
コバラミンを製造し得る生合成方法
【課題】組換え菌株を利用して、コバラミンの前駆物質の添加によってコバラミン産生を増加する生合成方法を提供する。
【解決手段】O−ホスホ−L−トレオニンもしくは5,6−ジメチルベンズイミダゾールのin situ合成もしくはこのような合成の増進を達成するために、Rhodobacter capsulatusのDNAフラグメントを使用して、Pseudomonas denitrificansを宿主とする組換えDNA技術によりB12補酵素の産生を増加させる方法を用いる。
【解決手段】O−ホスホ−L−トレオニンもしくは5,6−ジメチルベンズイミダゾールのin situ合成もしくはこのような合成の増進を達成するために、Rhodobacter capsulatusのDNAフラグメントを使用して、Pseudomonas denitrificansを宿主とする組換えDNA技術によりB12補酵素の産生を増加させる方法を用いる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、コバラミンを製造し得る生合成方法に関する。より詳細には本発明は、組換えDNA技術及び/または新規なコバラミン前駆物質の添加によってコバラミンの産生を増加させる方法、特にB12補酵素の産生を増加させる方法に関する。最後に本発明は、本発明のコバラミンの製造方法に有用な組換え菌株の作製方法に関する。
【背景技術】
【0002】
ビタミンB12はコバラミンと呼ばれる分子のクラスに属しており、その構造は特に国際特許WO91/11518に開示されている。
【0003】
コバラミンはほぼ例外なく、同じく国際特許WO91/11518に記載された複合プロセスによって細菌から合成されている。生合成メカニズムが極めて複雑であるため、工業的レベルのコバラミンの産生、特にビタミンB12の産生は主として、Pseudomonas denitrificans、Propionobacterium shermanii及びPropionobacterium freudenreichiiのような細菌の大量培養によって行われている。
【0004】
また、いくつかの微生物が、アミノレブリン酸、S−アデノシル−L−メチオニン、コバルト、グルタミン、R1−アミノ−2−プロパノール及び5,6−ジメチルベンズイミダゾールの基質からコバラミンを合成することも公知である。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】国際公開第WO91/11518号
【発明の概要】
【0006】
上記に挙げた前駆物質のうちで、5,6−ジメチルベンズイミダゾールは、コバラミン産生性微生物によって合成される。5,6−ジメチルベンズイミダゾールの生合成には2つの経路が存在すると考えられる。一方の経路は、分子酸素を利用する好気性微生物に特有の経路である。他方の経路は、嫌気性微生物によって使用される経路である。嫌気性経路に関与する遺伝子だけが単離されている。この遺伝子は、ネズミチフス菌(Salmonella thyphimurium)のcobT遺伝子である(Trzebiatowskiら,1994)。5,6−ジメチルベンズイミダゾールを合成する好気性微生物の遺伝子は現在まで全く同定されていない。多くの場合、微生物によって合成される5,6−ジメチルベンズイミダゾールの量には限界がある。
【0007】
このため、5,6−ジメチルベンズイミダゾールを化学的に製造して産生用培地に添加している。従って、培地に対するこのような添加の必要をなくすことができれば有利になることは確かである。
【0008】
従来のどのようなコバラミンの工業的製造方法も、コバルト及び5,6−ジメチルベンズイミダゾール以外の前駆物質の添加に言及したことはない。最近になって、培地がR1−アミノ−2−プロパノールを含有するときにだけコバラミンを産生するいくつかの菌株が記載された(Crouzetら,1990、Grabauら,1992)。従って、R1−アミノ−2−プロパノールもコバラミンの産生を増進するために使用し得るであろう。しかしながら、この物質は、工業的発酵に使用できるとしても、その使用が難しく費用も高い。その理由の1つは、R1−アミノ−2−プロパノールが刺激性及び揮発性の物質であるからであり、いま1つの理由は、この物質が微生物の増殖を阻害するからである。従って、R1−アミノ−2−プロパノール残基以外の、上記の欠点を有していないコバラミンの前駆物質を見つけることが特に有益であると考えられる。この観点から、幾つかの微生物ではL−トレオニンからアミノアセトンを経由してR1−アミノ−2−プロパノールを生合成する経路が存在することは開示されている。しかしながら、L−トレオニンは上記に引用した菌株を補完することができない。
【0009】
より一般的に、コバラミンの産生を増進するためには、培地中のコバラミンの前駆物質の量を増加させるのが有利であろう。コバラミンの前駆物質が制限的である場合には特にこのような増加が有利である。この方法は、制限的前駆物質またはその誘導体もしくはその類似体の1つを培地に直接添加することによって、あるいは、遺伝子技術、特に組換えDNA技術を使用して産生性菌株中でこの前駆物質のin situ合成を増進することによって行う。
【0010】
従って、コバラミンの産生を増進するために重要な段階は、コバラミン及びその前駆物質の生合成経路を明らかにすることである。
【0011】
最近には、ビタミンB12の生合成経路の大部分の段階がPseudomonas denitrificansで解明された(Blancheら,1995)。コバラミンの生合成に関与する22個以上のcob遺伝子が単離され、これらの遺伝子によってコードされたポリペプチドの殆どについてその機能が同定された。
【0012】
別の微生物においても、コバラミンまたはその前駆物質の生合成に関与すると推測される別の遺伝子が単離された。これらの遺伝子の殆どについてその機能は未だ解明されていない。これらの遺伝子の機能またはその効果を正確に決定することさえできれば、これらの遺伝子の応用が可能になるであろう。例えば、通性光合成細菌Rhodobacter capsulatus中で、光合成器官の形成に必要な少なくとも1つの遺伝子を含むDNAフラグメントが最近になって配列決定された(Pollichら,1993、Pollichら,1995a)。単離された8個の遺伝子のうちの5個の遺伝子が、前述のP.denitrificansの22個のcob遺伝子のうちの5個の遺伝子に高度な相同性を有するという理由でコバラミンの生合成遺伝子であると示唆された。逆に、残りの3つの遺伝子、即ちbluB遺伝子、bluE遺伝子及びbluF遺伝子に関しては、これらの遺伝子が直接に担当する正確な機能を明らかにすることができなかった。それらのプロモーター配列を含むbluB遺伝子、bluE遺伝子及びbluF遺伝子は、Pollichら,1995aによってその配列が決定され開示された。
【0013】
本発明によって、コバラミンの新規な前駆物質が知見された。その結果として本発明は、O−ホスホ−L−トレオニンを含有する培地によってコバラミンの産生を増進することに成功した。これまでに記載されたことのないこのコバラミンの前駆物質は、既に公知であった別の前駆物質R1−アミノ−2−プロパノールと同等の機能を有している。しかしながら、O−ホスホ−L−トレオニンは、無毒の物質である、及び、取り扱い容易な物質である、という点でR1−アミノ−2−プロパノールよりも有利である。更に、コバラミンの産生増進に対してO−ホスホ−L−トレオニンが示す効率は、R1−アミノ−2−プロパノールが示す効率の1,000倍以上である。
【0014】
本発明の目的はまた、コバラミンの産生増進を達成するためまたは所与の細胞のO−ホスホ−L−トレオニンもしくは5,6−ジメチルベンズイミダゾールのin situ合成もしくはこのような合成の増進を達成するために、Rhodobacter capsulatusのDNAフラグメントを使用することである。
【0015】
本発明によれば、Rhodobacter capsulatusの、特にbluE遺伝子及びbluF遺伝子を含むDNAフラグメントを使用することによって、R1−アミノ−2−プロパノールまたはO−ホスホ−L−トレオニンを含有しない培地においてコバラミンの産生を増進することが可能であった。
【0016】
最後に、本発明によれば、Rhodobacter capsulatusの、特にbluB遺伝子を含むDNAフラグメントを導入することによって、5,6−ジメチルベンズイミダゾールを含有しない培地においてコバラミンの産生を増進することが可能であった。
【0017】
本発明の1つの目的は、コバラミン産生性原核細胞微生物の発酵によるコバラミンの生合成方法を提供することである。本発明によるこの方法の特徴は、
−O−ホスホ−L−トレオニンの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって形質転換された微生物を使用し、前記酵素が発現しコバラミンが産生され得る条件下で前記微生物を培養する段階、及び/または、
−前記微生物の培養培地にO−ホスホ−L−トレオニンを添加する段階、または、
−5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって形質転換された好気性微生物を使用し、前記酵素が発現しコバラミンが産生され得る条件下で前記好気性微生物を培養する段階、
を含むことである。
【0018】
微生物は、上記に特定した遺伝子を内在的に含んでいてもよい。この場合には、本発明方法によって酵素が超発現し得る。しかしながらまた、微生物がこの種の遺伝子を内在的に含んでいなくてもよい。
【0019】
“O−ホスホ−L−トレオニンまたは5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードするDNAフラグメント”なる表現は、このDNAフラグメントの発現によってO−ホスホ−L−トレオニンまたは5,6−ジメチルベンズイミダゾールが細胞内で合成され、場合によってはその後に培養培地中に遊離されることを意味する。
【0020】
培養は、バッチ培養でもよく、または、連続培養でもよい。また、コバラミンの精製は既存の工業的方法によって行うことができる(Florent,1986)。
【0021】
1つの実施態様においては、Rhodobacter capsulatusのbluE遺伝子及びbluF遺伝子を含みO−ホスホ−L−トレオニンの生合成経路に関与するポリペプチドをコードするDNAフラグメントによって形質転換されたか、または、相同フラグメント、即ち、これらのbluE遺伝子及びbluF遺伝子とハイブリダイズし且つこれらの遺伝子と同様にO−ホスホ−L−トレオニンの生合成経路に関与する酵素をコードする機能を有しているフラグメントによって形質転換された微生物が使用される。
【0022】
別の実施態様においては、Rhodobacter capsulatusのbluB遺伝子を含み5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードするDNAフラグメントによって形質転換されたか、または、相同フラグメント、即ち、このbluB遺伝子とハイブリダイズし且つこの遺伝子と同様に5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする機能を有しているフラグメントによって形質転換された微生物が使用される。
【0023】
本発明は、天然、合成または組換えによって得られたDNAフラグメント及び相同フラグメントの使用を含む。フラグメントなる用語は、遺伝コードの縮重性に由来するフラグメント、または、少なくとも25%の配列相同性を有しており同じ機能のポリペプチドをコードするフラグメントを意味する。
【0024】
好気性条件下で培養される微生物を使用し、5,6−ジメチルベンズイミダゾールの好気性条件下の生合成経路に関与する酵素をコードするDNAフラグメントを使用するのが好ましい。
【0025】
本発明の目的はまた、遺伝子工学技術を用い、O−ホスホ−L−トレオニンまたは5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする少なくとも1つの上記のようなDNAフラグメントによって微生物を形質転換させることを特徴とするコバラミン産生性原核細胞微生物の組換え菌株の製造方法を提供することである。
【0026】
また、本発明の菌株の製造方法によって得られた組換え菌株を提供することも本発明の目的の1つである。
【0027】
ポリペプチドをコードする少なくとも1つのDNA配列を含み、1つまたは複数のこれらの配列が発現シグナルのコントロール下に配置されている組換えDNAを本発明方法に使用することも本発明に包含される。
【0028】
この観点から、特に、DNA配列の5′にプロモーター領域を配置し得る。このような領域は、DNA配列に同種(homologous)の領域でもよくまたは異種(heterologous)の領域でもよい。特に、強い細菌プロモーター、例えば、大腸菌のトリプトファンオペロンのプロモーターPtrpもしくはラクトースオペロンのプロモーターPlac、ラムダバクテリオファージの左もしくは右のプロモーター、コリネバクテリウム(corynebacterium)のような細菌のファージの強いプロモーター、大腸菌のプロモーターPtacのようなグラム陰性菌中で機能性のプロモーター、プラスミドTOLのキシレンの異化遺伝子のプロモーターPxylS、枯草菌(Bacillus subtilis)のアミラーゼのプロモーターPamyを使用し得る。また、ホスホグリセリン酸キナーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ラクターゼまたはエノラーゼをコードする遺伝子のプロモーターのような、酵母の解糖遺伝子に由来のプロモーターも例示し得る。これらのプロモーターは、組換えDNAを真核細胞宿主に導入するときに使用できる。また、DNA配列の5′端にリボソーム結合部位を配置してもよい。この部位は、同種部位でもよく、または、ラムダバクテリオファージのcII遺伝子のリボソーム結合部位のような異種部位でもよい。
【0029】
転写の終結に必要なシグナルはDNA配列の3′に配置され得る。
【0030】
本発明方法で使用される組換えDNAは、選択された発現シグナルに適合性の宿主細胞に直接導入されてもよく、または、問題のDNA配列を宿主細胞に安定に導入できるようにプラスミドベクターにクローニングされてもよい。
【0031】
本発明方法は公知の方法と同様に、ポリペプチドをコードするDNA配列を含むプラスミドの使用を包含する。更にこれらのプラスミドは公知のプラスミドと同様に、機能性複製系と選択マーカーとを含む。
【0032】
多様な種類のベクターを使用し得る。本発明の範囲内では、RK2型のベクター、即ち、RK2複製起点を有するベクターを使用するのが好ましい。特定例としては、ベクターRK2(Sauruggerら,1986)、ベクターpXL435(Cameronら,1989)、ベクターpRK290(米国特許第4,590,163号、Dittaら,1985)、及び、ベクターpXL1635(国際特許WO91/16439)を挙げることができる。特に有利なベクターはベクターpXL1635である。その他のベクターは国際特許出願WO91/16439に記載されている。
【0033】
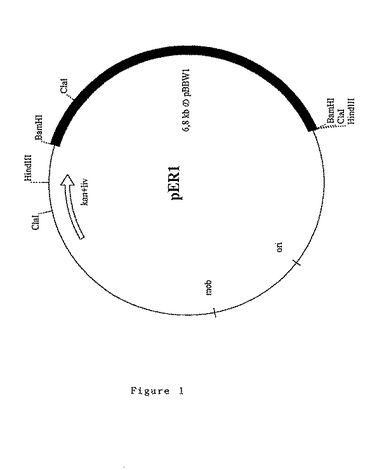
変形実施態様においては、後出の実施例に記載したように、2種類の前駆物質、即ち、O−ホスホ−L−トレオニン及び5,6−ジメチルベンズイミダゾールの合成をコードするプラスミドpER1(図1)の6.8kbのBamHIフラグメントから成るDNAフラグメントによって形質転換された微生物が使用される。
【0034】
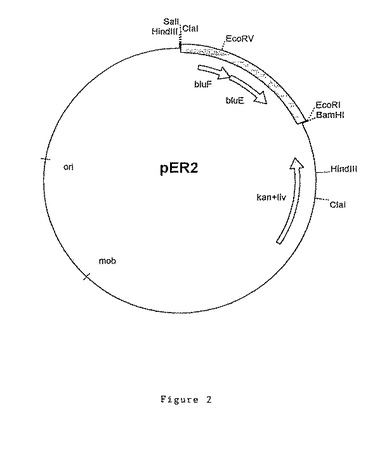
O−ホスホ−L−トレオニンの合成に関与するポリペプチドの発現に特に好適な実施態様では、後出の実施例に記載したようなプラスミドpER2(図2)の2.1kbのEcoRI/ClaIフラグメントから成るDNAフラグメントが使用される。
【0035】
O−ホスホ−L−トレオニンの合成に関与するポリペプチドの発現に特に好適な実施態様では、後出の実施例に記載したようなプラスミドpER2(図2)の1.6kbのEcoRI/EcoRVフラグメントから成るDNAフラグメントが使用される。
【0036】
5,6−ジメチルベンズイミダゾールの合成に関与するポリペプチドの発現に特に好適な別の実施態様では、後出の実施例に記載したようなプラスミドpER1の6.8kbのBamHIフラグメントから成るDNAフラグメントが使用される。
【0037】
本発明によって使用され得る宿主原核細胞微生物は、E.coli、Pseudomonas denitrificans、Agrobacterium radiobacter、Agrobacterium tumefaciensまたはRhizobium melitotiあるいはRhodobacter capsulatus属の細菌である。他の細菌は国際特許WO91/11518に記載されている。
【0038】
いずれにしても、P.denitrificansまたはA.radiobacter属の細菌の使用が特に有利である。
【0039】
本発明の別の特徴及び利点は以下の詳細な記載より明らかにされるであろう。
【0040】
実施例1及び2は、Pseudomonas denitrificansまたはRhodobacter capsulatusの菌株の培養培地にO−ホスホ−L−トレオニンを添加することによってコバラミンを産生させるのが可能であることを示している。実施例2は、産生培地中のR1−アミノ−2−プロパノールをO−ホスホ−L−トレオニンで代替することがどのように有利であるかを示している。同量のコバラミンを産生するために必要なR1−アミノ−2−プロパノールの量はO−ホスホ−L−トレオニンの1,000倍である。
【0041】
実施例2はまた、Rhodobacter capsulatusのbluE遺伝子及びbluF遺伝子を含む染色体の領域が、O−ホスホ−L−トレオニンの合成に関与することを示している。実施例3は、Rhodobacter capsulatusのDNAフラグメントを出発材料としたbluE遺伝子及びbluF遺伝子を含むプラスミドの構築を記載している。この実施例3は特に、これらのプラスミドがR1−アミノ−2−プロパノールまたはO−ホスホ−L−トレオニンの添加に依存してコバラミンを産生する菌株中に導入されたときに、R1−アミノ−2−プラスミドまたはO−ホスホ−L−トレオニン非含有の培地でどのようにしてコバラミンを産生し得るかを示している。実施例4は、Rhodobacter capsulatusのbluB遺伝子を含む染色体の領域が、5,6−ジメチルベンズイミダゾールの合成に関与することを示している。
【図面の簡単な説明】
【0042】
【図1】プラスミドpER1の制限地図を表す。
【図2】プラスミドpER2の制限地図を表す。
【図3】プラスミドpER3の制限地図を表す。
【0043】
図1から図3は夫々、プラスミドpER1、pER2及びpER3を表す。
【発明を実施するための形態】
【0044】
1.菌株及びプラスミド
Rhodobacter capsulatus菌のAH2株及びBB1株(Pollichら,1995a)を、37b4株(DSM938)から構築した。Pseudomonas denitrificans菌G2650株は、SBL 27 Rifr株を出発材料とし、トランスポゾンTn5の挿入によって構築した(Crouzetら,1990)。先ず、テトラサイクリン耐性遺伝子を含むトランスポゾンTn5を用いてG2650〔Tet〕株を構築した。次に、G2650〔Tet〕株を出発材料とし、トランスポゾンTn5のテトラサイクリン耐性遺伝子をスペクチノマイシン耐性遺伝子で置換することによってG2650〔Sp〕株を構築した。SBL 27 Rifr株はMB580株に由来する(米国特許US3,018,225)。
【0045】
プラスミドpBBW1(Pollichら,1995a)をRhodobacter capsulatusのDNAフラグメントから構築した。プラスミドpBBW1からプラスミドpAHW25(Pollich & Klug,1995a)を構築した。
【0046】
2.分子技術
DNA操作の汎用技術に関しては実験概論書(Sambrookら,1989)を参考として使用する。
【0047】
酵素はNew England Biolabs及びBoehringer Mannheimの研究所から入手し、製作者の指示通りに使用する。
【0048】
使用した技術は本質的に以下の2段階から成る。
【0049】
−制限酵素による消化、及び、
−T4バクテリオファージのリガーゼによるDNA分子の結合。
【0050】
3.形質転換技術
大腸菌の形質転換を電気穿孔によって行う(Dowerら,1988)。
【0051】
4.結合技術
大腸菌S17−1株とP.denitrificansの種々の菌株との結合は、Simonらによって記載されたプロトコル(Simonら,1986)を応用して行う。Pseudomonasの菌株の形質転換は遺伝子工学の他の任意の技術によって行う。
【0052】
5.コバラミン産生培地の調製
P.denitrificansの菌株によるコバラミンの産生に使用した培地は、Cameronら,1989、によって記載されたPS4培地である。
【0053】
Rhodobacter capsulatusの菌株によるコバラミンの産生に使用した培地は、Pollich,1995b、によって記載されたRA培地である。
【0054】
6.産生されたコバラミンの定量アッセイ
産生されたコバラミンの量を、微生物学的アッセイまたは高性能液体クロマトグラフィー(HPLC)によって測定する。
【0055】
−微生物学的アッセイ
ビタミンB12要求性の大腸菌指示株113−3(Davis & Mingioli,1950)を用い、半定量的方法によってコバラミンの産生量を測定する。
【0056】
この指示株は、B12依存性のホモシステインメチルトランスフェラーゼ(EC2.1.1.13)だけを有している大腸菌のmetE変異株である。この菌株が最小培地中で増殖するためにはビタミンB12の存在だけが必要である。(この菌株の増殖に必要なビタミンB12が欠乏した)ゲル状のM9最少培地(Miller,1972)の上層にこの菌株が含まれているときにビタミンB12を定量することが可能である。即ち、上層の表面にビタミンB12を含む溶液のサンプルを滴下し、37℃で16時間インキュベーション後、滴下した箇所に円形増殖斑が出現する。サンプルに含まれているビタミンB12が拡散して、寒天に含まれている細菌を増殖させ得るからである。円形増殖斑の直径はサンプル中のビタミンB12の濃度に比例する。
【0057】
サンプルは以下のプロトコルで細胞を溶解させることによって得られる。
【0058】
24mg/mlのリゾチーム(Boehringer Mannheim)を含む0.1mlの溶液(100mMのトリス−HCl,pH=8、20mMのEDTA、200g/リットルのショ糖)を、0.5mlの定量すべき細胞培養物と混合する。37℃で30分間インキュベーション後、30g/リットルの濃度のドデシル硫酸ナトリウムの溶液を60μl添加し、混合物の渦流を数秒間維持する。得られた10μlの細胞溶解液、あるいはこれを任意に1/50に希釈した液を、上層の表面に導入する。
【0059】
−HPLCアッセイ
高性能液体クロマトグラフィーによるコバラミンの定量方法としては、Blancheら,1990、に記載された方法を使用する。
【実施例1】
【0060】
Pseudomonas denitrificansの菌株によるコバラミンの産生に対するO−ホスホ−L−トレオニンの効果
100mlのエルレンマイヤーフラスコを用い、2μg/mlのテトラサイクリンを含む25mlのPS4培地でPseudomonas denitrificans菌G2650〔Tet〕株を培養する。撹拌下(250rpm)、30℃で24時間発酵後、10g/リットルのO−ホスホ−L−トレオニンの溶液を、0.16ml、0.32mlまたは1.5mlの量で培地に添加する。これらは夫々、66mg/リットル、132mg/リットル及び600mg/リットルの最終濃度に対応する。これらの各条件下で産生されたコバラミンの量を、148時間発酵後にHPLCによって定量する。結果(表1)は、G2650〔Tet〕株がPS4培地中でビタミンB12を産生しないことを示す。これに反して、O−ホスホ−L−トレオニンが培地中に存在すると、同じこの菌株によってB12が産生される。添加するO−ホスホ−L−トレオニンの量の増加に伴って産生されるビタミンB12の量が増加する。
【0061】
【表1】

【実施例2】
【0062】
Rhodobacter capsulatusの菌株によるコバラミンの産生に対するO−ホスホ−L−トレオニン及びR1−アミノ−2−プロパノールの効果の比較
100mlのエルレンマイヤーフラスコを用い、10μg/mlのカナマイシンと種々の濃度のO−ホスホ−L−トレオニン及びR1−アミノ−2−プロパノールを存在させた70mlのRA培地でRhodobacter capsulatus菌AH2株を培養する。撹拌下(100rpm)、30℃で24〜48時間発酵後、種々の条件下のコバラミンの産生量を微生物学的アッセイによって測定する。
【0063】
結果を表2に示す。表中の、“−”は、指示株の円形増殖斑が存在しないことを表し、“+”は円形増殖斑が存在することを表し、“+”の数が多いほど増殖斑の直径が大きいことを表している。これらの結果は、RA培地で培養したR.capsulatus菌AH2株がビタミンB12を産生しないことを示す。これに反して、O−ホスホ−L−トレオニンまたはR1−アミノ−2−プロパノールの存在下では、この菌株がビタミンB12を産生し得る。これらの結果はまた、R1−アミノ−2−プロパノールによってO−ホスホ−L−トレオニンと同じ結果を得るためには約6×103の添加量が必要であることを示している。
【0064】
【表2】
【実施例3】
【0065】
コバラミン産生性でないG2650株中のプラスミドpBBW1に由来のDNAフラグメントの存在の効果
3.1.プラスミドpER1の構築(図1)
ベクターpXL435(Cameronら,1989)のBamHI部位に、プラスミドpBBW1(Pollich & Klug,1995)から精製した6.8kbのBamHI DNAフラグメントをクローニングすることによって、17.4kbのプラスミドpER1を構築した。
【0066】
3.2.P.denitrificans菌G2650株へのプラスミドpER1の導入
第一段階で、大腸菌S17−1株にプラスミドpER1を電気穿孔によって導入し、第二段階で、プラスミドpER1を含む大腸菌S17−1株に結合させることによって、プラスミドpER1をP.denitrificans菌G2650〔Tet〕株に導入した。50μg/mlのリファンピシン及び100μg/lのリビドマイシンに耐性のトランスコンジュガントを選択した。分析した9個のクローンがプラスミドpER1を含んでいた。
【0067】
同様の手順で、ベクターpXL435だけを含む対照G2650〔Tet〕株を構築した。
【0068】
3.3.プラスミドpER1を含むG2650株によるコバラミンの産生
プラスミドpER1を含むクローンG2650〔Tet〕とプラスミドpXL435を含む2つのクローンとを、PS4培地中、リファンピシン、テトラサイクリン及びリビドマイシンの存在下で培養した。撹拌下(250rpm)、30℃で140時間発酵後、微生物学的アッセイ及びHPLCによってコバラミンの産生量を測定した。結果を表3に示す。
【0069】
【表3】
G2650〔Tet〕株及びプラスミドpXL435を含む菌株の双方がPS4培地中でビタミンB12を産生しないが、pER1を含む同じ菌株はB12を約3.5mg/リットルの濃度で産生し得る。プラスミドpXL435を含むG2650〔Tet〕株、即ちプラスミドpER1の構築に使用されるクローニングベクターだけを含むG2650〔Tet〕株はB12を産生しない。従って、B12が産生される理由は、プラスミドpBBW1に由来の6.8kbのDNAフラグメントの存在であることが判明する。
【0070】
即ち、プラスミドpBBW1から精製された6.8kbのBamHI DNAフラグメントは、P.denitrificans菌G2650〔Tet〕株に、PS4培地中でビタミンB12を産生する能力を与える。
【0071】
3.4.サブクローニング
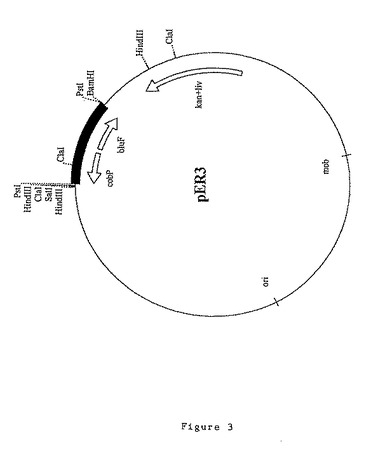
bluE遺伝子及びbluF遺伝子を含む6.8kbのBamHI DNAフラグメントの領域を、クローニングベクターpXL435にサブクローニングしてプラスミドを作製し、中間構築物に基づいてプラスミドpER2及びpER3と命名した。
【0072】
12.9kbのプラスミドpER2(図2)は、プラスミドpBBW1から精製されベクターpXL435にクローニングされた2.1kbのEcoRI/ClaIフラグメントを含む。このDNAフラグメントは、プラスミドpBluescript II SK+(Stratagene)のEcoRI/ClaI部位に予めクローニングされ、次いでこの組換えプラスミドからBamHI/SalI DNAフラグメントの形態で精製されてベクターpXL435にクローニングできるようにしたものである。
【0073】
11.9kbのプラスミドpER3(図3)は、プラスミドpBBW1から精製されベクターpXL435にクローニングされた1.2kbのPstIフラグメントを含む。このフラグメントは、プラスミドpBluescript II SK+のPstI部位に予めクローニングされ、次いでこの組換えプラスミドからBamHI/SalI制限フラグメントの形態で精製されてベクターpXL435にクローニングできるようにしたものである。
【0074】
プラスミドpAHW25(Pollich & Klug,1995a)は、プラスミドpBBW1から精製され、ベクターpRK415(Keenら,1988)にクローニングされた1.6kbのEcoRI/EcoRVフラグメントを含む。bluE遺伝子はベクターのlacプロモーターから転写される。
【0075】
プラスミドpER2またはpER3を、これらのプラスミドを含む大腸菌S17−1株に結合させることによってP.denitrificans菌G2650〔Tet〕株に導入した。プラスミドpER2、pER3またはpXL435を含むG2650〔Tet〕株のクローンを、5mlのPS4培地中、リファンピシン、テトラサイクリン及びリビドマイシンの存在下で培養した。プラスミドpAHW25及びpRK415を、これらのプラスミドを含む大腸菌S17−1株に結合させることによってP.denitrificans菌G2650〔Sp〕株に導入した。プラスミドpAHW25またはpRK415を含むG2650〔Sp〕株のクローンを25mlのPS4培地中、リファンピシン、リビドマイシン及びスペクチノマイシンの存在下で培養した。撹拌下(250rpm)、30℃で140時間発酵後、コバラミンの産生量を微生物学的アッセイ及びHPLCによって測定する。
【0076】
表4に示した結果は、プラスミドpER2またはpAHW25を含むG2650株のクローンだけがPS4培地中でビタミンB12を産生し得ることを示す。
【0077】
【表4】
【実施例4】
【0078】
bluB遺伝子は、B12の既知の前駆物質である5,6−ジメチルベンズイミダゾール(DBI)の生合成に関与する。
【0079】
Rhodobacter capsulatusは、bluB遺伝子にインターポゾンを挿入することによって得られた突然変異株である。この株はbluB−である(Pollichら,1995)。100mlのエルレンマイヤーフラスコを用い、70mlのRA培地中、10μg/mlのカナマイシン及び種々の濃度のDBIの存在下でBB1株を培養する。種々の条件下でこの菌株によって産生されるコバラミンの量を、撹拌下(100rpm)、30℃で24〜48時間発酵後に、微生物学的アッセイによって測定する。表5にまとめた結果は、突然変異株BB1がRA単独培地中ではB12産生性でないが、培地中に14nM以上のDBIが存在するときにはこの分子を合成することを示す。従って、bluB遺伝子は5,6−ジメチルベンズイミダゾールの生合成に関与する。
【0080】
【表5】
【0081】
【表6】
【技術分野】
【0001】
本発明は、コバラミンを製造し得る生合成方法に関する。より詳細には本発明は、組換えDNA技術及び/または新規なコバラミン前駆物質の添加によってコバラミンの産生を増加させる方法、特にB12補酵素の産生を増加させる方法に関する。最後に本発明は、本発明のコバラミンの製造方法に有用な組換え菌株の作製方法に関する。
【背景技術】
【0002】
ビタミンB12はコバラミンと呼ばれる分子のクラスに属しており、その構造は特に国際特許WO91/11518に開示されている。
【0003】
コバラミンはほぼ例外なく、同じく国際特許WO91/11518に記載された複合プロセスによって細菌から合成されている。生合成メカニズムが極めて複雑であるため、工業的レベルのコバラミンの産生、特にビタミンB12の産生は主として、Pseudomonas denitrificans、Propionobacterium shermanii及びPropionobacterium freudenreichiiのような細菌の大量培養によって行われている。
【0004】
また、いくつかの微生物が、アミノレブリン酸、S−アデノシル−L−メチオニン、コバルト、グルタミン、R1−アミノ−2−プロパノール及び5,6−ジメチルベンズイミダゾールの基質からコバラミンを合成することも公知である。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】国際公開第WO91/11518号
【発明の概要】
【0006】
上記に挙げた前駆物質のうちで、5,6−ジメチルベンズイミダゾールは、コバラミン産生性微生物によって合成される。5,6−ジメチルベンズイミダゾールの生合成には2つの経路が存在すると考えられる。一方の経路は、分子酸素を利用する好気性微生物に特有の経路である。他方の経路は、嫌気性微生物によって使用される経路である。嫌気性経路に関与する遺伝子だけが単離されている。この遺伝子は、ネズミチフス菌(Salmonella thyphimurium)のcobT遺伝子である(Trzebiatowskiら,1994)。5,6−ジメチルベンズイミダゾールを合成する好気性微生物の遺伝子は現在まで全く同定されていない。多くの場合、微生物によって合成される5,6−ジメチルベンズイミダゾールの量には限界がある。
【0007】
このため、5,6−ジメチルベンズイミダゾールを化学的に製造して産生用培地に添加している。従って、培地に対するこのような添加の必要をなくすことができれば有利になることは確かである。
【0008】
従来のどのようなコバラミンの工業的製造方法も、コバルト及び5,6−ジメチルベンズイミダゾール以外の前駆物質の添加に言及したことはない。最近になって、培地がR1−アミノ−2−プロパノールを含有するときにだけコバラミンを産生するいくつかの菌株が記載された(Crouzetら,1990、Grabauら,1992)。従って、R1−アミノ−2−プロパノールもコバラミンの産生を増進するために使用し得るであろう。しかしながら、この物質は、工業的発酵に使用できるとしても、その使用が難しく費用も高い。その理由の1つは、R1−アミノ−2−プロパノールが刺激性及び揮発性の物質であるからであり、いま1つの理由は、この物質が微生物の増殖を阻害するからである。従って、R1−アミノ−2−プロパノール残基以外の、上記の欠点を有していないコバラミンの前駆物質を見つけることが特に有益であると考えられる。この観点から、幾つかの微生物ではL−トレオニンからアミノアセトンを経由してR1−アミノ−2−プロパノールを生合成する経路が存在することは開示されている。しかしながら、L−トレオニンは上記に引用した菌株を補完することができない。
【0009】
より一般的に、コバラミンの産生を増進するためには、培地中のコバラミンの前駆物質の量を増加させるのが有利であろう。コバラミンの前駆物質が制限的である場合には特にこのような増加が有利である。この方法は、制限的前駆物質またはその誘導体もしくはその類似体の1つを培地に直接添加することによって、あるいは、遺伝子技術、特に組換えDNA技術を使用して産生性菌株中でこの前駆物質のin situ合成を増進することによって行う。
【0010】
従って、コバラミンの産生を増進するために重要な段階は、コバラミン及びその前駆物質の生合成経路を明らかにすることである。
【0011】
最近には、ビタミンB12の生合成経路の大部分の段階がPseudomonas denitrificansで解明された(Blancheら,1995)。コバラミンの生合成に関与する22個以上のcob遺伝子が単離され、これらの遺伝子によってコードされたポリペプチドの殆どについてその機能が同定された。
【0012】
別の微生物においても、コバラミンまたはその前駆物質の生合成に関与すると推測される別の遺伝子が単離された。これらの遺伝子の殆どについてその機能は未だ解明されていない。これらの遺伝子の機能またはその効果を正確に決定することさえできれば、これらの遺伝子の応用が可能になるであろう。例えば、通性光合成細菌Rhodobacter capsulatus中で、光合成器官の形成に必要な少なくとも1つの遺伝子を含むDNAフラグメントが最近になって配列決定された(Pollichら,1993、Pollichら,1995a)。単離された8個の遺伝子のうちの5個の遺伝子が、前述のP.denitrificansの22個のcob遺伝子のうちの5個の遺伝子に高度な相同性を有するという理由でコバラミンの生合成遺伝子であると示唆された。逆に、残りの3つの遺伝子、即ちbluB遺伝子、bluE遺伝子及びbluF遺伝子に関しては、これらの遺伝子が直接に担当する正確な機能を明らかにすることができなかった。それらのプロモーター配列を含むbluB遺伝子、bluE遺伝子及びbluF遺伝子は、Pollichら,1995aによってその配列が決定され開示された。
【0013】
本発明によって、コバラミンの新規な前駆物質が知見された。その結果として本発明は、O−ホスホ−L−トレオニンを含有する培地によってコバラミンの産生を増進することに成功した。これまでに記載されたことのないこのコバラミンの前駆物質は、既に公知であった別の前駆物質R1−アミノ−2−プロパノールと同等の機能を有している。しかしながら、O−ホスホ−L−トレオニンは、無毒の物質である、及び、取り扱い容易な物質である、という点でR1−アミノ−2−プロパノールよりも有利である。更に、コバラミンの産生増進に対してO−ホスホ−L−トレオニンが示す効率は、R1−アミノ−2−プロパノールが示す効率の1,000倍以上である。
【0014】
本発明の目的はまた、コバラミンの産生増進を達成するためまたは所与の細胞のO−ホスホ−L−トレオニンもしくは5,6−ジメチルベンズイミダゾールのin situ合成もしくはこのような合成の増進を達成するために、Rhodobacter capsulatusのDNAフラグメントを使用することである。
【0015】
本発明によれば、Rhodobacter capsulatusの、特にbluE遺伝子及びbluF遺伝子を含むDNAフラグメントを使用することによって、R1−アミノ−2−プロパノールまたはO−ホスホ−L−トレオニンを含有しない培地においてコバラミンの産生を増進することが可能であった。
【0016】
最後に、本発明によれば、Rhodobacter capsulatusの、特にbluB遺伝子を含むDNAフラグメントを導入することによって、5,6−ジメチルベンズイミダゾールを含有しない培地においてコバラミンの産生を増進することが可能であった。
【0017】
本発明の1つの目的は、コバラミン産生性原核細胞微生物の発酵によるコバラミンの生合成方法を提供することである。本発明によるこの方法の特徴は、
−O−ホスホ−L−トレオニンの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって形質転換された微生物を使用し、前記酵素が発現しコバラミンが産生され得る条件下で前記微生物を培養する段階、及び/または、
−前記微生物の培養培地にO−ホスホ−L−トレオニンを添加する段階、または、
−5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって形質転換された好気性微生物を使用し、前記酵素が発現しコバラミンが産生され得る条件下で前記好気性微生物を培養する段階、
を含むことである。
【0018】
微生物は、上記に特定した遺伝子を内在的に含んでいてもよい。この場合には、本発明方法によって酵素が超発現し得る。しかしながらまた、微生物がこの種の遺伝子を内在的に含んでいなくてもよい。
【0019】
“O−ホスホ−L−トレオニンまたは5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードするDNAフラグメント”なる表現は、このDNAフラグメントの発現によってO−ホスホ−L−トレオニンまたは5,6−ジメチルベンズイミダゾールが細胞内で合成され、場合によってはその後に培養培地中に遊離されることを意味する。
【0020】
培養は、バッチ培養でもよく、または、連続培養でもよい。また、コバラミンの精製は既存の工業的方法によって行うことができる(Florent,1986)。
【0021】
1つの実施態様においては、Rhodobacter capsulatusのbluE遺伝子及びbluF遺伝子を含みO−ホスホ−L−トレオニンの生合成経路に関与するポリペプチドをコードするDNAフラグメントによって形質転換されたか、または、相同フラグメント、即ち、これらのbluE遺伝子及びbluF遺伝子とハイブリダイズし且つこれらの遺伝子と同様にO−ホスホ−L−トレオニンの生合成経路に関与する酵素をコードする機能を有しているフラグメントによって形質転換された微生物が使用される。
【0022】
別の実施態様においては、Rhodobacter capsulatusのbluB遺伝子を含み5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードするDNAフラグメントによって形質転換されたか、または、相同フラグメント、即ち、このbluB遺伝子とハイブリダイズし且つこの遺伝子と同様に5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする機能を有しているフラグメントによって形質転換された微生物が使用される。
【0023】
本発明は、天然、合成または組換えによって得られたDNAフラグメント及び相同フラグメントの使用を含む。フラグメントなる用語は、遺伝コードの縮重性に由来するフラグメント、または、少なくとも25%の配列相同性を有しており同じ機能のポリペプチドをコードするフラグメントを意味する。
【0024】
好気性条件下で培養される微生物を使用し、5,6−ジメチルベンズイミダゾールの好気性条件下の生合成経路に関与する酵素をコードするDNAフラグメントを使用するのが好ましい。
【0025】
本発明の目的はまた、遺伝子工学技術を用い、O−ホスホ−L−トレオニンまたは5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする少なくとも1つの上記のようなDNAフラグメントによって微生物を形質転換させることを特徴とするコバラミン産生性原核細胞微生物の組換え菌株の製造方法を提供することである。
【0026】
また、本発明の菌株の製造方法によって得られた組換え菌株を提供することも本発明の目的の1つである。
【0027】
ポリペプチドをコードする少なくとも1つのDNA配列を含み、1つまたは複数のこれらの配列が発現シグナルのコントロール下に配置されている組換えDNAを本発明方法に使用することも本発明に包含される。
【0028】
この観点から、特に、DNA配列の5′にプロモーター領域を配置し得る。このような領域は、DNA配列に同種(homologous)の領域でもよくまたは異種(heterologous)の領域でもよい。特に、強い細菌プロモーター、例えば、大腸菌のトリプトファンオペロンのプロモーターPtrpもしくはラクトースオペロンのプロモーターPlac、ラムダバクテリオファージの左もしくは右のプロモーター、コリネバクテリウム(corynebacterium)のような細菌のファージの強いプロモーター、大腸菌のプロモーターPtacのようなグラム陰性菌中で機能性のプロモーター、プラスミドTOLのキシレンの異化遺伝子のプロモーターPxylS、枯草菌(Bacillus subtilis)のアミラーゼのプロモーターPamyを使用し得る。また、ホスホグリセリン酸キナーゼ、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ、ラクターゼまたはエノラーゼをコードする遺伝子のプロモーターのような、酵母の解糖遺伝子に由来のプロモーターも例示し得る。これらのプロモーターは、組換えDNAを真核細胞宿主に導入するときに使用できる。また、DNA配列の5′端にリボソーム結合部位を配置してもよい。この部位は、同種部位でもよく、または、ラムダバクテリオファージのcII遺伝子のリボソーム結合部位のような異種部位でもよい。
【0029】
転写の終結に必要なシグナルはDNA配列の3′に配置され得る。
【0030】
本発明方法で使用される組換えDNAは、選択された発現シグナルに適合性の宿主細胞に直接導入されてもよく、または、問題のDNA配列を宿主細胞に安定に導入できるようにプラスミドベクターにクローニングされてもよい。
【0031】
本発明方法は公知の方法と同様に、ポリペプチドをコードするDNA配列を含むプラスミドの使用を包含する。更にこれらのプラスミドは公知のプラスミドと同様に、機能性複製系と選択マーカーとを含む。
【0032】
多様な種類のベクターを使用し得る。本発明の範囲内では、RK2型のベクター、即ち、RK2複製起点を有するベクターを使用するのが好ましい。特定例としては、ベクターRK2(Sauruggerら,1986)、ベクターpXL435(Cameronら,1989)、ベクターpRK290(米国特許第4,590,163号、Dittaら,1985)、及び、ベクターpXL1635(国際特許WO91/16439)を挙げることができる。特に有利なベクターはベクターpXL1635である。その他のベクターは国際特許出願WO91/16439に記載されている。
【0033】
変形実施態様においては、後出の実施例に記載したように、2種類の前駆物質、即ち、O−ホスホ−L−トレオニン及び5,6−ジメチルベンズイミダゾールの合成をコードするプラスミドpER1(図1)の6.8kbのBamHIフラグメントから成るDNAフラグメントによって形質転換された微生物が使用される。
【0034】
O−ホスホ−L−トレオニンの合成に関与するポリペプチドの発現に特に好適な実施態様では、後出の実施例に記載したようなプラスミドpER2(図2)の2.1kbのEcoRI/ClaIフラグメントから成るDNAフラグメントが使用される。
【0035】
O−ホスホ−L−トレオニンの合成に関与するポリペプチドの発現に特に好適な実施態様では、後出の実施例に記載したようなプラスミドpER2(図2)の1.6kbのEcoRI/EcoRVフラグメントから成るDNAフラグメントが使用される。
【0036】
5,6−ジメチルベンズイミダゾールの合成に関与するポリペプチドの発現に特に好適な別の実施態様では、後出の実施例に記載したようなプラスミドpER1の6.8kbのBamHIフラグメントから成るDNAフラグメントが使用される。
【0037】
本発明によって使用され得る宿主原核細胞微生物は、E.coli、Pseudomonas denitrificans、Agrobacterium radiobacter、Agrobacterium tumefaciensまたはRhizobium melitotiあるいはRhodobacter capsulatus属の細菌である。他の細菌は国際特許WO91/11518に記載されている。
【0038】
いずれにしても、P.denitrificansまたはA.radiobacter属の細菌の使用が特に有利である。
【0039】
本発明の別の特徴及び利点は以下の詳細な記載より明らかにされるであろう。
【0040】
実施例1及び2は、Pseudomonas denitrificansまたはRhodobacter capsulatusの菌株の培養培地にO−ホスホ−L−トレオニンを添加することによってコバラミンを産生させるのが可能であることを示している。実施例2は、産生培地中のR1−アミノ−2−プロパノールをO−ホスホ−L−トレオニンで代替することがどのように有利であるかを示している。同量のコバラミンを産生するために必要なR1−アミノ−2−プロパノールの量はO−ホスホ−L−トレオニンの1,000倍である。
【0041】
実施例2はまた、Rhodobacter capsulatusのbluE遺伝子及びbluF遺伝子を含む染色体の領域が、O−ホスホ−L−トレオニンの合成に関与することを示している。実施例3は、Rhodobacter capsulatusのDNAフラグメントを出発材料としたbluE遺伝子及びbluF遺伝子を含むプラスミドの構築を記載している。この実施例3は特に、これらのプラスミドがR1−アミノ−2−プロパノールまたはO−ホスホ−L−トレオニンの添加に依存してコバラミンを産生する菌株中に導入されたときに、R1−アミノ−2−プラスミドまたはO−ホスホ−L−トレオニン非含有の培地でどのようにしてコバラミンを産生し得るかを示している。実施例4は、Rhodobacter capsulatusのbluB遺伝子を含む染色体の領域が、5,6−ジメチルベンズイミダゾールの合成に関与することを示している。
【図面の簡単な説明】
【0042】
【図1】プラスミドpER1の制限地図を表す。
【図2】プラスミドpER2の制限地図を表す。
【図3】プラスミドpER3の制限地図を表す。
【0043】
図1から図3は夫々、プラスミドpER1、pER2及びpER3を表す。
【発明を実施するための形態】
【0044】
1.菌株及びプラスミド
Rhodobacter capsulatus菌のAH2株及びBB1株(Pollichら,1995a)を、37b4株(DSM938)から構築した。Pseudomonas denitrificans菌G2650株は、SBL 27 Rifr株を出発材料とし、トランスポゾンTn5の挿入によって構築した(Crouzetら,1990)。先ず、テトラサイクリン耐性遺伝子を含むトランスポゾンTn5を用いてG2650〔Tet〕株を構築した。次に、G2650〔Tet〕株を出発材料とし、トランスポゾンTn5のテトラサイクリン耐性遺伝子をスペクチノマイシン耐性遺伝子で置換することによってG2650〔Sp〕株を構築した。SBL 27 Rifr株はMB580株に由来する(米国特許US3,018,225)。
【0045】
プラスミドpBBW1(Pollichら,1995a)をRhodobacter capsulatusのDNAフラグメントから構築した。プラスミドpBBW1からプラスミドpAHW25(Pollich & Klug,1995a)を構築した。
【0046】
2.分子技術
DNA操作の汎用技術に関しては実験概論書(Sambrookら,1989)を参考として使用する。
【0047】
酵素はNew England Biolabs及びBoehringer Mannheimの研究所から入手し、製作者の指示通りに使用する。
【0048】
使用した技術は本質的に以下の2段階から成る。
【0049】
−制限酵素による消化、及び、
−T4バクテリオファージのリガーゼによるDNA分子の結合。
【0050】
3.形質転換技術
大腸菌の形質転換を電気穿孔によって行う(Dowerら,1988)。
【0051】
4.結合技術
大腸菌S17−1株とP.denitrificansの種々の菌株との結合は、Simonらによって記載されたプロトコル(Simonら,1986)を応用して行う。Pseudomonasの菌株の形質転換は遺伝子工学の他の任意の技術によって行う。
【0052】
5.コバラミン産生培地の調製
P.denitrificansの菌株によるコバラミンの産生に使用した培地は、Cameronら,1989、によって記載されたPS4培地である。
【0053】
Rhodobacter capsulatusの菌株によるコバラミンの産生に使用した培地は、Pollich,1995b、によって記載されたRA培地である。
【0054】
6.産生されたコバラミンの定量アッセイ
産生されたコバラミンの量を、微生物学的アッセイまたは高性能液体クロマトグラフィー(HPLC)によって測定する。
【0055】
−微生物学的アッセイ
ビタミンB12要求性の大腸菌指示株113−3(Davis & Mingioli,1950)を用い、半定量的方法によってコバラミンの産生量を測定する。
【0056】
この指示株は、B12依存性のホモシステインメチルトランスフェラーゼ(EC2.1.1.13)だけを有している大腸菌のmetE変異株である。この菌株が最小培地中で増殖するためにはビタミンB12の存在だけが必要である。(この菌株の増殖に必要なビタミンB12が欠乏した)ゲル状のM9最少培地(Miller,1972)の上層にこの菌株が含まれているときにビタミンB12を定量することが可能である。即ち、上層の表面にビタミンB12を含む溶液のサンプルを滴下し、37℃で16時間インキュベーション後、滴下した箇所に円形増殖斑が出現する。サンプルに含まれているビタミンB12が拡散して、寒天に含まれている細菌を増殖させ得るからである。円形増殖斑の直径はサンプル中のビタミンB12の濃度に比例する。
【0057】
サンプルは以下のプロトコルで細胞を溶解させることによって得られる。
【0058】
24mg/mlのリゾチーム(Boehringer Mannheim)を含む0.1mlの溶液(100mMのトリス−HCl,pH=8、20mMのEDTA、200g/リットルのショ糖)を、0.5mlの定量すべき細胞培養物と混合する。37℃で30分間インキュベーション後、30g/リットルの濃度のドデシル硫酸ナトリウムの溶液を60μl添加し、混合物の渦流を数秒間維持する。得られた10μlの細胞溶解液、あるいはこれを任意に1/50に希釈した液を、上層の表面に導入する。
【0059】
−HPLCアッセイ
高性能液体クロマトグラフィーによるコバラミンの定量方法としては、Blancheら,1990、に記載された方法を使用する。
【実施例1】
【0060】
Pseudomonas denitrificansの菌株によるコバラミンの産生に対するO−ホスホ−L−トレオニンの効果
100mlのエルレンマイヤーフラスコを用い、2μg/mlのテトラサイクリンを含む25mlのPS4培地でPseudomonas denitrificans菌G2650〔Tet〕株を培養する。撹拌下(250rpm)、30℃で24時間発酵後、10g/リットルのO−ホスホ−L−トレオニンの溶液を、0.16ml、0.32mlまたは1.5mlの量で培地に添加する。これらは夫々、66mg/リットル、132mg/リットル及び600mg/リットルの最終濃度に対応する。これらの各条件下で産生されたコバラミンの量を、148時間発酵後にHPLCによって定量する。結果(表1)は、G2650〔Tet〕株がPS4培地中でビタミンB12を産生しないことを示す。これに反して、O−ホスホ−L−トレオニンが培地中に存在すると、同じこの菌株によってB12が産生される。添加するO−ホスホ−L−トレオニンの量の増加に伴って産生されるビタミンB12の量が増加する。
【0061】
【表1】
【実施例2】
【0062】
Rhodobacter capsulatusの菌株によるコバラミンの産生に対するO−ホスホ−L−トレオニン及びR1−アミノ−2−プロパノールの効果の比較
100mlのエルレンマイヤーフラスコを用い、10μg/mlのカナマイシンと種々の濃度のO−ホスホ−L−トレオニン及びR1−アミノ−2−プロパノールを存在させた70mlのRA培地でRhodobacter capsulatus菌AH2株を培養する。撹拌下(100rpm)、30℃で24〜48時間発酵後、種々の条件下のコバラミンの産生量を微生物学的アッセイによって測定する。
【0063】
結果を表2に示す。表中の、“−”は、指示株の円形増殖斑が存在しないことを表し、“+”は円形増殖斑が存在することを表し、“+”の数が多いほど増殖斑の直径が大きいことを表している。これらの結果は、RA培地で培養したR.capsulatus菌AH2株がビタミンB12を産生しないことを示す。これに反して、O−ホスホ−L−トレオニンまたはR1−アミノ−2−プロパノールの存在下では、この菌株がビタミンB12を産生し得る。これらの結果はまた、R1−アミノ−2−プロパノールによってO−ホスホ−L−トレオニンと同じ結果を得るためには約6×103の添加量が必要であることを示している。
【0064】
【表2】
【実施例3】
【0065】
コバラミン産生性でないG2650株中のプラスミドpBBW1に由来のDNAフラグメントの存在の効果
3.1.プラスミドpER1の構築(図1)
ベクターpXL435(Cameronら,1989)のBamHI部位に、プラスミドpBBW1(Pollich & Klug,1995)から精製した6.8kbのBamHI DNAフラグメントをクローニングすることによって、17.4kbのプラスミドpER1を構築した。
【0066】
3.2.P.denitrificans菌G2650株へのプラスミドpER1の導入
第一段階で、大腸菌S17−1株にプラスミドpER1を電気穿孔によって導入し、第二段階で、プラスミドpER1を含む大腸菌S17−1株に結合させることによって、プラスミドpER1をP.denitrificans菌G2650〔Tet〕株に導入した。50μg/mlのリファンピシン及び100μg/lのリビドマイシンに耐性のトランスコンジュガントを選択した。分析した9個のクローンがプラスミドpER1を含んでいた。
【0067】
同様の手順で、ベクターpXL435だけを含む対照G2650〔Tet〕株を構築した。
【0068】
3.3.プラスミドpER1を含むG2650株によるコバラミンの産生
プラスミドpER1を含むクローンG2650〔Tet〕とプラスミドpXL435を含む2つのクローンとを、PS4培地中、リファンピシン、テトラサイクリン及びリビドマイシンの存在下で培養した。撹拌下(250rpm)、30℃で140時間発酵後、微生物学的アッセイ及びHPLCによってコバラミンの産生量を測定した。結果を表3に示す。
【0069】
【表3】
G2650〔Tet〕株及びプラスミドpXL435を含む菌株の双方がPS4培地中でビタミンB12を産生しないが、pER1を含む同じ菌株はB12を約3.5mg/リットルの濃度で産生し得る。プラスミドpXL435を含むG2650〔Tet〕株、即ちプラスミドpER1の構築に使用されるクローニングベクターだけを含むG2650〔Tet〕株はB12を産生しない。従って、B12が産生される理由は、プラスミドpBBW1に由来の6.8kbのDNAフラグメントの存在であることが判明する。
【0070】
即ち、プラスミドpBBW1から精製された6.8kbのBamHI DNAフラグメントは、P.denitrificans菌G2650〔Tet〕株に、PS4培地中でビタミンB12を産生する能力を与える。
【0071】
3.4.サブクローニング
bluE遺伝子及びbluF遺伝子を含む6.8kbのBamHI DNAフラグメントの領域を、クローニングベクターpXL435にサブクローニングしてプラスミドを作製し、中間構築物に基づいてプラスミドpER2及びpER3と命名した。
【0072】
12.9kbのプラスミドpER2(図2)は、プラスミドpBBW1から精製されベクターpXL435にクローニングされた2.1kbのEcoRI/ClaIフラグメントを含む。このDNAフラグメントは、プラスミドpBluescript II SK+(Stratagene)のEcoRI/ClaI部位に予めクローニングされ、次いでこの組換えプラスミドからBamHI/SalI DNAフラグメントの形態で精製されてベクターpXL435にクローニングできるようにしたものである。
【0073】
11.9kbのプラスミドpER3(図3)は、プラスミドpBBW1から精製されベクターpXL435にクローニングされた1.2kbのPstIフラグメントを含む。このフラグメントは、プラスミドpBluescript II SK+のPstI部位に予めクローニングされ、次いでこの組換えプラスミドからBamHI/SalI制限フラグメントの形態で精製されてベクターpXL435にクローニングできるようにしたものである。
【0074】
プラスミドpAHW25(Pollich & Klug,1995a)は、プラスミドpBBW1から精製され、ベクターpRK415(Keenら,1988)にクローニングされた1.6kbのEcoRI/EcoRVフラグメントを含む。bluE遺伝子はベクターのlacプロモーターから転写される。
【0075】
プラスミドpER2またはpER3を、これらのプラスミドを含む大腸菌S17−1株に結合させることによってP.denitrificans菌G2650〔Tet〕株に導入した。プラスミドpER2、pER3またはpXL435を含むG2650〔Tet〕株のクローンを、5mlのPS4培地中、リファンピシン、テトラサイクリン及びリビドマイシンの存在下で培養した。プラスミドpAHW25及びpRK415を、これらのプラスミドを含む大腸菌S17−1株に結合させることによってP.denitrificans菌G2650〔Sp〕株に導入した。プラスミドpAHW25またはpRK415を含むG2650〔Sp〕株のクローンを25mlのPS4培地中、リファンピシン、リビドマイシン及びスペクチノマイシンの存在下で培養した。撹拌下(250rpm)、30℃で140時間発酵後、コバラミンの産生量を微生物学的アッセイ及びHPLCによって測定する。
【0076】
表4に示した結果は、プラスミドpER2またはpAHW25を含むG2650株のクローンだけがPS4培地中でビタミンB12を産生し得ることを示す。
【0077】
【表4】
【実施例4】
【0078】
bluB遺伝子は、B12の既知の前駆物質である5,6−ジメチルベンズイミダゾール(DBI)の生合成に関与する。
【0079】
Rhodobacter capsulatusは、bluB遺伝子にインターポゾンを挿入することによって得られた突然変異株である。この株はbluB−である(Pollichら,1995)。100mlのエルレンマイヤーフラスコを用い、70mlのRA培地中、10μg/mlのカナマイシン及び種々の濃度のDBIの存在下でBB1株を培養する。種々の条件下でこの菌株によって産生されるコバラミンの量を、撹拌下(100rpm)、30℃で24〜48時間発酵後に、微生物学的アッセイによって測定する。表5にまとめた結果は、突然変異株BB1がRA単独培地中ではB12産生性でないが、培地中に14nM以上のDBIが存在するときにはこの分子を合成することを示す。従って、bluB遺伝子は5,6−ジメチルベンズイミダゾールの生合成に関与する。
【0080】
【表5】
【0081】
【表6】
【特許請求の範囲】
【請求項1】
コバラミン産生性原核細胞微生物の発酵によるコバラミンの生合成方法であって、
5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって形質転換された好気性微生物を使用し、前記酵素が発現しコバラミンが産生され得る条件下で前記好気性微生物を培養することを特徴とするコバラミンの生合成方法。
【請求項2】
ロドバクター・カプスラタス(Rhodobacter capsulatus)のbluB遺伝子を含み5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードするDNAフラグメントによって形質転換されるか、または、相同フラグメント、及び/または、前記bluB遺伝子とハイブリダイズし且つ前記遺伝子と同様に5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする機能を有しているフラグメントによって形質転換された微生物を使用することを特徴とする請求項1に記載の方法。
【請求項3】
前記DNAフラグメントが、プラスミドpER1の6.8kbのBamHIフラグメントから成ることを特徴とする請求項2に記載の方法。
【請求項4】
請求項1から3のいずれか一項に記載のビタミンB12の製造方法。
【請求項5】
前記微生物がシュードモナス・デニトリフィカンス(Pseudomonas denitrificans)またはアグロバクテリウム・ラジオバクター(Agrobacterium radiobacter)の菌株であることを特徴とする請求項1から4のいずれか一項に記載の方法。
【請求項6】
遺伝子工学技術を用い、請求項1に記載のO−ホスホ−L−トレオニンの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって微生物を形質転換させることを特徴とするコバラミン産生性原核細胞微生物の組換え菌株の製造方法。
【請求項7】
遺伝子工学技術を用い、請求項1,2または3のいずれか一項に記載の5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって微生物を形質転換させることを特徴とするコバラミン産生性原核細胞微生物の組換え菌株の製造方法。
【請求項8】
請求項6または7に記載の方法によって得られた組換え微生物。
【請求項9】
シュードモナス・デニトリフィカンス(Pseudomonas denitrificans)またはアグロバクテリウム・ラジオバクター(Agrobacterium radiobacter)の菌株であることを特徴とする請求項8に記載の微生物。
【請求項1】
コバラミン産生性原核細胞微生物の発酵によるコバラミンの生合成方法であって、
5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって形質転換された好気性微生物を使用し、前記酵素が発現しコバラミンが産生され得る条件下で前記好気性微生物を培養することを特徴とするコバラミンの生合成方法。
【請求項2】
ロドバクター・カプスラタス(Rhodobacter capsulatus)のbluB遺伝子を含み5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードするDNAフラグメントによって形質転換されるか、または、相同フラグメント、及び/または、前記bluB遺伝子とハイブリダイズし且つ前記遺伝子と同様に5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする機能を有しているフラグメントによって形質転換された微生物を使用することを特徴とする請求項1に記載の方法。
【請求項3】
前記DNAフラグメントが、プラスミドpER1の6.8kbのBamHIフラグメントから成ることを特徴とする請求項2に記載の方法。
【請求項4】
請求項1から3のいずれか一項に記載のビタミンB12の製造方法。
【請求項5】
前記微生物がシュードモナス・デニトリフィカンス(Pseudomonas denitrificans)またはアグロバクテリウム・ラジオバクター(Agrobacterium radiobacter)の菌株であることを特徴とする請求項1から4のいずれか一項に記載の方法。
【請求項6】
遺伝子工学技術を用い、請求項1に記載のO−ホスホ−L−トレオニンの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって微生物を形質転換させることを特徴とするコバラミン産生性原核細胞微生物の組換え菌株の製造方法。
【請求項7】
遺伝子工学技術を用い、請求項1,2または3のいずれか一項に記載の5,6−ジメチルベンズイミダゾールの生合成経路に関与する酵素をコードする少なくとも1つのDNAフラグメントによって微生物を形質転換させることを特徴とするコバラミン産生性原核細胞微生物の組換え菌株の製造方法。
【請求項8】
請求項6または7に記載の方法によって得られた組換え微生物。
【請求項9】
シュードモナス・デニトリフィカンス(Pseudomonas denitrificans)またはアグロバクテリウム・ラジオバクター(Agrobacterium radiobacter)の菌株であることを特徴とする請求項8に記載の微生物。
【図1】

【図2】
【図3】
【図2】
【図3】
【公開番号】特開2009−142289(P2009−142289A)
【公開日】平成21年7月2日(2009.7.2)
【国際特許分類】
【出願番号】特願2009−29392(P2009−29392)
【出願日】平成21年2月12日(2009.2.12)
【分割の表示】特願平9−540573の分割
【原出願日】平成9年5月5日(1997.5.5)
【出願人】(508007569)
【Fターム(参考)】
【公開日】平成21年7月2日(2009.7.2)
【国際特許分類】
【出願日】平成21年2月12日(2009.2.12)
【分割の表示】特願平9−540573の分割
【原出願日】平成9年5月5日(1997.5.5)
【出願人】(508007569)
【Fターム(参考)】
[ Back to top ]