
グロボトリアオシルセラミドを特異的に認識するモノクローナル抗体及びその作製法
【課題】Gb3/CD77を標的とした有能な抗体を効率的に作製する手段を提供すること。
【解決手段】以下のステップ(1)〜(5)、即ち(1)α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物をGb3/CD77で免疫するステップ;(2)免疫後の非ヒト哺乳動物から抗体産生細胞を回収するステップ;(3)回収した抗体産生細胞を不死化させるステップ;(4)不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化するステップ;及び(5)モノクローナル化によって得られたクローンが産生する抗体を回収するステップを含む、抗Gb3/CD77モノクローナル抗体の作製法が提供される。
【解決手段】以下のステップ(1)〜(5)、即ち(1)α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物をGb3/CD77で免疫するステップ;(2)免疫後の非ヒト哺乳動物から抗体産生細胞を回収するステップ;(3)回収した抗体産生細胞を不死化させるステップ;(4)不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化するステップ;及び(5)モノクローナル化によって得られたクローンが産生する抗体を回収するステップを含む、抗Gb3/CD77モノクローナル抗体の作製法が提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はグロボ系糖脂質であるグロボトリアオシルセラミド(Gb3/CD77)を特異的に認識する抗体(抗Gb3/CD77抗体)の作製法及びその用途に関する。詳しくは、抗Gb3/CD77モノクローナル抗体の効率的な作製法、抗Gb3/CD77モノクローナル抗体及びその用途を提供する。
【背景技術】
【0002】
KohlerとMilsteinがモノクローナル抗体作製のためのハイブリドーマ樹立技術を確立して以来(非特許文献1)、モノクローナル技術は劇的に広まり、世界中の様々な生物学研究分野で広範に利用されている。B細胞ハイブリドーマに限らず、T細胞ハイブリドーマもまた開発され、免疫学的解析に幅広く利用されている(非特許文献2)。モノクローナル抗体は、現代生物学と医学の研究を推進するための、最も有用な技術の一つになってきている(非特許文献3)。モノクローナル抗体の作製過程で、動物の免疫とハイブリドーマのスクリーニングが最も重要である。疾患の診断や癌の免疫治療のために、抗体結合の特異性やCDC活性(補体依存性細胞傷害活性)あるいはADCC活性(抗体依存性細胞傷害活性)のような抗体の生物学的機能が極めて重要である。免疫の効果を向上させるために、免疫原の調製に関して様々な試行が行われてきた。例えば、ハプテンの結合、抗原へのアジュバントの添加、リポソームへの封埋、各々のタンパク質の立体配置から推定されるペプチドの利用などである。しかしながら、効率的なモノクローナル抗体の作製に有用な特定のマウスなどに関してはいまだ報告がない。
【0003】
糖脂質に反応するモノクローナル抗体の作製のために、通常のマウスに腫瘍細胞を注入する方法が、多くの研究室に広く流布した免疫方法であった。EBウイルスによるB細胞の不死化は、融合のための良好なパートナー細胞がなかったヒト型モノクローナル抗体の作製において特に効果的であった(非特許文献4、5)。その後、精製あるいはリコンビナントタンパク質がモノクローナル抗体の作製に広く用いられた。種々の機能性分子のペプチドも、様々な生体分子の免疫原として使用されてきた。ガングリオシドに対して有用なモノクローナル抗体の一式を作製するために精製ガングリオシドが免疫原として用いられた(非特許文献6〜8)が、その際には過去の報告に基づいて(非特許文献9)、特定のマウス系統が用いられた。抗ガングリオシドモノクローナル抗体の作製のために、複合型ガングリオシドを欠損するノックアウトマウスが利用されて、良好な抗体産生が示された(非特許文献10)。本発明者らの研究グループも、GM2/GD2合成酵素欠損(ノックアウト)マウスあるいはGD3合成酵素(ノックアウト)マウスを用いることにより、GD1a(非特許文献11)又はGD3及びGD1b(非特許文献12)に反応する有用な抗体を作製できたことを報告した。これらの抗ガングリオシド抗体は、免疫に使用された変異マウスにおいて欠損する構造に対してより高い特異性と親和性を示した。しかし、グロボ系糖脂質などの中性糖脂質を欠損するノックアウトマウスの利用に関しては、いまだ報告がない。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】G. Kohler, C. Milstein, Continuous cultures of fused cells secreting antibody of predefined specificity, Nature, 256 (1975) 495-497.
【非特許文献2】P.N. Nelson, G.M. Reynolds, E.E. Waldron, E. Ward, K. Giannopoulos, P.G. Murray, Monoclonal antibodies, Mol. Pathol. 53 (2000) 111-117.
【非特許文献3】R.H. Kennett, Hybridomas: a new dimension in biological analyses, In Vitro 17 (1981) 1036-1050.
【非特許文献4】Y. Nagatsuka, S. Hanazawa, T. Yasuda, Y. Ono, Anti-glycolipid antibodies produced by Epstein-Barr virus (EBV)-transformed oligoclonal B cell lines obtained from normal persons and autoimmune disease patients, Hum. Antibodies Hybridomas 5 (1994) 183-186.
【非特許文献5】H. Yamaguchi, K. Furukawa, S.R. Fortunato, P.O. Livingston, K.O. Lloyd, H.F. Oettgen, L.J. Old, Cell-surface antigens of melanoma recognized by human monoclonal antibodies, Proc. Natl. Acad. Sci. U. S. A. 84 (1987) 2416-2420.
【非特許文献6】I. Kawashima, N. Tada, S. Ikegami, S. Nakamura, R. Ueda, T. Tai, Mouse monoclonal antibodies detecting disialogangliosides on mouse and human T lymphomas, Int. J. Cancer 41 (1988) 267-274.
【非特許文献7】M. Kotani, H. Ozawa, I. Kawashima, S. Ando, T. Tai, Generation of one set of monoclonal antibodies specific for a-pathway ganglio-series gangliosides, Biochim. Biophys. Acta 1117 (1992) 97-103.
【非特許文献8】H. Ozawa, M. Kotani, I. Kawashima, T. Tai, Generation of one set of monoclonal antibodies specific for b-pathway ganglio-series gangliosides, Biochim. Biophys. Acta. 1123 (1992) 184-190.
【非特許文献9】I. Kawashima, O. Nakamura, T. Tai, Antibody responses to ganglio-series gangliosides in different strains of inbred mice, Mol. Immunol. 29 (1992) 625-632.
【非特許文献10】M.P. Lunn, L.A. Johnson, S.E. Fromholt, S. Itonori, J. Huang, A.A. Vyas, J.E. Hildreth, J.W. Griffin, R.L. Schnaar, K.A. Sheikh, High-affinity anti-ganglioside IgG antibodies raised in complex ganglioside knockout mice: reexamination of GD1a immunolocalization. J. Neurochem. 75 (2000) 404-412.
【非特許文献11】B.J. Willison, D. Nicholl, E.R. Wagner, K. Townson, C. Goodyear, K. Furukawa, K. Furukawa, J. Conner, H.J. Willison, Innate murine B cells produce anti-disialosyl antibodies reactive with Campylobacter jejuni LPS and gangliosides that are polyreactive and encoded by a restricted set of unmutated V genes, J. Neuroimmunol. 152 (2004) 98-111.
【非特許文献12】J. Boffey, M. Odaka, D. Nicoll, E.R. Wagner, K. Townson, T. Bowes, J. Conner, K. Furukawa, H.J. Willison, Characterization of the immunoglobulin variable region gene usage encoding the murine anti-ganglioside antibody repertoire, J. Neuroimmunol. 165 (2005) 92-103.
【発明の概要】
【発明が解決しようとする課題】
【0005】
Gb3/CD77は、ヒト(参考文献6)及びマウス(参考文献7)の胚中心B細胞の一部、腎臓、小腸、血管内皮細胞に発現するとともに、バーキットリンパ腫(参考文献6)及び大腸癌(参考文献9、10)などの腫瘍細胞において過剰発現することが知られている。多くの研究で、Gb3/CD77が細菌毒素の受容体として機能する(参考文献11)ことが報じられているが、その生理的な役割に関してはほとんど分かっていない。従って、Gb3/CD77の機能解明に有用な手段が切望されるところである。また、これまでの報告を総合すれば、医療目的においてGb3/CD77が重要な標的になることは明らかであり、Gb3/CD77を標的とした診断技術や治療技術の創出が期待される。このような状況下、本発明はGb3/CD77を標的とした有能な抗体を効率的に作製する手段を提供することを課題とする。また、当該手法によって得られる抗体の利用を図ること(用途の提供)も課題とする。
【課題を解決するための手段】
【0006】
上記課題を解決すべく本発明者らは、グロボ系列の糖脂質を欠損する変異マウスを利用すれば、当該マウスにおいて消失した既知あるいは未知の糖鎖構造に対するモノクローナル抗体を効率よく作製できるとの期待の下、検討を進めた。具体的には、グロボ系列の糖脂質の合成に必須のα-1.4ガラクトース転移酵素遺伝子を欠損するノックアウトマウスを用いて、スフィンゴ糖脂質グロボトリアオシルセラミド(Gb3)と反応するモノクローナル抗体の作製を試みた。その結果、野生型マウスの免疫による従来法に比べて、明らかに良好なモノクローナル抗体を作製できることが判明した。詳しくは、従来法よりも多数のクローンが得られ、しかも、得られたクローンの半数がIgGクラスであった(従来法で得られた3クローンはすべてIgMクラスであった)。しかも、得られた抗体の中には、CDC活性に加えて、ADCC活性を示すものが含まれていた。さらに、c.52と名付けた抗体はバーキットリンパ腫細胞株に対する凝集活性を示した。興味深いことに、凝集の過程で、c.52抗体の結合により、抗B細胞受容体抗体の結合時に見られるのと類似の数個のタンパク質のチロシンリン酸化が誘導された。特に、c.52抗体の結合により、E3ユビキチンリガーゼとして知られるc-Cblという名の120kDaのタンパク質の強いリン酸化が惹起された。抗B細胞受容体抗体による刺激を行う前に、B細胞を抗Gb3モノクローナル抗体で前処理した場合、Gb3を介するシグナルがB細胞受容体からの抗原疑似シグナルを抑制することが判明した。
【0007】
以上のように、期待を超える成果がもたらされ、α-1.4ガラクトース転移酵素遺伝子欠損マウスがグロボ系糖脂質に対するモノクローナル抗体の作製に極めて有効であることが示された。また、抗Gb3/CD77モノクローナル抗体の利用を図る上で有益な興味深い知見が得られた。当該知見は、抗Gb/CD77モノクローナル抗体の有用性を裏付けるとともに、本モノクローナル作製法によれば、生理機能を良好に発揮する抗Gb/CD77モノクローナル抗体を効率的に作製できることを示す。以下に列挙する本発明は、主として上記の成果に基づく。
[1]以下のステップ(1)〜(5)を含む、抗Gb3/CD77モノクローナル抗体の作製法:
(1)α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物をGb3/CD77で免疫するステップ;
(2)免疫後の非ヒト哺乳動物から抗体産生細胞を回収するステップ;
(3)回収した抗体産生細胞を不死化させるステップ;
(4)不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化するステップ;及び
(5)モノクローナル化によって得られたクローンが産生する抗体を回収するステップ。
[2]非ヒト哺乳動物がマウスである、[1]に記載の作製法。
[3]マウスがA/J系統である、[2]に記載の作製法。
[4]Gb3/CD77、リピッドA、コレステロール及びジパラミトイルホスファチジルコリンを含むリポソームを免疫原としてステップ(1)における免疫を行う、[1]〜[3]のいずれか一項に記載の作製法。
[5]抗体産生細胞が脾臓細胞である、[1]〜[4]のいずれか一項に記載の作製法。
[6]ステップ(3)が、抗体産生細胞と骨髄腫細胞との融合によって行われる、[1]〜[5]のいずれか一項に記載の作製法。
[7]ステップ(4)において、IgGクラスの抗Gb3/CD77抗体を産生する細胞をモノクローナル化する、[1]〜[6]のいずれか一項に記載の作製法。
[8]Gb3/CD77を特異的に認識し、補体依存性細胞傷害活性及び/又は抗体依存性細胞傷害活性を示す、単離されたモノクローナル抗体。
[9]バーキットリンパ腫細胞株に対する凝集活性を示す、[8]に記載の単離されたモノクローナル抗体。
[10][1]〜[7]のいずれか一項に記載の作製法によって得られる、[8]又は[9]に記載の単離されたモノクローナル抗体。
[11]ヒト抗体又はヒト化抗体である、[9]又は[10]に記載の単離されたモノクローナル抗体。
[12]IgG、Fab、Fab'、F(ab')2、scFv、又はdsFv抗体である、[9]〜[11]のいずれか一項に記載の単離されたモノクローナル抗体。
[13][9]〜[12]のいずれか一項に記載の単離されたモノクローナル抗体を含む、Gb3/CD77を発現する細胞を標的とした研究用試薬。
[14][9]〜[12]のいずれか一項に記載の単離されたモノクローナル抗体を含む、Gb3/CD77を過剰発現する悪性腫瘍を標的とした診断又は治療用の組成物。
[15]悪性腫瘍がバーキットリンパ腫、大腸癌、奇形腫又は巨核芽球性白血病である、[14]に記載の組成物。
【図面の簡単な説明】
【0008】
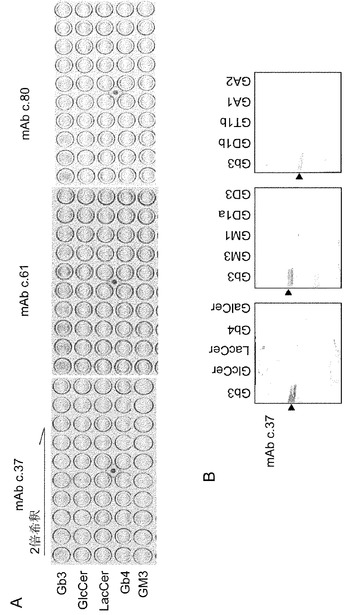
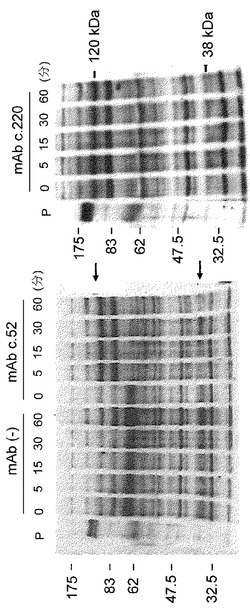
【図1】A/JマウスをGb3/CD77によって免疫して得られたモノクローナル抗体の特異性と反応性を示す図。(A) 精製糖脂質(20 ng/ウェル)に対して、2段階希釈のモノクローナル抗体(ハイブリドーマ上清)を用いて行ったELISAの結果の一例。上清を洗浄した後、HRP標識2次抗体を加え一時間静置した。洗浄後、基質溶液を添加し、モノクローナル抗体の結合を検出した。10〜20分後に、発色の程度をスキャナーにより記録した。(B) A/Jマウスから得られたモノクローナル抗体の、TLC免疫染色による様々な糖脂質に対する特異性。TLC免疫染色は、得られたモノクローナル抗体(ハイブリドーマ上清1:2 希釈)によって<材料と方法>に記載したように行った。TLCは、C:M:DW (60:35:8)の展開用溶媒にて行った。糖脂質(0.5μg/ウェル)を用いた。抗体の結合は、ABC complexTM キットとコニカイムノステインHRP 1000TMによって検出した。
【図2】Gb3合成酵素ノックアウトマウスから得られたモノクローナル抗体の特異性と反応性を示す図。TLC免疫染色は、得られたモノクローナル抗体(ハイブリドーマ上清1 : 2 希釈)を用いて、<材料と方法>に記載したように行った。TLCは、展開溶媒系C:M:DW (60:35:8)にて行った。糖脂質(0.5μg/ウェル)を用いた。抗体の結合をABC complexTM及びコニカイムノステインHRP 1000TMにて検出した。モノクローナル抗体c.76はこのアッセイ系に使用できなかったので、7種の抗体に関する検討結果を示す。
【図3】Gb3合成酵素ノックアウトマウスから得られたモノクローナル抗体の種々糖脂質に対する特異性と反応性を示す図。得られたモノクローナル抗体(ハイブリドーマ上清)を連続2倍希釈してELISAを行った。洗浄後、HRP標識2次抗体を添加した。洗浄後、基質溶液を加えることによってモノクローナル抗体の結合を検出した。
【図4】Ramos細胞に対するモノクローナル抗体の抗腫瘍活性を示す図。補体依存性細胞傷害活性。Ramos細胞を得られたモノクローナル抗体一式と静置した後(x20〜x10240希釈の腹水)、ウサギ血清を補体源として添加し、静置した。補体依存性細胞傷害活性は、<材料と方法>に記載したようにWST-1アッセイにより行った。実線:血清なし、広間隔の点線:血清(1:10希釈)、狭間隔の点線:血清(1:2 希釈)。
【図5】抗体依存性細胞傷害活性を示す図。Ramos細胞またはSK-MEL-28細胞をモノクローナル抗体c.52、c.95、c.37又はR24と各々静置した後、末梢血単核球を加えて一時間静置した。コントロールのIgGをコントロールとして用いた。ADCC活性は、放出されたLDHをCytoTox 96 Non-Radioactive Cytotoxicity AssayTM を用いて<材料と方法>に記載したように測定することで検出した。○:コントロールIgG、●:モノクローナル抗体c.52又はモノクローナルR24。
【図6】ベロ毒素によるアポトーシス誘導に対する、抗Gb3/CD77モノクローナル抗体の細胞前処理の効果を示す図。(A)カスパーゼ3のウエスタンイムノブロッテイングの結果。Ramos細胞をVT-2と様々な時間静置して、その細胞融解液を<材料と方法>に記したごとくイムノブロッテイングに使用した。元のカスパーゼ3(32 kDa)の分解産物が、ベロ毒素を加えてから3時間で出現した。(B) 抗Gb3/CD77抗体の前処置のアポトーシスに対する効果。Ramos細胞を抗Gb3/CD77抗体により4℃、2時間反応させた後、VT2(200 ng/ml)の存在下で3時間静置した。イムノブロッテイングの結果、モノクローナル抗体c.168、c.52及びc.95によって処理された時には、元のカスパーゼ3のバンドが少々残留することが分かった。カスパーゼ3のバンドの相対的強さを示す。即ち、非分解および分解カスパーゼ3の和に対する非分解カスパーゼ3の比率をBのバンド強度から計算してプロットした。繰り返し実験においても同様の結果が得られた。
【図7】Ramos細胞の形態に対する抗Gb3/CD77モノクローナル抗体の影響を示す図。 (A, B) Ramos細胞をモノクローナル抗体c.52、c.95又はc.220と静置した。PBSを陰性コントロールとして用いた。モノクローナル抗体c.52と一分間静置した細胞のみが凝集を起こした。(C) Ramos細胞(1x107細胞/ml)をモノクローナル抗体c.52、c.220又はc.95(20μg/ml)と反応させた。図に示された時間毎にWST-1アッセイを行った。同じ実験を何度か繰り返して、同様の結果が得られた。
【図8】モノクローナル抗体c.52によって誘導されたタンパク質のチロシンリン酸化を示す図。モノクローナル抗体c.52によって誘導されるタンパク質のチロシンリン酸化を検討した。Ramos細胞(107細胞/ml)を、モノクローナル抗体c.52(20μg/ml)又はc.220(20μg/ml)により37℃で図に示した時間、刺激した。細胞融解液を調製してPY20によるイムノブロッテイングの解析に用いた。
【図9】モノクローナル抗体c.52によってチロシンリン酸化されるタンパク質は、抗μ抗体によりリン酸化されるタンパク質に類似している。(A)モノクローナル抗体c.52の処置によって惹起されるチロシンリン酸化タンパク質のウエスタンイムノブロッテイングの結果。Ramos細胞をモノクローナル抗体c.52(0〜40μg/ml)と5分間反応させた後、細胞融解液をPY20によるイムノブロッテイングに用いた。(B) 抗μAbによるチロシンリン酸化タンパク質のウエスタンイムノブロッテイング。Ramos細胞を抗μ抗体(0〜40μg/ml)と反応させた後、その融解液をPY20によるウエスタンイムノブロッテイングに用いた。同様の実験を、より低濃度の抗μ抗体を用いて行った(B-2)。
【図10】p120がc-Cblであることの同定とリン酸化のキネテイクスを示す図。(A) モノクローナル抗体c.52で5分間処置した前後に、PY20あるいは抗c-Cbl抗体を用いて免疫沈降を行った。その免疫沈降物を用いて、PY20あるいは抗c-Cb抗体によるイムノブロッテイングを行った。(B) モノクローナル抗体c.52の刺激によるc-Cblのリン酸化の時間経過。モノクローナル抗体c.52により刺激されたRamos細胞の溶解液(1x107細胞由来)を用いて、抗c-Cbl抗体による免疫沈降を行った。その後、免疫沈降物に対して、HRP-PY20又は抗c-Cbl抗体とHRP標識抗ウサギIgG によるイムノブロッテイングを行った。
【図11】モノクローナル抗体c.52による前処置の、抗μ抗体によるタンパク質リン酸化パターンに対する影響を示す図。(A) 細胞融解液のウエスタンイムノブロッテイング。Ramos細胞をモノクローナル抗体c.52またはPBSと図に示された時間だけ反応させた後、抗μ抗体と5分間反応させた。細胞溶解液を調製してPY20によるイムノブロッテイングに用いた。(B) Aのp120のバンドの相対的強さを、NIHイメージ1.61によって測定して、βアクチンの値で補正した後、グラフにプロットした。繰り返し行った実験でも同様の結果を得た。
【図12】Gb3/CD77とBCR複合体の細胞内局在を示す図。(A) 分子の浮遊パターンにつき、Triton X-100による抽出物のショ糖密度勾配超遠心法で分画された画分を用いたイムノブロッテイングによって検討した。GM1、脂質ラフトのマーカーは、脂質ラフトに相当する第3−第4画分にすべて存在した。Gb3もまた、以前に報告されたように脂質ラフトに局在した。(B) モノクローナル抗体c.52または抗μ抗体の処置によるRamos細胞上のBCRの局在変化。各画分のイムノブロッテイングは、Aに示されたように、無処置(上)、モノクローナル抗体c.52による処置(中)、抗μ抗体による処置(下)を行ったRamos細胞の溶解液を用いて、抗BCR抗体によって行われた。
【発明を実施するための形態】
【0009】
(用語)
説明の便宜上、本明細書中で使用する用語の一部についてその定義・意味を以下にまとめて記す。
「モノクローナル抗体」は、単一の抗体産生細胞に由来するクローンから得られる抗体であり、複数種の混合物であるポリクローナル抗体に比較して格段に高い特異性を示す。
用語「単離された」とは、その本来の環境(例えば天然の物質の場合は天然の環境)から取り出された状態、即ち人為的操作によって本来の存在状態と異なる状態で存在していることを意味する。従って、「単離された抗体」には、天然であって且つ何ら外的操作(人為的操作)が施されていない抗体、即ちある個体の体内で産生され、そこに留まっている状態の抗体は含まれない。
グロボトリアオシルセラミドは、スフィンゴ糖脂質の一種であるグロボ系列の糖脂質(グロボ系糖脂質)でありGb3と略称される。Gb3は別名CD77抗原とも呼ばれることから、本明細書においては、グロボトリアオシルセラミドを「Gb3/CD77」と表記する場合がある。
【0010】
1.抗Gb3/CD77モノクローナル抗体の作製法
本発明の第1の局面は、Gb3/CD77を特異的に認識するモノクローナル抗体、即ち「抗Gb3/CD77モノクローナル抗体」の作製法を提供する。本発明では免疫学的手法を利用する。具体的には、以下のステップ(1)〜(5)が行われる。
(1)α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物をGb3/CD77で免疫するステップ
(2)免疫後の非ヒト哺乳動物から抗体産生細胞を回収するステップ
(3)回収した抗体産生細胞を不死化させるステップ
(4)不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化するステップ
(5)モノクローナル化によって得られたクローンが産生する抗体を回収するステップ
【0011】
ステップ(1)ではまず、α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物を用意する。当該非ヒト哺乳動物は、既報の方法(参考文献11)に準じて作製することができる。作製法(マウスを用いた場合の一例)の概要を説明すると、α1,4-ガラクトース転移酵素の遺伝子断片を含むゲノム断片の、酵素活性にとって重要な部位をコードする領域に挿入または置換によるneo耐性遺伝子を組込んだターゲッテイングベクターを作成する。培養ES細胞にこのベクターを電気穿孔法にて導入し、neo耐性クローンを得た後、相同遺伝子組換え体を得て、胚盤胞に注入する。その結果得られたキメラマウスから、へテロ接合体、ホモ接合体マウスを作製する。免疫動物として利用可能な非ヒト哺乳動物が用いられる。例えばマウス、ラット、ウサギ、ヤギなどをここでの非ヒト哺乳動物として採用する。好ましくはマウスを用いる。特に、糖脂質(特にガングリオシド)に対する反応性がよいことから、A/J系統のマウスを用いるとよい。A/J系統のマウスは日本チャールズリバーや日本エスエルシーなどから購入できる。
【0012】
一方、免疫用の抗原(免疫原)を用意する。抗原にはGb3/CD77が用いられる。Gb3/CD77は、生体由来の精製品であっても、合成品であってもよい。例えば、生体由来製品のGb3/CD77はLarodan AB社などから入手可能である。抗原として、好ましくは、Gb3/CD77、リピッドA、コレステロール及びジパラミトイルホスファチジルコリン(DPPC)を含むリポソームを免疫原とする。その理由は、抗原の存在様式を自然状態に近づけるためである。尚、当該リポソームの調製は、例えば既報(参考文献13)の方法に従えばよい。調製法(一例)の概要を説明すると、0.5μmolのDPPC、コレステロール、10μgのリピッドA、ガングリオシド100μgをC(クロロホルム):M(メタノール) (2:1)に溶解してエバポレートする。続いて、0.5 mlのPBSに溶かして50℃、1分間の処置後、よく攪拌する。
【0013】
必要に応じて免疫を繰り返し、抗体価の十分な上昇を認めた時点で免疫動物から抗体産生細胞を摘出、回収する。典型的には、脾臓又はその一部を摘出し、抗体産生細胞を回収する。但し、以降の操作が可能である限りにおいて、抗体産生細胞のソースは脾臓に限定されるものではない。
【0014】
次に、得られた抗体産生細胞を不死化させる。典型的には、抗体産生細胞を骨髄腫細胞と融合させハイブリドーマを得る。抗体産生細胞の動物種がマウスであれば、例えばNS-1骨髄腫細胞、P3X63-Ag8653、Sp2/0-Ag14、PAI等を用いることができる。
【0015】
続いて、不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化する。モノクローナル化にはHAT培地(ヒポキサンチン、アミノプテリン、チミジンを含む培地)を用いた選択培養及び限界希釈法が一般的である。この手法では、HAT培地による培養によって、抗体産生細胞と腫瘍細胞が融合したハイブリドーマを選択するとともに、限界希釈法を利用して目的の抗体(抗Gb3/CD77抗体)を産生するハイブリドーマをクローン化する。本発明の一態様では、有用性がより高いといえるIgGクラスの抗体を産生する細胞をモノクローナル化する。換言すれば、IgGクラスの抗Gb3/CD77抗体を産生するか否かを指標としてスクリーニングを行い、不死化した抗体産生細胞クローンを得る。本発明の作製法によれば、IgMクラスのみならずIgGクラスの抗体を産生するクローンも高率で得られることから(後述の実施例を参照)、このようなスクリーニングが可能であり且つ有効である。
【0016】
モノクローナル化によって得られたクローンが産生する抗体を回収することによって目的の抗体が得られる。例えば、選択されたクローンの培養液を精製することによって抗体を回収する。一方、選択されたクローンを所望数以上に増殖させた後、これを動物(例えばマウス)の腹腔内に移植し、腹水内で増殖させて腹水を精製することにより目的の抗体を取得することもできる。上記培養液の精製又は腹水の精製には、プロテインG、プロテインA等を用いたアフィニティークロマトグラフィーが好適に用いられる。また、抗原を固相化したアフィニティークロマトグラフィーを用いることもできる。更には、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、硫安分画、及び遠心分離等の方法を用いることもできる。これらの方法は単独ないし任意に組み合わされて用いられる。
【0017】
Gb3/CD77への特異的結合性を保持することを条件として、得られた抗体に種々の改変を施すことができる。例えば、低分子化合物、タンパク質、標識物質などを融合又は結合させることにより、融合抗体又は標識化抗体を構成することができる。標識物質としては例えば、フルオレセイン、ローダミン、テキサスレッド、オレゴングリーン等の蛍光色素、ホースラディッシュペルオキシダーゼ、マイクロペルオキシダーゼ、アルカリ性ホスファターゼ、β−D−ガラクトシダーゼ等の酵素、ルミノール、アクリジン色素等の化学又は生物発光化合物、32P、131I、125I等の放射性同位体、及びビオチンを挙げることができる。また、化学修飾(例えば、一部のアミノ酸をアセチル化、PEG化、リン酸化、アミド化する)を施すことも可能である。
【0018】
2.抗Gb3/CD77モノクローナル抗体及びその用途
本発明の第2の局面は、CDC活性を示す複数の抗Gb3/CD77モノクローナル抗体の取得に成功した事実、及びCDC活性に加えてはADCC活性をも示す抗Gb3/CD77モノクローナル抗体の取得に成功した事実に基づき、Gb3/CD77を特異的に認識し、CDC活性及び/又はADCC活性を示す、単離された抗体を提供する。本発明の抗体は、例えば、Gb3/CD77を発現する細胞の研究用試薬の成分として、或いはGb3/CD77を過剰発現する悪性腫瘍を標的とした診断又は治療用試薬の有効成分として有用である。
【0019】
本発明の抗体は、典型的には、上記本発明の抗体作製法により調製される。本発明の抗体にはマウス、ラットなどの非ヒト動物の抗体、一部の領域を他の動物(ヒトを含む)のものに置換したキメラ抗体、ヒト化抗体、及びヒト抗体が含まれる。本発明の抗体の由来、種類、クラス、形態などは特に限定されないが、好ましくはIgGクラスの抗体である。例えば、マウス抗体のサブクラスIgG1、IgG2a、IgG2b又はIgG3、或いはヒト抗体のサブクラスIgG1、IgG2、IgG3又はIgG4に属する抗体である。
【0020】
本発明の抗体の一形態はヒト化抗体である。「ヒト化抗体」とは、ヒトの抗体に構造を類似させた抗体のことをいい、ヒト型キメラ抗体(例えば抗体の一部がヒト化された抗体、CH2領域がヒト化された抗体、Fc領域がヒト化された抗体、定常領域がヒト化された抗体)、及び定常領域及び可変領域に存在するCDR(相補性決定領域)以外の部分がヒト化されたヒト型CDR移植(CDR-grafted)抗体(P.T.Johons et al., Nature 321,522(1986))、完全ヒト化抗体などが含まれる。ヒト型CDR移植抗体の抗原結合活性を高めるため、マウス抗体と相同性の高いヒト抗体FRを選択する方法、相同性の高いヒト化抗体を作製する方法、ヒト抗体にマウスCDRを移植した後さらにFRのアミノ酸を置換する方法の改良技術もすでに開発され(米国特許第5585089号、米国特許第5693761号、米国特許第5693762号、米国特許第6180370号、欧州特許第451216号、欧州特許第682040号、特許第2828340号などを参照)、本発明のヒト型CDR移植抗体の作製に利用することもできる。
【0021】
Fab、Fab'、F(ab')2、scFv、又はdsFv抗体などの抗体断片として本発明の抗体を調製することもできる。Fabは、IgG抗体をシステイン存在下パパイン消化することにより得られる、L鎖とH鎖可変領域、並びにCH1ドメイン及びヒンジ部の一部からなるH鎖フラグメントとから構成される分子量約5万の断片である。所望のIgG抗体があれば、これをパパイン消化することによりFabを得ることができる。また、H鎖の一部及びL鎖をコードするDNAを適当なベクターに組み込み、当該ベクターを用いて形質転換した形質転換体よりFabを調製することもできる。
【0022】
Fab'は、後述のF(ab')2のH鎖間のジスルフィド結合を切断することにより得られる分子量が約5万の断片である。所望のIgG抗体があれば、これをペプシン消化し、還元剤を用いてジスルフィド結合を切断することによりFab'を得ることができる。また、Fab同様に、Fab'をコードするDNAを用いて遺伝子工学的に調製することもできる。
【0023】
F(ab')2は、IgG抗体をペプシン消化することにより得られる、L鎖とH鎖可変領域、並びにCH1ドメイン及びヒンジ部の一部からなるH鎖フラグメントとから構成される断片(Fab')がジスルフィド結合で結合した分子量約10万の断片である。所望のIgG抗体があれば、これをペプシン消化することによりF(ab')2を得ることができる。また、Fab同様に、F(ab')2をコードするDNAを用いて遺伝子工学的に調製することもできる。
【0024】
scFvは、H鎖可変領域とL鎖可変領域とからなるFvを、片方の鎖のC末端と他方のN末端とを適当なペプチドリンカーで連結し一本鎖化した抗体断片である。ペプチドリンカーとしては、例えば柔軟性の高い(GGGGS)3などを用いることができる。例えば、H鎖可変領域及びL鎖可変領域をコードするDNAとペプチドリンカーをコードするDNAを用いてscFv抗体をコードするDNAを構築し、これを適当なベクターに組み込み、当該ベクターを用いて形質転換した形質転換体よりscFvを調製することができる。
【0025】
dsFvは、H鎖可変領域及びL鎖可変領域の適切な位置にCys残基を導入し、H鎖可変領域とL鎖可変領域とをジスルフィド結合により安定化させたFv断片である。各鎖におけるCys残基の導入位置は分子モデリングにより予測される立体構造に基づき決定することができる。例えばH鎖可変領域及びL鎖可変領域のアミノ酸配列から立体構造を予測し、かかる予測に基づき変異を導入したH鎖可変領域及びL鎖可変領域をそれぞれコードするDNAを構築し、これを適当なベクターに組み込み、そして当該ベクターを用いて形質転換した形質転換体よりdsFvを調製することができる。
【0026】
(研究用途)
本発明は更に、本発明の抗体の用途として、Gb3/CD77を発現する細胞を標的とした、本発明の抗体を含む研究用試薬を提供する。本発明の研究用試薬は、Gb3/CD77を発現する細胞(例えばB細胞の一部、腎臓細胞、小腸細胞、血管内皮細胞など)を標的とする或いは使用する研究に広く適用可能である。特に、バーキットリンパ腫や大腸癌など、Gb3/CD77の過剰発現を認める悪性腫瘍の研究や、Gb3/CD77の生理的役割の解明を目指した研究において有用である。標識化された本発明の抗体は、当該研究用途に好適な形態の一つである。本発明の研究用試薬には、本発明の抗体の他、溶媒、緩衝液、安定剤、防腐剤などを含有させることができる。
【0027】
(診断・治療用途)
本発明の抗体を用いれば、Gb3/CD77を発現する細胞を特異的に検出したり識別したりすることが可能である。従って、Gb3/CD77の過剰発現を認めるバーキットリンパ腫や大腸癌等の悪性腫瘍の診断、治療への本発明の抗体の適用も期待される。そこで本発明は更に、Gb3/CD77を過剰発現する悪性腫瘍を標的とした診断又は治療に利用可能な、本発明の抗体を含む組成物を提供する。標的となる悪性腫瘍の例は、バーキットリンパ腫、大腸癌、奇形腫、巨核芽球性白血病である。本発明の治療用組成物の一態様では、CDC活性又はADCC活性を示す抗Gb3/CD77モノクローナル抗体が有効成分として含有される。好ましくは、ADCC活性を示す抗Gb3/CD77モノクローナル抗体を用いる。この態様の治療用組成物では、ADCC活性を利用した細胞障害によって治療効果を得ることができる。
【0028】
本発明の治療用組成物の別の態様では、抗Gb3/CD77モノクローナル抗体をDDS用の運搬体として利用する。つまりこの態様は抗Gb3/CD77モノクローナル抗体に薬物(細胞毒など)又は放射性同位元素など(これらをまとめて「活性成分」ともいう)を結合して得られる免疫複合体を提供する。殺細胞活性又は細胞傷害活性を持つ薬物(細胞毒)を含有する免疫複合体は一般にイムノトキシンと呼ばれる。細胞毒の例には、タキソール、シトカラシンB、グラミシジンD、臭化エチジウム、エメチン、ミトマイシン、エトポシド、テノポシド、ビンクリスチン、ビンブラスチン、コルヒチン、ドキソルビシン、ダウノルビシン、ジヒドロキシアントラシンジオン、ミトキサントロン、ミトラマイシン、アクチノマイシンD、1-デヒドロテストステロン、糖質コルチコイド、プロカイン、テトラカイン、リドカイン、プロプラノロール、及びプロマイシン並びにこれらの類似体又は同族体を挙げることができる。免疫複合体に含有させる活性成分として、所望の生物活性を有するタンパク質又はペプチドを使用してもよい。このような目的において使用可能なタンパク質等の候補として、アブリン、リシンA、シュードモナス・エキソトキシン、ジフテリア毒素、腫瘍壊死因子、インターフェロン-γ、インターロイキン-1(IL-1)、インターロイキン-2(IL-2)、インターロイキン-6(IL-6)、顆粒球マクロファージコロニー刺激因子(GM-CSF)、顆粒球コロニー刺激因子(G-CSF)リンホカインを例示できる。活性部分を抗体に結合させる技術は公知であり、例えばMonoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985)、Controlled Drug Delivery (2nd edition.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987)、Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985)、Thorpe et al., "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-58 (1982)を参照することができる。
【0029】
治療用組成物の製剤化は常法に従って行うことができる。製剤化する場合には、製剤上許容される他の成分(例えば、担体、賦形剤、崩壊剤、緩衝剤、乳化剤、懸濁剤、無痛化剤、安定剤、保存剤、防腐剤、生理食塩水など)を含有させることができる。賦形剤としては乳糖、デンプン、ソルビトール、D-マンニトール、白糖等を用いることができる。崩壊剤としてはデンプン、カルボキシメチルセルロース、炭酸カルシウム等を用いることができる。緩衝剤としてはリン酸塩、クエン酸塩、酢酸塩等を用いることができる。乳化剤としてはアラビアゴム、アルギン酸ナトリウム、トラガント等を用いることができる。懸濁剤としてはモノステアリン酸グリセリン、モノステアリン酸アルミニウム、メチルセルロース、カルボキシメチルセルロース、ヒドロキシメチルセルロース、ラウリル硫酸ナトリウム等を用いることができる。無痛化剤としてはベンジルアルコール、クロロブタノール、ソルビトール等を用いることができる。安定剤としてはプロピレングリコール、ジエチリン亜硫酸塩、アスコルビン酸等を用いることができる。保存剤としてはフェノール、塩化ベンザルコニウム、ベンジルアルコール、クロロブタノール、メチルパラベン等を用いることができる。防腐剤としては塩化ベンザルコニウム、パラオキシ安息香酸、クロロブタノール等と用いることができる。製剤化する場合の剤型も特に限定されず、例えば錠剤、散剤、細粒剤、顆粒剤、カプセル剤、シロップ剤、注射剤、外用剤、及び座剤などとして調製できる。
【0030】
本発明の治療用組成物を用いた治療においては、標的となる悪性腫瘍を保有する対象(患者)に本発明の治療用組成物が投与される。本発明の治療用組成物はその形態に応じて経口投与又は非経口投与(静脈内、動脈内、皮下、筋肉、腹腔内注射、標的細胞への直接導入など)によって対象(患者)に適用され得る。投与量は症状、患者の年齢、性別、及び体重などによって異なるが、当業者であれば適宜適当な投与量を設定することが可能である。投与スケジュールとしては例えば一日一回〜数回、二日に一回、或いは三日に一回などを採用できる。投与スケジュールの設定においては、患者の病状や有効成分の効果持続時間などを考慮することができる。
【実施例】
【0031】
<材料と方法>
(1)動物
A/Jマウス、BALB/cnu/nuマウス及びKSNnu/nuマウスは日本チャールズリバーから購入した。α-1,4ガラクトース転移酵素遺伝子(A4galt-)欠損マウスは我々の研究室で樹立された(参考文献11)。動物実験に関するすべてのプロトコールは、「実験動物の管理と使用に関する指針(National Institutes of Health Guide for the Care and Use of Laboratory Animals(1966))に沿ったものであり、名古屋大学医学系研究科の動物実験委員会の承認を得た。すべての動物は特定病原微生物のいない条件下(specific pathogen-free)で維持された。
【0032】
(2)細胞培養
本研究で使用した骨髄腫細胞株NS-1、バーキットリンパ腫細胞Ramosは10% 牛胎仔血清(FCS)を含むRPMI 1640により培養した。cDNAをトランスフェクトしたL細胞は、ダルベッコ修飾イーグル培地(Dulbecco's modified eagle's medium;DMEM)に7.5% FCS及びG418(300μg/ml)を加えた培養液により維持された。バーキットリンパ腫細胞株であるRamos、HS-Sultan、Daudi及びRajiは日本健康科学財団(Japan Health Science Foundation)から入手した。A4galt-導入L細胞(LVTR-1)及びベクターコントロール細胞株(Lneo-2)は以前の報告の通り樹立された(参考文献12)。
【0033】
(3)Gb3/CD77に対するモノクローナル抗体の作製
3匹の8週齢A/Jマウスに対して、Gb3/CD77糖脂質(100μg)、リピッドA(10μg)、コレステロール(0.5μmol)、ジパラミトイルホスファチジルコリン(dipalmitoyl- phosphatidylcholine; DPPC)(0.5μmol)を含むリポソーム(参考文献13)を用いて2週間毎に皮下に免疫を行った。マウス血清中の抗体価を免疫蛍光法(IF)によって決定した。5回目の免疫から3日後に脾細胞を取り出し、ポリエチレングリコール(PEG)を用いてNS-1骨髄腫細胞との融合を行った。HAT選択培地(Sigma, St Louis, MO)により選択後、免疫蛍光法(IF)及び酵素結合免疫吸着法(enzyme-linked-immuno-solvent assay; ELISA)によって、培養上清中の抗体を検出した。陽性クローンを限界希釈法によりサブクローニングし、TLC-免疫染色法(TLC-immunostaining)によってその特異性を更に確認した。A4galt-欠損変異マウスについては、8週齢のマウスを用い、第0日、4日、7日、11日目に上記構成のリポソームを皮下免疫して、さらに第21日目に尾静脈からブースター免疫を実施した。各マウスの抗体価は、FITC標識抗マウスIgG抗体(Zymed Lab. Inc., SanFrancisco, CA)を用いたIFにより測定した。ハイブリドーマの培養上清をサンプルとしてELISA及び/又はIF法を実施し、陽性クローンについてはサブクローニングを行い、さらにTLC-免疫染色により特異性を確認した。陽性クローンの培養上清中の抗体は、プロテインA-アガロース4ファーストフローを用いたアフィニテイカラムにより、添付のプロトコール(Amersham Biosciences, Bucks, UK)に従って精製した。
【0034】
(4)ELISA
メタノールに溶かした市販の糖脂質(20 ng/10μl)を60穴Terasakiプレート(Greiner BioOne, Frickenhausen, Germany)にプレーテイングした。20分かけて空気乾燥した後、プレートに5% BSA含有リン酸緩衝液(PBS)を添加して2時間室温にてブロッキングを行った。2回洗浄後、段階希釈した抗体をプレートに加えて2時間室温で静置した。5回洗浄した後、HRP-標識抗マウスIgGs(Amersham Biosciences)を2次抗体として添加した。5回洗浄後、o-フェニレンジアミン(ortho-phenylene diamine)(2 mg) (Wako Junyaku, Osaka, Japan)とH2O2(8μl)(Wako)を5 mlのクエン酸緩衝液(citrate-phosphate buffer)に溶解した基質溶液(10μl)をプレートに添加した。適当な時間暗所で静置してから、発色度をスキャナーを用いて記録した。
【0035】
(5)IFアッセイ
LVTR-1あるいはLneo-2細胞をTerasakiプレートに加え(500細胞/穴)、一夜静置した。培養上清を除去した後、段階希釈した培養上清又は腹水をプレートに加え室温で1時間静置した。2回洗浄後、FITC標識抗マウスIgG (H+L)抗体(Cappel, Durham, NC)を加えて、1時間室温暗所にて静置した。2回洗浄後、抗体の結合を蛍光顕微鏡(BX51, OLYMPUS, Tokyo, Japan)下で観察した。
【0036】
(6)TLC免疫染色
TLCは高性能TLCプレート(Merck, Darmstadt, Germany)を用いて、クロロフォルム/メタノール/0.22%塩化カルシウム(chloroform/ methanol/0.22% CaCl2)(60:35:8)を展開溶媒として実施した。Gb3/CD77に対する特異性と反応性は、以前報告したように(参考文献14)、アルミニウム板のシリカプレート(Merck)を用いて検討した。TLCの後、プレートは既報のごとくPVDF(polyvinylidene difluoride)膜に対してブロットした。5% BSAを含むPBSによるブロッキングの後、プレートを培養上清と附置し、抗体の結合をABC kitTM(Vector Laboratories, Burlingame, CA)及びコニカ免疫染色HRP-1000TM(Konica, Tokyo)を用いて検出した。
【0037】
(7)フローサイトメトリー
細胞表面のGb3/CD77について、FACSCaliverTM(Becton Dickinson, Mountain View, CA)にて検討した。要約すると、細胞を希釈した腹水もしくは精製抗体と60分間氷上で静置した後、FITC標識ヤギ抗マウスIgG(H+L)(Cappel, Durham, NC)抗体と氷上で45分反応させた。フローサイトメトリーの対照細胞として2次抗体のみを使用して調製した細胞を用いた。陽性細胞の定量のためにCELLQuestTMプログラムを用いた。
【0038】
(8)増殖試験及び増殖抑制試験
作製した抗体の増殖抑制効果を検討するために、WST-1試験を行った。既報のごとく(参考文献12)、450 nmでの吸光度をELISAリーダー(Immuno Mini NJ-2300 (System Instruments, Tokyo))で検出することによって、WST-1のフォルマザンへの還元を測定した。要約すると、細胞(1x104)を48穴プレートに播き、10% FCSを含むRPMI 1640で一昼夜培養した。その後、希釈した腹水あるいは精製抗体をプレートに添加して培養した。WST-1試験はRamos細胞を用い、三連(triplicate)で24時間毎に行った。抗Gb3抗体の増殖抑制効果は、対照抗体のそれと比較して検討した。
【0039】
(9)ベロ毒素の毒性に対する抗体の阻害効果
細胞(1x106)を48穴プレートに播き、10% FCS含有RPMI 1640にて一晩培養した。その後、希釈した抗体をプレートに添加して氷上で1時間静置した。続いて、細胞を十分に殺しうる最小量のベロ毒素を洗浄なしに添加し、静置した。その後、細胞融解液を調製し、毒素の毒性に対する阻害効果を検討するためのイムノブロッテイングに用いた。
【0040】
(10)CDC試験
RPMIを除去するために細胞を5% FCS含有PBSで洗浄した後、48穴プレートに播いた(1x104/50μl(5% FCS含有PBS))。各ウェルに5% FCS及び種々の量の腹水を含むPBSを50μl添加して、1時間室温にて静置した。その後、5% FCS及びラビット血清(30μl:1/10又は150μl: 1/2)を含んだ200μlのPBSを各ウェルに添加して、最終的に300μl/ウェルの量にて37℃、1時間静置した。CDC反応の後、細胞増殖測定試薬WST-1(Roche, Manheim, Germany)を各々のウェルに添加した(30μl/ウェル)。2時間の静置後、プレートを1分間十分に振盪して、サンプルの450nmでの吸光度をマイクロプレートリーダー(ImmunoMini NJ-2300, Immuno Mini NJ-2300, InterMed, Tokyo, Japan)を用いて測定した。
【0041】
(11)ADCC試験
標的細胞(1x106)を希釈した抗体とファルコンチューブ内で氷上1時間静置し、10% FCS含有RPMIで2回洗浄した後、各々のウェルにプレーテイングした。正常人のヘパリン血(25 ml)よりヒトの末梢単核球を製造元の指示書に従ってフィコール・ハイパック(Ficoll-Hypaque)グラジエント(Amersham Biosciences Inc.)遠心により分離した。これらの細胞を所定の比(標的(target)/エフェクター(effector))になるようにミクロタイタープレートに添加して37℃で4時間静置した。ADCCにより放出されたLDHの量を測定するために、CytoTox 96 Non-Radioactive Cytotoxicity AssayTM(Promega, Madison, WI)を用い、製造元の指示書に沿って検討した。
【0042】
(12)イムノブロッテイングと免疫沈降
細胞を溶解バッファー(20 mM Tris-HCl, pH 7.4。以下の試薬を含む、即ち0.15 M NaCl, 1% Nonidet P-40, 1 mM phenylmethylsulfonyl fluoride (Sigma)及び5μg/ml aprotinin) により溶解した。細胞溶解液(4.0x106細胞に由来する)をSDS-PAGEに供し、その後PVDF膜(Millipore, Bedford, MA)に転写した。5% BSA含有PBSにより1時間室温にてブロッキングした後、PVDF膜を室温で1時間、一次抗体と反応させた。その後、0.05% Tween 20添加PBSにより3回洗浄し、ホースラディッシュペルオキシダーゼ標識抗ラビットIgG(H+L)(BD transduction Laboratories, San Jose, CA)を添加後1時間静置してカスパーゼ3(caspase-3)、p38及びリン酸化p38を検出した。ホースラディッシュペルオキシダーゼ標識した抗マウスIgG(H+L)抗体(Zymed Laboratories Inc. San Francisco, CA)を用いてFAK、Lyn及びc-Cblを検出した。検出はECL systemTM (Amersham Biosciences Inc.)を用いて、製造元の指示書に従った。免疫沈降では、細胞溶解液(4x106細胞に由来する)をPY20、FAK、Lynあるいはc-Cblに対するポリクローナル抗体とプロテインGセファローズ4 first flowTM (Amersham Biosciences)によって免疫沈降させた。免疫複合体を洗浄後、SDSサンプル緩衝液を添加して、3分間煮沸した。
【0043】
(13)抗B細胞受容体(BCR)抗体あるいはmAb c.52によるB細胞刺激
Ramos細胞を10% FCS含有RPMIに107/mLの濃度で懸濁した。刺激前に細胞を37℃で1時間静置した。その後、ヤギ抗ヒトIgM(10μg/ml)(Fc5μ断片特異的抗体、Jackson Immuno-Laboratory, West Grove, PA)又はモノクローナル抗体c.52(10μg/ml)を用いて所定時間刺激した。
【0044】
<結果>
最初に、Gb3を封埋したリポソームによりA/Jマウスを免疫した。免疫したマウスの中で、Gb3に対して最も高い抗体価を示したマウスの脾細胞を用いて、骨髄腫細胞株NS1との融合を行った。ELISA及び蛍光抗体法によるスクリーニングを行った。400クローンのスクリーニングの結果、c.37、c.61、そしてc.80という3種のハイブリドーマを樹立した。これらのハイブリドーマクローンが産生する抗体のGb3/CD77に対する特異性をELISAとTLC免疫染色で解析した(図1A及び1B)。13種の異なる糖脂質に対して、3クローンすべてがGb3/CD77に限って反応した。3クローンはいずれもラクトシルセラミド(lactosylceramide)、GlcCer、GM3及びGb4とは反応せず、得られた抗体が反応するために非還元末端のα4-ガラクトースが必須であることが示唆された。その他の糖脂質に関しては、GalCer、GD1b、GD3、GA1、GT1b、GA2、GM1及びGD1a等への反応性も認められなかった(データ示さず)。同様の反応特異性がTLC免疫染色法によっても観察された(図1B)。mAbs 61及び80もまた、Gb3とのみ反応した(データ示さず)。3クローンの抗体のアイソタイプをELISAにより決定したところ、IgMクラスであることが判明した(データ示さず)。
【0045】
Gb3/CD77に反応性を示し、抗腫瘍効果の点でより高性能である抗体の樹立を目指し、すべてのグロボ系糖脂質を欠損するA4galt欠損マウスを、Gb3封埋リポソームを免疫原として、野生型マウスの場合と同様に免疫した。免疫したマウスの中でGb3/CD77に対して最も高い抗体価を示したマウスを用いて、その脾細胞とマウスNS-1骨髄腫細胞との融合を行った。ELISA及び免疫蛍光法によるスクリーニングの結果、Gb3/CD7に対する反応性に基づいて323クローンから8クローンのハイブリドーマが樹立された。得られたモノクローナル抗体のGb3/CD77に対する特異性をELISA及びTLC免疫染色によって決定した(図2及び3)。13種の糖脂質の中、すべてのクローンがGb3/CD77に限って反応した。種々の糖脂質に対するELISAの結果を図3に示した(GalCer、GD1b、GD3、GA1、GA2、GT1b、GM1及びGD1aに関する結果は省略)。8クローンの抗体のアイソタイプをELISAにより決定したところ、IgM(4クローン)、IgG3(3クローン)及びIgG2b (1クローン)であった(データ示さず)。
【0046】
次に、得られたクローンの抗腫瘍活性を解析するために、バーキットリンパ腫Ramos細胞を標的に用いてCDC活性を検討した。A/Jマウスを用いて得られた3クローンのIgMクラスのモノクローナル抗体、及びA4galt欠損マウスを用いて得られたIgMクラスのモノクローナル抗体は強いCDC活性を示したが、抗体のみでは細胞傷害を誘導しなかった(図4)。A4galt欠損マウスを用いて得られた4クローンのIgGクラス抗体は同様に明らかなCDC活性を誘導したが、その強さはIgM型モノクローナル抗体のそれよりは弱かった。次に、これらの抗体のADCC活性を検討した。モノクローナル抗体c.52はRamos細胞に対してヒトの末梢血単核球をエフェクターとして明らかなADCC活性を誘導した(図5)。IgMあるいはIgG2bクラスのモノクローナル抗体はADCC活性がないか又はごく僅かしか示さなかった(図5)。
【0047】
A4galt欠損マウスからIgGクラスの抗体が複数得られたという事実から、例えばベロ毒素受容体に対する高性能の競合的な役割など、抗体の持つ他の機能の探索を行うことが必要と思われた。図6に示したように、Ramos細胞をベロ毒素2と3時間静置した時に、カスパーゼ3(caspase-3)の分解が惹起され、ベロ毒素による細胞傷害が示された。ベロ毒素2(2 ng/ml)(細胞死を誘導するために必要な最小量)の添加前にRamos細胞を抗Gb3/CD77抗体の存在下で氷上2時間処置した場合には、カスパーゼ3の分解がモノクローナル抗体c.168(IgM)、c.52(IgG3)及びc.95(IgG2b)によって部分的に抑制された。しかし、モノクローナル抗体c.220(IgG3)あるいはR24(コントロール)によっては抑制されなかった(図6B)。モノクローナル抗体によりカスパーゼ3の分解が部分的であれ確かに抑制されたにもかかわらず、細胞死の方は明らかには抑制されなかった。
【0048】
Gb3/CD77は胚中心Bリンパ球の亜集団に発現し、抗体の親和性成熟の過程に必要とされるのかもしれない。そこで、Gb3/CD77に対する未知のリガンドが存在し、胚中心B細胞においてシグナルを惹起するかもしれないと考えた。この仮説を検討するために、Ramos細胞をモノクローナル抗体で処理して形態学的な変化と細胞内のタンパク質リン酸化を解析した。検討したクローンの中、モノクローナル抗体c.52はカルシウム依存性の細胞凝集を惹起したが、モノクローナル抗体c.220は惹起しなかった(図7A及び7B)。モノクローナル抗体c.52はまた、Ramos細胞に対して軽度ではあるが確実な細胞増殖抑制活性を示した(図7C)。これらの抗体を加えた後の細胞を用いたリン酸化タンパク質のイムノブロッテイングの結果、細胞内タンパク質のリン酸化パターンに劇的な変化が起こったことが示された(図8)。これらの変化は、モノクローナル抗体c.220によって処置された細胞でも認められた。但し、リン酸化の程度はモノクローナル抗体c.52による場合に比して軽度であった(図8右)。モノクローナル抗体c.220が細胞凝集を誘導しなかったことからすると、モノクローナル抗体c.52によるリン酸化は細胞凝集によるものではないであろう。抗μ抗体によるB細胞受容体を介するシグナルと比べた時、モノクローナル抗体c.52によって惹起されるタンパク質リン酸化は、抗BCR抗体のクロスリンクによるリン酸化と若干異なりつつも類似していた(図9A及び9B)。数個のリン酸化タンパク質の中で120 kDaのタンパク質はどちらの刺激によっても共通にリン酸化される分子と思われた(図9)。上記したごとく、モノクローナル抗体c.52はRamos細胞に対して軽度であるが明らかな増殖抑制活性を示し、Gb3/CD77を介するシグナルがBCRを介するシグナルや生存シグナルなどの既存のシグナルに対して負の効果をもたらす可能性が示唆された。
【0049】
120 kDaのバンドの分子を同定するために、また、c-Cbl(参考文献15)がモノクローナル抗体c.52によるGb3/CD77のクロスリンクに基づいてリン酸化を受けたかどうかを明らかにするために、免疫沈降(IP)-イムノブロッテイング(IB)解析を行った。その結果、120 kDaのバンドは少なくともその一部がc-Cblであり、それがモノクローナル抗体c.52によるGb3/CD77のクロスリンクによってリン酸化されることが示された(図10A)。c-Cblリン酸化の時間経過を示した(図10B)。Gb3/CD77がBCRシグナルの負の調節役として働くか否かを検討するために、抗μ抗体を加える前にRamos細胞をモノクローナル抗体c.52で前処置した時のタンパク質リン酸化パターンに与える影響を検討した。予想されたように、モノクローナル抗体c.52による前処置により、コントロールIgGと比べて抗μ抗体によるタンパク質リン酸化が有意に抑制された(図11A及び11B)。
【0050】
<考察>
本研究では、Gb3/CD77合成酵素欠損(ノックアウト)マウスを免疫に用いると、抗Gb3抗体を効率よく作製できることが示された。モノクローナル抗体の作製効率及び樹立された抗体の機能の質は、通常のマウスを用いた場合に比して明らかに良好であった。過去の試みにおいては、精製Gb3を免疫原として用いて、もっぱらIgMモノクローナル抗体が産生された(参考文献23)。従来の方法ではIgMクラスのモノクローナル抗体が得られるのが普通であるが、本研究においてはGb3に反応するIgGクラスのモノクローナル抗体が比較的高率に得られた。
【0051】
Gb3はすべてのグロボ系糖脂質の合成の出発分子であり、バーキットリンパ腫の腫瘍マーカーとして(参考文献8)、ベロ毒素の受容体として(参考文献11、24)、あるいはB細胞系列の細胞死マーカーとして(参考文献25)、研究者から注目されてきた。生理的及び病理的な状況下でのグロボ系糖脂質の役割に関しては、現在、グロボ系糖脂質を欠損する変異マウスを用いて、我々の研究室で解析中である(参考文献11)。いずれにしても、様々な機能を有する抗Gb3抗体は、糖脂質の機能解明にとって極めて有用である。
【0052】
実際、カスパーゼ3の分解がこれらの抗体によって部分的に阻害された。しかしながら細胞死の方は、カスパーゼ3の分解が有意に抑制されたのにもかかわらず、明らかには抑制されなかった。これらの結果は、これらの抗体の結合がアポトーシスシグナルの遅延を招いたにせよ、死のシグナルを抑えるほどには十分強くないことを示唆している。
【0053】
モノクローナル抗体の生物学的な機能という点では、本研究で得られた抗Gb3抗体は非常に興味深い。というのは、それらのいくつかは、抗μ抗体によるクロスリンクの場合と同様の方法で、細胞表面に結合してB細胞のシグナル伝達経路の活性化を誘導したからである(参考文献26)。モノクローナル抗体c.52で処理された際に、チロシンリン酸化を受ける成分が、c-Cblを含めて数種存在する。c-CblはE3ユビキチンリガーゼとして知られており(参考文献27)、Sykのような分子をプロテアゾーム系での分解に誘導することによって、BCRを介するシグナルを減弱せしめる(参考文献28)。BCRを介するシグナルにおけるGb3/抗Gb3抗体からのシグナルの役割を解析する中で、Ramos細胞をモノクローナル抗体c.52で前処置することにより、抗μ抗体によるチロシンリン酸化のレベルが軽度に減弱した。この結果から、c-Cbl活性化がBCRを介するシグナルの調節に実際に働いていることが分かった。従って、ここで最も興味深い問題は、Gb3と相互作用する内因性因子が存在するか否か、存在するのであればそれは何か、という点である。
【0054】
Gb3/CD77が胚中心のBリンパ球の亜集団に発現しており、バーキットリンパ腫細胞にモノクローナル抗体が結合することによって細胞凝集やタンパク質のチロシンリン酸化が惹起されることから、我々はGb3/CD77の内因性リガンドが存在して、胚中心B細胞のシグナル制御に働いているのではないかと推測した。Ramos細胞をモノクローナル抗体で前処置した時に、細胞増殖の抑制が見られた事実は、我々の、Gb3/CD77がBCRのクロスリンクによるBCRシグナルなどの既存のシグナルに対して負の効果をもたらすシグナルを導入するのではないか、という想定を支持するものであった。c-Cbl、E3ユビキチンリガーゼがSykやSFK、TecなどBCRを介するシグナル伝達経路の上流に位置するシグナル分子のユビキチンプロテアソーム系による分解に基づいて、BCRシグナルの負の調節役を演じることはよく知られていた(参考文献28、29)。BCRのクロスリンクによって活性化を受けると、それらはリン酸化される。本研究で示されたように、c-Cblはモノクローナル抗体c.52によってリン酸化され、またRamos細胞をc.52で前処置すると、BCRシグナルの減弱とともに細胞増殖も抑制された。これらの結果は、発生過程におけるc-Cblの一般的機能(参考文献30)及び病態における意義(参考文献31)にも関連している。
【0055】
BCR複合体が、同様の抗原あるいは抗μ抗体によってBCRをクロスリンク(参考文献32、33)した後に、非脂質ラフトから脂質ラフトに移行することがよく知られていた(図12)。Gb3/CD77もまた脂質ラフトに局在することから、我々はGb3/CD77が脂質ラフトにおいてBCRシグナルの負の調節に働くとの仮説を立てた。実際、モノクローナル抗体c.52は、抗μ抗体によって誘導されるリン酸化を有意に抑制した。さらに、モノクローナル抗体c.52が結合することで、Ramos細胞の脂質ラフトへのBCRの移行が惹起された(図12)。この事実は、BCRの非脂質ラフトから脂質ラフトへの移行が、BCRの効果的なシグナル伝達を招くことを示唆するだけでなく、Gb3/CD77を介するシグナルの制御に基づいた、ユビキチンプロテアソームを介する分解やキー分子の脱リン酸化などの負のシグナル発信の開始の可能性をも示唆する。
【産業上の利用可能性】
【0056】
本発明のモノクローナル抗体作製法によれば、グロボ系糖脂質であるGb3/CD77を特異的に認識するモノクローナル抗体を効率的に作製できる。即ち、本発明のモノクローナル抗体作製法は、抗Gb3/CD77モノクローナル抗体を効率的に作製する手段として有用である。本発明のモノクローナル抗体作製法はIgGクラスのモノクローナル抗体を取得するための手段として特に有用である。本発明によってもたらされる抗Gb3/CD77モノクローナル抗体には、研究用途に留まらず、診断用途や治療用途など、様々な分野で利用・活用されることが期待される。
【0057】
この発明は、上記発明の実施の形態及び実施例の説明に何ら限定されるものではない。特許請求の範囲の記載を逸脱せず、当業者が容易に想到できる範囲で種々の変形態様もこの発明に含まれる。
本明細書の中で明示した論文、公開特許公報、及び特許公報などの内容は、その全ての内容を援用によって引用することとする。
【0058】
<参考論文>
[1] R.H. Kennett, Hybridomas: a new dimension in biological analyses, In Vitro 17 (1981) 1036-1050.
[2] P.N. Nelson, G.M. Reynolds, E.E. Waldron, E. Ward, K. Giannopoulos, P.G. Murray, Monoclonal antibodies, Mol. Pathol. 53 (2000) 111-117.
[3] B.J. Willison, D. Nicholl, E.R. Wagner, K. Townson, C. Goodyear, K. Furukawa, K. Furukawa, J. Conner, H.J. Willison, Innate murine B cells produce anti-disialosyl antibodies reactive with Campylobacter jejuni LPS and gangliosides that are polyreactive and encoded by a restricted set of unmutated V genes, J. Neuroimmunol. 152 (2004) 98-111.
[4] M.P. Lunn, L.A. Johnson, S.E. Fromholt, S. Itonori, J. Huang, A.A. Vyas, J.E. Hildreth, J.W. Griffin, R.L. Schnaar, K.A. Sheikh, High-affinity anti-ganglioside IgG antibodies raised in complex ganglioside knockout mice: reexamination of GD1a immunolocalization. J. Neurochem. 75 (2000) 404-412.
[5] J. Boffey, M. Odaka, D. Nicoll, E.R. Wagner, K. Townson, T. Bowes, J. Conner, K. Furukawa, H.J. Willison, Characterization of the immunoglobulin variable region gene usage encoding the murine anti-ganglioside antibody repertoire, J. Neuroimmunol. 165 (2005) 92-103.
[6] K.J. Kasai, J. Galton, P.I. Terasaki, A. Wakisaka, M. Kawahara, T. Root, S.I. Hakomori, Tissue distribution of the Pk antigen as determined by a monoclonal antibody, 12 (1985) J. Immunogenet. 213-220
[7] Y. Fujii, S. Numata, Y. Nakamura, T. Honda, K. Furukawa, T. Urano, J. Wiels, M. Uchikawa, N. Ozaki, S. Matsuo, Y. Sugiura, K. Furukawa, Murine glycosyltransferases responsible for the expression of globo-series glycolipids: cDNA structures, mRNA expression, and distribution of their products, Glycobiology 15 (2005) 1257-1267.
[8] E. Nudelman, R. Kannagi, S. Hakomori, M. Parsons, M. Lipinski, J. Wiels, M. Fellous, T.A. Tursz, glycolipid antigen associated with Burkitt's lymphoma defined by a monoclonal antibody, Science 220 (1983) 509-511.
[9] T. Falguieres, M. Maak, C. von Weyhern, M. Sarr, X. Sastre, M.F. Poupon, S. Robine, L. Johannes, K.P. Janssen, Human colorectal tumors and metastases express Gb3 and can be targeted by an intestinal pathogen-based delivery tool, Mol. Cancer Ther. 7 (2008)2498-2508.
[10] O. Kovbasnjuk, R. Mourtazina, B. Baibakov, T. Wang, C. Elowsky, M.A. Choti, A. Kane, M. Donowitz, The glycosphingolipid globotriaosylceramide in the metastatic transformation of colon cancers, Proc. Natl. Acad. Sci. U. S. A. 102 (2005) 19087-19092.
[11] T. Okuda, N. Tokuda, S. Numata, M. Ito, M. Ohta, K. Kawamura, J. Wiels, T. Urano, O. Tajima, K. Furukawa, K. Furukawa, Targeted disruption of Gb3/CD77 synthase gene resulted in the complete deletion of globo-series glyco- sphingolipids and loss of sensitivity to verotoxins, J. Biol. Chem. 281 (2006) 10230-10235.
[12] Y. Kojima, S. Fukumoto, K. Furukawa, T. Okajima, J. Wiels, K. Yokoyama, Y. Suzuki, M. Ohta, K. Furukawa, Molecular cloning of Gb3/CD77 synthase, a glycosyl- transferase that initiate the synthesis of globo-series glycosphingolipids, J. Biol. Chem. 275 (2000) 15152-15156.
[13] S. Watarai, K. Kiura, R. Shigeto, T. Shibayama, I. Kimura, T. Yasuda, Establishment of monoclonal antibodies specific for ganglioside GM1: detection of ganglioside GM1 in small cell lung carcinoma cell lines and tissues, J. Biochem. 116 (1994) 948-954.
[14] K. Furukawa, H. Clausen, S. Hakomori, J. Sakamoto, K. Look, A. Lundblad, M.J. Mattes, K.O. Lloyd, Analysis of the specificity of five murine anti-blood group A monoclonal antibodies, including one that identifies type 3 and type 4 determinants, Biochemistry 24 (1985) 7820-7826.
[15] M.L.J.Lupher, N. Rao, M.J. Eck, H. Band, The Cbl protooncoprotein: a negative regulator of immune receptor signal transduction, Immunol Today, 20 (1999) 375-382.
[16] G. Kohler, C. Milstein, Continuous cultures of fused cells secreting antibody of predefined specificity, Nature, 256 (1975) 495-497.
[17] Y. Nagatsuka, S. Hanazawa, T. Yasuda, Y. Ono, Anti-glycolipid antibodies produced by Epstein-Barr virus (EBV)-transformed oligoclonal B cell lines obtained from normal persons and autoimmune disease patients, Hum. Antibodies Hybridomas 5 (1994) 183-186.
[18] H. Yamaguchi, K. Furukawa, S.R. Fortunato, P.O. Livingston, K.O. Lloyd, H.F. Oettgen, L.J. Old, Cell-surface antigens of melanoma recognized by human monoclonal antibodies, Proc. Natl. Acad. Sci. U. S. A. 84 (1987) 2416-2420.
[19] I. Kawashima, N. Tada, S. Ikegami, S. Nakamura, R. Ueda, T. Tai, Mouse monoclonal antibodies detecting disialogangliosides on mouse and human T lymphomas, Int. J. Cancer 41 (1988) 267-274.
[20] M. Kotani, H. Ozawa, I. Kawashima, S. Ando, T. Tai, Generation of one set of monoclonal antibodies specific for a-pathway ganglio-series gangliosides, Biochim. Biophys. Acta 1117 (1992) 97-103.
[21] H. Ozawa, M. Kotani, I. Kawashima, T. Tai, Generation of one set of monoclonal antibodies specific for b-pathway ganglio-series gangliosides, Biochim. Biophys. Acta. 1123 (1992) 184-190.
[22] I. Kawashima, O. Nakamura, T. Tai, Antibody responses to ganglio-series gangliosides in different strains of inbred mice, Mol. Immunol. 29 (1992) 625-632.
[23] D. Miyamoto, T. Ueno, S. Takashima, K. Ohta, T. Miyawaki, T. Suzuki, Y. Suzuki, Establishment of a monoclonal antibody directed against Gb3Cer/CD77: a useful immunochemical reagent for a differentiation marker in Burkitt's lymphoma and germinal centre B cells, Glycoconj. J. 14 (1997) 379-388
[24] M. Jacewicz, H. Clausen, E. Nudelman, A. Donohue-Rolfe, G.T. Keusch, Pathogenesis of shigella diarrhea. XI. Isolation of a shigella toxin-binding glycolipid from rabbit jejunum and HeLa cells and its identification as globotriaosylceramide, J. Exp. Med. 163 (1986) 1391-1404.
[25] M. Mangeney, Y. Richard, D. Coulaud, T. Tursz, J. Wiels, CD77: an antigen of germinal center B cells entering apoptosis. Eur J Immunol. 21 (1991) 1131-1140.
[26] H.L. Weiner, J.W. Moorhead, K. Yamaga, R.T. Kubo, Anti-Immunoglobulin Stimulation of Murine Lymphocytes: II. Identification of Cell Surface Target Molecules and Requirements for Cross-Linkage, J. Immunol. 117 (1976) 1527-1531.
[27] D.L. Mueller, E3 ubiquitin ligases as T cell anergy factors, Nat. Immunol. 5 (2004) 883 - 890.
[28] C.B. Thien, W.Y. Langdon, c-Cbl and Cbl-b ubiquitin ligases: substrate diversity and the negative regulation of signaling responses, Biochem. J. 391 (2005) 153-166.
[29] L. Duan, A.L. Reddi, A. Ghosh, M. Dimri, H. Band, The Cbl family and other ubiquitin ligases: destructive forces in control of antigen receptor signaling, Immunity 21 (2004) 7-17.
[30] F. Huang, G. Hua, Negative regulation of lymphocyte development and function by the Cbl family of proteins, Immunol. Rev. 224 (2008) 1229-1238.
[31] C.E. Rudd, H. Schneider, Lymphocyte signaling: Cbl sets the threshold for autoimmunity, Curr. Biol. 10 (2000) R344-347.
[32] P.C. Cheng, M.L. Dykstra, R.N. Mitchell, S.K. Pierce, A role for lipid rafts in B cell antigen receptor signaling and antigen targeting, J. Exp. Med. 190 (1999) 1549-1560.
[33] R.J. Petrie, P.P. Schnetkamp, K.D. Patel, M. Awasthi-Kalia, J.P. Deans, Transient translocation of the B cell receptor and Src homology 2 domain-containing inositol phosphatase to lipid rafts: evidence toward a role in calcium regulation, J. Immunol. 165 (2000) 1220-1227.
【技術分野】
【0001】
本発明はグロボ系糖脂質であるグロボトリアオシルセラミド(Gb3/CD77)を特異的に認識する抗体(抗Gb3/CD77抗体)の作製法及びその用途に関する。詳しくは、抗Gb3/CD77モノクローナル抗体の効率的な作製法、抗Gb3/CD77モノクローナル抗体及びその用途を提供する。
【背景技術】
【0002】
KohlerとMilsteinがモノクローナル抗体作製のためのハイブリドーマ樹立技術を確立して以来(非特許文献1)、モノクローナル技術は劇的に広まり、世界中の様々な生物学研究分野で広範に利用されている。B細胞ハイブリドーマに限らず、T細胞ハイブリドーマもまた開発され、免疫学的解析に幅広く利用されている(非特許文献2)。モノクローナル抗体は、現代生物学と医学の研究を推進するための、最も有用な技術の一つになってきている(非特許文献3)。モノクローナル抗体の作製過程で、動物の免疫とハイブリドーマのスクリーニングが最も重要である。疾患の診断や癌の免疫治療のために、抗体結合の特異性やCDC活性(補体依存性細胞傷害活性)あるいはADCC活性(抗体依存性細胞傷害活性)のような抗体の生物学的機能が極めて重要である。免疫の効果を向上させるために、免疫原の調製に関して様々な試行が行われてきた。例えば、ハプテンの結合、抗原へのアジュバントの添加、リポソームへの封埋、各々のタンパク質の立体配置から推定されるペプチドの利用などである。しかしながら、効率的なモノクローナル抗体の作製に有用な特定のマウスなどに関してはいまだ報告がない。
【0003】
糖脂質に反応するモノクローナル抗体の作製のために、通常のマウスに腫瘍細胞を注入する方法が、多くの研究室に広く流布した免疫方法であった。EBウイルスによるB細胞の不死化は、融合のための良好なパートナー細胞がなかったヒト型モノクローナル抗体の作製において特に効果的であった(非特許文献4、5)。その後、精製あるいはリコンビナントタンパク質がモノクローナル抗体の作製に広く用いられた。種々の機能性分子のペプチドも、様々な生体分子の免疫原として使用されてきた。ガングリオシドに対して有用なモノクローナル抗体の一式を作製するために精製ガングリオシドが免疫原として用いられた(非特許文献6〜8)が、その際には過去の報告に基づいて(非特許文献9)、特定のマウス系統が用いられた。抗ガングリオシドモノクローナル抗体の作製のために、複合型ガングリオシドを欠損するノックアウトマウスが利用されて、良好な抗体産生が示された(非特許文献10)。本発明者らの研究グループも、GM2/GD2合成酵素欠損(ノックアウト)マウスあるいはGD3合成酵素(ノックアウト)マウスを用いることにより、GD1a(非特許文献11)又はGD3及びGD1b(非特許文献12)に反応する有用な抗体を作製できたことを報告した。これらの抗ガングリオシド抗体は、免疫に使用された変異マウスにおいて欠損する構造に対してより高い特異性と親和性を示した。しかし、グロボ系糖脂質などの中性糖脂質を欠損するノックアウトマウスの利用に関しては、いまだ報告がない。
【先行技術文献】
【非特許文献】
【0004】
【非特許文献1】G. Kohler, C. Milstein, Continuous cultures of fused cells secreting antibody of predefined specificity, Nature, 256 (1975) 495-497.
【非特許文献2】P.N. Nelson, G.M. Reynolds, E.E. Waldron, E. Ward, K. Giannopoulos, P.G. Murray, Monoclonal antibodies, Mol. Pathol. 53 (2000) 111-117.
【非特許文献3】R.H. Kennett, Hybridomas: a new dimension in biological analyses, In Vitro 17 (1981) 1036-1050.
【非特許文献4】Y. Nagatsuka, S. Hanazawa, T. Yasuda, Y. Ono, Anti-glycolipid antibodies produced by Epstein-Barr virus (EBV)-transformed oligoclonal B cell lines obtained from normal persons and autoimmune disease patients, Hum. Antibodies Hybridomas 5 (1994) 183-186.
【非特許文献5】H. Yamaguchi, K. Furukawa, S.R. Fortunato, P.O. Livingston, K.O. Lloyd, H.F. Oettgen, L.J. Old, Cell-surface antigens of melanoma recognized by human monoclonal antibodies, Proc. Natl. Acad. Sci. U. S. A. 84 (1987) 2416-2420.
【非特許文献6】I. Kawashima, N. Tada, S. Ikegami, S. Nakamura, R. Ueda, T. Tai, Mouse monoclonal antibodies detecting disialogangliosides on mouse and human T lymphomas, Int. J. Cancer 41 (1988) 267-274.
【非特許文献7】M. Kotani, H. Ozawa, I. Kawashima, S. Ando, T. Tai, Generation of one set of monoclonal antibodies specific for a-pathway ganglio-series gangliosides, Biochim. Biophys. Acta 1117 (1992) 97-103.
【非特許文献8】H. Ozawa, M. Kotani, I. Kawashima, T. Tai, Generation of one set of monoclonal antibodies specific for b-pathway ganglio-series gangliosides, Biochim. Biophys. Acta. 1123 (1992) 184-190.
【非特許文献9】I. Kawashima, O. Nakamura, T. Tai, Antibody responses to ganglio-series gangliosides in different strains of inbred mice, Mol. Immunol. 29 (1992) 625-632.
【非特許文献10】M.P. Lunn, L.A. Johnson, S.E. Fromholt, S. Itonori, J. Huang, A.A. Vyas, J.E. Hildreth, J.W. Griffin, R.L. Schnaar, K.A. Sheikh, High-affinity anti-ganglioside IgG antibodies raised in complex ganglioside knockout mice: reexamination of GD1a immunolocalization. J. Neurochem. 75 (2000) 404-412.
【非特許文献11】B.J. Willison, D. Nicholl, E.R. Wagner, K. Townson, C. Goodyear, K. Furukawa, K. Furukawa, J. Conner, H.J. Willison, Innate murine B cells produce anti-disialosyl antibodies reactive with Campylobacter jejuni LPS and gangliosides that are polyreactive and encoded by a restricted set of unmutated V genes, J. Neuroimmunol. 152 (2004) 98-111.
【非特許文献12】J. Boffey, M. Odaka, D. Nicoll, E.R. Wagner, K. Townson, T. Bowes, J. Conner, K. Furukawa, H.J. Willison, Characterization of the immunoglobulin variable region gene usage encoding the murine anti-ganglioside antibody repertoire, J. Neuroimmunol. 165 (2005) 92-103.
【発明の概要】
【発明が解決しようとする課題】
【0005】
Gb3/CD77は、ヒト(参考文献6)及びマウス(参考文献7)の胚中心B細胞の一部、腎臓、小腸、血管内皮細胞に発現するとともに、バーキットリンパ腫(参考文献6)及び大腸癌(参考文献9、10)などの腫瘍細胞において過剰発現することが知られている。多くの研究で、Gb3/CD77が細菌毒素の受容体として機能する(参考文献11)ことが報じられているが、その生理的な役割に関してはほとんど分かっていない。従って、Gb3/CD77の機能解明に有用な手段が切望されるところである。また、これまでの報告を総合すれば、医療目的においてGb3/CD77が重要な標的になることは明らかであり、Gb3/CD77を標的とした診断技術や治療技術の創出が期待される。このような状況下、本発明はGb3/CD77を標的とした有能な抗体を効率的に作製する手段を提供することを課題とする。また、当該手法によって得られる抗体の利用を図ること(用途の提供)も課題とする。
【課題を解決するための手段】
【0006】
上記課題を解決すべく本発明者らは、グロボ系列の糖脂質を欠損する変異マウスを利用すれば、当該マウスにおいて消失した既知あるいは未知の糖鎖構造に対するモノクローナル抗体を効率よく作製できるとの期待の下、検討を進めた。具体的には、グロボ系列の糖脂質の合成に必須のα-1.4ガラクトース転移酵素遺伝子を欠損するノックアウトマウスを用いて、スフィンゴ糖脂質グロボトリアオシルセラミド(Gb3)と反応するモノクローナル抗体の作製を試みた。その結果、野生型マウスの免疫による従来法に比べて、明らかに良好なモノクローナル抗体を作製できることが判明した。詳しくは、従来法よりも多数のクローンが得られ、しかも、得られたクローンの半数がIgGクラスであった(従来法で得られた3クローンはすべてIgMクラスであった)。しかも、得られた抗体の中には、CDC活性に加えて、ADCC活性を示すものが含まれていた。さらに、c.52と名付けた抗体はバーキットリンパ腫細胞株に対する凝集活性を示した。興味深いことに、凝集の過程で、c.52抗体の結合により、抗B細胞受容体抗体の結合時に見られるのと類似の数個のタンパク質のチロシンリン酸化が誘導された。特に、c.52抗体の結合により、E3ユビキチンリガーゼとして知られるc-Cblという名の120kDaのタンパク質の強いリン酸化が惹起された。抗B細胞受容体抗体による刺激を行う前に、B細胞を抗Gb3モノクローナル抗体で前処理した場合、Gb3を介するシグナルがB細胞受容体からの抗原疑似シグナルを抑制することが判明した。
【0007】
以上のように、期待を超える成果がもたらされ、α-1.4ガラクトース転移酵素遺伝子欠損マウスがグロボ系糖脂質に対するモノクローナル抗体の作製に極めて有効であることが示された。また、抗Gb3/CD77モノクローナル抗体の利用を図る上で有益な興味深い知見が得られた。当該知見は、抗Gb/CD77モノクローナル抗体の有用性を裏付けるとともに、本モノクローナル作製法によれば、生理機能を良好に発揮する抗Gb/CD77モノクローナル抗体を効率的に作製できることを示す。以下に列挙する本発明は、主として上記の成果に基づく。
[1]以下のステップ(1)〜(5)を含む、抗Gb3/CD77モノクローナル抗体の作製法:
(1)α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物をGb3/CD77で免疫するステップ;
(2)免疫後の非ヒト哺乳動物から抗体産生細胞を回収するステップ;
(3)回収した抗体産生細胞を不死化させるステップ;
(4)不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化するステップ;及び
(5)モノクローナル化によって得られたクローンが産生する抗体を回収するステップ。
[2]非ヒト哺乳動物がマウスである、[1]に記載の作製法。
[3]マウスがA/J系統である、[2]に記載の作製法。
[4]Gb3/CD77、リピッドA、コレステロール及びジパラミトイルホスファチジルコリンを含むリポソームを免疫原としてステップ(1)における免疫を行う、[1]〜[3]のいずれか一項に記載の作製法。
[5]抗体産生細胞が脾臓細胞である、[1]〜[4]のいずれか一項に記載の作製法。
[6]ステップ(3)が、抗体産生細胞と骨髄腫細胞との融合によって行われる、[1]〜[5]のいずれか一項に記載の作製法。
[7]ステップ(4)において、IgGクラスの抗Gb3/CD77抗体を産生する細胞をモノクローナル化する、[1]〜[6]のいずれか一項に記載の作製法。
[8]Gb3/CD77を特異的に認識し、補体依存性細胞傷害活性及び/又は抗体依存性細胞傷害活性を示す、単離されたモノクローナル抗体。
[9]バーキットリンパ腫細胞株に対する凝集活性を示す、[8]に記載の単離されたモノクローナル抗体。
[10][1]〜[7]のいずれか一項に記載の作製法によって得られる、[8]又は[9]に記載の単離されたモノクローナル抗体。
[11]ヒト抗体又はヒト化抗体である、[9]又は[10]に記載の単離されたモノクローナル抗体。
[12]IgG、Fab、Fab'、F(ab')2、scFv、又はdsFv抗体である、[9]〜[11]のいずれか一項に記載の単離されたモノクローナル抗体。
[13][9]〜[12]のいずれか一項に記載の単離されたモノクローナル抗体を含む、Gb3/CD77を発現する細胞を標的とした研究用試薬。
[14][9]〜[12]のいずれか一項に記載の単離されたモノクローナル抗体を含む、Gb3/CD77を過剰発現する悪性腫瘍を標的とした診断又は治療用の組成物。
[15]悪性腫瘍がバーキットリンパ腫、大腸癌、奇形腫又は巨核芽球性白血病である、[14]に記載の組成物。
【図面の簡単な説明】
【0008】
【図1】A/JマウスをGb3/CD77によって免疫して得られたモノクローナル抗体の特異性と反応性を示す図。(A) 精製糖脂質(20 ng/ウェル)に対して、2段階希釈のモノクローナル抗体(ハイブリドーマ上清)を用いて行ったELISAの結果の一例。上清を洗浄した後、HRP標識2次抗体を加え一時間静置した。洗浄後、基質溶液を添加し、モノクローナル抗体の結合を検出した。10〜20分後に、発色の程度をスキャナーにより記録した。(B) A/Jマウスから得られたモノクローナル抗体の、TLC免疫染色による様々な糖脂質に対する特異性。TLC免疫染色は、得られたモノクローナル抗体(ハイブリドーマ上清1:2 希釈)によって<材料と方法>に記載したように行った。TLCは、C:M:DW (60:35:8)の展開用溶媒にて行った。糖脂質(0.5μg/ウェル)を用いた。抗体の結合は、ABC complexTM キットとコニカイムノステインHRP 1000TMによって検出した。
【図2】Gb3合成酵素ノックアウトマウスから得られたモノクローナル抗体の特異性と反応性を示す図。TLC免疫染色は、得られたモノクローナル抗体(ハイブリドーマ上清1 : 2 希釈)を用いて、<材料と方法>に記載したように行った。TLCは、展開溶媒系C:M:DW (60:35:8)にて行った。糖脂質(0.5μg/ウェル)を用いた。抗体の結合をABC complexTM及びコニカイムノステインHRP 1000TMにて検出した。モノクローナル抗体c.76はこのアッセイ系に使用できなかったので、7種の抗体に関する検討結果を示す。
【図3】Gb3合成酵素ノックアウトマウスから得られたモノクローナル抗体の種々糖脂質に対する特異性と反応性を示す図。得られたモノクローナル抗体(ハイブリドーマ上清)を連続2倍希釈してELISAを行った。洗浄後、HRP標識2次抗体を添加した。洗浄後、基質溶液を加えることによってモノクローナル抗体の結合を検出した。
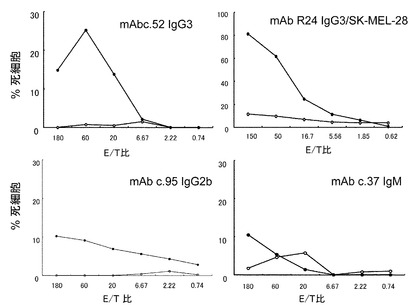
【図4】Ramos細胞に対するモノクローナル抗体の抗腫瘍活性を示す図。補体依存性細胞傷害活性。Ramos細胞を得られたモノクローナル抗体一式と静置した後(x20〜x10240希釈の腹水)、ウサギ血清を補体源として添加し、静置した。補体依存性細胞傷害活性は、<材料と方法>に記載したようにWST-1アッセイにより行った。実線:血清なし、広間隔の点線:血清(1:10希釈)、狭間隔の点線:血清(1:2 希釈)。
【図5】抗体依存性細胞傷害活性を示す図。Ramos細胞またはSK-MEL-28細胞をモノクローナル抗体c.52、c.95、c.37又はR24と各々静置した後、末梢血単核球を加えて一時間静置した。コントロールのIgGをコントロールとして用いた。ADCC活性は、放出されたLDHをCytoTox 96 Non-Radioactive Cytotoxicity AssayTM を用いて<材料と方法>に記載したように測定することで検出した。○:コントロールIgG、●:モノクローナル抗体c.52又はモノクローナルR24。
【図6】ベロ毒素によるアポトーシス誘導に対する、抗Gb3/CD77モノクローナル抗体の細胞前処理の効果を示す図。(A)カスパーゼ3のウエスタンイムノブロッテイングの結果。Ramos細胞をVT-2と様々な時間静置して、その細胞融解液を<材料と方法>に記したごとくイムノブロッテイングに使用した。元のカスパーゼ3(32 kDa)の分解産物が、ベロ毒素を加えてから3時間で出現した。(B) 抗Gb3/CD77抗体の前処置のアポトーシスに対する効果。Ramos細胞を抗Gb3/CD77抗体により4℃、2時間反応させた後、VT2(200 ng/ml)の存在下で3時間静置した。イムノブロッテイングの結果、モノクローナル抗体c.168、c.52及びc.95によって処理された時には、元のカスパーゼ3のバンドが少々残留することが分かった。カスパーゼ3のバンドの相対的強さを示す。即ち、非分解および分解カスパーゼ3の和に対する非分解カスパーゼ3の比率をBのバンド強度から計算してプロットした。繰り返し実験においても同様の結果が得られた。
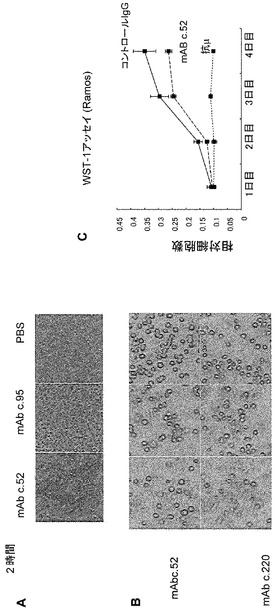
【図7】Ramos細胞の形態に対する抗Gb3/CD77モノクローナル抗体の影響を示す図。 (A, B) Ramos細胞をモノクローナル抗体c.52、c.95又はc.220と静置した。PBSを陰性コントロールとして用いた。モノクローナル抗体c.52と一分間静置した細胞のみが凝集を起こした。(C) Ramos細胞(1x107細胞/ml)をモノクローナル抗体c.52、c.220又はc.95(20μg/ml)と反応させた。図に示された時間毎にWST-1アッセイを行った。同じ実験を何度か繰り返して、同様の結果が得られた。
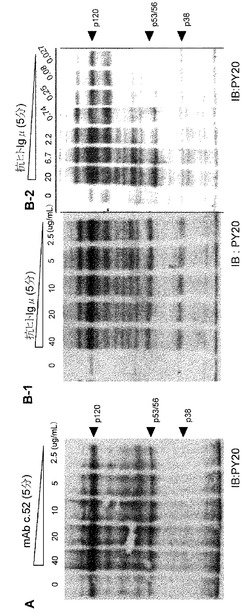
【図8】モノクローナル抗体c.52によって誘導されたタンパク質のチロシンリン酸化を示す図。モノクローナル抗体c.52によって誘導されるタンパク質のチロシンリン酸化を検討した。Ramos細胞(107細胞/ml)を、モノクローナル抗体c.52(20μg/ml)又はc.220(20μg/ml)により37℃で図に示した時間、刺激した。細胞融解液を調製してPY20によるイムノブロッテイングの解析に用いた。
【図9】モノクローナル抗体c.52によってチロシンリン酸化されるタンパク質は、抗μ抗体によりリン酸化されるタンパク質に類似している。(A)モノクローナル抗体c.52の処置によって惹起されるチロシンリン酸化タンパク質のウエスタンイムノブロッテイングの結果。Ramos細胞をモノクローナル抗体c.52(0〜40μg/ml)と5分間反応させた後、細胞融解液をPY20によるイムノブロッテイングに用いた。(B) 抗μAbによるチロシンリン酸化タンパク質のウエスタンイムノブロッテイング。Ramos細胞を抗μ抗体(0〜40μg/ml)と反応させた後、その融解液をPY20によるウエスタンイムノブロッテイングに用いた。同様の実験を、より低濃度の抗μ抗体を用いて行った(B-2)。
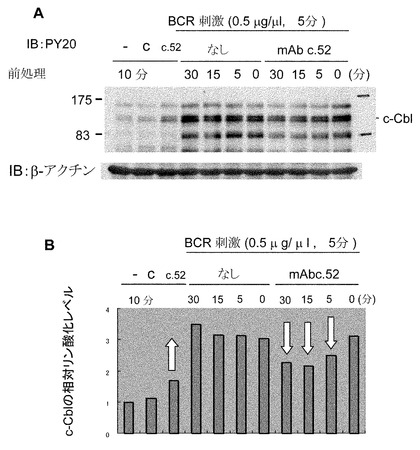
【図10】p120がc-Cblであることの同定とリン酸化のキネテイクスを示す図。(A) モノクローナル抗体c.52で5分間処置した前後に、PY20あるいは抗c-Cbl抗体を用いて免疫沈降を行った。その免疫沈降物を用いて、PY20あるいは抗c-Cb抗体によるイムノブロッテイングを行った。(B) モノクローナル抗体c.52の刺激によるc-Cblのリン酸化の時間経過。モノクローナル抗体c.52により刺激されたRamos細胞の溶解液(1x107細胞由来)を用いて、抗c-Cbl抗体による免疫沈降を行った。その後、免疫沈降物に対して、HRP-PY20又は抗c-Cbl抗体とHRP標識抗ウサギIgG によるイムノブロッテイングを行った。
【図11】モノクローナル抗体c.52による前処置の、抗μ抗体によるタンパク質リン酸化パターンに対する影響を示す図。(A) 細胞融解液のウエスタンイムノブロッテイング。Ramos細胞をモノクローナル抗体c.52またはPBSと図に示された時間だけ反応させた後、抗μ抗体と5分間反応させた。細胞溶解液を調製してPY20によるイムノブロッテイングに用いた。(B) Aのp120のバンドの相対的強さを、NIHイメージ1.61によって測定して、βアクチンの値で補正した後、グラフにプロットした。繰り返し行った実験でも同様の結果を得た。
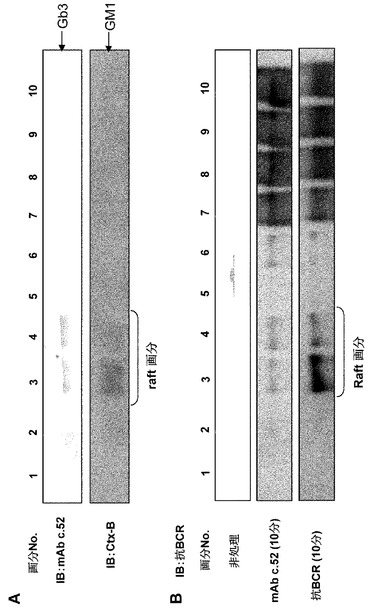
【図12】Gb3/CD77とBCR複合体の細胞内局在を示す図。(A) 分子の浮遊パターンにつき、Triton X-100による抽出物のショ糖密度勾配超遠心法で分画された画分を用いたイムノブロッテイングによって検討した。GM1、脂質ラフトのマーカーは、脂質ラフトに相当する第3−第4画分にすべて存在した。Gb3もまた、以前に報告されたように脂質ラフトに局在した。(B) モノクローナル抗体c.52または抗μ抗体の処置によるRamos細胞上のBCRの局在変化。各画分のイムノブロッテイングは、Aに示されたように、無処置(上)、モノクローナル抗体c.52による処置(中)、抗μ抗体による処置(下)を行ったRamos細胞の溶解液を用いて、抗BCR抗体によって行われた。
【発明を実施するための形態】
【0009】
(用語)
説明の便宜上、本明細書中で使用する用語の一部についてその定義・意味を以下にまとめて記す。
「モノクローナル抗体」は、単一の抗体産生細胞に由来するクローンから得られる抗体であり、複数種の混合物であるポリクローナル抗体に比較して格段に高い特異性を示す。
用語「単離された」とは、その本来の環境(例えば天然の物質の場合は天然の環境)から取り出された状態、即ち人為的操作によって本来の存在状態と異なる状態で存在していることを意味する。従って、「単離された抗体」には、天然であって且つ何ら外的操作(人為的操作)が施されていない抗体、即ちある個体の体内で産生され、そこに留まっている状態の抗体は含まれない。
グロボトリアオシルセラミドは、スフィンゴ糖脂質の一種であるグロボ系列の糖脂質(グロボ系糖脂質)でありGb3と略称される。Gb3は別名CD77抗原とも呼ばれることから、本明細書においては、グロボトリアオシルセラミドを「Gb3/CD77」と表記する場合がある。
【0010】
1.抗Gb3/CD77モノクローナル抗体の作製法
本発明の第1の局面は、Gb3/CD77を特異的に認識するモノクローナル抗体、即ち「抗Gb3/CD77モノクローナル抗体」の作製法を提供する。本発明では免疫学的手法を利用する。具体的には、以下のステップ(1)〜(5)が行われる。
(1)α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物をGb3/CD77で免疫するステップ
(2)免疫後の非ヒト哺乳動物から抗体産生細胞を回収するステップ
(3)回収した抗体産生細胞を不死化させるステップ
(4)不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化するステップ
(5)モノクローナル化によって得られたクローンが産生する抗体を回収するステップ
【0011】
ステップ(1)ではまず、α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物を用意する。当該非ヒト哺乳動物は、既報の方法(参考文献11)に準じて作製することができる。作製法(マウスを用いた場合の一例)の概要を説明すると、α1,4-ガラクトース転移酵素の遺伝子断片を含むゲノム断片の、酵素活性にとって重要な部位をコードする領域に挿入または置換によるneo耐性遺伝子を組込んだターゲッテイングベクターを作成する。培養ES細胞にこのベクターを電気穿孔法にて導入し、neo耐性クローンを得た後、相同遺伝子組換え体を得て、胚盤胞に注入する。その結果得られたキメラマウスから、へテロ接合体、ホモ接合体マウスを作製する。免疫動物として利用可能な非ヒト哺乳動物が用いられる。例えばマウス、ラット、ウサギ、ヤギなどをここでの非ヒト哺乳動物として採用する。好ましくはマウスを用いる。特に、糖脂質(特にガングリオシド)に対する反応性がよいことから、A/J系統のマウスを用いるとよい。A/J系統のマウスは日本チャールズリバーや日本エスエルシーなどから購入できる。
【0012】
一方、免疫用の抗原(免疫原)を用意する。抗原にはGb3/CD77が用いられる。Gb3/CD77は、生体由来の精製品であっても、合成品であってもよい。例えば、生体由来製品のGb3/CD77はLarodan AB社などから入手可能である。抗原として、好ましくは、Gb3/CD77、リピッドA、コレステロール及びジパラミトイルホスファチジルコリン(DPPC)を含むリポソームを免疫原とする。その理由は、抗原の存在様式を自然状態に近づけるためである。尚、当該リポソームの調製は、例えば既報(参考文献13)の方法に従えばよい。調製法(一例)の概要を説明すると、0.5μmolのDPPC、コレステロール、10μgのリピッドA、ガングリオシド100μgをC(クロロホルム):M(メタノール) (2:1)に溶解してエバポレートする。続いて、0.5 mlのPBSに溶かして50℃、1分間の処置後、よく攪拌する。
【0013】
必要に応じて免疫を繰り返し、抗体価の十分な上昇を認めた時点で免疫動物から抗体産生細胞を摘出、回収する。典型的には、脾臓又はその一部を摘出し、抗体産生細胞を回収する。但し、以降の操作が可能である限りにおいて、抗体産生細胞のソースは脾臓に限定されるものではない。
【0014】
次に、得られた抗体産生細胞を不死化させる。典型的には、抗体産生細胞を骨髄腫細胞と融合させハイブリドーマを得る。抗体産生細胞の動物種がマウスであれば、例えばNS-1骨髄腫細胞、P3X63-Ag8653、Sp2/0-Ag14、PAI等を用いることができる。
【0015】
続いて、不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化する。モノクローナル化にはHAT培地(ヒポキサンチン、アミノプテリン、チミジンを含む培地)を用いた選択培養及び限界希釈法が一般的である。この手法では、HAT培地による培養によって、抗体産生細胞と腫瘍細胞が融合したハイブリドーマを選択するとともに、限界希釈法を利用して目的の抗体(抗Gb3/CD77抗体)を産生するハイブリドーマをクローン化する。本発明の一態様では、有用性がより高いといえるIgGクラスの抗体を産生する細胞をモノクローナル化する。換言すれば、IgGクラスの抗Gb3/CD77抗体を産生するか否かを指標としてスクリーニングを行い、不死化した抗体産生細胞クローンを得る。本発明の作製法によれば、IgMクラスのみならずIgGクラスの抗体を産生するクローンも高率で得られることから(後述の実施例を参照)、このようなスクリーニングが可能であり且つ有効である。
【0016】
モノクローナル化によって得られたクローンが産生する抗体を回収することによって目的の抗体が得られる。例えば、選択されたクローンの培養液を精製することによって抗体を回収する。一方、選択されたクローンを所望数以上に増殖させた後、これを動物(例えばマウス)の腹腔内に移植し、腹水内で増殖させて腹水を精製することにより目的の抗体を取得することもできる。上記培養液の精製又は腹水の精製には、プロテインG、プロテインA等を用いたアフィニティークロマトグラフィーが好適に用いられる。また、抗原を固相化したアフィニティークロマトグラフィーを用いることもできる。更には、イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー、硫安分画、及び遠心分離等の方法を用いることもできる。これらの方法は単独ないし任意に組み合わされて用いられる。
【0017】
Gb3/CD77への特異的結合性を保持することを条件として、得られた抗体に種々の改変を施すことができる。例えば、低分子化合物、タンパク質、標識物質などを融合又は結合させることにより、融合抗体又は標識化抗体を構成することができる。標識物質としては例えば、フルオレセイン、ローダミン、テキサスレッド、オレゴングリーン等の蛍光色素、ホースラディッシュペルオキシダーゼ、マイクロペルオキシダーゼ、アルカリ性ホスファターゼ、β−D−ガラクトシダーゼ等の酵素、ルミノール、アクリジン色素等の化学又は生物発光化合物、32P、131I、125I等の放射性同位体、及びビオチンを挙げることができる。また、化学修飾(例えば、一部のアミノ酸をアセチル化、PEG化、リン酸化、アミド化する)を施すことも可能である。
【0018】
2.抗Gb3/CD77モノクローナル抗体及びその用途
本発明の第2の局面は、CDC活性を示す複数の抗Gb3/CD77モノクローナル抗体の取得に成功した事実、及びCDC活性に加えてはADCC活性をも示す抗Gb3/CD77モノクローナル抗体の取得に成功した事実に基づき、Gb3/CD77を特異的に認識し、CDC活性及び/又はADCC活性を示す、単離された抗体を提供する。本発明の抗体は、例えば、Gb3/CD77を発現する細胞の研究用試薬の成分として、或いはGb3/CD77を過剰発現する悪性腫瘍を標的とした診断又は治療用試薬の有効成分として有用である。
【0019】
本発明の抗体は、典型的には、上記本発明の抗体作製法により調製される。本発明の抗体にはマウス、ラットなどの非ヒト動物の抗体、一部の領域を他の動物(ヒトを含む)のものに置換したキメラ抗体、ヒト化抗体、及びヒト抗体が含まれる。本発明の抗体の由来、種類、クラス、形態などは特に限定されないが、好ましくはIgGクラスの抗体である。例えば、マウス抗体のサブクラスIgG1、IgG2a、IgG2b又はIgG3、或いはヒト抗体のサブクラスIgG1、IgG2、IgG3又はIgG4に属する抗体である。
【0020】
本発明の抗体の一形態はヒト化抗体である。「ヒト化抗体」とは、ヒトの抗体に構造を類似させた抗体のことをいい、ヒト型キメラ抗体(例えば抗体の一部がヒト化された抗体、CH2領域がヒト化された抗体、Fc領域がヒト化された抗体、定常領域がヒト化された抗体)、及び定常領域及び可変領域に存在するCDR(相補性決定領域)以外の部分がヒト化されたヒト型CDR移植(CDR-grafted)抗体(P.T.Johons et al., Nature 321,522(1986))、完全ヒト化抗体などが含まれる。ヒト型CDR移植抗体の抗原結合活性を高めるため、マウス抗体と相同性の高いヒト抗体FRを選択する方法、相同性の高いヒト化抗体を作製する方法、ヒト抗体にマウスCDRを移植した後さらにFRのアミノ酸を置換する方法の改良技術もすでに開発され(米国特許第5585089号、米国特許第5693761号、米国特許第5693762号、米国特許第6180370号、欧州特許第451216号、欧州特許第682040号、特許第2828340号などを参照)、本発明のヒト型CDR移植抗体の作製に利用することもできる。
【0021】
Fab、Fab'、F(ab')2、scFv、又はdsFv抗体などの抗体断片として本発明の抗体を調製することもできる。Fabは、IgG抗体をシステイン存在下パパイン消化することにより得られる、L鎖とH鎖可変領域、並びにCH1ドメイン及びヒンジ部の一部からなるH鎖フラグメントとから構成される分子量約5万の断片である。所望のIgG抗体があれば、これをパパイン消化することによりFabを得ることができる。また、H鎖の一部及びL鎖をコードするDNAを適当なベクターに組み込み、当該ベクターを用いて形質転換した形質転換体よりFabを調製することもできる。
【0022】
Fab'は、後述のF(ab')2のH鎖間のジスルフィド結合を切断することにより得られる分子量が約5万の断片である。所望のIgG抗体があれば、これをペプシン消化し、還元剤を用いてジスルフィド結合を切断することによりFab'を得ることができる。また、Fab同様に、Fab'をコードするDNAを用いて遺伝子工学的に調製することもできる。
【0023】
F(ab')2は、IgG抗体をペプシン消化することにより得られる、L鎖とH鎖可変領域、並びにCH1ドメイン及びヒンジ部の一部からなるH鎖フラグメントとから構成される断片(Fab')がジスルフィド結合で結合した分子量約10万の断片である。所望のIgG抗体があれば、これをペプシン消化することによりF(ab')2を得ることができる。また、Fab同様に、F(ab')2をコードするDNAを用いて遺伝子工学的に調製することもできる。
【0024】
scFvは、H鎖可変領域とL鎖可変領域とからなるFvを、片方の鎖のC末端と他方のN末端とを適当なペプチドリンカーで連結し一本鎖化した抗体断片である。ペプチドリンカーとしては、例えば柔軟性の高い(GGGGS)3などを用いることができる。例えば、H鎖可変領域及びL鎖可変領域をコードするDNAとペプチドリンカーをコードするDNAを用いてscFv抗体をコードするDNAを構築し、これを適当なベクターに組み込み、当該ベクターを用いて形質転換した形質転換体よりscFvを調製することができる。
【0025】
dsFvは、H鎖可変領域及びL鎖可変領域の適切な位置にCys残基を導入し、H鎖可変領域とL鎖可変領域とをジスルフィド結合により安定化させたFv断片である。各鎖におけるCys残基の導入位置は分子モデリングにより予測される立体構造に基づき決定することができる。例えばH鎖可変領域及びL鎖可変領域のアミノ酸配列から立体構造を予測し、かかる予測に基づき変異を導入したH鎖可変領域及びL鎖可変領域をそれぞれコードするDNAを構築し、これを適当なベクターに組み込み、そして当該ベクターを用いて形質転換した形質転換体よりdsFvを調製することができる。
【0026】
(研究用途)
本発明は更に、本発明の抗体の用途として、Gb3/CD77を発現する細胞を標的とした、本発明の抗体を含む研究用試薬を提供する。本発明の研究用試薬は、Gb3/CD77を発現する細胞(例えばB細胞の一部、腎臓細胞、小腸細胞、血管内皮細胞など)を標的とする或いは使用する研究に広く適用可能である。特に、バーキットリンパ腫や大腸癌など、Gb3/CD77の過剰発現を認める悪性腫瘍の研究や、Gb3/CD77の生理的役割の解明を目指した研究において有用である。標識化された本発明の抗体は、当該研究用途に好適な形態の一つである。本発明の研究用試薬には、本発明の抗体の他、溶媒、緩衝液、安定剤、防腐剤などを含有させることができる。
【0027】
(診断・治療用途)
本発明の抗体を用いれば、Gb3/CD77を発現する細胞を特異的に検出したり識別したりすることが可能である。従って、Gb3/CD77の過剰発現を認めるバーキットリンパ腫や大腸癌等の悪性腫瘍の診断、治療への本発明の抗体の適用も期待される。そこで本発明は更に、Gb3/CD77を過剰発現する悪性腫瘍を標的とした診断又は治療に利用可能な、本発明の抗体を含む組成物を提供する。標的となる悪性腫瘍の例は、バーキットリンパ腫、大腸癌、奇形腫、巨核芽球性白血病である。本発明の治療用組成物の一態様では、CDC活性又はADCC活性を示す抗Gb3/CD77モノクローナル抗体が有効成分として含有される。好ましくは、ADCC活性を示す抗Gb3/CD77モノクローナル抗体を用いる。この態様の治療用組成物では、ADCC活性を利用した細胞障害によって治療効果を得ることができる。
【0028】
本発明の治療用組成物の別の態様では、抗Gb3/CD77モノクローナル抗体をDDS用の運搬体として利用する。つまりこの態様は抗Gb3/CD77モノクローナル抗体に薬物(細胞毒など)又は放射性同位元素など(これらをまとめて「活性成分」ともいう)を結合して得られる免疫複合体を提供する。殺細胞活性又は細胞傷害活性を持つ薬物(細胞毒)を含有する免疫複合体は一般にイムノトキシンと呼ばれる。細胞毒の例には、タキソール、シトカラシンB、グラミシジンD、臭化エチジウム、エメチン、ミトマイシン、エトポシド、テノポシド、ビンクリスチン、ビンブラスチン、コルヒチン、ドキソルビシン、ダウノルビシン、ジヒドロキシアントラシンジオン、ミトキサントロン、ミトラマイシン、アクチノマイシンD、1-デヒドロテストステロン、糖質コルチコイド、プロカイン、テトラカイン、リドカイン、プロプラノロール、及びプロマイシン並びにこれらの類似体又は同族体を挙げることができる。免疫複合体に含有させる活性成分として、所望の生物活性を有するタンパク質又はペプチドを使用してもよい。このような目的において使用可能なタンパク質等の候補として、アブリン、リシンA、シュードモナス・エキソトキシン、ジフテリア毒素、腫瘍壊死因子、インターフェロン-γ、インターロイキン-1(IL-1)、インターロイキン-2(IL-2)、インターロイキン-6(IL-6)、顆粒球マクロファージコロニー刺激因子(GM-CSF)、顆粒球コロニー刺激因子(G-CSF)リンホカインを例示できる。活性部分を抗体に結合させる技術は公知であり、例えばMonoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985)、Controlled Drug Delivery (2nd edition.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987)、Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985)、Thorpe et al., "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-58 (1982)を参照することができる。
【0029】
治療用組成物の製剤化は常法に従って行うことができる。製剤化する場合には、製剤上許容される他の成分(例えば、担体、賦形剤、崩壊剤、緩衝剤、乳化剤、懸濁剤、無痛化剤、安定剤、保存剤、防腐剤、生理食塩水など)を含有させることができる。賦形剤としては乳糖、デンプン、ソルビトール、D-マンニトール、白糖等を用いることができる。崩壊剤としてはデンプン、カルボキシメチルセルロース、炭酸カルシウム等を用いることができる。緩衝剤としてはリン酸塩、クエン酸塩、酢酸塩等を用いることができる。乳化剤としてはアラビアゴム、アルギン酸ナトリウム、トラガント等を用いることができる。懸濁剤としてはモノステアリン酸グリセリン、モノステアリン酸アルミニウム、メチルセルロース、カルボキシメチルセルロース、ヒドロキシメチルセルロース、ラウリル硫酸ナトリウム等を用いることができる。無痛化剤としてはベンジルアルコール、クロロブタノール、ソルビトール等を用いることができる。安定剤としてはプロピレングリコール、ジエチリン亜硫酸塩、アスコルビン酸等を用いることができる。保存剤としてはフェノール、塩化ベンザルコニウム、ベンジルアルコール、クロロブタノール、メチルパラベン等を用いることができる。防腐剤としては塩化ベンザルコニウム、パラオキシ安息香酸、クロロブタノール等と用いることができる。製剤化する場合の剤型も特に限定されず、例えば錠剤、散剤、細粒剤、顆粒剤、カプセル剤、シロップ剤、注射剤、外用剤、及び座剤などとして調製できる。
【0030】
本発明の治療用組成物を用いた治療においては、標的となる悪性腫瘍を保有する対象(患者)に本発明の治療用組成物が投与される。本発明の治療用組成物はその形態に応じて経口投与又は非経口投与(静脈内、動脈内、皮下、筋肉、腹腔内注射、標的細胞への直接導入など)によって対象(患者)に適用され得る。投与量は症状、患者の年齢、性別、及び体重などによって異なるが、当業者であれば適宜適当な投与量を設定することが可能である。投与スケジュールとしては例えば一日一回〜数回、二日に一回、或いは三日に一回などを採用できる。投与スケジュールの設定においては、患者の病状や有効成分の効果持続時間などを考慮することができる。
【実施例】
【0031】
<材料と方法>
(1)動物
A/Jマウス、BALB/cnu/nuマウス及びKSNnu/nuマウスは日本チャールズリバーから購入した。α-1,4ガラクトース転移酵素遺伝子(A4galt-)欠損マウスは我々の研究室で樹立された(参考文献11)。動物実験に関するすべてのプロトコールは、「実験動物の管理と使用に関する指針(National Institutes of Health Guide for the Care and Use of Laboratory Animals(1966))に沿ったものであり、名古屋大学医学系研究科の動物実験委員会の承認を得た。すべての動物は特定病原微生物のいない条件下(specific pathogen-free)で維持された。
【0032】
(2)細胞培養
本研究で使用した骨髄腫細胞株NS-1、バーキットリンパ腫細胞Ramosは10% 牛胎仔血清(FCS)を含むRPMI 1640により培養した。cDNAをトランスフェクトしたL細胞は、ダルベッコ修飾イーグル培地(Dulbecco's modified eagle's medium;DMEM)に7.5% FCS及びG418(300μg/ml)を加えた培養液により維持された。バーキットリンパ腫細胞株であるRamos、HS-Sultan、Daudi及びRajiは日本健康科学財団(Japan Health Science Foundation)から入手した。A4galt-導入L細胞(LVTR-1)及びベクターコントロール細胞株(Lneo-2)は以前の報告の通り樹立された(参考文献12)。
【0033】
(3)Gb3/CD77に対するモノクローナル抗体の作製
3匹の8週齢A/Jマウスに対して、Gb3/CD77糖脂質(100μg)、リピッドA(10μg)、コレステロール(0.5μmol)、ジパラミトイルホスファチジルコリン(dipalmitoyl- phosphatidylcholine; DPPC)(0.5μmol)を含むリポソーム(参考文献13)を用いて2週間毎に皮下に免疫を行った。マウス血清中の抗体価を免疫蛍光法(IF)によって決定した。5回目の免疫から3日後に脾細胞を取り出し、ポリエチレングリコール(PEG)を用いてNS-1骨髄腫細胞との融合を行った。HAT選択培地(Sigma, St Louis, MO)により選択後、免疫蛍光法(IF)及び酵素結合免疫吸着法(enzyme-linked-immuno-solvent assay; ELISA)によって、培養上清中の抗体を検出した。陽性クローンを限界希釈法によりサブクローニングし、TLC-免疫染色法(TLC-immunostaining)によってその特異性を更に確認した。A4galt-欠損変異マウスについては、8週齢のマウスを用い、第0日、4日、7日、11日目に上記構成のリポソームを皮下免疫して、さらに第21日目に尾静脈からブースター免疫を実施した。各マウスの抗体価は、FITC標識抗マウスIgG抗体(Zymed Lab. Inc., SanFrancisco, CA)を用いたIFにより測定した。ハイブリドーマの培養上清をサンプルとしてELISA及び/又はIF法を実施し、陽性クローンについてはサブクローニングを行い、さらにTLC-免疫染色により特異性を確認した。陽性クローンの培養上清中の抗体は、プロテインA-アガロース4ファーストフローを用いたアフィニテイカラムにより、添付のプロトコール(Amersham Biosciences, Bucks, UK)に従って精製した。
【0034】
(4)ELISA
メタノールに溶かした市販の糖脂質(20 ng/10μl)を60穴Terasakiプレート(Greiner BioOne, Frickenhausen, Germany)にプレーテイングした。20分かけて空気乾燥した後、プレートに5% BSA含有リン酸緩衝液(PBS)を添加して2時間室温にてブロッキングを行った。2回洗浄後、段階希釈した抗体をプレートに加えて2時間室温で静置した。5回洗浄した後、HRP-標識抗マウスIgGs(Amersham Biosciences)を2次抗体として添加した。5回洗浄後、o-フェニレンジアミン(ortho-phenylene diamine)(2 mg) (Wako Junyaku, Osaka, Japan)とH2O2(8μl)(Wako)を5 mlのクエン酸緩衝液(citrate-phosphate buffer)に溶解した基質溶液(10μl)をプレートに添加した。適当な時間暗所で静置してから、発色度をスキャナーを用いて記録した。
【0035】
(5)IFアッセイ
LVTR-1あるいはLneo-2細胞をTerasakiプレートに加え(500細胞/穴)、一夜静置した。培養上清を除去した後、段階希釈した培養上清又は腹水をプレートに加え室温で1時間静置した。2回洗浄後、FITC標識抗マウスIgG (H+L)抗体(Cappel, Durham, NC)を加えて、1時間室温暗所にて静置した。2回洗浄後、抗体の結合を蛍光顕微鏡(BX51, OLYMPUS, Tokyo, Japan)下で観察した。
【0036】
(6)TLC免疫染色
TLCは高性能TLCプレート(Merck, Darmstadt, Germany)を用いて、クロロフォルム/メタノール/0.22%塩化カルシウム(chloroform/ methanol/0.22% CaCl2)(60:35:8)を展開溶媒として実施した。Gb3/CD77に対する特異性と反応性は、以前報告したように(参考文献14)、アルミニウム板のシリカプレート(Merck)を用いて検討した。TLCの後、プレートは既報のごとくPVDF(polyvinylidene difluoride)膜に対してブロットした。5% BSAを含むPBSによるブロッキングの後、プレートを培養上清と附置し、抗体の結合をABC kitTM(Vector Laboratories, Burlingame, CA)及びコニカ免疫染色HRP-1000TM(Konica, Tokyo)を用いて検出した。
【0037】
(7)フローサイトメトリー
細胞表面のGb3/CD77について、FACSCaliverTM(Becton Dickinson, Mountain View, CA)にて検討した。要約すると、細胞を希釈した腹水もしくは精製抗体と60分間氷上で静置した後、FITC標識ヤギ抗マウスIgG(H+L)(Cappel, Durham, NC)抗体と氷上で45分反応させた。フローサイトメトリーの対照細胞として2次抗体のみを使用して調製した細胞を用いた。陽性細胞の定量のためにCELLQuestTMプログラムを用いた。
【0038】
(8)増殖試験及び増殖抑制試験
作製した抗体の増殖抑制効果を検討するために、WST-1試験を行った。既報のごとく(参考文献12)、450 nmでの吸光度をELISAリーダー(Immuno Mini NJ-2300 (System Instruments, Tokyo))で検出することによって、WST-1のフォルマザンへの還元を測定した。要約すると、細胞(1x104)を48穴プレートに播き、10% FCSを含むRPMI 1640で一昼夜培養した。その後、希釈した腹水あるいは精製抗体をプレートに添加して培養した。WST-1試験はRamos細胞を用い、三連(triplicate)で24時間毎に行った。抗Gb3抗体の増殖抑制効果は、対照抗体のそれと比較して検討した。
【0039】
(9)ベロ毒素の毒性に対する抗体の阻害効果
細胞(1x106)を48穴プレートに播き、10% FCS含有RPMI 1640にて一晩培養した。その後、希釈した抗体をプレートに添加して氷上で1時間静置した。続いて、細胞を十分に殺しうる最小量のベロ毒素を洗浄なしに添加し、静置した。その後、細胞融解液を調製し、毒素の毒性に対する阻害効果を検討するためのイムノブロッテイングに用いた。
【0040】
(10)CDC試験
RPMIを除去するために細胞を5% FCS含有PBSで洗浄した後、48穴プレートに播いた(1x104/50μl(5% FCS含有PBS))。各ウェルに5% FCS及び種々の量の腹水を含むPBSを50μl添加して、1時間室温にて静置した。その後、5% FCS及びラビット血清(30μl:1/10又は150μl: 1/2)を含んだ200μlのPBSを各ウェルに添加して、最終的に300μl/ウェルの量にて37℃、1時間静置した。CDC反応の後、細胞増殖測定試薬WST-1(Roche, Manheim, Germany)を各々のウェルに添加した(30μl/ウェル)。2時間の静置後、プレートを1分間十分に振盪して、サンプルの450nmでの吸光度をマイクロプレートリーダー(ImmunoMini NJ-2300, Immuno Mini NJ-2300, InterMed, Tokyo, Japan)を用いて測定した。
【0041】
(11)ADCC試験
標的細胞(1x106)を希釈した抗体とファルコンチューブ内で氷上1時間静置し、10% FCS含有RPMIで2回洗浄した後、各々のウェルにプレーテイングした。正常人のヘパリン血(25 ml)よりヒトの末梢単核球を製造元の指示書に従ってフィコール・ハイパック(Ficoll-Hypaque)グラジエント(Amersham Biosciences Inc.)遠心により分離した。これらの細胞を所定の比(標的(target)/エフェクター(effector))になるようにミクロタイタープレートに添加して37℃で4時間静置した。ADCCにより放出されたLDHの量を測定するために、CytoTox 96 Non-Radioactive Cytotoxicity AssayTM(Promega, Madison, WI)を用い、製造元の指示書に沿って検討した。
【0042】
(12)イムノブロッテイングと免疫沈降
細胞を溶解バッファー(20 mM Tris-HCl, pH 7.4。以下の試薬を含む、即ち0.15 M NaCl, 1% Nonidet P-40, 1 mM phenylmethylsulfonyl fluoride (Sigma)及び5μg/ml aprotinin) により溶解した。細胞溶解液(4.0x106細胞に由来する)をSDS-PAGEに供し、その後PVDF膜(Millipore, Bedford, MA)に転写した。5% BSA含有PBSにより1時間室温にてブロッキングした後、PVDF膜を室温で1時間、一次抗体と反応させた。その後、0.05% Tween 20添加PBSにより3回洗浄し、ホースラディッシュペルオキシダーゼ標識抗ラビットIgG(H+L)(BD transduction Laboratories, San Jose, CA)を添加後1時間静置してカスパーゼ3(caspase-3)、p38及びリン酸化p38を検出した。ホースラディッシュペルオキシダーゼ標識した抗マウスIgG(H+L)抗体(Zymed Laboratories Inc. San Francisco, CA)を用いてFAK、Lyn及びc-Cblを検出した。検出はECL systemTM (Amersham Biosciences Inc.)を用いて、製造元の指示書に従った。免疫沈降では、細胞溶解液(4x106細胞に由来する)をPY20、FAK、Lynあるいはc-Cblに対するポリクローナル抗体とプロテインGセファローズ4 first flowTM (Amersham Biosciences)によって免疫沈降させた。免疫複合体を洗浄後、SDSサンプル緩衝液を添加して、3分間煮沸した。
【0043】
(13)抗B細胞受容体(BCR)抗体あるいはmAb c.52によるB細胞刺激
Ramos細胞を10% FCS含有RPMIに107/mLの濃度で懸濁した。刺激前に細胞を37℃で1時間静置した。その後、ヤギ抗ヒトIgM(10μg/ml)(Fc5μ断片特異的抗体、Jackson Immuno-Laboratory, West Grove, PA)又はモノクローナル抗体c.52(10μg/ml)を用いて所定時間刺激した。
【0044】
<結果>
最初に、Gb3を封埋したリポソームによりA/Jマウスを免疫した。免疫したマウスの中で、Gb3に対して最も高い抗体価を示したマウスの脾細胞を用いて、骨髄腫細胞株NS1との融合を行った。ELISA及び蛍光抗体法によるスクリーニングを行った。400クローンのスクリーニングの結果、c.37、c.61、そしてc.80という3種のハイブリドーマを樹立した。これらのハイブリドーマクローンが産生する抗体のGb3/CD77に対する特異性をELISAとTLC免疫染色で解析した(図1A及び1B)。13種の異なる糖脂質に対して、3クローンすべてがGb3/CD77に限って反応した。3クローンはいずれもラクトシルセラミド(lactosylceramide)、GlcCer、GM3及びGb4とは反応せず、得られた抗体が反応するために非還元末端のα4-ガラクトースが必須であることが示唆された。その他の糖脂質に関しては、GalCer、GD1b、GD3、GA1、GT1b、GA2、GM1及びGD1a等への反応性も認められなかった(データ示さず)。同様の反応特異性がTLC免疫染色法によっても観察された(図1B)。mAbs 61及び80もまた、Gb3とのみ反応した(データ示さず)。3クローンの抗体のアイソタイプをELISAにより決定したところ、IgMクラスであることが判明した(データ示さず)。
【0045】
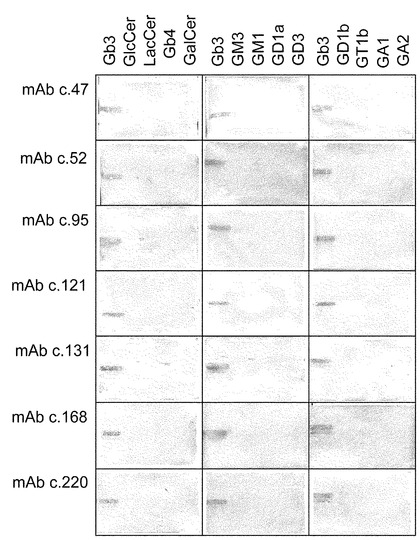
Gb3/CD77に反応性を示し、抗腫瘍効果の点でより高性能である抗体の樹立を目指し、すべてのグロボ系糖脂質を欠損するA4galt欠損マウスを、Gb3封埋リポソームを免疫原として、野生型マウスの場合と同様に免疫した。免疫したマウスの中でGb3/CD77に対して最も高い抗体価を示したマウスを用いて、その脾細胞とマウスNS-1骨髄腫細胞との融合を行った。ELISA及び免疫蛍光法によるスクリーニングの結果、Gb3/CD7に対する反応性に基づいて323クローンから8クローンのハイブリドーマが樹立された。得られたモノクローナル抗体のGb3/CD77に対する特異性をELISA及びTLC免疫染色によって決定した(図2及び3)。13種の糖脂質の中、すべてのクローンがGb3/CD77に限って反応した。種々の糖脂質に対するELISAの結果を図3に示した(GalCer、GD1b、GD3、GA1、GA2、GT1b、GM1及びGD1aに関する結果は省略)。8クローンの抗体のアイソタイプをELISAにより決定したところ、IgM(4クローン)、IgG3(3クローン)及びIgG2b (1クローン)であった(データ示さず)。
【0046】
次に、得られたクローンの抗腫瘍活性を解析するために、バーキットリンパ腫Ramos細胞を標的に用いてCDC活性を検討した。A/Jマウスを用いて得られた3クローンのIgMクラスのモノクローナル抗体、及びA4galt欠損マウスを用いて得られたIgMクラスのモノクローナル抗体は強いCDC活性を示したが、抗体のみでは細胞傷害を誘導しなかった(図4)。A4galt欠損マウスを用いて得られた4クローンのIgGクラス抗体は同様に明らかなCDC活性を誘導したが、その強さはIgM型モノクローナル抗体のそれよりは弱かった。次に、これらの抗体のADCC活性を検討した。モノクローナル抗体c.52はRamos細胞に対してヒトの末梢血単核球をエフェクターとして明らかなADCC活性を誘導した(図5)。IgMあるいはIgG2bクラスのモノクローナル抗体はADCC活性がないか又はごく僅かしか示さなかった(図5)。
【0047】
A4galt欠損マウスからIgGクラスの抗体が複数得られたという事実から、例えばベロ毒素受容体に対する高性能の競合的な役割など、抗体の持つ他の機能の探索を行うことが必要と思われた。図6に示したように、Ramos細胞をベロ毒素2と3時間静置した時に、カスパーゼ3(caspase-3)の分解が惹起され、ベロ毒素による細胞傷害が示された。ベロ毒素2(2 ng/ml)(細胞死を誘導するために必要な最小量)の添加前にRamos細胞を抗Gb3/CD77抗体の存在下で氷上2時間処置した場合には、カスパーゼ3の分解がモノクローナル抗体c.168(IgM)、c.52(IgG3)及びc.95(IgG2b)によって部分的に抑制された。しかし、モノクローナル抗体c.220(IgG3)あるいはR24(コントロール)によっては抑制されなかった(図6B)。モノクローナル抗体によりカスパーゼ3の分解が部分的であれ確かに抑制されたにもかかわらず、細胞死の方は明らかには抑制されなかった。
【0048】
Gb3/CD77は胚中心Bリンパ球の亜集団に発現し、抗体の親和性成熟の過程に必要とされるのかもしれない。そこで、Gb3/CD77に対する未知のリガンドが存在し、胚中心B細胞においてシグナルを惹起するかもしれないと考えた。この仮説を検討するために、Ramos細胞をモノクローナル抗体で処理して形態学的な変化と細胞内のタンパク質リン酸化を解析した。検討したクローンの中、モノクローナル抗体c.52はカルシウム依存性の細胞凝集を惹起したが、モノクローナル抗体c.220は惹起しなかった(図7A及び7B)。モノクローナル抗体c.52はまた、Ramos細胞に対して軽度ではあるが確実な細胞増殖抑制活性を示した(図7C)。これらの抗体を加えた後の細胞を用いたリン酸化タンパク質のイムノブロッテイングの結果、細胞内タンパク質のリン酸化パターンに劇的な変化が起こったことが示された(図8)。これらの変化は、モノクローナル抗体c.220によって処置された細胞でも認められた。但し、リン酸化の程度はモノクローナル抗体c.52による場合に比して軽度であった(図8右)。モノクローナル抗体c.220が細胞凝集を誘導しなかったことからすると、モノクローナル抗体c.52によるリン酸化は細胞凝集によるものではないであろう。抗μ抗体によるB細胞受容体を介するシグナルと比べた時、モノクローナル抗体c.52によって惹起されるタンパク質リン酸化は、抗BCR抗体のクロスリンクによるリン酸化と若干異なりつつも類似していた(図9A及び9B)。数個のリン酸化タンパク質の中で120 kDaのタンパク質はどちらの刺激によっても共通にリン酸化される分子と思われた(図9)。上記したごとく、モノクローナル抗体c.52はRamos細胞に対して軽度であるが明らかな増殖抑制活性を示し、Gb3/CD77を介するシグナルがBCRを介するシグナルや生存シグナルなどの既存のシグナルに対して負の効果をもたらす可能性が示唆された。
【0049】
120 kDaのバンドの分子を同定するために、また、c-Cbl(参考文献15)がモノクローナル抗体c.52によるGb3/CD77のクロスリンクに基づいてリン酸化を受けたかどうかを明らかにするために、免疫沈降(IP)-イムノブロッテイング(IB)解析を行った。その結果、120 kDaのバンドは少なくともその一部がc-Cblであり、それがモノクローナル抗体c.52によるGb3/CD77のクロスリンクによってリン酸化されることが示された(図10A)。c-Cblリン酸化の時間経過を示した(図10B)。Gb3/CD77がBCRシグナルの負の調節役として働くか否かを検討するために、抗μ抗体を加える前にRamos細胞をモノクローナル抗体c.52で前処置した時のタンパク質リン酸化パターンに与える影響を検討した。予想されたように、モノクローナル抗体c.52による前処置により、コントロールIgGと比べて抗μ抗体によるタンパク質リン酸化が有意に抑制された(図11A及び11B)。
【0050】
<考察>
本研究では、Gb3/CD77合成酵素欠損(ノックアウト)マウスを免疫に用いると、抗Gb3抗体を効率よく作製できることが示された。モノクローナル抗体の作製効率及び樹立された抗体の機能の質は、通常のマウスを用いた場合に比して明らかに良好であった。過去の試みにおいては、精製Gb3を免疫原として用いて、もっぱらIgMモノクローナル抗体が産生された(参考文献23)。従来の方法ではIgMクラスのモノクローナル抗体が得られるのが普通であるが、本研究においてはGb3に反応するIgGクラスのモノクローナル抗体が比較的高率に得られた。
【0051】
Gb3はすべてのグロボ系糖脂質の合成の出発分子であり、バーキットリンパ腫の腫瘍マーカーとして(参考文献8)、ベロ毒素の受容体として(参考文献11、24)、あるいはB細胞系列の細胞死マーカーとして(参考文献25)、研究者から注目されてきた。生理的及び病理的な状況下でのグロボ系糖脂質の役割に関しては、現在、グロボ系糖脂質を欠損する変異マウスを用いて、我々の研究室で解析中である(参考文献11)。いずれにしても、様々な機能を有する抗Gb3抗体は、糖脂質の機能解明にとって極めて有用である。
【0052】
実際、カスパーゼ3の分解がこれらの抗体によって部分的に阻害された。しかしながら細胞死の方は、カスパーゼ3の分解が有意に抑制されたのにもかかわらず、明らかには抑制されなかった。これらの結果は、これらの抗体の結合がアポトーシスシグナルの遅延を招いたにせよ、死のシグナルを抑えるほどには十分強くないことを示唆している。
【0053】
モノクローナル抗体の生物学的な機能という点では、本研究で得られた抗Gb3抗体は非常に興味深い。というのは、それらのいくつかは、抗μ抗体によるクロスリンクの場合と同様の方法で、細胞表面に結合してB細胞のシグナル伝達経路の活性化を誘導したからである(参考文献26)。モノクローナル抗体c.52で処理された際に、チロシンリン酸化を受ける成分が、c-Cblを含めて数種存在する。c-CblはE3ユビキチンリガーゼとして知られており(参考文献27)、Sykのような分子をプロテアゾーム系での分解に誘導することによって、BCRを介するシグナルを減弱せしめる(参考文献28)。BCRを介するシグナルにおけるGb3/抗Gb3抗体からのシグナルの役割を解析する中で、Ramos細胞をモノクローナル抗体c.52で前処置することにより、抗μ抗体によるチロシンリン酸化のレベルが軽度に減弱した。この結果から、c-Cbl活性化がBCRを介するシグナルの調節に実際に働いていることが分かった。従って、ここで最も興味深い問題は、Gb3と相互作用する内因性因子が存在するか否か、存在するのであればそれは何か、という点である。
【0054】
Gb3/CD77が胚中心のBリンパ球の亜集団に発現しており、バーキットリンパ腫細胞にモノクローナル抗体が結合することによって細胞凝集やタンパク質のチロシンリン酸化が惹起されることから、我々はGb3/CD77の内因性リガンドが存在して、胚中心B細胞のシグナル制御に働いているのではないかと推測した。Ramos細胞をモノクローナル抗体で前処置した時に、細胞増殖の抑制が見られた事実は、我々の、Gb3/CD77がBCRのクロスリンクによるBCRシグナルなどの既存のシグナルに対して負の効果をもたらすシグナルを導入するのではないか、という想定を支持するものであった。c-Cbl、E3ユビキチンリガーゼがSykやSFK、TecなどBCRを介するシグナル伝達経路の上流に位置するシグナル分子のユビキチンプロテアソーム系による分解に基づいて、BCRシグナルの負の調節役を演じることはよく知られていた(参考文献28、29)。BCRのクロスリンクによって活性化を受けると、それらはリン酸化される。本研究で示されたように、c-Cblはモノクローナル抗体c.52によってリン酸化され、またRamos細胞をc.52で前処置すると、BCRシグナルの減弱とともに細胞増殖も抑制された。これらの結果は、発生過程におけるc-Cblの一般的機能(参考文献30)及び病態における意義(参考文献31)にも関連している。
【0055】
BCR複合体が、同様の抗原あるいは抗μ抗体によってBCRをクロスリンク(参考文献32、33)した後に、非脂質ラフトから脂質ラフトに移行することがよく知られていた(図12)。Gb3/CD77もまた脂質ラフトに局在することから、我々はGb3/CD77が脂質ラフトにおいてBCRシグナルの負の調節に働くとの仮説を立てた。実際、モノクローナル抗体c.52は、抗μ抗体によって誘導されるリン酸化を有意に抑制した。さらに、モノクローナル抗体c.52が結合することで、Ramos細胞の脂質ラフトへのBCRの移行が惹起された(図12)。この事実は、BCRの非脂質ラフトから脂質ラフトへの移行が、BCRの効果的なシグナル伝達を招くことを示唆するだけでなく、Gb3/CD77を介するシグナルの制御に基づいた、ユビキチンプロテアソームを介する分解やキー分子の脱リン酸化などの負のシグナル発信の開始の可能性をも示唆する。
【産業上の利用可能性】
【0056】
本発明のモノクローナル抗体作製法によれば、グロボ系糖脂質であるGb3/CD77を特異的に認識するモノクローナル抗体を効率的に作製できる。即ち、本発明のモノクローナル抗体作製法は、抗Gb3/CD77モノクローナル抗体を効率的に作製する手段として有用である。本発明のモノクローナル抗体作製法はIgGクラスのモノクローナル抗体を取得するための手段として特に有用である。本発明によってもたらされる抗Gb3/CD77モノクローナル抗体には、研究用途に留まらず、診断用途や治療用途など、様々な分野で利用・活用されることが期待される。
【0057】
この発明は、上記発明の実施の形態及び実施例の説明に何ら限定されるものではない。特許請求の範囲の記載を逸脱せず、当業者が容易に想到できる範囲で種々の変形態様もこの発明に含まれる。
本明細書の中で明示した論文、公開特許公報、及び特許公報などの内容は、その全ての内容を援用によって引用することとする。
【0058】
<参考論文>
[1] R.H. Kennett, Hybridomas: a new dimension in biological analyses, In Vitro 17 (1981) 1036-1050.
[2] P.N. Nelson, G.M. Reynolds, E.E. Waldron, E. Ward, K. Giannopoulos, P.G. Murray, Monoclonal antibodies, Mol. Pathol. 53 (2000) 111-117.
[3] B.J. Willison, D. Nicholl, E.R. Wagner, K. Townson, C. Goodyear, K. Furukawa, K. Furukawa, J. Conner, H.J. Willison, Innate murine B cells produce anti-disialosyl antibodies reactive with Campylobacter jejuni LPS and gangliosides that are polyreactive and encoded by a restricted set of unmutated V genes, J. Neuroimmunol. 152 (2004) 98-111.
[4] M.P. Lunn, L.A. Johnson, S.E. Fromholt, S. Itonori, J. Huang, A.A. Vyas, J.E. Hildreth, J.W. Griffin, R.L. Schnaar, K.A. Sheikh, High-affinity anti-ganglioside IgG antibodies raised in complex ganglioside knockout mice: reexamination of GD1a immunolocalization. J. Neurochem. 75 (2000) 404-412.
[5] J. Boffey, M. Odaka, D. Nicoll, E.R. Wagner, K. Townson, T. Bowes, J. Conner, K. Furukawa, H.J. Willison, Characterization of the immunoglobulin variable region gene usage encoding the murine anti-ganglioside antibody repertoire, J. Neuroimmunol. 165 (2005) 92-103.
[6] K.J. Kasai, J. Galton, P.I. Terasaki, A. Wakisaka, M. Kawahara, T. Root, S.I. Hakomori, Tissue distribution of the Pk antigen as determined by a monoclonal antibody, 12 (1985) J. Immunogenet. 213-220
[7] Y. Fujii, S. Numata, Y. Nakamura, T. Honda, K. Furukawa, T. Urano, J. Wiels, M. Uchikawa, N. Ozaki, S. Matsuo, Y. Sugiura, K. Furukawa, Murine glycosyltransferases responsible for the expression of globo-series glycolipids: cDNA structures, mRNA expression, and distribution of their products, Glycobiology 15 (2005) 1257-1267.
[8] E. Nudelman, R. Kannagi, S. Hakomori, M. Parsons, M. Lipinski, J. Wiels, M. Fellous, T.A. Tursz, glycolipid antigen associated with Burkitt's lymphoma defined by a monoclonal antibody, Science 220 (1983) 509-511.
[9] T. Falguieres, M. Maak, C. von Weyhern, M. Sarr, X. Sastre, M.F. Poupon, S. Robine, L. Johannes, K.P. Janssen, Human colorectal tumors and metastases express Gb3 and can be targeted by an intestinal pathogen-based delivery tool, Mol. Cancer Ther. 7 (2008)2498-2508.
[10] O. Kovbasnjuk, R. Mourtazina, B. Baibakov, T. Wang, C. Elowsky, M.A. Choti, A. Kane, M. Donowitz, The glycosphingolipid globotriaosylceramide in the metastatic transformation of colon cancers, Proc. Natl. Acad. Sci. U. S. A. 102 (2005) 19087-19092.
[11] T. Okuda, N. Tokuda, S. Numata, M. Ito, M. Ohta, K. Kawamura, J. Wiels, T. Urano, O. Tajima, K. Furukawa, K. Furukawa, Targeted disruption of Gb3/CD77 synthase gene resulted in the complete deletion of globo-series glyco- sphingolipids and loss of sensitivity to verotoxins, J. Biol. Chem. 281 (2006) 10230-10235.
[12] Y. Kojima, S. Fukumoto, K. Furukawa, T. Okajima, J. Wiels, K. Yokoyama, Y. Suzuki, M. Ohta, K. Furukawa, Molecular cloning of Gb3/CD77 synthase, a glycosyl- transferase that initiate the synthesis of globo-series glycosphingolipids, J. Biol. Chem. 275 (2000) 15152-15156.
[13] S. Watarai, K. Kiura, R. Shigeto, T. Shibayama, I. Kimura, T. Yasuda, Establishment of monoclonal antibodies specific for ganglioside GM1: detection of ganglioside GM1 in small cell lung carcinoma cell lines and tissues, J. Biochem. 116 (1994) 948-954.
[14] K. Furukawa, H. Clausen, S. Hakomori, J. Sakamoto, K. Look, A. Lundblad, M.J. Mattes, K.O. Lloyd, Analysis of the specificity of five murine anti-blood group A monoclonal antibodies, including one that identifies type 3 and type 4 determinants, Biochemistry 24 (1985) 7820-7826.
[15] M.L.J.Lupher, N. Rao, M.J. Eck, H. Band, The Cbl protooncoprotein: a negative regulator of immune receptor signal transduction, Immunol Today, 20 (1999) 375-382.
[16] G. Kohler, C. Milstein, Continuous cultures of fused cells secreting antibody of predefined specificity, Nature, 256 (1975) 495-497.
[17] Y. Nagatsuka, S. Hanazawa, T. Yasuda, Y. Ono, Anti-glycolipid antibodies produced by Epstein-Barr virus (EBV)-transformed oligoclonal B cell lines obtained from normal persons and autoimmune disease patients, Hum. Antibodies Hybridomas 5 (1994) 183-186.
[18] H. Yamaguchi, K. Furukawa, S.R. Fortunato, P.O. Livingston, K.O. Lloyd, H.F. Oettgen, L.J. Old, Cell-surface antigens of melanoma recognized by human monoclonal antibodies, Proc. Natl. Acad. Sci. U. S. A. 84 (1987) 2416-2420.
[19] I. Kawashima, N. Tada, S. Ikegami, S. Nakamura, R. Ueda, T. Tai, Mouse monoclonal antibodies detecting disialogangliosides on mouse and human T lymphomas, Int. J. Cancer 41 (1988) 267-274.
[20] M. Kotani, H. Ozawa, I. Kawashima, S. Ando, T. Tai, Generation of one set of monoclonal antibodies specific for a-pathway ganglio-series gangliosides, Biochim. Biophys. Acta 1117 (1992) 97-103.
[21] H. Ozawa, M. Kotani, I. Kawashima, T. Tai, Generation of one set of monoclonal antibodies specific for b-pathway ganglio-series gangliosides, Biochim. Biophys. Acta. 1123 (1992) 184-190.
[22] I. Kawashima, O. Nakamura, T. Tai, Antibody responses to ganglio-series gangliosides in different strains of inbred mice, Mol. Immunol. 29 (1992) 625-632.
[23] D. Miyamoto, T. Ueno, S. Takashima, K. Ohta, T. Miyawaki, T. Suzuki, Y. Suzuki, Establishment of a monoclonal antibody directed against Gb3Cer/CD77: a useful immunochemical reagent for a differentiation marker in Burkitt's lymphoma and germinal centre B cells, Glycoconj. J. 14 (1997) 379-388
[24] M. Jacewicz, H. Clausen, E. Nudelman, A. Donohue-Rolfe, G.T. Keusch, Pathogenesis of shigella diarrhea. XI. Isolation of a shigella toxin-binding glycolipid from rabbit jejunum and HeLa cells and its identification as globotriaosylceramide, J. Exp. Med. 163 (1986) 1391-1404.
[25] M. Mangeney, Y. Richard, D. Coulaud, T. Tursz, J. Wiels, CD77: an antigen of germinal center B cells entering apoptosis. Eur J Immunol. 21 (1991) 1131-1140.
[26] H.L. Weiner, J.W. Moorhead, K. Yamaga, R.T. Kubo, Anti-Immunoglobulin Stimulation of Murine Lymphocytes: II. Identification of Cell Surface Target Molecules and Requirements for Cross-Linkage, J. Immunol. 117 (1976) 1527-1531.
[27] D.L. Mueller, E3 ubiquitin ligases as T cell anergy factors, Nat. Immunol. 5 (2004) 883 - 890.
[28] C.B. Thien, W.Y. Langdon, c-Cbl and Cbl-b ubiquitin ligases: substrate diversity and the negative regulation of signaling responses, Biochem. J. 391 (2005) 153-166.
[29] L. Duan, A.L. Reddi, A. Ghosh, M. Dimri, H. Band, The Cbl family and other ubiquitin ligases: destructive forces in control of antigen receptor signaling, Immunity 21 (2004) 7-17.
[30] F. Huang, G. Hua, Negative regulation of lymphocyte development and function by the Cbl family of proteins, Immunol. Rev. 224 (2008) 1229-1238.
[31] C.E. Rudd, H. Schneider, Lymphocyte signaling: Cbl sets the threshold for autoimmunity, Curr. Biol. 10 (2000) R344-347.
[32] P.C. Cheng, M.L. Dykstra, R.N. Mitchell, S.K. Pierce, A role for lipid rafts in B cell antigen receptor signaling and antigen targeting, J. Exp. Med. 190 (1999) 1549-1560.
[33] R.J. Petrie, P.P. Schnetkamp, K.D. Patel, M. Awasthi-Kalia, J.P. Deans, Transient translocation of the B cell receptor and Src homology 2 domain-containing inositol phosphatase to lipid rafts: evidence toward a role in calcium regulation, J. Immunol. 165 (2000) 1220-1227.
【特許請求の範囲】
【請求項1】
以下のステップ(1)〜(5)を含む、抗Gb3/CD77モノクローナル抗体の作製法:
(1)α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物をGb3/CD77で免疫するステップ;
(2)免疫後の非ヒト哺乳動物から抗体産生細胞を回収するステップ;
(3)回収した抗体産生細胞を不死化させるステップ;
(4)不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化するステップ;及び
(5)モノクローナル化によって得られたクローンが産生する抗体を回収するステップ。
【請求項2】
非ヒト哺乳動物がマウスである、請求項1に記載の作製法。
【請求項3】
マウスがA/J系統である、請求項2に記載の作製法。
【請求項4】
Gb3/CD77、リピッドA、コレステロール及びジパラミトイルホスファチジルコリンを含むリポソームを免疫原としてステップ(1)における免疫を行う、請求項1〜3のいずれか一項に記載の作製法。
【請求項5】
抗体産生細胞が脾臓細胞である、請求項1〜4のいずれか一項に記載の作製法。
【請求項6】
ステップ(3)が、抗体産生細胞と骨髄腫細胞との融合によって行われる、請求項1〜5のいずれか一項に記載の作製法。
【請求項7】
ステップ(4)において、IgGクラスの抗Gb3/CD77抗体を産生する細胞をモノクローナル化する、請求項1〜6のいずれか一項に記載の作製法。
【請求項8】
Gb3/CD77を特異的に認識し、補体依存性細胞傷害活性及び/又は抗体依存性細胞傷害活性を示す、単離されたモノクローナル抗体。
【請求項9】
バーキットリンパ腫細胞株に対する凝集活性を示す、請求項8に記載の単離されたモノクローナル抗体。
【請求項10】
請求項1〜7のいずれか一項に記載の作製法によって得られる、請求項8又は9に記載の単離されたモノクローナル抗体。
【請求項11】
ヒト抗体又はヒト化抗体である、請求項9又は10に記載の単離されたモノクローナル抗体。
【請求項12】
IgG、Fab、Fab'、F(ab')2、scFv、又はdsFv抗体である、請求項9〜11のいずれか一項に記載の単離されたモノクローナル抗体。
【請求項13】
請求項9〜12のいずれか一項に記載の単離されたモノクローナル抗体を含む、Gb3/CD77を発現する細胞を標的とした研究用試薬。
【請求項14】
請求項9〜12のいずれか一項に記載の単離されたモノクローナル抗体を含む、Gb3/CD77を過剰発現する悪性腫瘍を標的とした診断又は治療用の組成物。
【請求項15】
悪性腫瘍がバーキットリンパ腫、大腸癌、奇形腫又は巨核芽球性白血病である、請求項14に記載の組成物。
【請求項1】
以下のステップ(1)〜(5)を含む、抗Gb3/CD77モノクローナル抗体の作製法:
(1)α1,4-ガラクトース転移酵素遺伝子を欠損した非ヒト哺乳動物をGb3/CD77で免疫するステップ;
(2)免疫後の非ヒト哺乳動物から抗体産生細胞を回収するステップ;
(3)回収した抗体産生細胞を不死化させるステップ;
(4)不死化した抗体産生細胞の内、抗Gb3/CD77抗体を産生する細胞をモノクローナル化するステップ;及び
(5)モノクローナル化によって得られたクローンが産生する抗体を回収するステップ。
【請求項2】
非ヒト哺乳動物がマウスである、請求項1に記載の作製法。
【請求項3】
マウスがA/J系統である、請求項2に記載の作製法。
【請求項4】
Gb3/CD77、リピッドA、コレステロール及びジパラミトイルホスファチジルコリンを含むリポソームを免疫原としてステップ(1)における免疫を行う、請求項1〜3のいずれか一項に記載の作製法。
【請求項5】
抗体産生細胞が脾臓細胞である、請求項1〜4のいずれか一項に記載の作製法。
【請求項6】
ステップ(3)が、抗体産生細胞と骨髄腫細胞との融合によって行われる、請求項1〜5のいずれか一項に記載の作製法。
【請求項7】
ステップ(4)において、IgGクラスの抗Gb3/CD77抗体を産生する細胞をモノクローナル化する、請求項1〜6のいずれか一項に記載の作製法。
【請求項8】
Gb3/CD77を特異的に認識し、補体依存性細胞傷害活性及び/又は抗体依存性細胞傷害活性を示す、単離されたモノクローナル抗体。
【請求項9】
バーキットリンパ腫細胞株に対する凝集活性を示す、請求項8に記載の単離されたモノクローナル抗体。
【請求項10】
請求項1〜7のいずれか一項に記載の作製法によって得られる、請求項8又は9に記載の単離されたモノクローナル抗体。
【請求項11】
ヒト抗体又はヒト化抗体である、請求項9又は10に記載の単離されたモノクローナル抗体。
【請求項12】
IgG、Fab、Fab'、F(ab')2、scFv、又はdsFv抗体である、請求項9〜11のいずれか一項に記載の単離されたモノクローナル抗体。
【請求項13】
請求項9〜12のいずれか一項に記載の単離されたモノクローナル抗体を含む、Gb3/CD77を発現する細胞を標的とした研究用試薬。
【請求項14】
請求項9〜12のいずれか一項に記載の単離されたモノクローナル抗体を含む、Gb3/CD77を過剰発現する悪性腫瘍を標的とした診断又は治療用の組成物。
【請求項15】
悪性腫瘍がバーキットリンパ腫、大腸癌、奇形腫又は巨核芽球性白血病である、請求項14に記載の組成物。
【図1】


【図2】
【図3】

【図4】

【図5】
【図6】

【図7】
【図8】
【図9】
【図10】

【図11】
【図12】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【公開番号】特開2012−50407(P2012−50407A)
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【出願番号】特願2010−197639(P2010−197639)
【出願日】平成22年9月3日(2010.9.3)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人新エネルギー・産業技術総合開発機構、「糖鎖機能活用技術開発プロジェクト」、 産業技術力強化法19条の適用を受ける特許出願
【出願人】(504139662)国立大学法人名古屋大学 (996)
【出願人】(509131742)株式会社GPバイオサイエンス (5)
【Fターム(参考)】
【公開日】平成24年3月15日(2012.3.15)
【国際特許分類】
【出願日】平成22年9月3日(2010.9.3)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度、独立行政法人新エネルギー・産業技術総合開発機構、「糖鎖機能活用技術開発プロジェクト」、 産業技術力強化法19条の適用を受ける特許出願
【出願人】(504139662)国立大学法人名古屋大学 (996)
【出願人】(509131742)株式会社GPバイオサイエンス (5)
【Fターム(参考)】
[ Back to top ]