
コラーゲン生産植物及びその作成方法及びその使用
【課題】コラーゲン生産植物及びその作成方法の提供。
【解決手段】植物又は単離された植物細胞中で生産された組み換えコラーゲンであって、植物又は単離された植物細胞の液胞に少なくとも一種のコラーゲン鎖及び外因性プロリル−4−ヒドロキシラーゼ(P4H)を標的化して、前記外因性P4Hによるが植物又は単離された植物細胞の内因性P4Hによらない前記少なくとも一種のコラーゲン鎖のヒドロキシル化を可能にすることによって生産された組み換えコラーゲン。
【解決手段】植物又は単離された植物細胞中で生産された組み換えコラーゲンであって、植物又は単離された植物細胞の液胞に少なくとも一種のコラーゲン鎖及び外因性プロリル−4−ヒドロキシラーゼ(P4H)を標的化して、前記外因性P4Hによるが植物又は単離された植物細胞の内因性P4Hによらない前記少なくとも一種のコラーゲン鎖のヒドロキシル化を可能にすることによって生産された組み換えコラーゲン。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、コラーゲン生産植物及びその作成方法及びその使用に関する。特に、本発明は、生来的な三重らせんI型コラーゲン線維を形成することができる高レベルにヒドロキシル化されたコラーゲン鎖を生産することができる植物を作成するための新規のアプローチに関する。
【背景技術】
【0002】
コラーゲンは、脊椎動物及び多くの他の多細胞生物の構造的完全性を担う主要な構造タンパク質である。I型コラーゲンは原型線維状コラーゲンを表し、大部分の組織における主要なコラーゲン型である。
【0003】
I型コラーゲンは、骨及び腱の優勢なコラーゲン成分であり、皮膚、大動脈及び肺に多量に見出される。I型コラーゲン線維は、大きな引張り強度及び限定された伸度を与える。I型コラーゲンの最も豊富な分子形態は、二つの異なるアルファ鎖[アルファ1(I)]2及びアルファ2(I)からなるヘテロトリマーである(Inkinen、2003年)。すべての線維状コラーゲン分子は、繰返しGly−X−Yトリプレットから構築される三つのポリペプチド鎖を含み、ここでX及びYはいかなるアミノ酸であることができるが、イミノ酸プロリン及びヒドロキシプロリンであることが多い。
【0004】
原線維形成コラーゲン(線維状コラーゲン)は、球状N及びC末端延長プロペプチドを含む前駆体プロコラーゲンとして合成される。プロコラーゲンの生合成は、プロリン及びリシンのヒドロキシル化、N結合及びO結合グリコシル化及び鎖内及び鎖間ジスルフィド結合の形成を含む多数の異なる翻訳後修飾が関与する複雑な過程である。これらの修飾を行う酵素は、協調した様式で作用し、正確にアラインメントされた熱的に安定な三重らせん分子の折畳み及び会合を確実にする。
【0005】
各プロコラーゲン分子は、三つの構成的ポリペプチド鎖から大まかな小胞体内で会合する。ポリペプチド鎖が小胞体の膜を横切って共翻訳的に転位されるにつれて、プロリン及びリシン残基のヒドロキシル化がGly−X−Y繰返し領域内で起こる。一旦ポリペプチド鎖が小胞体の管腔中に十分に転位されたら、Cプロペプチドが折畳まれる。次に三つのプロアルファ鎖がそれらのCプロペプチドを介して会合し、Gly−X−Y繰返し領域がそのC末端に核形成点を形成して鎖の正確なアラインメントを確実にすることを可能にするトリマー分子を形成する。次にGly−X−Y領域はCからN方向に折畳まれ、三重らせんを形成する。
【0006】
ポリペプチド鎖修飾と三重らせん形成との間の時間的な関係は重大である。何故なら、プロリン残基のヒドロキシル化は体温での三重らせんの安定性を確実にするために必要であり、一旦形成されると三重らせんはヒドロキシル化酵素のための基質としてもはや作用しないからである。Cプロペプチド(及びより少ない程度のNプロペプチド)は、プロコラーゲンが細胞を通過する間、プロコラーゲンを可溶性に維持する(Bulleidら、2000年)。細胞外マトリックス中へのプロコラーゲン分子の分泌後又は分泌中、プロペプチドはプロコラーゲンN及びCプロテイナーゼによって除去され、それにより原線維へのコラーゲン分子の自発的な自己会合を引き起す(Hulmes、2002年)。プロコラーゲンN及びCプロテイナーゼによるプロペプチドの除去は、プロコラーゲンの可溶性を10000倍以上低下させ、コラーゲンの線維への自己会合を開始するのに必要かつ十分である。この会合過程にとって重要なのは、三重らせんドメインの終わりに位置する短い非三重らせんペプチド(テロペプチドと称される)であり、これは、原線維構造内のコラーゲン分子の正確な位置決めを確実にし、自己会合のための臨界濃度を低下させる(Bulleidら、2000年)。天然では、コラーゲンの三重らせん構造の安定性は、酵素プロリル−4−ヒドロキシラーゼ(P4H)によりプロリンがヒドロキシル化されてコラーゲン鎖内にヒドロキシプロリン残基を形成することを必要とする。
【0007】
コラーゲン鎖を発現する植物は当該技術分野で知られており、例えば米国特許第6617431号及び(Merleら、2002年、Ruggieroら、2000年)を参照できる。植物はヒドロキシプロリン含有タンパク質を合成することができるが、植物細胞中のヒドロキシプロリンの合成のために必要なプロリルヒドロキシラーゼは、哺乳類のP4Hと比較して比較的ゆるい基質配列特異性を示し、従って、Gly−X−YトリプレットのY位置のみにヒドロキシプロリンを含むコラーゲンの生産は、コラーゲン及びP4H遺伝子の植物中での共発現を必要とする(Olsenら、2003年)。
【0008】
植物中に天然に存在するヒドロキシル化作用機構に依存してヒトコラーゲンを生産する試みは、プロリンのヒドロキシル化が不十分なコラーゲンを生じた(Merleら、2002年)。かかるコラーゲンは、30℃より下の温度で溶解するか又はその三重らせん構造を失う。コラーゲン及びプロリルヒドロキシラーゼの共発現は、体温での用途のために生物学的に意味のある安定なヒドロキシル化コラーゲンを生じる(Merleら、20002年)。
【0009】
リシルヒロドキシラーゼ(LH,EC 1.14.11.4)、ガラクトシルトランスフェラーゼ(EC 2.4.1.50)及びグルコシルトランスフェラーゼ(EC 2.4.1.66)は、コラーゲンの翻訳後修飾に関与する酵素である。これらは特定位置のリシル残基をヒドロキシシリル、ガラクトシルヒドロキシリシル及びグルコシルガラクトシルヒドロキシリシル残基へと連続的に修飾する。これらの構造は、コラーゲンにとって特異的であり、それらの機能的活性のために必須である(Wangら、2002年)。単一のヒト酵素であるリシルヒドロキシラーゼ3(LH3)は、ヒドロキシリシンに結合された炭水化物形成において三つのすべての連続的ステップを触媒することができる(Wangら、2002年)。
【0010】
タバコ中で発現されるヒトコラーゲンのヒドロキシリシンは、ウシコラーゲン中で見出されるヒドロキシリシンの2%未満を形成する(0.04%の残基/1.88%の残基)。これは、植物に内在するリシルヒドロキシラーゼはコラーゲン中のリシンを十分にヒドロキシル化することができないということを示唆する。
【0011】
本発明を実施に移す際に、本発明者らは、コラーゲン鎖の効果的なヒドロキシル化は、このポリペプチドを正確に修飾することができる酵素に沿ってコラーゲン鎖を封鎖することに依存するということを発見した。
【発明の概要】
【0012】
本発明の一つの側面によれば、植物又は単離された植物細胞中でコラーゲンを生産する方法であって、内因性P4H活性を欠く細胞内区画中に少なくとも一種のコラーゲンアルファ鎖及び外因性P4Hが蓄積することを可能にする態様で、前記少なくとも一種のコラーゲンアルファ鎖及び前記外因性P4Hを植物又は単離された植物細胞中で発現させ、それにより植物中でコラーゲンを生産することを含む方法が提供される。
【0013】
以下に記述される本発明の好ましい実施態様におけるさらなる特徴によれば、方法は、内因性P4H活性を欠く前記細胞内区画中で外因性LH3を発現させることをさらに含む。
【0014】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0015】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、ER標的化又は残留配列を欠く。
【0016】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、植物のDNA含有オルガネラ中で発現される。
【0017】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0018】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ER標的化又は残留配列を欠く。
【0019】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、植物のDNA含有オルガネラ中で発現される。
【0020】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、アルファ1鎖である。
【0021】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、アルファ2鎖である。
【0022】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、C末端及び/又はN末端プロペプチドを含む。
【0023】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、植物は、タバコ、トウモロコシ、アルファルファ、イネ、ジャガイモ、ダイズ、トマト、小麦、大麦、カノーラ、ニンジン及び綿からなる群から選択される。
【0024】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖又は前記外因性P4Hは、前記植物の一部分のみで発現される。
【0025】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記植物の前記一部分は、葉、種子、根、塊茎又は茎である。
【0026】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、前記少なくとも一種のコラーゲンアルファ鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができる。
【0027】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ヒトのP4Hである。
【0028】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記植物は、ストレス条件に供される。
【0029】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記ストレス条件は、渇水、塩分、傷害、寒さ及びストレス誘導性化合物の噴霧からなる群から選択される。
【0030】
本発明の別の側面によれば、コラーゲンアルファ鎖がヒト細胞中で発現される場合に生産されるヒドロキシル化パターンと同一のヒドロキシルパターンを有するコラーゲンアルファ鎖を蓄積することができる遺伝的に改変された植物又は単離された植物細胞が提供される。
【0031】
本発明のなお別の側面によれば、内因性P4H活性を欠く細胞内区画中にコラーゲンアルファ鎖を蓄積することができる遺伝的に改変された植物又は単離された植物細胞が提供される。
【0032】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、遺伝的に改変された植物は、外因性P4Hをさらに含む。
【0033】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0034】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、ER標的化又は残留配列を欠く。
【0035】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、植物のDNA含有オルガネラ中で発現される。
【0036】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0037】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ER標的化又は残留配列を欠く。
【0038】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、植物のDNA含有オルガネラ中で発現される。
【0039】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ鎖は、アルファ1鎖である。
【0040】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ鎖は、アルファ2鎖である。
【0041】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ鎖は、C末端及び/又はN末端プロペプチドを含む。
【0042】
本発明のなお別の側面によれば、コラーゲンアルファ1鎖を蓄積することができる第一の遺伝的に改変された植物と、コラーゲンアルファ2鎖を蓄積することができる第二の遺伝的に改変された植物とを含む植物システムが提供される。
【0043】
本発明のさらに別の側面によれば、コラーゲンアルファ1鎖及びコラーゲンアルファ2鎖を蓄積することができる第一の遺伝的に改変された植物と、P4Hを蓄積することができる第二の遺伝的に改変された植物とを含む植物システムが提供される。
【0044】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記第一の遺伝的に改変された植物及び前記第二の遺伝的に改変された植物の少なくとも一つは外因性P4Hをさらに含む。
【0045】
本発明のさらに別の側面によれば、線維状コラーゲンの生産方法であって、(a)第一植物中でコラーゲンアルファ1鎖を発現させ;(b)第二植物中でコラーゲンアルファ2鎖を発現させ、ただし、前記第一植物及び前記第二植物中での発現は、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖がそれぞれ、内因性P4H活性を欠く細胞内区画中に蓄積することができるように構成されており、そして(c)前記第一植物と前記第二植物を交配させ、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖を発現する子孫を選択し、それにより線維状コラーゲンを生産することを含む方法が提供される。
【0046】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、方法は前記第一植物及び前記第二植物のそれぞれにおいて外因性P4Hを発現させることをさらに含む。
【0047】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖のそれぞれは、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0048】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖のそれぞれは、ER標的化又は残留配列を欠く。
【0049】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、ステップ(a)及び(b)は、植物のDNA含有オルガネラ中の発現を介して行われる。
【0050】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0051】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ER標的化又は残留配列を欠く。
【0052】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、植物のDNA含有オルガネラ中で発現される。
【0053】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖のそれぞれは、C末端及び/又はN末端プロペプチドを含む。
【0054】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、前記少なくとも一種のコラーゲンアルファ鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができる。
【0055】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ヒトのP4Hである。
【0056】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記植第一植物及び前記第二植物は、ストレス条件に供される。
【0057】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記ストレス条件は、渇水、塩分、傷害、重金属毒性及び寒さストレスからなる群から選択される。
【0058】
本発明のさらに別の側面によれば、線維状コラーゲンの生産方法であって、(a)第一植物中でコラーゲンアルファ1鎖及びコラーゲンアルファ2鎖を発現させ、ただし、前記第一植物中での発現は、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖がそれぞれ、内因性P4H活性を欠く細胞内区画中に蓄積することができるように構成されており;(b)第二植物中で、内因性P4H活性を欠く前記細胞内区画中に蓄積することができる外因性P4Hを発現させ;そして(c)前記第一植物と前記第二植物を交配させ、前記コラーゲンアルファ1鎖、前記コラーゲンアルファ2鎖及び前記P4Hを発現する子孫を選択し、それにより線維状コラーゲンを生産することを含む方法が提供される。
【0059】
本発明のさらに別の側面によれば、植物細胞中で機能的なプロモーターの転写制御下に配置されたヒトP4Hをコードするポリヌクレオチドを含む核酸構築物が提供される。
【0060】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記プロモーターは、CaMV 35Sプロモーター、Ubiquitinプロモーター、rbcSプロモーター及びSVBVプロモーターからなる群から選択される。
【0061】
本発明のさらに別の側面によれば、コラーゲンアルファ1鎖、コラーゲンアルファ2鎖、P4H、LH3及びプロテアーゼC及び/又はプロテアーゼNを発現することができる遺伝的に改変された植物又は単離された植物細胞が提供される。
【0062】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖はそれぞれ、内因性の植物P4H活性を欠く細胞内区画中に蓄積することができる。
【0063】
本発明のさらに別の側面によれば、哺乳類のコラーゲンの温度安定性特性と同一の温度安定性特性を有するコラーゲンを蓄積することができる遺伝的に改変された植物又は単離された植物細胞が提供される。
【0064】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンは、I型コラーゲンである。
【0065】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンは、ヒトのコラーゲンである。
【0066】
本発明のさらに別の側面によれば、植物中での発現のために最適化されたコラーゲンコード配列が提供される。
【0067】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンコード配列は配列番号1によって規定されるものである。
【0068】
本発明は、ヒトコラーゲンの特性と同様の特性を有するコラーゲンへと会合することができる正確にヒドロキシル化されたコラーゲン鎖を発現することができる植物を提供することによって、現在知られている構成の欠点を成功裏に克服する。
【0069】
本明細書で使用される技術用語と科学用語はすべて、特に断らない限り、本発明の属する技術分野の当業者が共通して理解しているのと同じ意味を持っている。本明細書に記載されているのと類似の又は均等の方法と材料は本発明を実施又は試験するのに使用できるが、適切な方法と材料は以下に述べる。争いが生じた場合、定義を含めて本特許明細書が基準である。さらに、本明細書の材料、方法及び実施例は例示することだけを目的とし本発明を限定するものではない。
【図面の簡単な説明】
【0070】
本明細書では本発明を単に例示し図面を参照して説明する。特に詳細に図面を参照して、示されている詳細が例示として本発明の好ましい実施態様を例示考察することだけを目的としており、本発明の原理や概念の側面の最も有用でかつ容易に理解される説明であると考えられるものを提供するために提示していることを強調するものである。この点について、本発明を基本的に理解するのに必要である以上に詳細に本発明の構造の詳細は示さないが、図面について行う説明によって本発明のいくつもの形態を実施する方法は当業者には明らかになるであろう。
【0071】
【図1a】図1aは、試験植物を形質転換するために使用される様々な発現カセット及びベクターの構造を示す。本研究の一部として合成されたすべてのコード配列は、タバコ中での発現のために最適化された。
【図1b】図1bは、試験植物を形質転換するために使用される様々な発現カセット及びベクターの構造を示す。本研究の一部として合成されたすべてのコード配列は、タバコ中での発現のために最適化された。
【図1c】図1cは、試験植物を形質転換するために使用される様々な発現カセット及びベクターの構造を示す。本研究の一部として合成されたすべてのコード配列は、タバコ中での発現のために最適化された。
【図1d】図1dは、試験植物を形質転換するために使用される様々な発現カセット及びベクターの構造を示す。本研究の一部として合成されたすべてのコード配列は、タバコ中での発現のために最適化された。
【図2】図2は、様々な共形質転換アプローチを示す。それぞれの発現カセットは、コード配列の短い名称によって表される。コード配列は、表1中で特定される。それぞれの共形質転換は、二つのpBINPLUSバイナリーベクターによって行われた。それぞれの長方形は、一つ、二つ又は三つの発現カセットを担持する単一のpBINPLUSベクターを表す。プロモーター及びターミネーターは、実施例1で特定される。
【図3】図3は、コラーゲンアルファ1(324bpフラグメント)又はコラーゲンアルファ2(537bpフラグメント)又は両者について陽性である植物を示す形質転換体のマルチプレックスPCRスクリーニングである。
【図4】図4は、共形質転換2,3及び4によって作成されたトランスジェニック植物のウェスタンブロット分析である。全可溶性タンパク質は、タバコ形質転換体#2、#3及び#4から抽出され、抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を用いて試験された。サイズマーカーは、Fermentas Inc.からの#SM0671であった。W.T.は野生型タバコである。陽性のコラーゲンバンドは、コラーゲンI型アルファ1又はアルファ2又は両者についてPCR陽性である植物において見ることができる。ヒト胎盤(ペプシン消化によってヒトの胎盤から抽出されたChemicon Inc.からの#CC050)からの500ngのコラーゲンI型の陽性コントロールバンドは、トランスジェニック植物からのサンプル中の全可溶性タンパク質の約0.3%(約150μg)を表す。ヒトコラーゲンサンプル中の約140kDaの大きなバンドは、コラーゲンI型抗体(Chemicon Inc.からの#MAB1913)の抗カルボキシ末端プロペプチドによって検出されるようにそのCプロペプチドを有するプロコラーゲンである。ヒトコラーゲンサンプル中の約120kDaの小さなバンドは、プロペプチドを有さないコラーゲンである。それらの異常な組成のため、プロリンリッチタンパク質(コラーゲンを含む)は、予測されたより高い分子量を有するバンドとしてポリアクリルアミドゲル上を一貫して移動する。それ故、約95kDaの分子量のプロペプチドを有さないコラーゲン鎖は、約120kDaのバンドとして移動する。
【図5】図5は、共形質転換によって作成されたトランスジェニック植物#8(コラーゲン鎖に翻訳的に融合されたアポプラストシグナルを担持する)のウェスタンブロット分析である。全可溶性タンパク質は、トランスジェニックタバコの葉から抽出され、抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を用いて試験された。陽性のコラーゲンアルファ2バンドが植物8−141で見ることができる。ヒト胎盤(Chemicon Inc.からの#CC050)からのコラーゲンI型は、コントロールとして使用された。
【図6a】図6aは、加熱処理及びトリプシン又はペプシン消化によって定性決定されるようなコラーゲン三重らせん会合体及び熱安定性を示す。図6aでは、タバコ2−9(コラーゲンアルファ1鎖のみを発現し、P4Hを発現しない)及び3−5(コラーゲンアルファ1鎖及びアルファ2鎖並びにヒトP4Hアルファ及びベータサブユニットの両方を発現する)からの全可溶性タンパク質は、加熱処理(38℃又は43℃で15分間)に供され、続いてトリプシン消化(室温で20分間)に供され、抗コラーゲンI抗体を用いてウェスタンブロット手順で試験された。陽性コントロールは、500ngのヒトコラーゲンI+w.t.タバコの全可溶性タンパク質のサンプルであった。
【図6b】図6bは、加熱処理及びトリプシン又はペプシン消化によって定性決定されるようなコラーゲン三重らせん会合体及び熱安定性を示す。図6bでは、全可溶性タンパク質はトランスジェニックタバコ13−6(矢印によって示されるコラーゲンIアルファ1及びアルファ2鎖、ヒトP4Hアルファ及びベータサブユニット及びヒトLH3を発現する)から抽出され、加熱処理(33℃、38℃又は42℃で20分間)に供され、直ちに氷上で冷却されて三重らせんの再会合を防止され、そしてペプシンと共に30分間室温(約22℃)でインキュベートされ、続いて抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を用いて標準的なウェスタンブロット手順で試験された。陽性コントロールは、w.t.タバコから抽出された全可溶性タンパク質に添加された約50ngのヒトコラーゲンI(ペプシン消化によってヒトの胎盤から抽出されたChemicon Inc.からの#CC050)のサンプルであった。
【図7】図7は、野生型タバコで行われたノーザンブロット分析を示す。ブロットは、タバコP4HのcDNAを用いてプローブされた。
【図8】図8は、共形質転換2,3及び13によって作成されたトランスジェニック植物のウェスタンブロット分析である。全可溶性タンパク質は、タバコ共形質転換体から抽出され、抗ヒトP4Hアルファ及びベータ並びに抗コラーゲンI抗体を用いて試験された。
【発明を実施するための形態】
【0072】
本発明は、哺乳類のコラーゲンの特徴を示すコラーゲン及びコラーゲン線維を生産するために使用されることができるコラーゲンを発現して蓄積する植物の発明である。
【0073】
本発明の原理および作用は、図面および付随する説明を参照してより十分に理解することができる。
【0074】
本発明の少なくとも1つの実施形態を詳しく説明する前に、本発明は、その適用において、下記の説明において示される細部、または、実施例によって例示される細部に限定されないことを理解しなければならない。本発明は他の実施形態が可能であり、または、様々な方法で実施することができ、または、様々な方法で実施される。また、本明細書中で用いられる表現法および用語法は記述のためであって、限定であると見なしてはならないことを理解しなければならない。
【0075】
コラーゲン生産植物は、当該技術分野で知られている。かかる植物は、コラーゲン鎖ならびにコラーゲンを生産するために使用されることができるが、かかる鎖は不正確にヒドロキシル化されており、従って、その自己会合は、植物中で生じるか否かにかかわらず、本来不安定なコラーゲンに導く。
【0076】
本発明を実施に移す際に、本発明者らは、コラーゲン鎖の正確なヒドロキシル化を確実にし、従ってヒトI型コラーゲンの特徴(例えば温度安定性)を良く模倣するコラーゲンの植物中での生産を可能にする植物発現アプローチを案出した。
【0077】
従って、本発明の一つの側面によれば、少なくとも一種のコラーゲンアルファ鎖を発現することができかつそれを内因性P4H活性を欠く細胞内区画中に蓄積することができる遺伝的に改変された植物が提供される。
【0078】
本明細書で使用される通り、「遺伝的に改変された植物(genetically modified plant)」は、外因性ポリヌクレオチド配列で安定に又は一過的に形質転換されるいかなる下等植物(例えば苔)又は高等植物(維管束植物)、又はその組織もしくは単離された細胞(例えば細胞懸濁液)に言及する。植物の例は、タバコ、トウモロコシ、アルファルファ、イネ、ジャガイモ、ダイズ、トマト、小麦、大麦、カノーラ、綿並びに苔の如き下等植物を含む。
【0079】
本明細書で使用される通り、句「コラーゲン鎖」は、コラーゲン線維のアルファ1又は2鎖の如きコラーゲンサブユニット、好ましくはI型線維に言及する。本明細書で使用される通り、句「コラーゲン」は、会合したコラーゲントリマーに言及し、これはI型コラーゲンの場合、二つのアルファ1鎖及び一つのアルファ2鎖を含む。コラーゲン線維は、末端プロペプチドC及びNを欠くコラーゲンである。
【0080】
本明細書で使用される通り、句「内因性P4H活性を欠く細胞内区画」は、植物のP4H又は植物様P4H活性を有する酵素を含まない細胞のいかなる区画化された領域に言及する。かかる細胞内区画の例は、液胞、アポプラスト及び細胞質並びにクロロプラスト、ミトコンドリアなどのオルガネラを含む。
【0081】
いかなる種類のコラーゲン鎖も本発明の遺伝的に改変された植物によって発現されることができる。その例は、原線維形成コラーゲン(I,II,III,V及びXI型)、網状組織形成コラーゲン(IV,VIII及びX型)、原線維表面に結合するコラーゲン(IX,XII及びXIV型)、膜貫通タンパク質として生じるコラーゲン(XIII及びXVII型)、又は11nmの周期でビーズを有するフィラメント(VI型)を含む。さらなる記述については、Hulmes、2002年を参照されたい。
【0082】
好ましくは、発現されるコラーゲン鎖は、I型コラーゲンのアルファ1及び/又は2鎖である。発現されるコラーゲンアルファ鎖は、いかなる哺乳類から由来するいかなるポリヌクレオチド配列によってコードされることができる。好ましくは、コラーゲンアルファ鎖をコードする配列は、ヒトのものであり、配列番号1及び4によって規定されるものである。
【0083】
典型的には、植物中で発現されるアルファコラーゲン鎖は、それらの末端ポリペプチド(即ち、プロペプチドC及びプロペプチドN)を含んでいてもよく、又は含まなくてもよい。
【0084】
Ruggieroら(2000年)は、植物のタンパク質分解活性によるプロコラーゲンの処理は、ヒトにおける正常な処理とは異なること、及び開裂部位は不明であるが、プロペプチドCは、植物のタンパク質分解活性によって除去されることに気付いた。Cプロペプチドの開裂は、トリマーの会合の前にプロコラーゲンペプチドに対して生じるであろう(三つのCプロペプチドの会合がトリマーの会合を開始するために必須である)。
【0085】
植物のタンパク質分解活性によるNプロペプチドの開裂は、成熟した植物では生じるが苗木では生じない。かかる開裂は、Nテロペプチドから2個のアミノ酸を除去する(17個のうちの2個)。
【0086】
Cプロペプチド(及びより少ない程度のNプロペプチド)は、プロコラーゲンが動物細胞を通過する間、プロコラーゲンを可溶性に維持し(Bulleidら、2000年)、植物細胞でも同様の効果を有すると予測される。細胞外マトリックス中へのプロコラーゲン分子の分泌後又は分泌中、プロペプチドはプロコラーゲンN及びCプロテイナーゼによって除去され、それにより原線維へのコラーゲン分子の自発的な自己会合を引き起す(Hulmes、2002年)。プロコラーゲンN及びCプロテイナーゼによるプロペプチドの除去は、プロコラーゲンの可溶性を10000倍以上低下させ、コラーゲンの線維への自己会合を開始するのに必要かつ十分である。この会合過程にとって重要なのは、三重らせんドメインの終わりに位置する短い非三重らせんペプチド(テロペプチドと称される)であり、これは、原線維構造内のコラーゲン分子の正確な位置決めを確実にし、自己会合のための臨界濃度を低下させる(Bulleidら、2000年)。従来技術は、コラーゲンの生産中にプロペプチドを開裂させるためのペプシンの使用を記述する(Bulleidら、2000年)。しかし、ペプシンはテロペプチドを損傷させ、結果としてペプシン抽出されたコラーゲンは、整列された線維状構造を形成することができない(Bulleidら、2000年)。
【0087】
ヒトP4Hのベータサブユニットを形成するタンパク質ジスルフィドイソメラーゼ(PDI)は、トリマー会合に先立ってCプロペプチドに結合し、それにより鎖会合の間の分子シャペロンとしても作用することが示されている(Ruggieroら、2000年)。様々な植物中で発現されるヒトI型プロコラーゲンNプロテイナーゼ及びプロコラーゲンCプロテイナーゼの使用は、生来的なヒトのコラーゲンに一層類似しかつ整列された原線維構造を形成することができるコラーゲンを生産することができるであろう。
【0088】
NもしくはCプロペプチド又はこれらの両方が、発現されるコラーゲン鎖に含まれる場合、本発明の遺伝的に改変された植物はそれぞれのプロテアーゼ(即ちCもしくはN又はこれらの両方)を発現することもできる。かかるプロテアーゼをコードするポリヌクレオチド配列は、配列番号18(プロテアーゼC)及び20(プロテアーゼN)によって例示される。かかるプロテアーゼは、それらがコラーゲン鎖と同じ細胞内区画中に蓄積されるように発現されることができる。
【0089】
内因性P4H活性を欠く細胞内区画中での発現されるコラーゲン鎖の蓄積は、いくつかのアプローチのいずれか一つによって実行されることができる。
【0090】
例えば、発現されるコラーゲン鎖は、発現されるタンパク質をアポプラスト又はオルガネラ(例えば葉緑体)の如き細胞内区画へと標的化するためのシグナル配列を含むことができる。好適なシグナル配列の例は、葉緑体輸送ペプチド(Swiss−ProtエントリーP07689、アミノ酸1〜57に含まれる)及びミトコンドリア輸送ペプチド(Swiss−ProtエントリーP46643、アミノ酸1〜28に含まれる)を含む。以下の実施例セクションは、好適なシグナル配列の追加の例示並びにかかるシグナル配列を植物細胞中でのコラーゲン鎖の発現において使用するためのガイドラインを与える。
【0091】
代わりに、コラーゲン鎖の配列は、植物中で発現されたときにコラーゲンの細胞内局在化を変化させるような方法で改変されることができる。
【0092】
上述の通り、植物のERは、コラーゲン鎖を正確にヒドロキシル化することができないP4Hを含む。コラーゲンアルファ鎖は、発現されるコラーゲンをER中に指向させるER標的化配列を生来的に含み、発現されるコラーゲンはER中で翻訳後修飾される(不正確なヒドロキシル化を含む)。従って、ER標的化配列の除去は、いかなるヒドロキシル化も含む翻訳後修飾を欠くコラーゲン鎖の細胞質内蓄積に導くであろう。
【0093】
以下の実施例セクションの実施例1は、ER配列を欠くコラーゲン配列の生成を記述する。
【0094】
また代わりに、コラーゲン鎖は、葉緑体又はミトコンドリアの如きDNA含有オルガネラ中に発現されて蓄積されることができる。葉緑体発現のさらなる記述は以下に与えられる。
【0095】
上述の通り、アルファ鎖のヒドロキシル化は、安定なI型コラーゲンの会合のために要求される。本発明の遺伝的に改変された植物によって発現されるアルファ鎖は、内因性P4H活性を欠く、細胞内区画中に蓄積するので、かかる鎖は、植物、植物組織又は細胞から単離されなければならず、インビトロでヒドロキシル化されなければならない。かかるヒドロキシル化は、Turpeenniemi−Hujanen及びMyllylaによって記述される方法(Concomitant hydroxylation of proline and lysine residues in collagen using purified enzymes in vitro.Biochim Biophys Acta.1984 Jul 16:800(1)59−65)によって達成されることができる。
【0096】
かかるインビトロヒドロキシル化は、正確にヒドロキシル化されたコラーゲン鎖に導くことができるが、達成するのが困難である場合があり費用がかかる場合がある。
【0097】
インビトロヒドロキシル化の制限を克服するため、本発明の遺伝的に改変された植物は、コラーゲンアルファ鎖を正確にヒドロキシル化する(即ち、Gly−X−Yトリプレットのプロリン(Y)位置のみをヒドロキシル化する)ことができるP4Hを共発現することが好ましい。P4Hは、二つのサブユニットアルファ及びベータからなる酵素である。これら両方が活性酵素を形成するのに必要であるが、ベータサブユニットはシャペロン機能をも担持する。
【0098】
本発明の遺伝的に改変された植物によって発現されるP4Hは、例えば配列番号12及び14によってコードされるヒトP4Hであることが好ましい。加えて、増大された基質特異性を示すP4H突然変異体又はP4H相同体も使用されることができる。
【0099】
好適なP4H相同体は、NCBIアクセッションNP 179363によって同定されるアラビドプシスのオキシドレダクターゼによって例示される。本発明者らによって行われたこのタンパク質の配列とヒトP4Hアルファサブユニットの対様式アラインメントは、植物のいかなる公知のP4H相同体の機能性ドメイン間の最高の相同性を明らかにした。
【0100】
P4Hは、発現されたコラーゲン鎖と共蓄積される必要があるので、そのコード配列は、それに応じて改変されることが好ましい(シグナル配列の付加、ER標的化を防止することができる欠失など)。
【0101】
哺乳類細胞では、コラーゲンはリシルヒドロキシラーゼ、ガラクトシルトランスフェラーゼ及びグルコシルトランスフェラーゼによっても修飾される。これらの酵素は、特定位置のリシル残基をヒドロキシリシル、ガラクトシルヒドロキシリシル及びグルコシルガラクトシルヒドロキシリシル残基へと連続的に修飾する。単一のヒト酵素であるリシルヒドロキシラーゼ3(LH3)は、ヒドロキシリシンに関連する炭水化物形成における三つの連続的ステップすべてを触媒することができる。
【0102】
従って、本発明の遺伝的に改変された植物は、哺乳類のLH3も発現することが好ましい。配列番号22によって規定されるようなLH3コード配列が、かかる目的のために使用されることができる。
【0103】
上述のコラーゲン鎖及び修飾酵素は、植物の機能的プロモーターの転写制御下に配置されたアルファ鎖及び/又は修飾酵素(例えばP4H及びLH3)をコードするポリヌクレオチド配列を含む、安定に組込まれた又は一過的に発現される核酸構築物から発現されることができる。かかる核酸構築物(本明細書では発現構築物とも称される)は、植物全体での発現のために、又は限定された植物組織もしくは限定された植物細胞での発現のために、又は植物の限定された発生段階での発現のために構成されることができる。かかる構築物は、選択マーカー(例えば抗生物質耐性)、エンハンサーエレメント及び細菌での複製のための複製起点も含むことができる。
【0104】
二つの発現性挿入物(例えば二つのアルファ鎖型又はアルファ鎖及びP4H)を含む構築物は、それぞれの挿入物について個別のプロモーターを含むことが好ましいことは理解されるであろう。又は代わりにかかる構築物は、単一のプロモーターから両方の挿入物配列を含む単一の転写キメラを発現することができる。かかる場合、キメラ転写物は、下流の挿入物が翻訳されることができるように二つの挿入物の間にIRES配列を含む。
【0105】
組織特異的、発生段階特異的、構成的、又は誘導性であることができる多数の植物機能的発現プロモーター及びエンハンサーが本発明の構築物によって利用されることができ、いくつかの例が以下に与えられる。
【0106】
本明細書及び特許請求の範囲で使用される通り、句「植物プロモーター」又は「プロモーター」は、植物細胞(DNA含有オルガネラを含む)中での遺伝子発現を指図することができるプロモーターを含む。かかるプロモーターは、植物、細菌、ウイルス、真菌又は動物起源のものであることができる。かかるプロモーターは、構成的、即ち複数の植物組織中での高レベルの遺伝子発現を指図することができるか、又は組織特異的、即ち特定の植物組織での遺伝子発現を指図することができるか、又は誘導性、即ち刺激の下で遺伝子発現を指図することができるか、又はキメラ、即ち少なくとも二つの異なるプロモーターの部分から形成されるものであることができる。
【0107】
従って、使用される植物プロモーターは、構成的プロモーター、組織特異的プロモーター、誘導性プロモーター、又はキメラプロモーターであることができる。
【0108】
構成的植物プロモーターの例は、CaMV35S及びCaMV19Sプロモーター、FMV34Sプロモーター、サトウキビ桿状バドナウイルスプロモーター、CsVMVプロモーター、アラビドプシスACT2/ACT8アクチンプロモーター、アラビドプシスユビキチンUBQ1プロモーター、大麦葉チオニンBTH6プロモーター、及びイネアクチンプロモーターを含むがこれらに限定されない。
【0109】
組織特異的プロモーターの例は、マメファゼオリン貯蔵タンパク質プロモーター、DLECプロモーター、PHSプロモーター、ゼイン貯蔵タンパク質プロモーター、ダイズからのコングルチンガンマプロモーター、AT2S1遺伝子プロモーター、アラビドプシスからのACT11アクチンプロモーター、ブラシカ ナプスからのnapAプロモーター、及びジャガイモパタチン遺伝子プロモーターを含むがこれらに限定されない。
【0110】
誘導性プロモーターは、例えば光、温度、化学物質、渇水、高塩分、浸透圧ショック、酸化的条件又は病原体を含むストレス条件の如き特定の刺激によって誘導されるプロモーターであり、エンドウマメrbcS遺伝子由来の光誘導性プロモーター、アルファルファrbcS遺伝子由来のプロモーター、渇水時に活性となるプロモーターDRE,MYC及びMYB、高塩分及び浸透圧ストレス時に活性となるプロモーターINT,INPS,prxEa,Ha hsp17.7G4及びRD21、及び病原体ストレス時に活性となるプロモーターhsr203J及びstr246Cを含むがこれらに限定されない。
【0111】
好ましくは、本発明によって利用されるプロモーターは、挿入された構築物の過剰発現が植物の形質転換後に行われるような強い構成的プロモーターである。
【0112】
本発明で使用されるいかなる構築物型も、各構築物型において同一又は異なる選択マーカーを使用して同一植物中へと共形質転換されることができることは理解されるであろう。代わりに、第一の構築物型が第一植物中に導入され、第二の構築物型が第二同系植物中に導入され、その後、生じたトランスジェニック植物が交配され、子孫が二重形質転換体について選択されることができる。かかる子孫のさらなる自家交配は、両方の構築物について同型接合体である遺伝系統を作成するために使用されることができる。
【0113】
単子葉植物及び双子葉植物の両方に核酸構築物を導入する様々な方法がある(Potrykus,L.,Annu.Rev.Plant.Physiol.,Plant,Mol.Biol.(1991)42:205−225;Shimamoto et al.,Nature(1989)338:274−276)。かかる方法は、核酸構築物又はその一部の植物ゲノム中への安定な組込み又は核酸構築物の一過性発現に依存しており、後者の場合、これらの配列は植物の子孫には遺伝しない。
【0114】
加えて、核酸構築物が、葉緑体の如きDNA含有オルガネラのDNA中に直接導入されることができるいくつかの方法がある。
【0115】
本発明の核酸構築物の中に含まれるような外因性配列を植物ゲノム中に安定に組込むためには二つの原理的な方法がある:
【0116】
(i)アグロバクテリウムによって媒介される遺伝子移入:Klee et al.(1987)Annu.Rev.Plant Physiol.38:467−486;Klee and Rogers in Cell Culture and Somatic Cell Genetics of Plants,Vol.6,Molecular Biology of Plant Nuclear Genes,eds.Schell,J.,and Vasil,L.K.,Academic Publishers,San Diego,Calif.(1989)p.2−25;Gatenby,in Plant Biotechnology,eds.Kung,S.and Arntzen,C.J.,Butterworth Publishers,Boston,Mass.(1989)p.93−112。
【0117】
(ii)直接DNA取込み:Paszkowski et al.,in Cell Culture and Somatic Cell Genetics of Plants,Vol.6,Molecular Biology of Plant Nuclear Genes eds.Schell,J.,and Vasil,L.K.,Academic Publishers,San Diego,Calif.(1989)p.52−68;この方法は、プロトプラストへDNAを直接取込むための方法を含む:Toriyama,K.et al.(1988)Bio/Technology 6:1072−1074。植物細胞の短い電気ショックによって誘導されるDNA取込み:Zhang et al.Plant Cell Rep.(1988)7:379−384.Fromm et al.Nature(1986)319:791−793。粒子衝撃による植物細胞又は組織中へのDNA注入:Klein et al.Bio/Technology(1988)6:559−563;McCabe et al.Bio/Technology(1988)6:923−926;Sanford,Physiol.Plant.(1990)79:206−209。マイクロピペット系の使用による植物細胞又は組織中へのDNA注入:Neuhaus et al.,Theor.Appl.Genet.(1987)75:30−36;Neuhaus and Spangenberg,Physiol.Plant.(1990)79:213−217。又は発芽花粉とのDNAの直接培養による植物細胞又は組織中へのDNA注入:DeWet et al.in Experimental Manipulation of Ovule Tissue,eds.Chapman,G.P.and Mantell,S.H.and Daniels,W.Longman,London,(1985)p.197−209;and Ohta,Proc.Natl.Acad.Sci.USA(1986)83:715−719。
【0118】
アグロバクテリウム系は、植物ゲノムDNA中に組込まれる限定されたDNAセグメントを含むプラスミドベクターの使用を含む。植物組織の接種方法は、植物種及びアグロバクテリウム送達系によって異なる。幅広く使用されているアプローチは、リーフディスク手順であり、これは植物全体への分化を開始するのに良好な源を提供するいかなる組織外植体を用いて行われることができる。Horsch et al.in Plant Molecular Biology Manual A5,Kluwer Academic Publishers,Dordrecht(1988)p.1−9参照。補助的なアプローチは、アグロバクテリウム送達系を減圧浸潤との組合せで使用する。アグロバクテリウム系は、トランスジェニック双子葉植物の作成において特に実行可能である。
【0119】
植物細胞中への直接DNA移入には様々な方法がある。エレクトロポレーションでは、プロトプラストは強い電場に短時間さらされる。マイクロ注入では、DNAは極めて小さいマイクロピペットを使用して細胞中へ直接機械的に注入される。微小粒子衝撃では、DNAは硫酸マグネシウムの結晶、タングステン粒子又は金粒子の如き微小投射物に吸着され、この微小投射物が細胞又は植物組織中へと物理的に加速される。
【0120】
形質転換後、植物の繁殖が行われる。植物繁殖の最も一般的な方法は種子による方法である。しかし、種子繁殖による再生は、異型接合性のため作物には均一性の欠失があるという欠点を有する。何故なら種子は、メンデルの法則によって支配される遺伝的変異に従って植物によって生産されるからである。基本的に、各種子は遺伝的に異なり、各種子はそれ自身の特異的な形質を有して成長するであろう。従って、形質転換植物は、再生された植物が親のトランスジェニック植物と同一の形質及び特徴を有するように生産されることが好ましい。それ故、形質転換植物は、形質転換植物の迅速で一貫した再生を与える微小繁殖によって再生されることが好ましい。
【0121】
本発明の核酸構築物内に含まれる単離された核酸を一過的に発現させるために利用されることができる一過的発現方法は、上述のマイクロ注入及び衝撃(しかし一過的発現が好ましい条件下で)、及びウイルスによって媒介される発現を含むがこれらに限定されない。ウイルスによって媒介される発現では、核酸構築物を含むパッケージ化された又はパッケージ化されていない組換えウイルスベクターが植物組織又は細胞を感染させるために利用され、そこに確立された繁殖する組換えウイルスが非ウイルス性核酸配列を発現する。
【0122】
植物宿主の形質転換のために有用であることが示されているウイルスはCaMV,TMV及びBVを含む。植物ウイルスを使用した植物の形質転換は、米国特許第4885237号(BGV),EP−A67553(TMV)、日本国公開特許公報第63−14693号(TMV),EPA第194809号(BV),EPA第278667号(BV)及びGluzman,Y.et al.,Communications in Molecular Biology:Viral Vectors,Cold Spring Harbor Laboratory,New York,pp.172−189(1988)に記述されている。植物を含む多くの宿主中で外来DNAを発現させるのに使用するためのシュードウイルス粒子は、WO 87/06261に記述されている。
【0123】
植物中に非ウイスル性外因性核酸配列を導入して発現させるための植物RNAウイルスの構築は、上述の文献並びにDawson,W.O.er al.,Virology(1989)172:285−292;Takamatsu et al.EMBO J.(1987)6:307−311;French et al.Science(1986)231:1294−1297;and Takamatsu et al.FEBS Letters(1990)269:73−76によって示されている。
【0124】
ウイルスがDNAウイルスである場合、構築はウイルス自体に対してなされることができる。代わりに、ウイルスは、外来DNAを有する望ましいウイルスベクターを構築するのが容易なように細菌プラスミド中へとまずクローニングされることができる。次にウイルスはプラスミドから切り出されることができる。もしウイルスがDNAウイルスであるなら、細菌の複製起点がウイルスDNAに取付けられることができ、ウイルスDNAは細菌によって複製されることができる。このDNAの転写及び翻訳は、ウイルスDNAを包囲するコートタンパク質を生産するであろう。もしウイルスがRNAウイルスであるなら、ウイルスは一般的にcDNAとしてクローニングされ、プラスミド中に挿入される。次にプラスミドは構築物のすべてを作るために使用される。次にRNAウイルスは、プラスミドのウイルス配列を転写することよって及びウイルス遺伝子を翻訳してウイルスRNAを包囲するコートタンパク質を生産することによって生産される。
【0125】
本発明の構築物中に含まれるような非ウイルス性外因性核酸配列の植物中への導入及び発現のための植物RNAウイルスの構築は、上述の文献並びに米国特許第5316931号によって示されている。
【0126】
一つの実施態様では、生来的コートタンパク質コード配列がウイルス核酸から欠失されており、かつ植物宿主中での発現、組換え植物ウイルス核酸のパッケージング及び組換え植物ウイルス核酸による宿主の全身的感染を確実にすることができる非生来的プロモーター、好ましくは非生来的コートタンパク質コード配列のサブゲノムプロモーターが挿入されている植物ウイルス核酸が提供される。代わりに、コートタンパク質遺伝子は、タンパク質が生産されるようにその中に非生来的核酸を挿入することによって不活性化されることができる。組換え植物ウイルス核酸は一以上の追加の非生来的サブゲノムプロモーターを含むことができる。それぞれの非生来的サブゲノムプロモーターは、植物宿主中の隣接遺伝子又は核酸配列を転写又は発現させることができるが、相互に及び生来的サブゲノムプロモーターと組換えられることができない。非生来的(外来性)核酸配列は、一以上の核酸配列が含められる場合、生来的な植物ウイルスサブゲノムプロモーター又は生来的な及び非生来的な植物ウイルスサブゲノムプロモーターに隣接して挿入されることができる。非生来的核酸配列はサブゲノムプロモーターの制御下で宿主植物中で転写又は発現され、望ましい生成物を生産する。
【0127】
第二の実施態様では、生来的なコートタンパク質コード配列が非生来的なコートタンパク質コード配列の代わりに非生来的なコートタンパク質サブゲノムプロモーターの一つに隣接して配置されていることを除いては第一の実施態様と同様の組換え植物ウイルス核酸が提供される。
【0128】
第三の実施態様では、生来的なコートタンパク質遺伝子がそのサブゲノムプロモーターに隣接されておりかつ一以上の非生来的サブゲノムプロモーターがウイルス核酸中に挿入されている組換え植物ウイルス核酸が提供される。挿入された非生来サブゲノムプロモーターは、隣接遺伝子を転写又は発現させることができるが、相互に及び生来的サブゲノムプロモーターと組換えられることができない。非生来的な核酸配列は、前記配列がサブゲノムプロモーターの制御下で宿主植物中で転写又は発現され、望ましい生成物を生産するように非生来的なサブゲノム植物ウイルスプロモーターに隣接して挿入されることができる。
【0129】
第四の実施態様では、生来的なコートタンパク質コード配列が非生来的なコートタンパク質コード配列によって置換されていることを除いては第三の実施態様と同様の組換え植物ウイルス核酸が提供される。
【0130】
ウイルスベクターは、組換え植物ウイルス核酸によってコードされるコートタンパク質によって包囲されて組換え植物ウイルスを生産する。組換え植物ウイルス核酸又は組換え植物ウイルスは、適切な宿主植物を感染させるために使用される。組換え植物ウイルス核酸は、宿主中で複製し、宿主中に全身的に広がり、宿主中で外来遺伝子(単離された核酸)を転写又は発現させて望ましいタンパク質を生産することができる。
【0131】
外因性核酸配列を葉緑体のゲノムに導入するための技術が知られている。この技術は下記の手順を伴う。第1に、植物細胞が、細胞あたりの葉緑体の数を約1個に減らすように化学的に処理される。その後、外因性核酸が、少なくとも1つの外因性核酸分子を葉緑体に導入することを目指して、粒子衝撃によって細胞に導入される。外因性核酸は、相同的組換え(これは葉緑体に固有の酵素によって容易に行われる)による葉緑体ゲノムへの組込みを可能にするように選択される。この目的のために、外因性核酸は、目的とする遺伝子に加えて、葉緑体ゲノムに由来する少なくとも1つの核酸配列を含む。加えて、外因性核酸は選択マーカーを含み、そのような選択マーカーは、逐次的な選抜手法によって、そのような選抜の後の葉緑体ゲノムのすべてまたは実質的にすべてのコピー体が外因性核酸を含むことを確認するために役立つ。この技術に関するさらなる詳細が、米国特許第4945050号および同第5693507号に見出される(これらは参考として本明細書中に組み込まれる)。従って、ポリペプチドを葉緑体のタンパク質発現系によって産生させることができ、かつ、葉緑体の内膜に一体化させることができる。
【0132】
上述の形質転換アプローチは、いかなる種の植物又はそこから由来する植物組織もしくは単離された植物細胞においてコラーゲン鎖及び/又は修飾酵素並びに会合されたコラーゲン(プロペプチドあり又はなし)を生産するために使用されることができる。
【0133】
好ましい植物は、本明細書に記述されたコラーゲン鎖、コラーゲン及び/又は処理酵素を多量に蓄積することができるものである。かかる植物は、ストレス条件に対するそれらの抵抗性及び発現された成分又は会合されたコラーゲンが抽出される容易さに従って選択されることもできる。好ましい植物の例は、タバコ、トウモロコシ、アルファルファ、イネ、ジャガイモ、ダイズ、トマト、小麦、大麦、カノーラ、ニンジン及び綿を含む。
【0134】
コラーゲン線維は、食品及び化粧品産業で広範囲に使用される。従って、植物によって発現されるコラーゲン線維成分(アルファ鎖)及び修飾酵素はコラーゲンの産業的合成において有用性を見出し、植物中での完全なコラーゲンの生産は、その簡単さ及び費用効率のために好ましい。
【0135】
植物中でI型コラーゲンを生成するためにいくつかのアプローチが使用されることができる。例えば、コラーゲンアルファ1鎖は、コラーゲンアルファ1及びP4H(及び所望によりLH3)を発現する植物から単離されることができ、コラーゲンアルファ2及びP4H(及び所望によりLH3及びプロテアーゼC及び/又はN)を発現する植物から単離されるコラーゲンアルファ2鎖と混合されることができる。コラーゲンアルファ1鎖はそれ自身によって三重らせんへと自己会合するため、混合前にかかるホモトリマーを変性させてコラーゲンアルファ2鎖と共に再生することが必要であるかもしれない。
【0136】
好ましくは、コラーゲンアルファ1及びP4H(及び所望によりLH3及びプロテアーゼC及び/又はN)を発現する第一植物は、コラーゲンアルファ2を発現する第二(及び好ましくは同系の)植物と交配されることができるか、又は代わりに両アルファ鎖を発現する第一植物は、P4H及び所望によりLH3及びプロテアーゼC及び/又はNを発現する第二植物と交配されることができる。
【0137】
上述の植物育種アプローチは二つの個別に形質転換された植物を利用するが、それぞれが一つ又は二つの成分を発現する三つ以上の個別に形質転換された植物を利用するアプローチも利用できるということは注意されるべきである。
【0138】
当業者は様々な植物育種技術に精通しているであろう。従って、かかる技術のさらなる記述は本明細書中では与えられない。
【0139】
植物育種アプローチが好ましいが、コラーゲンアルファ1及び2、P4H及びLH3(及び所望によりプロテアーゼC及び/又はN)を発現する単一の植物が、それぞれが一以上の発現可能な成分を細胞中に導入するために設計されているいくつかの形質転換事象を介して作成されることができることに注意すべきである。かかる場合には、各形質転換事象の安定性は特異的な選択マーカーを使用して確認されることができる。
【0140】
いかなる場合でも、形質転換及び植物育種アプローチはいかなる数の成分を発現するいかなる植物を作成するために使用されることができる。現在好まれているものはコラーゲンアルファ1及び2鎖、P4H,LH3及び少なくとも一つのプロテアーゼ(例えばプロテアーゼC及び/又はN)を発現する植物である。以下の実施例の節でさらに記述されるように、かかる植物は42℃までの温度で安定性を示すコラーゲンを蓄積する。
【0141】
育種から生じた子孫又は代わりに多数回形質転換植物は、核酸又はタンパク質プローブ(例えば抗体)を使用して外因性mRNA及び/又はポリペプチドの存在を確認することによって選択されることができる。後者のアプローチが好ましい。何故なら、それは発現されたポリペプチド成分の局在化を可能にし(例えば分別された植物抽出物をプロービングすることによって)、従って、正確な処理及び会合についての可能性も確認するからである。好適なプローブの例は以下の実施例の節に与えられる。
【0142】
一旦コラーゲンを発現する子孫が同定されたら、かかる植物はコラーゲン鎖ならびに修飾酵素の発現を最大にする条件下でさらに栽培される。
【0143】
遊離プロリンの蓄積は、本発明の遺伝的に改変された植物によって発現されるコラーゲン鎖を含む様々なプロリンリッチタンパク質の過剰生産を容易にすることができ、好ましい栽培条件は栽培された植物中での遊離プロリンの蓄積を増大させるようなものである。
【0144】
遊離プロリンの蓄積は、渇水、塩分、低温、高温、病原体感染、重金属毒性、嫌気生活、栄養欠乏、大気汚染及びUV照射を含む広範囲の環境ストレスに対する応答において様々な植物中に蓄積する(Hare and Cress,1997年)。
【0145】
遊離プロリンは、ABAの如き化合物での植物又は土壌の処理に対する応答において又は銅、塩、パラコート、サリチル酸などの如き化合物を含むストレスに対する応答においても蓄積することがあり得る。
【0146】
従って、コラーゲンを発現する子孫は様々なストレス条件(例えば50mMから250mMまでの範囲のNaClの異なる濃度)下で生長させられることができる。コラーゲン生産をさらに増大させるために、コラーゲン発現に対する様々なストレス条件の影響が調査され、植物の生存性、バイオマス及びコラーゲン蓄積に関して最適化されるであろう。
【0147】
植物組織/細胞は成熟時に収穫されることが好ましく、コラーゲン線維は周知の従来技術の抽出アプローチを使用して単離され、一つのかかるアプローチが以下に詳述される。
【0148】
形質転換植物の葉は液体窒素下で粉末へと磨砕され、ホモジネートは0.2MのNaClを含む0.5M酢酸中で60時間4℃で抽出される。不溶性材料は遠心分離によって除去される。組換えコラーゲンを含む上澄みは0.4M及び0.7MのNaClで塩分画される。組換えヘテロトリマーコラーゲンを含む0.7M NaCl沈殿は0.1Mの酢酸中に溶解されてこれに対して透析され、−20℃で貯蔵される(Ruggieroら、2000年に従う)。
【0149】
本発明のさらなる目的、利点および新規な特徴が、限定であることが意図されない下記の実施例を検討したとき、当業者には明らかになる。加えて、本明細書中上記に描かれるような、また、下記の請求項の節において特許請求されるような本発明の様々な実施形態および態様のそれぞれは、実験的裏付けが下記の実施例において見出される。
【実施例】
【0150】
次に下記の実施例が参照されるが、下記の実施例は、上記の説明と一緒に、本発明を非限定様式で例示する。
【0151】
本願で使用される用語と、本発明で利用される実験方法には、分子生化学、微生物学及び組み換えDNAの技法が広く含まれている。これらの技法は文献に詳細に説明されている[例えば以下の諸文献を参照されたい。「Molecular Cloning:A laboratory Manual」Sambrookら1989年;Ausubel,R.M.編1994年「Current Protocols in Molecular Biology」I〜III巻;Ausubelら著1989年「Current Protocols in Molecular Biology」John Wiley and Sons,米国メリーランド州バルチモア;Perbal著「A Practical Guide to Molecular Cloning」John Wiley & Sons,米国ニューヨーク1988年;Watsonら、「Recombinant DNA」Scientific American Books、米国ニューヨーク;Birrenら編「Genome Analysis:A Laboratory Manual Series」1〜4巻、Cold Spring Harbor Laboratory Press、米国ニューヨーク1998年;米国特許の4666828号、4683202号、4801531号、5192659号及び5272057号に記載される方法;Cellis,J.E.編「Cell Biology:A Laboratory Handbook」I〜III巻1994年;Freshney,Wiley−Liss著、「Culture of Animal Cells−A Manual of Basic Technique」(第3版)N.Y.(1994年);Coligan,J.E.編「Current Protocols in Immunology」I〜III巻1994年;Stitesら編「Basic and Clinical Immunology」(第8版)、Appleton & Lange、米国コネティカット州ノーウォーク1994年;MishellとShiigi編「Selected Methods in Cellular Immunology」、W.H.Freeman and Co.、米国ニューヨーク1980年;また利用可能な免疫検定法は、例えば以下の特許と科学文献に広範囲にわたって記載されている。米国特許の3791932号、3839153号、3850752号、3850578号、3853987号、3867517号、3879262号、3901654号、3935074号、3984533号、3996345号、4034074号、4098876号、4879219号、5011771号及び5281521号;Gait,M.J.編「Oligonucleotide Synthesis」1984年;Hames,B.D.及びHiggins S.J.編「Nucleic Acid Hybridization」1985年;Hames,B.D.及びHiggins S.J.編「Transcription and Translation」1984年;Freshney,R.I.編「Animal Cell Culture」1986年;「Immobilized Cells and Enzymes」IRL Press 1986年;Perbal,B.著「A Practical Guide to Molecular Cloning」1984年及び「Methods in Enzymology」1〜317巻、Academic Press;「PCR Protocols:A Guide To Methods And Applications」、Academic Press、米国カリフォルニア州サンディエゴ1990年;Marshakら、「Strategies for Protein Purification and Characterization−A Laboratory Course Manual」、CSHL Press、1996年;なおこれらの文献類は、あたかも本願に完全に記載されているように援用するものである]。その外の一般的な文献は、本明細書を通じて提供される。本明細書に記載の方法は当業技術界で周知であると考えられ、読者の便宜のために提供される。本明細書に含まれるすべての情報は本願に援用するものである。
【0152】
実施例1
構築及び形質転換スキーム
本研究で使用された発現カセットの構築は図1a−dに示されている。本研究におけるコード配列のすべては、タバコ中での発現のために最適化され、望ましいフランキング領域(配列番号1,4,7,12,14,16,18,20,22)を有するように化学的に合成された。図1a:液胞シグナルもしくはアポプラストシグナル(配列番号7によってコードされる)に融合された又はシグナルなしのCol1及びCol2(配列番号1,4)をコードする合成遺伝子は、キクrbcS1プロモーター及び5′UTR(配列番号10)並びにキクrbcS1 3′UTR及びターミネーター(配列番号11)からなる発現カセット中にクローニングされた。完全な発現カセットはpBINPLUS植物形質転換ベクターのマルチクローニングサイト中にクローニングされた(van Engelenら、1995年、Transgenic Res 4:288−290)。図1b:液胞シグナルもしくはアポプラストシグナル(配列番号7によってコードされる)に融合された又はシグナルなしのP4Hベータヒト、P4Hアルファヒト及びP4H植物(配列番号12,14及び16)をコードする合成遺伝子は、ベクターpJD330によって担持されるCaMV 35Sプロモーター及びTMVオメガ配列及びアグロバクテリウムノパリン合成酵素(NOS)ターミネーターからなる発現カセット中にクローニングされた。完全な発現カセットは、Col1又はCol2の発現ベクターを担持するpBINPLUSベクターのマルチクローニングサイト中にクローニングされた。図1c:液胞シグナルもしくはアポプラストシグナル(配列番号7によってコードされる)に融合されたプロテイナーゼC及びプロテイナーゼN(配列番号18,20)をコードする合成遺伝子は、キクrbcS1プロモーター及び5′UTR(配列番号10)並びにキクrbcS1 3′UTR及びターミネーター(配列番号11)からなる発現カセット中にクローニングされた。完全な発現カセットはpBINPLUS植物形質転換ベクターのマルチクローニングサイト中にクローニングされた。図1d:液胞シグナルもしくはアポプラストシグナル(配列番号7によってコードされる)に融合された又はシグナルなしのLH3(配列番号22)をコードする合成遺伝子(フランキングイチゴツル結合ウイルス(SVBV)プロモーター(NCBIアクセッションAF331666 REGION:623..950バージョンAF331666.1 GI:13345788)を有しかつアグロバクテリウムオクトピン合成酵素(OCS)ターミネーター(NCBIアクセッションZ37515 REGION:1344..1538バージョンZ37515.1 GI:886843)で末端停止されている)は、Col1及びP4Hベータの発現カセットを担持するpBINPLUSベクターのマルチクローニングサイト中にクローニングされた。
【0153】
図1に記述される発現カセットを利用した宿主植物中への共形質転換スキームは図2に図示される。各発現カセット挿入物はコード配列の短い名前によって表される。コード配列及び関連する配列番号は表1に記載されている。それぞれの共形質転換は二つのpBINPLUSバイナリーベクターによって行われる。それぞれの長方形は一つ、二つ又は三つの発現カセットを担持する単一のpBINPLUSベクターを表す。プロモーター及びターミネーターは図1で特定される。
【0154】
実施例2
植物コラーゲン発現
以下の表1に提示されるタンパク質をコードする合成ポリヌクレオチド配列が設計され、タバコ植物中での発現のために最適化された。
【0155】

【0156】
シグナルペプチド
(i)チオールプロテアーゼアレウレイン(aleurain)前駆体についての大麦遺伝子の液胞シグナル配列(NCBIアクセッションP05167 GI:113603)
(ii)アラビドプシスタリアーナのエンド1,4−ベータ−グルカナーゼのアポプラストシグナル(Cel1、NCBIアクセッションCAA67156.1 GI:2440033);配列番号9、配列番号7によってコードされる
【0157】
プラスミドの構築
植物発現ベクターは実施例1に教示されるようにして構築された。それぞれの構築された発現ベクターの組成は制限酵素分析及び配列決定を介して確認された。
【0158】
以下の発現カセットを含む発現ベクターが構築された:
1.コラーゲンアルファ1
2.コラーゲンアルファ1+ヒトP4Hベータサブユニット
3.コラーゲンアルファ1+ヒトP4Hベータサブユニット+ヒトLH3
4.コラーゲンアルファ2
5.コラーゲンアルファ2+ヒトP4Hアルファサブユニット
6.コラーゲンアルファ2+アラビドプシスP4H
7.ヒトP4Hベータサブユニット+ヒトLH3
8.ヒトP4Hアルファサブユニット
上述のコード配列のそれぞれは、液胞輸送ペプチドもしくはアポプラスト輸送ペプチドに翻訳的に融合されたか、又はいかなる輸送ペプチド配列も欠いており、その場合、細胞質蓄積が期待される。
【0159】
植物形質転換及びPCRスクリーニング
タバコ植物(ニコチアーナ・タバクム、Samsun NN)は、図2に教示される形質転換スキームに従って上述の発現ベクターで形質転換された。
【0160】
生じた形質転換植物は、コラーゲンアルファ1の324bpフラグメント及びコラーゲンアルファ2の537bpフラグメントを増幅することができるように設計された四つのプライマー(表2)を使用したマルチプレックスPCRを介してスクリーニングされた。図3は一つのマルチプレックスPCRスクリーニングの結果を示す。
【0161】
【0162】
実施例3
トランスジェニックタバコ植物中でのヒトコラーゲンの検出
全可溶性タンパク質は、「完全な」プロテアーゼ阻害剤カクテル(Roche Diagnostics GmbHからの製造#1836145、50mlの緩衝液あたり一錠)を使用して0.5mlの50mM Tris−HCl pH7.5中で500mgの葉を粉砕することによってタバコ形質転換体2,3及び4から抽出された。粗抽出物は、10%ベータ−メルカプト−エタノール及び8%SDSを含む250μlの4Xサンプル適用緩衝液と混合され、サンプルは7分間沸騰され、13000rpmで8分間遠心分離された。20μlの上澄みは10%ポリアクリルアミドゲル中に添加され、抗コラーゲンI(変性)抗体(Chemicon Inc.からの#AB745)を使用して標準的なウェスタンブロット手順でテストされた(図4)。W.T.は野生型タバコである。陽性コラーゲンバンドは、コラーゲンI型アルファ1又はアルファ2又は両方についてPCR陽性である植物において見ることができる。ヒト胎盤(Chemicon Inc.からの#CC050)からの500ngのコラーゲンI型の陽性コントロールバンドは、トランスジェニック植物からのサンプル中の約0.3%の全可溶性タンパク質(約150μg)を表す。
【0163】
全可溶性タンパク質の1%までの予測される分子量でコラーゲンを発現する植物は、コラーゲンが液胞に対して標準化された場合に検出された(図4)。アポプラストに対する全長コラーゲンの細胞内標的化は成功裏に達成された(図5)。細胞質中でコラーゲンを発現する植物(即ち標的化ペプチドなし)は、コラーゲンを検出可能なレベルに蓄積せず、植物中でのコラーゲンの細胞内標的化は成功のために重要であることを示した。
【0164】
N−プロペプチドを欠くコラーゲンが有意なタンパク質分解に供されることを示すRuggieroら2000年及びMerleら2002年の研究に加えて及びこれらの研究とは対照的に、現在のアプローチを使用すると、C−プロペプチド及びN−プロペプチドを有する全長コラーゲンタンパク質は高レベルで細胞内区画中に蓄積した。
【0165】
本データはまた、それぞれが異なるコラーゲン鎖型を発現する二つの植物を交配することは、それぞれの鎖型を最適レベルで発現する植物の選択及び望ましいコラーゲン生産植物を達成するための続く植物交配を可能にする点で有利であるということを明らかに示す。
【0166】
本発明の植物によって生産されるコラーゲンは、生来的なプロペプチドを含み、従って、タンパク質分解によって精製されたヒトのコントロールタンパク質より大きなタンパク質を形成すると予測される。ヒドロキシル化又はグリコシル化を有さないコラーゲンアルファ1及びアルファ2鎖の計算される分子量は以下の通りである:プロペプチドを有するCol1−136kDa、プロペプチドを有さないCol1−95kDa、プロペプチドを有するCol2−127kDa、プロペプチドを有さないCol2−92kDa。
【0167】
図4からわかるように、形質転換体3−5及び3−49中のCol1バンドは他の植物中のCol1バンドより大きく見える。これは、これらの植物中で共発現されてヒトコラーゲン鎖と同じ細胞内区画(例えば液胞)に対して標的化されたアルファ及びベータサブユニットからなるヒトプロリン−4−ヒドロキシラーゼホロ酵素によってコラーゲン鎖中でプロリンがヒドロキシル化されることを示す。
【0168】
実施例4
コラーゲン三重らせん会合及びトランスジェニック植物中での熱安定性
コラーゲン三重らせん会合及びトランスジェニック植物中でのらせん熱安定性は、トランスジェニック植物の全粗タンパク質抽出物の熱変性及び続くトリプシン又はペプシン消化によってテストされた(図6a〜b)。第一の実験では、タバコ2−9(Colアルファ1のみを発現し、P4Hを発現しない)及びタバコ3−5(Colアルファ+2及びP4Hの両方を発現する)からの全可溶性タンパク質は、0.5mlの50mM Tris−HCl pH=7.5中で500mgの葉を粉砕して13000rpmで10分間遠心分離し、上澄みを収集することによって抽出された。50μlの上澄みは熱処理(33℃又は43℃で15分間)に供され、次に直ちに氷上に置かれた。トリプシン消化は、各サンプルに50mM Tris−HCl pH=7.5中の1mg/mlトリプシン6μlを添加することによって開始された。サンプルは室温(約22℃)で20分間インキュベートされた。消化は、10%ベータメルカプトエタノール及び8%SDSを含む20μlの4Xサンプル適用緩衝液の添加によって停止され、サンプルは7分間沸騰され、13000rpmで7分間遠心分離された。50μlの上澄みは10%ポリアクリルアミドゲル中に添加され、抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を使用して標準的なウェスタンブロット手順でテストされた。陽性コントロールは、w.t.タバコから抽出された50μlの全可溶性タンパク質に添加された約500ngのヒトコラーゲンI(Chemicon Inc.からの#CC050であり、ペプシン消化によってヒト胎盤から抽出されたもの)のサンプルであった。
【0169】
図6aに示されるように、植物#3−5中で形成されたコラーゲン三重らせん並びにコントロールのヒトコラーゲンは33℃での変性に対して抵抗性であった。対照的に、植物#2−9によって形成されたコラーゲンは33℃で変性した。熱安定性におけるこの相違は、コラーゲンアルファ1及びコラーゲンアルファ2並びにP4Hベータ及びアルファサブユニットの両方を発現する形質転換体#3−5中での成功した三重らせん会合及び翻訳後のプロリンヒドロキシル化を示唆する。
【0170】
形質転換体#2−9における二つのバンドはダイマー又はトリマーを表すかもしれず、これらはSDS及びメルカプトエタノールを使用した7分間の沸騰後も安定である。同様のバンドがヒトのコラーゲン(上部パネル)において及び形質転換体#3−5において見ることができる。可能な説明は、異なる三重らせん中の二つのペプチドの間の等価なバンド(クロスリンク)がリシンオキシダーゼによる二つのリシンの酸化的脱アミノ化後に形成されたということである。第二の実験では、トランスジェニックタバコ13−6(コラーゲンIアルファ1及びアルファ2鎖(矢印によって示される)、ヒトP4Hアルファ及びベータサブユニット及びヒトLH3を発現する)からの全可溶性タンパク質は、0.5mlの100mM Tris−HCl pH=7.5及び300mM NaCl中で500mgの葉を粉砕して10000rpmで7分間遠心分離し、上澄みを収集することによって抽出された。50μlの上澄みは熱処理(33℃、38℃又は42℃で20分間)に供され、次に直ちに氷上に置かれた。ペプシン消化は、各サンプルに0.1M HCl4.5μl及び10mM酢酸中の2.5mg/mlペプシン4μlを添加することによって開始された。サンプルは室温(約22℃)で30分間インキュベートされた。消化は、5μlの非緩衝化1M Trisを添加することによって停止された。各サンプルは、10%ベータメルカプトエタノール及び8%SDSを含む22μlの4Xサンプル適用緩衝液と混合され、7分間沸騰され、13000rpmで7分間遠心分離された。40μlの上澄みは10%ポリアクリルアミドゲル中に添加され、抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を使用して標準的なウェスタンブロット手順でテストされた。陽性コントロールは、w.t.タバコから抽出された全可溶性タンパク質に添加された約50ngのヒトコラーゲン(Chemicon Inc.からの#CC050であり、ペプシン消化によってヒト胎盤から抽出されたもの)のサンプルであった。
【0171】
図6bに示されるように、植物#13−6中で形成されたコラーゲン三重らせんは42℃での変性に対して抵抗性であった。プロペプチドの開裂は、33℃でまず見ることができ、温度が38℃に上昇したとき及び42℃に上昇したとき再び効果的に徐々に増大する。開裂されたコラーゲン三重らせんドメインは、ペプシン処理されたヒトコラーゲンのゲル上での移動と同様の移動を示す。この実験で使用されたヒトコラーゲンは、ペプシンタンパク質分解によってヒト胎盤から抽出されたものであり、従ってプロペプチド及びいくつかのテロペプチドを欠いている。
【0172】
実施例5
植物のP4H発現
生来的な植物P4Hの誘導
タバコP4H cDNAは、クローニングされ、内因性P4H発現を誘導する条件及び処理を決定するためのプローブとして使用された。ノーザンブロット分析(図7)は、P4Hは茎頂では比較的高レベルでそして葉では低レベルで発現されることを明らかに示す。P4Hレベルは擦傷処理(下方パネルの「傷つけられた」)の4時間後に葉で有意に誘導された。同様の結果が他のストレス条件を使用して達成された(図示せず)。
【0173】
トランスジェニックタバコ植物中でのヒトP4Hアルファ及びベータサブユニット及びコラーゲンアルファ1及びアルファ2の検出
トランスジェニックタバコ植物中でのヒトP4Hアルファ及びベータサブユニット及びコラーゲンアルファ1及びアルファ2鎖の検出は、抗ヒトP4Hアルファサブユニット抗体(ICN Biomedicals Inc.からの#63−163)、抗ヒトP4Hベータサブユニット抗体(Chemicon Inc.からの#MAB2701)及び抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を使用して行われた。これらの抗体でプローブされたウェスタンブロットの結果は図8に示される。
【0174】
P4Hアルファ、P4Hベータ及びコラーゲンIアルファ1及びアルファ2バンドの発現は植物13−6(ヒトLH3でも形質転換されている)中で確認された。液胞シグナルペプチドを含むP4Hアルファ及びベータの計算された分子量はそれぞれ、65.5kDa及び53.4kDaである。プロペプチドを有する、ヒドロキシル化又はグリコシル化を有さないコラーゲンアルファ1及びアルファ2鎖の計算された分子量はそれぞれ、136kDa及び127kDaである。
【0175】
明確にするため別個の実施態様で説明されている本発明の特定の特徴は単一の実施態様に組み合わせて提供することもできることは分かるであろう。逆に、簡潔にするため単一の実施態様で説明されている本発明の各種の特徴は別個にまたは適切なサブコンビネーションで提供することもできる。
【0176】
本発明はその特定の実施態様によって説明してきたが、多くの別法、変更及び変形があることは当業者には明らかであることは明白である。従って、本発明は、本願の請求項の精神と広い範囲の中に入るこのような別法、変更及び変形すべてを包含するものである。本願で挙げた刊行物、特許、特許願及びGenBankアクセッション番号はすべて、個々の刊行物、特許、特許願及びGenBankアクセッション番号が各々あたかも具体的にかつ個々に引用提示されているのと同程度に、全体を本明細書に援用するものである。さらに、本願で引用又は確認したことは本発明の先行技術として利用できるという自白とみなすべきではない。
【0177】
【配列表フリーテキスト】
【0178】
配列番号1は、ヒトコラーゲンアルファ1(I)鎖に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号2及び3は、ヒトコラーゲンアルファ1(I)鎖に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域の合成配列である。
配列番号4及び6は、ヒトコラーゲンアルファ2(I)鎖に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号5は、ヒトコラーゲンアルファ2(I)鎖に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域の合成配列である。
配列番号7は、アラビドプシスタリアーナのエンド−1,4−ベータ−グルカナーゼのアポプラストシグナルのコード領域及びフランキング領域を含む合成配列である。
配列番号8及び9は、アラビドプシスタリアーナのエンド−1,4−ベータ−グルカナーゼのアポプラストシグナル及びフランキング領域の合成配列である。
配列番号10は、キクrbcS1プロモーター及び5′UTRの配列である。
配列番号11は、キクrbcS1 3′UTR及びターミネーターの配列である。
配列番号12は、ヒトプロリル4−ヒドロキシラーゼベータサブユニットに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号13は、ヒトプロリル4−ヒドロキシラーゼベータサブユニットに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域を含む合成配列である。
配列番号14は、ヒトプロリル4−ヒドロキシラーゼアルファ−1サブユニットに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号15は、ヒトプロリル4−ヒドロキシラーゼアルファ−1サブユニットに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域を含む合成配列である。
配列番号16は、植物プロリル4−ヒドロキシラーゼに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号17は、植物プロリル4−ヒドロキシラーゼに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域を含む合成配列である。
配列番号18及び19は、ヒトプロコラーゲンC−プロテイナーゼのコード領域及びフランキング領域を含む合成配列である。
配列番号20及び21は、ヒトプロコラーゲンI N−プロテイナーゼのコード領域及びフランキング領域を含む合成配列である。
配列番号22は、ヒトリシルヒドロキシラーゼ3に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号23は、ヒトリシルヒドロキシラーゼ3に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域を含む合成配列である。
配列番号24は、チオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列である。
配列番号25〜28は、一本鎖DNAオリゴヌクレオチドの配列である。
【技術分野】
【0001】
本発明は、コラーゲン生産植物及びその作成方法及びその使用に関する。特に、本発明は、生来的な三重らせんI型コラーゲン線維を形成することができる高レベルにヒドロキシル化されたコラーゲン鎖を生産することができる植物を作成するための新規のアプローチに関する。
【背景技術】
【0002】
コラーゲンは、脊椎動物及び多くの他の多細胞生物の構造的完全性を担う主要な構造タンパク質である。I型コラーゲンは原型線維状コラーゲンを表し、大部分の組織における主要なコラーゲン型である。
【0003】
I型コラーゲンは、骨及び腱の優勢なコラーゲン成分であり、皮膚、大動脈及び肺に多量に見出される。I型コラーゲン線維は、大きな引張り強度及び限定された伸度を与える。I型コラーゲンの最も豊富な分子形態は、二つの異なるアルファ鎖[アルファ1(I)]2及びアルファ2(I)からなるヘテロトリマーである(Inkinen、2003年)。すべての線維状コラーゲン分子は、繰返しGly−X−Yトリプレットから構築される三つのポリペプチド鎖を含み、ここでX及びYはいかなるアミノ酸であることができるが、イミノ酸プロリン及びヒドロキシプロリンであることが多い。
【0004】
原線維形成コラーゲン(線維状コラーゲン)は、球状N及びC末端延長プロペプチドを含む前駆体プロコラーゲンとして合成される。プロコラーゲンの生合成は、プロリン及びリシンのヒドロキシル化、N結合及びO結合グリコシル化及び鎖内及び鎖間ジスルフィド結合の形成を含む多数の異なる翻訳後修飾が関与する複雑な過程である。これらの修飾を行う酵素は、協調した様式で作用し、正確にアラインメントされた熱的に安定な三重らせん分子の折畳み及び会合を確実にする。
【0005】
各プロコラーゲン分子は、三つの構成的ポリペプチド鎖から大まかな小胞体内で会合する。ポリペプチド鎖が小胞体の膜を横切って共翻訳的に転位されるにつれて、プロリン及びリシン残基のヒドロキシル化がGly−X−Y繰返し領域内で起こる。一旦ポリペプチド鎖が小胞体の管腔中に十分に転位されたら、Cプロペプチドが折畳まれる。次に三つのプロアルファ鎖がそれらのCプロペプチドを介して会合し、Gly−X−Y繰返し領域がそのC末端に核形成点を形成して鎖の正確なアラインメントを確実にすることを可能にするトリマー分子を形成する。次にGly−X−Y領域はCからN方向に折畳まれ、三重らせんを形成する。
【0006】
ポリペプチド鎖修飾と三重らせん形成との間の時間的な関係は重大である。何故なら、プロリン残基のヒドロキシル化は体温での三重らせんの安定性を確実にするために必要であり、一旦形成されると三重らせんはヒドロキシル化酵素のための基質としてもはや作用しないからである。Cプロペプチド(及びより少ない程度のNプロペプチド)は、プロコラーゲンが細胞を通過する間、プロコラーゲンを可溶性に維持する(Bulleidら、2000年)。細胞外マトリックス中へのプロコラーゲン分子の分泌後又は分泌中、プロペプチドはプロコラーゲンN及びCプロテイナーゼによって除去され、それにより原線維へのコラーゲン分子の自発的な自己会合を引き起す(Hulmes、2002年)。プロコラーゲンN及びCプロテイナーゼによるプロペプチドの除去は、プロコラーゲンの可溶性を10000倍以上低下させ、コラーゲンの線維への自己会合を開始するのに必要かつ十分である。この会合過程にとって重要なのは、三重らせんドメインの終わりに位置する短い非三重らせんペプチド(テロペプチドと称される)であり、これは、原線維構造内のコラーゲン分子の正確な位置決めを確実にし、自己会合のための臨界濃度を低下させる(Bulleidら、2000年)。天然では、コラーゲンの三重らせん構造の安定性は、酵素プロリル−4−ヒドロキシラーゼ(P4H)によりプロリンがヒドロキシル化されてコラーゲン鎖内にヒドロキシプロリン残基を形成することを必要とする。
【0007】
コラーゲン鎖を発現する植物は当該技術分野で知られており、例えば米国特許第6617431号及び(Merleら、2002年、Ruggieroら、2000年)を参照できる。植物はヒドロキシプロリン含有タンパク質を合成することができるが、植物細胞中のヒドロキシプロリンの合成のために必要なプロリルヒドロキシラーゼは、哺乳類のP4Hと比較して比較的ゆるい基質配列特異性を示し、従って、Gly−X−YトリプレットのY位置のみにヒドロキシプロリンを含むコラーゲンの生産は、コラーゲン及びP4H遺伝子の植物中での共発現を必要とする(Olsenら、2003年)。
【0008】
植物中に天然に存在するヒドロキシル化作用機構に依存してヒトコラーゲンを生産する試みは、プロリンのヒドロキシル化が不十分なコラーゲンを生じた(Merleら、2002年)。かかるコラーゲンは、30℃より下の温度で溶解するか又はその三重らせん構造を失う。コラーゲン及びプロリルヒドロキシラーゼの共発現は、体温での用途のために生物学的に意味のある安定なヒドロキシル化コラーゲンを生じる(Merleら、20002年)。
【0009】
リシルヒロドキシラーゼ(LH,EC 1.14.11.4)、ガラクトシルトランスフェラーゼ(EC 2.4.1.50)及びグルコシルトランスフェラーゼ(EC 2.4.1.66)は、コラーゲンの翻訳後修飾に関与する酵素である。これらは特定位置のリシル残基をヒドロキシシリル、ガラクトシルヒドロキシリシル及びグルコシルガラクトシルヒドロキシリシル残基へと連続的に修飾する。これらの構造は、コラーゲンにとって特異的であり、それらの機能的活性のために必須である(Wangら、2002年)。単一のヒト酵素であるリシルヒドロキシラーゼ3(LH3)は、ヒドロキシリシンに結合された炭水化物形成において三つのすべての連続的ステップを触媒することができる(Wangら、2002年)。
【0010】
タバコ中で発現されるヒトコラーゲンのヒドロキシリシンは、ウシコラーゲン中で見出されるヒドロキシリシンの2%未満を形成する(0.04%の残基/1.88%の残基)。これは、植物に内在するリシルヒドロキシラーゼはコラーゲン中のリシンを十分にヒドロキシル化することができないということを示唆する。
【0011】
本発明を実施に移す際に、本発明者らは、コラーゲン鎖の効果的なヒドロキシル化は、このポリペプチドを正確に修飾することができる酵素に沿ってコラーゲン鎖を封鎖することに依存するということを発見した。
【発明の概要】
【0012】
本発明の一つの側面によれば、植物又は単離された植物細胞中でコラーゲンを生産する方法であって、内因性P4H活性を欠く細胞内区画中に少なくとも一種のコラーゲンアルファ鎖及び外因性P4Hが蓄積することを可能にする態様で、前記少なくとも一種のコラーゲンアルファ鎖及び前記外因性P4Hを植物又は単離された植物細胞中で発現させ、それにより植物中でコラーゲンを生産することを含む方法が提供される。
【0013】
以下に記述される本発明の好ましい実施態様におけるさらなる特徴によれば、方法は、内因性P4H活性を欠く前記細胞内区画中で外因性LH3を発現させることをさらに含む。
【0014】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0015】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、ER標的化又は残留配列を欠く。
【0016】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、植物のDNA含有オルガネラ中で発現される。
【0017】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0018】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ER標的化又は残留配列を欠く。
【0019】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、植物のDNA含有オルガネラ中で発現される。
【0020】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、アルファ1鎖である。
【0021】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、アルファ2鎖である。
【0022】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、C末端及び/又はN末端プロペプチドを含む。
【0023】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、植物は、タバコ、トウモロコシ、アルファルファ、イネ、ジャガイモ、ダイズ、トマト、小麦、大麦、カノーラ、ニンジン及び綿からなる群から選択される。
【0024】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖又は前記外因性P4Hは、前記植物の一部分のみで発現される。
【0025】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記植物の前記一部分は、葉、種子、根、塊茎又は茎である。
【0026】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、前記少なくとも一種のコラーゲンアルファ鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができる。
【0027】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ヒトのP4Hである。
【0028】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記植物は、ストレス条件に供される。
【0029】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記ストレス条件は、渇水、塩分、傷害、寒さ及びストレス誘導性化合物の噴霧からなる群から選択される。
【0030】
本発明の別の側面によれば、コラーゲンアルファ鎖がヒト細胞中で発現される場合に生産されるヒドロキシル化パターンと同一のヒドロキシルパターンを有するコラーゲンアルファ鎖を蓄積することができる遺伝的に改変された植物又は単離された植物細胞が提供される。
【0031】
本発明のなお別の側面によれば、内因性P4H活性を欠く細胞内区画中にコラーゲンアルファ鎖を蓄積することができる遺伝的に改変された植物又は単離された植物細胞が提供される。
【0032】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、遺伝的に改変された植物は、外因性P4Hをさらに含む。
【0033】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0034】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、ER標的化又は残留配列を欠く。
【0035】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記少なくとも一種のコラーゲンアルファ鎖は、植物のDNA含有オルガネラ中で発現される。
【0036】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0037】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ER標的化又は残留配列を欠く。
【0038】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、植物のDNA含有オルガネラ中で発現される。
【0039】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ鎖は、アルファ1鎖である。
【0040】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ鎖は、アルファ2鎖である。
【0041】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ鎖は、C末端及び/又はN末端プロペプチドを含む。
【0042】
本発明のなお別の側面によれば、コラーゲンアルファ1鎖を蓄積することができる第一の遺伝的に改変された植物と、コラーゲンアルファ2鎖を蓄積することができる第二の遺伝的に改変された植物とを含む植物システムが提供される。
【0043】
本発明のさらに別の側面によれば、コラーゲンアルファ1鎖及びコラーゲンアルファ2鎖を蓄積することができる第一の遺伝的に改変された植物と、P4Hを蓄積することができる第二の遺伝的に改変された植物とを含む植物システムが提供される。
【0044】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記第一の遺伝的に改変された植物及び前記第二の遺伝的に改変された植物の少なくとも一つは外因性P4Hをさらに含む。
【0045】
本発明のさらに別の側面によれば、線維状コラーゲンの生産方法であって、(a)第一植物中でコラーゲンアルファ1鎖を発現させ;(b)第二植物中でコラーゲンアルファ2鎖を発現させ、ただし、前記第一植物及び前記第二植物中での発現は、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖がそれぞれ、内因性P4H活性を欠く細胞内区画中に蓄積することができるように構成されており、そして(c)前記第一植物と前記第二植物を交配させ、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖を発現する子孫を選択し、それにより線維状コラーゲンを生産することを含む方法が提供される。
【0046】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、方法は前記第一植物及び前記第二植物のそれぞれにおいて外因性P4Hを発現させることをさらに含む。
【0047】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖のそれぞれは、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0048】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖のそれぞれは、ER標的化又は残留配列を欠く。
【0049】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、ステップ(a)及び(b)は、植物のDNA含有オルガネラ中の発現を介して行われる。
【0050】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、アポプラスト又は液胞に対して標的化するためのシグナルペプチドを含む。
【0051】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ER標的化又は残留配列を欠く。
【0052】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、植物のDNA含有オルガネラ中で発現される。
【0053】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖のそれぞれは、C末端及び/又はN末端プロペプチドを含む。
【0054】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、前記少なくとも一種のコラーゲンアルファ鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができる。
【0055】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記外因性P4Hは、ヒトのP4Hである。
【0056】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記植第一植物及び前記第二植物は、ストレス条件に供される。
【0057】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記ストレス条件は、渇水、塩分、傷害、重金属毒性及び寒さストレスからなる群から選択される。
【0058】
本発明のさらに別の側面によれば、線維状コラーゲンの生産方法であって、(a)第一植物中でコラーゲンアルファ1鎖及びコラーゲンアルファ2鎖を発現させ、ただし、前記第一植物中での発現は、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖がそれぞれ、内因性P4H活性を欠く細胞内区画中に蓄積することができるように構成されており;(b)第二植物中で、内因性P4H活性を欠く前記細胞内区画中に蓄積することができる外因性P4Hを発現させ;そして(c)前記第一植物と前記第二植物を交配させ、前記コラーゲンアルファ1鎖、前記コラーゲンアルファ2鎖及び前記P4Hを発現する子孫を選択し、それにより線維状コラーゲンを生産することを含む方法が提供される。
【0059】
本発明のさらに別の側面によれば、植物細胞中で機能的なプロモーターの転写制御下に配置されたヒトP4Hをコードするポリヌクレオチドを含む核酸構築物が提供される。
【0060】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記プロモーターは、CaMV 35Sプロモーター、Ubiquitinプロモーター、rbcSプロモーター及びSVBVプロモーターからなる群から選択される。
【0061】
本発明のさらに別の側面によれば、コラーゲンアルファ1鎖、コラーゲンアルファ2鎖、P4H、LH3及びプロテアーゼC及び/又はプロテアーゼNを発現することができる遺伝的に改変された植物又は単離された植物細胞が提供される。
【0062】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖はそれぞれ、内因性の植物P4H活性を欠く細胞内区画中に蓄積することができる。
【0063】
本発明のさらに別の側面によれば、哺乳類のコラーゲンの温度安定性特性と同一の温度安定性特性を有するコラーゲンを蓄積することができる遺伝的に改変された植物又は単離された植物細胞が提供される。
【0064】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンは、I型コラーゲンである。
【0065】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンは、ヒトのコラーゲンである。
【0066】
本発明のさらに別の側面によれば、植物中での発現のために最適化されたコラーゲンコード配列が提供される。
【0067】
以下に記述される本発明の好ましい実施態様におけるなおさらなる特徴によれば、前記コラーゲンコード配列は配列番号1によって規定されるものである。
【0068】
本発明は、ヒトコラーゲンの特性と同様の特性を有するコラーゲンへと会合することができる正確にヒドロキシル化されたコラーゲン鎖を発現することができる植物を提供することによって、現在知られている構成の欠点を成功裏に克服する。
【0069】
本明細書で使用される技術用語と科学用語はすべて、特に断らない限り、本発明の属する技術分野の当業者が共通して理解しているのと同じ意味を持っている。本明細書に記載されているのと類似の又は均等の方法と材料は本発明を実施又は試験するのに使用できるが、適切な方法と材料は以下に述べる。争いが生じた場合、定義を含めて本特許明細書が基準である。さらに、本明細書の材料、方法及び実施例は例示することだけを目的とし本発明を限定するものではない。
【図面の簡単な説明】
【0070】
本明細書では本発明を単に例示し図面を参照して説明する。特に詳細に図面を参照して、示されている詳細が例示として本発明の好ましい実施態様を例示考察することだけを目的としており、本発明の原理や概念の側面の最も有用でかつ容易に理解される説明であると考えられるものを提供するために提示していることを強調するものである。この点について、本発明を基本的に理解するのに必要である以上に詳細に本発明の構造の詳細は示さないが、図面について行う説明によって本発明のいくつもの形態を実施する方法は当業者には明らかになるであろう。
【0071】
【図1a】図1aは、試験植物を形質転換するために使用される様々な発現カセット及びベクターの構造を示す。本研究の一部として合成されたすべてのコード配列は、タバコ中での発現のために最適化された。
【図1b】図1bは、試験植物を形質転換するために使用される様々な発現カセット及びベクターの構造を示す。本研究の一部として合成されたすべてのコード配列は、タバコ中での発現のために最適化された。
【図1c】図1cは、試験植物を形質転換するために使用される様々な発現カセット及びベクターの構造を示す。本研究の一部として合成されたすべてのコード配列は、タバコ中での発現のために最適化された。
【図1d】図1dは、試験植物を形質転換するために使用される様々な発現カセット及びベクターの構造を示す。本研究の一部として合成されたすべてのコード配列は、タバコ中での発現のために最適化された。
【図2】図2は、様々な共形質転換アプローチを示す。それぞれの発現カセットは、コード配列の短い名称によって表される。コード配列は、表1中で特定される。それぞれの共形質転換は、二つのpBINPLUSバイナリーベクターによって行われた。それぞれの長方形は、一つ、二つ又は三つの発現カセットを担持する単一のpBINPLUSベクターを表す。プロモーター及びターミネーターは、実施例1で特定される。
【図3】図3は、コラーゲンアルファ1(324bpフラグメント)又はコラーゲンアルファ2(537bpフラグメント)又は両者について陽性である植物を示す形質転換体のマルチプレックスPCRスクリーニングである。
【図4】図4は、共形質転換2,3及び4によって作成されたトランスジェニック植物のウェスタンブロット分析である。全可溶性タンパク質は、タバコ形質転換体#2、#3及び#4から抽出され、抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を用いて試験された。サイズマーカーは、Fermentas Inc.からの#SM0671であった。W.T.は野生型タバコである。陽性のコラーゲンバンドは、コラーゲンI型アルファ1又はアルファ2又は両者についてPCR陽性である植物において見ることができる。ヒト胎盤(ペプシン消化によってヒトの胎盤から抽出されたChemicon Inc.からの#CC050)からの500ngのコラーゲンI型の陽性コントロールバンドは、トランスジェニック植物からのサンプル中の全可溶性タンパク質の約0.3%(約150μg)を表す。ヒトコラーゲンサンプル中の約140kDaの大きなバンドは、コラーゲンI型抗体(Chemicon Inc.からの#MAB1913)の抗カルボキシ末端プロペプチドによって検出されるようにそのCプロペプチドを有するプロコラーゲンである。ヒトコラーゲンサンプル中の約120kDaの小さなバンドは、プロペプチドを有さないコラーゲンである。それらの異常な組成のため、プロリンリッチタンパク質(コラーゲンを含む)は、予測されたより高い分子量を有するバンドとしてポリアクリルアミドゲル上を一貫して移動する。それ故、約95kDaの分子量のプロペプチドを有さないコラーゲン鎖は、約120kDaのバンドとして移動する。
【図5】図5は、共形質転換によって作成されたトランスジェニック植物#8(コラーゲン鎖に翻訳的に融合されたアポプラストシグナルを担持する)のウェスタンブロット分析である。全可溶性タンパク質は、トランスジェニックタバコの葉から抽出され、抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を用いて試験された。陽性のコラーゲンアルファ2バンドが植物8−141で見ることができる。ヒト胎盤(Chemicon Inc.からの#CC050)からのコラーゲンI型は、コントロールとして使用された。
【図6a】図6aは、加熱処理及びトリプシン又はペプシン消化によって定性決定されるようなコラーゲン三重らせん会合体及び熱安定性を示す。図6aでは、タバコ2−9(コラーゲンアルファ1鎖のみを発現し、P4Hを発現しない)及び3−5(コラーゲンアルファ1鎖及びアルファ2鎖並びにヒトP4Hアルファ及びベータサブユニットの両方を発現する)からの全可溶性タンパク質は、加熱処理(38℃又は43℃で15分間)に供され、続いてトリプシン消化(室温で20分間)に供され、抗コラーゲンI抗体を用いてウェスタンブロット手順で試験された。陽性コントロールは、500ngのヒトコラーゲンI+w.t.タバコの全可溶性タンパク質のサンプルであった。
【図6b】図6bは、加熱処理及びトリプシン又はペプシン消化によって定性決定されるようなコラーゲン三重らせん会合体及び熱安定性を示す。図6bでは、全可溶性タンパク質はトランスジェニックタバコ13−6(矢印によって示されるコラーゲンIアルファ1及びアルファ2鎖、ヒトP4Hアルファ及びベータサブユニット及びヒトLH3を発現する)から抽出され、加熱処理(33℃、38℃又は42℃で20分間)に供され、直ちに氷上で冷却されて三重らせんの再会合を防止され、そしてペプシンと共に30分間室温(約22℃)でインキュベートされ、続いて抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を用いて標準的なウェスタンブロット手順で試験された。陽性コントロールは、w.t.タバコから抽出された全可溶性タンパク質に添加された約50ngのヒトコラーゲンI(ペプシン消化によってヒトの胎盤から抽出されたChemicon Inc.からの#CC050)のサンプルであった。
【図7】図7は、野生型タバコで行われたノーザンブロット分析を示す。ブロットは、タバコP4HのcDNAを用いてプローブされた。
【図8】図8は、共形質転換2,3及び13によって作成されたトランスジェニック植物のウェスタンブロット分析である。全可溶性タンパク質は、タバコ共形質転換体から抽出され、抗ヒトP4Hアルファ及びベータ並びに抗コラーゲンI抗体を用いて試験された。
【発明を実施するための形態】
【0072】
本発明は、哺乳類のコラーゲンの特徴を示すコラーゲン及びコラーゲン線維を生産するために使用されることができるコラーゲンを発現して蓄積する植物の発明である。
【0073】
本発明の原理および作用は、図面および付随する説明を参照してより十分に理解することができる。
【0074】
本発明の少なくとも1つの実施形態を詳しく説明する前に、本発明は、その適用において、下記の説明において示される細部、または、実施例によって例示される細部に限定されないことを理解しなければならない。本発明は他の実施形態が可能であり、または、様々な方法で実施することができ、または、様々な方法で実施される。また、本明細書中で用いられる表現法および用語法は記述のためであって、限定であると見なしてはならないことを理解しなければならない。
【0075】
コラーゲン生産植物は、当該技術分野で知られている。かかる植物は、コラーゲン鎖ならびにコラーゲンを生産するために使用されることができるが、かかる鎖は不正確にヒドロキシル化されており、従って、その自己会合は、植物中で生じるか否かにかかわらず、本来不安定なコラーゲンに導く。
【0076】
本発明を実施に移す際に、本発明者らは、コラーゲン鎖の正確なヒドロキシル化を確実にし、従ってヒトI型コラーゲンの特徴(例えば温度安定性)を良く模倣するコラーゲンの植物中での生産を可能にする植物発現アプローチを案出した。
【0077】
従って、本発明の一つの側面によれば、少なくとも一種のコラーゲンアルファ鎖を発現することができかつそれを内因性P4H活性を欠く細胞内区画中に蓄積することができる遺伝的に改変された植物が提供される。
【0078】
本明細書で使用される通り、「遺伝的に改変された植物(genetically modified plant)」は、外因性ポリヌクレオチド配列で安定に又は一過的に形質転換されるいかなる下等植物(例えば苔)又は高等植物(維管束植物)、又はその組織もしくは単離された細胞(例えば細胞懸濁液)に言及する。植物の例は、タバコ、トウモロコシ、アルファルファ、イネ、ジャガイモ、ダイズ、トマト、小麦、大麦、カノーラ、綿並びに苔の如き下等植物を含む。
【0079】
本明細書で使用される通り、句「コラーゲン鎖」は、コラーゲン線維のアルファ1又は2鎖の如きコラーゲンサブユニット、好ましくはI型線維に言及する。本明細書で使用される通り、句「コラーゲン」は、会合したコラーゲントリマーに言及し、これはI型コラーゲンの場合、二つのアルファ1鎖及び一つのアルファ2鎖を含む。コラーゲン線維は、末端プロペプチドC及びNを欠くコラーゲンである。
【0080】
本明細書で使用される通り、句「内因性P4H活性を欠く細胞内区画」は、植物のP4H又は植物様P4H活性を有する酵素を含まない細胞のいかなる区画化された領域に言及する。かかる細胞内区画の例は、液胞、アポプラスト及び細胞質並びにクロロプラスト、ミトコンドリアなどのオルガネラを含む。
【0081】
いかなる種類のコラーゲン鎖も本発明の遺伝的に改変された植物によって発現されることができる。その例は、原線維形成コラーゲン(I,II,III,V及びXI型)、網状組織形成コラーゲン(IV,VIII及びX型)、原線維表面に結合するコラーゲン(IX,XII及びXIV型)、膜貫通タンパク質として生じるコラーゲン(XIII及びXVII型)、又は11nmの周期でビーズを有するフィラメント(VI型)を含む。さらなる記述については、Hulmes、2002年を参照されたい。
【0082】
好ましくは、発現されるコラーゲン鎖は、I型コラーゲンのアルファ1及び/又は2鎖である。発現されるコラーゲンアルファ鎖は、いかなる哺乳類から由来するいかなるポリヌクレオチド配列によってコードされることができる。好ましくは、コラーゲンアルファ鎖をコードする配列は、ヒトのものであり、配列番号1及び4によって規定されるものである。
【0083】
典型的には、植物中で発現されるアルファコラーゲン鎖は、それらの末端ポリペプチド(即ち、プロペプチドC及びプロペプチドN)を含んでいてもよく、又は含まなくてもよい。
【0084】
Ruggieroら(2000年)は、植物のタンパク質分解活性によるプロコラーゲンの処理は、ヒトにおける正常な処理とは異なること、及び開裂部位は不明であるが、プロペプチドCは、植物のタンパク質分解活性によって除去されることに気付いた。Cプロペプチドの開裂は、トリマーの会合の前にプロコラーゲンペプチドに対して生じるであろう(三つのCプロペプチドの会合がトリマーの会合を開始するために必須である)。
【0085】
植物のタンパク質分解活性によるNプロペプチドの開裂は、成熟した植物では生じるが苗木では生じない。かかる開裂は、Nテロペプチドから2個のアミノ酸を除去する(17個のうちの2個)。
【0086】
Cプロペプチド(及びより少ない程度のNプロペプチド)は、プロコラーゲンが動物細胞を通過する間、プロコラーゲンを可溶性に維持し(Bulleidら、2000年)、植物細胞でも同様の効果を有すると予測される。細胞外マトリックス中へのプロコラーゲン分子の分泌後又は分泌中、プロペプチドはプロコラーゲンN及びCプロテイナーゼによって除去され、それにより原線維へのコラーゲン分子の自発的な自己会合を引き起す(Hulmes、2002年)。プロコラーゲンN及びCプロテイナーゼによるプロペプチドの除去は、プロコラーゲンの可溶性を10000倍以上低下させ、コラーゲンの線維への自己会合を開始するのに必要かつ十分である。この会合過程にとって重要なのは、三重らせんドメインの終わりに位置する短い非三重らせんペプチド(テロペプチドと称される)であり、これは、原線維構造内のコラーゲン分子の正確な位置決めを確実にし、自己会合のための臨界濃度を低下させる(Bulleidら、2000年)。従来技術は、コラーゲンの生産中にプロペプチドを開裂させるためのペプシンの使用を記述する(Bulleidら、2000年)。しかし、ペプシンはテロペプチドを損傷させ、結果としてペプシン抽出されたコラーゲンは、整列された線維状構造を形成することができない(Bulleidら、2000年)。
【0087】
ヒトP4Hのベータサブユニットを形成するタンパク質ジスルフィドイソメラーゼ(PDI)は、トリマー会合に先立ってCプロペプチドに結合し、それにより鎖会合の間の分子シャペロンとしても作用することが示されている(Ruggieroら、2000年)。様々な植物中で発現されるヒトI型プロコラーゲンNプロテイナーゼ及びプロコラーゲンCプロテイナーゼの使用は、生来的なヒトのコラーゲンに一層類似しかつ整列された原線維構造を形成することができるコラーゲンを生産することができるであろう。
【0088】
NもしくはCプロペプチド又はこれらの両方が、発現されるコラーゲン鎖に含まれる場合、本発明の遺伝的に改変された植物はそれぞれのプロテアーゼ(即ちCもしくはN又はこれらの両方)を発現することもできる。かかるプロテアーゼをコードするポリヌクレオチド配列は、配列番号18(プロテアーゼC)及び20(プロテアーゼN)によって例示される。かかるプロテアーゼは、それらがコラーゲン鎖と同じ細胞内区画中に蓄積されるように発現されることができる。
【0089】
内因性P4H活性を欠く細胞内区画中での発現されるコラーゲン鎖の蓄積は、いくつかのアプローチのいずれか一つによって実行されることができる。
【0090】
例えば、発現されるコラーゲン鎖は、発現されるタンパク質をアポプラスト又はオルガネラ(例えば葉緑体)の如き細胞内区画へと標的化するためのシグナル配列を含むことができる。好適なシグナル配列の例は、葉緑体輸送ペプチド(Swiss−ProtエントリーP07689、アミノ酸1〜57に含まれる)及びミトコンドリア輸送ペプチド(Swiss−ProtエントリーP46643、アミノ酸1〜28に含まれる)を含む。以下の実施例セクションは、好適なシグナル配列の追加の例示並びにかかるシグナル配列を植物細胞中でのコラーゲン鎖の発現において使用するためのガイドラインを与える。
【0091】
代わりに、コラーゲン鎖の配列は、植物中で発現されたときにコラーゲンの細胞内局在化を変化させるような方法で改変されることができる。
【0092】
上述の通り、植物のERは、コラーゲン鎖を正確にヒドロキシル化することができないP4Hを含む。コラーゲンアルファ鎖は、発現されるコラーゲンをER中に指向させるER標的化配列を生来的に含み、発現されるコラーゲンはER中で翻訳後修飾される(不正確なヒドロキシル化を含む)。従って、ER標的化配列の除去は、いかなるヒドロキシル化も含む翻訳後修飾を欠くコラーゲン鎖の細胞質内蓄積に導くであろう。
【0093】
以下の実施例セクションの実施例1は、ER配列を欠くコラーゲン配列の生成を記述する。
【0094】
また代わりに、コラーゲン鎖は、葉緑体又はミトコンドリアの如きDNA含有オルガネラ中に発現されて蓄積されることができる。葉緑体発現のさらなる記述は以下に与えられる。
【0095】
上述の通り、アルファ鎖のヒドロキシル化は、安定なI型コラーゲンの会合のために要求される。本発明の遺伝的に改変された植物によって発現されるアルファ鎖は、内因性P4H活性を欠く、細胞内区画中に蓄積するので、かかる鎖は、植物、植物組織又は細胞から単離されなければならず、インビトロでヒドロキシル化されなければならない。かかるヒドロキシル化は、Turpeenniemi−Hujanen及びMyllylaによって記述される方法(Concomitant hydroxylation of proline and lysine residues in collagen using purified enzymes in vitro.Biochim Biophys Acta.1984 Jul 16:800(1)59−65)によって達成されることができる。
【0096】
かかるインビトロヒドロキシル化は、正確にヒドロキシル化されたコラーゲン鎖に導くことができるが、達成するのが困難である場合があり費用がかかる場合がある。
【0097】
インビトロヒドロキシル化の制限を克服するため、本発明の遺伝的に改変された植物は、コラーゲンアルファ鎖を正確にヒドロキシル化する(即ち、Gly−X−Yトリプレットのプロリン(Y)位置のみをヒドロキシル化する)ことができるP4Hを共発現することが好ましい。P4Hは、二つのサブユニットアルファ及びベータからなる酵素である。これら両方が活性酵素を形成するのに必要であるが、ベータサブユニットはシャペロン機能をも担持する。
【0098】
本発明の遺伝的に改変された植物によって発現されるP4Hは、例えば配列番号12及び14によってコードされるヒトP4Hであることが好ましい。加えて、増大された基質特異性を示すP4H突然変異体又はP4H相同体も使用されることができる。
【0099】
好適なP4H相同体は、NCBIアクセッションNP 179363によって同定されるアラビドプシスのオキシドレダクターゼによって例示される。本発明者らによって行われたこのタンパク質の配列とヒトP4Hアルファサブユニットの対様式アラインメントは、植物のいかなる公知のP4H相同体の機能性ドメイン間の最高の相同性を明らかにした。
【0100】
P4Hは、発現されたコラーゲン鎖と共蓄積される必要があるので、そのコード配列は、それに応じて改変されることが好ましい(シグナル配列の付加、ER標的化を防止することができる欠失など)。
【0101】
哺乳類細胞では、コラーゲンはリシルヒドロキシラーゼ、ガラクトシルトランスフェラーゼ及びグルコシルトランスフェラーゼによっても修飾される。これらの酵素は、特定位置のリシル残基をヒドロキシリシル、ガラクトシルヒドロキシリシル及びグルコシルガラクトシルヒドロキシリシル残基へと連続的に修飾する。単一のヒト酵素であるリシルヒドロキシラーゼ3(LH3)は、ヒドロキシリシンに関連する炭水化物形成における三つの連続的ステップすべてを触媒することができる。
【0102】
従って、本発明の遺伝的に改変された植物は、哺乳類のLH3も発現することが好ましい。配列番号22によって規定されるようなLH3コード配列が、かかる目的のために使用されることができる。
【0103】
上述のコラーゲン鎖及び修飾酵素は、植物の機能的プロモーターの転写制御下に配置されたアルファ鎖及び/又は修飾酵素(例えばP4H及びLH3)をコードするポリヌクレオチド配列を含む、安定に組込まれた又は一過的に発現される核酸構築物から発現されることができる。かかる核酸構築物(本明細書では発現構築物とも称される)は、植物全体での発現のために、又は限定された植物組織もしくは限定された植物細胞での発現のために、又は植物の限定された発生段階での発現のために構成されることができる。かかる構築物は、選択マーカー(例えば抗生物質耐性)、エンハンサーエレメント及び細菌での複製のための複製起点も含むことができる。
【0104】
二つの発現性挿入物(例えば二つのアルファ鎖型又はアルファ鎖及びP4H)を含む構築物は、それぞれの挿入物について個別のプロモーターを含むことが好ましいことは理解されるであろう。又は代わりにかかる構築物は、単一のプロモーターから両方の挿入物配列を含む単一の転写キメラを発現することができる。かかる場合、キメラ転写物は、下流の挿入物が翻訳されることができるように二つの挿入物の間にIRES配列を含む。
【0105】
組織特異的、発生段階特異的、構成的、又は誘導性であることができる多数の植物機能的発現プロモーター及びエンハンサーが本発明の構築物によって利用されることができ、いくつかの例が以下に与えられる。
【0106】
本明細書及び特許請求の範囲で使用される通り、句「植物プロモーター」又は「プロモーター」は、植物細胞(DNA含有オルガネラを含む)中での遺伝子発現を指図することができるプロモーターを含む。かかるプロモーターは、植物、細菌、ウイルス、真菌又は動物起源のものであることができる。かかるプロモーターは、構成的、即ち複数の植物組織中での高レベルの遺伝子発現を指図することができるか、又は組織特異的、即ち特定の植物組織での遺伝子発現を指図することができるか、又は誘導性、即ち刺激の下で遺伝子発現を指図することができるか、又はキメラ、即ち少なくとも二つの異なるプロモーターの部分から形成されるものであることができる。
【0107】
従って、使用される植物プロモーターは、構成的プロモーター、組織特異的プロモーター、誘導性プロモーター、又はキメラプロモーターであることができる。
【0108】
構成的植物プロモーターの例は、CaMV35S及びCaMV19Sプロモーター、FMV34Sプロモーター、サトウキビ桿状バドナウイルスプロモーター、CsVMVプロモーター、アラビドプシスACT2/ACT8アクチンプロモーター、アラビドプシスユビキチンUBQ1プロモーター、大麦葉チオニンBTH6プロモーター、及びイネアクチンプロモーターを含むがこれらに限定されない。
【0109】
組織特異的プロモーターの例は、マメファゼオリン貯蔵タンパク質プロモーター、DLECプロモーター、PHSプロモーター、ゼイン貯蔵タンパク質プロモーター、ダイズからのコングルチンガンマプロモーター、AT2S1遺伝子プロモーター、アラビドプシスからのACT11アクチンプロモーター、ブラシカ ナプスからのnapAプロモーター、及びジャガイモパタチン遺伝子プロモーターを含むがこれらに限定されない。
【0110】
誘導性プロモーターは、例えば光、温度、化学物質、渇水、高塩分、浸透圧ショック、酸化的条件又は病原体を含むストレス条件の如き特定の刺激によって誘導されるプロモーターであり、エンドウマメrbcS遺伝子由来の光誘導性プロモーター、アルファルファrbcS遺伝子由来のプロモーター、渇水時に活性となるプロモーターDRE,MYC及びMYB、高塩分及び浸透圧ストレス時に活性となるプロモーターINT,INPS,prxEa,Ha hsp17.7G4及びRD21、及び病原体ストレス時に活性となるプロモーターhsr203J及びstr246Cを含むがこれらに限定されない。
【0111】
好ましくは、本発明によって利用されるプロモーターは、挿入された構築物の過剰発現が植物の形質転換後に行われるような強い構成的プロモーターである。
【0112】
本発明で使用されるいかなる構築物型も、各構築物型において同一又は異なる選択マーカーを使用して同一植物中へと共形質転換されることができることは理解されるであろう。代わりに、第一の構築物型が第一植物中に導入され、第二の構築物型が第二同系植物中に導入され、その後、生じたトランスジェニック植物が交配され、子孫が二重形質転換体について選択されることができる。かかる子孫のさらなる自家交配は、両方の構築物について同型接合体である遺伝系統を作成するために使用されることができる。
【0113】
単子葉植物及び双子葉植物の両方に核酸構築物を導入する様々な方法がある(Potrykus,L.,Annu.Rev.Plant.Physiol.,Plant,Mol.Biol.(1991)42:205−225;Shimamoto et al.,Nature(1989)338:274−276)。かかる方法は、核酸構築物又はその一部の植物ゲノム中への安定な組込み又は核酸構築物の一過性発現に依存しており、後者の場合、これらの配列は植物の子孫には遺伝しない。
【0114】
加えて、核酸構築物が、葉緑体の如きDNA含有オルガネラのDNA中に直接導入されることができるいくつかの方法がある。
【0115】
本発明の核酸構築物の中に含まれるような外因性配列を植物ゲノム中に安定に組込むためには二つの原理的な方法がある:
【0116】
(i)アグロバクテリウムによって媒介される遺伝子移入:Klee et al.(1987)Annu.Rev.Plant Physiol.38:467−486;Klee and Rogers in Cell Culture and Somatic Cell Genetics of Plants,Vol.6,Molecular Biology of Plant Nuclear Genes,eds.Schell,J.,and Vasil,L.K.,Academic Publishers,San Diego,Calif.(1989)p.2−25;Gatenby,in Plant Biotechnology,eds.Kung,S.and Arntzen,C.J.,Butterworth Publishers,Boston,Mass.(1989)p.93−112。
【0117】
(ii)直接DNA取込み:Paszkowski et al.,in Cell Culture and Somatic Cell Genetics of Plants,Vol.6,Molecular Biology of Plant Nuclear Genes eds.Schell,J.,and Vasil,L.K.,Academic Publishers,San Diego,Calif.(1989)p.52−68;この方法は、プロトプラストへDNAを直接取込むための方法を含む:Toriyama,K.et al.(1988)Bio/Technology 6:1072−1074。植物細胞の短い電気ショックによって誘導されるDNA取込み:Zhang et al.Plant Cell Rep.(1988)7:379−384.Fromm et al.Nature(1986)319:791−793。粒子衝撃による植物細胞又は組織中へのDNA注入:Klein et al.Bio/Technology(1988)6:559−563;McCabe et al.Bio/Technology(1988)6:923−926;Sanford,Physiol.Plant.(1990)79:206−209。マイクロピペット系の使用による植物細胞又は組織中へのDNA注入:Neuhaus et al.,Theor.Appl.Genet.(1987)75:30−36;Neuhaus and Spangenberg,Physiol.Plant.(1990)79:213−217。又は発芽花粉とのDNAの直接培養による植物細胞又は組織中へのDNA注入:DeWet et al.in Experimental Manipulation of Ovule Tissue,eds.Chapman,G.P.and Mantell,S.H.and Daniels,W.Longman,London,(1985)p.197−209;and Ohta,Proc.Natl.Acad.Sci.USA(1986)83:715−719。
【0118】
アグロバクテリウム系は、植物ゲノムDNA中に組込まれる限定されたDNAセグメントを含むプラスミドベクターの使用を含む。植物組織の接種方法は、植物種及びアグロバクテリウム送達系によって異なる。幅広く使用されているアプローチは、リーフディスク手順であり、これは植物全体への分化を開始するのに良好な源を提供するいかなる組織外植体を用いて行われることができる。Horsch et al.in Plant Molecular Biology Manual A5,Kluwer Academic Publishers,Dordrecht(1988)p.1−9参照。補助的なアプローチは、アグロバクテリウム送達系を減圧浸潤との組合せで使用する。アグロバクテリウム系は、トランスジェニック双子葉植物の作成において特に実行可能である。
【0119】
植物細胞中への直接DNA移入には様々な方法がある。エレクトロポレーションでは、プロトプラストは強い電場に短時間さらされる。マイクロ注入では、DNAは極めて小さいマイクロピペットを使用して細胞中へ直接機械的に注入される。微小粒子衝撃では、DNAは硫酸マグネシウムの結晶、タングステン粒子又は金粒子の如き微小投射物に吸着され、この微小投射物が細胞又は植物組織中へと物理的に加速される。
【0120】
形質転換後、植物の繁殖が行われる。植物繁殖の最も一般的な方法は種子による方法である。しかし、種子繁殖による再生は、異型接合性のため作物には均一性の欠失があるという欠点を有する。何故なら種子は、メンデルの法則によって支配される遺伝的変異に従って植物によって生産されるからである。基本的に、各種子は遺伝的に異なり、各種子はそれ自身の特異的な形質を有して成長するであろう。従って、形質転換植物は、再生された植物が親のトランスジェニック植物と同一の形質及び特徴を有するように生産されることが好ましい。それ故、形質転換植物は、形質転換植物の迅速で一貫した再生を与える微小繁殖によって再生されることが好ましい。
【0121】
本発明の核酸構築物内に含まれる単離された核酸を一過的に発現させるために利用されることができる一過的発現方法は、上述のマイクロ注入及び衝撃(しかし一過的発現が好ましい条件下で)、及びウイルスによって媒介される発現を含むがこれらに限定されない。ウイルスによって媒介される発現では、核酸構築物を含むパッケージ化された又はパッケージ化されていない組換えウイルスベクターが植物組織又は細胞を感染させるために利用され、そこに確立された繁殖する組換えウイルスが非ウイルス性核酸配列を発現する。
【0122】
植物宿主の形質転換のために有用であることが示されているウイルスはCaMV,TMV及びBVを含む。植物ウイルスを使用した植物の形質転換は、米国特許第4885237号(BGV),EP−A67553(TMV)、日本国公開特許公報第63−14693号(TMV),EPA第194809号(BV),EPA第278667号(BV)及びGluzman,Y.et al.,Communications in Molecular Biology:Viral Vectors,Cold Spring Harbor Laboratory,New York,pp.172−189(1988)に記述されている。植物を含む多くの宿主中で外来DNAを発現させるのに使用するためのシュードウイルス粒子は、WO 87/06261に記述されている。
【0123】
植物中に非ウイスル性外因性核酸配列を導入して発現させるための植物RNAウイルスの構築は、上述の文献並びにDawson,W.O.er al.,Virology(1989)172:285−292;Takamatsu et al.EMBO J.(1987)6:307−311;French et al.Science(1986)231:1294−1297;and Takamatsu et al.FEBS Letters(1990)269:73−76によって示されている。
【0124】
ウイルスがDNAウイルスである場合、構築はウイルス自体に対してなされることができる。代わりに、ウイルスは、外来DNAを有する望ましいウイルスベクターを構築するのが容易なように細菌プラスミド中へとまずクローニングされることができる。次にウイルスはプラスミドから切り出されることができる。もしウイルスがDNAウイルスであるなら、細菌の複製起点がウイルスDNAに取付けられることができ、ウイルスDNAは細菌によって複製されることができる。このDNAの転写及び翻訳は、ウイルスDNAを包囲するコートタンパク質を生産するであろう。もしウイルスがRNAウイルスであるなら、ウイルスは一般的にcDNAとしてクローニングされ、プラスミド中に挿入される。次にプラスミドは構築物のすべてを作るために使用される。次にRNAウイルスは、プラスミドのウイルス配列を転写することよって及びウイルス遺伝子を翻訳してウイルスRNAを包囲するコートタンパク質を生産することによって生産される。
【0125】
本発明の構築物中に含まれるような非ウイルス性外因性核酸配列の植物中への導入及び発現のための植物RNAウイルスの構築は、上述の文献並びに米国特許第5316931号によって示されている。
【0126】
一つの実施態様では、生来的コートタンパク質コード配列がウイルス核酸から欠失されており、かつ植物宿主中での発現、組換え植物ウイルス核酸のパッケージング及び組換え植物ウイルス核酸による宿主の全身的感染を確実にすることができる非生来的プロモーター、好ましくは非生来的コートタンパク質コード配列のサブゲノムプロモーターが挿入されている植物ウイルス核酸が提供される。代わりに、コートタンパク質遺伝子は、タンパク質が生産されるようにその中に非生来的核酸を挿入することによって不活性化されることができる。組換え植物ウイルス核酸は一以上の追加の非生来的サブゲノムプロモーターを含むことができる。それぞれの非生来的サブゲノムプロモーターは、植物宿主中の隣接遺伝子又は核酸配列を転写又は発現させることができるが、相互に及び生来的サブゲノムプロモーターと組換えられることができない。非生来的(外来性)核酸配列は、一以上の核酸配列が含められる場合、生来的な植物ウイルスサブゲノムプロモーター又は生来的な及び非生来的な植物ウイルスサブゲノムプロモーターに隣接して挿入されることができる。非生来的核酸配列はサブゲノムプロモーターの制御下で宿主植物中で転写又は発現され、望ましい生成物を生産する。
【0127】
第二の実施態様では、生来的なコートタンパク質コード配列が非生来的なコートタンパク質コード配列の代わりに非生来的なコートタンパク質サブゲノムプロモーターの一つに隣接して配置されていることを除いては第一の実施態様と同様の組換え植物ウイルス核酸が提供される。
【0128】
第三の実施態様では、生来的なコートタンパク質遺伝子がそのサブゲノムプロモーターに隣接されておりかつ一以上の非生来的サブゲノムプロモーターがウイルス核酸中に挿入されている組換え植物ウイルス核酸が提供される。挿入された非生来サブゲノムプロモーターは、隣接遺伝子を転写又は発現させることができるが、相互に及び生来的サブゲノムプロモーターと組換えられることができない。非生来的な核酸配列は、前記配列がサブゲノムプロモーターの制御下で宿主植物中で転写又は発現され、望ましい生成物を生産するように非生来的なサブゲノム植物ウイルスプロモーターに隣接して挿入されることができる。
【0129】
第四の実施態様では、生来的なコートタンパク質コード配列が非生来的なコートタンパク質コード配列によって置換されていることを除いては第三の実施態様と同様の組換え植物ウイルス核酸が提供される。
【0130】
ウイルスベクターは、組換え植物ウイルス核酸によってコードされるコートタンパク質によって包囲されて組換え植物ウイルスを生産する。組換え植物ウイルス核酸又は組換え植物ウイルスは、適切な宿主植物を感染させるために使用される。組換え植物ウイルス核酸は、宿主中で複製し、宿主中に全身的に広がり、宿主中で外来遺伝子(単離された核酸)を転写又は発現させて望ましいタンパク質を生産することができる。
【0131】
外因性核酸配列を葉緑体のゲノムに導入するための技術が知られている。この技術は下記の手順を伴う。第1に、植物細胞が、細胞あたりの葉緑体の数を約1個に減らすように化学的に処理される。その後、外因性核酸が、少なくとも1つの外因性核酸分子を葉緑体に導入することを目指して、粒子衝撃によって細胞に導入される。外因性核酸は、相同的組換え(これは葉緑体に固有の酵素によって容易に行われる)による葉緑体ゲノムへの組込みを可能にするように選択される。この目的のために、外因性核酸は、目的とする遺伝子に加えて、葉緑体ゲノムに由来する少なくとも1つの核酸配列を含む。加えて、外因性核酸は選択マーカーを含み、そのような選択マーカーは、逐次的な選抜手法によって、そのような選抜の後の葉緑体ゲノムのすべてまたは実質的にすべてのコピー体が外因性核酸を含むことを確認するために役立つ。この技術に関するさらなる詳細が、米国特許第4945050号および同第5693507号に見出される(これらは参考として本明細書中に組み込まれる)。従って、ポリペプチドを葉緑体のタンパク質発現系によって産生させることができ、かつ、葉緑体の内膜に一体化させることができる。
【0132】
上述の形質転換アプローチは、いかなる種の植物又はそこから由来する植物組織もしくは単離された植物細胞においてコラーゲン鎖及び/又は修飾酵素並びに会合されたコラーゲン(プロペプチドあり又はなし)を生産するために使用されることができる。
【0133】
好ましい植物は、本明細書に記述されたコラーゲン鎖、コラーゲン及び/又は処理酵素を多量に蓄積することができるものである。かかる植物は、ストレス条件に対するそれらの抵抗性及び発現された成分又は会合されたコラーゲンが抽出される容易さに従って選択されることもできる。好ましい植物の例は、タバコ、トウモロコシ、アルファルファ、イネ、ジャガイモ、ダイズ、トマト、小麦、大麦、カノーラ、ニンジン及び綿を含む。
【0134】
コラーゲン線維は、食品及び化粧品産業で広範囲に使用される。従って、植物によって発現されるコラーゲン線維成分(アルファ鎖)及び修飾酵素はコラーゲンの産業的合成において有用性を見出し、植物中での完全なコラーゲンの生産は、その簡単さ及び費用効率のために好ましい。
【0135】
植物中でI型コラーゲンを生成するためにいくつかのアプローチが使用されることができる。例えば、コラーゲンアルファ1鎖は、コラーゲンアルファ1及びP4H(及び所望によりLH3)を発現する植物から単離されることができ、コラーゲンアルファ2及びP4H(及び所望によりLH3及びプロテアーゼC及び/又はN)を発現する植物から単離されるコラーゲンアルファ2鎖と混合されることができる。コラーゲンアルファ1鎖はそれ自身によって三重らせんへと自己会合するため、混合前にかかるホモトリマーを変性させてコラーゲンアルファ2鎖と共に再生することが必要であるかもしれない。
【0136】
好ましくは、コラーゲンアルファ1及びP4H(及び所望によりLH3及びプロテアーゼC及び/又はN)を発現する第一植物は、コラーゲンアルファ2を発現する第二(及び好ましくは同系の)植物と交配されることができるか、又は代わりに両アルファ鎖を発現する第一植物は、P4H及び所望によりLH3及びプロテアーゼC及び/又はNを発現する第二植物と交配されることができる。
【0137】
上述の植物育種アプローチは二つの個別に形質転換された植物を利用するが、それぞれが一つ又は二つの成分を発現する三つ以上の個別に形質転換された植物を利用するアプローチも利用できるということは注意されるべきである。
【0138】
当業者は様々な植物育種技術に精通しているであろう。従って、かかる技術のさらなる記述は本明細書中では与えられない。
【0139】
植物育種アプローチが好ましいが、コラーゲンアルファ1及び2、P4H及びLH3(及び所望によりプロテアーゼC及び/又はN)を発現する単一の植物が、それぞれが一以上の発現可能な成分を細胞中に導入するために設計されているいくつかの形質転換事象を介して作成されることができることに注意すべきである。かかる場合には、各形質転換事象の安定性は特異的な選択マーカーを使用して確認されることができる。
【0140】
いかなる場合でも、形質転換及び植物育種アプローチはいかなる数の成分を発現するいかなる植物を作成するために使用されることができる。現在好まれているものはコラーゲンアルファ1及び2鎖、P4H,LH3及び少なくとも一つのプロテアーゼ(例えばプロテアーゼC及び/又はN)を発現する植物である。以下の実施例の節でさらに記述されるように、かかる植物は42℃までの温度で安定性を示すコラーゲンを蓄積する。
【0141】
育種から生じた子孫又は代わりに多数回形質転換植物は、核酸又はタンパク質プローブ(例えば抗体)を使用して外因性mRNA及び/又はポリペプチドの存在を確認することによって選択されることができる。後者のアプローチが好ましい。何故なら、それは発現されたポリペプチド成分の局在化を可能にし(例えば分別された植物抽出物をプロービングすることによって)、従って、正確な処理及び会合についての可能性も確認するからである。好適なプローブの例は以下の実施例の節に与えられる。
【0142】
一旦コラーゲンを発現する子孫が同定されたら、かかる植物はコラーゲン鎖ならびに修飾酵素の発現を最大にする条件下でさらに栽培される。
【0143】
遊離プロリンの蓄積は、本発明の遺伝的に改変された植物によって発現されるコラーゲン鎖を含む様々なプロリンリッチタンパク質の過剰生産を容易にすることができ、好ましい栽培条件は栽培された植物中での遊離プロリンの蓄積を増大させるようなものである。
【0144】
遊離プロリンの蓄積は、渇水、塩分、低温、高温、病原体感染、重金属毒性、嫌気生活、栄養欠乏、大気汚染及びUV照射を含む広範囲の環境ストレスに対する応答において様々な植物中に蓄積する(Hare and Cress,1997年)。
【0145】
遊離プロリンは、ABAの如き化合物での植物又は土壌の処理に対する応答において又は銅、塩、パラコート、サリチル酸などの如き化合物を含むストレスに対する応答においても蓄積することがあり得る。
【0146】
従って、コラーゲンを発現する子孫は様々なストレス条件(例えば50mMから250mMまでの範囲のNaClの異なる濃度)下で生長させられることができる。コラーゲン生産をさらに増大させるために、コラーゲン発現に対する様々なストレス条件の影響が調査され、植物の生存性、バイオマス及びコラーゲン蓄積に関して最適化されるであろう。
【0147】
植物組織/細胞は成熟時に収穫されることが好ましく、コラーゲン線維は周知の従来技術の抽出アプローチを使用して単離され、一つのかかるアプローチが以下に詳述される。
【0148】
形質転換植物の葉は液体窒素下で粉末へと磨砕され、ホモジネートは0.2MのNaClを含む0.5M酢酸中で60時間4℃で抽出される。不溶性材料は遠心分離によって除去される。組換えコラーゲンを含む上澄みは0.4M及び0.7MのNaClで塩分画される。組換えヘテロトリマーコラーゲンを含む0.7M NaCl沈殿は0.1Mの酢酸中に溶解されてこれに対して透析され、−20℃で貯蔵される(Ruggieroら、2000年に従う)。
【0149】
本発明のさらなる目的、利点および新規な特徴が、限定であることが意図されない下記の実施例を検討したとき、当業者には明らかになる。加えて、本明細書中上記に描かれるような、また、下記の請求項の節において特許請求されるような本発明の様々な実施形態および態様のそれぞれは、実験的裏付けが下記の実施例において見出される。
【実施例】
【0150】
次に下記の実施例が参照されるが、下記の実施例は、上記の説明と一緒に、本発明を非限定様式で例示する。
【0151】
本願で使用される用語と、本発明で利用される実験方法には、分子生化学、微生物学及び組み換えDNAの技法が広く含まれている。これらの技法は文献に詳細に説明されている[例えば以下の諸文献を参照されたい。「Molecular Cloning:A laboratory Manual」Sambrookら1989年;Ausubel,R.M.編1994年「Current Protocols in Molecular Biology」I〜III巻;Ausubelら著1989年「Current Protocols in Molecular Biology」John Wiley and Sons,米国メリーランド州バルチモア;Perbal著「A Practical Guide to Molecular Cloning」John Wiley & Sons,米国ニューヨーク1988年;Watsonら、「Recombinant DNA」Scientific American Books、米国ニューヨーク;Birrenら編「Genome Analysis:A Laboratory Manual Series」1〜4巻、Cold Spring Harbor Laboratory Press、米国ニューヨーク1998年;米国特許の4666828号、4683202号、4801531号、5192659号及び5272057号に記載される方法;Cellis,J.E.編「Cell Biology:A Laboratory Handbook」I〜III巻1994年;Freshney,Wiley−Liss著、「Culture of Animal Cells−A Manual of Basic Technique」(第3版)N.Y.(1994年);Coligan,J.E.編「Current Protocols in Immunology」I〜III巻1994年;Stitesら編「Basic and Clinical Immunology」(第8版)、Appleton & Lange、米国コネティカット州ノーウォーク1994年;MishellとShiigi編「Selected Methods in Cellular Immunology」、W.H.Freeman and Co.、米国ニューヨーク1980年;また利用可能な免疫検定法は、例えば以下の特許と科学文献に広範囲にわたって記載されている。米国特許の3791932号、3839153号、3850752号、3850578号、3853987号、3867517号、3879262号、3901654号、3935074号、3984533号、3996345号、4034074号、4098876号、4879219号、5011771号及び5281521号;Gait,M.J.編「Oligonucleotide Synthesis」1984年;Hames,B.D.及びHiggins S.J.編「Nucleic Acid Hybridization」1985年;Hames,B.D.及びHiggins S.J.編「Transcription and Translation」1984年;Freshney,R.I.編「Animal Cell Culture」1986年;「Immobilized Cells and Enzymes」IRL Press 1986年;Perbal,B.著「A Practical Guide to Molecular Cloning」1984年及び「Methods in Enzymology」1〜317巻、Academic Press;「PCR Protocols:A Guide To Methods And Applications」、Academic Press、米国カリフォルニア州サンディエゴ1990年;Marshakら、「Strategies for Protein Purification and Characterization−A Laboratory Course Manual」、CSHL Press、1996年;なおこれらの文献類は、あたかも本願に完全に記載されているように援用するものである]。その外の一般的な文献は、本明細書を通じて提供される。本明細書に記載の方法は当業技術界で周知であると考えられ、読者の便宜のために提供される。本明細書に含まれるすべての情報は本願に援用するものである。
【0152】
実施例1
構築及び形質転換スキーム
本研究で使用された発現カセットの構築は図1a−dに示されている。本研究におけるコード配列のすべては、タバコ中での発現のために最適化され、望ましいフランキング領域(配列番号1,4,7,12,14,16,18,20,22)を有するように化学的に合成された。図1a:液胞シグナルもしくはアポプラストシグナル(配列番号7によってコードされる)に融合された又はシグナルなしのCol1及びCol2(配列番号1,4)をコードする合成遺伝子は、キクrbcS1プロモーター及び5′UTR(配列番号10)並びにキクrbcS1 3′UTR及びターミネーター(配列番号11)からなる発現カセット中にクローニングされた。完全な発現カセットはpBINPLUS植物形質転換ベクターのマルチクローニングサイト中にクローニングされた(van Engelenら、1995年、Transgenic Res 4:288−290)。図1b:液胞シグナルもしくはアポプラストシグナル(配列番号7によってコードされる)に融合された又はシグナルなしのP4Hベータヒト、P4Hアルファヒト及びP4H植物(配列番号12,14及び16)をコードする合成遺伝子は、ベクターpJD330によって担持されるCaMV 35Sプロモーター及びTMVオメガ配列及びアグロバクテリウムノパリン合成酵素(NOS)ターミネーターからなる発現カセット中にクローニングされた。完全な発現カセットは、Col1又はCol2の発現ベクターを担持するpBINPLUSベクターのマルチクローニングサイト中にクローニングされた。図1c:液胞シグナルもしくはアポプラストシグナル(配列番号7によってコードされる)に融合されたプロテイナーゼC及びプロテイナーゼN(配列番号18,20)をコードする合成遺伝子は、キクrbcS1プロモーター及び5′UTR(配列番号10)並びにキクrbcS1 3′UTR及びターミネーター(配列番号11)からなる発現カセット中にクローニングされた。完全な発現カセットはpBINPLUS植物形質転換ベクターのマルチクローニングサイト中にクローニングされた。図1d:液胞シグナルもしくはアポプラストシグナル(配列番号7によってコードされる)に融合された又はシグナルなしのLH3(配列番号22)をコードする合成遺伝子(フランキングイチゴツル結合ウイルス(SVBV)プロモーター(NCBIアクセッションAF331666 REGION:623..950バージョンAF331666.1 GI:13345788)を有しかつアグロバクテリウムオクトピン合成酵素(OCS)ターミネーター(NCBIアクセッションZ37515 REGION:1344..1538バージョンZ37515.1 GI:886843)で末端停止されている)は、Col1及びP4Hベータの発現カセットを担持するpBINPLUSベクターのマルチクローニングサイト中にクローニングされた。
【0153】
図1に記述される発現カセットを利用した宿主植物中への共形質転換スキームは図2に図示される。各発現カセット挿入物はコード配列の短い名前によって表される。コード配列及び関連する配列番号は表1に記載されている。それぞれの共形質転換は二つのpBINPLUSバイナリーベクターによって行われる。それぞれの長方形は一つ、二つ又は三つの発現カセットを担持する単一のpBINPLUSベクターを表す。プロモーター及びターミネーターは図1で特定される。
【0154】
実施例2
植物コラーゲン発現
以下の表1に提示されるタンパク質をコードする合成ポリヌクレオチド配列が設計され、タバコ植物中での発現のために最適化された。
【0155】
【0156】
シグナルペプチド
(i)チオールプロテアーゼアレウレイン(aleurain)前駆体についての大麦遺伝子の液胞シグナル配列(NCBIアクセッションP05167 GI:113603)
(ii)アラビドプシスタリアーナのエンド1,4−ベータ−グルカナーゼのアポプラストシグナル(Cel1、NCBIアクセッションCAA67156.1 GI:2440033);配列番号9、配列番号7によってコードされる
【0157】
プラスミドの構築
植物発現ベクターは実施例1に教示されるようにして構築された。それぞれの構築された発現ベクターの組成は制限酵素分析及び配列決定を介して確認された。
【0158】
以下の発現カセットを含む発現ベクターが構築された:
1.コラーゲンアルファ1
2.コラーゲンアルファ1+ヒトP4Hベータサブユニット
3.コラーゲンアルファ1+ヒトP4Hベータサブユニット+ヒトLH3
4.コラーゲンアルファ2
5.コラーゲンアルファ2+ヒトP4Hアルファサブユニット
6.コラーゲンアルファ2+アラビドプシスP4H
7.ヒトP4Hベータサブユニット+ヒトLH3
8.ヒトP4Hアルファサブユニット
上述のコード配列のそれぞれは、液胞輸送ペプチドもしくはアポプラスト輸送ペプチドに翻訳的に融合されたか、又はいかなる輸送ペプチド配列も欠いており、その場合、細胞質蓄積が期待される。
【0159】
植物形質転換及びPCRスクリーニング
タバコ植物(ニコチアーナ・タバクム、Samsun NN)は、図2に教示される形質転換スキームに従って上述の発現ベクターで形質転換された。
【0160】
生じた形質転換植物は、コラーゲンアルファ1の324bpフラグメント及びコラーゲンアルファ2の537bpフラグメントを増幅することができるように設計された四つのプライマー(表2)を使用したマルチプレックスPCRを介してスクリーニングされた。図3は一つのマルチプレックスPCRスクリーニングの結果を示す。
【0161】
【0162】
実施例3
トランスジェニックタバコ植物中でのヒトコラーゲンの検出
全可溶性タンパク質は、「完全な」プロテアーゼ阻害剤カクテル(Roche Diagnostics GmbHからの製造#1836145、50mlの緩衝液あたり一錠)を使用して0.5mlの50mM Tris−HCl pH7.5中で500mgの葉を粉砕することによってタバコ形質転換体2,3及び4から抽出された。粗抽出物は、10%ベータ−メルカプト−エタノール及び8%SDSを含む250μlの4Xサンプル適用緩衝液と混合され、サンプルは7分間沸騰され、13000rpmで8分間遠心分離された。20μlの上澄みは10%ポリアクリルアミドゲル中に添加され、抗コラーゲンI(変性)抗体(Chemicon Inc.からの#AB745)を使用して標準的なウェスタンブロット手順でテストされた(図4)。W.T.は野生型タバコである。陽性コラーゲンバンドは、コラーゲンI型アルファ1又はアルファ2又は両方についてPCR陽性である植物において見ることができる。ヒト胎盤(Chemicon Inc.からの#CC050)からの500ngのコラーゲンI型の陽性コントロールバンドは、トランスジェニック植物からのサンプル中の約0.3%の全可溶性タンパク質(約150μg)を表す。
【0163】
全可溶性タンパク質の1%までの予測される分子量でコラーゲンを発現する植物は、コラーゲンが液胞に対して標準化された場合に検出された(図4)。アポプラストに対する全長コラーゲンの細胞内標的化は成功裏に達成された(図5)。細胞質中でコラーゲンを発現する植物(即ち標的化ペプチドなし)は、コラーゲンを検出可能なレベルに蓄積せず、植物中でのコラーゲンの細胞内標的化は成功のために重要であることを示した。
【0164】
N−プロペプチドを欠くコラーゲンが有意なタンパク質分解に供されることを示すRuggieroら2000年及びMerleら2002年の研究に加えて及びこれらの研究とは対照的に、現在のアプローチを使用すると、C−プロペプチド及びN−プロペプチドを有する全長コラーゲンタンパク質は高レベルで細胞内区画中に蓄積した。
【0165】
本データはまた、それぞれが異なるコラーゲン鎖型を発現する二つの植物を交配することは、それぞれの鎖型を最適レベルで発現する植物の選択及び望ましいコラーゲン生産植物を達成するための続く植物交配を可能にする点で有利であるということを明らかに示す。
【0166】
本発明の植物によって生産されるコラーゲンは、生来的なプロペプチドを含み、従って、タンパク質分解によって精製されたヒトのコントロールタンパク質より大きなタンパク質を形成すると予測される。ヒドロキシル化又はグリコシル化を有さないコラーゲンアルファ1及びアルファ2鎖の計算される分子量は以下の通りである:プロペプチドを有するCol1−136kDa、プロペプチドを有さないCol1−95kDa、プロペプチドを有するCol2−127kDa、プロペプチドを有さないCol2−92kDa。
【0167】
図4からわかるように、形質転換体3−5及び3−49中のCol1バンドは他の植物中のCol1バンドより大きく見える。これは、これらの植物中で共発現されてヒトコラーゲン鎖と同じ細胞内区画(例えば液胞)に対して標的化されたアルファ及びベータサブユニットからなるヒトプロリン−4−ヒドロキシラーゼホロ酵素によってコラーゲン鎖中でプロリンがヒドロキシル化されることを示す。
【0168】
実施例4
コラーゲン三重らせん会合及びトランスジェニック植物中での熱安定性
コラーゲン三重らせん会合及びトランスジェニック植物中でのらせん熱安定性は、トランスジェニック植物の全粗タンパク質抽出物の熱変性及び続くトリプシン又はペプシン消化によってテストされた(図6a〜b)。第一の実験では、タバコ2−9(Colアルファ1のみを発現し、P4Hを発現しない)及びタバコ3−5(Colアルファ+2及びP4Hの両方を発現する)からの全可溶性タンパク質は、0.5mlの50mM Tris−HCl pH=7.5中で500mgの葉を粉砕して13000rpmで10分間遠心分離し、上澄みを収集することによって抽出された。50μlの上澄みは熱処理(33℃又は43℃で15分間)に供され、次に直ちに氷上に置かれた。トリプシン消化は、各サンプルに50mM Tris−HCl pH=7.5中の1mg/mlトリプシン6μlを添加することによって開始された。サンプルは室温(約22℃)で20分間インキュベートされた。消化は、10%ベータメルカプトエタノール及び8%SDSを含む20μlの4Xサンプル適用緩衝液の添加によって停止され、サンプルは7分間沸騰され、13000rpmで7分間遠心分離された。50μlの上澄みは10%ポリアクリルアミドゲル中に添加され、抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を使用して標準的なウェスタンブロット手順でテストされた。陽性コントロールは、w.t.タバコから抽出された50μlの全可溶性タンパク質に添加された約500ngのヒトコラーゲンI(Chemicon Inc.からの#CC050であり、ペプシン消化によってヒト胎盤から抽出されたもの)のサンプルであった。
【0169】
図6aに示されるように、植物#3−5中で形成されたコラーゲン三重らせん並びにコントロールのヒトコラーゲンは33℃での変性に対して抵抗性であった。対照的に、植物#2−9によって形成されたコラーゲンは33℃で変性した。熱安定性におけるこの相違は、コラーゲンアルファ1及びコラーゲンアルファ2並びにP4Hベータ及びアルファサブユニットの両方を発現する形質転換体#3−5中での成功した三重らせん会合及び翻訳後のプロリンヒドロキシル化を示唆する。
【0170】
形質転換体#2−9における二つのバンドはダイマー又はトリマーを表すかもしれず、これらはSDS及びメルカプトエタノールを使用した7分間の沸騰後も安定である。同様のバンドがヒトのコラーゲン(上部パネル)において及び形質転換体#3−5において見ることができる。可能な説明は、異なる三重らせん中の二つのペプチドの間の等価なバンド(クロスリンク)がリシンオキシダーゼによる二つのリシンの酸化的脱アミノ化後に形成されたということである。第二の実験では、トランスジェニックタバコ13−6(コラーゲンIアルファ1及びアルファ2鎖(矢印によって示される)、ヒトP4Hアルファ及びベータサブユニット及びヒトLH3を発現する)からの全可溶性タンパク質は、0.5mlの100mM Tris−HCl pH=7.5及び300mM NaCl中で500mgの葉を粉砕して10000rpmで7分間遠心分離し、上澄みを収集することによって抽出された。50μlの上澄みは熱処理(33℃、38℃又は42℃で20分間)に供され、次に直ちに氷上に置かれた。ペプシン消化は、各サンプルに0.1M HCl4.5μl及び10mM酢酸中の2.5mg/mlペプシン4μlを添加することによって開始された。サンプルは室温(約22℃)で30分間インキュベートされた。消化は、5μlの非緩衝化1M Trisを添加することによって停止された。各サンプルは、10%ベータメルカプトエタノール及び8%SDSを含む22μlの4Xサンプル適用緩衝液と混合され、7分間沸騰され、13000rpmで7分間遠心分離された。40μlの上澄みは10%ポリアクリルアミドゲル中に添加され、抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を使用して標準的なウェスタンブロット手順でテストされた。陽性コントロールは、w.t.タバコから抽出された全可溶性タンパク質に添加された約50ngのヒトコラーゲン(Chemicon Inc.からの#CC050であり、ペプシン消化によってヒト胎盤から抽出されたもの)のサンプルであった。
【0171】
図6bに示されるように、植物#13−6中で形成されたコラーゲン三重らせんは42℃での変性に対して抵抗性であった。プロペプチドの開裂は、33℃でまず見ることができ、温度が38℃に上昇したとき及び42℃に上昇したとき再び効果的に徐々に増大する。開裂されたコラーゲン三重らせんドメインは、ペプシン処理されたヒトコラーゲンのゲル上での移動と同様の移動を示す。この実験で使用されたヒトコラーゲンは、ペプシンタンパク質分解によってヒト胎盤から抽出されたものであり、従ってプロペプチド及びいくつかのテロペプチドを欠いている。
【0172】
実施例5
植物のP4H発現
生来的な植物P4Hの誘導
タバコP4H cDNAは、クローニングされ、内因性P4H発現を誘導する条件及び処理を決定するためのプローブとして使用された。ノーザンブロット分析(図7)は、P4Hは茎頂では比較的高レベルでそして葉では低レベルで発現されることを明らかに示す。P4Hレベルは擦傷処理(下方パネルの「傷つけられた」)の4時間後に葉で有意に誘導された。同様の結果が他のストレス条件を使用して達成された(図示せず)。
【0173】
トランスジェニックタバコ植物中でのヒトP4Hアルファ及びベータサブユニット及びコラーゲンアルファ1及びアルファ2の検出
トランスジェニックタバコ植物中でのヒトP4Hアルファ及びベータサブユニット及びコラーゲンアルファ1及びアルファ2鎖の検出は、抗ヒトP4Hアルファサブユニット抗体(ICN Biomedicals Inc.からの#63−163)、抗ヒトP4Hベータサブユニット抗体(Chemicon Inc.からの#MAB2701)及び抗コラーゲンI抗体(Chemicon Inc.からの#AB745)を使用して行われた。これらの抗体でプローブされたウェスタンブロットの結果は図8に示される。
【0174】
P4Hアルファ、P4Hベータ及びコラーゲンIアルファ1及びアルファ2バンドの発現は植物13−6(ヒトLH3でも形質転換されている)中で確認された。液胞シグナルペプチドを含むP4Hアルファ及びベータの計算された分子量はそれぞれ、65.5kDa及び53.4kDaである。プロペプチドを有する、ヒドロキシル化又はグリコシル化を有さないコラーゲンアルファ1及びアルファ2鎖の計算された分子量はそれぞれ、136kDa及び127kDaである。
【0175】
明確にするため別個の実施態様で説明されている本発明の特定の特徴は単一の実施態様に組み合わせて提供することもできることは分かるであろう。逆に、簡潔にするため単一の実施態様で説明されている本発明の各種の特徴は別個にまたは適切なサブコンビネーションで提供することもできる。
【0176】
本発明はその特定の実施態様によって説明してきたが、多くの別法、変更及び変形があることは当業者には明らかであることは明白である。従って、本発明は、本願の請求項の精神と広い範囲の中に入るこのような別法、変更及び変形すべてを包含するものである。本願で挙げた刊行物、特許、特許願及びGenBankアクセッション番号はすべて、個々の刊行物、特許、特許願及びGenBankアクセッション番号が各々あたかも具体的にかつ個々に引用提示されているのと同程度に、全体を本明細書に援用するものである。さらに、本願で引用又は確認したことは本発明の先行技術として利用できるという自白とみなすべきではない。
【0177】
【配列表フリーテキスト】
【0178】
配列番号1は、ヒトコラーゲンアルファ1(I)鎖に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号2及び3は、ヒトコラーゲンアルファ1(I)鎖に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域の合成配列である。
配列番号4及び6は、ヒトコラーゲンアルファ2(I)鎖に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号5は、ヒトコラーゲンアルファ2(I)鎖に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域の合成配列である。
配列番号7は、アラビドプシスタリアーナのエンド−1,4−ベータ−グルカナーゼのアポプラストシグナルのコード領域及びフランキング領域を含む合成配列である。
配列番号8及び9は、アラビドプシスタリアーナのエンド−1,4−ベータ−グルカナーゼのアポプラストシグナル及びフランキング領域の合成配列である。
配列番号10は、キクrbcS1プロモーター及び5′UTRの配列である。
配列番号11は、キクrbcS1 3′UTR及びターミネーターの配列である。
配列番号12は、ヒトプロリル4−ヒドロキシラーゼベータサブユニットに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号13は、ヒトプロリル4−ヒドロキシラーゼベータサブユニットに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域を含む合成配列である。
配列番号14は、ヒトプロリル4−ヒドロキシラーゼアルファ−1サブユニットに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号15は、ヒトプロリル4−ヒドロキシラーゼアルファ−1サブユニットに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域を含む合成配列である。
配列番号16は、植物プロリル4−ヒドロキシラーゼに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号17は、植物プロリル4−ヒドロキシラーゼに融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域を含む合成配列である。
配列番号18及び19は、ヒトプロコラーゲンC−プロテイナーゼのコード領域及びフランキング領域を含む合成配列である。
配列番号20及び21は、ヒトプロコラーゲンI N−プロテイナーゼのコード領域及びフランキング領域を含む合成配列である。
配列番号22は、ヒトリシルヒドロキシラーゼ3に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列のコード領域及びフランキング領域を含む合成配列である。
配列番号23は、ヒトリシルヒドロキシラーゼ3に融合されたチオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列及びフランキング領域を含む合成配列である。
配列番号24は、チオールプロテアーゼアレウレイン前駆体についての大麦遺伝子の液胞シグナル配列である。
配列番号25〜28は、一本鎖DNAオリゴヌクレオチドの配列である。
【特許請求の範囲】
【請求項1】
植物又は単離された植物細胞中で生産された組み換えコラーゲンであって、植物又は単離された植物細胞の液胞に少なくとも一種のコラーゲン鎖及び外因性プロリル−4−ヒドロキシラーゼ(P4H)を標的化して、前記外因性P4Hによるが植物又は単離された植物細胞の内因性P4Hによらない前記少なくとも一種のコラーゲン鎖のヒドロキシル化を可能にすることによって生産された組み換えコラーゲン。
【請求項2】
ウェスタンブロットによってアッセイされた場合に33℃で15分間安定である請求項1に記載の組み換えコラーゲン。
【請求項3】
前記植物又は植物細胞は、リシルヒドロキシラーゼ3(LH3)、プロテアーゼN及びプロテアーゼCからなる群から選択される外因性ポリペプチドを発現する請求項1に記載の組み換えコラーゲン。
【請求項4】
I型コラーゲンである請求項1に記載の組み換えコラーゲン。
【請求項5】
前記P4Hは、ヒトのP4Hである請求項1に記載の組み換えコラーゲン。
【請求項6】
前記コラーゲンは、哺乳類のコラーゲンである請求項1に記載の組み換えコラーゲン。
【請求項7】
前記哺乳類のコラーゲンは、ヒトのコラーゲンである請求項1に記載の組み換えコラーゲン。
【請求項8】
アルファ1鎖及びアルファ2鎖を含む請求項1に記載の組み換えコラーゲン。
【請求項9】
前記少なくとも一種のコラーゲン鎖は、アルファ鎖を含む請求項1に記載の組み換えコラーゲン。
【請求項10】
前記少なくとも一種のコラーゲン鎖は、C末端及び/又はN末端プロペプチドを含む請求項1に記載の組み換えコラーゲン。
【請求項11】
組み換えコラーゲンのコラーゲン鎖がGly−X−YトリプレットのY位置でヒドロキシル化されている請求項1に記載の組み換えコラーゲン。
【請求項12】
前記哺乳類のP4Hは、ヒトのP4Hである請求項5に記載の組み換えコラーゲン。
【請求項13】
植物又は単離された植物細胞中でコラーゲンを生産する方法であって、少なくとも一種のコラーゲンアルファ鎖及び外因性P4Hを植物又は単離された植物細胞中で発現させ、それにより植物中でコラーゲンを生産させることを含み、前記少なくとも一種のコラーゲンアルファ鎖は、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、ER残留配列を欠いており、前記外因性P4Hは、前記少なくとも一種のコラーゲンアルファ鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができ、前記外因性P4Hは、ヒトのP4H又はその相同体である、方法。
【請求項14】
少なくとも一種のコラーゲンアルファ鎖をコードする外因性ポリヌクレオチド及びP4Hをコードする外因性ポリヌクレオチドを含む、遺伝的に改変された植物又は単離された植物細胞であって、前記少なくとも一種のコラーゲンアルファ鎖は、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、ER残留配列を欠いており、前記外因性P4Hは、前記少なくとも一種のコラーゲンアルファ鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができ、前記外因性P4Hは、ヒトのP4H又はその相同体である、植物又は単離された植物細胞。
【請求項15】
以下のものを含む植物システム:
(i)コラーゲンアルファ1鎖をコードする外因性ポリヌクレオチド及び(ii)P4Hをコードする外因性ポリヌクレオチドを含む、第一の遺伝的に改変された植物;及び
(i)コラーゲンアルファ2鎖をコードする外因性ポリヌクレオチド及び(ii)P4Hをコードする外因性ポリヌクレオチドを含む、第二の遺伝的に改変された植物
ただし、前記コラーゲンアルファ1鎖及びコラーゲンアルファ2鎖のそれぞれは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、ER残留配列を欠いており、前記外因性P4Hは、前記少なくとも一種のコラーゲン鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができ、前記外因性P4Hは、ヒトのP4H又はその相同体である。
【請求項16】
(a)請求項15の植物システムを得ること;
(b)前記第一植物と前記第二植物を交配させること;及び
(c)前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖を発現する子孫を選択し、それによりコラーゲンを生産すること
を含む、請求項13に記載の方法。
【請求項17】
以下のものを含む植物システム:
(i)コラーゲンアルファ1鎖をコードする外因性ポリヌクレオチド及び(ii)コラーゲンアルファ2鎖をコードする外因性ポリヌクレオチドを含む、第一の遺伝的に改変された植物;及び
P4Hをコードする外因性ポリヌクレオチドを含む、第二の遺伝的に改変された植物
ただし、前記コラーゲンアルファ1鎖及びコラーゲンアルファ2鎖のそれぞれは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、ER残留配列を欠いており、前記外因性P4Hは、前記少なくとも一種のコラーゲン鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができ、前記外因性P4Hは、ヒトのP4H又はその相同体である。
【請求項18】
ヒトのP4H及び液胞に標的化するためのシグナルペプチドをコードし、かつER残留配列を欠いているポリヌクレオチドを含む核酸構築物であって、前記ポリヌクレオチドが、植物細胞中で機能的なプロモーターの転写制御下に配置されている、核酸構築物。
【請求項19】
前記コラーゲンは、I型コラーゲンである請求項14に記載の植物又は単離された植物細胞。
【請求項20】
外因性LH3を発現させることをさらに含む請求項13に記載の方法。
【請求項21】
前記少なくとも一種のコラーゲンアルファ鎖は、アルファ1鎖を含む請求項13に記載の方法。
【請求項22】
前記少なくとも一種のコラーゲンアルファ鎖は、アルファ2鎖を含む請求項13に記載の方法。
【請求項23】
前記少なくとも一種のコラーゲンアルファ鎖は、C末端及び/又はN末端プロペプチドを含む請求項13に記載の方法。
【請求項24】
前記植物は、ストレス条件に供される請求項13に記載の方法。
【請求項25】
前記ストレス条件は、渇水、塩分、傷害、寒さ、及びストレス誘導化合物の噴霧からなる群から選択される請求項24に記載の方法。
【請求項26】
前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖のそれぞれは、C末端及び/又はN末端プロペプチドを含む請求項15に記載の方法。
【請求項27】
外因性LH3を発現させることをさらに含む請求項14に記載の植物又は植物細胞。
【請求項28】
前記少なくとも一種のコラーゲン鎖の配列は、配列番号1又は4に規定される配列である請求項1に記載の組み換えコラーゲン。
【請求項29】
前記コラーゲンアルファ鎖は、コラーゲンERシグナルを欠く請求項13に記載の方法。
【請求項30】
前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖は、コラーゲンERシグナルを欠く請求項15又は17に記載の植物システム。
【請求項1】
植物又は単離された植物細胞中で生産された組み換えコラーゲンであって、植物又は単離された植物細胞の液胞に少なくとも一種のコラーゲン鎖及び外因性プロリル−4−ヒドロキシラーゼ(P4H)を標的化して、前記外因性P4Hによるが植物又は単離された植物細胞の内因性P4Hによらない前記少なくとも一種のコラーゲン鎖のヒドロキシル化を可能にすることによって生産された組み換えコラーゲン。
【請求項2】
ウェスタンブロットによってアッセイされた場合に33℃で15分間安定である請求項1に記載の組み換えコラーゲン。
【請求項3】
前記植物又は植物細胞は、リシルヒドロキシラーゼ3(LH3)、プロテアーゼN及びプロテアーゼCからなる群から選択される外因性ポリペプチドを発現する請求項1に記載の組み換えコラーゲン。
【請求項4】
I型コラーゲンである請求項1に記載の組み換えコラーゲン。
【請求項5】
前記P4Hは、ヒトのP4Hである請求項1に記載の組み換えコラーゲン。
【請求項6】
前記コラーゲンは、哺乳類のコラーゲンである請求項1に記載の組み換えコラーゲン。
【請求項7】
前記哺乳類のコラーゲンは、ヒトのコラーゲンである請求項1に記載の組み換えコラーゲン。
【請求項8】
アルファ1鎖及びアルファ2鎖を含む請求項1に記載の組み換えコラーゲン。
【請求項9】
前記少なくとも一種のコラーゲン鎖は、アルファ鎖を含む請求項1に記載の組み換えコラーゲン。
【請求項10】
前記少なくとも一種のコラーゲン鎖は、C末端及び/又はN末端プロペプチドを含む請求項1に記載の組み換えコラーゲン。
【請求項11】
組み換えコラーゲンのコラーゲン鎖がGly−X−YトリプレットのY位置でヒドロキシル化されている請求項1に記載の組み換えコラーゲン。
【請求項12】
前記哺乳類のP4Hは、ヒトのP4Hである請求項5に記載の組み換えコラーゲン。
【請求項13】
植物又は単離された植物細胞中でコラーゲンを生産する方法であって、少なくとも一種のコラーゲンアルファ鎖及び外因性P4Hを植物又は単離された植物細胞中で発現させ、それにより植物中でコラーゲンを生産させることを含み、前記少なくとも一種のコラーゲンアルファ鎖は、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、ER残留配列を欠いており、前記外因性P4Hは、前記少なくとも一種のコラーゲンアルファ鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができ、前記外因性P4Hは、ヒトのP4H又はその相同体である、方法。
【請求項14】
少なくとも一種のコラーゲンアルファ鎖をコードする外因性ポリヌクレオチド及びP4Hをコードする外因性ポリヌクレオチドを含む、遺伝的に改変された植物又は単離された植物細胞であって、前記少なくとも一種のコラーゲンアルファ鎖は、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、ER残留配列を欠いており、前記外因性P4Hは、前記少なくとも一種のコラーゲンアルファ鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができ、前記外因性P4Hは、ヒトのP4H又はその相同体である、植物又は単離された植物細胞。
【請求項15】
以下のものを含む植物システム:
(i)コラーゲンアルファ1鎖をコードする外因性ポリヌクレオチド及び(ii)P4Hをコードする外因性ポリヌクレオチドを含む、第一の遺伝的に改変された植物;及び
(i)コラーゲンアルファ2鎖をコードする外因性ポリヌクレオチド及び(ii)P4Hをコードする外因性ポリヌクレオチドを含む、第二の遺伝的に改変された植物
ただし、前記コラーゲンアルファ1鎖及びコラーゲンアルファ2鎖のそれぞれは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、ER残留配列を欠いており、前記外因性P4Hは、前記少なくとも一種のコラーゲン鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができ、前記外因性P4Hは、ヒトのP4H又はその相同体である。
【請求項16】
(a)請求項15の植物システムを得ること;
(b)前記第一植物と前記第二植物を交配させること;及び
(c)前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖を発現する子孫を選択し、それによりコラーゲンを生産すること
を含む、請求項13に記載の方法。
【請求項17】
以下のものを含む植物システム:
(i)コラーゲンアルファ1鎖をコードする外因性ポリヌクレオチド及び(ii)コラーゲンアルファ2鎖をコードする外因性ポリヌクレオチドを含む、第一の遺伝的に改変された植物;及び
P4Hをコードする外因性ポリヌクレオチドを含む、第二の遺伝的に改変された植物
ただし、前記コラーゲンアルファ1鎖及びコラーゲンアルファ2鎖のそれぞれは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、液胞に標的化させるためのシグナルペプチドを含み、前記外因性P4Hは、ER残留配列を欠いており、前記外因性P4Hは、前記少なくとも一種のコラーゲン鎖のGly−X−YトリプレットのY位置を特異的にヒドロキシル化することができ、前記外因性P4Hは、ヒトのP4H又はその相同体である。
【請求項18】
ヒトのP4H及び液胞に標的化するためのシグナルペプチドをコードし、かつER残留配列を欠いているポリヌクレオチドを含む核酸構築物であって、前記ポリヌクレオチドが、植物細胞中で機能的なプロモーターの転写制御下に配置されている、核酸構築物。
【請求項19】
前記コラーゲンは、I型コラーゲンである請求項14に記載の植物又は単離された植物細胞。
【請求項20】
外因性LH3を発現させることをさらに含む請求項13に記載の方法。
【請求項21】
前記少なくとも一種のコラーゲンアルファ鎖は、アルファ1鎖を含む請求項13に記載の方法。
【請求項22】
前記少なくとも一種のコラーゲンアルファ鎖は、アルファ2鎖を含む請求項13に記載の方法。
【請求項23】
前記少なくとも一種のコラーゲンアルファ鎖は、C末端及び/又はN末端プロペプチドを含む請求項13に記載の方法。
【請求項24】
前記植物は、ストレス条件に供される請求項13に記載の方法。
【請求項25】
前記ストレス条件は、渇水、塩分、傷害、寒さ、及びストレス誘導化合物の噴霧からなる群から選択される請求項24に記載の方法。
【請求項26】
前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖のそれぞれは、C末端及び/又はN末端プロペプチドを含む請求項15に記載の方法。
【請求項27】
外因性LH3を発現させることをさらに含む請求項14に記載の植物又は植物細胞。
【請求項28】
前記少なくとも一種のコラーゲン鎖の配列は、配列番号1又は4に規定される配列である請求項1に記載の組み換えコラーゲン。
【請求項29】
前記コラーゲンアルファ鎖は、コラーゲンERシグナルを欠く請求項13に記載の方法。
【請求項30】
前記コラーゲンアルファ1鎖及び前記コラーゲンアルファ2鎖は、コラーゲンERシグナルを欠く請求項15又は17に記載の植物システム。
【図1a】

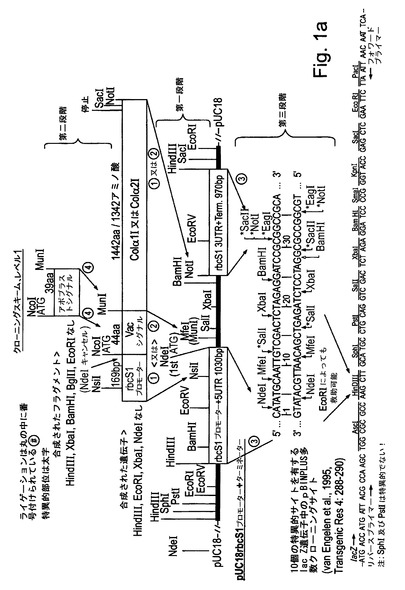
【図1b】
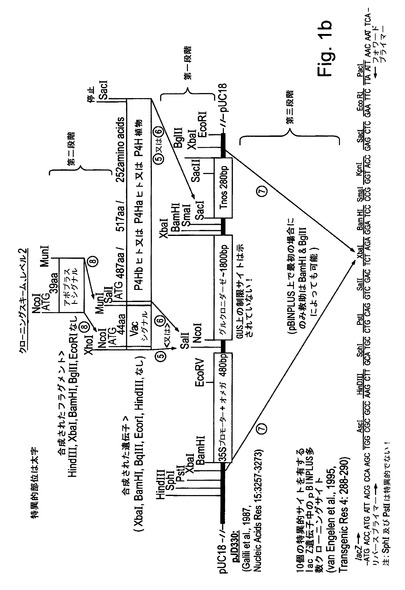
【図1c】
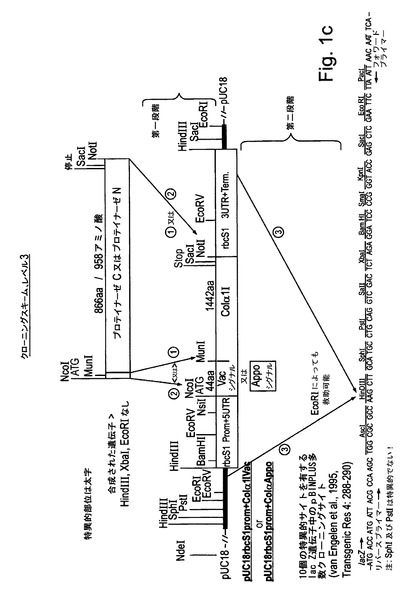
【図1d】
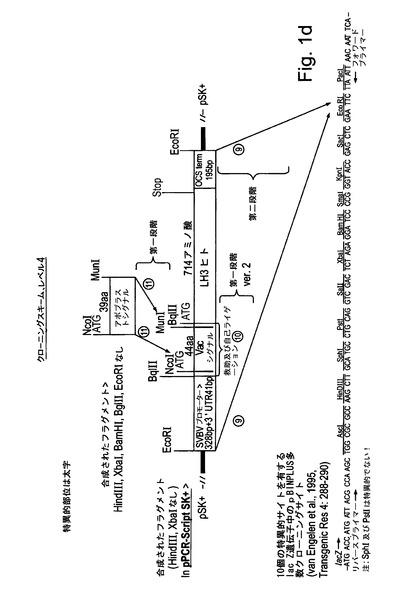
【図2】
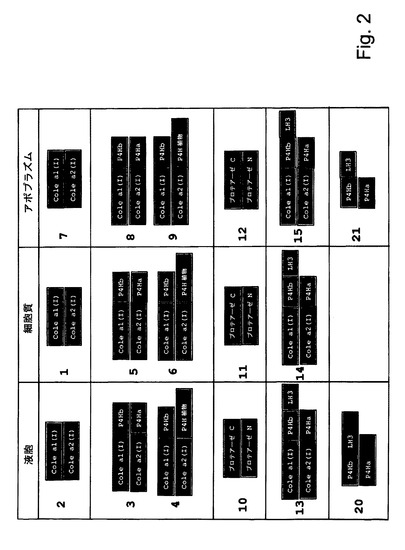
【図3】
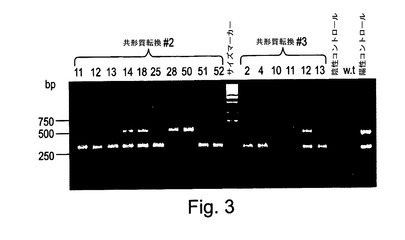
【図4】
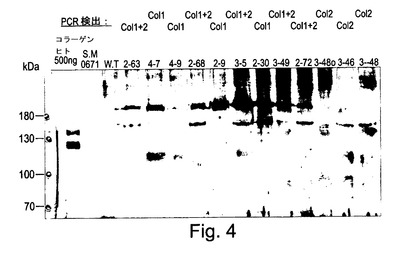
【図5】
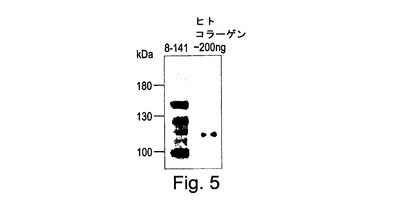
【図6a】
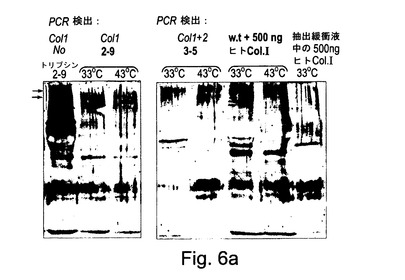
【図6b】
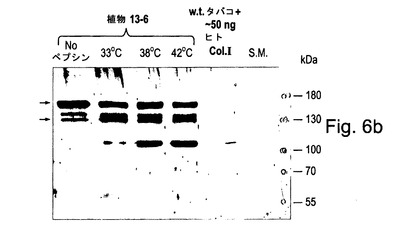
【図7】
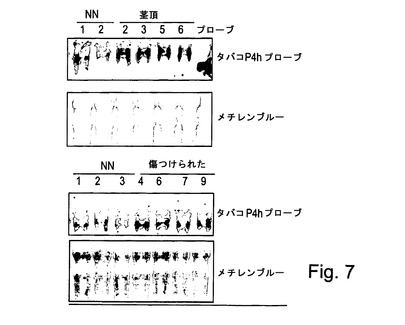
【図8】

【図1b】
【図1c】
【図1d】
【図2】
【図3】
【図4】
【図5】
【図6a】
【図6b】
【図7】
【図8】
【公開番号】特開2012−85644(P2012−85644A)
【公開日】平成24年5月10日(2012.5.10)
【国際特許分類】
【出願番号】特願2011−259678(P2011−259678)
【出願日】平成23年11月29日(2011.11.29)
【分割の表示】特願2007−534176(P2007−534176)の分割
【原出願日】平成17年9月28日(2005.9.28)
【出願人】(507101749)コルプラント リミテッド (2)
【Fターム(参考)】
【公開日】平成24年5月10日(2012.5.10)
【国際特許分類】
【出願日】平成23年11月29日(2011.11.29)
【分割の表示】特願2007−534176(P2007−534176)の分割
【原出願日】平成17年9月28日(2005.9.28)
【出願人】(507101749)コルプラント リミテッド (2)
【Fターム(参考)】
[ Back to top ]