
グレリン結合核酸
本発明は、好ましくはグレリンに結合する核酸に関し、この場合、核酸は、
第一のストレッチBoxAおよび第二のストレッチBoxBを含み、ここで、第一のストレッチBoxAは約25個の連続ヌクレオチドを含み、第二のストレッチBoxBは約6〜8個の連続ヌクレオチドを含み、ここで、第一のストレッチBoxAのヌクレオチドの3’末端ストレッチは、第二のストレッチBoxBとハイブリダイズし、このハイブリダイゼーションでは、第一の二本鎖構造が形成され、そしてそのような第一の二本鎖構造がバルジを含む。

第一のストレッチBoxAおよび第二のストレッチBoxBを含み、ここで、第一のストレッチBoxAは約25個の連続ヌクレオチドを含み、第二のストレッチBoxBは約6〜8個の連続ヌクレオチドを含み、ここで、第一のストレッチBoxAのヌクレオチドの3’末端ストレッチは、第二のストレッチBoxBとハイブリダイズし、このハイブリダイゼーションでは、第一の二本鎖構造が形成され、そしてそのような第一の二本鎖構造がバルジを含む。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、グレリン(ghrelin)に結合する核酸、および薬物の製造におけるそれらの使用、および診断作用物の製造におけるそれらの使用に関する。
【背景技術】
【0002】
グレリンは、成長ホルモン分泌促進物質受容体1a(GHSR1a)の天然のリガンドとして同定された。この受容体は、下垂体および脳の視床下部部分にもっとも豊富に存在するが、低濃度では他の組織においてもまた検出することができる。70年代末期以来、分泌促進物質と命名された合成ペプチドおよび他の化合物は、成長ホルモンの放出を刺激することが示された。しかしながら、成長ホルモンの放出に関与する天然のリガンドは、1999年におけるグレリンの発見まで知られていなかった。グレリンは、そのN末端の3番目のアミノ酸(セリン3)においてオクタン酸(octanoyl acid)側鎖をもつ高度に塩基性の28アミノ酸ペプチドホルモンである。この異例の修飾は、GHS受容体における相互作用およびその活性のために必要とされる。しかしながら、生物サンプルにおいては、生物活性グレリンの形態物であるオクタノイルグレリン、および未修飾またはデス−オクタノイルグレリンの両方が存在している。精製されたラット・グレリンのアミノ酸配列は、GSSFLSPEHQKAQQRKESKKPPAKLQPR(配列番号:2)であると決定され;対応するヒトの配列は、2つの位置においてのみ逸脱するが、アミノ酸位置セリン3において同じn−オクタノイル側鎖を保持し、そしてGSSFLSPEHQRVQQRKESKKPPAKLQPR(配列番号:1)であると決定された。
【0003】
天然に存在するn−オクタノイル残基の他に、グレリンの位置3において導入された不飽和または分枝オクタノイル基およびより長い脂肪族鎖が同様に受容体認識を媒介する。受容体相互作用ドメインはグレリンのまさにN末端に置かれ;欠失研究では、アミノ酸1〜5の最小モチーフ(グレリン(1〜5)[GSSFL])がGHSR1aの刺激のためには十分であるが、n−オクタノイル残基によるペプチド修飾に対する強力な必要性が観察されることが示された。
【0004】
グレリンは嫌気状態に対して適切な生理学的機能を媒介することが示されている。それは下垂体からの成長ホルモン(GH)の放出を直接刺激し、したがって、先端巨大症の処置における適当な標的になり得る。また齧歯類における実験は、グレリンが視床下部ニューロンへの作用によってGHに依存しない様式で栄養補給(feeding)を誘導することが示された。興味あることに、グレリン産生の一次部位は胃における酸分泌腺であり、それが、胃、下垂体および視床下部間のホルモン連鎖として働くことが示唆される。ラットにおけるグレリン投与がエネルギー獲得(intake)および/または燃料利用における変化の結果として体重獲得をもたらすという観察は、そのような役割を支持することになる。さらに、ヒトでの全身的グレリン投与は、試験の被験者における空腹感を惹起し、過食を誘導する。これらの知見に基づいて、グレリンは、食欲および体重の調節において決定的な役割を有して、食物が十分与えられない状態の急性ならびに慢性的シグナルとして働くと考えられる。この仮説のさらなる支持は、グレリンのレベルならびに食欲が、胃バイパス後の個人において低下され、体重減少を実施する手法の効力に少なくとも一部は貢献するという観察からも得られる。また、Prader−Willi症候群を有する患者からの臨床データは、この疾病に関係する過食および肥満が、大変な高グレリン血症(hyperghrelinemia)の結果であることを示唆する。さらに、グレリンは高血糖およびインスリン放出の抑制を誘導することが見出されて、グルコース代謝における関与が示された。エネルギー代謝におけるこれらの機能の他に、グレリンはまた、胃腸疾患の分野における多くの他のプロセス、例えば胃を空にしたり、腸の運動を調節することに関与している。さらに、グレリンはまた、多数の神経内分泌腫瘍において発現され、そして下垂体からのGH放出の他に、ACTH、PRLおよびコルチソルの放出を刺激することが見出された。健康な個体へのグレリンの単一注射は、心拍出量を増加し、血圧を低下させることが見出された。かくして、グレリンの作用は種々の異なる仕事に関与していると考えられる。これらに関するさらなる背景情報は、非特許文献1;非特許文献2;非特許文献3;非特許文献4;非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;非特許文献10において見出すことができる。
【非特許文献1】M.Kojima,H.Hosoda,Y.Date,M.Nakazato,H.Matsu,K.Kangawa,“Ghrelin is a growth−hormone−releasing acylated peptide from stomach”,Nature 402:656−60,1999
【非特許文献2】M.Tsch p,D.L.Smiley,M.L.Heiman,“Ghrelin induces adiposity in rodents”,Nature 407:908−13、2000
【非特許文献3】A.M.Wren et al.,”Ghrelin enhances appetite and increases food intake in humans”,Jounal of Clinical Endocrinology Metabolism 86:5992−6,2001
【非特許文献4】M.Nakazato et al.,”A role for ghrelin in the central regulation of feeding”,Nature 409:194−8,2001
【非特許文献5】N.Nagaya et al.,Am J Physiol Regul Integr Comp Physiol.2001 May;280(5):R1483−7;Hemodynamic and hormonal effects of human ghrelin in healthy volunteers
【非特許文献6】Volante M,et al.,J Clin Endocrinol Metab.2002 Mar;87(3):1300−8.Expression of ghrelin and of the GH secretagogue receptor by pancreatic islet cells and related endocrine tumors
【非特許文献7】Jeffery PL,et al.,J Endocrinol.2002 Mar;172(3):R7−11 Expression and action of the growth hormone releasing peptide ghrelin and its receptor in prostate cancer cell lines
【非特許文献8】Egido EM,et al.,Eur J Endocrinol.2002 Feb;146(2):241−4 Inhibitory effect of ghrelin on insulin and pancreatic somatostatin secretion
【非特許文献9】Broglio F,et al.,J Clin Endocrinol Metab.2001 Oct;86(10)5083−6,Ghrelin, a natural GH secretagogue produced by the stomach,induces hyperglycemia and reduces insulin secretion in humans
【非特許文献10】Bednarek MA,et al.,J Med Chem.2000 Oct;43:4370−6 Structure−function studies on the new growth hormone−releasing peptide,ghrelin:minimal sequence of ghrelin necessary for activation of growth hormone secretagogue receptor 1a.
【発明の開示】
【0005】
本発明の基礎をなす課題は、グレリンに対する特異的アンタゴニストを提供することである。本発明の基礎をなす課題のさらなる態様は、成長ホルモン分泌促進物質受容体1a(GHSR1a)に対する特異的アンタゴニストを提供することである。本発明の基礎をなす課題のその他の態様は、グレリンおよびGHSR1a受容体それぞれが関与する疾病および障害の処置のための化合物を提供することである。
【0006】
本発明の基礎をなすさらなる課題は、生物活性グレリンの結合のための手段を提供すること、より具体的には生物活性グレリンによって媒介される疾病および障害の処置のための方法ならびに生物活性グレリンの特異的検出のための方法を提供することである。
【0007】
本発明の基礎をなす課題は、好ましくはグレリンに結合する、核酸による第一の態様によって解決され、
こうして、核酸は、
第一のストレッチBoxAおよび
第二のストレッチBoxB
を含み、
ここで、
第一のストレッチBoxAは約25個の連続ヌクレオチドを含み、
第二のストレッチBoxBは約6〜8個の連続ヌクレオチドを含み、
ここで、
第一のストレッチBoxAのヌクレオチドの3’末端ストレッチは、第二のストレッチBoxBとハイブリダイズし、このハイブリダイゼーションでは、第一の二本鎖構造が形成され、そしてそのような第一の二本鎖構造はバルジを含む。
【0008】
本発明の各々およびいずれかの態様の実施態様では、グレリンは、生物活性グレリン、より好ましくはオクタノイルグレリン、そしてもっとも好ましくはn−オクタノイルグレリンである。
【0009】
1つの実施態様では、二本鎖構造は、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドおよび第二のストレッチBoxBのヌクレオチドの一部または全部、好ましくは第二のストレッチBoxBの6〜8個の連続ヌクレオチドによって形成される。
【0010】
1つの実施態様では、バルジは、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドと塩基対合しない、第二のストレッチBoxBの1〜3個のヌクレオチド、好ましくは第二のストレッチBoxBの1個のヌクレオチドによって形成される。
【0011】
1つの実施態様では、バルジは非塩基対合プリンによって形成され、ここで、プリンは好ましくはグアノシンである。
【0012】
好適な実施態様では、非塩基対合プリンは、第二のストレッチBoxBによって提供される。
【0013】
1つの実施態様では、核酸は、第三のストレッチBoxC1および第四のストレッチBoxC2をさらに含み、
ここで、第三のストレッチBoxC1は少なくとも1個のヌクレオチドを含み、そして
第四のストレッチBoxC2は少なくとも1個のヌクレオチドを含み、そして
ここで、第三のストレッチBoxC1は、その3’末端によって第二のストレッチBox
Bの5’末端に連結されており、そして第四のストレッチBoxC2は、その5’末端によって第一のストレッチBoxAの3’末端に連結されている。
【0014】
好適な実施態様では、第三のストレッチBoxC1および第四のストレッチBoxC2は、ハイブリダイゼーション可能であり、ここで、ハイブリダイゼーションでは、第二の二本鎖構造が形成される。
【0015】
1つの実施態様では、第一の二本鎖構造は第一のらせん構造を形成する。
【0016】
好適な実施態様では、第二の二本鎖構造は第二のらせん構造を形成する。
【0017】
より好適な実施態様では、第二のらせん構造は、1〜10個の塩基対、好ましくは1〜3個の塩基対、より好ましくは2〜3個の塩基対を含んでなる、ヘリックスまたはヘリックス様構造である。
【0018】
好適な実施態様では、第一のらせん構造は第二のらせん構造によって延長される。
【0019】
好適な実施態様では、第三のストレッチBoxC1は約1〜10個の連続ヌクレオチド、好ましくは1〜3個の連続ヌクレオチド、より好ましくは2または3個の連続ヌクレオチドを含む。
【0020】
好適な実施態様では、第四のストレッチBoxC2は約1〜10個の連続ヌクレオチド、好ましくは1〜3個の連続ヌクレオチド、より好ましくは2または3個の連続ヌクレオチドを含む。
【0021】
好適な実施態様では、核酸は、第五のストレッチBoxDをさらに含み、ここで、第五のストレッチBoxDは少なくとも2個の連続ヌクレオチドを含む。
【0022】
より好適な実施態様では、第五のストレッチBoxDは配列5’−CAを含む。
【0023】
より好適な実施態様では、第五のストレッチBoxDは、いずれかの長さの連続ヌクレオチドを含み、ここで、長さは2、3、4、5および6個の連続ヌクレオチドからなる群から選ばれる。
【0024】
好適な実施態様では、第五のストレッチBoxDは、
5’CA(X)n3’
[式中、Xは、好ましくはA,G,T,C,UおよびIを含んでなる群から選ばれるいずれかのヌクレオチドであり、そしてnは、0,1,2,3および4からなる群から選ばれるいずれかの整数である]
の配列を含む。
【0025】
より好適な実施態様では、第五のストレッチBoxDは、配列
5’CA(X)n3’
[式中、n=4]
からなる。
【0026】
好適な実施態様では、第五のストレッチBoxDは、その5’末端によって第二のストレッチBoxBの3’末端に連結されている。
【0027】
好適な実施態様では、核酸は、第六のストレッチBoxEをさらに含み、ここで、第六
のストレッチBoxEは少なくとも1個のヌクレオチドを含む。
【0028】
より好適な実施態様では、第六のストレッチBoxEは、約1〜10個の連続ヌクレオチド、好ましくは1〜4個の連続ヌクレオチド、より好ましくは3個の連続ヌクレオチドを含む。
【0029】
さらにより好適な実施態様では、第六のストレッチBoxEの少なくとも1個のヌクレオチドはUおよびGからなる群から選ばれる。
【0030】
特に好適な実施態様では、UまたはGヌクレオチドは、第一のストレッチBoxAの5’末端の直ぐ隣に配置されている。
【0031】
好適な実施態様では、第六のストレッチBoxEは、その3’末端によって第一のストレッチBoxAの5’末端に連結されている。
【0032】
好適な実施態様では、第五のストレッチBoxDの3’末端は、第一のスペーサーによって第六のストレッチBoxEの5’末端に連結されている。
【0033】
好適な実施態様では、第四のストレッチBoxC2の3’末端は、第二のスペーサーによって第三のストレッチBoxC1の5’末端に連結されている。
【0034】
好適な実施態様では、第一および第二のスペーサーは、各々別々に、独立して、親水性スペーサーを含んでなる群から選ばれる。
【0035】
より好適な実施態様では、親水性スペーサーは、核酸スペーサーおよび非核酸スペーサーを含んでなる群から選ばれる。
【0036】
より好適な実施態様では、第一のスペーサーは、約1〜20個の連続ヌクレオチド、好ましくは1〜5個の連続ヌクレオチド、より好ましくは2個の連続ヌクレオチドを含んでなる核酸スペーサーである。
【0037】
その他のより好適な実施態様では、第二のスペーサーは、約3〜20個の連続ヌクレオチド、好ましくは3〜5個の連続ヌクレオチド、より好ましくは3個の連続ヌクレオチドを含んでなる核酸スペーサーである。なおより好適な実施態様では、第二のスペーサーはACAまたはCAAからなる。
【0038】
さらなるその他のより好適な実施態様では、スペーサーは非核酸である。その特に好適な実施態様では、第一のスペーサーおよび/または第二のスペーサーは、少なくとも1個のエチレングリコール部分または複数個のそのようなエチレングリコール部分を含む。
【0039】
1つの実施態様では、スペーサーは約172〜688Da、好ましくは344Daの分子量を有する。
【0040】
1つの実施態様では、核酸は環状核酸である。
【0041】
1つの実施態様では、核酸は、
【0042】
【化1】
【0043】
の構造または
【0044】
【化2】
【0045】
の構造を有する。
【0046】
1つの実施態様では、第一のストレッチBoxAは、
【0047】
【化3】
【0048】
[式中、
X1=GまたはA;
X2=AまたはU;
X3=GまたはA;
X4=AまたはCまたはU;および
X5=GまたはA]
、好ましくは
【0049】
【化4】
【0050】
の配列を含んでなる。
【0051】
1つの実施態様では、第二のストレッチBoxBは5’GUGAGG3’の配列を含む。
【0052】
1つの実施態様では、核酸の配列は、次による配列を含んでなる群から選ばれる:
【0053】
【表1】
【0054】
【表2】
【0055】
好適な実施態様では、核酸の配列は、配列番号:29、配列番号:30、配列番号:33、配列番号:36、配列番号:38、配列番号:39、配列番号:40、配列番号:41、配列番号:42、配列番号:46、配列番号:47、配列番号:51および配列番号:52による配列を含んでなる群から選ばれる。
【0056】
1つの実施態様では、核酸はグレリン、好ましくはヒト・グレリンを結合することができる。
【0057】
好適な実施態様では、グレリンは配列番号:1によるアミノ酸配列を有する。
【0058】
1つの実施態様では、核酸は修飾(modification)を含む。
【0059】
好適な実施態様では、修飾は、HES部分およびPEG部分を含んでなる群から選ばれ
る。
【0060】
より好適な実施態様では、修飾体は直鎖または分枝状PEGからなるPEG部分であり、ここで、PEG部分の分子量は、好ましくは約20〜120kD、より好ましくは約30〜80kD、もっとも好ましくは約40kDである。
【0061】
その他のより好適な実施態様では、修飾体はHES部分であり、ここで、HES部分の分子量は、好ましくは約10〜130kD、より好ましくは約30〜80kD、もっとも好ましくは約50kDである。
【0062】
1つの実施態様では、核酸のヌクレオチドはL−ヌクレオチドである。
【0063】
さらなる実施態様では、核酸は完全にL−ヌクレオチドからなる。
【0064】
本発明の基礎をなす課題は、第一の態様による核酸および場合によってはさらなる成分を含んでなる製薬学的組成物によって第二の態様において解決され、この場合、さらなる成分は製薬学的に許容できる添加物および製薬学的活性作用物を含んでなる群から選ばれる。
【0065】
本発明の基礎をなす課題は、薬物の製造のための第一の態様による核酸の使用によって第三の態様において解決される。
【0066】
本発明の基礎をなす課題は、診断手段の製造のための第一の態様による核酸の使用によって第四の態様において解決される。
【0067】
第三の態様の実施態様では、薬物は、肥満、摂食障害、糖尿病、グルコース代謝障害、腫瘍、血圧障害、心臓血管病、先端巨大症、およびエネルギーバランス、食欲、体重の調節、および胃腸疾患を含んでなる群から選ばれる疾病または障害の処置および/または予防のために存在する。
【0068】
本発明の基礎をなす課題は、グレリンおよび第一の態様による核酸を含んでなる複合体によって第五の態様において解決され、ここで、好ましくは複合体は結晶性複合体である。
【0069】
本発明の基礎をなす課題は、グレリンの検出のための第一の態様による核酸の使用によって第六の態様において解決される。
【0070】
本発明の基礎をなす課題は、次の段階:
− 候補グレリンアンタゴニストおよび/または候補グレリンアゴニストを提供し、
− 第一の態様による核酸を提供し、
− グレリンアンタゴニストおよび/またはグレリンアゴニストの存在においてシグナルを提供する試験系を提供し、そして
− 候補グレリンアンタゴニストがグレリンアンタゴニストであるか、そして/または候補グレリンアゴニストがグレリンアゴニストであるか否かを決定すること、
を含んでなる、グレリンアンタゴニストまたはグレリンアゴニストのスクリーニングのための方法によって第7の態様において解決される。
【0071】
本発明の基礎をなす課題は、次の段階:
− 相、好ましくは固定相に固定化されたグレリンを提供し、
− 第一の態様による核酸、好ましくは標識されている第一の態様による核酸を提供し、
− 候補グレリンアゴニストおよび/または候補グレリンアンタゴニストを添加し、そして
− 候補グレリンアゴニストがグレリンアゴニストであるか、そして/または候補グレリンアンタゴニストがグレリンアンタゴニストであるか否かを決定すること、
を含んでなる、グレリンアゴニストおよび/またはグレリンアンタゴニストのスクリーニングのための方法によって第8の態様において解決される。
【0072】
1つの実施態様では、決定は、核酸が候補グレリンアゴニストによるか、または候補グレリンアンタゴニストによって置換されるか否かが調査されるように実施される。
【0073】
本発明の基礎をなす問題は、第一の態様による核酸を含んでなる、グレリンの検出のためのキットによって第9の態様において解決される。
【0074】
本発明の基礎をなす問題は、第8の態様による方法によって得ることができるグレリンアンタゴニストによって第十の態様において解決される。
【0075】
本発明の基礎をなす問題は、第8の態様による方法によって得ることができるグレリンアゴニストによって第十一の態様において解決される。
【0076】
本発明は、グレリンに特異的に結合し、かつグレリンに対して高い親和力を有する核酸を生成することができるという驚くべき発見に基づいている。より具体的には、本発明者らは、驚くべきことに、生物活性のあるグレリン、より好ましくはオクタノイルグレリン、もっとも好ましくはn−オクタノイルグレリンに特異的に結合する核酸を生成することができた。
【0077】
グレリンは配列番号:1によるアミノ酸配列を有する塩基性ペプチドであり、そして好ましくはオクタノイル側鎖、より好ましくはn−オクタノイル側鎖である脂肪酸側鎖により好ましくは修飾されている。グレリンの算出されたpIは、ヒト・グレリンについては11,07,およびラット・グレリンについては10,56である。好適な実施態様では、本発明による核酸が結合するグレリンは、脂肪酸側鎖により修飾されているグレリンである。その他の実施態様では、グレリンは、脂肪酸側鎖を有しないグレリンである。本明細書において使用されるような用語グレリンは、限定されるものではないが哺乳動物のグレリンを含むすべてのグレリンを指す。好ましくは、哺乳動物グレリンは、マウス、ラット、ウサギ、ハムスターおよびヒトのグレリンを含んでなる群から選ばれる。もっとも好ましくは、グレリンはヒト・グレリンである。
【0078】
グレリンに核酸を結合する高い親和力が同定できたという発見は、塩基性タンパク質に対向されるアプタマー(aptamer)、すなわち標的分子に結合するD−核酸の生成は、この種の標的が高いが非特異的なシグナル対ノイズ比を生成するので、一般に非常に困難であることをEatonら(Eaton,B.E.;Gold,L.;Hicke,B.J.;Janjic,N.;Jucker,F.M.;Sebosta,D.P.;Tarasow,T.M.;Willis,M.C.;Zichi,D.A.;Bioorganic & Medicinal Chemistry,Vol 5,No.6;pp1087−1096,1997)が観察した限りにおいて驚くべきことである。この高いシグナル対ノイズ比は、グレリンのような塩基性標的に対して核酸によって示される高い非特異的な親和力からもたらされる。
【0079】
非常に塩基性の全体的なグレリンのpIおよび5.5という算出されたpIをもつむしろ酸性のドメインであるグレリンの受容体結合モチーフGSSFL[グレリン(1−5)]にも拘わらず、本発明者らが、酸性の受容体結合ドメインを認識するが、ペプチドの塩基性の中央およびカルボキシ末端ドメインを認識しない全長グレリンを用いて核酸を同定できたことは、なお一層驚くべき発見である。このことは、標的分子、すなわちグレリンの電荷および核酸の電荷の両方の静電効果に関して驚くべきことである。標的分子の塩基性ドメインに対する負電荷の核酸の結合は、標的分子の酸性ドメインに対する核酸の結合に較べてはるかに得策であるに違いない。かくして、当業者が、グレリンの塩基性部分に対して結合性ではないが、標的分子の酸性ドメインに結合性である核酸リガンドを成功裏に選択する合理的な期待を有しなかったことを指摘しなければならない。
【0080】
アミノ末端の受容体結合モチーフGSSFLを有する他に、本明細書で生物活性グレリンとも呼ばれる生物学的に活性なグレリンは、好ましくは、アミノ酸セリン3においてn−オクタノイル基によるそのアシル化を特徴とする。好ましくは本明細書で開示されるアミノ末端モチーフGSSFLのリガンドである本発明による核酸分子は、好ましくは、グレリンの生物学的に不活性または生物活性のない形態と生物学的に活性な形態との区別を可能にする。結合は2つの部分、オクタノイル基とペプチドの存在に厳密に依存するので、このことは驚くべきことである:オクタノイル−グレリンへの核酸の結合は、デスオクタノイル−グレリンの1000倍過剰量の存在下で、より好ましくはデスオクタノイル−グレリンの100倍過剰量の存在下で、もっとも好ましくはデスオクタノイル−グレリンの10倍過剰量の存在下で特異的である。
【0081】
本明細書における好適な実施態様で使用されるように、生物活性グレリンは、天然に存在するグレリンの特徴のすべてを本質的に好適な実施態様において表すグレリンである。具体的には、好適な実施態様において本明細書で使用される生物活性グレリンは、より好ましくはGHS受容体との相互作用を介して、成長ホルモンの放出に関与するか、またはそれを惹起できるすべてのグレリンおよびグレリン誘導体である。好適な実施態様におけるこれとは反対に、非生物活性グレリンは、生物活性グレリンとは異なる、より好ましくは、より好ましくはGHS受容体との相互作用を介して、成長ホルモンの放出を惹起しないグレリンである。
【0082】
好適な実施態様では、本発明者らは、驚くべきことに、グレリン結合核酸を生成することができたが、ここで、グレリンは生物活性グレリンまたは生物学的に活性なグレリンであり、これらは、そのN末端の3番目アミノ酸(セリン3)においてオクタン酸側鎖を有するグレリンを識別し、一方、それらは、そのようなオクタン酸側鎖を欠如するグレリンには結合しない。
【0083】
本明細書で記述される本発明による核酸の特徴は、単独でも、またいかなる組み合わせ物においても、核酸が使用される本発明のいかなる態様においても実現することができる。
【0084】
いかなる理論によっても拘束されることを願わずに、本発明は、本発明によるグレリン結合核酸の観察された特異性が、引用が図20Aに対してなされる次において議論されるであろう若干の構造的特徴を共有すると推測する。しかしながら、図20Aが、本発明による核酸の各々およびいずれかにおいて必ずしも実現されるとは限らない該構造的特徴のいくつかを組み入れていることを理解すべきである。
【0085】
基本的な構造的特徴は、本明細書ではBoxAまたは第一のストレッチBoxAとして本明細書でまた言及される連続ヌクレオチドの第一のストレッチ、および本明細書ではBoxBまたは第二のストレッチBoxBとしてまた言及される連続ヌクレオチドの第二のストレッチである。第一のストレッチBoxAは約25個の連続ヌクレオチドを含み、これに対して第二のストレッチBoxBは約6〜8個の連続ヌクレオチドを含む。第一のストレッチBoxAの3’末端ストレッチは、第二のストレッチBoxBとハイブリダイズ
し、このハイブリダイゼーションでは、第一の二本鎖構造が形成され、そのような第一の二本鎖構造はバルジを含む。二本鎖構造は、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドおよび第二のストレッチBoxBの若干の6〜8個の連続ヌクレオチドによって形成される。バルジは、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドと塩基対合しない1個位の少ないおよび3個位の多いヌクレオチドによって形成される。したがって、バルジは、好ましくは第二のストレッチBoxBによって提供される、1,2または3個の非塩基対合ヌクレオチドからなってもよい。このバルジのサイズに鑑みて、第二のストレッチBoxBは6〜8個の連続ヌクレオチドを含んでもよい。より好ましくは、バルジは、好ましくは6個の連続ヌクレオチドを含んでなる第二のストレッチBoxBによって、第二のストレッチBoxBの5’末端から見られる第二のストレッチBoxB内の非塩基対合第三ヌクレオチドによって創成される。
【0086】
好適な実施態様では、本発明による核酸は単一の核酸分子である。さらなる実施態様では、単一の核酸分子は、多数の単一の核酸分子として存在する。好ましくは、用語核酸および核酸分子は、それとは違って示されない限り本明細書では互換可能な様式で使用される。
【0087】
本発明による核酸分子が、好ましくはホスホジエステル連結または結合をとおして、好ましくは、互いに共有結合されるヌクレオチドからなることは、当業者によって理解することができる。
【0088】
さらなる重要な特色は、第二の二本鎖構造である。そのような第二の二本鎖構造は、BoxC1または第三のストレッチBoxC1としてまた言及される連続ヌクレオチドの第三のストレッチ、およびBoxC2または第四のストレッチBoxC2としてまた言及される連続ヌクレオチドの第四のストレッチによって形成される。第三のストレッチBoxC1は、第二のストレッチBoxBの5’末端にその3’末端により連結され、そして第四のストレッチBoxC2は、第一のストレッチBoxAの3’末端にその5’末端により連結される。この第二の二本鎖構造は、典型的には、第二のらせん構造として本明細書でまた言及されるらせん構造を形成する。そのような第二のらせん構造は、好ましくは、第一の二本鎖構造によって典型的には形成されるらせん構造の伸長物である。好ましくは、第一のらせん構造として本明細書でまた言及されるらせん構造である該第一の二本鎖構造の長さは、第一のストレッチBoxAおよび第二のストレッチBoxBの長さによって、より正確には互いにハイブリダイズする該2種のBoxのストレッチによって定義される。第二の二本鎖構造によって該第一のらせん構造に提供される伸長物は、本発明者らの現在の理解によれば、本発明者らはこの理論に束縛されることを願わないけれども、より安定化する種類のものである。第二のらせん構造は、1〜10個の塩基対、好ましくは1〜3個の塩基対、より好ましくは2または3個の塩基対からなる。1〜3個位の少ない塩基対が、らせんを生成するのに必ずしも適当ではないことは当業者によって理解できる。この種の構造は、かくして、らせん様構造として本明細書でまた言及され、好ましくは、そのようならせん様構造は、1または数個の塩基対によって伸長される場合は、らせん構造をもたらすであろう構造物である。
【0089】
さらなる重要な特色は、BoxDまたは第五のストレッチBoxDとして本明細書でまた言及される連続ヌクレオチドの第五のストレッチである。このさらなるストレッチは、核酸の全体的結合において改良を与える。第五のストレッチは少なくともただ2個のヌクレオチドを含むけれども、グレリンの結合におけるその有益な劇的効果は、第五ストレッチの長さを好ましくは6個の連続ヌクレオチドまで増加することによってさらに改良することができる。また、第五のストレッチは、もし含有される場合は、好ましくはその5’末端CAジヌクレオチドおよび
5’CA(X)n3’
[式中、Xは、好ましくはA,G,T,C,UおよびIを含んでなる群から選ばれるいずれかのヌクレオチドである]
の配列を有する第五ストレッチにおいて特に効果的であることがわかった。
【0090】
本発明による核酸のさらなる特色は、BoxEまたは第六のストレッチBoxEとして本明細書でまた言及される隣接ヌクレオチドの第六のストレッチである。好ましくは、第六のストレッチは少なくとも1個のヌクレオチドからなる。
【0091】
種々のストレッチは図20Aから見られるように互いに連結される。そこに描かれるように、好ましくは、第三のストレッチBoxC1および第四のストレッチBoxC2の各々は、1または2個のヌクレオチド、より好ましくは2個のヌクレオチドを含み、第五のストレッチBoxDは少なくとも2個のヌクレオチド、好ましくは4個のヌクレオチド、もっとも好ましくは6個のヌクレオチドを含む。
【0092】
第三のストレッチBoxC1および第四のストレッチBoxC2の長さが0である実施態様では、第一のストレッチBoxAおよび第二のストレッチBoxBは場合によっては、リンカーまたはスペーサーを介して連結されてもよく、ここで、そのようなスペーサーは本明細書で開示されるスペーサーのいずれであってもよい。本明細書で使用されるように用語「スペーサー」および「リンカー」は、他に指示されない場合は互換可能な様式で使用される。
【0093】
本発明に関連して、第一のストレッチおよび第二のストレッチは、好ましくは共有結合を介して互いに連結される。好適な実施態様では、共有結合はホスホジエステル結合である。なおさらなる好適な実施態様では、第一のストレッチの3’末端は第二のストレッチの5’末端に連結される。
【0094】
その他の実施態様では、第六のストレッチBoxEは1〜10個の連続ヌクレオチド、好ましくは1〜4個の連続ヌクレオチド、もっとも好ましくは3個の連続ヌクレオチドを含む。
【0095】
これらの種々の配置から分るように、第五のストレッチと第六のストレッチは互いに連結されるか、または連結されず、第三と第四ストレッチは互いに連結されるか、または連結されず、そして第一のストレッチと第二のストレッチは互いに連結されるか、または連結されないことが可能である。連結は好ましくは共有結合を介してなされることは認識できる。より好ましくは連結は、少なくとも1個の、好ましくは多数のエチレングリコール部分を含んでなる親水性スペーサーを介してなされる。種々のリンカーおよびスペーサーは、それぞれ当業者には既知であり、そして次の基準、例えば、PilsおよびMicura(Nucleic Acid Research(2000),28(9),1859−1863)によって記述されたような基準を用いて選択することができる。リンカーは塩基対自体を妨害してはならない。芳香族炭素環を含有するリンカーの種類は末端塩基対を攻撃するので、適当ではない(J.Am.Chem.Soc.(1999),121,9905−9906;J.Am.Chem.Soc.(1998),120,11004−11005)。しかしながら、エチレングリコールに基づくかエチレングリコールから誘導されるリンカーは、それらが良好な水溶解度と高いコンホメーションの柔軟性という利点を有するので、これらの要件に適合する(J.Am.Chem.Soc.(1993),115,8483−8484;Nucleic Acids Research(1993)、21、5600−5603;Biochemistry(1993),32,1751−1758;Nucleic Acid Research(1990),18,6353−6359;J.Am.Chem.Soc.(1997),119,11591−11597)。好ましくは、スペーサーは、1または数個のエチレングリコール部分を含むか、またはそれからなり、ここで、酸素がCH3、リン酸または硫黄によって置換されている。
【0096】
これらの連結の選択に基づいて、次の構造物が実現できる。
【0097】
【化5】
【0098】
最後に、本発明による核酸のための完全に閉ざされた、すなわち環状の構造物が図20Dにおいて描かれるように実現されることも、また本発明内にはいる。
【0099】
本発明による核酸は、また、本明細書に開示される特定の配列に本質的に相同である核酸を含まねばならない。用語「本質的に相同」は、相同性が少なくとも75%、好ましくは85%、より好ましくは90%、もっとも好ましくは95%、96%、97%、98%または99%以上であるように理解されるべきである。
【0100】
用語「本発明の核酸」または「本発明による核酸」は、好ましくは核酸または該一部がグレリンへの結合に関与するほどに、より好ましくは生物活性グレリンを非生物活性グレリンと区別する、すなわち、特にオクタノイル−グレリンをデスオクタノイル−グレリンと区別するほどに、本明細書に開示される核酸またはその一部を含むそれらの核酸を含むべきである。そのような核酸は、例えば、切り取り(truncation)によって本明細書に開示される核酸から誘導されてもよい。切り取りは、本明細書に開示される核酸の末端のいずれか、または両方に関してもよい。また、切り取りは、ヌクレオチドの内部配列に関してもよい、すなわち、それぞれ5’および3’末端ヌクレオチドの間のヌクレオチドに関してもよい。さらに、切り取りは、本明細書に開示される核酸の配列から単一ヌクレオチド位の短い欠失を含んでもよい。また、切り取りは、本発明の核酸の1個以上のストレッチに関してもよく、ここで、ストレッチは1個位の短いヌクレオチド長であってもよい。
【0101】
本発明による核酸は、D−核酸でも、またL−核酸であってもよい。好ましくは、本発明の核酸はL−核酸である。さらに、核酸の一部または数部分がD−核酸として存在するか、または核酸の少なくとも一部または数部分がL−核酸であることも可能である。用語核酸の「一部」は、1ヌクレオチド位の小さいものを意味すべきである。そのような核酸は、一般にD−およびL−核酸とそれぞれ本明細書では言及される。したがって、特に好適な実施態様では、本発明による核酸はL−ヌクレオチドからなり、そして少なくとも1個のD−ヌクレオチドを含む。そのようなD−ヌクレオチドは、好ましくは、本発明による核酸を定義するストレッチとは異なる部分、好ましくは核酸の他の部分との相互作用が必要であるそれらのその部分に連結される。好ましくは、そのようなD−ヌクレオチドは、いずれかのストレッチおよび本発明によるいずれかの核酸のそれぞれの末端に連結される。さらなる好適な実施態様では、そのようなD−ヌクレオチドは、スペーサーまたはリンカー、好ましくは本発明による核酸に対するPEGおよびHESのような連結修飾体として働いてもよい。
【0102】
本発明による核酸がより長い核酸の一部であることもまた本発明の内にはいり、ここで
、このより長い核酸は、少なくとも一部分は、本発明による核酸またはその一部である数部分を含む。これらのより長い核酸の他の部分は、D−核酸でもL−核酸であってもよい。いずれの組み合わせ物が本発明に関して使用されてもよい。これらのより長い核酸の他の部分は、結合、好ましくはグレリンへの結合とは異なる機能を示すことができる。1つの可能性のある機能は、他の分子と相互作用させることであり、そのような他の分子は、例えば、固定化、架橋、検出または増幅のためのように、好ましくはグレリンとは異なる。
【0103】
本明細書で使用されるようなL−核酸は、L−ヌクレオチドからなる、好ましくは完全にL−ヌクレオチドからなる核酸である。
【0104】
本明細書で使用されるようなD−核酸は、D−ヌクレオチドからなる、好ましくは完全にD−ヌクレオチドからなる核酸である。
【0105】
本発明の核酸が、D−ヌクレオチド、L−ヌクレオチドあるいは例えば少なくとも1個のL−ヌクレオチドおよび少なくとも1個のD−ヌクレオチドからなるストレッチのランダム組み合わせ物または一定の配列である組み合わせによる両方の組み合わせ物からなるか否かに関係なく、核酸はデオキシリボヌクレオチド、リボヌクレオチドまたはその組み合わせ物からなってもよい。
【0106】
本発明の核酸をL−核酸として設計することは、いくつかの理由で得策である。L−核酸は天然に存在する核酸の鏡像異性体である。しかしながら、D−核酸は、水溶液中、特に、ヌクレアーゼの広範な存在によって生物系または生物サンプルにおいて非常に不安定である。天然に存在するヌクレアーゼ、特に動物細胞からのヌクレアーゼはL−核酸を分解することはできない。このために、L−核酸の生物学的半減期は、動物およびヒトの身体を含む、そのような系では有意に増大する。L−核酸の分解性の欠如により、ヌクレアーゼ分解産物は生成されず、したがってそれから起きる副作用は観察されない。この態様は、グレリンの存在に係わる疾病および/または障害の治療において使用される実際にすべての他の化合物のL−核酸の限界を定める。ワトソン・クリック塩基対合とは異なるメカニズムにより標的分子に特異的に結合するL−核酸、あるいは特に、標的分子へのアプタマーの結合に関与するアプタマーのそれらの部分をもつ、L−ヌクレオチドから部分的または完全になるアプタマーは、またspiegelmerと呼ばれる。
【0107】
本発明の核酸は、それらがD−核酸、L−核酸またはD,L−核酸として存在するか否か、あるいはそれらがDNAまたはRNAであるか否かに関係なく、一本鎖または二本鎖核酸として存在してもよいことは、また本発明内にはいる。典型的には、本発明の核酸は、一次配列により一定の二次構造を示す一本鎖核酸であり、したがってまた三次構造を形成してもよい。しかしながら、本発明の核酸は、互いに相補的または部分的に相補的である2本の鎖が互いにハイブリダイズされるという意味において二本鎖であってもよい。このことは、特に、核酸がL−型よりもむしろ天然に存在するD−型で存在する場合には有利であろう核酸への安定性を与える。
【0108】
本発明の核酸は修飾されてもよい。そのような修飾は核酸の単一ヌクレオチドに関してもよく、そして当該技術分野において周知である。そのような修飾の例は、なかんずく、Venkatesan N.et al.(2003)Curr Med Chem.Oct;10(19):1973−91;Kusser,W.(2000) J Biotechnol,74:27−38;Aurup,H.et al.(1994)Nucleic Acids Res,22,20−4;Cummins,L.L.et al.(1995) Nucleic Acids Res,23,2019−24;Eaton,B.E.et al.(1995)Chem Biol,2,633−8;Gree
n,L.S.et al.(1995)Chem Biol,2,683−95;Kawasaki,A.M.et al.(1993)J Med Chem、36,831−41;Lesnik,E.A.et al.(1993)Biochemistry,32,7832−8;Miller,L.E.et al.(1993)J Physiol,469,213−43において記述されている。そのような修飾は、核酸がなりたっている個々のヌクレオチドの2’位におけるH原子、F原子またはO−CH3基またはNH2基であってもよい。また、本発明による核酸は少なくとも1個のLNAヌクレオチドを含んでもよい。1つの実施態様では、本発明による核酸はLNAヌクレオチドからなる。
【0109】
1つの実施態様では、本発明による核酸は多くの部分に分かれた核酸であってもよい。本明細書で使用される多くの部分に分かれた核酸は、少なくとも2本の核酸鎖からなる核酸である。これらの少なくとも2本の核酸鎖は機能的単位を形成し、ここでの機能的単位は標的分子に対するリガンドである。少なくとも2本の核酸鎖は、核酸を切断して2本の鎖を生成するか、あるいは本発明の、すなわち核酸全体の第一の部分に対応する1つの核酸および核酸全体の第二の部分に対応するその他の核酸を合成するか、いずれかによって本発明の核酸のいずれから誘導されてもよい。両、切断および合成が、先に例示された2本以上の鎖である多くの部分に分かれた核酸を生成するために応用できることは認識されるべきである。言い換えれば、少なくとも2本の核酸鎖は、典型的には、種々の核酸部分の間で、ある程度の相補性が存在するかも知れないが、互いに相補的であり、かつハイブリダイズする2本の鎖とは異なる。
【0110】
本発明は、本発明による核酸が非常に好ましいKd値の範囲を示すことを発見した。より具体的には、完全なBoxAおよびBoxBの他に−最小2個のヌクレオチドを各々有するBoxC1およびC2、最小6個のヌクレオチドを有するBoxDおよび最小3個のヌクレオチドを有するBoxE−を含んでなるオリゴヌクレオチド(例えば、SOT−E)は、SOT−Cよりも6倍良好なグレリンに対する結合親和力(図18)を示す。
【0111】
結合定数の決定を可能にするのは、所謂バイオコア(biacore)・デバイスの使用であり、これはまた当業者には既知である。本明細書で使用される親和力は、実施例において記述されるような「平衡アッセイ」の使用によってまた測定できた。本事例ではグレリンである標的による核酸間の結合の強度を表すための適当な測定値は、その決定方法が当業者において十分に知られている所謂Kd値である。
【0112】
本発明による核酸はあるKd値を特徴とする。好ましくは、本発明による核酸によって示されるKd値は1μM以下である。約1μMのKd値は、標的に対する核酸の非特異的結合について特徴的であると言われる。当業者によって認識できるように、本発明による核酸のような1群の化合物のKd値は一定の範囲内にある。約1μMという上記KdはKd値について好適な上限である。核酸を結合する標的のKdの好適な下限は約10ピコモルまたはそれ以上であってもよい。グレリンに結合する個々の核酸のKd値が、好ましくは、この範囲内であることは本発明の内にはいる。好適な範囲は、この範囲内のいずれか第一の数とこの範囲内のいずれか第二の数を選ぶことによって定められてもよい。好適な上限値は0.25μM、0.1μMおよび0.01μMであり、好適な下限値は100nM、10nM、1nMおよび0.05nMである。
【0113】
本発明による核酸分子は、なおそれらが標的分子に結合でき、かつ生物活性グレリンを非生物活性グレリンと区別できる、すなわち好ましくはオクタノイル−グレリンをデスオクタノイル−グレリンと区別できる場合には、いかなる長さを有してもよい。本発明による核酸の好適な長さが存在することは当該技術分野において認識できる。典型的には、この長さは15〜120ヌクレオチドである。15〜120間のいかなる整数も本発明によ
る核酸について可能な長さであることは当業者によって認識できる。本発明による核酸の長さのより好適な範囲は、約20〜100ヌクレオチド、約20〜80ヌクレオチド、約20〜60ヌクレオチド、約20〜50ヌクレオチドおよび約30〜50ヌクレオチドの長さである。
【0114】
本明細書で開示される核酸が、好ましくは高分子量部分であり、かつ/または、なかんずく、動物体内、好ましくはヒト体内における滞留時間という用語で好ましくは核酸の特徴を修飾させる部分を含むことは本発明内にはいる。そのような修飾の特に好適な実施態様は、本発明による核酸のPEG化およびHES化である。本明細書で使用されるようなポリ(エチレングリコール)についてのPEG鎖およびヒドロキシエチル澱粉についてのHES。好ましくは本明細書で使用されるPEG化は本発明による核酸の修飾であり、そのような修飾体は、本発明による核酸に連結されるPEG部分からなる。本明細書で使用されるHES化は本発明による核酸の修飾であり、そのような修飾体は、本発明による核酸に連結されるHES部分からなる。これらの修飾ならびにそのような修飾を用いる核酸の修飾方法は、欧州特許第一 306 382号に記述されており、これの開示は引用によって完全に本明細書に組み入れられている。
【0115】
好ましくは、高分子量部分からなる、またはそれを含んでなる修飾体の分子量は、特にそのような高分子量部分であるPEGの場合には、約2,000〜200,000Da、好ましくは40,000〜120,000Daであり、そして特にそのような高分子量部分であるHESの場合には、好ましくは約3,000〜100,000Da、より好ましくは5,000〜60,000Daである。HES修飾の方法は、例えば、ドイツ特許出願第一 2004 006 249.8号に記述されており、これの開示は引用によって完全に本明細書に組み入れられている。
【0116】
いかなる理論によっても束縛されないことを願えば、好ましくは生理学的に許容できるポリマー、より具体的には本明細書で開示されるポリマーのような高分子量部分を用いる本発明による核酸の修飾によって排出動態が変化されると考えられる。より具体的には、そのような修飾された本発明の核酸の分子量の増加により、そして特にL型における場合の被験者が代謝することのない核酸により、動物の身体、好ましくは哺乳動物の身体、より好ましくはヒトの身体からの排出が減少されると思われる。排出は、典型的には腎臓を介して起きるので、本発明は、このように修飾された核酸の糸球体濾過速度が、体内の滞留時間の増加をもたらすこの種の高分子量修飾を有しない核酸に較べて有意に減少されると推測する。これに関連して、そのような高分子量修飾にもかかわらず、本発明による核酸の特異性が有害な様式で影響されないことは特に注目される。その限りにおいて、本発明による核酸は、持続放出を与える製薬学的調合物が、持続放出を与えるために必ずしも必要とされないような−通常には製薬学的活性化合物から期待することができないー驚くべき特徴を有する。むしろ、高分子量部分を含んでなるそれらの修飾形態物における本発明による核酸は、既にそれ自体、持続放出調合物として使用することができる。
【0117】
しかしながら、本明細書に開示される核酸が、いかなる修飾も、特にPEG化またはHES化のような高分子量修飾も含まないこともまた本発明の内にはいる。そのような実施態様は、投与後、身体からの核酸の速いクリアランスが所望される場合に特に好適である。そのような速いクリアランスは、本発明による核酸か、または同物質を含んでなる薬物を使用してインビボの想像性または一時的な食欲抑制の場合に望ましいであろう。
【0118】
本発明による核酸としてまた本明細書で言及される本発明の核酸および/または本発明によるアンタゴニストは、薬物の創成または製造のために使用されてもよい。そのような薬物は、少なくとも1種の本発明の核酸を、場合によってはさらなる製薬学的活性化合物とともに含有し、この場合、本発明の核酸は好ましくは製薬学的活性化合物自体として働
く。そのような薬物は好適な実施態様では少なくとも1種の製薬学的に許容できる担体を含む。そのような担体は、例えば、水、バッファー、PBS、グルコース溶液、好ましくは5%グルコース塩均衡溶液、澱粉、糖、ゼラチンまたはいずれか他の許容できる担体物質であってもよい。そのような担体は一般に当業者には既知である。
【0119】
さらなる実施態様では、薬物はさらなる製薬学的活性作用物を含む。そのようなさらなる製薬学的活性化合物は、食欲を減退することが知られているそれらの化合物であってもよく、そして好ましくは、PYY3−45、CCK、レプチンおよびインスリンを含んでなる群から選ばれる。あるいは、またはさらに、そのようなさらなる製薬学的活性作用物はさらなる本発明による核酸である。あるいはまた、薬物は、グレリンとは異なる標的分子に結合するか、または本発明による核酸の1種とは異なる機能を発揮する少なくとも1種以上の核酸を含む。1つの実施態様では、そのような核酸はオクタン酸部分を欠如するグレリンに結合している。
【0120】
そのような薬物が使用されてもよい処置および/または予防のための疾病および/または障害および/または疾病症状は、限定されるものではないが、肥満、エネルギーバランス、食欲および体重の調節、摂食障害、胃腸疾患、糖尿病、グルコース代謝、腫瘍、血圧、心臓血管病、先端巨大症、および他のGHインバランスを含む。当業者によって認識できるように、本発明の核酸は、事実上、グレリンに対するアンタゴニストがそのようなアンタゴニストを必要とする患者に投与でき、そしてそのようなアンタゴニストが疾病または障害の原因を除去できるか、または少なくとも疾病または障害からの影響を低下するために適当である、いかなる疾病において使用されてもよい。そのような影響は、限定されるものではないが、肥満、エネルギーバランス、食欲および体重の調節、摂食障害、胃腸疾患、糖尿病、グルコース代謝、腫瘍、血圧、心臓血管病、先端巨大症および他のGHインバランスを含む。これらおよび他の疾病または障害に関連する本発明による核酸の適応性は、なかんずく、本明細書の序論部分において概説されたようなグレリンの関与からもたらされる。本発明の目的では、エネルギーバランスの調節は疾病とみなされる。より具体的には、この使用は、エネルギーバランスの調節が直接にも間接にもグレリンによって影響され、グレリンの生物学的利用性の低下が望まれるすべての疾病の処置のためにある。同じことが、糖代謝、血圧および食欲と体重に適合する。本発明による核酸を用いて処置されてもよいさらなる疾病は、可能ならば全身的または局所的適用において、下垂体腫瘍、先端巨大症、中枢性Cushing症候群、副腎性Cushing症候群、腫瘍随伴性Cushing症候群、異所性Cushing症候群、副腎腫瘍、ストレス、副腎皮質亢進症、心不全、心筋梗塞、卒中、副腎皮質不全症、低血圧、大動脈狭窄、肺動脈緊張過度、梗塞性心外膜炎、感染疾患、感染毒性低血圧、血液量減少および低ナトリウム血症を含んでなる群から選択することができる疾病である。
【0121】
本明細書で使用されるように、用語、本発明による核酸を用いて処置できる胃腸疾患は、胃の疾病および障害、腸の疾病および障害、結腸の疾病および障害、および胃および結腸運動の調節を含む。好適な実施態様では、用語、腸の障害および腸の疾病は、炎症性腸疾患を含む。より好適な炎症性腸疾患は、潰瘍性大腸炎およびクローン病である。この種の疾病の処置のための本発明による核酸の適合性は、Karmirsら(Karmirs K,Koutroubakis IE,Xidakis C,Polychronaki M,Voudouri T,Kouroumalis EA,Circulatinglevels of leptin,adiponectin,resistin,and ghrelin in inflammatory bowl disease.Inflamm Bowel Dis.2006 Feb;12(2):100−5)、Tebbe JJら(Tebbe JJ,Mronga S,Tebbe CG,Ortmann E,Arnold R,Schafer MK.Ghrelin−induced stimulation of colonic propulsion isdependent on hypothalamic neuropeptide Y1− and corticotrophin−releasing factor 1 receptor activation.J Neuroendocrinol.2005 Sep;17(9):570−6)およびFukuda H.ら(Fukuda H,Mizuta Y,Isomoto H,Takeshima F,Ohnita K, Ohba K,Omagari K,Taniyama K,Kohno S.Ghrelin enhances gastric motility through direct stimulation of intrinsic neural pathways and capsaicin−sensitive afferent neurones in rats.Scand J Gastroenterol.2004 Dec;39(12):1209−14)により記述されたようなグレリンの関与から生じる。
【0122】
Kobeltらの公表(Kobelt P,Helmling S,Stengel A,Wlotzka B,Andresen V,Klapp BF,Wiedenmann B,Klussmann S,Monnikes H.Anti−ghrelin
SPIEGELMER NOX−B11 inhibits neurostimulatory and orexigenic effects of peripheral ghrelin in rats.Gut.2005 Jun 30,Epub ahead of print)は、抗グレリンSpiegelmer NOX−B11の投与において末梢グレリンにより誘発された短期間の食物摂取の用量依存性減退を報告している。より具体的には、PBSおよび3nmolグレリンによるポジティブ対照群(媒質/グレリン群)処置では、媒質/媒質群(1.13±0.59g/kg−BW,p<0.0002)に較べて腹腔内注射後初めの半時間内に食物摂取が有意に増加した(4.94±0.63g/kg−BW)(Kobeltら、前出の図2A)。Spiegelmer NOX−B11(=SOT−C=B11trc)の30nmolによる前処置は食物摂取に及ぼすグレリンの促進効果を阻止した(0.58±0.58g/kg−BW,p<0.0001)(Kobeltら、前出の図2A)。これに対して、ランダム配列からなる対照Spiegelmerの投与は、そのような抑制効果を有せず、食物摂取におけるグレリン誘導の促進は元のままであった(4.77±0.66g/kg−BW;p>0.864)(Kobeltら、前出の図2A)。
【0123】
食物摂取に及ぼすNOX−B11(=SOT−C=B11trc)の抑制効果は、厳密に用量依存性であることが証明された。Spiegelmer NOX−B11(=SOT−C=B11trc)の1nmolの用量は、食物摂取のグレリン刺激には効果を有しなかった(Kobeltら、前出の図2A)。中間的効果はNOX−B11(=SOT−C=B11trc)の10nmolの用量について観察された:この用量レベルでは、最初の30分間における3nmolグレリンの刺激効果は緩和された(3.51±0.66g/kg−BW対4.94±0.63g/kg−BW;p>0.159対グレリン単独)(Kobeltら、前出の図2A)。
【0124】
Shearmanらの公表(Shearman LP,Wang SP,Helmling S,Stribling DS,Mazur P,Ge L,Wang L,Klussmann S,Macintyre DE,Howard AD,Strack AM.Ghrelin Neutralisation by a Ribonucleic Acid−SPM Ameliorates Obesity in Diet−Induced Obese Mic.;Endocrinology.2006 Mar;147(3):1517−26.Epub 2005 Dec 8)は、食餌誘導の肥満(DIO)マウスにおける抗グレリンSpiegelmer NOX−B11−2(SOT−D−109)の投与において、体重の低下および食物摂取の減少を報告している。より具体的には、NOX−B11−2(SOT−D−109)注入は、対照に較べて体重喪失を惹起した(Shearmanら、前出の図4A、参照)。有意な体重喪失は、媒質処置のマウスに較べて1〜10日目および12日目において;対照Spiegelmer(対照SPM)注入群に較べて1〜13日目において、NOX−B11−11(SOT−D−109)注入により観察された(P<0.05vs.媒質または対照SPM)。13日目では、NOX−B11−2注入マウスは、体重の平均0.32gを増加したが、対照Spiegelmerを注入されたマウスは、平均1.85gを増加し、そして媒質を注入されたマウスは平均0.91g増加した。また、NOX−B11−2(SOT−D−109)注入は有意に食物摂取を減退させた(Shearmanら、前出の図4B、参照)。累積の食物摂取に及ぼす有意な効果は、媒質群に較べて1〜8日目に、対照Spiegelmer群に較べて1〜13日目に観察された(13日目に39.33g対42.61g;p<0.05)(Shearmanら、前出の図4C)。さらに、食物エネルギーが体質量の増大のためにいかに効率的に使用されたかを指示する摂食効率(消化された1kcal当たりの重量増加)が1〜5日目および6〜13日目から計算された(Shearmanら、前出の図4D,参照)。摂食効率は1〜5日目のNOX−B11−2(SOT−D−109)注入によって低下され、そしてこの効果は6〜13日目からは観察されず、このことは重量増加における一過性低下が、単に食物摂取における減退によるものではなかったことを示唆する。さらに、NOX−B11−2(SOT−D−109)による処置はDIOマウスの体組成を変えた(Shearmanら、前出の図4E)。赤身量含量(lean mass content)では変化がなかったのに対して、NOX−B11−2(SOT−D−109)注入されたマウスの脂肪量含量は、総体重についての校正後でさえ減少されなかった(Shearmanら、前出の図4F)。白色脂肪組織貯蔵物重量はNOX−B11−2(SOT−D−109)注入によって変わらず、対照Spiegelmer注入は体組成または白色脂肪組織重量を変えなかった。
【0125】
DIOグレリン欠乏および野生型の両マウスを使用するNOX−B11−2(SOT−D−109)による常習的注入研究において、有意な体重喪失が、1〜6日目の野生型マウスにおけるNOX−B11−2(SOT−D−109)注入において観察された(Shearmanら、前出の図5A,参照)。これに対して、NOX−B11−2(SOT−D−109)は、Ghrl−l−マウスでは体重を変えなかった(Shearmanら、前出の図5B、参照)。さらに、NOX−B11−2(SOT−D−109)注入は、野生型マウスでは1日目に一日の食物摂取を減退した(Shearmanら、前出の図5C,参照)。NOX−B11−2(SOT−D−109)注入は、Ghrl−l−マウスの食物摂取は変えなかった(Shearmanら、前出の図5D、参照)。
【0126】
薬物が、原則として、疾病の処置のための薬物の使用に関連して開示された該疾病のすべての予防のために代替的または追加的に使用されることは、本発明の内にはいる。したがって、それぞれのマーカーは、心臓血管の危険因子、例えば、コレステロールおよび低い好気的活性ならびに体重管理を必要とする一般因子を含んでなる群から選ばれる。
【0127】
本発明による薬物は、原則として、当業者には既知のいかなる形態物において投与されてもよい。好適な投与経路は全身的投与、より好ましくは注射による投与である。あるいはまた、薬物は局所的に投与されてもよい。他の投与経路は、筋肉内、腹腔内、および皮下、好ましくはもっとも侵入性が少ないが有効性が確保される投与経路であるper orumまたは鼻内を含む。
【0128】
さらなる態様では、本発明は製薬学的組成物に関する。そのような製薬学的組成物は、少なくとも1種の本発明による核酸および好ましくは製薬学的に許容できる結合剤を含む。そのような結合剤は当該技術分野において使用され、そして/または既知であるいかなる結合剤であってもよい。より具体的には、そのような結合剤は、本明細書で開示される薬物の製造に関連して議論されるすべての結合剤である。さらなる実施態様では、製薬学的組成物はさらなる製薬学的活性作用物を含む。
【0129】
本明細書で記述されるような薬物が本明細書で開示される製薬学的組成物を構成することは本発明内にはいる。
【0130】
さらなる態様では、本発明は、方法が少なくとも1種の本発明による核酸の製薬学的に活性な量の投与を含む、そのような処置を必要としている被験者の処置方法に関する。1つの実施態様では、被験者は、疾病を蒙っているか、またはそのような疾病を発生する危険性にあり、ここで、疾病は本明細書で開示されるすべての疾病、特に薬物の製造のためのすべての本発明による核酸の使用に関連して開示されるすべてのそれらの疾病である。
【0131】
本発明による核酸ならびにアンタゴニストが、特に肥満におけるグレリンの関与に関して、薬物としてまたは薬物の製造のためだけでなく、また化粧目的のために使用できることを理解すべきである。同じ目的および/または同じ理由のために、本発明による核酸ならびにアンタゴニストは、食品添加物、体重制御の手段、食欲制御の手段としておよび/または診断剤として使用することができる。本発明による核酸ならびにアンタゴニストを含んでなる組成物はすべての上記目的のために使用することができる。
【0132】
好ましくは本明細書で使用されるように、診断剤または診断作用物または診断手段は、直接的にもまた間接的にも、グレリン、好ましくは本明細書で記述されるグレリン、より好ましくは本明細書に記述される種々の障害および疾病に関連して本明細書で記述されるグレリンを検出するために適当である。診断剤は、本明細書に記述される障害および疾病それぞれすべての検出および/または追跡のために適当である。そのような検出はグレリンに対する本発明による核酸の結合をとおして可能である。そのような結合は直接的にも間接的にも検出することができる。各方法および手段は当業者には既知である。なかんずく、本発明による核酸は、本発明による核酸、好ましくはグレリンに結合される核酸の検出を可能にする標識を含む。そのような標識は、好ましくは放射能、酵素および蛍光標識を含んでなる群から選ばれる。原則として、抗体のために開発されたすべての既知アッセイが、本発明による核酸のために採用されてもよく、それにより、標的結合抗体が標的結合性核酸に置換される。無標識の標的結合抗体を用いる抗体アッセイでは、検出は、好ましくは、放射能、酵素および蛍光標識により修飾され、そしてそのFcフラグメントにおいて標的結合抗体に結合する二次抗体によって実施される。核酸、好ましくは本発明による核酸の場合には、核酸がそのような標識により修飾されるが、好ましくはそのような標識は、ビオチン、Cy−3およびCy−5を含んでなる群から選ばれ、そしてそのような標識は、そのような標識に対向される抗体、例えば、抗ビオチン抗体、抗Cy−3抗体または抗Cy−5抗体によって検出されるか、または−標識がビオチンの場合にはー標識は、ビオチンに自然に結合するストレプトアビジンまたはアビジンによって検出される。そのような抗体、ストレプトアビジンまたはアビジンは、順に、好ましくは、それぞれの標識、例えば放射能、酵素または蛍光標識により修飾される(二次抗体のように)。
【0133】
さらなる実施態様では、本発明による核酸は、検出手段が分子ビーコン(beacon)である第二の検出手段によって検出または分析される。分子ビーコンの方法は当業者には既知である。簡単に言えば、分子ビーコンとも呼ばれる核酸プローブは、検出されるべき核酸サンプルに対する逆相補物であり、このために、検出される核酸サンプルの一部にハイブリダイズする。核酸サンプルへの結合により、分子ビーコンの蛍光団基が分離されて、蛍光シグナルの変化、好ましくは強度における変化をもたらす。この変化は存在する核酸サンプルの量と相関する。
【0134】
本発明による生物活性および非生物活性グレリンの識別のためのアッセイは当業者には
既知の標準技術を用いて実施されてもよい。そのようなアッセイがまた、次に概説されるようなグレリン、好ましくは生物活性グレリンの検出のために使用できることは理解されるにちがいない。
【0135】
本発明による核酸を用いるグレリンの検出が、特に、本明細書で定義される生物活性グレリンの検出を可能にすることは認識できる。さらに、生物活性グレリンは、なかんずく、他の操作も当業者には明であるが、次の操作によってデスオクタノイルグレリンを別々に検出でき、したがって区別することができる。
【0136】
生物活性グレリンの検出に関連して、好適な方法は、次の段階:
(a)生物活性グレリンの存在について試験されるべきサンプルを提供し、
(b)本発明による核酸を提供し、
(c)好ましくは反応容器中で、サンプルを核酸と反応させること、
を含み、この場合、段階(a)は段階(b)の前に実施されてもよく、または段階(b)は段階(a)の前に実施されてもよい。
【0137】
好適な実施態様では、サンプルと核酸との反応の検出にある、さらなる段階d)が提供される。好ましくは、段階b)の核酸は、表面に固定化される。表面は、反応チューブ、プレートのウェルのような反応容器の表面、またはそのような反応容器中に含有されるデバイス、例えばビーズの表面であってもよい。表面への核酸の固定化は、限定されるものではないが、非共有または共有結合を含む、当業者には既知のいかなる手段によってなされてもよい。好ましくは、連結は、表面と核酸との間の共有化学結合により確立される。しかしながら、核酸が間接的に表面に固定化され、そのような間接的固定化がさらなる成分または相互作用パートナーのペアの使用を伴うことも本発明の内にはいる。そのようなさらなる成分は、好ましくは、相互作用パートナーとも呼ばれる、固定化される核酸と特異的に相互作用し、その結果表面への核酸の付着を媒介する化合物である。相互作用パートナーは、好ましくは、核酸、ポリペプチド、タンパク質および抗体を含んでなる群から選ばれる。好ましくは、相互作用パートナーは抗体、より好ましくはモノクローナル抗体である。あるいはまた、相互作用パートナーは、核酸、好ましくは機能的核酸である。より好ましくは、そのような機能的核酸は、アプタマー、spiegelmer、および核酸に対して少なくとも部分的に相補的である核酸を含んでなる群から選ばれる。さらなるその他の実施態様では、表面への核酸の結合は多分離した(multi−partite)相互作用パートナーによって媒介される。そのような多分離相互作用パートナーは、好ましくは、相互作用パートナーのペアまたは第一のメンバーと第二のメンバーからなる相互作用パートナーであり、ここでは、第一のメンバーは核酸によって含まれるか、または核酸に連結され、そして第二のメンバーは表面に付着されるか、または表面によって含まれる。多分離相互作用パートナーは、好ましくは、ビオチンとアビジン、ビオチンとストレプトアビジン、およびビオチンとノイトラビジン(neutravidin)を含んでなる相互作用パートナーのペアの群から選ばれる。好ましくは、相互作用パートナーのペアの第一のメンバーはビオチンである。
【0138】
そのような方法の好適な結果は、生物活性グレリンおよび核酸の非固定化複合体の形成であり、この場合、より好ましくは、該複合体が検出される。この複合体から生物活性グレリンが検出されることは実施態様内にはいる。より好ましくは、生物活性グレリンは、生物活性グレリンに特異的である検出手段によって検出される。特に好適な実施態様では、生物活性グレリンは、両、生物活性グレリンおよび非生物活性グレリンを検出する検出手段によって検出される。
【0139】
この要件に従うそれぞれの検出手段は、例えば、両、生物活性グレリンおよびデス−オクタノイルグレリンにおいて同一であるグレリンの、その/それらの部分に対して特異的
であるすべての検出手段である。好ましくは、そのような検出手段は、かくしてグレリンのC末端に結合するか、あるいは少なくとも、N末端およびn−オクタノイル側鎖によって形成されるドメインに結合することはない。特に好適な検出手段は、核酸、ポリペプチド、タンパク質および抗体を含んでなる群から選ばれる検出手段であり、これらの生成は当業者には既知である。
【0140】
グレリンの検出方法はまた、サンプルが、好ましくは段階c)を実施するために使用された反応容器から除去されることを含む。
【0141】
さらなる実施態様では、方法はまた、生物活性および/またはデス−オクタノイルグレリンの相互作用パートナーを表面、好ましくは先に定義された表面上に固定化する段階を含み、ここでの相互作用パートナーは、本明細書、好ましくはそれぞれの方法に関連して先のように定義されており、より好ましくは、それらの種々の実施態様における核酸、ポリペプチド、タンパク質および抗体を含む。この実施態様では、特に好適な検出手段は、本発明による核酸であり、そのような核酸は、好ましくは標識されても、または標識されていなくてもよい。そのような核酸が標識される場合には、それが、直接的にまたは間接的に検出できる。そのような検出はまた、好ましくは、核酸、ポリペプチド、タンパク質および本明細書で記述される種々の実施態様における実施態様を含んでなる群からまた選ばれる第二の検出手段の使用を伴ってもよい。そのような検出手段は、好ましくは、本発明による核酸に特異的である。より好適な実施態様では、第二の検出手段は分子ビーコンである。核酸または第二の検出手段または両方のいずれも、好適な実施態様では、検出標識を含んでもよい。検出標識は、好ましくは、ビオチン、ブロモ−デオキシウリジン標識、ジゴキシゲニン標識、蛍光標識、UV−標識、放射能標識およびキレーター分子を含んでなる群から選ばれる。あるいはまた、第二の検出手段は、好ましくは、核酸によって含有されるか、またはそれによって含まれるか、またはそれに連結される検出標識と相互作用する。特に好適な組み合わせは、次のとおりである:
検出標識がビオチンであり、そして第二の検出手段がビオチンに対向された抗体であるか、あるいは
検出標識がビオチンであり、そして第二の検出手段がアビジンまたはアビジン保持分子であるか、あるいは
検出標識がビオチンであり、そして第二の検出手段がストレプトアビジンまたはストレプトアビジン担持分子であるか、あるいは
検出標識がビオチンであり、そして第二の検出手段がノイトラビジンまたはノイトラビジン担持分子であるか、あるいは
検出標識がブロモ−デオキシウリジンであり、そして第二の検出手段がブロモ−デオキシウリジンに対向された抗体であるか、あるいは
検出標識がジゴキシゲニンであり、そして第二の検出手段がジゴキシゲニンに対向された抗体であるか、あるいは
検出標識がキレーターであり、そして第二の検出手段が放射性核種であり、
これらの場合、該検出標識は核酸に連結されていることが好ましい。この種の組み合わせがまた核酸が表面に付着される実施態様にも応用できることが理解されるべきである。そのような実施態様では、検出標識が相互作用パートナーに連結されることが好ましい。
【0142】
最後に、第二の検出手段が第三の検出手段を用いて検出される、好ましくは第三の検出手段が酵素、より好ましくは第二の検出手段の検出において酵素反応を示す酵素であるか、あるいは第三の検出手段が放射線、より好ましくは放射性核種によって放出される放射線を検出する手段であることは、また本発明内にはいる。好ましくは、第三の検出手段は具体的には第二の検出手段を検出することおよび/または第二の検出手段と相互作用することである。
【0143】
また、表面に固定されている生物活性および/またはデスーオクタノイルグレリンの相互作用パートナーおよび本発明による核種が、好ましくは相互作用パートナーとグレリン間に形成される複合体に付加される実施態様では、サンプルは、反応から、より好ましくは段階c)および/またはd)が実施される反応容器から除去されてもよい。
【0144】
1つの実施態様では、本発明による核種は蛍光部分を含み、ここで、蛍光部分の蛍光は、核種と生物活性グレリン間の複合体形成および遊離の生物活性グレリンにおいて異なる。
【0145】
さらなる実施態様では、核酸は本発明による核酸の誘導体であり、ここで、核酸の誘導体は、アデノシン置換アデノシンの少なくとも1種の蛍光誘導体を含む。好適な実施態様では、アデノシンの蛍光誘導体はエテノアデノシンである。
【0146】
さらなる実施態様では、本発明による核酸の誘導体および生物活性グレリンからなる複合体は蛍光を用いて検出される。
【0147】
本方法の実施態様では、シグナルは段階(c)または段階(d)において生成され、好ましくはシグナルはサンプル中の生物活性グレリンの濃度と相関される。
【0148】
好適な態様では、アッセイは96穴プレートにおいて実施されてもよく、ここでの成分は前述の反応容器および反応容器として働くウェル中に固定化される。
【0149】
反応段階の上記系列およびそれらに関連して記述される種々の実施態様は、原則として、両生物活性グレリン、すなわち、オクタノイルグレリン、より好ましくはn−オクタノイルグレリン、およびデス−オクタノイルグレリンを検出するために適当である。このことは、生物活性グレリンの検出について、少なくとも1種の相互作用パートナーおよび本発明による核酸が生物活性グレリンを特異的に検出するのに適当である場合には可能である。原則として、生物活性グレリンに特異的である本発明による核酸が使用されることが十分である。サンプル中に含有されるグレリン量を特定するこの種の方法の読み取りは、生物活性グレリンについての分析の結果としてそれ自体使用することができる。しかしながら、また結果は、グレリン、好ましくは両生物活性グレリンおよびデス−オクタノイルグレリンの総含量を決定する方法において使用されてもよい。そのような目的では、デス−オクタノイルグレリンに特異的な、あるいは両生物活性およびデス−オクタノイルグレリン、すなわち総グレリン含量またはその量を検出するのに適当な検出手段または相互作用パートナーのいずれかを使用する、上記と好ましくは同一な方法が実施される。該相互作用パートナーまたは手段が、生物活性グレリンであるか、またデス−オクタノイルグレリンであるかに拘わらず、いずれかのグレリンを検出するのに適当である場合は、サンプル中に含有されるグレリンの総量は、生物活性グレリンについて得られる値および総グレリン含量の商形成によってサンプル中の生物活性グレリンのパーセンテージを決定することを可能なような方法の読み取りによって計算することができる。さらなる実施態様では、方法はデス−オクタノイルグレリンを特異的に検出するために使用される。そのようなデス−オクタノイルグレリンは、例えば、n−オクタノイル部分を欠如しているグレリンのC末端またはN末端に対向されることによるような、デス−オクタノイルグレリンに特異的である手段または相互作用パートナーの検出によって検出することができる。かくして検出されるグレリンの量、すなわちデスーオクタノイルグレリンの量は、次いで、サンプル中の総含量を与える生物活性グレリンの量に加えることができる。
【0150】
本発明の核酸は、薬物設計のための出発材料としてさらに使用されてもよい。1つのアプローチは化合物ライブラリーのスクリーニングであり、そのような化合物ライブラリーは、好ましくは低分子量化合物ライブラリーである。1つの実施態様では、スクリーニン
グは高処理スクリーニングである。好ましくは、高処理スクリーニングは、標的に基づくアッセイにおける化合物の迅速、効率的、試行錯誤評価である。最良の場合には、分析は比色(colormatic)測定によって実施される。これに関連して使用されるライブラリーは当業者には既知である。
【0151】
あるいはまた、本発明による核酸は薬物の合理的設計のために使用されてもよい。好ましくは、合理的薬物設計は製薬学的先導(lead)構造の設計である。X線結晶学または核磁気共鳴分光法のような方法によって典型的には同定される標的の三次元構造から出発して、コンピュータープログラムが、多くの異なる化学化合物の構造を含有しているデータベースをとおして検索するために使用される。選択はコンピューターによって実施され、同定された化合物が続いて実験室において試験できる。
【0152】
薬物の合理的設計は、本発明による核酸のいずれから出発してもよく、そして本発明の核酸の構造に類似しているか、または本発明の核酸の構造の結合媒介部分に同一である構造、好ましくは三次元構造を必要とする。薬物の合理的設計におけるさらなる段階でもまた代替的段階においても、神経伝達物質に結合する核酸のそれらの部分の好ましくは三次元構造は、ヌクレオチドおよび核酸とは異なる化学基によって模倣される。この模倣によって、核酸とは異なる化合物が設計できる。そのような化合物は、好ましくは低分子またはペプチドである。
【0153】
当業者には既知である競合アッセイを使用するような、化合物ライブラリーのスクリーニングの場合では、適当なグレリン類似体、グレリンアゴニストまたはグレリンアンタゴニストが発見することができる。そのような競合アッセイは次のように設定されてもよい。本発明の核酸、好ましくは標的結合L−核酸であるspiegelmerが固相に結合される。グレリン類似体を同定するために、標識されたグレリンがアッセイに添加されてもよい。潜在的な類似体は、それぞれの標識によって得られるシグナルにおける減少とともに進行するspiegelmerに結合するグレリン分子と競合する。アゴニストまたはアンタゴニストのスクリーニングは、当業者には既知の細胞培養アッセイの使用を伴ってもよい。
【0154】
本発明によるキットは、少なくとも1種または数種の本発明の核酸を含んでもよい。さらに、キットは少なくとも1種または数種のポジティブまたはネガティブ対照を含んでもよい。ポジティブ対照は、例えば、グレリン、特にこれに対して本発明の核酸が選ばれるか、またはこれに対して、好ましくは液体形態においてそれが結合するものであってもよい。ネガティブ対照は、例えば、グレリンに類似する生物物理学的性質の見地から定義されるが、本発明の核酸によって認識されないペプチドであってもよい。さらにまた、該キットは1種以上のバッファーを含んでもよい。種々の成分は、乾燥または凍結乾燥形態においてキット中に含有されても、または液体に溶解されてもよい。キットは、キットの1種以上の成分を含有してもよい1種以上の容器を含んでもよい。さらなる実施態様では、キットは、キットおよびその種々の成分を使用する方法に関する情報を使用者に与える指示または指示紙片を含む。
【0155】
好ましくは本明細書で使用されるように、用語「処置」は、好適な実施態様では、追加的または代替的に予防および/または追跡処置を含む。
【0156】
好ましくは本明細書で使用されるように、用語「疾病および障害」は、改めて指示されない限り、互換可能な様式で使用されるべきである。
【0157】
本明細書で使用されるように、用語「含む」は、好ましくは、そのような用語によって追跡または記述される主題を限定することを意図していない。しかしながら、その他の実
施態様では、用語「含む」は、含有するという意味において理解されるべきであり、かくして、そのような用語によって追跡または記述される主題を限定するとして理解されるべきである。
【0158】
種々の配列番号、本発明による核酸分子の化学的性質および本明細書で使用される標的分子グレリン、その実際の配列および内部参照番号が、次の表において総括される。
【0159】
【表3】
【0160】
【表4】
【0161】
【表5】
【0162】
【表6】
【0163】
本発明は、図面、実施例および配列リストによってさらに具体的に説明され、これらからさらに、特徴、実施態様および利点が取り上げられてもよい。
【実施例】
【0164】
例1:グレリン結合配列
明確に別に指示されない限り、これらの実施例をとおして使用されるグレリンは、そのオクタノイル化形態におけるグレリンであった。
【0165】
ドイツ特許第一034944.3号に記述される技術から得られる技術を用いて、ビオチン化ヒト・D−グレリンに対向されるRNAインビトロ選択が実施された。dsDNA分子の富有化集団がクローン化され、配列決定された。配列分析の結果は図1において見ることができる。
【0166】
全配列は、既知のグレリン結合配列SOT−C(B11trc;特許出願WO2004/013274A2,参照)のグレリン結合モチーフA(25ヌクレオチド)を含む。これらの23クローンからの4個のみにおいてモチーフAが、配列の3’末端に置かれている。19配列内では、モチーフAはクローンの5’−末端に置かれている。モチーフDと呼ばれるさらなるモチーフはモチーフAの3’末端に置かれている。
【0167】
配列の結合特性
異なる位置において変異を示す15クローン(図1)が、実施例2に記述される「競合アッセイ」を用いる37℃での格付け実験のために選ばれた。参考として、放射能標識アプタマーSOT−C(B11trc)が使用された。数種の候補がSOT−C(B11trc)よりも強くD−グレリンに結合し、より低いKDまたは「競合アッセイ」では分別できない活性コンホメーションのより高い量を示した。クローンMS−P2−G2、MS−P2−D2、MS−P2−A2、MS−P2−H3は対照クローンSOT−C(B11trc)よりも良好な結合を有するように見え、クローンMS−P2−E1、MS−P2−B1、MS−P2−F1、MS−P2−C3、MS−P2−C2,MS−P2−A4およびMS−P2−B2はさらに良好な結合を示した。SOT−C(B11trc)に較べて類似の結合結果は、クローンMS−P2−E3、MS−P2−A3、MS−P2−H2およびMS−P2−D1について決定できた(図2;評価は図1参照)。したがって、最良クローンが、平衡アッセイにおいてアプタマーとして、そして37℃における細胞培養アッセイにおいてSpiegelmerとしてそれらの活性について試験された(プロトコルは実施例2参照)。結果は図1において総括されている。クローンMS−P2−D2、MS−P2−F1、MS−P2−C2、MS−P2−A4およびMS−P2−B2は、平衡アッセイにおいて37℃で22〜34nMのKD(約60%活性分子)と測定され、そして3,0〜8,0nMのIC50値が37℃の細胞培養実験において検出できた。これに較べて、SOT−C(B11trc)のIC50は37℃で20nMと検出され、そしてKDは37℃で100nM(47%活性分子)と決定された(特許出願WO2004/013274A2に記述されるように、室温におけるSOT−CのIC50は約5nMである)。結果は、SpiegelmerMS−P2−F1、MS−P2−C2およびMS−P2−D2の生物学的活性は、37℃においてSOT−C(B11trc)よりも約5倍も良好であり得ることを示している。連続的な先端切り取り実験では、クローンMS−P2−F1(4,0nMのIC50)が選択された(二次構造予想、図12参照)。
【0168】
図1に示される配列のいずれもが、その先端を切り取られた形態物を含む本発明による核酸であるが、それらはなお標的に結合することができる。
【0169】
例2: 配列の結合特性
2.1 「平衡結合アッセイ」の使用による放射能標識アプタマーの格付け
本明細書で開示され、そしてより具体的には図1において列挙されるほとんどのクローンが、「平衡結合アッセイ」の使用によって、ビオチン化ヒト・D−グレリンに対するそれらの結合挙動に関して放射能標識アプタマー(D−RNA)として格付けされた。
【0170】
ヒト・D−グレリンに対する分子の結合挙動の格付けが実施された。この目的のために、本明細書で記述される標準プロトコルを用いて、同定されたアプタマーが図1に描かれるような先端を切り取られたアプタマー(プライマー結合部位なし)として合成された。
【0171】
次に示すように、アプタマー配列が次のプロトコルを用いてγ−P32−ATPにより5’末端において放射能標識された。
成分 [最終]
オリゴヌクレオチド 5μM
T4 Forward反応バッファー(Invitrogen) 1x
T4 ポリヌクレオチドキナーゼ(Invitrogen) 10U/10μl反応容量[γ−32P]−ATP 1μl/10μl反応容量
【0172】
反応は37℃で1時間インキュベートされた。次いで、放射能標識アプタマーはゲル精製された。
【0173】
放射能標識RNAの2〜5pmoleが、Ca++およびMg++を含有しない選択バッファー(ヒト血液における生理学的条件にしたがう:20mM Tris、150mMNaCl、5mMKCl,pH7.4は37℃で調節された)において95℃で3分間変性され、37℃において1mMの最終濃度までのこれらのイオンの添加によって折り畳まれ、そして0,4〜3000nMの範囲の濃度のビオチン化ヒトD−グレリンとともに37℃で1時間インキュベートされた。続いて、NeutrAvidinアガロースの一定量がマトリックスとして添加され、そしてRNA:ペプチド複合体が30分以上37℃で振盪された。次いで、結合されたペプチドをもつマトリックスおよびチューブが沈降され、上澄液が除去され、マトリックスが100μl選択バッファーにより洗浄され、そして結合RNAと非結合RNAとの差異がBeckman Coulterを使用する放射能の測定によって決定された。算出された数から、対照(0nMビオチン化ヒトD−グレリン)がバックグラウンドとして差し引かれた。平衡定数はソフトウエアプログラム「GraFIT」(Version 4.0.10.,Erithacus Software Ltd.,Surrey,UK)を使用して計算された。
【0174】
2.2 競合アッセイの使用によるアプタマーの格付け
既知のグレリン結合配列SOT−C(B11trc;特許出願WO2004/013274A2参照)に対してクローンを比較するために、SOT−Cが、本明細書で記述される標準プロトコル(実施例4)を用いてアプタマー(D−RNA)として合成された。アプタマー配列は本明細書で記述される標準プロトコルを用いてγ−P32−ATPにより5’末端において放射能標識された。
【0175】
放射能標識SOT−Cおよび同定されたクローン(D−RNA)が平衡アッセイのために前述のように調製された。アッセイは20nMのペプチド濃度(ビオチン化ヒトD−グレリン)において実施された。その後、等モル量の放射能標識SOT−Cおよび2つの異なる濃度(40nMおよび200nM)のアプタマーが試験された(結果は図2に示される)。アッセイは平衡アッセイと同様に実施された。
【0176】
2.3 グレリン結合Spiegelmerによるグレリン誘導カルシウム放出の抑制
グレリン結合Spiegelmerの機能的特性決定が、ヒト(L−)グレリンとヒト
成長ホルモン分泌促進物質受容体(GHS−R)との相互作用を追跡する細胞アッセイ系において実施された。受容体−リガンド相互作用からもたらされる細胞内カルシウム放出は、蛍光カルシウム指示薬の手段によって可視化される。
【0177】
ヒト・グレリン受容体(GHS−R1a)を発現する安定なトランスフェクトされたCHO細胞(Euroscreen,Gosselies,Belgiumから得られた)が、透明な底を有する黒色の96穴プレート(Greiner)において5〜7x104細胞/ウェルを接種され、そして100単位/mlペニシリン、100μg/mlストレプトマイシン、400μg/mlゲネチシンおよび2.5μg/mlフンギゾン(fungizone)をさらに含有するUltraCHO培地(Cambrex)において37℃および5%CO2下で一夜培養された。
【0178】
カルシウム指示薬染料fluo−4を負荷する前に、細胞は200μlのCHO−U+により一回洗浄される。次いで、指示薬染料溶液(CHO−U+中、10μMfluo−4(Molecular Probes)、0.08%pluronic127(Molecular Probes))の50μlが添加され、そして細胞は37℃で60分間培養される。その後、細胞は180μlのCHO−U+により3回洗浄される。最後に、90μlのCHO−U+が各ウェルに添加される。
【0179】
種々の量のSpiegelmerが、0.2mlの低プロフィル(low profile)96穴プレートにおいて室温または37℃で15〜60分間、5mMprobenecidおよび20mM HEPES(CHO−U+)を含有するUltraCHO培地において、ヒト(L−)グレリン(Bachemから購入)とともにインキュベートされる。対照として、ペプチドのみを含有するサンプル(最大カルシウム放出)およびペプチドを含有しないサンプル(最小カルシウム放出)が分析される。これらの刺激溶液においては、ペプチドおよびSpiegelmer(添加される場合)は、このアッセイに較べて10倍濃縮されている。
【0180】
カルシウム放出の検出では、刺激溶液が細胞に添加され(10μl/ウェル)、そして蛍光シグナルの変化がモニターされる。蛍光シグナルの測定は、注入ポンプを備えたFluostar Optima多検出プレートリーダー(BMG)において485nmの励起波長および520nmの発光波長において実施される。
【0181】
数個のサンプルの並行測定では、96穴プレートの1つの(垂直の)列のウェルが一緒に記録される。4秒のタイムラグをもつ最初の3回の読み取りが、ベースラインの決定のために実施される。次いで、読み取りが中断され、プレートが装置から除かれる。多チャンネルピペットを用いて、刺激溶液の10μlがウェルに添加され、次いで、プレートが装置中に再び移動され、そして測定が継続される。4秒間隔での全20回の読み取りにおいて実施される。
【0182】
各ウェルでは、最大蛍光とベースライン値との間の差異が決定され、そしてヒト(L−)グレリン(オクタノイル化された)濃度に対してプロットされるか、あるいはSpiegelmerによるカルシウム放出の抑制における実験では、半最大抑制定数(IC50)の決定を可能にするSpiegelmerの濃度に対してプロットされる。
【0183】
2.4 表面プラズモン共鳴(SPR)測定
ビオチン化ヒトD−グレリンに結合するアプタマーの特性決定が、前記BIAcore2000装置(BIAcore AB,Uppsala,Sweden)を用いるSPR即時速度分析によって決定された。C末端ビオチン化ペプチドの100RU(フローセル2)および300RU(フローセル3)が、Streptavidin共役センサーチッ
プ(BIAcore AB,Freiburg,Germany)に固定化され、そして0,1μM〜1μMの濃度範囲におけるサンプルが、会合時間360(s)および解離時間360(s)を定めるKinject指令を用いて注入された。フローセル1はバッファーおよびデキストラン基質対照(Biacore SA−Chip surface)として使用され、フローセル4には、非特異的D−ペプチドが固定化されてアプタマーの非特異的結合を決定した。反応は37℃で実施された。BIAevaluation3.0 ソフトウエア(BIAcore AB,Uppsala,Sweden)によるデータ解析では、本研究者らはLangmuir1:1化学量論的フィッティング・アルゴリズムを使用した。
【0184】
例3: グレリン結合Spiegelmerモチーフの定義
3.1 SOT−D−000の切り取り
3.1.1 予め選ばれたSOT−D−000の末端切り取り
図3に示されたグレリン結合Spiegelmerは、特許出願WO2004/013274A2において既に示されるように予め得られた。二次構造の予測(最小自由エネルギーコンホメーション[Hofacker et al.,1994,Monatsh.Chem 125:167−188])では、5’−および3’末端の間の標準的分子内塩基対合が全Spiegelmerについて明白になる(図4−7;図3において下線を付した塩基)。Spiegelmerにおけるそのような構造的要素の存在は、活性のある三次元構造の正確な折り畳みのためには極めて重要であろうが、経済的なSpiegelmerの合成のためには、分子は可能な限り短くあるべきである。SpiegelmerSOT−D−000(別名、プライマー結合部位を含まないSOT−R04−DR14−F7の先端切り取り物)は、それが、最長の末端らせんを形成する可能性を有する最短の選ばれたSpiegelmerであるので、末端切り取りのための基本として選ばれた。
【0185】
10個の代わりに8個(SOT−D−108)、7個(SOT−D−100)および6個(SOT−D−104)の塩基対のらせんの長さをもつSOT−D−000の先端切り取り変異体が合成され(図8)、そして実施例2における記述のように細胞培養において試験された(図9)。SOT−D−104(39量体)は有意に結合活性を喪失したが、SOT−D−100(41量体)および−108(43量体)は完全なグレリン・アンタゴニスト性能を維持した。変異体SOT−D−106、G4が除去されたSOT−D−104のさらなる先端切り取り物は実質的に不活性であった。驚くべきことに、この不適合G−全選択グレリンSpiegelmerのらせんに存在する−は容易には除去されないであろう。むしろ、グレリン結合のためには必須であると思われる。
【0186】
3.1.2 SOT−D−100および−108の内部欠失変異体
図3における選ばれたグレリン結合物(binder)の整列を考慮する場合、強度に可変性の領域は、5’末端における末端らせんに直接隣接するすべての整列された分子において存在すると思われる。グレリン結合物のさらなる先端切り取りについてのこの明白な可変性を精査するために、最小の完全活性SOT−D変異体SOT−D−100およびー108がさらなる切り取りのための基礎として使用された。内部のSOT−D−100変異体の場合は、それぞれの塩基が単純に除かれたが、SOT−D−108変異体では、それらが、合成中に柔軟性の親水性スペーサーによって置換された。次に、SOT−D−100およびその誘導体SOT−D−101および−102ならびにSOT−D−108誘導体SOT−Dー109、SOT−D−110およびー111が、実施例2における記述のように細胞培養において試験された(図9)。図8に示されるように、3個ではないが2個の内部ヌクレオチドの欠失は、活性喪失なしに可能であった(SOT−D−101;−102)。これに対して、スペーサーによって置換された場合(SOT−D−109、−110、−111)には、3個のヌクレオチドが除くことができた。スペーサーの最適位置はSOT−D−109において実現される(図10)。
【0187】
40kDa部分による5’末端におけるSOT−D−109の修飾はグレリン結合活性の喪失なしに可能であった(図11)。
【0188】
3.2 SOT−Eの切り取り
次のように、先導配列として選ばれたグレリン結合配列MS−P2−F1(図1および図12)はSOT−Eと呼ばれる。
【0189】
二次構造の予測(最小自由エネルギーコンホメーション[Hofacker et al.,1994,Monatsh.Chem 125:167−188])では、C30およびG50の間の標準的分子内塩基対合の可能性が、SpiegelmerSOT−Eについて明白になる(図12)。SpiegelmerSOT−Eの切り取りをもたらす次の実験が記述される。SpiegelmerSOT−Eのすべての切り取り型が細胞培養においてそれらの活性に関して試験された(実施例2、参照)。
【0190】
3.2.1 SpiegelmerSOT−Eの末端切り取り
3’末端における5個のヌクレオチドは分子の他の部分にはハイブリダイズされないと思われる。反対に、5’末端では、ヌクレオチドは部分的に対合できる。驚くべきことに、3’末端は結合の低下なしには切り取りすることができない(SpiegelmerSOT−E−012)が、一方SOT−E−014(5’末端における6個のヌクレオチドの切り取り)は完全なグレリン・アンタゴニスト性能を維持した(図13)。
【0191】
3.2.2 SpiegelmerSOT−Eの内部欠失変異体
図1における選ばれたグレリン結合物MS−P2−E3、MS−P2−G2、MS−P2−D2、MS−P2−A3、MS−P2−E1、MS−P2−B1、MS−P2−C3、MS−P2−C2、MS−P2−H2、MS−P2−A4、およびSOT−E(配列ファミリーI)の整列を考慮する場合、可変性の領域は、末端において、かつ、らせんに直接隣接するすべての整列された分子において存在すると思われる(SOT−EにおけるG36−C43)。SpiegelmerSOT−Eの二次構造予測は、4個のヌクレオチドのループを示す(A38−U41;図12)。グレリン結合物のさらなる切り取りのためにこの明白な可変性を探査するために、最初に、SpiegelmerSOT−Eのそれぞれの塩基が、合成中に柔軟性の親水性スペーサーによって置換された。8個のヌクレオチド(G36−C43;SOT−E−008)ではない6個(U37−A42;SOT−E−011)の欠失が、活性の喪失なしに可能であった。5’末端における切り取り(G1−A6)の結果および柔軟性の親水性スペーサーによる6個のヌクレオチド(U37−A42)の置換の組み合わせは、56−ヌクレオチドSpiegelmerSOT−E(図12)として細胞培養において同一のIC50を示す44−ヌクレオチドのSpiegelmerSOT−E−019(図14)をもたらす。
【0192】
3.2.3 SOT−E−019内の配列セグメントの転位
配列MS−P2−A2、MS−P2−H3、MS−P2−D1(図1)によって表される配列ファミリーIIは配列ファミリーI(SOT−Eを含む)に対して類似性を担持する。これらの配列相同性は図1におけるboxA(UAAGACCGAAGGUAACCAAUCCUAC)として強調される。配列ファミリーIIでは、boxAは、むしろ3’末端に置かれるが、配列ファミリーIでは、boxAは5’末端に接近している。SpiegelmerSOT−Eのインビトロの選択および二次構造予測のこの結果により、boxAが5’末端に置かれるSOT−E−019の変異体が計画された。その目的のために、SOT−E−019の本来の5’末端および3’末端が、柔軟性の親水性スペーサーと連結され、そしてループが除去されて、新しい5’末端および3’末端を生成した(SOT−E−021;図15および21)。驚くべきことに、SOT−E−021では、活性の喪失は観察できなかった。SOT−E−019およびSOT−E−021(SOT−E−33(図16および21)およびSOT−E−25(図17および21))に基づくさらなる切り取り物は、グレリン・アンタゴニスト機能の低下なしには可能ではなかった(図13)。
【0193】
3.3 SOT−C、SOT−D−109およびSOT−Eの比較
SpiegelmerSOT−Eが、SOT−Cに比較して表面プラズモン共鳴(実施例2)によって測定され、6倍良好なKDが決定できた(図18)。同じ改良された結合性(5倍良好)が、D−109に較べてSpiegelmerSOT−Eを用いる細胞培養実験(実施例2)において検出することができた(図19)。動物体内での滞留時間を改良するために、SpiegelmerSOT−E−19が、40kDa−PEG部分によってその5’末端において修飾された(SOT−E19−5’−PEG)。SOT−E19−5’−PEGは、細胞培養実験においてSOT−Eに較べて類似の結果を示し(図19)、さらに、インビボ活性を例証した(実施例5;図22)。
【0194】
3.4 グレリン結合Spiegelmerモチーフの定義
グレリン、特に本明細書で定義されるオクタン酸側鎖を有するグレリンを結合させるすべての分子は、第一に、グレリン結合のために必須である25個のヌクレオチド(「BoxA」)のモチーフの存在を特徴とする。「BoxA」内の25個のヌクレオチドのあるものは他の塩基と相互に交換可能であると思われる(図20A)。
【0195】
グレリン結合Spiegelmerの「BoxA」の機能のための必須要件は、非対合の中央に置かれたグアノシンを含んでらせん構造を形成する配列5’−GUGAGG(BoxB)の相補鎖と、「BoxAの」3’末端の5個のヌクレオチド(5’−CCUAC)とのハイブリダイゼーションである。BoxBにおけるこの中央のグアノシンの欠失は、有意な結合の喪失をもたらす(「BoxAにより定義されるらせん」、非対合G;図20A)。「BoxA」、「BoxAにより定義されるらせん」に対する3’−隣接物は、1個、優先的には2個以上のさらなる定まっていない塩基対(「BoxC1」および「BoxC2」、図20A)によって最小に伸長されるべきである。
【0196】
「BoxB」の3’末端では、さらなるヌクレオチドが結合のために必須である。少なくとも2個のさらなるヌクレオチド(優先的には5’−CA)が結合のために必要である。結合性の有意な改良は、この位置において(「BoxD」、図20A)、さらなる2個、場合によっては4個のヌクレオチドを付加して2、3、4、5または6個のヌクレオチドを含んでなるBoxDをもたらすことによって達成できる。
【0197】
「BoxA」の5’末端では、さらなるヌクレオチドが結合のために必須である。少なくとも1個のさらなるヌクレオチド(特にUまたはG)が結合のために必要である。結合性の有意な改良は、この位置において(「BoxE」、図20A)、さらなる2個のヌクレオチドを付加して1、2、3または4個のヌクレオチドを含んでなるBoxEをもたらすことによって達成できる。
【0198】
BoxDおよびEが柔軟性の親水性スペーサーを介して連結される場合、該スペーサーはまた、種々の位置に挿入することができる(参照、図8;SOT−D−109、SOT−D−110およびSOT−D−111)。
【0199】
驚くべきことに、BoxDおよびEの代わりに「BoxC1」および「BoxC2」が柔軟性の親水性スペーサーに連結した場合に、一定の配列要素が機能する。
【0200】
変異体1:
【0201】
【化6】
【0202】
変異体2:
【0203】
【化7】
【0204】
本明細書、請求項および/または図面において開示される本発明の特徴は、別々に、およびいずれかのその組み合わせにおいて、その種々の形態において本発明を実現するための材料となり得る。
【0205】
例4: アプタマーおよびSpiegelmerの化学的合成
化学的固相合成
小規模合成
アプタマーは、2’TBDMS RNAホスホルアミダイト化学を使用してABI 394シンセサイザー(Applied Biosystems,Foster City,CA,USA)による固相合成によって製造された(M.J.Damha,K.K.Ogivie,Methods in Molecular Biology,Vol.20 Protocols for oligonucleotides and analogs,ed.S.Agrawal,p.81−114,Humana Press Inc.1993)。D−rA(N−Bz)−、D−rC(Ac)−、D−rG(N−ibu)−、D−rU−およびヘキサエチレングリコールホスホルアミダイトは、ChemGenes,Wilmington,MAから購入された。アプタマーはゲル電気泳動によって精製された。
【0206】
未修飾(PEG化のない)Spiegelmerは、2’TBDMS RNAホスホルアミダイト化学を使用してABI 394シンセサイザー(Applied Biosystems,Foster City,CA,USA)による固相合成によって製造された(M.J.Damha,K.K.Ogivie,Methods in Molecular Biology,Vol.20 Protocols for oligonucleotides and analogs,ed.S.Agrawal,p.81−114,Humana Press Inc.1993)。L−rA(N−Bz)−、L−rC(Ac)−、L−rG(N−ibu)−、L−rU−およびヘキサエチレングリコールホスホルアミダイトは、ChemGenes,Wilmington,MAから購入された。アプタマーはゲル電気泳動によって精製された。
【0207】
大規模合成および修飾
修飾SpiegelmerSOT−E−19は、2’TBDMS RNAホスホルアミダイト化学を使用して ktaPilot100シンセサイザー(Amersham Bioscience;General Electric Healthcare,Freiburg)による固相合成によって製造された(M.J.Damha,K.K.Ogilvie,Methods in Molecular Biology,Vol.20 Protocols for oligonucleotides and analogs,ed.S.Agrawal,p.81−114,Humana Press Inc.1993)。L−rA(N−Bz)−、L−rC(Ac)−、L−rG(N−ibu)−、L−rU−およびヘキサエチレングリコールホスホルアミダイトは、ChemGenes,Wilmington,MAから購入された。合成は、L−riboC修飾されたCPG孔径1000(Link Technology,Glasgow,UK)において開始された。カップリング(1サイクルについて15分)では、アセトニトリル中0.3Mベンジルチオテトラゾール(CMS−Chemicals,Abingdon,UK)、およびアセトニトリル中それぞれ0.1Mホスホルアミダイト溶液の3.5当量が使用された。Spiegelmerの5’末端におけるアミノ基は、アミノヘキシルホスホルアミダイト(ChemGenes)をカップリングすることによって連結された。酸化−キャッピング(capping)サイクルが使用された。オリゴヌクレオチド合成のためのさらなる標準溶媒および試薬はBiosolve(Valkenswaard,NL)から購入された。SpiegelmerはDMT−ON合成された;脱保護の後、それは、Source15RPC媒質(Amersham)を用いて調製用RP−HPLC(Wincott et al 1995 Nucleic Acids Res.23:2677)により精製された。5’DMT−基は80%酢酸を用いて除去された(RTで30分)。続いて、2MNaOAc水溶液が添加され、そしてSpiegelmerが、5K再生セルロース膜(Millipore,Bedford,MA)を用いる接線流動濾過によって脱塩された。
【0208】
PEG化
インビボの腎臓クリアランスを低下させるために、SpiegelmerSOT−E−19は5’末端において40kDaポリエチレングリコール(PEG)部分に共有結合された。そのような増加された分子質量をもつSpiegelmerを用いることは、血漿における保持を有意に延長させ、そのために、より長い有効性が達成できた。
【0209】
PEG化(欧州特許出願第一 306 382号)では、精製された5’−アミノ修飾Spiegelmerは、H2O(2.5ml)、DMF(5ml)およびバッファーA(5ml;クエン酸・H2O[7g]、ホウ酸[3.54g]、リン酸[2.26ml]および1MNaOH[343ml]を混合し、そして最終容量1 lまでH2Oを添加することによって調製された;pH=8.4は1MHClにより調節された)の混合液中に溶解された。
【0210】
Spiegelmer溶液のpHは1MNaOHにより8.4にもたらされた。次いで、40kDaのPEG−NHSエステル(Nektar Therapeutics,Huntsville,AL)が、75〜85%の最大収率が達成されるまで、0.6当量の4分量において30分毎に37℃で添加された。反応混合液のpHは、PEG−NHSエステルの添加中1MNaOHにより8〜8.5に維持された。
【0211】
反応混合液は、4ml尿素溶液(8M)、4mlバッファーA、および4mlバッファーB(H2O中0.1M酢酸トリエチルアンモニウム)と混合され、95℃に15分間加熱された。PEG化Spiegelmerは次に、アセトニトリル濃度勾配(バッファーB;バッファーC:アセトニトリル中0.1M酢酸トリエチルアンモニウム)を用いて、Source15RPC媒質(Amersham)によりRP−HPLCによって精製された。過剰のPEGは5%バッファーCにおいて、PEG化Spiegelmerは10〜15%バッファーCにおいて溶出した。>95%の純度(HPLCによって調査)をもつ生成物画分が合わされ、40mlの3M NaOAcと混合された。PEG化Spiegelmerは、接線流動濾過(5K再生セルロース膜、Millipore,Bedford,MA)によって脱塩された。
【0212】
例5: 抗グレリン−SpiegelmerSOT−E19−5’−PEGによる外因性グレリン投与後の成長ホルモン放出の抑制
外因性グレリンの投与は、ラットにおける成長ホルモン(GH)の放出を惹起することが知られている。グレリン結合Spiegelmerの前以ての投与は、GHのグレリン誘導放出を抑制する。
【0213】
Sprague Dawleyラットが、7日間それらの新しい環境に適応された。実験の設定は、各々5匹の動物をもつ5群からなった:1つのポジティブ対照群および4つの異なる用量のSpiegelmerSOT−E19−5’−PEG群。全動物は、実験期間中Ketamine/Xylazineで麻酔され、尾静脈中に2回の静脈注射を受けた。第一回の注射は第二回注射の30分前に与えられ、そしてPBS(ポジティブ対照群)または図22に指示されるSpiegelmerSOT−E19−5’−PEGの用量からなった。第二回の投与は全動物に対して3nmolラット・グレリンからなり、そして実験について時間0とマークされた。第二回の投与前に、最初の血液サンプルが眼窩の血脈洞から抜き取られた。さらなるサンプルは、ラット・グレリン投与後の5、15、30および45分に採取された。得られる血漿サンプルは、製造者の指示にしたがって市販の酵素免役アッセイ(Growth hormon,Rat,Biotrak Assay Kit,RPN2561,Amersham Biosciences Europe GmbH,Freiburg)により成長ホルモン濃度について分析された。
【0214】
図22において分るように、GHの放出はラット・グレリンの注射によって強力に刺激されるが、SOT−E19−5’−PEGの前以ての投与によって抑制することができた。最大の抑制は75nmolSOT−E19−5’−PEG/kg体重において達成される。150nmolSOT−E19−5’−PEG/kg体重までの投与用量におけるさらなる増加は、それ以上のGH放出を抑制しない。この結果は、抗グレリンSpiegelmerSOT−E19−5’−PEGのインビボ活性を例証している。
【0215】
例6: 内部リンカーを含まないSOT−E−19の誘導体
実施例3に記述されたように、抗グレリンSpiegelmerSOT−E−19は、L−ヌクレオチド、および6個のヌクレオチドを置換するために元の分子SOT−Eに組み入れられた内部リンカーからなる。6個のヌクレオチドからの4個はSpiegelmerSOT−Eにおいてループを形成し、残りの2ヌクレオチド(6ヌクレオチドからの)は互いにハイブリダイズできる(SOT−E、参照図23、第一列、ハイブリダイズされるヌクレオチドは下線を付される)。
【0216】
内部リンカーを含まないSOT−E−19の可能な限り短い誘導体を実現するために、リンカーは3個のヌクレオチドの種々の建造ブロックによって置換された。全誘導体がSpiegelmer(プロトコル、実施例4参照)として合成され、そして細胞培養実験(プロトコル、実施例2参照)において分析された。結合および抑制効力(IC50)の喪失のないSOT−E−19の内部リンカーの置換は、次の建造ブロックについて成功裏に示された:ACA(SOT−E−19−L3)およびCAA(SOT−E−19−L4)。SOT−E−19の種々の誘導体が図23に示される。
【0217】
例7: グレリン結合Spiegelmerによるオクタノイル−グレリンおよびデスオクタノイル−グレリンの識別
グレリンに対するSpiegelmerSOT−E−19の結合の特徴は、さらに、実施例2に記述された方法に基づいて、競合アッセイにおいて分析された。これらのアッセイにおいて、Spiegelmerは、細胞の刺激前の刺激溶液においてグレリンペプチド(オクタノイル−およびデスオクタノイル−グレリン)の種々の組み合わせ物とともにインキュベートされた。
【0218】
ペプチド組み合わせ物のスキームおよび全長オクタノイル−グレリン(ヒト・グレリン
=huグレリン)が図24(左から右へ数えられるバー)において総括される:いずれのオクタノイル−グレリン(ヒト・グレリン=huグレリン)も含まないか、または300nMの最終濃度においてデスオクタノイル−グレリンを含む場合は、細胞の刺激は検出できない(バー1および2)が、一方、13nMの濃度におけるオクタノイル−グレリン(ヒト・グレリン=huグレリン)は既にカルシウム放出を媒介するために十分であり(バー3);300nMデスオクタノイル−グレリン(バー4)のさらなる添加は細胞刺激を妨げず、これは、生物学的に不活性なデスオクタノイル−グレリンが受容体アンタゴニストではないことを示している。3nMオクタノイル−グレリン(ヒト・グレリン=huグレリン)によって媒介されるカルシウム放出はSOT−E−19の10倍過剰量によって抑制でき(バー5)、オクタノイル−グレリン(ヒト・グレリン=huグレリン)を超える100倍過剰量(300nM)でのデスオクタノイル−グレリンの存在でさえ抑制に対して競合しない(バー6)。これに対して、300nMオクタノイル−グレリンおよび30nMSpiegelmerのアッセイ濃度はカルシウム放出の増加を示し(バー7)、アッセイ条件下で、オクタノイル−グレリン(ヒト・グレリン=huグレリン)による刺激増進が達成できるという証拠を与える。この実験は、SOT−E−19がオクタノイル形態とデスオクタノイル形態におけるグレリン間を特異的に識別することを例証する。
【0219】
グレリンに対するSpiegelmerNOX−B11の結合特性(オクタノイル−グレリンには結合するが、デスオクタノイル−グレリンには結合しないか、または弱く結合する)は、SOT−E−19に類似している(同じ実験が既にWO2005/049828に記述されるように実施された)。
【0220】
両分子は、主としてboxAに基づく高い構造的相関関係(実施例3、参照)を有するので、この結果は、モチーフAが、グレリンのオクタノイル化形態に関して、特にオクタノイル基を含むN末端における5個のアミノ酸に関して、本発明による核酸分子の高い特異性のために必須であるという仮説を支持する。
【0221】
例8: グレリン結合SpiegelmerNOX−B11を使用するオクタノイル−グレリンの定量
グレリン結合SpiegelmerNOX−B11は、オクタノイル化されたヒトおよびラットのグレリンの非放射能定量のために、酵素免疫アッセイ(EIA)のアッセイ形式に類似する形式で使用することができる。
【0222】
原理
このアッセイでは、標準、対照および未知の処理血漿が、グレリン結合Spiegelmer、例えば、そのN末端においてオクタノイル−グレリンを認識するSpiegelmerNOX−B11によりコートされた96穴ミクロタイター・プレートにおいてインキュベートされる。インキュベーションおよび洗浄後、ウェルが、抗グレリン抗体(第一の抗体)またはグレリンのC末端においてグレリンに結合する核酸により処理される。この抗体または核酸は標識されても、されなくてもよいが、核酸は好ましくは標識される。(第一の)抗体が標識されない場合、(第一の)抗グレリン抗体を除去するインキュベーションおよび数回の洗浄段階後、第二の抗体(抗体は第一の抗体のFcフラグメントに対向され、そして標識されている)が添加される。第二の抗体の標識は、酵素セイヨウワサビ・ペルオキシダーゼ(HRP)であってもよい。第二の抗体とともにウェルをインキュベートし、未結合フラクションを除去した後、ウェルはHRP基質テトラメチルベンジジン(TMB)とともにインキュベートされる。次いで、酸性停止液が添加され、そして基質の酵素的ターンオーバーの度合が450nmにおける吸光度測定によって決定される。測定された吸光度はサンプル中に存在するオクタノイル−グレリンの濃度に直接比例している。1セットのグレリン標準が使用されて、グレリン濃度に対する吸光度の標準曲線がプロットされ、これから未知のグレリンが計算できる。
【0223】
プロトコル
最初に、ストレプトアビジンでコートされた96穴プレート(Reacti−Bind
Streptavidin Coated High Bind Capacity Black 96−well Plates,Perbio Science,Bonn,Germany)が、0.1%Tweenを含むPBS−Dulbecco(Mg2+およびCa2+を含む、Biochrom AG,Berlin,Germany)により3回洗浄された。各ウェルは、室温で1時間、100μlの50μMビオチン化SpiegelmerNOX−B11(PBSに溶解;配列番号:72)とともにインキュベートされた。ビオチン化SpiegelmerNOX−B11の固定化の後、未結合のSpiegelmerが1回の洗浄段階(100μl PBS)によって除去された。これは図26Aに図示される。
【0224】
洗浄バッファーを除去後、一定濃度のオクタノイル−グレリンを含む保存液が添加され、そして室温で1時間インキュベートされた。上澄液が除去され、ウェルが1回(100μl PBS)洗浄された。これは図26Bに図示される。
【0225】
抗グレリン抗体(Phoenix Peptides,Belmont,CA,USA;10ab)がブロッキング溶液(Phoenix Peptides,Belmont,CA,USA)において1時間インキュベートされ、未結合の抗体が3回の洗浄段階(各100μl PBS)によって除去された。これは図26Cに図示される。
【0226】
結合された抗グレリン抗体を検出するために、抗グレリン抗体(10ab)のFcフラグメントを特異的に認識し、そしてセイヨウワサビ・ペルオキシダーゼにより修飾されている第二の抗体(Phoenix Peptides,Belmont,CA,USA;20ab−HRP)が使用された。第二の抗体(20ab−HRP)の未結合フラクションは5回の洗浄段階(100μl PBS)によって除去された。これは図26Dに図示される。
【0227】
定量では、TMB基質(Amersham Biosciences,Little Chalfont,UK)の100μlが添加され、プレートが密封され、そして30分間暗室においてインキュベートされた。定義50μlが添加された。基質の酵素的ターンオーバーの度合が450nmにおける吸光度測定によって決定され、そして吸光度単位の差異が記録された。
【0228】
図25に示されるように、1セットのグレリン標準品が使用されて、グレリン濃度に対する吸光度の標準曲線をプロットした。
【0229】
結果
原則として、実験データは、SpiegelmerNOX−B11が濃度依存方式でオクタノイル−グレリンを固定化するために使用でき、したがって、EIA型検出アッセイにおいて試薬としての潜在力を有することを例証する。
【0230】
本明細書、請求項および/または図面において開示される本発明の特徴は、別々に、およびいずれかのその組み合わせにおいて、その種々の形態において本発明を実現するための材料となり得る。
【図面の簡単な説明】
【0231】
【図1】図1は、ヒト・グレリンに結合するRNAリガンドの配列の整列を示す;
【図2】図2は、先端を切り取られたアプタマー対既知の配列SOT−C(B11trc)によるグレリンへの結合の競合アッセイを示す;
【図3】図3は、RNAリガンドの選ばれた配列の整列を示し、これらは特許出願WO2004/013274A2において公表された;
【図4】図4は、グレリン結合RNAspiegelmerクローンSOT−Cの計算された二次構造を示し、二次構造はプログラム「RNAフォールド(fold)」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図5】図5は、グレリン結合RNAspiegelmerクローンSOT−D−000の計算された二次構造を示し、二次構造はプログラム「RNAフォールド」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図6】図6は、グレリン結合RNAspiegelmerクローンSOT−Dの計算された二次構造を示し、二次構造はプログラム「RNAフォールド」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図7】図7は,グレリン結合RNAspiegelmerクローンF−12の計算された二次構造を示し、二次構造はプログラム「RNAフォールド」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図8】図8は、SOT−D−000誘導体の整列を示し、これらは先端切り取りおよび合理的設計実験の結果である;
【図9】図9は、室温におけるSpiegelmerSOT−C、SOT−D−000およびそれらの変異体によるグレリン誘導Ca++−放出の抑制についてのワンポイント測定を示し;細胞は、種々の量のSpiegelmerSOT−C、SOT−D−000またはD−000の変異体とともに室温でプレインキュベートされた5nMグレリンにより刺激され、結果はSpiegelmerを無しで得られたシグナルに対して標準化された蛍光シグナルのパーセンテージを示す;
【図10】図10は、グレリン結合RNAspiegelmerクローンSOT−D−109の推定二次構造を示す;
【図11】図11は、室温におけるSpiegelmerD−109および5’−PEG化D−109によるグレリン誘導Ca++−放出の抑制についての用量−応答曲線を示し;細胞は、種々の量のSpiegelmerD−109および5’−PEG化D−109とともに室温でプレインキュベートされた5nMグレリンにより刺激され;結果はSpiegelmerを無しで得られたシグナルに対して標準化された蛍光シグナルのパーセンテージを示し;SpiegelmerD−109およびその修飾体は、同じIC50をもってグレリン誘導Ca++−放出を抑制することが見出された;
【図12】図12は,グレリン結合RNASpiegelmerクローンSOT−Eの計算された二次構造を示し、二次構造はプログラム「RNAフォールド」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図13】図13は、細胞培養実験によって試験されたSpiegelmerSOT−Eの先端を切り取られた変異体の結合分析を示す;
【図14】図14は、グレリン結合RNASpiegelmerSOT−E−19の推定二次構造を示す;
【図15】図15は、グレリン結合RNAspiegelmerクローンSOT−E−21の推定二次構造を示す;
【図16】図16は、グレリン結合RNAspiegelmerクローンSOT−E−33の推定二次構造を示す;
【図17】図17は、グレリン結合RNAspiegelmerクローンSOT−E−25の推定二次構造を示す;
【図18】図18は、D−グレリン結合RNAクローンSOT−CおよびSOT−EのKD値を指示するBiocore2000センサーグラム(sensorgram)を示す;
【図19】図19は、37℃におけるSpiegelmerSOT−E、SOT−E−19−5’−アミノ、SOT−E−19−5’−PEGまたはSOT−D−109によるグレリン誘導Ca++−放出の抑制についての用量−応答曲線を示し;細胞は、種々の量のSpiegelmerSOT−E、SOT−E−19−5’−アミノ、SOT−E−19−5’−PEGまたはSOT−D−109とともに37℃でプレインキュベートされた2nMヒト・グレリンにより刺激され;結果はSpiegelmerを無しで得られたシグナルに対して標準化された蛍光シグナルのパーセンテージを示し;SpiegelmerSOT−E−19およびその修飾体は、約4nMのIC50をもってグレリン誘導Ca++−放出を抑制することが見出された;
【図20A】図20Aは、グレリン結合Spiegelmerについて特徴的である配列boxの定義を示す;
【図20B】図20Bは、図20Aの1変異体;
【図20C】図20Cは、図20Aの1変異体;
【図20D】図20Dは、図20Aの1変異体;
【図21】図21は、SOT−D−109、SOT−E、SOT−E−19、SOT−E−21、SOT−E−33およびSOT−E−25の配列を示す;
【図22】図22は、抗グレリンSpiegelmerSOT−E−19−5’−PEGによる外因性グレリンの投与後の成長ホルモン放出の抑制を示す;
【図23】図23は、内部リンカーの代わりにさらなるヌクレオチドを含むSOT−E−19の異なる誘導体の概観、およびnMで表される観察されたIC50値を示す;
【図24】図24は、線の下に総括される成分の組み合わせおよび濃度をもつ、オクタノイルグレリン、デス−オクタノイルグレリンおよびSpiegelmerSOT−E−19による細胞競合アッセイの結果を示す;
【図25】図25は、EIA型検出アッセイによって記録されたヒト・グレリン(オクタノイル化)濃度に対する吸光度の標準曲線を示し、ここで、グレリン結合SpiegelmerNOX−B11はヒト・グレリン(オクタノイル化)を固定化するために使用されて、ヒト・グレリン(オクタノイル化)の定量を可能にした;
【図26A−D】図26A〜Dは、本発明によるグレリン結合核酸を用いるオクタノイルグレリンの定量方法の種々の段階を示す。
【技術分野】
【0001】
本発明は、グレリン(ghrelin)に結合する核酸、および薬物の製造におけるそれらの使用、および診断作用物の製造におけるそれらの使用に関する。
【背景技術】
【0002】
グレリンは、成長ホルモン分泌促進物質受容体1a(GHSR1a)の天然のリガンドとして同定された。この受容体は、下垂体および脳の視床下部部分にもっとも豊富に存在するが、低濃度では他の組織においてもまた検出することができる。70年代末期以来、分泌促進物質と命名された合成ペプチドおよび他の化合物は、成長ホルモンの放出を刺激することが示された。しかしながら、成長ホルモンの放出に関与する天然のリガンドは、1999年におけるグレリンの発見まで知られていなかった。グレリンは、そのN末端の3番目のアミノ酸(セリン3)においてオクタン酸(octanoyl acid)側鎖をもつ高度に塩基性の28アミノ酸ペプチドホルモンである。この異例の修飾は、GHS受容体における相互作用およびその活性のために必要とされる。しかしながら、生物サンプルにおいては、生物活性グレリンの形態物であるオクタノイルグレリン、および未修飾またはデス−オクタノイルグレリンの両方が存在している。精製されたラット・グレリンのアミノ酸配列は、GSSFLSPEHQKAQQRKESKKPPAKLQPR(配列番号:2)であると決定され;対応するヒトの配列は、2つの位置においてのみ逸脱するが、アミノ酸位置セリン3において同じn−オクタノイル側鎖を保持し、そしてGSSFLSPEHQRVQQRKESKKPPAKLQPR(配列番号:1)であると決定された。
【0003】
天然に存在するn−オクタノイル残基の他に、グレリンの位置3において導入された不飽和または分枝オクタノイル基およびより長い脂肪族鎖が同様に受容体認識を媒介する。受容体相互作用ドメインはグレリンのまさにN末端に置かれ;欠失研究では、アミノ酸1〜5の最小モチーフ(グレリン(1〜5)[GSSFL])がGHSR1aの刺激のためには十分であるが、n−オクタノイル残基によるペプチド修飾に対する強力な必要性が観察されることが示された。
【0004】
グレリンは嫌気状態に対して適切な生理学的機能を媒介することが示されている。それは下垂体からの成長ホルモン(GH)の放出を直接刺激し、したがって、先端巨大症の処置における適当な標的になり得る。また齧歯類における実験は、グレリンが視床下部ニューロンへの作用によってGHに依存しない様式で栄養補給(feeding)を誘導することが示された。興味あることに、グレリン産生の一次部位は胃における酸分泌腺であり、それが、胃、下垂体および視床下部間のホルモン連鎖として働くことが示唆される。ラットにおけるグレリン投与がエネルギー獲得(intake)および/または燃料利用における変化の結果として体重獲得をもたらすという観察は、そのような役割を支持することになる。さらに、ヒトでの全身的グレリン投与は、試験の被験者における空腹感を惹起し、過食を誘導する。これらの知見に基づいて、グレリンは、食欲および体重の調節において決定的な役割を有して、食物が十分与えられない状態の急性ならびに慢性的シグナルとして働くと考えられる。この仮説のさらなる支持は、グレリンのレベルならびに食欲が、胃バイパス後の個人において低下され、体重減少を実施する手法の効力に少なくとも一部は貢献するという観察からも得られる。また、Prader−Willi症候群を有する患者からの臨床データは、この疾病に関係する過食および肥満が、大変な高グレリン血症(hyperghrelinemia)の結果であることを示唆する。さらに、グレリンは高血糖およびインスリン放出の抑制を誘導することが見出されて、グルコース代謝における関与が示された。エネルギー代謝におけるこれらの機能の他に、グレリンはまた、胃腸疾患の分野における多くの他のプロセス、例えば胃を空にしたり、腸の運動を調節することに関与している。さらに、グレリンはまた、多数の神経内分泌腫瘍において発現され、そして下垂体からのGH放出の他に、ACTH、PRLおよびコルチソルの放出を刺激することが見出された。健康な個体へのグレリンの単一注射は、心拍出量を増加し、血圧を低下させることが見出された。かくして、グレリンの作用は種々の異なる仕事に関与していると考えられる。これらに関するさらなる背景情報は、非特許文献1;非特許文献2;非特許文献3;非特許文献4;非特許文献5;非特許文献6;非特許文献7;非特許文献8;非特許文献9;非特許文献10において見出すことができる。
【非特許文献1】M.Kojima,H.Hosoda,Y.Date,M.Nakazato,H.Matsu,K.Kangawa,“Ghrelin is a growth−hormone−releasing acylated peptide from stomach”,Nature 402:656−60,1999
【非特許文献2】M.Tsch p,D.L.Smiley,M.L.Heiman,“Ghrelin induces adiposity in rodents”,Nature 407:908−13、2000
【非特許文献3】A.M.Wren et al.,”Ghrelin enhances appetite and increases food intake in humans”,Jounal of Clinical Endocrinology Metabolism 86:5992−6,2001
【非特許文献4】M.Nakazato et al.,”A role for ghrelin in the central regulation of feeding”,Nature 409:194−8,2001
【非特許文献5】N.Nagaya et al.,Am J Physiol Regul Integr Comp Physiol.2001 May;280(5):R1483−7;Hemodynamic and hormonal effects of human ghrelin in healthy volunteers
【非特許文献6】Volante M,et al.,J Clin Endocrinol Metab.2002 Mar;87(3):1300−8.Expression of ghrelin and of the GH secretagogue receptor by pancreatic islet cells and related endocrine tumors
【非特許文献7】Jeffery PL,et al.,J Endocrinol.2002 Mar;172(3):R7−11 Expression and action of the growth hormone releasing peptide ghrelin and its receptor in prostate cancer cell lines
【非特許文献8】Egido EM,et al.,Eur J Endocrinol.2002 Feb;146(2):241−4 Inhibitory effect of ghrelin on insulin and pancreatic somatostatin secretion
【非特許文献9】Broglio F,et al.,J Clin Endocrinol Metab.2001 Oct;86(10)5083−6,Ghrelin, a natural GH secretagogue produced by the stomach,induces hyperglycemia and reduces insulin secretion in humans
【非特許文献10】Bednarek MA,et al.,J Med Chem.2000 Oct;43:4370−6 Structure−function studies on the new growth hormone−releasing peptide,ghrelin:minimal sequence of ghrelin necessary for activation of growth hormone secretagogue receptor 1a.
【発明の開示】
【0005】
本発明の基礎をなす課題は、グレリンに対する特異的アンタゴニストを提供することである。本発明の基礎をなす課題のさらなる態様は、成長ホルモン分泌促進物質受容体1a(GHSR1a)に対する特異的アンタゴニストを提供することである。本発明の基礎をなす課題のその他の態様は、グレリンおよびGHSR1a受容体それぞれが関与する疾病および障害の処置のための化合物を提供することである。
【0006】
本発明の基礎をなすさらなる課題は、生物活性グレリンの結合のための手段を提供すること、より具体的には生物活性グレリンによって媒介される疾病および障害の処置のための方法ならびに生物活性グレリンの特異的検出のための方法を提供することである。
【0007】
本発明の基礎をなす課題は、好ましくはグレリンに結合する、核酸による第一の態様によって解決され、
こうして、核酸は、
第一のストレッチBoxAおよび
第二のストレッチBoxB
を含み、
ここで、
第一のストレッチBoxAは約25個の連続ヌクレオチドを含み、
第二のストレッチBoxBは約6〜8個の連続ヌクレオチドを含み、
ここで、
第一のストレッチBoxAのヌクレオチドの3’末端ストレッチは、第二のストレッチBoxBとハイブリダイズし、このハイブリダイゼーションでは、第一の二本鎖構造が形成され、そしてそのような第一の二本鎖構造はバルジを含む。
【0008】
本発明の各々およびいずれかの態様の実施態様では、グレリンは、生物活性グレリン、より好ましくはオクタノイルグレリン、そしてもっとも好ましくはn−オクタノイルグレリンである。
【0009】
1つの実施態様では、二本鎖構造は、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドおよび第二のストレッチBoxBのヌクレオチドの一部または全部、好ましくは第二のストレッチBoxBの6〜8個の連続ヌクレオチドによって形成される。
【0010】
1つの実施態様では、バルジは、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドと塩基対合しない、第二のストレッチBoxBの1〜3個のヌクレオチド、好ましくは第二のストレッチBoxBの1個のヌクレオチドによって形成される。
【0011】
1つの実施態様では、バルジは非塩基対合プリンによって形成され、ここで、プリンは好ましくはグアノシンである。
【0012】
好適な実施態様では、非塩基対合プリンは、第二のストレッチBoxBによって提供される。
【0013】
1つの実施態様では、核酸は、第三のストレッチBoxC1および第四のストレッチBoxC2をさらに含み、
ここで、第三のストレッチBoxC1は少なくとも1個のヌクレオチドを含み、そして
第四のストレッチBoxC2は少なくとも1個のヌクレオチドを含み、そして
ここで、第三のストレッチBoxC1は、その3’末端によって第二のストレッチBox
Bの5’末端に連結されており、そして第四のストレッチBoxC2は、その5’末端によって第一のストレッチBoxAの3’末端に連結されている。
【0014】
好適な実施態様では、第三のストレッチBoxC1および第四のストレッチBoxC2は、ハイブリダイゼーション可能であり、ここで、ハイブリダイゼーションでは、第二の二本鎖構造が形成される。
【0015】
1つの実施態様では、第一の二本鎖構造は第一のらせん構造を形成する。
【0016】
好適な実施態様では、第二の二本鎖構造は第二のらせん構造を形成する。
【0017】
より好適な実施態様では、第二のらせん構造は、1〜10個の塩基対、好ましくは1〜3個の塩基対、より好ましくは2〜3個の塩基対を含んでなる、ヘリックスまたはヘリックス様構造である。
【0018】
好適な実施態様では、第一のらせん構造は第二のらせん構造によって延長される。
【0019】
好適な実施態様では、第三のストレッチBoxC1は約1〜10個の連続ヌクレオチド、好ましくは1〜3個の連続ヌクレオチド、より好ましくは2または3個の連続ヌクレオチドを含む。
【0020】
好適な実施態様では、第四のストレッチBoxC2は約1〜10個の連続ヌクレオチド、好ましくは1〜3個の連続ヌクレオチド、より好ましくは2または3個の連続ヌクレオチドを含む。
【0021】
好適な実施態様では、核酸は、第五のストレッチBoxDをさらに含み、ここで、第五のストレッチBoxDは少なくとも2個の連続ヌクレオチドを含む。
【0022】
より好適な実施態様では、第五のストレッチBoxDは配列5’−CAを含む。
【0023】
より好適な実施態様では、第五のストレッチBoxDは、いずれかの長さの連続ヌクレオチドを含み、ここで、長さは2、3、4、5および6個の連続ヌクレオチドからなる群から選ばれる。
【0024】
好適な実施態様では、第五のストレッチBoxDは、
5’CA(X)n3’
[式中、Xは、好ましくはA,G,T,C,UおよびIを含んでなる群から選ばれるいずれかのヌクレオチドであり、そしてnは、0,1,2,3および4からなる群から選ばれるいずれかの整数である]
の配列を含む。
【0025】
より好適な実施態様では、第五のストレッチBoxDは、配列
5’CA(X)n3’
[式中、n=4]
からなる。
【0026】
好適な実施態様では、第五のストレッチBoxDは、その5’末端によって第二のストレッチBoxBの3’末端に連結されている。
【0027】
好適な実施態様では、核酸は、第六のストレッチBoxEをさらに含み、ここで、第六
のストレッチBoxEは少なくとも1個のヌクレオチドを含む。
【0028】
より好適な実施態様では、第六のストレッチBoxEは、約1〜10個の連続ヌクレオチド、好ましくは1〜4個の連続ヌクレオチド、より好ましくは3個の連続ヌクレオチドを含む。
【0029】
さらにより好適な実施態様では、第六のストレッチBoxEの少なくとも1個のヌクレオチドはUおよびGからなる群から選ばれる。
【0030】
特に好適な実施態様では、UまたはGヌクレオチドは、第一のストレッチBoxAの5’末端の直ぐ隣に配置されている。
【0031】
好適な実施態様では、第六のストレッチBoxEは、その3’末端によって第一のストレッチBoxAの5’末端に連結されている。
【0032】
好適な実施態様では、第五のストレッチBoxDの3’末端は、第一のスペーサーによって第六のストレッチBoxEの5’末端に連結されている。
【0033】
好適な実施態様では、第四のストレッチBoxC2の3’末端は、第二のスペーサーによって第三のストレッチBoxC1の5’末端に連結されている。
【0034】
好適な実施態様では、第一および第二のスペーサーは、各々別々に、独立して、親水性スペーサーを含んでなる群から選ばれる。
【0035】
より好適な実施態様では、親水性スペーサーは、核酸スペーサーおよび非核酸スペーサーを含んでなる群から選ばれる。
【0036】
より好適な実施態様では、第一のスペーサーは、約1〜20個の連続ヌクレオチド、好ましくは1〜5個の連続ヌクレオチド、より好ましくは2個の連続ヌクレオチドを含んでなる核酸スペーサーである。
【0037】
その他のより好適な実施態様では、第二のスペーサーは、約3〜20個の連続ヌクレオチド、好ましくは3〜5個の連続ヌクレオチド、より好ましくは3個の連続ヌクレオチドを含んでなる核酸スペーサーである。なおより好適な実施態様では、第二のスペーサーはACAまたはCAAからなる。
【0038】
さらなるその他のより好適な実施態様では、スペーサーは非核酸である。その特に好適な実施態様では、第一のスペーサーおよび/または第二のスペーサーは、少なくとも1個のエチレングリコール部分または複数個のそのようなエチレングリコール部分を含む。
【0039】
1つの実施態様では、スペーサーは約172〜688Da、好ましくは344Daの分子量を有する。
【0040】
1つの実施態様では、核酸は環状核酸である。
【0041】
1つの実施態様では、核酸は、
【0042】
【化1】
【0043】
の構造または
【0044】
【化2】
【0045】
の構造を有する。
【0046】
1つの実施態様では、第一のストレッチBoxAは、
【0047】
【化3】
【0048】
[式中、
X1=GまたはA;
X2=AまたはU;
X3=GまたはA;
X4=AまたはCまたはU;および
X5=GまたはA]
、好ましくは
【0049】
【化4】
【0050】
の配列を含んでなる。
【0051】
1つの実施態様では、第二のストレッチBoxBは5’GUGAGG3’の配列を含む。
【0052】
1つの実施態様では、核酸の配列は、次による配列を含んでなる群から選ばれる:
【0053】
【表1】
【0054】
【表2】
【0055】
好適な実施態様では、核酸の配列は、配列番号:29、配列番号:30、配列番号:33、配列番号:36、配列番号:38、配列番号:39、配列番号:40、配列番号:41、配列番号:42、配列番号:46、配列番号:47、配列番号:51および配列番号:52による配列を含んでなる群から選ばれる。
【0056】
1つの実施態様では、核酸はグレリン、好ましくはヒト・グレリンを結合することができる。
【0057】
好適な実施態様では、グレリンは配列番号:1によるアミノ酸配列を有する。
【0058】
1つの実施態様では、核酸は修飾(modification)を含む。
【0059】
好適な実施態様では、修飾は、HES部分およびPEG部分を含んでなる群から選ばれ
る。
【0060】
より好適な実施態様では、修飾体は直鎖または分枝状PEGからなるPEG部分であり、ここで、PEG部分の分子量は、好ましくは約20〜120kD、より好ましくは約30〜80kD、もっとも好ましくは約40kDである。
【0061】
その他のより好適な実施態様では、修飾体はHES部分であり、ここで、HES部分の分子量は、好ましくは約10〜130kD、より好ましくは約30〜80kD、もっとも好ましくは約50kDである。
【0062】
1つの実施態様では、核酸のヌクレオチドはL−ヌクレオチドである。
【0063】
さらなる実施態様では、核酸は完全にL−ヌクレオチドからなる。
【0064】
本発明の基礎をなす課題は、第一の態様による核酸および場合によってはさらなる成分を含んでなる製薬学的組成物によって第二の態様において解決され、この場合、さらなる成分は製薬学的に許容できる添加物および製薬学的活性作用物を含んでなる群から選ばれる。
【0065】
本発明の基礎をなす課題は、薬物の製造のための第一の態様による核酸の使用によって第三の態様において解決される。
【0066】
本発明の基礎をなす課題は、診断手段の製造のための第一の態様による核酸の使用によって第四の態様において解決される。
【0067】
第三の態様の実施態様では、薬物は、肥満、摂食障害、糖尿病、グルコース代謝障害、腫瘍、血圧障害、心臓血管病、先端巨大症、およびエネルギーバランス、食欲、体重の調節、および胃腸疾患を含んでなる群から選ばれる疾病または障害の処置および/または予防のために存在する。
【0068】
本発明の基礎をなす課題は、グレリンおよび第一の態様による核酸を含んでなる複合体によって第五の態様において解決され、ここで、好ましくは複合体は結晶性複合体である。
【0069】
本発明の基礎をなす課題は、グレリンの検出のための第一の態様による核酸の使用によって第六の態様において解決される。
【0070】
本発明の基礎をなす課題は、次の段階:
− 候補グレリンアンタゴニストおよび/または候補グレリンアゴニストを提供し、
− 第一の態様による核酸を提供し、
− グレリンアンタゴニストおよび/またはグレリンアゴニストの存在においてシグナルを提供する試験系を提供し、そして
− 候補グレリンアンタゴニストがグレリンアンタゴニストであるか、そして/または候補グレリンアゴニストがグレリンアゴニストであるか否かを決定すること、
を含んでなる、グレリンアンタゴニストまたはグレリンアゴニストのスクリーニングのための方法によって第7の態様において解決される。
【0071】
本発明の基礎をなす課題は、次の段階:
− 相、好ましくは固定相に固定化されたグレリンを提供し、
− 第一の態様による核酸、好ましくは標識されている第一の態様による核酸を提供し、
− 候補グレリンアゴニストおよび/または候補グレリンアンタゴニストを添加し、そして
− 候補グレリンアゴニストがグレリンアゴニストであるか、そして/または候補グレリンアンタゴニストがグレリンアンタゴニストであるか否かを決定すること、
を含んでなる、グレリンアゴニストおよび/またはグレリンアンタゴニストのスクリーニングのための方法によって第8の態様において解決される。
【0072】
1つの実施態様では、決定は、核酸が候補グレリンアゴニストによるか、または候補グレリンアンタゴニストによって置換されるか否かが調査されるように実施される。
【0073】
本発明の基礎をなす問題は、第一の態様による核酸を含んでなる、グレリンの検出のためのキットによって第9の態様において解決される。
【0074】
本発明の基礎をなす問題は、第8の態様による方法によって得ることができるグレリンアンタゴニストによって第十の態様において解決される。
【0075】
本発明の基礎をなす問題は、第8の態様による方法によって得ることができるグレリンアゴニストによって第十一の態様において解決される。
【0076】
本発明は、グレリンに特異的に結合し、かつグレリンに対して高い親和力を有する核酸を生成することができるという驚くべき発見に基づいている。より具体的には、本発明者らは、驚くべきことに、生物活性のあるグレリン、より好ましくはオクタノイルグレリン、もっとも好ましくはn−オクタノイルグレリンに特異的に結合する核酸を生成することができた。
【0077】
グレリンは配列番号:1によるアミノ酸配列を有する塩基性ペプチドであり、そして好ましくはオクタノイル側鎖、より好ましくはn−オクタノイル側鎖である脂肪酸側鎖により好ましくは修飾されている。グレリンの算出されたpIは、ヒト・グレリンについては11,07,およびラット・グレリンについては10,56である。好適な実施態様では、本発明による核酸が結合するグレリンは、脂肪酸側鎖により修飾されているグレリンである。その他の実施態様では、グレリンは、脂肪酸側鎖を有しないグレリンである。本明細書において使用されるような用語グレリンは、限定されるものではないが哺乳動物のグレリンを含むすべてのグレリンを指す。好ましくは、哺乳動物グレリンは、マウス、ラット、ウサギ、ハムスターおよびヒトのグレリンを含んでなる群から選ばれる。もっとも好ましくは、グレリンはヒト・グレリンである。
【0078】
グレリンに核酸を結合する高い親和力が同定できたという発見は、塩基性タンパク質に対向されるアプタマー(aptamer)、すなわち標的分子に結合するD−核酸の生成は、この種の標的が高いが非特異的なシグナル対ノイズ比を生成するので、一般に非常に困難であることをEatonら(Eaton,B.E.;Gold,L.;Hicke,B.J.;Janjic,N.;Jucker,F.M.;Sebosta,D.P.;Tarasow,T.M.;Willis,M.C.;Zichi,D.A.;Bioorganic & Medicinal Chemistry,Vol 5,No.6;pp1087−1096,1997)が観察した限りにおいて驚くべきことである。この高いシグナル対ノイズ比は、グレリンのような塩基性標的に対して核酸によって示される高い非特異的な親和力からもたらされる。
【0079】
非常に塩基性の全体的なグレリンのpIおよび5.5という算出されたpIをもつむしろ酸性のドメインであるグレリンの受容体結合モチーフGSSFL[グレリン(1−5)]にも拘わらず、本発明者らが、酸性の受容体結合ドメインを認識するが、ペプチドの塩基性の中央およびカルボキシ末端ドメインを認識しない全長グレリンを用いて核酸を同定できたことは、なお一層驚くべき発見である。このことは、標的分子、すなわちグレリンの電荷および核酸の電荷の両方の静電効果に関して驚くべきことである。標的分子の塩基性ドメインに対する負電荷の核酸の結合は、標的分子の酸性ドメインに対する核酸の結合に較べてはるかに得策であるに違いない。かくして、当業者が、グレリンの塩基性部分に対して結合性ではないが、標的分子の酸性ドメインに結合性である核酸リガンドを成功裏に選択する合理的な期待を有しなかったことを指摘しなければならない。
【0080】
アミノ末端の受容体結合モチーフGSSFLを有する他に、本明細書で生物活性グレリンとも呼ばれる生物学的に活性なグレリンは、好ましくは、アミノ酸セリン3においてn−オクタノイル基によるそのアシル化を特徴とする。好ましくは本明細書で開示されるアミノ末端モチーフGSSFLのリガンドである本発明による核酸分子は、好ましくは、グレリンの生物学的に不活性または生物活性のない形態と生物学的に活性な形態との区別を可能にする。結合は2つの部分、オクタノイル基とペプチドの存在に厳密に依存するので、このことは驚くべきことである:オクタノイル−グレリンへの核酸の結合は、デスオクタノイル−グレリンの1000倍過剰量の存在下で、より好ましくはデスオクタノイル−グレリンの100倍過剰量の存在下で、もっとも好ましくはデスオクタノイル−グレリンの10倍過剰量の存在下で特異的である。
【0081】
本明細書における好適な実施態様で使用されるように、生物活性グレリンは、天然に存在するグレリンの特徴のすべてを本質的に好適な実施態様において表すグレリンである。具体的には、好適な実施態様において本明細書で使用される生物活性グレリンは、より好ましくはGHS受容体との相互作用を介して、成長ホルモンの放出に関与するか、またはそれを惹起できるすべてのグレリンおよびグレリン誘導体である。好適な実施態様におけるこれとは反対に、非生物活性グレリンは、生物活性グレリンとは異なる、より好ましくは、より好ましくはGHS受容体との相互作用を介して、成長ホルモンの放出を惹起しないグレリンである。
【0082】
好適な実施態様では、本発明者らは、驚くべきことに、グレリン結合核酸を生成することができたが、ここで、グレリンは生物活性グレリンまたは生物学的に活性なグレリンであり、これらは、そのN末端の3番目アミノ酸(セリン3)においてオクタン酸側鎖を有するグレリンを識別し、一方、それらは、そのようなオクタン酸側鎖を欠如するグレリンには結合しない。
【0083】
本明細書で記述される本発明による核酸の特徴は、単独でも、またいかなる組み合わせ物においても、核酸が使用される本発明のいかなる態様においても実現することができる。
【0084】
いかなる理論によっても拘束されることを願わずに、本発明は、本発明によるグレリン結合核酸の観察された特異性が、引用が図20Aに対してなされる次において議論されるであろう若干の構造的特徴を共有すると推測する。しかしながら、図20Aが、本発明による核酸の各々およびいずれかにおいて必ずしも実現されるとは限らない該構造的特徴のいくつかを組み入れていることを理解すべきである。
【0085】
基本的な構造的特徴は、本明細書ではBoxAまたは第一のストレッチBoxAとして本明細書でまた言及される連続ヌクレオチドの第一のストレッチ、および本明細書ではBoxBまたは第二のストレッチBoxBとしてまた言及される連続ヌクレオチドの第二のストレッチである。第一のストレッチBoxAは約25個の連続ヌクレオチドを含み、これに対して第二のストレッチBoxBは約6〜8個の連続ヌクレオチドを含む。第一のストレッチBoxAの3’末端ストレッチは、第二のストレッチBoxBとハイブリダイズ
し、このハイブリダイゼーションでは、第一の二本鎖構造が形成され、そのような第一の二本鎖構造はバルジを含む。二本鎖構造は、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドおよび第二のストレッチBoxBの若干の6〜8個の連続ヌクレオチドによって形成される。バルジは、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドと塩基対合しない1個位の少ないおよび3個位の多いヌクレオチドによって形成される。したがって、バルジは、好ましくは第二のストレッチBoxBによって提供される、1,2または3個の非塩基対合ヌクレオチドからなってもよい。このバルジのサイズに鑑みて、第二のストレッチBoxBは6〜8個の連続ヌクレオチドを含んでもよい。より好ましくは、バルジは、好ましくは6個の連続ヌクレオチドを含んでなる第二のストレッチBoxBによって、第二のストレッチBoxBの5’末端から見られる第二のストレッチBoxB内の非塩基対合第三ヌクレオチドによって創成される。
【0086】
好適な実施態様では、本発明による核酸は単一の核酸分子である。さらなる実施態様では、単一の核酸分子は、多数の単一の核酸分子として存在する。好ましくは、用語核酸および核酸分子は、それとは違って示されない限り本明細書では互換可能な様式で使用される。
【0087】
本発明による核酸分子が、好ましくはホスホジエステル連結または結合をとおして、好ましくは、互いに共有結合されるヌクレオチドからなることは、当業者によって理解することができる。
【0088】
さらなる重要な特色は、第二の二本鎖構造である。そのような第二の二本鎖構造は、BoxC1または第三のストレッチBoxC1としてまた言及される連続ヌクレオチドの第三のストレッチ、およびBoxC2または第四のストレッチBoxC2としてまた言及される連続ヌクレオチドの第四のストレッチによって形成される。第三のストレッチBoxC1は、第二のストレッチBoxBの5’末端にその3’末端により連結され、そして第四のストレッチBoxC2は、第一のストレッチBoxAの3’末端にその5’末端により連結される。この第二の二本鎖構造は、典型的には、第二のらせん構造として本明細書でまた言及されるらせん構造を形成する。そのような第二のらせん構造は、好ましくは、第一の二本鎖構造によって典型的には形成されるらせん構造の伸長物である。好ましくは、第一のらせん構造として本明細書でまた言及されるらせん構造である該第一の二本鎖構造の長さは、第一のストレッチBoxAおよび第二のストレッチBoxBの長さによって、より正確には互いにハイブリダイズする該2種のBoxのストレッチによって定義される。第二の二本鎖構造によって該第一のらせん構造に提供される伸長物は、本発明者らの現在の理解によれば、本発明者らはこの理論に束縛されることを願わないけれども、より安定化する種類のものである。第二のらせん構造は、1〜10個の塩基対、好ましくは1〜3個の塩基対、より好ましくは2または3個の塩基対からなる。1〜3個位の少ない塩基対が、らせんを生成するのに必ずしも適当ではないことは当業者によって理解できる。この種の構造は、かくして、らせん様構造として本明細書でまた言及され、好ましくは、そのようならせん様構造は、1または数個の塩基対によって伸長される場合は、らせん構造をもたらすであろう構造物である。
【0089】
さらなる重要な特色は、BoxDまたは第五のストレッチBoxDとして本明細書でまた言及される連続ヌクレオチドの第五のストレッチである。このさらなるストレッチは、核酸の全体的結合において改良を与える。第五のストレッチは少なくともただ2個のヌクレオチドを含むけれども、グレリンの結合におけるその有益な劇的効果は、第五ストレッチの長さを好ましくは6個の連続ヌクレオチドまで増加することによってさらに改良することができる。また、第五のストレッチは、もし含有される場合は、好ましくはその5’末端CAジヌクレオチドおよび
5’CA(X)n3’
[式中、Xは、好ましくはA,G,T,C,UおよびIを含んでなる群から選ばれるいずれかのヌクレオチドである]
の配列を有する第五ストレッチにおいて特に効果的であることがわかった。
【0090】
本発明による核酸のさらなる特色は、BoxEまたは第六のストレッチBoxEとして本明細書でまた言及される隣接ヌクレオチドの第六のストレッチである。好ましくは、第六のストレッチは少なくとも1個のヌクレオチドからなる。
【0091】
種々のストレッチは図20Aから見られるように互いに連結される。そこに描かれるように、好ましくは、第三のストレッチBoxC1および第四のストレッチBoxC2の各々は、1または2個のヌクレオチド、より好ましくは2個のヌクレオチドを含み、第五のストレッチBoxDは少なくとも2個のヌクレオチド、好ましくは4個のヌクレオチド、もっとも好ましくは6個のヌクレオチドを含む。
【0092】
第三のストレッチBoxC1および第四のストレッチBoxC2の長さが0である実施態様では、第一のストレッチBoxAおよび第二のストレッチBoxBは場合によっては、リンカーまたはスペーサーを介して連結されてもよく、ここで、そのようなスペーサーは本明細書で開示されるスペーサーのいずれであってもよい。本明細書で使用されるように用語「スペーサー」および「リンカー」は、他に指示されない場合は互換可能な様式で使用される。
【0093】
本発明に関連して、第一のストレッチおよび第二のストレッチは、好ましくは共有結合を介して互いに連結される。好適な実施態様では、共有結合はホスホジエステル結合である。なおさらなる好適な実施態様では、第一のストレッチの3’末端は第二のストレッチの5’末端に連結される。
【0094】
その他の実施態様では、第六のストレッチBoxEは1〜10個の連続ヌクレオチド、好ましくは1〜4個の連続ヌクレオチド、もっとも好ましくは3個の連続ヌクレオチドを含む。
【0095】
これらの種々の配置から分るように、第五のストレッチと第六のストレッチは互いに連結されるか、または連結されず、第三と第四ストレッチは互いに連結されるか、または連結されず、そして第一のストレッチと第二のストレッチは互いに連結されるか、または連結されないことが可能である。連結は好ましくは共有結合を介してなされることは認識できる。より好ましくは連結は、少なくとも1個の、好ましくは多数のエチレングリコール部分を含んでなる親水性スペーサーを介してなされる。種々のリンカーおよびスペーサーは、それぞれ当業者には既知であり、そして次の基準、例えば、PilsおよびMicura(Nucleic Acid Research(2000),28(9),1859−1863)によって記述されたような基準を用いて選択することができる。リンカーは塩基対自体を妨害してはならない。芳香族炭素環を含有するリンカーの種類は末端塩基対を攻撃するので、適当ではない(J.Am.Chem.Soc.(1999),121,9905−9906;J.Am.Chem.Soc.(1998),120,11004−11005)。しかしながら、エチレングリコールに基づくかエチレングリコールから誘導されるリンカーは、それらが良好な水溶解度と高いコンホメーションの柔軟性という利点を有するので、これらの要件に適合する(J.Am.Chem.Soc.(1993),115,8483−8484;Nucleic Acids Research(1993)、21、5600−5603;Biochemistry(1993),32,1751−1758;Nucleic Acid Research(1990),18,6353−6359;J.Am.Chem.Soc.(1997),119,11591−11597)。好ましくは、スペーサーは、1または数個のエチレングリコール部分を含むか、またはそれからなり、ここで、酸素がCH3、リン酸または硫黄によって置換されている。
【0096】
これらの連結の選択に基づいて、次の構造物が実現できる。
【0097】
【化5】
【0098】
最後に、本発明による核酸のための完全に閉ざされた、すなわち環状の構造物が図20Dにおいて描かれるように実現されることも、また本発明内にはいる。
【0099】
本発明による核酸は、また、本明細書に開示される特定の配列に本質的に相同である核酸を含まねばならない。用語「本質的に相同」は、相同性が少なくとも75%、好ましくは85%、より好ましくは90%、もっとも好ましくは95%、96%、97%、98%または99%以上であるように理解されるべきである。
【0100】
用語「本発明の核酸」または「本発明による核酸」は、好ましくは核酸または該一部がグレリンへの結合に関与するほどに、より好ましくは生物活性グレリンを非生物活性グレリンと区別する、すなわち、特にオクタノイル−グレリンをデスオクタノイル−グレリンと区別するほどに、本明細書に開示される核酸またはその一部を含むそれらの核酸を含むべきである。そのような核酸は、例えば、切り取り(truncation)によって本明細書に開示される核酸から誘導されてもよい。切り取りは、本明細書に開示される核酸の末端のいずれか、または両方に関してもよい。また、切り取りは、ヌクレオチドの内部配列に関してもよい、すなわち、それぞれ5’および3’末端ヌクレオチドの間のヌクレオチドに関してもよい。さらに、切り取りは、本明細書に開示される核酸の配列から単一ヌクレオチド位の短い欠失を含んでもよい。また、切り取りは、本発明の核酸の1個以上のストレッチに関してもよく、ここで、ストレッチは1個位の短いヌクレオチド長であってもよい。
【0101】
本発明による核酸は、D−核酸でも、またL−核酸であってもよい。好ましくは、本発明の核酸はL−核酸である。さらに、核酸の一部または数部分がD−核酸として存在するか、または核酸の少なくとも一部または数部分がL−核酸であることも可能である。用語核酸の「一部」は、1ヌクレオチド位の小さいものを意味すべきである。そのような核酸は、一般にD−およびL−核酸とそれぞれ本明細書では言及される。したがって、特に好適な実施態様では、本発明による核酸はL−ヌクレオチドからなり、そして少なくとも1個のD−ヌクレオチドを含む。そのようなD−ヌクレオチドは、好ましくは、本発明による核酸を定義するストレッチとは異なる部分、好ましくは核酸の他の部分との相互作用が必要であるそれらのその部分に連結される。好ましくは、そのようなD−ヌクレオチドは、いずれかのストレッチおよび本発明によるいずれかの核酸のそれぞれの末端に連結される。さらなる好適な実施態様では、そのようなD−ヌクレオチドは、スペーサーまたはリンカー、好ましくは本発明による核酸に対するPEGおよびHESのような連結修飾体として働いてもよい。
【0102】
本発明による核酸がより長い核酸の一部であることもまた本発明の内にはいり、ここで
、このより長い核酸は、少なくとも一部分は、本発明による核酸またはその一部である数部分を含む。これらのより長い核酸の他の部分は、D−核酸でもL−核酸であってもよい。いずれの組み合わせ物が本発明に関して使用されてもよい。これらのより長い核酸の他の部分は、結合、好ましくはグレリンへの結合とは異なる機能を示すことができる。1つの可能性のある機能は、他の分子と相互作用させることであり、そのような他の分子は、例えば、固定化、架橋、検出または増幅のためのように、好ましくはグレリンとは異なる。
【0103】
本明細書で使用されるようなL−核酸は、L−ヌクレオチドからなる、好ましくは完全にL−ヌクレオチドからなる核酸である。
【0104】
本明細書で使用されるようなD−核酸は、D−ヌクレオチドからなる、好ましくは完全にD−ヌクレオチドからなる核酸である。
【0105】
本発明の核酸が、D−ヌクレオチド、L−ヌクレオチドあるいは例えば少なくとも1個のL−ヌクレオチドおよび少なくとも1個のD−ヌクレオチドからなるストレッチのランダム組み合わせ物または一定の配列である組み合わせによる両方の組み合わせ物からなるか否かに関係なく、核酸はデオキシリボヌクレオチド、リボヌクレオチドまたはその組み合わせ物からなってもよい。
【0106】
本発明の核酸をL−核酸として設計することは、いくつかの理由で得策である。L−核酸は天然に存在する核酸の鏡像異性体である。しかしながら、D−核酸は、水溶液中、特に、ヌクレアーゼの広範な存在によって生物系または生物サンプルにおいて非常に不安定である。天然に存在するヌクレアーゼ、特に動物細胞からのヌクレアーゼはL−核酸を分解することはできない。このために、L−核酸の生物学的半減期は、動物およびヒトの身体を含む、そのような系では有意に増大する。L−核酸の分解性の欠如により、ヌクレアーゼ分解産物は生成されず、したがってそれから起きる副作用は観察されない。この態様は、グレリンの存在に係わる疾病および/または障害の治療において使用される実際にすべての他の化合物のL−核酸の限界を定める。ワトソン・クリック塩基対合とは異なるメカニズムにより標的分子に特異的に結合するL−核酸、あるいは特に、標的分子へのアプタマーの結合に関与するアプタマーのそれらの部分をもつ、L−ヌクレオチドから部分的または完全になるアプタマーは、またspiegelmerと呼ばれる。
【0107】
本発明の核酸は、それらがD−核酸、L−核酸またはD,L−核酸として存在するか否か、あるいはそれらがDNAまたはRNAであるか否かに関係なく、一本鎖または二本鎖核酸として存在してもよいことは、また本発明内にはいる。典型的には、本発明の核酸は、一次配列により一定の二次構造を示す一本鎖核酸であり、したがってまた三次構造を形成してもよい。しかしながら、本発明の核酸は、互いに相補的または部分的に相補的である2本の鎖が互いにハイブリダイズされるという意味において二本鎖であってもよい。このことは、特に、核酸がL−型よりもむしろ天然に存在するD−型で存在する場合には有利であろう核酸への安定性を与える。
【0108】
本発明の核酸は修飾されてもよい。そのような修飾は核酸の単一ヌクレオチドに関してもよく、そして当該技術分野において周知である。そのような修飾の例は、なかんずく、Venkatesan N.et al.(2003)Curr Med Chem.Oct;10(19):1973−91;Kusser,W.(2000) J Biotechnol,74:27−38;Aurup,H.et al.(1994)Nucleic Acids Res,22,20−4;Cummins,L.L.et al.(1995) Nucleic Acids Res,23,2019−24;Eaton,B.E.et al.(1995)Chem Biol,2,633−8;Gree
n,L.S.et al.(1995)Chem Biol,2,683−95;Kawasaki,A.M.et al.(1993)J Med Chem、36,831−41;Lesnik,E.A.et al.(1993)Biochemistry,32,7832−8;Miller,L.E.et al.(1993)J Physiol,469,213−43において記述されている。そのような修飾は、核酸がなりたっている個々のヌクレオチドの2’位におけるH原子、F原子またはO−CH3基またはNH2基であってもよい。また、本発明による核酸は少なくとも1個のLNAヌクレオチドを含んでもよい。1つの実施態様では、本発明による核酸はLNAヌクレオチドからなる。
【0109】
1つの実施態様では、本発明による核酸は多くの部分に分かれた核酸であってもよい。本明細書で使用される多くの部分に分かれた核酸は、少なくとも2本の核酸鎖からなる核酸である。これらの少なくとも2本の核酸鎖は機能的単位を形成し、ここでの機能的単位は標的分子に対するリガンドである。少なくとも2本の核酸鎖は、核酸を切断して2本の鎖を生成するか、あるいは本発明の、すなわち核酸全体の第一の部分に対応する1つの核酸および核酸全体の第二の部分に対応するその他の核酸を合成するか、いずれかによって本発明の核酸のいずれから誘導されてもよい。両、切断および合成が、先に例示された2本以上の鎖である多くの部分に分かれた核酸を生成するために応用できることは認識されるべきである。言い換えれば、少なくとも2本の核酸鎖は、典型的には、種々の核酸部分の間で、ある程度の相補性が存在するかも知れないが、互いに相補的であり、かつハイブリダイズする2本の鎖とは異なる。
【0110】
本発明は、本発明による核酸が非常に好ましいKd値の範囲を示すことを発見した。より具体的には、完全なBoxAおよびBoxBの他に−最小2個のヌクレオチドを各々有するBoxC1およびC2、最小6個のヌクレオチドを有するBoxDおよび最小3個のヌクレオチドを有するBoxE−を含んでなるオリゴヌクレオチド(例えば、SOT−E)は、SOT−Cよりも6倍良好なグレリンに対する結合親和力(図18)を示す。
【0111】
結合定数の決定を可能にするのは、所謂バイオコア(biacore)・デバイスの使用であり、これはまた当業者には既知である。本明細書で使用される親和力は、実施例において記述されるような「平衡アッセイ」の使用によってまた測定できた。本事例ではグレリンである標的による核酸間の結合の強度を表すための適当な測定値は、その決定方法が当業者において十分に知られている所謂Kd値である。
【0112】
本発明による核酸はあるKd値を特徴とする。好ましくは、本発明による核酸によって示されるKd値は1μM以下である。約1μMのKd値は、標的に対する核酸の非特異的結合について特徴的であると言われる。当業者によって認識できるように、本発明による核酸のような1群の化合物のKd値は一定の範囲内にある。約1μMという上記KdはKd値について好適な上限である。核酸を結合する標的のKdの好適な下限は約10ピコモルまたはそれ以上であってもよい。グレリンに結合する個々の核酸のKd値が、好ましくは、この範囲内であることは本発明の内にはいる。好適な範囲は、この範囲内のいずれか第一の数とこの範囲内のいずれか第二の数を選ぶことによって定められてもよい。好適な上限値は0.25μM、0.1μMおよび0.01μMであり、好適な下限値は100nM、10nM、1nMおよび0.05nMである。
【0113】
本発明による核酸分子は、なおそれらが標的分子に結合でき、かつ生物活性グレリンを非生物活性グレリンと区別できる、すなわち好ましくはオクタノイル−グレリンをデスオクタノイル−グレリンと区別できる場合には、いかなる長さを有してもよい。本発明による核酸の好適な長さが存在することは当該技術分野において認識できる。典型的には、この長さは15〜120ヌクレオチドである。15〜120間のいかなる整数も本発明によ
る核酸について可能な長さであることは当業者によって認識できる。本発明による核酸の長さのより好適な範囲は、約20〜100ヌクレオチド、約20〜80ヌクレオチド、約20〜60ヌクレオチド、約20〜50ヌクレオチドおよび約30〜50ヌクレオチドの長さである。
【0114】
本明細書で開示される核酸が、好ましくは高分子量部分であり、かつ/または、なかんずく、動物体内、好ましくはヒト体内における滞留時間という用語で好ましくは核酸の特徴を修飾させる部分を含むことは本発明内にはいる。そのような修飾の特に好適な実施態様は、本発明による核酸のPEG化およびHES化である。本明細書で使用されるようなポリ(エチレングリコール)についてのPEG鎖およびヒドロキシエチル澱粉についてのHES。好ましくは本明細書で使用されるPEG化は本発明による核酸の修飾であり、そのような修飾体は、本発明による核酸に連結されるPEG部分からなる。本明細書で使用されるHES化は本発明による核酸の修飾であり、そのような修飾体は、本発明による核酸に連結されるHES部分からなる。これらの修飾ならびにそのような修飾を用いる核酸の修飾方法は、欧州特許第一 306 382号に記述されており、これの開示は引用によって完全に本明細書に組み入れられている。
【0115】
好ましくは、高分子量部分からなる、またはそれを含んでなる修飾体の分子量は、特にそのような高分子量部分であるPEGの場合には、約2,000〜200,000Da、好ましくは40,000〜120,000Daであり、そして特にそのような高分子量部分であるHESの場合には、好ましくは約3,000〜100,000Da、より好ましくは5,000〜60,000Daである。HES修飾の方法は、例えば、ドイツ特許出願第一 2004 006 249.8号に記述されており、これの開示は引用によって完全に本明細書に組み入れられている。
【0116】
いかなる理論によっても束縛されないことを願えば、好ましくは生理学的に許容できるポリマー、より具体的には本明細書で開示されるポリマーのような高分子量部分を用いる本発明による核酸の修飾によって排出動態が変化されると考えられる。より具体的には、そのような修飾された本発明の核酸の分子量の増加により、そして特にL型における場合の被験者が代謝することのない核酸により、動物の身体、好ましくは哺乳動物の身体、より好ましくはヒトの身体からの排出が減少されると思われる。排出は、典型的には腎臓を介して起きるので、本発明は、このように修飾された核酸の糸球体濾過速度が、体内の滞留時間の増加をもたらすこの種の高分子量修飾を有しない核酸に較べて有意に減少されると推測する。これに関連して、そのような高分子量修飾にもかかわらず、本発明による核酸の特異性が有害な様式で影響されないことは特に注目される。その限りにおいて、本発明による核酸は、持続放出を与える製薬学的調合物が、持続放出を与えるために必ずしも必要とされないような−通常には製薬学的活性化合物から期待することができないー驚くべき特徴を有する。むしろ、高分子量部分を含んでなるそれらの修飾形態物における本発明による核酸は、既にそれ自体、持続放出調合物として使用することができる。
【0117】
しかしながら、本明細書に開示される核酸が、いかなる修飾も、特にPEG化またはHES化のような高分子量修飾も含まないこともまた本発明の内にはいる。そのような実施態様は、投与後、身体からの核酸の速いクリアランスが所望される場合に特に好適である。そのような速いクリアランスは、本発明による核酸か、または同物質を含んでなる薬物を使用してインビボの想像性または一時的な食欲抑制の場合に望ましいであろう。
【0118】
本発明による核酸としてまた本明細書で言及される本発明の核酸および/または本発明によるアンタゴニストは、薬物の創成または製造のために使用されてもよい。そのような薬物は、少なくとも1種の本発明の核酸を、場合によってはさらなる製薬学的活性化合物とともに含有し、この場合、本発明の核酸は好ましくは製薬学的活性化合物自体として働
く。そのような薬物は好適な実施態様では少なくとも1種の製薬学的に許容できる担体を含む。そのような担体は、例えば、水、バッファー、PBS、グルコース溶液、好ましくは5%グルコース塩均衡溶液、澱粉、糖、ゼラチンまたはいずれか他の許容できる担体物質であってもよい。そのような担体は一般に当業者には既知である。
【0119】
さらなる実施態様では、薬物はさらなる製薬学的活性作用物を含む。そのようなさらなる製薬学的活性化合物は、食欲を減退することが知られているそれらの化合物であってもよく、そして好ましくは、PYY3−45、CCK、レプチンおよびインスリンを含んでなる群から選ばれる。あるいは、またはさらに、そのようなさらなる製薬学的活性作用物はさらなる本発明による核酸である。あるいはまた、薬物は、グレリンとは異なる標的分子に結合するか、または本発明による核酸の1種とは異なる機能を発揮する少なくとも1種以上の核酸を含む。1つの実施態様では、そのような核酸はオクタン酸部分を欠如するグレリンに結合している。
【0120】
そのような薬物が使用されてもよい処置および/または予防のための疾病および/または障害および/または疾病症状は、限定されるものではないが、肥満、エネルギーバランス、食欲および体重の調節、摂食障害、胃腸疾患、糖尿病、グルコース代謝、腫瘍、血圧、心臓血管病、先端巨大症、および他のGHインバランスを含む。当業者によって認識できるように、本発明の核酸は、事実上、グレリンに対するアンタゴニストがそのようなアンタゴニストを必要とする患者に投与でき、そしてそのようなアンタゴニストが疾病または障害の原因を除去できるか、または少なくとも疾病または障害からの影響を低下するために適当である、いかなる疾病において使用されてもよい。そのような影響は、限定されるものではないが、肥満、エネルギーバランス、食欲および体重の調節、摂食障害、胃腸疾患、糖尿病、グルコース代謝、腫瘍、血圧、心臓血管病、先端巨大症および他のGHインバランスを含む。これらおよび他の疾病または障害に関連する本発明による核酸の適応性は、なかんずく、本明細書の序論部分において概説されたようなグレリンの関与からもたらされる。本発明の目的では、エネルギーバランスの調節は疾病とみなされる。より具体的には、この使用は、エネルギーバランスの調節が直接にも間接にもグレリンによって影響され、グレリンの生物学的利用性の低下が望まれるすべての疾病の処置のためにある。同じことが、糖代謝、血圧および食欲と体重に適合する。本発明による核酸を用いて処置されてもよいさらなる疾病は、可能ならば全身的または局所的適用において、下垂体腫瘍、先端巨大症、中枢性Cushing症候群、副腎性Cushing症候群、腫瘍随伴性Cushing症候群、異所性Cushing症候群、副腎腫瘍、ストレス、副腎皮質亢進症、心不全、心筋梗塞、卒中、副腎皮質不全症、低血圧、大動脈狭窄、肺動脈緊張過度、梗塞性心外膜炎、感染疾患、感染毒性低血圧、血液量減少および低ナトリウム血症を含んでなる群から選択することができる疾病である。
【0121】
本明細書で使用されるように、用語、本発明による核酸を用いて処置できる胃腸疾患は、胃の疾病および障害、腸の疾病および障害、結腸の疾病および障害、および胃および結腸運動の調節を含む。好適な実施態様では、用語、腸の障害および腸の疾病は、炎症性腸疾患を含む。より好適な炎症性腸疾患は、潰瘍性大腸炎およびクローン病である。この種の疾病の処置のための本発明による核酸の適合性は、Karmirsら(Karmirs K,Koutroubakis IE,Xidakis C,Polychronaki M,Voudouri T,Kouroumalis EA,Circulatinglevels of leptin,adiponectin,resistin,and ghrelin in inflammatory bowl disease.Inflamm Bowel Dis.2006 Feb;12(2):100−5)、Tebbe JJら(Tebbe JJ,Mronga S,Tebbe CG,Ortmann E,Arnold R,Schafer MK.Ghrelin−induced stimulation of colonic propulsion isdependent on hypothalamic neuropeptide Y1− and corticotrophin−releasing factor 1 receptor activation.J Neuroendocrinol.2005 Sep;17(9):570−6)およびFukuda H.ら(Fukuda H,Mizuta Y,Isomoto H,Takeshima F,Ohnita K, Ohba K,Omagari K,Taniyama K,Kohno S.Ghrelin enhances gastric motility through direct stimulation of intrinsic neural pathways and capsaicin−sensitive afferent neurones in rats.Scand J Gastroenterol.2004 Dec;39(12):1209−14)により記述されたようなグレリンの関与から生じる。
【0122】
Kobeltらの公表(Kobelt P,Helmling S,Stengel A,Wlotzka B,Andresen V,Klapp BF,Wiedenmann B,Klussmann S,Monnikes H.Anti−ghrelin
SPIEGELMER NOX−B11 inhibits neurostimulatory and orexigenic effects of peripheral ghrelin in rats.Gut.2005 Jun 30,Epub ahead of print)は、抗グレリンSpiegelmer NOX−B11の投与において末梢グレリンにより誘発された短期間の食物摂取の用量依存性減退を報告している。より具体的には、PBSおよび3nmolグレリンによるポジティブ対照群(媒質/グレリン群)処置では、媒質/媒質群(1.13±0.59g/kg−BW,p<0.0002)に較べて腹腔内注射後初めの半時間内に食物摂取が有意に増加した(4.94±0.63g/kg−BW)(Kobeltら、前出の図2A)。Spiegelmer NOX−B11(=SOT−C=B11trc)の30nmolによる前処置は食物摂取に及ぼすグレリンの促進効果を阻止した(0.58±0.58g/kg−BW,p<0.0001)(Kobeltら、前出の図2A)。これに対して、ランダム配列からなる対照Spiegelmerの投与は、そのような抑制効果を有せず、食物摂取におけるグレリン誘導の促進は元のままであった(4.77±0.66g/kg−BW;p>0.864)(Kobeltら、前出の図2A)。
【0123】
食物摂取に及ぼすNOX−B11(=SOT−C=B11trc)の抑制効果は、厳密に用量依存性であることが証明された。Spiegelmer NOX−B11(=SOT−C=B11trc)の1nmolの用量は、食物摂取のグレリン刺激には効果を有しなかった(Kobeltら、前出の図2A)。中間的効果はNOX−B11(=SOT−C=B11trc)の10nmolの用量について観察された:この用量レベルでは、最初の30分間における3nmolグレリンの刺激効果は緩和された(3.51±0.66g/kg−BW対4.94±0.63g/kg−BW;p>0.159対グレリン単独)(Kobeltら、前出の図2A)。
【0124】
Shearmanらの公表(Shearman LP,Wang SP,Helmling S,Stribling DS,Mazur P,Ge L,Wang L,Klussmann S,Macintyre DE,Howard AD,Strack AM.Ghrelin Neutralisation by a Ribonucleic Acid−SPM Ameliorates Obesity in Diet−Induced Obese Mic.;Endocrinology.2006 Mar;147(3):1517−26.Epub 2005 Dec 8)は、食餌誘導の肥満(DIO)マウスにおける抗グレリンSpiegelmer NOX−B11−2(SOT−D−109)の投与において、体重の低下および食物摂取の減少を報告している。より具体的には、NOX−B11−2(SOT−D−109)注入は、対照に較べて体重喪失を惹起した(Shearmanら、前出の図4A、参照)。有意な体重喪失は、媒質処置のマウスに較べて1〜10日目および12日目において;対照Spiegelmer(対照SPM)注入群に較べて1〜13日目において、NOX−B11−11(SOT−D−109)注入により観察された(P<0.05vs.媒質または対照SPM)。13日目では、NOX−B11−2注入マウスは、体重の平均0.32gを増加したが、対照Spiegelmerを注入されたマウスは、平均1.85gを増加し、そして媒質を注入されたマウスは平均0.91g増加した。また、NOX−B11−2(SOT−D−109)注入は有意に食物摂取を減退させた(Shearmanら、前出の図4B、参照)。累積の食物摂取に及ぼす有意な効果は、媒質群に較べて1〜8日目に、対照Spiegelmer群に較べて1〜13日目に観察された(13日目に39.33g対42.61g;p<0.05)(Shearmanら、前出の図4C)。さらに、食物エネルギーが体質量の増大のためにいかに効率的に使用されたかを指示する摂食効率(消化された1kcal当たりの重量増加)が1〜5日目および6〜13日目から計算された(Shearmanら、前出の図4D,参照)。摂食効率は1〜5日目のNOX−B11−2(SOT−D−109)注入によって低下され、そしてこの効果は6〜13日目からは観察されず、このことは重量増加における一過性低下が、単に食物摂取における減退によるものではなかったことを示唆する。さらに、NOX−B11−2(SOT−D−109)による処置はDIOマウスの体組成を変えた(Shearmanら、前出の図4E)。赤身量含量(lean mass content)では変化がなかったのに対して、NOX−B11−2(SOT−D−109)注入されたマウスの脂肪量含量は、総体重についての校正後でさえ減少されなかった(Shearmanら、前出の図4F)。白色脂肪組織貯蔵物重量はNOX−B11−2(SOT−D−109)注入によって変わらず、対照Spiegelmer注入は体組成または白色脂肪組織重量を変えなかった。
【0125】
DIOグレリン欠乏および野生型の両マウスを使用するNOX−B11−2(SOT−D−109)による常習的注入研究において、有意な体重喪失が、1〜6日目の野生型マウスにおけるNOX−B11−2(SOT−D−109)注入において観察された(Shearmanら、前出の図5A,参照)。これに対して、NOX−B11−2(SOT−D−109)は、Ghrl−l−マウスでは体重を変えなかった(Shearmanら、前出の図5B、参照)。さらに、NOX−B11−2(SOT−D−109)注入は、野生型マウスでは1日目に一日の食物摂取を減退した(Shearmanら、前出の図5C,参照)。NOX−B11−2(SOT−D−109)注入は、Ghrl−l−マウスの食物摂取は変えなかった(Shearmanら、前出の図5D、参照)。
【0126】
薬物が、原則として、疾病の処置のための薬物の使用に関連して開示された該疾病のすべての予防のために代替的または追加的に使用されることは、本発明の内にはいる。したがって、それぞれのマーカーは、心臓血管の危険因子、例えば、コレステロールおよび低い好気的活性ならびに体重管理を必要とする一般因子を含んでなる群から選ばれる。
【0127】
本発明による薬物は、原則として、当業者には既知のいかなる形態物において投与されてもよい。好適な投与経路は全身的投与、より好ましくは注射による投与である。あるいはまた、薬物は局所的に投与されてもよい。他の投与経路は、筋肉内、腹腔内、および皮下、好ましくはもっとも侵入性が少ないが有効性が確保される投与経路であるper orumまたは鼻内を含む。
【0128】
さらなる態様では、本発明は製薬学的組成物に関する。そのような製薬学的組成物は、少なくとも1種の本発明による核酸および好ましくは製薬学的に許容できる結合剤を含む。そのような結合剤は当該技術分野において使用され、そして/または既知であるいかなる結合剤であってもよい。より具体的には、そのような結合剤は、本明細書で開示される薬物の製造に関連して議論されるすべての結合剤である。さらなる実施態様では、製薬学的組成物はさらなる製薬学的活性作用物を含む。
【0129】
本明細書で記述されるような薬物が本明細書で開示される製薬学的組成物を構成することは本発明内にはいる。
【0130】
さらなる態様では、本発明は、方法が少なくとも1種の本発明による核酸の製薬学的に活性な量の投与を含む、そのような処置を必要としている被験者の処置方法に関する。1つの実施態様では、被験者は、疾病を蒙っているか、またはそのような疾病を発生する危険性にあり、ここで、疾病は本明細書で開示されるすべての疾病、特に薬物の製造のためのすべての本発明による核酸の使用に関連して開示されるすべてのそれらの疾病である。
【0131】
本発明による核酸ならびにアンタゴニストが、特に肥満におけるグレリンの関与に関して、薬物としてまたは薬物の製造のためだけでなく、また化粧目的のために使用できることを理解すべきである。同じ目的および/または同じ理由のために、本発明による核酸ならびにアンタゴニストは、食品添加物、体重制御の手段、食欲制御の手段としておよび/または診断剤として使用することができる。本発明による核酸ならびにアンタゴニストを含んでなる組成物はすべての上記目的のために使用することができる。
【0132】
好ましくは本明細書で使用されるように、診断剤または診断作用物または診断手段は、直接的にもまた間接的にも、グレリン、好ましくは本明細書で記述されるグレリン、より好ましくは本明細書に記述される種々の障害および疾病に関連して本明細書で記述されるグレリンを検出するために適当である。診断剤は、本明細書に記述される障害および疾病それぞれすべての検出および/または追跡のために適当である。そのような検出はグレリンに対する本発明による核酸の結合をとおして可能である。そのような結合は直接的にも間接的にも検出することができる。各方法および手段は当業者には既知である。なかんずく、本発明による核酸は、本発明による核酸、好ましくはグレリンに結合される核酸の検出を可能にする標識を含む。そのような標識は、好ましくは放射能、酵素および蛍光標識を含んでなる群から選ばれる。原則として、抗体のために開発されたすべての既知アッセイが、本発明による核酸のために採用されてもよく、それにより、標的結合抗体が標的結合性核酸に置換される。無標識の標的結合抗体を用いる抗体アッセイでは、検出は、好ましくは、放射能、酵素および蛍光標識により修飾され、そしてそのFcフラグメントにおいて標的結合抗体に結合する二次抗体によって実施される。核酸、好ましくは本発明による核酸の場合には、核酸がそのような標識により修飾されるが、好ましくはそのような標識は、ビオチン、Cy−3およびCy−5を含んでなる群から選ばれ、そしてそのような標識は、そのような標識に対向される抗体、例えば、抗ビオチン抗体、抗Cy−3抗体または抗Cy−5抗体によって検出されるか、または−標識がビオチンの場合にはー標識は、ビオチンに自然に結合するストレプトアビジンまたはアビジンによって検出される。そのような抗体、ストレプトアビジンまたはアビジンは、順に、好ましくは、それぞれの標識、例えば放射能、酵素または蛍光標識により修飾される(二次抗体のように)。
【0133】
さらなる実施態様では、本発明による核酸は、検出手段が分子ビーコン(beacon)である第二の検出手段によって検出または分析される。分子ビーコンの方法は当業者には既知である。簡単に言えば、分子ビーコンとも呼ばれる核酸プローブは、検出されるべき核酸サンプルに対する逆相補物であり、このために、検出される核酸サンプルの一部にハイブリダイズする。核酸サンプルへの結合により、分子ビーコンの蛍光団基が分離されて、蛍光シグナルの変化、好ましくは強度における変化をもたらす。この変化は存在する核酸サンプルの量と相関する。
【0134】
本発明による生物活性および非生物活性グレリンの識別のためのアッセイは当業者には
既知の標準技術を用いて実施されてもよい。そのようなアッセイがまた、次に概説されるようなグレリン、好ましくは生物活性グレリンの検出のために使用できることは理解されるにちがいない。
【0135】
本発明による核酸を用いるグレリンの検出が、特に、本明細書で定義される生物活性グレリンの検出を可能にすることは認識できる。さらに、生物活性グレリンは、なかんずく、他の操作も当業者には明であるが、次の操作によってデスオクタノイルグレリンを別々に検出でき、したがって区別することができる。
【0136】
生物活性グレリンの検出に関連して、好適な方法は、次の段階:
(a)生物活性グレリンの存在について試験されるべきサンプルを提供し、
(b)本発明による核酸を提供し、
(c)好ましくは反応容器中で、サンプルを核酸と反応させること、
を含み、この場合、段階(a)は段階(b)の前に実施されてもよく、または段階(b)は段階(a)の前に実施されてもよい。
【0137】
好適な実施態様では、サンプルと核酸との反応の検出にある、さらなる段階d)が提供される。好ましくは、段階b)の核酸は、表面に固定化される。表面は、反応チューブ、プレートのウェルのような反応容器の表面、またはそのような反応容器中に含有されるデバイス、例えばビーズの表面であってもよい。表面への核酸の固定化は、限定されるものではないが、非共有または共有結合を含む、当業者には既知のいかなる手段によってなされてもよい。好ましくは、連結は、表面と核酸との間の共有化学結合により確立される。しかしながら、核酸が間接的に表面に固定化され、そのような間接的固定化がさらなる成分または相互作用パートナーのペアの使用を伴うことも本発明の内にはいる。そのようなさらなる成分は、好ましくは、相互作用パートナーとも呼ばれる、固定化される核酸と特異的に相互作用し、その結果表面への核酸の付着を媒介する化合物である。相互作用パートナーは、好ましくは、核酸、ポリペプチド、タンパク質および抗体を含んでなる群から選ばれる。好ましくは、相互作用パートナーは抗体、より好ましくはモノクローナル抗体である。あるいはまた、相互作用パートナーは、核酸、好ましくは機能的核酸である。より好ましくは、そのような機能的核酸は、アプタマー、spiegelmer、および核酸に対して少なくとも部分的に相補的である核酸を含んでなる群から選ばれる。さらなるその他の実施態様では、表面への核酸の結合は多分離した(multi−partite)相互作用パートナーによって媒介される。そのような多分離相互作用パートナーは、好ましくは、相互作用パートナーのペアまたは第一のメンバーと第二のメンバーからなる相互作用パートナーであり、ここでは、第一のメンバーは核酸によって含まれるか、または核酸に連結され、そして第二のメンバーは表面に付着されるか、または表面によって含まれる。多分離相互作用パートナーは、好ましくは、ビオチンとアビジン、ビオチンとストレプトアビジン、およびビオチンとノイトラビジン(neutravidin)を含んでなる相互作用パートナーのペアの群から選ばれる。好ましくは、相互作用パートナーのペアの第一のメンバーはビオチンである。
【0138】
そのような方法の好適な結果は、生物活性グレリンおよび核酸の非固定化複合体の形成であり、この場合、より好ましくは、該複合体が検出される。この複合体から生物活性グレリンが検出されることは実施態様内にはいる。より好ましくは、生物活性グレリンは、生物活性グレリンに特異的である検出手段によって検出される。特に好適な実施態様では、生物活性グレリンは、両、生物活性グレリンおよび非生物活性グレリンを検出する検出手段によって検出される。
【0139】
この要件に従うそれぞれの検出手段は、例えば、両、生物活性グレリンおよびデス−オクタノイルグレリンにおいて同一であるグレリンの、その/それらの部分に対して特異的
であるすべての検出手段である。好ましくは、そのような検出手段は、かくしてグレリンのC末端に結合するか、あるいは少なくとも、N末端およびn−オクタノイル側鎖によって形成されるドメインに結合することはない。特に好適な検出手段は、核酸、ポリペプチド、タンパク質および抗体を含んでなる群から選ばれる検出手段であり、これらの生成は当業者には既知である。
【0140】
グレリンの検出方法はまた、サンプルが、好ましくは段階c)を実施するために使用された反応容器から除去されることを含む。
【0141】
さらなる実施態様では、方法はまた、生物活性および/またはデス−オクタノイルグレリンの相互作用パートナーを表面、好ましくは先に定義された表面上に固定化する段階を含み、ここでの相互作用パートナーは、本明細書、好ましくはそれぞれの方法に関連して先のように定義されており、より好ましくは、それらの種々の実施態様における核酸、ポリペプチド、タンパク質および抗体を含む。この実施態様では、特に好適な検出手段は、本発明による核酸であり、そのような核酸は、好ましくは標識されても、または標識されていなくてもよい。そのような核酸が標識される場合には、それが、直接的にまたは間接的に検出できる。そのような検出はまた、好ましくは、核酸、ポリペプチド、タンパク質および本明細書で記述される種々の実施態様における実施態様を含んでなる群からまた選ばれる第二の検出手段の使用を伴ってもよい。そのような検出手段は、好ましくは、本発明による核酸に特異的である。より好適な実施態様では、第二の検出手段は分子ビーコンである。核酸または第二の検出手段または両方のいずれも、好適な実施態様では、検出標識を含んでもよい。検出標識は、好ましくは、ビオチン、ブロモ−デオキシウリジン標識、ジゴキシゲニン標識、蛍光標識、UV−標識、放射能標識およびキレーター分子を含んでなる群から選ばれる。あるいはまた、第二の検出手段は、好ましくは、核酸によって含有されるか、またはそれによって含まれるか、またはそれに連結される検出標識と相互作用する。特に好適な組み合わせは、次のとおりである:
検出標識がビオチンであり、そして第二の検出手段がビオチンに対向された抗体であるか、あるいは
検出標識がビオチンであり、そして第二の検出手段がアビジンまたはアビジン保持分子であるか、あるいは
検出標識がビオチンであり、そして第二の検出手段がストレプトアビジンまたはストレプトアビジン担持分子であるか、あるいは
検出標識がビオチンであり、そして第二の検出手段がノイトラビジンまたはノイトラビジン担持分子であるか、あるいは
検出標識がブロモ−デオキシウリジンであり、そして第二の検出手段がブロモ−デオキシウリジンに対向された抗体であるか、あるいは
検出標識がジゴキシゲニンであり、そして第二の検出手段がジゴキシゲニンに対向された抗体であるか、あるいは
検出標識がキレーターであり、そして第二の検出手段が放射性核種であり、
これらの場合、該検出標識は核酸に連結されていることが好ましい。この種の組み合わせがまた核酸が表面に付着される実施態様にも応用できることが理解されるべきである。そのような実施態様では、検出標識が相互作用パートナーに連結されることが好ましい。
【0142】
最後に、第二の検出手段が第三の検出手段を用いて検出される、好ましくは第三の検出手段が酵素、より好ましくは第二の検出手段の検出において酵素反応を示す酵素であるか、あるいは第三の検出手段が放射線、より好ましくは放射性核種によって放出される放射線を検出する手段であることは、また本発明内にはいる。好ましくは、第三の検出手段は具体的には第二の検出手段を検出することおよび/または第二の検出手段と相互作用することである。
【0143】
また、表面に固定されている生物活性および/またはデスーオクタノイルグレリンの相互作用パートナーおよび本発明による核種が、好ましくは相互作用パートナーとグレリン間に形成される複合体に付加される実施態様では、サンプルは、反応から、より好ましくは段階c)および/またはd)が実施される反応容器から除去されてもよい。
【0144】
1つの実施態様では、本発明による核種は蛍光部分を含み、ここで、蛍光部分の蛍光は、核種と生物活性グレリン間の複合体形成および遊離の生物活性グレリンにおいて異なる。
【0145】
さらなる実施態様では、核酸は本発明による核酸の誘導体であり、ここで、核酸の誘導体は、アデノシン置換アデノシンの少なくとも1種の蛍光誘導体を含む。好適な実施態様では、アデノシンの蛍光誘導体はエテノアデノシンである。
【0146】
さらなる実施態様では、本発明による核酸の誘導体および生物活性グレリンからなる複合体は蛍光を用いて検出される。
【0147】
本方法の実施態様では、シグナルは段階(c)または段階(d)において生成され、好ましくはシグナルはサンプル中の生物活性グレリンの濃度と相関される。
【0148】
好適な態様では、アッセイは96穴プレートにおいて実施されてもよく、ここでの成分は前述の反応容器および反応容器として働くウェル中に固定化される。
【0149】
反応段階の上記系列およびそれらに関連して記述される種々の実施態様は、原則として、両生物活性グレリン、すなわち、オクタノイルグレリン、より好ましくはn−オクタノイルグレリン、およびデス−オクタノイルグレリンを検出するために適当である。このことは、生物活性グレリンの検出について、少なくとも1種の相互作用パートナーおよび本発明による核酸が生物活性グレリンを特異的に検出するのに適当である場合には可能である。原則として、生物活性グレリンに特異的である本発明による核酸が使用されることが十分である。サンプル中に含有されるグレリン量を特定するこの種の方法の読み取りは、生物活性グレリンについての分析の結果としてそれ自体使用することができる。しかしながら、また結果は、グレリン、好ましくは両生物活性グレリンおよびデス−オクタノイルグレリンの総含量を決定する方法において使用されてもよい。そのような目的では、デス−オクタノイルグレリンに特異的な、あるいは両生物活性およびデス−オクタノイルグレリン、すなわち総グレリン含量またはその量を検出するのに適当な検出手段または相互作用パートナーのいずれかを使用する、上記と好ましくは同一な方法が実施される。該相互作用パートナーまたは手段が、生物活性グレリンであるか、またデス−オクタノイルグレリンであるかに拘わらず、いずれかのグレリンを検出するのに適当である場合は、サンプル中に含有されるグレリンの総量は、生物活性グレリンについて得られる値および総グレリン含量の商形成によってサンプル中の生物活性グレリンのパーセンテージを決定することを可能なような方法の読み取りによって計算することができる。さらなる実施態様では、方法はデス−オクタノイルグレリンを特異的に検出するために使用される。そのようなデス−オクタノイルグレリンは、例えば、n−オクタノイル部分を欠如しているグレリンのC末端またはN末端に対向されることによるような、デス−オクタノイルグレリンに特異的である手段または相互作用パートナーの検出によって検出することができる。かくして検出されるグレリンの量、すなわちデスーオクタノイルグレリンの量は、次いで、サンプル中の総含量を与える生物活性グレリンの量に加えることができる。
【0150】
本発明の核酸は、薬物設計のための出発材料としてさらに使用されてもよい。1つのアプローチは化合物ライブラリーのスクリーニングであり、そのような化合物ライブラリーは、好ましくは低分子量化合物ライブラリーである。1つの実施態様では、スクリーニン
グは高処理スクリーニングである。好ましくは、高処理スクリーニングは、標的に基づくアッセイにおける化合物の迅速、効率的、試行錯誤評価である。最良の場合には、分析は比色(colormatic)測定によって実施される。これに関連して使用されるライブラリーは当業者には既知である。
【0151】
あるいはまた、本発明による核酸は薬物の合理的設計のために使用されてもよい。好ましくは、合理的薬物設計は製薬学的先導(lead)構造の設計である。X線結晶学または核磁気共鳴分光法のような方法によって典型的には同定される標的の三次元構造から出発して、コンピュータープログラムが、多くの異なる化学化合物の構造を含有しているデータベースをとおして検索するために使用される。選択はコンピューターによって実施され、同定された化合物が続いて実験室において試験できる。
【0152】
薬物の合理的設計は、本発明による核酸のいずれから出発してもよく、そして本発明の核酸の構造に類似しているか、または本発明の核酸の構造の結合媒介部分に同一である構造、好ましくは三次元構造を必要とする。薬物の合理的設計におけるさらなる段階でもまた代替的段階においても、神経伝達物質に結合する核酸のそれらの部分の好ましくは三次元構造は、ヌクレオチドおよび核酸とは異なる化学基によって模倣される。この模倣によって、核酸とは異なる化合物が設計できる。そのような化合物は、好ましくは低分子またはペプチドである。
【0153】
当業者には既知である競合アッセイを使用するような、化合物ライブラリーのスクリーニングの場合では、適当なグレリン類似体、グレリンアゴニストまたはグレリンアンタゴニストが発見することができる。そのような競合アッセイは次のように設定されてもよい。本発明の核酸、好ましくは標的結合L−核酸であるspiegelmerが固相に結合される。グレリン類似体を同定するために、標識されたグレリンがアッセイに添加されてもよい。潜在的な類似体は、それぞれの標識によって得られるシグナルにおける減少とともに進行するspiegelmerに結合するグレリン分子と競合する。アゴニストまたはアンタゴニストのスクリーニングは、当業者には既知の細胞培養アッセイの使用を伴ってもよい。
【0154】
本発明によるキットは、少なくとも1種または数種の本発明の核酸を含んでもよい。さらに、キットは少なくとも1種または数種のポジティブまたはネガティブ対照を含んでもよい。ポジティブ対照は、例えば、グレリン、特にこれに対して本発明の核酸が選ばれるか、またはこれに対して、好ましくは液体形態においてそれが結合するものであってもよい。ネガティブ対照は、例えば、グレリンに類似する生物物理学的性質の見地から定義されるが、本発明の核酸によって認識されないペプチドであってもよい。さらにまた、該キットは1種以上のバッファーを含んでもよい。種々の成分は、乾燥または凍結乾燥形態においてキット中に含有されても、または液体に溶解されてもよい。キットは、キットの1種以上の成分を含有してもよい1種以上の容器を含んでもよい。さらなる実施態様では、キットは、キットおよびその種々の成分を使用する方法に関する情報を使用者に与える指示または指示紙片を含む。
【0155】
好ましくは本明細書で使用されるように、用語「処置」は、好適な実施態様では、追加的または代替的に予防および/または追跡処置を含む。
【0156】
好ましくは本明細書で使用されるように、用語「疾病および障害」は、改めて指示されない限り、互換可能な様式で使用されるべきである。
【0157】
本明細書で使用されるように、用語「含む」は、好ましくは、そのような用語によって追跡または記述される主題を限定することを意図していない。しかしながら、その他の実
施態様では、用語「含む」は、含有するという意味において理解されるべきであり、かくして、そのような用語によって追跡または記述される主題を限定するとして理解されるべきである。
【0158】
種々の配列番号、本発明による核酸分子の化学的性質および本明細書で使用される標的分子グレリン、その実際の配列および内部参照番号が、次の表において総括される。
【0159】
【表3】
【0160】
【表4】
【0161】
【表5】
【0162】
【表6】
【0163】
本発明は、図面、実施例および配列リストによってさらに具体的に説明され、これらからさらに、特徴、実施態様および利点が取り上げられてもよい。
【実施例】
【0164】
例1:グレリン結合配列
明確に別に指示されない限り、これらの実施例をとおして使用されるグレリンは、そのオクタノイル化形態におけるグレリンであった。
【0165】
ドイツ特許第一034944.3号に記述される技術から得られる技術を用いて、ビオチン化ヒト・D−グレリンに対向されるRNAインビトロ選択が実施された。dsDNA分子の富有化集団がクローン化され、配列決定された。配列分析の結果は図1において見ることができる。
【0166】
全配列は、既知のグレリン結合配列SOT−C(B11trc;特許出願WO2004/013274A2,参照)のグレリン結合モチーフA(25ヌクレオチド)を含む。これらの23クローンからの4個のみにおいてモチーフAが、配列の3’末端に置かれている。19配列内では、モチーフAはクローンの5’−末端に置かれている。モチーフDと呼ばれるさらなるモチーフはモチーフAの3’末端に置かれている。
【0167】
配列の結合特性
異なる位置において変異を示す15クローン(図1)が、実施例2に記述される「競合アッセイ」を用いる37℃での格付け実験のために選ばれた。参考として、放射能標識アプタマーSOT−C(B11trc)が使用された。数種の候補がSOT−C(B11trc)よりも強くD−グレリンに結合し、より低いKDまたは「競合アッセイ」では分別できない活性コンホメーションのより高い量を示した。クローンMS−P2−G2、MS−P2−D2、MS−P2−A2、MS−P2−H3は対照クローンSOT−C(B11trc)よりも良好な結合を有するように見え、クローンMS−P2−E1、MS−P2−B1、MS−P2−F1、MS−P2−C3、MS−P2−C2,MS−P2−A4およびMS−P2−B2はさらに良好な結合を示した。SOT−C(B11trc)に較べて類似の結合結果は、クローンMS−P2−E3、MS−P2−A3、MS−P2−H2およびMS−P2−D1について決定できた(図2;評価は図1参照)。したがって、最良クローンが、平衡アッセイにおいてアプタマーとして、そして37℃における細胞培養アッセイにおいてSpiegelmerとしてそれらの活性について試験された(プロトコルは実施例2参照)。結果は図1において総括されている。クローンMS−P2−D2、MS−P2−F1、MS−P2−C2、MS−P2−A4およびMS−P2−B2は、平衡アッセイにおいて37℃で22〜34nMのKD(約60%活性分子)と測定され、そして3,0〜8,0nMのIC50値が37℃の細胞培養実験において検出できた。これに較べて、SOT−C(B11trc)のIC50は37℃で20nMと検出され、そしてKDは37℃で100nM(47%活性分子)と決定された(特許出願WO2004/013274A2に記述されるように、室温におけるSOT−CのIC50は約5nMである)。結果は、SpiegelmerMS−P2−F1、MS−P2−C2およびMS−P2−D2の生物学的活性は、37℃においてSOT−C(B11trc)よりも約5倍も良好であり得ることを示している。連続的な先端切り取り実験では、クローンMS−P2−F1(4,0nMのIC50)が選択された(二次構造予想、図12参照)。
【0168】
図1に示される配列のいずれもが、その先端を切り取られた形態物を含む本発明による核酸であるが、それらはなお標的に結合することができる。
【0169】
例2: 配列の結合特性
2.1 「平衡結合アッセイ」の使用による放射能標識アプタマーの格付け
本明細書で開示され、そしてより具体的には図1において列挙されるほとんどのクローンが、「平衡結合アッセイ」の使用によって、ビオチン化ヒト・D−グレリンに対するそれらの結合挙動に関して放射能標識アプタマー(D−RNA)として格付けされた。
【0170】
ヒト・D−グレリンに対する分子の結合挙動の格付けが実施された。この目的のために、本明細書で記述される標準プロトコルを用いて、同定されたアプタマーが図1に描かれるような先端を切り取られたアプタマー(プライマー結合部位なし)として合成された。
【0171】
次に示すように、アプタマー配列が次のプロトコルを用いてγ−P32−ATPにより5’末端において放射能標識された。
成分 [最終]
オリゴヌクレオチド 5μM
T4 Forward反応バッファー(Invitrogen) 1x
T4 ポリヌクレオチドキナーゼ(Invitrogen) 10U/10μl反応容量[γ−32P]−ATP 1μl/10μl反応容量
【0172】
反応は37℃で1時間インキュベートされた。次いで、放射能標識アプタマーはゲル精製された。
【0173】
放射能標識RNAの2〜5pmoleが、Ca++およびMg++を含有しない選択バッファー(ヒト血液における生理学的条件にしたがう:20mM Tris、150mMNaCl、5mMKCl,pH7.4は37℃で調節された)において95℃で3分間変性され、37℃において1mMの最終濃度までのこれらのイオンの添加によって折り畳まれ、そして0,4〜3000nMの範囲の濃度のビオチン化ヒトD−グレリンとともに37℃で1時間インキュベートされた。続いて、NeutrAvidinアガロースの一定量がマトリックスとして添加され、そしてRNA:ペプチド複合体が30分以上37℃で振盪された。次いで、結合されたペプチドをもつマトリックスおよびチューブが沈降され、上澄液が除去され、マトリックスが100μl選択バッファーにより洗浄され、そして結合RNAと非結合RNAとの差異がBeckman Coulterを使用する放射能の測定によって決定された。算出された数から、対照(0nMビオチン化ヒトD−グレリン)がバックグラウンドとして差し引かれた。平衡定数はソフトウエアプログラム「GraFIT」(Version 4.0.10.,Erithacus Software Ltd.,Surrey,UK)を使用して計算された。
【0174】
2.2 競合アッセイの使用によるアプタマーの格付け
既知のグレリン結合配列SOT−C(B11trc;特許出願WO2004/013274A2参照)に対してクローンを比較するために、SOT−Cが、本明細書で記述される標準プロトコル(実施例4)を用いてアプタマー(D−RNA)として合成された。アプタマー配列は本明細書で記述される標準プロトコルを用いてγ−P32−ATPにより5’末端において放射能標識された。
【0175】
放射能標識SOT−Cおよび同定されたクローン(D−RNA)が平衡アッセイのために前述のように調製された。アッセイは20nMのペプチド濃度(ビオチン化ヒトD−グレリン)において実施された。その後、等モル量の放射能標識SOT−Cおよび2つの異なる濃度(40nMおよび200nM)のアプタマーが試験された(結果は図2に示される)。アッセイは平衡アッセイと同様に実施された。
【0176】
2.3 グレリン結合Spiegelmerによるグレリン誘導カルシウム放出の抑制
グレリン結合Spiegelmerの機能的特性決定が、ヒト(L−)グレリンとヒト
成長ホルモン分泌促進物質受容体(GHS−R)との相互作用を追跡する細胞アッセイ系において実施された。受容体−リガンド相互作用からもたらされる細胞内カルシウム放出は、蛍光カルシウム指示薬の手段によって可視化される。
【0177】
ヒト・グレリン受容体(GHS−R1a)を発現する安定なトランスフェクトされたCHO細胞(Euroscreen,Gosselies,Belgiumから得られた)が、透明な底を有する黒色の96穴プレート(Greiner)において5〜7x104細胞/ウェルを接種され、そして100単位/mlペニシリン、100μg/mlストレプトマイシン、400μg/mlゲネチシンおよび2.5μg/mlフンギゾン(fungizone)をさらに含有するUltraCHO培地(Cambrex)において37℃および5%CO2下で一夜培養された。
【0178】
カルシウム指示薬染料fluo−4を負荷する前に、細胞は200μlのCHO−U+により一回洗浄される。次いで、指示薬染料溶液(CHO−U+中、10μMfluo−4(Molecular Probes)、0.08%pluronic127(Molecular Probes))の50μlが添加され、そして細胞は37℃で60分間培養される。その後、細胞は180μlのCHO−U+により3回洗浄される。最後に、90μlのCHO−U+が各ウェルに添加される。
【0179】
種々の量のSpiegelmerが、0.2mlの低プロフィル(low profile)96穴プレートにおいて室温または37℃で15〜60分間、5mMprobenecidおよび20mM HEPES(CHO−U+)を含有するUltraCHO培地において、ヒト(L−)グレリン(Bachemから購入)とともにインキュベートされる。対照として、ペプチドのみを含有するサンプル(最大カルシウム放出)およびペプチドを含有しないサンプル(最小カルシウム放出)が分析される。これらの刺激溶液においては、ペプチドおよびSpiegelmer(添加される場合)は、このアッセイに較べて10倍濃縮されている。
【0180】
カルシウム放出の検出では、刺激溶液が細胞に添加され(10μl/ウェル)、そして蛍光シグナルの変化がモニターされる。蛍光シグナルの測定は、注入ポンプを備えたFluostar Optima多検出プレートリーダー(BMG)において485nmの励起波長および520nmの発光波長において実施される。
【0181】
数個のサンプルの並行測定では、96穴プレートの1つの(垂直の)列のウェルが一緒に記録される。4秒のタイムラグをもつ最初の3回の読み取りが、ベースラインの決定のために実施される。次いで、読み取りが中断され、プレートが装置から除かれる。多チャンネルピペットを用いて、刺激溶液の10μlがウェルに添加され、次いで、プレートが装置中に再び移動され、そして測定が継続される。4秒間隔での全20回の読み取りにおいて実施される。
【0182】
各ウェルでは、最大蛍光とベースライン値との間の差異が決定され、そしてヒト(L−)グレリン(オクタノイル化された)濃度に対してプロットされるか、あるいはSpiegelmerによるカルシウム放出の抑制における実験では、半最大抑制定数(IC50)の決定を可能にするSpiegelmerの濃度に対してプロットされる。
【0183】
2.4 表面プラズモン共鳴(SPR)測定
ビオチン化ヒトD−グレリンに結合するアプタマーの特性決定が、前記BIAcore2000装置(BIAcore AB,Uppsala,Sweden)を用いるSPR即時速度分析によって決定された。C末端ビオチン化ペプチドの100RU(フローセル2)および300RU(フローセル3)が、Streptavidin共役センサーチッ
プ(BIAcore AB,Freiburg,Germany)に固定化され、そして0,1μM〜1μMの濃度範囲におけるサンプルが、会合時間360(s)および解離時間360(s)を定めるKinject指令を用いて注入された。フローセル1はバッファーおよびデキストラン基質対照(Biacore SA−Chip surface)として使用され、フローセル4には、非特異的D−ペプチドが固定化されてアプタマーの非特異的結合を決定した。反応は37℃で実施された。BIAevaluation3.0 ソフトウエア(BIAcore AB,Uppsala,Sweden)によるデータ解析では、本研究者らはLangmuir1:1化学量論的フィッティング・アルゴリズムを使用した。
【0184】
例3: グレリン結合Spiegelmerモチーフの定義
3.1 SOT−D−000の切り取り
3.1.1 予め選ばれたSOT−D−000の末端切り取り
図3に示されたグレリン結合Spiegelmerは、特許出願WO2004/013274A2において既に示されるように予め得られた。二次構造の予測(最小自由エネルギーコンホメーション[Hofacker et al.,1994,Monatsh.Chem 125:167−188])では、5’−および3’末端の間の標準的分子内塩基対合が全Spiegelmerについて明白になる(図4−7;図3において下線を付した塩基)。Spiegelmerにおけるそのような構造的要素の存在は、活性のある三次元構造の正確な折り畳みのためには極めて重要であろうが、経済的なSpiegelmerの合成のためには、分子は可能な限り短くあるべきである。SpiegelmerSOT−D−000(別名、プライマー結合部位を含まないSOT−R04−DR14−F7の先端切り取り物)は、それが、最長の末端らせんを形成する可能性を有する最短の選ばれたSpiegelmerであるので、末端切り取りのための基本として選ばれた。
【0185】
10個の代わりに8個(SOT−D−108)、7個(SOT−D−100)および6個(SOT−D−104)の塩基対のらせんの長さをもつSOT−D−000の先端切り取り変異体が合成され(図8)、そして実施例2における記述のように細胞培養において試験された(図9)。SOT−D−104(39量体)は有意に結合活性を喪失したが、SOT−D−100(41量体)および−108(43量体)は完全なグレリン・アンタゴニスト性能を維持した。変異体SOT−D−106、G4が除去されたSOT−D−104のさらなる先端切り取り物は実質的に不活性であった。驚くべきことに、この不適合G−全選択グレリンSpiegelmerのらせんに存在する−は容易には除去されないであろう。むしろ、グレリン結合のためには必須であると思われる。
【0186】
3.1.2 SOT−D−100および−108の内部欠失変異体
図3における選ばれたグレリン結合物(binder)の整列を考慮する場合、強度に可変性の領域は、5’末端における末端らせんに直接隣接するすべての整列された分子において存在すると思われる。グレリン結合物のさらなる先端切り取りについてのこの明白な可変性を精査するために、最小の完全活性SOT−D変異体SOT−D−100およびー108がさらなる切り取りのための基礎として使用された。内部のSOT−D−100変異体の場合は、それぞれの塩基が単純に除かれたが、SOT−D−108変異体では、それらが、合成中に柔軟性の親水性スペーサーによって置換された。次に、SOT−D−100およびその誘導体SOT−D−101および−102ならびにSOT−D−108誘導体SOT−Dー109、SOT−D−110およびー111が、実施例2における記述のように細胞培養において試験された(図9)。図8に示されるように、3個ではないが2個の内部ヌクレオチドの欠失は、活性喪失なしに可能であった(SOT−D−101;−102)。これに対して、スペーサーによって置換された場合(SOT−D−109、−110、−111)には、3個のヌクレオチドが除くことができた。スペーサーの最適位置はSOT−D−109において実現される(図10)。
【0187】
40kDa部分による5’末端におけるSOT−D−109の修飾はグレリン結合活性の喪失なしに可能であった(図11)。
【0188】
3.2 SOT−Eの切り取り
次のように、先導配列として選ばれたグレリン結合配列MS−P2−F1(図1および図12)はSOT−Eと呼ばれる。
【0189】
二次構造の予測(最小自由エネルギーコンホメーション[Hofacker et al.,1994,Monatsh.Chem 125:167−188])では、C30およびG50の間の標準的分子内塩基対合の可能性が、SpiegelmerSOT−Eについて明白になる(図12)。SpiegelmerSOT−Eの切り取りをもたらす次の実験が記述される。SpiegelmerSOT−Eのすべての切り取り型が細胞培養においてそれらの活性に関して試験された(実施例2、参照)。
【0190】
3.2.1 SpiegelmerSOT−Eの末端切り取り
3’末端における5個のヌクレオチドは分子の他の部分にはハイブリダイズされないと思われる。反対に、5’末端では、ヌクレオチドは部分的に対合できる。驚くべきことに、3’末端は結合の低下なしには切り取りすることができない(SpiegelmerSOT−E−012)が、一方SOT−E−014(5’末端における6個のヌクレオチドの切り取り)は完全なグレリン・アンタゴニスト性能を維持した(図13)。
【0191】
3.2.2 SpiegelmerSOT−Eの内部欠失変異体
図1における選ばれたグレリン結合物MS−P2−E3、MS−P2−G2、MS−P2−D2、MS−P2−A3、MS−P2−E1、MS−P2−B1、MS−P2−C3、MS−P2−C2、MS−P2−H2、MS−P2−A4、およびSOT−E(配列ファミリーI)の整列を考慮する場合、可変性の領域は、末端において、かつ、らせんに直接隣接するすべての整列された分子において存在すると思われる(SOT−EにおけるG36−C43)。SpiegelmerSOT−Eの二次構造予測は、4個のヌクレオチドのループを示す(A38−U41;図12)。グレリン結合物のさらなる切り取りのためにこの明白な可変性を探査するために、最初に、SpiegelmerSOT−Eのそれぞれの塩基が、合成中に柔軟性の親水性スペーサーによって置換された。8個のヌクレオチド(G36−C43;SOT−E−008)ではない6個(U37−A42;SOT−E−011)の欠失が、活性の喪失なしに可能であった。5’末端における切り取り(G1−A6)の結果および柔軟性の親水性スペーサーによる6個のヌクレオチド(U37−A42)の置換の組み合わせは、56−ヌクレオチドSpiegelmerSOT−E(図12)として細胞培養において同一のIC50を示す44−ヌクレオチドのSpiegelmerSOT−E−019(図14)をもたらす。
【0192】
3.2.3 SOT−E−019内の配列セグメントの転位
配列MS−P2−A2、MS−P2−H3、MS−P2−D1(図1)によって表される配列ファミリーIIは配列ファミリーI(SOT−Eを含む)に対して類似性を担持する。これらの配列相同性は図1におけるboxA(UAAGACCGAAGGUAACCAAUCCUAC)として強調される。配列ファミリーIIでは、boxAは、むしろ3’末端に置かれるが、配列ファミリーIでは、boxAは5’末端に接近している。SpiegelmerSOT−Eのインビトロの選択および二次構造予測のこの結果により、boxAが5’末端に置かれるSOT−E−019の変異体が計画された。その目的のために、SOT−E−019の本来の5’末端および3’末端が、柔軟性の親水性スペーサーと連結され、そしてループが除去されて、新しい5’末端および3’末端を生成した(SOT−E−021;図15および21)。驚くべきことに、SOT−E−021では、活性の喪失は観察できなかった。SOT−E−019およびSOT−E−021(SOT−E−33(図16および21)およびSOT−E−25(図17および21))に基づくさらなる切り取り物は、グレリン・アンタゴニスト機能の低下なしには可能ではなかった(図13)。
【0193】
3.3 SOT−C、SOT−D−109およびSOT−Eの比較
SpiegelmerSOT−Eが、SOT−Cに比較して表面プラズモン共鳴(実施例2)によって測定され、6倍良好なKDが決定できた(図18)。同じ改良された結合性(5倍良好)が、D−109に較べてSpiegelmerSOT−Eを用いる細胞培養実験(実施例2)において検出することができた(図19)。動物体内での滞留時間を改良するために、SpiegelmerSOT−E−19が、40kDa−PEG部分によってその5’末端において修飾された(SOT−E19−5’−PEG)。SOT−E19−5’−PEGは、細胞培養実験においてSOT−Eに較べて類似の結果を示し(図19)、さらに、インビボ活性を例証した(実施例5;図22)。
【0194】
3.4 グレリン結合Spiegelmerモチーフの定義
グレリン、特に本明細書で定義されるオクタン酸側鎖を有するグレリンを結合させるすべての分子は、第一に、グレリン結合のために必須である25個のヌクレオチド(「BoxA」)のモチーフの存在を特徴とする。「BoxA」内の25個のヌクレオチドのあるものは他の塩基と相互に交換可能であると思われる(図20A)。
【0195】
グレリン結合Spiegelmerの「BoxA」の機能のための必須要件は、非対合の中央に置かれたグアノシンを含んでらせん構造を形成する配列5’−GUGAGG(BoxB)の相補鎖と、「BoxAの」3’末端の5個のヌクレオチド(5’−CCUAC)とのハイブリダイゼーションである。BoxBにおけるこの中央のグアノシンの欠失は、有意な結合の喪失をもたらす(「BoxAにより定義されるらせん」、非対合G;図20A)。「BoxA」、「BoxAにより定義されるらせん」に対する3’−隣接物は、1個、優先的には2個以上のさらなる定まっていない塩基対(「BoxC1」および「BoxC2」、図20A)によって最小に伸長されるべきである。
【0196】
「BoxB」の3’末端では、さらなるヌクレオチドが結合のために必須である。少なくとも2個のさらなるヌクレオチド(優先的には5’−CA)が結合のために必要である。結合性の有意な改良は、この位置において(「BoxD」、図20A)、さらなる2個、場合によっては4個のヌクレオチドを付加して2、3、4、5または6個のヌクレオチドを含んでなるBoxDをもたらすことによって達成できる。
【0197】
「BoxA」の5’末端では、さらなるヌクレオチドが結合のために必須である。少なくとも1個のさらなるヌクレオチド(特にUまたはG)が結合のために必要である。結合性の有意な改良は、この位置において(「BoxE」、図20A)、さらなる2個のヌクレオチドを付加して1、2、3または4個のヌクレオチドを含んでなるBoxEをもたらすことによって達成できる。
【0198】
BoxDおよびEが柔軟性の親水性スペーサーを介して連結される場合、該スペーサーはまた、種々の位置に挿入することができる(参照、図8;SOT−D−109、SOT−D−110およびSOT−D−111)。
【0199】
驚くべきことに、BoxDおよびEの代わりに「BoxC1」および「BoxC2」が柔軟性の親水性スペーサーに連結した場合に、一定の配列要素が機能する。
【0200】
変異体1:
【0201】
【化6】
【0202】
変異体2:
【0203】
【化7】
【0204】
本明細書、請求項および/または図面において開示される本発明の特徴は、別々に、およびいずれかのその組み合わせにおいて、その種々の形態において本発明を実現するための材料となり得る。
【0205】
例4: アプタマーおよびSpiegelmerの化学的合成
化学的固相合成
小規模合成
アプタマーは、2’TBDMS RNAホスホルアミダイト化学を使用してABI 394シンセサイザー(Applied Biosystems,Foster City,CA,USA)による固相合成によって製造された(M.J.Damha,K.K.Ogivie,Methods in Molecular Biology,Vol.20 Protocols for oligonucleotides and analogs,ed.S.Agrawal,p.81−114,Humana Press Inc.1993)。D−rA(N−Bz)−、D−rC(Ac)−、D−rG(N−ibu)−、D−rU−およびヘキサエチレングリコールホスホルアミダイトは、ChemGenes,Wilmington,MAから購入された。アプタマーはゲル電気泳動によって精製された。
【0206】
未修飾(PEG化のない)Spiegelmerは、2’TBDMS RNAホスホルアミダイト化学を使用してABI 394シンセサイザー(Applied Biosystems,Foster City,CA,USA)による固相合成によって製造された(M.J.Damha,K.K.Ogivie,Methods in Molecular Biology,Vol.20 Protocols for oligonucleotides and analogs,ed.S.Agrawal,p.81−114,Humana Press Inc.1993)。L−rA(N−Bz)−、L−rC(Ac)−、L−rG(N−ibu)−、L−rU−およびヘキサエチレングリコールホスホルアミダイトは、ChemGenes,Wilmington,MAから購入された。アプタマーはゲル電気泳動によって精製された。
【0207】
大規模合成および修飾
修飾SpiegelmerSOT−E−19は、2’TBDMS RNAホスホルアミダイト化学を使用して ktaPilot100シンセサイザー(Amersham Bioscience;General Electric Healthcare,Freiburg)による固相合成によって製造された(M.J.Damha,K.K.Ogilvie,Methods in Molecular Biology,Vol.20 Protocols for oligonucleotides and analogs,ed.S.Agrawal,p.81−114,Humana Press Inc.1993)。L−rA(N−Bz)−、L−rC(Ac)−、L−rG(N−ibu)−、L−rU−およびヘキサエチレングリコールホスホルアミダイトは、ChemGenes,Wilmington,MAから購入された。合成は、L−riboC修飾されたCPG孔径1000(Link Technology,Glasgow,UK)において開始された。カップリング(1サイクルについて15分)では、アセトニトリル中0.3Mベンジルチオテトラゾール(CMS−Chemicals,Abingdon,UK)、およびアセトニトリル中それぞれ0.1Mホスホルアミダイト溶液の3.5当量が使用された。Spiegelmerの5’末端におけるアミノ基は、アミノヘキシルホスホルアミダイト(ChemGenes)をカップリングすることによって連結された。酸化−キャッピング(capping)サイクルが使用された。オリゴヌクレオチド合成のためのさらなる標準溶媒および試薬はBiosolve(Valkenswaard,NL)から購入された。SpiegelmerはDMT−ON合成された;脱保護の後、それは、Source15RPC媒質(Amersham)を用いて調製用RP−HPLC(Wincott et al 1995 Nucleic Acids Res.23:2677)により精製された。5’DMT−基は80%酢酸を用いて除去された(RTで30分)。続いて、2MNaOAc水溶液が添加され、そしてSpiegelmerが、5K再生セルロース膜(Millipore,Bedford,MA)を用いる接線流動濾過によって脱塩された。
【0208】
PEG化
インビボの腎臓クリアランスを低下させるために、SpiegelmerSOT−E−19は5’末端において40kDaポリエチレングリコール(PEG)部分に共有結合された。そのような増加された分子質量をもつSpiegelmerを用いることは、血漿における保持を有意に延長させ、そのために、より長い有効性が達成できた。
【0209】
PEG化(欧州特許出願第一 306 382号)では、精製された5’−アミノ修飾Spiegelmerは、H2O(2.5ml)、DMF(5ml)およびバッファーA(5ml;クエン酸・H2O[7g]、ホウ酸[3.54g]、リン酸[2.26ml]および1MNaOH[343ml]を混合し、そして最終容量1 lまでH2Oを添加することによって調製された;pH=8.4は1MHClにより調節された)の混合液中に溶解された。
【0210】
Spiegelmer溶液のpHは1MNaOHにより8.4にもたらされた。次いで、40kDaのPEG−NHSエステル(Nektar Therapeutics,Huntsville,AL)が、75〜85%の最大収率が達成されるまで、0.6当量の4分量において30分毎に37℃で添加された。反応混合液のpHは、PEG−NHSエステルの添加中1MNaOHにより8〜8.5に維持された。
【0211】
反応混合液は、4ml尿素溶液(8M)、4mlバッファーA、および4mlバッファーB(H2O中0.1M酢酸トリエチルアンモニウム)と混合され、95℃に15分間加熱された。PEG化Spiegelmerは次に、アセトニトリル濃度勾配(バッファーB;バッファーC:アセトニトリル中0.1M酢酸トリエチルアンモニウム)を用いて、Source15RPC媒質(Amersham)によりRP−HPLCによって精製された。過剰のPEGは5%バッファーCにおいて、PEG化Spiegelmerは10〜15%バッファーCにおいて溶出した。>95%の純度(HPLCによって調査)をもつ生成物画分が合わされ、40mlの3M NaOAcと混合された。PEG化Spiegelmerは、接線流動濾過(5K再生セルロース膜、Millipore,Bedford,MA)によって脱塩された。
【0212】
例5: 抗グレリン−SpiegelmerSOT−E19−5’−PEGによる外因性グレリン投与後の成長ホルモン放出の抑制
外因性グレリンの投与は、ラットにおける成長ホルモン(GH)の放出を惹起することが知られている。グレリン結合Spiegelmerの前以ての投与は、GHのグレリン誘導放出を抑制する。
【0213】
Sprague Dawleyラットが、7日間それらの新しい環境に適応された。実験の設定は、各々5匹の動物をもつ5群からなった:1つのポジティブ対照群および4つの異なる用量のSpiegelmerSOT−E19−5’−PEG群。全動物は、実験期間中Ketamine/Xylazineで麻酔され、尾静脈中に2回の静脈注射を受けた。第一回の注射は第二回注射の30分前に与えられ、そしてPBS(ポジティブ対照群)または図22に指示されるSpiegelmerSOT−E19−5’−PEGの用量からなった。第二回の投与は全動物に対して3nmolラット・グレリンからなり、そして実験について時間0とマークされた。第二回の投与前に、最初の血液サンプルが眼窩の血脈洞から抜き取られた。さらなるサンプルは、ラット・グレリン投与後の5、15、30および45分に採取された。得られる血漿サンプルは、製造者の指示にしたがって市販の酵素免役アッセイ(Growth hormon,Rat,Biotrak Assay Kit,RPN2561,Amersham Biosciences Europe GmbH,Freiburg)により成長ホルモン濃度について分析された。
【0214】
図22において分るように、GHの放出はラット・グレリンの注射によって強力に刺激されるが、SOT−E19−5’−PEGの前以ての投与によって抑制することができた。最大の抑制は75nmolSOT−E19−5’−PEG/kg体重において達成される。150nmolSOT−E19−5’−PEG/kg体重までの投与用量におけるさらなる増加は、それ以上のGH放出を抑制しない。この結果は、抗グレリンSpiegelmerSOT−E19−5’−PEGのインビボ活性を例証している。
【0215】
例6: 内部リンカーを含まないSOT−E−19の誘導体
実施例3に記述されたように、抗グレリンSpiegelmerSOT−E−19は、L−ヌクレオチド、および6個のヌクレオチドを置換するために元の分子SOT−Eに組み入れられた内部リンカーからなる。6個のヌクレオチドからの4個はSpiegelmerSOT−Eにおいてループを形成し、残りの2ヌクレオチド(6ヌクレオチドからの)は互いにハイブリダイズできる(SOT−E、参照図23、第一列、ハイブリダイズされるヌクレオチドは下線を付される)。
【0216】
内部リンカーを含まないSOT−E−19の可能な限り短い誘導体を実現するために、リンカーは3個のヌクレオチドの種々の建造ブロックによって置換された。全誘導体がSpiegelmer(プロトコル、実施例4参照)として合成され、そして細胞培養実験(プロトコル、実施例2参照)において分析された。結合および抑制効力(IC50)の喪失のないSOT−E−19の内部リンカーの置換は、次の建造ブロックについて成功裏に示された:ACA(SOT−E−19−L3)およびCAA(SOT−E−19−L4)。SOT−E−19の種々の誘導体が図23に示される。
【0217】
例7: グレリン結合Spiegelmerによるオクタノイル−グレリンおよびデスオクタノイル−グレリンの識別
グレリンに対するSpiegelmerSOT−E−19の結合の特徴は、さらに、実施例2に記述された方法に基づいて、競合アッセイにおいて分析された。これらのアッセイにおいて、Spiegelmerは、細胞の刺激前の刺激溶液においてグレリンペプチド(オクタノイル−およびデスオクタノイル−グレリン)の種々の組み合わせ物とともにインキュベートされた。
【0218】
ペプチド組み合わせ物のスキームおよび全長オクタノイル−グレリン(ヒト・グレリン
=huグレリン)が図24(左から右へ数えられるバー)において総括される:いずれのオクタノイル−グレリン(ヒト・グレリン=huグレリン)も含まないか、または300nMの最終濃度においてデスオクタノイル−グレリンを含む場合は、細胞の刺激は検出できない(バー1および2)が、一方、13nMの濃度におけるオクタノイル−グレリン(ヒト・グレリン=huグレリン)は既にカルシウム放出を媒介するために十分であり(バー3);300nMデスオクタノイル−グレリン(バー4)のさらなる添加は細胞刺激を妨げず、これは、生物学的に不活性なデスオクタノイル−グレリンが受容体アンタゴニストではないことを示している。3nMオクタノイル−グレリン(ヒト・グレリン=huグレリン)によって媒介されるカルシウム放出はSOT−E−19の10倍過剰量によって抑制でき(バー5)、オクタノイル−グレリン(ヒト・グレリン=huグレリン)を超える100倍過剰量(300nM)でのデスオクタノイル−グレリンの存在でさえ抑制に対して競合しない(バー6)。これに対して、300nMオクタノイル−グレリンおよび30nMSpiegelmerのアッセイ濃度はカルシウム放出の増加を示し(バー7)、アッセイ条件下で、オクタノイル−グレリン(ヒト・グレリン=huグレリン)による刺激増進が達成できるという証拠を与える。この実験は、SOT−E−19がオクタノイル形態とデスオクタノイル形態におけるグレリン間を特異的に識別することを例証する。
【0219】
グレリンに対するSpiegelmerNOX−B11の結合特性(オクタノイル−グレリンには結合するが、デスオクタノイル−グレリンには結合しないか、または弱く結合する)は、SOT−E−19に類似している(同じ実験が既にWO2005/049828に記述されるように実施された)。
【0220】
両分子は、主としてboxAに基づく高い構造的相関関係(実施例3、参照)を有するので、この結果は、モチーフAが、グレリンのオクタノイル化形態に関して、特にオクタノイル基を含むN末端における5個のアミノ酸に関して、本発明による核酸分子の高い特異性のために必須であるという仮説を支持する。
【0221】
例8: グレリン結合SpiegelmerNOX−B11を使用するオクタノイル−グレリンの定量
グレリン結合SpiegelmerNOX−B11は、オクタノイル化されたヒトおよびラットのグレリンの非放射能定量のために、酵素免疫アッセイ(EIA)のアッセイ形式に類似する形式で使用することができる。
【0222】
原理
このアッセイでは、標準、対照および未知の処理血漿が、グレリン結合Spiegelmer、例えば、そのN末端においてオクタノイル−グレリンを認識するSpiegelmerNOX−B11によりコートされた96穴ミクロタイター・プレートにおいてインキュベートされる。インキュベーションおよび洗浄後、ウェルが、抗グレリン抗体(第一の抗体)またはグレリンのC末端においてグレリンに結合する核酸により処理される。この抗体または核酸は標識されても、されなくてもよいが、核酸は好ましくは標識される。(第一の)抗体が標識されない場合、(第一の)抗グレリン抗体を除去するインキュベーションおよび数回の洗浄段階後、第二の抗体(抗体は第一の抗体のFcフラグメントに対向され、そして標識されている)が添加される。第二の抗体の標識は、酵素セイヨウワサビ・ペルオキシダーゼ(HRP)であってもよい。第二の抗体とともにウェルをインキュベートし、未結合フラクションを除去した後、ウェルはHRP基質テトラメチルベンジジン(TMB)とともにインキュベートされる。次いで、酸性停止液が添加され、そして基質の酵素的ターンオーバーの度合が450nmにおける吸光度測定によって決定される。測定された吸光度はサンプル中に存在するオクタノイル−グレリンの濃度に直接比例している。1セットのグレリン標準が使用されて、グレリン濃度に対する吸光度の標準曲線がプロットされ、これから未知のグレリンが計算できる。
【0223】
プロトコル
最初に、ストレプトアビジンでコートされた96穴プレート(Reacti−Bind
Streptavidin Coated High Bind Capacity Black 96−well Plates,Perbio Science,Bonn,Germany)が、0.1%Tweenを含むPBS−Dulbecco(Mg2+およびCa2+を含む、Biochrom AG,Berlin,Germany)により3回洗浄された。各ウェルは、室温で1時間、100μlの50μMビオチン化SpiegelmerNOX−B11(PBSに溶解;配列番号:72)とともにインキュベートされた。ビオチン化SpiegelmerNOX−B11の固定化の後、未結合のSpiegelmerが1回の洗浄段階(100μl PBS)によって除去された。これは図26Aに図示される。
【0224】
洗浄バッファーを除去後、一定濃度のオクタノイル−グレリンを含む保存液が添加され、そして室温で1時間インキュベートされた。上澄液が除去され、ウェルが1回(100μl PBS)洗浄された。これは図26Bに図示される。
【0225】
抗グレリン抗体(Phoenix Peptides,Belmont,CA,USA;10ab)がブロッキング溶液(Phoenix Peptides,Belmont,CA,USA)において1時間インキュベートされ、未結合の抗体が3回の洗浄段階(各100μl PBS)によって除去された。これは図26Cに図示される。
【0226】
結合された抗グレリン抗体を検出するために、抗グレリン抗体(10ab)のFcフラグメントを特異的に認識し、そしてセイヨウワサビ・ペルオキシダーゼにより修飾されている第二の抗体(Phoenix Peptides,Belmont,CA,USA;20ab−HRP)が使用された。第二の抗体(20ab−HRP)の未結合フラクションは5回の洗浄段階(100μl PBS)によって除去された。これは図26Dに図示される。
【0227】
定量では、TMB基質(Amersham Biosciences,Little Chalfont,UK)の100μlが添加され、プレートが密封され、そして30分間暗室においてインキュベートされた。定義50μlが添加された。基質の酵素的ターンオーバーの度合が450nmにおける吸光度測定によって決定され、そして吸光度単位の差異が記録された。
【0228】
図25に示されるように、1セットのグレリン標準品が使用されて、グレリン濃度に対する吸光度の標準曲線をプロットした。
【0229】
結果
原則として、実験データは、SpiegelmerNOX−B11が濃度依存方式でオクタノイル−グレリンを固定化するために使用でき、したがって、EIA型検出アッセイにおいて試薬としての潜在力を有することを例証する。
【0230】
本明細書、請求項および/または図面において開示される本発明の特徴は、別々に、およびいずれかのその組み合わせにおいて、その種々の形態において本発明を実現するための材料となり得る。
【図面の簡単な説明】
【0231】
【図1】図1は、ヒト・グレリンに結合するRNAリガンドの配列の整列を示す;
【図2】図2は、先端を切り取られたアプタマー対既知の配列SOT−C(B11trc)によるグレリンへの結合の競合アッセイを示す;
【図3】図3は、RNAリガンドの選ばれた配列の整列を示し、これらは特許出願WO2004/013274A2において公表された;
【図4】図4は、グレリン結合RNAspiegelmerクローンSOT−Cの計算された二次構造を示し、二次構造はプログラム「RNAフォールド(fold)」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図5】図5は、グレリン結合RNAspiegelmerクローンSOT−D−000の計算された二次構造を示し、二次構造はプログラム「RNAフォールド」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図6】図6は、グレリン結合RNAspiegelmerクローンSOT−Dの計算された二次構造を示し、二次構造はプログラム「RNAフォールド」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図7】図7は,グレリン結合RNAspiegelmerクローンF−12の計算された二次構造を示し、二次構造はプログラム「RNAフォールド」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図8】図8は、SOT−D−000誘導体の整列を示し、これらは先端切り取りおよび合理的設計実験の結果である;
【図9】図9は、室温におけるSpiegelmerSOT−C、SOT−D−000およびそれらの変異体によるグレリン誘導Ca++−放出の抑制についてのワンポイント測定を示し;細胞は、種々の量のSpiegelmerSOT−C、SOT−D−000またはD−000の変異体とともに室温でプレインキュベートされた5nMグレリンにより刺激され、結果はSpiegelmerを無しで得られたシグナルに対して標準化された蛍光シグナルのパーセンテージを示す;
【図10】図10は、グレリン結合RNAspiegelmerクローンSOT−D−109の推定二次構造を示す;
【図11】図11は、室温におけるSpiegelmerD−109および5’−PEG化D−109によるグレリン誘導Ca++−放出の抑制についての用量−応答曲線を示し;細胞は、種々の量のSpiegelmerD−109および5’−PEG化D−109とともに室温でプレインキュベートされた5nMグレリンにより刺激され;結果はSpiegelmerを無しで得られたシグナルに対して標準化された蛍光シグナルのパーセンテージを示し;SpiegelmerD−109およびその修飾体は、同じIC50をもってグレリン誘導Ca++−放出を抑制することが見出された;
【図12】図12は,グレリン結合RNASpiegelmerクローンSOT−Eの計算された二次構造を示し、二次構造はプログラム「RNAフォールド」(Hofacker et al.,1994,Monatsh.Chem 125:167−188)を用いて計算された;
【図13】図13は、細胞培養実験によって試験されたSpiegelmerSOT−Eの先端を切り取られた変異体の結合分析を示す;
【図14】図14は、グレリン結合RNASpiegelmerSOT−E−19の推定二次構造を示す;
【図15】図15は、グレリン結合RNAspiegelmerクローンSOT−E−21の推定二次構造を示す;
【図16】図16は、グレリン結合RNAspiegelmerクローンSOT−E−33の推定二次構造を示す;
【図17】図17は、グレリン結合RNAspiegelmerクローンSOT−E−25の推定二次構造を示す;
【図18】図18は、D−グレリン結合RNAクローンSOT−CおよびSOT−EのKD値を指示するBiocore2000センサーグラム(sensorgram)を示す;
【図19】図19は、37℃におけるSpiegelmerSOT−E、SOT−E−19−5’−アミノ、SOT−E−19−5’−PEGまたはSOT−D−109によるグレリン誘導Ca++−放出の抑制についての用量−応答曲線を示し;細胞は、種々の量のSpiegelmerSOT−E、SOT−E−19−5’−アミノ、SOT−E−19−5’−PEGまたはSOT−D−109とともに37℃でプレインキュベートされた2nMヒト・グレリンにより刺激され;結果はSpiegelmerを無しで得られたシグナルに対して標準化された蛍光シグナルのパーセンテージを示し;SpiegelmerSOT−E−19およびその修飾体は、約4nMのIC50をもってグレリン誘導Ca++−放出を抑制することが見出された;
【図20A】図20Aは、グレリン結合Spiegelmerについて特徴的である配列boxの定義を示す;
【図20B】図20Bは、図20Aの1変異体;
【図20C】図20Cは、図20Aの1変異体;
【図20D】図20Dは、図20Aの1変異体;
【図21】図21は、SOT−D−109、SOT−E、SOT−E−19、SOT−E−21、SOT−E−33およびSOT−E−25の配列を示す;
【図22】図22は、抗グレリンSpiegelmerSOT−E−19−5’−PEGによる外因性グレリンの投与後の成長ホルモン放出の抑制を示す;
【図23】図23は、内部リンカーの代わりにさらなるヌクレオチドを含むSOT−E−19の異なる誘導体の概観、およびnMで表される観察されたIC50値を示す;
【図24】図24は、線の下に総括される成分の組み合わせおよび濃度をもつ、オクタノイルグレリン、デス−オクタノイルグレリンおよびSpiegelmerSOT−E−19による細胞競合アッセイの結果を示す;
【図25】図25は、EIA型検出アッセイによって記録されたヒト・グレリン(オクタノイル化)濃度に対する吸光度の標準曲線を示し、ここで、グレリン結合SpiegelmerNOX−B11はヒト・グレリン(オクタノイル化)を固定化するために使用されて、ヒト・グレリン(オクタノイル化)の定量を可能にした;
【図26A−D】図26A〜Dは、本発明によるグレリン結合核酸を用いるオクタノイルグレリンの定量方法の種々の段階を示す。
【特許請求の範囲】
【請求項1】
選択的にグレリンに結合する核酸であって、
第一のストレッチBoxAおよび
第二のストレッチBoxB
を含み、
ここで、
第一のストレッチBoxAは約25個の連続ヌクレオチドを含み、
第二のストレッチBoxBは約6〜8個の連続ヌクレオチドを含み、
ここで、
第一のストレッチBoxAのヌクレオチドの3’末端ストレッチは、第二のストレッチBoxBとハイブリダイズし、このハイブリダイゼーションでは、第一の二本鎖構造が形成され、そしてそのような第一の二本鎖構造はバルジを含む、上記核酸。
【請求項2】
二本鎖構造が、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドおよび第二のストレッチBoxBのヌクレオチドの一部または全部、好ましくは第二のストレッチBoxBの6〜8個の連続ヌクレオチドによって形成される、請求項1に記載の核酸。
【請求項3】
バルジが、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドと塩基対合しない、第二のストレッチBoxBの1〜3個のヌクレオチド、好ましくは第二のストレッチBoxBの1個のヌクレオチドによって形成される、請求項1または2に記載の核酸。
【請求項4】
バルジが非塩基対合プリンによって形成され、ここで、プリンは好ましくはグアノシンである、請求項1〜3のいずれかに記載の核酸。
【請求項5】
非塩基対合プリンが、第二のストレッチBoxBによって提供される、請求項4に記載の核酸。
【請求項6】
核酸が、第三のストレッチBoxC1および第四のストレッチBoxC2をさらに含み、
ここで、第三のストレッチBoxC1は少なくとも1個のヌクレオチドを含み、そして
第四のストレッチBoxC2は少なくとも1個のヌクレオチドを含み、そして
ここで、第三のストレッチBoxC1は、その3’末端によって第二のストレッチBoxBの5’末端に連結されており、そして第四のストレッチBoxC2は、その5’末端によって第一のストレッチBoxAの3’末端に連結されている、請求項1〜5のいずれかに記載の核酸。
【請求項7】
第三のストレッチBoxC1および第四のストレッチBoxC2はハイブリダイゼーション可能であり、ハイブリダイゼーションにより、第二の二本鎖構造が形成される、請求項6に記載の核酸。
【請求項8】
第一の二本鎖構造が第一のらせん構造を形成する、請求項1〜7のいずれかに記載の核酸。
【請求項9】
第二の二本鎖構造が第二のらせん構造を形成する、請求項7に記載の核酸。
【請求項10】
第二のらせん構造が、1〜10個の塩基対、好ましくは1〜3個の塩基対、より好ましくは2〜3個の塩基対を含んでなる、ヘリックスまたはヘリックス様構造である、請求項9に記載の核酸。
【請求項11】
第一のらせん構造が第二のらせん構造によって延長されている、請求項9〜10に記載の核酸。
【請求項12】
第三のストレッチBoxC1が、約1〜10個の連続ヌクレオチド、好ましくは1〜3個の連続ヌクレオチド、より好ましくは2または3個の連続ヌクレオチドを含む、請求項6〜11のいずれかに記載の核酸。
【請求項13】
第四のストレッチBoxC2が、約1〜10個の連続ヌクレオチド、好ましくは1〜3個の連続ヌクレオチド、より好ましくは2または3個の連続ヌクレオチドを含む、請求項6〜12のいずれかに記載の核酸。
【請求項14】
核酸が第五のストレッチBoxDをさらに含み、ここで、第五のストレッチBoxDは少なくとも2個の連続ヌクレオチドを含む、請求項6〜13のいずれかに記載の核酸。
【請求項15】
第五のストレッチBoxDが配列5’−CAを含む、請求項14に記載の核酸。
【請求項16】
第五のストレッチBoxDがいずれかの長さの連続ヌクレオチドを含み、ここで、長さは2、3、4、5および6個の連続ヌクレオチドからなる群から選ばれる、請求項14または15に記載の核酸。
【請求項17】
第五のストレッチBoxDが、
5’CA(X)n3’
[式中、Xは、好ましくはA,G,T,C,UおよびIを含んでなる群から選ばれるいずれかのヌクレオチドであり、そしてnは、0,1,2,3および4からなる群から選ばれるいずれかの整数である]
の配列を含む、請求項13〜16のいずれかに記載の核酸。
【請求項18】
第五のストレッチBoxDが、配列
5’CA(X)n3’
[式中、n=4]
からなる、請求項17に記載の核酸。
【請求項19】
第五のストレッチBoxDが、その5’末端によって第二のストレッチBoxBの3’末端に連結される、請求項14〜18のいずれかに記載の核酸。
【請求項20】
核酸が第六のストレッチBoxEをさらに含み、ここで、第六のストレッチBoxEは少なくとも1個のヌクレオチドを含む、請求項14〜19のいずれかに記載の核酸。
【請求項21】
第六のストレッチBoxEが、約1〜10個の連続ヌクレオチド、好ましくは1〜4個の連続ヌクレオチド、より好ましくは3個の連続ヌクレオチドを含む、請求項20に記載の核酸。
【請求項22】
第六のストレッチBoxEの少なくとも1個のヌクレオチドが、UおよびGからなる群から選ばれる、請求項20または21に記載の核酸。
【請求項23】
UまたはGヌクレオチドが、第一のストレッチBoxAの5’末端の直ぐ隣に配置されている、請求項22に記載の核酸。
【請求項24】
第六のストレッチBoxEが、その3’末端によって第一のストレッチBoxAの5’末端に連結されている、請求項20〜23のいずれかに記載の核酸。
【請求項25】
第五のストレッチBoxDの3’末端が、第一のスペーサーによって第六のストレッチBoxEの5’末端に連結される、請求項20〜24のいずれかに記載の核酸。
【請求項26】
第四のストレッチBoxC2の3’末端が、第二のスペーサーによって第三のストレッチBoxC1の5’末端に連結される、請求項20〜25のいずれかに記載の核酸。
【請求項27】
第一および第二のスペーサーが、各々別々に、かつ独立して、親水性スペーサーを含んでなる群から選ばれている、請求項25または26に記載の核酸。
【請求項28】
親水性スペーサーが、核酸スペーサーおよび非核酸スペーサーを含んでなる群から選ばれる、請求項27に記載の核酸。
【請求項29】
第一のスペーサーが、約1〜20個の連続ヌクレオチド、好ましくは1〜5個の連続ヌクレオチド、より好ましくは2個の連続ヌクレオチドを含んでなる核酸スペーサーである、請求項28に記載の核酸。
【請求項30】
第二のスペーサーが、約3〜20個の連続ヌクレオチド、好ましくは3〜5個の連続ヌクレオチド、より好ましくは3個の連続ヌクレオチドを含んでなる核酸スペーサーである、請求項28に記載の核酸。
【請求項31】
第二のスペーサーがACAまたはCAAからなる、請求項30に記載の核酸。
【請求項32】
スペーサーが非核酸である、請求項28に記載の核酸。
【請求項33】
第一のスペーサーおよび/または第二のスペーサーが、少なくとも1個のエチレングリコール部分または複数個のそのようなエチレングリコール部分を含む、請求項32に記載の核酸。
【請求項34】
スペーサーが約172〜688Da、好ましくは344Daの分子量を有する、請求項33に記載の核酸。
【請求項35】
核酸が環状核酸である、請求項1〜34のいずれかに記載の核酸。
【請求項36】
【化1】
の構造または
【化2】
の構造を有する、請求項1〜35のいずれかに記載の核酸。
【請求項37】
第一のストレッチBoxAが、
【化3】
[式中、
X1=GまたはA;
X2=AまたはU;
X3=GまたはA;
X4=AまたはCまたはU;および
X5=GまたはA]
、好ましくは
【化4】
の配列を含んでなる、請求項1〜36のいずれかに記載の核酸。
【請求項38】
第二のストレッチBoxBが5’GUGAGG3’の配列を含んでなる、請求項1〜37、特に請求項37のいずれかに記載の核酸。
【請求項39】
核酸の配列が、
【表1】
【表2】
に記載の配列を含んでなる群から選ばれる、請求項1〜38のいずれかに記載の核酸。
【請求項40】
核酸が、
【表3】
に記載の配列を含んでなる群から選ばれる、請求項1〜38のいずれかに記載の核酸。
【請求項41】
核酸の配列が、配列番号:29、配列番号:32、配列番号:38、配列番号:39、配列番号:40および配列番号:41による配列を含んでなる群から選ばれる、請求項39に記載の核酸。
【請求項42】
核酸の配列が、配列番号:30、配列番号:33、配列番号:36、配列番号:42、配列番号:46、配列番号:47、配列番号:51および配列番号:52による配列を含んでなる群から選ばれる、請求項39又は40に記載の核酸。
【請求項43】
核酸が、グレリン、好ましくはヒト・グレリンを結合することができる、請求項1〜42のいずれかに記載の核酸。
【請求項44】
グレリンが配列番号:1によるアミノ酸配列を有する、請求項43に記載の核酸。
【請求項45】
核酸が修飾体(modification)を含んでなる、請求項1〜44のいずれかに記載の核酸。
【請求項46】
修飾体が、HES部分およびPEG部分を含んでなる群から選ばれる、請求項45に記載の核酸。
【請求項47】
修飾体が直鎖または分枝状PEGからなるPEG部分であり、ここで、PEG部分の分子量は、好ましくは約20〜120kD、より好ましくは約30〜80kD、もっとも好ましくは約40kDである、請求項46に記載の核酸。
【請求項48】
修飾体がHES部分であり、ここで、HES部分の分子量は、好ましくは約10〜130kD、より好ましくは約30〜80kD、もっとも好ましくは約50kDである、請求項46に記載の核酸。
【請求項49】
核酸のヌクレオチドがL−ヌクレオチドである、請求項1〜48のいずれかに記載の核酸。
【請求項50】
核酸が完全にL−ヌクレオチドからなる、請求項1〜48のいずれかに記載の核酸。
【請求項51】
さらなる成分が製薬学的に許容できる添加物および製薬学的活性作用物を含んでなる群から選ばれる、請求項1〜50のいずれかに記載の核酸および場合によってはさらなる成分を含んでなる製薬学的組成物。
【請求項52】
薬物の製造のための請求項1〜50のいずれかに記載の核酸の使用。
【請求項53】
診断手段の製造のための請求項1〜50のいずれかに記載の核酸の使用。
【請求項54】
薬物が、肥満、摂食障害、糖尿病、グルコース代謝障害、腫瘍、血圧障害、心臓血管病、先端巨大症、およびエネルギーバランス、食欲、体重の調節を含んでなる群から選ばれる疾病または障害の処置および/または予防のために存在する、請求項52に記載の使用。
【請求項55】
薬物が、胃腸疾患を含んでなる群から選ばれる疾病または障害の処置および/または予防のために存在する、請求項52に記載の使用。
【請求項56】
グレリンおよび請求項1〜50のいずれかに記載の核酸を含んでなる複合体であって、好ましくは結晶性複合体である上記複合体。
【請求項57】
グレリンの検出のための請求項1〜50のいずれかに記載の核酸の使用。
【請求項58】
次の段階:
− 候補グレリンアンタゴニストおよび/または候補グレリンアゴニストを提供し、
− 請求項1〜50のいずれかに記載の核酸を提供し、
− グレリンアンタゴニストおよび/またはグレリンアゴニストの存在においてシグナルを提供する試験系を提供し、そして
− 候補グレリンアンタゴニストがグレリンアンタゴニストであるか否か、そして/または候補グレリンアゴニストがグレリンアゴニストであるか否かを決定すること、
を含んでなる、グレリンアンタゴニストまたはグレリンアゴニストのスクリーニング方法。
【請求項59】
次の段階:
− 相、好ましくは固体相に固定化されたグレリンを提供し、
− 請求項1〜50のいずれかに記載の核酸、好ましくは標識されている請求項1〜50
のいずれかに記載の核酸を提供し、
− 候補グレリンアゴニストおよび/または候補グレリンアンタゴニストを添加し、そして
− 候補グレリンアゴニストがグレリンアゴニストであるか否か、そして/または候補グレリンアンタゴニストがグレリンアンタゴニストであるか否かを決定すること、
を含んでなる、グレリンアゴニストおよび/またはグレリンアンタゴニストのスクリーニングのための方法。
【請求項60】
決定は、核酸が候補グレリンアゴニストによるか、または候補グレリンアンタゴニストによって置換されるか否かが調査されるように実施されることを特徴とする、請求項59に記載の方法。
【請求項61】
請求項1〜50のいずれかに記載の核酸を含んでなる、グレリンの検出のためのキット。
【請求項62】
請求項59および60に記載の方法によって得ることができるグレリンアンタゴニスト。
【請求項63】
請求項59および60に記載の方法によって得ることができるグレリンアゴニスト。
【請求項1】
選択的にグレリンに結合する核酸であって、
第一のストレッチBoxAおよび
第二のストレッチBoxB
を含み、
ここで、
第一のストレッチBoxAは約25個の連続ヌクレオチドを含み、
第二のストレッチBoxBは約6〜8個の連続ヌクレオチドを含み、
ここで、
第一のストレッチBoxAのヌクレオチドの3’末端ストレッチは、第二のストレッチBoxBとハイブリダイズし、このハイブリダイゼーションでは、第一の二本鎖構造が形成され、そしてそのような第一の二本鎖構造はバルジを含む、上記核酸。
【請求項2】
二本鎖構造が、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドおよび第二のストレッチBoxBのヌクレオチドの一部または全部、好ましくは第二のストレッチBoxBの6〜8個の連続ヌクレオチドによって形成される、請求項1に記載の核酸。
【請求項3】
バルジが、第一のストレッチBoxAの5個の3’末端連続ヌクレオチドと塩基対合しない、第二のストレッチBoxBの1〜3個のヌクレオチド、好ましくは第二のストレッチBoxBの1個のヌクレオチドによって形成される、請求項1または2に記載の核酸。
【請求項4】
バルジが非塩基対合プリンによって形成され、ここで、プリンは好ましくはグアノシンである、請求項1〜3のいずれかに記載の核酸。
【請求項5】
非塩基対合プリンが、第二のストレッチBoxBによって提供される、請求項4に記載の核酸。
【請求項6】
核酸が、第三のストレッチBoxC1および第四のストレッチBoxC2をさらに含み、
ここで、第三のストレッチBoxC1は少なくとも1個のヌクレオチドを含み、そして
第四のストレッチBoxC2は少なくとも1個のヌクレオチドを含み、そして
ここで、第三のストレッチBoxC1は、その3’末端によって第二のストレッチBoxBの5’末端に連結されており、そして第四のストレッチBoxC2は、その5’末端によって第一のストレッチBoxAの3’末端に連結されている、請求項1〜5のいずれかに記載の核酸。
【請求項7】
第三のストレッチBoxC1および第四のストレッチBoxC2はハイブリダイゼーション可能であり、ハイブリダイゼーションにより、第二の二本鎖構造が形成される、請求項6に記載の核酸。
【請求項8】
第一の二本鎖構造が第一のらせん構造を形成する、請求項1〜7のいずれかに記載の核酸。
【請求項9】
第二の二本鎖構造が第二のらせん構造を形成する、請求項7に記載の核酸。
【請求項10】
第二のらせん構造が、1〜10個の塩基対、好ましくは1〜3個の塩基対、より好ましくは2〜3個の塩基対を含んでなる、ヘリックスまたはヘリックス様構造である、請求項9に記載の核酸。
【請求項11】
第一のらせん構造が第二のらせん構造によって延長されている、請求項9〜10に記載の核酸。
【請求項12】
第三のストレッチBoxC1が、約1〜10個の連続ヌクレオチド、好ましくは1〜3個の連続ヌクレオチド、より好ましくは2または3個の連続ヌクレオチドを含む、請求項6〜11のいずれかに記載の核酸。
【請求項13】
第四のストレッチBoxC2が、約1〜10個の連続ヌクレオチド、好ましくは1〜3個の連続ヌクレオチド、より好ましくは2または3個の連続ヌクレオチドを含む、請求項6〜12のいずれかに記載の核酸。
【請求項14】
核酸が第五のストレッチBoxDをさらに含み、ここで、第五のストレッチBoxDは少なくとも2個の連続ヌクレオチドを含む、請求項6〜13のいずれかに記載の核酸。
【請求項15】
第五のストレッチBoxDが配列5’−CAを含む、請求項14に記載の核酸。
【請求項16】
第五のストレッチBoxDがいずれかの長さの連続ヌクレオチドを含み、ここで、長さは2、3、4、5および6個の連続ヌクレオチドからなる群から選ばれる、請求項14または15に記載の核酸。
【請求項17】
第五のストレッチBoxDが、
5’CA(X)n3’
[式中、Xは、好ましくはA,G,T,C,UおよびIを含んでなる群から選ばれるいずれかのヌクレオチドであり、そしてnは、0,1,2,3および4からなる群から選ばれるいずれかの整数である]
の配列を含む、請求項13〜16のいずれかに記載の核酸。
【請求項18】
第五のストレッチBoxDが、配列
5’CA(X)n3’
[式中、n=4]
からなる、請求項17に記載の核酸。
【請求項19】
第五のストレッチBoxDが、その5’末端によって第二のストレッチBoxBの3’末端に連結される、請求項14〜18のいずれかに記載の核酸。
【請求項20】
核酸が第六のストレッチBoxEをさらに含み、ここで、第六のストレッチBoxEは少なくとも1個のヌクレオチドを含む、請求項14〜19のいずれかに記載の核酸。
【請求項21】
第六のストレッチBoxEが、約1〜10個の連続ヌクレオチド、好ましくは1〜4個の連続ヌクレオチド、より好ましくは3個の連続ヌクレオチドを含む、請求項20に記載の核酸。
【請求項22】
第六のストレッチBoxEの少なくとも1個のヌクレオチドが、UおよびGからなる群から選ばれる、請求項20または21に記載の核酸。
【請求項23】
UまたはGヌクレオチドが、第一のストレッチBoxAの5’末端の直ぐ隣に配置されている、請求項22に記載の核酸。
【請求項24】
第六のストレッチBoxEが、その3’末端によって第一のストレッチBoxAの5’末端に連結されている、請求項20〜23のいずれかに記載の核酸。
【請求項25】
第五のストレッチBoxDの3’末端が、第一のスペーサーによって第六のストレッチBoxEの5’末端に連結される、請求項20〜24のいずれかに記載の核酸。
【請求項26】
第四のストレッチBoxC2の3’末端が、第二のスペーサーによって第三のストレッチBoxC1の5’末端に連結される、請求項20〜25のいずれかに記載の核酸。
【請求項27】
第一および第二のスペーサーが、各々別々に、かつ独立して、親水性スペーサーを含んでなる群から選ばれている、請求項25または26に記載の核酸。
【請求項28】
親水性スペーサーが、核酸スペーサーおよび非核酸スペーサーを含んでなる群から選ばれる、請求項27に記載の核酸。
【請求項29】
第一のスペーサーが、約1〜20個の連続ヌクレオチド、好ましくは1〜5個の連続ヌクレオチド、より好ましくは2個の連続ヌクレオチドを含んでなる核酸スペーサーである、請求項28に記載の核酸。
【請求項30】
第二のスペーサーが、約3〜20個の連続ヌクレオチド、好ましくは3〜5個の連続ヌクレオチド、より好ましくは3個の連続ヌクレオチドを含んでなる核酸スペーサーである、請求項28に記載の核酸。
【請求項31】
第二のスペーサーがACAまたはCAAからなる、請求項30に記載の核酸。
【請求項32】
スペーサーが非核酸である、請求項28に記載の核酸。
【請求項33】
第一のスペーサーおよび/または第二のスペーサーが、少なくとも1個のエチレングリコール部分または複数個のそのようなエチレングリコール部分を含む、請求項32に記載の核酸。
【請求項34】
スペーサーが約172〜688Da、好ましくは344Daの分子量を有する、請求項33に記載の核酸。
【請求項35】
核酸が環状核酸である、請求項1〜34のいずれかに記載の核酸。
【請求項36】
【化1】
の構造または
【化2】
の構造を有する、請求項1〜35のいずれかに記載の核酸。
【請求項37】
第一のストレッチBoxAが、
【化3】
[式中、
X1=GまたはA;
X2=AまたはU;
X3=GまたはA;
X4=AまたはCまたはU;および
X5=GまたはA]
、好ましくは
【化4】
の配列を含んでなる、請求項1〜36のいずれかに記載の核酸。
【請求項38】
第二のストレッチBoxBが5’GUGAGG3’の配列を含んでなる、請求項1〜37、特に請求項37のいずれかに記載の核酸。
【請求項39】
核酸の配列が、
【表1】
【表2】
に記載の配列を含んでなる群から選ばれる、請求項1〜38のいずれかに記載の核酸。
【請求項40】
核酸が、
【表3】
に記載の配列を含んでなる群から選ばれる、請求項1〜38のいずれかに記載の核酸。
【請求項41】
核酸の配列が、配列番号:29、配列番号:32、配列番号:38、配列番号:39、配列番号:40および配列番号:41による配列を含んでなる群から選ばれる、請求項39に記載の核酸。
【請求項42】
核酸の配列が、配列番号:30、配列番号:33、配列番号:36、配列番号:42、配列番号:46、配列番号:47、配列番号:51および配列番号:52による配列を含んでなる群から選ばれる、請求項39又は40に記載の核酸。
【請求項43】
核酸が、グレリン、好ましくはヒト・グレリンを結合することができる、請求項1〜42のいずれかに記載の核酸。
【請求項44】
グレリンが配列番号:1によるアミノ酸配列を有する、請求項43に記載の核酸。
【請求項45】
核酸が修飾体(modification)を含んでなる、請求項1〜44のいずれかに記載の核酸。
【請求項46】
修飾体が、HES部分およびPEG部分を含んでなる群から選ばれる、請求項45に記載の核酸。
【請求項47】
修飾体が直鎖または分枝状PEGからなるPEG部分であり、ここで、PEG部分の分子量は、好ましくは約20〜120kD、より好ましくは約30〜80kD、もっとも好ましくは約40kDである、請求項46に記載の核酸。
【請求項48】
修飾体がHES部分であり、ここで、HES部分の分子量は、好ましくは約10〜130kD、より好ましくは約30〜80kD、もっとも好ましくは約50kDである、請求項46に記載の核酸。
【請求項49】
核酸のヌクレオチドがL−ヌクレオチドである、請求項1〜48のいずれかに記載の核酸。
【請求項50】
核酸が完全にL−ヌクレオチドからなる、請求項1〜48のいずれかに記載の核酸。
【請求項51】
さらなる成分が製薬学的に許容できる添加物および製薬学的活性作用物を含んでなる群から選ばれる、請求項1〜50のいずれかに記載の核酸および場合によってはさらなる成分を含んでなる製薬学的組成物。
【請求項52】
薬物の製造のための請求項1〜50のいずれかに記載の核酸の使用。
【請求項53】
診断手段の製造のための請求項1〜50のいずれかに記載の核酸の使用。
【請求項54】
薬物が、肥満、摂食障害、糖尿病、グルコース代謝障害、腫瘍、血圧障害、心臓血管病、先端巨大症、およびエネルギーバランス、食欲、体重の調節を含んでなる群から選ばれる疾病または障害の処置および/または予防のために存在する、請求項52に記載の使用。
【請求項55】
薬物が、胃腸疾患を含んでなる群から選ばれる疾病または障害の処置および/または予防のために存在する、請求項52に記載の使用。
【請求項56】
グレリンおよび請求項1〜50のいずれかに記載の核酸を含んでなる複合体であって、好ましくは結晶性複合体である上記複合体。
【請求項57】
グレリンの検出のための請求項1〜50のいずれかに記載の核酸の使用。
【請求項58】
次の段階:
− 候補グレリンアンタゴニストおよび/または候補グレリンアゴニストを提供し、
− 請求項1〜50のいずれかに記載の核酸を提供し、
− グレリンアンタゴニストおよび/またはグレリンアゴニストの存在においてシグナルを提供する試験系を提供し、そして
− 候補グレリンアンタゴニストがグレリンアンタゴニストであるか否か、そして/または候補グレリンアゴニストがグレリンアゴニストであるか否かを決定すること、
を含んでなる、グレリンアンタゴニストまたはグレリンアゴニストのスクリーニング方法。
【請求項59】
次の段階:
− 相、好ましくは固体相に固定化されたグレリンを提供し、
− 請求項1〜50のいずれかに記載の核酸、好ましくは標識されている請求項1〜50
のいずれかに記載の核酸を提供し、
− 候補グレリンアゴニストおよび/または候補グレリンアンタゴニストを添加し、そして
− 候補グレリンアゴニストがグレリンアゴニストであるか否か、そして/または候補グレリンアンタゴニストがグレリンアンタゴニストであるか否かを決定すること、
を含んでなる、グレリンアゴニストおよび/またはグレリンアンタゴニストのスクリーニングのための方法。
【請求項60】
決定は、核酸が候補グレリンアゴニストによるか、または候補グレリンアンタゴニストによって置換されるか否かが調査されるように実施されることを特徴とする、請求項59に記載の方法。
【請求項61】
請求項1〜50のいずれかに記載の核酸を含んでなる、グレリンの検出のためのキット。
【請求項62】
請求項59および60に記載の方法によって得ることができるグレリンアンタゴニスト。
【請求項63】
請求項59および60に記載の方法によって得ることができるグレリンアゴニスト。
【図1】

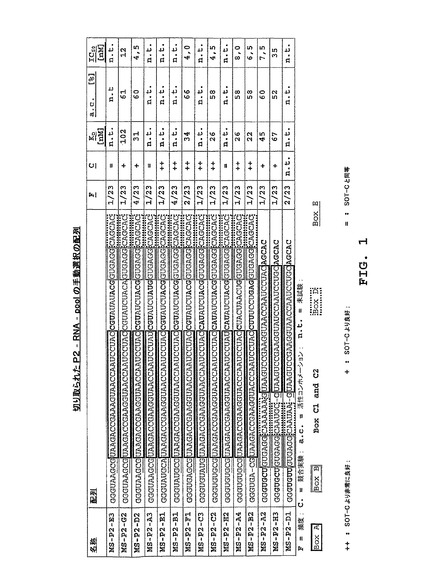
【図2】
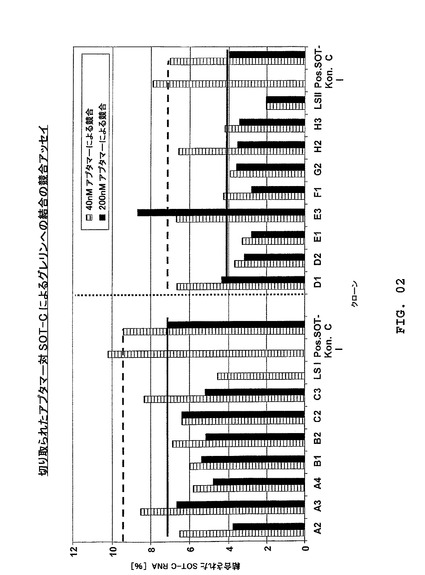
【図3】

【図4】
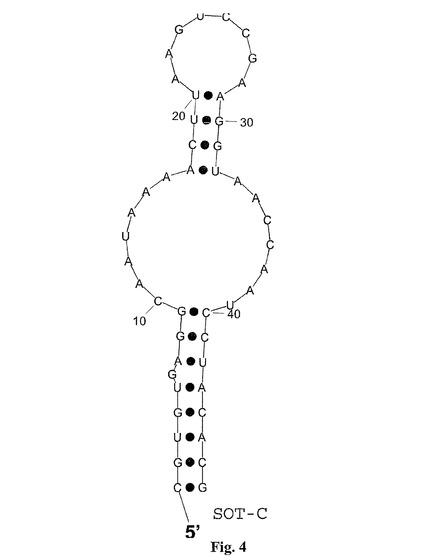
【図5】
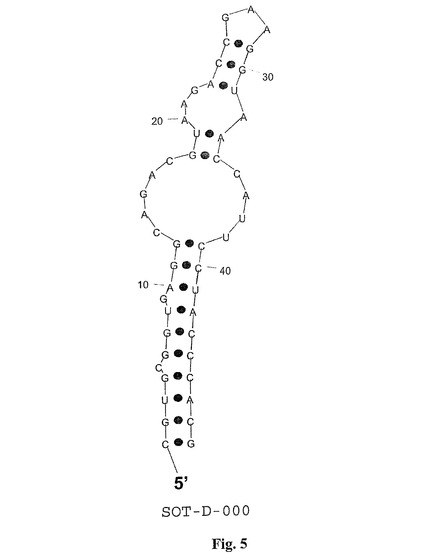
【図6】
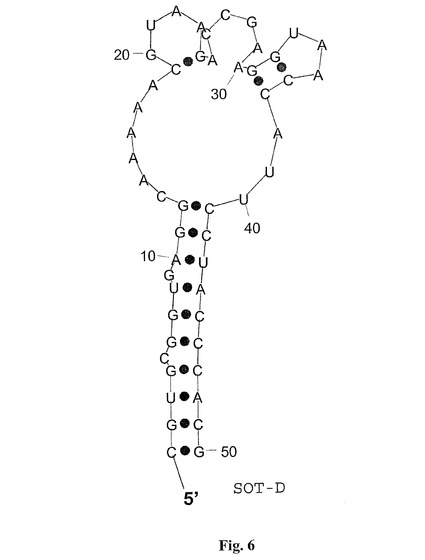
【図7】
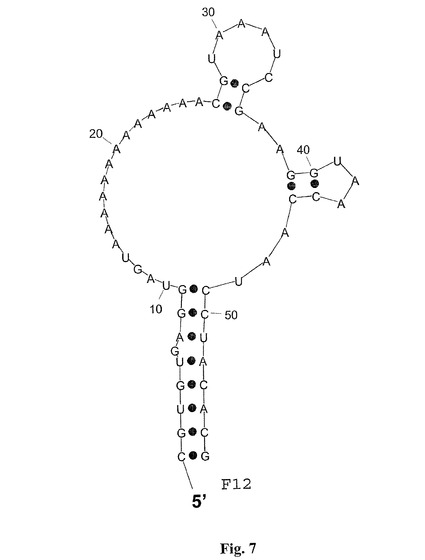
【図8】

【図9】
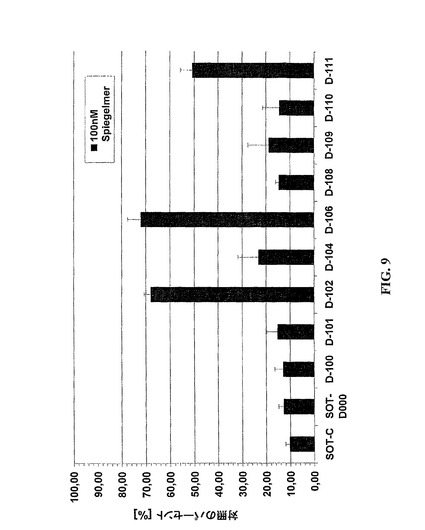
【図10】
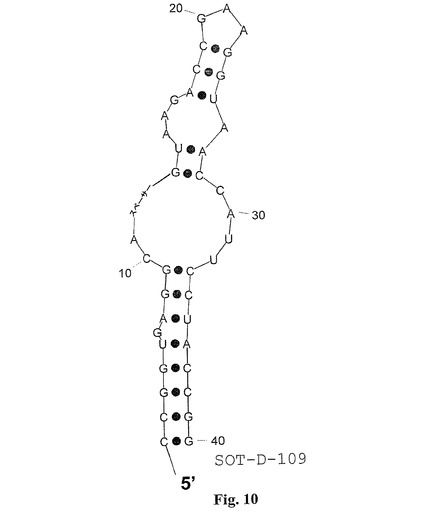
【図11】
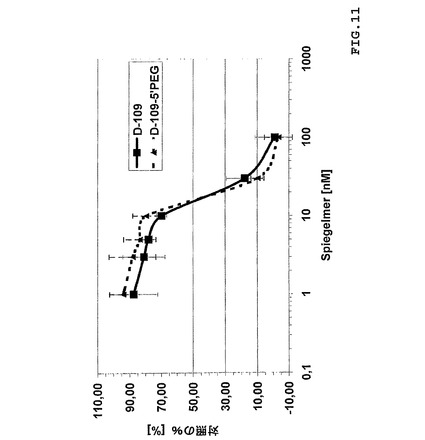
【図12】
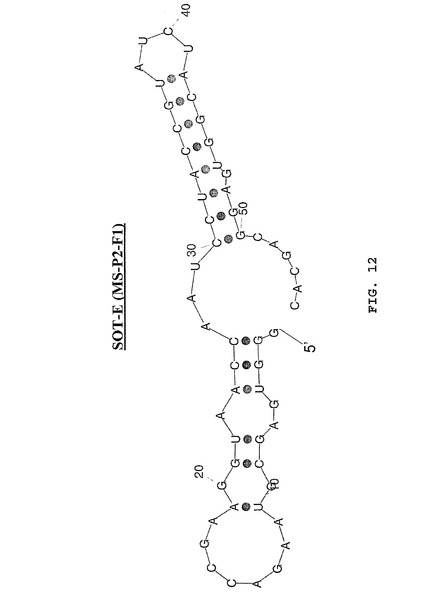
【図13】
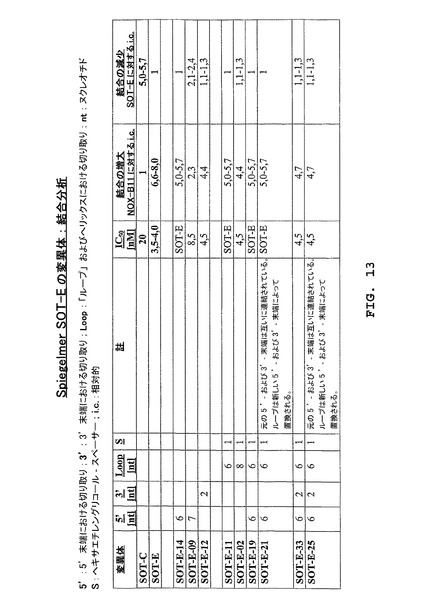
【図14】
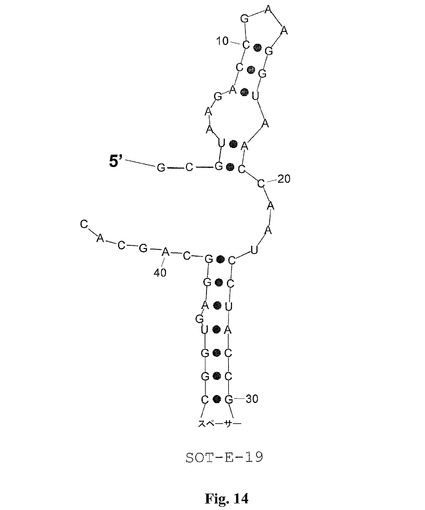
【図15】
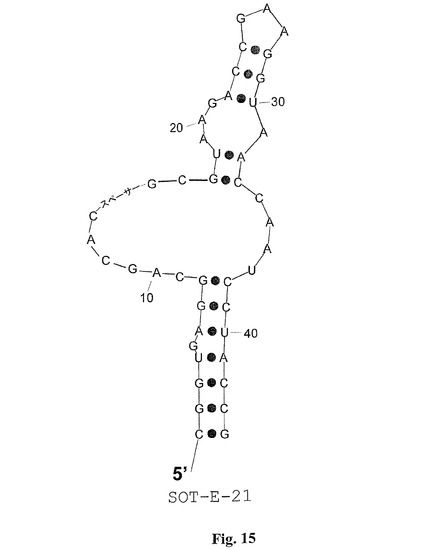
【図16】
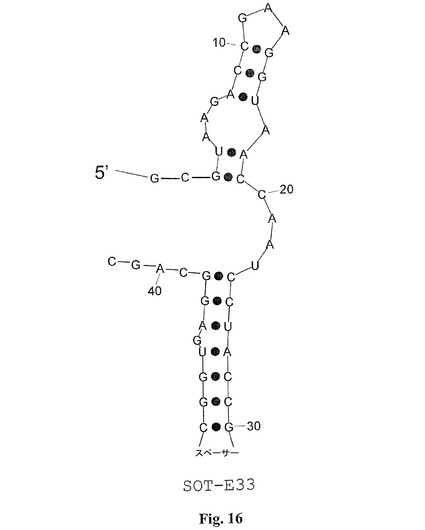
【図17】
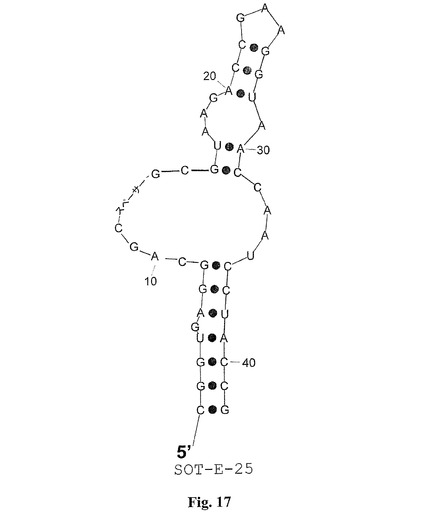
【図18】
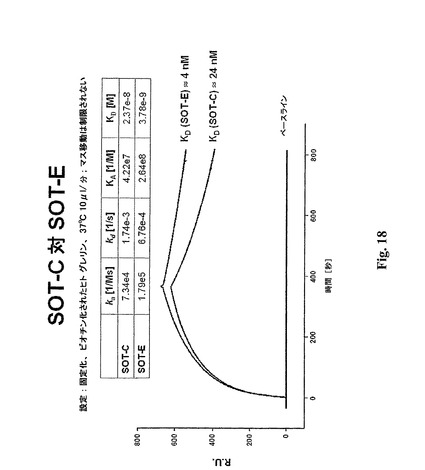
【図19】
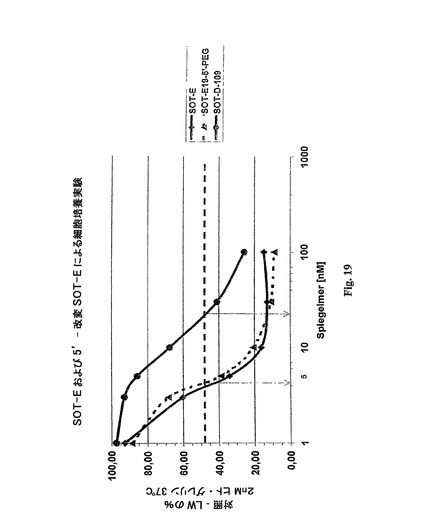
【図20A】
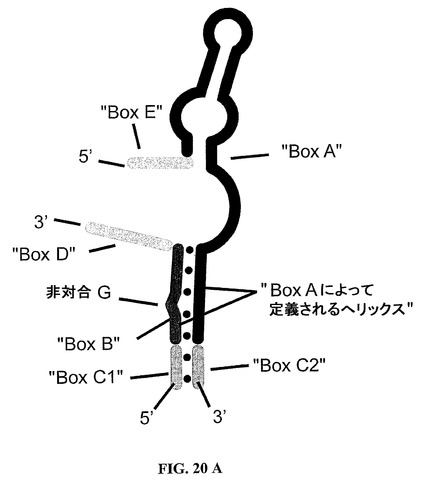
【図20B】
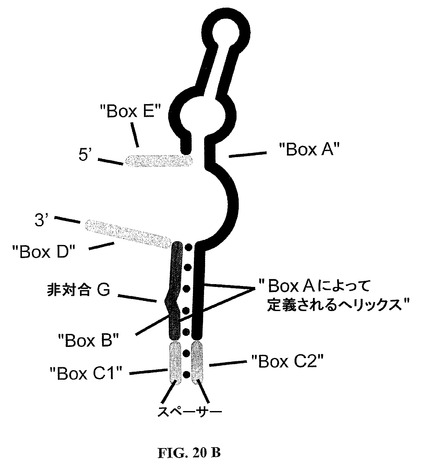
【図20C】
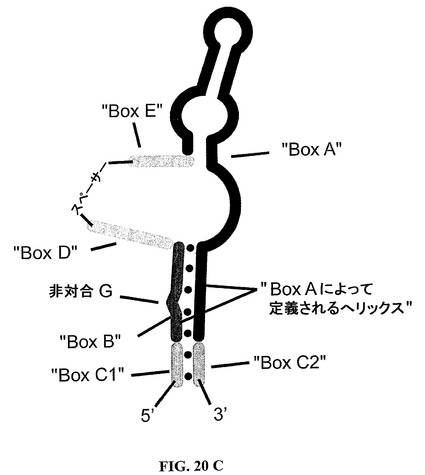
【図20D】
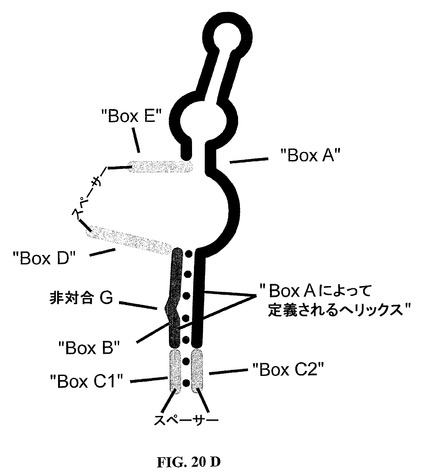
【図21】
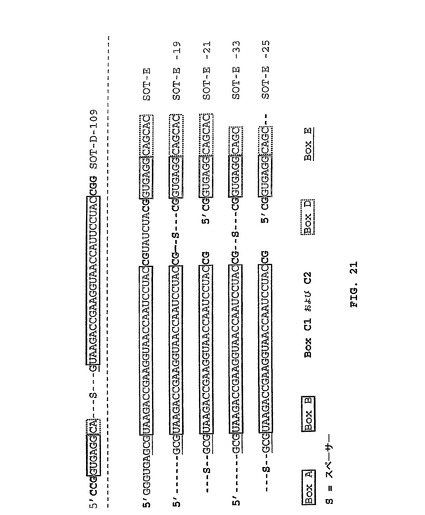
【図22】
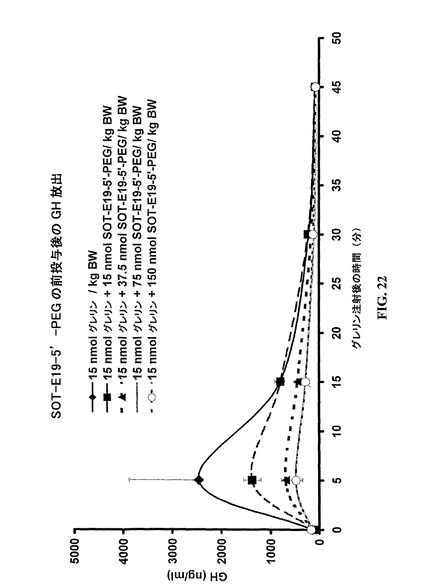
【図23】
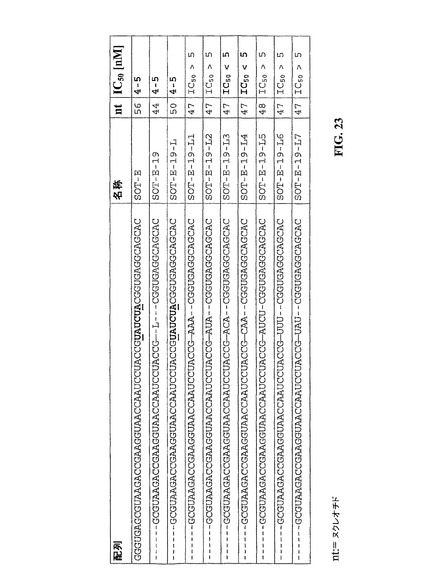
【図24】
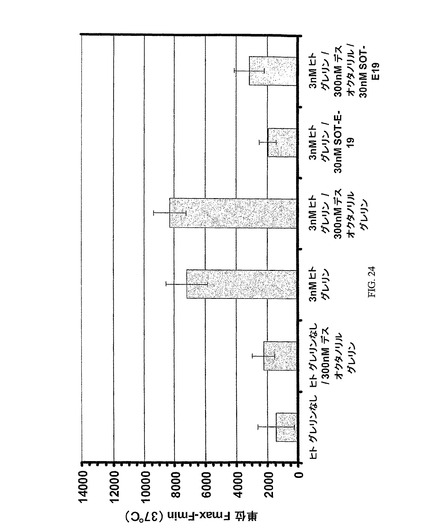
【図25】
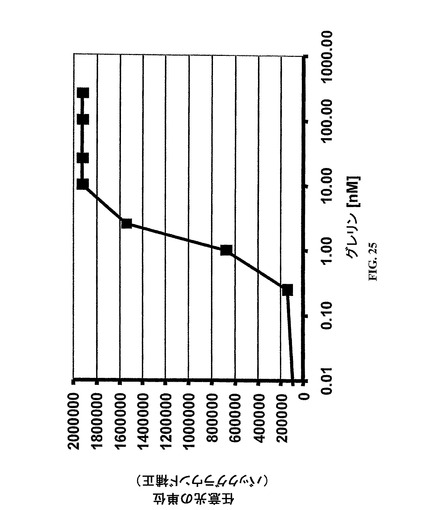
【図26A】
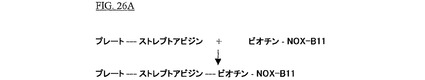
【図26B】
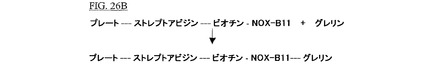
【図26C】
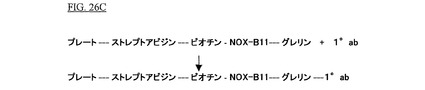
【図26D】
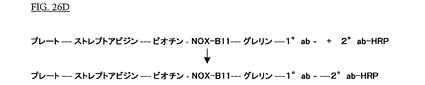
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20A】
【図20B】
【図20C】
【図20D】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26A】
【図26B】
【図26C】
【図26D】
【公表番号】特表2008−534024(P2008−534024A)
【公表日】平成20年8月28日(2008.8.28)
【国際特許分類】
【出願番号】特願2008−504702(P2008−504702)
【出願日】平成18年4月10日(2006.4.10)
【国際出願番号】PCT/EP2006/003276
【国際公開番号】WO2006/108599
【国際公開日】平成18年10月19日(2006.10.19)
【出願人】(504411214)ノクソン・フアルマ・アクチエンゲゼルシヤフト (8)
【Fターム(参考)】
【公表日】平成20年8月28日(2008.8.28)
【国際特許分類】
【出願日】平成18年4月10日(2006.4.10)
【国際出願番号】PCT/EP2006/003276
【国際公開番号】WO2006/108599
【国際公開日】平成18年10月19日(2006.10.19)
【出願人】(504411214)ノクソン・フアルマ・アクチエンゲゼルシヤフト (8)
【Fターム(参考)】
[ Back to top ]