
サポシンA特異的抗体及びその作成方法
【課題】スフィンゴ脂質活性化タンパク質であるサポシンAに特異的な抗体の提供。
【解決手段】不活性型プロサポシンの143位のリジン残基において切断されることで生成される、当該143位のリジン残基をC末端として有するサポシンAに特異的な抗体とその作成方法であって、サポシンAのアミノ酸残基139位〜143位のアミノ酸配列:Gly-Ser-Leu-Gln-Lys
を含んで成り、上記配列のC末端のLys残基をC末端とするペプチドを抗原として用いる方法。
【解決手段】不活性型プロサポシンの143位のリジン残基において切断されることで生成される、当該143位のリジン残基をC末端として有するサポシンAに特異的な抗体とその作成方法であって、サポシンAのアミノ酸残基139位〜143位のアミノ酸配列:Gly-Ser-Leu-Gln-Lys
を含んで成り、上記配列のC末端のLys残基をC末端とするペプチドを抗原として用いる方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、サポシンAに特異的な抗体及びその作成方法に関する。
【背景技術】
【0002】
ケラチノサイトは終末分化することで、有害な環境に対する「角層」と称される保護バリアーを形成する。角層細胞間脂質はこの保護バリアーにおいて重要な役割を担っている。角層細胞間脂質は、表皮ケラチノサイトにおいて分化とともに生合成され、オドランド小体に蓄積される。角層細胞間脂質の主成分であるセラミドは、オドランド小体の開口分泌により角層細胞間に放出されたグルコシルセラミドの糖鎖が、グルコセレブロシダーゼにより切断されることで生じる。また、別の経路として、スフィンゴミエリナーゼによるスフィンゴミエリンからセラミドへの分解経路も知られている。
【0003】
スフィンゴ脂質活性化タンパク質であるサポシンは、スフィンゴ脂質の加水分解に必須のタンパク質であり、4種類のサポシン(サポシンA、サポシンB、サポシンC、サポシンD)の存在が知られている。サポシンA、B、C、及びDは極めて相同性の高い糖タンパク質で、前駆タンパク質であるプロサポシンがライソゾームで分解されて生産される。サポシンA、B、C、Dはそれぞれプロサポシンのアミノ酸60〜143位、195〜275位、311〜390位、405〜487位に相当する(Kishimoto et al., Journal of Lipid Research (1992) 33, 1255-67)。サポシンは、角層間脂質の生成に必要なセラミド合成酵素、例えばグルコセレブロシダーゼ、スフィンゴミエリナーゼ等の補酵素として働く因子であることから、サポシンは表皮においては角層間脂質の合成や、皮膚バリアー機能にとって重要であると考えられる。
【0004】
これまで、サポシンA、C、Dが臓器に集積するとリソソーム蓄積病(例えば、ゴーシュ病、ニーマンピック病)が生じることや、プロサポシン遺伝子の突然変異が神経障害性の表現型をもたらすことが確認されている。皮膚との関連では、アトピー性皮膚炎や乾癬に罹患している患者において、プロサポシンが低下しているとの報告がある(それぞれ、Tezuka et al., Journal of Investigative Dermatology (1997) 109, 319-323(非特許文献1)及びAlessandrini et al., Journal of Investigative Dermatology (2001) 116, 394-400(非特許文献2)を参照のこと)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Tezuka et al., Journal of Investigative Dermatology (1997) 109, 319-323
【非特許文献2】Alessandrini et al., Journal of Investigative Dermatology (2001) 116, 394-400
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかしながら、プロサポシンの活性化の機構や活性化サポシンの機能に関しては明らかではなく、各サポシンが実際にどのように表皮において分布し、機能しているのかに解明が待たれる。プロサポシンの活性化の機構及び機能、更には活性型の生体内分布及びその生理学的役割の解明には、その活性化形態に特異的な抗体の利用が不可欠である。
【課題を解決するための手段】
【0007】
本発明者はサポシンAに特異的に結合する抗体の作製に成功した。
【0008】
第一の態様において、本発明はサポシンAに特異的な抗体の作成方法を提供する。サポシンAは、不活性型プロサポシンの143位のリジン残基において切断されることで生成される、当該143位のリジン残基をC末端として有するサポシンAに特異的な抗体である。この抗体作成方法は、不活性型プロサポシンのアミノ酸残基139位〜143位のアミノ酸配列:
Gly-Ser-Leu-Gln-Lys (配列番号1)
を含んで成り、上記配列のC末端のLys残基をC末端とするペプチドを抗原として用いることを特徴とする。好ましくは、上記ペプチドは配列番号1から成るペプチドである。
本発明はさらに、上記方法により作成された、不活性型プロサポシンの143位のリジン残基において切断されることで生成される、143位のリジン残基をC末端として有するサポシンAに特異的な抗体を提供する。
【0009】
別の観点において、本発明は組織、もしくは細胞、特に表皮中のサポシンAを検出するため、上記方法により作成された抗体を使用することを特徴とする方法を提供する。
【発明の効果】
【0010】
サポシンAは他のサポシンと比較して顕著に異なる挙動を示し、皮膚バリアー機能に関与している可能性が高く、その作用機能の解明は極めて有意義なものと考えられる。本願発明にかかるサポシンA特異的抗体はこのようなサポシンAの同定に有用であり、その結果カスパーゼ−14の生体内での役割を解明するのに極めて有用である。
【図面の簡単な説明】
【0011】
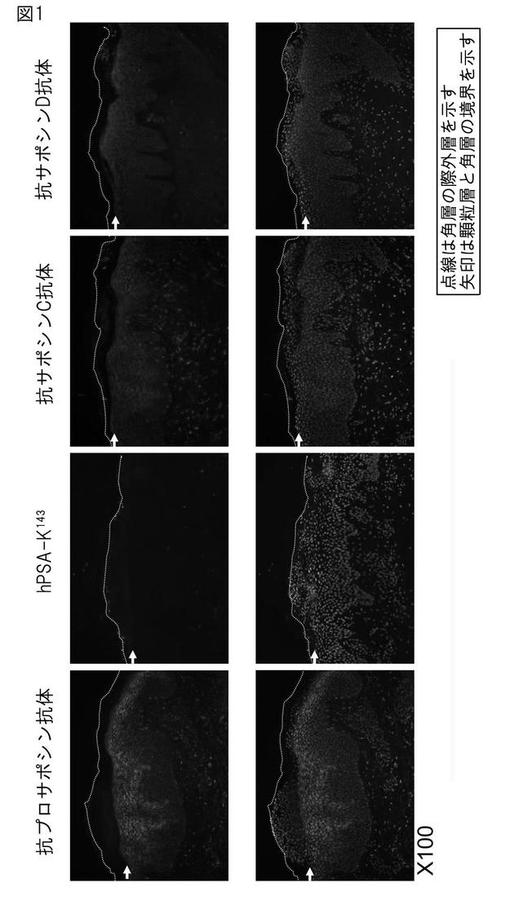
【図1】抗ヒトサポシンA特異的抗体(hSPA−K143抗体)によるヒト表皮の免疫組織化学染色。図中、点線は角層の最外層を示す。矢印は顆粒層と角層の境界を示す。
【発明を実施するための形態】
【0012】
本発明者は、プロサポシンのアミノ酸残基139位〜143位に相当するペンタペプチドを化学合成してハプテンとしてウサギの免疫に用いた。数回の感作ののち抗血清を得、抗血清中で免疫に用いたペプチドに強く結合する抗体が生成された。生成された抗体を用いて表皮細胞の免疫染色を行ったところ、驚くべきことに、この抗体は表皮の顆粒層を極めて特異的に染色した。このような特性を有する抗体は従来技術の抗サポシン抗体とは顕著に区別される。市販のサポシン抗体は活性化体である各サポシンを認識するが、その切断部位を認識するように設計されたものではなく、したがって活性化体のみならず、不活性型のプロサポシンも認識してしまう。従って、本発明の抗体は従来の抗サポシン抗体に比べ、特異性は極めて高いものと考えられる。
【0013】
本発明の抗体作製方法で使用する抗原は下記のペプチドが好ましい。
Gly-Ser-Leu-Gln-Lys
抗原は好ましくは免疫力を強化するため、適当な担体、例えばキーホールリンペットヘモシアニン(KLH)、ウシ胎児血清アルブミン(BSA)等に結合させるのが好ましい。ハプテンの免疫力を強化する担体及びその結合方法は当業者に周知である。好ましくは、ペプチドにかかる担体を結合させるため、Cys残基をペプチドのN末端に付加する。かかるシステイン残基の付加は、MBS(m-マレイミドベンゾイル-N-ヒドロキシスクシニミド-エステル)等の架橋剤を使用するのが好ましい。
【0014】
抗原として利用されるペプチドは、上記ペンタペプチドの他に、プロサポシンには結合せず、サポシンAにのみ特異的な抗体を生起する限りにおいて、プロサポシン内の143位のリジン残基を基準にN末端側の5個超の連続アミノ酸残基を含むものであってもよい。例えば、下記のペプチド配列であってよい:
Ala−Leu−Asn−Leu−Cys−Gly−Ser−Leu−Gln−Lys
Leu−Asn−Leu−Cys−Gly−Ser−Leu−Gln−Lys
Asn−Leu−Cys−Gly−Ser−Leu−Gln−Lys
Leu−Cys−Gly−Ser−Leu−Gln−Lys
Cys−Gly−Ser−Leu−Gln−Lys
【0015】
本発明に係るペプチドは例えば5〜50個のアミノ酸、好ましくは5〜20個のアミノ酸、より好ましくは5〜10個のアミノ酸、最も好ましくは5個のアミノ酸から成る。
【0016】
本発明において「抗体」とは、サポシンAにのみ特異的に結合し得る抗体分子全体又はその断片(例えば、Fab又はF(ab')2断片)を意味し、ポリクローナル抗体であってもモノクローナル抗体であってもよい。本発明の抗体は、種々の方法のいずれかによって製造することができる。このような抗体の製造法は当該分野において周知である[例えばHarlow E. & Lane D., Antibody, Cold Spring Harbor Laboratory Press(1988)を参照]。
【0017】
本発明に係るサポシンA特異的抗体をポリクローナル抗体として作製する場合、上記抗原、好ましくは適当な担体に結合させた抗原複合体を哺乳動物、例えばラット、マウス、ウサギなどに投与する。抗原又は抗原複合体の動物1匹当たりの投与量は、アジュバントを用いないときは0.1〜100mgであり、アジュバントを用いるときは10〜1000μgであるが、それらに限定されることはない。アジュバントとしては、フロイント完全アジュバント(FCA)、フロイント不完全アジュバント(FIA)、水酸化アルミニウムアジュバント等が挙げられる。免疫は、主として静脈内、皮下又は腹腔内等に注入することにより行われる。また、免疫の間隔は特に限定されず、数日から数週間間隔、好ましくは2〜5週間間隔で、1〜10回、好ましくは2〜5回免疫を行う。そして、最終の免疫日から6〜60日後に、ウェスタンブロット法、酵素免疫測定法(ELISA又は EIA)、放射性免疫測定法(RIA)等で抗体価を測定し、最大の抗体価を示した日に採血し、抗血清を得るのが好ましい。
【0018】
ポリクローナル抗体の精製は、抗原ペプチドの結合したカラムを用いるアフィニティークロマトグラフィー、その他の当業者周知の精製方法、例えばイオン交換クロマロトグラフィー、ゲル濾過クロマトグラフィー、高性能液体クロマトグラフィー等により精製することができる。
【0019】
本発明に係るサポシンA特異的抗体をモノクローナル抗体として作製する場合、上記抗原、好ましくは適当な担体に結合させた抗原複合体を、哺乳動物、例えばラット、マウス、ウサギなどに投与する。抗原の動物1匹当たりの投与量は、例えばアジュバントを用いないときは0.1〜100mgであり、アジュバントを用いるときは1〜100μgであるが、それらに限定されることはない。アジュバントとしては、フロイント完全アジュバント(FCA)、フロイント不完全アジュバント(FIA)、水酸化アルミニウムアジュバント等が挙げられる。免疫は、主として静脈内、皮下又は腹腔内等に注入することにより行われる。また、免疫の間隔は特に限定されず、数日から数週間間隔、好ましくは2〜5週間間隔で、1〜10回、好ましくは2〜5回免疫を行う。そして、最終の免疫日から1〜60日後、好ましくは1〜14日後に抗体産生細胞を採集する。抗体産生細胞としては、脾臓細胞、リンパ節細胞、末梢血細胞等が挙げられるが、脾臓細胞又は局所リンパ節細胞が好ましい。
【0020】
細胞融合ハイブリドーマを得るため、抗体産生細胞とミエローマ細胞との細胞融合を行う。抗体産生細胞と融合させるミエローマ細胞として、マウスなどの動物の一般に入手可能な株化細胞を使用することができる。使用する細胞株としては、薬剤選択性を有し、未融合の状態ではHAT選択培地(ヒポキサンチン、アミノプテリン、チミジンを含む)で生存できず、抗体産生細胞と融合した状態でのみ生存できる性質を有するものが好ましい。ミエローマ細胞としては、例えば X63Ag.8.653、NSI/1-Ag4-1、NS0/1などのマウスミエローマ細胞株、YB 2/0などのラットミエローマ細胞株が挙げられる。
【0021】
次に、上記ミエローマ細胞と抗体産生細胞とを細胞融合させる。細胞融合は、血清を含まないDMEM、RPMI-1640培地などの動物細胞培養用培地中で、1×106〜1×107個/mlの抗体産生細胞と2×105〜2×106個/mlのミエローマ細胞とを混合し(抗体産生細胞とミエローマ細胞との細胞比2:1〜3:1が好ましい)、細胞融合促進剤存在のもとで融合反応を行う。細胞融合促進剤として、平均分子量1000〜6000ダルトンのポリエチレングリコール等を使用することができる。また、電気刺激(例えばエレクトロポレーション)を利用した市販の細胞融合装置を用いて抗体産生細胞とミエローマ細胞とを融合させることもできる。
【0022】
ハイブリドーマの選別及びクローニング細胞融合処理後の細胞から目的とするハイブリドーマを選別する。その方法として、細胞懸濁液を例えばウシ胎児血清含有RPMI-1640培地などで適当に希釈後、マイクロタイタープレート上に3×105個/well程度まき、各ウエルに選択培地を加え、以後適当に選択培地を交換して培養を行う。その結果、選択培地で培養開始後、14日前後から生育してくる細胞をハイブリドーマとして得ることができる。
【0023】
次に、増殖してきたハイブリドーマの培養上清中に、サポシンAに特異的な抗体が存在するか否かをスクリーニングする。ハイブリドーマのスクリーニングは、通常の方法に従えばよく、特に限定されるものではない。例えば、ハイブリドーマとして生育したウエルに含まれる培養上清の一部を採集し、酵素免疫測定法、放射性免疫測定法等によってスクリーニングすることができる。融合細胞のクローニングは、限界希釈法等により行う。そして、最終的に、サポシンAとは反応し、全長不活性型プロカスパーゼ−14とは反応しないモノクローナル抗体を産生する細胞であるハイブリドーマを樹立する。
【0024】
モノクローナル抗体の採取樹立したハイブリドーマからモノクローナル抗体を採取する方法として、通常の細胞培養法又は腹水形成法等を採用することができる。細胞培養法においては、ハイブリドーマを10%ウシ胎児血清含有RPMI-1640培地、MEM培地又は無血清培地等の動物細胞培養培地中で、通常の培養条件(例えば37℃、5% CO2濃度)で7〜14日間培養し、その培養上清から抗体を取得する。腹水形成法の場合は、ミエローマ細胞由来の哺乳動物と同種系動物の腹腔内にハイブリドーマを約1×107個投与し、ハイブリドーマを大量に増殖させる。そして、1〜2週間後に腹水を採取する。上記抗体の採取方法において抗体の精製が必要とされる場合は、硫安塩析法、イオン交換クロマトグラフィー、ゲル濾過、アフィニティークロマトグラフィーなどの公知の方法を適宜選択して、又はこれらを組み合わせることにより精製することができる。
【0025】
本発明でいうプロサポシンは、NCBI NM_002778に掲載された全長524アミン酸のヒトプロサポシンである。
【0026】
本発明に係るサポシンAの検出方法は、本発明に係るサポシンA特異的抗体の検出を通じて行うことができ、その方法には例えば蛍光物質、色素、酵素等を利用する免疫染色法、ウェスタンブロット法、免疫測定方法、例えばELISA法、RIA法等、様々な方法が挙げられる。
【0027】
以下に本発明の限定でない実施例を提供する。
【実施例】
【0028】
実施例1
ヒトプロサポシンの上記切断点を認識する抗体(hSAP−K143抗体)野作成
hSAP−K143抗体は、簡単に説明すると、ヒトプロサポシン配列内の活性化に関与するものと推定される切断部位を含む領域(Gly-Ser-Leu-Gln-Lys)に対してN末端に担体を結合させるためのシステインを付加した計6個のアミノ酸から成るペプチド(Cys- Gly-Ser-Leu-Gln-Lys)を合成し、ペプチドをキーホールリンペットヘモシアニン(KLH:シグマ社製)に結合させたものをウサギに免疫して作成し、アフィニティークロマトグラフィーにより精製した。
【0029】
まず、ペプチドの担体タンパク質への結合については以下の通りに行った。10mgのKLHを2mlの0.2Mのリン酸カリウム緩衝液(pH7.0)に溶解した。これに1mgのMBS(m-マレイミドベンゾイル-N-ヒドロキシスクシニミド-エステル)をN,N'-ジメチルホルムアミド(岩井化学社製)67μlに溶解しておいたMBS溶液(用時調製)をゆっくり加え、室温で30分以上振とうした。予め作成しておいた0.2Mのリン酸カリウム緩衝液(pH7.0)で平衡化したセファデックスG−25ゲル濾過カラム(φ0.8×30cm:バイオラッド社製)にアプライし、同緩衝液にて流出画分を分取した。KLH−MB複合体を形成している画分について集めた(-80℃)で保存。尚、カラムを用いる一連の操作は4℃で行なった。
【0030】
更に、このKLH−MB複合体溶液0.4mlに2.5mg/0.1mlの合成ペプチド(サワデーテクノロジー社)を加え、室温にて4時間以上振とうし、KLH−MB−合成ペプチド結合物とした。分注して-20℃で保存した。
【0031】
免疫操作は以下の通りに行った。初回免疫では結核菌を含むフロインド完全アジュバント(ギブコ社製)を用いた。これを十分振って均等にし、23G注射針(テルモ社製)をつけた注射筒を用いて1ml測り取った。更に同針でKLH−MB−ペプチド結合物を吸い込んだ後、均一なエマルジョンを作製し、ウサギ(ニュージーランドホワイト種)の皮下に注射した。追加免疫として、2回目以降は結核菌を含まないフロインドの不完全アジュバント(ギブコ社製)を用いて約2週間毎に、計3回にわたりウサギに免疫を行った。初回免疫より3回目及び4回目の免疫後にウサギより血液を採取した。血液は37℃、2時間置いた後、4℃で一晩静置した。血清の回収は2,000×g、4℃で5分間遠心分離を行い、回収した血清を分注し、-30℃で保存した。
【0032】
2)アフィニティーカラムの作製及び精製方法
AH−セファロース4B(ファルマシア社製)約1.5gを0.5MのNaCl溶液と蒸留水で洗浄し、10mMのリン酸カリウム緩衝液(pH7.0)にて平衡化した後、N,N'-ジメチルホルムアミド1.4mlで溶解したMBS(20mg)をゆっくり加え、室温で80分間振とうした。これをガラス濾過器を用いて0.1Mのリン酸カリウム緩衝液(pH7.0)で洗浄し、再び同緩衝液4mlに膨潤させ、蒸留水に溶解した合成ペプチド5mgを加えた。水酸化カリウム溶液を用いてpH6.5に調整し、4℃で一晩振とうした。その後、2-メルカプトエタノール(終濃度75mM)(シグマ社製)を加え、更に4℃にて2.5時間振とうし、ガラス濾過器を用いて50mMのTris-HCl(pH7.4)で洗浄し、結合していないペプチド及び試薬を洗浄した。この樹脂をカラムに充填した。
【0033】
抗血清は、上記のペプチドを結合させたアフィニティーカラムに、3、4回繰り返しアプライした抗体を結合させた後、0.5MのNaClを含む50mMのTris-HCl(pH7.4)で洗浄し、更に50mMのTris-HCl(pH7.4)で洗浄し、夾雑物を取り除いた。0.1Mのグリシン−HCl(pH2.5)により溶出し、酸性による活性低下を防ぐため、予め冷却した2MのTris-HCl(pH8.0)に滴下し、分取した。波長280nmにて吸光度を測定し、タンパク質量の多い画分を実験に使用した。防腐剤として終濃度0.02%になるようにアジ化ナトリウムを加え、4℃に保存した。このようにして得られたhSAP−K143抗体はサポシンAを特異的に認識した(データーは示さない)。
【0034】
実施例2
hSAP−K143抗体によるヒト表皮の免疫組織化学染色方法
乾癬やアトピー性皮膚炎に罹患した皮膚ではプロサポシンのプロセシングが起きず、サポシンAの発現が認めらない知見がある。アトピー性皮膚を罹患した皮膚凍結切片を4%のPFA(パラホルムアルデヒド)で室温、20分固定し、ブリーチ(10%のメタノール、3%の過酸化水素/蒸留水)により、内因性のペルオキシダーゼを除去した後、ENVISIONキット(ダコ)を用いて染色を行った。具体的には、10%のヤギ正常血清(ニチレイ社製)にて室温、2時間ブロッキング処理し、実施例3のhSAP-K143抗体(1/200)と、又は対照として抗プロサポシン抗体(PSAP polyclonal antibody(A01):Abnova社)、抗サポシンB抗体(E−15, sc−27014: Santa Cruz社)、抗サポシンC抗体(H-81, sc-32875: Santa Cruz社)、抗サポシンD抗体(E-14, sc-27024R: Santa Cruz社)と4℃、一晩反応させた後、PBSで室温にて15分×3回、余剰抗体を洗浄した。次に、ペルオキシダーゼ標識抗ウサギ二次抗体(ダコ)と室温で1時間反応させ、再度PBSで室温にて15分×3回洗浄し、余剰抗体を洗浄し、DABを用いて発色させた。最後にヘマトキシニンを用いて核染色を行った。その結果を図1に示す。
【0035】
図1から、抗サポシンA特異抗体による染色ではアトピーの不全角化部位に染色はほとんど認められず、サポシンAが作られていないことがわかる。これに対し、市販のサポシンC,Dに対する抗体を用いた場合は、プロサポシンも染めてしまうため、これらのサポシンが作られているかどうかは判定できない。
【技術分野】
【0001】
本発明は、サポシンAに特異的な抗体及びその作成方法に関する。
【背景技術】
【0002】
ケラチノサイトは終末分化することで、有害な環境に対する「角層」と称される保護バリアーを形成する。角層細胞間脂質はこの保護バリアーにおいて重要な役割を担っている。角層細胞間脂質は、表皮ケラチノサイトにおいて分化とともに生合成され、オドランド小体に蓄積される。角層細胞間脂質の主成分であるセラミドは、オドランド小体の開口分泌により角層細胞間に放出されたグルコシルセラミドの糖鎖が、グルコセレブロシダーゼにより切断されることで生じる。また、別の経路として、スフィンゴミエリナーゼによるスフィンゴミエリンからセラミドへの分解経路も知られている。
【0003】
スフィンゴ脂質活性化タンパク質であるサポシンは、スフィンゴ脂質の加水分解に必須のタンパク質であり、4種類のサポシン(サポシンA、サポシンB、サポシンC、サポシンD)の存在が知られている。サポシンA、B、C、及びDは極めて相同性の高い糖タンパク質で、前駆タンパク質であるプロサポシンがライソゾームで分解されて生産される。サポシンA、B、C、Dはそれぞれプロサポシンのアミノ酸60〜143位、195〜275位、311〜390位、405〜487位に相当する(Kishimoto et al., Journal of Lipid Research (1992) 33, 1255-67)。サポシンは、角層間脂質の生成に必要なセラミド合成酵素、例えばグルコセレブロシダーゼ、スフィンゴミエリナーゼ等の補酵素として働く因子であることから、サポシンは表皮においては角層間脂質の合成や、皮膚バリアー機能にとって重要であると考えられる。
【0004】
これまで、サポシンA、C、Dが臓器に集積するとリソソーム蓄積病(例えば、ゴーシュ病、ニーマンピック病)が生じることや、プロサポシン遺伝子の突然変異が神経障害性の表現型をもたらすことが確認されている。皮膚との関連では、アトピー性皮膚炎や乾癬に罹患している患者において、プロサポシンが低下しているとの報告がある(それぞれ、Tezuka et al., Journal of Investigative Dermatology (1997) 109, 319-323(非特許文献1)及びAlessandrini et al., Journal of Investigative Dermatology (2001) 116, 394-400(非特許文献2)を参照のこと)。
【先行技術文献】
【非特許文献】
【0005】
【非特許文献1】Tezuka et al., Journal of Investigative Dermatology (1997) 109, 319-323
【非特許文献2】Alessandrini et al., Journal of Investigative Dermatology (2001) 116, 394-400
【発明の概要】
【発明が解決しようとする課題】
【0006】
しかしながら、プロサポシンの活性化の機構や活性化サポシンの機能に関しては明らかではなく、各サポシンが実際にどのように表皮において分布し、機能しているのかに解明が待たれる。プロサポシンの活性化の機構及び機能、更には活性型の生体内分布及びその生理学的役割の解明には、その活性化形態に特異的な抗体の利用が不可欠である。
【課題を解決するための手段】
【0007】
本発明者はサポシンAに特異的に結合する抗体の作製に成功した。
【0008】
第一の態様において、本発明はサポシンAに特異的な抗体の作成方法を提供する。サポシンAは、不活性型プロサポシンの143位のリジン残基において切断されることで生成される、当該143位のリジン残基をC末端として有するサポシンAに特異的な抗体である。この抗体作成方法は、不活性型プロサポシンのアミノ酸残基139位〜143位のアミノ酸配列:
Gly-Ser-Leu-Gln-Lys (配列番号1)
を含んで成り、上記配列のC末端のLys残基をC末端とするペプチドを抗原として用いることを特徴とする。好ましくは、上記ペプチドは配列番号1から成るペプチドである。
本発明はさらに、上記方法により作成された、不活性型プロサポシンの143位のリジン残基において切断されることで生成される、143位のリジン残基をC末端として有するサポシンAに特異的な抗体を提供する。
【0009】
別の観点において、本発明は組織、もしくは細胞、特に表皮中のサポシンAを検出するため、上記方法により作成された抗体を使用することを特徴とする方法を提供する。
【発明の効果】
【0010】
サポシンAは他のサポシンと比較して顕著に異なる挙動を示し、皮膚バリアー機能に関与している可能性が高く、その作用機能の解明は極めて有意義なものと考えられる。本願発明にかかるサポシンA特異的抗体はこのようなサポシンAの同定に有用であり、その結果カスパーゼ−14の生体内での役割を解明するのに極めて有用である。
【図面の簡単な説明】
【0011】
【図1】抗ヒトサポシンA特異的抗体(hSPA−K143抗体)によるヒト表皮の免疫組織化学染色。図中、点線は角層の最外層を示す。矢印は顆粒層と角層の境界を示す。
【発明を実施するための形態】
【0012】
本発明者は、プロサポシンのアミノ酸残基139位〜143位に相当するペンタペプチドを化学合成してハプテンとしてウサギの免疫に用いた。数回の感作ののち抗血清を得、抗血清中で免疫に用いたペプチドに強く結合する抗体が生成された。生成された抗体を用いて表皮細胞の免疫染色を行ったところ、驚くべきことに、この抗体は表皮の顆粒層を極めて特異的に染色した。このような特性を有する抗体は従来技術の抗サポシン抗体とは顕著に区別される。市販のサポシン抗体は活性化体である各サポシンを認識するが、その切断部位を認識するように設計されたものではなく、したがって活性化体のみならず、不活性型のプロサポシンも認識してしまう。従って、本発明の抗体は従来の抗サポシン抗体に比べ、特異性は極めて高いものと考えられる。
【0013】
本発明の抗体作製方法で使用する抗原は下記のペプチドが好ましい。
Gly-Ser-Leu-Gln-Lys
抗原は好ましくは免疫力を強化するため、適当な担体、例えばキーホールリンペットヘモシアニン(KLH)、ウシ胎児血清アルブミン(BSA)等に結合させるのが好ましい。ハプテンの免疫力を強化する担体及びその結合方法は当業者に周知である。好ましくは、ペプチドにかかる担体を結合させるため、Cys残基をペプチドのN末端に付加する。かかるシステイン残基の付加は、MBS(m-マレイミドベンゾイル-N-ヒドロキシスクシニミド-エステル)等の架橋剤を使用するのが好ましい。
【0014】
抗原として利用されるペプチドは、上記ペンタペプチドの他に、プロサポシンには結合せず、サポシンAにのみ特異的な抗体を生起する限りにおいて、プロサポシン内の143位のリジン残基を基準にN末端側の5個超の連続アミノ酸残基を含むものであってもよい。例えば、下記のペプチド配列であってよい:
Ala−Leu−Asn−Leu−Cys−Gly−Ser−Leu−Gln−Lys
Leu−Asn−Leu−Cys−Gly−Ser−Leu−Gln−Lys
Asn−Leu−Cys−Gly−Ser−Leu−Gln−Lys
Leu−Cys−Gly−Ser−Leu−Gln−Lys
Cys−Gly−Ser−Leu−Gln−Lys
【0015】
本発明に係るペプチドは例えば5〜50個のアミノ酸、好ましくは5〜20個のアミノ酸、より好ましくは5〜10個のアミノ酸、最も好ましくは5個のアミノ酸から成る。
【0016】
本発明において「抗体」とは、サポシンAにのみ特異的に結合し得る抗体分子全体又はその断片(例えば、Fab又はF(ab')2断片)を意味し、ポリクローナル抗体であってもモノクローナル抗体であってもよい。本発明の抗体は、種々の方法のいずれかによって製造することができる。このような抗体の製造法は当該分野において周知である[例えばHarlow E. & Lane D., Antibody, Cold Spring Harbor Laboratory Press(1988)を参照]。
【0017】
本発明に係るサポシンA特異的抗体をポリクローナル抗体として作製する場合、上記抗原、好ましくは適当な担体に結合させた抗原複合体を哺乳動物、例えばラット、マウス、ウサギなどに投与する。抗原又は抗原複合体の動物1匹当たりの投与量は、アジュバントを用いないときは0.1〜100mgであり、アジュバントを用いるときは10〜1000μgであるが、それらに限定されることはない。アジュバントとしては、フロイント完全アジュバント(FCA)、フロイント不完全アジュバント(FIA)、水酸化アルミニウムアジュバント等が挙げられる。免疫は、主として静脈内、皮下又は腹腔内等に注入することにより行われる。また、免疫の間隔は特に限定されず、数日から数週間間隔、好ましくは2〜5週間間隔で、1〜10回、好ましくは2〜5回免疫を行う。そして、最終の免疫日から6〜60日後に、ウェスタンブロット法、酵素免疫測定法(ELISA又は EIA)、放射性免疫測定法(RIA)等で抗体価を測定し、最大の抗体価を示した日に採血し、抗血清を得るのが好ましい。
【0018】
ポリクローナル抗体の精製は、抗原ペプチドの結合したカラムを用いるアフィニティークロマトグラフィー、その他の当業者周知の精製方法、例えばイオン交換クロマロトグラフィー、ゲル濾過クロマトグラフィー、高性能液体クロマトグラフィー等により精製することができる。
【0019】
本発明に係るサポシンA特異的抗体をモノクローナル抗体として作製する場合、上記抗原、好ましくは適当な担体に結合させた抗原複合体を、哺乳動物、例えばラット、マウス、ウサギなどに投与する。抗原の動物1匹当たりの投与量は、例えばアジュバントを用いないときは0.1〜100mgであり、アジュバントを用いるときは1〜100μgであるが、それらに限定されることはない。アジュバントとしては、フロイント完全アジュバント(FCA)、フロイント不完全アジュバント(FIA)、水酸化アルミニウムアジュバント等が挙げられる。免疫は、主として静脈内、皮下又は腹腔内等に注入することにより行われる。また、免疫の間隔は特に限定されず、数日から数週間間隔、好ましくは2〜5週間間隔で、1〜10回、好ましくは2〜5回免疫を行う。そして、最終の免疫日から1〜60日後、好ましくは1〜14日後に抗体産生細胞を採集する。抗体産生細胞としては、脾臓細胞、リンパ節細胞、末梢血細胞等が挙げられるが、脾臓細胞又は局所リンパ節細胞が好ましい。
【0020】
細胞融合ハイブリドーマを得るため、抗体産生細胞とミエローマ細胞との細胞融合を行う。抗体産生細胞と融合させるミエローマ細胞として、マウスなどの動物の一般に入手可能な株化細胞を使用することができる。使用する細胞株としては、薬剤選択性を有し、未融合の状態ではHAT選択培地(ヒポキサンチン、アミノプテリン、チミジンを含む)で生存できず、抗体産生細胞と融合した状態でのみ生存できる性質を有するものが好ましい。ミエローマ細胞としては、例えば X63Ag.8.653、NSI/1-Ag4-1、NS0/1などのマウスミエローマ細胞株、YB 2/0などのラットミエローマ細胞株が挙げられる。
【0021】
次に、上記ミエローマ細胞と抗体産生細胞とを細胞融合させる。細胞融合は、血清を含まないDMEM、RPMI-1640培地などの動物細胞培養用培地中で、1×106〜1×107個/mlの抗体産生細胞と2×105〜2×106個/mlのミエローマ細胞とを混合し(抗体産生細胞とミエローマ細胞との細胞比2:1〜3:1が好ましい)、細胞融合促進剤存在のもとで融合反応を行う。細胞融合促進剤として、平均分子量1000〜6000ダルトンのポリエチレングリコール等を使用することができる。また、電気刺激(例えばエレクトロポレーション)を利用した市販の細胞融合装置を用いて抗体産生細胞とミエローマ細胞とを融合させることもできる。
【0022】
ハイブリドーマの選別及びクローニング細胞融合処理後の細胞から目的とするハイブリドーマを選別する。その方法として、細胞懸濁液を例えばウシ胎児血清含有RPMI-1640培地などで適当に希釈後、マイクロタイタープレート上に3×105個/well程度まき、各ウエルに選択培地を加え、以後適当に選択培地を交換して培養を行う。その結果、選択培地で培養開始後、14日前後から生育してくる細胞をハイブリドーマとして得ることができる。
【0023】
次に、増殖してきたハイブリドーマの培養上清中に、サポシンAに特異的な抗体が存在するか否かをスクリーニングする。ハイブリドーマのスクリーニングは、通常の方法に従えばよく、特に限定されるものではない。例えば、ハイブリドーマとして生育したウエルに含まれる培養上清の一部を採集し、酵素免疫測定法、放射性免疫測定法等によってスクリーニングすることができる。融合細胞のクローニングは、限界希釈法等により行う。そして、最終的に、サポシンAとは反応し、全長不活性型プロカスパーゼ−14とは反応しないモノクローナル抗体を産生する細胞であるハイブリドーマを樹立する。
【0024】
モノクローナル抗体の採取樹立したハイブリドーマからモノクローナル抗体を採取する方法として、通常の細胞培養法又は腹水形成法等を採用することができる。細胞培養法においては、ハイブリドーマを10%ウシ胎児血清含有RPMI-1640培地、MEM培地又は無血清培地等の動物細胞培養培地中で、通常の培養条件(例えば37℃、5% CO2濃度)で7〜14日間培養し、その培養上清から抗体を取得する。腹水形成法の場合は、ミエローマ細胞由来の哺乳動物と同種系動物の腹腔内にハイブリドーマを約1×107個投与し、ハイブリドーマを大量に増殖させる。そして、1〜2週間後に腹水を採取する。上記抗体の採取方法において抗体の精製が必要とされる場合は、硫安塩析法、イオン交換クロマトグラフィー、ゲル濾過、アフィニティークロマトグラフィーなどの公知の方法を適宜選択して、又はこれらを組み合わせることにより精製することができる。
【0025】
本発明でいうプロサポシンは、NCBI NM_002778に掲載された全長524アミン酸のヒトプロサポシンである。
【0026】
本発明に係るサポシンAの検出方法は、本発明に係るサポシンA特異的抗体の検出を通じて行うことができ、その方法には例えば蛍光物質、色素、酵素等を利用する免疫染色法、ウェスタンブロット法、免疫測定方法、例えばELISA法、RIA法等、様々な方法が挙げられる。
【0027】
以下に本発明の限定でない実施例を提供する。
【実施例】
【0028】
実施例1
ヒトプロサポシンの上記切断点を認識する抗体(hSAP−K143抗体)野作成
hSAP−K143抗体は、簡単に説明すると、ヒトプロサポシン配列内の活性化に関与するものと推定される切断部位を含む領域(Gly-Ser-Leu-Gln-Lys)に対してN末端に担体を結合させるためのシステインを付加した計6個のアミノ酸から成るペプチド(Cys- Gly-Ser-Leu-Gln-Lys)を合成し、ペプチドをキーホールリンペットヘモシアニン(KLH:シグマ社製)に結合させたものをウサギに免疫して作成し、アフィニティークロマトグラフィーにより精製した。
【0029】
まず、ペプチドの担体タンパク質への結合については以下の通りに行った。10mgのKLHを2mlの0.2Mのリン酸カリウム緩衝液(pH7.0)に溶解した。これに1mgのMBS(m-マレイミドベンゾイル-N-ヒドロキシスクシニミド-エステル)をN,N'-ジメチルホルムアミド(岩井化学社製)67μlに溶解しておいたMBS溶液(用時調製)をゆっくり加え、室温で30分以上振とうした。予め作成しておいた0.2Mのリン酸カリウム緩衝液(pH7.0)で平衡化したセファデックスG−25ゲル濾過カラム(φ0.8×30cm:バイオラッド社製)にアプライし、同緩衝液にて流出画分を分取した。KLH−MB複合体を形成している画分について集めた(-80℃)で保存。尚、カラムを用いる一連の操作は4℃で行なった。
【0030】
更に、このKLH−MB複合体溶液0.4mlに2.5mg/0.1mlの合成ペプチド(サワデーテクノロジー社)を加え、室温にて4時間以上振とうし、KLH−MB−合成ペプチド結合物とした。分注して-20℃で保存した。
【0031】
免疫操作は以下の通りに行った。初回免疫では結核菌を含むフロインド完全アジュバント(ギブコ社製)を用いた。これを十分振って均等にし、23G注射針(テルモ社製)をつけた注射筒を用いて1ml測り取った。更に同針でKLH−MB−ペプチド結合物を吸い込んだ後、均一なエマルジョンを作製し、ウサギ(ニュージーランドホワイト種)の皮下に注射した。追加免疫として、2回目以降は結核菌を含まないフロインドの不完全アジュバント(ギブコ社製)を用いて約2週間毎に、計3回にわたりウサギに免疫を行った。初回免疫より3回目及び4回目の免疫後にウサギより血液を採取した。血液は37℃、2時間置いた後、4℃で一晩静置した。血清の回収は2,000×g、4℃で5分間遠心分離を行い、回収した血清を分注し、-30℃で保存した。
【0032】
2)アフィニティーカラムの作製及び精製方法
AH−セファロース4B(ファルマシア社製)約1.5gを0.5MのNaCl溶液と蒸留水で洗浄し、10mMのリン酸カリウム緩衝液(pH7.0)にて平衡化した後、N,N'-ジメチルホルムアミド1.4mlで溶解したMBS(20mg)をゆっくり加え、室温で80分間振とうした。これをガラス濾過器を用いて0.1Mのリン酸カリウム緩衝液(pH7.0)で洗浄し、再び同緩衝液4mlに膨潤させ、蒸留水に溶解した合成ペプチド5mgを加えた。水酸化カリウム溶液を用いてpH6.5に調整し、4℃で一晩振とうした。その後、2-メルカプトエタノール(終濃度75mM)(シグマ社製)を加え、更に4℃にて2.5時間振とうし、ガラス濾過器を用いて50mMのTris-HCl(pH7.4)で洗浄し、結合していないペプチド及び試薬を洗浄した。この樹脂をカラムに充填した。
【0033】
抗血清は、上記のペプチドを結合させたアフィニティーカラムに、3、4回繰り返しアプライした抗体を結合させた後、0.5MのNaClを含む50mMのTris-HCl(pH7.4)で洗浄し、更に50mMのTris-HCl(pH7.4)で洗浄し、夾雑物を取り除いた。0.1Mのグリシン−HCl(pH2.5)により溶出し、酸性による活性低下を防ぐため、予め冷却した2MのTris-HCl(pH8.0)に滴下し、分取した。波長280nmにて吸光度を測定し、タンパク質量の多い画分を実験に使用した。防腐剤として終濃度0.02%になるようにアジ化ナトリウムを加え、4℃に保存した。このようにして得られたhSAP−K143抗体はサポシンAを特異的に認識した(データーは示さない)。
【0034】
実施例2
hSAP−K143抗体によるヒト表皮の免疫組織化学染色方法
乾癬やアトピー性皮膚炎に罹患した皮膚ではプロサポシンのプロセシングが起きず、サポシンAの発現が認めらない知見がある。アトピー性皮膚を罹患した皮膚凍結切片を4%のPFA(パラホルムアルデヒド)で室温、20分固定し、ブリーチ(10%のメタノール、3%の過酸化水素/蒸留水)により、内因性のペルオキシダーゼを除去した後、ENVISIONキット(ダコ)を用いて染色を行った。具体的には、10%のヤギ正常血清(ニチレイ社製)にて室温、2時間ブロッキング処理し、実施例3のhSAP-K143抗体(1/200)と、又は対照として抗プロサポシン抗体(PSAP polyclonal antibody(A01):Abnova社)、抗サポシンB抗体(E−15, sc−27014: Santa Cruz社)、抗サポシンC抗体(H-81, sc-32875: Santa Cruz社)、抗サポシンD抗体(E-14, sc-27024R: Santa Cruz社)と4℃、一晩反応させた後、PBSで室温にて15分×3回、余剰抗体を洗浄した。次に、ペルオキシダーゼ標識抗ウサギ二次抗体(ダコ)と室温で1時間反応させ、再度PBSで室温にて15分×3回洗浄し、余剰抗体を洗浄し、DABを用いて発色させた。最後にヘマトキシニンを用いて核染色を行った。その結果を図1に示す。
【0035】
図1から、抗サポシンA特異抗体による染色ではアトピーの不全角化部位に染色はほとんど認められず、サポシンAが作られていないことがわかる。これに対し、市販のサポシンC,Dに対する抗体を用いた場合は、プロサポシンも染めてしまうため、これらのサポシンが作られているかどうかは判定できない。
【特許請求の範囲】
【請求項1】
不活性型プロサポシンの143位のリジン残基において切断されることで生成される、当該143位のリジン残基をC末端として有するサポシンAに特異的な抗体の作成方法であって、不活性型プロサポシンのアミノ酸残基139位〜143位のアミノ酸配列:
Gly-Ser-Leu-Gln-Lys
を含んで成り、上記配列のC末端のLys残基をC末端とするペプチドを抗原として用いることを特徴とする、方法。
【請求項2】
請求項1記載の方法により作成された、不活性型プロサポシンの143位のリジン残基において切断されることで生成される、143位のリジン残基をC末端として有するサポシンAに特異的な抗体。
【請求項1】
不活性型プロサポシンの143位のリジン残基において切断されることで生成される、当該143位のリジン残基をC末端として有するサポシンAに特異的な抗体の作成方法であって、不活性型プロサポシンのアミノ酸残基139位〜143位のアミノ酸配列:
Gly-Ser-Leu-Gln-Lys
を含んで成り、上記配列のC末端のLys残基をC末端とするペプチドを抗原として用いることを特徴とする、方法。
【請求項2】
請求項1記載の方法により作成された、不活性型プロサポシンの143位のリジン残基において切断されることで生成される、143位のリジン残基をC末端として有するサポシンAに特異的な抗体。
【図1】


【公開番号】特開2011−190202(P2011−190202A)
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願番号】特願2010−56573(P2010−56573)
【出願日】平成22年3月12日(2010.3.12)
【出願人】(000001959)株式会社 資生堂 (1,748)
【Fターム(参考)】
【公開日】平成23年9月29日(2011.9.29)
【国際特許分類】
【出願日】平成22年3月12日(2010.3.12)
【出願人】(000001959)株式会社 資生堂 (1,748)
【Fターム(参考)】
[ Back to top ]