
シュードモナスにおける組換え二十面体ウイルス様粒子の生産
本発明は、組換えペプチドと二十面体ウイルスカプシドの融合物及びシュードモナス起源の細菌細胞におけるこの融合物の発現による組換えペプチドの生産のための改善された方法を提供する。シュードモナス細胞は、インビボで二十面体ウイルスカプシドからのウイルス様粒子の形成を支持し、より大きな組換えペプチドを単量体又はコンカテマーとしてウイルス様粒子内に取り込むことを可能にする。本発明は、特に、ウイルスカプシド融合物を発現する細胞、毒性タンパク質と二十面体ウイルスカプシドの融合物をエンコードする核酸及び組換えタンパク質の製造のための方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、「High Efficiency Peptide Production in Pseudomonads」と題する2003年12月1日出願の米国特許仮出願第60/525,982号の優先権を主張する。
【0002】
本発明は、組換えペプチドの生産のための改善された方法を提供する。特に、本発明は、二十面体ウイルスからのウイルス様粒子を使用した細菌細胞における組換えペプチドの生産又は提示のための改善された方法を提供する。
【背景技術】
【0003】
遺伝子工学の革命は、ヒト及び動物治療薬としての使用のための組換えペプチドの開発へ発展した。現在、医学用途のために米国FDAによって承認された100以上のバイオテクノロジー由来の治療薬及びワクチンが存在し、1000を超えるさらなる薬剤及びワクチンが様々な相の臨床試験下にある。(M. Rai & H. Padh, (2001) "Expression systems for production of heterologous proteins, "Cur. Science 80 (9): 1121-1128参照)。
【0004】
細菌、酵母、ディクチオステリウム・ディスコイデウム、昆虫、及び哺乳動物細胞発現系が、組換えペプチドを生産するために現在使用されており、成功の程度は様々である。異種ペプチドの生産のための発現系を作成する上での1つの目標は、商業、治療、及びワクチン適用において利用できる、広範囲で、柔軟性があり、効率的、経済的及び実用的なプラットフォームを提供することである。例えば特定のペプチドの生産のためには、下流の再構築(reassembly)コストを取り除く又は低減するために、インビボで効率よく且つ安価に、大量の所望の最終産物を生産することができる発現系を得ることが理想である。
【0005】
現在、細菌は、豊富な量の組換えペプチドを生産するそれらの潜在能力の故に、組換えペプチドの生産のために最も広く使用される発現系である。しかしながら、細菌はしばしば特定のペプチドを生産する能力が限られており、代替的な、より費用のかかる発現系の使用を必要とする。例えば細菌系は、単量体抗菌ペプチドに関しては、しばしばペプチドの発現時に細胞死を導く、細菌に対するかかるペプチドの毒性の故に、生産能力が限られている。非細菌系の、コストと生産収率という観点における特有の不都合の故に、広い範囲の商業的及び治療的に有用なペプチドを生産する細菌系の能力を改善するために多大の時間と資源が費やされてきた。この領域で進歩はあったが、細菌系における異種ペプチドの生産のための付加的な方法及びプラットフォームは有益である。
【0006】
(ウイルス)
宿主細胞発現系におけるペプチド生産を改善するための1つのアプローチは、対象組換えペプチドを生産するために複製可能ウイルスの性質を利用することである。しかし、複製可能な完全長ウイルスの使用は、組換えペプチド生産戦略における使用において数多くの難点がある。例えば発酵状態の間は組換えペプチドの生産を制御することが困難であると考えられるが、発酵工程の効率を最大化するためには発現の厳密な調節が必要であり得る。さらに、組換えペプチドを生産するための複製可能ウイルスの使用は、高度な下流精製工程につながる、規制基準の負担をもたらし得る。
【0007】
特に発酵中の生産上の問題を克服するために、1つの研究領域は、特定のウイルスにとって天然の宿主ではない細胞(非親和性宿主細胞)におけるウイルスの発現と構築に焦点を当ててきた。非親和性細胞は、ウイルスカプシドと宿主受容体分子の不適合性、又はウイルスがその生活環を完了するのを妨げる、ウイルスの生化学と細胞の生化学の間の不適合性の故に、ウイルスがうまく入り込むことができない細胞である。例えばPriceらへの米国特許第5,869,287号は、ウイルスカプシド又はカプシド内に含まれるRNAがノダウイルス科又はブロモウイルス科の非酵母ウイルス種由来である場合に、酵母細胞において、RNAを含む複製可能又は感染性ウイルスを合成し、構築するための方法を述べている。しかしこのアプローチは、複製可能ウイルスにおけるタンパク質生産に関連する潜在的な規制上の障害を克服していない。
【0008】
(ウイルス様粒子)
組換えペプチドの生産を改善するためのもう1つのアプローチは、ウイルス様粒子(VLP)を使用することであった。一般に、カプシドに包まれたウイルスは、ウイルス核酸を含むように集合されるタンパク質コート又は「カプシド」を含む。多くのウイルスは、カプシドがVLPを形成するときに発現される細胞の内部で(「インビボ集合(in vivo assembly)」)及び単離及び精製後に細胞の外部で(「インビトロ集合(in vitro assembly)」)、個々に発現されたカプシドから「自己集合」され得るカプシドを有する。理想的には、カプシドは、標的組換えペプチドを含み、組換えウイルスカプシド−ペプチド融合物を生成するように修飾される。融合ペプチドを、次に、細胞において発現し、理想的には、組換えウイルス又はウイルス様粒子を形成するようにインビボで集合することができる。
【0009】
このアプローチは様々な程度の成功を収めている。例えばC Marusic et al., J Virol. 75 (18): 8434-39 (Sep 2001) (組換えウイルス粒子のインビボ形成を伴う、末端で抗原性HIVペプチドに融合した組換えらせんジャガイモXウイルスカプシドの植物における発現); FR Brennan et al., Vaccine 17 (15-16): 1846-57 (09 Apr 1999)(組換えウイルス粒子のインビボ形成を伴う、末端で抗原性黄色ブドウ球菌ペプチドに融合した組換え二十面体ササゲモザイクウイルス又はらせんジャガイモXウイルスカプシドの植物における発現)参照。
【0010】
Lomonossoff & Johnsonへの米国特許第5,874,087号は、 ウイルスカプシドが、生物活性ペプチド、例えばホルモン、成長因子又は抗原ペプチドを含むように加工されたカプシドを含む、植物細胞における組換え植物ウイルスの生産を述べている。コモウイルス、トンブスウイルス、ソベモウイルス、ネポウイルス属から選択されるウイルスを、外来性ペプチドをエンコードする配列を含むように加工し、加工したウイルスのゲノム全体を発現させて組換えウイルスを生産する。外来性ペプチドコード配列を、1又はそれ以上のカプシド表面ループモチーフをエンコードする配列内に挿入する。
【0011】
特定のウイルス様粒子を生産するために非親和性細胞を利用するための試みがなされてきた。例えばウイルス様粒子のインビボ形成を伴う、末端でヘプタデカペプチドに融合した組換え二十面体ジャガイモ巻葉ウイルスカプシドの昆虫細胞における発現を述べる、JW Lamb et al., J Gen. Virol. 77 (Pt. 7): 1349-58 (Jul 1996)参照のこと。ある状況では、非親和性VLPは好ましいと考えられる。例えば非親和性ウイルスカプシドは、天然のウイルスカプシドよりも、ウイルス様粒子へと集合する能力を破壊することのない異種ペプチドの挿入に対して適応しやすいと考えられる。あるいは、非親和性ウイルスカプシドは、天然の親和性ウイルスからのカプシドよりも詳細に特性が決定され、理解され得る。加えて、特定の適用、例えばワクチン生産は、特定の宿主細胞発現系における親和性ウイルスの使用を許容しないことがある。Chapmanらへの米国特許第6,232,099号は、植物においてウイルスカプシドサブユニットに結合された異種タンパク質を生産するための桿状ウイルスの使用を述べている。らせんウイルスとしても分類される桿状ウイルス、例えばジャガイモXウイルス(PVX)は、組換えウイルスカプシド−ペプチド融合物を作成するためにウイルスのゲノム内に挿入された対象組換えペプチドを有する。次に組換えウイルスを使用して宿主細胞に感染させると、ウイルスは宿主細胞において活発に複製し、さらに他の細胞に感染する。最終的に、組換えウイルスカプシド−ペプチド融合物を植物宿主細胞から精製する。
【0012】
(細菌発現系におけるウイルス様粒子の使用)
組換えペプチドの迅速で効率的且つ安価で豊富な収量の潜在的可能性の故に、細菌は、組換えウイルスカプシド−ペプチド融合ウイルス様粒子の生産のための発現系における宿主細胞として検討されてきた。
【0013】
研究者達は、組換えペプチド挿入物を含まない特定の野生型ウイルスカプシドが、非親和性腸内細菌においてトランスジェニック発現できることを示した。研究者達はまた、これらのカプシドが、インビボ及びインビトロの両方で、ウイルス様粒子を形成するように集合できることを示した。例えばSJ Shire et al., Biochemistry 29 (21): 5119-26 (29 May 1990) (大腸菌において発現されるらせんタバコモザイクウイルスカプシドからのウイルス様粒子のインビトロ集合) ; X Zhao et al., Virology 207 (2): 486-94 (10 Mar 1995) (大腸菌において発現される二十面体ササゲ退緑斑紋ウイルスカプシドからのウイルス様粒子のインビトロ集合); Y Stram et al., Virus Res. 28(1) : 29-35 (Apr 1993)(ウイルス様粒子のインビボ形成を伴う、大腸菌におけるひも状ジャガイモYウイルスカプシドの発現); J Joseph & HS Savithri, Arch. Virol. 144 (9): 1679-87 (1999) (ウイルス様粒子のインビボ形成を伴う、大腸菌におけるひも状トウガラシ葉脈緑帯ウイルスカプシドの発現); DJ Hwang et al., Proc. Nat'I Acad. Sci. USA 91 (19): 9067-71 (13 Sep 1994) (ウイルス様粒子のインビボ形成を伴う、大腸菌におけるらせんタバコモザイクウイルスカプシドの発現); M Sastri et al., J Mol. Biol. 272 (4): 541-52 (03 Oct 1997)(ウイルス様粒子のインビボ形成を伴う、大腸菌における二十面体ホオズキ斑紋ウイルスカプシドの発現)参照。
【0014】
現在まで、非親和性細菌細胞における組換えウイルスカプシド−ペプチド融合物の発現及びインビボ集合の成功は様々であった。一般に、これらの粒子のインビボ集合の成功は、非二十面体ウイルスカプシド−標的ペプチド融合物粒子に限られてきた。例えばMN Jagadish et al.,Intervirology 39 (1-2): 85-92 (1996) (ウイルス様粒子のインビボ形成を伴う、末端で抗原ペプチドに融合した組換えひも状非二十面体セイバンモロコシモザイクウイルスカプシドの非植物細胞における発現)参照。
【0015】
二十面体カプシドに連結されたペプチドの発現は、成功していないか又は限られた有用性しかなかった。例えばV Yusibov et al., J Gen. Virol. 77 (Pt. 4): 567-73 (Apr 1996)は、末端でヘキサヒスチジンペプチドに融合した、大腸菌で発現された組換え二十面体アルファルファモザイクウイルスカプシドからのウイルス様粒子のインビトロ集合を述べた。
【0016】
Brumfieldらは、二十面体カプシドに挿入した組換えペプチドをインビボ集合したウイルス様粒子として発現することを試みたが、成功しなかった。Brumfield et al. , (2004)「ササゲ退緑斑紋ブロモウイルスの修飾カプシドの異種発現は、変化した構造及び機能を有するタンパク質ケージの集合をもたらす」J. Gen. Vir. 85: 1049-1053参照。観察された、二十面体ウイルスカプシド−ペプチド融合物粒子を大腸菌においてインビボでウイルス様粒子として集合することの不可能性の理由はよく理解されていない。Brumfieldらはこの集合の失敗を、大腸菌が不溶性カプシドを生産するという事実に結び付けている。
【0017】
Chapmanは、米国特許第6,232,099号において、限られた挿入物サイズが二十面体ウイルスによって許容されることを指摘する。Chapmanは、自らの主張を裏付けるのに、植物宿主細胞における発現のための二十面体ウイルス内の組換えペプチドの大きさを26アミノ酸の長さに限定する、国際公開広報第WO92/18618を引用している。Chapmanは、二十面体ウイルスのカプシドの内部挿入部位に存在するより大きなペプチドは、タンパク質の形状及び/又は他のカプシドと成功裏に相互作用するその能力の破壊をもたらし、それがキメラウイルスの構築の失敗を導き得ると理論付けている。この文献はまた、大腸菌を含み得る細胞においてカプシド−組換えペプチド融合ペプチドを生産するための非複製桿状ウイルスの使用を述べている。
【発明の開示】
【発明が解決しようとする課題】
【0018】
したがって、二十面体ウイルスに由来するウイルス様粒子の生産のための改善された細菌発現系を提供することが本発明の1つの目的である。
【0019】
ウイルス様粒子の生産のための改善された発現系において宿主細胞として使用するための細菌生物を提供することが本発明のもう1つの目的である。
【0020】
細菌におけるウイルス様粒子の改善された生産のための方法を提供することが本発明のさらにもう1つの目的である。
【0021】
ウイルス様粒子の生産のための改善された細菌発現系において使用するための新規構築物及び核酸を提供することが本発明のさらにもう1つの目的である。
【課題を解決するための手段】
【0022】
二十面体カプシド−組換えペプチド融合物粒子は、シュードモナス生物において発現されたときインビボでウイルス様粒子又は可溶性ケージ構造へと集合する。さらに、50アミノ酸以上の大きな組換えペプチド又はペプチドコンカテマーを、二十面体カプシドに挿入し、シュードモナス生物においてインビボで集合することができる。
【0023】
本発明の1つの態様では、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を含むシュードモナス生物を提供する。本発明の1つの特定の実施形態では、シュードモナス細胞はシュードモナス・フルオレッセンスである。1つの実施形態では、前記細胞はウイルス様粒子又は可溶性ケージ構造を生産する。
【0024】
この細胞において生産されるウイルス様粒子は、典型的には細胞に感染することができない。ウイルスカプシド配列は、細胞に親和性でないウイルスに由来し得る。1つの実施形態では、細胞は、所望の二十面体カプシド以外のウイルスタンパク質を含まない。1つの実施形態では、ウイルスカプシドは、この細胞とは異なる科の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、この細胞とは異なる属の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、この細胞とは異なる種の生物に対して親和性を有するウイルスに由来する。本発明の1つの実施形態では、二十面体カプシドは植物二十面体ウイルスに由来する。特定の実施形態では、二十面体カプシドは、ササゲ退緑斑紋ウイルス(Cowpea Chlorotic Mottle Virus)、ササゲモザイクウイルス(Cowpea Mosaic Virus)及びアルファルファモザイクウイルス(Alfalfa Mosaic Virus)から選択される群に由来する。
【0025】
本発明の1つの実施形態では、二十面体カプシドに融合した組換えペプチドは、ヒト又は動物治療のために有用な治療ペプチドである。1つの特定の実施形態では、組換えペプチドは抗原である。1つの実施形態では、カプシド−組換えペプチドウイルス様粒子はヒト又は動物適用においてワクチンとして投与することができる。もう1つの特定の実施形態では、組換えペプチドは、遊離単量体形態であるとき宿主細胞に対して毒性であるペプチドである。より特定の実施形態では、毒性ペプチドは抗菌ペプチドである。
【0026】
1つの実施形態では、二十面体カプシドに融合した組換えペプチドは、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも35、少なくとも40、少なくとも45、少なくとも50、少なくとも55、少なくとも60、少なくとも65、少なくとも75、少なくとも85、少なくとも95、少なくとも99又は少なくとも100アミノ酸である。
【0027】
本発明の1つの実施形態では、二十面体カプシドに融合した組換えペプチドは、所望の標的ペプチドの少なくとも1個の単量体を含む。選択的実施形態では、組換えペプチドは、所望の標的ペプチドの2個以上の単量体を含む。特定の実施形態では、このペプチドは、コンカテマーペプチドとしてカプシドに作動可能に連結された少なくとも2個、少なくとも5個、少なくとも10個、少なくとも15個又は少なくとも20個の別々の単量体からなる。もう1つの実施形態では、コンカテマーペプチド内の個々の単量体は、切断可能なリンカー領域によって連結される。さらにもう1つの実施形態では、組換えペプチドを二十面体ウイルスカプシドの少なくとも1つの表面ループに挿入する。1つの実施形態では、少なくとも1個の単量体を二十面体ウイルスカプシドの2つ以上の表面ループに挿入する。
【0028】
ウイルス様粒子の2つ以上のループを修飾することができる。1つの特定の実施形態では、組換えペプチドを、二十面体ウイルス様粒子の少なくとも2つの表面ループ上で発現する。もう1つの実施形態では、少なくとも2個の異なるペプチドをウイルスカプシド、ケージ又はウイルス様粒子の少なくとも2つの表面ループに挿入する。もう1つの実施形態では、少なくとも3個の組換えペプチドをウイルス様粒子の少なくとも3つの表面ループに挿入する。表面ループ内の組換えペプチドは同じアミノ酸配列を有し得る。別の実施形態では、表面ループ内の組換えペプチドのアミノ酸配列は異なる。
【0029】
もう1つの実施形態では、この細胞は、第二の野生型カプシド又はカプシド−組換えペプチド融合ペプチドのいずれかをエンコードする少なくとも1個の付加的な核酸を含み、多数のカプシドがインビボでキメラウイルス様粒子を生成するように集合する。
【0030】
本発明の1つの態様では、二十面体カプシドと組換えペプチドの融合ペプチドを含むシュードモナス生物を提供する。本発明の1つの詳細な実施形態では、シュードモナス細胞はシュードモナス・フルオレッセンスである。1つの実施形態では、カプシド−組換えペプチド融合ペプチドは、インビボでウイルス様粒子を形成するように集合する。
【0031】
本発明のもう1つの態様では、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を提供する。本発明の1つの実施形態では、二十面体カプシドは植物二十面体ウイルスに由来する。特定の実施形態では、二十面体カプシドは、ササゲモザイクウイルス、ササゲ退緑斑紋ウイルス及びアルファルファモザイクウイルスから選択される群に由来する。
【0032】
1つの実施形態では、組換えペプチドは、遊離単量体形態であるとき宿主細胞に対して毒性であるペプチドである。より特定の実施形態では、毒性ペプチドは抗菌ペプチドである。
【0033】
本発明の1つの実施形態では、組換えペプチドは、所望の標的ペプチドの少なくとも1個の単量体を含む。別の実施形態では、組換えペプチドは、所望の標的ペプチドの2個以上の単量体を含む。さらにもう1つの実施形態では、組換えペプチドを二十面体ウイルスカプシドの少なくとも1つの表面ループに挿入する。
【0034】
もう1つの実施形態では、核酸構築物は、少なくとも1個のプロモーター、少なくとも1個の選択マーカー、少なくとも1個のオペレーター配列、少なくとも1個の複製起点、及び少なくとも1個のリボソーム結合部位を含む付加的な核酸配列を含み得る。
【0035】
1つの態様では、本発明は、
a)シュードモナス細胞を提供すること;
b)組換えペプチドと二十面体カプシドからなる融合ペプチドをエンコードする核酸を提供すること;
c)このシュードモナス細胞においてこの核酸を発現すること、ここで細胞における発現がこの融合ペプチドのウイルス様粒子へのインビボ集合を提供する;及び
d)ウイルス様粒子を単離すること
を含む、組換えペプチドを生産するための方法を提供する。
1つの実施形態では、この工程は、e)この融合生成物を切断して、カプシドから組換えペプチドを分離すること、をさらに含む。本発明の1つの実施形態では、シュードモナス細胞はシュードモナス・フルオレッセンスである。
【0036】
1つの実施形態では、この方法は、カプシドがインビボでキメラウイルス様粒子を生成するように集合する、野生型カプシド又はカプシド−組換えペプチド融合ペプチドをエンコードするもう1つ別の核酸を共発現することを含む。
【0037】
本発明のもう1つの態様では、
a)シュードモナス細胞;
b)少なくとも1個の組換えペプチドと少なくとも1個の二十面体ウイルスカプシドを含む融合ペプチドをエンコードする核酸;及び
c)培養培地
を含む組換えペプチドの生産のための発現系を提供する。
【0038】
発現されたとき融合ペプチドは、細胞内でウイルス様粒子へと集合することができる。
【発明を実施するための最良の形態】
【0039】
本発明は、二十面体ウイルスカプシドと対象組換えペプチドを含む融合ペプチドの細菌における発現のための方法を提供する。ここで使用する「ペプチド」という用語は、いかなる特定の分子量にも限定されず、タンパク質又はポリペプチドも包含し得る。本発明はさらに、この方法における使用のための細菌細胞及び核酸構築物を提供する。特に、本発明は、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を有するシュードモナス生物を提供する。本発明の1つの特定の実施形態では、シュードモナス細胞はシュードモナス・フルオレッセンスである。1つの実施形態では、この細胞は、ウイルス様粒子又は可溶性ケージ構造を生産する。本発明はまた、1つの実施形態ではヒト及び動物治療のために有用な治療ペプチドであり得る、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を提供する。
【0040】
本発明はまた、組換えペプチドと二十面体カプシドの融合ペプチドをエンコードする核酸を提供すること;この核酸をシュードモナス細胞において発現し、この細胞における発現が融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及びウイルス様粒子を単離すること、によってシュードモナス細胞において組換えペプチドを生産するための方法を提供する。
【0041】
(I.組換えシュードモナス細胞)
本発明は、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を含むシュードモナス細胞を提供する。この細胞は、組換えペプチドを生産するための方法において利用することができる。
【0042】
(ウイルスカプシド)
1つの実施形態では、本発明は、二十面体ウイルスカプシドに融合したペプチドの発現によってペプチドを生産するための方法における使用のためのシュードモナス細胞を提供する。この発現は、典型的には細胞内で少なくとも1個のウイルス様粒子(VLP)を生じさせる。
【0043】
ウイルスは、らせん対称又は二十面体対称を有するものに分類できる。一般に認識されるカプシド形態は、二十面体(正二十面体(icosahedral proper)、等軸晶系、準等軸晶系、及び双生又は「双晶(twinned)」を含む)、多面体(球状、卵形及びレモン形を含む)、桿菌状(棒又は弾丸形、及び紡錘状又は葉巻形を含む)、及びらせん状(桿状、円柱状及びひも状を含む)を含み、いずれも尾部を有していてもよく及び/又は表面突起、例えば棘又はこぶを含んでいてもよい。
【0044】
(形態)
本発明の1つの実施形態では、カプシドのアミノ酸配列は、何らかの二十面体形態を有すると分類されるウイルスのカプシドから選択される。1つの実施形態では、カプシドアミノ酸配列は、二十面体である実在のカプシドから選択される。もう1つの実施形態では、カプシドアミノ酸配列は、二十面体ウイルスのカプシドから選択される。1つの特定の実施形態では、カプシドアミノ酸配列は、二十面体植物ウイルスのカプシドから選択される。しかし、もう1つの実施形態では、ウイルスカプシドは、植物に対して感染性でない二十面体ウイルスに由来する。例えば1つの実施形態では、ウイルスは、哺乳動物に対して感染性のウイルスである。
【0045】
一般に、二十面体ウイルスのウイルスカプシドは、二十面体(立方)対称に配列された多数のタンパク質サブユニットからなる。天然の二十面体カプシドは、例えば3つのサブユニットが各々カプシドの各々の三角形の面を形成することによって構築することができ、60サブユニットが完全なカプシドを形成することになる。この小さなウイルス構造の典型は、例えばバクテリオファージφX174である。多くの二十面体ウイルスカプシドは60を超えるサブユニットを含む。二十面体ウイルスの多くのカプシドは、逆平行の8本鎖β−バレル型折りたたみモチーフを含む。このモチーフは、一方の側に4本のβ鎖(BIDGと称される)及び他方の側に4本のβ鎖(CHEFと称される)を備えたくさび形ブロックを有する。また2個の保存されたα−ヘリックスが存在し、1個はβCとβDの間、他の1個はβEとβFの間にある。
【0046】
エンベロープを持つウイルスは、膜を通して粒子が排出(出芽)することによる全面的な破壊を受けずに感染細胞を出て行くことができ、その間に粒子は細胞膜に由来する脂質エンベロープ内に被包される(例えばAJ Cann (ed. ) (2001)Principles of Molecular Virology (Academic Press); A Granoff and RG Webster (eds. ) (1999) Encyclopedia of Virology (Academic Press); DLD Caspar (1980) Bioplzys. J. 32: 103; DLD Caspar and A Klug (1962) Cold Spring Harbor Symp. Quant. Biol. 27: 1; J Grimes et al. (1988) Nature 395: 470; JE Johnson (1996) Proc. Nat'l Acad. Sci. USA 93: 27; and J Johnson and J Speir (1997) J. Mol. Biol. 269: 665参照)。
【0047】
(ウイルス)
ウイルス分類学は、以下の分類群のカプシド被包粒子の存在を認識する:
グループIウイルス、すなわちdsDNAウイルス;
・グループIIウイルス、すなわちssDNAウイルス;
・グループIIIウイルス、すなわちdsRNAウイルス;
・グループIVウイルス、すなわちDNA段階を持たないssRNA(+)鎖ウイルス;
・グループVウイルス、すなわちssRNA(−)鎖ウイルス;
・グループVIウイルス、すなわちssRNA逆転写ウイルスである、RNAレトロイドウイルス;
・グループVIIウイルス、すなわちdsDNA逆転写ウイルスである、DNAレトロイドウイルス;
・デルタウイルス;
・ウイロイド;及び
・サテライト核酸及びプリオンを除く、サテライトファージ及びサテライトウイルス。
【0048】
これらの分類群のメンバーは当業者に周知であり、H. V. Van Regenmortel et al. (eds.), Virus Taxonomy: Seventh Report of the International Committee on Taxonomy of Viruses (2000) (Academic Press/Elsevier, Burlington Mass. , USA); the University of Leicester (UK) Microbiology & Immunology Departmentのhttp : //wwwmicro. msb. le. ac. uk/3035/Virusgroups. htmlにおけるVirus Taxonomyウェブページ; およびthe National Center for Biotechnology Information(NCBI) of the National Library of Medicine of the National Institutes of Health of the US Department of Health & Human Services (Washington, D. C. , USA)のhttp : //www. ncbi. nlm. nih. gov/Taxonomy/tax. Html.におけるTaxonomy Browserのオンライン"Virus"および"Viroid"セクションにおいて総説されている。
【0049】
カプシドのアミノ酸配列は、これらの分類群のいずれかのメンバーのカプシドから選択され得る。これらの分類群のメンバーのカプシドについてのアミノ酸配列は、例えば、the NCBIによってhttp://www.ncbi. nlm. nih.gov/entrez/query. fcgi.において提供されているPubMed search facilityのオンライン"Nucleotide" (Genbank), "Protein,"および"Structure"セクションを含むがこれらに限定されないソースから入手し得る。
【0050】
1つの実施形態では、カプシドアミノ酸配列は、以下の宿主の少なくとも1つに特異的な分類群のメンバーから選択される:酵母を含む真菌、植物、藻類を含む原生生物、無脊椎動物、脊椎動物、及びヒト。1つの実施形態では、カプシドアミノ酸配列は、以下の分類群のいずれかのメンバーから選択される:グループI、グループII、グループIII、グループIV、グループV、グループVI、グループVII、ウイロイド、及びサテライトウイルス。1つの実施形態では、カプシドアミノ酸配列は、上述した6つの型の宿主の少なくとも1つに特異的なこれら7つの分類群のいずれかのメンバーから選択される。より特定の実施形態では、カプシドアミノ酸配列は、グループII、グループIII、グループIV、グループVII、及びサテライトウイルスのいずれかのメンバーから;又はグループII、グループIV、グループVII、及びサテライトウイルスのいずれかから選択される。もう1つの実施形態では、ウイルスカプシドはグループIV又はグループVIIから選択される。
【0051】
ウイルスカプシド配列は、当該細胞に対して親和性でないウイルスに由来し得る。1つの実施形態では、当該細胞は、所望の二十面体カプシド以外の特定の選択されたウイルスからのウイルスタンパク質を含まない。1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる科の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる属の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる種の生物に対して親和性を有するウイルスに由来する。
【0052】
特定の実施形態では、ウイルスカプシドはグループIVのウイルスから選択される。
【0053】
1つの実施形態では、ウイルスカプシドは二十面体ウイルスから選択される。二十面体ウイルスは、乳頭腫ウイルス科、トティウイルス科、ジシストロウイルス科、ヘパドナウイルス科、トガウイルス科、ポリオーマウイルス科、ノダウイルス科、テクティウイルス科、レヴィウイルス科、ミクロウイルス科、シフォビラーダ科、ノダウイルス科、ピコルナウイルス科、パルボウイルス科、カルシウイルス科、テトラウイルス科、及びサテライトウイルスのいずれかのメンバーから選択され得る。
【0054】
特定の実施形態では、この配列は、少なくとも1つの植物宿主に特異的な分類群のいずれかのメンバーから選択される。1つの実施形態では、二十面体植物ウイルス種は、ブンヤウイルス科、レオウイルス科、ラブドウイルス科、ルテオウイルス科、ナノウイルス科、パルティティウイルス科、セキウイルス科、ティモウイルス科、ウルミアウイルス属、タバコ壊死サテライトウイルス、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科分類群のいずれかである、又はそのいずれかのメンバーである、植物感染性ウイルス種である。1つの実施形態では、二十面体植物ウイルス種は、ルテオウイルス科、ナノウイルス科、パルティティウイルス科、セキウイルス科、ティモウイルス科、ウルミアウイルス属、タバコ壊死サテライトウイルス、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科分類群のいずれかである、又はそのいずれかのメンバーである、植物感染性ウイルス種である。特定の実施形態では、二十面体植物ウイルス種は、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科のいずれかである、又はそのいずれかのメンバーである、植物感染性ウイルス種である。より特定の実施形態では、二十面体植物ウイルス種は、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科のいずれかである、又はそのいずれかのメンバーである、植物感染性ウイルス種である。より特定の実施形態では、二十面体植物ウイルス種は、コモウイルス科又はブロモウイルス科のメンバーである植物感染性ウイルス種である。特定の実施形態では、ウイルスカプシドは、ササゲモザイクウイルス又はササゲ退緑斑紋ウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、ブロモウイルス科分類群の種に由来する。特定の実施形態では、カプシドは、イラルウイルス属又はアルファモウイルス属に由来する。より特定の実施形態では、カプシドは、タバコ条斑病ウイルス又はアルファルファモザイクウイルス(AMV)(AMV1又はAMV2を含む)に由来する。
【0055】
(VLP)
本発明の二十面体ウイルスカプシドは、ここで述べる宿主細胞において非感染性である。1つの実施形態では、ウイルスカプシドの発現の間又は発現後に宿主細胞においてウイルス様粒子(VLP)又はケージ構造を形成する。1つの実施形態では、VLP又はケージ構造はまた、対象とするペプチドを含み、特定の実施形態では、対象ペプチドをVLPの表面に発現する。この発現系は、典型的にはウイルス感染性を可能にする付加的なウイルスタンパク質を含まない。典型的実施形態では、発現系は、宿主細胞及び1又はそれ以上のウイルスカプシド及び作動可能に連結された対象ペプチドをコードするベクターを含む。このベクターは、典型的には付加的なウイルス集合タンパク質を含まない。本発明は、ウイルスカプシドが特定の宿主細胞においてより大きな度合で形成され、組換えペプチドのより効率的な回収を可能にするという発見に由来する。
【0056】
1つの実施形態では、VLP又はケージ構造は、3−約200カプシドを含む、カプシドの多量体構築物である。1つの実施形態では、VLP又はケージ構造は、少なくとも30、少なくとも50、少なくとも60、少なくとも90又は少なくとも120カプシドを含む。もう1つの実施形態では、各々のVLP又はケージ構造は、少なくとも150カプシド、少なくとも160カプシド、少なくとも170カプシド又は少なくとも180カプシドを含む。
【0057】
1つの実施形態では、VLPは二十面体構造として発現される。もう1つの実施形態では、VLPは、そのカプシド配列が由来する天然ウイルスと同じ形状で発現される。別の実施形態では、しかしながら、VLPは天然ウイルスと同じ形状を持たない。特定の実施形態では、例えば、構造は、多数のカプシドで形成されるが、天然型VLPを形成しない粒子中で生産される。例えば約6、9、12、15、18、21、24、27、30、33、36、39、42、45、48、51、54、57又は60カプシドのケージ構造が形成され得る。
【0058】
1つの実施形態では、カプシドの少なくとも1個は、少なくとも1個の対象ペプチドを含む。1つの実施形態では、ペプチドは、VLPの少なくとも1つの内部ループ又は少なくとも1つの外部表面ループ内で発現される。
【0059】
ウイルスカプシドの2つ以上のループを修飾することができる。1つの特定の実施形態では、組換えペプチドは、二十面体ウイルス様粒子の少なくとも2つの表面ループ上で発現される。もう1つの実施形態では、少なくとも2個の異なるペプチドを、ウイルスカプシド、ケージ又はウイルス様粒子の少なくとも2つの表面ループに挿入する。もう1つの実施形態では、少なくとも3個の組換えペプチドをウイルス様粒子の少なくとも3つの表面ループに挿入する。表面ループ内の組換えペプチドは、同じアミノ酸配列を有し得る。別の実施形態では、表面ループ内の組換えペプチドのアミノ酸配列は異なる。
【0060】
特定の実施形態では、宿主細胞を、VLPの集合を改善するように修飾することができる。宿主細胞を、例えば発現されたウイルスカプシドからのVLPの形成を促進するシャペロンタンパク質を含むように改変することができる。もう1つの実施形態では、宿主細胞を、カプシドの発現をより効率よく調節して、VLPの形成調節を促進するためのリプレッサータンパク質を含むように改変する。
【0061】
ウイルスカプシド又はタンパク質をエンコードする核酸配列はまた、VLPの形成を変化させるように付加的に修飾することができる(例えばBrumfield, et al. (2004) J. Gen. Virol. 85: 1049-1053参照)。例えば修飾の3つの一般的なクラスは、最も典型的にはVLPの発現及び集合を修飾するために行われる。これらの修飾は、集合されたタンパク質ケージ内の隣接サブユニットの内部、外部又はその間の界面を変化させるように設計される。これを実現するために、突然変異誘発プライマーを使用して、(i)N末端の塩基性残基(例えばK、R)を酸性グルタミン酸に置き換えることによってウイルス核酸結合領域の内表面電荷を変化させる(Douglas et al., 2002b);(ii)N末端から内部残基を欠失させる(CCMVにおいて、通常は残基4−37);(iii)11アミノ酸ペプチド細胞標的配列をエンコードするcDNA(Graf et al., 1987)を表面暴露ループに挿入する;及び(iv)金属結合部位を変化させることによってウイルスサブユニットの間の相互作用を改変する(CCMVにおいて、残基81/148突然変異体)ことができる。
【0062】
(組換えペプチド)
(大きさ)
1つの実施形態では、ウイルスカプシド配列に作動可能に連結されたペプチドは、少なくとも2個のアミノ酸を含む。もう1つの実施形態では、このペプチドは、少なくとも3、少なくとも4、少なくとも5又は少なくとも6アミノ酸の長さである。別の実施形態では、ペプチドは少なくとも7アミノ酸の長さである。ペプチドはまた、少なくとも8、少なくとも9、少なくとも10、少なくとも11、12、13、14、15、16、17、18、19、20、30、45、50、60、65、75、85、95、96、99又はそれ以上のアミノ酸の長さである。1つの実施形態では、コードされるペプチドは少なくとも25kDである。
【0063】
1つの実施形態では、ペプチドは、2−約300アミノ酸、又は約5−約250アミノ酸、又は約5−約200アミノ酸、又は約5−約150アミノ酸、又は約5−約100アミノ酸を含む。もう1つの実施形態では、ペプチドは、約10−約140アミノ酸、又は約10−約120アミノ酸、又は約10−約100アミノ酸を含む。
【0064】
1つの実施形態では、ウイルスカプシド配列に作動可能に連結されたペプチド又はタンパク質は約500アミノ酸を含む。1つの実施形態では、ペプチドは500個未満のアミノ酸を含む。もう1つの実施形態では、ペプチドは、約300個までのアミノ酸、又は約250個まで、又は約200個まで、又は約180個まで、又は約160個まで、又は約150個まで、又は約140個まで、又は約120個まで、又は約110個まで、又は約100個まで、又は約90個まで、又は約80個まで、又は約70個まで、又は約60個まで、又は約50個まで、又は約40個まで、又は約30個までのアミノ酸を含む。
【0065】
1つの実施形態では、二十面体カプシドに融合した組換えペプチドは、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも35、少なくとも40、少なくとも45、少なくとも50、少なくとも55、少なくとも60、少なくとも65、少なくとも75、少なくとも85、少なくとも95、少なくとも99、又は少なくとも100アミノ酸である。
【0066】
本発明の1つの実施形態では、組換えペプチドは、所望の標的ペプチドの少なくとも1個の単量体を含む。選択的実施形態では、組換えペプチドは、所望の標的ペプチドの2個以上の単量体を含む。特定の実施形態では、ペプチドは、コンカテマーペプチドとしてカプシドに作動可能に連結された、少なくとも2個、少なくとも5個、少なくとも10個、少なくとも15個又は少なくとも20個の別個の単量体からなる。もう1つの実施形態では、コンカテマーペプチド内の個々の単量体は、切断可能なリンカー領域によって連結される。さらにもう1つの実施形態では、組換えペプチドを、二十面体ウイルス様粒子の少なくとも1つの表面ループに挿入する。1つの実施形態では、少なくとも1個の単量体をウイルス様粒子の表面ループに挿入する。
【0067】
(分類)
ウイルスカプシドに融合する対象ペプチドは、ウイルスに由来しない、及び場合により、細胞と同じ宿主細胞に由来しない、異種タンパク質であり得る。
【0068】
ウイルスカプシドに融合する対象ペプチドは、機能性ペプチド:構造ペプチド;抗原ペプチド;毒性ペプチド、抗菌ペプチド、それらのフラグメント;それらの前駆体;これらのいずれかの組合せ;及び/又はこれらのいずれかのコンカテマーであり得る。本発明の1つの実施形態では、組換えペプチドは、ヒト及び動物の治療のために有用な治療ペプチドである。
【0069】
機能性ペプチドは、例えば:生物活性ペプチド(すなわち生物学的実在物(entity)、例えば生物、細胞、培養物、組織、器官又は細胞小器官、における又はその生物学的機能又は活性の開始、増強、延長、減衰、終結又は防止を及ぼす、惹起する又はさもなければ生じさせるペプチド);触媒ペプチド;ミクロ構造及びナノ構造活性ペプチド(すなわちその中で又はそれに関連して作用、例えば運動、エネルギー変換、を行う構築されたミクロ又はナノ構造の一部を形成するペプチド);及び刺激剤ペプチド(例えばペプチド着香料、着色料、着臭剤、フェロモン、誘引物質、抑制物質及び忌避物質)を含むが、これらに限定されない。
【0070】
生物活性ペプチドは、例えば:免疫活性ペプチド(例えば抗原ペプチド、アレルゲンペプチド、ペプチド免疫調節剤(immunoregulator)、ペプチド免疫調節剤(immunomodulator):シグナル伝達ペプチド(例えばペプチドホルモン、サイトカイン及び神経伝達物質;受容体;アゴニスト及びアンタゴニストペプチド;ペプチドターゲティング及び分泌シグナルペプチド);及び生物阻害性ペプチド(例えば毒性、生物致死性又は生物抑制性ペプチド、例えばペプチド毒素及び抗菌ペプチド)を含むが、これらに限定されない。
【0071】
構造ペプチドは、例えば:ペプチドアプタマー、折りたたみペプチド(例えばもう1つ別の分子の物理的立体配座の形成又は維持を促進する又は誘導するペプチド);接着促進ペプチド(例えば接着ペプチド、細胞接着促進ペプチド);界面ペプチド(例えばペプチド界面活性剤及び乳化剤);ミクロ構造及びナノ構造構築ペプチド(すなわち加工されたミクロ又はナノ構造の一部を形成する構造ペプチド);及びプレ活性化ペプチド(例えばプレ、プロ及びプレ−プロタンパク質のリーダーペプチド及びペプチド;インテイン)を含むが、これらに限定されない。
【0072】
触媒ペプチドは、例えばアポB RNAエディティングシチジンデアミナーゼペプチド;グルタミニル−tRNAシンテターゼの触媒ペプチド:アスパラギン酸トランスカルバモイラーゼの触媒ペプチド;植物1型リボソーム不活性化ペプチド;ウイルス触媒ペプチド、例えば口蹄疫ウイルス[FMDV−2A]触媒ペプチド;マトリックスメタロプロテイナーゼペプチド;及び触媒メタロオリゴペプチドを含む。
【0073】
ペプチドはまた、ペプチドエピトープ、ハプテン又は関連ペプチド(例えば抗原ウイルスペプチド;ウイルス関連ペプチド、例えばHIV関連ペプチド、肝炎関連ペプチド;抗体イディオタイプドメイン;細胞表面ペプチド;抗原性ヒト、動物、原生生物、植物、真菌、細菌及び/又は古細菌ペプチド;アレルゲンペプチド及びアレルゲン脱感作ペプチド)であり得る。
【0074】
ペプチドはまた、ペプチド免疫調節剤(immunoregulatorまたはimmunomodulator)(例えばインターフェロン、インターロイキン、ペプチド免疫抑制剤及び免疫増強剤);抗体ペプチド(例えば一本鎖抗体;一本鎖抗体フラグメント及び構築物、例えば一本鎖Fv分子;抗体軽鎖分子、抗体重鎖分子、ドメイン欠失抗体軽鎖又は重鎖分子;一本鎖抗体ドメイン及び分子、例えばCH1、CH1−3、CH3、CH1−4、CH4、VHCH1、CL、CDR1又はFR−CDR1−FR2ドメイン;パラトープペプチド;ミクロ抗体);別の結合ペプチド(例えばペプチドアプタマー、細胞内及び細胞表面受容体タンパク質、受容体フラグメント;抗腫瘍壊死因子ペプチド)であり得る。
【0075】
ペプチドはまた、酵素基質ペプチド又は酵素阻害剤ペプチド(例えばカスパーゼ基質及び阻害剤、プロテインキナーゼ基質及び阻害剤、蛍光共鳴エネルギー転移ペプチド酵素基質)であり得る。
【0076】
ペプチドはまた、細胞表面受容体ペプチドリガンド、アゴニスト及びアンタゴニスト(例えばセルレイン、ダイノルフィン、オレキシン、下垂体アデニル酸シクラーゼ活性化ペプチド、腫瘍壊死因子ペプチド;合成ペプチドリガンド、アゴニスト及びアンタゴニスト);ペプチドホルモン(例えば内分泌、パラ分泌及び自己分泌ホルモン、例えばアミリン、アンギオテンシン、ブラジキニン、カルシトニン、心臓興奮性神経ペプチド、カゾモルフィン、コレシストキニン、コルチコトロピン及びコルチコトロピン関連ペプチド、分化因子、エンドルフィン、エンドセリン、エンケファリン、エリスロポエチン、エキセンディン、卵胞刺激ホルモン、ガラニン、ガストリン、グルカゴン及びグルカゴン様ペプチド、ゴナドトロピン、成長ホルモン及び増殖因子、インスリン、カリジン、キニン、レプチン、向脂肪ホルモン、黄体形成ホルモン、メラニン細胞刺激ホルモン、メラトニン、ナトリウム排泄増加性ペプチド、ニューロキニン、ニューロメジン、ノシセプチン、オステオカルシン、オキシトシン(すなわちオシトシン)、副甲状腺ホルモン、プレイオトロフィン、プロラクチン、レラキシン、セクレチン、セロトニン、睡眠誘発ペプチド、ソマトメイジン、サイモポイエチン、甲状腺刺激ホルモン、チロトロピン、ウロテンシン、血管作用性小腸ペプチド、バソプレシン);ペプチドサイトカイン、ケモカイン、ウイロカイン(virokine)及びウイロセプター(viroceptor)ホルモン放出及び放出抑制ペプチド(例えばコルチコトロピン放出ホルモン、コルチスタチン、卵胞刺激ホルモン放出因子、胃抑制性ペプチド、ガストリン放出ペプチド、ゴナドトロピン放出ホルモン、成長ホルモン放出ホルモン、黄体形成ホルモン放出ホルモン、メラノトロピン放出ホルモン、メラノトロピン放出抑制因子;ノシスタチン、パンクリアスタチン、プロラクチン放出ペプチド、プロラクチン放出抑制因子;ソマトスタチン;甲状腺刺激ホルモン放出ホルモン);ペプチド神経伝達物質又はチャネルブロッカー(例えばボンベシン、神経ペプチドY、ニューロテンシン、サブスタンスP);ペプチド毒素、毒素前駆体ペプチド又は毒素ペプチド部分であり得る。特定の実施形態では、ペプチド毒素はD−アミノ酸を含まない。毒素前駆体ペプチドは、D−アミノ酸を含まないもの及び/又は翻訳後修飾によって、例えばアミノ酸にD−立体配置を導入することができるD立体配置誘導剤(例えばペプチドイソメラーゼ又はエピメラーゼ又はラセマーゼ又はトランスアミノアーゼ)の作用によって、及び/又は環状ペプチド構造を形成することができる環化剤(例えばペプチドチオエステラーゼ又はペプチドリガーゼ、例えばトランススプライシングタンパク質又はインテイン)の作用によって、天然の毒素構造に変換されていないものであり得る。
【0077】
毒素ペプチド部分は、ペプチド含有毒素の線状又は前環化オリゴ及びポリペプチド部分であり得る。ペプチド毒素の例は、例えばアガトキシン、アマトキシン、カリブドトキシン、クロロトキシン、コノトキシン、デンドロトキシン、インセクトトキシン、マルガトキシン、マスト細胞脱顆粒ペプチド、サポリン、サラフォトキシン;及び細菌体外毒素、例えば炭疽毒素、ボツリヌス毒素、ジフテリア毒素及び破傷風毒素を含む。
【0078】
ペプチドはまた、代謝及び消化関連ペプチド(例えばコレシストキニン−パンクレオザイミンペプチド、ペプチドyy、膵ペプチド、モチリン);細胞接着調節又は仲介ペプチド、細胞外マトリックスペプチド(例えば付着因子、セレクチン、ラミニン);神経保護又は髄鞘形成促進ペプチド;凝集阻害ペプチド(例えば細胞又は血小板凝集阻害剤ペプチド、アミロイド形成又は沈着阻害剤ペプチド);連結ペプチド(例えば心臓血管連結神経ペプチド、iga連結ペプチド);又はその他のペプチド(例えばアグーチ関連ペプチド、アミロイドペプチド、骨関連ペプチド、細胞透過性ペプチド、コナントキシン、コントリファン、コンチュラキン、ミエリン塩基性タンパク質等)であり得る。
【0079】
特定の実施形態では、対象ペプチドは、分泌されるウイルスカプシドに対して外因性である。ペプチドは、天然又は合成配列のいずれでもよい(及びそれらのコード配列は天然又は合成ヌクレオチド配列のいずれでもよい)。したがって、例えばアミノ酸の天然、修飾天然及び完全人工配列が包含される。これらのアミノ酸配列をエンコードする核酸分子の配列は、同様に、天然、修飾天然及び完全人工核酸配列であり得、例えばその核酸分子を得るために使用される(すなわちヒトの行為によって適用される)1又はそれ以上の論理的又はランダム突然変異及び/又は組換え及び/又は合成及び/又は選択工程の結果であり得る。
【0080】
コード配列は、使用可能な場合は、標的ペプチドについての天然のコード配列であり得るが、より典型的には、例えば宿主の種のコドン使用頻度を反映する遺伝子を合成することによって、選択した発現宿主細胞における使用のために選択された、改善された又は最適化されたコード配列である。本発明の1つの実施形態では、宿主種はP.フルオレッセンスであり、シグナル配列及びペプチド配列の両方を設計するときにP.フルオレッセンスのコドン使用頻度を考慮に入れる。
【0081】
(抗原ペプチド(ペプチドエピトープ))
1つの実施形態では、抗原ペプチドはウイルスカプシドによる発現を通して生産される。抗原ペプチドは、感染因子、寄生生物、癌細胞及び他の病原体を含む、ヒト又は動物病原体の抗原ペプチドであるものから選択することができる。かかる病原体はまた、毒性因子及び病原因子、例えばそれらの因子の体外毒素、内毒素等を含む。病原体は何らかのレベルの毒力を示し得る、すなわちそれらは、例えば毒性、無毒性、偽毒性、半毒性等であり得る。1つの実施形態では、抗原ペプチドは病原体からのエピトープアミノ酸配列を含む。1つの実施形態では、エピトープアミノ酸配列は、少なくとも1個のかかる因子の表面ペプチドの少なくとも一部であるものを含む。1つの実施形態では、カプシド−組換えペプチドウイルス様粒子は、ヒト又は動物適用におけるワクチンとして使用することができる。
【0082】
2個以上の抗原ペプチドを選択してもよく、その場合、生じるウイルス様粒子は多数の異なる抗原ペプチドを提示し得る。多抗原ペプチド方式の特定の実施形態では、様々な抗原ペプチドは全て同じ病原体からの複数のエピトープから選択される。多抗原ペプチド方式の特定の実施形態では、様々な抗原ペプチドは全て、密接に関連する病原体、例えば同じ種又は同じ属の異なる種の株、亜種、次亜種、病原型、血清型又は遺伝子型(genovar)から選択される。
【0083】
1つの実施形態では、病原体は以下の群の少なくとも1つに属する:病原性の:バチルス属種、例えば炭疽菌;バルトネラ属種、例えばB.クインタナ;ブルセラ属種;ブルクホルデリア属種、例えばB.シュードマレイ(類鼻疽菌);カンピロバクター種;クロストリジウム属種、例えば破傷風菌、ボツリヌス菌;コクシエラ属種、例えばC.バーネティー;エドワードシエラ属種、例えばE.ターダ;エンテロバクター属種、例えばE.クロアカ;エンテロコッカス属種、例えばE.フェカリス、E.フェシウム;エシェリキア属種、例えば大腸菌;フランシセラ属種、例えば野兎病菌(F. tularensis);ヘモフィルス属種、例えばインフルエンザ菌;クレブシエラ属種、例えば肺炎杆菌;レジオネラ属種;リステリア属種、例えばリステリア菌(L. monocytogenes);髄膜炎菌及び淋菌、例えばナイセリア属種;モラクセラ属種;ミコバクテリウム属種、例えばらい菌、ヒト型結核菌;肺炎球菌、例えば肺炎双球菌;シュードモナス属種、例えば緑膿菌;リケッチア属種、例えば発疹チフスリケッチア、斑点熱リケッチア、発疹熱リケッチア;サルモネラ属菌、例えば腸チフス菌;ブドウ球菌属種、例えば黄色ブドウ球菌;A型連鎖球菌属及び溶血連鎖球菌属を含む、連鎖球菌属種、例えば肺炎連鎖球菌、化膿連鎖球菌;ストレプトミセス属種;赤痢菌属種;ビブリオ属種、例えばコレラ菌;及びエルシニア属種、例えばペスト菌、腸炎エルシニア菌を含むが、これらに限定されない細菌及びマイコプラスマ因子。病原性の:アルテルナリア属種;アスペルギルス属種;ブラストミセス属種、例えばB.デルマチチジス;カンジダ属種、例えばカンジダアルビカンス;クラドスポリウム属種;コクシジオイデス属種、例えばC,イミチス;クリプトコッカス属種、例えばC.ネオフォルマンス;ヒストプラスマ属種、例えばH.カプスラーツム;及びスポロトリクス属種、例えばS.シェンキイ、を含むが、これらに限定されない真菌及び酵母因子。
【0084】
1つの実施形態では、病原体は、病原性の:アカントアメーバ属種、アメーバ属種、ネグレリア属種、エントアメーバ属種、例えば赤痢アメーバを含むアメーバ;クリプトスポリジウム属種、例えばC.パルブム;シクロスポラ属種;エンセファリトゾーン属種、例えばE.インテスティナリス;エンテロシトゾーン属種;ジアルジア属種、例えばランブル鞭毛虫;イソスポラ属種;ミクロスポリジウム(Microsporidium)種;プラスモディウム属種、例えば熱帯熱マラリア原虫、四日熱マラリア原虫、卵形マラリア原虫、三日熱マラリア原虫;トキソプラスマ属種、例えばトキソプラスマ原虫;及びトリパノソーマ属種、例えばトリパノソーマブルセイ、を含むが、これらに限定されない原生生物因子由来である。
【0085】
1つの実施形態では、病原体は、病原性の:回虫属種、例えば回虫;ドラクンクルス属種、例えばメジナ虫;オンコセルカ属種、例えば回旋糸状虫;住血吸虫属種;旋毛虫属種、例えば旋毛虫;及び鞭虫属種、例えばヒト鞭虫、を含むが、これらに限定されない寄生生物因子(例えば蠕虫寄生生物)由来である。
【0086】
もう1つの実施形態では、病原体は、病原性の:アデノウイルス;アレナウイルス、例えばラッサ熱ウイルス;アストロウイルス;ブンヤウイルス、例えばハンターンウイルス、リフトバレー熱ウイルス;コロナウイルス、デルタウイルス;サイトメガロウイルス、エプスタイン−バーウイルス、ヘルペスウイルス、水痘ウイルス;フィロウイルス、例えばエボラウイルス、マルブルクウイルス;フラビウイルス、例えばデング熱ウイルス、西ナイル熱ウイルス、黄熱病ウイルス;肝炎ウイルス;インフルエンザウイルス;レンチウイルス、T細胞向性ウイルス、他の白血病ウイルス;ノーウォークウイルス;パピローマウイルス、他の腫瘍ウイルス;パラミクソウイルス、例えば麻疹ウイルス、流行性耳下腺炎ウイルス、パラインフルエンザウイルス、肺炎ウイルス、センダイウイルス;パルボウイルス;ピコルナウイルス、例えばカーディオウイルス、コクサッキーウイルス、エコーウイルス、ポリオウイルス、ライノウイルス、他のエンテロウイルス;ポックスウイルス、例えば痘瘡ウイルス、ワクシニアウイルス、パラポックスウイルス;レオウイルス、例えばコルティウイルス、オルビウイルス、ロタウイルス;ラブドウイルス、例えばリッサウイルス、水泡性口内炎ウイルス;及びトガウイルス、例えば風疹ウイルス、シンドビスウイルス、西部脳炎ウイルス、を含むが、これらに限定されないウイルス因子由来である。
【0087】
1つの特定の実施形態では、抗原ペプチドは、イヌパルボウイルスペプチド、炭疽菌保護抗原(PA)抗原ペプチド及び東部ウマ脳脊髄炎ウイルス抗原ペプチドからなる群より選択される。特定の実施形態では、抗原ペプチドは、配列番号:7のアミノ酸配列を有するイヌパルボウイルス由来ペプチドである。もう1つの特定の実施形態では、抗原ペプチドは、配列番号:9、11、13又は15のアミノ酸配列のいずれか1つを有する炭疽菌保護抗原(PA)抗原ペプチドである。さらにもう1つの特定の実施形態では、抗原ペプチドは、配列番号:25又は27のいずれかのアミノ酸配列を有する東部ウマ脳脊髄炎ウイルス抗原ペプチドである。
【0088】
(宿主細胞毒性ペプチド)
もう1つの特定の実施形態では、組換えペプチドは、遊離単量体形態であるとき宿主細胞に対して毒性であるペプチドである。より特定の実施形態では、毒性ペプチドは抗菌ペプチドである。
【0089】
特定の実施形態では、ウイルスカプシドに関連して発現される対象ペプチドは、宿主細胞毒性ペプチドである。特定の実施形態では、このタンパク質は抗菌ペプチドである。宿主細胞毒性ペプチドは、それが発現される宿主細胞に対して、又は宿主細胞がそのメンバーである細胞培養中又は生物中の他の細胞に対して、又は宿主細胞を提供する生物又は種の細胞に対して生物抑制性、生物致死性又は毒性である、生物阻害性ペプチドを指示する。1つの実施形態では、宿主細胞毒性ペプチドは、それが発現される宿主細胞に対して生物抑制性、生物致死性又は毒性である生物阻害性ペプチドである。宿主細胞毒性ペプチドの一部の例は、ペプチド毒素、抗菌ペプチド及び他の抗生物質ペプチドを含むが、これらに限定されない。
【0090】
抗菌ペプチドは、例えば抗細菌性ペプチド、例えばマガイニン、βデフェンシン、一部のαデフェンシン;カテリシジン;ヒスタチン;抗真菌ペプチド;抗原生動物ペプチド;合成AMP;ペプチド抗生物質又はその線状又は前環化オリゴ又はポリペプチド部分;他の抗生物質ペプチド(例えば駆虫性ペプチド、溶血ペプチド、殺腫瘍性ペプチド);及び抗ウイルスペプチド(例えば一部のαデフェンシン;殺ウイルス性ペプチド;ウイルス感染を阻止するペプチド)を含む。1つの特定の実施形態では、抗菌ペプチドは、配列番号:20のアミノ酸配列を有するD2A21ペプチドである。もう1つの実施形態では、抗菌ペプチドは、配列番号:24に実質的に対応するアミノ酸配列を有する抗菌ペプチドPBF20である。
【0091】
(VLPを発現するときに使用するための細胞)
本発明のウイルスカプシド又はウイルスカプシド融合ペプチドの発現のための宿主として使用する細胞(「宿主細胞」とも称する)は、ウイルスカプシドが細胞の複製又は感染を許容しないものである。1つの実施形態では、ウイルスカプシドは、宿主細胞が由来する細胞の種に感染しないウイルスに由来する。例えば、1つの実施形態では、ウイルスカプシドは二十面体植物ウイルスに由来し、細菌種の宿主細胞において発現される。もう1つの実施形態では、ウイルス種は哺乳動物に感染し、発現系は細菌宿主細胞を含む。
【0092】
1つの実施形態では、宿主細胞は原核生物、例えばシュードモナス種を含むがこれに限定されない細菌細胞であり得る。典型的な細菌細胞は、例えばDr MJ Farabee of the Estrella Mountain Community College, Arizonaにより、URL:http://www. emc.maricopa.edu/faculty/farabee/BIOBK/BioBookDiversity#2. htmlにおいて提供される、「Biological Diversity: Bacteria and Archaean」, a chapter of the On-Line Biology Bookに述べられている。特定の実施形態では、宿主細胞はシュードモナス細胞であり得、典型的にはP.フルオレッセンス細胞であり得る。
【0093】
1つの実施形態では、宿主細胞は、真性細菌のいずれかの種のメンバーであり得る。宿主は、以下の分類群:アシドバクテリア門、アクチノバクテリア門、アクイフェックス門、バクテロイデス門、クロロビウム門、クラミジア門、クロロフレキシ門、クリシオジェネス門、藍細菌門、デフェリバクター門、ディノコッカス門、ディクティオグロムス門、フィブロバクター門、グラム陽性細菌門、フソバクテリア門、ゲマティモナス門、レンティスファエラ門、ニトロスピラ門、プラクトミセス門、プロテオバクテリア門、スピロヘータ門、サーモデスルフォバクテリア門、サーモミクロビア門、サーモトガ門、テルムス門(Thermales)、又はベルコミクロビア門、のいずれか1つのメンバーであり得る。真性細菌宿主細胞の実施形態では、細胞は、藍細菌門を除く真性細菌のいずれかの種のメンバーであり得る。
【0094】
細菌宿主はまた、プロテオバクテリア門のいずれかの種のメンバーであり得る。プロテオバクテリア宿主細胞は、以下の分類群:アルファプロテオバクテリア綱、ベータプロテオバクテリア綱、ガンマプロテオバクテリア綱、デルタプロテオバクテリア綱、又はイプシロンプロテオバクテリア綱のいずれかの1つのメンバーであり得る。加えて、宿主は、以下の分類群:アルファプロテオバクテリア綱、ベータプロテオバクテリア綱、又はガンマプロテオバクテリア綱のいずれかの1つのメンバーであり得、及びガンマプロテオバクテリア綱のいずれかの種のメンバーであり得る。
【0095】
ガンマプロテオバクテリア綱宿主細胞の1つの実施形態では、宿主は、以下の分類群:エアロモナス目、アルテロモナス目、エンテロバクター目、シュードモナス目、又はキサントモナス目のいずれか1つのメンバー;又はエンテロバクター目又はシュードモナス目のいずれかの種のメンバーである。1つの実施形態では、宿主細胞はエンテロバクター目であり得、宿主細胞は、腸内細菌科のメンバーであるか、又はエルウィニア属、エシェリキア属、又はセラチア属のいずれか1つのメンバー;又はエシェリキア属のメンバーである。シュードモナス目の宿主細胞の1つの実施形態では、宿主細胞は、シュードモナス科、さらにはシュードモナス属のメンバーである。ガンマプロテオバクテリア綱宿主は、大腸菌種のメンバー及びシュードモナス・フルオレッセンス種のメンバーを含む。
【0096】
他のシュードモナス属生物も使用し得る。シュードモナス及び密接に関連する種は、 R. E. Buchanan and N. E. Gibbons (eds.), Bergey's Manual of Determinative Bacteriology, pp. 217-289 (8th ed. , 1974) (The Williams & Wilkins Co. , Baltimore, MD, USA) (以下「Bergey(1974)」)により「グラム陰性好気性桿菌及び球菌」として述べられている科及び属に属するグラム(−)プロテオバクテリアの群を包含する、プロテオバクテリア亜群1を含む。表1はこれらの科及び属の生物を示す。
表1.「グラム陰性好気性桿菌及び球菌」の部分に列挙されている科及び属(Bergey(1974)」より)
【表1】

【0097】
「グラム(−)プロテオバクテリア亜群1」はまた、分類において使用される判定基準に従ってこの表題に分類されるプロテオバクテリアを包含する。この表題はまた、以前はこの部門に分類されたが現在はもはや分類されない群、例えばアシドボラクス属、ブレブンディモナス属、ブルクホルデリア属、ヒドロゲノファガ属、オセアニモナス属、ラルストニア属及びステノトロフォモナス属、キサントモナス属に属する(以前はキサントモナス属の種と呼ばれた)生物を再分類することによって作成されたスフィンゴモナス属(及びそれに由来する、ブラストモナス属)、Bergey(1974)において定義されるようにアセトバクター属に属する生物を再分類することによって作成されたアシドモナス属を包含する。加えて、宿主は、シュードモナス属由来の細胞、それぞれアルテロモナス・ハロプラクティス、アルテロモナス・ニグリファシエンス、及びアルテロモナス・プトレファシエンスとして再分類された、シュードモナス・エナリア(ATCC14393)、シュードモナス・ニグリファシエンス(ATCC19375)、及びシュードモナス・プトレファシエンス(ATCC8071)を含み得る。同様に、例えばシュードモナス・アシドボランス(ATCC15668)及びシュードモナス・テストステローニ(ATCC11996)は、その後それぞれコマモナス・アシドボランス及びコマモナス・テストステローニに再分類された;及びシュードモナス・ニグリファシエンス(ATCC19375)及びシュードモナス・ピスキキダ(ATCC15057)は、それぞれシュードアルテロモナス・ニグリファシエンス及びシュードモナス・ピスキキダに再分類された。「グラム(−)プロテオバクテリア亜群1」はまた、以下の科:シュードモナス科、アゾトバクター科(現在はしばしば同義語、シュードモナス科の「アゾトバクター群」と呼ばれる)、リゾビウム科、及びメチロモナス科(現在はしばしば同義語、「メチロコッカス科」と呼ばれる)、のいずれかに属すると分類されるプロテオバクテリアを包含する。その結果として、ここで述べた属に加えて、「グラム(−)プロテオバクテリア亜群1」に属するプロテオバクテリア属は:1)アゾリゾフィルス属のアゾトバクター群細菌;2)セルビブリロ属、オリゲラ属及びテレディニバクター属のシュードモナス科細菌;3)セラトバクター属、エンサイファ属、リベリバクター属(「カンジダ・リベリバクター」とも呼ばれる)、及びシノリゾビウム属のリゾビウム科細菌;及び4)メチロバクター属、メチロカルダム属、メチロミクロビウム属、メチロサルシナ属及びメチロスフェラ属のメチロコッカス科細菌が挙げられる。
【0098】
もう1つの実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群2」から選択される。「グラム(−)プロテオバクテリア亜群2」は、以下の属(特に異なる記載があるものを除き、全てATCCに寄託されている、カタログに列挙され、公的に入手可能なその寄託菌株の総数を括弧内に示す):アシドモナス(2);アセトバクター(93);グルコノバクター(37);ブレブンディモナス(23);ベイエリンキア(13);デルキシア(2);ブルセラ(4);アグロバクテリウム(79);ケラトバクター(Chelatobacter)(2);エンサイファ(3);リゾビウム(144);シノリゾビウム(24);ブラストモナス(1);スフィンゴモナス(27);アルカリゲネス(88);ボルデテラ(43);ブルクホルデリア(73);ラルストニア(33);アソドボラクス(20);ヒドロゲノファガ(9);ゾーグレア(9);メチロバクター(2);メチロカルダム(NCIMBに1);メチロコッカス(2);メチロミクロビウム(2);メチロモナス(9);メチロサルシナ(1);メチロスフェラ;アゾモナス(9);アゾリゾフィルス(5);アゾトバクター(64);セルビブリオ(3);オリゲラ(5);シュードモナス(1139);フランシセラ(4);キサントモナス(229);ステノトロフォモナス(50);及びオセアニモナス(4)、のプロテオバクテリアの群と定義される。
【0099】
「グラム(−)プロテオバクテリア亜群2」の例示的な宿主細胞種は、以下の細菌(その例示的な菌株のATCC又は他の寄託番号を括弧内に示す)を含むが、これらに限定されない:アシドモナス・メタノリカ(ATCC43581);アセトバクター・アセティ(ATCC15973);グルコノバクター・オキシダンス(ATCC19357);ブレブンディモナス・ディミヌタ(ATCC11568);ベイエリンキア・インディカ(ATCC9039及びATCC19361);デルキシア・グンモサ(ATCC15994);ブルセラ・メリテンシス(ATCC23456)、ブルセラ・アボルツス(ATCC23448);アグロバクテリウム・トゥメファシエンス(ATCC23308)、アグロバクテリウム・ラディオバクター(ATCC19358)、アグロバクテリウム・リゾゲネス(ATCC11325);ケラトバクター・ヘインツィー(ATCC29600);エンサイファ・アドヘレンス(ATCC33212);リゾビウム・レグミノサルム(ATCC10004);シノリゾビウム・フレディ(ATCC35423);ブラストモナス・ナタトリア(ATCC35951);スフィンゴモナス・パウチモビリス(ATCC29837);アルカリゲネス・フェカリス(ATCC8750);百日咳菌(ATCC9797);ブルクホルデリア・セパシア(ATCC25416);ラルストニア・ピックティー(ATCC27511);アシドボラクス・ファシリス(ATCC11228);ヒドロゲノファガ・フラバ(ATCC33667);ゾーグレア・ラミゲラ(ATCC19544);メチロバクター・ルテウス(ATCC49878);メチロカルダム・グラシル(NCIMB11912);メチロコッカス・カプシュラツス(ATCC19069);メチロミクロビウム・アジル(ATCC35068);メチロモナス・メタニカ(ATCC35067);メチロサルシナ・フィブラタ(ATCC700909);メチロスフェラ・ハンソニイ(ACAM549);アゾモナス・アジリス(ATCC7494);アゾリゾフィルス・パスパリ(ATCC23833);アゾトバクター・クロオコカム(ATCC9043);セルビブリオ・ミクツス(UQM2601);オリゲラ・ウレトラリス(ATCC17960);緑膿菌(ATCC10145);シュードモナス・フルオレッセンス(ATCC35858);野兎病菌(ATCC6223);ステノトロフォモナス・マルトフィリア(ATCC13637);キサントモナス・カンペストリス(ATCC33913);及びオセアニモナス・ドウドロフィ(ATCC27123)。
【0100】
もう1つの実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群3」から選択される。「グラム(−)プロテオバクテリア亜群3」は、以下の属:ブレブンディモナス;アグロバクテリウム;リゾビウム;シノリゾビウム;ブラストモナス;スフィンゴモナス;アルカリゲネス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;メチロバクター;メチロカルダム;メチロコッカス;メチロミクロビウム;メチロモナス;メチロサルシナ;メチロスフェラ;アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;フランシセラ;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0101】
もう1つの実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群4」から選択される。「グラム(−)プロテオバクテリア亜群4」は、以下の属:ブレブンディモナス;ブラストモナス;スフィンゴモナス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;メチロバクター;メチロカルダム;メチロコッカス;メチロミクロビウム;メチロモナス;メチロサルシナ;メチロスフェラ;アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;フランシセラ;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0102】
1つの実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群5」から選択される。「グラム(−)プロテオバクテリア亜群5」は、以下の属:メチロバクター;メチロカルダム;メチロコッカス;メチロミクロビウム;メチロモナス;メチロサルシナ;メチロスフェラ;アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;フランシセラ;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0103】
宿主細胞は、「グラム(−)プロテオバクテリア亜群6」から選択することができる。「グラム(−)プロテオバクテリア亜群6」は、以下の属:ブレブンディモナス;ブラストモナス;スフィンゴモナス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0104】
宿主細胞は、「グラム(−)プロテオバクテリア亜群7」から選択することができる。「グラム(−)プロテオバクテリア亜群7」は、以下の属:アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0105】
宿主細胞は、「グラム(−)プロテオバクテリア亜群8」から選択することができる。「グラム(−)プロテオバクテリア亜群8」は、以下の属:ブレブンディモナス;ブラストモナス;スフィンゴモナス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;シュードモナス;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0106】
宿主細胞は、「グラム(−)プロテオバクテリア亜群9」から選択することができる。「グラム(−)プロテオバクテリア亜群9」は、以下の属:ブレブンディモナス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;シュードモナス;ステノトロフォモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0107】
宿主細胞は、「グラム(−)プロテオバクテリア亜群10」から選択することができる。「グラム(−)プロテオバクテリア亜群10」は、以下の属:ブルクホルデリア;ラルストニア;シュードモナス;ステノトロフォモナス;及びキサントモナス、のプロテオバクテリアの群と定義される。
【0108】
宿主細胞は、「グラム(−)プロテオバクテリア亜群11」から選択することができる。「グラム(−)プロテオバクテリア亜群11」は、以下の属:シュードモナス;ステノトロフォモナス;及びキサントモナス、のプロテオバクテリアの群と定義される。宿主細胞は、「グラム(−)プロテオバクテリア亜群12」から選択することができる。「グラム(−)プロテオバクテリア亜群12」は、以下の属:ブルクホルデリア;ラルストニア;シュードモナス、のプロテオバクテリアの群と定義される。宿主細胞は、「グラム(−)プロテオバクテリア亜群13」から選択することができる。「グラム(−)プロテオバクテリア亜群13」は、以下の属:ブルクホルデリア;ラルストニア;シュードモナス;及びキサントモナス、のプロテオバクテリアの群と定義される。宿主細胞は、「グラム(−)プロテオバクテリア亜群14」から選択することができる。「グラム(−)プロテオバクテリア亜群14」は、以下の属:シュードモナス及びキサントモナス、のプロテオバクテリアの群と定義される。宿主細胞は、「グラム(−)プロテオバクテリア亜群15」から選択することができる。「グラム(−)プロテオバクテリア亜群15」は、シュードモナス属のプロテオバクテリアの群と定義される。
【0109】
宿主細胞は、「グラム(−)プロテオバクテリア亜群16」から選択することができる。「グラム(−)プロテオバクテリア亜群16」は、以下のシュードモナス種(例示的な菌株のATCC又は他の寄託番号を括弧内に示す)のプロテオバクテリアの群と定義される:シュードモナス・アビエタニフィラ(ATCC700689);緑膿菌(ATCC10145);シュードモナス・アルカリゲネス(ATCC14909);シュードモナス・アンギリセプティカ(ATCC33660);シュードモナス・シトロネロリス(ATCC13674);シュードモナス・フラベセンス(ATCC51555);シュードモナス・メンドシナ(ATCC25411);シュードモナス・ニトロレドゥセンス(ATCC33634);シュードモナス・オレオボランス(ATCC8062);シュードモナス・シュードアルカリゲネス(ATCC17440);シュードモナス・レシノボランス(ATCC14235);シュードモナス・ストラミネア(ATCC33636);シュードモナス・アガリシ(ATCC25941);シュードモナス・アルカリフィラ;シュードモナス・アルギノボラ;シュードモナス・アンデルソニ;シュードモナス・アスプレニ(ATCC23835);シュードモナス・アゼライカ(ATCC27162);シュードモナス・ベイエリンキ(ATCC19372);シュードモナス・ボレアリス;シュードモナス・ボレオポリス(ATCC33662);シュードモナス・ブラシカセルム;シュードモナス・ブタノボラ(ATCC43655);シュードモナス・セルロサ(ATCC55703);シュードモナス・オーランティアカ(ATCC33663);シュードモナス・クロロラフィス(ATCC9446、ATCC13985、ATCC17418、ATCC17461);シュードモナス・フラギ(ATCC4973);シュードモナス・ルンデンシス(ATCC49968);シュードモナス・テトロレンス(ATCC4683);シュードモナス・シシコラ(ATCC33616);シュードモナス・コロナファシエンス;シュードモナス・ディテルペニフィラ;シュードモナス・エロンガタ(ATCC10144);シュードモナス・フレクテンス(ATCC12775);シュードモナス・アゾトフォルマンス;シュードモナス・ブレネリ;シュードモナス・セドレラ;シュードモナス・コルガタ(ATCC29736);シュードモナス・エクストレモリエンタリス;シュードモナス・フルオレッセンス(ATCC35858);シュードモナス・ゲッサルディ;シュードモナス・リバネンシス;シュードモナス・マンデリ(ATCC700871);シュードモナス・マージナリス(ATCC10844);シュードモナス・ミグラ;シュードモナス・ムシドレンス(ATCC4685);シュードモナス・オリエンタリス;シュードモナス・ローデシア;シュードモナス・シンキサンタ(ATCC9890);シュードモナス・トラアシ(ATCC33618);シュードモナス・ベロニ(ATCC700474);シュードモナス・フレデリクスベルゲンシス;シュードモナス・ジェニクラタ(ATCC19374);シュードモナス・ギンゲリ;シュードモナス・グラミニス;シュードモナス・グリモンティ;シュードモナス・ハロデニトリフィカンス;シュードモナス・ハロフィラ;シュードモナス・ヒビシコラ(ATCC19867);シュードモナス・ハティエンシス(ATCC14670);シュードモナス・ヒドロゲノボラ;シュードモナス・ジェセニ(ATCC700870);シュードモナス・キロネンシス;シュードモナス・ランセオラタ(ATCC14669);シュードモナス・リニ;シュードモナス・マルギナタ(ATCC25417);シュードモナス・メフィティカ(ATCC33665);シュードモナス・デニトリフィカンス(ATCC19244);シュードモナス・ペルタシノゲナ(ATCC190);シュードモナス・ピクトラム(ATCC23328);シュードモナス・サイクロフィラ;シュードモナス・フルヴァ(ATCC31418);シュードモナス・モンテイリ(ATCC700476);シュードモナス・モセリ:シュードモナス・オリジハビタンス(ATCC43272);シュードモナス・プレコグロシシダ(ATCC700383);シュードモナス・プティダ(ATCC12633);シュードモナス・レアクタンス;シュードモナス・スピノサ(ATCC14606);シュードモナス・バレアリカ;シュードモナス・ルテオラ(ATCC43273);シュードモナス・スタツェリ(ATCC17588);シュードモナス・アミグダリ(ATCC33614);シュードモナス・アヴェラナ(ATCC700331);シュードモナス・カルシパパヤ(ATCC33615);シュードモナス・シコリ(ATCC10857);シュードモナス・フィクセレクタ(ATCC35104);シュードモナス・フスコヴァギナ;シュードモナス・メリア(ATCC33050);シュードモナス・シリンガ(ATCC19310);シュードモナス・ヴィリディフラバ(ATCC13223);シュードモナス・サーモカルボキシドヴォランス(ATCC35961);シュードモナス・サーモトレランス;シュードモナス・チヴェルヴァレンシス;シュードモナス・ヴァンクヴェレンシス(ATCC700688);シュードモナス・ウィスコンシネンシス;及びシュードモナス・キシアメネンシス。
【0110】
宿主細胞は、「グラム(−)プロテオバクテリア亜群17」から選択することができる。「グラム(−)プロテオバクテリア亜群17」は、例えば以下のシュードモナス種:シュードモナス・アゾトフォルマンス;シュードモナス・ブレネリ;シュードモナス・セドレラ;シュードモナス・コルガタ;シュードモナス・エクストレモリエンタリス;シュードモナス・フルオレッセンス;シュードモナス・ゲッサルディ;シュードモナス・リバネンシス;シュードモナス・マンデリ;シュードモナス・マージナリス;シュードモナス・ミグラ;シュードモナス・ムシドレンス;シュードモナス・オリエンタリス;シュードモナス・ローデシア;シュードモナス・シンキサンタ;シュードモナス・トラアシ;及びシュードモナス・ヴェロニ、に属するものを含む、「蛍光シュードモナス」として当技術分野において公知のプロテオバクテリアの群と定義される。
【0111】
この実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群18」から選択することができる。「グラム(−)プロテオバクテリア亜群18」は、例えば以下(例示的な菌株のATCC又は他の寄託番号を括弧内に示す)に属するものを含む、シュードモナス・フルオレッセンス種の全ての亜種、変種、菌株、及び他の亜種単位の群と定義される:次亜種1又は次亜種Iとも呼ばれる、シュードモナス・フルオレッセンス生物型A(ATCC13525);次亜種2又は次亜種IIとも呼ばれる、シュードモナス・フルオレッセンス生物型B(ATCC17816);次亜種3又は次亜種IIIとも呼ばれる、シュードモナス・フルオレッセンス生物型C(ATCC17400);次亜種4又は次亜種IVとも呼ばれる、シュードモナス・フルオレッセンス生物型F(ATCC12983);次亜種5又は次亜種Vとも呼ばれる、シュードモナス・フルオレッセンス生物型G(ATCC17518);シュードモナス・フルオレッセンス次亜種VI;シュードモナス・フルオレッセンスPf0−1;シュードモナス・フルオレッセンスPf−5(ATCC BAA−477);シュードモナス・フルオレッセンスSBW25;及びシュードモナス・フルオレッセンス亜種セルロサ(NCIMB10462)。
【0112】
宿主細胞は、「グラム(−)プロテオバクテリア亜群19」から選択することができる。「グラム(−)プロテオバクテリア亜群19」は、シュードモナス・フルオレッセンス生物型Aの全ての菌株の群と定義される。この生物型の1つの特定の菌株は、P.フルオレッセンスMB101株(Wilcoxへの米国特許第5,169,760号参照)及びその誘導体である。その誘導体の一例は、MB101染色体asd(アスパラギン酸デヒドロゲナーゼ遺伝子)遺伝子座に天然大腸菌PlacI−lacI−lacZYA構築物を挿入することによって構築される(すなわちPlacZが欠失した)P.フルオレッセンスMB214株である。
【0113】
本発明において使用できる付加的なP.フルオレッセンス株は、以下のATCC名称を有する、シュードモナス・フルオレッセンス・ミグラ及びシュードモナス・フルオレッセンス・ロイトキトクを含む:[NCIB8286];NRRL B−1244;NCIB 8865 CO1株;NCIB 8866 CO2株;1291[ATCC 17458;IFO 15837;NCIB 8917;LA;NRRL B−1864;ピロリジン;PW2[ICMP 3966;NCPPB 967;NRRL B−899];13475; NCTC 10038;NRRL B−1603[6;IFO 15840];52−1C;CCEB 488−A[BU140];CCEB 553[IEM15/47];IAM 1008 [AHH−27];IAM 1055[AHH−23];1[IFO 15842]; 12[ATCC25323;NIH 11;den Dooren de Jong 216];18[IFO15833;WRRL P−7];93[TR−10];108[52−22;IFO 15832];143[IFO 15836;PL];149[2−40−40;IFO 15838];182[IFO 3081;PJ 73];184[IFO 15830];185[W2 L−1];186[IFO 15829;PJ 79];187[NCPPB 263];188[NCPPB 316];189[PJ227;1208];191[IFO 15834;PJ 236;22/1];194[Klinge R−60;PJ 253];196[PJ 288];197[PJ 290];198 [PJ 302];201[PJ368];202[PJ 372];203[PJ 376];204[IFO 15835;PJ682];205[PJ 686];206[PJ 692];207[PJ 693];208[PJ722];212[PJ 832];215 [PJ 849];216[PJ 885];267[B−9];271[B−1612];401[C71A;IFO 15831;PJ187];NRRLB−3178[4;IFO15841];KY 8521;3081;30−21;[IFO 3081];N;PYR;PW;D946−B83[BU 2183;FERM−P 3328];P−2563 [FERM−P 2894;IFO 13658];IAM−1126[43F];M−1; A506[A5−06];A505[A5−05−1];A526[A5−26];B69;72;NRRL B−4290;PMW6[NCIB 11615];SC 12936;Al[1F0 15839];F 1847[CDC−EB];F1848[CDC 93]; NCIB 10586;P17;F−12;AmMS 257;PRA25;6133D02;6519E01;N1;SC15208;BNL−WVC;NCTC 2583[NCIB8194];H13;1013[ATCC 11251;CCEB 295];IFO 3903;1062;又はPf−5。
【0114】
(II.核酸構築物)
本発明はさらに、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を提供する。1つの実施形態では、a)組換えペプチドをエンコードする核酸及びb)二十面体カプシドをエンコードする核酸配列を含み、a)の核酸とb)の核酸は、細胞において発現されたとき融合タンパク質を形成するように作動可能に連結されている、シュードモナス宿主細胞を形質転換するのに使用するための核酸構築物を提供する。
【0115】
特定の実施形態では、ベクターは、多数のカプシド又は多数の対象ペプチドについての配列を含み得る。1つの実施形態では、ベクターは、少なくとも2個の異なるカプシド−ペプチドコード配列を含み得る。1つの実施形態では、コード配列は同じプロモーターに連結される。特定の実施形態では、コード配列は、内部リボソーム結合部位によって分離される。他の実施形態では、コード配列は、細胞内でのウイルス様粒子の形成を可能にするリンカー配列によって連結される。もう1つの実施形態では、コード配列は異なるプロモーターに連結される。これらのプロモーターは同じ誘導条件によって駆動され得る。もう1つの実施形態では、異なるカプシド−ペプチドの組合せをエンコードする多数のベクターを提供する。多数のベクターは、同じ誘導条件によって駆動されるか又は異なる誘導条件によって駆動されるプロモーターを含み得る。1つの実施形態では、プロモーターは、lacプロモーター又はlacプロモーターの誘導体、例えばtacプロモーターである。
【0116】
対象ペプチドについてのコード配列は、あらかじめ定められた部位でウイルスカプシド又はカプシドのコード配列に挿入することができる。このペプチドはまた、あらかじめ定められていない部位で挿入することもでき、細胞をVLPの生産に関してスクリーニングすることができる。1つの実施形態では、VLPの形成の間にループとして発現されるように、ペプチドをカプシドコード配列に挿入する。1つの実施形態では、1個のペプチドコード配列がベクター内に含まれるが、他の実施形態では、多数の配列が含まれる。多数の配列は、コンカテマーの形態、例えば切断可能なリンカー配列によって連結されたコンカテマーの形態であり得る。
【0117】
ペプチドは、カプシド内の2つまたはそれ以上の挿入部位に挿入し得る。したがってペプチドは、カプシドの2個またはそれ以上の表面ループモチーフに挿入し得る;ペプチドはまた、所与のループモチーフ内の多数の部位に挿入し得る。挿入物の個々の機能性及び/又は構造ペプチド、及び/又はペプチド挿入物全体は、切断可能部位、すなわちタンパク質を切断する又は加水分解する因子がカプシド構造又は集合物の残りの部分からペプチドを分離することができる部位によって分けられ得る。
【0118】
ペプチドは、外側に面したループ及び/又は内側に面したループ、すなわちそれぞれカプシドの中心から離れた方向又は中心に向かう方向に面したカプシドのループ内に挿入し得る。カプシドの表面ループ内のいかなるアミノ酸又はペプチド結合もペプチドの挿入部位として役立ち得る。典型的には、挿入部位は、ループのほぼ中央、すなわち折りたたまれたカプシドペプチドの三次構造の中心からほぼ最も遠位の位置で選択される。ペプチドコード配列は、選択されたループのこのほぼ中央に対応するカプシドコード配列の位置に作動可能に挿入され得る。これは、ペプチド挿入部位から下流に合成されるカプシドのペプチド配列のその部分についての読み枠の維持を含む。
【0119】
もう1つの実施形態では、ペプチドは、カプシドのアミノ末端に挿入することができる。ペプチドは、上述した切断可能リンカーを含む、1又はそれ以上のリンカー配列を通してカプシドに連結され得る。さらにもう1つの実施形態では、ペプチドは、カプシドのカルボキシ末端に挿入することができる。ペプチドは、化学的又は酵素的加水分解によって切断することができる1又はそれ以上のリンカー配列を通してカルボキシ末端に連結され得る。1つの実施形態では、ペプチド配列は、アミノ末端とカルボキシ末端の両方で、又は1つの末端と少なくとも1つの内部位置で、例えばその三次元立体配座においてカプシドの表面に発現される位置で、連結される。
【0120】
1つの実施形態では、ペプチドは、ササゲクロロティックモザイクウイルス由来のカプシドに挿入することができる。1つの特定の実施形態では、ペプチドは、CCMVウイルスのアミノ酸129で挿入できる。もう1つの実施形態では、ペプチド配列は、CCMVウイルスのアミノ酸60、61、62又は63で挿入できる。さらにもう1つの実施形態では、ペプチドは、CCMVウイルスのアミノ酸129とアミノ酸60−63の両方で挿入できる。
【0121】
特定の実施形態では、本発明は、a)抗菌ペプチドをエンコードする核酸及びb)二十面体カプシドをエンコードする核酸を含み、a)の核酸とb)の核酸は、細胞において発現されたとき融合タンパク質を形成するように作動可能に連結されている、核酸構築物を提供する。核酸構築物を構築する上で有用な他のカプシド及び組換えペプチドは上記に開示されている。
【0122】
(プロモーター)
1つの実施形態では、核酸構築物は、カプシド−組換えペプチド融合ペプチドをエンコードする核酸配列に作動可能に結合されたプロモーター配列を含む。作動可能な結合又は連結は、転写及び何らかの翻訳調節エレメントが、宿主細胞の作用により、調節エレメントが対象配列の発現を指令することができるようにこの配列に共有結合している立体配置を指す。
【0123】
発酵工程では、ひとたび標的組換えペプチドの発現が誘導されれば、発現系の効率を最大化するために高レベルの生産を得ることが理想的である。プロモーターは転写を開始させ、一般にリボソーム結合部位の10−100ヌクレオチド上流に位置する。理想的には、プロモーターは、宿主細胞の総細胞タンパク質の約50%の組換えペプチド蓄積を可能にするのに十分なほど強力であり、厳密な調節を受け、容易に(及び安価に)誘導される。
【0124】
本発明に従って使用されるプロモーターは、構成的プロモーター又は調節されるプロモーターであり得る。一般に使用される誘導的プロモーター及びそれらのその後の誘導物質の例は、lac(IPTG)、lacUV5(IPTG)、tac(IPTG)、trc(IPTG)、Psyn(IPTG)、trp(トリプトファン飢餓状態)、araBAD(1−アラビノース)、lppa(IPTG)、lpp−lac(IPTG)、phoA(リン酸飢餓状態)、recA(ナリジクス酸)、proU(浸透圧)、cst−1(グルコース飢餓状態)、tetA(テトラサイクリン)、cadA(pH)、nar(嫌気的条件)、PL(42℃への温度シフト)、cspA(20℃への温度シフト)、T7(熱誘導)、T7−lacオペレーター(IPTG)、T3−lacオペレーター(IPTG)、T5−lacオペレーター(IPTG)、T4遺伝子32(T4感染)、nprM−lacオペレーター(IPTG)、Pm(アルキル又はハロベンゾエート)、Pu(アルキル又はハロトルエン)、Psal(サリシレート)、及びVHb(酸素)を含む。例えばMakrides, S. C. (1996) Microbiol. Rev. 60,512-538 ; Hannig G. & Makrides, S. C. (1998) TIBTECH 16,54-60 ; Stevens, R. C. (2000) Structures 8, R177- R185参照。例えばJ. Sanchez-Romero & V. De Lorenzo, Genetic Engineering of Nonpathogenic Pseudomonas strains as Biocatalysts for Industrial and Environmental Processes, in Manual of Industrial Microbiology and Biotechnology (A. Demain & J. Davies, eds. ) pp. 460-74 (1999) (ASM Press, Washington, D. C. ) ; H. Schweizer, Vectors to express foreign genes and techniques to monitor gene expression for Pseudomonads, Current Opinion in Biotechnology, 12: 439-445 (2001); およびR. Slater & R. Williams, The Expression of Foreign DNA in Bacteria, in Molecular Biology and Biotechnology (J. Walker & R. Rapley, eds. ) pp.125-54 (2000) (The Royal Society of Chemistry, Cambridge, UK)参照。
【0125】
選択した細菌宿主細胞にネイティブなプロモーターのヌクレオチド配列を有するプロモーター、例えばシュードモナスアントラニレート又はベンゾエートオペロンプロモーター(Pant、Pben)も、標的ペプチドをエンコードする導入遺伝子の発現を制御するために使用できる。2個以上のプロモーターが、同じか又は異なる配列の、もう1つ別のプロモーターに共有結合しているタンデムプロモーター、例えばPant−Pbenタンデムプロモーター(プロモーター間ハイブリッド)又はPlac−Placタンデムプロモーター、も使用し得る。
【0126】
調節されるプロモーターは、プロモーターがその一部である遺伝子の転写を制御するためにプロモーター調節タンパク質を利用する。調節されるプロモーターをここで使用する場合は、対応するプロモーター調節タンパク質も本発明に従った発現系の一部である。プロモーター調節タンパク質の例は、アクチベータータンパク質、例えば大腸菌カタボライトアクチベータータンパク質、MalTタンパク質;AraCファミリー転写アクチベーター;リプレッサータンパク質、例えば大腸菌LacIタンパク質;及び二元機能調節タンパク質、例えば大腸菌NagCタンパク質を含む。多くの調節されるプロモーター/プロモーター調節タンパク質の対が当技術分野において公知である。
【0127】
プロモーター調節タンパク質は、エフェクター化合物、すなわち調節タンパク質が、放出されるか又はプロモーターの制御下にある遺伝子の少なくとも1つのDNA転写調節領域に結合し、それによって遺伝子の転写を開始させる転写酵素の作用を許容する又はブロックすることができるように、調節タンパク質と可逆的に又は不可逆的に結合する化合物と相互作用する。エフェクター化合物は誘導物質又はコリプレッサーのいずれかに分類され、これらの化合物は、天然エフェクター化合物及び非代謝性誘導物質化合物を含む。多くの調節されるプロモーター/プロモーター調節タンパク質/エフェクター化合物の三つ組が当技術分野において公知である。エフェクター化合物は細胞培養又は発酵全体を通じて使用できるが、調節されるプロモーターを使用する特定の実施形態では、所望量又は密度の宿主細胞バイオマスの増殖後、所望の標的遺伝子の発現を直接又は間接的に生じさせるために、適切なエフェクター化合物を培養に添加する。
【0128】
例として、lacファミリーのプロモーターを使用する場合は、lacI遺伝子又はその誘導体、例えばlacIQ又はlacIQ1遺伝子も系に存在し得る。(通常は)構成的に発現される遺伝子であるlacI遺伝子は、これらのプロモーターのlacオペレーターに結合するLacリプレッサータンパク質(LacIタンパク質)をエンコードする。そこで、lacファミリーのプロモーターを使用する場合は、lacI遺伝子も発現系に含まれ、発現され得る。lacプロモーターファミリーメンバー、例えばtacプロモーターの場合、エフェクター化合物は誘導物質、好ましくは非代謝性誘導物質、例えばIPTG(イソプロピル−β−D−1−チオガラクトピラノシド、「イソプロピルチオガラクトシド」とも呼ばれる)である。
【0129】
特定の実施形態では、Plac、Ptac、Ptrc、PtacII、PlacUV5、lpp−PlacUV5、lpp−lac、nprM−lac、T71ac、T51ac、 T31ac及びPmacを含む、lac又はtacファミリープロモーターを本発明において使用する。
【0130】
(他のエレメント)
lacO配列を含む、他の調節エレメントを発現構築物に含めることができる。かかるエレメントは、例えば転写エンハンサー配列、翻訳エンハンサー配列、他のプロモーター、アクチベーター、翻訳開始及び終結シグナル、転写ターミネーター、シストロニック調節因子、ポリシストロニック調節因子、Hisタグ、Flagタグ、T7タグ、Sタグ、HSVタグ、Bタグ、Strepタグ、ポリアルギニン、ポリシステイン、ポリフェニルアラニン、ポリアスパラギン酸、(Ala−Trp−Trp−Pro)n、チオレドキシン、β−ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、シクロマルトデキストリングルコノトランスフェラーゼ、CTP:CMP−3−デオキシ−D−マンノ−オクツロソネートシチジルトランスフェラーゼ、trpE又はtrpLE、アビジン、ストレプトアビジン、T7遺伝子10、T4gp55、ブドウ球菌プロテインA、連鎖球菌プロテインG、GST、DHFR、CBP、MBP、ガラクトース結合ドメイン、カルモジュリン結合ドメイン、GFP、KSI、c−myc、ompT、ompA、pelB、NusA、ユビキチン、及びヘモシリンAを含む、発現されるペプチドの特定、分離、精製又は単離を容易にするタグ配列、例えばヌクレオチド配列「タグ」及び「タグ」ペプチドコード配列を含むが、これらに限定されない。
【0131】
1つの実施形態では、核酸構築物は、対象組換えペプチドについてのコード配列に隣接する、又はウイルスカプシドについてのコード配列に連結された、タグ配列をさらに含む。1つの実施形態では、タグ配列はタンパク質の精製を可能にする。タグ配列は、アフィニティータグ、例えばヘキサヒスチジンアフィニティータグであり得る。もう1つの実施形態では、アフィニティータグは、グルタチオン−S−トランスフェラーゼ分子であり得る。タグはまた、蛍光分子、例えばYFP又はGFP、又はかかる蛍光タンパク質の類似体であり得る。タグはまた、抗体分子の一部、又は精製のために有用な公知の結合パートナーについての公知の抗原又はリガンドであり得る。
【0132】
本発明は、カプシド−組換えペプチドコード配列に加えて、それに作動可能に連結された以下の調節エレメント:プロモーター、リボソーム結合部位(RBS)、転写ターミネーター、翻訳開始及び終結シグナルを含み得る。有用なRBSは、本発明に従った発現系における宿主細胞として有用な種のいずれかから、好ましくは選択される宿主細胞から入手できる。多くの特定の及び多様なコンセンサスRBSが公知であり、例えばD.Frishman et al., Starts of bacterial genes: estimating the reliability of computer predictions, Gene 234 (2): 257-65 (8 Jul 1999); およびB. E. Suzek et al. , A probabilistic method for identifying start codons in bacterial genomes, Bioinformatics 17 (12): 1123-30 (Dec 2001)の中で記述され、言及されている。加えて、天然又は合成RBSが使用でき、例えば欧州特許第0207459号(合成RBS);O. Ikehata et al., Primary structure of nitrile hydratase deduced from the nucleotide sequence of a Rhodococcus species and its expression in Escherichia coli, Eur. J. Biochem. 181 (3): 563-70 (1989) (AAGGAAGの天然RBS配列)に述べられているものが使用できる。本発明において有用な方法、ベクター、及び翻訳及び転写エレメント、及び他のエレメントのさらなる例は、例えばGilroyへの米国特許第5,055,294号及びGilroyらへの米国特許第5,128,130号;Rammlerらへの米国特許第5,281,532号;Barnesらへの米国特許第4,695,455号及び同第4,861,595号;Grayらへの米国特許第4,755,465号;及びWilcoxへの米国特許第5,169,760号に述べられている。
【0133】
(ベクター)
本発明の酵素をエンコードするDNAのシュードモナス宿主による転写は、ベクター又はプラスミドにエンハンサー配列を挿入することによってさらに上昇させ得る。典型的なエンハンサーは、転写を上昇させるようにプロモーターに作用する、通常は約10−300bpの大きさの、DNAのシス作用性エレメントである。
【0134】
一般に、組換え発現ベクターは、シュードモナス宿主細胞、例えば本発明のカプシド−組換えペプチド融合ペプチド、の形質転換を可能にする複製起点及び選択マーカー、及び下流構造配列の転写を指令する高度発現遺伝子に由来するプロモーターを含む。かかるプロモーターは上記で説明した。異種構造配列は、翻訳開始及び終結配列と共に適切な段階で集合される。場合により及び本発明に従って、異種配列は、所望の特徴、例えば発現される組換え産物の安定化又は簡単な精製、を提供するN−末端特定ペプチドを含む融合ペプチドをコードし得る。
【0135】
カプシド−組換えペプチド融合ペプチドを発現するときP.フルオレッセンスと共に使用するための有用な発現ベクターは、カプシドペプチドと融合した所望の標的ペプチドをエンコードする構造DNA配列を、適切な翻訳開始及び終結シグナルと共に、機能性プロモーターを有する作動可能な読み枠(reading phase)に挿入することによって構築される。ベクターは、1又はそれ以上の表現型選択マーカー及びベクターの維持を確実にするため及び、所望する場合は、宿主内での増幅を提供するための複製起点を含む。本開示に従った形質転換のための適切な宿主は、シュードモナス属の中の様々な種、特にシュードモナス・フルオレッセンスの宿主細胞株を含む。
【0136】
宿主細胞内で組換えタンパク質を発現するために有用なベクターは当技術分野において公知であり、本発明に従った融合産物を発現するためにこれらのいずれかを修飾し、使用し得る。かかるベクターは、例えばプラスミド、コスミド及びファージ発現ベクターを含む。本発明に関する使用のために修飾することができる有用なプラスミドベクターの例は、発現プラスミドpBBRlMCS、pDSK519、pKT240、pML122、pPS10、RK2、RK6、pRO1600及びRSF1010を含むが、これらに限定されない。さらなる例は、pALTER−Exl、pALTER−Ex2、pBAD/His、pBAD/Myc−His、pBAD/gIII、pCal−n、pCal−n−EK、pCal−c、pCal−Kc、pcDNA2.1、pDUAL、pET−3a−c、pET 9a−d、pET−l la−d、pET−12a−c、pET−14b、pET15b、pET−16b、pET−17b、pET−19b、pET−20b(+)、pET−21a−d(+)、pET−22b(+)、pET−23a−d(+)、pET24a−d(+)、pET−25b(+)、pET−26b(+)、pET−27b(+)、pET28a−c(+)、pET−29a−c(+)、pET−30a−c(+)、pET31b (+)、pET−32a−c(+)、pET−33b(+)、pET−34b(+)、pET35b(+)、pET−36b(+)、pET−37b(+)、pET−38b(+)、pET−39b(+)、pET−40b(+)、pET−41a−c(+)、pET−42a−c(+)、pET−43a−c(+)、pETBlue−1、pETBlue−2、pETBlue−3、pGEMEX−1、pGEMEX−2、pGEXlλT、pGEX−2T、pGEX−2TK、pGEX−3X、pGEX−4T、pGEX−5X、pGEX−6P、pHAT10/11/12、pHAT20、pHAT−GFPuv、pKK223−3、pLEX、pMAL−c2X、pMAL−c2E、pMAL−c2g、pMAL−p2X、 pMAL−p2E、pMAL−p2G、pProEX HT、pPROLar.A、pPROTet.E、pQE−9、pQE−16、pQE−30/31/32、pQE−40、pQE−50、pQE−70、pQE−80/81/82L、pQE−100、pRSET、及びpSE280、pSE380、pSE420、pThioHis、pTrc99A、pTrcHis、pTrcHis2、pTriEx−1、pTriEx−2、pTrxFusを含み得る。かかる有用なベクターの他の例は、例えばN. Hayase, in Appl. Envir. Microbiol. 60 (9): 3336-42 (Sep 1994); A. A. Lushnikov et al. , in Basic Life Sci. 30: 657-62 (1985); S. Graupner & W. Wackernagel, in Biomolec. Eng. 17(1) : 11-16. (Oct 2000); H. P. Schweizer, in Curr. Opin. Biotech. 12 (5): 439-45 (Oct 2001); M. Bagdasarian & K. N. Timmis, in Curr. Topics Microbiol. Immunol. 96: 47-67 (1982); T. Ishii et al., in FEMS Microbiol. Lett. 116 (3): 307-13 (Mar 1,1994) ; I. N. Olekhnovich & Y. K. Fomichev, in Gene 140(1) : 63-65 (Mar 11,1994) ; M. Tsuda & T. Nakazawa, in Gene 136 (1-2): 257-62 (Dec 22,1993) ; C. Nieto et al. , in Gene 87(1) : 145-49 (Mar 1,1990) ; J. D. Jones & N. Gutterson, in Gene 61 (3): 299-306 (1987); M. Bagdasarian et al. , in Gene 16 (1-3): 237-47 (Dec 1981); H. P. Schweizer et al. , in Genet. Eng. (NY) 23: 69- 81 (2001); P.Mukhopadhyay et al., in J. Bact. 172(1) :477-80 (Jan 1990); D. O. Wood et al. , in J. Bact. 145 (3): 1448-51 (Mar 1981); およびR. Holtwick et al. , in Microbiology 147 (Pt 2): 337-44 (Feb 2001)によって記述されているものを含む。
【0137】
シュードモナス宿主細胞において有用であり得る発現ベクターのさらなる例は、表示されているレプリコンに由来する、表2に列挙されているものを含む。
表2.有用な発現ベクターの一部の例
【表2】
【0138】
発現プラスミド、RSF1010は、例えば F. Heffron et al., in Proc. Nat'1 Acad. Sci. USA 72 (9): 3623-27 (Sep 1975) 及びK. Nagahari & K. Sakaguchi, in J. Bact. 133 (3): 1527-29 (Mar 1978) によって述べられている。プラスミドRSF1010及びその誘導体は、本発明において特に有用なベクターである。当技術分野において公知である、RSF1010の有用な誘導体の例は、例えばpKT212、pKT214、pKT231及び関連プラスミド、及びpMYC1050及び関連プラスミド(例えばThompsonらへの米国特許第5,527,883号及び同第5,840,554号参照)、例えばpMYC1803を含む。プラスミドpMYC1803は、調節されるテトラサイクリン耐性マーカー及びRSF1010プラスミドからの複製及び動員遺伝子座を担持する、RSF1010に基づくプラスミドpTJS260(Wilcoxへの 米国特許第5,169,760号参照)に由来する。他の例示的な有用ベクターは、Puhlerらへの米国特許第4,680,264号に述べられているものを含む。
【0139】
1つの実施形態では、発現プラスミドを発現ベクターとして使用する。もう1つの実施形態では、RSF1010又はその誘導体を発現ベクターとして使用する。さらにもう1つの実施形態では、pMYC1050又はその誘導体、又はpMYC1803又はその誘導体を発現ベクターとして使用する。
【0140】
Champion(商標)pET発現系は高レベルのタンパク質生産を提供する。発現は、強力なT7lacプロモーターから誘導される。この系は、対象遺伝子の高レベル転写のためにバクテリオファージT7 RNAポリメラーゼの高い活性と特異性を利用する。プロモーター領域に位置するlacオペレーターは、伝統的なT7ベースのベクターよりも厳密な調節を提供し、プラスミドの安定性と細胞の生存度を改善する(Studier, F. W. and B. A. Moffatt (1986) J Molecular Biology 189(1) : 113-30; Rosenberg, et al. (1987)Gene 56(1) : 125-35)。T7発現系は、対象遺伝子の高レベル転写のためにT7プロモーターとT7 RNAポリメラーゼ(T7 RNAP)を使用する。T7 RNAPは天然大腸菌RNAPよりもプロセッシブであり、対象遺伝子の転写に専用されるので、T7発現系において高レベル発現が達成される。特定された遺伝子の発現は、宿主細胞においてT7 RNAPのソースを提供することによって誘導される。これは、T7 RNAP遺伝子の染色体コピーを含むBL21大腸菌宿主を使用することによって達成される。T7 RNAP遺伝子は、IPTGによって誘導することができるlacUV5プロモーターの制御下にある。誘導時にT7 RNAPが発現され、対象遺伝子を転写する。
【0141】
pBAD発現系は、特定の炭素源、例えばグルコース、グリセロール及びアラビノースの存在を通して組換えタンパク質の厳密に制御された滴定可能な発現を可能にする(Guzman, et al. (1995) J Bacteriology 177 (14): 4121- 30)。pBADベクターは、発現レベルに対する厳密な制御を与えるように独自に設計される。pBADベクターからの異種遺伝子発現はaraBADプロモーターで開始される。このプロモーターは、araC遺伝子の産物によって正及び負の両方に調節される。AraCは、L−アラビノースと複合体を形成する転写調節因子である。L−アラビノースの不在下では、AraC二量体は転写をブロックする。最大転写活性化のためには2つの事象が必要である:(i)L−アラビノースがAraCに結合して転写を開始させる、(ii)cAMPアクチベータータンパク質(CAP)−cAMP複合体がDNAに結合して、AraCの、プロモーター領域の正しい位置への結合を刺激する。
【0142】
trc発現系は、trcプロモーターからの大腸菌における高レベルの調節された発現を可能にする。trc発現ベクターは、大腸菌における真核生物遺伝子の発現に関して最適化されている。trcプロモーターは、トリプトファン(trp)及びラクトース(lac)プロモーターに由来する強力なハイブリッドプロモーターである。lacOオペレーター及びlacIQ遺伝子の産物によって調節される(Brosius, J. (1984) Gene 27 (2): 161-72)。
【0143】
(III.シュードモナスにおけるウイルス様粒子の発現)
本発明はまた、組換えペプチドを生産するための方法を提供する。この方法は、
a)シュードモナス細胞を提供すること;
b)組換えペプチドと二十面体カプシドの融合ペプチドをエンコードする核酸を提供すること;
c)このシュードモナス細胞においてこの核酸を発現すること、ここでこの細胞における発現がこの融合ペプチドのウイルス様粒子へのインビボ構築を提供する;及び
d)このウイルス様粒子を単離すること
を含む。
【0144】
ペプチドは、カプシドペプチド内の1コピーペプチド挿入物として発現され得る(すなわちペプチドに関してモノシストロニックである組換えカプシドペプチドコード配列から個別挿入物として発現される)か、又は2、3又は多コピー挿入物として発現され得る(すなわちペプチドに関してポリシストロニックである組換えカプシドペプチドコード配列からコンカテマー挿入物として発現される;コンカテマー挿入物は、同じ外来性対象ペプチドの多数のコピーを含み得るか又は異なる外来性対象ペプチドのコピーを含み得る)。コンカテマーは、ホモ又はヘテロコンカテマーであり得る。
【0145】
1つの実施形態では、単離されたウイルス様粒子を、ワクチン戦略においてヒト又は動物に投与することができる。
【0146】
もう1つの実施形態では、核酸構築物を、野生型カプシドをエンコードするもう1つ別の核酸と共発現することができる。特定の実施形態では、共発現されたカプシド/カプシド−組換えペプチド融合粒子は、インビボで集合してキメラウイルス様粒子を形成する。キメラVLPは、少なくとも2個の異なる核酸構築物によってエンコードされるカプシド又はカプシド−ペプチド融合物を含むウイルス様粒子である。
【0147】
さらにもう1つの実施形態では、核酸構築物を、異なるカプシド−組換えペプチド融合粒子をエンコードするもう1つ別の核酸と共発現することができる。特定の実施形態では、共発現されたカプシド融合粒子は、インビボで集合してキメラウイルス様粒子を形成する。
【0148】
さらにもう1つの実施形態では、異なるペプチド、例えばシャペロンタンパク質を発現するように設計された第二の核酸を、融合ペプチドをエンコードする核酸と同時に発現することができる。
【0149】
本発明のために有用なシュードモナス細胞、カプシド及び組換えペプチドは上記で論じている。
【0150】
1つの実施形態では、この方法は、VLPの形態の少なくとも0.1g/Lタンパク質を生産する。もう1つの実施形態では、この方法は、VLPの形態の0.1−10g/Lタンパク質を生産する。下位実施形態では、この方法は、VLP又はケージ構造の形態の少なくとも約0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9又は1.0g/Lタンパク質を生産する。1つの実施形態では、生産される総組換えタンパク質は少なくとも1.0g/Lである。一部の実施形態では、生産されるVLPタンパク質の量は、生産される総組換えタンパク質の少なくとも約5%、約10%、約15%、約20%、約25%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%又はそれ以上である。
【0151】
1つの実施形態では、この方法は、少なくとも0.1g/Lの前形成された(pre-formed)VLP又はケージ構造を生産する。もう1つの実施形態では、この方法は、細胞内で0.1−10g/Lの前形成VLPを生産する。もう1つの実施形態では、この方法は、細胞内で0.1−10g/Lの前形成ケージ構造を生産する。下位実施形態では、この方法は、少なくとも約0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9又は1.0g/Lの前形成VLPを生産する。1つの実施形態では、生産される総前形成VLPタンパク質は少なくとも1.0g/Lである。下位実施形態では、生産される総VLPタンパク質は少なくとも約2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、15.0、20.0又は50.0g/Lであり得る。一部の実施形態では、生産されるVLPタンパク質の量は、生産される総組換えタンパク質の少なくとも約5%、約10%、約15%、約20%、約25%又はそれ以上である。
【0152】
もう1つの実施形態では、生産される、発現されたトランスジェニックペプチド、ペプチド、タンパク質又はそのフラグメントの50%以上を、宿主細胞において復元可能な形態で生産することができる。もう1つの実施形態では、発現されるタンパク質の約60%、70%、75%、80%、85%、90%、95%が活性形態で得られるか又は活性形態に復元することができる。
【0153】
本発明の方法はまた、組換えタンパク質の収率増大を導き得る。1つの実施形態では、この方法は、組換えタンパク質を総細胞タンパク質(tcp)の5、10、15、20、25、30、40又は50、55、60、65、70又は75%として生産する。「パーセント総細胞タンパク質」は、集合細胞タンパク質のパーセンテージとしての宿主細胞内のペプチドの量である。パーセント総細胞タンパク質の測定は当技術分野において周知である。
【0154】
特定の実施形態では、宿主細胞は、無機塩類培地中で増殖させたとき(すなわち約4℃−約55℃(両端を含む)の温度範囲内で)、約1%tcpの組換えペプチド、ペプチド、タンパク質又はそのフラグメントの発現レベル及び少なくとも40g/Lの細胞密度を有し得る。特定の実施形態では、この発現系は、少なくとも10リットルの発酵スケールで無機塩類培地中において増殖させたとき(すなわち約4℃−約55℃(両端を含む)の温度範囲内で)、少なくとも5%tcpの組換えタンパク質又はペプチド発現レベル及び少なくとも40g/Lの細胞密度を有する。
【0155】
別の実施形態では、対象ペプチドに作動可能に連結された発現ウイルスカプシドの部分は、細胞内の不溶性凝集体中で形成される。1つの実施形態では、対象ペプチドを不溶性凝集体から復元することができる。
(対象ペプチドの切断)
【0156】
1つの実施形態では、この方法は、e)この融合産物を切断して、カプシドから組換えペプチドを分離すること、をさらに提供する。
【0157】
切断可能な連結配列は、ウイルスタンパク質と組換えペプチドの間に含まれ得る。かかる配列を切断することができる物質の例は、化学試薬、例えば酸(塩酸、ギ酸)、CNBr、ヒドロキシルアミン(アスパラギン−グリシンに関して)、2−ニトロ−5−チオシアノベンゾエート、O−ヨードソベンゾエート、及び酵素物質、例えばエンドペプチダーゼ、エンドプロテアーゼ、トリプシン、クロストリパイン及びスタフィロコッカスプロテアーゼを含むが、これらに限定されない。
【0158】
切断可能な連結配列は当技術分野において周知である。本発明では、ジペプチド切断配列、例えばAsp−Proを含む、切断物質によって認識されるいかなる切断可能連結配列も使用できる。
【0159】
(発現)
本発明の方法は、最適には、宿主細胞における組換えペプチドの生産増大を導く。生産増大は、選択的に、生産されるタンパク質のグラム当り又は宿主タンパク質のグラム当りの活性ペプチドの濃度増大であり得る。生産増大はまた、組換え体のグラム当り又は宿主細胞タンパク質のグラム当りで生産される回収可能なペプチド、例えば可溶性タンパク質の濃度上昇であり得る。生産増大はまた、総濃度上昇とタンパク質の活性又は可溶性濃度上昇の何らかの組合せであり得る。
【0160】
組換えタンパク質の改善された発現は、VLPにおける挿入タンパク質の発現を通してであり得る。特定の実施形態では、少なくとも60、少なくとも70、少なくとも80、少なくとも90、少なくとも100、少なくとも110、少なくとも120、少なくとも130、少なくとも140、少なくとも150、少なくとも160、少なくとも170、又は少なくとも180コピーの対象ペプチドが各々のVLPにおいて発現される。VLPを生産し、宿主細胞の細胞質、ペリプラズム又は細胞外液から回収することができる。
【0161】
もう1つの実施形態では、ペプチドは細胞において不溶性であり得る。特定の実施形態では、不溶性ペプチドは、多数のカプシドから形成されるが天然型VLPを形成しない粒子において生産される。例えばわずか3個のウイルスカプシドのケージ構造が形成され得る。特定の実施形態では、カプシド構造は2コピー以上の対象ペプチドを含み、特定の実施形態では、少なくとも10、少なくとも20又は少なくとも30コピーを含む。
【0162】
ペプチド又はウイルスカプシド配列はまた、1又はそれ以上のターゲティング配列又は精製を助けるための配列を含み得る。これらはアフィニティータグであり得る。これらはまた、カプシドのVLPへの構築を指令するターゲティング配列であり得る。
【0163】
(細胞増殖)
ベクターによるシュードモナス宿主細胞の形質転換は、当技術分野で公知のいかなる形質転換法を用いて実施してもよく、細菌宿主細胞を無傷細胞として又はプロトプラストとして(すなわち細胞質体を含む)形質転換し得る。例示的な形質転換法は、穿孔法、例えば電気穿孔法、プロトプラスト融合、細菌接合、及び二価陽イオン処理、例えば塩化カルシウム処理又はCaCl/Mg2+処理、又は当技術分野で周知の他の方法を含む。例えばMorrison, J. Bact., 132 : 349-351 (1977); Clark-Curtiss & Curtiss, Methods in Enzymology, 101: 347-362 (Wu et al., eds, 1983), Sambrook et al. , Molecular Cloning, A Laboratory Manual (2nd ed. 1989); Kriegler, Gene Transfer and Expression: A Laboratory Manual (1990); and Current Protocols in Molecular Biology (Ausubel et al. , eds., 1994)参照)。
【0164】
ここで使用する、「発酵」という用語は、文字通りの発酵を用いる実施形態と、他の非発酵培養法を用いる実施形態の両方を包含する。発酵はいかなる規模で実施してもよい。1つの実施形態では、発酵培地は、富栄養培地、最少培地及び無機塩類培地の中から選択し得る;富栄養培地を使用してもよいが、好ましくは回避される。もう1つの実施形態では、最少培地又は無機塩類培地のいずれかを選択する。
【0165】
さらにもう1つの実施形態では、最少培地を選択する。さらにもう1つの実施形態では、無機塩類培地を選択する。無機塩類培地は特に好ましい。
【0166】
無機塩類培地は、無機塩類と炭素源、例えばグルコース、スクロース又はグリセロールからなる。無機塩類培地の例は、例えばM9培地、シュードモナス培地(ATCC179)、DavisとMingioliの培地(BD Davis & ES Mingioli (1950) in J. Bact. 60: 17-28参照)。無機塩類培地を作製するために使用する無機塩類は、例えばリン酸カリウム、硫酸又は塩化アンモニウム、硫酸又は塩化マグネシウム、及び微量ミネラル、例えば塩化カルシウム、鉄、銅、マンガン及び亜鉛のホウ酸塩及び硫酸塩の中から選択されるものを含む。有機窒素源、例えばペプトン、トリプトン、アミノ酸、又は酵母抽出物は、無機塩類培地には含まれない。その代わりに、無機窒素源が使用され、これは、例えばアンモニウム塩、アンモニア水、及び気体アンモニアから選択し得る。好ましい無機塩類培地は、グルコースを炭素源として含む。無機塩類培地と比較して、最少培地も無機塩類と炭素源を含み得るが、例えば低濃度のアミノ酸、ビタミン、ペプトン又は他の成分を添加することができ、但しこれらは極小濃度で添加される。
【0167】
高細胞密度培養は、バッチ工程として開始し、次に二相フェド−バッチ培養を実施することができる。バッチ部分での無制限増殖後、増殖を、バイオマス濃度が数倍上昇できる3倍加時間にわたって低い特定の増殖速度に制御することができる。かかる培養手順についてのさらなる詳細は、Riesenberg, D.; Schulz, V.; Knorre, W. A.; Pohl, H. D.; Korz, D.; Sanders, E. A.; Ross,A. ; Deckwer, W. D. (1991) "High cell density cultivation of Escherichia coli at controlled specific growth rate" J Biotechnol: 20(1) 17-27によって述べられている。
【0168】
本発明に従った発現系はいかなる発酵方式でも培養することができる。例えばバッチ、フェドバッチ、半連続、及び連続発酵方式がここで使用し得る。
【0169】
本発明に従った発現系は、いかなる規模(すなわち容量)の発酵においても導入遺伝子発現のために有用である。したがって、例えばマイクロリットル規模、センチリットル規模及びデシリットル規模の発酵容量を使用し得る;及び1リットル又はそれ以上の規模の発酵容量が使用できる。1つの実施形態では、発酵容量は1リットル又はそれ以上である。もう1つの実施形態では、発酵容量は、5リットル、10リットル、15リットル、20リットル、25リットル、50リットル、75リットル、100リットル、200リットル、500リットル、1,000リットル、2,000リットル、5,000リットル、10,000リットル又は50,000リットル又はそれ以上である。
【0170】
本発明では、形質転換宿主細胞の増殖、培養及び/又は発酵は、宿主細胞の生存を許容する温度範囲内、好ましくは約4℃−約55℃(両端を含む)の範囲内の温度で実施する。そこで、本発明の宿主細胞に関してここで使用する、例えば「増殖」(及び「増殖する」、「増殖すること」)、「培養すること」(及び「培養」)、及び「発酵」(及び「発酵する」、「発酵すること」)という用語は、本質的に、約4℃−約55℃(両端を含む)の温度範囲内での「増殖」、「培養」及び「発酵」を意味する。加えて、「増殖」は、活発な細胞分裂及び/又は拡張(enlargement)の生物学的状態、並びに非分裂及び/又は非拡張細胞が代謝的に維持されている生物学的状態の両方を指示するために使用され、「増殖」という用語の後者の使用は「維持」という用語と同義である。
【0171】
(細胞密度)
VLPに入った組換えペプチドを発現するときにシュードモナス・フルオレッセンスを使用することの付加的な利点は、大腸菌又は他の細菌発現系に比して高い細胞密度で増殖するシュードモナス・フルオレッセンスの能力を含む。このために、本発明に従ったシュードモナス・フルオレッセンス発現系は約20g/L又はそれ以上の細胞密度を提供することができる。本発明に従ったシュードモナス・フルオレッセンス発現系は、同様に、容量当りバイオマスに関して述べたように、バイオマスを乾燥細胞重量として測定したとき、少なくとも約70g/Lの細胞密度を提供することができる。
【0172】
1つの実施形態では、細胞密度は少なくとも約20g/Lである。もう1つの実施形態では、細胞密度は、少なくとも約25g/L、30g/L、35g/L、40g/L、45g/L、50g/L、60g/L、70g/L、80g/L、90g/L、100g/L、110g/L、120g/L、130g/L、140g/L、又は少なくとも150g/Lである。
【0173】
もう1つの実施形態では、誘導時の細胞密度は、20g/L−150g/L;20g/L−120g/L;20g/L−80g/L;25g/L−80g/L;30g/L−80g/L;35g/L−80g/L;40g/L−80g/L;45g/L−80g/L;50g/L−80g/L;50g/L−75g/L;50g/L−70g/L;40g/L−80g/Lである。
【0174】
(VLP又は対象ペプチドの単離)
特定の実施形態では、本発明は、ウイルスカプシドとの連結及び共発現を通して発現の間ペプチドを保護することにより、対象ペプチドの回収率を改善するための方法を提供する。特定の実施形態では、ウイルスカプシド融合は、細胞溶解産物から容易に分離することができるVLPを形成する。
【0175】
本発明のタンパク質は、硫酸アンモニウム又はエタノール沈殿、酸抽出、陰イオン又は陽イオン交換クロマトグラフィ、ホスホセルロースクロマトグラフィ、疎水性相互作用クロマトグラフィ、アフィニティークロマトグラフィ、ニッケルクロマトグラフィ、ヒドロキシルアパタイトクロマトグラフィ、逆相クロマトグラフィ、レクチンクロマトグラフィ、分取電気泳動、界面活性剤可溶化、カラムクロマトグラフィのような物質による選択的沈殿、免疫精製法等を含むが、これらに限定されない、当技術分野において周知の標準手法によって単離し、実質的に純粋に精製し得る。例えば確立された分子接着特性を有するタンパク質は、リガンドに可逆的に融合することができる。適切なリガンドを用いて、タンパク質を選択的に精製カラムに吸着させ、その後比較的純粋な形態でカラムから遊離することができる。次に、酵素活性によって融合タンパク質を取り出す。加えて、免疫アフィニティーカラム又はNi−NTAカラムを用いてタンパク質を精製することができる。一般的な手法は、例えばR. Scopes, Protein Purification: Principles and Practice, Springer-Verlag: N. Y. (1982); Deutscher, Guide to Protein Purification, Academic Press (1990);米国特許第4,511,503号; S. Roe, Protein Purification Techniques: A Practical Approach (Practical Approach Series), Oxford Press (2001); D. Bollag, etal., Protein Methods, Wiley-Lisa, Inc. (1996); AK Patra et al., Protein
Expr Purif, 18 (2): p/182-92 (2000); and R. Mukhija, etal., Gene 165 (2): p. 303-6 (1995)において詳述されている。また、例えばAusubel, et al. (1987 and periodic supplements); Deutscher (1990) "Guide to Protein Purification, " Methods in Enzymology vol.182及びこのシリーズの他の巻; Coligan, et al. (1996 and periodic Supplements) Current Protocols in Protein Science Wiley/Greene, NY;及びタンパク質精製産物の使用に関する製造者の文献、例えばPharmacia,Piscataway,N.J.、 又はBio−Rad,Richmond,Calif.も参照のこと。組換え手法との組み合わせは、適切なセグメント、例えばFLAG配列又はプロテアーゼ−除去可能配列によって融合することができる等価物、への融合を可能にする。また、例えばHochuli (1989) Chemische Industrie 12: 69-70; Hochuli (1990) "Purification of Recombinant Proteins with Metal Chelate Absorbent" in Setlow (ed. ) Genetic Engineering, Principle and Methods 12: 87-98, Plenum Press, NY;およびCrowe, et al. (1992) QIAexpress : The High Level Expression & Protein Purification System QUIAGEN,Inc., Chatsworth, Calif.も参照のこと。
【0176】
同様に、ウイルス様粒子又はケージ様構造も、当技術分野で周知の標準手法によって単離する及び/又は実質的に純粋に精製することができる。VLPの単離のための手法は、上述したものに加えて、沈殿法、例えばポリエチレングリコール又は塩沈殿法を含む。分離手法は、陰イオン又は陽イオン交換クロマトグラフィ、サイズ排除クロマトグラフィ、ホスホセルロースクロマトグラフィ、疎水性相互作用クロマトグラフィ、アフィニティークロマトグラフィ、ニッケルクロマトグラフィ、ヒドロキシルアパタイトクロマトグラフィ、逆相クロマトグラフィ、レクチンクロマトグラフィ、分取電気泳動、免疫精製法、遠心分離、超遠心分離、密度勾配遠心分離(例えばショ糖又は塩化セシウム(CsCl)勾配で)、サイズ排除フィルターを通しての限外ろ過、及び当技術分野で公知の何らかの他のタンパク質単離法を含む。
【0177】
本発明はまた、活性組換えペプチドの回収率を改善することができる。活性タンパク質の濃度は、例えば特定されたペプチドと親ペプチド、ペプチド変異体、セグメント置換ペプチド及び/又は残基置換ペプチドの間の相互作用を従来の何らかのインビトロ又はインビボアッセイによって測定することによって測定できる。そこで、インビトロアッセイは、特定されたタンパク質と対象ペプチドの間、例えば酵素と基質の間、ホルモンとホルモン受容体の間、抗体と抗原の間等の、何らかの検出可能な相互作用を測定することによって使用できる。かかる検出は、比色変化、放射能の変化、溶解度の変化、ゲル電気泳動及び/又はゲル排除工程によって測定した際の分子量変化等の測定を含み得る。インビボアッセイは、生理的作用、例えば体重増加、電解質バランスの変化、血液凝固時間の変化、血餅溶解及び抗原応答の誘導の変化、を検出するアッセイを含むが、これらに限定されない。一般に、特定されたペプチドと対象ペプチドの間の相互作用の変化を検出するための可変パラメータが存在する限り、いかなるインビボアッセイも使用することができる。例えば米国特許第5,834,250号参照。
【0178】
ペリプラズムから組換えタンパク質を放出するには、化学物質、例えばクロロホルム(Ames et al. (1984) J.BacteiioL, 160: 1181-1183)、グアニジン−HCl、及びTriton X−100 (Naglak and Wang (1990)Enzyme Microb. Technol., 12: 603-611) を含む処理が使用されてきた。しかし、これらの化学物質は不活性ではなく、多くの組換えタンパク質産物又はその後の精製手順に有害作用を及ぼし得る。外膜の透過性上昇を生じさせる、大腸菌細胞のグリシン処理も、ペリプラズム内容物を放出することが報告されている(Ariga et al. (1989) J. Ferm. Bioeng, 68 : 243-246)。組換えタンパク質のペリプラズム放出の最も広く使用されている方法は、浸透圧衝撃(Nosal and Heppel (1966) J. Biol.Chem., 241: 3055-3062; Neu and Heppel (1965)J. Biol.Clam., 240: 3685-3692)、ニワトリ卵白(HEW)−リゾチーム/エチレンジアミン四酢酸(EDTA)処理 (Neu and Heppel (1964) J.Biol. Chem., 239: 3893-3900; Witholt et al. (1976) Biochim. Biophys. Acta, 443: 534-544; Pierce et al. (1995) ICheme Research. Event, 2: 995-997)、及びHEW−リゾチーム/浸透圧衝撃処理の組合せ (French et al. (1996) Enzyme and Microb. Tech., 19: 332-338)である。このFrenchの方法は、分画緩衝液中に細胞を再懸濁して、その後ペリプラズム画分を回収することを含み、リゾチーム処理の直後に浸透圧衝撃を実施する。回収率に対する組換えタンパク質の過剰発現、S.サーモビオラセウス(S. thermoviolaceus)α−アミラーゼ、及び宿主生物の増殖期の影響も論じられている。
【0179】
典型的には、これらの手順は、浸透圧安定化培地中での初期破壊とそれに続く非安定化培地中での選択的放出を含む。これらの培地の組成物(pH、保護剤)及び使用する破壊方法(クロロホルム、HEW−リゾチーム、EDTA、超音波処理)は、報告されている特定の手順によって異なる。EDTAの代わりに双極子イオン界面活性剤を使用する、HEW−リゾチーム/EDTA処理の変法が、Stabel et al. (1994)Veterinary Microbiol., 38: 307-314によって述べられている。大腸菌を破壊するための細胞内溶菌酵素系の使用の総説については、Dabora and Cooney (1990) in Advances in Biochemical Engineering/Biotechnology, Vol. 43, A. Fiechter, ed. (Springer-Verlag: Berlin), pp. 11-30参照。
【0180】
可溶性タンパク質又は屈折性粒子としての、サイトプラズムからの組換えタンパク質の回収のための従来の方法は、機械的破壊による細菌細胞の崩壊を含んだ。機械的破壊は、典型的には、液体懸濁液中での局所キャビテーションの生成、剛性ビーズとの急速攪拌、超音波処理、又は細胞懸濁液の摩砕を含む(Bacterial Cell Surface Techniques, Hancock and Poxton (John Wiley & Sons Ltd,1988), Chapter 3, p. 55)。
【0181】
HEW−リゾチームは、細胞壁のペプチドグリカン骨格を加水分解するように生化学的に作用する。その方法は、後にスフェロプラストとして知られる球状の細胞球体を生成するために大腸菌を卵アルブミン(HEW−リゾチームを含有する)で処理した、 Zinder and Arndt (1956) Proc. Natl. Acad. Sci. USA, 42: 586-590によって最初に開発された。これらの構造は、一部の細胞壁成分を保持するが、サイトプラズム膜が露出された大きな表面積を有していた。米国特許第5,169,772号は、浸透圧安定化培地、例えば20%スクロース溶液中で、例えばEDTA、リゾチーム又は有機化合物を使用して細菌のエンベロープを破壊すること、細菌を低イオン強度緩衝液に暴露することによって破壊された細菌のペリプラズムスペースから非ヘパリナーゼ様タンパク質を放出すること、及び低イオン強度洗浄した細菌を緩衝塩溶液に暴露することによってヘパリナーゼ様タンパク質を放出することを含む、細菌からヘパリナーゼを精製するための方法を開示する。
【0182】
これらの方法の多くの異なる修正が広い範囲の発現系で使用され、様々な程度の成功を収めている(Joseph-Liazun et al. (1990) Gene, 86: 291-295; Carter et al. (1992)BiolTechnology, 10: 163-167)。リゾチームを生産するために組換え細胞培養を誘導するための試みが報告されてきた。欧州特許第0 155 189号は、組換え細胞培養を、通常はかかる宿主細胞を死滅させると予想される細胞壁構造を破壊する又は溶解する手段によって、リゾチームを生産するように誘導するための手段を開示する。
【0183】
米国特許第4,595,658号は、大腸菌のペリプラズムスペースに輸送されたタンパク質の外在化(externalization)を促進するための方法を開示する。この方法は、細胞のリゾチーム処理、機械的摩砕又は浸透圧衝撃を必要とせずに、ペリプラズム内に位置するタンパク質の選択的単離を可能にする。米国特許第4,637,980号は、温度感受性溶原菌を、細菌産物を直接又は間接的にエンコードするDNA分子で形質転換し、細胞内で遺伝子産物を発現することを許容する条件下で形質転換体を培養して、ファージにコードされた機能を誘導するために温度を上昇させることによってこの産物を外在化させることにより、細菌産物を生産することを開示する。Asami et al. (1997) J. Ferment. and Bioeng., 83: 511-516 は、T4ファージ感染による大腸菌細胞の同調破壊を開示し、Tanji et al. (1998)J. Ferment. and Bioeng., 85 : 74-78 は、大腸菌細胞の穏やかな破壊のためにT4ファージ内にエンコードされる溶菌遺伝子の制御された発現を開示する。
【0184】
細胞溶解時に、ゲノムDNAがサイトプラズムから培地中に漏出し、遠心力場において固体の沈降を妨げ得る液体粘度の有意の上昇をもたらす。剪断力、例えばDNAポリマーを壊すための機械的破壊の間に及ぼされる剪断力、が存在しない場合は、粘性液体を通しての固体のより緩やかな沈降速度が、遠心分離において固体と液体の分離不良を生じさせる。機械的剪断力以外に、DNAポリマーを分解する核酸分解酵素が存在する。大腸菌において、内在性遺伝子endAは、正常にはペリプラズムに分泌されて、DNAをオリゴデオキシリボヌクレオチドへとエンドヌクレアーゼ的に切断する、エンドヌクレアーゼをエンコードする(成熟タンパク質の分子量は約24.5kDである)。endAは大腸菌によって比較的弱く発現されることが示唆された(Wackemagel et al. (1995) Gene 154: 55-59)。
【0185】
発現されたタンパク質の検出は、当技術分野で公知の方法によって達成され、例えば放射免疫測定法、ウエスタンブロット手法又は免疫沈降法を含む。
【0186】
本発明において発現される特定のタンパク質は不溶性凝集体(「封入体」)を形成し得る。いくつかのプロトコールが封入体からのタンパク質の精製に適する。例えば封入体の精製は、典型的には、例えば50mM TRIS/HCl pH7.5、50mM NaCl、5mM MgCl2、1mM DTT、0.1mM ATP、及び1mM PMSFの緩衝液中でのインキュベーションによる、宿主細胞の破壊による封入体の抽出、分離及び/又は精製を含む。細胞懸濁液は、典型的にはフレンチプレスの2−3回の通過を用いて溶菌される。細胞懸濁液はまた、Polytron(Brinkrnan Instruments)を用いて又は氷上で超音波処理して均一化することができる。細菌を溶解する代替的な方法は当業者に明白である(例えば Sambrook et al. 前出; Ausubel et al.前出参照)。
【0187】
必要に応じて、封入体を可溶化することができ、溶菌細胞懸濁液は、典型的には遠心分離して望ましくない不溶性物質を除去することができる。封入体を形成したタンパク質は、適合性緩衝液による希釈又は透析によって復元し得る。適切な溶媒は、尿素(約4M−約8M)、ホルムアミド(少なくとも80%、容量/容量ベース)、及び塩酸グアニジン(約4M−約8M)を含むが、これらに限定されない。塩酸グアニジン及び類似試薬は変性剤であるが、この変性は不可逆的ではなく、変性剤の除去(例えば透析による)又は希釈後に復元が起こり得、免疫学的及び/又は生物学的に活性なタンパク質の再形成が可能である。他の適切な緩衝液は当業者に公知である。
【0188】
あるいは、宿主ペリプラズムから組換えペプチドを精製することが可能である。宿主細胞の溶解後、組換えタンパク質が宿主細胞のペリプラズム内に輸送されたとき、当業者に公知の他の方法に加えて低温浸透圧衝撃によって細菌のペリプラズム画分を単離することができる。ペリプラズムから組換えタンパク質を単離するため、例えば細菌細胞を遠心分離してペレットを形成することができる。ペレットを、20%スクロースを含む緩衝液に再懸濁することができる。細胞を溶解するため、細菌を遠心分離し、ペレットを氷冷5mM MgSO4に再懸濁して、約10分間氷浴中に保持することができる。細胞懸濁液を遠心分離し、上清をデカントして保存することができる。上清中に存在する組換えタンパク質を、当業者に周知の標準分離手法によって宿主タンパク質から分離することができる。
【0189】
初期塩分画は、望ましくない宿主細胞タンパク質(又は細胞培養に由来するタンパク質)の多くを対象組換えタンパク質から分離することができる。1つのかかる例は硫酸アンモニウムであり得る。硫酸アンモニウムは、タンパク質混合物中の水分量を有効に減少させることによってタンパク質を沈殿させる。その後タンパク質は、それらの溶解度に基づいて沈殿する。タンパク質がより疎水性であるほど、より低い硫酸アンモニウム濃度で沈殿する可能性が高い。典型的なプロトコールは、生じる硫酸アンモニウム濃度が20−30%になるように飽和硫酸アンモニウムをタンパク質溶液に添加することを含む。この濃度は最も疎水性のタンパク質を沈殿させる。次に沈殿物を廃棄し(対象タンパク質が疎水性でない限り)、対象タンパク質を沈殿させることが既知である濃度まで硫酸アンモニウムを上清に添加する。次に沈殿物を緩衝液に溶解し、必要に応じて、透析又はダイアフィルトレーションのいずれかを通して過剰の塩を除去する。タンパク質の溶解度に基づく他の方法、例えば低温エタノール沈殿は当業者に周知であり、複雑なタンパク質混合物を分画するために使用できる。
【0190】
組換えタンパク質の分子量を利用して、異なる細孔径の膜(例えばアミコン又はミリポア膜)を通す限外ろ過を用いてより大きい及びより小さいサイズのタンパク質から分離することができる。第一段階として、タンパク質混合物を、対象タンパク質の分子量よりも低い分子量カットオフ値を有する細孔径の膜を通して限外ろ過することができる。限外ろ過の保持物質を、次に、対象タンパク質の分子量よりも大きい分子量カットオフ値を有する膜に対して限外ろ過することができる。組換えタンパク質は膜を通過してろ液中に入る。その後ろ液を以下で述べるようにクロマトグラフィに供することができる。
【0191】
組換えタンパク質はまた、その大きさ、正味表面電荷、疎水性、及びリガンドに対する親和性に基づいて他のタンパク質から分離することができる。加えて、タンパク質に対して惹起される抗体をカラムマトリックスに結合し、タンパク質を免疫精製することができる。これらの方法は全て当技術分野において周知である。クロマトグラフィ手法が、多くの異なる製造者(例えばPharmacia Biotech)からの装置を使用していかなる規模でも実施できることは、当業者には明白である。
【0192】
(復元及びリフォールディング)
不溶性タンパク質を復元又は再生して、二次及び三次タンパク質構造立体配座を形成することができる。タンパク質リフォールディング工程は、必要に応じて、組換え産物の立体配置を完成させるときに使用できる。リフォールディング及び復元は、タンパク質の解離/会合を促進する当技術分野で公知の物質を用いて実施することができる。例えばタンパク質をジチオトレイトールと共にインキュベートし、次に酸化グルタチオン二ナトリウム塩と共にインキュベートして、さらにその後リフォールディング剤、例えば尿素を含む緩衝液と共にインキュベートすることができる。
【0193】
組換えタンパク質はまた、例えばリン酸緩衝食塩水(PBS)又は50mM 酢酸ナトリウム、pH6緩衝液プラス200mM NaClに対して透析することによっても復元できる。あるいは、タンパク質をカラムに、例えばNi NTAカラムに固定化して、プロテアーゼ阻害因子を含有する、500mM NaCl、20%グリセロール、20mM Tris/HCl pH7.4中、線形6M−1M尿素勾配を使用することによって再生できる。復元は1.5時間又はそれ以上の期間で実施できる。復元後、250mM イミダゾールの添加によってタンパク質を溶出することができる。イミダゾールは、PBS又は50mM 酢酸ナトリウム、pH6緩衝液プラス200mM NaClに対する最終透析工程によって除去できる。精製タンパク質は、4℃で保存するか又は−80℃で凍結することができる。
【0194】
他の方法は、例えばMH Lee et al., Protein Expr. Purif., 25(1) : p. 166-73 (2002), W. K. Cho et al. , J. Biotechnology, 77 (2-3): p. 169-78 (2000), Ausubel, et al. (1987 and periodic supplements), Deutscher (1990) "Guide to Protein Purification, "Methods in Enzymology vol. 182, and other volumes in this series, Coligan, et al. (1996 and periodic Supplements) Current Protocols in Protein Science Wiley/Greene, NY, S. Roe, Protein Purification Techniques: A Practical Approach (Practical Approach Series), Oxford Press (2001); D. Bollag, et al. , Protein Methods, Wiley-Lisa, Inc. (1996)に述べられているものを含む。
【0195】
(活性ペプチド分析)
活性タンパク質は、その配列が由来する天然ペプチドの活性の少なくとも20%、30%又は40%、好ましくは少なくとも50%、60%又は70%、最も好ましくは少なくとも80%、90%又は95%の比活性を有し得る。さらに、基質特異性(Kcat/Km)は、場合により天然ペプチドと実質的に同様である。典型的には、Kcat/Kmは、天然ペプチドの少なくとも30%、40%又は50%、より好ましくは少なくとも60%、70%、80%又は90%である。タンパク質及びペプチドの活性及び基質特異性(Kcat/Km)を検定する方法及び定量手段は当業者に周知である。
【0196】
本発明に従って生産される組換えペプチドの活性は、当技術分野で公知のタンパク質特異的な従来の又は標準インビトロ又はインビボアッセイによって測定することができる。シュードモナス生産組換えペプチドの活性を対応する天然タンパク質の活性と比較して、組換えタンパク質が、同じ又は類似の生理的条件下で天然ペプチドにおいて一般に認められる活性と実質的に類似又は同等の活性を示すかどうかを判定することができる。
【0197】
組換えペプチドの活性を、これまでに確立された天然ペプチド標準活性と比較することができる。あるいは、組換えペプチドの活性を、天然ペプチドと同時又は実質的に同時の比較アッセイにおいて測定することができる。例えばインビトロアッセイを使用して、組換えペプチドと標的の間、例えば発現される酵素と基質の間、発現されるホルモンとホルモン受容体の間、発現される抗体と抗原の間等の、検出可能な相互作用を測定することができる。かかる検出は、比色変化、増殖変化、細胞死、細胞反発、放射能の変化、溶解度の変化、ゲル電気泳動及び/又はゲル排除法によって測定したときの分子量の変化、リン酸化能力、抗体特異性アッセイ、例えばELISAアッセイ等の測定を含み得る。加えて、インビボアッセイは、天然ペプチドの生理的作用と比較したシュードモナス生産ペプチドの生理的作用、例えば体重増加、電解質バランスの変化、血液凝固時間の変化、血餅溶解及び抗原応答の誘導の変化、を検出するアッセイを含むが、これらに限定されない。一般に、いかなるインビトロ又はインビボアッセイも、活性が検定可能である限り、天然ペプチドとの比較分析を可能にするシュードモナス生産組換えペプチドの活性を測定するために使用できる。あるいは、本発明において生産されるペプチドは、ペプチドと、通常そのペプチドと相互作用する分子、例えば基質又は天然タンパク質が通常相互作用するシグナル経路の成分、との間の相互作用を刺激する又は阻害する能力に関して検定することができる。かかるアッセイは、典型的には、ペプチドが標的分子と相互作用することを許容する条件下でタンパク質と基質分子を組み合わせる工程を含み、タンパク質と標的分子の相互作用の生化学的結果を検出する。ペプチド活性を測定するために使用できるアッセイは、例えばRalph, P. J. , et al. (1984) J. Immunol. 132:1858 or Saiki et al. (1981) J. Immunol. 127: 1044,. Steward, W. E.II (1980) The Interferon Systems. Springer-Verlag, Vienna and New York, Broxmeyer, H. E. , et al. (1982) Blood 60: 595, "Molecular Cloning: A Laboratory Manual", 2d ed. , Cold Spring Harbor Laboratory Press, Sambrook, J. , E. F. Fritsch and T. Maniatis eds. , 1989,および"Methods in Enzymology: Guide to Molecular Cloning Techniques", Academic Press, Berger, S. L. and A. R. Kimmel eds. , 1987, AK Patra et al., Protein Expr Purif, 18 (2):p/182-92 (2000), Kodama et al. , J. Biochem. 99: 1465-1472 (1986); Stewart et al. , Proc.Nat'1 Acad. Sci. USA 90: 5209- 5213 (1993); (Lombillo et al. , J. Cell Biol. 128: 107-115 (1995); (Vale et al. , Cell 42: 39-50 (1985) に述べられている。
【実施例】
【0198】
これらの実施形態では、ササゲ退緑斑紋ウイルス(CCMV)をペプチド担体として使用し、シュードモナス・フルオレッセンスを発現宿主として使用した。CCMVはブロモウイルス科のブロモウイルス群のメンバーである。ブロモウイルスは、4成分のポジティブセンス一本鎖RNAゲノムを有する直径25−28nmの二十面体ウイルスである。RNA1及びRNA2はレプリカーゼ酵素をエンコードする。RNA3は、植物宿主内のウイルス運動に関与するタンパク質をエンコードする。RNA4(RNA3に由来するサブゲノムRNA)、すなわちsgRNA4は、20kDaのカプシド(CP)、配列番号:1をエンコードする。
【0199】
【表3】
【0200】
各々のCCMV粒子は、約180コピーまでのCCMV CPを含む。CCMV CPをエンコードする例示的DNA配列を配列番号:21に示す。
【表4】
【0201】
CCMVの結晶構造を解明した。この構造は、粒子の安定性と動力学にとって決定的に重要であると思われるカプシド相互作用のより明瞭な像を提供し、挿入部位の合理的設計を導くのに有用であった。これまでの試験は、CCMVカプシドを、粒子を形成するそれらの能力を妨げることなく異種ペプチドを担持するように遺伝的に修飾できることを明らかにした。多くの適切な挿入部位が特定されている。
【0202】
P.フルオレッセンスにおけるカプシド−ペプチド融合VLPの生産のために従うべき一般的戦略を図2に示す。CCMV CP内の単一の挿入部位を使用する場合、合計約180コピーまでの異種ペプチド単位(個々のペプチド又はコンカテマー)をCCMV粒子に挿入することができる。現在までにCCMV CP内で特定された挿入部位は、様々な長さのペプチドを収容することができる。加えて、多量体形態のペプチドを挿入部位に挿入することができる。さらに、同じ粒子内/粒子上に同じか又は異なるペプチドを発現するために多数の挿入部位を同時に使用することができる。ペプチド挿入物は、約200アミノ酸残基又はそれ以下の長さ、より好ましくは約180又はそれ以下、さらに一層好ましくは約150又はそれ以下、さらに一層好ましくは約120又はそれ以下、さらに一層好ましくは約100アミノ酸残基又はそれ以下の長さであり得る。好ましい実施形態では、ペプチド挿入物は約5又はそれ以上のアミノ酸残基の長さである。好ましい実施形態では、ペプチド挿入物は、約5−約120、より好ましくは約5−約100アミノ酸残基の長さである。
【0203】
(実験材料及び方法)
異なる記載がない限り、分子生物学の分野において公知の標準手法、ベクター、制御配列エレメント及び他の発現系エレメントを核酸操作、形質転換及び発現のために使用する。かかる標準手法、ベクター及びエレメントは、例えばAusubel et al. (eds. ), Current Protocols in Molecular Biology (1995) (John Wiley & Sons); Sambrook, Fritsch, & Maniatis (eds.), Molecular Cloning (1989) (Cold Spring Harbor Laboratory Press, NY) ; Berger & Kimmel, Methods in Enzymology 152: Guide to Molecular Cloning Techniques(1987) (Academic Press); and Bukhari et al. (eds. ), DNA Insertion Elements, Plasmids and Episomes (1977) (Cold Spring Harbor Laboratory Press, NY)に見出すことができる。
【0204】
プラスミド地図の作成
全てのプラスミド地図は、VECTORNTI(InforMax Inc.,Frederick,MD,USA)を用いて作成した。
【0205】
DNA抽出
大腸菌からの全てのプラスミドで抽出は、Qiagen(Germany)からのミニ、ミディ及びマキシキットを製造者の指示に従って使用して実施した。
【0206】
実験戦略
以下の手順に従った。P.フルオレッセンス宿主細胞を、キメラウイルスカプシド−標的ペプチドインサート融合物をエンコードする発現プラスミドで形質転換した。形質転換細胞を所望の密度に増殖させ、キメラウイルスカプシド−ペプチド融合物を発現するように誘導した。次に細胞を溶解し、それらの内容物を分析した。
【0207】
加工した挿入部位を付加するための修飾CCMV−CP DNAの構築
CCMV CPコード配列を含むDNA分子を、読み枠内に、BamHI制限酵素認識及び切断部位(gggatcctn)を挿入することによって修飾し、これは、天然のCCMV−CPアミノ酸配列のAsn129とSer130の間にトリペプチド(Gly−Ile−Leu)を導入した。従って、天然のCCMV−CPアミノ酸配列(配列番号:1)を、CCMV129−CP(配列番号:2)を形成するように修飾した。
【0208】
【表5】
【0209】
CCMV−CPコード配列にACTAGT SpeI制限部位を付加するため及びコンセンサスシャイン-ダルガーノ配列を付加するために、プライマーCCMV−For(核酸配列:5’−gactagtagg aggaaagaga tgtctacagt cgg−3’(配列番号:3))を設計した。CCMV−CPコード配列にCTCGAG XhoI制限部位を付加するため及び2個の停止コドンを導入するためにプライマーCCMV−Rev(核酸配列:5’−ccgctcgagt cattactaat acaccgg−3’(配列番号:4))を設計した。これら2個のプライマーを、CCMV−CPのDNAコード配列に関する第一PCR反応において使用した。
【0210】
加工した挿入部位を付加するためのCCMV63−CP DNAの構築:
制限部位AscI及びNotIを、挿入部位として使用するためにCCMV−CP(配列番号:1)上に加工した。AscI(ggcgcgcc)、NotI(gcggccgc)についての認識及び切断部位、及び付加的なヌクレオチドは、CCMV−CP上の残基Ala60とAla61の間にヘプタペプチド(Glu−Ala−Trp−Arg−Ala−Ala−Ala)を導入した。したがって、CCMV−CPを、CCMV63−CPを形成するように修飾した。加えて、構築されたVLPに安定性を付加するために残基Arg26をCys26に突然変異させた。pCCMV63−CPのプラスミド地図を図4に示す。
【0211】
【表6】
【0212】
二重挿入物R26C−CCMV63/129−CPの構築:
制限部位AscI及びNotIを、第二の挿入部位として使用するためにCCMV129−CP(配列番号:2)上に加工した。AscI(ggcgcgcc)、NotI(gcggccgc)についての認識及び切断部位、及び付加的なヌクレオチドは、CCMV129−CP上の残基Ala60とAla61の間にヘプタペプチド(Glu−Ala−Trp−Arg−Ala−Ala−Ala)を導入した。したがって、CCMV129−CPを、CCMV63/129−CPを形成するように修飾した。加えて、構築されたVLPに安定性を付加するように残基Arg26をCys26に突然変異させ、R26C−CCMV63/129−CPを作製した。pR26C−CCMV63/129−CPのプラスミド地図を図5に示す。
【0213】
【表7】
【0214】
実施例1.シュードモナスのCCMV VLPにおけるペプチドPD1の生産
1.A.キメラCCMV−PD1遺伝子の構築
20アミノ酸の抗原ペプチドを、CCMVウイルスカプシドにおいて挿入物として発現するために選択した。この抗原ペプチドは、CCMV及びシュードモナス・フルオレッセンスと無関係であった。このペプチドをエンコードするオリゴヌクレオチドを、プライマーParvo−BamHI−F(核酸配列:5’−cgggatcctg gacccggatg−3’(配列番号:16))及びParvo−BamHI−R(核酸配列:5’−cgggatcccc gggtctcttt c−3’(配列番号:17))を用いてプラスミドpCP7Parvo1 DNAから増幅した。(これらのプライマーは、Integrated DNA Technologies,Inc.,Coralville,IA,USA、以下「IdtDNA」より入手した。)これらのプライマーは、そのBamHI制限部位をCCMV129コード配列に挿入するために両方の末端でBamHI制限部位を付加して、イヌパルボウイルスペプチドコード配列を増幅した。
【0215】
以下のプロトコールに従ってPTC225サーモサイクラー(MJ Research, South San Francisco,CA,USA)を使用してPCR反応を実施した:
【表8】
【0216】
DNA配列を、SpeI及びXhoI制限酵素を使用して、CCMV129 CP DNA配列を含む核酸を挿入することによってプラスミドpESC(Stratagene Corp.,LaJolla,CA,USAより入手した)から構築したプラスミド、CCMV129シャトルプラスミドに挿入した。PD1ペプチドをエンコードする核酸を、BamHI制限部位でCCMV129 CDSに挿入し、CCMV129−PD1シャトルプラスミドを作製した。
【0217】
PD1 CDSもCCMV129 CDSに挿入した。その結果として、挿入されたPDIコード配列は、そのアミノ酸配列が:Trp Ala Cys Arg Gly Thr Ala Gly Trp Pro Pro Ser Gly Cys Thr Ala Pro Ser Gly Ser(配列番号:7)であるPD1ペプチドをエンコードする、5’−tgg gcc tgc cgc ggc acg gcc ggc tgg ccg ccg tcc ggc tgc acg gcg ccg tcc ggg tcg−3’(配列番号:18)である。PD1をコードするヌクレオチド配列は、イヌパルボウイルスには無関係である。
【0218】
1.B.CCMV−PD1発現プラスミドの構築
CCMV129−PD1シャトルプラスミドをSpeI及びXhoI制限酵素で消化した。キメラCCMV129−PD1 DNA配列を含むフラグメントをゲル精製によって単離した。次にそれを、pMYC1803発現プラスミドのSpeI及びXhoI制限部位で、tacプロモーターに作動可能に連結して、ブイブイ毒素遺伝子の代わりにpMYC1803発現プラスミドに挿入した。図1参照。生じた発現プラスミドを、挿入物の存在を確認するためにSpeI及びXhoIでの制限酵素消化によってスクリーニングした。
【0219】
1.C.シュードモナス宿主細胞へのプラスミド形質転換
CCMV129−PD1発現プラスミドを、以下のプロトコールに従ってシュードモナス・フルオレッセンスMB214宿主細胞に形質転換した。宿主細胞を、氷上に保持したバイアル中で徐々に解凍した。各々の形質転換のために、1μLの精製発現プラスミドDNAを宿主細胞に添加し、生じた混合物をピペットの先端で静かに渦を描いて混合し、その後氷上で30分間インキュベートした。混合物を電気穿孔用使い捨てキュベット(BioRad Gene Pulser Cuvette,電極ギャップ0.2cm、カタログ番号165−2086)に移した。キュベットを、あらかじめ200Ω、25μファラッド、2.25kVに設定したBiorad Gene Pulserに入れた。細胞は、短時間(約1−2秒)パルス細胞であった。その後直ちに低温LB培地を添加し、生じた懸濁液を30℃で2時間インキュベートした。次に細胞をLB tet15(テトラサイクリン添加LB培地)寒天にプレートし、30℃で一晩増殖させた。
【0220】
1.D.CCMV−PD1構築物の振とうフラスコ発現
各々のプレートから1個のコロニーを採取し、採取した試料をバッフル付き振とうフラスコ中の50mL LB種培養に接種した。懸濁液培養物を、250rpmで振とうしながら30℃で一晩増殖させた。各々の生じた種培養10mLを使用して、次に、1リットルバッフル付き振とうフラスコ中の振とうフラスコ培地(すなわち微量元素、クエン酸ナトリウム及びグリセロールを含む酵母抽出物及び塩、pH6.8)200mLに接種した。選択のためにテトラサイクリンを添加した。接種した培養物を、250rpmで振とうしながら30℃で一晩増殖させ、CCMV−PD1キメラカプシドの発現のためにIPTGで誘導した。
【0221】
1.E.細胞培養溶解産物の可溶性画分と不溶性画分への分離
各々の振とうフラスコ培養物から1mLアリコートを遠心分離して、細胞をペレット化した。細胞ペレットを、2mM EDTAを含む低温50mM Tirs−HCl、pH8.2 0.75mLに再懸濁した。0.1%容量の10% TritonX−100界面活性剤を加え、次いでリゾチームを0.2mg/mLの最終濃度まで添加した。その後細胞を氷上で2時間インキュベートし、その時点で透明な粘性細胞溶解産物が見えるはずである。
【0222】
この溶解産物に、1/200容量の1M MgCl2を加え、次いで1/200容量の2mg/mL DNAseIを添加して、氷上で1時間インキュベートし、その時点までに溶解産物ははるかに粘性の低い液体になっているはずである。処理した溶解産物を、次に、卓上遠心分離機において最大速度、4℃で30分間遠心し、上清を透明なチューブにデカントした。デカントした上清は「可溶性」タンパク質画分である。その後残りのペレットを0.75mL TE緩衝液(10mM Tris−HCl、pH7.5、1mM EDTA)に再懸濁した。再懸濁したペレットは「不溶性」画分である。
【0223】
1.F.可溶性及び不溶性タンパク質画分のSDS−PAGE及びウエスタンブロット分析
これらの「可溶性」及び「不溶性」画分を、次に、製造者の仕様書に従って、1.0mm×15穴を有するNuPAGE4−12% Bis−Trisゲル(Invitrogenより、カタログ番号NP0323)で電気泳動した。ゲルをSimplyBlue Safe Stain(Invitrogenより、カタログ番号LC6060)で染色し、水で一晩脱染した。ウエスタンブロット検出は、CCMV IgG(アクセッション番号AS0011、DSMZ、Germanyより)及びWESTERN BREEZEキット(Invitrogenより、カタログ番号WB7105)を製造者のプロトコールに従って使用した。結果は、CCMVの生産に関して、特にキメラCCMV CPの生産に関して陽性であった(図6及び7参照)。
【0224】
1.G.キメラVLPのPEG沈殿
キメラ、すなわち組換えVLPを、別々の振とうフラスコ培養試料の溶解によって沈殿させ、次いで、生じた細胞溶解産物を以下のプロトコールに従ってPEG(ポリエチレングリコール)処理した。各々の振とうフラスコ培養物の5mLアリコートを遠心分離して、細胞をペレット化した。ペレット化した細胞を、緩衝液2容対ペレット1容の比率で0.1M リン酸緩衝液(好ましくはリン酸二水素カリウムとリン酸二カリウムの組合せ)、pH7.0に再懸濁した。その後細胞を10秒間ずつ4回超音波処理し、各々の処理の間2分間氷上に置いた。この超音波処理の間に、細胞溶解産物は幾分清澄化するはずである。超音波処理後、リゾチームを0.5mg/mLの最終濃度まで添加した。リゾチーム消化を室温で30分間進行させた。
【0225】
生じた処理溶解産物を、次に、4℃、15000×Gで5分間遠心分離した。生じた上清を取り出し、その容積を測定した。各々の上清に、PEG6000を4%の最終濃度まで添加した;次いでNaClを0.2Mの最終濃度まで添加し、4℃で1時間又は一晩、氷上でインキュベートした。次に、これらを4℃、20000×Gで15分間遠心分離した。沈殿したペレットを、1/10初期上清容量のリン酸緩衝液に再懸濁し、4℃で保存した。
【0226】
1.H.ショ糖勾配遠心分離
リン酸緩衝液中のショ糖(Sigma、カタログ番号S−5390)でショ糖溶液を作製した。ショ糖勾配を手動で上部から底部へ10%、20%、30%及び40%注いだ。再懸濁した沈殿ペレット試料を、Beckman−Coulter Optima XL 100K UltracentrifugeのBeckman−Coulter SW41−Tiローターにおいてブレーキをかけずに1時間遠心した。ショ糖勾配の各々1mL画分を別々に溶出し、さらに遠沈してVLPペレットを得た。VLPペレットをリン酸緩衝液に再懸濁し、SDS−PAGEゲルで電気泳動して、上記プロトコールに従ってCCMV IgGを用いてウエスタンブロット分析した。ウエスタンブロット分析はVLP形成に関して陽性であった(図8)。各々の生じたVLP試料の一部を電子顕微鏡検査のために使用した。
【0227】
1.I.電子顕微鏡分析
VLP試料を、コロジオン/カーボン又はフォルムバール/カーボン被覆グリッドにスポットした。試料を2%ホスホタングステン酸(PTA)で染色し、120kVの加速電圧で操作するPhilips CM−12 TEM透過型電子顕微鏡(シリアル番号D769)で画像化した。画像をMultiScan CCDカメラ(Gatan,Inc., Pleasanton,CA,USAより;749型、シリアル番号971119010)でデジタル記録した。VLPの形成を確認した(図9)。
【0228】
実施例2.シュードモナスのCCMV VLPにおけるD2A21 AMP三量体の生産及びそこからのAMPの回収
2.A.D2A21挿入物の合成:
抗菌ペプチド(「AMP」)三量体(「D2A21三量体」、すなわちD2A21単量体AMPを3個含む)をコードするヌクレオチド配列を、プライマーD2A21−BamHI−F(核酸配列:5’−cgggatcctg ggacagcaaa tgggtcgcga tccg−3’(配列番号:5))及びD2A21−BamHI−R(核酸配列:5’−cgggatcccg tcgacggagc tcgaattcgg atcacc−3’(配列番号:6))を用いてプラスミドpET−(D2A21)3から増幅した。上記実施例1.A.で述べたのと同じプロトコールに従ってPCR反応を実施した。
【0229】
生じた増幅挿入物は、加工したBamHI部位でCCMV129 CDSにD2A21三量体CDSを挿入するときに使用するための、各々の末端に付加されたBamHI制限部位を含んでいた。D2A21三量体をエンコードするヌクレオチド配列及びD2A21三量体のアミノ酸配列をそれぞれ配列番号:19及び20に示す。
【0230】
【表9】
【0231】
【表10】
【0232】
三量体CDSは、図3に示すように、ジペプチドAsp−Pro酸不安定切断部位CDSによって分けられた3個のAMP単量体CDSを含んでいた。三量体CDS全体も、ジペプチドAsp−Pro酸不安定切断部位CDSによって各々の末端で境界付けられた。増幅した挿入物をBamHI制限酵素で消化して、CCMV129 CDS内のBamHI部位でpESC−CCMV129BamHIシャトルプラスミドにクローニングするための付着末端を作成した。生じたシャトルプラスミドをSpeI及びXhoI制限酵素で消化した。所望のキメラRBS/CDSフラグメントをゲル精製によって単離した。
【0233】
2.B.発現プラスミド構築
生じたキメラCCMV129−(D2A21)3ポリヌクレオチドを、次に、tacプロモーターに作動可能に連結して、ブイブイコード配列の代わりにpMYC1803発現プラスミドに挿入した。生じた発現プラスミドを、挿入物の存在に関してSpeI及びXhoIでの制限酵素消化によってスクリーニングした。実施例1.B.に関して上述したのと同じプロトコールを使用した。
【0234】
2.C.形質転換及び発現
生じた発現プラスミドを、実施例1.C.に関して上述したのと同じプロトコールを用いてP.フルオレッセンスMB214に形質転換した。プレートでコロニー形成した形質転換体を採取し、上記実施例1.D.に関して述べたのと同じプロトコールに従って、発現のために振とうフラスコに移した。
【0235】
2.D.タンパク質及びVLPの回収と分析
振とうフラスコ培養細胞を、実施例1.E.の手順に従って溶解し、分画した。生じた画分を、実施例1.F.に関して述べたようにSDS−PAGE及びウエスタンブロット法によって分析した。キメラVLPをPEG沈殿及びショ糖勾配遠心分離によって回収し、実施例1.G.−1.I.で上述したように電子顕微鏡検査によって分析した。キメラCCMV VLP集合を確認した。結果はCCMVの生産に関して陽性であった。キメラCP発現についてのSDS−PAGEは96アミノ酸挿入物を示し(図10)、これは、VLP形成を示すためのショ糖勾配での分画後のウエスタンブロット(図11)及びVLP形成を確認するための電子顕微鏡写真(図12)によって確認された。
【0236】
2.E.D2A21抗菌ペプチド生産の分析
可溶性及び不溶性タンパク質画分を、以下のように、キメラVLPにおいて生産されたD2A21ペプチドを特性決定するためにさらに処理した。
【0237】
2.E.1.D2A21の酸切断
不溶性画分を15%v/vアセトニトリル水溶液及び約40−50%v/vギ酸水溶液に溶解した;可溶性画分を約45−50%v/vギ酸水溶液に再懸濁した。試料を、その後、60℃で24時間インキュベートして、酸切断を進行させた。−20℃に冷凍することによって反応を停止させ、この温度で、処理した試料をHPLC分析まで保存した。
2.E.2.HPLCによるD2A21分析
【0238】
可溶性画分を0.22μm膜でろ過した;不溶性画分を遠心分離して細胞デブリを沈殿させ、0.22μm膜でろ過した。各々の試料50μLを25%アセトニトリル水溶液950μLに添加した。D2A21’の内部対照ペプチド(合計10μgの対照ペプチド)を含む、各々の試料250μL容量を、Beckman高速液体クロマトグラフィ(HPLC)システムに据え付けた、内径6.4mmのVYDAC 250mm逆相C18カラム(Grace Vydac,Hesperia,CA,USAより入手可能)に注入した。30分間で25%アセトニトリル/0.1%トリフルオロ酢酸(TFA)から75%アセトニトリル/0.01%TFAまでの水性勾配を用いて溶出を実施した。溶出物をクロマトグラフィ画分に滴下収集した。加工ペプチドを含むVLPに由来する試料においてのみ適切なペプチドピークを認め、非加工VLPに由来する試料では認められなかった(図13)。
【0239】
2.E.3.D2A21ペプチドの質量分析
ペプチド対照及びクロマトグラフィ画分の質量分析を、Micromass M@LDI線形マトリックス支援レーザー脱離イオン化飛行時間型(MALDI−TOF)質量分析システム(Micromass UK Ltd.,Manchester,UKより)を用いて実施した。MS分析の前に、Speed Vacシステム(Thermo Savant, Milford,MA,USAより入手可能;250DDA型)を使用して、遠心蒸発によってHPLC画分を濃縮した。結果は、D2A21 AMPの正確な生産及び放出されるペプチドがD2A21ペプチドであることを明らかにした(図14)。これらの結果は、シュードモナスにおけるペプチド発現のためのVLP融合物の使用が、通常の宿主細胞毒性を有効に回避することを示した。(A)キメラVLPの生産が明らかにされ、また(B)ペプチド多量体の生産がVLP方式で明らかにされた(合計96アミノ酸まで)。従って、この実施例は:
(1)CCMV CP発現及び粒子集合を支持するP.フルオレッセンスの能力を試験し、
(2)簡単な方法(PEG沈殿)によってキメラVLPを精製し、
(3)あらかじめ試験した方法(酸加水分解)によって対象ペプチドを切断し、及び
(4)ペプチドの同一性と完全性を確認した。
【0240】
実施例3.シュードモナスのCCMV VLPにおける炭疽抗原の生産
3.A.PAペプチド挿入物の合成
4個の異なる炭疽菌防御抗原(「PA」)ペプチド(PA1−PA4)をCCMV VLPにおいて独立して発現した。PA1−PA4をエンコードする核酸を合成オリゴヌクレオチドのSOE(オーバーラップ伸長によるスプライシング)(splicing-by-overlap-extension)によって合成した。生じた核酸はBamHI認識部位末端を含んだ。これらのPAペプチドをエンコードするヌクレオチド配列及びこれらのPAペプチドのアミノ酸配列はそれぞれ以下の通りであった:1)PA1については、配列番号:8及び9;2)PA2については、配列番号:10及び11;3)PA3については、配列番号:12及び13;及び4)PA4については、配列番号:14及び15。生じた核酸をBamHIで消化して、シャトルベクターへのクローニングのための付着末端を作成した。生じたPA挿入物の各々を、CCMV129 CDSのBamHI部位でpESC−CCMV129BamHIシャトルプラスミドにクローニングした。各々の生じたシャトルプラスミドをSpeI及びXhoI制限酵素で消化した。各々の所望のキメラCCMV129−PAエンコードフラグメントをゲル精製によって単離した。
【表11】
【表12】
【表13】
【表14】
【0241】
3.B.発現プラスミド構築
生じたキメラCCMV129−PAポリヌクレオチドを各々、tacプロモーターに作動可能に連結して、ブイブイコード配列の代わりにpMYC1803発現プラスミドに挿入した。生じた発現プラスミドを、挿入物の存在に関してSpeI及びXhoIでの制限消化によってスクリーニングした。実施例1.B.について上述したのと同じプロトコールを使用した。
【0242】
3.C.形質転換及び発現
生じた発現プラスミドを、実施例1.C.に関して上述したプロトコールを用いてP.フルオレッセンスMB214に形質転換した。プレートでコロニー形成した形質転換体を採取し、上記実施例1.D.に関して述べたのと同じプロトコールに従って、発現のために振とうフラスコに移した。
【0243】
3.D.タンパク質及びVLPの回収と分析
振とうフラスコ培養細胞を、実施例1.E.の手順に従って溶解し、分画した。生じた画分を、実施例1.F.に関して述べたようにSDS−PAGE及びウエスタンブロット法によって分析した。結果はCCMVの生産に関して陽性であった。
【0244】
結果は、キメラCCMV CP生産に関して陽性であった(図15参照)。VLPを、実施例1.G.及び1.H.で述べたようにPEG沈殿及びショ糖勾配分画によって回収した。ショ糖勾配画分のウエスタンブロットを1.H.で述べたように実施した。結果はVLPの生産に関して陽性であった(図16)。
【0245】
実施例4.シュードモナスのCCMV VLPへの単一及び二重挿入によるPBF20 AMP単量体の生産
実施例1、2及び3で述べた手順に従った。PBF20単量体ペプチドをエンコードする(アミノ酸配列Asp Pro Lys Phe Ala Lys Lys Phe Ala Lys Lys Phe Ala Lys Lys Phe Ala Lys Lys Phe Ala Lys Asp Pro(配列番号:24)のアミノ酸配列3−22を含むAMPをエンコードする)核酸及び配列番号:24のアミノ酸配列1−2及び23−24を含む酸切断部位を、AscI/NotI部位でCCMV63−CPに及びBamHI部位でCCMV129−CPに独立して挿入した。ペプチドをまた、AscI/NotI部位及びBamHI部位の両方で同時にR26C−CCMV63/129−CPに挿入した。
【0246】
生じたキメラポリヌクレオチドを各々、tacプロモーターに作動可能に連結して、ブイブイコード配列の代わりにpMYC1803発現プラスミドに挿入した。生じた発現プラスミドを、実施例1.C.について上述したプロトコールを使用して、挿入物の存在に関してSpeI及びXhoIでの制限消化によってスクリーニングし、P.フルオレッセンスMB214に形質転換した。プレートでコロニー形成した形質転換体を採取し、上記実施例1.D.に関して述べたのと同じプロトコールに従って、発現のために振とうフラスコに移した。
【0247】
振とうフラスコ培養細胞を、実施例1.E.の手順に従って溶解し、分画した。生じた画分を、実施例1.F.に関して述べたようにSDS−PAGE及びウエスタンブロット法によって分析した。結果はVLPの生産に関して陽性であった。
【0248】
シュードモナス・フルオレッセンスにおいて酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を発現するように加工したキメラCCMV63−CPの発現を示すSDS−PAGEを図17に示す。キメラCCMV63−CP−PBF20は、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。CCMV63−CPに由来し、酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を提示するキメラCCMV VLPの電子顕微鏡(EM)画像を図18に示す。
【0249】
シュードモナス・フルオレッセンスにおいて酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を発現するように加工したキメラCCMV129−CPの発現を示すSDS−PAGEを図19に示す。キメラCCMV129−CP−PBF20は、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。CCMV129−CPに由来し、酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を提示するキメラCCMV VLPの電子顕微鏡(EM)画像を図20に示す。
【0250】
シュードモナス・フルオレッセンスにおいてCP内の2つの異なる挿入部位で酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を発現するように加工したキメラCCMV63/129 CPの発現を示すSDS−PAGEを図21に示す。二重挿入物(CP+2×20アミノ酸)を含むキメラCPは、同じペプチドの単一挿入物(CP+1×20アミノ酸)を発現するように加工したカプシドと比較して、SDS−PAGEゲルでより緩慢な移動度を有する。カプシド当り2つの挿入部位で酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を提示する、CCMV63/129−CP由来のキメラCCMV VLPの電子顕微鏡(EM)画像を図22に示す。各々のVLPは、粒子当り360までのPBF20単量体を含むことが認められた。
【0251】
実施例5.シュードモナスのCCMV VLPにおける東部ウマ脳脊髄炎ウイルス(EEE)抗原の生産
5.A.EEEペプチド挿入物の合成
2つの異なるEEEペプチド(EEE−1及びEEE−2)をCCMV VLPにおいて独立して発現した。
EEE−1ペプチド配列:
DLDTHFTQYKLARPYIADCPNCGHS(配列番号:25)
EEE−1核酸配列:
5’−gacctggacacccacttcacccagtacaagctggcccgcccgtacatcgccgactgcccgaactgcggccacagc−3’(配列番号:26)
EEE−2ペプチド配列:
GRLPRGEGDTFKGKLHVPFVPVKAK(配列番号:27)
EEE−2核酸配列:
5’−ggccgcctgccgcgcggcgaaggcgacaccttcaagggcaagctgcacgtgccgttcgtgccggtgaaggccaag−3’(配列番号:28)
【0252】
EEE−1及びEEE−2をエンコードする核酸を合成オリゴヌクレオチドのSOEによって合成した。生じた核酸は、BamHI認識部位末端を含んでいた。挿入物の合成のためのセンス及びアンチセンスオリゴヌクレオチドプライマーはBamHI認識部位を含み、以下の通りであった:
EEE1.センスプライマー:
5’−cgg gga tcc tgg acc tgg aca ccc act tca ccc agt aca agc tgg ccc gcc cgt ac−3’(配列番号:29)
EEE1.アンチセンスプライマー:
5’−cgc agg atc ccg ctg tgg ccg cag ttc ggg cag tcg gcg atg tac ggg cgg gcc agc−3’(配列番号:30)
EEE2.センスプライマー:
5’−cgg gga tcc tgg gcc gcc tgc cgc gcg gcg aag gcg aca cct tca agg gca agc−3’(配列番号:31)
EEE2.アンチセンスプライマー:
5’−cgc agg atc ccc ttg gcc ttc acc ggc acg aac ggc acg tgc agc ttg ccc ttg−3’(配列番号:32)
【0253】
生じた核酸をBamHIで消化して、pESC−CCMV129BamHIシャトルプラスミドにクローニングするための付着末端を作成した。
【0254】
生じたEEE挿入物の各々を、CCMV129 CDSのBamHI部位でpESC−CCMV129BamHIシャトルプラスミドにクローニングした。各々の生じたシャトルプラスミドをSpeI及びXhoI制限酵素で消化した。所望のキメラCCMV−129−EEEをエンコードするフラグメントの各々をゲル精製によって単離した。
【0255】
5.B.発現プラスミド構築
生じたキメラCCMV129−EEEポリヌクレオチドフラグメントを各々、次に、tacプロモーターに作動可能に連結して、ブイブイコード配列の代わりにSpeI及びXhoIで制限消化したpMYC1803発現プラスミドに挿入した。生じた発現プラスミドを、挿入物の存在に関してSpeI及びXhoIでの制限消化によってスクリーニングした。実施例1.B.に関して上述したのと同じプロトコールを使用した。
【0256】
5.C.形質転換及び発現
生じた発現プラスミドを、実施例1.C.において上述したプロトコールを用いてP.フルオレッセンスMB214に形質転換する。実施例1.D.に関して上述したのと同じプロトコールを、EEE抗原を提示するキメラVLPの発現のために使用する。
【0257】
5.D.タンパク質及びVLPの回収と分析
振とうフラスコ培養細胞を、実施例1.E.の手順に従って溶解し、分画する。生じた画分を、実施例1.F.に関して述べたようにSDS−PAGE及びウエスタンブロット法によって分析する。
【図面の簡単な説明】
【0258】
【図1】図1は、シュードモナス宿主細胞における組換えVLPの発現のために有用なCCMV129−CP発現プラスミドのプラスミド地図を示す。
【図2】図2は、宿主細胞、例えばシュードモナス宿主細胞におけるウイルス様粒子(VLP)内のペプチド単量体の生産のための概要図を示す。組換えウイルスカプシド遺伝子(「rCP」)を構築するとき、所望の標的ペプチド挿入物コード配列(「I」)をインフレームでウイルスカプシドコード配列(「CP」)に挿入し、それが、ベクターの一部として、宿主細胞に形質転換され、組換えカプシド(「rCP」)を形成するように発現される。これらは、次に、CCMVの場合には、各々180個までのrCPを含むVLPを形成するように集合される。カプシドの外部ループにおいて発現される標的ペプチド挿入物(「I」)を有するVLPを例示する。集合されたVLPは各々、粒子当り多数のペプチド挿入物、例えば180個まで又はその倍数のペプチド挿入物を含む。VLPは、その後、例えばPEG沈降法によって、回収のために細胞溶解産物から容易に沈殿される。カプシド表面ループ及び/又は末端において発現された組換えペプチド挿入物は、沈殿したVLPから高度に純粋な形態で単離され得る。
【図3】図3は、宿主細胞、例えばシュードモナス宿主細胞におけるVLP内のペプチド多量体の生産のための概要図を示す。ペプチド挿入物は、そのコード配列(「i」)が、組換えウイルスカプシド遺伝子(「rCP」)を構築するときウイルスカプシドコード配列(「CP」)に挿入される、所望の標的ペプチドの多量体(三量体を示す)である。標的ペプチドコード配列の各々は、切断部位(「*」)についてのコード配列によって境界づけられ、核酸挿入物全体が標識「I」である。図では、CCMVカプシド当り1個の三量体挿入だけが行われ、生じるVLPは各々180個までのペプチド挿入物(「I」)を含み、合計540個までの標的ペプチド(「i」)を含む。標的ペプチドは、その後、切断剤、例えば酸又は酵素でのVLPの処理によって、VLPの沈殿後、高度に純粋な形態で容易に単離される。
【図4】図4は、組換えVLPの発現のために有用なCCMV63−CP発現プラスミドのプラスミド地図を示す。制限部位AscI及びNotIを、ペプチドのための挿入部位として働くようにCCMV−CP(配列番号:1)上に作製した。
【図5】図5は、組換えVLPの発現のために有用なR26C−CCMV63/129−CP発現プラスミドのプラスミド地図を示す。2つの挿入部位(AscI−NotI及びBamHI)を、2個の同じ又は異なるペプチドの挿入のためにCP内に作製した。
【図6】図6は、誘導後24時間目のシュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEゲルの画像である。キメラCPは、20アミノ酸の抗原ペプチドPD1を発現するように加工した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目の野生型CPであり、レーン3は誘導後24時間目の野生型CPであり、レーン4は誘導後0時間目のCCMV129−PD1であり、及びレーン5は誘導後24時間目のCCMV129−PD1である。
【図7】図7は、シュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すウエスタンブロットの画像である。キメラCPは、20アミノ酸の抗原ペプチドPD1を発現するように加工した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目の野生型CPであり、レーン3は誘導後24時間目の野生型CPであり、レーン4は誘導後0時間目のCCMV129−PD1であり、及びレーン5は誘導後24時間目のCCMV129−PD1である。
【図8】図8は、CCMV129−PD1 VLPショ糖勾配画分のウエスタンブロットの画像である。20アミノ酸の抗原ペプチドPD1を発現するように加工したキメラCCMV CPをシュードモナス・フルオレッセンスにおいて発現した。キメラVLPをPEG沈降法によって誘導後24時間目に単離し、ショ糖密度勾配で分画した。VLP画分はキメラCPに関して陽性であった。レーン1はCCMV129−PD1 VLPショ糖勾配画分であり、レーン2はCCMV129−PD1 VLPショ糖勾配画分であり、レーン3はCCMV129−PD1 VLPショ糖勾配画分であり、及びレーン4はサイズラダーである。
【図9】図9は、20アミノ酸の抗原ペプチドPD1を示すキメラCCMV VLPの電子顕微鏡(EM)画像である。VLPは、PEG沈降法及びショ糖密度勾配を用いてP.フルオレッセンスから単離した。
【図10】図10は、誘導後12、24及び48時間目のシュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEゲルの画像である。キメラCPは、酸加水分解部位によって分離される抗菌ペプチドD2A21三量体を発現するように作製した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目の野生型CPであり、レーン3は誘導後12時間目の野生型CPであり、レーン4は誘導後24時間目の野生型CPであり、レーン5は誘導後48時間目の野生型CPであり、レーン6は誘導後0時間目のCCMV129−(D2A21)3であり、レーン7は誘導後12時間目のCCMV129−(D2A21)3であり、レーン8は誘導後24時間目のCCMV129−(D2A21)3であり、及びレーン9は誘導後48時間目のCCMV129−(D2A21)3である。
【図11】図11は、CMV129−(D2A21)3 VLPショ糖勾配画分のウエスタンブロットの画像である。酸加水分解部位によって分離される96アミノ酸の抗菌ペプチドD2A21三量体を発現するように加工したキメラCCMV CPをシュードモナス・フルオレッセンスにおいて発現した。キメラVLPをPEG沈降法によって誘導後24時間目に単離し、ショ糖密度勾配で分画した。VLP画分はキメラCPに関して陽性であった。レーン1はサイズラダーであり、レーン2−4はCMV129−(D2A21)3 VLPショ糖勾配画分である。
【図12】図12は、酸加水分解部位によって分離される抗菌ペプチドD2A21三量体を示すキメラCCMV VLPの電子顕微鏡(EM)画像である。VLPは、PEG沈降法及びショ糖密度分画を用いてP.フルオレッセンスから単離した。
【図13】図13は、酸での処理により、酸切断部位によって分離される抗菌ペプチドD2A21三量体を提示するように加工したキメラVLPからのAMP D2A21ペプチド単量体の放出を示すHPLCクロマトグラムである。加工されていない(空の)VLPでは、AMPペプチドのピークは検出されなかった。
【図14】図14は、酸での処理により、酸切断部位によって分離される抗菌ペプチドD2A21三量体を提示するように加工したキメラVLPから放出されるAMP D2A21ペプチド単量体の同一性を示すMALDI−MSグラフである。分子量は、D2A21ペプチド単量体に関して予測されるとおりである。
【図15】図15は、誘導後12及び24時間目のシュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEゲルの画像である。キメラCPは、4つの異なる25アミノ酸の抗原ペプチドPA1、PA2、PA3及びPA4を発現するように作製した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目のCCMV129−PA1であり、レーン3は誘導後12時間目のCCMV129−PA1であり、レーン4は誘導後24時間目のCCMV129−PA1であり、レーン5は誘導後0時間目のCCMV129−PA2であり、レーン6は誘導後12時間目のCCMV129−PA2であり、レーン7は誘導後24時間目のCCMV129−PA2であり、レーン8は誘導後0時間目のCCMV129−PA3であり、レーン9は誘導後12時間目のCCMV129−PA3であり、レーン10は誘導後24時間目のCCMV129−PA3であり、レーン11は誘導後0時間目のCCMV129−PA4であり、レーン12は誘導後12時間目のCCMV129−PA4であり、レーン13は誘導後24時間目のCCMV129−PA4である。
【図16】図16は、CCMV129−PA1、CCMV129−PA2、CCMV129−PA3、CCMV129−PA4 VLPショ糖勾配画分のウエスタンブロットの画像である。25アミノ酸の抗原ペプチドPAを発現するように加工したキメラCCMV CPをシュードモナス・フルオレッセンスにおいて発現した。キメラVLPをPEG沈降法によって誘導後24時間目に単離し、ショ糖密度勾配で分画した。VLP画分はキメラCPに関して陽性であった。レーン1はサイズラダーであり、レーン2−4はCCMV129−PA1 VLPショ糖勾配画分であり、レーン5−7はCCMV129−PA2 VLPショ糖勾配画分であり、レーン8−10はCCMV129−PA3 VLPショ糖勾配画分であり、及びレーン11−13はCCMV129−PA4 VLPショ糖勾配画分である。
【図17】図17は、シュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEの画像である。キメラCCMV63−CPは、酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を発現するように作製した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目の野生型CPであり、レーン3は誘導後24時間目の野生型CPであり、レーン4は誘導後0時間目のCCMV63−PBF20であり、レーン5は誘導後24時間目のCCMV63−PBF20である。
【図18】図18は、CCMV63−CPに由来し、酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を提示するキメラCCMV VLPの電子顕微鏡(EM)画像である。キメラVLPは、PEG沈降法及びショ糖密度分画を用いてP.フルオレッセンスから単離した。
【図19】図19は、シュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEの画像である。キメラCCMV129−CPは、酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を発現するように作製した。レーン1はサイズラダーであり、レーン2は誘導後0時間目のCCMV129−PBF20であり、及びレーン3は誘導後24時間目のCCMV129−PBF20である。
【図20】図20は、CCMV129−CPに由来し、酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を提示するキメラCCMV VLPの電子顕微鏡(EM)画像である。キメラVLPは、PEG沈降法及びショ糖密度分画を用いてP.フルオレッセンスから単離した。
【図21】図21は、シュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEの画像である。キメラCCMV63/129−CPは、CP内の2つの異なる挿入部位(63及び129)の酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を発現するように作製した。二重挿入物(CP+2×20AA)を含むキメラCPは、同じペプチドの単一挿入物(CP+1×20AA)を発現するように加工したカプシドと比較して、SDS−PAGEゲルでより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目のCCMV63−PBF20であり、レーン3は誘導後24時間目のCCMV63−PBF20であり、レーン4は誘導後0時間目のCCMV63/129−2×(PBF20)であり、レーン5は誘導後24時間目のCCMV63/129−2×(PBF20)であり、レーン6は誘導後0時間目のCCMV63/129−2×(PBF20)であり、レーン7は誘導後24時間目のCCMV63/129−2×(PBF20)であり、レーン8は誘導後0時間目のCCMV63/129−2×(PBF20)であり、レーン9は誘導後24時間目のCCMV63/129−2×(PBF20)である。
【図22】図22は、カプシド当り2つの挿入部位(63及び129)の酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を提示する、CCMV63/129−CPに由来するキメラCCMV VLPの電子顕微鏡(EM)画像である。キメラVLPは、PEG沈降法及びショ糖密度分画を用いてP.フルオレッセンスから単離した。
【技術分野】
【0001】
(関連出願の相互参照)
本出願は、「High Efficiency Peptide Production in Pseudomonads」と題する2003年12月1日出願の米国特許仮出願第60/525,982号の優先権を主張する。
【0002】
本発明は、組換えペプチドの生産のための改善された方法を提供する。特に、本発明は、二十面体ウイルスからのウイルス様粒子を使用した細菌細胞における組換えペプチドの生産又は提示のための改善された方法を提供する。
【背景技術】
【0003】
遺伝子工学の革命は、ヒト及び動物治療薬としての使用のための組換えペプチドの開発へ発展した。現在、医学用途のために米国FDAによって承認された100以上のバイオテクノロジー由来の治療薬及びワクチンが存在し、1000を超えるさらなる薬剤及びワクチンが様々な相の臨床試験下にある。(M. Rai & H. Padh, (2001) "Expression systems for production of heterologous proteins, "Cur. Science 80 (9): 1121-1128参照)。
【0004】
細菌、酵母、ディクチオステリウム・ディスコイデウム、昆虫、及び哺乳動物細胞発現系が、組換えペプチドを生産するために現在使用されており、成功の程度は様々である。異種ペプチドの生産のための発現系を作成する上での1つの目標は、商業、治療、及びワクチン適用において利用できる、広範囲で、柔軟性があり、効率的、経済的及び実用的なプラットフォームを提供することである。例えば特定のペプチドの生産のためには、下流の再構築(reassembly)コストを取り除く又は低減するために、インビボで効率よく且つ安価に、大量の所望の最終産物を生産することができる発現系を得ることが理想である。
【0005】
現在、細菌は、豊富な量の組換えペプチドを生産するそれらの潜在能力の故に、組換えペプチドの生産のために最も広く使用される発現系である。しかしながら、細菌はしばしば特定のペプチドを生産する能力が限られており、代替的な、より費用のかかる発現系の使用を必要とする。例えば細菌系は、単量体抗菌ペプチドに関しては、しばしばペプチドの発現時に細胞死を導く、細菌に対するかかるペプチドの毒性の故に、生産能力が限られている。非細菌系の、コストと生産収率という観点における特有の不都合の故に、広い範囲の商業的及び治療的に有用なペプチドを生産する細菌系の能力を改善するために多大の時間と資源が費やされてきた。この領域で進歩はあったが、細菌系における異種ペプチドの生産のための付加的な方法及びプラットフォームは有益である。
【0006】
(ウイルス)
宿主細胞発現系におけるペプチド生産を改善するための1つのアプローチは、対象組換えペプチドを生産するために複製可能ウイルスの性質を利用することである。しかし、複製可能な完全長ウイルスの使用は、組換えペプチド生産戦略における使用において数多くの難点がある。例えば発酵状態の間は組換えペプチドの生産を制御することが困難であると考えられるが、発酵工程の効率を最大化するためには発現の厳密な調節が必要であり得る。さらに、組換えペプチドを生産するための複製可能ウイルスの使用は、高度な下流精製工程につながる、規制基準の負担をもたらし得る。
【0007】
特に発酵中の生産上の問題を克服するために、1つの研究領域は、特定のウイルスにとって天然の宿主ではない細胞(非親和性宿主細胞)におけるウイルスの発現と構築に焦点を当ててきた。非親和性細胞は、ウイルスカプシドと宿主受容体分子の不適合性、又はウイルスがその生活環を完了するのを妨げる、ウイルスの生化学と細胞の生化学の間の不適合性の故に、ウイルスがうまく入り込むことができない細胞である。例えばPriceらへの米国特許第5,869,287号は、ウイルスカプシド又はカプシド内に含まれるRNAがノダウイルス科又はブロモウイルス科の非酵母ウイルス種由来である場合に、酵母細胞において、RNAを含む複製可能又は感染性ウイルスを合成し、構築するための方法を述べている。しかしこのアプローチは、複製可能ウイルスにおけるタンパク質生産に関連する潜在的な規制上の障害を克服していない。
【0008】
(ウイルス様粒子)
組換えペプチドの生産を改善するためのもう1つのアプローチは、ウイルス様粒子(VLP)を使用することであった。一般に、カプシドに包まれたウイルスは、ウイルス核酸を含むように集合されるタンパク質コート又は「カプシド」を含む。多くのウイルスは、カプシドがVLPを形成するときに発現される細胞の内部で(「インビボ集合(in vivo assembly)」)及び単離及び精製後に細胞の外部で(「インビトロ集合(in vitro assembly)」)、個々に発現されたカプシドから「自己集合」され得るカプシドを有する。理想的には、カプシドは、標的組換えペプチドを含み、組換えウイルスカプシド−ペプチド融合物を生成するように修飾される。融合ペプチドを、次に、細胞において発現し、理想的には、組換えウイルス又はウイルス様粒子を形成するようにインビボで集合することができる。
【0009】
このアプローチは様々な程度の成功を収めている。例えばC Marusic et al., J Virol. 75 (18): 8434-39 (Sep 2001) (組換えウイルス粒子のインビボ形成を伴う、末端で抗原性HIVペプチドに融合した組換えらせんジャガイモXウイルスカプシドの植物における発現); FR Brennan et al., Vaccine 17 (15-16): 1846-57 (09 Apr 1999)(組換えウイルス粒子のインビボ形成を伴う、末端で抗原性黄色ブドウ球菌ペプチドに融合した組換え二十面体ササゲモザイクウイルス又はらせんジャガイモXウイルスカプシドの植物における発現)参照。
【0010】
Lomonossoff & Johnsonへの米国特許第5,874,087号は、 ウイルスカプシドが、生物活性ペプチド、例えばホルモン、成長因子又は抗原ペプチドを含むように加工されたカプシドを含む、植物細胞における組換え植物ウイルスの生産を述べている。コモウイルス、トンブスウイルス、ソベモウイルス、ネポウイルス属から選択されるウイルスを、外来性ペプチドをエンコードする配列を含むように加工し、加工したウイルスのゲノム全体を発現させて組換えウイルスを生産する。外来性ペプチドコード配列を、1又はそれ以上のカプシド表面ループモチーフをエンコードする配列内に挿入する。
【0011】
特定のウイルス様粒子を生産するために非親和性細胞を利用するための試みがなされてきた。例えばウイルス様粒子のインビボ形成を伴う、末端でヘプタデカペプチドに融合した組換え二十面体ジャガイモ巻葉ウイルスカプシドの昆虫細胞における発現を述べる、JW Lamb et al., J Gen. Virol. 77 (Pt. 7): 1349-58 (Jul 1996)参照のこと。ある状況では、非親和性VLPは好ましいと考えられる。例えば非親和性ウイルスカプシドは、天然のウイルスカプシドよりも、ウイルス様粒子へと集合する能力を破壊することのない異種ペプチドの挿入に対して適応しやすいと考えられる。あるいは、非親和性ウイルスカプシドは、天然の親和性ウイルスからのカプシドよりも詳細に特性が決定され、理解され得る。加えて、特定の適用、例えばワクチン生産は、特定の宿主細胞発現系における親和性ウイルスの使用を許容しないことがある。Chapmanらへの米国特許第6,232,099号は、植物においてウイルスカプシドサブユニットに結合された異種タンパク質を生産するための桿状ウイルスの使用を述べている。らせんウイルスとしても分類される桿状ウイルス、例えばジャガイモXウイルス(PVX)は、組換えウイルスカプシド−ペプチド融合物を作成するためにウイルスのゲノム内に挿入された対象組換えペプチドを有する。次に組換えウイルスを使用して宿主細胞に感染させると、ウイルスは宿主細胞において活発に複製し、さらに他の細胞に感染する。最終的に、組換えウイルスカプシド−ペプチド融合物を植物宿主細胞から精製する。
【0012】
(細菌発現系におけるウイルス様粒子の使用)
組換えペプチドの迅速で効率的且つ安価で豊富な収量の潜在的可能性の故に、細菌は、組換えウイルスカプシド−ペプチド融合ウイルス様粒子の生産のための発現系における宿主細胞として検討されてきた。
【0013】
研究者達は、組換えペプチド挿入物を含まない特定の野生型ウイルスカプシドが、非親和性腸内細菌においてトランスジェニック発現できることを示した。研究者達はまた、これらのカプシドが、インビボ及びインビトロの両方で、ウイルス様粒子を形成するように集合できることを示した。例えばSJ Shire et al., Biochemistry 29 (21): 5119-26 (29 May 1990) (大腸菌において発現されるらせんタバコモザイクウイルスカプシドからのウイルス様粒子のインビトロ集合) ; X Zhao et al., Virology 207 (2): 486-94 (10 Mar 1995) (大腸菌において発現される二十面体ササゲ退緑斑紋ウイルスカプシドからのウイルス様粒子のインビトロ集合); Y Stram et al., Virus Res. 28(1) : 29-35 (Apr 1993)(ウイルス様粒子のインビボ形成を伴う、大腸菌におけるひも状ジャガイモYウイルスカプシドの発現); J Joseph & HS Savithri, Arch. Virol. 144 (9): 1679-87 (1999) (ウイルス様粒子のインビボ形成を伴う、大腸菌におけるひも状トウガラシ葉脈緑帯ウイルスカプシドの発現); DJ Hwang et al., Proc. Nat'I Acad. Sci. USA 91 (19): 9067-71 (13 Sep 1994) (ウイルス様粒子のインビボ形成を伴う、大腸菌におけるらせんタバコモザイクウイルスカプシドの発現); M Sastri et al., J Mol. Biol. 272 (4): 541-52 (03 Oct 1997)(ウイルス様粒子のインビボ形成を伴う、大腸菌における二十面体ホオズキ斑紋ウイルスカプシドの発現)参照。
【0014】
現在まで、非親和性細菌細胞における組換えウイルスカプシド−ペプチド融合物の発現及びインビボ集合の成功は様々であった。一般に、これらの粒子のインビボ集合の成功は、非二十面体ウイルスカプシド−標的ペプチド融合物粒子に限られてきた。例えばMN Jagadish et al.,Intervirology 39 (1-2): 85-92 (1996) (ウイルス様粒子のインビボ形成を伴う、末端で抗原ペプチドに融合した組換えひも状非二十面体セイバンモロコシモザイクウイルスカプシドの非植物細胞における発現)参照。
【0015】
二十面体カプシドに連結されたペプチドの発現は、成功していないか又は限られた有用性しかなかった。例えばV Yusibov et al., J Gen. Virol. 77 (Pt. 4): 567-73 (Apr 1996)は、末端でヘキサヒスチジンペプチドに融合した、大腸菌で発現された組換え二十面体アルファルファモザイクウイルスカプシドからのウイルス様粒子のインビトロ集合を述べた。
【0016】
Brumfieldらは、二十面体カプシドに挿入した組換えペプチドをインビボ集合したウイルス様粒子として発現することを試みたが、成功しなかった。Brumfield et al. , (2004)「ササゲ退緑斑紋ブロモウイルスの修飾カプシドの異種発現は、変化した構造及び機能を有するタンパク質ケージの集合をもたらす」J. Gen. Vir. 85: 1049-1053参照。観察された、二十面体ウイルスカプシド−ペプチド融合物粒子を大腸菌においてインビボでウイルス様粒子として集合することの不可能性の理由はよく理解されていない。Brumfieldらはこの集合の失敗を、大腸菌が不溶性カプシドを生産するという事実に結び付けている。
【0017】
Chapmanは、米国特許第6,232,099号において、限られた挿入物サイズが二十面体ウイルスによって許容されることを指摘する。Chapmanは、自らの主張を裏付けるのに、植物宿主細胞における発現のための二十面体ウイルス内の組換えペプチドの大きさを26アミノ酸の長さに限定する、国際公開広報第WO92/18618を引用している。Chapmanは、二十面体ウイルスのカプシドの内部挿入部位に存在するより大きなペプチドは、タンパク質の形状及び/又は他のカプシドと成功裏に相互作用するその能力の破壊をもたらし、それがキメラウイルスの構築の失敗を導き得ると理論付けている。この文献はまた、大腸菌を含み得る細胞においてカプシド−組換えペプチド融合ペプチドを生産するための非複製桿状ウイルスの使用を述べている。
【発明の開示】
【発明が解決しようとする課題】
【0018】
したがって、二十面体ウイルスに由来するウイルス様粒子の生産のための改善された細菌発現系を提供することが本発明の1つの目的である。
【0019】
ウイルス様粒子の生産のための改善された発現系において宿主細胞として使用するための細菌生物を提供することが本発明のもう1つの目的である。
【0020】
細菌におけるウイルス様粒子の改善された生産のための方法を提供することが本発明のさらにもう1つの目的である。
【0021】
ウイルス様粒子の生産のための改善された細菌発現系において使用するための新規構築物及び核酸を提供することが本発明のさらにもう1つの目的である。
【課題を解決するための手段】
【0022】
二十面体カプシド−組換えペプチド融合物粒子は、シュードモナス生物において発現されたときインビボでウイルス様粒子又は可溶性ケージ構造へと集合する。さらに、50アミノ酸以上の大きな組換えペプチド又はペプチドコンカテマーを、二十面体カプシドに挿入し、シュードモナス生物においてインビボで集合することができる。
【0023】
本発明の1つの態様では、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を含むシュードモナス生物を提供する。本発明の1つの特定の実施形態では、シュードモナス細胞はシュードモナス・フルオレッセンスである。1つの実施形態では、前記細胞はウイルス様粒子又は可溶性ケージ構造を生産する。
【0024】
この細胞において生産されるウイルス様粒子は、典型的には細胞に感染することができない。ウイルスカプシド配列は、細胞に親和性でないウイルスに由来し得る。1つの実施形態では、細胞は、所望の二十面体カプシド以外のウイルスタンパク質を含まない。1つの実施形態では、ウイルスカプシドは、この細胞とは異なる科の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、この細胞とは異なる属の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、この細胞とは異なる種の生物に対して親和性を有するウイルスに由来する。本発明の1つの実施形態では、二十面体カプシドは植物二十面体ウイルスに由来する。特定の実施形態では、二十面体カプシドは、ササゲ退緑斑紋ウイルス(Cowpea Chlorotic Mottle Virus)、ササゲモザイクウイルス(Cowpea Mosaic Virus)及びアルファルファモザイクウイルス(Alfalfa Mosaic Virus)から選択される群に由来する。
【0025】
本発明の1つの実施形態では、二十面体カプシドに融合した組換えペプチドは、ヒト又は動物治療のために有用な治療ペプチドである。1つの特定の実施形態では、組換えペプチドは抗原である。1つの実施形態では、カプシド−組換えペプチドウイルス様粒子はヒト又は動物適用においてワクチンとして投与することができる。もう1つの特定の実施形態では、組換えペプチドは、遊離単量体形態であるとき宿主細胞に対して毒性であるペプチドである。より特定の実施形態では、毒性ペプチドは抗菌ペプチドである。
【0026】
1つの実施形態では、二十面体カプシドに融合した組換えペプチドは、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも35、少なくとも40、少なくとも45、少なくとも50、少なくとも55、少なくとも60、少なくとも65、少なくとも75、少なくとも85、少なくとも95、少なくとも99又は少なくとも100アミノ酸である。
【0027】
本発明の1つの実施形態では、二十面体カプシドに融合した組換えペプチドは、所望の標的ペプチドの少なくとも1個の単量体を含む。選択的実施形態では、組換えペプチドは、所望の標的ペプチドの2個以上の単量体を含む。特定の実施形態では、このペプチドは、コンカテマーペプチドとしてカプシドに作動可能に連結された少なくとも2個、少なくとも5個、少なくとも10個、少なくとも15個又は少なくとも20個の別々の単量体からなる。もう1つの実施形態では、コンカテマーペプチド内の個々の単量体は、切断可能なリンカー領域によって連結される。さらにもう1つの実施形態では、組換えペプチドを二十面体ウイルスカプシドの少なくとも1つの表面ループに挿入する。1つの実施形態では、少なくとも1個の単量体を二十面体ウイルスカプシドの2つ以上の表面ループに挿入する。
【0028】
ウイルス様粒子の2つ以上のループを修飾することができる。1つの特定の実施形態では、組換えペプチドを、二十面体ウイルス様粒子の少なくとも2つの表面ループ上で発現する。もう1つの実施形態では、少なくとも2個の異なるペプチドをウイルスカプシド、ケージ又はウイルス様粒子の少なくとも2つの表面ループに挿入する。もう1つの実施形態では、少なくとも3個の組換えペプチドをウイルス様粒子の少なくとも3つの表面ループに挿入する。表面ループ内の組換えペプチドは同じアミノ酸配列を有し得る。別の実施形態では、表面ループ内の組換えペプチドのアミノ酸配列は異なる。
【0029】
もう1つの実施形態では、この細胞は、第二の野生型カプシド又はカプシド−組換えペプチド融合ペプチドのいずれかをエンコードする少なくとも1個の付加的な核酸を含み、多数のカプシドがインビボでキメラウイルス様粒子を生成するように集合する。
【0030】
本発明の1つの態様では、二十面体カプシドと組換えペプチドの融合ペプチドを含むシュードモナス生物を提供する。本発明の1つの詳細な実施形態では、シュードモナス細胞はシュードモナス・フルオレッセンスである。1つの実施形態では、カプシド−組換えペプチド融合ペプチドは、インビボでウイルス様粒子を形成するように集合する。
【0031】
本発明のもう1つの態様では、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を提供する。本発明の1つの実施形態では、二十面体カプシドは植物二十面体ウイルスに由来する。特定の実施形態では、二十面体カプシドは、ササゲモザイクウイルス、ササゲ退緑斑紋ウイルス及びアルファルファモザイクウイルスから選択される群に由来する。
【0032】
1つの実施形態では、組換えペプチドは、遊離単量体形態であるとき宿主細胞に対して毒性であるペプチドである。より特定の実施形態では、毒性ペプチドは抗菌ペプチドである。
【0033】
本発明の1つの実施形態では、組換えペプチドは、所望の標的ペプチドの少なくとも1個の単量体を含む。別の実施形態では、組換えペプチドは、所望の標的ペプチドの2個以上の単量体を含む。さらにもう1つの実施形態では、組換えペプチドを二十面体ウイルスカプシドの少なくとも1つの表面ループに挿入する。
【0034】
もう1つの実施形態では、核酸構築物は、少なくとも1個のプロモーター、少なくとも1個の選択マーカー、少なくとも1個のオペレーター配列、少なくとも1個の複製起点、及び少なくとも1個のリボソーム結合部位を含む付加的な核酸配列を含み得る。
【0035】
1つの態様では、本発明は、
a)シュードモナス細胞を提供すること;
b)組換えペプチドと二十面体カプシドからなる融合ペプチドをエンコードする核酸を提供すること;
c)このシュードモナス細胞においてこの核酸を発現すること、ここで細胞における発現がこの融合ペプチドのウイルス様粒子へのインビボ集合を提供する;及び
d)ウイルス様粒子を単離すること
を含む、組換えペプチドを生産するための方法を提供する。
1つの実施形態では、この工程は、e)この融合生成物を切断して、カプシドから組換えペプチドを分離すること、をさらに含む。本発明の1つの実施形態では、シュードモナス細胞はシュードモナス・フルオレッセンスである。
【0036】
1つの実施形態では、この方法は、カプシドがインビボでキメラウイルス様粒子を生成するように集合する、野生型カプシド又はカプシド−組換えペプチド融合ペプチドをエンコードするもう1つ別の核酸を共発現することを含む。
【0037】
本発明のもう1つの態様では、
a)シュードモナス細胞;
b)少なくとも1個の組換えペプチドと少なくとも1個の二十面体ウイルスカプシドを含む融合ペプチドをエンコードする核酸;及び
c)培養培地
を含む組換えペプチドの生産のための発現系を提供する。
【0038】
発現されたとき融合ペプチドは、細胞内でウイルス様粒子へと集合することができる。
【発明を実施するための最良の形態】
【0039】
本発明は、二十面体ウイルスカプシドと対象組換えペプチドを含む融合ペプチドの細菌における発現のための方法を提供する。ここで使用する「ペプチド」という用語は、いかなる特定の分子量にも限定されず、タンパク質又はポリペプチドも包含し得る。本発明はさらに、この方法における使用のための細菌細胞及び核酸構築物を提供する。特に、本発明は、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を有するシュードモナス生物を提供する。本発明の1つの特定の実施形態では、シュードモナス細胞はシュードモナス・フルオレッセンスである。1つの実施形態では、この細胞は、ウイルス様粒子又は可溶性ケージ構造を生産する。本発明はまた、1つの実施形態ではヒト及び動物治療のために有用な治療ペプチドであり得る、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を提供する。
【0040】
本発明はまた、組換えペプチドと二十面体カプシドの融合ペプチドをエンコードする核酸を提供すること;この核酸をシュードモナス細胞において発現し、この細胞における発現が融合ペプチドのウイルス様粒子へのインビボ集合を提供すること;及びウイルス様粒子を単離すること、によってシュードモナス細胞において組換えペプチドを生産するための方法を提供する。
【0041】
(I.組換えシュードモナス細胞)
本発明は、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を含むシュードモナス細胞を提供する。この細胞は、組換えペプチドを生産するための方法において利用することができる。
【0042】
(ウイルスカプシド)
1つの実施形態では、本発明は、二十面体ウイルスカプシドに融合したペプチドの発現によってペプチドを生産するための方法における使用のためのシュードモナス細胞を提供する。この発現は、典型的には細胞内で少なくとも1個のウイルス様粒子(VLP)を生じさせる。
【0043】
ウイルスは、らせん対称又は二十面体対称を有するものに分類できる。一般に認識されるカプシド形態は、二十面体(正二十面体(icosahedral proper)、等軸晶系、準等軸晶系、及び双生又は「双晶(twinned)」を含む)、多面体(球状、卵形及びレモン形を含む)、桿菌状(棒又は弾丸形、及び紡錘状又は葉巻形を含む)、及びらせん状(桿状、円柱状及びひも状を含む)を含み、いずれも尾部を有していてもよく及び/又は表面突起、例えば棘又はこぶを含んでいてもよい。
【0044】
(形態)
本発明の1つの実施形態では、カプシドのアミノ酸配列は、何らかの二十面体形態を有すると分類されるウイルスのカプシドから選択される。1つの実施形態では、カプシドアミノ酸配列は、二十面体である実在のカプシドから選択される。もう1つの実施形態では、カプシドアミノ酸配列は、二十面体ウイルスのカプシドから選択される。1つの特定の実施形態では、カプシドアミノ酸配列は、二十面体植物ウイルスのカプシドから選択される。しかし、もう1つの実施形態では、ウイルスカプシドは、植物に対して感染性でない二十面体ウイルスに由来する。例えば1つの実施形態では、ウイルスは、哺乳動物に対して感染性のウイルスである。
【0045】
一般に、二十面体ウイルスのウイルスカプシドは、二十面体(立方)対称に配列された多数のタンパク質サブユニットからなる。天然の二十面体カプシドは、例えば3つのサブユニットが各々カプシドの各々の三角形の面を形成することによって構築することができ、60サブユニットが完全なカプシドを形成することになる。この小さなウイルス構造の典型は、例えばバクテリオファージφX174である。多くの二十面体ウイルスカプシドは60を超えるサブユニットを含む。二十面体ウイルスの多くのカプシドは、逆平行の8本鎖β−バレル型折りたたみモチーフを含む。このモチーフは、一方の側に4本のβ鎖(BIDGと称される)及び他方の側に4本のβ鎖(CHEFと称される)を備えたくさび形ブロックを有する。また2個の保存されたα−ヘリックスが存在し、1個はβCとβDの間、他の1個はβEとβFの間にある。
【0046】
エンベロープを持つウイルスは、膜を通して粒子が排出(出芽)することによる全面的な破壊を受けずに感染細胞を出て行くことができ、その間に粒子は細胞膜に由来する脂質エンベロープ内に被包される(例えばAJ Cann (ed. ) (2001)Principles of Molecular Virology (Academic Press); A Granoff and RG Webster (eds. ) (1999) Encyclopedia of Virology (Academic Press); DLD Caspar (1980) Bioplzys. J. 32: 103; DLD Caspar and A Klug (1962) Cold Spring Harbor Symp. Quant. Biol. 27: 1; J Grimes et al. (1988) Nature 395: 470; JE Johnson (1996) Proc. Nat'l Acad. Sci. USA 93: 27; and J Johnson and J Speir (1997) J. Mol. Biol. 269: 665参照)。
【0047】
(ウイルス)
ウイルス分類学は、以下の分類群のカプシド被包粒子の存在を認識する:
グループIウイルス、すなわちdsDNAウイルス;
・グループIIウイルス、すなわちssDNAウイルス;
・グループIIIウイルス、すなわちdsRNAウイルス;
・グループIVウイルス、すなわちDNA段階を持たないssRNA(+)鎖ウイルス;
・グループVウイルス、すなわちssRNA(−)鎖ウイルス;
・グループVIウイルス、すなわちssRNA逆転写ウイルスである、RNAレトロイドウイルス;
・グループVIIウイルス、すなわちdsDNA逆転写ウイルスである、DNAレトロイドウイルス;
・デルタウイルス;
・ウイロイド;及び
・サテライト核酸及びプリオンを除く、サテライトファージ及びサテライトウイルス。
【0048】
これらの分類群のメンバーは当業者に周知であり、H. V. Van Regenmortel et al. (eds.), Virus Taxonomy: Seventh Report of the International Committee on Taxonomy of Viruses (2000) (Academic Press/Elsevier, Burlington Mass. , USA); the University of Leicester (UK) Microbiology & Immunology Departmentのhttp : //wwwmicro. msb. le. ac. uk/3035/Virusgroups. htmlにおけるVirus Taxonomyウェブページ; およびthe National Center for Biotechnology Information(NCBI) of the National Library of Medicine of the National Institutes of Health of the US Department of Health & Human Services (Washington, D. C. , USA)のhttp : //www. ncbi. nlm. nih. gov/Taxonomy/tax. Html.におけるTaxonomy Browserのオンライン"Virus"および"Viroid"セクションにおいて総説されている。
【0049】
カプシドのアミノ酸配列は、これらの分類群のいずれかのメンバーのカプシドから選択され得る。これらの分類群のメンバーのカプシドについてのアミノ酸配列は、例えば、the NCBIによってhttp://www.ncbi. nlm. nih.gov/entrez/query. fcgi.において提供されているPubMed search facilityのオンライン"Nucleotide" (Genbank), "Protein,"および"Structure"セクションを含むがこれらに限定されないソースから入手し得る。
【0050】
1つの実施形態では、カプシドアミノ酸配列は、以下の宿主の少なくとも1つに特異的な分類群のメンバーから選択される:酵母を含む真菌、植物、藻類を含む原生生物、無脊椎動物、脊椎動物、及びヒト。1つの実施形態では、カプシドアミノ酸配列は、以下の分類群のいずれかのメンバーから選択される:グループI、グループII、グループIII、グループIV、グループV、グループVI、グループVII、ウイロイド、及びサテライトウイルス。1つの実施形態では、カプシドアミノ酸配列は、上述した6つの型の宿主の少なくとも1つに特異的なこれら7つの分類群のいずれかのメンバーから選択される。より特定の実施形態では、カプシドアミノ酸配列は、グループII、グループIII、グループIV、グループVII、及びサテライトウイルスのいずれかのメンバーから;又はグループII、グループIV、グループVII、及びサテライトウイルスのいずれかから選択される。もう1つの実施形態では、ウイルスカプシドはグループIV又はグループVIIから選択される。
【0051】
ウイルスカプシド配列は、当該細胞に対して親和性でないウイルスに由来し得る。1つの実施形態では、当該細胞は、所望の二十面体カプシド以外の特定の選択されたウイルスからのウイルスタンパク質を含まない。1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる科の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる属の生物に対して親和性を有するウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、当該細胞とは異なる種の生物に対して親和性を有するウイルスに由来する。
【0052】
特定の実施形態では、ウイルスカプシドはグループIVのウイルスから選択される。
【0053】
1つの実施形態では、ウイルスカプシドは二十面体ウイルスから選択される。二十面体ウイルスは、乳頭腫ウイルス科、トティウイルス科、ジシストロウイルス科、ヘパドナウイルス科、トガウイルス科、ポリオーマウイルス科、ノダウイルス科、テクティウイルス科、レヴィウイルス科、ミクロウイルス科、シフォビラーダ科、ノダウイルス科、ピコルナウイルス科、パルボウイルス科、カルシウイルス科、テトラウイルス科、及びサテライトウイルスのいずれかのメンバーから選択され得る。
【0054】
特定の実施形態では、この配列は、少なくとも1つの植物宿主に特異的な分類群のいずれかのメンバーから選択される。1つの実施形態では、二十面体植物ウイルス種は、ブンヤウイルス科、レオウイルス科、ラブドウイルス科、ルテオウイルス科、ナノウイルス科、パルティティウイルス科、セキウイルス科、ティモウイルス科、ウルミアウイルス属、タバコ壊死サテライトウイルス、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科分類群のいずれかである、又はそのいずれかのメンバーである、植物感染性ウイルス種である。1つの実施形態では、二十面体植物ウイルス種は、ルテオウイルス科、ナノウイルス科、パルティティウイルス科、セキウイルス科、ティモウイルス科、ウルミアウイルス属、タバコ壊死サテライトウイルス、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科分類群のいずれかである、又はそのいずれかのメンバーである、植物感染性ウイルス種である。特定の実施形態では、二十面体植物ウイルス種は、カリモウイルス科、ジェミニウイルス科、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科のいずれかである、又はそのいずれかのメンバーである、植物感染性ウイルス種である。より特定の実施形態では、二十面体植物ウイルス種は、コモウイルス科、ソベモウイルス属、トンブスウイルス科、又はブロモウイルス科のいずれかである、又はそのいずれかのメンバーである、植物感染性ウイルス種である。より特定の実施形態では、二十面体植物ウイルス種は、コモウイルス科又はブロモウイルス科のメンバーである植物感染性ウイルス種である。特定の実施形態では、ウイルスカプシドは、ササゲモザイクウイルス又はササゲ退緑斑紋ウイルスに由来する。もう1つの実施形態では、ウイルスカプシドは、ブロモウイルス科分類群の種に由来する。特定の実施形態では、カプシドは、イラルウイルス属又はアルファモウイルス属に由来する。より特定の実施形態では、カプシドは、タバコ条斑病ウイルス又はアルファルファモザイクウイルス(AMV)(AMV1又はAMV2を含む)に由来する。
【0055】
(VLP)
本発明の二十面体ウイルスカプシドは、ここで述べる宿主細胞において非感染性である。1つの実施形態では、ウイルスカプシドの発現の間又は発現後に宿主細胞においてウイルス様粒子(VLP)又はケージ構造を形成する。1つの実施形態では、VLP又はケージ構造はまた、対象とするペプチドを含み、特定の実施形態では、対象ペプチドをVLPの表面に発現する。この発現系は、典型的にはウイルス感染性を可能にする付加的なウイルスタンパク質を含まない。典型的実施形態では、発現系は、宿主細胞及び1又はそれ以上のウイルスカプシド及び作動可能に連結された対象ペプチドをコードするベクターを含む。このベクターは、典型的には付加的なウイルス集合タンパク質を含まない。本発明は、ウイルスカプシドが特定の宿主細胞においてより大きな度合で形成され、組換えペプチドのより効率的な回収を可能にするという発見に由来する。
【0056】
1つの実施形態では、VLP又はケージ構造は、3−約200カプシドを含む、カプシドの多量体構築物である。1つの実施形態では、VLP又はケージ構造は、少なくとも30、少なくとも50、少なくとも60、少なくとも90又は少なくとも120カプシドを含む。もう1つの実施形態では、各々のVLP又はケージ構造は、少なくとも150カプシド、少なくとも160カプシド、少なくとも170カプシド又は少なくとも180カプシドを含む。
【0057】
1つの実施形態では、VLPは二十面体構造として発現される。もう1つの実施形態では、VLPは、そのカプシド配列が由来する天然ウイルスと同じ形状で発現される。別の実施形態では、しかしながら、VLPは天然ウイルスと同じ形状を持たない。特定の実施形態では、例えば、構造は、多数のカプシドで形成されるが、天然型VLPを形成しない粒子中で生産される。例えば約6、9、12、15、18、21、24、27、30、33、36、39、42、45、48、51、54、57又は60カプシドのケージ構造が形成され得る。
【0058】
1つの実施形態では、カプシドの少なくとも1個は、少なくとも1個の対象ペプチドを含む。1つの実施形態では、ペプチドは、VLPの少なくとも1つの内部ループ又は少なくとも1つの外部表面ループ内で発現される。
【0059】
ウイルスカプシドの2つ以上のループを修飾することができる。1つの特定の実施形態では、組換えペプチドは、二十面体ウイルス様粒子の少なくとも2つの表面ループ上で発現される。もう1つの実施形態では、少なくとも2個の異なるペプチドを、ウイルスカプシド、ケージ又はウイルス様粒子の少なくとも2つの表面ループに挿入する。もう1つの実施形態では、少なくとも3個の組換えペプチドをウイルス様粒子の少なくとも3つの表面ループに挿入する。表面ループ内の組換えペプチドは、同じアミノ酸配列を有し得る。別の実施形態では、表面ループ内の組換えペプチドのアミノ酸配列は異なる。
【0060】
特定の実施形態では、宿主細胞を、VLPの集合を改善するように修飾することができる。宿主細胞を、例えば発現されたウイルスカプシドからのVLPの形成を促進するシャペロンタンパク質を含むように改変することができる。もう1つの実施形態では、宿主細胞を、カプシドの発現をより効率よく調節して、VLPの形成調節を促進するためのリプレッサータンパク質を含むように改変する。
【0061】
ウイルスカプシド又はタンパク質をエンコードする核酸配列はまた、VLPの形成を変化させるように付加的に修飾することができる(例えばBrumfield, et al. (2004) J. Gen. Virol. 85: 1049-1053参照)。例えば修飾の3つの一般的なクラスは、最も典型的にはVLPの発現及び集合を修飾するために行われる。これらの修飾は、集合されたタンパク質ケージ内の隣接サブユニットの内部、外部又はその間の界面を変化させるように設計される。これを実現するために、突然変異誘発プライマーを使用して、(i)N末端の塩基性残基(例えばK、R)を酸性グルタミン酸に置き換えることによってウイルス核酸結合領域の内表面電荷を変化させる(Douglas et al., 2002b);(ii)N末端から内部残基を欠失させる(CCMVにおいて、通常は残基4−37);(iii)11アミノ酸ペプチド細胞標的配列をエンコードするcDNA(Graf et al., 1987)を表面暴露ループに挿入する;及び(iv)金属結合部位を変化させることによってウイルスサブユニットの間の相互作用を改変する(CCMVにおいて、残基81/148突然変異体)ことができる。
【0062】
(組換えペプチド)
(大きさ)
1つの実施形態では、ウイルスカプシド配列に作動可能に連結されたペプチドは、少なくとも2個のアミノ酸を含む。もう1つの実施形態では、このペプチドは、少なくとも3、少なくとも4、少なくとも5又は少なくとも6アミノ酸の長さである。別の実施形態では、ペプチドは少なくとも7アミノ酸の長さである。ペプチドはまた、少なくとも8、少なくとも9、少なくとも10、少なくとも11、12、13、14、15、16、17、18、19、20、30、45、50、60、65、75、85、95、96、99又はそれ以上のアミノ酸の長さである。1つの実施形態では、コードされるペプチドは少なくとも25kDである。
【0063】
1つの実施形態では、ペプチドは、2−約300アミノ酸、又は約5−約250アミノ酸、又は約5−約200アミノ酸、又は約5−約150アミノ酸、又は約5−約100アミノ酸を含む。もう1つの実施形態では、ペプチドは、約10−約140アミノ酸、又は約10−約120アミノ酸、又は約10−約100アミノ酸を含む。
【0064】
1つの実施形態では、ウイルスカプシド配列に作動可能に連結されたペプチド又はタンパク質は約500アミノ酸を含む。1つの実施形態では、ペプチドは500個未満のアミノ酸を含む。もう1つの実施形態では、ペプチドは、約300個までのアミノ酸、又は約250個まで、又は約200個まで、又は約180個まで、又は約160個まで、又は約150個まで、又は約140個まで、又は約120個まで、又は約110個まで、又は約100個まで、又は約90個まで、又は約80個まで、又は約70個まで、又は約60個まで、又は約50個まで、又は約40個まで、又は約30個までのアミノ酸を含む。
【0065】
1つの実施形態では、二十面体カプシドに融合した組換えペプチドは、少なくとも7、少なくとも8、少なくとも9、少なくとも10、少なくとも12、少なくとも15、少なくとも20、少なくとも25、少なくとも30、少なくとも35、少なくとも40、少なくとも45、少なくとも50、少なくとも55、少なくとも60、少なくとも65、少なくとも75、少なくとも85、少なくとも95、少なくとも99、又は少なくとも100アミノ酸である。
【0066】
本発明の1つの実施形態では、組換えペプチドは、所望の標的ペプチドの少なくとも1個の単量体を含む。選択的実施形態では、組換えペプチドは、所望の標的ペプチドの2個以上の単量体を含む。特定の実施形態では、ペプチドは、コンカテマーペプチドとしてカプシドに作動可能に連結された、少なくとも2個、少なくとも5個、少なくとも10個、少なくとも15個又は少なくとも20個の別個の単量体からなる。もう1つの実施形態では、コンカテマーペプチド内の個々の単量体は、切断可能なリンカー領域によって連結される。さらにもう1つの実施形態では、組換えペプチドを、二十面体ウイルス様粒子の少なくとも1つの表面ループに挿入する。1つの実施形態では、少なくとも1個の単量体をウイルス様粒子の表面ループに挿入する。
【0067】
(分類)
ウイルスカプシドに融合する対象ペプチドは、ウイルスに由来しない、及び場合により、細胞と同じ宿主細胞に由来しない、異種タンパク質であり得る。
【0068】
ウイルスカプシドに融合する対象ペプチドは、機能性ペプチド:構造ペプチド;抗原ペプチド;毒性ペプチド、抗菌ペプチド、それらのフラグメント;それらの前駆体;これらのいずれかの組合せ;及び/又はこれらのいずれかのコンカテマーであり得る。本発明の1つの実施形態では、組換えペプチドは、ヒト及び動物の治療のために有用な治療ペプチドである。
【0069】
機能性ペプチドは、例えば:生物活性ペプチド(すなわち生物学的実在物(entity)、例えば生物、細胞、培養物、組織、器官又は細胞小器官、における又はその生物学的機能又は活性の開始、増強、延長、減衰、終結又は防止を及ぼす、惹起する又はさもなければ生じさせるペプチド);触媒ペプチド;ミクロ構造及びナノ構造活性ペプチド(すなわちその中で又はそれに関連して作用、例えば運動、エネルギー変換、を行う構築されたミクロ又はナノ構造の一部を形成するペプチド);及び刺激剤ペプチド(例えばペプチド着香料、着色料、着臭剤、フェロモン、誘引物質、抑制物質及び忌避物質)を含むが、これらに限定されない。
【0070】
生物活性ペプチドは、例えば:免疫活性ペプチド(例えば抗原ペプチド、アレルゲンペプチド、ペプチド免疫調節剤(immunoregulator)、ペプチド免疫調節剤(immunomodulator):シグナル伝達ペプチド(例えばペプチドホルモン、サイトカイン及び神経伝達物質;受容体;アゴニスト及びアンタゴニストペプチド;ペプチドターゲティング及び分泌シグナルペプチド);及び生物阻害性ペプチド(例えば毒性、生物致死性又は生物抑制性ペプチド、例えばペプチド毒素及び抗菌ペプチド)を含むが、これらに限定されない。
【0071】
構造ペプチドは、例えば:ペプチドアプタマー、折りたたみペプチド(例えばもう1つ別の分子の物理的立体配座の形成又は維持を促進する又は誘導するペプチド);接着促進ペプチド(例えば接着ペプチド、細胞接着促進ペプチド);界面ペプチド(例えばペプチド界面活性剤及び乳化剤);ミクロ構造及びナノ構造構築ペプチド(すなわち加工されたミクロ又はナノ構造の一部を形成する構造ペプチド);及びプレ活性化ペプチド(例えばプレ、プロ及びプレ−プロタンパク質のリーダーペプチド及びペプチド;インテイン)を含むが、これらに限定されない。
【0072】
触媒ペプチドは、例えばアポB RNAエディティングシチジンデアミナーゼペプチド;グルタミニル−tRNAシンテターゼの触媒ペプチド:アスパラギン酸トランスカルバモイラーゼの触媒ペプチド;植物1型リボソーム不活性化ペプチド;ウイルス触媒ペプチド、例えば口蹄疫ウイルス[FMDV−2A]触媒ペプチド;マトリックスメタロプロテイナーゼペプチド;及び触媒メタロオリゴペプチドを含む。
【0073】
ペプチドはまた、ペプチドエピトープ、ハプテン又は関連ペプチド(例えば抗原ウイルスペプチド;ウイルス関連ペプチド、例えばHIV関連ペプチド、肝炎関連ペプチド;抗体イディオタイプドメイン;細胞表面ペプチド;抗原性ヒト、動物、原生生物、植物、真菌、細菌及び/又は古細菌ペプチド;アレルゲンペプチド及びアレルゲン脱感作ペプチド)であり得る。
【0074】
ペプチドはまた、ペプチド免疫調節剤(immunoregulatorまたはimmunomodulator)(例えばインターフェロン、インターロイキン、ペプチド免疫抑制剤及び免疫増強剤);抗体ペプチド(例えば一本鎖抗体;一本鎖抗体フラグメント及び構築物、例えば一本鎖Fv分子;抗体軽鎖分子、抗体重鎖分子、ドメイン欠失抗体軽鎖又は重鎖分子;一本鎖抗体ドメイン及び分子、例えばCH1、CH1−3、CH3、CH1−4、CH4、VHCH1、CL、CDR1又はFR−CDR1−FR2ドメイン;パラトープペプチド;ミクロ抗体);別の結合ペプチド(例えばペプチドアプタマー、細胞内及び細胞表面受容体タンパク質、受容体フラグメント;抗腫瘍壊死因子ペプチド)であり得る。
【0075】
ペプチドはまた、酵素基質ペプチド又は酵素阻害剤ペプチド(例えばカスパーゼ基質及び阻害剤、プロテインキナーゼ基質及び阻害剤、蛍光共鳴エネルギー転移ペプチド酵素基質)であり得る。
【0076】
ペプチドはまた、細胞表面受容体ペプチドリガンド、アゴニスト及びアンタゴニスト(例えばセルレイン、ダイノルフィン、オレキシン、下垂体アデニル酸シクラーゼ活性化ペプチド、腫瘍壊死因子ペプチド;合成ペプチドリガンド、アゴニスト及びアンタゴニスト);ペプチドホルモン(例えば内分泌、パラ分泌及び自己分泌ホルモン、例えばアミリン、アンギオテンシン、ブラジキニン、カルシトニン、心臓興奮性神経ペプチド、カゾモルフィン、コレシストキニン、コルチコトロピン及びコルチコトロピン関連ペプチド、分化因子、エンドルフィン、エンドセリン、エンケファリン、エリスロポエチン、エキセンディン、卵胞刺激ホルモン、ガラニン、ガストリン、グルカゴン及びグルカゴン様ペプチド、ゴナドトロピン、成長ホルモン及び増殖因子、インスリン、カリジン、キニン、レプチン、向脂肪ホルモン、黄体形成ホルモン、メラニン細胞刺激ホルモン、メラトニン、ナトリウム排泄増加性ペプチド、ニューロキニン、ニューロメジン、ノシセプチン、オステオカルシン、オキシトシン(すなわちオシトシン)、副甲状腺ホルモン、プレイオトロフィン、プロラクチン、レラキシン、セクレチン、セロトニン、睡眠誘発ペプチド、ソマトメイジン、サイモポイエチン、甲状腺刺激ホルモン、チロトロピン、ウロテンシン、血管作用性小腸ペプチド、バソプレシン);ペプチドサイトカイン、ケモカイン、ウイロカイン(virokine)及びウイロセプター(viroceptor)ホルモン放出及び放出抑制ペプチド(例えばコルチコトロピン放出ホルモン、コルチスタチン、卵胞刺激ホルモン放出因子、胃抑制性ペプチド、ガストリン放出ペプチド、ゴナドトロピン放出ホルモン、成長ホルモン放出ホルモン、黄体形成ホルモン放出ホルモン、メラノトロピン放出ホルモン、メラノトロピン放出抑制因子;ノシスタチン、パンクリアスタチン、プロラクチン放出ペプチド、プロラクチン放出抑制因子;ソマトスタチン;甲状腺刺激ホルモン放出ホルモン);ペプチド神経伝達物質又はチャネルブロッカー(例えばボンベシン、神経ペプチドY、ニューロテンシン、サブスタンスP);ペプチド毒素、毒素前駆体ペプチド又は毒素ペプチド部分であり得る。特定の実施形態では、ペプチド毒素はD−アミノ酸を含まない。毒素前駆体ペプチドは、D−アミノ酸を含まないもの及び/又は翻訳後修飾によって、例えばアミノ酸にD−立体配置を導入することができるD立体配置誘導剤(例えばペプチドイソメラーゼ又はエピメラーゼ又はラセマーゼ又はトランスアミノアーゼ)の作用によって、及び/又は環状ペプチド構造を形成することができる環化剤(例えばペプチドチオエステラーゼ又はペプチドリガーゼ、例えばトランススプライシングタンパク質又はインテイン)の作用によって、天然の毒素構造に変換されていないものであり得る。
【0077】
毒素ペプチド部分は、ペプチド含有毒素の線状又は前環化オリゴ及びポリペプチド部分であり得る。ペプチド毒素の例は、例えばアガトキシン、アマトキシン、カリブドトキシン、クロロトキシン、コノトキシン、デンドロトキシン、インセクトトキシン、マルガトキシン、マスト細胞脱顆粒ペプチド、サポリン、サラフォトキシン;及び細菌体外毒素、例えば炭疽毒素、ボツリヌス毒素、ジフテリア毒素及び破傷風毒素を含む。
【0078】
ペプチドはまた、代謝及び消化関連ペプチド(例えばコレシストキニン−パンクレオザイミンペプチド、ペプチドyy、膵ペプチド、モチリン);細胞接着調節又は仲介ペプチド、細胞外マトリックスペプチド(例えば付着因子、セレクチン、ラミニン);神経保護又は髄鞘形成促進ペプチド;凝集阻害ペプチド(例えば細胞又は血小板凝集阻害剤ペプチド、アミロイド形成又は沈着阻害剤ペプチド);連結ペプチド(例えば心臓血管連結神経ペプチド、iga連結ペプチド);又はその他のペプチド(例えばアグーチ関連ペプチド、アミロイドペプチド、骨関連ペプチド、細胞透過性ペプチド、コナントキシン、コントリファン、コンチュラキン、ミエリン塩基性タンパク質等)であり得る。
【0079】
特定の実施形態では、対象ペプチドは、分泌されるウイルスカプシドに対して外因性である。ペプチドは、天然又は合成配列のいずれでもよい(及びそれらのコード配列は天然又は合成ヌクレオチド配列のいずれでもよい)。したがって、例えばアミノ酸の天然、修飾天然及び完全人工配列が包含される。これらのアミノ酸配列をエンコードする核酸分子の配列は、同様に、天然、修飾天然及び完全人工核酸配列であり得、例えばその核酸分子を得るために使用される(すなわちヒトの行為によって適用される)1又はそれ以上の論理的又はランダム突然変異及び/又は組換え及び/又は合成及び/又は選択工程の結果であり得る。
【0080】
コード配列は、使用可能な場合は、標的ペプチドについての天然のコード配列であり得るが、より典型的には、例えば宿主の種のコドン使用頻度を反映する遺伝子を合成することによって、選択した発現宿主細胞における使用のために選択された、改善された又は最適化されたコード配列である。本発明の1つの実施形態では、宿主種はP.フルオレッセンスであり、シグナル配列及びペプチド配列の両方を設計するときにP.フルオレッセンスのコドン使用頻度を考慮に入れる。
【0081】
(抗原ペプチド(ペプチドエピトープ))
1つの実施形態では、抗原ペプチドはウイルスカプシドによる発現を通して生産される。抗原ペプチドは、感染因子、寄生生物、癌細胞及び他の病原体を含む、ヒト又は動物病原体の抗原ペプチドであるものから選択することができる。かかる病原体はまた、毒性因子及び病原因子、例えばそれらの因子の体外毒素、内毒素等を含む。病原体は何らかのレベルの毒力を示し得る、すなわちそれらは、例えば毒性、無毒性、偽毒性、半毒性等であり得る。1つの実施形態では、抗原ペプチドは病原体からのエピトープアミノ酸配列を含む。1つの実施形態では、エピトープアミノ酸配列は、少なくとも1個のかかる因子の表面ペプチドの少なくとも一部であるものを含む。1つの実施形態では、カプシド−組換えペプチドウイルス様粒子は、ヒト又は動物適用におけるワクチンとして使用することができる。
【0082】
2個以上の抗原ペプチドを選択してもよく、その場合、生じるウイルス様粒子は多数の異なる抗原ペプチドを提示し得る。多抗原ペプチド方式の特定の実施形態では、様々な抗原ペプチドは全て同じ病原体からの複数のエピトープから選択される。多抗原ペプチド方式の特定の実施形態では、様々な抗原ペプチドは全て、密接に関連する病原体、例えば同じ種又は同じ属の異なる種の株、亜種、次亜種、病原型、血清型又は遺伝子型(genovar)から選択される。
【0083】
1つの実施形態では、病原体は以下の群の少なくとも1つに属する:病原性の:バチルス属種、例えば炭疽菌;バルトネラ属種、例えばB.クインタナ;ブルセラ属種;ブルクホルデリア属種、例えばB.シュードマレイ(類鼻疽菌);カンピロバクター種;クロストリジウム属種、例えば破傷風菌、ボツリヌス菌;コクシエラ属種、例えばC.バーネティー;エドワードシエラ属種、例えばE.ターダ;エンテロバクター属種、例えばE.クロアカ;エンテロコッカス属種、例えばE.フェカリス、E.フェシウム;エシェリキア属種、例えば大腸菌;フランシセラ属種、例えば野兎病菌(F. tularensis);ヘモフィルス属種、例えばインフルエンザ菌;クレブシエラ属種、例えば肺炎杆菌;レジオネラ属種;リステリア属種、例えばリステリア菌(L. monocytogenes);髄膜炎菌及び淋菌、例えばナイセリア属種;モラクセラ属種;ミコバクテリウム属種、例えばらい菌、ヒト型結核菌;肺炎球菌、例えば肺炎双球菌;シュードモナス属種、例えば緑膿菌;リケッチア属種、例えば発疹チフスリケッチア、斑点熱リケッチア、発疹熱リケッチア;サルモネラ属菌、例えば腸チフス菌;ブドウ球菌属種、例えば黄色ブドウ球菌;A型連鎖球菌属及び溶血連鎖球菌属を含む、連鎖球菌属種、例えば肺炎連鎖球菌、化膿連鎖球菌;ストレプトミセス属種;赤痢菌属種;ビブリオ属種、例えばコレラ菌;及びエルシニア属種、例えばペスト菌、腸炎エルシニア菌を含むが、これらに限定されない細菌及びマイコプラスマ因子。病原性の:アルテルナリア属種;アスペルギルス属種;ブラストミセス属種、例えばB.デルマチチジス;カンジダ属種、例えばカンジダアルビカンス;クラドスポリウム属種;コクシジオイデス属種、例えばC,イミチス;クリプトコッカス属種、例えばC.ネオフォルマンス;ヒストプラスマ属種、例えばH.カプスラーツム;及びスポロトリクス属種、例えばS.シェンキイ、を含むが、これらに限定されない真菌及び酵母因子。
【0084】
1つの実施形態では、病原体は、病原性の:アカントアメーバ属種、アメーバ属種、ネグレリア属種、エントアメーバ属種、例えば赤痢アメーバを含むアメーバ;クリプトスポリジウム属種、例えばC.パルブム;シクロスポラ属種;エンセファリトゾーン属種、例えばE.インテスティナリス;エンテロシトゾーン属種;ジアルジア属種、例えばランブル鞭毛虫;イソスポラ属種;ミクロスポリジウム(Microsporidium)種;プラスモディウム属種、例えば熱帯熱マラリア原虫、四日熱マラリア原虫、卵形マラリア原虫、三日熱マラリア原虫;トキソプラスマ属種、例えばトキソプラスマ原虫;及びトリパノソーマ属種、例えばトリパノソーマブルセイ、を含むが、これらに限定されない原生生物因子由来である。
【0085】
1つの実施形態では、病原体は、病原性の:回虫属種、例えば回虫;ドラクンクルス属種、例えばメジナ虫;オンコセルカ属種、例えば回旋糸状虫;住血吸虫属種;旋毛虫属種、例えば旋毛虫;及び鞭虫属種、例えばヒト鞭虫、を含むが、これらに限定されない寄生生物因子(例えば蠕虫寄生生物)由来である。
【0086】
もう1つの実施形態では、病原体は、病原性の:アデノウイルス;アレナウイルス、例えばラッサ熱ウイルス;アストロウイルス;ブンヤウイルス、例えばハンターンウイルス、リフトバレー熱ウイルス;コロナウイルス、デルタウイルス;サイトメガロウイルス、エプスタイン−バーウイルス、ヘルペスウイルス、水痘ウイルス;フィロウイルス、例えばエボラウイルス、マルブルクウイルス;フラビウイルス、例えばデング熱ウイルス、西ナイル熱ウイルス、黄熱病ウイルス;肝炎ウイルス;インフルエンザウイルス;レンチウイルス、T細胞向性ウイルス、他の白血病ウイルス;ノーウォークウイルス;パピローマウイルス、他の腫瘍ウイルス;パラミクソウイルス、例えば麻疹ウイルス、流行性耳下腺炎ウイルス、パラインフルエンザウイルス、肺炎ウイルス、センダイウイルス;パルボウイルス;ピコルナウイルス、例えばカーディオウイルス、コクサッキーウイルス、エコーウイルス、ポリオウイルス、ライノウイルス、他のエンテロウイルス;ポックスウイルス、例えば痘瘡ウイルス、ワクシニアウイルス、パラポックスウイルス;レオウイルス、例えばコルティウイルス、オルビウイルス、ロタウイルス;ラブドウイルス、例えばリッサウイルス、水泡性口内炎ウイルス;及びトガウイルス、例えば風疹ウイルス、シンドビスウイルス、西部脳炎ウイルス、を含むが、これらに限定されないウイルス因子由来である。
【0087】
1つの特定の実施形態では、抗原ペプチドは、イヌパルボウイルスペプチド、炭疽菌保護抗原(PA)抗原ペプチド及び東部ウマ脳脊髄炎ウイルス抗原ペプチドからなる群より選択される。特定の実施形態では、抗原ペプチドは、配列番号:7のアミノ酸配列を有するイヌパルボウイルス由来ペプチドである。もう1つの特定の実施形態では、抗原ペプチドは、配列番号:9、11、13又は15のアミノ酸配列のいずれか1つを有する炭疽菌保護抗原(PA)抗原ペプチドである。さらにもう1つの特定の実施形態では、抗原ペプチドは、配列番号:25又は27のいずれかのアミノ酸配列を有する東部ウマ脳脊髄炎ウイルス抗原ペプチドである。
【0088】
(宿主細胞毒性ペプチド)
もう1つの特定の実施形態では、組換えペプチドは、遊離単量体形態であるとき宿主細胞に対して毒性であるペプチドである。より特定の実施形態では、毒性ペプチドは抗菌ペプチドである。
【0089】
特定の実施形態では、ウイルスカプシドに関連して発現される対象ペプチドは、宿主細胞毒性ペプチドである。特定の実施形態では、このタンパク質は抗菌ペプチドである。宿主細胞毒性ペプチドは、それが発現される宿主細胞に対して、又は宿主細胞がそのメンバーである細胞培養中又は生物中の他の細胞に対して、又は宿主細胞を提供する生物又は種の細胞に対して生物抑制性、生物致死性又は毒性である、生物阻害性ペプチドを指示する。1つの実施形態では、宿主細胞毒性ペプチドは、それが発現される宿主細胞に対して生物抑制性、生物致死性又は毒性である生物阻害性ペプチドである。宿主細胞毒性ペプチドの一部の例は、ペプチド毒素、抗菌ペプチド及び他の抗生物質ペプチドを含むが、これらに限定されない。
【0090】
抗菌ペプチドは、例えば抗細菌性ペプチド、例えばマガイニン、βデフェンシン、一部のαデフェンシン;カテリシジン;ヒスタチン;抗真菌ペプチド;抗原生動物ペプチド;合成AMP;ペプチド抗生物質又はその線状又は前環化オリゴ又はポリペプチド部分;他の抗生物質ペプチド(例えば駆虫性ペプチド、溶血ペプチド、殺腫瘍性ペプチド);及び抗ウイルスペプチド(例えば一部のαデフェンシン;殺ウイルス性ペプチド;ウイルス感染を阻止するペプチド)を含む。1つの特定の実施形態では、抗菌ペプチドは、配列番号:20のアミノ酸配列を有するD2A21ペプチドである。もう1つの実施形態では、抗菌ペプチドは、配列番号:24に実質的に対応するアミノ酸配列を有する抗菌ペプチドPBF20である。
【0091】
(VLPを発現するときに使用するための細胞)
本発明のウイルスカプシド又はウイルスカプシド融合ペプチドの発現のための宿主として使用する細胞(「宿主細胞」とも称する)は、ウイルスカプシドが細胞の複製又は感染を許容しないものである。1つの実施形態では、ウイルスカプシドは、宿主細胞が由来する細胞の種に感染しないウイルスに由来する。例えば、1つの実施形態では、ウイルスカプシドは二十面体植物ウイルスに由来し、細菌種の宿主細胞において発現される。もう1つの実施形態では、ウイルス種は哺乳動物に感染し、発現系は細菌宿主細胞を含む。
【0092】
1つの実施形態では、宿主細胞は原核生物、例えばシュードモナス種を含むがこれに限定されない細菌細胞であり得る。典型的な細菌細胞は、例えばDr MJ Farabee of the Estrella Mountain Community College, Arizonaにより、URL:http://www. emc.maricopa.edu/faculty/farabee/BIOBK/BioBookDiversity#2. htmlにおいて提供される、「Biological Diversity: Bacteria and Archaean」, a chapter of the On-Line Biology Bookに述べられている。特定の実施形態では、宿主細胞はシュードモナス細胞であり得、典型的にはP.フルオレッセンス細胞であり得る。
【0093】
1つの実施形態では、宿主細胞は、真性細菌のいずれかの種のメンバーであり得る。宿主は、以下の分類群:アシドバクテリア門、アクチノバクテリア門、アクイフェックス門、バクテロイデス門、クロロビウム門、クラミジア門、クロロフレキシ門、クリシオジェネス門、藍細菌門、デフェリバクター門、ディノコッカス門、ディクティオグロムス門、フィブロバクター門、グラム陽性細菌門、フソバクテリア門、ゲマティモナス門、レンティスファエラ門、ニトロスピラ門、プラクトミセス門、プロテオバクテリア門、スピロヘータ門、サーモデスルフォバクテリア門、サーモミクロビア門、サーモトガ門、テルムス門(Thermales)、又はベルコミクロビア門、のいずれか1つのメンバーであり得る。真性細菌宿主細胞の実施形態では、細胞は、藍細菌門を除く真性細菌のいずれかの種のメンバーであり得る。
【0094】
細菌宿主はまた、プロテオバクテリア門のいずれかの種のメンバーであり得る。プロテオバクテリア宿主細胞は、以下の分類群:アルファプロテオバクテリア綱、ベータプロテオバクテリア綱、ガンマプロテオバクテリア綱、デルタプロテオバクテリア綱、又はイプシロンプロテオバクテリア綱のいずれかの1つのメンバーであり得る。加えて、宿主は、以下の分類群:アルファプロテオバクテリア綱、ベータプロテオバクテリア綱、又はガンマプロテオバクテリア綱のいずれかの1つのメンバーであり得、及びガンマプロテオバクテリア綱のいずれかの種のメンバーであり得る。
【0095】
ガンマプロテオバクテリア綱宿主細胞の1つの実施形態では、宿主は、以下の分類群:エアロモナス目、アルテロモナス目、エンテロバクター目、シュードモナス目、又はキサントモナス目のいずれか1つのメンバー;又はエンテロバクター目又はシュードモナス目のいずれかの種のメンバーである。1つの実施形態では、宿主細胞はエンテロバクター目であり得、宿主細胞は、腸内細菌科のメンバーであるか、又はエルウィニア属、エシェリキア属、又はセラチア属のいずれか1つのメンバー;又はエシェリキア属のメンバーである。シュードモナス目の宿主細胞の1つの実施形態では、宿主細胞は、シュードモナス科、さらにはシュードモナス属のメンバーである。ガンマプロテオバクテリア綱宿主は、大腸菌種のメンバー及びシュードモナス・フルオレッセンス種のメンバーを含む。
【0096】
他のシュードモナス属生物も使用し得る。シュードモナス及び密接に関連する種は、 R. E. Buchanan and N. E. Gibbons (eds.), Bergey's Manual of Determinative Bacteriology, pp. 217-289 (8th ed. , 1974) (The Williams & Wilkins Co. , Baltimore, MD, USA) (以下「Bergey(1974)」)により「グラム陰性好気性桿菌及び球菌」として述べられている科及び属に属するグラム(−)プロテオバクテリアの群を包含する、プロテオバクテリア亜群1を含む。表1はこれらの科及び属の生物を示す。
表1.「グラム陰性好気性桿菌及び球菌」の部分に列挙されている科及び属(Bergey(1974)」より)
【表1】
【0097】
「グラム(−)プロテオバクテリア亜群1」はまた、分類において使用される判定基準に従ってこの表題に分類されるプロテオバクテリアを包含する。この表題はまた、以前はこの部門に分類されたが現在はもはや分類されない群、例えばアシドボラクス属、ブレブンディモナス属、ブルクホルデリア属、ヒドロゲノファガ属、オセアニモナス属、ラルストニア属及びステノトロフォモナス属、キサントモナス属に属する(以前はキサントモナス属の種と呼ばれた)生物を再分類することによって作成されたスフィンゴモナス属(及びそれに由来する、ブラストモナス属)、Bergey(1974)において定義されるようにアセトバクター属に属する生物を再分類することによって作成されたアシドモナス属を包含する。加えて、宿主は、シュードモナス属由来の細胞、それぞれアルテロモナス・ハロプラクティス、アルテロモナス・ニグリファシエンス、及びアルテロモナス・プトレファシエンスとして再分類された、シュードモナス・エナリア(ATCC14393)、シュードモナス・ニグリファシエンス(ATCC19375)、及びシュードモナス・プトレファシエンス(ATCC8071)を含み得る。同様に、例えばシュードモナス・アシドボランス(ATCC15668)及びシュードモナス・テストステローニ(ATCC11996)は、その後それぞれコマモナス・アシドボランス及びコマモナス・テストステローニに再分類された;及びシュードモナス・ニグリファシエンス(ATCC19375)及びシュードモナス・ピスキキダ(ATCC15057)は、それぞれシュードアルテロモナス・ニグリファシエンス及びシュードモナス・ピスキキダに再分類された。「グラム(−)プロテオバクテリア亜群1」はまた、以下の科:シュードモナス科、アゾトバクター科(現在はしばしば同義語、シュードモナス科の「アゾトバクター群」と呼ばれる)、リゾビウム科、及びメチロモナス科(現在はしばしば同義語、「メチロコッカス科」と呼ばれる)、のいずれかに属すると分類されるプロテオバクテリアを包含する。その結果として、ここで述べた属に加えて、「グラム(−)プロテオバクテリア亜群1」に属するプロテオバクテリア属は:1)アゾリゾフィルス属のアゾトバクター群細菌;2)セルビブリロ属、オリゲラ属及びテレディニバクター属のシュードモナス科細菌;3)セラトバクター属、エンサイファ属、リベリバクター属(「カンジダ・リベリバクター」とも呼ばれる)、及びシノリゾビウム属のリゾビウム科細菌;及び4)メチロバクター属、メチロカルダム属、メチロミクロビウム属、メチロサルシナ属及びメチロスフェラ属のメチロコッカス科細菌が挙げられる。
【0098】
もう1つの実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群2」から選択される。「グラム(−)プロテオバクテリア亜群2」は、以下の属(特に異なる記載があるものを除き、全てATCCに寄託されている、カタログに列挙され、公的に入手可能なその寄託菌株の総数を括弧内に示す):アシドモナス(2);アセトバクター(93);グルコノバクター(37);ブレブンディモナス(23);ベイエリンキア(13);デルキシア(2);ブルセラ(4);アグロバクテリウム(79);ケラトバクター(Chelatobacter)(2);エンサイファ(3);リゾビウム(144);シノリゾビウム(24);ブラストモナス(1);スフィンゴモナス(27);アルカリゲネス(88);ボルデテラ(43);ブルクホルデリア(73);ラルストニア(33);アソドボラクス(20);ヒドロゲノファガ(9);ゾーグレア(9);メチロバクター(2);メチロカルダム(NCIMBに1);メチロコッカス(2);メチロミクロビウム(2);メチロモナス(9);メチロサルシナ(1);メチロスフェラ;アゾモナス(9);アゾリゾフィルス(5);アゾトバクター(64);セルビブリオ(3);オリゲラ(5);シュードモナス(1139);フランシセラ(4);キサントモナス(229);ステノトロフォモナス(50);及びオセアニモナス(4)、のプロテオバクテリアの群と定義される。
【0099】
「グラム(−)プロテオバクテリア亜群2」の例示的な宿主細胞種は、以下の細菌(その例示的な菌株のATCC又は他の寄託番号を括弧内に示す)を含むが、これらに限定されない:アシドモナス・メタノリカ(ATCC43581);アセトバクター・アセティ(ATCC15973);グルコノバクター・オキシダンス(ATCC19357);ブレブンディモナス・ディミヌタ(ATCC11568);ベイエリンキア・インディカ(ATCC9039及びATCC19361);デルキシア・グンモサ(ATCC15994);ブルセラ・メリテンシス(ATCC23456)、ブルセラ・アボルツス(ATCC23448);アグロバクテリウム・トゥメファシエンス(ATCC23308)、アグロバクテリウム・ラディオバクター(ATCC19358)、アグロバクテリウム・リゾゲネス(ATCC11325);ケラトバクター・ヘインツィー(ATCC29600);エンサイファ・アドヘレンス(ATCC33212);リゾビウム・レグミノサルム(ATCC10004);シノリゾビウム・フレディ(ATCC35423);ブラストモナス・ナタトリア(ATCC35951);スフィンゴモナス・パウチモビリス(ATCC29837);アルカリゲネス・フェカリス(ATCC8750);百日咳菌(ATCC9797);ブルクホルデリア・セパシア(ATCC25416);ラルストニア・ピックティー(ATCC27511);アシドボラクス・ファシリス(ATCC11228);ヒドロゲノファガ・フラバ(ATCC33667);ゾーグレア・ラミゲラ(ATCC19544);メチロバクター・ルテウス(ATCC49878);メチロカルダム・グラシル(NCIMB11912);メチロコッカス・カプシュラツス(ATCC19069);メチロミクロビウム・アジル(ATCC35068);メチロモナス・メタニカ(ATCC35067);メチロサルシナ・フィブラタ(ATCC700909);メチロスフェラ・ハンソニイ(ACAM549);アゾモナス・アジリス(ATCC7494);アゾリゾフィルス・パスパリ(ATCC23833);アゾトバクター・クロオコカム(ATCC9043);セルビブリオ・ミクツス(UQM2601);オリゲラ・ウレトラリス(ATCC17960);緑膿菌(ATCC10145);シュードモナス・フルオレッセンス(ATCC35858);野兎病菌(ATCC6223);ステノトロフォモナス・マルトフィリア(ATCC13637);キサントモナス・カンペストリス(ATCC33913);及びオセアニモナス・ドウドロフィ(ATCC27123)。
【0100】
もう1つの実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群3」から選択される。「グラム(−)プロテオバクテリア亜群3」は、以下の属:ブレブンディモナス;アグロバクテリウム;リゾビウム;シノリゾビウム;ブラストモナス;スフィンゴモナス;アルカリゲネス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;メチロバクター;メチロカルダム;メチロコッカス;メチロミクロビウム;メチロモナス;メチロサルシナ;メチロスフェラ;アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;フランシセラ;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0101】
もう1つの実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群4」から選択される。「グラム(−)プロテオバクテリア亜群4」は、以下の属:ブレブンディモナス;ブラストモナス;スフィンゴモナス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;メチロバクター;メチロカルダム;メチロコッカス;メチロミクロビウム;メチロモナス;メチロサルシナ;メチロスフェラ;アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;フランシセラ;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0102】
1つの実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群5」から選択される。「グラム(−)プロテオバクテリア亜群5」は、以下の属:メチロバクター;メチロカルダム;メチロコッカス;メチロミクロビウム;メチロモナス;メチロサルシナ;メチロスフェラ;アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;フランシセラ;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0103】
宿主細胞は、「グラム(−)プロテオバクテリア亜群6」から選択することができる。「グラム(−)プロテオバクテリア亜群6」は、以下の属:ブレブンディモナス;ブラストモナス;スフィンゴモナス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0104】
宿主細胞は、「グラム(−)プロテオバクテリア亜群7」から選択することができる。「グラム(−)プロテオバクテリア亜群7」は、以下の属:アゾモナス;アゾリゾフィルス;アゾトバクター;セルビブリオ;オリゲラ;シュードモナス;テレディニバクター;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0105】
宿主細胞は、「グラム(−)プロテオバクテリア亜群8」から選択することができる。「グラム(−)プロテオバクテリア亜群8」は、以下の属:ブレブンディモナス;ブラストモナス;スフィンゴモナス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;シュードモナス;ステノトロフォモナス;キサントモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0106】
宿主細胞は、「グラム(−)プロテオバクテリア亜群9」から選択することができる。「グラム(−)プロテオバクテリア亜群9」は、以下の属:ブレブンディモナス;ブルクホルデリア;ラルストニア;アシドボラクス;ヒドロゲノファガ;シュードモナス;ステノトロフォモナス;及びオセアニモナス、のプロテオバクテリアの群と定義される。
【0107】
宿主細胞は、「グラム(−)プロテオバクテリア亜群10」から選択することができる。「グラム(−)プロテオバクテリア亜群10」は、以下の属:ブルクホルデリア;ラルストニア;シュードモナス;ステノトロフォモナス;及びキサントモナス、のプロテオバクテリアの群と定義される。
【0108】
宿主細胞は、「グラム(−)プロテオバクテリア亜群11」から選択することができる。「グラム(−)プロテオバクテリア亜群11」は、以下の属:シュードモナス;ステノトロフォモナス;及びキサントモナス、のプロテオバクテリアの群と定義される。宿主細胞は、「グラム(−)プロテオバクテリア亜群12」から選択することができる。「グラム(−)プロテオバクテリア亜群12」は、以下の属:ブルクホルデリア;ラルストニア;シュードモナス、のプロテオバクテリアの群と定義される。宿主細胞は、「グラム(−)プロテオバクテリア亜群13」から選択することができる。「グラム(−)プロテオバクテリア亜群13」は、以下の属:ブルクホルデリア;ラルストニア;シュードモナス;及びキサントモナス、のプロテオバクテリアの群と定義される。宿主細胞は、「グラム(−)プロテオバクテリア亜群14」から選択することができる。「グラム(−)プロテオバクテリア亜群14」は、以下の属:シュードモナス及びキサントモナス、のプロテオバクテリアの群と定義される。宿主細胞は、「グラム(−)プロテオバクテリア亜群15」から選択することができる。「グラム(−)プロテオバクテリア亜群15」は、シュードモナス属のプロテオバクテリアの群と定義される。
【0109】
宿主細胞は、「グラム(−)プロテオバクテリア亜群16」から選択することができる。「グラム(−)プロテオバクテリア亜群16」は、以下のシュードモナス種(例示的な菌株のATCC又は他の寄託番号を括弧内に示す)のプロテオバクテリアの群と定義される:シュードモナス・アビエタニフィラ(ATCC700689);緑膿菌(ATCC10145);シュードモナス・アルカリゲネス(ATCC14909);シュードモナス・アンギリセプティカ(ATCC33660);シュードモナス・シトロネロリス(ATCC13674);シュードモナス・フラベセンス(ATCC51555);シュードモナス・メンドシナ(ATCC25411);シュードモナス・ニトロレドゥセンス(ATCC33634);シュードモナス・オレオボランス(ATCC8062);シュードモナス・シュードアルカリゲネス(ATCC17440);シュードモナス・レシノボランス(ATCC14235);シュードモナス・ストラミネア(ATCC33636);シュードモナス・アガリシ(ATCC25941);シュードモナス・アルカリフィラ;シュードモナス・アルギノボラ;シュードモナス・アンデルソニ;シュードモナス・アスプレニ(ATCC23835);シュードモナス・アゼライカ(ATCC27162);シュードモナス・ベイエリンキ(ATCC19372);シュードモナス・ボレアリス;シュードモナス・ボレオポリス(ATCC33662);シュードモナス・ブラシカセルム;シュードモナス・ブタノボラ(ATCC43655);シュードモナス・セルロサ(ATCC55703);シュードモナス・オーランティアカ(ATCC33663);シュードモナス・クロロラフィス(ATCC9446、ATCC13985、ATCC17418、ATCC17461);シュードモナス・フラギ(ATCC4973);シュードモナス・ルンデンシス(ATCC49968);シュードモナス・テトロレンス(ATCC4683);シュードモナス・シシコラ(ATCC33616);シュードモナス・コロナファシエンス;シュードモナス・ディテルペニフィラ;シュードモナス・エロンガタ(ATCC10144);シュードモナス・フレクテンス(ATCC12775);シュードモナス・アゾトフォルマンス;シュードモナス・ブレネリ;シュードモナス・セドレラ;シュードモナス・コルガタ(ATCC29736);シュードモナス・エクストレモリエンタリス;シュードモナス・フルオレッセンス(ATCC35858);シュードモナス・ゲッサルディ;シュードモナス・リバネンシス;シュードモナス・マンデリ(ATCC700871);シュードモナス・マージナリス(ATCC10844);シュードモナス・ミグラ;シュードモナス・ムシドレンス(ATCC4685);シュードモナス・オリエンタリス;シュードモナス・ローデシア;シュードモナス・シンキサンタ(ATCC9890);シュードモナス・トラアシ(ATCC33618);シュードモナス・ベロニ(ATCC700474);シュードモナス・フレデリクスベルゲンシス;シュードモナス・ジェニクラタ(ATCC19374);シュードモナス・ギンゲリ;シュードモナス・グラミニス;シュードモナス・グリモンティ;シュードモナス・ハロデニトリフィカンス;シュードモナス・ハロフィラ;シュードモナス・ヒビシコラ(ATCC19867);シュードモナス・ハティエンシス(ATCC14670);シュードモナス・ヒドロゲノボラ;シュードモナス・ジェセニ(ATCC700870);シュードモナス・キロネンシス;シュードモナス・ランセオラタ(ATCC14669);シュードモナス・リニ;シュードモナス・マルギナタ(ATCC25417);シュードモナス・メフィティカ(ATCC33665);シュードモナス・デニトリフィカンス(ATCC19244);シュードモナス・ペルタシノゲナ(ATCC190);シュードモナス・ピクトラム(ATCC23328);シュードモナス・サイクロフィラ;シュードモナス・フルヴァ(ATCC31418);シュードモナス・モンテイリ(ATCC700476);シュードモナス・モセリ:シュードモナス・オリジハビタンス(ATCC43272);シュードモナス・プレコグロシシダ(ATCC700383);シュードモナス・プティダ(ATCC12633);シュードモナス・レアクタンス;シュードモナス・スピノサ(ATCC14606);シュードモナス・バレアリカ;シュードモナス・ルテオラ(ATCC43273);シュードモナス・スタツェリ(ATCC17588);シュードモナス・アミグダリ(ATCC33614);シュードモナス・アヴェラナ(ATCC700331);シュードモナス・カルシパパヤ(ATCC33615);シュードモナス・シコリ(ATCC10857);シュードモナス・フィクセレクタ(ATCC35104);シュードモナス・フスコヴァギナ;シュードモナス・メリア(ATCC33050);シュードモナス・シリンガ(ATCC19310);シュードモナス・ヴィリディフラバ(ATCC13223);シュードモナス・サーモカルボキシドヴォランス(ATCC35961);シュードモナス・サーモトレランス;シュードモナス・チヴェルヴァレンシス;シュードモナス・ヴァンクヴェレンシス(ATCC700688);シュードモナス・ウィスコンシネンシス;及びシュードモナス・キシアメネンシス。
【0110】
宿主細胞は、「グラム(−)プロテオバクテリア亜群17」から選択することができる。「グラム(−)プロテオバクテリア亜群17」は、例えば以下のシュードモナス種:シュードモナス・アゾトフォルマンス;シュードモナス・ブレネリ;シュードモナス・セドレラ;シュードモナス・コルガタ;シュードモナス・エクストレモリエンタリス;シュードモナス・フルオレッセンス;シュードモナス・ゲッサルディ;シュードモナス・リバネンシス;シュードモナス・マンデリ;シュードモナス・マージナリス;シュードモナス・ミグラ;シュードモナス・ムシドレンス;シュードモナス・オリエンタリス;シュードモナス・ローデシア;シュードモナス・シンキサンタ;シュードモナス・トラアシ;及びシュードモナス・ヴェロニ、に属するものを含む、「蛍光シュードモナス」として当技術分野において公知のプロテオバクテリアの群と定義される。
【0111】
この実施形態では、宿主細胞は、「グラム(−)プロテオバクテリア亜群18」から選択することができる。「グラム(−)プロテオバクテリア亜群18」は、例えば以下(例示的な菌株のATCC又は他の寄託番号を括弧内に示す)に属するものを含む、シュードモナス・フルオレッセンス種の全ての亜種、変種、菌株、及び他の亜種単位の群と定義される:次亜種1又は次亜種Iとも呼ばれる、シュードモナス・フルオレッセンス生物型A(ATCC13525);次亜種2又は次亜種IIとも呼ばれる、シュードモナス・フルオレッセンス生物型B(ATCC17816);次亜種3又は次亜種IIIとも呼ばれる、シュードモナス・フルオレッセンス生物型C(ATCC17400);次亜種4又は次亜種IVとも呼ばれる、シュードモナス・フルオレッセンス生物型F(ATCC12983);次亜種5又は次亜種Vとも呼ばれる、シュードモナス・フルオレッセンス生物型G(ATCC17518);シュードモナス・フルオレッセンス次亜種VI;シュードモナス・フルオレッセンスPf0−1;シュードモナス・フルオレッセンスPf−5(ATCC BAA−477);シュードモナス・フルオレッセンスSBW25;及びシュードモナス・フルオレッセンス亜種セルロサ(NCIMB10462)。
【0112】
宿主細胞は、「グラム(−)プロテオバクテリア亜群19」から選択することができる。「グラム(−)プロテオバクテリア亜群19」は、シュードモナス・フルオレッセンス生物型Aの全ての菌株の群と定義される。この生物型の1つの特定の菌株は、P.フルオレッセンスMB101株(Wilcoxへの米国特許第5,169,760号参照)及びその誘導体である。その誘導体の一例は、MB101染色体asd(アスパラギン酸デヒドロゲナーゼ遺伝子)遺伝子座に天然大腸菌PlacI−lacI−lacZYA構築物を挿入することによって構築される(すなわちPlacZが欠失した)P.フルオレッセンスMB214株である。
【0113】
本発明において使用できる付加的なP.フルオレッセンス株は、以下のATCC名称を有する、シュードモナス・フルオレッセンス・ミグラ及びシュードモナス・フルオレッセンス・ロイトキトクを含む:[NCIB8286];NRRL B−1244;NCIB 8865 CO1株;NCIB 8866 CO2株;1291[ATCC 17458;IFO 15837;NCIB 8917;LA;NRRL B−1864;ピロリジン;PW2[ICMP 3966;NCPPB 967;NRRL B−899];13475; NCTC 10038;NRRL B−1603[6;IFO 15840];52−1C;CCEB 488−A[BU140];CCEB 553[IEM15/47];IAM 1008 [AHH−27];IAM 1055[AHH−23];1[IFO 15842]; 12[ATCC25323;NIH 11;den Dooren de Jong 216];18[IFO15833;WRRL P−7];93[TR−10];108[52−22;IFO 15832];143[IFO 15836;PL];149[2−40−40;IFO 15838];182[IFO 3081;PJ 73];184[IFO 15830];185[W2 L−1];186[IFO 15829;PJ 79];187[NCPPB 263];188[NCPPB 316];189[PJ227;1208];191[IFO 15834;PJ 236;22/1];194[Klinge R−60;PJ 253];196[PJ 288];197[PJ 290];198 [PJ 302];201[PJ368];202[PJ 372];203[PJ 376];204[IFO 15835;PJ682];205[PJ 686];206[PJ 692];207[PJ 693];208[PJ722];212[PJ 832];215 [PJ 849];216[PJ 885];267[B−9];271[B−1612];401[C71A;IFO 15831;PJ187];NRRLB−3178[4;IFO15841];KY 8521;3081;30−21;[IFO 3081];N;PYR;PW;D946−B83[BU 2183;FERM−P 3328];P−2563 [FERM−P 2894;IFO 13658];IAM−1126[43F];M−1; A506[A5−06];A505[A5−05−1];A526[A5−26];B69;72;NRRL B−4290;PMW6[NCIB 11615];SC 12936;Al[1F0 15839];F 1847[CDC−EB];F1848[CDC 93]; NCIB 10586;P17;F−12;AmMS 257;PRA25;6133D02;6519E01;N1;SC15208;BNL−WVC;NCTC 2583[NCIB8194];H13;1013[ATCC 11251;CCEB 295];IFO 3903;1062;又はPf−5。
【0114】
(II.核酸構築物)
本発明はさらに、二十面体カプシドと組換えペプチドの融合ペプチドをエンコードする核酸構築物を提供する。1つの実施形態では、a)組換えペプチドをエンコードする核酸及びb)二十面体カプシドをエンコードする核酸配列を含み、a)の核酸とb)の核酸は、細胞において発現されたとき融合タンパク質を形成するように作動可能に連結されている、シュードモナス宿主細胞を形質転換するのに使用するための核酸構築物を提供する。
【0115】
特定の実施形態では、ベクターは、多数のカプシド又は多数の対象ペプチドについての配列を含み得る。1つの実施形態では、ベクターは、少なくとも2個の異なるカプシド−ペプチドコード配列を含み得る。1つの実施形態では、コード配列は同じプロモーターに連結される。特定の実施形態では、コード配列は、内部リボソーム結合部位によって分離される。他の実施形態では、コード配列は、細胞内でのウイルス様粒子の形成を可能にするリンカー配列によって連結される。もう1つの実施形態では、コード配列は異なるプロモーターに連結される。これらのプロモーターは同じ誘導条件によって駆動され得る。もう1つの実施形態では、異なるカプシド−ペプチドの組合せをエンコードする多数のベクターを提供する。多数のベクターは、同じ誘導条件によって駆動されるか又は異なる誘導条件によって駆動されるプロモーターを含み得る。1つの実施形態では、プロモーターは、lacプロモーター又はlacプロモーターの誘導体、例えばtacプロモーターである。
【0116】
対象ペプチドについてのコード配列は、あらかじめ定められた部位でウイルスカプシド又はカプシドのコード配列に挿入することができる。このペプチドはまた、あらかじめ定められていない部位で挿入することもでき、細胞をVLPの生産に関してスクリーニングすることができる。1つの実施形態では、VLPの形成の間にループとして発現されるように、ペプチドをカプシドコード配列に挿入する。1つの実施形態では、1個のペプチドコード配列がベクター内に含まれるが、他の実施形態では、多数の配列が含まれる。多数の配列は、コンカテマーの形態、例えば切断可能なリンカー配列によって連結されたコンカテマーの形態であり得る。
【0117】
ペプチドは、カプシド内の2つまたはそれ以上の挿入部位に挿入し得る。したがってペプチドは、カプシドの2個またはそれ以上の表面ループモチーフに挿入し得る;ペプチドはまた、所与のループモチーフ内の多数の部位に挿入し得る。挿入物の個々の機能性及び/又は構造ペプチド、及び/又はペプチド挿入物全体は、切断可能部位、すなわちタンパク質を切断する又は加水分解する因子がカプシド構造又は集合物の残りの部分からペプチドを分離することができる部位によって分けられ得る。
【0118】
ペプチドは、外側に面したループ及び/又は内側に面したループ、すなわちそれぞれカプシドの中心から離れた方向又は中心に向かう方向に面したカプシドのループ内に挿入し得る。カプシドの表面ループ内のいかなるアミノ酸又はペプチド結合もペプチドの挿入部位として役立ち得る。典型的には、挿入部位は、ループのほぼ中央、すなわち折りたたまれたカプシドペプチドの三次構造の中心からほぼ最も遠位の位置で選択される。ペプチドコード配列は、選択されたループのこのほぼ中央に対応するカプシドコード配列の位置に作動可能に挿入され得る。これは、ペプチド挿入部位から下流に合成されるカプシドのペプチド配列のその部分についての読み枠の維持を含む。
【0119】
もう1つの実施形態では、ペプチドは、カプシドのアミノ末端に挿入することができる。ペプチドは、上述した切断可能リンカーを含む、1又はそれ以上のリンカー配列を通してカプシドに連結され得る。さらにもう1つの実施形態では、ペプチドは、カプシドのカルボキシ末端に挿入することができる。ペプチドは、化学的又は酵素的加水分解によって切断することができる1又はそれ以上のリンカー配列を通してカルボキシ末端に連結され得る。1つの実施形態では、ペプチド配列は、アミノ末端とカルボキシ末端の両方で、又は1つの末端と少なくとも1つの内部位置で、例えばその三次元立体配座においてカプシドの表面に発現される位置で、連結される。
【0120】
1つの実施形態では、ペプチドは、ササゲクロロティックモザイクウイルス由来のカプシドに挿入することができる。1つの特定の実施形態では、ペプチドは、CCMVウイルスのアミノ酸129で挿入できる。もう1つの実施形態では、ペプチド配列は、CCMVウイルスのアミノ酸60、61、62又は63で挿入できる。さらにもう1つの実施形態では、ペプチドは、CCMVウイルスのアミノ酸129とアミノ酸60−63の両方で挿入できる。
【0121】
特定の実施形態では、本発明は、a)抗菌ペプチドをエンコードする核酸及びb)二十面体カプシドをエンコードする核酸を含み、a)の核酸とb)の核酸は、細胞において発現されたとき融合タンパク質を形成するように作動可能に連結されている、核酸構築物を提供する。核酸構築物を構築する上で有用な他のカプシド及び組換えペプチドは上記に開示されている。
【0122】
(プロモーター)
1つの実施形態では、核酸構築物は、カプシド−組換えペプチド融合ペプチドをエンコードする核酸配列に作動可能に結合されたプロモーター配列を含む。作動可能な結合又は連結は、転写及び何らかの翻訳調節エレメントが、宿主細胞の作用により、調節エレメントが対象配列の発現を指令することができるようにこの配列に共有結合している立体配置を指す。
【0123】
発酵工程では、ひとたび標的組換えペプチドの発現が誘導されれば、発現系の効率を最大化するために高レベルの生産を得ることが理想的である。プロモーターは転写を開始させ、一般にリボソーム結合部位の10−100ヌクレオチド上流に位置する。理想的には、プロモーターは、宿主細胞の総細胞タンパク質の約50%の組換えペプチド蓄積を可能にするのに十分なほど強力であり、厳密な調節を受け、容易に(及び安価に)誘導される。
【0124】
本発明に従って使用されるプロモーターは、構成的プロモーター又は調節されるプロモーターであり得る。一般に使用される誘導的プロモーター及びそれらのその後の誘導物質の例は、lac(IPTG)、lacUV5(IPTG)、tac(IPTG)、trc(IPTG)、Psyn(IPTG)、trp(トリプトファン飢餓状態)、araBAD(1−アラビノース)、lppa(IPTG)、lpp−lac(IPTG)、phoA(リン酸飢餓状態)、recA(ナリジクス酸)、proU(浸透圧)、cst−1(グルコース飢餓状態)、tetA(テトラサイクリン)、cadA(pH)、nar(嫌気的条件)、PL(42℃への温度シフト)、cspA(20℃への温度シフト)、T7(熱誘導)、T7−lacオペレーター(IPTG)、T3−lacオペレーター(IPTG)、T5−lacオペレーター(IPTG)、T4遺伝子32(T4感染)、nprM−lacオペレーター(IPTG)、Pm(アルキル又はハロベンゾエート)、Pu(アルキル又はハロトルエン)、Psal(サリシレート)、及びVHb(酸素)を含む。例えばMakrides, S. C. (1996) Microbiol. Rev. 60,512-538 ; Hannig G. & Makrides, S. C. (1998) TIBTECH 16,54-60 ; Stevens, R. C. (2000) Structures 8, R177- R185参照。例えばJ. Sanchez-Romero & V. De Lorenzo, Genetic Engineering of Nonpathogenic Pseudomonas strains as Biocatalysts for Industrial and Environmental Processes, in Manual of Industrial Microbiology and Biotechnology (A. Demain & J. Davies, eds. ) pp. 460-74 (1999) (ASM Press, Washington, D. C. ) ; H. Schweizer, Vectors to express foreign genes and techniques to monitor gene expression for Pseudomonads, Current Opinion in Biotechnology, 12: 439-445 (2001); およびR. Slater & R. Williams, The Expression of Foreign DNA in Bacteria, in Molecular Biology and Biotechnology (J. Walker & R. Rapley, eds. ) pp.125-54 (2000) (The Royal Society of Chemistry, Cambridge, UK)参照。
【0125】
選択した細菌宿主細胞にネイティブなプロモーターのヌクレオチド配列を有するプロモーター、例えばシュードモナスアントラニレート又はベンゾエートオペロンプロモーター(Pant、Pben)も、標的ペプチドをエンコードする導入遺伝子の発現を制御するために使用できる。2個以上のプロモーターが、同じか又は異なる配列の、もう1つ別のプロモーターに共有結合しているタンデムプロモーター、例えばPant−Pbenタンデムプロモーター(プロモーター間ハイブリッド)又はPlac−Placタンデムプロモーター、も使用し得る。
【0126】
調節されるプロモーターは、プロモーターがその一部である遺伝子の転写を制御するためにプロモーター調節タンパク質を利用する。調節されるプロモーターをここで使用する場合は、対応するプロモーター調節タンパク質も本発明に従った発現系の一部である。プロモーター調節タンパク質の例は、アクチベータータンパク質、例えば大腸菌カタボライトアクチベータータンパク質、MalTタンパク質;AraCファミリー転写アクチベーター;リプレッサータンパク質、例えば大腸菌LacIタンパク質;及び二元機能調節タンパク質、例えば大腸菌NagCタンパク質を含む。多くの調節されるプロモーター/プロモーター調節タンパク質の対が当技術分野において公知である。
【0127】
プロモーター調節タンパク質は、エフェクター化合物、すなわち調節タンパク質が、放出されるか又はプロモーターの制御下にある遺伝子の少なくとも1つのDNA転写調節領域に結合し、それによって遺伝子の転写を開始させる転写酵素の作用を許容する又はブロックすることができるように、調節タンパク質と可逆的に又は不可逆的に結合する化合物と相互作用する。エフェクター化合物は誘導物質又はコリプレッサーのいずれかに分類され、これらの化合物は、天然エフェクター化合物及び非代謝性誘導物質化合物を含む。多くの調節されるプロモーター/プロモーター調節タンパク質/エフェクター化合物の三つ組が当技術分野において公知である。エフェクター化合物は細胞培養又は発酵全体を通じて使用できるが、調節されるプロモーターを使用する特定の実施形態では、所望量又は密度の宿主細胞バイオマスの増殖後、所望の標的遺伝子の発現を直接又は間接的に生じさせるために、適切なエフェクター化合物を培養に添加する。
【0128】
例として、lacファミリーのプロモーターを使用する場合は、lacI遺伝子又はその誘導体、例えばlacIQ又はlacIQ1遺伝子も系に存在し得る。(通常は)構成的に発現される遺伝子であるlacI遺伝子は、これらのプロモーターのlacオペレーターに結合するLacリプレッサータンパク質(LacIタンパク質)をエンコードする。そこで、lacファミリーのプロモーターを使用する場合は、lacI遺伝子も発現系に含まれ、発現され得る。lacプロモーターファミリーメンバー、例えばtacプロモーターの場合、エフェクター化合物は誘導物質、好ましくは非代謝性誘導物質、例えばIPTG(イソプロピル−β−D−1−チオガラクトピラノシド、「イソプロピルチオガラクトシド」とも呼ばれる)である。
【0129】
特定の実施形態では、Plac、Ptac、Ptrc、PtacII、PlacUV5、lpp−PlacUV5、lpp−lac、nprM−lac、T71ac、T51ac、 T31ac及びPmacを含む、lac又はtacファミリープロモーターを本発明において使用する。
【0130】
(他のエレメント)
lacO配列を含む、他の調節エレメントを発現構築物に含めることができる。かかるエレメントは、例えば転写エンハンサー配列、翻訳エンハンサー配列、他のプロモーター、アクチベーター、翻訳開始及び終結シグナル、転写ターミネーター、シストロニック調節因子、ポリシストロニック調節因子、Hisタグ、Flagタグ、T7タグ、Sタグ、HSVタグ、Bタグ、Strepタグ、ポリアルギニン、ポリシステイン、ポリフェニルアラニン、ポリアスパラギン酸、(Ala−Trp−Trp−Pro)n、チオレドキシン、β−ガラクトシダーゼ、クロラムフェニコールアセチルトランスフェラーゼ、シクロマルトデキストリングルコノトランスフェラーゼ、CTP:CMP−3−デオキシ−D−マンノ−オクツロソネートシチジルトランスフェラーゼ、trpE又はtrpLE、アビジン、ストレプトアビジン、T7遺伝子10、T4gp55、ブドウ球菌プロテインA、連鎖球菌プロテインG、GST、DHFR、CBP、MBP、ガラクトース結合ドメイン、カルモジュリン結合ドメイン、GFP、KSI、c−myc、ompT、ompA、pelB、NusA、ユビキチン、及びヘモシリンAを含む、発現されるペプチドの特定、分離、精製又は単離を容易にするタグ配列、例えばヌクレオチド配列「タグ」及び「タグ」ペプチドコード配列を含むが、これらに限定されない。
【0131】
1つの実施形態では、核酸構築物は、対象組換えペプチドについてのコード配列に隣接する、又はウイルスカプシドについてのコード配列に連結された、タグ配列をさらに含む。1つの実施形態では、タグ配列はタンパク質の精製を可能にする。タグ配列は、アフィニティータグ、例えばヘキサヒスチジンアフィニティータグであり得る。もう1つの実施形態では、アフィニティータグは、グルタチオン−S−トランスフェラーゼ分子であり得る。タグはまた、蛍光分子、例えばYFP又はGFP、又はかかる蛍光タンパク質の類似体であり得る。タグはまた、抗体分子の一部、又は精製のために有用な公知の結合パートナーについての公知の抗原又はリガンドであり得る。
【0132】
本発明は、カプシド−組換えペプチドコード配列に加えて、それに作動可能に連結された以下の調節エレメント:プロモーター、リボソーム結合部位(RBS)、転写ターミネーター、翻訳開始及び終結シグナルを含み得る。有用なRBSは、本発明に従った発現系における宿主細胞として有用な種のいずれかから、好ましくは選択される宿主細胞から入手できる。多くの特定の及び多様なコンセンサスRBSが公知であり、例えばD.Frishman et al., Starts of bacterial genes: estimating the reliability of computer predictions, Gene 234 (2): 257-65 (8 Jul 1999); およびB. E. Suzek et al. , A probabilistic method for identifying start codons in bacterial genomes, Bioinformatics 17 (12): 1123-30 (Dec 2001)の中で記述され、言及されている。加えて、天然又は合成RBSが使用でき、例えば欧州特許第0207459号(合成RBS);O. Ikehata et al., Primary structure of nitrile hydratase deduced from the nucleotide sequence of a Rhodococcus species and its expression in Escherichia coli, Eur. J. Biochem. 181 (3): 563-70 (1989) (AAGGAAGの天然RBS配列)に述べられているものが使用できる。本発明において有用な方法、ベクター、及び翻訳及び転写エレメント、及び他のエレメントのさらなる例は、例えばGilroyへの米国特許第5,055,294号及びGilroyらへの米国特許第5,128,130号;Rammlerらへの米国特許第5,281,532号;Barnesらへの米国特許第4,695,455号及び同第4,861,595号;Grayらへの米国特許第4,755,465号;及びWilcoxへの米国特許第5,169,760号に述べられている。
【0133】
(ベクター)
本発明の酵素をエンコードするDNAのシュードモナス宿主による転写は、ベクター又はプラスミドにエンハンサー配列を挿入することによってさらに上昇させ得る。典型的なエンハンサーは、転写を上昇させるようにプロモーターに作用する、通常は約10−300bpの大きさの、DNAのシス作用性エレメントである。
【0134】
一般に、組換え発現ベクターは、シュードモナス宿主細胞、例えば本発明のカプシド−組換えペプチド融合ペプチド、の形質転換を可能にする複製起点及び選択マーカー、及び下流構造配列の転写を指令する高度発現遺伝子に由来するプロモーターを含む。かかるプロモーターは上記で説明した。異種構造配列は、翻訳開始及び終結配列と共に適切な段階で集合される。場合により及び本発明に従って、異種配列は、所望の特徴、例えば発現される組換え産物の安定化又は簡単な精製、を提供するN−末端特定ペプチドを含む融合ペプチドをコードし得る。
【0135】
カプシド−組換えペプチド融合ペプチドを発現するときP.フルオレッセンスと共に使用するための有用な発現ベクターは、カプシドペプチドと融合した所望の標的ペプチドをエンコードする構造DNA配列を、適切な翻訳開始及び終結シグナルと共に、機能性プロモーターを有する作動可能な読み枠(reading phase)に挿入することによって構築される。ベクターは、1又はそれ以上の表現型選択マーカー及びベクターの維持を確実にするため及び、所望する場合は、宿主内での増幅を提供するための複製起点を含む。本開示に従った形質転換のための適切な宿主は、シュードモナス属の中の様々な種、特にシュードモナス・フルオレッセンスの宿主細胞株を含む。
【0136】
宿主細胞内で組換えタンパク質を発現するために有用なベクターは当技術分野において公知であり、本発明に従った融合産物を発現するためにこれらのいずれかを修飾し、使用し得る。かかるベクターは、例えばプラスミド、コスミド及びファージ発現ベクターを含む。本発明に関する使用のために修飾することができる有用なプラスミドベクターの例は、発現プラスミドpBBRlMCS、pDSK519、pKT240、pML122、pPS10、RK2、RK6、pRO1600及びRSF1010を含むが、これらに限定されない。さらなる例は、pALTER−Exl、pALTER−Ex2、pBAD/His、pBAD/Myc−His、pBAD/gIII、pCal−n、pCal−n−EK、pCal−c、pCal−Kc、pcDNA2.1、pDUAL、pET−3a−c、pET 9a−d、pET−l la−d、pET−12a−c、pET−14b、pET15b、pET−16b、pET−17b、pET−19b、pET−20b(+)、pET−21a−d(+)、pET−22b(+)、pET−23a−d(+)、pET24a−d(+)、pET−25b(+)、pET−26b(+)、pET−27b(+)、pET28a−c(+)、pET−29a−c(+)、pET−30a−c(+)、pET31b (+)、pET−32a−c(+)、pET−33b(+)、pET−34b(+)、pET35b(+)、pET−36b(+)、pET−37b(+)、pET−38b(+)、pET−39b(+)、pET−40b(+)、pET−41a−c(+)、pET−42a−c(+)、pET−43a−c(+)、pETBlue−1、pETBlue−2、pETBlue−3、pGEMEX−1、pGEMEX−2、pGEXlλT、pGEX−2T、pGEX−2TK、pGEX−3X、pGEX−4T、pGEX−5X、pGEX−6P、pHAT10/11/12、pHAT20、pHAT−GFPuv、pKK223−3、pLEX、pMAL−c2X、pMAL−c2E、pMAL−c2g、pMAL−p2X、 pMAL−p2E、pMAL−p2G、pProEX HT、pPROLar.A、pPROTet.E、pQE−9、pQE−16、pQE−30/31/32、pQE−40、pQE−50、pQE−70、pQE−80/81/82L、pQE−100、pRSET、及びpSE280、pSE380、pSE420、pThioHis、pTrc99A、pTrcHis、pTrcHis2、pTriEx−1、pTriEx−2、pTrxFusを含み得る。かかる有用なベクターの他の例は、例えばN. Hayase, in Appl. Envir. Microbiol. 60 (9): 3336-42 (Sep 1994); A. A. Lushnikov et al. , in Basic Life Sci. 30: 657-62 (1985); S. Graupner & W. Wackernagel, in Biomolec. Eng. 17(1) : 11-16. (Oct 2000); H. P. Schweizer, in Curr. Opin. Biotech. 12 (5): 439-45 (Oct 2001); M. Bagdasarian & K. N. Timmis, in Curr. Topics Microbiol. Immunol. 96: 47-67 (1982); T. Ishii et al., in FEMS Microbiol. Lett. 116 (3): 307-13 (Mar 1,1994) ; I. N. Olekhnovich & Y. K. Fomichev, in Gene 140(1) : 63-65 (Mar 11,1994) ; M. Tsuda & T. Nakazawa, in Gene 136 (1-2): 257-62 (Dec 22,1993) ; C. Nieto et al. , in Gene 87(1) : 145-49 (Mar 1,1990) ; J. D. Jones & N. Gutterson, in Gene 61 (3): 299-306 (1987); M. Bagdasarian et al. , in Gene 16 (1-3): 237-47 (Dec 1981); H. P. Schweizer et al. , in Genet. Eng. (NY) 23: 69- 81 (2001); P.Mukhopadhyay et al., in J. Bact. 172(1) :477-80 (Jan 1990); D. O. Wood et al. , in J. Bact. 145 (3): 1448-51 (Mar 1981); およびR. Holtwick et al. , in Microbiology 147 (Pt 2): 337-44 (Feb 2001)によって記述されているものを含む。
【0137】
シュードモナス宿主細胞において有用であり得る発現ベクターのさらなる例は、表示されているレプリコンに由来する、表2に列挙されているものを含む。
表2.有用な発現ベクターの一部の例
【表2】
【0138】
発現プラスミド、RSF1010は、例えば F. Heffron et al., in Proc. Nat'1 Acad. Sci. USA 72 (9): 3623-27 (Sep 1975) 及びK. Nagahari & K. Sakaguchi, in J. Bact. 133 (3): 1527-29 (Mar 1978) によって述べられている。プラスミドRSF1010及びその誘導体は、本発明において特に有用なベクターである。当技術分野において公知である、RSF1010の有用な誘導体の例は、例えばpKT212、pKT214、pKT231及び関連プラスミド、及びpMYC1050及び関連プラスミド(例えばThompsonらへの米国特許第5,527,883号及び同第5,840,554号参照)、例えばpMYC1803を含む。プラスミドpMYC1803は、調節されるテトラサイクリン耐性マーカー及びRSF1010プラスミドからの複製及び動員遺伝子座を担持する、RSF1010に基づくプラスミドpTJS260(Wilcoxへの 米国特許第5,169,760号参照)に由来する。他の例示的な有用ベクターは、Puhlerらへの米国特許第4,680,264号に述べられているものを含む。
【0139】
1つの実施形態では、発現プラスミドを発現ベクターとして使用する。もう1つの実施形態では、RSF1010又はその誘導体を発現ベクターとして使用する。さらにもう1つの実施形態では、pMYC1050又はその誘導体、又はpMYC1803又はその誘導体を発現ベクターとして使用する。
【0140】
Champion(商標)pET発現系は高レベルのタンパク質生産を提供する。発現は、強力なT7lacプロモーターから誘導される。この系は、対象遺伝子の高レベル転写のためにバクテリオファージT7 RNAポリメラーゼの高い活性と特異性を利用する。プロモーター領域に位置するlacオペレーターは、伝統的なT7ベースのベクターよりも厳密な調節を提供し、プラスミドの安定性と細胞の生存度を改善する(Studier, F. W. and B. A. Moffatt (1986) J Molecular Biology 189(1) : 113-30; Rosenberg, et al. (1987)Gene 56(1) : 125-35)。T7発現系は、対象遺伝子の高レベル転写のためにT7プロモーターとT7 RNAポリメラーゼ(T7 RNAP)を使用する。T7 RNAPは天然大腸菌RNAPよりもプロセッシブであり、対象遺伝子の転写に専用されるので、T7発現系において高レベル発現が達成される。特定された遺伝子の発現は、宿主細胞においてT7 RNAPのソースを提供することによって誘導される。これは、T7 RNAP遺伝子の染色体コピーを含むBL21大腸菌宿主を使用することによって達成される。T7 RNAP遺伝子は、IPTGによって誘導することができるlacUV5プロモーターの制御下にある。誘導時にT7 RNAPが発現され、対象遺伝子を転写する。
【0141】
pBAD発現系は、特定の炭素源、例えばグルコース、グリセロール及びアラビノースの存在を通して組換えタンパク質の厳密に制御された滴定可能な発現を可能にする(Guzman, et al. (1995) J Bacteriology 177 (14): 4121- 30)。pBADベクターは、発現レベルに対する厳密な制御を与えるように独自に設計される。pBADベクターからの異種遺伝子発現はaraBADプロモーターで開始される。このプロモーターは、araC遺伝子の産物によって正及び負の両方に調節される。AraCは、L−アラビノースと複合体を形成する転写調節因子である。L−アラビノースの不在下では、AraC二量体は転写をブロックする。最大転写活性化のためには2つの事象が必要である:(i)L−アラビノースがAraCに結合して転写を開始させる、(ii)cAMPアクチベータータンパク質(CAP)−cAMP複合体がDNAに結合して、AraCの、プロモーター領域の正しい位置への結合を刺激する。
【0142】
trc発現系は、trcプロモーターからの大腸菌における高レベルの調節された発現を可能にする。trc発現ベクターは、大腸菌における真核生物遺伝子の発現に関して最適化されている。trcプロモーターは、トリプトファン(trp)及びラクトース(lac)プロモーターに由来する強力なハイブリッドプロモーターである。lacOオペレーター及びlacIQ遺伝子の産物によって調節される(Brosius, J. (1984) Gene 27 (2): 161-72)。
【0143】
(III.シュードモナスにおけるウイルス様粒子の発現)
本発明はまた、組換えペプチドを生産するための方法を提供する。この方法は、
a)シュードモナス細胞を提供すること;
b)組換えペプチドと二十面体カプシドの融合ペプチドをエンコードする核酸を提供すること;
c)このシュードモナス細胞においてこの核酸を発現すること、ここでこの細胞における発現がこの融合ペプチドのウイルス様粒子へのインビボ構築を提供する;及び
d)このウイルス様粒子を単離すること
を含む。
【0144】
ペプチドは、カプシドペプチド内の1コピーペプチド挿入物として発現され得る(すなわちペプチドに関してモノシストロニックである組換えカプシドペプチドコード配列から個別挿入物として発現される)か、又は2、3又は多コピー挿入物として発現され得る(すなわちペプチドに関してポリシストロニックである組換えカプシドペプチドコード配列からコンカテマー挿入物として発現される;コンカテマー挿入物は、同じ外来性対象ペプチドの多数のコピーを含み得るか又は異なる外来性対象ペプチドのコピーを含み得る)。コンカテマーは、ホモ又はヘテロコンカテマーであり得る。
【0145】
1つの実施形態では、単離されたウイルス様粒子を、ワクチン戦略においてヒト又は動物に投与することができる。
【0146】
もう1つの実施形態では、核酸構築物を、野生型カプシドをエンコードするもう1つ別の核酸と共発現することができる。特定の実施形態では、共発現されたカプシド/カプシド−組換えペプチド融合粒子は、インビボで集合してキメラウイルス様粒子を形成する。キメラVLPは、少なくとも2個の異なる核酸構築物によってエンコードされるカプシド又はカプシド−ペプチド融合物を含むウイルス様粒子である。
【0147】
さらにもう1つの実施形態では、核酸構築物を、異なるカプシド−組換えペプチド融合粒子をエンコードするもう1つ別の核酸と共発現することができる。特定の実施形態では、共発現されたカプシド融合粒子は、インビボで集合してキメラウイルス様粒子を形成する。
【0148】
さらにもう1つの実施形態では、異なるペプチド、例えばシャペロンタンパク質を発現するように設計された第二の核酸を、融合ペプチドをエンコードする核酸と同時に発現することができる。
【0149】
本発明のために有用なシュードモナス細胞、カプシド及び組換えペプチドは上記で論じている。
【0150】
1つの実施形態では、この方法は、VLPの形態の少なくとも0.1g/Lタンパク質を生産する。もう1つの実施形態では、この方法は、VLPの形態の0.1−10g/Lタンパク質を生産する。下位実施形態では、この方法は、VLP又はケージ構造の形態の少なくとも約0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9又は1.0g/Lタンパク質を生産する。1つの実施形態では、生産される総組換えタンパク質は少なくとも1.0g/Lである。一部の実施形態では、生産されるVLPタンパク質の量は、生産される総組換えタンパク質の少なくとも約5%、約10%、約15%、約20%、約25%、約30%、約40%、約50%、約60%、約70%、約80%、約90%、約95%又はそれ以上である。
【0151】
1つの実施形態では、この方法は、少なくとも0.1g/Lの前形成された(pre-formed)VLP又はケージ構造を生産する。もう1つの実施形態では、この方法は、細胞内で0.1−10g/Lの前形成VLPを生産する。もう1つの実施形態では、この方法は、細胞内で0.1−10g/Lの前形成ケージ構造を生産する。下位実施形態では、この方法は、少なくとも約0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9又は1.0g/Lの前形成VLPを生産する。1つの実施形態では、生産される総前形成VLPタンパク質は少なくとも1.0g/Lである。下位実施形態では、生産される総VLPタンパク質は少なくとも約2.0、3.0、4.0、5.0、6.0、7.0、8.0、9.0、10.0、15.0、20.0又は50.0g/Lであり得る。一部の実施形態では、生産されるVLPタンパク質の量は、生産される総組換えタンパク質の少なくとも約5%、約10%、約15%、約20%、約25%又はそれ以上である。
【0152】
もう1つの実施形態では、生産される、発現されたトランスジェニックペプチド、ペプチド、タンパク質又はそのフラグメントの50%以上を、宿主細胞において復元可能な形態で生産することができる。もう1つの実施形態では、発現されるタンパク質の約60%、70%、75%、80%、85%、90%、95%が活性形態で得られるか又は活性形態に復元することができる。
【0153】
本発明の方法はまた、組換えタンパク質の収率増大を導き得る。1つの実施形態では、この方法は、組換えタンパク質を総細胞タンパク質(tcp)の5、10、15、20、25、30、40又は50、55、60、65、70又は75%として生産する。「パーセント総細胞タンパク質」は、集合細胞タンパク質のパーセンテージとしての宿主細胞内のペプチドの量である。パーセント総細胞タンパク質の測定は当技術分野において周知である。
【0154】
特定の実施形態では、宿主細胞は、無機塩類培地中で増殖させたとき(すなわち約4℃−約55℃(両端を含む)の温度範囲内で)、約1%tcpの組換えペプチド、ペプチド、タンパク質又はそのフラグメントの発現レベル及び少なくとも40g/Lの細胞密度を有し得る。特定の実施形態では、この発現系は、少なくとも10リットルの発酵スケールで無機塩類培地中において増殖させたとき(すなわち約4℃−約55℃(両端を含む)の温度範囲内で)、少なくとも5%tcpの組換えタンパク質又はペプチド発現レベル及び少なくとも40g/Lの細胞密度を有する。
【0155】
別の実施形態では、対象ペプチドに作動可能に連結された発現ウイルスカプシドの部分は、細胞内の不溶性凝集体中で形成される。1つの実施形態では、対象ペプチドを不溶性凝集体から復元することができる。
(対象ペプチドの切断)
【0156】
1つの実施形態では、この方法は、e)この融合産物を切断して、カプシドから組換えペプチドを分離すること、をさらに提供する。
【0157】
切断可能な連結配列は、ウイルスタンパク質と組換えペプチドの間に含まれ得る。かかる配列を切断することができる物質の例は、化学試薬、例えば酸(塩酸、ギ酸)、CNBr、ヒドロキシルアミン(アスパラギン−グリシンに関して)、2−ニトロ−5−チオシアノベンゾエート、O−ヨードソベンゾエート、及び酵素物質、例えばエンドペプチダーゼ、エンドプロテアーゼ、トリプシン、クロストリパイン及びスタフィロコッカスプロテアーゼを含むが、これらに限定されない。
【0158】
切断可能な連結配列は当技術分野において周知である。本発明では、ジペプチド切断配列、例えばAsp−Proを含む、切断物質によって認識されるいかなる切断可能連結配列も使用できる。
【0159】
(発現)
本発明の方法は、最適には、宿主細胞における組換えペプチドの生産増大を導く。生産増大は、選択的に、生産されるタンパク質のグラム当り又は宿主タンパク質のグラム当りの活性ペプチドの濃度増大であり得る。生産増大はまた、組換え体のグラム当り又は宿主細胞タンパク質のグラム当りで生産される回収可能なペプチド、例えば可溶性タンパク質の濃度上昇であり得る。生産増大はまた、総濃度上昇とタンパク質の活性又は可溶性濃度上昇の何らかの組合せであり得る。
【0160】
組換えタンパク質の改善された発現は、VLPにおける挿入タンパク質の発現を通してであり得る。特定の実施形態では、少なくとも60、少なくとも70、少なくとも80、少なくとも90、少なくとも100、少なくとも110、少なくとも120、少なくとも130、少なくとも140、少なくとも150、少なくとも160、少なくとも170、又は少なくとも180コピーの対象ペプチドが各々のVLPにおいて発現される。VLPを生産し、宿主細胞の細胞質、ペリプラズム又は細胞外液から回収することができる。
【0161】
もう1つの実施形態では、ペプチドは細胞において不溶性であり得る。特定の実施形態では、不溶性ペプチドは、多数のカプシドから形成されるが天然型VLPを形成しない粒子において生産される。例えばわずか3個のウイルスカプシドのケージ構造が形成され得る。特定の実施形態では、カプシド構造は2コピー以上の対象ペプチドを含み、特定の実施形態では、少なくとも10、少なくとも20又は少なくとも30コピーを含む。
【0162】
ペプチド又はウイルスカプシド配列はまた、1又はそれ以上のターゲティング配列又は精製を助けるための配列を含み得る。これらはアフィニティータグであり得る。これらはまた、カプシドのVLPへの構築を指令するターゲティング配列であり得る。
【0163】
(細胞増殖)
ベクターによるシュードモナス宿主細胞の形質転換は、当技術分野で公知のいかなる形質転換法を用いて実施してもよく、細菌宿主細胞を無傷細胞として又はプロトプラストとして(すなわち細胞質体を含む)形質転換し得る。例示的な形質転換法は、穿孔法、例えば電気穿孔法、プロトプラスト融合、細菌接合、及び二価陽イオン処理、例えば塩化カルシウム処理又はCaCl/Mg2+処理、又は当技術分野で周知の他の方法を含む。例えばMorrison, J. Bact., 132 : 349-351 (1977); Clark-Curtiss & Curtiss, Methods in Enzymology, 101: 347-362 (Wu et al., eds, 1983), Sambrook et al. , Molecular Cloning, A Laboratory Manual (2nd ed. 1989); Kriegler, Gene Transfer and Expression: A Laboratory Manual (1990); and Current Protocols in Molecular Biology (Ausubel et al. , eds., 1994)参照)。
【0164】
ここで使用する、「発酵」という用語は、文字通りの発酵を用いる実施形態と、他の非発酵培養法を用いる実施形態の両方を包含する。発酵はいかなる規模で実施してもよい。1つの実施形態では、発酵培地は、富栄養培地、最少培地及び無機塩類培地の中から選択し得る;富栄養培地を使用してもよいが、好ましくは回避される。もう1つの実施形態では、最少培地又は無機塩類培地のいずれかを選択する。
【0165】
さらにもう1つの実施形態では、最少培地を選択する。さらにもう1つの実施形態では、無機塩類培地を選択する。無機塩類培地は特に好ましい。
【0166】
無機塩類培地は、無機塩類と炭素源、例えばグルコース、スクロース又はグリセロールからなる。無機塩類培地の例は、例えばM9培地、シュードモナス培地(ATCC179)、DavisとMingioliの培地(BD Davis & ES Mingioli (1950) in J. Bact. 60: 17-28参照)。無機塩類培地を作製するために使用する無機塩類は、例えばリン酸カリウム、硫酸又は塩化アンモニウム、硫酸又は塩化マグネシウム、及び微量ミネラル、例えば塩化カルシウム、鉄、銅、マンガン及び亜鉛のホウ酸塩及び硫酸塩の中から選択されるものを含む。有機窒素源、例えばペプトン、トリプトン、アミノ酸、又は酵母抽出物は、無機塩類培地には含まれない。その代わりに、無機窒素源が使用され、これは、例えばアンモニウム塩、アンモニア水、及び気体アンモニアから選択し得る。好ましい無機塩類培地は、グルコースを炭素源として含む。無機塩類培地と比較して、最少培地も無機塩類と炭素源を含み得るが、例えば低濃度のアミノ酸、ビタミン、ペプトン又は他の成分を添加することができ、但しこれらは極小濃度で添加される。
【0167】
高細胞密度培養は、バッチ工程として開始し、次に二相フェド−バッチ培養を実施することができる。バッチ部分での無制限増殖後、増殖を、バイオマス濃度が数倍上昇できる3倍加時間にわたって低い特定の増殖速度に制御することができる。かかる培養手順についてのさらなる詳細は、Riesenberg, D.; Schulz, V.; Knorre, W. A.; Pohl, H. D.; Korz, D.; Sanders, E. A.; Ross,A. ; Deckwer, W. D. (1991) "High cell density cultivation of Escherichia coli at controlled specific growth rate" J Biotechnol: 20(1) 17-27によって述べられている。
【0168】
本発明に従った発現系はいかなる発酵方式でも培養することができる。例えばバッチ、フェドバッチ、半連続、及び連続発酵方式がここで使用し得る。
【0169】
本発明に従った発現系は、いかなる規模(すなわち容量)の発酵においても導入遺伝子発現のために有用である。したがって、例えばマイクロリットル規模、センチリットル規模及びデシリットル規模の発酵容量を使用し得る;及び1リットル又はそれ以上の規模の発酵容量が使用できる。1つの実施形態では、発酵容量は1リットル又はそれ以上である。もう1つの実施形態では、発酵容量は、5リットル、10リットル、15リットル、20リットル、25リットル、50リットル、75リットル、100リットル、200リットル、500リットル、1,000リットル、2,000リットル、5,000リットル、10,000リットル又は50,000リットル又はそれ以上である。
【0170】
本発明では、形質転換宿主細胞の増殖、培養及び/又は発酵は、宿主細胞の生存を許容する温度範囲内、好ましくは約4℃−約55℃(両端を含む)の範囲内の温度で実施する。そこで、本発明の宿主細胞に関してここで使用する、例えば「増殖」(及び「増殖する」、「増殖すること」)、「培養すること」(及び「培養」)、及び「発酵」(及び「発酵する」、「発酵すること」)という用語は、本質的に、約4℃−約55℃(両端を含む)の温度範囲内での「増殖」、「培養」及び「発酵」を意味する。加えて、「増殖」は、活発な細胞分裂及び/又は拡張(enlargement)の生物学的状態、並びに非分裂及び/又は非拡張細胞が代謝的に維持されている生物学的状態の両方を指示するために使用され、「増殖」という用語の後者の使用は「維持」という用語と同義である。
【0171】
(細胞密度)
VLPに入った組換えペプチドを発現するときにシュードモナス・フルオレッセンスを使用することの付加的な利点は、大腸菌又は他の細菌発現系に比して高い細胞密度で増殖するシュードモナス・フルオレッセンスの能力を含む。このために、本発明に従ったシュードモナス・フルオレッセンス発現系は約20g/L又はそれ以上の細胞密度を提供することができる。本発明に従ったシュードモナス・フルオレッセンス発現系は、同様に、容量当りバイオマスに関して述べたように、バイオマスを乾燥細胞重量として測定したとき、少なくとも約70g/Lの細胞密度を提供することができる。
【0172】
1つの実施形態では、細胞密度は少なくとも約20g/Lである。もう1つの実施形態では、細胞密度は、少なくとも約25g/L、30g/L、35g/L、40g/L、45g/L、50g/L、60g/L、70g/L、80g/L、90g/L、100g/L、110g/L、120g/L、130g/L、140g/L、又は少なくとも150g/Lである。
【0173】
もう1つの実施形態では、誘導時の細胞密度は、20g/L−150g/L;20g/L−120g/L;20g/L−80g/L;25g/L−80g/L;30g/L−80g/L;35g/L−80g/L;40g/L−80g/L;45g/L−80g/L;50g/L−80g/L;50g/L−75g/L;50g/L−70g/L;40g/L−80g/Lである。
【0174】
(VLP又は対象ペプチドの単離)
特定の実施形態では、本発明は、ウイルスカプシドとの連結及び共発現を通して発現の間ペプチドを保護することにより、対象ペプチドの回収率を改善するための方法を提供する。特定の実施形態では、ウイルスカプシド融合は、細胞溶解産物から容易に分離することができるVLPを形成する。
【0175】
本発明のタンパク質は、硫酸アンモニウム又はエタノール沈殿、酸抽出、陰イオン又は陽イオン交換クロマトグラフィ、ホスホセルロースクロマトグラフィ、疎水性相互作用クロマトグラフィ、アフィニティークロマトグラフィ、ニッケルクロマトグラフィ、ヒドロキシルアパタイトクロマトグラフィ、逆相クロマトグラフィ、レクチンクロマトグラフィ、分取電気泳動、界面活性剤可溶化、カラムクロマトグラフィのような物質による選択的沈殿、免疫精製法等を含むが、これらに限定されない、当技術分野において周知の標準手法によって単離し、実質的に純粋に精製し得る。例えば確立された分子接着特性を有するタンパク質は、リガンドに可逆的に融合することができる。適切なリガンドを用いて、タンパク質を選択的に精製カラムに吸着させ、その後比較的純粋な形態でカラムから遊離することができる。次に、酵素活性によって融合タンパク質を取り出す。加えて、免疫アフィニティーカラム又はNi−NTAカラムを用いてタンパク質を精製することができる。一般的な手法は、例えばR. Scopes, Protein Purification: Principles and Practice, Springer-Verlag: N. Y. (1982); Deutscher, Guide to Protein Purification, Academic Press (1990);米国特許第4,511,503号; S. Roe, Protein Purification Techniques: A Practical Approach (Practical Approach Series), Oxford Press (2001); D. Bollag, etal., Protein Methods, Wiley-Lisa, Inc. (1996); AK Patra et al., Protein
Expr Purif, 18 (2): p/182-92 (2000); and R. Mukhija, etal., Gene 165 (2): p. 303-6 (1995)において詳述されている。また、例えばAusubel, et al. (1987 and periodic supplements); Deutscher (1990) "Guide to Protein Purification, " Methods in Enzymology vol.182及びこのシリーズの他の巻; Coligan, et al. (1996 and periodic Supplements) Current Protocols in Protein Science Wiley/Greene, NY;及びタンパク質精製産物の使用に関する製造者の文献、例えばPharmacia,Piscataway,N.J.、 又はBio−Rad,Richmond,Calif.も参照のこと。組換え手法との組み合わせは、適切なセグメント、例えばFLAG配列又はプロテアーゼ−除去可能配列によって融合することができる等価物、への融合を可能にする。また、例えばHochuli (1989) Chemische Industrie 12: 69-70; Hochuli (1990) "Purification of Recombinant Proteins with Metal Chelate Absorbent" in Setlow (ed. ) Genetic Engineering, Principle and Methods 12: 87-98, Plenum Press, NY;およびCrowe, et al. (1992) QIAexpress : The High Level Expression & Protein Purification System QUIAGEN,Inc., Chatsworth, Calif.も参照のこと。
【0176】
同様に、ウイルス様粒子又はケージ様構造も、当技術分野で周知の標準手法によって単離する及び/又は実質的に純粋に精製することができる。VLPの単離のための手法は、上述したものに加えて、沈殿法、例えばポリエチレングリコール又は塩沈殿法を含む。分離手法は、陰イオン又は陽イオン交換クロマトグラフィ、サイズ排除クロマトグラフィ、ホスホセルロースクロマトグラフィ、疎水性相互作用クロマトグラフィ、アフィニティークロマトグラフィ、ニッケルクロマトグラフィ、ヒドロキシルアパタイトクロマトグラフィ、逆相クロマトグラフィ、レクチンクロマトグラフィ、分取電気泳動、免疫精製法、遠心分離、超遠心分離、密度勾配遠心分離(例えばショ糖又は塩化セシウム(CsCl)勾配で)、サイズ排除フィルターを通しての限外ろ過、及び当技術分野で公知の何らかの他のタンパク質単離法を含む。
【0177】
本発明はまた、活性組換えペプチドの回収率を改善することができる。活性タンパク質の濃度は、例えば特定されたペプチドと親ペプチド、ペプチド変異体、セグメント置換ペプチド及び/又は残基置換ペプチドの間の相互作用を従来の何らかのインビトロ又はインビボアッセイによって測定することによって測定できる。そこで、インビトロアッセイは、特定されたタンパク質と対象ペプチドの間、例えば酵素と基質の間、ホルモンとホルモン受容体の間、抗体と抗原の間等の、何らかの検出可能な相互作用を測定することによって使用できる。かかる検出は、比色変化、放射能の変化、溶解度の変化、ゲル電気泳動及び/又はゲル排除工程によって測定した際の分子量変化等の測定を含み得る。インビボアッセイは、生理的作用、例えば体重増加、電解質バランスの変化、血液凝固時間の変化、血餅溶解及び抗原応答の誘導の変化、を検出するアッセイを含むが、これらに限定されない。一般に、特定されたペプチドと対象ペプチドの間の相互作用の変化を検出するための可変パラメータが存在する限り、いかなるインビボアッセイも使用することができる。例えば米国特許第5,834,250号参照。
【0178】
ペリプラズムから組換えタンパク質を放出するには、化学物質、例えばクロロホルム(Ames et al. (1984) J.BacteiioL, 160: 1181-1183)、グアニジン−HCl、及びTriton X−100 (Naglak and Wang (1990)Enzyme Microb. Technol., 12: 603-611) を含む処理が使用されてきた。しかし、これらの化学物質は不活性ではなく、多くの組換えタンパク質産物又はその後の精製手順に有害作用を及ぼし得る。外膜の透過性上昇を生じさせる、大腸菌細胞のグリシン処理も、ペリプラズム内容物を放出することが報告されている(Ariga et al. (1989) J. Ferm. Bioeng, 68 : 243-246)。組換えタンパク質のペリプラズム放出の最も広く使用されている方法は、浸透圧衝撃(Nosal and Heppel (1966) J. Biol.Chem., 241: 3055-3062; Neu and Heppel (1965)J. Biol.Clam., 240: 3685-3692)、ニワトリ卵白(HEW)−リゾチーム/エチレンジアミン四酢酸(EDTA)処理 (Neu and Heppel (1964) J.Biol. Chem., 239: 3893-3900; Witholt et al. (1976) Biochim. Biophys. Acta, 443: 534-544; Pierce et al. (1995) ICheme Research. Event, 2: 995-997)、及びHEW−リゾチーム/浸透圧衝撃処理の組合せ (French et al. (1996) Enzyme and Microb. Tech., 19: 332-338)である。このFrenchの方法は、分画緩衝液中に細胞を再懸濁して、その後ペリプラズム画分を回収することを含み、リゾチーム処理の直後に浸透圧衝撃を実施する。回収率に対する組換えタンパク質の過剰発現、S.サーモビオラセウス(S. thermoviolaceus)α−アミラーゼ、及び宿主生物の増殖期の影響も論じられている。
【0179】
典型的には、これらの手順は、浸透圧安定化培地中での初期破壊とそれに続く非安定化培地中での選択的放出を含む。これらの培地の組成物(pH、保護剤)及び使用する破壊方法(クロロホルム、HEW−リゾチーム、EDTA、超音波処理)は、報告されている特定の手順によって異なる。EDTAの代わりに双極子イオン界面活性剤を使用する、HEW−リゾチーム/EDTA処理の変法が、Stabel et al. (1994)Veterinary Microbiol., 38: 307-314によって述べられている。大腸菌を破壊するための細胞内溶菌酵素系の使用の総説については、Dabora and Cooney (1990) in Advances in Biochemical Engineering/Biotechnology, Vol. 43, A. Fiechter, ed. (Springer-Verlag: Berlin), pp. 11-30参照。
【0180】
可溶性タンパク質又は屈折性粒子としての、サイトプラズムからの組換えタンパク質の回収のための従来の方法は、機械的破壊による細菌細胞の崩壊を含んだ。機械的破壊は、典型的には、液体懸濁液中での局所キャビテーションの生成、剛性ビーズとの急速攪拌、超音波処理、又は細胞懸濁液の摩砕を含む(Bacterial Cell Surface Techniques, Hancock and Poxton (John Wiley & Sons Ltd,1988), Chapter 3, p. 55)。
【0181】
HEW−リゾチームは、細胞壁のペプチドグリカン骨格を加水分解するように生化学的に作用する。その方法は、後にスフェロプラストとして知られる球状の細胞球体を生成するために大腸菌を卵アルブミン(HEW−リゾチームを含有する)で処理した、 Zinder and Arndt (1956) Proc. Natl. Acad. Sci. USA, 42: 586-590によって最初に開発された。これらの構造は、一部の細胞壁成分を保持するが、サイトプラズム膜が露出された大きな表面積を有していた。米国特許第5,169,772号は、浸透圧安定化培地、例えば20%スクロース溶液中で、例えばEDTA、リゾチーム又は有機化合物を使用して細菌のエンベロープを破壊すること、細菌を低イオン強度緩衝液に暴露することによって破壊された細菌のペリプラズムスペースから非ヘパリナーゼ様タンパク質を放出すること、及び低イオン強度洗浄した細菌を緩衝塩溶液に暴露することによってヘパリナーゼ様タンパク質を放出することを含む、細菌からヘパリナーゼを精製するための方法を開示する。
【0182】
これらの方法の多くの異なる修正が広い範囲の発現系で使用され、様々な程度の成功を収めている(Joseph-Liazun et al. (1990) Gene, 86: 291-295; Carter et al. (1992)BiolTechnology, 10: 163-167)。リゾチームを生産するために組換え細胞培養を誘導するための試みが報告されてきた。欧州特許第0 155 189号は、組換え細胞培養を、通常はかかる宿主細胞を死滅させると予想される細胞壁構造を破壊する又は溶解する手段によって、リゾチームを生産するように誘導するための手段を開示する。
【0183】
米国特許第4,595,658号は、大腸菌のペリプラズムスペースに輸送されたタンパク質の外在化(externalization)を促進するための方法を開示する。この方法は、細胞のリゾチーム処理、機械的摩砕又は浸透圧衝撃を必要とせずに、ペリプラズム内に位置するタンパク質の選択的単離を可能にする。米国特許第4,637,980号は、温度感受性溶原菌を、細菌産物を直接又は間接的にエンコードするDNA分子で形質転換し、細胞内で遺伝子産物を発現することを許容する条件下で形質転換体を培養して、ファージにコードされた機能を誘導するために温度を上昇させることによってこの産物を外在化させることにより、細菌産物を生産することを開示する。Asami et al. (1997) J. Ferment. and Bioeng., 83: 511-516 は、T4ファージ感染による大腸菌細胞の同調破壊を開示し、Tanji et al. (1998)J. Ferment. and Bioeng., 85 : 74-78 は、大腸菌細胞の穏やかな破壊のためにT4ファージ内にエンコードされる溶菌遺伝子の制御された発現を開示する。
【0184】
細胞溶解時に、ゲノムDNAがサイトプラズムから培地中に漏出し、遠心力場において固体の沈降を妨げ得る液体粘度の有意の上昇をもたらす。剪断力、例えばDNAポリマーを壊すための機械的破壊の間に及ぼされる剪断力、が存在しない場合は、粘性液体を通しての固体のより緩やかな沈降速度が、遠心分離において固体と液体の分離不良を生じさせる。機械的剪断力以外に、DNAポリマーを分解する核酸分解酵素が存在する。大腸菌において、内在性遺伝子endAは、正常にはペリプラズムに分泌されて、DNAをオリゴデオキシリボヌクレオチドへとエンドヌクレアーゼ的に切断する、エンドヌクレアーゼをエンコードする(成熟タンパク質の分子量は約24.5kDである)。endAは大腸菌によって比較的弱く発現されることが示唆された(Wackemagel et al. (1995) Gene 154: 55-59)。
【0185】
発現されたタンパク質の検出は、当技術分野で公知の方法によって達成され、例えば放射免疫測定法、ウエスタンブロット手法又は免疫沈降法を含む。
【0186】
本発明において発現される特定のタンパク質は不溶性凝集体(「封入体」)を形成し得る。いくつかのプロトコールが封入体からのタンパク質の精製に適する。例えば封入体の精製は、典型的には、例えば50mM TRIS/HCl pH7.5、50mM NaCl、5mM MgCl2、1mM DTT、0.1mM ATP、及び1mM PMSFの緩衝液中でのインキュベーションによる、宿主細胞の破壊による封入体の抽出、分離及び/又は精製を含む。細胞懸濁液は、典型的にはフレンチプレスの2−3回の通過を用いて溶菌される。細胞懸濁液はまた、Polytron(Brinkrnan Instruments)を用いて又は氷上で超音波処理して均一化することができる。細菌を溶解する代替的な方法は当業者に明白である(例えば Sambrook et al. 前出; Ausubel et al.前出参照)。
【0187】
必要に応じて、封入体を可溶化することができ、溶菌細胞懸濁液は、典型的には遠心分離して望ましくない不溶性物質を除去することができる。封入体を形成したタンパク質は、適合性緩衝液による希釈又は透析によって復元し得る。適切な溶媒は、尿素(約4M−約8M)、ホルムアミド(少なくとも80%、容量/容量ベース)、及び塩酸グアニジン(約4M−約8M)を含むが、これらに限定されない。塩酸グアニジン及び類似試薬は変性剤であるが、この変性は不可逆的ではなく、変性剤の除去(例えば透析による)又は希釈後に復元が起こり得、免疫学的及び/又は生物学的に活性なタンパク質の再形成が可能である。他の適切な緩衝液は当業者に公知である。
【0188】
あるいは、宿主ペリプラズムから組換えペプチドを精製することが可能である。宿主細胞の溶解後、組換えタンパク質が宿主細胞のペリプラズム内に輸送されたとき、当業者に公知の他の方法に加えて低温浸透圧衝撃によって細菌のペリプラズム画分を単離することができる。ペリプラズムから組換えタンパク質を単離するため、例えば細菌細胞を遠心分離してペレットを形成することができる。ペレットを、20%スクロースを含む緩衝液に再懸濁することができる。細胞を溶解するため、細菌を遠心分離し、ペレットを氷冷5mM MgSO4に再懸濁して、約10分間氷浴中に保持することができる。細胞懸濁液を遠心分離し、上清をデカントして保存することができる。上清中に存在する組換えタンパク質を、当業者に周知の標準分離手法によって宿主タンパク質から分離することができる。
【0189】
初期塩分画は、望ましくない宿主細胞タンパク質(又は細胞培養に由来するタンパク質)の多くを対象組換えタンパク質から分離することができる。1つのかかる例は硫酸アンモニウムであり得る。硫酸アンモニウムは、タンパク質混合物中の水分量を有効に減少させることによってタンパク質を沈殿させる。その後タンパク質は、それらの溶解度に基づいて沈殿する。タンパク質がより疎水性であるほど、より低い硫酸アンモニウム濃度で沈殿する可能性が高い。典型的なプロトコールは、生じる硫酸アンモニウム濃度が20−30%になるように飽和硫酸アンモニウムをタンパク質溶液に添加することを含む。この濃度は最も疎水性のタンパク質を沈殿させる。次に沈殿物を廃棄し(対象タンパク質が疎水性でない限り)、対象タンパク質を沈殿させることが既知である濃度まで硫酸アンモニウムを上清に添加する。次に沈殿物を緩衝液に溶解し、必要に応じて、透析又はダイアフィルトレーションのいずれかを通して過剰の塩を除去する。タンパク質の溶解度に基づく他の方法、例えば低温エタノール沈殿は当業者に周知であり、複雑なタンパク質混合物を分画するために使用できる。
【0190】
組換えタンパク質の分子量を利用して、異なる細孔径の膜(例えばアミコン又はミリポア膜)を通す限外ろ過を用いてより大きい及びより小さいサイズのタンパク質から分離することができる。第一段階として、タンパク質混合物を、対象タンパク質の分子量よりも低い分子量カットオフ値を有する細孔径の膜を通して限外ろ過することができる。限外ろ過の保持物質を、次に、対象タンパク質の分子量よりも大きい分子量カットオフ値を有する膜に対して限外ろ過することができる。組換えタンパク質は膜を通過してろ液中に入る。その後ろ液を以下で述べるようにクロマトグラフィに供することができる。
【0191】
組換えタンパク質はまた、その大きさ、正味表面電荷、疎水性、及びリガンドに対する親和性に基づいて他のタンパク質から分離することができる。加えて、タンパク質に対して惹起される抗体をカラムマトリックスに結合し、タンパク質を免疫精製することができる。これらの方法は全て当技術分野において周知である。クロマトグラフィ手法が、多くの異なる製造者(例えばPharmacia Biotech)からの装置を使用していかなる規模でも実施できることは、当業者には明白である。
【0192】
(復元及びリフォールディング)
不溶性タンパク質を復元又は再生して、二次及び三次タンパク質構造立体配座を形成することができる。タンパク質リフォールディング工程は、必要に応じて、組換え産物の立体配置を完成させるときに使用できる。リフォールディング及び復元は、タンパク質の解離/会合を促進する当技術分野で公知の物質を用いて実施することができる。例えばタンパク質をジチオトレイトールと共にインキュベートし、次に酸化グルタチオン二ナトリウム塩と共にインキュベートして、さらにその後リフォールディング剤、例えば尿素を含む緩衝液と共にインキュベートすることができる。
【0193】
組換えタンパク質はまた、例えばリン酸緩衝食塩水(PBS)又は50mM 酢酸ナトリウム、pH6緩衝液プラス200mM NaClに対して透析することによっても復元できる。あるいは、タンパク質をカラムに、例えばNi NTAカラムに固定化して、プロテアーゼ阻害因子を含有する、500mM NaCl、20%グリセロール、20mM Tris/HCl pH7.4中、線形6M−1M尿素勾配を使用することによって再生できる。復元は1.5時間又はそれ以上の期間で実施できる。復元後、250mM イミダゾールの添加によってタンパク質を溶出することができる。イミダゾールは、PBS又は50mM 酢酸ナトリウム、pH6緩衝液プラス200mM NaClに対する最終透析工程によって除去できる。精製タンパク質は、4℃で保存するか又は−80℃で凍結することができる。
【0194】
他の方法は、例えばMH Lee et al., Protein Expr. Purif., 25(1) : p. 166-73 (2002), W. K. Cho et al. , J. Biotechnology, 77 (2-3): p. 169-78 (2000), Ausubel, et al. (1987 and periodic supplements), Deutscher (1990) "Guide to Protein Purification, "Methods in Enzymology vol. 182, and other volumes in this series, Coligan, et al. (1996 and periodic Supplements) Current Protocols in Protein Science Wiley/Greene, NY, S. Roe, Protein Purification Techniques: A Practical Approach (Practical Approach Series), Oxford Press (2001); D. Bollag, et al. , Protein Methods, Wiley-Lisa, Inc. (1996)に述べられているものを含む。
【0195】
(活性ペプチド分析)
活性タンパク質は、その配列が由来する天然ペプチドの活性の少なくとも20%、30%又は40%、好ましくは少なくとも50%、60%又は70%、最も好ましくは少なくとも80%、90%又は95%の比活性を有し得る。さらに、基質特異性(Kcat/Km)は、場合により天然ペプチドと実質的に同様である。典型的には、Kcat/Kmは、天然ペプチドの少なくとも30%、40%又は50%、より好ましくは少なくとも60%、70%、80%又は90%である。タンパク質及びペプチドの活性及び基質特異性(Kcat/Km)を検定する方法及び定量手段は当業者に周知である。
【0196】
本発明に従って生産される組換えペプチドの活性は、当技術分野で公知のタンパク質特異的な従来の又は標準インビトロ又はインビボアッセイによって測定することができる。シュードモナス生産組換えペプチドの活性を対応する天然タンパク質の活性と比較して、組換えタンパク質が、同じ又は類似の生理的条件下で天然ペプチドにおいて一般に認められる活性と実質的に類似又は同等の活性を示すかどうかを判定することができる。
【0197】
組換えペプチドの活性を、これまでに確立された天然ペプチド標準活性と比較することができる。あるいは、組換えペプチドの活性を、天然ペプチドと同時又は実質的に同時の比較アッセイにおいて測定することができる。例えばインビトロアッセイを使用して、組換えペプチドと標的の間、例えば発現される酵素と基質の間、発現されるホルモンとホルモン受容体の間、発現される抗体と抗原の間等の、検出可能な相互作用を測定することができる。かかる検出は、比色変化、増殖変化、細胞死、細胞反発、放射能の変化、溶解度の変化、ゲル電気泳動及び/又はゲル排除法によって測定したときの分子量の変化、リン酸化能力、抗体特異性アッセイ、例えばELISAアッセイ等の測定を含み得る。加えて、インビボアッセイは、天然ペプチドの生理的作用と比較したシュードモナス生産ペプチドの生理的作用、例えば体重増加、電解質バランスの変化、血液凝固時間の変化、血餅溶解及び抗原応答の誘導の変化、を検出するアッセイを含むが、これらに限定されない。一般に、いかなるインビトロ又はインビボアッセイも、活性が検定可能である限り、天然ペプチドとの比較分析を可能にするシュードモナス生産組換えペプチドの活性を測定するために使用できる。あるいは、本発明において生産されるペプチドは、ペプチドと、通常そのペプチドと相互作用する分子、例えば基質又は天然タンパク質が通常相互作用するシグナル経路の成分、との間の相互作用を刺激する又は阻害する能力に関して検定することができる。かかるアッセイは、典型的には、ペプチドが標的分子と相互作用することを許容する条件下でタンパク質と基質分子を組み合わせる工程を含み、タンパク質と標的分子の相互作用の生化学的結果を検出する。ペプチド活性を測定するために使用できるアッセイは、例えばRalph, P. J. , et al. (1984) J. Immunol. 132:1858 or Saiki et al. (1981) J. Immunol. 127: 1044,. Steward, W. E.II (1980) The Interferon Systems. Springer-Verlag, Vienna and New York, Broxmeyer, H. E. , et al. (1982) Blood 60: 595, "Molecular Cloning: A Laboratory Manual", 2d ed. , Cold Spring Harbor Laboratory Press, Sambrook, J. , E. F. Fritsch and T. Maniatis eds. , 1989,および"Methods in Enzymology: Guide to Molecular Cloning Techniques", Academic Press, Berger, S. L. and A. R. Kimmel eds. , 1987, AK Patra et al., Protein Expr Purif, 18 (2):p/182-92 (2000), Kodama et al. , J. Biochem. 99: 1465-1472 (1986); Stewart et al. , Proc.Nat'1 Acad. Sci. USA 90: 5209- 5213 (1993); (Lombillo et al. , J. Cell Biol. 128: 107-115 (1995); (Vale et al. , Cell 42: 39-50 (1985) に述べられている。
【実施例】
【0198】
これらの実施形態では、ササゲ退緑斑紋ウイルス(CCMV)をペプチド担体として使用し、シュードモナス・フルオレッセンスを発現宿主として使用した。CCMVはブロモウイルス科のブロモウイルス群のメンバーである。ブロモウイルスは、4成分のポジティブセンス一本鎖RNAゲノムを有する直径25−28nmの二十面体ウイルスである。RNA1及びRNA2はレプリカーゼ酵素をエンコードする。RNA3は、植物宿主内のウイルス運動に関与するタンパク質をエンコードする。RNA4(RNA3に由来するサブゲノムRNA)、すなわちsgRNA4は、20kDaのカプシド(CP)、配列番号:1をエンコードする。
【0199】
【表3】
【0200】
各々のCCMV粒子は、約180コピーまでのCCMV CPを含む。CCMV CPをエンコードする例示的DNA配列を配列番号:21に示す。
【表4】
【0201】
CCMVの結晶構造を解明した。この構造は、粒子の安定性と動力学にとって決定的に重要であると思われるカプシド相互作用のより明瞭な像を提供し、挿入部位の合理的設計を導くのに有用であった。これまでの試験は、CCMVカプシドを、粒子を形成するそれらの能力を妨げることなく異種ペプチドを担持するように遺伝的に修飾できることを明らかにした。多くの適切な挿入部位が特定されている。
【0202】
P.フルオレッセンスにおけるカプシド−ペプチド融合VLPの生産のために従うべき一般的戦略を図2に示す。CCMV CP内の単一の挿入部位を使用する場合、合計約180コピーまでの異種ペプチド単位(個々のペプチド又はコンカテマー)をCCMV粒子に挿入することができる。現在までにCCMV CP内で特定された挿入部位は、様々な長さのペプチドを収容することができる。加えて、多量体形態のペプチドを挿入部位に挿入することができる。さらに、同じ粒子内/粒子上に同じか又は異なるペプチドを発現するために多数の挿入部位を同時に使用することができる。ペプチド挿入物は、約200アミノ酸残基又はそれ以下の長さ、より好ましくは約180又はそれ以下、さらに一層好ましくは約150又はそれ以下、さらに一層好ましくは約120又はそれ以下、さらに一層好ましくは約100アミノ酸残基又はそれ以下の長さであり得る。好ましい実施形態では、ペプチド挿入物は約5又はそれ以上のアミノ酸残基の長さである。好ましい実施形態では、ペプチド挿入物は、約5−約120、より好ましくは約5−約100アミノ酸残基の長さである。
【0203】
(実験材料及び方法)
異なる記載がない限り、分子生物学の分野において公知の標準手法、ベクター、制御配列エレメント及び他の発現系エレメントを核酸操作、形質転換及び発現のために使用する。かかる標準手法、ベクター及びエレメントは、例えばAusubel et al. (eds. ), Current Protocols in Molecular Biology (1995) (John Wiley & Sons); Sambrook, Fritsch, & Maniatis (eds.), Molecular Cloning (1989) (Cold Spring Harbor Laboratory Press, NY) ; Berger & Kimmel, Methods in Enzymology 152: Guide to Molecular Cloning Techniques(1987) (Academic Press); and Bukhari et al. (eds. ), DNA Insertion Elements, Plasmids and Episomes (1977) (Cold Spring Harbor Laboratory Press, NY)に見出すことができる。
【0204】
プラスミド地図の作成
全てのプラスミド地図は、VECTORNTI(InforMax Inc.,Frederick,MD,USA)を用いて作成した。
【0205】
DNA抽出
大腸菌からの全てのプラスミドで抽出は、Qiagen(Germany)からのミニ、ミディ及びマキシキットを製造者の指示に従って使用して実施した。
【0206】
実験戦略
以下の手順に従った。P.フルオレッセンス宿主細胞を、キメラウイルスカプシド−標的ペプチドインサート融合物をエンコードする発現プラスミドで形質転換した。形質転換細胞を所望の密度に増殖させ、キメラウイルスカプシド−ペプチド融合物を発現するように誘導した。次に細胞を溶解し、それらの内容物を分析した。
【0207】
加工した挿入部位を付加するための修飾CCMV−CP DNAの構築
CCMV CPコード配列を含むDNA分子を、読み枠内に、BamHI制限酵素認識及び切断部位(gggatcctn)を挿入することによって修飾し、これは、天然のCCMV−CPアミノ酸配列のAsn129とSer130の間にトリペプチド(Gly−Ile−Leu)を導入した。従って、天然のCCMV−CPアミノ酸配列(配列番号:1)を、CCMV129−CP(配列番号:2)を形成するように修飾した。
【0208】
【表5】
【0209】
CCMV−CPコード配列にACTAGT SpeI制限部位を付加するため及びコンセンサスシャイン-ダルガーノ配列を付加するために、プライマーCCMV−For(核酸配列:5’−gactagtagg aggaaagaga tgtctacagt cgg−3’(配列番号:3))を設計した。CCMV−CPコード配列にCTCGAG XhoI制限部位を付加するため及び2個の停止コドンを導入するためにプライマーCCMV−Rev(核酸配列:5’−ccgctcgagt cattactaat acaccgg−3’(配列番号:4))を設計した。これら2個のプライマーを、CCMV−CPのDNAコード配列に関する第一PCR反応において使用した。
【0210】
加工した挿入部位を付加するためのCCMV63−CP DNAの構築:
制限部位AscI及びNotIを、挿入部位として使用するためにCCMV−CP(配列番号:1)上に加工した。AscI(ggcgcgcc)、NotI(gcggccgc)についての認識及び切断部位、及び付加的なヌクレオチドは、CCMV−CP上の残基Ala60とAla61の間にヘプタペプチド(Glu−Ala−Trp−Arg−Ala−Ala−Ala)を導入した。したがって、CCMV−CPを、CCMV63−CPを形成するように修飾した。加えて、構築されたVLPに安定性を付加するために残基Arg26をCys26に突然変異させた。pCCMV63−CPのプラスミド地図を図4に示す。
【0211】
【表6】
【0212】
二重挿入物R26C−CCMV63/129−CPの構築:
制限部位AscI及びNotIを、第二の挿入部位として使用するためにCCMV129−CP(配列番号:2)上に加工した。AscI(ggcgcgcc)、NotI(gcggccgc)についての認識及び切断部位、及び付加的なヌクレオチドは、CCMV129−CP上の残基Ala60とAla61の間にヘプタペプチド(Glu−Ala−Trp−Arg−Ala−Ala−Ala)を導入した。したがって、CCMV129−CPを、CCMV63/129−CPを形成するように修飾した。加えて、構築されたVLPに安定性を付加するように残基Arg26をCys26に突然変異させ、R26C−CCMV63/129−CPを作製した。pR26C−CCMV63/129−CPのプラスミド地図を図5に示す。
【0213】
【表7】
【0214】
実施例1.シュードモナスのCCMV VLPにおけるペプチドPD1の生産
1.A.キメラCCMV−PD1遺伝子の構築
20アミノ酸の抗原ペプチドを、CCMVウイルスカプシドにおいて挿入物として発現するために選択した。この抗原ペプチドは、CCMV及びシュードモナス・フルオレッセンスと無関係であった。このペプチドをエンコードするオリゴヌクレオチドを、プライマーParvo−BamHI−F(核酸配列:5’−cgggatcctg gacccggatg−3’(配列番号:16))及びParvo−BamHI−R(核酸配列:5’−cgggatcccc gggtctcttt c−3’(配列番号:17))を用いてプラスミドpCP7Parvo1 DNAから増幅した。(これらのプライマーは、Integrated DNA Technologies,Inc.,Coralville,IA,USA、以下「IdtDNA」より入手した。)これらのプライマーは、そのBamHI制限部位をCCMV129コード配列に挿入するために両方の末端でBamHI制限部位を付加して、イヌパルボウイルスペプチドコード配列を増幅した。
【0215】
以下のプロトコールに従ってPTC225サーモサイクラー(MJ Research, South San Francisco,CA,USA)を使用してPCR反応を実施した:
【表8】
【0216】
DNA配列を、SpeI及びXhoI制限酵素を使用して、CCMV129 CP DNA配列を含む核酸を挿入することによってプラスミドpESC(Stratagene Corp.,LaJolla,CA,USAより入手した)から構築したプラスミド、CCMV129シャトルプラスミドに挿入した。PD1ペプチドをエンコードする核酸を、BamHI制限部位でCCMV129 CDSに挿入し、CCMV129−PD1シャトルプラスミドを作製した。
【0217】
PD1 CDSもCCMV129 CDSに挿入した。その結果として、挿入されたPDIコード配列は、そのアミノ酸配列が:Trp Ala Cys Arg Gly Thr Ala Gly Trp Pro Pro Ser Gly Cys Thr Ala Pro Ser Gly Ser(配列番号:7)であるPD1ペプチドをエンコードする、5’−tgg gcc tgc cgc ggc acg gcc ggc tgg ccg ccg tcc ggc tgc acg gcg ccg tcc ggg tcg−3’(配列番号:18)である。PD1をコードするヌクレオチド配列は、イヌパルボウイルスには無関係である。
【0218】
1.B.CCMV−PD1発現プラスミドの構築
CCMV129−PD1シャトルプラスミドをSpeI及びXhoI制限酵素で消化した。キメラCCMV129−PD1 DNA配列を含むフラグメントをゲル精製によって単離した。次にそれを、pMYC1803発現プラスミドのSpeI及びXhoI制限部位で、tacプロモーターに作動可能に連結して、ブイブイ毒素遺伝子の代わりにpMYC1803発現プラスミドに挿入した。図1参照。生じた発現プラスミドを、挿入物の存在を確認するためにSpeI及びXhoIでの制限酵素消化によってスクリーニングした。
【0219】
1.C.シュードモナス宿主細胞へのプラスミド形質転換
CCMV129−PD1発現プラスミドを、以下のプロトコールに従ってシュードモナス・フルオレッセンスMB214宿主細胞に形質転換した。宿主細胞を、氷上に保持したバイアル中で徐々に解凍した。各々の形質転換のために、1μLの精製発現プラスミドDNAを宿主細胞に添加し、生じた混合物をピペットの先端で静かに渦を描いて混合し、その後氷上で30分間インキュベートした。混合物を電気穿孔用使い捨てキュベット(BioRad Gene Pulser Cuvette,電極ギャップ0.2cm、カタログ番号165−2086)に移した。キュベットを、あらかじめ200Ω、25μファラッド、2.25kVに設定したBiorad Gene Pulserに入れた。細胞は、短時間(約1−2秒)パルス細胞であった。その後直ちに低温LB培地を添加し、生じた懸濁液を30℃で2時間インキュベートした。次に細胞をLB tet15(テトラサイクリン添加LB培地)寒天にプレートし、30℃で一晩増殖させた。
【0220】
1.D.CCMV−PD1構築物の振とうフラスコ発現
各々のプレートから1個のコロニーを採取し、採取した試料をバッフル付き振とうフラスコ中の50mL LB種培養に接種した。懸濁液培養物を、250rpmで振とうしながら30℃で一晩増殖させた。各々の生じた種培養10mLを使用して、次に、1リットルバッフル付き振とうフラスコ中の振とうフラスコ培地(すなわち微量元素、クエン酸ナトリウム及びグリセロールを含む酵母抽出物及び塩、pH6.8)200mLに接種した。選択のためにテトラサイクリンを添加した。接種した培養物を、250rpmで振とうしながら30℃で一晩増殖させ、CCMV−PD1キメラカプシドの発現のためにIPTGで誘導した。
【0221】
1.E.細胞培養溶解産物の可溶性画分と不溶性画分への分離
各々の振とうフラスコ培養物から1mLアリコートを遠心分離して、細胞をペレット化した。細胞ペレットを、2mM EDTAを含む低温50mM Tirs−HCl、pH8.2 0.75mLに再懸濁した。0.1%容量の10% TritonX−100界面活性剤を加え、次いでリゾチームを0.2mg/mLの最終濃度まで添加した。その後細胞を氷上で2時間インキュベートし、その時点で透明な粘性細胞溶解産物が見えるはずである。
【0222】
この溶解産物に、1/200容量の1M MgCl2を加え、次いで1/200容量の2mg/mL DNAseIを添加して、氷上で1時間インキュベートし、その時点までに溶解産物ははるかに粘性の低い液体になっているはずである。処理した溶解産物を、次に、卓上遠心分離機において最大速度、4℃で30分間遠心し、上清を透明なチューブにデカントした。デカントした上清は「可溶性」タンパク質画分である。その後残りのペレットを0.75mL TE緩衝液(10mM Tris−HCl、pH7.5、1mM EDTA)に再懸濁した。再懸濁したペレットは「不溶性」画分である。
【0223】
1.F.可溶性及び不溶性タンパク質画分のSDS−PAGE及びウエスタンブロット分析
これらの「可溶性」及び「不溶性」画分を、次に、製造者の仕様書に従って、1.0mm×15穴を有するNuPAGE4−12% Bis−Trisゲル(Invitrogenより、カタログ番号NP0323)で電気泳動した。ゲルをSimplyBlue Safe Stain(Invitrogenより、カタログ番号LC6060)で染色し、水で一晩脱染した。ウエスタンブロット検出は、CCMV IgG(アクセッション番号AS0011、DSMZ、Germanyより)及びWESTERN BREEZEキット(Invitrogenより、カタログ番号WB7105)を製造者のプロトコールに従って使用した。結果は、CCMVの生産に関して、特にキメラCCMV CPの生産に関して陽性であった(図6及び7参照)。
【0224】
1.G.キメラVLPのPEG沈殿
キメラ、すなわち組換えVLPを、別々の振とうフラスコ培養試料の溶解によって沈殿させ、次いで、生じた細胞溶解産物を以下のプロトコールに従ってPEG(ポリエチレングリコール)処理した。各々の振とうフラスコ培養物の5mLアリコートを遠心分離して、細胞をペレット化した。ペレット化した細胞を、緩衝液2容対ペレット1容の比率で0.1M リン酸緩衝液(好ましくはリン酸二水素カリウムとリン酸二カリウムの組合せ)、pH7.0に再懸濁した。その後細胞を10秒間ずつ4回超音波処理し、各々の処理の間2分間氷上に置いた。この超音波処理の間に、細胞溶解産物は幾分清澄化するはずである。超音波処理後、リゾチームを0.5mg/mLの最終濃度まで添加した。リゾチーム消化を室温で30分間進行させた。
【0225】
生じた処理溶解産物を、次に、4℃、15000×Gで5分間遠心分離した。生じた上清を取り出し、その容積を測定した。各々の上清に、PEG6000を4%の最終濃度まで添加した;次いでNaClを0.2Mの最終濃度まで添加し、4℃で1時間又は一晩、氷上でインキュベートした。次に、これらを4℃、20000×Gで15分間遠心分離した。沈殿したペレットを、1/10初期上清容量のリン酸緩衝液に再懸濁し、4℃で保存した。
【0226】
1.H.ショ糖勾配遠心分離
リン酸緩衝液中のショ糖(Sigma、カタログ番号S−5390)でショ糖溶液を作製した。ショ糖勾配を手動で上部から底部へ10%、20%、30%及び40%注いだ。再懸濁した沈殿ペレット試料を、Beckman−Coulter Optima XL 100K UltracentrifugeのBeckman−Coulter SW41−Tiローターにおいてブレーキをかけずに1時間遠心した。ショ糖勾配の各々1mL画分を別々に溶出し、さらに遠沈してVLPペレットを得た。VLPペレットをリン酸緩衝液に再懸濁し、SDS−PAGEゲルで電気泳動して、上記プロトコールに従ってCCMV IgGを用いてウエスタンブロット分析した。ウエスタンブロット分析はVLP形成に関して陽性であった(図8)。各々の生じたVLP試料の一部を電子顕微鏡検査のために使用した。
【0227】
1.I.電子顕微鏡分析
VLP試料を、コロジオン/カーボン又はフォルムバール/カーボン被覆グリッドにスポットした。試料を2%ホスホタングステン酸(PTA)で染色し、120kVの加速電圧で操作するPhilips CM−12 TEM透過型電子顕微鏡(シリアル番号D769)で画像化した。画像をMultiScan CCDカメラ(Gatan,Inc., Pleasanton,CA,USAより;749型、シリアル番号971119010)でデジタル記録した。VLPの形成を確認した(図9)。
【0228】
実施例2.シュードモナスのCCMV VLPにおけるD2A21 AMP三量体の生産及びそこからのAMPの回収
2.A.D2A21挿入物の合成:
抗菌ペプチド(「AMP」)三量体(「D2A21三量体」、すなわちD2A21単量体AMPを3個含む)をコードするヌクレオチド配列を、プライマーD2A21−BamHI−F(核酸配列:5’−cgggatcctg ggacagcaaa tgggtcgcga tccg−3’(配列番号:5))及びD2A21−BamHI−R(核酸配列:5’−cgggatcccg tcgacggagc tcgaattcgg atcacc−3’(配列番号:6))を用いてプラスミドpET−(D2A21)3から増幅した。上記実施例1.A.で述べたのと同じプロトコールに従ってPCR反応を実施した。
【0229】
生じた増幅挿入物は、加工したBamHI部位でCCMV129 CDSにD2A21三量体CDSを挿入するときに使用するための、各々の末端に付加されたBamHI制限部位を含んでいた。D2A21三量体をエンコードするヌクレオチド配列及びD2A21三量体のアミノ酸配列をそれぞれ配列番号:19及び20に示す。
【0230】
【表9】
【0231】
【表10】
【0232】
三量体CDSは、図3に示すように、ジペプチドAsp−Pro酸不安定切断部位CDSによって分けられた3個のAMP単量体CDSを含んでいた。三量体CDS全体も、ジペプチドAsp−Pro酸不安定切断部位CDSによって各々の末端で境界付けられた。増幅した挿入物をBamHI制限酵素で消化して、CCMV129 CDS内のBamHI部位でpESC−CCMV129BamHIシャトルプラスミドにクローニングするための付着末端を作成した。生じたシャトルプラスミドをSpeI及びXhoI制限酵素で消化した。所望のキメラRBS/CDSフラグメントをゲル精製によって単離した。
【0233】
2.B.発現プラスミド構築
生じたキメラCCMV129−(D2A21)3ポリヌクレオチドを、次に、tacプロモーターに作動可能に連結して、ブイブイコード配列の代わりにpMYC1803発現プラスミドに挿入した。生じた発現プラスミドを、挿入物の存在に関してSpeI及びXhoIでの制限酵素消化によってスクリーニングした。実施例1.B.に関して上述したのと同じプロトコールを使用した。
【0234】
2.C.形質転換及び発現
生じた発現プラスミドを、実施例1.C.に関して上述したのと同じプロトコールを用いてP.フルオレッセンスMB214に形質転換した。プレートでコロニー形成した形質転換体を採取し、上記実施例1.D.に関して述べたのと同じプロトコールに従って、発現のために振とうフラスコに移した。
【0235】
2.D.タンパク質及びVLPの回収と分析
振とうフラスコ培養細胞を、実施例1.E.の手順に従って溶解し、分画した。生じた画分を、実施例1.F.に関して述べたようにSDS−PAGE及びウエスタンブロット法によって分析した。キメラVLPをPEG沈殿及びショ糖勾配遠心分離によって回収し、実施例1.G.−1.I.で上述したように電子顕微鏡検査によって分析した。キメラCCMV VLP集合を確認した。結果はCCMVの生産に関して陽性であった。キメラCP発現についてのSDS−PAGEは96アミノ酸挿入物を示し(図10)、これは、VLP形成を示すためのショ糖勾配での分画後のウエスタンブロット(図11)及びVLP形成を確認するための電子顕微鏡写真(図12)によって確認された。
【0236】
2.E.D2A21抗菌ペプチド生産の分析
可溶性及び不溶性タンパク質画分を、以下のように、キメラVLPにおいて生産されたD2A21ペプチドを特性決定するためにさらに処理した。
【0237】
2.E.1.D2A21の酸切断
不溶性画分を15%v/vアセトニトリル水溶液及び約40−50%v/vギ酸水溶液に溶解した;可溶性画分を約45−50%v/vギ酸水溶液に再懸濁した。試料を、その後、60℃で24時間インキュベートして、酸切断を進行させた。−20℃に冷凍することによって反応を停止させ、この温度で、処理した試料をHPLC分析まで保存した。
2.E.2.HPLCによるD2A21分析
【0238】
可溶性画分を0.22μm膜でろ過した;不溶性画分を遠心分離して細胞デブリを沈殿させ、0.22μm膜でろ過した。各々の試料50μLを25%アセトニトリル水溶液950μLに添加した。D2A21’の内部対照ペプチド(合計10μgの対照ペプチド)を含む、各々の試料250μL容量を、Beckman高速液体クロマトグラフィ(HPLC)システムに据え付けた、内径6.4mmのVYDAC 250mm逆相C18カラム(Grace Vydac,Hesperia,CA,USAより入手可能)に注入した。30分間で25%アセトニトリル/0.1%トリフルオロ酢酸(TFA)から75%アセトニトリル/0.01%TFAまでの水性勾配を用いて溶出を実施した。溶出物をクロマトグラフィ画分に滴下収集した。加工ペプチドを含むVLPに由来する試料においてのみ適切なペプチドピークを認め、非加工VLPに由来する試料では認められなかった(図13)。
【0239】
2.E.3.D2A21ペプチドの質量分析
ペプチド対照及びクロマトグラフィ画分の質量分析を、Micromass M@LDI線形マトリックス支援レーザー脱離イオン化飛行時間型(MALDI−TOF)質量分析システム(Micromass UK Ltd.,Manchester,UKより)を用いて実施した。MS分析の前に、Speed Vacシステム(Thermo Savant, Milford,MA,USAより入手可能;250DDA型)を使用して、遠心蒸発によってHPLC画分を濃縮した。結果は、D2A21 AMPの正確な生産及び放出されるペプチドがD2A21ペプチドであることを明らかにした(図14)。これらの結果は、シュードモナスにおけるペプチド発現のためのVLP融合物の使用が、通常の宿主細胞毒性を有効に回避することを示した。(A)キメラVLPの生産が明らかにされ、また(B)ペプチド多量体の生産がVLP方式で明らかにされた(合計96アミノ酸まで)。従って、この実施例は:
(1)CCMV CP発現及び粒子集合を支持するP.フルオレッセンスの能力を試験し、
(2)簡単な方法(PEG沈殿)によってキメラVLPを精製し、
(3)あらかじめ試験した方法(酸加水分解)によって対象ペプチドを切断し、及び
(4)ペプチドの同一性と完全性を確認した。
【0240】
実施例3.シュードモナスのCCMV VLPにおける炭疽抗原の生産
3.A.PAペプチド挿入物の合成
4個の異なる炭疽菌防御抗原(「PA」)ペプチド(PA1−PA4)をCCMV VLPにおいて独立して発現した。PA1−PA4をエンコードする核酸を合成オリゴヌクレオチドのSOE(オーバーラップ伸長によるスプライシング)(splicing-by-overlap-extension)によって合成した。生じた核酸はBamHI認識部位末端を含んだ。これらのPAペプチドをエンコードするヌクレオチド配列及びこれらのPAペプチドのアミノ酸配列はそれぞれ以下の通りであった:1)PA1については、配列番号:8及び9;2)PA2については、配列番号:10及び11;3)PA3については、配列番号:12及び13;及び4)PA4については、配列番号:14及び15。生じた核酸をBamHIで消化して、シャトルベクターへのクローニングのための付着末端を作成した。生じたPA挿入物の各々を、CCMV129 CDSのBamHI部位でpESC−CCMV129BamHIシャトルプラスミドにクローニングした。各々の生じたシャトルプラスミドをSpeI及びXhoI制限酵素で消化した。各々の所望のキメラCCMV129−PAエンコードフラグメントをゲル精製によって単離した。
【表11】
【表12】
【表13】
【表14】
【0241】
3.B.発現プラスミド構築
生じたキメラCCMV129−PAポリヌクレオチドを各々、tacプロモーターに作動可能に連結して、ブイブイコード配列の代わりにpMYC1803発現プラスミドに挿入した。生じた発現プラスミドを、挿入物の存在に関してSpeI及びXhoIでの制限消化によってスクリーニングした。実施例1.B.について上述したのと同じプロトコールを使用した。
【0242】
3.C.形質転換及び発現
生じた発現プラスミドを、実施例1.C.に関して上述したプロトコールを用いてP.フルオレッセンスMB214に形質転換した。プレートでコロニー形成した形質転換体を採取し、上記実施例1.D.に関して述べたのと同じプロトコールに従って、発現のために振とうフラスコに移した。
【0243】
3.D.タンパク質及びVLPの回収と分析
振とうフラスコ培養細胞を、実施例1.E.の手順に従って溶解し、分画した。生じた画分を、実施例1.F.に関して述べたようにSDS−PAGE及びウエスタンブロット法によって分析した。結果はCCMVの生産に関して陽性であった。
【0244】
結果は、キメラCCMV CP生産に関して陽性であった(図15参照)。VLPを、実施例1.G.及び1.H.で述べたようにPEG沈殿及びショ糖勾配分画によって回収した。ショ糖勾配画分のウエスタンブロットを1.H.で述べたように実施した。結果はVLPの生産に関して陽性であった(図16)。
【0245】
実施例4.シュードモナスのCCMV VLPへの単一及び二重挿入によるPBF20 AMP単量体の生産
実施例1、2及び3で述べた手順に従った。PBF20単量体ペプチドをエンコードする(アミノ酸配列Asp Pro Lys Phe Ala Lys Lys Phe Ala Lys Lys Phe Ala Lys Lys Phe Ala Lys Lys Phe Ala Lys Asp Pro(配列番号:24)のアミノ酸配列3−22を含むAMPをエンコードする)核酸及び配列番号:24のアミノ酸配列1−2及び23−24を含む酸切断部位を、AscI/NotI部位でCCMV63−CPに及びBamHI部位でCCMV129−CPに独立して挿入した。ペプチドをまた、AscI/NotI部位及びBamHI部位の両方で同時にR26C−CCMV63/129−CPに挿入した。
【0246】
生じたキメラポリヌクレオチドを各々、tacプロモーターに作動可能に連結して、ブイブイコード配列の代わりにpMYC1803発現プラスミドに挿入した。生じた発現プラスミドを、実施例1.C.について上述したプロトコールを使用して、挿入物の存在に関してSpeI及びXhoIでの制限消化によってスクリーニングし、P.フルオレッセンスMB214に形質転換した。プレートでコロニー形成した形質転換体を採取し、上記実施例1.D.に関して述べたのと同じプロトコールに従って、発現のために振とうフラスコに移した。
【0247】
振とうフラスコ培養細胞を、実施例1.E.の手順に従って溶解し、分画した。生じた画分を、実施例1.F.に関して述べたようにSDS−PAGE及びウエスタンブロット法によって分析した。結果はVLPの生産に関して陽性であった。
【0248】
シュードモナス・フルオレッセンスにおいて酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を発現するように加工したキメラCCMV63−CPの発現を示すSDS−PAGEを図17に示す。キメラCCMV63−CP−PBF20は、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。CCMV63−CPに由来し、酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を提示するキメラCCMV VLPの電子顕微鏡(EM)画像を図18に示す。
【0249】
シュードモナス・フルオレッセンスにおいて酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を発現するように加工したキメラCCMV129−CPの発現を示すSDS−PAGEを図19に示す。キメラCCMV129−CP−PBF20は、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。CCMV129−CPに由来し、酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を提示するキメラCCMV VLPの電子顕微鏡(EM)画像を図20に示す。
【0250】
シュードモナス・フルオレッセンスにおいてCP内の2つの異なる挿入部位で酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を発現するように加工したキメラCCMV63/129 CPの発現を示すSDS−PAGEを図21に示す。二重挿入物(CP+2×20アミノ酸)を含むキメラCPは、同じペプチドの単一挿入物(CP+1×20アミノ酸)を発現するように加工したカプシドと比較して、SDS−PAGEゲルでより緩慢な移動度を有する。カプシド当り2つの挿入部位で酸加水分解部位によって分けられた20アミノ酸抗菌ペプチドPBF20を提示する、CCMV63/129−CP由来のキメラCCMV VLPの電子顕微鏡(EM)画像を図22に示す。各々のVLPは、粒子当り360までのPBF20単量体を含むことが認められた。
【0251】
実施例5.シュードモナスのCCMV VLPにおける東部ウマ脳脊髄炎ウイルス(EEE)抗原の生産
5.A.EEEペプチド挿入物の合成
2つの異なるEEEペプチド(EEE−1及びEEE−2)をCCMV VLPにおいて独立して発現した。
EEE−1ペプチド配列:
DLDTHFTQYKLARPYIADCPNCGHS(配列番号:25)
EEE−1核酸配列:
5’−gacctggacacccacttcacccagtacaagctggcccgcccgtacatcgccgactgcccgaactgcggccacagc−3’(配列番号:26)
EEE−2ペプチド配列:
GRLPRGEGDTFKGKLHVPFVPVKAK(配列番号:27)
EEE−2核酸配列:
5’−ggccgcctgccgcgcggcgaaggcgacaccttcaagggcaagctgcacgtgccgttcgtgccggtgaaggccaag−3’(配列番号:28)
【0252】
EEE−1及びEEE−2をエンコードする核酸を合成オリゴヌクレオチドのSOEによって合成した。生じた核酸は、BamHI認識部位末端を含んでいた。挿入物の合成のためのセンス及びアンチセンスオリゴヌクレオチドプライマーはBamHI認識部位を含み、以下の通りであった:
EEE1.センスプライマー:
5’−cgg gga tcc tgg acc tgg aca ccc act tca ccc agt aca agc tgg ccc gcc cgt ac−3’(配列番号:29)
EEE1.アンチセンスプライマー:
5’−cgc agg atc ccg ctg tgg ccg cag ttc ggg cag tcg gcg atg tac ggg cgg gcc agc−3’(配列番号:30)
EEE2.センスプライマー:
5’−cgg gga tcc tgg gcc gcc tgc cgc gcg gcg aag gcg aca cct tca agg gca agc−3’(配列番号:31)
EEE2.アンチセンスプライマー:
5’−cgc agg atc ccc ttg gcc ttc acc ggc acg aac ggc acg tgc agc ttg ccc ttg−3’(配列番号:32)
【0253】
生じた核酸をBamHIで消化して、pESC−CCMV129BamHIシャトルプラスミドにクローニングするための付着末端を作成した。
【0254】
生じたEEE挿入物の各々を、CCMV129 CDSのBamHI部位でpESC−CCMV129BamHIシャトルプラスミドにクローニングした。各々の生じたシャトルプラスミドをSpeI及びXhoI制限酵素で消化した。所望のキメラCCMV−129−EEEをエンコードするフラグメントの各々をゲル精製によって単離した。
【0255】
5.B.発現プラスミド構築
生じたキメラCCMV129−EEEポリヌクレオチドフラグメントを各々、次に、tacプロモーターに作動可能に連結して、ブイブイコード配列の代わりにSpeI及びXhoIで制限消化したpMYC1803発現プラスミドに挿入した。生じた発現プラスミドを、挿入物の存在に関してSpeI及びXhoIでの制限消化によってスクリーニングした。実施例1.B.に関して上述したのと同じプロトコールを使用した。
【0256】
5.C.形質転換及び発現
生じた発現プラスミドを、実施例1.C.において上述したプロトコールを用いてP.フルオレッセンスMB214に形質転換する。実施例1.D.に関して上述したのと同じプロトコールを、EEE抗原を提示するキメラVLPの発現のために使用する。
【0257】
5.D.タンパク質及びVLPの回収と分析
振とうフラスコ培養細胞を、実施例1.E.の手順に従って溶解し、分画する。生じた画分を、実施例1.F.に関して述べたようにSDS−PAGE及びウエスタンブロット法によって分析する。
【図面の簡単な説明】
【0258】
【図1】図1は、シュードモナス宿主細胞における組換えVLPの発現のために有用なCCMV129−CP発現プラスミドのプラスミド地図を示す。
【図2】図2は、宿主細胞、例えばシュードモナス宿主細胞におけるウイルス様粒子(VLP)内のペプチド単量体の生産のための概要図を示す。組換えウイルスカプシド遺伝子(「rCP」)を構築するとき、所望の標的ペプチド挿入物コード配列(「I」)をインフレームでウイルスカプシドコード配列(「CP」)に挿入し、それが、ベクターの一部として、宿主細胞に形質転換され、組換えカプシド(「rCP」)を形成するように発現される。これらは、次に、CCMVの場合には、各々180個までのrCPを含むVLPを形成するように集合される。カプシドの外部ループにおいて発現される標的ペプチド挿入物(「I」)を有するVLPを例示する。集合されたVLPは各々、粒子当り多数のペプチド挿入物、例えば180個まで又はその倍数のペプチド挿入物を含む。VLPは、その後、例えばPEG沈降法によって、回収のために細胞溶解産物から容易に沈殿される。カプシド表面ループ及び/又は末端において発現された組換えペプチド挿入物は、沈殿したVLPから高度に純粋な形態で単離され得る。
【図3】図3は、宿主細胞、例えばシュードモナス宿主細胞におけるVLP内のペプチド多量体の生産のための概要図を示す。ペプチド挿入物は、そのコード配列(「i」)が、組換えウイルスカプシド遺伝子(「rCP」)を構築するときウイルスカプシドコード配列(「CP」)に挿入される、所望の標的ペプチドの多量体(三量体を示す)である。標的ペプチドコード配列の各々は、切断部位(「*」)についてのコード配列によって境界づけられ、核酸挿入物全体が標識「I」である。図では、CCMVカプシド当り1個の三量体挿入だけが行われ、生じるVLPは各々180個までのペプチド挿入物(「I」)を含み、合計540個までの標的ペプチド(「i」)を含む。標的ペプチドは、その後、切断剤、例えば酸又は酵素でのVLPの処理によって、VLPの沈殿後、高度に純粋な形態で容易に単離される。
【図4】図4は、組換えVLPの発現のために有用なCCMV63−CP発現プラスミドのプラスミド地図を示す。制限部位AscI及びNotIを、ペプチドのための挿入部位として働くようにCCMV−CP(配列番号:1)上に作製した。
【図5】図5は、組換えVLPの発現のために有用なR26C−CCMV63/129−CP発現プラスミドのプラスミド地図を示す。2つの挿入部位(AscI−NotI及びBamHI)を、2個の同じ又は異なるペプチドの挿入のためにCP内に作製した。
【図6】図6は、誘導後24時間目のシュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEゲルの画像である。キメラCPは、20アミノ酸の抗原ペプチドPD1を発現するように加工した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目の野生型CPであり、レーン3は誘導後24時間目の野生型CPであり、レーン4は誘導後0時間目のCCMV129−PD1であり、及びレーン5は誘導後24時間目のCCMV129−PD1である。
【図7】図7は、シュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すウエスタンブロットの画像である。キメラCPは、20アミノ酸の抗原ペプチドPD1を発現するように加工した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目の野生型CPであり、レーン3は誘導後24時間目の野生型CPであり、レーン4は誘導後0時間目のCCMV129−PD1であり、及びレーン5は誘導後24時間目のCCMV129−PD1である。
【図8】図8は、CCMV129−PD1 VLPショ糖勾配画分のウエスタンブロットの画像である。20アミノ酸の抗原ペプチドPD1を発現するように加工したキメラCCMV CPをシュードモナス・フルオレッセンスにおいて発現した。キメラVLPをPEG沈降法によって誘導後24時間目に単離し、ショ糖密度勾配で分画した。VLP画分はキメラCPに関して陽性であった。レーン1はCCMV129−PD1 VLPショ糖勾配画分であり、レーン2はCCMV129−PD1 VLPショ糖勾配画分であり、レーン3はCCMV129−PD1 VLPショ糖勾配画分であり、及びレーン4はサイズラダーである。
【図9】図9は、20アミノ酸の抗原ペプチドPD1を示すキメラCCMV VLPの電子顕微鏡(EM)画像である。VLPは、PEG沈降法及びショ糖密度勾配を用いてP.フルオレッセンスから単離した。
【図10】図10は、誘導後12、24及び48時間目のシュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEゲルの画像である。キメラCPは、酸加水分解部位によって分離される抗菌ペプチドD2A21三量体を発現するように作製した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目の野生型CPであり、レーン3は誘導後12時間目の野生型CPであり、レーン4は誘導後24時間目の野生型CPであり、レーン5は誘導後48時間目の野生型CPであり、レーン6は誘導後0時間目のCCMV129−(D2A21)3であり、レーン7は誘導後12時間目のCCMV129−(D2A21)3であり、レーン8は誘導後24時間目のCCMV129−(D2A21)3であり、及びレーン9は誘導後48時間目のCCMV129−(D2A21)3である。
【図11】図11は、CMV129−(D2A21)3 VLPショ糖勾配画分のウエスタンブロットの画像である。酸加水分解部位によって分離される96アミノ酸の抗菌ペプチドD2A21三量体を発現するように加工したキメラCCMV CPをシュードモナス・フルオレッセンスにおいて発現した。キメラVLPをPEG沈降法によって誘導後24時間目に単離し、ショ糖密度勾配で分画した。VLP画分はキメラCPに関して陽性であった。レーン1はサイズラダーであり、レーン2−4はCMV129−(D2A21)3 VLPショ糖勾配画分である。
【図12】図12は、酸加水分解部位によって分離される抗菌ペプチドD2A21三量体を示すキメラCCMV VLPの電子顕微鏡(EM)画像である。VLPは、PEG沈降法及びショ糖密度分画を用いてP.フルオレッセンスから単離した。
【図13】図13は、酸での処理により、酸切断部位によって分離される抗菌ペプチドD2A21三量体を提示するように加工したキメラVLPからのAMP D2A21ペプチド単量体の放出を示すHPLCクロマトグラムである。加工されていない(空の)VLPでは、AMPペプチドのピークは検出されなかった。
【図14】図14は、酸での処理により、酸切断部位によって分離される抗菌ペプチドD2A21三量体を提示するように加工したキメラVLPから放出されるAMP D2A21ペプチド単量体の同一性を示すMALDI−MSグラフである。分子量は、D2A21ペプチド単量体に関して予測されるとおりである。
【図15】図15は、誘導後12及び24時間目のシュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEゲルの画像である。キメラCPは、4つの異なる25アミノ酸の抗原ペプチドPA1、PA2、PA3及びPA4を発現するように作製した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目のCCMV129−PA1であり、レーン3は誘導後12時間目のCCMV129−PA1であり、レーン4は誘導後24時間目のCCMV129−PA1であり、レーン5は誘導後0時間目のCCMV129−PA2であり、レーン6は誘導後12時間目のCCMV129−PA2であり、レーン7は誘導後24時間目のCCMV129−PA2であり、レーン8は誘導後0時間目のCCMV129−PA3であり、レーン9は誘導後12時間目のCCMV129−PA3であり、レーン10は誘導後24時間目のCCMV129−PA3であり、レーン11は誘導後0時間目のCCMV129−PA4であり、レーン12は誘導後12時間目のCCMV129−PA4であり、レーン13は誘導後24時間目のCCMV129−PA4である。
【図16】図16は、CCMV129−PA1、CCMV129−PA2、CCMV129−PA3、CCMV129−PA4 VLPショ糖勾配画分のウエスタンブロットの画像である。25アミノ酸の抗原ペプチドPAを発現するように加工したキメラCCMV CPをシュードモナス・フルオレッセンスにおいて発現した。キメラVLPをPEG沈降法によって誘導後24時間目に単離し、ショ糖密度勾配で分画した。VLP画分はキメラCPに関して陽性であった。レーン1はサイズラダーであり、レーン2−4はCCMV129−PA1 VLPショ糖勾配画分であり、レーン5−7はCCMV129−PA2 VLPショ糖勾配画分であり、レーン8−10はCCMV129−PA3 VLPショ糖勾配画分であり、及びレーン11−13はCCMV129−PA4 VLPショ糖勾配画分である。
【図17】図17は、シュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEの画像である。キメラCCMV63−CPは、酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を発現するように作製した。キメラCPは、加工されていない野生型(wt)CCMV CPと比べてより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目の野生型CPであり、レーン3は誘導後24時間目の野生型CPであり、レーン4は誘導後0時間目のCCMV63−PBF20であり、レーン5は誘導後24時間目のCCMV63−PBF20である。
【図18】図18は、CCMV63−CPに由来し、酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を提示するキメラCCMV VLPの電子顕微鏡(EM)画像である。キメラVLPは、PEG沈降法及びショ糖密度分画を用いてP.フルオレッセンスから単離した。
【図19】図19は、シュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEの画像である。キメラCCMV129−CPは、酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を発現するように作製した。レーン1はサイズラダーであり、レーン2は誘導後0時間目のCCMV129−PBF20であり、及びレーン3は誘導後24時間目のCCMV129−PBF20である。
【図20】図20は、CCMV129−CPに由来し、酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を提示するキメラCCMV VLPの電子顕微鏡(EM)画像である。キメラVLPは、PEG沈降法及びショ糖密度分画を用いてP.フルオレッセンスから単離した。
【図21】図21は、シュードモナス・フルオレッセンスにおけるキメラCCMV CPの発現を示すSDS−PAGEの画像である。キメラCCMV63/129−CPは、CP内の2つの異なる挿入部位(63及び129)の酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を発現するように作製した。二重挿入物(CP+2×20AA)を含むキメラCPは、同じペプチドの単一挿入物(CP+1×20AA)を発現するように加工したカプシドと比較して、SDS−PAGEゲルでより緩慢な移動度を有する。レーン1はサイズラダーであり、レーン2は誘導後0時間目のCCMV63−PBF20であり、レーン3は誘導後24時間目のCCMV63−PBF20であり、レーン4は誘導後0時間目のCCMV63/129−2×(PBF20)であり、レーン5は誘導後24時間目のCCMV63/129−2×(PBF20)であり、レーン6は誘導後0時間目のCCMV63/129−2×(PBF20)であり、レーン7は誘導後24時間目のCCMV63/129−2×(PBF20)であり、レーン8は誘導後0時間目のCCMV63/129−2×(PBF20)であり、レーン9は誘導後24時間目のCCMV63/129−2×(PBF20)である。
【図22】図22は、カプシド当り2つの挿入部位(63及び129)の酸加水分解部位によって分離される20アミノ酸の抗菌ペプチドPBF20を提示する、CCMV63/129−CPに由来するキメラCCMV VLPの電子顕微鏡(EM)画像である。キメラVLPは、PEG沈降法及びショ糖密度分画を用いてP.フルオレッセンスから単離した。
【特許請求の範囲】
【請求項1】
a)二十面体ウイルスカプシドをエンコードする少なくとも1個の核酸配列;及び
b)組換えペプチドをエンコードする少なくとも1個の核酸配列
を含む第一の核酸構築物からなるシュードモナス(Pseudomonad)細胞。
【請求項2】
前記シュードモナスが、シュードモナス・フルオレッセンスである、請求項1に記載の細胞。
【請求項3】
前記二十面体ウイルスカプシドが、シュードモナス細胞に対する天然の親和性を示さないウイルス由来である、請求項1に記載の細胞。
【請求項4】
前記二十面体ウイルスカプシドが、植物二十面体ウイルス由来である、請求項3に記載の細胞。
【請求項5】
前記植物二十面体ウイルスが、ササゲ退緑斑紋ウイルス(Cowpea Chlorotic Mottle Virus)、ササゲモザイクウイルス(Cowpea Mosaic Virus)及びアルファルファモザイクウイルス(Alfalfa Mosaic Virus)からなる群より選択される、請求項4に記載の細胞。
【請求項6】
前記核酸が、少なくとも2個の異なる二十面体ウイルスカプシドをエンコードする、請求項1に記載の細胞。
【請求項7】
前記二十面体ウイルスカプシドの少なくとも1個が植物二十面体ウイルス由来である、請求項6に記載の細胞。
【請求項8】
前記組換えペプチドをエンコードする核酸が2個以上の単量体を含む、請求項1に記載の細胞。
【請求項9】
前記組換えペプチドをエンコードする核酸が少なくとも3個の単量体を含む、請求項8に記載の細胞。
【請求項10】
前記単量体がコンカテマーとして作動可能に連結されている、請求項8に記載の細胞。
【請求項11】
前記二十面体カプシドに融合された組換えペプチドが治療ペプチドである、請求項1に記載の細胞。
【請求項12】
前記組換えペプチドが抗原である、請求項1に記載の細胞。
【請求項13】
前記抗原が、イヌパルボウイルス抗原、炭疽菌抗原及び東部ウマ脳脊髄炎ウイルス抗原からなる群より選択される、請求項12に記載の細胞。
【請求項14】
前記組換えペプチドが抗菌ペプチドである、請求項1に記載の細胞。
【請求項15】
前記抗菌ペプチドが、D2A21及びPBF20からなる群より選択される、請求項14に記載の細胞。
【請求項16】
前記組換えペプチドが、少なくとも7アミノ酸の長さである、請求項1に記載の細胞。
【請求項17】
前記組換えペプチドが、少なくとも15アミノ酸の長さである、請求項16に記載の細胞。
【請求項18】
前記細胞が、野生型二十面体ウイルスタンパク質をエンコードする第二の核酸をさらに含む、請求項1に記載の細胞。
【請求項19】
前記細胞が、
c)第二の二十面体ウイルスカプシドをエンコードする少なくとも1個の核酸配列;及び
d)第二の組換えペプチドをエンコードする少なくとも1個の核酸配列
からなる第二の核酸をさらに含む、請求項1に記載の細胞。
【請求項20】
第一と第二の二十面体ウイルスカプシドが異なる、請求項19に記載の細胞。
【請求項21】
a)少なくとも1個の二十面体ウイルスカプシド;及び
b)少なくとも1個の組換えペプチド
を含む融合ペプチドを含むシュードモナス細胞。
【請求項22】
前記融合ペプチドが細胞内でウイルス様粒子を形成するように集合する、請求項21に記載の細胞。
【請求項23】
前記融合ペプチドが細胞内で可溶性ケージ構造を形成するように集合する、請求項21に記載の細胞。
【請求項24】
前記ウイルス様粒子が複製することができない、請求項22に記載の細胞。
【請求項25】
前記ウイルス様粒子が細胞に感染することができない、請求項22に記載の細胞。
【請求項26】
前記組換えペプチドが、二十面体カプシドの少なくとも1つの表面ループに挿入される、請求項21に記載の細胞。
【請求項27】
前記組換えペプチドが、二十面体カプシドの2つ以上の表面ループに挿入される、請求項21に記載の細胞。
【請求項28】
前記融合ペプチドが、二十面体ウイルスカプシドに融合された2個以上の組換えペプチドを含む、請求項21に記載の細胞。
【請求項29】
前記組換えペプチドが異なる、請求項28に記載の細胞。
【請求項30】
前記組換えペプチドが治療ペプチドである、請求項21に記載の細胞。
【請求項31】
前記組換えペプチドが抗原である、請求項21に記載の細胞。
【請求項32】
前記抗原が、イヌパルボウイルス抗原、炭疽菌抗原及び東部ウマ脳脊髄炎ウイルス抗原からなる群より選択される、請求項31に記載の細胞。
【請求項33】
前記ウイルス様粒子がワクチンとして使用できる、請求項22に記載の細胞。
【請求項34】
前記組換えペプチドが抗菌ペプチドである、請求項21に記載の細胞。
【請求項35】
前記抗菌ペプチドが、D2A21及びPBF20からなる群より選択される、請求項34に記載の細胞。
【請求項36】
前記組換えペプチドが、少なくとも7アミノ酸の長さである、請求項21に記載の細胞。
【請求項37】
前記組換えペプチドが、少なくとも15アミノ酸の長さである、請求項21に記載の細胞。
【請求項38】
前記細胞が、野生型二十面体ウイルスカプシドをさらに含む、請求項21に記載の細胞。
【請求項39】
前記細胞が、
a)少なくとも1個の第二の二十面体ウイルスカプシド;及び
b)少なくとも1個の第二の組換えペプチド
からなる第二の融合ペプチドをさらに含む、請求項21に記載の細胞。
【請求項40】
前記第二の融合ペプチドが細胞内でウイルス様粒子又は可溶性ケージ構造を形成するように集合する、請求項39に記載の細胞。
【請求項41】
前記第二の融合ペプチドが第一の融合ペプチドとは異なるアミノ酸配列を含む、請求項39に記載の細胞。
【請求項42】
前記ウイルスカプシド及び組換えペプチドが、リンカーを含むアミノ酸配列によって連結されている、請求項21に記載の細胞。
【請求項43】
前記リンカーアミノ酸配列が切断可能配列を含む、請求項42に記載の細胞。
【請求項44】
前記シュードモナスがシュードモナス・フルオレッセンスである、請求項21に記載の細胞。
【請求項45】
微生物細胞に対して毒性であるペプチドをエンコードする第二の核酸配列に作動可能に連結された二十面体ウイルスカプシドをエンコードする第一の核酸配列を含む核酸構築物。
【請求項46】
前記二十面体ウイルスカプシドが植物二十面体ウイルス由来である、請求項45に記載の構築物。
【請求項47】
前記植物二十面体ウイルスが、ササゲ退緑斑紋ウイルス、ササゲモザイクウイルス及びアルファルファモザイクウイルスからなる群より選択される、請求項46に記載の構築物。
【請求項48】
前記毒性ペプチドが2個以上のペプチド単量体配列を含む、請求項46に記載の構築物。
【請求項49】
前記毒性ペプチドが少なくとも3個のペプチド単量体配列を含む、請求項46に記載の構築物。
【請求項50】
前記単量体がコンカテマーに作動可能に連結されている、請求項48に記載の構築物。
【請求項51】
前記作動可能な連結が、前記カプシドをエンコードする第一の核酸配列の内部にある、請求項45に記載の構築物。
【請求項52】
前記毒性ペプチドをエンコードする第二の核酸配列が、カプシドの少なくとも1つの表面ループをエンコードする位置のカプシド配列に作動可能に連結されている、請求項45に記載の構築物。
【請求項53】
前記構築物が、カプシドの2つ以上の表面ループをエンコードするカプシド配列位置に作動可能に連結された2個以上の毒性ペプチド配列をエンコードする、請求項45に記載の構築物。
【請求項54】
前記組換えペプチドが抗菌ペプチドである、請求項45に記載の構築物。
【請求項55】
前記抗菌ペプチドが、D2A21及びPBF20からなる群より選択される、請求項54に記載の構築物。
【請求項56】
a)シュードモナス細胞を提供すること;
b)少なくとも1個の組換えペプチドと少なくとも1個の二十面体カプシドを含む融合ペプチドをエンコードする核酸を提供すること;
c)前記シュードモナス細胞において前記核酸を発現すること、ここで前記融合ペプチドがウイルス様粒子へと集合する;及び
d)前記ウイルス様粒子を単離すること
を含む、組換えペプチドを生産するための方法。
【請求項57】
e)前記融合ペプチドを切断して、二十面体ウイルスカプシドから組換えペプチドを分離すること
をさらに含む、請求項56に記載の方法。
【請求項58】
前記シュードモナスがシュードモナス・フルオレッセンスである、請求項56に記載の方法。
【請求項59】
前記ウイルス様粒子が複製することができない、請求項56に記載の方法。
【請求項60】
前記ウイルス様粒子が細胞に感染することができない、請求項56に記載の方法。
【請求項61】
前記二十面体ウイルスカプシドが、シュードモナス細胞に対する天然の親和性を示さないウイルス由来である、請求項56に記載の方法。
【請求項62】
前記二十面体ウイルスカプシドが植物二十面体ウイルス由来である、請求項56に記載の方法。
【請求項63】
前記植物二十面体ウイルスが、ササゲ退緑斑紋ウイルス、ササゲモザイクウイルス及びアルファルファモザイクウイルスからなる群より選択される、請求項62に記載の方法。
【請求項64】
前記核酸が、少なくとも2個の異なる二十面体ウイルスカプシドをエンコードする核酸配列を含む、請求項56に記載の方法。
【請求項65】
前記二十面体ウイルスカプシドの少なくとも1個が植物二十面体ウイルス由来である、請求項64に記載の方法。
【請求項66】
前記組換えペプチドが2個以上のペプチド単量体を含む、請求項56に記載の方法。
【請求項67】
前記組換えペプチドが少なくとも3個の単量体を含む、請求項56に記載の方法。
【請求項68】
前記単量体がコンカテマーとして作動可能に連結されている、請求項66に記載の方法。
【請求項69】
前記組換えペプチドが、二十面体カプシドの少なくとも1つの表面ループに作動可能に連結される、請求項56に記載の方法。
【請求項70】
前記組換えペプチドが、二十面体カプシドの2つ以上の表面ループに作動可能に連結される、請求項69に記載の方法。
【請求項71】
前記融合ペプチドが2個以上の組換えペプチドを含み、前記組換えペプチドが異なる、請求項56に記載の方法。
【請求項72】
前記組換えペプチドが治療ペプチドである、請求項56に記載の方法。
【請求項73】
前記組換えペプチドが抗原である、請求項56に記載の方法。
【請求項74】
前記抗原が、イヌパルボウイルス抗原、炭疽菌抗原及び東部ウマ脳脊髄炎ウイルス抗原からなる群より選択される、請求項73に記載の方法。
【請求項75】
前記ウイルス様粒子がワクチンとして使用できる、請求項56に記載の方法。
【請求項76】
前記組換えペプチドが抗菌ペプチドである、請求項56に記載の方法。
【請求項77】
前記抗菌ペプチドが、D2A21及びPBF20からなる群より選択される、請求項76に記載の方法。
【請求項78】
前記組換えペプチドが、少なくとも7アミノ酸の長さである、請求項56に記載の方法。
【請求項79】
前記組換えペプチドが、少なくとも15アミノ酸の長さである、請求項56に記載の方法。
【請求項80】
前記細胞が、野生型二十面体ウイルスカプシドをエンコードする第二の核酸をさらに含む、請求項56に記載の方法。
【請求項81】
前記細胞が、
a)少なくとも1個の第二の二十面体ウイルスカプシド;及び
b)少なくとも1個の第二の組換えペプチド
を含有する第二の融合ペプチドをエンコードする第二の核酸をさらに含む、請求項56に記載の方法。
【請求項82】
前記細胞において前記第二の核酸を発現することを含む、請求項81に記載の方法。
【請求項83】
前記第二の融合ペプチドが細胞内でウイルス様粒子又は可溶性ケージ構造を形成するように集合する、請求項81に記載の方法。
【請求項84】
前記第一の二十面体ウイルスカプシドが第一のアミノ酸配列を含み、第二の二十面体ウイルスカプシドが第二のアミノ酸配列を含み、第一と第二のカプシド配列が異なる、請求項81に記載の方法。
【請求項85】
前記第一の組換えペプチドが第一のアミノ酸配列を含み、第二の組換えペプチドが第二のアミノ酸配列を含み、第一と第二の組換えペプチド配列が異なる、請求項81に記載の方法。
【請求項1】
a)二十面体ウイルスカプシドをエンコードする少なくとも1個の核酸配列;及び
b)組換えペプチドをエンコードする少なくとも1個の核酸配列
を含む第一の核酸構築物からなるシュードモナス(Pseudomonad)細胞。
【請求項2】
前記シュードモナスが、シュードモナス・フルオレッセンスである、請求項1に記載の細胞。
【請求項3】
前記二十面体ウイルスカプシドが、シュードモナス細胞に対する天然の親和性を示さないウイルス由来である、請求項1に記載の細胞。
【請求項4】
前記二十面体ウイルスカプシドが、植物二十面体ウイルス由来である、請求項3に記載の細胞。
【請求項5】
前記植物二十面体ウイルスが、ササゲ退緑斑紋ウイルス(Cowpea Chlorotic Mottle Virus)、ササゲモザイクウイルス(Cowpea Mosaic Virus)及びアルファルファモザイクウイルス(Alfalfa Mosaic Virus)からなる群より選択される、請求項4に記載の細胞。
【請求項6】
前記核酸が、少なくとも2個の異なる二十面体ウイルスカプシドをエンコードする、請求項1に記載の細胞。
【請求項7】
前記二十面体ウイルスカプシドの少なくとも1個が植物二十面体ウイルス由来である、請求項6に記載の細胞。
【請求項8】
前記組換えペプチドをエンコードする核酸が2個以上の単量体を含む、請求項1に記載の細胞。
【請求項9】
前記組換えペプチドをエンコードする核酸が少なくとも3個の単量体を含む、請求項8に記載の細胞。
【請求項10】
前記単量体がコンカテマーとして作動可能に連結されている、請求項8に記載の細胞。
【請求項11】
前記二十面体カプシドに融合された組換えペプチドが治療ペプチドである、請求項1に記載の細胞。
【請求項12】
前記組換えペプチドが抗原である、請求項1に記載の細胞。
【請求項13】
前記抗原が、イヌパルボウイルス抗原、炭疽菌抗原及び東部ウマ脳脊髄炎ウイルス抗原からなる群より選択される、請求項12に記載の細胞。
【請求項14】
前記組換えペプチドが抗菌ペプチドである、請求項1に記載の細胞。
【請求項15】
前記抗菌ペプチドが、D2A21及びPBF20からなる群より選択される、請求項14に記載の細胞。
【請求項16】
前記組換えペプチドが、少なくとも7アミノ酸の長さである、請求項1に記載の細胞。
【請求項17】
前記組換えペプチドが、少なくとも15アミノ酸の長さである、請求項16に記載の細胞。
【請求項18】
前記細胞が、野生型二十面体ウイルスタンパク質をエンコードする第二の核酸をさらに含む、請求項1に記載の細胞。
【請求項19】
前記細胞が、
c)第二の二十面体ウイルスカプシドをエンコードする少なくとも1個の核酸配列;及び
d)第二の組換えペプチドをエンコードする少なくとも1個の核酸配列
からなる第二の核酸をさらに含む、請求項1に記載の細胞。
【請求項20】
第一と第二の二十面体ウイルスカプシドが異なる、請求項19に記載の細胞。
【請求項21】
a)少なくとも1個の二十面体ウイルスカプシド;及び
b)少なくとも1個の組換えペプチド
を含む融合ペプチドを含むシュードモナス細胞。
【請求項22】
前記融合ペプチドが細胞内でウイルス様粒子を形成するように集合する、請求項21に記載の細胞。
【請求項23】
前記融合ペプチドが細胞内で可溶性ケージ構造を形成するように集合する、請求項21に記載の細胞。
【請求項24】
前記ウイルス様粒子が複製することができない、請求項22に記載の細胞。
【請求項25】
前記ウイルス様粒子が細胞に感染することができない、請求項22に記載の細胞。
【請求項26】
前記組換えペプチドが、二十面体カプシドの少なくとも1つの表面ループに挿入される、請求項21に記載の細胞。
【請求項27】
前記組換えペプチドが、二十面体カプシドの2つ以上の表面ループに挿入される、請求項21に記載の細胞。
【請求項28】
前記融合ペプチドが、二十面体ウイルスカプシドに融合された2個以上の組換えペプチドを含む、請求項21に記載の細胞。
【請求項29】
前記組換えペプチドが異なる、請求項28に記載の細胞。
【請求項30】
前記組換えペプチドが治療ペプチドである、請求項21に記載の細胞。
【請求項31】
前記組換えペプチドが抗原である、請求項21に記載の細胞。
【請求項32】
前記抗原が、イヌパルボウイルス抗原、炭疽菌抗原及び東部ウマ脳脊髄炎ウイルス抗原からなる群より選択される、請求項31に記載の細胞。
【請求項33】
前記ウイルス様粒子がワクチンとして使用できる、請求項22に記載の細胞。
【請求項34】
前記組換えペプチドが抗菌ペプチドである、請求項21に記載の細胞。
【請求項35】
前記抗菌ペプチドが、D2A21及びPBF20からなる群より選択される、請求項34に記載の細胞。
【請求項36】
前記組換えペプチドが、少なくとも7アミノ酸の長さである、請求項21に記載の細胞。
【請求項37】
前記組換えペプチドが、少なくとも15アミノ酸の長さである、請求項21に記載の細胞。
【請求項38】
前記細胞が、野生型二十面体ウイルスカプシドをさらに含む、請求項21に記載の細胞。
【請求項39】
前記細胞が、
a)少なくとも1個の第二の二十面体ウイルスカプシド;及び
b)少なくとも1個の第二の組換えペプチド
からなる第二の融合ペプチドをさらに含む、請求項21に記載の細胞。
【請求項40】
前記第二の融合ペプチドが細胞内でウイルス様粒子又は可溶性ケージ構造を形成するように集合する、請求項39に記載の細胞。
【請求項41】
前記第二の融合ペプチドが第一の融合ペプチドとは異なるアミノ酸配列を含む、請求項39に記載の細胞。
【請求項42】
前記ウイルスカプシド及び組換えペプチドが、リンカーを含むアミノ酸配列によって連結されている、請求項21に記載の細胞。
【請求項43】
前記リンカーアミノ酸配列が切断可能配列を含む、請求項42に記載の細胞。
【請求項44】
前記シュードモナスがシュードモナス・フルオレッセンスである、請求項21に記載の細胞。
【請求項45】
微生物細胞に対して毒性であるペプチドをエンコードする第二の核酸配列に作動可能に連結された二十面体ウイルスカプシドをエンコードする第一の核酸配列を含む核酸構築物。
【請求項46】
前記二十面体ウイルスカプシドが植物二十面体ウイルス由来である、請求項45に記載の構築物。
【請求項47】
前記植物二十面体ウイルスが、ササゲ退緑斑紋ウイルス、ササゲモザイクウイルス及びアルファルファモザイクウイルスからなる群より選択される、請求項46に記載の構築物。
【請求項48】
前記毒性ペプチドが2個以上のペプチド単量体配列を含む、請求項46に記載の構築物。
【請求項49】
前記毒性ペプチドが少なくとも3個のペプチド単量体配列を含む、請求項46に記載の構築物。
【請求項50】
前記単量体がコンカテマーに作動可能に連結されている、請求項48に記載の構築物。
【請求項51】
前記作動可能な連結が、前記カプシドをエンコードする第一の核酸配列の内部にある、請求項45に記載の構築物。
【請求項52】
前記毒性ペプチドをエンコードする第二の核酸配列が、カプシドの少なくとも1つの表面ループをエンコードする位置のカプシド配列に作動可能に連結されている、請求項45に記載の構築物。
【請求項53】
前記構築物が、カプシドの2つ以上の表面ループをエンコードするカプシド配列位置に作動可能に連結された2個以上の毒性ペプチド配列をエンコードする、請求項45に記載の構築物。
【請求項54】
前記組換えペプチドが抗菌ペプチドである、請求項45に記載の構築物。
【請求項55】
前記抗菌ペプチドが、D2A21及びPBF20からなる群より選択される、請求項54に記載の構築物。
【請求項56】
a)シュードモナス細胞を提供すること;
b)少なくとも1個の組換えペプチドと少なくとも1個の二十面体カプシドを含む融合ペプチドをエンコードする核酸を提供すること;
c)前記シュードモナス細胞において前記核酸を発現すること、ここで前記融合ペプチドがウイルス様粒子へと集合する;及び
d)前記ウイルス様粒子を単離すること
を含む、組換えペプチドを生産するための方法。
【請求項57】
e)前記融合ペプチドを切断して、二十面体ウイルスカプシドから組換えペプチドを分離すること
をさらに含む、請求項56に記載の方法。
【請求項58】
前記シュードモナスがシュードモナス・フルオレッセンスである、請求項56に記載の方法。
【請求項59】
前記ウイルス様粒子が複製することができない、請求項56に記載の方法。
【請求項60】
前記ウイルス様粒子が細胞に感染することができない、請求項56に記載の方法。
【請求項61】
前記二十面体ウイルスカプシドが、シュードモナス細胞に対する天然の親和性を示さないウイルス由来である、請求項56に記載の方法。
【請求項62】
前記二十面体ウイルスカプシドが植物二十面体ウイルス由来である、請求項56に記載の方法。
【請求項63】
前記植物二十面体ウイルスが、ササゲ退緑斑紋ウイルス、ササゲモザイクウイルス及びアルファルファモザイクウイルスからなる群より選択される、請求項62に記載の方法。
【請求項64】
前記核酸が、少なくとも2個の異なる二十面体ウイルスカプシドをエンコードする核酸配列を含む、請求項56に記載の方法。
【請求項65】
前記二十面体ウイルスカプシドの少なくとも1個が植物二十面体ウイルス由来である、請求項64に記載の方法。
【請求項66】
前記組換えペプチドが2個以上のペプチド単量体を含む、請求項56に記載の方法。
【請求項67】
前記組換えペプチドが少なくとも3個の単量体を含む、請求項56に記載の方法。
【請求項68】
前記単量体がコンカテマーとして作動可能に連結されている、請求項66に記載の方法。
【請求項69】
前記組換えペプチドが、二十面体カプシドの少なくとも1つの表面ループに作動可能に連結される、請求項56に記載の方法。
【請求項70】
前記組換えペプチドが、二十面体カプシドの2つ以上の表面ループに作動可能に連結される、請求項69に記載の方法。
【請求項71】
前記融合ペプチドが2個以上の組換えペプチドを含み、前記組換えペプチドが異なる、請求項56に記載の方法。
【請求項72】
前記組換えペプチドが治療ペプチドである、請求項56に記載の方法。
【請求項73】
前記組換えペプチドが抗原である、請求項56に記載の方法。
【請求項74】
前記抗原が、イヌパルボウイルス抗原、炭疽菌抗原及び東部ウマ脳脊髄炎ウイルス抗原からなる群より選択される、請求項73に記載の方法。
【請求項75】
前記ウイルス様粒子がワクチンとして使用できる、請求項56に記載の方法。
【請求項76】
前記組換えペプチドが抗菌ペプチドである、請求項56に記載の方法。
【請求項77】
前記抗菌ペプチドが、D2A21及びPBF20からなる群より選択される、請求項76に記載の方法。
【請求項78】
前記組換えペプチドが、少なくとも7アミノ酸の長さである、請求項56に記載の方法。
【請求項79】
前記組換えペプチドが、少なくとも15アミノ酸の長さである、請求項56に記載の方法。
【請求項80】
前記細胞が、野生型二十面体ウイルスカプシドをエンコードする第二の核酸をさらに含む、請求項56に記載の方法。
【請求項81】
前記細胞が、
a)少なくとも1個の第二の二十面体ウイルスカプシド;及び
b)少なくとも1個の第二の組換えペプチド
を含有する第二の融合ペプチドをエンコードする第二の核酸をさらに含む、請求項56に記載の方法。
【請求項82】
前記細胞において前記第二の核酸を発現することを含む、請求項81に記載の方法。
【請求項83】
前記第二の融合ペプチドが細胞内でウイルス様粒子又は可溶性ケージ構造を形成するように集合する、請求項81に記載の方法。
【請求項84】
前記第一の二十面体ウイルスカプシドが第一のアミノ酸配列を含み、第二の二十面体ウイルスカプシドが第二のアミノ酸配列を含み、第一と第二のカプシド配列が異なる、請求項81に記載の方法。
【請求項85】
前記第一の組換えペプチドが第一のアミノ酸配列を含み、第二の組換えペプチドが第二のアミノ酸配列を含み、第一と第二の組換えペプチド配列が異なる、請求項81に記載の方法。
【図1】

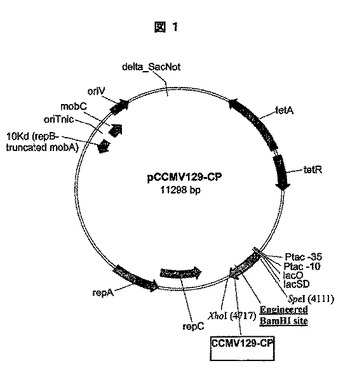
【図2】
【図3】
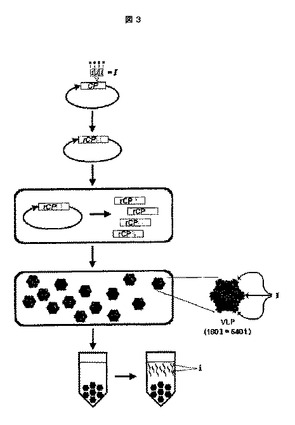
【図4】
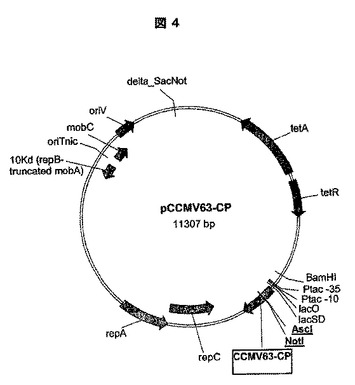
【図5】
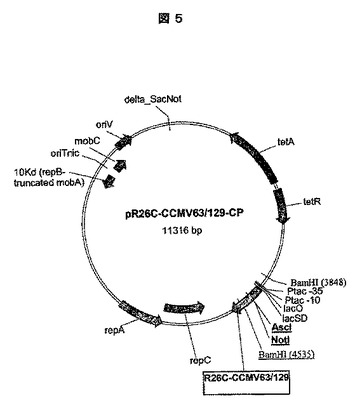
【図6】
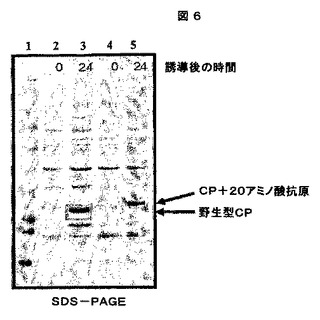
【図7】
【図8】
【図9】

【図10】
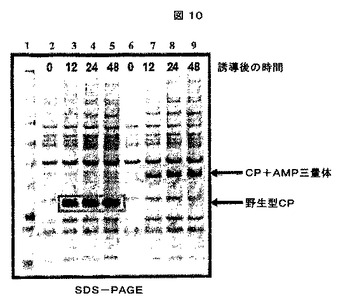
【図11】
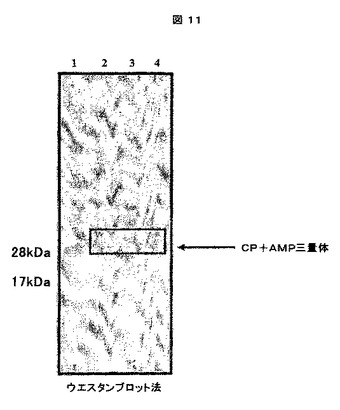
【図12】
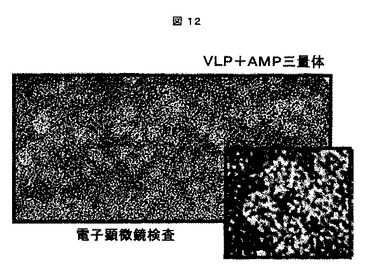
【図13】
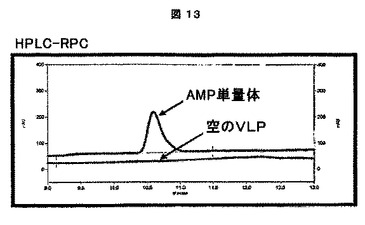
【図14】
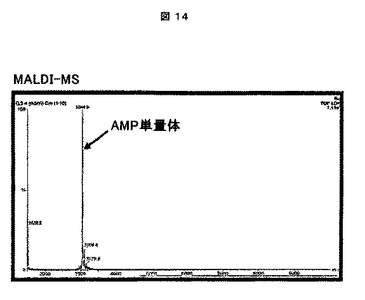
【図15】
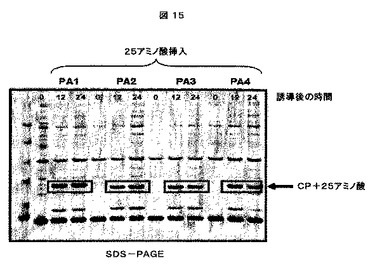
【図16】
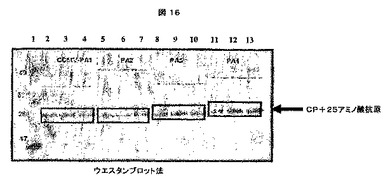
【図17】
【図18】
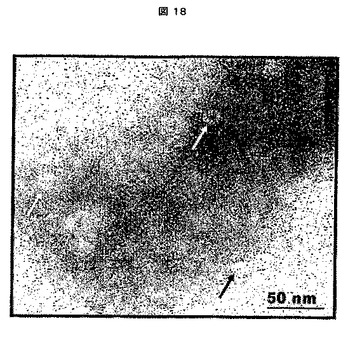
【図19】
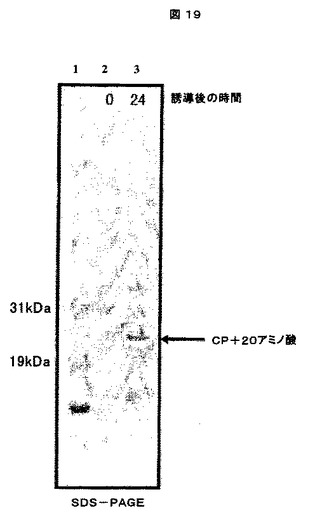
【図20】

【図21】
【図22】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【公表番号】特表2007−512842(P2007−512842A)
【公表日】平成19年5月24日(2007.5.24)
【国際特許分類】
【出願番号】特願2006−542692(P2006−542692)
【出願日】平成16年12月1日(2004.12.1)
【国際出願番号】PCT/US2004/040117
【国際公開番号】WO2005/067478
【国際公開日】平成17年7月28日(2005.7.28)
【出願人】(502141050)ダウ グローバル テクノロジーズ インコーポレイティド (1,383)
【Fターム(参考)】
【公表日】平成19年5月24日(2007.5.24)
【国際特許分類】
【出願日】平成16年12月1日(2004.12.1)
【国際出願番号】PCT/US2004/040117
【国際公開番号】WO2005/067478
【国際公開日】平成17年7月28日(2005.7.28)
【出願人】(502141050)ダウ グローバル テクノロジーズ インコーポレイティド (1,383)
【Fターム(参考)】
[ Back to top ]