
シイタケ菌糸のウイルスフリー化方法

【課題】ウイルス感染したシイタケの菌糸からウイルスを除去して、伸長可能なウイルスを含まない菌糸を確実に得るシイタケ菌糸のウイルスフリー化方法を提供する。
【解決手段】本発明のウイルスフリー化方法は、ウイルス感染したシイタケの菌糸を耐熱限界温度で培養する加熱処理工程と、加熱処理中新たに伸長した菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、分離した菌糸先端部を適温で培養し菌糸を伸長させて菌株を得る菌糸増殖工程と、から成ることを特徴とする。
【解決手段】本発明のウイルスフリー化方法は、ウイルス感染したシイタケの菌糸を耐熱限界温度で培養する加熱処理工程と、加熱処理中新たに伸長した菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、分離した菌糸先端部を適温で培養し菌糸を伸長させて菌株を得る菌糸増殖工程と、から成ることを特徴とする。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ウイルス感染したシイタケ菌株から、ウイルスフリー株を得る方法に関する。
【背景技術】
【0002】
現在、生シイタケ生産は菌床栽培が主流であり、収益性の高い施設栽培化が進んでいる。しかしその反面、栽培規模拡大にともない原因不明の栽培不良症状の発生による減収が深刻な問題となる状況も増えている。病害の主な原因になるものとして細菌や糸状菌、ウイルスや害虫等が挙げられ、シイタケ生産、栽培におけるこれら病原に対する研究の発展は防除の必要性からも重要度を増している。
【0003】
一方、菌類を宿主とする菌類ウイルスについては、1962年HollingsがLa France病のツクリタケから病原体として検出したのが最初の発見である。日本でも1970年代にシイタケからウイルスが検出されており、また、ウイルス感染したシイタケ菌からウイルスを除去する方法についても、加熱と菌糸先端分離培養を繰り返し長期間行う処理によって一部ウイルスフリー化株が得られた研究報告があった。しかしながら、このウイルスフリー化方法は長期間(320日)を所要する上に、ウイルスフリー化できる確率が低い(3割に満たない)ため安定的要素に欠けており、一般的に実用化される技術には至っていない。しかも当時、正常なシイタケであってもウイルス感染が認められることからウイルスが病害因子とは見なされず、その後の長い間研究の進歩はなかった。
【0004】
近年、2004年にエノキタケのウイルス病が登録されるなど、シイタケやエノキタケ、ハタケシメジなど栽培キノコにおけるウイルス病害を示唆する研究報告例があがっている。また、今まで知見の少なかった菌類ウイルスのゲノム解析などの情報も増えつつあり、RT−PCR法を用いた高感度なウイルス検出方法も開発された。現在、栽培キノコの病害因子としてさらなるウイルスの解明と同時に、防除の面から、確実なウイルスフリー化技術が必要とされている。
【0005】
植物ウイルスについては、茎頂培養が一般的なウイルスフリー化技術として実用化されている。また、下記特許文献1には、活性酸素を用い、ウイルスを不活性化させて損傷の少ない植物の成長点のみ生かすという選択的手法によって植物をウイルスフリーにする方法が提案されている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開平10−249364号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、本発明者等が実験をしたところ、シイタケ菌糸については菌糸先端培養や抗生物質などの薬品を用いる方法ではウイルスを除去することができなかった。
そこで、本発明の課題は、ウイルス感染したシイタケ菌株からウイルス感染がなく且つ正常な伸長が可能となる菌株を確実に得るウイルスフリー化方法を提供することにある。
【課題を解決するための手段】
【0008】
上記課題を解決するために、本発明者等は、ウイルスが高温に弱い性質があり、またキノコ類も高温で菌糸が衰弱や死滅する性質があるので、両者の温度に対する生存限界の違いによって、温度制御をすれば選択的に菌糸は生かしてウイルスは死滅させることができるのではないかと考えた。そして、工夫と実験を重ねた結果、本発明のシイタケ菌糸のウイルスフリー化方法を完成させることができた。
【0009】
本発明における請求項1に係るシイタケ菌糸のウイルスフリー化方法は、ウイルス感染したシイタケの菌糸を、シイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度で培養する加熱処理工程と、加熱処理中新たに伸びてきた菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、から成る。
【0010】
また請求項2に係るシイタケ菌糸のウイルスフリー化方法は、ウイルス感染したシイタケの菌糸を、シイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度で培養する加熱処理工程と、加熱処理中新たに伸びてきた菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、分離した菌糸先端部を適温で培養し菌糸を伸長させて菌株を得る菌糸増殖工程と、から成る。
【発明の効果】
【0011】
本発明のシイタケ菌糸のウイルスフリー化方法は、菌糸を加熱する加熱処理工程において、ウイルス感染している菌糸をシイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度(以下耐熱限界温度と呼ぶ)で培養することで、その間にウイルス感染したシイタケの菌糸の先端は生き延びさせつつ培養し、ウイルスは弱らせて菌糸の先端にまでには繁殖が進まない段階で、その菌糸の先端にあるウイルスがなく伸長が可能となる2核の先端細胞を先端部分離工程で切り出し、これを元の菌糸から別の場所に隔離してウイルスフリーの状態で菌糸を培養することが可能となった。
元のシイタケ菌糸についてはウイルス感染していても、耐熱限界温度で培養すると2核を有した先端細胞にはウイルスの感染が起こらないことが、RT−PCR法を用いたアガロースゲル電気泳動で現れるバンドの有無で確認できた。
なお、RT−PCR法は、Reverse Transcription Polymerase Chain Reactionの略で「逆転写酵素ポリメラーゼ連鎖反応」のことであり、ウイルスの検出に有効な手法である
【0012】
請求項2に係るシイタケ菌糸のウイルスフリー化方法では、上記方法の先端部分離工程で得られたウイルス感染していない2核の先端細胞を、さらに菌糸増殖工程において隔離した別の場所で適度な温度環境下に置くことで菌糸が大きく伸長した正常な菌株をウイルスが存在しない状態で得ることが可能となる。
【図面の簡単な説明】
【0013】
【図1】本発明のシイタケ菌糸のウイルスフリー化方法のフロー図である。
【図2】別形態のシイタケ菌糸のウイルスフリー化方法のフロー図である。
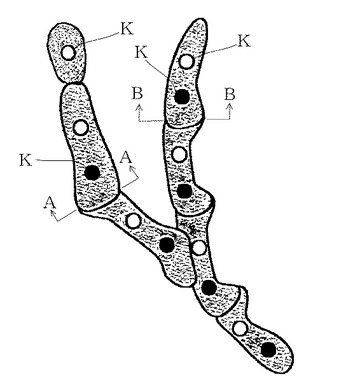
【図3】菌糸先端細胞の拡大図である。
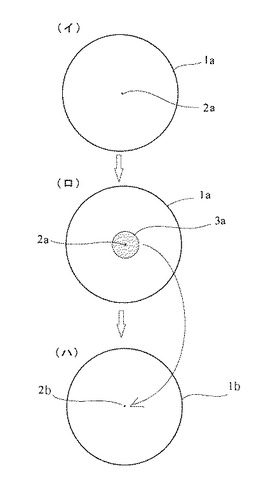
【図4】(イ)はシャーレの培地に菌糸片を置いた状態を示し、(ロ)は菌糸の伸長した状態を示し、(ハ)は加熱処理中新たに伸長した菌糸の先端部を別のシャーレの培地に移した状態を示す各平面図である。
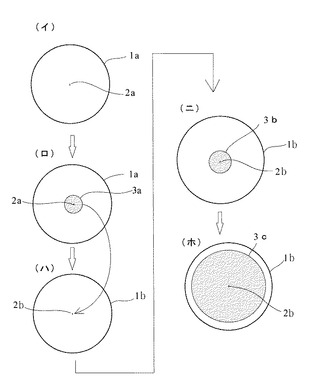
【図5】(イ)はシャーレの培地に菌糸片を置いた状態を示し、(ロ)は菌糸の伸長した状態を示し、(ハ)は加熱処理中新たに伸長した菌糸の先端部を別のシャーレの培地に移した状態を示し、(ニ)は菌糸が伸長した状態を示し、(ホ)は菌糸が増殖した状態を示す各平面図である。
【発明を実施するための形態】
【0014】
以下本発明の実施の形態を説明する。
【0015】
本発明のシイタケ菌糸のウイルスフリー化方法は、図1に示すように、ウイルス感染したシイタケの菌糸をPDA培地に、ウイルス感染している菌糸をシイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度(耐熱限界温度)で培養する加熱処理工程と、該加熱処理工程中新たに伸びてきた菌糸の2核の菌糸先端部を、ウイルス感染している元の菌糸から切り離す先端部分離工程とから成る。
【0016】
また、図2に示すように、前記先端部分離工程の後に、分離した菌糸先端部を適温で培養して菌糸を伸長させて菌株を得る菌糸増殖工程を有する形態が可能である。
【0017】
本発明を以下さらに詳しく説明する。
図3に示すように、ウイルス感染している菌株の中から菌糸片2aを切り出し、図4の(イ)に示すように、シャーレ1aの中のPDA(寒天)培地に移す。このとき菌糸はまだウイルス感染している。
【0018】
次に、ウイルス感染しているシイタケの菌糸片2aを耐熱限界温度で培養する。すると、図4(ロ)に示すように、新たに菌糸が周囲に少し伸長して菌株3aが得られる。
次に、顕微鏡で菌糸先端部を確認し、前記菌株3aの中から図3中のA−A、B−B部分から先部を含む先端部、即ち菌糸先端の1つ以上の2核細胞又は1つ以上の2核細胞と1核細胞を切り出す。
そして、切り出した該菌糸先端部2bを、図4の(ハ)に示すように、シャーレ1bの中のPDA培地に移す。
この菌糸先端部2bにはウイルスが存在しない。したがって、この時点で実質的にウイルスフリー化がなされている。
【0019】
また、前記先端部分離工程以降は、図5の(ニ)に示すように、シャーレ1bの中でPDA培地にウイルスのいない前記菌糸先端部2bを置き、耐熱限界温度で1ヶ月培養すると、図5の(ニ)に示すように、菌糸先端部2bの周囲に菌糸が少し伸長して菌株3bが得られる。このとき伸長した菌株3bはまだ小さい。
そこで、そのシャーレ1bの中で23°Cの温度で7日間培養する。すると、図5の(ホ)に示すように、菌糸が周囲に広く増殖する。そして全ての菌糸細胞にウイルスが感染していない菌株3cが得られる。
【0020】
以上のように、耐熱限界温度で培養する加熱処理した菌株3aから、菌糸先端部2bを分離して取り出した菌糸には、ウイルス感染していないが、ウイルスフリーをより確実にするために、前記先端部分離工程以降に、さらに前記先端部分離工程と菌糸増殖工程とを数回繰り返せば、全くウイルスのないウイルスフリーの菌株を得ることが可能となる。
【0021】
本発明では、加熱処理工程後は、2核の菌糸先端部を切り離す先端部分離工程を行うが、この工程で切り取って使用する菌糸先端部は必ず2核を含む細胞が含まれなければならない。
菌糸先端部は、図3に示すように、2核菌糸が伸長するとき、先ず新たな細胞ができ、その後、前の細胞の核が分裂して2個の核がクランプを通り新たな細胞へ移動することで新たな2核細胞となる。新たな細胞への核移動の途中で存在する可能性のある1個の核のみを持った先端細胞では、子実体を作る菌株は得られない。
したがって、元のシイタケ菌株の性質をそのまま受け継いだ子実体を収穫するためには、2核の細胞が必要となる。また、2核を持った細胞を多く使用すると、時間とともにウイルス感染している可能性が増大するので、先端側の1個以上の2核細胞を使用する。
したがって、図3中のA−A、B−B部分から先部分、即ち菌糸先端の1つ以上の2核細胞又は1つ以上の2核細胞と1核細胞を切り出して使用する。
【0022】
次に、植物のウイルスフリー化方法等を参考に各種の方法を試みた実験の結果を説明する。
【0023】
本実験では事前に、保有していたシイタケ菌46菌株を供試菌株として、2本鎖RNAウイルスの検出を試みた。このうち2株に2本鎖RNAウイルス感染が確認された。
この2株は「Lentinula edodes mycovirus HKAウイルスに感染したシイタケ菌株(LeAウイルス感染シイタケ菌株Aと呼ぶ)」と「Lentinula edodes mycovirus HKBウイルスに感染したシイタケ菌株(LeBウイルス感染シイタケ菌株Bと呼ぶ)」である。
そして、上記2株の内LeAウイルス感染シイタケ菌株Aは、熟成させても白色のままで正常な褐変を起こさない「褐変化不良症状」が確認された。
その内LeBウイルス感染シイタケ菌株Bは、正常な菌株である。
【0024】
次に、本実験で行った2本鎖RNAウイルスの検出方法について説明する。
先ず、供試菌株にウイルスフリー化を目的とした処理をした後、菌糸をバイオマッシャー(フナコシ)で破砕し、セパゾール(ナカライテスク)を用いてRNA抽出した。
得られたRNAを熱変性した後、RT−PCR法を用いたアガロースゲル電気泳動を行い、バンドの有無を確認した。
【0025】
本実験では、LeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bの2種類のウイルス感染シイタケ菌糸を用いて実験を行った。
その実験の内容は以下の菌糸先端分離、加熱処理、低温処理等の下記(1)〜(8)の8通りであり、その結果を示すのが下記「表1」及び「表2」である。
【表1】

【0026】
【表2】
【0027】
(1)無処理
他の手法と比較するための基準として無処理の場合を確認した。
LeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株BをPDA培地20°Cで培養し、2本鎖RNAウイルス検出試験を行った結果、ウイルスが存在していることが確認された。
【0028】
(2)菌糸先端分離
LeAウイルス感染シイタケ菌株AをPDA培地18°Cで培養して、その伸長した菌糸の先端を切り出した菌糸の2本鎖RNAウイルス検出試験を行った結果、ウイルスが除去されていないことが確認された。
【0029】
(3)加熱処理
PDA培地18°Cで7日間培養したLeAウイルス感染シイタケ菌株Aを、35、40、50、60、70、80、90、100°Cの各温度に設定したインキュベータに処理時間内静置した。
その後18°Cの温度でインキュベータに移して培養した。
各温度の中でそれぞれ最も長い時間の加熱処理に耐えて発菌してきた菌糸について、2本鎖RNAウイルス検出試験を行った。
下記表3中の数値は発菌数であり、「−」は未試験であることを表している。
【0030】
【表3】
【0031】
上記「表3」では、温度が高くなると短い加熱時間で菌糸の発菌が不能(発菌数が「0」となる)となるここが確認できる。発菌数が「10」以下時間は正常に発菌し、発菌数が「0」以上の時間では発菌しない発菌限界であることが把握できた。
また、上記の各温度で最も長い時間の加熱処理に耐えて発菌してきた菌糸について、2本鎖RNAウイルス検出試験を行ったが、すべてウイルスが除去されていないことが確認された。
【0032】
(4)低温処理
LeAウイルス感染シイタケ菌株Aを、−85°Cの超低温フリーザーで冷凍し、解凍後PDA培地18°Cで培養し、発菌してきた菌糸について2本鎖RNAウイルス検出試験を行った。ウイルスが除去されていないことが確認された。
【0033】
(5)抗生物質(シクロヘキシミド;Ch)と菌糸先端分離
Ch添加したPDA培地(Ch0.05μg/ml)でLeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bを23°Cの温度でそれぞれ培養し、伸長した菌糸の先端部分を顕微鏡で確認して切り出し、培養を繰り返した。その結果、いずれもウイルスが除去されていないことが確認された。
【0034】
(6)栄養条件(素寒天培地)と菌糸先端分離
素寒天培地(1.5%寒天)23°CでLeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bをそれぞれ培養し、伸長した菌糸の先端部分を顕微鏡で確認して切り出し、培養を繰り返した。その結果、いずれもウイルスが除去されていないことが確認された。
【0035】
(7)栄養条件(木粉培地)継代
木粉培地(ブナ材使用・含水率約65%に調節)23°CでLeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bをそれぞれ培養し、継代を繰り返した。いずれもウイルスが除去されていないことが確認された。
【0036】
(8)加熱処理と菌糸先端分離
LeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bについて、先ず「耐熱限界温度」の確認試験をした。「耐熱限界温度」とは、シイタケ菌糸が伸長し且つその菌糸先端部が正常に発菌できる最も高い温度を示す。下記表4に示すように、32°Cではいずれも先端菌糸の発菌が不能となることが確認され、31°Cが耐熱限界温度であった。
【0037】
【表4】
【0038】
上記表4の実験では、加熱期間が同じ10日間であっても、加熱する温度が31°Cの場合と32°Cの場合とでは全く異なる結果となっている。
このことから、加熱処理後の菌糸先端部の伸長にとって、31°Cでは伸長できて32°Cでは伸長できないことから31°Cが菌糸の「耐熱限界温度」であると考えられた。
【0039】
確認された「耐熱限界温度」31°Cで10日間培養し、ウイルス感染する前の2核の菌糸先端部を切り離してこれを培養することで、いずれの菌糸についてもウイルスが除去されたことが確認され、ウイルスフリー化が成功した。
温度が32°C以上ではウイルスフリーの菌株は得られない。
なお、31°C以下の温度では菌糸先端部以外の伸長前の菌糸に感染したウイルスは死滅したわけではなく、31°Cの温度で10日間に伸長した菌糸先端細胞にはウイルスの感染が及んでいない状態となっている。
【産業上の利用可能性】
【0040】
本発明は、シイタケ菌株以外のキノコ類の菌株についても利用できる可能性がある。
【符号の説明】
【0041】
1a シャーレ
1b シャーレ
2a 菌糸片
2b 菌糸先端部
3a ウイルスを含む伸長した菌株
3b ウイルスを含まない伸長した菌株
3c ウイルスを含まない増殖した菌株
【技術分野】
【0001】
本発明は、ウイルス感染したシイタケ菌株から、ウイルスフリー株を得る方法に関する。
【背景技術】
【0002】
現在、生シイタケ生産は菌床栽培が主流であり、収益性の高い施設栽培化が進んでいる。しかしその反面、栽培規模拡大にともない原因不明の栽培不良症状の発生による減収が深刻な問題となる状況も増えている。病害の主な原因になるものとして細菌や糸状菌、ウイルスや害虫等が挙げられ、シイタケ生産、栽培におけるこれら病原に対する研究の発展は防除の必要性からも重要度を増している。
【0003】
一方、菌類を宿主とする菌類ウイルスについては、1962年HollingsがLa France病のツクリタケから病原体として検出したのが最初の発見である。日本でも1970年代にシイタケからウイルスが検出されており、また、ウイルス感染したシイタケ菌からウイルスを除去する方法についても、加熱と菌糸先端分離培養を繰り返し長期間行う処理によって一部ウイルスフリー化株が得られた研究報告があった。しかしながら、このウイルスフリー化方法は長期間(320日)を所要する上に、ウイルスフリー化できる確率が低い(3割に満たない)ため安定的要素に欠けており、一般的に実用化される技術には至っていない。しかも当時、正常なシイタケであってもウイルス感染が認められることからウイルスが病害因子とは見なされず、その後の長い間研究の進歩はなかった。
【0004】
近年、2004年にエノキタケのウイルス病が登録されるなど、シイタケやエノキタケ、ハタケシメジなど栽培キノコにおけるウイルス病害を示唆する研究報告例があがっている。また、今まで知見の少なかった菌類ウイルスのゲノム解析などの情報も増えつつあり、RT−PCR法を用いた高感度なウイルス検出方法も開発された。現在、栽培キノコの病害因子としてさらなるウイルスの解明と同時に、防除の面から、確実なウイルスフリー化技術が必要とされている。
【0005】
植物ウイルスについては、茎頂培養が一般的なウイルスフリー化技術として実用化されている。また、下記特許文献1には、活性酸素を用い、ウイルスを不活性化させて損傷の少ない植物の成長点のみ生かすという選択的手法によって植物をウイルスフリーにする方法が提案されている。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】特開平10−249364号公報
【発明の概要】
【発明が解決しようとする課題】
【0007】
しかしながら、本発明者等が実験をしたところ、シイタケ菌糸については菌糸先端培養や抗生物質などの薬品を用いる方法ではウイルスを除去することができなかった。
そこで、本発明の課題は、ウイルス感染したシイタケ菌株からウイルス感染がなく且つ正常な伸長が可能となる菌株を確実に得るウイルスフリー化方法を提供することにある。
【課題を解決するための手段】
【0008】
上記課題を解決するために、本発明者等は、ウイルスが高温に弱い性質があり、またキノコ類も高温で菌糸が衰弱や死滅する性質があるので、両者の温度に対する生存限界の違いによって、温度制御をすれば選択的に菌糸は生かしてウイルスは死滅させることができるのではないかと考えた。そして、工夫と実験を重ねた結果、本発明のシイタケ菌糸のウイルスフリー化方法を完成させることができた。
【0009】
本発明における請求項1に係るシイタケ菌糸のウイルスフリー化方法は、ウイルス感染したシイタケの菌糸を、シイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度で培養する加熱処理工程と、加熱処理中新たに伸びてきた菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、から成る。
【0010】
また請求項2に係るシイタケ菌糸のウイルスフリー化方法は、ウイルス感染したシイタケの菌糸を、シイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度で培養する加熱処理工程と、加熱処理中新たに伸びてきた菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、分離した菌糸先端部を適温で培養し菌糸を伸長させて菌株を得る菌糸増殖工程と、から成る。
【発明の効果】
【0011】
本発明のシイタケ菌糸のウイルスフリー化方法は、菌糸を加熱する加熱処理工程において、ウイルス感染している菌糸をシイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度(以下耐熱限界温度と呼ぶ)で培養することで、その間にウイルス感染したシイタケの菌糸の先端は生き延びさせつつ培養し、ウイルスは弱らせて菌糸の先端にまでには繁殖が進まない段階で、その菌糸の先端にあるウイルスがなく伸長が可能となる2核の先端細胞を先端部分離工程で切り出し、これを元の菌糸から別の場所に隔離してウイルスフリーの状態で菌糸を培養することが可能となった。
元のシイタケ菌糸についてはウイルス感染していても、耐熱限界温度で培養すると2核を有した先端細胞にはウイルスの感染が起こらないことが、RT−PCR法を用いたアガロースゲル電気泳動で現れるバンドの有無で確認できた。
なお、RT−PCR法は、Reverse Transcription Polymerase Chain Reactionの略で「逆転写酵素ポリメラーゼ連鎖反応」のことであり、ウイルスの検出に有効な手法である
【0012】
請求項2に係るシイタケ菌糸のウイルスフリー化方法では、上記方法の先端部分離工程で得られたウイルス感染していない2核の先端細胞を、さらに菌糸増殖工程において隔離した別の場所で適度な温度環境下に置くことで菌糸が大きく伸長した正常な菌株をウイルスが存在しない状態で得ることが可能となる。
【図面の簡単な説明】
【0013】
【図1】本発明のシイタケ菌糸のウイルスフリー化方法のフロー図である。
【図2】別形態のシイタケ菌糸のウイルスフリー化方法のフロー図である。
【図3】菌糸先端細胞の拡大図である。
【図4】(イ)はシャーレの培地に菌糸片を置いた状態を示し、(ロ)は菌糸の伸長した状態を示し、(ハ)は加熱処理中新たに伸長した菌糸の先端部を別のシャーレの培地に移した状態を示す各平面図である。
【図5】(イ)はシャーレの培地に菌糸片を置いた状態を示し、(ロ)は菌糸の伸長した状態を示し、(ハ)は加熱処理中新たに伸長した菌糸の先端部を別のシャーレの培地に移した状態を示し、(ニ)は菌糸が伸長した状態を示し、(ホ)は菌糸が増殖した状態を示す各平面図である。
【発明を実施するための形態】
【0014】
以下本発明の実施の形態を説明する。
【0015】
本発明のシイタケ菌糸のウイルスフリー化方法は、図1に示すように、ウイルス感染したシイタケの菌糸をPDA培地に、ウイルス感染している菌糸をシイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度(耐熱限界温度)で培養する加熱処理工程と、該加熱処理工程中新たに伸びてきた菌糸の2核の菌糸先端部を、ウイルス感染している元の菌糸から切り離す先端部分離工程とから成る。
【0016】
また、図2に示すように、前記先端部分離工程の後に、分離した菌糸先端部を適温で培養して菌糸を伸長させて菌株を得る菌糸増殖工程を有する形態が可能である。
【0017】
本発明を以下さらに詳しく説明する。
図3に示すように、ウイルス感染している菌株の中から菌糸片2aを切り出し、図4の(イ)に示すように、シャーレ1aの中のPDA(寒天)培地に移す。このとき菌糸はまだウイルス感染している。
【0018】
次に、ウイルス感染しているシイタケの菌糸片2aを耐熱限界温度で培養する。すると、図4(ロ)に示すように、新たに菌糸が周囲に少し伸長して菌株3aが得られる。
次に、顕微鏡で菌糸先端部を確認し、前記菌株3aの中から図3中のA−A、B−B部分から先部を含む先端部、即ち菌糸先端の1つ以上の2核細胞又は1つ以上の2核細胞と1核細胞を切り出す。
そして、切り出した該菌糸先端部2bを、図4の(ハ)に示すように、シャーレ1bの中のPDA培地に移す。
この菌糸先端部2bにはウイルスが存在しない。したがって、この時点で実質的にウイルスフリー化がなされている。
【0019】
また、前記先端部分離工程以降は、図5の(ニ)に示すように、シャーレ1bの中でPDA培地にウイルスのいない前記菌糸先端部2bを置き、耐熱限界温度で1ヶ月培養すると、図5の(ニ)に示すように、菌糸先端部2bの周囲に菌糸が少し伸長して菌株3bが得られる。このとき伸長した菌株3bはまだ小さい。
そこで、そのシャーレ1bの中で23°Cの温度で7日間培養する。すると、図5の(ホ)に示すように、菌糸が周囲に広く増殖する。そして全ての菌糸細胞にウイルスが感染していない菌株3cが得られる。
【0020】
以上のように、耐熱限界温度で培養する加熱処理した菌株3aから、菌糸先端部2bを分離して取り出した菌糸には、ウイルス感染していないが、ウイルスフリーをより確実にするために、前記先端部分離工程以降に、さらに前記先端部分離工程と菌糸増殖工程とを数回繰り返せば、全くウイルスのないウイルスフリーの菌株を得ることが可能となる。
【0021】
本発明では、加熱処理工程後は、2核の菌糸先端部を切り離す先端部分離工程を行うが、この工程で切り取って使用する菌糸先端部は必ず2核を含む細胞が含まれなければならない。
菌糸先端部は、図3に示すように、2核菌糸が伸長するとき、先ず新たな細胞ができ、その後、前の細胞の核が分裂して2個の核がクランプを通り新たな細胞へ移動することで新たな2核細胞となる。新たな細胞への核移動の途中で存在する可能性のある1個の核のみを持った先端細胞では、子実体を作る菌株は得られない。
したがって、元のシイタケ菌株の性質をそのまま受け継いだ子実体を収穫するためには、2核の細胞が必要となる。また、2核を持った細胞を多く使用すると、時間とともにウイルス感染している可能性が増大するので、先端側の1個以上の2核細胞を使用する。
したがって、図3中のA−A、B−B部分から先部分、即ち菌糸先端の1つ以上の2核細胞又は1つ以上の2核細胞と1核細胞を切り出して使用する。
【0022】
次に、植物のウイルスフリー化方法等を参考に各種の方法を試みた実験の結果を説明する。
【0023】
本実験では事前に、保有していたシイタケ菌46菌株を供試菌株として、2本鎖RNAウイルスの検出を試みた。このうち2株に2本鎖RNAウイルス感染が確認された。
この2株は「Lentinula edodes mycovirus HKAウイルスに感染したシイタケ菌株(LeAウイルス感染シイタケ菌株Aと呼ぶ)」と「Lentinula edodes mycovirus HKBウイルスに感染したシイタケ菌株(LeBウイルス感染シイタケ菌株Bと呼ぶ)」である。
そして、上記2株の内LeAウイルス感染シイタケ菌株Aは、熟成させても白色のままで正常な褐変を起こさない「褐変化不良症状」が確認された。
その内LeBウイルス感染シイタケ菌株Bは、正常な菌株である。
【0024】
次に、本実験で行った2本鎖RNAウイルスの検出方法について説明する。
先ず、供試菌株にウイルスフリー化を目的とした処理をした後、菌糸をバイオマッシャー(フナコシ)で破砕し、セパゾール(ナカライテスク)を用いてRNA抽出した。
得られたRNAを熱変性した後、RT−PCR法を用いたアガロースゲル電気泳動を行い、バンドの有無を確認した。
【0025】
本実験では、LeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bの2種類のウイルス感染シイタケ菌糸を用いて実験を行った。
その実験の内容は以下の菌糸先端分離、加熱処理、低温処理等の下記(1)〜(8)の8通りであり、その結果を示すのが下記「表1」及び「表2」である。
【表1】
【0026】
【表2】
【0027】
(1)無処理
他の手法と比較するための基準として無処理の場合を確認した。
LeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株BをPDA培地20°Cで培養し、2本鎖RNAウイルス検出試験を行った結果、ウイルスが存在していることが確認された。
【0028】
(2)菌糸先端分離
LeAウイルス感染シイタケ菌株AをPDA培地18°Cで培養して、その伸長した菌糸の先端を切り出した菌糸の2本鎖RNAウイルス検出試験を行った結果、ウイルスが除去されていないことが確認された。
【0029】
(3)加熱処理
PDA培地18°Cで7日間培養したLeAウイルス感染シイタケ菌株Aを、35、40、50、60、70、80、90、100°Cの各温度に設定したインキュベータに処理時間内静置した。
その後18°Cの温度でインキュベータに移して培養した。
各温度の中でそれぞれ最も長い時間の加熱処理に耐えて発菌してきた菌糸について、2本鎖RNAウイルス検出試験を行った。
下記表3中の数値は発菌数であり、「−」は未試験であることを表している。
【0030】
【表3】
【0031】
上記「表3」では、温度が高くなると短い加熱時間で菌糸の発菌が不能(発菌数が「0」となる)となるここが確認できる。発菌数が「10」以下時間は正常に発菌し、発菌数が「0」以上の時間では発菌しない発菌限界であることが把握できた。
また、上記の各温度で最も長い時間の加熱処理に耐えて発菌してきた菌糸について、2本鎖RNAウイルス検出試験を行ったが、すべてウイルスが除去されていないことが確認された。
【0032】
(4)低温処理
LeAウイルス感染シイタケ菌株Aを、−85°Cの超低温フリーザーで冷凍し、解凍後PDA培地18°Cで培養し、発菌してきた菌糸について2本鎖RNAウイルス検出試験を行った。ウイルスが除去されていないことが確認された。
【0033】
(5)抗生物質(シクロヘキシミド;Ch)と菌糸先端分離
Ch添加したPDA培地(Ch0.05μg/ml)でLeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bを23°Cの温度でそれぞれ培養し、伸長した菌糸の先端部分を顕微鏡で確認して切り出し、培養を繰り返した。その結果、いずれもウイルスが除去されていないことが確認された。
【0034】
(6)栄養条件(素寒天培地)と菌糸先端分離
素寒天培地(1.5%寒天)23°CでLeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bをそれぞれ培養し、伸長した菌糸の先端部分を顕微鏡で確認して切り出し、培養を繰り返した。その結果、いずれもウイルスが除去されていないことが確認された。
【0035】
(7)栄養条件(木粉培地)継代
木粉培地(ブナ材使用・含水率約65%に調節)23°CでLeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bをそれぞれ培養し、継代を繰り返した。いずれもウイルスが除去されていないことが確認された。
【0036】
(8)加熱処理と菌糸先端分離
LeAウイルス感染シイタケ菌株AとLeBウイルス感染シイタケ菌株Bについて、先ず「耐熱限界温度」の確認試験をした。「耐熱限界温度」とは、シイタケ菌糸が伸長し且つその菌糸先端部が正常に発菌できる最も高い温度を示す。下記表4に示すように、32°Cではいずれも先端菌糸の発菌が不能となることが確認され、31°Cが耐熱限界温度であった。
【0037】
【表4】
【0038】
上記表4の実験では、加熱期間が同じ10日間であっても、加熱する温度が31°Cの場合と32°Cの場合とでは全く異なる結果となっている。
このことから、加熱処理後の菌糸先端部の伸長にとって、31°Cでは伸長できて32°Cでは伸長できないことから31°Cが菌糸の「耐熱限界温度」であると考えられた。
【0039】
確認された「耐熱限界温度」31°Cで10日間培養し、ウイルス感染する前の2核の菌糸先端部を切り離してこれを培養することで、いずれの菌糸についてもウイルスが除去されたことが確認され、ウイルスフリー化が成功した。
温度が32°C以上ではウイルスフリーの菌株は得られない。
なお、31°C以下の温度では菌糸先端部以外の伸長前の菌糸に感染したウイルスは死滅したわけではなく、31°Cの温度で10日間に伸長した菌糸先端細胞にはウイルスの感染が及んでいない状態となっている。
【産業上の利用可能性】
【0040】
本発明は、シイタケ菌株以外のキノコ類の菌株についても利用できる可能性がある。
【符号の説明】
【0041】
1a シャーレ
1b シャーレ
2a 菌糸片
2b 菌糸先端部
3a ウイルスを含む伸長した菌株
3b ウイルスを含まない伸長した菌株
3c ウイルスを含まない増殖した菌株
【特許請求の範囲】
【請求項1】
ウイルス感染したシイタケ菌糸を、シイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度で培養する加熱処理工程と、加熱処理中新たに伸長した菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、から成るシイタケ菌糸のウイルスフリー化方法。
【請求項2】
ウイルスが感染したシイタケの菌糸を、シイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度で培養する加熱処理工程と、加熱処理中新たに伸長した菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、分離した菌糸先端部を適温で培養し菌糸を伸長させて菌株を得る菌糸増殖工程と、から成るシイタケ菌糸のウイルスフリー化方法。
【請求項1】
ウイルス感染したシイタケ菌糸を、シイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度で培養する加熱処理工程と、加熱処理中新たに伸長した菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、から成るシイタケ菌糸のウイルスフリー化方法。
【請求項2】
ウイルスが感染したシイタケの菌糸を、シイタケ菌糸が伸長し且つその分離した菌糸先端部が正常に発育できる最も高い温度で培養する加熱処理工程と、加熱処理中新たに伸長した菌糸の2核細胞を1つ以上含む先端細胞を切り離す先端部分離工程と、分離した菌糸先端部を適温で培養し菌糸を伸長させて菌株を得る菌糸増殖工程と、から成るシイタケ菌糸のウイルスフリー化方法。
【図1】


【図2】

【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−178992(P2012−178992A)
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願番号】特願2011−43146(P2011−43146)
【出願日】平成23年2月28日(2011.2.28)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 研究集会名 日本きのこ学会第14回大会 主催者名 日本きのこ学会 開催日 平成22年9月15日〜17日(講演要旨集の発行: 平成22年9月1日)
【出願人】(000242024)株式会社北研 (17)
【Fターム(参考)】
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願日】平成23年2月28日(2011.2.28)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り 研究集会名 日本きのこ学会第14回大会 主催者名 日本きのこ学会 開催日 平成22年9月15日〜17日(講演要旨集の発行: 平成22年9月1日)
【出願人】(000242024)株式会社北研 (17)
【Fターム(参考)】
[ Back to top ]