
シグナル配列非依存性pIXファージディスプレイ
本発明は、主にpIXに由来する新規融合タンパク質を通した、繊維状ファージ上に提示されるペプチドのための代替足場を提供する。繊維状ファージのライブラリーが、融合タンパク質から作出され得、ファージミドとヘルパーファージとを含むファージディスプレイシステムは、本発明の一部である。本発明の一局面は、本発明の融合タンパク質をコードする核酸を含有しているファージミドとヘルパーファージとを含むファージディスプレイシステムを含有しているキットである。
【発明の詳細な説明】
【背景技術】
【0001】
発明の背景
タンパク質の同定、特徴決定、および修飾のためのコンビナトリアルアプローチの使用は、学術的な研究開発においても商業的な研究開発においても高度に成功している。この点で、繊維状バクテリオファージディスプレイまたは繊維状ファージディスプレイのテクノロジーは、最初のライブラリープラットフォームとしてその道を開き、今なお優勢なテクノロジーとして王座についている。従って、ファージディスプレイは、基礎的なタンパク質発見および応用的なタンパク質発見の両方において、そして世界的に最も急成長中の化合物クラスである新規のタンパク質に基づく診断薬および治療薬の両方の開発において、広く適用されている。
【0002】
コンビナトリアルファージディスプレイテクノロジーの原理は、各ビリオンが、タンパク質コートに封入されたゲノムによってコードされるまさに同一のタンパク質を、表面上に単に提示する、という特性により与えられる、遺伝子型-表現型連鎖に基づく。ファージ粒子自体は、多様な生理化学的条件に対して高度に抵抗性である;従って、ファージディスプレイは、競合するコンビナトリアルテクノロジーと比較して、多くの選択計画における優れた多用途性を示す。
【0003】
異種ポリペプチドのファージディスプレイは、繊維状ファージのコートの5種の構造タンパク質全てを使用して達成されているが、pIIIディスプレイのみが広範に使用されており、pVIIIディスプレイもある程度は使用されている(図1)。
【0004】
異種融合体が短いペプチドのみである場合には、ファージゲノムに基づくベクターを使用した多価ディスプレイシステムが好ましいが、折り畳まれたドメインを必要とする、より大きな融合体については、大部分の適用がファージミドシステムより利益を得るであろう。後者の場合、抗体-pIIIファージディスプレイが、この領域において極めて優勢であるが、代替足場が出現しつつあり、将来のタンパク質工学ツールの増大の必要性は続いている。高度に望まれる適用は、単に、ファージビリオンを細菌に感染させることにより、ファージディスプレイライブラリーから、高親和性の特異的なペプチド結合剤またはタンパク質結合剤を、標的に結合したまま効率的に入手することである。
【0005】
Endemann and Model, 1995(PMID:7616570)は、マイナーなコートpIXは、N末端にもう一つのタンパク質が融合すると機能しないことを報告した。従って、この報告は、pIXはファージディスプレイのために使用され得ないとの結論を下した。
【0006】
Gaoら(PMID:10339535、12239343、およびWO0071694)、ならびにKhalilら(PMID:17360403)の両方は、その後、ファージミドから発現させ、かつシグナル配列依存性周辺質ターゲティングと組み合わせて使用した場合には、N末端pIX融合ディスプレイが可能であることを示した。これらのシステムにおいては、ファージミドレスキューによりヘルパーファージゲノムからwt pIXが供与され、補完が起こる。
【0007】
(融合タンパク質が挿入されていない)WO0071694の図2Aに開示されたようなファージミドおよび(融合タンパク質が挿入されている)WO0071694の図2Bに開示されたようなファージミドは、明白に、pelBシグナル配列を含む(図面の説明は7頁2〜14行目)。
【0008】
前述のように、pIXは、N末端にもう一つのタンパク質が融合すると機能しないことが、以前に示唆され、Gaoらが、単独の、または両方の組み合わせによる、成功のための二つの可能性のある理由を与えた。
【0009】
一つの可能性のある理由は、原核生物リーダー配列(シグナル配列)が融合タンパク質のN末端に付着させられ、従って、組換えタンパク質の周辺質空間へのターゲティングが確実になり、それにより、細胞質における蓄積が防止されたというものである。
【0010】
もう一つの可能性のある理由は、EndemannおよびModelでのようなファージゲノムではなく、ファージミドから組換えタンパク質が発現され、従って、ファージミドレスキューのために必然的に必要とされるヘルパーファージ由来の野生型pIXが、組換えpIX融合タンパク質を補完し、従って、さもなければ、組換え修飾のために失われていたかもしれない野生型機能性が保存されたというものである。即ち、ファージは、野生型タンパク質と融合タンパク質とのミックスを含んでいたであろう。
【0011】
Khalilら(PMID:17360403)は、まさに同一のビリオンの各遠位先端に外来性ペプチドが提示される二重特異的な繊維状ファージビリオンの特色を活用する適用を記載している。彼らは、原核生物シグナル配列依存性pIXディスプレイファージミドを補完する一般的なpIIIファージゲノムベクターの組み合わせを使用することにより、これを達成した。この環境において、ファージゲノムベクターは、ファージミドレスキューにおけるヘルパーファージとして役立った。
【発明の概要】
【0012】
本発明の目的は、繊維状ファージ上に提示されたペプチドのための代替足場を提供することである。
【0013】
本発明の第1の局面は、原核生物N末端シグナル配列を含まず、従って、外来性ペプチドとの直接融合体である、繊維状ファージに由来するpIX融合タンパク質である。
【0014】
本発明のもう一つの局面は、本発明の融合タンパク質をコードする核酸に関する。
【0015】
本発明の一つの局面は、本発明の融合タンパク質を含む繊維状ファージに関する。
【0016】
本発明のもう一つの局面は、繊維状ファージのライブラリーに関する。
【0017】
本発明のもう一つの局面は、本発明のpIX融合タンパク質をコードする核酸を含むファージミドとヘルパーファージとを含むファージディスプレイシステムに関する。
【0018】
一つの局面は、本発明のpIX融合タンパク質をコードする核酸を含むファージミドとヘルパーファージとを含むファージディスプレイシステムを含むキットに関する。
【図面の簡単な説明】
【0019】
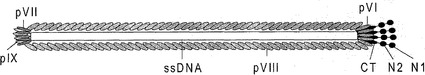
【図1】繊維状ファージ構造の概略図である。ビリオンは、一本鎖DNA分子をコートする5種の構造タンパク質により構築される。野生型(wt)ファージには、pVIIIが約2700コピー存在し、ビリオンの各先端に見出される4種のタンパク質pIII、pVI、pVII、およびpIXがいずれもおよそ3〜5コピー存在する。ビリオンサイズは、pVIIIコートタンパク質1個当たりおよそ2.3ヌクレオチドで、ゲノムサイズに依存し、従って、粒子の長さは、挿入されるpVIIIのコピーの増加または減少に適合する。特に、pIIIおよびpVIIIの構造は、X線繊維回折、結晶学、およびNMRにより特徴決定されている。マイナーなコートタンパク質pIIIは、グリシンリッチ領域により分離された3個の別個のドメイン:N1(TolAに結合する)、N2(F繊毛に結合する)、およびCT(ビリオンに組み込まれ、正常なビリオン組み立てのために重要である)を含有している。
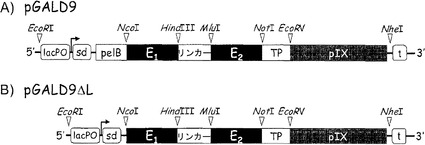
【図2】新規のpGALD9 pIXディスプレイファージミド(A)およびpGALD97ΔL pIXディスプレイファージミド(B)の概略図である。両ファージミドのベクター骨格は、GenBankアクセッション番号:Y14584からアクセス可能な配列を有するpIIIディスプレイファージミドpSEX81(SEQ ID NO:2)に基づいており、構築に関する詳細は材料および方法に記載される。両ファージミドは、それぞれ、NcoI/HindIII部分およびMluI/NotI部分の容易なカセット交換を通して、インフレームの(E1およびE2と呼ばれる)外来性配列のセグメントを収容することができる。カセットは、本明細書に記載された異なる構築物によって変動する合成リンカー配列により接続されている。略語:lacPO、lacプロモーター;sd、シャインダルガーノ配列;pelB、細菌ペクチン酸リアーゼのシグナル配列;TP、トリプシンプロテアーゼ部位;t、T7転写ターミネーター。
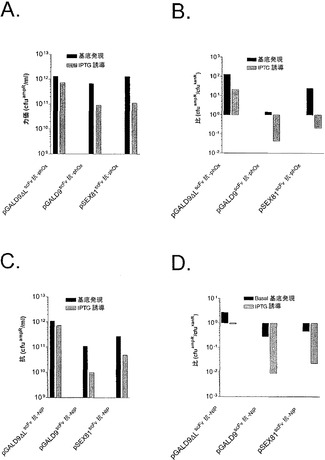
【図3】pGALD9ΔL、pGALD9、pSEX81から提示された(A)scFv抗phOx(SEQ ID NO:11)および(C)scFv抗NIPのファージミド力価。全てのファージミドが、アンピシリン耐性マーカーを保有しており;従って、力価は、溶液1ミリリットル当たりのアンピシリン耐性コロニー形成単位(cfuampR/ml)として示される。ファージミド力価(cfuampR/ml)をヘルパーファージ力価(cfukanR/ml)により割った比として示された、pGALD9ΔL、pGALD9、pSEX81から提示された(B)scFv抗phOx(SEQ ID NO:11)および(D)scFv抗NIPのファージミド対ヘルパーファージ比。ビリオンパッケージングは、IPTGなしで(基底発現)、または重感染後に0.1mM IPTGの最終濃度を存在させて(IPTG誘導)、材料および方法に記載されるような標準的なファージミドレスキューにより行われた。
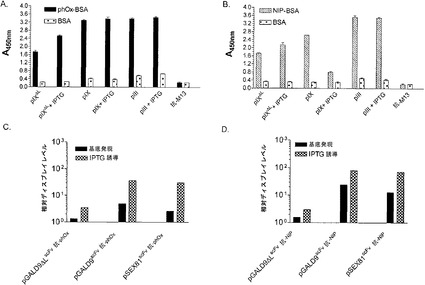
【図4】pIXとpIIIとの間の、シグナル配列がある場合とシグナル配列がない場合(ΔL)との間の、機能的な(A)scFv抗phOx(SEQ ID NO:11)および(B)scFv抗NIP(SEQ ID NO:3)のディスプレイを比較する抗原特異的ELISA。ELISAは、100μl/ウェルの精製されたビリオン含有上清を使用して、材料および方法に記載されるように実施された。抗M13HRPは、ビリオン検出MAbの抗原およびブロックへの非特異的な吸着についての陰性対照である。(CおよびD)AおよびBのELISAを繰り返したが、シグナル飽和の前に発色を中止し、力価の関数として抗原反応性を使用して相対ディスプレイレベルを決定した。
【図5】アフィニティ選択における明白な標的特異的濃縮は、カプシドディスプレイ足場、関心対象のタンパク質(Protein of Interest/POI)の発現、および溶出条件に依る。選択ラウンド1および2の後の、phOx/NIPをスパイクされた8つのライブラリーの各々からの等しい容量の未滴定のビリオン含有上清を、ELISAにより抗原反応性について査定した。ラウンド0は、1×1010cfuampRのスパイクされたインプットに相当する。最大可能応答を推定するため、ラウンド0におけるスパイクされた濃縮物が由来するクローン上清が含まれ、図示された結果は、円錐形により示された最大可能応答の分画として、バックグラウンドを差し引いたシグナルにより与えられている。
【発明を実施するための形態】
【0020】
発明の詳細な説明
本発明の局面のうちの一つに関して記載された態様および特色が、本発明のその他の局面にも当てはまることに、注意するべきである。
【0021】
本願において引用された全ての特許参照および非特許参照は、参照によりその全体が本明細書に組み入れられる。
【0022】
本発明者らは、修飾されたバージョンがN末端のペプチドドメインまたはタンパク質ドメインをコードするよう、繊維状ファージビリオンの構造コートタンパク質pIXが遺伝学的に改変される、新規の概念を、ここに提示する。
【0023】
pIX融合タンパク質
一つの局面において、本発明は、pIXのN末端への外来性ペプチドの融合を含む、繊維状ファージに由来するpIX融合タンパク質を提供する。そのような融合タンパク質は、例えば、ファージディスプレイに関して有用である。
【0024】
外来性ペプチドとの言及により意味されるのは、リンカーアミノ酸により、またはリンカーアミノ酸なしで、融合タンパク質のpIXアミノ酸部分のN末端へ融合させられる、元々はpIXタンパク質の一部でないペプチドである。
【0025】
好ましい態様において、融合タンパク質をコードする核酸は、原核生物N末端シグナル配列を含まない。
【0026】
本明細書において使用されるように、ペプチドという用語には、短いペプチド、ポリペプチド、タンパク質、およびそれらの断片の両方が包含される。
【0027】
pIXタンパク質という用語は、SEQ ID NO 1に開示されたアミノ酸配列(MSVLVYSFASFVLGWCLRSGITYFTRLMETSS)をさす。
【0028】
一態様において、pIXタンパク質は、SEQ ID NO 1のものとの少なくとも70%の配列同一性、例えば、75%の同一性、80%の同一性、81%の同一性、82%の同一性、83%の同一性、84%の同一性、85%の同一性、86%の同一性、87%の同一性、88%の同一性、89%の同一性、90%の同一性、91%の同一性、92%の同一性、93%の同一性、94%の同一性、95%の同一性、96%の同一性、97%の同一性、98%の同一性、99%の同一性を有するアミノ酸を含む。
【0029】
配列同一性
一般的に定義されるように、「同一性」とは、ここで、それぞれヌクレオチドレベルまたはアミノ酸レベルでの遺伝子間またはタンパク質間の配列同一性として定義される。
【0030】
従って、本発明において、「配列同一性」とは、アミノ酸レベルでのタンパク質間の同一性の尺度、およびヌクレオチドレベルでの核酸間の同一性の尺度である。タンパク質配列同一性は、配列を整列化した場合の、各配列の所定の位置のアミノ酸配列を比較することにより決定され得る。同様に、核酸配列同一性は、配列を整列化した場合の、各配列の所定の位置のヌクレオチド配列を比較することにより決定され得る。
【0031】
二つのアミノ酸配列または二つの核酸配列の同一率を決定するためには、配列が最適の比較目的のため整列化される(例えば、第2のアミノ酸配列または核酸配列との最適の整列化のため、第1のアミノ酸配列または核酸配列にギャップが導入されてもよい)。次いで、対応するアミノ酸位置またはヌクレオチド位置のアミノ酸残基またはヌクレオチドが比較される。第1の配列のある位置が、第2の配列の対応する位置と同一のアミノ酸残基またはヌクレオチドにより占有されている場合、その分子はその位置において同一である。二つの配列の間の同一率は、それらの配列が共有している同一である位置の数の関数である(即ち、同一%=同一である位置の数/位置の総数(例えば、重複している位置)×100)。一つの態様において、二つの配列は同一の長さである。
【0032】
手動で配列を整列化し、同一のアミノ酸の数を計数してもよい。あるいは、同一率の決定のための二つの配列の整列化は、数学的アルゴリズムを使用して達成されてもよい。そのようなアルゴリズムは、(Altschul et al. 1990)のNBLASTプログラムおよびXBLASTプログラムに組み入れられている。本発明の核酸分子と相同なヌクレオチド配列を入手するためには、NBLASTプログラム(score=100、wordlength=12)により、BLASTヌクレオチド検索が実施され得る。本発明のタンパク質分子と相同なアミノ酸配列を入手するためには、XBLASTプログラム(score=50、wordlength=3)により、BLASTタンパク質検索が実施され得る。比較目的のため、ギャップを挿入された整列化を入手するためには、Gapped BLASTが利用され得る。あるいは、PSI-Blastが、分子間の遠い関係を検出する反復検索を実施するために使用され得る。NBLASTプログラム、XBLASTプログラム、およびGapped BLASTプログラムを利用する場合、それぞれのプログラムのデフォルトパラメーターが使用され得る。http://www.ncbi.nlm.nih.govを参照のこと。あるいは、配列同一性は、例えば、EMBLデータベース内のBLASTプログラム(www.ncbi.nlm.gov/cgi-bin/BLAST)により、配列を整列化した後、計算され得る。一般に、例えば、「スコアリングマトリックス」および「ギャップペナルティ」に関しては、デフォルト設定が整列化のために使用され得る。本発明に関して、BLASTNおよびPSI BLASTのデフォルト設定が有利であり得る。
【0033】
二つの配列の間の同一率は、ギャップを許容し、または許容せず、上記のものと類似の技術を使用して決定され得る。同一率の計算においては、正確なマッチのみが計数される。
【0034】
折り畳まれたタンパク質
好ましい態様において、ペプチドという用語は、抗体に由来するドメインのような折り畳まれたタンパク質を排他的にさす。折り畳まれたタンパク質は、Fv、scFv、Fab、単一ドメインを含む抗体またはその断片、プロテインAのZドメインまたはその断片(アフィボディ)、アンキリンまたはその断片、DARPinまたはその断片、T細胞受容体またはその断片、MHCクラスIもしくはIIまたはそれらの断片、フィブロネクチンまたはその断片、アンチカリンまたはその断片、PDZドメインまたはその断片、IgNARまたはその断片、CTLA4またはその断片、ImmE7またはその断片、ノッチンまたはその断片、アビマーまたはその断片、GFPまたはその断片、およびその他の遺伝子によりコードされた生物学的フルオロフォアであり得ることを、当業者は認識するであろう。
【0035】
原則的には、それが提示される限り任意のライブラリーを作成することが可能であり、従って、最も単純な形態において、折り畳まれた秩序正しい構造と比較して、組織化されていない配置を有するもののみを分離することができる。
【0036】
もう一つの好ましい態様において、ペプチドという用語は、2〜50aaの短いペプチドを排他的にさす。ある長さにおいては、短いランダムコイルペプチドは、明確な二次または三次の折り畳みを採るために十分に長く、従って、折り畳まれたドメインの定義に入るであろう。明らかに、これは化学的組成に依り、従って、ある20aaのペプチドはやはりランダムコイルであろうが、もう一つの20aaペプチドは折り畳まれていて、折り畳まれたドメインの定義に分類されるかもしれない。
【0037】
もう一つの好ましい態様において、本発明のpIX融合タンパク質は、SEQ ID NO:1の1〜32位、2〜32位、3〜32位、4〜32位、5〜32位、6〜32位、7〜32位、8〜32位、9〜32位、10〜32位、11〜32位、および12〜32位からなる群より選択される配列を含む。原則的に、ファージミド環境において、膜貫通部分が保持され、正常なビリオンの取り込みおよび組み立てが可能であるならば、pIXの任意のN末端修飾が構想され得る。
【0038】

は、繊維状ファージの構造コートタンパク質pIX(野生型pIX)のアミノ酸配列である。最も好ましくは、pIX融合タンパク質は、SEQ ID NO:1の1〜32位を含む。
【0039】
SEQ ID NO:1は、以下に説明されるシグナル/リーダー配列と混同されるべきではない。
【0040】
シグナル配列
好ましくは、外来性ペプチドは、融合タンパク質のpIX配列のN末端へ、リンカーアミノ酸により、またはリンカーアミノ酸なしに、直接融合される。
【0041】
さらにもう一つの好ましい態様において、pIX融合タンパク質は原核生物N末端リーダー配列を含まない。
【0042】
「リーダー配列」という用語は、「シグナルペプチド」および「シグナル配列」という用語と交換可能に使用され、グラム陰性菌の周辺質膜空間へと(そのリーダー配列を含む)タンパク質をターゲティングするアミノ酸配列をさす。しばしば使用されるリーダー配列の例は、pelBss、OmpAss、TorAss、malEss、phoAss、lamBss、Blass、DspAss、mglBss、sfmCss、tolBss、およびTorTssである。そのようなシグナル配列は、少なくともSRP依存性、SEC依存性、TatABC依存性、またはYidC依存性の細胞質から周辺質空間への転移を含むことが公知の大腸菌の分泌機構へ、完全タンパク質をターゲティングすることが公知である(Baneyx et al. PMID:15529165)。従って、N末端シグナル配列という用語は、タンパク質のN末端部分にあるシグナル配列をさす。
【0043】
(それを含む)タンパク質を大腸菌の分泌機構へターゲティングし、それにより、細胞質コンパートメントから周辺質コンパートメントへと転移させる特性を保有しているシグナル配列は、部分的には、アミノ酸組成の化学的特性により定義されるサインまたはモチーフを通して同定され得る。
【0044】
しかしながら、既存の機能性シグナル配列の多様性は、それらの同定における現在の知識をまだ越えており、従って、同族シグナル配列としてのペプチドの定義における現在の最先端技術は、典型的には、例えば、ニューラルネットワークまたは発見的方法論により、知識に基づくデータベースをテンプレートとして使用して、データ発掘を通して行われている。SignalP、PROSITEのPPSEARCH(EMBL-EBI)、SecretomeP、TatPのような、いくつかのそのようなツールが、現時点で、オープンアクセスのチャネルを通して公に入手可能になっている。
【0045】
分泌タンパク質のクラスでは、それらが細胞質コンパートメントから輸送され、ルールから逸脱するため、シグナル配列モチーフが同定され得ないという意味で、問題がより大きいが、ここでも、データ発掘を通して、シグナル配列特色を定義するか、または問題の真核生物タンパク質の分泌能の可能性を得ることが可能である。原核生物分類群のためのそのようなツールは、まだ存在しない。
【0046】
従って、ペプチドをシグナル配列として決定的に同定した現在利用可能な唯一の方法は、それが現実のシグナル配列であるか否かを確立するためにペプチドの特性をバリデートするための実験的な手段による。シグナル配列内の所定のアミノ酸位置を改変し、それでもネイティブの機能性、または増加した輸送能力のような改変された機能性により、シグナルペプチドとしての機能は保持されるよう、そのようなペプチドを操作してもよいことも明白である。アミノ酸の欠失または付加も利用され得る。そのような分析および操作は、実際、Ff pVIIIシグナル配列、Sec経路をターゲティングするg8pss、およびTat経路をターゲティングするTorAssについて行われている。特に、Shenらの結果は、pIIIシグナル配列および細菌ペクチン酸リアーゼシグナル配列の機能性ではあるが改変されている変異体の操作のための、根拠の確かなガイドラインとして役立つかもしれない。
【0047】
シグナル配列の機能性は、以下の二つの特性にさらに分解され得る:
1. (それを含む)タンパク質を大腸菌の分泌機構へとターゲティングし、それにより、細胞質コンパートメントから周辺質コンパートメントへと転移させ、この過程の途中で、リポタンパク質シグナルペプチターゼまたはリーダーペプチダーゼのような特定のプロテアーゼによるタンパク質分解によって残りのタンパク質から分離される。
2. (それを含む)タンパク質を大腸菌の分泌機構へとターゲティングし、それにより、細胞質コンパートメントから周辺質コンパートメントへと移転させ、転移後もやはりタンパク質の一部として残存する。
【0048】
シグナル配列の大多数は、上に与えられた情況(1)にあるが、これらのタンパク質が情況(2)になるよう容易に操作され得ることは明白である。従って、元々は情況(1)に属するが、情況(2)へと改変された現在公知のシグナル配列、例えば、変異型pelBssおよびその他も、やはり、同族のシグナル配列と見なされる。
【0049】
さらに、情況(1)のシグナル配列を情況(2)に改変するか、または情況(2)にあるシグナル配列を直接選び、次いで、転移後にシグナル配列を除去することも構想され得る。これは、宿主の内在性プロテアーゼにより、かつ/または、例えばファージディスプレイの場合には、タンパク質がカプシドタンパク質と融合する時に、なされる。その場合には、明確な切断が達成され得るよう、シグナル配列またはそれを含むタンパク質の適切な領域に、人工的なプロテアーゼ部位が組み込まれるであろう。ここで、選ばれる二つの異なる型のプロテアーゼ部位が構想され得る:
A. プロテアーゼ部位は関心対象のタンパク質を切断しない。抗体、または主要組織適合性抗原分子もしくはT細胞受容体のようなその他の関心対象の足場と組み合わせられた、例えば、カルボキシペプチダーゼA部位または3Cライノウイルスプロテアーゼ部位のような予測された部位のみ。このアプローチを使用することにより、例えば、上記の情況(2)にあるシグナル配列を使用して、関心対象のタンパク質をファージディスプレイし、選択等において使用する前に、カプシド融合体へ機能性および均質性を入手するために、人工的にシグナルペプチドを除去することが構想され得る。
B. プロテアーゼ部位は、例えば、トリプシンのような操作された部位に加えて、興味のあるタンパク質を切断する。
【0050】
いずれの情況も、やはり、シグナル配列依存性ファージディスプレイと見なされるであろう。
【0051】
外来性ペプチド
好ましい態様において、pIX融合タンパク質の外来性ペプチドは、Fv、scFv、Fab、単一ドメインを含む抗体またはその断片、プロテインAのZドメインまたはその断片(アフィボディ)、アンキリンまたはその断片、DARPinまたはその断片、T細胞受容体またはその断片、MHCクラスIもしくはIIまたはそれらの断片、フィブロネクチンまたはその断片、アンチカリンまたはその断片、PDZドメインまたはその断片、IgNARまたはその断片、CTLA4またはその断片、ImmE7またはその断片、ノッチンまたはその断片、アビマーまたはその断片、GFPまたはその断片、およびその他の遺伝子によりコードされた生物学的フルオロフォアからなる群より選択される。
【0052】
好ましい態様において、pIX融合タンパク質の外来性ペプチドは、ライブラリーメンバーである。
【0053】
ライブラリーとは、本発明に関して使用されるように、異なるペプチドの集合体をさす。ペプチドは、折り畳まれたドメイン、または例えば2〜50アミノ酸の短いペプチドであり得る。そのようなライブラリーは、所定の標的への新たなリガンドの結合を同定するために使用され得るため、興味深い。
【0054】
ライブラリーを提示するためのpIXの使用には、pIIIまたはpVIIIを使用して提示されるライブラリーと比較して、いくつかの利点がある。pIXディスプレイは、方向性および価数に関してpIIIディスプレイと同一の長所を含有しているが、感染性に影響を与えないであろう。その現象は、pIIIディスプレイで起こることが公知であり、例えば、アフィニティ選択後のレスキューの際に、制御されない望まれない不均一性をシステムに導入する。
【0055】
さらに、pIXディスプレイは、pIIIディスプレイおよびpVIIIディスプレイの両方についての必要条件である原核生物N末端シグナル配列の必要性なしに達成され得る。最後に、pIIIディスプレイにおける標的に固定化された種は、通常、この標的-ファージ結合の(通常、競合的溶出または高pHもしくは低pHによる溶出による)破壊を必要とする。これは、例えば、pIIIディスプレイにおける高親和性のまたは安定的な結合剤の回収を著しく妨害することが公知である。感染のために必要とされるpIIIは、ファージ-標的相互作用の後ですら、未改変であり、pIXディスプレイにおける代替的な相互作用のために容易に利用可能であるため、これは、結合破壊、例えば、酸性溶出の必要性を完全に排除し、固定化されたファージは完全な感染性を保持し、従って、単なる感染により、標的と結合したまま回収されるかもしれない。
【0056】
核酸
本発明の第2の局面は、本発明の融合タンパク質をコードする核酸である。本発明の核酸は、プラスミド、ベクター、ファージゲノム、ファージミド、またはファスミドの一部であり得る。
【0057】
核酸という用語は、モノマーヌクレオチドの鎖から構成される高分子をさす。生化学において、これらの分子は、遺伝情報を保持するか、または細胞内の構造を形成する。最も一般的な核酸は、デオキシリボ核酸(DNA)およびリボ核酸(RNA)である。さらに、核酸という用語には、ペプチド核酸(PNA)、モルホリノ核酸、ロックド核酸(LNA)、グリコール核酸(GNA)、およびトレオース核酸(TNA)のような人工核酸が含まれる。これらの各々は、分子の骨格の変化により、天然に存在するDNAまたはRNAと区別される。
【0058】
ファージミドまたはファスミドは、プラスミドとして繁殖することができ、かつウイルス粒子内に一本鎖DNAとしてパッケージングされることもできるベクターを作製するため、繊維状ファージFfとプラスミドとのハイブリッドとして開発されたクローニングベクターの一つの型である。プラスミドと同様に、ファージミドは、DNA断片をクローニングするために使用され得、かつ一連の技術(形質転換、電気穿孔)により細菌宿主に導入され得る。しかしながら、ファージミドを含有している細菌宿主への、「ヘルパー」ファージ、例えば、VCSM13またはM13K07の感染が、一本鎖DNAの複製およびファージミドDNAのファージ粒子へのパッケージングを可能にするために必要なウイルス成分を提供する。
【0059】
従って、本発明の一つの局面は、原核生物N末端シグナル配列を含まない、繊維状ファージに由来するpIX融合タンパク質をコードする核酸を含む、ファージゲノムまたはファージミドに関する。
【0060】
本発明の一態様において、ファージゲノムまたはファージミドは、原核生物N末端シグナル配列を含まず、かつ
の1〜32位、2〜32位、3〜32位、4〜32位、および5〜32位からなる群より選択される配列を含む、繊維状ファージに由来するpIX融合タンパク質をコードする核酸を含む。
。
【0061】
本発明のもう一つの態様において、外来性ペプチドはpIX配列のN末端に直接融合している。
【0062】
本発明の一態様は、pIXに融合した外来性ペプチドが、Fv、scFv、Fab、単一ドメインを含む抗体またはその断片、プロテインAのZドメインまたはその断片(アフィボディ)、アンキリンまたはその断片、DARPinまたはその断片、T細胞受容体またはその断片、MHCクラスIもしくはIIまたはそれらの断片、フィブロネクチンまたはその断片、アンチカリンまたはその断片、PDZドメインまたはその断片、IgNARまたはその断片、CTLA4またはその断片、ImmE7またはその断片、ノッチンまたはその断片、アビマーまたはその断片、GFPまたはその断片、およびその他の遺伝子によりコードされた生物学的フルオロフォアからなる群より選択される、本発明のファージゲノムまたはファージミドに関する。
【0063】
本発明のもう一つの態様において、pIXに融合した外来性ペプチドはライブラリーメンバーである。
【0064】
繊維状ファージ
本発明の第3の局面は、本発明の融合タンパク質を含む繊維状ファージである。繊維状ファージビリオンは、ファージミドを保有し得る。
【0065】
しばしばバクテリオファージと呼ばれるファージは、ここで、感染性、複製性であり、細菌から分泌されるウイルスを意味する。繊維状バクテリオファージまたは繊維状ファージとは、ファージコートタンパク質によりパッケージングされる一本鎖DNA分子(ssDNA)を有するファージである。分泌された繊維状ファージ粒子は、表現型的には繊維状の構造を有する。
【0066】
繊維状ファージという用語には、本明細書において使用されるように、ファージゲノムに由来するビリオンおよびファージミドに由来するビリオンの両方が包含される。
【0067】
ヘルパーファージという用語は、例えば、欠陥ウイルスが既に占有している同じ宿主細胞に感染し、ファージミドを含有しているビリオンを形成するために必要な、欠陥ウイルスが欠いているタンパク質を提供することにより、繁殖するための、ファージゲノムに由来する一種または数種の要素を含有している、それ自体はファージゲノムでもなく機能性ウイルスでもなく、単なるプラスミドであるファージミドとして定義される、別の無関係な欠陥ウイルスを補助するウイルスをさす。
【0068】
一つの態様において、繊維状ファージは、本発明の融合タンパク質をコードする核酸を含む。特に好ましいのは、本発明の融合タンパク質をコードする核酸を含むファージミドを含むファージである。
【0069】
ファージライブラリーとは、繊維状ファージコートタンパク質のうちの一つまたは複数の一部としてペプチドまたはタンパク質を提示している繊維状ファージの集合体である。そのようなライブラリーは、異なるペプチドまたはタンパク質を提示する二種以上のファージを含むことができる。
【0070】
従って、本発明の一態様において、繊維状ファージは、繊維状ファージコートタンパク質のうちの一つまたは複数の一部としてペプチドまたはタンパク質を提示している。
【0071】
一態様において、繊維状ファージは、pIII融合タンパク質、pVII融合タンパク質、またはpVIII融合タンパク質をさらに含む。
【0072】
本発明の一局面は、pIII、pVII、pVIII、またはpIXのうちの一つまたは複数との融合体として外来性のペプチドまたはタンパク質を提示している本発明の繊維状ファージのライブラリーである。
【0073】
ライブラリーとは、繊維状ファージコートタンパク質のうちの一つまたは複数の一部としてペプチドまたはタンパク質を提示している繊維状ファージの集合体である。
【0074】
そのようなライブラリーは、異なるペプチドまたはタンパク質を提示している二種以上のファージを含むことができる。
【0075】
本発明の一局面は、異なるタンパク質を提示している二種以上の繊維状ファージを含み、これらのタンパク質のうちの少なくとも一種が本発明のファージゲノムまたはファージミドから発現されたpIX融合タンパク質である、ファージライブラリーに関する。本発明の一態様は、異なるペプチドまたはタンパク質を提示している二種以上の繊維状ファージを含むファージライブラリーに関する。
【0076】
特定の態様において、これらのペプチドまたはタンパク質のうちの少なくとも一つは、本発明のpIX融合タンパク質である。一態様において、ペプチドは、pIXと、pIII、pVII、またはpVIIIのいずれかとで、同時に提示される。
【0077】
もう一つの態様において、ペプチドは、pIXと、pIII、pVII、またはpVIIIからなる群より選択される二つまたは三つとで、同時に提示される。
【0078】
野生型(wildtype)、野生型(wild-type)、またはwtと記載されることもある野生型(wild type)という用語は、自然界に存在するような、生物、株、遺伝子、または特徴の典型的な型である。野生型とは、天然集団において最も一般的な表現型をさす。野生型とは、野生型表現型を生ずるのに必要とされる各遺伝子座における対立遺伝子もさす。野生型は、遺伝子型および表現型についての参照の標準である。生物学において、それは、特に、天然に存在する生物と意図的に変異させられた生物との間の差に関する。部位特異的変異誘発は、野生型遺伝子の遺伝子配列内の特定のヌクレオチドの変異を可能にする研究技術である。野生型タンパク質は、wt(タンパク質の名称)と記載され、例えば、野生型pIXタンパク質は、wt pIX、wt-pIX、または野生型pIXと記載される。
【0079】
従って、本発明の一つの局面は、wt pIXをコードする遺伝子および/またはwt pIXタンパク質を含まない本発明の繊維状ファージに関する。
【0080】
本発明のもう一つの局面は、wt pIXをコードする遺伝子および/またはwt pIXタンパク質をさらに含む本発明の繊維状ファージに関する。
【0081】
本発明のもう一つの局面は、本発明のファージゲノムまたはファージミドを含む繊維状ファージに関する。
【0082】
本発明の一態様において、本発明のファージゲノムまたはファージミドを含む繊維状ファージは、wt pIXをコードする遺伝子および/またはwt pIXタンパク質をさらに含む。
【0083】
本発明のもう一つの態様において、本発明のファージゲノムまたはファージミドを含む繊維状ファージは、wt pIXをコードする遺伝子および/またはwt pIXタンパク質を含まない。
【0084】
本発明のさらなる局面は、wt pIXタンパク質による補完なしにファージディスプレイにおいて機能するpIX融合タンパク質に関する。本発明の一局面は、pIII融合タンパク質、pVII融合タンパク質、およびpVIII融合タンパク質より選択される群一つまたは複数をさらに含む、本発明のファージゲノムまたはファージミドを含む繊維状ファージに関する。
【0085】
ファージディスプレイシステム
本発明の第5の局面は、ファージミドと、本発明のpIX融合タンパク質をコードする核酸を含むヘルパーファージとを含むファージディスプレイシステムである。
【0086】
ファージディスプレイシステム、ファージディスプレイ技術、ファージディスプレイテクノロジー、または単にファージディスプレイとは、タンパク質を、それをコードする遺伝情報と結び付けるためにバクテリオファージを利用する、タンパク質-タンパク質、タンパク質-ペプチド、およびタンパク質-DNAの相互作用の発見および研究のための方法をさす。
【0087】
タンパク質の提示または提示されたタンパク質とは、リガンドによる検出または固定化のためにアクセス可能なファージコートタンパク質に融合したタンパク質をさす。
【0088】
本発明の第6の局面は、本発明のpIX融合タンパク質をコードする核酸を含むファージミドと、ヘルパーファージとを含むファージディスプレイシステムである。
【0089】
本発明の一局面は、本発明のファージゲノムまたはファージミドを含むファージディスプレイシステムに関する。
【0090】
本発明のもう一つの局面は、本発明のファージゲノムまたはファージミドとヘルパーファージとを含むファージディスプレイシステムに関する。
【0091】
本発明の一局面は、異なるタンパク質を提示している二種以上の繊維状ファージを含み、これらのタンパク質のうちの少なくとも一つが本発明のファージゲノムまたはファージミドから発現されたpIX融合タンパク質である、ファージライブラリーに関する。
【0092】
本発明の一態様において、本発明のファージライブラリーは、pIII融合タンパク質、pVII融合タンパク質、およびpVIII融合タンパク質より選択される一つまたは複数の付加的な融合タンパク質を含む。
【0093】
キット
本発明の第7の局面は、本発明のpIX融合タンパク質をコードする核酸を含むファージミドと、ヘルパーファージとから構成されるファージディスプレイシステムを含むキットである。キットは、コーディング領域のN末端にマルチクローニング部位を有する、pIXをコードする遺伝子を含むファージミドと、ヘルパーファージ(例えば、M13K07、VCSM13、またはその他)とを含んでいるべきである。キットには、ファージクローンの感染、発現、固定化、選択、および検出のためのプロトコルが補足されるべきである。キットには、特定のアッセイを実施するための緩衝液および培地についての必要なレシピも添付されるべきである。
【0094】
キットとは、ここで、ファージディスプレイライブラリーとして、または単一のファージ粒子として、単一特異的または二重特異的な融合タンパク質を含むファージ粒子を生成するための試薬の集合体をさす。キットは、ファージミド、ヘルパーファージ、細菌株、ならびに試薬のレシピおよびアッセイの説明を含むプロトコルを含み得る。キットは、研究用、診断用、および治療用の試薬の開発のために使用され得る。
【0095】
本発明の一局面は、本発明のファージゲノムまたはファージミドとヘルパーファージとを含むファージディスプレイを含むキットに関する。
【0096】
本発明のもう一つの局面は、本発明のファージゲノムまたはファージミドを含むキットに関する。本発明のさらにもう一つの局面は、本発明のファージゲノムまたはファージミドを含む繊維状ファージを含むキットに関する。
【0097】
以下の非限定的な例において、本発明をさらに詳細に説明する。
【実施例】
【0098】
実施例1:pIX上のファージミドディスプレイ
試薬
全ての培地および緩衝液を、本質的に、Sambrook et al. (2001) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Pressに記載されたようにして調製した。抗M13-HRP抗体はGE Healthcare Bio-Sciences AB(Uppsala, Sweden)から購入した。制限酵素(RE)は、Stratagene(LaJolla, CA, USA)から入手したDpnIを除き、New England Biolabs(Ipswich, MA, USA)から購入した。DNAオリゴは、MWG Biotech AG(Ebersberg, Germany)から購入した。ウシ血清アルブミン(BSA)およびトゥイーン20は、Sigma-Aldrich(Oslo, Norway)から購入した。Pfu Turbo DNAポリメラーゼは、Stratagene(LaJolla, CA, USA)から購入した。BSAにコンジュゲートしたハプテン、2-フェニルオキサゾール-5-オン(phOx)および5-ニトロフェンアセチル(NIP)は、本質的に、他に記載されたようにして調製した(Nakela et al, PMID;722243およびMichaelsen et al, PMID:2125362)。イソプロピル-β-D-チオガラクトピラノシド(IPTG)は、Fermentas(Burlington, Canada)から購入した。トリエチルアミン(TEA)およびトリプシン/EDTAは、それぞれ、Sigma-Aldrich(Oslo, Norway)およびBioWhittaker(Lonza Group Ltd., Visp, Switzerland)から購入した。大腸菌株XL1-Blueは、Stratagene(LaJolla, CA, USA)から購入した。M13K07ヘルパーファージは、GE Healthcare Bio-Sciences AB(Uppsala, Sweden)から購入した。phOx-BSAに対する特異性を有する単鎖Fv(scFv)を保有しているpSEX81(SEQ ID NO: 2)ファージミド(pIIIディスプレイ)は、Affitech AS(Oslo, Norway)から厚意により提供を受けた。scFv抗NIP(SEQ ID NO:3)を保有している原核生物発現ベクターpSG1(未発表)は、pHOG21(Kiprianov et al, PMID:9005945)に基づいており、pLNOH2およびpLNOK(Norderhaug et al, PMID:9202712)に由来する抗体可変遺伝子から社内で作成した。
【0099】
新規のpIXディスプレイファージミドベクターpGALD9およびpGALD9ΔLの構築
ベクター骨格の出発テンプレートとして、上記のpSEX81(SEQ ID NO:2)ファージミドを選んだ(GenBankアクセッション番号:Y14584)。まず、このベクター内の原核生物pelBシグナル配列
をコードするストレッチを除去するため、プライマー対
を使用したQuikChange(商標)インビトロ変異誘発により、NcoI RE部位を最もN末端に導入し、それにより、pelB ORFの第2コドンの最初のヌクレオチドをAからGに変化させた。変異誘発の後、ベクターをNcoI消化し、再ライゲートさせ、プライマー対
を使用した、ベクターの関連部分を回収する二次PCRにおいて、テンプレートとして使用した。次いで、このPCR断片を、標準的な技術を使用して、適合性のEcoRI/HindIII RE部位で、元のpSEX81(SEQ ID NO:2)に移し、DNA配列決定により確認した。この工程により、pelBシグナル配列コーディング部分が完全に除去されたが、正常な転写および翻訳のために重要な開始コドンならびにlacPOおよびシャインダルガーノ配列(SD)に対するその相対位置は保存され、さらに、元のpSEX81(SEQ ID NO:2)に見出されたNcoI/NotI RE部位により画定された外来性配列の前にAla残基が1個だけ付加された。新たな構築物をpSEX81ΔLと名付けた。
【0100】
第2に、pXIコーディング配列を、5'末REタグ付きプライマー対
を使用して、M13K07から増幅した。次いで、このPCR断片を、適合性のRE部位で、pSEX81(SEQ ID NO:2)ファージミドおよびpSEX81ΔLファージミドの両方に移し、それにより、両方のpIIIコーディング領域を交換し、元のpSEX81(SEQ ID NO:2)のNcoI/NotIにより画定されたカセットのN末端インフレームpIX融合体を得た。新たな構築物をDNA配列決定により確認し、それぞれ、pGALD9およびpGALD9ΔLと名付けた。上記の様々なファージミドのscFv抗phOx(SEQ ID NO:11)単位を、pSG1由来のscFv抗NIP(SEQ ID NO:3)単位に切り替えるため、標準的な技術を使用して、NcoI/NotI REにより画定されたカセットの交換として、これを行った。本明細書に記載された全てのファージミドを、標準的な技術を使用して、電気穿孔により大腸菌XL1-Blueに導入した。
【0101】
ファージ粒子の調製
M13K07ヘルパーファージを使用した大腸菌XL1-Blueからのファージミドレスキューおよびビリオン組み立てを、記載されたようなスポット滴定によりモニタリングした(Welschof et al, PMID:9050877およびKoch et al, PMID:11126120)。
【0102】
ファージ捕獲酵素結合免疫吸着アッセイ(ELISA)
phOx-BSAまたはNIP-BSAを、4℃で一夜、PBS(pH7.4)中5μg/mlで、MaxiSorp(商標)マイクロタイタープレートウェル(Nunc, Roskilde, Denmark)に吸着させた。RTで1時間、PBS(w/v)中2%BSAによりウェルをブロッキングし、次いで、ビリオン調製物を添加し、RTで1〜2時間、反応させた後、捕獲されたビリオンを、RTで1時間、抗M13-HRP(5,000倍)により検出した。各工程の間に、ウェルをPBST(PBS/0.05%トゥイーン20)で3回洗浄した。ウェルをTMB可溶性基質により発色させ、30分後に1M HClにより中止し、A450nmで吸光度を読み取った。1ビリオン当たりの提示された融合タンパク質の関数として、Ag反応性を定量化するため、任意の試料についてシグナル飽和が観察される前に、1M HClを添加することによりELISA発色を中止し、吸光度読み取り値を、以下の式に従って変換した:相対ディスプレイレベル=(A450nm/ファージミド力価)×1012。
【0103】
スパイクされたphOx-BSA/NIP-BSA選択
新鮮なビリオン試料を、1mM IPTG誘導により、または誘導なしに調製し、PEG沈殿させ、記載されたようにして滴定した。
【0104】
次いで、中サイズのコンビナトリアルライブラリーに相当する107という既知の多様性を与える1:107レベルで、標的特異的な実体を、無関係のバックグラウンドにスパイクした。
【0105】
NIP-BSA選択については、scFv抗NIPを、scFv抗phOxカウンターパートにスパイクし、その逆も同様に行った。初期インプットは、12のモデルライブラリー全てについてパニングラウンド1において103という複雑さレベルをもたらす1×1010cfuampRであった。
【0106】
簡単に説明すると、ラウンド1については1μg/ml、ラウンド2については0.1μg/mlの100μl容量を使用して、同一のプレート上にトリプリケートで、MaxiSorp(商標)マイクロタイタープレートウェル(Nunc, Roskilde, Denmark)に標的を固定化した。
【0107】
パニング前に、RTで1〜2時間、ウェルをPBSTMによりブロッキングした後、それぞれの(PBSTMで)予めブロッキングされたビリオン調製物100μlを添加し、RTで1.5時間、撹拌しながら反応させた。
【0108】
マイクロタイター洗浄装置を使用して、PBSTで9回、続いてdH2Oで5回、ウェルを洗浄した後、(トリプリケートウェル内の)標的に結合したビリオンを、以下のいずれかにより溶出させた。
(1)RTで5分間、100μl/ウェルの100mM TEA(pH12)を添加し、続いて、100μl/ウェルのトリスHCl(pH6.8)を含有している新鮮なウェルに移すことにより中和する;
(2)RTで10分間、100μl/ウェルのトリプシン/EDTAを添加し、続いて、新鮮なウェルに移す;
(3)37℃で30分間、撹拌しながら、200μl/ウェルの対数期(A600nmおよそ0.5、≧5×107細胞に相当)の大腸菌XL1-Blueを添加し、続いて、MOI10でM13K07ヘルパーファージが補足された10mlの予め加温されたYT-TAG(30μg/mlテトラサイクリン、100μg/mlアンピシリン、および0.1Mグルコースを含有している2×YT)に移す。
【0109】
弱く撹拌しながら37℃で15分間、続いて強く撹拌しながら37℃で30分間、インキュベーションを続けた。平行して、TEAおよびトリプシンにより溶出させた試料を、9ml YT-TAG中の対数期大腸菌XL1-Blue培養物に感染させるために使用し、37℃で15分間弱く撹拌しながらインキュベートした後、MOI10でM13K07ヘルパーファージが補足された1ml YT-TAGを添加した。
【0110】
弱く撹拌しながら37℃で15分間、続いて激しく撹拌しながら37℃で30分間、インキュベーションを続けた。次いで、全ての試料を3000g/10分/RTで遠心分離し、上清を廃棄し、100μg/mlアンピシリンおよび50μg/mlカナマイシンを含有している10mlの予め加温された2×YTにペレットを穏和に再懸濁させた。
【0111】
適切な試料に1mM IPTGを補足し、全ての試料を厳しく撹拌しながら30℃で一夜、インキュベートした。翌日、培養物を4000g/10分/RTで遠心分離し、上清を0.2μmフィルターで新鮮な15mlチューブへと滅菌濾過した。
【0112】
次いで、これらの上清を、少なくとも各試料109cfuampRのインプットに相当する、各試料50μlの容量を使用して、記載されたようなパニングの次のラウンドに供した。選択の第2ラウンドの後、ビリオンを含有している上清を、上記のような抗原特異的ELISAに供した。
【0113】
結果
繊維状ファージ(fd、M13、およびf1)の5種の構造コートタンパク質は、全て、突出するビリオンに取り込まれる前は、グラム陰性宿主の内膜に見出される膜内在性タンパク質であることが公知である(Endeman and Model, PMID:7616570)。この報告は、pIXはN末端の融合修飾を可能にしないが、カプシドタンパク質自体は、完全ビリオンにおいて溶媒に露出しているとの結論も下した。Gaoら(PMID:10339535および12239343)ならびにKhalilら(PMID:17360403)の両方は、その後、ファージミドから発現させ、かつシグナル配列依存性周辺質ターゲティングと組み合わせて使用した場合には、N末端pIX融合ディスプレイが可能であることを示した。これらのシステムにおいては、ファージミドレスキューによりヘルパーファージゲノムからwt pIXが供与され、補完が起こる。そのようなディスプレイが、周辺質へのシグナル配列依存性ターゲティングなしに可能であるか否かを試験するため、本発明者らは、そのようなシグナル配列を含むか、または含まない、N末端pIXディスプレイを可能にするpGALD9およびpGALD9ΔLと名付けられた二つの新規ファージミドを構築した(図2)。
【0114】
二つの異なるpIX融合体を、これらの新たなファージミドを使用した分析のために選び、標準的なpIIIディスプレイを使用したカウンターパートと比較した。両融合体は、ヒト抗体可変遺伝子セグメントに基づくハプテンコンジュゲートphOx-BSAに特異的な抗体断片scFv(scFv抗phOx)、またはマウス抗体可変遺伝子セグメントに基づくハプテンコンジュゲートNIP-BSAに特異的な抗体断片scFv(scFv抗NIP)であった。特に、scFv抗phOx(SEQ ID NO:11)は、ヒト抗体scFvライブラリーから選択されており、大腸菌において極めてよく発現することが公知である(Marks et al., PMID:1748994)。対照的に、多くのマウスハイブリドーマ可変遺伝子は、大腸菌において、ファージディスプレイされた場合にも、よく発現しないことが周知である(Krebber et al, PMID:9032408)。
【0115】
scFvのpIXディスプレイは、正常なビリオン組み立てに干渉しないはずである。従って、本発明者らは、材料および方法に記載されるような標準的なファージミドレスキューおよび滴定を使用して、これらのシグナル配列を含むscFvディスプレイファージミドおよびシグナル配列を含まないscFvディスプレイファージミドの性能を比較し、(シグナル配列依存性周辺質ターゲティングを絶対的に必要とする)標準的なpIIIディスプレイとも比較した(図3)。
【0116】
滴定結果は、ファージミドを含有しているビリオンが全てのケースで作成されたことを実際示した(図3AおよびC)。scFv融合体をプロモーター誘導なしにlac POから発現させる場合(基底発現)、両方のpIXディスプレイバージョンおよびpIII対照が、scFv抗phOxについて比較可能な力価を与える(図2A)。対照的に、シグナル配列非依存性scFv抗NIP pIXディスプレイの力価は、pIIIディスプレイおよびシグナル配列依存性pIXディスプレイと比較して、それぞれ5〜10倍高い(図3C)。lacPOのIPTG誘導によりscFv融合体発現を増加させると、シグナル配列依存性pIXディスプレイおよびpIIIディスプレイの両方の力価が10倍低下した。対照的に、両方のシグナル配列非依存性pIXディスプレイバリアントに対しては軽微な効果しかなかった(図3AおよびC)。このシステムにおいてはヘルパーファージからのpIXのwt補完が存在するため、この所見は驚くべきものでありかつ重要であった。なぜなら、シグナル配列依存性pIXディスプレイおよびpIIIディスプレイは、ビリオン組み立て過程に干渉するが、シグナル配列非依存性pIXディスプレイの場合には、IPTG誘導時ですら、この効果が軽微に過ぎないことを、それが示すためである。シグナル配列を含むか、またはシグナル配列を含まないpIX上に提示されたscFv抗NIPからの結果を比較した場合に、効果は最も顕著であって、前者は力価の100倍の低下を示す。両方のscFvについて、シグナル配列非依存性pIXバージョンで行われた場合、ビリオン組み立てが、pIIIと同等であるかまたはpIIIより良好であることも注目に値する。
【0117】
コンビナトリアル選択プラットフォームのコアは、表現型の選択を通した遺伝子型の回収を可能にする、物理的な表現型-遺伝子型カップリングである。この物理的な関係が損なわれるかまたは失われた場合、システムは非機能性となる。ファージミドディスプレイに翻訳すると、これは、ヘルパーファージレスキューによりビリオンに封入されるのが、ヘルパーファージゲノムではなくファージミドであることが、選択のために不可欠であることを意味する。ファージミドおよびヘルパーファージは異なる抗生物質選択マーカーを保有するため、この概念は、適切な選択増殖(アンピシリン(ampR)、またはカナマイシン(kanR))に起因する、それぞれのコロニー形成単位(cfu)に基づき、ファージミド対ヘルパーファージ比を計算することにより、感染性滴定において容易に査定され得る。その際、その比は、効率的な下流の選択が実行可能であるよう、1より大きいべきである。
【0118】
上記のビリオン調製物のファージミド対ヘルパーファージ比を査定したところ(図3BおよびD)、3種のディスプレイ経路の間の有意な差が明らかになった。プロモーター誘導なしの標準的なビリオンパッケージング(基底発現)を使用すると、scFv抗phOxについては3種の経路全てが実施可能であるが(図2B)、scFv抗NIPについて実行可能であったのは、配列非依存性pIXバリアントのみであった(図3D)。プロモーター誘導(IPTG誘導)時、pIIIバリアントおよびシグナル依存性pIXバリアントの両方が、表現型-遺伝子型連鎖の著しい損失を示し、その効果はscFv抗NIP融合体で最も顕著であった。対照的に、この効果は、両方のシグナル配列非依存性pIXバリアントについては、存在しないか(scFv抗phOx)または軽度であった(scFv抗NIP)。従って、結果は、シグナル配列非依存性pIXディスプレイが、標準的なpIIIディスプレイおよびシグナル配列依存性pIXディスプレイの両方と比較して、優れた表現型-遺伝子型連鎖表現型を保有していることを明白に示している。
【0119】
次いで、上記試料に基づき、本発明者らは、抗原特異的ELISAでこれらのビリオン上の機能的なscFvディスプレイを査定した(図4)。結果は、3種のディスプレイ経路全てからの機能的なscFvディスプレイを明白に示した(図4AおよびB)。試料はファージミド力価によってノーマライズされていないため、シグナル強度は直接比較可能ではない。さらに、シグナル飽和が、いくつかの試料において観察された。3種のディスプレイ経路の間の機能的な差を十分に査定するため、相対ディスプレイレベル(1ビリオン当たりの機能性ディスプレイ単位)を、材料および方法に記載されるような新たな抗原特異的ELISAにおいて決定した(図4CおよびD)。ディスプレイレベルは、pIIIバリアントとシグナル配列依存性pIXバリアントとの間で比較可能であったが、シグナル配列非依存性pIXディスプレイは有意に低かった。このことは、両方のscFvについて当てはまり、基底発現時にもIPTG誘導時にも当てはまった。予想通り、3種のディスプレイ経路全てが、両方のscFvについて、IPTG誘導時に、より高いディスプレイを示した。しかしながら、図3BおよびDに与えられた比を考慮して、図4CおよびDの結果を検討することが最も重要である。例えば、scFv抗NIP単位のpIIIディスプレイおよびシグナル配列依存性pIXディスプレイの両方が、シグナル配列非依存性pIXカウンターパートと比較して、10〜20倍(基底発現)、およそ25倍(IPTG誘導)高いディスプレイを示すことは明白である。しかしながら、前者の2種の経路は、いずれも、極めて弱いかまたは失われた表現型-遺伝子型関係を有し、表現型選択時に遺伝子型が失われるため、コンビナトリアル選択計画において非機能性であろう。この効果は、IPTG誘導時のみであるが、scFv抗phOx単位についても当てはまる。さらに、IPTGにより誘導されたシグナル配列非依存性pIXバリアントにおけるscFv抗phOx単位のディスプレイレベルは、基底発現時のpIIIバリアントおよびシグナル配列依存性pIXバリアントと比較可能である。これらのデータを考慮すると、scFvディスプレイのためにpIX(シグナル配列依存性)を活用している唯一の報告(Gao, PMID:12239343)が、ビリオンパッケージングの際に常にIPTG誘導を使用しているが、ファージミド対ヘルパーファージ比を報告していないことに注目することは興味深い。
【0120】
scFvフォーマットは、大腸菌における好都合な発現プロファイルのためしばしば使用されているが(Bradbury and Marks, PMID:15261570)、いくつかの報告は、アフィニティ選択による高親和性結合剤の回収においては、例えば、抗体Fab断片のような、より低レベルのディスプレイフォーマットが有利であることを指摘している(de Haard, et al, PMID:10373423およびHoogenboom, PMID:16151404)ならびにRotheら(PMID:18191144)。
【0121】
従って、たとえシグナル配列非依存性scFv-pIXディスプレイが低レベルのディスプレイを与えるようであっても、これは、実際、高親和性選択のために適用された場合、高度に有利であることが判明するかもしれない。
【0122】
従って、シグナル配列非依存性pIXディスプレイのアフィニティ選択における性能を査定するため、本発明者らは従来のpIIIディスプレイと比較した。
【0123】
IPTG誘導の存在下または非存在下でビリオンを作製した。関心対象のタンパク質はscFv抗phOxまたはscFv抗NIPのいずれかであった。
【0124】
従って、2種の標的phOx-BSAおよびNIP-BSAそれぞれについての計4つのファージ集団を評価した。各ケースにおいて、標的特異的scFvを1:107の比で特異性が無関係のscFvと混合した。重要なことに、2種のscFvは交差反応しない。次いで、2ラウンドのアフィニティ選択を実施した。
【0125】
3種の異なる溶出戦略;高pH、タンパク質分解、または直接感染のいずれかを利用した。選択の第2ラウンドの後、抗原特異的ポリクローナルファージELISAにより、濃縮を確証した(図5)。
【0126】
関心対象のタンパク質をpIX上に提示した場合、選択は、POIをpIII上に提示した場合とほぼ同等の効率であり、ほとんどのケースで、より効率的であった。特に、高pH(TEA)またはタンパク質分解(トリプシン)による溶出を利用した標準的なpIIIディスプレイ経路は、pIXと比較して、不十分な濃縮を示した。
【0127】
選択は、ディスプレイ経路および溶出条件とは無関係に、IPTG誘導時よりIPTG誘導なしの方が効率的であり、負の効果は、pIIIディスプレイ経路で最も大きいようであった。
【0128】
従って、シグナル配列非依存性pIXの低いディスプレイの傾向は、不十分な選択を意味しなかった。pIIIを改変せず、完全に溶媒に露出させておくことにより、ライブラリー選択工程後のビリオンレスキューが、ビリオン-標的結合を破壊することなく効率的に達成され得、従って、溶出工程が不要となり、ハイスループットプロトコルが加速されるかもしれない。
【0129】
後者は、溶出が多様な戦略に対して抵抗性であるかもしれない高親和性結合剤の単離も容易にするかもしれない(Balass et al, PMID:8954559)。
【0130】
参照
【背景技術】
【0001】
発明の背景
タンパク質の同定、特徴決定、および修飾のためのコンビナトリアルアプローチの使用は、学術的な研究開発においても商業的な研究開発においても高度に成功している。この点で、繊維状バクテリオファージディスプレイまたは繊維状ファージディスプレイのテクノロジーは、最初のライブラリープラットフォームとしてその道を開き、今なお優勢なテクノロジーとして王座についている。従って、ファージディスプレイは、基礎的なタンパク質発見および応用的なタンパク質発見の両方において、そして世界的に最も急成長中の化合物クラスである新規のタンパク質に基づく診断薬および治療薬の両方の開発において、広く適用されている。
【0002】
コンビナトリアルファージディスプレイテクノロジーの原理は、各ビリオンが、タンパク質コートに封入されたゲノムによってコードされるまさに同一のタンパク質を、表面上に単に提示する、という特性により与えられる、遺伝子型-表現型連鎖に基づく。ファージ粒子自体は、多様な生理化学的条件に対して高度に抵抗性である;従って、ファージディスプレイは、競合するコンビナトリアルテクノロジーと比較して、多くの選択計画における優れた多用途性を示す。
【0003】
異種ポリペプチドのファージディスプレイは、繊維状ファージのコートの5種の構造タンパク質全てを使用して達成されているが、pIIIディスプレイのみが広範に使用されており、pVIIIディスプレイもある程度は使用されている(図1)。
【0004】
異種融合体が短いペプチドのみである場合には、ファージゲノムに基づくベクターを使用した多価ディスプレイシステムが好ましいが、折り畳まれたドメインを必要とする、より大きな融合体については、大部分の適用がファージミドシステムより利益を得るであろう。後者の場合、抗体-pIIIファージディスプレイが、この領域において極めて優勢であるが、代替足場が出現しつつあり、将来のタンパク質工学ツールの増大の必要性は続いている。高度に望まれる適用は、単に、ファージビリオンを細菌に感染させることにより、ファージディスプレイライブラリーから、高親和性の特異的なペプチド結合剤またはタンパク質結合剤を、標的に結合したまま効率的に入手することである。
【0005】
Endemann and Model, 1995(PMID:7616570)は、マイナーなコートpIXは、N末端にもう一つのタンパク質が融合すると機能しないことを報告した。従って、この報告は、pIXはファージディスプレイのために使用され得ないとの結論を下した。
【0006】
Gaoら(PMID:10339535、12239343、およびWO0071694)、ならびにKhalilら(PMID:17360403)の両方は、その後、ファージミドから発現させ、かつシグナル配列依存性周辺質ターゲティングと組み合わせて使用した場合には、N末端pIX融合ディスプレイが可能であることを示した。これらのシステムにおいては、ファージミドレスキューによりヘルパーファージゲノムからwt pIXが供与され、補完が起こる。
【0007】
(融合タンパク質が挿入されていない)WO0071694の図2Aに開示されたようなファージミドおよび(融合タンパク質が挿入されている)WO0071694の図2Bに開示されたようなファージミドは、明白に、pelBシグナル配列を含む(図面の説明は7頁2〜14行目)。
【0008】
前述のように、pIXは、N末端にもう一つのタンパク質が融合すると機能しないことが、以前に示唆され、Gaoらが、単独の、または両方の組み合わせによる、成功のための二つの可能性のある理由を与えた。
【0009】
一つの可能性のある理由は、原核生物リーダー配列(シグナル配列)が融合タンパク質のN末端に付着させられ、従って、組換えタンパク質の周辺質空間へのターゲティングが確実になり、それにより、細胞質における蓄積が防止されたというものである。
【0010】
もう一つの可能性のある理由は、EndemannおよびModelでのようなファージゲノムではなく、ファージミドから組換えタンパク質が発現され、従って、ファージミドレスキューのために必然的に必要とされるヘルパーファージ由来の野生型pIXが、組換えpIX融合タンパク質を補完し、従って、さもなければ、組換え修飾のために失われていたかもしれない野生型機能性が保存されたというものである。即ち、ファージは、野生型タンパク質と融合タンパク質とのミックスを含んでいたであろう。
【0011】
Khalilら(PMID:17360403)は、まさに同一のビリオンの各遠位先端に外来性ペプチドが提示される二重特異的な繊維状ファージビリオンの特色を活用する適用を記載している。彼らは、原核生物シグナル配列依存性pIXディスプレイファージミドを補完する一般的なpIIIファージゲノムベクターの組み合わせを使用することにより、これを達成した。この環境において、ファージゲノムベクターは、ファージミドレスキューにおけるヘルパーファージとして役立った。
【発明の概要】
【0012】
本発明の目的は、繊維状ファージ上に提示されたペプチドのための代替足場を提供することである。
【0013】
本発明の第1の局面は、原核生物N末端シグナル配列を含まず、従って、外来性ペプチドとの直接融合体である、繊維状ファージに由来するpIX融合タンパク質である。
【0014】
本発明のもう一つの局面は、本発明の融合タンパク質をコードする核酸に関する。
【0015】
本発明の一つの局面は、本発明の融合タンパク質を含む繊維状ファージに関する。
【0016】
本発明のもう一つの局面は、繊維状ファージのライブラリーに関する。
【0017】
本発明のもう一つの局面は、本発明のpIX融合タンパク質をコードする核酸を含むファージミドとヘルパーファージとを含むファージディスプレイシステムに関する。
【0018】
一つの局面は、本発明のpIX融合タンパク質をコードする核酸を含むファージミドとヘルパーファージとを含むファージディスプレイシステムを含むキットに関する。
【図面の簡単な説明】
【0019】
【図1】繊維状ファージ構造の概略図である。ビリオンは、一本鎖DNA分子をコートする5種の構造タンパク質により構築される。野生型(wt)ファージには、pVIIIが約2700コピー存在し、ビリオンの各先端に見出される4種のタンパク質pIII、pVI、pVII、およびpIXがいずれもおよそ3〜5コピー存在する。ビリオンサイズは、pVIIIコートタンパク質1個当たりおよそ2.3ヌクレオチドで、ゲノムサイズに依存し、従って、粒子の長さは、挿入されるpVIIIのコピーの増加または減少に適合する。特に、pIIIおよびpVIIIの構造は、X線繊維回折、結晶学、およびNMRにより特徴決定されている。マイナーなコートタンパク質pIIIは、グリシンリッチ領域により分離された3個の別個のドメイン:N1(TolAに結合する)、N2(F繊毛に結合する)、およびCT(ビリオンに組み込まれ、正常なビリオン組み立てのために重要である)を含有している。
【図2】新規のpGALD9 pIXディスプレイファージミド(A)およびpGALD97ΔL pIXディスプレイファージミド(B)の概略図である。両ファージミドのベクター骨格は、GenBankアクセッション番号:Y14584からアクセス可能な配列を有するpIIIディスプレイファージミドpSEX81(SEQ ID NO:2)に基づいており、構築に関する詳細は材料および方法に記載される。両ファージミドは、それぞれ、NcoI/HindIII部分およびMluI/NotI部分の容易なカセット交換を通して、インフレームの(E1およびE2と呼ばれる)外来性配列のセグメントを収容することができる。カセットは、本明細書に記載された異なる構築物によって変動する合成リンカー配列により接続されている。略語:lacPO、lacプロモーター;sd、シャインダルガーノ配列;pelB、細菌ペクチン酸リアーゼのシグナル配列;TP、トリプシンプロテアーゼ部位;t、T7転写ターミネーター。
【図3】pGALD9ΔL、pGALD9、pSEX81から提示された(A)scFv抗phOx(SEQ ID NO:11)および(C)scFv抗NIPのファージミド力価。全てのファージミドが、アンピシリン耐性マーカーを保有しており;従って、力価は、溶液1ミリリットル当たりのアンピシリン耐性コロニー形成単位(cfuampR/ml)として示される。ファージミド力価(cfuampR/ml)をヘルパーファージ力価(cfukanR/ml)により割った比として示された、pGALD9ΔL、pGALD9、pSEX81から提示された(B)scFv抗phOx(SEQ ID NO:11)および(D)scFv抗NIPのファージミド対ヘルパーファージ比。ビリオンパッケージングは、IPTGなしで(基底発現)、または重感染後に0.1mM IPTGの最終濃度を存在させて(IPTG誘導)、材料および方法に記載されるような標準的なファージミドレスキューにより行われた。
【図4】pIXとpIIIとの間の、シグナル配列がある場合とシグナル配列がない場合(ΔL)との間の、機能的な(A)scFv抗phOx(SEQ ID NO:11)および(B)scFv抗NIP(SEQ ID NO:3)のディスプレイを比較する抗原特異的ELISA。ELISAは、100μl/ウェルの精製されたビリオン含有上清を使用して、材料および方法に記載されるように実施された。抗M13HRPは、ビリオン検出MAbの抗原およびブロックへの非特異的な吸着についての陰性対照である。(CおよびD)AおよびBのELISAを繰り返したが、シグナル飽和の前に発色を中止し、力価の関数として抗原反応性を使用して相対ディスプレイレベルを決定した。
【図5】アフィニティ選択における明白な標的特異的濃縮は、カプシドディスプレイ足場、関心対象のタンパク質(Protein of Interest/POI)の発現、および溶出条件に依る。選択ラウンド1および2の後の、phOx/NIPをスパイクされた8つのライブラリーの各々からの等しい容量の未滴定のビリオン含有上清を、ELISAにより抗原反応性について査定した。ラウンド0は、1×1010cfuampRのスパイクされたインプットに相当する。最大可能応答を推定するため、ラウンド0におけるスパイクされた濃縮物が由来するクローン上清が含まれ、図示された結果は、円錐形により示された最大可能応答の分画として、バックグラウンドを差し引いたシグナルにより与えられている。
【発明を実施するための形態】
【0020】
発明の詳細な説明
本発明の局面のうちの一つに関して記載された態様および特色が、本発明のその他の局面にも当てはまることに、注意するべきである。
【0021】
本願において引用された全ての特許参照および非特許参照は、参照によりその全体が本明細書に組み入れられる。
【0022】
本発明者らは、修飾されたバージョンがN末端のペプチドドメインまたはタンパク質ドメインをコードするよう、繊維状ファージビリオンの構造コートタンパク質pIXが遺伝学的に改変される、新規の概念を、ここに提示する。
【0023】
pIX融合タンパク質
一つの局面において、本発明は、pIXのN末端への外来性ペプチドの融合を含む、繊維状ファージに由来するpIX融合タンパク質を提供する。そのような融合タンパク質は、例えば、ファージディスプレイに関して有用である。
【0024】
外来性ペプチドとの言及により意味されるのは、リンカーアミノ酸により、またはリンカーアミノ酸なしで、融合タンパク質のpIXアミノ酸部分のN末端へ融合させられる、元々はpIXタンパク質の一部でないペプチドである。
【0025】
好ましい態様において、融合タンパク質をコードする核酸は、原核生物N末端シグナル配列を含まない。
【0026】
本明細書において使用されるように、ペプチドという用語には、短いペプチド、ポリペプチド、タンパク質、およびそれらの断片の両方が包含される。
【0027】
pIXタンパク質という用語は、SEQ ID NO 1に開示されたアミノ酸配列(MSVLVYSFASFVLGWCLRSGITYFTRLMETSS)をさす。
【0028】
一態様において、pIXタンパク質は、SEQ ID NO 1のものとの少なくとも70%の配列同一性、例えば、75%の同一性、80%の同一性、81%の同一性、82%の同一性、83%の同一性、84%の同一性、85%の同一性、86%の同一性、87%の同一性、88%の同一性、89%の同一性、90%の同一性、91%の同一性、92%の同一性、93%の同一性、94%の同一性、95%の同一性、96%の同一性、97%の同一性、98%の同一性、99%の同一性を有するアミノ酸を含む。
【0029】
配列同一性
一般的に定義されるように、「同一性」とは、ここで、それぞれヌクレオチドレベルまたはアミノ酸レベルでの遺伝子間またはタンパク質間の配列同一性として定義される。
【0030】
従って、本発明において、「配列同一性」とは、アミノ酸レベルでのタンパク質間の同一性の尺度、およびヌクレオチドレベルでの核酸間の同一性の尺度である。タンパク質配列同一性は、配列を整列化した場合の、各配列の所定の位置のアミノ酸配列を比較することにより決定され得る。同様に、核酸配列同一性は、配列を整列化した場合の、各配列の所定の位置のヌクレオチド配列を比較することにより決定され得る。
【0031】
二つのアミノ酸配列または二つの核酸配列の同一率を決定するためには、配列が最適の比較目的のため整列化される(例えば、第2のアミノ酸配列または核酸配列との最適の整列化のため、第1のアミノ酸配列または核酸配列にギャップが導入されてもよい)。次いで、対応するアミノ酸位置またはヌクレオチド位置のアミノ酸残基またはヌクレオチドが比較される。第1の配列のある位置が、第2の配列の対応する位置と同一のアミノ酸残基またはヌクレオチドにより占有されている場合、その分子はその位置において同一である。二つの配列の間の同一率は、それらの配列が共有している同一である位置の数の関数である(即ち、同一%=同一である位置の数/位置の総数(例えば、重複している位置)×100)。一つの態様において、二つの配列は同一の長さである。
【0032】
手動で配列を整列化し、同一のアミノ酸の数を計数してもよい。あるいは、同一率の決定のための二つの配列の整列化は、数学的アルゴリズムを使用して達成されてもよい。そのようなアルゴリズムは、(Altschul et al. 1990)のNBLASTプログラムおよびXBLASTプログラムに組み入れられている。本発明の核酸分子と相同なヌクレオチド配列を入手するためには、NBLASTプログラム(score=100、wordlength=12)により、BLASTヌクレオチド検索が実施され得る。本発明のタンパク質分子と相同なアミノ酸配列を入手するためには、XBLASTプログラム(score=50、wordlength=3)により、BLASTタンパク質検索が実施され得る。比較目的のため、ギャップを挿入された整列化を入手するためには、Gapped BLASTが利用され得る。あるいは、PSI-Blastが、分子間の遠い関係を検出する反復検索を実施するために使用され得る。NBLASTプログラム、XBLASTプログラム、およびGapped BLASTプログラムを利用する場合、それぞれのプログラムのデフォルトパラメーターが使用され得る。http://www.ncbi.nlm.nih.govを参照のこと。あるいは、配列同一性は、例えば、EMBLデータベース内のBLASTプログラム(www.ncbi.nlm.gov/cgi-bin/BLAST)により、配列を整列化した後、計算され得る。一般に、例えば、「スコアリングマトリックス」および「ギャップペナルティ」に関しては、デフォルト設定が整列化のために使用され得る。本発明に関して、BLASTNおよびPSI BLASTのデフォルト設定が有利であり得る。
【0033】
二つの配列の間の同一率は、ギャップを許容し、または許容せず、上記のものと類似の技術を使用して決定され得る。同一率の計算においては、正確なマッチのみが計数される。
【0034】
折り畳まれたタンパク質
好ましい態様において、ペプチドという用語は、抗体に由来するドメインのような折り畳まれたタンパク質を排他的にさす。折り畳まれたタンパク質は、Fv、scFv、Fab、単一ドメインを含む抗体またはその断片、プロテインAのZドメインまたはその断片(アフィボディ)、アンキリンまたはその断片、DARPinまたはその断片、T細胞受容体またはその断片、MHCクラスIもしくはIIまたはそれらの断片、フィブロネクチンまたはその断片、アンチカリンまたはその断片、PDZドメインまたはその断片、IgNARまたはその断片、CTLA4またはその断片、ImmE7またはその断片、ノッチンまたはその断片、アビマーまたはその断片、GFPまたはその断片、およびその他の遺伝子によりコードされた生物学的フルオロフォアであり得ることを、当業者は認識するであろう。
【0035】
原則的には、それが提示される限り任意のライブラリーを作成することが可能であり、従って、最も単純な形態において、折り畳まれた秩序正しい構造と比較して、組織化されていない配置を有するもののみを分離することができる。
【0036】
もう一つの好ましい態様において、ペプチドという用語は、2〜50aaの短いペプチドを排他的にさす。ある長さにおいては、短いランダムコイルペプチドは、明確な二次または三次の折り畳みを採るために十分に長く、従って、折り畳まれたドメインの定義に入るであろう。明らかに、これは化学的組成に依り、従って、ある20aaのペプチドはやはりランダムコイルであろうが、もう一つの20aaペプチドは折り畳まれていて、折り畳まれたドメインの定義に分類されるかもしれない。
【0037】
もう一つの好ましい態様において、本発明のpIX融合タンパク質は、SEQ ID NO:1の1〜32位、2〜32位、3〜32位、4〜32位、5〜32位、6〜32位、7〜32位、8〜32位、9〜32位、10〜32位、11〜32位、および12〜32位からなる群より選択される配列を含む。原則的に、ファージミド環境において、膜貫通部分が保持され、正常なビリオンの取り込みおよび組み立てが可能であるならば、pIXの任意のN末端修飾が構想され得る。
【0038】
は、繊維状ファージの構造コートタンパク質pIX(野生型pIX)のアミノ酸配列である。最も好ましくは、pIX融合タンパク質は、SEQ ID NO:1の1〜32位を含む。
【0039】
SEQ ID NO:1は、以下に説明されるシグナル/リーダー配列と混同されるべきではない。
【0040】
シグナル配列
好ましくは、外来性ペプチドは、融合タンパク質のpIX配列のN末端へ、リンカーアミノ酸により、またはリンカーアミノ酸なしに、直接融合される。
【0041】
さらにもう一つの好ましい態様において、pIX融合タンパク質は原核生物N末端リーダー配列を含まない。
【0042】
「リーダー配列」という用語は、「シグナルペプチド」および「シグナル配列」という用語と交換可能に使用され、グラム陰性菌の周辺質膜空間へと(そのリーダー配列を含む)タンパク質をターゲティングするアミノ酸配列をさす。しばしば使用されるリーダー配列の例は、pelBss、OmpAss、TorAss、malEss、phoAss、lamBss、Blass、DspAss、mglBss、sfmCss、tolBss、およびTorTssである。そのようなシグナル配列は、少なくともSRP依存性、SEC依存性、TatABC依存性、またはYidC依存性の細胞質から周辺質空間への転移を含むことが公知の大腸菌の分泌機構へ、完全タンパク質をターゲティングすることが公知である(Baneyx et al. PMID:15529165)。従って、N末端シグナル配列という用語は、タンパク質のN末端部分にあるシグナル配列をさす。
【0043】
(それを含む)タンパク質を大腸菌の分泌機構へターゲティングし、それにより、細胞質コンパートメントから周辺質コンパートメントへと転移させる特性を保有しているシグナル配列は、部分的には、アミノ酸組成の化学的特性により定義されるサインまたはモチーフを通して同定され得る。
【0044】
しかしながら、既存の機能性シグナル配列の多様性は、それらの同定における現在の知識をまだ越えており、従って、同族シグナル配列としてのペプチドの定義における現在の最先端技術は、典型的には、例えば、ニューラルネットワークまたは発見的方法論により、知識に基づくデータベースをテンプレートとして使用して、データ発掘を通して行われている。SignalP、PROSITEのPPSEARCH(EMBL-EBI)、SecretomeP、TatPのような、いくつかのそのようなツールが、現時点で、オープンアクセスのチャネルを通して公に入手可能になっている。
【0045】
分泌タンパク質のクラスでは、それらが細胞質コンパートメントから輸送され、ルールから逸脱するため、シグナル配列モチーフが同定され得ないという意味で、問題がより大きいが、ここでも、データ発掘を通して、シグナル配列特色を定義するか、または問題の真核生物タンパク質の分泌能の可能性を得ることが可能である。原核生物分類群のためのそのようなツールは、まだ存在しない。
【0046】
従って、ペプチドをシグナル配列として決定的に同定した現在利用可能な唯一の方法は、それが現実のシグナル配列であるか否かを確立するためにペプチドの特性をバリデートするための実験的な手段による。シグナル配列内の所定のアミノ酸位置を改変し、それでもネイティブの機能性、または増加した輸送能力のような改変された機能性により、シグナルペプチドとしての機能は保持されるよう、そのようなペプチドを操作してもよいことも明白である。アミノ酸の欠失または付加も利用され得る。そのような分析および操作は、実際、Ff pVIIIシグナル配列、Sec経路をターゲティングするg8pss、およびTat経路をターゲティングするTorAssについて行われている。特に、Shenらの結果は、pIIIシグナル配列および細菌ペクチン酸リアーゼシグナル配列の機能性ではあるが改変されている変異体の操作のための、根拠の確かなガイドラインとして役立つかもしれない。
【0047】
シグナル配列の機能性は、以下の二つの特性にさらに分解され得る:
1. (それを含む)タンパク質を大腸菌の分泌機構へとターゲティングし、それにより、細胞質コンパートメントから周辺質コンパートメントへと転移させ、この過程の途中で、リポタンパク質シグナルペプチターゼまたはリーダーペプチダーゼのような特定のプロテアーゼによるタンパク質分解によって残りのタンパク質から分離される。
2. (それを含む)タンパク質を大腸菌の分泌機構へとターゲティングし、それにより、細胞質コンパートメントから周辺質コンパートメントへと移転させ、転移後もやはりタンパク質の一部として残存する。
【0048】
シグナル配列の大多数は、上に与えられた情況(1)にあるが、これらのタンパク質が情況(2)になるよう容易に操作され得ることは明白である。従って、元々は情況(1)に属するが、情況(2)へと改変された現在公知のシグナル配列、例えば、変異型pelBssおよびその他も、やはり、同族のシグナル配列と見なされる。
【0049】
さらに、情況(1)のシグナル配列を情況(2)に改変するか、または情況(2)にあるシグナル配列を直接選び、次いで、転移後にシグナル配列を除去することも構想され得る。これは、宿主の内在性プロテアーゼにより、かつ/または、例えばファージディスプレイの場合には、タンパク質がカプシドタンパク質と融合する時に、なされる。その場合には、明確な切断が達成され得るよう、シグナル配列またはそれを含むタンパク質の適切な領域に、人工的なプロテアーゼ部位が組み込まれるであろう。ここで、選ばれる二つの異なる型のプロテアーゼ部位が構想され得る:
A. プロテアーゼ部位は関心対象のタンパク質を切断しない。抗体、または主要組織適合性抗原分子もしくはT細胞受容体のようなその他の関心対象の足場と組み合わせられた、例えば、カルボキシペプチダーゼA部位または3Cライノウイルスプロテアーゼ部位のような予測された部位のみ。このアプローチを使用することにより、例えば、上記の情況(2)にあるシグナル配列を使用して、関心対象のタンパク質をファージディスプレイし、選択等において使用する前に、カプシド融合体へ機能性および均質性を入手するために、人工的にシグナルペプチドを除去することが構想され得る。
B. プロテアーゼ部位は、例えば、トリプシンのような操作された部位に加えて、興味のあるタンパク質を切断する。
【0050】
いずれの情況も、やはり、シグナル配列依存性ファージディスプレイと見なされるであろう。
【0051】
外来性ペプチド
好ましい態様において、pIX融合タンパク質の外来性ペプチドは、Fv、scFv、Fab、単一ドメインを含む抗体またはその断片、プロテインAのZドメインまたはその断片(アフィボディ)、アンキリンまたはその断片、DARPinまたはその断片、T細胞受容体またはその断片、MHCクラスIもしくはIIまたはそれらの断片、フィブロネクチンまたはその断片、アンチカリンまたはその断片、PDZドメインまたはその断片、IgNARまたはその断片、CTLA4またはその断片、ImmE7またはその断片、ノッチンまたはその断片、アビマーまたはその断片、GFPまたはその断片、およびその他の遺伝子によりコードされた生物学的フルオロフォアからなる群より選択される。
【0052】
好ましい態様において、pIX融合タンパク質の外来性ペプチドは、ライブラリーメンバーである。
【0053】
ライブラリーとは、本発明に関して使用されるように、異なるペプチドの集合体をさす。ペプチドは、折り畳まれたドメイン、または例えば2〜50アミノ酸の短いペプチドであり得る。そのようなライブラリーは、所定の標的への新たなリガンドの結合を同定するために使用され得るため、興味深い。
【0054】
ライブラリーを提示するためのpIXの使用には、pIIIまたはpVIIIを使用して提示されるライブラリーと比較して、いくつかの利点がある。pIXディスプレイは、方向性および価数に関してpIIIディスプレイと同一の長所を含有しているが、感染性に影響を与えないであろう。その現象は、pIIIディスプレイで起こることが公知であり、例えば、アフィニティ選択後のレスキューの際に、制御されない望まれない不均一性をシステムに導入する。
【0055】
さらに、pIXディスプレイは、pIIIディスプレイおよびpVIIIディスプレイの両方についての必要条件である原核生物N末端シグナル配列の必要性なしに達成され得る。最後に、pIIIディスプレイにおける標的に固定化された種は、通常、この標的-ファージ結合の(通常、競合的溶出または高pHもしくは低pHによる溶出による)破壊を必要とする。これは、例えば、pIIIディスプレイにおける高親和性のまたは安定的な結合剤の回収を著しく妨害することが公知である。感染のために必要とされるpIIIは、ファージ-標的相互作用の後ですら、未改変であり、pIXディスプレイにおける代替的な相互作用のために容易に利用可能であるため、これは、結合破壊、例えば、酸性溶出の必要性を完全に排除し、固定化されたファージは完全な感染性を保持し、従って、単なる感染により、標的と結合したまま回収されるかもしれない。
【0056】
核酸
本発明の第2の局面は、本発明の融合タンパク質をコードする核酸である。本発明の核酸は、プラスミド、ベクター、ファージゲノム、ファージミド、またはファスミドの一部であり得る。
【0057】
核酸という用語は、モノマーヌクレオチドの鎖から構成される高分子をさす。生化学において、これらの分子は、遺伝情報を保持するか、または細胞内の構造を形成する。最も一般的な核酸は、デオキシリボ核酸(DNA)およびリボ核酸(RNA)である。さらに、核酸という用語には、ペプチド核酸(PNA)、モルホリノ核酸、ロックド核酸(LNA)、グリコール核酸(GNA)、およびトレオース核酸(TNA)のような人工核酸が含まれる。これらの各々は、分子の骨格の変化により、天然に存在するDNAまたはRNAと区別される。
【0058】
ファージミドまたはファスミドは、プラスミドとして繁殖することができ、かつウイルス粒子内に一本鎖DNAとしてパッケージングされることもできるベクターを作製するため、繊維状ファージFfとプラスミドとのハイブリッドとして開発されたクローニングベクターの一つの型である。プラスミドと同様に、ファージミドは、DNA断片をクローニングするために使用され得、かつ一連の技術(形質転換、電気穿孔)により細菌宿主に導入され得る。しかしながら、ファージミドを含有している細菌宿主への、「ヘルパー」ファージ、例えば、VCSM13またはM13K07の感染が、一本鎖DNAの複製およびファージミドDNAのファージ粒子へのパッケージングを可能にするために必要なウイルス成分を提供する。
【0059】
従って、本発明の一つの局面は、原核生物N末端シグナル配列を含まない、繊維状ファージに由来するpIX融合タンパク質をコードする核酸を含む、ファージゲノムまたはファージミドに関する。
【0060】
本発明の一態様において、ファージゲノムまたはファージミドは、原核生物N末端シグナル配列を含まず、かつ
の1〜32位、2〜32位、3〜32位、4〜32位、および5〜32位からなる群より選択される配列を含む、繊維状ファージに由来するpIX融合タンパク質をコードする核酸を含む。
。
【0061】
本発明のもう一つの態様において、外来性ペプチドはpIX配列のN末端に直接融合している。
【0062】
本発明の一態様は、pIXに融合した外来性ペプチドが、Fv、scFv、Fab、単一ドメインを含む抗体またはその断片、プロテインAのZドメインまたはその断片(アフィボディ)、アンキリンまたはその断片、DARPinまたはその断片、T細胞受容体またはその断片、MHCクラスIもしくはIIまたはそれらの断片、フィブロネクチンまたはその断片、アンチカリンまたはその断片、PDZドメインまたはその断片、IgNARまたはその断片、CTLA4またはその断片、ImmE7またはその断片、ノッチンまたはその断片、アビマーまたはその断片、GFPまたはその断片、およびその他の遺伝子によりコードされた生物学的フルオロフォアからなる群より選択される、本発明のファージゲノムまたはファージミドに関する。
【0063】
本発明のもう一つの態様において、pIXに融合した外来性ペプチドはライブラリーメンバーである。
【0064】
繊維状ファージ
本発明の第3の局面は、本発明の融合タンパク質を含む繊維状ファージである。繊維状ファージビリオンは、ファージミドを保有し得る。
【0065】
しばしばバクテリオファージと呼ばれるファージは、ここで、感染性、複製性であり、細菌から分泌されるウイルスを意味する。繊維状バクテリオファージまたは繊維状ファージとは、ファージコートタンパク質によりパッケージングされる一本鎖DNA分子(ssDNA)を有するファージである。分泌された繊維状ファージ粒子は、表現型的には繊維状の構造を有する。
【0066】
繊維状ファージという用語には、本明細書において使用されるように、ファージゲノムに由来するビリオンおよびファージミドに由来するビリオンの両方が包含される。
【0067】
ヘルパーファージという用語は、例えば、欠陥ウイルスが既に占有している同じ宿主細胞に感染し、ファージミドを含有しているビリオンを形成するために必要な、欠陥ウイルスが欠いているタンパク質を提供することにより、繁殖するための、ファージゲノムに由来する一種または数種の要素を含有している、それ自体はファージゲノムでもなく機能性ウイルスでもなく、単なるプラスミドであるファージミドとして定義される、別の無関係な欠陥ウイルスを補助するウイルスをさす。
【0068】
一つの態様において、繊維状ファージは、本発明の融合タンパク質をコードする核酸を含む。特に好ましいのは、本発明の融合タンパク質をコードする核酸を含むファージミドを含むファージである。
【0069】
ファージライブラリーとは、繊維状ファージコートタンパク質のうちの一つまたは複数の一部としてペプチドまたはタンパク質を提示している繊維状ファージの集合体である。そのようなライブラリーは、異なるペプチドまたはタンパク質を提示する二種以上のファージを含むことができる。
【0070】
従って、本発明の一態様において、繊維状ファージは、繊維状ファージコートタンパク質のうちの一つまたは複数の一部としてペプチドまたはタンパク質を提示している。
【0071】
一態様において、繊維状ファージは、pIII融合タンパク質、pVII融合タンパク質、またはpVIII融合タンパク質をさらに含む。
【0072】
本発明の一局面は、pIII、pVII、pVIII、またはpIXのうちの一つまたは複数との融合体として外来性のペプチドまたはタンパク質を提示している本発明の繊維状ファージのライブラリーである。
【0073】
ライブラリーとは、繊維状ファージコートタンパク質のうちの一つまたは複数の一部としてペプチドまたはタンパク質を提示している繊維状ファージの集合体である。
【0074】
そのようなライブラリーは、異なるペプチドまたはタンパク質を提示している二種以上のファージを含むことができる。
【0075】
本発明の一局面は、異なるタンパク質を提示している二種以上の繊維状ファージを含み、これらのタンパク質のうちの少なくとも一種が本発明のファージゲノムまたはファージミドから発現されたpIX融合タンパク質である、ファージライブラリーに関する。本発明の一態様は、異なるペプチドまたはタンパク質を提示している二種以上の繊維状ファージを含むファージライブラリーに関する。
【0076】
特定の態様において、これらのペプチドまたはタンパク質のうちの少なくとも一つは、本発明のpIX融合タンパク質である。一態様において、ペプチドは、pIXと、pIII、pVII、またはpVIIIのいずれかとで、同時に提示される。
【0077】
もう一つの態様において、ペプチドは、pIXと、pIII、pVII、またはpVIIIからなる群より選択される二つまたは三つとで、同時に提示される。
【0078】
野生型(wildtype)、野生型(wild-type)、またはwtと記載されることもある野生型(wild type)という用語は、自然界に存在するような、生物、株、遺伝子、または特徴の典型的な型である。野生型とは、天然集団において最も一般的な表現型をさす。野生型とは、野生型表現型を生ずるのに必要とされる各遺伝子座における対立遺伝子もさす。野生型は、遺伝子型および表現型についての参照の標準である。生物学において、それは、特に、天然に存在する生物と意図的に変異させられた生物との間の差に関する。部位特異的変異誘発は、野生型遺伝子の遺伝子配列内の特定のヌクレオチドの変異を可能にする研究技術である。野生型タンパク質は、wt(タンパク質の名称)と記載され、例えば、野生型pIXタンパク質は、wt pIX、wt-pIX、または野生型pIXと記載される。
【0079】
従って、本発明の一つの局面は、wt pIXをコードする遺伝子および/またはwt pIXタンパク質を含まない本発明の繊維状ファージに関する。
【0080】
本発明のもう一つの局面は、wt pIXをコードする遺伝子および/またはwt pIXタンパク質をさらに含む本発明の繊維状ファージに関する。
【0081】
本発明のもう一つの局面は、本発明のファージゲノムまたはファージミドを含む繊維状ファージに関する。
【0082】
本発明の一態様において、本発明のファージゲノムまたはファージミドを含む繊維状ファージは、wt pIXをコードする遺伝子および/またはwt pIXタンパク質をさらに含む。
【0083】
本発明のもう一つの態様において、本発明のファージゲノムまたはファージミドを含む繊維状ファージは、wt pIXをコードする遺伝子および/またはwt pIXタンパク質を含まない。
【0084】
本発明のさらなる局面は、wt pIXタンパク質による補完なしにファージディスプレイにおいて機能するpIX融合タンパク質に関する。本発明の一局面は、pIII融合タンパク質、pVII融合タンパク質、およびpVIII融合タンパク質より選択される群一つまたは複数をさらに含む、本発明のファージゲノムまたはファージミドを含む繊維状ファージに関する。
【0085】
ファージディスプレイシステム
本発明の第5の局面は、ファージミドと、本発明のpIX融合タンパク質をコードする核酸を含むヘルパーファージとを含むファージディスプレイシステムである。
【0086】
ファージディスプレイシステム、ファージディスプレイ技術、ファージディスプレイテクノロジー、または単にファージディスプレイとは、タンパク質を、それをコードする遺伝情報と結び付けるためにバクテリオファージを利用する、タンパク質-タンパク質、タンパク質-ペプチド、およびタンパク質-DNAの相互作用の発見および研究のための方法をさす。
【0087】
タンパク質の提示または提示されたタンパク質とは、リガンドによる検出または固定化のためにアクセス可能なファージコートタンパク質に融合したタンパク質をさす。
【0088】
本発明の第6の局面は、本発明のpIX融合タンパク質をコードする核酸を含むファージミドと、ヘルパーファージとを含むファージディスプレイシステムである。
【0089】
本発明の一局面は、本発明のファージゲノムまたはファージミドを含むファージディスプレイシステムに関する。
【0090】
本発明のもう一つの局面は、本発明のファージゲノムまたはファージミドとヘルパーファージとを含むファージディスプレイシステムに関する。
【0091】
本発明の一局面は、異なるタンパク質を提示している二種以上の繊維状ファージを含み、これらのタンパク質のうちの少なくとも一つが本発明のファージゲノムまたはファージミドから発現されたpIX融合タンパク質である、ファージライブラリーに関する。
【0092】
本発明の一態様において、本発明のファージライブラリーは、pIII融合タンパク質、pVII融合タンパク質、およびpVIII融合タンパク質より選択される一つまたは複数の付加的な融合タンパク質を含む。
【0093】
キット
本発明の第7の局面は、本発明のpIX融合タンパク質をコードする核酸を含むファージミドと、ヘルパーファージとから構成されるファージディスプレイシステムを含むキットである。キットは、コーディング領域のN末端にマルチクローニング部位を有する、pIXをコードする遺伝子を含むファージミドと、ヘルパーファージ(例えば、M13K07、VCSM13、またはその他)とを含んでいるべきである。キットには、ファージクローンの感染、発現、固定化、選択、および検出のためのプロトコルが補足されるべきである。キットには、特定のアッセイを実施するための緩衝液および培地についての必要なレシピも添付されるべきである。
【0094】
キットとは、ここで、ファージディスプレイライブラリーとして、または単一のファージ粒子として、単一特異的または二重特異的な融合タンパク質を含むファージ粒子を生成するための試薬の集合体をさす。キットは、ファージミド、ヘルパーファージ、細菌株、ならびに試薬のレシピおよびアッセイの説明を含むプロトコルを含み得る。キットは、研究用、診断用、および治療用の試薬の開発のために使用され得る。
【0095】
本発明の一局面は、本発明のファージゲノムまたはファージミドとヘルパーファージとを含むファージディスプレイを含むキットに関する。
【0096】
本発明のもう一つの局面は、本発明のファージゲノムまたはファージミドを含むキットに関する。本発明のさらにもう一つの局面は、本発明のファージゲノムまたはファージミドを含む繊維状ファージを含むキットに関する。
【0097】
以下の非限定的な例において、本発明をさらに詳細に説明する。
【実施例】
【0098】
実施例1:pIX上のファージミドディスプレイ
試薬
全ての培地および緩衝液を、本質的に、Sambrook et al. (2001) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Pressに記載されたようにして調製した。抗M13-HRP抗体はGE Healthcare Bio-Sciences AB(Uppsala, Sweden)から購入した。制限酵素(RE)は、Stratagene(LaJolla, CA, USA)から入手したDpnIを除き、New England Biolabs(Ipswich, MA, USA)から購入した。DNAオリゴは、MWG Biotech AG(Ebersberg, Germany)から購入した。ウシ血清アルブミン(BSA)およびトゥイーン20は、Sigma-Aldrich(Oslo, Norway)から購入した。Pfu Turbo DNAポリメラーゼは、Stratagene(LaJolla, CA, USA)から購入した。BSAにコンジュゲートしたハプテン、2-フェニルオキサゾール-5-オン(phOx)および5-ニトロフェンアセチル(NIP)は、本質的に、他に記載されたようにして調製した(Nakela et al, PMID;722243およびMichaelsen et al, PMID:2125362)。イソプロピル-β-D-チオガラクトピラノシド(IPTG)は、Fermentas(Burlington, Canada)から購入した。トリエチルアミン(TEA)およびトリプシン/EDTAは、それぞれ、Sigma-Aldrich(Oslo, Norway)およびBioWhittaker(Lonza Group Ltd., Visp, Switzerland)から購入した。大腸菌株XL1-Blueは、Stratagene(LaJolla, CA, USA)から購入した。M13K07ヘルパーファージは、GE Healthcare Bio-Sciences AB(Uppsala, Sweden)から購入した。phOx-BSAに対する特異性を有する単鎖Fv(scFv)を保有しているpSEX81(SEQ ID NO: 2)ファージミド(pIIIディスプレイ)は、Affitech AS(Oslo, Norway)から厚意により提供を受けた。scFv抗NIP(SEQ ID NO:3)を保有している原核生物発現ベクターpSG1(未発表)は、pHOG21(Kiprianov et al, PMID:9005945)に基づいており、pLNOH2およびpLNOK(Norderhaug et al, PMID:9202712)に由来する抗体可変遺伝子から社内で作成した。
【0099】
新規のpIXディスプレイファージミドベクターpGALD9およびpGALD9ΔLの構築
ベクター骨格の出発テンプレートとして、上記のpSEX81(SEQ ID NO:2)ファージミドを選んだ(GenBankアクセッション番号:Y14584)。まず、このベクター内の原核生物pelBシグナル配列
をコードするストレッチを除去するため、プライマー対
を使用したQuikChange(商標)インビトロ変異誘発により、NcoI RE部位を最もN末端に導入し、それにより、pelB ORFの第2コドンの最初のヌクレオチドをAからGに変化させた。変異誘発の後、ベクターをNcoI消化し、再ライゲートさせ、プライマー対
を使用した、ベクターの関連部分を回収する二次PCRにおいて、テンプレートとして使用した。次いで、このPCR断片を、標準的な技術を使用して、適合性のEcoRI/HindIII RE部位で、元のpSEX81(SEQ ID NO:2)に移し、DNA配列決定により確認した。この工程により、pelBシグナル配列コーディング部分が完全に除去されたが、正常な転写および翻訳のために重要な開始コドンならびにlacPOおよびシャインダルガーノ配列(SD)に対するその相対位置は保存され、さらに、元のpSEX81(SEQ ID NO:2)に見出されたNcoI/NotI RE部位により画定された外来性配列の前にAla残基が1個だけ付加された。新たな構築物をpSEX81ΔLと名付けた。
【0100】
第2に、pXIコーディング配列を、5'末REタグ付きプライマー対
を使用して、M13K07から増幅した。次いで、このPCR断片を、適合性のRE部位で、pSEX81(SEQ ID NO:2)ファージミドおよびpSEX81ΔLファージミドの両方に移し、それにより、両方のpIIIコーディング領域を交換し、元のpSEX81(SEQ ID NO:2)のNcoI/NotIにより画定されたカセットのN末端インフレームpIX融合体を得た。新たな構築物をDNA配列決定により確認し、それぞれ、pGALD9およびpGALD9ΔLと名付けた。上記の様々なファージミドのscFv抗phOx(SEQ ID NO:11)単位を、pSG1由来のscFv抗NIP(SEQ ID NO:3)単位に切り替えるため、標準的な技術を使用して、NcoI/NotI REにより画定されたカセットの交換として、これを行った。本明細書に記載された全てのファージミドを、標準的な技術を使用して、電気穿孔により大腸菌XL1-Blueに導入した。
【0101】
ファージ粒子の調製
M13K07ヘルパーファージを使用した大腸菌XL1-Blueからのファージミドレスキューおよびビリオン組み立てを、記載されたようなスポット滴定によりモニタリングした(Welschof et al, PMID:9050877およびKoch et al, PMID:11126120)。
【0102】
ファージ捕獲酵素結合免疫吸着アッセイ(ELISA)
phOx-BSAまたはNIP-BSAを、4℃で一夜、PBS(pH7.4)中5μg/mlで、MaxiSorp(商標)マイクロタイタープレートウェル(Nunc, Roskilde, Denmark)に吸着させた。RTで1時間、PBS(w/v)中2%BSAによりウェルをブロッキングし、次いで、ビリオン調製物を添加し、RTで1〜2時間、反応させた後、捕獲されたビリオンを、RTで1時間、抗M13-HRP(5,000倍)により検出した。各工程の間に、ウェルをPBST(PBS/0.05%トゥイーン20)で3回洗浄した。ウェルをTMB可溶性基質により発色させ、30分後に1M HClにより中止し、A450nmで吸光度を読み取った。1ビリオン当たりの提示された融合タンパク質の関数として、Ag反応性を定量化するため、任意の試料についてシグナル飽和が観察される前に、1M HClを添加することによりELISA発色を中止し、吸光度読み取り値を、以下の式に従って変換した:相対ディスプレイレベル=(A450nm/ファージミド力価)×1012。
【0103】
スパイクされたphOx-BSA/NIP-BSA選択
新鮮なビリオン試料を、1mM IPTG誘導により、または誘導なしに調製し、PEG沈殿させ、記載されたようにして滴定した。
【0104】
次いで、中サイズのコンビナトリアルライブラリーに相当する107という既知の多様性を与える1:107レベルで、標的特異的な実体を、無関係のバックグラウンドにスパイクした。
【0105】
NIP-BSA選択については、scFv抗NIPを、scFv抗phOxカウンターパートにスパイクし、その逆も同様に行った。初期インプットは、12のモデルライブラリー全てについてパニングラウンド1において103という複雑さレベルをもたらす1×1010cfuampRであった。
【0106】
簡単に説明すると、ラウンド1については1μg/ml、ラウンド2については0.1μg/mlの100μl容量を使用して、同一のプレート上にトリプリケートで、MaxiSorp(商標)マイクロタイタープレートウェル(Nunc, Roskilde, Denmark)に標的を固定化した。
【0107】
パニング前に、RTで1〜2時間、ウェルをPBSTMによりブロッキングした後、それぞれの(PBSTMで)予めブロッキングされたビリオン調製物100μlを添加し、RTで1.5時間、撹拌しながら反応させた。
【0108】
マイクロタイター洗浄装置を使用して、PBSTで9回、続いてdH2Oで5回、ウェルを洗浄した後、(トリプリケートウェル内の)標的に結合したビリオンを、以下のいずれかにより溶出させた。
(1)RTで5分間、100μl/ウェルの100mM TEA(pH12)を添加し、続いて、100μl/ウェルのトリスHCl(pH6.8)を含有している新鮮なウェルに移すことにより中和する;
(2)RTで10分間、100μl/ウェルのトリプシン/EDTAを添加し、続いて、新鮮なウェルに移す;
(3)37℃で30分間、撹拌しながら、200μl/ウェルの対数期(A600nmおよそ0.5、≧5×107細胞に相当)の大腸菌XL1-Blueを添加し、続いて、MOI10でM13K07ヘルパーファージが補足された10mlの予め加温されたYT-TAG(30μg/mlテトラサイクリン、100μg/mlアンピシリン、および0.1Mグルコースを含有している2×YT)に移す。
【0109】
弱く撹拌しながら37℃で15分間、続いて強く撹拌しながら37℃で30分間、インキュベーションを続けた。平行して、TEAおよびトリプシンにより溶出させた試料を、9ml YT-TAG中の対数期大腸菌XL1-Blue培養物に感染させるために使用し、37℃で15分間弱く撹拌しながらインキュベートした後、MOI10でM13K07ヘルパーファージが補足された1ml YT-TAGを添加した。
【0110】
弱く撹拌しながら37℃で15分間、続いて激しく撹拌しながら37℃で30分間、インキュベーションを続けた。次いで、全ての試料を3000g/10分/RTで遠心分離し、上清を廃棄し、100μg/mlアンピシリンおよび50μg/mlカナマイシンを含有している10mlの予め加温された2×YTにペレットを穏和に再懸濁させた。
【0111】
適切な試料に1mM IPTGを補足し、全ての試料を厳しく撹拌しながら30℃で一夜、インキュベートした。翌日、培養物を4000g/10分/RTで遠心分離し、上清を0.2μmフィルターで新鮮な15mlチューブへと滅菌濾過した。
【0112】
次いで、これらの上清を、少なくとも各試料109cfuampRのインプットに相当する、各試料50μlの容量を使用して、記載されたようなパニングの次のラウンドに供した。選択の第2ラウンドの後、ビリオンを含有している上清を、上記のような抗原特異的ELISAに供した。
【0113】
結果
繊維状ファージ(fd、M13、およびf1)の5種の構造コートタンパク質は、全て、突出するビリオンに取り込まれる前は、グラム陰性宿主の内膜に見出される膜内在性タンパク質であることが公知である(Endeman and Model, PMID:7616570)。この報告は、pIXはN末端の融合修飾を可能にしないが、カプシドタンパク質自体は、完全ビリオンにおいて溶媒に露出しているとの結論も下した。Gaoら(PMID:10339535および12239343)ならびにKhalilら(PMID:17360403)の両方は、その後、ファージミドから発現させ、かつシグナル配列依存性周辺質ターゲティングと組み合わせて使用した場合には、N末端pIX融合ディスプレイが可能であることを示した。これらのシステムにおいては、ファージミドレスキューによりヘルパーファージゲノムからwt pIXが供与され、補完が起こる。そのようなディスプレイが、周辺質へのシグナル配列依存性ターゲティングなしに可能であるか否かを試験するため、本発明者らは、そのようなシグナル配列を含むか、または含まない、N末端pIXディスプレイを可能にするpGALD9およびpGALD9ΔLと名付けられた二つの新規ファージミドを構築した(図2)。
【0114】
二つの異なるpIX融合体を、これらの新たなファージミドを使用した分析のために選び、標準的なpIIIディスプレイを使用したカウンターパートと比較した。両融合体は、ヒト抗体可変遺伝子セグメントに基づくハプテンコンジュゲートphOx-BSAに特異的な抗体断片scFv(scFv抗phOx)、またはマウス抗体可変遺伝子セグメントに基づくハプテンコンジュゲートNIP-BSAに特異的な抗体断片scFv(scFv抗NIP)であった。特に、scFv抗phOx(SEQ ID NO:11)は、ヒト抗体scFvライブラリーから選択されており、大腸菌において極めてよく発現することが公知である(Marks et al., PMID:1748994)。対照的に、多くのマウスハイブリドーマ可変遺伝子は、大腸菌において、ファージディスプレイされた場合にも、よく発現しないことが周知である(Krebber et al, PMID:9032408)。
【0115】
scFvのpIXディスプレイは、正常なビリオン組み立てに干渉しないはずである。従って、本発明者らは、材料および方法に記載されるような標準的なファージミドレスキューおよび滴定を使用して、これらのシグナル配列を含むscFvディスプレイファージミドおよびシグナル配列を含まないscFvディスプレイファージミドの性能を比較し、(シグナル配列依存性周辺質ターゲティングを絶対的に必要とする)標準的なpIIIディスプレイとも比較した(図3)。
【0116】
滴定結果は、ファージミドを含有しているビリオンが全てのケースで作成されたことを実際示した(図3AおよびC)。scFv融合体をプロモーター誘導なしにlac POから発現させる場合(基底発現)、両方のpIXディスプレイバージョンおよびpIII対照が、scFv抗phOxについて比較可能な力価を与える(図2A)。対照的に、シグナル配列非依存性scFv抗NIP pIXディスプレイの力価は、pIIIディスプレイおよびシグナル配列依存性pIXディスプレイと比較して、それぞれ5〜10倍高い(図3C)。lacPOのIPTG誘導によりscFv融合体発現を増加させると、シグナル配列依存性pIXディスプレイおよびpIIIディスプレイの両方の力価が10倍低下した。対照的に、両方のシグナル配列非依存性pIXディスプレイバリアントに対しては軽微な効果しかなかった(図3AおよびC)。このシステムにおいてはヘルパーファージからのpIXのwt補完が存在するため、この所見は驚くべきものでありかつ重要であった。なぜなら、シグナル配列依存性pIXディスプレイおよびpIIIディスプレイは、ビリオン組み立て過程に干渉するが、シグナル配列非依存性pIXディスプレイの場合には、IPTG誘導時ですら、この効果が軽微に過ぎないことを、それが示すためである。シグナル配列を含むか、またはシグナル配列を含まないpIX上に提示されたscFv抗NIPからの結果を比較した場合に、効果は最も顕著であって、前者は力価の100倍の低下を示す。両方のscFvについて、シグナル配列非依存性pIXバージョンで行われた場合、ビリオン組み立てが、pIIIと同等であるかまたはpIIIより良好であることも注目に値する。
【0117】
コンビナトリアル選択プラットフォームのコアは、表現型の選択を通した遺伝子型の回収を可能にする、物理的な表現型-遺伝子型カップリングである。この物理的な関係が損なわれるかまたは失われた場合、システムは非機能性となる。ファージミドディスプレイに翻訳すると、これは、ヘルパーファージレスキューによりビリオンに封入されるのが、ヘルパーファージゲノムではなくファージミドであることが、選択のために不可欠であることを意味する。ファージミドおよびヘルパーファージは異なる抗生物質選択マーカーを保有するため、この概念は、適切な選択増殖(アンピシリン(ampR)、またはカナマイシン(kanR))に起因する、それぞれのコロニー形成単位(cfu)に基づき、ファージミド対ヘルパーファージ比を計算することにより、感染性滴定において容易に査定され得る。その際、その比は、効率的な下流の選択が実行可能であるよう、1より大きいべきである。
【0118】
上記のビリオン調製物のファージミド対ヘルパーファージ比を査定したところ(図3BおよびD)、3種のディスプレイ経路の間の有意な差が明らかになった。プロモーター誘導なしの標準的なビリオンパッケージング(基底発現)を使用すると、scFv抗phOxについては3種の経路全てが実施可能であるが(図2B)、scFv抗NIPについて実行可能であったのは、配列非依存性pIXバリアントのみであった(図3D)。プロモーター誘導(IPTG誘導)時、pIIIバリアントおよびシグナル依存性pIXバリアントの両方が、表現型-遺伝子型連鎖の著しい損失を示し、その効果はscFv抗NIP融合体で最も顕著であった。対照的に、この効果は、両方のシグナル配列非依存性pIXバリアントについては、存在しないか(scFv抗phOx)または軽度であった(scFv抗NIP)。従って、結果は、シグナル配列非依存性pIXディスプレイが、標準的なpIIIディスプレイおよびシグナル配列依存性pIXディスプレイの両方と比較して、優れた表現型-遺伝子型連鎖表現型を保有していることを明白に示している。
【0119】
次いで、上記試料に基づき、本発明者らは、抗原特異的ELISAでこれらのビリオン上の機能的なscFvディスプレイを査定した(図4)。結果は、3種のディスプレイ経路全てからの機能的なscFvディスプレイを明白に示した(図4AおよびB)。試料はファージミド力価によってノーマライズされていないため、シグナル強度は直接比較可能ではない。さらに、シグナル飽和が、いくつかの試料において観察された。3種のディスプレイ経路の間の機能的な差を十分に査定するため、相対ディスプレイレベル(1ビリオン当たりの機能性ディスプレイ単位)を、材料および方法に記載されるような新たな抗原特異的ELISAにおいて決定した(図4CおよびD)。ディスプレイレベルは、pIIIバリアントとシグナル配列依存性pIXバリアントとの間で比較可能であったが、シグナル配列非依存性pIXディスプレイは有意に低かった。このことは、両方のscFvについて当てはまり、基底発現時にもIPTG誘導時にも当てはまった。予想通り、3種のディスプレイ経路全てが、両方のscFvについて、IPTG誘導時に、より高いディスプレイを示した。しかしながら、図3BおよびDに与えられた比を考慮して、図4CおよびDの結果を検討することが最も重要である。例えば、scFv抗NIP単位のpIIIディスプレイおよびシグナル配列依存性pIXディスプレイの両方が、シグナル配列非依存性pIXカウンターパートと比較して、10〜20倍(基底発現)、およそ25倍(IPTG誘導)高いディスプレイを示すことは明白である。しかしながら、前者の2種の経路は、いずれも、極めて弱いかまたは失われた表現型-遺伝子型関係を有し、表現型選択時に遺伝子型が失われるため、コンビナトリアル選択計画において非機能性であろう。この効果は、IPTG誘導時のみであるが、scFv抗phOx単位についても当てはまる。さらに、IPTGにより誘導されたシグナル配列非依存性pIXバリアントにおけるscFv抗phOx単位のディスプレイレベルは、基底発現時のpIIIバリアントおよびシグナル配列依存性pIXバリアントと比較可能である。これらのデータを考慮すると、scFvディスプレイのためにpIX(シグナル配列依存性)を活用している唯一の報告(Gao, PMID:12239343)が、ビリオンパッケージングの際に常にIPTG誘導を使用しているが、ファージミド対ヘルパーファージ比を報告していないことに注目することは興味深い。
【0120】
scFvフォーマットは、大腸菌における好都合な発現プロファイルのためしばしば使用されているが(Bradbury and Marks, PMID:15261570)、いくつかの報告は、アフィニティ選択による高親和性結合剤の回収においては、例えば、抗体Fab断片のような、より低レベルのディスプレイフォーマットが有利であることを指摘している(de Haard, et al, PMID:10373423およびHoogenboom, PMID:16151404)ならびにRotheら(PMID:18191144)。
【0121】
従って、たとえシグナル配列非依存性scFv-pIXディスプレイが低レベルのディスプレイを与えるようであっても、これは、実際、高親和性選択のために適用された場合、高度に有利であることが判明するかもしれない。
【0122】
従って、シグナル配列非依存性pIXディスプレイのアフィニティ選択における性能を査定するため、本発明者らは従来のpIIIディスプレイと比較した。
【0123】
IPTG誘導の存在下または非存在下でビリオンを作製した。関心対象のタンパク質はscFv抗phOxまたはscFv抗NIPのいずれかであった。
【0124】
従って、2種の標的phOx-BSAおよびNIP-BSAそれぞれについての計4つのファージ集団を評価した。各ケースにおいて、標的特異的scFvを1:107の比で特異性が無関係のscFvと混合した。重要なことに、2種のscFvは交差反応しない。次いで、2ラウンドのアフィニティ選択を実施した。
【0125】
3種の異なる溶出戦略;高pH、タンパク質分解、または直接感染のいずれかを利用した。選択の第2ラウンドの後、抗原特異的ポリクローナルファージELISAにより、濃縮を確証した(図5)。
【0126】
関心対象のタンパク質をpIX上に提示した場合、選択は、POIをpIII上に提示した場合とほぼ同等の効率であり、ほとんどのケースで、より効率的であった。特に、高pH(TEA)またはタンパク質分解(トリプシン)による溶出を利用した標準的なpIIIディスプレイ経路は、pIXと比較して、不十分な濃縮を示した。
【0127】
選択は、ディスプレイ経路および溶出条件とは無関係に、IPTG誘導時よりIPTG誘導なしの方が効率的であり、負の効果は、pIIIディスプレイ経路で最も大きいようであった。
【0128】
従って、シグナル配列非依存性pIXの低いディスプレイの傾向は、不十分な選択を意味しなかった。pIIIを改変せず、完全に溶媒に露出させておくことにより、ライブラリー選択工程後のビリオンレスキューが、ビリオン-標的結合を破壊することなく効率的に達成され得、従って、溶出工程が不要となり、ハイスループットプロトコルが加速されるかもしれない。
【0129】
後者は、溶出が多様な戦略に対して抵抗性であるかもしれない高親和性結合剤の単離も容易にするかもしれない(Balass et al, PMID:8954559)。
【0130】
参照
【特許請求の範囲】
【請求項1】
繊維状ファージに由来するpIX融合タンパク質をコードする核酸を含むファージゲノムまたはファージミドであって、該融合タンパク質が原核生物N末端シグナル配列を含まない、ファージゲノムまたはファージミド。
【請求項2】
pIX融合タンパク質が、
の1〜32位、2〜32位、3〜32位、4〜32位、および5〜32位からなる群より選択される配列を含む、請求項1記載のファージゲノムまたはファージミド。
【請求項3】
外来性ペプチドが、pIX配列のN末端に直接融合している、前記請求項のいずれか一項記載のファージゲノムまたはファージミド。
【請求項4】
外来性ペプチドが、Fv、scFv、Fab、単一ドメインを含む抗体またはその断片、プロテインAのZドメインまたはその断片(アフィボディ)、アンキリンまたはその断片、DARPinまたはその断片、T細胞受容体またはその断片、MHCクラスIもしくはIIまたはそれらの断片、フィブロネクチンまたはその断片、アンチカリンまたはその断片、PDZドメインまたはその断片、IgNARまたはその断片、CTLA4またはその断片、ImmE7またはその断片、ノッチンまたはその断片、アビマーまたはその断片、GFPまたはその断片、およびその他の遺伝子によりコードされた生物学的フルオロフォアからなる群より選択される、前記請求項のいずれか一項記載のファージゲノムまたはファージミド。
【請求項5】
外来性ペプチドがライブラリーメンバーである、前記請求項のいずれか一項記載のファージゲノムまたはファージミド。
【請求項6】
請求項1〜5のいずれか一項記載のファージゲノムまたはファージミドを含む繊維状ファージ。
【請求項7】
wt pIXをコードする遺伝子および/またはwt pIXタンパク質をさらに含む、請求項6記載の繊維状ファージ。
【請求項8】
wt pIXをコードする遺伝子および/またはwt pIXタンパク質を含まない、請求項6記載の繊維状ファージ。
【請求項9】
pIII融合タンパク質、pVII融合タンパク質、およびpVIII融合タンパク質より選択される群一つまたは複数をさらに含む、請求項6〜8のいずれか一項記載の繊維状ファージ。
【請求項10】
請求項1〜5のいずれか一項記載のファージゲノムまたはファージミドとヘルパーファージとを含むファージディスプレイシステム。
【請求項10】
請求項10記載のファージディスプレイシステムを含むキット。
【請求項11】
異なるタンパク質を提示する2種以上の繊維状ファージを含むファージライブラリーであって、これらのタンパク質のうちの少なくとも1種が、請求項1記載のファージゲノムまたはファージミドから発現されたpIX融合タンパク質である、ファージライブラリー。
【請求項1】
繊維状ファージに由来するpIX融合タンパク質をコードする核酸を含むファージゲノムまたはファージミドであって、該融合タンパク質が原核生物N末端シグナル配列を含まない、ファージゲノムまたはファージミド。
【請求項2】
pIX融合タンパク質が、
の1〜32位、2〜32位、3〜32位、4〜32位、および5〜32位からなる群より選択される配列を含む、請求項1記載のファージゲノムまたはファージミド。
【請求項3】
外来性ペプチドが、pIX配列のN末端に直接融合している、前記請求項のいずれか一項記載のファージゲノムまたはファージミド。
【請求項4】
外来性ペプチドが、Fv、scFv、Fab、単一ドメインを含む抗体またはその断片、プロテインAのZドメインまたはその断片(アフィボディ)、アンキリンまたはその断片、DARPinまたはその断片、T細胞受容体またはその断片、MHCクラスIもしくはIIまたはそれらの断片、フィブロネクチンまたはその断片、アンチカリンまたはその断片、PDZドメインまたはその断片、IgNARまたはその断片、CTLA4またはその断片、ImmE7またはその断片、ノッチンまたはその断片、アビマーまたはその断片、GFPまたはその断片、およびその他の遺伝子によりコードされた生物学的フルオロフォアからなる群より選択される、前記請求項のいずれか一項記載のファージゲノムまたはファージミド。
【請求項5】
外来性ペプチドがライブラリーメンバーである、前記請求項のいずれか一項記載のファージゲノムまたはファージミド。
【請求項6】
請求項1〜5のいずれか一項記載のファージゲノムまたはファージミドを含む繊維状ファージ。
【請求項7】
wt pIXをコードする遺伝子および/またはwt pIXタンパク質をさらに含む、請求項6記載の繊維状ファージ。
【請求項8】
wt pIXをコードする遺伝子および/またはwt pIXタンパク質を含まない、請求項6記載の繊維状ファージ。
【請求項9】
pIII融合タンパク質、pVII融合タンパク質、およびpVIII融合タンパク質より選択される群一つまたは複数をさらに含む、請求項6〜8のいずれか一項記載の繊維状ファージ。
【請求項10】
請求項1〜5のいずれか一項記載のファージゲノムまたはファージミドとヘルパーファージとを含むファージディスプレイシステム。
【請求項10】
請求項10記載のファージディスプレイシステムを含むキット。
【請求項11】
異なるタンパク質を提示する2種以上の繊維状ファージを含むファージライブラリーであって、これらのタンパク質のうちの少なくとも1種が、請求項1記載のファージゲノムまたはファージミドから発現されたpIX融合タンパク質である、ファージライブラリー。
【図1】

【図2】
【図3】
【図4】
【図5】

【図2】
【図3】
【図4】
【図5】
【公表番号】特表2012−518423(P2012−518423A)
【公表日】平成24年8月16日(2012.8.16)
【国際特許分類】
【出願番号】特願2011−551486(P2011−551486)
【出願日】平成22年2月24日(2010.2.24)
【国際出願番号】PCT/EP2010/052344
【国際公開番号】WO2010/097411
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(511206397)
【Fターム(参考)】
【公表日】平成24年8月16日(2012.8.16)
【国際特許分類】
【出願日】平成22年2月24日(2010.2.24)
【国際出願番号】PCT/EP2010/052344
【国際公開番号】WO2010/097411
【国際公開日】平成22年9月2日(2010.9.2)
【出願人】(511206397)
【Fターム(参考)】
[ Back to top ]