
シガトキシン類を認識するヒト化抗体
【課題】ヒト抗マウス応答を最小限にとどめ、かつシガトキシンに対する強い親和性を示すヒト化抗シガトキシン抗体を提供する。
【解決手段】ヒト免疫グロブリンのフレームワーク領域(FW)と、FERM BP−8292のハイブリドーマにより産生されるシガトキシン類を認識するマウス抗体10C9の相補性決定領域(CDR)とを含む可変領域をそれぞれ有する重鎖及び軽鎖を少なくとも含み、軽鎖及び重鎖のFWの特定の位置のアミノ酸が、前記マウス抗体10C9の軽鎖の対応する位置に存在するアミノ酸で置き換えられていることを特徴とする、シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識するヒト化抗体又はそのフラグメント。
【解決手段】ヒト免疫グロブリンのフレームワーク領域(FW)と、FERM BP−8292のハイブリドーマにより産生されるシガトキシン類を認識するマウス抗体10C9の相補性決定領域(CDR)とを含む可変領域をそれぞれ有する重鎖及び軽鎖を少なくとも含み、軽鎖及び重鎖のFWの特定の位置のアミノ酸が、前記マウス抗体10C9の軽鎖の対応する位置に存在するアミノ酸で置き換えられていることを特徴とする、シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識するヒト化抗体又はそのフラグメント。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は一般に、シガトキシンCTX3C及び51−ヒドロキシ−CTX3Cを特異的に認識できるヒト化抗体に関する。
【背景技術】
【0002】
食中毒シガテラは、ポリネシア、ハワイ、沖縄、カリブ海などの広いサンゴ礁海域で頻発し、年間2万人以上の中毒患者が発生している。シガテラ毒素は植物プランクトンが生産し、食物連鎖を介して魚介類に含まれることとなる。したがって、一旦発生すると多数の食用魚が広範に毒化して深刻な社会問題となることから、シガテラ毒素を迅速に検出することが食中毒シガテラの予防にとって重要である。
【0003】
食中毒シガテラの主要成分であるシガトキシン類(ciguatoxin、CTX)の免疫学的手法を用いた検出に関する研究は、ラジオイムノアッセイの発展に伴い、1977年頃から始まった。免疫学的手法による研究においては、シガトキシン類を認識する抗体が必要であるが、シガトキシン類は天然からごく微量しか採集されず、培養による生産も困難なことから、抗体を製造することも困難である。ハワイ大学ホカマらは、貴重な毒本体シガトキシンを用いて、モノクローナル抗体を調製し(非特許文献1参照)、この抗体を用いて、シガトキシン類を検出するための試薬、キットなどが開発されている。
【0004】
ところで、シガトキシン類は、単一の化合物ではなく、多種の毒素の混合物であり、次の一般式:
【0005】
【化1】

【0006】
で表される4種(化合物1〜4)が主に知られている。
【0007】
このような状況のなかで、本発明者らは、以前にシガトキシンCTX3Cの右末端の部分構造であるIJKLM環部を含む合成ハプテンを設計、合成し、この合成ハプテンのタンパク質コンジュゲートでマウスを免疫する工程を含む方法により、受託番号FERM BP‐8293のハイブリドーマ3D11を作製し、該ハイブリドーマを用いて、上記の化合物1及び3のシガトキシン類に特異性の高いモノクローナル抗体3D11を製造することに成功した(特許文献1参照)。
【0008】
さらに、本発明者らは、以前にシガトキシンCTX3Cの左末端の部分構造であるABCDE環部を含む合成ハプテンを設計、合成し、この合成ハプテンのタンパク質コンジュゲートでマウスを免疫する工程を含む方法により、受託番号FERM BP‐8292のハイブリドーマ10C9を作製し、該ハイブリドーマを用いて、上記の化合物1及び2のシガトキシン類に特異性の高いモノクローナル抗体10C9を調製することに成功した(特許文献2参照)。モノクローナル抗体10C9のシガトキシンCTX3Cに対する解離定数(Kd)は、2.8nMであった。
【0009】
また、本発明者らは、上記の2種のモノクローナル抗体を組み合わせて、検出特性をより改善したサンドイッチ法により、シガトキシンCTX3Cを検出するキットを作製した(特許文献3及び非特許文献2参照)。
【0010】
シガトキシン類は、神経細胞膜上のナトリウムチャネルに作用するので、神経細胞にナトリウムイオンの流入を引き起こす。この結果、軸索で脱分極が起こり、アセチルコリン等の神経伝達物質放出を促進すると考えられている。一方、シガトキシンの作用機構の研究が進められるにつれて、いくつかのアンタゴニストが発見されてきた。現在、シガトキシン類の解毒剤として、シガトキシン類のアンタゴニストである抗コリン性のアトロピンや、マンニトールの有効性が報告されている(非特許文献3参照)が、治療薬の開発には至っていない。シガテラ中毒を発症した場合、自然回復を待つしかないのが現状であり、現時点では有効な治療方法が存在しない。
【0011】
これまでに本発明者らは、上記の2種のモノクローナル抗体(3D11、10C9)について、細胞を用いたin vivoでの試験及びマウスを用いたin vitroでの試験を行い、2種の抗体を用いてシガテラ中毒に対する中和が有効であることを確認している(非特許文献4参照)。
【0012】
1975年にハイブリドーマ技術が開発されて以来(非特許文献5参照)、モノクローナル抗体の産生が可能となり、マウスモノクローナル抗体は現在、様々な疾病の診断的手法として広く利用されている。しかしマウス抗体を治療薬としてヒト体内に投与すると、ヒト抗マウス抗体(human anti−mouse antibodies;HAMA)応答が引き起こされ、この反応は時にヒトにとって致命的なものになりうる。このように、ヒト体内でマウス抗体は免疫原性を持つので、マウス抗体をヒトに直接投与することは回避することが望ましい。しかし、動物由来抗体の免疫原性を軽減するために、ヒトのモノクローナル抗体を作製することは、技術的及び倫理的問題により困難である。
【0013】
そこで、マウス抗体の抗原結合部位を含む可変領域をヒト抗体の定常領域に接合したキメラ抗体を作製することが行われている。しかし、キメラ抗体もなお、その抗原性が高く、ヒト抗キメラ抗体(human anti−chimeric antibodies;HACA)応答を引き起こすと考えられる(非特許文献6参照)。
これを改善するため、マウス抗体の抗原結合に最も重要な相補性決定領域(CDR)のみをヒト抗体フレームワークに移植した(CDR−grafting)ヒト化抗体が開発されている。このようなヒト化抗体作製技術は1990年代から発展し、現在ではがん、白血病、関節リウマチなど多くの疾病にヒト化抗体が医薬品として用いられている。
【0014】
ヒト化抗体の作製技術は、キメラ抗体作製よりも複雑な技術である。抗原に対する特異性や親和性を維持するためには、CDRの移植のみでは不十分な場合が多い。抗原との結合に重要なアミノ酸残基は、マウス抗体のフレームワーク領域にも存在するので、ヒト抗体フレームワークのアミノ酸残基をマウス抗体アミノ酸に戻す必要がある。このような残基を同定するには、X線結晶解析などによる構造情報があると理想的であることが知られている。
【先行技術文献】
【特許文献】
【0015】
【特許文献1】特開2003−55400号公報
【特許文献2】特許第3845796号公報
【特許文献3】特許第3820519号公報
【非特許文献】
【0016】
【非特許文献1】Toxicon 第15巻、(1977年)、第317頁
【非特許文献2】Journal of American Chemical Society 第125巻、(2003年)、第7608頁
【非特許文献3】Laurentら編、Ciguatera: field reference guide/un guide pratique、Secretariat of the Pacific Community出版、(2005年)
【非特許文献4】Toxicon 第53巻、(2009年)、第802頁
【非特許文献5】Nature 第256巻、(1975年)、第495頁
【非特許文献6】Dubel編、Handbook of Therapeutic Antibodies、Wiley−VCH出版、(2007年)、第119頁
【発明の概要】
【発明が解決しようとする課題】
【0017】
本発明の課題は、食中毒シガテラの治療薬開発のために、ヒト抗マウス応答を最小限にとどめ、かつシガトキシン類に対する強い親和性を示すヒト化抗シガトキシン抗体を提供することにある。
本発明者らは、上記の課題を解決するために、従来のマウス抗体をヒト化及び最適化して、従来のマウス抗体と同等の強い親和性をシガトキシン類に対して示すヒト化抗シガトキシン抗体を取得することに成功した。
【課題を解決するための手段】
【0018】
すなわち、本発明は、
ヒト免疫グロブリンのフレームワーク領域(FW)と、
FERM BP−8292のハイブリドーマにより産生されるシガトキシン類を認識するマウス抗体10C9の相補性決定領域(CDR)と
を含む可変領域をそれぞれ有する重鎖及び軽鎖を少なくとも含み、
軽鎖の46位のアミノ酸と、任意に、1位、2位、7位、60位及び71位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の軽鎖の対応する位置に存在するアミノ酸で置き換えられ、及び/又は
重鎖の94位のアミノ酸と、任意に、5位、71位及び73位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の重鎖の対応する位置に存在するアミノ酸で置き換えられており、
シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識する、ヒト化抗体又はそのフラグメント(以下、単に「ヒト化抗体」ともいう)を提供する。
【0019】
なお、本明細書において、免疫グロブリンの軽鎖及び重鎖のアミノ酸配列の番号付けは、Kabatら、Sequences of Proteins of Immunological Interest、第5版、Bethesda,MD:U.S.Dept.of Health and Human Services,Public Health Service,National Institutes of Health,1991で定義された免疫グロブリンの軽鎖及び重鎖の番号付けの定義に従う。
任意の免疫グロブリンのアミノ酸配列の番号付けは、http://www.bioinf.org.uk/abs/abnumのウェブサイトで行うこともできる。
【0020】
また、本発明は、上記のヒト化抗体の重鎖又はそのフラグメントをコードする配列を含むポリヌクレオチド、上記のヒト化抗体の軽鎖又はそのフラグメントをコードする配列を含むポリヌクレオチド、上記のポリヌクレオチドを含むベクター、上記のベクターが導入された宿主細胞、並びに上記の宿主細胞を培養して、上記のヒト化抗体又はそのフラグメントを発現させる工程を含むヒト化抗体又はそのフラグメントを生産する方法も提供する。
【発明の効果】
【0021】
本発明によって、シガトキシンに対して強い親和性を示すヒト化抗シガトキシン抗体が提供される。このような特質を有する本発明の抗体は、ヒトに投与したときに、マウス抗体と比較して免疫原性が低下していると考えられるので、食中毒シガテラの予防又は治療用の医薬品の開発に有用である。
【図面の簡単な説明】
【0022】
【図1−1】マウス抗体10C9及び作製したヒト化抗体の軽鎖可変領域のアミノ酸配列を示す。m10C9:マウス10C9抗体;1GKV1D−39*01:ヒトgermline配列の軽鎖V領域;1GKJ2*01:ヒトgermline配列の軽鎖J領域;h10C9g−VL:グラフト化抗体;各h10C9d−VL及びh10C9o−VL:設計したヒト化抗体;1GKC*01:ヒトgermline配列の軽鎖定常領域。
【図1−2】マウス抗体10C9及び作製したヒト化抗体の重鎖可変領域のアミノ酸配列を示す。m10C9:マウス10C9抗体;1GKV4−31*03:ヒトgermline配列の重鎖V領域;1GHJ4*01:ヒトgermline配列の重鎖J領域;h10C9g−VH:グラフト化抗体;各h10C9d−VH及びh10C9o−VH:設計したヒト化抗体;1GHG1*01:ヒトgermline配列の重鎖定常領域。
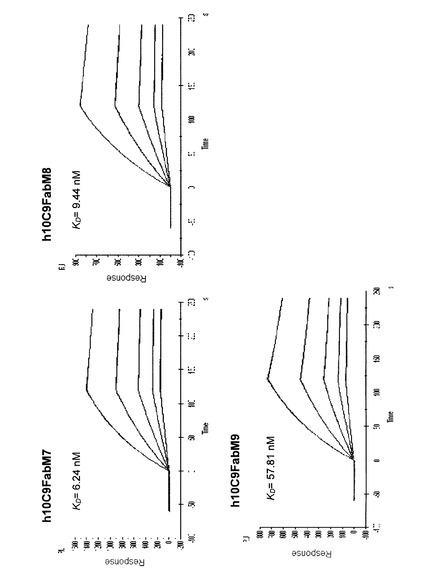
【図2】大腸菌内でマウス抗体及びヒト化抗体を産生するために使用されるFab発現ベクターの例を示す図である。
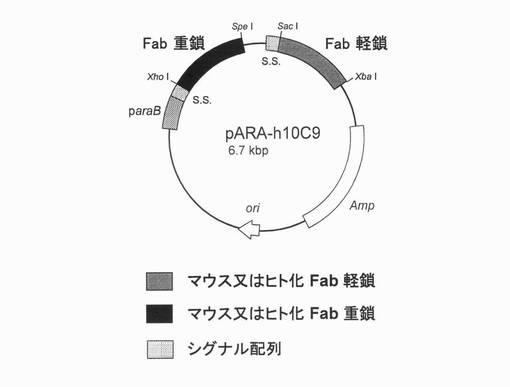
【図3】精製したマウス10C9抗体及び記載されるヒト化抗体のSDS−PAGE結果を示す図である。
【図4】マウス10C9抗体及び記載されるヒト化抗体を、抗マウス抗体及び/又は抗ヒト抗体で検出したウェスタンブロッティングの結果を示す図である。
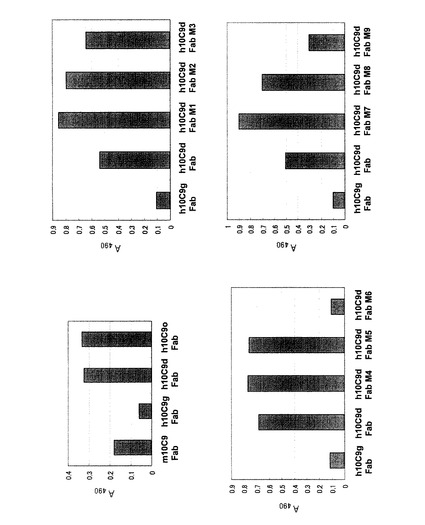
【図5】マウス10C9抗体及び各ヒト化抗体のCTX3C−ABCDEに対する抗原結合性をELISA法で調べた結果を示すグラフである。
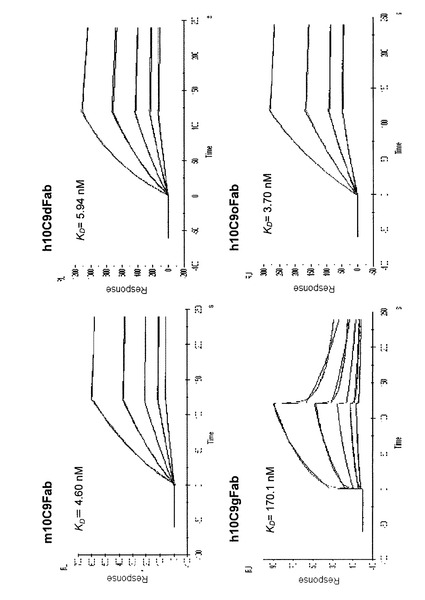
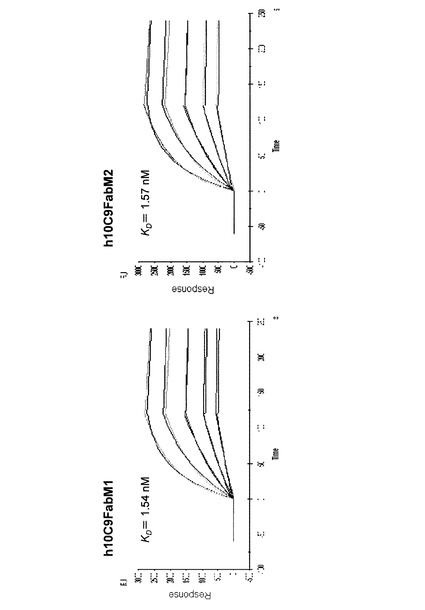
【図6】精製したマウス10C9抗体及び各ヒト化抗体のCTX3C−ABCDEに対する結合性をBiacoreで調べた結果を示すグラフである。
【発明を実施するための形態】
【0023】
本明細書において、「シガトキシン類」とは、マウス抗体10C9が認識して結合できるシガトキシンCTX3C及び51−ヒドロキシ−CTX3Cから選択されるシガトキシンのことをいう。
【0024】
本明細書において、「免疫グロブリン」及び「抗体」との用語は、同等の意味を有すると解される。「抗体」との用語は、一般的に、それに対する特定の「抗原」との関係において、機能に注目した名称であると解される。
【0025】
本発明のヒト化抗体又はそのフラグメントは、シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識して、これと結合できる。本発明のヒト化抗体のフラグメントとは、抗原に結合できるフラグメントであればよく、Fab、F(ab’)2フラグメントを含む。
【0026】
一般に、抗体分子の基本構造は、各クラス共通で、分子量5万〜7万の重鎖と2〜3万の軽鎖から構成される。重鎖は、通常約440個のアミノ酸を含むポリペプチド鎖からなり、クラスごとに特徴的な構造をもち、IgG、IgM、IgA、IgD、IgEに対応してγ、μ、α、δ、ε鎖とよばれる。さらにIgGには、IgG1、IgG2、IgG3、IgG4が存在し、それぞれγ1、γ2、γ3、γ4とよばれている。軽鎖は、通常約220個のアミノ酸を含むポリペプチド鎖からなり、L型とK型の2種が知られており、それぞれλ、κ鎖とよばれる。抗体分子の基本構造のペプチド構成は、それぞれ相同な2本の重鎖及び2本の軽鎖が、ジスルフィド結合(S−S結合)及び非共有結合によって結合され、分子量15万〜19万である。2種の軽鎖は、どの重鎖とも対をなすことができる。個々の抗体分子は、常に同一の軽鎖2本と同一の重鎖2本からできている。
【0027】
鎖内S−S結合は、重鎖に四つ(μ、ε鎖には五つ)、軽鎖には二つあって、アミノ酸100〜110残基ごとに一つのループを成し、この立体構造は各ループ間で類似していて、構造単位あるいはドメインとよばれる。重鎖、軽鎖ともにN末端に位置するドメインは、同種動物の同一クラス(サブクラス)からの標品であっても、そのアミノ酸配列が一定せず、可変領域(V領域、variable region、可変部)とよばれている(各ドメインは、それぞれ、VH及びVLと表される)。これよりC末端側のアミノ酸配列は、各クラスあるいはサブクラスごとにほぼ一定で定常領域(C領域、constant region、定常部)とよばれている(各ドメインは、それぞれ、CH1、CH2、CH3あるいはCLと表される)。軽鎖及び重鎖の中で、可変及び定常領域は、重鎖も同じく約10個以上のアミノ酸の「D」領域を内含している状態で、約12個以上のアミノ酸の「J」領域によって連結されている(一般に、Fundamental Immunology(基礎免疫学)(Paul,W.編,第2版,Raven Press,N.Y.,1989、第7章を参照のこと)。
【0028】
抗体の抗原決定部位はVH及びVLによって構成され、結合の特異性はこの部位のアミノ酸配列によっている。一方、補体や各種細胞との結合といった生物学的活性は各クラスIgのC領域の構造の差を反映している。重鎖と軽鎖の可変領域の可変性は、どちらの鎖にも存在する3つの小さな超可変領域(hypervariable region)にほぼ限られることがわかっており、これらの領域を相補性決定領域(CDR、complementarity determining region)と呼んでいる。可変領域の残りの部分はフレームワーク領域(FW、framework region)とよばれ、比較的一定である。通常、各可変領域の相補性決定領域の5〜10個のアミノ酸だけが抗原結合部位を形成している。
通常、抗体の重鎖及び軽鎖の可変領域には、3つのCDR(CDR1〜3)と、4つのFW(FW1〜4)が含まれる。
【0029】
本明細書において、「キメラ抗体」とは、可変領域がマウス抗体(ドナー異種抗体とも称する)由来であり、定常領域がヒト免疫グロブリン由来である抗体のことである。
本明細書において、「ヒト化抗体」とは、抗原特異的なマウス抗体分子の相補性決定領域(抗原結合部位)以外がヒト免疫グロブリンに由来する組換え抗体のことである。ヒト化抗体は、その機能の向上のためにアミノ酸の改変(置換、挿入、欠失及び/又は付加)を含むことができる。
【0030】
一般に、ヒト化抗体の作製においては、相補性決定領域のアミノ酸配列のみを鋳型のヒト抗体フレームワークに移植しただけ(グラフト化抗体)では、多くの場合、オリジナルのマウス抗体よりも抗原結合活性が低下することが知られている。
本発明に用いられるマウス抗体10C9のCDRを、ヒト免疫グロブリンのフレームワークに移植することのみにより得られるグラフト化抗体も、シガトキシンCTX3Cに対する結合性は有するものの、抗原親和性が極めて低く、抗体医薬としての使用には好ましくないことが確認された(後記実施例D−5及びD−6を参照)。
【0031】
本発明者らは、このようなヒト化抗体の活性低下を改善し、かつ、抗体医薬として使用するためにより優れたヒト化抗体を得るために検討を重ねた結果、上記の本発明のヒト化抗シガトキシン類抗体が、従来のマウス抗シガトキシン類抗体と比較して、抗原親和性を維持しつつ、免疫原性が低下され得るという有意に改善された特性を有することを見出した。
【0032】
本発明のヒト化抗体に含まれるヒト免疫グロブリン(以下、「Ig」ともいう)のFW領域は、マウス抗体10C9のFWと同じ又は類似のコンホメーションをとることが好ましい。このことにより、マウス抗体のCDRと融合させたときに、抗原との結合のための正しい空間的配向が保持できる確率が高くなる。
このために、本発明では、軽鎖のFWの46位のアミノ酸と、任意に、1位、2位、7位、60位及び71位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の軽鎖の対応する位置に存在するアミノ酸で置き換えられ、及び/又は
重鎖のFWの94位のアミノ酸と、任意に、5位、71位及び73位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の重鎖の対応する位置に存在するアミノ酸で置き換えられている。
【0033】
上記の軽鎖及び重鎖のFW領域内のマウス抗体10C9の対応する位置に存在するアミノ酸で置き換えるアミノ酸は、軽鎖及び重鎖においてそれぞれ少なくとも1つである。軽鎖では、46位のアミノ酸と、上記の任意に選択される位置のアミノ酸の1つ、2つ、3つ、4つ又は5つの位置のさらなるアミノ酸を置き換えることができ、重鎖では、94位のアミノ酸と、上記の任意に選択される位置のアミノ酸の1つ、2つ又は3つの位置のさらなるアミノ酸を置き換えることができる。
【0034】
本発明のヒト化抗体の重鎖及び軽鎖のFW領域が由来するヒト免疫グロブリンは、天然に存在するヒト免疫グロブリン(IgG、IgM、IgA、IgD又はIgE)のFW領域の配列であってもよいし、いくつかのサブクラスのヒト免疫グロブリンのFW領域に共通するコンセンサス配列でもあり得る。Dubel編,Handbook of Therpeutic Antibodies、Wiley−VCH出版(2007)を参照。
重鎖及び軽鎖のそれぞれに含まれる複数のFW領域(FW1〜FW4)は、同じサブクラスのヒトIgに由来してもよいし、異なるサブクラスのヒトIgに由来してもよい。また、これらの複数のFW領域は、同じ免疫グロブリンに由来してもよいし、異なる免疫グロブリンに由来してもよい。
【0035】
ヒト免疫グロブリンのFW領域のアミノ酸配列は公知であり、例えばEntrez Protein(http://www.ncbi.nlm.nih.gov/sites/entrez?db=protein)などのデータベースから入手できる。
【0036】
上記のFW領域として、ヒト免疫グロブリンのgermline配列を利用することもできる(Dubel編,Handbook of Therapeutics Antibodies,Wiley−VCH出版,p.119,2007;Hwangら,Methods,vol.36,p.35,2005)。germline配列とは、体細胞変異が生じる前の生物が元来有する生殖細胞の遺伝子系列であり、ヒト免疫グロブリンについてもデータベースが公開されている(http://imgt.cines.fr//IMGT#GENE-DB/GENElect?query=4.2+&species=Homo+sapiens)。
ヒトgermline配列は、特異的なモチーフの発現が最小であり、免疫原性(免疫システムが非自己として認識する可能性)を低く抑えることができると考えられている。
【0037】
本発明のヒト化抗体のFW領域は、マウス抗体10C9の可変領域全体の配列と相同性が高くなるように選択したgermline配列に由来することが好ましい。このようにFW領域を選択することにより、マウス抗体の重要なアミノ酸残基と同じ位置で等しいアミノ酸残基を有するFW領域を用いることができる。また、germline配列を用いると、他の免疫グロブリン配列を用いるよりも、抗原結合活性の損失率が低いという利点もある。
【0038】
本発明のヒト化抗体に用いるためのヒト免疫グロブリン配列は、既知のヒト免疫グロブリンgermline配列のデータベースを利用して、軽鎖及び重鎖それぞれについてマウス抗体可変領域のアミノ酸配列と相同性が高い配列を、コンピュータにより(例えばBLASTにより)比較することによって同定できる。この比較により、マウス抗体10C9の可変領域と高い配列同一性を示すヒトgermlineからの軽鎖及び重鎖フレームワーク領域を選択できる。
【0039】
本発明のヒト化抗体は、実質的にヒトIgからのFW領域と、実質的にマウス抗体10C9からのCDRとを含む。このようなアミノ酸配列の並置は、天然には見られないので、ある種のアミノ酸の改変を加えることにより、より天然に近いコンホメーションを有し、抗原結合親和性の向上を導くことができる。
よって、得られるヒト化抗体の特性を最適化するために、フレームワーク領域内でどのアミノ酸残基を置換すればよいかを決定する(バックミューテーション残基)。一般に、マウス抗体に由来する残基をヒト化抗体に導入することは、ヒトの体内でのHAMA応答を引き起こす危険性を増大させる可能性が高いので、最小限に抑えることが好ましい。
【0040】
CDRコンホメーション及び/又は抗原への結合に対して考えられるその影響に基づいて、FW領域内で置換すべきアミノ酸を同定できる。
置換すべきアミノ酸残基は、(1)抗原に直接、非共有的に結合するアミノ酸残基、(2)CDR領域に隣接するか、Chothiaら(J.Mol.Biol.,Vol.196,p.901,1987;Nature,Vol.342,p.878,1989)により提案される代替的定義づけの下でCDR領域の一部であるか、CDRループの立体配座の支持に関与するか(Footeら,J.Mol.Biol.,Vol.224,p.487,1992)、又はその他の形でCDR領域と相互作用するアミノ酸残基、又は(3)VL−VHインタフェイスに参与するアミノ酸(Chothiaら,J.Mol.Biol.,Vol.186,p.651,1985)であることが好ましい。
【0041】
或いは、FW領域内で置換すべきアミノ酸残基は、マウス免疫グロブリン配列中でマウスの体細胞変異(ソマティック変異)が導入されたと考えられる位置に対応する位置のヒトFW領域内のアミノ酸残基が好ましい。マウス免疫グロブリン中で体細胞変異が導入されたと考えられるアミノ酸残基は、マウス抗体10C9のアミノ酸配列を、マウス免疫グロブリンgermline配列と、ClustalWなどのアルゴリズムをデフォルト設定で用いて相同性がなるべく高くなるように整列させ、マウス抗体10C9中の配列がgermline配列とは異なる位置のアミノ酸残基を選択することにより決定できる。体細胞変異が導入されたと考えられる位置のアミノ酸残基は、ヒト抗体にとっては普通でないアミノ酸であるので、マウス免疫グロブリン中で体細胞変異が導入された位置に対応する位置のヒト免疫グロブリンのアミノ酸をマウス抗体10C9の対応する位置のアミノ酸残基で置換することにより、抗体の抗原結合性の低下を最小限に抑えて、ヒト化抗体を作製できる。
【0042】
マウス10C9抗体の立体構造は、X線結晶構造解析によって以前に報告されている(Uiら,J.Biol.Chem.,Vol.283,p.19440,2008)。この立体構造情報に基づいて、抗原に直接結合するアミノ酸残基、及び抗原−抗体結合に間接的に関わる(結合部位の構造を維持する上で重要な)アミノ酸残基を同定できる。
【0043】
上記の情報に基づいて鋭意検討を重ねた結果、本発明のヒト化抗体は、軽鎖の46位と、任意に、1位、2位、7位、60位及び71位の少なくとも1つ、例えば1つ、2つ、3つ、4つ又は5つの位置のアミノ酸残基が、マウス抗体10C9の対応する位置のアミノ酸残基で置き換えられ、かつ重鎖の94位と、任意に、5位、71位及び73位の少なくとも1つ、例えば1つ、2つ又は3つの位置のアミノ酸残基が、マウス抗体10C9の対応する位置のアミノ酸残基で置き換えられたものであることにより、シガトキシン類との良好な抗原結合特性を示すことが見出された。
これらの置換され得るアミノ酸は、図1に下線を付して示す。
【0044】
本明細書において、「マウス抗体10C9の対応する位置のアミノ酸残基」とは、上記のKabatらによる免疫グロブリンの番号付けに従って番号を付したときに、ヒト免疫グロブリン中のアミノ酸残基と同じ番号を付与される位置のアミノ酸残基のことである。
【0045】
本発明のヒト化抗体に用いられるマウス抗体10C9のCDRは、独立行政法人産業技術総合研究所特許生物寄託センターに2003年2月13日に、ブダペスト条約に基づく寄託の移管申請により受託番号FERM BP−8292として寄託されたハイブリドーマ10C9から生成されるマウス抗体10C9のCDRである。このマウス抗体10C9は、シガトキシンCTX3C及び51−OH−CTX3CのABCDE環部を特異的に認識する。この抗体については、特許第3845796号及び特許第3820519号を参照されたい。
【0046】
マウス抗体10C9のアミノ酸配列は、当該技術において公知の方法により決定できる。マウス抗体10C9の軽鎖可変領域VL、軽鎖定常領域CL、重鎖可変領域VH及び重鎖定常領域CH1のcDNA配列及びそれらから推測されるアミノ酸配列を、それぞれ配列番号1、3、5及び7(cDNA配列)及び配列番号2、4、6及び8(アミノ酸配列)に示す。
【0047】
本発明のヒト化抗体は、より好ましくは、配列番号15〜19及び21のいずれか1つに示すアミノ酸配列を有する軽鎖と、配列番号22〜25及び27のいずれか1つに示すアミノ酸配列を有する重鎖とを含む。
上記のヒト化抗体は、最も好ましくは、配列番号21に示すアミノ酸配列を有する軽鎖と、配列番号27に示すアミノ酸配列を有する重鎖とを含む。
【0048】
本発明のヒト化抗体の定常領域としては、従来公知のヒト免疫グロブリンの定常領域を用いることができる。ヒトIgの定常領域は、軽鎖定常領域(CL領域)及び重鎖定常領域(CH1、CH2及びCH3)からなるが、本発明のヒト化抗体又はそのフラグメントは、シガトキシン類との結合性を有するように、これらの定常領域のいずれかを適切に有することができる。
ヒト免疫グロブリンの可変領域について述べたことと同様に、上記の定常領域も、IgG、IgM、IgA、IgD又はIgEのサブクラスのヒト免疫グロブリンのいずれに基づくものであってもよい。好ましくは重鎖定常領域としてはヒトIgγ1を、また、軽鎖定常領域としてはヒトIgκを用いることができる。
また、軽鎖及び重鎖のそれぞれの定常領域は、同じサブクラスのヒトIgに基づいてもよいし、異なるサブクラスのヒトIgに基づいてもよい。
【0049】
本発明のヒト化抗体は、本願明細書に開示される、本発明のヒト化抗体のアミノ酸配列をコードする塩基配列を有するポリヌクレオチドを当該分野で公知の方法を使用して作製し、該ポリヌクレオチドを発現ベクターに導入し、得られた発現ベクターを宿主細胞に導入し、宿主細胞を培養することを含む方法により得ることができる。
培養後の培養液から、ヒト化抗体を精製してもよい。
上記の各工程は、当該分野で公知の種々の方法を使用して行うことができる。
【0050】
上記の本発明の抗体の重鎖及び軽鎖のアミノ酸配列をコードするポリヌクレオチドは、例えば、当業者に公知の核酸合成方法に従って、マウス10C9抗体の重鎖及び軽鎖の各々のCDRをコードする遺伝子断片を調製し、ヒト免疫グロブリンのフレームワーク領域をコードする遺伝子断片を調製し、所定の位置に所定の変異を導入し、軽鎖及び重鎖のそれぞれのCDRとFW領域と定常領域の配列とを連結することを含む方法によって作製することができる。
核酸の合成は、化学的な核酸合成法を用いるか、適切な鋳型及びプライマーを用いてポリメラーゼ連鎖反応(PCR)を行うことにより増幅させる方法を用いることができる。
フレームワーク領域の所定の部位に所定の変異を導入する方法としては、部位特異的変異誘発法(Ausebelら編,Current Protocols in Molecular Biology,Publish.John Wiley & Sons,8.1〜8.5章,1987)などの当業者に公知の種々の方法が使用され得る。
【0051】
上記の重鎖及び軽鎖をコードするポリヌクレオチドは、それぞれ同じ発現ベクターに導入してもよいし、別々の発現ベクターに導入してもよい。
本明細書において、「発現ベクター」とは、遺伝子からのタンパク質の宿主細胞での発現に必要な配列(コードされる発現させるポリヌクレオチドに機能可能に連結されたプロモーター、リボソーム結合部位、ポリアデニル化部位、転写終結部位など)を備えたベクターのことをいう。
【0052】
ヒト化抗体遺伝子を連結させる発現ベクターとしては、ヒト化抗体遺伝子を発現できるものであれば特に制限されず、細菌由来、ウイルス由来(例えばアデノウイルス、レトロウイルス)などの公知の発現ベクターを用いることができる。
なお、発現ベクターとして予めヒトIg定常領域遺伝子を保持するものを利用すれば、これに所望の配列を有する可変領域遺伝子を挿入するだけでヒト化抗体を発現できる発現ベクターとなるため好ましい。このような発現ベクターとしては、pFUSE2−CLIg−hk、pFUSE−CHIg−hG1(InvivoGen社)などが知られている。
【0053】
本発明の発現ベクターが含み得るプロモーターとしては、宿主細胞が大腸菌である場合、例えば、Trpプロモーター、lacプロモーター、T7プロモーター、recA プロモーター、λPLプロモーター、lppプロモーター、tacプロモーターなどが挙げられる。宿主細胞がバチルス属菌の場合、プロモーターとしては、SL01プロモーター、SP02プロモーター、penP プロモーターなどが挙げられる。宿主細胞が酵母である場合、プロモーターとしては、例えば、PH05プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーターが挙げられる。また、宿主が哺乳動物細胞等の真核細胞である場合、SV40由来のプロモーター、ヒトEF−1α由来プロモーター、レトロウイルスのプロモーター、ヒートショックプロモーターなどが挙げられる。
【0054】
上記の発現ベクターを導入する宿主細胞は、原核細胞又は真核細胞のいずれであってもよく、発現ベクターの種類に応じて適宜選択できる。例えば、大腸菌(エシェリキア属菌)、バチルス属などの細菌細胞、サッカロマイセス属、ピキア属などの酵母細胞、Sf9などの昆虫細胞、哺乳動物細胞などを用いることができる。なかでも、簡便にヒト化抗体を生産できるので、大腸菌が好ましい。
上記の発現ベクターを宿主細胞に導入する方法は、宿主細胞の種類に応じて適宜選択でき、例えば、宿主細胞が大腸菌である場合、塩化カルシウム法を用いることができる。
【0055】
発現ベクターを導入した宿主細胞は、宿主細胞の種類に応じた適切な培地中で、従来公知の方法に従って培養することにより、本発明のヒト化抗体を作製できる。
【0056】
上記の培養は、特に宿主細胞が細菌である場合、栄養培地中で行うことが好ましい。栄養培地は、形質転換体の生育に必要な炭素源、無機窒素源もしくは有機窒素源を含んでいることが好ましい。炭素源としては、例えばグルコース、デキストラン、可溶性デンプン、ショ糖など、無機窒素源もしくは有機窒素源としては、例えばアンモニウム塩類、硝酸塩類、アミノ酸、コーンスチープ・リカー、ペプトン、カゼイン、肉エキス、大豆粕、バレイショ抽出液などが挙げられる。また、所望により他の栄養素(例えば、無機塩(例えば塩化カルシウム、リン酸二水素ナトリウム、塩化マグネシウム)、ビタミン類、抗生物質(例えばテトラサイクリン、ネオマイシン、アンピシリン、カナマイシン等)など)を含んでいてもよい。
【0057】
上記の宿主細胞が大腸菌である場合、好ましい培地としてLB培地、M9培地(Millerら、Exp.Mol.Genet,Cold Spring Harbor Laboratory,p.431,1972)などが例示される。この場合、培養は、必要により通気、撹拌しながら、通常14〜43℃、約3〜24時間行うことができる。
宿主細胞がバチルス属菌の場合、必要により通気、撹拌をしながら、通常30〜40℃、約16〜96時間培養を行うことができる。
【0058】
宿主細胞が酵母である場合、培地としては、例えばBurkholder最小培地(Bostian,Proc.Natl.Acad.Sci.USA,Vol.77,p.4505,1980)が挙げられ、pHは5〜8であることが望ましい。培養は、通常、約20〜35℃で約14〜144時間行なわれ、必要により通気や撹拌を行うこともできる。
宿主細胞が哺乳動物細胞の場合、培地としては、約5〜20%の胎児牛血清を含むMEM培地(Eagle,Science,Vol.122,p.501,1952)、DMEM培地(Dulbecco,Virology,Vol.8,p.396,1959)、RPMI1640培地(George E.ら,J.Am.Med.Assoc.,Vol.199,p.519,1967)、199培地(Morganら,Proc.Soc.Exp.Biol.Med.,Vol.73,p.1,1950)等を用いることができる。培地のpHは約6〜8であるのが好ましく、培養は通常約30〜40℃で約15〜72時間行なわれ、必要により通気や撹拌を行うこともできる。
【0059】
宿主細胞が昆虫細胞の場合、例えば胎児牛血清を含むGrace’s培地(Smithら,Proc.Natl.Acad.Sci.USA,Vol.82,p.8404,1985)等が挙げられ、そのpHは約5〜8であるのが好ましい。培養は通常約20〜40℃で15〜100時間行なわれ、必要により通気や撹拌を行うこともできる。
【0060】
上記の培養後、培養した宿主細胞及び/又は培養液を回収し、宿主細胞内及び/又は培養上清中に蓄積された抗体を単離して精製できる。
ヒト化抗体を単離、精製する方法としては、例えば塩析、溶媒沈澱法等の溶解度を利用する方法、透析、限外濾過、ゲル濾過、ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動など分子量の差を利用する方法、イオン交換クロマトグラフィーやヒドロキシルアパタイトクロマトグラフィーなどの荷電を利用する方法、プロテインAなどのアフィニティークロマトグラフィーなどの特異的親和性を利用する方法、逆相高速液体クロマトグラフィーなどの疎水性の差を利用する方法、等電点電気泳動などの等電点の差を利用する方法などが挙げられる。
【0061】
こうして得られたヒト化抗シガトキシン抗体の抗原活性(抗原親和性)は、例えば、後記実施例に記載されるようなシガトキシンCTX−3Cを用いたELISAや、Biacore(GEヘルスケア・ジャパン社)などにより測定可能である。
【0062】
本発明は、本発明のヒト化抗体又はそのフラグメントをコードする配列を含むポリヌクレオチドも提供する。
本明細書において、「コードする」とは、mRNAに転写され、タンパク質に翻訳されたときに、コードされるアミノ酸配列を有するタンパク質を生成し得る遺伝子情報を、該ポリヌクレオチドが有することを意味する。
上記のポリヌクレオチドは、より好ましくは配列番号30、32〜35及び41のいずれか1つで表されるポリヌクレオチドを含むか、及び/又は配列番号31、37〜39及び42のいずれか1つで表されるポリヌクレオチドを含む。
上記のポリヌクレオチドは、最も好ましくは、配列番号41及び/又は42のポリヌクレオチドを含む。
【0063】
本発明は、本発明のヒト化抗体又はそのフラグメントをコードするポリヌクレオチドを含むベクターも提供する。
本発明のベクターは、上述した当該技術において公知の方法により作製された本発明のヒト化抗体又はそのフラグメントをコードするポリヌクレオチドを、適切なベクターに挿入することにより得ることができる。本発明のベクターに用いることができるベクターは、上記の発現ベクターについて述べたものと同様である。
本発明のベクターは、上記のポリヌクレオチドの遺伝子産物の発現に必要な配列をさらに有することが好ましい。このような配列は、当該技術において公知であり、例えばコードされるポリヌクレオチドに機能可能に連結されたプロモーター、リボソーム結合部位、ポリアデニル化部位、転写終結部位などを含む。
上記のベクターは、複製起点、エンハンサー、及び目的に応じて、通常用いられる選択マーカー(例えば、テトラサイクリン、アンピシリン又はカナマイシンに対する耐性遺伝子)を含んでいてもよい。
【0064】
本発明はまた、上記のベクターが導入された宿主細胞も提供する。好ましい宿主細胞は、上記のとおりである。
【0065】
本発明はまた、本発明のヒト化抗体又はそのフラグメントをコードするポリヌクレオチドを適切な発現ベクターに導入し、得られた発現ベクターを適切な宿主細胞に導入し、得られた宿主細胞を培養することを含む、本発明のヒト化抗体又はそのフラグメントの生産方法も提供する。
本発明の生産方法は、宿主細胞及び/又はその培養液からヒト化抗体又はそのフラグメントを精製することをさらに含んでもよい。
本発明の生産方法の各工程は、上記の本発明のヒト化抗体又はそのフラグメントの作製について述べたことと同様である。
【0066】
本発明のヒト化抗体又はそのフラグメントは、シガトキシン類を認識して結合することができるので、食中毒シガテラにより体内にシガトキシン類が存在する対象者(ヒト)に投与すれば、シガトキシン類を捕捉して、その毒性を低減できる可能性がある。
よって、本発明は、上記のヒト化抗体又はそのフラグメントを含む医薬組成物も提供する。該組成物は、食中毒シガテラの治療又は予防に用いることができる。
【0067】
上記の組成物は、いずれの投与形態であってもよいが、静脈内投与用の形態が好ましい。
【0068】
上記の組成物は、ヒト化抗体又はそのフラグメントとともに、少なくとも1種の医薬的に許容される医薬品添加物を含む。用い得る医薬品添加物は、投与形態に応じて、当該技術において公知の適切なものを用いることができる。
例えば静脈内投与用の医薬組成物の場合、医薬的に許容される医薬品添加物としては、溶剤(例えばオリーブ油、大豆油のような油性溶剤、水、アルコール、プロピレングリコールのような親水性溶剤など)、溶解補助剤(例えばポリエチレングリコール、コレステロールなど)、乳化剤(例えば界面活性剤など)、懸濁化剤(ポリビニル系化合物、セルロース類のような親水性高分子、界面活性剤など)、保存剤(例えばパラベン、ソルビン酸など)、着色剤などを用いることができる。
【0069】
上記の医薬組成物は、対象者に治療又は予防上有効な量で投与することができる。治療又は予防上有効な量は、臨床試験などにより適宜決定できる。
【実施例】
【0070】
以下に、本発明についてより具体的に理解するために、特定の実施例を示すが、これらは例示目的とするものであって、本発明を限定するものではない。
【0071】
以下の記載について、キット等を使用した部分については特にことわりの無い限り添付のプロトコールに従った。
【0072】
A.マウス10C9Fab断片のクローニング及び配列決定
マウス抗シガトキシン抗体10C9については、特許第3845796号公報に記載されている。
マウス10C9抗体を産生するハイブリドーマ細胞(FERM BP−8292)から、全RNAを分離した。ProSTAR FirstStrand RT−PCR Kit(STRATAGENE社)を用いて、逆転写により第1ストランドのcDNAを合成した。マウス抗体Fab断片のコーディング配列のクローニングを可能にするように設計されたPCRプライマー(Lernerら,Science,Vol.246,p.1275,1989)を用いて、重鎖及び軽鎖Fab領域を得た。マウス抗体10C9の軽鎖はL5とL9、重鎖はH3とH12のPCRプライマーを組み合わせて用いた(表1)。
【0073】
【表1】
【0074】
上記のプライマーを用いてPCR反応により増幅したcDNAフラグメントは、約700bp長の主要生成物を有した。cDNAフラグメントをpDriveベクター(QIAGEN社)にクローニングした。適正なサイズのインサートを含むベクターを、DTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)を用いて配列決定した。
決定したマウス抗体10C9の軽鎖可変領域VL、軽鎖定常領域CL、重鎖可変領域VH及び重鎖定常領域CH1のcDNA配列及びそれらから推測されるアミノ酸配列を、それぞれ配列番号1、3、5及び7(cDNA配列)及び配列番号2、4、6及び8(アミノ酸配列)に示す。
【0075】
B.シガトキシン類を認識するヒト化抗体の設計
(1)フレームワーク構造配列のための相同ヒト抗体の選択
マウス10C9軽鎖及び重鎖の可変領域に対して高い同一性を示すヒトgermline配列(V、J遺伝子領域)を、ClustalWを用いたアミノ酸配列の比較により同定した。
マウス10C9のVL領域に対して、61.5%という同一性百分率をもつヒトgermlineのκ鎖V領域(1GKV1D−39*01;配列番号9)及び70.4%という同一百分率をもつJ領域(1GKJ2*01;配列番号10)が同定された。
【0076】
マウス10C9のVH領域に対して、70.4%という同一性百分率をもつヒトgermlineのV領域(1GHV4−31*03;配列番号11)及び86.7%という同一百分率をもつJ領域(1GHJ4*01;配列番号12)が同定された。
マウス抗体10C9の可変領域と高い同一性を有するこれらのヒトgermline配列の各々からのフレームワーク構造領域(FW)のV領域及びJ領域をそれぞれ軽鎖及び重鎖について連結したアミノ酸配列を基本として、本発明のヒト化抗体の軽鎖及び重鎖を設計することにした。
【0077】
(2)相補性決定領域とフレームワーク構造領域の連結
ヒト化抗体のVL及びVH領域の作製のための次の段階として、マウス10C9のVL及びVH領域からのCDRを、(1)で設計したヒトgermline配列からのFWに連結させた。このマウス抗体のCDRとヒト免疫グロブリンのFW領域とを連結しただけのVL領域及びVH領域のバージョン(グラフト化抗体)を、h10C9g−VL及びh10C9g−VHと呼ぶこととした。h10C9g−VL及びh10C9g−VHのアミノ酸配列を、それぞれ配列番号13及び14に示す。
【0078】
(3)フレームワーク構造領域内のアミノ酸の置換
(a)軽鎖
上記のようにして設計されたh10C9g−VLのアミノ酸配列において、以下の表2に示すように、軽鎖のFWの1位、2位、7位、46位、60位及び71位の6個のアミノ酸の少なくとも1つを、マウス抗体10C9の対応する位置に存在するアミノ酸に置換した。
【0079】
【表2】
【0080】
具体的には、FW1の位置1、2及び7のアミノ酸(ヒトgermline配列内に通常存在するアスパラギン酸、イソロイシン及びセリン)を、マウス抗体10C9のVL領域内の対応する位置に見られるグルタミン酸、ロイシン及びトレオニンに変更した。この位置のアミノ酸残基は、マウス免疫グロブリンgermline配列をマウス抗体10C9の配列と比較したときに変異されている配列であり、マウス10C9のVL領域のソマティック変異の位置であると考えられ、抗体−抗原結合や抗体の構造を保持する上で間接的に関係があると考えられる。
【0081】
FW2の位置46のアミノ酸(ヒトgermline配列内に通常存在するロイシン)を、マウス抗体10C9のVL領域内の対応する位置に見られるアルギニンに変更した。マウス抗体10C9のVL領域内のこの位置にあるアルギニンは、結合部位の中央に位置づけされているものとして、既に決定されているマウス10C9の立体構造(Uiら,J.Biol.Chem.,Vol.283,p.19440,2008)内で観察され、抗体−抗原結合の間に抗原と直接相互作用している重要な残基であると考えられる。
【0082】
FW3内の位置60のアミノ酸(ヒトgermline配列内に通常存在するセリン)を、マウス抗体10C9のVL領域内の対応する位置に見られるグリシンに変更した。この位置にあるアミノ酸残基は、マウス抗体10C9のVL領域のソマティック変異の位置で、抗体−抗原結合や構造を保持する上で間接的に関係があると考えられる。
【0083】
FW3の位置71のアミノ酸(ヒトgermline配列内に通常存在するフェニルアラニン)を、マウス抗体10C9のVL領域内の対応する位置に見られるチロシンに変更した。マウス抗体10C9のVL領域内のこの位置にあるチロシンは、マウス10C9VL領域のCDR2ループを支持する上で重要であると考えられる。この位置におけるフェニルアラニンの存在が、CDR1ループの配置を分断する可能性がある。
【0084】
(b)重鎖
上記のようにして設計されたh10C9g−VHのアミノ酸配列において、以下の表3に示すように、重鎖のFWの5位、71位、73位及び94位の4個のアミノ酸の少なくとも1つを、マウス抗体10C9の対応する位置に存在するアミノ酸に改変した。
【0085】
【表3】
【0086】
具体的には、FW1内の位置5のアミノ酸(ヒトgermline配列内に通常存在するグルタミン)を、マウス抗体10C9VH領域内の対応する位置で見られるロイシンに変更した。マウス抗体10C9VH領域内のこの位置にあるロイシンは、マウス10C9のVH領域のソマティック変異の位置であると考えられ、またCDR1ループを支持する上で重要であると考えられる。
【0087】
FW3の位置71のアミノ酸(ヒトgermline配列内に通常存在するバリン)を、マウス抗体10C9のVH領域内の対応する位置に見られるアルギニンに変更した。この位置にあるアルギニンは、マウス抗体10C9のVH領域のCDR2ループを支持する上で重要であると考えられる。この位置におけるバリンの存在が、VH領域のCDR2ループの配置を分断する可能性がある。
【0088】
FW3の位置73のアミノ酸(ヒトgermline配列内に通常存在するトレオニン)を、マウス抗体10C9のVH領域内の対応する位置で見られるセリンに変更した。マウス抗体10C9のVH領域内のこの位置にあるセリンは、マウス抗体10C9のVH領域のCDR2ループを支持する上で重要であると考えられる。
【0089】
FW3内の位置94のアミノ酸(ヒトgermline内に通常存在するアルギニン)を、マウス抗体10C9のVH領域内の対応する位置に見られるシステインに変更した。マウス抗体10C9のVH領域内のこの位置にあるシステインは、マウス抗体10C9のVH領域のソマティック変異の位置であると考えられ、またCDR3ループを支持する上で重要であると考えられる。
【0090】
(4)定常領域の選択
マウス抗体10C9の軽鎖のCL及び重鎖のCH1に対して高い同一性を示すヒトgermline配列のC遺伝子領域を、ClustalWを用いたアミノ酸配列の比較により同定した。
マウス抗体10C9の軽鎖C領域に対して、70.4%という同一性百分率をもつヒトgermlineのκ鎖C領域(1GKC*01;配列番号28)が同定された。
マウス抗体10C9の重鎖CH1領域に対して、61.9%という同一性百分率をもつヒトgermlineのγ1鎖C領域(1GHG1*01;配列番号29)が同定された。
マウス抗体10C9の定常領域と高い同一性を示すこれらのヒトgermline配列のCL及びCH1領域を、(3)で設計した可変領域に連結して、ヒト化抗体のFabフラグメントの作製に使用した。
【0091】
C.ヒト化抗体をコードするポリヌクレオチドの構築
C−1.h10C9d−VL及びh10C9d−VHを含む軽鎖及び重鎖をコードするDNAの作製
(3)で設計した軽鎖及び重鎖の可変領域(h10C9d−VL及びh10C9d−VH)を、(4)で選択した軽鎖及び重鎖の定常領域(CL(1GKC*01)及びCH1(1GHG1*01))とそれぞれ連結したヒト化抗体のFabフラグメントの軽鎖及び重鎖をそれぞれコードするDNAを構築した。DNAの合成は、TAKARA社に委託した。
ヒト化抗体軽鎖FabフラグメントはSacI及びXbaIの制限酵素認識部位をもつように作製し、pMD19内へクローニングさせた。適正な配列をもつクローンを、h10C9dFab-L(配列番号30)と命名した。
ヒト化抗体重鎖FabフラグメントはXhoI及びSpeIの制限酵素認識部位をもつように作製し、pMD19内へクローニングさせた。適正な配列をもつクローンを、h10C9dFab-H(配列番号31)と命名した。
【0092】
C−2.h10C9d−VL−a〜e又はh10C9o−VL、及びh10C9d−VH−a〜d又はh10C9o−VHを含む軽鎖及び重鎖をコードするDNAの作製
上記のh10C9dFab-L及びh10C9dFab-Hの所望の位置のアミノ酸をマウス抗体10C9の対応する位置のアミノ酸に変換するために、シグマジェノシス社に依頼して、表4に示すPCRプライマーを合成した。
変換するアミノ酸の位置に従って選択される2組のプライマーを用いて、部位特異的変異誘発法を応用した市販のキット(STRATAGENE社;QuickChange Site−Directed Mutagenesis Kit)を用いてDNAに変異導入を行った。DTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)によりDNA配列を決定し、所望の配列をもつクローンを選択した。
【0093】
【表4】
【0094】
C−3.グラフト化抗体の作製
マウス抗体10C9のCDRと、ヒト免疫グロブリンのFW領域とを連結したVL領域及びVH領域のバージョン(グラフト化抗体;h10C9g−VL及びh10C9g−VH)と、(4)で選択した定常領域とを連結したFabフラグメントは、h10C9dFab−VL及びh10C9dFab−VHを鋳型として表1のPCRプライマーを用いてPCRを行うことにより構築された。DTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)により配列決定して、適正な配列をもつクローンを、h10C9Fabg−L(配列番号43)及びh10C9Fabg−H(配列番号44)と命名した。
【0095】
D.マウス抗体及びヒト化抗体の発現及び分析
D−1.発現プラスミドの構築
マウス抗体及びヒト化抗体の発現には、抗体Fab遺伝子を高効率に発現できることが報告されているpARA7(Miyashitaら,J.Mol.Biol.,Vol.267,p.1247,1997)を発現ベクターとして用いた(図2)。pARA7ベクターは、アラビノースにより誘導されるaraBプロモーターを含む。さらに、シグナル配列をコードする塩基配列が付加されていて、発現されるポリペプチドは、大腸菌培養上清中に分泌される。
【0096】
まず、マウス抗体の発現プラスミドは、次のようにして作製した。A.でクローニングしたマウスFab重鎖をコードするDNA配列中には、pARAに挿入する際に利用する制限酵素XhoI認識部位が含まれる。そこで、表5に示した2組のプライマーを用いて、QuickChange Site−Directed Mutagenesis Kit(STRATAGENE社)を用いてDNAに変異導入を行った。DTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)によりDNA配列を決定し、適正な配列をもつクローンをm10C9Fab−Hm(配列番号65)と命名した。
【0097】
【表5】
【0098】
上記で作製したマウスFab重鎖をコードするDNA(m10C9Fab−Hm)を、制限酵素XhoI認識部位及びSpeI認識部位を利用して、pARA7に挿入した。重鎖をコードするDNAが挿入されたプラスミドに、A.でクローニングしたマウスFab軽鎖をコードするDNA(m10C9Fab−L;配列番号68)を、制限酵素SacI認識部位及びXbaI認識部位を利用して挿入し、発現プラスミドを作製した。
【0099】
ヒト化抗体の発現プラスミドは、以下のようにして作製した。C.で作製した各々のヒト化Fab重鎖をコードするDNAのそれぞれを、制限酵素XhoI認識部位及びSpeI認識部位を利用して、pARA7に挿入した。重鎖をコードするDNAが挿入されたプラスミドに、C.で作製した各々のヒト化Fab軽鎖をコードするDNAのそれぞれを、制限酵素SacI認識部位及びXbaI認識部位を利用して挿入し、発現プラスミドを作製した。なお、軽鎖及び重鎖の可変領域の組み合わせは、以下の表6に示すとおりである。
【0100】
【表6】
【0101】
これらの発現プラスミドを、大腸菌に導入して増殖させ、市販のプラスミド精製キット(QIAGEN社;QIAprep Spin Miniprep Kit)により精製し、DNA塩基配列解析によりFabのコード領域の配列を確認した。DNA塩基配列解析にはDTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)を使用した。
【0102】
ヒト化10C9のうち、h10C9gFab及びh10C9oFabについては、マウスアミノ酸に置換した軽鎖内の位置1及び位置2と重鎖内の位置5がpARA7に挿入するための制限酵素認識部位に含まれるので、pARA7に挿入後、合成プライマー(表7)を用いてヒトgermline由来アミノ酸に戻すためのDNA変異導入を行った。
【0103】
【表7】
【0104】
D−2.発現プラスミドによる大腸菌の形質転換及び培養
次に、上記のD−1のようにして精製した発現プラスミドを、常法に従って大腸菌MC1061株に導入し、LB寒天プレート(100μg/mlのアンピシリン含有)に播いて形質転換株を得た。この形質転換株を、100μg/mlのアンピシリンを含むSB培地(Miyashitaら、上記文献)で増殖させ、発現誘導剤であるアラビノースを0.2%になるように添加して30℃で3日間培養を行った。培養終了後、遠心分離を行い、抗体を含む培養上清を回収した。
【0105】
D−3.マウス抗体及びヒト化抗体の精製
上記の培養上清に硫酸アンモニウムを65%飽和になるように加え、硫安沈殿を行った。遠心分離後の沈殿を10mMリン酸ナトリウム緩衝液に再懸濁し、同緩衝液に対して透析を行った。遠心分離後、上清を0.45μmのフィルターでろ過した。この上清を材料として、プロテインGカラム(GEヘルスケア・ジャパン社)を用いて精製した。その後、各分画フラクションについてSDS−PAGEを行うことによって、純度の高いFabが得られたことを確認した(図3;マウス抗体10C9Fab(m10C9Fab)、h10C9dFab、h10C9gFab及びh10C9oFabについてのみ結果を示す)。
【0106】
D−4.マウス抗体及びヒト化抗体の検出
D−2で作製した大腸菌で発現させたマウス抗体10C9のFab及びそれぞれのヒト化抗体のFabについて、ウェスタンブロッティングにより、アルカリフォスファターゼ標識抗マウスIgG(Millipore社)又はアルカリフォスファターゼ標識抗ヒトIgG(Millipore社)を用いて、抗体の検出を行った。
【0107】
図4の結果から明らかなように、マウス抗体10C9(m10C9Fab)は抗マウス抗体によって検出されるが、抗ヒト抗体によっては検出されない。一方、作製したヒト化抗体h10C9dFabは抗マウス抗体によって検出されないが、抗ヒト抗体によって検出された。その他のヒト化抗体(h10C9gFab、h10C9oFab、h10C9dFabM1〜M9)についても抗ヒト抗体によって検出された。
よって、本発明のヒト化抗体は、抗ヒト抗体により認識されるが、抗マウス抗体によっては認識されないので、ヒトに投与した場合に、免疫原性が低いことが期待される。
【0108】
D−5.ELISAによる抗原結合性の確認
シガトキシンCTX3CのABCDE環部の構造を有する抗原(CTX3C−ABCDEという)に対する各ヒト化抗体の結合活性を、マウス10C9抗体と比較してELISAによって検定した。ELISAの方法は、特許第3845796号公報に記載された方法に従う。
96ウェルELISA用プレート(COSTAR社)の各ウェルにCTX3C−ABCDE−BSAコンジュゲートを50μL入れ、4℃で一晩放置して、プレートに固定化した。プレートを洗浄後、1%スキムミルクでブロッキングした。プレートをさらに洗浄した後、連続希釈したマウス抗体又はヒト化抗体を、ウェルに50μLずつ添加して、室温で1時間反応させた。
検出は、ペルオキシダーゼ(HRP)標識抗ヒトIgG(Millipore社)又はHRP標識抗マウスIgG(GEヘルスケア・ジャパン社)を用いて行った。波長490nmにおける吸光度を、マイクロプレートリーダー(BioRad社)を用いて測定した。
【0109】
図5は、マウス抗体10C9(m10C9Fab)とヒト化抗体(h10C9gFab、h10C9dFab、h10C9oFab、h10C9dFabM1〜M9)の抗原との結合を比較している。その結果、マウスCDRとヒトIgのFW領域とを含むグラフト化抗体h10C9gFabは、マウス抗体10C9よりも、抗原に対する結合性が低かった。h10C9dFab及びh10C9oFabは、マウス10C9抗体と同程度又はそれ以上の結合活性を示すことが確認された。h10C9dFabM1〜M5、M7及びM8は、h10C9dFabと比較して同程度又はそれ以上の結合活性を示すことが確認された。
しかし、軽鎖の46位のアミノ酸がヒト免疫グロブリン由来のアミノ酸であるh10C9dFabM6、及び重鎖の94位のアミノ酸がヒト免疫グロブリン由来のアミノ酸であるh10C9dFabM9は、結合活性が低下していることがわかった。
【0110】
D−6.Biacoreによる結合親和力の測定
マウス抗体10C9(m10C9Fab)又はヒト化抗体のシガトキシンCTX3C−ABCDEに対する結合活性を、表面プラズモン共鳴測定法を用いて確認した。
CTX3C−ABCDE−BSAコンジュゲートをSensor Chip CM5(GEヘルスケア・ジャパン社)に固定化し、HBS−EPバッファー(GEヘルスケア・ジャパン社)で連続希釈した各抗体溶液を用いて結合活性を確認した結果を、図6に示す。
Kd値を求めたところ、マウス抗体10C9と比較してh10C9gFabは明らかに親和性が低下していた。
これに対して、h10C9dFab、h10C9oFab並びにh10C9dFabM1〜M5、M7及びM8は、CTX3C−ABCDEに対するマウス抗体10C9と同様又はわずかに良い結合親和力を示すことが確認された。
しかし、軽鎖の46位のアミノ酸がヒト免疫グロブリン由来のアミノ酸であるh10C9dFabM6、及び重鎖の94位のアミノ酸がヒト免疫グロブリン由来のアミノ酸であるh10C9dFabM9は、Kd値が非常に大きく、シガトキシン類との結合親和性が低いことがわかった。
【0111】
従って、本発明のヒト化抗体は、マウス抗体10C9とほぼ同様の抗原親和性を有することがわかった。さらに、ヒトFW内のわずかな修正(VLの位置46及びVHの位置94)だけを有するヒト化抗体(h10C9oFab)であっても、マウス抗体と同様の抗原親和性でシガトキシンを認識できることがわかった。
【0112】
添付の配列表に示す配列は、以下のものに相当する。
【0113】
【表8−1】
【0114】
【表8−2】
【技術分野】
【0001】
本発明は一般に、シガトキシンCTX3C及び51−ヒドロキシ−CTX3Cを特異的に認識できるヒト化抗体に関する。
【背景技術】
【0002】
食中毒シガテラは、ポリネシア、ハワイ、沖縄、カリブ海などの広いサンゴ礁海域で頻発し、年間2万人以上の中毒患者が発生している。シガテラ毒素は植物プランクトンが生産し、食物連鎖を介して魚介類に含まれることとなる。したがって、一旦発生すると多数の食用魚が広範に毒化して深刻な社会問題となることから、シガテラ毒素を迅速に検出することが食中毒シガテラの予防にとって重要である。
【0003】
食中毒シガテラの主要成分であるシガトキシン類(ciguatoxin、CTX)の免疫学的手法を用いた検出に関する研究は、ラジオイムノアッセイの発展に伴い、1977年頃から始まった。免疫学的手法による研究においては、シガトキシン類を認識する抗体が必要であるが、シガトキシン類は天然からごく微量しか採集されず、培養による生産も困難なことから、抗体を製造することも困難である。ハワイ大学ホカマらは、貴重な毒本体シガトキシンを用いて、モノクローナル抗体を調製し(非特許文献1参照)、この抗体を用いて、シガトキシン類を検出するための試薬、キットなどが開発されている。
【0004】
ところで、シガトキシン類は、単一の化合物ではなく、多種の毒素の混合物であり、次の一般式:
【0005】
【化1】
【0006】
で表される4種(化合物1〜4)が主に知られている。
【0007】
このような状況のなかで、本発明者らは、以前にシガトキシンCTX3Cの右末端の部分構造であるIJKLM環部を含む合成ハプテンを設計、合成し、この合成ハプテンのタンパク質コンジュゲートでマウスを免疫する工程を含む方法により、受託番号FERM BP‐8293のハイブリドーマ3D11を作製し、該ハイブリドーマを用いて、上記の化合物1及び3のシガトキシン類に特異性の高いモノクローナル抗体3D11を製造することに成功した(特許文献1参照)。
【0008】
さらに、本発明者らは、以前にシガトキシンCTX3Cの左末端の部分構造であるABCDE環部を含む合成ハプテンを設計、合成し、この合成ハプテンのタンパク質コンジュゲートでマウスを免疫する工程を含む方法により、受託番号FERM BP‐8292のハイブリドーマ10C9を作製し、該ハイブリドーマを用いて、上記の化合物1及び2のシガトキシン類に特異性の高いモノクローナル抗体10C9を調製することに成功した(特許文献2参照)。モノクローナル抗体10C9のシガトキシンCTX3Cに対する解離定数(Kd)は、2.8nMであった。
【0009】
また、本発明者らは、上記の2種のモノクローナル抗体を組み合わせて、検出特性をより改善したサンドイッチ法により、シガトキシンCTX3Cを検出するキットを作製した(特許文献3及び非特許文献2参照)。
【0010】
シガトキシン類は、神経細胞膜上のナトリウムチャネルに作用するので、神経細胞にナトリウムイオンの流入を引き起こす。この結果、軸索で脱分極が起こり、アセチルコリン等の神経伝達物質放出を促進すると考えられている。一方、シガトキシンの作用機構の研究が進められるにつれて、いくつかのアンタゴニストが発見されてきた。現在、シガトキシン類の解毒剤として、シガトキシン類のアンタゴニストである抗コリン性のアトロピンや、マンニトールの有効性が報告されている(非特許文献3参照)が、治療薬の開発には至っていない。シガテラ中毒を発症した場合、自然回復を待つしかないのが現状であり、現時点では有効な治療方法が存在しない。
【0011】
これまでに本発明者らは、上記の2種のモノクローナル抗体(3D11、10C9)について、細胞を用いたin vivoでの試験及びマウスを用いたin vitroでの試験を行い、2種の抗体を用いてシガテラ中毒に対する中和が有効であることを確認している(非特許文献4参照)。
【0012】
1975年にハイブリドーマ技術が開発されて以来(非特許文献5参照)、モノクローナル抗体の産生が可能となり、マウスモノクローナル抗体は現在、様々な疾病の診断的手法として広く利用されている。しかしマウス抗体を治療薬としてヒト体内に投与すると、ヒト抗マウス抗体(human anti−mouse antibodies;HAMA)応答が引き起こされ、この反応は時にヒトにとって致命的なものになりうる。このように、ヒト体内でマウス抗体は免疫原性を持つので、マウス抗体をヒトに直接投与することは回避することが望ましい。しかし、動物由来抗体の免疫原性を軽減するために、ヒトのモノクローナル抗体を作製することは、技術的及び倫理的問題により困難である。
【0013】
そこで、マウス抗体の抗原結合部位を含む可変領域をヒト抗体の定常領域に接合したキメラ抗体を作製することが行われている。しかし、キメラ抗体もなお、その抗原性が高く、ヒト抗キメラ抗体(human anti−chimeric antibodies;HACA)応答を引き起こすと考えられる(非特許文献6参照)。
これを改善するため、マウス抗体の抗原結合に最も重要な相補性決定領域(CDR)のみをヒト抗体フレームワークに移植した(CDR−grafting)ヒト化抗体が開発されている。このようなヒト化抗体作製技術は1990年代から発展し、現在ではがん、白血病、関節リウマチなど多くの疾病にヒト化抗体が医薬品として用いられている。
【0014】
ヒト化抗体の作製技術は、キメラ抗体作製よりも複雑な技術である。抗原に対する特異性や親和性を維持するためには、CDRの移植のみでは不十分な場合が多い。抗原との結合に重要なアミノ酸残基は、マウス抗体のフレームワーク領域にも存在するので、ヒト抗体フレームワークのアミノ酸残基をマウス抗体アミノ酸に戻す必要がある。このような残基を同定するには、X線結晶解析などによる構造情報があると理想的であることが知られている。
【先行技術文献】
【特許文献】
【0015】
【特許文献1】特開2003−55400号公報
【特許文献2】特許第3845796号公報
【特許文献3】特許第3820519号公報
【非特許文献】
【0016】
【非特許文献1】Toxicon 第15巻、(1977年)、第317頁
【非特許文献2】Journal of American Chemical Society 第125巻、(2003年)、第7608頁
【非特許文献3】Laurentら編、Ciguatera: field reference guide/un guide pratique、Secretariat of the Pacific Community出版、(2005年)
【非特許文献4】Toxicon 第53巻、(2009年)、第802頁
【非特許文献5】Nature 第256巻、(1975年)、第495頁
【非特許文献6】Dubel編、Handbook of Therapeutic Antibodies、Wiley−VCH出版、(2007年)、第119頁
【発明の概要】
【発明が解決しようとする課題】
【0017】
本発明の課題は、食中毒シガテラの治療薬開発のために、ヒト抗マウス応答を最小限にとどめ、かつシガトキシン類に対する強い親和性を示すヒト化抗シガトキシン抗体を提供することにある。
本発明者らは、上記の課題を解決するために、従来のマウス抗体をヒト化及び最適化して、従来のマウス抗体と同等の強い親和性をシガトキシン類に対して示すヒト化抗シガトキシン抗体を取得することに成功した。
【課題を解決するための手段】
【0018】
すなわち、本発明は、
ヒト免疫グロブリンのフレームワーク領域(FW)と、
FERM BP−8292のハイブリドーマにより産生されるシガトキシン類を認識するマウス抗体10C9の相補性決定領域(CDR)と
を含む可変領域をそれぞれ有する重鎖及び軽鎖を少なくとも含み、
軽鎖の46位のアミノ酸と、任意に、1位、2位、7位、60位及び71位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の軽鎖の対応する位置に存在するアミノ酸で置き換えられ、及び/又は
重鎖の94位のアミノ酸と、任意に、5位、71位及び73位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の重鎖の対応する位置に存在するアミノ酸で置き換えられており、
シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識する、ヒト化抗体又はそのフラグメント(以下、単に「ヒト化抗体」ともいう)を提供する。
【0019】
なお、本明細書において、免疫グロブリンの軽鎖及び重鎖のアミノ酸配列の番号付けは、Kabatら、Sequences of Proteins of Immunological Interest、第5版、Bethesda,MD:U.S.Dept.of Health and Human Services,Public Health Service,National Institutes of Health,1991で定義された免疫グロブリンの軽鎖及び重鎖の番号付けの定義に従う。
任意の免疫グロブリンのアミノ酸配列の番号付けは、http://www.bioinf.org.uk/abs/abnumのウェブサイトで行うこともできる。
【0020】
また、本発明は、上記のヒト化抗体の重鎖又はそのフラグメントをコードする配列を含むポリヌクレオチド、上記のヒト化抗体の軽鎖又はそのフラグメントをコードする配列を含むポリヌクレオチド、上記のポリヌクレオチドを含むベクター、上記のベクターが導入された宿主細胞、並びに上記の宿主細胞を培養して、上記のヒト化抗体又はそのフラグメントを発現させる工程を含むヒト化抗体又はそのフラグメントを生産する方法も提供する。
【発明の効果】
【0021】
本発明によって、シガトキシンに対して強い親和性を示すヒト化抗シガトキシン抗体が提供される。このような特質を有する本発明の抗体は、ヒトに投与したときに、マウス抗体と比較して免疫原性が低下していると考えられるので、食中毒シガテラの予防又は治療用の医薬品の開発に有用である。
【図面の簡単な説明】
【0022】
【図1−1】マウス抗体10C9及び作製したヒト化抗体の軽鎖可変領域のアミノ酸配列を示す。m10C9:マウス10C9抗体;1GKV1D−39*01:ヒトgermline配列の軽鎖V領域;1GKJ2*01:ヒトgermline配列の軽鎖J領域;h10C9g−VL:グラフト化抗体;各h10C9d−VL及びh10C9o−VL:設計したヒト化抗体;1GKC*01:ヒトgermline配列の軽鎖定常領域。
【図1−2】マウス抗体10C9及び作製したヒト化抗体の重鎖可変領域のアミノ酸配列を示す。m10C9:マウス10C9抗体;1GKV4−31*03:ヒトgermline配列の重鎖V領域;1GHJ4*01:ヒトgermline配列の重鎖J領域;h10C9g−VH:グラフト化抗体;各h10C9d−VH及びh10C9o−VH:設計したヒト化抗体;1GHG1*01:ヒトgermline配列の重鎖定常領域。
【図2】大腸菌内でマウス抗体及びヒト化抗体を産生するために使用されるFab発現ベクターの例を示す図である。
【図3】精製したマウス10C9抗体及び記載されるヒト化抗体のSDS−PAGE結果を示す図である。
【図4】マウス10C9抗体及び記載されるヒト化抗体を、抗マウス抗体及び/又は抗ヒト抗体で検出したウェスタンブロッティングの結果を示す図である。
【図5】マウス10C9抗体及び各ヒト化抗体のCTX3C−ABCDEに対する抗原結合性をELISA法で調べた結果を示すグラフである。
【図6】精製したマウス10C9抗体及び各ヒト化抗体のCTX3C−ABCDEに対する結合性をBiacoreで調べた結果を示すグラフである。
【発明を実施するための形態】
【0023】
本明細書において、「シガトキシン類」とは、マウス抗体10C9が認識して結合できるシガトキシンCTX3C及び51−ヒドロキシ−CTX3Cから選択されるシガトキシンのことをいう。
【0024】
本明細書において、「免疫グロブリン」及び「抗体」との用語は、同等の意味を有すると解される。「抗体」との用語は、一般的に、それに対する特定の「抗原」との関係において、機能に注目した名称であると解される。
【0025】
本発明のヒト化抗体又はそのフラグメントは、シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識して、これと結合できる。本発明のヒト化抗体のフラグメントとは、抗原に結合できるフラグメントであればよく、Fab、F(ab’)2フラグメントを含む。
【0026】
一般に、抗体分子の基本構造は、各クラス共通で、分子量5万〜7万の重鎖と2〜3万の軽鎖から構成される。重鎖は、通常約440個のアミノ酸を含むポリペプチド鎖からなり、クラスごとに特徴的な構造をもち、IgG、IgM、IgA、IgD、IgEに対応してγ、μ、α、δ、ε鎖とよばれる。さらにIgGには、IgG1、IgG2、IgG3、IgG4が存在し、それぞれγ1、γ2、γ3、γ4とよばれている。軽鎖は、通常約220個のアミノ酸を含むポリペプチド鎖からなり、L型とK型の2種が知られており、それぞれλ、κ鎖とよばれる。抗体分子の基本構造のペプチド構成は、それぞれ相同な2本の重鎖及び2本の軽鎖が、ジスルフィド結合(S−S結合)及び非共有結合によって結合され、分子量15万〜19万である。2種の軽鎖は、どの重鎖とも対をなすことができる。個々の抗体分子は、常に同一の軽鎖2本と同一の重鎖2本からできている。
【0027】
鎖内S−S結合は、重鎖に四つ(μ、ε鎖には五つ)、軽鎖には二つあって、アミノ酸100〜110残基ごとに一つのループを成し、この立体構造は各ループ間で類似していて、構造単位あるいはドメインとよばれる。重鎖、軽鎖ともにN末端に位置するドメインは、同種動物の同一クラス(サブクラス)からの標品であっても、そのアミノ酸配列が一定せず、可変領域(V領域、variable region、可変部)とよばれている(各ドメインは、それぞれ、VH及びVLと表される)。これよりC末端側のアミノ酸配列は、各クラスあるいはサブクラスごとにほぼ一定で定常領域(C領域、constant region、定常部)とよばれている(各ドメインは、それぞれ、CH1、CH2、CH3あるいはCLと表される)。軽鎖及び重鎖の中で、可変及び定常領域は、重鎖も同じく約10個以上のアミノ酸の「D」領域を内含している状態で、約12個以上のアミノ酸の「J」領域によって連結されている(一般に、Fundamental Immunology(基礎免疫学)(Paul,W.編,第2版,Raven Press,N.Y.,1989、第7章を参照のこと)。
【0028】
抗体の抗原決定部位はVH及びVLによって構成され、結合の特異性はこの部位のアミノ酸配列によっている。一方、補体や各種細胞との結合といった生物学的活性は各クラスIgのC領域の構造の差を反映している。重鎖と軽鎖の可変領域の可変性は、どちらの鎖にも存在する3つの小さな超可変領域(hypervariable region)にほぼ限られることがわかっており、これらの領域を相補性決定領域(CDR、complementarity determining region)と呼んでいる。可変領域の残りの部分はフレームワーク領域(FW、framework region)とよばれ、比較的一定である。通常、各可変領域の相補性決定領域の5〜10個のアミノ酸だけが抗原結合部位を形成している。
通常、抗体の重鎖及び軽鎖の可変領域には、3つのCDR(CDR1〜3)と、4つのFW(FW1〜4)が含まれる。
【0029】
本明細書において、「キメラ抗体」とは、可変領域がマウス抗体(ドナー異種抗体とも称する)由来であり、定常領域がヒト免疫グロブリン由来である抗体のことである。
本明細書において、「ヒト化抗体」とは、抗原特異的なマウス抗体分子の相補性決定領域(抗原結合部位)以外がヒト免疫グロブリンに由来する組換え抗体のことである。ヒト化抗体は、その機能の向上のためにアミノ酸の改変(置換、挿入、欠失及び/又は付加)を含むことができる。
【0030】
一般に、ヒト化抗体の作製においては、相補性決定領域のアミノ酸配列のみを鋳型のヒト抗体フレームワークに移植しただけ(グラフト化抗体)では、多くの場合、オリジナルのマウス抗体よりも抗原結合活性が低下することが知られている。
本発明に用いられるマウス抗体10C9のCDRを、ヒト免疫グロブリンのフレームワークに移植することのみにより得られるグラフト化抗体も、シガトキシンCTX3Cに対する結合性は有するものの、抗原親和性が極めて低く、抗体医薬としての使用には好ましくないことが確認された(後記実施例D−5及びD−6を参照)。
【0031】
本発明者らは、このようなヒト化抗体の活性低下を改善し、かつ、抗体医薬として使用するためにより優れたヒト化抗体を得るために検討を重ねた結果、上記の本発明のヒト化抗シガトキシン類抗体が、従来のマウス抗シガトキシン類抗体と比較して、抗原親和性を維持しつつ、免疫原性が低下され得るという有意に改善された特性を有することを見出した。
【0032】
本発明のヒト化抗体に含まれるヒト免疫グロブリン(以下、「Ig」ともいう)のFW領域は、マウス抗体10C9のFWと同じ又は類似のコンホメーションをとることが好ましい。このことにより、マウス抗体のCDRと融合させたときに、抗原との結合のための正しい空間的配向が保持できる確率が高くなる。
このために、本発明では、軽鎖のFWの46位のアミノ酸と、任意に、1位、2位、7位、60位及び71位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の軽鎖の対応する位置に存在するアミノ酸で置き換えられ、及び/又は
重鎖のFWの94位のアミノ酸と、任意に、5位、71位及び73位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の重鎖の対応する位置に存在するアミノ酸で置き換えられている。
【0033】
上記の軽鎖及び重鎖のFW領域内のマウス抗体10C9の対応する位置に存在するアミノ酸で置き換えるアミノ酸は、軽鎖及び重鎖においてそれぞれ少なくとも1つである。軽鎖では、46位のアミノ酸と、上記の任意に選択される位置のアミノ酸の1つ、2つ、3つ、4つ又は5つの位置のさらなるアミノ酸を置き換えることができ、重鎖では、94位のアミノ酸と、上記の任意に選択される位置のアミノ酸の1つ、2つ又は3つの位置のさらなるアミノ酸を置き換えることができる。
【0034】
本発明のヒト化抗体の重鎖及び軽鎖のFW領域が由来するヒト免疫グロブリンは、天然に存在するヒト免疫グロブリン(IgG、IgM、IgA、IgD又はIgE)のFW領域の配列であってもよいし、いくつかのサブクラスのヒト免疫グロブリンのFW領域に共通するコンセンサス配列でもあり得る。Dubel編,Handbook of Therpeutic Antibodies、Wiley−VCH出版(2007)を参照。
重鎖及び軽鎖のそれぞれに含まれる複数のFW領域(FW1〜FW4)は、同じサブクラスのヒトIgに由来してもよいし、異なるサブクラスのヒトIgに由来してもよい。また、これらの複数のFW領域は、同じ免疫グロブリンに由来してもよいし、異なる免疫グロブリンに由来してもよい。
【0035】
ヒト免疫グロブリンのFW領域のアミノ酸配列は公知であり、例えばEntrez Protein(http://www.ncbi.nlm.nih.gov/sites/entrez?db=protein)などのデータベースから入手できる。
【0036】
上記のFW領域として、ヒト免疫グロブリンのgermline配列を利用することもできる(Dubel編,Handbook of Therapeutics Antibodies,Wiley−VCH出版,p.119,2007;Hwangら,Methods,vol.36,p.35,2005)。germline配列とは、体細胞変異が生じる前の生物が元来有する生殖細胞の遺伝子系列であり、ヒト免疫グロブリンについてもデータベースが公開されている(http://imgt.cines.fr//IMGT#GENE-DB/GENElect?query=4.2+&species=Homo+sapiens)。
ヒトgermline配列は、特異的なモチーフの発現が最小であり、免疫原性(免疫システムが非自己として認識する可能性)を低く抑えることができると考えられている。
【0037】
本発明のヒト化抗体のFW領域は、マウス抗体10C9の可変領域全体の配列と相同性が高くなるように選択したgermline配列に由来することが好ましい。このようにFW領域を選択することにより、マウス抗体の重要なアミノ酸残基と同じ位置で等しいアミノ酸残基を有するFW領域を用いることができる。また、germline配列を用いると、他の免疫グロブリン配列を用いるよりも、抗原結合活性の損失率が低いという利点もある。
【0038】
本発明のヒト化抗体に用いるためのヒト免疫グロブリン配列は、既知のヒト免疫グロブリンgermline配列のデータベースを利用して、軽鎖及び重鎖それぞれについてマウス抗体可変領域のアミノ酸配列と相同性が高い配列を、コンピュータにより(例えばBLASTにより)比較することによって同定できる。この比較により、マウス抗体10C9の可変領域と高い配列同一性を示すヒトgermlineからの軽鎖及び重鎖フレームワーク領域を選択できる。
【0039】
本発明のヒト化抗体は、実質的にヒトIgからのFW領域と、実質的にマウス抗体10C9からのCDRとを含む。このようなアミノ酸配列の並置は、天然には見られないので、ある種のアミノ酸の改変を加えることにより、より天然に近いコンホメーションを有し、抗原結合親和性の向上を導くことができる。
よって、得られるヒト化抗体の特性を最適化するために、フレームワーク領域内でどのアミノ酸残基を置換すればよいかを決定する(バックミューテーション残基)。一般に、マウス抗体に由来する残基をヒト化抗体に導入することは、ヒトの体内でのHAMA応答を引き起こす危険性を増大させる可能性が高いので、最小限に抑えることが好ましい。
【0040】
CDRコンホメーション及び/又は抗原への結合に対して考えられるその影響に基づいて、FW領域内で置換すべきアミノ酸を同定できる。
置換すべきアミノ酸残基は、(1)抗原に直接、非共有的に結合するアミノ酸残基、(2)CDR領域に隣接するか、Chothiaら(J.Mol.Biol.,Vol.196,p.901,1987;Nature,Vol.342,p.878,1989)により提案される代替的定義づけの下でCDR領域の一部であるか、CDRループの立体配座の支持に関与するか(Footeら,J.Mol.Biol.,Vol.224,p.487,1992)、又はその他の形でCDR領域と相互作用するアミノ酸残基、又は(3)VL−VHインタフェイスに参与するアミノ酸(Chothiaら,J.Mol.Biol.,Vol.186,p.651,1985)であることが好ましい。
【0041】
或いは、FW領域内で置換すべきアミノ酸残基は、マウス免疫グロブリン配列中でマウスの体細胞変異(ソマティック変異)が導入されたと考えられる位置に対応する位置のヒトFW領域内のアミノ酸残基が好ましい。マウス免疫グロブリン中で体細胞変異が導入されたと考えられるアミノ酸残基は、マウス抗体10C9のアミノ酸配列を、マウス免疫グロブリンgermline配列と、ClustalWなどのアルゴリズムをデフォルト設定で用いて相同性がなるべく高くなるように整列させ、マウス抗体10C9中の配列がgermline配列とは異なる位置のアミノ酸残基を選択することにより決定できる。体細胞変異が導入されたと考えられる位置のアミノ酸残基は、ヒト抗体にとっては普通でないアミノ酸であるので、マウス免疫グロブリン中で体細胞変異が導入された位置に対応する位置のヒト免疫グロブリンのアミノ酸をマウス抗体10C9の対応する位置のアミノ酸残基で置換することにより、抗体の抗原結合性の低下を最小限に抑えて、ヒト化抗体を作製できる。
【0042】
マウス10C9抗体の立体構造は、X線結晶構造解析によって以前に報告されている(Uiら,J.Biol.Chem.,Vol.283,p.19440,2008)。この立体構造情報に基づいて、抗原に直接結合するアミノ酸残基、及び抗原−抗体結合に間接的に関わる(結合部位の構造を維持する上で重要な)アミノ酸残基を同定できる。
【0043】
上記の情報に基づいて鋭意検討を重ねた結果、本発明のヒト化抗体は、軽鎖の46位と、任意に、1位、2位、7位、60位及び71位の少なくとも1つ、例えば1つ、2つ、3つ、4つ又は5つの位置のアミノ酸残基が、マウス抗体10C9の対応する位置のアミノ酸残基で置き換えられ、かつ重鎖の94位と、任意に、5位、71位及び73位の少なくとも1つ、例えば1つ、2つ又は3つの位置のアミノ酸残基が、マウス抗体10C9の対応する位置のアミノ酸残基で置き換えられたものであることにより、シガトキシン類との良好な抗原結合特性を示すことが見出された。
これらの置換され得るアミノ酸は、図1に下線を付して示す。
【0044】
本明細書において、「マウス抗体10C9の対応する位置のアミノ酸残基」とは、上記のKabatらによる免疫グロブリンの番号付けに従って番号を付したときに、ヒト免疫グロブリン中のアミノ酸残基と同じ番号を付与される位置のアミノ酸残基のことである。
【0045】
本発明のヒト化抗体に用いられるマウス抗体10C9のCDRは、独立行政法人産業技術総合研究所特許生物寄託センターに2003年2月13日に、ブダペスト条約に基づく寄託の移管申請により受託番号FERM BP−8292として寄託されたハイブリドーマ10C9から生成されるマウス抗体10C9のCDRである。このマウス抗体10C9は、シガトキシンCTX3C及び51−OH−CTX3CのABCDE環部を特異的に認識する。この抗体については、特許第3845796号及び特許第3820519号を参照されたい。
【0046】
マウス抗体10C9のアミノ酸配列は、当該技術において公知の方法により決定できる。マウス抗体10C9の軽鎖可変領域VL、軽鎖定常領域CL、重鎖可変領域VH及び重鎖定常領域CH1のcDNA配列及びそれらから推測されるアミノ酸配列を、それぞれ配列番号1、3、5及び7(cDNA配列)及び配列番号2、4、6及び8(アミノ酸配列)に示す。
【0047】
本発明のヒト化抗体は、より好ましくは、配列番号15〜19及び21のいずれか1つに示すアミノ酸配列を有する軽鎖と、配列番号22〜25及び27のいずれか1つに示すアミノ酸配列を有する重鎖とを含む。
上記のヒト化抗体は、最も好ましくは、配列番号21に示すアミノ酸配列を有する軽鎖と、配列番号27に示すアミノ酸配列を有する重鎖とを含む。
【0048】
本発明のヒト化抗体の定常領域としては、従来公知のヒト免疫グロブリンの定常領域を用いることができる。ヒトIgの定常領域は、軽鎖定常領域(CL領域)及び重鎖定常領域(CH1、CH2及びCH3)からなるが、本発明のヒト化抗体又はそのフラグメントは、シガトキシン類との結合性を有するように、これらの定常領域のいずれかを適切に有することができる。
ヒト免疫グロブリンの可変領域について述べたことと同様に、上記の定常領域も、IgG、IgM、IgA、IgD又はIgEのサブクラスのヒト免疫グロブリンのいずれに基づくものであってもよい。好ましくは重鎖定常領域としてはヒトIgγ1を、また、軽鎖定常領域としてはヒトIgκを用いることができる。
また、軽鎖及び重鎖のそれぞれの定常領域は、同じサブクラスのヒトIgに基づいてもよいし、異なるサブクラスのヒトIgに基づいてもよい。
【0049】
本発明のヒト化抗体は、本願明細書に開示される、本発明のヒト化抗体のアミノ酸配列をコードする塩基配列を有するポリヌクレオチドを当該分野で公知の方法を使用して作製し、該ポリヌクレオチドを発現ベクターに導入し、得られた発現ベクターを宿主細胞に導入し、宿主細胞を培養することを含む方法により得ることができる。
培養後の培養液から、ヒト化抗体を精製してもよい。
上記の各工程は、当該分野で公知の種々の方法を使用して行うことができる。
【0050】
上記の本発明の抗体の重鎖及び軽鎖のアミノ酸配列をコードするポリヌクレオチドは、例えば、当業者に公知の核酸合成方法に従って、マウス10C9抗体の重鎖及び軽鎖の各々のCDRをコードする遺伝子断片を調製し、ヒト免疫グロブリンのフレームワーク領域をコードする遺伝子断片を調製し、所定の位置に所定の変異を導入し、軽鎖及び重鎖のそれぞれのCDRとFW領域と定常領域の配列とを連結することを含む方法によって作製することができる。
核酸の合成は、化学的な核酸合成法を用いるか、適切な鋳型及びプライマーを用いてポリメラーゼ連鎖反応(PCR)を行うことにより増幅させる方法を用いることができる。
フレームワーク領域の所定の部位に所定の変異を導入する方法としては、部位特異的変異誘発法(Ausebelら編,Current Protocols in Molecular Biology,Publish.John Wiley & Sons,8.1〜8.5章,1987)などの当業者に公知の種々の方法が使用され得る。
【0051】
上記の重鎖及び軽鎖をコードするポリヌクレオチドは、それぞれ同じ発現ベクターに導入してもよいし、別々の発現ベクターに導入してもよい。
本明細書において、「発現ベクター」とは、遺伝子からのタンパク質の宿主細胞での発現に必要な配列(コードされる発現させるポリヌクレオチドに機能可能に連結されたプロモーター、リボソーム結合部位、ポリアデニル化部位、転写終結部位など)を備えたベクターのことをいう。
【0052】
ヒト化抗体遺伝子を連結させる発現ベクターとしては、ヒト化抗体遺伝子を発現できるものであれば特に制限されず、細菌由来、ウイルス由来(例えばアデノウイルス、レトロウイルス)などの公知の発現ベクターを用いることができる。
なお、発現ベクターとして予めヒトIg定常領域遺伝子を保持するものを利用すれば、これに所望の配列を有する可変領域遺伝子を挿入するだけでヒト化抗体を発現できる発現ベクターとなるため好ましい。このような発現ベクターとしては、pFUSE2−CLIg−hk、pFUSE−CHIg−hG1(InvivoGen社)などが知られている。
【0053】
本発明の発現ベクターが含み得るプロモーターとしては、宿主細胞が大腸菌である場合、例えば、Trpプロモーター、lacプロモーター、T7プロモーター、recA プロモーター、λPLプロモーター、lppプロモーター、tacプロモーターなどが挙げられる。宿主細胞がバチルス属菌の場合、プロモーターとしては、SL01プロモーター、SP02プロモーター、penP プロモーターなどが挙げられる。宿主細胞が酵母である場合、プロモーターとしては、例えば、PH05プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーターが挙げられる。また、宿主が哺乳動物細胞等の真核細胞である場合、SV40由来のプロモーター、ヒトEF−1α由来プロモーター、レトロウイルスのプロモーター、ヒートショックプロモーターなどが挙げられる。
【0054】
上記の発現ベクターを導入する宿主細胞は、原核細胞又は真核細胞のいずれであってもよく、発現ベクターの種類に応じて適宜選択できる。例えば、大腸菌(エシェリキア属菌)、バチルス属などの細菌細胞、サッカロマイセス属、ピキア属などの酵母細胞、Sf9などの昆虫細胞、哺乳動物細胞などを用いることができる。なかでも、簡便にヒト化抗体を生産できるので、大腸菌が好ましい。
上記の発現ベクターを宿主細胞に導入する方法は、宿主細胞の種類に応じて適宜選択でき、例えば、宿主細胞が大腸菌である場合、塩化カルシウム法を用いることができる。
【0055】
発現ベクターを導入した宿主細胞は、宿主細胞の種類に応じた適切な培地中で、従来公知の方法に従って培養することにより、本発明のヒト化抗体を作製できる。
【0056】
上記の培養は、特に宿主細胞が細菌である場合、栄養培地中で行うことが好ましい。栄養培地は、形質転換体の生育に必要な炭素源、無機窒素源もしくは有機窒素源を含んでいることが好ましい。炭素源としては、例えばグルコース、デキストラン、可溶性デンプン、ショ糖など、無機窒素源もしくは有機窒素源としては、例えばアンモニウム塩類、硝酸塩類、アミノ酸、コーンスチープ・リカー、ペプトン、カゼイン、肉エキス、大豆粕、バレイショ抽出液などが挙げられる。また、所望により他の栄養素(例えば、無機塩(例えば塩化カルシウム、リン酸二水素ナトリウム、塩化マグネシウム)、ビタミン類、抗生物質(例えばテトラサイクリン、ネオマイシン、アンピシリン、カナマイシン等)など)を含んでいてもよい。
【0057】
上記の宿主細胞が大腸菌である場合、好ましい培地としてLB培地、M9培地(Millerら、Exp.Mol.Genet,Cold Spring Harbor Laboratory,p.431,1972)などが例示される。この場合、培養は、必要により通気、撹拌しながら、通常14〜43℃、約3〜24時間行うことができる。
宿主細胞がバチルス属菌の場合、必要により通気、撹拌をしながら、通常30〜40℃、約16〜96時間培養を行うことができる。
【0058】
宿主細胞が酵母である場合、培地としては、例えばBurkholder最小培地(Bostian,Proc.Natl.Acad.Sci.USA,Vol.77,p.4505,1980)が挙げられ、pHは5〜8であることが望ましい。培養は、通常、約20〜35℃で約14〜144時間行なわれ、必要により通気や撹拌を行うこともできる。
宿主細胞が哺乳動物細胞の場合、培地としては、約5〜20%の胎児牛血清を含むMEM培地(Eagle,Science,Vol.122,p.501,1952)、DMEM培地(Dulbecco,Virology,Vol.8,p.396,1959)、RPMI1640培地(George E.ら,J.Am.Med.Assoc.,Vol.199,p.519,1967)、199培地(Morganら,Proc.Soc.Exp.Biol.Med.,Vol.73,p.1,1950)等を用いることができる。培地のpHは約6〜8であるのが好ましく、培養は通常約30〜40℃で約15〜72時間行なわれ、必要により通気や撹拌を行うこともできる。
【0059】
宿主細胞が昆虫細胞の場合、例えば胎児牛血清を含むGrace’s培地(Smithら,Proc.Natl.Acad.Sci.USA,Vol.82,p.8404,1985)等が挙げられ、そのpHは約5〜8であるのが好ましい。培養は通常約20〜40℃で15〜100時間行なわれ、必要により通気や撹拌を行うこともできる。
【0060】
上記の培養後、培養した宿主細胞及び/又は培養液を回収し、宿主細胞内及び/又は培養上清中に蓄積された抗体を単離して精製できる。
ヒト化抗体を単離、精製する方法としては、例えば塩析、溶媒沈澱法等の溶解度を利用する方法、透析、限外濾過、ゲル濾過、ドデシル硫酸ナトリウム−ポリアクリルアミドゲル電気泳動など分子量の差を利用する方法、イオン交換クロマトグラフィーやヒドロキシルアパタイトクロマトグラフィーなどの荷電を利用する方法、プロテインAなどのアフィニティークロマトグラフィーなどの特異的親和性を利用する方法、逆相高速液体クロマトグラフィーなどの疎水性の差を利用する方法、等電点電気泳動などの等電点の差を利用する方法などが挙げられる。
【0061】
こうして得られたヒト化抗シガトキシン抗体の抗原活性(抗原親和性)は、例えば、後記実施例に記載されるようなシガトキシンCTX−3Cを用いたELISAや、Biacore(GEヘルスケア・ジャパン社)などにより測定可能である。
【0062】
本発明は、本発明のヒト化抗体又はそのフラグメントをコードする配列を含むポリヌクレオチドも提供する。
本明細書において、「コードする」とは、mRNAに転写され、タンパク質に翻訳されたときに、コードされるアミノ酸配列を有するタンパク質を生成し得る遺伝子情報を、該ポリヌクレオチドが有することを意味する。
上記のポリヌクレオチドは、より好ましくは配列番号30、32〜35及び41のいずれか1つで表されるポリヌクレオチドを含むか、及び/又は配列番号31、37〜39及び42のいずれか1つで表されるポリヌクレオチドを含む。
上記のポリヌクレオチドは、最も好ましくは、配列番号41及び/又は42のポリヌクレオチドを含む。
【0063】
本発明は、本発明のヒト化抗体又はそのフラグメントをコードするポリヌクレオチドを含むベクターも提供する。
本発明のベクターは、上述した当該技術において公知の方法により作製された本発明のヒト化抗体又はそのフラグメントをコードするポリヌクレオチドを、適切なベクターに挿入することにより得ることができる。本発明のベクターに用いることができるベクターは、上記の発現ベクターについて述べたものと同様である。
本発明のベクターは、上記のポリヌクレオチドの遺伝子産物の発現に必要な配列をさらに有することが好ましい。このような配列は、当該技術において公知であり、例えばコードされるポリヌクレオチドに機能可能に連結されたプロモーター、リボソーム結合部位、ポリアデニル化部位、転写終結部位などを含む。
上記のベクターは、複製起点、エンハンサー、及び目的に応じて、通常用いられる選択マーカー(例えば、テトラサイクリン、アンピシリン又はカナマイシンに対する耐性遺伝子)を含んでいてもよい。
【0064】
本発明はまた、上記のベクターが導入された宿主細胞も提供する。好ましい宿主細胞は、上記のとおりである。
【0065】
本発明はまた、本発明のヒト化抗体又はそのフラグメントをコードするポリヌクレオチドを適切な発現ベクターに導入し、得られた発現ベクターを適切な宿主細胞に導入し、得られた宿主細胞を培養することを含む、本発明のヒト化抗体又はそのフラグメントの生産方法も提供する。
本発明の生産方法は、宿主細胞及び/又はその培養液からヒト化抗体又はそのフラグメントを精製することをさらに含んでもよい。
本発明の生産方法の各工程は、上記の本発明のヒト化抗体又はそのフラグメントの作製について述べたことと同様である。
【0066】
本発明のヒト化抗体又はそのフラグメントは、シガトキシン類を認識して結合することができるので、食中毒シガテラにより体内にシガトキシン類が存在する対象者(ヒト)に投与すれば、シガトキシン類を捕捉して、その毒性を低減できる可能性がある。
よって、本発明は、上記のヒト化抗体又はそのフラグメントを含む医薬組成物も提供する。該組成物は、食中毒シガテラの治療又は予防に用いることができる。
【0067】
上記の組成物は、いずれの投与形態であってもよいが、静脈内投与用の形態が好ましい。
【0068】
上記の組成物は、ヒト化抗体又はそのフラグメントとともに、少なくとも1種の医薬的に許容される医薬品添加物を含む。用い得る医薬品添加物は、投与形態に応じて、当該技術において公知の適切なものを用いることができる。
例えば静脈内投与用の医薬組成物の場合、医薬的に許容される医薬品添加物としては、溶剤(例えばオリーブ油、大豆油のような油性溶剤、水、アルコール、プロピレングリコールのような親水性溶剤など)、溶解補助剤(例えばポリエチレングリコール、コレステロールなど)、乳化剤(例えば界面活性剤など)、懸濁化剤(ポリビニル系化合物、セルロース類のような親水性高分子、界面活性剤など)、保存剤(例えばパラベン、ソルビン酸など)、着色剤などを用いることができる。
【0069】
上記の医薬組成物は、対象者に治療又は予防上有効な量で投与することができる。治療又は予防上有効な量は、臨床試験などにより適宜決定できる。
【実施例】
【0070】
以下に、本発明についてより具体的に理解するために、特定の実施例を示すが、これらは例示目的とするものであって、本発明を限定するものではない。
【0071】
以下の記載について、キット等を使用した部分については特にことわりの無い限り添付のプロトコールに従った。
【0072】
A.マウス10C9Fab断片のクローニング及び配列決定
マウス抗シガトキシン抗体10C9については、特許第3845796号公報に記載されている。
マウス10C9抗体を産生するハイブリドーマ細胞(FERM BP−8292)から、全RNAを分離した。ProSTAR FirstStrand RT−PCR Kit(STRATAGENE社)を用いて、逆転写により第1ストランドのcDNAを合成した。マウス抗体Fab断片のコーディング配列のクローニングを可能にするように設計されたPCRプライマー(Lernerら,Science,Vol.246,p.1275,1989)を用いて、重鎖及び軽鎖Fab領域を得た。マウス抗体10C9の軽鎖はL5とL9、重鎖はH3とH12のPCRプライマーを組み合わせて用いた(表1)。
【0073】
【表1】
【0074】
上記のプライマーを用いてPCR反応により増幅したcDNAフラグメントは、約700bp長の主要生成物を有した。cDNAフラグメントをpDriveベクター(QIAGEN社)にクローニングした。適正なサイズのインサートを含むベクターを、DTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)を用いて配列決定した。
決定したマウス抗体10C9の軽鎖可変領域VL、軽鎖定常領域CL、重鎖可変領域VH及び重鎖定常領域CH1のcDNA配列及びそれらから推測されるアミノ酸配列を、それぞれ配列番号1、3、5及び7(cDNA配列)及び配列番号2、4、6及び8(アミノ酸配列)に示す。
【0075】
B.シガトキシン類を認識するヒト化抗体の設計
(1)フレームワーク構造配列のための相同ヒト抗体の選択
マウス10C9軽鎖及び重鎖の可変領域に対して高い同一性を示すヒトgermline配列(V、J遺伝子領域)を、ClustalWを用いたアミノ酸配列の比較により同定した。
マウス10C9のVL領域に対して、61.5%という同一性百分率をもつヒトgermlineのκ鎖V領域(1GKV1D−39*01;配列番号9)及び70.4%という同一百分率をもつJ領域(1GKJ2*01;配列番号10)が同定された。
【0076】
マウス10C9のVH領域に対して、70.4%という同一性百分率をもつヒトgermlineのV領域(1GHV4−31*03;配列番号11)及び86.7%という同一百分率をもつJ領域(1GHJ4*01;配列番号12)が同定された。
マウス抗体10C9の可変領域と高い同一性を有するこれらのヒトgermline配列の各々からのフレームワーク構造領域(FW)のV領域及びJ領域をそれぞれ軽鎖及び重鎖について連結したアミノ酸配列を基本として、本発明のヒト化抗体の軽鎖及び重鎖を設計することにした。
【0077】
(2)相補性決定領域とフレームワーク構造領域の連結
ヒト化抗体のVL及びVH領域の作製のための次の段階として、マウス10C9のVL及びVH領域からのCDRを、(1)で設計したヒトgermline配列からのFWに連結させた。このマウス抗体のCDRとヒト免疫グロブリンのFW領域とを連結しただけのVL領域及びVH領域のバージョン(グラフト化抗体)を、h10C9g−VL及びh10C9g−VHと呼ぶこととした。h10C9g−VL及びh10C9g−VHのアミノ酸配列を、それぞれ配列番号13及び14に示す。
【0078】
(3)フレームワーク構造領域内のアミノ酸の置換
(a)軽鎖
上記のようにして設計されたh10C9g−VLのアミノ酸配列において、以下の表2に示すように、軽鎖のFWの1位、2位、7位、46位、60位及び71位の6個のアミノ酸の少なくとも1つを、マウス抗体10C9の対応する位置に存在するアミノ酸に置換した。
【0079】
【表2】
【0080】
具体的には、FW1の位置1、2及び7のアミノ酸(ヒトgermline配列内に通常存在するアスパラギン酸、イソロイシン及びセリン)を、マウス抗体10C9のVL領域内の対応する位置に見られるグルタミン酸、ロイシン及びトレオニンに変更した。この位置のアミノ酸残基は、マウス免疫グロブリンgermline配列をマウス抗体10C9の配列と比較したときに変異されている配列であり、マウス10C9のVL領域のソマティック変異の位置であると考えられ、抗体−抗原結合や抗体の構造を保持する上で間接的に関係があると考えられる。
【0081】
FW2の位置46のアミノ酸(ヒトgermline配列内に通常存在するロイシン)を、マウス抗体10C9のVL領域内の対応する位置に見られるアルギニンに変更した。マウス抗体10C9のVL領域内のこの位置にあるアルギニンは、結合部位の中央に位置づけされているものとして、既に決定されているマウス10C9の立体構造(Uiら,J.Biol.Chem.,Vol.283,p.19440,2008)内で観察され、抗体−抗原結合の間に抗原と直接相互作用している重要な残基であると考えられる。
【0082】
FW3内の位置60のアミノ酸(ヒトgermline配列内に通常存在するセリン)を、マウス抗体10C9のVL領域内の対応する位置に見られるグリシンに変更した。この位置にあるアミノ酸残基は、マウス抗体10C9のVL領域のソマティック変異の位置で、抗体−抗原結合や構造を保持する上で間接的に関係があると考えられる。
【0083】
FW3の位置71のアミノ酸(ヒトgermline配列内に通常存在するフェニルアラニン)を、マウス抗体10C9のVL領域内の対応する位置に見られるチロシンに変更した。マウス抗体10C9のVL領域内のこの位置にあるチロシンは、マウス10C9VL領域のCDR2ループを支持する上で重要であると考えられる。この位置におけるフェニルアラニンの存在が、CDR1ループの配置を分断する可能性がある。
【0084】
(b)重鎖
上記のようにして設計されたh10C9g−VHのアミノ酸配列において、以下の表3に示すように、重鎖のFWの5位、71位、73位及び94位の4個のアミノ酸の少なくとも1つを、マウス抗体10C9の対応する位置に存在するアミノ酸に改変した。
【0085】
【表3】
【0086】
具体的には、FW1内の位置5のアミノ酸(ヒトgermline配列内に通常存在するグルタミン)を、マウス抗体10C9VH領域内の対応する位置で見られるロイシンに変更した。マウス抗体10C9VH領域内のこの位置にあるロイシンは、マウス10C9のVH領域のソマティック変異の位置であると考えられ、またCDR1ループを支持する上で重要であると考えられる。
【0087】
FW3の位置71のアミノ酸(ヒトgermline配列内に通常存在するバリン)を、マウス抗体10C9のVH領域内の対応する位置に見られるアルギニンに変更した。この位置にあるアルギニンは、マウス抗体10C9のVH領域のCDR2ループを支持する上で重要であると考えられる。この位置におけるバリンの存在が、VH領域のCDR2ループの配置を分断する可能性がある。
【0088】
FW3の位置73のアミノ酸(ヒトgermline配列内に通常存在するトレオニン)を、マウス抗体10C9のVH領域内の対応する位置で見られるセリンに変更した。マウス抗体10C9のVH領域内のこの位置にあるセリンは、マウス抗体10C9のVH領域のCDR2ループを支持する上で重要であると考えられる。
【0089】
FW3内の位置94のアミノ酸(ヒトgermline内に通常存在するアルギニン)を、マウス抗体10C9のVH領域内の対応する位置に見られるシステインに変更した。マウス抗体10C9のVH領域内のこの位置にあるシステインは、マウス抗体10C9のVH領域のソマティック変異の位置であると考えられ、またCDR3ループを支持する上で重要であると考えられる。
【0090】
(4)定常領域の選択
マウス抗体10C9の軽鎖のCL及び重鎖のCH1に対して高い同一性を示すヒトgermline配列のC遺伝子領域を、ClustalWを用いたアミノ酸配列の比較により同定した。
マウス抗体10C9の軽鎖C領域に対して、70.4%という同一性百分率をもつヒトgermlineのκ鎖C領域(1GKC*01;配列番号28)が同定された。
マウス抗体10C9の重鎖CH1領域に対して、61.9%という同一性百分率をもつヒトgermlineのγ1鎖C領域(1GHG1*01;配列番号29)が同定された。
マウス抗体10C9の定常領域と高い同一性を示すこれらのヒトgermline配列のCL及びCH1領域を、(3)で設計した可変領域に連結して、ヒト化抗体のFabフラグメントの作製に使用した。
【0091】
C.ヒト化抗体をコードするポリヌクレオチドの構築
C−1.h10C9d−VL及びh10C9d−VHを含む軽鎖及び重鎖をコードするDNAの作製
(3)で設計した軽鎖及び重鎖の可変領域(h10C9d−VL及びh10C9d−VH)を、(4)で選択した軽鎖及び重鎖の定常領域(CL(1GKC*01)及びCH1(1GHG1*01))とそれぞれ連結したヒト化抗体のFabフラグメントの軽鎖及び重鎖をそれぞれコードするDNAを構築した。DNAの合成は、TAKARA社に委託した。
ヒト化抗体軽鎖FabフラグメントはSacI及びXbaIの制限酵素認識部位をもつように作製し、pMD19内へクローニングさせた。適正な配列をもつクローンを、h10C9dFab-L(配列番号30)と命名した。
ヒト化抗体重鎖FabフラグメントはXhoI及びSpeIの制限酵素認識部位をもつように作製し、pMD19内へクローニングさせた。適正な配列をもつクローンを、h10C9dFab-H(配列番号31)と命名した。
【0092】
C−2.h10C9d−VL−a〜e又はh10C9o−VL、及びh10C9d−VH−a〜d又はh10C9o−VHを含む軽鎖及び重鎖をコードするDNAの作製
上記のh10C9dFab-L及びh10C9dFab-Hの所望の位置のアミノ酸をマウス抗体10C9の対応する位置のアミノ酸に変換するために、シグマジェノシス社に依頼して、表4に示すPCRプライマーを合成した。
変換するアミノ酸の位置に従って選択される2組のプライマーを用いて、部位特異的変異誘発法を応用した市販のキット(STRATAGENE社;QuickChange Site−Directed Mutagenesis Kit)を用いてDNAに変異導入を行った。DTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)によりDNA配列を決定し、所望の配列をもつクローンを選択した。
【0093】
【表4】
【0094】
C−3.グラフト化抗体の作製
マウス抗体10C9のCDRと、ヒト免疫グロブリンのFW領域とを連結したVL領域及びVH領域のバージョン(グラフト化抗体;h10C9g−VL及びh10C9g−VH)と、(4)で選択した定常領域とを連結したFabフラグメントは、h10C9dFab−VL及びh10C9dFab−VHを鋳型として表1のPCRプライマーを用いてPCRを行うことにより構築された。DTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)により配列決定して、適正な配列をもつクローンを、h10C9Fabg−L(配列番号43)及びh10C9Fabg−H(配列番号44)と命名した。
【0095】
D.マウス抗体及びヒト化抗体の発現及び分析
D−1.発現プラスミドの構築
マウス抗体及びヒト化抗体の発現には、抗体Fab遺伝子を高効率に発現できることが報告されているpARA7(Miyashitaら,J.Mol.Biol.,Vol.267,p.1247,1997)を発現ベクターとして用いた(図2)。pARA7ベクターは、アラビノースにより誘導されるaraBプロモーターを含む。さらに、シグナル配列をコードする塩基配列が付加されていて、発現されるポリペプチドは、大腸菌培養上清中に分泌される。
【0096】
まず、マウス抗体の発現プラスミドは、次のようにして作製した。A.でクローニングしたマウスFab重鎖をコードするDNA配列中には、pARAに挿入する際に利用する制限酵素XhoI認識部位が含まれる。そこで、表5に示した2組のプライマーを用いて、QuickChange Site−Directed Mutagenesis Kit(STRATAGENE社)を用いてDNAに変異導入を行った。DTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)によりDNA配列を決定し、適正な配列をもつクローンをm10C9Fab−Hm(配列番号65)と命名した。
【0097】
【表5】
【0098】
上記で作製したマウスFab重鎖をコードするDNA(m10C9Fab−Hm)を、制限酵素XhoI認識部位及びSpeI認識部位を利用して、pARA7に挿入した。重鎖をコードするDNAが挿入されたプラスミドに、A.でクローニングしたマウスFab軽鎖をコードするDNA(m10C9Fab−L;配列番号68)を、制限酵素SacI認識部位及びXbaI認識部位を利用して挿入し、発現プラスミドを作製した。
【0099】
ヒト化抗体の発現プラスミドは、以下のようにして作製した。C.で作製した各々のヒト化Fab重鎖をコードするDNAのそれぞれを、制限酵素XhoI認識部位及びSpeI認識部位を利用して、pARA7に挿入した。重鎖をコードするDNAが挿入されたプラスミドに、C.で作製した各々のヒト化Fab軽鎖をコードするDNAのそれぞれを、制限酵素SacI認識部位及びXbaI認識部位を利用して挿入し、発現プラスミドを作製した。なお、軽鎖及び重鎖の可変領域の組み合わせは、以下の表6に示すとおりである。
【0100】
【表6】
【0101】
これらの発現プラスミドを、大腸菌に導入して増殖させ、市販のプラスミド精製キット(QIAGEN社;QIAprep Spin Miniprep Kit)により精製し、DNA塩基配列解析によりFabのコード領域の配列を確認した。DNA塩基配列解析にはDTCS−Quick Start Kit及びCEQ2000XL DNA Analysis System(ともにベックマンコールター社)を使用した。
【0102】
ヒト化10C9のうち、h10C9gFab及びh10C9oFabについては、マウスアミノ酸に置換した軽鎖内の位置1及び位置2と重鎖内の位置5がpARA7に挿入するための制限酵素認識部位に含まれるので、pARA7に挿入後、合成プライマー(表7)を用いてヒトgermline由来アミノ酸に戻すためのDNA変異導入を行った。
【0103】
【表7】
【0104】
D−2.発現プラスミドによる大腸菌の形質転換及び培養
次に、上記のD−1のようにして精製した発現プラスミドを、常法に従って大腸菌MC1061株に導入し、LB寒天プレート(100μg/mlのアンピシリン含有)に播いて形質転換株を得た。この形質転換株を、100μg/mlのアンピシリンを含むSB培地(Miyashitaら、上記文献)で増殖させ、発現誘導剤であるアラビノースを0.2%になるように添加して30℃で3日間培養を行った。培養終了後、遠心分離を行い、抗体を含む培養上清を回収した。
【0105】
D−3.マウス抗体及びヒト化抗体の精製
上記の培養上清に硫酸アンモニウムを65%飽和になるように加え、硫安沈殿を行った。遠心分離後の沈殿を10mMリン酸ナトリウム緩衝液に再懸濁し、同緩衝液に対して透析を行った。遠心分離後、上清を0.45μmのフィルターでろ過した。この上清を材料として、プロテインGカラム(GEヘルスケア・ジャパン社)を用いて精製した。その後、各分画フラクションについてSDS−PAGEを行うことによって、純度の高いFabが得られたことを確認した(図3;マウス抗体10C9Fab(m10C9Fab)、h10C9dFab、h10C9gFab及びh10C9oFabについてのみ結果を示す)。
【0106】
D−4.マウス抗体及びヒト化抗体の検出
D−2で作製した大腸菌で発現させたマウス抗体10C9のFab及びそれぞれのヒト化抗体のFabについて、ウェスタンブロッティングにより、アルカリフォスファターゼ標識抗マウスIgG(Millipore社)又はアルカリフォスファターゼ標識抗ヒトIgG(Millipore社)を用いて、抗体の検出を行った。
【0107】
図4の結果から明らかなように、マウス抗体10C9(m10C9Fab)は抗マウス抗体によって検出されるが、抗ヒト抗体によっては検出されない。一方、作製したヒト化抗体h10C9dFabは抗マウス抗体によって検出されないが、抗ヒト抗体によって検出された。その他のヒト化抗体(h10C9gFab、h10C9oFab、h10C9dFabM1〜M9)についても抗ヒト抗体によって検出された。
よって、本発明のヒト化抗体は、抗ヒト抗体により認識されるが、抗マウス抗体によっては認識されないので、ヒトに投与した場合に、免疫原性が低いことが期待される。
【0108】
D−5.ELISAによる抗原結合性の確認
シガトキシンCTX3CのABCDE環部の構造を有する抗原(CTX3C−ABCDEという)に対する各ヒト化抗体の結合活性を、マウス10C9抗体と比較してELISAによって検定した。ELISAの方法は、特許第3845796号公報に記載された方法に従う。
96ウェルELISA用プレート(COSTAR社)の各ウェルにCTX3C−ABCDE−BSAコンジュゲートを50μL入れ、4℃で一晩放置して、プレートに固定化した。プレートを洗浄後、1%スキムミルクでブロッキングした。プレートをさらに洗浄した後、連続希釈したマウス抗体又はヒト化抗体を、ウェルに50μLずつ添加して、室温で1時間反応させた。
検出は、ペルオキシダーゼ(HRP)標識抗ヒトIgG(Millipore社)又はHRP標識抗マウスIgG(GEヘルスケア・ジャパン社)を用いて行った。波長490nmにおける吸光度を、マイクロプレートリーダー(BioRad社)を用いて測定した。
【0109】
図5は、マウス抗体10C9(m10C9Fab)とヒト化抗体(h10C9gFab、h10C9dFab、h10C9oFab、h10C9dFabM1〜M9)の抗原との結合を比較している。その結果、マウスCDRとヒトIgのFW領域とを含むグラフト化抗体h10C9gFabは、マウス抗体10C9よりも、抗原に対する結合性が低かった。h10C9dFab及びh10C9oFabは、マウス10C9抗体と同程度又はそれ以上の結合活性を示すことが確認された。h10C9dFabM1〜M5、M7及びM8は、h10C9dFabと比較して同程度又はそれ以上の結合活性を示すことが確認された。
しかし、軽鎖の46位のアミノ酸がヒト免疫グロブリン由来のアミノ酸であるh10C9dFabM6、及び重鎖の94位のアミノ酸がヒト免疫グロブリン由来のアミノ酸であるh10C9dFabM9は、結合活性が低下していることがわかった。
【0110】
D−6.Biacoreによる結合親和力の測定
マウス抗体10C9(m10C9Fab)又はヒト化抗体のシガトキシンCTX3C−ABCDEに対する結合活性を、表面プラズモン共鳴測定法を用いて確認した。
CTX3C−ABCDE−BSAコンジュゲートをSensor Chip CM5(GEヘルスケア・ジャパン社)に固定化し、HBS−EPバッファー(GEヘルスケア・ジャパン社)で連続希釈した各抗体溶液を用いて結合活性を確認した結果を、図6に示す。
Kd値を求めたところ、マウス抗体10C9と比較してh10C9gFabは明らかに親和性が低下していた。
これに対して、h10C9dFab、h10C9oFab並びにh10C9dFabM1〜M5、M7及びM8は、CTX3C−ABCDEに対するマウス抗体10C9と同様又はわずかに良い結合親和力を示すことが確認された。
しかし、軽鎖の46位のアミノ酸がヒト免疫グロブリン由来のアミノ酸であるh10C9dFabM6、及び重鎖の94位のアミノ酸がヒト免疫グロブリン由来のアミノ酸であるh10C9dFabM9は、Kd値が非常に大きく、シガトキシン類との結合親和性が低いことがわかった。
【0111】
従って、本発明のヒト化抗体は、マウス抗体10C9とほぼ同様の抗原親和性を有することがわかった。さらに、ヒトFW内のわずかな修正(VLの位置46及びVHの位置94)だけを有するヒト化抗体(h10C9oFab)であっても、マウス抗体と同様の抗原親和性でシガトキシンを認識できることがわかった。
【0112】
添付の配列表に示す配列は、以下のものに相当する。
【0113】
【表8−1】
【0114】
【表8−2】
【特許請求の範囲】
【請求項1】
ヒト免疫グロブリンのフレームワーク領域(FW)と、
FERM BP−8292のハイブリドーマにより産生されるシガトキシン類を認識するマウス抗体10C9の相補性決定領域(CDR)と
を含む可変領域をそれぞれ有する重鎖及び軽鎖を少なくとも含み、
軽鎖の46位のアミノ酸と、任意に、1位、2位、7位、60位及び71位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の軽鎖の対応する位置に存在するアミノ酸で置き換えられ、及び/又は
重鎖の94位のアミノ酸と、任意に、5位、71位及び73位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の重鎖の対応する位置に存在するアミノ酸で置き換えられており、
シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識することを特徴とする、ヒト化抗体又はそのフラグメント。
【請求項2】
前記軽鎖が配列番号15〜19及び21のいずれか1つのアミノ酸配列を有し、前記重鎖が配列番号22〜25及び27のいずれか1つのアミノ酸配列を有する請求項1に記載のヒト化抗体又はそのフラグメント。
【請求項3】
請求項1又は2に記載のヒト化抗体又はそのフラグメントをコードする配列を含むポリヌクレオチド。
【請求項4】
請求項3に記載のポリヌクレオチドを含むベクター。
【請求項5】
請求項4に記載のベクターが導入された宿主細胞。
【請求項6】
請求項5に記載の宿主細胞を培養し、シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識するヒト化抗体又はそのフラグメントを発現させる工程を含む前記ヒト化抗体又はそのフラグメントを生産する方法。
【請求項1】
ヒト免疫グロブリンのフレームワーク領域(FW)と、
FERM BP−8292のハイブリドーマにより産生されるシガトキシン類を認識するマウス抗体10C9の相補性決定領域(CDR)と
を含む可変領域をそれぞれ有する重鎖及び軽鎖を少なくとも含み、
軽鎖の46位のアミノ酸と、任意に、1位、2位、7位、60位及び71位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の軽鎖の対応する位置に存在するアミノ酸で置き換えられ、及び/又は
重鎖の94位のアミノ酸と、任意に、5位、71位及び73位から選択される少なくとも1つの位置のアミノ酸とが、前記マウス抗体10C9の重鎖の対応する位置に存在するアミノ酸で置き換えられており、
シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識することを特徴とする、ヒト化抗体又はそのフラグメント。
【請求項2】
前記軽鎖が配列番号15〜19及び21のいずれか1つのアミノ酸配列を有し、前記重鎖が配列番号22〜25及び27のいずれか1つのアミノ酸配列を有する請求項1に記載のヒト化抗体又はそのフラグメント。
【請求項3】
請求項1又は2に記載のヒト化抗体又はそのフラグメントをコードする配列を含むポリヌクレオチド。
【請求項4】
請求項3に記載のポリヌクレオチドを含むベクター。
【請求項5】
請求項4に記載のベクターが導入された宿主細胞。
【請求項6】
請求項5に記載の宿主細胞を培養し、シガトキシンCTX3C又は51−ヒドロキシ−CTX3Cであるシガトキシン類を認識するヒト化抗体又はそのフラグメントを発現させる工程を含む前記ヒト化抗体又はそのフラグメントを生産する方法。
【図1−1】


【図1−2】

【図5】
【図6−1】
【図6−2】
【図6−3】
【図6−4】
【図2】
【図3】
【図4】

【図1−2】
【図5】
【図6−1】
【図6−2】
【図6−3】
【図6−4】
【図2】
【図3】
【図4】
【公開番号】特開2011−200164(P2011−200164A)
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願番号】特願2010−70349(P2010−70349)
【出願日】平成22年3月25日(2010.3.25)
【出願人】(505127721)公立大学法人大阪府立大学 (688)
【Fターム(参考)】
【公開日】平成23年10月13日(2011.10.13)
【国際特許分類】
【出願日】平成22年3月25日(2010.3.25)
【出願人】(505127721)公立大学法人大阪府立大学 (688)
【Fターム(参考)】
[ Back to top ]