
シイタケの保存過程で発現する細胞壁分解酵素関連遺伝子群、及びその利用方法
【課題】シイタケ菌の保存性に影響する遺伝子を特定し、シイタケの保存性をはじめとする、これら遺伝子や遺伝子産物の利用方法を提供すること。
【解決手段】特定の塩基配列を持つシイタケ菌の細胞壁分解関連酵素遺伝子(キチナーゼ、グルカナーゼ等)の発現を、特異的に抑制することにより、保存性が親株よりも改善されたシイタケ菌を作出する。また、前記遺伝子産物を用いて糸状菌のプロトプラストを作製する。
【解決手段】特定の塩基配列を持つシイタケ菌の細胞壁分解関連酵素遺伝子(キチナーゼ、グルカナーゼ等)の発現を、特異的に抑制することにより、保存性が親株よりも改善されたシイタケ菌を作出する。また、前記遺伝子産物を用いて糸状菌のプロトプラストを作製する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、シイタケ菌の保存過程で発現する細胞壁分解酵素関連遺伝子群とこれを利用したシイタケ菌の保存性の改善、及び当該遺伝子産物を用いたプロトプラストの作製方法に関する。
【背景技術】
【0002】
シイタケは、担子菌類のキシメジ科シイタケ属に属し、古くから現在に至るまで、優秀な食用菌として用いられ、日本においては古来より栽培されている。シイタケは生シイタケとしてだけではなく、乾シイタケとしても利用されている。しかし、近年、中国産のシイタケの輸入が増加して、国内生産が圧迫され、2001年には輸入制限措置も取られた。こうした状況のなかで、現在国内のシイタケ栽培では、国際競争力の高い、新品種の開発が求められている。
【0003】
シイタケ菌の育種のほとんどは交配育種に頼っており、変異原処理等による育種は行われていない。しかし、交配による育種は胞子形成を伴うことから非常に時間がかかるうえに、形質が変化した株を選抜するのに、多大な労力がかかるという問題点があった。
【0004】
近年、シイタケ由来の遺伝子プロモーターを用いたベクター及びシイタケの形質転換方法が報告されている(特許文献1〜4、及び非特許文献1〜3)。これらのベクターを用いて、アンチセンス発現ベクターあるいは、RNAiベクターを導入することにより、シイタケ由来の遺伝子発現抑制が可能と考えられる。しかしながら、この方法を行うには、対象となる遺伝子の単離が必要であるが、シイタケ菌の遺伝子解析研究は発展途上であり、単離されている遺伝子の数は、非常に少ない。
【0005】
レンチナンは免疫機能を活性化させることにより、抗癌活性を示す物質として、シイタケから抽出、精製されたβ-1,3-1,6-グルカンである(非特許文献4)。レンチナンはすでに抗ガン剤として認可されており、実際に臨床現場で使用されている。また、生シイタケから精製されたレンチナンも市販されているが、レンチナンは収穫後の保存過程において分解されるために、収量が一定しないという問題点がある(非特許文献5)。また、これまでの研究により、シイタケ保存過程で、exo-グルカナーゼ活性が上昇することでレンチナンが分解されることも明らかになっている(非特許文献6)。
【0006】
これまで、シイタケのプロトプラスト作製に関しては、他の菌で産生された細胞壁分解酵素が用いられているが、高価なうえに、lot間で効率が異なるなどの問題が生じることがある。またシイタケの保存過程に発現するグルカナーゼ遺伝子としてexg2, tlg1が単離されているが、細胞壁の分解との関連は明らかになっておらず、また、細胞壁の分解にはその他多くの細胞壁分解関連酵素が関わっていることが予想される。
【特許文献1】特開平11-15556号公報
【特許文献2】特開2000-069975号公報
【特許文献3】特開2001-321182号公報
【特許文献4】特開2003-189855号公報
【非特許文献1】Sato et al., Biosci. Biotech. Biochem., 62(12), 2346-2350 (1998)
【非特許文献2】Hirano et.al., Mol. Gen. Genet., 263, 1047-1052 (2000)
【非特許文献3】Irie et.al., Biosci. Biotech. Biochem., 67(9), 2006-2009 (2003)
【非特許文献4】千原吾郎、2.レンチナン. キノコの化学、生化学. 水野卓、川合 正允 編 学会出版センター 東京. 323-333 (1992)
【非特許文献5】Minato et. al., Int. J. Med. Mushroom 1, 265-272 (1999)
【非特許文献6】Minato et. al., Carbohydr. Polym. 56, 279-286 (2004)
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明は、シイタケ菌の保存性に影響する細胞壁分解関連酵素遺伝子を特定し、該遺伝子の発現を抑制することにより、保存性が改善させたシイタケ菌を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者らは、シイタケ菌の保存過程で発現量が変化する遺伝子群を単離し、その中に細胞壁の分解に関与する酵素の遺伝子が多数存在することを明らかにした。そして、シイタケの保存性にはこれら細胞壁分解酵素の作用が影響していることを見出し、本発明を完成させるに至った。
【0009】
すなわち、本発明は、以下の(a)〜(d)のいずれかの核酸分子のシイタケ菌における発現を抑制することにより、親株よりも保存性が改善された形質転換シイタケ菌を作出する方法を提供する。
(a)配列番号1〜12のいずれかに示される塩基配列を含む核酸分子、
(b)配列番号1〜12のいずれかに示される塩基配列からなる核酸分子と相補的な塩基配列からなる核酸分子とストリンジェントな条件下でハイブリダイズし、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子、
(c)配列番号13〜24のいずれかに示されるアミノ酸配列をコードする核酸分子、
(d)配列番号13〜24のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子。
【0010】
前記発現抑制は、アンチセンス法、RNA干渉法、遺伝子破壊法、共抑制法等、公知の方法を用いて実施することができる。
【0011】
本発明はまた、以下の(a)又は(b)のタンパク質:を糸状菌の培養菌糸に作用させることにより、当該糸状菌のプロトプラストを作製する方法も提供する:
(a)配列番号13〜24のいずれかに示されるアミノ酸配列を含むタンパク質、
(b)配列番号13〜24のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質。
【0012】
本発明はまた、以下の(a)〜(d)のいずれかの核酸分子を提供する:
(a)配列番号1〜10のいずれかに示される塩基配列を含む核酸分子、
(b)配列番号1〜10のいずれかに示される塩基配列からなる核酸分子と相補的な塩基配列からなる核酸分子とストリンジェントな条件下でハイブリダイズし、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子、
(c)配列番号13〜20のいずれかに示されるアミノ酸配列をコードする核酸分子、
(d)配列番号13〜20のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子。
【0013】
本発明はまた、前記核酸分子の発現を抑制するベクターを提供する。そのようなベクターとしては、配列番号1〜10のいずれかに示される配列中の連続した100塩基以上からなる塩基配列をアンチセンス方向に含む発現抑制ベクターや、配列番号1〜10のいずれかに示される配列中の連続した19塩基以上からなる塩基配列及びその相補配列をそれぞれ1回以上含み、前記19塩基以上からなる塩基配列に対しそのインバーテッドリピート配列を有するRNAを発現させる、発現抑制ベクターを挙げることができる。
【0014】
前記ベクターは、形質転換マーカー遺伝子をさらに含んでいてもよい。形質転換マーカーとしては、栄養要求性マーカーまたは薬剤耐性マーカーが挙げられる。具体的には、たとえば、ハイグロマイシンB耐性遺伝子(hph)、カルボキシン耐性遺伝子(cbx)、ビアラホス耐性遺伝子(bar)等を用いることができる。
【0015】
本発明はまた、前記した発現抑制ベクターをシイタケ菌に導入して得られる、親株よりも保存性が改善された形質転換シイタケ菌を提供する。
【0016】
本発明はまた、本発明の核酸分子含む発現ベクターや、該ベクターを宿主に導入して得られる形質転換体、該形質転換体を利用した組換え細胞壁分解関連酵素の製造方法も提供する。
【0017】
前記発現ベクターは、形質転換マーカー遺伝子をさらに含んでいてもよい。形質転換マーカーとしては、栄養要求性マーカーまたは薬剤耐性マーカーが挙げられる。具体的には、たとえば、ハイグロマイシンB耐性遺伝子(hph)、カルボキシン耐性遺伝子(cbx)、ビアラホス耐性遺伝子(bar)等を用いることができる。
【0018】
本発明はまた、以下の(a)又は(b)のタンパク質も提供する:
(a)配列番号13〜20のいずれかに示されるアミノ酸配列を含むタンパク質、
(b)配列番号13〜20のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質。
【発明の効果】
【0019】
本発明によれば、保存過程における細胞壁の破壊を抑制して、シイタケの保存性を改善することができる。さらに、本発明によれば、糸状菌のプロトプラストを簡便に作製することができる。
【発明を実施するための最良の形態】
【0020】
1.本発明の遺伝子
本発明にかかる遺伝子:chi1(配列番号1)、chi29(配列番号2)、mlg1(配列番号3)、ghf16.1(配列番号4)、ghf16.2(配列番号5)、mlg2(配列番号6)、ghf30 chd1(配列番号7)、(配列番号8)、pro1(配列番号9)、pep1(配列番号10)、tlg1(配列番号11)、exg2(配列番号12)は、シイタケの保存過程において発現が大きく変動する遺伝子として特定されたもので、キチナーゼ、グルカナーゼ等の細胞壁溶解活性を有する酵素をコードする。
【0021】
このうちchi1とchi29は、その推定ORFからいずれもキチナーゼ活性を有することが示唆される。
mlg1とmlg2は、その推定ORFからいずれもmixed-linked glucanase活性を有することが示唆される。
ghf16.1とghf16.2とghf30は、その推定ORFからいずれも糖鎖分解活性を有することが示唆される。
chd1は、その推定ORFからキチン脱アセチル化酵素活性を有することが示唆される。
【0022】
pro1とpep1は、その推定ORFからアスパラギン酸プロテアーゼ活性を有することが示唆され、胞子形成促進活性を有することが予測される。
tlg1は、エンドグルカナーゼ活性を有し、レンチナン分解活性を有することを発明者らは見出している(特願2005-091557, Sakamoto et al. 2006, Plant Physiol. in press)。
exg2は、その推定ORFからエキソグルカナーゼ活性を有することが示唆される(Sakamoto et al., (2005) Characterization of the Lentinula edodes exg2 gene encoding a lentinan-degrading exo-b-1, 3-glucanase. Current Genetics、 Vol. 48:195-203)。
【0023】
発明者らは、前記遺伝子をクローニングし、その全長cDNA配列を決定した。本明細書の配列表の配列番号1〜12に、chi1, chi29, mlg1, ghf16.1, ghf16.2, mlg2, chd1, ghf30, pro1, pep1, tlg1, exg2のcDNA配列をそれぞれ示す。また、その推定ORFを配列番号13〜24にそれぞれ示す。
【0024】
本発明の遺伝子は、それが細胞壁溶解活性を有する本発明のタンパク質をコードする限り、配列番号1〜12で示される配列に限定されず、これらの配列に相補的な塩基配列を有する核酸分子とストリンジェントな条件下でハイブリダイズしうる配列を有する核酸分子であってもよい。なお、ストリンジェントな条件とは、例えば、ナトリウム濃度が300-2000mMで温度が40-75℃、好ましくはナトリウム濃度が600-900mMで温度が65℃の条件をいう。当業者であれば、Molecular Cloning(Sambrook, J. et al., Molecular Cloning :a Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, 10 Skyline Drive Plainview, NY (1989))等を参照することにより、こうした本発明の遺伝子ホモログを容易に取得することができる。
【0025】
本発明のタンパク質も、それが細胞壁溶解活性を有する限り、配列番号13〜24に示されるアミノ酸配列に限定されず、これらの配列において1または数個のアミノ酸が欠失、置換あるいは付加した配列であってもよい。なお、「数個」とは、好ましくは2〜7個、より好ましくは2〜5個、最も好ましくは2〜3個のアミノ酸を意味する。
【0026】
また配列番号13〜24に表されるアミノ酸配列と、BLAST等を用いて計算したときに(例えば、BLASTのデフォルトすなわち初期条件のパラメーターを用いた場合に)、少なくとも70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%、98%若しくは99%以上の相同性を有しているタンパク質も、それが細胞壁溶解活性を有する限り、本発明のタンパク質に包含される。
【0027】
2.シイタケの保存性の改善
2.1 発現抑制ベクター
本発明にかかる、本発明の遺伝子発現抑制用ベクターは、本発明の遺伝子の発現を抑制するためのアンチセンスベクターあるいはRNAiベクターである。
【0028】
前記アンチセンスベクターは、本発明の遺伝子のcDNA配列(配列番号1〜12に示される配列)中の連続した100塩基以上、好ましくは100〜1000塩基、より好ましくは300〜800塩基)からなる塩基配列を適当なベクターにアンチセンス方向に連結して作製される。前記アンチセンスベクターは、導入された宿主内で前記100塩基以上からなる塩基配列に対するアンチセンスRNAを発現させることにより、本発明の遺伝子の発現を抑制する。アンチセンスベクターの構築は、基本的に前項の記載に従えばよい。
【0029】
前記RNAiベクターは、RNA干渉(RNAi)により、標的とする本発明の遺伝子の宿主における発現を抑制しうるベクターである。具体的には、前記RNAiベクターは本発明の遺伝子のcDNA配列中の連続した少なくとも19塩基以上からなる塩基配列及びその相補配列を各々1以上含み、前記19塩基以上からなる塩基配列に対しそのインバーテッドリピート配列を有するRNAを発現させるように作製される。
【0030】
RNAiベクターの遺伝子抑制効果は、一般にその二重鎖領域の長さに依存するといわれ(Yang, D. et al., 2000年, Curr.Biol., 10,1191-1200)、したがって前記インバーテッドリピート配列は100塩基対以上となるように作製されることが好ましい。このようなRNAiベクターは、当該分野で周知の方法に従い、あるいは市販のRNAi用ベクターやシステム(例えば、psiRNA(Invitrogen)、pSUPER RNAi SystemTM(OligoEngine)等)を利用して容易に構築することができる。
【0031】
前記ベクターは、形質転換マーカー遺伝子をさらに含んでいてもよい。形質転換マーカーとしては、栄養要求性マーカー又は薬剤耐性マーカーが挙げられる。具体的には、たとえば、ジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子、カナマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子、ビアラホス耐性遺伝子、カルボキシン耐性遺伝子、フレオマイシン耐性遺伝子等を用いることができる。
【0032】
なお、ベクターとしては、発明者らがすでに開発し、特開2000-069975号や特開2001-157586号で開示しているpLGベクター(Hirano, T. et al., Mol. Gen. Genet.. 263, 1047-1052、2000)、pLTベクター、pCHSベクター、及びpChGベクター(特開2001-321182)などを好適に利用できる。
【0033】
2.2 形質転換シイタケ菌
本発明は、本発明の遺伝子の発現を特異的に抑制することにより、当該遺伝子によってコードされる細胞壁溶解酵素(本発明のタンパク質)の発現量を低減させ、あるいは胞子形成を抑制させることにより、親株よりも保存性が改善された形質転換シイタケを提供する。
【0034】
本発明の遺伝子の発現抑制方法としては、例えば、アンチセンス法、RNA干渉法、遺伝子破壊法、及び共抑制法(Napoli, C. et al.(1990), Plant Cell 2, 279-289、Van Der Krol, A. R. et al (1990), Plant Cell 2, 291-299参照)等の方法を挙げることができる。
【0035】
アンチセンス法、及びRNA干渉法は、それぞれ前項に記載したアンチセンスベクター、及びRNAiベクターを宿主シイタケ菌のプロトプラストに導入することにより実施できる。シイタケ菌へのベクターの導入は、前述したとおり、ポリエチレングリコール法、エレクトロポレーション法、REMI法等を用いて実施できる。形質転換されたシイタケ菌は、ベクター内に連結された形質転換マーカー遺伝子により容易に選抜される。
【0036】
本発明の形質転換シイタケでは、細胞壁の溶解に関わるタンパク質の発現が通常のシイタケ菌(すなわち、親株)よりも抑制されている。したがって、この形質転換シイタケ菌を用いれば、収穫後の保存過程における細胞壁の溶解や胞子形成を防止し、シイタケの保存性を改善することができる。
【0037】
上記はシイタケの保存性について述べたが、本発明の遺伝子のホモログは他の糸状菌にも存在することが予測され、これらホモログを抑制することで、同様に他の(子実体を形成する)糸状菌類についてもその保存性を改善できることが期待される。
【0038】
3.プロトプラストの作製
3.1 発現ベクター
本発明の遺伝子を発現させるためのベクター(発現ベクター)は、公知のベクターに本発明の遺伝子を連結(挿入)して得ることができる。前記ベクターは宿主中で複製可能なものであれば特に限定されず、例えば、プラスミドDNA、ファージDNA等が挙げられる。
【0039】
前記プラスミドDNAとしては、大腸菌由来のプラスミド(例えば、pBR322,pBR325,pUC18, pUC119,pTrcHis,pBlueBacHis等)、枯草菌由来のプラスミド(例えば、pUB110,pTP5等)、酵母由来のプラスミド(例えば、YEp13,YEp24,YCp50,pYE52等)、植物細胞宿主用プラスミド(pBI221、pBI121)等が挙げられ、ファージ DNAとしてはλファージ等が挙げられる。さらに、レトロウイルス又はワクシニアウイルス等の動物ウイルス、バキュロウイルス等の昆虫ウイルスベクターを用いることもできる。
【0040】
ベクターに本発明の遺伝子を挿入するには、まず、精製されたDNAを適当な制限酵素で切断し、適当なベクター DNAの制限酵素部位又はマルチクローニングサイトに挿入してベクターに連結する方法等が採用される。本発明の遺伝子は、その遺伝子が機能しうる態様で、宿主に応じたプロモーターに連結して導入される必要がある。ここで「機能しうる態様」とは、プロモーター活性によって、その下流に配置された本発明の遺伝子が宿主中で適切に発現され、その機能を発揮することをいう。使用されるプロモーターの種類は、宿主細胞によって適宜決定されるが、その詳細は次項で説明する。
【0041】
本発明のベクターは、プロモーター、本発明の遺伝子のほか、所望によりエンハンサー等のシスエレメント、スプライシングシグナル、ポリA付加シグナル、形質転換マーカー遺伝子(例えば、ジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子、カナマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子、ビアラホス耐性遺伝子、カルボキシン耐性遺伝子、フレオマイシン耐性遺伝子等)、リボソーム結合配列(SD配列)等を含んでいてもよい。
【0042】
特にシイタケ菌を宿主とする場合、シイタケ発現ベクターとして、発明者らがすでに開発し、特開2000-069975号や特開2001-157586号で開示しているpLGベクター(Hirano, T. et al., Mol. Gen. Genet.. 263, 1047-1052、2000)、pLTベクター、pCHSベクター、及びpChGベクター(特開2001-321182)などを好適に利用できる。
【0043】
3.2 形質転換体(組換えタンパク質生産用宿主細胞)
組換えタンパク質を生産するための形質転換体は、前記ベクターを適当な宿主に導入することにより得ることができる。宿主は、本発明の本発明の遺伝子が発現できるものであれば特に限定されない。例えば、エッシェリヒア・コリ(Escherichia coli)等のエッシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、シュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属、リゾビウム・メリロティ(Rhizobium meliloti)等のリゾビウム属に属する細菌、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)等の酵母、麹菌(Aspergillus oryzae)、シロイヌナズナ、タバコ、トウモロコシ、イネ、ニンジン等から株化した植物細胞やプロトプラスト、COS細胞、CHO細胞等の動物細胞、あるいはSf9、Sf21等の昆虫細胞等が挙げられる。
【0044】
大腸菌等の細菌を宿主とする場合は、本発明のベクターが該細菌中で自律複製可能であると同時に、プロモーター、リボゾーム結合配列、本発明の遺伝子、転写終結配列により構成されていることが好ましい。また、プロモーターを制御する遺伝子が含まれていてもよい。大腸菌としては、例えば、エッシェリヒア・コリ(Escherichia coli)HMS174(DE3)、K12、DH1等が挙げられ、枯草菌としては、例えば、バチルス・ズブチリス(Bacillus subtilis)MI 114、207-21等が挙げられる。プロモーターとしては、大腸菌等の上記宿主中で発現できるものであれば特に限定されず、例えば、trpプロモーター、lacプロモーター、PLプロモーター、PRプロモーター等の、大腸菌やファージに由来するプロモーターが挙げられる。また、tacプロモーター等のように、人為的に設計改変されたプロモーターを用いてもよい。細菌へのベクターの導入方法は、特に限定されず、例えば、カルシウムイオンを用いる方法[Cohen, S.N. et al.:Proc. Natl. Acad. Sci., USA, 69:2110-2114 (1972)]や、エレクトロポレーション法等が挙げることができる。
【0045】
酵母を宿主とする場合は、例えば、サッカロミセス・セレビシエ、シゾサッカロミセス・ポンベ、ピヒア・パストリス等が用いられる。プロモーターとしては、酵母中で発現できるものであれば特に限定されず、例えば、gal1プロモーター、gal10プロモーター、ヒートショックタンパク質プロモーター、MFα1プロモーター、PHO5プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーター、AOX1プロモーター等を挙げることができる。酵母へのベクターの導入方法は、特に限定されず、例えば、エレクトロポレーション法[Becker, D.M. et al.:Methods. Enzymol., 194: 182-187 (1990)]、スフェロプラスト法[Hinnen, A.et al.:Proc. Natl. Acad. Sci., USA, 75: 1929-1933 (1978)]、酢酸リチウム法[Itoh, H.:J. Bacteriol., 153:163-168 (1983)]等を挙げることができる。
【0046】
麹菌を宿主とする場合、プロモーターとしては、例えば、GlaA プロモーター(Hata et al. Curr. Genet., Vol 22, 85-91, 1992)、AmyB プロモーター(Tuchiya et al. Biosci. Biotechnol. Biochem., Vol 46, 1849-1853, 1992)、No. 8 プロモーター(Ozeki et al. Biosci. Biotech. Biochem., Vol 60, 383-389, 1996)が挙げられる。麹菌へのベクターの導入方法は、特に限定されず、例えば、エレクトロポレーション法、カルシウムイオン法等を用いることができる。
【0047】
植物細胞を宿主とする場合は、例えば、イネ、トウモロコシ、コムギ、シロイヌナズナ、タバコ、ニンジン等から株化した細胞や該植物から調製したプロトプラストが用いられる。この場合、プロモーターとしては植物中で発現できるものであれば特に限定されず、例えば、カリフラワーモザイクウイルスの35S RNAプロモーター、rd29A遺伝子プロモーター、rbcSプロモーター等が挙げられる。植物へのベクターの導入方法としては、アグロバクテリウム感染法等の間接導入法や、パーティクルガン法、ポリエチレングリコール法、リポソーム法、マイクロインジェクション法等の直接導入法等が挙げられる。
【0048】
シイタケ菌を宿主とする場合は、ポリエチレングリコール法、エレクトロポレーション法、REMI法等を用いてシイタケ菌のプロトプラストに前記ベクターを導入することが好ましい。REMI法とは、発明者らによって開発されたシイタケ菌へのベクターの導入方法で、その詳細は、Sato, T. et. al., Biosci. Biotech. Biochem., 62, 2346-2350, (1998)、及び特開平11-155568号等に記載されている。
【0049】
3.3 組換えタンパク質の製造
本発明のタンパク質は、前述の形質転換体(宿主細胞)を適当な培地で培養し、その培養物から細胞壁溶解活性を有するタンパク質を採取することによって得ることができる。本発明の形質転換体の培養は、常法に従って行えばよい。例えば、大腸菌や酵母等の微生物を宿主とする形質転換体の場合は、微生物が資化しうる炭素源、窒素源、無機塩類等を含有し、形質転換体を効率的に培養しうる天然培地、あるいは合成培地で培養すればよい。また、植物細胞を宿主として用いている場合には、チアミン、ピリドキシン等のビタミン類を添加した植物細胞用の培地で培養すればよい。
【0050】
炭素源としては、グルコース、フラクトース、スクロース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類が用いられる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸若しくは有機酸のアンモニウム塩又はその他の含窒素化合物のほか、ペプトン、肉エキス、コーンスティープリカー等が用いられる。無機物としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等が用いられる。
【0051】
培地中は必要に応じてアンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。プロモーターとして誘導性のプロモーターを用いたベクターで形質転換した微生物を培養する場合は、必要に応じてインデューサーを培地に添加してもよい。例えば、Lacプロモーターを用いたベクターで形質転換した微生物を培養するときにはイソプロピル-β-D-チオガラクトピラノシド(IPTG)等を、trpプロモーターを用いたベクターで形質転換した微生物を培養するときにはインドールアクリル酸(IAA)等を培地に添加してもよい。
【0052】
培養は、通常、振盪培養又は通気攪拌培養等の好気的条件下、30〜37℃位で6時間〜3日間程度行う。培養期間中、pHは7.0〜7.5程度に保持する。pHの調整は、無機又は有機酸、アルカリ溶液等を用いて行う。培養後、本発明のタンパク質が菌体内又は細胞内に生産される場合には、菌体又は細胞を破砕することにより該タンパク質を抽出する。また、本発明のタンパク質が菌体外又は細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離等により菌体又は細胞を除去する。その後、タンパク質の単離精製に用いられる一般的な生化学的方法、例えば、硫酸アンモニウム沈殿、SDS-PAGE、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、疎水クロマトグラフィー、アフィニティークロマトグラフィー等を単独で又は適宜組み合わせて用いることにより、前記培養物中から本発明のタンパク質を単離精製することができる。
【0053】
3.4 糸状菌のプロトプラスト化
本発明の組換えタンパク質は細胞壁溶解活性を有するため、これをシイタケ菌をはじめとする糸状菌に作用させることにより、当該糸状菌のプロトプラストを容易に調製することができる。保存後期の子実体抽出液には本発明のタンパク質が豊富に含まれているが、実際この抽出液をシイタケ菌をはじめとする糸状菌に作用させると、容易にプロトプラストを調製できる。なお、保存後期とは、菌種によっても異なるが、シイタケ菌の場合であれば、一般に収穫したときから25℃で保存した場合3日目〜5日目を意味する。
【実施例】
【0054】
実施例1:シイタケ菌を由来とする細胞壁分解関連遺伝子群のクローニング
遺伝子資源としては、レンチヌラ・エドデス(H600)株を用い、菌傘の膜が切れた段階で収穫した子実体を、湿度約80%、25℃のデシケーター内で保存試験を行った。サンプリングは収穫した直後をD0(0日目)とし、D0〜D4(0日〜4日目)まで毎日サンプリングを行った(数字は保存日数を示す)。サンプリングは、柄、傘、ひだに分けた後、液体窒素で凍結させて行った。発現比較を行うサンプルとして、D0子実体とD3子実体のひだを用いた。各サンプル1gより、ISOGEN(日本ジーン)を用いてRNAを抽出し、PCR-select subtraction 用のサンプルとした。PCR-select subtraction はクロンテックのカスタムサービスにより行った。表1にサブトラクションの結果を示す。
【0055】
【表1】

【0056】
得られたサブトラクション済みのクローンは、D3子実体をテスターとしたものをフォワード、D0子実体をテスターとしたものをリバースライブラリーとし、各576クローンの大腸菌ストックとしてライブラリー化した。
【0057】
得られた大腸菌ストックをテンプレートにして、Nested primer 1(TCGAGCGGCCGCCCGGGCAGGT:配列番号103)とNested primer 2R(AGCGTGGTCGCGGCCGAGGT:配列番号104)を用いてPCRを行い、得られたDNA断片をテンプレートとして、前述のNested Primer 1を用いてシークエンス反応を行い、ABI3100により配列を決定した。得られた配列はBlast検索、InterProScan(http://www.ebi.ac.uk/InterProScan/)によるモチーフ検索を行い、遺伝子の機能の推定を行った。
【0058】
その結果、フォワードサブトラクションで得られたクローンの配列中には、キチナーゼと相同性の高い配列を持つクローン2種類(chi1, chi29)、mixed-linked glucanaseと相同性の高いクローン(mlg1)、キチン脱アセチル化酵素と相同性の高い配列(chd1)、Glycosyl hydrolase(GH)family 30のドメインを持つクローン(ghf30)等の細胞壁分解関連酵素と相同性の高いクローンが見つかった。
【0059】
見つかったクローンのうち、細胞壁の分解に関わる可能性のある遺伝子について、遺伝子全長のクローニングを行った。まず、PCR-subtractionで得られた配列をもとに3’及び5'RACEを行った。3'RACE用のcDNAは、SMART RACEキット(BD Biosciences)を用いて合成した。プライマーとして、表2に記載した各遺伝子に特異的なプライマー及びUPM(BD Biosciences :universal primer mix:CTAATACGACTCACTATA-GGGCAAGCAGTGGTATCAACGCAGAGT(配列番号105)=0.4μM, CTAATACGACTCAC-TATAGGGC(配列番号106)=2μM)を用い、変性94℃30秒,アニーリング55℃30秒、伸長反応72℃1分の条件でPCRを行った。5’RACE用のcDNAはGeneRACERキット(Invitrogen)を用いて合成した。cDNAの合成は各キットのマニュアルに従った。1st PCRではGeneRace 5'primerと各遺伝子に特異的なプライマーを用いた。PCRの条件は変性94℃30秒、アニーリング55℃30秒、伸長反応72℃1分で、Ex-Taqを用いて行った。得られたPCR反応液を50倍希釈し、2nd PCRのテンプレートとした。2nd PCRは、GeneRacer 5' Nested Primer(GGACACTGACATGGACTGAAGGAGTA(配列番号107):BD Biosciences)と各遺伝子に特異的なプライマーを用い,2nd PCRは、1st PCRと同じ条件で、Ex-Taqを用いて行った。
【0060】
【表2】
【0061】
糸状菌類のGH16ファミリーと相同性を示した配列の保存配列をもとに縮重プライマーを作製した。H600株D4のSMART cDNAをテンプレートとして用い、プライマーghf16.1-1Uとghf16.1-2U、ghf16.2-1Uとghf16.2-2Uを用いて3’RACEを行った。3'RACEの条件はSakamotoら(Sakamoto Y, Irie T, Sato T (2005a) Isolation and characterization of a fruiting body-specific exo-b-1,3-glucanase-encoding gene, exg1, from Lentinula edodes. Curr Genet 47: 244-252)の方法に従った。5’RACEは表2に記載のプライマー:ghf16.1-544L-RACE, ghf16.1-393L-RACE, ghf.16.2-519L-RACE, ghf.16.2-178L-RACEを用い、テンプレートはD4のRNAを用い、GeneRacer(Invitrogen)により合成したcDNAを用いた。5'RACEの条件は上記の方法に従った。
【0062】
exg2遺伝子は、麹菌や、トリコデルマなどの糸状菌の保存配列をもとに、縮重プライマーを作製し、クローニングを行った。H600株保存4日目のSMART cDNAをテンプレートとして用い、プライマーexga-8Uとexga-7Lにより、変性94℃30秒、アニーリング45℃30秒、伸長反応72℃2分のPCRの条件でEx-Taqを用いて行った。得られたPCR断片は、前述のNested Primer 1を用いてシークエンスを行った。3' RACEのテンプレートはH600株D4のSMART cDNAを用い,プライマーには、exga-8UとUPMを用いた。1st PCRの条件は変性94℃30秒,アニーリング55℃30秒、伸長反応72℃1分で行った。得られた配列は、同様にしてシークエンスを行った。得られた配列をもとに5' RACE用のプライマーを作製した。テンプレートはD1のGeneRacer cDNAを用いた。PCRの条件は変性94℃30秒,アニーリング55℃30秒、伸長反応72℃1分で、Ex-Taqを用いて行った。得られた配列は同様にしてシークエンスを行った。
【0063】
tlg1遺伝子は、シイタケにおいて報告されているThaumatin-like proteinのN末端の配列を元に(Grenier J, Potvin C, Asselin A. (2000) Some fungi express b-1,3-glucanases similar to thaumatin-like proteins Mycologia 92: 841-848)、プライマーを作製し、クローニングを行った。1st PCRにはtlg-1U (TAYAAYGGNTGYCCNTTYAC:配列番号108)とUPM, 2nd PCRにはtlg-2U (ATHTGGCCNGCNATGTTYAC:配列番号109)とSMART PCR Primer (AAGCAGTGGTATCAACGCAGAGT:配列番号110)を用いてPCRを行った後、シークエンスを行った。
【0064】
同様にして、保存過程において発現変化が大きい遺伝子についてその全長クローニングを行なった。以下、各遺伝子の全長クローニング結果について記載する。
(1)chi1及びchi29のクローニング
まず、発現変化が大きいキチナーゼ2種類:chi1及びchi29の全長クローニングを行った。サブトラクションで得られた配列をもとに、3'及び5'RACE を行い、それぞれ1,311 bpと960 bpのcDNA 配列を得ることが出来た。chi1 のORFは436アミノ酸、分子量 45kDa、pI値 4.47 であった。推定アミノ酸配列をホモロジー検索にかけたところ、Cryptococcus neoformansの推定キチナーゼ遺伝子と約46%の相同性を示し、Glycosyl hydrolase(GH)family 18のキチナーゼモチーフを有することが明らかとなった。chi29のORFは319アミノ酸、分子量 34kDa、pI値 6.62であった。推定アミノ酸配列をホモロジー検索にかけたところ、C. neoformansの推定キチナーゼ遺伝子と約43%の相同性を示し、Glycosyl hydrolase(GH)family 18のキチナーゼモチーフを有することが明らかとなった。また、キチナーゼの活性部位に保存されている酸性アミノ酸が保存されていることが明らかとなった。
【0065】
(2)mlg1のクローニング
5' RACE及び3' RACEを行い、1223bpのcDNA配列を得た。ORFは、977bp、325アミノ酸、分子量35kDa、pI値4.55と推定された。推定アミノ酸配列をホモロジー検索にかけたところ、C. carbonumのmixed-linked グルカナーゼと約40%の相同性を示すことが明らかになった。また、推定アミノ酸配列中には、endo-グルカナーゼで保存されている領域が含まれており、特にグルカンとの結合に重要なGEDIの配列(Gorlach, J. M.、Van Der Knaap, E., and Walton, J. D.(1998) Cloning and targeted disruption of MLG1, a gene encoding two of three extracellular mixed-linked glucanases of Cochliobolus carbonum. Appl Environ Microbiol 64(2): 385-91.)が完全に保存されていた。
【0066】
(3)mlg2のクローニング
ghf16.2の3’RACEを行った際に、ghf16.2とは異なり、mixed-linked glucanaseと相同性を示す配列が見つかった(mlg2)。その配列をもとに5' RACEを行い、1,352 bpのmlg2遺伝子のcDNA配列を得ることが出来た。予想されるORFは620アミノ酸、分子量 36kDa、pI値 4.45であった。推定アミノ酸配列をホモロジー検索にかけたところ、endo-beta-1,3(4)-グルカナーゼ及びシイタケのmlg1と約30%の相同性を示し、Glycosyl hydrolase(GH)family 17のモチーフを有することが明らかとなった。また、グルカナーゼの活性部位に保存されている酸性アミノ酸が保存されていることが明らかとなった。
【0067】
(4)ghf16.1のクローニング
縮重プライマーを用いた3’RACEにより、約750 bpの断片を得た。得られた断片の配列をホモロジー検索にかけたところ、細菌のendo-グルカナーゼ遺伝子と相同性を示すことが明らかになり、目的の断片を得ることが出来たと考えられた。得られた配列を元に、5' RACEを行い、1,310 bpのghf16.1のcDNA配列を得ることが出来た。推定アミノ酸配列をPSROTIIで検索したところ、21残基目のところで切断され、細胞外に分泌される可能性が示唆された。また予想されるORFは372アミノ酸、分子量37 kDa、pI値 4.2であった。推定アミノ酸配列をホモロジー検索にかけたところ、糸状菌類のhypothetical proteinと約50%の相同性を示し、Glycosyl hydrolase(GH)family 16のモチーフを有することが明らかとなった。また、グルカナーゼの活性部位に保存されている酸性アミノ酸が保存されていることが明らかとなった。
【0068】
(5)ghf16.2のクローニング
縮重プライマーを用いた3’RACEにより、約700 bpの断片を得た。得られた断片の配列をホモロジー検索にかけたところ、糸状菌類のendo-グルカナーゼ遺伝子と相同性を示し、目的の断片を得ることが出来たと考えられた。得られた配列を元に、5' RACEを行い、2,093 bpのghf16.2のcDNA配列を得ることが出来た。予想されるORFは620アミノ酸、分子量 63kDa、pI値5.5であった。推定アミノ酸配列をホモロジー検索にかけたところ、glucan binding proteinと約35%の相同性を示し、Glycosyl hydrolase(GH)family 16のモチーフを有することが明らかとなった。また、グルカナーゼの活性部位に保存されている酸性アミノ酸が保存されていることが明らかとなった。
【0069】
(6)chd1のクローニング
キチン脱アセチル化酵素と相同性の高かったクローンの配列をもとに3'及び5'RACEを行い、chd1遺伝子のクローニングを行った。その結果、1,230 bpのchd1遺伝子のcDNA配列を得ることが出来た。chd1のORFは409アミノ酸、分子量 45kDa、pI値 4.26であった。推定アミノ酸配列をホモロジー検索にかけたところ、C. neoformansのキチン脱アセチル化酵素遺伝子と約55%の相同性を示し、多糖類脱アセチル化酵素のモチーフを有すること、及びC末端側にセリンリッチ領域が存在することが明らかとなった。
【0070】
(7)ghf30のクローニング
Glycosyl hydrolase(GH)family30と相同性の高かったクローンの配列をもとに3'及び5'RACEを行い、ghf30遺伝子のクローニングを行った。その結果、559 bpのghf30遺伝子のcDNA配列を得ることが出来た。Ghf30のORFは159アミノ酸、分子量 17.5kDa、pI値 4.66であった。推定アミノ酸配列をホモロジー検索にかけたところ、Caulobacter crescentusのGlycosyl hydrolase family30遺伝子と約37%の相同性を示した。
【0071】
(8)pro1のクローニング
リバースライブラリーで得られた配列をもとに、pro1の全長クローニングを行った。アスパラギン酸プロテアーゼと相同性の高いクローンの配列をもとに3'及び5'RACEを行い、1406bpのpro1遺伝子のcDNA配列を得ることが出来た。pro1のORFは423アミノ酸、分子量 45kDa、pI値 5.19であった。推定アミノ酸配列をホモロジー検索にかけたところ、Amanita muscariaのアスパラギン酸プロテアーゼと49%の相同性を示し、アスパラギン酸プロテアーゼのモチーフを有することが明らかとなった。
【0072】
(9)pep1のクローニング
リバースライブラリーで得られた配列をもとに、pep1の全長クローニングを行った。アスパラギン酸プロテアーゼと相同性の高いクローンの配列をもとに3'及び5'RACEを行い、1377bpのpep1遺伝子のcDNA配列を得ることが出来た。pep1のORFは415アミノ酸、分子量 44kDa、pI値 4.84であった。推定アミノ酸配列をホモロジー検索にかけたところ、Irpex lacteusのアスパラギン酸プロテアーゼと50%の相同性を示し、アスパラギン酸プロテアーゼのモチーフを有することが明らかとなった。シイタケ保存過程では2種類のアスパラギン酸プロテアーゼが存在し、そのうちの一つは胞子形成に関わることが報告されていることから(Takao Terashita, Kohei Oda, Mihoko Tsuji, Kentaro Yoshikawa, Jiko Shishiyama and Sawao Murao The role of carboxyl proteinasa in the maturation of Basidiospores of Lentinus edodes furuiting bodies. (1993) Mokuzai Gakkaishi 39(8): 951-957)、pro1,もしくはpep1にコードされているアスパラギン酸プロテアーゼが胞子形成に関わる可能性が示唆された。
【0073】
(10)exg2のクローニング
degenerate PCRにより、約1.2kbpの断片を得た。得られた断片の配列をホモロジー検索にかけたところ、麹菌のexo-グルカナーゼと相同性が高いことが明らかになり、目的の断片を得ることが出来た。5' RACE及び3' RACEをおこない、2447bpのcDNA配列を得ることが出来た。ORFは、2341bp、779アミノ酸、分子量81kDa、pI値4.66と推定された。推定アミノ酸配列をホモロジー検索にかけたところ、A. saitoiのexo-グルカナーゼと約40%の相同性を示すことが明らかになった。また、exo-グルカナーゼで保存されている繰り返しモチーフ(Nikolskaya AN, Pitkin JW, Schaeffer HJ, Ahn J-H, Walton JD (1998) EXG1p, a novel exo-β-1,3-glucanase from the fungus Cochliobolus carbonum, contains a repeated motif present in other proteins that interact with polysaccharides. Biochem Biophys Acta 1425: 632-636)は、シイタケのexo-グルカナーゼにおいても保存されていることが明らかになった。
【0074】
(11)tlg1のクローニング
縮重プライマーを用いた3'RACEにより、約800bpの断片を得た。得られた断片の配列をホモロジー検索にかけたところ、植物のThaumatin-like proteinと相同性が高いことが明らかになり、目的の断片を得ることが出来た。5' RACEを行い981bpのcDNA配列を得ることが出来た。ORFは、782bp、259アミノ酸、分子量25kDa、pI値3.84と推定された。推定アミノ酸配列をホモロジー検索にかけたところ、A. thalianaのthaumatin-like proteinと約40%の相同性を示すことが明らかになった。また、既報のシイタケThaumatin-like proteinの、N末端配列(Grenier J, Potvin C, Asselin A. (2000) Some fungi express b-1,3-glucanases similar to thaumatin-like proteins Mycologia 92: 841-848)と完全に一致した配列を含んでいることが明らかになった。
【0075】
実施例2:各遺伝子の発現様式の解析
実施例1で特定された各遺伝子について、その発現様式の解析をリアルタイムPCRにより行った。
【0076】
RNAサンプルはPCR-subtractionに用いたものと同じものを使用した。リアルタイムPCR用のcDNAの合成はQuantiTect(登録商標:QIAGEN)を用い、添付のマニュアルに従って行った。リアルタイムPCRの反応試薬にはTaKaRaのSYBR(登録商標)Premix ExTaqTM を用い、7500 Real Time PCR System(Applied Biosystems)にてPCR反応を行った。使用したプライマーは、表3に示すとおりである。
【0077】
【表3】
【0078】
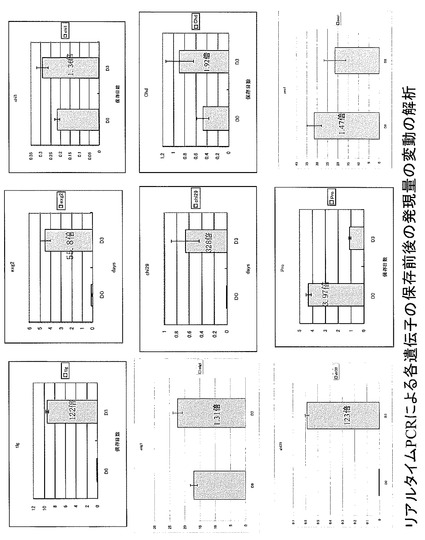
サブトラクションで得られた遺伝子の中で子実体の老化に関わると考えられる遺伝子について、リアルタイムPCR法で発現量の確認を行った。各遺伝子の発現量は、一定量構成的に発現していると考えられる gpdに対する発現量の比で求めた。はじめにノーザン解析によって、保存過程で発現量が増加することがわかっている exg2, tlg1 遺伝子で発現量の差をみたところ、両遺伝子ともにリアルタイムPCR法による解析においてもD3で発現量が多いことが確認できた(図1A,B)。そこで、フォワードサブトラクションで得られた各遺伝子で発現量の差をみたところ、chi1, chi29, chd1で発現量が増加していることが確認できた(図1C,D,E)。mlg1では、ノーザン解析では発現量が保存過程で増加することが明らかになっている。chi1に関してはD0-D4間での発現量を比較したところ、D3, D4で発現が認められ、D4で最も高い発現量であった。リバースライブラリーで得られた遺伝子の中から、pro1の発現量を比較したところ、保存過程で発現量が1/4程度に減少することが明らかになった(図1F)。
【0079】
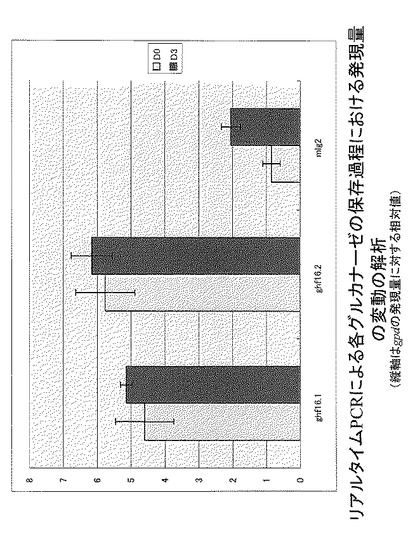
ghf16.1, ghf16.2, mlg2各遺伝子のリアルタイムPCRを行い、保存過程での発現パターンを解析した(図2)。各遺伝子ともに、保存過程の子実体において、発現量の顕著な増加は認められなかったが、gpdに対して数倍程度の発現量を示していることから、恒常的に高発現しており、保存過程においても若干発現量が増加していることが示唆された。
【0080】
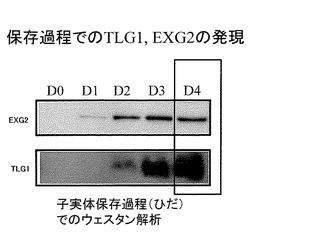
保存過程におけるシイタケタンパク質抽出液を用いてウェスタンブロッティングを行ったところ、EXG2, TLG2ともに保存過程においてタンパク質の発現量が増加していることが明らかになった(図3)。
【0081】
実施例3:酵素活性の測定
グルカナーゼ活性の測定:
グルカナーゼ活性測定用のサンプルは、ノーザン解析で用いたものと同じものを用いた。液体窒素中で粉砕したサンプル1gを、抽出バッファー(200mM 酢酸ナトリウム pH4.2)5mlに懸濁し、タンパク質を抽出した。抽出液5μlに、0.1%ラミナリン溶液を加えて、37℃30分インキュベートした後に、ソモギ-ネルソン法で還元糖の測定を行った。一時間に1μmol還元糖が放出される活性を1ユニットと定義した。タンパク質濃度の測定は、Bio-Rad Protein Assay(Bio-Rad)により行い、BSAをスタンダードとして使用した。
【0082】
キチナーゼ活性の測定:
キチナーゼ活性測定用のサンプルは、保存過程の子実体ひだ(D0-D4)を用いた。液体窒素中で粉砕したサンプル1gを、抽出バッファー(200mM 酢酸ナトリウム pH4.2)5mlに懸濁し、タンパク質を抽出した。抽出液10μlに、蛍光ラベルされたN-アセチルグルコサミン3量体(4-methylumbelliferyl-β-D-N,N',N”-triacetylchitotrioside)溶液を10μl、10mMリン酸バッファー(pH 7.0)180mlを加えて、37℃1時間インキュベートした後に、蛍光強度(励起波長366nm, 吸収波長456nm)を測定した。一時間に1μmolの蛍光N-アセチルグルコサミンが放出される活性を1ユニットと定義した。蛍光スタンダードとして、4-Methylumbelliferoneを用いた。また、タンパク質濃度の測定は、Bio-Rad Protein Assay(Bio-Rad)により行い、BSAをスタンダードとして使用した。
【0083】
結果:
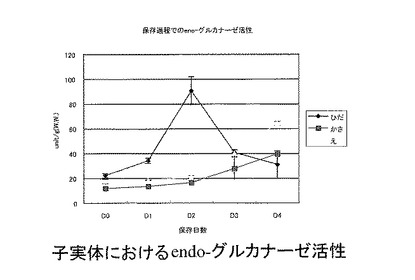
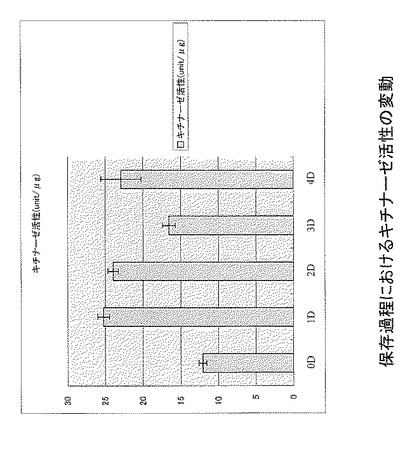
子実体保存過程におけるグルカナーゼ活性の変動を測定したところ、保存に伴い、各部位ともに活性が上昇することが明らかになった(図4)。ひだではすみやかに発現が上昇するが、D3以降では柄や傘で活性が強いことが明らかになった。蛍光ラベルされたキチンを基質としてキチナーゼ活性を測定した。保存過程の子実体からタンパク質を抽出し、キチナーゼ活性を測定したところ、D0でも比較的高い活性が認められ、保存過程でキチナーゼ活性が増加した。保存後期でのD3, D4においては、D1, D2と比較するとやや低い傾向にあった(図5)。
【0084】
実施例4:tlg1遺伝子の異種発現
ペリプラズム移行シグナルを持つpET26ベクター(Novagen社)に挿入するために、tlg1遺伝子をプライマーtlg1-Nt, mlg1-TGA-Not Iを用いて増幅した。得られたtlg1遺伝子断片を平滑化した後Not Iで消化した。pET26ベクターをEcoR V, Not Iで消化後、消化しtlg1断片とライゲーションした。キット添付のマニュアルに基づいて発現を行い、ペリプラズム画分からタンパク質を抽出した。ペリプラズム画分のタンパク質を抽出するために、遠心して回収した大腸菌ペレットをペリプラズム分画バッファー(100mM Tris-Hcl pH8.0, 500mM sucrose, 1mM EDTA)に懸濁後、30分氷上で処理後、13000rpmで遠心し、上清をペリプラズム画分として得た。麹菌を用いたtlg1の発現用ベクターの構築用に、プライマー、tlg1-ATGU及びtlg1-TAAL用いて遺伝子断片を増幅し、PmaC Iにより切断した麹菌発現用ベクターpPAN8142にライゲーションした。得られたベクターをGomiら(Gomi K, Iimura Y, Hara S (1987) Integrative transformation of Aspergillus oryzae with a plasmid containing the Aspergillus nidulans argB gene. Agric Biol Chem 51: 2549-2555)の方法により麹菌RIB40に形質転換し、得られた組換え体及び親株をDPY培地25℃で培養してタンパク質の抽出を行った。
【0085】
還元化状態にあるペリプラズムでのタンパク合成を行うためのベクターpET26を用いたところ、ペリプラズム画分に可溶化されたタンパクが検出できた(図6A)。得られたタンパク質は弱いながらも活性が認められたが、大量に発現している細胞質画分のタンパク質からは活性が認められなかった。次に麹菌による発現を行った。麹菌で発現させたところ、麹菌においTLG1の発現が認められた(図6C)。親株の内在性のグルカナーゼ活性が高いことから明確な差は認められなかったが、組換え体のグルカナーゼ活性(総活性)が高い傾向が認められた(図6B)。また、イオン交換カラムにて分画したところ、シイタケのTLG1と同じフラクションに分画され、そのフラクションにはグルカナーゼ活性が認められた。
【0086】
実施例5:保存後期の子実体抽出液を用いたシイタケ菌糸のプロトプラスト化
保存後期のD4子実体を液体窒素中で粉砕し、子実体重量に対して2倍量の抽出バッファー(200mM 酢酸ナトリウム pH4.2)に懸濁し、タンパク質を抽出した。タンパク質の抽出方法はSakamotoら(前掲)の方法に従った。MYPG培地で2週間培養したシイタケ菌糸1gに対して子実体抽出液10mlを加え、28℃で4時間インキュベートした。また、プロトプラスト化するために、子実体抽出液に0.6Mになるようにマンニトールを加え、同様の反応条件でインキュベートした。
【0087】
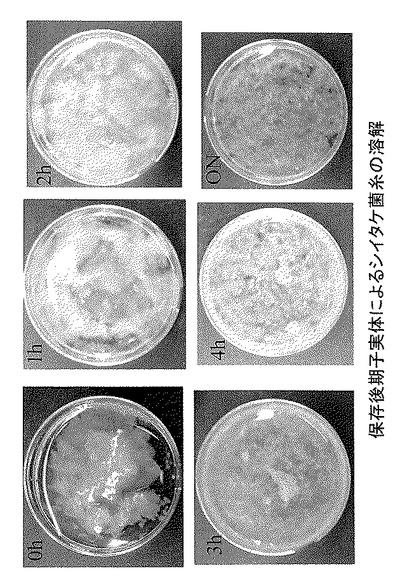
保存後期の子実体抽出液を用いて、シイタケ菌糸を処理したところ、1.4gの菌糸が4時間のインキュベートで0.4gに減少することが明らかになった(図7)。このことから、シイタケ菌糸が子実体抽出液中のグルカナーゼやキチナーゼなどの細胞壁分解酵素により溶解していることが示唆された。そこで、子実体抽出液に浸透圧調整剤として、0.6Mになるようにマンニトールを加えたものを用い、シイタケ菌糸を上記の条件で処理した。結果、1gの菌糸から4時間の処理で2.1×107個のプロトプラストが得られた。
【産業上の利用可能性】
【0088】
本発明の遺伝子の発現を抑制することにより、シイタケ菌をはじめとする糸状菌の保存性を向上させることができる。また、当該遺伝子産物を用いることにより、シイタケ菌をはじめとする糸状菌のプロトプラストを容易に作製することができる。したがって、本発明は糸状菌の品種改良や研究等において有用である。
【図面の簡単な説明】
【0089】
【図1】図1は、リアルタイムPCRによる各遺伝子の保存前後の発現量の変動の解析結果を示す。
【図2】図2は、リアルタイムPCRによる各グルカナーゼの保存過程における発現量の変動の解析結果を示す。
【図3】図3は、保存過程でのTLG1, EXG2の発現(ウェスタン解析)結果を示す。
【図4】図4は、子実体におけるendo-グルカナーゼ活性の変動を示す。
【図5】図5は、保存過程におけるキチナーゼ活性の変動を示す。
【図6】図6は、大腸菌、麹菌によるtlg1の異種発現の結果を示す。A:大腸菌での発現(ウェスタン解析)結果、B:麹菌におけるグルカナーゼ活性、C:麹菌での発現(ウェスタン解析)結果。
【図7】図7は、保存後期子実体によるシイタケ菌糸の溶解を経時的に示したものである(図中、ON(overnight)は一晩のインキュベーションを示す)。
【配列表フリーテキスト】
【0090】
配列番号1:chi1 cDNA
配列番号2:chi29 cDNA
配列番号3:mlg1 cDNA
配列番号4:ghf16.1 cDNA
配列番号5:ghf16.2 cDNA
配列番号6:mlg2 cDNA
配列番号7:chd1 cDNA
配列番号8:ghf30 cDNA
配列番号9:pro1 cDNA
配列番号10:pep1 cDNA
配列番号11:tlg1 cDNA
配列番号12:exg2 cDNA
配列番号13:CHI1
配列番号14:CHI29
配列番号15:MLG1
配列番号16:GHF16.1
配列番号17:GHF16.2
配列番号18:MLG2
配列番号19:CHD1
配列番号20:GHF30
配列番号21:PRO1
配列番号22:PEP1
配列番号23:TLG1
配列番号24:EXG2
配列番号25〜110:人工配列の説明−プライマー
【技術分野】
【0001】
本発明は、シイタケ菌の保存過程で発現する細胞壁分解酵素関連遺伝子群とこれを利用したシイタケ菌の保存性の改善、及び当該遺伝子産物を用いたプロトプラストの作製方法に関する。
【背景技術】
【0002】
シイタケは、担子菌類のキシメジ科シイタケ属に属し、古くから現在に至るまで、優秀な食用菌として用いられ、日本においては古来より栽培されている。シイタケは生シイタケとしてだけではなく、乾シイタケとしても利用されている。しかし、近年、中国産のシイタケの輸入が増加して、国内生産が圧迫され、2001年には輸入制限措置も取られた。こうした状況のなかで、現在国内のシイタケ栽培では、国際競争力の高い、新品種の開発が求められている。
【0003】
シイタケ菌の育種のほとんどは交配育種に頼っており、変異原処理等による育種は行われていない。しかし、交配による育種は胞子形成を伴うことから非常に時間がかかるうえに、形質が変化した株を選抜するのに、多大な労力がかかるという問題点があった。
【0004】
近年、シイタケ由来の遺伝子プロモーターを用いたベクター及びシイタケの形質転換方法が報告されている(特許文献1〜4、及び非特許文献1〜3)。これらのベクターを用いて、アンチセンス発現ベクターあるいは、RNAiベクターを導入することにより、シイタケ由来の遺伝子発現抑制が可能と考えられる。しかしながら、この方法を行うには、対象となる遺伝子の単離が必要であるが、シイタケ菌の遺伝子解析研究は発展途上であり、単離されている遺伝子の数は、非常に少ない。
【0005】
レンチナンは免疫機能を活性化させることにより、抗癌活性を示す物質として、シイタケから抽出、精製されたβ-1,3-1,6-グルカンである(非特許文献4)。レンチナンはすでに抗ガン剤として認可されており、実際に臨床現場で使用されている。また、生シイタケから精製されたレンチナンも市販されているが、レンチナンは収穫後の保存過程において分解されるために、収量が一定しないという問題点がある(非特許文献5)。また、これまでの研究により、シイタケ保存過程で、exo-グルカナーゼ活性が上昇することでレンチナンが分解されることも明らかになっている(非特許文献6)。
【0006】
これまで、シイタケのプロトプラスト作製に関しては、他の菌で産生された細胞壁分解酵素が用いられているが、高価なうえに、lot間で効率が異なるなどの問題が生じることがある。またシイタケの保存過程に発現するグルカナーゼ遺伝子としてexg2, tlg1が単離されているが、細胞壁の分解との関連は明らかになっておらず、また、細胞壁の分解にはその他多くの細胞壁分解関連酵素が関わっていることが予想される。
【特許文献1】特開平11-15556号公報
【特許文献2】特開2000-069975号公報
【特許文献3】特開2001-321182号公報
【特許文献4】特開2003-189855号公報
【非特許文献1】Sato et al., Biosci. Biotech. Biochem., 62(12), 2346-2350 (1998)
【非特許文献2】Hirano et.al., Mol. Gen. Genet., 263, 1047-1052 (2000)
【非特許文献3】Irie et.al., Biosci. Biotech. Biochem., 67(9), 2006-2009 (2003)
【非特許文献4】千原吾郎、2.レンチナン. キノコの化学、生化学. 水野卓、川合 正允 編 学会出版センター 東京. 323-333 (1992)
【非特許文献5】Minato et. al., Int. J. Med. Mushroom 1, 265-272 (1999)
【非特許文献6】Minato et. al., Carbohydr. Polym. 56, 279-286 (2004)
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明は、シイタケ菌の保存性に影響する細胞壁分解関連酵素遺伝子を特定し、該遺伝子の発現を抑制することにより、保存性が改善させたシイタケ菌を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者らは、シイタケ菌の保存過程で発現量が変化する遺伝子群を単離し、その中に細胞壁の分解に関与する酵素の遺伝子が多数存在することを明らかにした。そして、シイタケの保存性にはこれら細胞壁分解酵素の作用が影響していることを見出し、本発明を完成させるに至った。
【0009】
すなわち、本発明は、以下の(a)〜(d)のいずれかの核酸分子のシイタケ菌における発現を抑制することにより、親株よりも保存性が改善された形質転換シイタケ菌を作出する方法を提供する。
(a)配列番号1〜12のいずれかに示される塩基配列を含む核酸分子、
(b)配列番号1〜12のいずれかに示される塩基配列からなる核酸分子と相補的な塩基配列からなる核酸分子とストリンジェントな条件下でハイブリダイズし、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子、
(c)配列番号13〜24のいずれかに示されるアミノ酸配列をコードする核酸分子、
(d)配列番号13〜24のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子。
【0010】
前記発現抑制は、アンチセンス法、RNA干渉法、遺伝子破壊法、共抑制法等、公知の方法を用いて実施することができる。
【0011】
本発明はまた、以下の(a)又は(b)のタンパク質:を糸状菌の培養菌糸に作用させることにより、当該糸状菌のプロトプラストを作製する方法も提供する:
(a)配列番号13〜24のいずれかに示されるアミノ酸配列を含むタンパク質、
(b)配列番号13〜24のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質。
【0012】
本発明はまた、以下の(a)〜(d)のいずれかの核酸分子を提供する:
(a)配列番号1〜10のいずれかに示される塩基配列を含む核酸分子、
(b)配列番号1〜10のいずれかに示される塩基配列からなる核酸分子と相補的な塩基配列からなる核酸分子とストリンジェントな条件下でハイブリダイズし、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子、
(c)配列番号13〜20のいずれかに示されるアミノ酸配列をコードする核酸分子、
(d)配列番号13〜20のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子。
【0013】
本発明はまた、前記核酸分子の発現を抑制するベクターを提供する。そのようなベクターとしては、配列番号1〜10のいずれかに示される配列中の連続した100塩基以上からなる塩基配列をアンチセンス方向に含む発現抑制ベクターや、配列番号1〜10のいずれかに示される配列中の連続した19塩基以上からなる塩基配列及びその相補配列をそれぞれ1回以上含み、前記19塩基以上からなる塩基配列に対しそのインバーテッドリピート配列を有するRNAを発現させる、発現抑制ベクターを挙げることができる。
【0014】
前記ベクターは、形質転換マーカー遺伝子をさらに含んでいてもよい。形質転換マーカーとしては、栄養要求性マーカーまたは薬剤耐性マーカーが挙げられる。具体的には、たとえば、ハイグロマイシンB耐性遺伝子(hph)、カルボキシン耐性遺伝子(cbx)、ビアラホス耐性遺伝子(bar)等を用いることができる。
【0015】
本発明はまた、前記した発現抑制ベクターをシイタケ菌に導入して得られる、親株よりも保存性が改善された形質転換シイタケ菌を提供する。
【0016】
本発明はまた、本発明の核酸分子含む発現ベクターや、該ベクターを宿主に導入して得られる形質転換体、該形質転換体を利用した組換え細胞壁分解関連酵素の製造方法も提供する。
【0017】
前記発現ベクターは、形質転換マーカー遺伝子をさらに含んでいてもよい。形質転換マーカーとしては、栄養要求性マーカーまたは薬剤耐性マーカーが挙げられる。具体的には、たとえば、ハイグロマイシンB耐性遺伝子(hph)、カルボキシン耐性遺伝子(cbx)、ビアラホス耐性遺伝子(bar)等を用いることができる。
【0018】
本発明はまた、以下の(a)又は(b)のタンパク質も提供する:
(a)配列番号13〜20のいずれかに示されるアミノ酸配列を含むタンパク質、
(b)配列番号13〜20のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質。
【発明の効果】
【0019】
本発明によれば、保存過程における細胞壁の破壊を抑制して、シイタケの保存性を改善することができる。さらに、本発明によれば、糸状菌のプロトプラストを簡便に作製することができる。
【発明を実施するための最良の形態】
【0020】
1.本発明の遺伝子
本発明にかかる遺伝子:chi1(配列番号1)、chi29(配列番号2)、mlg1(配列番号3)、ghf16.1(配列番号4)、ghf16.2(配列番号5)、mlg2(配列番号6)、ghf30 chd1(配列番号7)、(配列番号8)、pro1(配列番号9)、pep1(配列番号10)、tlg1(配列番号11)、exg2(配列番号12)は、シイタケの保存過程において発現が大きく変動する遺伝子として特定されたもので、キチナーゼ、グルカナーゼ等の細胞壁溶解活性を有する酵素をコードする。
【0021】
このうちchi1とchi29は、その推定ORFからいずれもキチナーゼ活性を有することが示唆される。
mlg1とmlg2は、その推定ORFからいずれもmixed-linked glucanase活性を有することが示唆される。
ghf16.1とghf16.2とghf30は、その推定ORFからいずれも糖鎖分解活性を有することが示唆される。
chd1は、その推定ORFからキチン脱アセチル化酵素活性を有することが示唆される。
【0022】
pro1とpep1は、その推定ORFからアスパラギン酸プロテアーゼ活性を有することが示唆され、胞子形成促進活性を有することが予測される。
tlg1は、エンドグルカナーゼ活性を有し、レンチナン分解活性を有することを発明者らは見出している(特願2005-091557, Sakamoto et al. 2006, Plant Physiol. in press)。
exg2は、その推定ORFからエキソグルカナーゼ活性を有することが示唆される(Sakamoto et al., (2005) Characterization of the Lentinula edodes exg2 gene encoding a lentinan-degrading exo-b-1, 3-glucanase. Current Genetics、 Vol. 48:195-203)。
【0023】
発明者らは、前記遺伝子をクローニングし、その全長cDNA配列を決定した。本明細書の配列表の配列番号1〜12に、chi1, chi29, mlg1, ghf16.1, ghf16.2, mlg2, chd1, ghf30, pro1, pep1, tlg1, exg2のcDNA配列をそれぞれ示す。また、その推定ORFを配列番号13〜24にそれぞれ示す。
【0024】
本発明の遺伝子は、それが細胞壁溶解活性を有する本発明のタンパク質をコードする限り、配列番号1〜12で示される配列に限定されず、これらの配列に相補的な塩基配列を有する核酸分子とストリンジェントな条件下でハイブリダイズしうる配列を有する核酸分子であってもよい。なお、ストリンジェントな条件とは、例えば、ナトリウム濃度が300-2000mMで温度が40-75℃、好ましくはナトリウム濃度が600-900mMで温度が65℃の条件をいう。当業者であれば、Molecular Cloning(Sambrook, J. et al., Molecular Cloning :a Laboratory Manual 2nd ed., Cold Spring Harbor Laboratory Press, 10 Skyline Drive Plainview, NY (1989))等を参照することにより、こうした本発明の遺伝子ホモログを容易に取得することができる。
【0025】
本発明のタンパク質も、それが細胞壁溶解活性を有する限り、配列番号13〜24に示されるアミノ酸配列に限定されず、これらの配列において1または数個のアミノ酸が欠失、置換あるいは付加した配列であってもよい。なお、「数個」とは、好ましくは2〜7個、より好ましくは2〜5個、最も好ましくは2〜3個のアミノ酸を意味する。
【0026】
また配列番号13〜24に表されるアミノ酸配列と、BLAST等を用いて計算したときに(例えば、BLASTのデフォルトすなわち初期条件のパラメーターを用いた場合に)、少なくとも70%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは97%、98%若しくは99%以上の相同性を有しているタンパク質も、それが細胞壁溶解活性を有する限り、本発明のタンパク質に包含される。
【0027】
2.シイタケの保存性の改善
2.1 発現抑制ベクター
本発明にかかる、本発明の遺伝子発現抑制用ベクターは、本発明の遺伝子の発現を抑制するためのアンチセンスベクターあるいはRNAiベクターである。
【0028】
前記アンチセンスベクターは、本発明の遺伝子のcDNA配列(配列番号1〜12に示される配列)中の連続した100塩基以上、好ましくは100〜1000塩基、より好ましくは300〜800塩基)からなる塩基配列を適当なベクターにアンチセンス方向に連結して作製される。前記アンチセンスベクターは、導入された宿主内で前記100塩基以上からなる塩基配列に対するアンチセンスRNAを発現させることにより、本発明の遺伝子の発現を抑制する。アンチセンスベクターの構築は、基本的に前項の記載に従えばよい。
【0029】
前記RNAiベクターは、RNA干渉(RNAi)により、標的とする本発明の遺伝子の宿主における発現を抑制しうるベクターである。具体的には、前記RNAiベクターは本発明の遺伝子のcDNA配列中の連続した少なくとも19塩基以上からなる塩基配列及びその相補配列を各々1以上含み、前記19塩基以上からなる塩基配列に対しそのインバーテッドリピート配列を有するRNAを発現させるように作製される。
【0030】
RNAiベクターの遺伝子抑制効果は、一般にその二重鎖領域の長さに依存するといわれ(Yang, D. et al., 2000年, Curr.Biol., 10,1191-1200)、したがって前記インバーテッドリピート配列は100塩基対以上となるように作製されることが好ましい。このようなRNAiベクターは、当該分野で周知の方法に従い、あるいは市販のRNAi用ベクターやシステム(例えば、psiRNA(Invitrogen)、pSUPER RNAi SystemTM(OligoEngine)等)を利用して容易に構築することができる。
【0031】
前記ベクターは、形質転換マーカー遺伝子をさらに含んでいてもよい。形質転換マーカーとしては、栄養要求性マーカー又は薬剤耐性マーカーが挙げられる。具体的には、たとえば、ジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子、カナマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子、ビアラホス耐性遺伝子、カルボキシン耐性遺伝子、フレオマイシン耐性遺伝子等を用いることができる。
【0032】
なお、ベクターとしては、発明者らがすでに開発し、特開2000-069975号や特開2001-157586号で開示しているpLGベクター(Hirano, T. et al., Mol. Gen. Genet.. 263, 1047-1052、2000)、pLTベクター、pCHSベクター、及びpChGベクター(特開2001-321182)などを好適に利用できる。
【0033】
2.2 形質転換シイタケ菌
本発明は、本発明の遺伝子の発現を特異的に抑制することにより、当該遺伝子によってコードされる細胞壁溶解酵素(本発明のタンパク質)の発現量を低減させ、あるいは胞子形成を抑制させることにより、親株よりも保存性が改善された形質転換シイタケを提供する。
【0034】
本発明の遺伝子の発現抑制方法としては、例えば、アンチセンス法、RNA干渉法、遺伝子破壊法、及び共抑制法(Napoli, C. et al.(1990), Plant Cell 2, 279-289、Van Der Krol, A. R. et al (1990), Plant Cell 2, 291-299参照)等の方法を挙げることができる。
【0035】
アンチセンス法、及びRNA干渉法は、それぞれ前項に記載したアンチセンスベクター、及びRNAiベクターを宿主シイタケ菌のプロトプラストに導入することにより実施できる。シイタケ菌へのベクターの導入は、前述したとおり、ポリエチレングリコール法、エレクトロポレーション法、REMI法等を用いて実施できる。形質転換されたシイタケ菌は、ベクター内に連結された形質転換マーカー遺伝子により容易に選抜される。
【0036】
本発明の形質転換シイタケでは、細胞壁の溶解に関わるタンパク質の発現が通常のシイタケ菌(すなわち、親株)よりも抑制されている。したがって、この形質転換シイタケ菌を用いれば、収穫後の保存過程における細胞壁の溶解や胞子形成を防止し、シイタケの保存性を改善することができる。
【0037】
上記はシイタケの保存性について述べたが、本発明の遺伝子のホモログは他の糸状菌にも存在することが予測され、これらホモログを抑制することで、同様に他の(子実体を形成する)糸状菌類についてもその保存性を改善できることが期待される。
【0038】
3.プロトプラストの作製
3.1 発現ベクター
本発明の遺伝子を発現させるためのベクター(発現ベクター)は、公知のベクターに本発明の遺伝子を連結(挿入)して得ることができる。前記ベクターは宿主中で複製可能なものであれば特に限定されず、例えば、プラスミドDNA、ファージDNA等が挙げられる。
【0039】
前記プラスミドDNAとしては、大腸菌由来のプラスミド(例えば、pBR322,pBR325,pUC18, pUC119,pTrcHis,pBlueBacHis等)、枯草菌由来のプラスミド(例えば、pUB110,pTP5等)、酵母由来のプラスミド(例えば、YEp13,YEp24,YCp50,pYE52等)、植物細胞宿主用プラスミド(pBI221、pBI121)等が挙げられ、ファージ DNAとしてはλファージ等が挙げられる。さらに、レトロウイルス又はワクシニアウイルス等の動物ウイルス、バキュロウイルス等の昆虫ウイルスベクターを用いることもできる。
【0040】
ベクターに本発明の遺伝子を挿入するには、まず、精製されたDNAを適当な制限酵素で切断し、適当なベクター DNAの制限酵素部位又はマルチクローニングサイトに挿入してベクターに連結する方法等が採用される。本発明の遺伝子は、その遺伝子が機能しうる態様で、宿主に応じたプロモーターに連結して導入される必要がある。ここで「機能しうる態様」とは、プロモーター活性によって、その下流に配置された本発明の遺伝子が宿主中で適切に発現され、その機能を発揮することをいう。使用されるプロモーターの種類は、宿主細胞によって適宜決定されるが、その詳細は次項で説明する。
【0041】
本発明のベクターは、プロモーター、本発明の遺伝子のほか、所望によりエンハンサー等のシスエレメント、スプライシングシグナル、ポリA付加シグナル、形質転換マーカー遺伝子(例えば、ジヒドロ葉酸還元酵素遺伝子、アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子、カナマイシン耐性遺伝子、ハイグロマイシン耐性遺伝子、ビアラホス耐性遺伝子、カルボキシン耐性遺伝子、フレオマイシン耐性遺伝子等)、リボソーム結合配列(SD配列)等を含んでいてもよい。
【0042】
特にシイタケ菌を宿主とする場合、シイタケ発現ベクターとして、発明者らがすでに開発し、特開2000-069975号や特開2001-157586号で開示しているpLGベクター(Hirano, T. et al., Mol. Gen. Genet.. 263, 1047-1052、2000)、pLTベクター、pCHSベクター、及びpChGベクター(特開2001-321182)などを好適に利用できる。
【0043】
3.2 形質転換体(組換えタンパク質生産用宿主細胞)
組換えタンパク質を生産するための形質転換体は、前記ベクターを適当な宿主に導入することにより得ることができる。宿主は、本発明の本発明の遺伝子が発現できるものであれば特に限定されない。例えば、エッシェリヒア・コリ(Escherichia coli)等のエッシェリヒア属、バチルス・ズブチリス(Bacillus subtilis)等のバチルス属、シュードモナス・プチダ(Pseudomonas putida)等のシュードモナス属、リゾビウム・メリロティ(Rhizobium meliloti)等のリゾビウム属に属する細菌、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロミセス・ポンベ(Schizosaccharomyces pombe)等の酵母、麹菌(Aspergillus oryzae)、シロイヌナズナ、タバコ、トウモロコシ、イネ、ニンジン等から株化した植物細胞やプロトプラスト、COS細胞、CHO細胞等の動物細胞、あるいはSf9、Sf21等の昆虫細胞等が挙げられる。
【0044】
大腸菌等の細菌を宿主とする場合は、本発明のベクターが該細菌中で自律複製可能であると同時に、プロモーター、リボゾーム結合配列、本発明の遺伝子、転写終結配列により構成されていることが好ましい。また、プロモーターを制御する遺伝子が含まれていてもよい。大腸菌としては、例えば、エッシェリヒア・コリ(Escherichia coli)HMS174(DE3)、K12、DH1等が挙げられ、枯草菌としては、例えば、バチルス・ズブチリス(Bacillus subtilis)MI 114、207-21等が挙げられる。プロモーターとしては、大腸菌等の上記宿主中で発現できるものであれば特に限定されず、例えば、trpプロモーター、lacプロモーター、PLプロモーター、PRプロモーター等の、大腸菌やファージに由来するプロモーターが挙げられる。また、tacプロモーター等のように、人為的に設計改変されたプロモーターを用いてもよい。細菌へのベクターの導入方法は、特に限定されず、例えば、カルシウムイオンを用いる方法[Cohen, S.N. et al.:Proc. Natl. Acad. Sci., USA, 69:2110-2114 (1972)]や、エレクトロポレーション法等が挙げることができる。
【0045】
酵母を宿主とする場合は、例えば、サッカロミセス・セレビシエ、シゾサッカロミセス・ポンベ、ピヒア・パストリス等が用いられる。プロモーターとしては、酵母中で発現できるものであれば特に限定されず、例えば、gal1プロモーター、gal10プロモーター、ヒートショックタンパク質プロモーター、MFα1プロモーター、PHO5プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーター、AOX1プロモーター等を挙げることができる。酵母へのベクターの導入方法は、特に限定されず、例えば、エレクトロポレーション法[Becker, D.M. et al.:Methods. Enzymol., 194: 182-187 (1990)]、スフェロプラスト法[Hinnen, A.et al.:Proc. Natl. Acad. Sci., USA, 75: 1929-1933 (1978)]、酢酸リチウム法[Itoh, H.:J. Bacteriol., 153:163-168 (1983)]等を挙げることができる。
【0046】
麹菌を宿主とする場合、プロモーターとしては、例えば、GlaA プロモーター(Hata et al. Curr. Genet., Vol 22, 85-91, 1992)、AmyB プロモーター(Tuchiya et al. Biosci. Biotechnol. Biochem., Vol 46, 1849-1853, 1992)、No. 8 プロモーター(Ozeki et al. Biosci. Biotech. Biochem., Vol 60, 383-389, 1996)が挙げられる。麹菌へのベクターの導入方法は、特に限定されず、例えば、エレクトロポレーション法、カルシウムイオン法等を用いることができる。
【0047】
植物細胞を宿主とする場合は、例えば、イネ、トウモロコシ、コムギ、シロイヌナズナ、タバコ、ニンジン等から株化した細胞や該植物から調製したプロトプラストが用いられる。この場合、プロモーターとしては植物中で発現できるものであれば特に限定されず、例えば、カリフラワーモザイクウイルスの35S RNAプロモーター、rd29A遺伝子プロモーター、rbcSプロモーター等が挙げられる。植物へのベクターの導入方法としては、アグロバクテリウム感染法等の間接導入法や、パーティクルガン法、ポリエチレングリコール法、リポソーム法、マイクロインジェクション法等の直接導入法等が挙げられる。
【0048】
シイタケ菌を宿主とする場合は、ポリエチレングリコール法、エレクトロポレーション法、REMI法等を用いてシイタケ菌のプロトプラストに前記ベクターを導入することが好ましい。REMI法とは、発明者らによって開発されたシイタケ菌へのベクターの導入方法で、その詳細は、Sato, T. et. al., Biosci. Biotech. Biochem., 62, 2346-2350, (1998)、及び特開平11-155568号等に記載されている。
【0049】
3.3 組換えタンパク質の製造
本発明のタンパク質は、前述の形質転換体(宿主細胞)を適当な培地で培養し、その培養物から細胞壁溶解活性を有するタンパク質を採取することによって得ることができる。本発明の形質転換体の培養は、常法に従って行えばよい。例えば、大腸菌や酵母等の微生物を宿主とする形質転換体の場合は、微生物が資化しうる炭素源、窒素源、無機塩類等を含有し、形質転換体を効率的に培養しうる天然培地、あるいは合成培地で培養すればよい。また、植物細胞を宿主として用いている場合には、チアミン、ピリドキシン等のビタミン類を添加した植物細胞用の培地で培養すればよい。
【0050】
炭素源としては、グルコース、フラクトース、スクロース、デンプン等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類が用いられる。窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸若しくは有機酸のアンモニウム塩又はその他の含窒素化合物のほか、ペプトン、肉エキス、コーンスティープリカー等が用いられる。無機物としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等が用いられる。
【0051】
培地中は必要に応じてアンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。プロモーターとして誘導性のプロモーターを用いたベクターで形質転換した微生物を培養する場合は、必要に応じてインデューサーを培地に添加してもよい。例えば、Lacプロモーターを用いたベクターで形質転換した微生物を培養するときにはイソプロピル-β-D-チオガラクトピラノシド(IPTG)等を、trpプロモーターを用いたベクターで形質転換した微生物を培養するときにはインドールアクリル酸(IAA)等を培地に添加してもよい。
【0052】
培養は、通常、振盪培養又は通気攪拌培養等の好気的条件下、30〜37℃位で6時間〜3日間程度行う。培養期間中、pHは7.0〜7.5程度に保持する。pHの調整は、無機又は有機酸、アルカリ溶液等を用いて行う。培養後、本発明のタンパク質が菌体内又は細胞内に生産される場合には、菌体又は細胞を破砕することにより該タンパク質を抽出する。また、本発明のタンパク質が菌体外又は細胞外に生産される場合には、培養液をそのまま使用するか、遠心分離等により菌体又は細胞を除去する。その後、タンパク質の単離精製に用いられる一般的な生化学的方法、例えば、硫酸アンモニウム沈殿、SDS-PAGE、ゲルクロマトグラフィー、イオン交換クロマトグラフィー、疎水クロマトグラフィー、アフィニティークロマトグラフィー等を単独で又は適宜組み合わせて用いることにより、前記培養物中から本発明のタンパク質を単離精製することができる。
【0053】
3.4 糸状菌のプロトプラスト化
本発明の組換えタンパク質は細胞壁溶解活性を有するため、これをシイタケ菌をはじめとする糸状菌に作用させることにより、当該糸状菌のプロトプラストを容易に調製することができる。保存後期の子実体抽出液には本発明のタンパク質が豊富に含まれているが、実際この抽出液をシイタケ菌をはじめとする糸状菌に作用させると、容易にプロトプラストを調製できる。なお、保存後期とは、菌種によっても異なるが、シイタケ菌の場合であれば、一般に収穫したときから25℃で保存した場合3日目〜5日目を意味する。
【実施例】
【0054】
実施例1:シイタケ菌を由来とする細胞壁分解関連遺伝子群のクローニング
遺伝子資源としては、レンチヌラ・エドデス(H600)株を用い、菌傘の膜が切れた段階で収穫した子実体を、湿度約80%、25℃のデシケーター内で保存試験を行った。サンプリングは収穫した直後をD0(0日目)とし、D0〜D4(0日〜4日目)まで毎日サンプリングを行った(数字は保存日数を示す)。サンプリングは、柄、傘、ひだに分けた後、液体窒素で凍結させて行った。発現比較を行うサンプルとして、D0子実体とD3子実体のひだを用いた。各サンプル1gより、ISOGEN(日本ジーン)を用いてRNAを抽出し、PCR-select subtraction 用のサンプルとした。PCR-select subtraction はクロンテックのカスタムサービスにより行った。表1にサブトラクションの結果を示す。
【0055】
【表1】
【0056】
得られたサブトラクション済みのクローンは、D3子実体をテスターとしたものをフォワード、D0子実体をテスターとしたものをリバースライブラリーとし、各576クローンの大腸菌ストックとしてライブラリー化した。
【0057】
得られた大腸菌ストックをテンプレートにして、Nested primer 1(TCGAGCGGCCGCCCGGGCAGGT:配列番号103)とNested primer 2R(AGCGTGGTCGCGGCCGAGGT:配列番号104)を用いてPCRを行い、得られたDNA断片をテンプレートとして、前述のNested Primer 1を用いてシークエンス反応を行い、ABI3100により配列を決定した。得られた配列はBlast検索、InterProScan(http://www.ebi.ac.uk/InterProScan/)によるモチーフ検索を行い、遺伝子の機能の推定を行った。
【0058】
その結果、フォワードサブトラクションで得られたクローンの配列中には、キチナーゼと相同性の高い配列を持つクローン2種類(chi1, chi29)、mixed-linked glucanaseと相同性の高いクローン(mlg1)、キチン脱アセチル化酵素と相同性の高い配列(chd1)、Glycosyl hydrolase(GH)family 30のドメインを持つクローン(ghf30)等の細胞壁分解関連酵素と相同性の高いクローンが見つかった。
【0059】
見つかったクローンのうち、細胞壁の分解に関わる可能性のある遺伝子について、遺伝子全長のクローニングを行った。まず、PCR-subtractionで得られた配列をもとに3’及び5'RACEを行った。3'RACE用のcDNAは、SMART RACEキット(BD Biosciences)を用いて合成した。プライマーとして、表2に記載した各遺伝子に特異的なプライマー及びUPM(BD Biosciences :universal primer mix:CTAATACGACTCACTATA-GGGCAAGCAGTGGTATCAACGCAGAGT(配列番号105)=0.4μM, CTAATACGACTCAC-TATAGGGC(配列番号106)=2μM)を用い、変性94℃30秒,アニーリング55℃30秒、伸長反応72℃1分の条件でPCRを行った。5’RACE用のcDNAはGeneRACERキット(Invitrogen)を用いて合成した。cDNAの合成は各キットのマニュアルに従った。1st PCRではGeneRace 5'primerと各遺伝子に特異的なプライマーを用いた。PCRの条件は変性94℃30秒、アニーリング55℃30秒、伸長反応72℃1分で、Ex-Taqを用いて行った。得られたPCR反応液を50倍希釈し、2nd PCRのテンプレートとした。2nd PCRは、GeneRacer 5' Nested Primer(GGACACTGACATGGACTGAAGGAGTA(配列番号107):BD Biosciences)と各遺伝子に特異的なプライマーを用い,2nd PCRは、1st PCRと同じ条件で、Ex-Taqを用いて行った。
【0060】
【表2】
【0061】
糸状菌類のGH16ファミリーと相同性を示した配列の保存配列をもとに縮重プライマーを作製した。H600株D4のSMART cDNAをテンプレートとして用い、プライマーghf16.1-1Uとghf16.1-2U、ghf16.2-1Uとghf16.2-2Uを用いて3’RACEを行った。3'RACEの条件はSakamotoら(Sakamoto Y, Irie T, Sato T (2005a) Isolation and characterization of a fruiting body-specific exo-b-1,3-glucanase-encoding gene, exg1, from Lentinula edodes. Curr Genet 47: 244-252)の方法に従った。5’RACEは表2に記載のプライマー:ghf16.1-544L-RACE, ghf16.1-393L-RACE, ghf.16.2-519L-RACE, ghf.16.2-178L-RACEを用い、テンプレートはD4のRNAを用い、GeneRacer(Invitrogen)により合成したcDNAを用いた。5'RACEの条件は上記の方法に従った。
【0062】
exg2遺伝子は、麹菌や、トリコデルマなどの糸状菌の保存配列をもとに、縮重プライマーを作製し、クローニングを行った。H600株保存4日目のSMART cDNAをテンプレートとして用い、プライマーexga-8Uとexga-7Lにより、変性94℃30秒、アニーリング45℃30秒、伸長反応72℃2分のPCRの条件でEx-Taqを用いて行った。得られたPCR断片は、前述のNested Primer 1を用いてシークエンスを行った。3' RACEのテンプレートはH600株D4のSMART cDNAを用い,プライマーには、exga-8UとUPMを用いた。1st PCRの条件は変性94℃30秒,アニーリング55℃30秒、伸長反応72℃1分で行った。得られた配列は、同様にしてシークエンスを行った。得られた配列をもとに5' RACE用のプライマーを作製した。テンプレートはD1のGeneRacer cDNAを用いた。PCRの条件は変性94℃30秒,アニーリング55℃30秒、伸長反応72℃1分で、Ex-Taqを用いて行った。得られた配列は同様にしてシークエンスを行った。
【0063】
tlg1遺伝子は、シイタケにおいて報告されているThaumatin-like proteinのN末端の配列を元に(Grenier J, Potvin C, Asselin A. (2000) Some fungi express b-1,3-glucanases similar to thaumatin-like proteins Mycologia 92: 841-848)、プライマーを作製し、クローニングを行った。1st PCRにはtlg-1U (TAYAAYGGNTGYCCNTTYAC:配列番号108)とUPM, 2nd PCRにはtlg-2U (ATHTGGCCNGCNATGTTYAC:配列番号109)とSMART PCR Primer (AAGCAGTGGTATCAACGCAGAGT:配列番号110)を用いてPCRを行った後、シークエンスを行った。
【0064】
同様にして、保存過程において発現変化が大きい遺伝子についてその全長クローニングを行なった。以下、各遺伝子の全長クローニング結果について記載する。
(1)chi1及びchi29のクローニング
まず、発現変化が大きいキチナーゼ2種類:chi1及びchi29の全長クローニングを行った。サブトラクションで得られた配列をもとに、3'及び5'RACE を行い、それぞれ1,311 bpと960 bpのcDNA 配列を得ることが出来た。chi1 のORFは436アミノ酸、分子量 45kDa、pI値 4.47 であった。推定アミノ酸配列をホモロジー検索にかけたところ、Cryptococcus neoformansの推定キチナーゼ遺伝子と約46%の相同性を示し、Glycosyl hydrolase(GH)family 18のキチナーゼモチーフを有することが明らかとなった。chi29のORFは319アミノ酸、分子量 34kDa、pI値 6.62であった。推定アミノ酸配列をホモロジー検索にかけたところ、C. neoformansの推定キチナーゼ遺伝子と約43%の相同性を示し、Glycosyl hydrolase(GH)family 18のキチナーゼモチーフを有することが明らかとなった。また、キチナーゼの活性部位に保存されている酸性アミノ酸が保存されていることが明らかとなった。
【0065】
(2)mlg1のクローニング
5' RACE及び3' RACEを行い、1223bpのcDNA配列を得た。ORFは、977bp、325アミノ酸、分子量35kDa、pI値4.55と推定された。推定アミノ酸配列をホモロジー検索にかけたところ、C. carbonumのmixed-linked グルカナーゼと約40%の相同性を示すことが明らかになった。また、推定アミノ酸配列中には、endo-グルカナーゼで保存されている領域が含まれており、特にグルカンとの結合に重要なGEDIの配列(Gorlach, J. M.、Van Der Knaap, E., and Walton, J. D.(1998) Cloning and targeted disruption of MLG1, a gene encoding two of three extracellular mixed-linked glucanases of Cochliobolus carbonum. Appl Environ Microbiol 64(2): 385-91.)が完全に保存されていた。
【0066】
(3)mlg2のクローニング
ghf16.2の3’RACEを行った際に、ghf16.2とは異なり、mixed-linked glucanaseと相同性を示す配列が見つかった(mlg2)。その配列をもとに5' RACEを行い、1,352 bpのmlg2遺伝子のcDNA配列を得ることが出来た。予想されるORFは620アミノ酸、分子量 36kDa、pI値 4.45であった。推定アミノ酸配列をホモロジー検索にかけたところ、endo-beta-1,3(4)-グルカナーゼ及びシイタケのmlg1と約30%の相同性を示し、Glycosyl hydrolase(GH)family 17のモチーフを有することが明らかとなった。また、グルカナーゼの活性部位に保存されている酸性アミノ酸が保存されていることが明らかとなった。
【0067】
(4)ghf16.1のクローニング
縮重プライマーを用いた3’RACEにより、約750 bpの断片を得た。得られた断片の配列をホモロジー検索にかけたところ、細菌のendo-グルカナーゼ遺伝子と相同性を示すことが明らかになり、目的の断片を得ることが出来たと考えられた。得られた配列を元に、5' RACEを行い、1,310 bpのghf16.1のcDNA配列を得ることが出来た。推定アミノ酸配列をPSROTIIで検索したところ、21残基目のところで切断され、細胞外に分泌される可能性が示唆された。また予想されるORFは372アミノ酸、分子量37 kDa、pI値 4.2であった。推定アミノ酸配列をホモロジー検索にかけたところ、糸状菌類のhypothetical proteinと約50%の相同性を示し、Glycosyl hydrolase(GH)family 16のモチーフを有することが明らかとなった。また、グルカナーゼの活性部位に保存されている酸性アミノ酸が保存されていることが明らかとなった。
【0068】
(5)ghf16.2のクローニング
縮重プライマーを用いた3’RACEにより、約700 bpの断片を得た。得られた断片の配列をホモロジー検索にかけたところ、糸状菌類のendo-グルカナーゼ遺伝子と相同性を示し、目的の断片を得ることが出来たと考えられた。得られた配列を元に、5' RACEを行い、2,093 bpのghf16.2のcDNA配列を得ることが出来た。予想されるORFは620アミノ酸、分子量 63kDa、pI値5.5であった。推定アミノ酸配列をホモロジー検索にかけたところ、glucan binding proteinと約35%の相同性を示し、Glycosyl hydrolase(GH)family 16のモチーフを有することが明らかとなった。また、グルカナーゼの活性部位に保存されている酸性アミノ酸が保存されていることが明らかとなった。
【0069】
(6)chd1のクローニング
キチン脱アセチル化酵素と相同性の高かったクローンの配列をもとに3'及び5'RACEを行い、chd1遺伝子のクローニングを行った。その結果、1,230 bpのchd1遺伝子のcDNA配列を得ることが出来た。chd1のORFは409アミノ酸、分子量 45kDa、pI値 4.26であった。推定アミノ酸配列をホモロジー検索にかけたところ、C. neoformansのキチン脱アセチル化酵素遺伝子と約55%の相同性を示し、多糖類脱アセチル化酵素のモチーフを有すること、及びC末端側にセリンリッチ領域が存在することが明らかとなった。
【0070】
(7)ghf30のクローニング
Glycosyl hydrolase(GH)family30と相同性の高かったクローンの配列をもとに3'及び5'RACEを行い、ghf30遺伝子のクローニングを行った。その結果、559 bpのghf30遺伝子のcDNA配列を得ることが出来た。Ghf30のORFは159アミノ酸、分子量 17.5kDa、pI値 4.66であった。推定アミノ酸配列をホモロジー検索にかけたところ、Caulobacter crescentusのGlycosyl hydrolase family30遺伝子と約37%の相同性を示した。
【0071】
(8)pro1のクローニング
リバースライブラリーで得られた配列をもとに、pro1の全長クローニングを行った。アスパラギン酸プロテアーゼと相同性の高いクローンの配列をもとに3'及び5'RACEを行い、1406bpのpro1遺伝子のcDNA配列を得ることが出来た。pro1のORFは423アミノ酸、分子量 45kDa、pI値 5.19であった。推定アミノ酸配列をホモロジー検索にかけたところ、Amanita muscariaのアスパラギン酸プロテアーゼと49%の相同性を示し、アスパラギン酸プロテアーゼのモチーフを有することが明らかとなった。
【0072】
(9)pep1のクローニング
リバースライブラリーで得られた配列をもとに、pep1の全長クローニングを行った。アスパラギン酸プロテアーゼと相同性の高いクローンの配列をもとに3'及び5'RACEを行い、1377bpのpep1遺伝子のcDNA配列を得ることが出来た。pep1のORFは415アミノ酸、分子量 44kDa、pI値 4.84であった。推定アミノ酸配列をホモロジー検索にかけたところ、Irpex lacteusのアスパラギン酸プロテアーゼと50%の相同性を示し、アスパラギン酸プロテアーゼのモチーフを有することが明らかとなった。シイタケ保存過程では2種類のアスパラギン酸プロテアーゼが存在し、そのうちの一つは胞子形成に関わることが報告されていることから(Takao Terashita, Kohei Oda, Mihoko Tsuji, Kentaro Yoshikawa, Jiko Shishiyama and Sawao Murao The role of carboxyl proteinasa in the maturation of Basidiospores of Lentinus edodes furuiting bodies. (1993) Mokuzai Gakkaishi 39(8): 951-957)、pro1,もしくはpep1にコードされているアスパラギン酸プロテアーゼが胞子形成に関わる可能性が示唆された。
【0073】
(10)exg2のクローニング
degenerate PCRにより、約1.2kbpの断片を得た。得られた断片の配列をホモロジー検索にかけたところ、麹菌のexo-グルカナーゼと相同性が高いことが明らかになり、目的の断片を得ることが出来た。5' RACE及び3' RACEをおこない、2447bpのcDNA配列を得ることが出来た。ORFは、2341bp、779アミノ酸、分子量81kDa、pI値4.66と推定された。推定アミノ酸配列をホモロジー検索にかけたところ、A. saitoiのexo-グルカナーゼと約40%の相同性を示すことが明らかになった。また、exo-グルカナーゼで保存されている繰り返しモチーフ(Nikolskaya AN, Pitkin JW, Schaeffer HJ, Ahn J-H, Walton JD (1998) EXG1p, a novel exo-β-1,3-glucanase from the fungus Cochliobolus carbonum, contains a repeated motif present in other proteins that interact with polysaccharides. Biochem Biophys Acta 1425: 632-636)は、シイタケのexo-グルカナーゼにおいても保存されていることが明らかになった。
【0074】
(11)tlg1のクローニング
縮重プライマーを用いた3'RACEにより、約800bpの断片を得た。得られた断片の配列をホモロジー検索にかけたところ、植物のThaumatin-like proteinと相同性が高いことが明らかになり、目的の断片を得ることが出来た。5' RACEを行い981bpのcDNA配列を得ることが出来た。ORFは、782bp、259アミノ酸、分子量25kDa、pI値3.84と推定された。推定アミノ酸配列をホモロジー検索にかけたところ、A. thalianaのthaumatin-like proteinと約40%の相同性を示すことが明らかになった。また、既報のシイタケThaumatin-like proteinの、N末端配列(Grenier J, Potvin C, Asselin A. (2000) Some fungi express b-1,3-glucanases similar to thaumatin-like proteins Mycologia 92: 841-848)と完全に一致した配列を含んでいることが明らかになった。
【0075】
実施例2:各遺伝子の発現様式の解析
実施例1で特定された各遺伝子について、その発現様式の解析をリアルタイムPCRにより行った。
【0076】
RNAサンプルはPCR-subtractionに用いたものと同じものを使用した。リアルタイムPCR用のcDNAの合成はQuantiTect(登録商標:QIAGEN)を用い、添付のマニュアルに従って行った。リアルタイムPCRの反応試薬にはTaKaRaのSYBR(登録商標)Premix ExTaqTM を用い、7500 Real Time PCR System(Applied Biosystems)にてPCR反応を行った。使用したプライマーは、表3に示すとおりである。
【0077】
【表3】
【0078】
サブトラクションで得られた遺伝子の中で子実体の老化に関わると考えられる遺伝子について、リアルタイムPCR法で発現量の確認を行った。各遺伝子の発現量は、一定量構成的に発現していると考えられる gpdに対する発現量の比で求めた。はじめにノーザン解析によって、保存過程で発現量が増加することがわかっている exg2, tlg1 遺伝子で発現量の差をみたところ、両遺伝子ともにリアルタイムPCR法による解析においてもD3で発現量が多いことが確認できた(図1A,B)。そこで、フォワードサブトラクションで得られた各遺伝子で発現量の差をみたところ、chi1, chi29, chd1で発現量が増加していることが確認できた(図1C,D,E)。mlg1では、ノーザン解析では発現量が保存過程で増加することが明らかになっている。chi1に関してはD0-D4間での発現量を比較したところ、D3, D4で発現が認められ、D4で最も高い発現量であった。リバースライブラリーで得られた遺伝子の中から、pro1の発現量を比較したところ、保存過程で発現量が1/4程度に減少することが明らかになった(図1F)。
【0079】
ghf16.1, ghf16.2, mlg2各遺伝子のリアルタイムPCRを行い、保存過程での発現パターンを解析した(図2)。各遺伝子ともに、保存過程の子実体において、発現量の顕著な増加は認められなかったが、gpdに対して数倍程度の発現量を示していることから、恒常的に高発現しており、保存過程においても若干発現量が増加していることが示唆された。
【0080】
保存過程におけるシイタケタンパク質抽出液を用いてウェスタンブロッティングを行ったところ、EXG2, TLG2ともに保存過程においてタンパク質の発現量が増加していることが明らかになった(図3)。
【0081】
実施例3:酵素活性の測定
グルカナーゼ活性の測定:
グルカナーゼ活性測定用のサンプルは、ノーザン解析で用いたものと同じものを用いた。液体窒素中で粉砕したサンプル1gを、抽出バッファー(200mM 酢酸ナトリウム pH4.2)5mlに懸濁し、タンパク質を抽出した。抽出液5μlに、0.1%ラミナリン溶液を加えて、37℃30分インキュベートした後に、ソモギ-ネルソン法で還元糖の測定を行った。一時間に1μmol還元糖が放出される活性を1ユニットと定義した。タンパク質濃度の測定は、Bio-Rad Protein Assay(Bio-Rad)により行い、BSAをスタンダードとして使用した。
【0082】
キチナーゼ活性の測定:
キチナーゼ活性測定用のサンプルは、保存過程の子実体ひだ(D0-D4)を用いた。液体窒素中で粉砕したサンプル1gを、抽出バッファー(200mM 酢酸ナトリウム pH4.2)5mlに懸濁し、タンパク質を抽出した。抽出液10μlに、蛍光ラベルされたN-アセチルグルコサミン3量体(4-methylumbelliferyl-β-D-N,N',N”-triacetylchitotrioside)溶液を10μl、10mMリン酸バッファー(pH 7.0)180mlを加えて、37℃1時間インキュベートした後に、蛍光強度(励起波長366nm, 吸収波長456nm)を測定した。一時間に1μmolの蛍光N-アセチルグルコサミンが放出される活性を1ユニットと定義した。蛍光スタンダードとして、4-Methylumbelliferoneを用いた。また、タンパク質濃度の測定は、Bio-Rad Protein Assay(Bio-Rad)により行い、BSAをスタンダードとして使用した。
【0083】
結果:
子実体保存過程におけるグルカナーゼ活性の変動を測定したところ、保存に伴い、各部位ともに活性が上昇することが明らかになった(図4)。ひだではすみやかに発現が上昇するが、D3以降では柄や傘で活性が強いことが明らかになった。蛍光ラベルされたキチンを基質としてキチナーゼ活性を測定した。保存過程の子実体からタンパク質を抽出し、キチナーゼ活性を測定したところ、D0でも比較的高い活性が認められ、保存過程でキチナーゼ活性が増加した。保存後期でのD3, D4においては、D1, D2と比較するとやや低い傾向にあった(図5)。
【0084】
実施例4:tlg1遺伝子の異種発現
ペリプラズム移行シグナルを持つpET26ベクター(Novagen社)に挿入するために、tlg1遺伝子をプライマーtlg1-Nt, mlg1-TGA-Not Iを用いて増幅した。得られたtlg1遺伝子断片を平滑化した後Not Iで消化した。pET26ベクターをEcoR V, Not Iで消化後、消化しtlg1断片とライゲーションした。キット添付のマニュアルに基づいて発現を行い、ペリプラズム画分からタンパク質を抽出した。ペリプラズム画分のタンパク質を抽出するために、遠心して回収した大腸菌ペレットをペリプラズム分画バッファー(100mM Tris-Hcl pH8.0, 500mM sucrose, 1mM EDTA)に懸濁後、30分氷上で処理後、13000rpmで遠心し、上清をペリプラズム画分として得た。麹菌を用いたtlg1の発現用ベクターの構築用に、プライマー、tlg1-ATGU及びtlg1-TAAL用いて遺伝子断片を増幅し、PmaC Iにより切断した麹菌発現用ベクターpPAN8142にライゲーションした。得られたベクターをGomiら(Gomi K, Iimura Y, Hara S (1987) Integrative transformation of Aspergillus oryzae with a plasmid containing the Aspergillus nidulans argB gene. Agric Biol Chem 51: 2549-2555)の方法により麹菌RIB40に形質転換し、得られた組換え体及び親株をDPY培地25℃で培養してタンパク質の抽出を行った。
【0085】
還元化状態にあるペリプラズムでのタンパク合成を行うためのベクターpET26を用いたところ、ペリプラズム画分に可溶化されたタンパクが検出できた(図6A)。得られたタンパク質は弱いながらも活性が認められたが、大量に発現している細胞質画分のタンパク質からは活性が認められなかった。次に麹菌による発現を行った。麹菌で発現させたところ、麹菌においTLG1の発現が認められた(図6C)。親株の内在性のグルカナーゼ活性が高いことから明確な差は認められなかったが、組換え体のグルカナーゼ活性(総活性)が高い傾向が認められた(図6B)。また、イオン交換カラムにて分画したところ、シイタケのTLG1と同じフラクションに分画され、そのフラクションにはグルカナーゼ活性が認められた。
【0086】
実施例5:保存後期の子実体抽出液を用いたシイタケ菌糸のプロトプラスト化
保存後期のD4子実体を液体窒素中で粉砕し、子実体重量に対して2倍量の抽出バッファー(200mM 酢酸ナトリウム pH4.2)に懸濁し、タンパク質を抽出した。タンパク質の抽出方法はSakamotoら(前掲)の方法に従った。MYPG培地で2週間培養したシイタケ菌糸1gに対して子実体抽出液10mlを加え、28℃で4時間インキュベートした。また、プロトプラスト化するために、子実体抽出液に0.6Mになるようにマンニトールを加え、同様の反応条件でインキュベートした。
【0087】
保存後期の子実体抽出液を用いて、シイタケ菌糸を処理したところ、1.4gの菌糸が4時間のインキュベートで0.4gに減少することが明らかになった(図7)。このことから、シイタケ菌糸が子実体抽出液中のグルカナーゼやキチナーゼなどの細胞壁分解酵素により溶解していることが示唆された。そこで、子実体抽出液に浸透圧調整剤として、0.6Mになるようにマンニトールを加えたものを用い、シイタケ菌糸を上記の条件で処理した。結果、1gの菌糸から4時間の処理で2.1×107個のプロトプラストが得られた。
【産業上の利用可能性】
【0088】
本発明の遺伝子の発現を抑制することにより、シイタケ菌をはじめとする糸状菌の保存性を向上させることができる。また、当該遺伝子産物を用いることにより、シイタケ菌をはじめとする糸状菌のプロトプラストを容易に作製することができる。したがって、本発明は糸状菌の品種改良や研究等において有用である。
【図面の簡単な説明】
【0089】
【図1】図1は、リアルタイムPCRによる各遺伝子の保存前後の発現量の変動の解析結果を示す。
【図2】図2は、リアルタイムPCRによる各グルカナーゼの保存過程における発現量の変動の解析結果を示す。
【図3】図3は、保存過程でのTLG1, EXG2の発現(ウェスタン解析)結果を示す。
【図4】図4は、子実体におけるendo-グルカナーゼ活性の変動を示す。
【図5】図5は、保存過程におけるキチナーゼ活性の変動を示す。
【図6】図6は、大腸菌、麹菌によるtlg1の異種発現の結果を示す。A:大腸菌での発現(ウェスタン解析)結果、B:麹菌におけるグルカナーゼ活性、C:麹菌での発現(ウェスタン解析)結果。
【図7】図7は、保存後期子実体によるシイタケ菌糸の溶解を経時的に示したものである(図中、ON(overnight)は一晩のインキュベーションを示す)。
【配列表フリーテキスト】
【0090】
配列番号1:chi1 cDNA
配列番号2:chi29 cDNA
配列番号3:mlg1 cDNA
配列番号4:ghf16.1 cDNA
配列番号5:ghf16.2 cDNA
配列番号6:mlg2 cDNA
配列番号7:chd1 cDNA
配列番号8:ghf30 cDNA
配列番号9:pro1 cDNA
配列番号10:pep1 cDNA
配列番号11:tlg1 cDNA
配列番号12:exg2 cDNA
配列番号13:CHI1
配列番号14:CHI29
配列番号15:MLG1
配列番号16:GHF16.1
配列番号17:GHF16.2
配列番号18:MLG2
配列番号19:CHD1
配列番号20:GHF30
配列番号21:PRO1
配列番号22:PEP1
配列番号23:TLG1
配列番号24:EXG2
配列番号25〜110:人工配列の説明−プライマー
【特許請求の範囲】
【請求項1】
以下の(a)〜(d)のいずれかの核酸分子のシイタケ菌における発現を抑制することにより、親株よりも保存性が改善された形質転換シイタケ菌を作出する方法:
(a)配列番号1〜12のいずれかに示される塩基配列を含む核酸分子、
(b)配列番号1〜12のいずれかに示される塩基配列からなる核酸分子と相補的な塩基配列からなる核酸分子とストリンジェントな条件下でハイブリダイズし、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子、
(c)配列番号13〜24のいずれかに示されるアミノ酸配列をコードする核酸分子、
(d)配列番号13〜24のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子。
【請求項2】
前記発現抑制が、アンチセンス法、RNA干渉法、遺伝子破壊法、及び共抑制法から選ばれるいずれかの手法を用いて行われる、請求項1記載の方法。
【請求項3】
以下の(a)又は(b)のタンパク質を糸状菌の培養菌糸に作用させることにより、当該糸状菌のプロトプラストを作製する方法:
(a)配列番号13〜24のいずれかに示されるアミノ酸配列を含むタンパク質、
(b)配列番号13〜24のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質。
【請求項4】
以下の(a)〜(d)のいずれかの核酸分子:
(a)配列番号1〜10のいずれかに示される塩基配列を含む核酸分子、
(b)配列番号1〜10のいずれかに示される塩基配列からなる核酸分子と相補的な塩基配列からなる核酸分子とストリンジェントな条件下でハイブリダイズし、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子、
(c)配列番号13〜22のいずれかに示されるアミノ酸配列をコードする核酸分子、
(d)配列番号13〜22のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子。
【請求項5】
配列番号1〜10のいずれかに示される配列中の連続した100塩基以上からなる塩基配列をアンチセンス方向に含む、発現抑制ベクター。
【請求項6】
配列番号1〜10のいずれかに示される配列中の連続した19塩基以上からなる塩基配列及びその相補配列をそれぞれ1回以上含み、前記19塩基以上からなる塩基配列に対しそのインバーテッドリピート配列を有するRNAを発現させる、発現抑制ベクター。
【請求項7】
形質転換マーカー遺伝子をさらに含む、請求項5又は6記載の発現抑制ベクター。
【請求項8】
請求項5〜7のいずれか1項に記載の発現抑制ベクターをシイタケ菌に導入して得られる、親株よりも保存性が改善された形質転換シイタケ菌。
【請求項9】
請求項4記載の核酸分子を含む発現ベクター。
【請求項10】
形質転換マーカー遺伝子をさらに含む、請求項9に記載の発現ベクター。
【請求項11】
請求項9又は10記載の発現ベクターを宿主に導入して得られる形質転換体。
【請求項12】
請求項11記載の形質転換体を培養し、得られる培養液から細胞壁溶解活性を有するタンパク質を回収することを特徴とする、組換え細胞壁分解関連酵素の製造方法。
【請求項13】
以下の(a)又は(b)のタンパク質:
(a)配列番号13〜22のいずれかに示されるアミノ酸配列を含むタンパク質、
(b)配列番号13〜22のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質。
【請求項1】
以下の(a)〜(d)のいずれかの核酸分子のシイタケ菌における発現を抑制することにより、親株よりも保存性が改善された形質転換シイタケ菌を作出する方法:
(a)配列番号1〜12のいずれかに示される塩基配列を含む核酸分子、
(b)配列番号1〜12のいずれかに示される塩基配列からなる核酸分子と相補的な塩基配列からなる核酸分子とストリンジェントな条件下でハイブリダイズし、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子、
(c)配列番号13〜24のいずれかに示されるアミノ酸配列をコードする核酸分子、
(d)配列番号13〜24のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子。
【請求項2】
前記発現抑制が、アンチセンス法、RNA干渉法、遺伝子破壊法、及び共抑制法から選ばれるいずれかの手法を用いて行われる、請求項1記載の方法。
【請求項3】
以下の(a)又は(b)のタンパク質を糸状菌の培養菌糸に作用させることにより、当該糸状菌のプロトプラストを作製する方法:
(a)配列番号13〜24のいずれかに示されるアミノ酸配列を含むタンパク質、
(b)配列番号13〜24のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質。
【請求項4】
以下の(a)〜(d)のいずれかの核酸分子:
(a)配列番号1〜10のいずれかに示される塩基配列を含む核酸分子、
(b)配列番号1〜10のいずれかに示される塩基配列からなる核酸分子と相補的な塩基配列からなる核酸分子とストリンジェントな条件下でハイブリダイズし、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子、
(c)配列番号13〜22のいずれかに示されるアミノ酸配列をコードする核酸分子、
(d)配列番号13〜22のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質をコードするシイタケ菌由来の核酸分子。
【請求項5】
配列番号1〜10のいずれかに示される配列中の連続した100塩基以上からなる塩基配列をアンチセンス方向に含む、発現抑制ベクター。
【請求項6】
配列番号1〜10のいずれかに示される配列中の連続した19塩基以上からなる塩基配列及びその相補配列をそれぞれ1回以上含み、前記19塩基以上からなる塩基配列に対しそのインバーテッドリピート配列を有するRNAを発現させる、発現抑制ベクター。
【請求項7】
形質転換マーカー遺伝子をさらに含む、請求項5又は6記載の発現抑制ベクター。
【請求項8】
請求項5〜7のいずれか1項に記載の発現抑制ベクターをシイタケ菌に導入して得られる、親株よりも保存性が改善された形質転換シイタケ菌。
【請求項9】
請求項4記載の核酸分子を含む発現ベクター。
【請求項10】
形質転換マーカー遺伝子をさらに含む、請求項9に記載の発現ベクター。
【請求項11】
請求項9又は10記載の発現ベクターを宿主に導入して得られる形質転換体。
【請求項12】
請求項11記載の形質転換体を培養し、得られる培養液から細胞壁溶解活性を有するタンパク質を回収することを特徴とする、組換え細胞壁分解関連酵素の製造方法。
【請求項13】
以下の(a)又は(b)のタンパク質:
(a)配列番号13〜22のいずれかに示されるアミノ酸配列を含むタンパク質、
(b)配列番号13〜22のいずれかに示されるアミノ酸配列において1または数個のアミノ酸が欠失、置換もしくは付加されたアミノ酸配列からなり、かつ細胞壁溶解活性を有するタンパク質。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】

【図7】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【公開番号】特開2007−312618(P2007−312618A)
【公開日】平成19年12月6日(2007.12.6)
【国際特許分類】
【出願番号】特願2006−142930(P2006−142930)
【出願日】平成18年5月23日(2006.5.23)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 1. 第28回日本分子生物学会年会(2006)にて発表 主催者名:第28回日本分子生物学会年会 組織委員会 開催日:平成17年12月7日〜平成17年12月10日 発表日:平成17年12月7日 演題番号:1P0019 講演要旨集発行日:平成17年11月25日 2. 刊行物名:日本菌学会50周年記念大会 講演予稿集 講演番号:132−C 発行所:日本菌学会 発行日:平成18年5月10日
【出願人】(390025793)岩手県 (38)
【Fターム(参考)】
【公開日】平成19年12月6日(2007.12.6)
【国際特許分類】
【出願日】平成18年5月23日(2006.5.23)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 1. 第28回日本分子生物学会年会(2006)にて発表 主催者名:第28回日本分子生物学会年会 組織委員会 開催日:平成17年12月7日〜平成17年12月10日 発表日:平成17年12月7日 演題番号:1P0019 講演要旨集発行日:平成17年11月25日 2. 刊行物名:日本菌学会50周年記念大会 講演予稿集 講演番号:132−C 発行所:日本菌学会 発行日:平成18年5月10日
【出願人】(390025793)岩手県 (38)
【Fターム(参考)】
[ Back to top ]