
シナプトフィジンに対する特異的結合要素
【課題】シナプトフィジンに結合する特異的結合要素を提供する。
【解決手段】前記結合要素は、C1-3 VHドメインと特定のアミノ酸配列を有するVH CDR3を含むVHドメインと場合により特定のアミノ酸配列を有する1つまたは複数のVH CDRとからなる群から選択された抗体VHドメイン;および/またはC1-3 VLドメインと特定のアミノ酸配列を有する1つまたは複数のVL CDRを含むVLドメインとからなる群から選択された抗体VLドメインを含む。核酸、キットおよび組成物などの関連材料、ならびに結合要素の使用方法、例えば、肝線維症に関連している肝星細胞に薬剤実体を向けることにおける使用方法。
【解決手段】前記結合要素は、C1-3 VHドメインと特定のアミノ酸配列を有するVH CDR3を含むVHドメインと場合により特定のアミノ酸配列を有する1つまたは複数のVH CDRとからなる群から選択された抗体VHドメイン;および/またはC1-3 VLドメインと特定のアミノ酸配列を有する1つまたは複数のVL CDRを含むVLドメインとからなる群から選択された抗体VLドメインを含む。核酸、キットおよび組成物などの関連材料、ならびに結合要素の使用方法、例えば、肝線維症に関連している肝星細胞に薬剤実体を向けることにおける使用方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はシナプトフィジンに対する特異的結合要素に関する。本発明の好ましい実施形態は、本明細書においてC1-3と称するscFvフラグメントの抗体VHおよび/またはVLドメインを使用する。さらに好ましい実施形態では、C1-3の重鎖可変(VH)および/または軽鎖可変(VL)ドメイン、特にVH C1-3の1つまたは複数の相補性決定領域(CDR)を他の抗体のフレームワーク領域中に使用する。本発明者らは、有利な性質、特に肝星細胞の外表面を標的にする能力を有する多数の抗体分子を同定している。
【背景技術】
【0002】
肝線維症は、肝硬変および肝不全の発生に先立つ、肝臓での細胞外基質タンパク質の蓄積により特徴付けられる可逆的過程である(Friedman S.L.、J Biol Chem、2000年、275巻、2247〜50頁、Batallex R.ら、Semin Liver Dis、2001年、21巻、437〜51頁)。ウィルス感染(例えばC型肝炎)およびアルコール乱用を含む多くの条件が、線維症に至る肝臓への損傷を惹起し得る。線維症は長年の間検知されないままで、致命的になることのある重度の損傷を与えることがある。肝線維症を患う人々は、世界中では2億人を超える可能性があるにもかかわらず、この疾患を治療するために臨床医が使用できる治療の選択肢はない。
【0003】
肝線維症は、慢性的な肝損傷に反応して肝星細胞(HSC)によって惹き起こされる。HSCは、肝線維症の発生および回復において主要かつ中心的な役割を演ずる。HSCは、正常肝では静止状態で存在し、ビタミンAを貯蔵するように機能している(Geerts A.Semin、Liver Dis、2001年、21巻、311〜35頁)。慢性肝損傷に反応して、静止状態のHSCが、筋線維芽細胞様表現型に「活性化」する。活性化したHSCは増殖し、肝線維症で観察される瘢痕を構成する細胞外基質タンパク質の大部分を発現すると考えられている。活性化したHSCおよび線維発生は、何らかの原因による慢性的肝損傷に対する有害な反応であること、および増大した活性化HSCアポトーシスが肝線維症を消散させ、慢性的損傷に対する肝臓の反応を増大させ得ることを示唆する強い証拠がある(Iredale J.P.ら、J Clin Invest、1998年、102巻、538〜49頁、Wright M.C.ら、Gastroenterology 2001年、121巻、685〜98頁、Orr J.G.ら、Hepatology 2004年、40巻、232〜42頁、Issa R.ら、Gut、2001年、48巻、548〜57頁)。
【0004】
過去における多くの抗線維化薬候補の主な問題は、おそらく外来性化合物を代謝するように機能する肝細胞の近傍に肝星細胞があるために、薬剤が肝星細胞内で治療濃度に達しなかったことである。これらの薬剤には、in vitroおよびin vivoで抗炎症性活性を有し、星細胞の活性化を低下させることができるいくつかの作用薬が含まれる。これらの薬剤には、コルチコステロイド、TNFα拮抗剤、抗酸化剤、サイトカイン(γインターフェロン)および肝細胞成長因子(HGF)PPARγリガンド(チアゾリジンジオン)、エンドテリン-1拮抗剤、ハロフジノン(抗コクシジウム薬)および遺伝子治療(動物モデルにおける遺伝子治療によるメタロプロテイナーゼmRNAの投与)が含まれる。成功したものがないということは、任意の抗線維化療法を肝臓中の肝星細胞に向けることが有用であろうということを示している。
【0005】
線維症の間、活性化肝星細胞中でマンノース6-リン酸/インスリン様成長因子II(M6P/IGF-II)受容体が高レベルで発現していることおよびマンノース6-リン酸(M6P)で修飾された血清アルブミン(SA)をラットに投与すると(M6P:SAのモル比は28:1)、肝細胞内の投与量の70%が肝星細胞に見出されるように肝臓に分布することが、最近Polestraおよび共同研究者によって示された(Beljaarsら、Hepatology、1999年、29巻、1486〜93頁)。VI型コラーゲン受容体を認識する10個の環状ペプチド部分(C*GRGDSPC*、ただしC*は環化しているシステイン残基を示す)で修飾したSAもまた、静脈注射後10分以内にラット肝臓に優先的に分布する結果となる(Beljaarsら、J Biol Chem、2000年、275巻、12743〜51頁)。線維症の肝臓においては、肝臓に投与したペプチド修飾アルブミンの70%が、活性化した肝星細胞に結合した。しかし、ヒト肝組織灌流において、これらの作用薬は、大分子量分子および異物粒子の除去に関与する型の肝細胞であるクップファー細胞に取り込まれて、星細胞には取り込まれなかった。
【0006】
シナプトフィジンは神経細胞および肝星細胞だけに発現するタンパク質である(Cassiman D.ら、Am J Pathol、1999年、155巻、1831〜1839頁、Bargou R.C.ら、Gene、1991年、99巻、197〜204頁)。これは膜結合タンパク質であり、機能性のある精製形態では入手できない。この大きな制限にもかかわらず、本出願人らは、肝星細胞上に存在するシナプトフィジンの細胞外ドメインに対して特異性のある初めての完全なヒトモノクローナル抗体フラグメントを単離することに成功した。この抗体はファージディスプレイ法およびMRC(Cambridge、UK所在)によって使用可能になったヒト抗体ライブラリーを使用して単離した。この抗体はあるペプチドに対して作られたもので、星細胞膜中で自然のままの立体配座にある天然のシナプトフィジンタンパク質全体を認識するのも、この抗ペプチド抗体である。シナプスフィジンを認識して結合する抗体は、下で論ずるように多くの治療応用に適した試薬を初めて提供する可能性を有する。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】欧州特許A184187号
【特許文献2】英国特許2188638A号
【特許文献3】欧州特許A239400号
【特許文献4】欧州特許A0120694号
【特許文献5】欧州特許A0125023号
【特許文献6】国際出願PCT/US92/09965
【特許文献7】国際公開WO94/13804
【特許文献8】国際公開WO92/01047
【特許文献9】国際公開WO99/58572
【非特許文献】
【0008】
【非特許文献1】Friedman S.L.、J Biol Chem、2000年、275巻、2247〜50頁
【非特許文献2】Batallex R.ら、Semin Liver Dis、2001年、21巻、437〜51頁
【非特許文献3】Geerts A.Semin、Liver Dis、2001年、21巻、311〜35頁
【非特許文献4】Iredale J.P.ら、J Clin Invest、1998年、102巻、538〜49頁
【非特許文献5】Wright M.C.ら、Gastroenterology 2001年、121巻、685〜98頁
【非特許文献6】Orr J.G.ら、Hepatology 2004年、40巻、232〜42頁
【非特許文献7】Issa R.ら、Gut、2001年、48巻、548〜57頁
【非特許文献8】Beljaarsら、Hepatology、1999年、29巻、1486〜93頁
【非特許文献9】Beljaarsら、J Biol Chem、2000年、275巻、12743〜51頁
【非特許文献10】Cassiman D.ら、Am J Pathol、1999年、155巻、1831〜1839頁
【非特許文献11】Bargou R.C.ら、Gene、1991年、99巻、197〜204頁
【非特許文献12】Ward、E.S.ら、Nature 341巻、544〜546頁、1989年
【非特許文献13】Birdら、Science、242巻、423-426頁、1988年
【非特許文献14】Hustonら、PNAS USA、85巻、5879〜5883頁、1988年
【非特許文献15】P.Holligerら、Proc.Natl.Acad.Sci.USA、90巻、6444〜6448頁、1993年
【非特許文献16】Y.Reiterら、Nature Biotech、14巻、1239〜1245頁、1996年
【非特許文献17】S.Huら、Cancer Res.、56巻、3055〜3061頁、1996年
【非特許文献18】Holliger,P.、Winter G.、Current Opinion Biotechnol.、4巻、446〜449頁、1993年
【非特許文献19】J.B.B.Ridgewayら、Protein Eng.、9巻、616〜621頁、1996年
【非特許文献20】Kabat、E.A.ら、「Sequences of Proteins of Immunological Interest」第4版、US Department of Health and Human Services、1987年、および現在インターネット上で使用できるその更新版(http://immuno.bme.nwu.eduまたは任意の検索エンジンを使用して「Kabat」)
【非特許文献21】Markら、Bio/Technology、1992年、10巻、779〜783頁
【非特許文献22】Stemmer、Nature、1994年、370巻、389〜391頁
【非特許文献23】Gramら、1992年、Proc.Natl.Acad.Sci.、USA、89巻、3576〜3580頁
【非特許文献24】Barbasら、1994年、Proc.Natl.Acad.Sci.、USA、91巻、3809〜3813頁
【非特許文献25】Schierら、1996年、J.Mol.Biol.、263巻、551〜567頁
【非特許文献26】Ledermann J.A.ら、(1991年)、Int.J.Cancer 47巻、659〜664頁、
【非特許文献27】Bagshawe K.D.ら、(1991年)、Antibody,Immunoconjugates and Radiopharmaceuticals 4巻、915〜922頁
【非特許文献28】Pluckthun、A.Bio/Technology 9巻、545〜551頁、1991年
【非特許文献29】Ref、M.E.、(1993年)Curr.Opinion Biotech.4巻、573〜576頁、
【非特許文献30】Trill J.J.ら、(1995年)Curr.Opinion Biotech 6巻、553〜560頁
【非特許文献31】SambrookおよびRussell、「Molecular Cloning:a Laboratory Manual」第3版、2001年、Cold Spring Harbor Laboratory Press
【非特許文献32】「Current Protocols in Molecular Biology」第2版、Ausubelら(編)、John Wiley & Sons、1992年
【非特許文献33】Leel V.ら、Biochem Biophys Res Commun 2004年、316巻、872〜7頁
【非特許文献34】Hayhurst A.ら、Protein Expr Purif 1999年、15巻、336〜43頁
【非特許文献35】Wright M.C.ら、Mol Pharmacol 1996年、50巻、856〜63頁
【非特許文献36】Wright M.C.ら、Hum Exp Toxicol、1996年、15巻、203〜4頁
【非特許文献37】Harvey J.L.ら、Drug Metab Dispos、2000年、28巻、96〜101頁
【非特許文献38】Marek C.J.ら、Biochem J、2003年、370巻、763〜9頁
【非特許文献39】Eastwood S.L.ら、Brain Res Bull、2001年、55巻、569〜78頁
【非特許文献40】McElhiney J.ら、Appl Environ Microbiol、2002年、68巻、5288〜95頁
【非特許文献41】Foxら、J Microbiol Methods、2004年、56巻、221〜230頁
【発明の開示】
【課題を解決するための手段】
【0009】
本明細書において次の配列を開示する。
配列番号1 C1-3 VHをコードするヌクレオチド配列
配列番号2 C1-3 VHのアミノ酸配列
配列番号3 C1-3 VLをコードするヌクレオチド配列
配列番号4 C1-3 VLのアミノ酸配列
配列番号5 C1-3 リンカーをコードするヌクレオチド配列
配列番号6 C1-3 リンカーのアミノ酸配列
配列番号7 C1-3をコードするヌクレオチド配列
配列番号8 C1-3のアミノ酸配列
配列番号9 C1-3抗原
配列番号10 VHアミノ酸配列(配列番号2)内のC1-3 VH CDR1
配列番号11 VHアミノ酸配列(配列番号2)内のC1-3 VH CDR2
配列番号12 VHアミノ酸配列(配列番号2)内のC1-3 VH CDR3
配列番号13 VLアミノ酸配列(配列番号4)内のC1-3 VH CDR1
配列番号14 VLアミノ酸配列(配列番号4)内のC1-3 VH CDR2
配列番号15 VLアミノ酸配列(配列番号4)内のC1-3 VH CDR3
配列番号16 ヒト定常カッパドメイン(Hu Cκ)および6ヒスチジン残基精製タグの全アミノ酸配列(1文字コード)- NotIサイトの末端でL鎖フレームワーク4(LFW4)が終り、Hu Cκが始まる。
配列番号17 C1-3 VLをコードするヌクレオチド配列の初期バージョン
配列番号18 C1-3 VLのアミノ酸配列の初期バージョン
配列番号19 C1-3をコードするヌクレオチド配列の初期バージョン
配列番号20 C1-3のアミノ酸配列の初期バージョン
上記の配列において、図4におけるように、抗原結合部位を構成する超可変相補性決定領域(CDR)は太字で示している。HおよびL鎖に結合する柔軟なアミノ酸リンカー(Gly4、Ser)3は下線を引いてある(______)。フレームワーク領域(FW)の開始点およびヒト定常カッパ領域(Cκ)の開始点も示す。クローニング部位を下線(___)で示し、対応する制限酵素をアミノ酸配列の上に示した。
1つの態様において、本発明は、シナプトフィジンに結合し、C1-3 VHドメイン(配列番号2)および/またはC1-3 VLドメイン(配列番号4)を含む特異的結合要素を提供する。
【0010】
一般的に、VHドメインはVLドメインと対になって抗体の抗原結合部位を提供するが、以下にさらに論ずるように、VHドメインのみでも抗原に結合するために使用することができる。好ましい一実施形態においては、C1-3 VHドメイン(配列番号2)はC1-3 VLドメイン(配列番号4)と対になり、その結果抗体の抗原結合部位が、C1-3 VHおよびC1-3 VLドメイン両方を含んで形成される。他の実施形態においては、C1-3 VHはC1-3 VL以外のVLドメインと対になる。軽鎖の乱交は当業者においては十分に確かめられている。
【0011】
1つまたは複数のCDRをC1-3 VHまたはVLドメインから取って、適当なフレームワーク中に導入することができる。このことは後でさらに論ずる。C1-3 VH CDR1、2および3は、それぞれ配列番号10、11および12で示される。C1-3 VL CDR1、2および3は、それぞれ配列番号13、14および15で示される。
【0012】
配列を本明細書で提示した、シナプトフィジンに対する特異的結合体に使用できる変異体のVHおよびVLドメインは、配列の改変または変異およびスクリーニングという方法によって得ることができる。そのような方法もまた本発明によって提供される。
【0013】
本明細書で配列を具体的に開示するVHおよびVLドメインのいずれの可変領域アミノ酸配列の変異体も、説明するように本発明に従って使用することができる。特定の変異体には、1つまたは複数のアミノ酸配列の改変、おそらく20個未満、約15個未満、約10個未満または約5個未満、4、3、2または1個のアミノ酸配列の改変(アミノ酸残基の付加、欠失、置換および/または挿入)が含まれ得る。改変は1つまたは複数のフレームワーク領域および/または1つまたは複数のCDR中になすことができる。
【0014】
本発明による特異的結合要素は、抗原に結合することについて、任意の特異的結合要素と競合するものであってよく、これらのいずれも抗原に結合し、特異的結合要素、本明細書で開示したVHおよび/もしくはVLドメイン、または本明細書で開示したVH CDR3、またはこれらいずれの変異体をも含むものである。結合要素間の競合はin vitroで容易に測定することができ、例えばELISAを使用するか、および/または一方の結合要素、他方のタグ付けしていない結合要素の存在下に検出できる特異的なレポーター分子をタグ付けすることにより、同一のエピトープまたは重複するエピトープに結合する特異的結合要素の同定が可能になる。
【0015】
それ故、本発明のさらなる態様は、シナプトフィジンとの結合についてC1-3と競合する、ヒト抗体の抗原結合部位を含む特異的結合要素を提供する。
【0016】
シナプトフィジンに対する抗体を得るために、当業者は種々の方法を使用でき、その抗体はシナプトフィジンへの結合についてC1-3と競合し得るであろう。
【0017】
さらに他の態様において、本発明は抗原に結合し得る1つまたは複数の特異的結合要素を得る方法を提供する。該方法は本発明による特異的結合要素のライブラリーと前記抗原とを接触させて、前記抗原に結合し得るライブラリーの1つまたは複数の特異的結合要素を選択することを含む。
【0018】
好ましい実施形態において、特異的結合要素はアミノ酸配列YPFRLHQVYFDAPSC(配列番号9)内のエピトープに結合する。
【0019】
ライブラリーはバクテリオファージ粒子表面に提示されることがあり、各粒子は表面に提示された抗体VH可変ドメインをコードする核酸を含み、場合により提示されたVLドメインがあればそれも含む。
【0020】
前記抗原に結合できてバクテリオファージ粒子表面に提示された特異的結合要素の選択に続いて、前記選択された特異的結合要素を提示するバクテリオファージ粒子から核酸を取り出すことができる。そのような核酸は、それの後に続く、前記の選択された特異的結合要素を提示するバクテリオファージ粒子から取った核酸の配列を用いる核酸の発現による特異的結合要素または抗体VH可変ドメイン(場合によっては抗体VL可変ドメイン)の生産に使用できる。
【0021】
前記の選択された特異的結合要素の抗体VH可変ドメインのアミノ酸配列を有する抗体VH可変ドメインは、単離された形態で提供することができ、そのようなVHドメインを含む特異的結合要素も同様である。
【0022】
さらにシナプトフィジンに結合する能力を試験することができ、シナプトフィジンへの結合でC1-3と競争する能力も同様である。シナプトフィジンの作用に拮抗する能力は、後でさらに説明するようにして試験できる。
【0023】
本発明による特異的結合要素はC1-3の親和性によりシナプトフィジンに結合できる。好ましくは、特異的結合要素はシナプトフィジンのエピトープYPFRLHQVYFDAPSCに結合する。
【0024】
特異的結合要素はマウス、ラットおよび/またはヒトのシナプトフィジンに結合できる。好ましくは、特異的結合要素はヒトシナプトフィジンに結合する。
【0025】
異なる特異的結合要素の結合親和性および中和能力は、適当な条件で比較できる。
【0026】
抗体配列に加えて、本発明の特異的結合要素は、例えばペプチドもしくは折りたたまれたドメインなどのポリペプチドを形成する、または抗原に結合する能力に加えて他の機能的特性を分子に賦与する他のアミノ酸を含むことができる。
【0027】
本発明の特異的結合要素は、検出可能な標識を有することができ、または毒素もしくは酵素に(例えばペプチド結合またはリンカーにより)結合させることができる。
【0028】
当業者はタンパク質に分子を化学的に結合する多くの手法に通じている。特異的結合要素が医薬用のものであるとき、結合体の結合は、好ましくは、血液循環中では安定であるが、ひとたび結合体が細胞内に隔離されると変化しやすくなるものである。
【0029】
本発明の好ましい実施形態において、特異的結合要素は検出可能な、蛍光性標識のフルオレッセインイソチオシアネート(FITC)に結合させることができる。
【0030】
さらに他の態様において、本発明は、本発明による特異的結合要素、VHドメインおよび/またはVLドメインをコードする配列を含む単離した核酸を提供し、ならびに前記本発明による特異的結合要素、VHドメインおよび/またはVLドメインの生成を起こす条件下で前記核酸を発現させ、生成物を回収することを含む、本発明による特異的結合要素、VHドメインおよび/またはVLドメインを調製する方法を提供する。
【0031】
本発明による特異的結合要素は、ヒト患者に本発明の特異的結合要素の有効量を投与することを含む、前記患者における疾患または障害を治療(予防的治療を含む)する方法などのヒトまたは動物の体の治療または診断に使用できる。本発明により治療可能な疾患は、本明細書の他の箇所で説明するものを含む。
【0032】
本発明による特異的結合要素は、画像法、例えば、特異的結合要素が結合する細胞の存在または位置を決定する方法に使用できる。
【0033】
さらに他の態様において、本発明は、本発明による特異的結合要素および抗原に対する特異的結合要素の結合を測定する1つまたは複数の試薬を含む診断キットを提供する。
【0034】
本発明のさらに他の態様では、通常は単離された本明細書で開示した抗体VH可変ドメイン(配列番号1)および/またはVL可変ドメイン(配列番号3)をコードする核酸が、提供される。
【0035】
本発明の別の態様では、通常は単離された本明細書において開示したVH CDRまたはVL CDR配列、特に配列番号10、11および12から選択されたVH CDRまたは配列番号13、14および15から選択されたVL CDR、最も好ましくはC1-3 VH CDR3(配列番号12)をコードする核酸が、提供される。
【0036】
さらに他の態様では、本発明の核酸で形質転換した宿主細胞が提供される。
【0037】
さらに他の態様では、抗体VH可変ドメインを生成する方法が提供され、前記方法はコードする核酸からの発現を惹起することを含む。そのような方法は前記抗体VH可変ドメインの生成のための条件下で宿主細胞を培養することを含むことができる。
【0038】
VL可変ドメイン生成する類似の方法ならびにVHおよび/またはVLドメインを含む特異的結合要素が、本発明のさらに他の態様として提供される。
【0039】
製造方法は生成物の単離および/または精製のステップを含むことができる。
【0040】
製造方法は、生成物を、薬学的に許容可能な賦形剤などの少なくとも1つの追加成分を含む組成物に配合することを含むことができる。
【0041】
本発明のこれらのおよび他の態様を以下にさらに詳細に説明する。
【0042】
用語
特異的結合要素
この用語は、互いに結合する特異性を有する一対の分子の1つの構成要素を表す。特異的に結合する一対の構成要素は、天然に由来してもまたは全体もしくは一部が合成的に製造されてもよい。分子対の一方は、その表面または空孔に、分子対の他方の特定の空間的および極性的組織に特異的に結合し、かつそれ故相補的である区域を有する。このように、前記分子対の構成要素は互いに特異的に結合する性質を有する。特異的結合対の種類の例は、抗原-抗体、ビオチン-アビジン、ホルモン-ホルモン受容体、受容体-リガンド、酵素-基質である。本発明の応用は抗原-抗体タイプの反応に関係している。
【0043】
抗体分子
この用語は、天然または全体もしくは一部が合成的に製造された免疫グロブリンを表す。この用語はまた、抗体の結合ドメインを含む如何なるポリペプチドまたはタンパク質をも包含する。抗体結合ドメインを含む抗体フラグメントはFab、scFv、Fv、dAb、Fd;およびダイアボディなどである。
【0044】
モノクローナル抗体および他の抗体を取り、組換えDNA技法を使用して、元の抗体の特異性を保持する他の抗体またはキメラ分子を製造することが可能である。そのような技法は、免疫グロブリンの可変領域、すなわち抗体の相補性決定領域(CDR)をコードするDNAを、異なる免疫グロブリンの定常部、または定常部とフレームワーク領域とを合わせた領域に導入することを含むことができる。例えば、欧州特許A184187号、英国特許2188638A号または欧州特許A239400号を参照されたい。ハイブリドーマまたは抗体を生成する他の細胞は、生成された抗体の結合特異性が変化するかまたは変化しないことがある遺伝的変異または他の変化を受けやすいことがある。
【0045】
抗体は多くの方法で改変できるので、「抗体分子」という用語は、抗体の要求される特性を有する抗原結合ドメインを有する、如何なる特異的結合要素または物質をも包含すると解釈すべきである。このように、この用語は、天然のものか全体的もしくは一部的に合成であるかを問わず、免疫グロブリン結合ドメインを含む如何なるポリペプチドをも含む、抗体フラグメントおよび誘導体を包含する。それ故、他のポリペプチドに融合した、免疫グロブリンの結合ドメイン、またはその同等物を含むキメラ分子が含まれる。キメラ抗体のクローニングと発現は欧州特許A0120694号および欧州特許A0125023号に記載されている。
【0046】
全抗体のフラグメントが抗原に結合する機能を果たすことができることが示されている。結合するフラグメントの例は、(i)VL、VH、CLおよびCH1ドメインにより構成されるFabフラグメント;(ii)VHおよびCH1ドメインにより構成されるFdフラグメント;(iii)単一抗体のVLおよびVHドメインにより構成されるFvフラグメント;(iv)VHドメインにより構成されるdAbフラグメント(Ward、E.S.ら、Nature 341巻、544〜546頁(1989年));(v)単離したCDR領域;(vi)F(ab')2フラグメント、2つの連結したFabフラグメントにより構成される2価のフラグメント;(vii)2ドメインが共同して抗原結合部位を形成することを許容するペプチドリンカーによりVHドメインおよびVLドメインが連結した単鎖Fv分子(scFv)(Birdら、Science、242巻、423-426頁、1988年;Hustonら、PNAS USA、85巻、5879〜5883頁、1988年);(viii)二重特異性単鎖Fv2量体(国際出願PCT/US92/09965)および(ix)「ダイアボディ」、遺伝子融合により構築された多価または多重特異性フラグメント(国際公開WO94/13804;P.Holligerら、Proc.Natl.Acad.Sci.USA、90巻、6444〜6448頁、1993年)である。Fv、scFvまたはダイアボディ分子はVHおよびVLドメインを連結するジスルフィド架橋の導入により安定化できる(Y.Reiterら、Nature Biotech、14巻、1239〜1245頁、1996年)。CH3領域に接続したscFvを含むミニボディも作製できる(S.Huら、Cancer Res.、56巻、3055〜3061頁、1996年)。
【0047】
二重特異性抗体が使用されるべきところでは、これらは、種々の方法(Holliger,P.、Winter G.、Current Opinion Biotechnol.、4巻、446〜449頁(1993年))で作製され、例えば、化学的にまたは混成ハイブリドーマから調製できる従来の二重特異性抗体であってよく、あるいは上記の二重特異性抗体フラグメントのいずれであってもよい。ダイアボディおよびscFvは、Fc領域なしで、可変領域のみを使用して、抗イディオタイプ反応の効果を減少させる可能性をもって構築できる。
【0048】
二重特異性のダイアボディも、二重特異性の全抗体とは対照的に、大腸菌中で容易に構築および発現できるので、特に有用である。適当な結合特異性のダイアボディ(および抗体フラグメントなど多くの他のポリペプチド)は、ライブラリーからファージディスプレイ(国際公開W094/13804号)を使用して容易に選択できる。ダイアボディの一方のアームが、例えば、シナプトフィジンに対して向けられた特異性をもって、一定に保たれるべきであれば、その際には、他方のアームが変化したライブラリーを作製することができて、適当な特異性の抗体が選択される。二重特異性の全抗体は、窪みに突起を入れる技術(knobs-into-holes engineering)により作製できる(J.B.B.Ridgewayら、Protein Eng.、9巻、616〜621頁、1996年)。
【0049】
抗原結合ドメイン
この用語は、抗体分子の、抗原の一部または全部に特異的に結合しかつ相補的である区域を含む部分を表す。抗原が大きい場合、抗体は、エピトープと名付けられた部分にのみ結合できる。抗原結合ドメインは1つまたは複数の抗体可変ドメインにより提供される(例えば、VHドメインにより構成される、いわゆるFd抗体フラグメント)。好ましくは、抗原結合ドメインは抗体L鎖可変領域(VL)および抗体H鎖可変領域(VH)を含む。
【0050】
特異的
この用語は、特異的結合対の1構成要素が、その特異的結合の相手以外の分子に如何なる有効な結合も示さない状態を指すことに使用できる。この用語は、例えば抗原結合ドメインが、多くの抗原中に存在する特定のエピトープに対して特異的であるときにも適用でき、その場合には、抗原結合ドメインを担う特異的結合要素が、エピトープを担う種々の抗原に結合できるであろう。
【0051】
典型的には、抗原のリストを使用してELISAなどの結合測定により特異性を決定できる。本発明による特異的結合要素は肝星細胞のシナプトフィジンを認識できるが、神経細胞のものは認識しない。
【0052】
含む(comprise)
この用語は、一般的に「含む(include)」の意味で、すなわち1つまたは複数の特徴の存在を認める意味で使用する。
【0053】
単離した
この用語は、本発明の特異的結合要素、またはそのような結合要素をコードする核酸が、本発明に一般的に合致するであろうという状態を指す。結合要素および核酸は、それらの天然の環境で、またはそれらがin vitroもしくはin vivoで実施された組換えDNA技法により作製されたとき、それらが作製された環境(例えば細胞培養)で、共存していた他のポリペプチドもしくは核酸など元来付随していた物質を含まないかまたは実質的に含まないであろう。結合要素および核酸は希釈剤または補助剤と共に配合することができ、それでもなお実用目的にとっては単離された状態である。例えば、前記結合要素は、ゼラチンまたは他の担体が免疫測定で使用するためのマイクロタイタープレートのコーティングに使用されれば、当然これらと混合されるであろうし、あるいは診断または治療に使用されるときには、薬理学的に許容できる担体または希釈剤と混合されるであろう。特異的結合要素は、自然に、または異種真核細胞(例えばCHOもしくはNSO(ECACC 85110503)細胞)系によりグリコシル化されてもよく、あるいは(例えば原核細胞中での発現により生成されるなら)グリコシル化されなくてもよい。
【0054】
「実質的に述べたように」という表現により、本発明の関連するCDRまたはVHもしくはVLドメインが、本明細書において配列を述べた特定の領域と同一であるかまたは高度に類似であるかのいずれかであろうということを意味する。「高度に類似」という表現により、CDRおよび/またはVHもしくはVLドメインにおいて、1から5、好ましくは1から3または1または2、または3または4など1から4のアミノ酸置換がなされてよいということを予期している。
【0055】
本発明のCDRを担う構造は、通常抗体HもしくはL鎖配列の、または、それらの実質的部分の構造、すなわちそれらの中で再配列免疫グロブリン遺伝子のコードする天然のVHおよびVL抗体可変ドメインのCDRに対応する位置に前記CDRが位置するそれらの実質的部分の構造であろう。免疫グロブリン可変ドメインの構造および位置は、Kabat、E.A.ら、「Sequences of Proteins of Immunological Interest」 第4版、US Department of Health and Human Services、1987年、および現在インターネット上で使用できるその更新版(http://immuno.bme.nwu.eduまたは任意の検索エンジンを使用して「Kabat」を見出されたい)を参照して決定できる。
【0056】
本発明で使用する可変ドメインは、任意の生殖細胞系または再配列ヒト可変ドメインから得ることができ、あるいは知られているヒト可変ドメインの共通配列に基づく合成可変ドメインであってもよい。本発明のCDR配列(例えばCDR3)を、組換えDNA技法を使用してCDR欠損可変ドメインのレパートリーに導入することができる。
【0057】
例えば、Markら(Bio/Technology、1992年、10巻、779〜783頁)は抗体可変ドメインのレパートリーを作製する方法を述べていて、その方法では、可変ドメイン区域の5'末端に向けられたまたは隣接した共通プライマーが、ヒトVH遺伝子の第3フレームワーク領域への共通プライマーと共に使用されて、CDRを欠損するVH可変ドメインのレパートリーが提供される。Markらは、いかにしてこのレパートリーを特定の抗体のCDR3と組み合わせられるかをさらに述べている。類似の技法を使用して本発明のCDR3由来の配列をCDR3の欠損するVHまたはVLドメインのレパートリーと入れ替えることができ、入れ替えた完全なVHまたはVLドメインを同属のVHまたはVLドメインと組み合わせて、本発明の特異的結合要素を提供する。それから前記レパートリーを、WO92/01047のファージディスプレイ系などの適当な宿主系で提示させて、その結果適当な特異的結合要素が選択される。レパートリーは、104以上の独立の構成要素のいずれか、例えば、106から108または1010の構成要素により構成され得る。
【0058】
類似の入れ替えまたは組合せ技法は、Stemmerによっても開示されており(Nature、1994年、370巻、389〜391頁)、彼はβ-ラクタマーゼ遺伝子に関する技法を述べているが、この手法が抗体の生成のために使用できることを認めている。
【0059】
さらに他の別法は、全可変ドメイン内に変異を生じさせるために、1つまたは複数の選択されたVHおよび/またはVL遺伝子のランダム変異誘発を使用して、本発明のCDR由来配列を担う新規なVHまたはVL領域を生成させる。そのような技法は、Gramらによって述べられており(1992年、Proc.Natl.Acad.Sci.、USA、89巻、3576〜3580頁)、彼らは誤りがちなPCRを使用した。
【0060】
使用してよい他の方法は、VHまたはVLの遺伝子CDR領域に定方向突然変異誘発を起こさせることである。そのような技法はBarbasら(1994年、Proc.Natl.Acad.Sci.、USA、91巻、3809〜3813頁)およびSchierら(1996年、J.Mol.Biol.、263巻、551〜567頁)によって開示されている。
【0061】
上記の技法はすべて当業者に知られたものであり、それら自体では本発明の一部を形成しない。当業者は、当業者に定常的な手法を使用して本発明の特異的結合要素を提供するためにそのような技法を使用できるであろう。
【0062】
本発明のさらに他の態様は、シナプトフィジンのエピトープYPFRLHQVYFDAPSCに特異的な抗体の抗原結合ドメインを得る方法を提供し、その方法は、本明細書で述べたVHドメインのアミノ酸配列中の1つまたは複数のアミノ酸の付加、欠失、置換または挿入という手段により、VHドメインのアミノ酸配列変異体であるVHドメインを提供すること、場合によりこのようにして提供されたVHドメインを1つまたは複数のVLドメインと組み合わせること、およびシナプトフィジンに対して特異的な特異的結合要素または抗体の抗原結合ドメインを同定するためにVHドメインまたはVH/VLの組合せを試験することを含む。前記VLドメインは本明細書で実質的に述べたアミノ酸配列を有することができる。
【0063】
本明細書で開示したVLドメインの1つまたは複数の配列変異体を1つまたは複数のVHドメインと組み合わせる類似の方法が使用できる。
【0064】
本発明のさらに他の態様は、シナプトフィジンに対して特異的な特異的結合要素を調製する方法を提供し、その方法は、
(a)置換すべきCDR3を含むかまたはCDR3をコードする領域を欠くVHドメインをコードする核酸の出発レパートリーを提供すること、
(b)前記レパートリーと本明細書でVH CDR3のものとして実質的に述べたアミノ酸配列をコードするドナー核酸とを組み合わせ、その結果前記ドナー核酸がレパートリーのCDR3領域に挿入されて、VHドメインをコードする核酸の生成物レパートリーを提供すること、
(c)前記生成物レパートリーの核酸を発現させること、
(d)シナプトフィジンに対して特異的な特異的結合要素を選択すること、および
(e)前記特異的結合要素またはそれをコードする核酸を回収すること
を含む。
【0065】
この場合も同様に、本発明のVL CDR3と、置換すべきCDR3を含むかまたはCDR3をコードする領域を欠くVLドメインをコードする核酸のレパートリーとを組み合わせる類似の方法を使用することができる。
【0066】
同様に、1つまたは2つ以上、またはすべての3つのCDRをVHまたはVLドメインのレパートリーに移植でき、次にそれらをシナプトフィジンに対する特異的結合要素としてスクリーニングする。
【0067】
免疫グロブリンの可変ドメインの実質的部分は、少なくともその3つのCDR領域を、それらの間に介在するフレームワーク領域と共に含むであろう。好ましくは、前記部分も第1および第4フレームワーク領域のどちらかまたは両方の少なくとも約50%を含むであろう。その50%は第1フレームワーク領域のC末端50%、および第4フレームワーク領域のN末端50%である。可変ドメインの実質部分のN末端またはC末端に付加する残基は、天然の可変ドメイン区域とは本来なら関係がないものでよい。例えば、組換えDNA技法によりなされた本発明の特異的結合要素の構築は、クローニングまたは他の操作段階を容易にするために導入されたリンカーによりコードされるN-またはC-末端残基を導入する結果になる。他の操作段階には、本発明の可変ドメインを免疫グロブリンH鎖、他の可変ドメイン(例えばダイアボディの生成における)または以下でさらに詳細に論ずるタンパク質標識を含むさらなるタンパク質配列に接続するためのリンカーの導入が含まれる。
【0068】
本発明の好ましい態様において、VHおよびVLドメインの一対を含む特異的結合要素が好ましいが、VHまたはVLドメイン配列のいずれかに基づく単結合ドメインは、本発明のさらに他の態様を形成する。単免疫グロブリンドメイン、特にVHドメインは特異的に標的抗原に結合し得ることが知られている。
【0069】
単鎖特異的結合ドメインのいずれの場合においても、これらのドメインは、シナプトフィジンに結合できる2ドメイン特異的結合要素を形成し得る相補性ドメインをスクリーニングするために使用できる。
【0070】
これは国際公開WO92/01047に開示された、いわゆる階層的二重コンビナトリアル手法を使用するファージディスプレイスクリーニング法によって達成できる。前記スクリーニング法では、HまたはL鎖クローンのいずれかを含む個々のコロニーが他鎖(LまたはH)をコードするクローンの全ライブラリーを感染させるために使用され、その結果生ずる2鎖特異的結合要素が、前記文献に記載されたようなファージディスプレイ技法に従って選択される。この技法はMarkらの前掲の文献にも開示されている。
【0071】
本発明の特異的結合要素はさらに抗体定常部またはその部分を含むことができる。例えば、VLドメインはそのC末端で、ヒトCκまたはCλ鎖、好ましくはCκを含む抗体L鎖の定常ドメインに結合できる。同様に、VHドメインに基づく特異的結合要素はそのC末端で、如何なる抗体アイソタイプ、例えばIgG、IgA、IgEおよびIgMならびに任意のアイソタイプのサブクラスに由来する免疫グロブリのH鎖の全体または部分に結合できる。WO99/58572に開示されたΔnabおよびΔnacなどのFc領域が使用できる。
【0072】
本発明の特異的結合要素は検出可能なまたは機能性の標識で標識できる。検出可能な標識には、131Iまたは99Tcなどの放射性標識が含まれ、それらは抗体イメージングの当業者に知られた常套的な化学的方法を使用して抗体に付けることができる。標識には、西洋ワサビペルオキシダーゼなどの酵素標識も含まれる。標識にはさらに、特異的同族検出可能成分例えば標識アビジンへの結合により検出できるビオチンなどの化学的成分が含まれる。好ましくは、標識はFITCなどの蛍光性標識を含む。
【0073】
本発明の特異的結合要素は、ヒトまたは動物の患者、好ましくはヒトの診断または治療方法に使用されるようにデザインされる。
【0074】
したがって、本発明のさらに他の態様は、提供された特異的結合要素をそのままで、または例えばFITCなどの検出可能な標識に結合した1つまたは複数の他の試薬と共に投与することを含む診断方法を提供する。提供された特異的結合要素は、生検組織由来の肝線維症細胞の迅速かつ信頼性ある検査の開発に使用できる。
【0075】
本発明のさらに他の態様は、提供された特異的結合要素の投与を含む治療方法、そのような特異的結合要素を含む薬剤組成物、および投与のための医薬の製造における、例えば薬理学的に許容できる賦形剤と共に特異的結合要素を配合することを含む医薬または薬剤組成物の製法におけるそのような特異的結合要素の使用を提供する。
【0076】
肝星細胞に対する抗体を使用して治療に役立ち得る臨床的適応症には、肝線維症の結果病気になる如何なる疾患、例えばウィルス感染例えば肝炎およびアルコール乱用などの肝疾患も含まれる。提供された特異的結合要素はそのままヒト抗星細胞抗体または抗体様構造の受動免疫による肝線維症の直接的治療にも使用できる。
【0077】
本発明による抗線維症治療は肝線維症を有する患者にとって明らかな恩恵を提供するために使用できる。抗線維症治療は注射(例えば静脈に)または局所送達法によって与えることができる。提供された特異的結合要素は薬剤組成物の送達を標的肝星細胞へ向かわせるために使用できる。
【0078】
別の配合戦略は、経口または座剤経路に適した製剤を提供できる。投与経路は、治療剤の物理化学的性質により、疾患に対する特別の考慮により、有効性を最適化するように、または副作用を最小化するように決定することができる。
【0079】
本発明に従い、提供された組成物は個々人に投与できる。投与は好ましくは「治療的有効量」で行われ、治療的有効量は患者の利益になるのに十分な量である。そのような利益とは、少なくとも1つの症状の少なくとも1つ改善があればよい。実際に投与する量、および投与の速度と時間経過は治療されているものの性質および重症度に依存するであろう。治療の処方、例えば、投与量の決定その他は一般診療医およびその他の医師の責任範囲になる。抗体の適当な投与量は当業者にはよく知られている。Ledermann J.A.ら、(1991年)、Int.J.Cancer 47巻、659〜664頁、Bagshawe K.D.ら、(1991年)、Antibody,Immunoconjugates and Radiopharmaceuticals 4巻、915〜922頁を参照されたい。
【0080】
正確な投与量は、抗体が診断用か治療用か、治療すべき区域のサイズおよび位置、抗体の詳細な性質(例えば全抗体かフラグメントかダイアボディか)、および何らかの抗体に付けた検出可能な標識または他の分子の性質を含む多くの要因に依存するであろう。典型的な抗体投与量は0.5mg〜1.0gの範囲にあり、静脈への大量瞬時投与として投与できる。他の様式の投与には、同様の累積投与量を達成するために数時間かける静脈注射が含まれる。これは成人患者の単回治療のための投与量であって、少児および幼児のためには比例的に調節してよく、また他の抗体型に対しても分子量に比例して調節できる。治療は、医師の判断で、毎日、週2回、毎週または毎月の間隔で繰り返すことができる。
【0081】
他の様式の投与は、留置するデバイスのプレコートか、そうでなければその中への取込みを使用するもので、そのための最適抗体量は適当な実験によって決定されるであろう。
【0082】
本発明のある好ましい実施形態における抗体分子は、F(ab)またはscFvなどの単量体フラグメントである。そのような抗体フラグメントは半減期が比較的短いという利点を有する。
【0083】
本発明の特異的結合要素は、通常、薬剤組成物の形態で投与されるもので、特異的結合要素に加えて少なくとも1つの成分を含むことができる。
【0084】
したがって、本発明による、および本発明による使用のための薬剤組成物は、活性成分に加えて薬理学的に許容できる賦形剤、担体、緩衝剤、安定剤または当業者によく知られた他の材料を含むことができる。そのような材料は、無毒性であるべきで、かつ活性成分の有効性に干渉すべきでない。担体または他の材料の詳細な性質は投与経路に依存するであろう。投与経路は経口または注射、例えば静脈注射によるものでもよい。
【0085】
経口投与のための薬剤組成物は、錠剤、カプセル、粉末または液体の形態であってよい。錠剤はゼラチンなどの固体担体または助剤を含むことができる。液状薬剤組成物は通常、水、石油、動物もしくは植物油、鉱油または合成油などの液体担体を含む。生理食塩水、デキストロースまたは他の糖類溶液またはエチレングリコール、プロピレングリコールもしくはポリエチレングリコールなどのグリコールが含まれてよい。
【0086】
静脈注射または患部への注射のためには、活性成分は、発熱物質を含まず適当なpH、等張性および安定性を有する非経口的に許容できる水溶液の形態であろう。当業者は、例えば、塩化ナトリウム注射液、リンゲル注射液、乳酸リンゲル注射液などの等張溶媒を使用して適当な溶液を十分に調製できる。保存料、安定化剤、緩衝剤、抗酸化剤および/または他の添加剤が必要に応じて含まれてよい。
【0087】
組成物は、治療する疾患に依存して、単独でまたは他の治療と同時にもしくは順次に併用して投与できる。他の治療には、非ステロイド抗炎症薬(例えばアスピリン、イブプロフェンもしくはケトプロフェン)などの鎮痛薬またはモルヒネなどのアヘン剤、または抗嘔吐剤の適当な投与量の投与が含まれる。
【0088】
本発明は、本明細書で提供したシナプトフィジンに対する特異的結合要素の結合を惹起するかまたは許容することを含む方法を提供する。指摘したように、そのような結合はin vivoで、例えば、特異的結合要素、または特異的結合要素をコードする核酸の投与に続いて起こることができ、あるいはin vitroで、例えば、ELISA、ウェスタンブロット、免疫細胞化学、免疫沈降またはアフィニティークロマトグラフィーで起こることができる。
【0089】
特異的結合要素のシナプトフィジンに対する結合の量を測定することができる。定量は試験試料中の抗原量に関係づけることができ、試験試料は診断上興味があるものであってよい。
【0090】
抗体の試料への反応性は何らかの適当な手段によって測定できる。放射免疫測定(RIA)は1つの可能性である。放射活性標識抗原を非標識抗原(試験試料)と混合して、抗体に結合させる。結合していない抗原から結合した抗原を物理的に分離して、抗体に結合した放射性抗原の量を測定する。試験試料中に抗原が多くあればあるほど、抗体に結合する放射性抗原は少なくなるであろう。競争的結合測定はまた、レポーター分子に結合した抗原または類似体を使用して、非放射性抗原でも使用できる。前記レポーター分子は、蛍光体、リン光体または分光的に分離した吸収もしくは発光特性を有するレーザー色素であることができる。適当な蛍光体には、フルオレッセイン、ローダミン、フィコエリトリンおよびテキサスレッドが含まれる。適当な発色色素にはジアミノベンジジンが含まれる。
【0091】
他のレポーターには、着色しているか、磁性もしくは常磁性である高分子コロイド粒子もしくはラテックスビーズなどの粒子状材料、および視覚的に観察されるか、電子的に検出されるかもしくは他の方法で記録される検出可能なシグナルを直接的もしくは間接的に生じさせ得る、生物学的にまたは化学的に活性な作用物質が含まれる。これらの分子は、例えば、色を発せさせるかもしくは変化させるかまたは電気的性質に変化を起こさせる反応の触媒になる酵素であってよい。それらは分子的に励起可能であることができ、そのために電子のエネルギー状態間遷移の結果が特徴的な分光学的吸収または発光を生ずる。それらはバイオセンサーと併用する化学的要素を含むことができる。ビオチン/アビジンもしくはビオチン/ストレプトアビジンおよびアルカリホスファターゼ検出系が使用できる。
【0092】
個々の抗体-レポーター結合体により生じたシグナルは、試料中(正常試料および被検試料)における関連抗体結合の測定可能な絶対または相対データを引き出すのに使用できる。
【0093】
本発明は競争的測定において抗原レベルを測定するための上記のような特異的結合要素の使用、すなわち競争的測定において本発明により提供される特異的結合要素を使用することによる試料中の抗原レベルの測定方法も提供する。これは、結合していない抗原からの結合している抗原の物理的単離が必要ない場合であるだろう。結合したときに物理的または光学的変化が起こるようにレポーター分子を特異的結合要素に連結することは、1つの可能性である。前記レポーター分子は、検出可能な、好ましくは測定可能なシグナルを直接的または間接的に生ずることができる。レポーター分子の連結は直接的でもまたは間接的でも、共有結合でも、例えばペプチド結合でもまたは非共有結合でもよい。ペプチド結合による連結は、抗体およびレポーター分子をコードする遺伝子融合の組換え発現の結果として可能である。
【0094】
本発明は、例えばバイオセンサー系において本発明による特異的結合要素を使用することにより抗原レベルを直接的に測定することも提供する。
【0095】
結合を測定する様式は本発明の特徴ではなく、当業者はその好みおよび一般的知識により適当な様式を選択できる。
【0096】
本発明はさらに、シナプトフィジンへの結合に対して如何なる特異的結合要素にも劣らない、抗原に結合しかつ本明細書で実質的に開示したアミノ酸を有するCDRを含むVドメインまたは本明細書で実質的に開示したアミノ酸配列を有するVドメインを含む特異的結合要素に及ぶ。結合要素間の競争は、例えば、タグを付けていない一方の結合要素の存在下で検出できる特異的レポーター分子を、タグとして他方の結合要素に付けて、同一のエピトープまたは重複しているエピトープに結合する特異的結合要素の同定を可能にすることにより、in vitroで容易に測定できる。競争は例えばELISAまたはフローサイトメトリーを使用して測定できる。
【0097】
競争の試験において、抗原のペプチドフラグメント、特に興味のあるエピトープを含むペプチドを使用できる。エピトープ配列に加えて1つまたは複数のアミノ酸をいずれかの末端に有するペプチドが使用できる。そのようなペプチドは特定された配列により「本質的に構成されている」といわれる。本発明による特異的結合要素は、それらの抗原に対する結合が、特定の配列を有するかまたは含むペプチドによって阻止されるようなものである。これを試験するために、1つまたは複数のいずれかのアミノ酸を加えた配列を有するペプチドが使用できる。
【0098】
特定のペプチドに結合する特異的結合要素を、例えば、ファージディスプレイライブラリーから前記ペプチドでのパニングにより単離できる。
【0099】
本発明はさらに、本発明の特異的結合要素をコードする単離した核酸を提供する。核酸はDNAおよびRNAを含む。好ましい態様において、本発明は上で定義した本発明のCDR、VHまたはVLドメインをコードする核酸を提供する。
【0100】
本発明はまた、構築体を、上記の少なくとも1つのポリヌクレオチドを含むプラスミド、ベクター、転写または発現カセットの形態で提供する。
【0101】
本発明はまた、上記の1つまたは複数の構築体を含む組換え宿主細胞も提供する。任意のCDR、VHまたはVLドメインをコードする核酸または提供された特異的結合要素それ自体が本発明の態様を形成し、コードされた生成物の製造方法も同様であって、その方法はその生成物をコードする核酸からの発現を含む。発現は核酸を含む組換え宿主細胞を適当な条件下で培養することにより簡便に達成できる。発現による生成に続いて、VHもしくはVLドメインまたは特異的結合要素を、任意の適当な技法を使用して単離および/または精製し、それから適宜使用する。
【0102】
本発明による特異的結合要素、VHおよび/またはVLドメイン、およびコードする核酸分子およびベクターは、例えばそれらの自然の環境から、実質的に純粋なまたは均質な形態で単離しおよび/または精製して、あるいは核酸の場合には、求められている機能を有するポリペプチドをコードする配列以外に原料由来の核酸または遺伝子を含まずにまたは実質的に含まずに提供できる。本発明による核酸はDNAまたはRNAを含むことができ、かつ、全体または部分が合成によることができる。本明細書で述べたヌクレオチド配列への言及は、特に断らない限り、前記の特定した配列を有するDNA分子を包含し、前記特定した配列においてTの代わりにUで置換したRNA分子をも包含する。
【0103】
種々の異なった宿主細胞におけるポリペプチドのクローニングおよび発現のための系がよく知られている。適当な宿主細胞には、細菌、哺乳動物細胞、酵母およびバキュロウイルス系が含まれる。異種ポリペプチドの発現に当業者の使用できる哺乳動物細胞ラインには、チャイニーズハムスター卵巣細胞、HeLa細胞、仔ハムスター腎細胞、NSOマウス黒色腫細胞、YB2/0ラット骨髄腫細胞およびその他多くのものが含まれる。一般的で、好ましい細菌宿主は大腸菌である。
【0104】
大腸菌などの原核細胞における抗体および抗体フラグメントの発現は、当業者においては十分確立されている。総説としては、Pluckthun、A.、Bio/Technology 9巻、545〜551頁(1991年)を参照されたい。培養における真核細胞中の発現も、特異的結合要素の生成のための選択肢として当業者に使用できる。最近の総説として、例えば、Ref、M.E.、(1993年)Curr.Opinion Biotech.4巻、573〜576頁、Trill J.J.ら、(1995年)Curr.Opinion Biotech 6巻、553〜560頁を参照されたい。
【0105】
適当なベクターは、プロモーター配列、ターミネーター配列、ポリアデニル化配列、エンハンサー配列、マーカー遺伝子および他の配列を適宜含む適当な制御配列を含んで、選択または構築できる。ベクターは適宜、プラスミド、ウィルス性の例えばファージまたはファージミドにすることができる。「Molecular Cloning:a Laboratory Manual」第3版、SambrookおよびRussell、2001年、Cold Spring Harbor Laboratory Press。核酸の操作、例えば、核酸構築体の調製、変異誘発、塩基配列決定、DNAの細胞への導入および遺伝子発現、およびタンパク質の分析のために多くの知られた技法およびプロトコルが、「Current Protocols in Molecular Biology」第2版、Ausubelら(編)、John Wiley & Sons、(1992年)に詳細に説明されている。
【0106】
したがって、本発明のさらなる態様は、本明細書で開示したように核酸を含む宿主細胞を提供する。さらに他の態様は、そのような核酸を宿主細胞へ導入することを含む方法を提供する。前記導入には如何なる使用可能な技法も使用できる。真核細胞に対して、適当な技法は、リン酸カルシウムトランスフェクション、DEAE-デキストラン、エレクトロポレーション、リポソーム媒介トランスフェクションおよびレトロウィルスまたは他のウィルス例えばワクシニアまたは昆虫細胞に対してはバキュロウイルスを使用する形質導入を含むことができる。細菌細胞に対して、適当な技法は、塩化カルシウム形質転換、エレクトロポレーションおよびバクテリオファージを使用するトランスフェクションを含むことができる。
【0107】
前記導入に続いて、例えば、遺伝子の発現のための条件下で宿主細胞を培養することにより、核酸の発現を起こさせるまたは起こすに任せることができる。
【0108】
一実施形態において、本発明の核酸は宿主細胞のゲノム(例えば染色体)中に組込まれる。組込みは、標準的技法に従って、ゲノムとの組換えを促進する配列を含めることにより促進することができる。
【0109】
本発明は、上記の特異的結合要素またはポリペプチドを発現させる目的で、発現系に上で述べたような構築体を使用することを含む方法も提供する。
【0110】
本発明の態様および実施形態を、ここで次の実験を参照して実施例により例示することにする。
【0111】
本明細書において如何なる箇所で引用した文献もすべて参照により組み込む。
【発明を実施するための最良の形態】
【0112】
材料および方法
ペプチド-ウシ血清アルブミン結合体
シナプトフィジンタンパク質(図1)の外形質側に存在するアミノ酸に対応するペプチドYPFRLHQVYFDAPSCをFmoc法の使用により合成し(Proteomics Group、University of Aberdeen)、HPLCにより純粋と判定した。ペプチドの分子量は質量分析法によって確かめた(データは示していない)。前記ペプチドを、3-マレイミド酢酸N-ヒドロキシスクシンイミド(MBS)を使用して、ウシ血清アルブミンに化学的に結合した(Leel V.ら、Biochem Biophys Res Commun 2004年、316巻、872〜7頁)。SDS-PAGEおよび質量分析法による分析(MALDI-TOF、Applied Biosystems Voyager-DE STR)により前記ペプチドがBSAに結合したことを確認した。一般に平均して、アルブミン1分子当りおよそ4ペプチド分子であった(データは示していない)。
【0113】
ペプチド配列に対する可溶性単鎖抗体フラグメント(scAb)のファージディスプレイ生成
ヒト単倍(single-fold)scFvファージミド(pIT2)ライブラリー(Tomlinson I + J、MRC、UK)を、本質的にはLeel V.らがBiochem Biophys Res Commun 2004年、316巻、872〜7頁で概略を説明しているようにして、ペプチド配列に特異性のあるファージ抗体をスクリーニングするために使用した。ペプチド配列に対してパニングを合計3回実施した。ペプチドへの特異性のあるファージ抗体のBSAに対する比を上げるために、パニングを2%(w/v)ドライミルク(MPBS)および2ml/mlBSAを含有する4mlのPBS(137mM NaCl、2.7mM KC1、10mMリン酸塩、pH7.4)中で実施した。
【0114】
抗ペプチドファージ抗体ELISA
PBS中のタンパク質(peptide-BSAまたはBSA)2μg/ウェルを4℃で一晩かけてコートした平底96ウェルのImmulon-4マイクロタイタープレート(Dynex、Sussex、UK)を使用して、ペプチドに対する結合要素を増やした精製ポリクローナルファージをELISAにより測定した。結合したファージを、100μl/ウェルの西洋ワサビペルオキシダーゼ(HRP)結合抗M13抗体(AmershamPharmacia)と1時間インキュベートし、洗浄して、また100μl/ウェルのテトラメチルベンジジン二塩酸塩溶液(KPL Laboratories、Gaithersburg、メリーランド州)とインキュベートすることにより検出した。反応を50μl/ウェルの1M H2SO4で停止して、450nmの吸収をマイクロプレートリーダーを使用して読み取った。
【0115】
ペプチドに対するモノクローナルファージ抗体の同定および発現ベクターpIMS147中へのサブクローニング
第3パニングからの個々のコロニーを96ウェルのプレート(Greiner)中で成長させ、M13 K07でファージ抗体を回収した。
【0116】
ペプチド-BSAおよびBSA単独に対する結合について、ファージ上清の特異性をELISAにより測定した。陽性クローンのscFvコード領域を発現ベクターpIMS147中にサブクローニングした。これは、IPTG(イソプロピル-β-D-チオガラクトピラノシド)誘導pUC系ベクターpIMS100(Hayhurst A.ら、Protein Expr Purif 1999年、15巻、336〜43頁)の改質である。ヘキサヒスチジンのタグ付加により、固定化金属イオンキレートアフィニティークロマトグラフィーによる、発現したscAbの精製が可能になる。ベクターは大腸菌 TG1またはXL 1ブルー細胞に形質転換した。
【0117】
scAbの発現、精製およびキャラクタリゼーション
Leel V.ら、Biochem Biophys Res Commun 2004年、316巻、872〜7頁で概略が説明されているようにして、IPTG処理した細胞中でscAbを発現させ、Ni2+負荷IMACFast Flow Sepharose樹脂(AmershamPharmacia)をメーカーの指示に従って使用して、ヘキサヒスチジンC末端タグ尾部により精製した。PBSに対して4℃で十分透析した後、scAbを-20℃で保存した。精製したscAbは、Leel V.ら、Biochem Biophys Res Commun 2004年、316巻、872〜7頁で概略が説明されているようにして、全ヒトIgG測定用標準品を使用して、ヒトCκドメインによる捕捉ELISAによって測定した。
【0118】
発現し精製された抗ペプチドscAB ELISA
発現したscAbは、本質的にはポリクローナル抗ペプチドファージ抗体ELISAに対して概略を説明したようにして、ペプチドと相互作用する能力をELISAにより試験した。結合したscAbは、西洋ワサビペルオキシダーゼ結合抗ヒトCκL鎖抗体(Sigma Chem.Co.Poole、UK)を使用して検出した。
【0119】
SDS-PAGEおよびウェスタンブロット
ウェスタンブロットは、本質的にはWright M.C.ら、Mol Pharmacol 1996年、50巻、856〜63頁に述べられているようにして実施した。
【0120】
タンパク質のFITC標識
ScAbは、FluoroporterFITCタンパク質標識キット(Molecular Probes)を、メーカーの指示に従って使用して、蛍光で標識した。この方法は、トリブチル錫イソチオシアナート(Aldrich Chemicals、Poole、UK)を使用して、C1-3scAbをトリブチル錫に結合させるためにも使用した。
【0121】
動物および細胞の調製
ヒト肝細胞はUK Human Tissue Bank(DeMontfort University、Leicester、UK)から入手した。ヒトHSCは腫瘍が存在するという理由で患者から除去した肝臓の正常組織の縁から単離された。これらの研究におけるヒト組織の使用はGrampian Regional Ethics Committeeにより許可され、供与者の全面的同意を得た。ヒト肝細胞培養のプロトコルはWright M.C.ら、Hum Exp Toxicol、1996年、15巻、203〜4頁およびHarvey J.L.ら、Drug Metab Dispos、2000年、28巻、96〜101頁に掲載されている。ラットHSCおよびB-13細胞の培養はWright M.C.ら、Gastroenterology、2001年、121巻、685〜98頁およびMarek C.J.ら、Biochem J、2003年、370巻、763〜9頁に掲載されている。
【0122】
FITC標識scAbの細胞とのインキュベーション
FITC-C1-3scAbを、蛍光顕微鏡により、培養中の肝星細胞と相互作用する能力について試験した。ヒト肝星細胞は、実験に先立って完全培地中のチャンバースライド(Nunclon、Naperville)上に播種した(チャンバー当り1×103細胞)。細胞はリン酸緩衝生理食塩水(PBS)で2度洗浄し、10μgのscABまたはFITC-C1-3scAbを含有する500μlのHepes/HBSS緩衝液(0.14M NaCl、5.4mM KC1、0.34mM Na2HPO4、0.44mM KH2PO4、5.6mMグルコース、1mM CaCl2、6mM HEPES、4mM NaHCO3 pH7.4)中、室温暗所で1時間インキュベートした。次いで、細胞をPBSで洗浄し、4',6'-ジアミジノ-2-フェニルインドール(DAPI)を含有するVectashield封入剤(Vector laboratories、Burlingame、カリフォルニア州)で顕微鏡標本を作製して、細胞核を染色しカバーガラスをかけた。顕微鏡観察はPhotometrics Sensisカメラ付きZeiss Axeola II顕微鏡を使用して実施し、データはSmart Captureプログラムを使用して取り込んだ。ヒト肝細胞(UK Human Tissue Bank)は、最初コラーゲンをコートした24ウェルプレートで培養し(1×105/ウェル)、80単位/mlのペニシリンおよび80μg/mlのストレプトマイシンを補充したWilliamの培地E中に維持した。
【0123】
共培養実験のためには、肝星細胞を肝細胞培養物(ウェル当り1×103細胞)中にシードして、実験に先立って24時間培養した。純肝星細胞の培養について簡単に説明したのと同様にして、C1-3またはFITC-C1-3scAbをインキュベートし、1時間のインキュベーションおよび洗浄後、細胞をトリプシン処理および1000rpmで5分間遠心することにより回収した。次に細胞ペレットを0.4%ホルムアルデヒド/106細胞で再懸濁して、細胞遠心およびDAPI含有Vectashieldで顕微鏡標本を作製することによりガラススライドに付けた。スライドは前記のようにして蛍光顕微鏡により観察した。
【0124】
DAPI染色は核のFITC-C1-3染色を隠蔽するので、FITC-C1-3のHSCへの結合に対するププチドおよびモネンシンの影響を比較するために、細胞固定なしの蛍光顕微鏡観察を使用した。24ウェルプレートにシードしたHSCをHepes/HBSSで洗浄してから、l0μgのscAbを含有し添加化合物を含むかまたは含まない500μl/well Hepes/HBSS中37℃でインキュベートした。1時間後、細胞を500μlのHepes/HBSSで3回洗浄した。フィルターセット#9(BP 450〜490nm;LP 515nm)を使用するAxiovert100顕微鏡(Zeiss、ドイツ)を使用して細胞を分析した。
【0125】
FACS分析
FITC-C1-3scAbを培養液に加え(200μg/10m1)、HSCの集密的な(直径10cmのシャーレ)培養物と共に37℃で1時間インキュベートした。次に媒質を除去して細胞をPBSで洗浄した。次に細胞単離溶液(Sigma、Poole、UK)を使用して細胞を非酵素的に培養基体から剥離し、遠心によりペレット化して、BD cytofix/cytoperm(BD Biosciences、Oxford、UK)を使用して固定/透過処理した。15分後、細胞を0.5mlの1×wash/perm buffer(BD Biosciences、Oxford、UK)で洗浄し、l×wash/perm bufferで希釈したマウス抗α-平滑筋アクチンモノクローナル抗体(Sigma、Poole、UK)とインキュベートした。次に細胞をl×wash/perm bufferで洗浄し、ビオチン結合抗マウスIgG(1:200、DakoCytomatron、Cambridgeshire、UK)と氷上で30分間インキュベートし、2回洗浄してから、ストレプトアビジン-アロフィコシアニン(APC)結合体(1:200、BD Biosciences、Oxford、UK)と氷上で30分間インキュベートした。細胞はフローサイトメトリー(LSR I、BD、Oxford、UK)で分析した。細胞をアイソタイプ特異的対照抗体とインキュベートすることにより、非特異的蛍光の対照とした。
【0126】
結果
標的ペプチドYPFRLHQVYFDAPSCに対する親和性のあるTomlinson I+Jライブラリーから組換えファージを探索するために、ファージディスプレイの技法を使用した。図2は標的ペプチドに親和性のあるファージ抗体が選択的に免疫チューブに結合し、3回の別々の探索(pan)を通して増幅されたことを示す。第3パニングライブラリーからの96の別々のクローンを、BSAおよび標的ペプチド-BSAに結合するファージ抗体を求めて、個々にスクリーニングした。この測定においては、57クローンが標的ペプチド-BSAに対する高い親和性を示し、BSAに対して何らかの親和性を示したクローンはなかった(データは示していない)。モノクローナルファージ-抗体ELISAにおいて最高の反応を与えた12クローンを選択して、scABコード領域の配列を決定した。
【0127】
配列決定は、多くのクローンがscAbコード領域内にインフレーム停止コドンを含むことを示した。構築体をpIMS147にサブクローニングして、サプレッサtRNAを含むE.cloi菌株(例えばTG-1)に形質転換した。これらの構築体はTG-1でペプチドに高い親和性を有するscAbを生成したが、高レベルのscAb発現は得られなかった(データは示していない)。しかしながら、ファージ抗体クローンC1は、scAbをコードする領域内に停止コドンを含まず、モノクローナルファージ抗体ELISAスクリーニングにおいて良好な結合反応を与えた(図3)。クローンC1はpIMS147ベクターにサブクローンして、XL-1ブルー大腸菌に形質転換した。クローンpIMS147 C1-3(図4)は、ELISAで前記ペプチドを特異的に認識する(図6)scAbの高レベルの発現(図5)を指令した。発現したscAbのためのELISAアッセイにより、ペプチドYPFRLHQVYFDAPSCに対して特異的であるscAbの生成が確認された。図7は、発現したscAbがBSA単独に対してよりもペプチド-BSA(すなわち、YPFRLHQVYFDAPSC-BSA)に対して約8倍多く結合したことを示す。scAbを欠損する対照のペプチド-BSAまたはBSAのいずれに対する結合もわずかであった。いくつかの異なったタイプの細胞抽出物に対するウェスタン分析は、C1-3scAbが、ヒト抗体のターゲティングであるHSC12の非グリコシル化シナプトフィジンタンパク質(Eastwood S.L.ら、Brain Res Bull、2001年、55巻、569〜78頁)の予測サイズである約32kDのヒトHSC中のタンパク質に結合することを示す。興味深いことに、C1-3scAbはラットHSC中のシナプトフィジンと交差反応しない(図8)。
【0128】
C1-3scAbが生きているHSC C1-3の表面に結合できるか否かを決めるために、C1-3scAbをFITCで標識し、SDS-PAGEにより、図9に示すようにFITC-C1-3scAbの移動距離の減少により判定して確認した。透析の前後および濃縮後のC1-3Abは約40Kdaの分子量を有していた。FITCで標識したC1-3scAbの分子量は約45Kdaであった。
【0129】
次にFITC-C1-3 scAbを活性化した培養中のヒトHSCに加えた。ヒトHSC培養物の免疫細胞化学染色(示していない)およびFACS分析は、細胞がまったくα平滑筋アクチン陽性のものだけで、したがって活性化したHSCおよび/または肝臓由来の筋線維芽細胞であることを示した(両方の細胞のタイプは線維発生を助長すると信じられている(Ramadori G.ら、Liver、2002年、22巻、283〜94頁)。図10を参照されたい)。固定に先立つ自然条件下におけるFITC-C1-3での共染色細胞は、すべての細胞がC1-3 scAbに結合したことを示したが、α平滑筋アクチンに比較してFITC-C1-3染色の強度にはより大きな変動があった(図10)。このことは、シナプトフィジンのレベルが上がった、および/またはFITC-Cl-3 scAb取込み機序が貪欲化したα平滑筋アクチン発現細胞の亜集団があり得ることを示唆している。HSCとC1-3、FITC-C1-3またはトリブチル錫結合C1-3 scAb(TBT-C1-3)との会合は培養媒質濃度の低下に対応していた(図11)。HSCをアポトーシス(グリオトキシン)または壊死(クロルプロマジン)により殺すことが知られている化合物と対照的に、C1-3またはFITC-C1-3について毒性の証拠はない(図12)。興味深いことに、TBT-C1-3 scAbはHSCに有毒であったので、C1-3 scAbが内部移行することおよび薬剤をC1-3に結合して薬剤の薬理作用を保持することが可能であることが示唆される。
【0130】
蛍光実験は、(フルオレッセインによる)緑色蛍光がFITC-C1-3 scAbとインキュベートした肝星細胞に付随するので、FITC-C1-3 scAbが培養中の肝星細胞と特異的に相互作用することを示唆する。光漂白を阻止しかつ核をブルーに染色した封入剤を使用して、大部分の場合、蛍光が核膜および細胞膜の付近に局在することが見出された。染色はしばしば点状に見え、それは局所化された構造との関連を反映しているのかもしれない。この点において、シナプトフィジンはニューロン中のシナプス小胞に濃縮されて、その場所でサブユニット数可変の単独オリゴマーを形成していることが知られている。染色は核膜付近でも観察されて、C1-3 scAbが細胞内に蓄積しやすいという提案を支持している。(フルオレッセインと結合していない)C1-3Abとインキュベートした肝星細胞培養物では検出可能な緑色蛍光は観察されない。C1-3 scAbと肝星細胞との間の相互作用の特異性を決定するために、ヒト肝細胞もFITC-C1-3 scAbとインキュベートした。C1-3 scAbまたはFITC-C1-3 scAbのいずれとインキュベートした肝細胞も、関連する緑色蛍光を示さず、C1-3 scAb抗体は肝細胞に結合しないことを示唆した。さらに、C1-3 scAbの能力の細胞タイプ間での差を知るために、肝星細胞と肝細胞とを共培養して、FITC C1-3 scAbとインキュベートした。蛍光は星細胞上に観察され、一方肝細胞は検出可能なレベルの関連する緑色蛍光を示さないことで一貫していた。共培養物を(フルオレッセインを結合していない)C1-3 scAbとインキュベートした場合、有意な緑色蛍光は実験では観察されなかった。肝星細胞への結合についてのC1-3scAbの作用の特異性をFITC標識BSAを使用して検討した。ヒト肝星細胞および肝細胞の両方をBSAおよびFITC-BSAとインキュベートした。ヒトHSCおよび肝細胞の両方とも、FITC-BSAとインキュベートしたときに蛍光の細胞質への集中を示し、蛍光は肝細胞へのC1-3scAb取込みを非特異的に阻止するタンパク質(すなわちC1-3)上の蛍光の存在ではなく、C1-3scAbは肝細胞とは相互作用しないことを示唆した。COS-7またはHepG2細胞では、C1-3scAbまたはC1-3scAb
-FITCとインキュベートしたとき、蛍光の結合は検出されなかった。
【0131】
ペプチドまたはモネンシンのFITC-C1-3 HSC結合/取込みへの影響を比較するために、実験に先立つ固定およびDAPI共染色なしの蛍光顕微鏡観察を使用した。図13は、対照のFITC標識3A8scAb(毒素のミクロシスチンに対して前に調製した(McElhiney J.ら、Appl Environ Microbiol、2002年、68巻、5288〜95頁))と対照的に、FITC-C1-3 scAbがHSCと結合したことを示す。シナプトフィジンタンパク質の外形質側に存在する他のアミノ酸に対応するペプチドATDPENIIKEMPMCの培養への添加は、FITC-C1-3のHSCとの結合を有意に阻止しなかったが、一方標的ペプチドYPFRLHQVYFDAPSCは、HSCへのFITC-C1-3の結合を、投与量に依存して減少させた。エンドサイトーシス阻止剤であるモネンシンの添加は、FITC-C13で染色された細胞の総数を抑えなかったが、核のFITC-C1-3染色の減少によって示されるように取込みを減少させた。
【0132】
本質的には、Foxら、J Microbiol Methods、2004年、56巻、221〜230頁により述べられているようにして、C1-3 scAbを1mg/mlの濃度でグリオトキシンと結合した。SDS-PAGEにより、前記タンパク質が改質されたことが確認された(データは示していない)。グリオトキシンに対する抗体(Foxらによって得られた)は、グリオトキシンがC1-3scAbに共有結合で結合したことを確認した。MALDI-TOF分析は前記タンパク質の質量が1683ドルトンだけ増加することが最も多いことを示した。結合方法を考慮に入れると、大部分のC1-3タンパク質が4分子のグリオトキシンと結合したことが示唆される。図14は、グリオトキシンがヒトHSCを殺し、グリオトキシンと結合したC1-3もまたHSCを殺したことを示す。このデータは薬剤をC1-3scAbに結合してかつターゲティングおよび薬効を保持することが可能であることを示している。
【図面の簡単な説明】
【0133】
【図1】シナプトフィジンの模式図である。ヒトのシナプトフィジンの理論分子量は33.8kDaで、概略を描いたように細胞の形質膜の厚さにわたると予測される。このタンパク質はグリコシル化されていると報告されていて、実験的にはおよそ38〜40kDaの分子量を有する(Eastwood S.L.ら、Brain Res Bull 2001年、55巻、569〜78頁)。
【図2】標的ペプチド-BSAに対する親和性を有するファージ抗体(ポリクローナル)の単離および増幅を示す図である。バイオパニング中における標的ペプチド-BSA抗原を認識するファージ抗体(ポリクローナル)の濃縮をELISAによって評価した。各回の選択から回収したポリクローナルファージ抗体(〜1×1010)を標的ペプチド-BSAへの結合について測定した。結合したファージは、方法の項で概説したようにHRP-標識抗M13抗体(Pharmacia)を使用して検出した。各パニングでのファージ力価を示す(---□---)。
【図3】図2のために概略を示したのと同様にして標的ペプチド-BSAへの結合を測定したC1-ファージ抗体クローン(モノクローナル)を示す図である。
【図4A】pIMS147ベクター(配列番号7および配列番号8の配列)内のC1-3scAbのヌクレオチドおよびアミノ酸配列(前部)を示す図である。抗原結合部位を構成する超可変相補性決定領域(CDR)は太字である。HおよびL鎖に結合する柔軟性のアミノ酸リンカー(Gly4、Ser)3は下線を引いてある(______)。フレームワーク領域(FW)の開始点およびヒト定常カッパドメイン(Cκ)の開始点も示す。クローニング部位を、アミノ酸配列の上に示した対応する制限酵素と共に下線(___)で示した。HuCκ定常ドメインの開始および6ヒスチジン残基精製タグを示す。全アミノ酸配列は以下の通りである。
【図4B】pIMS147ベクター(配列番号7および配列番号8の配列)内のC1-3scAbのヌクレオチドおよびアミノ酸配列(後部)を示す図である。抗原結合部位を構成する超可変相補性決定領域(CDR)は太字である。HおよびL鎖に結合する柔軟性のアミノ酸リンカー(Gly4、Ser)3は下線を引いてある(______)。フレームワーク領域(FW)の開始点およびヒト定常カッパドメイン(Cκ)の開始点も示す。クローニング部位を、アミノ酸配列の上に示した対応する制限酵素と共に下線(___)で示した。HuCκ定常ドメインの開始および6ヒスチジン残基精製タグを示す。全アミノ酸配列は以下の通りである。 AAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQES VTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGESHHHHHH
【図5】XL-1ブルー細胞におけるC1-3scAb発現の誘起およびNiアフィニティークロマトグラフィーによる精製を示す図である。上方のブロットは、示したタンパク質を負荷したSDS-PAGE後のクーマジーブルー染色を示し、下方のブロットは抗ヒトCκL鎖抗体による試料のウェスタン分析を示す。
【図6】C1-3scAbの抗ヒトCκL鎖捕捉ELISA測定を示す図である。ヤギ抗ヒトCκL鎖抗体を96ウェルプレートの表面にコートした。ヒトIgGまたはC1-3scAb標品の希釈溶液を個々のウェルでインキュベートした。十分に洗浄した後捕捉されたIgG(-□-)またはscAb(-○-)を方法の項で概説したようにして測定した。データは別々の7標品の代表的なものの、3回の別々の測定の平均および標準偏差である。
【図7】発現したscAbを、ペプチドYPFRLHQVYFDAPSC(配列番号9)と相互作用する能力について、抗BSA-標的ペプチドscAbを使用するELISAによって試験した図である。96ウェルプレートのウェルをBSA-標的ペプチドまたはBSAでコートして、C1-3scAb(グレイの縦線)または対照scAbインキュベーション緩衝液(無色の縦線)とインキュベートし、続いて十分に洗浄した。結合したscAbはHRP結合抗ヒトCκL鎖抗体を使用して検出し、方法の項で概説したようにして測定した。データは3回の別々の測定の平均および標準偏差である。「BSA-ペプチド2」は標的ペプチドである。
【図8】C1-3 scAbの標的ペプチドへの結合を示すウェスタンブロットの図である。細胞抽出物(5μg/lane)をウェスタンブロットにかけて、膜をC1-3scAbのプローブで調べ、続いてHRP結合抗ヒトCκL鎖抗体とインキュベートし、ECL試薬を使用して検出した。rHSC - ラット肝星細胞; hHSC - ヒト肝星細胞(AH4=匿名の患者コード);rHSC-ラット肝細胞;B13-ラット膵幹細胞;B13-H-肝細胞に分化転換後のラット膵幹細胞。
【図9】C1-3 scFvをFITCで標識して、SDS-PAGEにより確認した図である。Ni2+をチャージしたIMAC Fast Flow Sepharose樹脂で精製したC1-3 acAbを透析にかけ、次にcentricon YM-3遠心濃縮機を使用して濃縮した。次に濃縮したC1-3 scAbをFITC標識し、各画分の一定分量(約0.1μg/レーン)をSDS-PAGEにかけ、続いてゲルのクーマジーブルー(全タンパク質)染色を行った。
【図10】1次抗体なしで(上方パネル)またはありで(下方パネル)インキュベートしたヒトHSCのFACS分析、すなわちFITC-C13 scAb(FITC)およびα平滑筋アクチン(α-APC)に対するマウスモノクローナル抗体の図である。抗α平滑筋アクチン抗体は、方法の項で概説したようにして、ビオチン結合2次抗体を使用し、続いて蛍光体結合ストレプトアビジンとインキュベートすることにより検出した。数字は各象限内の細胞の百分率を表す。データは3つの別々の細胞標本の代表的なものである。
【図11】培養中のヒトHSCによるCl-3 scAbおよびC1-3結合体の取込みを示す図である。24ウェルのプレートにシードしたHSCを、5μgのscAbを含む0.3mlの培地中でインキュベートした。提示した時間に培地試料を取り、ウェスタンブロットにかけ、HRP結合抗ヒトCκL鎖抗体を使用してscAb(上方パネル)および負荷対照の血清アルブミン(下方パネル)を検出した。4つの別々の実験の代表的な結果である。
【図12】scAbインキュベートのHSC生存度への効果を示す図である。24ウェルのプレートにシードしたHSCを、5μgのscAbまたは提示した化学薬品を含む0.3mlの培地中でインキュベートした。細胞を3時間培養した後、培地を除去して1×PBSで洗浄した。付着率(生存度の尺度)を培養ウェル中のタンパク質を直接測定して測定した。データは同一の実験の3つの別々のウェル、別々の3回の実験の代表的なものの、平均および標準偏差である。*スチューデントT検定(両側)を使用すると、対照から有意の差がある。P>95%。
【図13】FITC-C1-3 scAbのヒトHSCへの結合に対するシナプトフィジンペプチドおよびモネンシンの効果を示す図である。24ウェルのプレートにシードしたヒトHSCを洗浄し、10μgのC1-3 scAb;10μgのFITC-C1-3 scAb;10μgのFITC-C1-3 scAbおよび0.25ナノモルの標的ペプチド;10μgのFITC-C1-3 scAbおよび2.5ナノモルの標的ペプチド;10μgのFITC-C1-3 scAbおよび25ナノモルの標的ペプチド;10μgのFITC-C1-3 scAbおよび25ナノモルの標的ペプチドP1;10μgのFITC-C1-3 scAbおよび2.5μMのモネンシン;10μgのFITC-C1-3 scAbおよび15μMのモネンシン;10μgのFITC-3A8 scAb(scAbはミクロシスチンに対して生成されたもので、HSCには結合しないと予想される)のいずれかを含むウェル当り500μlのHepes/HBSS中で1時間インキュベートした。モネンシンとインキュベートした細胞は10分間プレインキュベートした。scABを添加する直前に、培地を交換して細胞にモネンシンを再投与した。図には、無作為に選択した20視野からの1視野当りの蛍光性細胞の百分率の平均±標準偏差の測定分析を示す。バー=50μm。Momen=モネンシン;P1=ATDPENIIKEMPMCペプチド;P2=標的ペプチド。
【図14】HSC生存度を示す図である。24ウェルのプレートで培養したヒトHSCを、賦形剤DMSO;1.5μMグリオトキシン(750ピコモル);20μlの1mg/mlC1-3scAb(20μgのC1-3scAb);または20μlの0.5mg/mlC1-3-グリオトキシン結合体(10μgC1-3scAb-250ピコモルC1-3scAb*/1000ピコモルグリオトキシン)のいずれかを含む500μlの媒質で処理した。*0.5mg C1-3 タンパク質/ml貯蔵液=12.5μM=12.5ナノモル/ml=12.5ピコモル/μl。
【技術分野】
【0001】
本発明はシナプトフィジンに対する特異的結合要素に関する。本発明の好ましい実施形態は、本明細書においてC1-3と称するscFvフラグメントの抗体VHおよび/またはVLドメインを使用する。さらに好ましい実施形態では、C1-3の重鎖可変(VH)および/または軽鎖可変(VL)ドメイン、特にVH C1-3の1つまたは複数の相補性決定領域(CDR)を他の抗体のフレームワーク領域中に使用する。本発明者らは、有利な性質、特に肝星細胞の外表面を標的にする能力を有する多数の抗体分子を同定している。
【背景技術】
【0002】
肝線維症は、肝硬変および肝不全の発生に先立つ、肝臓での細胞外基質タンパク質の蓄積により特徴付けられる可逆的過程である(Friedman S.L.、J Biol Chem、2000年、275巻、2247〜50頁、Batallex R.ら、Semin Liver Dis、2001年、21巻、437〜51頁)。ウィルス感染(例えばC型肝炎)およびアルコール乱用を含む多くの条件が、線維症に至る肝臓への損傷を惹起し得る。線維症は長年の間検知されないままで、致命的になることのある重度の損傷を与えることがある。肝線維症を患う人々は、世界中では2億人を超える可能性があるにもかかわらず、この疾患を治療するために臨床医が使用できる治療の選択肢はない。
【0003】
肝線維症は、慢性的な肝損傷に反応して肝星細胞(HSC)によって惹き起こされる。HSCは、肝線維症の発生および回復において主要かつ中心的な役割を演ずる。HSCは、正常肝では静止状態で存在し、ビタミンAを貯蔵するように機能している(Geerts A.Semin、Liver Dis、2001年、21巻、311〜35頁)。慢性肝損傷に反応して、静止状態のHSCが、筋線維芽細胞様表現型に「活性化」する。活性化したHSCは増殖し、肝線維症で観察される瘢痕を構成する細胞外基質タンパク質の大部分を発現すると考えられている。活性化したHSCおよび線維発生は、何らかの原因による慢性的肝損傷に対する有害な反応であること、および増大した活性化HSCアポトーシスが肝線維症を消散させ、慢性的損傷に対する肝臓の反応を増大させ得ることを示唆する強い証拠がある(Iredale J.P.ら、J Clin Invest、1998年、102巻、538〜49頁、Wright M.C.ら、Gastroenterology 2001年、121巻、685〜98頁、Orr J.G.ら、Hepatology 2004年、40巻、232〜42頁、Issa R.ら、Gut、2001年、48巻、548〜57頁)。
【0004】
過去における多くの抗線維化薬候補の主な問題は、おそらく外来性化合物を代謝するように機能する肝細胞の近傍に肝星細胞があるために、薬剤が肝星細胞内で治療濃度に達しなかったことである。これらの薬剤には、in vitroおよびin vivoで抗炎症性活性を有し、星細胞の活性化を低下させることができるいくつかの作用薬が含まれる。これらの薬剤には、コルチコステロイド、TNFα拮抗剤、抗酸化剤、サイトカイン(γインターフェロン)および肝細胞成長因子(HGF)PPARγリガンド(チアゾリジンジオン)、エンドテリン-1拮抗剤、ハロフジノン(抗コクシジウム薬)および遺伝子治療(動物モデルにおける遺伝子治療によるメタロプロテイナーゼmRNAの投与)が含まれる。成功したものがないということは、任意の抗線維化療法を肝臓中の肝星細胞に向けることが有用であろうということを示している。
【0005】
線維症の間、活性化肝星細胞中でマンノース6-リン酸/インスリン様成長因子II(M6P/IGF-II)受容体が高レベルで発現していることおよびマンノース6-リン酸(M6P)で修飾された血清アルブミン(SA)をラットに投与すると(M6P:SAのモル比は28:1)、肝細胞内の投与量の70%が肝星細胞に見出されるように肝臓に分布することが、最近Polestraおよび共同研究者によって示された(Beljaarsら、Hepatology、1999年、29巻、1486〜93頁)。VI型コラーゲン受容体を認識する10個の環状ペプチド部分(C*GRGDSPC*、ただしC*は環化しているシステイン残基を示す)で修飾したSAもまた、静脈注射後10分以内にラット肝臓に優先的に分布する結果となる(Beljaarsら、J Biol Chem、2000年、275巻、12743〜51頁)。線維症の肝臓においては、肝臓に投与したペプチド修飾アルブミンの70%が、活性化した肝星細胞に結合した。しかし、ヒト肝組織灌流において、これらの作用薬は、大分子量分子および異物粒子の除去に関与する型の肝細胞であるクップファー細胞に取り込まれて、星細胞には取り込まれなかった。
【0006】
シナプトフィジンは神経細胞および肝星細胞だけに発現するタンパク質である(Cassiman D.ら、Am J Pathol、1999年、155巻、1831〜1839頁、Bargou R.C.ら、Gene、1991年、99巻、197〜204頁)。これは膜結合タンパク質であり、機能性のある精製形態では入手できない。この大きな制限にもかかわらず、本出願人らは、肝星細胞上に存在するシナプトフィジンの細胞外ドメインに対して特異性のある初めての完全なヒトモノクローナル抗体フラグメントを単離することに成功した。この抗体はファージディスプレイ法およびMRC(Cambridge、UK所在)によって使用可能になったヒト抗体ライブラリーを使用して単離した。この抗体はあるペプチドに対して作られたもので、星細胞膜中で自然のままの立体配座にある天然のシナプトフィジンタンパク質全体を認識するのも、この抗ペプチド抗体である。シナプスフィジンを認識して結合する抗体は、下で論ずるように多くの治療応用に適した試薬を初めて提供する可能性を有する。
【先行技術文献】
【特許文献】
【0007】
【特許文献1】欧州特許A184187号
【特許文献2】英国特許2188638A号
【特許文献3】欧州特許A239400号
【特許文献4】欧州特許A0120694号
【特許文献5】欧州特許A0125023号
【特許文献6】国際出願PCT/US92/09965
【特許文献7】国際公開WO94/13804
【特許文献8】国際公開WO92/01047
【特許文献9】国際公開WO99/58572
【非特許文献】
【0008】
【非特許文献1】Friedman S.L.、J Biol Chem、2000年、275巻、2247〜50頁
【非特許文献2】Batallex R.ら、Semin Liver Dis、2001年、21巻、437〜51頁
【非特許文献3】Geerts A.Semin、Liver Dis、2001年、21巻、311〜35頁
【非特許文献4】Iredale J.P.ら、J Clin Invest、1998年、102巻、538〜49頁
【非特許文献5】Wright M.C.ら、Gastroenterology 2001年、121巻、685〜98頁
【非特許文献6】Orr J.G.ら、Hepatology 2004年、40巻、232〜42頁
【非特許文献7】Issa R.ら、Gut、2001年、48巻、548〜57頁
【非特許文献8】Beljaarsら、Hepatology、1999年、29巻、1486〜93頁
【非特許文献9】Beljaarsら、J Biol Chem、2000年、275巻、12743〜51頁
【非特許文献10】Cassiman D.ら、Am J Pathol、1999年、155巻、1831〜1839頁
【非特許文献11】Bargou R.C.ら、Gene、1991年、99巻、197〜204頁
【非特許文献12】Ward、E.S.ら、Nature 341巻、544〜546頁、1989年
【非特許文献13】Birdら、Science、242巻、423-426頁、1988年
【非特許文献14】Hustonら、PNAS USA、85巻、5879〜5883頁、1988年
【非特許文献15】P.Holligerら、Proc.Natl.Acad.Sci.USA、90巻、6444〜6448頁、1993年
【非特許文献16】Y.Reiterら、Nature Biotech、14巻、1239〜1245頁、1996年
【非特許文献17】S.Huら、Cancer Res.、56巻、3055〜3061頁、1996年
【非特許文献18】Holliger,P.、Winter G.、Current Opinion Biotechnol.、4巻、446〜449頁、1993年
【非特許文献19】J.B.B.Ridgewayら、Protein Eng.、9巻、616〜621頁、1996年
【非特許文献20】Kabat、E.A.ら、「Sequences of Proteins of Immunological Interest」第4版、US Department of Health and Human Services、1987年、および現在インターネット上で使用できるその更新版(http://immuno.bme.nwu.eduまたは任意の検索エンジンを使用して「Kabat」)
【非特許文献21】Markら、Bio/Technology、1992年、10巻、779〜783頁
【非特許文献22】Stemmer、Nature、1994年、370巻、389〜391頁
【非特許文献23】Gramら、1992年、Proc.Natl.Acad.Sci.、USA、89巻、3576〜3580頁
【非特許文献24】Barbasら、1994年、Proc.Natl.Acad.Sci.、USA、91巻、3809〜3813頁
【非特許文献25】Schierら、1996年、J.Mol.Biol.、263巻、551〜567頁
【非特許文献26】Ledermann J.A.ら、(1991年)、Int.J.Cancer 47巻、659〜664頁、
【非特許文献27】Bagshawe K.D.ら、(1991年)、Antibody,Immunoconjugates and Radiopharmaceuticals 4巻、915〜922頁
【非特許文献28】Pluckthun、A.Bio/Technology 9巻、545〜551頁、1991年
【非特許文献29】Ref、M.E.、(1993年)Curr.Opinion Biotech.4巻、573〜576頁、
【非特許文献30】Trill J.J.ら、(1995年)Curr.Opinion Biotech 6巻、553〜560頁
【非特許文献31】SambrookおよびRussell、「Molecular Cloning:a Laboratory Manual」第3版、2001年、Cold Spring Harbor Laboratory Press
【非特許文献32】「Current Protocols in Molecular Biology」第2版、Ausubelら(編)、John Wiley & Sons、1992年
【非特許文献33】Leel V.ら、Biochem Biophys Res Commun 2004年、316巻、872〜7頁
【非特許文献34】Hayhurst A.ら、Protein Expr Purif 1999年、15巻、336〜43頁
【非特許文献35】Wright M.C.ら、Mol Pharmacol 1996年、50巻、856〜63頁
【非特許文献36】Wright M.C.ら、Hum Exp Toxicol、1996年、15巻、203〜4頁
【非特許文献37】Harvey J.L.ら、Drug Metab Dispos、2000年、28巻、96〜101頁
【非特許文献38】Marek C.J.ら、Biochem J、2003年、370巻、763〜9頁
【非特許文献39】Eastwood S.L.ら、Brain Res Bull、2001年、55巻、569〜78頁
【非特許文献40】McElhiney J.ら、Appl Environ Microbiol、2002年、68巻、5288〜95頁
【非特許文献41】Foxら、J Microbiol Methods、2004年、56巻、221〜230頁
【発明の開示】
【課題を解決するための手段】
【0009】
本明細書において次の配列を開示する。
配列番号1 C1-3 VHをコードするヌクレオチド配列
配列番号2 C1-3 VHのアミノ酸配列
配列番号3 C1-3 VLをコードするヌクレオチド配列
配列番号4 C1-3 VLのアミノ酸配列
配列番号5 C1-3 リンカーをコードするヌクレオチド配列
配列番号6 C1-3 リンカーのアミノ酸配列
配列番号7 C1-3をコードするヌクレオチド配列
配列番号8 C1-3のアミノ酸配列
配列番号9 C1-3抗原
配列番号10 VHアミノ酸配列(配列番号2)内のC1-3 VH CDR1
配列番号11 VHアミノ酸配列(配列番号2)内のC1-3 VH CDR2
配列番号12 VHアミノ酸配列(配列番号2)内のC1-3 VH CDR3
配列番号13 VLアミノ酸配列(配列番号4)内のC1-3 VH CDR1
配列番号14 VLアミノ酸配列(配列番号4)内のC1-3 VH CDR2
配列番号15 VLアミノ酸配列(配列番号4)内のC1-3 VH CDR3
配列番号16 ヒト定常カッパドメイン(Hu Cκ)および6ヒスチジン残基精製タグの全アミノ酸配列(1文字コード)- NotIサイトの末端でL鎖フレームワーク4(LFW4)が終り、Hu Cκが始まる。
配列番号17 C1-3 VLをコードするヌクレオチド配列の初期バージョン
配列番号18 C1-3 VLのアミノ酸配列の初期バージョン
配列番号19 C1-3をコードするヌクレオチド配列の初期バージョン
配列番号20 C1-3のアミノ酸配列の初期バージョン
上記の配列において、図4におけるように、抗原結合部位を構成する超可変相補性決定領域(CDR)は太字で示している。HおよびL鎖に結合する柔軟なアミノ酸リンカー(Gly4、Ser)3は下線を引いてある(______)。フレームワーク領域(FW)の開始点およびヒト定常カッパ領域(Cκ)の開始点も示す。クローニング部位を下線(___)で示し、対応する制限酵素をアミノ酸配列の上に示した。
1つの態様において、本発明は、シナプトフィジンに結合し、C1-3 VHドメイン(配列番号2)および/またはC1-3 VLドメイン(配列番号4)を含む特異的結合要素を提供する。
【0010】
一般的に、VHドメインはVLドメインと対になって抗体の抗原結合部位を提供するが、以下にさらに論ずるように、VHドメインのみでも抗原に結合するために使用することができる。好ましい一実施形態においては、C1-3 VHドメイン(配列番号2)はC1-3 VLドメイン(配列番号4)と対になり、その結果抗体の抗原結合部位が、C1-3 VHおよびC1-3 VLドメイン両方を含んで形成される。他の実施形態においては、C1-3 VHはC1-3 VL以外のVLドメインと対になる。軽鎖の乱交は当業者においては十分に確かめられている。
【0011】
1つまたは複数のCDRをC1-3 VHまたはVLドメインから取って、適当なフレームワーク中に導入することができる。このことは後でさらに論ずる。C1-3 VH CDR1、2および3は、それぞれ配列番号10、11および12で示される。C1-3 VL CDR1、2および3は、それぞれ配列番号13、14および15で示される。
【0012】
配列を本明細書で提示した、シナプトフィジンに対する特異的結合体に使用できる変異体のVHおよびVLドメインは、配列の改変または変異およびスクリーニングという方法によって得ることができる。そのような方法もまた本発明によって提供される。
【0013】
本明細書で配列を具体的に開示するVHおよびVLドメインのいずれの可変領域アミノ酸配列の変異体も、説明するように本発明に従って使用することができる。特定の変異体には、1つまたは複数のアミノ酸配列の改変、おそらく20個未満、約15個未満、約10個未満または約5個未満、4、3、2または1個のアミノ酸配列の改変(アミノ酸残基の付加、欠失、置換および/または挿入)が含まれ得る。改変は1つまたは複数のフレームワーク領域および/または1つまたは複数のCDR中になすことができる。
【0014】
本発明による特異的結合要素は、抗原に結合することについて、任意の特異的結合要素と競合するものであってよく、これらのいずれも抗原に結合し、特異的結合要素、本明細書で開示したVHおよび/もしくはVLドメイン、または本明細書で開示したVH CDR3、またはこれらいずれの変異体をも含むものである。結合要素間の競合はin vitroで容易に測定することができ、例えばELISAを使用するか、および/または一方の結合要素、他方のタグ付けしていない結合要素の存在下に検出できる特異的なレポーター分子をタグ付けすることにより、同一のエピトープまたは重複するエピトープに結合する特異的結合要素の同定が可能になる。
【0015】
それ故、本発明のさらなる態様は、シナプトフィジンとの結合についてC1-3と競合する、ヒト抗体の抗原結合部位を含む特異的結合要素を提供する。
【0016】
シナプトフィジンに対する抗体を得るために、当業者は種々の方法を使用でき、その抗体はシナプトフィジンへの結合についてC1-3と競合し得るであろう。
【0017】
さらに他の態様において、本発明は抗原に結合し得る1つまたは複数の特異的結合要素を得る方法を提供する。該方法は本発明による特異的結合要素のライブラリーと前記抗原とを接触させて、前記抗原に結合し得るライブラリーの1つまたは複数の特異的結合要素を選択することを含む。
【0018】
好ましい実施形態において、特異的結合要素はアミノ酸配列YPFRLHQVYFDAPSC(配列番号9)内のエピトープに結合する。
【0019】
ライブラリーはバクテリオファージ粒子表面に提示されることがあり、各粒子は表面に提示された抗体VH可変ドメインをコードする核酸を含み、場合により提示されたVLドメインがあればそれも含む。
【0020】
前記抗原に結合できてバクテリオファージ粒子表面に提示された特異的結合要素の選択に続いて、前記選択された特異的結合要素を提示するバクテリオファージ粒子から核酸を取り出すことができる。そのような核酸は、それの後に続く、前記の選択された特異的結合要素を提示するバクテリオファージ粒子から取った核酸の配列を用いる核酸の発現による特異的結合要素または抗体VH可変ドメイン(場合によっては抗体VL可変ドメイン)の生産に使用できる。
【0021】
前記の選択された特異的結合要素の抗体VH可変ドメインのアミノ酸配列を有する抗体VH可変ドメインは、単離された形態で提供することができ、そのようなVHドメインを含む特異的結合要素も同様である。
【0022】
さらにシナプトフィジンに結合する能力を試験することができ、シナプトフィジンへの結合でC1-3と競争する能力も同様である。シナプトフィジンの作用に拮抗する能力は、後でさらに説明するようにして試験できる。
【0023】
本発明による特異的結合要素はC1-3の親和性によりシナプトフィジンに結合できる。好ましくは、特異的結合要素はシナプトフィジンのエピトープYPFRLHQVYFDAPSCに結合する。
【0024】
特異的結合要素はマウス、ラットおよび/またはヒトのシナプトフィジンに結合できる。好ましくは、特異的結合要素はヒトシナプトフィジンに結合する。
【0025】
異なる特異的結合要素の結合親和性および中和能力は、適当な条件で比較できる。
【0026】
抗体配列に加えて、本発明の特異的結合要素は、例えばペプチドもしくは折りたたまれたドメインなどのポリペプチドを形成する、または抗原に結合する能力に加えて他の機能的特性を分子に賦与する他のアミノ酸を含むことができる。
【0027】
本発明の特異的結合要素は、検出可能な標識を有することができ、または毒素もしくは酵素に(例えばペプチド結合またはリンカーにより)結合させることができる。
【0028】
当業者はタンパク質に分子を化学的に結合する多くの手法に通じている。特異的結合要素が医薬用のものであるとき、結合体の結合は、好ましくは、血液循環中では安定であるが、ひとたび結合体が細胞内に隔離されると変化しやすくなるものである。
【0029】
本発明の好ましい実施形態において、特異的結合要素は検出可能な、蛍光性標識のフルオレッセインイソチオシアネート(FITC)に結合させることができる。
【0030】
さらに他の態様において、本発明は、本発明による特異的結合要素、VHドメインおよび/またはVLドメインをコードする配列を含む単離した核酸を提供し、ならびに前記本発明による特異的結合要素、VHドメインおよび/またはVLドメインの生成を起こす条件下で前記核酸を発現させ、生成物を回収することを含む、本発明による特異的結合要素、VHドメインおよび/またはVLドメインを調製する方法を提供する。
【0031】
本発明による特異的結合要素は、ヒト患者に本発明の特異的結合要素の有効量を投与することを含む、前記患者における疾患または障害を治療(予防的治療を含む)する方法などのヒトまたは動物の体の治療または診断に使用できる。本発明により治療可能な疾患は、本明細書の他の箇所で説明するものを含む。
【0032】
本発明による特異的結合要素は、画像法、例えば、特異的結合要素が結合する細胞の存在または位置を決定する方法に使用できる。
【0033】
さらに他の態様において、本発明は、本発明による特異的結合要素および抗原に対する特異的結合要素の結合を測定する1つまたは複数の試薬を含む診断キットを提供する。
【0034】
本発明のさらに他の態様では、通常は単離された本明細書で開示した抗体VH可変ドメイン(配列番号1)および/またはVL可変ドメイン(配列番号3)をコードする核酸が、提供される。
【0035】
本発明の別の態様では、通常は単離された本明細書において開示したVH CDRまたはVL CDR配列、特に配列番号10、11および12から選択されたVH CDRまたは配列番号13、14および15から選択されたVL CDR、最も好ましくはC1-3 VH CDR3(配列番号12)をコードする核酸が、提供される。
【0036】
さらに他の態様では、本発明の核酸で形質転換した宿主細胞が提供される。
【0037】
さらに他の態様では、抗体VH可変ドメインを生成する方法が提供され、前記方法はコードする核酸からの発現を惹起することを含む。そのような方法は前記抗体VH可変ドメインの生成のための条件下で宿主細胞を培養することを含むことができる。
【0038】
VL可変ドメイン生成する類似の方法ならびにVHおよび/またはVLドメインを含む特異的結合要素が、本発明のさらに他の態様として提供される。
【0039】
製造方法は生成物の単離および/または精製のステップを含むことができる。
【0040】
製造方法は、生成物を、薬学的に許容可能な賦形剤などの少なくとも1つの追加成分を含む組成物に配合することを含むことができる。
【0041】
本発明のこれらのおよび他の態様を以下にさらに詳細に説明する。
【0042】
用語
特異的結合要素
この用語は、互いに結合する特異性を有する一対の分子の1つの構成要素を表す。特異的に結合する一対の構成要素は、天然に由来してもまたは全体もしくは一部が合成的に製造されてもよい。分子対の一方は、その表面または空孔に、分子対の他方の特定の空間的および極性的組織に特異的に結合し、かつそれ故相補的である区域を有する。このように、前記分子対の構成要素は互いに特異的に結合する性質を有する。特異的結合対の種類の例は、抗原-抗体、ビオチン-アビジン、ホルモン-ホルモン受容体、受容体-リガンド、酵素-基質である。本発明の応用は抗原-抗体タイプの反応に関係している。
【0043】
抗体分子
この用語は、天然または全体もしくは一部が合成的に製造された免疫グロブリンを表す。この用語はまた、抗体の結合ドメインを含む如何なるポリペプチドまたはタンパク質をも包含する。抗体結合ドメインを含む抗体フラグメントはFab、scFv、Fv、dAb、Fd;およびダイアボディなどである。
【0044】
モノクローナル抗体および他の抗体を取り、組換えDNA技法を使用して、元の抗体の特異性を保持する他の抗体またはキメラ分子を製造することが可能である。そのような技法は、免疫グロブリンの可変領域、すなわち抗体の相補性決定領域(CDR)をコードするDNAを、異なる免疫グロブリンの定常部、または定常部とフレームワーク領域とを合わせた領域に導入することを含むことができる。例えば、欧州特許A184187号、英国特許2188638A号または欧州特許A239400号を参照されたい。ハイブリドーマまたは抗体を生成する他の細胞は、生成された抗体の結合特異性が変化するかまたは変化しないことがある遺伝的変異または他の変化を受けやすいことがある。
【0045】
抗体は多くの方法で改変できるので、「抗体分子」という用語は、抗体の要求される特性を有する抗原結合ドメインを有する、如何なる特異的結合要素または物質をも包含すると解釈すべきである。このように、この用語は、天然のものか全体的もしくは一部的に合成であるかを問わず、免疫グロブリン結合ドメインを含む如何なるポリペプチドをも含む、抗体フラグメントおよび誘導体を包含する。それ故、他のポリペプチドに融合した、免疫グロブリンの結合ドメイン、またはその同等物を含むキメラ分子が含まれる。キメラ抗体のクローニングと発現は欧州特許A0120694号および欧州特許A0125023号に記載されている。
【0046】
全抗体のフラグメントが抗原に結合する機能を果たすことができることが示されている。結合するフラグメントの例は、(i)VL、VH、CLおよびCH1ドメインにより構成されるFabフラグメント;(ii)VHおよびCH1ドメインにより構成されるFdフラグメント;(iii)単一抗体のVLおよびVHドメインにより構成されるFvフラグメント;(iv)VHドメインにより構成されるdAbフラグメント(Ward、E.S.ら、Nature 341巻、544〜546頁(1989年));(v)単離したCDR領域;(vi)F(ab')2フラグメント、2つの連結したFabフラグメントにより構成される2価のフラグメント;(vii)2ドメインが共同して抗原結合部位を形成することを許容するペプチドリンカーによりVHドメインおよびVLドメインが連結した単鎖Fv分子(scFv)(Birdら、Science、242巻、423-426頁、1988年;Hustonら、PNAS USA、85巻、5879〜5883頁、1988年);(viii)二重特異性単鎖Fv2量体(国際出願PCT/US92/09965)および(ix)「ダイアボディ」、遺伝子融合により構築された多価または多重特異性フラグメント(国際公開WO94/13804;P.Holligerら、Proc.Natl.Acad.Sci.USA、90巻、6444〜6448頁、1993年)である。Fv、scFvまたはダイアボディ分子はVHおよびVLドメインを連結するジスルフィド架橋の導入により安定化できる(Y.Reiterら、Nature Biotech、14巻、1239〜1245頁、1996年)。CH3領域に接続したscFvを含むミニボディも作製できる(S.Huら、Cancer Res.、56巻、3055〜3061頁、1996年)。
【0047】
二重特異性抗体が使用されるべきところでは、これらは、種々の方法(Holliger,P.、Winter G.、Current Opinion Biotechnol.、4巻、446〜449頁(1993年))で作製され、例えば、化学的にまたは混成ハイブリドーマから調製できる従来の二重特異性抗体であってよく、あるいは上記の二重特異性抗体フラグメントのいずれであってもよい。ダイアボディおよびscFvは、Fc領域なしで、可変領域のみを使用して、抗イディオタイプ反応の効果を減少させる可能性をもって構築できる。
【0048】
二重特異性のダイアボディも、二重特異性の全抗体とは対照的に、大腸菌中で容易に構築および発現できるので、特に有用である。適当な結合特異性のダイアボディ(および抗体フラグメントなど多くの他のポリペプチド)は、ライブラリーからファージディスプレイ(国際公開W094/13804号)を使用して容易に選択できる。ダイアボディの一方のアームが、例えば、シナプトフィジンに対して向けられた特異性をもって、一定に保たれるべきであれば、その際には、他方のアームが変化したライブラリーを作製することができて、適当な特異性の抗体が選択される。二重特異性の全抗体は、窪みに突起を入れる技術(knobs-into-holes engineering)により作製できる(J.B.B.Ridgewayら、Protein Eng.、9巻、616〜621頁、1996年)。
【0049】
抗原結合ドメイン
この用語は、抗体分子の、抗原の一部または全部に特異的に結合しかつ相補的である区域を含む部分を表す。抗原が大きい場合、抗体は、エピトープと名付けられた部分にのみ結合できる。抗原結合ドメインは1つまたは複数の抗体可変ドメインにより提供される(例えば、VHドメインにより構成される、いわゆるFd抗体フラグメント)。好ましくは、抗原結合ドメインは抗体L鎖可変領域(VL)および抗体H鎖可変領域(VH)を含む。
【0050】
特異的
この用語は、特異的結合対の1構成要素が、その特異的結合の相手以外の分子に如何なる有効な結合も示さない状態を指すことに使用できる。この用語は、例えば抗原結合ドメインが、多くの抗原中に存在する特定のエピトープに対して特異的であるときにも適用でき、その場合には、抗原結合ドメインを担う特異的結合要素が、エピトープを担う種々の抗原に結合できるであろう。
【0051】
典型的には、抗原のリストを使用してELISAなどの結合測定により特異性を決定できる。本発明による特異的結合要素は肝星細胞のシナプトフィジンを認識できるが、神経細胞のものは認識しない。
【0052】
含む(comprise)
この用語は、一般的に「含む(include)」の意味で、すなわち1つまたは複数の特徴の存在を認める意味で使用する。
【0053】
単離した
この用語は、本発明の特異的結合要素、またはそのような結合要素をコードする核酸が、本発明に一般的に合致するであろうという状態を指す。結合要素および核酸は、それらの天然の環境で、またはそれらがin vitroもしくはin vivoで実施された組換えDNA技法により作製されたとき、それらが作製された環境(例えば細胞培養)で、共存していた他のポリペプチドもしくは核酸など元来付随していた物質を含まないかまたは実質的に含まないであろう。結合要素および核酸は希釈剤または補助剤と共に配合することができ、それでもなお実用目的にとっては単離された状態である。例えば、前記結合要素は、ゼラチンまたは他の担体が免疫測定で使用するためのマイクロタイタープレートのコーティングに使用されれば、当然これらと混合されるであろうし、あるいは診断または治療に使用されるときには、薬理学的に許容できる担体または希釈剤と混合されるであろう。特異的結合要素は、自然に、または異種真核細胞(例えばCHOもしくはNSO(ECACC 85110503)細胞)系によりグリコシル化されてもよく、あるいは(例えば原核細胞中での発現により生成されるなら)グリコシル化されなくてもよい。
【0054】
「実質的に述べたように」という表現により、本発明の関連するCDRまたはVHもしくはVLドメインが、本明細書において配列を述べた特定の領域と同一であるかまたは高度に類似であるかのいずれかであろうということを意味する。「高度に類似」という表現により、CDRおよび/またはVHもしくはVLドメインにおいて、1から5、好ましくは1から3または1または2、または3または4など1から4のアミノ酸置換がなされてよいということを予期している。
【0055】
本発明のCDRを担う構造は、通常抗体HもしくはL鎖配列の、または、それらの実質的部分の構造、すなわちそれらの中で再配列免疫グロブリン遺伝子のコードする天然のVHおよびVL抗体可変ドメインのCDRに対応する位置に前記CDRが位置するそれらの実質的部分の構造であろう。免疫グロブリン可変ドメインの構造および位置は、Kabat、E.A.ら、「Sequences of Proteins of Immunological Interest」 第4版、US Department of Health and Human Services、1987年、および現在インターネット上で使用できるその更新版(http://immuno.bme.nwu.eduまたは任意の検索エンジンを使用して「Kabat」を見出されたい)を参照して決定できる。
【0056】
本発明で使用する可変ドメインは、任意の生殖細胞系または再配列ヒト可変ドメインから得ることができ、あるいは知られているヒト可変ドメインの共通配列に基づく合成可変ドメインであってもよい。本発明のCDR配列(例えばCDR3)を、組換えDNA技法を使用してCDR欠損可変ドメインのレパートリーに導入することができる。
【0057】
例えば、Markら(Bio/Technology、1992年、10巻、779〜783頁)は抗体可変ドメインのレパートリーを作製する方法を述べていて、その方法では、可変ドメイン区域の5'末端に向けられたまたは隣接した共通プライマーが、ヒトVH遺伝子の第3フレームワーク領域への共通プライマーと共に使用されて、CDRを欠損するVH可変ドメインのレパートリーが提供される。Markらは、いかにしてこのレパートリーを特定の抗体のCDR3と組み合わせられるかをさらに述べている。類似の技法を使用して本発明のCDR3由来の配列をCDR3の欠損するVHまたはVLドメインのレパートリーと入れ替えることができ、入れ替えた完全なVHまたはVLドメインを同属のVHまたはVLドメインと組み合わせて、本発明の特異的結合要素を提供する。それから前記レパートリーを、WO92/01047のファージディスプレイ系などの適当な宿主系で提示させて、その結果適当な特異的結合要素が選択される。レパートリーは、104以上の独立の構成要素のいずれか、例えば、106から108または1010の構成要素により構成され得る。
【0058】
類似の入れ替えまたは組合せ技法は、Stemmerによっても開示されており(Nature、1994年、370巻、389〜391頁)、彼はβ-ラクタマーゼ遺伝子に関する技法を述べているが、この手法が抗体の生成のために使用できることを認めている。
【0059】
さらに他の別法は、全可変ドメイン内に変異を生じさせるために、1つまたは複数の選択されたVHおよび/またはVL遺伝子のランダム変異誘発を使用して、本発明のCDR由来配列を担う新規なVHまたはVL領域を生成させる。そのような技法は、Gramらによって述べられており(1992年、Proc.Natl.Acad.Sci.、USA、89巻、3576〜3580頁)、彼らは誤りがちなPCRを使用した。
【0060】
使用してよい他の方法は、VHまたはVLの遺伝子CDR領域に定方向突然変異誘発を起こさせることである。そのような技法はBarbasら(1994年、Proc.Natl.Acad.Sci.、USA、91巻、3809〜3813頁)およびSchierら(1996年、J.Mol.Biol.、263巻、551〜567頁)によって開示されている。
【0061】
上記の技法はすべて当業者に知られたものであり、それら自体では本発明の一部を形成しない。当業者は、当業者に定常的な手法を使用して本発明の特異的結合要素を提供するためにそのような技法を使用できるであろう。
【0062】
本発明のさらに他の態様は、シナプトフィジンのエピトープYPFRLHQVYFDAPSCに特異的な抗体の抗原結合ドメインを得る方法を提供し、その方法は、本明細書で述べたVHドメインのアミノ酸配列中の1つまたは複数のアミノ酸の付加、欠失、置換または挿入という手段により、VHドメインのアミノ酸配列変異体であるVHドメインを提供すること、場合によりこのようにして提供されたVHドメインを1つまたは複数のVLドメインと組み合わせること、およびシナプトフィジンに対して特異的な特異的結合要素または抗体の抗原結合ドメインを同定するためにVHドメインまたはVH/VLの組合せを試験することを含む。前記VLドメインは本明細書で実質的に述べたアミノ酸配列を有することができる。
【0063】
本明細書で開示したVLドメインの1つまたは複数の配列変異体を1つまたは複数のVHドメインと組み合わせる類似の方法が使用できる。
【0064】
本発明のさらに他の態様は、シナプトフィジンに対して特異的な特異的結合要素を調製する方法を提供し、その方法は、
(a)置換すべきCDR3を含むかまたはCDR3をコードする領域を欠くVHドメインをコードする核酸の出発レパートリーを提供すること、
(b)前記レパートリーと本明細書でVH CDR3のものとして実質的に述べたアミノ酸配列をコードするドナー核酸とを組み合わせ、その結果前記ドナー核酸がレパートリーのCDR3領域に挿入されて、VHドメインをコードする核酸の生成物レパートリーを提供すること、
(c)前記生成物レパートリーの核酸を発現させること、
(d)シナプトフィジンに対して特異的な特異的結合要素を選択すること、および
(e)前記特異的結合要素またはそれをコードする核酸を回収すること
を含む。
【0065】
この場合も同様に、本発明のVL CDR3と、置換すべきCDR3を含むかまたはCDR3をコードする領域を欠くVLドメインをコードする核酸のレパートリーとを組み合わせる類似の方法を使用することができる。
【0066】
同様に、1つまたは2つ以上、またはすべての3つのCDRをVHまたはVLドメインのレパートリーに移植でき、次にそれらをシナプトフィジンに対する特異的結合要素としてスクリーニングする。
【0067】
免疫グロブリンの可変ドメインの実質的部分は、少なくともその3つのCDR領域を、それらの間に介在するフレームワーク領域と共に含むであろう。好ましくは、前記部分も第1および第4フレームワーク領域のどちらかまたは両方の少なくとも約50%を含むであろう。その50%は第1フレームワーク領域のC末端50%、および第4フレームワーク領域のN末端50%である。可変ドメインの実質部分のN末端またはC末端に付加する残基は、天然の可変ドメイン区域とは本来なら関係がないものでよい。例えば、組換えDNA技法によりなされた本発明の特異的結合要素の構築は、クローニングまたは他の操作段階を容易にするために導入されたリンカーによりコードされるN-またはC-末端残基を導入する結果になる。他の操作段階には、本発明の可変ドメインを免疫グロブリンH鎖、他の可変ドメイン(例えばダイアボディの生成における)または以下でさらに詳細に論ずるタンパク質標識を含むさらなるタンパク質配列に接続するためのリンカーの導入が含まれる。
【0068】
本発明の好ましい態様において、VHおよびVLドメインの一対を含む特異的結合要素が好ましいが、VHまたはVLドメイン配列のいずれかに基づく単結合ドメインは、本発明のさらに他の態様を形成する。単免疫グロブリンドメイン、特にVHドメインは特異的に標的抗原に結合し得ることが知られている。
【0069】
単鎖特異的結合ドメインのいずれの場合においても、これらのドメインは、シナプトフィジンに結合できる2ドメイン特異的結合要素を形成し得る相補性ドメインをスクリーニングするために使用できる。
【0070】
これは国際公開WO92/01047に開示された、いわゆる階層的二重コンビナトリアル手法を使用するファージディスプレイスクリーニング法によって達成できる。前記スクリーニング法では、HまたはL鎖クローンのいずれかを含む個々のコロニーが他鎖(LまたはH)をコードするクローンの全ライブラリーを感染させるために使用され、その結果生ずる2鎖特異的結合要素が、前記文献に記載されたようなファージディスプレイ技法に従って選択される。この技法はMarkらの前掲の文献にも開示されている。
【0071】
本発明の特異的結合要素はさらに抗体定常部またはその部分を含むことができる。例えば、VLドメインはそのC末端で、ヒトCκまたはCλ鎖、好ましくはCκを含む抗体L鎖の定常ドメインに結合できる。同様に、VHドメインに基づく特異的結合要素はそのC末端で、如何なる抗体アイソタイプ、例えばIgG、IgA、IgEおよびIgMならびに任意のアイソタイプのサブクラスに由来する免疫グロブリのH鎖の全体または部分に結合できる。WO99/58572に開示されたΔnabおよびΔnacなどのFc領域が使用できる。
【0072】
本発明の特異的結合要素は検出可能なまたは機能性の標識で標識できる。検出可能な標識には、131Iまたは99Tcなどの放射性標識が含まれ、それらは抗体イメージングの当業者に知られた常套的な化学的方法を使用して抗体に付けることができる。標識には、西洋ワサビペルオキシダーゼなどの酵素標識も含まれる。標識にはさらに、特異的同族検出可能成分例えば標識アビジンへの結合により検出できるビオチンなどの化学的成分が含まれる。好ましくは、標識はFITCなどの蛍光性標識を含む。
【0073】
本発明の特異的結合要素は、ヒトまたは動物の患者、好ましくはヒトの診断または治療方法に使用されるようにデザインされる。
【0074】
したがって、本発明のさらに他の態様は、提供された特異的結合要素をそのままで、または例えばFITCなどの検出可能な標識に結合した1つまたは複数の他の試薬と共に投与することを含む診断方法を提供する。提供された特異的結合要素は、生検組織由来の肝線維症細胞の迅速かつ信頼性ある検査の開発に使用できる。
【0075】
本発明のさらに他の態様は、提供された特異的結合要素の投与を含む治療方法、そのような特異的結合要素を含む薬剤組成物、および投与のための医薬の製造における、例えば薬理学的に許容できる賦形剤と共に特異的結合要素を配合することを含む医薬または薬剤組成物の製法におけるそのような特異的結合要素の使用を提供する。
【0076】
肝星細胞に対する抗体を使用して治療に役立ち得る臨床的適応症には、肝線維症の結果病気になる如何なる疾患、例えばウィルス感染例えば肝炎およびアルコール乱用などの肝疾患も含まれる。提供された特異的結合要素はそのままヒト抗星細胞抗体または抗体様構造の受動免疫による肝線維症の直接的治療にも使用できる。
【0077】
本発明による抗線維症治療は肝線維症を有する患者にとって明らかな恩恵を提供するために使用できる。抗線維症治療は注射(例えば静脈に)または局所送達法によって与えることができる。提供された特異的結合要素は薬剤組成物の送達を標的肝星細胞へ向かわせるために使用できる。
【0078】
別の配合戦略は、経口または座剤経路に適した製剤を提供できる。投与経路は、治療剤の物理化学的性質により、疾患に対する特別の考慮により、有効性を最適化するように、または副作用を最小化するように決定することができる。
【0079】
本発明に従い、提供された組成物は個々人に投与できる。投与は好ましくは「治療的有効量」で行われ、治療的有効量は患者の利益になるのに十分な量である。そのような利益とは、少なくとも1つの症状の少なくとも1つ改善があればよい。実際に投与する量、および投与の速度と時間経過は治療されているものの性質および重症度に依存するであろう。治療の処方、例えば、投与量の決定その他は一般診療医およびその他の医師の責任範囲になる。抗体の適当な投与量は当業者にはよく知られている。Ledermann J.A.ら、(1991年)、Int.J.Cancer 47巻、659〜664頁、Bagshawe K.D.ら、(1991年)、Antibody,Immunoconjugates and Radiopharmaceuticals 4巻、915〜922頁を参照されたい。
【0080】
正確な投与量は、抗体が診断用か治療用か、治療すべき区域のサイズおよび位置、抗体の詳細な性質(例えば全抗体かフラグメントかダイアボディか)、および何らかの抗体に付けた検出可能な標識または他の分子の性質を含む多くの要因に依存するであろう。典型的な抗体投与量は0.5mg〜1.0gの範囲にあり、静脈への大量瞬時投与として投与できる。他の様式の投与には、同様の累積投与量を達成するために数時間かける静脈注射が含まれる。これは成人患者の単回治療のための投与量であって、少児および幼児のためには比例的に調節してよく、また他の抗体型に対しても分子量に比例して調節できる。治療は、医師の判断で、毎日、週2回、毎週または毎月の間隔で繰り返すことができる。
【0081】
他の様式の投与は、留置するデバイスのプレコートか、そうでなければその中への取込みを使用するもので、そのための最適抗体量は適当な実験によって決定されるであろう。
【0082】
本発明のある好ましい実施形態における抗体分子は、F(ab)またはscFvなどの単量体フラグメントである。そのような抗体フラグメントは半減期が比較的短いという利点を有する。
【0083】
本発明の特異的結合要素は、通常、薬剤組成物の形態で投与されるもので、特異的結合要素に加えて少なくとも1つの成分を含むことができる。
【0084】
したがって、本発明による、および本発明による使用のための薬剤組成物は、活性成分に加えて薬理学的に許容できる賦形剤、担体、緩衝剤、安定剤または当業者によく知られた他の材料を含むことができる。そのような材料は、無毒性であるべきで、かつ活性成分の有効性に干渉すべきでない。担体または他の材料の詳細な性質は投与経路に依存するであろう。投与経路は経口または注射、例えば静脈注射によるものでもよい。
【0085】
経口投与のための薬剤組成物は、錠剤、カプセル、粉末または液体の形態であってよい。錠剤はゼラチンなどの固体担体または助剤を含むことができる。液状薬剤組成物は通常、水、石油、動物もしくは植物油、鉱油または合成油などの液体担体を含む。生理食塩水、デキストロースまたは他の糖類溶液またはエチレングリコール、プロピレングリコールもしくはポリエチレングリコールなどのグリコールが含まれてよい。
【0086】
静脈注射または患部への注射のためには、活性成分は、発熱物質を含まず適当なpH、等張性および安定性を有する非経口的に許容できる水溶液の形態であろう。当業者は、例えば、塩化ナトリウム注射液、リンゲル注射液、乳酸リンゲル注射液などの等張溶媒を使用して適当な溶液を十分に調製できる。保存料、安定化剤、緩衝剤、抗酸化剤および/または他の添加剤が必要に応じて含まれてよい。
【0087】
組成物は、治療する疾患に依存して、単独でまたは他の治療と同時にもしくは順次に併用して投与できる。他の治療には、非ステロイド抗炎症薬(例えばアスピリン、イブプロフェンもしくはケトプロフェン)などの鎮痛薬またはモルヒネなどのアヘン剤、または抗嘔吐剤の適当な投与量の投与が含まれる。
【0088】
本発明は、本明細書で提供したシナプトフィジンに対する特異的結合要素の結合を惹起するかまたは許容することを含む方法を提供する。指摘したように、そのような結合はin vivoで、例えば、特異的結合要素、または特異的結合要素をコードする核酸の投与に続いて起こることができ、あるいはin vitroで、例えば、ELISA、ウェスタンブロット、免疫細胞化学、免疫沈降またはアフィニティークロマトグラフィーで起こることができる。
【0089】
特異的結合要素のシナプトフィジンに対する結合の量を測定することができる。定量は試験試料中の抗原量に関係づけることができ、試験試料は診断上興味があるものであってよい。
【0090】
抗体の試料への反応性は何らかの適当な手段によって測定できる。放射免疫測定(RIA)は1つの可能性である。放射活性標識抗原を非標識抗原(試験試料)と混合して、抗体に結合させる。結合していない抗原から結合した抗原を物理的に分離して、抗体に結合した放射性抗原の量を測定する。試験試料中に抗原が多くあればあるほど、抗体に結合する放射性抗原は少なくなるであろう。競争的結合測定はまた、レポーター分子に結合した抗原または類似体を使用して、非放射性抗原でも使用できる。前記レポーター分子は、蛍光体、リン光体または分光的に分離した吸収もしくは発光特性を有するレーザー色素であることができる。適当な蛍光体には、フルオレッセイン、ローダミン、フィコエリトリンおよびテキサスレッドが含まれる。適当な発色色素にはジアミノベンジジンが含まれる。
【0091】
他のレポーターには、着色しているか、磁性もしくは常磁性である高分子コロイド粒子もしくはラテックスビーズなどの粒子状材料、および視覚的に観察されるか、電子的に検出されるかもしくは他の方法で記録される検出可能なシグナルを直接的もしくは間接的に生じさせ得る、生物学的にまたは化学的に活性な作用物質が含まれる。これらの分子は、例えば、色を発せさせるかもしくは変化させるかまたは電気的性質に変化を起こさせる反応の触媒になる酵素であってよい。それらは分子的に励起可能であることができ、そのために電子のエネルギー状態間遷移の結果が特徴的な分光学的吸収または発光を生ずる。それらはバイオセンサーと併用する化学的要素を含むことができる。ビオチン/アビジンもしくはビオチン/ストレプトアビジンおよびアルカリホスファターゼ検出系が使用できる。
【0092】
個々の抗体-レポーター結合体により生じたシグナルは、試料中(正常試料および被検試料)における関連抗体結合の測定可能な絶対または相対データを引き出すのに使用できる。
【0093】
本発明は競争的測定において抗原レベルを測定するための上記のような特異的結合要素の使用、すなわち競争的測定において本発明により提供される特異的結合要素を使用することによる試料中の抗原レベルの測定方法も提供する。これは、結合していない抗原からの結合している抗原の物理的単離が必要ない場合であるだろう。結合したときに物理的または光学的変化が起こるようにレポーター分子を特異的結合要素に連結することは、1つの可能性である。前記レポーター分子は、検出可能な、好ましくは測定可能なシグナルを直接的または間接的に生ずることができる。レポーター分子の連結は直接的でもまたは間接的でも、共有結合でも、例えばペプチド結合でもまたは非共有結合でもよい。ペプチド結合による連結は、抗体およびレポーター分子をコードする遺伝子融合の組換え発現の結果として可能である。
【0094】
本発明は、例えばバイオセンサー系において本発明による特異的結合要素を使用することにより抗原レベルを直接的に測定することも提供する。
【0095】
結合を測定する様式は本発明の特徴ではなく、当業者はその好みおよび一般的知識により適当な様式を選択できる。
【0096】
本発明はさらに、シナプトフィジンへの結合に対して如何なる特異的結合要素にも劣らない、抗原に結合しかつ本明細書で実質的に開示したアミノ酸を有するCDRを含むVドメインまたは本明細書で実質的に開示したアミノ酸配列を有するVドメインを含む特異的結合要素に及ぶ。結合要素間の競争は、例えば、タグを付けていない一方の結合要素の存在下で検出できる特異的レポーター分子を、タグとして他方の結合要素に付けて、同一のエピトープまたは重複しているエピトープに結合する特異的結合要素の同定を可能にすることにより、in vitroで容易に測定できる。競争は例えばELISAまたはフローサイトメトリーを使用して測定できる。
【0097】
競争の試験において、抗原のペプチドフラグメント、特に興味のあるエピトープを含むペプチドを使用できる。エピトープ配列に加えて1つまたは複数のアミノ酸をいずれかの末端に有するペプチドが使用できる。そのようなペプチドは特定された配列により「本質的に構成されている」といわれる。本発明による特異的結合要素は、それらの抗原に対する結合が、特定の配列を有するかまたは含むペプチドによって阻止されるようなものである。これを試験するために、1つまたは複数のいずれかのアミノ酸を加えた配列を有するペプチドが使用できる。
【0098】
特定のペプチドに結合する特異的結合要素を、例えば、ファージディスプレイライブラリーから前記ペプチドでのパニングにより単離できる。
【0099】
本発明はさらに、本発明の特異的結合要素をコードする単離した核酸を提供する。核酸はDNAおよびRNAを含む。好ましい態様において、本発明は上で定義した本発明のCDR、VHまたはVLドメインをコードする核酸を提供する。
【0100】
本発明はまた、構築体を、上記の少なくとも1つのポリヌクレオチドを含むプラスミド、ベクター、転写または発現カセットの形態で提供する。
【0101】
本発明はまた、上記の1つまたは複数の構築体を含む組換え宿主細胞も提供する。任意のCDR、VHまたはVLドメインをコードする核酸または提供された特異的結合要素それ自体が本発明の態様を形成し、コードされた生成物の製造方法も同様であって、その方法はその生成物をコードする核酸からの発現を含む。発現は核酸を含む組換え宿主細胞を適当な条件下で培養することにより簡便に達成できる。発現による生成に続いて、VHもしくはVLドメインまたは特異的結合要素を、任意の適当な技法を使用して単離および/または精製し、それから適宜使用する。
【0102】
本発明による特異的結合要素、VHおよび/またはVLドメイン、およびコードする核酸分子およびベクターは、例えばそれらの自然の環境から、実質的に純粋なまたは均質な形態で単離しおよび/または精製して、あるいは核酸の場合には、求められている機能を有するポリペプチドをコードする配列以外に原料由来の核酸または遺伝子を含まずにまたは実質的に含まずに提供できる。本発明による核酸はDNAまたはRNAを含むことができ、かつ、全体または部分が合成によることができる。本明細書で述べたヌクレオチド配列への言及は、特に断らない限り、前記の特定した配列を有するDNA分子を包含し、前記特定した配列においてTの代わりにUで置換したRNA分子をも包含する。
【0103】
種々の異なった宿主細胞におけるポリペプチドのクローニングおよび発現のための系がよく知られている。適当な宿主細胞には、細菌、哺乳動物細胞、酵母およびバキュロウイルス系が含まれる。異種ポリペプチドの発現に当業者の使用できる哺乳動物細胞ラインには、チャイニーズハムスター卵巣細胞、HeLa細胞、仔ハムスター腎細胞、NSOマウス黒色腫細胞、YB2/0ラット骨髄腫細胞およびその他多くのものが含まれる。一般的で、好ましい細菌宿主は大腸菌である。
【0104】
大腸菌などの原核細胞における抗体および抗体フラグメントの発現は、当業者においては十分確立されている。総説としては、Pluckthun、A.、Bio/Technology 9巻、545〜551頁(1991年)を参照されたい。培養における真核細胞中の発現も、特異的結合要素の生成のための選択肢として当業者に使用できる。最近の総説として、例えば、Ref、M.E.、(1993年)Curr.Opinion Biotech.4巻、573〜576頁、Trill J.J.ら、(1995年)Curr.Opinion Biotech 6巻、553〜560頁を参照されたい。
【0105】
適当なベクターは、プロモーター配列、ターミネーター配列、ポリアデニル化配列、エンハンサー配列、マーカー遺伝子および他の配列を適宜含む適当な制御配列を含んで、選択または構築できる。ベクターは適宜、プラスミド、ウィルス性の例えばファージまたはファージミドにすることができる。「Molecular Cloning:a Laboratory Manual」第3版、SambrookおよびRussell、2001年、Cold Spring Harbor Laboratory Press。核酸の操作、例えば、核酸構築体の調製、変異誘発、塩基配列決定、DNAの細胞への導入および遺伝子発現、およびタンパク質の分析のために多くの知られた技法およびプロトコルが、「Current Protocols in Molecular Biology」第2版、Ausubelら(編)、John Wiley & Sons、(1992年)に詳細に説明されている。
【0106】
したがって、本発明のさらなる態様は、本明細書で開示したように核酸を含む宿主細胞を提供する。さらに他の態様は、そのような核酸を宿主細胞へ導入することを含む方法を提供する。前記導入には如何なる使用可能な技法も使用できる。真核細胞に対して、適当な技法は、リン酸カルシウムトランスフェクション、DEAE-デキストラン、エレクトロポレーション、リポソーム媒介トランスフェクションおよびレトロウィルスまたは他のウィルス例えばワクシニアまたは昆虫細胞に対してはバキュロウイルスを使用する形質導入を含むことができる。細菌細胞に対して、適当な技法は、塩化カルシウム形質転換、エレクトロポレーションおよびバクテリオファージを使用するトランスフェクションを含むことができる。
【0107】
前記導入に続いて、例えば、遺伝子の発現のための条件下で宿主細胞を培養することにより、核酸の発現を起こさせるまたは起こすに任せることができる。
【0108】
一実施形態において、本発明の核酸は宿主細胞のゲノム(例えば染色体)中に組込まれる。組込みは、標準的技法に従って、ゲノムとの組換えを促進する配列を含めることにより促進することができる。
【0109】
本発明は、上記の特異的結合要素またはポリペプチドを発現させる目的で、発現系に上で述べたような構築体を使用することを含む方法も提供する。
【0110】
本発明の態様および実施形態を、ここで次の実験を参照して実施例により例示することにする。
【0111】
本明細書において如何なる箇所で引用した文献もすべて参照により組み込む。
【発明を実施するための最良の形態】
【0112】
材料および方法
ペプチド-ウシ血清アルブミン結合体
シナプトフィジンタンパク質(図1)の外形質側に存在するアミノ酸に対応するペプチドYPFRLHQVYFDAPSCをFmoc法の使用により合成し(Proteomics Group、University of Aberdeen)、HPLCにより純粋と判定した。ペプチドの分子量は質量分析法によって確かめた(データは示していない)。前記ペプチドを、3-マレイミド酢酸N-ヒドロキシスクシンイミド(MBS)を使用して、ウシ血清アルブミンに化学的に結合した(Leel V.ら、Biochem Biophys Res Commun 2004年、316巻、872〜7頁)。SDS-PAGEおよび質量分析法による分析(MALDI-TOF、Applied Biosystems Voyager-DE STR)により前記ペプチドがBSAに結合したことを確認した。一般に平均して、アルブミン1分子当りおよそ4ペプチド分子であった(データは示していない)。
【0113】
ペプチド配列に対する可溶性単鎖抗体フラグメント(scAb)のファージディスプレイ生成
ヒト単倍(single-fold)scFvファージミド(pIT2)ライブラリー(Tomlinson I + J、MRC、UK)を、本質的にはLeel V.らがBiochem Biophys Res Commun 2004年、316巻、872〜7頁で概略を説明しているようにして、ペプチド配列に特異性のあるファージ抗体をスクリーニングするために使用した。ペプチド配列に対してパニングを合計3回実施した。ペプチドへの特異性のあるファージ抗体のBSAに対する比を上げるために、パニングを2%(w/v)ドライミルク(MPBS)および2ml/mlBSAを含有する4mlのPBS(137mM NaCl、2.7mM KC1、10mMリン酸塩、pH7.4)中で実施した。
【0114】
抗ペプチドファージ抗体ELISA
PBS中のタンパク質(peptide-BSAまたはBSA)2μg/ウェルを4℃で一晩かけてコートした平底96ウェルのImmulon-4マイクロタイタープレート(Dynex、Sussex、UK)を使用して、ペプチドに対する結合要素を増やした精製ポリクローナルファージをELISAにより測定した。結合したファージを、100μl/ウェルの西洋ワサビペルオキシダーゼ(HRP)結合抗M13抗体(AmershamPharmacia)と1時間インキュベートし、洗浄して、また100μl/ウェルのテトラメチルベンジジン二塩酸塩溶液(KPL Laboratories、Gaithersburg、メリーランド州)とインキュベートすることにより検出した。反応を50μl/ウェルの1M H2SO4で停止して、450nmの吸収をマイクロプレートリーダーを使用して読み取った。
【0115】
ペプチドに対するモノクローナルファージ抗体の同定および発現ベクターpIMS147中へのサブクローニング
第3パニングからの個々のコロニーを96ウェルのプレート(Greiner)中で成長させ、M13 K07でファージ抗体を回収した。
【0116】
ペプチド-BSAおよびBSA単独に対する結合について、ファージ上清の特異性をELISAにより測定した。陽性クローンのscFvコード領域を発現ベクターpIMS147中にサブクローニングした。これは、IPTG(イソプロピル-β-D-チオガラクトピラノシド)誘導pUC系ベクターpIMS100(Hayhurst A.ら、Protein Expr Purif 1999年、15巻、336〜43頁)の改質である。ヘキサヒスチジンのタグ付加により、固定化金属イオンキレートアフィニティークロマトグラフィーによる、発現したscAbの精製が可能になる。ベクターは大腸菌 TG1またはXL 1ブルー細胞に形質転換した。
【0117】
scAbの発現、精製およびキャラクタリゼーション
Leel V.ら、Biochem Biophys Res Commun 2004年、316巻、872〜7頁で概略が説明されているようにして、IPTG処理した細胞中でscAbを発現させ、Ni2+負荷IMACFast Flow Sepharose樹脂(AmershamPharmacia)をメーカーの指示に従って使用して、ヘキサヒスチジンC末端タグ尾部により精製した。PBSに対して4℃で十分透析した後、scAbを-20℃で保存した。精製したscAbは、Leel V.ら、Biochem Biophys Res Commun 2004年、316巻、872〜7頁で概略が説明されているようにして、全ヒトIgG測定用標準品を使用して、ヒトCκドメインによる捕捉ELISAによって測定した。
【0118】
発現し精製された抗ペプチドscAB ELISA
発現したscAbは、本質的にはポリクローナル抗ペプチドファージ抗体ELISAに対して概略を説明したようにして、ペプチドと相互作用する能力をELISAにより試験した。結合したscAbは、西洋ワサビペルオキシダーゼ結合抗ヒトCκL鎖抗体(Sigma Chem.Co.Poole、UK)を使用して検出した。
【0119】
SDS-PAGEおよびウェスタンブロット
ウェスタンブロットは、本質的にはWright M.C.ら、Mol Pharmacol 1996年、50巻、856〜63頁に述べられているようにして実施した。
【0120】
タンパク質のFITC標識
ScAbは、FluoroporterFITCタンパク質標識キット(Molecular Probes)を、メーカーの指示に従って使用して、蛍光で標識した。この方法は、トリブチル錫イソチオシアナート(Aldrich Chemicals、Poole、UK)を使用して、C1-3scAbをトリブチル錫に結合させるためにも使用した。
【0121】
動物および細胞の調製
ヒト肝細胞はUK Human Tissue Bank(DeMontfort University、Leicester、UK)から入手した。ヒトHSCは腫瘍が存在するという理由で患者から除去した肝臓の正常組織の縁から単離された。これらの研究におけるヒト組織の使用はGrampian Regional Ethics Committeeにより許可され、供与者の全面的同意を得た。ヒト肝細胞培養のプロトコルはWright M.C.ら、Hum Exp Toxicol、1996年、15巻、203〜4頁およびHarvey J.L.ら、Drug Metab Dispos、2000年、28巻、96〜101頁に掲載されている。ラットHSCおよびB-13細胞の培養はWright M.C.ら、Gastroenterology、2001年、121巻、685〜98頁およびMarek C.J.ら、Biochem J、2003年、370巻、763〜9頁に掲載されている。
【0122】
FITC標識scAbの細胞とのインキュベーション
FITC-C1-3scAbを、蛍光顕微鏡により、培養中の肝星細胞と相互作用する能力について試験した。ヒト肝星細胞は、実験に先立って完全培地中のチャンバースライド(Nunclon、Naperville)上に播種した(チャンバー当り1×103細胞)。細胞はリン酸緩衝生理食塩水(PBS)で2度洗浄し、10μgのscABまたはFITC-C1-3scAbを含有する500μlのHepes/HBSS緩衝液(0.14M NaCl、5.4mM KC1、0.34mM Na2HPO4、0.44mM KH2PO4、5.6mMグルコース、1mM CaCl2、6mM HEPES、4mM NaHCO3 pH7.4)中、室温暗所で1時間インキュベートした。次いで、細胞をPBSで洗浄し、4',6'-ジアミジノ-2-フェニルインドール(DAPI)を含有するVectashield封入剤(Vector laboratories、Burlingame、カリフォルニア州)で顕微鏡標本を作製して、細胞核を染色しカバーガラスをかけた。顕微鏡観察はPhotometrics Sensisカメラ付きZeiss Axeola II顕微鏡を使用して実施し、データはSmart Captureプログラムを使用して取り込んだ。ヒト肝細胞(UK Human Tissue Bank)は、最初コラーゲンをコートした24ウェルプレートで培養し(1×105/ウェル)、80単位/mlのペニシリンおよび80μg/mlのストレプトマイシンを補充したWilliamの培地E中に維持した。
【0123】
共培養実験のためには、肝星細胞を肝細胞培養物(ウェル当り1×103細胞)中にシードして、実験に先立って24時間培養した。純肝星細胞の培養について簡単に説明したのと同様にして、C1-3またはFITC-C1-3scAbをインキュベートし、1時間のインキュベーションおよび洗浄後、細胞をトリプシン処理および1000rpmで5分間遠心することにより回収した。次に細胞ペレットを0.4%ホルムアルデヒド/106細胞で再懸濁して、細胞遠心およびDAPI含有Vectashieldで顕微鏡標本を作製することによりガラススライドに付けた。スライドは前記のようにして蛍光顕微鏡により観察した。
【0124】
DAPI染色は核のFITC-C1-3染色を隠蔽するので、FITC-C1-3のHSCへの結合に対するププチドおよびモネンシンの影響を比較するために、細胞固定なしの蛍光顕微鏡観察を使用した。24ウェルプレートにシードしたHSCをHepes/HBSSで洗浄してから、l0μgのscAbを含有し添加化合物を含むかまたは含まない500μl/well Hepes/HBSS中37℃でインキュベートした。1時間後、細胞を500μlのHepes/HBSSで3回洗浄した。フィルターセット#9(BP 450〜490nm;LP 515nm)を使用するAxiovert100顕微鏡(Zeiss、ドイツ)を使用して細胞を分析した。
【0125】
FACS分析
FITC-C1-3scAbを培養液に加え(200μg/10m1)、HSCの集密的な(直径10cmのシャーレ)培養物と共に37℃で1時間インキュベートした。次に媒質を除去して細胞をPBSで洗浄した。次に細胞単離溶液(Sigma、Poole、UK)を使用して細胞を非酵素的に培養基体から剥離し、遠心によりペレット化して、BD cytofix/cytoperm(BD Biosciences、Oxford、UK)を使用して固定/透過処理した。15分後、細胞を0.5mlの1×wash/perm buffer(BD Biosciences、Oxford、UK)で洗浄し、l×wash/perm bufferで希釈したマウス抗α-平滑筋アクチンモノクローナル抗体(Sigma、Poole、UK)とインキュベートした。次に細胞をl×wash/perm bufferで洗浄し、ビオチン結合抗マウスIgG(1:200、DakoCytomatron、Cambridgeshire、UK)と氷上で30分間インキュベートし、2回洗浄してから、ストレプトアビジン-アロフィコシアニン(APC)結合体(1:200、BD Biosciences、Oxford、UK)と氷上で30分間インキュベートした。細胞はフローサイトメトリー(LSR I、BD、Oxford、UK)で分析した。細胞をアイソタイプ特異的対照抗体とインキュベートすることにより、非特異的蛍光の対照とした。
【0126】
結果
標的ペプチドYPFRLHQVYFDAPSCに対する親和性のあるTomlinson I+Jライブラリーから組換えファージを探索するために、ファージディスプレイの技法を使用した。図2は標的ペプチドに親和性のあるファージ抗体が選択的に免疫チューブに結合し、3回の別々の探索(pan)を通して増幅されたことを示す。第3パニングライブラリーからの96の別々のクローンを、BSAおよび標的ペプチド-BSAに結合するファージ抗体を求めて、個々にスクリーニングした。この測定においては、57クローンが標的ペプチド-BSAに対する高い親和性を示し、BSAに対して何らかの親和性を示したクローンはなかった(データは示していない)。モノクローナルファージ-抗体ELISAにおいて最高の反応を与えた12クローンを選択して、scABコード領域の配列を決定した。
【0127】
配列決定は、多くのクローンがscAbコード領域内にインフレーム停止コドンを含むことを示した。構築体をpIMS147にサブクローニングして、サプレッサtRNAを含むE.cloi菌株(例えばTG-1)に形質転換した。これらの構築体はTG-1でペプチドに高い親和性を有するscAbを生成したが、高レベルのscAb発現は得られなかった(データは示していない)。しかしながら、ファージ抗体クローンC1は、scAbをコードする領域内に停止コドンを含まず、モノクローナルファージ抗体ELISAスクリーニングにおいて良好な結合反応を与えた(図3)。クローンC1はpIMS147ベクターにサブクローンして、XL-1ブルー大腸菌に形質転換した。クローンpIMS147 C1-3(図4)は、ELISAで前記ペプチドを特異的に認識する(図6)scAbの高レベルの発現(図5)を指令した。発現したscAbのためのELISAアッセイにより、ペプチドYPFRLHQVYFDAPSCに対して特異的であるscAbの生成が確認された。図7は、発現したscAbがBSA単独に対してよりもペプチド-BSA(すなわち、YPFRLHQVYFDAPSC-BSA)に対して約8倍多く結合したことを示す。scAbを欠損する対照のペプチド-BSAまたはBSAのいずれに対する結合もわずかであった。いくつかの異なったタイプの細胞抽出物に対するウェスタン分析は、C1-3scAbが、ヒト抗体のターゲティングであるHSC12の非グリコシル化シナプトフィジンタンパク質(Eastwood S.L.ら、Brain Res Bull、2001年、55巻、569〜78頁)の予測サイズである約32kDのヒトHSC中のタンパク質に結合することを示す。興味深いことに、C1-3scAbはラットHSC中のシナプトフィジンと交差反応しない(図8)。
【0128】
C1-3scAbが生きているHSC C1-3の表面に結合できるか否かを決めるために、C1-3scAbをFITCで標識し、SDS-PAGEにより、図9に示すようにFITC-C1-3scAbの移動距離の減少により判定して確認した。透析の前後および濃縮後のC1-3Abは約40Kdaの分子量を有していた。FITCで標識したC1-3scAbの分子量は約45Kdaであった。
【0129】
次にFITC-C1-3 scAbを活性化した培養中のヒトHSCに加えた。ヒトHSC培養物の免疫細胞化学染色(示していない)およびFACS分析は、細胞がまったくα平滑筋アクチン陽性のものだけで、したがって活性化したHSCおよび/または肝臓由来の筋線維芽細胞であることを示した(両方の細胞のタイプは線維発生を助長すると信じられている(Ramadori G.ら、Liver、2002年、22巻、283〜94頁)。図10を参照されたい)。固定に先立つ自然条件下におけるFITC-C1-3での共染色細胞は、すべての細胞がC1-3 scAbに結合したことを示したが、α平滑筋アクチンに比較してFITC-C1-3染色の強度にはより大きな変動があった(図10)。このことは、シナプトフィジンのレベルが上がった、および/またはFITC-Cl-3 scAb取込み機序が貪欲化したα平滑筋アクチン発現細胞の亜集団があり得ることを示唆している。HSCとC1-3、FITC-C1-3またはトリブチル錫結合C1-3 scAb(TBT-C1-3)との会合は培養媒質濃度の低下に対応していた(図11)。HSCをアポトーシス(グリオトキシン)または壊死(クロルプロマジン)により殺すことが知られている化合物と対照的に、C1-3またはFITC-C1-3について毒性の証拠はない(図12)。興味深いことに、TBT-C1-3 scAbはHSCに有毒であったので、C1-3 scAbが内部移行することおよび薬剤をC1-3に結合して薬剤の薬理作用を保持することが可能であることが示唆される。
【0130】
蛍光実験は、(フルオレッセインによる)緑色蛍光がFITC-C1-3 scAbとインキュベートした肝星細胞に付随するので、FITC-C1-3 scAbが培養中の肝星細胞と特異的に相互作用することを示唆する。光漂白を阻止しかつ核をブルーに染色した封入剤を使用して、大部分の場合、蛍光が核膜および細胞膜の付近に局在することが見出された。染色はしばしば点状に見え、それは局所化された構造との関連を反映しているのかもしれない。この点において、シナプトフィジンはニューロン中のシナプス小胞に濃縮されて、その場所でサブユニット数可変の単独オリゴマーを形成していることが知られている。染色は核膜付近でも観察されて、C1-3 scAbが細胞内に蓄積しやすいという提案を支持している。(フルオレッセインと結合していない)C1-3Abとインキュベートした肝星細胞培養物では検出可能な緑色蛍光は観察されない。C1-3 scAbと肝星細胞との間の相互作用の特異性を決定するために、ヒト肝細胞もFITC-C1-3 scAbとインキュベートした。C1-3 scAbまたはFITC-C1-3 scAbのいずれとインキュベートした肝細胞も、関連する緑色蛍光を示さず、C1-3 scAb抗体は肝細胞に結合しないことを示唆した。さらに、C1-3 scAbの能力の細胞タイプ間での差を知るために、肝星細胞と肝細胞とを共培養して、FITC C1-3 scAbとインキュベートした。蛍光は星細胞上に観察され、一方肝細胞は検出可能なレベルの関連する緑色蛍光を示さないことで一貫していた。共培養物を(フルオレッセインを結合していない)C1-3 scAbとインキュベートした場合、有意な緑色蛍光は実験では観察されなかった。肝星細胞への結合についてのC1-3scAbの作用の特異性をFITC標識BSAを使用して検討した。ヒト肝星細胞および肝細胞の両方をBSAおよびFITC-BSAとインキュベートした。ヒトHSCおよび肝細胞の両方とも、FITC-BSAとインキュベートしたときに蛍光の細胞質への集中を示し、蛍光は肝細胞へのC1-3scAb取込みを非特異的に阻止するタンパク質(すなわちC1-3)上の蛍光の存在ではなく、C1-3scAbは肝細胞とは相互作用しないことを示唆した。COS-7またはHepG2細胞では、C1-3scAbまたはC1-3scAb
-FITCとインキュベートしたとき、蛍光の結合は検出されなかった。
【0131】
ペプチドまたはモネンシンのFITC-C1-3 HSC結合/取込みへの影響を比較するために、実験に先立つ固定およびDAPI共染色なしの蛍光顕微鏡観察を使用した。図13は、対照のFITC標識3A8scAb(毒素のミクロシスチンに対して前に調製した(McElhiney J.ら、Appl Environ Microbiol、2002年、68巻、5288〜95頁))と対照的に、FITC-C1-3 scAbがHSCと結合したことを示す。シナプトフィジンタンパク質の外形質側に存在する他のアミノ酸に対応するペプチドATDPENIIKEMPMCの培養への添加は、FITC-C1-3のHSCとの結合を有意に阻止しなかったが、一方標的ペプチドYPFRLHQVYFDAPSCは、HSCへのFITC-C1-3の結合を、投与量に依存して減少させた。エンドサイトーシス阻止剤であるモネンシンの添加は、FITC-C13で染色された細胞の総数を抑えなかったが、核のFITC-C1-3染色の減少によって示されるように取込みを減少させた。
【0132】
本質的には、Foxら、J Microbiol Methods、2004年、56巻、221〜230頁により述べられているようにして、C1-3 scAbを1mg/mlの濃度でグリオトキシンと結合した。SDS-PAGEにより、前記タンパク質が改質されたことが確認された(データは示していない)。グリオトキシンに対する抗体(Foxらによって得られた)は、グリオトキシンがC1-3scAbに共有結合で結合したことを確認した。MALDI-TOF分析は前記タンパク質の質量が1683ドルトンだけ増加することが最も多いことを示した。結合方法を考慮に入れると、大部分のC1-3タンパク質が4分子のグリオトキシンと結合したことが示唆される。図14は、グリオトキシンがヒトHSCを殺し、グリオトキシンと結合したC1-3もまたHSCを殺したことを示す。このデータは薬剤をC1-3scAbに結合してかつターゲティングおよび薬効を保持することが可能であることを示している。
【図面の簡単な説明】
【0133】
【図1】シナプトフィジンの模式図である。ヒトのシナプトフィジンの理論分子量は33.8kDaで、概略を描いたように細胞の形質膜の厚さにわたると予測される。このタンパク質はグリコシル化されていると報告されていて、実験的にはおよそ38〜40kDaの分子量を有する(Eastwood S.L.ら、Brain Res Bull 2001年、55巻、569〜78頁)。
【図2】標的ペプチド-BSAに対する親和性を有するファージ抗体(ポリクローナル)の単離および増幅を示す図である。バイオパニング中における標的ペプチド-BSA抗原を認識するファージ抗体(ポリクローナル)の濃縮をELISAによって評価した。各回の選択から回収したポリクローナルファージ抗体(〜1×1010)を標的ペプチド-BSAへの結合について測定した。結合したファージは、方法の項で概説したようにHRP-標識抗M13抗体(Pharmacia)を使用して検出した。各パニングでのファージ力価を示す(---□---)。
【図3】図2のために概略を示したのと同様にして標的ペプチド-BSAへの結合を測定したC1-ファージ抗体クローン(モノクローナル)を示す図である。
【図4A】pIMS147ベクター(配列番号7および配列番号8の配列)内のC1-3scAbのヌクレオチドおよびアミノ酸配列(前部)を示す図である。抗原結合部位を構成する超可変相補性決定領域(CDR)は太字である。HおよびL鎖に結合する柔軟性のアミノ酸リンカー(Gly4、Ser)3は下線を引いてある(______)。フレームワーク領域(FW)の開始点およびヒト定常カッパドメイン(Cκ)の開始点も示す。クローニング部位を、アミノ酸配列の上に示した対応する制限酵素と共に下線(___)で示した。HuCκ定常ドメインの開始および6ヒスチジン残基精製タグを示す。全アミノ酸配列は以下の通りである。
【図4B】pIMS147ベクター(配列番号7および配列番号8の配列)内のC1-3scAbのヌクレオチドおよびアミノ酸配列(後部)を示す図である。抗原結合部位を構成する超可変相補性決定領域(CDR)は太字である。HおよびL鎖に結合する柔軟性のアミノ酸リンカー(Gly4、Ser)3は下線を引いてある(______)。フレームワーク領域(FW)の開始点およびヒト定常カッパドメイン(Cκ)の開始点も示す。クローニング部位を、アミノ酸配列の上に示した対応する制限酵素と共に下線(___)で示した。HuCκ定常ドメインの開始および6ヒスチジン残基精製タグを示す。全アミノ酸配列は以下の通りである。 AAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQES VTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGESHHHHHH
【図5】XL-1ブルー細胞におけるC1-3scAb発現の誘起およびNiアフィニティークロマトグラフィーによる精製を示す図である。上方のブロットは、示したタンパク質を負荷したSDS-PAGE後のクーマジーブルー染色を示し、下方のブロットは抗ヒトCκL鎖抗体による試料のウェスタン分析を示す。
【図6】C1-3scAbの抗ヒトCκL鎖捕捉ELISA測定を示す図である。ヤギ抗ヒトCκL鎖抗体を96ウェルプレートの表面にコートした。ヒトIgGまたはC1-3scAb標品の希釈溶液を個々のウェルでインキュベートした。十分に洗浄した後捕捉されたIgG(-□-)またはscAb(-○-)を方法の項で概説したようにして測定した。データは別々の7標品の代表的なものの、3回の別々の測定の平均および標準偏差である。
【図7】発現したscAbを、ペプチドYPFRLHQVYFDAPSC(配列番号9)と相互作用する能力について、抗BSA-標的ペプチドscAbを使用するELISAによって試験した図である。96ウェルプレートのウェルをBSA-標的ペプチドまたはBSAでコートして、C1-3scAb(グレイの縦線)または対照scAbインキュベーション緩衝液(無色の縦線)とインキュベートし、続いて十分に洗浄した。結合したscAbはHRP結合抗ヒトCκL鎖抗体を使用して検出し、方法の項で概説したようにして測定した。データは3回の別々の測定の平均および標準偏差である。「BSA-ペプチド2」は標的ペプチドである。
【図8】C1-3 scAbの標的ペプチドへの結合を示すウェスタンブロットの図である。細胞抽出物(5μg/lane)をウェスタンブロットにかけて、膜をC1-3scAbのプローブで調べ、続いてHRP結合抗ヒトCκL鎖抗体とインキュベートし、ECL試薬を使用して検出した。rHSC - ラット肝星細胞; hHSC - ヒト肝星細胞(AH4=匿名の患者コード);rHSC-ラット肝細胞;B13-ラット膵幹細胞;B13-H-肝細胞に分化転換後のラット膵幹細胞。
【図9】C1-3 scFvをFITCで標識して、SDS-PAGEにより確認した図である。Ni2+をチャージしたIMAC Fast Flow Sepharose樹脂で精製したC1-3 acAbを透析にかけ、次にcentricon YM-3遠心濃縮機を使用して濃縮した。次に濃縮したC1-3 scAbをFITC標識し、各画分の一定分量(約0.1μg/レーン)をSDS-PAGEにかけ、続いてゲルのクーマジーブルー(全タンパク質)染色を行った。
【図10】1次抗体なしで(上方パネル)またはありで(下方パネル)インキュベートしたヒトHSCのFACS分析、すなわちFITC-C13 scAb(FITC)およびα平滑筋アクチン(α-APC)に対するマウスモノクローナル抗体の図である。抗α平滑筋アクチン抗体は、方法の項で概説したようにして、ビオチン結合2次抗体を使用し、続いて蛍光体結合ストレプトアビジンとインキュベートすることにより検出した。数字は各象限内の細胞の百分率を表す。データは3つの別々の細胞標本の代表的なものである。
【図11】培養中のヒトHSCによるCl-3 scAbおよびC1-3結合体の取込みを示す図である。24ウェルのプレートにシードしたHSCを、5μgのscAbを含む0.3mlの培地中でインキュベートした。提示した時間に培地試料を取り、ウェスタンブロットにかけ、HRP結合抗ヒトCκL鎖抗体を使用してscAb(上方パネル)および負荷対照の血清アルブミン(下方パネル)を検出した。4つの別々の実験の代表的な結果である。
【図12】scAbインキュベートのHSC生存度への効果を示す図である。24ウェルのプレートにシードしたHSCを、5μgのscAbまたは提示した化学薬品を含む0.3mlの培地中でインキュベートした。細胞を3時間培養した後、培地を除去して1×PBSで洗浄した。付着率(生存度の尺度)を培養ウェル中のタンパク質を直接測定して測定した。データは同一の実験の3つの別々のウェル、別々の3回の実験の代表的なものの、平均および標準偏差である。*スチューデントT検定(両側)を使用すると、対照から有意の差がある。P>95%。
【図13】FITC-C1-3 scAbのヒトHSCへの結合に対するシナプトフィジンペプチドおよびモネンシンの効果を示す図である。24ウェルのプレートにシードしたヒトHSCを洗浄し、10μgのC1-3 scAb;10μgのFITC-C1-3 scAb;10μgのFITC-C1-3 scAbおよび0.25ナノモルの標的ペプチド;10μgのFITC-C1-3 scAbおよび2.5ナノモルの標的ペプチド;10μgのFITC-C1-3 scAbおよび25ナノモルの標的ペプチド;10μgのFITC-C1-3 scAbおよび25ナノモルの標的ペプチドP1;10μgのFITC-C1-3 scAbおよび2.5μMのモネンシン;10μgのFITC-C1-3 scAbおよび15μMのモネンシン;10μgのFITC-3A8 scAb(scAbはミクロシスチンに対して生成されたもので、HSCには結合しないと予想される)のいずれかを含むウェル当り500μlのHepes/HBSS中で1時間インキュベートした。モネンシンとインキュベートした細胞は10分間プレインキュベートした。scABを添加する直前に、培地を交換して細胞にモネンシンを再投与した。図には、無作為に選択した20視野からの1視野当りの蛍光性細胞の百分率の平均±標準偏差の測定分析を示す。バー=50μm。Momen=モネンシン;P1=ATDPENIIKEMPMCペプチド;P2=標的ペプチド。
【図14】HSC生存度を示す図である。24ウェルのプレートで培養したヒトHSCを、賦形剤DMSO;1.5μMグリオトキシン(750ピコモル);20μlの1mg/mlC1-3scAb(20μgのC1-3scAb);または20μlの0.5mg/mlC1-3-グリオトキシン結合体(10μgC1-3scAb-250ピコモルC1-3scAb*/1000ピコモルグリオトキシン)のいずれかを含む500μlの媒質で処理した。*0.5mg C1-3 タンパク質/ml貯蔵液=12.5μM=12.5ナノモル/ml=12.5ピコモル/μl。
【特許請求の範囲】
【請求項1】
アミノ酸配列YPFRLHQVYFDAPSC(配列番号9)内のシナプトフィジンエピトープに結合する、活性化肝星細胞に結合した際に取り込まれる抗体。
【請求項1】
アミノ酸配列YPFRLHQVYFDAPSC(配列番号9)内のシナプトフィジンエピトープに結合する、活性化肝星細胞に結合した際に取り込まれる抗体。
【図1】


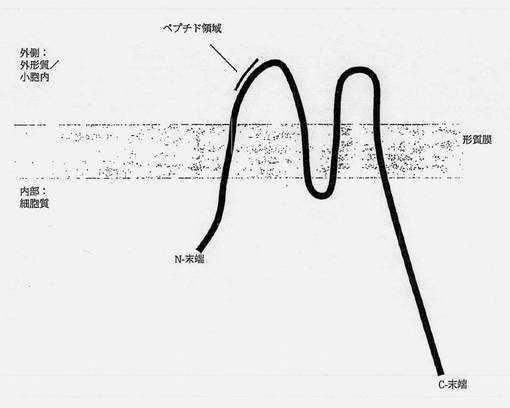
【図2】
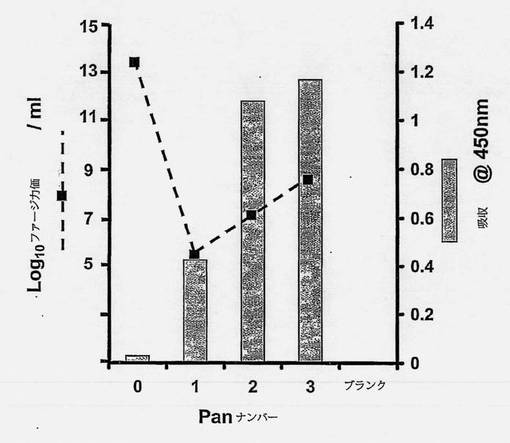
【図3】
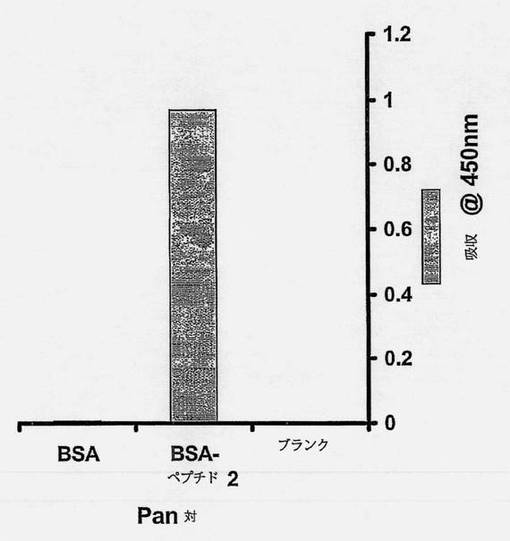
【図4A】

【図4B】

【図5】
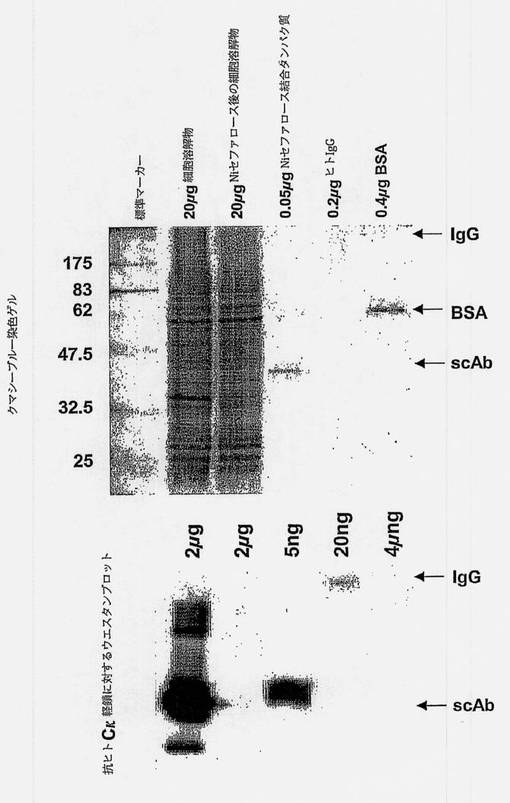
【図6】
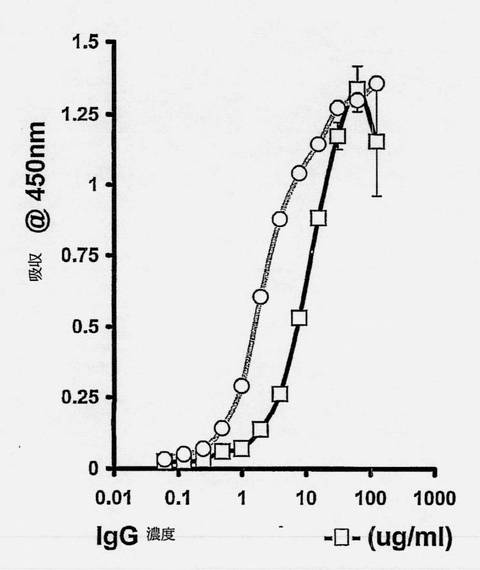
【図7】
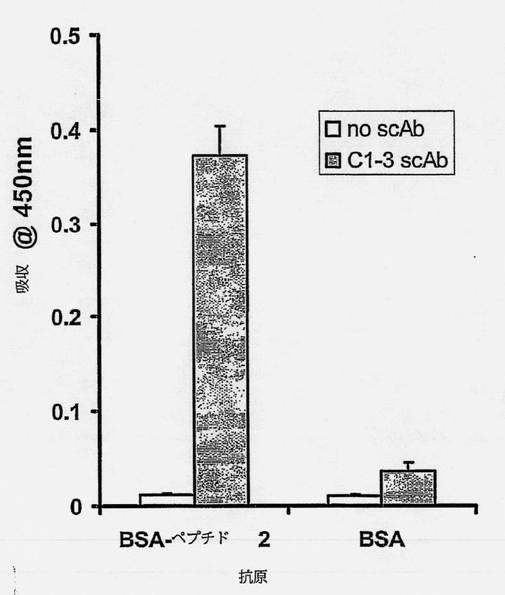
【図8】
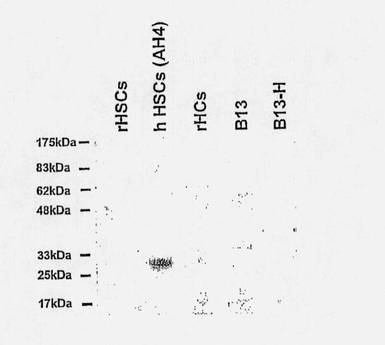
【図9】
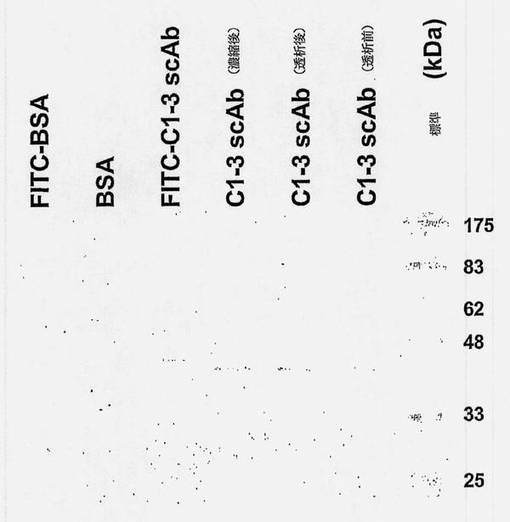
【図10】
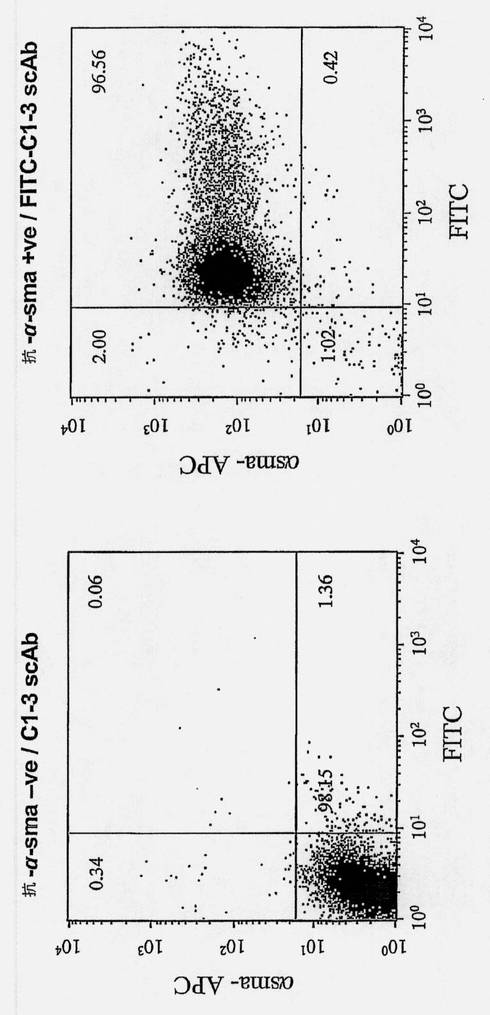
【図11】
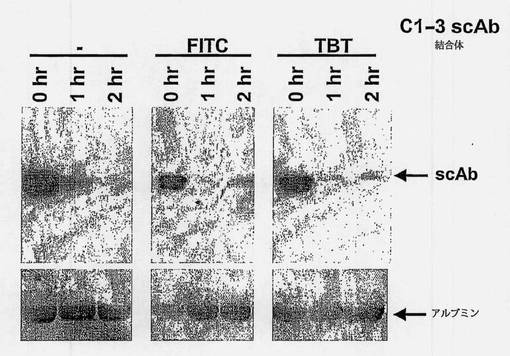
【図12】
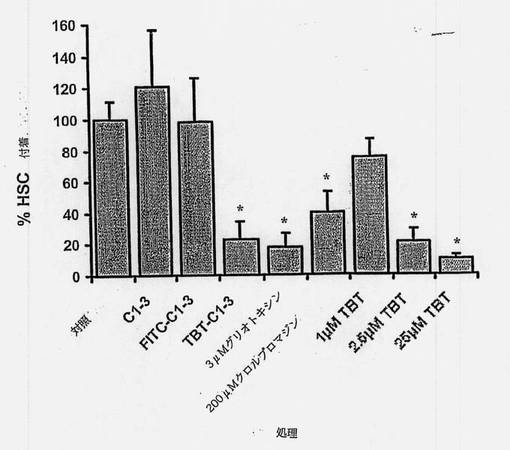
【図13】
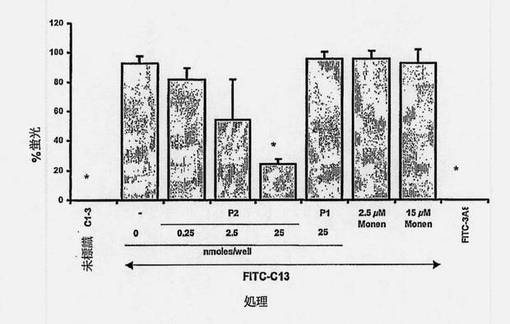
【図14】
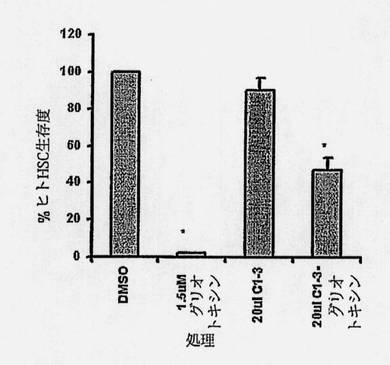
【図2】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【公開番号】特開2012−162551(P2012−162551A)
【公開日】平成24年8月30日(2012.8.30)
【国際特許分類】
【出願番号】特願2012−87427(P2012−87427)
【出願日】平成24年4月6日(2012.4.6)
【分割の表示】特願2007−505627(P2007−505627)の分割
【原出願日】平成17年3月29日(2005.3.29)
【出願人】(506330922)ザ・ユニバーシティ・コート・オブ・ザ・ユニバーシティ・オブ・アバディーン (2)
【Fターム(参考)】
【公開日】平成24年8月30日(2012.8.30)
【国際特許分類】
【出願日】平成24年4月6日(2012.4.6)
【分割の表示】特願2007−505627(P2007−505627)の分割
【原出願日】平成17年3月29日(2005.3.29)
【出願人】(506330922)ザ・ユニバーシティ・コート・オブ・ザ・ユニバーシティ・オブ・アバディーン (2)
【Fターム(参考)】
[ Back to top ]