
サトウキビの茎径関連マーカーとその利用
【課題】サトウキビの量的形質の中でも茎径に関連するマーカーを提供する。
【解決手段】サトウキビの染色体における特定の塩基配列及び特定の塩基配列により挟まれる領域、特定の塩基配列及び特定の塩基配列により挟まれる領域、特定の塩基配列及び特定の塩基配列により挟まれる領域、又は特定の塩基配列及び特定の塩基配列により挟まれる領域から選ばれる連続する核酸領域からなる、サトウキビの茎径関連マーカー。少なくとも一方の親がサトウキビである後代植物の染色体を抽出する工程と、上記で得られた染色体におけるサトウキビの茎径関連マーカーの存在・非存在を測定する工程とを含む、茎径が増大したサトウキビ系統の製造方法。
【解決手段】サトウキビの染色体における特定の塩基配列及び特定の塩基配列により挟まれる領域、特定の塩基配列及び特定の塩基配列により挟まれる領域、特定の塩基配列及び特定の塩基配列により挟まれる領域、又は特定の塩基配列及び特定の塩基配列により挟まれる領域から選ばれる連続する核酸領域からなる、サトウキビの茎径関連マーカー。少なくとも一方の親がサトウキビである後代植物の染色体を抽出する工程と、上記で得られた染色体におけるサトウキビの茎径関連マーカーの存在・非存在を測定する工程とを含む、茎径が増大したサトウキビ系統の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、茎径が増大した特性を有するサトウキビ系統を選抜することができる茎径関連マーカー及びその利用方法に関する。
【背景技術】
【0002】
サトウキビは、砂糖の原料、酒類原料など、食用に栽培されている他、バイオ燃料原料としての利用を含む様々な産業分野で利用されている。このような状況下、所望の特性(例えば、糖含有量、生長力の増強、新芽形成能、耐病性及び虫害抵抗性、耐寒性、葉長の増大、茎径や茎長の増大など)を有するサトウキビの新品種を開発するニーズがある。
【0003】
一般に、植物品種・系統の識別として、特性データを比較する「特性比較」、同一条件で栽培し比較する「比較栽培」、DNAを解析する「DNA分析」の3つの方法がある。特性比較および比較栽培による系統識別は、栽培条件の違いによる精度低下や多大な工数が必要とされる長期間の圃場調査など多くの問題を抱える。特に、サトウキビは、イネやトウモロコシなど、他のイネ科作物と比べ植物体が極めて大きく、圃場調査による系統識別の実施が困難である。また、葉長や茎径、茎長といった形質に特徴的な特性を有する品種・系統を識別するには、サトウキビを長期間栽培した後、これら特性データを収集する必要がある。また、サトウキビを長期間栽培したとしても、これら形質は環境の影響を受けやすいため精度良く識別することが困難である。
【0004】
さらに、サトウキビの新品種作出には、先ず、交配により数万種類の交配種を作製し、そこから実生選抜、さらに優良な系統を段階的に選抜し、最終的に所望な特性を有する2〜3種の新品種を得ることができる。このように、サトウキビの新品種作出には、非常に多くの系統を栽培・評価する必要があり、大規模な圃場を準備して多大な手間をかける必要がある。
【0005】
したがって、所望の特性を有するサトウキビ系統を、ゲノムに存在するマーカーを用いて識別する方法の開発が求められている。特に、サトウキビにおける新品種作出に際して、種々の特性について優れたマーカーが使用できれば、上述したようなサトウキビに特有の諸問題を回避でき、非常に有効なツールとなりうる。しかしながら、サトウキビは、高次倍数性で染色体数が多いため(約100〜130)、マーカー技術開発が遅れている。サトウキビでは、USDAにおいてSSRマーカーを用いた遺伝子型決定に関する報告があるものの(非特許文献1)、マーカー数及び各マーカーからの多型数が少ないことに起因して精度が低く、適用範囲がアメリカ・オーストラリア品種に限られるため、日本国内および台湾・インドなどの主要品種および有用な遺伝資源の系統識別に利用できない。
【0006】
また、非特許文献2は、マーカー数を増やし、各々のマーカーの特性関係を比較し、検証することでサトウキビにおける遺伝子地図を作製する可能性を示唆している。しかしながら、非特許文献2には、十分な数のマーカーが開示されておらず、目的とする特性に連鎖したマーカーも見つかっていない。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Maydica 48(2003)319-329 “Molecular genotyping of sugarcane clones with microsatellite DNA markers”
【非特許文献2】Nathalie Piperidis et al., Molecular Breeding, 2008, Vol 21, 233-247
【発明の概要】
【発明が解決しようとする課題】
【0008】
そこで、本発明は、サトウキビの量的形質の中でも茎径に関連するマーカーを提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題を解決するために鋭意検討を重ねた結果、サトウキビにおける多数のマーカーを準備し、交雑後代系統における量的形質とマーカーとの連鎖解析によって、茎径が増大するといった量的形質に連鎖するマーカーを見いだし、本発明を完成するに至った。
【0010】
本発明は以下を包含する。
[1] サトウキビの染色体における配列番号1に示す塩基配列及び配列番号20に示す塩基配列により挟まれる領域、配列番号21に示す塩基配列及び配列番号34に示す塩基配列により挟まれる領域、配列番号35に示す塩基配列及び配列番号40に示す塩基配列により挟まれる領域、又は配列番号41に示す塩基配列及び配列番号52に示す塩基配列により挟まれる領域から選ばれる連続する核酸領域からなる、サトウキビの茎径関連マーカー。
[2] 上記核酸領域は、配列番号1〜52からなる群から選ばれるいずれか1つの塩基配列を含むことを特徴とする[1]のサトウキビの茎径関連マーカー。
[3] 上記核酸領域は、サトウキビの染色体における配列番号5に示す塩基配列と配列番号7に示す塩基配列とにより挟み込まれる領域、配列番号21に示す塩基配列と配列番号22に示す塩基配列とにより挟み込まれる領域、配列番号36に示す塩基配列と配列番号38に示す塩基配列とにより挟み込まれる領域、又は配列番号41に示す塩基配列と配列番号43に示す塩基配列とにより挟み込まれる領域に位置することを特徴とする[1]の茎径関連マーカー。
[4] 少なくとも一方の親がサトウキビである後代植物の染色体を抽出する工程と、
上記で得られた染色体における[1]乃至[3]のいずれかのサトウキビの茎径関連マーカーの存在・非存在を測定する工程とを含む、茎径が増大したサトウキビ系統の製造方法。
[5] 上記測定する工程では、上記サトウキビの茎径関連マーカーに対応するプローブを備えるDNAチップを使用することを特徴とする[4]のサトウキビ系統の製造方法。
[6] 上記後代植物は種子又は幼苗であり、当該種子又は幼苗から染色体を抽出することを特徴とする[4]のサトウキビ系統の製造方法。
【発明の効果】
【0011】
本発明によれば、サトウキビにおける量的形質の中でも茎径が増大するといった特性に連鎖する新規なサトウキビの茎径関連マーカーを提供することができる。本発明に係るサトウキビの茎径関連マーカーを利用することによって、サトウキビの交配系統における茎径を識別することができる。これにより、茎径が増大した特性を有するサトウキビ系統を非常に低コストに識別することができる。
【図面の簡単な説明】
【0012】
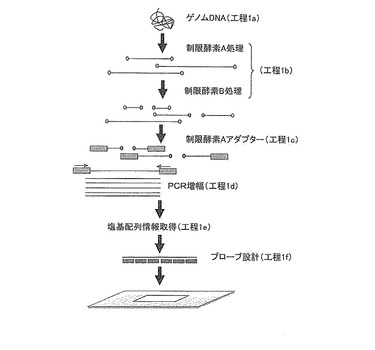
【図1】サトウキビ染色体のマーカーを得る際に使用したDNAマイクロアレイの製造フローを示す模式図である。
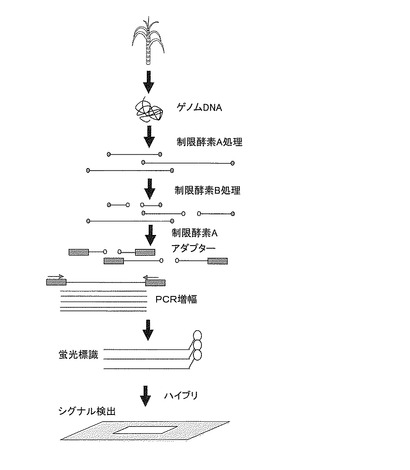
【図2】DNAマイクロアレイを使用したシグナル検出の工程を示す模式図である。
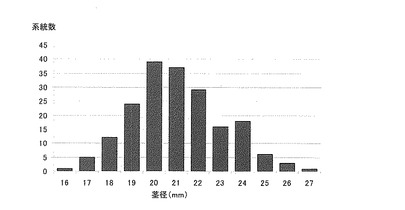
【図3】実施例で使用したサトウキビ品種・系統群について茎径データを示す特性図である。
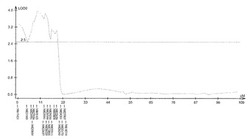
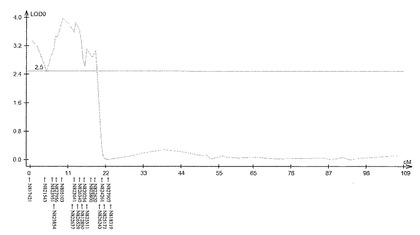
【図4】サトウキビ品種NiF8における茎径に関するQTL解析の結果(第2連鎖群)を示す特性図である。
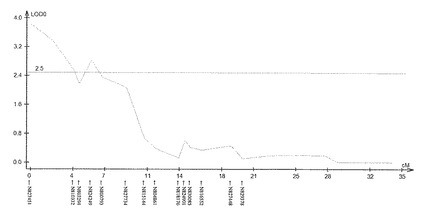
【図5】サトウキビ品種NiF8における茎径に関するQTL解析の結果(第40連鎖群)を示す特性図である。
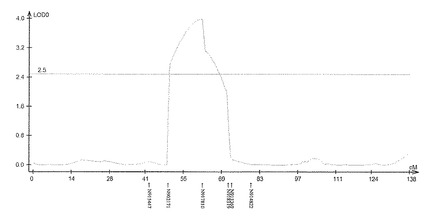
【図6】サトウキビ品種Ni9における茎径に関するQTL解析の結果(第21連鎖群)を示す特性図である。
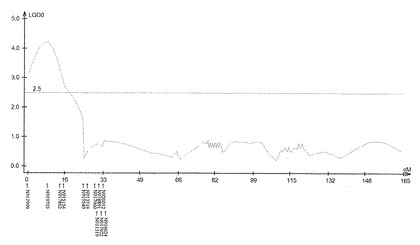
【図7】サトウキビ品種Ni9における茎径に関するQTL解析の結果(第46連鎖群)を示す特性図である。
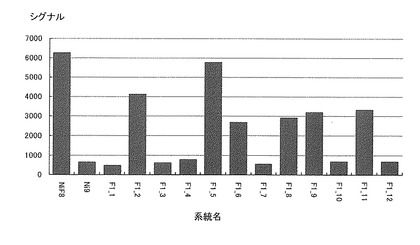
【図8】各系統毎におけるN812115(NiF8第2連鎖群のマーカー)のシグナル値を示す特性図である。
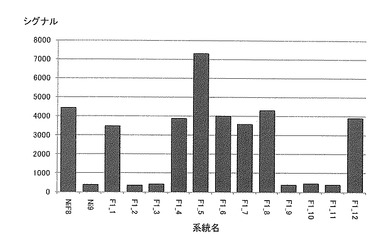
【図9】各系統毎におけるN815144(NiF8第40連鎖群のマーカー)のシグナル値を示す特性図である。
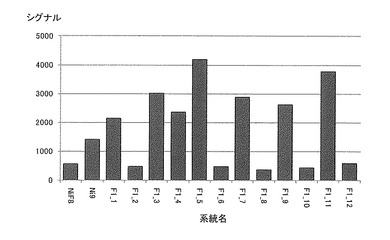
【図10】各系統毎におけるN917810(Ni9第21連鎖群のマーカー)のシグナル値を示す特性図である。
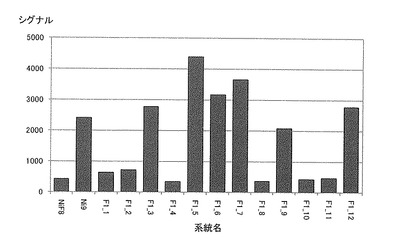
【図11】各系統毎におけるN919549(Ni9第46連鎖群のマーカー)のシグナル値を示す特性図である。
【発明を実施するための形態】
【0013】
以下、本発明に係るサトウキビの茎径関連マーカー及びその利用方法、特にサトウキビの茎径関連マーカーを用いたサトウキビ系統の製造方法について説明する。
【0014】
サトウキビの茎径関連マーカー
本発明に係るサトウキビの茎径関連マーカーとは、サトウキビの染色体上に存在する特定の領域であり、サトウキビの茎径を増大させる形質の原因遺伝子(群)に連鎖して、サトウキビ茎径の増大という形質を判別できる機能を有する。すなわち、既知のサトウキビ系統を用いて得られた後代系統において、サトウキビの茎径関連マーカーの存在を確認することで茎径の増大という形質を有する系統であると判断することができる。
【0015】
ここで、茎径とは茎一本あたりの直径を意味し、当該分野において一般的な手法を用いて計測することができる。特に限定されないが、例えばサトウキビ茎を原料茎に調整し、茎中央部に相当する節間のさらに中央部の短径をノギスなどを用いて測定する。ここで「原料茎」とは粗糖などを生産する際の原料として利用される茎の部分であり、無処理のサトウキビ茎から糖度の低い梢頭部、葉、根等を除去した後の茎で、一般的に1m以上のものを指す。
【0016】
サトウキビとは、イネ科サトウキビ属に属する植物を意味する。また、サトウキビとは、所謂、高貴種(学名:Saccharum officinarum)と野生種(学名:Saccharum spontaneum)のいずれも含むことを意味する。既知のサトウキビ品種・系統としては、特に限定されず、日本国内にて使用可能なあらゆる品種・系統、日本国外において使用されている品種・系統等を含む意味である。例えば、サトウキビ日本国内育成品種としては、特に限定されないが、Ni1、NiN2、NiF3、NiF4、NiF5、Ni6、NiN7、NiF8、Ni9、NiTn10、Ni11、Ni12、Ni14、Ni15、Ni16、Ni17、NiTn19、NiTn20、Ni22及びNi23等を挙げることができる。また、サトウキビ日本国内主要品種としては、特に限定されないが、NiF8、Ni9、NiTn10及びNi15等を挙げることができる。さらに、サトウキビ日本国内導入主要品種としては、特に限定されないが、F177、Nco310及びF172等を挙げることができる。
【0017】
また、後代系統は、母本及び父本の両方がサトウキビ品種・系統である同種交配による系統であっても良いし、いずれか一方がサトウキビ品種・系統であり他方が近縁のErianthus arundinaceusであるような交雑系統であっても良い。また、後代品種は、いわゆる戻し交雑によって得られたものでも良い。
【0018】
サトウキビの茎径関連マーカーは、サトウキビ品種NiF8の染色体から独自に取得した3004個のマーカーを含む遺伝子連鎖地図およびサトウキビ品種Ni9の染色体から独自に取得した4569個のマーカーを含む遺伝子連鎖地図とサトウキビ茎径データとを用いたQTL(Quantitative Trait Loci)解析によって新たに同定されたものである。なお、サトウキビ茎径は、多数の遺伝子が関与していると考えられ連続分布をとる量的形質である。QTL解析には、遺伝解析ソフトQTL Cartographer(Wang S., C. J. Basten, and Z.-B. Zeng (2010). Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC)を使用し、Composite interval mapping(CIM)法を適用している。
【0019】
具体的に、上述したQTL解析により、ロッドスコア(LOD score)が所定の閾値(例えば3.0)以上となるピークを上記それぞれの遺伝子連鎖地図に含まれる4つの領域で見いだした。すなわち、当該ピークを含む約23.4cM(センチモルガン)領域及び約20.0cM領域(サトウキビ品種NiF8)、及並びに当該ピークを含む約37.2cM領域及び約33.7cM領域(サトウキビ品種Ni9)の4領域を特定した。ここで、「モルガン(M)」は、染色体上の遺伝子間の距離を相対的に示した単位であり、交叉価をパーセントにした値である。サトウキビの染色体において、1cMは、約2000kbに相当する。なお、このピーク位置又はその近傍には、茎径を増大させる形質の原因遺伝子(群)が存在することが示唆される。
【0020】
上記ピークを含む23.4cMの領域は、上記マーカーのうち表1に示す20種類のマーカーがこの順で含まれる領域である。
【0021】
【表1】

【0022】
上記ピークを含む20.0cMの領域は、上記マーカーのうち表2に示す14種類のマーカーがこの順で含まれる領域である。
【0023】
【表2】
【0024】
上記ピークを含む37.2cMの領域は、上記マーカーのうち表3に示す6種類のマーカーがこの順で含まれる領域である。
【0025】
【表3】
【0026】
上記ピークを含む33.7cMの領域は、上記マーカーのうち表4に示す12種類のマーカーがこの順で含まれる領域である。
【0027】
【表4】
【0028】
なお、表1−4において、連鎖群とは、QTL解析において特定された複数の連鎖群についてそれぞれ付された番号である。表1−4においてマーカー名とは、本発明で独自に取得したマーカーに付された名称である。表1−4においてシグナル閾値とは、マーカーの有無を判定するための閾値である。
【0029】
サトウキビ品種NiF8の上記23.4cMの領域に含まれるピークは、配列番号5に示す塩基配列からなるマーカー(N827956)及び配列番号7に示す塩基配列からなるマーカー(N822637)により挟み込まれる領域に存在している。
【0030】
また、サトウキビ品種NiF8の上記20.0cMの領域に含まれるピークは、配列番号21に示す塩基配列からなるマーカー(N827451)及び配列番号22に示す塩基配列からなるマーカー(N810332)により挟み込まれる領域に存在している。
【0031】
さらに、サトウキビ品種Ni9の上記37.2cMの領域に含まれるピークは、配列番号36に示す塩基配列からなるマーカー(N902171)及び配列番号38に示す塩基配列からなるマーカー(N918319)により挟み込まれる領域に存在している。
【0032】
またさらに、サトウキビ品種Ni9の上記33.7cMの領域に含まれるピークは、配列番号41に示す塩基配列からなるマーカー(N917999)及び配列番号43に示す塩基配列からなるマーカー(N917862)により挟み込まれる領域に存在している。
【0033】
表1−4に示したマーカーが含まれる4つの領域から選ばれる連続した核酸領域を、サトウキビの茎径関連マーカーとして使用することができる。ここで、核酸領域とは、サトウキビの染色体に存在する他の領域との同一性が95%以下、好ましくは90%以下、より好ましくは80%以下、最も好ましくは70%以下となるような塩基配列からなる領域を意味する。サトウキビの茎径関連マーカーとなる核酸領域と他の領域との同一性が上記範囲であれば、定法に従って、当該核酸領域を特異的に検出することができる。ここで、同一性の値は、例えばBLASTを用いてデフォルトパラメータを用いて算出することができる。
【0034】
また、サトウキビの茎径関連マーカーとなる核酸領域の塩基長は、少なくとも8塩基長以上、好ましくは15塩基長以上、より好ましくは20塩基長以上、最も好ましくは30塩基長とすることができる。サトウキビの茎径関連マーカーとなる核酸領域の塩基長が上記範囲であれば、定法に従って、当該核酸領域を特異的に検出することができる。
【0035】
特に、サトウキビの茎径関連マーカーとしては、サトウキビ品種NiF8の23.4cMの領域に含まれる20種類のマーカーのうち、配列番号5に示す塩基配列と配列番号7に示す塩基配列とにより挟み込まれる領域から選ばれることが好ましい。上記ピークが配列番号5に示す塩基配列と配列番号7に示す塩基配列とにより挟み込まれる領域に存在するためである。また、サトウキビの茎径関連マーカーとしては、サトウキビ品種NiF8の20.0cMの領域に含まれる14種類のマーカーのうち、配列番号21に示す塩基配列と配列番号22に示す塩基配列とにより挟み込まれる領域から選ばれることが好ましい。上記ピークが配列番号21に示す塩基配列と配列番号22に示す塩基配列とにより挟み込まれる領域に存在するためである。さらに、サトウキビの茎径関連マーカーとしては、サトウキビ品種Ni9の37.2cMの領域に含まれる6種類のマーカーのうち、配列番号36に示す塩基配列と配列番号38に示す塩基配列とにより挟み込まれる領域から選ばれることが好ましい。上記ピークが配列番号36に示す塩基配列と配列番号38に示す塩基配列とにより挟み込まれる領域に存在するためである。またさらに、サトウキビの茎径関連マーカーとしては、サトウキビ品種Ni9の33.7cMの領域に含まれる12種類のマーカーのうち、配列番号41に示す塩基配列と配列番号43に示す塩基配列とにより挟み込まれる領域から選ばれることが好ましい。上記ピークが配列番号41に示す塩基配列と配列番号43に示す塩基配列とにより挟み込まれる領域に存在するためである。
【0036】
また、サトウキビの茎径関連マーカーとしては、上記表1−4に示した52種類のマーカーから選ばれる1種類のマーカーを含む核酸領域とすることもできる。例えば、サトウキビの茎径関連マーカーとしては、サトウキビ品種NiF8の23.4cMの領域に含まれるピークの位置に最も近い配列番号6に示す塩基配列からなるマーカー(N805103)を含む核酸領域、サトウキビ品種NiF8の20.0cMの領域に含まれるピークの位置に最も近い配列番号21に示す塩基配列からなるマーカー(N827451)を含む核酸領域、サトウキビ品種Ni9に由来する37.2cMの領域に含まれるピークの位置に最も近い配列番号37に示す塩基配列からなるマーカー(N917810)、サトウキビ品種Ni9に由来する33.7cMの領域に含まれるピークの位置に最も近い配列番号42に示す塩基配列からなるマーカー(N919703)を含む核酸領域を使用することが好ましい。このとき、マーカーを含む核酸領域の塩基配列は、当該マーカーの塩基配列に基づいて設計したプライマーを用いたインバースPCRによって特定することができる。
【0037】
さらに、サトウキビの茎径関連マーカーとしては、上記52種類のマーカーそのものを使用することができる。すなわち、これら52種類のマーカーのうち1種類以上のマーカーをサトウキビの茎径関連マーカーとして使用することができる。例えば、サトウキビ品種NiF8の23.4cMの領域に含まれるピークの位置に最も近い配列番号6に示す塩基配列からなるマーカー(N805103)、サトウキビ品種NiF8の20.0cMの領域に含まれるピークの位置に最も近い配列番号21に示す塩基配列からなるマーカー(N827451)、サトウキビ品種Ni9に由来する37.2cMの領域に含まれるピークの位置に最も近い配列番号37に示す塩基配列からなるマーカー(N917810)、サトウキビ品種Ni9に由来する33.7cMの領域に含まれるピークの位置に最も近い配列番号42に示す塩基配列からなるマーカー(N919703)を使用することが好ましい。
【0038】
サトウキビにおけるマーカーの同定
本発明では、上述したように、サトウキビ品種NiF8の染色体から独自に取得した3004個のマーカーおよびサトウキビ品種Ni9の染色体から独自に取得した4569個のマーカーからサトウキビの茎径関連マーカーを特定した。ここでは、これらのマーカーについて説明する。これらのマーカーを同定する際には、特願2009-283430号に開示された方法を適用したDNAマイクロアレイを使用することができる。
【0039】
具体的に、サトウキビの染色体から独自に取得したこれらのマーカーは、特願2009-283430号に開示された方法により設計されたプローブを有するDNAマイクロアレイを使用する。プローブの設計方法は、図1に示すように、先ず、サトウキビからゲノムDNAを抽出する(工程1a)。次に、抽出したゲノムDNAを1又は複数の制限酵素により消化する(工程1b)。なお、図1に示した例では、制限酵素A及び制限酵素Bの2種類の制限酵素をこの順で用いてゲノムDNAを消化している。ここで、制限酵素としては、特に限定されないが、例えば、PstI、EcoRI、HindIII、BstNI、HpaII、HaeIII等を使用することができる。特に制限酵素としては、ゲノムDNAを完全に消化した際に20〜10000塩基長のゲノムDNA断片となるよう、認識配列の出現頻度等を考慮して適宜選択することができる。また、複数の制限酵素を使用する場合、全ての制限酵素を使用した後のゲノムDNA断片が200〜6000塩基長となっていることが好ましい。さらに、複数の制限酵素を使用する場合、処理に供する制限酵素の順序は特に限定されず、また、処理条件(溶液組成や温度等)が共通する場合には複数の制限酵素を同一の反応系で使用しても良い。すなわち、図1に示した例においては、制限酵素A及び制限酵素Bをこの順で使用してゲノムDNAを消化しているが、制限酵素A及び制限酵素Bを同じ反応系で同時に使用してゲノムDNAを消化しても良いし、制限酵素B及び制限酵素Aをこの順で使用してゲノムDNAを消化してもよい。さらに、使用する制限酵素の数は3以上であってもよい。
【0040】
次に、制限酵素処理後のゲノムDNA断片に対してアダプターを結合する(工程1c)。ここで、アダプターとは、上述した制限酵素処理によって得られたゲノムDNA断片の両端に結合できるものであれば特に限定されない。アダプターとしては、例えば、制限酵素処理によってゲノムDNAの両末端に形成される突出末端(粘着末端)に対して相補的な一本鎖を有し、詳細を後述する増幅処理の際に使用するプライマーがハイブリダイズしうるプライマー結合配列を有するものを使用することができる。また、アダプターとしては、上記突出末端(粘着末端)に対して相補的な一本鎖を有し、クローニングする際のベクターに組み入れるための制限酵素認識部位を有するものを使用することもできる。
【0041】
また、複数の制限酵素を使用してゲノムDNAを消化した場合には、各制限酵素に対応する複数のアダプターを準備して使用することができる。すなわち、複数の制限酵素でゲノムDNAを消化した場合に生ずる複数種類の突出末端のそれぞれに対して、相補的な一本鎖を有する複数のアダプターを使用することができる。このとき、複数の制限酵素に対応する複数のアダプターは、共通するプライマーがハイブリダイズできるように共通するプライマー結合配列を有しているものであっても良いし、それぞれ異なるプライマーがハイブリダイズできるように異なるプライマー結合配列を有するものであっても良い。
【0042】
さらに、複数の制限酵素を使用してゲノムDNAを消化した場合、アダプターとしては、使用した複数の制限酵素のなかから選ばれる1つの制限酵素若しくは、使用した制限酵素のうち一部の制限酵素に対応するアダプターを準備して使用することもできる。
【0043】
次に、両末端にアダプターが付加されたゲノムDNA断片を増幅する(工程1d)。プライマー結合配列を有するアダプターを使用した場合には、当該プライマー結合配列にハイブリダイズできるプライマーを使用することで上記ゲノムDNA断片を増幅することができる。或いは、アダプターを付加したゲノムDNA断片を、アダプター配列を利用してベクターにクローニングし、当該ベクターにおける所定の領域にハイブリダイズできるプライマーを用いてゲノムDNA断片を増幅することができる。なお、プライマーを用いたゲノムDNA断片の増幅反応としては、一例としてPCRを使用することができる。
【0044】
また、複数の制限酵素を使用してゲノムDNAを消化するとともに、各制限酵素に対応する複数のアダプターをゲノムDNA断片に連結した場合、複数の制限酵素を用いた処理によって得られたゲノムDNA断片の全てにアダプターが連結されることとなる。この場合、アダプターに含まれるプライマー結合配列を用いて核酸増幅反応を行うことで、得られた全てのゲノムDNA断片を増幅することができる。
【0045】
或いは、複数の制限酵素を使用してゲノムDNAを消化するとともに、使用した複数の制限酵素のなかから選ばれる1つ制限酵素若しくは、使用した制限酵素のうち一部の制限酵素に対応するアダプターをゲノムDNA断片に連結した場合、得られたゲノムDNA断片のうち、選ばれた制限酵素の認識配列を両末端に有するゲノムDNA断片のみを増幅することができる。
【0046】
次に、増幅されたゲノムDNA断片の塩基配列を決定し(工程1e)、当該ゲノムDNA断片より短い塩基長を有し、ゲノムDNA断片内の少なくとも一部をカバーする1又は複数の領域を特定し、特定した1又は複数の領域を、サトウキビにおけるプローブとして設計する(工程1f)。ゲノムDNA断片の塩基配列を決定する方法は、特に限定されず、サンガー法等を適用したDNAシークエンサーを利用した従来公知の方法を使用することができる。ここで、設計する領域としては、上述したように、例えば20〜100塩基長、好ましくは30〜90塩基長、より好ましくは50〜75塩基長とする。
【0047】
以上のように、サトウキビから抽出したゲノムDNAを使用して多数のプローブを設計し、設計したプローブの塩基配列に基づいて、担体上にて目的の塩基配列を有するオリゴヌクレオチドを合成することでDNAマイクロアレイを作製することができる。このように作製したDNAマイクロアレイを使用することで、上述した配列番号1〜52に示した52種類のサトウキビの茎径関連マーカーを含む3004個および4569個のマーカーを、サトウキビ品種NiF8及びNi9よりそれぞれ同定することができる。
【0048】
より具体的に、本発明者らは、既知のサトウキビ品種NiF8、Ni9及びこれらの交配後代系統(191系統)について、上述したDNAマイクロアレイを用いてシグナルデータを取得した。そして、得られたシグナルデータから遺伝子型データを取得し、この遺伝子型データを元にして、遺伝地図作成ソフトウェアAntMap(Iwata H, Ninomiya S (2006) AntMap: constructing genetic linkage maps using an ant colony optimization algorithm. Breed Sci 56: 371-378)を使用し、遺伝距離計算式Kosambiにより染色体におけるマーカーの位置情報を算出した。さらに、取得したマーカーの位置情報をもとに、Mapmaker/EXP ver.3.0(A Whitehead Institute for Biomedical Research Technical Report, Third Edition, January, 1993)により遺伝地図データシートを作成した。その結果、上述した配列番号1〜52に示した52種類のサトウキビの茎径関連マーカーを含む3004個及び4569個のマーカーを、サトウキビ品種NiF8及びNi9よりそれぞれ同定している。
【0049】
サトウキビの茎径関連マーカーの利用
サトウキビの茎径関連マーカーを利用することで、後代系統等に茎径の表現型が未知のサトウキビ系統について茎径の増大という表現型を示す系統であるか判断することができる。ここで、サトウキビの茎径関連マーカーを利用するとは、当該マーカーに対応するプローブを有するDNAマイクロアレイを利用する形態を含む意味である。サトウキビの茎径関連マーカーに対応するプローブとは、上述のように定義されたサトウキビの茎径関連マーカーに対して、ストリンジェントな条件下で特異的にハイブリダイズできるオリゴヌクレオチドを意味する。このようなオリゴヌクレオチドは、例えば、上述のように定義されたサトウキビの茎径関連マーカーの塩基配列又はその相補鎖の少なくとも連続する10塩基、15塩基、20塩基、25塩基、30塩基、35塩基、40塩基、45塩基、50塩基又はそれ以上の塩基長の部分領域若しくは全領域として設計することができる。なお、このプローブを有するDNAマイクロアレイとしては、ガラスやシリコーン等の平面基板を担体とするマイクロアレイや、マイクロビーズを担体とするビーズアレイ、或いは中空繊維の内壁にプローブを固定する3次元マイクロアレイ等の如何なるタイプのマイクロアレイであってもよい。
【0050】
以上のように作製されたDNAマイクロアレイを使用することで、後代系統等に代表される茎径の表現型が未知のサトウキビ系統について、茎径の増大という表現型を示す系統であるか判断することができる。なお、上述したDNAマイクロアレイを使用する方法以外であっても、従来公知の手法を用いて上述したサトウキビの茎径関連マーカーを検出して、茎径の表現型が未知のサトウキビ系統について茎径の増大という形質を有する系統であるか判断してもよい。
【0051】
DNAマイクロアレイを使用する方法をより詳細に説明する。図2に示すように、先ず供試サトウキビからゲノムDNAを抽出する。この供試サトウキビとは、後代系統等の茎径の表現型が未知のサトウキビ系統のことであり、茎径が増大する形質を有するか判定する対象となるサトウキビ系統である。次に、抽出したゲノムDNAを、DNAマイクロアレイを作製する際に使用した制限酵素で消化して複数のゲノムDNA断片を調整する。次に、得られたゲノムDNA断片と、DNAマイクロアレイを作製する際に使用したアダプターとを連結する。次に、両末端にアダプターが付加されたゲノムDNA断片を、DNAマイクロアレイを作製する際に使用したプライマーを用いて増幅する。これにより、DNAマイクロアレイを作製する際の工程1dで増幅したゲノムDNA断片に対応する、供試サトウキビ由来のゲノムDNA断片を増幅することができる。
【0052】
この工程においては、アダプターが付加されたゲノムDNA断片のうち、所定のゲノムDNA断片を選択的に増幅してもよい。例えば、複数の制限酵素に対応する複数のアダプターを使用した場合には、特定のアダプターが付加されたゲノムDNA断片を選択的に増幅することができる。また、複数の制限酵素でゲノムDNAを消化した場合、得られたゲノムDNA断片のうち、所定の制限酵素に対応する突出末端を有するゲノムDNA断片のみにアダプターを付加することで、アダプターが付加されたゲノムDNA断片を選択的に増幅することができる。このように、所定のゲノムDNA断片を選択的に増幅することで濃縮することができる。
【0053】
次に、増幅したゲノムDNA断片に標識を付加する。標識としては、従来公知の如何なる物質を使用しても良い。標識としては、例えば蛍光分子、色素分子、放射性分子等を使用することができる。なお、本工程は、ゲノムDNA断片を増幅する工程において標識を有するヌクレオチドを用いることで省略することができる。上記工程において標識を有するヌクレオチドを用いてゲノムDNA断片を増幅することで、増幅されたDNA断片が標識化されるためである。
【0054】
次に、標識を有するゲノムDNA断片を所定の条件下でDNAマイクロアレイに接触させ、DNAマイクロアレイに固定されたプローブと標識を有するゲノムDNA断片とをハイブリダイズさせる。このとき、ハイブリダイズさせる際には高いストリンジェンシー条件とすることが好ましい。このような高いストリンジェンシー条件とすることによって、供試サトウキビにサトウキビの茎径関連マーカーが存在しているか否かを、より高精度に判定することができる。なお、ストリンジェンシー条件は、反応温度及び塩濃度で調節することができる。すなわち、より高温とすることでより高いストリンジェンシー条件となり、またより低い塩濃度でより高いストリンジェンシー条件となる。例えば、50〜75塩基長のプローブを使用する場合、ハイブリダイゼーション条件としては、40〜44℃、0.2SDS、6×SSCの条件とすることでより高いストリンジェンシー条件とすることができる。
【0055】
また、プローブと標識を有するゲノムDNA断片とのハイブリダイズは、標識に基づいて検出することができる。すなわち、上述した標識を有するゲノムDNA断片とプローブのハイブリダイズ反応の後、未反応のゲノムDNA断片等を洗浄し、その後、プローブに対して特異的にハイブリダイズしたゲノムDNA断片の標識を観察する。例えば、標識が蛍光物質である場合にはその蛍光波長を検出し、標識が色素分子であればその色素波長を検出する。より具体的には、通常のDNAマイクロアレイ解析に使用している、蛍光検出装置やイメージアナライザー等の装置を使用することができる。
【0056】
以上のように、上述したDNAマイクロアレイを使用することにより、供試サトウキビが上述したサトウキビの茎径関連マーカーを有するか否か判断することができる。特に、上述した方法では、供試サトウキビを実際の茎径が測定可能な程度まで成長させる必要はなく、例えば後代系統の種子や当該種子を発芽させた幼苗を使用することができる。したがって、サトウキビの茎径関連マーカーを利用することによって、供試サトウキビを生育させるための圃場やその他、生育のためのコストを大幅に削減することができる。
【0057】
特に、サトウキビの新品種作出に際して、先ず、交配により数万種類の交配種を作製した後、実生選抜に先立って若しくは実生選抜に代えて、サトウキビの茎径関連マーカーを利用した判断を行うことが好ましい。これにより、実際の圃場において、優良な系統を栽培する数を大幅に削減することができ、サトウキビの新品種作出に係る手間やコストを大幅に抑制することができる。
【0058】
上記サトウキビの茎径関連マーカーを利用することで、茎径を増大させる形質の原因遺伝子又は原因遺伝子群を単離することができる。単離方法としては、従来公知の手法を用いることができる(「バイオ実験イラストレイテッド4 苦労なしのクローニング」真壁和裕著(1997)、秀潤社)。例えば、上記定義のサトウキビの茎径関連マーカーに対応するプライマー又はプローブを作製し、サトウキビのゲノムDNA又はcDNAをスクリーニングすることによって、茎径を増大させる形質の原因遺伝子を単離することができる。また、上記サトウキビのゲノムDNA又はcDNAに代えて、その他のイネ科植物由来のゲノムDNA又はcDNAを、当該サトウキビの茎径関連マーカーに対応するプライマー又はプローブを用いてスクリーニングすることによって、サトウキビ以外のイネ科植物の茎径を増大させる形質の原因遺伝子又は原因遺伝子群を単離することも可能である。
【0059】
また、上記で得られた茎径を増大させる形質の原因遺伝子が組み込まれた組換えベクターを用いて植物細胞を形質転換することによって、茎径が増大した形質を有する形質転換植物体を製造することもできる。
【実施例】
【0060】
以下、実施例により本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0061】
1.DNAマイクロアレイ用プローブの作成
(1)材料
サトウキビ品種:NiF8、Ni9、US56-15-8、POJ2878、Q165、R570、Co290及びB3439、サトウキビ近縁野生種:Glagah Kloet、Chunee、Natal Uba及びRobustum9、並びにエリアンサス:IJ76-349及びJW630を用いた。
【0062】
(2)制限酵素処理
これらサトウキビ品種、サトウキビ近縁野生種及びエリアンサスからそれぞれゲノムDNAを、DNeasy Plant Mini Kit(QIAGEN社製)を用いて抽出した。ゲノムDNA(750ng)を制限酵素PstI(NEB社、25unit)で37℃、2時間処理後、制限酵素BstNI(NEB社、25unit)を添加、60℃、2時間処理した。
【0063】
(3)アダプターライゲーション
(2)で処理したゲノムDNA断片(120ng)にPstI配列アダプター(5’-CACGATGGATCCAGTGCA-3’(配列番号53)、5’-CTGGATCCATCGTGCA-3’ (配列番号54))とT4 DNA Ligase(NEB社、800 unit)を加え、16℃、4時間以上処理した。これにより、(2)で処理したゲノムDNA断片のうち、両末端にPstI認識配列を有するゲノムDNA断片に対して選択的にアダプターを付加した。
【0064】
(4)PCR増幅
(3)で得られたアダプターを有するゲノムDNA断片(15ng)にPstI配列アダプター認識プライマー(5’-GATGGATCCAGTGCAG-3’(配列番号55))とTaq polymerase(タカラバイオ社PrimeSTAR、1.25unit)を加え、PCR(98℃を10秒間、55℃を15秒間、72℃を1分間、30サイクル後、72℃で3分間処理後、4℃で保存)でゲノムDNA断片を増幅した。
【0065】
(5)ゲノムシークエンス取得
(4)においてPCR増幅したゲノムDNA断片についてFLX454(Roche社)又はサンガー法により塩基配列を決定した。また、ゲノムデータベース(Gramene:http://www.gramene.org/)に格納されたソルガム全ゲノム配列情報から、PstI認識配列により挟み込まれる塩基配列情報を取得した。
【0066】
(6)プローブ設計及びDNAマイクロアレイの作成
(5)のゲノムシークエンス情報をもとに50〜75bpのプローブを設計した。設計したプローブの塩基配列情報をもとに、これらプローブを有するDNAマイクロアレイを作製した。
【0067】
2.DNAマイクロアレイを用いたシグナルデータの取得
(1)材料
サトウキビ品種・系統:NiF8及びNi9、並びに、これらの交配後代191系統を用いた。
【0068】
(2)制限酵素処理
これらNiF8及びNi9、並びに191系統の後代系統からそれぞれゲノムDNAを、DNeasy Plant Mini Kit(QIAGEN社製)を用いて抽出した。ゲノムDNA(750ng)を制限酵素PstI(NEB社、25unit)で37℃、2時間処理後、制限酵素BstNI(NEB社、25unit)を添加、60℃、2時間処理した。
【0069】
(3)アダプターライゲーション
(2)で処理したゲノムDNA断片(120ng)にPstI配列アダプター(5’-CACGATGGATCCAGTGCA-3’(配列番号53)、5’-CTGGATCCATCGTGCA-3’ (配列番号54))とT4 DNA Ligase(NEB社、800 unit)を加え、16℃、4時間以上処理した。これにより、(2)で処理したゲノムDNA断片のうち、両末端にPstI認識配列を有するゲノムDNA断片に対して選択的にアダプターを付加した。
【0070】
(4)PCR増幅
(3)で得られたアダプターを有するゲノムDNA断片(15ng)にPstI配列アダプター認識プライマー(5’-GATGGATCCAGTGCAG-3’(配列番号55))とTaq polymerase(タカラバイオ社PrimeSTAR、1.25unit)を加え、PCR(98℃を10秒間、55℃を15秒間、72℃を1分間、30サイクル後、72℃で3分間処理後、4℃で保存)でゲノムDNA断片を増幅した。
【0071】
(5)ラベル化
上述した(4)で得られたPCR増幅断片をカラム(QIAGEN社)で精製後、Cy3 9mer wobble(TriLink社、1O.D.)を加え、98℃、10分間処理後、氷上で10分間静置した。その後、Klenow(NEB社、100unit)を加え37℃、2時間処理した。そして、イソプロパノール沈殿によりラベル化サンプルを調整した。
【0072】
(6)ハイブリ・シグナル検出
(5)のラベル化サンプルを用い、NimbleGen Arrays User's Guideに従い、上記1.で作製したDNAマイクロアレイを用いてハイブリダイズを行い、ラベルに基づくシグナルを検出した。
【0073】
3.サトウキビ茎径のQTLの同定及びマーカーの開発
(1)遺伝地図データシート作成
上記2.にて検出したサトウキビ品種NiF8、Ni9及びこれらの交配後代系統(191系統)のシグナルデータから、NiF8由来の3004個のマーカー及びNi9由来の4569個のマーカーとなりうる遺伝子型データを取得した。この遺伝子型データを元にして、遺伝地図作成ソフトウェアAntMap(Iwata H, Ninomiya S (2006) AntMap: constructing genetic linkage maps using an ant colony optimization algorithm. Breed Sci 56: 371-378)を使用し、遺伝距離計算式Kosambiにより染色体におけるマーカーの位置情報を算出した。さらに、取得したマーカーの位置情報をもとに、Mapmaker/EXP ver.3.0(A Whitehead Institute for Biomedical Research Technical Report, Third Edition, January, 1993)により遺伝地図データシートを作成した。
【0074】
(2)茎径データの取得
供試したサトウキビ品種:NiF8及びNi9、並びに、交配後代191系統を、2009年4月、一区画(2.2m2)に13個体ずつ植え付けした。2010年3月、各区画5個体について茎を収穫した。収穫した茎の原料茎中央部に相当する節間のさらに中央部の短径を対象として直径計測された各系統の茎径の平均値を図3にまとめた。NiF8は21(mm)、Ni9は23(mm)のデータ区間にそれぞれ含まれる。
【0075】
(3)量的形質(Quantitative trait loci: QTL)の解析
上記(1)で得られた遺伝地図データシート及び上記(2)で得られた茎径データを元にして、遺伝解析ソフトQTL Cartographer(Wang S., C. J. Basten, and Z.-B. Zeng (2010). Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC; http://statgen.ncsu.edu/qtlcart/cartographer.html)を使用し、Composite interval mapping(CIM)法によりQTL解析を行った。このときLODの閾値を3.0とした。その結果、図4−7に示すように、サトウキビ品種NiF8の第2連鎖群に存在するマーカーN817421からN818319の区間内及び第40連鎖群に存在するマーカーN827451からN829378の区間内、並びにサトウキビ品種Ni9の第21連鎖群に存在するマーカーN915447からN914822の区間内及び第46連鎖群に存在するマーカーN917999からN916624の区間内に、LODの閾値を超えるピークが得られた。得られたピークは表5に示すように特定することができ、当該ピークの位置に茎径を増大させる機能を有する原因遺伝子(群)が存在することが示唆された。
【0076】
【表5】
【0077】
そして、図4−7に示すように、当該ピークの近傍に位置するマーカーは、茎径を増大させる機能を有する原因遺伝子(群)と連鎖して遺伝するためサトウキビの茎径関連マーカーとして使用できることが示された。すなわち、図4−7に示した52種類のマーカーは、サトウキビの茎径関連マーカーとして使用できることが明らかとなった。なお、上記2.における(6)シグナル検出の一例として、NiF8及びNi9、並びに12の後代系統(F1_1〜F1_12)における、サトウキビ品種NiF8の第2連鎖群に存在するマーカーN817421からN818319、サトウキビ品種NiF8の第40連鎖群に存在するマーカーN827451からN829378、サトウキビ品種Ni9の第21連鎖群に存在するマーカーN915447からN914822及びサトウキビ品種Ni9の第46連鎖群に存在するマーカーN917999からN916624に含まれる52種類のマーカーのシグナル値を表6−9に示し、なかでもN812115、N815144、N917810及びN919549については当該シグナル値をそれぞれ図8−11に示した。
【0078】
【表6】
【0079】
【表7】
【0080】
【表8】
【0081】
【表9】
【0082】
茎径が比較的に大である後代系統F1_5、F1_6、F1_7、F1_9等については、これら52種類のマーカーのシグナル値が非常に高いことが示された。これらの結果からも、サトウキビ品種NiF8の第2連鎖群に存在するマーカーN817421からN818319、サトウキビ品種NiF8の第40連鎖群に存在するマーカーN827451からN829378、サトウキビ品種Ni9の第21連鎖群に存在するマーカーN915447からN914822及びサトウキビ品種Ni9の第46連鎖群に存在するマーカーN917999からN916624に含まれる52種類のマーカーは、サトウキビの茎径関連マーカーとして使用できることが明らかとなった。
【技術分野】
【0001】
本発明は、茎径が増大した特性を有するサトウキビ系統を選抜することができる茎径関連マーカー及びその利用方法に関する。
【背景技術】
【0002】
サトウキビは、砂糖の原料、酒類原料など、食用に栽培されている他、バイオ燃料原料としての利用を含む様々な産業分野で利用されている。このような状況下、所望の特性(例えば、糖含有量、生長力の増強、新芽形成能、耐病性及び虫害抵抗性、耐寒性、葉長の増大、茎径や茎長の増大など)を有するサトウキビの新品種を開発するニーズがある。
【0003】
一般に、植物品種・系統の識別として、特性データを比較する「特性比較」、同一条件で栽培し比較する「比較栽培」、DNAを解析する「DNA分析」の3つの方法がある。特性比較および比較栽培による系統識別は、栽培条件の違いによる精度低下や多大な工数が必要とされる長期間の圃場調査など多くの問題を抱える。特に、サトウキビは、イネやトウモロコシなど、他のイネ科作物と比べ植物体が極めて大きく、圃場調査による系統識別の実施が困難である。また、葉長や茎径、茎長といった形質に特徴的な特性を有する品種・系統を識別するには、サトウキビを長期間栽培した後、これら特性データを収集する必要がある。また、サトウキビを長期間栽培したとしても、これら形質は環境の影響を受けやすいため精度良く識別することが困難である。
【0004】
さらに、サトウキビの新品種作出には、先ず、交配により数万種類の交配種を作製し、そこから実生選抜、さらに優良な系統を段階的に選抜し、最終的に所望な特性を有する2〜3種の新品種を得ることができる。このように、サトウキビの新品種作出には、非常に多くの系統を栽培・評価する必要があり、大規模な圃場を準備して多大な手間をかける必要がある。
【0005】
したがって、所望の特性を有するサトウキビ系統を、ゲノムに存在するマーカーを用いて識別する方法の開発が求められている。特に、サトウキビにおける新品種作出に際して、種々の特性について優れたマーカーが使用できれば、上述したようなサトウキビに特有の諸問題を回避でき、非常に有効なツールとなりうる。しかしながら、サトウキビは、高次倍数性で染色体数が多いため(約100〜130)、マーカー技術開発が遅れている。サトウキビでは、USDAにおいてSSRマーカーを用いた遺伝子型決定に関する報告があるものの(非特許文献1)、マーカー数及び各マーカーからの多型数が少ないことに起因して精度が低く、適用範囲がアメリカ・オーストラリア品種に限られるため、日本国内および台湾・インドなどの主要品種および有用な遺伝資源の系統識別に利用できない。
【0006】
また、非特許文献2は、マーカー数を増やし、各々のマーカーの特性関係を比較し、検証することでサトウキビにおける遺伝子地図を作製する可能性を示唆している。しかしながら、非特許文献2には、十分な数のマーカーが開示されておらず、目的とする特性に連鎖したマーカーも見つかっていない。
【先行技術文献】
【非特許文献】
【0007】
【非特許文献1】Maydica 48(2003)319-329 “Molecular genotyping of sugarcane clones with microsatellite DNA markers”
【非特許文献2】Nathalie Piperidis et al., Molecular Breeding, 2008, Vol 21, 233-247
【発明の概要】
【発明が解決しようとする課題】
【0008】
そこで、本発明は、サトウキビの量的形質の中でも茎径に関連するマーカーを提供することを目的とする。
【課題を解決するための手段】
【0009】
本発明者らは、上記課題を解決するために鋭意検討を重ねた結果、サトウキビにおける多数のマーカーを準備し、交雑後代系統における量的形質とマーカーとの連鎖解析によって、茎径が増大するといった量的形質に連鎖するマーカーを見いだし、本発明を完成するに至った。
【0010】
本発明は以下を包含する。
[1] サトウキビの染色体における配列番号1に示す塩基配列及び配列番号20に示す塩基配列により挟まれる領域、配列番号21に示す塩基配列及び配列番号34に示す塩基配列により挟まれる領域、配列番号35に示す塩基配列及び配列番号40に示す塩基配列により挟まれる領域、又は配列番号41に示す塩基配列及び配列番号52に示す塩基配列により挟まれる領域から選ばれる連続する核酸領域からなる、サトウキビの茎径関連マーカー。
[2] 上記核酸領域は、配列番号1〜52からなる群から選ばれるいずれか1つの塩基配列を含むことを特徴とする[1]のサトウキビの茎径関連マーカー。
[3] 上記核酸領域は、サトウキビの染色体における配列番号5に示す塩基配列と配列番号7に示す塩基配列とにより挟み込まれる領域、配列番号21に示す塩基配列と配列番号22に示す塩基配列とにより挟み込まれる領域、配列番号36に示す塩基配列と配列番号38に示す塩基配列とにより挟み込まれる領域、又は配列番号41に示す塩基配列と配列番号43に示す塩基配列とにより挟み込まれる領域に位置することを特徴とする[1]の茎径関連マーカー。
[4] 少なくとも一方の親がサトウキビである後代植物の染色体を抽出する工程と、
上記で得られた染色体における[1]乃至[3]のいずれかのサトウキビの茎径関連マーカーの存在・非存在を測定する工程とを含む、茎径が増大したサトウキビ系統の製造方法。
[5] 上記測定する工程では、上記サトウキビの茎径関連マーカーに対応するプローブを備えるDNAチップを使用することを特徴とする[4]のサトウキビ系統の製造方法。
[6] 上記後代植物は種子又は幼苗であり、当該種子又は幼苗から染色体を抽出することを特徴とする[4]のサトウキビ系統の製造方法。
【発明の効果】
【0011】
本発明によれば、サトウキビにおける量的形質の中でも茎径が増大するといった特性に連鎖する新規なサトウキビの茎径関連マーカーを提供することができる。本発明に係るサトウキビの茎径関連マーカーを利用することによって、サトウキビの交配系統における茎径を識別することができる。これにより、茎径が増大した特性を有するサトウキビ系統を非常に低コストに識別することができる。
【図面の簡単な説明】
【0012】
【図1】サトウキビ染色体のマーカーを得る際に使用したDNAマイクロアレイの製造フローを示す模式図である。
【図2】DNAマイクロアレイを使用したシグナル検出の工程を示す模式図である。
【図3】実施例で使用したサトウキビ品種・系統群について茎径データを示す特性図である。
【図4】サトウキビ品種NiF8における茎径に関するQTL解析の結果(第2連鎖群)を示す特性図である。
【図5】サトウキビ品種NiF8における茎径に関するQTL解析の結果(第40連鎖群)を示す特性図である。
【図6】サトウキビ品種Ni9における茎径に関するQTL解析の結果(第21連鎖群)を示す特性図である。
【図7】サトウキビ品種Ni9における茎径に関するQTL解析の結果(第46連鎖群)を示す特性図である。
【図8】各系統毎におけるN812115(NiF8第2連鎖群のマーカー)のシグナル値を示す特性図である。
【図9】各系統毎におけるN815144(NiF8第40連鎖群のマーカー)のシグナル値を示す特性図である。
【図10】各系統毎におけるN917810(Ni9第21連鎖群のマーカー)のシグナル値を示す特性図である。
【図11】各系統毎におけるN919549(Ni9第46連鎖群のマーカー)のシグナル値を示す特性図である。
【発明を実施するための形態】
【0013】
以下、本発明に係るサトウキビの茎径関連マーカー及びその利用方法、特にサトウキビの茎径関連マーカーを用いたサトウキビ系統の製造方法について説明する。
【0014】
サトウキビの茎径関連マーカー
本発明に係るサトウキビの茎径関連マーカーとは、サトウキビの染色体上に存在する特定の領域であり、サトウキビの茎径を増大させる形質の原因遺伝子(群)に連鎖して、サトウキビ茎径の増大という形質を判別できる機能を有する。すなわち、既知のサトウキビ系統を用いて得られた後代系統において、サトウキビの茎径関連マーカーの存在を確認することで茎径の増大という形質を有する系統であると判断することができる。
【0015】
ここで、茎径とは茎一本あたりの直径を意味し、当該分野において一般的な手法を用いて計測することができる。特に限定されないが、例えばサトウキビ茎を原料茎に調整し、茎中央部に相当する節間のさらに中央部の短径をノギスなどを用いて測定する。ここで「原料茎」とは粗糖などを生産する際の原料として利用される茎の部分であり、無処理のサトウキビ茎から糖度の低い梢頭部、葉、根等を除去した後の茎で、一般的に1m以上のものを指す。
【0016】
サトウキビとは、イネ科サトウキビ属に属する植物を意味する。また、サトウキビとは、所謂、高貴種(学名:Saccharum officinarum)と野生種(学名:Saccharum spontaneum)のいずれも含むことを意味する。既知のサトウキビ品種・系統としては、特に限定されず、日本国内にて使用可能なあらゆる品種・系統、日本国外において使用されている品種・系統等を含む意味である。例えば、サトウキビ日本国内育成品種としては、特に限定されないが、Ni1、NiN2、NiF3、NiF4、NiF5、Ni6、NiN7、NiF8、Ni9、NiTn10、Ni11、Ni12、Ni14、Ni15、Ni16、Ni17、NiTn19、NiTn20、Ni22及びNi23等を挙げることができる。また、サトウキビ日本国内主要品種としては、特に限定されないが、NiF8、Ni9、NiTn10及びNi15等を挙げることができる。さらに、サトウキビ日本国内導入主要品種としては、特に限定されないが、F177、Nco310及びF172等を挙げることができる。
【0017】
また、後代系統は、母本及び父本の両方がサトウキビ品種・系統である同種交配による系統であっても良いし、いずれか一方がサトウキビ品種・系統であり他方が近縁のErianthus arundinaceusであるような交雑系統であっても良い。また、後代品種は、いわゆる戻し交雑によって得られたものでも良い。
【0018】
サトウキビの茎径関連マーカーは、サトウキビ品種NiF8の染色体から独自に取得した3004個のマーカーを含む遺伝子連鎖地図およびサトウキビ品種Ni9の染色体から独自に取得した4569個のマーカーを含む遺伝子連鎖地図とサトウキビ茎径データとを用いたQTL(Quantitative Trait Loci)解析によって新たに同定されたものである。なお、サトウキビ茎径は、多数の遺伝子が関与していると考えられ連続分布をとる量的形質である。QTL解析には、遺伝解析ソフトQTL Cartographer(Wang S., C. J. Basten, and Z.-B. Zeng (2010). Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC)を使用し、Composite interval mapping(CIM)法を適用している。
【0019】
具体的に、上述したQTL解析により、ロッドスコア(LOD score)が所定の閾値(例えば3.0)以上となるピークを上記それぞれの遺伝子連鎖地図に含まれる4つの領域で見いだした。すなわち、当該ピークを含む約23.4cM(センチモルガン)領域及び約20.0cM領域(サトウキビ品種NiF8)、及並びに当該ピークを含む約37.2cM領域及び約33.7cM領域(サトウキビ品種Ni9)の4領域を特定した。ここで、「モルガン(M)」は、染色体上の遺伝子間の距離を相対的に示した単位であり、交叉価をパーセントにした値である。サトウキビの染色体において、1cMは、約2000kbに相当する。なお、このピーク位置又はその近傍には、茎径を増大させる形質の原因遺伝子(群)が存在することが示唆される。
【0020】
上記ピークを含む23.4cMの領域は、上記マーカーのうち表1に示す20種類のマーカーがこの順で含まれる領域である。
【0021】
【表1】
【0022】
上記ピークを含む20.0cMの領域は、上記マーカーのうち表2に示す14種類のマーカーがこの順で含まれる領域である。
【0023】
【表2】
【0024】
上記ピークを含む37.2cMの領域は、上記マーカーのうち表3に示す6種類のマーカーがこの順で含まれる領域である。
【0025】
【表3】
【0026】
上記ピークを含む33.7cMの領域は、上記マーカーのうち表4に示す12種類のマーカーがこの順で含まれる領域である。
【0027】
【表4】
【0028】
なお、表1−4において、連鎖群とは、QTL解析において特定された複数の連鎖群についてそれぞれ付された番号である。表1−4においてマーカー名とは、本発明で独自に取得したマーカーに付された名称である。表1−4においてシグナル閾値とは、マーカーの有無を判定するための閾値である。
【0029】
サトウキビ品種NiF8の上記23.4cMの領域に含まれるピークは、配列番号5に示す塩基配列からなるマーカー(N827956)及び配列番号7に示す塩基配列からなるマーカー(N822637)により挟み込まれる領域に存在している。
【0030】
また、サトウキビ品種NiF8の上記20.0cMの領域に含まれるピークは、配列番号21に示す塩基配列からなるマーカー(N827451)及び配列番号22に示す塩基配列からなるマーカー(N810332)により挟み込まれる領域に存在している。
【0031】
さらに、サトウキビ品種Ni9の上記37.2cMの領域に含まれるピークは、配列番号36に示す塩基配列からなるマーカー(N902171)及び配列番号38に示す塩基配列からなるマーカー(N918319)により挟み込まれる領域に存在している。
【0032】
またさらに、サトウキビ品種Ni9の上記33.7cMの領域に含まれるピークは、配列番号41に示す塩基配列からなるマーカー(N917999)及び配列番号43に示す塩基配列からなるマーカー(N917862)により挟み込まれる領域に存在している。
【0033】
表1−4に示したマーカーが含まれる4つの領域から選ばれる連続した核酸領域を、サトウキビの茎径関連マーカーとして使用することができる。ここで、核酸領域とは、サトウキビの染色体に存在する他の領域との同一性が95%以下、好ましくは90%以下、より好ましくは80%以下、最も好ましくは70%以下となるような塩基配列からなる領域を意味する。サトウキビの茎径関連マーカーとなる核酸領域と他の領域との同一性が上記範囲であれば、定法に従って、当該核酸領域を特異的に検出することができる。ここで、同一性の値は、例えばBLASTを用いてデフォルトパラメータを用いて算出することができる。
【0034】
また、サトウキビの茎径関連マーカーとなる核酸領域の塩基長は、少なくとも8塩基長以上、好ましくは15塩基長以上、より好ましくは20塩基長以上、最も好ましくは30塩基長とすることができる。サトウキビの茎径関連マーカーとなる核酸領域の塩基長が上記範囲であれば、定法に従って、当該核酸領域を特異的に検出することができる。
【0035】
特に、サトウキビの茎径関連マーカーとしては、サトウキビ品種NiF8の23.4cMの領域に含まれる20種類のマーカーのうち、配列番号5に示す塩基配列と配列番号7に示す塩基配列とにより挟み込まれる領域から選ばれることが好ましい。上記ピークが配列番号5に示す塩基配列と配列番号7に示す塩基配列とにより挟み込まれる領域に存在するためである。また、サトウキビの茎径関連マーカーとしては、サトウキビ品種NiF8の20.0cMの領域に含まれる14種類のマーカーのうち、配列番号21に示す塩基配列と配列番号22に示す塩基配列とにより挟み込まれる領域から選ばれることが好ましい。上記ピークが配列番号21に示す塩基配列と配列番号22に示す塩基配列とにより挟み込まれる領域に存在するためである。さらに、サトウキビの茎径関連マーカーとしては、サトウキビ品種Ni9の37.2cMの領域に含まれる6種類のマーカーのうち、配列番号36に示す塩基配列と配列番号38に示す塩基配列とにより挟み込まれる領域から選ばれることが好ましい。上記ピークが配列番号36に示す塩基配列と配列番号38に示す塩基配列とにより挟み込まれる領域に存在するためである。またさらに、サトウキビの茎径関連マーカーとしては、サトウキビ品種Ni9の33.7cMの領域に含まれる12種類のマーカーのうち、配列番号41に示す塩基配列と配列番号43に示す塩基配列とにより挟み込まれる領域から選ばれることが好ましい。上記ピークが配列番号41に示す塩基配列と配列番号43に示す塩基配列とにより挟み込まれる領域に存在するためである。
【0036】
また、サトウキビの茎径関連マーカーとしては、上記表1−4に示した52種類のマーカーから選ばれる1種類のマーカーを含む核酸領域とすることもできる。例えば、サトウキビの茎径関連マーカーとしては、サトウキビ品種NiF8の23.4cMの領域に含まれるピークの位置に最も近い配列番号6に示す塩基配列からなるマーカー(N805103)を含む核酸領域、サトウキビ品種NiF8の20.0cMの領域に含まれるピークの位置に最も近い配列番号21に示す塩基配列からなるマーカー(N827451)を含む核酸領域、サトウキビ品種Ni9に由来する37.2cMの領域に含まれるピークの位置に最も近い配列番号37に示す塩基配列からなるマーカー(N917810)、サトウキビ品種Ni9に由来する33.7cMの領域に含まれるピークの位置に最も近い配列番号42に示す塩基配列からなるマーカー(N919703)を含む核酸領域を使用することが好ましい。このとき、マーカーを含む核酸領域の塩基配列は、当該マーカーの塩基配列に基づいて設計したプライマーを用いたインバースPCRによって特定することができる。
【0037】
さらに、サトウキビの茎径関連マーカーとしては、上記52種類のマーカーそのものを使用することができる。すなわち、これら52種類のマーカーのうち1種類以上のマーカーをサトウキビの茎径関連マーカーとして使用することができる。例えば、サトウキビ品種NiF8の23.4cMの領域に含まれるピークの位置に最も近い配列番号6に示す塩基配列からなるマーカー(N805103)、サトウキビ品種NiF8の20.0cMの領域に含まれるピークの位置に最も近い配列番号21に示す塩基配列からなるマーカー(N827451)、サトウキビ品種Ni9に由来する37.2cMの領域に含まれるピークの位置に最も近い配列番号37に示す塩基配列からなるマーカー(N917810)、サトウキビ品種Ni9に由来する33.7cMの領域に含まれるピークの位置に最も近い配列番号42に示す塩基配列からなるマーカー(N919703)を使用することが好ましい。
【0038】
サトウキビにおけるマーカーの同定
本発明では、上述したように、サトウキビ品種NiF8の染色体から独自に取得した3004個のマーカーおよびサトウキビ品種Ni9の染色体から独自に取得した4569個のマーカーからサトウキビの茎径関連マーカーを特定した。ここでは、これらのマーカーについて説明する。これらのマーカーを同定する際には、特願2009-283430号に開示された方法を適用したDNAマイクロアレイを使用することができる。
【0039】
具体的に、サトウキビの染色体から独自に取得したこれらのマーカーは、特願2009-283430号に開示された方法により設計されたプローブを有するDNAマイクロアレイを使用する。プローブの設計方法は、図1に示すように、先ず、サトウキビからゲノムDNAを抽出する(工程1a)。次に、抽出したゲノムDNAを1又は複数の制限酵素により消化する(工程1b)。なお、図1に示した例では、制限酵素A及び制限酵素Bの2種類の制限酵素をこの順で用いてゲノムDNAを消化している。ここで、制限酵素としては、特に限定されないが、例えば、PstI、EcoRI、HindIII、BstNI、HpaII、HaeIII等を使用することができる。特に制限酵素としては、ゲノムDNAを完全に消化した際に20〜10000塩基長のゲノムDNA断片となるよう、認識配列の出現頻度等を考慮して適宜選択することができる。また、複数の制限酵素を使用する場合、全ての制限酵素を使用した後のゲノムDNA断片が200〜6000塩基長となっていることが好ましい。さらに、複数の制限酵素を使用する場合、処理に供する制限酵素の順序は特に限定されず、また、処理条件(溶液組成や温度等)が共通する場合には複数の制限酵素を同一の反応系で使用しても良い。すなわち、図1に示した例においては、制限酵素A及び制限酵素Bをこの順で使用してゲノムDNAを消化しているが、制限酵素A及び制限酵素Bを同じ反応系で同時に使用してゲノムDNAを消化しても良いし、制限酵素B及び制限酵素Aをこの順で使用してゲノムDNAを消化してもよい。さらに、使用する制限酵素の数は3以上であってもよい。
【0040】
次に、制限酵素処理後のゲノムDNA断片に対してアダプターを結合する(工程1c)。ここで、アダプターとは、上述した制限酵素処理によって得られたゲノムDNA断片の両端に結合できるものであれば特に限定されない。アダプターとしては、例えば、制限酵素処理によってゲノムDNAの両末端に形成される突出末端(粘着末端)に対して相補的な一本鎖を有し、詳細を後述する増幅処理の際に使用するプライマーがハイブリダイズしうるプライマー結合配列を有するものを使用することができる。また、アダプターとしては、上記突出末端(粘着末端)に対して相補的な一本鎖を有し、クローニングする際のベクターに組み入れるための制限酵素認識部位を有するものを使用することもできる。
【0041】
また、複数の制限酵素を使用してゲノムDNAを消化した場合には、各制限酵素に対応する複数のアダプターを準備して使用することができる。すなわち、複数の制限酵素でゲノムDNAを消化した場合に生ずる複数種類の突出末端のそれぞれに対して、相補的な一本鎖を有する複数のアダプターを使用することができる。このとき、複数の制限酵素に対応する複数のアダプターは、共通するプライマーがハイブリダイズできるように共通するプライマー結合配列を有しているものであっても良いし、それぞれ異なるプライマーがハイブリダイズできるように異なるプライマー結合配列を有するものであっても良い。
【0042】
さらに、複数の制限酵素を使用してゲノムDNAを消化した場合、アダプターとしては、使用した複数の制限酵素のなかから選ばれる1つの制限酵素若しくは、使用した制限酵素のうち一部の制限酵素に対応するアダプターを準備して使用することもできる。
【0043】
次に、両末端にアダプターが付加されたゲノムDNA断片を増幅する(工程1d)。プライマー結合配列を有するアダプターを使用した場合には、当該プライマー結合配列にハイブリダイズできるプライマーを使用することで上記ゲノムDNA断片を増幅することができる。或いは、アダプターを付加したゲノムDNA断片を、アダプター配列を利用してベクターにクローニングし、当該ベクターにおける所定の領域にハイブリダイズできるプライマーを用いてゲノムDNA断片を増幅することができる。なお、プライマーを用いたゲノムDNA断片の増幅反応としては、一例としてPCRを使用することができる。
【0044】
また、複数の制限酵素を使用してゲノムDNAを消化するとともに、各制限酵素に対応する複数のアダプターをゲノムDNA断片に連結した場合、複数の制限酵素を用いた処理によって得られたゲノムDNA断片の全てにアダプターが連結されることとなる。この場合、アダプターに含まれるプライマー結合配列を用いて核酸増幅反応を行うことで、得られた全てのゲノムDNA断片を増幅することができる。
【0045】
或いは、複数の制限酵素を使用してゲノムDNAを消化するとともに、使用した複数の制限酵素のなかから選ばれる1つ制限酵素若しくは、使用した制限酵素のうち一部の制限酵素に対応するアダプターをゲノムDNA断片に連結した場合、得られたゲノムDNA断片のうち、選ばれた制限酵素の認識配列を両末端に有するゲノムDNA断片のみを増幅することができる。
【0046】
次に、増幅されたゲノムDNA断片の塩基配列を決定し(工程1e)、当該ゲノムDNA断片より短い塩基長を有し、ゲノムDNA断片内の少なくとも一部をカバーする1又は複数の領域を特定し、特定した1又は複数の領域を、サトウキビにおけるプローブとして設計する(工程1f)。ゲノムDNA断片の塩基配列を決定する方法は、特に限定されず、サンガー法等を適用したDNAシークエンサーを利用した従来公知の方法を使用することができる。ここで、設計する領域としては、上述したように、例えば20〜100塩基長、好ましくは30〜90塩基長、より好ましくは50〜75塩基長とする。
【0047】
以上のように、サトウキビから抽出したゲノムDNAを使用して多数のプローブを設計し、設計したプローブの塩基配列に基づいて、担体上にて目的の塩基配列を有するオリゴヌクレオチドを合成することでDNAマイクロアレイを作製することができる。このように作製したDNAマイクロアレイを使用することで、上述した配列番号1〜52に示した52種類のサトウキビの茎径関連マーカーを含む3004個および4569個のマーカーを、サトウキビ品種NiF8及びNi9よりそれぞれ同定することができる。
【0048】
より具体的に、本発明者らは、既知のサトウキビ品種NiF8、Ni9及びこれらの交配後代系統(191系統)について、上述したDNAマイクロアレイを用いてシグナルデータを取得した。そして、得られたシグナルデータから遺伝子型データを取得し、この遺伝子型データを元にして、遺伝地図作成ソフトウェアAntMap(Iwata H, Ninomiya S (2006) AntMap: constructing genetic linkage maps using an ant colony optimization algorithm. Breed Sci 56: 371-378)を使用し、遺伝距離計算式Kosambiにより染色体におけるマーカーの位置情報を算出した。さらに、取得したマーカーの位置情報をもとに、Mapmaker/EXP ver.3.0(A Whitehead Institute for Biomedical Research Technical Report, Third Edition, January, 1993)により遺伝地図データシートを作成した。その結果、上述した配列番号1〜52に示した52種類のサトウキビの茎径関連マーカーを含む3004個及び4569個のマーカーを、サトウキビ品種NiF8及びNi9よりそれぞれ同定している。
【0049】
サトウキビの茎径関連マーカーの利用
サトウキビの茎径関連マーカーを利用することで、後代系統等に茎径の表現型が未知のサトウキビ系統について茎径の増大という表現型を示す系統であるか判断することができる。ここで、サトウキビの茎径関連マーカーを利用するとは、当該マーカーに対応するプローブを有するDNAマイクロアレイを利用する形態を含む意味である。サトウキビの茎径関連マーカーに対応するプローブとは、上述のように定義されたサトウキビの茎径関連マーカーに対して、ストリンジェントな条件下で特異的にハイブリダイズできるオリゴヌクレオチドを意味する。このようなオリゴヌクレオチドは、例えば、上述のように定義されたサトウキビの茎径関連マーカーの塩基配列又はその相補鎖の少なくとも連続する10塩基、15塩基、20塩基、25塩基、30塩基、35塩基、40塩基、45塩基、50塩基又はそれ以上の塩基長の部分領域若しくは全領域として設計することができる。なお、このプローブを有するDNAマイクロアレイとしては、ガラスやシリコーン等の平面基板を担体とするマイクロアレイや、マイクロビーズを担体とするビーズアレイ、或いは中空繊維の内壁にプローブを固定する3次元マイクロアレイ等の如何なるタイプのマイクロアレイであってもよい。
【0050】
以上のように作製されたDNAマイクロアレイを使用することで、後代系統等に代表される茎径の表現型が未知のサトウキビ系統について、茎径の増大という表現型を示す系統であるか判断することができる。なお、上述したDNAマイクロアレイを使用する方法以外であっても、従来公知の手法を用いて上述したサトウキビの茎径関連マーカーを検出して、茎径の表現型が未知のサトウキビ系統について茎径の増大という形質を有する系統であるか判断してもよい。
【0051】
DNAマイクロアレイを使用する方法をより詳細に説明する。図2に示すように、先ず供試サトウキビからゲノムDNAを抽出する。この供試サトウキビとは、後代系統等の茎径の表現型が未知のサトウキビ系統のことであり、茎径が増大する形質を有するか判定する対象となるサトウキビ系統である。次に、抽出したゲノムDNAを、DNAマイクロアレイを作製する際に使用した制限酵素で消化して複数のゲノムDNA断片を調整する。次に、得られたゲノムDNA断片と、DNAマイクロアレイを作製する際に使用したアダプターとを連結する。次に、両末端にアダプターが付加されたゲノムDNA断片を、DNAマイクロアレイを作製する際に使用したプライマーを用いて増幅する。これにより、DNAマイクロアレイを作製する際の工程1dで増幅したゲノムDNA断片に対応する、供試サトウキビ由来のゲノムDNA断片を増幅することができる。
【0052】
この工程においては、アダプターが付加されたゲノムDNA断片のうち、所定のゲノムDNA断片を選択的に増幅してもよい。例えば、複数の制限酵素に対応する複数のアダプターを使用した場合には、特定のアダプターが付加されたゲノムDNA断片を選択的に増幅することができる。また、複数の制限酵素でゲノムDNAを消化した場合、得られたゲノムDNA断片のうち、所定の制限酵素に対応する突出末端を有するゲノムDNA断片のみにアダプターを付加することで、アダプターが付加されたゲノムDNA断片を選択的に増幅することができる。このように、所定のゲノムDNA断片を選択的に増幅することで濃縮することができる。
【0053】
次に、増幅したゲノムDNA断片に標識を付加する。標識としては、従来公知の如何なる物質を使用しても良い。標識としては、例えば蛍光分子、色素分子、放射性分子等を使用することができる。なお、本工程は、ゲノムDNA断片を増幅する工程において標識を有するヌクレオチドを用いることで省略することができる。上記工程において標識を有するヌクレオチドを用いてゲノムDNA断片を増幅することで、増幅されたDNA断片が標識化されるためである。
【0054】
次に、標識を有するゲノムDNA断片を所定の条件下でDNAマイクロアレイに接触させ、DNAマイクロアレイに固定されたプローブと標識を有するゲノムDNA断片とをハイブリダイズさせる。このとき、ハイブリダイズさせる際には高いストリンジェンシー条件とすることが好ましい。このような高いストリンジェンシー条件とすることによって、供試サトウキビにサトウキビの茎径関連マーカーが存在しているか否かを、より高精度に判定することができる。なお、ストリンジェンシー条件は、反応温度及び塩濃度で調節することができる。すなわち、より高温とすることでより高いストリンジェンシー条件となり、またより低い塩濃度でより高いストリンジェンシー条件となる。例えば、50〜75塩基長のプローブを使用する場合、ハイブリダイゼーション条件としては、40〜44℃、0.2SDS、6×SSCの条件とすることでより高いストリンジェンシー条件とすることができる。
【0055】
また、プローブと標識を有するゲノムDNA断片とのハイブリダイズは、標識に基づいて検出することができる。すなわち、上述した標識を有するゲノムDNA断片とプローブのハイブリダイズ反応の後、未反応のゲノムDNA断片等を洗浄し、その後、プローブに対して特異的にハイブリダイズしたゲノムDNA断片の標識を観察する。例えば、標識が蛍光物質である場合にはその蛍光波長を検出し、標識が色素分子であればその色素波長を検出する。より具体的には、通常のDNAマイクロアレイ解析に使用している、蛍光検出装置やイメージアナライザー等の装置を使用することができる。
【0056】
以上のように、上述したDNAマイクロアレイを使用することにより、供試サトウキビが上述したサトウキビの茎径関連マーカーを有するか否か判断することができる。特に、上述した方法では、供試サトウキビを実際の茎径が測定可能な程度まで成長させる必要はなく、例えば後代系統の種子や当該種子を発芽させた幼苗を使用することができる。したがって、サトウキビの茎径関連マーカーを利用することによって、供試サトウキビを生育させるための圃場やその他、生育のためのコストを大幅に削減することができる。
【0057】
特に、サトウキビの新品種作出に際して、先ず、交配により数万種類の交配種を作製した後、実生選抜に先立って若しくは実生選抜に代えて、サトウキビの茎径関連マーカーを利用した判断を行うことが好ましい。これにより、実際の圃場において、優良な系統を栽培する数を大幅に削減することができ、サトウキビの新品種作出に係る手間やコストを大幅に抑制することができる。
【0058】
上記サトウキビの茎径関連マーカーを利用することで、茎径を増大させる形質の原因遺伝子又は原因遺伝子群を単離することができる。単離方法としては、従来公知の手法を用いることができる(「バイオ実験イラストレイテッド4 苦労なしのクローニング」真壁和裕著(1997)、秀潤社)。例えば、上記定義のサトウキビの茎径関連マーカーに対応するプライマー又はプローブを作製し、サトウキビのゲノムDNA又はcDNAをスクリーニングすることによって、茎径を増大させる形質の原因遺伝子を単離することができる。また、上記サトウキビのゲノムDNA又はcDNAに代えて、その他のイネ科植物由来のゲノムDNA又はcDNAを、当該サトウキビの茎径関連マーカーに対応するプライマー又はプローブを用いてスクリーニングすることによって、サトウキビ以外のイネ科植物の茎径を増大させる形質の原因遺伝子又は原因遺伝子群を単離することも可能である。
【0059】
また、上記で得られた茎径を増大させる形質の原因遺伝子が組み込まれた組換えベクターを用いて植物細胞を形質転換することによって、茎径が増大した形質を有する形質転換植物体を製造することもできる。
【実施例】
【0060】
以下、実施例により本発明をより詳細に説明するが、本発明の技術的範囲は以下の実施例に限定されるものではない。
【0061】
1.DNAマイクロアレイ用プローブの作成
(1)材料
サトウキビ品種:NiF8、Ni9、US56-15-8、POJ2878、Q165、R570、Co290及びB3439、サトウキビ近縁野生種:Glagah Kloet、Chunee、Natal Uba及びRobustum9、並びにエリアンサス:IJ76-349及びJW630を用いた。
【0062】
(2)制限酵素処理
これらサトウキビ品種、サトウキビ近縁野生種及びエリアンサスからそれぞれゲノムDNAを、DNeasy Plant Mini Kit(QIAGEN社製)を用いて抽出した。ゲノムDNA(750ng)を制限酵素PstI(NEB社、25unit)で37℃、2時間処理後、制限酵素BstNI(NEB社、25unit)を添加、60℃、2時間処理した。
【0063】
(3)アダプターライゲーション
(2)で処理したゲノムDNA断片(120ng)にPstI配列アダプター(5’-CACGATGGATCCAGTGCA-3’(配列番号53)、5’-CTGGATCCATCGTGCA-3’ (配列番号54))とT4 DNA Ligase(NEB社、800 unit)を加え、16℃、4時間以上処理した。これにより、(2)で処理したゲノムDNA断片のうち、両末端にPstI認識配列を有するゲノムDNA断片に対して選択的にアダプターを付加した。
【0064】
(4)PCR増幅
(3)で得られたアダプターを有するゲノムDNA断片(15ng)にPstI配列アダプター認識プライマー(5’-GATGGATCCAGTGCAG-3’(配列番号55))とTaq polymerase(タカラバイオ社PrimeSTAR、1.25unit)を加え、PCR(98℃を10秒間、55℃を15秒間、72℃を1分間、30サイクル後、72℃で3分間処理後、4℃で保存)でゲノムDNA断片を増幅した。
【0065】
(5)ゲノムシークエンス取得
(4)においてPCR増幅したゲノムDNA断片についてFLX454(Roche社)又はサンガー法により塩基配列を決定した。また、ゲノムデータベース(Gramene:http://www.gramene.org/)に格納されたソルガム全ゲノム配列情報から、PstI認識配列により挟み込まれる塩基配列情報を取得した。
【0066】
(6)プローブ設計及びDNAマイクロアレイの作成
(5)のゲノムシークエンス情報をもとに50〜75bpのプローブを設計した。設計したプローブの塩基配列情報をもとに、これらプローブを有するDNAマイクロアレイを作製した。
【0067】
2.DNAマイクロアレイを用いたシグナルデータの取得
(1)材料
サトウキビ品種・系統:NiF8及びNi9、並びに、これらの交配後代191系統を用いた。
【0068】
(2)制限酵素処理
これらNiF8及びNi9、並びに191系統の後代系統からそれぞれゲノムDNAを、DNeasy Plant Mini Kit(QIAGEN社製)を用いて抽出した。ゲノムDNA(750ng)を制限酵素PstI(NEB社、25unit)で37℃、2時間処理後、制限酵素BstNI(NEB社、25unit)を添加、60℃、2時間処理した。
【0069】
(3)アダプターライゲーション
(2)で処理したゲノムDNA断片(120ng)にPstI配列アダプター(5’-CACGATGGATCCAGTGCA-3’(配列番号53)、5’-CTGGATCCATCGTGCA-3’ (配列番号54))とT4 DNA Ligase(NEB社、800 unit)を加え、16℃、4時間以上処理した。これにより、(2)で処理したゲノムDNA断片のうち、両末端にPstI認識配列を有するゲノムDNA断片に対して選択的にアダプターを付加した。
【0070】
(4)PCR増幅
(3)で得られたアダプターを有するゲノムDNA断片(15ng)にPstI配列アダプター認識プライマー(5’-GATGGATCCAGTGCAG-3’(配列番号55))とTaq polymerase(タカラバイオ社PrimeSTAR、1.25unit)を加え、PCR(98℃を10秒間、55℃を15秒間、72℃を1分間、30サイクル後、72℃で3分間処理後、4℃で保存)でゲノムDNA断片を増幅した。
【0071】
(5)ラベル化
上述した(4)で得られたPCR増幅断片をカラム(QIAGEN社)で精製後、Cy3 9mer wobble(TriLink社、1O.D.)を加え、98℃、10分間処理後、氷上で10分間静置した。その後、Klenow(NEB社、100unit)を加え37℃、2時間処理した。そして、イソプロパノール沈殿によりラベル化サンプルを調整した。
【0072】
(6)ハイブリ・シグナル検出
(5)のラベル化サンプルを用い、NimbleGen Arrays User's Guideに従い、上記1.で作製したDNAマイクロアレイを用いてハイブリダイズを行い、ラベルに基づくシグナルを検出した。
【0073】
3.サトウキビ茎径のQTLの同定及びマーカーの開発
(1)遺伝地図データシート作成
上記2.にて検出したサトウキビ品種NiF8、Ni9及びこれらの交配後代系統(191系統)のシグナルデータから、NiF8由来の3004個のマーカー及びNi9由来の4569個のマーカーとなりうる遺伝子型データを取得した。この遺伝子型データを元にして、遺伝地図作成ソフトウェアAntMap(Iwata H, Ninomiya S (2006) AntMap: constructing genetic linkage maps using an ant colony optimization algorithm. Breed Sci 56: 371-378)を使用し、遺伝距離計算式Kosambiにより染色体におけるマーカーの位置情報を算出した。さらに、取得したマーカーの位置情報をもとに、Mapmaker/EXP ver.3.0(A Whitehead Institute for Biomedical Research Technical Report, Third Edition, January, 1993)により遺伝地図データシートを作成した。
【0074】
(2)茎径データの取得
供試したサトウキビ品種:NiF8及びNi9、並びに、交配後代191系統を、2009年4月、一区画(2.2m2)に13個体ずつ植え付けした。2010年3月、各区画5個体について茎を収穫した。収穫した茎の原料茎中央部に相当する節間のさらに中央部の短径を対象として直径計測された各系統の茎径の平均値を図3にまとめた。NiF8は21(mm)、Ni9は23(mm)のデータ区間にそれぞれ含まれる。
【0075】
(3)量的形質(Quantitative trait loci: QTL)の解析
上記(1)で得られた遺伝地図データシート及び上記(2)で得られた茎径データを元にして、遺伝解析ソフトQTL Cartographer(Wang S., C. J. Basten, and Z.-B. Zeng (2010). Windows QTL Cartographer 2.5. Department of Statistics, North Carolina State University, Raleigh, NC; http://statgen.ncsu.edu/qtlcart/cartographer.html)を使用し、Composite interval mapping(CIM)法によりQTL解析を行った。このときLODの閾値を3.0とした。その結果、図4−7に示すように、サトウキビ品種NiF8の第2連鎖群に存在するマーカーN817421からN818319の区間内及び第40連鎖群に存在するマーカーN827451からN829378の区間内、並びにサトウキビ品種Ni9の第21連鎖群に存在するマーカーN915447からN914822の区間内及び第46連鎖群に存在するマーカーN917999からN916624の区間内に、LODの閾値を超えるピークが得られた。得られたピークは表5に示すように特定することができ、当該ピークの位置に茎径を増大させる機能を有する原因遺伝子(群)が存在することが示唆された。
【0076】
【表5】
【0077】
そして、図4−7に示すように、当該ピークの近傍に位置するマーカーは、茎径を増大させる機能を有する原因遺伝子(群)と連鎖して遺伝するためサトウキビの茎径関連マーカーとして使用できることが示された。すなわち、図4−7に示した52種類のマーカーは、サトウキビの茎径関連マーカーとして使用できることが明らかとなった。なお、上記2.における(6)シグナル検出の一例として、NiF8及びNi9、並びに12の後代系統(F1_1〜F1_12)における、サトウキビ品種NiF8の第2連鎖群に存在するマーカーN817421からN818319、サトウキビ品種NiF8の第40連鎖群に存在するマーカーN827451からN829378、サトウキビ品種Ni9の第21連鎖群に存在するマーカーN915447からN914822及びサトウキビ品種Ni9の第46連鎖群に存在するマーカーN917999からN916624に含まれる52種類のマーカーのシグナル値を表6−9に示し、なかでもN812115、N815144、N917810及びN919549については当該シグナル値をそれぞれ図8−11に示した。
【0078】
【表6】
【0079】
【表7】
【0080】
【表8】
【0081】
【表9】
【0082】
茎径が比較的に大である後代系統F1_5、F1_6、F1_7、F1_9等については、これら52種類のマーカーのシグナル値が非常に高いことが示された。これらの結果からも、サトウキビ品種NiF8の第2連鎖群に存在するマーカーN817421からN818319、サトウキビ品種NiF8の第40連鎖群に存在するマーカーN827451からN829378、サトウキビ品種Ni9の第21連鎖群に存在するマーカーN915447からN914822及びサトウキビ品種Ni9の第46連鎖群に存在するマーカーN917999からN916624に含まれる52種類のマーカーは、サトウキビの茎径関連マーカーとして使用できることが明らかとなった。
【特許請求の範囲】
【請求項1】
サトウキビの染色体における配列番号1に示す塩基配列及び配列番号20に示す塩基配列により挟まれる領域、配列番号21に示す塩基配列及び配列番号34に示す塩基配列により挟まれる領域、配列番号35に示す塩基配列及び配列番号40に示す塩基配列により挟まれる領域、又は配列番号41に示す塩基配列及び配列番号52に示す塩基配列により挟まれる領域から選ばれる連続する核酸領域からなる、サトウキビの茎径関連マーカー。
【請求項2】
上記核酸領域は、配列番号1〜52からなる群から選ばれるいずれか1つの塩基配列を含むことを特徴とする請求項1記載のサトウキビの茎径関連マーカー。
【請求項3】
上記核酸領域は、サトウキビの染色体における配列番号5に示す塩基配列と配列番号7に示す塩基配列とにより挟み込まれる領域、配列番号21に示す塩基配列と配列番号22に示す塩基配列とにより挟み込まれる領域、配列番号36に示す塩基配列と配列番号38に示す塩基配列とにより挟み込まれる領域、又は配列番号41に示す塩基配列と配列番号43に示す塩基配列とにより挟み込まれる領域に位置することを特徴とする請求項1記載の茎径関連マーカー。
【請求項4】
少なくとも一方の親がサトウキビである後代植物の染色体を抽出する工程と、
上記で得られた染色体における請求項1乃至3いずれか一項に記載のサトウキビの茎径関連マーカーの存在・非存在を測定する工程とを含む、茎径が増大したサトウキビ系統の製造方法。
【請求項5】
上記測定する工程では、上記サトウキビの茎径関連マーカーに対応するプローブを備えるDNAチップを使用することを特徴とする請求項4記載のサトウキビ系統の製造方法。
【請求項6】
上記後代植物は種子又は幼苗であり、当該種子又は幼苗から染色体を抽出することを特徴とする請求項4記載のサトウキビ系統の製造方法。
【請求項1】
サトウキビの染色体における配列番号1に示す塩基配列及び配列番号20に示す塩基配列により挟まれる領域、配列番号21に示す塩基配列及び配列番号34に示す塩基配列により挟まれる領域、配列番号35に示す塩基配列及び配列番号40に示す塩基配列により挟まれる領域、又は配列番号41に示す塩基配列及び配列番号52に示す塩基配列により挟まれる領域から選ばれる連続する核酸領域からなる、サトウキビの茎径関連マーカー。
【請求項2】
上記核酸領域は、配列番号1〜52からなる群から選ばれるいずれか1つの塩基配列を含むことを特徴とする請求項1記載のサトウキビの茎径関連マーカー。
【請求項3】
上記核酸領域は、サトウキビの染色体における配列番号5に示す塩基配列と配列番号7に示す塩基配列とにより挟み込まれる領域、配列番号21に示す塩基配列と配列番号22に示す塩基配列とにより挟み込まれる領域、配列番号36に示す塩基配列と配列番号38に示す塩基配列とにより挟み込まれる領域、又は配列番号41に示す塩基配列と配列番号43に示す塩基配列とにより挟み込まれる領域に位置することを特徴とする請求項1記載の茎径関連マーカー。
【請求項4】
少なくとも一方の親がサトウキビである後代植物の染色体を抽出する工程と、
上記で得られた染色体における請求項1乃至3いずれか一項に記載のサトウキビの茎径関連マーカーの存在・非存在を測定する工程とを含む、茎径が増大したサトウキビ系統の製造方法。
【請求項5】
上記測定する工程では、上記サトウキビの茎径関連マーカーに対応するプローブを備えるDNAチップを使用することを特徴とする請求項4記載のサトウキビ系統の製造方法。
【請求項6】
上記後代植物は種子又は幼苗であり、当該種子又は幼苗から染色体を抽出することを特徴とする請求項4記載のサトウキビ系統の製造方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【公開番号】特開2012−115245(P2012−115245A)
【公開日】平成24年6月21日(2012.6.21)
【国際特許分類】
【出願番号】特願2010−270822(P2010−270822)
【出願日】平成22年12月3日(2010.12.3)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
【公開日】平成24年6月21日(2012.6.21)
【国際特許分類】
【出願日】平成22年12月3日(2010.12.3)
【出願人】(000003207)トヨタ自動車株式会社 (59,920)
【出願人】(501203344)独立行政法人農業・食品産業技術総合研究機構 (827)
【Fターム(参考)】
[ Back to top ]