
サイトカインレセプターファミリーのメンバーであるMU−1
【課題】造血素レセプタースーパーファミリーの従来未知であったメンバーに関するDNAおよびタンパク質の配列を提供すること。
【解決手段】MU−1造血素レセプタースーパーファミリー鎖をコードするマウス由来のポリヌクレオチド、およびMU−1造血素レセプタースーパーファミリー鎖をコードするヒト由来のポリヌクレオチド、ならびにマウスMU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有するアミノ酸配列を含む単離されたMU−1タンパク質、およびヒトMU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有するアミノ酸配列を含む単離されたMU−1タンパク質。
【解決手段】MU−1造血素レセプタースーパーファミリー鎖をコードするマウス由来のポリヌクレオチド、およびMU−1造血素レセプタースーパーファミリー鎖をコードするヒト由来のポリヌクレオチド、ならびにマウスMU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有するアミノ酸配列を含む単離されたMU−1タンパク質、およびヒトMU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有するアミノ酸配列を含む単離されたMU−1タンパク質。
【発明の詳細な説明】
【技術分野】
【0001】
(発明の分野)
本発明は、哺乳動物サイトカインレセプターファミリーのタンパク質の新規なメンバー(限定はしないが、ヒトレセプタータンパク質およびマウスレセプタータンパク質が、挙げられる)、そのフラグメント、ならびにそのようなタンパク質を発現するために有用な組換えポリヌクレオチドおよび細胞に関する。
【背景技術】
【0002】
(発明の背景)
種々の造血細胞集団または血球集団の発生および増殖に関与する、種々の調節分子(造血素として公知)が、同定されている。ほとんどの造血素は、標的細胞の表面上のレセプターと相互作用することにより、特定の生物学的活性を示す。サイトカインレセプターは、1つ、2つ、または3つの鎖から一般に構成される。多くのサイトカインレセプターおよびいくつかのサイトカイン(例えば、IL−12 p40)が、造血素レセプタースーパーファミリータンパク質のメンバーである。この造血素レセプタースーパーファミリーの新規なメンバーを同定することは、造血の調節、免疫応答の調節、および造血素スーパーファミリーの他のメンバー(サイトカインおよびレセプターを含む)の同定において、有用であり得る。
【発明の開示】
【発明が解決しようとする課題】
【0003】
この造血素レセプタースーパーファミリーの従来未知であったメンバーに関するDNAおよびタンパク質の配列を、同定および決定することが望ましい。
【課題を解決するための手段】
【0004】
(発明の要旨)
本発明に従って、MU−1造血素レセプタースーパーファミリー鎖をコードするポリヌクレオチド(マウス供給源およびヒト供給源由来のものが挙げられるが、限定はしない)が、開示される。
【0005】
特定の実施形態において、本発明は、ヌクレオチド配列を含む単離されたポリヌクレオチドを提供し、このヌクレオチド配列は、以下:
(a)配列番号1のヌクレオチド配列;
(b)配列番号1のヌクレオチド238〜ヌクレオチド1852のヌクレオチド配列;
(c)配列番号1のヌクレオチド301〜ヌクレオチド1852のヌクレオチド配列;
(d)配列番号1のヌクレオチド301〜ヌクレオチド945のヌクレオチド配列;
(e)(a)〜(d)のいずれかにおいて特定されるヌクレオチド配列の配列と、遺伝暗号の縮重の結果として異なる、ヌクレオチド配列;
(f)(a)〜(d)のいずれかにおいて特定されるヌクレオチドに、ストリンジェントな条件下でハイブリダイズし得る、ヌクレオチド配列;
(g)配列番号2の配列の種ホモログをコードするヌクレオチド配列;および
(h)(a)〜(d)のいずれかにおいて特定されるヌクレオチド配列の対立遺伝子改変体、
からなる群より選択される。好ましくは、このヌクレオチド配列は、MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有するタンパク質をコードする。このヌクレオチド配列は、発現制御配列に作動可能に連結され得る。
【0006】
本発明はまた、アミノ酸配列を含むペプチドまたはタンパク質をコードするヌクレオチド配列を含む単離されたポリヌクレオチドも提供し、このアミノ酸配列は、以下:
(a)配列番号2のアミノ酸配列;
(b)配列番号2のアミノ酸22〜538のアミノ酸配列;
(c)配列番号2のアミノ酸22〜236のアミノ酸配列;
(d)配列番号2のアミノ酸1〜236のアミノ酸配列;および
(e)MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有する、(a)〜(d)のフラグメント、
からなる群より選択される。
【0007】
別の実施形態において、本発明は、ヌクレオチド配列を含む単離されたポリヌクレオチドを提供し、このヌクレオチド配列は、以下:
(a)配列番号9のヌクレオチド配列;
(b)(a)において特定されるヌクレオチド配列と、遺伝暗号の縮重の結果として異なる、ヌクレオチド配列;
(c)(a)において特定されるヌクレオチドに、ストリンジェントな条件下でハイブリダイズし得る、ヌクレオチド配列;
(d)(a)において特定されるヌクレオチド配列の対立遺伝子改変体、
からなる群より選択される。
【0008】
本発明はまた、アミノ酸配列を含むペプチドまたはタンパク質をコードするヌクレオチド配列を含む単離されたポリヌクレオチドを提供し、このアミノ酸配列は、以下:
(a)配列番号10のアミノ酸配列;
(b)MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有する、(a)のフラグメント、
からなる群より選択される。
【0009】
これらのポリヌクレオチドで形質転換された宿主細胞(好ましくは、哺乳動物細胞)もまた、提供される。
【0010】
他の実施形態において、本発明は、MU−1タンパク質を産生するためのプロセスを提供する。このプロセスは、以下:
(a)適切な培養培地において本発明の宿主細胞の培養物を増殖させる工程;および
(b)その培養物からヒトMU−1タンパク質を精製する工程、
を包含する。これらの方法により産生されたタンパク質もまた、提供される。
【0011】
本発明はまた、アミノ酸配列を含む単離されたMU−1タンパク質を提供し、このアミノ酸配列は、以下:
(a)配列番号2のアミノ酸配列;
(b)配列番号2のアミノ酸22〜538のアミノ酸配列;
(c)配列番号2のアミノ酸22〜236のアミノ酸配列;
(d)配列番号2のアミノ酸1〜236のアミノ酸配列;および
(e)MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有する、(a)〜(d)のフラグメント、
からなる群より選択される。
【0012】
本発明はまた、アミノ酸配列を含む単離されたMU−1タンパク質を提供し、このアミノ酸配列は、以下:
(a)配列番号10のアミノ酸配列;
(b)MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有する、(a)のフラグメント、
からなる群より選択される。
【0013】
マウスMU−1(配列番号10)のアミノ酸配列は、ヒトMU−1のアミノ酸配列と約65%同一である。
【0014】
他の好ましい実施形態において、特定のアミノ酸配列は、融合タンパク質の一部(MU−1に由来しないさらなるアミノ酸配列)である。好ましい融合タンパク質は、抗体フラグメント(例えば、Fcフラグメント)を含む。
【0015】
本発明のタンパク質および薬学的に受容可能なキャリアを含む薬学的組成物もまた、提供される。
【0016】
本発明はさらに、本発明のタンパク質と特異的に反応する抗体を含む組成物を提供する。
【0017】
1つの実施形態において、本発明のMU−1核酸分子は、配列番号1または9に示されるヌクレオチド配列に対して(例えば、全長ヌクレオチド配列に対して)、少なくとも65%、70%、75%、80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%またはそれ以上同一である。
【0018】
別の好ましい実施形態において、核酸分子は、配列番号1または9に示されるヌクレオチド配列からなる。別の好ましい実施形態において、核酸分子は、配列番号1または9のヌクレオチド配列のうちの少なくとも50ヌクレオチド、100ヌクレオチド、200ヌクレオチド、300ヌクレオチド、400ヌクレオチド、500ヌクレオチド、600ヌクレオチド、700ヌクレオチド、800ヌクレオチド、900ヌクレオチド、1000ヌクレオチド、1100ヌクレオチド、1200ヌクレオチド、1300ヌクレオチド、1400ヌクレオチド、1500ヌクレオチド、1600ヌクレオチド、1700ヌクレオチド、1800ヌクレオチドまたはそれ以上のヌクレオチド(例えば、連続するヌクレオチド)のフラグメント、あるいはその相補体を含む。
【0019】
好ましい実施形態において、MU−1タンパク質ファミリーメンバーは、配列番号2または10のアミノ酸配列に対して、少なくとも約65%、70%、75%、80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%またはそれ以上同一である、アミノ酸配列を有する。
【0020】
別の実施形態において、本発明は、配列番号2または10のアミノ酸配列を有するタンパク質のフラグメントを特徴とし、ここで、このフラグメントは、配列番号2または10のアミノ酸配列のうちの少なくとも15アミノ酸、20アミノ酸、30アミノ酸、40アミノ酸、50アミノ酸、60アミノ酸、70アミノ酸、80アミノ酸、90アミノ酸または100アミノ酸(例えば、連続するアミノ酸)を含む。
【0021】
別の局面において、本発明は、生物学的サンプルにおけるMU−1核酸分子、タンパク質またはポリペプチドの存在を検出するための方法を提供し、この方法は、MU−1核酸分子、MU−1タンパク質またはMU−1ポリペプチドがこの生物学的サンプルにおいて検出されるように、この生物学的サンプルに、MU−1核酸分子、MU−1タンパク質またはMU−1ポリペプチドを検出し得る因子を接触させることによる。
【0022】
別の局面において、本発明は、生物学的サンプルにおけるMU−1活性の存在を検出するための方法を提供し、この方法は、MU−1活性の存在がこの生物学的サンプルにおいて検出されるように、この生物学的サンプルに、MU−1活性の指標を検出し得る因子を接触させることによる。
【0023】
別の局面において、本発明は、MU−1活性を調節するための方法を提供し、この方法は、MU−1を発現し得る細胞においてMU−1活性が調節されるように、この細胞に、MU−1活性を調節する因子を接触させる工程を包含する。1つの実施形態において、この因子は、MU−1活性を阻害する。別の実施形態において、この因子は、MU−1活性を刺激する。1つの実施形態において、この因子は、MU−1タンパク質に特異的に結合する抗体である。別の実施形態において、この因子は、MU−1遺伝子の転写またはMU−1 mRNAの翻訳を調節することによって、MU−1の発現を調節する。なお別の実施形態において、この因子は、MU−1 mRNAまたはMU−1遺伝子のコード鎖に対してアンチセンスであるヌクレオチド配列を有する核酸分子である。
【0024】
1つの実施形態において、本発明の方法は、異常であるかまたは所望されない、MU−1タンパク質またはMU−1核酸の発現または活性によって特徴付けられる障害(例えば、MU−1関連障害)を有する被験体を、その被験体にMU−1モジュレーターである因子を投与することによって処置するために使用される。1つの実施形態において、MU−1モジュレーターは、MU−1タンパク質である。別の実施形態において、MU−1モジュレーターは、MU−1核酸分子である。なお別の実施形態において、MU−1モジュレーターは、ペプチド、ペプチド模倣物、抗体または他の低分子である。
【0025】
本発明はまた、以下の少なくとも1つによって特徴付けられる遺伝的変化の存在または非存在を同定するための診断アッセイを提供する:(i)MU−1タンパク質をコードする遺伝子の異常な修飾または変異;(ii)MU−1遺伝子の調節不全(mis-regulation);および(iii)MU−1タンパク質の異常な翻訳後修飾(ここで、この遺伝子の野生型形態は、MU−1活性を有するタンパク質をコードする)。
【0026】
別の局面において、本発明は、MU−1タンパク質に結合するかまたはMU−1タンパク質の活性を調節する、化合物を同定するための方法を提供し、この方法は、MU−1活性を有するMU−1タンパク質を含む指標組成物を提供する工程、この指標組成物に試験化合物を接触させる工程、および指標組成物におけるMU−1活性に対する試験化合物の効果(例えば、STATリン酸化(例えば、STAT3またはSTAT5リン酸化)の調節)を決定し、MU−1タンパク質の活性を調節する化合物を同定する工程による。
・本発明はまた、以下を提供し得る:
・(項目1) ヌクレオチド配列を含む単離されたポリヌクレオチドであって、上記ヌクレオチド配列は、以下:
(a)配列番号9のヌクレオチド配列;
(b)(a)において特定されるヌクレオチド配列の配列と、遺伝暗号の縮重の結果として異なる、ヌクレオチド配列;
(c)(a)において特定されるヌクレオチド配列に、ストリンジェントな条件下でハイブリダイズし得る、ヌクレオチド配列;
(d)配列番号10の配列の種ホモログをコードする、ヌクレオチド配列;および
(e)(a)において特定されるヌクレオチド配列の対立遺伝子改変体、
からなる群より選択される、ポリヌクレオチド。
・(項目2) 項目1に記載のポリヌクレオチドであって、上記ヌクレオチド配列が、MU−1の生物学的活性を有するタンパク質をコードする、ポリヌクレオチド。
・(項目3) 項目2に記載のポリヌクレオチドであって、上記生物学的活性が、STAT分子との相互作用である、ポリヌクレオチド。
・(項目4) 項目1に記載のポリヌクレオチドであって、上記ヌクレオチド配列が、発現制御配列に作動可能に連結されている、ポリヌクレオチド。
・(項目5) 項目1に記載のポリヌクレオチドであって、配列番号9のヌクレオチド配列を含む、ポリヌクレオチド。
・(項目6) 項目4に記載のポリヌクレオチドで形質転換された、宿主細胞。
・(項目7) 哺乳動物細胞である、項目6に記載の宿主細胞。
・(項目8) MU−1タンパク質を産生するためのプロセスであって、上記プロセスは、以下:
(a)適切な培養培地において項目6に記載の宿主細胞の培養物を増殖させる工程;および
(b)上記培養物から上記MU−1タンパク質を精製する工程、
を包含する、プロセス。
・(項目9) アミノ酸配列を含む単離されたMU−1タンパク質であって、上記アミノ酸配列は、以下:
(a)配列番号10のアミノ酸配列;および
(b)MU−1の生物学的活性を有する、(a)のフラグメント、
からなる群より選択される、タンパク質。
・(項目10) 配列番号10のアミノ酸配列を含む、項目9に記載のタンパク質。
・(項目11) 薬学的組成物であって、
項目9に記載のタンパク質と、
薬学的に受容可能なキャリアと、
を含む、組成物。
・(項目12) 項目8に記載のプロセスにより産生された、タンパク質。
・(項目13) 抗体を含む組成物であって、上記抗体は、項目9に記載のタンパク質と特異的に反応する、組成物。
・(項目14) アミノ酸配列を含むペプチドまたはタンパク質をコードするヌクレオチド配列を含む単離されたポリヌクレオチドであって、上記アミノ酸配列は、以下:
(a)配列番号10のアミノ酸配列;および
(b)MU−1の生物学的活性を有する、(a)のフラグメント、
からなる群より選択される、ポリヌクレオチド。
・(項目15) 項目9に記載のタンパク質であって、上記アミノ酸配列が融合タンパク質の一部である、タンパク質。
・(項目16) Fcフラグメントを含む、項目15に記載のタンパク質。
・(項目17) 項目14に記載のポリヌクレオチドであって、配列番号10のアミノ酸配列を含むペプチドまたはタンパク質をコードする、ポリヌクレオチド。
・(項目18) 配列番号9のヌクレオチド配列を含む単離されたポリヌクレオチドであって、STAT分子と相互作用するMU−1タンパク質をコードする、ポリヌクレオチド。
・(項目19) STATシグナル伝達経路を調節するための方法であって、
STATシグナル伝達に関与する細胞を、MU−1活性を調節し得る因子と接触させ、それによりSTATシグナル伝達経路を調節する工程、
を包含する、方法。
・(項目20) 項目19に記載の方法であって、上記STATシグナル伝達経路が、STAT 3シグナル伝達経路であるか、またはSTAT 5シグナル伝達経路である、方法。
・(項目21) 免疫障害を処置し得る化合物を同定するための方法であって、
STAT分子とMU−1ポリペプチドとの相互作用を上記化合物が調節する能力をアッセイし、それにより免疫障害を処置し得る化合物を同定する工程、
を包含する、方法。
・(項目22) 項目21に記載の方法であって、上記STATシグナル伝達経路が、STAT 3シグナル伝達経路であるか、またはSTAT 5シグナル伝達経路である、方法。
【発明を実施するための最良の形態】
【0027】
(好ましい実施形態の詳細な説明)
本出願の発明者は、MU−1造血素レセプタースーパーファミリー鎖(本明細書中以後、「MU−1」または「MU−1タンパク質」)をコードするポリヌクレオチド(ヒトおよびマウスMU−1をコードするポリヌクレオチドが挙げられるが、これらに限定されない)を、初めて同定および提供した。
【0028】
特に好ましい実施形態において、本発明のMU−1タンパク質およびMU−1核酸分子は、ヒトMU−1分子である。ヒトIL5レセプターの70アミノ酸領域
【0029】
【化1】

を用いて、TBLASTNアルゴリズムを使用してGenBank ESTデータベースを検索した。ヒト染色体16p12由来のゲノムBACクローンAC002303中の配列は、この領域に対する相同性を有することが同定され、これは、このセグメントが新規造血素レセプターの遺伝子をコードすることを示唆する。ヌクレオチド40,886の1000bp中のオープンリーディングフレームの試験は、GenPeptのBLASTP検索を使用した場合にサイトカインレセプターファミリーのメンバーを専ら同定した、270bpのオープンリーディングフレームを明らかにした。このリーディングフレームの末端の終止コドンは、エキソン/イントロン境界にまたがるトランジションの表れであると解釈された。
【0030】
次いで、RNAが、第16p12染色体由来のこのBACクローン内に含まれる遺伝子から転写されるか否かを決定した。PCRプライマーを、サイトカインレセプターファミリー内で保存されるペプチド配列を含む最大のORFセグメントに基づいて合成した。プライマーGAGTCCGAGGAGAAAGCTGATCTCA(5p)(配列番号4)およびプライマーGAAAGATGACCGGGTCACTCCATT(3p)(配列番号5)をPCRにおいて用いて、種々のヒト組織由来のファージライブラリー(Clontech)をスクリーニングした。配列ACTCGAGCTATGAGCTGCAGGTGCGGGCA(配列番号6)の32−P標識オリゴヌクレオチドに特異的にハイブリダイズする、予測された164bpサイズのPCR産物が、肺、腎臓、胎盤および心臓由来のファージにおいて観察された。オリゴヌクレオチドACTCGAGCTATGAGCTGCAGGTGCGGGCA(配列番号7)を用いて、全長のcDNAクローンNN14−1b(MU−1)が同定され、これを精製し、そして配列決定した。DNA配列および推定アミノ酸配列をそれぞれ、配列番号1および配列番号2に示す。ヒトMU−1レセプター鎖の推定アミノ酸配列は、アミノ酸1〜21の推定シグナル配列を含む。成熟ヒトMU−1は、配列番号2のアミノ酸24〜538の配列を有すると考えられる。膜貫通ドメインは、アミノ酸237〜254に見出される。
【0031】
別の特に好ましい実施形態では、本発明のMU−1タンパク質分子およびMU−1核酸分子は、マウスMU−1分子である。マウスMU−1タンパク質をコードするポリヌクレオチド配列を同定するために、MU−1レセプターのマウスホモログの部分フラグメントを、実施例1に記載されるように、ヒト配列から誘導されたオリゴヌクレオチドを用いるPCRによってマウスcDNAから単離した。このフラグメントのDNA配列を決定し、そして2つのオリゴヌクレオチドを、以下の配列を用いて、このフラグメントの内部部分から誘導した:
TTGAACGTGACTGTGGCCTT(5p)(配列番号13)
TGAATGAAGTGCCTGGCTGA(3p)(配列番号14)。
これらのオリゴヌクレオチドを用いて、元のPCR産物(図1および配列番号9のマウスcDNA配列のヌクレオチド781〜1043に対応する)の内部の262ヌクレオチドフラグメントを増幅し、2D6 T細胞株から単離されたcDNAライブラリーをスクリーニングするためのハイブリダイゼーションプローブとして用いた。DNA配列を、2つの独立したクローンから決定した。クローン6を配列決定し、そしてヒトMU−1の全長マウスホモログであることを確認した。
【0032】
マウスMU−1の全長ヌクレオチド配列を、図1(配列番号9のヌクレオチド1〜2628に対応する)に示す。このヌクレオチド配列は、推定リーダー配列をヌクレオチド407〜464に、コード配列をヌクレオチド407〜1993に、そして終結コドンをヌクレオチド1994〜1997に有する。ヌクレオチド1〜406は、5’非翻訳領域に対応し、そしてヌクレオチド1998〜2628は3’非翻訳領域に対応する。マウスMU−1の推定タンパク質配列を図2(配列番号10のアミノ酸1〜529に対応する)に示す。
【0033】
このマウスMU−1タンパク質は、SPScan(スコア=10.1)によって決定したところ、推定リーダー配列(配列番号10のアミノ酸1〜19に対応する)、および推定膜貫通ドメイン(配列番号10のアミノ酸237〜253に対応する)を含む。推定シグナルモチーフは、以下の領域を含む:ボックス1:配列番号10のアミノ酸265〜274、ボックス2:配列番号10のアミノ酸310〜324、配列番号10の281位、319位、361位、368位、397位および510位の6個のチロシン残基。潜在的なSTATドッキング部位は、STAT 5:EDDGYPA、STAT 3:YLQRを含むがこれらに限定されない。
【0034】
MU−1のオープンリーディングフレームは、造血素レセプターファミリーの新規メンバーをコードする。好ましい実施形態では、MU−1は、リーダー配列、保存されたシステイン対、このファミリーに特有のPPモチーフおよびWSXWS(配列番号8)モチーフ、ならびに膜貫通ドメインおよび広範囲にわたる細胞質ドメインを有する。MU−1はまた、細胞質ドメイン中に、保存されたPXPPならびにボックスIおよびボックスIIのシグナル伝達モチーフを含む(図5を参照のこと)。これらのドメインは、マウスMU−1とヒトMU−1との間で保存される。その後のGenPeptを用いたMU−1配列のFASTA整列は、ヒトIL−2Rbと最大の相同性を示した(図5を参照のこと)。
【0035】
ヒトMU−1 cDNAを、登録番号ATCC 98687として1998年3月10日にAmerican Type Culture Collectionに寄託した。
【0036】
ノーザン分析によって、実施例4に記載されるように、マウスMU−1は、成体マウスの脾臓組織、肺組織および心臓組織において検出された。ヒトMU−1は、成体ヒトのリンパ組織、PBL、胸腺、脾臓およびリンパ節において、ならびに胎児肺において検出された。
【0037】
全長よりも短い任意の形態のMU−1タンパク質は、本発明の範囲内に包含され、そして全長および成熟形態とともに本明細書中で集合的に「MU−1」または「MU−1タンパク質」と呼ばれる。全長よりも短いMU−1タンパク質は、全長MU−1タンパク質(配列番号4または配列番号6)をコードするポリヌクレオチドの対応するフラグメントを発現させることによって産生され得る。これらの対応するポリヌクレオチドフラグメントもまた、本発明の一部である。上記の通りの改変されたポリヌクレオチドは、標準的な分子生物学的技術(適切な所望の欠失変異体の構築を含む)によって、部位特異的変異誘発法、または適切なオリゴヌクレオチドプライマーを使用するポリメラーゼ連鎖反応によって作製され得る。
【0038】
本発明の目的のために、タンパク質は、対応する成熟MU−1タンパク質の1以上の生物学的活性を保有する場合、「MU−1造血素レセプタースーパーファミリー鎖の生物学的活性」を有する。1つの実施形態では、MU−1活性は、STAT分子(例えば、STAT 5、STAT 3)との相互作用を含む。別の実施形態では、MU−1タンパク質の活性は、いかなる公知のサイトカインへの結合も含まない。
【0039】
MU−1またはその活性なフラグメント(MU−1タンパク質)は、キャリア分子(例えば、免疫グロブリンまたは免疫グロブリンフラグメント)に融合され得る。例えば、可溶性形態のMU−1は、「リンカー」配列を通して、免疫グロブリンのFc部分へと融合され得る。他の融合タンパク質(例えば、GST、Lex−AまたはMBPを有する融合タンパク質)もまた用いられ得る。
【0040】
本発明はまた、配列番号1および配列番号9に示すようなヌクレオチド配列の対立遺伝子改変体(すなわち、MU−1タンパク質(好ましくはMU−1の生物学的活性を有するタンパク質)をまたコードする、配列番号1または配列番号9の天然に存在する選択的形態の単離されたポリヌクレオチド)を包含する。本発明にはまた包含されるのは、配列番号1または配列番号9に示されるヌクレオチド配列に、非常にストリンジェントな条件(例えば、65℃で0.1×SSC)下でハイブリダイズする、単離されたポリヌクレオチドである。MU−1タンパク質をコードするが、遺伝暗号の縮重によって、配列番号1または配列番号9に示されるヌクレオチド配列とは異なる、単離されたポリヌクレオチドもまた本発明によって包含される。点変異または誘導改変によって引き起こされる、配列番号1または配列番号9に示されるようなヌクレオチド配列におけるバリエーションもまた、本発明に包含される。
【0041】
本明細書中で使用される場合、用語「ストリンジェントな条件下でハイブリダイズする」は、互いに少なくとも60%同一なヌクレオチド配列が代表的に、互いにハイブリダイズしたままである、ハイブリダイゼーションおよび洗浄についての条件を記載することを意図する。好ましくは、この条件は、互いに少なくとも約70%、より好ましくは少なくとも約80%、さらにより好ましくは少なくとも約85%または90%同一な配列が代表的に、互いにハイブリダイズしたままである条件である。このようなストリンジェントな条件は当業者に公知であり、そしてCurrent Protocols in Molecular Biology,John Wiley & Sons,N.Y.(1989),6.3.1−6.3.6に見出され得る。ストリンジェントなハイブリダイゼーション条件の好ましい非限定的な例は、約45℃での6×塩化ナトリウム/クエン酸ナトリウム(SSC)中でのハイブリダイゼーション、続いて0.2×SSC、0.1% SDS中での50℃(好ましくは55℃、より好ましくは60℃、そしてさらにより好ましくは65℃)での1回以上の洗浄である。上記で記載した値にわたる範囲(例えば、55℃〜60℃または50℃〜65℃)は、本発明によって包含される。好ましくは、配列番号1または配列番号9の配列に、ストリンジェントな条件下でハイブリダイズする、本発明の単離された核酸分子は、天然に存在する核酸分子に対応する。本明細書中で使用される場合、「天然に存在する」核酸分子とは、天然に存在する(例えば、天然タンパク質をコードする)ヌクレオチド配列を有する、RNA分子またはDNA分子をいう。
【0042】
2つのアミノ酸配列または2つの核酸配列の同一性%を決定するために、これらの配列は、最適な比較目的のために整列される(例えば、第一および第二のアミノ酸配列または核酸配列の一方または両方に、最適な整列のためにギャップが導入され得、そして同一でない配列は、比較目的のために無視され得る)。好ましい実施形態では、比較目的のために整列された参照配列の長さは、参照配列の長さの少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、さらにより好ましくは少なくとも60%、そしてさらにより好ましくは少なくとも70%、80%、または90%である。次いで、対応するアミノ酸位置もしくはヌクレオチド位置でのアミノ酸残基またはヌクレオチドが比較される。第一の配列における位置が、第二の配列における対応する位置と同じアミノ酸残基またはヌクレオチドで占められていたならば、この分子はその位置で同一である(本明細書中で使用される場合、アミノ酸または核酸の「同一性」は、アミノ酸または核酸の「相同性」と等価である)。2つの配列間の同一性パーセントは、2つの配列の最適な整列のために導入する必要があるギャップの数および各ギャップの長さを考慮に入れた、これらの配列によって共有される同一位置数の関数である。
【0043】
2つの配列間の配列比較および同一性パーセントの決定は、数理的アルゴリズムを使用して達成され得る。好ましい実施形態において、2つのアミノ酸配列間の同一性パーセントは、GCGソフトウェアパッケージ(http://www.gcg.comにおいて入手可能)中のGAPプログラム中に組込まれたNeedlemanおよびWunsch(J.Mol.Biol.(48):444−453(1970))アルゴリズムを用い、Blosum 62マトリックスまたはPAM250マトリックスのいずれか、ならびにギャップウェイト(gap weight)=16、14、12、10、8、6、または4、およびレングスウェイト(length weight)=1、2、3、4、5、または6を使用して決定される。さらに別の好ましい実施形態において、2つのヌクレオチド配列間の同一性パーセントは、GCGソフトウェアパッケージ(http://www.gcg.comにおいて入手可能)中のGAPプログラムを用い、NWSgapdna.CMPマトリックス、およびギャップウェイト(gap weight)=40、50、60、70、または80、およびレングスウェイト(length weight)=1、2、3、4、5、または6を使用して決定される。別の実施形態において、2つのアミノ酸配列間または2つのヌクレオチド配列間の同一性パーセントは、ALIGNプログラム(バージョン2.0)中に組込まれたE.MeyersおよびW.Miller(MyersおよびMiller,1988,Comput.Appl.Biosci.4:11−17)のアルゴリズムを用い、PAM120ウェイト残基表(weight resiue table)、ギャップレングスペナルティ(gap length penalty)=12、およびギャップペナルティ(gap penalty)=4を使用して決定される。
【0044】
本発明の核酸配列およびタンパク質配列はさらに、公共のデータベースに対する検索を実行するための「問い合わせ配列」としてさらに使用されて、例えば、他のファミリーのメンバーまたは関連する配列を同定し得る。このような検索はAltschulら((1990)J.Mol.Biol.215:403−10)のNBLASTプログラムおよびXBLASTプログラム(バージョン2.0)を用いて実行され得る。BLASTヌクレオチド検索は、NBLASTプログラム、スコア(score)=100、ワードレングス(wordlength)=12を用いて実行されて、本発明のAdhr−1核酸分子に相同性のヌクレオチド配列を獲得し得る。BLASTタンパク質検索は、XBLASTプログラム、スコア(score)=100、ワードレングス(wordlength)=3を用いて実行されて、本発明のAdhr−1タンパク質分子に相同性のアミノ酸配列を獲得し得る。比較目的のためにギャップの入った整列を獲得するために、Gapped BLASTを、Altschulら(1997)Nucleic Acids Res.25(17):3389−3402に記載されるように利用し得る。BLASTプログラムおよびGapped BLASTプログラムを利用する場合、それぞれのプログラム(例えば、XBLASTおよびNBLAST)のデフォルトパラメーターを使用し得る。http://www.ncbi.nlm.nih.gov.を参照のこと。
【0045】
本発明はまた、他の動物種(特に、他の哺乳動物種)由来のヒトMU−1のホモログをコードするポリヌクレオチドを提供する。種ホモログは、本明細書中に開示されるマウス配列またはヒト配列からプローブまたはプライマーを作製し、そして適切な種由来のライブラリー(例えば、関連する種のPBMC、胸腺または精巣から構築されたライブラリーなど)をスクリーニングすることによって同定および単離され得る。
【0046】
本発明の単離されたポリヌクレオチドは、MU−1タンパク質を組み換え産生するために、Kaufmanら、Nucleic Acids Res.19,4485−4490(1991)に開示されるpMT2発現ベクターまたはpED発現ベクターのような発現制御配列に作動可能に連結され得る。多くの適切な発現制御配列が、当該分野で公知である。組換えタンパク質を発現させる一般的な方法もまた公知であり、そしてR.Kaufman、Methods in Enzymology 185,537−566(1990)に例示される。本明細書中に規定されるように、「作動可能に連結された」は、連結されたポリヌクレオチド/発現制御配列を用いて形質転換(トランスフェクト)された宿主細胞によってMU−1タンパク質が発現されるような様式で、本発明の単離されたポリヌクレオチドと発現制御配列との間に共有結合を形成するように酵素的または化学的に連結されることを意味する。
【0047】
多数の型の細胞が、MU−1タンパク質の発現のための適切な宿主細胞として作用し得る。機能的なMU−1タンパク質を発現し得る任意の細胞型が、使用され得る。適切な哺乳動物宿主細胞としては、例えば、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、ヒト腎臓293細胞、ヒト上皮A431細胞、ヒトColo205細胞、3T3細胞、CV−1細胞、他の形質転換された霊長類細胞株、正常複相体細胞、インビトロ初代培養の組織由来の細胞株、初代外植片、HeLa細胞、マウスL細胞、BHK細胞、HL−60細胞、U937細胞、HaK細胞、Rat2細胞、BaF3細胞、32D細胞、FDCP−1細胞、PC12細胞、M1x細胞、C2C12細胞が挙げられる。
【0048】
MU−1タンパク質はまた、1つ以上の昆虫発現ベクターにおいて本発明の単離されたポリヌクレオチドを適切な制御配列に作動可能に連結し、そして昆虫発現系を使用することによって、産生され得る。バキュロウイルス/昆虫細胞発現系についての材料および方法は、例えば、Invitrogen,San Diego,California,U.S.A.からのキットの形態(MaxBac(登録商標)キット)で市販され、そしてそのような方法は、SummersおよびSmith,Texas Agricultural Experiment Station Bulletin No.1555(1987)(本明細書中に参考として援用される)に記載されるように当該分野で周知である。MU−1タンパク質の可溶性形態がまた、上記のような適切な単離ポリヌクレオチドを用いて昆虫細胞中で産生され得る。
【0049】
あるいは、MU−1タンパク質は、酵母のような下等真核生物または細菌のような原核生物中で産生され得る。適切な酵母株としては、Saccharomyces cerevisiae、Schizosaccharomyces pombe、Kluyveromyces株、Candida、または異種タンパク質を発現し得る任意の酵母株が挙げられる。適切な細菌株としては、Escherichia coli、Bacillus subtilis、Salmonella typhimurium、または異種タンパク質を発現し得る任意の細菌株が挙げられる。
【0050】
細菌中での発現は、組換えタンパク質を組込んだ封入体の形成をもたらし得る。従って、活性な材料またはより活性な材料を産生するために、組換えタンパク質の再折り畳みが、必要であり得る。正確に折り畳まれた異種タンパク質を細菌封入体から得るためのいくつかの方法が、当該分野で公知である。これらの方法は一般的に、タンパク質を封入体から可溶化する工程、次いで、カオトロピック薬剤を使用してこのタンパク質を完全に変性する工程を包含する。システイン残基がタンパク質の一次アミノ酸配列中に存在する場合、ジスルフィド結合の正確な形成を可能にする環境下(酸化還元系)で再折り畳みを達成することが、しばしば必要である。再折り畳みの一般的な方法は、Kohno,Meth.Enzym.,185:187−195(1990)に開示される。EP 0433225および同時係属出願USSN 08/163,877は、他の適切な方法を記載する。
【0051】
本発明のMU−1タンパク質はまた、トランスジェニック動物の生成物として、例えば、MU−1タンパク質をコードするポリヌクレオチド配列を含む体細胞または生殖細胞によって特徴付けられる、トランスジェニックウシ、トランスジェニックヤギ、トランスジェニックブタ、またはトランスジェニックヒツジの乳汁成分として発現され得る。
【0052】
本発明のMU−1タンパク質は、所望のタンパク質を発現するのに必要な培養条件下で形質転換された培養宿主細胞を増殖させることによって調製され得る。次いで、生じる発現タンパク質は、この培養培地または細胞抽出物から精製され得る。本発明のMU−1タンパク質の可溶性形態は、馴化培地から精製され得る。本発明のMU−1タンパク質の膜結合形態は、発現している細胞から総膜画分を調製し、そして非イオン性変性剤(例えば、Triton X−100)を用いてこの膜を抽出することによって精製され得る。
【0053】
MU−1タンパク質は、当業者に公知の方法を用いて精製され得る。例えば、本発明のMU−1タンパク質は、市販のタンパク質濃縮フィルター(例えば、AmiconまたはMillipore Pellicon限外濾過ユニット)を用いて濃縮され得る。濃縮工程後、濃縮物は、ゲル濾過媒体のような精製マトリックスにアプライされ得る。あるいは、陰イオン交換樹脂(例えば、ペンダントジエチルアミノエチル(DEAE)基もしくはポリエチレンイミン(PEI)基を有するマトリックスまたは基材)が、使用され得る。マトリックスは、アクリルアミド、アガロース、デキストラン、セルロース、またはタンパク質精製に一般的に使用される他の型であり得る。あるいは、陽イオン交換工程が、使用され得る。適切な陽イオン交換体としては、スルホプロピル基またはカルボキシメチル基を含む種々の不溶性マトリックスが挙げられる。スルホプロピル基(例えば、S−Sepharose(登録商標)カラム)が、好ましい。培養上清からのMU−1タンパク質の精製はまた、コンカナバリンA−アガロース、heparin−toyopearl(登録商標)またはCibacrom blue 3GA Sepharose(登録商標)のようなアフィニティー樹脂で;あるいは、フェニルエーテル、ブチルエーテル、またはプロピルエーテルのような樹脂を用いた疎水性相互作用クロマトグラフィーによる;あるいは、免疫アフィニティークロマトグラフィーによる、1つ以上のカラム工程が挙げられ得る。最後に、疎水性RP−HPLC媒体(例えば、ペンダントメチル基または他の脂肪族基を有するシリカゲル)を使用する1つ以上の逆相高速液体クロマトグラフィー(RP−HPLC)工程が、MU−1タンパク質をさらに精製するために使用され得る。MU−1タンパク質に対する抗体を含むアフィニティーカラムもまた、公知の方法に従って精製に使用され得る。種々の組合わせまたは他の公知の方法における、上記の精製工程のうちのいくつかまたは全てがまた使用されて、実質的に精製された単離組み換えタンパク質を提供し得る。好ましくは、単離されたMU−1タンパク質は、他の哺乳動物タンパク質を実質的に含まないように、精製される。
【0054】
本発明のMU−1タンパク質はまた、MU−1に結合し得る因子をスクリーニングするために使用され得る。所望の結合タンパク質(固定されているかまたは固定されていない)を用いた結合アッセイが、当該分野で周知であり、そして本発明のMU−1タンパク質を使用する本目的のために使用され得る。精製された細胞に基づくスクリーニングアッセイまたは精製されたタンパク質に基づく(無細胞)スクリーニングアッセイが、このような因子を同定するために使用され得る。例えば、MU−1タンパク質は、精製形態でキャリアに固定され得、そして精製されたMU−1タンパク質に対する結合または潜在的なリガンドが、測定され得る。
【0055】
MU−1タンパク質(細胞から精製されたか、または組み換え生成された)は、薬学的に受容可能なキャリアと組み合わせた場合に、薬学的組成物として用いられ得る。このような組成物は、MU−1またはインヒビターおよびキャリアに加えて、種々の希釈剤、充填剤、塩、緩衝液、安定化剤、可溶化剤、および当該分野で周知の他の物質を含み得る。用語、「薬学的に受容可能な」とは、活性成分の生物学的活性の有効性を妨害しない非毒性の物質を意味する。キャリアの特徴は、投与の経路に依存する。
【0056】
本発明の薬学的組成物はまた、サイトカイン、リンホカイン、またはM−CSF、GM−CSF、IL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12、IL−14、IL−15、G−CSF、幹細胞因子、およびエリスロポエチンのような他の造血因子を含み得る。この薬学的組成物はまた、抗サイトカイン抗体も含み得る。この薬学的組成物は、プラスミノゲン活性化因子および第VIII因子のような血栓溶解性因子または抗血栓性因子を含み得る。この薬学的組成物はさらに、他の抗炎症性因子を含み得る。このようなさらなる因子(factor)および/または物質(agent)は、この薬学的組成物中に含まれて、単離されたMU−1タンパク質と相乗効果を生じ得るか、または単離されたMU−1タンパク質によって生じる副作用を最小にし得る。逆に、単離されたMU−1タンパク質は、特定のサイトカイン、リンホカイン、他の造血因子、血栓溶解因子もしくは抗血栓性因子、または抗炎症剤の処方物に含まれて、サイトカイン、リンホカイン、他の造血因子、血栓溶解因子もしくは抗血栓性因子、または抗炎症剤の副作用を最小限にし得る。
【0057】
本発明の薬学的組成物は、単離されたMU−1タンパク質が、他の薬学的に受容可能なキャリアに加えて、水溶液中に、ミセルとして凝集形態で存在する脂質のような両親媒性因子と組み合わせられるリポソーム、不溶性単層、液晶、または層状層(lamellar layer)の形態中に存在し得る。リポソーム処方物のために適切な脂質としては、限定はしないが、モノグリセリド、ジグリセリド、スルファチド、リゾレシチン、リン脂質、サポニン、胆汁酸などが挙げられるがこれらに限定されない。このようなリポソーム処方物の調製は、例えば、米国特許第4,235,871号;米国特許第4,501,728号;米国特許第4,837,028号;および米国特許第4,737,323号(これらの全てが、本明細書において参考として援用されている)に記載のように、当業者のレベルの範囲内である。
【0058】
本明細書において用いる場合、用語「治療的有効量」とは、意味のある患者の利点(例えば、このような状態の症状の改善、このような状態の治癒、またはこのような状態の治癒率の上昇)を示すのに十分である、この薬学的組成物の各々の活性成分の総量または方法を意味する。単独で投与された、個々の活性成分に対して適用される場合、この用語は、その成分単独をいう。組み合わせ(併用)に対して適用される場合、この用語は、組み合わせて投与されても、連続して投与されても、または同時に投与されても、治療効果を生じる活性成分の合わせた量をいう。
【0059】
本発明の処置または使用の方法の実施において、治療的有効量の単離されたMU−Iタンパク質が哺乳動物に投与される。単離されたMU−1タンパク質は、単独でか、または他の治療(例えば、サイトカイン、リンホカイン、または他の造血因子を使用する処置)と組み合わせてのいずれかで、本発明の方法に従って投与され得る。1つ以上のサイトカイン、リンホカイン、または他の造血因子とともに同時投与される場合、MU−1タンパク質は、サイトカイン、リンホカイン、他の造血因子、血栓溶解因子もしくは抗血栓性因子と同時にか、または連続してのいずれかで投与され得る。連続的に投与される場合、主治医は、サイトカイン、リンホカイン、他の造血因子、血栓溶解因子もしくは抗血栓性因子と組み合わせた、MU−1タンパク質投与の適切な順序を決定する。
【0060】
薬学的組成物において用いられるMU−1タンパク質の投与、または本発明の本発明法を実施するために用いられるMU−1タンパク質の投与は、従来の種々の方法(例えば、経口摂取、吸引、または皮膚注射、皮下注射、もしくは静脈注射)で実行され得る。患者に対する静脈投与が好ましい。
【0061】
治療上有効な量のMU−1タンパク質を経口的に投与する場合、MU−1タンパク質は、錠剤、カプセル、粉末、溶液、またはエリキシルの形態である。錠剤形態で投与される場合、本発明の薬学的組成物は、ゼラチンまたはアジュバントのような固体キャリアをさらに含み得る。この錠剤、カプセル、および粉末は、約5〜95%のMU−1タンパク質、そして好ましくは、約25〜90%のMU−1タンパク質を含む。液体形態で投与される場合、液体キャリア(例えば、水、石油、動物もしくは植物起源の油脂(例えば、ピーナッツオイル、鉱油、ダイズ油、またはゴマ油)、または合成油)が添加され得る。この薬学的組成物の液体形態は、さらに、生理食塩水溶液、デキストロースもしくは他の糖溶液、またはグリコール(例えば、エチレングリコール、プロピレングリコール、またはポリエチレングリコール)を含み得る。液体形態で投与される場合、この薬学的組成物は、約0.5〜90重量%のMU−1タンパク質、そして好ましくは約1〜50%のMU−1タンパク質を含む。
【0062】
治療上有効な量のMU−1タンパク質を静脈注射、皮膚注射、または皮下注射によって投与する場合、MU−1タンパク質は、パイロジェン(発熱物質)のない、非経口的に受容可能な水溶液の形態である。このような非経口的に受容可能なタンパク質溶液(pHによって、等張性、安定性などを有する)の調製は、当該分野の範囲内である。静脈注射、皮膚注射、または皮下注射のために好ましい薬学的組成物は、MU−1タンパク質に加えて、等張性ビヒクル(例えば、注射用塩化ナトリウム、注射用リンゲル液、注射用デキストロース、注射用デキストロースおよび塩化ナトリウム、注射用乳酸リンゲル液、または当該分野で公知の他のビヒクル)を含むべきである。本発明の薬学的組成物はまた、安定化剤、保存剤、緩衝液、抗酸化剤、または当業者に公知の他の添加物を含み得る。
【0063】
本発明の薬学的組成物中のMU−1タンパク質の量は、処置される状態の性質および重篤度、ならびに患者が以前に受けた事前処置の性質に依存する。結局、主治医が、各々の個々の患者を処置するMU−1タンパク質の量を決定する。最初に、主治医は、低用量のMU−1タンパク質を投与して、患者の反応を観察する。MU−1タンパク質のさらに高い用量を、患者に至適の治療効果が得られるまで投与し得るが、その時点でこの投薬量は、さらに漸増されることはない。本発明の方法を実施するために用いられる種々の薬学的組成物は、体重1kgあたり、約0.1μg〜約100mgのMU−1タンパク質を含むべきことが意図される。
【0064】
本発明の薬学的組成物を用いる静脈内治療の期間は、処置される疾患の重篤度、ならびに各々の個々の患者の状態および可能性のある特有の応答に依存して変化する。MU−1タンパク質の各出願の期間は、12〜24時間の範囲の連続的静脈投与であることが意図される。結局、主治医は、本発明の薬学的組成物を用いて静脈治療の適切な期間を決定する。
【0065】
本発明のポリヌクレオチドおよびタンパク質は、以下に同定された、1つ以上の用途または生物学的活性(本明細書に言及されるアッセイに関連するものを含む)を示すと期待される。本発明のタンパク質について記載された用途または活性は、このようなタンパク質の投与もしくは使用によって、またはこのようなタンパク質をコードするポリヌクレオチドの投与もしくは使用によって(例えば、遺伝子治療において、またはDNAの導入に適切なベクターにおいて)、提供され得る。
【0066】
(サイトカインおよび細胞増殖/細胞分化活性)
本発明のタンパク質は、サイトカイン活性、細胞増殖(誘導または阻害のいずれか)活性、もしくは細胞分化(誘導または阻害のいずれか)活性を示し得るか、または特定の細胞集団における他のサイトカインの産生を誘導し得る。
【0067】
今日までに発見された多くのタンパク質因子(全ての公知のサイトカインを含む)は、1つ以上の因子依存性細胞増殖アッセイにおいて活性を示しており、従って、このアッセイは、サイトカイン活性の都合のよい確証として機能する。本発明のタンパク質の活性は、以下:32D、DA2、DA1G、T10、B9、B9/11、BaF3、MC9/G、M+(プレB M+)、2E8、RB5、DA1、123、T1165、HT2、CTLL2、TF−1、Mo7e、およびCMKを含むがこれらに限定されない細胞株についての多数の慣用的な因子依存性細胞増殖アッセイのいずれか1つによって証明されている。
【0068】
huEPOR−huMUキメラレセプターを発現するBAF−3細胞は、huEPOに応答して増殖する。MUレセプターが、その細胞質ドメインを通じてシグナル伝達する能力を試験するために、BAF−3細胞を操作して、EPOr/MU(cyto)キメラレセプターを発現し、そしてEPOの存在下における3Hチミジンの取り込みについてアッセイした。インタクトなEPOr分子を発現するBAF−3細胞は、EPOに応答して増殖するが、親のBAF−3細胞は増殖しない。A5クローン(キメラEPOr/MU(cyto)を保有する)は、EPOに応答して増殖し、このことは、MU−1の細胞質部分が、増殖シグナルを保持し得ることを実証する。BAF−3細胞(その表面上でEPOrを発現する)はまた、EPOに応答する。
【0069】
本発明のタンパク質の活性は、他にも方法はあるが、以下の方法によって測定され得る:
T細胞または胸腺細胞の増殖についてのアッセイとしては、以下に記載されるアッセイが、これらに制限することなく挙げられる:Current Protocols in Immunology,J.E.Coligan,A.M.Kruisbeek,D.H.Margulies,E.M.Shevach,W Strober編,Pub.Greene Publishing Associates and Wiley−Interscience(第3章、In Vitro assays for Mouse Lymphocyte Function 3.1−3.19;第7章、Immunologic studies in Humans);Takaiら、J.Immunol.137:3494−3500,1986;Bertagnolliら、J.Immunol.145:1706−1712,1990;Bertagnolliら、Cellular Immunology 133:327−341,1991;Bertagnolliら、J.Immunol.149:3778−3783,1992;Bowmanら、J.Immunol.152:1756−1761,1994。
【0070】
脾細胞、リンパ節細胞、または胸腺細胞のサイトカイン産生および/または増殖についてのアッセイとしては、以下に記載されるアッセイが、これらに制限することなく挙げられる:Polyclonal T cell stimulation,Kruisbeek,A.M.およびShevach,E.M.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、3.12.1−3.12.14頁、John Wiley and Sons,Toronto.1994;ならびにMeasurement of mouse and human Interferon γ,Schreiber,R.D.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、6.8.1−6.8.8頁、John Wiley and Sons,Toronto.1994。
【0071】
造血細胞およびリンパ球産生細胞の増殖および分化についてのアッセイとしては、以下に記載されるアッセイが、これらに制限することなく挙げられる:Measurement of Human and Murine Interleukin 2 and Interleukin 4,Bottomly,K.,Davis,L.S.およびLipsky,P.E.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、6.3.1−6.3.12頁、John Wiley and Sons,Toronto.1991;deVriesら、J.Exp.Med.173 :1205−1211,1991;Moreauら、Nature 336:690−692,1988;Greenbergerら、Proc.Natl.Acad.Sci.U.S.A.80:2931−2938,1983;Measurement of mouse and human interleukin 6−Nordan,R.Current Protocols
in Immunology.J.E.e.a.Coligan編、第1巻、6.6.1−6.6.5頁、John Wiley and Sons,Toronto.1991;Smithら、Proc.Natl.Acad.Sci.U.S.A.83:1857−1861,1986;Measurement of human Interleukin 11−Bennett,F.,Giannotti,J.,Clark,S.C.およびTurner,K.J.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、6.15.1頁、John Wiley and Sons,Toronto.1991;Measurement of
mouse and human Interleukin 9−Ciarletta,A.,Giannotti,J.,Clark,S.C.およびTurner,K.J.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、6.13.1頁、John Wiley and Sons,Toronto.1991。
【0072】
抗原に対するT細胞クローンの応答についてのアッセイ(とりわけ、APC−T細胞相互作用に影響する、ならびにT細胞効果を指向するタンパク質を、増殖およびサイトカイン産生を測定することにより同定する)としては、以下に記載されるアッセイが、これらに制限することなく挙げられる:Current Protocols in Immunology,J.E.Coligan,A.M.Kruisbeek,D.H.Margulies,E.M.Shevach,W Strober編、Pub.Greene Publishing Associates and Wiley−Interscience(第3章、In Vitro assays for Mouse Lymphocyte Function;第6章、Cytokines and their cellular receptors;第7章、Immunologic studies in Humans);Weinbergerら、Proc.Natl.Acad.Sci.USA 77:6091−6095,1980;Weinbergerら、Eur.J.Immun.11:405−411,1981;Takaiら、J.Immunol.137:3494−3500,1986;Takaiら、J.Immunol.140:508−512,1988。
【0073】
(免疫刺激活性または免疫抑制活性)
本発明のタンパク質はまた、免疫刺激活性または免疫抑制活性(その活性に対するアッセイが本明細書中に記載される活性が挙げられるが、これらに限定されない)を示し得る。タンパク質は、種々の免疫不全および障害(重症複合型免疫不全(SCID)を含む)の処置において(例えば、Tリンパ球および/またはBリンパ球の成長および増殖の調節(上方または下方)、ならびにNK細胞および他の細胞集団の細胞溶解活性の誘導において)有用であり得る。これらの免疫不全は、遺伝的であり得るか、またはウイルス(例えば、HIV)および細菌もしくは真菌の感染によって引き起こされ得るか、あるいは自己免疫障害から生じ得る。より具体的には、ウイルス、細菌、真菌または他の感染(HIV、肝炎ウイルス、ヘルペスウイルス、ミコバクテリア、Leishmania spp.、マラリアspp.による感染、およびカンジダ症のような種々の真菌感染が挙げられる)によって引き起こされる感染性疾患は、本発明のタンパク質を使用して、処置可能であり得る。もちろん、これに関して、本発明のタンパク質はまた、免疫系に対するブーストが一般的に所望であり得る場合に(すなわち、癌の処置において)有用であり得る。
【0074】
本発明のタンパク質を使用して処置され得る自己免疫障害としては、例えば、結合組織疾患、多発性硬化症、全身性エリテマトーデス、慢性関節リウマチ、自己免疫肺炎症、ギヤン−バレー症候群、自己免疫性甲状腺炎、インスリン依存性糖尿病、筋無力症、対宿主性移植片病および自己免疫炎症性眼疾患が挙げられる。本発明のこのようなタンパク質はまた、アレルギー性の反応および状態(例えば、喘息(特に、アレルギー性喘息))または他の呼吸の問題の処置において、有用であり得る。免疫抑制が所望である他の状態(例えば、器官移植が挙げられる)もまた、本発明のタンパク質を使用して処置可能であり得る。
【0075】
MU−1 DNAはまた、クローン病に対する染色体遺伝子座をマッピングする。その結果、本発明のタンパク質は、クローン病および他の炎症性腸疾患を処置するために、使用され得る。
【0076】
本発明のタンパク質を使用して、免疫応答を多数の様式で調節することもまた可能であり得る。ダウンレギュレーションは、既に進行している免疫応答を阻害またはブロックする形態であり得るか、あるいは免疫応答の誘導の防止を包含し得る。活性化T細胞の機能は、T細胞応答を抑制することによってか、またはT細胞における特異的耐性を誘導することによってか、あるいはその両方によって、阻害され得る。T細胞応答の免疫抑制は、一般に、活性な、非抗原特異的なプロセスであり、このプロセスは、抑制因子に対するT細胞の連続的な曝露を必要とする。耐性(これは、T細胞における非応答性またはアネルギーの誘導を伴う)は、一般的に抗原特異性である点、および耐性化因子への曝露が終了した後にも持続する点で、免疫抑制から区別可能である。機能上、耐性は、耐性化因子の非存在下での特異的抗原への再曝露の際の、T細胞応答の欠如によって、実証され得る。
【0077】
1つ以上の抗原機能(Bリンパ球抗原機能(例えば、B7のような)が挙げられるが、これらに限定されない)のダウンレギュレーションまたは防止(例えば、活性化T細胞による高レベルのリンホカイン合成の防止)は、組織、皮膚および器官の移植の状況において、ならびに対宿主性移植片病(GVHD)において、有用である。例えば、T細胞機能のブロックは、組織移植において、組織破壊の低下を生じるはずである。代表的に、組織移植において、移植物の拒絶は、T細胞によるその移植物の外来物質としての認識を介して開始され、続いて、この移植物を破壊する免疫反応が起こる。B7リンパ球抗原の、その天然のリガンドとの相互作用を阻害またはブロックする分子を(例えば、B7−2活性を有するペプチドの可溶性の単量体形態を、単独でか、あるいは別のBリンパ球抗原(例えば、B7−1、B7−3)の活性を有するペプチドの単量体形態または遮断抗体と組合せて)免疫細胞に、移植前に投与することによって、その天然のリガンドに対するその分子の結合が、その免疫細胞において、対応する同時刺激信号を伝達することなくもたらされ得る。この様式で、Bリンパ球抗原機能をブロックすることにより、免疫細胞(例えば、T細胞)によるサイトカイン合成が防止され、従って、免疫抑制剤として作用する。さらに、同時刺激の欠如はまた、T細胞をアネルギー化し、これによって被験体において耐性を誘導するに十分であり得る。Bリンパ球抗原ブロック試薬による、長期間の耐性の誘導は、これらのブロッキング試薬のたびたびの投与の必要性を回避し得る。十分な免疫抑制または耐性を被験体において達成するためには、Bリンパ球抗原の組み合わせの機能をブロックすることもまた必要であり得る。
【0078】
器官移植拒絶またはGVHDの予防における、特定のブロッキング試薬の効力は、ヒトにおける効力を予測可能な動物モデルを使用して、評価され得る。使用され得る適切な系の例としては、ラットにおける同種異型の心臓移植片およびマウスにおける異種のランゲルハンス島細胞移植片が挙げられ、これらの両方は、Lenschowら、Science 257:789−792(1992)およびTurkaら、Proc.Natl.Acad.Sci USA,89:11102−11105(1992)に記載されるように、インビボでのCTLA4Ig融合タンパク質の免疫抑制効果を試験するために使用された。さらに、GVHDのマウスモデル(Paul編、Fundamental Immunology,Raven Press,New York,1989,846−847頁を参照のこと)は、インビボでのBリンパ球抗原機能のブロックの、この疾患の発達に対する影響を決定するために、使用され得る。
【0079】
抗原機能をブロックすることはまた、自己免疫疾患を処置するために治療的に有用であり得る。多くの自己免疫障害は、自己組織に対して反応性であり、そしてこれらの疾患の病理に関与するサイトカインおよび自己抗原の産生を促進する、T細胞の不適切な活性化の結果である。自己反応性T細胞の活性化の防止は、疾患の症状を低下または排除し得る。Bリンパ球抗原のレセプター:リガンド相互作用を妨害することによってT細胞の同時刺激をブロックする試薬の投与を使用して、T細胞の活性化を阻害し得、そしてこの疾患プロセスに関与し得る自己抗原またはT細胞由来のサイトカインの産生を防止し得る。さらに、ブロッキング試薬は、自己反応性T細胞の抗原特異的耐性を誘導し得、このことは、疾患の長期間の軽減をもたらし得る。自己免疫障害の予防または軽減におけるブロッキング試薬の効力は、ヒト自己免疫疾患の十分に特徴付けられた多数の動物モデルを使用して、決定され得る。例としては、マウス実験的自己免疫性脳脊髄炎、MRL/lpr/lprマウスまたはNZBハイブリッドマウスにおける全身性エリテマトーデス、マウス自己免疫性コラーゲン関節炎、NODマウスおよびBBラットにおける真性糖尿病、ならびにマウス実験的重症筋無力症が挙げられる(Paul編、Fundamental Immunology,Raven Press,New York,1989,840−856頁を参照のこと)。
【0080】
免疫応答をアップレギュレートする手段としての抗原機能(好ましくは、Bリンパ球抗原機能)のアップレギュレーションはまた、治療において有用であり得る。免疫応答のアップレギュレーションは、既存の免疫応答の増大または最初の免疫応答の誘発の形態であり得る。例えば、Bリンパ球抗原機能の刺激を介する免疫応答の増大は、ウイルス感染の場合に有用であり得る。さらに、全身性ウイルス疾患(例えば、インフルエンザ、感冒、および脳炎)は、Bリンパ球抗原の刺激性形態を全身的に投与することによって緩和され得る。
【0081】
あるいは、抗ウイルス免疫応答は、患者からのT細胞の除去、本発明のペプチドを発現するウイルス抗原パルスAPCで、または本発明の可溶性ペプチドの刺激性形態と共にウイルス抗原パルスAPCでのいずれかでの、インビトロでのT細胞の同時刺激、およびインビトロで活性化されたT細胞のこの患者への再導入によって、感染した患者において増大され得る。抗ウイルス免疫応答を増大する別の方法は、感染した細胞を患者から単離し、これらの細胞を、本明細書中に記載される本発明のタンパク質をコードする核酸で、この細胞がその表面上にこのタンパク質の全てまたは一部を発現するようにトランスフェクトし、そしてこのトランスフェクトした細胞を患者に再導入することである。感染した細胞は、ここで同時刺激シグナルをインビボでT細胞に送達し得、そしてこれによってT細胞を活性化し得る。
【0082】
別の適用において、抗原機能(好ましくは、Bリンパ球抗原機能)のアップレギュレーションまたは増大は、腫瘍免疫の誘導において有用であり得る。本発明の少なくとも1つのペプチドをコードする核酸でトランスフェクトされた腫瘍細胞(例えば、肉腫、黒色腫、リンパ腫、白血病、神経芽細胞腫、癌)は、被験体における腫瘍特異的耐性を克服するために、被験体に投与され得る。所望の場合、この腫瘍細胞は、ペプチドの組み合わせを発現するためにトランスフェクトされ得る。例えば、患者から得られる腫瘍細胞は、B7−2様活性を単独で有するペプチドの発現、またはB7−1様活性および/もしくはB7−3様活性を有するペプチドと関連した発現を指向する発現ベクターを用いて、エキソビボでトランスフェクトされ得る。このトランスフェクトされた腫瘍細胞は患者に戻されて、このトランスフェクトされた細胞の表面上でのこのペプチドの発現を生じる。あるいは、遺伝子治療技術が、インビボでのトランスフェクションのための腫瘍細胞を標的するために使用され得る。
【0083】
腫瘍細胞の表面上での、Bリンパ球抗原の活性を有する本発明のペプチドの存在は、トランスフェクトされた腫瘍細胞に対するT細胞媒介免疫応答を誘導するために必要な同時刺激シグナルを、T細胞に提供する。さらに、MHCクラスI分子またはMHCクラスII分子を欠いた腫瘍細胞、あるいは十分な量のMHCクラスI分子またはMHCクラスII分子を再発現できない腫瘍細胞は、MHCクラスIα鎖タンパク質およびβ2ミクログロブリンタンパク質またはMHCクラスIIα鎖タンパク質およびMHCクラスIIβ鎖タンパク質の全てまたは一部(例えば、細胞質ドメインの短縮部分)をコードする核酸でトランスフェクトされ得、これによってMHCクラスIまたはMHCクラスIIタンパク質を細胞表面に発現する。Bリンパ球抗原(例えば、B7−1、B7−2、B7−3)の活性を有するペプチドと関連しての、適切なクラスIまたはクラスII MHCの発現は、トランスフェクトされた腫瘍細胞に対するT細胞媒介免疫応答を誘導する。必要に応じて、MHCクラスII関連タンパク質(例えば、不変の鎖)の発現をブロックするアンチセンス構築物をコードする遺伝子はまた、Bリンパ球抗原の活性を有するペプチドをコードするDNAで同時トランスフェクトされ得、腫瘍関連抗原の提示を促進し、そして腫瘍特異的免疫を誘導する。従って、ヒト被験体におけるT細胞媒介免疫応答の誘導は、この被験体における腫瘍特異的耐性を克服するために十分であり得る。
【0084】
本発明のタンパク質の活性は、他の手段の中でも、以下の方法によって測定され得る:
胸腺細胞または脾細胞の細胞傷害性についての適切なアッセイとしては、以下に記載されるものが挙げられるが、これらに限定されない:Current Protocols in Immunology,J.E.Coligan編,A.M.Kruisbeek,D.H.Margulies,E.M.Shevach,W Strober,Pub.Greene Publishing Associates and Wiley−Interscience(Chapter 3,In Vitro assays for Mouse Lymphocyte Function 3.1−3.19;Chapter 7,Immunologic studies in Humans);Herrmannら,Proc.Natl.Acad.Sci.USA 78:2488−2492,1981;Herrmannら,J.Immunol.128:1968−1974,1982;Handaら,J.Immunol.135:1564−1572,1985;Takaiら,J.Immunol.137:3494−3500,1986;Takaiら,J.Immunol.140:508−512,1988;Herrmannら,Proc.Natl.Acad.Sci.USA 78:2488−2492,1981;Herrmannら,J.Immunol.128:1968−1974,1982;Handaら,J.Immunol.135:1564−1572,1985;Takaiら,J.Immunol.137:3494−3500,1986;Bowmanら,J.Virology 61:1992−1998;Takaiら,J.Immunol.140:508−512,1988;Bertagnolliら,Cellular Immunology 133:327−341,1991;Brownら,J.Immunol.153:3079−3092,1994。
【0085】
T細胞依存性免疫グロブリン応答およびアイソタイプのスイッチングについてのアッセイ(これは、とりわけ、T細胞依存性抗体応答を調節しそしてTh1/Th2プロフィールに影響するタンパク質を同定する)としては、以下に記載されるものが挙げられるが、これらに限定されない:Maliszewski,J.Immunol.144:3028−3033,1990;ならびにAssays for B cell function:In vitro antibody production,Mond,J.J.およびBrunswick,M.、Current Protocols in Immunology.J.E.e.a.Coligan編,Vol 1,pp.3.8.1−3.8.16,John Wiley and Sons,Toronto.1994。
【0086】
混合リンパ球反応(MLR)アッセイ(これは、とりわけ、主にTh1およびCTL応答を生成するタンパク質を同定する)としては、以下に記載されるものが挙げられるが、これらに限定されない:Current Protocols
in Immunology,J.E.Coligan編,A.M.Kruisbeek,D.H.Margulies,E.M.Shevach,W Strober,Pub.Greene Publishing Associates and Wiley−Interscience(Chapter 3,In Vitro assays for Mouse Lymphocyte
Function 3.1−3.19;Chapter 7,Immunologic studies in Humans);Takaiら,J.Immunol.137:3494−3500,1986;Takaiら,J.Immunol.140:508−512,1988;Bertagnolliら,J.Immunol.149:3778−3783,1992。
【0087】
樹状細胞依存性アッセイ(これは、とりわけ、ネイティブのT細胞を活性化する樹状細胞によって発現されるタンパク質を同定する)としては、以下に記載されるものが挙げられるが、これらに限定されない:Gueryら,J.Immunol.134:536−544,1995;Inabaら,Journal of Experimental Medicine 173:549−559,1991;Macatoniaら,Journal of Immunology 154:5071−5079,1995;Porgadorら,Journal of Experimental Medicine 182:255−260,1995;Nairら,Journal of Virology 67:4062−4069,1993;Huangら,Science 264:961−965,1994;Macatoniaら,Journal of Experimental Medicine 169:1255−1264,1989;Bhardwajら,Journal of Clinical Investigation 94:797−807,1994;およびInabaら,Journal of Experimental Medicine 172:631−640,1990。
【0088】
リンパ球の生存/アポトーシスについてのアッセイ(これは、とりわけ、スーパー抗原誘導後にアポトーシスを防止するタンパク質およびリンパ球のホメオスタシスを調節するタンパク質を同定する)としては、以下に記載されるものが挙げられるが、これらに限定されない:Darzynkiewiczら,Cytometry 13:795−808,1992;Gorczycaら,Leukemia 7:659−670,1993;Gorczycaら,Cancer
Research 53:1945−1951,1993;Itohら,Cell 66:233−243,1991;Zacharchuk,Journal of Immunology 145:4037−4045,1990;Zamaiら,Cytometry 14:891−897,1993;Gorczycaら,International Journal of Oncology 1:639−648,1992。
【0089】
T細胞の拘束および発生の初期段階に影響するタンパク質についてのアッセイとしては、以下に記載されるものが挙げられるが、これらに限定されない:Anticaら,Blood 84:111−117,1994;Fineら,Celluler Immunology 155:111−122,1994;Galyら,Blood 85:2770−2778,1995;Tokiら,Proc.Nat.Acad Sci.USA 88:7548−7551,1991。
【0090】
本発明のMU−1分子はまた、STAT活性(例えば、シグナル伝達)の調節(例えば、STAT分子(例えば、STAT3および/またはSTAT5)との相互作用を介する調節、およびSTATリン酸化の調節)に関与する。STATのシグナル伝達のモジュレーター、およびSTAT分子とのMU−1の相互作用のモジュレーターは、本明細書中に記載されるスクリーニングアッセイを介して同定され得る。
【0091】
MU−1活性のモジュレーターを同定するために使用され得るアッセイとしては、STAT活性(例えば、STATシグナル伝達)についてのアッセイ(例えば、当該分野で公知のアッセイおよび本明細書中に記載されるアッセイ)が挙げられる。STAT活性についてのアッセイはまた、例えば、Friedrich
K.ら(2001)Biol Chem 382(2):343−51に記載される。STAT活性についての他のアッセイとしては、細胞増殖およびSTATリン酸化についてのアッセイが挙げられる。
【0092】
1つの局面において、MU−1活性のモジュレーターについてのアッセイは、細胞ベースのアッセイ(ここで、MU−1タンパク質またはその生物学的に活性な部分を発現する細胞が、試験化合物と接触され、そしてこの試験化合物がMU−1活性を調節する能力(例えば、STAT分子との相互作用またはSTATシグナル伝達の調節)が決定される)である。試験化合物がMU−1活性を調節する能力の決定は、例えば、STATシグナル伝達、STAT分子のリン酸化、細胞増殖、細胞の分化、またはサイトカインの産生をモニターすることによって達成され得る。この細胞は、例えば、哺乳動物起源の細胞(例えば、活性化T細胞)であり得る。
【0093】
(造血調節活性)
本発明のタンパク質は、造血の調節において、従って骨髄性細胞またはリンパ球様細胞の欠失の処置において有用であり得る。コロニー形成細胞または因子依存性細胞株の支持における辺縁の生物学的な活性でさえ、以下の造血の調節への関与を示す:例えば、赤血球系前駆細胞の成長および増殖を単独または他のサイトカインと組み合わせて支持し、これにより、例えば、種々の貧血症の処置、または赤血球系前駆体細胞および/または赤血球系細胞の産生を刺激するための照射/化学療法と組み合わせた使用の有用性を示すこと;例えば、結果的に骨髄の抑制を予防または処置するための化学療法と組み合わせて有用な、骨髄性細胞(例えば、有用な顆粒球および単球/マクロファージ)の成長および増殖を支持すること(すなわち、古典的なCSF活性);巨核球および結果的に血小板の成長および増殖を支持し、これにより、血小板減少症のような種々の血小板障害の予防または処置、および一般に、血小板の輸血の代わりにまたは補充での使用を可能にすること;ならびに/または上記の任意および全ての造血細胞に成熟し得る造血幹細胞の成長および増殖を支持すること。これにより種々の幹細胞の障害(例えば、移植を用いて処置される障害(これには、再生不良性貧血および発作性夜間血色素尿症が挙げられるが、これらに限定されない))、および正常な細胞または遺伝子治療のために遺伝子操作された細胞として、インビボまたはエキソビボのいずれかで(すなわち、骨髄移植と組み合わせるかまたは辺縁の前駆体細胞(相同性または非相同性)の移植を用いて)、照射/化学療法後に幹細胞のコンパートメントを再増殖することでの治療利用を見出す。
【0094】
本発明のタンパク質の活性は、とりわけ、以下の方法によって測定され得る。
【0095】
種々の造血株の増殖および分化のための適切なアッセイは、上に挙げられる。
【0096】
胚幹細胞の分化についてのアッセイ(このアッセイにより、とりわけ、胚性分化造血に影響を及ぼすタンパク質を同定する)としては、以下に記載されるアッセイが挙げられるが、これらに限定されない:Johanssonら、Cellular Biology 15:141−151、1995;Kellerら、Molecular and Cellular Biology 13:473−486、1993;McClanahanら、Blood 81:2903−2915、1993。
【0097】
幹細胞の生存および分化についてのアッセイ(このアッセイにより、とりわけ、リンパ−造血を調節するタンパク質を同定する)としては、以下に記載されるアッセイが挙げられるが、これらに限定されない:Methylcellulose colony forming assays、Freshney、M.G.、Culture of Hematopoietic Cells.R.I.Freshneyら編、第265〜268頁、Wiely−Liss,Inc.、New York、NY.1994;Hirayamaら、Proc.Natl.Acad.Sci.USA 89:5907−5911、1992;Primitive hematopoietic colony forming cells with high proliferative potential、McNiece、I.K.およびBriddell,R.A.、Culture of Hematopoietic Cells、R.I.Freshneyら編、第23〜39頁、Wiley−Liss,Inc.、New
York,NY.1994;Nebenら、Experimental Hematology 22:353−359、1994;Cobblestone
area forming cell assay、Ploemacher、R.E.、Culture of Hematopoietic Cells.R.I.Freshneyら編、第1〜21頁、Wiley−Liss,Inc.、New York,NY.1994;Long term bone marrow cultures in the presence of stromal cells、Spooncer,E.、Dexter,M.およびAllen,T、Culture of Hematopoietic Cells、R.I.Freshneyら編、第163〜179頁、Wiley−Liss,Inc.、New York,NY.1994;Long term culture initiating cell assay、Sutherland,H.J.、Culture of Hematopoietic Cells、R.I.Freshneyら編、第139〜162頁、Wiley−Liss,Inc.、New York,NY.1994。
【0098】
(リサーチの使用および有用性)
本発明により提供されるポリヌクレオチドは、種々の目的のためにリサーチコミュニティによって使用され得る。ポリヌクレオチドは、分析、特徴付け、または治療に使用するための組換えタンパク質を発現するために;対応するタンパク質が優先的に発現される組織(構成的に、あるいは組織の分化もしくは発達または疾患状態の特定の段階のいずれか)のためのマーカーとして;サザンゲルにおける分子量マーカーとして;染色体を同定するためもしくは関連する遺伝子座マッピングするための染色体マーカーもしくはタグ(標識された場合)として;可能性のある遺伝子障害を同定するために、患者の内因性DNA配列と比較するため;ハイブリダイズするため、従って新規な関連するDNA配列を発見するためのプローブとして;遺伝子フィンガープリントのためのPCRプライマーを誘導するための情報の供給源として;他の新規なポリヌクレオチドを発見するプロセスにおいて、既知の配列を「差引きする(subtract−out)」ためのプローブとして;「遺伝子チップ」または他の支持体への連結のためにオリゴマーを選択し、そして作製するため(発現パターンの試験を含む);DNA免疫化技術を使用して抗タンパク質抗体を産生させるため;ならびに抗DNA抗体を産生するため、または別の免疫応答を惹起するための抗原として、使用され得る。ポリヌクレオチドが、別のタンパク質に(例えば、レセプター−リガンドの相互作用において)結合または部分的に結合するタンパク質をコードする場合、このポリヌクレオチドはまた、相互作用トラップアッセイ(例えば、Gyurisら、Cell 75:791−803(1993)に記載されるアッセイ)において、結合が生じる他のタンパク質をコードするポリヌクレオチドを同定するため、または結合相互作用のインヒビターを同定するために使用され得る。
【0099】
本発明によって提供されるタンパク質は、以下に挙げられる生物学的な活性を決定するためのアッセイにおいて同様に使用され得る:ハイスループットスクリーニングのための複数のタンパク質のパネル;抗体を産生するか、または別の免疫応答を惹起するため;生物学的流体中でのタンパク質(またはそのレセプター)のレベルを定量的に決定するように設計されたアッセイにおける試薬(標識化された試薬を含む)として;対応するタンパク質が優先的に発現される組織(構成的に、あるいは組織の分化もしくは発達または疾患状態の特定の段階でのいずれか)のためのマーカーとして;ならびに当然、相関するレセプターまたはリガンドを単離するため。タンパク質が、別のタンパク質に(例えば、レセプター−リガンド相互作用により)結合または部分的に結合する場合、このタンパク質は、結合が生じる他のタンパク質を同定するためか、または結合相互作用のインヒビターを同定するために使用され得る。これらの結合相互作用に関与するタンパク質はまた、この結合相互作用のペプチドまたは小分子のインヒビターもしくはアゴニストをスクリーニングするために使用され得る。
【0100】
これらのリサーチのいずれかまたは全ての有用性は、リサーチ製品として市販するための、試薬等級またはキットフォーマットに発展され得る。
【0101】
上に列挙される使用を実施するための方法は、当業者に周知である。このような方法を開示する参考文献としては、以下が挙げられるが、これらに限定されない:「Molecular Cloning:A Laboratory Manual」、第2版、Cold Spring Harbor Laboratory Press、Sambrook,J.、E.F.FritschおよびT.Maniatis編、1989、ならびに「Methods in Enzymology:Guide to Molecular Cloning Techniques」,Academic Press,Berger,S.L.およびA.R.Kimmel編、1987。
【0102】
(栄養性使用)
本発明のポリヌクレオチドおよびタンパク質はまた、栄養性供給源または補充物として使用され得る。このような使用は、以下が挙げられるが、これらに限定されない:タンパク質またはアミノ酸サプリメントとしての使用、炭素供給源としての使用、窒素供給源としての使用および炭化水素の供給源としての使用。このような場合において、本発明のタンパク質またはポリヌクレオチドは、特定の生物の飼料(フィード)に添加され得、あるいは例えば、粉末、ピル、溶液、懸濁物またはカプセルの形態のような、別の固体調製物または液体調製物として投与され得る。微生物の場合において、本発明のタンパク質およびポリヌクレオチドは、培地中に添加され得、この培地中または培地上で、微生物は培養される。
【0103】
本発明のMU−1タンパク質はまた、ポリクローナル抗体およびモノクローナル抗体を得るために動物を免疫するために使用され得、これらの抗体は、MU−1タンパク質と特異的に反応し、そしてこれらの抗体は、リガンドのレセプターに対する結合を阻害し得る。このような抗体は、免疫原として全MU−1を使用することによるか、またはMU−1のフラグメントを使用することによって得られ得る。このMU−1のより小さなフラグメントはまた、動物を免疫するために使用され得る。このペプチド免疫原は、カルボキシル末端にシステイン残基をされに含み得、そしてキーホールリンペットヘモシアニン(keyhole limpet hemocyanin(KLH))のようなハプテンに結合される。さらなるペプチド免疫原は、チロシン残基を硫酸化されたチロシン残基を用いて置換することによって生成され得る。このようなペプチドを合成するための方法は、例えばR.P.Merrifield,J.Amer.Chem.Soc.85,2149−2154(1963);J.L.Krstenanskyら、FEBS Lett.211、10(1987)にあるように当該分野で公知である。
【0104】
MU−1タンパク質に結合する、中和抗体または非中和抗体(好ましくは、モノクローナル抗体)はまた、特定の腫瘍のための有用な治療剤であり得、そして上記の状態の処置においてまた有用であり得る。これらの中和モノクローナル抗体は、MU−1レセプター鎖に結合するリガンドをブロックし得る。
【0105】
本明細書中で引用される全ての特許および参考文献は、完全に記載されているかのように参考として援用されている。本発明は、以下の実施例によってさらに例示され、これらの実施例は、限定するものとして構成されるべきではない。
【実施例】
【0106】
(実施例)
(実施例1:マウスMU−1 cDNAの単離および特徴付け)
Mu−1レセプターのマウスホモログの部分的フラグメントを、ヒト配列に由来するオリゴヌクレオチドを使用するPCRによって単離した。cDNAを、17日齢のマウス胸腺およびマウス2D6T細胞株から単離したRNAから調製した。約300ヌクレオチドのDNAフラグメントを、以下のオリゴヌクレオチド(図1のヒトcDNA配列(配列番号1に対応する)のそれぞれ領域584〜603および876〜896に対応する)を用いるPCRによってcDNAから増幅した。
【0107】
【化2】
増幅は、94℃で1分間、50℃で1分間、および72℃で1分間の30サイクルの間、1.5mMの塩化マグネシウムを含む1×Taq緩衝液中でTaqポリメラーゼを使用して行なった。このフラグメントのDNA配列を決定し、そして2つのオリゴヌクレオチドは、以下の配列を有するこのフラグメントの内部部分に由来した:
【0108】
【化3】
オリゴヌクレオチドを使用して、2D6 T細胞株から単離したcDNAライブラリーをスクリーニングためのハイブリダイゼーションプローブとして使用するために、本来のPCR産物(図1のマウスcDNA配列のヌクレオチド781〜1043、および配列番号9に対応する)の内部262ヌクレオチドフラグメントを増幅した。フィルターを、標準5×SSCハイブリダイゼーション条件を使用して、65℃でハイブリダイズし、そして65℃でSSC中に洗浄した。426,000個のクローンのスクリーニングにおいてプローブにハイブリダイズした20個のクローンを単離した。DNA配列を、2つの独立したクローンから決定した。クローン番号6の全長配列は、これがヒトMU−1(配列番号9)の全長マウスホモログであることを確認した。
【0109】
マウスMU−1の全長ヌクレオチド配列(配列番号9に対応する)を、図1に示す。ヌクレオチド配列は、ヌクレオチド407〜764の推定リーダー配列、ヌクレオチド407〜1993のコード配列、ヌクレオチド1994〜1997の終止コドンを有する。ヌクレオチド1〜406は、5’非翻訳領域に対応し、そしてヌクレオチド1998〜2628は、3’非翻訳領域に対応する。
【0110】
マウスMU−1の推定タンパク質配列(配列番号10に対応する)を、図2に示す。このマウスMU−1タンパク質は、SPScan(スコア=10.1)によって決定された推定リーダー配列(配列番号10のアミノ酸1〜19に対応する)および推定膜貫通ドメイン(配列番号10のアミノ酸237〜253に対応する)を含む。推定シグナル伝達モチーフは、以下の領域を含む:ボックス1:配列番号10のアミノ酸265〜274、ボックス2:配列番号10のアミノ酸310〜324、配列番号10の281位、319位、361位、368位、397位、および510位の6つのチロシン残基。潜在的なSTATドッキング部位は、以下を含む:STAT5:EDDGYPA、STAT3:YLQR。
【0111】
(実施例2:ヒトMU−1およびマウスMU−1の比較)
GAPアルゴニズムを使用して、ヒトMU−1およびマウスMU−1のアミノ酸を比較する。マウスおよびヒト推定タンパク質配列の比較を、図4に示す。アミノ酸は、GAPアルゴリズムを使用して、65.267%同一であった。整列を、BLOSUM62アミノ酸置換マトリクスによって作成した(Henikoff,S.およびHenikoff,J.G.(1992))。タンパク質ブロック質由来のアミノ酸置換マトリクス(Proc.Natl.Acad.Sci.USA 89:10915−10919)。Gapパラメーター:Gap Weight=8、Average Match=2.912、Length Weight=2、Average Mismatch=−2.003。Percent Similarity=69.466。
【0112】
ヒトおよびマウスcDNAヌクレオチド配列の比較を図3に示す。DNA配列は、GAPアルゴリズムを使用して整列される場合、66.116%同一である。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。Percent Similarity=66.198。
【0113】
ヒトMU−1タンパク質およびマウスMU−1タンパク質の両方は、1型サイトカインレセプタースーパーファミリーのメンバーである。マウスMU−1およびヒトMU−1の両方の配列の評価は、潜在的なボックス1およびボックス2シグナル伝達モチーフの存在を明らかにする。6つのチロシン残基が、細胞質ドメインに存在し、そしてMU−1のシグナル伝達機能において重要であり得る。MU−1の他のファミリーのメンバーとの配列の比較は、STAT5およびSTAT3についての潜在的なドッキング部位の存在を示唆した。
【0114】
(実施例3:ヒトMU−1によって使用されるSTATシグナル伝達経路の決定)
BAF−3細胞を操作して、ヒトEPOレセプターの細胞外ドメインおよびMU−1のレセプターの細胞内ドメインからなるキメラサイトカインレセプターを発現した。huEPOR/MU−1(細胞質)キメラレセプターを発現したBAF−3細胞は、ヒト可溶性EPOに応答して増殖した。これらの細胞を分析して、どのSTAT分子がEPOシグナル伝達に応答してリン酸化されているかを決定した。手短にいうと、コントロール未改変親BAF−3細胞およびEPOR/MUキメラBAF−3細胞を、IL−3含有増殖培養から静止させ、そしてIL−3またはEPOのいずれかを用いて、0、15、30および60分間再刺激した。リン酸化されたチロシンを保存するために、細胞をペレット化し、そしてオルトバナジン酸塩を含む氷冷した溶解緩衝液中に再懸濁した。等量の細胞溶解物を、SDS−PAGEによって電気泳動して、そしてウエスタン分析についてニトロセルロース膜にブロットした。二連のブロットを、STAT分子の各形態に特異的な抗体を使用することによって、STAT1、3、5、および6のリン酸化形態および非リン酸化形態について染色した。活性化していないHELA細胞、α−インターフェロンで活性化したHELA細胞を陽性コントロールとして使用した。
【0115】
これらの結果は、これらの特定の条件化で、MU−1を介するシグナル伝達が、試験された全ての時点(T=0、T=15’、T=30’、T=60’)においてSTAT5のリン酸化を生じることを示した。コントロールまたはキメラBAF−3細胞のIL−3を用いる処理は、STAT1または5ではなく、STAT3のリン酸化を生じた。
【0116】
(実施例4:マウスMU−1およびヒトMU−1の組織発現)
(ノーザン分析)
種々の組織(Clonetech,Palo Alto,CA)由来のポリA+RNAのノーザンブロットを、製造業者によって推奨されるようにして行なった。マウスブロットについて、図1のヌクレオチド781〜1043および配列番号9に対応する262ヌクレオチドフラグメントを、ハイブリダイゼーションのために使用した。
【0117】
マウスMU−1の単一の転写物を、成体マウスの脾臓、肺および心臓組織において検出した。ヒト組織において観察されるより大きい転写物は、マウス組織において観察されなかった。
【0118】
ヒトMU−1の2つの転写物を、成体ヒトリンパ様組織、PBL、胸腺、脾臓およびリンパ節、ならびに胎児肺において検出した。
【0119】
(インサイチュハイブリダイゼーション)
インサイチュハイブリダイゼーションにおいて、研究は、Columbus,OHのPhylogency Inc.によって行なわれた(Lyonsら、1990、J.Cell.Biol:111:2427−2436の方法に従う)。手短にいうと、連続的な5〜7ミクロンのパラフィン切片を、脱パラフィン化し、固定し、プロテイナーゼKを用いて消化し、トリエタノールアミンを用いて処理し、そして脱水した。cRNAを、アンチセンスおよびセンスプローブを作成するために、直鎖状cDNAテンプレートから調製した。cRNA転写物を、製造業者の条件(Ambion)に従って合成し、そして35S−UTPで標識した。切片を、一晩ハイブリダイズし、ストリンジェントに洗浄し、そしてRNAase Aを用いて処理し、そして核トラックエマルジョンに浸漬し、そして2〜3週間露光した。コントロール切片を、手順のバックグラウンドレベルを示すために、センスプローブを用いてハイブリダイズした。マウスプローブは、ヌクレオチド860〜1064(配列番号9)に対応する186bpフラグメントからなった。ヒトプローブは、ヒトMU−1 DNAから作成された231bp
PCR産物であった。
【0120】
マウスMU−1発現は、胚中心および外部筋層(muscularis externa)における成体小腸のリンパ節において観察された。分化したリンパ節およびパイアー斑はまた、マウスMU−1発現を示した。
【0121】
ヒトMU−1発現は、皮質のリンパ小節の胚中心において検出された。髄質(これは、マクロファージを含む)は、ヒトMU−1について陰性であった。ヒト脾臓において、ヒトMU−1発現は、赤脾髄ではなく、白脾髄の領域において検出された。
【0122】
(実施例5:細胞および細胞株におけるヒトMU−1の発現)
RNアーセプロテクション分析を、休止しているおよび活性化したヒトT細胞株およびB細胞株、RajiおよびRPMI 8866、ならびにT細胞株ジャーカットについて行った。ヒトT細胞を、抗CD3および抗CD28を用いて活性化した。細胞株をホルボールエステルおよびイオノマイシンで活性化した。231bpのPCR産物(PCRを、5’プライマー:
【0123】
【化4】
(配列番号15)
および3’プライマー:
【0124】
【化5】
(配列番号16)
を使用することによって行った)をpGEM3zf(−)(Promega,Madison,WI)ベクターのBamH1部位およびHindIII部位に挿入することによって、MU−1リボプローブ産生プラスミドを構築した。このリボプローブを作製するために、リボプローブ産生プラスミドをHindIIIで直線化した。得られたDNAを、フェノール/クロロホルム抽出し、そしてエタノールを用いて沈殿させた。T7 RNAポリメラーゼを使用して、メーカー(PharMingen,San Diego,CA)により指示されたプロトコルに従って、リボプローブを作製した。PharMingenのRiboQuant Multi−Probe Ribonuclease Protection Assayシステムを使用することによって、RNAse保護アッセイを行った。RNAse消化の後、2.0μgの総RNAが、各RPA反応物に含まれ、保護されたリボプローブを、QuickPoint高速核酸分離システム(Novex,San Diego,CA)で実行した。ゲルを、メーカーの指示に従って乾燥し、そして暴露した。
【0125】
ヒトMU−1 RNAは、非刺激集団と比較する場合、抗CD3+抗CD28刺激ヒト精製CD3+細胞においてアップレギュレートされる。MU−1はまた、ThおよびTh2非対称T細胞集団において再刺激される際に、アップレギュレートされる。B細胞株、RPMI 8866およびRajiは、MU−1を構成的に発現し、一方、ジャーカットT細胞株はMU−1を発現しない。
【0126】
(実施例6:既知のサイトカインへのヒトMU−1の結合)
ヒトIg融合タンパク質およびマウスIg融合タンパク質の両方は、MU−1に対するリガンドを同定する試みにおいて、Biacoreチップ上で構築されこのチップ上に固定される。様々な細胞培養物の馴化培地ならびに既知のサイトカインのパネルを、MU−1に対する結合について評価した。いくつかのサイトカインをまた、このファミリーの他のレセプター鎖と組み合わせて試験して、MU−1がリガンド結合のために第2のレセプター鎖を必要とし得る可能性を考察した。以下のサイトカインを試験し、そしてMU−1結合に対して陰性であることを見出した:mIL−2、hIL−2、hIL−15、mIL−7、TSLP、TSLP+IL7、TSLP+IL7R、TSLP+IL7g、TSLP+IL−2、TSLP+IL2+IL2Rβ、IL2Rβ、IL2Rγ、IL7R、IL2+2Rβ、IL2+2Rγ、IL15+IL2Rβ、Il15+2Rγ、IL7+2Rγ、IL2+IL7R、IL15+IL7R、IL7+IL7R。既知のレセプターを同様に固定し、そしてMUFc結合について試験し、陰性結果を得た。IL−15はIL2Rbに結合するが、IL2RgにもMUFcにも結合しない。
【0127】
(等価物)
当業者は、単に慣用的な実験を使用して、本明細書中に記載される本発明の特定の実施形態の多くの等価物を認識するか、または確認し得る。このような等価物は、上記の特許請求の範囲に包含されることが意図される。
【図面の簡単な説明】
【0128】
【図1−1】図1は、マウスMU−1の全長cDNA配列を示す。このヌクレオチド配列は、配列番号9のヌクレオチド1〜2528に対応する。
【図1−2】図1は、マウスMU−1の全長cDNA配列を示す。このヌクレオチド配列は、配列番号9のヌクレオチド1〜2528に対応する。
【図2】図2は、マウスMU−1のアミノ酸配列(配列番号10のアミノ酸1〜529に対応する)を示す。SPScanによって10.1のスコアで推定された、アミノ酸1〜19に推定リーダー配列が存在する(太字)。アミノ酸237〜257に推定膜貫通ドメインが存在する(下線)。推定シグナル伝達モチーフは、以下の領域を含む:ボックス1:アミノ酸265〜274およびボックス2:アミノ酸310〜324(太字および下線)。6つのチロシンが、アミノ酸第281位、第319位、第361位、第368位、第397位および第510位に位置する。WSXWSモチーフ(配列番号8)は、アミノ酸残基214〜アミノ酸残基218に位置付する(大きい太字)。可能性のあるSTAT結合(docking)部位は、アミノ酸393〜398およびアミノ酸510〜513を含む。
【図3−1】図3は、ヒトMU−1およびマウスMU−1のcDNA配列(それぞれ、配列番号1の核酸1〜2665および配列番号9の1〜2628に対応する)のGAP比較を示す。HuMU−1=ヒトMU−1、murMU−1=マウスMU−1。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。同一性%=66.116。
【図3−2】図3は、ヒトMU−1およびマウスMU−1のcDNA配列(それぞれ、配列番号1の核酸1〜2665および配列番号9の1〜2628に対応する)のGAP比較を示す。HuMU−1=ヒトMU−1、murMU−1=マウスMU−1。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。同一性%=66.116。
【図3−3】図3は、ヒトMU−1およびマウスMU−1のcDNA配列(それぞれ、配列番号1の核酸1〜2665および配列番号9の1〜2628に対応する)のGAP比較を示す。HuMU−1=ヒトMU−1、murMU−1=マウスMU−1。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。同一性%=66.116。
【図3−4】図3は、ヒトMU−1およびマウスMU−1のcDNA配列(それぞれ、配列番号1の核酸1〜2665および配列番号9の1〜2628に対応する)のGAP比較を示す。HuMU−1=ヒトMU−1、murMU−1=マウスMU−1。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。同一性%=66.116。
【図4】図4は、ヒトMU−1タンパク質(配列番号2のアミノ酸1〜538に対応する)およびマウスMU−1タンパク質(配列番号10のアミノ酸1〜529に対応する)のGAP比較を示す。BLOSUM62アミノ酸置換マトリクス(Henikoff,S.およびHenikoff,J.G.(1992))。タンパク質ブロックからのアミノ酸置換マトリクス(Proc.Natl.Acad.Sci.USA 89:10915−10919)。Gapパラメーター:Gap Weight=8、Average Match=2.912、Length Weight=2、Average Mismatch=−2.003。同一性%=65.267。
【図5】図5は、ヒトMU−1(humu)(配列番号2に対応する)、マウスMU−1(mousemu)(配列番号10)およびヒトIL2β鎖(humil2rbc)(GENbank登録番号M26062)のアミノ酸の多重配列アライメントを示す。リーダーおよび膜貫通ドメインに下線を付す。保存サイトカインレセプターモジュールモチーフを、太字で示す。可能性のあるシグナル伝達領域を、下線および太字で示す。
【図6】図6は、MU−1を介するシグナル伝達を示す。MU−1は、クローンE7 EPO−MU−1キメラにおいてSTAT5をリン酸化する。実施例3に示される条件下で、MU−1を介するシグナル伝達は、試験された全ての時点でSTAT5のリン酸化を生じる。IL−3でのコントロールまたはキメラBAF−3細胞の処理は、STAT3のリン酸化を生じたが、STAT1およびSTAT5のいずれのリン酸化も生じなかった。
【技術分野】
【0001】
(発明の分野)
本発明は、哺乳動物サイトカインレセプターファミリーのタンパク質の新規なメンバー(限定はしないが、ヒトレセプタータンパク質およびマウスレセプタータンパク質が、挙げられる)、そのフラグメント、ならびにそのようなタンパク質を発現するために有用な組換えポリヌクレオチドおよび細胞に関する。
【背景技術】
【0002】
(発明の背景)
種々の造血細胞集団または血球集団の発生および増殖に関与する、種々の調節分子(造血素として公知)が、同定されている。ほとんどの造血素は、標的細胞の表面上のレセプターと相互作用することにより、特定の生物学的活性を示す。サイトカインレセプターは、1つ、2つ、または3つの鎖から一般に構成される。多くのサイトカインレセプターおよびいくつかのサイトカイン(例えば、IL−12 p40)が、造血素レセプタースーパーファミリータンパク質のメンバーである。この造血素レセプタースーパーファミリーの新規なメンバーを同定することは、造血の調節、免疫応答の調節、および造血素スーパーファミリーの他のメンバー(サイトカインおよびレセプターを含む)の同定において、有用であり得る。
【発明の開示】
【発明が解決しようとする課題】
【0003】
この造血素レセプタースーパーファミリーの従来未知であったメンバーに関するDNAおよびタンパク質の配列を、同定および決定することが望ましい。
【課題を解決するための手段】
【0004】
(発明の要旨)
本発明に従って、MU−1造血素レセプタースーパーファミリー鎖をコードするポリヌクレオチド(マウス供給源およびヒト供給源由来のものが挙げられるが、限定はしない)が、開示される。
【0005】
特定の実施形態において、本発明は、ヌクレオチド配列を含む単離されたポリヌクレオチドを提供し、このヌクレオチド配列は、以下:
(a)配列番号1のヌクレオチド配列;
(b)配列番号1のヌクレオチド238〜ヌクレオチド1852のヌクレオチド配列;
(c)配列番号1のヌクレオチド301〜ヌクレオチド1852のヌクレオチド配列;
(d)配列番号1のヌクレオチド301〜ヌクレオチド945のヌクレオチド配列;
(e)(a)〜(d)のいずれかにおいて特定されるヌクレオチド配列の配列と、遺伝暗号の縮重の結果として異なる、ヌクレオチド配列;
(f)(a)〜(d)のいずれかにおいて特定されるヌクレオチドに、ストリンジェントな条件下でハイブリダイズし得る、ヌクレオチド配列;
(g)配列番号2の配列の種ホモログをコードするヌクレオチド配列;および
(h)(a)〜(d)のいずれかにおいて特定されるヌクレオチド配列の対立遺伝子改変体、
からなる群より選択される。好ましくは、このヌクレオチド配列は、MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有するタンパク質をコードする。このヌクレオチド配列は、発現制御配列に作動可能に連結され得る。
【0006】
本発明はまた、アミノ酸配列を含むペプチドまたはタンパク質をコードするヌクレオチド配列を含む単離されたポリヌクレオチドも提供し、このアミノ酸配列は、以下:
(a)配列番号2のアミノ酸配列;
(b)配列番号2のアミノ酸22〜538のアミノ酸配列;
(c)配列番号2のアミノ酸22〜236のアミノ酸配列;
(d)配列番号2のアミノ酸1〜236のアミノ酸配列;および
(e)MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有する、(a)〜(d)のフラグメント、
からなる群より選択される。
【0007】
別の実施形態において、本発明は、ヌクレオチド配列を含む単離されたポリヌクレオチドを提供し、このヌクレオチド配列は、以下:
(a)配列番号9のヌクレオチド配列;
(b)(a)において特定されるヌクレオチド配列と、遺伝暗号の縮重の結果として異なる、ヌクレオチド配列;
(c)(a)において特定されるヌクレオチドに、ストリンジェントな条件下でハイブリダイズし得る、ヌクレオチド配列;
(d)(a)において特定されるヌクレオチド配列の対立遺伝子改変体、
からなる群より選択される。
【0008】
本発明はまた、アミノ酸配列を含むペプチドまたはタンパク質をコードするヌクレオチド配列を含む単離されたポリヌクレオチドを提供し、このアミノ酸配列は、以下:
(a)配列番号10のアミノ酸配列;
(b)MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有する、(a)のフラグメント、
からなる群より選択される。
【0009】
これらのポリヌクレオチドで形質転換された宿主細胞(好ましくは、哺乳動物細胞)もまた、提供される。
【0010】
他の実施形態において、本発明は、MU−1タンパク質を産生するためのプロセスを提供する。このプロセスは、以下:
(a)適切な培養培地において本発明の宿主細胞の培養物を増殖させる工程;および
(b)その培養物からヒトMU−1タンパク質を精製する工程、
を包含する。これらの方法により産生されたタンパク質もまた、提供される。
【0011】
本発明はまた、アミノ酸配列を含む単離されたMU−1タンパク質を提供し、このアミノ酸配列は、以下:
(a)配列番号2のアミノ酸配列;
(b)配列番号2のアミノ酸22〜538のアミノ酸配列;
(c)配列番号2のアミノ酸22〜236のアミノ酸配列;
(d)配列番号2のアミノ酸1〜236のアミノ酸配列;および
(e)MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有する、(a)〜(d)のフラグメント、
からなる群より選択される。
【0012】
本発明はまた、アミノ酸配列を含む単離されたMU−1タンパク質を提供し、このアミノ酸配列は、以下:
(a)配列番号10のアミノ酸配列;
(b)MU−1造血素レセプタースーパーファミリー鎖の生物学的活性を有する、(a)のフラグメント、
からなる群より選択される。
【0013】
マウスMU−1(配列番号10)のアミノ酸配列は、ヒトMU−1のアミノ酸配列と約65%同一である。
【0014】
他の好ましい実施形態において、特定のアミノ酸配列は、融合タンパク質の一部(MU−1に由来しないさらなるアミノ酸配列)である。好ましい融合タンパク質は、抗体フラグメント(例えば、Fcフラグメント)を含む。
【0015】
本発明のタンパク質および薬学的に受容可能なキャリアを含む薬学的組成物もまた、提供される。
【0016】
本発明はさらに、本発明のタンパク質と特異的に反応する抗体を含む組成物を提供する。
【0017】
1つの実施形態において、本発明のMU−1核酸分子は、配列番号1または9に示されるヌクレオチド配列に対して(例えば、全長ヌクレオチド配列に対して)、少なくとも65%、70%、75%、80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%またはそれ以上同一である。
【0018】
別の好ましい実施形態において、核酸分子は、配列番号1または9に示されるヌクレオチド配列からなる。別の好ましい実施形態において、核酸分子は、配列番号1または9のヌクレオチド配列のうちの少なくとも50ヌクレオチド、100ヌクレオチド、200ヌクレオチド、300ヌクレオチド、400ヌクレオチド、500ヌクレオチド、600ヌクレオチド、700ヌクレオチド、800ヌクレオチド、900ヌクレオチド、1000ヌクレオチド、1100ヌクレオチド、1200ヌクレオチド、1300ヌクレオチド、1400ヌクレオチド、1500ヌクレオチド、1600ヌクレオチド、1700ヌクレオチド、1800ヌクレオチドまたはそれ以上のヌクレオチド(例えば、連続するヌクレオチド)のフラグメント、あるいはその相補体を含む。
【0019】
好ましい実施形態において、MU−1タンパク質ファミリーメンバーは、配列番号2または10のアミノ酸配列に対して、少なくとも約65%、70%、75%、80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%またはそれ以上同一である、アミノ酸配列を有する。
【0020】
別の実施形態において、本発明は、配列番号2または10のアミノ酸配列を有するタンパク質のフラグメントを特徴とし、ここで、このフラグメントは、配列番号2または10のアミノ酸配列のうちの少なくとも15アミノ酸、20アミノ酸、30アミノ酸、40アミノ酸、50アミノ酸、60アミノ酸、70アミノ酸、80アミノ酸、90アミノ酸または100アミノ酸(例えば、連続するアミノ酸)を含む。
【0021】
別の局面において、本発明は、生物学的サンプルにおけるMU−1核酸分子、タンパク質またはポリペプチドの存在を検出するための方法を提供し、この方法は、MU−1核酸分子、MU−1タンパク質またはMU−1ポリペプチドがこの生物学的サンプルにおいて検出されるように、この生物学的サンプルに、MU−1核酸分子、MU−1タンパク質またはMU−1ポリペプチドを検出し得る因子を接触させることによる。
【0022】
別の局面において、本発明は、生物学的サンプルにおけるMU−1活性の存在を検出するための方法を提供し、この方法は、MU−1活性の存在がこの生物学的サンプルにおいて検出されるように、この生物学的サンプルに、MU−1活性の指標を検出し得る因子を接触させることによる。
【0023】
別の局面において、本発明は、MU−1活性を調節するための方法を提供し、この方法は、MU−1を発現し得る細胞においてMU−1活性が調節されるように、この細胞に、MU−1活性を調節する因子を接触させる工程を包含する。1つの実施形態において、この因子は、MU−1活性を阻害する。別の実施形態において、この因子は、MU−1活性を刺激する。1つの実施形態において、この因子は、MU−1タンパク質に特異的に結合する抗体である。別の実施形態において、この因子は、MU−1遺伝子の転写またはMU−1 mRNAの翻訳を調節することによって、MU−1の発現を調節する。なお別の実施形態において、この因子は、MU−1 mRNAまたはMU−1遺伝子のコード鎖に対してアンチセンスであるヌクレオチド配列を有する核酸分子である。
【0024】
1つの実施形態において、本発明の方法は、異常であるかまたは所望されない、MU−1タンパク質またはMU−1核酸の発現または活性によって特徴付けられる障害(例えば、MU−1関連障害)を有する被験体を、その被験体にMU−1モジュレーターである因子を投与することによって処置するために使用される。1つの実施形態において、MU−1モジュレーターは、MU−1タンパク質である。別の実施形態において、MU−1モジュレーターは、MU−1核酸分子である。なお別の実施形態において、MU−1モジュレーターは、ペプチド、ペプチド模倣物、抗体または他の低分子である。
【0025】
本発明はまた、以下の少なくとも1つによって特徴付けられる遺伝的変化の存在または非存在を同定するための診断アッセイを提供する:(i)MU−1タンパク質をコードする遺伝子の異常な修飾または変異;(ii)MU−1遺伝子の調節不全(mis-regulation);および(iii)MU−1タンパク質の異常な翻訳後修飾(ここで、この遺伝子の野生型形態は、MU−1活性を有するタンパク質をコードする)。
【0026】
別の局面において、本発明は、MU−1タンパク質に結合するかまたはMU−1タンパク質の活性を調節する、化合物を同定するための方法を提供し、この方法は、MU−1活性を有するMU−1タンパク質を含む指標組成物を提供する工程、この指標組成物に試験化合物を接触させる工程、および指標組成物におけるMU−1活性に対する試験化合物の効果(例えば、STATリン酸化(例えば、STAT3またはSTAT5リン酸化)の調節)を決定し、MU−1タンパク質の活性を調節する化合物を同定する工程による。
・本発明はまた、以下を提供し得る:
・(項目1) ヌクレオチド配列を含む単離されたポリヌクレオチドであって、上記ヌクレオチド配列は、以下:
(a)配列番号9のヌクレオチド配列;
(b)(a)において特定されるヌクレオチド配列の配列と、遺伝暗号の縮重の結果として異なる、ヌクレオチド配列;
(c)(a)において特定されるヌクレオチド配列に、ストリンジェントな条件下でハイブリダイズし得る、ヌクレオチド配列;
(d)配列番号10の配列の種ホモログをコードする、ヌクレオチド配列;および
(e)(a)において特定されるヌクレオチド配列の対立遺伝子改変体、
からなる群より選択される、ポリヌクレオチド。
・(項目2) 項目1に記載のポリヌクレオチドであって、上記ヌクレオチド配列が、MU−1の生物学的活性を有するタンパク質をコードする、ポリヌクレオチド。
・(項目3) 項目2に記載のポリヌクレオチドであって、上記生物学的活性が、STAT分子との相互作用である、ポリヌクレオチド。
・(項目4) 項目1に記載のポリヌクレオチドであって、上記ヌクレオチド配列が、発現制御配列に作動可能に連結されている、ポリヌクレオチド。
・(項目5) 項目1に記載のポリヌクレオチドであって、配列番号9のヌクレオチド配列を含む、ポリヌクレオチド。
・(項目6) 項目4に記載のポリヌクレオチドで形質転換された、宿主細胞。
・(項目7) 哺乳動物細胞である、項目6に記載の宿主細胞。
・(項目8) MU−1タンパク質を産生するためのプロセスであって、上記プロセスは、以下:
(a)適切な培養培地において項目6に記載の宿主細胞の培養物を増殖させる工程;および
(b)上記培養物から上記MU−1タンパク質を精製する工程、
を包含する、プロセス。
・(項目9) アミノ酸配列を含む単離されたMU−1タンパク質であって、上記アミノ酸配列は、以下:
(a)配列番号10のアミノ酸配列;および
(b)MU−1の生物学的活性を有する、(a)のフラグメント、
からなる群より選択される、タンパク質。
・(項目10) 配列番号10のアミノ酸配列を含む、項目9に記載のタンパク質。
・(項目11) 薬学的組成物であって、
項目9に記載のタンパク質と、
薬学的に受容可能なキャリアと、
を含む、組成物。
・(項目12) 項目8に記載のプロセスにより産生された、タンパク質。
・(項目13) 抗体を含む組成物であって、上記抗体は、項目9に記載のタンパク質と特異的に反応する、組成物。
・(項目14) アミノ酸配列を含むペプチドまたはタンパク質をコードするヌクレオチド配列を含む単離されたポリヌクレオチドであって、上記アミノ酸配列は、以下:
(a)配列番号10のアミノ酸配列;および
(b)MU−1の生物学的活性を有する、(a)のフラグメント、
からなる群より選択される、ポリヌクレオチド。
・(項目15) 項目9に記載のタンパク質であって、上記アミノ酸配列が融合タンパク質の一部である、タンパク質。
・(項目16) Fcフラグメントを含む、項目15に記載のタンパク質。
・(項目17) 項目14に記載のポリヌクレオチドであって、配列番号10のアミノ酸配列を含むペプチドまたはタンパク質をコードする、ポリヌクレオチド。
・(項目18) 配列番号9のヌクレオチド配列を含む単離されたポリヌクレオチドであって、STAT分子と相互作用するMU−1タンパク質をコードする、ポリヌクレオチド。
・(項目19) STATシグナル伝達経路を調節するための方法であって、
STATシグナル伝達に関与する細胞を、MU−1活性を調節し得る因子と接触させ、それによりSTATシグナル伝達経路を調節する工程、
を包含する、方法。
・(項目20) 項目19に記載の方法であって、上記STATシグナル伝達経路が、STAT 3シグナル伝達経路であるか、またはSTAT 5シグナル伝達経路である、方法。
・(項目21) 免疫障害を処置し得る化合物を同定するための方法であって、
STAT分子とMU−1ポリペプチドとの相互作用を上記化合物が調節する能力をアッセイし、それにより免疫障害を処置し得る化合物を同定する工程、
を包含する、方法。
・(項目22) 項目21に記載の方法であって、上記STATシグナル伝達経路が、STAT 3シグナル伝達経路であるか、またはSTAT 5シグナル伝達経路である、方法。
【発明を実施するための最良の形態】
【0027】
(好ましい実施形態の詳細な説明)
本出願の発明者は、MU−1造血素レセプタースーパーファミリー鎖(本明細書中以後、「MU−1」または「MU−1タンパク質」)をコードするポリヌクレオチド(ヒトおよびマウスMU−1をコードするポリヌクレオチドが挙げられるが、これらに限定されない)を、初めて同定および提供した。
【0028】
特に好ましい実施形態において、本発明のMU−1タンパク質およびMU−1核酸分子は、ヒトMU−1分子である。ヒトIL5レセプターの70アミノ酸領域
【0029】
【化1】
を用いて、TBLASTNアルゴリズムを使用してGenBank ESTデータベースを検索した。ヒト染色体16p12由来のゲノムBACクローンAC002303中の配列は、この領域に対する相同性を有することが同定され、これは、このセグメントが新規造血素レセプターの遺伝子をコードすることを示唆する。ヌクレオチド40,886の1000bp中のオープンリーディングフレームの試験は、GenPeptのBLASTP検索を使用した場合にサイトカインレセプターファミリーのメンバーを専ら同定した、270bpのオープンリーディングフレームを明らかにした。このリーディングフレームの末端の終止コドンは、エキソン/イントロン境界にまたがるトランジションの表れであると解釈された。
【0030】
次いで、RNAが、第16p12染色体由来のこのBACクローン内に含まれる遺伝子から転写されるか否かを決定した。PCRプライマーを、サイトカインレセプターファミリー内で保存されるペプチド配列を含む最大のORFセグメントに基づいて合成した。プライマーGAGTCCGAGGAGAAAGCTGATCTCA(5p)(配列番号4)およびプライマーGAAAGATGACCGGGTCACTCCATT(3p)(配列番号5)をPCRにおいて用いて、種々のヒト組織由来のファージライブラリー(Clontech)をスクリーニングした。配列ACTCGAGCTATGAGCTGCAGGTGCGGGCA(配列番号6)の32−P標識オリゴヌクレオチドに特異的にハイブリダイズする、予測された164bpサイズのPCR産物が、肺、腎臓、胎盤および心臓由来のファージにおいて観察された。オリゴヌクレオチドACTCGAGCTATGAGCTGCAGGTGCGGGCA(配列番号7)を用いて、全長のcDNAクローンNN14−1b(MU−1)が同定され、これを精製し、そして配列決定した。DNA配列および推定アミノ酸配列をそれぞれ、配列番号1および配列番号2に示す。ヒトMU−1レセプター鎖の推定アミノ酸配列は、アミノ酸1〜21の推定シグナル配列を含む。成熟ヒトMU−1は、配列番号2のアミノ酸24〜538の配列を有すると考えられる。膜貫通ドメインは、アミノ酸237〜254に見出される。
【0031】
別の特に好ましい実施形態では、本発明のMU−1タンパク質分子およびMU−1核酸分子は、マウスMU−1分子である。マウスMU−1タンパク質をコードするポリヌクレオチド配列を同定するために、MU−1レセプターのマウスホモログの部分フラグメントを、実施例1に記載されるように、ヒト配列から誘導されたオリゴヌクレオチドを用いるPCRによってマウスcDNAから単離した。このフラグメントのDNA配列を決定し、そして2つのオリゴヌクレオチドを、以下の配列を用いて、このフラグメントの内部部分から誘導した:
TTGAACGTGACTGTGGCCTT(5p)(配列番号13)
TGAATGAAGTGCCTGGCTGA(3p)(配列番号14)。
これらのオリゴヌクレオチドを用いて、元のPCR産物(図1および配列番号9のマウスcDNA配列のヌクレオチド781〜1043に対応する)の内部の262ヌクレオチドフラグメントを増幅し、2D6 T細胞株から単離されたcDNAライブラリーをスクリーニングするためのハイブリダイゼーションプローブとして用いた。DNA配列を、2つの独立したクローンから決定した。クローン6を配列決定し、そしてヒトMU−1の全長マウスホモログであることを確認した。
【0032】
マウスMU−1の全長ヌクレオチド配列を、図1(配列番号9のヌクレオチド1〜2628に対応する)に示す。このヌクレオチド配列は、推定リーダー配列をヌクレオチド407〜464に、コード配列をヌクレオチド407〜1993に、そして終結コドンをヌクレオチド1994〜1997に有する。ヌクレオチド1〜406は、5’非翻訳領域に対応し、そしてヌクレオチド1998〜2628は3’非翻訳領域に対応する。マウスMU−1の推定タンパク質配列を図2(配列番号10のアミノ酸1〜529に対応する)に示す。
【0033】
このマウスMU−1タンパク質は、SPScan(スコア=10.1)によって決定したところ、推定リーダー配列(配列番号10のアミノ酸1〜19に対応する)、および推定膜貫通ドメイン(配列番号10のアミノ酸237〜253に対応する)を含む。推定シグナルモチーフは、以下の領域を含む:ボックス1:配列番号10のアミノ酸265〜274、ボックス2:配列番号10のアミノ酸310〜324、配列番号10の281位、319位、361位、368位、397位および510位の6個のチロシン残基。潜在的なSTATドッキング部位は、STAT 5:EDDGYPA、STAT 3:YLQRを含むがこれらに限定されない。
【0034】
MU−1のオープンリーディングフレームは、造血素レセプターファミリーの新規メンバーをコードする。好ましい実施形態では、MU−1は、リーダー配列、保存されたシステイン対、このファミリーに特有のPPモチーフおよびWSXWS(配列番号8)モチーフ、ならびに膜貫通ドメインおよび広範囲にわたる細胞質ドメインを有する。MU−1はまた、細胞質ドメイン中に、保存されたPXPPならびにボックスIおよびボックスIIのシグナル伝達モチーフを含む(図5を参照のこと)。これらのドメインは、マウスMU−1とヒトMU−1との間で保存される。その後のGenPeptを用いたMU−1配列のFASTA整列は、ヒトIL−2Rbと最大の相同性を示した(図5を参照のこと)。
【0035】
ヒトMU−1 cDNAを、登録番号ATCC 98687として1998年3月10日にAmerican Type Culture Collectionに寄託した。
【0036】
ノーザン分析によって、実施例4に記載されるように、マウスMU−1は、成体マウスの脾臓組織、肺組織および心臓組織において検出された。ヒトMU−1は、成体ヒトのリンパ組織、PBL、胸腺、脾臓およびリンパ節において、ならびに胎児肺において検出された。
【0037】
全長よりも短い任意の形態のMU−1タンパク質は、本発明の範囲内に包含され、そして全長および成熟形態とともに本明細書中で集合的に「MU−1」または「MU−1タンパク質」と呼ばれる。全長よりも短いMU−1タンパク質は、全長MU−1タンパク質(配列番号4または配列番号6)をコードするポリヌクレオチドの対応するフラグメントを発現させることによって産生され得る。これらの対応するポリヌクレオチドフラグメントもまた、本発明の一部である。上記の通りの改変されたポリヌクレオチドは、標準的な分子生物学的技術(適切な所望の欠失変異体の構築を含む)によって、部位特異的変異誘発法、または適切なオリゴヌクレオチドプライマーを使用するポリメラーゼ連鎖反応によって作製され得る。
【0038】
本発明の目的のために、タンパク質は、対応する成熟MU−1タンパク質の1以上の生物学的活性を保有する場合、「MU−1造血素レセプタースーパーファミリー鎖の生物学的活性」を有する。1つの実施形態では、MU−1活性は、STAT分子(例えば、STAT 5、STAT 3)との相互作用を含む。別の実施形態では、MU−1タンパク質の活性は、いかなる公知のサイトカインへの結合も含まない。
【0039】
MU−1またはその活性なフラグメント(MU−1タンパク質)は、キャリア分子(例えば、免疫グロブリンまたは免疫グロブリンフラグメント)に融合され得る。例えば、可溶性形態のMU−1は、「リンカー」配列を通して、免疫グロブリンのFc部分へと融合され得る。他の融合タンパク質(例えば、GST、Lex−AまたはMBPを有する融合タンパク質)もまた用いられ得る。
【0040】
本発明はまた、配列番号1および配列番号9に示すようなヌクレオチド配列の対立遺伝子改変体(すなわち、MU−1タンパク質(好ましくはMU−1の生物学的活性を有するタンパク質)をまたコードする、配列番号1または配列番号9の天然に存在する選択的形態の単離されたポリヌクレオチド)を包含する。本発明にはまた包含されるのは、配列番号1または配列番号9に示されるヌクレオチド配列に、非常にストリンジェントな条件(例えば、65℃で0.1×SSC)下でハイブリダイズする、単離されたポリヌクレオチドである。MU−1タンパク質をコードするが、遺伝暗号の縮重によって、配列番号1または配列番号9に示されるヌクレオチド配列とは異なる、単離されたポリヌクレオチドもまた本発明によって包含される。点変異または誘導改変によって引き起こされる、配列番号1または配列番号9に示されるようなヌクレオチド配列におけるバリエーションもまた、本発明に包含される。
【0041】
本明細書中で使用される場合、用語「ストリンジェントな条件下でハイブリダイズする」は、互いに少なくとも60%同一なヌクレオチド配列が代表的に、互いにハイブリダイズしたままである、ハイブリダイゼーションおよび洗浄についての条件を記載することを意図する。好ましくは、この条件は、互いに少なくとも約70%、より好ましくは少なくとも約80%、さらにより好ましくは少なくとも約85%または90%同一な配列が代表的に、互いにハイブリダイズしたままである条件である。このようなストリンジェントな条件は当業者に公知であり、そしてCurrent Protocols in Molecular Biology,John Wiley & Sons,N.Y.(1989),6.3.1−6.3.6に見出され得る。ストリンジェントなハイブリダイゼーション条件の好ましい非限定的な例は、約45℃での6×塩化ナトリウム/クエン酸ナトリウム(SSC)中でのハイブリダイゼーション、続いて0.2×SSC、0.1% SDS中での50℃(好ましくは55℃、より好ましくは60℃、そしてさらにより好ましくは65℃)での1回以上の洗浄である。上記で記載した値にわたる範囲(例えば、55℃〜60℃または50℃〜65℃)は、本発明によって包含される。好ましくは、配列番号1または配列番号9の配列に、ストリンジェントな条件下でハイブリダイズする、本発明の単離された核酸分子は、天然に存在する核酸分子に対応する。本明細書中で使用される場合、「天然に存在する」核酸分子とは、天然に存在する(例えば、天然タンパク質をコードする)ヌクレオチド配列を有する、RNA分子またはDNA分子をいう。
【0042】
2つのアミノ酸配列または2つの核酸配列の同一性%を決定するために、これらの配列は、最適な比較目的のために整列される(例えば、第一および第二のアミノ酸配列または核酸配列の一方または両方に、最適な整列のためにギャップが導入され得、そして同一でない配列は、比較目的のために無視され得る)。好ましい実施形態では、比較目的のために整列された参照配列の長さは、参照配列の長さの少なくとも30%、好ましくは少なくとも40%、より好ましくは少なくとも50%、さらにより好ましくは少なくとも60%、そしてさらにより好ましくは少なくとも70%、80%、または90%である。次いで、対応するアミノ酸位置もしくはヌクレオチド位置でのアミノ酸残基またはヌクレオチドが比較される。第一の配列における位置が、第二の配列における対応する位置と同じアミノ酸残基またはヌクレオチドで占められていたならば、この分子はその位置で同一である(本明細書中で使用される場合、アミノ酸または核酸の「同一性」は、アミノ酸または核酸の「相同性」と等価である)。2つの配列間の同一性パーセントは、2つの配列の最適な整列のために導入する必要があるギャップの数および各ギャップの長さを考慮に入れた、これらの配列によって共有される同一位置数の関数である。
【0043】
2つの配列間の配列比較および同一性パーセントの決定は、数理的アルゴリズムを使用して達成され得る。好ましい実施形態において、2つのアミノ酸配列間の同一性パーセントは、GCGソフトウェアパッケージ(http://www.gcg.comにおいて入手可能)中のGAPプログラム中に組込まれたNeedlemanおよびWunsch(J.Mol.Biol.(48):444−453(1970))アルゴリズムを用い、Blosum 62マトリックスまたはPAM250マトリックスのいずれか、ならびにギャップウェイト(gap weight)=16、14、12、10、8、6、または4、およびレングスウェイト(length weight)=1、2、3、4、5、または6を使用して決定される。さらに別の好ましい実施形態において、2つのヌクレオチド配列間の同一性パーセントは、GCGソフトウェアパッケージ(http://www.gcg.comにおいて入手可能)中のGAPプログラムを用い、NWSgapdna.CMPマトリックス、およびギャップウェイト(gap weight)=40、50、60、70、または80、およびレングスウェイト(length weight)=1、2、3、4、5、または6を使用して決定される。別の実施形態において、2つのアミノ酸配列間または2つのヌクレオチド配列間の同一性パーセントは、ALIGNプログラム(バージョン2.0)中に組込まれたE.MeyersおよびW.Miller(MyersおよびMiller,1988,Comput.Appl.Biosci.4:11−17)のアルゴリズムを用い、PAM120ウェイト残基表(weight resiue table)、ギャップレングスペナルティ(gap length penalty)=12、およびギャップペナルティ(gap penalty)=4を使用して決定される。
【0044】
本発明の核酸配列およびタンパク質配列はさらに、公共のデータベースに対する検索を実行するための「問い合わせ配列」としてさらに使用されて、例えば、他のファミリーのメンバーまたは関連する配列を同定し得る。このような検索はAltschulら((1990)J.Mol.Biol.215:403−10)のNBLASTプログラムおよびXBLASTプログラム(バージョン2.0)を用いて実行され得る。BLASTヌクレオチド検索は、NBLASTプログラム、スコア(score)=100、ワードレングス(wordlength)=12を用いて実行されて、本発明のAdhr−1核酸分子に相同性のヌクレオチド配列を獲得し得る。BLASTタンパク質検索は、XBLASTプログラム、スコア(score)=100、ワードレングス(wordlength)=3を用いて実行されて、本発明のAdhr−1タンパク質分子に相同性のアミノ酸配列を獲得し得る。比較目的のためにギャップの入った整列を獲得するために、Gapped BLASTを、Altschulら(1997)Nucleic Acids Res.25(17):3389−3402に記載されるように利用し得る。BLASTプログラムおよびGapped BLASTプログラムを利用する場合、それぞれのプログラム(例えば、XBLASTおよびNBLAST)のデフォルトパラメーターを使用し得る。http://www.ncbi.nlm.nih.gov.を参照のこと。
【0045】
本発明はまた、他の動物種(特に、他の哺乳動物種)由来のヒトMU−1のホモログをコードするポリヌクレオチドを提供する。種ホモログは、本明細書中に開示されるマウス配列またはヒト配列からプローブまたはプライマーを作製し、そして適切な種由来のライブラリー(例えば、関連する種のPBMC、胸腺または精巣から構築されたライブラリーなど)をスクリーニングすることによって同定および単離され得る。
【0046】
本発明の単離されたポリヌクレオチドは、MU−1タンパク質を組み換え産生するために、Kaufmanら、Nucleic Acids Res.19,4485−4490(1991)に開示されるpMT2発現ベクターまたはpED発現ベクターのような発現制御配列に作動可能に連結され得る。多くの適切な発現制御配列が、当該分野で公知である。組換えタンパク質を発現させる一般的な方法もまた公知であり、そしてR.Kaufman、Methods in Enzymology 185,537−566(1990)に例示される。本明細書中に規定されるように、「作動可能に連結された」は、連結されたポリヌクレオチド/発現制御配列を用いて形質転換(トランスフェクト)された宿主細胞によってMU−1タンパク質が発現されるような様式で、本発明の単離されたポリヌクレオチドと発現制御配列との間に共有結合を形成するように酵素的または化学的に連結されることを意味する。
【0047】
多数の型の細胞が、MU−1タンパク質の発現のための適切な宿主細胞として作用し得る。機能的なMU−1タンパク質を発現し得る任意の細胞型が、使用され得る。適切な哺乳動物宿主細胞としては、例えば、サルCOS細胞、チャイニーズハムスター卵巣(CHO)細胞、ヒト腎臓293細胞、ヒト上皮A431細胞、ヒトColo205細胞、3T3細胞、CV−1細胞、他の形質転換された霊長類細胞株、正常複相体細胞、インビトロ初代培養の組織由来の細胞株、初代外植片、HeLa細胞、マウスL細胞、BHK細胞、HL−60細胞、U937細胞、HaK細胞、Rat2細胞、BaF3細胞、32D細胞、FDCP−1細胞、PC12細胞、M1x細胞、C2C12細胞が挙げられる。
【0048】
MU−1タンパク質はまた、1つ以上の昆虫発現ベクターにおいて本発明の単離されたポリヌクレオチドを適切な制御配列に作動可能に連結し、そして昆虫発現系を使用することによって、産生され得る。バキュロウイルス/昆虫細胞発現系についての材料および方法は、例えば、Invitrogen,San Diego,California,U.S.A.からのキットの形態(MaxBac(登録商標)キット)で市販され、そしてそのような方法は、SummersおよびSmith,Texas Agricultural Experiment Station Bulletin No.1555(1987)(本明細書中に参考として援用される)に記載されるように当該分野で周知である。MU−1タンパク質の可溶性形態がまた、上記のような適切な単離ポリヌクレオチドを用いて昆虫細胞中で産生され得る。
【0049】
あるいは、MU−1タンパク質は、酵母のような下等真核生物または細菌のような原核生物中で産生され得る。適切な酵母株としては、Saccharomyces cerevisiae、Schizosaccharomyces pombe、Kluyveromyces株、Candida、または異種タンパク質を発現し得る任意の酵母株が挙げられる。適切な細菌株としては、Escherichia coli、Bacillus subtilis、Salmonella typhimurium、または異種タンパク質を発現し得る任意の細菌株が挙げられる。
【0050】
細菌中での発現は、組換えタンパク質を組込んだ封入体の形成をもたらし得る。従って、活性な材料またはより活性な材料を産生するために、組換えタンパク質の再折り畳みが、必要であり得る。正確に折り畳まれた異種タンパク質を細菌封入体から得るためのいくつかの方法が、当該分野で公知である。これらの方法は一般的に、タンパク質を封入体から可溶化する工程、次いで、カオトロピック薬剤を使用してこのタンパク質を完全に変性する工程を包含する。システイン残基がタンパク質の一次アミノ酸配列中に存在する場合、ジスルフィド結合の正確な形成を可能にする環境下(酸化還元系)で再折り畳みを達成することが、しばしば必要である。再折り畳みの一般的な方法は、Kohno,Meth.Enzym.,185:187−195(1990)に開示される。EP 0433225および同時係属出願USSN 08/163,877は、他の適切な方法を記載する。
【0051】
本発明のMU−1タンパク質はまた、トランスジェニック動物の生成物として、例えば、MU−1タンパク質をコードするポリヌクレオチド配列を含む体細胞または生殖細胞によって特徴付けられる、トランスジェニックウシ、トランスジェニックヤギ、トランスジェニックブタ、またはトランスジェニックヒツジの乳汁成分として発現され得る。
【0052】
本発明のMU−1タンパク質は、所望のタンパク質を発現するのに必要な培養条件下で形質転換された培養宿主細胞を増殖させることによって調製され得る。次いで、生じる発現タンパク質は、この培養培地または細胞抽出物から精製され得る。本発明のMU−1タンパク質の可溶性形態は、馴化培地から精製され得る。本発明のMU−1タンパク質の膜結合形態は、発現している細胞から総膜画分を調製し、そして非イオン性変性剤(例えば、Triton X−100)を用いてこの膜を抽出することによって精製され得る。
【0053】
MU−1タンパク質は、当業者に公知の方法を用いて精製され得る。例えば、本発明のMU−1タンパク質は、市販のタンパク質濃縮フィルター(例えば、AmiconまたはMillipore Pellicon限外濾過ユニット)を用いて濃縮され得る。濃縮工程後、濃縮物は、ゲル濾過媒体のような精製マトリックスにアプライされ得る。あるいは、陰イオン交換樹脂(例えば、ペンダントジエチルアミノエチル(DEAE)基もしくはポリエチレンイミン(PEI)基を有するマトリックスまたは基材)が、使用され得る。マトリックスは、アクリルアミド、アガロース、デキストラン、セルロース、またはタンパク質精製に一般的に使用される他の型であり得る。あるいは、陽イオン交換工程が、使用され得る。適切な陽イオン交換体としては、スルホプロピル基またはカルボキシメチル基を含む種々の不溶性マトリックスが挙げられる。スルホプロピル基(例えば、S−Sepharose(登録商標)カラム)が、好ましい。培養上清からのMU−1タンパク質の精製はまた、コンカナバリンA−アガロース、heparin−toyopearl(登録商標)またはCibacrom blue 3GA Sepharose(登録商標)のようなアフィニティー樹脂で;あるいは、フェニルエーテル、ブチルエーテル、またはプロピルエーテルのような樹脂を用いた疎水性相互作用クロマトグラフィーによる;あるいは、免疫アフィニティークロマトグラフィーによる、1つ以上のカラム工程が挙げられ得る。最後に、疎水性RP−HPLC媒体(例えば、ペンダントメチル基または他の脂肪族基を有するシリカゲル)を使用する1つ以上の逆相高速液体クロマトグラフィー(RP−HPLC)工程が、MU−1タンパク質をさらに精製するために使用され得る。MU−1タンパク質に対する抗体を含むアフィニティーカラムもまた、公知の方法に従って精製に使用され得る。種々の組合わせまたは他の公知の方法における、上記の精製工程のうちのいくつかまたは全てがまた使用されて、実質的に精製された単離組み換えタンパク質を提供し得る。好ましくは、単離されたMU−1タンパク質は、他の哺乳動物タンパク質を実質的に含まないように、精製される。
【0054】
本発明のMU−1タンパク質はまた、MU−1に結合し得る因子をスクリーニングするために使用され得る。所望の結合タンパク質(固定されているかまたは固定されていない)を用いた結合アッセイが、当該分野で周知であり、そして本発明のMU−1タンパク質を使用する本目的のために使用され得る。精製された細胞に基づくスクリーニングアッセイまたは精製されたタンパク質に基づく(無細胞)スクリーニングアッセイが、このような因子を同定するために使用され得る。例えば、MU−1タンパク質は、精製形態でキャリアに固定され得、そして精製されたMU−1タンパク質に対する結合または潜在的なリガンドが、測定され得る。
【0055】
MU−1タンパク質(細胞から精製されたか、または組み換え生成された)は、薬学的に受容可能なキャリアと組み合わせた場合に、薬学的組成物として用いられ得る。このような組成物は、MU−1またはインヒビターおよびキャリアに加えて、種々の希釈剤、充填剤、塩、緩衝液、安定化剤、可溶化剤、および当該分野で周知の他の物質を含み得る。用語、「薬学的に受容可能な」とは、活性成分の生物学的活性の有効性を妨害しない非毒性の物質を意味する。キャリアの特徴は、投与の経路に依存する。
【0056】
本発明の薬学的組成物はまた、サイトカイン、リンホカイン、またはM−CSF、GM−CSF、IL−1、IL−2、IL−3、IL−4、IL−5、IL−6、IL−7、IL−8、IL−9、IL−10、IL−11、IL−12、IL−14、IL−15、G−CSF、幹細胞因子、およびエリスロポエチンのような他の造血因子を含み得る。この薬学的組成物はまた、抗サイトカイン抗体も含み得る。この薬学的組成物は、プラスミノゲン活性化因子および第VIII因子のような血栓溶解性因子または抗血栓性因子を含み得る。この薬学的組成物はさらに、他の抗炎症性因子を含み得る。このようなさらなる因子(factor)および/または物質(agent)は、この薬学的組成物中に含まれて、単離されたMU−1タンパク質と相乗効果を生じ得るか、または単離されたMU−1タンパク質によって生じる副作用を最小にし得る。逆に、単離されたMU−1タンパク質は、特定のサイトカイン、リンホカイン、他の造血因子、血栓溶解因子もしくは抗血栓性因子、または抗炎症剤の処方物に含まれて、サイトカイン、リンホカイン、他の造血因子、血栓溶解因子もしくは抗血栓性因子、または抗炎症剤の副作用を最小限にし得る。
【0057】
本発明の薬学的組成物は、単離されたMU−1タンパク質が、他の薬学的に受容可能なキャリアに加えて、水溶液中に、ミセルとして凝集形態で存在する脂質のような両親媒性因子と組み合わせられるリポソーム、不溶性単層、液晶、または層状層(lamellar layer)の形態中に存在し得る。リポソーム処方物のために適切な脂質としては、限定はしないが、モノグリセリド、ジグリセリド、スルファチド、リゾレシチン、リン脂質、サポニン、胆汁酸などが挙げられるがこれらに限定されない。このようなリポソーム処方物の調製は、例えば、米国特許第4,235,871号;米国特許第4,501,728号;米国特許第4,837,028号;および米国特許第4,737,323号(これらの全てが、本明細書において参考として援用されている)に記載のように、当業者のレベルの範囲内である。
【0058】
本明細書において用いる場合、用語「治療的有効量」とは、意味のある患者の利点(例えば、このような状態の症状の改善、このような状態の治癒、またはこのような状態の治癒率の上昇)を示すのに十分である、この薬学的組成物の各々の活性成分の総量または方法を意味する。単独で投与された、個々の活性成分に対して適用される場合、この用語は、その成分単独をいう。組み合わせ(併用)に対して適用される場合、この用語は、組み合わせて投与されても、連続して投与されても、または同時に投与されても、治療効果を生じる活性成分の合わせた量をいう。
【0059】
本発明の処置または使用の方法の実施において、治療的有効量の単離されたMU−Iタンパク質が哺乳動物に投与される。単離されたMU−1タンパク質は、単独でか、または他の治療(例えば、サイトカイン、リンホカイン、または他の造血因子を使用する処置)と組み合わせてのいずれかで、本発明の方法に従って投与され得る。1つ以上のサイトカイン、リンホカイン、または他の造血因子とともに同時投与される場合、MU−1タンパク質は、サイトカイン、リンホカイン、他の造血因子、血栓溶解因子もしくは抗血栓性因子と同時にか、または連続してのいずれかで投与され得る。連続的に投与される場合、主治医は、サイトカイン、リンホカイン、他の造血因子、血栓溶解因子もしくは抗血栓性因子と組み合わせた、MU−1タンパク質投与の適切な順序を決定する。
【0060】
薬学的組成物において用いられるMU−1タンパク質の投与、または本発明の本発明法を実施するために用いられるMU−1タンパク質の投与は、従来の種々の方法(例えば、経口摂取、吸引、または皮膚注射、皮下注射、もしくは静脈注射)で実行され得る。患者に対する静脈投与が好ましい。
【0061】
治療上有効な量のMU−1タンパク質を経口的に投与する場合、MU−1タンパク質は、錠剤、カプセル、粉末、溶液、またはエリキシルの形態である。錠剤形態で投与される場合、本発明の薬学的組成物は、ゼラチンまたはアジュバントのような固体キャリアをさらに含み得る。この錠剤、カプセル、および粉末は、約5〜95%のMU−1タンパク質、そして好ましくは、約25〜90%のMU−1タンパク質を含む。液体形態で投与される場合、液体キャリア(例えば、水、石油、動物もしくは植物起源の油脂(例えば、ピーナッツオイル、鉱油、ダイズ油、またはゴマ油)、または合成油)が添加され得る。この薬学的組成物の液体形態は、さらに、生理食塩水溶液、デキストロースもしくは他の糖溶液、またはグリコール(例えば、エチレングリコール、プロピレングリコール、またはポリエチレングリコール)を含み得る。液体形態で投与される場合、この薬学的組成物は、約0.5〜90重量%のMU−1タンパク質、そして好ましくは約1〜50%のMU−1タンパク質を含む。
【0062】
治療上有効な量のMU−1タンパク質を静脈注射、皮膚注射、または皮下注射によって投与する場合、MU−1タンパク質は、パイロジェン(発熱物質)のない、非経口的に受容可能な水溶液の形態である。このような非経口的に受容可能なタンパク質溶液(pHによって、等張性、安定性などを有する)の調製は、当該分野の範囲内である。静脈注射、皮膚注射、または皮下注射のために好ましい薬学的組成物は、MU−1タンパク質に加えて、等張性ビヒクル(例えば、注射用塩化ナトリウム、注射用リンゲル液、注射用デキストロース、注射用デキストロースおよび塩化ナトリウム、注射用乳酸リンゲル液、または当該分野で公知の他のビヒクル)を含むべきである。本発明の薬学的組成物はまた、安定化剤、保存剤、緩衝液、抗酸化剤、または当業者に公知の他の添加物を含み得る。
【0063】
本発明の薬学的組成物中のMU−1タンパク質の量は、処置される状態の性質および重篤度、ならびに患者が以前に受けた事前処置の性質に依存する。結局、主治医が、各々の個々の患者を処置するMU−1タンパク質の量を決定する。最初に、主治医は、低用量のMU−1タンパク質を投与して、患者の反応を観察する。MU−1タンパク質のさらに高い用量を、患者に至適の治療効果が得られるまで投与し得るが、その時点でこの投薬量は、さらに漸増されることはない。本発明の方法を実施するために用いられる種々の薬学的組成物は、体重1kgあたり、約0.1μg〜約100mgのMU−1タンパク質を含むべきことが意図される。
【0064】
本発明の薬学的組成物を用いる静脈内治療の期間は、処置される疾患の重篤度、ならびに各々の個々の患者の状態および可能性のある特有の応答に依存して変化する。MU−1タンパク質の各出願の期間は、12〜24時間の範囲の連続的静脈投与であることが意図される。結局、主治医は、本発明の薬学的組成物を用いて静脈治療の適切な期間を決定する。
【0065】
本発明のポリヌクレオチドおよびタンパク質は、以下に同定された、1つ以上の用途または生物学的活性(本明細書に言及されるアッセイに関連するものを含む)を示すと期待される。本発明のタンパク質について記載された用途または活性は、このようなタンパク質の投与もしくは使用によって、またはこのようなタンパク質をコードするポリヌクレオチドの投与もしくは使用によって(例えば、遺伝子治療において、またはDNAの導入に適切なベクターにおいて)、提供され得る。
【0066】
(サイトカインおよび細胞増殖/細胞分化活性)
本発明のタンパク質は、サイトカイン活性、細胞増殖(誘導または阻害のいずれか)活性、もしくは細胞分化(誘導または阻害のいずれか)活性を示し得るか、または特定の細胞集団における他のサイトカインの産生を誘導し得る。
【0067】
今日までに発見された多くのタンパク質因子(全ての公知のサイトカインを含む)は、1つ以上の因子依存性細胞増殖アッセイにおいて活性を示しており、従って、このアッセイは、サイトカイン活性の都合のよい確証として機能する。本発明のタンパク質の活性は、以下:32D、DA2、DA1G、T10、B9、B9/11、BaF3、MC9/G、M+(プレB M+)、2E8、RB5、DA1、123、T1165、HT2、CTLL2、TF−1、Mo7e、およびCMKを含むがこれらに限定されない細胞株についての多数の慣用的な因子依存性細胞増殖アッセイのいずれか1つによって証明されている。
【0068】
huEPOR−huMUキメラレセプターを発現するBAF−3細胞は、huEPOに応答して増殖する。MUレセプターが、その細胞質ドメインを通じてシグナル伝達する能力を試験するために、BAF−3細胞を操作して、EPOr/MU(cyto)キメラレセプターを発現し、そしてEPOの存在下における3Hチミジンの取り込みについてアッセイした。インタクトなEPOr分子を発現するBAF−3細胞は、EPOに応答して増殖するが、親のBAF−3細胞は増殖しない。A5クローン(キメラEPOr/MU(cyto)を保有する)は、EPOに応答して増殖し、このことは、MU−1の細胞質部分が、増殖シグナルを保持し得ることを実証する。BAF−3細胞(その表面上でEPOrを発現する)はまた、EPOに応答する。
【0069】
本発明のタンパク質の活性は、他にも方法はあるが、以下の方法によって測定され得る:
T細胞または胸腺細胞の増殖についてのアッセイとしては、以下に記載されるアッセイが、これらに制限することなく挙げられる:Current Protocols in Immunology,J.E.Coligan,A.M.Kruisbeek,D.H.Margulies,E.M.Shevach,W Strober編,Pub.Greene Publishing Associates and Wiley−Interscience(第3章、In Vitro assays for Mouse Lymphocyte Function 3.1−3.19;第7章、Immunologic studies in Humans);Takaiら、J.Immunol.137:3494−3500,1986;Bertagnolliら、J.Immunol.145:1706−1712,1990;Bertagnolliら、Cellular Immunology 133:327−341,1991;Bertagnolliら、J.Immunol.149:3778−3783,1992;Bowmanら、J.Immunol.152:1756−1761,1994。
【0070】
脾細胞、リンパ節細胞、または胸腺細胞のサイトカイン産生および/または増殖についてのアッセイとしては、以下に記載されるアッセイが、これらに制限することなく挙げられる:Polyclonal T cell stimulation,Kruisbeek,A.M.およびShevach,E.M.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、3.12.1−3.12.14頁、John Wiley and Sons,Toronto.1994;ならびにMeasurement of mouse and human Interferon γ,Schreiber,R.D.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、6.8.1−6.8.8頁、John Wiley and Sons,Toronto.1994。
【0071】
造血細胞およびリンパ球産生細胞の増殖および分化についてのアッセイとしては、以下に記載されるアッセイが、これらに制限することなく挙げられる:Measurement of Human and Murine Interleukin 2 and Interleukin 4,Bottomly,K.,Davis,L.S.およびLipsky,P.E.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、6.3.1−6.3.12頁、John Wiley and Sons,Toronto.1991;deVriesら、J.Exp.Med.173 :1205−1211,1991;Moreauら、Nature 336:690−692,1988;Greenbergerら、Proc.Natl.Acad.Sci.U.S.A.80:2931−2938,1983;Measurement of mouse and human interleukin 6−Nordan,R.Current Protocols
in Immunology.J.E.e.a.Coligan編、第1巻、6.6.1−6.6.5頁、John Wiley and Sons,Toronto.1991;Smithら、Proc.Natl.Acad.Sci.U.S.A.83:1857−1861,1986;Measurement of human Interleukin 11−Bennett,F.,Giannotti,J.,Clark,S.C.およびTurner,K.J.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、6.15.1頁、John Wiley and Sons,Toronto.1991;Measurement of
mouse and human Interleukin 9−Ciarletta,A.,Giannotti,J.,Clark,S.C.およびTurner,K.J.Current Protocols in Immunology.J.E.e.a.Coligan編、第1巻、6.13.1頁、John Wiley and Sons,Toronto.1991。
【0072】
抗原に対するT細胞クローンの応答についてのアッセイ(とりわけ、APC−T細胞相互作用に影響する、ならびにT細胞効果を指向するタンパク質を、増殖およびサイトカイン産生を測定することにより同定する)としては、以下に記載されるアッセイが、これらに制限することなく挙げられる:Current Protocols in Immunology,J.E.Coligan,A.M.Kruisbeek,D.H.Margulies,E.M.Shevach,W Strober編、Pub.Greene Publishing Associates and Wiley−Interscience(第3章、In Vitro assays for Mouse Lymphocyte Function;第6章、Cytokines and their cellular receptors;第7章、Immunologic studies in Humans);Weinbergerら、Proc.Natl.Acad.Sci.USA 77:6091−6095,1980;Weinbergerら、Eur.J.Immun.11:405−411,1981;Takaiら、J.Immunol.137:3494−3500,1986;Takaiら、J.Immunol.140:508−512,1988。
【0073】
(免疫刺激活性または免疫抑制活性)
本発明のタンパク質はまた、免疫刺激活性または免疫抑制活性(その活性に対するアッセイが本明細書中に記載される活性が挙げられるが、これらに限定されない)を示し得る。タンパク質は、種々の免疫不全および障害(重症複合型免疫不全(SCID)を含む)の処置において(例えば、Tリンパ球および/またはBリンパ球の成長および増殖の調節(上方または下方)、ならびにNK細胞および他の細胞集団の細胞溶解活性の誘導において)有用であり得る。これらの免疫不全は、遺伝的であり得るか、またはウイルス(例えば、HIV)および細菌もしくは真菌の感染によって引き起こされ得るか、あるいは自己免疫障害から生じ得る。より具体的には、ウイルス、細菌、真菌または他の感染(HIV、肝炎ウイルス、ヘルペスウイルス、ミコバクテリア、Leishmania spp.、マラリアspp.による感染、およびカンジダ症のような種々の真菌感染が挙げられる)によって引き起こされる感染性疾患は、本発明のタンパク質を使用して、処置可能であり得る。もちろん、これに関して、本発明のタンパク質はまた、免疫系に対するブーストが一般的に所望であり得る場合に(すなわち、癌の処置において)有用であり得る。
【0074】
本発明のタンパク質を使用して処置され得る自己免疫障害としては、例えば、結合組織疾患、多発性硬化症、全身性エリテマトーデス、慢性関節リウマチ、自己免疫肺炎症、ギヤン−バレー症候群、自己免疫性甲状腺炎、インスリン依存性糖尿病、筋無力症、対宿主性移植片病および自己免疫炎症性眼疾患が挙げられる。本発明のこのようなタンパク質はまた、アレルギー性の反応および状態(例えば、喘息(特に、アレルギー性喘息))または他の呼吸の問題の処置において、有用であり得る。免疫抑制が所望である他の状態(例えば、器官移植が挙げられる)もまた、本発明のタンパク質を使用して処置可能であり得る。
【0075】
MU−1 DNAはまた、クローン病に対する染色体遺伝子座をマッピングする。その結果、本発明のタンパク質は、クローン病および他の炎症性腸疾患を処置するために、使用され得る。
【0076】
本発明のタンパク質を使用して、免疫応答を多数の様式で調節することもまた可能であり得る。ダウンレギュレーションは、既に進行している免疫応答を阻害またはブロックする形態であり得るか、あるいは免疫応答の誘導の防止を包含し得る。活性化T細胞の機能は、T細胞応答を抑制することによってか、またはT細胞における特異的耐性を誘導することによってか、あるいはその両方によって、阻害され得る。T細胞応答の免疫抑制は、一般に、活性な、非抗原特異的なプロセスであり、このプロセスは、抑制因子に対するT細胞の連続的な曝露を必要とする。耐性(これは、T細胞における非応答性またはアネルギーの誘導を伴う)は、一般的に抗原特異性である点、および耐性化因子への曝露が終了した後にも持続する点で、免疫抑制から区別可能である。機能上、耐性は、耐性化因子の非存在下での特異的抗原への再曝露の際の、T細胞応答の欠如によって、実証され得る。
【0077】
1つ以上の抗原機能(Bリンパ球抗原機能(例えば、B7のような)が挙げられるが、これらに限定されない)のダウンレギュレーションまたは防止(例えば、活性化T細胞による高レベルのリンホカイン合成の防止)は、組織、皮膚および器官の移植の状況において、ならびに対宿主性移植片病(GVHD)において、有用である。例えば、T細胞機能のブロックは、組織移植において、組織破壊の低下を生じるはずである。代表的に、組織移植において、移植物の拒絶は、T細胞によるその移植物の外来物質としての認識を介して開始され、続いて、この移植物を破壊する免疫反応が起こる。B7リンパ球抗原の、その天然のリガンドとの相互作用を阻害またはブロックする分子を(例えば、B7−2活性を有するペプチドの可溶性の単量体形態を、単独でか、あるいは別のBリンパ球抗原(例えば、B7−1、B7−3)の活性を有するペプチドの単量体形態または遮断抗体と組合せて)免疫細胞に、移植前に投与することによって、その天然のリガンドに対するその分子の結合が、その免疫細胞において、対応する同時刺激信号を伝達することなくもたらされ得る。この様式で、Bリンパ球抗原機能をブロックすることにより、免疫細胞(例えば、T細胞)によるサイトカイン合成が防止され、従って、免疫抑制剤として作用する。さらに、同時刺激の欠如はまた、T細胞をアネルギー化し、これによって被験体において耐性を誘導するに十分であり得る。Bリンパ球抗原ブロック試薬による、長期間の耐性の誘導は、これらのブロッキング試薬のたびたびの投与の必要性を回避し得る。十分な免疫抑制または耐性を被験体において達成するためには、Bリンパ球抗原の組み合わせの機能をブロックすることもまた必要であり得る。
【0078】
器官移植拒絶またはGVHDの予防における、特定のブロッキング試薬の効力は、ヒトにおける効力を予測可能な動物モデルを使用して、評価され得る。使用され得る適切な系の例としては、ラットにおける同種異型の心臓移植片およびマウスにおける異種のランゲルハンス島細胞移植片が挙げられ、これらの両方は、Lenschowら、Science 257:789−792(1992)およびTurkaら、Proc.Natl.Acad.Sci USA,89:11102−11105(1992)に記載されるように、インビボでのCTLA4Ig融合タンパク質の免疫抑制効果を試験するために使用された。さらに、GVHDのマウスモデル(Paul編、Fundamental Immunology,Raven Press,New York,1989,846−847頁を参照のこと)は、インビボでのBリンパ球抗原機能のブロックの、この疾患の発達に対する影響を決定するために、使用され得る。
【0079】
抗原機能をブロックすることはまた、自己免疫疾患を処置するために治療的に有用であり得る。多くの自己免疫障害は、自己組織に対して反応性であり、そしてこれらの疾患の病理に関与するサイトカインおよび自己抗原の産生を促進する、T細胞の不適切な活性化の結果である。自己反応性T細胞の活性化の防止は、疾患の症状を低下または排除し得る。Bリンパ球抗原のレセプター:リガンド相互作用を妨害することによってT細胞の同時刺激をブロックする試薬の投与を使用して、T細胞の活性化を阻害し得、そしてこの疾患プロセスに関与し得る自己抗原またはT細胞由来のサイトカインの産生を防止し得る。さらに、ブロッキング試薬は、自己反応性T細胞の抗原特異的耐性を誘導し得、このことは、疾患の長期間の軽減をもたらし得る。自己免疫障害の予防または軽減におけるブロッキング試薬の効力は、ヒト自己免疫疾患の十分に特徴付けられた多数の動物モデルを使用して、決定され得る。例としては、マウス実験的自己免疫性脳脊髄炎、MRL/lpr/lprマウスまたはNZBハイブリッドマウスにおける全身性エリテマトーデス、マウス自己免疫性コラーゲン関節炎、NODマウスおよびBBラットにおける真性糖尿病、ならびにマウス実験的重症筋無力症が挙げられる(Paul編、Fundamental Immunology,Raven Press,New York,1989,840−856頁を参照のこと)。
【0080】
免疫応答をアップレギュレートする手段としての抗原機能(好ましくは、Bリンパ球抗原機能)のアップレギュレーションはまた、治療において有用であり得る。免疫応答のアップレギュレーションは、既存の免疫応答の増大または最初の免疫応答の誘発の形態であり得る。例えば、Bリンパ球抗原機能の刺激を介する免疫応答の増大は、ウイルス感染の場合に有用であり得る。さらに、全身性ウイルス疾患(例えば、インフルエンザ、感冒、および脳炎)は、Bリンパ球抗原の刺激性形態を全身的に投与することによって緩和され得る。
【0081】
あるいは、抗ウイルス免疫応答は、患者からのT細胞の除去、本発明のペプチドを発現するウイルス抗原パルスAPCで、または本発明の可溶性ペプチドの刺激性形態と共にウイルス抗原パルスAPCでのいずれかでの、インビトロでのT細胞の同時刺激、およびインビトロで活性化されたT細胞のこの患者への再導入によって、感染した患者において増大され得る。抗ウイルス免疫応答を増大する別の方法は、感染した細胞を患者から単離し、これらの細胞を、本明細書中に記載される本発明のタンパク質をコードする核酸で、この細胞がその表面上にこのタンパク質の全てまたは一部を発現するようにトランスフェクトし、そしてこのトランスフェクトした細胞を患者に再導入することである。感染した細胞は、ここで同時刺激シグナルをインビボでT細胞に送達し得、そしてこれによってT細胞を活性化し得る。
【0082】
別の適用において、抗原機能(好ましくは、Bリンパ球抗原機能)のアップレギュレーションまたは増大は、腫瘍免疫の誘導において有用であり得る。本発明の少なくとも1つのペプチドをコードする核酸でトランスフェクトされた腫瘍細胞(例えば、肉腫、黒色腫、リンパ腫、白血病、神経芽細胞腫、癌)は、被験体における腫瘍特異的耐性を克服するために、被験体に投与され得る。所望の場合、この腫瘍細胞は、ペプチドの組み合わせを発現するためにトランスフェクトされ得る。例えば、患者から得られる腫瘍細胞は、B7−2様活性を単独で有するペプチドの発現、またはB7−1様活性および/もしくはB7−3様活性を有するペプチドと関連した発現を指向する発現ベクターを用いて、エキソビボでトランスフェクトされ得る。このトランスフェクトされた腫瘍細胞は患者に戻されて、このトランスフェクトされた細胞の表面上でのこのペプチドの発現を生じる。あるいは、遺伝子治療技術が、インビボでのトランスフェクションのための腫瘍細胞を標的するために使用され得る。
【0083】
腫瘍細胞の表面上での、Bリンパ球抗原の活性を有する本発明のペプチドの存在は、トランスフェクトされた腫瘍細胞に対するT細胞媒介免疫応答を誘導するために必要な同時刺激シグナルを、T細胞に提供する。さらに、MHCクラスI分子またはMHCクラスII分子を欠いた腫瘍細胞、あるいは十分な量のMHCクラスI分子またはMHCクラスII分子を再発現できない腫瘍細胞は、MHCクラスIα鎖タンパク質およびβ2ミクログロブリンタンパク質またはMHCクラスIIα鎖タンパク質およびMHCクラスIIβ鎖タンパク質の全てまたは一部(例えば、細胞質ドメインの短縮部分)をコードする核酸でトランスフェクトされ得、これによってMHCクラスIまたはMHCクラスIIタンパク質を細胞表面に発現する。Bリンパ球抗原(例えば、B7−1、B7−2、B7−3)の活性を有するペプチドと関連しての、適切なクラスIまたはクラスII MHCの発現は、トランスフェクトされた腫瘍細胞に対するT細胞媒介免疫応答を誘導する。必要に応じて、MHCクラスII関連タンパク質(例えば、不変の鎖)の発現をブロックするアンチセンス構築物をコードする遺伝子はまた、Bリンパ球抗原の活性を有するペプチドをコードするDNAで同時トランスフェクトされ得、腫瘍関連抗原の提示を促進し、そして腫瘍特異的免疫を誘導する。従って、ヒト被験体におけるT細胞媒介免疫応答の誘導は、この被験体における腫瘍特異的耐性を克服するために十分であり得る。
【0084】
本発明のタンパク質の活性は、他の手段の中でも、以下の方法によって測定され得る:
胸腺細胞または脾細胞の細胞傷害性についての適切なアッセイとしては、以下に記載されるものが挙げられるが、これらに限定されない:Current Protocols in Immunology,J.E.Coligan編,A.M.Kruisbeek,D.H.Margulies,E.M.Shevach,W Strober,Pub.Greene Publishing Associates and Wiley−Interscience(Chapter 3,In Vitro assays for Mouse Lymphocyte Function 3.1−3.19;Chapter 7,Immunologic studies in Humans);Herrmannら,Proc.Natl.Acad.Sci.USA 78:2488−2492,1981;Herrmannら,J.Immunol.128:1968−1974,1982;Handaら,J.Immunol.135:1564−1572,1985;Takaiら,J.Immunol.137:3494−3500,1986;Takaiら,J.Immunol.140:508−512,1988;Herrmannら,Proc.Natl.Acad.Sci.USA 78:2488−2492,1981;Herrmannら,J.Immunol.128:1968−1974,1982;Handaら,J.Immunol.135:1564−1572,1985;Takaiら,J.Immunol.137:3494−3500,1986;Bowmanら,J.Virology 61:1992−1998;Takaiら,J.Immunol.140:508−512,1988;Bertagnolliら,Cellular Immunology 133:327−341,1991;Brownら,J.Immunol.153:3079−3092,1994。
【0085】
T細胞依存性免疫グロブリン応答およびアイソタイプのスイッチングについてのアッセイ(これは、とりわけ、T細胞依存性抗体応答を調節しそしてTh1/Th2プロフィールに影響するタンパク質を同定する)としては、以下に記載されるものが挙げられるが、これらに限定されない:Maliszewski,J.Immunol.144:3028−3033,1990;ならびにAssays for B cell function:In vitro antibody production,Mond,J.J.およびBrunswick,M.、Current Protocols in Immunology.J.E.e.a.Coligan編,Vol 1,pp.3.8.1−3.8.16,John Wiley and Sons,Toronto.1994。
【0086】
混合リンパ球反応(MLR)アッセイ(これは、とりわけ、主にTh1およびCTL応答を生成するタンパク質を同定する)としては、以下に記載されるものが挙げられるが、これらに限定されない:Current Protocols
in Immunology,J.E.Coligan編,A.M.Kruisbeek,D.H.Margulies,E.M.Shevach,W Strober,Pub.Greene Publishing Associates and Wiley−Interscience(Chapter 3,In Vitro assays for Mouse Lymphocyte
Function 3.1−3.19;Chapter 7,Immunologic studies in Humans);Takaiら,J.Immunol.137:3494−3500,1986;Takaiら,J.Immunol.140:508−512,1988;Bertagnolliら,J.Immunol.149:3778−3783,1992。
【0087】
樹状細胞依存性アッセイ(これは、とりわけ、ネイティブのT細胞を活性化する樹状細胞によって発現されるタンパク質を同定する)としては、以下に記載されるものが挙げられるが、これらに限定されない:Gueryら,J.Immunol.134:536−544,1995;Inabaら,Journal of Experimental Medicine 173:549−559,1991;Macatoniaら,Journal of Immunology 154:5071−5079,1995;Porgadorら,Journal of Experimental Medicine 182:255−260,1995;Nairら,Journal of Virology 67:4062−4069,1993;Huangら,Science 264:961−965,1994;Macatoniaら,Journal of Experimental Medicine 169:1255−1264,1989;Bhardwajら,Journal of Clinical Investigation 94:797−807,1994;およびInabaら,Journal of Experimental Medicine 172:631−640,1990。
【0088】
リンパ球の生存/アポトーシスについてのアッセイ(これは、とりわけ、スーパー抗原誘導後にアポトーシスを防止するタンパク質およびリンパ球のホメオスタシスを調節するタンパク質を同定する)としては、以下に記載されるものが挙げられるが、これらに限定されない:Darzynkiewiczら,Cytometry 13:795−808,1992;Gorczycaら,Leukemia 7:659−670,1993;Gorczycaら,Cancer
Research 53:1945−1951,1993;Itohら,Cell 66:233−243,1991;Zacharchuk,Journal of Immunology 145:4037−4045,1990;Zamaiら,Cytometry 14:891−897,1993;Gorczycaら,International Journal of Oncology 1:639−648,1992。
【0089】
T細胞の拘束および発生の初期段階に影響するタンパク質についてのアッセイとしては、以下に記載されるものが挙げられるが、これらに限定されない:Anticaら,Blood 84:111−117,1994;Fineら,Celluler Immunology 155:111−122,1994;Galyら,Blood 85:2770−2778,1995;Tokiら,Proc.Nat.Acad Sci.USA 88:7548−7551,1991。
【0090】
本発明のMU−1分子はまた、STAT活性(例えば、シグナル伝達)の調節(例えば、STAT分子(例えば、STAT3および/またはSTAT5)との相互作用を介する調節、およびSTATリン酸化の調節)に関与する。STATのシグナル伝達のモジュレーター、およびSTAT分子とのMU−1の相互作用のモジュレーターは、本明細書中に記載されるスクリーニングアッセイを介して同定され得る。
【0091】
MU−1活性のモジュレーターを同定するために使用され得るアッセイとしては、STAT活性(例えば、STATシグナル伝達)についてのアッセイ(例えば、当該分野で公知のアッセイおよび本明細書中に記載されるアッセイ)が挙げられる。STAT活性についてのアッセイはまた、例えば、Friedrich
K.ら(2001)Biol Chem 382(2):343−51に記載される。STAT活性についての他のアッセイとしては、細胞増殖およびSTATリン酸化についてのアッセイが挙げられる。
【0092】
1つの局面において、MU−1活性のモジュレーターについてのアッセイは、細胞ベースのアッセイ(ここで、MU−1タンパク質またはその生物学的に活性な部分を発現する細胞が、試験化合物と接触され、そしてこの試験化合物がMU−1活性を調節する能力(例えば、STAT分子との相互作用またはSTATシグナル伝達の調節)が決定される)である。試験化合物がMU−1活性を調節する能力の決定は、例えば、STATシグナル伝達、STAT分子のリン酸化、細胞増殖、細胞の分化、またはサイトカインの産生をモニターすることによって達成され得る。この細胞は、例えば、哺乳動物起源の細胞(例えば、活性化T細胞)であり得る。
【0093】
(造血調節活性)
本発明のタンパク質は、造血の調節において、従って骨髄性細胞またはリンパ球様細胞の欠失の処置において有用であり得る。コロニー形成細胞または因子依存性細胞株の支持における辺縁の生物学的な活性でさえ、以下の造血の調節への関与を示す:例えば、赤血球系前駆細胞の成長および増殖を単独または他のサイトカインと組み合わせて支持し、これにより、例えば、種々の貧血症の処置、または赤血球系前駆体細胞および/または赤血球系細胞の産生を刺激するための照射/化学療法と組み合わせた使用の有用性を示すこと;例えば、結果的に骨髄の抑制を予防または処置するための化学療法と組み合わせて有用な、骨髄性細胞(例えば、有用な顆粒球および単球/マクロファージ)の成長および増殖を支持すること(すなわち、古典的なCSF活性);巨核球および結果的に血小板の成長および増殖を支持し、これにより、血小板減少症のような種々の血小板障害の予防または処置、および一般に、血小板の輸血の代わりにまたは補充での使用を可能にすること;ならびに/または上記の任意および全ての造血細胞に成熟し得る造血幹細胞の成長および増殖を支持すること。これにより種々の幹細胞の障害(例えば、移植を用いて処置される障害(これには、再生不良性貧血および発作性夜間血色素尿症が挙げられるが、これらに限定されない))、および正常な細胞または遺伝子治療のために遺伝子操作された細胞として、インビボまたはエキソビボのいずれかで(すなわち、骨髄移植と組み合わせるかまたは辺縁の前駆体細胞(相同性または非相同性)の移植を用いて)、照射/化学療法後に幹細胞のコンパートメントを再増殖することでの治療利用を見出す。
【0094】
本発明のタンパク質の活性は、とりわけ、以下の方法によって測定され得る。
【0095】
種々の造血株の増殖および分化のための適切なアッセイは、上に挙げられる。
【0096】
胚幹細胞の分化についてのアッセイ(このアッセイにより、とりわけ、胚性分化造血に影響を及ぼすタンパク質を同定する)としては、以下に記載されるアッセイが挙げられるが、これらに限定されない:Johanssonら、Cellular Biology 15:141−151、1995;Kellerら、Molecular and Cellular Biology 13:473−486、1993;McClanahanら、Blood 81:2903−2915、1993。
【0097】
幹細胞の生存および分化についてのアッセイ(このアッセイにより、とりわけ、リンパ−造血を調節するタンパク質を同定する)としては、以下に記載されるアッセイが挙げられるが、これらに限定されない:Methylcellulose colony forming assays、Freshney、M.G.、Culture of Hematopoietic Cells.R.I.Freshneyら編、第265〜268頁、Wiely−Liss,Inc.、New York、NY.1994;Hirayamaら、Proc.Natl.Acad.Sci.USA 89:5907−5911、1992;Primitive hematopoietic colony forming cells with high proliferative potential、McNiece、I.K.およびBriddell,R.A.、Culture of Hematopoietic Cells、R.I.Freshneyら編、第23〜39頁、Wiley−Liss,Inc.、New
York,NY.1994;Nebenら、Experimental Hematology 22:353−359、1994;Cobblestone
area forming cell assay、Ploemacher、R.E.、Culture of Hematopoietic Cells.R.I.Freshneyら編、第1〜21頁、Wiley−Liss,Inc.、New York,NY.1994;Long term bone marrow cultures in the presence of stromal cells、Spooncer,E.、Dexter,M.およびAllen,T、Culture of Hematopoietic Cells、R.I.Freshneyら編、第163〜179頁、Wiley−Liss,Inc.、New York,NY.1994;Long term culture initiating cell assay、Sutherland,H.J.、Culture of Hematopoietic Cells、R.I.Freshneyら編、第139〜162頁、Wiley−Liss,Inc.、New York,NY.1994。
【0098】
(リサーチの使用および有用性)
本発明により提供されるポリヌクレオチドは、種々の目的のためにリサーチコミュニティによって使用され得る。ポリヌクレオチドは、分析、特徴付け、または治療に使用するための組換えタンパク質を発現するために;対応するタンパク質が優先的に発現される組織(構成的に、あるいは組織の分化もしくは発達または疾患状態の特定の段階のいずれか)のためのマーカーとして;サザンゲルにおける分子量マーカーとして;染色体を同定するためもしくは関連する遺伝子座マッピングするための染色体マーカーもしくはタグ(標識された場合)として;可能性のある遺伝子障害を同定するために、患者の内因性DNA配列と比較するため;ハイブリダイズするため、従って新規な関連するDNA配列を発見するためのプローブとして;遺伝子フィンガープリントのためのPCRプライマーを誘導するための情報の供給源として;他の新規なポリヌクレオチドを発見するプロセスにおいて、既知の配列を「差引きする(subtract−out)」ためのプローブとして;「遺伝子チップ」または他の支持体への連結のためにオリゴマーを選択し、そして作製するため(発現パターンの試験を含む);DNA免疫化技術を使用して抗タンパク質抗体を産生させるため;ならびに抗DNA抗体を産生するため、または別の免疫応答を惹起するための抗原として、使用され得る。ポリヌクレオチドが、別のタンパク質に(例えば、レセプター−リガンドの相互作用において)結合または部分的に結合するタンパク質をコードする場合、このポリヌクレオチドはまた、相互作用トラップアッセイ(例えば、Gyurisら、Cell 75:791−803(1993)に記載されるアッセイ)において、結合が生じる他のタンパク質をコードするポリヌクレオチドを同定するため、または結合相互作用のインヒビターを同定するために使用され得る。
【0099】
本発明によって提供されるタンパク質は、以下に挙げられる生物学的な活性を決定するためのアッセイにおいて同様に使用され得る:ハイスループットスクリーニングのための複数のタンパク質のパネル;抗体を産生するか、または別の免疫応答を惹起するため;生物学的流体中でのタンパク質(またはそのレセプター)のレベルを定量的に決定するように設計されたアッセイにおける試薬(標識化された試薬を含む)として;対応するタンパク質が優先的に発現される組織(構成的に、あるいは組織の分化もしくは発達または疾患状態の特定の段階でのいずれか)のためのマーカーとして;ならびに当然、相関するレセプターまたはリガンドを単離するため。タンパク質が、別のタンパク質に(例えば、レセプター−リガンド相互作用により)結合または部分的に結合する場合、このタンパク質は、結合が生じる他のタンパク質を同定するためか、または結合相互作用のインヒビターを同定するために使用され得る。これらの結合相互作用に関与するタンパク質はまた、この結合相互作用のペプチドまたは小分子のインヒビターもしくはアゴニストをスクリーニングするために使用され得る。
【0100】
これらのリサーチのいずれかまたは全ての有用性は、リサーチ製品として市販するための、試薬等級またはキットフォーマットに発展され得る。
【0101】
上に列挙される使用を実施するための方法は、当業者に周知である。このような方法を開示する参考文献としては、以下が挙げられるが、これらに限定されない:「Molecular Cloning:A Laboratory Manual」、第2版、Cold Spring Harbor Laboratory Press、Sambrook,J.、E.F.FritschおよびT.Maniatis編、1989、ならびに「Methods in Enzymology:Guide to Molecular Cloning Techniques」,Academic Press,Berger,S.L.およびA.R.Kimmel編、1987。
【0102】
(栄養性使用)
本発明のポリヌクレオチドおよびタンパク質はまた、栄養性供給源または補充物として使用され得る。このような使用は、以下が挙げられるが、これらに限定されない:タンパク質またはアミノ酸サプリメントとしての使用、炭素供給源としての使用、窒素供給源としての使用および炭化水素の供給源としての使用。このような場合において、本発明のタンパク質またはポリヌクレオチドは、特定の生物の飼料(フィード)に添加され得、あるいは例えば、粉末、ピル、溶液、懸濁物またはカプセルの形態のような、別の固体調製物または液体調製物として投与され得る。微生物の場合において、本発明のタンパク質およびポリヌクレオチドは、培地中に添加され得、この培地中または培地上で、微生物は培養される。
【0103】
本発明のMU−1タンパク質はまた、ポリクローナル抗体およびモノクローナル抗体を得るために動物を免疫するために使用され得、これらの抗体は、MU−1タンパク質と特異的に反応し、そしてこれらの抗体は、リガンドのレセプターに対する結合を阻害し得る。このような抗体は、免疫原として全MU−1を使用することによるか、またはMU−1のフラグメントを使用することによって得られ得る。このMU−1のより小さなフラグメントはまた、動物を免疫するために使用され得る。このペプチド免疫原は、カルボキシル末端にシステイン残基をされに含み得、そしてキーホールリンペットヘモシアニン(keyhole limpet hemocyanin(KLH))のようなハプテンに結合される。さらなるペプチド免疫原は、チロシン残基を硫酸化されたチロシン残基を用いて置換することによって生成され得る。このようなペプチドを合成するための方法は、例えばR.P.Merrifield,J.Amer.Chem.Soc.85,2149−2154(1963);J.L.Krstenanskyら、FEBS Lett.211、10(1987)にあるように当該分野で公知である。
【0104】
MU−1タンパク質に結合する、中和抗体または非中和抗体(好ましくは、モノクローナル抗体)はまた、特定の腫瘍のための有用な治療剤であり得、そして上記の状態の処置においてまた有用であり得る。これらの中和モノクローナル抗体は、MU−1レセプター鎖に結合するリガンドをブロックし得る。
【0105】
本明細書中で引用される全ての特許および参考文献は、完全に記載されているかのように参考として援用されている。本発明は、以下の実施例によってさらに例示され、これらの実施例は、限定するものとして構成されるべきではない。
【実施例】
【0106】
(実施例)
(実施例1:マウスMU−1 cDNAの単離および特徴付け)
Mu−1レセプターのマウスホモログの部分的フラグメントを、ヒト配列に由来するオリゴヌクレオチドを使用するPCRによって単離した。cDNAを、17日齢のマウス胸腺およびマウス2D6T細胞株から単離したRNAから調製した。約300ヌクレオチドのDNAフラグメントを、以下のオリゴヌクレオチド(図1のヒトcDNA配列(配列番号1に対応する)のそれぞれ領域584〜603および876〜896に対応する)を用いるPCRによってcDNAから増幅した。
【0107】
【化2】
増幅は、94℃で1分間、50℃で1分間、および72℃で1分間の30サイクルの間、1.5mMの塩化マグネシウムを含む1×Taq緩衝液中でTaqポリメラーゼを使用して行なった。このフラグメントのDNA配列を決定し、そして2つのオリゴヌクレオチドは、以下の配列を有するこのフラグメントの内部部分に由来した:
【0108】
【化3】
オリゴヌクレオチドを使用して、2D6 T細胞株から単離したcDNAライブラリーをスクリーニングためのハイブリダイゼーションプローブとして使用するために、本来のPCR産物(図1のマウスcDNA配列のヌクレオチド781〜1043、および配列番号9に対応する)の内部262ヌクレオチドフラグメントを増幅した。フィルターを、標準5×SSCハイブリダイゼーション条件を使用して、65℃でハイブリダイズし、そして65℃でSSC中に洗浄した。426,000個のクローンのスクリーニングにおいてプローブにハイブリダイズした20個のクローンを単離した。DNA配列を、2つの独立したクローンから決定した。クローン番号6の全長配列は、これがヒトMU−1(配列番号9)の全長マウスホモログであることを確認した。
【0109】
マウスMU−1の全長ヌクレオチド配列(配列番号9に対応する)を、図1に示す。ヌクレオチド配列は、ヌクレオチド407〜764の推定リーダー配列、ヌクレオチド407〜1993のコード配列、ヌクレオチド1994〜1997の終止コドンを有する。ヌクレオチド1〜406は、5’非翻訳領域に対応し、そしてヌクレオチド1998〜2628は、3’非翻訳領域に対応する。
【0110】
マウスMU−1の推定タンパク質配列(配列番号10に対応する)を、図2に示す。このマウスMU−1タンパク質は、SPScan(スコア=10.1)によって決定された推定リーダー配列(配列番号10のアミノ酸1〜19に対応する)および推定膜貫通ドメイン(配列番号10のアミノ酸237〜253に対応する)を含む。推定シグナル伝達モチーフは、以下の領域を含む:ボックス1:配列番号10のアミノ酸265〜274、ボックス2:配列番号10のアミノ酸310〜324、配列番号10の281位、319位、361位、368位、397位、および510位の6つのチロシン残基。潜在的なSTATドッキング部位は、以下を含む:STAT5:EDDGYPA、STAT3:YLQR。
【0111】
(実施例2:ヒトMU−1およびマウスMU−1の比較)
GAPアルゴニズムを使用して、ヒトMU−1およびマウスMU−1のアミノ酸を比較する。マウスおよびヒト推定タンパク質配列の比較を、図4に示す。アミノ酸は、GAPアルゴリズムを使用して、65.267%同一であった。整列を、BLOSUM62アミノ酸置換マトリクスによって作成した(Henikoff,S.およびHenikoff,J.G.(1992))。タンパク質ブロック質由来のアミノ酸置換マトリクス(Proc.Natl.Acad.Sci.USA 89:10915−10919)。Gapパラメーター:Gap Weight=8、Average Match=2.912、Length Weight=2、Average Mismatch=−2.003。Percent Similarity=69.466。
【0112】
ヒトおよびマウスcDNAヌクレオチド配列の比較を図3に示す。DNA配列は、GAPアルゴリズムを使用して整列される場合、66.116%同一である。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。Percent Similarity=66.198。
【0113】
ヒトMU−1タンパク質およびマウスMU−1タンパク質の両方は、1型サイトカインレセプタースーパーファミリーのメンバーである。マウスMU−1およびヒトMU−1の両方の配列の評価は、潜在的なボックス1およびボックス2シグナル伝達モチーフの存在を明らかにする。6つのチロシン残基が、細胞質ドメインに存在し、そしてMU−1のシグナル伝達機能において重要であり得る。MU−1の他のファミリーのメンバーとの配列の比較は、STAT5およびSTAT3についての潜在的なドッキング部位の存在を示唆した。
【0114】
(実施例3:ヒトMU−1によって使用されるSTATシグナル伝達経路の決定)
BAF−3細胞を操作して、ヒトEPOレセプターの細胞外ドメインおよびMU−1のレセプターの細胞内ドメインからなるキメラサイトカインレセプターを発現した。huEPOR/MU−1(細胞質)キメラレセプターを発現したBAF−3細胞は、ヒト可溶性EPOに応答して増殖した。これらの細胞を分析して、どのSTAT分子がEPOシグナル伝達に応答してリン酸化されているかを決定した。手短にいうと、コントロール未改変親BAF−3細胞およびEPOR/MUキメラBAF−3細胞を、IL−3含有増殖培養から静止させ、そしてIL−3またはEPOのいずれかを用いて、0、15、30および60分間再刺激した。リン酸化されたチロシンを保存するために、細胞をペレット化し、そしてオルトバナジン酸塩を含む氷冷した溶解緩衝液中に再懸濁した。等量の細胞溶解物を、SDS−PAGEによって電気泳動して、そしてウエスタン分析についてニトロセルロース膜にブロットした。二連のブロットを、STAT分子の各形態に特異的な抗体を使用することによって、STAT1、3、5、および6のリン酸化形態および非リン酸化形態について染色した。活性化していないHELA細胞、α−インターフェロンで活性化したHELA細胞を陽性コントロールとして使用した。
【0115】
これらの結果は、これらの特定の条件化で、MU−1を介するシグナル伝達が、試験された全ての時点(T=0、T=15’、T=30’、T=60’)においてSTAT5のリン酸化を生じることを示した。コントロールまたはキメラBAF−3細胞のIL−3を用いる処理は、STAT1または5ではなく、STAT3のリン酸化を生じた。
【0116】
(実施例4:マウスMU−1およびヒトMU−1の組織発現)
(ノーザン分析)
種々の組織(Clonetech,Palo Alto,CA)由来のポリA+RNAのノーザンブロットを、製造業者によって推奨されるようにして行なった。マウスブロットについて、図1のヌクレオチド781〜1043および配列番号9に対応する262ヌクレオチドフラグメントを、ハイブリダイゼーションのために使用した。
【0117】
マウスMU−1の単一の転写物を、成体マウスの脾臓、肺および心臓組織において検出した。ヒト組織において観察されるより大きい転写物は、マウス組織において観察されなかった。
【0118】
ヒトMU−1の2つの転写物を、成体ヒトリンパ様組織、PBL、胸腺、脾臓およびリンパ節、ならびに胎児肺において検出した。
【0119】
(インサイチュハイブリダイゼーション)
インサイチュハイブリダイゼーションにおいて、研究は、Columbus,OHのPhylogency Inc.によって行なわれた(Lyonsら、1990、J.Cell.Biol:111:2427−2436の方法に従う)。手短にいうと、連続的な5〜7ミクロンのパラフィン切片を、脱パラフィン化し、固定し、プロテイナーゼKを用いて消化し、トリエタノールアミンを用いて処理し、そして脱水した。cRNAを、アンチセンスおよびセンスプローブを作成するために、直鎖状cDNAテンプレートから調製した。cRNA転写物を、製造業者の条件(Ambion)に従って合成し、そして35S−UTPで標識した。切片を、一晩ハイブリダイズし、ストリンジェントに洗浄し、そしてRNAase Aを用いて処理し、そして核トラックエマルジョンに浸漬し、そして2〜3週間露光した。コントロール切片を、手順のバックグラウンドレベルを示すために、センスプローブを用いてハイブリダイズした。マウスプローブは、ヌクレオチド860〜1064(配列番号9)に対応する186bpフラグメントからなった。ヒトプローブは、ヒトMU−1 DNAから作成された231bp
PCR産物であった。
【0120】
マウスMU−1発現は、胚中心および外部筋層(muscularis externa)における成体小腸のリンパ節において観察された。分化したリンパ節およびパイアー斑はまた、マウスMU−1発現を示した。
【0121】
ヒトMU−1発現は、皮質のリンパ小節の胚中心において検出された。髄質(これは、マクロファージを含む)は、ヒトMU−1について陰性であった。ヒト脾臓において、ヒトMU−1発現は、赤脾髄ではなく、白脾髄の領域において検出された。
【0122】
(実施例5:細胞および細胞株におけるヒトMU−1の発現)
RNアーセプロテクション分析を、休止しているおよび活性化したヒトT細胞株およびB細胞株、RajiおよびRPMI 8866、ならびにT細胞株ジャーカットについて行った。ヒトT細胞を、抗CD3および抗CD28を用いて活性化した。細胞株をホルボールエステルおよびイオノマイシンで活性化した。231bpのPCR産物(PCRを、5’プライマー:
【0123】
【化4】
(配列番号15)
および3’プライマー:
【0124】
【化5】
(配列番号16)
を使用することによって行った)をpGEM3zf(−)(Promega,Madison,WI)ベクターのBamH1部位およびHindIII部位に挿入することによって、MU−1リボプローブ産生プラスミドを構築した。このリボプローブを作製するために、リボプローブ産生プラスミドをHindIIIで直線化した。得られたDNAを、フェノール/クロロホルム抽出し、そしてエタノールを用いて沈殿させた。T7 RNAポリメラーゼを使用して、メーカー(PharMingen,San Diego,CA)により指示されたプロトコルに従って、リボプローブを作製した。PharMingenのRiboQuant Multi−Probe Ribonuclease Protection Assayシステムを使用することによって、RNAse保護アッセイを行った。RNAse消化の後、2.0μgの総RNAが、各RPA反応物に含まれ、保護されたリボプローブを、QuickPoint高速核酸分離システム(Novex,San Diego,CA)で実行した。ゲルを、メーカーの指示に従って乾燥し、そして暴露した。
【0125】
ヒトMU−1 RNAは、非刺激集団と比較する場合、抗CD3+抗CD28刺激ヒト精製CD3+細胞においてアップレギュレートされる。MU−1はまた、ThおよびTh2非対称T細胞集団において再刺激される際に、アップレギュレートされる。B細胞株、RPMI 8866およびRajiは、MU−1を構成的に発現し、一方、ジャーカットT細胞株はMU−1を発現しない。
【0126】
(実施例6:既知のサイトカインへのヒトMU−1の結合)
ヒトIg融合タンパク質およびマウスIg融合タンパク質の両方は、MU−1に対するリガンドを同定する試みにおいて、Biacoreチップ上で構築されこのチップ上に固定される。様々な細胞培養物の馴化培地ならびに既知のサイトカインのパネルを、MU−1に対する結合について評価した。いくつかのサイトカインをまた、このファミリーの他のレセプター鎖と組み合わせて試験して、MU−1がリガンド結合のために第2のレセプター鎖を必要とし得る可能性を考察した。以下のサイトカインを試験し、そしてMU−1結合に対して陰性であることを見出した:mIL−2、hIL−2、hIL−15、mIL−7、TSLP、TSLP+IL7、TSLP+IL7R、TSLP+IL7g、TSLP+IL−2、TSLP+IL2+IL2Rβ、IL2Rβ、IL2Rγ、IL7R、IL2+2Rβ、IL2+2Rγ、IL15+IL2Rβ、Il15+2Rγ、IL7+2Rγ、IL2+IL7R、IL15+IL7R、IL7+IL7R。既知のレセプターを同様に固定し、そしてMUFc結合について試験し、陰性結果を得た。IL−15はIL2Rbに結合するが、IL2RgにもMUFcにも結合しない。
【0127】
(等価物)
当業者は、単に慣用的な実験を使用して、本明細書中に記載される本発明の特定の実施形態の多くの等価物を認識するか、または確認し得る。このような等価物は、上記の特許請求の範囲に包含されることが意図される。
【図面の簡単な説明】
【0128】
【図1−1】図1は、マウスMU−1の全長cDNA配列を示す。このヌクレオチド配列は、配列番号9のヌクレオチド1〜2528に対応する。
【図1−2】図1は、マウスMU−1の全長cDNA配列を示す。このヌクレオチド配列は、配列番号9のヌクレオチド1〜2528に対応する。
【図2】図2は、マウスMU−1のアミノ酸配列(配列番号10のアミノ酸1〜529に対応する)を示す。SPScanによって10.1のスコアで推定された、アミノ酸1〜19に推定リーダー配列が存在する(太字)。アミノ酸237〜257に推定膜貫通ドメインが存在する(下線)。推定シグナル伝達モチーフは、以下の領域を含む:ボックス1:アミノ酸265〜274およびボックス2:アミノ酸310〜324(太字および下線)。6つのチロシンが、アミノ酸第281位、第319位、第361位、第368位、第397位および第510位に位置する。WSXWSモチーフ(配列番号8)は、アミノ酸残基214〜アミノ酸残基218に位置付する(大きい太字)。可能性のあるSTAT結合(docking)部位は、アミノ酸393〜398およびアミノ酸510〜513を含む。
【図3−1】図3は、ヒトMU−1およびマウスMU−1のcDNA配列(それぞれ、配列番号1の核酸1〜2665および配列番号9の1〜2628に対応する)のGAP比較を示す。HuMU−1=ヒトMU−1、murMU−1=マウスMU−1。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。同一性%=66.116。
【図3−2】図3は、ヒトMU−1およびマウスMU−1のcDNA配列(それぞれ、配列番号1の核酸1〜2665および配列番号9の1〜2628に対応する)のGAP比較を示す。HuMU−1=ヒトMU−1、murMU−1=マウスMU−1。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。同一性%=66.116。
【図3−3】図3は、ヒトMU−1およびマウスMU−1のcDNA配列(それぞれ、配列番号1の核酸1〜2665および配列番号9の1〜2628に対応する)のGAP比較を示す。HuMU−1=ヒトMU−1、murMU−1=マウスMU−1。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。同一性%=66.116。
【図3−4】図3は、ヒトMU−1およびマウスMU−1のcDNA配列(それぞれ、配列番号1の核酸1〜2665および配列番号9の1〜2628に対応する)のGAP比較を示す。HuMU−1=ヒトMU−1、murMU−1=マウスMU−1。Gapパラメーター:Gap Weight=50、Average Match=10.000、Length Weight=3、Average Mismatch=0.000。同一性%=66.116。
【図4】図4は、ヒトMU−1タンパク質(配列番号2のアミノ酸1〜538に対応する)およびマウスMU−1タンパク質(配列番号10のアミノ酸1〜529に対応する)のGAP比較を示す。BLOSUM62アミノ酸置換マトリクス(Henikoff,S.およびHenikoff,J.G.(1992))。タンパク質ブロックからのアミノ酸置換マトリクス(Proc.Natl.Acad.Sci.USA 89:10915−10919)。Gapパラメーター:Gap Weight=8、Average Match=2.912、Length Weight=2、Average Mismatch=−2.003。同一性%=65.267。
【図5】図5は、ヒトMU−1(humu)(配列番号2に対応する)、マウスMU−1(mousemu)(配列番号10)およびヒトIL2β鎖(humil2rbc)(GENbank登録番号M26062)のアミノ酸の多重配列アライメントを示す。リーダーおよび膜貫通ドメインに下線を付す。保存サイトカインレセプターモジュールモチーフを、太字で示す。可能性のあるシグナル伝達領域を、下線および太字で示す。
【図6】図6は、MU−1を介するシグナル伝達を示す。MU−1は、クローンE7 EPO−MU−1キメラにおいてSTAT5をリン酸化する。実施例3に示される条件下で、MU−1を介するシグナル伝達は、試験された全ての時点でSTAT5のリン酸化を生じる。IL−3でのコントロールまたはキメラBAF−3細胞の処理は、STAT3のリン酸化を生じたが、STAT1およびSTAT5のいずれのリン酸化も生じなかった。
【特許請求の範囲】
【請求項1】
本明細書に記載されるような、タンパク質。
【請求項1】
本明細書に記載されるような、タンパク質。
【図1−1】


【図1−2】

【図2】

【図3−1】

【図3−2】

【図3−3】

【図3−4】
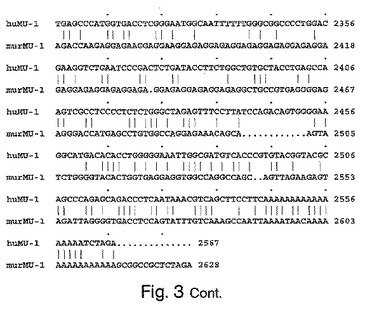
【図4】

【図5】

【図6】
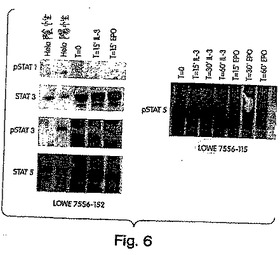
【図1−2】
【図2】
【図3−1】
【図3−2】
【図3−3】
【図3−4】
【図4】
【図5】
【図6】
【公開番号】特開2009−19037(P2009−19037A)
【公開日】平成21年1月29日(2009.1.29)
【国際特許分類】
【出願番号】特願2008−161010(P2008−161010)
【出願日】平成20年6月19日(2008.6.19)
【分割の表示】特願2001−582391(P2001−582391)の分割
【原出願日】平成13年5月11日(2001.5.11)
【出願人】(501418214)ジェネティクス インスティテュート,エルエルシー (35)
【Fターム(参考)】
【公開日】平成21年1月29日(2009.1.29)
【国際特許分類】
【出願日】平成20年6月19日(2008.6.19)
【分割の表示】特願2001−582391(P2001−582391)の分割
【原出願日】平成13年5月11日(2001.5.11)
【出願人】(501418214)ジェネティクス インスティテュート,エルエルシー (35)
【Fターム(参考)】
[ Back to top ]