる。
【0072】
受精促進ペプチド(FPP)は、サイロトロピン放出ホルモン(TRH)に関連するトリペプ
チドであり、精漿に見い出される。インビトロ、及びインビボから得られた最近の証拠は
、FPPが精子受精能力の調整において重要な役割を果たしていることを示した。特に、FPP
は初めに受精能力のない(受精不能な)精子を“スイッチオン”にし、かつより速やかに
受精可能にするよう刺激促進し、その後、精子が自発アクロソーム損失を起こし、それゆ
え受精能力を喪失しないように、受精能獲得を抑制する。これらの反応は、アデニリルシ
クラーゼ(AC)/cAMPシグナル伝達経路を調節することが公知の、アデノシンによって模
倣され、かつ実際、増大される。FPP、及びアデノシンは両方とも、FPP受容体がどういう
わけかアデノシン受容体、及びGタンパク質と相互作用しACの調節をもたらし、受精不能
な細胞でcAMP産生を刺激促進し、受精可能な細胞ではそれを阻害することが示された。こ
れらの事象は、様々な細胞のチロシンリン酸化状態に影響し、初期の“スイッチオン”に
おいて重要なものもあれば、おそらく先体反応自身に関与しているものもある。また、精
漿に存在するカルシトニン、及びアンギオテンシンIIは、インビボで受精不能な精子に同
様の効果をもち、FPPの反応を増大し得る。これらの分子は、インビボで同様の効果をも
ち、受精能の刺激促進、及び維持により、受精に影響する。FPP、アデノシン、カルシト
ニン、及びアンギオテンシンIIの供給の低下、又はそのレセプターの異常は、男性不妊に
寄与する(Fraser, L.R.、及びAdeoya-Osiguwa, S. A.の論文、2001 Vitam Horm 63, 1-2
8)。
【0073】
第八の実施態様において、本発明は、哺乳動物における受精抑制薬の調剤、及び/又は
、受精の低下のための、QCの活性低下エフェクターの使用を提供する。該QCの活性低
下エフェクターは、活性型[pGlu1]FPP濃度を低下させ、精子の受精能獲得を抑制、及び精
子細胞の不活性化をもたらす。これに対し、QCの活性上昇エフェクターにより、男性生
殖能力を刺激を促進し、かつ不妊を治療することが可能であることが示され得る。
【0074】
第九の実施態様において、さらなるQCの生理学的基質が、本発明の範囲内で同定され
た。これらは、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 18
、及び[Gln1]フラクタルカインである。詳細は、表2を参照されたい。これらのポリペプ
チドは、骨髄性前駆細胞増殖の抑制、新生組織形成、炎症性宿主反応、癌、乾癬、関節リ
ウマチ、アステローム性動脈硬化、体液性及び細胞性免疫反応、内皮における白血球接着
及び遊走プロセスのような病態生理学的状態において、重要な役割を果たす。
【0075】
最近、B型肝炎、HIV、及びメラノーマに対する、いくつかの細胞傷害性Tリンパ球ペ
プチドを基にしたワクチンが、臨床試験で研究されている。デカペプチドELAは、単独で
、或いは腫瘍抗原と組合せる、1種の興味深いメラノーマワクチン候補である。このペプ
チドは、N末端グルタミン酸をもつ、Melan-A/MART-1抗原免疫優勢ペプチドアナログで
ある。グルタミンのアミノ基、及びガンマカルボン酸基は、グルタミン酸のアミノ基、及
びガンマカルボキサミド基と同様に、容易に縮合し、ピログルタミン誘導体を産生する。
医薬上興味深いいくつかのペプチドは、N末端グルタミン、及びグルタミン酸の代わりに
ピログルタミン誘導体で開発し、医薬上の特性を損失することなくこの安定性の問題を克
服した。残念なことに、ELAに比べ、該ピログルタミン誘導体(PyrELA)、及びN末端ア
セチル修飾誘導体(AcELA)は、細胞傷害性Tリンパ球(CTL)の活性を引き出せなかった
。PyrELA、及びAcELAにおける外見上の些少な修飾にかかわらず、おそらく該二誘導体は
該特異クラスI主要組織適合複合体に対して、ELAより低い親和性を示す。それゆえ、ELA
の最大の活性を維持するために、PyrELAの産生を回避しなければならない(Beck A.らの
論文、2001, J Pept Res 57(6):528-38)。また、最近、メラノーマにおいて、グルタ
ミニルシクラーゼ(QC)酵素が過剰発現していることが見い出された(Ross D. T.らの
論文、2000, Nat Genet 24:227-35)。
【0076】
第十の実施態様において、本発明は、骨髄性前駆細胞増殖の抑制、新生組織形成、炎症
性宿主反応、癌、悪性転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化
、体液性及び細胞性免疫反応障害、内皮における白血球接着及び遊走プロセスのような病
態生理学的状態の治療に用いる薬剤の製造のための、QCのエフェクターの使用を提供す
る。
【0077】
第十一の実施態様において、[Gln1]オレキシンAは、本発明の範囲内で、QCの生理学
的基質と同定した。摂食、睡眠覚醒の調整に有意な影響を及ぼす神経ペプチドであり、お
そらく、これらの相補的な恒常性機能の複雑な行動、及び生理学的応答を調整している。
また、それは、エネルギー代謝の恒常性調節、自律機能、ホルモンバランス、及び体液調
節において影響を及ぼす。
第十二の実施態様において、本発明は、摂食障害、及び睡眠覚醒、エネルギー代謝の恒
常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害の治療に用いる
薬剤の製造のための、QCのエフェクターの使用を提供する。
【0078】
様々なタンパク質におけるプログルタミン伸長は、パーキンソン病、及びケネディ病の
ような神経変性疾患を生じる。それゆえ、該機構は、ほとんどわかっていない。ポリグル
タミン繰り返し配列の生化学的特性は、1つの考えられる説明を示唆し:グルタミニル-
グルタミニル結合の切断(endolytic cleavage)後の、ピログルタミン酸産生が、ポリグ
ルタミンタンパク質の異化安定性、疎水性、アミロイド凝集性、及び神経傷害性に寄与し
得るということである(Saido, Tの論文、Med Hypotheses(2000)Mar;54(3):427-9
)。したがって、第十三の実施態様において、本発明は、パーキンソン病、及びハンチン
トン病の治療に用いる薬剤の製造のための、QCのエフェクターの使用を提供する。
【0079】
第十四の実施態様において、本発明は、QCの酵素活性を低下、又は阻害させる一般的
な方法を提供する。また、阻害化合物の例を提供する。
当初、哺乳動物QCの阻害は、1,10-フェナントロリン、及び還元型 6-メチルプテリン
でのみ検出された(Busby, W. H. J.らの論文、1987 J Biol Chem 262, 8532-8536)。ED
TAはQCを阻害せず、故にQCは金属依存性酵素ではないと結論された(Busby, W. H. J
.らの論文、1987 J Biol Chem 262, 8532-8536, Bateman, R.C.J.らの論文、2001 Bioche
mistry 40, 11246-11250、Booth, R.E.らの論文、2004 BMC Biology 2)。しかしながら
、本発明では、1,10-フェナントロリン、ジピコリン酸、8-ヒドロキシ-キノリン、及び他
のキレーターによるQCの阻害特性(図18、19)、及び遷移金属イオンによるQCの
再活性化(図20)によって明らかであるように、ヒトQC、及び他の動物QCが金属依
存性酵素であることを示した。最後に、該金属依存性を他の金属依存性酵素との配列比較
をまとめ、ヒトQCにも、キレートするアミノ酸残基が保存されていることを示した(図
21)。化合物の、金属イオン結合部位との相互作用は、QC活性を低下、又は阻害する
一般的な方法である。
【0080】
本発明では、イミダゾール誘導体がQCの有力なインヒビターであることを示す。連続
分析(詳細は実施例2参照)を用い、多数のイミダゾール誘導体を、高度に保存された哺
乳動物QCの1種であるヒトQCを阻害する能力に関して分析した。
このように、本発明は、QCの活性低下エフェクターとして、イミダゾール誘導体、及
びヒスチジン、及びその誘導体、及びその特性を阻害型、及び有効性の面から提供する。
構造、及びKi値を、表3、及び4に示す。その結果は、実施例7に詳しく記載した。
【0081】
【表3】
【0082】
【表4】
【0083】
また、驚いたことに、酵素活性の解析中、N末端グルタミニル残基以外にN末端β-ホ
モグルタミル残基が、植物、又は動物由来QCの基質としての特性を満たすことを発見し
た。該N末端β-ホモグルタミル残基は、ヒト、及びパパイヤ由来QCにより、それぞれ
五員のラクタム環に転換された。該結果を、実施例5に示した。適用した方法は実施例2
に示し、ペプチド合成は、実施例6に記載したように行った。
本発明のもう一つの好ましい実施態様は、QCのエフェクターのスクリーニング方法を
含む。
【0084】
化合物群からQCの活性を調整するエフェクターを同定する、好ましいスクリーニング
方法は、下記工程を含む前記方法である:
a)前記の化合物とQCとを、それらの間で結合が可能な条件下で接触させること;
b)QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残存QC活性を測定すること;及び
d)QCの該基質転換、及び/又は、酵素活性の変化を計算し、QCの活性調整フェクタ
ーを同定することである。
【0085】
もう一つの好ましいスクリーニング方法は、QCの金属イオン結合活性部位と、直接、
又は間接的に相互作用するエフェクターの同定、及び選定に用いる方法に関連し、下記の
工程を含む方法である:
a)前記の化合物とQCとを、それらの間で結合が可能な条件下で接触させること;
b)QCにより転換可能な、QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残存QC活性を測定すること;及び
d)QCの該基質転換、及び/又は酵素活性の変化を計算し、QCの活性調整エフェクタ
ーを同定することである。
【0086】
前記スクリーニング方法での使用が好ましいのは、哺乳動物QC、又はパパイヤQCで
ある。特に好ましいのは哺乳動物QCであり、それはこれらのスクリーニング方法で同定
したエフェクターが哺乳動物、特にヒトの疾患の治療に使用され得るからである。
前記スクリーニング方法で選定された薬剤は、少なくとも1種のQC基質の転換を低下
させる(負のエフェクター、インヒビター)、又は、少なくとも1種のQC基質の転換を
増加させる(正のエフェクター、アクチベーター)ことにより機能することが可能である
。
本発明の化合物は、酸付加塩、特に医薬として許容し得る酸付加塩へ転換することがで
きる。
該発明の化合物の該塩は、無機、又は有機の形態とし得る。
【0087】
本発明の該化合物は、転換し、酸付加塩、特に医薬として許容し得る酸付加塩として使
用することが可能である。医薬として許容し得る塩は、一般に、塩基性側鎖が無機、又は
有機酸でプロトン化されているような形態をとる。典型的な有機、又は無機酸は、塩酸、
臭化水素酸、過塩素酸、硫酸、硝酸、リン酸、酢酸、プロピオン酸、グリコール酸、乳酸
、コハク酸、マレイン酸、フマル酸、リンゴ酸、酒石酸、クエン酸、安息香酸、マンデル
酸、メタンスルホン酸、ヒドロキシエタンスルホン酸、ベンゼンスルホン酸、シュウ酸、
パモ酸、2-ナフタレンスルホン酸、p-トルエンスルホン酸、サイクラミン酸、サリチル酸
、サッカリン酸、又はトリフルオロ酢酸を含む。すべての本発明の該化合物の医薬として
許容し得る塩は、本発明の範囲内に取り込むことを意図している。
【0088】
化合物自身と、塩の形態の該化合物の密接な関係から、本明細書の文脈で化合物に言及
するときは常に、その状況下でそれが考え得るか、又は適切であるならば、相当する塩も
対象とする。
本発明の該化合物が少なくとも1種の不斉中心を持つ場合は、それ相応に光学異性体が
存在し得る。該化合物が2種以上の不斉中心を持つ場合は、さらにジアステレオマーが存
在し得る。その異性体、及び混合物のすべては、本発明の範囲内に含まれると理解すべき
である。さらに、該化合物の結晶性形態は多形体として存在し得て、それらはさらに、本
発明に含まれる対象である。該化合物によっては、水(すなわち、水和物)、又は慣用の
有機溶媒との溶媒和化合物を形成し得る。
また、その塩を含む該化合物は、水和物の形態で調達することが可能であるか、或いは
その結晶化に用いる他の溶媒を含む。
【0089】
さらなる実施態様において、本発明は、必要としている個体における、QC酵素活性の
調整を介した疾患状態の予防、又は治療法を提供し、それは、本発明の該化合物、又はそ
の医薬組成物のいずれかを、治療上効果的な量、及び投与法で投与し、病態を治療するこ
とを含む。さらに、本発明は、個体におけるQC活性調整を介した病態の予防、又は治療
に用いる薬剤の調製のための、この発明における化合物、及びそれに相当する医薬として
許容される酸付加塩の形態の使用を含む。該化合物は、静脈注射、経口、皮下注射、筋肉
注射、皮内注射、非経口、及びその組み合わせ含むが、これに限定されない、慣用の投与
経路で患者に投与され得る。
さらなる好ましい実施態様において、本発明は、少なくとも1種の本発明の化合物、又
はその塩を、任意に、少なくとも1種の医薬として許容し得る担体、及び/又は溶媒と組
合せて含有する医薬組成物、すなわち、薬剤に関するものである。
【0090】
例えば、医薬組成物は、非経口、又は経腸製剤の形態であり得て、適切な担体を含むか
、又は経口投与に適した適切な担体を含む、経口製剤の形態である。好ましくは、経口製
剤の形態である。
本発明の記載に従って投与されるQC活性のエフェクターは、インヒビター、又はイン
ヒビター、基質、擬似基質、QC発現のインヒビター、哺乳動物のQCタンパク質濃度を
低下させる酵素の結合タンパク質、又は抗体と組合せて、医薬上投与できる製剤、又は製
剤複合物の形で用いられる。本発明の該化合物により、患者、及び疾患個々に治療を調節
することが可能になり、特に、個々の不耐性、アレルギー、及び副作用を回避することが
可能である。
【0091】
また、該化合物は、時間の関数として、異なる活性の程度を示す。それによって、治療
を施す医師は、患者の個々の状況に応じて対処する機会を与えられ:医師は、一方では作
用開始の速度、他方では、作用の持続期間、及び特に作用強度を正確に調節することが可
能である。
本発明の好ましい治療方法は、哺乳動物のQC酵素活性の調整を介した病態の予防、又
は治療に用いる新規のアプローチを示す。それは、有利に単純で、商業的適用の余地があ
り、特に、哺乳動物、及び特にヒト医薬において、例えば、表1、及び2に記載された、
生理活性QC基質のアンバランスに基づく疾患の治療における使用に適している。
【0092】
例えば、該化合物は、有効成分を希釈剤、賦形剤、及び/又は、先行技術から公知の担
体のような添加剤を組合せて含む調剤の形態で、有利に投与し得る。例えば、それらは、
非経口的(例えば、生理食塩水の点滴で)、又は経腸的に(例えば、慣用の担体と処方さ
れ、経口的に)投与することが可能である。
その内生安定性、及びその生体利用効率に依存して、該化合物は、所望する血糖グルコ
ース値の正常化を達成するために、1日に1回、又は複数回投与することが可能である。
例えば、ヒトにおけるそのような用量域は、1日あたり約0.01 mgから250.0 mgの範囲で
、好ましくは、体重キログラムあたり化合物約0.01 mgから100 mgの範囲であり得る。
【0093】
QC活性のエフェクターを哺乳動物に投与することによって、アルツハイマー病、ダウ
ン症、ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は伴わない、潰瘍性疾患及び胃
癌、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、乾
癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性免疫反応障害、内皮にお
ける白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネルギー代謝の恒常性調節障
害、自律機能障害、ホルモンバランス障害、及び体液調節障害から、選択される病態を、
予防、又は緩和、又は治療し得る。
【0094】
さらに、QC活性のエフェクターを哺乳動物に投与することによって、胃腸管細胞増殖
、好ましくは、胃粘膜細胞、上皮細胞の増殖、急性の酸分泌、及び酸産生壁細胞、及びヒ
スタミン分泌クロム親和性様細胞分化を刺激、促進し得る。
さらに、哺乳動物へのQCインヒビターの投与は、精子細胞機能の喪失、したがって男
性生殖能力の抑制を可能にし得る。従って、本発明は、男性生殖能力の調整、及びコント
ロールに用いる方法、及び男性用避妊薬剤の製造のための、QCの活性低下エフェクター
の使用を提供する。
さらに、QC活性のエフェクターを哺乳動物への投与することにより、骨髄性前駆細胞
増殖の抑制を可能にし得る。
【0095】
本発明で使用される化合物は、本来の公知の方法で、例えば、不活性無毒の、医薬上適
切な担体、及び添加剤、又は溶媒を用いて、錠剤、カプセル剤、糖衣剤、丸剤、顆粒剤、
噴霧剤、シロップ剤、液剤、固形剤、及びクリーム状乳剤、及び懸濁剤、及び溶液剤のよ
うな慣用調剤形態へ、それ相応に転換することが可能である。その各々の調剤形態におい
て、該医薬有効化合物は、好ましくは、全混合物の重量に対して約0.1〜80%、より好ま
しくは、1〜50%の濃度で、すなわち、言及した投薬量域を得るのに十分な量で存在する
。
【0096】
また、該物質は、糖衣剤、カプセル剤、可噛性カプセル剤、錠剤、ドロップ剤、シロッ
プ剤の形態の薬剤として、又は座薬として、又は鼻腔用スプレーとして使用することがで
きる。
例えば、該調剤形態は、有効成分を溶媒、及び/又は担体と共に、任意に乳化剤、及び
/又は分散剤の使用と共に供与され、有利に製造することができ、例えば、水が希釈剤と
して使用される場合は、有機溶媒は、任意に補助溶剤として使用することが可能である。
【0097】
本発明に関わる有用な賦形剤の例を挙げると:水、パラフィン(例えば天然油画分)、
植物油(例えば、ナタネ油、ラッカセイ油、ゴマ油)、グリコール(例えば、プロピレン
グリコール、ポリエチレングリコール)のような無毒性有機溶媒;例えば、天然粉末ミネ
ラル(例えば、抗分散シリカ、ケイ酸塩)、糖類(例えば、粗糖、ラクトース、デキスト
ロース)のような固形担体;非イオン性、及び陰イオン性乳化剤(例えば、ポリオキシエ
チレン脂肪酸エステル、ポリオキシエチレン脂肪アルコールエステル、アルキルスルホン
酸、アリールスルホン酸)のような乳化剤、分散剤(例えば、リグニン、亜硫酸溶液、メ
チルセルロース、澱粉、及びポリビニルピロリドン)、潤滑剤(例えば、ステアリン酸マ
グネシウム、滑石粉、ステアリン酸、及びラウリル硫酸ナトリウム)、及び任意の香料が
ある。
【0098】
投与は、従来の方法で、好ましくは経腸的、又は非経口的に、特に経口的に行われ得る
。経腸投与の場合に、錠剤は上記の担体に加えて、さらにクエン酸ナトリウム、炭酸カル
シウム、及びリン酸カルシウムのような添加剤を、澱粉、好ましくは片栗粉、ゼラチン、
及びそのような多様な添加剤と共に含有し得る。さらに、ステアリン酸マグネイウム、ラ
ウリル硫酸ナトリウム、及び滑石粉のような潤滑剤は、錠剤製造に付随して使用すること
が可能である。経口投与を目的とした水性懸濁液、及び/又はエリキシル剤の場合は、多
様な矯味料、又は着色料が、上記記載の賦形剤に加えて、有効成分に付加することが可能
である。
【0099】
非経口投与の場合、適切な液体担体を用いた有効成分の溶液を使用することができる。
一般に、静脈注射の場合、一日当たり体重に対して約0.01〜2.0 mg/kg、好ましくは、0.
01〜1.0 mg/kgの量を投与すると、効果的な結果が得られる利点が見いだされ、かつ経腸
投与の場合、該投与量は一日当たり体重に対して約0.01〜2 mg/kg、好ましくは、0.01〜
1 mg/kgである。
しかしながら、実験動物、及び患者の体重、又は投与経路の種類だげでなく、動物の種
類、又は薬剤に対する個々の反応、又は投与が行われる間隔などによって、該記載量から
外れることが不可欠である場合もあり得る。したがって、上記記載の最低量より少ない使
用で十分な場合もある一方、記載の上限を超過しなければならない場合もある。比較的大
量に投与される場合は、該投与量を一日に複数回の投薬に分割することが望ましいかもし
れない。ヒト薬剤の投与には、同様の投与許容度を提供する。その場合、上記の所見が同
様に適用される。
【0100】
(医薬調剤形態の例)
1.カプセル剤当り、本発明の化合物100 mgを含有するカプセル剤
およそ10,000カプセル剤に対して、下記組成の溶液を調製する:
本発明の化合物 1.0 kg
グリセロール 0.5 kg
ポリエチレングリコール 3.0 kg
水 5.0 kg
該溶液は、本質的に公知の方法で、軟ゼラチンカプセルに封入する。該カプセルは、咀
嚼、及び嚥下に適している。
【0101】
2.本発明の化合物100 mgを含有する錠剤、又はコーティングされた錠剤、又は糖衣剤
下記の量は錠剤100,000粒の製造に関するものである:
細かく粉砕した本発明の化合物 10.0 kg
グルコース 4.35 kg
ラクトース 4.35 kg
澱粉 4.50 kg
細かく粉砕したセルロース 4.50 kg
【0102】
上記組成物を、混合し、下記成分から製造される溶液と共に提供し、該湿塊をおろし、
かつステアリン酸マグネシウム0.2 kg付加後乾燥させる、本質的に公知の方法で粒状にさ
れる。
ポリビニルピロリドン 2.0 kg
ポリソルベート 0.1 kg
及び水 約5.0 kg
【0103】
最終錠剤混合物30.0 kgを加工し、300 mgの重さの突型錠剤を形成する。理想的には、
該錠剤を、本質的に公知の方法でコーティング、又は糖コーティングすることが可能であ
ることである。
有利には、本明細書中で定義された医薬組成物は、少なくとも1種のQC活性のエフェ
クター、及び少なくとも1種のDP IVインヒビターを含有する。そのような医薬組成物は、
特にアルツハイマー病、及びダウン症の治療に有用である。
【実施例】
【0104】
(実施例1):ヒト、及びパパイヤQCの調製法
(宿主株、及び培地)
ヒトQCの発現に使用される酵母(Pichia pastoris)X33株(AOX1、AOX2)を、製造会
社の取扱説明書(Invitrogen社)に従って、培養し、形質転換し、かつ解析した。酵母(
P.pastoris)が必要とする培地、例えば、緩衝化したグリセロール(BMGY)複合体、又は
メタノール(BMMY)複合体培地、及び、発酵基礎塩培地は、製造会社の推奨にしたがって
調製した。
【0105】
(ヒトQCをコードするプラスミドベクターの分子クローニング)
すべてのクローニング手順は、標準的分子生物学的技法を適用して行なった。酵母にお
ける発現のために、ベクターpPICZαB(Invitrogen社)を使用した。pQE-31ベクター(Qi
agen社)を使用し、ヒトQCを大腸菌において発現させた。第38番目のコドンより始ま
る成熟型QCのcDNAと、6xヒスチジン標識をコードするプラスミドとを、読み枠内で融合
した。pQCyc-1、及びpQCyc-2プライマー(表1)を使用し、増幅、かつサブクローニング
の後、該フラグメントを、SphI及び、HindIIIの制限酵素切断部位を用いて、発現ベクタ
ーに挿入した。
【0106】
(P. pastorisの形質転換、及び小量発現実験)
プラスミドDNAは、大腸菌JM109内で増幅され、製造会社(Qiagen社)の推奨に従って精
製した。使用した発現プラスミドpPICZαBにおいては、3つの直線化に用いる制限酵素切
断部位が提供される。SacI、及びBstXIは、該QCcDNAの内部で切断するため、PmeIを選
択し、直線化した。20〜30μgのプラスミドDNAをPmeIで切断し、エタノールにより沈澱さ
せ、滅菌脱イオン水に溶かした。その後、10μgのDNAを、製造会社(BioRad社)の取扱説
明書に従って、電気穿孔法による酵母(P.pastoris)コンピテント細胞の形質転換に使用
した。選択は、150μg/ml Zeocinを含むプレートにおいて行なった。直線化されたプラ
スミドを使用した一回の形質転換実験から、数百の形質転換体を得た。
【0107】
組み換え酵母クローンをQCの発現に関して調べるために、組み換え体を、2 mlのBMGY
培地を含む10 mlコニカルチューブにおいて、24時間培養した。その後、該酵母を遠心
分離し、0.5%メタノールを含む2 mlのBMMYに再懸濁した。この濃度は、72時間まで、
24時間毎にメタノールを加えることによって維持された。その後、上清のQC活性が決
定した。融合タンパク質の存在を、6xヒスチジン標識に対する抗体(Qiagen社)を使用し
たウェスタンブロット解析によって確認した。最も高いQC活性を示した酵母のクローン
を、さらなる実験、及び発酵のために選択した。
【0108】
(発酵槽における大量発現実験)
QCの発現は、5 l(リットル)のリアクター(Biostat B,B.Braun biotech社)におい
て、基本的に"酵母Pichia発酵操作ガイドライン(Pichia fermentation process guideli
nes)"(Invitrogen社)に記載されているように行なった。簡潔に述べると、酵母細胞は
、微量の塩を添加し、かつグリセロールを唯一の炭素源とする、発酵基礎塩培地(pH 5.5
)において培養した。約24時間の初期バッチ段階、及びその後の約5時間のフェドバッ
チ(fed-batch)段階の間、酵母細胞塊が蓄積された。細胞湿重量が200 g/lに達した時
点で、約60時間の全発酵時間に三段階の栄養添加プロフィールを適用し、メタノールを
用いたQC発現の誘導を行なった。その後、細胞を、6000x g、4℃における15分間の遠
心によって、QC含有上清から除いた。そのpHは、NaOHの添加により、6.8に調整し、そ
の結果生じた懸濁溶液は、再び、37000x g、4℃において、40分間遠心した。遠心後も
液の濁りが継続した場合は、セルロース膜(孔径0.45μm)を用いた、さらなる濾過工程
を適用した。
【0109】
(P. pastorisにおいて発現された6xヒスチジン標識QCの精製)
最初に、His標識QCは、固定化金属アフィニティークロマトグラフィー(IMAC)によ
って精製した。典型的な精製においては、1000 mlの培養上清を、750 mMのNaClを含む、5
0 mMのリン酸緩衝液(pH 6.8)で平衡化したNi2+結合キレーティングセファロースFFカラ
ム(Chelating Sepharose FF column)(1.6 cm x 20 cm、Pharmacia社)に、流速5 ml/
分でかけた。10カラム容量の平衡化緩衝液、及び5カラム容量の5 mMヒスチジンを含む
平衡化緩衝液で洗浄した後、150 mM NaCl、及び100 mMヒスチジンを含む50 mMリン酸緩衝
液(pH6.8)へ変更することによって、結合したタンパク質を溶出した。その結果生じた
溶出液を、20 mMのビス-トリス/HCl緩衝液(pH 6.8)に対し、4℃において一晩透析し
た。その後さらに、該QCは、透析緩衝液によって平衡化されたMono Q6カラム(BioRad
社)における陰イオン交換クロマトグラフィーにより、精製した。QC含有画分は、流速
5 ml/分で、Mono Q6カラムへかけた。その後、該Mono Q6カラムを、100 mMのNaClを含む
平衡化緩衝液で洗浄した。溶出は、240 mM、及び360 mMのNaClを含む平衡化緩衝液を、各
々、30カラム容量、又は5カラム容量もたらす、2通りの濃度勾配によって行なった。
6 mlずつの画分を集め、各画分のタンパク質の純度を、SDS-PAGEにより解析した。均質の
QC含有画分を集め、限外濾過により濃縮した。長期保存(-20℃)のため、グルセロー
ルを最終濃度50%まで加えた。タンパク質は、Bradford、又は、Gill、及びvon Hippelの
方法に従い、定量した(Bradford, M. M.の論文、1976 Anal Biochem 72, 248-254;Gill
, S.C.、及びvon Hippel, P.H.の論文、1989 Anal Biochem 182, 319-326)。
【0110】
(大腸菌におけるQCの発現と精製)
QCをコードするコンストラクトで、M15細胞(Qiagen社)を形質転換し、選択的LB寒
天プレートにおいて、37℃で培養した。タンパク質の発現は、1%グルコース、及び1%の
エタノールを含むLB培地において、室温にて行なった。菌培養液が、OD600約0.8に達した
とき、0.1 mMのIPTGにより、発現を一晩誘導した。一回の凍結溶融のサイクルの後、300
mMのNaCl、及び2mMのヒスチジンを含む、50 mMのリン酸緩衝液、pH 8.0中の、2.5 mg/ml
リゾチーム溶液の添加により、4℃にて、約30分間、細胞を溶解した。該溶液は、37000x
g、4℃、30分間の遠心分離によって、不純物を除去し、その後、ガラスフリットを用いた
濾過(DNAの分離)、及び粗い沈殿物、及び細かい沈殿物用のセルロース・フィルターを
用いた、さらなる2段階の濾過工程を行なった。その上清(約500 ml)は、Ni2+アフィニ
ティーカラム(1.6 x 20 cm)に、流速毎分1 mlでかけた。QCの溶出は、150 mMの塩化
ナトリム、及び100 mMのヒスチジンを含む50 mMリン酸緩衝液によって行なった。QC含
有画分は、限外濾過により濃縮した。
【0111】
(パパイヤラテックス由来QCの精製)
パパイヤラテックス由来QCは、BioCAD 700E(Perseptive Biosystems、Wiesbaden社
、ドイツ)を用いて、以前に報告された方法(Zerhouni, S.らの論文、1989 Biochim Bio
phys Acta 138, 275-290)の修正版で調製した。50 gのラテックスを水に溶解し、その中
に記載されているように遠心した。プロテアーゼの不活化は、S-メチルメタンチオスルフ
ォネートによって行い、その結果得られた粗抽出液を、透析した。透析後、上清全体は、
100 mMの酢酸ナトリウム緩衝液、pH 5.0で平衡化したSPセファロースファーストフロウ(
Fast Flow)カラム(21 x内径2.5 cm)にかけた(流速3 ml/分)。溶出は、流速2 ml/
分で、酢酸ナトリム緩衝液の濃度を上げていく3工程において行なった。第一工程は、0.
5カラム容量の、0.1 Mから0.5 Mへの酢酸ナトリム緩衝液濃度の直線勾配による溶出であ
った。第二工程は、4カラム容量の、0.5 Mから0.68 Mへの酢酸ナトリム緩衝液濃度の直線
勾配による溶出であった。そして、最終溶出工程の間は、1カラム容量の、0.85 Mの緩衝
液を用いた。最も高い酵素活性を含む画分(6 ml)を溜めた。濃縮と、0.02 Mトリス/HC
l緩衝液(pH 8.0)への緩衝液交換は、限外濾過(アミコン(Amicon);透過分子量10 kD
aの膜を使用)により行なった。
【0112】
イオン交換クロマトグラフィー工程から得られた、濃縮されたパパイヤの酵素に、最終
濃度2 Mになるように硫酸アンモニウムを加えた。この溶液を、2 M硫酸アンモニウムを含
む0.02 Mのトリス/HCl溶液(pH8.0)によって平衡化されたブチルセファロース4 ファー
ストフロウ(Fast Flow)カラム(21 x内径2.5 cm)にかけた(流速1.3 ml/分)。溶出
は、硫酸アンモニウム濃度を下げて行く3工程で行なった。第一工程の間は、2 Mから0.6
Mへの硫酸アンモニウムの直線勾配を、0.5カラム容量の0.02 Mトリス/HCl溶液(pH 8.0
)で用いた(流速1.3 ml/分)。第二工程は、0.6 Mから0 M硫酸アンモニウムの直線勾配
を5カラム容量の0.02 Mのトリス/HCl溶液(pH8.0)により、流速毎分1.5 mlでの溶出で
あった。0.02 Mのトリス/HCl溶液(pH 8.0)を、流速1.5 ml/分で用いることにより、
最終溶出工程を行なった。すべてのQC活性を含む画分を溜め、限外濾過により濃縮した
。その結果得られた均質のQCは、-70℃に保存した。最終タンパク質濃度は、Bradford
の方法を用い、ウシ血清アルブミンで得られた標準曲線と比較して決定した。
【0113】
(実施例2):グルタミニルシクラーゼ活性の測定
(蛍光測定法)
すべての測定は、30℃で、マイクロプレート用のバイオアッセイリーダーHTS-7000プラ
ス(BioAssay Reader HTS-7000Plus)(Perkin Elmer社)で行った。QC活性は、H-Gln-
βNAを用い、蛍光測定的に評価した。試料は、20 mM EDTAを含む0.2 Mトリス/HCl溶液(
pH 8.0)中の0.2 mM蛍光基質、0.25 Uピログルタミニルアミノペプチダーゼ(Unizyme, H
orsholm社、Denmark)、及び適切に希釈されたQCの分注液を、最終容量250μlに調整し
た。励起/発光波長は、320/410 nmであった。測定反応は、グルタミニルシクラーゼの
添加により開始した。QC活性は、測定条件下のβ-ナフチルアミンの標準曲線から決定
した。1ユニットは、記載された条件下で、1分当たりH-Gln-βNAから1μmol pGlu-βNA
の産生を触媒するQCの量として定義した。
第二の蛍光測定法において、QCの活性は、H-Gln-AMCを基質に用いて決定した。反応
は、ノボスター(NOVOStar)マイクロプレートリーダー(BMG labtechnologies社)を用
い、30℃で行った。試料は、5 mMのEDTAを含む0.05 Mトリス/HCl溶液(pH 8.0)中の多
様な濃度の該蛍光基質、0.1 Uピログルタミルアミノペプチダーゼ(Qiagen社)、及び適
切に希釈されたQCの分注液を、最終容量を250μlに調整した。励起/発光波長は、380
/460 nmであった。測定反応は、グルタミニルシクラーゼの添加により開始した。QC活
性は、測定条件下の7-アミノ-4-メチルクマリンの標準曲線から決定した。速度論的デー
タは、グラフィト(GraFit)ソフトウェアを用いて査定した。
【0114】
(QCの分光学的測定法)
この新規測定法を使い、ほとんどのQC基質の速度論的パラメータを決定した。QC活
性は、グルタミン酸デヒドロゲナーゼを補助酵素として使用した、以前の非連続測定法(
Bateman, R. C. J.の論文、1989 J Neurosci Methods 30, 23-28)を適応させることによ
り得られた、連続法を使って、分光光学的に分析した。最終容量250μl中の試料は、各Q
C基質、0.3 mMの NADH、14 mMのα-ケトグルタル酸、及び30 U/mlのグルタミン酸デヒ
ドロゲナーゼから成った。測定反応は、グルタミニルシクラーゼの添加により開始し、34
0 nmでの吸光度の減少を8〜15分間モニターすることにより遂行した。産物産生の典型的
経時変化を、図1に示す。
初期の速度を測定し、測定条件下のアンモニアの標準曲線から、該酵素活性を決定した
。全試料は、30℃でスペクトラフルオープラス(SPECTRAFluor Plus)、又はサンライズ
(Sunrise)(両者ともTECAN社)マイクロプレートリーダーを用い、測定した。速度論的
データは、グラフィト(GraFit)ソフトウェアを用いて査定した。
【0115】
(インヒビター測定)
インヒビター試験において、試料組成は、推定上のインヒビター化合物を添加加した以
外は、前記のものと同様であった。短時間QC阻害試験では、試料は4 mMの各インヒビタ
ーを含み、かつ1 KMの基質濃度であった。阻害の詳細な研究、及びKi値の決定においては
、最初に、インヒビターの補助酵素における影響を調べた。どの場合も、どちらかの酵素
に対する影響は検出されず、従って、信頼性のあるQC阻害測定を可能にした。阻害定数
は、グラフィト(GraFit)ソフトウェアを用いて、該時間経過の曲線を競合阻害の一般式
に当てはめることにより決定した。
【0116】
(実施例3):MALDI-TOF質量分析
ヒューレットパッカード(Hewlett-Packard)G2025 LD-TOFシステムをリニア飛行時間
型分析計と共に用いて、マトリックス支援イオン化−飛行時間型質量分析を行なった。機
器には、337 nm窒素レーザー、加速電位源(5)、及び1.0 mの飛行管を装着した。検出器
操作は、陽イオンモードで、かつシグナルはパーソナルコンピューターに接続したレクロ
イ9350 Mデジタルストレージオシロスコープ(LeCroy 9350M digital storage oscillosc
ope)で記録、及びフィルターをかけた。試料(5μl)は、同量のマトリックス溶液と混
合した。マトリックス溶液には、1 mlアセトニトリル/0.1% TFA水溶液(1/1、v/v)
に、30 mgの2',6'-ジヒドロキシアセトフェノン(Aldrich社)、及び44 mgのクエン酸水
素二アンモニウム(Fluka社)を溶解して調製したDHAP/DAHCを用いた。少量(≒1μl)
のマトリックス−検体混合物は、プローブ先端に移動させ、かつ速やかに真空管(Hewlet
t-Packard G2024A 試料調製付属品)の中で蒸発させ、急速、かつ均一な試料結晶化を確
実にした。
【0117】
長期間Glu1環化反応試験においては、Aβ由来ペプチドは、30℃の100μl 0.1 M酢酸ナ
トリウム緩衝液(pH 5.2)、または0.1 Mビス-トリス緩衝液(pH 6.5)中に準備した。ペ
プチドは、0.5 mM [Aβ3-11a]、又は0.15 mM [Aβ3-21a]濃度で使用し、0.2 U QCは、全2
4時間加えた。Aβ3-21aの場合は、1% DMSOを加え測定した。様々な時間で、試料は測定
管から採取され、ペプチドは、製造会社推奨に従いジップチップ(ZipTip)(Millipore
社)で抽出し、マトリックス溶液と混合(1/1、v/v)し、かつその後、マススペクトル
を記録した。ネガティブコントロールは、QCを加えないか、又は加熱不活化酵素を加え
るかのいずれかであった。インヒビターの研究において、該試料組成は、阻害化合物(5
mMベンズイミダゾール、又は2 mM 1,10-フェナントロリン)を添加する以外は、前記と同
様であった。
【0118】
(実施例4):pH依存性
ヒト、及びパパイヤQCの触媒作用のpH依存性を、一次反応条件下で研究し、それ故に
特異性定数kcat/KMにおけるプロトン濃度の影響を示した。この目的のために、ピログル
タミルアミノペプチダーゼを補助酵素、及びGln-βNAを器質として使用した、結合酵素測
定(the coupled enzymatic assay)を使用した。ピログルタミルアミノペプチダーゼは
、pH 5.5〜8.5で活性を有し、かつ安定であることが示された(Tsuru, D.らの論文、1978
J Biochem(Tokyo)84, 467-476)。それ故に、該測定によって、このpH範囲内でQC触
媒の研究が可能になった。得られた速度プロファイルは、図2に示されるように、典型的
ベル型曲線に適合した。ヒトQCは、至適pHがおよそ7.8〜8.0の、大変狭いpH依存性を有
する。該速度は、より塩基性のpHで、降下する傾向があった。これは、pH 8.5まで活性の
降下を示さなかった(図2、差し込み図)、パパイヤQCで見られた速度プロファイルと
対照的である。しかしながら、両酵素は、pH 8で至適特異性を示した。意外にも、曲線の
査定により、ヒト、及びパパイヤの酸性範囲でのpKa値は、各々7.17±0.02、及び7.15±0
.02と、同一であることが明らかになった。
【0119】
明らかに、ヒトQCの、塩基pH値での活性の減少は、pKa値およそ8.5を有する原子団の
解離に起因した。パパイヤQCの場合は、その第二pKa値の信用できる測定を可能にする
ような、塩基性pHにおけるデータポイント収集はなかった。これは、該データの単独解離
モデルへの適合が、該データの二重解離モデルへの適合と比べて、ほぼ同一のpKa値(pKa
7.13±0.03)をもたらすことにより、支持される。これは、双方のpKa値が、かなり離れ
ていることを示唆する。
【0120】
(pH安定性)
グルタミニルシクラーゼの安定性は、該植物、及び動物酵素をpH 4〜10間の異なるpH値
において、30℃で30分間インキュベートすることにより調べた。その後、QC活性は、標
準的条件下で調べた。その結果を、図3に示す。
パパイヤラテックス由来の該QCは、酸性、又は塩基性の範囲での顕著な不安定性への
傾向はなく、研究したpH範囲内で安定であった。その一方、ヒトQCは、7〜8.5間のpH範
囲内で同程度に安定であった。従って、pH 8のあたりの領域が、植物、及びヒトQCの活
性、及び安定性にとって至適で、かつ該QCの基質特異性の比較を行うのに適しているよ
うである。
【0121】
(実施例5):QCの基質特異性の決定
(分光学的測定)
実施例2に記載したように、連続分光学的測定を行った。従って、QCの活性は、α-
ケトグルタル酸からのグルタミン酸の産生のための、アンモニアの遊離、及びその後のNA
DH/H+の消費に起因する340 nmでの吸光度の減少に反映する。図1に示すように、リニア
プログレス曲線をモニターしたところ、測定した活性、及びQC濃度の間には、直線関係
があった。さらに、ここ(表1)に示す連続測定を使用し、得られたH-Gln-Gln-OHの速度
パラメータは、断続法を使用して得られたものとよく一致した(KM=175±18μM、kcat=
21.3±0.6 s-1)。その上、表1に示したパパイヤQCによる、基質H-Gln-Ala-OH、H-Gln
-Glu-OH、H-Gln-Gln-OH、H-Gln-OtBu、及びH-Gln-NH2の転換速度パラメータは、pH 8.8、
及び37℃の条件で、直接法を使用して決定されたものとよく一致した(Gololobov, M. Y.
らの論文、1996 Biol Chem Hoppe Seyler 377, 395-398)。それ故、新規連続アッセイ法
が、信用できる結果を提供することは明らかである。
【0122】
(ジ、トリ、及びジペプチド代用物)
先に記載した該新規連続アッセイ法を使用し、約30種の化合物をパパイヤ(C. papay
a)、及びヒト由来QCの潜在基質として調べた。結果を、表5に示した。特異性の比較
により、ほぼ全部の短いペプチドがパパイヤQCにより、該ヒト酵素に比べてより効果的
に転換されることが示された。興味深いことに、他のトリペプチドと比較したH-Gln-Tyr-
Ala-OH、H-Gln-Phe-Ala-NH2、及びH-Gln-Trp-Ala-NH2の特異性、又はジペプチド基質と比
較した発色基質H-Gln-AMC、H-Gln-βNA、及びH-Gln-Tyr-OHの反応性により示されるよう
に、両酵素において、2位に疎水性の残基を有する基質が、最も有効な基質である。パパ
イヤQCにおいて、この知見は、その特異性が2位のアミノ酸残基の大きさに相関関係が
あるという初期の結果と一致する(Gololobov, M. Y.らの論文、1996 Biol Chem Hoppe S
eyler 377, 395-398)。植物、及び動物QCの特異性における唯一顕著な相違点は、H-Gl
n-OtBuにおいて観察された。エステルはパパイヤQCにより、ジペプチド基質と同様の特
異性で転換されるのに対し、ヒトQCでは、ほぼ一桁遅く転換された。
【0123】
(オリゴペプチド)
様々なジペプチド、及びトリペプチドに加えて、パパイヤ、及びヒトQCによる転換に
より、多くのオリゴペプチドを調べた(図5)。興味深いことに、テトラペプチドグルー
プにおけるヒト、及びパパイヤQCにおける特異性の相違点は、ジペプチ、及びトリペプ
チドで観察されたほど大きくなかった。これにより、3位及び4位のアミノ酸でも、速度
論的挙動に影響することが示唆された。しかしながら、H-Gln-Xaa-Tyr-Phe-NH2の構造の
テトラペプチドグループにおいて著しく減少したkcat/KM値を示す、2位のアミノ酸がプ
ロリンのペプチドは例外である(表5)。その特異性の低下は、ヒトQCにおいてより顕
著であり、パパイヤQCと比較して該kcat/KM値においてほぼ8倍の相違をもたらした。
【0124】
また、ヒトQCの特異性の微低下は、H-Gln-Arg-Tyr-Phe-NH2、H-Gln-Arg-Tyr-Phe-NH2
、及びH-Gln-Lys-Arg-Leu-NH2の特異性を他のテトラペプチドと比較して示唆されるよう
に、アミノ酸C末端に正に荷電されたグルタミンを有する基質の転換において観察された
。明らかに、該特異性低下は、主としてより小さい代謝回転数に起因した。この効果は、
該植物酵素では見られなかった。
【0125】
【表5】
【0126】
また、該テトラペプチドで得られた結果は、もう一つの結論をもたらした。既に指摘し
たように、パパイヤQCは、ジペプチドにより高い特異性を示した。しかしながら、2位
のアミノ酸にグルタミン酸を含有するペプチドグループにおいて、表5に与えられたデー
タを曲線で表した図4で示されるように、テトラペプチドによっては、ヒトQCにおいて
より高い特異性定数が観察された。さらに、ペプチド鎖長が、ジから、テトラペプチドへ
伸張するにつれて、パパイヤQCで得られた結果と対照的に、ヒトQCの選択性は上昇し
た。さらに、ヒトQCは、3位、又は4位のアミノ酸に大型の疎水性残基を有するペプチ
ドにおいて、最も高い選択性を記録し、それにより、該酵素の疎水性相互作用を示唆した
。それぞれのペプチドの速度パラメータを比較すると、該変化は主に低いKM値に起因する
と考えられ、該ペプチドの代謝回転数は同様であることが見い出された。従って、ヒトQ
Cの長いペプチドに対する高選択性は、より疎水性の基質の該酵素へのより強い結合の結
果であると考えられる。
【0127】
また、3位と4位に疎水性アミノ酸を含有するペプチドにおいて観察されたヒト,及び
植物QCの相違点は、H-Gln-Arg-Gly-Ile-NH2、又はH-Gln-Arg-Tyr-Phe-NH2、又はH-Gln-
Gln-OH、又はH-Gln-Gln-Tyr-Phe-OHに対する該酵素の特異性定数を比較により明らかにな
る。
また、ヒトQCが、N末端にGlnを含有し、かつC末端Ala残基が増加する同族基質にお
いて、より選択的であるこを見い出した(表6)。ヒトQCの選択性が、基質の鎖長が伸
張するにつれ増加したのに対し、パパイヤQCにはそのような傾向はなかった。また、ヒ
トQCがSer残基を配列に含有するペプチドに対して、より低い特異性であったので、側
鎖の性質が重要であるように考えられる(表6)。
【0128】
【表6】
【0129】
(触媒反応におけるイオン強度の影響)
イオン強度は、基質特異性の影響に関して調べたもう1つのパラメータであった。その
目的において、様々な基質の環化反応における速度パラメータを、0.5 M KClの存在、又
は非存在下で決定した(表7)。驚いたことに、パパイヤラテックス由来QC、及びヒト
QCにおいて、荷電されていない基幹をもつ基質の選択性は、塩の付加により有意に変化
しなかったが、ヒトQCのH-Gln-Ala-OH、及びH-Gln-Glu-OHにおける特異性定数は、KCl
の添加により低下した。各々の速度パラメータにより示唆されるように、この効果は、上
昇したKM、及びごくわずかに減少したkcat値に起因した。パパイヤQCの場合は、どちら
のパラメータにおいても効果は検出されなかった。負に荷電されたH-Gln-Glu-Asp-Leu-NH
2において、パラメータの変化が見い出されなかったことから、該効果は、そのように負
に荷電された基質に起因しないように考えられる。塩付加の興味深い効果は、正に荷電さ
れたH-Gln-Arg-Gly-Ile-NH2、及びH-Gln-Lys-Arg-Leu-NH2において見い出された。植物、
及びヒトQCの場合、触媒反応における正の効果は、主に、より小さいKM値、及びわずか
に高い代謝回転数に起因すると決定した。
【0130】
【表7】
【0131】
(生理的基質)
既に、初期の研究において、QCによる[Gln1]-TRH、及び[Gln1]-GnRHの転換は、ウシ
、及びブタ脳下垂体由来QCにおいて示された(Busby, W. H. J.らの論文、1987 J Biol
Chem 262, 8532-8536;Fischer, W. H.、及びSpiess, J.の論文、1987 Proc Natl Acad
Sci U S A 84, 3628-3632)。既に調べられたこれらの脳下垂体ホルモンに加え、ヒトQ
Cの3つの潜在基質、すなわち[Gln1]ガストリン、[Gln1]ニューロテンシン、及び[Gln1]
FPPを合成し、調べた。それらの速度パラメータは表1に示した。興味深いことに、グル
タミニルペプチドは、その大きさに依存して特異性定数の上昇を伴いながら、各々のピロ
グルタミルペプチドへ転換される。すなわち、17アミノ酸の最も大きいペプチドガスト
リン前駆体が最初で、続いて、ニューロテンシン前駆体、GnRH前駆体、TRH前駆体、及びF
PP前駆体である。これらの知見は、合成ペプチドを用いて得られたデータに一致する。
また、驚いたことに、長い基質ほど、該植物酵素による高い選択性で転換され、それは
、より短いオリゴペプチドにおける知見と部分的に対照的である。おそらく、活性部位か
らはるかに離れた、基質、及び酵素間の二次的結合相互作用が存在する。
【0132】
(改質アミノ酸含有ペプチド)
さらに、該QCの該特異性、及び該選択性を調べるために、改質N末端グルタミニル残
基、及び2位に改質アミノ酸を含むペプチドを合成した。これらのペプチドの転換は、MA
LDI-TOF質量分析法を用いて、定性的に調べた(実施例3も参照)。グルタミニル残基、
又はその類似体のそれぞれの環化反応に起因して、該基質と該触媒反応生成物の質量差を
検出する。また、基質モル当たり、1モルのアンモニア遊離の場合は、分光学的定量法を
用いて定量的に分析した。
【0133】
(H-Gln-Lys(Gln)-Arg-Leu-Ala-NH2)
N末端において、ペプチド、及び部分イソペプチド結合でリシル残基に結合している2
グルタミニル残基含有する、N末端分鎖ペプチドは、ヒト(図5)、及びパパイヤQC(
図示せず)により明らかに同一の方法で転換された。一貫した基質転換で示唆される(図
5)ように、両グルタミニル残基は、個別の残基に対する検出可能な優先はなく、ピログ
ルタミン酸へ転換された。従って、QCの、異なった結合をしたグルタミニル残基におけ
る選択性は、根本的に相違しない。
【0134】
(H-Gln(NMe)-Phe-Lys-Ala-Glu-NH2)
唯一、メチル化されたグルタミニル残基は、パパイヤQCにより、ピログルタミル残基に
転換された(図6)。さらに、該ヒトQCの該ペプチドによる阻害は検出されなかったの
で、該メチル化された残基はヒトQCにより認識されないことが示唆された。
【0135】
(H-Glu(OMe)-βNA、及びH-Glu-βNA)
これらの化合物はどちらも、パパイヤ、及びヒトQCによって転換されなかった。これ
らの蛍光発生基質は、ピログルタミルアミノペプチダーゼを補助酵素として使用しながら
、蛍光分析学的に分析した。しかしながら、O-メチル化グルタミン酸残基は、非酵素的触
媒環化反応する傾向があるトリス、及びトリシン緩衝液両者において顕著な不安定性を示
した。さらに、両QCのH-Gln-AMCを基質とする活性は、H-Glu(OMe)-Phe-Lys-Arg-Leu-
Ala-NH2、又はH-Glu-Phe-Lys-Arg-Leu-Ala-NH2の長鎖ペプチドにより阻害されなかったこ
とは、グルタミン酸、又は誘導体が両QC型により認識されないことを示唆する。さらに
、該結果はグルタミン酸残基の負電荷のみが、該ペプチドの該活性部位からの反発力の原
因でないことを意味する。
【0136】
(H-Gln-cyclo(Nε-Lys-Arg-Pro-Ala-Gly-Phe))
分子内部分イソペプチド結合を含有するH-Gln-cyclo(Nε-Lys-Arg-Pro-Ala-Gly-Phe)
の転換を定量的に分析し、ヒト、及びパパイヤQCにおけるKM値が各々240±14μM、及び
133±5μMであることを明らかにした。ヒトQC(22.8±0.6 s-1)と比較して、パパイヤ
QC(49.4±0.6 s-1)による転換におけるより高い代謝回転数に起因して、該植物QC
は、ヒトQCのおよそ4倍高いkcat/KM値372±9 mM-1min-1を示した。従って、パパイヤ
QCの場合、特異性定数はH-Gln-Ala-Ala-Ser-Ala-Ala-NH2のような同様の大きさをもつ
基質と比べてごくわずかに小さい。しかしながら、ヒトQCのkcat/KM値は、95±3 mM-1
s-1で、同様の大きさの基質に比べておよそ一桁小さいことを見い出した(表5)。
【0137】
(H-βhomoGln-Phe-Lys-Arg-Leu-Ala-NH2)
N末端β-ホモグルタミニル残基は、ヒト、及びパパイヤQCにより、各々五員ラクタ
ム環へ転換された。アンモニアの付随遊離を、先に記載したように、分光学的、及びMALD
I-tof分析法により分析した。QCを除いたり、又は沸騰した場合、アンモニアの遊離が
検出されなかったことから、該環化反応の特異的な触媒反応であることが示唆された。興
味深いことに、パパイヤ(C. papaya)(KM=3.1±0.3 mM、kcat=4.0±0.4 s-1)、及び
ヒト(KM=2.5±0.2 mM、kcat=3.5±0.1 s-1)由来該QCは、各々1.4±0.1、及び1.3±
0.1 mM-1s-1とほぼ同一のkcat/KMで、該ペプチドの転換を触媒する。従って、β-グルタ
ミン残基の環化反応は、N末端にグルタミニル残基を含有する同様の大きさのペプチドに
比べて、およそ1000倍低下した効率で触媒される。これは、基質のα-炭素の構造が、Q
C型による基質認識において重要であるが、必須ではないことを示す。基質である必須条
件は、環化反応しやすい距離と角度の、γ-アミド基、及びプロトン化されていないN末
端アミノ基であり、N末端グルタミニル、及びγ-ホモ-グルタミニル残基は必須条件を満
たしている。
【0138】
(実施例6):QC基質の合成
(オリゴペプチド)
ペプチドを、既に記載されたように、ペプチド合成機(ラボーテックSP650(Labortec
SP650)、Bachem社、スイス)を使用し、0.5 mmolスケールで半自動的に合成した(Schil
ling, S.らの論文、2002 Biochemistry 41, 10849-10857)。長鎖ペプチドは、記載され
たように、自動化シンフォニーペプチド合成機(Symphony peptide synthesizer)(Rain
in Instrument社)を使用し、25μmolスケールで合成した(Manhart, S.らの論文、2003
Biochemistry 42, 3081-3088)。すべてのペプチド結合において、2-(1H-ベンゾトリア
ゾール-1-イル)-1,1,3,3,-テトラメチルウロニウムテトラフルオロホウ酸(TBTU;Novab
iochem社)/塩基(ジイソプロピルエチルアミン、又はN-メチル- モルホリン;Merck社
)を使用する、改質した固相ペプチド合成のFmocプロトコルを採用し、又は困難な結合の
場合は、N-[(ジメチルアミノ)-1H-1,2,3,-トリアゾロ[4,5-b]ピリジン-1-イルメチレン
]-N-メチルメタンアンモニウムヘキサフルオロリン酸N-オキシド(4,5)(HATU;Applied
Biosystems社)/ジイソプロピルエチルアミンを活性化剤として使用した。カクテルを
含むトリフルオロ酢酸(TFA;Merck社)によるレジンからの開裂の後、粗ペプチドは無酸
性溶媒の分取用HPLCにより精製し、N末端グルタミンのさらなる環化反応を回避した。25
0-21ルナ(Luna)RP18カラム(Phenomenex社)におけるアセトニトリル(Merck社)水溶
液リニアグラジエント(40分間を通して、5〜40%、又は65%アセトニトリル)で分取HPL
Cを行った。ペプチド精度と同定を裏付けるために、分析用HPLC、及びESI-MSを使用した
。
【0139】
(Glu(NH-NH2)-Ser-Pro-Thr-Ala-NH2)
直鎖前駆体ペプチド(Fmoc-Glu-Ser-Pro-Thr-Ala-NH2)は、標準的Fmoc方法(Schillin
g, S.らの論文、2002 Biochemistry 41, 10849-10857)に従って、リンクアミド(Rink a
mide)MBHAリジン(Novabiochem社)において合成した。Fmoc保護ペプチドのレジンから
の開裂後、該ペプチドを、ジエチルエーテル(Merck)で沈殿させ、濾過し、かつ乾燥さ
せた。ジクロロメタン(DCM、Merck社)中で、該前駆体のγ-カルボン酸基(0.3等量)を
結合させるのに、HMBA-AM レジン(1.16 mmol/g、Novabiochem社)を使用した。結合剤
として、ジシクロへキシルカルボジイミド(DCC、Serva社)(4等量)、及びジメチルア
ミノピリジン(DMAP、Aldrich社)(0.1等量)を使用した。12時間後、レジンを、濾過し
、DCMで洗い、反応を繰り返した。20%ピペリジンDMF溶液(5分間、3回)を用いたN末端
Fmoc基の脱保護後、該ペプチドリジンを、5%ヒドラジン溶液(20 ml/g)で1.5時間処理
した。該レジンを、濾過し、ジメチルホルムアミド(DMF、Roth社、ドイツ)、及びTFAで
洗った。蒸発後、該粗ペプチドをエーテルで沈殿し、76%の収率を得た。
【0140】
(H-Gln-Lys(Gln)-Arg-Leu-Ala-NH2)
直鎖ペプチドを、標準的Fmoc/tBu方法に従って、Fmoc-Lys(Fmoc)-OHを最後から2番
目のアミノ酸結合として使用して、リンクアミド(Rink amide)MBHAレジン(Novabioche
m社)において合成した(Schilling, S.らの論文、2002 Biochemistry 41, 10849-10857
)。20%ピペリジン(Merck社)DMF溶液で、レジンの2つのアミノ保護基を脱保護後、4等
量のFmoc-Gln(Trt)-OHを結合させた。標準的開裂法で、95%の収率を得た。
【0141】
(H-Gln(NMe)-Phe-Lys-Ala-Glu-NH2)
Fmoc-Gln(NMe)-OHを、Fmoc-MI-AM(Novabiochem社)レジンに充填したFmoc-Glu-OtBu
から開始して合成した。DCMで膨潤後、該レジン(0.5 g)をDMFで洗い、20%ピペリジン
(Merck社)DMF溶液で脱保護した。該レジンを5 ml DMF中に入れ、5等量Fmoc-Glu-OtBu、
5等量HATU、及び10等量DIPEAを次いで加え、6時間振とうした。濾過、及び洗浄後、生成
物を標準的TFA開裂条件に従って、開裂した。該H-Gln(NMe)-Phe-Lys-Ala-Glu-NH2ペプ
チドは、記載されたように合成した(Schilling, S.らの論文、2002 Biochemistry 41, 1
0849-10857)。Fmoc-Gln(NMe)-OHは、HATU/DIPEAと一晩結合させた。標準的開裂法で
、78%の収率を得た。
【0142】
(H-Glu(OMe)-β-ナフチルアミド、H-Gln-Val-OH、H-Gln-Tyr-OH)
Boc保護ジペプチドを、クロロ炭酸イソブチル(Merck社)を使用して、標準的無水物混
合法で合成した。C末端メチルエステルBoc-Gln-Tyr-OMe、及びBoc-Gln-Val-OMeを、1 N
NaOHジオキサン溶液により鹸化した。Boc保護ペプチドは、HCl/ジオキサン溶液により、
10分間脱保護した。蒸発後、残基を様々な溶媒で結晶化させ、固体化合物の60〜70%の収
率を得た。
【0143】
(H-Gln-cyclo(Nε-Lys-Arg-Pro-Ala-Gly-Phe))
直鎖前駆体Boc-Gln(Trt)-Lys-Arg(Pmc)-Ala-Gly-Phe-OHを、酸感受性2-クロロトリ
チルリジンにおいて合成した。結合を、Fmoc-Lys(Mtt)-OHを用いたFmoc/tBu方法の標
準的プロトコルを用いて行った。3%TFA DCM 溶液(10回、5分)で開裂後、該溶液は、10
%ピリジン(Merck社)メタノール溶液(MeOH;Merck社)で中和し、DCM、及びMeOHで3回
洗い、5%の容量に蒸発させ、粗ペプチドは氷冷水で沈殿させた。その後、該粗シペプチ
ドは、DCC/N-ヒドロキシベンゾトリアゾール(HOBt;Aldrich社)活性化を利用して、環
化した。該粗ペプチドは、乾燥ジクロロメタン溶液(0.2 mmol/50 ml)に、溶解し、0.2
mmolのN-メチルモルホリン、及び0.4 mmolの1-ヒドロキシベンゾトリアゾールを添加し
た。この溶液を、0℃において、250 mlの0.4 mmolジシクロヘキシルカルボジイミドのジ
クロロメタン溶液へ滴下した。該反応は、室温で、一晩撹拌して完了した。N,N'-ジシク
ロヘキシル尿素の濾過後、溶媒は、蒸発により除いた。残留物は、エチルアセテートに溶
解し、1規定HCl、NaHCO3飽和溶液、及び水で洗った。該溶液は、無水Na2SO4上で乾燥さ
せ、濾過し、真空で乾燥するまで蒸発させた。
【0144】
(実施例7):QCのエフェクターの特性
(イミダゾール誘導体)
五員環の異なった位置に置換基をもつイミダゾール、及びベンズイミダゾール誘導体を
、QCのインヒビターとして調べた(表3)。構造式の数は、イミダゾール環を示す。該
方法は、実施例2に示す。
【0145】
(C-4(5)、及びC-4,5誘導体)
イミダゾール環の構造上同等の4、又は5位のいずれか、又は両位置に置換基をもつ化
合物は、ヒトQCの阻害において有効性の減少を示した。しかしながら、最も効力のある
阻害化合物の1種であることが証明されたN-ω-アセチル化ヒスタミンは、唯一の例外であ
る。イミダゾールと比べて5-ヒドロキシメチル-4-メチル-イミダゾールが同様の阻害定数
を示したように、これらの位置の小さい置換基は、結合にほとんど効果がなかった。これ
らの位置についた大きく、かつ巨大な基は、酵素による化合物の結合を低下、又は消失し
た。他の調べた置換基の中には、イミダゾール環の電子密度を低下できる負の誘導、又は
メソメリー効果を及ぼすことが知られているものもあり、また、それはより低い結合定数
の原因となる。また、L-ヒスチジン、及びヒスチジンアミドのKi値の相違は、結合におけ
る荷電の若干の影響を示唆する。既に、荷電した基質の静電反発力の証拠は、該基質特異
性の研究において示され、例えば、グルタミンアミドは、ヒトQCにより容易に生成物に
転換されたが、遊離グルタミンにおける基質としての反応性は観察されなかった。
【0146】
(C-2誘導体)
調べたすべての誘導体は、イミダゾールのように、より弱くQCを阻害した。プロトン
より大きいすべての置換は、適切なQC結合を妨げる。唯一2-メチル-ベンズイミダゾー
ルのメチル基に起因して、該阻害定数は、およそ一桁の程度低下する。ベンズイミダゾー
ル、及び2-アミノ-ベンズイミダゾールにおけるKi値の比較により、ごく同様の関係が示
された。さらに、その結果により、影響が電子論的改質に関連しないことが示唆された。
【0147】
(N-1誘導体)
ヒトQCの阻害を調べたイミダゾール誘導体の中で、イミダゾールと比べて向上したKi
値を持つほとんどの化合物は、1窒素原子において改質が見られた。また、これらの化合
物には、最も有効なQCインヒビターの1種、1-ベンジルイミダゾールが含まれた。興味
深いことに、1-ベンゾイルイミダゾール、及びフェニルイミダゾールにおいて見られるよ
うに、この構造のごくわずかな改質が、阻害特性の消失を起こし、実験条件下では不活性
であった。また、この場合において、該観察された変化は、フェニル基の負のメソメリー
効果に起因するイミダゾール環の低下した電子密度よって引き起こされるだけでないよう
に思われ、それは、正の誘導効果を示す、巨大なトリメチルシリル基もまた、他の残基に
比べて結合の低下を示したからである。興味深いことに、1-アミノプロピル-イミダゾー
ルは、この族においてさほど有効性がない化合物の1種であった。立体的に同様の化合物
1-メチルイミダゾール、及び1-ビニルイミダゾールにおいては、活性部位への結合の向上
が見られたので、この化合物の該低有効性は、該塩基アミノ基によって引き起こされる。
従って、正に荷電されたアミノ基が、該低Ki値の原因となり、その結果はN-ω-アセチル
化ヒスタミン(表3)、及びヒスタミン(表4)により裏付けられる。
【0148】
(3,4、及び3,5誘導体化の効果)
4(5)位に、又は両位置に置環基を含有するイミダゾール誘導体は、該酵素の結合に
おいて限られた有効性をもつことを示した。該特異的置換基の効果を、L-ヒスタミン、及
びヒスタミンの生物分解の2中間体3-メチル-4-ヒスタミン、及び3-メチル-5-ヒスタミン
(表4)の阻害定数と比較することにより特定した。L-ヒスタミンは、そのアセチル化対
応物に比べて、およそ一桁小さいKi値を示した。3-メチル-4-ヒスタミンの場合において
、一窒素のメチル化は、有効性をかなり向上させた。しかしながら、3-メチル-5-ヒスタ
ミンをもたらすメチル化は、阻害活性の完全な消失を引き起こした。従って、該観察され
た効果は、主に、塩基窒素に隣接した炭素の誘導体化に起因した、結合の立体障害により
引き起こされるように考えられる。おそらく、該塩基窒素は、該酵素の結合において、重
要な役割を果たす。
【0149】
(実施例8):Aβ3-40/42誘導体の産生
Aβ3-40/42のN末端の配列で、その3位にグルタミン酸残基の代わりにグルタミンを
含有する短鎖ペプチド2種、[Gln3]-Aβ1-11(配列:DAQFRHDSGYE)、及び[Gln3]Aβ3-11
を用いて、測定を行った。2種のペプチドのDP IVによる開裂、及びQCによるN末端グル
タミン残基の環化反応を、MALDI-TOF質量分析で調べた。測定は、連続触媒反応測定用の
両酵素においてと同様、精製DP IV(ブタ腎臓)、又は粗ブタ脳下垂体ホモジネートをQ
C源として用いて行った。
【0150】
(結果)
1.DPIVにより触媒された[Gln3]Aβ1-11aから[Gln3]Aβ3-11aの産生、及びDP IVインヒ
ビター、Val-ピロリジド(Val-Pyrr)によるその抑制
DPIV、又はDPIV様活性は、[Gln3]Aβ3-11a産生下における、[Gln3]Aβ1-11aの開裂をす
る(図7)。その3位の残基は、この開裂により露出し、それゆえ他の酵素、例えば、Q
Cによる改質のために、利用されやすくなる。予想どおり、触媒作用は、Val-Pyrrにより
完全に抑制することが可能である(表8)。
【0151】
2.脳下垂体ホモジネートにおけるQC触媒作用による[Gln3]Aβ3-11aから[pGlu3]Aβ3-
11aの産生、及び1,10-フェナントロリンによる抑制
ブタ脳下垂体ホモジネートに存在するQCは、[Gln3]Aβ3-11aの[pGlu3]Aβ3-11aへの
転換を触媒する(図9)。[pGlu3]Aβ3-11aの産生は、1,10-フェナントロリンの添加によ
り阻害された(図10)。
【0152】
3.[pGlu3]Aβ3-11aの産生をもたらすDP IV、及びQCの連続触媒作用、及びVal-Pyrr、
及び1,10-フェナントロリンによる抑制
粗ブタ脳下垂体ホモジネートにブタ腎臓由来DP IVを付加して測定した、[Gln3]Aβ1-11
aから[pGlu3]Aβ3-11aの産生は、DP IV、及びQCによる連続触媒反応後、起こる(図1
1)。QCインヒビター1,10-フェナントロリン(図12)、又はDP IVインヒビターVal-
Pyrrを添加したとき(図13)は、[pGlu3]Aβ3-11aは産生されなかった。[pGlu3]Aβ3-1
1aのわずかな出現は、アミノペプチダーゼの開裂、次いでグルタミン残基の環化反応に起
因し、また、[Gln3]Aβ2-11aの産生により示唆された。
【0153】
4.アミノペプチダーゼ触媒作用による、粗脳下垂体ホモジネートにおける[pGlu3]Aβ3-
11aの産生
DPIV触媒作用に依存しない[pGlu3]Aβ3-11aに起因して、[Gln3]Aβ1-11aの分解を、DP
IVを添加しない粗脳下垂体ホモジネートにおいて調べた(図14)。4項におけるデータ
から予想されるように、[pGlu3]Aβ3-11aの産生が観察された。また、該データは、[Gln3
]Aβ1-11aの分解はアミノペプチダーゼによって触媒され、[pGlu3]Aβ3-11aを生じ得るこ
とを示す。それ故、該結果は、ピログルタミル産生は、この組織におけるN末端ペプチド
分解の指標であることを示し、さらに斑形成におけるQCの役割を支持した。
【0154】
(実施例9):組み換えヒトQCによる[Gln3]Aβ3-11a;3-21a、及び3-40の代謝回転
調べた全ての[Gln3]Aβ誘導ペプチドは、ヒトQCにより、対応するピログルタミル型
へ効率よく転換された(表8)。水溶液における[Gln3]Aβ3-21a、及び[Gln3]Aβ3-40の
難溶解性のため、該決定は、1% DMSOの存在下で行った。しかしながら、[Gln3]Aβ3-11a
では、良好な溶解性により、DMSO存在、及び非存在下におけるQC触媒作用の代謝回転の
動態解析が可能であった(表8)。まとめると、鎖長8、18、及び37アミノ酸の、Q
C基質としてのAβ-ペプチドの研究(表8参照)により、基質の鎖長が伸張するにつれヒ
トQC活性が上昇するという観察を裏付けた。従って、その特異性定数を考慮に入れると
、Gln1-ガストリン、Gln1-ニューロテンシン、Gln1-GnRHは、QC基質の中でも最良であ
る。同様に、これまで調べた最長のQC基質である[Gln3]Aβ3-40、及びグルカゴンは、1
% DMSO存在下においても、高二次速度定数(各々449 mM-1s-1、及び526 mM-1s-1)を示
した(表8)。
興味深いことに、調べたアミロイドペプチドにおける該転換の該速度パラメータが、鎖
長の伸張につれて劇的に変化しなかったことは、QC触媒おけるAβのC末端のごく軽度
の影響を示唆した。従って、より良溶解性、及び実験処理によって、Aβの小断片、[Gln3
]Aβ1-11a、[Gln3]Aβ3-11a、及びAβ3-11aを用いて、これらのペプチドN末端アミノペ
プチダーゼプロセズに関するさらなる研究を行った。
【0155】
【表8】
【0156】
(実施例10):組み換えヒトQCによるAβ3-11a、及びAβ3-21aの代謝回転
QC存在下でのAβ3-11a、及びAβ3-21aのインキュベートにより、以前の研究と対照的
に、グルタミン酸含有ペプチドがQCの基質になる得ることが示された(図15C、及び
D)。QC触媒による[pGlu3]Aβ3-11a、及び[pGlu3]Aβ3-21aの産生を、各々pH 5.2、及
び6.5で調べた。QCの添加による活性の開始前に、QCインヒビター、ベンズイミダゾ
ールを該溶液に添加した場合は、[pGlu3]Aβ3-11a、又は[pGlu3]Aβ3-21aを生じる基質転
換は抑制された(図15E、F)。QCを添加前に煮沸した場合は、pGluペプチドの産生
はごく僅かであった(図15A、及びB)。
【0157】
(実施例11):パパイヤQC触媒のGln-βNA、及びGlu-βNA の環化反応におけるpH依
存性
パパイヤQCは、ミカエリスメンテン速度論に従い、2 mM(基質溶解性により限定され
た)までの濃度範囲においてGlu-βNAを転換した(図16)。pH 6.1〜8.5の間で調べた
、QC触媒のGlu-βNAの転換における基質濃度に対する代謝回転の図を検討することによ
り、このGlu基質における、KM、及びkcatの両パラメータが、pHに依存することが示され
た(図16)。これは、以前に記載されたQC触媒のグルタミン環化反応とは対照的であ
り、その反応においては、該既定のpH範囲ではKMの変化のみが観察された(Gololobov, M
. Y.、Song, I.、Wang, W.、及びBateman, R. C.の論文、(1994)Arch Biochem Biophys
309, 300-307)。
続いて、Glu、及びGln環化反応中のプロトン濃度の影響を検討するために、低一次速度
条件下(すなわち、KM値をはるかに下回る基質濃度)における、Glu-βNA、及びGln-βNA
の環化反応のpH依存性を調べた(図17)。pH 6.0で至適pHを示したグルタミン酸の該環
化反応に対し、グルタミンの環化反応は、pH 8.0で至適pHを示す。各々の至適pHにおける
該特異性定数は、およそ80,000倍異なる一方、pH 6.0付近でのECに対するQCの活性は
8,000倍だけである。
pH 6.0において4週間調べた、Gln-βNAから非酵素的pGluの産生により、一次速度定数
を1.2*10-7 s-1と示された。しかしながら、同期間においてGlu-βNAからpGlu-βNAは産
生されなかったことから、代謝回転の律速速度定数は1.0*10-9 s-1と推定できた。
【0158】
(実施例12):酵素失活化/再活性化法
ヒトQC(0.1-0.5 mg、1 mg/ml)の分注液を、3000倍超過の5 mM 1,10-フェナントロ
リン、又は5 mMジピコリン酸含有0.05 Mビス-トリス/HCl溶液(pH 6.8)溶液に対して一
晩透析して、失活させた。続いて、試料を1 mM EDTA含有0.05 Mビス-トリス/HCl溶液(p
H 6.8)に対する透析(3回、2000倍超過)により、失活物質を注意深く除去した。再活
性実験は0.025 Mビス-トリス(pH 6.8)においてZn++、Mn++、Ni++、Ca++、K+、及びCo++
イオンを1.0、0.5、0.25 mMの濃度で用いて、室温で15分間行なった。QC活性測定は
、2 mM EDTA含有0.05 M トリス/HCl溶液(pH 8.0)で行い、緩衝溶液中に存在する微量
の金属イオンにより急速な再活性化を防いだ。
【0159】
既に、1,10-フェナントロリンによるブタQCの阻害は、記載されている(Busby, W. H
. J.らの論文、1987 J Biol Chem 262, 8532-8536、Bateman, R.C.J.らの論文、2001 Bio
chemistry 40)。しかしながら、EDTAがQC触媒反応において活性化する効果を持つこと
が示されていた事実は、フェナントロリンによる阻害が金属キレートに起因しないことを
示唆した(Busby, W. H. J.らの論文、1987 J Biol Chem 262, 8532-8536、Bateman, R.C
.J.らの論文、2001 Biochemistry 40)。また、1,10-フェナントロリンによる阻害に加え
て、ヒトQC触媒の基質環化反応は、ジピコリン酸、及び8-ヒドロキシキノリン、他の金
属酵素インヒビター存在下で消失した。これらのキレーターは、QCを競合的、及び時間
依存的に阻害し、すなわち、既に競合的に阻害された初期の活性は、該化合物の長時間イ
ンキュベート後、さらに低下することが見い出された(図18、19)。興味深いことに
、EDTAは、インキュベート時間、又は条件にかかわらず、顕著な阻害を示さなかった。
【0160】
ヒトQCは、5 mM 1,10-フェナントロリン、又は5 mMジピコリン酸に対する大規模な透
析後、ほぼ完全に阻害された。キレーター非存在緩衝溶液に対して一晩透析を繰り返した
後、QC活性は50〜60%まで部分的に再活性化された。しかしながら、1 mM EDTA含有緩
衝液に対して透析した場合は、再活性化は観察されなかった。
該タンパク質を0.5 mM EDTA 存在下で0.5 mM ZnSO4と10分間インキュベートすること
により、ジピコリン酸、又は1,10-フェナントロリンのいずれかによる不活性化後のQC
活性の、ほぼ完全な再活性化が実現された(図20)。同様に、再活性化にCo++、及びMn
++を用いて、QCの部分再活性化を得た。0.25 mM Zn++存在下ですら、本来の活性の25%
までの再活性化が可能だった。Ni++、Ca++、又はK+イオンを利用すると再活性化は観察さ
れなかった。同様に、完全な活性型QCと、これらのイオンとのインキュベートは、該酵
素活性に影響しなかった。
(図面の簡単な説明)
本発明のこれらおよび他の態様のさらなる理解は、以下の図を参照することより、得ら
れるであろう。
【図面の簡単な説明】
【0161】
【図1】図1は、ヒトQCにより触媒されるH-Gln-Ala-OHの環化反応のプログレス曲線を示し、340 nmにおける吸光度の低下をモニターする。該試料は、0.3 mM NADH/H+、14 mM α-ケトグルタル酸、30 U/mlグルタミン酸デヒドロゲナーゼ、及び1 mM H-Gln-Ala-OHを含んだ。曲線A〜Dには、様々な濃度のQCを加えて:その濃度はA、10mU/ml、B、5 mU/ml、C、2.5 mU/mlであった。曲線Dの場合は、QCは除いた。QC濃度、及び観察された活性との間に、直線関係を得た(挿入図)。
【図2】図2は、Gln-βNAを基質として用い、一次反応速度条件下で決定されたヒト、及びパパイヤ(挿入図)QCのpH依存性を示す。ヒトQCの場合、Ellis、及びMorrisonに従い、25 mM MES、25 mM 酢酸、及び50 mMトリスからなる一定イオン強度をもたらす緩衝系を用いた(Ellis, K. J.、及びMorrison, J. F.の論文、1982 Methods Enzymol. 87, 405-426)。トリスのわずかな阻害効果のため、パパイヤQCは、50 mM Mops緩衝液を用いて調べた。イオン強度はNaClの添加によって0.05 Mに調整した。速度プロファイルは、解離基に基づくモデルに適合させることにより評価した。パパイヤQCの場合、データを単一解離モデルに適合させることにより、pKa値7.13±0.03を得た。
【0162】
【図3】図3は、パパイヤラテックス、及びヒトQCの安定性における、pHの影響を示す。酵素原液は、様々なpH値の0.1 M緩衝液に20倍希釈した(pH 4〜7のクエン酸ナトリウム緩衝液、及びpH 7〜10のリン酸ナトリウム緩衝液)。酵素溶液は、30℃で30分間インキュベートし、その後、酵素活性を標準プロトコルに従って解析した。
【図4】図4は、2位のアミノ酸に、グルタミン酸を含有する一組の基質における、特異性定数kcat/KMの比較を示す。ジからテトラペプチドへと、ヒトQCの特異性上昇が検出された一方、パパイヤQCの場合は、変化は観察されなかった。ここに示されたデータは、表3で与えられたパラメータの再プロットである。
【図5】図5は、ヒトQCにより触媒される、H-Gln-Lys(Gln)-Arg-Leu-Ala-NH2からのpGlu-Lys(pGlu)-Arg-Leu-Ala-NH2の産生を示す。基質の転換は、アンモニア放出に起因する、m/z比における時間依存的変化によりモニターする。試料組成は、40 mMトリス/HCl溶液(pH 7.7)における0.5 mM基質、38 nM QCであった。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。パパイヤQCの場合にも、ごく同様の依存性が観察された。
【0163】
【図6】図6は、パパイヤQCにより触媒される、H-Gln(NMe)-Phe-Lys-Ala-Glu-NH2からのpGlu-Phe-Lys-Ala-Glu-NH2の産生を示す。基質の転換は、メチルアミンの放出による、m/z比の時間依存性変化によってモニターする。試料組成は、40 mMトリス/HCl溶液(pH 7.7)における0.5 mM基質、0.65μMパパイヤQCであった。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。パパイヤQCを含まない試料において、又は該基質に、1.5μMまでのヒトQCを添加することによって、基質の転換は観察されなかった(図示せず)。
【図7】図7は、DPIVにより触媒される、[Gln3]Aβ1-11から[Gln3]-Aβ3-11の産生を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図8】図8は、DP IVインヒビター、Val-ピロリジド(Val-Pyrr)による[Gln3]Aβ1-11の切断の抑制を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【0164】
【図9】図9は、QCにより触媒される、[Gln3]Aβ3-11から[pGlu3]Aβ3-11の産生を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図10】図10は、該QCインヒビター、1,10-フェナントロリンによる、[Gln3]Aβ3-11から[pGlu3]Aβ3-11の産生阻害を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図11】図11は、DP IV、及びQCによる連続触媒後の、[Gln3]Aβ1-11から[pGlu3]Aβ3-11の産生を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図12】図12は、触媒活性型DP IV、及びQCの存在下における、QCインヒビター、1,10-フェナントロリンによる、[Gln3]Aβ1-11から[pGlu3]Aβ3-11産生の阻害を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【0165】
【図13】図13は、触媒活性型DP IV、及びQCの存在下における、DP IVインヒビター、Val-Pyrrによる、[Gln3]Aβ1-11から[pGlu3]Aβ3-11産生の低下を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図14】図14は、ブタ脳下垂体ホモジネートに存在するアミノペプチダーゼ、及びQCによる連続触媒作用後の、[Gln3]Aβ1-11から[pGlu3]Aβペプチド3-11の産生を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図15】図15A、及びBは、使用前に10分間煮沸した、組み換えヒトQCとインキュベートされたAβ3-11a、及びAβ3-21aの質量スペクトルを示す。C、及びDは、活性型ヒトQC存在下における、Aβ3-11、及びAβ3-21aの質量スペクトルを示し、それらは各々、[pGlu3]Aβ3-11a、及び[pGlu3]Aβ3-21aの産生をもたらす。E、及びFは、活性型QC、及び[pGlu3]産生を抑制する5 mMベンズイミダゾール存在下における、Aβ3-11a、及びAβ3-21aの質量スペクトルを示す。
【0166】
【図16】図16は、基質の濃度に対してプロットされた、パパイヤQCに触媒されるGlu-βNA転換の反応速度を示す。初速度は、0.1 Mピロリン酸緩衝液(pH 6.1)(四角)、0.1 Mリン酸緩衝液(pH 7.5)(丸)、及び0.1 Mホウ酸緩衝液(pH 8.5)(三角)で測定した。速度パラメータは、下記のとおりであり:KM=1.13±0.07 mM、kcat=1.13±0.04 min-1(pH 6.1);KM=1.45±0.03 mM、kcat=0.92±0.01 min-1(pH 7.5);KM=1.76±0.06 mM、kcat=0.56±0.01 min-1(pH 8.5)であった。
【図17】図17は、低一次反応速度条件下(S<<KM)で決定された、Gln-βNA(丸)、及びGlu-βNA(四角)の転換のpH依存性を示す。基質濃度は、各々、0.01 mM、及び0.25 mMであった。両決定には、0.05 M酢酸、0.05 Mピロリン酸、及び0.05 Mトリシンを含む、三成分緩衝系を使用した。全ての緩衝液は、イオン強度の相違を防ぐため、NaClの添加により等電気伝導度に調整した。該データは、2種の解離基を説明する公式に適合し、pKa値は、Gln-βNAにおいては、6.91±0.02、及び9.5±0.1、及びGlu-βNAにおいては4.6±0.1、及び7.55±0.02であった。滴定により決定された、各々の基質アミノ基のpKa値は、6.97±0.01(Gln-βNA)、及び7.57±0.05(Glu-βNA)であった。全ての決定は、30℃で行った。
【図18】図18は、イミダゾール、及びジピコリン酸存在下、及び阻害化合物非存在下における、ヒトQCに触媒されるH-Gln-AMCの環化反応の経時変化曲線を示す。ジピコリン酸存在下のハイパーボリック(hyperbolic)型は、QC活性部位からの金属イオンの除去を示唆する。
【0167】
【図19】図19は、複素環キレーター、1,10-フェナントロリンによる、QCの時間依存的不活性化を示す。該QC酵素と、基質非存在下におけるインヒビターとのインキュベート後(実線)では、インヒビターと予めインキュべートしなかった試料(点跡)に比べて、酵素活性低下が観察され、これはQC活性部位からの金属イオンの除去を示唆する。
【図20】図20は、一価、及び一価金属イオンとの、ヒトQCの再活性化を示す。QCは、50 mMビス-トリス(pH 6.8)中の2 mMジピコリン酸の添加により、不活性化した。その後、該酵素を、1.0 mM EDTAを含む、50 mM ビス-トリス(pH 6.8)に対して透析させた。該酵素の再活性化は、該不活性化された酵素試料と、0.5 mM EDTA存在下における、0.5 mMの濃度の金属イオンとのインキュベートにより達成し、緩衝溶液に存在する微量の金属イオンにより、非特異的再活性化を防いだ。コントロールは、不活性化しなかったが、不活性化された酵素としてEDTA溶液に対して透析した酵素試料(+EDTA)、及びEDTAを添加しない緩衝溶液に対して透析した酵素試料(-EDTA)により示された。
【図21】図21は、ヒトQC(hQC)、及び他のメタロペプチダーゼClan MHのM28ファミリー員の配列アライメントを示す。多重配列アライメントは、ch. EMBnet. orgにおけるClustalWをデフォルト設定で使用し、行なった。ヒトQC(hQC;GenBank X71125)、放線菌(Streptomyces griseus)由来Zn依存性アミノペプチダーゼ(SGAP;Swiss-Prot P80561)、及びヒトグルタミン酸カルボキシペプチダーゼII(hGCP II;Swiss-Prot Q04609)のN-アセチル化-α-結合酸性ジペプチダーゼ(N-acetylated-alpha-linked acidic dipeptidase)(NAALADase I)ドメイン(残基274〜587)内おける、亜鉛イオン結合残基の保存を示す。金属結合に関与する該アミノ酸は、太字でタイプし、下線を引いた。ヒトQCの場合は、これらの残基は、該ペプチダーゼへの推定相応残基である。
【技術分野】
【0001】
(発明の分野)
本発明は、アンモニアの遊離下における、N末端グルタミン残基のピログルタミン酸(
5-オクソ-プロリン、pGlu*)への分子内環化反応、及び水の遊離下における、N末端グル
タミン酸残基のピログルタミン酸への分子内環化反応を触媒する、グルタミニルシクラー
ゼ(QC、EC 2.3.2.5)に関するものである。
本発明は、哺乳動物QCを金属酵素として同定し、哺乳動物における新規の生理学的基
質、及びQC活性の調整によって治療可能な病態の治療のための、QCのエフェクターの
使用、及びQCのエフェクターを含む医薬組成物を提供する。さらに、金属相互作用が、
QCインヒビターの開発に有用なアプローチであることが示される。
好ましい実施態様において、本発明は、QC、及び/又はDP IV活性の調整によって治
療可能な病態の治療、又は緩和のために、DP IV、又はDP IV様酵素のインヒビターを組合
せるQC活性のエフェクターの使用を提供する。
また、QC活性のエフェクターの同定、及び選定のためのスクリーニング方法を提供す
る。
【背景技術】
【0002】
(背景)
グルタミニルシクラーゼ(QC、EC 2.3.2.5)は、アンモニアの遊離を伴う、N末端グ
ルタミン残基のピログルタミン酸(pGlu*)への分子内環化反応を触媒する。QCは、196
3年にMesserが熱帯植物パパイヤ(Carica papaya)のラテックスからはじめて精製された
(Messer, Mの論文、1963 Nature 4874, 1299)。24年後、類似の酵素活性が、動物脳
下垂体において発見された(Busby, W. H. J.らの論文、1987 J Biol Chem 262, 8532-85
36;Fischer, W. H. 及びSpiess, J.の論文1987 Proc Natl Acad Sci U S A 84, 3628-36
32)。該哺乳動物QCにおいて、QCによる pGluへのGln の転換は、TRH 及びGnRHの前
駆体に見ることができる(Busby, W. H. J.らの論文1987 J Biol Chem 262, 8532-8536;
Fischer, W. H. 及びSpiess, J.の論文、1987 Proc Natl Acad Sci U S A 84, 3628-3632
)。その上、QCの初期局在実験によって、ウシ脳下垂体における触媒の推定的産物との
同一場所での局在が明らかになり、さらに、ペプチドホルモン合成における示唆された機
能が裏付けられた(Bockers, T. M.らの論文、1995 J Neuroendocrinol 7, 445-453)。
これに対し、植物QCの生理学的機能は、明らかでない。パパイヤ由来の該酵素の場合、
病原微生物に対する植物防御における役割が示唆された(El Moussaoui, A.らの論文、20
01 Cell Mol Life Sci 58, 556-570)。近年、配列比較によって、他の植物からの推定上
QCが同定された(Dahl, S. W.らの論文、2000 Protein Expr Purif 20, 27-36)。これ
らの酵素の生理学的機能は、依然として明らかとなっていない。
【0003】
植物、及び動物由来の公知のQCは、基質N末端のL-グルタミンに高い特異性を示し、
その速度論的挙動は、ミカエリス−メンテン式に従うことが見い出された(Pohl, T.らの
論文、1991 Proc Natl Acad Sci U S A 88, 10059-10063;Consalvo, A. P.らの論文、19
88 Anal Biochem 175, 131-138;Gololobov, M. Y.らの論文、1996 Biol Chem Hoppe Sey
ler 377, 395-398)。しかしながら、パパイヤ由来QC、及び保存性の高い哺乳動物由来
のQCの一次構造の比較では、配列の相同性がないことが明らかになった(Dahl, S. W.
らの論文、2000 Protein Expr Purif 20, 27-36)。該植物QCが、新しい酵素ファミリ
ーに属していると思われるのに対し(Dahl, S. W.らの論文、2000 Protein Expr Purif 2
0, 27-36)、哺乳動物QCは細菌アミノペプチダーゼと顕著な配列相同性をもつことが見
い出され(Bateman, R. C.らの論文、2001 Biochemistry 40, 11246-11250)、植物、及
び動物由来のQCは異なる進化上の起源をもつという結論が導かれた。
【0004】
欧州特許出願公開第02 011 349.4号には、昆虫グルタミニルシクラーゼをコードしてい
るポリヌクレオチド、それによってコードされるポリペプチドが開示されている。さらに
、この出願は、該発明のポリヌクレオチドを含む発現ベクターを含む宿主細胞を提供する
。昆虫QCを含む精製ポリペプチド、及び宿主細胞は、グルタミニルシクラーゼ活性を低
下させる薬剤のスクリーニング方法において有用である。そのような薬剤は、殺虫剤と同
様に有用であると記載されている。
【0005】
アルツハイマー病(AD)は、神経細胞変性、活性化アストロサイト、及び活性化マイク
ログリアが密接に関連する細胞外アミロイド斑の異常蓄積が特徴である(Terry, R. D.、
及びKatzman, R.の論文、1983 Ann Neurol 14, 497-506;Glenner, G. G.、及びWong, C.
W.の論文、1984 Biochem Biophys Res Comm 120, 885-890;Intagaki, S.らの論文、198
9 J Neuroimmunol 24, 173-182;Funato, H.らの論文、1998 Am J Pathol 152, 983-992
;Selkoe, D. J.の論文、2001 Physiol Rev 81, 741-766)。アミロイド-β(Aβ)ペプ
チドは老人斑の主要構成成分であり、ADの発病、及び進行、遺伝学的研究が支持する仮説
に直接関与していると考えられている(Glenner, G. G.、及びWong, C. W.の論文、1984
Biochem Biophys Res Comm 120, 885-890;Borchelt, D. R.らの論文、1996 Neuron 17,
1005-1013;Lemere, C. A.らの論文、1996 Nat Med 2, 1146-1150;Mann, D. M.、及びIw
atsubo, T.の論文、1996 Neurodegeneration 5, 115-120;Citron, M.らの論文、1997 Na
t Med 3, 67-72;Selkoe, D. J.の論文、2001 Physiol Rev 81, 741-766)。Aβは、β-
アミロイド前駆体タンパク質(APP)のタンパク質から(Kang, J.らの論文、1987 Nature
325, 733-736;Selkoe, D. J. 1998 Trends Cell Biol 8, 447-453)、AβのN末端でベ
ータセクレターゼ、及びC末端でのガンマセクレターゼにより配列切断されるタンパク質
分解プロセスによって生成される(Haass, C.、及びSelkoe, D. J.の論文 1993 Cell 75,
1039-1042;Simons, M.らの論文、1996 J Neurosci 16 899-908)。N末端がL-Aspで始
まる主要なAβペプチド(Aβ1-42/40)に加えて、N末端切断型の多異種混合が老人斑に
生じる。そのような短縮型ペプチドは、フルレングス型に比べてインビトロでより神経毒
性が高く、より急速に凝集することが報告された(Pike, C. J.らの論文、1995 J Biol C
hem 270 23895-23898)。N末端切断型ペプチドは、家族性アルツハイマー病(FAD)患者
において過剰生成されること(Saido, T. C.らの論文、1995 Neuron 14, 457-466;Russo
, C.らの論文、2000 Nature 405, 531-532)、ダウン症の脳で早期に出現し、加齢ととも
に増加することが知られている(Russo, C.らの論文、1997 FEBS Lett 409, 411-416, Ru
sso, C.らの論文、2001 Neurobiol Dis 8, 173-180;Tekirian, T. L.らの論文、1998 J
Neuropathol Exp Neurol 57, 76-94)。最後に、それらの量は該疾患の進行性重症度を反
映する(Russo, C.らの論文、1997 FEBS Lett 409, 411-416)。さらに、付加的翻訳後プ
ロセスは、1及び7位のアスパラギン酸の異性化、又はラセミ化によって、及び残基3及
び11位のグルタミン酸の環化によって、N末端を改質し得る。3位にピログルタミン酸
を含む異性体[pGlu3]Aβ3-40/42は、老人斑内のN末端切断種の顕著な型で、−全Aβ量
の約50%−を占め(Mori, H.らの論文、1992 J Biol Chem 267, 17082-17086;Saido,
T. C.らの論文、1995 Neuron 14, 457-466;Russo, C.らの論文、1997 FEBS Lett 409, 4
11-416;Tekirian, T. L.らの論文、1998 J Neuropathol Exp Neurol 57, 76-94;Geddes
, J. W.らの論文、1999 Neurobiol Aging 20, 75-79;Harigaya, Y.らの論文、2000 Bioc
hem Biophys Res Commun 276, 422-427)、かつ、それはアミロイド傷害以前にも存在す
る(Lalowski, M.らの論文、1996 J Biol Chem 271, 33623-33631)。AβN3(pE)ペプチ
ドの蓄積は、凝集を増進しほとんどのアミノペプチダーゼに対する耐性を付与する、構造
上の改質によるものと思われる(Saido, T. C.らの論文、1995 Neuron 14, 457-466 ;Te
kirian, T. L.らの論文、1999 J Neurochem 73, 1584-1589)。この証拠は、アルツハイ
マー病の発症機序におけるAβN3(pE)ペプチドのきわめて重要な役割の手掛かりを提供
する。しかしながら、それらの神経毒性、及び凝集特性は比較的ほとんど知られていない
(He, W.、及びBarrow, C. J.の論文、1999 Biochemistry 38, 10871-10877;Tekirian,
T. L.らの論文、1999 J Neurochem 73, 1584-1589)。さらに、活性化グリアは厳密に老
人斑に関連しており、アミロイド沈着の凝集に積極的に寄与しうるが、該イソ型のグリア
細胞への作用、及び該ペプチドへのグリア反応は全く知られていない。近年の研究では、
Aβ1-42、Aβ1-40、[pGlu3]Aβ3-42、及び[pGlu3]Aβ3-40ペプチドの毒性、凝集特性、及
び異化を神経細胞培養、及びグリア細胞培養で調べ、ピログルタミン酸改質がAβ-ペプチ
ドの毒性を高め、さらに培養アストロサイトによる該ペプチドの分解を阻害することが示
された。Shirotaniらは、インビトロでシンドビスウィルスにより感染した初代皮質神経
細胞における[pGlu3]Aβ-ペプチドの産生を調べた。彼らは、アミノ酸置換、及びアミノ
酸欠失により、[pGlu3]Aβの潜在前駆体をコードする相補的DNAを作成した。本来の前駆
体のグルタミン酸の代わりにN末端グルタミン残基ではじまる1種の人工前駆体では、グ
ルタミニルシクラーゼによるピログルタミン酸への自然転換、又は酵素的転換が示唆され
た。[pGlu3]Aβの本来の前駆体における3位のN末端グルタミン酸の環化機構は、インビ
ボでは確定されなかった(Shirotani, K.、Tsubuki, S.、Lee, H. J.、Maruyama, K.、及
びSaido, T. C.の論文、(2002)Neurosci Lett 327, 25-28)。
【0006】
ジペプチジルペプチダーゼIV(DP IV)は、腎臓、肝臓、及び腸を含む多様な人体組織
で見出される、プロリンの後(より少ない程度にアラニンの後、セリンの後、又はグリシ
ンの後)を開裂するセリンプロテアーゼであり、かつペプチド鎖からN末端のジペプチド
を開裂する。最近、DP IVが神経ペプチド代謝、T細胞活性化、癌細胞の内皮への接着、
及びHIVのリンパ球様細胞への侵入に重要な役割を果たすことが示された。それについて
は、国際公開第02/34242号、国際公開第02/34243号、国際公開第03/002595号、及び国
際公開第03/002596号を参照されたい。
【0007】
DP IVインヒビターは、グルコース耐性障害、及び糖尿病の治療に有用であり得ること
が知られている(国際特許出願、公開番号、国際公開第99/61431号、Pederson, R. A.ら
の論文(1998 Diabetes 47, 1253-1258)、及びPauly, R. P.らの論文(1999 Metabolism
48, 385-389))。特に、国際公開第99/61431号では、アミノ酸残基、及びチアゾリジ
ン、又はピロリジン群、及びその塩、特にL-threo-イソロイシルチアゾリン、L-allo-イ
ソロイシルチアゾリン、L-threo-イソロイシルピロリジン、L-allo-イソロイシルチアゾ
リン、L-allo-イソロイシルピロリジン、及びその塩を含むDP IVインヒビターを開示する
。
【0008】
さらなる低分子量ジペプチジルペプチダーゼIVインヒビターの例には、テトラヒドロイ
ソキノリン-3-カルボキサミド誘導体、N-置換2-シアノピロール、及びピロリジン、N-(N
'-置換グリシル)2-シアノピロリジン、N-(置換グリシル)-チアゾリジン、N-(置換グ
リシル)-4-シアノチアゾリジン、アミノ-アシル-ボロノ-プロピル-インヒビター、シク
ロプロピル-融合ピロリジン、及び六員環化合物のような薬剤がある。ジペプチジルペプ
チダーゼIVインヒビターは、下記の文献に記載されている:米国特許第6,380,398号、米
国特許第6,011,155号、米国特許第6,107,317号、米国特許第6,110,949号、米国特許第6,1
24,305号、米国特許第6,172,081号、国際公開第 95/15309号、国際公開第99/61431号、
国際公開第99/67278号、国際公開第99/67279号、ドイツ特許第198 34 591号公報、国際
公開第97/40832号、ドイツ特許第196 16 486 C 2号公報、国際公開第98/19998号、国際
公開第00/07617号、国際公開第99/38501号、国際公開第99/46272号、国際公開第99/3
8501号、国際公開第01/68603号、国際公開第01/40180号、国際公開第01/8133号、国際
公開第01/81304号、国際公開第01/55105、国際公開第02/02560号、及び国際公開第02
/14271号、国際公開第02/04610号、国際公開第02/051836号、国際公開第02/068420号
、国際公開第02/076450号、国際公開第02/083128号、国際公開第02/38541号、国際公
開第03/000180号、国際公開第03/000181号、国際公開第03/000250号、国際公開第03/
002530号、国際公開第03/002531号、国際公開第03/002553号、国際公開第03/002593号
、国際公開第03/004496号、国際公開第03/024942号、及び国際公開第03/024965号であ
る。これらの文献に教示されている内容は、その全体、特にインヒビター、その定義、使
用、及び産生に関して、引用により本明細書に取り込まれるものとする。
【発明の概要】
【発明が解決しようとする課題】
【0009】
(本発明の要旨)
本発明は、哺乳動物QCの新規生理学的基質、Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu
11]Aβ11-40/42、[Gln11]Aβ11-40/42、[Gln1]ガストリン、[Gln1]ニューロテンシン、
[Gln1]FPP、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 18、[G
ln1]フラクタルカイン、[Gln1]オレキシンA、[Gln3]グルカゴン3-29、及び[Gln5]サブス
タンスP5-11、及びQC活性の調節によって治療可能な病態の治療のための、QCエフェ
クターの使用、及びQCエフェクターを含む医薬組成物を提供する。
【0010】
阻害研究により、ヒトQCは、金属依存性トランスフェラーゼであることが示された。
QCのアポ酵素は、亜鉛イオンにより最も効果的に活性化され得て、かつ、亜鉛依存性ア
ミノペプチダーゼの金属結合モチーフは、ヒトQCにも存在する。金属と結合する活性部
位と相互作用する化合物は、有力なインヒビターである。
【0011】
予想外に、組み換えQCは、脳抽出物由来QCの活性と同様に、N末端グルタミニルと
同様グルタミン酸環化反応の両方を触媒することが示された。最も特筆すべきことは、シ
クラーゼ触媒のGlu1-転換がpH 6.0付近で活発なのに対し、pGlu-誘導体へのGln1-転換は
至適pH 8.0付近で起こる。pGlu-Aβに関連するペプチドの産生は、組み換えヒトQC、及
びブタ脳下垂体抽出物由来QCの活性阻害によって抑制できるので、該酵素QCは、アル
ツハイマー病治療に用いる薬剤開発の標的である。
【0012】
QC活性のエフェクターを哺乳動物に投与することによって、アルツハイマー病、ダウ
ン症、ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は伴わない、潰瘍性疾患及び胃
癌、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、悪
性転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性
免疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネル
ギー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害か
ら選択される病態を、予防、又は緩和、又は治療することが可能である。
【0013】
さらに、哺乳動物へのQC活性のエフェクターの投与により、胃腸管細胞増殖、好まし
くは、胃粘膜細胞、上皮細胞の増殖、急性の酸分泌、及び酸産生壁細胞、及びヒスタミン
分泌クロム親和性様細胞分化を刺激促進することが可能である。
さらに、哺乳動物へのQC活性のエフェクターの投与により、骨髄性前駆細胞増殖を抑
制することが可能である。
さらに、QCインヒビターの投与は、男性生殖能力の抑制をもたらすことができる。
【0014】
好ましい実施態様において、本発明は、QC、及び/又はDP IV活性を調整することに
よって治療可能な病態の治療、又は緩和のために、DP IV、又はDP IV様酵素のインヒビタ
ーと組合せる、QC活性のエフェクターの使用を提供する。
本発明は、任意に慣用の担体、及び/又は賦形剤と共に少なくとも1種のQCのエフェ
クターを含むか;又は任意に慣用の担体、及び/又は賦形剤と共に少なくとも1種のQC
のエフェクターを、少なくとも1種のDP IVインヒビターと共に含む、非経口、経腸、又
は経口投与に用いる医薬組成物を提供する。
また、QCエフェクターの同定、及び選定のためのスクリーニング方法を提供する。
【0015】
(ペプチド配列)
本明細書中で言及し、かつ使用したペプチドは、下記の配列を有する。
Aβ1-42:
Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-
Ala-Glu-Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val-
Ile-Ala
Aβ1-40:
Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-
Ala-Glu-Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val
Aβ3-42:
Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-Glu-
Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val-Ile-Ala
Aβ3-40:
Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-Glu-
Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val
Aβ1-11a:
Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-NH2
Aβ3-11a:
Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-NH2
【0016】
Aβ1-21a:
Asp-Ala-Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-
Ala-NH2
Aβ3-21a:
Glu-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-NH2
Gln3-Aβ3-40:
Gln-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-Glu-
Asp-Val-Gly-Ser-Asn-Lys-Gly-Ala-Ile-Ile-Gly-Leu-Met-Val-Gly-Gly-Val-Val
Gln3-Aβ3-21a:
Gln-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-Val-His-His-Gln-Lys-Leu-Val-Phe-Phe-Ala-NH2
Gln3-Aβ1-11a:
Asp-Ala-Gln-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-NH2
Gln3-Aβ3-11a:
Gln-Phe-Arg-His-Asp-Ser-Gly-Tyr-Glu-NH2
【課題を解決するための手段】
【0017】
(発明の詳細な説明)
本発明は、a)インビボQC活性の調整によって治療可能な、哺乳動物の疾患の治療、
及び/又は、b)QC活性の調整によって引き起こされる、pGlu含有ペプチドの活性に基
づく生理的プロセスの調整のための、グルタミニルシクラーゼ(QC)のエフェクターを
提供する。
さらに、本発明は哺乳動物におけるグルタミニルシクラーゼ(QC、EC 2.3.2.5)、及
び/又はQC様酵素の阻害に用いる化合物を提供し、かつQC活性に関わる病態の治療の
ための、QC活性のインヒビターの使用を提供する。
【0018】
また、本発明はアルツハイマー病、及びダウン症の新規な治療方法を提供する。アルツ
ハイマー病、及びダウン症の脳に沈着したアミロイドβ-ペプチドのN末端は、ピログル
タミン酸を有する。pGluの産生は、該疾患の発症、及び進行における重要な事象である。
その理由は、改質されたアミロイドβ-ペプチドは、β-アミロイドの凝集、及び毒性を増
大し、疾患の発病、及び進行を悪化させ得るからである(Russo, C.らの論文、2002 J Ne
urochem 82, 1480-1489)。
【0019】
これに対し、本来のAβ-ペプチド(3-40/42)においては、グルタミン酸がN末端アミ
ノ酸として存在する。今日までに公知のGluのpGluへの酵素転換は存在しない。さらに、G
luペプチドのpGluペプチドへの自然環化反応は、今までのところ観察されていない。した
がって、本発明の一態様は、アルツハイマー病、及びダウン症におけるQCの役割を決定
することであった。この一態様においては、3位のアミノ酸としてグルタミン酸の代わり
にグルタミンを含む、Aβ3-11、及びAβ1-11の合成、これらの改質アミロイドβ-ペプチ
ドのQC、DP IV、及びDP IV様酵素、及びアミノペプチダーゼに対する基質特性の決定、
及び、QCインヒビターを使用して、該アミロイドβ-誘導体ペプチド1-11、及び3-11の
N末端グルタミン残基からpGluの産生を抑制し、解決することに向けられている。該結果
を、実施例8に示す。適用された方法は、実施例3に記載されている。
【0020】
今日まで、該疾患の進行におけるQCの関与が全く示唆されていない。その理由は、グ
ルタミン酸がAβ(3-40/42、及び11-40/42)におけるN末端アミノ酸だからである。し
かし、QCはペプチドのN末端でpGluを産生できる唯一公知の酵素である。本発明の他の
形態は下記の知見、及び発見に関与している:
a)副反応として、QCはごく低速度でグルタミン酸のピログルタミン酸への環化反応を
触媒する、
b)APP、又はその後に形成されるアミロイドβ-ペプチドのグルタミン酸は、未知の酵素
活性により翻訳後にグルタミンに転換され、かつ第二工程として、QCはグルタミンのピ
ログルタミン酸への環化反応を、アミロイドβ-ペプチドN末端のプロセス後、触媒する
、
c)グルタミン酸は化学触媒、又は自己触媒により、翻訳後にグルタミンに転換され、か
つ続いて、該アミロイドβ-ペプチドN末端のプロセス後、QCは、グルタミンのピログ
ルタミン酸への環化反応を触媒する、
d)アミロイドβ-タンパク質をコードするAPP遺伝子において、3位のGluの代わりにGln
をもたらす突然変異がある。翻訳、及び該N末端のプロセシング後、QCはグルタミンの
ピログルタミン酸への該環化反応を触媒する、
e)グルタミンは、未知の酵素活性の異常により、APPの未完成のペプチド鎖に取り込ま
れ、かつ続いて、該アミロイドβ-ペプチドN末端のプロセシング後、QCは、N末端グ
ルタミンのピログルタミン酸への環化反応を触媒する。
【0021】
QCは上記全5事例における重要な工程、すなわちアミロイドβ-ペプチドの凝集を促
進するピログルタミン酸の産生に関与している。このように、QCの阻害は、環化反応が
起こる機構に関わらず、アルツハイマー病、及びダウン症の発症と進行を起こす、斑形成
型Aβ3-40/41/43、又はAβ11-40/41/43の沈着の予防を可能にする。
グルタミン酸は、アミロイドβ-ペプチドの3、11、及び22位に見い出される。そ
の中で、22位のグルタミン酸(E)からグルタミン(Q)への突然変異(アミロイド前駆
体タンパク質APP 693、スイスプロットP05067に相当)は、いわゆるオランダ型脳動脈ア
ミロイドーシス突然変異として記載されている。
3、11、及び22位にピログルタミン酸を有するβ-アミロイドペプチドは、A β1-4
0/42/43に比べて、細胞毒性、及び疎水性がより高いことが記載されている(Saido T.C
.の論文、2000 Medical Hypotheses 54(3):427-429)。
【0022】
多種のN末端変異は、異なる部位でのβ-セクレターゼ酵素、β-部位アミロイド前駆体
タンパク質-切断酵素(BACE)(Huse J.T.らの論文、2002 J. Biol. Chem. 277(18):1
6278-16284)、及び/又はアミノペプチダーゼのプロセシングによって、産生が可能であ
る。全ての場合において、先に記載したa)〜e)の経路によって環化反応が起こり得る
。
【0023】
今までのところ、経路a)に相当する、未知のグルタミルシクラーゼ(EC)による、
Glu1ペプチドのpGluペプチドへの酵素的転換を支持する実験的証拠はない(Garden, R. W
.、Moroz, T. P.、Gleeson, J. M.、Floyd, P. D.、Li, L. J.、Rubakhin, S. S.、及びS
weedler, J. V.の論文(1999)J Neurochem 72, 676-681;Hosoda R.らの論文(1998)J
Neuropathol Exp Neurol. 57, 1089-1095)。今日までに、弱アルカリpH条件下で、N末
端側がプロトン化され、かつ負電荷のGlu1γ-カルボン酸部分をもつGlu1ペプチドを、環
化できるような酵素活性は同定されていない。
【0024】
Gln1基質に対するQC活性は、pH 7.0以下で劇的に低下する。これに対し、Glu1転換は
酸性反応条件で起こり得る(Iwatsubo, T.、Saido, T. C.、Mann, D. M.、Lee, V. M.、
及びTrojanowski, J. Q.の論文、(1996)Am J Pathol 149, 1823-1830;Russo, C.、Sai
do, T. C.、DeBusk, L. M.、Tabaton, M.、Gambetti, P.、及びTeller, J. K.の論文、(
1977)FEBS Lett 409, 411-416;Russo, C.、Salis, S.、Dolcini, V.、Venezia, V.、So
ng, X. H.、Teller, J. K.、及びSchettini, G.の論文(2001)Neurobiol Dis 8, 173-1
80;Tekirian, T. L.、Saido, T. C.、Markesbery, W. R.、Russell, M. J.、Wekstein,
D. R.、Patel, E.、及びGeddes, J. W.の論文(1998)J Neuropathol Exp Neurol. 57, 7
6-94;Russo, C.、Violani, E.、Salis, S.、Venezia, V.、Dolcini, V.、Damonte, G.、
Benatti, U.、DArrigo, C.、Patrone, E.、Carlo, P.、及びSchettini, G.の論文(2002
)J Neurochem 82, 1480-1489;Hosoda, R.、Saido, T. C.、Otvos, L., Jr.、Arai, T.
、Mann, D. M.、Lee, V. M.、Trojanowski, J. Q.、及びIwatsubo, T.の論文(1998)J N
europathol Exp Neurol. 57, 1089-1095;Garden, R. W.、Moroz, T. P.、Gleeson, J. M
.、Floyd, P. D.、Li, L. J.、Rubakhin, S. S.、及びSweedler, J. V.の論文(1999)J
Neurochem 72, 676-681)。
【0025】
本発明に従って、QCが弱酸性条件下でアミロイド-β誘導ペプチドを認識し、かつ代
謝回転できるか否かを調べた。したがって、該酵素の潜在的基質として、[Gln3]Aβ1-11a
、Aβ3-11a、[Gln3]Aβ3-11a、Aβ3-21a、[Gln3]Aβ3-21a、及び[Gln3]Aβ3-40 を合成し
、かつ調べた。これらの配列は、翻訳後のGluアミド化が原因で起こり得る、本来のN末
端、及びC末端切断型[Glu3]Aβペプチド、及び[Gln3]Aβペプチドを模倣するように選択
された。
【0026】
本発明において、パパイヤ、及びヒトQCがグルタミニル、及びグルタミン酸環化反応
の両方を触媒することを示した。明らかに、QCの主要生理学的機能は、ホルモン分泌プ
ロセスに先立つ、又はその間の、グルタミン環化反応による内分泌細胞内のホルモン成熟
を完成させることである。そのような分泌小胞は、酸性pHであることが知られている。従
って、pH 5.0〜7.0の狭いpH範囲内での該酵素の副活性は、Glu-Aβペプチドをも転換する
、新規に発見されたグルタミルシクラーゼの活性であり得る。しかしながら、Gln環化に
比べてGlu環化はかなり遅く起こるので、該グルタミルシクラーゼが有意義な生理的役割
を果たすかどうかは、疑問の余地がある。
【0027】
しかしながら、神経変性疾患の病理においては、該グルタミル環化反応は有意義である
。該酵素反応のpH依存性を研究し、我々はプロトン化されていないN末端がGln1ペプチド
の環化反応に不可欠であり、かつ従って、該基質のpKa値は、QC触媒のpKa値と同一であ
ることを見いだした(図17参照)。このように、QCは、プロトン化されていないα-
アミノ部分の、アミド化により親電子的に活性化されたγ-カルボニル炭素への分子内求
核攻撃を安定化させる(スキーム1)。
【0028】
N末端グルタミン含有ペプチドに存在する一価の電荷に対して、Glu含有ペプチドにお
けるN末端Glu残基は、中性付近pHで、大部分は二価に荷電される。グルタミン酸は、γ-
カルボキシル、及びα-アミノ部位に対し、各々、約4.2、及び7.5のpKa値を示した。すな
わち、中性、及びそれ以上のpHでは、該α-アミノ窒素は、部分的に、又は全部プロトン
化されず、かつ求核的であるが、γ-カルボキシル基はプロトン化されず、かつそれによ
り親電子的カルボニル活性を発揮しない。したがって、分子内環化反応は不可能である。
【0029】
しかしながら、その各々のpKa値の間、pH約5.2〜6.5の範囲において、全N末端Glu含有
ペプチドの約1〜10%(-NH2)、又は10〜1%(-COOH)の濃度で、2官能基は両方非イオ
ン化型で存在する。その結果として、弱酸性pH範囲間で、電荷していない両基を持つN末
端Glu含有ペプチド種が存在し、かつ従って、QCがpGluペプチドへの分子内環化反応の
中間体を安定化させることが可能である。すなわち、もし、γ-カルボキシル基がプロト
ン化していると、カルボニル炭素は親電子的で、プロトン化されていないα-アミノ基に
よる親核攻撃が可能になる。このpHにおいて、水酸基イオンは脱離基として機能する(ス
キーム3)。これらの仮説は、QCが触媒するGlu-βNAの転換のために得られたpH依存性
のデータによって確認された(実施例11参照)。QCによるGln-βNAのグルタミン転換に
対し、該触媒の至適pHはpH 6.0付近の酸性範囲、すなわち、基質分子種がプロトン化され
たγ-カルボキシル基、及びプロトン化されていないα-アミノ基を同時に十分に持って存
在するようなpH範囲に移動する。さらに、速度論的に決定したpKa値7.55±0.02は、滴定
により決定されたGlu-βNAのα-アミノ基のpKa値に極めてよく一致している(7.57±0.05
)。
【0030】
生理学的に、pH 6.0で、QCが触媒するグルタミン酸環化反応の二次速度定数(又は特
異性定数、kcat/KM)は、グルタミン環化反応の該定数より、8,000倍程度遅くなり得る
(図17)。しかしながら、Glu-βNA、及びGln-βNAの両モデル基質の非酵素的転換がご
くわずかであることは、本発明において観察されたごくわずかなpGluペプチド産生と一致
する。それゆえ、QCによるpGluの産生において、酵素対非酵素の速度定数の比から、少
なくとも108の加速を推定することが可能である(該酵素触媒の該二次速度定数を、各非
酵素環化反応の一次速度定数と比較すると、Gln転換、及びGlu転換における触媒有効係数
は、各々109〜1010 M-1である)。これらのデータからの結論は、pGlu産生をもたらす酵
素経路は、インビボでのみあり得ると思われる。
【0031】
QCは脳内に非常に豊富に存在しており、かつ最近見い出された、30μM(Gln-)TRH様
ペプチドを成熟するための高代謝回転(Prokai, L.、Prokai-Tatrai, K.、Ouyang, X.、K
im, H. S.、Wu, W. M.、Zharikova, A.、及びBodor, N.の論文、(1999)J Med Chem 42,
4563-4571)であるから、同様の反応条件下での適切なグルタミン酸基質には、約100時
間の環化反応半減期を予測し得る。さらに、分泌経路における脳QC/ECの局在化、お
よび位置を考慮すると、実際のインビボ酵素、及び基質濃度、及び反応条件は、正常細胞
の酵素的環化反応において、なおさら好都合であり得る。かつ、N末端GluがGlnに転換さ
れているなら、QCにより調節される、かなり急速なpGluの産生を予想し得る。インビト
ロにおいて、両反応はQC/EC活性のインヒビターを適用することによって抑制された
(図9、10、及び15)。
【0032】
要約すると、本発明は、脳内にかなり豊富に存在するヒトQCが、Glu-Aβ、及びGln-A
β前駆体から、アルツハイマー病で見い出される斑沈着の50%以上を構成する、アミロ
イド生成性pGlu-Aβ-ペプチドを産生する有望な触媒であることを示した。これらの知見
は、QC/ECを老人斑形成に関わる一因子と同定し、かつ従って、アルツハイマー病の
治療における新規な薬剤標的と同定した。
【0033】
本発明の第二の実施態様において、アミロイドβ-誘導ペプチドが、ジルペプチダーゼI
V(DP IV)、又はDP IV様酵素、好ましくはジペプチジルペプチダーゼII(DP II)の基質
であることを見い出された。DP IV、DP II、又はDP IV様酵素は、改質したアミロイドβ-
ペプチド(1-11)のN末端からジペプチドを放出し、グルタミンをN末端アミノ酸残基に
持つアミロイドβ-ペプチド(3-11)を産生する。その結果は、実施例8に示した。
【0034】
DP II、DP IV、又は他のDP IV様酵素の切断に先立ち、アスパラギン酸(アミロイドβ-
ペプチドの1番めの残基)、及びアラニン(アミロイドβ-ペプチドの2番めの残基)間
のペプチド結合は、文献に記載されているように、異性化されイソアスパルチル残基を生
じ得る(Kuo, Y.-M.、Emmerling, M. R.、Woods, A. S.、Cotter, R. J.、及びRoher, A.
E.の論文、(1997)BBRC 237, 188-191;Shimizu, T.、Watanabe, A.、Ogawara, M.、Mo
ri, H.、及びShirasawa, T.の論文、(2000)Arch. Biochem. Biophys. 381, 225-234)
。
これらのイソアスパラチル残基は、該アミロイドβ-ペプチドにアミノペプチダーゼ分
解に対する耐性を与え、かつその結果として、核化老人斑は多量のisoAsp1アミロイドベ
ータペプチドを含んでおり、それはN末端における低下した代謝回転を示唆する。
【0035】
しかしながら、本発明において初めて、N末端ジペプチドH-isoAsp1-Ala2-OHが、特に
酸性条件下でジペプチダーゼにより放出されることを示した。さらにまた、異性化はβ-
セクレターゼによる切断に先行することが可能であり、かつ該異性化はタンパク質分解を
促進し、isoAsp1アミロイドβ-ペプチドのN末端イソアスパルチル結合の遊離を可能にし
、次いでDP II、DPIV、又は DP IV様酵素により代謝回転され得ることが示された(Moman
d, J.、及びClarke, S.(1987)Biochemistry 26, 7798-7805;Kuo, Y.-M.、Emmerling,
M. R.、Woods, A. S.、Cotter, R. J.、Roher, A. E.の論文、(1997)BBRC 237, 188-19
1)。従って、イソアスパルチル産生の阻害は、β-セクレターゼによる切断の減少を可能
にし、かつ順に、アミロイドβ-ペプチドの産生の減少を可能にし得る。その上、isoAsp1
アミロイドβ-ペプチドのDP II、DPIV、又は DP IV様酵素による代謝回転の阻害は、[Glu
3]Aβの露出を防げ、QC/EC触媒による[pGlu3]Aβ産生を抑制し得る。
本発明の第三の実施態様において、DP IV活性のインヒビター、及びQCのインヒビタ
ーとの組み合わせを、アルツハイマー病、及びダウン症の治療のために使用することがで
きる。
【0036】
DP IV、及び/又は DP IV様酵素、及びQCを組合せた効果は、以下に示すとおりであ
る:
a)DP IV、及び/又は DP IV様酵素はAβ1-40/42を切断し、H-Asp-Ala-OHを含むジペプ
チド、及びAβ3-40/42が放出される、
b)副反応として、QCは低速度でグルタミン酸のピログルタミン酸への環化反応を触媒
する、
c)グルタミン酸は、未知の酵素活性により翻訳後にグルタミンに転換され、かつ続いて
、アミロイドβ-ペプチドN末端のプロセス後、QCはグルタミンのピログルタミン酸へ
の環化反応を触媒する、
d)グルタミン酸は化学触媒、又は自己触媒により、翻訳後にグルタミンに転換され、か
つ第二工程として、アミロイドβ-ペプチドN末端のプロセシング後、QCはグルタミン
のピログルタミン酸への環化反応を触媒する、
e)アミロイドβ-タンパク質をコードするAPP遺伝子において、Aβの3位のGluの代わり
にGlnをもたらす突然変異がある。翻訳、及びN末端のプロセシング後、QCはグルタミ
ンのピログルタミン酸への環化反応を触媒する、
f)グルタミンは、未知の酵素活性の異常により未完成のペプチド鎖に取り込まれ、かつ
続いて、アミロイドβ-ペプチドN末端のプロセス後、QCはグルタミンのピログルタミ
ン酸への環化反応を触媒する。
【0037】
また、QC活性への該N末端Glnの露出は、異なるペプチダーゼ活性によって、引き起
こし得る。アミノペプチダーゼは、Aβ1-40/41/43のN末端から、Asp、及びAlaを順次
取り去ることが可能で、従って、環化反応され易い3番目のアミノ酸が露出される。DP I
、DP II、DP IV、DP 8、DP 9、及びDP 10のようなジペプチジルペプチダーゼは、一工程
でAsp-Alaを除去する。従って、アミノペプチダーゼ、又はジペプチジルペプチダーゼの
活性の阻害は、Aβ3-40/41/43の産生を防止するのに有用である。
【0038】
DP IV、及び/又はDP IV様酵素のインヒビター、及びQCの活性低下エフェクターとの
組合せ効果を、下記の経路で示す:
a)DP IV、及び/又はDP IV様酵素は、Aβ1-40/42のAβ3-40/42への転換を阻害する。
b)それにより、グルタミン酸のN末端露出を妨げ、酵素による、又は化学触媒によるグ
ルタミンへの転換はなく、続いてピログルタミン酸産生することは不可能である。
c)さらに、QCのインヒビターは、残基が改質されたAβ3-40/42分子、及びAPP遺伝子
の突然変異により産生されたAβ3-40/42分子からのピログルタミン酸形成を阻害する。
本発明の範囲内で、DP IV、又はDP IV様酵素、及びQCとの類似の組合せ効果は、さら
にグルカゴン、CCケモカイン、及びサブスタンスPのような、ペプチドホルモンで示さ
れた。
【0039】
グルカゴンは、すい臓のランゲルハンス島アルファ細胞から放出される、29アミノ酸ポ
リペプチドであり、肝臓でのグリコーゲン分解、及びグルコース新生を刺激促進すること
により、血糖値を維持するよう作用する。その重要性にかかわらず、体内でのグルカゴン
排除をつかさどる機構に関しては見解の一致を見ないままである。Pospisilikらは、高感
度の質量分光学的技術を用い、グルカゴンの酵素代謝を測定し、分子産物を同定した。グ
ルカゴンを精製されたブタジペプチジルペプチダーゼIV(DP IV)とインキュベートする
ことにより、グルカゴン3-29、及びグルカゴン5-29が生じた。ヒト血清では、グルカゴン
3-29への分解後速やかにグルカゴンのN末端環化反応が起こり、さらなる加水分解を抑制
した。グルカゴンのバイオアッセイにおいて、精製DP IV、又は、正常ラット血清とイン
キュベートの後、高血糖活性の有意な低下が示されたのに対し、DP IV欠損ラット血清と
の同様のインキュベートでは、グルカゴン生物活性の低下は示されなかった。質量分析、
およびバイオアッセイによってモニターすると、分解は、DP IV特異的インヒビター、イ
ソロイシルチアゾリジンにより阻害された。これらの結果により、DP IVがグルカゴンの
分解、及び不活性化に関係する主要酵素と同定した。これらの知見は、ヒト血漿中のグル
カゴンレベルの決定に、重要な影響をもたらす(Pospisilikらの論文、Regul Pept 2001
Jan 12;96(3):133-41)。
【0040】
当初、ヒト単球走化性タンパク質(MCP)-2は、MCP-1、及びMCP-3と共に生成されるケ
モカインとして、刺激促進された骨肉腫細胞から精製されてきた。Von Coillieら(Van C
oillie, E.らの論文、1998 Biochemistry 37, 12672-12680)は、ヒト精巣cDNAライブラ
リーから5'末端伸張型MCP-2 cDNAをクローニングした。それは76残基のMCP-2タンパク
質をコードしたが、報告されている骨髄由来MCP-2 cDNA配列とはコドン46で異なり、Gl
nの代わりにLysをコードした。一塩基変異多型(SNP)により生じた、該MCP-2Lys46変異
体をMCP-2Gln46と生物学的に比較した。該コード領域をバクテリア発現ベクターpHEN1に
サブクローンし、かつ大腸菌(Escherichia coli)の形質転換後、該2種のMCP-2変異体を
ペリプラズムから回収した。エドマン分解によって、N末端がpGluの代わりにGln残基で
あることが明らかになった。rMCP-2Gln46、及びrMCP-2Lys46、及びそのN末端が環化した
該対応型を、単球細胞上でカルシウム流動、及びアッセイで、分析した。rMCP-2Gln46、
及びrMCP-2Lys46 異性体間で、生物学活性には有意な差はなかった。しかしながら、両MC
P-2において、N末端ピログルタミン酸が遊走性に不可欠であるが、カルシウム流動には
不可欠でないことが示された。セリンプロテアーゼCD26/ジペプチジルペプチダーゼIV(
CD26/DPP IV)による、rMCP-2Lys46のN末端切断型は、アミノ末端Gln-Proジペプチドの
放出をもたらすが、アミノ末端pGluを含む人工MCP-2は、影響を受けなかった。CD26/DPP
IVで切断されたrMCP-2Lys46(3-76)は、遊走性、及びシグナル伝達アッセイの両方でほ
ぼ完全に不活性であった。これらの観察は、MCP-2のNH2末端pGluが遊走活性に不可欠であ
ること、また、それが該タンパク質をCD26/DPP IVによる分解から保護することを示唆し
た(van Coillie, E.らの論文、Biochemistry 1998 37, 12672-80)。
【0041】
本発明の範囲内で、LC/MS分析により、グルカゴン3-29(Pospisilikらの論文、2001)
、及びMCP-2異性体(van Coillieらの論文、1998)のN末端ピログルタミン酸残基の産生
がQCにより触媒されることが決定された。
さらに、LC/MS実験により、サブスタンスPからLys-Pro、及びArg-Proの2種のジペプ
チドのN末端DP IV触媒による除去後、残存[Gln5]サブスタンスP5-11は、QCにより[pGl
u5]サブスタンスP5-11へ転換されることを証明した。
【0042】
DP IVインヒビターは、国際公開第99/61431号において開示されている。特に、アミノ
酸残基、及びチアゾリジン、又はピロリジン群、及びその塩を含むDP IVインヒビターが
開示されており、特にL-threo-イソロイシルチアゾリン、L-allo-イソロイシルチアゾリ
ン、L-threo-イソロイシルピロリジン、L-allo-イソロイシルチアゾリン、L-allo-イソロ
イシルピロリジン、及びその塩である。
【0043】
低分子量ジペプチジルペプチダーゼIVインヒビターのさらなる例には、テトラヒドロイ
ソキノリン-3-カルボキサミド誘導体、N-置換 2-シアノピロール、及びピロリジン、N-(
N'-置換グリシル)2-シアノピロリジン、N-(置換グリシル)-チアゾリジン、N-(置換グ
リシル)-4-シアノチアゾリジン、アミノ-アシル-ボロノ-プロピル-インヒビター、シク
ロプロピル-融合ピロリジン、及び六員環化合物のような薬剤がある。ジペプチジルペプ
チダーゼIVインヒビターは、下記の文献に記載されている:米国特許第6,380,398号、米
国特許第6,011,155号、米国特許第6,107,317号、米国特許第6,110,949号、米国特許第6,1
24,305号、米国特許第6,172,081号、国際公開第95/15309号、国際公開第99/61431号、
国際公開第99/67278号、国際公開第99/67279号、ドイツ特許第198 34 591号公報、国際
公開第97/40832号、ドイツ特許第196 16 486 C 2号公報、国際公開第98/19998号、国際
公開第00/07617号、国際公開第99/38501号、国際公開第99/46272号、国際公開第99/3
8501号、国際公開第01/68603号、国際公開第01/40180号、国際公開第01/8133号、国際
公開第01/81304号、国際公開第01/55105、国際公開第02/02560号、及び国際公開第02
/14271号、国際公開第02/04610号、国際公開第02/051836号、国際公開第02/068420号
、国際公開第02/076450号、国際公開第02/083128号、国際公開第02/38541号、国際公
開第03/000180号、国際公開第03/000181号、国際公開第03/000250号、国際公開第03/
002530号、国際公開第03/002531号、国際公開第03/002553号、国際公開第03/002593号
、国際公開第03/004496号、国際公開第03/024942号、及び国際公開第03/024965号であ
る。これらの文献に教示されている内容は、その全体、特にインヒビター、その定義、使
用、及び産生に関して、引用により本明細書に取り込まれるものとする。
【0044】
QCのエフェクターと組合せる使用が好ましいのは、Hughesらの論文(1999 Biochemis
try 38 11597-11603)により開示されたNVP-DPP728A(1-[[[2-[{5-シアノピリジン-2-イ
ル}アミノ]エチル]アミノ]アセチル]-2-シアノ-(S)-ピロリジン)(Novartis社)、Hug
hesら(アメリカ糖尿病協会会議、2002年、要旨番号272)により開示されたLAF-237(1-[
(3-ヒドロキシ-アダマンチル-1-アミノ)-アセチル]-プロリジン-2(S)-カルボニトリ
ル)(Novartis社)、Yamadaらの論文(1998 Bioorg Med Chem Lett 8, 1537-1540)に
開示されたTSL-225(トリプトフィル-1,2,3,4-テトラヒドロイソキノリン-3-カルボン酸
)、Asworthらの論文(1996 Bioorg Med Chem Lett 6, 1163-1166、及び2745-2748)に
開示された2-シアノピロリジド、及び4-シアノピロリジド、Sudreらの論文(2002 Diabet
es 51, 1461-1469)によって開示されたFE-999011(Ferring社)、及び国際公開第01/34
594号で開示された化合物(Guilford)のようなDP IVインヒビターがあり、上記の文献で
設定した投与量を採用する。
【0045】
QCのエフェクターと組合せる使用がより好ましいDP IVインヒビターは、ジペプチド
化合物であり、そのアミノ酸は、例えば、ロイシン、バリン、グルタミン、グルタミン酸
、プロリン、イソロイシン、アスパラギン、及びアスパラギン酸のような天然アミノ酸、
から選択されるのが好ましい。本発明で使用されるジペプチド様化合物は、10μMの(ジ
ペプチド化合物の)濃度で、血漿ジペプチジルペプチダーゼIV、又はDP IVアナログ酵素
活性の少なくとも10%、特に少なくとも40%活性の低下を示す。また、インビボでは、多
くの場合少なくとも60%、又は少なくとも70%の活性の低下が所望される。また、好まし
い化合物では、最高20%、又は30%の活性の低下が示され得る。
【0046】
好ましいジペプチド化合物は、N-バリルプロリル、O-ベンゾイルヒドロキシルアミン、
アラニルピロリジン、L-allo-イソロイシルチアゾリジン、L-threo-イソロイシルピロリ
ジンのようなイソロイシルチアゾリジン、及びその塩、特にフマル酸塩、及び、L-allo-
イソロイシルピロリジン、及びその塩である。特に好ましい化合物は、グルタミニルピロ
リジン、及びグルタミニルチアゾリジン、H-Asn-ピロリジン、H-Asn-チアゾリジン、H-As
p-ピロリジン、H-Asp-チアゾリジン、H-Asp(NHOH)-ピロリジン、H-Asp(NHOH)-チアゾ
リジン、H-Glu-ピロリジン、H-Glu-チアゾリジン、H-Glu(NHOH)-ピロリジン、H-Glu(N
HOH)-チアゾリジン、H-His-ピロリジン、H-His-チアゾリジン、H-Pro-ピロリジン、H-Pr
o-チアゾリジン、H-Ile-アジジジン、H-Ile-ピロリジン、H-L-allo-Ile-チアゾリジン、H
-Val-ピロリジン、及びH-Val-チアゾリジン、及び医薬として許容し得るその塩である。
これらの化合物は、国際公開第99/61431号、及び欧州特許出願公開第1 304 327号に記載
されている。
【0047】
さらに本発明は、ジペプチジルペプチダーゼIV触媒の競合的調整に有用な基質様ペプチ
ド化合物と組合せるQCのエフェクターの使用を提供する。好ましいペプチド化合物は、
2-アミノオクタン酸-Pro-Ile、Abu-Pro-Ile、Aib-Pro-Ile、Aze-Pro-Ile、Cha-Pro-Ile、
Ile-Hyp-Ile、Ile-Pro-allo-Ile、Ile-Pro-t-ブチル-Gly、Ile-Pro-Val、Nle-Pro-Ile、N
va-Pro-Ile、Orn-Pro-Ile、Phe-Pro-Ile、Phg-Pro-Ile、Pip-Pro-Ile、Ser(Bzl)-Pro-I
le、Ser(P)-Pro-Ile、Ser-Pro-Ile、t-ブチル-Gly-Pro-D-Val、t-ブチル-Gly-Pro-Gly
、t-ブチル-Gly-Pro-Ile、t-ブチル-Gly-Pro-Ile-アミド、t-ブチル-Gly-Pro-t-ブチル-G
ly、t-ブチル-Gly-Pro-Val、Thr-Pro-Ile、Tic-Pro-Ile、Trp-Pro-Ile、Tyr(P)-Pro-Il
e、Tyr-Pro-allo-Ile、Val-Pro-allo-Ile、Val-Pro-t-ブチル-Gly、Val-Pro-Val、及びそ
の医薬として許容し得るその塩である。そのうち、t-ブチル-Glyは、下記式のように定義
し、かつ、Ser(Bzl)、及びSer(P)は、それぞれベンジル-セリン、及びホスホリル-セ
リンと定義する。Tyr(P)は、ホスホリル-チロシンと定義する。これらの化合物は、国
際公開第03/002593号に開示されている。
【0048】
【化1】
【0049】
本発明に従ってQCのエフェクターと組合せて使用でき、さらに好ましいDP IVインヒビ
ターは、ペプチジルケトンである。例を挙げると、下記化合物、及びその医薬として許容
し得るその塩である:
2-メチルカルボニル-1-N-[(L)-アラニル-(L)-バリニル]-(2S)-ピロリジン臭化水素
酸塩、2-メチル)カルボニル-1-N-[(L)-バリニル-(L)-プロリル-(L)-バリニル]-(
2S)-ピロリジン臭化水素酸塩、
-[(アセチル-オキシ-メチル)カルボニル]-1-N-[(L)-アラニル-(L)-バリニル]-(2S
)-ピロリジン臭化水素酸塩、
2-[ベンゾイル-オキシ-メチル)カルボニル]-1-N-[{(L)-アラニル}-(L)-バリニル]-
(2S)-ピロリジン臭化水素酸塩、
2-{[(2,6-ジクロロベンジル)チオメチル]カルボニル}-1-N-[{(L)-アラニル}-(L)-
バリニル]-(2S)-ピロリジン、
2-[ベンゾイ-ルオキシ-メチル)カルボニル]-1-N-[グリシル-(L)-バリニル]-(2S)-ピ
ロリジン臭化水素酸塩、
2-[([1,3]-チアゾールチアゾール-2-イル)カルボニル]-1-N-[{(L)-アラニル}-(L)-
バリニル]-(2S)-ピロリジントリフルオロ酢酸塩、
2-[(ベンゾチアゾールチアゾール-2-イル)カルボニル]-1-N-[N-{(L)-アラニル}-(L
)-バリニル]-(2S)-ピロリジントリフルオロ酢酸塩、
2-[(-ベンゾチアゾールチアゾール-2-イル)カルボニル]-1-N-[{(L)-アラニル}-グリ
シル]-(2S)-ピロリジントリフルオロ酢酸塩、及び
2-[(ピリジン-2-イル)カルボニル]-1-N-[N-{(L)-アラニル}-(L)-バリニル]-(2S)
-ピロリジントリフルオロ酢酸塩である。これらの化合物は、国際公開第03/033524号に
開示されている。
【0050】
さらに、本発明に従って、置換アミノケトンをQCエフェクターと組合せて使用するこ
とが可能である。好ましい置換ケトンを挙げると、
1-シクロペンチル-3-メチル-1-オクソ-2-ペンタンアミウムクロライド、
1-シクロペンチル-3-メチル-1-オクソ-2-ブタンアミウムクロライド、
1-シクロペンチル-3,3-ジメチル-1-オクソ-2-ブタンアミウムクロライド、
1-シクロヘキシル-3,3-ジメチル-1-オクソ-2-ブタンアミウムクロライド、
3-(シクロペンチルカルボニル)-1,2,3,4-テトラヒドロイソキノリニウムクロライド、
N-(2-シクロペンチル-2-オクソエチル)シクロヘキサンアミウムクロライド、及び
その医薬として許容し得るその塩がある。
【0051】
当初、希少なプロリン特異的プロテアーゼグループのうちで、DP IVは、唯一膜結合性
酵素で、ポリペプチド鎖のアミノ末端でその端から2番目残基のプロリンに特異性がある
と考えられていた。しかしながら、DP IVと構造的に相同性はないが、同様の酵素活性を
もつ、他の分子が同定された。今までに同定されたDP IV様酵素は、例えば、Sedo、及びM
alik(Sedo、及びMalikの論文、2001, Biochim Biophys Acta, 36506, 1-10)、及びAbbo
tt、及びGorrell(Abbott, C.A.、及びGorrell, M.Dの著書、エクトペプチダーゼ(Langn
e、及びAnsorge編集、Kluwer Academic/Plenum Publishers, New York中、171-195ペー
ジ))による総説に記載されている繊維芽活性化タンパク質α(fibroblast activation
protein α)、ジペプチジルペプチダーゼIV β、ジペプチジルアミノペプチダーゼ様タ
ンパク質、N-アセチル化α結合型酸性ジペプチダーゼ、休止細胞プロリンジペプチダーゼ
(quiescent cell proline dipeptidase)、ジペプチジルペプチダーゼII、アトラクチン
、及びジペプチジルペプチダーゼIV 関連タンパク質(DPP 8)、DPL1(DPX、DP6)、DPL2
、及びDPP 9 がある。最近、ジペプチジルペプチダーゼ10(DPP 10)のクローニング、及
び特性が報告された(Qi, S.Y.らの論文、Biochemical Journal Immediate Publication
において、2003年3月28日に原稿番号BJ20021914として発表)。
【0052】
本明細書中で使われる用語、エフェクターは、酵素に結合し、インビボ、及び/又はイ
ンビトロでその活性を上昇、又は低下させる分子として定義する。酵素によっては、その
触媒活性に影響する低分子の結合部位を有し;刺激促進分子はアクチベーターと呼ばれる
。酵素は1種以上のアクチベーター、又はインヒビターを認識するのための複数の結合部
位を有すことすらあり得る。酵素は様々な分子の濃度を感知し、かつその情報を使用し、
それ自身の活性を変えることが可能である。
【0053】
エフェクターが酵素活性を調整可能なのは、酵素が活性型と不活性型の両構造を呈する
からであり:アクチベーターは、正のエフェクターであり、インヒビターは負のエフェク
ターである。エフェクターは、酵素の活性部位においてだけでなく、調節部位、又はアロ
ステリック部位においても作用し、この用語は、調節部位が触媒部位から異なる酵素の要
素であることを強調し、触媒部位における基質、及びインヒビター間の競合による調節形
態と区別するために用いられる(Darnell, J.、Lodish, H.、及びBaltimore, D. の著書
、分子生物学第2版(1990, Molecular Cell Biology 2nd Edition, Scientific American
Books, New York)63ページ)。
【0054】
本発明のペプチドにおいて、各アミノ酸残基は、以下の慣用一覧表に従い、アミノ酸の
慣用名に相当する一文字、又は三文字表示で表す。
アミノ酸 一文字表記 三文字表記
アラニン A Ala
アルギニン R Arg
アスパラギン N Asn
アスパラギン酸 D Asp
システイン C Cys
グルタミン Q Gln
グルタミン酸 E Glu
グリシン G Gly
ヒスチジン H His
イソロイシン I Ile
ロイシン L Leu
リシン K Lys
メチオニン M Met
フェニルアラニン F Phe
プロリン P Pro
セリン S Ser
トレオニン T Thr
トリプトファン W Trp
チロシン Y Tyr
バリン V Val
【0055】
本明細書中で使用する用語“QC”とは、グルタミニルシクラーゼ(QC)、及びQC
様酵素を含む。さらに、QC、及びQC様酵素は同一の、又は同様の酵素活性を有し、Q
C活性と定義する。この点において、根本的に、QC様酵素は、分子構造上QCから異な
ることが可能である。
【0056】
本明細書中で使用する用語“QC活性”とは、アンモニア遊離下における、N末端グル
タミン残基のピログルタミン酸(pGlu*)への、又はN末端L-ホモグルタミン、又はL-β
ホモグルタミンの環状ピロ-ホモグルタミン誘導体への分子内環化反応と定義する。それ
については、スキーム1、及び2を参照されたい。
【0057】
スキーム1:QCによるグルタミンの環化反応
【化2】
【0058】
スキーム2:QCによるL-ホモグルタミンの環化反応
【化3】
【0059】
本明細書中で使用する用語“EC”とは、QC、及びQC様酵素のグルタミン酸シクラ
ーゼ(EC)ような副活性を含み、さらにEC活性として定義する。
本明細書中で使用する用語“EC活性”とは、QCによるN末端グルタミン酸残基の、
ピログルタミン酸(pGlu*)への分子間環化と定義する。それについては、スキーム3を
参照されたい。
本明細書中で使用する用語“金属依存性酵素”とは、その触媒機能を果たすために結合
した金属イオンとの結合を必要とする、かつ/又は、触媒活性の構造を形成するために結
合した金属イオンを必要とする酵素と定義する。
【0060】
酵素に結合して、かつその活性を上昇させたり、又は低下させる分子は、“エフェクタ
ー”と呼ばれる。エフェクターが酵素活性を調整可能なのは、酵素が活性型と不活性型の
両構造を呈するからであり:アクチベーターは、正のエフェクターであり;インヒビター
は負のエフェクターである。エフェクターは、調節部位、又はアロステリック部位(ギリ
シャ語の“もう一つの形”由来)において結合し、この用語は、調節部位が触媒部位から
異なる酵素の要素であることを強調し、触媒部位における基質、及びインヒビター間の競
合による調節形態と区別するために用いられる。本発明の個々の実施態様によって、アク
チベーター、又はインヒビターのどちらかが好まれる。
【0061】
スキーム3:QCによる、荷電していないグルタミルペプチドのN末端環化反応(EC)
【化4】
【0062】
本発明のもう一つの態様は、QCの新規の生理学的基質の同定である。これらは、実施
例5に記載した哺乳動物ペプチドでの環化反応実験を行い、同定した。それに先立ち、ヒ
トQC、及びパパイヤQCを実施例1に記載されるように精製した。該適用方法は実施例
2に記載し、かつ使用した該ペプチド合成は実施例6に概要を示す。該研究の結果は、表
1に示す。
【0063】
【表1】
【0064】
すべての分析は、実施例4に示されるように、ヒト、又は植物QCのいずれかの最適活
性、及び安定性の範囲で行った。N末端にグルタミン残基をもち、かつそれゆえQC酵素
の基質である、生理活性ペプチドのアミノ酸配列は表2に記載する:
【0065】
【表2】
【0066】
第四の実施態様において、[Gln1]ガストリン(長さ17、及び34アミノ酸)、[Gln1]ニュ
ーロテンシン、及び[Gln1]FPPペプチドを、QCの新規生理的基質として同定した。ガス
トリン、ニューロテンシン、及びFPPは、N末端にpGlu残基を含有する。すべてのペプチ
ドで、N末端pGlu残基が、QCによりN末端のグルタミンから産生されることを示した。
結果として、N末端グルタミン残基のpGluへの転換により、その生物学機能の点で活性化
される。
【0067】
経上皮変換細胞、特にガストリン(G)細胞は、胃への食物到来とともに胃酸分泌を調
節する。最近の研究により、複数の活性産物がガストリン前駆体より産生され、かつガス
トリン生合成には、複数のコントロール箇所があることが示された。生合成前駆体、及び
中間体(プロガストリン、及びGly-ガストリン)は、推定成長因子であり;その産物、ア
ミド化されたガストリンは、上皮細胞の増殖、酸産生壁細胞、及びヒスタミン分泌クロム
親和性様(ECL)細胞の分化、及びECL細胞でのヒスタミン合成、及び貯蔵に関わる遺伝子
の発現を、急性の酸分泌刺激促進と同様に調節する。また、ガストリンは、上皮成長因子
(EGF)ファミリーメンバーの産生を刺激促進し、順に壁細胞機能を阻害し、表面上皮細
胞の増殖を促進する。血漿ガストリン濃度は、ピロリ菌を保有し、十二指腸潰瘍、及び胃
癌のリスクが高いことが公知の個体で上昇する(Dockray, G.J.の論文、1999 J Physiol
15, 315-324)。
【0068】
管腔G細胞から放出されるペプチドホルモンガストリンは、CCK-2レセプターを介して、
酸分泌粘膜中のECL細胞由来のヒスタミン合成、及び放出を刺激促進することが知られて
いる。動員されたヒスタミンは、壁細胞にあるH(2)レセプターに結合することにより、
酸分泌を引き起こす。最近の研究により、ガストリンは、完全にアミド化された、及び、
プロセスが少ない型(プロガストリン、及びグリシン延長型ガストリン)の両方で、胃腸
管の成長因子であることが示唆された。アミド化されたガストリンの主要な栄養作用は、
胃の酸分泌粘膜に対してであり、胃幹細胞、及びECL細胞の細胞増殖の促進を起こし、壁
、及びECL細胞塊増大を生じることが確立している。一方、プロセスが少ないガストリン
(例えば、グリシン延長ガストリン)の主要栄養標的は、結腸粘膜であるように思われる
(Koh, T.J.、及びChen, D.の論文、2000 Regul Pept 93, 37-44)。
【0069】
第五の実施態様において、本発明は、胃腸管細胞増殖、特に、胃粘膜細胞及び上皮細胞
の増殖、酸産生壁細胞、及びヒスタミン分泌クロム親和性様(ECL)細胞の分化、及びECL
細胞内のヒスタミン生合成、及び貯蔵に関連した遺伝子の発現を刺激し、並びに、活性型
[pGlu1]-ガストリンの濃度を維持、又は上昇させることによる、哺乳動物における急性
の酸分泌をするために、QCの活性上昇エフェクターの使用を提供する。
第六の実施態様において、本発明は、不活性型[Gln1]のガストリンの活性型[pGlu1]ガ
ストリンへの転換率を低下させることによる、哺乳動物におけるピロリ菌を伴う、又は伴
なわない十二指腸潰瘍、及び胃癌の治療のために、QCの活性低下エフェクターの使用を
提供する。
【0070】
ニューロテンシン(NT)は、統合失調症の病態生理学に関わる、特に神経伝達物質シ
ステムを調整する神経ペプチドで、今までに、該疾患では調節障害が起こっていることが
示された。脳脊髄液(CSF)中のNT濃度を測定する臨床研究では、脳脊髄液(CSF)中の
NT濃度が低下している統合失調症患者の一部は、効果的な抗精神病剤治療で、回復する
ことを示した。また、抗精神病剤の作用機構におけるNTシステムの関与に一致した多数
の証拠がある。中枢系に投与されたNTの行動学的、及び生化学効果は、抗精神病剤を全
身に投与した該効果に著しく類似し、かつ抗精神病剤は、NT神経伝達を上昇させる。こ
の知見の連鎖から、NTが内在性の抗精神病剤として機能するという仮説が導かれた。さ
らに、典型、及び非典型抗精神病剤は、黒質線条体系、及び中脳辺縁系のドーパミン経路
の終末領域のNT神経伝達を異なった形で改質し、かつ該効果で、副作用障害、及び有効
性が予想可能である。
【0071】
第七の実施態様において、本発明は、抗精神病剤の調剤、及び/又は哺乳動物における
統合失調症治療のために、QCの活性上昇エフェクターの使用を提供する。該QCのエフ
ェクターは、活性型[pGlu1]ニューロテンシン濃度を維持、又は上昇させる。
【0072】
受精促進ペプチド(FPP)は、サイロトロピン放出ホルモン(TRH)に関連するトリペプ
チドであり、精漿に見い出される。インビトロ、及びインビボから得られた最近の証拠は
、FPPが精子受精能力の調整において重要な役割を果たしていることを示した。特に、FPP
は初めに受精能力のない(受精不能な)精子を“スイッチオン”にし、かつより速やかに
受精可能にするよう刺激促進し、その後、精子が自発アクロソーム損失を起こし、それゆ
え受精能力を喪失しないように、受精能獲得を抑制する。これらの反応は、アデニリルシ
クラーゼ(AC)/cAMPシグナル伝達経路を調節することが公知の、アデノシンによって模
倣され、かつ実際、増大される。FPP、及びアデノシンは両方とも、FPP受容体がどういう
わけかアデノシン受容体、及びGタンパク質と相互作用しACの調節をもたらし、受精不能
な細胞でcAMP産生を刺激促進し、受精可能な細胞ではそれを阻害することが示された。こ
れらの事象は、様々な細胞のチロシンリン酸化状態に影響し、初期の“スイッチオン”に
おいて重要なものもあれば、おそらく先体反応自身に関与しているものもある。また、精
漿に存在するカルシトニン、及びアンギオテンシンIIは、インビボで受精不能な精子に同
様の効果をもち、FPPの反応を増大し得る。これらの分子は、インビボで同様の効果をも
ち、受精能の刺激促進、及び維持により、受精に影響する。FPP、アデノシン、カルシト
ニン、及びアンギオテンシンIIの供給の低下、又はそのレセプターの異常は、男性不妊に
寄与する(Fraser, L.R.、及びAdeoya-Osiguwa, S. A.の論文、2001 Vitam Horm 63, 1-2
8)。
【0073】
第八の実施態様において、本発明は、哺乳動物における受精抑制薬の調剤、及び/又は
、受精の低下のための、QCの活性低下エフェクターの使用を提供する。該QCの活性低
下エフェクターは、活性型[pGlu1]FPP濃度を低下させ、精子の受精能獲得を抑制、及び精
子細胞の不活性化をもたらす。これに対し、QCの活性上昇エフェクターにより、男性生
殖能力を刺激を促進し、かつ不妊を治療することが可能であることが示され得る。
【0074】
第九の実施態様において、さらなるQCの生理学的基質が、本発明の範囲内で同定され
た。これらは、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 18
、及び[Gln1]フラクタルカインである。詳細は、表2を参照されたい。これらのポリペプ
チドは、骨髄性前駆細胞増殖の抑制、新生組織形成、炎症性宿主反応、癌、乾癬、関節リ
ウマチ、アステローム性動脈硬化、体液性及び細胞性免疫反応、内皮における白血球接着
及び遊走プロセスのような病態生理学的状態において、重要な役割を果たす。
【0075】
最近、B型肝炎、HIV、及びメラノーマに対する、いくつかの細胞傷害性Tリンパ球ペ
プチドを基にしたワクチンが、臨床試験で研究されている。デカペプチドELAは、単独で
、或いは腫瘍抗原と組合せる、1種の興味深いメラノーマワクチン候補である。このペプ
チドは、N末端グルタミン酸をもつ、Melan-A/MART-1抗原免疫優勢ペプチドアナログで
ある。グルタミンのアミノ基、及びガンマカルボン酸基は、グルタミン酸のアミノ基、及
びガンマカルボキサミド基と同様に、容易に縮合し、ピログルタミン誘導体を産生する。
医薬上興味深いいくつかのペプチドは、N末端グルタミン、及びグルタミン酸の代わりに
ピログルタミン誘導体で開発し、医薬上の特性を損失することなくこの安定性の問題を克
服した。残念なことに、ELAに比べ、該ピログルタミン誘導体(PyrELA)、及びN末端ア
セチル修飾誘導体(AcELA)は、細胞傷害性Tリンパ球(CTL)の活性を引き出せなかった
。PyrELA、及びAcELAにおける外見上の些少な修飾にかかわらず、おそらく該二誘導体は
該特異クラスI主要組織適合複合体に対して、ELAより低い親和性を示す。それゆえ、ELA
の最大の活性を維持するために、PyrELAの産生を回避しなければならない(Beck A.らの
論文、2001, J Pept Res 57(6):528-38)。また、最近、メラノーマにおいて、グルタ
ミニルシクラーゼ(QC)酵素が過剰発現していることが見い出された(Ross D. T.らの
論文、2000, Nat Genet 24:227-35)。
【0076】
第十の実施態様において、本発明は、骨髄性前駆細胞増殖の抑制、新生組織形成、炎症
性宿主反応、癌、悪性転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化
、体液性及び細胞性免疫反応障害、内皮における白血球接着及び遊走プロセスのような病
態生理学的状態の治療に用いる薬剤の製造のための、QCのエフェクターの使用を提供す
る。
【0077】
第十一の実施態様において、[Gln1]オレキシンAは、本発明の範囲内で、QCの生理学
的基質と同定した。摂食、睡眠覚醒の調整に有意な影響を及ぼす神経ペプチドであり、お
そらく、これらの相補的な恒常性機能の複雑な行動、及び生理学的応答を調整している。
また、それは、エネルギー代謝の恒常性調節、自律機能、ホルモンバランス、及び体液調
節において影響を及ぼす。
第十二の実施態様において、本発明は、摂食障害、及び睡眠覚醒、エネルギー代謝の恒
常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害の治療に用いる
薬剤の製造のための、QCのエフェクターの使用を提供する。
【0078】
様々なタンパク質におけるプログルタミン伸長は、パーキンソン病、及びケネディ病の
ような神経変性疾患を生じる。それゆえ、該機構は、ほとんどわかっていない。ポリグル
タミン繰り返し配列の生化学的特性は、1つの考えられる説明を示唆し:グルタミニル-
グルタミニル結合の切断(endolytic cleavage)後の、ピログルタミン酸産生が、ポリグ
ルタミンタンパク質の異化安定性、疎水性、アミロイド凝集性、及び神経傷害性に寄与し
得るということである(Saido, Tの論文、Med Hypotheses(2000)Mar;54(3):427-9
)。したがって、第十三の実施態様において、本発明は、パーキンソン病、及びハンチン
トン病の治療に用いる薬剤の製造のための、QCのエフェクターの使用を提供する。
【0079】
第十四の実施態様において、本発明は、QCの酵素活性を低下、又は阻害させる一般的
な方法を提供する。また、阻害化合物の例を提供する。
当初、哺乳動物QCの阻害は、1,10-フェナントロリン、及び還元型 6-メチルプテリン
でのみ検出された(Busby, W. H. J.らの論文、1987 J Biol Chem 262, 8532-8536)。ED
TAはQCを阻害せず、故にQCは金属依存性酵素ではないと結論された(Busby, W. H. J
.らの論文、1987 J Biol Chem 262, 8532-8536, Bateman, R.C.J.らの論文、2001 Bioche
mistry 40, 11246-11250、Booth, R.E.らの論文、2004 BMC Biology 2)。しかしながら
、本発明では、1,10-フェナントロリン、ジピコリン酸、8-ヒドロキシ-キノリン、及び他
のキレーターによるQCの阻害特性(図18、19)、及び遷移金属イオンによるQCの
再活性化(図20)によって明らかであるように、ヒトQC、及び他の動物QCが金属依
存性酵素であることを示した。最後に、該金属依存性を他の金属依存性酵素との配列比較
をまとめ、ヒトQCにも、キレートするアミノ酸残基が保存されていることを示した(図
21)。化合物の、金属イオン結合部位との相互作用は、QC活性を低下、又は阻害する
一般的な方法である。
【0080】
本発明では、イミダゾール誘導体がQCの有力なインヒビターであることを示す。連続
分析(詳細は実施例2参照)を用い、多数のイミダゾール誘導体を、高度に保存された哺
乳動物QCの1種であるヒトQCを阻害する能力に関して分析した。
このように、本発明は、QCの活性低下エフェクターとして、イミダゾール誘導体、及
びヒスチジン、及びその誘導体、及びその特性を阻害型、及び有効性の面から提供する。
構造、及びKi値を、表3、及び4に示す。その結果は、実施例7に詳しく記載した。
【0081】
【表3】
【0082】
【表4】
【0083】
また、驚いたことに、酵素活性の解析中、N末端グルタミニル残基以外にN末端β-ホ
モグルタミル残基が、植物、又は動物由来QCの基質としての特性を満たすことを発見し
た。該N末端β-ホモグルタミル残基は、ヒト、及びパパイヤ由来QCにより、それぞれ
五員のラクタム環に転換された。該結果を、実施例5に示した。適用した方法は実施例2
に示し、ペプチド合成は、実施例6に記載したように行った。
本発明のもう一つの好ましい実施態様は、QCのエフェクターのスクリーニング方法を
含む。
【0084】
化合物群からQCの活性を調整するエフェクターを同定する、好ましいスクリーニング
方法は、下記工程を含む前記方法である:
a)前記の化合物とQCとを、それらの間で結合が可能な条件下で接触させること;
b)QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残存QC活性を測定すること;及び
d)QCの該基質転換、及び/又は、酵素活性の変化を計算し、QCの活性調整フェクタ
ーを同定することである。
【0085】
もう一つの好ましいスクリーニング方法は、QCの金属イオン結合活性部位と、直接、
又は間接的に相互作用するエフェクターの同定、及び選定に用いる方法に関連し、下記の
工程を含む方法である:
a)前記の化合物とQCとを、それらの間で結合が可能な条件下で接触させること;
b)QCにより転換可能な、QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残存QC活性を測定すること;及び
d)QCの該基質転換、及び/又は酵素活性の変化を計算し、QCの活性調整エフェクタ
ーを同定することである。
【0086】
前記スクリーニング方法での使用が好ましいのは、哺乳動物QC、又はパパイヤQCで
ある。特に好ましいのは哺乳動物QCであり、それはこれらのスクリーニング方法で同定
したエフェクターが哺乳動物、特にヒトの疾患の治療に使用され得るからである。
前記スクリーニング方法で選定された薬剤は、少なくとも1種のQC基質の転換を低下
させる(負のエフェクター、インヒビター)、又は、少なくとも1種のQC基質の転換を
増加させる(正のエフェクター、アクチベーター)ことにより機能することが可能である
。
本発明の化合物は、酸付加塩、特に医薬として許容し得る酸付加塩へ転換することがで
きる。
該発明の化合物の該塩は、無機、又は有機の形態とし得る。
【0087】
本発明の該化合物は、転換し、酸付加塩、特に医薬として許容し得る酸付加塩として使
用することが可能である。医薬として許容し得る塩は、一般に、塩基性側鎖が無機、又は
有機酸でプロトン化されているような形態をとる。典型的な有機、又は無機酸は、塩酸、
臭化水素酸、過塩素酸、硫酸、硝酸、リン酸、酢酸、プロピオン酸、グリコール酸、乳酸
、コハク酸、マレイン酸、フマル酸、リンゴ酸、酒石酸、クエン酸、安息香酸、マンデル
酸、メタンスルホン酸、ヒドロキシエタンスルホン酸、ベンゼンスルホン酸、シュウ酸、
パモ酸、2-ナフタレンスルホン酸、p-トルエンスルホン酸、サイクラミン酸、サリチル酸
、サッカリン酸、又はトリフルオロ酢酸を含む。すべての本発明の該化合物の医薬として
許容し得る塩は、本発明の範囲内に取り込むことを意図している。
【0088】
化合物自身と、塩の形態の該化合物の密接な関係から、本明細書の文脈で化合物に言及
するときは常に、その状況下でそれが考え得るか、又は適切であるならば、相当する塩も
対象とする。
本発明の該化合物が少なくとも1種の不斉中心を持つ場合は、それ相応に光学異性体が
存在し得る。該化合物が2種以上の不斉中心を持つ場合は、さらにジアステレオマーが存
在し得る。その異性体、及び混合物のすべては、本発明の範囲内に含まれると理解すべき
である。さらに、該化合物の結晶性形態は多形体として存在し得て、それらはさらに、本
発明に含まれる対象である。該化合物によっては、水(すなわち、水和物)、又は慣用の
有機溶媒との溶媒和化合物を形成し得る。
また、その塩を含む該化合物は、水和物の形態で調達することが可能であるか、或いは
その結晶化に用いる他の溶媒を含む。
【0089】
さらなる実施態様において、本発明は、必要としている個体における、QC酵素活性の
調整を介した疾患状態の予防、又は治療法を提供し、それは、本発明の該化合物、又はそ
の医薬組成物のいずれかを、治療上効果的な量、及び投与法で投与し、病態を治療するこ
とを含む。さらに、本発明は、個体におけるQC活性調整を介した病態の予防、又は治療
に用いる薬剤の調製のための、この発明における化合物、及びそれに相当する医薬として
許容される酸付加塩の形態の使用を含む。該化合物は、静脈注射、経口、皮下注射、筋肉
注射、皮内注射、非経口、及びその組み合わせ含むが、これに限定されない、慣用の投与
経路で患者に投与され得る。
さらなる好ましい実施態様において、本発明は、少なくとも1種の本発明の化合物、又
はその塩を、任意に、少なくとも1種の医薬として許容し得る担体、及び/又は溶媒と組
合せて含有する医薬組成物、すなわち、薬剤に関するものである。
【0090】
例えば、医薬組成物は、非経口、又は経腸製剤の形態であり得て、適切な担体を含むか
、又は経口投与に適した適切な担体を含む、経口製剤の形態である。好ましくは、経口製
剤の形態である。
本発明の記載に従って投与されるQC活性のエフェクターは、インヒビター、又はイン
ヒビター、基質、擬似基質、QC発現のインヒビター、哺乳動物のQCタンパク質濃度を
低下させる酵素の結合タンパク質、又は抗体と組合せて、医薬上投与できる製剤、又は製
剤複合物の形で用いられる。本発明の該化合物により、患者、及び疾患個々に治療を調節
することが可能になり、特に、個々の不耐性、アレルギー、及び副作用を回避することが
可能である。
【0091】
また、該化合物は、時間の関数として、異なる活性の程度を示す。それによって、治療
を施す医師は、患者の個々の状況に応じて対処する機会を与えられ:医師は、一方では作
用開始の速度、他方では、作用の持続期間、及び特に作用強度を正確に調節することが可
能である。
本発明の好ましい治療方法は、哺乳動物のQC酵素活性の調整を介した病態の予防、又
は治療に用いる新規のアプローチを示す。それは、有利に単純で、商業的適用の余地があ
り、特に、哺乳動物、及び特にヒト医薬において、例えば、表1、及び2に記載された、
生理活性QC基質のアンバランスに基づく疾患の治療における使用に適している。
【0092】
例えば、該化合物は、有効成分を希釈剤、賦形剤、及び/又は、先行技術から公知の担
体のような添加剤を組合せて含む調剤の形態で、有利に投与し得る。例えば、それらは、
非経口的(例えば、生理食塩水の点滴で)、又は経腸的に(例えば、慣用の担体と処方さ
れ、経口的に)投与することが可能である。
その内生安定性、及びその生体利用効率に依存して、該化合物は、所望する血糖グルコ
ース値の正常化を達成するために、1日に1回、又は複数回投与することが可能である。
例えば、ヒトにおけるそのような用量域は、1日あたり約0.01 mgから250.0 mgの範囲で
、好ましくは、体重キログラムあたり化合物約0.01 mgから100 mgの範囲であり得る。
【0093】
QC活性のエフェクターを哺乳動物に投与することによって、アルツハイマー病、ダウ
ン症、ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は伴わない、潰瘍性疾患及び胃
癌、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、乾
癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性免疫反応障害、内皮にお
ける白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネルギー代謝の恒常性調節障
害、自律機能障害、ホルモンバランス障害、及び体液調節障害から、選択される病態を、
予防、又は緩和、又は治療し得る。
【0094】
さらに、QC活性のエフェクターを哺乳動物に投与することによって、胃腸管細胞増殖
、好ましくは、胃粘膜細胞、上皮細胞の増殖、急性の酸分泌、及び酸産生壁細胞、及びヒ
スタミン分泌クロム親和性様細胞分化を刺激、促進し得る。
さらに、哺乳動物へのQCインヒビターの投与は、精子細胞機能の喪失、したがって男
性生殖能力の抑制を可能にし得る。従って、本発明は、男性生殖能力の調整、及びコント
ロールに用いる方法、及び男性用避妊薬剤の製造のための、QCの活性低下エフェクター
の使用を提供する。
さらに、QC活性のエフェクターを哺乳動物への投与することにより、骨髄性前駆細胞
増殖の抑制を可能にし得る。
【0095】
本発明で使用される化合物は、本来の公知の方法で、例えば、不活性無毒の、医薬上適
切な担体、及び添加剤、又は溶媒を用いて、錠剤、カプセル剤、糖衣剤、丸剤、顆粒剤、
噴霧剤、シロップ剤、液剤、固形剤、及びクリーム状乳剤、及び懸濁剤、及び溶液剤のよ
うな慣用調剤形態へ、それ相応に転換することが可能である。その各々の調剤形態におい
て、該医薬有効化合物は、好ましくは、全混合物の重量に対して約0.1〜80%、より好ま
しくは、1〜50%の濃度で、すなわち、言及した投薬量域を得るのに十分な量で存在する
。
【0096】
また、該物質は、糖衣剤、カプセル剤、可噛性カプセル剤、錠剤、ドロップ剤、シロッ
プ剤の形態の薬剤として、又は座薬として、又は鼻腔用スプレーとして使用することがで
きる。
例えば、該調剤形態は、有効成分を溶媒、及び/又は担体と共に、任意に乳化剤、及び
/又は分散剤の使用と共に供与され、有利に製造することができ、例えば、水が希釈剤と
して使用される場合は、有機溶媒は、任意に補助溶剤として使用することが可能である。
【0097】
本発明に関わる有用な賦形剤の例を挙げると:水、パラフィン(例えば天然油画分)、
植物油(例えば、ナタネ油、ラッカセイ油、ゴマ油)、グリコール(例えば、プロピレン
グリコール、ポリエチレングリコール)のような無毒性有機溶媒;例えば、天然粉末ミネ
ラル(例えば、抗分散シリカ、ケイ酸塩)、糖類(例えば、粗糖、ラクトース、デキスト
ロース)のような固形担体;非イオン性、及び陰イオン性乳化剤(例えば、ポリオキシエ
チレン脂肪酸エステル、ポリオキシエチレン脂肪アルコールエステル、アルキルスルホン
酸、アリールスルホン酸)のような乳化剤、分散剤(例えば、リグニン、亜硫酸溶液、メ
チルセルロース、澱粉、及びポリビニルピロリドン)、潤滑剤(例えば、ステアリン酸マ
グネシウム、滑石粉、ステアリン酸、及びラウリル硫酸ナトリウム)、及び任意の香料が
ある。
【0098】
投与は、従来の方法で、好ましくは経腸的、又は非経口的に、特に経口的に行われ得る
。経腸投与の場合に、錠剤は上記の担体に加えて、さらにクエン酸ナトリウム、炭酸カル
シウム、及びリン酸カルシウムのような添加剤を、澱粉、好ましくは片栗粉、ゼラチン、
及びそのような多様な添加剤と共に含有し得る。さらに、ステアリン酸マグネイウム、ラ
ウリル硫酸ナトリウム、及び滑石粉のような潤滑剤は、錠剤製造に付随して使用すること
が可能である。経口投与を目的とした水性懸濁液、及び/又はエリキシル剤の場合は、多
様な矯味料、又は着色料が、上記記載の賦形剤に加えて、有効成分に付加することが可能
である。
【0099】
非経口投与の場合、適切な液体担体を用いた有効成分の溶液を使用することができる。
一般に、静脈注射の場合、一日当たり体重に対して約0.01〜2.0 mg/kg、好ましくは、0.
01〜1.0 mg/kgの量を投与すると、効果的な結果が得られる利点が見いだされ、かつ経腸
投与の場合、該投与量は一日当たり体重に対して約0.01〜2 mg/kg、好ましくは、0.01〜
1 mg/kgである。
しかしながら、実験動物、及び患者の体重、又は投与経路の種類だげでなく、動物の種
類、又は薬剤に対する個々の反応、又は投与が行われる間隔などによって、該記載量から
外れることが不可欠である場合もあり得る。したがって、上記記載の最低量より少ない使
用で十分な場合もある一方、記載の上限を超過しなければならない場合もある。比較的大
量に投与される場合は、該投与量を一日に複数回の投薬に分割することが望ましいかもし
れない。ヒト薬剤の投与には、同様の投与許容度を提供する。その場合、上記の所見が同
様に適用される。
【0100】
(医薬調剤形態の例)
1.カプセル剤当り、本発明の化合物100 mgを含有するカプセル剤
およそ10,000カプセル剤に対して、下記組成の溶液を調製する:
本発明の化合物 1.0 kg
グリセロール 0.5 kg
ポリエチレングリコール 3.0 kg
水 5.0 kg
該溶液は、本質的に公知の方法で、軟ゼラチンカプセルに封入する。該カプセルは、咀
嚼、及び嚥下に適している。
【0101】
2.本発明の化合物100 mgを含有する錠剤、又はコーティングされた錠剤、又は糖衣剤
下記の量は錠剤100,000粒の製造に関するものである:
細かく粉砕した本発明の化合物 10.0 kg
グルコース 4.35 kg
ラクトース 4.35 kg
澱粉 4.50 kg
細かく粉砕したセルロース 4.50 kg
【0102】
上記組成物を、混合し、下記成分から製造される溶液と共に提供し、該湿塊をおろし、
かつステアリン酸マグネシウム0.2 kg付加後乾燥させる、本質的に公知の方法で粒状にさ
れる。
ポリビニルピロリドン 2.0 kg
ポリソルベート 0.1 kg
及び水 約5.0 kg
【0103】
最終錠剤混合物30.0 kgを加工し、300 mgの重さの突型錠剤を形成する。理想的には、
該錠剤を、本質的に公知の方法でコーティング、又は糖コーティングすることが可能であ
ることである。
有利には、本明細書中で定義された医薬組成物は、少なくとも1種のQC活性のエフェ
クター、及び少なくとも1種のDP IVインヒビターを含有する。そのような医薬組成物は、
特にアルツハイマー病、及びダウン症の治療に有用である。
【実施例】
【0104】
(実施例1):ヒト、及びパパイヤQCの調製法
(宿主株、及び培地)
ヒトQCの発現に使用される酵母(Pichia pastoris)X33株(AOX1、AOX2)を、製造会
社の取扱説明書(Invitrogen社)に従って、培養し、形質転換し、かつ解析した。酵母(
P.pastoris)が必要とする培地、例えば、緩衝化したグリセロール(BMGY)複合体、又は
メタノール(BMMY)複合体培地、及び、発酵基礎塩培地は、製造会社の推奨にしたがって
調製した。
【0105】
(ヒトQCをコードするプラスミドベクターの分子クローニング)
すべてのクローニング手順は、標準的分子生物学的技法を適用して行なった。酵母にお
ける発現のために、ベクターpPICZαB(Invitrogen社)を使用した。pQE-31ベクター(Qi
agen社)を使用し、ヒトQCを大腸菌において発現させた。第38番目のコドンより始ま
る成熟型QCのcDNAと、6xヒスチジン標識をコードするプラスミドとを、読み枠内で融合
した。pQCyc-1、及びpQCyc-2プライマー(表1)を使用し、増幅、かつサブクローニング
の後、該フラグメントを、SphI及び、HindIIIの制限酵素切断部位を用いて、発現ベクタ
ーに挿入した。
【0106】
(P. pastorisの形質転換、及び小量発現実験)
プラスミドDNAは、大腸菌JM109内で増幅され、製造会社(Qiagen社)の推奨に従って精
製した。使用した発現プラスミドpPICZαBにおいては、3つの直線化に用いる制限酵素切
断部位が提供される。SacI、及びBstXIは、該QCcDNAの内部で切断するため、PmeIを選
択し、直線化した。20〜30μgのプラスミドDNAをPmeIで切断し、エタノールにより沈澱さ
せ、滅菌脱イオン水に溶かした。その後、10μgのDNAを、製造会社(BioRad社)の取扱説
明書に従って、電気穿孔法による酵母(P.pastoris)コンピテント細胞の形質転換に使用
した。選択は、150μg/ml Zeocinを含むプレートにおいて行なった。直線化されたプラ
スミドを使用した一回の形質転換実験から、数百の形質転換体を得た。
【0107】
組み換え酵母クローンをQCの発現に関して調べるために、組み換え体を、2 mlのBMGY
培地を含む10 mlコニカルチューブにおいて、24時間培養した。その後、該酵母を遠心
分離し、0.5%メタノールを含む2 mlのBMMYに再懸濁した。この濃度は、72時間まで、
24時間毎にメタノールを加えることによって維持された。その後、上清のQC活性が決
定した。融合タンパク質の存在を、6xヒスチジン標識に対する抗体(Qiagen社)を使用し
たウェスタンブロット解析によって確認した。最も高いQC活性を示した酵母のクローン
を、さらなる実験、及び発酵のために選択した。
【0108】
(発酵槽における大量発現実験)
QCの発現は、5 l(リットル)のリアクター(Biostat B,B.Braun biotech社)におい
て、基本的に"酵母Pichia発酵操作ガイドライン(Pichia fermentation process guideli
nes)"(Invitrogen社)に記載されているように行なった。簡潔に述べると、酵母細胞は
、微量の塩を添加し、かつグリセロールを唯一の炭素源とする、発酵基礎塩培地(pH 5.5
)において培養した。約24時間の初期バッチ段階、及びその後の約5時間のフェドバッ
チ(fed-batch)段階の間、酵母細胞塊が蓄積された。細胞湿重量が200 g/lに達した時
点で、約60時間の全発酵時間に三段階の栄養添加プロフィールを適用し、メタノールを
用いたQC発現の誘導を行なった。その後、細胞を、6000x g、4℃における15分間の遠
心によって、QC含有上清から除いた。そのpHは、NaOHの添加により、6.8に調整し、そ
の結果生じた懸濁溶液は、再び、37000x g、4℃において、40分間遠心した。遠心後も
液の濁りが継続した場合は、セルロース膜(孔径0.45μm)を用いた、さらなる濾過工程
を適用した。
【0109】
(P. pastorisにおいて発現された6xヒスチジン標識QCの精製)
最初に、His標識QCは、固定化金属アフィニティークロマトグラフィー(IMAC)によ
って精製した。典型的な精製においては、1000 mlの培養上清を、750 mMのNaClを含む、5
0 mMのリン酸緩衝液(pH 6.8)で平衡化したNi2+結合キレーティングセファロースFFカラ
ム(Chelating Sepharose FF column)(1.6 cm x 20 cm、Pharmacia社)に、流速5 ml/
分でかけた。10カラム容量の平衡化緩衝液、及び5カラム容量の5 mMヒスチジンを含む
平衡化緩衝液で洗浄した後、150 mM NaCl、及び100 mMヒスチジンを含む50 mMリン酸緩衝
液(pH6.8)へ変更することによって、結合したタンパク質を溶出した。その結果生じた
溶出液を、20 mMのビス-トリス/HCl緩衝液(pH 6.8)に対し、4℃において一晩透析し
た。その後さらに、該QCは、透析緩衝液によって平衡化されたMono Q6カラム(BioRad
社)における陰イオン交換クロマトグラフィーにより、精製した。QC含有画分は、流速
5 ml/分で、Mono Q6カラムへかけた。その後、該Mono Q6カラムを、100 mMのNaClを含む
平衡化緩衝液で洗浄した。溶出は、240 mM、及び360 mMのNaClを含む平衡化緩衝液を、各
々、30カラム容量、又は5カラム容量もたらす、2通りの濃度勾配によって行なった。
6 mlずつの画分を集め、各画分のタンパク質の純度を、SDS-PAGEにより解析した。均質の
QC含有画分を集め、限外濾過により濃縮した。長期保存(-20℃)のため、グルセロー
ルを最終濃度50%まで加えた。タンパク質は、Bradford、又は、Gill、及びvon Hippelの
方法に従い、定量した(Bradford, M. M.の論文、1976 Anal Biochem 72, 248-254;Gill
, S.C.、及びvon Hippel, P.H.の論文、1989 Anal Biochem 182, 319-326)。
【0110】
(大腸菌におけるQCの発現と精製)
QCをコードするコンストラクトで、M15細胞(Qiagen社)を形質転換し、選択的LB寒
天プレートにおいて、37℃で培養した。タンパク質の発現は、1%グルコース、及び1%の
エタノールを含むLB培地において、室温にて行なった。菌培養液が、OD600約0.8に達した
とき、0.1 mMのIPTGにより、発現を一晩誘導した。一回の凍結溶融のサイクルの後、300
mMのNaCl、及び2mMのヒスチジンを含む、50 mMのリン酸緩衝液、pH 8.0中の、2.5 mg/ml
リゾチーム溶液の添加により、4℃にて、約30分間、細胞を溶解した。該溶液は、37000x
g、4℃、30分間の遠心分離によって、不純物を除去し、その後、ガラスフリットを用いた
濾過(DNAの分離)、及び粗い沈殿物、及び細かい沈殿物用のセルロース・フィルターを
用いた、さらなる2段階の濾過工程を行なった。その上清(約500 ml)は、Ni2+アフィニ
ティーカラム(1.6 x 20 cm)に、流速毎分1 mlでかけた。QCの溶出は、150 mMの塩化
ナトリム、及び100 mMのヒスチジンを含む50 mMリン酸緩衝液によって行なった。QC含
有画分は、限外濾過により濃縮した。
【0111】
(パパイヤラテックス由来QCの精製)
パパイヤラテックス由来QCは、BioCAD 700E(Perseptive Biosystems、Wiesbaden社
、ドイツ)を用いて、以前に報告された方法(Zerhouni, S.らの論文、1989 Biochim Bio
phys Acta 138, 275-290)の修正版で調製した。50 gのラテックスを水に溶解し、その中
に記載されているように遠心した。プロテアーゼの不活化は、S-メチルメタンチオスルフ
ォネートによって行い、その結果得られた粗抽出液を、透析した。透析後、上清全体は、
100 mMの酢酸ナトリウム緩衝液、pH 5.0で平衡化したSPセファロースファーストフロウ(
Fast Flow)カラム(21 x内径2.5 cm)にかけた(流速3 ml/分)。溶出は、流速2 ml/
分で、酢酸ナトリム緩衝液の濃度を上げていく3工程において行なった。第一工程は、0.
5カラム容量の、0.1 Mから0.5 Mへの酢酸ナトリム緩衝液濃度の直線勾配による溶出であ
った。第二工程は、4カラム容量の、0.5 Mから0.68 Mへの酢酸ナトリム緩衝液濃度の直線
勾配による溶出であった。そして、最終溶出工程の間は、1カラム容量の、0.85 Mの緩衝
液を用いた。最も高い酵素活性を含む画分(6 ml)を溜めた。濃縮と、0.02 Mトリス/HC
l緩衝液(pH 8.0)への緩衝液交換は、限外濾過(アミコン(Amicon);透過分子量10 kD
aの膜を使用)により行なった。
【0112】
イオン交換クロマトグラフィー工程から得られた、濃縮されたパパイヤの酵素に、最終
濃度2 Mになるように硫酸アンモニウムを加えた。この溶液を、2 M硫酸アンモニウムを含
む0.02 Mのトリス/HCl溶液(pH8.0)によって平衡化されたブチルセファロース4 ファー
ストフロウ(Fast Flow)カラム(21 x内径2.5 cm)にかけた(流速1.3 ml/分)。溶出
は、硫酸アンモニウム濃度を下げて行く3工程で行なった。第一工程の間は、2 Mから0.6
Mへの硫酸アンモニウムの直線勾配を、0.5カラム容量の0.02 Mトリス/HCl溶液(pH 8.0
)で用いた(流速1.3 ml/分)。第二工程は、0.6 Mから0 M硫酸アンモニウムの直線勾配
を5カラム容量の0.02 Mのトリス/HCl溶液(pH8.0)により、流速毎分1.5 mlでの溶出で
あった。0.02 Mのトリス/HCl溶液(pH 8.0)を、流速1.5 ml/分で用いることにより、
最終溶出工程を行なった。すべてのQC活性を含む画分を溜め、限外濾過により濃縮した
。その結果得られた均質のQCは、-70℃に保存した。最終タンパク質濃度は、Bradford
の方法を用い、ウシ血清アルブミンで得られた標準曲線と比較して決定した。
【0113】
(実施例2):グルタミニルシクラーゼ活性の測定
(蛍光測定法)
すべての測定は、30℃で、マイクロプレート用のバイオアッセイリーダーHTS-7000プラ
ス(BioAssay Reader HTS-7000Plus)(Perkin Elmer社)で行った。QC活性は、H-Gln-
βNAを用い、蛍光測定的に評価した。試料は、20 mM EDTAを含む0.2 Mトリス/HCl溶液(
pH 8.0)中の0.2 mM蛍光基質、0.25 Uピログルタミニルアミノペプチダーゼ(Unizyme, H
orsholm社、Denmark)、及び適切に希釈されたQCの分注液を、最終容量250μlに調整し
た。励起/発光波長は、320/410 nmであった。測定反応は、グルタミニルシクラーゼの
添加により開始した。QC活性は、測定条件下のβ-ナフチルアミンの標準曲線から決定
した。1ユニットは、記載された条件下で、1分当たりH-Gln-βNAから1μmol pGlu-βNA
の産生を触媒するQCの量として定義した。
第二の蛍光測定法において、QCの活性は、H-Gln-AMCを基質に用いて決定した。反応
は、ノボスター(NOVOStar)マイクロプレートリーダー(BMG labtechnologies社)を用
い、30℃で行った。試料は、5 mMのEDTAを含む0.05 Mトリス/HCl溶液(pH 8.0)中の多
様な濃度の該蛍光基質、0.1 Uピログルタミルアミノペプチダーゼ(Qiagen社)、及び適
切に希釈されたQCの分注液を、最終容量を250μlに調整した。励起/発光波長は、380
/460 nmであった。測定反応は、グルタミニルシクラーゼの添加により開始した。QC活
性は、測定条件下の7-アミノ-4-メチルクマリンの標準曲線から決定した。速度論的デー
タは、グラフィト(GraFit)ソフトウェアを用いて査定した。
【0114】
(QCの分光学的測定法)
この新規測定法を使い、ほとんどのQC基質の速度論的パラメータを決定した。QC活
性は、グルタミン酸デヒドロゲナーゼを補助酵素として使用した、以前の非連続測定法(
Bateman, R. C. J.の論文、1989 J Neurosci Methods 30, 23-28)を適応させることによ
り得られた、連続法を使って、分光光学的に分析した。最終容量250μl中の試料は、各Q
C基質、0.3 mMの NADH、14 mMのα-ケトグルタル酸、及び30 U/mlのグルタミン酸デヒ
ドロゲナーゼから成った。測定反応は、グルタミニルシクラーゼの添加により開始し、34
0 nmでの吸光度の減少を8〜15分間モニターすることにより遂行した。産物産生の典型的
経時変化を、図1に示す。
初期の速度を測定し、測定条件下のアンモニアの標準曲線から、該酵素活性を決定した
。全試料は、30℃でスペクトラフルオープラス(SPECTRAFluor Plus)、又はサンライズ
(Sunrise)(両者ともTECAN社)マイクロプレートリーダーを用い、測定した。速度論的
データは、グラフィト(GraFit)ソフトウェアを用いて査定した。
【0115】
(インヒビター測定)
インヒビター試験において、試料組成は、推定上のインヒビター化合物を添加加した以
外は、前記のものと同様であった。短時間QC阻害試験では、試料は4 mMの各インヒビタ
ーを含み、かつ1 KMの基質濃度であった。阻害の詳細な研究、及びKi値の決定においては
、最初に、インヒビターの補助酵素における影響を調べた。どの場合も、どちらかの酵素
に対する影響は検出されず、従って、信頼性のあるQC阻害測定を可能にした。阻害定数
は、グラフィト(GraFit)ソフトウェアを用いて、該時間経過の曲線を競合阻害の一般式
に当てはめることにより決定した。
【0116】
(実施例3):MALDI-TOF質量分析
ヒューレットパッカード(Hewlett-Packard)G2025 LD-TOFシステムをリニア飛行時間
型分析計と共に用いて、マトリックス支援イオン化−飛行時間型質量分析を行なった。機
器には、337 nm窒素レーザー、加速電位源(5)、及び1.0 mの飛行管を装着した。検出器
操作は、陽イオンモードで、かつシグナルはパーソナルコンピューターに接続したレクロ
イ9350 Mデジタルストレージオシロスコープ(LeCroy 9350M digital storage oscillosc
ope)で記録、及びフィルターをかけた。試料(5μl)は、同量のマトリックス溶液と混
合した。マトリックス溶液には、1 mlアセトニトリル/0.1% TFA水溶液(1/1、v/v)
に、30 mgの2',6'-ジヒドロキシアセトフェノン(Aldrich社)、及び44 mgのクエン酸水
素二アンモニウム(Fluka社)を溶解して調製したDHAP/DAHCを用いた。少量(≒1μl)
のマトリックス−検体混合物は、プローブ先端に移動させ、かつ速やかに真空管(Hewlet
t-Packard G2024A 試料調製付属品)の中で蒸発させ、急速、かつ均一な試料結晶化を確
実にした。
【0117】
長期間Glu1環化反応試験においては、Aβ由来ペプチドは、30℃の100μl 0.1 M酢酸ナ
トリウム緩衝液(pH 5.2)、または0.1 Mビス-トリス緩衝液(pH 6.5)中に準備した。ペ
プチドは、0.5 mM [Aβ3-11a]、又は0.15 mM [Aβ3-21a]濃度で使用し、0.2 U QCは、全2
4時間加えた。Aβ3-21aの場合は、1% DMSOを加え測定した。様々な時間で、試料は測定
管から採取され、ペプチドは、製造会社推奨に従いジップチップ(ZipTip)(Millipore
社)で抽出し、マトリックス溶液と混合(1/1、v/v)し、かつその後、マススペクトル
を記録した。ネガティブコントロールは、QCを加えないか、又は加熱不活化酵素を加え
るかのいずれかであった。インヒビターの研究において、該試料組成は、阻害化合物(5
mMベンズイミダゾール、又は2 mM 1,10-フェナントロリン)を添加する以外は、前記と同
様であった。
【0118】
(実施例4):pH依存性
ヒト、及びパパイヤQCの触媒作用のpH依存性を、一次反応条件下で研究し、それ故に
特異性定数kcat/KMにおけるプロトン濃度の影響を示した。この目的のために、ピログル
タミルアミノペプチダーゼを補助酵素、及びGln-βNAを器質として使用した、結合酵素測
定(the coupled enzymatic assay)を使用した。ピログルタミルアミノペプチダーゼは
、pH 5.5〜8.5で活性を有し、かつ安定であることが示された(Tsuru, D.らの論文、1978
J Biochem(Tokyo)84, 467-476)。それ故に、該測定によって、このpH範囲内でQC触
媒の研究が可能になった。得られた速度プロファイルは、図2に示されるように、典型的
ベル型曲線に適合した。ヒトQCは、至適pHがおよそ7.8〜8.0の、大変狭いpH依存性を有
する。該速度は、より塩基性のpHで、降下する傾向があった。これは、pH 8.5まで活性の
降下を示さなかった(図2、差し込み図)、パパイヤQCで見られた速度プロファイルと
対照的である。しかしながら、両酵素は、pH 8で至適特異性を示した。意外にも、曲線の
査定により、ヒト、及びパパイヤの酸性範囲でのpKa値は、各々7.17±0.02、及び7.15±0
.02と、同一であることが明らかになった。
【0119】
明らかに、ヒトQCの、塩基pH値での活性の減少は、pKa値およそ8.5を有する原子団の
解離に起因した。パパイヤQCの場合は、その第二pKa値の信用できる測定を可能にする
ような、塩基性pHにおけるデータポイント収集はなかった。これは、該データの単独解離
モデルへの適合が、該データの二重解離モデルへの適合と比べて、ほぼ同一のpKa値(pKa
7.13±0.03)をもたらすことにより、支持される。これは、双方のpKa値が、かなり離れ
ていることを示唆する。
【0120】
(pH安定性)
グルタミニルシクラーゼの安定性は、該植物、及び動物酵素をpH 4〜10間の異なるpH値
において、30℃で30分間インキュベートすることにより調べた。その後、QC活性は、標
準的条件下で調べた。その結果を、図3に示す。
パパイヤラテックス由来の該QCは、酸性、又は塩基性の範囲での顕著な不安定性への
傾向はなく、研究したpH範囲内で安定であった。その一方、ヒトQCは、7〜8.5間のpH範
囲内で同程度に安定であった。従って、pH 8のあたりの領域が、植物、及びヒトQCの活
性、及び安定性にとって至適で、かつ該QCの基質特異性の比較を行うのに適しているよ
うである。
【0121】
(実施例5):QCの基質特異性の決定
(分光学的測定)
実施例2に記載したように、連続分光学的測定を行った。従って、QCの活性は、α-
ケトグルタル酸からのグルタミン酸の産生のための、アンモニアの遊離、及びその後のNA
DH/H+の消費に起因する340 nmでの吸光度の減少に反映する。図1に示すように、リニア
プログレス曲線をモニターしたところ、測定した活性、及びQC濃度の間には、直線関係
があった。さらに、ここ(表1)に示す連続測定を使用し、得られたH-Gln-Gln-OHの速度
パラメータは、断続法を使用して得られたものとよく一致した(KM=175±18μM、kcat=
21.3±0.6 s-1)。その上、表1に示したパパイヤQCによる、基質H-Gln-Ala-OH、H-Gln
-Glu-OH、H-Gln-Gln-OH、H-Gln-OtBu、及びH-Gln-NH2の転換速度パラメータは、pH 8.8、
及び37℃の条件で、直接法を使用して決定されたものとよく一致した(Gololobov, M. Y.
らの論文、1996 Biol Chem Hoppe Seyler 377, 395-398)。それ故、新規連続アッセイ法
が、信用できる結果を提供することは明らかである。
【0122】
(ジ、トリ、及びジペプチド代用物)
先に記載した該新規連続アッセイ法を使用し、約30種の化合物をパパイヤ(C. papay
a)、及びヒト由来QCの潜在基質として調べた。結果を、表5に示した。特異性の比較
により、ほぼ全部の短いペプチドがパパイヤQCにより、該ヒト酵素に比べてより効果的
に転換されることが示された。興味深いことに、他のトリペプチドと比較したH-Gln-Tyr-
Ala-OH、H-Gln-Phe-Ala-NH2、及びH-Gln-Trp-Ala-NH2の特異性、又はジペプチド基質と比
較した発色基質H-Gln-AMC、H-Gln-βNA、及びH-Gln-Tyr-OHの反応性により示されるよう
に、両酵素において、2位に疎水性の残基を有する基質が、最も有効な基質である。パパ
イヤQCにおいて、この知見は、その特異性が2位のアミノ酸残基の大きさに相関関係が
あるという初期の結果と一致する(Gololobov, M. Y.らの論文、1996 Biol Chem Hoppe S
eyler 377, 395-398)。植物、及び動物QCの特異性における唯一顕著な相違点は、H-Gl
n-OtBuにおいて観察された。エステルはパパイヤQCにより、ジペプチド基質と同様の特
異性で転換されるのに対し、ヒトQCでは、ほぼ一桁遅く転換された。
【0123】
(オリゴペプチド)
様々なジペプチド、及びトリペプチドに加えて、パパイヤ、及びヒトQCによる転換に
より、多くのオリゴペプチドを調べた(図5)。興味深いことに、テトラペプチドグルー
プにおけるヒト、及びパパイヤQCにおける特異性の相違点は、ジペプチ、及びトリペプ
チドで観察されたほど大きくなかった。これにより、3位及び4位のアミノ酸でも、速度
論的挙動に影響することが示唆された。しかしながら、H-Gln-Xaa-Tyr-Phe-NH2の構造の
テトラペプチドグループにおいて著しく減少したkcat/KM値を示す、2位のアミノ酸がプ
ロリンのペプチドは例外である(表5)。その特異性の低下は、ヒトQCにおいてより顕
著であり、パパイヤQCと比較して該kcat/KM値においてほぼ8倍の相違をもたらした。
【0124】
また、ヒトQCの特異性の微低下は、H-Gln-Arg-Tyr-Phe-NH2、H-Gln-Arg-Tyr-Phe-NH2
、及びH-Gln-Lys-Arg-Leu-NH2の特異性を他のテトラペプチドと比較して示唆されるよう
に、アミノ酸C末端に正に荷電されたグルタミンを有する基質の転換において観察された
。明らかに、該特異性低下は、主としてより小さい代謝回転数に起因した。この効果は、
該植物酵素では見られなかった。
【0125】
【表5】
【0126】
また、該テトラペプチドで得られた結果は、もう一つの結論をもたらした。既に指摘し
たように、パパイヤQCは、ジペプチドにより高い特異性を示した。しかしながら、2位
のアミノ酸にグルタミン酸を含有するペプチドグループにおいて、表5に与えられたデー
タを曲線で表した図4で示されるように、テトラペプチドによっては、ヒトQCにおいて
より高い特異性定数が観察された。さらに、ペプチド鎖長が、ジから、テトラペプチドへ
伸張するにつれて、パパイヤQCで得られた結果と対照的に、ヒトQCの選択性は上昇し
た。さらに、ヒトQCは、3位、又は4位のアミノ酸に大型の疎水性残基を有するペプチ
ドにおいて、最も高い選択性を記録し、それにより、該酵素の疎水性相互作用を示唆した
。それぞれのペプチドの速度パラメータを比較すると、該変化は主に低いKM値に起因する
と考えられ、該ペプチドの代謝回転数は同様であることが見い出された。従って、ヒトQ
Cの長いペプチドに対する高選択性は、より疎水性の基質の該酵素へのより強い結合の結
果であると考えられる。
【0127】
また、3位と4位に疎水性アミノ酸を含有するペプチドにおいて観察されたヒト,及び
植物QCの相違点は、H-Gln-Arg-Gly-Ile-NH2、又はH-Gln-Arg-Tyr-Phe-NH2、又はH-Gln-
Gln-OH、又はH-Gln-Gln-Tyr-Phe-OHに対する該酵素の特異性定数を比較により明らかにな
る。
また、ヒトQCが、N末端にGlnを含有し、かつC末端Ala残基が増加する同族基質にお
いて、より選択的であるこを見い出した(表6)。ヒトQCの選択性が、基質の鎖長が伸
張するにつれ増加したのに対し、パパイヤQCにはそのような傾向はなかった。また、ヒ
トQCがSer残基を配列に含有するペプチドに対して、より低い特異性であったので、側
鎖の性質が重要であるように考えられる(表6)。
【0128】
【表6】
【0129】
(触媒反応におけるイオン強度の影響)
イオン強度は、基質特異性の影響に関して調べたもう1つのパラメータであった。その
目的において、様々な基質の環化反応における速度パラメータを、0.5 M KClの存在、又
は非存在下で決定した(表7)。驚いたことに、パパイヤラテックス由来QC、及びヒト
QCにおいて、荷電されていない基幹をもつ基質の選択性は、塩の付加により有意に変化
しなかったが、ヒトQCのH-Gln-Ala-OH、及びH-Gln-Glu-OHにおける特異性定数は、KCl
の添加により低下した。各々の速度パラメータにより示唆されるように、この効果は、上
昇したKM、及びごくわずかに減少したkcat値に起因した。パパイヤQCの場合は、どちら
のパラメータにおいても効果は検出されなかった。負に荷電されたH-Gln-Glu-Asp-Leu-NH
2において、パラメータの変化が見い出されなかったことから、該効果は、そのように負
に荷電された基質に起因しないように考えられる。塩付加の興味深い効果は、正に荷電さ
れたH-Gln-Arg-Gly-Ile-NH2、及びH-Gln-Lys-Arg-Leu-NH2において見い出された。植物、
及びヒトQCの場合、触媒反応における正の効果は、主に、より小さいKM値、及びわずか
に高い代謝回転数に起因すると決定した。
【0130】
【表7】
【0131】
(生理的基質)
既に、初期の研究において、QCによる[Gln1]-TRH、及び[Gln1]-GnRHの転換は、ウシ
、及びブタ脳下垂体由来QCにおいて示された(Busby, W. H. J.らの論文、1987 J Biol
Chem 262, 8532-8536;Fischer, W. H.、及びSpiess, J.の論文、1987 Proc Natl Acad
Sci U S A 84, 3628-3632)。既に調べられたこれらの脳下垂体ホルモンに加え、ヒトQ
Cの3つの潜在基質、すなわち[Gln1]ガストリン、[Gln1]ニューロテンシン、及び[Gln1]
FPPを合成し、調べた。それらの速度パラメータは表1に示した。興味深いことに、グル
タミニルペプチドは、その大きさに依存して特異性定数の上昇を伴いながら、各々のピロ
グルタミルペプチドへ転換される。すなわち、17アミノ酸の最も大きいペプチドガスト
リン前駆体が最初で、続いて、ニューロテンシン前駆体、GnRH前駆体、TRH前駆体、及びF
PP前駆体である。これらの知見は、合成ペプチドを用いて得られたデータに一致する。
また、驚いたことに、長い基質ほど、該植物酵素による高い選択性で転換され、それは
、より短いオリゴペプチドにおける知見と部分的に対照的である。おそらく、活性部位か
らはるかに離れた、基質、及び酵素間の二次的結合相互作用が存在する。
【0132】
(改質アミノ酸含有ペプチド)
さらに、該QCの該特異性、及び該選択性を調べるために、改質N末端グルタミニル残
基、及び2位に改質アミノ酸を含むペプチドを合成した。これらのペプチドの転換は、MA
LDI-TOF質量分析法を用いて、定性的に調べた(実施例3も参照)。グルタミニル残基、
又はその類似体のそれぞれの環化反応に起因して、該基質と該触媒反応生成物の質量差を
検出する。また、基質モル当たり、1モルのアンモニア遊離の場合は、分光学的定量法を
用いて定量的に分析した。
【0133】
(H-Gln-Lys(Gln)-Arg-Leu-Ala-NH2)
N末端において、ペプチド、及び部分イソペプチド結合でリシル残基に結合している2
グルタミニル残基含有する、N末端分鎖ペプチドは、ヒト(図5)、及びパパイヤQC(
図示せず)により明らかに同一の方法で転換された。一貫した基質転換で示唆される(図
5)ように、両グルタミニル残基は、個別の残基に対する検出可能な優先はなく、ピログ
ルタミン酸へ転換された。従って、QCの、異なった結合をしたグルタミニル残基におけ
る選択性は、根本的に相違しない。
【0134】
(H-Gln(NMe)-Phe-Lys-Ala-Glu-NH2)
唯一、メチル化されたグルタミニル残基は、パパイヤQCにより、ピログルタミル残基に
転換された(図6)。さらに、該ヒトQCの該ペプチドによる阻害は検出されなかったの
で、該メチル化された残基はヒトQCにより認識されないことが示唆された。
【0135】
(H-Glu(OMe)-βNA、及びH-Glu-βNA)
これらの化合物はどちらも、パパイヤ、及びヒトQCによって転換されなかった。これ
らの蛍光発生基質は、ピログルタミルアミノペプチダーゼを補助酵素として使用しながら
、蛍光分析学的に分析した。しかしながら、O-メチル化グルタミン酸残基は、非酵素的触
媒環化反応する傾向があるトリス、及びトリシン緩衝液両者において顕著な不安定性を示
した。さらに、両QCのH-Gln-AMCを基質とする活性は、H-Glu(OMe)-Phe-Lys-Arg-Leu-
Ala-NH2、又はH-Glu-Phe-Lys-Arg-Leu-Ala-NH2の長鎖ペプチドにより阻害されなかったこ
とは、グルタミン酸、又は誘導体が両QC型により認識されないことを示唆する。さらに
、該結果はグルタミン酸残基の負電荷のみが、該ペプチドの該活性部位からの反発力の原
因でないことを意味する。
【0136】
(H-Gln-cyclo(Nε-Lys-Arg-Pro-Ala-Gly-Phe))
分子内部分イソペプチド結合を含有するH-Gln-cyclo(Nε-Lys-Arg-Pro-Ala-Gly-Phe)
の転換を定量的に分析し、ヒト、及びパパイヤQCにおけるKM値が各々240±14μM、及び
133±5μMであることを明らかにした。ヒトQC(22.8±0.6 s-1)と比較して、パパイヤ
QC(49.4±0.6 s-1)による転換におけるより高い代謝回転数に起因して、該植物QC
は、ヒトQCのおよそ4倍高いkcat/KM値372±9 mM-1min-1を示した。従って、パパイヤ
QCの場合、特異性定数はH-Gln-Ala-Ala-Ser-Ala-Ala-NH2のような同様の大きさをもつ
基質と比べてごくわずかに小さい。しかしながら、ヒトQCのkcat/KM値は、95±3 mM-1
s-1で、同様の大きさの基質に比べておよそ一桁小さいことを見い出した(表5)。
【0137】
(H-βhomoGln-Phe-Lys-Arg-Leu-Ala-NH2)
N末端β-ホモグルタミニル残基は、ヒト、及びパパイヤQCにより、各々五員ラクタ
ム環へ転換された。アンモニアの付随遊離を、先に記載したように、分光学的、及びMALD
I-tof分析法により分析した。QCを除いたり、又は沸騰した場合、アンモニアの遊離が
検出されなかったことから、該環化反応の特異的な触媒反応であることが示唆された。興
味深いことに、パパイヤ(C. papaya)(KM=3.1±0.3 mM、kcat=4.0±0.4 s-1)、及び
ヒト(KM=2.5±0.2 mM、kcat=3.5±0.1 s-1)由来該QCは、各々1.4±0.1、及び1.3±
0.1 mM-1s-1とほぼ同一のkcat/KMで、該ペプチドの転換を触媒する。従って、β-グルタ
ミン残基の環化反応は、N末端にグルタミニル残基を含有する同様の大きさのペプチドに
比べて、およそ1000倍低下した効率で触媒される。これは、基質のα-炭素の構造が、Q
C型による基質認識において重要であるが、必須ではないことを示す。基質である必須条
件は、環化反応しやすい距離と角度の、γ-アミド基、及びプロトン化されていないN末
端アミノ基であり、N末端グルタミニル、及びγ-ホモ-グルタミニル残基は必須条件を満
たしている。
【0138】
(実施例6):QC基質の合成
(オリゴペプチド)
ペプチドを、既に記載されたように、ペプチド合成機(ラボーテックSP650(Labortec
SP650)、Bachem社、スイス)を使用し、0.5 mmolスケールで半自動的に合成した(Schil
ling, S.らの論文、2002 Biochemistry 41, 10849-10857)。長鎖ペプチドは、記載され
たように、自動化シンフォニーペプチド合成機(Symphony peptide synthesizer)(Rain
in Instrument社)を使用し、25μmolスケールで合成した(Manhart, S.らの論文、2003
Biochemistry 42, 3081-3088)。すべてのペプチド結合において、2-(1H-ベンゾトリア
ゾール-1-イル)-1,1,3,3,-テトラメチルウロニウムテトラフルオロホウ酸(TBTU;Novab
iochem社)/塩基(ジイソプロピルエチルアミン、又はN-メチル- モルホリン;Merck社
)を使用する、改質した固相ペプチド合成のFmocプロトコルを採用し、又は困難な結合の
場合は、N-[(ジメチルアミノ)-1H-1,2,3,-トリアゾロ[4,5-b]ピリジン-1-イルメチレン
]-N-メチルメタンアンモニウムヘキサフルオロリン酸N-オキシド(4,5)(HATU;Applied
Biosystems社)/ジイソプロピルエチルアミンを活性化剤として使用した。カクテルを
含むトリフルオロ酢酸(TFA;Merck社)によるレジンからの開裂の後、粗ペプチドは無酸
性溶媒の分取用HPLCにより精製し、N末端グルタミンのさらなる環化反応を回避した。25
0-21ルナ(Luna)RP18カラム(Phenomenex社)におけるアセトニトリル(Merck社)水溶
液リニアグラジエント(40分間を通して、5〜40%、又は65%アセトニトリル)で分取HPL
Cを行った。ペプチド精度と同定を裏付けるために、分析用HPLC、及びESI-MSを使用した
。
【0139】
(Glu(NH-NH2)-Ser-Pro-Thr-Ala-NH2)
直鎖前駆体ペプチド(Fmoc-Glu-Ser-Pro-Thr-Ala-NH2)は、標準的Fmoc方法(Schillin
g, S.らの論文、2002 Biochemistry 41, 10849-10857)に従って、リンクアミド(Rink a
mide)MBHAリジン(Novabiochem社)において合成した。Fmoc保護ペプチドのレジンから
の開裂後、該ペプチドを、ジエチルエーテル(Merck)で沈殿させ、濾過し、かつ乾燥さ
せた。ジクロロメタン(DCM、Merck社)中で、該前駆体のγ-カルボン酸基(0.3等量)を
結合させるのに、HMBA-AM レジン(1.16 mmol/g、Novabiochem社)を使用した。結合剤
として、ジシクロへキシルカルボジイミド(DCC、Serva社)(4等量)、及びジメチルア
ミノピリジン(DMAP、Aldrich社)(0.1等量)を使用した。12時間後、レジンを、濾過し
、DCMで洗い、反応を繰り返した。20%ピペリジンDMF溶液(5分間、3回)を用いたN末端
Fmoc基の脱保護後、該ペプチドリジンを、5%ヒドラジン溶液(20 ml/g)で1.5時間処理
した。該レジンを、濾過し、ジメチルホルムアミド(DMF、Roth社、ドイツ)、及びTFAで
洗った。蒸発後、該粗ペプチドをエーテルで沈殿し、76%の収率を得た。
【0140】
(H-Gln-Lys(Gln)-Arg-Leu-Ala-NH2)
直鎖ペプチドを、標準的Fmoc/tBu方法に従って、Fmoc-Lys(Fmoc)-OHを最後から2番
目のアミノ酸結合として使用して、リンクアミド(Rink amide)MBHAレジン(Novabioche
m社)において合成した(Schilling, S.らの論文、2002 Biochemistry 41, 10849-10857
)。20%ピペリジン(Merck社)DMF溶液で、レジンの2つのアミノ保護基を脱保護後、4等
量のFmoc-Gln(Trt)-OHを結合させた。標準的開裂法で、95%の収率を得た。
【0141】
(H-Gln(NMe)-Phe-Lys-Ala-Glu-NH2)
Fmoc-Gln(NMe)-OHを、Fmoc-MI-AM(Novabiochem社)レジンに充填したFmoc-Glu-OtBu
から開始して合成した。DCMで膨潤後、該レジン(0.5 g)をDMFで洗い、20%ピペリジン
(Merck社)DMF溶液で脱保護した。該レジンを5 ml DMF中に入れ、5等量Fmoc-Glu-OtBu、
5等量HATU、及び10等量DIPEAを次いで加え、6時間振とうした。濾過、及び洗浄後、生成
物を標準的TFA開裂条件に従って、開裂した。該H-Gln(NMe)-Phe-Lys-Ala-Glu-NH2ペプ
チドは、記載されたように合成した(Schilling, S.らの論文、2002 Biochemistry 41, 1
0849-10857)。Fmoc-Gln(NMe)-OHは、HATU/DIPEAと一晩結合させた。標準的開裂法で
、78%の収率を得た。
【0142】
(H-Glu(OMe)-β-ナフチルアミド、H-Gln-Val-OH、H-Gln-Tyr-OH)
Boc保護ジペプチドを、クロロ炭酸イソブチル(Merck社)を使用して、標準的無水物混
合法で合成した。C末端メチルエステルBoc-Gln-Tyr-OMe、及びBoc-Gln-Val-OMeを、1 N
NaOHジオキサン溶液により鹸化した。Boc保護ペプチドは、HCl/ジオキサン溶液により、
10分間脱保護した。蒸発後、残基を様々な溶媒で結晶化させ、固体化合物の60〜70%の収
率を得た。
【0143】
(H-Gln-cyclo(Nε-Lys-Arg-Pro-Ala-Gly-Phe))
直鎖前駆体Boc-Gln(Trt)-Lys-Arg(Pmc)-Ala-Gly-Phe-OHを、酸感受性2-クロロトリ
チルリジンにおいて合成した。結合を、Fmoc-Lys(Mtt)-OHを用いたFmoc/tBu方法の標
準的プロトコルを用いて行った。3%TFA DCM 溶液(10回、5分)で開裂後、該溶液は、10
%ピリジン(Merck社)メタノール溶液(MeOH;Merck社)で中和し、DCM、及びMeOHで3回
洗い、5%の容量に蒸発させ、粗ペプチドは氷冷水で沈殿させた。その後、該粗シペプチ
ドは、DCC/N-ヒドロキシベンゾトリアゾール(HOBt;Aldrich社)活性化を利用して、環
化した。該粗ペプチドは、乾燥ジクロロメタン溶液(0.2 mmol/50 ml)に、溶解し、0.2
mmolのN-メチルモルホリン、及び0.4 mmolの1-ヒドロキシベンゾトリアゾールを添加し
た。この溶液を、0℃において、250 mlの0.4 mmolジシクロヘキシルカルボジイミドのジ
クロロメタン溶液へ滴下した。該反応は、室温で、一晩撹拌して完了した。N,N'-ジシク
ロヘキシル尿素の濾過後、溶媒は、蒸発により除いた。残留物は、エチルアセテートに溶
解し、1規定HCl、NaHCO3飽和溶液、及び水で洗った。該溶液は、無水Na2SO4上で乾燥さ
せ、濾過し、真空で乾燥するまで蒸発させた。
【0144】
(実施例7):QCのエフェクターの特性
(イミダゾール誘導体)
五員環の異なった位置に置換基をもつイミダゾール、及びベンズイミダゾール誘導体を
、QCのインヒビターとして調べた(表3)。構造式の数は、イミダゾール環を示す。該
方法は、実施例2に示す。
【0145】
(C-4(5)、及びC-4,5誘導体)
イミダゾール環の構造上同等の4、又は5位のいずれか、又は両位置に置換基をもつ化
合物は、ヒトQCの阻害において有効性の減少を示した。しかしながら、最も効力のある
阻害化合物の1種であることが証明されたN-ω-アセチル化ヒスタミンは、唯一の例外であ
る。イミダゾールと比べて5-ヒドロキシメチル-4-メチル-イミダゾールが同様の阻害定数
を示したように、これらの位置の小さい置換基は、結合にほとんど効果がなかった。これ
らの位置についた大きく、かつ巨大な基は、酵素による化合物の結合を低下、又は消失し
た。他の調べた置換基の中には、イミダゾール環の電子密度を低下できる負の誘導、又は
メソメリー効果を及ぼすことが知られているものもあり、また、それはより低い結合定数
の原因となる。また、L-ヒスチジン、及びヒスチジンアミドのKi値の相違は、結合におけ
る荷電の若干の影響を示唆する。既に、荷電した基質の静電反発力の証拠は、該基質特異
性の研究において示され、例えば、グルタミンアミドは、ヒトQCにより容易に生成物に
転換されたが、遊離グルタミンにおける基質としての反応性は観察されなかった。
【0146】
(C-2誘導体)
調べたすべての誘導体は、イミダゾールのように、より弱くQCを阻害した。プロトン
より大きいすべての置換は、適切なQC結合を妨げる。唯一2-メチル-ベンズイミダゾー
ルのメチル基に起因して、該阻害定数は、およそ一桁の程度低下する。ベンズイミダゾー
ル、及び2-アミノ-ベンズイミダゾールにおけるKi値の比較により、ごく同様の関係が示
された。さらに、その結果により、影響が電子論的改質に関連しないことが示唆された。
【0147】
(N-1誘導体)
ヒトQCの阻害を調べたイミダゾール誘導体の中で、イミダゾールと比べて向上したKi
値を持つほとんどの化合物は、1窒素原子において改質が見られた。また、これらの化合
物には、最も有効なQCインヒビターの1種、1-ベンジルイミダゾールが含まれた。興味
深いことに、1-ベンゾイルイミダゾール、及びフェニルイミダゾールにおいて見られるよ
うに、この構造のごくわずかな改質が、阻害特性の消失を起こし、実験条件下では不活性
であった。また、この場合において、該観察された変化は、フェニル基の負のメソメリー
効果に起因するイミダゾール環の低下した電子密度よって引き起こされるだけでないよう
に思われ、それは、正の誘導効果を示す、巨大なトリメチルシリル基もまた、他の残基に
比べて結合の低下を示したからである。興味深いことに、1-アミノプロピル-イミダゾー
ルは、この族においてさほど有効性がない化合物の1種であった。立体的に同様の化合物
1-メチルイミダゾール、及び1-ビニルイミダゾールにおいては、活性部位への結合の向上
が見られたので、この化合物の該低有効性は、該塩基アミノ基によって引き起こされる。
従って、正に荷電されたアミノ基が、該低Ki値の原因となり、その結果はN-ω-アセチル
化ヒスタミン(表3)、及びヒスタミン(表4)により裏付けられる。
【0148】
(3,4、及び3,5誘導体化の効果)
4(5)位に、又は両位置に置環基を含有するイミダゾール誘導体は、該酵素の結合に
おいて限られた有効性をもつことを示した。該特異的置換基の効果を、L-ヒスタミン、及
びヒスタミンの生物分解の2中間体3-メチル-4-ヒスタミン、及び3-メチル-5-ヒスタミン
(表4)の阻害定数と比較することにより特定した。L-ヒスタミンは、そのアセチル化対
応物に比べて、およそ一桁小さいKi値を示した。3-メチル-4-ヒスタミンの場合において
、一窒素のメチル化は、有効性をかなり向上させた。しかしながら、3-メチル-5-ヒスタ
ミンをもたらすメチル化は、阻害活性の完全な消失を引き起こした。従って、該観察され
た効果は、主に、塩基窒素に隣接した炭素の誘導体化に起因した、結合の立体障害により
引き起こされるように考えられる。おそらく、該塩基窒素は、該酵素の結合において、重
要な役割を果たす。
【0149】
(実施例8):Aβ3-40/42誘導体の産生
Aβ3-40/42のN末端の配列で、その3位にグルタミン酸残基の代わりにグルタミンを
含有する短鎖ペプチド2種、[Gln3]-Aβ1-11(配列:DAQFRHDSGYE)、及び[Gln3]Aβ3-11
を用いて、測定を行った。2種のペプチドのDP IVによる開裂、及びQCによるN末端グル
タミン残基の環化反応を、MALDI-TOF質量分析で調べた。測定は、連続触媒反応測定用の
両酵素においてと同様、精製DP IV(ブタ腎臓)、又は粗ブタ脳下垂体ホモジネートをQ
C源として用いて行った。
【0150】
(結果)
1.DPIVにより触媒された[Gln3]Aβ1-11aから[Gln3]Aβ3-11aの産生、及びDP IVインヒ
ビター、Val-ピロリジド(Val-Pyrr)によるその抑制
DPIV、又はDPIV様活性は、[Gln3]Aβ3-11a産生下における、[Gln3]Aβ1-11aの開裂をす
る(図7)。その3位の残基は、この開裂により露出し、それゆえ他の酵素、例えば、Q
Cによる改質のために、利用されやすくなる。予想どおり、触媒作用は、Val-Pyrrにより
完全に抑制することが可能である(表8)。
【0151】
2.脳下垂体ホモジネートにおけるQC触媒作用による[Gln3]Aβ3-11aから[pGlu3]Aβ3-
11aの産生、及び1,10-フェナントロリンによる抑制
ブタ脳下垂体ホモジネートに存在するQCは、[Gln3]Aβ3-11aの[pGlu3]Aβ3-11aへの
転換を触媒する(図9)。[pGlu3]Aβ3-11aの産生は、1,10-フェナントロリンの添加によ
り阻害された(図10)。
【0152】
3.[pGlu3]Aβ3-11aの産生をもたらすDP IV、及びQCの連続触媒作用、及びVal-Pyrr、
及び1,10-フェナントロリンによる抑制
粗ブタ脳下垂体ホモジネートにブタ腎臓由来DP IVを付加して測定した、[Gln3]Aβ1-11
aから[pGlu3]Aβ3-11aの産生は、DP IV、及びQCによる連続触媒反応後、起こる(図1
1)。QCインヒビター1,10-フェナントロリン(図12)、又はDP IVインヒビターVal-
Pyrrを添加したとき(図13)は、[pGlu3]Aβ3-11aは産生されなかった。[pGlu3]Aβ3-1
1aのわずかな出現は、アミノペプチダーゼの開裂、次いでグルタミン残基の環化反応に起
因し、また、[Gln3]Aβ2-11aの産生により示唆された。
【0153】
4.アミノペプチダーゼ触媒作用による、粗脳下垂体ホモジネートにおける[pGlu3]Aβ3-
11aの産生
DPIV触媒作用に依存しない[pGlu3]Aβ3-11aに起因して、[Gln3]Aβ1-11aの分解を、DP
IVを添加しない粗脳下垂体ホモジネートにおいて調べた(図14)。4項におけるデータ
から予想されるように、[pGlu3]Aβ3-11aの産生が観察された。また、該データは、[Gln3
]Aβ1-11aの分解はアミノペプチダーゼによって触媒され、[pGlu3]Aβ3-11aを生じ得るこ
とを示す。それ故、該結果は、ピログルタミル産生は、この組織におけるN末端ペプチド
分解の指標であることを示し、さらに斑形成におけるQCの役割を支持した。
【0154】
(実施例9):組み換えヒトQCによる[Gln3]Aβ3-11a;3-21a、及び3-40の代謝回転
調べた全ての[Gln3]Aβ誘導ペプチドは、ヒトQCにより、対応するピログルタミル型
へ効率よく転換された(表8)。水溶液における[Gln3]Aβ3-21a、及び[Gln3]Aβ3-40の
難溶解性のため、該決定は、1% DMSOの存在下で行った。しかしながら、[Gln3]Aβ3-11a
では、良好な溶解性により、DMSO存在、及び非存在下におけるQC触媒作用の代謝回転の
動態解析が可能であった(表8)。まとめると、鎖長8、18、及び37アミノ酸の、Q
C基質としてのAβ-ペプチドの研究(表8参照)により、基質の鎖長が伸張するにつれヒ
トQC活性が上昇するという観察を裏付けた。従って、その特異性定数を考慮に入れると
、Gln1-ガストリン、Gln1-ニューロテンシン、Gln1-GnRHは、QC基質の中でも最良であ
る。同様に、これまで調べた最長のQC基質である[Gln3]Aβ3-40、及びグルカゴンは、1
% DMSO存在下においても、高二次速度定数(各々449 mM-1s-1、及び526 mM-1s-1)を示
した(表8)。
興味深いことに、調べたアミロイドペプチドにおける該転換の該速度パラメータが、鎖
長の伸張につれて劇的に変化しなかったことは、QC触媒おけるAβのC末端のごく軽度
の影響を示唆した。従って、より良溶解性、及び実験処理によって、Aβの小断片、[Gln3
]Aβ1-11a、[Gln3]Aβ3-11a、及びAβ3-11aを用いて、これらのペプチドN末端アミノペ
プチダーゼプロセズに関するさらなる研究を行った。
【0155】
【表8】
【0156】
(実施例10):組み換えヒトQCによるAβ3-11a、及びAβ3-21aの代謝回転
QC存在下でのAβ3-11a、及びAβ3-21aのインキュベートにより、以前の研究と対照的
に、グルタミン酸含有ペプチドがQCの基質になる得ることが示された(図15C、及び
D)。QC触媒による[pGlu3]Aβ3-11a、及び[pGlu3]Aβ3-21aの産生を、各々pH 5.2、及
び6.5で調べた。QCの添加による活性の開始前に、QCインヒビター、ベンズイミダゾ
ールを該溶液に添加した場合は、[pGlu3]Aβ3-11a、又は[pGlu3]Aβ3-21aを生じる基質転
換は抑制された(図15E、F)。QCを添加前に煮沸した場合は、pGluペプチドの産生
はごく僅かであった(図15A、及びB)。
【0157】
(実施例11):パパイヤQC触媒のGln-βNA、及びGlu-βNA の環化反応におけるpH依
存性
パパイヤQCは、ミカエリスメンテン速度論に従い、2 mM(基質溶解性により限定され
た)までの濃度範囲においてGlu-βNAを転換した(図16)。pH 6.1〜8.5の間で調べた
、QC触媒のGlu-βNAの転換における基質濃度に対する代謝回転の図を検討することによ
り、このGlu基質における、KM、及びkcatの両パラメータが、pHに依存することが示され
た(図16)。これは、以前に記載されたQC触媒のグルタミン環化反応とは対照的であ
り、その反応においては、該既定のpH範囲ではKMの変化のみが観察された(Gololobov, M
. Y.、Song, I.、Wang, W.、及びBateman, R. C.の論文、(1994)Arch Biochem Biophys
309, 300-307)。
続いて、Glu、及びGln環化反応中のプロトン濃度の影響を検討するために、低一次速度
条件下(すなわち、KM値をはるかに下回る基質濃度)における、Glu-βNA、及びGln-βNA
の環化反応のpH依存性を調べた(図17)。pH 6.0で至適pHを示したグルタミン酸の該環
化反応に対し、グルタミンの環化反応は、pH 8.0で至適pHを示す。各々の至適pHにおける
該特異性定数は、およそ80,000倍異なる一方、pH 6.0付近でのECに対するQCの活性は
8,000倍だけである。
pH 6.0において4週間調べた、Gln-βNAから非酵素的pGluの産生により、一次速度定数
を1.2*10-7 s-1と示された。しかしながら、同期間においてGlu-βNAからpGlu-βNAは産
生されなかったことから、代謝回転の律速速度定数は1.0*10-9 s-1と推定できた。
【0158】
(実施例12):酵素失活化/再活性化法
ヒトQC(0.1-0.5 mg、1 mg/ml)の分注液を、3000倍超過の5 mM 1,10-フェナントロ
リン、又は5 mMジピコリン酸含有0.05 Mビス-トリス/HCl溶液(pH 6.8)溶液に対して一
晩透析して、失活させた。続いて、試料を1 mM EDTA含有0.05 Mビス-トリス/HCl溶液(p
H 6.8)に対する透析(3回、2000倍超過)により、失活物質を注意深く除去した。再活
性実験は0.025 Mビス-トリス(pH 6.8)においてZn++、Mn++、Ni++、Ca++、K+、及びCo++
イオンを1.0、0.5、0.25 mMの濃度で用いて、室温で15分間行なった。QC活性測定は
、2 mM EDTA含有0.05 M トリス/HCl溶液(pH 8.0)で行い、緩衝溶液中に存在する微量
の金属イオンにより急速な再活性化を防いだ。
【0159】
既に、1,10-フェナントロリンによるブタQCの阻害は、記載されている(Busby, W. H
. J.らの論文、1987 J Biol Chem 262, 8532-8536、Bateman, R.C.J.らの論文、2001 Bio
chemistry 40)。しかしながら、EDTAがQC触媒反応において活性化する効果を持つこと
が示されていた事実は、フェナントロリンによる阻害が金属キレートに起因しないことを
示唆した(Busby, W. H. J.らの論文、1987 J Biol Chem 262, 8532-8536、Bateman, R.C
.J.らの論文、2001 Biochemistry 40)。また、1,10-フェナントロリンによる阻害に加え
て、ヒトQC触媒の基質環化反応は、ジピコリン酸、及び8-ヒドロキシキノリン、他の金
属酵素インヒビター存在下で消失した。これらのキレーターは、QCを競合的、及び時間
依存的に阻害し、すなわち、既に競合的に阻害された初期の活性は、該化合物の長時間イ
ンキュベート後、さらに低下することが見い出された(図18、19)。興味深いことに
、EDTAは、インキュベート時間、又は条件にかかわらず、顕著な阻害を示さなかった。
【0160】
ヒトQCは、5 mM 1,10-フェナントロリン、又は5 mMジピコリン酸に対する大規模な透
析後、ほぼ完全に阻害された。キレーター非存在緩衝溶液に対して一晩透析を繰り返した
後、QC活性は50〜60%まで部分的に再活性化された。しかしながら、1 mM EDTA含有緩
衝液に対して透析した場合は、再活性化は観察されなかった。
該タンパク質を0.5 mM EDTA 存在下で0.5 mM ZnSO4と10分間インキュベートすること
により、ジピコリン酸、又は1,10-フェナントロリンのいずれかによる不活性化後のQC
活性の、ほぼ完全な再活性化が実現された(図20)。同様に、再活性化にCo++、及びMn
++を用いて、QCの部分再活性化を得た。0.25 mM Zn++存在下ですら、本来の活性の25%
までの再活性化が可能だった。Ni++、Ca++、又はK+イオンを利用すると再活性化は観察さ
れなかった。同様に、完全な活性型QCと、これらのイオンとのインキュベートは、該酵
素活性に影響しなかった。
(図面の簡単な説明)
本発明のこれらおよび他の態様のさらなる理解は、以下の図を参照することより、得ら
れるであろう。
【図面の簡単な説明】
【0161】
【図1】図1は、ヒトQCにより触媒されるH-Gln-Ala-OHの環化反応のプログレス曲線を示し、340 nmにおける吸光度の低下をモニターする。該試料は、0.3 mM NADH/H+、14 mM α-ケトグルタル酸、30 U/mlグルタミン酸デヒドロゲナーゼ、及び1 mM H-Gln-Ala-OHを含んだ。曲線A〜Dには、様々な濃度のQCを加えて:その濃度はA、10mU/ml、B、5 mU/ml、C、2.5 mU/mlであった。曲線Dの場合は、QCは除いた。QC濃度、及び観察された活性との間に、直線関係を得た(挿入図)。
【図2】図2は、Gln-βNAを基質として用い、一次反応速度条件下で決定されたヒト、及びパパイヤ(挿入図)QCのpH依存性を示す。ヒトQCの場合、Ellis、及びMorrisonに従い、25 mM MES、25 mM 酢酸、及び50 mMトリスからなる一定イオン強度をもたらす緩衝系を用いた(Ellis, K. J.、及びMorrison, J. F.の論文、1982 Methods Enzymol. 87, 405-426)。トリスのわずかな阻害効果のため、パパイヤQCは、50 mM Mops緩衝液を用いて調べた。イオン強度はNaClの添加によって0.05 Mに調整した。速度プロファイルは、解離基に基づくモデルに適合させることにより評価した。パパイヤQCの場合、データを単一解離モデルに適合させることにより、pKa値7.13±0.03を得た。
【0162】
【図3】図3は、パパイヤラテックス、及びヒトQCの安定性における、pHの影響を示す。酵素原液は、様々なpH値の0.1 M緩衝液に20倍希釈した(pH 4〜7のクエン酸ナトリウム緩衝液、及びpH 7〜10のリン酸ナトリウム緩衝液)。酵素溶液は、30℃で30分間インキュベートし、その後、酵素活性を標準プロトコルに従って解析した。
【図4】図4は、2位のアミノ酸に、グルタミン酸を含有する一組の基質における、特異性定数kcat/KMの比較を示す。ジからテトラペプチドへと、ヒトQCの特異性上昇が検出された一方、パパイヤQCの場合は、変化は観察されなかった。ここに示されたデータは、表3で与えられたパラメータの再プロットである。
【図5】図5は、ヒトQCにより触媒される、H-Gln-Lys(Gln)-Arg-Leu-Ala-NH2からのpGlu-Lys(pGlu)-Arg-Leu-Ala-NH2の産生を示す。基質の転換は、アンモニア放出に起因する、m/z比における時間依存的変化によりモニターする。試料組成は、40 mMトリス/HCl溶液(pH 7.7)における0.5 mM基質、38 nM QCであった。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。パパイヤQCの場合にも、ごく同様の依存性が観察された。
【0163】
【図6】図6は、パパイヤQCにより触媒される、H-Gln(NMe)-Phe-Lys-Ala-Glu-NH2からのpGlu-Phe-Lys-Ala-Glu-NH2の産生を示す。基質の転換は、メチルアミンの放出による、m/z比の時間依存性変化によってモニターする。試料組成は、40 mMトリス/HCl溶液(pH 7.7)における0.5 mM基質、0.65μMパパイヤQCであった。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。パパイヤQCを含まない試料において、又は該基質に、1.5μMまでのヒトQCを添加することによって、基質の転換は観察されなかった(図示せず)。
【図7】図7は、DPIVにより触媒される、[Gln3]Aβ1-11から[Gln3]-Aβ3-11の産生を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図8】図8は、DP IVインヒビター、Val-ピロリジド(Val-Pyrr)による[Gln3]Aβ1-11の切断の抑制を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【0164】
【図9】図9は、QCにより触媒される、[Gln3]Aβ3-11から[pGlu3]Aβ3-11の産生を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図10】図10は、該QCインヒビター、1,10-フェナントロリンによる、[Gln3]Aβ3-11から[pGlu3]Aβ3-11の産生阻害を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図11】図11は、DP IV、及びQCによる連続触媒後の、[Gln3]Aβ1-11から[pGlu3]Aβ3-11の産生を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図12】図12は、触媒活性型DP IV、及びQCの存在下における、QCインヒビター、1,10-フェナントロリンによる、[Gln3]Aβ1-11から[pGlu3]Aβ3-11産生の阻害を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【0165】
【図13】図13は、触媒活性型DP IV、及びQCの存在下における、DP IVインヒビター、Val-Pyrrによる、[Gln3]Aβ1-11から[pGlu3]Aβ3-11産生の低下を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図14】図14は、ブタ脳下垂体ホモジネートに存在するアミノペプチダーゼ、及びQCによる連続触媒作用後の、[Gln3]Aβ1-11から[pGlu3]Aβペプチド3-11の産生を示す。表示された時間に、試料を測定管から取り出し、マトリックス溶液と混合(1:1 v/v)し、その後、質量スペクトルを記録した。
【図15】図15A、及びBは、使用前に10分間煮沸した、組み換えヒトQCとインキュベートされたAβ3-11a、及びAβ3-21aの質量スペクトルを示す。C、及びDは、活性型ヒトQC存在下における、Aβ3-11、及びAβ3-21aの質量スペクトルを示し、それらは各々、[pGlu3]Aβ3-11a、及び[pGlu3]Aβ3-21aの産生をもたらす。E、及びFは、活性型QC、及び[pGlu3]産生を抑制する5 mMベンズイミダゾール存在下における、Aβ3-11a、及びAβ3-21aの質量スペクトルを示す。
【0166】
【図16】図16は、基質の濃度に対してプロットされた、パパイヤQCに触媒されるGlu-βNA転換の反応速度を示す。初速度は、0.1 Mピロリン酸緩衝液(pH 6.1)(四角)、0.1 Mリン酸緩衝液(pH 7.5)(丸)、及び0.1 Mホウ酸緩衝液(pH 8.5)(三角)で測定した。速度パラメータは、下記のとおりであり:KM=1.13±0.07 mM、kcat=1.13±0.04 min-1(pH 6.1);KM=1.45±0.03 mM、kcat=0.92±0.01 min-1(pH 7.5);KM=1.76±0.06 mM、kcat=0.56±0.01 min-1(pH 8.5)であった。
【図17】図17は、低一次反応速度条件下(S<<KM)で決定された、Gln-βNA(丸)、及びGlu-βNA(四角)の転換のpH依存性を示す。基質濃度は、各々、0.01 mM、及び0.25 mMであった。両決定には、0.05 M酢酸、0.05 Mピロリン酸、及び0.05 Mトリシンを含む、三成分緩衝系を使用した。全ての緩衝液は、イオン強度の相違を防ぐため、NaClの添加により等電気伝導度に調整した。該データは、2種の解離基を説明する公式に適合し、pKa値は、Gln-βNAにおいては、6.91±0.02、及び9.5±0.1、及びGlu-βNAにおいては4.6±0.1、及び7.55±0.02であった。滴定により決定された、各々の基質アミノ基のpKa値は、6.97±0.01(Gln-βNA)、及び7.57±0.05(Glu-βNA)であった。全ての決定は、30℃で行った。
【図18】図18は、イミダゾール、及びジピコリン酸存在下、及び阻害化合物非存在下における、ヒトQCに触媒されるH-Gln-AMCの環化反応の経時変化曲線を示す。ジピコリン酸存在下のハイパーボリック(hyperbolic)型は、QC活性部位からの金属イオンの除去を示唆する。
【0167】
【図19】図19は、複素環キレーター、1,10-フェナントロリンによる、QCの時間依存的不活性化を示す。該QC酵素と、基質非存在下におけるインヒビターとのインキュベート後(実線)では、インヒビターと予めインキュべートしなかった試料(点跡)に比べて、酵素活性低下が観察され、これはQC活性部位からの金属イオンの除去を示唆する。
【図20】図20は、一価、及び一価金属イオンとの、ヒトQCの再活性化を示す。QCは、50 mMビス-トリス(pH 6.8)中の2 mMジピコリン酸の添加により、不活性化した。その後、該酵素を、1.0 mM EDTAを含む、50 mM ビス-トリス(pH 6.8)に対して透析させた。該酵素の再活性化は、該不活性化された酵素試料と、0.5 mM EDTA存在下における、0.5 mMの濃度の金属イオンとのインキュベートにより達成し、緩衝溶液に存在する微量の金属イオンにより、非特異的再活性化を防いだ。コントロールは、不活性化しなかったが、不活性化された酵素としてEDTA溶液に対して透析した酵素試料(+EDTA)、及びEDTAを添加しない緩衝溶液に対して透析した酵素試料(-EDTA)により示された。
【図21】図21は、ヒトQC(hQC)、及び他のメタロペプチダーゼClan MHのM28ファミリー員の配列アライメントを示す。多重配列アライメントは、ch. EMBnet. orgにおけるClustalWをデフォルト設定で使用し、行なった。ヒトQC(hQC;GenBank X71125)、放線菌(Streptomyces griseus)由来Zn依存性アミノペプチダーゼ(SGAP;Swiss-Prot P80561)、及びヒトグルタミン酸カルボキシペプチダーゼII(hGCP II;Swiss-Prot Q04609)のN-アセチル化-α-結合酸性ジペプチダーゼ(N-acetylated-alpha-linked acidic dipeptidase)(NAALADase I)ドメイン(残基274〜587)内おける、亜鉛イオン結合残基の保存を示す。金属結合に関与する該アミノ酸は、太字でタイプし、下線を引いた。ヒトQCの場合は、これらの残基は、該ペプチダーゼへの推定相応残基である。
【特許請求の範囲】
【請求項1】
a)インビボQC活性の調整によって治療可能な、哺乳動物の疾患の治療、及び/又は
、b)QC活性の調整によって引き起こされる、pGlu含有ペプチドの作用に基づく生理的
プロセスの調整、に用いる薬剤製造のための、グルタミニルシクラーゼ(QC)のエフェ
クターの使用。
【請求項2】
Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu11]Aβ11-40/42、[Gln11]Aβ11-40/42、[Gln
1]ガストリン(17及び34)、[Gln1]ニューロテンシン、[Gln1]FPP、[Gln1]TRH、[Gln1]Gn
RH、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 18、[Gln1]ELA
、[Gln1]フラクタルカイン、[Gln1]オレキシンA、[Gln3]グルカゴン3-29、及び[Gln5]サ
ブスタンスP 5-11から選択される、少なくとも1種のQC基質における、N末端のグル
タミン酸、又はグルタミン残基の、ピログルタミン酸残基への転換の改質のための、請求
項1記載の使用。
【請求項3】
アルツハイマー病、及びダウン症の治療のための、請求項1記載の使用。
【請求項4】
男性生殖能力の調節、及び/又は、コントロールのための、請求項1記載の使用。
【請求項5】
ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は、伴わない、潰瘍性疾患及び胃癌
、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、悪性
転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性免
疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネルギ
ー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害から
、選択される疾患治療のための、請求項1記載の使用。
【請求項6】
胃腸管細胞増殖、好ましくは、胃粘膜細胞及び上皮細胞の増殖、急性の酸分泌の刺激促
進のため、及び、酸産生壁細胞、及びヒスタミン分泌クロム親和性様細胞の分化のための
、請求項1記載の使用。
【請求項7】
骨髄性前駆細胞増殖の抑制のための、請求項1記載の使用。
【請求項8】
ハンチントン病、及びケネディ病から選択される疾患の治療のための、請求項1記載の
使用。
【請求項9】
QCの該エフェクターを、DP IV、又はDP IV様酵素のインヒビター、及び/又は、アミ
ノぺプチダーゼインヒビターと共に投与する、請求項1〜8のいずれか1項記載の使用。
【請求項10】
DP IV様酵素のH-isoAsp-Ala-OH生成活性を阻害するような、請求項9記載の使用。
【請求項11】
DP IV様酵素がDP IIである、請求項9、又は10記載の使用。
【請求項12】
少なくとも1種のQCのエフェクターを、任意に、慣用の担体、及び/又は賦形剤と共
に含有することを特徴とする、非経口、経腸、又は経口投与に用いる医薬組成物。
【請求項13】
少なくとも1種のQCのエフェクターを、少なくとも1種のDP IV、又はDP IV様酵素、
及び/又は少なくとも1種のアミノぺプチダーゼインヒビターと共に、任意に、慣用の担
体、及び/又は賦形剤と共に含有することを特徴とする、非経口、経腸、又は経口投与に
用いる医薬組成物。
【請求項14】
QC、及び/又は、DP IV、及びDP IV様酵素、及び/又はアミノぺプチダーゼの酵素活
性のインビボ調整のための、請求項12、又は13記載の医薬組成物の使用。
【請求項15】
DP IV様酵素のH-isoAsp-Ala-OH生成活性を阻害する、請求項14記載の使用。
【請求項16】
DP IV様酵素がDP IIである、請求項14、又は15記載の使用。
【請求項17】
Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu11]Aβ11-40/42、[Gln11]Aβ11-40/42、[Gln
1]ガストリン(17及び34)、[Gln1]ニューロテンシン、[Gln1]FPP、[Gln1]TRH、[Gln1]Gn
RH、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 18、[Gln1]ELA
、[Gln1]フラクタルカイン、[Gln1]オレキシンAから、選択される、少なくとも1種のQ
C基質における、N末端のグルタミン酸、又はグルタミン残基の、ピログルタミル(5-オ
クソ-プロリル)残基への転換の改質のための、請求項14〜16のいずれか1項記載の
使用。
【請求項18】
アルツハイマー病、及びダウン症の治療のための、請求項14〜16記載のいずれか1
項記載の使用。
【請求項19】
QCエフェクター投与による男性生殖能力の調整のための、請求項14〜16のいずれ
か1項記載の使用。
【請求項20】
ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は、伴わない、潰瘍性疾患及び胃癌
、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、悪性
転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性免
疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネルギ
ー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害から
なる群から選択される、疾患治療のための、請求項14〜16のいずれか1項記載の使用
。
【請求項21】
胃腸管細胞増殖、好ましくは、胃粘膜細胞及び上皮細胞の増殖、急性の酸分泌の刺激促
進のため、及び、酸産生壁細胞、及びヒスタミン分泌クロム親和性様細胞の分化のための
、請求項14〜16のいずれか1項記載の使用。
【請求項22】
骨髄性前駆細胞増殖の抑制のための、請求項14〜16のいずれか1項記載の使用。
【請求項23】
下記工程を含む、QCのエフェクターの同定、及び選定のためのスクリーニング方法で
あって:
a)前記の化合物QCを、それらの間で結合が可能な条件下で接触させること;
b)QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残留QC活性を測定すること;及び
d)該基質転換、及び/又は、QCの酵素活性の変化を計算し、QC活性調節エフェクタ
ーを同定することを含む、前記方法。
【請求項24】
下記工程を含む、QCの金属イオン結合活性部位と、直接、又は間接的に相互作用する
エフェクターの同定、及び選定のためのスクリーニング方法であって:
a)前記の化合物QCを、それらの間で結合が可能な条件下で接触させること;
b)QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残留QC活性を測定すること;及び
d)QCの該基質転換、及び/又は、酵素活性の変化を計算し、QC活性調節エフェクタ
ーを同定することを含む、前記方法。
【請求項25】
請求項23、又は24のスクリーニング方法で同定された、QCのエフェクター。
【請求項26】
前記哺乳動物に、少なくとも1種のグルタミニルシクラーゼ(QC)の治療上有効量を
投与することによって、グルタミン、又はグルタミン酸ペプチドのQC活性を調整するこ
とを含む、
a)インビボQC活性の調整によって、及び/又は
b)QC活性の調整によって引き起こされる、pGlu含有ペプチドの作用に基づく生理的プ
ロセスの調整によって、治療可能な、哺乳動物の疾患の治療方法。
【請求項27】
前記QC活性が、Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu11]Aβ11-40/42、[Gln11]A
β11-40/42、[Gln1]ガストリン(17及び34)、[Gln1]ニューロテンシン、[Gln1]FPP、[G
ln1]TRH、[Gln1]GnRH、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]
CCL 18、[Gln1]ELA、[Gln1]フラクタルカイン、[Gln1]オレキシンA、[Gln3]グルカゴン3
-29、及び[Gln5]サブスタンスP 5-11から選択される、少なくとも1種のQC基質におけ
る、N末端のグルタミン酸、又はグルタミン残基の、ピログルタミン酸残基への転換の改
質をもたらす、請求項26記載の方法。
【請求項28】
アルツハイマー病、及びダウン症の治療のための、請求項26記載の方法。
【請求項29】
前記QC活性の調整が、男性生殖能力の調節、及び/又は、コントロールを含む、請求
項26記載の方法。
【請求項30】
前記疾患が、ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は、伴わない、潰瘍性
疾患及び胃癌、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反
応、癌、悪性転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性
及び細胞性免疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚
醒、エネルギー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液
調節障害からなる群より選択される、請求項26記載の方法。
【請求項31】
前記治療が、胃腸管細胞増殖、好ましくは、胃粘膜細胞及び上皮細胞の増殖、急性の酸
分泌の刺激促進、及び、酸産生壁細胞、及びヒスタミン分泌クロム親和性様細胞の分化を
もたらす、請求項26記載の方法。
【請求項32】
前記治療が、骨髄性前駆細胞増殖の抑制をもたらす、請求項26記載の使用。
【請求項33】
QCの該エフェクターを、DP IV、又はDP IV様酵素のインヒビター、及び/又はアミノ
ぺプチダーゼインヒビターと共に投与する、請求項26〜32のいずれか1項記載の方法
。
【請求項34】
DP IV様酵素のH-isoAsp-Ala-OH生成活性を阻害する、請求項33記載の方法。
【請求項35】
DP IV様酵素がDP IIである、請求項34記載の治療方法。
【請求項36】
少なくとも1種のQCのエフェクターを、任意に、治療上許容し得る担体、及び/又は
賦形剤と共に含む、非経口、経腸、又は経口投与に用いる医薬組成物。
【請求項37】
さらに、少なくとも1種のDP IV、又はDP IV様酵素、及び/又は、少なくとも1種のア
ミノぺプチダーゼインヒビターを含む、請求項36記載の医薬組成物。
【請求項38】
前記哺乳動物に請求項36、又は37記載の医薬組成物を投与することを含む、
a)インビボQC活性の調整によって、及び/又は、
b)QC活性、及び/又はDP IV及びDP IV様酵素、及び/又はインビボアミノペプチダー
ゼ活性の調整によって引き起こされる、pGlu含有ペプチドの作用に基づく生理的プロセス
の調整によって、治療可能な哺乳動物の疾患の治療方法。
【請求項39】
前記基質が、Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu11]Aβ11-40/42、[Gln11]Aβ11-
40/42、[Gln1]ガストリン(17及び34)、[Gln1]ニューロテンシン、[Gln1]FPP、[Gln1]T
RH、[Gln1]GnRH、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 1
8、[Gln1]ELA、[Gln1]フラクタルカイン、[Gln1]オレキシンA、からなる群から選択され
る、少なくとも1種のQC基質における、N末端のグルタミン酸、又はグルタミン残基の
、ピログルタミル(5-オクソ-プロリル)残基への転換の改質のための、請求項38記載
の方法。
【請求項40】
アルツハイマー病、及びダウン症の治療のための、請求項38記載の方法。
【請求項41】
男性生殖能力を調節する、請求項38記載の方法。
【請求項42】
ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は、伴わない、潰瘍性疾患及び胃癌
、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、悪性
転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性免
疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネルギ
ー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害から
なる群から選択される、疾患治療のための、請求項38記載の方法。
【請求項43】
胃腸管細胞増殖の刺激促進のため、胃粘膜細胞及び上皮細胞の増殖、急性の酸分泌の刺
激促進のため、及び、酸産生壁細胞、及びヒスタミン分泌クロム親和性様細胞の分化の刺
激促進のための、請求項38記載の方法。
【請求項44】
骨髄性前駆細胞増殖の抑制のための、請求項38記載の方法。
【請求項45】
下記工程を含む、QCのエフェクターの同定、及び選定のためのスクリーニング方法で
あって:
a)前記化合物とQCとを、それらの間で結合が可能な条件下で接触させること;
b)QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残留QC活性を測定すること;及び
d)QCの該基質転換、及び/又は、酵素活性の変化を計算し、QC活性調節エフェクタ
ーを同定することを含む、前記方法。
【請求項46】
下記工程を含む、QCの金属イオン結合活性部位と、直接、又は間接的に相互作用する
エフェクターの同定、及び選定のためのスクリーニング方法であって:
a)前記化合物とQCとを、それらの間で結合が可能な条件下で接触させること;
b)QCによって転換され得る、QCの基質を加えること;
c)該基質の該転換をモニターすること、又は任意に該残留QC活性を測定すること;及
び
d)QCの該基質転換、及び/又は、酵素活性の変化を計算し、QC活性調節エフェクタ
ーを同定することを含む、前記方法。
【請求項47】
請求項45、又は46のスクリーニング方法で同定される、該QCエフェクター。
【請求項1】
a)インビボQC活性の調整によって治療可能な、哺乳動物の疾患の治療、及び/又は
、b)QC活性の調整によって引き起こされる、pGlu含有ペプチドの作用に基づく生理的
プロセスの調整、に用いる薬剤製造のための、グルタミニルシクラーゼ(QC)のエフェ
クターの使用。
【請求項2】
Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu11]Aβ11-40/42、[Gln11]Aβ11-40/42、[Gln
1]ガストリン(17及び34)、[Gln1]ニューロテンシン、[Gln1]FPP、[Gln1]TRH、[Gln1]Gn
RH、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 18、[Gln1]ELA
、[Gln1]フラクタルカイン、[Gln1]オレキシンA、[Gln3]グルカゴン3-29、及び[Gln5]サ
ブスタンスP 5-11から選択される、少なくとも1種のQC基質における、N末端のグル
タミン酸、又はグルタミン残基の、ピログルタミン酸残基への転換の改質のための、請求
項1記載の使用。
【請求項3】
アルツハイマー病、及びダウン症の治療のための、請求項1記載の使用。
【請求項4】
男性生殖能力の調節、及び/又は、コントロールのための、請求項1記載の使用。
【請求項5】
ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は、伴わない、潰瘍性疾患及び胃癌
、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、悪性
転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性免
疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネルギ
ー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害から
、選択される疾患治療のための、請求項1記載の使用。
【請求項6】
胃腸管細胞増殖、好ましくは、胃粘膜細胞及び上皮細胞の増殖、急性の酸分泌の刺激促
進のため、及び、酸産生壁細胞、及びヒスタミン分泌クロム親和性様細胞の分化のための
、請求項1記載の使用。
【請求項7】
骨髄性前駆細胞増殖の抑制のための、請求項1記載の使用。
【請求項8】
ハンチントン病、及びケネディ病から選択される疾患の治療のための、請求項1記載の
使用。
【請求項9】
QCの該エフェクターを、DP IV、又はDP IV様酵素のインヒビター、及び/又は、アミ
ノぺプチダーゼインヒビターと共に投与する、請求項1〜8のいずれか1項記載の使用。
【請求項10】
DP IV様酵素のH-isoAsp-Ala-OH生成活性を阻害するような、請求項9記載の使用。
【請求項11】
DP IV様酵素がDP IIである、請求項9、又は10記載の使用。
【請求項12】
少なくとも1種のQCのエフェクターを、任意に、慣用の担体、及び/又は賦形剤と共
に含有することを特徴とする、非経口、経腸、又は経口投与に用いる医薬組成物。
【請求項13】
少なくとも1種のQCのエフェクターを、少なくとも1種のDP IV、又はDP IV様酵素、
及び/又は少なくとも1種のアミノぺプチダーゼインヒビターと共に、任意に、慣用の担
体、及び/又は賦形剤と共に含有することを特徴とする、非経口、経腸、又は経口投与に
用いる医薬組成物。
【請求項14】
QC、及び/又は、DP IV、及びDP IV様酵素、及び/又はアミノぺプチダーゼの酵素活
性のインビボ調整のための、請求項12、又は13記載の医薬組成物の使用。
【請求項15】
DP IV様酵素のH-isoAsp-Ala-OH生成活性を阻害する、請求項14記載の使用。
【請求項16】
DP IV様酵素がDP IIである、請求項14、又は15記載の使用。
【請求項17】
Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu11]Aβ11-40/42、[Gln11]Aβ11-40/42、[Gln
1]ガストリン(17及び34)、[Gln1]ニューロテンシン、[Gln1]FPP、[Gln1]TRH、[Gln1]Gn
RH、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 18、[Gln1]ELA
、[Gln1]フラクタルカイン、[Gln1]オレキシンAから、選択される、少なくとも1種のQ
C基質における、N末端のグルタミン酸、又はグルタミン残基の、ピログルタミル(5-オ
クソ-プロリル)残基への転換の改質のための、請求項14〜16のいずれか1項記載の
使用。
【請求項18】
アルツハイマー病、及びダウン症の治療のための、請求項14〜16記載のいずれか1
項記載の使用。
【請求項19】
QCエフェクター投与による男性生殖能力の調整のための、請求項14〜16のいずれ
か1項記載の使用。
【請求項20】
ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は、伴わない、潰瘍性疾患及び胃癌
、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、悪性
転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性免
疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネルギ
ー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害から
なる群から選択される、疾患治療のための、請求項14〜16のいずれか1項記載の使用
。
【請求項21】
胃腸管細胞増殖、好ましくは、胃粘膜細胞及び上皮細胞の増殖、急性の酸分泌の刺激促
進のため、及び、酸産生壁細胞、及びヒスタミン分泌クロム親和性様細胞の分化のための
、請求項14〜16のいずれか1項記載の使用。
【請求項22】
骨髄性前駆細胞増殖の抑制のための、請求項14〜16のいずれか1項記載の使用。
【請求項23】
下記工程を含む、QCのエフェクターの同定、及び選定のためのスクリーニング方法で
あって:
a)前記の化合物QCを、それらの間で結合が可能な条件下で接触させること;
b)QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残留QC活性を測定すること;及び
d)該基質転換、及び/又は、QCの酵素活性の変化を計算し、QC活性調節エフェクタ
ーを同定することを含む、前記方法。
【請求項24】
下記工程を含む、QCの金属イオン結合活性部位と、直接、又は間接的に相互作用する
エフェクターの同定、及び選定のためのスクリーニング方法であって:
a)前記の化合物QCを、それらの間で結合が可能な条件下で接触させること;
b)QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残留QC活性を測定すること;及び
d)QCの該基質転換、及び/又は、酵素活性の変化を計算し、QC活性調節エフェクタ
ーを同定することを含む、前記方法。
【請求項25】
請求項23、又は24のスクリーニング方法で同定された、QCのエフェクター。
【請求項26】
前記哺乳動物に、少なくとも1種のグルタミニルシクラーゼ(QC)の治療上有効量を
投与することによって、グルタミン、又はグルタミン酸ペプチドのQC活性を調整するこ
とを含む、
a)インビボQC活性の調整によって、及び/又は
b)QC活性の調整によって引き起こされる、pGlu含有ペプチドの作用に基づく生理的プ
ロセスの調整によって、治療可能な、哺乳動物の疾患の治療方法。
【請求項27】
前記QC活性が、Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu11]Aβ11-40/42、[Gln11]A
β11-40/42、[Gln1]ガストリン(17及び34)、[Gln1]ニューロテンシン、[Gln1]FPP、[G
ln1]TRH、[Gln1]GnRH、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]
CCL 18、[Gln1]ELA、[Gln1]フラクタルカイン、[Gln1]オレキシンA、[Gln3]グルカゴン3
-29、及び[Gln5]サブスタンスP 5-11から選択される、少なくとも1種のQC基質におけ
る、N末端のグルタミン酸、又はグルタミン残基の、ピログルタミン酸残基への転換の改
質をもたらす、請求項26記載の方法。
【請求項28】
アルツハイマー病、及びダウン症の治療のための、請求項26記載の方法。
【請求項29】
前記QC活性の調整が、男性生殖能力の調節、及び/又は、コントロールを含む、請求
項26記載の方法。
【請求項30】
前記疾患が、ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は、伴わない、潰瘍性
疾患及び胃癌、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反
応、癌、悪性転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性
及び細胞性免疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚
醒、エネルギー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液
調節障害からなる群より選択される、請求項26記載の方法。
【請求項31】
前記治療が、胃腸管細胞増殖、好ましくは、胃粘膜細胞及び上皮細胞の増殖、急性の酸
分泌の刺激促進、及び、酸産生壁細胞、及びヒスタミン分泌クロム親和性様細胞の分化を
もたらす、請求項26記載の方法。
【請求項32】
前記治療が、骨髄性前駆細胞増殖の抑制をもたらす、請求項26記載の使用。
【請求項33】
QCの該エフェクターを、DP IV、又はDP IV様酵素のインヒビター、及び/又はアミノ
ぺプチダーゼインヒビターと共に投与する、請求項26〜32のいずれか1項記載の方法
。
【請求項34】
DP IV様酵素のH-isoAsp-Ala-OH生成活性を阻害する、請求項33記載の方法。
【請求項35】
DP IV様酵素がDP IIである、請求項34記載の治療方法。
【請求項36】
少なくとも1種のQCのエフェクターを、任意に、治療上許容し得る担体、及び/又は
賦形剤と共に含む、非経口、経腸、又は経口投与に用いる医薬組成物。
【請求項37】
さらに、少なくとも1種のDP IV、又はDP IV様酵素、及び/又は、少なくとも1種のア
ミノぺプチダーゼインヒビターを含む、請求項36記載の医薬組成物。
【請求項38】
前記哺乳動物に請求項36、又は37記載の医薬組成物を投与することを含む、
a)インビボQC活性の調整によって、及び/又は、
b)QC活性、及び/又はDP IV及びDP IV様酵素、及び/又はインビボアミノペプチダー
ゼ活性の調整によって引き起こされる、pGlu含有ペプチドの作用に基づく生理的プロセス
の調整によって、治療可能な哺乳動物の疾患の治療方法。
【請求項39】
前記基質が、Aβ3-40/42、[Gln3]Aβ3-40/42、[Glu11]Aβ11-40/42、[Gln11]Aβ11-
40/42、[Gln1]ガストリン(17及び34)、[Gln1]ニューロテンシン、[Gln1]FPP、[Gln1]T
RH、[Gln1]GnRH、[Gln1]CCL 2、[Gln1]CCL 7、[Gln1]CCL 8、[Gln1]CCL 16、[Gln1]CCL 1
8、[Gln1]ELA、[Gln1]フラクタルカイン、[Gln1]オレキシンA、からなる群から選択され
る、少なくとも1種のQC基質における、N末端のグルタミン酸、又はグルタミン残基の
、ピログルタミル(5-オクソ-プロリル)残基への転換の改質のための、請求項38記載
の方法。
【請求項40】
アルツハイマー病、及びダウン症の治療のための、請求項38記載の方法。
【請求項41】
男性生殖能力を調節する、請求項38記載の方法。
【請求項42】
ピロリ菌(ヘリコバクターピロリ)感染を伴う、又は、伴わない、潰瘍性疾患及び胃癌
、病原性精神病的状態、統合失調病、不妊症、新生組織形成、炎症性宿主反応、癌、悪性
転移、メラノーマ、乾癬、関節リウマチ、アステローム性動脈硬化、体液性及び細胞性免
疫反応障害、内皮における白血球接着及び遊走プロセス、摂食障害、睡眠覚醒、エネルギ
ー代謝の恒常性調節障害、自律機能障害、ホルモンバランス障害、及び体液調節障害から
なる群から選択される、疾患治療のための、請求項38記載の方法。
【請求項43】
胃腸管細胞増殖の刺激促進のため、胃粘膜細胞及び上皮細胞の増殖、急性の酸分泌の刺
激促進のため、及び、酸産生壁細胞、及びヒスタミン分泌クロム親和性様細胞の分化の刺
激促進のための、請求項38記載の方法。
【請求項44】
骨髄性前駆細胞増殖の抑制のための、請求項38記載の方法。
【請求項45】
下記工程を含む、QCのエフェクターの同定、及び選定のためのスクリーニング方法で
あって:
a)前記化合物とQCとを、それらの間で結合が可能な条件下で接触させること;
b)QCの基質を加えること;
c)該基質の転換をモニターすること、又は任意に該残留QC活性を測定すること;及び
d)QCの該基質転換、及び/又は、酵素活性の変化を計算し、QC活性調節エフェクタ
ーを同定することを含む、前記方法。
【請求項46】
下記工程を含む、QCの金属イオン結合活性部位と、直接、又は間接的に相互作用する
エフェクターの同定、及び選定のためのスクリーニング方法であって:
a)前記化合物とQCとを、それらの間で結合が可能な条件下で接触させること;
b)QCによって転換され得る、QCの基質を加えること;
c)該基質の該転換をモニターすること、又は任意に該残留QC活性を測定すること;及
び
d)QCの該基質転換、及び/又は、酵素活性の変化を計算し、QC活性調節エフェクタ
ーを同定することを含む、前記方法。
【請求項47】
請求項45、又は46のスクリーニング方法で同定される、該QCエフェクター。
【図1】

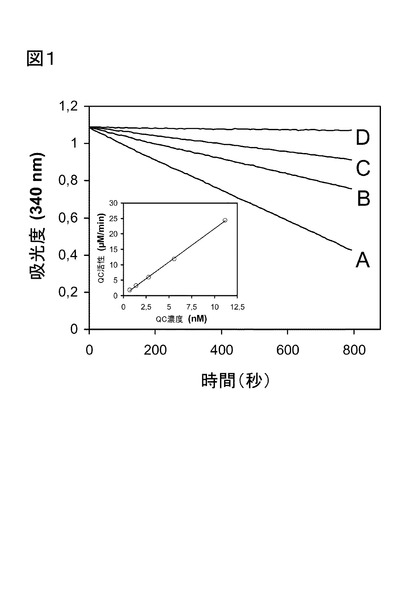
【図2】
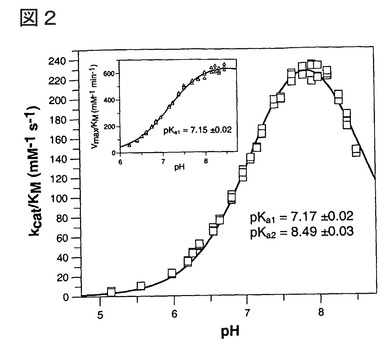
【図3】
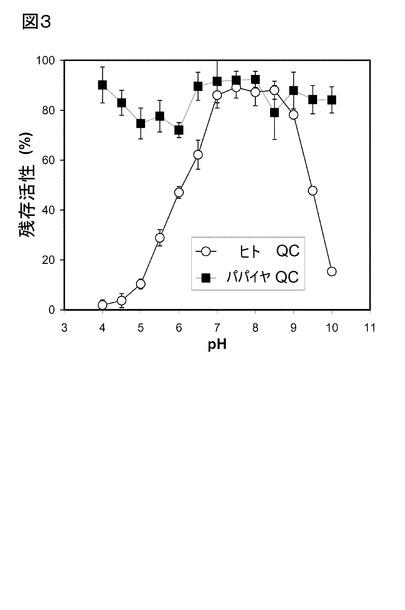
【図4】
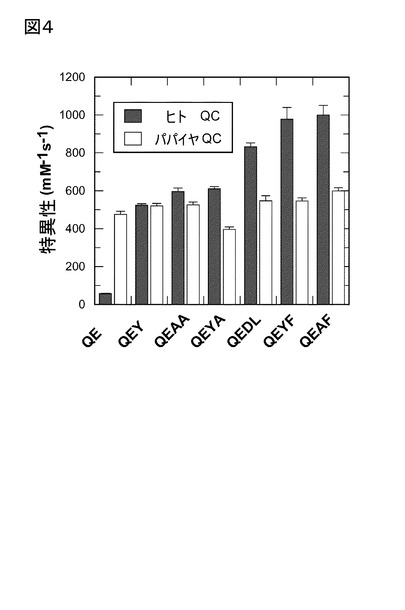
【図5】
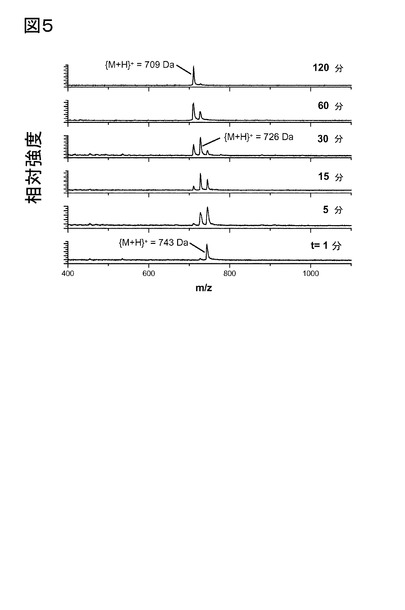
【図6】
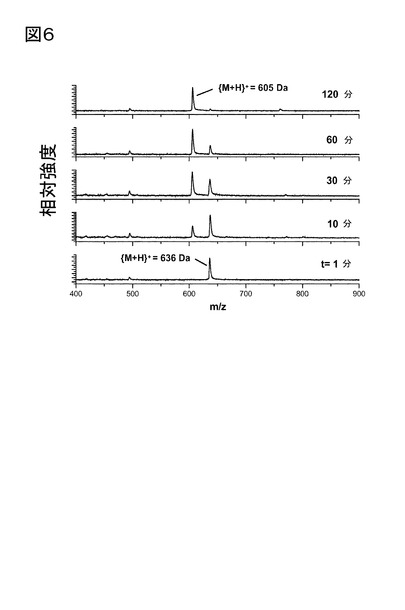
【図7】
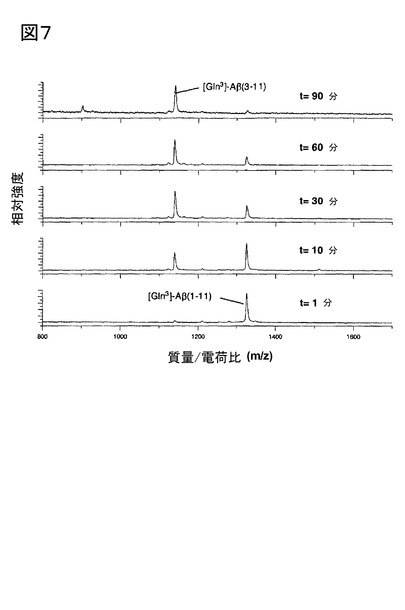
【図8】
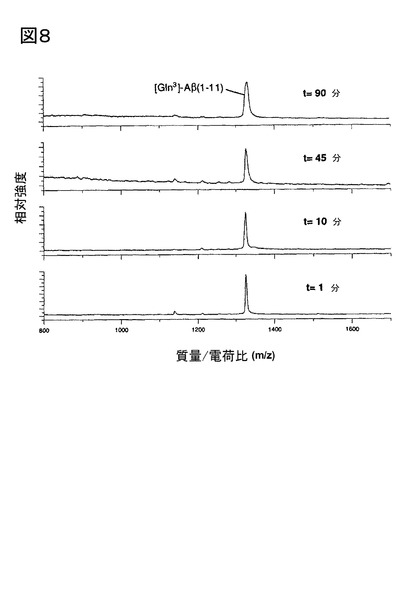
【図9】
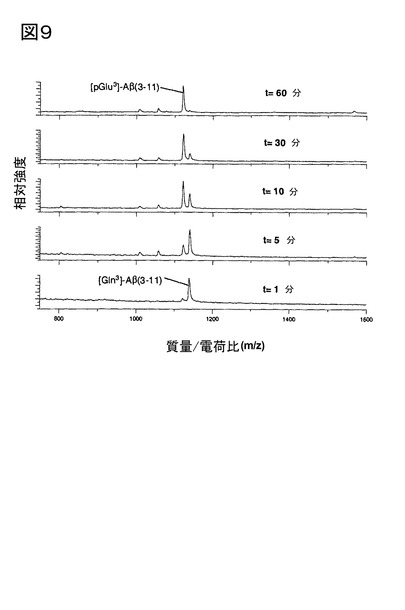
【図10】
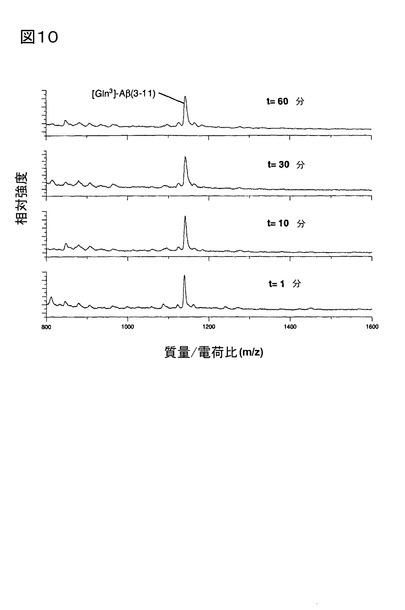
【図11】
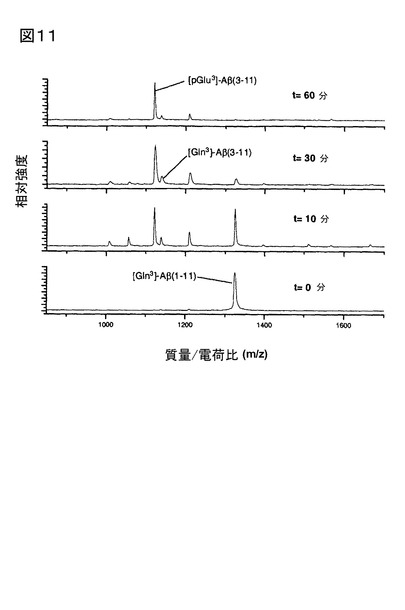
【図12】
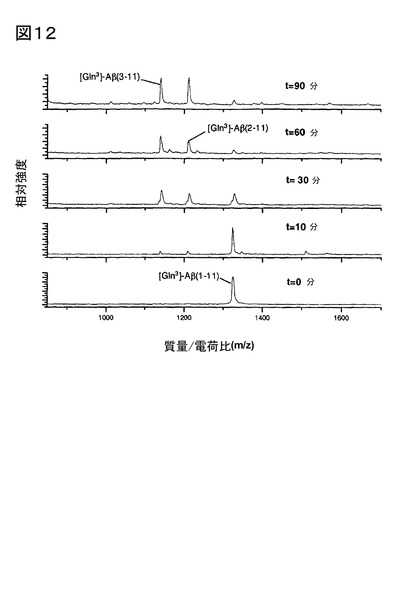
【図13】
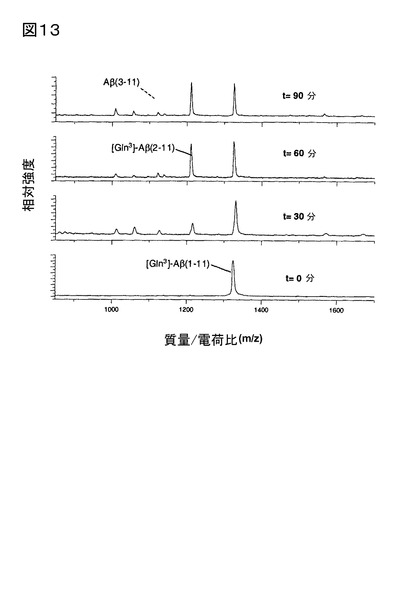
【図14】
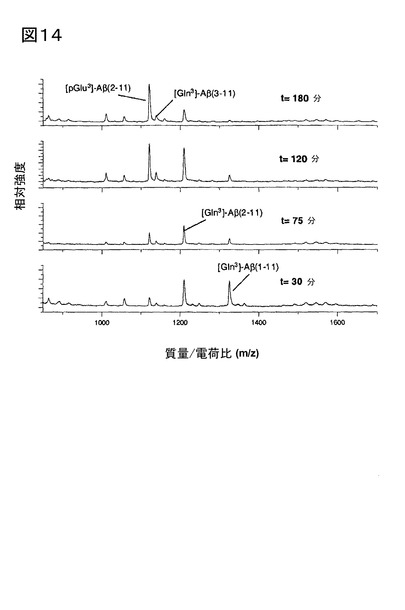
【図15】
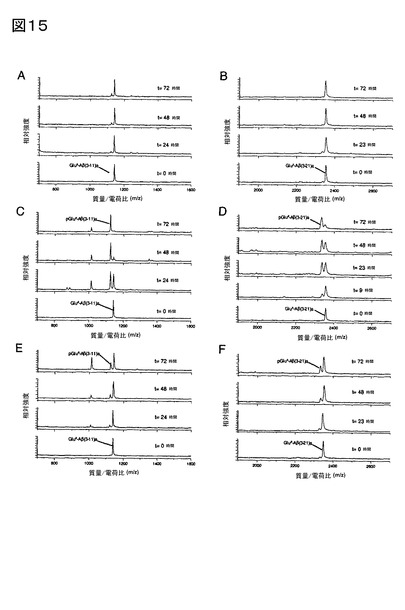
【図16】
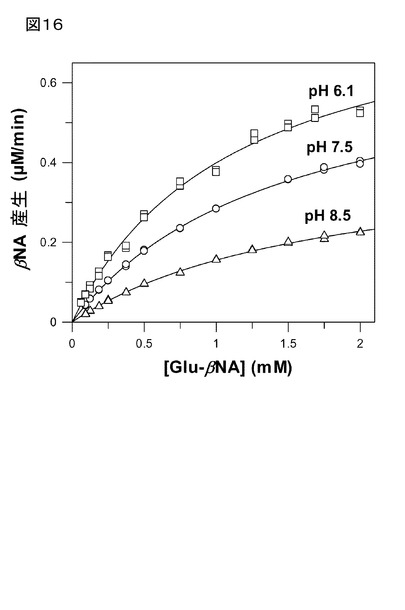
【図17】
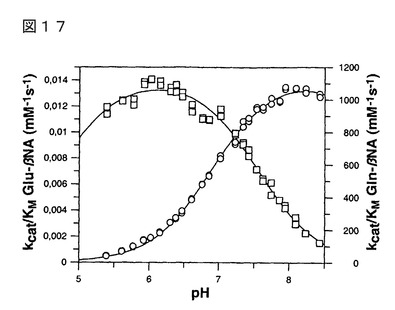
【図18】
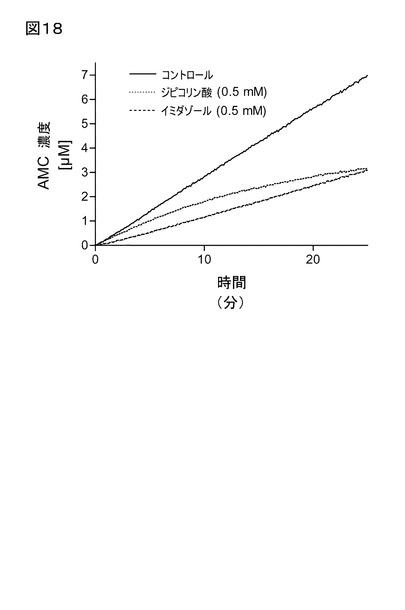
【図19】
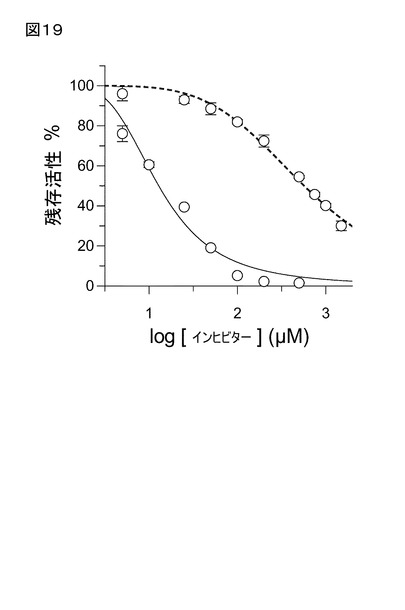
【図20】
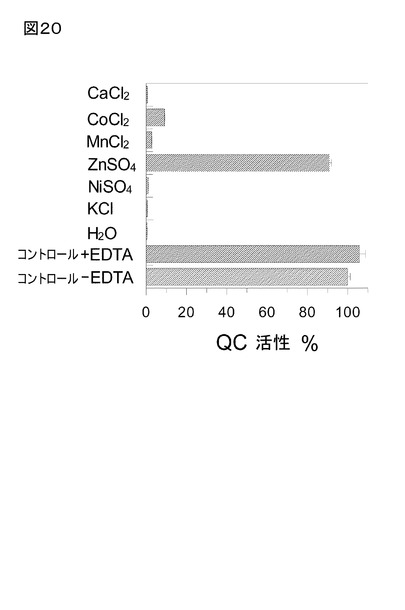
【図21】
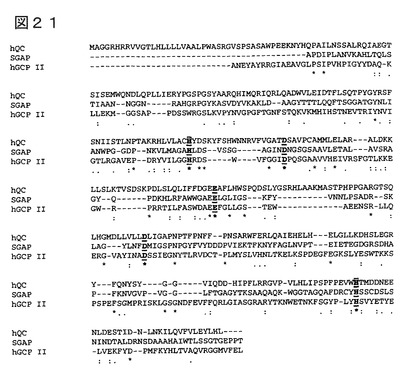
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【公開番号】特開2012−184237(P2012−184237A)
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2012−100566(P2012−100566)
【出願日】平成24年4月26日(2012.4.26)
【分割の表示】特願2006−505377(P2006−505377)の分割
【原出願日】平成16年5月5日(2004.5.5)
【出願人】(505403119)プロビオドルグ エージー (39)
【Fターム(参考)】
【公開日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願日】平成24年4月26日(2012.4.26)
【分割の表示】特願2006−505377(P2006−505377)の分割
【原出願日】平成16年5月5日(2004.5.5)
【出願人】(505403119)プロビオドルグ エージー (39)
【Fターム(参考)】
[ Back to top ]