
シャペロン融合タンパク質としての風疹E1エンベロープタンパク質変異体の組換え発現、可溶性で天然様かつ免疫反応性の抗原コンホメーションへのそのリフォールディングならびに抗風疹抗体の検出におけるその用途
【課題】風疹E1抗原変異体及び抗風疹抗体検出における用途の提供。
【解決手段】可溶性風疹E1抗原及びこのペプチドの変異体は、C末端における少なくとも膜貫通領域とアンカーセグメント、及びアミノ酸143-164を欠き、並びに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有し、更にこの領域のN末端(Cys349)は少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は隣接風疹E1抗原配列の少なくとも8アミノ酸を含有する。該抗原は、シャペロン融合タンパク質として組換え発現され、可溶性かつ免疫反応性のコンホメーションにリフォールディングされ、抗風疹抗体の血清学的検出に使用される。風疹E1抗原及び変異体をコードする組換えDNA分子も得られる。サンプル中のIgG及び/又はIgMサブクラスの抗風疹抗体の検出、測定及び定量の方法も得られる。
【解決手段】可溶性風疹E1抗原及びこのペプチドの変異体は、C末端における少なくとも膜貫通領域とアンカーセグメント、及びアミノ酸143-164を欠き、並びに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有し、更にこの領域のN末端(Cys349)は少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は隣接風疹E1抗原配列の少なくとも8アミノ酸を含有する。該抗原は、シャペロン融合タンパク質として組換え発現され、可溶性かつ免疫反応性のコンホメーションにリフォールディングされ、抗風疹抗体の血清学的検出に使用される。風疹E1抗原及び変異体をコードする組換えDNA分子も得られる。サンプル中のIgG及び/又はIgMサブクラスの抗風疹抗体の検出、測定及び定量の方法も得られる。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、可溶性風疹E1抗原及びこのペプチドの変異体に関する。これらは、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。この領域のN末端(Cys349)は更に少なくとも隣接風疹E1抗原配列の25、30、又は34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも10、11、15、25、又は35アミノ酸を含有する。
【背景技術】
【0002】
風疹ウイルス(RV)はトガウイルスであり、ルビウイルス亜群の唯一の種である。この小さな包膜ウイルスは65nmのサイズを有し、脂質エンベロープに包まれた正二十面体ヌクレオカプシド内に封入された10kbの一本鎖RNA分子よりなる。風疹ウイルスは、Tリンパ球及び抗体の両方により媒介されると考えられている永久免疫を通常は与える比較的軽度な小児疾患(三日はしか)を引き起こす。成人における風疹感染は、一過性/慢性関節炎、筋骨格症候群、インスリン依存性糖尿病、及び後になって発現する神経後遺症を引き起こす可能性がある。風疹ウイルスは妊娠初期における主要パラメーターである。抗風疹IgM及び/又は抗風疹IgGの特異的検出は、それぞれ急性感染が生じたのか又は激しい再感染が生じたのかを明らかにするために決定的に重要である。妊娠中の母体の風疹ウイルス感染は胎児の先天性風疹症候群(CRS)のリスクを伴い、妊娠の最初の12週のうちに一次感染が生じた場合に先天性奇形の頻度が最も高い。この理由により、妊娠初期におけるRV感染により引き起こされる先天性異常の予防は、血清学的方法により個体の免疫状態を測定することを要する。これは、風疹ウイルス抗原に選択的なIgG抗体及びIgM抗体の選択的測定を意味する。一次風疹感染は特異的IgM抗体応答に関連しており、一方、検出可能なIgMの非存在下のIgGレベルの上昇は、急性風疹ウイルス感染に対して防御的である免疫状態を示す。出生前スクリーニングが、女性が妊娠初期に一次風疹感染を受けたことを示した場合には、治療的流産が推奨されることが多い。したがって、その試験結果が正確であることが何よりも重要である。
【0003】
RVに対する抗体を検出するための通常の検査診断は、主として、ELISA(酵素結合イムノソルベントアッセイ)試験に基づくものであり、欧州中部においてRV免疫状態の測定のための最も広く受け入れられている方法は、RV-IgG ELISAの証明のための赤血球凝集阻止(HAI)試験である(Weber,1997)。ある免疫グロブリンクラスの特異的抗体の検出は、特異的抗原が固定化されている固相に該免疫グロブリンを結合させることにより行われうる。ついで、結合した免疫グロブリンを、あるクラスのヒト免疫グロブリンに特異的な標識抗体により検出する。このアッセイ形態は、検出前に未結合免疫グロブリンを除去する洗浄工程を含む2工程アッセイ形態によってのみ行われうる。自動イムノアッセイ分析装置においてしばしば実施される1工程アッセイ形態は二重抗原サンドイッチの直接アッセイ形態を要する。すなわち、特異的抗体が、固相に固定化された第1抗原に結合する免疫複合体を形成するか、あるいは固相への又は標識を含有する第2抗原への固定化を媒介して、該特異的結合抗体の定量的又は定性的検出を可能にする。1工程二重抗原サンドイッチ形態における同じ特異性のIgM抗体の存在下の特異的IgG抗体の選択的測定のためには、可溶性単量体又は既定オリゴマー抗原の使用が厳しく要求され(EP 0 944 838)、これにより、免疫反応性コンホメーションが明らかにされる。
【0004】
風疹ウイルスは、動物及びヒトにおいて抗原性であることが示されている4つの構造タンパク質を含有する。これらは、3つのエンベロープ糖タンパク質E1(58kDa)、E2a(47kDa)及びE2b(42kDa)、ならびにウイルスゲノムを構成する一本鎖RNAプラス鎖を装飾する非グリコシル化カプシド(C)タンパク質(33kDa)である(Waxham及びWolinsky, 1985a; Oker-Blomら, 1984)。E2a及びE2bは同一遺伝子産物の変異体であることが示されており、ポリアクリルアミドゲル内での泳動における相違は、それらのタンパク質の不均一なグリコシル化によるものである。E1及びE2は単量体又はジスルフィド結合複合体(E1-E1及びE1-E2)を形成することが判明しており、一方、Cは専らホモ二量体として存在する(Waxham及びWolinsky, 1983; Wolinsky, 1996)。RVの生物学的、物理学的及び生化学的特性ならびにその感染の臨床的特徴はWolinsky, (1996) に公開されている。
【0005】
現在のところ、風疹感染の検出のためには抗原が使用されており、これは、安定に感染した/トランスフェクトされた細胞系及び一般には真核生物過剰産生からの幹細胞(stem)から誘導される。Seppanenら (1991)は、バキュロウイルス発現系を使用することによるスポドプテラ・フルジペルダ(Spodoptera frugiperda)Sf9昆虫細胞における風疹ウイルスのE1及びE2の発現を記載している。さらに、RV様粒子(RLP)の形態でRVの構造タンパク質E1、E2及びCを発現し分泌する安定にトランスフェクトされたCHO細胞系がHobmanら (1994)において開示されている。風疹様粒子は風疹主要抗原E1、E2及びCから構成され、これらはウイルスポリペプチド前駆体タンパク質として発現される。シグナル配列により、この未プロセシング前駆体タンパク質は培地内に分泌され、ウイルス様粒子が形成される。これらのRLPの表面は、風疹陽性血清中の抗RV抗体の免疫学的検出に適したエピトープを提示する。BHK(子ハムスター腎細胞)細胞系における、RVの3つの構造タンパク質を含有する非感染性RV様粒子(VLP)の発現が、Quiら, (1994)に概説されている。しかし、真核細胞系におけるRV抗原の製造は労働集約的であり、長時間を要し、一方、収率は比較的低い。抗風疹ウイルス抗体の検出のin vitro診断方法の確立のためには、可溶性で機能的で再現可能な一定の質で大腸菌(E. coli)のような原核生物から組換えRV抗原を製造する方法が必要であり、そのような方法は、確立されたRLPと比べて明らかな利点を伴うことが必要である。タンパク質の設計及び操作の分野におけるノウハウは著しく増加しているが、風疹抗原E1、E2及びCは大腸菌(E. coli)宿主細胞内で非常に少量でしか発現されず、さらに、それらは難溶性である。
【0006】
免疫優性風疹抗原(すなわち、ヒト血清からの抗体の免疫学的検出のための選択された抗原)はエンベロープタンパク質E1である。E1タンパク質の全体的なトポロジーは、α-らせん領域(438-452)を含む大きな細胞外エクトドメイン(1-452)及び膜貫通ドメイン(453-468)及びそれに続く短いC末端アンカーセグメント(481まで)を含む。RVのE1の不溶性の1つの理由は、10個のジスルフィド架橋により安定化されると考えられるエクトドメインのコンホメーションによるものであろう。風疹E1ペプチドはアミノ末端(N末端)領域及びカルボキシ末端(C末端)領域内に構築され、合計24個のシステイン残基を含有する。一般に、N末端は、遊離アミノ基(NH2)を有するアミノ酸で終結するタンパク質又はポリペプチドの端を意味し、C末端は、遊離カルボキシル基(COOH)を有するアミノ酸で終結するタンパク質又はポリペプチドの端を意味する。
【0007】
風疹E1タンパク質のN末端領域はシステイン残基C(8)からC(287)までの8個のジスルフィドを含有し、C末端領域は、システイン残基C(249)からC(401)までに生じる2個のジスルフィドを含有することを、Grosら, (1997)は開示している。残基C(456)、C(466)及びC(468)は推定膜貫通セグメント内に位置し、残基C(470)は該ウイルスの内部に位置すると予想される。これらのデータは、残基C(456)-C(470)がジスルフィド形成に関与していないことを示している。
【0008】
システイン残基が多数存在すること、及びそれに伴う、誤った架橋事象の可能性(分子内及び分子間SHシャフリングの生成は十分に考えられうる)は、E1のコンホメーション的及び酸化的リフォールディングが、in vitroで制御することが困難な非常に複雑な過程であることを示唆している。したがって、免疫反応性(すなわち、抗原性)コンホメーションへの風疹抗原の高収率リフォールディング及び精製の両方を促進する簡便かつ信頼しうるプロトコールが緊急に必要とされている。天然のようにフォールディングした組換えE1エンベロープ変異体は、RVに対する抗体に関する信頼しうるイムノアッセイを達成するための必須要件である。
【0009】
風疹E1エクトドメイン(1-452)は20個のシステイン残基を含有する。それらの正しい架橋はその三次元構造を決定し、正しい天然様コンホメーションエピトープの露出のために決定的に重要である。正しい分子内及び/又は分子間ジスルフィド架橋の形成である、システインに富むタンパク質の酸化的リフォールディングは、複雑な過程であり、過度な実験を要する過程である。新たに合成された又は輸送されたタンパク質内へのin vivoでのジスルフィド架橋の挿入のためには、細胞区画(真核細胞における小胞体、原核細胞におけるペリプラズム)内の多数のフォールディング補助体(シャペロン、オキシドレダクターゼ、プロリルイソメラーゼ)が利用可能である。細菌サイトゾル内のレドックス電位は強還元性(-270mV)であるため、異種標的タンパク質のシステインは天然においてはチオール部分として生じる。発現が非常に高い場合には、標的タンパク質は不溶性凝集物(いわゆる、封入体、IB)として宿主細胞内に貯蔵される。通常、部分的にリフォールディングした及びアンフォールディング(変性)したタンパク質画分は、カオトロピック溶液(7.0M GdmCl、8.0M尿素)中の還元条件(例えば、5mM TCEP)下で可溶化されて、アンフォールディングしたタンパク質鎖を与え、これは再生(すなわち、天然又は天然様コンホメーションにリフォールディング)されなければならない。一般に、天然様にフォールディングしたタンパク質の収率は、正しくないジスルフィド架橋によって著しく減少する。
【0010】
コンホメーションの観点を軽視すれば、限られた数のシステイン(及び厳密に分子内架橋事象が生じると仮定した場合)でさえも、莫大な数の誤った架橋事象を招いて、凝集過程のような非生産的な副反応という結果に終わる。試験管内のin vitroリフォールディングとは異なり、in vivoでのde novoフォールディングには、フォールディングを助ける最適化レドックス環境中の非常に多くのフォールディング補助体が利用可能である。それでも、大腸菌(E. coli)のような原核宿主細胞における、システインに富む機能性タンパク質の可溶性発現は、尚も非常に複雑である。RVの免疫反応性抗原の過剰生産率を改善し、リフォールディングしたタンパク質の溶解度を増加させる目的は、限られた程度でしか達成されていない。
【0011】
Newcombeら, (1994)においては、風疹E1抗原断片を可溶性形態で大腸菌(E. coli)内で発現させるために9個のグルタチオン-S-トランスフェラーゼ(GST)E1融合タンパク質が使用された。E1配列の相当な末端切断(トランケート化)を行った後に初めて、システイン非含有領域243-286(44アミノ酸残基)に関する可溶性発現の成功が可能となった。EP0299673は、風疹Ig特異的結合特性を保有するアミノ酸残基207-353のペプチドを開示している。さらに、Starkeyら, (1995)は、GST-E1融合タンパク質の44から75までのアミノ酸残基の非常に狭い領域のみが可溶性であったことを開示している。全E1コード配列及びより大きな亜断片を含有するGST融合タンパク質は、精製され得ない形態で発現された。したがって、それらは不溶性封入体として発現されたと考えられる。
【0012】
RV抗原内の免疫反応性決定基を同定するために、合成ペプチドを使用してE1及びE2エピトープが過去に詳細にマッピングされている(Mitchellら, (1993))。また、ウイルス感染性及び赤血球凝集に重要なドメインを含む、RV E1タンパク質内の、異なる無関係なエピトープが位置決定されている(Waxham及びWolinsky, (1985), Green及びDorsett, (1986), Ho-Terryら, (1985))。Terryら, (1989)に記載されている組換えタンパク質A-E1融合体は直鎖状エピトープのみにより特徴づけられている。RV E1糖タンパク質のエピトープの、より詳細な位置決定は、Terryら, (1988)、Wolinskyら, (1991) 及びChayeら, (1993)に記載されている。さらに、Giessaufら, (2004)は、風疹に対する免疫の測定を改善するために、E1のコート化ペプチドのいくつかを使用することによりELISAを評価している。これらのペプチドのうちの1つ(BCH-178ペプチド)だけが、低陽性又はボーダーラインHAI力価又はRV-IgG値を確認するための追加的な方法として、中和抗体をスクリーニングするのに成功裏に使用されるようであった。小さなBCH-178ペプチド(アミノ酸残基213-239)が天然E1分子の抗原エピトープのほとんどを含有するという更なる証拠は存在しない。しかし、免疫優性風疹E1エンベロープタンパク質の主要反応性の局在化に関しては、現在までに言及されていない。
【0013】
血清学的アッセイにおける高い免疫反応性と高い溶解度とを兼ね備えた、原核細胞系由来の可溶性風疹E1変異体を提供することが、本発明の目的であった。
【発明の開示】
【0014】
発明の概要
本発明の対象は可溶性風疹E1抗原及びこのペプチドの変異体であり、これらは、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。ジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域は、この領域のN末端(Cys349)において、更に少なくとも隣接風疹E1抗原配列の25、30、34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも10、11、15、25、35アミノ酸を含有する。
【0015】
可溶性風疹E1抗原は更に、C末端において、アミノ酸残基438-452の間のα-らせん領域を更に欠くことにより特徴づけられる。
【0016】
さらに、可溶性風疹E1抗原は、少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130の組合せにわたる領域を含有するN末端領域により特徴づけられる。さらに、可溶性風疹E1抗原は、少なくともジスルフィド架橋Cys349-Cys352及びCys368-Cys401、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せにわたる領域を含有するC末端領域により特徴づけられる。
【0017】
また、本発明は、シャペロン融合タンパク質として組換え発現され、可溶性かつ免疫反応性のコンホメーションにリフォールディングされ、さらに、抗風疹抗体の血清学的検出に使用される、風疹E1抗原及び変異体をコードする組換えDNA分子に関する。
【0018】
本発明は、サンプル中のIgG及び/又はIgMサブクラスの抗風疹抗体の検出、測定及び定量のための方法を開示する。この場合、該抗体に対する捕捉試薬及び/又は結合相手として、風疹E1抗原を使用する。本発明は更に、風疹E1抗原の少なくとも1つの抗原を含有する、抗風疹抗体の検出のための診断試験及び試薬キットを含む。
【0019】
発明の詳細な説明
本発明は、可溶性風疹E1抗原及びこのペプチドの変異体を開示する。これらは、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。
【0020】
ジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域は、この領域のCys349位のN末端において、更に少なくとも隣接風疹E1抗原配列の25、30、34アミノ酸を含有し、及び/又は、この領域のCys401位のC末端は更に隣接風疹E1抗原配列の少なくとも10、11、15、25、35アミノ酸を含有する。風疹E1抗原の最低限の要件に必要なアミノ酸の前記の定義は絶対的なものではなく、当業者により実験により容易に確認されうる。
【0021】
可溶性風疹E1抗原及びこのペプチドの変異体は、好ましくは、アミノ酸143-164を欠く。より好ましくは、この領域のN末端(アミノ酸143)は更に、少なくとも1、2、5及び/もしくは少なくとも10アミノ酸を欠き、ならびに/又は、この領域のC末端(アミノ酸164)は更に隣接風疹E1抗原配列の少なくとも1、2、5及び/又は少なくとも10アミノ酸を欠く。当業者は、前記の境界は、風疹E1タンパク質の配列内で欠けているアミノ酸残基の或る範囲を示すために用いられていると理解するであろう。該境界の前記定義は絶対的なものではなく、この領域内で欠けているアミノ酸残基の量は可変的なものである。
【0022】
さらに、本発明は更に、アミノ酸残基438-452の間のα-らせん領域を更に欠く可溶性組換え風疹E1抗原及びこのペプチドの変異体に関する。
【0023】
或るシステイン残基の置換及び非常に疎水性のセグメントを有する或る配列の除去により風疹E1タンパク質の酸化的及びコンホメーション的リフォールディングの複雑性を軽減することが可能かどうかを評価するために、欠失分析を行った。
【0024】
本発明において、「ジスルフィド架橋」という用語は、タンパクの3次元構造に隣接し、酸化されてジスルフィド結合を形成しうる2つのシステイン残基(Freedman, Curr. Op. Struct. Biol. (1995) 5, 85-91; Creighton et al., TIBTECH 1995 (13), 18-23; Raina, Annu. Rev. Microbiol. (1997) 51, 179-202)に関する。ジスルフィド結合形成の割合は、2つのシステイン残基の近接性によって決まり、それはそれらの硫黄原子がチオール/ジスルフィド変換に必要な距離に到達する確率として規定される。ジスルフィド架橋とも称されるジスルフィド結合(同義語は、それぞれSS 結合及びSS 架橋である)は共有結合的な三次元関係を構成し、通常フォールディングされた構造の安定化に寄与する。それらは、フォールディングされていないポリペプチド鎖の構造的フレキシビリティーを制限することによりそのような安定化に寄与する。すなわちSS結合のタンパクの安定性への寄与は自然界においてかなりエントロピックである。ジスルフィド結合の形成は酸化されやすい環境を必要とする。それゆえ、細菌の細胞質や真核細胞の細胞質ゾルのような細胞内コンパートメントは本質的に還元されやすい環境のため、細胞内タンパク質はほとんどジスルフィド結合を含まない。しかしながら、ジスルフィド架橋は、各々HIV-1及びHIV-2由来のgp41及び gp36 エクトドメインのような分泌あるいは移行性タンパク質、及び風疹エンベロープタンパク質 E1及びE2でよく見られる。
【0025】
驚くべきことに、可溶性風疹E1抗原変異体は、膜貫通領域とC末端アンカーセグメント(好ましくはアミノ酸残基453-481)の欠失により得られることが判明した。最も好ましくは、さらに、アミノ酸453-約468のα-らせん領域も欠失される。N末端領域内には、少なくともアミノ酸残基143-164、より好ましくは、アミノ酸134-約168も欠失されなければならない。
【0026】
それぞれジスルフィド架橋Cys8-Cys13、Cys59-Cys71、Cys117-Cys130及びジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せは、膜貫通領域とC末端アンカーセグメント、及び少なくともアミノ酸143-164、好ましくは134-約168が欠けている場合には、血清学的アッセイにおける好ましい高い免疫反応性と共に可能な限り多数のエピトープを含有し尚も可溶性である、最も伸張した本発明の風疹E1抗原を与える。
【0027】
本発明の風疹E1エンベロープタンパク質及びその変異体中のN末端領域は、好ましくは、少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71を含有し、より好ましくは、ジスルフィドCys8-Cys13及びCys59-Cys71及びCys117-Cys130の組合せを含有する。ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130の組合せは本発明の風疹E1断片に対して有益な効果をもたらす。なぜなら、それは、高い免疫反応性抗原との或る風疹E1エピトープの三次的固定を保証するからである。
【0028】
本発明の風疹E1抗原及びその変異体中のC末端領域は、少なくともジスルフィド架橋Cys349-Cys352及びCys368-Cys401、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せを含む領域を含有する。ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せは風疹E1抗原に対する追加的な効果をもたらす。なぜなら、それは、高い免疫反応性を有する主要エピトープの接近を可能にするからである。
【0029】
風疹E1抗原の主要免疫反応性の測定のために、本発明者らの目的においては、該タンパク質を、まず、アミノ酸残基1-314を含むN末端部分、及びアミノ酸残基315-432を含むC末端部分に分割した。両部分の配列を、ジスルフィドシャフリングによるリフォールディングアーチファクトを回避するためにシステインを含有しないように設計し、さらに、発現ベクター内にクローニングした。ついで、Grosら, (1997)に記載されている風疹E1ペプチドの各単一ジスルフィド架橋を該N末端及びC末端部分内に個々に又は組合せて挿入した。エピトープ固定のための、共有的三次的接触の重要性を、それぞれの免疫反応性により評価した。
【0030】
興味深いことに、N末端断片(1-314)は高分子量会合体として出現し、一方、C末端断片(315-432)は見掛け上は二量体として溶出することが、ゲル濾過分析から明らかである。これは、単量体、ホモ及びヘテロ二量体よりなる凝集体としての風疹E1/E2ペプチドを開示しているBaron及びForsell, (1991)に記載の最先端研究内容とは対照的である。さらに、本発明の結果は、異なる溶解度特性が発現活性と相関することを明らかにした。すなわち、N末端セグメント(1-314)の生産率はタンパク質約1mg/gバイオマスであるに過ぎなかったが、C末端セグメント(315-432)の生産率はタンパク質20mg/g(湿潤細胞重量)を著しく上回った。
【0031】
システイン非含有変異体は、自動Elecsys(登録商標)分析装置において風疹陽性血清でアッセイされた場合には、有意な免疫反応性を示さなかった。重要なことに、N末端断片(1-314)内の単一のジスルフィド架橋又はジスルフィド架橋の組合せは反応性における有意な上昇を示さなかった。しかし、C末端断片(アミノ酸残基315-432)の反応性は、ジスルフィド架橋Cys349-Cys352及びCys368-Cys401を挿入することにより有意に上昇した。N末端風疹E1タンパク質の反応性の欠如の理由は、凝集又は会合を媒介して主要エピトープの接近を損なう該配列内の構造要素によるものでありうるであろう。
【0032】
この仮定を更に証明するために、風疹E1エンベロープタンパク質のN末端断片(アミノ酸残基1-314)をそのC末端から段階的に末端切断した。同様に、C末端断片(315-432)をそのN末端から徐々に伸長させた。C末端断片は或る程度伸長されると不溶性になり、一方、N末端断片は或る程度末端切断されると可溶性になると予想された。
【0033】
発現及び特徴づけのために、C末端変異体E1-C(aa260-432)、E1-C(aa201-432)、E1-C(aa143-432)、E1(aa105-432)、E1(aa56-432)及びE1(aa34-432)を使用した。変異体E1(aa201-432)、E1(aa260-432)及びE1(aa315-432)は安定な二量体を形成し、一方、より長い構築物は凝集及び会合する著しい傾向を示した。したがって、E1エクトドメインのオリゴマー化モチーフを約143aa-約200aaの領域に限定した。より厳密なマッピングのために、断片E1(169-432)をクローニングし、発現させ、特徴づけした。驚くべきことに、この構築物は安定な且つ可溶性の二量体を形成し、したがって抗風疹IgG分子の検出のための優れた候補であった。この結果に基づき、該会合モチーフをセグメント143aa-168aaに限定することができた。より詳細なマッピングのために、断片E1(156-432)及びE1(163-432)を特徴づけした。どちらの構築物も、部分的に二量体として、部分的に高分子量凝集体として溶出し、したがって、1工程二重抗原サンドイッチ形態の抗風疹IgG抗体の検出のための抗原としては限られた価値しか有さなかった。
【0034】
本発明のこの一連の試験の主な結果として、溶解度、オリゴマー化状態及び安定性を損なうことなく、C末端E1(315-432)断片をアミノ酸残基315からアミノ酸残基169まで伸長させることが可能であることが、明らかに示されうるであろう。第2に、E1エクトドメインの最初の3分の1と次の3分の1との間の領域(143-168)は、高分子量凝集体を形成するよう該タンパク質の会合を引き起こす構造又は配列モチーフを含有する。この領域の内在的疎水性は謎である。なぜなら、それは、単純な一次構造分析からは演繹され得ないからである。該会合モチーフを含む領域は、IgG分子の血清学的検出のためのE1エンベロープタンパク質の使用を著しく限定する。
【0035】
N末端セグメントの溶解度の最適化のために、同等の方法を用いた。E1断片E1-N (1-200)、E1-N (1-142)、E1-N (1-104) 及びE1-N (1-55)を発現ベクター内にクローニングし、大腸菌(E. coli)宿主細胞内に導入し、発現させ、特徴づけした。予想どおり、最長N断片(エクトドメイン1-200)は高分子量凝集体として溶出し、したがってE1エクトドメイン変異体E1-N (1-314)に著しく類似している。有意に改善された溶解度特性は、部分的に可溶性二量体として、部分的に高分子量凝集体として溶出する末端切断断片E1-N (1-142)で達成された。最後に、末端切断変異体E1-N (1-104) 及びE1-N (1-55)は見掛け上は二量体として溶出した。断片E1-N-(1-117) 及び(1-133)を使用して、アミノ酸残基104と142との間の領域を更に分析した。どちらの変異体もFPLC分析において可溶性二量体として出現し、したがって、IgG分子の血清学的検出のための重要な前提条件を満たしている。しかし、変異体E1-N (1-117)はより安定であり、したがって、更なる最適化実験のための、より優れた候補であった。
【0036】
簡潔に説明すると、本発明において、長さの最適化の後、大腸菌(E. coli)において非常に良く発現され可溶性二量体を形成する風疹E1タンパク質の2つの領域、すなわち、アミノ酸残基1-117/1-133のN末端断片(E1-N)及び領域169-432/201-432を含むC末端断片(E1-C)を同定した。これらのE1断片を使用して、組換え風疹E1エクトドメインの抗原性のためにはどのジスルフィド架橋が重要であるかという問題について検討した。したがって該システイン対を個々に及び組合せて再構築し、対応するE1変異体を自動Elecsys(登録商標)2010分析装置(Roche Diagnostics GmbH)において抗風疹陽性血清で評価した。
【0037】
以下のシステイン変異体を風疹E1(201-432)において特徴づけした:単一架橋変異体Cys 225-Cys235、Cys349-Cys352及びCys368-Cys401、二重架橋の組合せCys349-Cys352/Cys368-Cys401、ならびに三重架橋の組合せCys225-Cys235/Cys349-Cys352/Cys368-Cys401。さらに、伸長構築物E1(169-432)において、二重組合せCys176-185/Cys225-Cys235を分析した。
【0038】
以下のシステインの組合せを風疹E1断片1-133において特徴づけした:単一架橋変異体Cys8-Cys13、Cys59-Cys71及びCys117-Cys130ならびに二重架橋の組合せCys8-Cys13/Cys59-Cys71、Cys8-Cys13/Cys117-130及びCys59-Cys71/Cys117-Cys130。さらに、二重架橋の組合せCys 8-Cys 13/Cys59-Cys71を融合構築物E1-N (1-117) 及びE1-N (1-104)において特徴づけした。自動Elecsys(登録商標)2010分析装置における風疹E1変異体の免疫学的評価は、抗原性が、ジスルフィド架橋によりもたらされる共有的三次的接触により固定される該エクトドメイン分子の天然様コンホメーションに依存することを示した。
【0039】
驚くべきことに、これらの特徴的知見は、免疫反応性に関して、風疹E1抗原のジスルフィド架橋に対して、異なった力点を置く必要があることを示している。ジスルフィド架橋Cys-349-Cys-352及びCys-368-Cys-401は決定的な役割を果たしている。興味深いことに、これらのジスルフィド架橋の組合せによる感受性の上昇は相加的なものではなく、協同的なものである。構築物E1-C(201-432)の評価は、各場合にCys349-Cys352及びCys368-Cys401の三次的固定のための有意なシグナルを示している。両方の架橋の組合せにより、シグナルのレベルは、単一架橋構築物と比べて、倍数的に驚くほどに上昇した。おそらく、風疹E1エクトドメインの天然様コンホメーションが、それらの2つの分子内組合せを介して固定され、風疹陽性血清中の、より大量の特異的IgG分子を検出したのであろう。共有的三次的接触の妥当性は、システイン非含有構築物E1-C(201-432)の免疫学的評価により、特に明らかである。驚くべきことに、単一架橋構築物の場合とは対照的に、システイン非含有構築物においては、陽性シグナルは全く検出できない。
【0040】
天然サンプル中の抗風疹IgG抗体の免疫学的検出は、最適化抗原E1-C(201-432, Cys349-Cys352, Cys368-Cys401)で更に改善された。抗風疹IgG陽性として分類されたこれらの研究において適用されたすべてのヒト血清は陽性であることが判明した。より複雑な三次架橋融合構築物-E1-C(169-432)、Cys225-Cys235、Cys349-Cys352、Cys368-Cys401)を使用することにより、抗風疹IgG抗体の検出に関して同等の結果を得ることができた。風疹E1抗原断片の最低限の要件は、少なくともジスルフィド架橋Cys349-Cys352及びCys368-Cys401により分子内で連結されたポリペプチドバックボーンにまで縮小することができた。E1断片E1-C(342-412、Cys349-Cys352、Cys368-Cys401)は不十分な免疫反応を示すため、E1-C(342−412)のN末端及び/又はC末端に隣接した配列要素は免疫反応性に重要だと考えられた。これを証明するために、構築物E1-C(315-412、Cys349-Cys352、Cys368-Cys401)をクローニングし、特徴づけした。この断片に関して、E1-C(201-432、Cys349-Cys352、Cys368-Cys401)と比較しうる免疫反応性を示すことができた。この結果は、構築物E1-C(アミノ酸315-412)のN末端領域(315-342)のアミノ酸残基が風疹E1抗原の免疫反応性に重要らしいことを示している。当業者は、E1-C(315-412)のN末端及び/又はC末端に隣接したどの配列要素が免疫反応性に重要なのかを、更なる実験により証明することが可能である。
【0041】
エクトドメインのC末端部分からのジスルフィド架橋を有する高反応性断片に加えて、いくつかのN末端断片をクローニングし、発現させた。それらは有意な免疫反応性を示した。免疫反応性は2つの二重架橋融合構築物E1-N(1-117、Cys8-Cys13/Cys59-Cys71)及びE1-N(1-133、Cys8-Cys13/Cys59-Cys71)から得られた。
【0042】
可溶性かつ免疫反応性のコンホメーションにリフォールディングしうる最も長い風疹E1断片は、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130を含むアミノ酸残基1-133、ならびにジスルフィド架橋Cys349-Cys352及びCys368-Cys401ならびにジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せを含むアミノ酸残基169-432にわたる(これは、C末端における少なくとも膜貫通領域と短いC末端アンカーセグメント、及び少なくともアミノ酸143-164、好ましくはアミノ酸134-168を欠くことによりもたらされる)。
【0043】
したがって、本発明の好ましい実施形態は、ジスルフィド架橋Cys349-Cys352及びCys368-Cys401にわたる領域ならびに更に少なくともジスルフィド架橋Cys225-Cys235にわたる領域を含有する可溶性風疹E1抗原ならびにこのペプチドの変異体である。
【0044】
本発明のもう1つの主題は、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401にわたる領域ならびに更に少なくともジスルフィド架橋Cys176-Cys185にわたる領域を含有する可溶性風疹E1抗原ならびにこのペプチドの変異体に関する。さらに、ジスルフィド架橋Cys349-Cys352及びCys368-Cys401の組合せ、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せが、本発明の好ましい実施形態である。これらの風疹E1抗原はエピトープの形成のための主要免疫反応性を含有する。
【0045】
本発明のもう1つの好ましい実施形態は、更にジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130にわたる領域を更に含有する可溶性風疹E1抗原及びこのタンパク質の変異体に関する。
【0046】
少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130にわたる風疹E1抗原の領域も、エピトープの形成のための或る免疫反応性を含有する。ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401を含有するC末端配列に連結させることなく、これらのN末端断片を使用することも可能である。好ましくは、これらのN末端風疹E1抗原は、前記のC末端風疹E1抗原と共に、アッセイにおいて使用される。
【0047】
本発明においては、大腸菌(E. coli)内で組換え発現されin vitroで再現可能な様態で機能性コンホメーションにリフォールディングされる風疹E1エクトドメインの包括的領域及び断片を同定した。長さ及びシステイン生成に関して最適化された本発明のE1断片は可溶性であり、特に、風疹陽性血清中の抗風疹IgGの検出に適している。
【0048】
この知見は公知風疹E1断片のもの(Newcombeら, (1994), Starkeyら, (1995), EP0299673)とは対照的である。これらの公知構築物はジスルフィド架橋Cys349-Cys352及びCys368-Cys401の組合せを含有せず、したがって、それらはエピトープの形成のための主要免疫反応性を欠いている。さらに、該公知構築物に関しては、高溶解性かつ免疫反応性コンホメーションにリフォールディングされうる、本発明において開示されている、C末端における少なくとも膜貫通領域及び少なくともアミノ酸143-164、好ましくはアミノ酸134-約168を欠くことによる、N末端領域のジスルフィド架橋Cys8-Cys13及びCys59-Cys 71及びCys117-Cys130ならびに/又はC末端領域及びN末端領域のジスルフィド架橋の組合せが開示されていない。さらに、本発明の好ましい融合構築物は、より多数のエピトープを有する、風疹E1タンパク質の、より長い部分を含有しうる。なぜなら、それらは可溶性であり、したがって、ほとんどの抗風疹抗体の検出に、より適しているからである。
【0049】
「変異体」なる語は、異なる量のシステイン対を含みうる、風疹E1エクトドメインの、異なる長さの配列の部分を意味する。これらの配列、挿入カセットの核酸配列、及び風疹E1の得られた融合ポリペプチドのアミノ酸配列は、本発明の配列プロトコールにおいて記載されている。
【0050】
さらに、この場合、「変異体」なる語は、該タンパク質に実質的に類似したタンパク質を意味する。特に、変異体は、最も一般的なタンパク質アイソフォームのアミノ酸配列と比べてアミノ酸の変化、欠失又は挿入を示すアイソフォーム又はアレルでありうる。好ましくは、そのような実質的に類似したタンパク質は、該タンパク質の最も一般的なアイソフォームに対して、少なくとも80%、好ましくは少なくとも85%、より好ましくは少なくとも90%、最も好ましくは少なくとも95%の配列類似性を有する。また、「変異体」なる語は、翻訳後修飾されたタンパク質、例えばグリコシル化タンパク質を意味する。また、「変異体」は、例えば、該タンパク質への標識(特に放射能蛍光標識)の共有結合又は非共有結合により、サンプルの収集後に修飾されたタンパク質である。他の考えられうる標識としては、放射能、蛍光、化学発光、電気化学発光、酵素などが挙げられる。さらに、「変異体」は、例えばビオチン化タンパク質のような固相結合基である。標識の詳細な説明は本明細書中、後記において記載されている。
【0051】
風疹E1抗原は化学合成により製造されうる。該合成は均一溶液中又は固相中で行われうる。例えば、用いられうる均一溶液中での合成技術としては、Houbenweyl (1974)に記載されているものが挙げられる。本発明のタンパク質は、Atheton及びShepard (1989)に記載されている方法に従い、固相中でも製造されうる。
【0052】
可溶性及び一定の単量体又はオリゴマー状態に関して最適化された前記風疹E1タンパク質はIgM検出モジュールの可能性をも提供する。この目的には、見掛け上二量体であるE1融合タンパク質を、例えば化学的架橋により重合させることが可能であろう。本発明のもう1つの好ましい実施形態は、タンパク質E1、E2及びコアタンパク質Cから構成される混合重合体である。風疹E2及びコアタンパク質は当技術分野で公知である。
【0053】
本発明の風疹E1タンパク質は組換えDNA技術によっても製造されうる。「組換えDNA分子」なる語は、遺伝子工学技術又は化学合成により、単離されたポリヌクレオチド断片を人工的に操作して、2つの分離した配列断片を組合せることにより製造された分子を意味する。それを行う場合には、所望の機能を有するポリヌクレオチド断片同士を互いに連結させて、所望の機能の組合せを得ることが可能である。適当な宿主細胞内での複製により、大量の該ポリヌクレオチドを製造することが可能である。タンパク質又はその断片をコードする天然又は合成DNA断片を、例えばSambrookら, (1989)に記載されている原核生物細胞又は下等もしくは高等真核生物細胞内への導入及びそのような細胞内での複製が可能な組換えポリヌクレオチド構築物(典型的にはDNA構築物)内に組込む。「下等真核生物」なる語は、酵母、真菌などのような宿主細胞を意味する。下等真核生物は、一般には(必ずしもそうではないが)単細胞である。「原核生物」なる語は、大腸菌(E. coli)、ラクトバシラス(Lactobacillus)、ラクトコッカス(Lactococcus)、サルモネラ(Salmonella)、ストレプトコッカス(Streptococcus)、バシラス・サチリス(Bacillus subtillis)又はストレプトマイセス(Streptomyces)のような宿主を意味する。これらの宿主も本発明において予想される。好ましい下等真核生物は、酵母、特に、シゾサッカロミセス(Schizosaccharomyces)、サッカロミセス(Saccharomyces)、クルイベロミセス(Kluiveromyces)、ピチア(Pichia)(例えば、ピチア・パストリス(Pichia pastoris))、ハンゼヌラ(Hansenula)(例えば、ハンゼヌラ・ポリモルファ)、シュワニオミセス(Schwaniomyces)、シゾサッカロミセス(Schizosaccharomyces)、ヤロウィア(Yarowia)、ジゴサッカロミセス(Zygosaccharomyces)などの種である。サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、エス・カールスベルゲンシス(S. carlsbergensis)及びケイ・ラクチス(K. lactis)は、最もよく使用される酵母宿主であり、簡便な真菌宿主である。「高等真核生物」なる語は、動物、例えば哺乳動物、爬虫類動物、昆虫などに由来する宿主細胞を意味する。現在好ましい高等真核生物宿主細胞は、チャイニーズハムスター(例えば、CHO)、サル(例えば、COS及びVero細胞)、子ハムスター腎細胞(BHK)、ブタ腎臓(PK15)、ウサギ腎臓13細胞(RK13)、ヒト骨肉種細胞系143B、ヒト細胞系HeLa及びヒトヘパトーマ細胞系、例えばHep G2及び昆虫細胞系(例えば、スポドプテラ・フルジペルダ(Spodoptera frugiperda))に由来するものである。該宿主細胞は、懸濁又はフラスコ培養、組織培養、器官培養などにおいて供給されうる。
【0054】
本発明の好ましい実施形態においては、風疹E1抗原は組換え融合タンパク質として製造される。本発明において用いる「融合タンパク質」なる語は、風疹E1タンパク質に対応する少なくとも1つのタンパク質ドメインと、後記のとおりの発現手段として使用されるFKBPシャペロンに好適な別の融合タンパク質に対応する少なくとも1つタンパク質ドメインとを含むタンパク質を意味する。他のよく使用される融合タンパク質としては、例えば、融合タンパク質の高レベル発現ならびに細菌及び真核細胞ライセートからのその迅速な精製のためのグルタチオンS-トランスフェラーゼ(GST)融合タンパク質系が挙げられる。タンパク質のフォールディング及び精製は、しばしば、それらを、自ら活発にフォールディングするパートナータンパク質又はタグと共有結合により融合させることにより促進される。これらの融合モジュールには、マルトース結合タンパク質、グルタチオンS-トランスフェラーゼ、チオレドキシン、NusA、DsbA及びFkpAが含まれる。それらの使用は、通常、過剰生産性大腸菌(E. coli)宿主のサイトゾル又はペリプラズムにおけるそれぞれの標的タンパク質の可溶性発現(すなわち、天然様フォールディング)の向上を目的としている。可溶性かつ機能性のタンパク質の生物工学的製造においてシャペロンが果たす役割は益々重要なものとなっている。好ましくは、シャペロンは風疹E1融合ポリペプチドにおける融合タンパク質として使用される。シャペロンは、古典的な「フォールディング補助体(folding helper)」として公知であり、他のタンパク質のフォールディング及び構造的完全性維持を補助するタンパク質である。それらは、in vivo及びin vitroの両方においてタンパク質のフォールディングを促進する能力を有する。一般には、フォールディング補助体はフォールディング触媒及びシャペロンに細分される。フォールディング触媒は、その触媒機能により、タンパク質のフォールディングにおける律速段階を加速させる。シャペロンは、変性した又は部分的に変性したタンパク質に結合することが公知であり、したがって、タンパク質が再生するのを助ける。したがって、フォールディング触媒とは異なり、シャペロンは単なる結合機能を発揮するに過ぎない(Buchner, (1996))。触媒及びシャペロンの具体例はWO 03/000877に詳細に記載されている。
【0055】
現在までに、いくつかの異なるシャペロンファミリーが公知である。すべてのこれらのシャペロンは、アンフォールディングした又は部分的にアンフォールディングしたタンパク質に結合するそれらの能力により特徴づけられ、タンパク質の適切なフォールディング又は変性もしくは凝集タンパク質の除去に関連した生理的機能を有する。さらに、WO 94/08012には、シャペロンの発現の増強はタンパク質の組換え産生を促進しうることが示されている。また、タンパク質の産生の増加は、タンパク質コード配列とシャペロン配列とを含む遺伝子構築物を使用することにより達成されうることが公知である。天然のようにフォールディングしたタンパク質の産生の増加のためにシャペロンを使用するアプローチは、主として、シャペロンタンパク質結合機能及びそれによる可溶化機能によるものである。シャペロンと標的タンパク質とを含む融合タンパク質の組換え産生の後、通常、シャペロンを、生じたタンパク質から切り離して、所望のタンパク質を純粋な形態で得る。
【0056】
フォールディング補助体、例えば、ペプチジルプロピルイソメラーゼ(PPI)クラスのメンバー、特に、FKBPファミリーのメンバーは、触媒活性を示すだけでなく、凝集傾向にあるタンパク質の溶解度に対する劇的な有益な効果をも与えることが、WO 03/000877に示されている。それらは、そうでなければ(すなわち、未シャペロン化単離形態であれば)凝集傾向にあるそのようなタンパク質との可溶性複合体を形成することにより、それを達成する。そうでなければ生理的条件下で難溶性又は不溶性であるそのようなタンパク質は、それらが適当なPPIシャペロンとの複合体内に結合すると、穏やかな生理的条件下で(すなわち、界面活性剤又はカオトロピック剤のような可溶化添加剤を要することなく)可溶性となる。
【0057】
本発明者らは、本発明の組換え産生融合タンパク質が封入体から可溶性形態で容易に得られうることを見出した。組換え産生された風疹E1タンパク質に含まれる風疹E1タンパク質の顕著な特徴は生理的バッファー条件下でのその溶解性である。さらに、融合タンパク質に含まれる本発明の風疹E1タンパク質は天然様構造として容易に得られうる。
【0058】
また、本発明の風疹E1融合タンパク質は、取扱いが非常に容易である。例えば、そのような融合タンパク質を再生させることは非常に容易である。カオトロピック物質は、異なる様態でリフォールディングされて、すべて、熱力学的に安定かつ可溶性の天然様形態を与えうることは興味深い。リフォールディングは、透析及び急速希釈のどちらによっても並びにサイズ排除クロマトグラフィー又はマトリックス介助リフォールディングの再生によっても、高収率で達成される。これらの知見は、この共有結合形態においては、風疹E1抗原融合タンパク質が、準安定タンパク質ではなく熱力学的に安定であることを示唆している。
【0059】
好ましくは、本発明の可溶性タンパク質は、ペプチジル-プロピル-イソメラーゼクラスのシャペロンとの風疹E1抗原の融合により製造されうる。したがって、本発明の好ましい実施形態は、風疹E1抗原とペプチジル-プロピル-イソメラーゼクラスのシャペロンとの融合体に関する。
【0060】
本発明のもう1つの主題は、風疹E1抗原をコードする少なくとも1つのヌクレオチド配列と、その上流に位置する、FKBPシャペロンをコードする少なくとも1つのヌクレオチド配列とを含む、風疹E1抗原をコードする組換えDNA分子に関する。
【0061】
プロリルイソメラーゼは、種々の機能の種々のサブユニット又はモジュール(例えば、触媒活性を示すモジュール、及びシャペロン又は結合活性を示すモジュール)を含みうる。FKBPファミリーのそのようなモジュラーメンバーとしては、FkpA(Ramm, (2000))、SlyD(Hottenrottら, (1997))及び誘発因子(Scholzら, (1997))が挙げられる。好ましい実施形態においては、本発明は、FKBPシャペロンが、FkpA、SlyD及び誘発因子よりなる群から選ばれることにより特徴づけられる組換えDNA分子に関する。
【0062】
また、分子シャペロンの完全な配列を常に使用する必要はないことが、よく知られており理解されている。必要な能力及び機能を尚も有する、シャペロンの機能性断片(いわゆる、モジュール)も使用されうる(WO 98/13496を参照されたい)。本発明において発現手段として使用されるFkpAはN末端シグナル配列を欠く。FkpAの近縁体、すなわちSlyDは、触媒機能及びシャペロン機能をもたらす構造化N末端ドメインと、例外的にヒスチジン及びシステイン残基に富む、大部分が非構造化されたC末端とよりなる(Hottenrott, 1997)。WO 03/000878は、アミノ酸1-165を含むSlyDのC末端切断型変異体が標的タンパク質の効率的発現に対して例外的に正の効果を及ぼすことを開示している。野生型SlyDの場合とは異なり、使用される末端切断型SlyD変異体(1-165)においては、ジスルフィドシャフリングを損なう恐れが成功裏に排除されている。末端切断型SlyD(1-165)を含む組換えDNA分子は本発明の好ましい実施形態の1つである。
【0063】
本発明の風疹E1抗原を設計する好ましい態様においては、シグナルペプチドは含まれない。本発明の発現系は、サイトゾル発現系として働く場合に最も有利であることが判明している。このサイトゾル発現は封入体の形成をもたらす。封入体に通常は伴う顕著でよく知られた問題とは異なり、本発明者らは本発明において、格別に大量の風疹E1タンパク質が産生されるだけでなく、本発明の組換え風疹E1タンパク質が取り扱い易くもある(例えば、可溶化及びリフォールディングが容易である)ことを見出した。
【0064】
好ましくは、本発明の組換えDNA分子は更に、それが、風疹E1抗原をコードする配列とFKBPシャペロンをコードする配列との間に位置する10〜100アミノ酸のペプチド性リンカーをコードする少なくとも1つのヌクレオチド配列を含むことにより特徴づけられる。当業者に理解されるとおり、そのようなリンカーポリペプチドは、特に長さ、柔軟性、電荷及び親水性の点で、意図される用途に最も適したものとして設計される。さらに、リンカーをコードするそのようなDNA配列は、例えばタンパク質分解部位を与えることに加えて、ポリリンカーとしても働きうる。すなわち、それは、風疹E1タンパク質をコードするDNA断片とシャペロンドメインをコードするDNA断片との融合を促進する複数のDNA制限部位を与えうる。得られた融合タンパク質の発現及び精製ならびに更には可溶性かつ免疫反応性コンホメーションへのリフォールディングの後、該ポリリンカーは融合タンパク質複合体からの風疹E1タンパク質の遊離をも促進する。
【0065】
それにより、本発明の可溶性風疹E1抗原及びこのタンパク質の変異体は融合構築物から切り出されることが可能であり、この場合、該風疹E1抗原は、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくとも、ジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。さらに、風疹E1抗原の前記領域のN末端(Cys349)は更に少なくとも隣接風疹E1抗原配列の25、30、34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも10、11、15、25、35アミノ酸を含有する。
【0066】
本発明のもう1つの主題は、FKBPシャペロンをコードする単一のヌクレオチド配列と風疹E1タンパク質をコードする単一のヌクレオチド配列とを含む組換えDNAに関する。
【0067】
少なくとも2つのFKBPシャペロンドメインと1つの標的タンパク質ドメインとを含む融合タンパク質も非常に有利である。もう1つの好ましい実施形態においては、本発明の組換えDNA分子は、FKBPシャペロンをコードする2つの配列と風疹E1タンパク質をコードする1つの配列とを含む。Scholzら, (2005)は、第2のシャペロン単位が融合されると融合タンパク質の溶解度が顕著に改善されることを示している。天然において二量体であるPPIアーゼは2つのFKPBシャペロンドメインの融合により厳密に模擬され、それにより溶解度が改善されることを示唆する証拠が存在する。
【0068】
当業者に理解されるとおり、「少なくとも2つ」なる表現は、FKBPシャペロンドメインをコードする2以上のヌクレオチド配列が、本発明の範囲から逸脱するこなく組換えDNA分子の構築において使用されうることを示すために用いられる。好ましくは、風疹E1シャペロンタンパク質は、シャペロンを含有する少なくとも2つ且つ多くとも4つの配列を含有する。
【0069】
該DNA分子は、標的タンパク質の上流にFKBPシャペロンをコードする両方のDNA配列を含むように設計されうる。あるいは、それらの2つのFKBPドメインは、標的タンパク質を挟むように配置されうる。風疹E1抗原をコードする配列の上流に両方のFKBPドメインを含む組換えDNA分子は、本発明の好ましい実施形態の1つである。
【0070】
本発明のもう1つの実施形態においては、該組換えDNA分子は、PPIシャペロンをコードする一方の配列が風疹E1抗原の上流に位置し、PPIシャペロンをコードするその他の配列が、風疹E1抗原をコードする配列の下流に位置することにより特徴づけられる。
【0071】
2つのシャペロンドメイン及び風疹E1抗原をコードする配列を含むDNA構築物は、好ましくは、これらのドメインの間に10〜100アミノ酸の2つのリンカーペプチドをも含有する。組織的クローニングを可能にするために、これらの2つのリンカーペプチド配列をコードするヌクレオチド配列は、好ましくは、異なる。該ヌクレオチド配列におけるこの相違は、該リンカーペプチドのアミノ酸配列における相違を必ずしも与える必要はない。さらにもう1つの好ましい実施形態においては、それらの2つのリンカーペプチドのアミノ酸配列は同一である。そのような同一リンカーペプチド配列は、例えば、2つのFKBPシャペロンドメインと風疹E1タンパク質とを含む融合タンパク質をイムノアッセイにおいて使用したい場合に有利である。
【0072】
シャペロンの1つ又は全てを本発明の融合タンパク質から遊離させたい場合には、該リンカーペプチドを、タンパク質分解切断部位を含むように構築する。既に記載されているとおり、タンパク質分解切断部位はポリリンカーとしても働きうる。すなわち、それは、風疹E1タンパク質をコードするDNA断片とシャペロンドメインをコードするDNA断片との融合を促進する複数のDNA制限部位を与えうる。風疹E1タンパク質をコードする少なくとも1つのポリペプチド配列をコードする核酸配列と、その上流の、FkpA、SlyD及び誘発因子よりなる群から選ばれるFKBPシャペロンをコードする少なくとも1つのヌクレオチド配列と、さらに、タンパク質分解切断部位を含むペプチドリンカーをコードする核酸配列とを含む、融合タンパク質をコードする組換えDNA分子は、本発明のもう1つの実施形態である。本発明の組換えDNA分子(すなわち、風疹E1タンパク質をコードする少なくとも1つのポリヌクレオチド配列と、その上流の、FkpA、SlyD及び誘発因子から選ばれるFKBPシャペロンをコードする少なくとも1つのヌクレオチド配列とを含む、融合タンパク質をコードする組換えDNA分子)を、機能しうる形で連結された形態で含む発現ベクターは、非常に有利であることが判明している。
【0073】
本発明の組換えDNAを含む発現ベクターは、無細胞翻訳系において融合タンパク質を発現させるために使用されることが可能であり、あるいは宿主細胞を形質転換するために使用されうる。好ましい実施形態においては、本発明は、本発明の発現ベクターで形質転換された宿主細胞に関する。
【0074】
発現及びクローニングベクターは選択マーカーを含有しうる。該選択マーカーは、該ベクターで形質転換された宿主細胞の生存又は成長に必要なタンパク質をコードする遺伝子であるが、そのようなマーカー遺伝子は、宿主細胞内に共導入される別のポリヌクレオチド配列上に含有されうる。マーカー遺伝子を発現する宿主細胞のみが選択条件下で生存及び/又は成長する。典型的な選択遺伝子には、(a)抗生物質又は他の毒性物質、例えばアンピシリン、テトラサイクリンなどに対する耐性を付与するタンパク質をコードするもの、(b)栄養要求性欠損を相補するタンパク質をコードするもの、あるいは(c)複合培地からは利用可能でない決定的に重要な栄養素を供給するタンパク質をコードするものが含まれるが、これらに限定されるものではない。適当な選択マーカーの選択は宿主細胞に左右され、種々の宿主のための適当なマーカーが当技術分野で公知である。
【0075】
関心のある風疹E1タンパク質を含有するベクターは、当技術分野で公知の任意の方法により宿主細胞内に導入されうる。これらの方法は細胞宿主の型によって様々であり、塩化カルシウム、塩化ルビジウム、リン酸カルシウム、DEAE-デキストラン、他の物質を使用するトランスフェクション、及びウイルスによる感染を包含するが、これらに限定されるものではない。大量の本発明の風疹E1タンパク質は、和合性宿主内でベクター又は他の発現ビヒクル中の本発明のポリペプチドを発現させることにより製造されうる。本発明のベクターの構築は通常の連結技術を用いる。単離されたプラスミド又はDNA断片を切断し、操作処理し、所望の形態で再連結して、必要なプラスミドを得る。所望により、構築されたプラスミド内の配列が正しいことを確認するための分析を公知様態で行う。発現ベクターを構築し、in vitro転写産物を調製し、宿主細胞内にDNAを導入し、発現及び機能の評価のための分析を行うための適当な方法は、当業者に公知である。遺伝子の存在、増幅及び/又は発現は、例えば、本明細書に記載の配列に基づきうる適当に標識されたプローブを使用する、通常のサザンブロット法、mRNAの転写を定量するためのノーザンブロット法、ドットブロット法(DNA又はRNA分析)、又はin situハイブリダイゼーションにより、サンプル中で直接的に測定されうる。当業者は、所望によりこれらの方法がどのようにして修飾されうるかを容易に予想するであろう。
【0076】
本発明のもう1つの好ましい実施形態は、可溶性かつ免疫反応性の風疹E1抗原-シャペロン融合タンパク質の製造方法を含み、該方法は、
a.宿主細胞の培養、
b.該融合タンパク質の発現、
c.該融合タンパク質の精製、
d.可溶性かつ免疫反応性のコンホメーションへのリフォールディング
の工程を含む。
【0077】
本発明の方法は、ペプチジルプロリルイソメラーゼがFKBPシャペロンである、可溶性かつ免疫反応性の風疹E1抗原-シャペロン融合タンパク質の製造に関する。さらに、該方法は、SlyD、FkpA及び誘発因子よりなる群から選ばれるFKBPシャペロンに関する。
【0078】
本発明のもう1つの主題は、サンプル中のサブクラスIgG及び/又はIgMの抗風疹抗体の検出、測定及び定量のための方法であり、この場合、風疹E1抗原を該抗体に対する捕捉試薬及び/又は結合相手として使用する。当業者に公知のすべての生物学的液体が、抗風疹抗体の検出のためのサンプルとして使用されうる。好ましいサンプルは、全血、血清、血漿、尿、唾液などの体液である。診断方法に関しては、本発明の可溶性風疹E1抗原融合タンパク質、より好ましくは、可溶性風疹E1抗原-シャペロンタンパク質の明らかな利点は、例えば、生理的バッファー条件下の風疹E1タンパク質の安定性の増強、診断感度の増加、存在するコンホメーションエピトープの数の増加、及び適切にフォールディングした風疹E1タンパク質を容易に標識する可能性があることである。
【0079】
よく知られた標識は、マーカー基又はエフェクター基、例えば固相結合基である。標識された可溶性風疹E1抗原融合タンパク質、より好ましくは、標識された可溶性風疹E1抗原-シャペロンタンパク質は、本発明のもう1つの好ましい実施形態である。標識基は、任意の公知の検出可能なマーカー基、例えば色素、発光標識基、例えば化学発光基、例えばアクリジニウムエステルもしくはジオキセタン、又は蛍光色素、例えばフルオレセイン、クマリン、ローダミン、オキサジン、レゾルフィン、シアニン及びそれらの誘導体から選ばれうる。標識基の他の具体例としては、発光性金属錯体、例えばルテニウム又はユーロピウム錯体、酵素、例えばELISA又はCEDIA(クローン化酵素ドナーイムノアッセイ(Cloned Enzyme Donor Immunoassay)、例えばEP-A-0 061 888)に使用される酵素、及び放射性同位体が挙げられる。エフェクター基は、例えば、バイオアフィン(bioaffine)結合ペアのうちの一方のパートナーを含む。アッセイの実施中に、エフェクター基は、バイオアフィン結合ペアのもう一方のパートナーと特異的に、そして好ましくは非共有結合的に相互作用する。適当な結合ペアの具体例としては、ハプテン又は抗原/抗体、ビオチン又はビオチン類似体、例えばアミノビオチン、イミノビオチン又はデスチオビオチン/アビジン又はストレプトアビジン、糖/レクチン、核酸又は核酸類似体/相補的核酸、及び受容体/リガンド、例えばステロイドホルモン受容体/ステロイドホルモンが挙げられる。好ましい結合ペアメンバーはハプテン、抗原及びホルモンを含む。特に好ましいのは、ジゴキシンのようなハプテンならびにビオチン及びその類似体である。好ましくは、風疹E1タンパク質と好ましくはPPIシャペロンとを含む可溶性複合体が、風疹に対する抗体の検出のためのイムノアッセイにおいて使用される。もう1つの好ましい実施形態においては、風疹E1抗原と好ましくはPPIシャペロンとを含む標識された可溶性複合体が、風疹に対する抗体の検出のためのイムノアッセイにおいて使用される。最も好ましくは、その標識された複合体は、好ましくはPPIシャペロンと風疹E1タンパク質とを含む組換えポリペプチドにおける分子内複合体である。
【0080】
イムノアッセイは当業者によく知られている。そのようなアッセイを行うための方法ならびに実際の適用及び手順は関連参考書において要約されている。関連参考書の具体例としては、Tijssen, P., Preparation of enzym-antibody or other enzyme-macromolecule conjugates in "Practice and theory of enzyme immunoassays" (1990) 221-278, R. H. Burdon及びv. P. H. Knippenberg編, Elsevier, AmsterdamならびにTijssen, in "Methods in Enzymology" (1980), S. P. Colowick, N. O. Caplan及びS. P.編, Academic Pressの、免疫学的検出方法を扱った様々な巻、特に第70, 73, 74, 84, 92及び121巻が挙げられる。
【0081】
該新規可溶性風疹E1抗原融合タンパク質、より好ましくは、該新規可溶性風疹E1抗原-PPIシャペロンタンパク質は、検出の様式(例えば、放射性同位体アッセイ、酵素イムノアッセイ、電気化学発光アッセイなど)又はアッセイ原理(例えば、試験ストリップアッセイ、サンドイッチアッセイ又は均一系アッセイなど)には無関係に、抗風疹抗体の検出のためのアッセイを改良するために使用されうる。風疹感染の、信頼しうる且つ高感度の早期検出のためには、体液サンプル中の抗ウイルス抗体を測定することが必須である。本発明の可溶性複合体は生理的バッファー条件における抗風疹抗体の検出を可能にする。抗風疹抗体の検出はそのような複合的な風疹検出系の重要な部分である。したがって、好ましい実施形態においては、本発明は、風疹E1抗原融合タンパク質の使用、より好ましくは、風疹E1抗原-シャペロンタンパク質の使用に基づく抗風疹抗体の検出を含む風疹検出系に関する。
【0082】
当技術分野で公知のとおり、細菌、真菌又はウイルスのような感染因子に対する抗体は、好ましくは、二重抗原架橋の概念(時には、このアッセイ概念は、2つの抗原が抗体により架橋されるため、二重抗原架橋概念とも称される)に従うアッセイにより検出される。そのようなアッセイにおいては、与えられた抗原の、少なくとも2つの異なる分子に、抗体がその2個(IgG、IgA、IgE)又は10個(IgM)のパラトープで結合する能力が要求され、用いられる。
【0083】
該架橋概念による、体液からの抗体の検出は、多種多様なアッセイ設定で行われうる。単純な設定は、固相への抗原の直接的なコーティング、及び標識された形態の同じ抗原の使用を含む。適当なアッセイ条件下、サンプル中の抗体は該固相結合抗原と該標識抗原との間で架橋を形成する。したがって、検査中の抗体がサンプル内に存在する場合にのみ、架橋が形成され、シグナルが検出されうる。「固相抗原」及び「検出抗原」の基本的構造は、好ましくは同じである。例えば、1つ又はいくつかのエピトープを含むタンパク質が、固相にコートされて直接的又は間接的に使用されることが可能であり、標識又はマーカーに結合しているが同じ合成タンパク質が検出抗原として使用される。また、二重抗原架橋アッセイにおいて免疫学的に交差反応性である、類似しているが異なる風疹E1抗原を使用することも可能である。そのようなアッセイを行うための必須要件は、関連エピトープが両方の抗原上に存在することである。明らかに、二重抗原架橋アッセイ形態には多数の変法が存在する。そのような変法は、例えば、固相への風疹E1抗原の間接的コーティングを含む。好ましくは、特異的結合ペア、最も好ましくは、ビオチン-ストレプトアビジン(又はアビジン)系が、風疹抗原を固相に間接的に結合させるために使用される。一方、そのような系における検出に使用される風疹E1抗原は直接的にはマーカー(例えば、放射性同位体、酵素、蛍光分子など)を担持していないことも可能であり、その代わりに、例えばハプテン(例えばジゴキシン)を担持することにより間接的に検出されうることが可能である。そのような間接的検出は、例えば、標識された抗ジゴキシン抗体により行われうる。
【0084】
したがって、本発明の好ましい実施形態は、本発明の第1風疹E1抗原と本発明の第2風疹E1抗原とを含む、二重抗原架橋概念に従うイムノアッセイである。
【0085】
もう1つの好ましい実施形態においては、本発明は、第1風疹E1抗原融合タンパク質複合体を捕捉抗原として使用し、第2風疹E1抗原融合タンパク質を検出抗原として使用することにより特徴づけられる、二重抗原架橋概念に従うイムノアッセイに関する。
【0086】
本明細書に記載の風疹E1抗原融合タンパク質複合体、好ましくは、風疹E1抗原-シャペロン複合体は、それを使用しなければ取り扱いが困難な種々のタンパク質の可溶化をもたらすだけでなく、二重抗原架橋概念に従う非常に有利なイムノアッセイをも可能にする。それぞれ固相結合抗原との複合体形成のために及び検出抗原との複合体形成のために種々のシャペロンを使用することが今や可能になったことは、二重抗原架橋概念に従うそのようなイムノアッセイの特に魅力的な特徴である。アッセイのそのような修飾は更に、非特異的結合の問題を改善する。シャペロンに対して反応性であり従って偽陽性シグナルを生成しうる、サンプル中の抗体は、それぞれ固相抗原及び検出抗原を複合体形成させるために異なるシャペロンを使用した場合には、架橋を形成しないであろう。したがって、本発明のこの態様においては、非特異的結合による陽性シグナルの可能性が著しく減少する。したがって、好ましい実施形態において、本発明は、第1及び第2シャペロン-風疹E1抗原複合体の第1シャペロンと第2シャペロンとが互いに異なることにより特徴づけられる、二重抗原架橋概念に従うイムノアッセイに関する。
【0087】
最もよく特徴づけられているシャペロンのほとんどは、バイオテクノロジー研究において広く使用されている大腸菌(Escherichia coli)から単離されている。大腸菌(Escherichia coli)は広く分布する細菌種であるため、多数の哺乳動物は、この細菌に由来するタンパク質に対する抗体を持つようになっている。そのような抗体により引き起こされる偽陽性反応の可能性を減少させるためには、異なる細菌種(好ましくは好熱菌)に由来する少なくとも1つのPPIシャペロンを使用することが好ましい。好ましくは、シャペロンは、好極限性細菌、特に、テルマトガ・マリチマ(Thermatoga maritima)、アクイフェックス・エオリクス(Aquifex aeolicus)及びテルムス・サーモフィルス(Thermus thermophilus)を含む細菌群の好極限性細菌に由来する。イムノアッセイ全般及び好ましくは該架橋概念に従うイムノアッセイにおけるシャペロン-抗原複合体の使用は、そのような複合体のシャペロンを誘導体化する可能性をも提供し、抗原自体の修飾を要しない。第2化学部分によるタンパク質の修飾、例えば、その分子への標識のカップリングは、該ポリペプチドに負の影響を及ぼす危険性を含む、と一般に認識されている。例えば、そのような標識により、検査中のエピトープが損なわれることがあり、あるいは非特異的結合が引き起こされうる。本発明において、風疹E1抗原-シャペロン複合体内のシャペロンを特異的に誘導体化することが今や可能となった。好ましい実施形態においては、二重抗原架橋概念に従うイムノアッセイは更に、捕捉抗原として使用する第1風疹E1抗原-シャペロン複合体が固相結合基を含むことにより特徴づけられる。もう1つの好ましい実施形態においては、該架橋概念に従うイムノアッセイを行い、これは更に、検出抗原として使用する第2風疹E1抗原-シャペロン複合体がマーカー基を含むことにより特徴づけられる。
【0088】
本発明は更に、抗風疹抗体の検出のための診断試験における、風疹E1の少なくとも1つの抗原及び必要に応じて使用する他の通常の添加剤の使用に関する。
【0089】
本発明のもう1つの主題は、イムノアッセイのための通常の試験添加剤に加えて、測定すべき風疹抗体への特異的結合に適した風疹E1抗原の少なくとも1つの抗原を含有し、場合によっては標識及び必要に応じて他の通常の添加剤を含有する、風疹に対する抗体の検出のための試薬キットに関する。特に、該試験キットは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130にわたるN末端領域を含有する風疹E1抗原及びこのタンパク質の変異体を含有する。好ましくは、該試験キットは、ジスルフィド架橋Cys349-Cys352及びCys368-Cys401、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せにわたるC末端領域を含有する風疹E1抗原及びこのタンパク質の変異体を含有する。
【0090】
本発明のもう1つの実施形態の試験キットが含有する風疹E1抗原及びこのペプチドの変異体は、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164、好ましくは134-約168を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。
【0091】
さらに、この領域のN末端(Cys349)は更に、少なくとも25、30、34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に、隣接風疹E1抗原配列の少なくとも10、11、15、25、35アミノ酸を含有する。該試薬キットは更に、少なくともジスルフィド架橋Cys349-Cys352及びCys368-Cys401、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せにわたるC末端領域、ならびに少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130の組合せにわたるN末端領域を含有する風疹E1抗原の組合せを含有する。
【0092】
また、該試薬キットは、対照及び標準溶液、ならびに当業者により使用される通常の添加剤、バッファー、塩、界面活性剤などを含有する1以上の溶液中の試薬を含有する。
【0093】
実施例
【実施例1】
【0094】
縦列EcSlyD及び風疹E1エクトドメイン断片E1(201-432)を含む発現プラスミドの構築
Novagen(Madison, WI, USA)のpET24a発現プラスミドに基づき、以下のクローニング工程を行った。該ベクターをNdeI及びXhoIで消化し、縦列SlyD及び風疹E1断片201-432を含む半合成カセットを挿入した。この風疹E1断片はアミノ酸349、352、368及び401位にシステイン残基を含有する。
【化1】

【0095】
得られたプラスミドのインサートを配列決定したところ、それは所望の融合タンパク質をコードしていることが判明した。該挿入カセットの核酸配列及び得られた融合タンパク質のアミノ酸配列を本発明の配列プロトコールに示す。
【実施例2】
【0096】
SS-E1(201-432)融合タンパク質の、共役された精製及びリフォールディング
該発現プラスミドを含有する大腸菌(E. coli)BL21(DE3)細胞をLB培地+ カナマイシン(30μg/ml)内で、OD600が1になるまで増殖させ、37℃の増殖温度でイソプロピル-β-D-チオガラクトシド(IPTG)を最終濃度1mMまで加えることによりサイトゾル過剰発現を誘導した。誘導の4時間後、細胞を遠心分離(5000×gで20分間)し、凍結させ、-20℃で保存した。細胞溶解のために、該凍結ペレットを100mMリン酸ナトリウム(pH8.0)、7.0M GuHCl、10mM イミダゾールに室温で再懸濁させ、得られた懸濁液を2時間攪拌して細胞溶解を完了させた。遠心分離及び濾過の後、前記細胞溶解バッファー中で予め平衡化されたNi-NTA(ニッケル-ニトリロ-トリアセタート)カラム上にライセートをアプライした。未熟なジスルフィド架橋及びSSシャフリングを避けるために、金属キレート化カラムに適合しうる還元剤として5mM TCEPを洗浄バッファー中に加えた。過剰の洗浄工程(>20カラム容量の細胞溶解バッファー + TCEP)の後、マトリックス結合タンパク質のコンホメーションリフォールディングを誘導するために、該カオトロピック細胞溶解バッファーを50mMリン酸ナトリウム(pH7.8)、100mM塩化ナトリウム、5mM TCEPで置換した(残留GuHClがカオトロピック濃度で存在しないことが保証されるよう、少なくとも10カラム容量のリフォールディングバッファーをアプライした)。ついで、酸化的フォールディング(すなわち、システイン残基の酸化的架橋)を、50mMリン酸ナトリウム pH7,8、100mM塩化ナトリウムでの洗浄によりを誘導した。二価Ni2+イオンの有効濃度が高いため、マトリックス結合融合タンパク質内のジスルフィド架橋の形成は非常に速い過程である。溶出前に、〜50kDaの見掛け分子量を有する汚染(コンタミ)タンパク質を除去するために、該イミダゾール濃度を55mMまで上昇させた。ついで、50mMリン酸ナトリウム(pH7.8)、100mM塩化ナトリウム中の55mMから500mMまでのイミダゾール勾配を適用することにより、該天然融合タンパク質を溶出した。タンパク質含有画分を純度に関して評価し(SDS-PAGEによる判定で>95%)、プールした。最後に、該タンパク質をサイ
ズ排除クロマトグラフィーに付し、二量体画分をプールし、濃縮し、その分光学的特性に関して評価した。
【実施例3】
【0097】
風疹E1融合タンパク質の特徴づけ
a.UV分光学
SS-E1(201-432)は、可溶性かつ天然のようにフォールディングしたタンパク質として溶出する。組換え産生されマトリックスリフォールディングした融合タンパク質のUVスペクトルは凝集傾向を何ら示していない。図1に示すとおり、生理的バッファー条件中のSS-E1(201-432)のUV吸収スペクトルのベースラインは横軸にほぼ等しく(310nm以上)、したがってこのことは、自己会合又は凝集減少により生じる迷光粒子が存在しないことを示している。
【0098】
図1:マトリックス介助リフォールディング及びイミダゾール勾配溶出の後の融合タンパク質SS-E1(201-432)のUVスペクトル。該スペクトルは、1cmの光路長を用いたUvicon XS光度計上で記録された。バッファー条件は50mM リン酸ナトリウム(pH8.0)、100mM 塩化ナトリウム及び〜250mM イミダゾールであった。SS-E1(201-432)に関して48370 M-1cm-1のモル吸光係数εを使用して、タンパク質濃度は7.25μMと決定された。スペクトルの形状はシャペロン化風疹E1エクトドメイン断片(201-432)の溶解度を表す。
【0099】
b.SDSゲル-電気泳動
融合ポリペプチド風疹SS-E1アミノ酸残基201-432の純度をSDS-PAGEにより確認した(図2)。2工程のクロマトグラフィーの後、該融合ポリペプチドの純度は95%を超えている。
【0100】
図2:非還元性SDS-PAGEにより実証されたSS-E1(201-432)の精製。該クーマシー染色ゲルは、(左から右に)Invitrogenのタンパク質標準M12(レーン1)、大腸菌(E. coli)カオトロピック粗抽出物(不溶性画分レーン2、可溶性画分レーン3)、IMAC流動物(レーン5)、〜50kDaの見掛け分子量を有する顕著な混入物(コンタミ)を含有する洗浄画分(レーン7)、及びSS-E1(201-432)イミダゾール溶出画分(レーン9-11)を示している。SDS-PAGEからの判断によると、該融合タンパク質の純度は95%を超えている。混合ジスルフィド又は共有分子間SS付加体の徴候は何ら存在しない。これは、それぞれシステイン349&352及びシステイン368&401の定量的架橋を示している。
【0101】
C.FPLC分析(高速タンパク質液体クロマトグラフィー分析)
その抗原性のほかに、SS-E1融合タンパク質のオリゴマー状態、溶解度及び安定性が診断目的に対するその適合性を決定する。該組換え風疹エクトドメイン断片のオリゴマー状態を明らかにするために、システイン含量において異なるSS-E1(201-432)をSuperdex 200HR 10/30カラム上の分析用ゲル濾過に付した。泳動バッファーは50mM リン酸ナトリウム(pH7.8)、100mM NaClであった。150μlのSS-E1(201-432)溶液(タンパク質濃度〜1.0mg/ml)をSECカラム上にアプライし、溶出を280nmの吸光度によりモニターした(図3)。該実験の結果は、すべてのSS-E1(201-432)システイン変異体が〜12.8mlで定量的に溶出したことを示しており、このことは見掛け上二量体の融合タンパク質を示している。これは、ジスルフィド結合の導入が風疹エクトドメインのポリペプチドバックボーンを拘束し該分子の全体的な小型化を招くという予想と一致する。FPLC分析における高分子会合の明らかな欠如はSDS-PAGEの結果を証明しており、SS-E1(201-432)のマトリックス共役酸化的リフォールディングが、相当に安定で異性化傾向もシャフリング傾向も示さない天然様ジスルフィド架橋を与えるという仮定を裏付けている。SS-E1(201-432)のオリゴマー化は、おそらく、安定な二量体を形成すると報告されている(Mukherjeeら, Biotechnol. Appl. Biochem. (2003) 37, 183-186; Mitterauerら, Biochem. J. (1999) 342, 33-39)シャペロン融合相手SlyDにより媒介されるのであろう。
【0102】
図3:SS-E1(201-432)のシステイン変異体はすべて、FPLC分析により評価した場合、可溶性二量体を形成する。SS-E1(201-432)のシステイン非含有及びジスルフィド架橋変異体をSuperdex 200 HR 10/30分析用ゲル濾過カラム上で泳動させ、それらのオリゴマー状態に関して評価した。泳動バッファーは50mM リン酸ナトリウム(pH7.8)、100mM 塩化ナトリウム、1mM EDTAであり、タンパク質ローデング量は〜200μgであった。溶出を280nmの吸光度によりモニターした。より伸張したシステイン非含有SS-E1(201-432)は12.6ml(黒線)で溶出し、一方、より小型の二重(青線)及び三重(赤線)ジスルフィド架橋変異体は共に12.9mlで溶出する。Roche HPLCタンパク質標準(灰色)はβ-ガラクトシダーゼ(465kDa)、IgG(150kDa)、Fab(50kDa)、ミオグロビン(17kDa)及びジペプチドGly-Tyr(238Da)を含む。
【実施例4】
【0103】
SS-E1(201-432)へのビオチン及びルテニウム部分のカップリング
組換え風疹エクトドメインのリシンε-アミノ基を〜10mg/mlのタンパク質濃度で、それぞれN-ヒドロキシ-スクシンイミド活性化ビオチン及びルテニウム標識により修飾した。標識/タンパク質のモル比は、それぞれの融合タンパク質に応じて2:1から5:1まで様々となった。反応バッファーは150mM リン酸ナトリウム(pH8.0)、50mM NaCl、1mM EDTAであった。反応を室温で15分間行い、緩衝化L-リシンを最終濃度10mMまで加えることにより停止させた。該カップリング反応の後、該粗タンパク質コンジュゲートをゲル濾過カラム(Superdex 200 HI Load)上に通過させることにより未反応遊離標識を除去した。
【実施例5】
【0104】
免疫診断試験における組換え風疹E1融合タンパク質SS-E1(aa201-432)の免疫反応性の検査:天然血清中の抗風疹IgG抗体の検出
種々の該融合タンパク質の免疫反応性を自動Elecsys(登録商標)2010分析装置(Roche Diagnostics GmbH)において評価した。二重抗原サンドイッチ形態で測定を行った。それにより、該ビオチン-コンジュゲート(すなわち、捕捉抗原)をストレプトアビジンコート化磁気ビーズの表面上に固定化した。一方、検出抗原はシグナリング部分として錯化ルテニウムカチオンを含有する。Elecsys(登録商標)2010におけるシグナル検出は電気化学発光に基づくものである。特異的免疫グロブリンアナラアイトの存在下では、該発色性ルテニウム錯体は該固相へ架橋され、白金電極における励起の後に620nmの光を発する。ババリア赤十字の血清収集物からの抗風疹IgG陽性サンプルで測定を行った。
【表1】
表1:風疹SS-E1(aa201-432)抗原を使用することによる天然血清中の抗風疹IgG抗体の検出。
陽性として分類された全ての血清は、それで正しいと分析判定された。さらに、該結果は、陽性(COI* > 1)サンプルと陰性(COI* < 1)サンプルとの間の明らかな相違を示している。
【実施例6】
【0105】
風疹SS-E1(aa315-432)を使用することによる天然血清中の抗風疹IgG抗体の免疫学的検出。免疫学的活性に対する天然様三次構造の関連性
システイン349と352との間、システイン349とシステイン352との間及びシステイン368と401との間でジスルフィド架橋を形成する風疹E1変異体の3つの異なる組合せの測定。前記風疹変異体と比較して、システイン非含有抗原を使用した。サンプル(WHO標準から)、及びババリア赤十字の血清収集物からの抗風疹IgG陽性サンプルを使用して、測定を行った。測定は、二重抗原サンドイッチ形態を用いて自動Elecsys(登録商標)2010分析装置(Roche Diagnostics GmbH)において行った。
【表2】
表2:風疹SS-E1(aa315-432)抗原を使用することによる天然血清中の抗風疹IgG抗体の検出。
ジスルフィド架橋の形成において異なる種々の変異体の比較は風疹SS-E1(aa315-432)の免疫反応性を示している。風疹SS-E1(aa315-432)システイン349とシステイン352との間及びシステイン368とシステイン401との間のジスルフィド架橋は適切に形成されている。陽性として分類された全ての血清は、それで正しいと分析判定された。さらに、該結果は、陽性(COI* > 1)サンプルと陰性(COI* < 1)サンプルとの間の明らかな相違を示している。
【実施例7】
【0106】
風疹SS-E1(aa315-412)及び風疹SS-E1(aa201-432)を使用することによる天然血清中の抗風疹IgG抗体の免疫学的検出
ババリア赤十字の血清収集物からの抗風疹IgG陽性サンプルを使用して、測定を行った。測定は、二重抗原サンドイッチ形態を用いて自動Elecsys(登録商標)2010分析装置(Roche Diagnostics GmbH)において行った。
【表3】
表3:風疹SS-E1(aa315-412)及び風疹SS-E1(aa201-432)抗原を使用することによる天然血清中の抗風疹IgG抗体の検出。
【0107】
長さにおいて異なる種々の変異体の比較は風疹SS-E1(aa315-412)及び風疹SS-E1(aa201-432)の両方の比較しうる免疫反応性を示している。どちらの変異体に関しても、システイン349とシステイン352との間及びシステイン368とシステイン401との間のジスルフィド架橋は適切に形成されている。陽性として分類された全ての血清は、それで正しいと分析判定された。さらに、該結果は、陽性(COI* > 1)サンプルと陰性(COI* < 1)サンプルとの間の明らかな相違を示している。
【0108】
表に関する更なる説明
「カットオフ」なる語は、陽性結果と陰性結果とを識別するためのシグナルである。すなわち、「シグナル > カットオフ」は陽性サンプルを表し、「シグナル < カットオフ」は陰性サンプルを表す。「COI」なる語はカットオフ指数として定義される。COIは、(サンプルのシグナル)/(カットオフでのシグナル)として計算される。すなわち、COI > 1は陽性サンプルを表し、COI < 1は陰性サンプルを表す。
【0109】
参考文献の一覧
Artherton and Shepard in “Solid phase peptide synthesis” IRL Press, Oxford, UK, 1989.
Baron and Forsell, Virology. 1991 Dec;185(2):811-9
Buchner, J., Faseb J 10 (1996) 10-19
Chaye et al., J Clin Immunol. 1993 Mar;13(2) 93-100.
Giessauf et al., J. Immun. Meth. 287 (2004), 1-11
Green and Dorsett, J. Virol. (1986) 57, 893-898
Gros et al., 1997 Virology 230, 179-186
Hobman et al., 1994 (574-585) Virology
Ho-Terry et al., Arch Virol. 1985;84(3-4):207-15
Houbenweyl in “Methode der Organischen Chemie” edited by E. Wunsch, vol 15-I et II, Thiem, Stuttgart, Germany, 1974
Hottenrott, S., et al., J Biol Chem 272 (1997) 15697-701
Mitchell et al., Virus Research 29(1993), 33-54
Mitterauer et al., Biochem. J. (1999) 342, 33-39)
Mukherjee et al., Biotechnol. Appl. Biochem. (2003) 37, 183-186
Newcombe et al., Clin Diagn Virol. 1994 (3)149-63
Oker-Blom et al., J Virol. 1984 (2):403-8
Qui et al., Journal of virology 1994 (4086-4091)
Ramm, K. and Pluckthun, A., J Biol Chem 275 (2000) 17106-13
Sambrook, J., et al., in "Molecular Cloning: A Laboratory Manual" (1989) -, Eds. J. Sambrook,
Scholz, C., et al., Embo J 16 (1997) 54-8
Scholz, C., et al., J Mol Biol., 345, 1229-41, (2005)
Seppaenen et al., J. Clin. Microbiol (1991) 1877-1882
Starkey et al., J. Clin. Microbiol. (1995), 270-274)
Terry et al., Arch Virol. (1985) 86, 29-36
Terry et al Arch Virol. (1988);98, 189-97
Tijssen, P., Preparation of enzym-antibody or other enzyme-macromolecule conjugates in "Practice and theory of enzyme immunoassays" (1990) 221-278, Eds. R. H. Burdon and v. P. H. Knippenberg, Elsevier, Amsterdam)
Tijssen, P. "Methods in Enzymology" (1980), Eds. S. P. Colowick, N. O. Caplan and S. P., Academic Press
Waxham and Wolinsky, Virology. (1983)126(1),194-203
Waxham and Wolinsky, Rev Infect Dis. (1985) 133-9
Waxham and Wolinsky, Virology (1985) 153-65
Weber et al., Bull Soc Sci Med Grand Duche Luxemb. (1997), 31-41
Wolinsky et al., J Virol. (1991), 3986-94
Wolinsky et al., Rubella (1996) Fields Virology. Lippincott-Raven Publisher, Philadelpia, 899-929
EP 0299673
EP 0061888
EP0944838
WO 03/000877
WO 98/13496
WO 03/000878
【図面の簡単な説明】
【0110】
【図1】図1は、マトリックス介助リフォールディング及びイミダゾール勾配溶出の後の融合タンパク質SS-E1(201-432)のUVスペクトルを示す。該スペクトルは、1cmの光路長を用いたUvicon XS光度計上で記録された。バッファー条件は50mM リン酸ナトリウム(pH8.0)、100mM 塩化ナトリウム及び〜250mM イミダゾールであった。SS-E1(201-432)に関して48370 M-1cm-1のモル吸光係数εを使用して、タンパク質濃度は7.25μMと決定された。スペクトルの形状はシャペロン化風疹E1エクトドメイン断片(201-432)の溶解度を表す(実施例3aも参照されたい)。
【図2】図2は、非還元性SDS-PAGEにより実証されたSS-E1(201-432)の精製を示す。該クーマシー染色ゲルは、(左から右に)Invitrogenのタンパク質標準M12(レーン1)、大腸菌(E. coli)カオトロピック粗抽出物(不溶性画分レーン2、可溶性画分レーン3)、IMAC流動物(レーン5)、〜50kDaの見掛け分子量を有する顕著な混入物(コンタミ)を含有する洗浄画分(レーン7)、及びSS-E1(201-432)イミダゾール溶出画分(レーン9-11)を示している。SDS-PAGEからの判断によると、該融合タンパク質の純度は95%を超えている。混合ジスルフィド又は共有分子間SS付加体の徴候は何ら存在しない。これは、それぞれシステイン349&352及びシステイン368&401の定量的架橋を示している(実施例3bも参照されたい)。
【図3】図3は、SS-E1(201-432)のシステイン変異体がすべて、FPLC分析により評価した場合に可溶性二量体を形成することを示す。SS-E1(201-432)のシステイン非含有及びジスルフィド架橋変異体をSuperdex 200 HR 10/30分析用ゲル濾過カラム上で泳動させ、それらのオリゴマー状態に関して評価した。泳動バッファーは50mM リン酸ナトリウム(pH7.8)、100mM 塩化ナトリウム、1mM EDTAであり、タンパク質ローデング量は〜200μgであった。溶出を280nmの吸光度によりモニターした。より伸張したシステイン非含有SS-E1(201-432)は12.6ml(濃灰色実線)で溶出し、一方、より小型の二重(黒色実線)及び三重(黒色点線)ジスルフィド架橋変異体は共に12.9mlで溶出する。Roche HPLCタンパク質標準(淡灰色点線)はβ-ガラクトシダーゼ(465kDa)、IgG(150kDa)、Fab(50kDa)、ミオグロビン(17kDa)及びジペプチドGly-Tyr(238Da)を含む(実施例3aも参照されたい)。
【技術分野】
【0001】
本発明は、可溶性風疹E1抗原及びこのペプチドの変異体に関する。これらは、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。この領域のN末端(Cys349)は更に少なくとも隣接風疹E1抗原配列の25、30、又は34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも10、11、15、25、又は35アミノ酸を含有する。
【背景技術】
【0002】
風疹ウイルス(RV)はトガウイルスであり、ルビウイルス亜群の唯一の種である。この小さな包膜ウイルスは65nmのサイズを有し、脂質エンベロープに包まれた正二十面体ヌクレオカプシド内に封入された10kbの一本鎖RNA分子よりなる。風疹ウイルスは、Tリンパ球及び抗体の両方により媒介されると考えられている永久免疫を通常は与える比較的軽度な小児疾患(三日はしか)を引き起こす。成人における風疹感染は、一過性/慢性関節炎、筋骨格症候群、インスリン依存性糖尿病、及び後になって発現する神経後遺症を引き起こす可能性がある。風疹ウイルスは妊娠初期における主要パラメーターである。抗風疹IgM及び/又は抗風疹IgGの特異的検出は、それぞれ急性感染が生じたのか又は激しい再感染が生じたのかを明らかにするために決定的に重要である。妊娠中の母体の風疹ウイルス感染は胎児の先天性風疹症候群(CRS)のリスクを伴い、妊娠の最初の12週のうちに一次感染が生じた場合に先天性奇形の頻度が最も高い。この理由により、妊娠初期におけるRV感染により引き起こされる先天性異常の予防は、血清学的方法により個体の免疫状態を測定することを要する。これは、風疹ウイルス抗原に選択的なIgG抗体及びIgM抗体の選択的測定を意味する。一次風疹感染は特異的IgM抗体応答に関連しており、一方、検出可能なIgMの非存在下のIgGレベルの上昇は、急性風疹ウイルス感染に対して防御的である免疫状態を示す。出生前スクリーニングが、女性が妊娠初期に一次風疹感染を受けたことを示した場合には、治療的流産が推奨されることが多い。したがって、その試験結果が正確であることが何よりも重要である。
【0003】
RVに対する抗体を検出するための通常の検査診断は、主として、ELISA(酵素結合イムノソルベントアッセイ)試験に基づくものであり、欧州中部においてRV免疫状態の測定のための最も広く受け入れられている方法は、RV-IgG ELISAの証明のための赤血球凝集阻止(HAI)試験である(Weber,1997)。ある免疫グロブリンクラスの特異的抗体の検出は、特異的抗原が固定化されている固相に該免疫グロブリンを結合させることにより行われうる。ついで、結合した免疫グロブリンを、あるクラスのヒト免疫グロブリンに特異的な標識抗体により検出する。このアッセイ形態は、検出前に未結合免疫グロブリンを除去する洗浄工程を含む2工程アッセイ形態によってのみ行われうる。自動イムノアッセイ分析装置においてしばしば実施される1工程アッセイ形態は二重抗原サンドイッチの直接アッセイ形態を要する。すなわち、特異的抗体が、固相に固定化された第1抗原に結合する免疫複合体を形成するか、あるいは固相への又は標識を含有する第2抗原への固定化を媒介して、該特異的結合抗体の定量的又は定性的検出を可能にする。1工程二重抗原サンドイッチ形態における同じ特異性のIgM抗体の存在下の特異的IgG抗体の選択的測定のためには、可溶性単量体又は既定オリゴマー抗原の使用が厳しく要求され(EP 0 944 838)、これにより、免疫反応性コンホメーションが明らかにされる。
【0004】
風疹ウイルスは、動物及びヒトにおいて抗原性であることが示されている4つの構造タンパク質を含有する。これらは、3つのエンベロープ糖タンパク質E1(58kDa)、E2a(47kDa)及びE2b(42kDa)、ならびにウイルスゲノムを構成する一本鎖RNAプラス鎖を装飾する非グリコシル化カプシド(C)タンパク質(33kDa)である(Waxham及びWolinsky, 1985a; Oker-Blomら, 1984)。E2a及びE2bは同一遺伝子産物の変異体であることが示されており、ポリアクリルアミドゲル内での泳動における相違は、それらのタンパク質の不均一なグリコシル化によるものである。E1及びE2は単量体又はジスルフィド結合複合体(E1-E1及びE1-E2)を形成することが判明しており、一方、Cは専らホモ二量体として存在する(Waxham及びWolinsky, 1983; Wolinsky, 1996)。RVの生物学的、物理学的及び生化学的特性ならびにその感染の臨床的特徴はWolinsky, (1996) に公開されている。
【0005】
現在のところ、風疹感染の検出のためには抗原が使用されており、これは、安定に感染した/トランスフェクトされた細胞系及び一般には真核生物過剰産生からの幹細胞(stem)から誘導される。Seppanenら (1991)は、バキュロウイルス発現系を使用することによるスポドプテラ・フルジペルダ(Spodoptera frugiperda)Sf9昆虫細胞における風疹ウイルスのE1及びE2の発現を記載している。さらに、RV様粒子(RLP)の形態でRVの構造タンパク質E1、E2及びCを発現し分泌する安定にトランスフェクトされたCHO細胞系がHobmanら (1994)において開示されている。風疹様粒子は風疹主要抗原E1、E2及びCから構成され、これらはウイルスポリペプチド前駆体タンパク質として発現される。シグナル配列により、この未プロセシング前駆体タンパク質は培地内に分泌され、ウイルス様粒子が形成される。これらのRLPの表面は、風疹陽性血清中の抗RV抗体の免疫学的検出に適したエピトープを提示する。BHK(子ハムスター腎細胞)細胞系における、RVの3つの構造タンパク質を含有する非感染性RV様粒子(VLP)の発現が、Quiら, (1994)に概説されている。しかし、真核細胞系におけるRV抗原の製造は労働集約的であり、長時間を要し、一方、収率は比較的低い。抗風疹ウイルス抗体の検出のin vitro診断方法の確立のためには、可溶性で機能的で再現可能な一定の質で大腸菌(E. coli)のような原核生物から組換えRV抗原を製造する方法が必要であり、そのような方法は、確立されたRLPと比べて明らかな利点を伴うことが必要である。タンパク質の設計及び操作の分野におけるノウハウは著しく増加しているが、風疹抗原E1、E2及びCは大腸菌(E. coli)宿主細胞内で非常に少量でしか発現されず、さらに、それらは難溶性である。
【0006】
免疫優性風疹抗原(すなわち、ヒト血清からの抗体の免疫学的検出のための選択された抗原)はエンベロープタンパク質E1である。E1タンパク質の全体的なトポロジーは、α-らせん領域(438-452)を含む大きな細胞外エクトドメイン(1-452)及び膜貫通ドメイン(453-468)及びそれに続く短いC末端アンカーセグメント(481まで)を含む。RVのE1の不溶性の1つの理由は、10個のジスルフィド架橋により安定化されると考えられるエクトドメインのコンホメーションによるものであろう。風疹E1ペプチドはアミノ末端(N末端)領域及びカルボキシ末端(C末端)領域内に構築され、合計24個のシステイン残基を含有する。一般に、N末端は、遊離アミノ基(NH2)を有するアミノ酸で終結するタンパク質又はポリペプチドの端を意味し、C末端は、遊離カルボキシル基(COOH)を有するアミノ酸で終結するタンパク質又はポリペプチドの端を意味する。
【0007】
風疹E1タンパク質のN末端領域はシステイン残基C(8)からC(287)までの8個のジスルフィドを含有し、C末端領域は、システイン残基C(249)からC(401)までに生じる2個のジスルフィドを含有することを、Grosら, (1997)は開示している。残基C(456)、C(466)及びC(468)は推定膜貫通セグメント内に位置し、残基C(470)は該ウイルスの内部に位置すると予想される。これらのデータは、残基C(456)-C(470)がジスルフィド形成に関与していないことを示している。
【0008】
システイン残基が多数存在すること、及びそれに伴う、誤った架橋事象の可能性(分子内及び分子間SHシャフリングの生成は十分に考えられうる)は、E1のコンホメーション的及び酸化的リフォールディングが、in vitroで制御することが困難な非常に複雑な過程であることを示唆している。したがって、免疫反応性(すなわち、抗原性)コンホメーションへの風疹抗原の高収率リフォールディング及び精製の両方を促進する簡便かつ信頼しうるプロトコールが緊急に必要とされている。天然のようにフォールディングした組換えE1エンベロープ変異体は、RVに対する抗体に関する信頼しうるイムノアッセイを達成するための必須要件である。
【0009】
風疹E1エクトドメイン(1-452)は20個のシステイン残基を含有する。それらの正しい架橋はその三次元構造を決定し、正しい天然様コンホメーションエピトープの露出のために決定的に重要である。正しい分子内及び/又は分子間ジスルフィド架橋の形成である、システインに富むタンパク質の酸化的リフォールディングは、複雑な過程であり、過度な実験を要する過程である。新たに合成された又は輸送されたタンパク質内へのin vivoでのジスルフィド架橋の挿入のためには、細胞区画(真核細胞における小胞体、原核細胞におけるペリプラズム)内の多数のフォールディング補助体(シャペロン、オキシドレダクターゼ、プロリルイソメラーゼ)が利用可能である。細菌サイトゾル内のレドックス電位は強還元性(-270mV)であるため、異種標的タンパク質のシステインは天然においてはチオール部分として生じる。発現が非常に高い場合には、標的タンパク質は不溶性凝集物(いわゆる、封入体、IB)として宿主細胞内に貯蔵される。通常、部分的にリフォールディングした及びアンフォールディング(変性)したタンパク質画分は、カオトロピック溶液(7.0M GdmCl、8.0M尿素)中の還元条件(例えば、5mM TCEP)下で可溶化されて、アンフォールディングしたタンパク質鎖を与え、これは再生(すなわち、天然又は天然様コンホメーションにリフォールディング)されなければならない。一般に、天然様にフォールディングしたタンパク質の収率は、正しくないジスルフィド架橋によって著しく減少する。
【0010】
コンホメーションの観点を軽視すれば、限られた数のシステイン(及び厳密に分子内架橋事象が生じると仮定した場合)でさえも、莫大な数の誤った架橋事象を招いて、凝集過程のような非生産的な副反応という結果に終わる。試験管内のin vitroリフォールディングとは異なり、in vivoでのde novoフォールディングには、フォールディングを助ける最適化レドックス環境中の非常に多くのフォールディング補助体が利用可能である。それでも、大腸菌(E. coli)のような原核宿主細胞における、システインに富む機能性タンパク質の可溶性発現は、尚も非常に複雑である。RVの免疫反応性抗原の過剰生産率を改善し、リフォールディングしたタンパク質の溶解度を増加させる目的は、限られた程度でしか達成されていない。
【0011】
Newcombeら, (1994)においては、風疹E1抗原断片を可溶性形態で大腸菌(E. coli)内で発現させるために9個のグルタチオン-S-トランスフェラーゼ(GST)E1融合タンパク質が使用された。E1配列の相当な末端切断(トランケート化)を行った後に初めて、システイン非含有領域243-286(44アミノ酸残基)に関する可溶性発現の成功が可能となった。EP0299673は、風疹Ig特異的結合特性を保有するアミノ酸残基207-353のペプチドを開示している。さらに、Starkeyら, (1995)は、GST-E1融合タンパク質の44から75までのアミノ酸残基の非常に狭い領域のみが可溶性であったことを開示している。全E1コード配列及びより大きな亜断片を含有するGST融合タンパク質は、精製され得ない形態で発現された。したがって、それらは不溶性封入体として発現されたと考えられる。
【0012】
RV抗原内の免疫反応性決定基を同定するために、合成ペプチドを使用してE1及びE2エピトープが過去に詳細にマッピングされている(Mitchellら, (1993))。また、ウイルス感染性及び赤血球凝集に重要なドメインを含む、RV E1タンパク質内の、異なる無関係なエピトープが位置決定されている(Waxham及びWolinsky, (1985), Green及びDorsett, (1986), Ho-Terryら, (1985))。Terryら, (1989)に記載されている組換えタンパク質A-E1融合体は直鎖状エピトープのみにより特徴づけられている。RV E1糖タンパク質のエピトープの、より詳細な位置決定は、Terryら, (1988)、Wolinskyら, (1991) 及びChayeら, (1993)に記載されている。さらに、Giessaufら, (2004)は、風疹に対する免疫の測定を改善するために、E1のコート化ペプチドのいくつかを使用することによりELISAを評価している。これらのペプチドのうちの1つ(BCH-178ペプチド)だけが、低陽性又はボーダーラインHAI力価又はRV-IgG値を確認するための追加的な方法として、中和抗体をスクリーニングするのに成功裏に使用されるようであった。小さなBCH-178ペプチド(アミノ酸残基213-239)が天然E1分子の抗原エピトープのほとんどを含有するという更なる証拠は存在しない。しかし、免疫優性風疹E1エンベロープタンパク質の主要反応性の局在化に関しては、現在までに言及されていない。
【0013】
血清学的アッセイにおける高い免疫反応性と高い溶解度とを兼ね備えた、原核細胞系由来の可溶性風疹E1変異体を提供することが、本発明の目的であった。
【発明の開示】
【0014】
発明の概要
本発明の対象は可溶性風疹E1抗原及びこのペプチドの変異体であり、これらは、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。ジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域は、この領域のN末端(Cys349)において、更に少なくとも隣接風疹E1抗原配列の25、30、34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも10、11、15、25、35アミノ酸を含有する。
【0015】
可溶性風疹E1抗原は更に、C末端において、アミノ酸残基438-452の間のα-らせん領域を更に欠くことにより特徴づけられる。
【0016】
さらに、可溶性風疹E1抗原は、少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130の組合せにわたる領域を含有するN末端領域により特徴づけられる。さらに、可溶性風疹E1抗原は、少なくともジスルフィド架橋Cys349-Cys352及びCys368-Cys401、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せにわたる領域を含有するC末端領域により特徴づけられる。
【0017】
また、本発明は、シャペロン融合タンパク質として組換え発現され、可溶性かつ免疫反応性のコンホメーションにリフォールディングされ、さらに、抗風疹抗体の血清学的検出に使用される、風疹E1抗原及び変異体をコードする組換えDNA分子に関する。
【0018】
本発明は、サンプル中のIgG及び/又はIgMサブクラスの抗風疹抗体の検出、測定及び定量のための方法を開示する。この場合、該抗体に対する捕捉試薬及び/又は結合相手として、風疹E1抗原を使用する。本発明は更に、風疹E1抗原の少なくとも1つの抗原を含有する、抗風疹抗体の検出のための診断試験及び試薬キットを含む。
【0019】
発明の詳細な説明
本発明は、可溶性風疹E1抗原及びこのペプチドの変異体を開示する。これらは、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。
【0020】
ジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域は、この領域のCys349位のN末端において、更に少なくとも隣接風疹E1抗原配列の25、30、34アミノ酸を含有し、及び/又は、この領域のCys401位のC末端は更に隣接風疹E1抗原配列の少なくとも10、11、15、25、35アミノ酸を含有する。風疹E1抗原の最低限の要件に必要なアミノ酸の前記の定義は絶対的なものではなく、当業者により実験により容易に確認されうる。
【0021】
可溶性風疹E1抗原及びこのペプチドの変異体は、好ましくは、アミノ酸143-164を欠く。より好ましくは、この領域のN末端(アミノ酸143)は更に、少なくとも1、2、5及び/もしくは少なくとも10アミノ酸を欠き、ならびに/又は、この領域のC末端(アミノ酸164)は更に隣接風疹E1抗原配列の少なくとも1、2、5及び/又は少なくとも10アミノ酸を欠く。当業者は、前記の境界は、風疹E1タンパク質の配列内で欠けているアミノ酸残基の或る範囲を示すために用いられていると理解するであろう。該境界の前記定義は絶対的なものではなく、この領域内で欠けているアミノ酸残基の量は可変的なものである。
【0022】
さらに、本発明は更に、アミノ酸残基438-452の間のα-らせん領域を更に欠く可溶性組換え風疹E1抗原及びこのペプチドの変異体に関する。
【0023】
或るシステイン残基の置換及び非常に疎水性のセグメントを有する或る配列の除去により風疹E1タンパク質の酸化的及びコンホメーション的リフォールディングの複雑性を軽減することが可能かどうかを評価するために、欠失分析を行った。
【0024】
本発明において、「ジスルフィド架橋」という用語は、タンパクの3次元構造に隣接し、酸化されてジスルフィド結合を形成しうる2つのシステイン残基(Freedman, Curr. Op. Struct. Biol. (1995) 5, 85-91; Creighton et al., TIBTECH 1995 (13), 18-23; Raina, Annu. Rev. Microbiol. (1997) 51, 179-202)に関する。ジスルフィド結合形成の割合は、2つのシステイン残基の近接性によって決まり、それはそれらの硫黄原子がチオール/ジスルフィド変換に必要な距離に到達する確率として規定される。ジスルフィド架橋とも称されるジスルフィド結合(同義語は、それぞれSS 結合及びSS 架橋である)は共有結合的な三次元関係を構成し、通常フォールディングされた構造の安定化に寄与する。それらは、フォールディングされていないポリペプチド鎖の構造的フレキシビリティーを制限することによりそのような安定化に寄与する。すなわちSS結合のタンパクの安定性への寄与は自然界においてかなりエントロピックである。ジスルフィド結合の形成は酸化されやすい環境を必要とする。それゆえ、細菌の細胞質や真核細胞の細胞質ゾルのような細胞内コンパートメントは本質的に還元されやすい環境のため、細胞内タンパク質はほとんどジスルフィド結合を含まない。しかしながら、ジスルフィド架橋は、各々HIV-1及びHIV-2由来のgp41及び gp36 エクトドメインのような分泌あるいは移行性タンパク質、及び風疹エンベロープタンパク質 E1及びE2でよく見られる。
【0025】
驚くべきことに、可溶性風疹E1抗原変異体は、膜貫通領域とC末端アンカーセグメント(好ましくはアミノ酸残基453-481)の欠失により得られることが判明した。最も好ましくは、さらに、アミノ酸453-約468のα-らせん領域も欠失される。N末端領域内には、少なくともアミノ酸残基143-164、より好ましくは、アミノ酸134-約168も欠失されなければならない。
【0026】
それぞれジスルフィド架橋Cys8-Cys13、Cys59-Cys71、Cys117-Cys130及びジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せは、膜貫通領域とC末端アンカーセグメント、及び少なくともアミノ酸143-164、好ましくは134-約168が欠けている場合には、血清学的アッセイにおける好ましい高い免疫反応性と共に可能な限り多数のエピトープを含有し尚も可溶性である、最も伸張した本発明の風疹E1抗原を与える。
【0027】
本発明の風疹E1エンベロープタンパク質及びその変異体中のN末端領域は、好ましくは、少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71を含有し、より好ましくは、ジスルフィドCys8-Cys13及びCys59-Cys71及びCys117-Cys130の組合せを含有する。ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130の組合せは本発明の風疹E1断片に対して有益な効果をもたらす。なぜなら、それは、高い免疫反応性抗原との或る風疹E1エピトープの三次的固定を保証するからである。
【0028】
本発明の風疹E1抗原及びその変異体中のC末端領域は、少なくともジスルフィド架橋Cys349-Cys352及びCys368-Cys401、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せを含む領域を含有する。ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せは風疹E1抗原に対する追加的な効果をもたらす。なぜなら、それは、高い免疫反応性を有する主要エピトープの接近を可能にするからである。
【0029】
風疹E1抗原の主要免疫反応性の測定のために、本発明者らの目的においては、該タンパク質を、まず、アミノ酸残基1-314を含むN末端部分、及びアミノ酸残基315-432を含むC末端部分に分割した。両部分の配列を、ジスルフィドシャフリングによるリフォールディングアーチファクトを回避するためにシステインを含有しないように設計し、さらに、発現ベクター内にクローニングした。ついで、Grosら, (1997)に記載されている風疹E1ペプチドの各単一ジスルフィド架橋を該N末端及びC末端部分内に個々に又は組合せて挿入した。エピトープ固定のための、共有的三次的接触の重要性を、それぞれの免疫反応性により評価した。
【0030】
興味深いことに、N末端断片(1-314)は高分子量会合体として出現し、一方、C末端断片(315-432)は見掛け上は二量体として溶出することが、ゲル濾過分析から明らかである。これは、単量体、ホモ及びヘテロ二量体よりなる凝集体としての風疹E1/E2ペプチドを開示しているBaron及びForsell, (1991)に記載の最先端研究内容とは対照的である。さらに、本発明の結果は、異なる溶解度特性が発現活性と相関することを明らかにした。すなわち、N末端セグメント(1-314)の生産率はタンパク質約1mg/gバイオマスであるに過ぎなかったが、C末端セグメント(315-432)の生産率はタンパク質20mg/g(湿潤細胞重量)を著しく上回った。
【0031】
システイン非含有変異体は、自動Elecsys(登録商標)分析装置において風疹陽性血清でアッセイされた場合には、有意な免疫反応性を示さなかった。重要なことに、N末端断片(1-314)内の単一のジスルフィド架橋又はジスルフィド架橋の組合せは反応性における有意な上昇を示さなかった。しかし、C末端断片(アミノ酸残基315-432)の反応性は、ジスルフィド架橋Cys349-Cys352及びCys368-Cys401を挿入することにより有意に上昇した。N末端風疹E1タンパク質の反応性の欠如の理由は、凝集又は会合を媒介して主要エピトープの接近を損なう該配列内の構造要素によるものでありうるであろう。
【0032】
この仮定を更に証明するために、風疹E1エンベロープタンパク質のN末端断片(アミノ酸残基1-314)をそのC末端から段階的に末端切断した。同様に、C末端断片(315-432)をそのN末端から徐々に伸長させた。C末端断片は或る程度伸長されると不溶性になり、一方、N末端断片は或る程度末端切断されると可溶性になると予想された。
【0033】
発現及び特徴づけのために、C末端変異体E1-C(aa260-432)、E1-C(aa201-432)、E1-C(aa143-432)、E1(aa105-432)、E1(aa56-432)及びE1(aa34-432)を使用した。変異体E1(aa201-432)、E1(aa260-432)及びE1(aa315-432)は安定な二量体を形成し、一方、より長い構築物は凝集及び会合する著しい傾向を示した。したがって、E1エクトドメインのオリゴマー化モチーフを約143aa-約200aaの領域に限定した。より厳密なマッピングのために、断片E1(169-432)をクローニングし、発現させ、特徴づけした。驚くべきことに、この構築物は安定な且つ可溶性の二量体を形成し、したがって抗風疹IgG分子の検出のための優れた候補であった。この結果に基づき、該会合モチーフをセグメント143aa-168aaに限定することができた。より詳細なマッピングのために、断片E1(156-432)及びE1(163-432)を特徴づけした。どちらの構築物も、部分的に二量体として、部分的に高分子量凝集体として溶出し、したがって、1工程二重抗原サンドイッチ形態の抗風疹IgG抗体の検出のための抗原としては限られた価値しか有さなかった。
【0034】
本発明のこの一連の試験の主な結果として、溶解度、オリゴマー化状態及び安定性を損なうことなく、C末端E1(315-432)断片をアミノ酸残基315からアミノ酸残基169まで伸長させることが可能であることが、明らかに示されうるであろう。第2に、E1エクトドメインの最初の3分の1と次の3分の1との間の領域(143-168)は、高分子量凝集体を形成するよう該タンパク質の会合を引き起こす構造又は配列モチーフを含有する。この領域の内在的疎水性は謎である。なぜなら、それは、単純な一次構造分析からは演繹され得ないからである。該会合モチーフを含む領域は、IgG分子の血清学的検出のためのE1エンベロープタンパク質の使用を著しく限定する。
【0035】
N末端セグメントの溶解度の最適化のために、同等の方法を用いた。E1断片E1-N (1-200)、E1-N (1-142)、E1-N (1-104) 及びE1-N (1-55)を発現ベクター内にクローニングし、大腸菌(E. coli)宿主細胞内に導入し、発現させ、特徴づけした。予想どおり、最長N断片(エクトドメイン1-200)は高分子量凝集体として溶出し、したがってE1エクトドメイン変異体E1-N (1-314)に著しく類似している。有意に改善された溶解度特性は、部分的に可溶性二量体として、部分的に高分子量凝集体として溶出する末端切断断片E1-N (1-142)で達成された。最後に、末端切断変異体E1-N (1-104) 及びE1-N (1-55)は見掛け上は二量体として溶出した。断片E1-N-(1-117) 及び(1-133)を使用して、アミノ酸残基104と142との間の領域を更に分析した。どちらの変異体もFPLC分析において可溶性二量体として出現し、したがって、IgG分子の血清学的検出のための重要な前提条件を満たしている。しかし、変異体E1-N (1-117)はより安定であり、したがって、更なる最適化実験のための、より優れた候補であった。
【0036】
簡潔に説明すると、本発明において、長さの最適化の後、大腸菌(E. coli)において非常に良く発現され可溶性二量体を形成する風疹E1タンパク質の2つの領域、すなわち、アミノ酸残基1-117/1-133のN末端断片(E1-N)及び領域169-432/201-432を含むC末端断片(E1-C)を同定した。これらのE1断片を使用して、組換え風疹E1エクトドメインの抗原性のためにはどのジスルフィド架橋が重要であるかという問題について検討した。したがって該システイン対を個々に及び組合せて再構築し、対応するE1変異体を自動Elecsys(登録商標)2010分析装置(Roche Diagnostics GmbH)において抗風疹陽性血清で評価した。
【0037】
以下のシステイン変異体を風疹E1(201-432)において特徴づけした:単一架橋変異体Cys 225-Cys235、Cys349-Cys352及びCys368-Cys401、二重架橋の組合せCys349-Cys352/Cys368-Cys401、ならびに三重架橋の組合せCys225-Cys235/Cys349-Cys352/Cys368-Cys401。さらに、伸長構築物E1(169-432)において、二重組合せCys176-185/Cys225-Cys235を分析した。
【0038】
以下のシステインの組合せを風疹E1断片1-133において特徴づけした:単一架橋変異体Cys8-Cys13、Cys59-Cys71及びCys117-Cys130ならびに二重架橋の組合せCys8-Cys13/Cys59-Cys71、Cys8-Cys13/Cys117-130及びCys59-Cys71/Cys117-Cys130。さらに、二重架橋の組合せCys 8-Cys 13/Cys59-Cys71を融合構築物E1-N (1-117) 及びE1-N (1-104)において特徴づけした。自動Elecsys(登録商標)2010分析装置における風疹E1変異体の免疫学的評価は、抗原性が、ジスルフィド架橋によりもたらされる共有的三次的接触により固定される該エクトドメイン分子の天然様コンホメーションに依存することを示した。
【0039】
驚くべきことに、これらの特徴的知見は、免疫反応性に関して、風疹E1抗原のジスルフィド架橋に対して、異なった力点を置く必要があることを示している。ジスルフィド架橋Cys-349-Cys-352及びCys-368-Cys-401は決定的な役割を果たしている。興味深いことに、これらのジスルフィド架橋の組合せによる感受性の上昇は相加的なものではなく、協同的なものである。構築物E1-C(201-432)の評価は、各場合にCys349-Cys352及びCys368-Cys401の三次的固定のための有意なシグナルを示している。両方の架橋の組合せにより、シグナルのレベルは、単一架橋構築物と比べて、倍数的に驚くほどに上昇した。おそらく、風疹E1エクトドメインの天然様コンホメーションが、それらの2つの分子内組合せを介して固定され、風疹陽性血清中の、より大量の特異的IgG分子を検出したのであろう。共有的三次的接触の妥当性は、システイン非含有構築物E1-C(201-432)の免疫学的評価により、特に明らかである。驚くべきことに、単一架橋構築物の場合とは対照的に、システイン非含有構築物においては、陽性シグナルは全く検出できない。
【0040】
天然サンプル中の抗風疹IgG抗体の免疫学的検出は、最適化抗原E1-C(201-432, Cys349-Cys352, Cys368-Cys401)で更に改善された。抗風疹IgG陽性として分類されたこれらの研究において適用されたすべてのヒト血清は陽性であることが判明した。より複雑な三次架橋融合構築物-E1-C(169-432)、Cys225-Cys235、Cys349-Cys352、Cys368-Cys401)を使用することにより、抗風疹IgG抗体の検出に関して同等の結果を得ることができた。風疹E1抗原断片の最低限の要件は、少なくともジスルフィド架橋Cys349-Cys352及びCys368-Cys401により分子内で連結されたポリペプチドバックボーンにまで縮小することができた。E1断片E1-C(342-412、Cys349-Cys352、Cys368-Cys401)は不十分な免疫反応を示すため、E1-C(342−412)のN末端及び/又はC末端に隣接した配列要素は免疫反応性に重要だと考えられた。これを証明するために、構築物E1-C(315-412、Cys349-Cys352、Cys368-Cys401)をクローニングし、特徴づけした。この断片に関して、E1-C(201-432、Cys349-Cys352、Cys368-Cys401)と比較しうる免疫反応性を示すことができた。この結果は、構築物E1-C(アミノ酸315-412)のN末端領域(315-342)のアミノ酸残基が風疹E1抗原の免疫反応性に重要らしいことを示している。当業者は、E1-C(315-412)のN末端及び/又はC末端に隣接したどの配列要素が免疫反応性に重要なのかを、更なる実験により証明することが可能である。
【0041】
エクトドメインのC末端部分からのジスルフィド架橋を有する高反応性断片に加えて、いくつかのN末端断片をクローニングし、発現させた。それらは有意な免疫反応性を示した。免疫反応性は2つの二重架橋融合構築物E1-N(1-117、Cys8-Cys13/Cys59-Cys71)及びE1-N(1-133、Cys8-Cys13/Cys59-Cys71)から得られた。
【0042】
可溶性かつ免疫反応性のコンホメーションにリフォールディングしうる最も長い風疹E1断片は、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130を含むアミノ酸残基1-133、ならびにジスルフィド架橋Cys349-Cys352及びCys368-Cys401ならびにジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せを含むアミノ酸残基169-432にわたる(これは、C末端における少なくとも膜貫通領域と短いC末端アンカーセグメント、及び少なくともアミノ酸143-164、好ましくはアミノ酸134-168を欠くことによりもたらされる)。
【0043】
したがって、本発明の好ましい実施形態は、ジスルフィド架橋Cys349-Cys352及びCys368-Cys401にわたる領域ならびに更に少なくともジスルフィド架橋Cys225-Cys235にわたる領域を含有する可溶性風疹E1抗原ならびにこのペプチドの変異体である。
【0044】
本発明のもう1つの主題は、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401にわたる領域ならびに更に少なくともジスルフィド架橋Cys176-Cys185にわたる領域を含有する可溶性風疹E1抗原ならびにこのペプチドの変異体に関する。さらに、ジスルフィド架橋Cys349-Cys352及びCys368-Cys401の組合せ、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せが、本発明の好ましい実施形態である。これらの風疹E1抗原はエピトープの形成のための主要免疫反応性を含有する。
【0045】
本発明のもう1つの好ましい実施形態は、更にジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130にわたる領域を更に含有する可溶性風疹E1抗原及びこのタンパク質の変異体に関する。
【0046】
少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130にわたる風疹E1抗原の領域も、エピトープの形成のための或る免疫反応性を含有する。ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401を含有するC末端配列に連結させることなく、これらのN末端断片を使用することも可能である。好ましくは、これらのN末端風疹E1抗原は、前記のC末端風疹E1抗原と共に、アッセイにおいて使用される。
【0047】
本発明においては、大腸菌(E. coli)内で組換え発現されin vitroで再現可能な様態で機能性コンホメーションにリフォールディングされる風疹E1エクトドメインの包括的領域及び断片を同定した。長さ及びシステイン生成に関して最適化された本発明のE1断片は可溶性であり、特に、風疹陽性血清中の抗風疹IgGの検出に適している。
【0048】
この知見は公知風疹E1断片のもの(Newcombeら, (1994), Starkeyら, (1995), EP0299673)とは対照的である。これらの公知構築物はジスルフィド架橋Cys349-Cys352及びCys368-Cys401の組合せを含有せず、したがって、それらはエピトープの形成のための主要免疫反応性を欠いている。さらに、該公知構築物に関しては、高溶解性かつ免疫反応性コンホメーションにリフォールディングされうる、本発明において開示されている、C末端における少なくとも膜貫通領域及び少なくともアミノ酸143-164、好ましくはアミノ酸134-約168を欠くことによる、N末端領域のジスルフィド架橋Cys8-Cys13及びCys59-Cys 71及びCys117-Cys130ならびに/又はC末端領域及びN末端領域のジスルフィド架橋の組合せが開示されていない。さらに、本発明の好ましい融合構築物は、より多数のエピトープを有する、風疹E1タンパク質の、より長い部分を含有しうる。なぜなら、それらは可溶性であり、したがって、ほとんどの抗風疹抗体の検出に、より適しているからである。
【0049】
「変異体」なる語は、異なる量のシステイン対を含みうる、風疹E1エクトドメインの、異なる長さの配列の部分を意味する。これらの配列、挿入カセットの核酸配列、及び風疹E1の得られた融合ポリペプチドのアミノ酸配列は、本発明の配列プロトコールにおいて記載されている。
【0050】
さらに、この場合、「変異体」なる語は、該タンパク質に実質的に類似したタンパク質を意味する。特に、変異体は、最も一般的なタンパク質アイソフォームのアミノ酸配列と比べてアミノ酸の変化、欠失又は挿入を示すアイソフォーム又はアレルでありうる。好ましくは、そのような実質的に類似したタンパク質は、該タンパク質の最も一般的なアイソフォームに対して、少なくとも80%、好ましくは少なくとも85%、より好ましくは少なくとも90%、最も好ましくは少なくとも95%の配列類似性を有する。また、「変異体」なる語は、翻訳後修飾されたタンパク質、例えばグリコシル化タンパク質を意味する。また、「変異体」は、例えば、該タンパク質への標識(特に放射能蛍光標識)の共有結合又は非共有結合により、サンプルの収集後に修飾されたタンパク質である。他の考えられうる標識としては、放射能、蛍光、化学発光、電気化学発光、酵素などが挙げられる。さらに、「変異体」は、例えばビオチン化タンパク質のような固相結合基である。標識の詳細な説明は本明細書中、後記において記載されている。
【0051】
風疹E1抗原は化学合成により製造されうる。該合成は均一溶液中又は固相中で行われうる。例えば、用いられうる均一溶液中での合成技術としては、Houbenweyl (1974)に記載されているものが挙げられる。本発明のタンパク質は、Atheton及びShepard (1989)に記載されている方法に従い、固相中でも製造されうる。
【0052】
可溶性及び一定の単量体又はオリゴマー状態に関して最適化された前記風疹E1タンパク質はIgM検出モジュールの可能性をも提供する。この目的には、見掛け上二量体であるE1融合タンパク質を、例えば化学的架橋により重合させることが可能であろう。本発明のもう1つの好ましい実施形態は、タンパク質E1、E2及びコアタンパク質Cから構成される混合重合体である。風疹E2及びコアタンパク質は当技術分野で公知である。
【0053】
本発明の風疹E1タンパク質は組換えDNA技術によっても製造されうる。「組換えDNA分子」なる語は、遺伝子工学技術又は化学合成により、単離されたポリヌクレオチド断片を人工的に操作して、2つの分離した配列断片を組合せることにより製造された分子を意味する。それを行う場合には、所望の機能を有するポリヌクレオチド断片同士を互いに連結させて、所望の機能の組合せを得ることが可能である。適当な宿主細胞内での複製により、大量の該ポリヌクレオチドを製造することが可能である。タンパク質又はその断片をコードする天然又は合成DNA断片を、例えばSambrookら, (1989)に記載されている原核生物細胞又は下等もしくは高等真核生物細胞内への導入及びそのような細胞内での複製が可能な組換えポリヌクレオチド構築物(典型的にはDNA構築物)内に組込む。「下等真核生物」なる語は、酵母、真菌などのような宿主細胞を意味する。下等真核生物は、一般には(必ずしもそうではないが)単細胞である。「原核生物」なる語は、大腸菌(E. coli)、ラクトバシラス(Lactobacillus)、ラクトコッカス(Lactococcus)、サルモネラ(Salmonella)、ストレプトコッカス(Streptococcus)、バシラス・サチリス(Bacillus subtillis)又はストレプトマイセス(Streptomyces)のような宿主を意味する。これらの宿主も本発明において予想される。好ましい下等真核生物は、酵母、特に、シゾサッカロミセス(Schizosaccharomyces)、サッカロミセス(Saccharomyces)、クルイベロミセス(Kluiveromyces)、ピチア(Pichia)(例えば、ピチア・パストリス(Pichia pastoris))、ハンゼヌラ(Hansenula)(例えば、ハンゼヌラ・ポリモルファ)、シュワニオミセス(Schwaniomyces)、シゾサッカロミセス(Schizosaccharomyces)、ヤロウィア(Yarowia)、ジゴサッカロミセス(Zygosaccharomyces)などの種である。サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、エス・カールスベルゲンシス(S. carlsbergensis)及びケイ・ラクチス(K. lactis)は、最もよく使用される酵母宿主であり、簡便な真菌宿主である。「高等真核生物」なる語は、動物、例えば哺乳動物、爬虫類動物、昆虫などに由来する宿主細胞を意味する。現在好ましい高等真核生物宿主細胞は、チャイニーズハムスター(例えば、CHO)、サル(例えば、COS及びVero細胞)、子ハムスター腎細胞(BHK)、ブタ腎臓(PK15)、ウサギ腎臓13細胞(RK13)、ヒト骨肉種細胞系143B、ヒト細胞系HeLa及びヒトヘパトーマ細胞系、例えばHep G2及び昆虫細胞系(例えば、スポドプテラ・フルジペルダ(Spodoptera frugiperda))に由来するものである。該宿主細胞は、懸濁又はフラスコ培養、組織培養、器官培養などにおいて供給されうる。
【0054】
本発明の好ましい実施形態においては、風疹E1抗原は組換え融合タンパク質として製造される。本発明において用いる「融合タンパク質」なる語は、風疹E1タンパク質に対応する少なくとも1つのタンパク質ドメインと、後記のとおりの発現手段として使用されるFKBPシャペロンに好適な別の融合タンパク質に対応する少なくとも1つタンパク質ドメインとを含むタンパク質を意味する。他のよく使用される融合タンパク質としては、例えば、融合タンパク質の高レベル発現ならびに細菌及び真核細胞ライセートからのその迅速な精製のためのグルタチオンS-トランスフェラーゼ(GST)融合タンパク質系が挙げられる。タンパク質のフォールディング及び精製は、しばしば、それらを、自ら活発にフォールディングするパートナータンパク質又はタグと共有結合により融合させることにより促進される。これらの融合モジュールには、マルトース結合タンパク質、グルタチオンS-トランスフェラーゼ、チオレドキシン、NusA、DsbA及びFkpAが含まれる。それらの使用は、通常、過剰生産性大腸菌(E. coli)宿主のサイトゾル又はペリプラズムにおけるそれぞれの標的タンパク質の可溶性発現(すなわち、天然様フォールディング)の向上を目的としている。可溶性かつ機能性のタンパク質の生物工学的製造においてシャペロンが果たす役割は益々重要なものとなっている。好ましくは、シャペロンは風疹E1融合ポリペプチドにおける融合タンパク質として使用される。シャペロンは、古典的な「フォールディング補助体(folding helper)」として公知であり、他のタンパク質のフォールディング及び構造的完全性維持を補助するタンパク質である。それらは、in vivo及びin vitroの両方においてタンパク質のフォールディングを促進する能力を有する。一般には、フォールディング補助体はフォールディング触媒及びシャペロンに細分される。フォールディング触媒は、その触媒機能により、タンパク質のフォールディングにおける律速段階を加速させる。シャペロンは、変性した又は部分的に変性したタンパク質に結合することが公知であり、したがって、タンパク質が再生するのを助ける。したがって、フォールディング触媒とは異なり、シャペロンは単なる結合機能を発揮するに過ぎない(Buchner, (1996))。触媒及びシャペロンの具体例はWO 03/000877に詳細に記載されている。
【0055】
現在までに、いくつかの異なるシャペロンファミリーが公知である。すべてのこれらのシャペロンは、アンフォールディングした又は部分的にアンフォールディングしたタンパク質に結合するそれらの能力により特徴づけられ、タンパク質の適切なフォールディング又は変性もしくは凝集タンパク質の除去に関連した生理的機能を有する。さらに、WO 94/08012には、シャペロンの発現の増強はタンパク質の組換え産生を促進しうることが示されている。また、タンパク質の産生の増加は、タンパク質コード配列とシャペロン配列とを含む遺伝子構築物を使用することにより達成されうることが公知である。天然のようにフォールディングしたタンパク質の産生の増加のためにシャペロンを使用するアプローチは、主として、シャペロンタンパク質結合機能及びそれによる可溶化機能によるものである。シャペロンと標的タンパク質とを含む融合タンパク質の組換え産生の後、通常、シャペロンを、生じたタンパク質から切り離して、所望のタンパク質を純粋な形態で得る。
【0056】
フォールディング補助体、例えば、ペプチジルプロピルイソメラーゼ(PPI)クラスのメンバー、特に、FKBPファミリーのメンバーは、触媒活性を示すだけでなく、凝集傾向にあるタンパク質の溶解度に対する劇的な有益な効果をも与えることが、WO 03/000877に示されている。それらは、そうでなければ(すなわち、未シャペロン化単離形態であれば)凝集傾向にあるそのようなタンパク質との可溶性複合体を形成することにより、それを達成する。そうでなければ生理的条件下で難溶性又は不溶性であるそのようなタンパク質は、それらが適当なPPIシャペロンとの複合体内に結合すると、穏やかな生理的条件下で(すなわち、界面活性剤又はカオトロピック剤のような可溶化添加剤を要することなく)可溶性となる。
【0057】
本発明者らは、本発明の組換え産生融合タンパク質が封入体から可溶性形態で容易に得られうることを見出した。組換え産生された風疹E1タンパク質に含まれる風疹E1タンパク質の顕著な特徴は生理的バッファー条件下でのその溶解性である。さらに、融合タンパク質に含まれる本発明の風疹E1タンパク質は天然様構造として容易に得られうる。
【0058】
また、本発明の風疹E1融合タンパク質は、取扱いが非常に容易である。例えば、そのような融合タンパク質を再生させることは非常に容易である。カオトロピック物質は、異なる様態でリフォールディングされて、すべて、熱力学的に安定かつ可溶性の天然様形態を与えうることは興味深い。リフォールディングは、透析及び急速希釈のどちらによっても並びにサイズ排除クロマトグラフィー又はマトリックス介助リフォールディングの再生によっても、高収率で達成される。これらの知見は、この共有結合形態においては、風疹E1抗原融合タンパク質が、準安定タンパク質ではなく熱力学的に安定であることを示唆している。
【0059】
好ましくは、本発明の可溶性タンパク質は、ペプチジル-プロピル-イソメラーゼクラスのシャペロンとの風疹E1抗原の融合により製造されうる。したがって、本発明の好ましい実施形態は、風疹E1抗原とペプチジル-プロピル-イソメラーゼクラスのシャペロンとの融合体に関する。
【0060】
本発明のもう1つの主題は、風疹E1抗原をコードする少なくとも1つのヌクレオチド配列と、その上流に位置する、FKBPシャペロンをコードする少なくとも1つのヌクレオチド配列とを含む、風疹E1抗原をコードする組換えDNA分子に関する。
【0061】
プロリルイソメラーゼは、種々の機能の種々のサブユニット又はモジュール(例えば、触媒活性を示すモジュール、及びシャペロン又は結合活性を示すモジュール)を含みうる。FKBPファミリーのそのようなモジュラーメンバーとしては、FkpA(Ramm, (2000))、SlyD(Hottenrottら, (1997))及び誘発因子(Scholzら, (1997))が挙げられる。好ましい実施形態においては、本発明は、FKBPシャペロンが、FkpA、SlyD及び誘発因子よりなる群から選ばれることにより特徴づけられる組換えDNA分子に関する。
【0062】
また、分子シャペロンの完全な配列を常に使用する必要はないことが、よく知られており理解されている。必要な能力及び機能を尚も有する、シャペロンの機能性断片(いわゆる、モジュール)も使用されうる(WO 98/13496を参照されたい)。本発明において発現手段として使用されるFkpAはN末端シグナル配列を欠く。FkpAの近縁体、すなわちSlyDは、触媒機能及びシャペロン機能をもたらす構造化N末端ドメインと、例外的にヒスチジン及びシステイン残基に富む、大部分が非構造化されたC末端とよりなる(Hottenrott, 1997)。WO 03/000878は、アミノ酸1-165を含むSlyDのC末端切断型変異体が標的タンパク質の効率的発現に対して例外的に正の効果を及ぼすことを開示している。野生型SlyDの場合とは異なり、使用される末端切断型SlyD変異体(1-165)においては、ジスルフィドシャフリングを損なう恐れが成功裏に排除されている。末端切断型SlyD(1-165)を含む組換えDNA分子は本発明の好ましい実施形態の1つである。
【0063】
本発明の風疹E1抗原を設計する好ましい態様においては、シグナルペプチドは含まれない。本発明の発現系は、サイトゾル発現系として働く場合に最も有利であることが判明している。このサイトゾル発現は封入体の形成をもたらす。封入体に通常は伴う顕著でよく知られた問題とは異なり、本発明者らは本発明において、格別に大量の風疹E1タンパク質が産生されるだけでなく、本発明の組換え風疹E1タンパク質が取り扱い易くもある(例えば、可溶化及びリフォールディングが容易である)ことを見出した。
【0064】
好ましくは、本発明の組換えDNA分子は更に、それが、風疹E1抗原をコードする配列とFKBPシャペロンをコードする配列との間に位置する10〜100アミノ酸のペプチド性リンカーをコードする少なくとも1つのヌクレオチド配列を含むことにより特徴づけられる。当業者に理解されるとおり、そのようなリンカーポリペプチドは、特に長さ、柔軟性、電荷及び親水性の点で、意図される用途に最も適したものとして設計される。さらに、リンカーをコードするそのようなDNA配列は、例えばタンパク質分解部位を与えることに加えて、ポリリンカーとしても働きうる。すなわち、それは、風疹E1タンパク質をコードするDNA断片とシャペロンドメインをコードするDNA断片との融合を促進する複数のDNA制限部位を与えうる。得られた融合タンパク質の発現及び精製ならびに更には可溶性かつ免疫反応性コンホメーションへのリフォールディングの後、該ポリリンカーは融合タンパク質複合体からの風疹E1タンパク質の遊離をも促進する。
【0065】
それにより、本発明の可溶性風疹E1抗原及びこのタンパク質の変異体は融合構築物から切り出されることが可能であり、この場合、該風疹E1抗原は、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくとも、ジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。さらに、風疹E1抗原の前記領域のN末端(Cys349)は更に少なくとも隣接風疹E1抗原配列の25、30、34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも10、11、15、25、35アミノ酸を含有する。
【0066】
本発明のもう1つの主題は、FKBPシャペロンをコードする単一のヌクレオチド配列と風疹E1タンパク質をコードする単一のヌクレオチド配列とを含む組換えDNAに関する。
【0067】
少なくとも2つのFKBPシャペロンドメインと1つの標的タンパク質ドメインとを含む融合タンパク質も非常に有利である。もう1つの好ましい実施形態においては、本発明の組換えDNA分子は、FKBPシャペロンをコードする2つの配列と風疹E1タンパク質をコードする1つの配列とを含む。Scholzら, (2005)は、第2のシャペロン単位が融合されると融合タンパク質の溶解度が顕著に改善されることを示している。天然において二量体であるPPIアーゼは2つのFKPBシャペロンドメインの融合により厳密に模擬され、それにより溶解度が改善されることを示唆する証拠が存在する。
【0068】
当業者に理解されるとおり、「少なくとも2つ」なる表現は、FKBPシャペロンドメインをコードする2以上のヌクレオチド配列が、本発明の範囲から逸脱するこなく組換えDNA分子の構築において使用されうることを示すために用いられる。好ましくは、風疹E1シャペロンタンパク質は、シャペロンを含有する少なくとも2つ且つ多くとも4つの配列を含有する。
【0069】
該DNA分子は、標的タンパク質の上流にFKBPシャペロンをコードする両方のDNA配列を含むように設計されうる。あるいは、それらの2つのFKBPドメインは、標的タンパク質を挟むように配置されうる。風疹E1抗原をコードする配列の上流に両方のFKBPドメインを含む組換えDNA分子は、本発明の好ましい実施形態の1つである。
【0070】
本発明のもう1つの実施形態においては、該組換えDNA分子は、PPIシャペロンをコードする一方の配列が風疹E1抗原の上流に位置し、PPIシャペロンをコードするその他の配列が、風疹E1抗原をコードする配列の下流に位置することにより特徴づけられる。
【0071】
2つのシャペロンドメイン及び風疹E1抗原をコードする配列を含むDNA構築物は、好ましくは、これらのドメインの間に10〜100アミノ酸の2つのリンカーペプチドをも含有する。組織的クローニングを可能にするために、これらの2つのリンカーペプチド配列をコードするヌクレオチド配列は、好ましくは、異なる。該ヌクレオチド配列におけるこの相違は、該リンカーペプチドのアミノ酸配列における相違を必ずしも与える必要はない。さらにもう1つの好ましい実施形態においては、それらの2つのリンカーペプチドのアミノ酸配列は同一である。そのような同一リンカーペプチド配列は、例えば、2つのFKBPシャペロンドメインと風疹E1タンパク質とを含む融合タンパク質をイムノアッセイにおいて使用したい場合に有利である。
【0072】
シャペロンの1つ又は全てを本発明の融合タンパク質から遊離させたい場合には、該リンカーペプチドを、タンパク質分解切断部位を含むように構築する。既に記載されているとおり、タンパク質分解切断部位はポリリンカーとしても働きうる。すなわち、それは、風疹E1タンパク質をコードするDNA断片とシャペロンドメインをコードするDNA断片との融合を促進する複数のDNA制限部位を与えうる。風疹E1タンパク質をコードする少なくとも1つのポリペプチド配列をコードする核酸配列と、その上流の、FkpA、SlyD及び誘発因子よりなる群から選ばれるFKBPシャペロンをコードする少なくとも1つのヌクレオチド配列と、さらに、タンパク質分解切断部位を含むペプチドリンカーをコードする核酸配列とを含む、融合タンパク質をコードする組換えDNA分子は、本発明のもう1つの実施形態である。本発明の組換えDNA分子(すなわち、風疹E1タンパク質をコードする少なくとも1つのポリヌクレオチド配列と、その上流の、FkpA、SlyD及び誘発因子から選ばれるFKBPシャペロンをコードする少なくとも1つのヌクレオチド配列とを含む、融合タンパク質をコードする組換えDNA分子)を、機能しうる形で連結された形態で含む発現ベクターは、非常に有利であることが判明している。
【0073】
本発明の組換えDNAを含む発現ベクターは、無細胞翻訳系において融合タンパク質を発現させるために使用されることが可能であり、あるいは宿主細胞を形質転換するために使用されうる。好ましい実施形態においては、本発明は、本発明の発現ベクターで形質転換された宿主細胞に関する。
【0074】
発現及びクローニングベクターは選択マーカーを含有しうる。該選択マーカーは、該ベクターで形質転換された宿主細胞の生存又は成長に必要なタンパク質をコードする遺伝子であるが、そのようなマーカー遺伝子は、宿主細胞内に共導入される別のポリヌクレオチド配列上に含有されうる。マーカー遺伝子を発現する宿主細胞のみが選択条件下で生存及び/又は成長する。典型的な選択遺伝子には、(a)抗生物質又は他の毒性物質、例えばアンピシリン、テトラサイクリンなどに対する耐性を付与するタンパク質をコードするもの、(b)栄養要求性欠損を相補するタンパク質をコードするもの、あるいは(c)複合培地からは利用可能でない決定的に重要な栄養素を供給するタンパク質をコードするものが含まれるが、これらに限定されるものではない。適当な選択マーカーの選択は宿主細胞に左右され、種々の宿主のための適当なマーカーが当技術分野で公知である。
【0075】
関心のある風疹E1タンパク質を含有するベクターは、当技術分野で公知の任意の方法により宿主細胞内に導入されうる。これらの方法は細胞宿主の型によって様々であり、塩化カルシウム、塩化ルビジウム、リン酸カルシウム、DEAE-デキストラン、他の物質を使用するトランスフェクション、及びウイルスによる感染を包含するが、これらに限定されるものではない。大量の本発明の風疹E1タンパク質は、和合性宿主内でベクター又は他の発現ビヒクル中の本発明のポリペプチドを発現させることにより製造されうる。本発明のベクターの構築は通常の連結技術を用いる。単離されたプラスミド又はDNA断片を切断し、操作処理し、所望の形態で再連結して、必要なプラスミドを得る。所望により、構築されたプラスミド内の配列が正しいことを確認するための分析を公知様態で行う。発現ベクターを構築し、in vitro転写産物を調製し、宿主細胞内にDNAを導入し、発現及び機能の評価のための分析を行うための適当な方法は、当業者に公知である。遺伝子の存在、増幅及び/又は発現は、例えば、本明細書に記載の配列に基づきうる適当に標識されたプローブを使用する、通常のサザンブロット法、mRNAの転写を定量するためのノーザンブロット法、ドットブロット法(DNA又はRNA分析)、又はin situハイブリダイゼーションにより、サンプル中で直接的に測定されうる。当業者は、所望によりこれらの方法がどのようにして修飾されうるかを容易に予想するであろう。
【0076】
本発明のもう1つの好ましい実施形態は、可溶性かつ免疫反応性の風疹E1抗原-シャペロン融合タンパク質の製造方法を含み、該方法は、
a.宿主細胞の培養、
b.該融合タンパク質の発現、
c.該融合タンパク質の精製、
d.可溶性かつ免疫反応性のコンホメーションへのリフォールディング
の工程を含む。
【0077】
本発明の方法は、ペプチジルプロリルイソメラーゼがFKBPシャペロンである、可溶性かつ免疫反応性の風疹E1抗原-シャペロン融合タンパク質の製造に関する。さらに、該方法は、SlyD、FkpA及び誘発因子よりなる群から選ばれるFKBPシャペロンに関する。
【0078】
本発明のもう1つの主題は、サンプル中のサブクラスIgG及び/又はIgMの抗風疹抗体の検出、測定及び定量のための方法であり、この場合、風疹E1抗原を該抗体に対する捕捉試薬及び/又は結合相手として使用する。当業者に公知のすべての生物学的液体が、抗風疹抗体の検出のためのサンプルとして使用されうる。好ましいサンプルは、全血、血清、血漿、尿、唾液などの体液である。診断方法に関しては、本発明の可溶性風疹E1抗原融合タンパク質、より好ましくは、可溶性風疹E1抗原-シャペロンタンパク質の明らかな利点は、例えば、生理的バッファー条件下の風疹E1タンパク質の安定性の増強、診断感度の増加、存在するコンホメーションエピトープの数の増加、及び適切にフォールディングした風疹E1タンパク質を容易に標識する可能性があることである。
【0079】
よく知られた標識は、マーカー基又はエフェクター基、例えば固相結合基である。標識された可溶性風疹E1抗原融合タンパク質、より好ましくは、標識された可溶性風疹E1抗原-シャペロンタンパク質は、本発明のもう1つの好ましい実施形態である。標識基は、任意の公知の検出可能なマーカー基、例えば色素、発光標識基、例えば化学発光基、例えばアクリジニウムエステルもしくはジオキセタン、又は蛍光色素、例えばフルオレセイン、クマリン、ローダミン、オキサジン、レゾルフィン、シアニン及びそれらの誘導体から選ばれうる。標識基の他の具体例としては、発光性金属錯体、例えばルテニウム又はユーロピウム錯体、酵素、例えばELISA又はCEDIA(クローン化酵素ドナーイムノアッセイ(Cloned Enzyme Donor Immunoassay)、例えばEP-A-0 061 888)に使用される酵素、及び放射性同位体が挙げられる。エフェクター基は、例えば、バイオアフィン(bioaffine)結合ペアのうちの一方のパートナーを含む。アッセイの実施中に、エフェクター基は、バイオアフィン結合ペアのもう一方のパートナーと特異的に、そして好ましくは非共有結合的に相互作用する。適当な結合ペアの具体例としては、ハプテン又は抗原/抗体、ビオチン又はビオチン類似体、例えばアミノビオチン、イミノビオチン又はデスチオビオチン/アビジン又はストレプトアビジン、糖/レクチン、核酸又は核酸類似体/相補的核酸、及び受容体/リガンド、例えばステロイドホルモン受容体/ステロイドホルモンが挙げられる。好ましい結合ペアメンバーはハプテン、抗原及びホルモンを含む。特に好ましいのは、ジゴキシンのようなハプテンならびにビオチン及びその類似体である。好ましくは、風疹E1タンパク質と好ましくはPPIシャペロンとを含む可溶性複合体が、風疹に対する抗体の検出のためのイムノアッセイにおいて使用される。もう1つの好ましい実施形態においては、風疹E1抗原と好ましくはPPIシャペロンとを含む標識された可溶性複合体が、風疹に対する抗体の検出のためのイムノアッセイにおいて使用される。最も好ましくは、その標識された複合体は、好ましくはPPIシャペロンと風疹E1タンパク質とを含む組換えポリペプチドにおける分子内複合体である。
【0080】
イムノアッセイは当業者によく知られている。そのようなアッセイを行うための方法ならびに実際の適用及び手順は関連参考書において要約されている。関連参考書の具体例としては、Tijssen, P., Preparation of enzym-antibody or other enzyme-macromolecule conjugates in "Practice and theory of enzyme immunoassays" (1990) 221-278, R. H. Burdon及びv. P. H. Knippenberg編, Elsevier, AmsterdamならびにTijssen, in "Methods in Enzymology" (1980), S. P. Colowick, N. O. Caplan及びS. P.編, Academic Pressの、免疫学的検出方法を扱った様々な巻、特に第70, 73, 74, 84, 92及び121巻が挙げられる。
【0081】
該新規可溶性風疹E1抗原融合タンパク質、より好ましくは、該新規可溶性風疹E1抗原-PPIシャペロンタンパク質は、検出の様式(例えば、放射性同位体アッセイ、酵素イムノアッセイ、電気化学発光アッセイなど)又はアッセイ原理(例えば、試験ストリップアッセイ、サンドイッチアッセイ又は均一系アッセイなど)には無関係に、抗風疹抗体の検出のためのアッセイを改良するために使用されうる。風疹感染の、信頼しうる且つ高感度の早期検出のためには、体液サンプル中の抗ウイルス抗体を測定することが必須である。本発明の可溶性複合体は生理的バッファー条件における抗風疹抗体の検出を可能にする。抗風疹抗体の検出はそのような複合的な風疹検出系の重要な部分である。したがって、好ましい実施形態においては、本発明は、風疹E1抗原融合タンパク質の使用、より好ましくは、風疹E1抗原-シャペロンタンパク質の使用に基づく抗風疹抗体の検出を含む風疹検出系に関する。
【0082】
当技術分野で公知のとおり、細菌、真菌又はウイルスのような感染因子に対する抗体は、好ましくは、二重抗原架橋の概念(時には、このアッセイ概念は、2つの抗原が抗体により架橋されるため、二重抗原架橋概念とも称される)に従うアッセイにより検出される。そのようなアッセイにおいては、与えられた抗原の、少なくとも2つの異なる分子に、抗体がその2個(IgG、IgA、IgE)又は10個(IgM)のパラトープで結合する能力が要求され、用いられる。
【0083】
該架橋概念による、体液からの抗体の検出は、多種多様なアッセイ設定で行われうる。単純な設定は、固相への抗原の直接的なコーティング、及び標識された形態の同じ抗原の使用を含む。適当なアッセイ条件下、サンプル中の抗体は該固相結合抗原と該標識抗原との間で架橋を形成する。したがって、検査中の抗体がサンプル内に存在する場合にのみ、架橋が形成され、シグナルが検出されうる。「固相抗原」及び「検出抗原」の基本的構造は、好ましくは同じである。例えば、1つ又はいくつかのエピトープを含むタンパク質が、固相にコートされて直接的又は間接的に使用されることが可能であり、標識又はマーカーに結合しているが同じ合成タンパク質が検出抗原として使用される。また、二重抗原架橋アッセイにおいて免疫学的に交差反応性である、類似しているが異なる風疹E1抗原を使用することも可能である。そのようなアッセイを行うための必須要件は、関連エピトープが両方の抗原上に存在することである。明らかに、二重抗原架橋アッセイ形態には多数の変法が存在する。そのような変法は、例えば、固相への風疹E1抗原の間接的コーティングを含む。好ましくは、特異的結合ペア、最も好ましくは、ビオチン-ストレプトアビジン(又はアビジン)系が、風疹抗原を固相に間接的に結合させるために使用される。一方、そのような系における検出に使用される風疹E1抗原は直接的にはマーカー(例えば、放射性同位体、酵素、蛍光分子など)を担持していないことも可能であり、その代わりに、例えばハプテン(例えばジゴキシン)を担持することにより間接的に検出されうることが可能である。そのような間接的検出は、例えば、標識された抗ジゴキシン抗体により行われうる。
【0084】
したがって、本発明の好ましい実施形態は、本発明の第1風疹E1抗原と本発明の第2風疹E1抗原とを含む、二重抗原架橋概念に従うイムノアッセイである。
【0085】
もう1つの好ましい実施形態においては、本発明は、第1風疹E1抗原融合タンパク質複合体を捕捉抗原として使用し、第2風疹E1抗原融合タンパク質を検出抗原として使用することにより特徴づけられる、二重抗原架橋概念に従うイムノアッセイに関する。
【0086】
本明細書に記載の風疹E1抗原融合タンパク質複合体、好ましくは、風疹E1抗原-シャペロン複合体は、それを使用しなければ取り扱いが困難な種々のタンパク質の可溶化をもたらすだけでなく、二重抗原架橋概念に従う非常に有利なイムノアッセイをも可能にする。それぞれ固相結合抗原との複合体形成のために及び検出抗原との複合体形成のために種々のシャペロンを使用することが今や可能になったことは、二重抗原架橋概念に従うそのようなイムノアッセイの特に魅力的な特徴である。アッセイのそのような修飾は更に、非特異的結合の問題を改善する。シャペロンに対して反応性であり従って偽陽性シグナルを生成しうる、サンプル中の抗体は、それぞれ固相抗原及び検出抗原を複合体形成させるために異なるシャペロンを使用した場合には、架橋を形成しないであろう。したがって、本発明のこの態様においては、非特異的結合による陽性シグナルの可能性が著しく減少する。したがって、好ましい実施形態において、本発明は、第1及び第2シャペロン-風疹E1抗原複合体の第1シャペロンと第2シャペロンとが互いに異なることにより特徴づけられる、二重抗原架橋概念に従うイムノアッセイに関する。
【0087】
最もよく特徴づけられているシャペロンのほとんどは、バイオテクノロジー研究において広く使用されている大腸菌(Escherichia coli)から単離されている。大腸菌(Escherichia coli)は広く分布する細菌種であるため、多数の哺乳動物は、この細菌に由来するタンパク質に対する抗体を持つようになっている。そのような抗体により引き起こされる偽陽性反応の可能性を減少させるためには、異なる細菌種(好ましくは好熱菌)に由来する少なくとも1つのPPIシャペロンを使用することが好ましい。好ましくは、シャペロンは、好極限性細菌、特に、テルマトガ・マリチマ(Thermatoga maritima)、アクイフェックス・エオリクス(Aquifex aeolicus)及びテルムス・サーモフィルス(Thermus thermophilus)を含む細菌群の好極限性細菌に由来する。イムノアッセイ全般及び好ましくは該架橋概念に従うイムノアッセイにおけるシャペロン-抗原複合体の使用は、そのような複合体のシャペロンを誘導体化する可能性をも提供し、抗原自体の修飾を要しない。第2化学部分によるタンパク質の修飾、例えば、その分子への標識のカップリングは、該ポリペプチドに負の影響を及ぼす危険性を含む、と一般に認識されている。例えば、そのような標識により、検査中のエピトープが損なわれることがあり、あるいは非特異的結合が引き起こされうる。本発明において、風疹E1抗原-シャペロン複合体内のシャペロンを特異的に誘導体化することが今や可能となった。好ましい実施形態においては、二重抗原架橋概念に従うイムノアッセイは更に、捕捉抗原として使用する第1風疹E1抗原-シャペロン複合体が固相結合基を含むことにより特徴づけられる。もう1つの好ましい実施形態においては、該架橋概念に従うイムノアッセイを行い、これは更に、検出抗原として使用する第2風疹E1抗原-シャペロン複合体がマーカー基を含むことにより特徴づけられる。
【0088】
本発明は更に、抗風疹抗体の検出のための診断試験における、風疹E1の少なくとも1つの抗原及び必要に応じて使用する他の通常の添加剤の使用に関する。
【0089】
本発明のもう1つの主題は、イムノアッセイのための通常の試験添加剤に加えて、測定すべき風疹抗体への特異的結合に適した風疹E1抗原の少なくとも1つの抗原を含有し、場合によっては標識及び必要に応じて他の通常の添加剤を含有する、風疹に対する抗体の検出のための試薬キットに関する。特に、該試験キットは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130にわたるN末端領域を含有する風疹E1抗原及びこのタンパク質の変異体を含有する。好ましくは、該試験キットは、ジスルフィド架橋Cys349-Cys352及びCys368-Cys401、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せにわたるC末端領域を含有する風疹E1抗原及びこのタンパク質の変異体を含有する。
【0090】
本発明のもう1つの実施形態の試験キットが含有する風疹E1抗原及びこのペプチドの変異体は、C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164、好ましくは134-約168を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)は更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる。
【0091】
さらに、この領域のN末端(Cys349)は更に、少なくとも25、30、34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)は更に、隣接風疹E1抗原配列の少なくとも10、11、15、25、35アミノ酸を含有する。該試薬キットは更に、少なくともジスルフィド架橋Cys349-Cys352及びCys368-Cys401、より好ましくは、ジスルフィド架橋Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せ又はジスルフィド架橋Cys176-Cys185、Cys349-Cys352及びCys368-Cys401の組合せ、最も好ましくは、ジスルフィド架橋Cys176-Cys185、Cys225-Cys235、Cys349-Cys352及びCys368-Cys401の組合せにわたるC末端領域、ならびに少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71、より好ましくは、ジスルフィド架橋Cys8-Cys13及びCys59-Cys71及びCys117-Cys130の組合せにわたるN末端領域を含有する風疹E1抗原の組合せを含有する。
【0092】
また、該試薬キットは、対照及び標準溶液、ならびに当業者により使用される通常の添加剤、バッファー、塩、界面活性剤などを含有する1以上の溶液中の試薬を含有する。
【0093】
実施例
【実施例1】
【0094】
縦列EcSlyD及び風疹E1エクトドメイン断片E1(201-432)を含む発現プラスミドの構築
Novagen(Madison, WI, USA)のpET24a発現プラスミドに基づき、以下のクローニング工程を行った。該ベクターをNdeI及びXhoIで消化し、縦列SlyD及び風疹E1断片201-432を含む半合成カセットを挿入した。この風疹E1断片はアミノ酸349、352、368及び401位にシステイン残基を含有する。
【化1】
【0095】
得られたプラスミドのインサートを配列決定したところ、それは所望の融合タンパク質をコードしていることが判明した。該挿入カセットの核酸配列及び得られた融合タンパク質のアミノ酸配列を本発明の配列プロトコールに示す。
【実施例2】
【0096】
SS-E1(201-432)融合タンパク質の、共役された精製及びリフォールディング
該発現プラスミドを含有する大腸菌(E. coli)BL21(DE3)細胞をLB培地+ カナマイシン(30μg/ml)内で、OD600が1になるまで増殖させ、37℃の増殖温度でイソプロピル-β-D-チオガラクトシド(IPTG)を最終濃度1mMまで加えることによりサイトゾル過剰発現を誘導した。誘導の4時間後、細胞を遠心分離(5000×gで20分間)し、凍結させ、-20℃で保存した。細胞溶解のために、該凍結ペレットを100mMリン酸ナトリウム(pH8.0)、7.0M GuHCl、10mM イミダゾールに室温で再懸濁させ、得られた懸濁液を2時間攪拌して細胞溶解を完了させた。遠心分離及び濾過の後、前記細胞溶解バッファー中で予め平衡化されたNi-NTA(ニッケル-ニトリロ-トリアセタート)カラム上にライセートをアプライした。未熟なジスルフィド架橋及びSSシャフリングを避けるために、金属キレート化カラムに適合しうる還元剤として5mM TCEPを洗浄バッファー中に加えた。過剰の洗浄工程(>20カラム容量の細胞溶解バッファー + TCEP)の後、マトリックス結合タンパク質のコンホメーションリフォールディングを誘導するために、該カオトロピック細胞溶解バッファーを50mMリン酸ナトリウム(pH7.8)、100mM塩化ナトリウム、5mM TCEPで置換した(残留GuHClがカオトロピック濃度で存在しないことが保証されるよう、少なくとも10カラム容量のリフォールディングバッファーをアプライした)。ついで、酸化的フォールディング(すなわち、システイン残基の酸化的架橋)を、50mMリン酸ナトリウム pH7,8、100mM塩化ナトリウムでの洗浄によりを誘導した。二価Ni2+イオンの有効濃度が高いため、マトリックス結合融合タンパク質内のジスルフィド架橋の形成は非常に速い過程である。溶出前に、〜50kDaの見掛け分子量を有する汚染(コンタミ)タンパク質を除去するために、該イミダゾール濃度を55mMまで上昇させた。ついで、50mMリン酸ナトリウム(pH7.8)、100mM塩化ナトリウム中の55mMから500mMまでのイミダゾール勾配を適用することにより、該天然融合タンパク質を溶出した。タンパク質含有画分を純度に関して評価し(SDS-PAGEによる判定で>95%)、プールした。最後に、該タンパク質をサイ
ズ排除クロマトグラフィーに付し、二量体画分をプールし、濃縮し、その分光学的特性に関して評価した。
【実施例3】
【0097】
風疹E1融合タンパク質の特徴づけ
a.UV分光学
SS-E1(201-432)は、可溶性かつ天然のようにフォールディングしたタンパク質として溶出する。組換え産生されマトリックスリフォールディングした融合タンパク質のUVスペクトルは凝集傾向を何ら示していない。図1に示すとおり、生理的バッファー条件中のSS-E1(201-432)のUV吸収スペクトルのベースラインは横軸にほぼ等しく(310nm以上)、したがってこのことは、自己会合又は凝集減少により生じる迷光粒子が存在しないことを示している。
【0098】
図1:マトリックス介助リフォールディング及びイミダゾール勾配溶出の後の融合タンパク質SS-E1(201-432)のUVスペクトル。該スペクトルは、1cmの光路長を用いたUvicon XS光度計上で記録された。バッファー条件は50mM リン酸ナトリウム(pH8.0)、100mM 塩化ナトリウム及び〜250mM イミダゾールであった。SS-E1(201-432)に関して48370 M-1cm-1のモル吸光係数εを使用して、タンパク質濃度は7.25μMと決定された。スペクトルの形状はシャペロン化風疹E1エクトドメイン断片(201-432)の溶解度を表す。
【0099】
b.SDSゲル-電気泳動
融合ポリペプチド風疹SS-E1アミノ酸残基201-432の純度をSDS-PAGEにより確認した(図2)。2工程のクロマトグラフィーの後、該融合ポリペプチドの純度は95%を超えている。
【0100】
図2:非還元性SDS-PAGEにより実証されたSS-E1(201-432)の精製。該クーマシー染色ゲルは、(左から右に)Invitrogenのタンパク質標準M12(レーン1)、大腸菌(E. coli)カオトロピック粗抽出物(不溶性画分レーン2、可溶性画分レーン3)、IMAC流動物(レーン5)、〜50kDaの見掛け分子量を有する顕著な混入物(コンタミ)を含有する洗浄画分(レーン7)、及びSS-E1(201-432)イミダゾール溶出画分(レーン9-11)を示している。SDS-PAGEからの判断によると、該融合タンパク質の純度は95%を超えている。混合ジスルフィド又は共有分子間SS付加体の徴候は何ら存在しない。これは、それぞれシステイン349&352及びシステイン368&401の定量的架橋を示している。
【0101】
C.FPLC分析(高速タンパク質液体クロマトグラフィー分析)
その抗原性のほかに、SS-E1融合タンパク質のオリゴマー状態、溶解度及び安定性が診断目的に対するその適合性を決定する。該組換え風疹エクトドメイン断片のオリゴマー状態を明らかにするために、システイン含量において異なるSS-E1(201-432)をSuperdex 200HR 10/30カラム上の分析用ゲル濾過に付した。泳動バッファーは50mM リン酸ナトリウム(pH7.8)、100mM NaClであった。150μlのSS-E1(201-432)溶液(タンパク質濃度〜1.0mg/ml)をSECカラム上にアプライし、溶出を280nmの吸光度によりモニターした(図3)。該実験の結果は、すべてのSS-E1(201-432)システイン変異体が〜12.8mlで定量的に溶出したことを示しており、このことは見掛け上二量体の融合タンパク質を示している。これは、ジスルフィド結合の導入が風疹エクトドメインのポリペプチドバックボーンを拘束し該分子の全体的な小型化を招くという予想と一致する。FPLC分析における高分子会合の明らかな欠如はSDS-PAGEの結果を証明しており、SS-E1(201-432)のマトリックス共役酸化的リフォールディングが、相当に安定で異性化傾向もシャフリング傾向も示さない天然様ジスルフィド架橋を与えるという仮定を裏付けている。SS-E1(201-432)のオリゴマー化は、おそらく、安定な二量体を形成すると報告されている(Mukherjeeら, Biotechnol. Appl. Biochem. (2003) 37, 183-186; Mitterauerら, Biochem. J. (1999) 342, 33-39)シャペロン融合相手SlyDにより媒介されるのであろう。
【0102】
図3:SS-E1(201-432)のシステイン変異体はすべて、FPLC分析により評価した場合、可溶性二量体を形成する。SS-E1(201-432)のシステイン非含有及びジスルフィド架橋変異体をSuperdex 200 HR 10/30分析用ゲル濾過カラム上で泳動させ、それらのオリゴマー状態に関して評価した。泳動バッファーは50mM リン酸ナトリウム(pH7.8)、100mM 塩化ナトリウム、1mM EDTAであり、タンパク質ローデング量は〜200μgであった。溶出を280nmの吸光度によりモニターした。より伸張したシステイン非含有SS-E1(201-432)は12.6ml(黒線)で溶出し、一方、より小型の二重(青線)及び三重(赤線)ジスルフィド架橋変異体は共に12.9mlで溶出する。Roche HPLCタンパク質標準(灰色)はβ-ガラクトシダーゼ(465kDa)、IgG(150kDa)、Fab(50kDa)、ミオグロビン(17kDa)及びジペプチドGly-Tyr(238Da)を含む。
【実施例4】
【0103】
SS-E1(201-432)へのビオチン及びルテニウム部分のカップリング
組換え風疹エクトドメインのリシンε-アミノ基を〜10mg/mlのタンパク質濃度で、それぞれN-ヒドロキシ-スクシンイミド活性化ビオチン及びルテニウム標識により修飾した。標識/タンパク質のモル比は、それぞれの融合タンパク質に応じて2:1から5:1まで様々となった。反応バッファーは150mM リン酸ナトリウム(pH8.0)、50mM NaCl、1mM EDTAであった。反応を室温で15分間行い、緩衝化L-リシンを最終濃度10mMまで加えることにより停止させた。該カップリング反応の後、該粗タンパク質コンジュゲートをゲル濾過カラム(Superdex 200 HI Load)上に通過させることにより未反応遊離標識を除去した。
【実施例5】
【0104】
免疫診断試験における組換え風疹E1融合タンパク質SS-E1(aa201-432)の免疫反応性の検査:天然血清中の抗風疹IgG抗体の検出
種々の該融合タンパク質の免疫反応性を自動Elecsys(登録商標)2010分析装置(Roche Diagnostics GmbH)において評価した。二重抗原サンドイッチ形態で測定を行った。それにより、該ビオチン-コンジュゲート(すなわち、捕捉抗原)をストレプトアビジンコート化磁気ビーズの表面上に固定化した。一方、検出抗原はシグナリング部分として錯化ルテニウムカチオンを含有する。Elecsys(登録商標)2010におけるシグナル検出は電気化学発光に基づくものである。特異的免疫グロブリンアナラアイトの存在下では、該発色性ルテニウム錯体は該固相へ架橋され、白金電極における励起の後に620nmの光を発する。ババリア赤十字の血清収集物からの抗風疹IgG陽性サンプルで測定を行った。
【表1】
表1:風疹SS-E1(aa201-432)抗原を使用することによる天然血清中の抗風疹IgG抗体の検出。
陽性として分類された全ての血清は、それで正しいと分析判定された。さらに、該結果は、陽性(COI* > 1)サンプルと陰性(COI* < 1)サンプルとの間の明らかな相違を示している。
【実施例6】
【0105】
風疹SS-E1(aa315-432)を使用することによる天然血清中の抗風疹IgG抗体の免疫学的検出。免疫学的活性に対する天然様三次構造の関連性
システイン349と352との間、システイン349とシステイン352との間及びシステイン368と401との間でジスルフィド架橋を形成する風疹E1変異体の3つの異なる組合せの測定。前記風疹変異体と比較して、システイン非含有抗原を使用した。サンプル(WHO標準から)、及びババリア赤十字の血清収集物からの抗風疹IgG陽性サンプルを使用して、測定を行った。測定は、二重抗原サンドイッチ形態を用いて自動Elecsys(登録商標)2010分析装置(Roche Diagnostics GmbH)において行った。
【表2】
表2:風疹SS-E1(aa315-432)抗原を使用することによる天然血清中の抗風疹IgG抗体の検出。
ジスルフィド架橋の形成において異なる種々の変異体の比較は風疹SS-E1(aa315-432)の免疫反応性を示している。風疹SS-E1(aa315-432)システイン349とシステイン352との間及びシステイン368とシステイン401との間のジスルフィド架橋は適切に形成されている。陽性として分類された全ての血清は、それで正しいと分析判定された。さらに、該結果は、陽性(COI* > 1)サンプルと陰性(COI* < 1)サンプルとの間の明らかな相違を示している。
【実施例7】
【0106】
風疹SS-E1(aa315-412)及び風疹SS-E1(aa201-432)を使用することによる天然血清中の抗風疹IgG抗体の免疫学的検出
ババリア赤十字の血清収集物からの抗風疹IgG陽性サンプルを使用して、測定を行った。測定は、二重抗原サンドイッチ形態を用いて自動Elecsys(登録商標)2010分析装置(Roche Diagnostics GmbH)において行った。
【表3】
表3:風疹SS-E1(aa315-412)及び風疹SS-E1(aa201-432)抗原を使用することによる天然血清中の抗風疹IgG抗体の検出。
【0107】
長さにおいて異なる種々の変異体の比較は風疹SS-E1(aa315-412)及び風疹SS-E1(aa201-432)の両方の比較しうる免疫反応性を示している。どちらの変異体に関しても、システイン349とシステイン352との間及びシステイン368とシステイン401との間のジスルフィド架橋は適切に形成されている。陽性として分類された全ての血清は、それで正しいと分析判定された。さらに、該結果は、陽性(COI* > 1)サンプルと陰性(COI* < 1)サンプルとの間の明らかな相違を示している。
【0108】
表に関する更なる説明
「カットオフ」なる語は、陽性結果と陰性結果とを識別するためのシグナルである。すなわち、「シグナル > カットオフ」は陽性サンプルを表し、「シグナル < カットオフ」は陰性サンプルを表す。「COI」なる語はカットオフ指数として定義される。COIは、(サンプルのシグナル)/(カットオフでのシグナル)として計算される。すなわち、COI > 1は陽性サンプルを表し、COI < 1は陰性サンプルを表す。
【0109】
参考文献の一覧
Artherton and Shepard in “Solid phase peptide synthesis” IRL Press, Oxford, UK, 1989.
Baron and Forsell, Virology. 1991 Dec;185(2):811-9
Buchner, J., Faseb J 10 (1996) 10-19
Chaye et al., J Clin Immunol. 1993 Mar;13(2) 93-100.
Giessauf et al., J. Immun. Meth. 287 (2004), 1-11
Green and Dorsett, J. Virol. (1986) 57, 893-898
Gros et al., 1997 Virology 230, 179-186
Hobman et al., 1994 (574-585) Virology
Ho-Terry et al., Arch Virol. 1985;84(3-4):207-15
Houbenweyl in “Methode der Organischen Chemie” edited by E. Wunsch, vol 15-I et II, Thiem, Stuttgart, Germany, 1974
Hottenrott, S., et al., J Biol Chem 272 (1997) 15697-701
Mitchell et al., Virus Research 29(1993), 33-54
Mitterauer et al., Biochem. J. (1999) 342, 33-39)
Mukherjee et al., Biotechnol. Appl. Biochem. (2003) 37, 183-186
Newcombe et al., Clin Diagn Virol. 1994 (3)149-63
Oker-Blom et al., J Virol. 1984 (2):403-8
Qui et al., Journal of virology 1994 (4086-4091)
Ramm, K. and Pluckthun, A., J Biol Chem 275 (2000) 17106-13
Sambrook, J., et al., in "Molecular Cloning: A Laboratory Manual" (1989) -, Eds. J. Sambrook,
Scholz, C., et al., Embo J 16 (1997) 54-8
Scholz, C., et al., J Mol Biol., 345, 1229-41, (2005)
Seppaenen et al., J. Clin. Microbiol (1991) 1877-1882
Starkey et al., J. Clin. Microbiol. (1995), 270-274)
Terry et al., Arch Virol. (1985) 86, 29-36
Terry et al Arch Virol. (1988);98, 189-97
Tijssen, P., Preparation of enzym-antibody or other enzyme-macromolecule conjugates in "Practice and theory of enzyme immunoassays" (1990) 221-278, Eds. R. H. Burdon and v. P. H. Knippenberg, Elsevier, Amsterdam)
Tijssen, P. "Methods in Enzymology" (1980), Eds. S. P. Colowick, N. O. Caplan and S. P., Academic Press
Waxham and Wolinsky, Virology. (1983)126(1),194-203
Waxham and Wolinsky, Rev Infect Dis. (1985) 133-9
Waxham and Wolinsky, Virology (1985) 153-65
Weber et al., Bull Soc Sci Med Grand Duche Luxemb. (1997), 31-41
Wolinsky et al., J Virol. (1991), 3986-94
Wolinsky et al., Rubella (1996) Fields Virology. Lippincott-Raven Publisher, Philadelpia, 899-929
EP 0299673
EP 0061888
EP0944838
WO 03/000877
WO 98/13496
WO 03/000878
【図面の簡単な説明】
【0110】
【図1】図1は、マトリックス介助リフォールディング及びイミダゾール勾配溶出の後の融合タンパク質SS-E1(201-432)のUVスペクトルを示す。該スペクトルは、1cmの光路長を用いたUvicon XS光度計上で記録された。バッファー条件は50mM リン酸ナトリウム(pH8.0)、100mM 塩化ナトリウム及び〜250mM イミダゾールであった。SS-E1(201-432)に関して48370 M-1cm-1のモル吸光係数εを使用して、タンパク質濃度は7.25μMと決定された。スペクトルの形状はシャペロン化風疹E1エクトドメイン断片(201-432)の溶解度を表す(実施例3aも参照されたい)。
【図2】図2は、非還元性SDS-PAGEにより実証されたSS-E1(201-432)の精製を示す。該クーマシー染色ゲルは、(左から右に)Invitrogenのタンパク質標準M12(レーン1)、大腸菌(E. coli)カオトロピック粗抽出物(不溶性画分レーン2、可溶性画分レーン3)、IMAC流動物(レーン5)、〜50kDaの見掛け分子量を有する顕著な混入物(コンタミ)を含有する洗浄画分(レーン7)、及びSS-E1(201-432)イミダゾール溶出画分(レーン9-11)を示している。SDS-PAGEからの判断によると、該融合タンパク質の純度は95%を超えている。混合ジスルフィド又は共有分子間SS付加体の徴候は何ら存在しない。これは、それぞれシステイン349&352及びシステイン368&401の定量的架橋を示している(実施例3bも参照されたい)。
【図3】図3は、SS-E1(201-432)のシステイン変異体がすべて、FPLC分析により評価した場合に可溶性二量体を形成することを示す。SS-E1(201-432)のシステイン非含有及びジスルフィド架橋変異体をSuperdex 200 HR 10/30分析用ゲル濾過カラム上で泳動させ、それらのオリゴマー状態に関して評価した。泳動バッファーは50mM リン酸ナトリウム(pH7.8)、100mM 塩化ナトリウム、1mM EDTAであり、タンパク質ローデング量は〜200μgであった。溶出を280nmの吸光度によりモニターした。より伸張したシステイン非含有SS-E1(201-432)は12.6ml(濃灰色実線)で溶出し、一方、より小型の二重(黒色実線)及び三重(黒色点線)ジスルフィド架橋変異体は共に12.9mlで溶出する。Roche HPLCタンパク質標準(淡灰色点線)はβ-ガラクトシダーゼ(465kDa)、IgG(150kDa)、Fab(50kDa)、ミオグロビン(17kDa)及びジペプチドGly-Tyr(238Da)を含む(実施例3aも参照されたい)。
【特許請求の範囲】
【請求項1】
C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)が更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)が更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる可溶性風疹E1抗原ならびにこのペプチドの変異体。
【請求項2】
少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)が更に隣接風疹E1抗原配列の少なくとも25、30、又は34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)が更に隣接風疹E1抗原配列の少なくとも10、11、15、25、又は35アミノ酸を含有することにより特徴づけられる、請求項1記載の可溶性風疹E1抗原。
【請求項3】
C末端において、アミノ酸残基438-452の間のα-らせん領域を更に欠くことにより特徴づけられる、請求項1又は2記載の可溶性風疹E1抗原。
【請求項4】
少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71にわたる領域を含有するN末端領域により特徴づけられる、請求項1又は2記載の可溶性風疹E1抗原。
【請求項5】
少なくともジスルフィド架橋Cys117-Cys130にわたる領域を更に含有することにより特徴づけられる、請求項4記載の可溶性風疹E1抗原。
【請求項6】
少なくともジスルフィド架橋Cys225-Cys235にわたる領域を更に含有することにより特徴づけられる、請求項5記載の可溶性風疹E1抗原。
【請求項7】
少なくともジスルフィド架橋Cys176-Cys185にわたる領域を更に含有することにより特徴づけられる、請求項5又は6記載の可溶性風疹E1抗原。
【請求項8】
該タンパク質が合成的に製造される、請求項1〜7のいずれか1項記載の可溶性風疹E1抗原。
【請求項9】
該タンパク質が組換え的に製造される、請求項1〜7のいずれか1項記載の可溶性風疹E1抗原。
【請求項10】
該タンパク質が組換え融合タンパク質として製造される、請求項9記載の可溶性風疹E1抗原。
【請求項11】
該風疹E1抗原がペプチジル-プロリル-イソメラーゼクラスのシャペロンに融合されている、請求項10記載の可溶性風疹E1抗原。
【請求項12】
請求項1〜7のいずれか1項記載の風疹E1抗原をコードする少なくとも1つのヌクレオチド配列を含んでなり、その上流に、FKBPシャペロンをコードする少なくとも1つのヌクレオチド配列が位置する、風疹E1抗原をコードする組換えDNA分子。
【請求項13】
FKBPシャペロンが、FkpA、SlyD及び誘発因子よりなる群から選ばれることにより特徴づけられる、請求項12記載の組換えDNA分子。
【請求項14】
それが、風疹E1抗原をコードする配列とFKBPシャペロンをコードする配列との間に位置する10〜100アミノ酸のペプチドリンカーをコードする少なくとも1つのヌクレオチド配列を含むことにより更に特徴づけられる、請求項12又は13記載の組換えDNA分子。
【請求項15】
FKBPシャペロンをコードする1つのヌクレオチド配列を含む、請求項12〜14のいずれか1項記載の組換えDNA分子。
【請求項16】
FKBPシャペロンをコードする少なくとも2つのヌクレオチド配列を含む、請求項12〜14のいずれか1項記載の組換えDNA分子。
【請求項17】
FKBPシャペロンをコードする少なくとも2つの配列が、風疹E1抗原をコードする配列の上流に位置することにより更に特徴づけられる、請求項16記載の組換えDNA分子。
【請求項18】
PPIシャペロンをコードする1つの配列が風疹E1抗原の上流に位置し、PPIシャペロンをコードするその他の配列が、風疹E1抗原をコードする配列の下流に位置することにより更に特徴づけられる、請求項17記載の組換えDNA分子。
【請求項19】
それが、10〜100アミノ酸のリンカーポリペプチドをコードする2つの核酸配列を含むことにより更に特徴づけられる、請求項16〜18のいずれか1項記載の組換えDNA分子。
【請求項20】
10〜100アミノ酸のリンカーをコードする2つの核酸配列が互いに異なる、請求項19記載の組換えDNA分子。
【請求項21】
該リンカー配列の少なくとも1つが、タンパク質分解切断部位を含むポリペプチドリンカーをコードする、請求項12〜20のいずれか1項記載の組換えDNA分子。
【請求項22】
請求項12〜21のいずれか1項記載の組換えDNA分子を、機能しうる形で連結されて含んでなる発現ベクター。
【請求項23】
請求項22記載の発現ベクターで形質転換された宿主細胞。
【請求項24】
可溶性かつ免疫反応性の風疹E1抗原-シャペロン融合タンパク質の製造方法であって、
a.請求項23記載の宿主細胞の培養、
b.該融合タンパク質の発現、
c.該融合タンパク質の精製、
d.可溶性かつ免疫反応性のコンホメーションへのリフォールディング
の工程を含んでなる製造方法。
【請求項25】
ペプチジルプロリルイソメラーゼがFKBPシャペロンである、請求項24記載の製造方法。
【請求項26】
FKBPシャペロンが、SlyD、FkpA及び誘発因子よりなる群から選ばれる、請求項25記載の製造方法。
【請求項27】
サンプル中のIgG及び/又はIgMサブクラスの抗風疹抗体の検出及び測定及び定量のための方法であって、請求項1〜11のいずれか1項記載の風疹E1抗原を該抗体に対する捕捉試薬及び/又は結合相手として使用する、方法。
【請求項28】
請求項1〜12のいずれか1項記載の第1風疹E1抗原と第2風疹E1抗原とを含んでなる、二重抗原架橋概念に従うイムノアッセイ。
【請求項29】
抗風疹抗体の検出のための診断試験における、請求項1〜12のいずれか1項記載の風疹E1の少なくとも1つの抗原の使用。
【請求項30】
請求項1〜12のいずれか1項記載の風疹E1抗原の少なくとも1つの抗原を含有する、抗風疹抗体の検出のための試薬キット。
【請求項1】
C末端における少なくとも膜貫通領域とアンカーセグメント、及び少なくともアミノ酸143-164を欠いていること、ならびに少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)が更に少なくとも15アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)が更に隣接風疹E1抗原配列の少なくとも8アミノ酸を含有することにより特徴づけられる可溶性風疹E1抗原ならびにこのペプチドの変異体。
【請求項2】
少なくともジスルフィド架橋Cys349-Cys352及びCys368-401にわたる領域を含有することにより特徴づけられ、一方、この領域のN末端(Cys349)が更に隣接風疹E1抗原配列の少なくとも25、30、又は34アミノ酸を含有し、及び/又は、この領域のC末端(Cys401)が更に隣接風疹E1抗原配列の少なくとも10、11、15、25、又は35アミノ酸を含有することにより特徴づけられる、請求項1記載の可溶性風疹E1抗原。
【請求項3】
C末端において、アミノ酸残基438-452の間のα-らせん領域を更に欠くことにより特徴づけられる、請求項1又は2記載の可溶性風疹E1抗原。
【請求項4】
少なくともジスルフィド架橋Cys8-Cys13及びCys59-Cys71にわたる領域を含有するN末端領域により特徴づけられる、請求項1又は2記載の可溶性風疹E1抗原。
【請求項5】
少なくともジスルフィド架橋Cys117-Cys130にわたる領域を更に含有することにより特徴づけられる、請求項4記載の可溶性風疹E1抗原。
【請求項6】
少なくともジスルフィド架橋Cys225-Cys235にわたる領域を更に含有することにより特徴づけられる、請求項5記載の可溶性風疹E1抗原。
【請求項7】
少なくともジスルフィド架橋Cys176-Cys185にわたる領域を更に含有することにより特徴づけられる、請求項5又は6記載の可溶性風疹E1抗原。
【請求項8】
該タンパク質が合成的に製造される、請求項1〜7のいずれか1項記載の可溶性風疹E1抗原。
【請求項9】
該タンパク質が組換え的に製造される、請求項1〜7のいずれか1項記載の可溶性風疹E1抗原。
【請求項10】
該タンパク質が組換え融合タンパク質として製造される、請求項9記載の可溶性風疹E1抗原。
【請求項11】
該風疹E1抗原がペプチジル-プロリル-イソメラーゼクラスのシャペロンに融合されている、請求項10記載の可溶性風疹E1抗原。
【請求項12】
請求項1〜7のいずれか1項記載の風疹E1抗原をコードする少なくとも1つのヌクレオチド配列を含んでなり、その上流に、FKBPシャペロンをコードする少なくとも1つのヌクレオチド配列が位置する、風疹E1抗原をコードする組換えDNA分子。
【請求項13】
FKBPシャペロンが、FkpA、SlyD及び誘発因子よりなる群から選ばれることにより特徴づけられる、請求項12記載の組換えDNA分子。
【請求項14】
それが、風疹E1抗原をコードする配列とFKBPシャペロンをコードする配列との間に位置する10〜100アミノ酸のペプチドリンカーをコードする少なくとも1つのヌクレオチド配列を含むことにより更に特徴づけられる、請求項12又は13記載の組換えDNA分子。
【請求項15】
FKBPシャペロンをコードする1つのヌクレオチド配列を含む、請求項12〜14のいずれか1項記載の組換えDNA分子。
【請求項16】
FKBPシャペロンをコードする少なくとも2つのヌクレオチド配列を含む、請求項12〜14のいずれか1項記載の組換えDNA分子。
【請求項17】
FKBPシャペロンをコードする少なくとも2つの配列が、風疹E1抗原をコードする配列の上流に位置することにより更に特徴づけられる、請求項16記載の組換えDNA分子。
【請求項18】
PPIシャペロンをコードする1つの配列が風疹E1抗原の上流に位置し、PPIシャペロンをコードするその他の配列が、風疹E1抗原をコードする配列の下流に位置することにより更に特徴づけられる、請求項17記載の組換えDNA分子。
【請求項19】
それが、10〜100アミノ酸のリンカーポリペプチドをコードする2つの核酸配列を含むことにより更に特徴づけられる、請求項16〜18のいずれか1項記載の組換えDNA分子。
【請求項20】
10〜100アミノ酸のリンカーをコードする2つの核酸配列が互いに異なる、請求項19記載の組換えDNA分子。
【請求項21】
該リンカー配列の少なくとも1つが、タンパク質分解切断部位を含むポリペプチドリンカーをコードする、請求項12〜20のいずれか1項記載の組換えDNA分子。
【請求項22】
請求項12〜21のいずれか1項記載の組換えDNA分子を、機能しうる形で連結されて含んでなる発現ベクター。
【請求項23】
請求項22記載の発現ベクターで形質転換された宿主細胞。
【請求項24】
可溶性かつ免疫反応性の風疹E1抗原-シャペロン融合タンパク質の製造方法であって、
a.請求項23記載の宿主細胞の培養、
b.該融合タンパク質の発現、
c.該融合タンパク質の精製、
d.可溶性かつ免疫反応性のコンホメーションへのリフォールディング
の工程を含んでなる製造方法。
【請求項25】
ペプチジルプロリルイソメラーゼがFKBPシャペロンである、請求項24記載の製造方法。
【請求項26】
FKBPシャペロンが、SlyD、FkpA及び誘発因子よりなる群から選ばれる、請求項25記載の製造方法。
【請求項27】
サンプル中のIgG及び/又はIgMサブクラスの抗風疹抗体の検出及び測定及び定量のための方法であって、請求項1〜11のいずれか1項記載の風疹E1抗原を該抗体に対する捕捉試薬及び/又は結合相手として使用する、方法。
【請求項28】
請求項1〜12のいずれか1項記載の第1風疹E1抗原と第2風疹E1抗原とを含んでなる、二重抗原架橋概念に従うイムノアッセイ。
【請求項29】
抗風疹抗体の検出のための診断試験における、請求項1〜12のいずれか1項記載の風疹E1の少なくとも1つの抗原の使用。
【請求項30】
請求項1〜12のいずれか1項記載の風疹E1抗原の少なくとも1つの抗原を含有する、抗風疹抗体の検出のための試薬キット。
【図1】

【図2】
【図3】
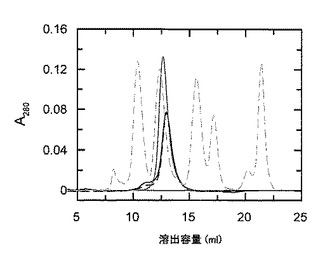
【図2】
【図3】
【公開番号】特開2007−117090(P2007−117090A)
【公開日】平成19年5月17日(2007.5.17)
【国際特許分類】
【外国語出願】
【出願番号】特願2006−290414(P2006−290414)
【出願日】平成18年10月25日(2006.10.25)
【出願人】(591003013)エフ.ホフマン−ラ ロシュ アーゲー (1,754)
【氏名又は名称原語表記】F. HOFFMANN−LA ROCHE AKTIENGESELLSCHAFT
【Fターム(参考)】
【公開日】平成19年5月17日(2007.5.17)
【国際特許分類】
【出願番号】特願2006−290414(P2006−290414)
【出願日】平成18年10月25日(2006.10.25)
【出願人】(591003013)エフ.ホフマン−ラ ロシュ アーゲー (1,754)
【氏名又は名称原語表記】F. HOFFMANN−LA ROCHE AKTIENGESELLSCHAFT
【Fターム(参考)】
[ Back to top ]