
サンプル分子を単離および標識するための方法
【課題】本発明は、サンプル分子についての反応性基へのサンプル分子の共有結合を可能にする条件下で、サンプル分子と、切断可能な官能基、1つ以上の官能基、および反応性基を含む化学基に結合された固体支持体とを接触させること;および切断可能な官能基を切断し、これにより1つ以上の官能基(これらは、タグであり得る)を含むサンプル分子を放出することによって分子を標識する方法を提供すること。
【解決手段】上記方法。本発明はまた、切断可能な官能基、質量分析タグおよびサンプル分子に共有結合するための反応性基を含む化学基に共有結合された固体支持体を提供し、ここで切断可能な官能基、タグおよび反応性基は、切断可能な官能基の切断の際に、サンプル分子へのタグの移動を可能にするように互いに対して配置されている。
【解決手段】上記方法。本発明はまた、切断可能な官能基、質量分析タグおよびサンプル分子に共有結合するための反応性基を含む化学基に共有結合された固体支持体を提供し、ここで切断可能な官能基、タグおよび反応性基は、切断可能な官能基の切断の際に、サンプル分子へのタグの移動を可能にするように互いに対して配置されている。
【発明の詳細な説明】
【背景技術】
【0001】
(発明の背景)
本発明は、国立癌研究所によって与えられた許可番号1R33CA84698−0下で政府に支援されてなされた。政府は、本発明に特定の権利を有する。
【0002】
本発明は、一般にプロテオーム分析に関し、より詳細には、分子の分析および定量のために、サンプル中の分子に官能基を移動させるための方法に関する。
【0003】
生物学的プロセスを研究するための伝統的な生化学的アプローチは、連続的な分画、およびプロセスを構築する比活性のアッセイサイクルによる均質化への精製、それぞれの単離された成分の詳細な構造分析、機能的分析および調節分析、ならびに単離された成分からのプロセスの再構築に基づく。ヒトゲノム計画および他のゲノム配列決定プログラムは、特定の種の完全ゲノム配列および本質的に、潜在的にその種によってコードされる全てのタンパク質のアミノ酸配列を、迅速に連続して作製している。生物学における歴史で先例のないこの情報資源が、伝統的な研究方法を拡大し、そして全く異なる研究パラダイム(その1つがプロテオミクスである)における進歩を触媒することが、期待される。
【0004】
多数の他の種のゲノムとともにヒトゲノムの全体を配列決定するための努力は、非常に成功している。46の微生物種のゲノム(TIGR Microbial Database;www.tigr.org)は、完全にされ、そして120を超える他の微生物種のゲノムが、現在配列決定の過程にある。さらに、より複雑な真核生物のゲノム(特に、遺伝的に十分に特徴づけられた単細胞生物Saccharomyces cerevisiaeおよび多細胞の種Caenorhabditis elegansおよびDrosophila melanogasterのゲノム)が、完全に配列決定されている。さらに、コメ、ヒトおよびArabidopsisゲノムの「ドラフト配列(draft sequence)」が公開されている。完全なゲノム配列の非存在下でさえも、豊富なDNA配列データベースが、公的に利用可能にされており、これらとしては、210万を超えるヒトおよび120万を超えるマウスの発現配列タグ(EST)を含むデータベースが挙げられる。
【0005】
ESTは、cDNAライブラリーにおけるクローンの系統的な単一通過配列決定によって生成される部分遺伝子配列を表す約300〜500の連続したヌクレオチドのストレッチである。進化を明らかに例外とするほとんどの生物学的プロセセスの時間尺度において、ゲノムDNA配列は静的と見なされ得、故にゲノム配列データベースは、ライブラリーに類似する情報資源を表す。激しい努力が、配列データベースにおける個々の配列に対して「機能」を割当するために進行中である。これは、既知の機能を有する配列のファミリーに対する配列の統計学的に有意な類似性を示す、線状配列モチーフまたはより高次の構造的モチーフのコンピューター分析によってか、または種に渡る相同性タンパク質の機能の比較のような他の手段によって試みられる。他の方法もまた、個々の配列の機能を決定するために使用されており、この方法としては、遺伝子ノックアウトおよびアンチセンスヌクレオチド技術を用いる遺伝子発現の抑制のような実験方法が挙げられ、これらは、時間を消費し得、そしていくつかの場合、配列によってコードされるポリペプチドに対する生物学的機能の割当を可能にするのになお不十分であり得る。
【0006】
プロテオームは、ゲノムによって発現されるタンパク質補体として規定されている。この幾分制限的な規定は、プロテオームの静的な性質を示唆する。実際、発現されたタンパク質の型、その存在量、改変の状態、亜細胞の位置、および他の生体分子(例えば、ポリペプチドおよび核酸)との相互作用は、細胞または組織の生理学的状態に依存するので、プロテオームは、高度に動的である。故に、プロテオームは、細胞の状態、または細胞によって遭遇する外部の状態を反映し得、そしてプロテオーム分析は、細胞の状態を区別および研究するためのゲノムにわたるアッセイ、ならびにこれらを制御する分子機構を決定するためのゲノムにわたるアッセイとして見なされ得る。分化した細胞のプロテオームは、何1000個〜何10000個もの異なる型のタンパク質からなると見積もられることを考慮して、少なくとも5桁の発現の見積もられたダイナミックレンジを用いて、プロテオーム分析のための見通しは煩わしいようである。しかし、全ての潜在的に発現されるタンパク質の配列を列挙するDNAデータベースの利用可能性は、実際に発現されるタンパク質を同定し得る技術における迅速な進展と結びついて、現在、プロテオミクスを現実的な仕事とする。質量分析法は、現在のプロテオミクス技術が位置する基本的な脚の1つである。
【0007】
定量的なプロテオミクスは、その量および同一性に関する、細胞または組織によって発現される全てのタンパク質の系統的な分析である。所定の時間で、細胞、組織、生物学的流体またはタンパク質複合体で発現されるタンパク質は、その時点での細胞または組織の状態を正確に規定する。異なる状態での同じ細胞型のタンパク質プロフィール間の量的差異および質的差異は、それぞれの状態の間の移行を理解するために使用され得る。慣習的に、プロテオーム分析は、タンパク質を分離するための、高い解像度のゲル電気泳動、特に2次元ゲル電気泳動と、タンパク質を同定するための質量分析法との組み合わせを用いて実行された。このアプローチは、連続的かつ単調であるが、より重要なことには、タンパク質の生物学的に重要なクラスが本質的に検出不可能である点で、基本的に制限されている(非特許文献1)。
【0008】
多数の種のゲノム配列の完成は、代表的に発見科学といわれる生物学への新しいアプローチを触媒している。発見科学の本質は、細胞または組織によって発現される特定のクラスの分子の全てのメンバーの系統的かつ定量的な分析である。発見科学の例示的な実施としては、遺伝子発現アレイによる細胞または組織によって発現されるmRNA分子の系統的分析、生物学的サンプル中に含まれるタンパク質の系統的分析が挙げられる。発見科学の主な観点は、生体分子の系統的な測定から得られるデータに基づく細胞または組織の状態(活性、病理学、ストレス)の説明、および2つの状態を表す細胞の分子組成物の比較分析による、1つの状態から他の状態への細胞の移行を制御する分子機構の同定である。細胞の状態およびそれを制御する機構の分子の的説明のためには、可能な限り多くのパラメーターが所望される。現在の発現アレイ方法は、細胞中のmRNA分子の系統的分析を可能にする。
【0009】
近年、同位体コード化親和性タグといわれる等級の試薬および質量分析法に基づく方法が、生物学的サンプル中に存在するタンパク質の系統的な同定および定量に適していることが記載されている。細胞の状態に関連する他の特性(例えば、タンパク質リン酸化および他の翻訳後修飾、ならびにタンパク質または核酸以外の生体分子(例えば、脂質、セカンドメッセンジャー、代謝産物))の定量的プロフィールは、現在の技術では系統的および定量的に測定することは困難である。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Gygiら、Proc.Natl.Acad.Sci.USA 97:9390〜9395(2000)
【発明の概要】
【発明が解決しようとする課題】
【0011】
従って、細胞中の分子の分析のための迅速、効果的そしてコストがかからない方法への要求が存在する。本発明は、この要求を満たし、そして関連の利点も提供する。
【課題を解決するための手段】
【0012】
(発明の要旨)
本発明は、サンプル分子が反応性基に共有結合することを可能にする条件下で、サンプル分子を、切断可能な官能基、1つ以上の官能基、およびサンプル分子のための反応性基を含む化学基と連結した固体支持体と接触させ;そして切断可能な官能基を切断し、これによってタグ化され得る1つ以上の官能基を含むサンプル分子を放出することによって、分子を標識するための方法を提供する。本発明はまた、切断可能な官能基、質量分析タグおよびサンプル分子に共有結合するための反応性基を含む化学基に共有結合した固体支持体を提供し、ここで、切断可能な官能基、タグおよび反応性基は、切断可能な官能基の切断の際のサンプル分子へのタグの移動を可能にするように、互いに関連して位置される。
【図面の簡単な説明】
【0013】
【図1A】図1は、ポリペプチドの光捕捉回収を可能にする光切断可能なリンカーを介してポリペプチドを捕捉するための固相ベースのアプローチの概略図を示す。図1Aは、以下の4つのエレメントを有する固相同位体タグ化試薬の概略図を示す:ビーズ、光切断可能なリンカー、安定な同位体タグ、および特異的な反応性基。
【図1B】図1は、ポリペプチドの光捕捉回収を可能にする光切断可能なリンカーを介してポリペプチドを捕捉するための固相ベースのアプローチの概略図を示す。図1Bは、スルフヒドリル−反応性固相同位体タグ化試薬を示す。o−ニトロベンジル−ベースの光切断可能なリンカーは、アミノプロピルガラスビーズに連結される。光切断可能なリンカーの末梢に、7個の水素(H)または7個の重水素(D)のいずれか(「X」によって示される)を含むロイシン分子として表される同位体タグが、付着され、続いてSH−反応性基としてヨードアセチル基が付着される。
【図1C】図1は、ポリペプチドの光捕捉回収を可能にする光切断可能なリンカーを介してポリペプチドを捕捉するための固相ベースのアプローチの概略図を示す。図1Cは、固相同位体タグ化方法を用いた捕捉後のポリペプチドへの同位体タグの移動および放出を示す。リンカーは、切断の際、特異的な官能基が、放出されたポリペプチドに移動されるように構築される。
【図2A】図2は、トリス(2−カルボキシエチル)ホスフィン(TCEP)を用いた還元後のペプチドラミニンB、およびコントロールペプチドホスホアンジオテンシンの液体クロマトグラフィーマスウペクトロメトリー(LC−MS)分析を示す。図2Aは、ラミニンBおよびコントロールホスホアンジオテンシンの混合物の逆相HPLCによるLC分析を示す(RT:8.95−13.04、NL:1.22E7 基底ピークMS tcep30分)。
【図2B】図2は、トリス(2−カルボキシエチル)ホスフィン(TCEP)を用いた還元後のペプチドラミニンB、およびコントロールペプチドホスホアンジオテンシンの液体クロマトグラフィーマススペクトロメトリー(LC−MS)分析を示す。図2Bは、TCEP処理ラミニンBのエレクトロスプレーイオン化MS分析を示す(tcep30分、#384、RT:9.55、AV:1、NL:6.50EB、T:+c ESI Q3MS[400.00−1600.00])。
【図3A】図3は、図1に示されるビーズと接触させた後の還元されたラミニンBおよびホスホアンジオテンシンの混合物のLC−MSを示す。図3Aは、図2で処置されたような上清のアリコートのLC分析を示す(RT:8.97−13.08,NL:1.84E7 基底ピーク MS 60分ビーズ)。
【図3B】図3は、図1に示されるビーズと接触させた後の還元されたラミニンBおよびホスホアンジオテンシンの混合物のLC−MSを示す。図3Bは、還元されたラミニンBと一致する保持時間を有する図3Aの9.62分のLC画分のMS分析を示す(60分ビーズ、#387 RT:9.62、AV:1、NL:1.14E5、T:+c ESI Q3MS[400.00−1600.00])。ラミニンBの量を、ビーズへの結合に起因して減少させた。
【図4A】図4は、図3で処置されたようなビーズから光切断されたラミニンBのLC−MS分析を示す。図4Aは、図2で使用される量と等量の濃度でコントロールホスホアンジオテンシンを添加することによる、光切断されたラミニンBのLC分析を示す(RT:9.19−15.18、NL:2.09E6 基底ピーク m/z=560.0−570.0 MS 1時間光_0104 15121039)。
【図4B】図4は、図3で処置されたようなビーズから光切断されたラミニンBのLC−MS分析を示す。図4Bは、改変に起因する質量の予測された増加を伴なう、図4Aで10.72分で溶出した光切断されたラミニンBのピークのMS分析を示す(1時間光_010415121039 #430、RT:10.70、AV:1、NL:9.17E5、T:+c ESI Q3MS[400.00−1600.00])。
【図5】図5は、スルフヒドリル基を組み込むためのポリペプチドの1級アミン基のタグ化についてのストラテジーを示す。ポリペプチドのアミノ基は、N−スクシンイミジルS−アセチルチオアセテート(acetylthioacctate)(SATA)によって改変される。ヒドロキシアミン処理に際して、トリス(2−カルボキシエチル)ホスフィン(TCEP)での還元後、ポリペプチドのアミノ基は、スルフヒドリル基に転換される。
【図6A】図6は、SATAで改変されるホスホアンジオテンシンのLC−MS分析を示す。図6Aは、SATA処理ホスホアンジオテンシンのLC分析を示す(RT:9.56−16.88、NL:3.29E7 基底ピーク MS sata15分)。
【図6B】図6は、SATAで改変されるホスホアンジオテンシンのLC−MS分析を示す。図6Bは、SATA処理ホスホアンジオテンシンのMS分析を示す(sata15分 #582、RT:14.48、AV:1、NL:2.75E7、T:+c ESI Q3MS[400.00−1600.00])。2つの主なシグナルは、1荷電の[M+H]+=1242.7質量単位、および2荷電の[M+2H]2+=622.3質量単位の形態を表す。
【図7A】図7は、図5に示されるように、ヒドロキシアミンでさらに処理され、そしてTCEPで還元されたSATA処理ホスホアンジオテンシンのLC−MS分析を示す。図7Aは、還元された改変ホスホアンジオテンシンのLC分析を示す(RT:12.08−14.93、NL:1.32E7 基底ピーク MS 30分10m MTCEP)。
【図7B】図7は、図5に示されるように、ヒドロキシアミンでさらに処理され、そしてTCEPで還元されたSATA処理ホスホアンジオテンシンのLC−MS分析を示す。図7Bは、還元された改変ホスホアンジオテンシンのMS分析を示し、改変に起因して予測される質量における変化を示す(30分10mMTCEP #557 RT:13.86、AV:1、NL:1.32E7、T:+c ESI Q3MS[400.00−1600.00])。2つの主なシグナルは、1荷電の[M+H]+=1200.7質量単位および2荷電の[M+2H]2+=601.1質量単位の形態を表す。
【図8】図8は、固相捕獲を使用する差次的改変による2つのサンプルの比較のための概略図を示す。比較される2つのタンパク質サンプルは、タンパク質分解(例えば、トリプシンを用いて)にかけられる。スルフヒドリル反応性試薬の場合において、Cys含有ペプチドは還元され、そして差次的に同位体的に標識された試薬(例えば、それぞれ0個または7個の重水素を含むd0−ロイシンタグまたはd7−ロイシンタグ)を保有するビーズによって捕獲される。このビーズは、合わされそして洗浄され、そしてタグ化されたペプチドは、光切断によって放出される。放出されたペプチドは、例えば、マイクロキャピラリー液体クロマトグラフィーおよびタンデム質量分析(μLC−MS/MS)のような質量分析(MS)を使用してさらに特徴付けられ得る。
【図9】図9は、固相法およびICAT法によって同定され、定量化されるタンパク質数の概要を示す。図9Aは、大規模実験(L)から同定されるタンパク質の数を示し、その中で100μgの全タンパク質サンプルが標識され、そして20μgがμLC−MS/MSによって分析された。固相法は、82個のタンパク質を定量し、そして33個のタンパク質は、ICAT法によって定量され、25個のタンパク質は共通であった。図9Bは、小規模実験(S)から同定されたタンパク質数を示し、この中で10μgの全サンプルが標識され、5μgが分析された。固相法は、57個のタンパク質を定量し、そして18個のタンパク質は、ICAT法によって定量され、13個のタンパク質は共通であった。図9Cは、大規模実験(L)および小規模実験(S)における固相法によって同定されたタンパク質の数を示す。ベン図の区画中の数は、定量されたタンパク質の数を示す。
【発明を実施するための形態】
【0014】
(発明の詳細な説明)
本発明は、所望の官能基(タグ化分子の検出を増強し、そして同定および定量を容易にするのに有用なタグを含む)の分子への輸送を可能にする化学基を有する固体支持体上に分子を捕獲することによって、サンプル中の分子を標識するための方法および組成物を提供する。この方法は、サンプル由来の分子を選択的に単離しそして標識するために使用され得、分析物中の複雑な混合物の定量的な分析(質量分析のような方法による分析を含む)を可能にするという点で有利である。従って、この方法は、基本的に全ての特定のクラスの分子または分子のサブセット(例えば、基本的に全てのポリペプチドまたはリンタンパク質、糖タンパク質、またはその他の改変されたポリペプチドのサブセット)を単離するために使用され得る。
【0015】
一般的な共有結合的捕獲−放出化学を使用することによって、特定の官能基が複雑なサンプルの成分に輸送され得る。さらに、捕獲された分子を放出するための能力を組み入れることによって、この方法はまた、サンプル分子を単離または精製するために有利に使用され得、このことは、分析されたサンプルの複雑さを低減するのに有用であり得る。この方法は、定量的なプロテオーム分析、タンパク質リン酸化および他の翻訳後修飾の系統的かつ定量的な分析に十分適切であり、そしてタンパク質およびペプチド以外の分子の系統的かつ定量的な分析まで拡張され得る。さらに、本発明の方法は、サンプル分子が、効率良く捕獲および放出され得、使用される開始サンプルの量をより少なくし、プロテオミクス分析のための複雑な生物学的サンプルを分析するのに特に有用であるという点で有利である。
【0016】
本発明の方法は特に、生物学的サンプル中に含まれる分子の同定および定量分析、特に定量的なプロテオンミクスについてのタンパク質の分析のために有用である。この方法はまた、他の改変されたタンパク質における、タンパク質リン酸化または他の改変の系統的、定量的な分析のために使用され得る。本発明はまた、分子を標識するために有用な試薬を提供する。プロテオミクス分析に加えて、この方法はまた、タンパク質に加えて、その他の生体分子の系統的かつ定量的な分析のために有用である。この方法は、特に標識またはタグを質量分析(MS)に適切な分子に輸送するのに特に有用である。
【0017】
本明細書中で使用される場合、用語「ポリペプチド」は、2つ以上のアミノ酸のペプチドまたはポリペプチドをいう。ポリペプチドはまた、翻訳後修飾または合成的改変(リン酸化、脂質化、プレニル化、パルミチル化、ミリスチル化、硫酸化、ヒドロキシル化、アセチル化、グリコシル化、ユビキチン化、補欠分子族または補因子の付加、ジスルフィド結合の形成、タンパク質分解、高分子複合体へのアセンブリなどを含む)のような天然に存在する改変によって改変され得る。
【0018】
ポリペプチドは、2、3またはいくつかのアミノ酸を有する小さいポリペプチドならびに数百またはそれ以上のアミノ酸を有する大きいポリペプチドを含む。通常、2つ以上のアミノ酸残基間の共有結合は、アミド結合である。しかし、これらのアミノ酸は、ペプチドおよび化学の分野における当業者に公知の種々の他の手段によって共に結合され得る。従って、用語ポリペプチドは、アミノ酸、アミノ酸アナログ、および模倣物の間の非アミド結合を全体的にまたは部分的に含む分子を含むことが意図される。同様に、この用語はまた、環状ポリペプチドおよび他の立体配置的に構成された構築物を含む。
【0019】
ポリペプチド、特にリガンドポリペプチドの改変はまた、化学合成によって作製された、天然に存在しない誘導体、そのアナログおよび機能的模倣物を含み得、ただしこのようなポリペプチド改変は、親ポリペプチドと比較して類似の機能的活性を示す。例えば、誘導体は、このポリペプチドの化学修飾(例えば、アルキル化、アクリル化、カルバミル化、ヨード化、またはポリペプチドを誘導体化する任意の修飾)を含み得る。このような誘導体化分子としては、例えば、分子中の遊離アミノ基が誘導体化されてアミン塩酸塩、p−トルエンスルホン基、カルボベンゾキシ基、t−ブチルオキシカルボニル基、クロロアセチル基またはホルミル基を形成する分子が挙げられる。遊離カルボキシル基は、誘導体化され、塩、メチルおよびエチルエステルまたはエステルもしくはヒドラジドの他の型を形成し得る。遊離ヒドロキシル基は、誘導体化され、O−アシル誘導体またはO−アルキル誘導体を形成し得る。ヒスチジンのイミダゾール窒素は、誘導体化され、N−im−ベンジルヒスチジンを形成し得る。また、20個の標準的なアミノ酸のうち天然に存在するアミノ酸誘導体(例えば、4−ヒドロキシプロリン、5−ヒドロキシリジン、3−メチルヒスチジン、ホモセリン、オルニチンまたはカルボキシグルタメート)を1つ以上含むポリペプチドがまた、誘導体またはアナログとして含まれ、そしてペプチド結合によって連結されないアミノ酸を含み得る。
【0020】
特に有用なポリペプチド誘導体は、本明細書中に開示された方法を用いて所望の機能的特徴を取り込むための改変を含む。このような改変は、標識またはタグ(特に、MS分析に有用な標識またはタグ)の取り込みを含む。
【0021】
本明細書中で使用される用語「核酸」は、生化学系の成分に関して用いられる場合、デオキシリボ核酸(DNA)またはリボ核酸(RNA)のような共に共有結合された2つ以上のヌクレオチドを意味すると意図され、そして例えば、一本鎖および二本鎖の核酸を含む。この用語は、同様に、ゲノムDNA、cDNA、mRNA、およびそれらに対応する合成オリゴヌクレオチドを含むことが意図され、これらは。センス鎖、アンチセンス鎖またはその両方を表し得る。
【0022】
本明細書中で使用される用語「アミノ酸」は、天然に存在するアミノ酸および天然に存在しないアミノ酸の両方、ならびにアミノ酸アナログおよび模倣体を意味することが意図される。天然に存在するアミノ酸は、タンパク質生合成の間に利用される20個の(L)−アミノ酸ならびに例えば、4−ヒドロキシプロリン、ヒドロキシリジン、デスモシン、イソデスモシン、ホモシステイン、シトルリン、およびオルニチンのような他のアミノ酸を含む。天然に存在しないアミノ酸は、例えば、(D)−アミノ酸、ノルロイシン、ノルバリン、p−フルオロフェニルアラニン、エチオニンなどを含む。アミノ酸アナログは、天然に存在するアミノ酸および天然に存在しないアミノ酸の改変形態を含む。このような改変は、例えば、アミノ酸上の化学基および化学部分の置換(substitution or replacement)またはアミノ酸の誘導体化によるものを含み得る。アミノ酸模倣物は、例えば、参照アミノ酸に特徴的な電荷および電荷間隔のような機能的に類似の特性を示す有機構造を含む。例えば、リジン(LysまたはK)を模倣する有機構造は、同様の分子領域に位置し、そして天然に存在するLysアミノ酸の側鎖のε−アミノ基と同じ程度の移動度を有する正の電荷部分を有する。模倣物はまた、アミノ酸またはアミノ酸官能基の最適な間隔および電荷相互作用を維持するように制限された構造を含む。
【0023】
本明細書中で使用される「官能基」は、所望の機能特性を有する化学基をいう。所望の機能特性は、分子に所望の化学特徴を付与する任意の特性である。官能基は、分子の物理化学特性を変更する(例えば、質量、電荷、疎水性などを変更する)基を含み得る。特に有用な官能基は、標識またはタグ(例えば、発蛍光団、発色団、スピン標識、アイソトープ、分布タグなど)である。
【0024】
本明細書中で使用される用語「標識」は、分子の質量の変化を生じる分子に付着され得る任意の部分を意味することが意図される。一般に、標識は、共有結合されるけれども、この標識は、共有結合または非共有結合のいずれかで分子に結合され得る。非共有結合相互作用が標識と分子との間で生じる場合、非共有結合相互作用は、標識を、本発明の方法において使用される化学的および/または物理的操作の間に分子に結合されたままとするのに十分に高い親和性を有することが理解される。
【0025】
特に有用な標識は、MSによるサンプル分析に有用な質量標識である。質量標識の取り込みに起因する分子の質量の変化は、質量決定について選択された機器の感受性範囲内にあるべきである。さらに、当業者は、異なる大きさおよび異なる組成の分子についての適当な質量の標識を認識し、そして決定し得る。さらに、例えば、分子の差次的な標識について重質量標識および軽質量標識を用いる場合、約1〜3質量単位の間の程度の小ささの質量差または約10質量単位より大きい程度の大きさが用いられ得、例えば、約4、約5、約6、約7、約8、約9、約10、約12、約15、または約20の質量単位またはそれ以上が所望の通りである。2つのサンプルを差次的に標識するために適した質量標識は、化学的に同一であるが質量は異なる。
【0026】
本明細書中で使用される「タグ」は、検出可能な標識をいう。タグは、分子が種々の分析法(MS、クロマトグラフィー、蛍光光度分析、分光光度法、免疫学的技術などを含む)のいずれかによって検出され得るように、その分子に特徴を付与する。タグは、例えば、アイソトープ、蛍光(fluor)、クロマゲン(chromagen)、強磁性物質、発光タグ、または抗体もしくは抗体フラグメントにより認識されるエピトープタグであり得る。特に有用なタグは、質量タグであり、これは、MSによる分子の検出および分析に適切な質量標識である。例示の質量タグは、例えば、適切なアイソトープタグ、アイソトープ分布タグ、荷電アミノ酸、差次的にアイソトープで標識されたタグなどを含む。タグは、気相塩基性基(例えば、ピリジル)または疎水性基であり得る。タグはまた、特徴的なアイソトープ分布を有する元素、例えば、塩素、臭素、または識別可能なアイソトープ分布を有する任意の元素であり得る。さらに、タグは、適切な条件下での質量分光計の衝突セルまたはイオン源において破壊しリポーターイオンを生じる結合を有し得る。
【0027】
タグはまた、アフィニティータグの同族結合パートナーへの結合によりアフィニティー試薬にカップリングされた分子の単離を可能にするアフィニティータグであり得る。ポリペプチドタグ化のために、サンプル中の1つまたは複数のポリペプチドは変性され得、必要に応じて還元され得、そしてポリペプチドの化学反応性基は、化学改変試薬で共有結合により誘導体化され得る。タグ化されたポリペプチドは、サンプル中でタグ化されていないポリペプチドおよび他の成分から容易に単離され得る。これは、質量分析により分析されるべきであるサンプルの複雑度を低減する。このようなアフィニティータグ化は、他の分子(例えば、核酸、脂質、炭水化物、第二メッセンジャー、代謝物など)に同様に適用され得る。さらに、タグは、化学反応または酵素触媒反応によって導入され得る。
【0028】
本明細書中で使用される「開裂可能な官能基」は、種々の方法(エネルギー、化学物質、酵素などの投入を含む)によって開裂され得る化学基である。本発明の方法において使用するために、開裂可能な官能基は、一般に特異的であり、すなわち、開裂される分子に変化も損傷も与えることなく特異的に開裂可能であり、または比較的に均一に、再現様式で分子を変化させる。例えば、開裂可能な官能基は、光開裂可能な基であり得る。このような場合、光開裂可能な基は、一般に、放出される分子を損傷しない光の波長で(例えば、紫外光から可視光範囲で)開裂される(実施例Iを参照のこと)。例示の光開裂可能なリンカーとしては、例えば、o−ニトロベンジル基、デシル基、トランス−o−シンナモイル基、m−ニトロフェニル基、ベンジルスルホニル基などを含むリンカーが挙げられる(例えば、DormanおよびPrestwich、Trens Biotech.18:64−77(2000);GreeneおよびWuts、Protective Groups in Organic Synthesis、第2版、John Wiley & Sons、New York(1991);米国特許第5,143,854号;米国特許第5,986,076号;米国特許第5,917,016号;米国特許第5,489,678号;米国特許第5,405,783号を参照のこと)。
【0029】
切断可能な官能基はまた、酸または塩基のような化学物質によって切断可能な、化学的切断可能な基であり得る。望ましい場合、化学的切断反応は、その化学的切断可能な基が本質的に唯一の切断される化学結合である、比較的穏やかな条件下で実行され得る。化学的切断可能な基はまた、CNBr(これは、メチオニン残基を切断し得る)のような化学物質によって切断可能な基であり得る。CNBrは、メチオニンのような化学的切断可能な基が、(特に、メチオニン残基を有さないポリペプチドにおいて)ある分子に付加されている場合、その分子を遊離させるために特に有用であり得る。適切な化学切断可能な基は、当業者に周知である(例えば、WilsonおよびCzarnik編、Combinatorial Chemistry:Synthesis and Application,John Wiley & Sons,New York(1997);Merrifield,J.Am.Chem.Soc.85:2149(1964);Bodanszky,M.,Principles of Peptide Synthesis(Springer−Verlag,1984);Houghten,Proc.Natl.Acad.Sci.USA 82:5131(1985)を参照のこと)。例示的な化学的切断可能なリンカーは、ジスルフィド(これは、還元剤により切断され得る);ジオール(これは、過ヨウ素酸塩)により切断され得る);ジアゾ結合(これは、ジチオネートにより切断され得る);エステル(これは、ヒドロキシルアミンにより切断され得る);スルホン(これは、塩基により切断され得る)などを含み得る(Hermanson,Bioconjugate Technique,Academic Press,San Diego(1996);Pierce Chemical Co.,Rockford,ILを参照のこと)。
【0030】
この切断可能な官能基はまた、酵素切断可能な基であり得る。例えば、プロテアーゼが、そのプロテアーゼにとって適切な認識配列を有する切断可能な官能基を切断するために、使用され得る。特に有用なプロテアーゼは、エンドペプチダーゼ(例えば、第Xa因子、タバコエッチ(etch)ウイルス(TEV)プロテアーゼ、トリプシン、キモトリプシン、Staphylococcus aureusプロテアーゼ、顎下腺(submaxillaris)プロテアーゼなど)である。そのプロテアーゼは、切断可能な特定の認識配列を官能基として組み込むことに基づいて選択され得る。プロテアーゼを選択するための他の考慮事項としては、捕捉されそして遊離される分子中に認識配列が存在するかまたは存在しないかが挙げられる。
【0031】
例えば、希に切断するプロテアーゼ(例えば、TEVプロテアーゼまたは第Xa因子)は、対応するプロテアーゼ認識配列を含む官能基を切断して捕捉された分子を遊離するために、使用され得る。このような希に切断するプロテアーゼは、インタクトなポリペプチド分子を遊離するために特に有用である。なぜなら、これらのプロテアーゼの認識配列は、大部分のポリペプチド中に存在しないからである。あるいは、ポリペプチドサンプルは、特定のプロテアーゼで処理され得、そして消化されたペプチドが、本明細書中に開示される方法によって単離され得る。このような場合、捕捉されたペプチドは、切断のために使用されるプロテアーゼの認識配列を含まない。なぜなら、そのポリペプチドは、すでに消化されているからである。さらに、望ましい場合、インタクトなポリペプチドは、固体支持体に結合された後に、捕捉されそしてプロテアーゼで消化され得、その固体支持体上に捕捉されたポリペプチドのペプチドフラグメント上の標識の組み込みおよび遊離を生じ得る。従って、プロテアーゼ消化は、望ましい場合、サンプル分子(特にポリペプチドサンプル分子)の捕捉の前にもまたは後にも使用され得る。
【0032】
プロテアーゼに加えて、切断可能な官能基は、エンドヌクレアーゼ(例えば、制限酵素)の認識配列であり得る。従って、制限酵素の適切な認識配列が、切断可能な官能基として組込まれ得、そしてその個々の制限酵素により切断され得る。そのようなヌクレオチド官能基は、核酸またはポリペプチドまたは望ましい他の任意の型の分子を捕捉および遊離させるために有用であり得ることが、理解される。同様に、プロテアーゼ認識配列が、核酸またはポリペプチドまたは望ましい他の任意の型の分子を捕捉および遊離させるために有用であり得る。
【0033】
本明細書中で使用される場合、用語「反応性基」は、分子(例えば、ポリペプチド、核酸、脂質、糖質、セカンドメッセンジャー、代謝物など)との反応およびその分子への共有結合に適切な有用な化学的特性を有する、種々の化学基のうちのいずれかを意味することが意図される。例えば、反応性基は、AspもしくはGluにおいて見出されるカルボキシル基と反応し得るか、またはその反応性基は、他のアミノ酸(例えば、His、Tyr、Arg、およびMet)と反応し得る。反応性基はまた、Lysのようなアミンと反応し得る(例えば、イミドエステルおよびN−ヒドロキシスクシンイミジルエステルである)。さらに、反応性基はまた、当該分野で周知の化学反応を使用して、酸素または硫黄と反応し得る。反応性基はまた、ホスホペプチドもしくは核酸の選択的標識のためにリン酸基と反応し得るし、または他の共有結合により改変されたペプチド(グリコペプチド、リポペプチド、または本明細書中に開示されるような共有結合ポリペプチド改変物のうちのいずれかを含む)と反応し得る。
【0034】
本明細書中で使用される場合、用語「同位体標識」または「同位体タグ」とは、2つの別個の同位体形態で生成され得る化学基(例えば、その化学基を構成する構成元素の重い同位体バージョンおよび軽い同位体バージョン)を指す。そのような構成元素としては、例えば、炭素、酸素、水素、窒素、および硫黄が挙げられる。さらに、化学的または機能的に類似する他の元素が、天然に存在する上記元素に代わって置き換わり得る。例えば、セレンが、硫黄に代わる置換基として使用され得る。特に有用な同位体標識または同位体タグは、MSによる簡便な分析を可能にする、同位体標識または同位体タグである。例えば、重い同位体バージョンおよび軽い同位体バージョンのアミノ酸が、ポリペプチドを差次的に同位体標識するために使用され得る(実施例Iを参照のこと)。
【0035】
本明細書中で使用される場合、「結合している(coupled)」またはその文法上の形態は、分子間の結合相互作用を指す。例えば、固体支持体は、その固体支持体の化学部分と、化学基の化学部分との間の結合相互作用を介して、その化学基に結合され得る。結合している分子間の結合相互作用は、共有結合性相互作用でもまたは非共有結合性相互作用でもあり得る。一般的に、化学基は、共有結合性相互作用を介して、固体支持体または他の分子に結合される。その固体支持体と、結合された分子との間に非共有結合性相互作用が生じる場合、その非共有結合性相互作用は、本発明の方法において使用される化学的操作および/または物理的操作(例えば、その固体支持体に結合している分子に対して実施される化学的改変工程または洗浄工程)の間に結合したままであるに十分に高親和性であることが、理解される。
【0036】
本発明は、分子を標識するための方法を提供し、この方法は、サンプル分子を、化学基(切断可能な官能基と、1つ以上の官能基と、そのサンプル分子についての反応性基とを含む)に結合されている固体支持体と、そのサンプル分子をその反応性基に共有結合させる条件下で接触させることによる。この方法は、その切断可能な官能基を切断し、それによって、1つ以上の結合した官能基を有するそのサンプル分子を遊離させる工程をさらに包含し得る。この方法は、複雑なサンプル混合物中の分子が、捕捉され得、そして簡便な分析のために標識され得、そしてその固体支持体から遊離される能力に起因して、精製もされ得る点で、有利である。
【0037】
本発明は、目的のポリペプチドまたは他の分子を、切断可能なリンカー(例えば、光切断可能なリンカー)を介して共有結合により捕捉するために、固相ベースのアプローチを使用する方法を提供し、この光切断可能なリンカーは、捕捉された分子の光触媒回復を可能にする。この切断可能なリンカーは、切断の際に、遊離される分子に特定の官能基が転移するように構築される。そのような官能基としては、例えば、安定な同位体タグ(同位体希釈理論に基づいて質量分光法によって正確なペプチド定量を可能にする)、同位体分布タグ(その同位体分布によってタグ化ペプチドまたはそのフラグメントを同定する)、荷電アミノ酸または他の化合物(質量分析計中での効率的なイオン化を媒介し、そしてタンデム質量分析計の衝突セルにおける断片化パターンをもたらす)が挙げられる。さらに、この方法は、その分子(例えば、ポリペプチド)が固体支持体上に固定されている間の、その分子の化学的改変または酵素的改変、脱改変(de−modification)、切断または他の操作を可能にする。光切断可能なリンカーが特に有用であるが、本明細書中に開示されるような特異的に切断され得る任意のリンカーが使用され得ることが、理解される。光切断可能なリンカーの代替案としては、酸切断可能なリンカーおよび塩基切断可能なリンカー、熱により切断可能なリンカー、ならびに本明細書中に記載されるような、酵素の標的切断部を含むリンカーが、挙げられる。
【0038】
本発明の方法は、サンプルから分子を固定し、そして捕捉されたサンプル分子に標識(例えば、タグ)を簡便に転移するのを可能にする、固相化学の有利な使用に基づく。この方法は、固体支持体に化学基を結合することに基づく。その化学基は、図1に例示される特徴を有する。その化学基の1つの特徴は、分子の可逆的な捕捉および遊離を可能にする、切断可能な官能基である。その化学基の第2の特徴は、望ましい化学的特性を有する1つ以上の官能基である。そのような官能基は、例えば、その分子または化学部分のその後の分析のために便利な、その分子に望ましい化学的特性(例えば、電荷の変化、疎水性の変化、または質量の変化)を付与する標識またはタグであり得る。その化学基の第3の特徴は、サンプル中の分子にその化学基を共有結合するのを可能にする、反応性基である。これらの3つの特徴は、サンプル分子がその反応性基を介して捕捉され得、切断の際に、遊離される分子にその官能基が転移するように、その化学基上で配置される(図1を参照のこと)。
【0039】
その化学基はまた、固体支持体にその化学基が結合されるのを可能にし、同時に上記特徴が活用されるのを可能にする、化学部分を含む。上記特徴を有する化学部分は、その特徴を付与する化学部分の連続付加によって、固体支持体上で合成され得るか、または望ましい場合は、化学基として合成され得、その後、その固体支持体に結合され得る。
【0040】
本発明の方法の特定の実施形態が、図1において例示される。切断可能な官能基と、1つ以上の望ましい官能基(例えば、タグ)と、反応性基とを有する化学基が、固体支持体に結合される。図1は、その固体支持体に共有結合されたアミノ官能基を含む、光切断可能なリンカーを示す。この光切断可能なリンカーは、分子が固相上に捕捉された後に、それらの分子の光開始切断を可能にする。望ましい特定の官能基を有するリンカー分子が、そのアミノ官能基を介して、その光切断可能なリンカーに結合される。光切断の際、その官能基を含むリンカー分子は、図1Cにペプチドとして示される捕捉された分子に転移され、望ましい官能基のその分子への転移をもたらす。例えば、本明細書中に開示されるように、転移される官能基は、定量的質量分光法に有用な、安定な同位体がコードするアミノ酸であり得る。1つ以上の官能基を有する機能的リンカーから延びているのは、その分子上の化学部分について特異性を有する反応性基(例えば、ポリペプチドのアミノ基と反応性の基、スルフヒドリル基と反応性の基、カルボキシ基と反応性の基または他の基と反応性の基)である。
【0041】
分子(例えば、サンプル中の分子)は、上記のように結合された化学基を有する固体支持体と、接触される。その分子は、そのサンプル分子がその化学基を介してその固体支持体に共有結合するのを可能にする条件下で、インキュベートされる。その化学基およびサンプル分子上の反応性基に基づいて、共有結合を可能にする適切な条件を、当業者は容易に決定し得る。同様に、本明細書中に開示されるように、非共有結合性相互作用のための適切な条件を、当業者は容易に決定し得る。
【0042】
実施例Iに例示され、図1に示される特定の実施形態において、固相支持体は、光切断性リンカーが、シラン結合を介して結合されている、制御された孔を有するガラスビーズである。図1に示される光切断性リンカーは、360nmのUV光で切断され得る。重水素化アミノ酸または非重水素化アミノ酸(例えば、ロイシン)は、図1に例示されたポリペプチドに転位される官能基として使用され得る。2つの異なるサンプル供給源が、比較分析または定量分析のために使用される場合、2つの同位体タグは、ロイシンの重水素化の状態に依存して、一般的に質量が、7または10質量単位異なる。本明細書中に開示されるように、異なる同位体分布を有する他のアミノ酸またはアミノ酸とは異なる分子もまた、安定な同位体タグとして使用され得る。図1に示されるペプチド反応性基は、スルフヒドリル基と特異的に反応するヨードアセチル基である。光切断の際に、改変されたロイシンに起因する質量タグの増加を有する、図1Cに示されるポリペプチドが、放出される。重水素化されたアミノ酸を用いて例示されるけれども、任意の適切な同位体形態(例えば、13C,15Nなどのような他の構成要素の同位体)が、本発明の方法において、使用され得ることが理解される。非放射性同位体が一般的に使用されるが、トリチウムのような放射性同位体でさえも使用され得る。
【0043】
本発明はまた、図1に例示される試薬のような試薬を提供する。この試薬は、サンプル分子(複合サンプルを含む)を単離し、タグ化するために使用され得る(実施例Vを参照のこと)。従って、本発明は、図1Aに概略的に示されるような、ビーズ、切断性官能基、タグのような官能基、および反応性基を含む試薬を提供する。
【0044】
このようにして、本発明はさらに、化学基に結合された固体支持体を含有する構成物(composition)を提供し、この化学基は、切断性官能基、タグのような官能基およびサンプル分子に共有結合された反応性基を含み、ここで、切断性官能基、タグおよび反応性基は互いに関して、切断性官能基の切断の際に、タグがサンプル分子に移動可能であるように配置される。本発明はさらに、化学基に共有結合された固体支持体を含有する構成物を提供し、この化学基は、切断性官能基、質量分析法タグおよびサンプル分子に共有結合された反応性基を含み、ここで、切断性官能基、タグおよび反応性基は互いに関して、切断性官能基の切断の際に、タグがサンプル分子に移動可能であるように配置される。
【0045】
本発明の方法は、分子を選択的に単離し、かつ放出の際に、標識またはタグを含有する1つ以上の官能基をこの分子上に転位する能力を提供するという点で有利である。従って、化学基(すなわち、切断性官能基、1つ以上の官能基および反応性基)の官能基は互いに関して、官能基(例えば、タグ)が分子に転位可能になるように配置される。従って、化学基の官能基は、官能基(例えば、タグ)が図1に示されるように切断性官能基と反応性基との間に配置され、切断性官能基の切断およびこの分子の放出の際に、この官能基が捕捉分子に転位し得るように、一般的に配置される。
【0046】
本発明の方法は、サンプル分子を捕捉する能力を使用し、官能基をこの分子に転位し、かつ結合官能基を有する分子を放出する能力を利用しているので、有利である。従って、本方法は、サンプル分子を標識し、そして単一工程でサンプル分子を同時に精製するために使用され得る。切断性官能基の組み込みは、結合官能基を有するサンプル分子の放出を容易にし、この結合官能基が次いで、分析され得る。本発明の方法は、切断性官能基を一般的に使用するけれども、本発明の方法が、さらなる分析の前に捕捉分子の放出を必要としない標識またはタグのような官能基を転位するためにもまた使用され得ることが理解される。化学反応または酵素反応によって切断可能な切断性化学基の非存在下での、本発明の方法の使用は、分析方法(例えば、MS、特に、MALDI−TOF)に適用可能であり、ここでは、レーザーが、結合分子の切断に使用され、同時に分子をイオン化する。
【0047】
本発明の方法は、広範囲の種々の分子に容易に適用され得る。上記に記載されるように、いくつかの場合、分子は、本明細書中に開示される捕捉方法に適切な反応性化学部分を有し得る。しかし、所望される場合、分子は、所望の官能基、特に、本明細書中に開示される捕捉方法に適切な反応性基を組み込むように改変され得る。
【0048】
例えば、システイン残基を含まないポリペプチド、つまりスルフヒドリル側鎖を含有する天然アミノ酸を含まないポリペプチドは、図1に示される固相試薬に結合しない。いくつかの場合、システイン含有ポリペプチドを選択的に単離することが望ましいが、他の場合、サンプル中に含有される他のポリペプチドまたはさらなるポリペプチドを、単離し、同定し、そして定量することが、望ましい。例えば、アミノ反応性基(例えば、固相試薬の反応性基としてのスクシンイミドエステル)を合成することが可能である。あるいは、捕捉されるべき分子は、化学的に改変され、特異的な官能基を組み込み得る。
【0049】
例えば、本明細書中に開示される特定の実施形態において、ポリペプチドの一級アミノ基は、スルフヒドリル基へと改変され、図1に示されるような、同一のSH−反応性固相ビーズを、ポリペプチドの捕捉に使用されるようにする。この戦略において、ポリペプチドのアミノ基は、図5に示されるワンポット化学によってスルフヒドリル基へと転換され得る。第1に、アミノ基は、N−スクシンイミジルS−アセチルチオアセテート(SATA)によって改変される。ヒドロキシルアミン処理の際に、トリス(2−カルボキシエチル)ホスフィン(TCEP)での還元の後に、このペプチドのアミノ基が、スルフヒドリル基へと転換される。本明細書中に例示されるように、サンプル分子中の実質的に全てのアミノ基が、スルフヒドリル基へと転換され得る(実施例IIを参照のこと)。改変されたペプチドは、必要に応じて、例えばC18逆相カートリッジ上で脱塩することによって精製され得、回収され、次いで、図1に示されるようなビーズに結合され得る。
【0050】
アミノ基のスルフヒドリル基への転換の際に、プロトン化部位は、消失する。従って、例えば、ロイシンの代わりの荷電性アミノ酸(例えば、リジン)が、ポリペプチドのタグ化に使用され得る。この方法において、リジン側鎖の遊離アミノ基は、代替のプロトン化部位を提供し得、ペプチドの荷電状態は、この手順によって変わらない。タグを付加するための他の適切な基(荷電性基を含む)としては、例えば、アルギニン、ピリジル、トリメチルアミンなどが挙げられ、これらは、溶液中または気相中で強塩基であり、つまりイオン化を促進する基である。代表的な質量分光法実験下では、ペプチドの荷電状態が、ペプチドの配列決定および検出のための質量分析器における、衝突により誘導されるフラグメント化に影響するので、このことは重要である。
【0051】
ポリペプチド中のアミノ酸側鎖を改変するための方法および化学は、当業者に周知である(例えば、Glazerら、Laboratory Techniques in Biochemistry and Molecular Biology:Chemical Modification of Proteins,Chapter 3,68頁〜120頁,Elsevier Biomedical Press,New York(1975)(これは参考として本明細書中に援用される);およびPierce Catalog(1994),Pierce,Rockford ILを参照のこと)。
【0052】
ポリペプチドのアミノ末端を改変するための方法もまた、使用され得る。本明細書中に例示される、(N末端を含む(実施例IIを参照のこと))アミノ基を改変するための方法に加えて、N末端を改変するための他の方法は、当業者に周知である(例えば、Branciaら、Electrophoresis 22:552−559(2001);Hovingら、Anal.Chem.72:1006−1014(2000);Munchbachら、Anal.Chem.72:4047−4057(2000)(各々は、参考として本明細書中に援用される)を参照のこと)。
【0053】
さらに、反応性基は、分子上に形成され得、続いてこの分子は、所望の化学部分を組み込むように改変され得る。例えば、CNBrによる切断は、ホモセリンラクトンを生じる。従って、メチオニンを含むポリペプチドは、CNBrによって化学的に切断され、ホモセリンラクトンを生じ得る。得られたホモセリンラクトンは、アミンによって改変され得、反応性アミンを有する化学基の組み込みを可能にする。
【0054】
分子は、所望される場合、化学的にかまたは酵素的に改変され得る。例えば、分子は、上記に記載されるような方法を使用して化学的に改変され得る。さらに、分子は、酵素的に改変され得る。捕捉された分子は、酵素的に改変され、この分子に基を組込み得るかまたはこの分子から基を除去し得る。例えば、ポリペプチドは、キナーゼによってリン酸化され得るかもしくはホスファターゼによって脱リン酸化され得るか、または、ポリペプチドを翻訳後修飾する能力を有する任意の他の酵素によって分子への化学部分の付加もしくは分子から化学部分の除去を生じ得る。同様に、核酸は、核酸へと化学部分を付加するかまたは核酸から化学部分を除去する、任意の周知の酵素によって捕捉された後、改変され得る(例えば、Sambrookら、Molecular Cloning:A Laboratory Manual,第2版、Cold Spring Harbor Press,Plainview,New York(1989);Ausubelら、Current Protocols in Molecular Biology(補遺47),John Wiley & Sons,New York(1999);SambrookおよびRussel,Molecular Cloning:A Laboratory Manual,第3版,Cold Spring Harbor Press,Cold Spring Harbor(2001)を参照のこと)。捕捉された分子(ポリペプチドまたは核酸を含む)を改変するために有用な例示的な酵素としては、キナーゼ、ホスファターゼ、メチラーゼ、およびデカルボキシラーゼなど、または捕捉された分子へと化学部分を付加し得るかまたは捕捉された分子から化学部分を除去し得る任意の酵素が挙げられる。
【0055】
任意の種々の反応性基は、この反応性基がポリペプチドのような分子に共有結合され得る限り、サンプル分子との反応のために化学基に組み込まれ得る。反応性基は、当業者に周知である(例えば、Hermanson,(上述)1996;Glazerら、Laboratory Techniques in Biochemistry and Molecular Biology:Chemical Modification of Proteins,第3章,68頁〜120頁,Elsevier Biomedical Press,New York(1975);Pierce Catalog(1994),Pierce,Rockford ILを参照のこと)。例えば、反応性基は、AspまたはGluに見出されるカルボキシル基と反応しえるかあるいは、この反応性基は、他のアミノ酸(例えば、His、Tyr、Arg、およびMet)と反応し得る。反応性基はまた、Lysのようなアミン(例えば、イミドエステルおよびN−ヒドロキシスクシンイミジルエステル)と反応し得る。さらに、反応性基はまた、当該分野で周知の化学を使用して、酸素または硫黄と反応し得る。反応性基はまた、ホスホペプチドの選択的標識のためのリン酸基、または他の共有結合的に改変されたペプチド(グリコペプチド、リポペプチド、または本明細書中に開示される任意の共有結合ポリペプチド改変体を含む)と反応し得る。さらに、当業者は、公知の試薬を使用してポリペプチドを改変するための条件、ポリペプチドまたは本発明の方法における使用のための他の分子の改変のために最適な条件を得るためのインキュベーション条件およびインキュベーションの時間を知るかあるいはそれらのことを容易に決定し得る。
【0056】
例示的なスルフヒドリル反応性基としては、ヨードアセトアミド基が挙げられる(Gygiら、Nature Biotechnol.17:994−999(1999)を参照のこと)。他の例示的なスルフヒドリル反応性基としては、マレイミド、アルキルハライドおよびアリールハライド、ハロアセチル、α−ハロアシル、ピリジルジスルフィド、アジリジン、アクリロリル、ならびにアリール化剤が挙げられる。所望の場合、ポリペプチドは、本発明の試薬と反応する前に還元され得、これは、この試薬が、スルフヒドリル反応性基を含む場合に、特に有用である。
【0057】
反応性基はまた、例えば、ペプチドのα−アミノ基またはLys側鎖のε−アミノ基のようなアミンと、例えば、イミドエステル、N−ヒドロキシスクシンイミジルエステル(NHS)、イソチオシアネート、イソシアネート、アシルアジド、スルホニルクロリド、アルデヒド、ケトン、グリオキサール、エポキシド、オキシラン、カーボネート、アリール化剤、カルボジイミド、無水物などとを反応し得る。反応性基はまた、AspもしくはGluまたはペプチドのC末端に見られるカルボキシル基と、例えば、ジアゾアルカン、ジアゾアセチル、カルボニルジイミダゾール、カルボジイミドなどとを反応し得る。ヒドロキシル基と反応する反応性基としては、例えば、エポキシド、オキシラン、カルボニルジイミダゾール、N,N’−ジスクシンイミジルカーボネート、N−ヒドロキシスクシンイミジルクロロホルメートなどが挙げられる。反応性基はまた、アミノ酸(例えば、His、Tyr、Arg、およびMet)と反応し得る。さらに、反応性基はまた、ホスホペプチドの選択的標識のためのリン酸基、または他の共有結合的に改変されたペプチド(グリコペプチド、リポペプチド、または任意の公知の共有結合ポリペプチド改変が挙げられる)と反応し得る。当業者は、本発明の方法における使用のためのサンプル分子の改変について最適な条件を得るための種々の試薬、インキュベーション条件およびインキュベーション時間を用いることによって、サンプル分子を改変するための条件を容易に決定し得る。
【0058】
サンプル分子を捕捉することによって、種々の化学修飾および/または酵素的改変が、結合した分子に対して実行され得るが、このサンプル分子が、固体支持体に結合したままであるという点で、本発明の方法は、有利である。サンプル分子が、物理的操作、化学的操作および/または酵素的操作の間、固体支持体に結合したままであるので、改変されたサンプル分子の収率は、溶液相方法よりも高い。従って、本発明の方法は、本発明のタグ試薬と固体相捕捉の反応性基と反応し得る化学特性を有する、特定のクラス(class or classes)の分子の少なくとも約5%、約10%、約20%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%、約98%、約99%でか、または本質的に全てを捕捉するために使用され得る。さらに、本発明の方法は、捕捉試薬およびタグ試薬を含む本発明の固体支持体と接触され、そしてこの支持体に結合されるサンプル中の特定のクラスの分子の少なくとも約5%、約10%、約20%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%、約98%、約99%でか、または本質的に全てを捕捉および放出するために使用され得る。例えば、本明細書中で開示されるように、実質的に100%の分子は、捕捉および放出され得る(実施例Iを参照のこと)。当業者は、固体支持体からサンプル分子を捕捉するため、および/またはサンプル分子を放出するために最適化された条件を容易に決定し得る。
【0059】
本発明はさらに、分子を標識するための方法を提供する。この方法は、サンプル分子が反応性基に共有結合し得る条件下で、サンプル分子を化学基が結合された固体支持体と接触させる工程であって、この化学基が、切断可能な官能基、1つ以上の官能基、およびサンプル分子に対する反応性基を含む工程;固体支持体に結合されるサンプル分子を改変する工程;および切断可能な官能基を切断する工程であって、これによって、1つ以上の官能基を含む改変されたサンプル分子を放出する工程を包含する。
【0060】
本発明の方法は、望ましい場合、捕捉された分子を改変するために使用され得る。例えば、この方法は、リン酸化ポリペプチドおよび非リン酸化ポリペプチドの両方を含むサンプルからポリペプチドを捕捉するために使用され得る。タンパク質リン酸化を系統的に分析する方法は、最近記述された(Zhouら、Nature Biotechnol.19:375〜378(2001))。Zhouらのように溶液中で化学反応を実行するというのでなく、ポリペプチドは、まず、本明細書中に開示される方法を用いて捕捉され得、そして固体支持体に結合される間に改変され得る(実施例IVを参照のこと)。あるいは、分子を捕捉および標識するための本明細書中に開示される方法は、Zhouら(前出、2001)に記載される反応の実施後、または任意の他の化学改変もしくは酵素的改変の後に適用され得る。
【0061】
本発明の構成物および方法は、種々の適用において有利に使用され得る。1つの特に有用な適用は、定量的タンパク質発現分析である。例えば、本明細書中に開示される方法および試薬は、複雑なサンプル中に含まれるポリペプチドのような分子に特定の同位体サインを与えるために使用され得る。定量分析の1つの特定の適用において、2つ以上のサンプルが、比較され得る(図8、実施例IIIを参照のこと)。2つの異なるサンプル由来のペプチドの定量分析は、タンパク質発現の相対的定量化についてのICAT技術と同様な様式で、安定な同位体の希釈の概念に基づいて達成され得る(Gygiら、NatureBiotechnol.17:994〜999(1999);実施例Vを参照のこと)。示差的に同位体で標識した分子は、比較のために2つのサンプル(例えば、アミノ酸(例えば、重水素化ロイシンまたは非重水素化ロイシン)、または2つの異なるサンプルに対して質量タグとして組込まれ得る他のアミノ酸)を標識するために使用され得る。本発明の方法は、分子の単離およびタグ(例えば、安定な同位体)の組込みの両方が、両方達成される点で有利である。従って、本発明の方法は、定量的な質量スペクトル分析について特に有用である。
【0062】
本明細書中に開示されるように、ポリペプチドへの官能基の組込みはまた、ペプチドのアミノ基を介して達成され得、この組込みは、ポリペプチド中のスルフヒドリル基の存在に基づく捕捉方法が適用されるとき、遊離のシステイン残基を含まないポリペプチドが分析される場合、特に有用である。ほとんどのポリペプチドは、そのN−末端に少なくとも1つのアミノ基を含むので、システイン残基を有さないポリペプチドでさえも、ポリペプチド中にスルフヒドリル基を組込む方法に基づく固相で標識され得る。ブロックされたアミノ末端を有するサンプル中のポリペプチドの場合にて、このようなポリペプチドは、フラグメント化されて、切断されたフラグメント上に遊離のアミノ末端を生成し得る。従って、本発明の方法は、特定の化学改変を使用する種々の分子の単離に適用され得る。
【0063】
さらに、移入されたタグの選択における大きな構造的柔軟性が存在する。従って、タグの構造は、特定の目標を達成するために意図的に選択され得る。例えば、非常に極性のペプチドは、より疎水性に作製され得、従って、疎水性タグの移入によって逆相カラム上に良好に保持され得るか、強力な気相塩基性基(例えば、ピリジル)が、質量分析計の衝突セルにおいて直接的にフラグメント化するために移動され得るか、または特徴的な同位体分布を有する元素(例えば、塩素または臭素)が、タグ化ペプチドに対して異なる同位体サインを提供するために付加され得る。
【0064】
本発明の方法はまた、改変分子(例えば、翻訳後修飾によって修飾されたポリペプチド)の分析に適用され得る。例えば、この方法は、タンパク質リン酸化の定量分析に適用され得る。タンパク質リン酸化の系統的な分析に関する方法は、すでに記載されている(Zhouら、Nature Biotechnol.19:375〜378(2001))。一連の化学反応は、溶液中でリン酸化ポリペプチドおよび非リン酸化ポリペプチドを含む、複雑なペプチド溶液からリン酸化ペプチドの選択的単離のために、溶液中で実行される(Zhouら、前出、2001)。固相の捕捉および放出を用いる本明細書中に開示される方法はまた、ホスホペプチドの選択的単離のために使用され得る。リン酸化ポリペプチドおよび非リン酸化ポリペプチドは、本明細書中に開示されるように、固相ビーズ上に捕捉される。一旦固定化されると、ホスホペプチドのリン特異的標識化を導く一連の化学反応が、実行される(Zhouら、前出、2001)。ペプチドは、切断によって放出され、正確な定量化のために各々のペプチドに安定な同位体サインを与える。最初からリン酸化されているポリペプチドは、スルフヒドリル基に新たに変換され、スルフヒドリル含有分子を単離するための本明細書中に開示される方法を用いて捕捉され得る。捕捉されたポリペプチドは、非リン酸化ペプチドがなくなるまで洗浄され得、そして例えば、質量スペクトル分析のために放出され得る。本明細書中に開示される本発明の方法は、各々の化学反応後に過剰の試薬を除去するために必要とされるサンプルを取扱う工程の数が、劇的に減少および単純化されるので、溶液化学よりも明らかに有利である。
【0065】
リン酸化に加えて、本発明の方法は、本明細書中に開示されるように、翻訳後修飾の多くの異なる形態(例えば、グリコシル化、ユビキチン化、アセチル化、パルミチル化、ミリスチル化など)を有するポリペプチドに容易に適用され得る。従って、本発明の方法は、ペプチドへの種々の官能基の同じ移入を伴うポリペプチドを含む、他の翻訳後修飾された分子を選択的に単離するために使用され得る。翻訳後修飾の特定の型の選択的な単離は、本発明の方法を用いて達成され得る。例えば、ユビキチン化に対する特異的結合活性を有する抗体は、ユビキチン化ポリペプチドを単離するために使用され得、タンパク質リン酸化の定量分析と同じ方法で、ポリペプチドのユビキチン化の定量分析が達成され得る。このような方法はまた、所望の場合、分子の他の修飾に適用され得る。
【0066】
抗体もまた、エピトープタグで修飾されたサンプル分子の引き続く分析および/または単離のために使用され得る。抗体を調製するための方法は、当業者に周知である。用語抗体は、その最も広範な意味で使用され、ポリクローナル抗体およびモノクローナル抗体、ならびにこのような抗体の抗原結合フラグメントを含む。本発明において有用な抗体、またはこのような抗体の抗原結合フラグメントは、少なくとも約1×105M−1のポリペプチドまたはペプチド部分に対して、特異的結合活性を有することによって特徴付けられる。従って、ポリペプチドに対して特異的結合活性を保持する、抗体のFabフラグメント、F(ab’)2フラグメント、Fdフラグメント、Fvフラグメント、単鎖Fv(scFv)フラグメントなどである。ポリペプチドに対する抗体の特異的結合活性は、例えば、特定の抗原 対 コントロール抗原に対する抗体の結合活性を比較することによって、当業者によって容易に決定され得る。ポリクローナル抗体またはモノクローナル抗体を調製する方法ならびに結合活性および/または特異性を決定する方法は、当業者に周知である(例えば、HarlowおよびLane、Antibodies:A Laboratory Manual,Cold Spring Harbor Laboratory Press(1988))。
【0067】
本発明の方法は、一般的に、ポリペプチドを用いて本明細書中に例証したが、サンプル中の任意の種々の分子が、本明細書中に開示される方法によって容易に標識され得ることが理解される。一般的には、多くのクラスの生体分子(例えば、オリゴヌクレオチド、代謝産物など)は、本明細書中に開示される方法によって官能化されて、改善された定性分析または定量分析のための所望の官能基を組込み得る。従って、分子への特定の官能基の可逆的な捕捉および移入を可能にする、本明細書中に開示される方法は、一般的には、プロテオミクス、他の型の発見科学および一般的な定量的生物学的分析の分野における多くの適用において有用である。
【0068】
本発明の方法は、あるクラスの分子(例えば、ポリペプチド、核酸、脂質、セカンドメッセンジャー、代謝産物など)、またはこのようなあるクラスの分子の一部を、効率的に捕捉および放出するために使用され得る。所望の場合、本発明の方法はまた、2つ以上のクラスの分子および/またはこれらのクラスの分子の一部を捕捉するために使用され得る。例えば、この方法は、ポリペプチドおよび核酸の両方、または所望の場合、2つ以上のクラスの分子の任意の組合せを捕捉するために使用され得る。
【0069】
本発明はまた、分子を分析するための方法を提供する。この方法は、サンプル分子が反応性基に共有結合し得る条件下で、サンプル分子を化学基が結合した固体支持体と接触させる工程であって、この化学基が、切断可能な官能基、1つ以上の官能基、およびサンプル分子に対する反応性基を含む工程;サンプル分子を固体支持体から切断する工程であって、1つ以上の特定の官能基が、放出されたサンプル分子に移入される工程;ならびに放出されたサンプル分子を分析する工程を包含する。任意の種々の分析方法が、使用され得、例えば、質量分析法、配列決定、液体クロマトグラフィー、分光光度法、蛍光光度法などが挙げられる。分子が、共有結合的に捕捉され、分析に先立って、非検体物質を除去するために広範に洗浄し得るという理由で、本発明の方法は、有利である。さらに、捕捉される分子に移入されるべき官能基は、分子のさらなる分析を容易にするために有用な官能性を、例えば、発色団、フルオロフォア、質量タグなどを付加することによって組込み得る。
【0070】
質量分析は、サンプル分子を分析するための特定の有用な方法である。本発明の方法を用いて、種々の質量分析システムを使用して捕捉されるサンプル分子を分析し得る。高度の質量正確性、高感度および高度の解像度を伴う質量分析器が用いられ得、そしてマトリクス支援レーザー吸着飛行時間型(MALDI−TOF)質量分析器、電子スプレー電離飛行時間型(ESI−TOF)質量分析器およびフーリエ変換イオンサイクトロトン質量分析器(FT−ICR−MS)が挙げられるがそれらに限定されない。他のモードのMSとしては、イオントラップおよび三重四極質量分析器が挙げられる。イオントラップMSにおいて、分析物は、電子スプレー電離またはMALDIによってイオン化され、ついでイオントラップへと入れられる。次いで、捕捉されたイオンは、そのイオントラップからの選択的放出に応じて別々にMSにより分析され得る。フラグメントもまた、このイオントラップにおいて生成され得、そして分析され得る。本発明の方法を用いて質量タグを伴う標識されるサンプル分子は、例えば、MALDI−TOFシステムまたはESI−TOFのシステムを用いて単一段階質量分析によって分析され得る。さらに、LC−MS/MSまたはLC−ESI−TOFもまた使用され得る。任意のMS方法およびMS方法の任意の組み合わせを使用してサンプル分子を分析し得ることが理解される。質量分析による分析の方法は、当業者に周知である(例えば、Yates,J.Mass Spect.33:1−19(1998);KinterおよびSherman,Protein Sequencing and Identification Using Tandem Mass Spectrometry,John Wiley&Sons,New York/(2000)を参照のこと)。
本発明の方法を用いてサンプル中の分子を分析し得る。このサンプルは、たとえば、生物学的検体に由来し得る。検体とは、具体的には、生物または個体から得られたサンプルをいう。検体は、流体または組織検体として個体から得られ得る。例えば、組織検体は、皮膚生検、組織生検または腫瘍生検のような生検として得られ得る。流体検体は、血液、血清、尿、唾液、脳脊髄液または他の体液であり得る。流体検体は、本発明の方法において特に有用である。なぜなら、流体検体は、個体から容易に入手されるからである。検体の収集のための方法は、当業者に周知である(例えば、Young and Bermes,in Tietz Textbook of Clinical Chemistry,第3版,BurtisおよびAshwood(編),W.B.Saunders,Philadelphia,第2章,42頁−72頁(1999)を参照のこと)。検体はまた、微生物検体であり得、この微生物検体は、微生物の培養物に由来し得、この微生物の培養物としては、個体からの検体から培養される培養物が挙げられる。従って、本発明の方法を用いて、生物学的サンプルにおける複合混合物を分析し得る。
【0071】
本発明の方法は、複雑な生物学的サンプルが直接分析され得るという点において有利であるが、サンプルはまた、所望の場合、処理され得る。例えば、血液サンプルを分画して特定の細胞型(例えば、赤血球、白血球など)を分離し得る。血清サンプルを分画して、例えば、構造または機能的な特性に基づいて、グリコシル化、リン酸化、および他の翻訳後修飾によって改変される血清タンパク質、または特定の親和性(例えば、核酸に対する親和性)を有するタンパク質のような特定の型のタンパク質を単離し得る。血清サンプルはまた、物理化学的特性(例えば、サイズ、pIなど)に基づいて分画され得る。血清サンプルをさらに分画して大量に存在する嵩高いタンパク質(例えば、アルブミン)を取り除いて、あまり多く存在しない血清ポリペプチドの分析を容易にし得る。さらに、細胞サンプルを分画して、細胞レベル下のオルガネラを単離し得る。さらに、細胞サンプルまたは組織サンプルは、任意の周知の分画方法(クロマトグラフィー技術(例えば、イオン交換、疎水性および逆相、サイズ排除、アフィニティー、疎水性荷電誘導のクロマトグラフィーなど)によって可溶化および分画され得る(Ausubelら,前出,1999;Scopes,Protein Purification: Principles and Practice,第3版,Springer−Verlag,New York (1993);Burton and Harding,J.Chromatogr.A 814:71−81(1998))。
【0072】
本発明の方法は、生物学的サンプルのような複雑なサンプルを分析するために特に有用であるが(実施例Vを参照のこと)、この方法はまた、複雑度が減少したサンプルにおいて使用され得る。例えば、このサンプルは、上記されるように分画されて(予備的なアフィニティークロマトグラフィーを含む)、固相に捕捉されるサンプル分子をより少なく提供し得る。さらに、このサンプルは、例えば組み換え方法によってサンプル中に高いレベルで発現されるポリペプチドまたは核酸または分子のような本質的に単一の精製された分子を含む、高度に精製されたサンプルであり得る。
【0073】
本発明の方法は、自動化に容易に適合させることができる。例えば、所望であれば、自動化されたサンプリング、ロボット工学、または任意の適切な自動化方法を本発明の方法に適用し得る。すべての反応は、自動化された様式で容易になし得ることから、この方法は、ハイスループットサンプル調製を可能にする。さらに、転移工程のようなサンプル処理が実質的にないことから、捕捉された分子の損失が最小化され、したがって、分子回収率を改善する。捕捉された分子はまた、徹底的に洗浄して捕捉されていないサンプル分子または任意の試薬(regent)を取り除き得る。なぜなら、捕捉されたサンプル分子は、この洗浄工程の間、固体支持体に結合したまま残るからである。本発明の方法を用いて所望に応じて、サンプルから実質的にすべてのクラスまたは複数のクラスの分子を、またはサンプルから分子の一部を捕捉し得る。
【0074】
本発明の方法を用いてサンプル中の分子の発現レベルを決定し得る。発現レベルとは、サンプル中の分子の量をいう。分子の発現レベルは、遺伝子によってコードされるメッセンジャーRNA(mRNA)、遺伝子によってコードされる所定のアミノ酸配列に対応するポリペプチドの量、または細胞中に蓄積する分子の生化学的形態の量(例えば、ポリペプチド、核酸または低分子のような分子の特定の合成後改変の量を含む)の代表例であり得る。発現レベルとは、サンプル中の分子の絶対量、または標準物に比較しての、その分子の相対量をいい得、これには、定常状態または非定常状態の条件下で決定される量が含まれる。分子の発現レベルは、サンプル中のコントロール成分分子に対して相対的に決定され得る。発現レベルは、本明細書に開示されるように、2つまたはそれを超えるサンプルの直接の比較によって決定され得る(実施例IIIを参照のこと)。
【0075】
本明細書において開示されるように、固体相は、ガラスビーズ(例えば、制御された孔ガラスビーズ(実施例Iを参照のこと)であり得る。しかし、サンプル分子を結合し、そして所望の化学を実行し、そして洗浄条件について有用な、任意の適切な固体支持体が使用され得る。従って、固体支持体は、ガラス、誘導体化ガラス、シリコン、プラスチックまたは他の支持体であり得る。固相化学合成に適切な任意の固相物質は、本発明の方法における固体支持体として有用である。固体支持体は、孔性または非孔性の物質、表面フィルム、磁性ビーズ、コロイド、膜などであり得る。固体支持体は、ビーズ、平坦表面または本明細書において開示される方法を用いた分子の捕捉のために適切な任意の構造の形態であり得る。固体支持体は、所望の場合、他の化学基に結合させるに適切な化学部分を組み込むように誘導体化され得る。
【0076】
本発明はまた、本発明の試薬または構成物を備えるキットを提供する。従って、本発明は、固相捕捉物およびタグ試薬を伴うキットを提供し、これは図1に例示される。本発明のキットの内容物(例えば、固相捕捉物およびタグ試薬または本発明の構成物)は、包装材料中に、所望の場合滅菌され、汚染がない環境で含まれる。さらに、包装材料は、キット内の物がどのようにサンプル分子を標識するために用いられ得るかを示す指示書を備える。使用のための指示書は、代表的に、試薬濃度または少なくとも1つのアッセイ方法パラメータ(例えば、試薬の相対量)および混合されるべきサンプル、試薬/サンプル混合のための維持時間、温度、緩衝条件などを記載する具体的な表現を含む。
【0077】
本発明の種々の実施形態の活動に実質的に影響を与えない改変もまた、本明細書において提供される発明の定義内に含まれることが理解される。従って、以下の実施例は、本発明の例示であり、限定を意図しない。
【実施例】
【0078】
(実施例I:システイン含有ペプチドの固体支持体への付着および回収)
本実施例は、システイン含有ペプチドのビーズへの付着および光切断による改変されたペプチドの回収を記載する。
【0079】
システイン含有ペプチドの固体支持体ビーズへの付着および光切断を介した改変されたペプチドの回収を例示する模式図を図1Cに示す。方法は、固相同位体タグ化試薬を用いてシステインペプチドの部位特異的で、安定同位体標識となるように開発した(図1)。o−ニトロベンジルベースの光切断可能なリンカーを第一にアミノプロピルコーティングしたガラスビーズを固相ペプチド合成によって付着させた(Holmes and Jones,J.Org.Chem.60: 2318−2319 (1995))。次に、同位体タグである7つの水素(d0)または7つの重水素(d7)を有するロイシン分子を、再び固相ペプチド合成によってこの光切断可能なリンカーに付着させた(Holmes and Jones,前出,1995)。最後に、スルフヒドリル特異的ヨードアセチル基を付着させた。
【0080】
システイン含有ペプチドの捕捉および改変を実証するために、配列CDPGYIGSR(配列番号1;分子量967)を有する市販のラミニンBペプチドを用いた。手短には、1molのシステイン含有ラミニンBペプチド(CDPGYIGSR;配列番号1)1nmolおよびシステインを含有しないホスホアンギオテンシン(DRVY*IHPF;配列番号2、アステリスクはリン酸化チロシンを示す)500pmolからなるサンプルを用いた。ペプチドを0.2M Tris(pH8.0)、10mMエチレンジアミンテトラ酢酸(EDTA)の100μl中の5mM tris(カルボキシエチル)ホスフィン(TCEP)で30分間室温にて還元した。
【0081】
このペプチドを逆相HPLC上で液体クロマトグラフィーおよび質量分析(LC−MS)によって分析した。このペプチドをtris(2−カルボキシエチル)ホスフィン(TCEP)で還元し、二重に荷電したイオンについて、9.55分においてm/z=484.6の質量荷電比(m/z)で検出される単一のピークが検出された(図2を参照のこと)。11.94分において溶出される第二のピークは、コントロールとしてサンプルに加えられた別の標準的なペプチド(すなわちホスホアンギオテンシン、配列DRVYIHPF (配列番号2、分子量1126)を伴う)である。ホスホアンギオテンシンは、システインを含まず、そして予測された質量を伴って観察された。
【0082】
制御された孔ガラスの固相支持体ビーズを、アミノ官能性を光分解可能なリンカーに共有結合させることによって改変した(図1)。光分解可能なリンカーをシラン結合を介して固体支持体に付着させた。この反応性基は、ヨードアセチル基であり、このヨードアセチル基は、スルフヒドリル基と特異的に反応する。アミノ基で官能化されたガラスビーズ(Sigma Aldrich;St Louis MO)を固体支持体として用いたFmoc保護された光リンカー(4− [4− (l− (Fmocアミノ)エチル)−2−メトキシ−5−ニトロフェノキシ)ブタン酸またはFmoc−アミノエチル−光リンカー;NovaBiochem,Merck KGaAの関連会社 ;Darmstadt Germany)およびロイシンを、標準的な固相ペプチド合成手順のもとで、カルボジイミド化学を介してアミノ基官能化されたビーズに特異的に付着させた。次いで、ロイシンにおけるFmoc保護は、ピペリジン処理によって取り除き、ロイシンの遊離αアミノ基を、無水ヨード酢酸と反応させて、反応性ヨードアセチル基を生成させた。
【0083】
手短には、固相同位体標識ビーズを、以下のように合成した。他に言及しない限り、化学物質は、Aldrich(Milwaukee,WI)から購入した。第一に0.5gのアミノプロピルコーティングされた制御された孔ガラスビーズ(アミン含量約400μmol/g;Sigma,St.Louis,MO)を無水ジメチルホルムアミド(DMP)で洗浄した。次いで、各々600μmolの1−ヒドロキシベンゾトリアゾール(HOBt;Nova Biochem,Laufelfingen,Switzerland)、Fmoc−アミノエチル光リンカー(Nova Biochem)およびジイソプロピルカルボジイミド(DIC)を、室温で30分にわたり混合した。そして、この混合物を、90分間ビーズに加えた。次いで、ビーズを順番にDMFおよびジクロロメタンを用いて洗浄し、ジクロロメタン中で2mlの40%無水酢酸/60%ピリジンを用いて30分間洗浄した。ビーズは、DMFで再び洗浄し、そして30分間DMF中の20%ピペリジンで処理してFmoc保護を取り除いた。このプロセスを反復してFmoc−ロイシン(Nova Biochem)を同位体タグとして付着した。最後に、ヨードアセチル基を、以前に記載されるように(Zhouら,Nat.Biotechnol.19: 375〜378(2001))ビーズに付着させた。ビーズを、DMF、水およびメタノールの順に洗浄し、減圧下で乾燥させ、そして暗所にて室温で保存した。重同位体を有するビーズの合成のために、Fmoc−d7−ロイシンをd7−ロイシン(Isotec,Miamisberg,OH)およびFmoc−N−ヒドロキシスクシンイミド(Nova Biochem)から、Fmoc−ロイシンを精製する再結晶化工程が省略されることを除き、本質的に以前に記載(Lapatsanisら,Synthesis 671−673 (1983))されるように調製した。
【0084】
上記のように調製された5μgのビーズ(2μmolの結合能力)を使用して、一定の撹拌下で100μlの体積中でペプチドを捕獲した。上清のアリコート(1μl)を、μLC−MSのためにビーズの添加前にそしてそれらの添加後の異なる時点で、反応混合物から除去した。15分のインキュベーションの後、ビーズを、2μlのβ−メルカプトエタノールの添加によって5分間不活性化し、そして2.0Mの塩化ナトリウム、メタノールおよび水で連続的に洗浄した。光切断のために、ビーズを、100μlの0.2M Tris(pH8.0)、10mM EDTA、2% β−メルカプトエタノールで再懸濁した。リン酸アンギオテンシン(500pmol)を、内部標準として添加した。Blak−Ray長波UVランプ(100W、VWR Scientific、West Chester、PA)からの光を、10%硫酸銅(II)溶液(1cmパッチ長(pach length))によって濾過し、そして10cmの距離から照射するために使用した。照射のことなる時点で、1μlのアリコートを、μLC−MS分析のために上清から取り出した。ビーズを、均一な光照明に曝露するために時折撹拌した。光切断の緩衝液でのβ−メルカプトエタノールの使用は、光切断の間の可能なメチオニン酸化を提供する。
【0085】
ビーズとペプチドをインキュベートした後、図1に示されるように、上清のアリコートを、再びLC−MSいよって分析した。データを図3に示す。予期されたように、非システイン含有ホスホアンギオテンシン(phosphoangiotensin)ペプチドのみが上清中に存在するが、システイン含有ラミニンBペプチドは、溶液から完全に消失し、これはおそらくビーズによる量的な捕獲に起因する。
【0086】
2Mの塩化ナトリウム、メタノールおよび水でビーズを洗浄した後、ビーズを0.2M Tris、pH8.0、10mM エチレンジアミン四酢酸(EDTA)を含む緩衝液に再懸濁し、同じ濃度のホスホアンギオテンシンペプチドで再びコントロールとしてスパイクした。光切断の後、予測された質量改変([M+2H]2+イオンに対してm/z=569)を有するラミニンBペプチドを、初期のラミニンBペプチドと比較して、わずかに遅い溶出時間で観測した。より長い保持時間は、転移したロイシンアミノ酸の付加に起因して、予測された光切断産物の減少した極性と一貫する。光切断産物の構造を、MS/MS配列分析によってさらに確認した。改変したラミニンBペプチドの相対的なピーク強度およびホスホアンギオテンシン標準は、光切断の有効性の概算を与えた。ラミニンBペプチドの本来の入力の少なくとも50%で、回収した。収率を、反応物の捕獲および放出のさらなる最適化によって改善し得る。
【0087】
反応物の捕獲および放出の有効性を説明するために、システイン含有ラミニンBペプチドおよび非システイン含有ホスホアンギオテンシンからなる混合物を使用した。ラミニンBを、固相上へと量的に捕獲した。光切断の1時間後、タグ化ラミニンBを回収した;予測された質量改変(+170Da)(これはまた、MS/MSによって確認された)は、システイン残基へのロイシンタグの添加に起因することが示された。ロイシンタグの疎水性の性質は、非タグ化形態と比較してタグ化ラミニンBの保持時間を増加する。ホスホアンギオテンシンの同量と対照する非タグ化ラミニンBおよびタグ化ラミニンBのシグナル強度は、特定の捕獲を示し、そしてタグ化ラミニンBの回収をほとんど回収した。より長い光切断時間は、収率またはタグ化ラミニンBの量のいずれにも影響せず、光触媒化副反応は、いずれの実質的な程度を生じないことが示された。
【0088】
これらの結果は、アミノ酸ロイシンの捕獲−放出化学を介するペプチドのサルファイドリル側鎖への成功した転移を照明する。これらは、選択的捕獲ペプチドおよび官能基(例えば、ペプチドに対するアミノ酸)の転移への例示的な一般的なアプローチを結果として生じる。
【0089】
(実施例II:スルフヒドリル基を組み込むためのポリペプチドのアミノ基の改変)
この実施例は、スルフヒドリル基を組み込むためのペプチドのN末端改変を記載する。
【0090】
ペプチドがシステイン残基を含んでいない場合、そのペプチドを改変して、スルフヒドリル基を組み込み得、このことによりアミノ基をスルフヒドリル基に変換し、組み込まれたスルフヒドリル基を介してペプチドを捕捉することが可能である。ポリペプチドのアミノ基の改変のためのストラテジーを、図5に示す。
【0091】
ホスホアンジオテンシンを使用して、アミノ基をスルフヒドリル基に改変する原理を示した。このペプチドを最初にN−スクシンイミジルS−アセチルチオアセテート(SATA)(これは、1級アミノ基に対して特異的に反応する)により改変した。ホスホアンジオテンシンを、図5に示すように改変した。簡潔には、ホスホアンジオテンシンを10mM SATAと30分間、0.1M K3PO4(pH8.0)中で室温にてインキュベートした。
【0092】
改変したホスホアンジオテンシンを、LC−MSにより分析した。このペプチドのN末端改変を観察した。図6に示されるデータは、1つの産物が、14.5〜14.55分([M+2H]2+イオンについてm/z=622.3)で溶出されることを示す。測定された質量は、SATA誘導体化ホスホアンジオテンシンペプチドについての計算された質量(m/z=622.3)と十分に相関した。
【0093】
改変したペプチドを、0.1M ヒドロキシルアミンを用いてpH8にて2時間処理し、続いて、5mM TCEPで30分間還元した。図5に示されるように、ヒドロキシルアミンによる処理およびTCEPによる還元は、SATA誘導体化ホスホアンジオテンシンにおいて遊離SH基を生成する。このように処置された化合物のLC−MS分析からのデータを、図7に示す。再び、13.83分に溶出する1つの産物を観察した。MS分析により、[M+2H]2+イオンについてm/z=601が示された。測定した質量は、予測されたペプチド産物についての計算された質量と一致する(m/z=601)。従って、ペプチドのアミノ基の遊離SH基への変換は、定量的に、最小量の副産物とともになされ得る。さらに、このようなスルフヒドリル基改変ポリペプチドは、実施例Iにおいて記載されるように、引き続いて捕捉され得る。
【0094】
これらの結果は、ポリペプチドのアミノ基がスルフヒドリル基を組み込むために、本質的に定量的に改変され得ることを示す。
【0095】
(実施例III:重い標識および軽い標識の組み込みによるタンパク質サンプルの分析)
この実施例は、質量分析法によるサンプルペプチドの定性的かつ定量的分析のための、2つのサンプルの差示的標識を記載する。
【0096】
ポリペプチドの差示的標識の模式図を、図8に示す。サンプルを、トリプシンのようなプロテアーゼで消化する。消化したポリペプチドを、実施例Iに記載のように処理する。所望であれば、そのサンプルポリペプチドを、実施例IIに記載の方法により改変して、システインを欠くポリペプチドを試験し得る。
【0097】
1つの消化サンプルを、ロイシンのような非変性アミノ酸または別の適切なアミノ酸を含む化学基に共有結合させたビーズに添加する。第2のサンプルを、その化学基が同じアミノ酸の変性された形態を含むことを除いて、化学的に同一の基に共有結合させたビーズに添加する。差示的に同位体標識したロイシンは、ロイシンの変性の状態に依存して、7または10質量単位異なる。
【0098】
サンプルポリペプチドを、差示的に同位体標識したアミノ酸を含むそれぞれのビーズに共有結合的にカップリングした後、そのビーズを合わせ、次いで、光切断により切断する。光切断したポリペプチド(ここでは、差示的に標識されている)を、質量分析法により同時に分析する。2つのサンプルからのMSシグナルの強度を直接比較することにより、サンプル定量のためにMSを使用し得る。この2つのサンプルは、所定の質量が異なり、所定の質量が異なる二重のピークとして、MSで容易に区別され得る。さらに、このサンプルを、MSにより配列決定し得る。
【0099】
この実施例は、質量分析法によりサンプル分子の差示的標識および分析を実証する。
【0100】
(実施例IV:捕捉されたホスホペプチドの改変)
この実施例は、ポリペプチドの捕捉および捕捉されたホスホペプチドの改変を記載する。
【0101】
サンプルからのポリペプチドを、本質的に実施例Iに記載されるようにビーズ上に捕捉する。この捕捉されたポリペプチドを、本質的にZhouら、Nature Biotechnol.19:375−378(2001)(本明細書中に参考として援用される)に記載されるように改変する。
【0102】
簡潔には、この捕捉されたポリペプチドを、以下の工程により改変する。(1)アミノ保護:ペプチドのアミノ基を、必要に応じて、t−ブチル−ジカーボネート(tBoc)化学物質を用いて保護し、その後の反応における分子内縮合および分子間縮合の可能性を排除する。(2)縮合反応:カルボジイミドは、ペプチドと過剰なアミンとの間の縮合反応を触媒して、それぞれ、ペプチドのカルボキシレート結合およびホスフェート結合にて、アミド結合およびホスホルアミダイト結合を形成する。(3)ホスフェート再形成:遊離のホスフェート基をホスホルアミダイト結合の短時間の酸加水分解により再形成する。(4)縮合および還元:カルボジイミド触媒縮合反応は、再形成されたホスフェート基にシスタミンを結合させる。シスタミンの内部ジスルフィドの還元は、次に、捕捉されたホスホペプチドのホスフェート全てに対して遊離のスルフヒドリル基を生成する。(5)改変ポリペプチドの放出:捕捉されたポリペプチドを、例えば、実施例Iに記載のように、光を用いて、化学的切断基の切断により、固体支持体から放出させる。(6)固相捕捉:この放出されたポリペプチド(遊離のスルフヒドリル基を含む改変ホスホポリペプチドを含む)を、このペプチド中の遊離のスルフヒドリル基とガラスビーズ上に固定化されたヨードアセチル基とを反応させることにより、第2の固相に結合させる。(7)ホスホペプチドの回収:樹脂のストリンジェントな洗浄後に、ホスホペプチドを、tBoc保護基をも除去する濃度にて、トリフルオロ酢酸(TFA)を使用してホスホルアミダイト結合を切断することにより回収する。従って、遊離のアミノ基とホスフェート基を用いてペプチドを再生する。カルボキシレート基は、工程(2)からブロックされたままである。
【0103】
化学反応を、以下に詳細に記載するように行う。捕捉されたポリペプチドを含む固相を、50%(vol/vol)の0.1M リン酸緩衝液(pH11)およびアセトニトリル中でインキュベートする。0.1M tBocを、室温にて4時間添加する。次いで、アセトニトリルを除去する。捕捉されたポリペプチドを含む固相を、1M エタノールアミン、25mM N−ヒドロキシルスクシンイミド(NHS)、および0.5M N,N’−ジメチルアミノプロピルエチルカルボジイミドHCL(EDC)中でインキュベートし、室温で2時間インキュベートする。10%TFAを添加し、室温にて30分間インキュベートする。この固相を洗浄して、過剰な試薬を除去し、脱塩し、1M イミダゾール(pH6.0)を添加する。0.5M EDCを室温にて3時間添加する。この固相を洗浄し、次いで、1M シスタミン(pH8.0)と50℃にて2時間インキュベートする。この固相を水で洗浄し、次いで、10mM ジチオスレイトール(DTT)で還元して、遊離のスルフヒドリル基を生成する。この固相を洗浄してDTTを除去し、次いで、捕捉された分子を切断可能な官能基の切断により、放出する。
【0104】
放出されたポリペプチド(スルフヒドリル基で改変されたホスホペプチドを含む)を、第2の型の固相ビーズ(このビーズは、ヨードアセチル基を有し、1M Tris(pH8.0)、50mM エチレンジアミン四酢酸(EDTA)でpH8.0まで滴定される)と少なくとも2時間インキュベートする。固定化されたヨードアセチル基を有するビーズは、3当量の無水ヨード酢酸と1当量のアミノビーズ(G4643;Sigma;St.Louis MO)との間の、ジメチルホルムアミド(dimethylformide)中の3.3当量のジイソプロピルエチレンアミンを用いた2時間の反応により調製される。カルボジイミドによるチロシン付加は、あり得る副反応であるので、スルフヒドリル基で改変された捕捉されたホスホペプチドを、1M ヒドロキシルアミン(pH10)中で室温にて2時間インキュベートして、全ての改変されたチロシンを元に戻す。次いで、2M NaCl、メタノールおよび水で連続してビーズを洗浄して、非特異的に結合した分子を除去する。このビーズを100% TFAと30分間インキュベートして、ホスホペプチドを回収すると同時に、tBoc改変基からtBoc保護を外す。次いで、例えば、質量分析法により、回収したホスホペプチドを分析する。
【0105】
第1固体支持体上に捕捉された分子は、一般的に、第2固相上に再捕捉される直前に放出され、この第2固相は、改変ホスホペプチドを選択的に捕捉し、このことにより、有効な洗浄、および捕捉されたポリペプチドを改変するために使用される化学物質の除去が可能となる。しかし、第1固相への結合が、標識またはタグを、捕捉されたポリペプチドに転移するようにのみ意図される場合、このペプチドは、より初期の段階(始めの捕捉の後でさえあれば)で放出され得る。あるいは、ホスホペプチドの改変を、本質的にZhouらに記載されるように実行し得、次いで回収されたホスホペプチドは、本質的に実施例Iに記載されるように、捕捉されそして標識される。
【0106】
この実施例は、ホスホペプチドの選択的な改変および単離を実証する。
【0107】
(実施例V:酵母タンパク質の単離および同位体標識)
この実施例は、固相の捕捉および放出のプロセスを介した、複雑なペプチド混合物中のシステイン性ペプチドの部位特異的な、安定同位体標識のための方法、および標識ペプチドの同時性の単離を記載する。
【0108】
酵母のタンパク質を、固相同位体タグ化試薬からの捕捉および放出によって、差次的に同位体標識した。回収されたペプチドを、マイクロキャピラリー液体クロマトグラフィーおよびタンデム質量分析(μLC−MS/MS)によって分析して、それらの配列および相対質量を決定した。この方法を使用して、酵母Saccharomyces cerevisiae中の、タンパク質存在比におけるガラクトースで誘導された変化を検出した。同位体コード親和性タグ(ICAT)方法(Gygiら、前出、1999)との横並び(side−by−side)比較は、ペプチドの安定同位体タグ化のための固相法が、比較的より簡単であり、より効果的であり、そしてより感度がよいことを実証した。
【0109】
比較される2つのサンプル由来のシステイン性ペプチドを、同位体的に重いタグまたは通常のタグを含有する固相上に、共有結合で捕捉した。次いで、このビーズを混ぜ合わせ、洗浄し、そしてUV光に曝した(360nm、任意の起こり得る光触媒性の副反応を最小限にするように選択される)。このことは、リンカーの光切断、およびシステイン残基の側鎖上への固相からの同位体タグの転移を生じた。最終的に、回収されたタグ化ペプチドを、本質的に以前に記載されるように、μLC−MS/MSによって分析して、各々のペプチドの配列および相対存在比を決定した(Gygiら、前出、1999)。
【0110】
安定同位体タグ化は、質量分析による、タンパク質の定量分析のための一般的なストラテジーである(Gygiら、前出、1999;Odaら、Proc.Natl.Acad.Sci.USA 96:6591−6596(1999);Idekerら、Science 292:929−934(2001);Hanら、Nat.Biotechnol.19:946−951(2001))。固相アプローチの性能を、標準的なICATアプローチ(Gygiら、前出、1999)の性能と比較するために、ガラクトースを用いた誘導への応答において、この2つの方法を使用して、酵母S.cerevisiae中のタンパク質発現の変化を検出する、横並び比較を実施した。同じの酵母のタンパク質の多量のサンプルの装填については100μg、少量のサンプルの装填については10μgの、2つの異なる量の開始時のタンパク質物質を評価した。この目的は、標識化ストラテジーの相対的な性能を比較することであり、そしてペプチドの分離ストラテジーでもタンパク質の同定ストラテジーでもないので、タンパク質同定のため、全てのサンプルに対して1回のLC−MS/MS実施を行った。このことは、サンプルの標識化よりもむしろサンプルの処理に起因して、結果についての多様性を最小限にしたが、おそらくは、より多量のサンプルサイズおよびLC−MS/MSより前の最適化されたペプチド分離を用いて得られ得るよりも、より少ないタンパク質の同定を生じた。
【0111】
酵母のタンパク質サンプルの調製のために、酵母BY4742株を100mlのYPR(1%酵母抽出物、2%ペプトン(Difco,Detroit,MI)および2%ラフィノース)またはYPR+2%ガラクトースのいずれかで、1のA600値まで増殖させた。以前に記載されるように(Ausubelら、前出)スフェロプラストを調製した。スフェロプラストを50mM Tris(pH8.0)、5mM EDTA、6M 尿素、0.5% SDS中で溶解した。細胞溶解物を15,000g(14,000rpm)で15分間遠心分離した。この上清を回収し、そして50mM Tris(pH8.0)、5mM EDTA中で、Econo−Pac 10DGカラム(Bio−Rad,Hercules,CA)で脱塩した。タンパク質濃度を、Bio−Radタンパク質アッセイによって決定した。
【0112】
固相の捕捉−放出方法による、酵母タンパク質のトリプシン消化物の単離および同位体標識化のために、多量スケールおよび小規模実験を実施した。多量スケールの実験について、ガラクトースの添加または無添加で増殖した酵母細胞からの50μgの各々のタンパク質抽出物(全部で100μg)を、100μlの200mM Tris(pH8.0)、5mM EDTA中に調製した。各々のタンパク質抽出物を、5μgのトリプシンによって37℃で3時間消化し、そして5mM TCEPで還元し、次いでシステイン性ペプチドを、実施例Iに記載されるように、d0−ロイシンタグまたはd7−ロイシンタグのいずれかを有するビーズによって15分間捕捉した。このビーズを混ぜ合わせて洗浄し、そして標識されたペプチドを2時間の光照射によって放出した。この放出されたペプチドを、MCXカラム(Waters,Milford,MA)に充填し、そして4mlの0.1%トリフルオロ酢酸(TFA)、4mlの80%アセトニトリル/0.1%TFA、および水(中性化のため)で連続的に洗浄した。ペプチドを、9容量のメタノールと1容量の28%アンモニアとの混合物1mlによって溶出し、そして減圧下で乾燥した。乾燥したペプチドを、μLC−MS/MS分析のために水に再懸濁した。次に、20%の回収されたペプチド(20μgの全タンパク質を示す)を、以前に記載されるように(Zhouら、Nat.Biotechnol.19:375−378(2001))、LCQイオントラップ質量分析(Finnigan,San Jose,CA)を使用するμLC−MS/MSによって分析した。タンパク質の同定および定量を、Sequestおよび既存のソフトウェア(Hanら、Nat.Biotechnol.19:946−951(2001);Engら、J.Am.Soc.Mass.Spectrom.5:976−989(1994))を用いて実施した。小規模実験のために、ガラクトースの添加または無添加で増殖した細胞からの、5μgの各々のタンパク質抽出物を、0.5μgのトリプシンによって消化し、そして上記のように処理し、次いで回収したペプチド混合物の50%(5μgの全タンパク質抽出物を示す)を、同様のμLC−MS/MS方法によって分析した。
【0113】
ICATによって酵母のタンパク質の同位体標識化およびペプチドの単離のために、ガラクトースの添加または無添加で増殖した細胞からの、100μgの各々のタンパク質抽出物を、200mM Tris(pH8)、5mM EDTA、6M 尿素、0.05% SDSを含む標識化緩衝液中に、出発物質として調製した。タンパク質を、5mM TCEPで30分間還元し、そして100μgのd0−ICATまたはd8−ICATを、それぞれ、ラフィノースで増殖した細胞、またはガラクトースで増殖した細胞からのタンパク質抽出液に添加した。室温で90分の標識化の後、この反応物をβ−メルカプトエタノールの10mMまでの添加によってクエンチし、そして混ぜ合わせた。各々のサンプルを、20mM Tris(pH8.3)、0.01% SDSで10倍に希釈した。トリプシン(10μg)を添加して、タンパク質を3時間37℃で消化した。このサンプルを等容量の緩衝液A(5mM KH2PHO4(pH3)、25% CH3CN)で希釈し、そしてそのpHを、TFAを用いてpH3に調整した。100μg(大規模実験)または10μg(小規模実験)のいずれかの混合したタンパク質消化物を、緩衝液Aで平衡化した陽イオン交換カートリッジ(Applied Biosystems,Foster City,CA)に適用した。このカートリッジを2mlの緩衝液A、その後2mlの緩衝液A+40mM KClで洗浄した。結合したペプチドを、600μlの緩衝液A+600mM KClで溶出した。サンプルの容量を減圧下で300μlに減らし、そして500μlの2×PBSおよび12μlの1M NH4HCO3を添加した。サンプルを、モノマーのアビジンカートリッジ(Applied Biosystems)に通し、そして2mlの2×PBS、1mlの1×PBS、および1mlの20%メタノールを含む50mM NH4HCO3で洗浄した。標識ペプチドを、1mlの30%アセトニトリルを含む0.4% TFAで溶出し、減圧下で乾燥し、そして10μlの5%アセトニトリルを含む0.4%酢酸に再懸濁した。大規模実験または小規模実験のどちらかについて、固相の方法において使用した方法と同一のμLC−MS/MS方法によって、同容量のサンプルを分析した。
【0114】
小規模実験および大規模実験の両方において、固相法によって同定および定量されたタンパク質の数は、ICATによる数よりも多かった(図9)。実際に、固相アプローチはより感度が高く、従来のICATによって識別されるタンパク質の大部分のタンパク質、さらにICATによっては同定されない他の多くを同定した(図9A、9B)。複数の実験において同定された同位置のタンパク質の定量もまた一致した。従って、タンパク質定量は、同位体タグの構造によっても固相法の捕捉および開放によっても影響されなかった。
【0115】
ガラクトースは、ガラクトース利用に関連するいくつかの遺伝子の発現を強力に誘導することが既知であり、これら遺伝子としては、ガラクトキナーゼ(GAL1)、ガラクトースパーミアーゼ(GAL2)、ガラクトトランスフェラーゼ。(GAL7)およびUDP−グルコース−4−エピメラーゼ(GALX)が挙げられる(JohnstonおよびCarlson、The Molecular and Cellular Biology of the Yeast Saccharomyces、Jonesら、編、193−281頁、Cold Spring Harbor Press、Cold Spring Harbor、NY(1992))。ガラクトースでの誘導後、GAL1、GAL2、GAL7およびGALXを含むタンパク質由来の複数のペプチドを固相法によって同定および定量した。対照的に、ICAT方法を使用してGAL1タンパク質由来の1つのペプチドのみを、小規模実験および大規模実験の両方において同定し、そして固相を用いるよりも、より低いシグナル対ノイズ比を検出した。これらのデータは、ICATアプローチを超える、固相の優勢なサンプル回収および感度を確認した。さらに、小規模実験および大規模実験によって識別されたタンパク質の実質的な重複によって、固相法の再現性を確認した(図9C)。
【0116】
これらの結果は、固相法が、定量性タンパク質分析について、簡単であり、再現性があり、有効であり、そして感度が高いことを示す。実際に、ICAT方法と比較して、この方法はいくつかの利点を有する。第1に、システイン含有ペプチドの単離、および同位体の安定な組み込みの両方が、本質的に1工程で達成される。従って、固相法は、ICATアプローチよりも、より迅速かつより簡単で、より少ない手動操作を必要とする。第2に、固相へのペプチドの共有結合性の捕捉により、非共有結合性に会合した分子を除去するための、ストリンジェントな洗浄条件の使用が可能となる。実際に、本明細書に提示される実験は、最小限の手動操作に起因して、ほぼ排他的なシステイン性ペプチドの回収を生じた。第3に、この手順は、トリプシンのようなタンパク質分解性酵素の存在によっても、尿素およびSDSのような強力な変性剤および界面活性剤の存在によっても、影響されない。従って、このような分子を除去するためのさらなる工程の必要性が存在しない。最低限のサンプル操作に起因して、この固相法は、ICATよりも感度が高い。多くの生物学的に有益な事象は、相対的に低い存在比の調節タンパク質に関するので、固相法は、このようなタンパク質に誘導される、変化の分析に有用である。第4に、カップリング工程に関係する標準的な固相ペプチド化学は、本明細書で使用されるd0/d7−ロイシンの場所に、同位体質量タグとして、ある範囲の天然または非天然のアミノ酸の使用を可能とする。このことは、1つの実験における複数のサンプル(2つより多い)の分析のために、ある範囲の質量タグを有するビーズの合成に役立ち得る。第5に、本明細書で使用されるシステイン上の質量タグは、d0−ロイシンタグについては170Daの重量である。ICATで標識されたペプチドとは対照的に、タグのサイズの小ささ、およびタグの化学的性質に起因して、MS/MSモードにおいて観察されるペプチドのフラグメント化は、標識自体の望ましくないフラグメント化によって複雑化されることがない(Hanら、前出、2001)。最後に、光切断の前に、共有結合で固定化されたペプチドは、所望される場合、さらなる化学反応および酵素反応のための理想的な基質を提供する(Zhouら、前出、2001)。
【0117】
固相法がICAT方法と異なる1つの重要な方法は、固相試薬がタンパク質分解の後にペプチドを標識するが、一方、ICATにおいては、タンパク質はタンパク質分解の前に標識されることである。従って、ICATアプローチは、標識されたタンパク質の分離(例えば、ゲル電気泳動)が必要とされる場合に好ましい。上記の固相法は、定量的プロテオミクスへの一般的な適用に適切なツールを提供し、そして自動化された装置に利用可能である。従って、固定化方法は、質量分析による定量的タンパク質分析のための安定同位体タグ化のさらにより広範な適用に向けられる工程を表す。
【0118】
これらの結果は、定量的プロテオーム分析に適切な複雑なタンパク質混合物の差次的な同位体標識化を実証する。
【0119】
この出願全体にわたって、種々の刊行物が引用されている。これらの刊行物の開示は、本明細書によって、本発明が関する技術分野の状態をより完全に記載するために、本明細書における参考としてその全体が援用される。
【0120】
本発明は、開示された実施形態に関して記載されたが、当業者は、詳説された特定の実験が本発明の単なる例示であることを容易に認識する。種々の改変が、本発明の精神から逸脱すること無くなされ得ることが、理解されるべきである。
【産業上の利用可能性】
【0121】
本発明により、例えば、以下が提供される:
(項1)
分子を標識する方法であって、以下の工程:
(a)サンプル分子の、該サンプル分子についての反応性基への共有結合を可能にする条件下で、該サンプル分子と、化学基に結合された固体支持体とを接触させる工程であって、該化学基は、切断可能な官能基、1つ以上の官能基、および該反応性基を含む、工程;ならびに
(b)該切断可能な官能基を切断する工程であって、それにより該1つ以上の官能基を含む該サンプル分子を放出する、工程、
を包含する、方法。
(項2)
前記サンプル分子が、ポリペプチド、核酸、脂質、セカンドメッセンジャー、および代謝産物からなる群より選択される、項1に記載の方法。
(項3)
前記サンプル分子が、ポリペプチドである、項1に記載の方法。
(項4)
項3に記載の方法であって、前記ポリペプチドが、リン酸化、グリコシル化、ユビキチン化、アセチル化、プレニル化、パルミチル化、ミリスチル化、硫酸化、およびヒドロキシル化からなる群より選択される改変を有する、方法。
(項5)
前記ポリペプチドが、ホスホポリペプチドである、項4に記載の方法。
(項6)
前記固体支持体が、ガラスビーズである、項1に記載の方法。
(項7)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項1に記載の方法。
(項8)
前記官能基の1つが、タグである、項1に記載の方法。
(項9)
前記タグが、質量分析タグである、項8に記載の方法。
(項10)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項8に記載の方法。
(項11)
前記タグが、安定同位体をコードするアミノ酸である、項10に記載の方法。
(項12)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項11に記載の方法。
(項13)
前記タグが、気相塩基性基または疎水性基である、項8に記載の方法。
(項14)
前記気相塩基性基が、ピリジルである、項13に記載の方法。
(項15)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項8に記載の方法。
(項16)
前記官能基の1つが、特徴的な同位体分配を有する元素を含む、項8に記載の方法。
(項17)
前記元素が、塩素または臭素である、項16に記載の方法。
(項18)
前記化学基の前記反応性基が、スクシンイミドエステル基およびヨードアセチル基からなる群より選択される、項3に記載の方法。
(項19)
前記ポリペプチドの第一級アミン基がN−スクシンイミジル S−アセチルチオアセテート、ヒドロキシルアミン、およびトリス(2−カルボキシエチル)ホスフィンを用いる処理によって改変されている、項3に記載の方法。
(項20)
前記ポリペプチドが、該ポリペプチドの前記改変に対する特異的な結合活性を有する抗体を用いて単離されている、項4に記載の方法。
(項21)
前記方法の工程が、自動化プロセスにより実施される、項1に記載の方法。
(項22)
前記固体支持体に接触された前記サンプル分子の少なくとも50%が、放出される、項1に記載の方法。
(項23)
サンプル分子を分析するための方法であって、以下の工程:
(a)サンプル分子の、該サンプル分子についての反応性基への共有結合を可能にする条件下で、該サンプル分子と、化学基に結合された固体支持体とを接触させる工程であって、該化学基は、切断可能な官能基、1つ以上の官能基、および該反応性基を含む、工程;
(b)該サンプル分子を該固体支持体から切断する工程であって、ここで1つ以上の特定の官能基が、該放出されたサンプル分子に移動する、工程;ならびに
(c)該放出されたサンプル分子を分析する工程、
を包含する、方法。
(項24)
前記放出されたサンプル分子が、質量分析法により分析される、項23に記載の方法。
(項25)
細胞または組織により発現された、複数のクラスの分子が分析される、項23に記載の方法。
(項26)
前記サンプル分子が、ポリペプチド、核酸、脂質、セカンドメッセンジャー、および代謝産物からなる群より選択される、項23に記載の方法。
(項27)
前記サンプル分子が、ポリペプチドである、項26に記載の方法。
(項28)
項27に記載の方法であって、前記ポリペプチドが、リン酸化、グリコシル化、ユビキチン化、アセチル化、パルミチル化、プレニル化、硫酸化、ヒドロキシル化およびミリスチル化からなる群より選択される改変を有する、方法。
(項29)
前記ポリペプチドが、ホスホポリペプチドである、項28に記載の方法。
(項30)
前記固体支持体が、ガラスビーズである、項23に記載の方法。
(項31)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項23に記載の方法。
(項32)
前記官能基の1つが、タグである、項23に記載の方法。
(項33)
前記タグが、質量分析タグである、項32に記載の方法。
(項34)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項32に記載の方法。
(項35)
前記タグが、安定同位体をコードするアミノ酸である、項34に記載の方法。
(項36)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項35に記載の方法。
(項37)
前記タグが、気相塩基性基または疎水性基である、項32に記載の方法。
(項38)
前記気相塩基性基が、ピリジルである、項37に記載の方法。
(項39)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項32に記載の方法。
(項40)
前記官能基の1つが、特徴的な同位体分配を有する元素を含む、項32に記載の方法。
(項41)
前記元素が、塩素または臭素である、項40に記載の方法。
(項42)
前記化学基の前記反応性基が、スクシンイミドエステル基およびヨードアセチル基からなる群より選択される、項23に記載の方法。
(項43)
前記ポリペプチドの第一級アミン基が、N−スクシンイミジル S−アセチルチオアセテート、ヒドロキシルアミン、およびトリス(2−カルボキシエチル)ホスフィンを用いる処理によって改変されている、項27に記載の方法。
(項44)
前記ポリペプチドが、該ポリペプチドの前記改変に対する特異的な結合活性を有する抗体を用いて単離されている、項28に記載の方法。
(項45)
前記方法の工程が、自動化プロセスにより実施される、項23に記載の方法。
(項46)
前記固体支持体に接触された前記サンプル分子の少なくとも50%が、放出される、項23に記載の方法。
(項47)
2つ以上のサンプルからの分子が、比較して分析される、項23に記載の方法。
(項48)
前記サンプルが示差的に標識されている、項47に記載の方法。
(項49)
前記2つ以上のサンプルが、質量分析タグを用いて示差的に標識されている、項48に記載の方法。
(項50)
前記サンプルが、安定同位体タグ、同位体分布タグ、または荷電アミノ酸を用いて示差的に標識されている、項48に記載の方法。
(項51)
前記サンプルが、発蛍光団、発光団、またはスピン標識を用いて標識されている、項48に記載の方法。
(項52)
分子を標識する方法であって、以下の工程:
(a)サンプル分子の、該サンプル分子についての反応性基への共有結合を可能にする条件下で、該サンプル分子と、化学基に結合された固体支持体とを接触させる工程であって、該化学基は、切断可能な官能基、1つ以上の官能基、および該反応性基を含む、工程;
(b)該固体支持体に結合した該サンプル分子を改変する工程;ならびに
(c)該切断可能な官能基を切断する工程であって、それにより該1つ以上の官能基を含む該改変されたサンプル分子を放出する、工程、
を包含する、方法。
(項53)
前記改変する工程が、化学的改変または酵素的改変である、項52に記載の方法。
(項54)
前記サンプル分子が、ポリペプチド、核酸、脂質、セカンドメッセンジャー、および代謝産物からなる群より選択される、項52に記載の方法。
(項55)
前記サンプル分子が、ポリペプチドである、項52に記載の方法。
(項56)
項55に記載の方法であって、前記ポリペプチドが、リン酸化、グリコシル化、ユビキチン化、アセチル化、プレニル化、パルミチル化、ミリスチル化、硫酸化、およびヒドロキシル化からなる群より選択される改変を有する、方法。
(項57)
前記ポリペプチドが、ホスホポリペプチドである、項56に記載の方法。
(項58)
前記改変する工程が、前記ホスホポリペプチド上のホスフェート基を改変する、項57に記載の方法。
(項59)
前記固体支持体が、ガラスビーズである、項52に記載の方法。
(項60)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項52に記載の方法。
(項61)
前記官能基の1つが、タグである、項52に記載の方法。
(項62)
前記タグが、質量分析タグである、項61に記載の方法。
(項63)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項61に記載の方法。
(項64)
前記タグが、安定同位体をコードするアミノ酸である、項63に記載の方法。
(項65)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項64に記載の方法。
(項66)
前記タグが、気相塩基性基または疎水性基である、項61に記載の方法。
(項67)
前記気相塩基性基が、ピリジルである、項66に記載の方法。
(項68)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項61に記載の方法。
(項69)
前記官能基の1つが、特徴的な同位体分配を有する元素を含む、項61に記載の方法。
(項70)
前記元素が、塩素または臭素である、項69に記載の方法。
(項71)
前記化学基の前記反応性基が、スクシンイミドエステル基およびヨードアセチル基からなる群より選択される、項55に記載の方法。
(項72)
前記ポリペプチドの第一級アミン基がN−スクシンイミジル S−アセチルチオアセテート、ヒドロキシルアミン、およびトリス(2−カルボキシエチル)ホスフィンを用いる処理によって改変されている、項55に記載の方法。
(項73)
前記ポリペプチドが、該ポリペプチドの前記改変に対する特異的な結合活性を有する抗体を用いて単離されている、項56に記載の方法。
(項74)
前記方法の工程が、自動化プロセスにより実施される、項52に記載の方法。
(項75)
前記固体支持体に接触された前記サンプル分子の少なくとも50%が、放出される、項52に記載の方法。
(項76)
化学基に結合された支持体を含む構成物であって、該構成物は、切断可能な官能基、タグおよび反応性基を含み、該反応性基は、サンプル分子に共有結合され、ここで該切断可能な官能基、該タグおよび該反応性基は、該切断可能な官能基の切断の際に、該サンプル分子への該タグの移動を可能にするように互いに対して配置されている、構成物。
(項77)
項76に記載の構成物であって、前記サンプル分子が、ポリペプチド、核酸、脂質、セカンドメッセンジャー、および代謝産物より選択される、構成物。
(項78)
前記サンプル分子が、ホスホポリペプチドである、項77に記載の構成物。
(項79)
項78に記載の構成物であって、前記ポリペプチドが、リン酸化、グリコシル化、ユビキチン化、アセチル化、パルミチル化、プレニル化、硫酸化、ヒドロキシル化、およびミリスチル化からなる群より選択される改変を有する、構成物。
(項80)
前記ポリペプチドが、ホスホポリペプチドである、項79に記載の構成物。
(項81)
前記固体支持体が、ガラスビーズである、項76に記載の構成物。
(項82)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項76に記載の構成物。
(項83)
前記タグが、質量分析タグである、項76に記載の構成物。
(項84)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項76に記載の構成物。
(項85)
前記タグが、安定同位体をコードするアミノ酸である、項84に記載の構成物。
(項86)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項85に記載の構成物。
(項87)
前記タグが、気相塩基性基または疎水性基である、項76に記載の構成物。
(項88)
前記気相塩基性基が、ピリジルである、項87に記載の構成物。
(項89)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項76に記載の構成物。
(項90)
前記タグが、特徴的な同位体分配を有する元素を含む、項76に記載の構成物。
(項91)
前記元素が、塩素または臭素である、項90に記載の構成物。
(項92)
前記共有結合された反応性基が、スクシンイミドエステル基およびヨードアセチル基から誘導される、項76に記載の構成物。
(項93)
前記ポリペプチドの第一級アミン基がN−スクシンイミジル S−アセチルチオアセテート、ヒドロキシルアミン、およびトリス(2−カルボキシエチル)ホスフィンを用いる処理によって改変されている、項78に記載の構成物。
(項94)
化学基に共有結合された固体支持体を含む構成物であって、該化学基は、切断可能な官能基、質量分析タグおよびサンプル分子に共有結合するための反応性基を含み、該切断可能な官能基、該タグおよび該反応性基は、該切断可能な官能基の切断の際に、該サンプル分子への該タグの移動を可能にするように互いに対して配置されている、構成物。
(項95)
前記固体支持体が、ガラスビーズである、項94に記載の構成物。
(項96)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項94に記載の構成物。
(項97)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項94に記載の構成物。
(項98)
前記タグが、安定な同位体をコードするアミノ酸である、項97に記載の構成物。
(項99)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項98に記載の構成物。
(項100)
前記タグが、気相塩基性基または疎水性基である、項94に記載の構成物。
(項101)
前記気相塩基性基が、ピリジルである、項100に記載の構成物。
(項102)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項94に記載の構成物。
(項103)
前記タグが、特徴的な同位体分配を有する元素を含む、項94に記載の構成物。
(項104)
前記元素が、塩素または臭素である、項103に記載の構成物。
(項105)
前記化学基の前記反応性基が、スクシンイミドエステル基およびヨードアセチル基からなる群より選択される、項94に記載の構成物。
【背景技術】
【0001】
(発明の背景)
本発明は、国立癌研究所によって与えられた許可番号1R33CA84698−0下で政府に支援されてなされた。政府は、本発明に特定の権利を有する。
【0002】
本発明は、一般にプロテオーム分析に関し、より詳細には、分子の分析および定量のために、サンプル中の分子に官能基を移動させるための方法に関する。
【0003】
生物学的プロセスを研究するための伝統的な生化学的アプローチは、連続的な分画、およびプロセスを構築する比活性のアッセイサイクルによる均質化への精製、それぞれの単離された成分の詳細な構造分析、機能的分析および調節分析、ならびに単離された成分からのプロセスの再構築に基づく。ヒトゲノム計画および他のゲノム配列決定プログラムは、特定の種の完全ゲノム配列および本質的に、潜在的にその種によってコードされる全てのタンパク質のアミノ酸配列を、迅速に連続して作製している。生物学における歴史で先例のないこの情報資源が、伝統的な研究方法を拡大し、そして全く異なる研究パラダイム(その1つがプロテオミクスである)における進歩を触媒することが、期待される。
【0004】
多数の他の種のゲノムとともにヒトゲノムの全体を配列決定するための努力は、非常に成功している。46の微生物種のゲノム(TIGR Microbial Database;www.tigr.org)は、完全にされ、そして120を超える他の微生物種のゲノムが、現在配列決定の過程にある。さらに、より複雑な真核生物のゲノム(特に、遺伝的に十分に特徴づけられた単細胞生物Saccharomyces cerevisiaeおよび多細胞の種Caenorhabditis elegansおよびDrosophila melanogasterのゲノム)が、完全に配列決定されている。さらに、コメ、ヒトおよびArabidopsisゲノムの「ドラフト配列(draft sequence)」が公開されている。完全なゲノム配列の非存在下でさえも、豊富なDNA配列データベースが、公的に利用可能にされており、これらとしては、210万を超えるヒトおよび120万を超えるマウスの発現配列タグ(EST)を含むデータベースが挙げられる。
【0005】
ESTは、cDNAライブラリーにおけるクローンの系統的な単一通過配列決定によって生成される部分遺伝子配列を表す約300〜500の連続したヌクレオチドのストレッチである。進化を明らかに例外とするほとんどの生物学的プロセセスの時間尺度において、ゲノムDNA配列は静的と見なされ得、故にゲノム配列データベースは、ライブラリーに類似する情報資源を表す。激しい努力が、配列データベースにおける個々の配列に対して「機能」を割当するために進行中である。これは、既知の機能を有する配列のファミリーに対する配列の統計学的に有意な類似性を示す、線状配列モチーフまたはより高次の構造的モチーフのコンピューター分析によってか、または種に渡る相同性タンパク質の機能の比較のような他の手段によって試みられる。他の方法もまた、個々の配列の機能を決定するために使用されており、この方法としては、遺伝子ノックアウトおよびアンチセンスヌクレオチド技術を用いる遺伝子発現の抑制のような実験方法が挙げられ、これらは、時間を消費し得、そしていくつかの場合、配列によってコードされるポリペプチドに対する生物学的機能の割当を可能にするのになお不十分であり得る。
【0006】
プロテオームは、ゲノムによって発現されるタンパク質補体として規定されている。この幾分制限的な規定は、プロテオームの静的な性質を示唆する。実際、発現されたタンパク質の型、その存在量、改変の状態、亜細胞の位置、および他の生体分子(例えば、ポリペプチドおよび核酸)との相互作用は、細胞または組織の生理学的状態に依存するので、プロテオームは、高度に動的である。故に、プロテオームは、細胞の状態、または細胞によって遭遇する外部の状態を反映し得、そしてプロテオーム分析は、細胞の状態を区別および研究するためのゲノムにわたるアッセイ、ならびにこれらを制御する分子機構を決定するためのゲノムにわたるアッセイとして見なされ得る。分化した細胞のプロテオームは、何1000個〜何10000個もの異なる型のタンパク質からなると見積もられることを考慮して、少なくとも5桁の発現の見積もられたダイナミックレンジを用いて、プロテオーム分析のための見通しは煩わしいようである。しかし、全ての潜在的に発現されるタンパク質の配列を列挙するDNAデータベースの利用可能性は、実際に発現されるタンパク質を同定し得る技術における迅速な進展と結びついて、現在、プロテオミクスを現実的な仕事とする。質量分析法は、現在のプロテオミクス技術が位置する基本的な脚の1つである。
【0007】
定量的なプロテオミクスは、その量および同一性に関する、細胞または組織によって発現される全てのタンパク質の系統的な分析である。所定の時間で、細胞、組織、生物学的流体またはタンパク質複合体で発現されるタンパク質は、その時点での細胞または組織の状態を正確に規定する。異なる状態での同じ細胞型のタンパク質プロフィール間の量的差異および質的差異は、それぞれの状態の間の移行を理解するために使用され得る。慣習的に、プロテオーム分析は、タンパク質を分離するための、高い解像度のゲル電気泳動、特に2次元ゲル電気泳動と、タンパク質を同定するための質量分析法との組み合わせを用いて実行された。このアプローチは、連続的かつ単調であるが、より重要なことには、タンパク質の生物学的に重要なクラスが本質的に検出不可能である点で、基本的に制限されている(非特許文献1)。
【0008】
多数の種のゲノム配列の完成は、代表的に発見科学といわれる生物学への新しいアプローチを触媒している。発見科学の本質は、細胞または組織によって発現される特定のクラスの分子の全てのメンバーの系統的かつ定量的な分析である。発見科学の例示的な実施としては、遺伝子発現アレイによる細胞または組織によって発現されるmRNA分子の系統的分析、生物学的サンプル中に含まれるタンパク質の系統的分析が挙げられる。発見科学の主な観点は、生体分子の系統的な測定から得られるデータに基づく細胞または組織の状態(活性、病理学、ストレス)の説明、および2つの状態を表す細胞の分子組成物の比較分析による、1つの状態から他の状態への細胞の移行を制御する分子機構の同定である。細胞の状態およびそれを制御する機構の分子の的説明のためには、可能な限り多くのパラメーターが所望される。現在の発現アレイ方法は、細胞中のmRNA分子の系統的分析を可能にする。
【0009】
近年、同位体コード化親和性タグといわれる等級の試薬および質量分析法に基づく方法が、生物学的サンプル中に存在するタンパク質の系統的な同定および定量に適していることが記載されている。細胞の状態に関連する他の特性(例えば、タンパク質リン酸化および他の翻訳後修飾、ならびにタンパク質または核酸以外の生体分子(例えば、脂質、セカンドメッセンジャー、代謝産物))の定量的プロフィールは、現在の技術では系統的および定量的に測定することは困難である。
【先行技術文献】
【非特許文献】
【0010】
【非特許文献1】Gygiら、Proc.Natl.Acad.Sci.USA 97:9390〜9395(2000)
【発明の概要】
【発明が解決しようとする課題】
【0011】
従って、細胞中の分子の分析のための迅速、効果的そしてコストがかからない方法への要求が存在する。本発明は、この要求を満たし、そして関連の利点も提供する。
【課題を解決するための手段】
【0012】
(発明の要旨)
本発明は、サンプル分子が反応性基に共有結合することを可能にする条件下で、サンプル分子を、切断可能な官能基、1つ以上の官能基、およびサンプル分子のための反応性基を含む化学基と連結した固体支持体と接触させ;そして切断可能な官能基を切断し、これによってタグ化され得る1つ以上の官能基を含むサンプル分子を放出することによって、分子を標識するための方法を提供する。本発明はまた、切断可能な官能基、質量分析タグおよびサンプル分子に共有結合するための反応性基を含む化学基に共有結合した固体支持体を提供し、ここで、切断可能な官能基、タグおよび反応性基は、切断可能な官能基の切断の際のサンプル分子へのタグの移動を可能にするように、互いに関連して位置される。
【図面の簡単な説明】
【0013】
【図1A】図1は、ポリペプチドの光捕捉回収を可能にする光切断可能なリンカーを介してポリペプチドを捕捉するための固相ベースのアプローチの概略図を示す。図1Aは、以下の4つのエレメントを有する固相同位体タグ化試薬の概略図を示す:ビーズ、光切断可能なリンカー、安定な同位体タグ、および特異的な反応性基。
【図1B】図1は、ポリペプチドの光捕捉回収を可能にする光切断可能なリンカーを介してポリペプチドを捕捉するための固相ベースのアプローチの概略図を示す。図1Bは、スルフヒドリル−反応性固相同位体タグ化試薬を示す。o−ニトロベンジル−ベースの光切断可能なリンカーは、アミノプロピルガラスビーズに連結される。光切断可能なリンカーの末梢に、7個の水素(H)または7個の重水素(D)のいずれか(「X」によって示される)を含むロイシン分子として表される同位体タグが、付着され、続いてSH−反応性基としてヨードアセチル基が付着される。
【図1C】図1は、ポリペプチドの光捕捉回収を可能にする光切断可能なリンカーを介してポリペプチドを捕捉するための固相ベースのアプローチの概略図を示す。図1Cは、固相同位体タグ化方法を用いた捕捉後のポリペプチドへの同位体タグの移動および放出を示す。リンカーは、切断の際、特異的な官能基が、放出されたポリペプチドに移動されるように構築される。
【図2A】図2は、トリス(2−カルボキシエチル)ホスフィン(TCEP)を用いた還元後のペプチドラミニンB、およびコントロールペプチドホスホアンジオテンシンの液体クロマトグラフィーマスウペクトロメトリー(LC−MS)分析を示す。図2Aは、ラミニンBおよびコントロールホスホアンジオテンシンの混合物の逆相HPLCによるLC分析を示す(RT:8.95−13.04、NL:1.22E7 基底ピークMS tcep30分)。
【図2B】図2は、トリス(2−カルボキシエチル)ホスフィン(TCEP)を用いた還元後のペプチドラミニンB、およびコントロールペプチドホスホアンジオテンシンの液体クロマトグラフィーマススペクトロメトリー(LC−MS)分析を示す。図2Bは、TCEP処理ラミニンBのエレクトロスプレーイオン化MS分析を示す(tcep30分、#384、RT:9.55、AV:1、NL:6.50EB、T:+c ESI Q3MS[400.00−1600.00])。
【図3A】図3は、図1に示されるビーズと接触させた後の還元されたラミニンBおよびホスホアンジオテンシンの混合物のLC−MSを示す。図3Aは、図2で処置されたような上清のアリコートのLC分析を示す(RT:8.97−13.08,NL:1.84E7 基底ピーク MS 60分ビーズ)。
【図3B】図3は、図1に示されるビーズと接触させた後の還元されたラミニンBおよびホスホアンジオテンシンの混合物のLC−MSを示す。図3Bは、還元されたラミニンBと一致する保持時間を有する図3Aの9.62分のLC画分のMS分析を示す(60分ビーズ、#387 RT:9.62、AV:1、NL:1.14E5、T:+c ESI Q3MS[400.00−1600.00])。ラミニンBの量を、ビーズへの結合に起因して減少させた。
【図4A】図4は、図3で処置されたようなビーズから光切断されたラミニンBのLC−MS分析を示す。図4Aは、図2で使用される量と等量の濃度でコントロールホスホアンジオテンシンを添加することによる、光切断されたラミニンBのLC分析を示す(RT:9.19−15.18、NL:2.09E6 基底ピーク m/z=560.0−570.0 MS 1時間光_0104 15121039)。
【図4B】図4は、図3で処置されたようなビーズから光切断されたラミニンBのLC−MS分析を示す。図4Bは、改変に起因する質量の予測された増加を伴なう、図4Aで10.72分で溶出した光切断されたラミニンBのピークのMS分析を示す(1時間光_010415121039 #430、RT:10.70、AV:1、NL:9.17E5、T:+c ESI Q3MS[400.00−1600.00])。
【図5】図5は、スルフヒドリル基を組み込むためのポリペプチドの1級アミン基のタグ化についてのストラテジーを示す。ポリペプチドのアミノ基は、N−スクシンイミジルS−アセチルチオアセテート(acetylthioacctate)(SATA)によって改変される。ヒドロキシアミン処理に際して、トリス(2−カルボキシエチル)ホスフィン(TCEP)での還元後、ポリペプチドのアミノ基は、スルフヒドリル基に転換される。
【図6A】図6は、SATAで改変されるホスホアンジオテンシンのLC−MS分析を示す。図6Aは、SATA処理ホスホアンジオテンシンのLC分析を示す(RT:9.56−16.88、NL:3.29E7 基底ピーク MS sata15分)。
【図6B】図6は、SATAで改変されるホスホアンジオテンシンのLC−MS分析を示す。図6Bは、SATA処理ホスホアンジオテンシンのMS分析を示す(sata15分 #582、RT:14.48、AV:1、NL:2.75E7、T:+c ESI Q3MS[400.00−1600.00])。2つの主なシグナルは、1荷電の[M+H]+=1242.7質量単位、および2荷電の[M+2H]2+=622.3質量単位の形態を表す。
【図7A】図7は、図5に示されるように、ヒドロキシアミンでさらに処理され、そしてTCEPで還元されたSATA処理ホスホアンジオテンシンのLC−MS分析を示す。図7Aは、還元された改変ホスホアンジオテンシンのLC分析を示す(RT:12.08−14.93、NL:1.32E7 基底ピーク MS 30分10m MTCEP)。
【図7B】図7は、図5に示されるように、ヒドロキシアミンでさらに処理され、そしてTCEPで還元されたSATA処理ホスホアンジオテンシンのLC−MS分析を示す。図7Bは、還元された改変ホスホアンジオテンシンのMS分析を示し、改変に起因して予測される質量における変化を示す(30分10mMTCEP #557 RT:13.86、AV:1、NL:1.32E7、T:+c ESI Q3MS[400.00−1600.00])。2つの主なシグナルは、1荷電の[M+H]+=1200.7質量単位および2荷電の[M+2H]2+=601.1質量単位の形態を表す。
【図8】図8は、固相捕獲を使用する差次的改変による2つのサンプルの比較のための概略図を示す。比較される2つのタンパク質サンプルは、タンパク質分解(例えば、トリプシンを用いて)にかけられる。スルフヒドリル反応性試薬の場合において、Cys含有ペプチドは還元され、そして差次的に同位体的に標識された試薬(例えば、それぞれ0個または7個の重水素を含むd0−ロイシンタグまたはd7−ロイシンタグ)を保有するビーズによって捕獲される。このビーズは、合わされそして洗浄され、そしてタグ化されたペプチドは、光切断によって放出される。放出されたペプチドは、例えば、マイクロキャピラリー液体クロマトグラフィーおよびタンデム質量分析(μLC−MS/MS)のような質量分析(MS)を使用してさらに特徴付けられ得る。
【図9】図9は、固相法およびICAT法によって同定され、定量化されるタンパク質数の概要を示す。図9Aは、大規模実験(L)から同定されるタンパク質の数を示し、その中で100μgの全タンパク質サンプルが標識され、そして20μgがμLC−MS/MSによって分析された。固相法は、82個のタンパク質を定量し、そして33個のタンパク質は、ICAT法によって定量され、25個のタンパク質は共通であった。図9Bは、小規模実験(S)から同定されたタンパク質数を示し、この中で10μgの全サンプルが標識され、5μgが分析された。固相法は、57個のタンパク質を定量し、そして18個のタンパク質は、ICAT法によって定量され、13個のタンパク質は共通であった。図9Cは、大規模実験(L)および小規模実験(S)における固相法によって同定されたタンパク質の数を示す。ベン図の区画中の数は、定量されたタンパク質の数を示す。
【発明を実施するための形態】
【0014】
(発明の詳細な説明)
本発明は、所望の官能基(タグ化分子の検出を増強し、そして同定および定量を容易にするのに有用なタグを含む)の分子への輸送を可能にする化学基を有する固体支持体上に分子を捕獲することによって、サンプル中の分子を標識するための方法および組成物を提供する。この方法は、サンプル由来の分子を選択的に単離しそして標識するために使用され得、分析物中の複雑な混合物の定量的な分析(質量分析のような方法による分析を含む)を可能にするという点で有利である。従って、この方法は、基本的に全ての特定のクラスの分子または分子のサブセット(例えば、基本的に全てのポリペプチドまたはリンタンパク質、糖タンパク質、またはその他の改変されたポリペプチドのサブセット)を単離するために使用され得る。
【0015】
一般的な共有結合的捕獲−放出化学を使用することによって、特定の官能基が複雑なサンプルの成分に輸送され得る。さらに、捕獲された分子を放出するための能力を組み入れることによって、この方法はまた、サンプル分子を単離または精製するために有利に使用され得、このことは、分析されたサンプルの複雑さを低減するのに有用であり得る。この方法は、定量的なプロテオーム分析、タンパク質リン酸化および他の翻訳後修飾の系統的かつ定量的な分析に十分適切であり、そしてタンパク質およびペプチド以外の分子の系統的かつ定量的な分析まで拡張され得る。さらに、本発明の方法は、サンプル分子が、効率良く捕獲および放出され得、使用される開始サンプルの量をより少なくし、プロテオミクス分析のための複雑な生物学的サンプルを分析するのに特に有用であるという点で有利である。
【0016】
本発明の方法は特に、生物学的サンプル中に含まれる分子の同定および定量分析、特に定量的なプロテオンミクスについてのタンパク質の分析のために有用である。この方法はまた、他の改変されたタンパク質における、タンパク質リン酸化または他の改変の系統的、定量的な分析のために使用され得る。本発明はまた、分子を標識するために有用な試薬を提供する。プロテオミクス分析に加えて、この方法はまた、タンパク質に加えて、その他の生体分子の系統的かつ定量的な分析のために有用である。この方法は、特に標識またはタグを質量分析(MS)に適切な分子に輸送するのに特に有用である。
【0017】
本明細書中で使用される場合、用語「ポリペプチド」は、2つ以上のアミノ酸のペプチドまたはポリペプチドをいう。ポリペプチドはまた、翻訳後修飾または合成的改変(リン酸化、脂質化、プレニル化、パルミチル化、ミリスチル化、硫酸化、ヒドロキシル化、アセチル化、グリコシル化、ユビキチン化、補欠分子族または補因子の付加、ジスルフィド結合の形成、タンパク質分解、高分子複合体へのアセンブリなどを含む)のような天然に存在する改変によって改変され得る。
【0018】
ポリペプチドは、2、3またはいくつかのアミノ酸を有する小さいポリペプチドならびに数百またはそれ以上のアミノ酸を有する大きいポリペプチドを含む。通常、2つ以上のアミノ酸残基間の共有結合は、アミド結合である。しかし、これらのアミノ酸は、ペプチドおよび化学の分野における当業者に公知の種々の他の手段によって共に結合され得る。従って、用語ポリペプチドは、アミノ酸、アミノ酸アナログ、および模倣物の間の非アミド結合を全体的にまたは部分的に含む分子を含むことが意図される。同様に、この用語はまた、環状ポリペプチドおよび他の立体配置的に構成された構築物を含む。
【0019】
ポリペプチド、特にリガンドポリペプチドの改変はまた、化学合成によって作製された、天然に存在しない誘導体、そのアナログおよび機能的模倣物を含み得、ただしこのようなポリペプチド改変は、親ポリペプチドと比較して類似の機能的活性を示す。例えば、誘導体は、このポリペプチドの化学修飾(例えば、アルキル化、アクリル化、カルバミル化、ヨード化、またはポリペプチドを誘導体化する任意の修飾)を含み得る。このような誘導体化分子としては、例えば、分子中の遊離アミノ基が誘導体化されてアミン塩酸塩、p−トルエンスルホン基、カルボベンゾキシ基、t−ブチルオキシカルボニル基、クロロアセチル基またはホルミル基を形成する分子が挙げられる。遊離カルボキシル基は、誘導体化され、塩、メチルおよびエチルエステルまたはエステルもしくはヒドラジドの他の型を形成し得る。遊離ヒドロキシル基は、誘導体化され、O−アシル誘導体またはO−アルキル誘導体を形成し得る。ヒスチジンのイミダゾール窒素は、誘導体化され、N−im−ベンジルヒスチジンを形成し得る。また、20個の標準的なアミノ酸のうち天然に存在するアミノ酸誘導体(例えば、4−ヒドロキシプロリン、5−ヒドロキシリジン、3−メチルヒスチジン、ホモセリン、オルニチンまたはカルボキシグルタメート)を1つ以上含むポリペプチドがまた、誘導体またはアナログとして含まれ、そしてペプチド結合によって連結されないアミノ酸を含み得る。
【0020】
特に有用なポリペプチド誘導体は、本明細書中に開示された方法を用いて所望の機能的特徴を取り込むための改変を含む。このような改変は、標識またはタグ(特に、MS分析に有用な標識またはタグ)の取り込みを含む。
【0021】
本明細書中で使用される用語「核酸」は、生化学系の成分に関して用いられる場合、デオキシリボ核酸(DNA)またはリボ核酸(RNA)のような共に共有結合された2つ以上のヌクレオチドを意味すると意図され、そして例えば、一本鎖および二本鎖の核酸を含む。この用語は、同様に、ゲノムDNA、cDNA、mRNA、およびそれらに対応する合成オリゴヌクレオチドを含むことが意図され、これらは。センス鎖、アンチセンス鎖またはその両方を表し得る。
【0022】
本明細書中で使用される用語「アミノ酸」は、天然に存在するアミノ酸および天然に存在しないアミノ酸の両方、ならびにアミノ酸アナログおよび模倣体を意味することが意図される。天然に存在するアミノ酸は、タンパク質生合成の間に利用される20個の(L)−アミノ酸ならびに例えば、4−ヒドロキシプロリン、ヒドロキシリジン、デスモシン、イソデスモシン、ホモシステイン、シトルリン、およびオルニチンのような他のアミノ酸を含む。天然に存在しないアミノ酸は、例えば、(D)−アミノ酸、ノルロイシン、ノルバリン、p−フルオロフェニルアラニン、エチオニンなどを含む。アミノ酸アナログは、天然に存在するアミノ酸および天然に存在しないアミノ酸の改変形態を含む。このような改変は、例えば、アミノ酸上の化学基および化学部分の置換(substitution or replacement)またはアミノ酸の誘導体化によるものを含み得る。アミノ酸模倣物は、例えば、参照アミノ酸に特徴的な電荷および電荷間隔のような機能的に類似の特性を示す有機構造を含む。例えば、リジン(LysまたはK)を模倣する有機構造は、同様の分子領域に位置し、そして天然に存在するLysアミノ酸の側鎖のε−アミノ基と同じ程度の移動度を有する正の電荷部分を有する。模倣物はまた、アミノ酸またはアミノ酸官能基の最適な間隔および電荷相互作用を維持するように制限された構造を含む。
【0023】
本明細書中で使用される「官能基」は、所望の機能特性を有する化学基をいう。所望の機能特性は、分子に所望の化学特徴を付与する任意の特性である。官能基は、分子の物理化学特性を変更する(例えば、質量、電荷、疎水性などを変更する)基を含み得る。特に有用な官能基は、標識またはタグ(例えば、発蛍光団、発色団、スピン標識、アイソトープ、分布タグなど)である。
【0024】
本明細書中で使用される用語「標識」は、分子の質量の変化を生じる分子に付着され得る任意の部分を意味することが意図される。一般に、標識は、共有結合されるけれども、この標識は、共有結合または非共有結合のいずれかで分子に結合され得る。非共有結合相互作用が標識と分子との間で生じる場合、非共有結合相互作用は、標識を、本発明の方法において使用される化学的および/または物理的操作の間に分子に結合されたままとするのに十分に高い親和性を有することが理解される。
【0025】
特に有用な標識は、MSによるサンプル分析に有用な質量標識である。質量標識の取り込みに起因する分子の質量の変化は、質量決定について選択された機器の感受性範囲内にあるべきである。さらに、当業者は、異なる大きさおよび異なる組成の分子についての適当な質量の標識を認識し、そして決定し得る。さらに、例えば、分子の差次的な標識について重質量標識および軽質量標識を用いる場合、約1〜3質量単位の間の程度の小ささの質量差または約10質量単位より大きい程度の大きさが用いられ得、例えば、約4、約5、約6、約7、約8、約9、約10、約12、約15、または約20の質量単位またはそれ以上が所望の通りである。2つのサンプルを差次的に標識するために適した質量標識は、化学的に同一であるが質量は異なる。
【0026】
本明細書中で使用される「タグ」は、検出可能な標識をいう。タグは、分子が種々の分析法(MS、クロマトグラフィー、蛍光光度分析、分光光度法、免疫学的技術などを含む)のいずれかによって検出され得るように、その分子に特徴を付与する。タグは、例えば、アイソトープ、蛍光(fluor)、クロマゲン(chromagen)、強磁性物質、発光タグ、または抗体もしくは抗体フラグメントにより認識されるエピトープタグであり得る。特に有用なタグは、質量タグであり、これは、MSによる分子の検出および分析に適切な質量標識である。例示の質量タグは、例えば、適切なアイソトープタグ、アイソトープ分布タグ、荷電アミノ酸、差次的にアイソトープで標識されたタグなどを含む。タグは、気相塩基性基(例えば、ピリジル)または疎水性基であり得る。タグはまた、特徴的なアイソトープ分布を有する元素、例えば、塩素、臭素、または識別可能なアイソトープ分布を有する任意の元素であり得る。さらに、タグは、適切な条件下での質量分光計の衝突セルまたはイオン源において破壊しリポーターイオンを生じる結合を有し得る。
【0027】
タグはまた、アフィニティータグの同族結合パートナーへの結合によりアフィニティー試薬にカップリングされた分子の単離を可能にするアフィニティータグであり得る。ポリペプチドタグ化のために、サンプル中の1つまたは複数のポリペプチドは変性され得、必要に応じて還元され得、そしてポリペプチドの化学反応性基は、化学改変試薬で共有結合により誘導体化され得る。タグ化されたポリペプチドは、サンプル中でタグ化されていないポリペプチドおよび他の成分から容易に単離され得る。これは、質量分析により分析されるべきであるサンプルの複雑度を低減する。このようなアフィニティータグ化は、他の分子(例えば、核酸、脂質、炭水化物、第二メッセンジャー、代謝物など)に同様に適用され得る。さらに、タグは、化学反応または酵素触媒反応によって導入され得る。
【0028】
本明細書中で使用される「開裂可能な官能基」は、種々の方法(エネルギー、化学物質、酵素などの投入を含む)によって開裂され得る化学基である。本発明の方法において使用するために、開裂可能な官能基は、一般に特異的であり、すなわち、開裂される分子に変化も損傷も与えることなく特異的に開裂可能であり、または比較的に均一に、再現様式で分子を変化させる。例えば、開裂可能な官能基は、光開裂可能な基であり得る。このような場合、光開裂可能な基は、一般に、放出される分子を損傷しない光の波長で(例えば、紫外光から可視光範囲で)開裂される(実施例Iを参照のこと)。例示の光開裂可能なリンカーとしては、例えば、o−ニトロベンジル基、デシル基、トランス−o−シンナモイル基、m−ニトロフェニル基、ベンジルスルホニル基などを含むリンカーが挙げられる(例えば、DormanおよびPrestwich、Trens Biotech.18:64−77(2000);GreeneおよびWuts、Protective Groups in Organic Synthesis、第2版、John Wiley & Sons、New York(1991);米国特許第5,143,854号;米国特許第5,986,076号;米国特許第5,917,016号;米国特許第5,489,678号;米国特許第5,405,783号を参照のこと)。
【0029】
切断可能な官能基はまた、酸または塩基のような化学物質によって切断可能な、化学的切断可能な基であり得る。望ましい場合、化学的切断反応は、その化学的切断可能な基が本質的に唯一の切断される化学結合である、比較的穏やかな条件下で実行され得る。化学的切断可能な基はまた、CNBr(これは、メチオニン残基を切断し得る)のような化学物質によって切断可能な基であり得る。CNBrは、メチオニンのような化学的切断可能な基が、(特に、メチオニン残基を有さないポリペプチドにおいて)ある分子に付加されている場合、その分子を遊離させるために特に有用であり得る。適切な化学切断可能な基は、当業者に周知である(例えば、WilsonおよびCzarnik編、Combinatorial Chemistry:Synthesis and Application,John Wiley & Sons,New York(1997);Merrifield,J.Am.Chem.Soc.85:2149(1964);Bodanszky,M.,Principles of Peptide Synthesis(Springer−Verlag,1984);Houghten,Proc.Natl.Acad.Sci.USA 82:5131(1985)を参照のこと)。例示的な化学的切断可能なリンカーは、ジスルフィド(これは、還元剤により切断され得る);ジオール(これは、過ヨウ素酸塩)により切断され得る);ジアゾ結合(これは、ジチオネートにより切断され得る);エステル(これは、ヒドロキシルアミンにより切断され得る);スルホン(これは、塩基により切断され得る)などを含み得る(Hermanson,Bioconjugate Technique,Academic Press,San Diego(1996);Pierce Chemical Co.,Rockford,ILを参照のこと)。
【0030】
この切断可能な官能基はまた、酵素切断可能な基であり得る。例えば、プロテアーゼが、そのプロテアーゼにとって適切な認識配列を有する切断可能な官能基を切断するために、使用され得る。特に有用なプロテアーゼは、エンドペプチダーゼ(例えば、第Xa因子、タバコエッチ(etch)ウイルス(TEV)プロテアーゼ、トリプシン、キモトリプシン、Staphylococcus aureusプロテアーゼ、顎下腺(submaxillaris)プロテアーゼなど)である。そのプロテアーゼは、切断可能な特定の認識配列を官能基として組み込むことに基づいて選択され得る。プロテアーゼを選択するための他の考慮事項としては、捕捉されそして遊離される分子中に認識配列が存在するかまたは存在しないかが挙げられる。
【0031】
例えば、希に切断するプロテアーゼ(例えば、TEVプロテアーゼまたは第Xa因子)は、対応するプロテアーゼ認識配列を含む官能基を切断して捕捉された分子を遊離するために、使用され得る。このような希に切断するプロテアーゼは、インタクトなポリペプチド分子を遊離するために特に有用である。なぜなら、これらのプロテアーゼの認識配列は、大部分のポリペプチド中に存在しないからである。あるいは、ポリペプチドサンプルは、特定のプロテアーゼで処理され得、そして消化されたペプチドが、本明細書中に開示される方法によって単離され得る。このような場合、捕捉されたペプチドは、切断のために使用されるプロテアーゼの認識配列を含まない。なぜなら、そのポリペプチドは、すでに消化されているからである。さらに、望ましい場合、インタクトなポリペプチドは、固体支持体に結合された後に、捕捉されそしてプロテアーゼで消化され得、その固体支持体上に捕捉されたポリペプチドのペプチドフラグメント上の標識の組み込みおよび遊離を生じ得る。従って、プロテアーゼ消化は、望ましい場合、サンプル分子(特にポリペプチドサンプル分子)の捕捉の前にもまたは後にも使用され得る。
【0032】
プロテアーゼに加えて、切断可能な官能基は、エンドヌクレアーゼ(例えば、制限酵素)の認識配列であり得る。従って、制限酵素の適切な認識配列が、切断可能な官能基として組込まれ得、そしてその個々の制限酵素により切断され得る。そのようなヌクレオチド官能基は、核酸またはポリペプチドまたは望ましい他の任意の型の分子を捕捉および遊離させるために有用であり得ることが、理解される。同様に、プロテアーゼ認識配列が、核酸またはポリペプチドまたは望ましい他の任意の型の分子を捕捉および遊離させるために有用であり得る。
【0033】
本明細書中で使用される場合、用語「反応性基」は、分子(例えば、ポリペプチド、核酸、脂質、糖質、セカンドメッセンジャー、代謝物など)との反応およびその分子への共有結合に適切な有用な化学的特性を有する、種々の化学基のうちのいずれかを意味することが意図される。例えば、反応性基は、AspもしくはGluにおいて見出されるカルボキシル基と反応し得るか、またはその反応性基は、他のアミノ酸(例えば、His、Tyr、Arg、およびMet)と反応し得る。反応性基はまた、Lysのようなアミンと反応し得る(例えば、イミドエステルおよびN−ヒドロキシスクシンイミジルエステルである)。さらに、反応性基はまた、当該分野で周知の化学反応を使用して、酸素または硫黄と反応し得る。反応性基はまた、ホスホペプチドもしくは核酸の選択的標識のためにリン酸基と反応し得るし、または他の共有結合により改変されたペプチド(グリコペプチド、リポペプチド、または本明細書中に開示されるような共有結合ポリペプチド改変物のうちのいずれかを含む)と反応し得る。
【0034】
本明細書中で使用される場合、用語「同位体標識」または「同位体タグ」とは、2つの別個の同位体形態で生成され得る化学基(例えば、その化学基を構成する構成元素の重い同位体バージョンおよび軽い同位体バージョン)を指す。そのような構成元素としては、例えば、炭素、酸素、水素、窒素、および硫黄が挙げられる。さらに、化学的または機能的に類似する他の元素が、天然に存在する上記元素に代わって置き換わり得る。例えば、セレンが、硫黄に代わる置換基として使用され得る。特に有用な同位体標識または同位体タグは、MSによる簡便な分析を可能にする、同位体標識または同位体タグである。例えば、重い同位体バージョンおよび軽い同位体バージョンのアミノ酸が、ポリペプチドを差次的に同位体標識するために使用され得る(実施例Iを参照のこと)。
【0035】
本明細書中で使用される場合、「結合している(coupled)」またはその文法上の形態は、分子間の結合相互作用を指す。例えば、固体支持体は、その固体支持体の化学部分と、化学基の化学部分との間の結合相互作用を介して、その化学基に結合され得る。結合している分子間の結合相互作用は、共有結合性相互作用でもまたは非共有結合性相互作用でもあり得る。一般的に、化学基は、共有結合性相互作用を介して、固体支持体または他の分子に結合される。その固体支持体と、結合された分子との間に非共有結合性相互作用が生じる場合、その非共有結合性相互作用は、本発明の方法において使用される化学的操作および/または物理的操作(例えば、その固体支持体に結合している分子に対して実施される化学的改変工程または洗浄工程)の間に結合したままであるに十分に高親和性であることが、理解される。
【0036】
本発明は、分子を標識するための方法を提供し、この方法は、サンプル分子を、化学基(切断可能な官能基と、1つ以上の官能基と、そのサンプル分子についての反応性基とを含む)に結合されている固体支持体と、そのサンプル分子をその反応性基に共有結合させる条件下で接触させることによる。この方法は、その切断可能な官能基を切断し、それによって、1つ以上の結合した官能基を有するそのサンプル分子を遊離させる工程をさらに包含し得る。この方法は、複雑なサンプル混合物中の分子が、捕捉され得、そして簡便な分析のために標識され得、そしてその固体支持体から遊離される能力に起因して、精製もされ得る点で、有利である。
【0037】
本発明は、目的のポリペプチドまたは他の分子を、切断可能なリンカー(例えば、光切断可能なリンカー)を介して共有結合により捕捉するために、固相ベースのアプローチを使用する方法を提供し、この光切断可能なリンカーは、捕捉された分子の光触媒回復を可能にする。この切断可能なリンカーは、切断の際に、遊離される分子に特定の官能基が転移するように構築される。そのような官能基としては、例えば、安定な同位体タグ(同位体希釈理論に基づいて質量分光法によって正確なペプチド定量を可能にする)、同位体分布タグ(その同位体分布によってタグ化ペプチドまたはそのフラグメントを同定する)、荷電アミノ酸または他の化合物(質量分析計中での効率的なイオン化を媒介し、そしてタンデム質量分析計の衝突セルにおける断片化パターンをもたらす)が挙げられる。さらに、この方法は、その分子(例えば、ポリペプチド)が固体支持体上に固定されている間の、その分子の化学的改変または酵素的改変、脱改変(de−modification)、切断または他の操作を可能にする。光切断可能なリンカーが特に有用であるが、本明細書中に開示されるような特異的に切断され得る任意のリンカーが使用され得ることが、理解される。光切断可能なリンカーの代替案としては、酸切断可能なリンカーおよび塩基切断可能なリンカー、熱により切断可能なリンカー、ならびに本明細書中に記載されるような、酵素の標的切断部を含むリンカーが、挙げられる。
【0038】
本発明の方法は、サンプルから分子を固定し、そして捕捉されたサンプル分子に標識(例えば、タグ)を簡便に転移するのを可能にする、固相化学の有利な使用に基づく。この方法は、固体支持体に化学基を結合することに基づく。その化学基は、図1に例示される特徴を有する。その化学基の1つの特徴は、分子の可逆的な捕捉および遊離を可能にする、切断可能な官能基である。その化学基の第2の特徴は、望ましい化学的特性を有する1つ以上の官能基である。そのような官能基は、例えば、その分子または化学部分のその後の分析のために便利な、その分子に望ましい化学的特性(例えば、電荷の変化、疎水性の変化、または質量の変化)を付与する標識またはタグであり得る。その化学基の第3の特徴は、サンプル中の分子にその化学基を共有結合するのを可能にする、反応性基である。これらの3つの特徴は、サンプル分子がその反応性基を介して捕捉され得、切断の際に、遊離される分子にその官能基が転移するように、その化学基上で配置される(図1を参照のこと)。
【0039】
その化学基はまた、固体支持体にその化学基が結合されるのを可能にし、同時に上記特徴が活用されるのを可能にする、化学部分を含む。上記特徴を有する化学部分は、その特徴を付与する化学部分の連続付加によって、固体支持体上で合成され得るか、または望ましい場合は、化学基として合成され得、その後、その固体支持体に結合され得る。
【0040】
本発明の方法の特定の実施形態が、図1において例示される。切断可能な官能基と、1つ以上の望ましい官能基(例えば、タグ)と、反応性基とを有する化学基が、固体支持体に結合される。図1は、その固体支持体に共有結合されたアミノ官能基を含む、光切断可能なリンカーを示す。この光切断可能なリンカーは、分子が固相上に捕捉された後に、それらの分子の光開始切断を可能にする。望ましい特定の官能基を有するリンカー分子が、そのアミノ官能基を介して、その光切断可能なリンカーに結合される。光切断の際、その官能基を含むリンカー分子は、図1Cにペプチドとして示される捕捉された分子に転移され、望ましい官能基のその分子への転移をもたらす。例えば、本明細書中に開示されるように、転移される官能基は、定量的質量分光法に有用な、安定な同位体がコードするアミノ酸であり得る。1つ以上の官能基を有する機能的リンカーから延びているのは、その分子上の化学部分について特異性を有する反応性基(例えば、ポリペプチドのアミノ基と反応性の基、スルフヒドリル基と反応性の基、カルボキシ基と反応性の基または他の基と反応性の基)である。
【0041】
分子(例えば、サンプル中の分子)は、上記のように結合された化学基を有する固体支持体と、接触される。その分子は、そのサンプル分子がその化学基を介してその固体支持体に共有結合するのを可能にする条件下で、インキュベートされる。その化学基およびサンプル分子上の反応性基に基づいて、共有結合を可能にする適切な条件を、当業者は容易に決定し得る。同様に、本明細書中に開示されるように、非共有結合性相互作用のための適切な条件を、当業者は容易に決定し得る。
【0042】
実施例Iに例示され、図1に示される特定の実施形態において、固相支持体は、光切断性リンカーが、シラン結合を介して結合されている、制御された孔を有するガラスビーズである。図1に示される光切断性リンカーは、360nmのUV光で切断され得る。重水素化アミノ酸または非重水素化アミノ酸(例えば、ロイシン)は、図1に例示されたポリペプチドに転位される官能基として使用され得る。2つの異なるサンプル供給源が、比較分析または定量分析のために使用される場合、2つの同位体タグは、ロイシンの重水素化の状態に依存して、一般的に質量が、7または10質量単位異なる。本明細書中に開示されるように、異なる同位体分布を有する他のアミノ酸またはアミノ酸とは異なる分子もまた、安定な同位体タグとして使用され得る。図1に示されるペプチド反応性基は、スルフヒドリル基と特異的に反応するヨードアセチル基である。光切断の際に、改変されたロイシンに起因する質量タグの増加を有する、図1Cに示されるポリペプチドが、放出される。重水素化されたアミノ酸を用いて例示されるけれども、任意の適切な同位体形態(例えば、13C,15Nなどのような他の構成要素の同位体)が、本発明の方法において、使用され得ることが理解される。非放射性同位体が一般的に使用されるが、トリチウムのような放射性同位体でさえも使用され得る。
【0043】
本発明はまた、図1に例示される試薬のような試薬を提供する。この試薬は、サンプル分子(複合サンプルを含む)を単離し、タグ化するために使用され得る(実施例Vを参照のこと)。従って、本発明は、図1Aに概略的に示されるような、ビーズ、切断性官能基、タグのような官能基、および反応性基を含む試薬を提供する。
【0044】
このようにして、本発明はさらに、化学基に結合された固体支持体を含有する構成物(composition)を提供し、この化学基は、切断性官能基、タグのような官能基およびサンプル分子に共有結合された反応性基を含み、ここで、切断性官能基、タグおよび反応性基は互いに関して、切断性官能基の切断の際に、タグがサンプル分子に移動可能であるように配置される。本発明はさらに、化学基に共有結合された固体支持体を含有する構成物を提供し、この化学基は、切断性官能基、質量分析法タグおよびサンプル分子に共有結合された反応性基を含み、ここで、切断性官能基、タグおよび反応性基は互いに関して、切断性官能基の切断の際に、タグがサンプル分子に移動可能であるように配置される。
【0045】
本発明の方法は、分子を選択的に単離し、かつ放出の際に、標識またはタグを含有する1つ以上の官能基をこの分子上に転位する能力を提供するという点で有利である。従って、化学基(すなわち、切断性官能基、1つ以上の官能基および反応性基)の官能基は互いに関して、官能基(例えば、タグ)が分子に転位可能になるように配置される。従って、化学基の官能基は、官能基(例えば、タグ)が図1に示されるように切断性官能基と反応性基との間に配置され、切断性官能基の切断およびこの分子の放出の際に、この官能基が捕捉分子に転位し得るように、一般的に配置される。
【0046】
本発明の方法は、サンプル分子を捕捉する能力を使用し、官能基をこの分子に転位し、かつ結合官能基を有する分子を放出する能力を利用しているので、有利である。従って、本方法は、サンプル分子を標識し、そして単一工程でサンプル分子を同時に精製するために使用され得る。切断性官能基の組み込みは、結合官能基を有するサンプル分子の放出を容易にし、この結合官能基が次いで、分析され得る。本発明の方法は、切断性官能基を一般的に使用するけれども、本発明の方法が、さらなる分析の前に捕捉分子の放出を必要としない標識またはタグのような官能基を転位するためにもまた使用され得ることが理解される。化学反応または酵素反応によって切断可能な切断性化学基の非存在下での、本発明の方法の使用は、分析方法(例えば、MS、特に、MALDI−TOF)に適用可能であり、ここでは、レーザーが、結合分子の切断に使用され、同時に分子をイオン化する。
【0047】
本発明の方法は、広範囲の種々の分子に容易に適用され得る。上記に記載されるように、いくつかの場合、分子は、本明細書中に開示される捕捉方法に適切な反応性化学部分を有し得る。しかし、所望される場合、分子は、所望の官能基、特に、本明細書中に開示される捕捉方法に適切な反応性基を組み込むように改変され得る。
【0048】
例えば、システイン残基を含まないポリペプチド、つまりスルフヒドリル側鎖を含有する天然アミノ酸を含まないポリペプチドは、図1に示される固相試薬に結合しない。いくつかの場合、システイン含有ポリペプチドを選択的に単離することが望ましいが、他の場合、サンプル中に含有される他のポリペプチドまたはさらなるポリペプチドを、単離し、同定し、そして定量することが、望ましい。例えば、アミノ反応性基(例えば、固相試薬の反応性基としてのスクシンイミドエステル)を合成することが可能である。あるいは、捕捉されるべき分子は、化学的に改変され、特異的な官能基を組み込み得る。
【0049】
例えば、本明細書中に開示される特定の実施形態において、ポリペプチドの一級アミノ基は、スルフヒドリル基へと改変され、図1に示されるような、同一のSH−反応性固相ビーズを、ポリペプチドの捕捉に使用されるようにする。この戦略において、ポリペプチドのアミノ基は、図5に示されるワンポット化学によってスルフヒドリル基へと転換され得る。第1に、アミノ基は、N−スクシンイミジルS−アセチルチオアセテート(SATA)によって改変される。ヒドロキシルアミン処理の際に、トリス(2−カルボキシエチル)ホスフィン(TCEP)での還元の後に、このペプチドのアミノ基が、スルフヒドリル基へと転換される。本明細書中に例示されるように、サンプル分子中の実質的に全てのアミノ基が、スルフヒドリル基へと転換され得る(実施例IIを参照のこと)。改変されたペプチドは、必要に応じて、例えばC18逆相カートリッジ上で脱塩することによって精製され得、回収され、次いで、図1に示されるようなビーズに結合され得る。
【0050】
アミノ基のスルフヒドリル基への転換の際に、プロトン化部位は、消失する。従って、例えば、ロイシンの代わりの荷電性アミノ酸(例えば、リジン)が、ポリペプチドのタグ化に使用され得る。この方法において、リジン側鎖の遊離アミノ基は、代替のプロトン化部位を提供し得、ペプチドの荷電状態は、この手順によって変わらない。タグを付加するための他の適切な基(荷電性基を含む)としては、例えば、アルギニン、ピリジル、トリメチルアミンなどが挙げられ、これらは、溶液中または気相中で強塩基であり、つまりイオン化を促進する基である。代表的な質量分光法実験下では、ペプチドの荷電状態が、ペプチドの配列決定および検出のための質量分析器における、衝突により誘導されるフラグメント化に影響するので、このことは重要である。
【0051】
ポリペプチド中のアミノ酸側鎖を改変するための方法および化学は、当業者に周知である(例えば、Glazerら、Laboratory Techniques in Biochemistry and Molecular Biology:Chemical Modification of Proteins,Chapter 3,68頁〜120頁,Elsevier Biomedical Press,New York(1975)(これは参考として本明細書中に援用される);およびPierce Catalog(1994),Pierce,Rockford ILを参照のこと)。
【0052】
ポリペプチドのアミノ末端を改変するための方法もまた、使用され得る。本明細書中に例示される、(N末端を含む(実施例IIを参照のこと))アミノ基を改変するための方法に加えて、N末端を改変するための他の方法は、当業者に周知である(例えば、Branciaら、Electrophoresis 22:552−559(2001);Hovingら、Anal.Chem.72:1006−1014(2000);Munchbachら、Anal.Chem.72:4047−4057(2000)(各々は、参考として本明細書中に援用される)を参照のこと)。
【0053】
さらに、反応性基は、分子上に形成され得、続いてこの分子は、所望の化学部分を組み込むように改変され得る。例えば、CNBrによる切断は、ホモセリンラクトンを生じる。従って、メチオニンを含むポリペプチドは、CNBrによって化学的に切断され、ホモセリンラクトンを生じ得る。得られたホモセリンラクトンは、アミンによって改変され得、反応性アミンを有する化学基の組み込みを可能にする。
【0054】
分子は、所望される場合、化学的にかまたは酵素的に改変され得る。例えば、分子は、上記に記載されるような方法を使用して化学的に改変され得る。さらに、分子は、酵素的に改変され得る。捕捉された分子は、酵素的に改変され、この分子に基を組込み得るかまたはこの分子から基を除去し得る。例えば、ポリペプチドは、キナーゼによってリン酸化され得るかもしくはホスファターゼによって脱リン酸化され得るか、または、ポリペプチドを翻訳後修飾する能力を有する任意の他の酵素によって分子への化学部分の付加もしくは分子から化学部分の除去を生じ得る。同様に、核酸は、核酸へと化学部分を付加するかまたは核酸から化学部分を除去する、任意の周知の酵素によって捕捉された後、改変され得る(例えば、Sambrookら、Molecular Cloning:A Laboratory Manual,第2版、Cold Spring Harbor Press,Plainview,New York(1989);Ausubelら、Current Protocols in Molecular Biology(補遺47),John Wiley & Sons,New York(1999);SambrookおよびRussel,Molecular Cloning:A Laboratory Manual,第3版,Cold Spring Harbor Press,Cold Spring Harbor(2001)を参照のこと)。捕捉された分子(ポリペプチドまたは核酸を含む)を改変するために有用な例示的な酵素としては、キナーゼ、ホスファターゼ、メチラーゼ、およびデカルボキシラーゼなど、または捕捉された分子へと化学部分を付加し得るかまたは捕捉された分子から化学部分を除去し得る任意の酵素が挙げられる。
【0055】
任意の種々の反応性基は、この反応性基がポリペプチドのような分子に共有結合され得る限り、サンプル分子との反応のために化学基に組み込まれ得る。反応性基は、当業者に周知である(例えば、Hermanson,(上述)1996;Glazerら、Laboratory Techniques in Biochemistry and Molecular Biology:Chemical Modification of Proteins,第3章,68頁〜120頁,Elsevier Biomedical Press,New York(1975);Pierce Catalog(1994),Pierce,Rockford ILを参照のこと)。例えば、反応性基は、AspまたはGluに見出されるカルボキシル基と反応しえるかあるいは、この反応性基は、他のアミノ酸(例えば、His、Tyr、Arg、およびMet)と反応し得る。反応性基はまた、Lysのようなアミン(例えば、イミドエステルおよびN−ヒドロキシスクシンイミジルエステル)と反応し得る。さらに、反応性基はまた、当該分野で周知の化学を使用して、酸素または硫黄と反応し得る。反応性基はまた、ホスホペプチドの選択的標識のためのリン酸基、または他の共有結合的に改変されたペプチド(グリコペプチド、リポペプチド、または本明細書中に開示される任意の共有結合ポリペプチド改変体を含む)と反応し得る。さらに、当業者は、公知の試薬を使用してポリペプチドを改変するための条件、ポリペプチドまたは本発明の方法における使用のための他の分子の改変のために最適な条件を得るためのインキュベーション条件およびインキュベーションの時間を知るかあるいはそれらのことを容易に決定し得る。
【0056】
例示的なスルフヒドリル反応性基としては、ヨードアセトアミド基が挙げられる(Gygiら、Nature Biotechnol.17:994−999(1999)を参照のこと)。他の例示的なスルフヒドリル反応性基としては、マレイミド、アルキルハライドおよびアリールハライド、ハロアセチル、α−ハロアシル、ピリジルジスルフィド、アジリジン、アクリロリル、ならびにアリール化剤が挙げられる。所望の場合、ポリペプチドは、本発明の試薬と反応する前に還元され得、これは、この試薬が、スルフヒドリル反応性基を含む場合に、特に有用である。
【0057】
反応性基はまた、例えば、ペプチドのα−アミノ基またはLys側鎖のε−アミノ基のようなアミンと、例えば、イミドエステル、N−ヒドロキシスクシンイミジルエステル(NHS)、イソチオシアネート、イソシアネート、アシルアジド、スルホニルクロリド、アルデヒド、ケトン、グリオキサール、エポキシド、オキシラン、カーボネート、アリール化剤、カルボジイミド、無水物などとを反応し得る。反応性基はまた、AspもしくはGluまたはペプチドのC末端に見られるカルボキシル基と、例えば、ジアゾアルカン、ジアゾアセチル、カルボニルジイミダゾール、カルボジイミドなどとを反応し得る。ヒドロキシル基と反応する反応性基としては、例えば、エポキシド、オキシラン、カルボニルジイミダゾール、N,N’−ジスクシンイミジルカーボネート、N−ヒドロキシスクシンイミジルクロロホルメートなどが挙げられる。反応性基はまた、アミノ酸(例えば、His、Tyr、Arg、およびMet)と反応し得る。さらに、反応性基はまた、ホスホペプチドの選択的標識のためのリン酸基、または他の共有結合的に改変されたペプチド(グリコペプチド、リポペプチド、または任意の公知の共有結合ポリペプチド改変が挙げられる)と反応し得る。当業者は、本発明の方法における使用のためのサンプル分子の改変について最適な条件を得るための種々の試薬、インキュベーション条件およびインキュベーション時間を用いることによって、サンプル分子を改変するための条件を容易に決定し得る。
【0058】
サンプル分子を捕捉することによって、種々の化学修飾および/または酵素的改変が、結合した分子に対して実行され得るが、このサンプル分子が、固体支持体に結合したままであるという点で、本発明の方法は、有利である。サンプル分子が、物理的操作、化学的操作および/または酵素的操作の間、固体支持体に結合したままであるので、改変されたサンプル分子の収率は、溶液相方法よりも高い。従って、本発明の方法は、本発明のタグ試薬と固体相捕捉の反応性基と反応し得る化学特性を有する、特定のクラス(class or classes)の分子の少なくとも約5%、約10%、約20%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%、約98%、約99%でか、または本質的に全てを捕捉するために使用され得る。さらに、本発明の方法は、捕捉試薬およびタグ試薬を含む本発明の固体支持体と接触され、そしてこの支持体に結合されるサンプル中の特定のクラスの分子の少なくとも約5%、約10%、約20%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%、約98%、約99%でか、または本質的に全てを捕捉および放出するために使用され得る。例えば、本明細書中で開示されるように、実質的に100%の分子は、捕捉および放出され得る(実施例Iを参照のこと)。当業者は、固体支持体からサンプル分子を捕捉するため、および/またはサンプル分子を放出するために最適化された条件を容易に決定し得る。
【0059】
本発明はさらに、分子を標識するための方法を提供する。この方法は、サンプル分子が反応性基に共有結合し得る条件下で、サンプル分子を化学基が結合された固体支持体と接触させる工程であって、この化学基が、切断可能な官能基、1つ以上の官能基、およびサンプル分子に対する反応性基を含む工程;固体支持体に結合されるサンプル分子を改変する工程;および切断可能な官能基を切断する工程であって、これによって、1つ以上の官能基を含む改変されたサンプル分子を放出する工程を包含する。
【0060】
本発明の方法は、望ましい場合、捕捉された分子を改変するために使用され得る。例えば、この方法は、リン酸化ポリペプチドおよび非リン酸化ポリペプチドの両方を含むサンプルからポリペプチドを捕捉するために使用され得る。タンパク質リン酸化を系統的に分析する方法は、最近記述された(Zhouら、Nature Biotechnol.19:375〜378(2001))。Zhouらのように溶液中で化学反応を実行するというのでなく、ポリペプチドは、まず、本明細書中に開示される方法を用いて捕捉され得、そして固体支持体に結合される間に改変され得る(実施例IVを参照のこと)。あるいは、分子を捕捉および標識するための本明細書中に開示される方法は、Zhouら(前出、2001)に記載される反応の実施後、または任意の他の化学改変もしくは酵素的改変の後に適用され得る。
【0061】
本発明の構成物および方法は、種々の適用において有利に使用され得る。1つの特に有用な適用は、定量的タンパク質発現分析である。例えば、本明細書中に開示される方法および試薬は、複雑なサンプル中に含まれるポリペプチドのような分子に特定の同位体サインを与えるために使用され得る。定量分析の1つの特定の適用において、2つ以上のサンプルが、比較され得る(図8、実施例IIIを参照のこと)。2つの異なるサンプル由来のペプチドの定量分析は、タンパク質発現の相対的定量化についてのICAT技術と同様な様式で、安定な同位体の希釈の概念に基づいて達成され得る(Gygiら、NatureBiotechnol.17:994〜999(1999);実施例Vを参照のこと)。示差的に同位体で標識した分子は、比較のために2つのサンプル(例えば、アミノ酸(例えば、重水素化ロイシンまたは非重水素化ロイシン)、または2つの異なるサンプルに対して質量タグとして組込まれ得る他のアミノ酸)を標識するために使用され得る。本発明の方法は、分子の単離およびタグ(例えば、安定な同位体)の組込みの両方が、両方達成される点で有利である。従って、本発明の方法は、定量的な質量スペクトル分析について特に有用である。
【0062】
本明細書中に開示されるように、ポリペプチドへの官能基の組込みはまた、ペプチドのアミノ基を介して達成され得、この組込みは、ポリペプチド中のスルフヒドリル基の存在に基づく捕捉方法が適用されるとき、遊離のシステイン残基を含まないポリペプチドが分析される場合、特に有用である。ほとんどのポリペプチドは、そのN−末端に少なくとも1つのアミノ基を含むので、システイン残基を有さないポリペプチドでさえも、ポリペプチド中にスルフヒドリル基を組込む方法に基づく固相で標識され得る。ブロックされたアミノ末端を有するサンプル中のポリペプチドの場合にて、このようなポリペプチドは、フラグメント化されて、切断されたフラグメント上に遊離のアミノ末端を生成し得る。従って、本発明の方法は、特定の化学改変を使用する種々の分子の単離に適用され得る。
【0063】
さらに、移入されたタグの選択における大きな構造的柔軟性が存在する。従って、タグの構造は、特定の目標を達成するために意図的に選択され得る。例えば、非常に極性のペプチドは、より疎水性に作製され得、従って、疎水性タグの移入によって逆相カラム上に良好に保持され得るか、強力な気相塩基性基(例えば、ピリジル)が、質量分析計の衝突セルにおいて直接的にフラグメント化するために移動され得るか、または特徴的な同位体分布を有する元素(例えば、塩素または臭素)が、タグ化ペプチドに対して異なる同位体サインを提供するために付加され得る。
【0064】
本発明の方法はまた、改変分子(例えば、翻訳後修飾によって修飾されたポリペプチド)の分析に適用され得る。例えば、この方法は、タンパク質リン酸化の定量分析に適用され得る。タンパク質リン酸化の系統的な分析に関する方法は、すでに記載されている(Zhouら、Nature Biotechnol.19:375〜378(2001))。一連の化学反応は、溶液中でリン酸化ポリペプチドおよび非リン酸化ポリペプチドを含む、複雑なペプチド溶液からリン酸化ペプチドの選択的単離のために、溶液中で実行される(Zhouら、前出、2001)。固相の捕捉および放出を用いる本明細書中に開示される方法はまた、ホスホペプチドの選択的単離のために使用され得る。リン酸化ポリペプチドおよび非リン酸化ポリペプチドは、本明細書中に開示されるように、固相ビーズ上に捕捉される。一旦固定化されると、ホスホペプチドのリン特異的標識化を導く一連の化学反応が、実行される(Zhouら、前出、2001)。ペプチドは、切断によって放出され、正確な定量化のために各々のペプチドに安定な同位体サインを与える。最初からリン酸化されているポリペプチドは、スルフヒドリル基に新たに変換され、スルフヒドリル含有分子を単離するための本明細書中に開示される方法を用いて捕捉され得る。捕捉されたポリペプチドは、非リン酸化ペプチドがなくなるまで洗浄され得、そして例えば、質量スペクトル分析のために放出され得る。本明細書中に開示される本発明の方法は、各々の化学反応後に過剰の試薬を除去するために必要とされるサンプルを取扱う工程の数が、劇的に減少および単純化されるので、溶液化学よりも明らかに有利である。
【0065】
リン酸化に加えて、本発明の方法は、本明細書中に開示されるように、翻訳後修飾の多くの異なる形態(例えば、グリコシル化、ユビキチン化、アセチル化、パルミチル化、ミリスチル化など)を有するポリペプチドに容易に適用され得る。従って、本発明の方法は、ペプチドへの種々の官能基の同じ移入を伴うポリペプチドを含む、他の翻訳後修飾された分子を選択的に単離するために使用され得る。翻訳後修飾の特定の型の選択的な単離は、本発明の方法を用いて達成され得る。例えば、ユビキチン化に対する特異的結合活性を有する抗体は、ユビキチン化ポリペプチドを単離するために使用され得、タンパク質リン酸化の定量分析と同じ方法で、ポリペプチドのユビキチン化の定量分析が達成され得る。このような方法はまた、所望の場合、分子の他の修飾に適用され得る。
【0066】
抗体もまた、エピトープタグで修飾されたサンプル分子の引き続く分析および/または単離のために使用され得る。抗体を調製するための方法は、当業者に周知である。用語抗体は、その最も広範な意味で使用され、ポリクローナル抗体およびモノクローナル抗体、ならびにこのような抗体の抗原結合フラグメントを含む。本発明において有用な抗体、またはこのような抗体の抗原結合フラグメントは、少なくとも約1×105M−1のポリペプチドまたはペプチド部分に対して、特異的結合活性を有することによって特徴付けられる。従って、ポリペプチドに対して特異的結合活性を保持する、抗体のFabフラグメント、F(ab’)2フラグメント、Fdフラグメント、Fvフラグメント、単鎖Fv(scFv)フラグメントなどである。ポリペプチドに対する抗体の特異的結合活性は、例えば、特定の抗原 対 コントロール抗原に対する抗体の結合活性を比較することによって、当業者によって容易に決定され得る。ポリクローナル抗体またはモノクローナル抗体を調製する方法ならびに結合活性および/または特異性を決定する方法は、当業者に周知である(例えば、HarlowおよびLane、Antibodies:A Laboratory Manual,Cold Spring Harbor Laboratory Press(1988))。
【0067】
本発明の方法は、一般的に、ポリペプチドを用いて本明細書中に例証したが、サンプル中の任意の種々の分子が、本明細書中に開示される方法によって容易に標識され得ることが理解される。一般的には、多くのクラスの生体分子(例えば、オリゴヌクレオチド、代謝産物など)は、本明細書中に開示される方法によって官能化されて、改善された定性分析または定量分析のための所望の官能基を組込み得る。従って、分子への特定の官能基の可逆的な捕捉および移入を可能にする、本明細書中に開示される方法は、一般的には、プロテオミクス、他の型の発見科学および一般的な定量的生物学的分析の分野における多くの適用において有用である。
【0068】
本発明の方法は、あるクラスの分子(例えば、ポリペプチド、核酸、脂質、セカンドメッセンジャー、代謝産物など)、またはこのようなあるクラスの分子の一部を、効率的に捕捉および放出するために使用され得る。所望の場合、本発明の方法はまた、2つ以上のクラスの分子および/またはこれらのクラスの分子の一部を捕捉するために使用され得る。例えば、この方法は、ポリペプチドおよび核酸の両方、または所望の場合、2つ以上のクラスの分子の任意の組合せを捕捉するために使用され得る。
【0069】
本発明はまた、分子を分析するための方法を提供する。この方法は、サンプル分子が反応性基に共有結合し得る条件下で、サンプル分子を化学基が結合した固体支持体と接触させる工程であって、この化学基が、切断可能な官能基、1つ以上の官能基、およびサンプル分子に対する反応性基を含む工程;サンプル分子を固体支持体から切断する工程であって、1つ以上の特定の官能基が、放出されたサンプル分子に移入される工程;ならびに放出されたサンプル分子を分析する工程を包含する。任意の種々の分析方法が、使用され得、例えば、質量分析法、配列決定、液体クロマトグラフィー、分光光度法、蛍光光度法などが挙げられる。分子が、共有結合的に捕捉され、分析に先立って、非検体物質を除去するために広範に洗浄し得るという理由で、本発明の方法は、有利である。さらに、捕捉される分子に移入されるべき官能基は、分子のさらなる分析を容易にするために有用な官能性を、例えば、発色団、フルオロフォア、質量タグなどを付加することによって組込み得る。
【0070】
質量分析は、サンプル分子を分析するための特定の有用な方法である。本発明の方法を用いて、種々の質量分析システムを使用して捕捉されるサンプル分子を分析し得る。高度の質量正確性、高感度および高度の解像度を伴う質量分析器が用いられ得、そしてマトリクス支援レーザー吸着飛行時間型(MALDI−TOF)質量分析器、電子スプレー電離飛行時間型(ESI−TOF)質量分析器およびフーリエ変換イオンサイクトロトン質量分析器(FT−ICR−MS)が挙げられるがそれらに限定されない。他のモードのMSとしては、イオントラップおよび三重四極質量分析器が挙げられる。イオントラップMSにおいて、分析物は、電子スプレー電離またはMALDIによってイオン化され、ついでイオントラップへと入れられる。次いで、捕捉されたイオンは、そのイオントラップからの選択的放出に応じて別々にMSにより分析され得る。フラグメントもまた、このイオントラップにおいて生成され得、そして分析され得る。本発明の方法を用いて質量タグを伴う標識されるサンプル分子は、例えば、MALDI−TOFシステムまたはESI−TOFのシステムを用いて単一段階質量分析によって分析され得る。さらに、LC−MS/MSまたはLC−ESI−TOFもまた使用され得る。任意のMS方法およびMS方法の任意の組み合わせを使用してサンプル分子を分析し得ることが理解される。質量分析による分析の方法は、当業者に周知である(例えば、Yates,J.Mass Spect.33:1−19(1998);KinterおよびSherman,Protein Sequencing and Identification Using Tandem Mass Spectrometry,John Wiley&Sons,New York/(2000)を参照のこと)。
本発明の方法を用いてサンプル中の分子を分析し得る。このサンプルは、たとえば、生物学的検体に由来し得る。検体とは、具体的には、生物または個体から得られたサンプルをいう。検体は、流体または組織検体として個体から得られ得る。例えば、組織検体は、皮膚生検、組織生検または腫瘍生検のような生検として得られ得る。流体検体は、血液、血清、尿、唾液、脳脊髄液または他の体液であり得る。流体検体は、本発明の方法において特に有用である。なぜなら、流体検体は、個体から容易に入手されるからである。検体の収集のための方法は、当業者に周知である(例えば、Young and Bermes,in Tietz Textbook of Clinical Chemistry,第3版,BurtisおよびAshwood(編),W.B.Saunders,Philadelphia,第2章,42頁−72頁(1999)を参照のこと)。検体はまた、微生物検体であり得、この微生物検体は、微生物の培養物に由来し得、この微生物の培養物としては、個体からの検体から培養される培養物が挙げられる。従って、本発明の方法を用いて、生物学的サンプルにおける複合混合物を分析し得る。
【0071】
本発明の方法は、複雑な生物学的サンプルが直接分析され得るという点において有利であるが、サンプルはまた、所望の場合、処理され得る。例えば、血液サンプルを分画して特定の細胞型(例えば、赤血球、白血球など)を分離し得る。血清サンプルを分画して、例えば、構造または機能的な特性に基づいて、グリコシル化、リン酸化、および他の翻訳後修飾によって改変される血清タンパク質、または特定の親和性(例えば、核酸に対する親和性)を有するタンパク質のような特定の型のタンパク質を単離し得る。血清サンプルはまた、物理化学的特性(例えば、サイズ、pIなど)に基づいて分画され得る。血清サンプルをさらに分画して大量に存在する嵩高いタンパク質(例えば、アルブミン)を取り除いて、あまり多く存在しない血清ポリペプチドの分析を容易にし得る。さらに、細胞サンプルを分画して、細胞レベル下のオルガネラを単離し得る。さらに、細胞サンプルまたは組織サンプルは、任意の周知の分画方法(クロマトグラフィー技術(例えば、イオン交換、疎水性および逆相、サイズ排除、アフィニティー、疎水性荷電誘導のクロマトグラフィーなど)によって可溶化および分画され得る(Ausubelら,前出,1999;Scopes,Protein Purification: Principles and Practice,第3版,Springer−Verlag,New York (1993);Burton and Harding,J.Chromatogr.A 814:71−81(1998))。
【0072】
本発明の方法は、生物学的サンプルのような複雑なサンプルを分析するために特に有用であるが(実施例Vを参照のこと)、この方法はまた、複雑度が減少したサンプルにおいて使用され得る。例えば、このサンプルは、上記されるように分画されて(予備的なアフィニティークロマトグラフィーを含む)、固相に捕捉されるサンプル分子をより少なく提供し得る。さらに、このサンプルは、例えば組み換え方法によってサンプル中に高いレベルで発現されるポリペプチドまたは核酸または分子のような本質的に単一の精製された分子を含む、高度に精製されたサンプルであり得る。
【0073】
本発明の方法は、自動化に容易に適合させることができる。例えば、所望であれば、自動化されたサンプリング、ロボット工学、または任意の適切な自動化方法を本発明の方法に適用し得る。すべての反応は、自動化された様式で容易になし得ることから、この方法は、ハイスループットサンプル調製を可能にする。さらに、転移工程のようなサンプル処理が実質的にないことから、捕捉された分子の損失が最小化され、したがって、分子回収率を改善する。捕捉された分子はまた、徹底的に洗浄して捕捉されていないサンプル分子または任意の試薬(regent)を取り除き得る。なぜなら、捕捉されたサンプル分子は、この洗浄工程の間、固体支持体に結合したまま残るからである。本発明の方法を用いて所望に応じて、サンプルから実質的にすべてのクラスまたは複数のクラスの分子を、またはサンプルから分子の一部を捕捉し得る。
【0074】
本発明の方法を用いてサンプル中の分子の発現レベルを決定し得る。発現レベルとは、サンプル中の分子の量をいう。分子の発現レベルは、遺伝子によってコードされるメッセンジャーRNA(mRNA)、遺伝子によってコードされる所定のアミノ酸配列に対応するポリペプチドの量、または細胞中に蓄積する分子の生化学的形態の量(例えば、ポリペプチド、核酸または低分子のような分子の特定の合成後改変の量を含む)の代表例であり得る。発現レベルとは、サンプル中の分子の絶対量、または標準物に比較しての、その分子の相対量をいい得、これには、定常状態または非定常状態の条件下で決定される量が含まれる。分子の発現レベルは、サンプル中のコントロール成分分子に対して相対的に決定され得る。発現レベルは、本明細書に開示されるように、2つまたはそれを超えるサンプルの直接の比較によって決定され得る(実施例IIIを参照のこと)。
【0075】
本明細書において開示されるように、固体相は、ガラスビーズ(例えば、制御された孔ガラスビーズ(実施例Iを参照のこと)であり得る。しかし、サンプル分子を結合し、そして所望の化学を実行し、そして洗浄条件について有用な、任意の適切な固体支持体が使用され得る。従って、固体支持体は、ガラス、誘導体化ガラス、シリコン、プラスチックまたは他の支持体であり得る。固相化学合成に適切な任意の固相物質は、本発明の方法における固体支持体として有用である。固体支持体は、孔性または非孔性の物質、表面フィルム、磁性ビーズ、コロイド、膜などであり得る。固体支持体は、ビーズ、平坦表面または本明細書において開示される方法を用いた分子の捕捉のために適切な任意の構造の形態であり得る。固体支持体は、所望の場合、他の化学基に結合させるに適切な化学部分を組み込むように誘導体化され得る。
【0076】
本発明はまた、本発明の試薬または構成物を備えるキットを提供する。従って、本発明は、固相捕捉物およびタグ試薬を伴うキットを提供し、これは図1に例示される。本発明のキットの内容物(例えば、固相捕捉物およびタグ試薬または本発明の構成物)は、包装材料中に、所望の場合滅菌され、汚染がない環境で含まれる。さらに、包装材料は、キット内の物がどのようにサンプル分子を標識するために用いられ得るかを示す指示書を備える。使用のための指示書は、代表的に、試薬濃度または少なくとも1つのアッセイ方法パラメータ(例えば、試薬の相対量)および混合されるべきサンプル、試薬/サンプル混合のための維持時間、温度、緩衝条件などを記載する具体的な表現を含む。
【0077】
本発明の種々の実施形態の活動に実質的に影響を与えない改変もまた、本明細書において提供される発明の定義内に含まれることが理解される。従って、以下の実施例は、本発明の例示であり、限定を意図しない。
【実施例】
【0078】
(実施例I:システイン含有ペプチドの固体支持体への付着および回収)
本実施例は、システイン含有ペプチドのビーズへの付着および光切断による改変されたペプチドの回収を記載する。
【0079】
システイン含有ペプチドの固体支持体ビーズへの付着および光切断を介した改変されたペプチドの回収を例示する模式図を図1Cに示す。方法は、固相同位体タグ化試薬を用いてシステインペプチドの部位特異的で、安定同位体標識となるように開発した(図1)。o−ニトロベンジルベースの光切断可能なリンカーを第一にアミノプロピルコーティングしたガラスビーズを固相ペプチド合成によって付着させた(Holmes and Jones,J.Org.Chem.60: 2318−2319 (1995))。次に、同位体タグである7つの水素(d0)または7つの重水素(d7)を有するロイシン分子を、再び固相ペプチド合成によってこの光切断可能なリンカーに付着させた(Holmes and Jones,前出,1995)。最後に、スルフヒドリル特異的ヨードアセチル基を付着させた。
【0080】
システイン含有ペプチドの捕捉および改変を実証するために、配列CDPGYIGSR(配列番号1;分子量967)を有する市販のラミニンBペプチドを用いた。手短には、1molのシステイン含有ラミニンBペプチド(CDPGYIGSR;配列番号1)1nmolおよびシステインを含有しないホスホアンギオテンシン(DRVY*IHPF;配列番号2、アステリスクはリン酸化チロシンを示す)500pmolからなるサンプルを用いた。ペプチドを0.2M Tris(pH8.0)、10mMエチレンジアミンテトラ酢酸(EDTA)の100μl中の5mM tris(カルボキシエチル)ホスフィン(TCEP)で30分間室温にて還元した。
【0081】
このペプチドを逆相HPLC上で液体クロマトグラフィーおよび質量分析(LC−MS)によって分析した。このペプチドをtris(2−カルボキシエチル)ホスフィン(TCEP)で還元し、二重に荷電したイオンについて、9.55分においてm/z=484.6の質量荷電比(m/z)で検出される単一のピークが検出された(図2を参照のこと)。11.94分において溶出される第二のピークは、コントロールとしてサンプルに加えられた別の標準的なペプチド(すなわちホスホアンギオテンシン、配列DRVYIHPF (配列番号2、分子量1126)を伴う)である。ホスホアンギオテンシンは、システインを含まず、そして予測された質量を伴って観察された。
【0082】
制御された孔ガラスの固相支持体ビーズを、アミノ官能性を光分解可能なリンカーに共有結合させることによって改変した(図1)。光分解可能なリンカーをシラン結合を介して固体支持体に付着させた。この反応性基は、ヨードアセチル基であり、このヨードアセチル基は、スルフヒドリル基と特異的に反応する。アミノ基で官能化されたガラスビーズ(Sigma Aldrich;St Louis MO)を固体支持体として用いたFmoc保護された光リンカー(4− [4− (l− (Fmocアミノ)エチル)−2−メトキシ−5−ニトロフェノキシ)ブタン酸またはFmoc−アミノエチル−光リンカー;NovaBiochem,Merck KGaAの関連会社 ;Darmstadt Germany)およびロイシンを、標準的な固相ペプチド合成手順のもとで、カルボジイミド化学を介してアミノ基官能化されたビーズに特異的に付着させた。次いで、ロイシンにおけるFmoc保護は、ピペリジン処理によって取り除き、ロイシンの遊離αアミノ基を、無水ヨード酢酸と反応させて、反応性ヨードアセチル基を生成させた。
【0083】
手短には、固相同位体標識ビーズを、以下のように合成した。他に言及しない限り、化学物質は、Aldrich(Milwaukee,WI)から購入した。第一に0.5gのアミノプロピルコーティングされた制御された孔ガラスビーズ(アミン含量約400μmol/g;Sigma,St.Louis,MO)を無水ジメチルホルムアミド(DMP)で洗浄した。次いで、各々600μmolの1−ヒドロキシベンゾトリアゾール(HOBt;Nova Biochem,Laufelfingen,Switzerland)、Fmoc−アミノエチル光リンカー(Nova Biochem)およびジイソプロピルカルボジイミド(DIC)を、室温で30分にわたり混合した。そして、この混合物を、90分間ビーズに加えた。次いで、ビーズを順番にDMFおよびジクロロメタンを用いて洗浄し、ジクロロメタン中で2mlの40%無水酢酸/60%ピリジンを用いて30分間洗浄した。ビーズは、DMFで再び洗浄し、そして30分間DMF中の20%ピペリジンで処理してFmoc保護を取り除いた。このプロセスを反復してFmoc−ロイシン(Nova Biochem)を同位体タグとして付着した。最後に、ヨードアセチル基を、以前に記載されるように(Zhouら,Nat.Biotechnol.19: 375〜378(2001))ビーズに付着させた。ビーズを、DMF、水およびメタノールの順に洗浄し、減圧下で乾燥させ、そして暗所にて室温で保存した。重同位体を有するビーズの合成のために、Fmoc−d7−ロイシンをd7−ロイシン(Isotec,Miamisberg,OH)およびFmoc−N−ヒドロキシスクシンイミド(Nova Biochem)から、Fmoc−ロイシンを精製する再結晶化工程が省略されることを除き、本質的に以前に記載(Lapatsanisら,Synthesis 671−673 (1983))されるように調製した。
【0084】
上記のように調製された5μgのビーズ(2μmolの結合能力)を使用して、一定の撹拌下で100μlの体積中でペプチドを捕獲した。上清のアリコート(1μl)を、μLC−MSのためにビーズの添加前にそしてそれらの添加後の異なる時点で、反応混合物から除去した。15分のインキュベーションの後、ビーズを、2μlのβ−メルカプトエタノールの添加によって5分間不活性化し、そして2.0Mの塩化ナトリウム、メタノールおよび水で連続的に洗浄した。光切断のために、ビーズを、100μlの0.2M Tris(pH8.0)、10mM EDTA、2% β−メルカプトエタノールで再懸濁した。リン酸アンギオテンシン(500pmol)を、内部標準として添加した。Blak−Ray長波UVランプ(100W、VWR Scientific、West Chester、PA)からの光を、10%硫酸銅(II)溶液(1cmパッチ長(pach length))によって濾過し、そして10cmの距離から照射するために使用した。照射のことなる時点で、1μlのアリコートを、μLC−MS分析のために上清から取り出した。ビーズを、均一な光照明に曝露するために時折撹拌した。光切断の緩衝液でのβ−メルカプトエタノールの使用は、光切断の間の可能なメチオニン酸化を提供する。
【0085】
ビーズとペプチドをインキュベートした後、図1に示されるように、上清のアリコートを、再びLC−MSいよって分析した。データを図3に示す。予期されたように、非システイン含有ホスホアンギオテンシン(phosphoangiotensin)ペプチドのみが上清中に存在するが、システイン含有ラミニンBペプチドは、溶液から完全に消失し、これはおそらくビーズによる量的な捕獲に起因する。
【0086】
2Mの塩化ナトリウム、メタノールおよび水でビーズを洗浄した後、ビーズを0.2M Tris、pH8.0、10mM エチレンジアミン四酢酸(EDTA)を含む緩衝液に再懸濁し、同じ濃度のホスホアンギオテンシンペプチドで再びコントロールとしてスパイクした。光切断の後、予測された質量改変([M+2H]2+イオンに対してm/z=569)を有するラミニンBペプチドを、初期のラミニンBペプチドと比較して、わずかに遅い溶出時間で観測した。より長い保持時間は、転移したロイシンアミノ酸の付加に起因して、予測された光切断産物の減少した極性と一貫する。光切断産物の構造を、MS/MS配列分析によってさらに確認した。改変したラミニンBペプチドの相対的なピーク強度およびホスホアンギオテンシン標準は、光切断の有効性の概算を与えた。ラミニンBペプチドの本来の入力の少なくとも50%で、回収した。収率を、反応物の捕獲および放出のさらなる最適化によって改善し得る。
【0087】
反応物の捕獲および放出の有効性を説明するために、システイン含有ラミニンBペプチドおよび非システイン含有ホスホアンギオテンシンからなる混合物を使用した。ラミニンBを、固相上へと量的に捕獲した。光切断の1時間後、タグ化ラミニンBを回収した;予測された質量改変(+170Da)(これはまた、MS/MSによって確認された)は、システイン残基へのロイシンタグの添加に起因することが示された。ロイシンタグの疎水性の性質は、非タグ化形態と比較してタグ化ラミニンBの保持時間を増加する。ホスホアンギオテンシンの同量と対照する非タグ化ラミニンBおよびタグ化ラミニンBのシグナル強度は、特定の捕獲を示し、そしてタグ化ラミニンBの回収をほとんど回収した。より長い光切断時間は、収率またはタグ化ラミニンBの量のいずれにも影響せず、光触媒化副反応は、いずれの実質的な程度を生じないことが示された。
【0088】
これらの結果は、アミノ酸ロイシンの捕獲−放出化学を介するペプチドのサルファイドリル側鎖への成功した転移を照明する。これらは、選択的捕獲ペプチドおよび官能基(例えば、ペプチドに対するアミノ酸)の転移への例示的な一般的なアプローチを結果として生じる。
【0089】
(実施例II:スルフヒドリル基を組み込むためのポリペプチドのアミノ基の改変)
この実施例は、スルフヒドリル基を組み込むためのペプチドのN末端改変を記載する。
【0090】
ペプチドがシステイン残基を含んでいない場合、そのペプチドを改変して、スルフヒドリル基を組み込み得、このことによりアミノ基をスルフヒドリル基に変換し、組み込まれたスルフヒドリル基を介してペプチドを捕捉することが可能である。ポリペプチドのアミノ基の改変のためのストラテジーを、図5に示す。
【0091】
ホスホアンジオテンシンを使用して、アミノ基をスルフヒドリル基に改変する原理を示した。このペプチドを最初にN−スクシンイミジルS−アセチルチオアセテート(SATA)(これは、1級アミノ基に対して特異的に反応する)により改変した。ホスホアンジオテンシンを、図5に示すように改変した。簡潔には、ホスホアンジオテンシンを10mM SATAと30分間、0.1M K3PO4(pH8.0)中で室温にてインキュベートした。
【0092】
改変したホスホアンジオテンシンを、LC−MSにより分析した。このペプチドのN末端改変を観察した。図6に示されるデータは、1つの産物が、14.5〜14.55分([M+2H]2+イオンについてm/z=622.3)で溶出されることを示す。測定された質量は、SATA誘導体化ホスホアンジオテンシンペプチドについての計算された質量(m/z=622.3)と十分に相関した。
【0093】
改変したペプチドを、0.1M ヒドロキシルアミンを用いてpH8にて2時間処理し、続いて、5mM TCEPで30分間還元した。図5に示されるように、ヒドロキシルアミンによる処理およびTCEPによる還元は、SATA誘導体化ホスホアンジオテンシンにおいて遊離SH基を生成する。このように処置された化合物のLC−MS分析からのデータを、図7に示す。再び、13.83分に溶出する1つの産物を観察した。MS分析により、[M+2H]2+イオンについてm/z=601が示された。測定した質量は、予測されたペプチド産物についての計算された質量と一致する(m/z=601)。従って、ペプチドのアミノ基の遊離SH基への変換は、定量的に、最小量の副産物とともになされ得る。さらに、このようなスルフヒドリル基改変ポリペプチドは、実施例Iにおいて記載されるように、引き続いて捕捉され得る。
【0094】
これらの結果は、ポリペプチドのアミノ基がスルフヒドリル基を組み込むために、本質的に定量的に改変され得ることを示す。
【0095】
(実施例III:重い標識および軽い標識の組み込みによるタンパク質サンプルの分析)
この実施例は、質量分析法によるサンプルペプチドの定性的かつ定量的分析のための、2つのサンプルの差示的標識を記載する。
【0096】
ポリペプチドの差示的標識の模式図を、図8に示す。サンプルを、トリプシンのようなプロテアーゼで消化する。消化したポリペプチドを、実施例Iに記載のように処理する。所望であれば、そのサンプルポリペプチドを、実施例IIに記載の方法により改変して、システインを欠くポリペプチドを試験し得る。
【0097】
1つの消化サンプルを、ロイシンのような非変性アミノ酸または別の適切なアミノ酸を含む化学基に共有結合させたビーズに添加する。第2のサンプルを、その化学基が同じアミノ酸の変性された形態を含むことを除いて、化学的に同一の基に共有結合させたビーズに添加する。差示的に同位体標識したロイシンは、ロイシンの変性の状態に依存して、7または10質量単位異なる。
【0098】
サンプルポリペプチドを、差示的に同位体標識したアミノ酸を含むそれぞれのビーズに共有結合的にカップリングした後、そのビーズを合わせ、次いで、光切断により切断する。光切断したポリペプチド(ここでは、差示的に標識されている)を、質量分析法により同時に分析する。2つのサンプルからのMSシグナルの強度を直接比較することにより、サンプル定量のためにMSを使用し得る。この2つのサンプルは、所定の質量が異なり、所定の質量が異なる二重のピークとして、MSで容易に区別され得る。さらに、このサンプルを、MSにより配列決定し得る。
【0099】
この実施例は、質量分析法によりサンプル分子の差示的標識および分析を実証する。
【0100】
(実施例IV:捕捉されたホスホペプチドの改変)
この実施例は、ポリペプチドの捕捉および捕捉されたホスホペプチドの改変を記載する。
【0101】
サンプルからのポリペプチドを、本質的に実施例Iに記載されるようにビーズ上に捕捉する。この捕捉されたポリペプチドを、本質的にZhouら、Nature Biotechnol.19:375−378(2001)(本明細書中に参考として援用される)に記載されるように改変する。
【0102】
簡潔には、この捕捉されたポリペプチドを、以下の工程により改変する。(1)アミノ保護:ペプチドのアミノ基を、必要に応じて、t−ブチル−ジカーボネート(tBoc)化学物質を用いて保護し、その後の反応における分子内縮合および分子間縮合の可能性を排除する。(2)縮合反応:カルボジイミドは、ペプチドと過剰なアミンとの間の縮合反応を触媒して、それぞれ、ペプチドのカルボキシレート結合およびホスフェート結合にて、アミド結合およびホスホルアミダイト結合を形成する。(3)ホスフェート再形成:遊離のホスフェート基をホスホルアミダイト結合の短時間の酸加水分解により再形成する。(4)縮合および還元:カルボジイミド触媒縮合反応は、再形成されたホスフェート基にシスタミンを結合させる。シスタミンの内部ジスルフィドの還元は、次に、捕捉されたホスホペプチドのホスフェート全てに対して遊離のスルフヒドリル基を生成する。(5)改変ポリペプチドの放出:捕捉されたポリペプチドを、例えば、実施例Iに記載のように、光を用いて、化学的切断基の切断により、固体支持体から放出させる。(6)固相捕捉:この放出されたポリペプチド(遊離のスルフヒドリル基を含む改変ホスホポリペプチドを含む)を、このペプチド中の遊離のスルフヒドリル基とガラスビーズ上に固定化されたヨードアセチル基とを反応させることにより、第2の固相に結合させる。(7)ホスホペプチドの回収:樹脂のストリンジェントな洗浄後に、ホスホペプチドを、tBoc保護基をも除去する濃度にて、トリフルオロ酢酸(TFA)を使用してホスホルアミダイト結合を切断することにより回収する。従って、遊離のアミノ基とホスフェート基を用いてペプチドを再生する。カルボキシレート基は、工程(2)からブロックされたままである。
【0103】
化学反応を、以下に詳細に記載するように行う。捕捉されたポリペプチドを含む固相を、50%(vol/vol)の0.1M リン酸緩衝液(pH11)およびアセトニトリル中でインキュベートする。0.1M tBocを、室温にて4時間添加する。次いで、アセトニトリルを除去する。捕捉されたポリペプチドを含む固相を、1M エタノールアミン、25mM N−ヒドロキシルスクシンイミド(NHS)、および0.5M N,N’−ジメチルアミノプロピルエチルカルボジイミドHCL(EDC)中でインキュベートし、室温で2時間インキュベートする。10%TFAを添加し、室温にて30分間インキュベートする。この固相を洗浄して、過剰な試薬を除去し、脱塩し、1M イミダゾール(pH6.0)を添加する。0.5M EDCを室温にて3時間添加する。この固相を洗浄し、次いで、1M シスタミン(pH8.0)と50℃にて2時間インキュベートする。この固相を水で洗浄し、次いで、10mM ジチオスレイトール(DTT)で還元して、遊離のスルフヒドリル基を生成する。この固相を洗浄してDTTを除去し、次いで、捕捉された分子を切断可能な官能基の切断により、放出する。
【0104】
放出されたポリペプチド(スルフヒドリル基で改変されたホスホペプチドを含む)を、第2の型の固相ビーズ(このビーズは、ヨードアセチル基を有し、1M Tris(pH8.0)、50mM エチレンジアミン四酢酸(EDTA)でpH8.0まで滴定される)と少なくとも2時間インキュベートする。固定化されたヨードアセチル基を有するビーズは、3当量の無水ヨード酢酸と1当量のアミノビーズ(G4643;Sigma;St.Louis MO)との間の、ジメチルホルムアミド(dimethylformide)中の3.3当量のジイソプロピルエチレンアミンを用いた2時間の反応により調製される。カルボジイミドによるチロシン付加は、あり得る副反応であるので、スルフヒドリル基で改変された捕捉されたホスホペプチドを、1M ヒドロキシルアミン(pH10)中で室温にて2時間インキュベートして、全ての改変されたチロシンを元に戻す。次いで、2M NaCl、メタノールおよび水で連続してビーズを洗浄して、非特異的に結合した分子を除去する。このビーズを100% TFAと30分間インキュベートして、ホスホペプチドを回収すると同時に、tBoc改変基からtBoc保護を外す。次いで、例えば、質量分析法により、回収したホスホペプチドを分析する。
【0105】
第1固体支持体上に捕捉された分子は、一般的に、第2固相上に再捕捉される直前に放出され、この第2固相は、改変ホスホペプチドを選択的に捕捉し、このことにより、有効な洗浄、および捕捉されたポリペプチドを改変するために使用される化学物質の除去が可能となる。しかし、第1固相への結合が、標識またはタグを、捕捉されたポリペプチドに転移するようにのみ意図される場合、このペプチドは、より初期の段階(始めの捕捉の後でさえあれば)で放出され得る。あるいは、ホスホペプチドの改変を、本質的にZhouらに記載されるように実行し得、次いで回収されたホスホペプチドは、本質的に実施例Iに記載されるように、捕捉されそして標識される。
【0106】
この実施例は、ホスホペプチドの選択的な改変および単離を実証する。
【0107】
(実施例V:酵母タンパク質の単離および同位体標識)
この実施例は、固相の捕捉および放出のプロセスを介した、複雑なペプチド混合物中のシステイン性ペプチドの部位特異的な、安定同位体標識のための方法、および標識ペプチドの同時性の単離を記載する。
【0108】
酵母のタンパク質を、固相同位体タグ化試薬からの捕捉および放出によって、差次的に同位体標識した。回収されたペプチドを、マイクロキャピラリー液体クロマトグラフィーおよびタンデム質量分析(μLC−MS/MS)によって分析して、それらの配列および相対質量を決定した。この方法を使用して、酵母Saccharomyces cerevisiae中の、タンパク質存在比におけるガラクトースで誘導された変化を検出した。同位体コード親和性タグ(ICAT)方法(Gygiら、前出、1999)との横並び(side−by−side)比較は、ペプチドの安定同位体タグ化のための固相法が、比較的より簡単であり、より効果的であり、そしてより感度がよいことを実証した。
【0109】
比較される2つのサンプル由来のシステイン性ペプチドを、同位体的に重いタグまたは通常のタグを含有する固相上に、共有結合で捕捉した。次いで、このビーズを混ぜ合わせ、洗浄し、そしてUV光に曝した(360nm、任意の起こり得る光触媒性の副反応を最小限にするように選択される)。このことは、リンカーの光切断、およびシステイン残基の側鎖上への固相からの同位体タグの転移を生じた。最終的に、回収されたタグ化ペプチドを、本質的に以前に記載されるように、μLC−MS/MSによって分析して、各々のペプチドの配列および相対存在比を決定した(Gygiら、前出、1999)。
【0110】
安定同位体タグ化は、質量分析による、タンパク質の定量分析のための一般的なストラテジーである(Gygiら、前出、1999;Odaら、Proc.Natl.Acad.Sci.USA 96:6591−6596(1999);Idekerら、Science 292:929−934(2001);Hanら、Nat.Biotechnol.19:946−951(2001))。固相アプローチの性能を、標準的なICATアプローチ(Gygiら、前出、1999)の性能と比較するために、ガラクトースを用いた誘導への応答において、この2つの方法を使用して、酵母S.cerevisiae中のタンパク質発現の変化を検出する、横並び比較を実施した。同じの酵母のタンパク質の多量のサンプルの装填については100μg、少量のサンプルの装填については10μgの、2つの異なる量の開始時のタンパク質物質を評価した。この目的は、標識化ストラテジーの相対的な性能を比較することであり、そしてペプチドの分離ストラテジーでもタンパク質の同定ストラテジーでもないので、タンパク質同定のため、全てのサンプルに対して1回のLC−MS/MS実施を行った。このことは、サンプルの標識化よりもむしろサンプルの処理に起因して、結果についての多様性を最小限にしたが、おそらくは、より多量のサンプルサイズおよびLC−MS/MSより前の最適化されたペプチド分離を用いて得られ得るよりも、より少ないタンパク質の同定を生じた。
【0111】
酵母のタンパク質サンプルの調製のために、酵母BY4742株を100mlのYPR(1%酵母抽出物、2%ペプトン(Difco,Detroit,MI)および2%ラフィノース)またはYPR+2%ガラクトースのいずれかで、1のA600値まで増殖させた。以前に記載されるように(Ausubelら、前出)スフェロプラストを調製した。スフェロプラストを50mM Tris(pH8.0)、5mM EDTA、6M 尿素、0.5% SDS中で溶解した。細胞溶解物を15,000g(14,000rpm)で15分間遠心分離した。この上清を回収し、そして50mM Tris(pH8.0)、5mM EDTA中で、Econo−Pac 10DGカラム(Bio−Rad,Hercules,CA)で脱塩した。タンパク質濃度を、Bio−Radタンパク質アッセイによって決定した。
【0112】
固相の捕捉−放出方法による、酵母タンパク質のトリプシン消化物の単離および同位体標識化のために、多量スケールおよび小規模実験を実施した。多量スケールの実験について、ガラクトースの添加または無添加で増殖した酵母細胞からの50μgの各々のタンパク質抽出物(全部で100μg)を、100μlの200mM Tris(pH8.0)、5mM EDTA中に調製した。各々のタンパク質抽出物を、5μgのトリプシンによって37℃で3時間消化し、そして5mM TCEPで還元し、次いでシステイン性ペプチドを、実施例Iに記載されるように、d0−ロイシンタグまたはd7−ロイシンタグのいずれかを有するビーズによって15分間捕捉した。このビーズを混ぜ合わせて洗浄し、そして標識されたペプチドを2時間の光照射によって放出した。この放出されたペプチドを、MCXカラム(Waters,Milford,MA)に充填し、そして4mlの0.1%トリフルオロ酢酸(TFA)、4mlの80%アセトニトリル/0.1%TFA、および水(中性化のため)で連続的に洗浄した。ペプチドを、9容量のメタノールと1容量の28%アンモニアとの混合物1mlによって溶出し、そして減圧下で乾燥した。乾燥したペプチドを、μLC−MS/MS分析のために水に再懸濁した。次に、20%の回収されたペプチド(20μgの全タンパク質を示す)を、以前に記載されるように(Zhouら、Nat.Biotechnol.19:375−378(2001))、LCQイオントラップ質量分析(Finnigan,San Jose,CA)を使用するμLC−MS/MSによって分析した。タンパク質の同定および定量を、Sequestおよび既存のソフトウェア(Hanら、Nat.Biotechnol.19:946−951(2001);Engら、J.Am.Soc.Mass.Spectrom.5:976−989(1994))を用いて実施した。小規模実験のために、ガラクトースの添加または無添加で増殖した細胞からの、5μgの各々のタンパク質抽出物を、0.5μgのトリプシンによって消化し、そして上記のように処理し、次いで回収したペプチド混合物の50%(5μgの全タンパク質抽出物を示す)を、同様のμLC−MS/MS方法によって分析した。
【0113】
ICATによって酵母のタンパク質の同位体標識化およびペプチドの単離のために、ガラクトースの添加または無添加で増殖した細胞からの、100μgの各々のタンパク質抽出物を、200mM Tris(pH8)、5mM EDTA、6M 尿素、0.05% SDSを含む標識化緩衝液中に、出発物質として調製した。タンパク質を、5mM TCEPで30分間還元し、そして100μgのd0−ICATまたはd8−ICATを、それぞれ、ラフィノースで増殖した細胞、またはガラクトースで増殖した細胞からのタンパク質抽出液に添加した。室温で90分の標識化の後、この反応物をβ−メルカプトエタノールの10mMまでの添加によってクエンチし、そして混ぜ合わせた。各々のサンプルを、20mM Tris(pH8.3)、0.01% SDSで10倍に希釈した。トリプシン(10μg)を添加して、タンパク質を3時間37℃で消化した。このサンプルを等容量の緩衝液A(5mM KH2PHO4(pH3)、25% CH3CN)で希釈し、そしてそのpHを、TFAを用いてpH3に調整した。100μg(大規模実験)または10μg(小規模実験)のいずれかの混合したタンパク質消化物を、緩衝液Aで平衡化した陽イオン交換カートリッジ(Applied Biosystems,Foster City,CA)に適用した。このカートリッジを2mlの緩衝液A、その後2mlの緩衝液A+40mM KClで洗浄した。結合したペプチドを、600μlの緩衝液A+600mM KClで溶出した。サンプルの容量を減圧下で300μlに減らし、そして500μlの2×PBSおよび12μlの1M NH4HCO3を添加した。サンプルを、モノマーのアビジンカートリッジ(Applied Biosystems)に通し、そして2mlの2×PBS、1mlの1×PBS、および1mlの20%メタノールを含む50mM NH4HCO3で洗浄した。標識ペプチドを、1mlの30%アセトニトリルを含む0.4% TFAで溶出し、減圧下で乾燥し、そして10μlの5%アセトニトリルを含む0.4%酢酸に再懸濁した。大規模実験または小規模実験のどちらかについて、固相の方法において使用した方法と同一のμLC−MS/MS方法によって、同容量のサンプルを分析した。
【0114】
小規模実験および大規模実験の両方において、固相法によって同定および定量されたタンパク質の数は、ICATによる数よりも多かった(図9)。実際に、固相アプローチはより感度が高く、従来のICATによって識別されるタンパク質の大部分のタンパク質、さらにICATによっては同定されない他の多くを同定した(図9A、9B)。複数の実験において同定された同位置のタンパク質の定量もまた一致した。従って、タンパク質定量は、同位体タグの構造によっても固相法の捕捉および開放によっても影響されなかった。
【0115】
ガラクトースは、ガラクトース利用に関連するいくつかの遺伝子の発現を強力に誘導することが既知であり、これら遺伝子としては、ガラクトキナーゼ(GAL1)、ガラクトースパーミアーゼ(GAL2)、ガラクトトランスフェラーゼ。(GAL7)およびUDP−グルコース−4−エピメラーゼ(GALX)が挙げられる(JohnstonおよびCarlson、The Molecular and Cellular Biology of the Yeast Saccharomyces、Jonesら、編、193−281頁、Cold Spring Harbor Press、Cold Spring Harbor、NY(1992))。ガラクトースでの誘導後、GAL1、GAL2、GAL7およびGALXを含むタンパク質由来の複数のペプチドを固相法によって同定および定量した。対照的に、ICAT方法を使用してGAL1タンパク質由来の1つのペプチドのみを、小規模実験および大規模実験の両方において同定し、そして固相を用いるよりも、より低いシグナル対ノイズ比を検出した。これらのデータは、ICATアプローチを超える、固相の優勢なサンプル回収および感度を確認した。さらに、小規模実験および大規模実験によって識別されたタンパク質の実質的な重複によって、固相法の再現性を確認した(図9C)。
【0116】
これらの結果は、固相法が、定量性タンパク質分析について、簡単であり、再現性があり、有効であり、そして感度が高いことを示す。実際に、ICAT方法と比較して、この方法はいくつかの利点を有する。第1に、システイン含有ペプチドの単離、および同位体の安定な組み込みの両方が、本質的に1工程で達成される。従って、固相法は、ICATアプローチよりも、より迅速かつより簡単で、より少ない手動操作を必要とする。第2に、固相へのペプチドの共有結合性の捕捉により、非共有結合性に会合した分子を除去するための、ストリンジェントな洗浄条件の使用が可能となる。実際に、本明細書に提示される実験は、最小限の手動操作に起因して、ほぼ排他的なシステイン性ペプチドの回収を生じた。第3に、この手順は、トリプシンのようなタンパク質分解性酵素の存在によっても、尿素およびSDSのような強力な変性剤および界面活性剤の存在によっても、影響されない。従って、このような分子を除去するためのさらなる工程の必要性が存在しない。最低限のサンプル操作に起因して、この固相法は、ICATよりも感度が高い。多くの生物学的に有益な事象は、相対的に低い存在比の調節タンパク質に関するので、固相法は、このようなタンパク質に誘導される、変化の分析に有用である。第4に、カップリング工程に関係する標準的な固相ペプチド化学は、本明細書で使用されるd0/d7−ロイシンの場所に、同位体質量タグとして、ある範囲の天然または非天然のアミノ酸の使用を可能とする。このことは、1つの実験における複数のサンプル(2つより多い)の分析のために、ある範囲の質量タグを有するビーズの合成に役立ち得る。第5に、本明細書で使用されるシステイン上の質量タグは、d0−ロイシンタグについては170Daの重量である。ICATで標識されたペプチドとは対照的に、タグのサイズの小ささ、およびタグの化学的性質に起因して、MS/MSモードにおいて観察されるペプチドのフラグメント化は、標識自体の望ましくないフラグメント化によって複雑化されることがない(Hanら、前出、2001)。最後に、光切断の前に、共有結合で固定化されたペプチドは、所望される場合、さらなる化学反応および酵素反応のための理想的な基質を提供する(Zhouら、前出、2001)。
【0117】
固相法がICAT方法と異なる1つの重要な方法は、固相試薬がタンパク質分解の後にペプチドを標識するが、一方、ICATにおいては、タンパク質はタンパク質分解の前に標識されることである。従って、ICATアプローチは、標識されたタンパク質の分離(例えば、ゲル電気泳動)が必要とされる場合に好ましい。上記の固相法は、定量的プロテオミクスへの一般的な適用に適切なツールを提供し、そして自動化された装置に利用可能である。従って、固定化方法は、質量分析による定量的タンパク質分析のための安定同位体タグ化のさらにより広範な適用に向けられる工程を表す。
【0118】
これらの結果は、定量的プロテオーム分析に適切な複雑なタンパク質混合物の差次的な同位体標識化を実証する。
【0119】
この出願全体にわたって、種々の刊行物が引用されている。これらの刊行物の開示は、本明細書によって、本発明が関する技術分野の状態をより完全に記載するために、本明細書における参考としてその全体が援用される。
【0120】
本発明は、開示された実施形態に関して記載されたが、当業者は、詳説された特定の実験が本発明の単なる例示であることを容易に認識する。種々の改変が、本発明の精神から逸脱すること無くなされ得ることが、理解されるべきである。
【産業上の利用可能性】
【0121】
本発明により、例えば、以下が提供される:
(項1)
分子を標識する方法であって、以下の工程:
(a)サンプル分子の、該サンプル分子についての反応性基への共有結合を可能にする条件下で、該サンプル分子と、化学基に結合された固体支持体とを接触させる工程であって、該化学基は、切断可能な官能基、1つ以上の官能基、および該反応性基を含む、工程;ならびに
(b)該切断可能な官能基を切断する工程であって、それにより該1つ以上の官能基を含む該サンプル分子を放出する、工程、
を包含する、方法。
(項2)
前記サンプル分子が、ポリペプチド、核酸、脂質、セカンドメッセンジャー、および代謝産物からなる群より選択される、項1に記載の方法。
(項3)
前記サンプル分子が、ポリペプチドである、項1に記載の方法。
(項4)
項3に記載の方法であって、前記ポリペプチドが、リン酸化、グリコシル化、ユビキチン化、アセチル化、プレニル化、パルミチル化、ミリスチル化、硫酸化、およびヒドロキシル化からなる群より選択される改変を有する、方法。
(項5)
前記ポリペプチドが、ホスホポリペプチドである、項4に記載の方法。
(項6)
前記固体支持体が、ガラスビーズである、項1に記載の方法。
(項7)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項1に記載の方法。
(項8)
前記官能基の1つが、タグである、項1に記載の方法。
(項9)
前記タグが、質量分析タグである、項8に記載の方法。
(項10)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項8に記載の方法。
(項11)
前記タグが、安定同位体をコードするアミノ酸である、項10に記載の方法。
(項12)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項11に記載の方法。
(項13)
前記タグが、気相塩基性基または疎水性基である、項8に記載の方法。
(項14)
前記気相塩基性基が、ピリジルである、項13に記載の方法。
(項15)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項8に記載の方法。
(項16)
前記官能基の1つが、特徴的な同位体分配を有する元素を含む、項8に記載の方法。
(項17)
前記元素が、塩素または臭素である、項16に記載の方法。
(項18)
前記化学基の前記反応性基が、スクシンイミドエステル基およびヨードアセチル基からなる群より選択される、項3に記載の方法。
(項19)
前記ポリペプチドの第一級アミン基がN−スクシンイミジル S−アセチルチオアセテート、ヒドロキシルアミン、およびトリス(2−カルボキシエチル)ホスフィンを用いる処理によって改変されている、項3に記載の方法。
(項20)
前記ポリペプチドが、該ポリペプチドの前記改変に対する特異的な結合活性を有する抗体を用いて単離されている、項4に記載の方法。
(項21)
前記方法の工程が、自動化プロセスにより実施される、項1に記載の方法。
(項22)
前記固体支持体に接触された前記サンプル分子の少なくとも50%が、放出される、項1に記載の方法。
(項23)
サンプル分子を分析するための方法であって、以下の工程:
(a)サンプル分子の、該サンプル分子についての反応性基への共有結合を可能にする条件下で、該サンプル分子と、化学基に結合された固体支持体とを接触させる工程であって、該化学基は、切断可能な官能基、1つ以上の官能基、および該反応性基を含む、工程;
(b)該サンプル分子を該固体支持体から切断する工程であって、ここで1つ以上の特定の官能基が、該放出されたサンプル分子に移動する、工程;ならびに
(c)該放出されたサンプル分子を分析する工程、
を包含する、方法。
(項24)
前記放出されたサンプル分子が、質量分析法により分析される、項23に記載の方法。
(項25)
細胞または組織により発現された、複数のクラスの分子が分析される、項23に記載の方法。
(項26)
前記サンプル分子が、ポリペプチド、核酸、脂質、セカンドメッセンジャー、および代謝産物からなる群より選択される、項23に記載の方法。
(項27)
前記サンプル分子が、ポリペプチドである、項26に記載の方法。
(項28)
項27に記載の方法であって、前記ポリペプチドが、リン酸化、グリコシル化、ユビキチン化、アセチル化、パルミチル化、プレニル化、硫酸化、ヒドロキシル化およびミリスチル化からなる群より選択される改変を有する、方法。
(項29)
前記ポリペプチドが、ホスホポリペプチドである、項28に記載の方法。
(項30)
前記固体支持体が、ガラスビーズである、項23に記載の方法。
(項31)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項23に記載の方法。
(項32)
前記官能基の1つが、タグである、項23に記載の方法。
(項33)
前記タグが、質量分析タグである、項32に記載の方法。
(項34)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項32に記載の方法。
(項35)
前記タグが、安定同位体をコードするアミノ酸である、項34に記載の方法。
(項36)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項35に記載の方法。
(項37)
前記タグが、気相塩基性基または疎水性基である、項32に記載の方法。
(項38)
前記気相塩基性基が、ピリジルである、項37に記載の方法。
(項39)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項32に記載の方法。
(項40)
前記官能基の1つが、特徴的な同位体分配を有する元素を含む、項32に記載の方法。
(項41)
前記元素が、塩素または臭素である、項40に記載の方法。
(項42)
前記化学基の前記反応性基が、スクシンイミドエステル基およびヨードアセチル基からなる群より選択される、項23に記載の方法。
(項43)
前記ポリペプチドの第一級アミン基が、N−スクシンイミジル S−アセチルチオアセテート、ヒドロキシルアミン、およびトリス(2−カルボキシエチル)ホスフィンを用いる処理によって改変されている、項27に記載の方法。
(項44)
前記ポリペプチドが、該ポリペプチドの前記改変に対する特異的な結合活性を有する抗体を用いて単離されている、項28に記載の方法。
(項45)
前記方法の工程が、自動化プロセスにより実施される、項23に記載の方法。
(項46)
前記固体支持体に接触された前記サンプル分子の少なくとも50%が、放出される、項23に記載の方法。
(項47)
2つ以上のサンプルからの分子が、比較して分析される、項23に記載の方法。
(項48)
前記サンプルが示差的に標識されている、項47に記載の方法。
(項49)
前記2つ以上のサンプルが、質量分析タグを用いて示差的に標識されている、項48に記載の方法。
(項50)
前記サンプルが、安定同位体タグ、同位体分布タグ、または荷電アミノ酸を用いて示差的に標識されている、項48に記載の方法。
(項51)
前記サンプルが、発蛍光団、発光団、またはスピン標識を用いて標識されている、項48に記載の方法。
(項52)
分子を標識する方法であって、以下の工程:
(a)サンプル分子の、該サンプル分子についての反応性基への共有結合を可能にする条件下で、該サンプル分子と、化学基に結合された固体支持体とを接触させる工程であって、該化学基は、切断可能な官能基、1つ以上の官能基、および該反応性基を含む、工程;
(b)該固体支持体に結合した該サンプル分子を改変する工程;ならびに
(c)該切断可能な官能基を切断する工程であって、それにより該1つ以上の官能基を含む該改変されたサンプル分子を放出する、工程、
を包含する、方法。
(項53)
前記改変する工程が、化学的改変または酵素的改変である、項52に記載の方法。
(項54)
前記サンプル分子が、ポリペプチド、核酸、脂質、セカンドメッセンジャー、および代謝産物からなる群より選択される、項52に記載の方法。
(項55)
前記サンプル分子が、ポリペプチドである、項52に記載の方法。
(項56)
項55に記載の方法であって、前記ポリペプチドが、リン酸化、グリコシル化、ユビキチン化、アセチル化、プレニル化、パルミチル化、ミリスチル化、硫酸化、およびヒドロキシル化からなる群より選択される改変を有する、方法。
(項57)
前記ポリペプチドが、ホスホポリペプチドである、項56に記載の方法。
(項58)
前記改変する工程が、前記ホスホポリペプチド上のホスフェート基を改変する、項57に記載の方法。
(項59)
前記固体支持体が、ガラスビーズである、項52に記載の方法。
(項60)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項52に記載の方法。
(項61)
前記官能基の1つが、タグである、項52に記載の方法。
(項62)
前記タグが、質量分析タグである、項61に記載の方法。
(項63)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項61に記載の方法。
(項64)
前記タグが、安定同位体をコードするアミノ酸である、項63に記載の方法。
(項65)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項64に記載の方法。
(項66)
前記タグが、気相塩基性基または疎水性基である、項61に記載の方法。
(項67)
前記気相塩基性基が、ピリジルである、項66に記載の方法。
(項68)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項61に記載の方法。
(項69)
前記官能基の1つが、特徴的な同位体分配を有する元素を含む、項61に記載の方法。
(項70)
前記元素が、塩素または臭素である、項69に記載の方法。
(項71)
前記化学基の前記反応性基が、スクシンイミドエステル基およびヨードアセチル基からなる群より選択される、項55に記載の方法。
(項72)
前記ポリペプチドの第一級アミン基がN−スクシンイミジル S−アセチルチオアセテート、ヒドロキシルアミン、およびトリス(2−カルボキシエチル)ホスフィンを用いる処理によって改変されている、項55に記載の方法。
(項73)
前記ポリペプチドが、該ポリペプチドの前記改変に対する特異的な結合活性を有する抗体を用いて単離されている、項56に記載の方法。
(項74)
前記方法の工程が、自動化プロセスにより実施される、項52に記載の方法。
(項75)
前記固体支持体に接触された前記サンプル分子の少なくとも50%が、放出される、項52に記載の方法。
(項76)
化学基に結合された支持体を含む構成物であって、該構成物は、切断可能な官能基、タグおよび反応性基を含み、該反応性基は、サンプル分子に共有結合され、ここで該切断可能な官能基、該タグおよび該反応性基は、該切断可能な官能基の切断の際に、該サンプル分子への該タグの移動を可能にするように互いに対して配置されている、構成物。
(項77)
項76に記載の構成物であって、前記サンプル分子が、ポリペプチド、核酸、脂質、セカンドメッセンジャー、および代謝産物より選択される、構成物。
(項78)
前記サンプル分子が、ホスホポリペプチドである、項77に記載の構成物。
(項79)
項78に記載の構成物であって、前記ポリペプチドが、リン酸化、グリコシル化、ユビキチン化、アセチル化、パルミチル化、プレニル化、硫酸化、ヒドロキシル化、およびミリスチル化からなる群より選択される改変を有する、構成物。
(項80)
前記ポリペプチドが、ホスホポリペプチドである、項79に記載の構成物。
(項81)
前記固体支持体が、ガラスビーズである、項76に記載の構成物。
(項82)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項76に記載の構成物。
(項83)
前記タグが、質量分析タグである、項76に記載の構成物。
(項84)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項76に記載の構成物。
(項85)
前記タグが、安定同位体をコードするアミノ酸である、項84に記載の構成物。
(項86)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項85に記載の構成物。
(項87)
前記タグが、気相塩基性基または疎水性基である、項76に記載の構成物。
(項88)
前記気相塩基性基が、ピリジルである、項87に記載の構成物。
(項89)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項76に記載の構成物。
(項90)
前記タグが、特徴的な同位体分配を有する元素を含む、項76に記載の構成物。
(項91)
前記元素が、塩素または臭素である、項90に記載の構成物。
(項92)
前記共有結合された反応性基が、スクシンイミドエステル基およびヨードアセチル基から誘導される、項76に記載の構成物。
(項93)
前記ポリペプチドの第一級アミン基がN−スクシンイミジル S−アセチルチオアセテート、ヒドロキシルアミン、およびトリス(2−カルボキシエチル)ホスフィンを用いる処理によって改変されている、項78に記載の構成物。
(項94)
化学基に共有結合された固体支持体を含む構成物であって、該化学基は、切断可能な官能基、質量分析タグおよびサンプル分子に共有結合するための反応性基を含み、該切断可能な官能基、該タグおよび該反応性基は、該切断可能な官能基の切断の際に、該サンプル分子への該タグの移動を可能にするように互いに対して配置されている、構成物。
(項95)
前記固体支持体が、ガラスビーズである、項94に記載の構成物。
(項96)
前記切断可能な官能基が、光、酸、塩基、または酵素により切断可能な化学リンカーである、項94に記載の構成物。
(項97)
前記タグが、安定同位体タグ、同位体分布タグ、および荷電アミノ酸からなる群より選択される、項94に記載の構成物。
(項98)
前記タグが、安定な同位体をコードするアミノ酸である、項97に記載の構成物。
(項99)
前記タグが、重水素化アミノ酸、または重水素化されていないアミノ酸である、項98に記載の構成物。
(項100)
前記タグが、気相塩基性基または疎水性基である、項94に記載の構成物。
(項101)
前記気相塩基性基が、ピリジルである、項100に記載の構成物。
(項102)
前記タグが、発蛍光団、発光団、およびスピン標識より選択される、項94に記載の構成物。
(項103)
前記タグが、特徴的な同位体分配を有する元素を含む、項94に記載の構成物。
(項104)
前記元素が、塩素または臭素である、項103に記載の構成物。
(項105)
前記化学基の前記反応性基が、スクシンイミドエステル基およびヨードアセチル基からなる群より選択される、項94に記載の構成物。
【特許請求の範囲】
【請求項1】
本明細書に記載の方法および構成物。
【請求項1】
本明細書に記載の方法および構成物。
【図1A】



【図1B】
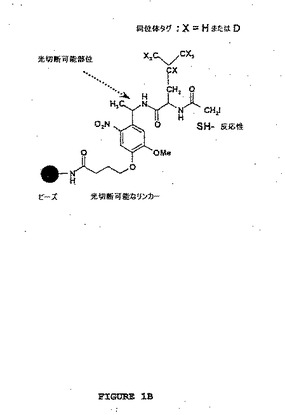
【図1C】
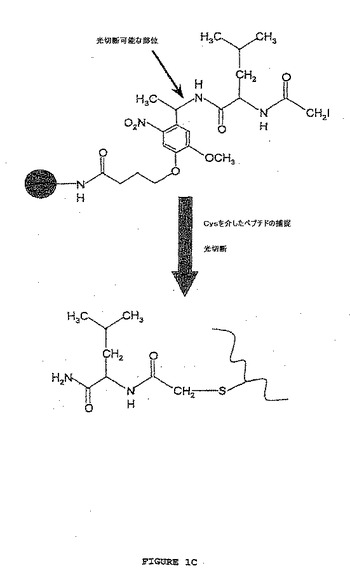
【図2A】
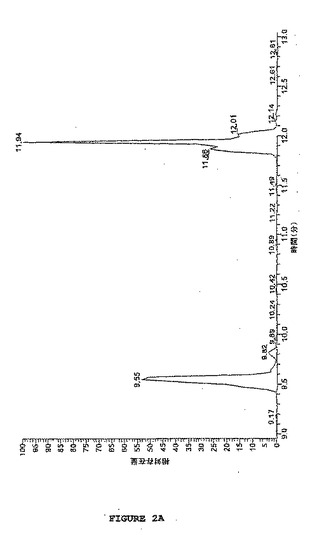
【図2B】

【図3A】

【図3B】

【図4A】
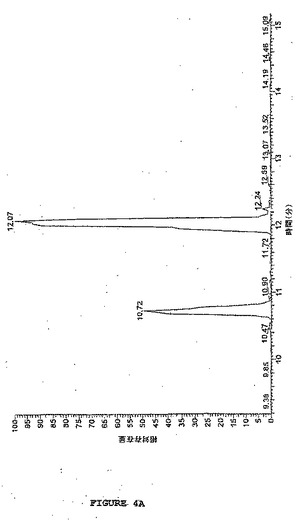
【図4B】
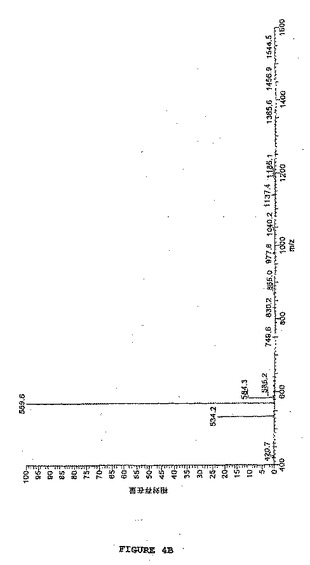
【図5】
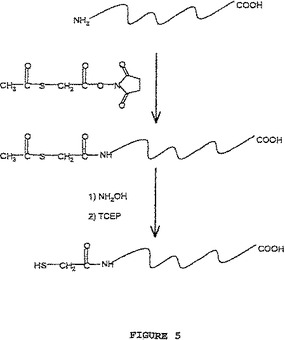
【図6A】
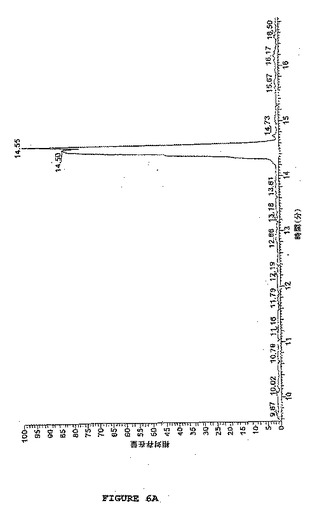
【図6B】
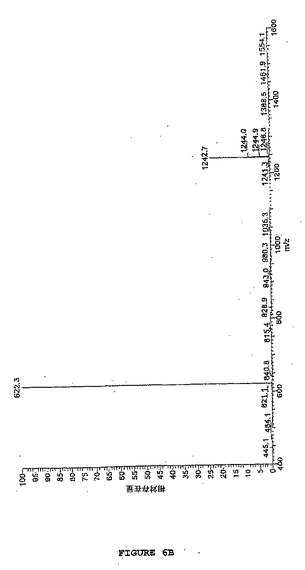
【図7A】
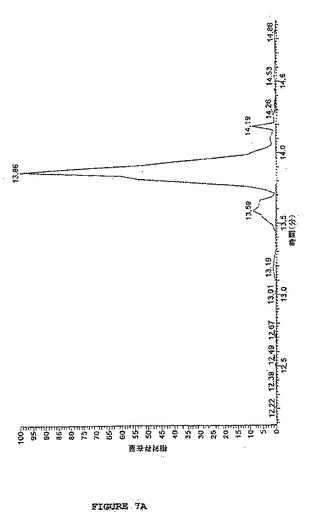
【図7B】
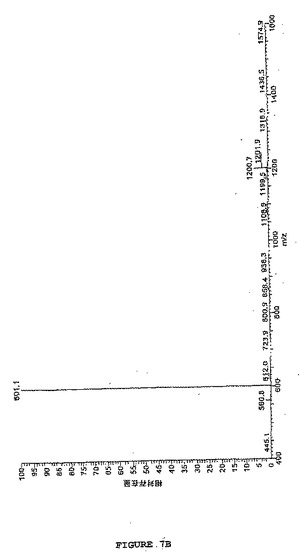
【図8】
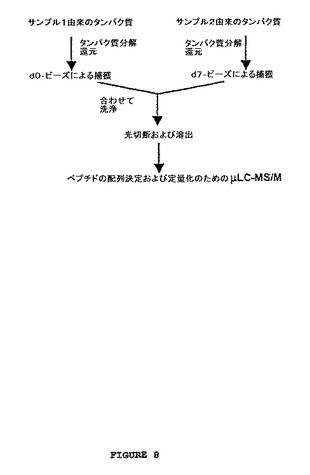
【図9】
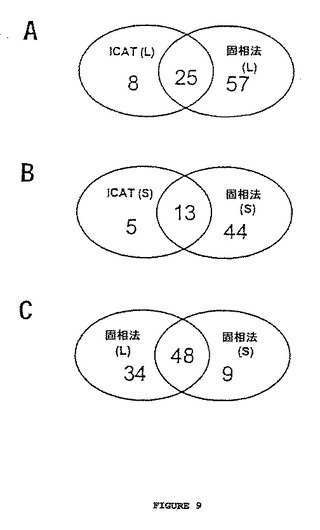
【図1B】
【図1C】
【図2A】
【図2B】
【図3A】
【図3B】
【図4A】
【図4B】
【図5】
【図6A】
【図6B】
【図7A】
【図7B】
【図8】
【図9】
【公開番号】特開2009−175157(P2009−175157A)
【公開日】平成21年8月6日(2009.8.6)
【国際特許分類】
【出願番号】特願2009−113092(P2009−113092)
【出願日】平成21年5月7日(2009.5.7)
【分割の表示】特願2002−589762(P2002−589762)の分割
【原出願日】平成14年5月14日(2002.5.14)
【出願人】(503230117)ザ インスティチュート フォー システムズ バイオロジー (5)
【出願人】(502457803)ユニヴァーシティ オブ ワシントン (93)
【出願人】(500069057)アプライド バイオシステムズ インコーポレイテッド (120)
【Fターム(参考)】
【公開日】平成21年8月6日(2009.8.6)
【国際特許分類】
【出願日】平成21年5月7日(2009.5.7)
【分割の表示】特願2002−589762(P2002−589762)の分割
【原出願日】平成14年5月14日(2002.5.14)
【出願人】(503230117)ザ インスティチュート フォー システムズ バイオロジー (5)
【出願人】(502457803)ユニヴァーシティ オブ ワシントン (93)
【出願人】(500069057)アプライド バイオシステムズ インコーポレイテッド (120)
【Fターム(参考)】
[ Back to top ]