
コレステロール関連障害の治療におけるMIR−33マイクロRNAの調節
MiR−33マイクロRNAを標的とする核酸配列を含む組成物が、コレステロール関連障害の治療におけるその組成物の使用と共に記載される。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願/特許
本出願は、代理人整理番号MGH20023として2009年3月31日に出願した米国仮出願第61/165,041号の利益を主張し、その内容は本明細書に参照として組み込まれている。
【0002】
連邦支援研究下でなされた発明に対する権利の声明
本研究は、国立衛生研究所助成番号R01GM71449及びR21DK084459により援助された。米国政府は本発明に一定の権利を有し得る。
【背景技術】
【0003】
異常なコレステロール及び脂質ホメオスタシスは、メタボリックシンドローム、アテローム性動脈硬化症/心血管疾患、及び2型糖尿病などの蔓延している疾患と関連している。コレステロール及び脂質は、それらの脂肪性のカーゴを異なる細胞及び組織に運ぶ、低比重リポタンパク質(LDL)及び高比重リポタンパク質(HDL)などのリポタンパク質粒子として血液中で輸送される。過剰な循環LDLは、酸化され、動脈内のマクロファージにより取り込まれる可能性があり、それらのマクロファージは、動脈硬化性プラークの形成に関与するコレステロール/脂質に満ちた「泡沫細胞」に変化する。反対に、HDL−コレステロールは、マクロファージを含む、末梢細胞から運び出され、コレステロールの逆輸送経路によって分解するために肝臓に輸送されるが、しかしながら、心臓疾患を患っている患者は、多くの場合、循環HDLが低く、コレステロールクリアランスが低下している。したがって、心血管疾患を寛解させるための治療努力は、コレステロール生合成及びLDL産生を低下させること(例えばスタチン治療による)、並びにHDLを介したコレステロールの逆輸送を増加させること(例えばナイアシン添加による)に重点を置かれている。これらの治療はかなりの効果を示しているが、心血管疾患は、米国において主要な死亡原因のままである。コレステロール及び脂質調節の分子機構の詳細な理解により、心血管疾患を治療するための新規治療戦略の開発が促進する可能性がある。
【0004】
ATP結合カセット輸送体ABCA1は、HDL合成及びマクロファージ/泡沫細胞を含む、末梢組織からのコレステロールの逆輸送に重要である。ABCA1遺伝子における変異/SNPがアテローム性動脈硬化に関係している。例えば、ABCA1レベル、HDL産生及び/又はコレステロールの逆輸送を増加できる新規の治療は、調節を改善でき、心血管疾患を寛解させる可能性がある。
【発明の概要】
【課題を解決するための手段】
【0005】
ステロール調節要素結合タンパク質(SREBP)転写因子は、コレステロール及び脂質の生合成及び取り込みの両方に関与する、公知の遺伝子調節因子である。現在、ヒトSREBP−2及びヒトSREBP−1遺伝子におけるイントロン配列内に埋め込まれている保存されたマイクロRNAをコードする配列の存在は発見されている。本明細書の以下に記載している予備研究により、本明細書において配列番号1及び2としてそれぞれ示しているmiR−33a及びmiR−33bは、翻訳抑制のためにATP結合カセット輸送体ABCA1を標的とするという驚くべき知見が得られた。哺乳動物におけるコレステロール恒常性の調節において、miR−33とSREBP宿主遺伝子産物との協調した役割は、今まで知られていなかった。更に、miR−33の阻害が、ABCA1レベル、HDL産生、及びコレステロールの逆輸送を増加させる新規の治療手段を表すことが、ここで発見された。
【0006】
一態様において、本発明は、配列番号1又は2と相補的である単離された核酸配列を含む組成物であって、その核酸配列は、哺乳動物細胞におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効である、組成物を提供する。一部の実施形態において、組成物は、コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で核酸を含む。
【0007】
一部の実施形態において、配列は配列番号7又は9と相補的である。
【0008】
一実施形態において、配列番号1又は2と相補的である核酸配列は、限定されないが、本明細書に記載されるアンチセンスオリゴヌクレオチド、例えば配列番号3、4、6、又は8を含む、アンチセンスオリゴヌクレオチドであり得る。一部の実施形態において、核酸は修飾され、例えば、1つ又は複数の天然に存在しないヌクレオシドを含有する。一部の実施形態において、核酸はロックされている。
【0009】
別の実施形態において、配列番号1又は2と相補的である核酸配列は、限定されないが、shRNA又はsiRNAを含む、干渉RNAであり得る。
【0010】
更に別の実施形態において、配列番号1又は2と相補的である核酸配列は、アンタゴマー(antagomir)である。
【0011】
更に別の実施形態において、配列番号1と相補的である核酸配列は、配列番号1の転写後プロセシングを阻害する。
【0012】
更に別の実施形態において、配列番号2と相補的である核酸配列は、配列番号2の転写後プロセシングを阻害する。
【0013】
別の態様において、本発明は、コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で、配列番号1又は2と相補的である核酸配列を発現できるベクターを含む組成物を提供する。
【0014】
一実施形態において、ベクターは、プラスミドベクター又はウイルスベクターである。
【0015】
更に別の態様において、本発明は、細胞、例えば、コレステロール恒常性を必要とする被験体の細胞におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させる方法であって、配列番号1又は2と相補的である核酸配列を細胞又は被験体に投与することによって、細胞又は被験体におけるABCA1タンパク質の発現を増加させることを含む、方法を提供する。
【0016】
更に別の態様において、本発明は、被験体の肝臓における細胞内コレステロールの流出及び/又はHDLの産生を増加させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体の肝臓における細胞内コレステロールの流出及び/又はHDLの産生を増加させることを含む、方法を提供する。
【0017】
更に別の態様において、本発明は、被験体の血液を循環するコレステロールの量を減少させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体の血液を循環するコレステロールの量を減少させることを含む、方法を提供する。
【0018】
本発明の他の特徴及び利点は、詳細な説明、及び特許請求の範囲から明らかとなろう。したがって、本発明の他の態様は以下の開示に説明され、それらは本発明の範囲内である。
【0019】
以下の詳細な図面は例として与えられ、記載される特定の実施形態に本発明を限定することを意図せず、本明細書に参照として組み込まれる添付の図面と併せて理解され得る。
【図面の簡単な説明】
【0020】
【図1】SREBPによって調節される遺伝子を示す。この図は、コレステロール、脂肪酸、及びトリグリセリドの合成経路における主要な代謝中間体を示す。インビボにおいて、SREBP−2は、コレステロール代謝の遺伝子を選択的に活性化するのに対して、SREBP−1cは、脂肪酸及びトリグリセリド代謝の遺伝子を選択的に活性化する。DHCR、7−デヒドロコレステロールレダクターゼ;FPP、ファルネシル二リン酸;GPP、ゲラニルゲラニルピロリン酸シンターゼ;CYP51、ラノステロール14α−デメチラーゼ;G6PD、グルコース−6−リン酸デヒドロゲナーゼ;PGDH、6−ホスホグルコン酸デヒドロゲナーゼ;GPAT、グリセロール−3−リン酸アシルトランスフェラーゼ。出典Hortonら,J Clin Invest.2002.109:1125−31。
【図2】CBP/p300ヒストンアセチルトランスフェラーゼ及びARC/メディエーター共活性化因子複合体のARC105/MED15サブユニットに存在する構造保存性のKIX領域(青色楕円形)と相互作用することによる標的遺伝子の転写のSREBP活性化を示す。CBP/p300の動員により、ヒストン/クロマチンの増加したアセチル化が生じるのに対して、ARC/メディエーター相互作用により、その大きさサブユニットでC末端領域(CTD)を経由するRNAポリメラーゼII(Pol II)の動員が生じる。TFIID及び他の基本転写因子(GTF)もまた、転写開始を促進する、プロモーターに動員される。
【図3】イントロンに位置するmiRNA(それぞれ、miR−33b及びmiR−33a)をコードする配列を保有する、ヒトSREBP−1及びSREBP−2遺伝子を示す。特に、マウスのみが、マウスSREBP−2遺伝子においてmiR−33a miRNAを保有するのに対して、対応するマウスSREBP−1イントロンはヒトSREBP−1遺伝子との相同性を欠き、miRNAコード配列を欠いている。
【図4】miRNAプロセシング酵素のドローシャによって認識されるヘアピン構造を形成する一次miRNA転写産物(pri−miRNA)の合成を生じる、miRNAコード配列を介するPol IIによる転写を示す。pri−miRNAは、ドローシャ(Drosha)によってpre−miRNAにプロセシングされ、次いで細胞質に送り出され、細胞質において、それらのpre−miRNAはダイサー(Dicer)によって成熟miRNA二本鎖までのプロセシングを更に受ける。それらの成熟miRNA二本鎖はRISC複合体によって損傷を受けず、mRNA標的鎖がRISCに組み込まれ、次いでそのmRNA標的鎖は、miRNA−mRNA配列相補性に応じて翻訳抑制又はmRNA分解のいずれかによってmRNAを標的とする。出典van den Bergら,Biochim.et Biophys.Acta 2008。
【図5】マクロファージ及びアテローム生成泡沫細胞などの末梢細胞からApoA−I及び高比重リポタンパク質(HDL)に遊離コレステロールを送り出す、ABC輸送体ABCA1(及び関連ABCG1)を示す。HDLは、肝細胞においてスカベンジャー受容体−B1(SR−B1)に結合し、それらの付随したコレステロール及びコレステロールエステル(CE)を肝臓に移す。コレステロールは、遊離コレステロールとして、又は胆汁塩に変換後のいずれかで胆汁に排出される。出典Maxfield及びTabas.Nature 2005.438:612−21。
【図6】miR−33及びその宿主遺伝子産物SREBPによるコレステロール恒常性の協調した調節についてのモデルを示す。miR−33は、その3’UTRを標的化することによりABCA1翻訳を抑制する。このステップは、コレステロール産生遺伝子(cholesterogenic gene)のトランス活性化を介するコレステロール生合成及び取り込みを活性化する、SREBPの発現と協調するように提案される。コレステロール蓄積は、SREBP経路を阻害し、ABCA1の発現を活性化する。そのABCA1は、細胞膜へ移行し、ApoA−I及びHDLへのコレステロール流出を媒介する。
【図7A】miRNAによるABCA1発現の調節を示す。図7Aにおいて、3つの細胞株(J774マウスマクロファージ、ヒトHepG2肝細胞癌細胞、及び正常なヒトIMR−90線維芽細胞)においてsiRNAを用いるmiRNAプロセシング酵素のドローシャ及びダイサーのRNAiに向けられたノックダウンにより、ABCA1タンパク質レベルの増加が生じた。タンパク質レベルはブラッドフォード分析により正規化した。β−チューブリンを負荷対照として使用した。
【図7B】miRNAによるABCA1発現の調節を示す。図7Bにおいて、抗miR−33アンチセンスオリゴヌクレオチドによるmiR−33機能の拮抗により、ABCA1のより高い発現が生じることが示される。
【図7C】miRNA前駆体Pre−miR−33でトランスフェクトした細胞によって模倣される、miR−33の効果を示す。予想されるように、ABCA1発現は、増加したレベルのmiR−33による翻訳抑制を介して阻害される。
【図8A】HEK293細胞におけるルシフェラーゼレポーター構築物によるABCA1 3’−UTRにおけるmiR−33についての予測標的部位の評価を示す。(A)2つの潜在的標的部位が、ABCA1 3’UTR(配列番号5)の最初の160塩基内に存在するmiR−33について予測される。ルシフェラーゼORFのABCA1 3’UTR下流の対応する配列(塩基18〜205)の挿入により、ルシフェラーゼ活性の下方調節が生じる(右のグラフ)。
【図8B】HEK293細胞におけるルシフェラーゼレポーター構築物によるABCA1 3’−UTRにおけるmiR−33についての予測標的部位の評価を示す。(B)(A)において観察された下方調節は、30nMの濃度にて抗マイクロRNAオリゴヌクレオチド(Ambion)A33a及びA33b(配列番号6及び8)を用いるmiR−33a/bアイソフォーム(配列番号7及び9)の配列特異的喪失により遮断される(AC:陰性対照としての関係のない抗miR)。
【図8C】HEK293細胞におけるルシフェラーゼレポーター構築物によるABCA1 3’−UTRにおけるmiR−33についての予測標的部位の評価を示す。(C)変異したmiR−33前駆体(30nMにおけるm33a(配列番号10)及びm33b(配列番号11))ではなく、過剰な外因性野生型miR−33前駆体(30nMにおける33a及び33b)の添加は、予想されるようにルシフェラーゼ活性を更に阻害した。
【図9A】コレステロール及びSREBP/miR−33による相互調節を示す。J774マウスマクロファージにおけるコレステロールのロバスタチン及びβ−シクロデキストリンを介する喪失により、(A)SREBP−2の増加した発現が生じる。エラーバーはs.dを表す。**はp<0.01を示す。
【図9B】コレステロール及びSREBP/miR−33による相互調節を示す。J774マウスマクロファージにおけるコレステロールのロバスタチン及びβ−シクロデキストリンを介する喪失により、(B)miR−33aの増加した発現が生じる。エラーバーはs.dを表す。**はp<0.01を示す。
【図9C】コレステロール及びSREBP/miR−33による相互調節を示す。J774マウスマクロファージにおけるコレステロールのロバスタチン及びβ−シクロデキストリンを介する喪失により、(C)ABCA1タンパク質レベルの減少が生じる。
【図9D】コレステロール及びSREBP/miR−33による相互調節を示す。(D)コレステロールが喪失したJ774マウスマクロファージは、偽トランスフェクション(UT)又はアンチセンス対照(AC)オリゴヌクレオチドと比較して、miR−33bアンチセンスオリゴヌクレオチドでのトランスフェクションに応答してABCA1タンパク質の上方調節を示す。β−チューブリンは、この免疫ブロット法において負荷対照としての機能を果たした。
【図9E】コレステロール及びSREBP/miR−33による相互調節を示す。(E)miR−33a前駆体を有する血清/コレステロールの存在下で培養したJ774マクロファージのトランスフェクションにより、前駆体対照(PC)と比較して、減少したレベルのABCA1が生じた。
【図9F】コレステロール及びSREBP/miR−33による相互調節を示す。(F)放射性標識したコレステロールを負荷したJ775マクロファージへのmiR−33aアンチセンスオリゴヌクレオチド(A)の導入により、増加したコレステロール流出が生じるのに対して、miR−33a前駆体(P)は、トランスフェクション対照(UT)又はスクランブルアンチセンス対照(AC)又は前駆体対照(PC)オリゴヌクレオチドと比較して、コレステロール流出を阻害する。エラーバーはs.dを表す。**はP<0.01を示す。
【図10A】インビボでのmiR−33aによるHDLの調節を示す。(A)マウスmiR−33aに対して向けられるロックド核酸(LNA)−アンチセンスオリゴヌクレオチドを有する西洋型食餌を与えたマウスの尾静脈注射により、ビヒクル(PBS)及びスクランブルLNA対照(LC)オリゴヌクレオチドと比較して、増大した総血清コレステロールが生じる。
【図10B】インビボでのmiR−33aによるHDLの調節を示す。(B)LNA−miR−33aアンチセンス(LNA)及びLNA(対照)(LC)で処置した西洋型食餌を与えた5匹のマウス由来のプールされた血清のFPLC分析により、miR−33aアンチセンスで処置した動物においてHDL−コレステロールピークの大きさの増加が明らかにされた。リポタンパク質標準物の溶出は標識により示される。
【図10C】インビボでのmiR−33aによるHDLの調節を示す。血漿トリグリセリド(C)は処置に影響を与えなかった。
【図10D】インビボでのmiR−33aによるHDLの調節を示す。グルコース(D)は処置に影響を与えなかった。
【図10E】インビボでのmiR−33aによるHDLの調節を示す。AST(E)は処置に影響を与えなかった。
【図10F】インビボでのmiR−33aによるHDLの調節を示す。ALTは処置に影響を与えなかった。
【図10G】インビボでのmiR−33aによるHDLの調節を示す。(G)は、miR−33及びその宿主遺伝子産物SREBPによるコレステロール恒常性の協調した調節についてのモデルを表す。
【図11A】ヒト及びマウス組織におけるSREBPの同時発現を示す。ヒトSREBP−1(A;配列番号12)遺伝子は、関連するイントロンmiRNA(miR−33b)を保有する。pri−miRNAをコードする配列を、成熟miRNA配列に下線を引いて示す。
【図11B】ヒト及びマウス組織におけるSREBPの同時発現を示す。ヒトSREBP−2(B;配列番号13)遺伝子は、関連するイントロンmiRNA(miR−33a)を保有する。pri−miRNAをコードする配列を、成熟miRNA配列に下線を引いて示す。
【図11C】ヒト及びマウス組織におけるSREBPの同時発現を示す。選択されたヒト及びマウス組織におけるmiR−33a/b及びSREBP宿主遺伝子の発現の相関関係:(C)hSREBP−1/−hsa−miR−33b。エラーバーはs.e.mを表す。
【図11D】ヒト及びマウス組織におけるSREBPの同時発現を示す。選択されたヒト及びマウス組織におけるmiR−33a/b及びSREBP宿主遺伝子の発現の相関関係:(D)hSREBP−2/hsa−miR−33a。エラーバーはs.e.mを表す。
【図11E】ヒト及びマウス組織におけるSREBPの同時発現を示す。選択されたヒト及びマウス組織におけるmiR−33a/b及びSREBP宿主遺伝子の発現の相関関係:(E)mSREBP−1/mmu−miR−33b。エラーバーはs.e.mを表す。
【図12A】選択された種におけるmiR−33a(A)アイソフォーム(ヒトにおけるmir−33a、配列番号14;マウスにおけるmir−33a、配列番号15;ニワトリにおけるmir−33a、配列番号16;推測上カエルにおけるmir−33a、配列番号17、及びショウジョウバエにおけるmir−33a、配列番号18)の配列及びゲノム構成を示す。miRNAステムループを示し、それらが存在するSREBP遺伝子に従って分類する。アスタリスクは完全に保存された残基の位置を表す。
【図12B】選択された種におけるmiR−33b(B)アイソフォーム(ヒトにおけるmir−33b、配列番号19、マウスにおけるmir−33b、配列番号20)の配列及びゲノム構成を示す。miRNAステムループを示し、それらが存在するSREBP遺伝子に従って分類する。アスタリスクは完全に保存された残基の位置を表す。
【図13A】ヒト組織におけるSREBP−1の相対的発現を示す。β−アクチンを組織間の発現を正規化するために使用した。各組織から使用した全RNAは、男性及び女性の患者を含む少なくとも3体のドナーからプールした。
【図13B】ヒト組織におけるhsa−miR−33bの相対的発現を示す。U6を組織間の発現を正規化するために使用した。各組織から使用した全RNAは、男性及び女性の患者を含む少なくとも3体のドナーからプールした。
【図14A】ヒト組織におけるSREBP−2の相対的発現を示す。β−アクチンを組織間の発現を正規化するために使用した。低い相対的発現を有する組織をグラフの左側において比較し、高い相対的発現を有する組織をグラフの右側において比較する。各組織から使用した全RNAは、男性及び女性の患者を含む少なくとも3体のドナーからプールした。
【図14B】ヒト組織におけるhsa−miR−33aの相対的発現を示す。U6を組織間の発現を正規化するために使用した。低い相対的発現を有する組織をグラフの左側において比較し、高い相対的発現を有する組織をグラフの右側において比較する。各組織から使用した全RNAは、男性及び女性の患者を含む少なくとも3体のドナーからプールした。
【図15A】マウス組織におけるSREBP−2の相対的発現を示す。β−アクチンを組織間の発現を正規化するために使用した。
【図15B】マウス組織におけるmmu−miR−33aの相対的発現を示す。U6を組織間の発現を正規化するために使用した。
【図16】ヒトNPC1 3’UTRにおいて予測されるmiRNA標的部位を示す(TargetScan 4.2)。miR−33標的部位は矢印によって示される。
【図17】配列番号1及び2として、それぞれ、miR−33a及びmir−33bを示す。また、配列番号3及び4として、それぞれ、抗miR−33a及び抗miR−33bを示す。
【発明を実施するための形態】
【0021】
定義
他に定義されない限り、本明細書に使用される全ての技術用語及び科学用語は、本発明が属する技術分野における当業者によって一般的に理解されているものと同じ意味を有する。矛盾する場合には、定義を含む本出願が適用される。
【0022】
本明細書で言及する場合、「相補的核酸配列」は、相補的ヌクレオチド塩基対からなる別の核酸配列とハイブリダイズできる核酸配列である。「ハイブリダイズする」とは、ストリンジェンシーの適切な条件下で、相補的ヌクレオチド塩基間で二本鎖分子を形成する対を意味する(例えば、DNAにおいて、アデニン(A)はチミン(T)と塩基対を形成し、同様にグアニン(G)はシトシン(C)と塩基対を形成する)(例えば、Wahl,G.M.及びS.L.Berger(1987)Methods Enzymol.152:399;Kimmel,A.R.(1987)Methods Enzymol.152:507を参照のこと)。
【0023】
本明細書で使用する場合、「アンチセンスオリゴヌクレオチド」とは、マイクロRNAの配列などのDNA又はmRNA配列と相補的である合成核酸配列をいう。
【0024】
「RNA」は、少なくとも1つ又は複数のリボヌクレオチド残基を含む分子である。「リボヌクレオチド」は、ベータ−D−リボフラノース部分の2’位にヒドロキシル基を有するヌクレオチドである。本明細書で使用する場合、用語RNAは、二本鎖RNA、一本鎖RNA、部分的に精製されたRNAなどの単離されたRNA、実質的に純粋なRNA、合成RNA、組換えで生成されたRNA、並びに1つ若しくは複数のヌクレオチドの付加、欠失、置換及び/又は変更によって天然に存在するRNAとは異なった改変RNAを含む。RNA分子のヌクレオチドはまた、天然に存在しないヌクレオチド又は化学的に合成されたヌクレオチド若しくはデオキシヌクレオチドなどの非標準的ヌクレオチドを含んでもよい。
【0025】
「マイクロRNA」(miRNA)は、典型的に、遺伝子発現を調節する、長さが約21〜23ヌクレオチドの一本鎖RNA分子をいう。miRNAは遺伝子によってコードされ、その遺伝子のDNAからmiRNAは転写されるが、miRNAはタンパク質に翻訳されず、代わりに、各々の一次転写産物は短いステムループ構造にプロセシングされ、その後、機能的miRNAへの更なるプロセシングを受ける。成熟miRNA分子は、1つ又は複数のメッセンジャーRNA(mRNA)分子と部分的に相補的であり、それらの主な機能は遺伝子発現を下方調節することである。
【0026】
本明細書で使用する場合、「干渉RNA」とは、直接又は間接(すなわち変換の際に)のいずれかで、RNA干渉を介することによって遺伝子発現を阻害又は下方調節できる、任意の二本鎖又は一本鎖RNA配列をいう。干渉RNAには、限定されないが、低分子干渉RNA(「siRNA」)及び低分子ヘアピン型RNA(「shRNA」)が含まれる。「RNA干渉」とは、配列適合性のメッセンジャーRNA転写産物の選択的分解をいう。
【0027】
本明細書で使用する場合、「shRNA」(低分子ヘアピン型RNA)とは、アンチセンス領域、ループ部分及びセンス領域を含むRNA分子であって、そのセンス領域が、二重ステムを形成するためにアンチセンス領域と塩基対合する相補的ヌクレオチドを有する、RNA分子をいう。転写後プロセシング後、低分子ヘアピン型RNAは、RNase IIIファミリーのメンバーである、酵素のダイサーを介して行われる切断事象によって低分子干渉RNAに変換される。
【0028】
本明細書で使用する場合、「低分子干渉RNA」又は「siRNA」とは、配列特異的にRNA干渉を介することによって遺伝子発現を阻害又は下方調節できる、任意の低分子RNA分子をいう。低分子RNAは、例えば、約18〜21ヌクレオチド長であり得る。
【0029】
本明細書で使用する場合、「アンタゴマー」とは、切断を阻害するために切断部位又は1つ若しくは複数の塩基修飾部位のいずれかに誤対合を有する、特定のマイクロRNA標的と相補性を有する低分子合成RNAをいう。
【0030】
本明細書で使用する場合、「転写後プロセシング」という語句は、転写後に起こり、例えば、酵素のダイサー及び/又はドローシャを介して行われる、mRNAプロセシングをいう。
【0031】
本明細書で使用する場合、「ABCA1」という用語は、例えば、Remaley,A.T.,Proc.Nat.Acad.Sci.96:12685−12690,1999に記載されている、ATP結合カセット、サブファミリーA、メンバー1、タンパク質輸送体をいう。
【0032】
本明細書で使用する場合、「ABCA1タンパク質発現の増加」とは、本明細書に記載される方法による治療の前の被験体におけるABCA1タンパク質の量より少なくとも約1倍以上(例えば1、2、3、4、5、10、20、30、40、50、60、70、80、90、100、1000、10,000倍又はそれ以上)のABCA1タンパク質の量をいう。被験体におけるABCA1タンパク質発現の量をいう場合、「増加」とは、また、本明細書に記載される方法による治療の前の被験体におけるABCA1タンパク質の量より少なくとも約5%以上(例えば5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、99又は100%)であることを意味する。タンパク質量は、当該技術分野において公知の方法によって測定できる。
【0033】
「有効量」とは、未治療の患者に対して疾患の症状を寛解させるのに必要とされる薬剤又は薬剤を含む組成物の量を意味する。疾患の治療的処置のために本発明を実施するのに使用される組成物(複数可)の有効量は、投与様式、被験体の年齢、体重、及び全体的な健康に応じて変わる。最終的に、主治医又は獣医が、適した量及び投薬レジメンを決定するだろう。かかる量を「有効」量」という。
【0034】
本明細書で使用する場合、「コレステロール恒常性」とは、コレステロール取り込み、コレステロール生合成、胆汁酸へのコレステロールの変換及び胆汁酸の排泄のような、健康レベルの血中のLDL、HDL及びコレステロール(例えば、本明細書においてかかる健康レベルは「参照基準」ともいわれる)を有する被験体において起こるプロセスの調節をいう。したがって、コレステロール恒常性を必要とする被験体は、健康レベルの血中のLDL、HDL及び/又はコレステロールへの回復を生じる、改善された調節を必要とする。
【0035】
「被験体」は、ヒト、家庭用家畜及び飼育場用家畜、並びに動物園の動物、スポーツ用の動物又はペット用の動物、例えばマウス、ウサギ、ブタ、ヒツジ、ヤギ、ウシ及び高等霊長類を含む、哺乳網の任意のメンバーを含む、脊椎動物である。
【0036】
本明細書で使用する場合、「ベクター」又は「発現ベクター」は、所望の調節配列及び遺伝子を含む、核酸ベースの送達ビヒクルであり、その内容物を細胞に移動させるために使用され得る。
【0037】
本明細書の開示において、「含む(comprise)」、「含んでいる(comprising)」、「含有している」及び「有している」などは、米国特許法でみなされている意味を有することができ、「包含する(include)」、「包含している(including)」などを意味することができ、同様に「から実質的になる」又は「実質的になる」は、米国特許法でみなされている意味を有していて、この用語には制約がなく、列挙されている基本的又は新規な特徴が、列挙されているものよりも多くの存在によって変えられない限り、列挙されているものより多くのものの存在を許容しているが、先行技術の実施形態は除外する。
【0038】
本開示全体にわたって状況に応じて、他の定義が現れる。
【0039】
本発明の組成物及び方法
マイクロRNA(miRNA)は、哺乳動物を含む、種々の生物に存在し、進化において保存されている低分子(例えば、18〜24ヌクレオチド)の非コードRNAのクラスである。miRNAは、RNAse III酵素のドローシャ及びダイサーによる配列切断を介する一次転写産物に由来する約70ヌクレオチドのヘアピン前駆体からプロセシングされる。多くのマイクロRNAは、プレmRNAのイントロン内又はncRNA遺伝子内に受け入れられる、遺伝子間領域においてコードされ得る。多くのmiRNAはまた、クラスター化され、ポリシストロンとして転写される傾向があり、多くの場合、類似した時空間的発現パターンを有する。miRNAは、発生のタイミング、分化、アポトーシス、細胞増殖、臓器発生、及び代謝を含む、種々の生物学的プロセスにおいて役割を有することが見出されている。
【0040】
本明細書において配列番号1及び2とそれぞれ称する、マイクロRNA−33a及びマイクロRNA−33b(miR−33a及びmiR−33b)は、翻訳抑制のためにATP結合カセット輸送体ABCA1を標的とする。したがって、miR−33の阻害は、ABCA1レベル、HDL産生、及びコレステロールの逆輸送を増加させるための新規の治療手段を表す。
【0041】
本発明の組成物及び方法は、コレステロール恒常性を必要とする被験体においてmiR−33a及び/又はmiR−33bを阻害するように設計される。一態様において、本発明は、コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で、配列番号1又は2と相補的である核酸配列を含む組成物を提供する。これは、例えば、限定されないが、配列番号3又は配列番号4のアンチセンスオリゴヌクレオチドを含む、miR−33a又はmiR−33bと相補的であるアンチセンスオリゴヌクレオチドを投与することによって達成され得る。本発明の方法を実施するのに使用するため、及びmiR−33a又はmiR−33bと相補的である、他の核酸配列は、限定されないが、shRNA若しくはsiRNAを含む干渉RNA、又はアンタゴマーなどのmiR−33a又はmiR−33bの転写後プロセシングを阻害するものであり得る。
【0042】
アンチセンスオリゴヌクレオチドは、通常、標的に結合し、転写、翻訳、又はスプライシングのレベルで発現を停止することによってDNA又はRNA標的の発現を遮断するように設計される。本発明のアンチセンスオリゴヌクレオチドは、ストリンジェントな条件下で、miR−33a及び/又はmiR−33bとハイブリダイズするように設計されている相補的核酸配列である。したがって、オリゴヌクレオチドは、標的と十分に相補的であるように、すなわち、所望の効果を与えるのに非常に十分に、かつ、十分な特異性でハイブリダイズするように選択される。
【0043】
本発明の文脈において、ハイブリダイゼーションとは、相補的ヌクレオシド又はヌクレオチド塩基間のワトソン−クリック水素結合、フーグスティーン水素結合、又は逆フーグスティーン水素結合であり得る、水素結合を意味する。例えば、アデニン及びチミンは、水素結合の形成を介して対になる相補的核酸塩基である。本明細書で使用する場合、相補的とは、2つのヌクレオチド間の正確な対合のための能力をいう。例えば、オリゴヌクレオチドの特定の位置におけるヌクレオチドが、DNA又はRNA分子の同じ位置におけるヌクレオチドと水素結合できる場合、オリゴヌクレオチド及びDNA又はRNAは、その位置において互いに相補的であるとみなされる。各分子における十分な数の対応する位置が、互いに水素結合できるヌクレオチドによって占められている場合、オリゴヌクレオチド及びDNA又はRNAは互いに相補的である。したがって、「特異的にハイブリダイズできる」及び「相補的」は、安定及び特異的な結合が、オリゴヌクレオチドと、DNA又はRNA標的との間で起こるような十分な程度の相補性又は正確な対合を示すために使用される用語である。
【0044】
相補的核酸配列は、特異的にハイブリダイズできるその標的核酸の配列と100%相補的であることを必要としないことは、当該技術分野において理解されている。標的DNA又はRNA分子に対する配列の結合が、活性を喪失させるために標的DNA又はRNAの正常な機能を妨げる場合、並びに特異的結合が所望される条件下、すなわち、インビボアッセイ又は治療的処置の場合、及びインビトロアッセイの場合における生理学的条件下、アッセイが適切なストリンジェンシーの条件下で実施される条件下で、非標的配列に対する配列の非特異的結合を回避するために十分な程度の相補性が存在する場合、本発明の相補的核酸配列は特定的にハイブリダイズできる。例えば、ストリンジェント塩濃度は、通常、約750mMのNaCl及び75mMのクエン酸三ナトリウムより少なく、好ましくは約500mMのNaCl及び50mMのクエン酸三ナトリウムより少なく、より好ましくは約250mMのNaCl及び25mMのクエン酸三ナトリウムより少ない。低ストリンジェンシーハイブリダイゼーションは、有機溶媒、例えばホルムアミドの非存在下で得ることができるのに対して、高ストリンジェンシーハイブリダイゼーションは、少なくとも約35%のホルムアミド、より好ましくは少なくとも約50%のホルムアミドの存在下で得ることができる。ストリンジェント温度条件としては、通常、少なくとも約30℃、より好ましくは少なくとも約37℃、最も好ましくは少なくとも約42℃の温度が挙げられる。ハイブリダイゼーション時間、界面活性剤、例えば、ドデシル硫酸ナトリウム(SDS)の濃度、及びキャリアDNAの包含又は排除などの付加パラメーターを変化させることは、当業者に周知である。必要に応じて、これらの種々の条件を組み合わせることで種々のレベルのストリンジェンシーが達成される。好ましい実施形態において、750mMのNaCl、75mMのクエン酸三ナトリウム、及び1%のSDS中で30℃にてハイブリダイゼーションが起こる。より好ましい実施形態において、500mMのNaCl、50mMのクエン酸三ナトリウム、1%のSDS、35%のホルムアミド、及び100μg/mlの変性サケ精子DNA(ssDNA)中で37℃にてハイブリダイゼーションが起こる。最も好ましい実施形態において、250mMのNaCl、25mMのクエン酸三ナトリウム、1%のSDS、50%のホルムアミド、及び200μg/mlのssDNA中で42℃にてハイブリダイゼーションが起こる。これらの条件における有用な変化は、当業者であれば容易に明らかとなる。
【0045】
ほとんどの用途において、ハイブリダイゼーション後の洗浄工程もストリンジェンシーを変化させる。洗浄ストリンジェンシー条件は、塩濃度及び温度で定義できる。前述のように、塩濃度を低下させるか又は温度を上昇させることで洗浄ストリンジェンシーを増加させることができる。例えば、洗浄工程用のストリンジェント塩濃度は、好ましくは約30mMのNaCl及び3mMのクエン酸三ナトリウムより少なく、最も好ましくは約15mMのNaCl及び1.5mMのクエン酸三ナトリウムより少ない。洗浄工程用のストリンジェント温度条件としては、通常、少なくとも約25℃、より好ましくは少なくとも約42℃、更により好ましくは少なくとも約68℃の温度が挙げられる。好ましい実施形態において、洗浄工程は、30mMのNaCl、3mMのクエン酸三ナトリウム、及び0.1%のSDS中で25℃にて起こる。より好ましい実施形態において、洗浄工程は、15mMのNaCl、1.5mMのクエン酸三ナトリウム、及び0.1%のSDS中で42℃にて起こる。より好ましい実施形態において、洗浄工程は、15mMのNaCl、1.5mMのクエン酸三ナトリウム、及び0.1%のSDS中で68℃にて起こる。これらの条件における更なる変化は、当業者であれば容易に明らかとなる。ハイブリダイゼーション技術は、当業者に周知であり、例えば、Benton及びDavis(Science 196:180,1977);Grunstein及びHogness(Proc.Natl.Acad.Sci.,USA 72:3961,1975);Ausubelら,(Current Protocols in Molecular Biology,Wiley Interscience,New York,2001);Berger及びKimmel(Guide to Molecular Cloning Techniques,1987,Academic Press,New York);並びにSambrookら,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,New Yorkに記載されている。
【0046】
本発明のアンチセンスオリゴヌクレオチドは、標的核酸内の標的領域と少なくとも80%の配列相補性を含むことが好ましく、更にそれらは90%の配列相補性を含むことが好ましく、標的とされる標的核酸配列内の標的領域と95%の配列相補性を含むことが更により好ましい。例えば、アンチセンスオリゴヌクレオチドの20核酸塩基のうちの18が相補的であると、標的領域と特異的にハイブリダイズするアンチセンス化合物は90パーセントの相補性を表す。標的核酸領域を有するアンチセンス化合物の相補性パーセントは、基本ローカルアラインメント検索ツール(BLASTプログラム)を用いて慣例的に決定することができる(Altschulら,J.Mol.Biol.1990,215,403−410:Zhang及びMadden,Genome Res.,1997,7,649−656)。miR−33a又はmiR−33bとハイブリダイズする、本発明のアンチセンス及び他の化合物は実験により確認され、それらの化合物の代表的な配列は、本発明の好ましい実施形態として本明細書の以下で確認される(例えば、限定されないが、配列番号3又は配列番号4のアンチセンスオリゴヌクレオチドを含む)。
【0047】
別の実施形態において、配列番号1又は2と相補的である核酸配列は、限定されないが、shRNA又はsiRNAを含む、干渉RNAであり得る。干渉RNAとしては、限定されないが、低分子干渉RNA(「siRNA」)及び低分子ヘアピン型RNA(「shRNA」)が挙げられる。干渉RNAを構築するための方法は当該技術分野において周知である。例えば、干渉RNAは、2つの別のオリゴヌクレオチドから構築することができ、一方の鎖はセンス鎖であり、他方の鎖はアンチセンス鎖であり、アンチセンス鎖及びセンス鎖は自己相補的であり(すなわち、例えばアンチセンス鎖及びセンス鎖が二本鎖又は二本鎖構造を形成する場合などに、各々の鎖は他方の鎖におけるヌクレオチド配列と相補的であるヌクレオチド配列を含む)、アンチセンス鎖は、標的核酸分子又はその一部(すなわち望まれていない遺伝子)におけるヌクレオチド配列と相補的であるヌクレオチド配列を含み、センス鎖は、標的核酸配列又はその一部に対応するヌクレオチド配列を含む。代替として、干渉RNAは単一のオリゴヌクレオチドから構築され、自己相補的センス領域及びアンチセンス領域は、核酸ベース又は非核酸ベースのリンカー(複数可)によって連結される。干渉RNAは、自己相補的センス領域及びアンチセンス領域を有する、二本鎖、非対称二本鎖、ヘアピン又は非対称ヘアピン二次構造を有するポリヌクレオチドであってもよく、アンチセンス領域は、別の標的核酸分子又はその一部におけるヌクレオチド配列と相補的であるヌクレオチド配列を含み、センス領域は、標的核酸配列又はその一部に対応するヌクレオチド配列を有する。干渉は、自己相補的センス領域及びアンチセンス領域を含む2つ以上のループ構造及び1つのステムを有する環状の一本鎖ポリヌクレオチドであってもよく、アンチセンス領域は、標的核酸分子又はその一部におけるヌクレオチド配列と相補的であるヌクレオチド配列を含み、センス領域は、標的核酸配列又はその一部に対応するヌクレオチド配列を有し、環状ポリヌクレオチドは、RNA干渉を介することができる活性siRNA分子を生成するためにインビボ又はインビトロのいずれかでプロセシングされ得る。
【0048】
本発明の特定の実施形態において、干渉RNAコード領域は、センス領域、アンチセンス領域及びループ領域を有する自己相補的RNA分子をコードする。望ましく発現される場合、かかるRNA分子は「ヘアピン」構造を形成し、本明細書で「shRNA」という。ループ領域は、通常、長さが約2から約10の間のヌクレオチドである。好ましい実施形態において、ループ領域は、長さが約6から約9のヌクレオチドである。本発明の1つのかかる実施形態において、センス領域及びアンチセンス領域は、長さが約15から約20の間のヌクレオチドである。転写後プロセシング後、低分子ヘアピン型RNAは、RNase IIIファミリーのメンバーである、酵素のダイサーを介して行われる切断事象によってsiRNAに変換される。次いで、siRNAは、相同性を共有する遺伝子の発現を阻害できる。詳細については、Brummelkampら,Science 296:550−553,(2002);Leeら,Nature Biotechnol.,20,500−505,(2002);Miyagishi及びTaira,Nature Biotechnol 20:497−500,(2002);Paddisonら,Genes & Dev.16:948−958,(2002);Paul,Nature Biotechnol,20,505−508,(2002);Sui,Proc.Natl.Acad.Sd.USA,99(6),5515−5520,(2002);Yuら,Proc NatlAcadSci USA 99:6047−6052,(2002)を参照のこと。
【0049】
siRNAによって誘導される標的RNA切断反応は高度に配列特異的である。一般に、標的遺伝子(すなわち、miR−33a及び/又はmiR−33b)の一部と同一であるヌクレオチド配列を含有するsiRNAが阻害のために好ましい。しかしながら、siRNAと標的遺伝子との間の100%の配列同一性は、本発明を実施するのに必要とされない。したがって、本発明は、遺伝子変異、株多型性、又は進化的分岐に起因して予測され得る配列変異を耐容できる利点を有する。例えば、標的配列に対する挿入、欠失、及び単一点突然変異を有するsiRNA配列もまた、阻害に効果的であることが見出されている。代替として、ヌクレオチド類似体置換又は挿入を有するsiRNA配列が阻害に効果的であってもよい。
【0050】
更に別の実施形態において、配列番号1又は2と相補的である核酸配列はアンタゴマーである。アンタゴマーは、マイクロRNAを標的とする一本鎖、二本鎖、部分的に二本鎖及びヘアピン構造化された化学的に修飾されたオリゴヌクレオチドである。好ましくは、本発明において特徴付けられたアンタゴマーは、約12〜25ヌクレオチド、好ましくは約15〜23ヌクレオチドのmiRNA標的配列とハイブリダイズするのに十分に相補的であるヌクレオチド配列を含む。
【0051】
特定の実施形態において、アンタゴマーは、RNaseプロテクション並びに増大した組織及び細胞取り込みなどの薬理学的特性についての種々の修飾を保有するRNA様オリゴヌクレオチドである。アンタゴマーは、糖の完全な2’−O−メチル化、ホスホロチオエート骨格及び3’末端におけるコレステロール部分を有することによって正常なRNAとは異なり得る。ホスホロチオエート修飾は、増大した組織取り込みの一因となるRNase活性及びそれらの親油性に対する保護を与える。好ましい実施形態において、アンタゴマーは6つのホスホロチオエート骨格修飾を含み、2つのホスホロチオエートは5’末端に位置し、4つは3’末端に位置する。本発明のアンタゴマーはまた、それらの長さ又はアンタゴマーを構成するヌクレオチドの数に関して修飾されてもよい。本発明のアンタゴマーは、最適機能のために長さが約20〜21ヌクレオチドであることが好ましい。なぜならこのサイズはmiR−33a及びmiR−33bについての成熟マイクロRNAのサイズと一致するからである。
【0052】
RNA、cDNA、ゲノムDNA、ベクター、ウイルス又はそのハイブリッドであろうとなかろうと、本発明を実施するために使用される核酸配列は、種々の源から単離でき、遺伝子操作でき、増幅でき、及び/又は発現/組換え的に生成できる。組換え核酸配列は、個々に単離又はクローニングでき、所望の活性について試験できる。例えばインビトロ、細菌、真菌、哺乳動物、酵母、昆虫又は植物細胞発現系を含む、任意の組換え発現系が使用されてもよい。
【0053】
本発明の核酸配列は、送達ベクターに挿入でき、そのベクター内の転写ユニットから発現できる。組換えベクターは、DNAプラスミド又はウイルスベクターであってもよい。ベクター構築物の生成は、限定されないが、例えば、Sambrookら,Molecular Cloning:A Laboratory Manual.(1989)),Coffinら,(Retroviruses.(1997))及び「RNA Viruses:A Practical Approach」(Alan J.Cann,Ed.,Oxford University Press,(2000))に記載されているようなPCR、オリゴヌクレオチド合成、制限エンドヌクレアーゼ消化、ライゲーション、形質転換、プラスミド精製、及びDNAシークエンシングの標準的技術を含む、当該技術分野において周知の任意の適切な遺伝子操作技術を用いて成され得る。当業者に明らかであるように、種々の適切なベクターが、本発明の核酸を細胞に移すのに利用可能である。核酸を送達するのに適したベクターの選択及び選択された発現ベクターの細胞への挿入のための条件の最適化は、過度の実験を必要とせずに、当業者の想定の範囲内である。ウイルスベクターは、パッケージング細胞において組換えウイルスの産生のための配列を有するヌクレオチド配列を含む。本発明の核酸を発現するウイルスベクターは、限定されないが、レトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、ポックスウイルス又はアルファウイルスを含む、ウイルス骨格に基づいて構築できる。本発明の核酸を発現できる組換えベクターは本明細書に記載されるように送達でき、標的細胞(例えば、安定な形質転換体)に存続する。
【0054】
本発明を実施するために使用される核酸配列は、例えば、Adams(1983)J.Am.Chem.Soc.105:661;Belousov(1997)Nucleic Acids Res.25:3440−3444;Frenkel(1995)Free Radic.Biol.Med.19:373−380;Blommers(1994)Biochemistry 33:7886−7896;Narang(1979)Meth.Enzymol.68:90;Brown(1979)Meth.Enzymol.68:109;Beaucage(1981)Tetra.Lett.22:1859;米国特許第4,458,066号に記載されているような周知の化学合成技術によってインビトロで合成できる。
【0055】
本発明の核酸配列は、修飾、例えばヌクレオチド修飾の導入などによって核酸分解に対して安定化できる。例えば、本発明の核酸配列は、ヌクレオチド配列の5’又は3’末端においてホスホロチオエートの少なくとも第1、第2又は第3のヌクレオチド間結合を含む。別の例として、核酸配列は、2’修飾ヌクレオチド、例えば、2’−デオキシ、2’−デオキシ−2’−フルオロ、2’−O−メチル、2’−O−メトキシエチル(2’−O−MOE)、2’−O−アミノプロピル(2’−O−AP)、2’−O−ジメチルアミノエチル(2’−O−DMAOE)、2’−O−ジメチルアミノプロピル(2’−O−DMAP)、2’−O−ジメチルアミノエチルオキシエチル(2’−O−DMAEOE)、又は2’−O−N−メチルアセトアミド(2’−O−NMA)を含んでもよい。別の例として、核酸配列は、少なくとも1つの2’−O−メチル修飾ヌクレオチド含んでもよく、一部の実施形態において、ヌクレオチドの全ては2’−O−メチル修飾を含む。一部の実施形態において、核酸は「ロック」されている。すなわち、核酸は、2’−O原子及び4’−C原子を結合するメチレン架橋によってリボース環が「ロック」されている核酸類似体を含む(例えば、Kaupinnenら,Drug Disc.Today 2(3):287−290(2005);Koshkinら,J.Am.Chem.Soc.,120(50):13252−13253(1998)を参照のこと)。
【0056】
例えば、サブクローニング、ラベリングプローブ(例えば、クレノウポリメラーゼを用いるランダムプライマーラベリング、ニックトランスレーション、増幅)、シークエンシング、ハイブリダイゼーションなどの本発明を実施するために使用される核酸操作のための技術は、科学文献及び特許文献に十分に記載されている。例えば、Sambrook,ed.,MOLECULAR CLONING:A LABORATORY MANUAL(2ND ED.),Vols.1−3,Cold Spring Harbor Laboratory,(1989);CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,Ausubel,ed.john Wiley & Sons,Inc.,New York(1997);LABORATORY TECHNIQUES IN BIOCHEMISTRY AND MOLECULAR BIOLOGY:HYBRIDIZATION WITH NUCLEIC ACID PROBES,Part I.Theory and Nucleic Acid Preparation,Tijssen,ed.Elsevier,N.Y.(1993)を参照のこと。
【0057】
本発明は、翻訳抑制のためにATP結合カセット輸送体ABCA1を標的とし、それによって、ABCA1レベル、HDL産生、及びコレステロールの逆輸送を増加させるように設計される核酸配列を含む医薬組成物及び製剤を提供する。
【0058】
代替の実施形態において、本発明の組成物は薬学的に許容される担体と共に製剤化される。代替の実施形態において、本発明の医薬組成物及び製剤は、非経口、局所的、経口、又はエアゾール若しくは経皮などによる局所投与によって投与できる。医薬組成物は任意の方法で製剤化でき、状態又は疾患(例えば、心血管障害の種類)及び病気の程度、各患者の全身の医学的状態、もたらされる好ましい投与方法に応じて、種々の単位剤形で投与できる。製剤化及び薬剤の投与についての技術に関する詳細は、科学文献及び特許文献に十分に記載されている。例えば、Remington’s Pharmaceutical Sciences,Maack Publishing Co,Easton PA(「Remington’s」)の最新版を参照のこと。
【0059】
本発明の核酸配列は、単独で、又は医薬製剤(組成物)の成分として投与できる。化合物は、ヒト又は獣医用の医薬に使用するための任意の簡便な方法で投与のために製剤化できる。湿潤剤、乳化剤及びラウリル硫酸ナトリウム及びステアリン酸マグネシウムなどの滑沢剤、並びに着色剤、離型剤、コーティング剤、甘味剤、香味剤及び芳香剤、防腐剤及び酸化防止剤も組成物中に存在してもよい。
【0060】
本発明の組成物の製剤は、皮内、吸入、経口/経鼻、局所、非経口、直腸、及び/又は膣内投与に適切なものを含む。製剤は簡便に単位剤形で存在してもよく、薬学の分野で周知の任意の方法によって調製されてもよい。単一の剤形を生成するために担体物質と組み合わされ得る活性成分(例えば、本発明の核酸配列)の量は、治療される宿主、特定の投与様式、例えば皮内又は吸入に応じて変更されるだろう。単一の剤形を生成するために担体物質と組み合わされ得る活性成分の量は、通常、治療的効果、例えば、抗原特異的T細胞又は体液性応答を生じる化合物の量である。
【0061】
本発明の医薬製剤は、製薬に関する分野で知られている任意の方法に従って調製できる。かかる薬物は、甘味剤、香味剤、着色剤及び保存剤を含有してもよい。製剤は、製造に適切である非毒性の薬学的に許容される添加剤と混合されてもよい。製剤は、1つ以上の賦形剤、乳化剤、防腐剤、緩衝剤、添加剤などを含んでもよく、液体剤、散剤、乳剤、凍結乾燥粉末、スプレー剤、クリーム剤、ローション剤、放出制御製剤、錠剤、丸剤、ゲル剤、パッチ剤上、インプラントにおいてなどの形態で提供されてもよい。
【0062】
経口投与のための医薬製剤は、適した及び適切な用量で、当該技術分野において周知の薬学的に許容される担体を用いて製剤化されてよい。かかる担体は、薬剤が、患者による摂取に適切な錠剤、丸剤、散剤、糖衣錠、カプセル剤、液体剤、ロゼンジ剤、ゲル剤、シロップ剤、スラリー剤、懸濁剤などとして単位剤形で製剤化されることを可能にする。経口使用のための医薬調製物は固形添加剤として製剤化でき、所望の場合、錠剤又は糖衣錠コアを得るために、適切な付加化合物を加えた後で、任意選択に、得られた混合物を粉砕し、顆粒の混合物を加工する。適切な固形添加剤は、例えば、ラクトース、スクロース、マンニトール、又はソルビトールを含む糖;トウモロコシ、コムギ、コメ、ジャガイモ、又は他の植物由来のデンプン;メチルセルロース、ヒドロキシプロピルメチル−セルロース、又はナトリウムカルボキシ−メチルセルロースなどのセルロース;並びにアラビア及びトラガントを含むガム;並びにタンパク質、例えばゼラチン及びコラーゲンを含む、糖質又はタンパク質の充填剤である。架橋ポリビニルピロリドン、寒天、アルギン酸、又はアルギン酸ナトリウムなどのその塩などの崩壊剤又は可溶化剤が加えられてもよい。押し込み式カプセル剤は、ラクトース又はデンプンなどの充填剤又は結合剤、タルク又はステアリン酸マグネシウムなどの滑沢剤、及び任意選択に安定剤と混合した活性剤を含有してもよい。軟カプセル剤において、活性剤は、安定剤を含んでも含まなくてもよく、脂肪油、液体パラフィン、又は液体ポリエチレングリコールなどの適切な液体に溶解又は懸濁されてもよい。
【0063】
水性懸濁剤は、例えば、水性皮内注射用の水性懸濁剤の製造に適切な添加剤と混合して活性剤(例えば、本発明の核酸配列)を含有してもよい。かかる添加剤としては、ナトリウムカルボキシメチルセルロース、メチルセルロース、ヒドロキシプロピルメチルセルロース、アルギン酸ナトリウム、ポリビニルピロリドン、トラガントガム及びアカシアガムなどの懸濁化剤、天然に存在するリン脂質(例えば、レシチン)などの分散化剤又は湿潤剤、アルキレンオキシドと脂肪酸との縮合生成物(例えば、ポリオキシエチレンステアレート)、エチレンオキシドと長鎖脂肪族アルコールとの縮合生成物(例えば、ヘプタデカエチレンオキシセタノール(heptadecaethylene oxycetanol))、エチレンオキシドと脂肪酸及びヘキシトールから得られる部分エステルとの縮合生成物(例えば、ポリオキシエチレンソルビトールモノ−オレエート)、又はエチレンオキシドと脂肪酸及びヘキシトール無水物から得られる部分エステルとの縮合生成物(例えば、ポリオキシエチレンソルビタンモノオレエート)が挙げられる。また、水性懸濁剤は、p−ヒドロキシ安息香酸エチル又はp−ヒドロキシ安息香酸−n−プロピルなどの1つ又は複数の防腐剤、1つ又は複数の着色剤、1つ又は複数の香味剤、及びスクロース、アスパルテーム又はサッカリンなどの1つ又は複数の甘味剤を含有してもよい。製剤はオスモル濃度で調整できる。
【0064】
一実施形態において、油ベースの薬剤が、本発明の核酸配列を投与するために使用される。油ベースの懸濁剤は、ラッカセイ油、オリーブ油、ゴマ油若しくはココナッツ油などの植物油中、又は流動パラフィンなどの鉱油中、或いはそれらの混合物中で活性剤を懸濁することにより製剤化できる。例えば、生物学的利用能を増加させ、経口投与した疎水性医薬化合物の個体間変動及び個体内変動を減少させるための精油又は精油成分を用いることを記載している米国特許第5,716,928号を参照のこと(米国特許第5,858,401号も参照のこと)。油懸濁剤は、蜜ろう、固形パラフィン又はセチルアルコールなどの増粘剤を含有してもよい。甘味剤が、グリセロール、ソルビトール又はスクロースなどの味の良い経口用調製物を提供するために加えられてもよい。これらの製剤は、アスコルビン酸などの酸化防止剤を加えることによって保存されてもよい。注射用油ビヒクルの例としては、Minto(1997)J.Pharmacol.Exp.Ther.281:93−102を参照のこと。
【0065】
本発明の医薬製剤はまた、水中油型エマルションの形態であってもよい。油性相は、上記の植物油若しくは鉱油又はそれらの混合物であってもよい。適切な乳化剤としては、アカシアガム及びトラガントガムなどの天然に存在するガム、大豆レシチンなどの天然に存在するリン脂質、ソルビタンモノオレエートなどの、脂肪酸及びヘキシトール無水物に由来するエステル又は部分エステル、並びにポリオキシエチレンソルビタンモノオレエートなどの、上記部分エステルとエチレンオキシドとの縮合生成物が挙げられる。エマルションはまた、シロップ剤及びエリキシル剤の製剤などで甘味剤及び香味剤を含有してもよい。かかる製剤はまた、鎮痛剤、防腐剤、又は着色剤を含有してもよい。代替の実施形態において、本発明のこれらの注射用水中油型エマルションは、パラフィン油、ソルビタンモノオレエート、エトキシ化ソルビタンモノオレエート及び/又はエトキシ化ソルビタントリオレートを含む。
【0066】
本発明を実施する際に、医薬化合物はまた、坐剤、インサフレーション、粉末及びエアゾール製剤(例えばステロイド吸入剤、例えば、Rohatagi(1995)J.Clin.Pharmacol.35:1187−1193;Tjwa(1995)Ann.Allergy Asthma Immunol.75:107−111を参照のこと)を含み、経鼻、眼内及び膣内経路により投与できる。坐剤製剤は、常温で固体であるが、体温で液体になるので、身体で溶解して薬物を放出する適切な非刺激性の添加剤と、薬物とを混合することにより調製できる。かかる物質はココアバター及びポリエチレングリコールである。
【0067】
本発明を実施する際に、医薬化合物は、塗布棒、溶液剤、懸濁剤、乳剤、ゲル剤、クリーム剤、軟膏剤、ペースト剤、ゼリー剤、塗布剤、粉末剤、及びエアゾール剤として製剤化され、局所経路により経皮的に送達できる。
【0068】
本発明を実施する際に、医薬化合物はまた、身体における持続放出のためにミクロスフェアとして送達できる。例えば、ミクロスフェアは、皮下に持続放出される薬物の皮内注射により投与できる;Rao(1995)J.Biomater Sci.Polym.Ed.7:623−645を参照のこと;生分解性及び注射用ゲル製剤として、例えば、Gao(1995)Pharm.Res.12:857−863(1995)を参照のこと;又は経口投与のためのミクロスフェアとして、例えば、Eyles(1997)J.Pharm.Pharmacol.49:669−674を参照のこと。
【0069】
本発明を実施する際に、医薬化合物は、静脈内(IV)投与又は体腔若しくは器官の内腔への投与などにより非経口で投与できる。それらの製剤は、薬学的に許容される担体に溶解される活性剤の溶液を含んでもよい。利用され得る許容されるビヒクル及び溶媒は、水及びリンガー溶液、等張食塩水である。加えて、滅菌不揮発性油が、溶媒又は懸濁媒として利用されてもよい。この目的のために、合成モノグリセリド又はジグリセリドを含む、任意の無菌不揮発性油が利用されてもよい。加えて、オレイン酸などの脂肪酸が、同様に、注射用調製物に使用されてもよい。それらの溶液は滅菌され、通常、望ましくない物質を含まない。それらの製剤は、従来の周知の滅菌技術により滅菌されてもよい。製剤は、pH調整剤及び緩衝剤、毒性調整剤、例えば、酢酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム、乳酸ナトリウムなどのほぼ生理的条件に必要とされる薬学的に許容される補助物質を含有してもよい。それらの製剤中の活性剤の濃度は、広範に変化されてもよく、選択される特定の投与様式及び患者の必要性に従って、液量、粘性、体重などに主に基づいて選択される。IV投与のために、製剤は、滅菌注射用水溶液又は油性懸濁液などの滅菌注射用調製物であってもよい。この懸濁液は、上記の適切な分散化剤又は湿潤剤及び懸濁化剤を用いて製剤化されてもよい。滅菌注射用調製物はまた、1,3−ブタンジオール溶液などの非毒性の非経口的に許容される賦形剤又は溶媒中の懸濁液であってもよい。投与は、ボーラス又は持続注入(例えば、特定期間、血管内への実質的に連続導入)によってなされてもよい。
【0070】
本発明の医薬化合物及び製剤は凍結乾燥されてもよい。本発明は、本発明の薬剤及び充填剤、例えば、マンニトール、トレハロース、ラフィノース、及びスクロース又はそれらの混合物を含む溶液を凍結乾燥することによって作製され得る、本発明の組成物を含む安定な凍結乾燥製剤を提供する。安定な凍結乾燥製剤を調製するためのプロセスは、約2.5mg/mLのタンパク質、約15mg/mLのスクロース、約19mg/mLのNaCl、及び5.5より高いが、6.5未満のpHを有するクエン酸ナトリウム緩衝剤の溶液を凍結乾燥することを含んでもよい。例えば、米国特許出願公開第20040028670号を参照のこと。
【0071】
本発明の組成物及び製剤は、リポソームを使用することにより送達できる。リポソームを使用することにより、特に、リポソーム表面が、標的細胞に特異的なリガンドを保持するか、又はそうでなければ特定の器官に選択的に向けられている場合、インビボにおける標的細胞への活性剤の送達に集中することができる。例えば、米国特許第6,063,400号;米国特許第6,007,839号;Al−Muhammed(1996)J.Microencapsul.13:293−306;Chonn(1995)Curr.Opin.Biotechnol.6:698−708;Ostro(1989)Am.J.Hosp.Pharm.46:1576−1587を参照のこと。
【0072】
本発明の製剤は、予防的及び/又は治療的処置のために投与できる。代替の実施形態において、治療的用途のために、組成物は、障害又はその合併症、例えば、心臓血管異常の臨床症状を治療、緩和又は部分的に停止するのに十分な量(これは治療有効量と呼ばれ得る)で、コレステロール恒常性を必要とする被験体に投与される。例えば、代替の実施形態において、本発明の医薬組成物は、ATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させ、細胞内コレステロールの流出及び/若しくは肝臓におけるHDLの産生を増加させ、並びに/又はコレステロール恒常性を必要とする被験体の血液を循環するコレステロールの量を減少させるのに十分な量で投与される。
【0073】
これを達成するのに適切な医薬組成物の量が治療有効量である。これに使用するための投薬スケジュール及び有効量、すなわち投与レジメンは、疾患又は状態の段階、疾患又は状態の重症度、患者の健康の一般状態、患者の身体状況、年齢などを含む、種々の要因に依存するだろう。患者のための投薬レジメンを判断する際に、投与様式も考慮される。
【0074】
投薬レジメンはまた、当該技術分野において周知の薬物動態パラメーター、すなわち、活性剤の吸収率、生物学的利用能、代謝、クリアランスなども考慮される(例えば、Hidalgo−Aragones(1996)J.Steroid Biochem.Mol.Biol.58:611−617;Groning(1996)Pharmazie 51:337−341;Fotherby(1996)Contraception 54:59−69;Johnson(1995)J.Pharm.Sci.84:1144−1146;Rohatagi(1995)Pharmazie 50:610−613;Brophy(1983)Eur.J.Clin.Pharmacol.24:103−108;最新のRemington’s,前出を参照のこと)。現在の技術水準により、臨床医は、個々の患者、活性剤及び治療される疾患又は状態の各々についての投薬レジメンを決定することができる。薬剤として使用される類似組成物のために提供されるガイドラインは、投薬レジメン、すなわち、投与スケジュール及び投薬レベルを決定するための指針として使用され、本発明の方法を正確かつ適正に実施することができる。
【0075】
製剤の単回又は多回投与は、例えば、患者に必要とされ、忍容される用量及び頻度、各投与後に生じるコレステロール恒常性の程度及び量などに応じて与えることができる。製剤は、状態、疾患又は症状を有効に治療、予防又は寛解する(例えば、ABCA1の発現を増加させ、細胞内コレステロールの流出及び/若しくは肝臓におけるHDLの産生を増加させ、並びに/又はコレステロール恒常性を必要とする被験体の血液を循環するコレステロールの量を減少させる)のに十分な量の活性剤を与えるべきである。
【0076】
代替の実施形態において、経口投与のための医薬製剤は、約1〜100mg/kg体重/日又はそれ以上の日用量である。経口投与と対照的に、血流、体腔又は器官の内腔内へのより低い用量が使用されてもよい。十分に高い用量が、局所若しくは経口投与又は粉末、スプレー若しくは吸入による投与において使用されてもよい。非経口又は経口投与可能な製剤を調製するための実際の方法は、当業者に公知であるか、又は明らかであり、Remington’s、前出などの文献により詳細に記載されている。
【0077】
種々の研究により、相補的核酸配列を用いた成功している哺乳動物の投薬が報告されている。例えば、Esau C.ら,(2006)Cell Metabolism,3(2):87−98は、4週間、週に2回の12.5〜75mg/kgの範囲のmiR−122アンチセンスオリゴヌクレオチドの腹腔内投与による正常なマウスの投薬を報告している。マウスは、体重を損失せず、又は食物摂取を減少せずに、治療の終わりに健康で、正常なように見えた。血漿アミノ基転移酵素レベルは、ALT及びASTレベルの非常に穏やかな増加を示した、75mg/kg用量のmiR−122 ASOを除いて、全ての用量について正常な範囲(AST3/4 45、ALT3/4 35)であった。これらにより、50mg/kgが有効で非毒性の用量であることが結論付けられた。Krutzfeldt Jら,(2005)Nature 438,685−689による別の研究は、80、160又は240mg/kg体重の総量を用いてマウスにおいtmiR−122をサイレンシングするためにアンタゴマーを注入した。最も高い用量はmiR−122シグナルの完全な損失を生じた。更に別の研究において、ロックド核酸(「LNA」)が、miR−122をサイレンシングするために霊長類において首尾よく適用された。Elmen J.ら,(2008)Nature 452,896−899は、miR−122の効果的なサイレンシングを、3用量の10mg/kgのLNA−抗miRによって霊長類において達成し、研究動物においてLNA関連毒性又は組織病理学的変化についてのいかなる形跡もなく、総血漿コレステロールの長期間存続し、可逆的な減少を導いたことを報告している。
【0078】
本発明の方法は、他の薬物又は薬剤、例えば、コレステロール恒常性を提供するための組成物との同時投与を更に含んでもよい。例えば、本発明の方法並びに/又は組成物及び製剤は、HMG CoA還元酵素阻害剤、例えば、アトルバスタチン、フルバスタチン、ロバスタチン、プラバスタチン、ロスバスタチン及びシンバスタチン、胆汁酸捕捉剤、例えば、コレセベラム、コレスチラミン及びコレスチポール、並びにニコチン酸(ナイアシン)などの増大したLDLコレステロールを治療するための薬物と同時投与できる。
【0079】
本発明は、本発明のより十分な理解及びその多くの利点を与える、以下に示す、非限定的な実施例によって更に説明される。
【実施例】
【0080】
本明細書中で、SREBPと協調してコレステロール恒常性を制御する際にmiR−33の役割を更に詳細に説明するための一連のインビトロ及びインビボでの研究を記載する。以下の実施例は例示の目的のみのために提示し、発明者らが彼らの発明とみなす発明の範囲を限定することを意図しない。
【実施例1】
【0081】
ヒト及びマウスにおけるmiR−33の予測される標的
miRNAによるmRNA標的の識別に対する利用可能なアプローチの中で、計算による予測研究が、miRNA標的の実験検証に対する第1のステップを確立する際の有用な方法を表すことが示された。miRNA標的を予測する際に考慮される最も重要なパラメーターは、「種対合」として知られている、その同種標的との特定の塩基対合を受けるmiRNA配列の能力である(Bartel DP(2004)Cell 116:281−297;van den Berg A,Mols J,Han J(2008)Biochem Biophys Acta)。これは、miRNA5’末端の7〜8塩基に関する。この文脈において、Miranda及びTargetScan 4.2標的予測データベース(microrna.sanger.ac.uk;www.targetscan.org/vert_42/)の両方から導かれた100より多い予測される標的の記載の中で、ヒト及びマウスの両方におけるmiR−33についての最も関連のある保存された標的はABCA1である(表1及び図7)。興味深いことに、これらの研究は、コレステロール輸送ネットワーク又は脂質ホメオスタシスにおける役割を果たし、ABCA1と機能的に関連することが知られている付加的タンパク質はまた、ヒト及びマウスの両方における標的として役立つことを明らかにした。それらとしては、ニーマンピック病C1型(NPC1)が挙げられる(表1)。
【0082】
【表1】
【実施例2】
【0083】
ヒト及びマウス細胞株におけるmiR−33によるABCA1の下方調節
バイオインフォマティクス予測研究からの結果を検証するために、一連の実験を、モデル系として、ヒトHepG2肝細胞癌細胞、IMR−90初代ヒト線維芽細胞、及びマウスマクロファージ細胞株J774を含む、ヒト及びマウス細胞株を用いて実施した。それらの細胞株は、低細胞密度でABCA1タンパク質の低い発現を示す。miR−33によるABCA1の翻訳阻害は、この低いレベルの発現に起因すると予測された。したがって、miRNA生物発生経路の成分のsiRNAノックダウンを、ABCA1タンパク質発現がmiRNAにより調節されるか否かを決定するために実施した。実際に、ドローシャ及びダイサーmiRNAプロセシング酵素に向けられたsiRNAでのトランスフェクションにより、試験した3つ全ての細胞株において(特にドローシャsiRNAでの)ABCA1タンパク質発現の十分な増加が生じた(図7A)。これらの結果はmiRNAによるABCA1の調節と一致している。
【0084】
この上方調節が、miR−33プロセシングの欠如から特異的に生じることを決定するために、3つの異なる細胞株を、抗miR−33a/bオリゴヌクレオチド(Ambion)でトランスフェクトした。抗miRは、塩基対合による配列特異的に内因性miRNAの機能を遮断する修飾したmiRNAアンチセンスオリゴヌクレオチドである。抗miR−33a及び抗miR−33bの両方のトランスフェクションは、ヒト細胞株において、及びJ774マウスマクロファージの抗miR−33a処置の際に、ABCA1タンパク質のかなり高い発現を生じた(図7B)。
【0085】
これらのデータは、内因性miR−33によるABCA1 mRNAの翻訳又は安定性の調節を示す。ABCA1発現に対するmiR−33の効果を更に評価するために、miR−33a及びmiR−33bのそれぞれの細胞内レベルを増加させるために、細胞を合成Pre−miR−33(Ambion)でトランスフェクトした。ABCA1タンパク質レベルは、細胞播種の24時間後に検出することが困難であるため、Pre−miR33の効果を、トランスフェクション及び播種の48時間後に評価した。この実験により、ABCA1発現が、特にマウスJ774マクロファージ及びヒトIMR−90線維芽細胞において、miR−33により特異的に抑制され得ることが明確に示される(図7C)。
【0086】
最近の研究により、ABCA1が、効果的なコレステロール流出及びHDL生合成を促進するためにマクロファージにおいて、別のコレステロール輸送体、ABCG1と共に作用することが示された(図5)(Tall ARら(2008)Cell Metab 7:365−375;Wang Xら(2007)J Clin Invest 117:2216−2224)。実際に、データは、一方の輸送体が欠損して、他方が誘導される場合、代償的調節を示唆することを示した(Yvan−Charvet Lら(2007)J Clin Invest 117:3900−3908;Ranalletta Mら(2006)Arterioscler Thromb Vasc Biol 26:2308−2315)。したがって、ABCG1発現が、miRNA経路及びmiR−33aの操作後、ABCA1レベルの変化の際に変わるか否かを調べた。興味深いことに、マウスマクロファージ細胞株J774において、ダイサー及びドローシャのRNAiによるABCG1発現、抗miR−33aトランスフェクション、又はPre−miR−33a発現に対する有意な効果は観察されなかった。これらの知見により、少なくともこれらの予備実験条件下で、ABCA1及びABCG1の発現がmiR−33操作に応答して相互に関連しないことが示唆される。
【実施例3】
【0087】
miR−33によるABCA1 3’UTRの標的化
miR−33が、ABCA1 mRNA及び/又はタンパク質レベルに影響を与えるか否かを決定するために、Pre−miR−33a/bを、miR−33のレベルを増加させるために、いくつかの細胞株(すなわち、IMR−90、HepG2、及びJ774)において発現させ、miR−33を拮抗するために抗miR−33オリゴヌクレオチドをトランスフェクトし、続いて、定量的RT−PCR及び免疫ブロット法のそれぞれによってABCA1 mRNA及びタンパク質レベルを分析する。アクチンmRNA(qRT−PCR)及びチューブリンタンパク質(免疫ブロット法)を対照として使用する。
【0088】
miR−33が、3’UTRの標的化を介してABCA1レベルに影響を与えるか否かを規定するために、予測されるmiR−33標的配列を保有するヒト及びマウスABCA1 3’UTRの断片を単離し、ルシフェラーゼレポータープラスミド(pMIR−REPORT、Ambion)の3’UTRクローニング部位にクローニングした。得られたABCA1 3’UTRルシフェラーゼレポーターを、miR−33a/b抗miR、前駆体、又は対照オリゴヌクレオチドの存在下で、ヒトHEK293細胞にトランスフェクトした。図8Aに示すように、ルシフェラーゼレポーターベクター内に予測されるmiR−33標的部位を保有するABCA1 3’UTRの断片の挿入により、挿入しないベクターと比較してレポーター活性の著しい減少が生じ、挿入された断片(例えばmiR−33認識部位)内の阻害配列の存在と一致する。重要なことに、ヒトmiR−33a/bと相補的である抗miRオリゴヌクレオチドでの同時トランスフェクションはレポーターの高い発現を引き起こし、内因性miR−33a/bが、実際に、ヒトHEK293細胞におけるABCA1 3’UTR断片を標的とすることを示唆する(図8B)。反対に、野生型ヒトmiR−33a/b前駆体オリゴヌクレオチド(Ambion)での同時トランスフェクションは、レポーター活性の更なる抑制を引き起こし、一方、種塩基対合領域において変異した前駆体オリゴヌクレオチドは、前駆体対照オリゴヌクレオチドと比較して効果を有さなかった(図8C)。相対ルシフェラーゼ活性を、ウミシイタケルシフェラーゼ活性によって正規化した絶対ルシフェラーゼ活性から測定した(PC:陰性対照として非関連マイクロRNA前駆体)。
【0089】
一緒に、これらのデータは、miR−33a/bが、翻訳抑制又はmRNA分解のためのABCA1 3’UTRを標的化することによりヒトABCA1の発現を阻害するという見解についての重要な支持を与える。
【実施例4】
【0090】
マウスにおけるマクロファージからのコレステロール輸送におけるmiR−33についての役割
SREBPは、コレステロールによって古典的な負のフィードバック形式で調節される(Brown MS,Goldstein JL(1997)Cell 89:331−340)。miR−33/SREBP/ABCA1コレステロール調節回路の生物医学及び生理学関連を、マウスJ774マクロファージモデルを用いて調べた(de la Llera−Moya Mら(2010,Epub ahead of print)Arterioscler.Thromb.Vasc.Biol)。J774マウスマクロファージを、以下のようにSREBP発現モジュレーションに関してコレステロール処置に供した:150K細胞を、24ウェルプレートフォーマットにおいてウェルごとに播種し、48時間、10μg/mlのコレステロール及び1μg/mlの25−ヒドロキシコレステロール(Sigma)で処理した。SREBP発現を活性化するために、次いで細胞をPBSで2回洗浄し、培地を、10%の脂質を含まないFBS、5mMの(2−ヒドロキシプロピル)−β−シクロデキストリン(Sigma)及び50μMのロバスタチン(Mevinolin,Sigma)を含有するDMEMに置き換え、示した時点で収集する。コレステロールで処理した細胞及びコレステロールを含まない細胞の両方を同じ時点で収集した。ロバスタチン/β−シクロデキストリン処理によるコレステロールの喪失は、J774マクロファージにおいてmiR−33a及びmSREBP−2宿主遺伝子の両方の増加した発現を生じ、ABCA1タンパク質発現の減少と一致した(図9A〜C)。コレステロール喪失に応答するABCA1タンパク質レベルの強力な抑制は、miR−33aアンチセンスオリゴヌクレオチドによって少なくとも部分的に反転し(図9D)、miR−33が、マウスマクロファージにおけるABCA1レベルのコレステロールにより調節される転写後制御を介するという考えと一致した。血清/コレステロールの存在下で培養したJ774マクロファージにおいて観察されたABCA1の高いタンパク質レベルは、更なるmiR−33a前駆体によって十分に抑制され(図9E)、ABCA1のmiR−33調節と一致する。加えて、コレステロールによるABCA1調節のmiRNAを介して行われるこの機構は、以前に記載した肝臓X受容体(LXR)を介するオキシステロールによるABCA1発現の転写制御であり(Tontonoz P,Mangelsdorf D,(2003)Mol.Endocrinol.17:985)、複数レベルのコレステロールによるABCA1の細胞調節を示す。したがって、マウスmiR−33a及びSREBP−2宿主遺伝子は、コレステロールにより同時調節され、マクロファージにおけるSREBP、miR−33、ABCA1、及びコレステロールの逆調節ネットワークの存在を示唆する。
【0091】
次に、J774マクロファージからのコレステロール流出に対するmiR−33操作の効果を試験した。まず、細胞を[3H]−コレステロールで標識し、次いでmiR−33a前駆体、miR−33aアンチセンス、又は対照オリゴヌクレオチドでトランスフェクトし、続いて血清/apoA−Iに対する[3H]−コレステロール流出のレベルを測定する。手短に述べると、低い継代のJ774細胞を、抗miR又はPre−マイクロRNA二本鎖によりトランスフェクトし、300K細胞/ウェルにて24ウェルプレートに播種し、[3H]−コレステロールを加えた。インキュベーションの20時間後、細胞を、室温にてPBSで2回洗浄し、培地を、15μg/mlの遊離apo−A−Iを含有する新鮮な培地と置き換えた。2又は4時間後、培地のアリコートを除去し、[3H]−コレステロールを、液体シンチレーション計数により測定した。細胞に存在する[3H]−コレステロールを、水酸化ナトリウム中で細胞を抽出することにより決定し、続いて液体シンチレーション計数により測定した。図9Fに示すように、miR−33aアンチセンスオリゴヌクレオチド(A)のマウスマクロファージへの導入により、[3H]−コレステロール流出の著しい(2倍)増加が生じたが、miR−33a前駆体(P)処理は、対照オリゴヌクレオチド(AC/PC)又は偽トランスフェクト細胞(UT)と比較して、[3H]−コレステロール流出の著しい減少を生じた。これらの知見により、ABCA1のmiR−33転写後抑制が、マクロファージにおいて細胞内コレステロールの保持を生じ、ABCA1レベルの制御を介するコレステロール輸送のmiR−33調節と一致することが実証される。
【実施例5】
【0092】
インビボモデルでの哺乳動物におけるmiR−33アンチセンスによるABCA1レベルの操作
miR−33アンチセンスアプローチによるABCA1レベルの操作もまた、インビボモデルでの哺乳動物において増大したHDLコレステロールレベルを生じる。尾静脈注射を、PBS(ビヒクル)、LNA−miR−33aアンチセンス、又はLNA対照オリゴヌクレオチドを用いる処置の6週間前の間、及び処置の間、西洋型食餌(すなわち、乳脂肪から42%kcalを補足した)を与えたマウスに実施した。血液を、治療の開始前にコレステロール/トリグリセリドプロファイルのために実験用マウスから顎下腺出血により得た。注射を行うために、20mg/kg/日のmiR−33−LNA又は対照LNAをPBS(全体積200μl)に溶解し、各日、同時間に毎日の尾静脈注射を介して3日連続投与した。マウスを、最後の尾静脈注射から48時間後に屠殺した。5日にわたる注射後、マウスを屠殺し、血清を以下のように回収した。屠殺の際に、総量1mLの血液を右側脳質穿刺によってマウスから得た。血液を5分間8,000rpmにて遠心分離して血清を得、−80℃にて凍結した。血清コレステロール、トリグリセリド、グルコース、アラニンアミノトランスフェラーゼ(ALT)及びアスパラギン酸アミノトランスフェラーゼ(AST)の全てを、マサチューセッツジェネラルホスピタル、比較医学のためのセンター、診断研究所においてHeska Dri−Chem 4000 Chemistry Analyzer(Heska,Loveland,CO)によってmiR−33−LNA又は対照−LNAでの処置前後に測定した。プールした血清のFPLC分析を記載(Hyogo Hら,(2002)J.Biol.Chem.277:34117)されるように実施した。血漿HDL−コレステロール濃度は、LNA−対照で処置したマウスと比較して、LNA−miR−33aアンチセンスで処置した動物において著しく増加した(図10A、B及び表2)。対照的に、LDL−コレステロール、トリグリセリド、又はグルコースレベルの血漿濃度で有意な効果は存在しなかった(図10B〜D及び表2)。これらの実験において、肝毒性(血漿AST/ALT)も見られなかった(図10E、F及び表2)。LNA−miR−33aアンチセンス処置に応答して血漿HDL−コレステロールにおいて観察された増加は、インビボでのmiR−33aによるABCA1依存性コレステロール流出の調節と一致する。
【0093】
これらの結果により更に、コレステロール生成及び脂肪合成遺伝子の重要な調節因子である、哺乳動物のSREBPファミリーの転写因子が、脊椎動物において細胞内コレステロールレベル及びコレステロール恒常性を決定するSREBP宿主遺伝子産物と協調して機能する、保存されたmiRNA(miR−33a/b)に対する宿主であることが確認される。図10は、miR−33及びその宿主遺伝子産物SREBPによるコレステロール恒常性の協調した調節についてのモデルを示す。miR−33は、その3’UTRを標的とすることによってABCA1翻訳を抑制する。このステップは、コレステロール産生遺伝子のトランス活性化を介してコレステロール生合成及び取り込みを活性化する、SREBPの発現と協調する。コレステロール蓄積は、SREBP経路を阻害し、ABCA1の発現を活性化する。そのABCA1は、細胞膜に移行し、apoA−I及びHDLへのコレステロール流出を媒介する。
【0094】
【表2】
表2.ロックド核酸(LNA)−miR−33aアンチマー(antimer)を有する西洋型食餌を与えたマウスの処置。血清コレステロール(A)、血漿トリグリセリド(B)、グルコース(C)、AST(D)、及びALT(E)の全てを、PBS(ビヒクル)、マウスmiR−33aに向けられたLNA−アンチセンスオリゴヌクレオチド、及びスクランブルLNA対照(LC)オリゴヌクレオチドの注入前後に西洋型食餌を与えたマウスにおいて測定した。
【0095】
これらの実施例に示すように、コレステロール生成及び脂肪合成遺伝子の重要な調節因子である、哺乳動物のSREBPファミリーのメンバーの転写因子は、脊椎動物において細胞内コレステロールレベル及びコレステロール恒常性を決定するSREBP宿主遺伝子産物と協調して機能する、保存されたmiRNA(miR−33a/b)に対する宿主である。miR−33は、インビトロでのコレステロール輸送及びインビボでのHDL合成にとって重要な結果を有する、ABCA1コレステロール輸送体の転写後制御に影響を及ぼす。
【0096】
(予測実施例6)
miR−33によるNPC1の標的化
ニーマンピックCI型タンパク質(NPC1)は、トランス−ゴルジネットワーク(TGN)を介して、後期のエンドソーム/リソソームから形質膜までのコレステロール/脂質の動員において重要な役割を果たす。このプロセスは、発生期のHDLを生成するためにABCA1を介して行われる細胞表面におけるapoA−I及び他のアポリポタンパク質に対するリン脂質及びコレステロールの動因と協調すると考えられる。NPC1の3’UTRは、1つの予測されるmiR−33標的部位を保有する(7−mer;TargetScan 4.2)。細胞内コレステロール輸送のその既知の役割並びに最近のABCA1及びコレステロール流出に対する機能的関連のために、そのNPC1の3’UTRはmiR−33に対する候補標的である。
【0097】
第1に、miRNAプロセシング酵素のドローシャ及びダイサーを標的とするsiRNAの使用によって、miRNAが、上記の3つの細胞株においてNPC1レベルを制御するか否かを決定する。第2に、Pre−miR−33及び抗miR−33オリゴヌクレオチドを、NPC1調節におけるmiR−33の特定の役割を扱うためにこれらの細胞株に導入する。NPC1タンパク質レベルを、対照としてβ−チューブリンを用いる免疫ブロット法によって決定する。NPC1 mRNAレベルに対するmiR−33操作の何らかの効果をqRT−PCRにより評価する。NCP1タンパク質及び/又はmRNAレベルに対する効果をmiR−33操作後に観察した場合、予測されるmiR−33標的配列を保有するNPC1 3’UTRを、pMIR−REPORTベクター内にクローニングし、その後、ABCA1に関して本明細書で上記したようにmiR−33標的部位を変異し、ルシフェラーゼ発現分析を行う。コレステロール輸送/流出におけるNPC1の潜在的な役割を確立するために、J774マウスマクロファージ細胞株におけるNPC1のsiRNA標的化の効果を、ABCA1に関して上記した方法を用いて試験する。
【0098】
前述の説明から、変更及び修飾が種々の用途及び状態を採用するために本明細書に記載される本発明に対してなされ得ることは明らかである。かかる実施形態はまた、添付の特許請求の範囲内である。本明細書の可変物の任意の定義における要素の記載の列挙は、記載した要素の任意の単一の要素又は組合せ(又は部分的組合せ)として可変のものの定義を含む。本明細書の実施形態の列挙は、任意の単一の実施形態又は任意の他の実施形態若しくはその一部と組み合わせた実施形態を含む。
【0099】
参考文献
本出願に記載した全ての特許、特許出願、データベース登録、及び刊行物は、各々の独立した特許、データベース登録、及び刊行物が、参照として組み込まれている具体的かつ個々に示されるものと同程度まで本明細書に参照として組み込まれている。本明細書に参照として組み込まれるものとしては、限定されないが、以下のものが挙げられる。
(参考文献)
【0100】
1. Maxfield FR, Tabas I (2005) Role of cholesterol and lipid organization in disease. Nature 438: 612-621.
2. Moller DE, Kaufman KD (2005) Metabolic syndrome: a clinical and molecular perspective. Annu Rev Med 56: 45-62.
3. Bengoechea-Alonso MT, Ericsson J (2007) SREBP in signal transduction: cholesterol metabolism and beyond. Curr Opin Cell Biol 19: 215-222.
4. Brown MS, Goldstein JL (1997) The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 89: 331-340.
5. Eberle D, et al. (2004) SREBP transcription factors: master regulators of lipid homeostasis. Biochimie 86: 839-848.
6. Horton JD, Goldstein JL, Brown MS (2002) SREBPs: activators of the complete program of cholesterol and fatty acid synthesis in the liver. J Clin Invest 109: 1125-1131.
7. Raghow R, et al. (2008) SREBPs: the crossroads of physiological and pathological lipid homeostasis. Trends Endocrinol Metab 19: 65-73.
8. Ericsson J, Edwards PA (1998) CBP is required for sterol-regulated and sterol regulatory element-binding protein-regulated transcription. J Biol Chem 273: 17865-17870.
9. Naar AM, et al. (1998) Chromatin, TAFs, and a novel multiprotein coactivator are required for synergistic activation by Sp1 and SREBP-1a in vitro. Genes Dev. 12: 3020-3031.
10. Naar AM, et al. (1999) Composite coactivator ARC mediates chromatin-directed transcriptional activation. Nature 398: 828-832.
11. Naar AM, Lemon BD, Tjian R (2001) Transcriptional coactivator complexes. Annu Rev Biochem 70: 475-501.
12. Naar AM, et al. (2002) Human CRSP interacts with RNA polymerase II CTD and adopts a specific CTD-bound conformation. Genes Dev 16: 1339-1344.
13. Oliner JD AJ, Hansen SK, Zhou S, Tjian R. (1996) SREBP transcriptional activity is mediated through an interaction with the CREB-binding protein. Genes Dev 10: 2903-2911.
14. Taatjes DJ, et al. (2002) Structure, function, and activator-induced conformations of the CRSP coactivator. Science 295: 1058-1062.
15. Yang F, et al. (2006) An ARC/Mediator subunit required for SREBP control of cholesterol and lipid homeostasis. Nature 442: 700-704.
16. Bartel DP (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281-297.
17. He L, Hannon GJ (2004) MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet 5: 522-531.
18. Krutzfeldt J, Stoffel M (2006) MicroRNAs: a new class of regulatory genes affecting metabolism. Cell Metab 4: 9-12.
19. Lewis BP, et al. (2003) Prediction of mammalian microRNA targets. Cell 115: 787-798.
20. Lee YS, Dutta A (2008) MicroRNAs in Cancer. Annu Rev Pathol.
21. Mattes J, Collison A, Foster PS (2008) Emerging role of microRNAs in disease pathogenesis and strategies for therapeutic modulation. Curr Opin Mol Ther 10: 150-157.
22. Thum T, Catalucci D, Bauersachs J (2008) MicroRNAs: novel regulators in cardiac development and disease. Cardiovasc Res 79: 562-570.
23. Stefani G, Slack FJ (2008) Small non-coding RNAs in animal development. Nat Rev Mol Cell Biol 9: 219-230.
24. Sonkoly E, Stahle M, Pivarcsi A (2008) MicroRNAs and immunity: novel players in the regulation of normal immune function and inflammation. Semin Cancer Biol 18: 131-140.
25. van den Berg A, Mols J, Han J (2008) RISC-target interaction: Cleavage and translational suppression. Biochim Biophys Acta.
26. Vasudevan S, Tong Y, Steitz JA (2007) Switching from repression to activation: microRNAs can up-regulate translation. Science 318: 1931-1934.
27. Lagos-Quintana M, Rauhut R, Lendeckel W, Tuschl T (2001) Identification of novel genes coding for small expressed RNAs. Science 294: 853-858.
28. Rodriguez A, Griffiths-Jones S, Ashurst JL, Bradley A (2004) Identification of mammalian microRNA host genes and transcription units. Genome Res 14: 1902-1910.
29. Elmen J, et al. (2008) LNA-mediated microRNA silencing in non-human primates. Nature 452: 896-899.
30. Elmen J, et al. (2008) Antagonism of microRNA-122 in mice by systemically administered LNA-antimiR leads to up-regulation of a large set of predicted target mRNAs in the liver. Nucleic Acids Res 36: 1153-1162.
31. Esau C, et al. (2006) miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab 3: 87-98.
32. Krutzfeldt J, et al. (2005) Silencing of microRNAs in vivo with 'antagomirs'. Nature 438: 685-689.
33. Tall AR, et al. (2008) HDL, ABC transporters, and cholesterol efflux: implications for the treatment of atherosclerosis. Cell Metab 7: 365-375.
34. Wang X, Rader DJ (2007) Molecular regulation of macrophage reverse cholesterol transport. Curr Opin Cardiol 22: 368-372.
35. Singaraja RR, et al. (2002) Increased ABCA1 activity protects against atherosclerosis. J Clin Invest 110: 35-42.
36. Wang X, et al. (2007) Macrophage ABCA1 and ABCG1, but not SR-BI, promote macrophage reverse cholesterol transport in vivo. J Clin Invest 117: 2216-2224.
37. Van Eck M, et al. (2006) Macrophage ATP-binding cassette transporter A1 overexpression inhibits atherosclerotic lesion progression in low-density lipoprotein receptor knockout mice. Arterioscler Thromb Vasc Biol 26: 929-934.
38. van Eck M, et al. (2002) Leukocyte ABCA1 controls susceptibility to atherosclerosis and macrophage recruitment into tissues. Proc Natl Acad Sci U S A 99: 6298-6303.
39. Koldamova R, Lefterov I (2007) Role of LXR and ABCA1 in the pathogenesis of Alzheimer's disease - implications for a new therapeutic approach. Curr Alzheimer Res 4: 171-178.
40. Wahrle SE, et al. (2008) Overexpression of ABCA1 reduces amyloid deposition in the PDAPP mouse model of Alzheimer disease. J Clin Invest 118: 671-682.
41. Brunham LR, Kruit JK, Verchere CB, Hayden MR (2008) Cholesterol in islet dysfunction and type 2 diabetes. J Clin Invest 118: 403-408.
42. Chawla A, et al. (2001) A PPAR gamma-LXR-ABCA1 pathway in macrophages is involved in cholesterol efflux and atherogenesis. Mol Cell 7: 161-171.
43. Venkateswaran A, et al. (2000) Control of cellular cholesterol efflux by the nuclear oxysterol receptor LXR alpha. Proc Natl Acad Sci U S A 97: 12097-12102.
44. Chisholm JW, Hong J, Mills SA, Lawn RM (2003) The LXR ligand T0901317 induces severe lipogenesis in the db/db diabetic mouse. J Lipid Res 44: 2039-2048.
45. Millatt LJ, Bocher V, Fruchart JC, Staels B (2003) Liver X receptors and the control of cholesterol homeostasis: potential therapeutic targets for the treatment of atherosclerosis. Biochim Biophys Acta 1631: 107-118.
46. Schmitz G, Langmann T (2005) Transcriptional regulatory networks in lipid metabolism control ABCA1 expression. Biochim Biophys Acta 1735: 1-19.
47. Tamehiro N, et al. (2007) Sterol regulatory element-binding protein-2- and liver X receptor-driven dual promoter regulation of hepatic ABC transporter A1 gene expression: mechanism underlying the unique response to cellular cholesterol status. J Biol Chem 282: 21090-21099.
48. Wong J, Quinn CM, Brown AJ (2006) SREBP-2 positively regulates transcription of the cholesterol efflux gene, ABCA1, by generating oxysterol ligands for LXR. Biochem J 400: 485-491.
49. Zeng L, et al. (2004) Sterol-responsive element-binding protein (SREBP) 2 down-regulates ATP-binding cassette transporter A1 in vascular endothelial cells: a novel role of SREBP in regulating cholesterol metabolism. J Biol Chem 279: 48801-48807.
50. Haidar B, et al. (2002) cAMP induces ABCA1 phosphorylation activity and promotes cholesterol efflux from fibroblasts. J Lipid Res 43: 2087-2094.
51. See RH, et al. (2002) Protein kinase A site-specific phosphorylation regulates ATP-binding cassette A1 (ABCA1)-mediated phospholipid efflux. J Biol Chem 277: 41835-41842.
52. Lu R, et al. (2008) ApoA-I facilitates ABCA1 recycle/accumulation to cell surface by inhibiting its intracellular degradation and increases HDL generation. Arterioscler Thromb Vasc Biol 28: 1820-1824.
53. Taylor AJ (2008) Evidence to support aggressive management of high-density lipoprotein cholesterol: implications of recent imaging trials. Am J Cardiol 101: 36B-43B.
54. Yvan-Charvet L, et al. (2007) Combined deficiency of ABCA1 and ABCG1 promotes foam cell accumulation and accelerates atherosclerosis in mice. J Clin Invest 117: 3900-3908.
55. Ranalletta M, et al. (2006) Decreased atherosclerosis in low-density lipoprotein receptor knockout mice transplanted with Abcg1-/- bone marrow. Arterioscler Thromb Vasc Biol 26: 2308-2315.
56. deGoma EM, deGoma RL, Rader DJ (2008) Beyond high-density lipoprotein cholesterol levels evaluating high-density lipoprotein function as influenced by novel therapeutic approaches. J Am Coll Cardiol 51: 2199-2211.
57. Neufeld EB, et al. (2004) The ABCA1 transporter modulates late endocytic trafficking: insights from the correction of the genetic defect in Tangier disease. J Biol Chem 279: 15571-15578.
58. Wang MD, et al. (2007) Differential regulation of ATP binding cassette protein A1 expression and ApoA-I lipidation by Niemann-Pick type C1 in murine hepatocytes and macrophages. J Biol Chem 282: 22525-22533.
59. Boadu E, Francis GA (2006) The role of vesicular transport in ABCA1-dependent lipid efflux and its connection with NPC pathways. J Mol Med 84: 266-275.
60. Baek D, et al. (2008) The impact of microRNAs on protein output. Nature 455: 64-71.
61. Baskerville S, Bartel DP (2005) Microarray profiling of microRNAs reveals frequent coexpression with neighboring miRNAs and host genes. Rna 11: 241-247.
62. Foufelle F, Ferre P (2002) New perspectives in the regulation of hepatic glycolytic and lipogenic genes by insulin and glucose: a role for the transcription factor sterol regulatory element binding protein-1c. Biochem J 366: 377-391.
63. Foretz M, et al. (1999) ADD1/SREBP-1c is required in the activation of hepatic lipogenic gene expression by glucose. Mol Cell Biol 19: 3760-3768.
64. Radhakrishnan A, et al. (2007) Sterol-regulated transport of SREBPs from endoplasmic reticulum to Golgi: oxysterols block transport by binding to Insig. Proc Natl Acad Sci U S A 104: 6511-6518.
65. Repa JJ, et al. (2000) Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRalpha and LXRbeta. Genes Dev 14: 2819-2830.
66. Hannah VC, et al. (2001) Unsaturated fatty acids down-regulate srebp isoforms 1a and 1c by two mechanisms in HEK-293 cells. J Biol Chem 276: 4365-4372.
67. Ou J, et al. (2001) Unsaturated fatty acids inhibit transcription of the sterol regulatory element-binding protein-1c (SREBP-1c) gene by antagonizing ligand-dependent activation of the LXR. Proc Natl Acad Sci U S A 98: 6027-6032.
68. Sheng Z, Otani H, Brown MS, Goldstein JL (1995) Independent regulation of sterol regulatory element-binding proteins 1 and 2 in hamster liver. Proc Natl Acad Sci U S A 92: 935-938.
69. Akinc A, et al. (2008) A combinatorial library of lipid-like materials for delivery of RNAi therapeutics. Nat Biotechnol 26: 561-569.
70. de la Llera-Moya M, et al. (2010, Epub ahead of print) Arterioscler. Thromb. Vasc. Biol.
72. Tontonoz P, Mangelsdorf D, (2003) Mol. Endocrinol. 17:985.
71. Hyogo H, Roy S, Paigen B, Cohen D, (2002) J. Biol. Chem. 277: 34117.
【技術分野】
【0001】
関連出願/特許
本出願は、代理人整理番号MGH20023として2009年3月31日に出願した米国仮出願第61/165,041号の利益を主張し、その内容は本明細書に参照として組み込まれている。
【0002】
連邦支援研究下でなされた発明に対する権利の声明
本研究は、国立衛生研究所助成番号R01GM71449及びR21DK084459により援助された。米国政府は本発明に一定の権利を有し得る。
【背景技術】
【0003】
異常なコレステロール及び脂質ホメオスタシスは、メタボリックシンドローム、アテローム性動脈硬化症/心血管疾患、及び2型糖尿病などの蔓延している疾患と関連している。コレステロール及び脂質は、それらの脂肪性のカーゴを異なる細胞及び組織に運ぶ、低比重リポタンパク質(LDL)及び高比重リポタンパク質(HDL)などのリポタンパク質粒子として血液中で輸送される。過剰な循環LDLは、酸化され、動脈内のマクロファージにより取り込まれる可能性があり、それらのマクロファージは、動脈硬化性プラークの形成に関与するコレステロール/脂質に満ちた「泡沫細胞」に変化する。反対に、HDL−コレステロールは、マクロファージを含む、末梢細胞から運び出され、コレステロールの逆輸送経路によって分解するために肝臓に輸送されるが、しかしながら、心臓疾患を患っている患者は、多くの場合、循環HDLが低く、コレステロールクリアランスが低下している。したがって、心血管疾患を寛解させるための治療努力は、コレステロール生合成及びLDL産生を低下させること(例えばスタチン治療による)、並びにHDLを介したコレステロールの逆輸送を増加させること(例えばナイアシン添加による)に重点を置かれている。これらの治療はかなりの効果を示しているが、心血管疾患は、米国において主要な死亡原因のままである。コレステロール及び脂質調節の分子機構の詳細な理解により、心血管疾患を治療するための新規治療戦略の開発が促進する可能性がある。
【0004】
ATP結合カセット輸送体ABCA1は、HDL合成及びマクロファージ/泡沫細胞を含む、末梢組織からのコレステロールの逆輸送に重要である。ABCA1遺伝子における変異/SNPがアテローム性動脈硬化に関係している。例えば、ABCA1レベル、HDL産生及び/又はコレステロールの逆輸送を増加できる新規の治療は、調節を改善でき、心血管疾患を寛解させる可能性がある。
【発明の概要】
【課題を解決するための手段】
【0005】
ステロール調節要素結合タンパク質(SREBP)転写因子は、コレステロール及び脂質の生合成及び取り込みの両方に関与する、公知の遺伝子調節因子である。現在、ヒトSREBP−2及びヒトSREBP−1遺伝子におけるイントロン配列内に埋め込まれている保存されたマイクロRNAをコードする配列の存在は発見されている。本明細書の以下に記載している予備研究により、本明細書において配列番号1及び2としてそれぞれ示しているmiR−33a及びmiR−33bは、翻訳抑制のためにATP結合カセット輸送体ABCA1を標的とするという驚くべき知見が得られた。哺乳動物におけるコレステロール恒常性の調節において、miR−33とSREBP宿主遺伝子産物との協調した役割は、今まで知られていなかった。更に、miR−33の阻害が、ABCA1レベル、HDL産生、及びコレステロールの逆輸送を増加させる新規の治療手段を表すことが、ここで発見された。
【0006】
一態様において、本発明は、配列番号1又は2と相補的である単離された核酸配列を含む組成物であって、その核酸配列は、哺乳動物細胞におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効である、組成物を提供する。一部の実施形態において、組成物は、コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で核酸を含む。
【0007】
一部の実施形態において、配列は配列番号7又は9と相補的である。
【0008】
一実施形態において、配列番号1又は2と相補的である核酸配列は、限定されないが、本明細書に記載されるアンチセンスオリゴヌクレオチド、例えば配列番号3、4、6、又は8を含む、アンチセンスオリゴヌクレオチドであり得る。一部の実施形態において、核酸は修飾され、例えば、1つ又は複数の天然に存在しないヌクレオシドを含有する。一部の実施形態において、核酸はロックされている。
【0009】
別の実施形態において、配列番号1又は2と相補的である核酸配列は、限定されないが、shRNA又はsiRNAを含む、干渉RNAであり得る。
【0010】
更に別の実施形態において、配列番号1又は2と相補的である核酸配列は、アンタゴマー(antagomir)である。
【0011】
更に別の実施形態において、配列番号1と相補的である核酸配列は、配列番号1の転写後プロセシングを阻害する。
【0012】
更に別の実施形態において、配列番号2と相補的である核酸配列は、配列番号2の転写後プロセシングを阻害する。
【0013】
別の態様において、本発明は、コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で、配列番号1又は2と相補的である核酸配列を発現できるベクターを含む組成物を提供する。
【0014】
一実施形態において、ベクターは、プラスミドベクター又はウイルスベクターである。
【0015】
更に別の態様において、本発明は、細胞、例えば、コレステロール恒常性を必要とする被験体の細胞におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させる方法であって、配列番号1又は2と相補的である核酸配列を細胞又は被験体に投与することによって、細胞又は被験体におけるABCA1タンパク質の発現を増加させることを含む、方法を提供する。
【0016】
更に別の態様において、本発明は、被験体の肝臓における細胞内コレステロールの流出及び/又はHDLの産生を増加させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体の肝臓における細胞内コレステロールの流出及び/又はHDLの産生を増加させることを含む、方法を提供する。
【0017】
更に別の態様において、本発明は、被験体の血液を循環するコレステロールの量を減少させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体の血液を循環するコレステロールの量を減少させることを含む、方法を提供する。
【0018】
本発明の他の特徴及び利点は、詳細な説明、及び特許請求の範囲から明らかとなろう。したがって、本発明の他の態様は以下の開示に説明され、それらは本発明の範囲内である。
【0019】
以下の詳細な図面は例として与えられ、記載される特定の実施形態に本発明を限定することを意図せず、本明細書に参照として組み込まれる添付の図面と併せて理解され得る。
【図面の簡単な説明】
【0020】
【図1】SREBPによって調節される遺伝子を示す。この図は、コレステロール、脂肪酸、及びトリグリセリドの合成経路における主要な代謝中間体を示す。インビボにおいて、SREBP−2は、コレステロール代謝の遺伝子を選択的に活性化するのに対して、SREBP−1cは、脂肪酸及びトリグリセリド代謝の遺伝子を選択的に活性化する。DHCR、7−デヒドロコレステロールレダクターゼ;FPP、ファルネシル二リン酸;GPP、ゲラニルゲラニルピロリン酸シンターゼ;CYP51、ラノステロール14α−デメチラーゼ;G6PD、グルコース−6−リン酸デヒドロゲナーゼ;PGDH、6−ホスホグルコン酸デヒドロゲナーゼ;GPAT、グリセロール−3−リン酸アシルトランスフェラーゼ。出典Hortonら,J Clin Invest.2002.109:1125−31。
【図2】CBP/p300ヒストンアセチルトランスフェラーゼ及びARC/メディエーター共活性化因子複合体のARC105/MED15サブユニットに存在する構造保存性のKIX領域(青色楕円形)と相互作用することによる標的遺伝子の転写のSREBP活性化を示す。CBP/p300の動員により、ヒストン/クロマチンの増加したアセチル化が生じるのに対して、ARC/メディエーター相互作用により、その大きさサブユニットでC末端領域(CTD)を経由するRNAポリメラーゼII(Pol II)の動員が生じる。TFIID及び他の基本転写因子(GTF)もまた、転写開始を促進する、プロモーターに動員される。
【図3】イントロンに位置するmiRNA(それぞれ、miR−33b及びmiR−33a)をコードする配列を保有する、ヒトSREBP−1及びSREBP−2遺伝子を示す。特に、マウスのみが、マウスSREBP−2遺伝子においてmiR−33a miRNAを保有するのに対して、対応するマウスSREBP−1イントロンはヒトSREBP−1遺伝子との相同性を欠き、miRNAコード配列を欠いている。
【図4】miRNAプロセシング酵素のドローシャによって認識されるヘアピン構造を形成する一次miRNA転写産物(pri−miRNA)の合成を生じる、miRNAコード配列を介するPol IIによる転写を示す。pri−miRNAは、ドローシャ(Drosha)によってpre−miRNAにプロセシングされ、次いで細胞質に送り出され、細胞質において、それらのpre−miRNAはダイサー(Dicer)によって成熟miRNA二本鎖までのプロセシングを更に受ける。それらの成熟miRNA二本鎖はRISC複合体によって損傷を受けず、mRNA標的鎖がRISCに組み込まれ、次いでそのmRNA標的鎖は、miRNA−mRNA配列相補性に応じて翻訳抑制又はmRNA分解のいずれかによってmRNAを標的とする。出典van den Bergら,Biochim.et Biophys.Acta 2008。
【図5】マクロファージ及びアテローム生成泡沫細胞などの末梢細胞からApoA−I及び高比重リポタンパク質(HDL)に遊離コレステロールを送り出す、ABC輸送体ABCA1(及び関連ABCG1)を示す。HDLは、肝細胞においてスカベンジャー受容体−B1(SR−B1)に結合し、それらの付随したコレステロール及びコレステロールエステル(CE)を肝臓に移す。コレステロールは、遊離コレステロールとして、又は胆汁塩に変換後のいずれかで胆汁に排出される。出典Maxfield及びTabas.Nature 2005.438:612−21。
【図6】miR−33及びその宿主遺伝子産物SREBPによるコレステロール恒常性の協調した調節についてのモデルを示す。miR−33は、その3’UTRを標的化することによりABCA1翻訳を抑制する。このステップは、コレステロール産生遺伝子(cholesterogenic gene)のトランス活性化を介するコレステロール生合成及び取り込みを活性化する、SREBPの発現と協調するように提案される。コレステロール蓄積は、SREBP経路を阻害し、ABCA1の発現を活性化する。そのABCA1は、細胞膜へ移行し、ApoA−I及びHDLへのコレステロール流出を媒介する。
【図7A】miRNAによるABCA1発現の調節を示す。図7Aにおいて、3つの細胞株(J774マウスマクロファージ、ヒトHepG2肝細胞癌細胞、及び正常なヒトIMR−90線維芽細胞)においてsiRNAを用いるmiRNAプロセシング酵素のドローシャ及びダイサーのRNAiに向けられたノックダウンにより、ABCA1タンパク質レベルの増加が生じた。タンパク質レベルはブラッドフォード分析により正規化した。β−チューブリンを負荷対照として使用した。
【図7B】miRNAによるABCA1発現の調節を示す。図7Bにおいて、抗miR−33アンチセンスオリゴヌクレオチドによるmiR−33機能の拮抗により、ABCA1のより高い発現が生じることが示される。
【図7C】miRNA前駆体Pre−miR−33でトランスフェクトした細胞によって模倣される、miR−33の効果を示す。予想されるように、ABCA1発現は、増加したレベルのmiR−33による翻訳抑制を介して阻害される。
【図8A】HEK293細胞におけるルシフェラーゼレポーター構築物によるABCA1 3’−UTRにおけるmiR−33についての予測標的部位の評価を示す。(A)2つの潜在的標的部位が、ABCA1 3’UTR(配列番号5)の最初の160塩基内に存在するmiR−33について予測される。ルシフェラーゼORFのABCA1 3’UTR下流の対応する配列(塩基18〜205)の挿入により、ルシフェラーゼ活性の下方調節が生じる(右のグラフ)。
【図8B】HEK293細胞におけるルシフェラーゼレポーター構築物によるABCA1 3’−UTRにおけるmiR−33についての予測標的部位の評価を示す。(B)(A)において観察された下方調節は、30nMの濃度にて抗マイクロRNAオリゴヌクレオチド(Ambion)A33a及びA33b(配列番号6及び8)を用いるmiR−33a/bアイソフォーム(配列番号7及び9)の配列特異的喪失により遮断される(AC:陰性対照としての関係のない抗miR)。
【図8C】HEK293細胞におけるルシフェラーゼレポーター構築物によるABCA1 3’−UTRにおけるmiR−33についての予測標的部位の評価を示す。(C)変異したmiR−33前駆体(30nMにおけるm33a(配列番号10)及びm33b(配列番号11))ではなく、過剰な外因性野生型miR−33前駆体(30nMにおける33a及び33b)の添加は、予想されるようにルシフェラーゼ活性を更に阻害した。
【図9A】コレステロール及びSREBP/miR−33による相互調節を示す。J774マウスマクロファージにおけるコレステロールのロバスタチン及びβ−シクロデキストリンを介する喪失により、(A)SREBP−2の増加した発現が生じる。エラーバーはs.dを表す。**はp<0.01を示す。
【図9B】コレステロール及びSREBP/miR−33による相互調節を示す。J774マウスマクロファージにおけるコレステロールのロバスタチン及びβ−シクロデキストリンを介する喪失により、(B)miR−33aの増加した発現が生じる。エラーバーはs.dを表す。**はp<0.01を示す。
【図9C】コレステロール及びSREBP/miR−33による相互調節を示す。J774マウスマクロファージにおけるコレステロールのロバスタチン及びβ−シクロデキストリンを介する喪失により、(C)ABCA1タンパク質レベルの減少が生じる。
【図9D】コレステロール及びSREBP/miR−33による相互調節を示す。(D)コレステロールが喪失したJ774マウスマクロファージは、偽トランスフェクション(UT)又はアンチセンス対照(AC)オリゴヌクレオチドと比較して、miR−33bアンチセンスオリゴヌクレオチドでのトランスフェクションに応答してABCA1タンパク質の上方調節を示す。β−チューブリンは、この免疫ブロット法において負荷対照としての機能を果たした。
【図9E】コレステロール及びSREBP/miR−33による相互調節を示す。(E)miR−33a前駆体を有する血清/コレステロールの存在下で培養したJ774マクロファージのトランスフェクションにより、前駆体対照(PC)と比較して、減少したレベルのABCA1が生じた。
【図9F】コレステロール及びSREBP/miR−33による相互調節を示す。(F)放射性標識したコレステロールを負荷したJ775マクロファージへのmiR−33aアンチセンスオリゴヌクレオチド(A)の導入により、増加したコレステロール流出が生じるのに対して、miR−33a前駆体(P)は、トランスフェクション対照(UT)又はスクランブルアンチセンス対照(AC)又は前駆体対照(PC)オリゴヌクレオチドと比較して、コレステロール流出を阻害する。エラーバーはs.dを表す。**はP<0.01を示す。
【図10A】インビボでのmiR−33aによるHDLの調節を示す。(A)マウスmiR−33aに対して向けられるロックド核酸(LNA)−アンチセンスオリゴヌクレオチドを有する西洋型食餌を与えたマウスの尾静脈注射により、ビヒクル(PBS)及びスクランブルLNA対照(LC)オリゴヌクレオチドと比較して、増大した総血清コレステロールが生じる。
【図10B】インビボでのmiR−33aによるHDLの調節を示す。(B)LNA−miR−33aアンチセンス(LNA)及びLNA(対照)(LC)で処置した西洋型食餌を与えた5匹のマウス由来のプールされた血清のFPLC分析により、miR−33aアンチセンスで処置した動物においてHDL−コレステロールピークの大きさの増加が明らかにされた。リポタンパク質標準物の溶出は標識により示される。
【図10C】インビボでのmiR−33aによるHDLの調節を示す。血漿トリグリセリド(C)は処置に影響を与えなかった。
【図10D】インビボでのmiR−33aによるHDLの調節を示す。グルコース(D)は処置に影響を与えなかった。
【図10E】インビボでのmiR−33aによるHDLの調節を示す。AST(E)は処置に影響を与えなかった。
【図10F】インビボでのmiR−33aによるHDLの調節を示す。ALTは処置に影響を与えなかった。
【図10G】インビボでのmiR−33aによるHDLの調節を示す。(G)は、miR−33及びその宿主遺伝子産物SREBPによるコレステロール恒常性の協調した調節についてのモデルを表す。
【図11A】ヒト及びマウス組織におけるSREBPの同時発現を示す。ヒトSREBP−1(A;配列番号12)遺伝子は、関連するイントロンmiRNA(miR−33b)を保有する。pri−miRNAをコードする配列を、成熟miRNA配列に下線を引いて示す。
【図11B】ヒト及びマウス組織におけるSREBPの同時発現を示す。ヒトSREBP−2(B;配列番号13)遺伝子は、関連するイントロンmiRNA(miR−33a)を保有する。pri−miRNAをコードする配列を、成熟miRNA配列に下線を引いて示す。
【図11C】ヒト及びマウス組織におけるSREBPの同時発現を示す。選択されたヒト及びマウス組織におけるmiR−33a/b及びSREBP宿主遺伝子の発現の相関関係:(C)hSREBP−1/−hsa−miR−33b。エラーバーはs.e.mを表す。
【図11D】ヒト及びマウス組織におけるSREBPの同時発現を示す。選択されたヒト及びマウス組織におけるmiR−33a/b及びSREBP宿主遺伝子の発現の相関関係:(D)hSREBP−2/hsa−miR−33a。エラーバーはs.e.mを表す。
【図11E】ヒト及びマウス組織におけるSREBPの同時発現を示す。選択されたヒト及びマウス組織におけるmiR−33a/b及びSREBP宿主遺伝子の発現の相関関係:(E)mSREBP−1/mmu−miR−33b。エラーバーはs.e.mを表す。
【図12A】選択された種におけるmiR−33a(A)アイソフォーム(ヒトにおけるmir−33a、配列番号14;マウスにおけるmir−33a、配列番号15;ニワトリにおけるmir−33a、配列番号16;推測上カエルにおけるmir−33a、配列番号17、及びショウジョウバエにおけるmir−33a、配列番号18)の配列及びゲノム構成を示す。miRNAステムループを示し、それらが存在するSREBP遺伝子に従って分類する。アスタリスクは完全に保存された残基の位置を表す。
【図12B】選択された種におけるmiR−33b(B)アイソフォーム(ヒトにおけるmir−33b、配列番号19、マウスにおけるmir−33b、配列番号20)の配列及びゲノム構成を示す。miRNAステムループを示し、それらが存在するSREBP遺伝子に従って分類する。アスタリスクは完全に保存された残基の位置を表す。
【図13A】ヒト組織におけるSREBP−1の相対的発現を示す。β−アクチンを組織間の発現を正規化するために使用した。各組織から使用した全RNAは、男性及び女性の患者を含む少なくとも3体のドナーからプールした。
【図13B】ヒト組織におけるhsa−miR−33bの相対的発現を示す。U6を組織間の発現を正規化するために使用した。各組織から使用した全RNAは、男性及び女性の患者を含む少なくとも3体のドナーからプールした。
【図14A】ヒト組織におけるSREBP−2の相対的発現を示す。β−アクチンを組織間の発現を正規化するために使用した。低い相対的発現を有する組織をグラフの左側において比較し、高い相対的発現を有する組織をグラフの右側において比較する。各組織から使用した全RNAは、男性及び女性の患者を含む少なくとも3体のドナーからプールした。
【図14B】ヒト組織におけるhsa−miR−33aの相対的発現を示す。U6を組織間の発現を正規化するために使用した。低い相対的発現を有する組織をグラフの左側において比較し、高い相対的発現を有する組織をグラフの右側において比較する。各組織から使用した全RNAは、男性及び女性の患者を含む少なくとも3体のドナーからプールした。
【図15A】マウス組織におけるSREBP−2の相対的発現を示す。β−アクチンを組織間の発現を正規化するために使用した。
【図15B】マウス組織におけるmmu−miR−33aの相対的発現を示す。U6を組織間の発現を正規化するために使用した。
【図16】ヒトNPC1 3’UTRにおいて予測されるmiRNA標的部位を示す(TargetScan 4.2)。miR−33標的部位は矢印によって示される。
【図17】配列番号1及び2として、それぞれ、miR−33a及びmir−33bを示す。また、配列番号3及び4として、それぞれ、抗miR−33a及び抗miR−33bを示す。
【発明を実施するための形態】
【0021】
定義
他に定義されない限り、本明細書に使用される全ての技術用語及び科学用語は、本発明が属する技術分野における当業者によって一般的に理解されているものと同じ意味を有する。矛盾する場合には、定義を含む本出願が適用される。
【0022】
本明細書で言及する場合、「相補的核酸配列」は、相補的ヌクレオチド塩基対からなる別の核酸配列とハイブリダイズできる核酸配列である。「ハイブリダイズする」とは、ストリンジェンシーの適切な条件下で、相補的ヌクレオチド塩基間で二本鎖分子を形成する対を意味する(例えば、DNAにおいて、アデニン(A)はチミン(T)と塩基対を形成し、同様にグアニン(G)はシトシン(C)と塩基対を形成する)(例えば、Wahl,G.M.及びS.L.Berger(1987)Methods Enzymol.152:399;Kimmel,A.R.(1987)Methods Enzymol.152:507を参照のこと)。
【0023】
本明細書で使用する場合、「アンチセンスオリゴヌクレオチド」とは、マイクロRNAの配列などのDNA又はmRNA配列と相補的である合成核酸配列をいう。
【0024】
「RNA」は、少なくとも1つ又は複数のリボヌクレオチド残基を含む分子である。「リボヌクレオチド」は、ベータ−D−リボフラノース部分の2’位にヒドロキシル基を有するヌクレオチドである。本明細書で使用する場合、用語RNAは、二本鎖RNA、一本鎖RNA、部分的に精製されたRNAなどの単離されたRNA、実質的に純粋なRNA、合成RNA、組換えで生成されたRNA、並びに1つ若しくは複数のヌクレオチドの付加、欠失、置換及び/又は変更によって天然に存在するRNAとは異なった改変RNAを含む。RNA分子のヌクレオチドはまた、天然に存在しないヌクレオチド又は化学的に合成されたヌクレオチド若しくはデオキシヌクレオチドなどの非標準的ヌクレオチドを含んでもよい。
【0025】
「マイクロRNA」(miRNA)は、典型的に、遺伝子発現を調節する、長さが約21〜23ヌクレオチドの一本鎖RNA分子をいう。miRNAは遺伝子によってコードされ、その遺伝子のDNAからmiRNAは転写されるが、miRNAはタンパク質に翻訳されず、代わりに、各々の一次転写産物は短いステムループ構造にプロセシングされ、その後、機能的miRNAへの更なるプロセシングを受ける。成熟miRNA分子は、1つ又は複数のメッセンジャーRNA(mRNA)分子と部分的に相補的であり、それらの主な機能は遺伝子発現を下方調節することである。
【0026】
本明細書で使用する場合、「干渉RNA」とは、直接又は間接(すなわち変換の際に)のいずれかで、RNA干渉を介することによって遺伝子発現を阻害又は下方調節できる、任意の二本鎖又は一本鎖RNA配列をいう。干渉RNAには、限定されないが、低分子干渉RNA(「siRNA」)及び低分子ヘアピン型RNA(「shRNA」)が含まれる。「RNA干渉」とは、配列適合性のメッセンジャーRNA転写産物の選択的分解をいう。
【0027】
本明細書で使用する場合、「shRNA」(低分子ヘアピン型RNA)とは、アンチセンス領域、ループ部分及びセンス領域を含むRNA分子であって、そのセンス領域が、二重ステムを形成するためにアンチセンス領域と塩基対合する相補的ヌクレオチドを有する、RNA分子をいう。転写後プロセシング後、低分子ヘアピン型RNAは、RNase IIIファミリーのメンバーである、酵素のダイサーを介して行われる切断事象によって低分子干渉RNAに変換される。
【0028】
本明細書で使用する場合、「低分子干渉RNA」又は「siRNA」とは、配列特異的にRNA干渉を介することによって遺伝子発現を阻害又は下方調節できる、任意の低分子RNA分子をいう。低分子RNAは、例えば、約18〜21ヌクレオチド長であり得る。
【0029】
本明細書で使用する場合、「アンタゴマー」とは、切断を阻害するために切断部位又は1つ若しくは複数の塩基修飾部位のいずれかに誤対合を有する、特定のマイクロRNA標的と相補性を有する低分子合成RNAをいう。
【0030】
本明細書で使用する場合、「転写後プロセシング」という語句は、転写後に起こり、例えば、酵素のダイサー及び/又はドローシャを介して行われる、mRNAプロセシングをいう。
【0031】
本明細書で使用する場合、「ABCA1」という用語は、例えば、Remaley,A.T.,Proc.Nat.Acad.Sci.96:12685−12690,1999に記載されている、ATP結合カセット、サブファミリーA、メンバー1、タンパク質輸送体をいう。
【0032】
本明細書で使用する場合、「ABCA1タンパク質発現の増加」とは、本明細書に記載される方法による治療の前の被験体におけるABCA1タンパク質の量より少なくとも約1倍以上(例えば1、2、3、4、5、10、20、30、40、50、60、70、80、90、100、1000、10,000倍又はそれ以上)のABCA1タンパク質の量をいう。被験体におけるABCA1タンパク質発現の量をいう場合、「増加」とは、また、本明細書に記載される方法による治療の前の被験体におけるABCA1タンパク質の量より少なくとも約5%以上(例えば5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、99又は100%)であることを意味する。タンパク質量は、当該技術分野において公知の方法によって測定できる。
【0033】
「有効量」とは、未治療の患者に対して疾患の症状を寛解させるのに必要とされる薬剤又は薬剤を含む組成物の量を意味する。疾患の治療的処置のために本発明を実施するのに使用される組成物(複数可)の有効量は、投与様式、被験体の年齢、体重、及び全体的な健康に応じて変わる。最終的に、主治医又は獣医が、適した量及び投薬レジメンを決定するだろう。かかる量を「有効」量」という。
【0034】
本明細書で使用する場合、「コレステロール恒常性」とは、コレステロール取り込み、コレステロール生合成、胆汁酸へのコレステロールの変換及び胆汁酸の排泄のような、健康レベルの血中のLDL、HDL及びコレステロール(例えば、本明細書においてかかる健康レベルは「参照基準」ともいわれる)を有する被験体において起こるプロセスの調節をいう。したがって、コレステロール恒常性を必要とする被験体は、健康レベルの血中のLDL、HDL及び/又はコレステロールへの回復を生じる、改善された調節を必要とする。
【0035】
「被験体」は、ヒト、家庭用家畜及び飼育場用家畜、並びに動物園の動物、スポーツ用の動物又はペット用の動物、例えばマウス、ウサギ、ブタ、ヒツジ、ヤギ、ウシ及び高等霊長類を含む、哺乳網の任意のメンバーを含む、脊椎動物である。
【0036】
本明細書で使用する場合、「ベクター」又は「発現ベクター」は、所望の調節配列及び遺伝子を含む、核酸ベースの送達ビヒクルであり、その内容物を細胞に移動させるために使用され得る。
【0037】
本明細書の開示において、「含む(comprise)」、「含んでいる(comprising)」、「含有している」及び「有している」などは、米国特許法でみなされている意味を有することができ、「包含する(include)」、「包含している(including)」などを意味することができ、同様に「から実質的になる」又は「実質的になる」は、米国特許法でみなされている意味を有していて、この用語には制約がなく、列挙されている基本的又は新規な特徴が、列挙されているものよりも多くの存在によって変えられない限り、列挙されているものより多くのものの存在を許容しているが、先行技術の実施形態は除外する。
【0038】
本開示全体にわたって状況に応じて、他の定義が現れる。
【0039】
本発明の組成物及び方法
マイクロRNA(miRNA)は、哺乳動物を含む、種々の生物に存在し、進化において保存されている低分子(例えば、18〜24ヌクレオチド)の非コードRNAのクラスである。miRNAは、RNAse III酵素のドローシャ及びダイサーによる配列切断を介する一次転写産物に由来する約70ヌクレオチドのヘアピン前駆体からプロセシングされる。多くのマイクロRNAは、プレmRNAのイントロン内又はncRNA遺伝子内に受け入れられる、遺伝子間領域においてコードされ得る。多くのmiRNAはまた、クラスター化され、ポリシストロンとして転写される傾向があり、多くの場合、類似した時空間的発現パターンを有する。miRNAは、発生のタイミング、分化、アポトーシス、細胞増殖、臓器発生、及び代謝を含む、種々の生物学的プロセスにおいて役割を有することが見出されている。
【0040】
本明細書において配列番号1及び2とそれぞれ称する、マイクロRNA−33a及びマイクロRNA−33b(miR−33a及びmiR−33b)は、翻訳抑制のためにATP結合カセット輸送体ABCA1を標的とする。したがって、miR−33の阻害は、ABCA1レベル、HDL産生、及びコレステロールの逆輸送を増加させるための新規の治療手段を表す。
【0041】
本発明の組成物及び方法は、コレステロール恒常性を必要とする被験体においてmiR−33a及び/又はmiR−33bを阻害するように設計される。一態様において、本発明は、コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で、配列番号1又は2と相補的である核酸配列を含む組成物を提供する。これは、例えば、限定されないが、配列番号3又は配列番号4のアンチセンスオリゴヌクレオチドを含む、miR−33a又はmiR−33bと相補的であるアンチセンスオリゴヌクレオチドを投与することによって達成され得る。本発明の方法を実施するのに使用するため、及びmiR−33a又はmiR−33bと相補的である、他の核酸配列は、限定されないが、shRNA若しくはsiRNAを含む干渉RNA、又はアンタゴマーなどのmiR−33a又はmiR−33bの転写後プロセシングを阻害するものであり得る。
【0042】
アンチセンスオリゴヌクレオチドは、通常、標的に結合し、転写、翻訳、又はスプライシングのレベルで発現を停止することによってDNA又はRNA標的の発現を遮断するように設計される。本発明のアンチセンスオリゴヌクレオチドは、ストリンジェントな条件下で、miR−33a及び/又はmiR−33bとハイブリダイズするように設計されている相補的核酸配列である。したがって、オリゴヌクレオチドは、標的と十分に相補的であるように、すなわち、所望の効果を与えるのに非常に十分に、かつ、十分な特異性でハイブリダイズするように選択される。
【0043】
本発明の文脈において、ハイブリダイゼーションとは、相補的ヌクレオシド又はヌクレオチド塩基間のワトソン−クリック水素結合、フーグスティーン水素結合、又は逆フーグスティーン水素結合であり得る、水素結合を意味する。例えば、アデニン及びチミンは、水素結合の形成を介して対になる相補的核酸塩基である。本明細書で使用する場合、相補的とは、2つのヌクレオチド間の正確な対合のための能力をいう。例えば、オリゴヌクレオチドの特定の位置におけるヌクレオチドが、DNA又はRNA分子の同じ位置におけるヌクレオチドと水素結合できる場合、オリゴヌクレオチド及びDNA又はRNAは、その位置において互いに相補的であるとみなされる。各分子における十分な数の対応する位置が、互いに水素結合できるヌクレオチドによって占められている場合、オリゴヌクレオチド及びDNA又はRNAは互いに相補的である。したがって、「特異的にハイブリダイズできる」及び「相補的」は、安定及び特異的な結合が、オリゴヌクレオチドと、DNA又はRNA標的との間で起こるような十分な程度の相補性又は正確な対合を示すために使用される用語である。
【0044】
相補的核酸配列は、特異的にハイブリダイズできるその標的核酸の配列と100%相補的であることを必要としないことは、当該技術分野において理解されている。標的DNA又はRNA分子に対する配列の結合が、活性を喪失させるために標的DNA又はRNAの正常な機能を妨げる場合、並びに特異的結合が所望される条件下、すなわち、インビボアッセイ又は治療的処置の場合、及びインビトロアッセイの場合における生理学的条件下、アッセイが適切なストリンジェンシーの条件下で実施される条件下で、非標的配列に対する配列の非特異的結合を回避するために十分な程度の相補性が存在する場合、本発明の相補的核酸配列は特定的にハイブリダイズできる。例えば、ストリンジェント塩濃度は、通常、約750mMのNaCl及び75mMのクエン酸三ナトリウムより少なく、好ましくは約500mMのNaCl及び50mMのクエン酸三ナトリウムより少なく、より好ましくは約250mMのNaCl及び25mMのクエン酸三ナトリウムより少ない。低ストリンジェンシーハイブリダイゼーションは、有機溶媒、例えばホルムアミドの非存在下で得ることができるのに対して、高ストリンジェンシーハイブリダイゼーションは、少なくとも約35%のホルムアミド、より好ましくは少なくとも約50%のホルムアミドの存在下で得ることができる。ストリンジェント温度条件としては、通常、少なくとも約30℃、より好ましくは少なくとも約37℃、最も好ましくは少なくとも約42℃の温度が挙げられる。ハイブリダイゼーション時間、界面活性剤、例えば、ドデシル硫酸ナトリウム(SDS)の濃度、及びキャリアDNAの包含又は排除などの付加パラメーターを変化させることは、当業者に周知である。必要に応じて、これらの種々の条件を組み合わせることで種々のレベルのストリンジェンシーが達成される。好ましい実施形態において、750mMのNaCl、75mMのクエン酸三ナトリウム、及び1%のSDS中で30℃にてハイブリダイゼーションが起こる。より好ましい実施形態において、500mMのNaCl、50mMのクエン酸三ナトリウム、1%のSDS、35%のホルムアミド、及び100μg/mlの変性サケ精子DNA(ssDNA)中で37℃にてハイブリダイゼーションが起こる。最も好ましい実施形態において、250mMのNaCl、25mMのクエン酸三ナトリウム、1%のSDS、50%のホルムアミド、及び200μg/mlのssDNA中で42℃にてハイブリダイゼーションが起こる。これらの条件における有用な変化は、当業者であれば容易に明らかとなる。
【0045】
ほとんどの用途において、ハイブリダイゼーション後の洗浄工程もストリンジェンシーを変化させる。洗浄ストリンジェンシー条件は、塩濃度及び温度で定義できる。前述のように、塩濃度を低下させるか又は温度を上昇させることで洗浄ストリンジェンシーを増加させることができる。例えば、洗浄工程用のストリンジェント塩濃度は、好ましくは約30mMのNaCl及び3mMのクエン酸三ナトリウムより少なく、最も好ましくは約15mMのNaCl及び1.5mMのクエン酸三ナトリウムより少ない。洗浄工程用のストリンジェント温度条件としては、通常、少なくとも約25℃、より好ましくは少なくとも約42℃、更により好ましくは少なくとも約68℃の温度が挙げられる。好ましい実施形態において、洗浄工程は、30mMのNaCl、3mMのクエン酸三ナトリウム、及び0.1%のSDS中で25℃にて起こる。より好ましい実施形態において、洗浄工程は、15mMのNaCl、1.5mMのクエン酸三ナトリウム、及び0.1%のSDS中で42℃にて起こる。より好ましい実施形態において、洗浄工程は、15mMのNaCl、1.5mMのクエン酸三ナトリウム、及び0.1%のSDS中で68℃にて起こる。これらの条件における更なる変化は、当業者であれば容易に明らかとなる。ハイブリダイゼーション技術は、当業者に周知であり、例えば、Benton及びDavis(Science 196:180,1977);Grunstein及びHogness(Proc.Natl.Acad.Sci.,USA 72:3961,1975);Ausubelら,(Current Protocols in Molecular Biology,Wiley Interscience,New York,2001);Berger及びKimmel(Guide to Molecular Cloning Techniques,1987,Academic Press,New York);並びにSambrookら,Molecular Cloning:A Laboratory Manual,Cold Spring Harbor Laboratory Press,New Yorkに記載されている。
【0046】
本発明のアンチセンスオリゴヌクレオチドは、標的核酸内の標的領域と少なくとも80%の配列相補性を含むことが好ましく、更にそれらは90%の配列相補性を含むことが好ましく、標的とされる標的核酸配列内の標的領域と95%の配列相補性を含むことが更により好ましい。例えば、アンチセンスオリゴヌクレオチドの20核酸塩基のうちの18が相補的であると、標的領域と特異的にハイブリダイズするアンチセンス化合物は90パーセントの相補性を表す。標的核酸領域を有するアンチセンス化合物の相補性パーセントは、基本ローカルアラインメント検索ツール(BLASTプログラム)を用いて慣例的に決定することができる(Altschulら,J.Mol.Biol.1990,215,403−410:Zhang及びMadden,Genome Res.,1997,7,649−656)。miR−33a又はmiR−33bとハイブリダイズする、本発明のアンチセンス及び他の化合物は実験により確認され、それらの化合物の代表的な配列は、本発明の好ましい実施形態として本明細書の以下で確認される(例えば、限定されないが、配列番号3又は配列番号4のアンチセンスオリゴヌクレオチドを含む)。
【0047】
別の実施形態において、配列番号1又は2と相補的である核酸配列は、限定されないが、shRNA又はsiRNAを含む、干渉RNAであり得る。干渉RNAとしては、限定されないが、低分子干渉RNA(「siRNA」)及び低分子ヘアピン型RNA(「shRNA」)が挙げられる。干渉RNAを構築するための方法は当該技術分野において周知である。例えば、干渉RNAは、2つの別のオリゴヌクレオチドから構築することができ、一方の鎖はセンス鎖であり、他方の鎖はアンチセンス鎖であり、アンチセンス鎖及びセンス鎖は自己相補的であり(すなわち、例えばアンチセンス鎖及びセンス鎖が二本鎖又は二本鎖構造を形成する場合などに、各々の鎖は他方の鎖におけるヌクレオチド配列と相補的であるヌクレオチド配列を含む)、アンチセンス鎖は、標的核酸分子又はその一部(すなわち望まれていない遺伝子)におけるヌクレオチド配列と相補的であるヌクレオチド配列を含み、センス鎖は、標的核酸配列又はその一部に対応するヌクレオチド配列を含む。代替として、干渉RNAは単一のオリゴヌクレオチドから構築され、自己相補的センス領域及びアンチセンス領域は、核酸ベース又は非核酸ベースのリンカー(複数可)によって連結される。干渉RNAは、自己相補的センス領域及びアンチセンス領域を有する、二本鎖、非対称二本鎖、ヘアピン又は非対称ヘアピン二次構造を有するポリヌクレオチドであってもよく、アンチセンス領域は、別の標的核酸分子又はその一部におけるヌクレオチド配列と相補的であるヌクレオチド配列を含み、センス領域は、標的核酸配列又はその一部に対応するヌクレオチド配列を有する。干渉は、自己相補的センス領域及びアンチセンス領域を含む2つ以上のループ構造及び1つのステムを有する環状の一本鎖ポリヌクレオチドであってもよく、アンチセンス領域は、標的核酸分子又はその一部におけるヌクレオチド配列と相補的であるヌクレオチド配列を含み、センス領域は、標的核酸配列又はその一部に対応するヌクレオチド配列を有し、環状ポリヌクレオチドは、RNA干渉を介することができる活性siRNA分子を生成するためにインビボ又はインビトロのいずれかでプロセシングされ得る。
【0048】
本発明の特定の実施形態において、干渉RNAコード領域は、センス領域、アンチセンス領域及びループ領域を有する自己相補的RNA分子をコードする。望ましく発現される場合、かかるRNA分子は「ヘアピン」構造を形成し、本明細書で「shRNA」という。ループ領域は、通常、長さが約2から約10の間のヌクレオチドである。好ましい実施形態において、ループ領域は、長さが約6から約9のヌクレオチドである。本発明の1つのかかる実施形態において、センス領域及びアンチセンス領域は、長さが約15から約20の間のヌクレオチドである。転写後プロセシング後、低分子ヘアピン型RNAは、RNase IIIファミリーのメンバーである、酵素のダイサーを介して行われる切断事象によってsiRNAに変換される。次いで、siRNAは、相同性を共有する遺伝子の発現を阻害できる。詳細については、Brummelkampら,Science 296:550−553,(2002);Leeら,Nature Biotechnol.,20,500−505,(2002);Miyagishi及びTaira,Nature Biotechnol 20:497−500,(2002);Paddisonら,Genes & Dev.16:948−958,(2002);Paul,Nature Biotechnol,20,505−508,(2002);Sui,Proc.Natl.Acad.Sd.USA,99(6),5515−5520,(2002);Yuら,Proc NatlAcadSci USA 99:6047−6052,(2002)を参照のこと。
【0049】
siRNAによって誘導される標的RNA切断反応は高度に配列特異的である。一般に、標的遺伝子(すなわち、miR−33a及び/又はmiR−33b)の一部と同一であるヌクレオチド配列を含有するsiRNAが阻害のために好ましい。しかしながら、siRNAと標的遺伝子との間の100%の配列同一性は、本発明を実施するのに必要とされない。したがって、本発明は、遺伝子変異、株多型性、又は進化的分岐に起因して予測され得る配列変異を耐容できる利点を有する。例えば、標的配列に対する挿入、欠失、及び単一点突然変異を有するsiRNA配列もまた、阻害に効果的であることが見出されている。代替として、ヌクレオチド類似体置換又は挿入を有するsiRNA配列が阻害に効果的であってもよい。
【0050】
更に別の実施形態において、配列番号1又は2と相補的である核酸配列はアンタゴマーである。アンタゴマーは、マイクロRNAを標的とする一本鎖、二本鎖、部分的に二本鎖及びヘアピン構造化された化学的に修飾されたオリゴヌクレオチドである。好ましくは、本発明において特徴付けられたアンタゴマーは、約12〜25ヌクレオチド、好ましくは約15〜23ヌクレオチドのmiRNA標的配列とハイブリダイズするのに十分に相補的であるヌクレオチド配列を含む。
【0051】
特定の実施形態において、アンタゴマーは、RNaseプロテクション並びに増大した組織及び細胞取り込みなどの薬理学的特性についての種々の修飾を保有するRNA様オリゴヌクレオチドである。アンタゴマーは、糖の完全な2’−O−メチル化、ホスホロチオエート骨格及び3’末端におけるコレステロール部分を有することによって正常なRNAとは異なり得る。ホスホロチオエート修飾は、増大した組織取り込みの一因となるRNase活性及びそれらの親油性に対する保護を与える。好ましい実施形態において、アンタゴマーは6つのホスホロチオエート骨格修飾を含み、2つのホスホロチオエートは5’末端に位置し、4つは3’末端に位置する。本発明のアンタゴマーはまた、それらの長さ又はアンタゴマーを構成するヌクレオチドの数に関して修飾されてもよい。本発明のアンタゴマーは、最適機能のために長さが約20〜21ヌクレオチドであることが好ましい。なぜならこのサイズはmiR−33a及びmiR−33bについての成熟マイクロRNAのサイズと一致するからである。
【0052】
RNA、cDNA、ゲノムDNA、ベクター、ウイルス又はそのハイブリッドであろうとなかろうと、本発明を実施するために使用される核酸配列は、種々の源から単離でき、遺伝子操作でき、増幅でき、及び/又は発現/組換え的に生成できる。組換え核酸配列は、個々に単離又はクローニングでき、所望の活性について試験できる。例えばインビトロ、細菌、真菌、哺乳動物、酵母、昆虫又は植物細胞発現系を含む、任意の組換え発現系が使用されてもよい。
【0053】
本発明の核酸配列は、送達ベクターに挿入でき、そのベクター内の転写ユニットから発現できる。組換えベクターは、DNAプラスミド又はウイルスベクターであってもよい。ベクター構築物の生成は、限定されないが、例えば、Sambrookら,Molecular Cloning:A Laboratory Manual.(1989)),Coffinら,(Retroviruses.(1997))及び「RNA Viruses:A Practical Approach」(Alan J.Cann,Ed.,Oxford University Press,(2000))に記載されているようなPCR、オリゴヌクレオチド合成、制限エンドヌクレアーゼ消化、ライゲーション、形質転換、プラスミド精製、及びDNAシークエンシングの標準的技術を含む、当該技術分野において周知の任意の適切な遺伝子操作技術を用いて成され得る。当業者に明らかであるように、種々の適切なベクターが、本発明の核酸を細胞に移すのに利用可能である。核酸を送達するのに適したベクターの選択及び選択された発現ベクターの細胞への挿入のための条件の最適化は、過度の実験を必要とせずに、当業者の想定の範囲内である。ウイルスベクターは、パッケージング細胞において組換えウイルスの産生のための配列を有するヌクレオチド配列を含む。本発明の核酸を発現するウイルスベクターは、限定されないが、レトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、ポックスウイルス又はアルファウイルスを含む、ウイルス骨格に基づいて構築できる。本発明の核酸を発現できる組換えベクターは本明細書に記載されるように送達でき、標的細胞(例えば、安定な形質転換体)に存続する。
【0054】
本発明を実施するために使用される核酸配列は、例えば、Adams(1983)J.Am.Chem.Soc.105:661;Belousov(1997)Nucleic Acids Res.25:3440−3444;Frenkel(1995)Free Radic.Biol.Med.19:373−380;Blommers(1994)Biochemistry 33:7886−7896;Narang(1979)Meth.Enzymol.68:90;Brown(1979)Meth.Enzymol.68:109;Beaucage(1981)Tetra.Lett.22:1859;米国特許第4,458,066号に記載されているような周知の化学合成技術によってインビトロで合成できる。
【0055】
本発明の核酸配列は、修飾、例えばヌクレオチド修飾の導入などによって核酸分解に対して安定化できる。例えば、本発明の核酸配列は、ヌクレオチド配列の5’又は3’末端においてホスホロチオエートの少なくとも第1、第2又は第3のヌクレオチド間結合を含む。別の例として、核酸配列は、2’修飾ヌクレオチド、例えば、2’−デオキシ、2’−デオキシ−2’−フルオロ、2’−O−メチル、2’−O−メトキシエチル(2’−O−MOE)、2’−O−アミノプロピル(2’−O−AP)、2’−O−ジメチルアミノエチル(2’−O−DMAOE)、2’−O−ジメチルアミノプロピル(2’−O−DMAP)、2’−O−ジメチルアミノエチルオキシエチル(2’−O−DMAEOE)、又は2’−O−N−メチルアセトアミド(2’−O−NMA)を含んでもよい。別の例として、核酸配列は、少なくとも1つの2’−O−メチル修飾ヌクレオチド含んでもよく、一部の実施形態において、ヌクレオチドの全ては2’−O−メチル修飾を含む。一部の実施形態において、核酸は「ロック」されている。すなわち、核酸は、2’−O原子及び4’−C原子を結合するメチレン架橋によってリボース環が「ロック」されている核酸類似体を含む(例えば、Kaupinnenら,Drug Disc.Today 2(3):287−290(2005);Koshkinら,J.Am.Chem.Soc.,120(50):13252−13253(1998)を参照のこと)。
【0056】
例えば、サブクローニング、ラベリングプローブ(例えば、クレノウポリメラーゼを用いるランダムプライマーラベリング、ニックトランスレーション、増幅)、シークエンシング、ハイブリダイゼーションなどの本発明を実施するために使用される核酸操作のための技術は、科学文献及び特許文献に十分に記載されている。例えば、Sambrook,ed.,MOLECULAR CLONING:A LABORATORY MANUAL(2ND ED.),Vols.1−3,Cold Spring Harbor Laboratory,(1989);CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,Ausubel,ed.john Wiley & Sons,Inc.,New York(1997);LABORATORY TECHNIQUES IN BIOCHEMISTRY AND MOLECULAR BIOLOGY:HYBRIDIZATION WITH NUCLEIC ACID PROBES,Part I.Theory and Nucleic Acid Preparation,Tijssen,ed.Elsevier,N.Y.(1993)を参照のこと。
【0057】
本発明は、翻訳抑制のためにATP結合カセット輸送体ABCA1を標的とし、それによって、ABCA1レベル、HDL産生、及びコレステロールの逆輸送を増加させるように設計される核酸配列を含む医薬組成物及び製剤を提供する。
【0058】
代替の実施形態において、本発明の組成物は薬学的に許容される担体と共に製剤化される。代替の実施形態において、本発明の医薬組成物及び製剤は、非経口、局所的、経口、又はエアゾール若しくは経皮などによる局所投与によって投与できる。医薬組成物は任意の方法で製剤化でき、状態又は疾患(例えば、心血管障害の種類)及び病気の程度、各患者の全身の医学的状態、もたらされる好ましい投与方法に応じて、種々の単位剤形で投与できる。製剤化及び薬剤の投与についての技術に関する詳細は、科学文献及び特許文献に十分に記載されている。例えば、Remington’s Pharmaceutical Sciences,Maack Publishing Co,Easton PA(「Remington’s」)の最新版を参照のこと。
【0059】
本発明の核酸配列は、単独で、又は医薬製剤(組成物)の成分として投与できる。化合物は、ヒト又は獣医用の医薬に使用するための任意の簡便な方法で投与のために製剤化できる。湿潤剤、乳化剤及びラウリル硫酸ナトリウム及びステアリン酸マグネシウムなどの滑沢剤、並びに着色剤、離型剤、コーティング剤、甘味剤、香味剤及び芳香剤、防腐剤及び酸化防止剤も組成物中に存在してもよい。
【0060】
本発明の組成物の製剤は、皮内、吸入、経口/経鼻、局所、非経口、直腸、及び/又は膣内投与に適切なものを含む。製剤は簡便に単位剤形で存在してもよく、薬学の分野で周知の任意の方法によって調製されてもよい。単一の剤形を生成するために担体物質と組み合わされ得る活性成分(例えば、本発明の核酸配列)の量は、治療される宿主、特定の投与様式、例えば皮内又は吸入に応じて変更されるだろう。単一の剤形を生成するために担体物質と組み合わされ得る活性成分の量は、通常、治療的効果、例えば、抗原特異的T細胞又は体液性応答を生じる化合物の量である。
【0061】
本発明の医薬製剤は、製薬に関する分野で知られている任意の方法に従って調製できる。かかる薬物は、甘味剤、香味剤、着色剤及び保存剤を含有してもよい。製剤は、製造に適切である非毒性の薬学的に許容される添加剤と混合されてもよい。製剤は、1つ以上の賦形剤、乳化剤、防腐剤、緩衝剤、添加剤などを含んでもよく、液体剤、散剤、乳剤、凍結乾燥粉末、スプレー剤、クリーム剤、ローション剤、放出制御製剤、錠剤、丸剤、ゲル剤、パッチ剤上、インプラントにおいてなどの形態で提供されてもよい。
【0062】
経口投与のための医薬製剤は、適した及び適切な用量で、当該技術分野において周知の薬学的に許容される担体を用いて製剤化されてよい。かかる担体は、薬剤が、患者による摂取に適切な錠剤、丸剤、散剤、糖衣錠、カプセル剤、液体剤、ロゼンジ剤、ゲル剤、シロップ剤、スラリー剤、懸濁剤などとして単位剤形で製剤化されることを可能にする。経口使用のための医薬調製物は固形添加剤として製剤化でき、所望の場合、錠剤又は糖衣錠コアを得るために、適切な付加化合物を加えた後で、任意選択に、得られた混合物を粉砕し、顆粒の混合物を加工する。適切な固形添加剤は、例えば、ラクトース、スクロース、マンニトール、又はソルビトールを含む糖;トウモロコシ、コムギ、コメ、ジャガイモ、又は他の植物由来のデンプン;メチルセルロース、ヒドロキシプロピルメチル−セルロース、又はナトリウムカルボキシ−メチルセルロースなどのセルロース;並びにアラビア及びトラガントを含むガム;並びにタンパク質、例えばゼラチン及びコラーゲンを含む、糖質又はタンパク質の充填剤である。架橋ポリビニルピロリドン、寒天、アルギン酸、又はアルギン酸ナトリウムなどのその塩などの崩壊剤又は可溶化剤が加えられてもよい。押し込み式カプセル剤は、ラクトース又はデンプンなどの充填剤又は結合剤、タルク又はステアリン酸マグネシウムなどの滑沢剤、及び任意選択に安定剤と混合した活性剤を含有してもよい。軟カプセル剤において、活性剤は、安定剤を含んでも含まなくてもよく、脂肪油、液体パラフィン、又は液体ポリエチレングリコールなどの適切な液体に溶解又は懸濁されてもよい。
【0063】
水性懸濁剤は、例えば、水性皮内注射用の水性懸濁剤の製造に適切な添加剤と混合して活性剤(例えば、本発明の核酸配列)を含有してもよい。かかる添加剤としては、ナトリウムカルボキシメチルセルロース、メチルセルロース、ヒドロキシプロピルメチルセルロース、アルギン酸ナトリウム、ポリビニルピロリドン、トラガントガム及びアカシアガムなどの懸濁化剤、天然に存在するリン脂質(例えば、レシチン)などの分散化剤又は湿潤剤、アルキレンオキシドと脂肪酸との縮合生成物(例えば、ポリオキシエチレンステアレート)、エチレンオキシドと長鎖脂肪族アルコールとの縮合生成物(例えば、ヘプタデカエチレンオキシセタノール(heptadecaethylene oxycetanol))、エチレンオキシドと脂肪酸及びヘキシトールから得られる部分エステルとの縮合生成物(例えば、ポリオキシエチレンソルビトールモノ−オレエート)、又はエチレンオキシドと脂肪酸及びヘキシトール無水物から得られる部分エステルとの縮合生成物(例えば、ポリオキシエチレンソルビタンモノオレエート)が挙げられる。また、水性懸濁剤は、p−ヒドロキシ安息香酸エチル又はp−ヒドロキシ安息香酸−n−プロピルなどの1つ又は複数の防腐剤、1つ又は複数の着色剤、1つ又は複数の香味剤、及びスクロース、アスパルテーム又はサッカリンなどの1つ又は複数の甘味剤を含有してもよい。製剤はオスモル濃度で調整できる。
【0064】
一実施形態において、油ベースの薬剤が、本発明の核酸配列を投与するために使用される。油ベースの懸濁剤は、ラッカセイ油、オリーブ油、ゴマ油若しくはココナッツ油などの植物油中、又は流動パラフィンなどの鉱油中、或いはそれらの混合物中で活性剤を懸濁することにより製剤化できる。例えば、生物学的利用能を増加させ、経口投与した疎水性医薬化合物の個体間変動及び個体内変動を減少させるための精油又は精油成分を用いることを記載している米国特許第5,716,928号を参照のこと(米国特許第5,858,401号も参照のこと)。油懸濁剤は、蜜ろう、固形パラフィン又はセチルアルコールなどの増粘剤を含有してもよい。甘味剤が、グリセロール、ソルビトール又はスクロースなどの味の良い経口用調製物を提供するために加えられてもよい。これらの製剤は、アスコルビン酸などの酸化防止剤を加えることによって保存されてもよい。注射用油ビヒクルの例としては、Minto(1997)J.Pharmacol.Exp.Ther.281:93−102を参照のこと。
【0065】
本発明の医薬製剤はまた、水中油型エマルションの形態であってもよい。油性相は、上記の植物油若しくは鉱油又はそれらの混合物であってもよい。適切な乳化剤としては、アカシアガム及びトラガントガムなどの天然に存在するガム、大豆レシチンなどの天然に存在するリン脂質、ソルビタンモノオレエートなどの、脂肪酸及びヘキシトール無水物に由来するエステル又は部分エステル、並びにポリオキシエチレンソルビタンモノオレエートなどの、上記部分エステルとエチレンオキシドとの縮合生成物が挙げられる。エマルションはまた、シロップ剤及びエリキシル剤の製剤などで甘味剤及び香味剤を含有してもよい。かかる製剤はまた、鎮痛剤、防腐剤、又は着色剤を含有してもよい。代替の実施形態において、本発明のこれらの注射用水中油型エマルションは、パラフィン油、ソルビタンモノオレエート、エトキシ化ソルビタンモノオレエート及び/又はエトキシ化ソルビタントリオレートを含む。
【0066】
本発明を実施する際に、医薬化合物はまた、坐剤、インサフレーション、粉末及びエアゾール製剤(例えばステロイド吸入剤、例えば、Rohatagi(1995)J.Clin.Pharmacol.35:1187−1193;Tjwa(1995)Ann.Allergy Asthma Immunol.75:107−111を参照のこと)を含み、経鼻、眼内及び膣内経路により投与できる。坐剤製剤は、常温で固体であるが、体温で液体になるので、身体で溶解して薬物を放出する適切な非刺激性の添加剤と、薬物とを混合することにより調製できる。かかる物質はココアバター及びポリエチレングリコールである。
【0067】
本発明を実施する際に、医薬化合物は、塗布棒、溶液剤、懸濁剤、乳剤、ゲル剤、クリーム剤、軟膏剤、ペースト剤、ゼリー剤、塗布剤、粉末剤、及びエアゾール剤として製剤化され、局所経路により経皮的に送達できる。
【0068】
本発明を実施する際に、医薬化合物はまた、身体における持続放出のためにミクロスフェアとして送達できる。例えば、ミクロスフェアは、皮下に持続放出される薬物の皮内注射により投与できる;Rao(1995)J.Biomater Sci.Polym.Ed.7:623−645を参照のこと;生分解性及び注射用ゲル製剤として、例えば、Gao(1995)Pharm.Res.12:857−863(1995)を参照のこと;又は経口投与のためのミクロスフェアとして、例えば、Eyles(1997)J.Pharm.Pharmacol.49:669−674を参照のこと。
【0069】
本発明を実施する際に、医薬化合物は、静脈内(IV)投与又は体腔若しくは器官の内腔への投与などにより非経口で投与できる。それらの製剤は、薬学的に許容される担体に溶解される活性剤の溶液を含んでもよい。利用され得る許容されるビヒクル及び溶媒は、水及びリンガー溶液、等張食塩水である。加えて、滅菌不揮発性油が、溶媒又は懸濁媒として利用されてもよい。この目的のために、合成モノグリセリド又はジグリセリドを含む、任意の無菌不揮発性油が利用されてもよい。加えて、オレイン酸などの脂肪酸が、同様に、注射用調製物に使用されてもよい。それらの溶液は滅菌され、通常、望ましくない物質を含まない。それらの製剤は、従来の周知の滅菌技術により滅菌されてもよい。製剤は、pH調整剤及び緩衝剤、毒性調整剤、例えば、酢酸ナトリウム、塩化ナトリウム、塩化カリウム、塩化カルシウム、乳酸ナトリウムなどのほぼ生理的条件に必要とされる薬学的に許容される補助物質を含有してもよい。それらの製剤中の活性剤の濃度は、広範に変化されてもよく、選択される特定の投与様式及び患者の必要性に従って、液量、粘性、体重などに主に基づいて選択される。IV投与のために、製剤は、滅菌注射用水溶液又は油性懸濁液などの滅菌注射用調製物であってもよい。この懸濁液は、上記の適切な分散化剤又は湿潤剤及び懸濁化剤を用いて製剤化されてもよい。滅菌注射用調製物はまた、1,3−ブタンジオール溶液などの非毒性の非経口的に許容される賦形剤又は溶媒中の懸濁液であってもよい。投与は、ボーラス又は持続注入(例えば、特定期間、血管内への実質的に連続導入)によってなされてもよい。
【0070】
本発明の医薬化合物及び製剤は凍結乾燥されてもよい。本発明は、本発明の薬剤及び充填剤、例えば、マンニトール、トレハロース、ラフィノース、及びスクロース又はそれらの混合物を含む溶液を凍結乾燥することによって作製され得る、本発明の組成物を含む安定な凍結乾燥製剤を提供する。安定な凍結乾燥製剤を調製するためのプロセスは、約2.5mg/mLのタンパク質、約15mg/mLのスクロース、約19mg/mLのNaCl、及び5.5より高いが、6.5未満のpHを有するクエン酸ナトリウム緩衝剤の溶液を凍結乾燥することを含んでもよい。例えば、米国特許出願公開第20040028670号を参照のこと。
【0071】
本発明の組成物及び製剤は、リポソームを使用することにより送達できる。リポソームを使用することにより、特に、リポソーム表面が、標的細胞に特異的なリガンドを保持するか、又はそうでなければ特定の器官に選択的に向けられている場合、インビボにおける標的細胞への活性剤の送達に集中することができる。例えば、米国特許第6,063,400号;米国特許第6,007,839号;Al−Muhammed(1996)J.Microencapsul.13:293−306;Chonn(1995)Curr.Opin.Biotechnol.6:698−708;Ostro(1989)Am.J.Hosp.Pharm.46:1576−1587を参照のこと。
【0072】
本発明の製剤は、予防的及び/又は治療的処置のために投与できる。代替の実施形態において、治療的用途のために、組成物は、障害又はその合併症、例えば、心臓血管異常の臨床症状を治療、緩和又は部分的に停止するのに十分な量(これは治療有効量と呼ばれ得る)で、コレステロール恒常性を必要とする被験体に投与される。例えば、代替の実施形態において、本発明の医薬組成物は、ATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させ、細胞内コレステロールの流出及び/若しくは肝臓におけるHDLの産生を増加させ、並びに/又はコレステロール恒常性を必要とする被験体の血液を循環するコレステロールの量を減少させるのに十分な量で投与される。
【0073】
これを達成するのに適切な医薬組成物の量が治療有効量である。これに使用するための投薬スケジュール及び有効量、すなわち投与レジメンは、疾患又は状態の段階、疾患又は状態の重症度、患者の健康の一般状態、患者の身体状況、年齢などを含む、種々の要因に依存するだろう。患者のための投薬レジメンを判断する際に、投与様式も考慮される。
【0074】
投薬レジメンはまた、当該技術分野において周知の薬物動態パラメーター、すなわち、活性剤の吸収率、生物学的利用能、代謝、クリアランスなども考慮される(例えば、Hidalgo−Aragones(1996)J.Steroid Biochem.Mol.Biol.58:611−617;Groning(1996)Pharmazie 51:337−341;Fotherby(1996)Contraception 54:59−69;Johnson(1995)J.Pharm.Sci.84:1144−1146;Rohatagi(1995)Pharmazie 50:610−613;Brophy(1983)Eur.J.Clin.Pharmacol.24:103−108;最新のRemington’s,前出を参照のこと)。現在の技術水準により、臨床医は、個々の患者、活性剤及び治療される疾患又は状態の各々についての投薬レジメンを決定することができる。薬剤として使用される類似組成物のために提供されるガイドラインは、投薬レジメン、すなわち、投与スケジュール及び投薬レベルを決定するための指針として使用され、本発明の方法を正確かつ適正に実施することができる。
【0075】
製剤の単回又は多回投与は、例えば、患者に必要とされ、忍容される用量及び頻度、各投与後に生じるコレステロール恒常性の程度及び量などに応じて与えることができる。製剤は、状態、疾患又は症状を有効に治療、予防又は寛解する(例えば、ABCA1の発現を増加させ、細胞内コレステロールの流出及び/若しくは肝臓におけるHDLの産生を増加させ、並びに/又はコレステロール恒常性を必要とする被験体の血液を循環するコレステロールの量を減少させる)のに十分な量の活性剤を与えるべきである。
【0076】
代替の実施形態において、経口投与のための医薬製剤は、約1〜100mg/kg体重/日又はそれ以上の日用量である。経口投与と対照的に、血流、体腔又は器官の内腔内へのより低い用量が使用されてもよい。十分に高い用量が、局所若しくは経口投与又は粉末、スプレー若しくは吸入による投与において使用されてもよい。非経口又は経口投与可能な製剤を調製するための実際の方法は、当業者に公知であるか、又は明らかであり、Remington’s、前出などの文献により詳細に記載されている。
【0077】
種々の研究により、相補的核酸配列を用いた成功している哺乳動物の投薬が報告されている。例えば、Esau C.ら,(2006)Cell Metabolism,3(2):87−98は、4週間、週に2回の12.5〜75mg/kgの範囲のmiR−122アンチセンスオリゴヌクレオチドの腹腔内投与による正常なマウスの投薬を報告している。マウスは、体重を損失せず、又は食物摂取を減少せずに、治療の終わりに健康で、正常なように見えた。血漿アミノ基転移酵素レベルは、ALT及びASTレベルの非常に穏やかな増加を示した、75mg/kg用量のmiR−122 ASOを除いて、全ての用量について正常な範囲(AST3/4 45、ALT3/4 35)であった。これらにより、50mg/kgが有効で非毒性の用量であることが結論付けられた。Krutzfeldt Jら,(2005)Nature 438,685−689による別の研究は、80、160又は240mg/kg体重の総量を用いてマウスにおいtmiR−122をサイレンシングするためにアンタゴマーを注入した。最も高い用量はmiR−122シグナルの完全な損失を生じた。更に別の研究において、ロックド核酸(「LNA」)が、miR−122をサイレンシングするために霊長類において首尾よく適用された。Elmen J.ら,(2008)Nature 452,896−899は、miR−122の効果的なサイレンシングを、3用量の10mg/kgのLNA−抗miRによって霊長類において達成し、研究動物においてLNA関連毒性又は組織病理学的変化についてのいかなる形跡もなく、総血漿コレステロールの長期間存続し、可逆的な減少を導いたことを報告している。
【0078】
本発明の方法は、他の薬物又は薬剤、例えば、コレステロール恒常性を提供するための組成物との同時投与を更に含んでもよい。例えば、本発明の方法並びに/又は組成物及び製剤は、HMG CoA還元酵素阻害剤、例えば、アトルバスタチン、フルバスタチン、ロバスタチン、プラバスタチン、ロスバスタチン及びシンバスタチン、胆汁酸捕捉剤、例えば、コレセベラム、コレスチラミン及びコレスチポール、並びにニコチン酸(ナイアシン)などの増大したLDLコレステロールを治療するための薬物と同時投与できる。
【0079】
本発明は、本発明のより十分な理解及びその多くの利点を与える、以下に示す、非限定的な実施例によって更に説明される。
【実施例】
【0080】
本明細書中で、SREBPと協調してコレステロール恒常性を制御する際にmiR−33の役割を更に詳細に説明するための一連のインビトロ及びインビボでの研究を記載する。以下の実施例は例示の目的のみのために提示し、発明者らが彼らの発明とみなす発明の範囲を限定することを意図しない。
【実施例1】
【0081】
ヒト及びマウスにおけるmiR−33の予測される標的
miRNAによるmRNA標的の識別に対する利用可能なアプローチの中で、計算による予測研究が、miRNA標的の実験検証に対する第1のステップを確立する際の有用な方法を表すことが示された。miRNA標的を予測する際に考慮される最も重要なパラメーターは、「種対合」として知られている、その同種標的との特定の塩基対合を受けるmiRNA配列の能力である(Bartel DP(2004)Cell 116:281−297;van den Berg A,Mols J,Han J(2008)Biochem Biophys Acta)。これは、miRNA5’末端の7〜8塩基に関する。この文脈において、Miranda及びTargetScan 4.2標的予測データベース(microrna.sanger.ac.uk;www.targetscan.org/vert_42/)の両方から導かれた100より多い予測される標的の記載の中で、ヒト及びマウスの両方におけるmiR−33についての最も関連のある保存された標的はABCA1である(表1及び図7)。興味深いことに、これらの研究は、コレステロール輸送ネットワーク又は脂質ホメオスタシスにおける役割を果たし、ABCA1と機能的に関連することが知られている付加的タンパク質はまた、ヒト及びマウスの両方における標的として役立つことを明らかにした。それらとしては、ニーマンピック病C1型(NPC1)が挙げられる(表1)。
【0082】
【表1】
【実施例2】
【0083】
ヒト及びマウス細胞株におけるmiR−33によるABCA1の下方調節
バイオインフォマティクス予測研究からの結果を検証するために、一連の実験を、モデル系として、ヒトHepG2肝細胞癌細胞、IMR−90初代ヒト線維芽細胞、及びマウスマクロファージ細胞株J774を含む、ヒト及びマウス細胞株を用いて実施した。それらの細胞株は、低細胞密度でABCA1タンパク質の低い発現を示す。miR−33によるABCA1の翻訳阻害は、この低いレベルの発現に起因すると予測された。したがって、miRNA生物発生経路の成分のsiRNAノックダウンを、ABCA1タンパク質発現がmiRNAにより調節されるか否かを決定するために実施した。実際に、ドローシャ及びダイサーmiRNAプロセシング酵素に向けられたsiRNAでのトランスフェクションにより、試験した3つ全ての細胞株において(特にドローシャsiRNAでの)ABCA1タンパク質発現の十分な増加が生じた(図7A)。これらの結果はmiRNAによるABCA1の調節と一致している。
【0084】
この上方調節が、miR−33プロセシングの欠如から特異的に生じることを決定するために、3つの異なる細胞株を、抗miR−33a/bオリゴヌクレオチド(Ambion)でトランスフェクトした。抗miRは、塩基対合による配列特異的に内因性miRNAの機能を遮断する修飾したmiRNAアンチセンスオリゴヌクレオチドである。抗miR−33a及び抗miR−33bの両方のトランスフェクションは、ヒト細胞株において、及びJ774マウスマクロファージの抗miR−33a処置の際に、ABCA1タンパク質のかなり高い発現を生じた(図7B)。
【0085】
これらのデータは、内因性miR−33によるABCA1 mRNAの翻訳又は安定性の調節を示す。ABCA1発現に対するmiR−33の効果を更に評価するために、miR−33a及びmiR−33bのそれぞれの細胞内レベルを増加させるために、細胞を合成Pre−miR−33(Ambion)でトランスフェクトした。ABCA1タンパク質レベルは、細胞播種の24時間後に検出することが困難であるため、Pre−miR33の効果を、トランスフェクション及び播種の48時間後に評価した。この実験により、ABCA1発現が、特にマウスJ774マクロファージ及びヒトIMR−90線維芽細胞において、miR−33により特異的に抑制され得ることが明確に示される(図7C)。
【0086】
最近の研究により、ABCA1が、効果的なコレステロール流出及びHDL生合成を促進するためにマクロファージにおいて、別のコレステロール輸送体、ABCG1と共に作用することが示された(図5)(Tall ARら(2008)Cell Metab 7:365−375;Wang Xら(2007)J Clin Invest 117:2216−2224)。実際に、データは、一方の輸送体が欠損して、他方が誘導される場合、代償的調節を示唆することを示した(Yvan−Charvet Lら(2007)J Clin Invest 117:3900−3908;Ranalletta Mら(2006)Arterioscler Thromb Vasc Biol 26:2308−2315)。したがって、ABCG1発現が、miRNA経路及びmiR−33aの操作後、ABCA1レベルの変化の際に変わるか否かを調べた。興味深いことに、マウスマクロファージ細胞株J774において、ダイサー及びドローシャのRNAiによるABCG1発現、抗miR−33aトランスフェクション、又はPre−miR−33a発現に対する有意な効果は観察されなかった。これらの知見により、少なくともこれらの予備実験条件下で、ABCA1及びABCG1の発現がmiR−33操作に応答して相互に関連しないことが示唆される。
【実施例3】
【0087】
miR−33によるABCA1 3’UTRの標的化
miR−33が、ABCA1 mRNA及び/又はタンパク質レベルに影響を与えるか否かを決定するために、Pre−miR−33a/bを、miR−33のレベルを増加させるために、いくつかの細胞株(すなわち、IMR−90、HepG2、及びJ774)において発現させ、miR−33を拮抗するために抗miR−33オリゴヌクレオチドをトランスフェクトし、続いて、定量的RT−PCR及び免疫ブロット法のそれぞれによってABCA1 mRNA及びタンパク質レベルを分析する。アクチンmRNA(qRT−PCR)及びチューブリンタンパク質(免疫ブロット法)を対照として使用する。
【0088】
miR−33が、3’UTRの標的化を介してABCA1レベルに影響を与えるか否かを規定するために、予測されるmiR−33標的配列を保有するヒト及びマウスABCA1 3’UTRの断片を単離し、ルシフェラーゼレポータープラスミド(pMIR−REPORT、Ambion)の3’UTRクローニング部位にクローニングした。得られたABCA1 3’UTRルシフェラーゼレポーターを、miR−33a/b抗miR、前駆体、又は対照オリゴヌクレオチドの存在下で、ヒトHEK293細胞にトランスフェクトした。図8Aに示すように、ルシフェラーゼレポーターベクター内に予測されるmiR−33標的部位を保有するABCA1 3’UTRの断片の挿入により、挿入しないベクターと比較してレポーター活性の著しい減少が生じ、挿入された断片(例えばmiR−33認識部位)内の阻害配列の存在と一致する。重要なことに、ヒトmiR−33a/bと相補的である抗miRオリゴヌクレオチドでの同時トランスフェクションはレポーターの高い発現を引き起こし、内因性miR−33a/bが、実際に、ヒトHEK293細胞におけるABCA1 3’UTR断片を標的とすることを示唆する(図8B)。反対に、野生型ヒトmiR−33a/b前駆体オリゴヌクレオチド(Ambion)での同時トランスフェクションは、レポーター活性の更なる抑制を引き起こし、一方、種塩基対合領域において変異した前駆体オリゴヌクレオチドは、前駆体対照オリゴヌクレオチドと比較して効果を有さなかった(図8C)。相対ルシフェラーゼ活性を、ウミシイタケルシフェラーゼ活性によって正規化した絶対ルシフェラーゼ活性から測定した(PC:陰性対照として非関連マイクロRNA前駆体)。
【0089】
一緒に、これらのデータは、miR−33a/bが、翻訳抑制又はmRNA分解のためのABCA1 3’UTRを標的化することによりヒトABCA1の発現を阻害するという見解についての重要な支持を与える。
【実施例4】
【0090】
マウスにおけるマクロファージからのコレステロール輸送におけるmiR−33についての役割
SREBPは、コレステロールによって古典的な負のフィードバック形式で調節される(Brown MS,Goldstein JL(1997)Cell 89:331−340)。miR−33/SREBP/ABCA1コレステロール調節回路の生物医学及び生理学関連を、マウスJ774マクロファージモデルを用いて調べた(de la Llera−Moya Mら(2010,Epub ahead of print)Arterioscler.Thromb.Vasc.Biol)。J774マウスマクロファージを、以下のようにSREBP発現モジュレーションに関してコレステロール処置に供した:150K細胞を、24ウェルプレートフォーマットにおいてウェルごとに播種し、48時間、10μg/mlのコレステロール及び1μg/mlの25−ヒドロキシコレステロール(Sigma)で処理した。SREBP発現を活性化するために、次いで細胞をPBSで2回洗浄し、培地を、10%の脂質を含まないFBS、5mMの(2−ヒドロキシプロピル)−β−シクロデキストリン(Sigma)及び50μMのロバスタチン(Mevinolin,Sigma)を含有するDMEMに置き換え、示した時点で収集する。コレステロールで処理した細胞及びコレステロールを含まない細胞の両方を同じ時点で収集した。ロバスタチン/β−シクロデキストリン処理によるコレステロールの喪失は、J774マクロファージにおいてmiR−33a及びmSREBP−2宿主遺伝子の両方の増加した発現を生じ、ABCA1タンパク質発現の減少と一致した(図9A〜C)。コレステロール喪失に応答するABCA1タンパク質レベルの強力な抑制は、miR−33aアンチセンスオリゴヌクレオチドによって少なくとも部分的に反転し(図9D)、miR−33が、マウスマクロファージにおけるABCA1レベルのコレステロールにより調節される転写後制御を介するという考えと一致した。血清/コレステロールの存在下で培養したJ774マクロファージにおいて観察されたABCA1の高いタンパク質レベルは、更なるmiR−33a前駆体によって十分に抑制され(図9E)、ABCA1のmiR−33調節と一致する。加えて、コレステロールによるABCA1調節のmiRNAを介して行われるこの機構は、以前に記載した肝臓X受容体(LXR)を介するオキシステロールによるABCA1発現の転写制御であり(Tontonoz P,Mangelsdorf D,(2003)Mol.Endocrinol.17:985)、複数レベルのコレステロールによるABCA1の細胞調節を示す。したがって、マウスmiR−33a及びSREBP−2宿主遺伝子は、コレステロールにより同時調節され、マクロファージにおけるSREBP、miR−33、ABCA1、及びコレステロールの逆調節ネットワークの存在を示唆する。
【0091】
次に、J774マクロファージからのコレステロール流出に対するmiR−33操作の効果を試験した。まず、細胞を[3H]−コレステロールで標識し、次いでmiR−33a前駆体、miR−33aアンチセンス、又は対照オリゴヌクレオチドでトランスフェクトし、続いて血清/apoA−Iに対する[3H]−コレステロール流出のレベルを測定する。手短に述べると、低い継代のJ774細胞を、抗miR又はPre−マイクロRNA二本鎖によりトランスフェクトし、300K細胞/ウェルにて24ウェルプレートに播種し、[3H]−コレステロールを加えた。インキュベーションの20時間後、細胞を、室温にてPBSで2回洗浄し、培地を、15μg/mlの遊離apo−A−Iを含有する新鮮な培地と置き換えた。2又は4時間後、培地のアリコートを除去し、[3H]−コレステロールを、液体シンチレーション計数により測定した。細胞に存在する[3H]−コレステロールを、水酸化ナトリウム中で細胞を抽出することにより決定し、続いて液体シンチレーション計数により測定した。図9Fに示すように、miR−33aアンチセンスオリゴヌクレオチド(A)のマウスマクロファージへの導入により、[3H]−コレステロール流出の著しい(2倍)増加が生じたが、miR−33a前駆体(P)処理は、対照オリゴヌクレオチド(AC/PC)又は偽トランスフェクト細胞(UT)と比較して、[3H]−コレステロール流出の著しい減少を生じた。これらの知見により、ABCA1のmiR−33転写後抑制が、マクロファージにおいて細胞内コレステロールの保持を生じ、ABCA1レベルの制御を介するコレステロール輸送のmiR−33調節と一致することが実証される。
【実施例5】
【0092】
インビボモデルでの哺乳動物におけるmiR−33アンチセンスによるABCA1レベルの操作
miR−33アンチセンスアプローチによるABCA1レベルの操作もまた、インビボモデルでの哺乳動物において増大したHDLコレステロールレベルを生じる。尾静脈注射を、PBS(ビヒクル)、LNA−miR−33aアンチセンス、又はLNA対照オリゴヌクレオチドを用いる処置の6週間前の間、及び処置の間、西洋型食餌(すなわち、乳脂肪から42%kcalを補足した)を与えたマウスに実施した。血液を、治療の開始前にコレステロール/トリグリセリドプロファイルのために実験用マウスから顎下腺出血により得た。注射を行うために、20mg/kg/日のmiR−33−LNA又は対照LNAをPBS(全体積200μl)に溶解し、各日、同時間に毎日の尾静脈注射を介して3日連続投与した。マウスを、最後の尾静脈注射から48時間後に屠殺した。5日にわたる注射後、マウスを屠殺し、血清を以下のように回収した。屠殺の際に、総量1mLの血液を右側脳質穿刺によってマウスから得た。血液を5分間8,000rpmにて遠心分離して血清を得、−80℃にて凍結した。血清コレステロール、トリグリセリド、グルコース、アラニンアミノトランスフェラーゼ(ALT)及びアスパラギン酸アミノトランスフェラーゼ(AST)の全てを、マサチューセッツジェネラルホスピタル、比較医学のためのセンター、診断研究所においてHeska Dri−Chem 4000 Chemistry Analyzer(Heska,Loveland,CO)によってmiR−33−LNA又は対照−LNAでの処置前後に測定した。プールした血清のFPLC分析を記載(Hyogo Hら,(2002)J.Biol.Chem.277:34117)されるように実施した。血漿HDL−コレステロール濃度は、LNA−対照で処置したマウスと比較して、LNA−miR−33aアンチセンスで処置した動物において著しく増加した(図10A、B及び表2)。対照的に、LDL−コレステロール、トリグリセリド、又はグルコースレベルの血漿濃度で有意な効果は存在しなかった(図10B〜D及び表2)。これらの実験において、肝毒性(血漿AST/ALT)も見られなかった(図10E、F及び表2)。LNA−miR−33aアンチセンス処置に応答して血漿HDL−コレステロールにおいて観察された増加は、インビボでのmiR−33aによるABCA1依存性コレステロール流出の調節と一致する。
【0093】
これらの結果により更に、コレステロール生成及び脂肪合成遺伝子の重要な調節因子である、哺乳動物のSREBPファミリーの転写因子が、脊椎動物において細胞内コレステロールレベル及びコレステロール恒常性を決定するSREBP宿主遺伝子産物と協調して機能する、保存されたmiRNA(miR−33a/b)に対する宿主であることが確認される。図10は、miR−33及びその宿主遺伝子産物SREBPによるコレステロール恒常性の協調した調節についてのモデルを示す。miR−33は、その3’UTRを標的とすることによってABCA1翻訳を抑制する。このステップは、コレステロール産生遺伝子のトランス活性化を介してコレステロール生合成及び取り込みを活性化する、SREBPの発現と協調する。コレステロール蓄積は、SREBP経路を阻害し、ABCA1の発現を活性化する。そのABCA1は、細胞膜に移行し、apoA−I及びHDLへのコレステロール流出を媒介する。
【0094】
【表2】
表2.ロックド核酸(LNA)−miR−33aアンチマー(antimer)を有する西洋型食餌を与えたマウスの処置。血清コレステロール(A)、血漿トリグリセリド(B)、グルコース(C)、AST(D)、及びALT(E)の全てを、PBS(ビヒクル)、マウスmiR−33aに向けられたLNA−アンチセンスオリゴヌクレオチド、及びスクランブルLNA対照(LC)オリゴヌクレオチドの注入前後に西洋型食餌を与えたマウスにおいて測定した。
【0095】
これらの実施例に示すように、コレステロール生成及び脂肪合成遺伝子の重要な調節因子である、哺乳動物のSREBPファミリーのメンバーの転写因子は、脊椎動物において細胞内コレステロールレベル及びコレステロール恒常性を決定するSREBP宿主遺伝子産物と協調して機能する、保存されたmiRNA(miR−33a/b)に対する宿主である。miR−33は、インビトロでのコレステロール輸送及びインビボでのHDL合成にとって重要な結果を有する、ABCA1コレステロール輸送体の転写後制御に影響を及ぼす。
【0096】
(予測実施例6)
miR−33によるNPC1の標的化
ニーマンピックCI型タンパク質(NPC1)は、トランス−ゴルジネットワーク(TGN)を介して、後期のエンドソーム/リソソームから形質膜までのコレステロール/脂質の動員において重要な役割を果たす。このプロセスは、発生期のHDLを生成するためにABCA1を介して行われる細胞表面におけるapoA−I及び他のアポリポタンパク質に対するリン脂質及びコレステロールの動因と協調すると考えられる。NPC1の3’UTRは、1つの予測されるmiR−33標的部位を保有する(7−mer;TargetScan 4.2)。細胞内コレステロール輸送のその既知の役割並びに最近のABCA1及びコレステロール流出に対する機能的関連のために、そのNPC1の3’UTRはmiR−33に対する候補標的である。
【0097】
第1に、miRNAプロセシング酵素のドローシャ及びダイサーを標的とするsiRNAの使用によって、miRNAが、上記の3つの細胞株においてNPC1レベルを制御するか否かを決定する。第2に、Pre−miR−33及び抗miR−33オリゴヌクレオチドを、NPC1調節におけるmiR−33の特定の役割を扱うためにこれらの細胞株に導入する。NPC1タンパク質レベルを、対照としてβ−チューブリンを用いる免疫ブロット法によって決定する。NPC1 mRNAレベルに対するmiR−33操作の何らかの効果をqRT−PCRにより評価する。NCP1タンパク質及び/又はmRNAレベルに対する効果をmiR−33操作後に観察した場合、予測されるmiR−33標的配列を保有するNPC1 3’UTRを、pMIR−REPORTベクター内にクローニングし、その後、ABCA1に関して本明細書で上記したようにmiR−33標的部位を変異し、ルシフェラーゼ発現分析を行う。コレステロール輸送/流出におけるNPC1の潜在的な役割を確立するために、J774マウスマクロファージ細胞株におけるNPC1のsiRNA標的化の効果を、ABCA1に関して上記した方法を用いて試験する。
【0098】
前述の説明から、変更及び修飾が種々の用途及び状態を採用するために本明細書に記載される本発明に対してなされ得ることは明らかである。かかる実施形態はまた、添付の特許請求の範囲内である。本明細書の可変物の任意の定義における要素の記載の列挙は、記載した要素の任意の単一の要素又は組合せ(又は部分的組合せ)として可変のものの定義を含む。本明細書の実施形態の列挙は、任意の単一の実施形態又は任意の他の実施形態若しくはその一部と組み合わせた実施形態を含む。
【0099】
参考文献
本出願に記載した全ての特許、特許出願、データベース登録、及び刊行物は、各々の独立した特許、データベース登録、及び刊行物が、参照として組み込まれている具体的かつ個々に示されるものと同程度まで本明細書に参照として組み込まれている。本明細書に参照として組み込まれるものとしては、限定されないが、以下のものが挙げられる。
(参考文献)
【0100】
1. Maxfield FR, Tabas I (2005) Role of cholesterol and lipid organization in disease. Nature 438: 612-621.
2. Moller DE, Kaufman KD (2005) Metabolic syndrome: a clinical and molecular perspective. Annu Rev Med 56: 45-62.
3. Bengoechea-Alonso MT, Ericsson J (2007) SREBP in signal transduction: cholesterol metabolism and beyond. Curr Opin Cell Biol 19: 215-222.
4. Brown MS, Goldstein JL (1997) The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 89: 331-340.
5. Eberle D, et al. (2004) SREBP transcription factors: master regulators of lipid homeostasis. Biochimie 86: 839-848.
6. Horton JD, Goldstein JL, Brown MS (2002) SREBPs: activators of the complete program of cholesterol and fatty acid synthesis in the liver. J Clin Invest 109: 1125-1131.
7. Raghow R, et al. (2008) SREBPs: the crossroads of physiological and pathological lipid homeostasis. Trends Endocrinol Metab 19: 65-73.
8. Ericsson J, Edwards PA (1998) CBP is required for sterol-regulated and sterol regulatory element-binding protein-regulated transcription. J Biol Chem 273: 17865-17870.
9. Naar AM, et al. (1998) Chromatin, TAFs, and a novel multiprotein coactivator are required for synergistic activation by Sp1 and SREBP-1a in vitro. Genes Dev. 12: 3020-3031.
10. Naar AM, et al. (1999) Composite coactivator ARC mediates chromatin-directed transcriptional activation. Nature 398: 828-832.
11. Naar AM, Lemon BD, Tjian R (2001) Transcriptional coactivator complexes. Annu Rev Biochem 70: 475-501.
12. Naar AM, et al. (2002) Human CRSP interacts with RNA polymerase II CTD and adopts a specific CTD-bound conformation. Genes Dev 16: 1339-1344.
13. Oliner JD AJ, Hansen SK, Zhou S, Tjian R. (1996) SREBP transcriptional activity is mediated through an interaction with the CREB-binding protein. Genes Dev 10: 2903-2911.
14. Taatjes DJ, et al. (2002) Structure, function, and activator-induced conformations of the CRSP coactivator. Science 295: 1058-1062.
15. Yang F, et al. (2006) An ARC/Mediator subunit required for SREBP control of cholesterol and lipid homeostasis. Nature 442: 700-704.
16. Bartel DP (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281-297.
17. He L, Hannon GJ (2004) MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev Genet 5: 522-531.
18. Krutzfeldt J, Stoffel M (2006) MicroRNAs: a new class of regulatory genes affecting metabolism. Cell Metab 4: 9-12.
19. Lewis BP, et al. (2003) Prediction of mammalian microRNA targets. Cell 115: 787-798.
20. Lee YS, Dutta A (2008) MicroRNAs in Cancer. Annu Rev Pathol.
21. Mattes J, Collison A, Foster PS (2008) Emerging role of microRNAs in disease pathogenesis and strategies for therapeutic modulation. Curr Opin Mol Ther 10: 150-157.
22. Thum T, Catalucci D, Bauersachs J (2008) MicroRNAs: novel regulators in cardiac development and disease. Cardiovasc Res 79: 562-570.
23. Stefani G, Slack FJ (2008) Small non-coding RNAs in animal development. Nat Rev Mol Cell Biol 9: 219-230.
24. Sonkoly E, Stahle M, Pivarcsi A (2008) MicroRNAs and immunity: novel players in the regulation of normal immune function and inflammation. Semin Cancer Biol 18: 131-140.
25. van den Berg A, Mols J, Han J (2008) RISC-target interaction: Cleavage and translational suppression. Biochim Biophys Acta.
26. Vasudevan S, Tong Y, Steitz JA (2007) Switching from repression to activation: microRNAs can up-regulate translation. Science 318: 1931-1934.
27. Lagos-Quintana M, Rauhut R, Lendeckel W, Tuschl T (2001) Identification of novel genes coding for small expressed RNAs. Science 294: 853-858.
28. Rodriguez A, Griffiths-Jones S, Ashurst JL, Bradley A (2004) Identification of mammalian microRNA host genes and transcription units. Genome Res 14: 1902-1910.
29. Elmen J, et al. (2008) LNA-mediated microRNA silencing in non-human primates. Nature 452: 896-899.
30. Elmen J, et al. (2008) Antagonism of microRNA-122 in mice by systemically administered LNA-antimiR leads to up-regulation of a large set of predicted target mRNAs in the liver. Nucleic Acids Res 36: 1153-1162.
31. Esau C, et al. (2006) miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab 3: 87-98.
32. Krutzfeldt J, et al. (2005) Silencing of microRNAs in vivo with 'antagomirs'. Nature 438: 685-689.
33. Tall AR, et al. (2008) HDL, ABC transporters, and cholesterol efflux: implications for the treatment of atherosclerosis. Cell Metab 7: 365-375.
34. Wang X, Rader DJ (2007) Molecular regulation of macrophage reverse cholesterol transport. Curr Opin Cardiol 22: 368-372.
35. Singaraja RR, et al. (2002) Increased ABCA1 activity protects against atherosclerosis. J Clin Invest 110: 35-42.
36. Wang X, et al. (2007) Macrophage ABCA1 and ABCG1, but not SR-BI, promote macrophage reverse cholesterol transport in vivo. J Clin Invest 117: 2216-2224.
37. Van Eck M, et al. (2006) Macrophage ATP-binding cassette transporter A1 overexpression inhibits atherosclerotic lesion progression in low-density lipoprotein receptor knockout mice. Arterioscler Thromb Vasc Biol 26: 929-934.
38. van Eck M, et al. (2002) Leukocyte ABCA1 controls susceptibility to atherosclerosis and macrophage recruitment into tissues. Proc Natl Acad Sci U S A 99: 6298-6303.
39. Koldamova R, Lefterov I (2007) Role of LXR and ABCA1 in the pathogenesis of Alzheimer's disease - implications for a new therapeutic approach. Curr Alzheimer Res 4: 171-178.
40. Wahrle SE, et al. (2008) Overexpression of ABCA1 reduces amyloid deposition in the PDAPP mouse model of Alzheimer disease. J Clin Invest 118: 671-682.
41. Brunham LR, Kruit JK, Verchere CB, Hayden MR (2008) Cholesterol in islet dysfunction and type 2 diabetes. J Clin Invest 118: 403-408.
42. Chawla A, et al. (2001) A PPAR gamma-LXR-ABCA1 pathway in macrophages is involved in cholesterol efflux and atherogenesis. Mol Cell 7: 161-171.
43. Venkateswaran A, et al. (2000) Control of cellular cholesterol efflux by the nuclear oxysterol receptor LXR alpha. Proc Natl Acad Sci U S A 97: 12097-12102.
44. Chisholm JW, Hong J, Mills SA, Lawn RM (2003) The LXR ligand T0901317 induces severe lipogenesis in the db/db diabetic mouse. J Lipid Res 44: 2039-2048.
45. Millatt LJ, Bocher V, Fruchart JC, Staels B (2003) Liver X receptors and the control of cholesterol homeostasis: potential therapeutic targets for the treatment of atherosclerosis. Biochim Biophys Acta 1631: 107-118.
46. Schmitz G, Langmann T (2005) Transcriptional regulatory networks in lipid metabolism control ABCA1 expression. Biochim Biophys Acta 1735: 1-19.
47. Tamehiro N, et al. (2007) Sterol regulatory element-binding protein-2- and liver X receptor-driven dual promoter regulation of hepatic ABC transporter A1 gene expression: mechanism underlying the unique response to cellular cholesterol status. J Biol Chem 282: 21090-21099.
48. Wong J, Quinn CM, Brown AJ (2006) SREBP-2 positively regulates transcription of the cholesterol efflux gene, ABCA1, by generating oxysterol ligands for LXR. Biochem J 400: 485-491.
49. Zeng L, et al. (2004) Sterol-responsive element-binding protein (SREBP) 2 down-regulates ATP-binding cassette transporter A1 in vascular endothelial cells: a novel role of SREBP in regulating cholesterol metabolism. J Biol Chem 279: 48801-48807.
50. Haidar B, et al. (2002) cAMP induces ABCA1 phosphorylation activity and promotes cholesterol efflux from fibroblasts. J Lipid Res 43: 2087-2094.
51. See RH, et al. (2002) Protein kinase A site-specific phosphorylation regulates ATP-binding cassette A1 (ABCA1)-mediated phospholipid efflux. J Biol Chem 277: 41835-41842.
52. Lu R, et al. (2008) ApoA-I facilitates ABCA1 recycle/accumulation to cell surface by inhibiting its intracellular degradation and increases HDL generation. Arterioscler Thromb Vasc Biol 28: 1820-1824.
53. Taylor AJ (2008) Evidence to support aggressive management of high-density lipoprotein cholesterol: implications of recent imaging trials. Am J Cardiol 101: 36B-43B.
54. Yvan-Charvet L, et al. (2007) Combined deficiency of ABCA1 and ABCG1 promotes foam cell accumulation and accelerates atherosclerosis in mice. J Clin Invest 117: 3900-3908.
55. Ranalletta M, et al. (2006) Decreased atherosclerosis in low-density lipoprotein receptor knockout mice transplanted with Abcg1-/- bone marrow. Arterioscler Thromb Vasc Biol 26: 2308-2315.
56. deGoma EM, deGoma RL, Rader DJ (2008) Beyond high-density lipoprotein cholesterol levels evaluating high-density lipoprotein function as influenced by novel therapeutic approaches. J Am Coll Cardiol 51: 2199-2211.
57. Neufeld EB, et al. (2004) The ABCA1 transporter modulates late endocytic trafficking: insights from the correction of the genetic defect in Tangier disease. J Biol Chem 279: 15571-15578.
58. Wang MD, et al. (2007) Differential regulation of ATP binding cassette protein A1 expression and ApoA-I lipidation by Niemann-Pick type C1 in murine hepatocytes and macrophages. J Biol Chem 282: 22525-22533.
59. Boadu E, Francis GA (2006) The role of vesicular transport in ABCA1-dependent lipid efflux and its connection with NPC pathways. J Mol Med 84: 266-275.
60. Baek D, et al. (2008) The impact of microRNAs on protein output. Nature 455: 64-71.
61. Baskerville S, Bartel DP (2005) Microarray profiling of microRNAs reveals frequent coexpression with neighboring miRNAs and host genes. Rna 11: 241-247.
62. Foufelle F, Ferre P (2002) New perspectives in the regulation of hepatic glycolytic and lipogenic genes by insulin and glucose: a role for the transcription factor sterol regulatory element binding protein-1c. Biochem J 366: 377-391.
63. Foretz M, et al. (1999) ADD1/SREBP-1c is required in the activation of hepatic lipogenic gene expression by glucose. Mol Cell Biol 19: 3760-3768.
64. Radhakrishnan A, et al. (2007) Sterol-regulated transport of SREBPs from endoplasmic reticulum to Golgi: oxysterols block transport by binding to Insig. Proc Natl Acad Sci U S A 104: 6511-6518.
65. Repa JJ, et al. (2000) Regulation of mouse sterol regulatory element-binding protein-1c gene (SREBP-1c) by oxysterol receptors, LXRalpha and LXRbeta. Genes Dev 14: 2819-2830.
66. Hannah VC, et al. (2001) Unsaturated fatty acids down-regulate srebp isoforms 1a and 1c by two mechanisms in HEK-293 cells. J Biol Chem 276: 4365-4372.
67. Ou J, et al. (2001) Unsaturated fatty acids inhibit transcription of the sterol regulatory element-binding protein-1c (SREBP-1c) gene by antagonizing ligand-dependent activation of the LXR. Proc Natl Acad Sci U S A 98: 6027-6032.
68. Sheng Z, Otani H, Brown MS, Goldstein JL (1995) Independent regulation of sterol regulatory element-binding proteins 1 and 2 in hamster liver. Proc Natl Acad Sci U S A 92: 935-938.
69. Akinc A, et al. (2008) A combinatorial library of lipid-like materials for delivery of RNAi therapeutics. Nat Biotechnol 26: 561-569.
70. de la Llera-Moya M, et al. (2010, Epub ahead of print) Arterioscler. Thromb. Vasc. Biol.
72. Tontonoz P, Mangelsdorf D, (2003) Mol. Endocrinol. 17:985.
71. Hyogo H, Roy S, Paigen B, Cohen D, (2002) J. Biol. Chem. 277: 34117.
【特許請求の範囲】
【請求項1】
コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で、配列番号1又は2と相補的である単離された核酸配列を含む、組成物。
【請求項2】
配列番号1又は2と相補的である核酸配列は、アンチセンスオリゴヌクレオチドである、請求項1に記載の組成物。
【請求項3】
アンチセンスオリゴヌクレオチドは、配列番号3である、請求項2に記載の組成物。
【請求項4】
アンチセンスオリゴヌクレオチドは、配列番号4である、請求項2に記載の組成物。
【請求項5】
配列番号1又は2と相補的である核酸配列は、干渉RNAである、請求項1に記載の組成物。
【請求項6】
干渉RNAは、shRNA又はsiRNAである、請求項5に記載の組成物。
【請求項7】
配列番号1又は2と相補的である核酸配列は、アンタゴマーである、請求項1に記載の組成物。
【請求項8】
配列番号1と相補的である核酸配列は、配列番号1の転写後プロセシングを阻害する、請求項1に記載の組成物。
【請求項9】
配列番号2と相補的である核酸配列は、配列番号2の転写後プロセシングを阻害する、請求項1に記載の組成物。
【請求項10】
コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で、配列番号1又は2と相補的である核酸配列を発現できるベクターを含む、組成物。
【請求項11】
前記ベクターは、プラスミドベクター又はウイルスベクターである、請求項10に記載の組成物。
【請求項12】
コレステロール恒常性を必要とする被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体におけるABCA1タンパク質の発現を増加させることを含む、方法。
【請求項13】
被験体の肝臓における細胞内コレステロールの流出及び/又はHDLの産生を増加させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体の肝臓における細胞内コレステロールの流出及び/又はHDLの産生を増加させることを含む、方法。
【請求項14】
被験体の血液を循環するコレステロールの量を減少させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体の血液を循環するコレステロールの量を減少させることを含む、方法。
【請求項15】
配列番号1又は2と相補的である核酸配列は、アンチセンスオリゴヌクレオチドである、請求項12、13、又は14に記載の方法。
【請求項16】
アンチセンスオリゴヌクレオチドは、配列番号3である、請求項15に記載の方法。
【請求項17】
アンチセンスオリゴヌクレオチドは、配列番号4である、請求項15に記載の方法。
【請求項18】
配列番号1又は2と相補的である核酸配列は、干渉RNAである、請求項12、13、又は14に記載の方法。
【請求項19】
干渉RNAは、shRNA又はsiRNAである、請求項18に記載の方法。
【請求項20】
配列番号1又は2と相補的である核酸配列は、アンタゴマーである、請求項12、13、又は14に記載の方法。
【請求項21】
配列番号1と相補的である核酸配列は、配列番号1の転写後プロセシングを阻害する、請求項12、13、又は14に記載の方法。
【請求項22】
配列番号2と相補的である核酸配列は、配列番号2の転写後プロセシングを阻害する、請求項12、13、又は14に記載の方法。
【請求項1】
コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で、配列番号1又は2と相補的である単離された核酸配列を含む、組成物。
【請求項2】
配列番号1又は2と相補的である核酸配列は、アンチセンスオリゴヌクレオチドである、請求項1に記載の組成物。
【請求項3】
アンチセンスオリゴヌクレオチドは、配列番号3である、請求項2に記載の組成物。
【請求項4】
アンチセンスオリゴヌクレオチドは、配列番号4である、請求項2に記載の組成物。
【請求項5】
配列番号1又は2と相補的である核酸配列は、干渉RNAである、請求項1に記載の組成物。
【請求項6】
干渉RNAは、shRNA又はsiRNAである、請求項5に記載の組成物。
【請求項7】
配列番号1又は2と相補的である核酸配列は、アンタゴマーである、請求項1に記載の組成物。
【請求項8】
配列番号1と相補的である核酸配列は、配列番号1の転写後プロセシングを阻害する、請求項1に記載の組成物。
【請求項9】
配列番号2と相補的である核酸配列は、配列番号2の転写後プロセシングを阻害する、請求項1に記載の組成物。
【請求項10】
コレステロール恒常性を必要とするヒト被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させるのに有効な量で、配列番号1又は2と相補的である核酸配列を発現できるベクターを含む、組成物。
【請求項11】
前記ベクターは、プラスミドベクター又はウイルスベクターである、請求項10に記載の組成物。
【請求項12】
コレステロール恒常性を必要とする被験体におけるATP結合カセット、サブファミリーA、メンバー1、(ABCA1)タンパク質の発現を増加させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体におけるABCA1タンパク質の発現を増加させることを含む、方法。
【請求項13】
被験体の肝臓における細胞内コレステロールの流出及び/又はHDLの産生を増加させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体の肝臓における細胞内コレステロールの流出及び/又はHDLの産生を増加させることを含む、方法。
【請求項14】
被験体の血液を循環するコレステロールの量を減少させる方法であって、配列番号1又は2と相補的である核酸配列を被験体に投与することによって、被験体の血液を循環するコレステロールの量を減少させることを含む、方法。
【請求項15】
配列番号1又は2と相補的である核酸配列は、アンチセンスオリゴヌクレオチドである、請求項12、13、又は14に記載の方法。
【請求項16】
アンチセンスオリゴヌクレオチドは、配列番号3である、請求項15に記載の方法。
【請求項17】
アンチセンスオリゴヌクレオチドは、配列番号4である、請求項15に記載の方法。
【請求項18】
配列番号1又は2と相補的である核酸配列は、干渉RNAである、請求項12、13、又は14に記載の方法。
【請求項19】
干渉RNAは、shRNA又はsiRNAである、請求項18に記載の方法。
【請求項20】
配列番号1又は2と相補的である核酸配列は、アンタゴマーである、請求項12、13、又は14に記載の方法。
【請求項21】
配列番号1と相補的である核酸配列は、配列番号1の転写後プロセシングを阻害する、請求項12、13、又は14に記載の方法。
【請求項22】
配列番号2と相補的である核酸配列は、配列番号2の転写後プロセシングを阻害する、請求項12、13、又は14に記載の方法。
【図1】

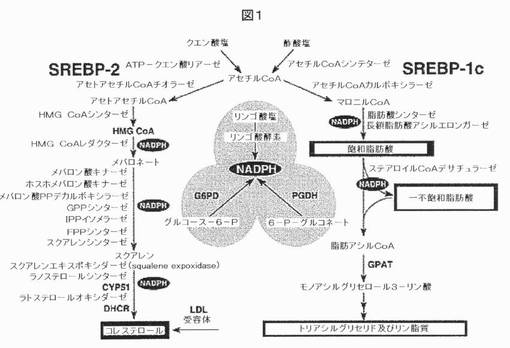
【図2】
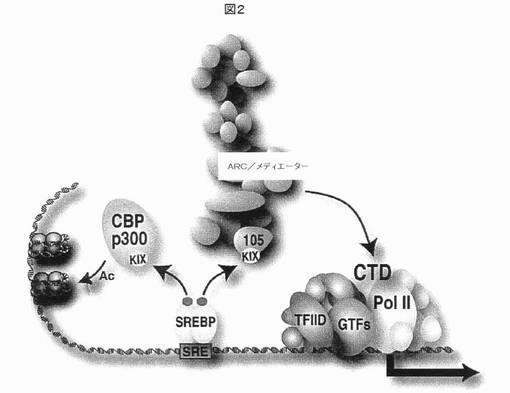
【図3】
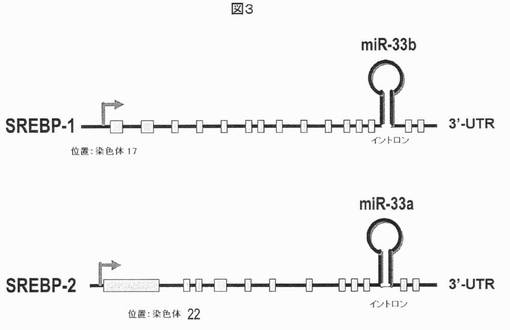
【図4】
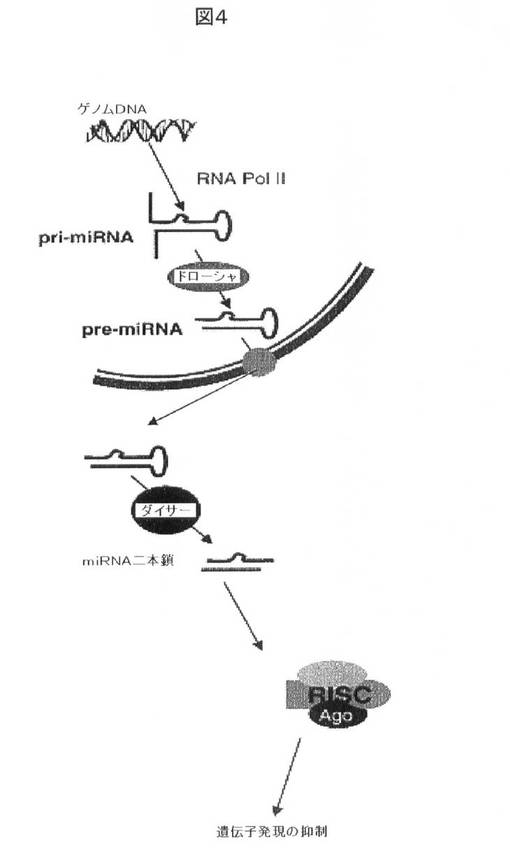
【図5】
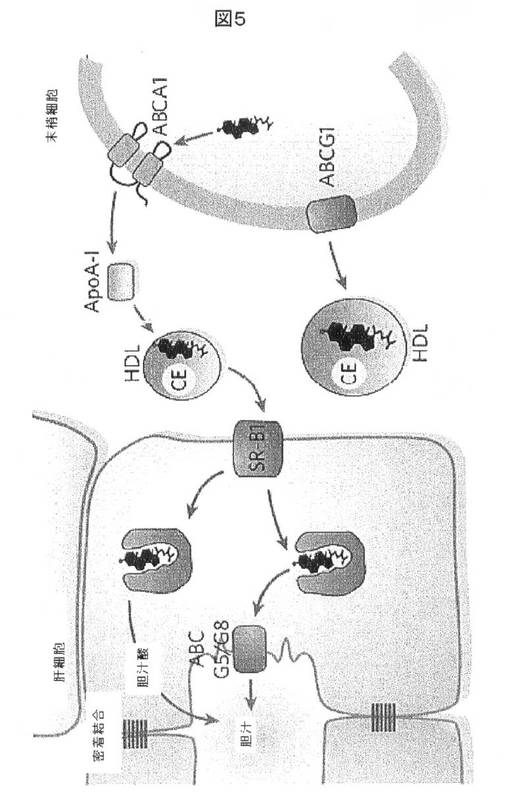
【図6】
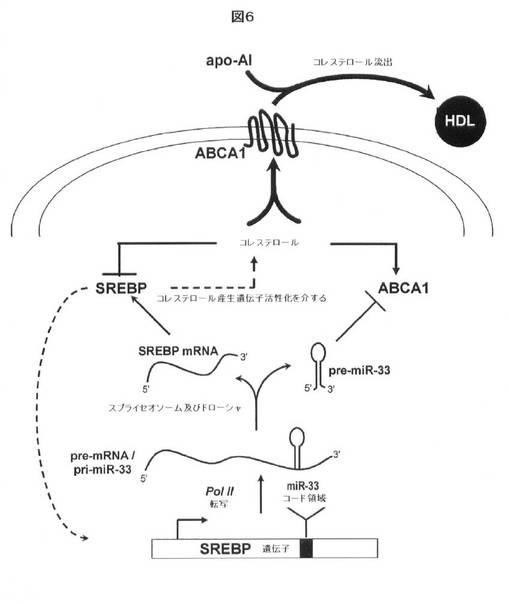
【図7A】
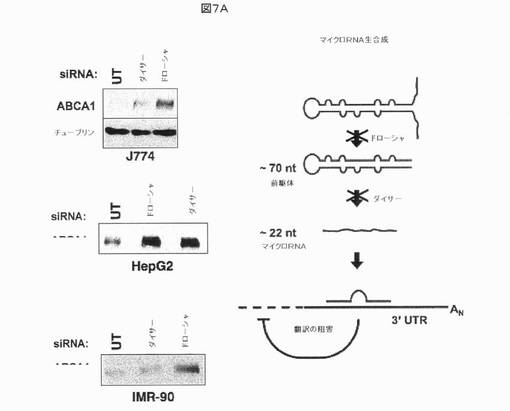
【図7B】
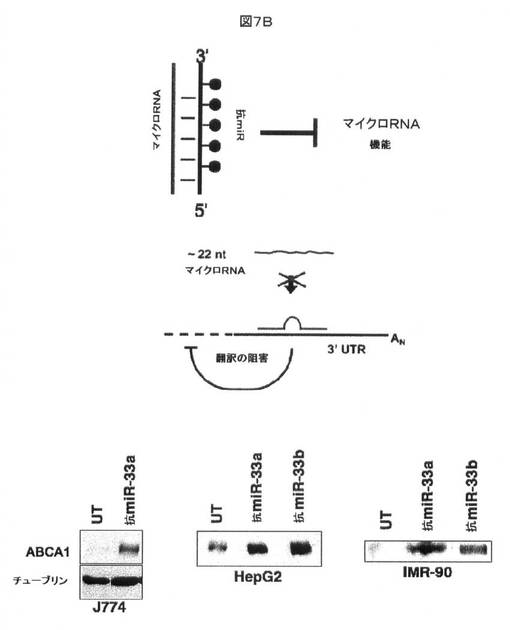
【図7C】
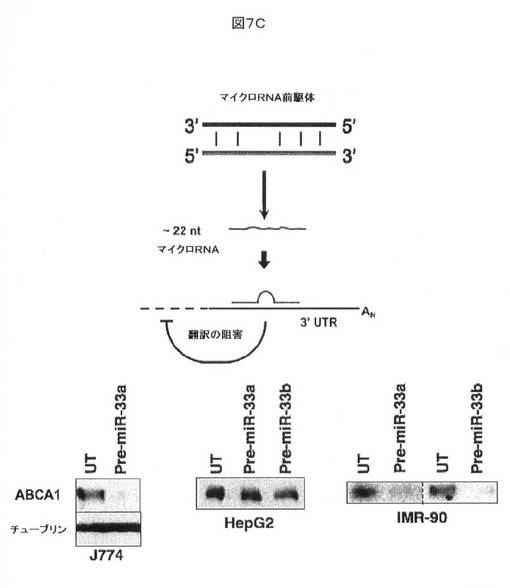
【図8A】
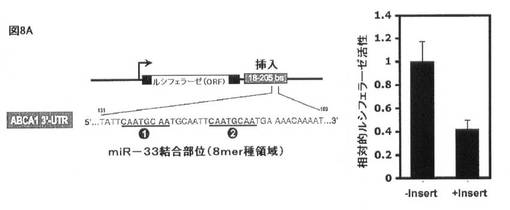
【図8B】
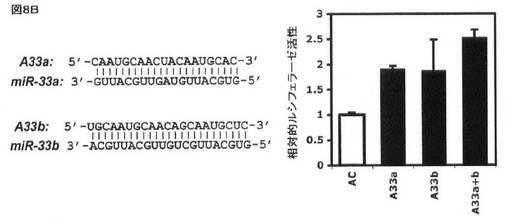
【図8C】
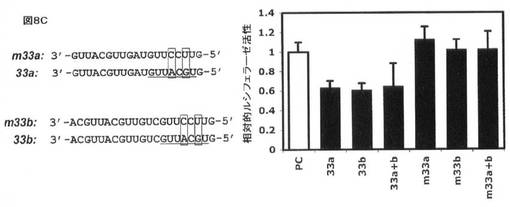
【図9A】
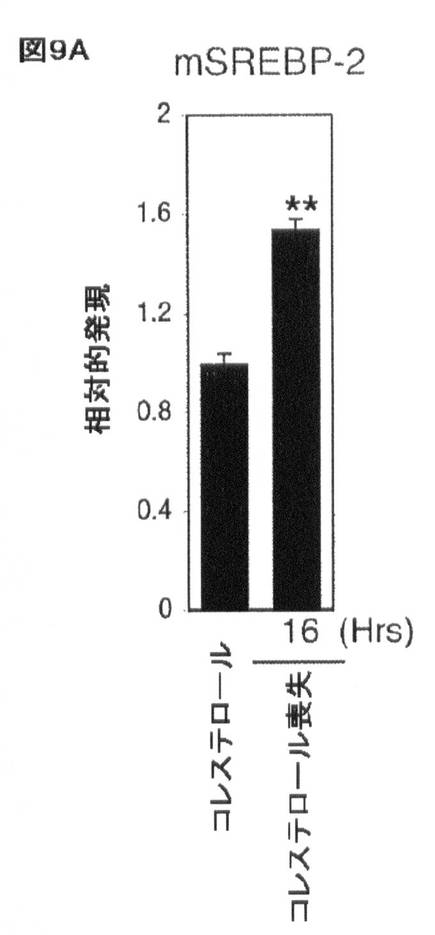
【図9B】
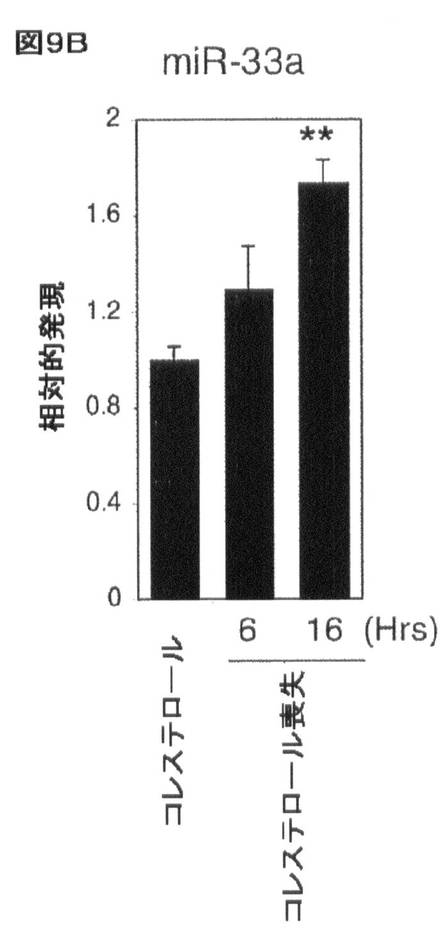
【図9C】
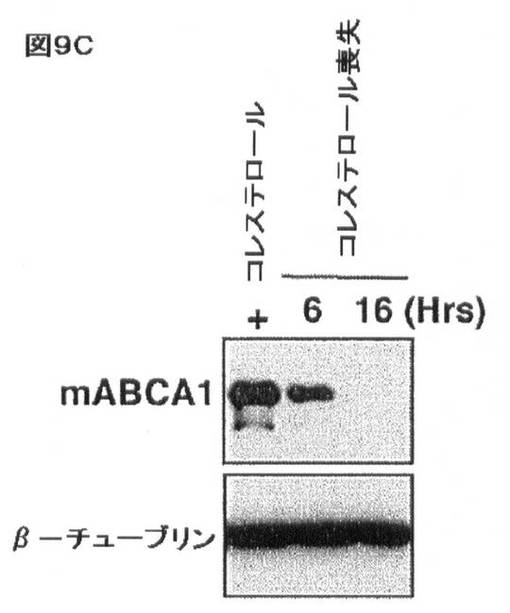
【図9D】
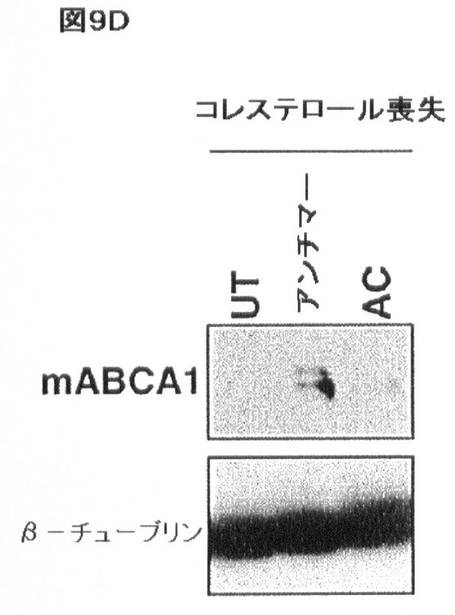
【図9E】
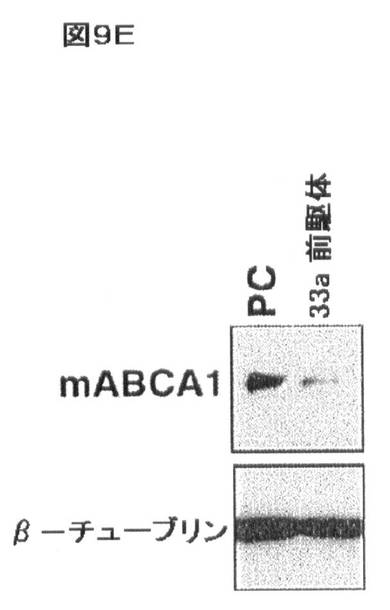
【図9F】
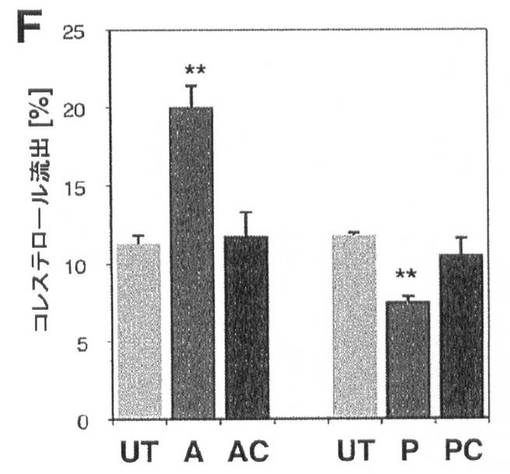
【図10A】
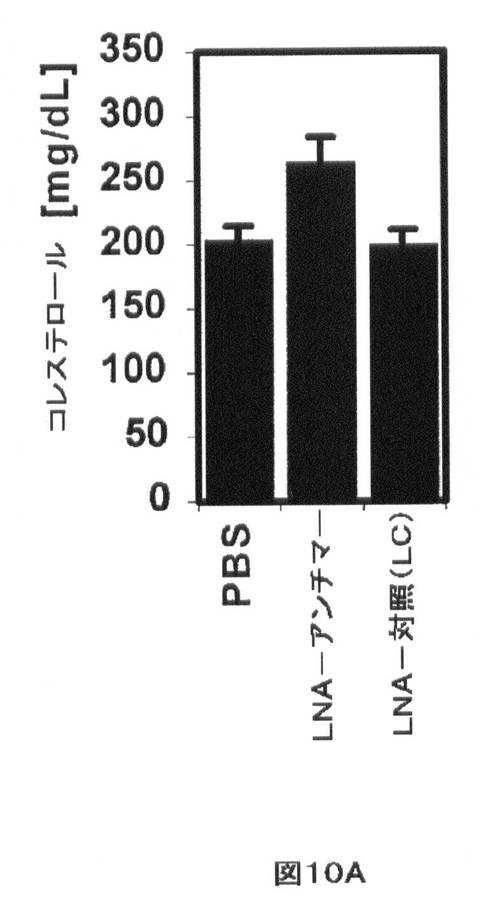
【図10B】
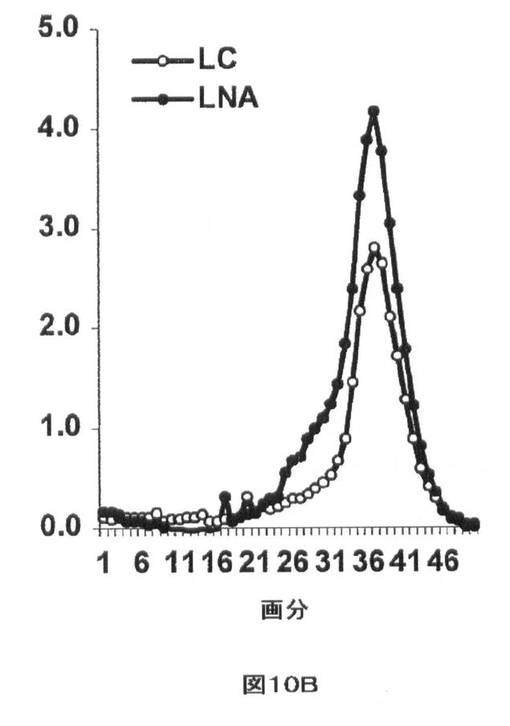
【図10C】
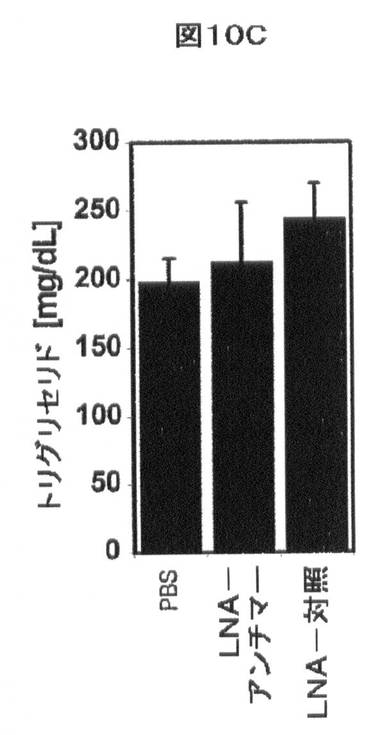
【図10D】
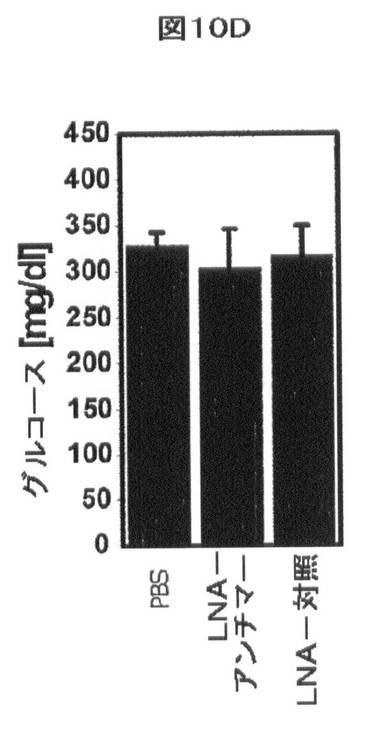
【図10E】
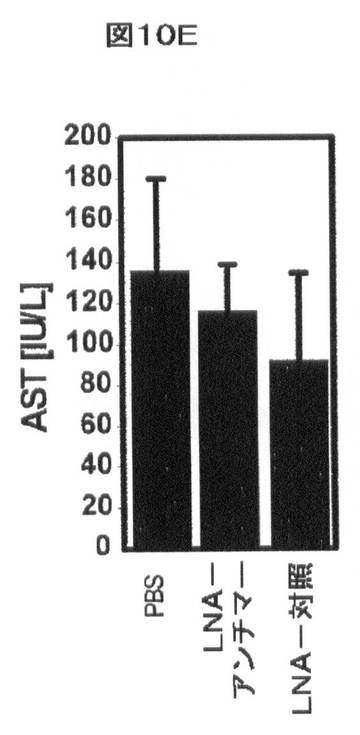
【図10F】
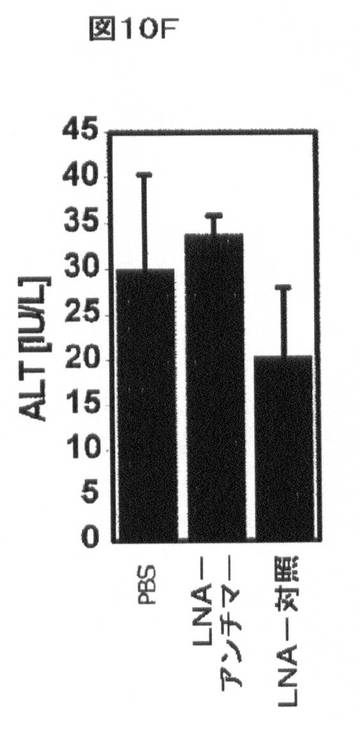
【図11A】
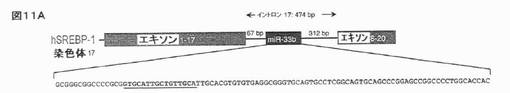
【図11B】
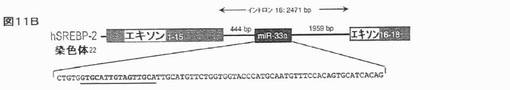
【図11C】
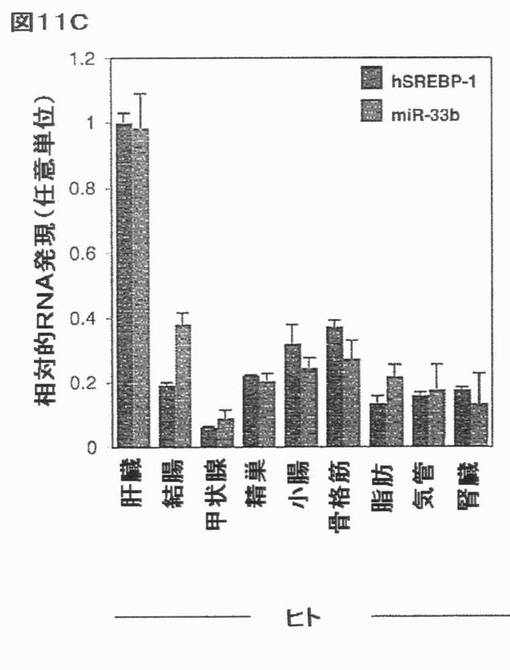
【図11D】
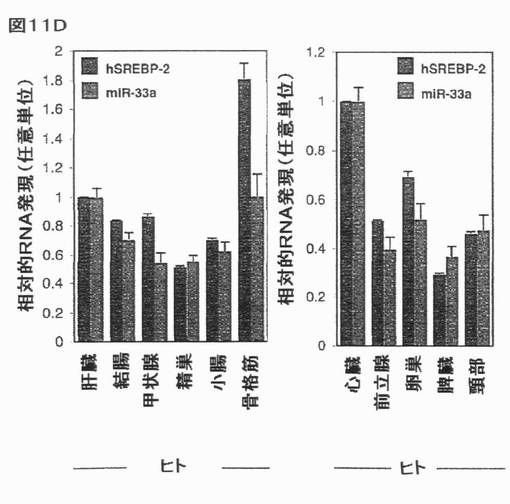
【図11E】
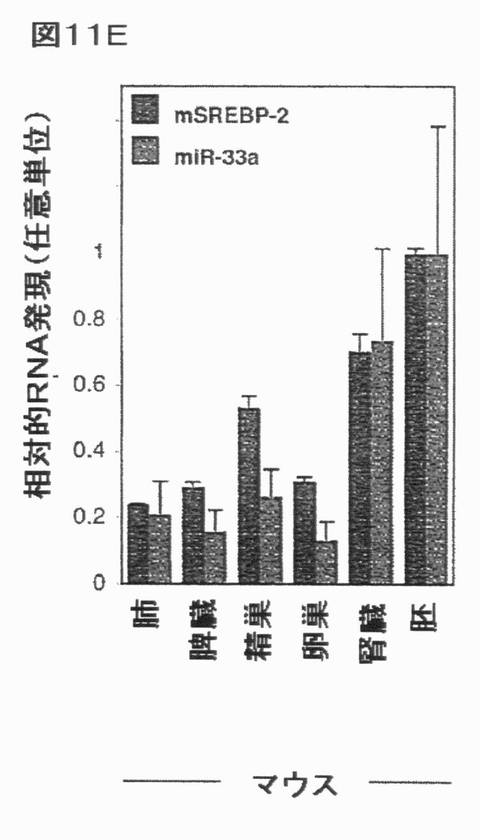
【図12A】
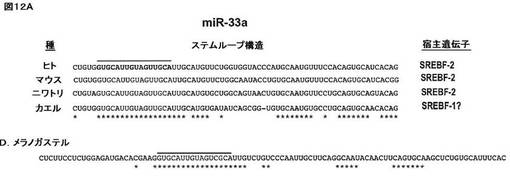
【図12B】

【図13A】
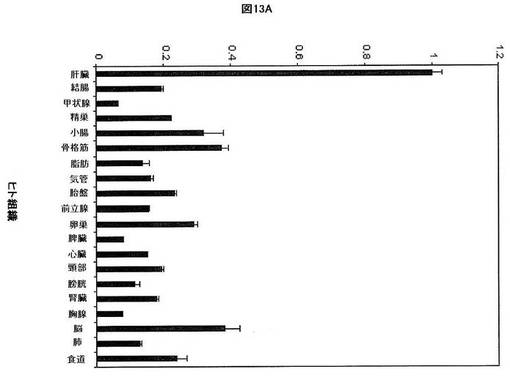
【図13B】
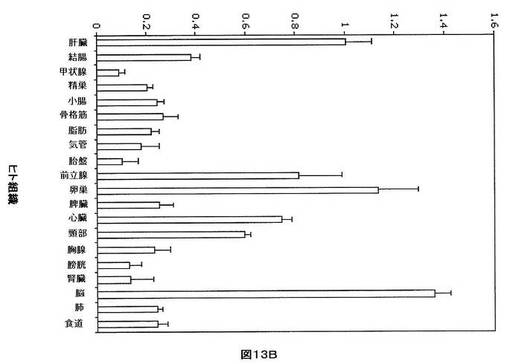
【図14A】
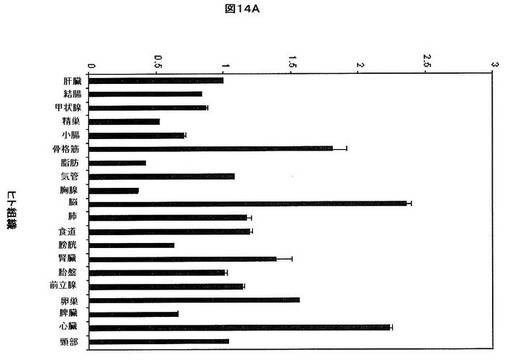
【図14B】
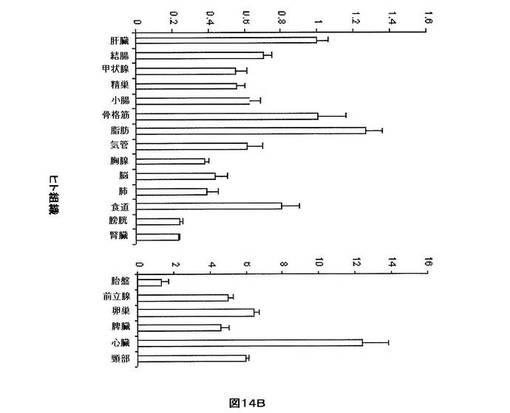
【図15A】
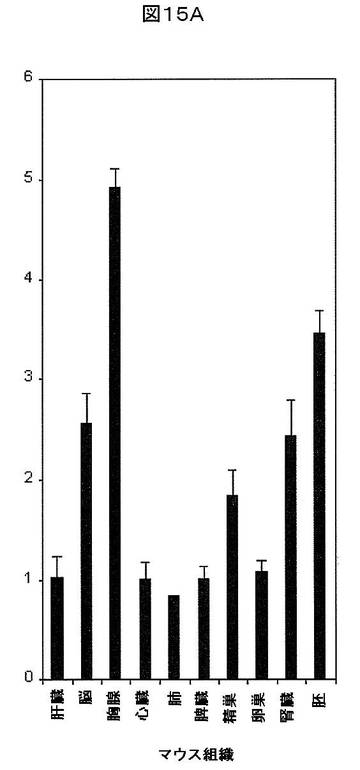
【図15B】
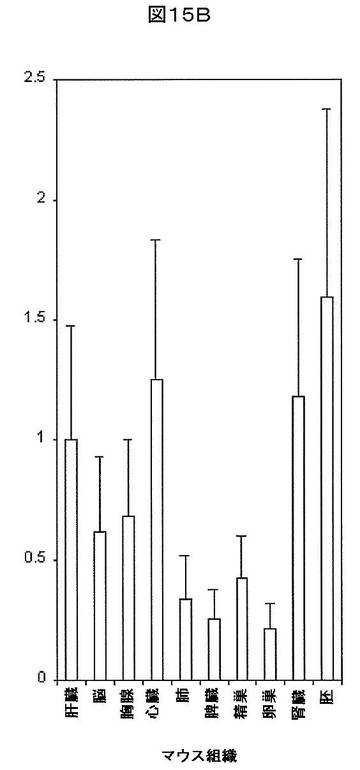
【図16】
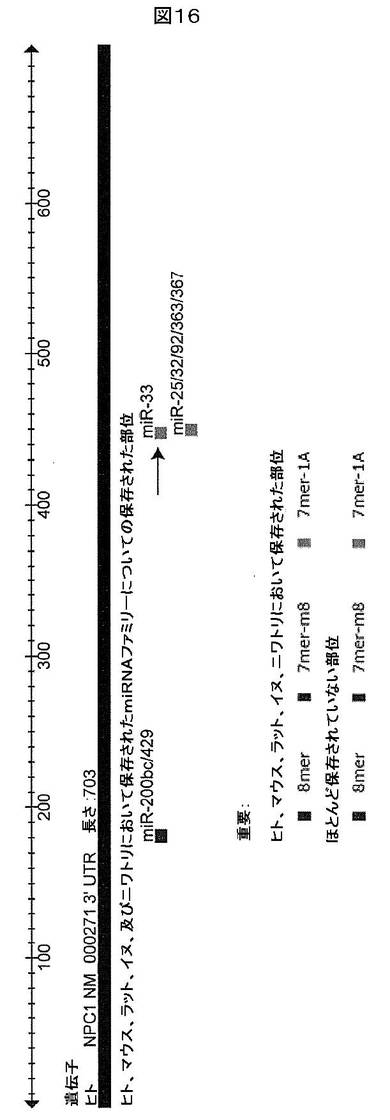
【図17】
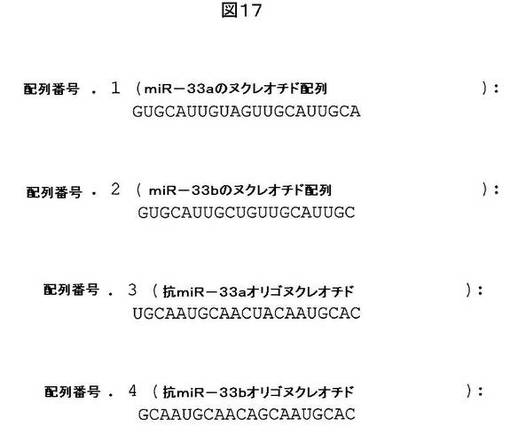
【図2】
【図3】
【図4】
【図5】
【図6】
【図7A】
【図7B】
【図7C】
【図8A】
【図8B】
【図8C】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【図9F】
【図10A】
【図10B】
【図10C】
【図10D】
【図10E】
【図10F】
【図11A】
【図11B】
【図11C】
【図11D】
【図11E】
【図12A】
【図12B】
【図13A】
【図13B】
【図14A】
【図14B】
【図15A】
【図15B】
【図16】
【図17】
【公表番号】特表2012−522509(P2012−522509A)
【公表日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願番号】特願2012−503656(P2012−503656)
【出願日】平成22年3月31日(2010.3.31)
【国際出願番号】PCT/US2010/029376
【国際公開番号】WO2010/120508
【国際公開日】平成22年10月21日(2010.10.21)
【出願人】(592017633)ザ ジェネラル ホスピタル コーポレイション (177)
【Fターム(参考)】
【公表日】平成24年9月27日(2012.9.27)
【国際特許分類】
【出願日】平成22年3月31日(2010.3.31)
【国際出願番号】PCT/US2010/029376
【国際公開番号】WO2010/120508
【国際公開日】平成22年10月21日(2010.10.21)
【出願人】(592017633)ザ ジェネラル ホスピタル コーポレイション (177)
【Fターム(参考)】
[ Back to top ]