
シンターゼ
【課題】テルペノイド反応中間体および反応産物を生成する、新規のシンターゼおよび該シンターゼをコードする核酸を提供する。
【解決手段】重要なアミノ酸残基を含む活性部位ポケットがあり、該アミノ酸残基が目的のテルペノイド反応中間体および反応産物を生成するように改変された、テルペンシンターゼと該シンターゼをコードする遺伝子。シンターゼの改変は、例えば、活性部位に基質が結合したまたは結合していない、タバコの5-エピ-アリストロケンシンターゼの三次元座標に基づいて設計される。
【解決手段】重要なアミノ酸残基を含む活性部位ポケットがあり、該アミノ酸残基が目的のテルペノイド反応中間体および反応産物を生成するように改変された、テルペンシンターゼと該シンターゼをコードする遺伝子。シンターゼの改変は、例えば、活性部位に基質が結合したまたは結合していない、タバコの5-エピ-アリストロケンシンターゼの三次元座標に基づいて設計される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、シンターゼタンパク質の活性部位やそこに位置するα-炭素原子やそれらの空間的関係などの情報を用いて新しい合成能を有する新規シンターゼを開発し生産する方法に関する。
【0002】
関連出願の相互参照
本出願は、1998年9月18日付けの仮出願第60/100,993号、1999年4月22日付けの仮出願第60/130,628号、および1999年8月23日付けの仮出願第60/150,262号の優先権の利益を主張するものである。
【0003】
本研究は、一部には、連邦政府機関からの基金によって支援されたものである。したがって、米国連邦政府は本発明に対して一定の権利を有する。
【背景技術】
【0004】
発明の背景
イソプレノイド化合物はさまざまな生物(例えば、植物、細菌、真菌など)によって産生される有機分子である。これまでに23,000種を超えるイソプレノイド分子が特徴づけられており、毎年何十、何百もの新しい構造が同定されている。これらの分子は種々の役割を果たすことができる。例えば、モノテルペン類は芳香剤や香味料として使用される。セスキテルペン類とジテルペン類はフェロモン、防御用薬剤、視覚色素、抗腫瘍薬、シグナル伝達経路の成分として作用しうる。トリテルペン類は膜の構成成分として、さらにステロイドホルモンと胆汁酸の前駆物質として重要な機能を果たしている。ポリプレノール類は光感受性物質およびコファクター側鎖として機能し、また天然ポリマーとしても存在しうる。
【0005】
イソプレノイド経路により産生される種々の分子化合物はモノ不飽和イソプレン単位の二リン酸エステルから作られる。イソプレンはプレニルトランスフェラーゼによって2、3、または4の倍数で一緒に付加されて、それぞれC10、C15、およびC20単位を作る。C10、C15、およびC20分子はそれぞれゲラニル二リン酸(GPP)、ファルネシル二リン酸(FPP)、およびゲラニルゲラニル二リン酸(GGPP)と呼ばれており、テルペンシンターゼの基質として作用する。
【0006】
テルペンシンターゼは化学または生物学の分野で知られている最も複雑な反応のうちの一つを介してイソプレノイド化合物の生成を触媒している。一般的に、テルペンシンターゼは約40〜100kDの分子量をもつ中程度の大きさの酵素である。酵素として、テルペンシンターゼは、絶妙な反応特異性およびキラリティーの保存と結びついた低度から中程度の代謝回転率をもつとして分類することができる。
【0007】
代謝回転は、基質の酵素への結合、基質コンホメーションの確立、基質の生成物への変換、および生成物の放出から成っている。反応は水性溶媒中でin vitroで行なうことができ、典型的にはコファクターとしてマグネシウムイオンを必要とし、得られる生成物はしばしば非常に疎水性であって、有機溶媒に分配することで回収できる。
【0008】
テルペンシンターゼ遺伝子は細菌、真菌、植物などの様々な生物中に見出される。異なるテルペンシンターゼ間でエキソンに近似する領域を交換することによって、最終的な酵素段階に関与する機能性ドメインが同定された。例えば、Nicotiana tabacum(タバコ)由来の5-エピ-アリストロケンシンターゼ(TEAS)およびヒヨス(henbane)由来のHyoscyamus muticusベチスピラジエンシンターゼ(HVS)について行なった研究により、エキソン4とエキソン6はそれぞれが反応生成物特異性に関与していることが明らかにされた。機能性ドメインを組み合わせることにより、新しい反応生成物を合成する能力がある新規の酵素が得られている(米国特許第5,824,774号)。
【0009】
多くの研究により、イソプレノイドを生成するための反応機構が提案されている。例えば、Caneら, 1985, Bioorg. Chem., 13:246-265; Wheeler and Croteau, 1987, Proc. Natl. Acad. Sci. USA, 84:4856-4859;およびPyunら, 1994, Arch. Biochem. Biophys., 308:488-496を参照されたい。これらの研究では、基質類似体および自己不活化阻害剤(Croteau, 1994, Arch. Biochem. Biophys., 251:777-782; Caneら, 1995, Biochemistry, 34:2471-2479;およびCroteau, 1993, Arch. Biochem. Biophys., 307:397-404)、ならびに触媒作用に不可欠なアミノ酸を同定しようとして化学修飾剤と部位特異的突然変異誘発(Caneら, 1995, Biochemistry, 34:2480-2488; Rajaonarivonyら, 1992, Arch. Biochem. Biophys., 296:49-57;およびRajaonarivonyら, 1992, Arch. Biochem. Biophys., 299:77-82)が採用された。しかしながら、これらの研究は、こうした技法に伴う固有の限界のため、活性部位を規定するうえで限られた成功を収めたにすぎなかった。
【発明の概要】
【0010】
本発明は、テルペンシンターゼの活性部位に存在するα-炭素原子を同定する方法、ならびに三次元空間におけるこれらの原子およびそれらの間の空間的関係を開示する。本発明はまた、そのようなα-炭素と関連したR基、および新しい反応産物を生成することができる新規テルペンシンターゼを作るためにこれらのR基を変更する方法を開示する。
【発明が解決しようとする課題】
【0011】
本発明が本出願において教示されるまで、シンターゼタンパク質の活性部位、そこに位置するアミノ酸、触媒作用に関与するアミノ酸残基、および活性部位内のα-炭素とR基の立体配置は知られていなかった。本発明は今回、シンターゼの構造を教示するのみならず、それから得られる情報を用いて新しい合成能を有する新規シンターゼを開発し生産する手段を提供する。ここに記載の方法を用いて得られたデータは、種々のイソプレノイド基質を使用して種々のイソプレノイド産物を生成することができるシンターゼ変異体の創製および生産に有用である。
【課題を解決するための手段】
【0012】
一実施形態において、本発明は、配列番号2の残基265〜535に対して約20%以上の配列同一性を有する単離されたテルペンシンターゼを特徴とする。該シンターゼは9個のα-炭素を含み、該9個のα-炭素は表6にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有する。該α-炭素のそれぞれの中心点は半径2.3Åの球内に位置する。該球のそれぞれの中心点は表5に示した構造座標を有する。該α-炭素のそれぞれは関連するR基を有しており、該シンターゼは表9に示したR基の順序配列とは異なるR基の順序配列を有する。該シンターゼは配列番号2の残基265〜535に対して約25%以上の配列同一性、または配列番号2の残基265〜535に対して約35%以上の配列同一性を有する。この種のシンターゼはモノテルペン基質、セスキテルペン基質、またはジテルペン基質からのテルペノイド産物の生成を触媒することができる。該産物は環式テルペノイド炭化水素または非環式テルペノイド炭化水素でありうる。どちらのタイプの産物もヒドロキシル化されていてもよい。α-炭素1と関連したR基は、次の群:Cys、SerおよびThrからなる群、Phe、TyrおよびTrpからなる群、Pro、GlyおよびAlaからなる群、GluおよびAspからなる群、Met、Ile、ValおよびLeuからなる群、ArgおよびLysからなる群、ならびにGln、AsnおよびHisからなる群のうちの一つより選択することができる。α-炭素2〜9と関連したR基は、表9の順序配列を有するものを除いた任意のアミノ酸でありうる。同様に、α-炭素2〜9のそれぞれと関連したR基は、Cys、SerおよびThrからなる群、Phe、TyrおよびTrpからなる群、Pro、GlyおよびAlaからなる群、GluおよびAspからなる群、Met、Ile、ValおよびLeuからなる群、ArgおよびLysからなる群、ならびにGln、AsnおよびHisからなる群から独立に選択することができる。これらの実施形態において、残りの8個のα-炭素と関連したR基は、表9の順序配列を有するものを除いた任意のアミノ酸でありうる。
【0013】
いくつかの実施形態において、α-炭素1〜9と関連したR基の順序配列は、それぞれTrp、Ile、Thr、Thr、Tyr、Leu、Cys、ThrおよびPhe、それぞれSer、Ile、Thr、Thr、Tyr、Leu、Cys、ThrおよびTyr、それぞれTrp、Ile、Thr、Thr、Tyr、Leu、Trp、ThrおよびTyr、それぞれSer、Ile、Thr、Thr、Tyr、Leu、Trp、ThrおよびTyr、またはそれぞれGlu、Ile、Thr、Thr、Tyr、Leu、Cys、ThrおよびTyrである。
【0014】
本発明はまた、所定のテルペンシンターゼポリペプチドの一次アミノ酸配列を、配列番号2の残基265〜535のアミノ酸配列にアライメントし、該所定のポリペプチドをコードする核酸に、配列番号2の残基265〜535に対して20%以上の配列同一性を有する該ポリペプチドの一次アミノ酸配列の領域中の9個のアミノ酸残基のうち1個以上のコドンで突然変異を起こさせ、ただし、該ポリペプチド中の該9個の残基は配列番号2の残基273、294、402、403、404、407、440、519および520とアライメントするものであり、そして変異型のテルペンシンターゼが産生されるように突然変異させた核酸を発現させることにより得られる、テルペンシンターゼを特徴とする。
【0015】
本発明はまた、配列番号2の残基265〜535に対して20%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、該シンターゼが16個のα-炭素を含み、該16個のα-炭素が表4にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有する、該テルペンシンターゼを特徴とする。該α-炭素のそれぞれの中心点は半径2.3Åの球内に位置する。該球のそれぞれの中心点は表3に示した構造座標を有する。該α-炭素のそれぞれは関連するR基を有しており、該シンターゼは表8に示したR基の順序配列とは異なるR基の順序配列を有する。該シンターゼは、配列番号2の残基265〜535に対して約25%以上の配列同一性、または配列番号2の残基265〜535に対して約35%以上の配列同一性を有しうる。該シンターゼはモノテルペン基質、セスキテルペン基質、またはジテルペン基質からのテルペノイド産物の生成を触媒することができる。前記産物は例えば環式テルペノイド炭化水素でありうる。α-炭素1〜16と関連した前記シンターゼ中のR基の順序配列は、それぞれCys、Trp、Ile、Ile、Ser、Thr、Thr、Tyr、Leu、Cys、Val、Thr、Tyr、Asp、PheおよびThrでありうる。
【0016】
本発明はまた、配列番号2の残基265〜535に対して20%以上の配列同一性を有する単離されたテルペンシンターゼであって、該シンターゼが19個のα-炭素を含み、該19個のα-炭素が表2にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有する、該テルペンシンターゼを特徴とする。該α-炭素のそれぞれの中心点は半径2.3Åの球内に位置する。該球のそれぞれの中心点は表1に示した構造座標を有する。該α-炭素のそれぞれは関連するR基を有しており、該シンターゼは表7に示したR基の順序配列とは異なるR基の順序配列を有する。該シンターゼは、配列番号2の残基265〜535に対して約25%以上の配列同一性、または配列番号2の残基265〜535に対して約35%以上の配列同一性を有しうる。該シンターゼはモノテルペン基質、セスキテルペン基質、またはジテルペン基質からのテルペノイド産物の生成を触媒することができる。前記産物は例えば環式テルペノイド炭化水素でありうる。
【0017】
本発明はまた、アミノ末端とカルボキシル末端とを有する第1のドメインを含む単離されたタンパク質を特徴とする。該第1のドメインは三次元空間においてグリコシルヒドロラーゼの触媒コアと構造的にアライメントするアミノ酸を含み、ここで、該グリコシルヒドロラーゼの触媒コアは、Aspergillus awamoriのグルコアミラーゼタンパク質データバンク(PDB)コード3GLYのアミノ酸36〜230、およびエンドグルカナーゼCelD PDBコード1CLCのアミノ酸36〜230からなる群より選択されるものである。該単離されたタンパク質はさらに、アミノ末端とカルボキシル末端とを有する第2のドメインを含む。該第2のドメインは三次元空間においてトリFPPシンターゼと構造的にアライメントするアミノ酸を含む。第1のドメインのカルボキシル末端は第2のドメインのアミノ末端に連結されている。第2のドメインの領域は配列番号2の残基265〜535に対して約20%以上の配列同一性を有し、かつ9個のα-炭素を含み、該9個のα-炭素は表6にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有する。該α-炭素のそれぞれの中心点は半径2.3Åの球内に位置し、該球のそれぞれの中心点は表5に示した構造座標を有する。該α-炭素のそれぞれは関連するR基を有しており、該シンターゼは表9に示したR基の順序配列とは異なるR基の順序配列を有する。前記タンパク質は配列番号2に対して約25%以上の配列同一性、または配列番号2に対して約35%以上の配列同一性を有する。前記シンターゼはモノテルペン基質、セスキテルペン基質、またはジテルペン基質からのテルペノイド産物の生成を触媒することができる。前記産物は例えば環式テルペノイド炭化水素でありうる。
【0018】
本発明はまた、配列番号20の残基343〜606に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号20の348、351、372、375、376、454、479、480、481、482、485、519、523、597、600、601、605、607および608位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸Y、L、C、I、T、Y、S、C、G、H、S、L、G、F、G、Y、D、YおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0019】
本発明はまた、配列番号22の残基316〜586に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号22の321、324、345、348、349、427、452、453、454、455、458、492、496、569、572、573、577、579および580位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、N、I、T、Y、S、I、S、G、M、L、D、A、M、Y、D、HおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0020】
本発明はまた、配列番号58の残基352〜622に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号58の357、360、381、384、385、463、487、488、489、490、493、528、532、606、609、610、614、616および617位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸Y、M、C、V、T、F、V、S、S、G、I、L、G、F、V、Y、D、YおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0021】
本発明はまた、配列番号33によりコードされるアミノ酸残基272〜540に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号33によりコードされる277、280、301、304、305、383、408、409、410、411、414、448、452、524、527、528、532、534および535位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸G、W、I、A、S、Y、T、S、G、Y、L、C、D、M、L、Y、D、YおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0022】
本発明はまた、配列番号42の残基319〜571に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号42の324、327、348、351、352、430、455、456、457、458、461、495、499、571、574、575、579、581および582位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸I、W、V、I、S、Y、T、T、G、L、V、I、N、T、S、Y、D、YおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0023】
本発明はまた、配列番号44の残基579〜847に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号44の584、587、606、609、610、688、713、714、715、716、719、753、757、831、834、835、839、841および842位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸V、S、G、Q、V、Y、S、V、G、L、C、W、N、V、F、Y、D、YおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0024】
本発明はまた、配列番号46の残基495〜767に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号46の500、503、524、527、528、606、631、632、633、634、637、674、678、751、754、755、759、761および762位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸F、L、A、Q、T、Y、S、I、G、Q、L、S、D、T、I、F、D、FおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0025】
本発明はまた、配列番号48の残基295〜564に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号48の300、303、324、327、328、406、431、432、433、434、437、471、475、548、551、552、556、558および559位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸Y、W、A、C、T、Y、S、S、G、M、L、G、D、L、I、Y、D、LおよびYとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0026】
本発明はまた、配列番号50の残基307〜578に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号50の312、315、336、339、340、419、444、445、446、447、450、484、488、562、565、566、570、572および573位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸F、W、A、M、T、Y、N、T、G、M、L、S、D、I、M、Y、D、FおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0027】
本発明はまた、配列番号52の残基264〜533に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号52の269、272、293、296、297、375、401、402、403、404、407、441、445、517、520、521、525、527および528位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、L、T、S、Y、S、A、G、Y、I、A、N、A、L、Y、D、YおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0028】
本発明はまた、配列番号56の残基585〜853に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号56の590、593、614、617、618、696、721、722、723、724、727、761、765、837、840、841、845、847および848位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸I、S、S、T、V、Y、S、I、A、L、V、G、N、M、F、Y、D、LおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0029】
本発明はまた、配列番号54の残基307〜574に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号54の312、315、336、339、340、418、443、444、445、446、449、483、487、560、563、564、566、568および569位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、I、I、T、Y、S、I、S、A、I、L、D、A、I、Y、D、DおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0030】
本発明はまた、配列番号24の残基309〜577に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号24の314、317、338、341、342、420、446、447、448、449、452、485、489、560、563、564、569、571および572位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、N、V、T、Y、I、G、G、I、L、L、D、A、I、Y、D、FおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0031】
本発明はまた、配列番号26の残基315〜584に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号26の320、323、344、347、348、426、451、452、453、454、457、492、496、568、571、572、576、578および579位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸S、W、I、A、T、Y、S、V、A、S、I、L、D、A、I、Y、D、FおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0032】
本発明はまた、配列番号28の残基265〜536に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号28の270、273、294、297、298、376、401、402、403、404、407、440、444、518、521、522、528、530および531位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸A、W、V、C、G、F、T、S、C、I、M、G、N、C、S、Y、D、YおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0033】
本発明はまた、配列番号30の残基342〜612に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号30の347、350、371、374、375、453、478、479、480、481、483、518、522、596、599、600、604、606および607位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸F、L、C、V、T、Y、S、S、A、Y、V、L、G、L、L、Y、D、FおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0034】
本発明はまた、配列番号32の残基273〜541に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号32の278、281、302、305、306、384、409、410、411、412、415、448、452、524、527、528、533、535および536位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、I、I、S、Y、T、S、T、Y、L、C、D、I、T、Y、D、YおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0035】
本発明はまた、配列番号2の残基265〜535に対して20%以上の配列同一性を有する領域をもつ所定のポリペプチドにおいて、9個のアミノ酸残基を同定すること、ただし該9個のアミノ酸残基のα-炭素は表6にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有し、該α-炭素のそれぞれの中心点が半径2.3Åの球内に位置し、該球のそれぞれの中心点が表5に示した構造座標を有するものであること;および所定のポリペプチドから改変されているポリペプチドを合成すること、ただし、該改変型ポリペプチドは、所定のポリペプチド中の該α-炭素と関連したR基とは異なる該9個のα-炭素と関連したR基を1個以上有すること;を含んでなる、テルペンシンターゼの製造方法を特徴とする。前記合成工程は、該9個のα-炭素に対応するアミノ酸1個以上のコード配列が所定のポリペプチド中に存在するアミノ酸とは異なるアミノ酸のコード配列で置換されている、所定のポリペプチドをコードする核酸を作製することを含む。所定のポリペプチドは例えば配列番号2、4、6,8、10、12、20、22,24、26、28、30、32,34-40、42、44、46、48、50、52、54、56または58に示したポリペプチドのいずれか1つでありうる。
【0036】
本発明はまた、配列番号2の残基265〜535に対して20%以上の配列同一性を有する領域をもつ所定のポリペプチドにおいて、配列番号2の273、294、402、403、404、407、440、519および520位のアミノ酸残基とアライメントする9つの位置のアミノ酸残基を同定すること;および所定のポリペプチドから改変されているポリペプチドを合成すること;を含んでなる、テルペンシンターゼの使用方法を特徴とする。該新規のポリペプチドは、該9つの位置のうち1つ以上に、所定のポリペプチド中に存在するアミノ酸残基とは異なるアミノ酸残基を有することにより改変される。いくつかの実施形態において、前記同定工程は、配列番号2の270、273、294、297、298、402、403、404、407、440、516、519、520、525、527および528位のアミノ酸残基とアライメントする所定のポリペプチド中の16個のアミノ酸残基を同定することを含み、前記合成工程は、所定のポリペプチドから改変されているポリペプチドを合成することを含み、該改変型ポリペプチドが、該16の位置のうち1つ以上に、所定のポリペプチド中に存在するアミノ酸残基とは異なるアミノ酸残基を有する。いくつかの実施形態において、前記同定工程は、配列番号2の270、273、294、297、298、376、401、402、403、404、407、440、444、516、519、520、525、527および528位のアミノ酸残基とアライメントする所定のポリペプチド中の19個のアミノ酸残基を同定することを含み、前記合成工程は、所定のポリペプチドから改変されているポリペプチドを合成することを含み、該改変型ポリペプチドが、該19の位置のうち1つ以上に、所定のポリペプチド中に存在するアミノ酸残基とは異なるアミノ酸残基を有する。前記合成工程は、同定されたアミノ酸残基のうち1個以上をコードする核酸中のコード配列が所定のポリペプチド中に存在するアミノ酸とは異なるアミノ酸をコードするコード配列で置換されている、所定のポリペプチドをコードする核酸を作製することを含む。所定のポリペプチドは例えば配列番号2、4、6,8、10、12、20、22,24、26、28、30、32,34-40、42、44、46、48、50、52、54、56または58に示したポリペプ
チドのいずれか1つでありうる。前記方法はさらに、該改変型ポリペプチドとイソプレノイド基質とを、該基質が該ポリペプチドに結合するのに有効な条件下で接触させること;および該改変型ポリペプチドの、該イソプレノイド基質からの反応産物の生成を触媒する能力を測定すること;を含む。イソプレノイド基質はモノテルペン、セスキテルペン、またはジデルペンでありうる。
【0037】
本発明はまた、ポリペプチド類をコードする核酸分子の集団を作製することを含んでなる、テルペンシンターゼの製造方法を特徴とする。前記集団は、配列番号2の残基265〜535に対して約20%以上の配列同一性を有する所定のテルペンシンターゼのアミノ酸を特定する9つのコドンのうち1つ以上において相互に異なっているメンバーを含み、該9個のアミノ酸のα-炭素は表6にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有し、該α-炭素のそれぞれの中心点が半径2.3Åの球内に位置し、該球のそれぞれの中心点が表5に示した構造座標を有するものである。いくつかの実施形態において、前記コドンは所定のテルペンシンターゼの表1〜2または3〜4に記載のアミノ酸を特定する。該核酸集団の一部または全部を発現させてポリペプチドの集団を作製するようにする。該ポリペプチド集団の少なくとも1つのメンバーは変異型テルペンシンターゼである。前記発現工程は該核酸集団のin vitro転写およびin vitro翻訳を含むことができる。いくつかの実施形態において、前記発現工程は、該核酸集団のメンバーを発現ベクターにクローニングすること;該発現ベクターを宿主細胞に導入し、該宿主細胞においてクローニングした核酸集団メンバーを発現させて該ポリペプチドの集団を作製すること;を含みうる。所定のテルペンシンターゼポリペプチドはモノテルペンシンターゼ、セスキテルペンシンターゼ、またはジテルペンシンターゼでありうる。宿主細胞は原核細胞または真核細胞であり、例えば、細菌細胞、真菌細胞および動物細胞(例:哺乳動物細胞または昆虫細胞)が含まれる。また、宿主細胞は植物細胞であってもよく、例えば、Graminaceae(イネ科)植物由来の細胞、Legumineae(マメ科)植物由来の細胞、Solanaceae(ナス科)植物由来の細胞、Brassicaeae(アブラナ科)植物由来の細胞、またはConifereae(針葉樹類)植物由来の細胞がある。
【0038】
本発明はまた、本明細書に記載のシンターゼをコードする核酸、および該核酸を含有する宿主細胞を特徴とする。本発明はまた、そのような核酸を含むトランスジェニック植物、またはそのような核酸を含むトランスジェニック動物細胞培養物を特徴とする。
【0039】
いくつかの実施形態において、本発明のシンターゼポリペプチドは、表5の座標を有する9個のα-炭素原子(該α-炭素間の原子間距離は表6に示した距離の±2.3Åである)を含む活性部位を有するドメインを含んでなる。これらのα-炭素原子は、三次元空間において、結合した基質または基質類似体の存在下または不在下でトリFPPシンターゼと構造的にアライメントする。別の実施形態において、本発明のシンターゼは、(i) 三次元空間において(溶解状態または結晶形態で、かつ基質が結合したまたは結合していない状態で)、(a) Aspergillus awamoriのグリコシルヒドロラーゼ(PDBコード3GLY)のアミノ酸36〜230、および(b) エンドグルカナーゼCellB(PDBコード1CLC)のアミノ酸36〜230からなる群より選択されるグリコシルヒドロラーゼ触媒コアとアライメントするアミノ酸残基を含む第1のドメイン、および(ii) 三次元空間において、基質または基質類似体が活性部位に結合したまたは結合していない状態で、トリFPPシンターゼと構造的にアライメントするアミノ酸残基を含む第2のドメイン、を含むものである。第2のドメインは表1〜2、3〜4、または5〜6の原子間距離および構造座標を有する9個、16個または19個のα-炭素原子を含む活性部位を含んでなる。これらのα-炭素原子にはイソプレノイド基質と直接または間接に相互作用しうるR基が結合されている。
【0040】
本発明はまた、触媒活性を有する変異型テルペンシンターゼの作製方法を特徴とする。この方法は、(a) 活性部位を有する触媒的に活性なテルペンシンターゼの結晶学的モデルを提供すること、および(b) 該モデルを用いて、所定のシンターゼに対して活性部位中の少なくとも1個のR基が改変されたテルペンシンターゼを設計すること、を含んでなる。本発明はまた、基質特異性が改変されているテルペンシンターゼ、該テルペンシンターゼの製造方法、およびその三次元構造の作製方法を特徴とする。
【0041】
本明細書に記載したものと同様のまたは等価の方法および材料を用いて本発明を実施することができるけれども、適当な方法および材料を以下に記載する。本明細書中で挙げた刊行物、特許出願、特許および他の文献はいずれもその全体を参照により本明細書中に組み入れるものとする。
【0042】
本発明の他の態様、実施形態、効果および特徴は本明細書から明らかになるであろう。
【図面の簡単な説明】
【0043】
【図1】図1は、Carson, M.およびBugg, C., J. Mol. Graphics 4:121(1986)のRIBBONSソフトウェアプログラムを用いて作成した、ファルネシルヒドロキシホスホネート(FHP)が結合した状態のタバコの5-エピ-アリストロケンシンターゼ(TEAS)の模式図である。
【図2A】図2Aは、α-炭素と関連するR基を示す20種の天然アミノ酸の構造である。
【図2B】図2Bは、α-炭素と関連するR基を示す20種の天然アミノ酸の構造である。
【図3】図3は、GGPPを基質として用いてTEASおよび変異体TEAS酵素により産生されたテルペノイド炭化水素産物の銀薄層クロマトグラムのオートラジオグラムである。DM:W273S/C440W変異体TEAS酵素。
【図4】図4は、FPPを基質として用いてTEASおよび変異体TEAS酵素により産生されたテルペノイド炭化水素産物の銀薄層クロマトグラムのオートラジオグラムである。
【0044】
表の説明
表1は、テルペンシンターゼの活性部位において見出された19個のα-炭素についてのX線結晶構造座標である。
【0045】
表2は、Åで表した表1の各α-炭素間の原子間距離である。各α-炭素は2.3Åの半径を有する球として空間を占有する。原子間距離は各球の中心から計算した。
【0046】
表3は、テルペンシンターゼの活性部位において見出された16個のα-炭素についてのX線結晶構造座標である。
【0047】
表4は、Åで表した表3の各α-炭素間の原子間距離である。各α-炭素は2.3Åの半径を有する球として空間を占有する。原子間距離は各球の中心から計算した。
【0048】
表5は、テルペンシンターゼの活性部位において見出された9個のα-炭素に
ついてのX線結晶構造座標である。
【0049】
表6は、Åで表した表5の各α-炭素間の原子間距離である。各α-炭素は2.3Åの半径を有する球として空間を占有する。原子間距離は各球の中心から計算した。
【0050】
表7は、表1のα-炭素に関連して見出されなかったR基の順序配列である。
【0051】
表8は、表3のα-炭素に関連して見出されなかったR基の順序配列である。
【0052】
表9は、表5のα-炭素に関連して見出されなかったR基の順序配列である。
【0053】
表10は、活性部位に基質類似体FHPが結合したTEASについてのX線構造座標である。
【0054】
表11は、基質が存在しない状態でのTEASについてのX線構造座標である。
【0055】
表12は、BLASTpアライメントプログラムを用いた、リモネンシンターゼ(配
列番号22)とTEASの残基265-535とのアライメントである。
【0056】
表13は、BLASTpアライメントプログラムを用いた、配列番号44の残基579-
847と配列番号26とのアライメントである。
【0057】
表14は、BLASTpアライメントプログラムを用いた、TEASの残基265-535と配列番号48とのアライメントである。
【0058】
表15は、BLASTpアライメントプログラムを用いた、配列番号50の残基307-593と配列番号56とのアライメントである。
【0059】
配列表の簡単な説明
配列番号1は、タバコの5-エピ-アリストロケンシンターゼ(TEAS)タンパク質のDNAコード配列である。Genbank受託番号:Q40577。
【0060】
配列番号2は、配列番号1のDNAによりコードされるタンパク質のアミノ酸配列である。
【0061】
配列番号3は、Trp273のコドンがGluのコドンに変わっているTEASタンパク質のDNAコード配列である。
【0062】
配列番号4は、配列番号3のTEAS DNAによりコードされるW273Eタンパク質のアミノ酸配列である。
【0063】
配列番号5は、Tyr520のコドンがPheのコドンに変わっているTEASタンパク質のDNAコード配列である。
【0064】
配列番号6は、配列番号5のTEAS DNAによりコードされるY520Fタンパク質のアミノ酸配列である。
【0065】
配列番号7は、Tyr527のコドンがPheのコドンに変わっているTEASタンパク質のDNAコード配列である。
【0066】
配列番号8は、配列番号7のTEAS DNAによりコードされるY527Fタンパク質のアミノ酸配列である。
【0067】
配列番号9は、Trp273のコドンがSerのコドンに変わっており、かつCys440のコドンがTrpのコドンに変わっているTEASタンパク質のDNAコード配列である。
【0068】
配列番号10は、配列番号9のTEAS DNAによりコードされるW273S/C440Wタンパク質のアミノ酸配列である。
【0069】
配列番号11は、Tyr406およびLeu407のコドンがそれぞれヌクレオチドNNSに変わっているTEASタンパク質のDNAコード配列である。
【0070】
配列番号12は、配列番号11のTEAS DNAによりコードされるY406X/L407Xタンパク質の集団のアミノ酸配列であり、但し、Xは任意の天然に存在するアミノ酸である。
【0071】
配列番号13は、DNAプライマー配列である。
【0072】
配列番号14は、DNAプライマー配列である。
【0073】
配列番号15は、DNAプライマー配列である。
【0074】
配列番号16は、DNAプライマー配列である。
【0075】
配列番号17は、DNAプライマー配列である。
【0076】
配列番号18は、DNAプライマー配列である。
【0077】
配列番号19は、アメリカオオモミ(grand fir)のピネンシンターゼのDNAコード配列である。Genbank受託番号:U87909。
【0078】
配列番号20は、配列番号19のDNAによりコードされるタンパク質のアミノ酸配列である。
【0079】
配列番号21は、オランダハッカ(spearmint)のリモネンシンターゼのDNAコード配列である。Genbank受託番号:L13459。
【0080】
配列番号22は、配列番号21のDNAによりコードされるタンパク質のアミノ酸配列である。
【0081】
配列番号23は、ヤクヨウサルビア(sage)の1,8シネオールシンターゼのDNAコード配列である。Genbank受託番号:AF051899。
【0082】
配列番号24は、配列番号23のDNAによりコードされるタンパク質のアミノ酸配列である。
【0083】
配列番号25は、ヤクヨウサルビアのボルニルジホスフェートシンターゼのDNA
コード配列である。Genbank受託番号:AF051900。
【0084】
配列番号26は、配列番号25のDNAによりコードされるタンパク質のアミノ酸配列である。
【0085】
配列番号27は、ハッカ(mint)のE-b-ファメセン(E-b-famesene)シンターゼのDNAコード配列である。Genbank受託番号:AF024615。
【0086】
配列番号28は、配列番号27のDNAによりコードされるタンパク質のアミノ酸配列である。
【0087】
配列番号29は、アメリカオオモミのミルセンシンターゼのDNAコード配列である。Genbank受託番号:U87908。
【0088】
配列番号30は、配列番号29のDNAによりコードされるタンパク質のアミノ酸配列である。
【0089】
配列番号31は、馬鈴薯のベタスピラジエン(vetaspiradiene)シンターゼのDNAコード配列である。Genbank受託番号:AB022598。
【0090】
配列番号32は、配列番号31のDNAによりコードされるタンパク質のアミノ酸配列である。
【0091】
配列番号33は、綿(cotton)のデルタ-カジネンシンターゼのゲノムDNAコード配列である。Genbank受託番号:Y18484。
【0092】
配列番号34〜40は、配列番号33のDNAによりコードされるエキソンのアミノ酸配列である。
【0093】
配列番号41は、トウゴマ(castor bean)のカスベン(casbene)シンターゼのDNAコード配列である。Genbank受託番号:L32134。
【0094】
配列番号42は、配列番号41のDNAによりコードされるタンパク質のアミノ酸配列である。
【0095】
配列番号43は、イチイ(yew)のタキサジエンシンターゼのDNAコード配列である。Genbank受託番号:U48796。
【0096】
配列番号44は、配列番号43のDNAによりコードされるタンパク質のアミノ酸配列である。
【0097】
配列番号45は、アメリカオオモミのE-アルファ-ビザボレンシンターゼのDNAコード配列である。Genbank受託番号:AF006194。
【0098】
配列番号46は、配列番号45のDNAによりコードされるタンパク質のアミノ酸配列である。
【0099】
配列番号47は、アメリカオオモミのデルタ-セリネン(delta-silenene)シンターゼのDNAコード配列である。Genbank受託番号:U92266。
【0100】
配列番号48は、配列番号47のDNAによりコードされるタンパク質のアミノ酸配列である。
【0101】
配列番号49は、アメリカオオモミのガンマ-フムレンシンターゼのDNAコード配列である。Genbank受託番号:U92267。
【0102】
配列番号50は、配列番号49のDNAによりコードされるタンパク質のアミノ酸配列である。
【0103】
配列番号51は、トマトのゲルマクレンCシンターゼのDNAコード配列である。Genbank受託番号:AF035631。
【0104】
配列番号52は、配列番号51のDNAによりコードされるタンパク質のアミノ酸配列である。
【0105】
配列番号53は、ヤクヨウサルビアの+サビネンシンターゼのDNAコード配列である。Genbank受託番号:AF051901。
【0106】
配列番号54は、配列番号53のDNAによりコードされるタンパク質のアミノ酸配列である。
【0107】
配列番号55は、アメリカオオモミのアビエタジエン(abietadiene)シンターゼのDNAコード配列である。Genbank受託番号:U50768。
【0108】
配列番号56は、配列番号55のDNAによりコードされるタンパク質のアミノ酸配列である。
【0109】
配列番号57は、アメリカオオモミのリモネンシンターゼのDNAコード配列である。Genbank受託番号:AF006193。
【0110】
配列番号58は、配列番号57のDNAによりコードされるタンパク質のアミノ酸配列である。
【0111】
詳細な説明
本明細書中では以下の用語が用いられる。
【0112】
「α-炭素」とは、アミノ酸残基中で見られるキラル炭素原子をいう。α-炭素には、アミノ基、カルボキシル基、水素原子およびR基を含む4つの置換基が共有結合している。
【0113】
「R基」とは、タンパク質内でのペプチド結合の形成に関与しないアミノ酸残基のα-炭素に結合している置換基をいう。R基は、アミノ酸の全体的な化学的特性の重要な決定要素(determinant)である。タンパク質中に見られる20の天然に存在するアミノ酸および各アミノ酸のα-炭素に結合するR基を図2A、Bに掲載する。本明細書中では、特定のアミノ酸に結合するR基について言及するために、天然に存在するアミノ酸についての3文字または1文字の略語を用いることがある。
【0114】
「天然に存在するアミノ酸」としては、タンパク質において天然に存在する20種のアミノ酸のL-異性体が含まれる。天然に存在するアミノ酸は、グリシン、
アラニン、バリン、ロイシン、イソロイシン、セリン、メチオニン、トレオニン、フェニルアラニン、チロシン、トリプトファン、システイン、プロリン、ヒスチジン、アスパラギン酸、アスパラギン、グルタミン酸、グルタミン、アルギニン、およびリシンである。特に指示しない限り、本出願中で言及する全てのアミノ酸はL形である。本明細書中では、天然に存在するアミノ酸について言及するために、3文字または1文字の略語を用いることがある。これらの略語は当業界で公知である。
【0115】
「非天然のアミノ酸」としては、タンパク質において天然には見られないアミノ酸が含まれる。本明細書中での非天然のアミノ酸の例としては、セレノシステインおよびセレノメチオニンのラセミ混合物が挙げられる。さらに、非天然のアミノ酸には、ノルロイシン、パラ-ニトロフェニルアラニン、ホモフェニルアラニン、パラ-フルオロフェニルアラニン、3-アミノ-2-ベンジルプロピオン酸、ホモアルギニン、D-フェニルアラニンなどのD形またはL形が含まれる。
【0116】
「正に荷電したアミノ酸」としては、通常の生理学的条件下で正電荷を保有するR基を有する任意の天然に存在するアミノ酸または非天然のアミノ酸が含まれる。正に荷電した天然に存在するアミノ酸の例としては、アルギニンおよびリシンが挙げられる。
【0117】
「負に荷電したアミノ酸」としては、通常の生理学的条件下で負電荷を保有するR基を有する任意の天然に存在するアミノ酸または非天然のアミノ酸が含まれる。負に荷電した天然に存在するアミノ酸の例としては、アスパラギン酸およびグルタミン酸が挙げられる。
【0118】
「疎水性のアミノ酸」としては、通常の生理学的条件下で荷電していない非極性の側鎖を有する任意の天然に存在するアミノ酸または非天然のアミノ酸が含まれる。天然に存在する疎水性のアミノ酸の例としては、ロイシン、イソロイシン、バリンおよびメチオニンが挙げられる。
【0119】
「親水性のアミノ酸」としては、荷電している極性の側鎖を有する任意の天然に存在するアミノ酸または非天然のアミノ酸が含まれる。天然に存在する親水性のアミノ酸の例としては、セリン、トレオニンおよびシステインが挙げられる。
【0120】
「突然変異テルペンシンターゼ」または「突然変異型テルペンシンターゼ」とは、一次アミノ酸配列を有するシンターゼポリペプチドをいう。該ポリペプチドの9つの残基のα-炭素の中心点は、半径が2.3Åの球内に位置する。その9つの球の中心点は、表5の構造座標、または表5の座標に合せて回転および/もしくは変換され得る座標を有する。該9つのα-炭素の間の相対的な原子間距離は、表6に示す原子間距離の±2.3Åである。各α-炭素は結合しているR基を有する。突然変異シンターゼは、非突然変異シンターゼとは、該9つのα-炭素に結合しているR基の順序配列が異なる。突然変異シンターゼは、該9つのα-炭素上に、表9に示すR基の順序配列以外のR基の順序配列を有する。該シンターゼの一次アミノ酸配列の他のα-炭素に結合しているR基は、非突然変異型シンターゼにおけるものと同じものであってもよいし、同じものでなくてもよい。
【0121】
幾つかの実施形態において、突然変異シンターゼとは、該ポリペプチドの16の残基のα-炭素の中心点が半径2.3Åの球内に位置するシンターゼをいう。該16の球の中心点は、表3の構造座標、または表3の座標に合せて回転および/もしくは変換され得る座標を有する。該9つのα-炭素の間の相対的な原子間距離は、表4に示す原子間距離の±2.3Åである。各α-炭素は結合しているR基を有する。突然変異シンターゼは、非突然変異シンターゼとは、その16のα-炭素に結合しているR基の順序配列が異なる。突然変異シンターゼは、該16のα-炭素上に、表8に示すR基の順序配列以外のR基の順序配列を有する。該シンターゼの一次アミノ酸配列の他のα-炭素に結合しているR基は、非突然変異型シンターゼにおけるものと同じものであってもよいし、同じものでなくてもよい。
【0122】
幾つかの実施形態において、突然変異シンターゼとは、該ポリペプチドの19の残基のα-炭素の中心点が半径2.3Åの球内に位置するシンターゼをいう。該19の球の中心点は、表1の三次元座標、または表1の座標に合せて回転および/もしくは変換され得る座標を有する。該19のα-炭素の間の相対的な原子間距離は、表2に示す原子間距離の±2.3Åである。各α-炭素は結合しているR基を有する。突然変異シンターゼは、非突然変異シンターゼとは、その19のα-炭素に結合しているR基の順序配列が異なる。突然変異シンターゼは、該19のα-炭素上に、表7に示すR基の順序配列以外のR基の順序配列を有する。該シンターゼの一次アミノ酸配列の他のα-炭素に結合しているR基は、非突然変異型シンターゼにおけるものと同じものであってもよいし、同じものでなくてもよい。
【0123】
「非突然変異型シンターゼ」または「非突然変異シンターゼ」としては、9、16または19のアミノ酸残基を含む一次アミノ酸配列を有するシンターゼが含まれる。これらの残基の各α-炭素の中心点は、半径2.3Åの球内に位置する。該球の中心点は、それぞれ表5、3もしくは1の三次元座標、または表5、3もしくは1の座標に合せて回転および/もしくは変換され得る座標を有する。該9、16または19のα-炭素の間の相対的な原子間距離は、それぞれ表6、4または2に示す原子間距離の±2.3Åである。各α-炭素は結合しているR基を有する。非突然変異シンターゼは、該9、16または19のα-炭素上に、それぞれ表9、8または7に示すようなR基の順序配列を有する。
【0124】
「その縮重変異体」とは、それにより、遺伝暗号の縮重のために異なるヌクレオチド配列により同じポリペプチドがコードされる遺伝子コード配列の変異体をいう。例えば、本発明のシンターゼは、一次アミノ酸配列を有する。縮重シンターゼ変異体は、異なる核酸コード配列であるが、それにもかかわらず、遺伝暗号の縮重のために同じ一次アミノ酸配列をコードする。
【0125】
「発現」とは、遺伝子または核酸分子の転写および該核酸のポリペプチドへの翻訳をいう。遺伝子の発現とはまた、真核系におけるRNAのmRNAへのプロセシングを含む。その遺伝子は、必ずしも、発現を達成するために細胞のゲノムに組み込まれる必要はない。この定義は、特定の系または特定の細胞型における発現に限られるものではなく、(限定しようとするものではないが)安定な、一過性の、in vitroでの、およびin vivoでの発現を含む。
【0126】
「プロモーター」および「プロモーター調節エレメント」とは、遺伝子の発現の調節に関与する核酸をいう。遺伝子発現を促進するために、種々の供給源に由来するプロモーター調節エレメントなどを効率的に用いることができる。プロモーター調節エレメントとしては、構成的、組織特異的、発生特異的(developmental-specific)、誘導的、サブゲノムプロモーターなどが含まれる。プロモーター調節エレメントはまた、転写効率を向上または調節する特定のエンハンサーエレメントまたはサイレンシングエレメントも含み得る。
【0127】
「活性部位」とは、テルペン基質であるGPP、FPPおよび/またはGGPPの疎水性部分を結合させるテルペンシンターゼ内の部位をいう。この活性部位は、特定の条件下で、1つ以上の反応産物が生成されるようにする生合成反応を触媒できる。
【0128】
「改変された酵素特異性」としては、対応する非突然変異型シンターゼの基質特異性およびそれにより作られる反応産物と比較した場合の、突然変異シンターゼの、特定のテルペン基質を使う能力の改変、または突然変異シンターゼからの反応産物のプロフィールの変化が含まれる。改変された特異性としては、シンターゼの、非突然変異型シンターゼと比較して異なる酵素パラメーター(Km、Vmax、など)を示す能力、および/または対応する非突然変異シンターゼにより生成されるものとは異なる産物を生成する能力を含み得る。
【0129】
「構造座標(structure coordinatesまたはstructural coordinates)」とは、結晶形態におけるシンターゼ分子の原子(散乱中心)によるX線の単色光線の回折により得られたパターンから求めた場合のフーリエ合成に関する数学的式から誘導された直交座標(x、yおよびz位置)をいう。回折データを用いて、該結晶(単位胞)中の繰り返しタンパク質単位の電子密度マップを算出する。電子密度マップを用いて、結晶の単位胞内での個々の原子の位置を確証する。本明細書中で示す構造座標の絶対値は、原子間の相対的な空間的関係を意味する。何故ならば、構造座標に帰せられる絶対値は、同じ該原子間の相対的な空間的関係を保ちながら、x、yおよび/またはz軸に沿って一緒に、または別々に回転運動および/または変換運動させることにより変えることができるからである。したがって、構造座標のセットの絶対値を表1、3または5に示す特定の値に合せて回転または変換により調節できるテルペンシンターゼは、表1、3または5のものと同じ構造座標を有すると考えられる。回転および/または変換させた後で本明細書中で示す絶対値と一致する構造座標の例は表11の座標である。
【0130】
「重原子誘導体化(heavy atom derivatization)」とは、シンターゼ結晶の化学的に改変された形態を作製する方法をいう。実際には、結晶を、該結晶中に拡散して該タンパク質の表面に結合できる重原子塩または有機金属化合物(例えば、塩化鉛、チオリンゴ酸金、チメロサール、酢酸ウラニルなど)を含む溶液に浸漬する。結合した重原子の位置は、浸漬した結晶のX線回折分析により求めることができる。次に、その情報を用いて位相の情報を構築し、次にBlundel, T.L.,およびJohnson, N.L., Protein Crystallography, Academic Press (1976)に記載のようにして、該情報を用いて該酵素の三次元構造を構築する。
【0131】
「単位胞」とは、基本的な平行六面体形のブロックをいう。そのようなブロックの規則的な集合体が結晶の全体積を構築している場合もある。各単位胞は、単位パターンの完全な表現(representation)を含んでおり、その繰返しが該結晶を構築している。
【0132】
「突然変異誘発」とは、タンパク質の一次アミノ酸配列内の特定の位置において異なるアミノ酸残基により置換して、それによりその位置に存在するR基を改変することをいう。突然変異誘発は、該タンパク質をコードする核酸のコード配列を変化させて、該核酸中の該コード配列がその位置に元々存在する残基とは異なるアミノ酸残基を特定するようにすることによって最も容易に行うことができる。
【0133】
「空間群」とは、結晶内における対称要素の配置をいう。
【0134】
「分子置換」とは、構造座標がわかっていないシンターゼの予備モデル(preliminary model)を、構造座標がわかっている分子を該未知の結晶の単位胞内で、該未知の結晶の観察される回折パターンを最もよく説明するように、配向および配置させることにより作製することをいう。次に、このモデルから位相を算出し、観察される振幅と組合せて、座標がわかっていない構造のおよそのフーリエ合成を得ることができる。次に、これを、幾つかの形態の精密化のいずれかに供して、該未知の結晶の最終的な正確な構造を得る(Lattman, E., 1985, Method in Enzymology中, 115:55-77; Rossmann, MG.,編, “The Molecular Replacement Method” 1972, Int. Sci. Rev. Ser., No. 13, Gorden & Breach, New York)。構造座標および原子間距離のマトリックスを用いれば、分子置換は、テルペンシンターゼの結晶性の突然変異体、相同体または異なる結晶形態の構造座標を求めるのに用いることができる。
【0135】
「組換えタンパク質」としては、化学的に合成された、または単離された遺伝子から生合成により誘導されたタンパク質が含まれる。
【0136】
「遺伝子」としては、ポリペプチドを作製するのに必要な情報を含む、自然に誘導された、または遺伝子操作された核酸が含まれる。
【0137】
「核酸」としては、ヌクレオチドであるグアニン、アデニン、チミン、シトシン、ウラシル、イノシンなどからなる任意の遺伝子物質が含まれる。核酸は一本鎖、二本鎖または三本鎖であってもよい。核酸は、デオキシリボ核酸であってもリボ核酸であってもよい。
【0138】
「遺伝子操作された」とは、所定の核酸中に存在するものとは異なるヌクレオチド配列を含むように改変されている遺伝子を含む。遺伝子は、合成により、または従来のクローニング、PCR、化学的遺伝子合成、直接もしくはランダムな突然変異誘発、および遺伝子シャッフリングにより操作できる。遺伝子操作された、とは、所定のタンパク質をコードする核酸の縮重変異体である遺伝子を作製するプロセスも含む。
【0139】
「第1のドメイン」とは、第1および第2の末端を有するポリペプチドを含み、そこにおいて、第1の末端は遊離アミノ基を有するアミノ末端アミノ酸を有してもよく、ペプチド結合により第2のアミノ酸に連結させてもよい。該第1の末端はまた、アセチル化などにより修飾されてもよい。該第1のドメインの第2の末端は遊離カルボキシル末端基を有してもよいし、有していなくてもよい。
【0140】
「第2のドメイン」とは、第1および第2の末端を有するポリペプチドを含み、そこにおいて、第1の末端はアミノ末端アミノ酸を有してもよく、ペプチド結合により第2のアミノ酸に連結されていてもよい。該第2のドメインの第2の末端はカルボキシル末端基を有してもよいし、有していなくてもよい。典型的には、該第2のドメインの第1の末端は、ペプチド結合を介して該第1のドメインの第2の末端に連結されている。
【0141】
「イソプレノイド基質」とは、それぞれゲラニルジホスフェート(GPP)、ファルネシルジホスフェート(FPP)およびゲラニルゲラニルジホスフェート(GGPP)と名付けられているC10、C15およびC20の分子をいう。
【0142】
「配列同一性」または「配列同一性の割合」とは、2つのタンパク質配列またはヌクレオチド配列[つまり、参照配列(query sequence)と対象配列(subject sequence)]のアライメントをとった場合の、同じ相対する位置を占めるアミノ酸またはヌクレオチドの割合(%)をいう。対象配列と参照配列との間で同一であるアミノ酸またはヌクレオチドの残基の数を数え、参照配列中の残基の数で割り、100をかける。最も高い割合の配列同一性をもたらすアライメントが見つかるまで、このプロセスを繰り返す。配列同一性の割合は、目視による確認により、および/またはMegAlign(DNASTAR, Inc., Madison, Wisconsin)またはNational Center for Biotechnology Information(NCBI)からワールドワイドウェブ上で入手可能なBLASTプログラムのような種々のコンピュータープログラムを用いて求めてもよい。配列アライメントを最大にするために、1つ以上の残基のギャップを参照配列の構造上保存されたドメイン(すなわち、α-ヘリックス、β-シートおよびループ)に挿入してもよい場合がある。
【0143】
「モノテルペン産物」とは、基質GPPからつくられる直鎖状、環状および/またはヒドロシキル化された反応産物をいう。「セスキテルペン産物」とは、基質FPPからつくられる直鎖状、環状および/またはヒドロシキル化された反応産物をいう。「ジテルペン産物」とは、基質GGPPからつくられる直鎖状、環状および/またはヒドロシキル化された反応産物をいう。
【0144】
本発明は、活性部位を含む、特定のα-炭素およびそれに結合しているR基の位置が、三次元空間において決定できるテルペンシンターゼおよびその突然変異体に関する。本発明はまた、該シンターゼの構造座標、シンターゼ相同体、突然変異体などに関する構造的情報を創り出すための該構造座標の使用、ならびにそのようなシンターゼの結晶形態に関する。さらに、本発明は、所定のテルペンシンターゼの活性部位を含む原子のα-炭素の構造座標を用いて、活性部位α-炭素原子に結合しているR基が該所定のテルペンシンターゼ中で見られるR基とは異なるシンターゼを開発する方法を提供する。さらに、本発明は、本明細書中で提供される構造的情報に基づく新規テルペンシンターゼの製造、および種々のイソプレノイド化合物を製造するためのそのようなシンターゼの使用を提供する。
【0145】
本発明はさらに、初めて、タバコの5-エピ-アリストロケンシンターゼ(TEAS)に例示されるような、基質または基質類似体の存在下または不在下で成長して、それに関連する構造座標の規定を可能にするシンターゼの結晶を提供する。該構造座標は、活性部位を含むα-炭素原子およびそれに結合しているR基の決定を可能にする。本発明の結晶は、正方晶系の空間群P41212に属し、単位胞の大きさは結晶間で数Åだけ変動するが、平均してa=126Å、c=122Å、a=b、α=90°、β=90°、およびγ=90°である。
【0146】
構造座標は、好ましくは、結合した基質または基質類似体の存在下および不在下でシンターゼについて約2.2〜約2.8Åの解像度(resolution)で得られる。活性部位に結合した基質類似体を有する場合のシンターゼの座標を表10に示す。活性部位に結合した基質類似体がない場合のシンターゼの座標は表11に示す。当業者であれば、X線結晶学により求めた構造座標のセットには標準誤差が含まれないことを理解する。したがって、本発明の目的のためには、シンターゼ、シンターゼ相同体またはその突然変異体の活性部位α-炭素が表1、3または5に示す構造座標を用いて重ねあわせた場合に±2.3Å未満の二乗平均平方根偏差を有する任意のセットの構造座標は同一であると考えられる。
【0147】
RIBBONS(CarsonおよびBugg, 1986, J. Mol. Graphics, 4:121)により作製されたシンターゼの三次元的形状の概略図を図1に示す。図1に示すシンターゼは、全体がα-ヘリックスならびに短い連結ループおよびターンからなり、第1および第2の構造ドメインに分かれて構成されている。
【0148】
1つの実施形態では、本発明の単離されたシンターゼは、表3の構造座標および表4に示す距離の±2.3Åの相対的距離を有する16の活性部位α-炭素を含む。表3の活性部位α-炭素は通常、種々の活性部位α-炭素間に介在するアミノ酸残基のために、全てが隣接しているわけではない(すなわち、シンターゼの一次アミノ酸配列において互いに隣接していない)。一方、幾つかの例においては、該活性部位α-炭素の幾つかは互いに隣接していてもよいと理解すべきである。例えば、TEAS Y527Fタンパク質(配列番号8)に示す実施形態では、一次アミノ酸配列において、それぞれ402位、403位および404位にて、活性部位α-炭素は互いに隣接しているが、残基273および294における活性部位α-炭素は分断されており、したがって隣接していない。したがって、表1、2、3、4、5または6に示す活性部位α-炭素の番号付けは単に便宜上のものにすぎず、そのようなα-炭素は、表1、3または5に示す構造座標および表2、4または6に示す±2.3Åの相対的原子間距離を達成する一次アミノ酸配列中の任意の位置に存在し得る。
【0149】
活性部位α-炭素に結合しているR基を適切に組み合わせれば、1種以上の目的の反応産物の生成を容易にし得る。本発明のテルペンシンターゼにおいて用いるために選ばれるR基の組合せは、表7、8または9に示すR基および対応する活性部位α-炭素の順序配列以外のどのような組合せであってもよい。R基およびα-炭素の適切な配列の代表例は、表3の活性部位α-炭素1〜16にそれぞれ結合しているCys、Trp、Ile、Ile、Ser、Thr、Thr、Tyr、Leu、Cys、Val、Thr、Phe、Asp、TyrおよびThrである。R基およびα-炭素の適切な配列の別の例は、表3の活性部位α-炭素1〜16のそれぞれにおけるCys、Trp、Ile、Ile、Ser、Thr、Thr、Tyr、Leu、Cys、Val、Thr、Tyr、Asp、PheおよびThrである。幾つかの実施形態では、本発明のシンターゼは、配列番号4、配列番号6、配列番号8および配列番号10に示す一次アミノ酸配列、それらをコードするDNA分子(それぞれ、配列番号3、配列番号5、配列番号7および配列番号9に示される)、ならびにそれらの縮重変異体を有する。典型的には、活性部位α-炭素で見られるR基は、天然に存在するアミノ酸において見られるものである。例えば図2A、Bを参照されたい。しかし、幾つかの実施形態では、天然に存在するアミノ酸以外のR基を用い得る。
【0150】
R基および活性部位α-炭素の配列には、反応産物を生成する突然変異テルペンシンターゼを生じさせるものもある。そのような酵素的に活性なシンターゼおよびそれらの対応する遺伝子は、例えばモノテルペン(例えばピネン)、セスキテルペン(例えばデルタ-カジネン)およびジテルペン(例えばアビエタジエン)のような既知のテルペノイド炭化水素の製造に有用である。他の酵素的に活性なシンターゼは、新規テルペノイド産物の製造に有用であり得る。
【0151】
R基および活性部位α-炭素の配列には、反応産物を所望の速度で生成しない突然変異テルペンシンターゼを生じさせるものもある。そのようなシンターゼおよびそれらの遺伝子は、酵素的に活性な突然変異シンターゼによる産物生成の分析における対照(コントロール)として有用である。そのようなシンターゼおよびそれらの遺伝子はまた、酵素的に活性な突然変異シンターゼ遺伝子の翻訳の分析においても、あるいは栄養補助剤としても有用である。そのようなシンターゼは、Sepharoseビーズに付着させて、粗調製物からのイソプレノイド化合物のアフィニティ精製に用いることができる。さらに、そのようなシンターゼおよびそれらの遺伝子はまた、種々の目的の試薬(例えばテルペンシンターゼタンパク質の発現をモニターするための免疫学的試薬)または植物育種プログラムにおけるテルペンシンターゼ遺伝子の遺伝をモニターするための核酸プローブもしくはプライマーの開発にも有用である。
【0152】
幾つかの実施形態では、シンターゼの第1のドメインのα-炭素骨格は、Aspergillus awamori由来のグルコアミラーゼ(Brookhaven Protein Database (PDB)コード3GLY)(Aleshinら, 1994, J. Mol. Biol. 238:575)およびClostridium thermocelum由来のエンドグルカナーゼCelD(PDBコードICLC)(Juyら, 1992, Nature, 357:89)により例示されるようはグリコシルヒドロラーゼの触媒コアと構造的に整列(align)し、シンターゼの第2のドメインのα-炭素骨格(活性部位を含む)は、鳥類ファルネシルジホスフェートシンターゼ(FPS)と構造的に整列し、そこにおいて、活性部位は、表1、3または5に示すα-炭素の構造座標および表2、4または6に記載の原子間距離を有する9、16または19のアミノ酸残基からなる。そのようなα-炭素は、非突然変異型シンターゼにおいて見られるものとは異なるR基の順序配列を有する。
【0153】
本発明において、第1のドメインは、水和された場合、きちんと並んだ水分子で満たされた表面の窪みを囲む8つの短い(10〜15アミノ酸残基の)ヘリックスからなる捻じれたα-バレルを形成する。第2のドメインは、疎水性で芳香族に富む活性部位ポケットを囲むα-ヘリックスの二層のバレルからなる。典型的には、この第2のドメインは基質結合部位を含む。図1に例示するように、ヘリックスHは、プロリンのようなアミノ酸によりセグメントH1およびH2の間で分断されているが、ヘリックスGによるそのヘリックス間パッキングは、ヘリックスG中でG1とG2との間で対応するキンク(kink)により収容されている。このキンク内では、トレオニンで見られるような水酸基と他のアミノ酸のカルボニル酸素との間の水素結合がヘリックスGの主鎖ヘリックス内水素結合を分断し、したがって所定の構造をとるのを助ける。
【0154】
TEASにより例示されるように、本発明のテルペンシンターゼは、ヘリックスA、BおよびC(A−Cループ)を含む第1のドメインセグメントと、ヘリックスJおよびK(J−Kループ)を含む第2のドメインとを有し得る(図1)。基質の結合の際にこれらのループが規則的に配列(ordering)することにより、閉じた、溶媒に接触不可能な活性部位ポケットが生じる。J−Kループが規則的に配列されると、基質の存在下で活性部位の入り口を締めつける蓋形の構造が形成され、活性部位ポケットに深く入り込んでいる伸びた芳香族パッチが形成される。A−Cループが規則的に配列されると、それは活性部位に向って内側に平行移動(translate)し、特定のR基をこのループ内で活性部位に、または活性部位付近に配置させる。このように、活性部位への基質の結合は、タンパク質のコンフォメーションの変化をもたらす。
【0155】
突然変異テルペンシンターゼを同定または作製するために、配列アライメントを行って、表1−2、3−4または5−6の構造座標および原子間距離を有する所定のポリペプチド中で特定の残基およびα-炭素の位置を特定し得る。この所定のポリペプチドは、該アライメントにおける対象配列として、例えば全長一次アミノ酸配列、長さ190残基の領域、長さ220残基の領域または長さ300残基の領域として用いる。このアライメントには、TEAS(配列番号2)の残基265〜535を用いることができ、これは該所定のポリペプチドとアライメントをとる参照配列として表1、3または5のα-炭素を含む。該所定のポリペプチドおよび該参照配列は、BLOSUM62スコアリングマトリックス、期待値(expect value)が10、ギャップオープン値が11、xドロップオフ値が50、ギャップイクステンション値が1、ワードサイズが3および低複雑性(low complexity)配列のフィルタリングなしを用いるBLASTp2.0.9コンピュータープログラムを用いてアライメントをとることができる。別法として、BLASTp2.0.9プログラムは、BLOSUM50スコアリングマトリックス、期待値が10、ギャップオープン値が13、xドロップオフ値が50、ギャップイクステンション値が2、ワードサイズが3および低複雑性配列のフィルタリングなしと共に用いることができる。他のパラメーター値(例えば0〜4のギャップイクステンション値)も用い得る。Altschulら, Nucl. Acids Res. 25:3389-3402を参照されたい。
【0156】
TEASの残基265〜535に対して有意な配列同一性、例えば20%以上の配列同一性、25%以上の配列同一性、35%以上の配列同一性、40%以上の配列同一性、50%以上の配列同一性、60%以上の配列同一性、70%以上の配列同一性、または80%以上の配列同一性、を有する所定のポリペプチドの領域を、表1、3または5に示すものに対応するTEASの残基と整列する特定の残基について調べる。幾つかの場合、コンピュータープログラムアライメントの出力は、アライメントプログラムによりギャップが導入されている、またはされていない表1−2、3−4または5−6の構造座標および原子間距離を有する参照配列中の該9、16または19の残基の各々に対して、所定のポリペプチド中の特定の残基を同定する。他の場合、参照配列または対象配列のいずれかにアライメントプログラムによりギャップが導入されて、関心のある参照配列中の該9、16または19の残基の1以上の間に直接的なアライメントまたはミスアライメントが起こらないようにする。いずれの場合も、出力は目視で確認でき、参照配列中のα-ヘリックスおよびβ-シートの領域が維持され、かつ対象配列中のギャップまたは挿入が参照配列のループ領域と整列するようにアライメントを調整した後で、対象配列中で特定の残基を選択できる。
【0157】
配列アライメントから、他のテルペンシンターゼがTEASの残基265〜535に対して20%以上の配列同一性を有する領域を有することが示唆される。したがって、配列番号2の残基265〜535に対して有意な配列同一性を有する配列番号4、6、8、10、12、20、22、24、26、28、30、32、34〜40、42、44、46、48、50、52、54、56または58に示すテルペンシンターゼの領域のような、TEAS以外のテルペンシンターゼの領域を参照配列として用いることができる。例えば、TEASと比較して、テキサジエンシンターゼ(配列番号44)においては大きな配列挿入物はアミノ末端に存在するか、あるいはアミノ末端ドメイン中の溶媒に暴露されるループ内に存在する。したがって、配列番号2に対して20%より高い配列同一性を有するテキサジエンシンターゼの領域は、カルボキシ末端により近接しており、例えば配列番号44の残基579〜残基847である。
【0158】
参照配列として用い得る他のテルペンシンターゼの有用な領域としては、配列番号20の残基343〜606、配列番号22の316〜586、配列番号58の残基352〜622、配列番号33の残基272〜540、配列番号42の残基319〜571、 配列番号44の残基579〜847、配列番号46の残基495〜767、配列番号48の残基295〜564、配列番号50の残基307〜578、配列番号52の残基264〜533、配列番号56の残基585〜853、配列番号54の残基307〜574、配列番号24の残基309〜577、配列番号26の残基315〜584、配列番号28の残基265〜536、配列番号30の残基342〜612、および配列番号32の残基273〜541が挙げられるが、それらに限定されない。
【0159】
参照配列中の残基と整列する対象配列中の特定の残基のうちの1つ以上は、所定のポリペプチド中では、例えば該ポリペプチドをコードする核酸中に突然変異を起こすことにより、突然変異されている。こうして作製された突然変異テルペンシンターゼは、次いで宿主細胞内で発現され、所望によりそのタンパク質を酵素活性について評価することができる。
【0160】
本発明の突然変異タンパク質は、オリゴヌクレオチド指定突然変異誘発、欠失、化学的突然変異誘発など(しかし、それらに限定されない)のような幾つかの方法により調製できる。テルペンシンターゼにおける活性部位α-炭素原子に結合している1つ以上のR基は、対応する遺伝子のヌクレオチド配列を改変することにより変えられる。例えば、突然変異は、配列番号1(TEASのヌクレオチド配列)に、次の16のα-炭素の1つ以上をコードするコドンにおいて導入することができる:α-炭素1=Cys270;α-炭素2=Trp273;α-炭素3=Ile294;α-炭素4=Ile297;α-炭素5=Ser298;α-炭素6=Thr402;α-炭素7=Thr403;α-炭素8=Tyr404;α-炭素9=Leu407;α-炭素10=Cys440;α-炭素11=Val516;α-炭素12=Thr519;α-炭素13=Tyr520;α-炭素14=Asp525;α-炭素15=Tyr527;α-炭素16=Thr528。次いで、該遺伝子を例えば細菌または植物発現系内で発現することにより、該突然変異遺伝子によりコードされるタンパク質が生産される。あるいはまた、シンターゼ突然変異体は、特定のアミノ酸を天然には存在しないアミノ酸で部位指定置換することにより作製してもよい。そのようにして、シンターゼ突然変異体は、アミノ酸残基または特定のシステインもしくはメチオニン残基をセレノシステインまたはセレノメチオニンで置換することにより作製できる。これは、野生型または突然変異型のポリペプチドを発現できる宿主生物を、天然のシステインもしくはメチオニンまたはそれらの双方が欠乏している増殖培地上で増殖させ、セレノシステイン、セレノメチオニンまたはそれらの両者のいずれかに富む増殖培地上で増殖させることにより達成できる。これらおよび類似の技法は、Sambrookら、[Molecular Cloning, A Laboratory Manual, 第2版,(1989) Cold Spring Harbor Laboratory Press]に記載されている。
【0161】
本発明の突然変異シンターゼを作製する別の適切な方法は、NoelおよびTsal(1989) J. Cell. Biochem., 40:309-320に記載の手順に基づくものである。その場合、該シンターゼをコードする核酸は、重複領域を有するオリゴヌクレオチドを用いて合成により作製でき、該オリゴヌクレオチドは、突然変異が誘導されるように特定の塩基において縮重している。
【0162】
本発明によれば、突然変異型シンターゼをコードする核酸配列は、本明細書中で記載の方法または当業者に利用可能な任意の別の方法により作製できる。関心のある核酸配列(遺伝子)の設計において、特定の発現系における発現を向上させるために、該遺伝子を再操作することが望ましいものであり得る。例えば、多くの細菌由来の遺伝子は植物系では十分に発現されないことが示されている。幾つかの場合、植物由来の遺伝子は細菌内では十分に発現しない。この現象は、用いる発現系に関して該遺伝子のG+C含量またはA+T含量が最適でないことによるものと思われる。例えば、多くの細菌遺伝子の非常に低いG+C含量により、高度にA+Tに富む植物遺伝子調節配列を模倣または複製する配列が作製されるようになる。植物に導入された遺伝子内でのA+Tに富む配列(例えば、一般に遺伝子プロモーター中で見られるTATAボックス領域)の存在は、該遺伝子の異常な転写を引き起こす可能性がある。さらに、転写されたmRNA中に存在する他の調節配列[例えば、ポリアデニル化シグナル配列(AAUAAA)またはプレmRNAスプライシングに関与する低分子の核RNAに相補的な配列]の存在は、RNAの不安定性につながり得る。したがって、遺伝子の設計における1つの目標は、特定の発現系のためのmRNAの安定性および転写精度を賦与するG+C含量を有する核酸配列を作製することである。
【0163】
遺伝暗号の重複性によりもたらされる適応性(plasticity)(すなわち、幾つかのアミノ酸は1以上のコドンにより特定されること)のために、異なる生物または生物クラスのゲノムの進化は、重複コドンの特異的な使用法をもたらしている。この「コドンバイアス」は、タンパク質コード領域の平均塩基組成において反映される。例えば、比較的低いG+C含量を有する生物は重複コドンの3番目の位置にAまたはTを有するコドンを利用し、一方高いG+C含量を有する生物は3番目の位置にGまたはCを有するコドンを利用する。したがって、発現のための遺伝子の再操作において、遺伝子を発現させようとする生物のコドンバイアスを測定したいと思うかもしれない。GenBankに寄託された特定の生物の遺伝子について調べたコドン利用を参照すれば、この情報が得られる。そのバイアスを調べた後、新規な遺伝子配列は、制限酵素部位、ならびにエキソン:イントロン接合部、ポリA付加シグナルまたはRNAポリメラーゼ終結シグナルのような転写に影響を及ぼし得る他の部位について分析することができる。
【0164】
シンターゼをコードする遺伝子は、当業者の関心に応じて適当なベクター内に配置することができ、適切な発現系を用いて発現させることができる。当業界では周知であるように、発現ベクターは、典型的には、宿主細胞内で該ベクターの複製を可能にするエレメントを含んでおり、該遺伝子を含む細胞の選択のための1つ以上の表現型マーカーを含んでもよい。発現ベクターは、典型的には、プロモーター配列、リボゾーム結合部位ならびに翻訳開始および終結配列などの発現を調節する配列を含む。発現ベクターはまた、サブゲノムプロモーター、リプレッサー遺伝子または種々のアクチベーター遺伝子のようなエレメントを含んでもよい。当業者であれば、遺伝子産物の分泌、植物色素体のような特定の細胞小器官への該産物の移動をもたらす核酸配列(米国特許第4,762,785号、同第5,451,513号および同第5,545,817号を参照)、またはアフィニティータグのようなペプチド精製をより容易にする他の配列を含ませることも選ぶであろう。
【0165】
多種多様な発現調節配列が、機能し得るように連結されれば突然変異型シンターゼの発現に有用である。そのような発現調節配列としては、例えば、動物細胞のためのSV40の初期および後期プロモーター、lac系、trp系、ファージλの主要なオペレーターおよびプロモーター系、ならびにコートタンパク質の調節領域(特に植物中のRNAウイルス由来のもの)が挙げられる。大腸菌において、有用な転写調節配列はT7 RNAポリメラーゼ結合プロモーターであり、これは、Studierら(1990)Methods Enzymology, 185:60-89に記載のようにしてpETベクターに組み込むことができる。
【0166】
発現のためには、所望の遺伝子は該発現調節配列に機能し得るように連結し、適切な読み取り枠を維持して、所望のシンターゼの産生を可能にしなければならない。多種多様な周知の発現ベクターのいずれもが、本発明では有用である。これらとしては、例えば、SV40に由来するもののような染色体DNA配列、非染色体DNA配列および合成DNA配列のセグメントからなるベクター、細菌プラスミド(大腸菌に由来するもの、例えばcol E1、pCR1、pBR322およびそれらの誘導体、pMB9)、より広い宿主範囲のプラスミド(例えばRP4)、ファージDNA(例えばファージλ、NM989、M13)、ならびにSambrookら[Molecular Cloning, A Laboratory Manual,第2版, (1989) Cold Spring Harbor Laboratory Press]により記載されているような他のそのような系が挙げられる。
【0167】
多種多様な宿主細胞が、本発明のシンターゼ突然変異体の発現に利用可能である。そのような宿主細胞としては、(限定しようとするものではないが)大腸菌、バシルス(Bacillus)およびストレプトマイセスのような細菌、真菌、酵母、動物細胞、植物細胞、昆虫細胞などが挙げられる。本発明の好ましい実施形態は、大腸菌または植物細胞において発現されるテルペンシンターゼ突然変異体を含む。該植物細胞は、懸濁液培養物または寒天ベースの培地のような固体支持体上での培養物のいずれかであり得る。
【0168】
本発明のシンターゼをコードする遺伝子はまた、トランスジェニック植物細胞内で発現させてもよい。トランスジェニック植物を作製するためには、突然変異テルペンシンターゼをコードする核酸構築物を含むベクターを植物ゲノムに挿入する。好ましくは、これらの組換えベクターは、植物ゲノムへ安定に組み込まれることができる。トランスジェニック植物の作製における1つの変化し得る要素は、選択マーカー遺伝子の選択である。選択マーカー遺伝子は、非形質転換細胞の高いバックグラウンドから形質転換細胞を同定するのに用いられる。そのような選択マーカー遺伝子としては、抗生物質カナマイシン、ネオマイシンおよびG418に対する耐性をコードするトランスポゾンTn5のアミノグルコシドホスホトランスフェラーゼ遺伝子(Aph II)、ならびにグリホセート(glyphosate)、ハイグロマイシン、メトトレキサート、ホスフィノトリシン(phosphinothricin)、イミダゾリノン、スルホニル尿素、およびトリアゾロピリミジン殺草剤(例えばクロロスルフロン、ブロモキシニル、ダラポンなど)に対する耐性または寛容性をコードする遺伝子が挙げられるが、それらに限定されない。選択マーカー遺伝子に加えて、レポーター遺伝子を用いることが好ましい場合がある。幾つかの例において、レポーター遺伝子は選択マーカーと共に用いることができる。レポーター遺伝子は、形質転換細胞の検出を可能にし、当業者の随意で用いることができる。これらのレポーター遺伝子の一覧は、K. Weisingら, 1988, Ann. Rev. Genetics, 22:421に提供されている。
【0169】
該遺伝子は、あらゆる組織においてあらゆる時期に発現するプロモーター(構成的プロモーター)、特定の組織において発現するプロモーター(組織特異的プロモーター)、特定の発生段階で発現するプロモーター(発生的プロモーター)、および/または刺激に応答して発現するプロモーター(誘導的プロモーター)により発現される。これらの選択は、当業者であれば随意に決定される。
【0170】
外来遺伝子を植物細胞に導入するため、および導入された遺伝子を安定に維持し発現する植物を得るための幾つかの技法が存在する。そのような技法としては、被覆された遺伝物質を細胞に直接的に促進すること(米国特許第4,945,050号)が挙げられる。植物はまた、アグロバクテリウム(Agrobacterium)法(米国特許第5,177,010号、同第5,104,310号、同第5,149,645号、同第5,469,976号、同第5,464,763号、同第4,940,838号、同第4,693,976号、同第5,591,616号、同第5,231,019号、同第5,463,174号、同第4,762,785号、同第5,004,863号および同第5,159,135号;欧州特許出願第116718号、同第290799号、同第320500号、同第604662号、同第627752号、同第0267159号および同第0292435号)を用いて形質転換してもよい。他の形質転換法としてはウィスカー(whiskers)法が挙げられる(米国特許第5,302,523号および同第5,464,765号を参照)。エレクトロポレーション法もまた、植物を形質転換するのに用いられている(WO87/06614、WO92/09696およびWO93/21335ならびに米国特許第5,472,869号および同第5,384,253号を参照)。米国特許第5,316,931号、同第5,589,367号、同第5,811,653号および同第5,866,785号に記載のもののようなウイルスベクター発現系も用い得る。
【0171】
植物を形質転換するための多くの技法に加えて、関心のある遺伝子と接触させる組織のタイプもまた様々なものであり得る。適切な組織としては、胚形成組織、カルス組織、胚軸組織、分裂組織などが挙げられるが、それらに限定されない。ほぼ全ての植物組織が、本明細書中に記載の適切な技法を用いて分化の間に形質転換できる。
【0172】
変異型シンターゼをコードする遺伝子は、使用する形質転換系とは関係なく、発現制御配列(植物プロモーター調節エレメント)をベクターに含ませることによって植物細胞中で該遺伝子を発現するように適合された遺伝子転移ベクターに導入されることが好ましい。植物プロモーター調節エレメントに加えて、様々な由来源から得たプロモーター調節エレメントも、植物細胞において外来遺伝子を発現するために有効に使用できる。例えば、オクトピンシンターゼプロモーター、ノパリンシンターゼプロモーター、およびマンノピン(mannopine)シンターゼプロモーター等の細菌由来のプロモーター調節エレメントを使用してもよい。カリフラワーモザイクウイルス(35Sおよび19S)等のウイルス由来のプロモーターも望ましい。植物プロモーター調節エレメントとしては、リブロース-1,6-ビスホスフェートカルボキシラーゼ小サブユニットプロモーター、β-コングリシニン(conglycinin)プロモーター、ファゼオリン(phaseolin)プロモーター、ADHプロモーター、熱ショックプロモーターおよび組織特異的プロモーター等も挙げられるがこれらに限定されない。多数のプロモーターが、入手可能であり、当業者の裁量
下で使用できる。
【0173】
全ての発現ベクターおよび発現系が、本発明の変異型遺伝子配列を発現するために同様に機能するわけではないことが理解されるべきである。宿主細胞も全てが同じ発現系に対して同等に上手く機能するわけではない。しかし、当業者は、これらのベクター、発現制御配列、および宿主を、過度の実験を要することなく、また本発明の範囲から逸脱することなく選択できるであろう。
【0174】
本発明のシンターゼを発現させた後は、それから得たタンパク質を精製して、本明細書に例示するように、構造分析、モデリング、および/または生化学的分析を行うことができる。得られるタンパク質の性質は、使用する発現系に依存しうる。例えば、遺伝子は、哺乳動物またはその他の真核生物細胞で発現された場合には、機能を変化するかもしれないしまたは変化しないかもしれないグリコシル化、リン酸化、またはその他の翻訳後修飾を生じうる潜在的シグナル配列を含み得る。タンパク質は、発現後、タンパク質生化学分野の当業者には常識的な技法を使用して、またはColliganら(1997), Current Protocols in Protein Science, Chanda, V.B., Ed., John Wiley & Sons, Inc.に記載されているように、容易に単離および精製できる。このような技法には、陽イオン交換または陰イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー等の使用が含まれることが多い。一般的に使用されうる別の技法は、アフィニティークロマトグラフィーである。アフィニティークロマトグラフィーは、抗体、基質類似体またはヒス
チジン残基(His-タグ技法)の使用を含みうる。
【0175】
本発明の変異体は、精製後、いくつかの異なる特性のいずれかによって特徴決定してもよい。例えば、このような変異体では、1つ以上の電荷ユニットの活性部位表面電荷が変化しているかもしれない。さらに、変異体は、非変異型シンターゼと比べて、基質特異性または反応産物のスペクトルが変化しているかもしれない。
【0176】
本発明は、結晶化およびその後のX線回折により変異型テルペンシンターゼの特徴決定を可能にする。ポリペプチド結晶化は、ポリペプチド濃度がその最大溶解度を超える溶液において(すなわち、ポリペプチド溶液が過飽和の状態で)生じる。このような溶液は、好ましくはポリペプチド結晶の沈殿によりポリペプチド濃度を低くすることによって、平衡状態に戻すことができる。ポリペプチド表面電荷を変化させるか、ポリペプチドとバルク水との相互作用を摂動させて結晶化をもたらす会合を促進する作用物質を添加することにより、過飽和溶液から、ポリペプチドを誘導して結晶化させることが多い。
【0177】
「沈殿剤」として知られる化合物は、ポリペプチド分子の周りにエネルギー学的に好ましくない沈殿する減損層(depleted layer)を形成することにより、濃縮溶液中のポリペプチドの溶解度を低くするためにしばしば用いられる(Weber, 1991, Advances in Protein Chemistry, 41:1-36)。沈殿剤に加えて、他の物質をポリペプチド結晶化溶液に添加する場合もある。これらには、溶液のpHを調節するための緩衝剤、およびポリペプチドの溶解度を下げるための塩が挙げられる。様々な沈殿剤が当該分野において公知であり、以下のものが挙げられる。すなわち、エタノール、3-エチル-2-4-ペンタンジオール、ならびにポリエチレングリコールなどの多くのポリグリコールである。
【0178】
よく使用されるポリペプチド結晶化方法としては、以下の技法が挙げられる。すなわち、バッチ法、懸滴法、結晶種開始法(seed initiation)および透析法である。これらの各方法では、過飽和溶液を維持することにより、核形成後の持続的な結晶化を促進することが重要である。バッチ法では、ポリペプチドを沈殿剤と混合して過飽和状態を得、容器を密封し、結晶が現れるまで置いておく。透析法では、ポリペプチドを密封した透析膜内に保持し、沈殿剤を含有する溶液に入れる。膜を介した平衡により、ポリペプチドおよび沈殿剤の濃度が高くなり、ポリペプチドを過飽和レベルに到達させる。
【0179】
好ましい懸滴技法(McPherson, 1976, J.Biol.Chem., 6300-6306)では、沈殿剤を濃縮ポリペプチド溶液に添加して、初期ポリペプチド混合液を作る。ポリペプチドおよび沈殿剤の濃度は、この初期状態においてポリペプチドが結晶化しない程度のものである。この混合液の小滴を、スライドガラスに滴下し、第2の溶液の貯蔵器上で反転させ、懸濁させる。次に、この系を密封する。典型的には、第2の溶液は、より高い濃度の沈殿剤またはその他の脱水剤を含む。沈殿剤濃度の差により、溶液よりも高い蒸気圧を有するタンパク質溶液が生じる。2つの溶液を含む系は密封されているため、平衡が確立され、ポリペプチド混合液由来の水は第2の溶液に移る。この平衡により、ポリペプチド溶液中のポリペプチドおよび沈殿剤の濃度が高くなる。ポリペプチドおよび沈殿剤の臨界濃度において、ポリペプチドの結晶が形成されうる。
【0180】
別の結晶化方法では、核形成部位を濃縮ポリペプチド溶液に導入する。通常、濃縮ポリペプチド溶液を用意し、ポリペプチドの結晶種をこの溶液に導入する。ポリペプチドおよび任意の沈殿剤の濃度が正しければ、結晶種は核形成部位を提供し、その周りにより大きい結晶が形成される。好ましい実施形態では、本発明の結晶は、沈殿剤として15%PEG 8000;200mM酢酸マグネシウムまたは塩化マグネシウム、100mM 3-(N-モルホリノ)-2-ヒドロキシプロパンスルホン酸(pH 7.0)、1mMジチオスレイトールを用いた懸滴で形成される。
【0181】
一部のタンパク質は、結晶化が困難であるかもしれない。しかし、結晶化を誘導するためにいくつかの技法を当業者は利用できる。タンパク質のアミノ末端またはカルボキシル末端におけるポリペプチドセグメントの除去により、結晶タンパク質サンプルの生成が容易になる。このようなセグメントの除去は、トリプシン、キモトリプシン、サブチリシン等のプロテアーゼを用いた、タンパク質の分子生物学的技法または処理を使用して行うことができる。このような処置は、結晶化に対して負に影響を及ぼしうるフレキシブルなポリペプチドセグメントを除去しうる。
【0182】
このように形成された結晶の用途は広範囲にわたる。例えば、高品質結晶は、変異型シンターゼの三次元構造を決定し、さらに別の変異体を設計するためのX線または中性子回折分析に適している。さらに、結晶化は、さらなる精製方法として作用しうる。いくつかの例では、ポリペプチドまたはタンパク質は、不均質混合物から結晶に結晶化する。濾過、遠心分離などによりこのような結晶を単離し、その後ポリペプチドを再溶解することにより、回折研究に必要な高品質結晶を成長させるのに適した精製溶液が得られる。この高品質結晶を、水に溶解した後に、処方して、所望に応じた他の使用用途を有する水性溶液を得ても良い。
【0183】
シンターゼは、1つ以上の結晶形態で結晶化しうるため、本発明により得られる、シンターゼまたはその一部から決定した活性部位のα-炭素の構造座標は、シンターゼの他の結晶形態の構造を解読するのに特に有用である。本明細書で得られる構造座標は、同一であるかまたは同一でないR基を有する野生型と同様に、活性部位内にα-炭素位置を有するシンターゼの構造を解読するためにも使用できる。さらに、本明細書に開示する構造座標を使用して、シンターゼの任意の機能ドメインに対して有意なアミノ酸相同性または構造的相同性を有する他のタンパク質の結晶形態の構造を決定できる。このような目的のために採用できる方法の1つとして分子置換(molecular replacement)がある。この方法では、未知の結晶構造は、それがシンターゼの別の結晶形態であるのか、変異活性部位を有するシンターゼなのか、またはシンターゼと有意な配列同一性および/もしくは構造相同性を有するなんらかの他のタンパク質の結晶なのかを、表10および/または表11に示す座標を用いて決定できる。この方法は、未知の結晶についての十分な構造形態を、最初からこのような情報を決定しようとする場合よりも効率的に提供する。さらに、本方法を用いて、問題となる所与のシンターゼが本発明の範囲に含まれるか否かを決定できる。
【0184】
本明細書にさらに開示するように、シンターゼおよびその変異体は、基質および基質類似体の存在下または不在下において結晶化できる。その後、一連の複合体の結晶構造を、分子置換により解読し、野生型のものと比較して、活性部位内のR基の適切な置換を決定するのを助けて、本発明のシンターゼ変異体を作成する。
【0185】
本発明の変異体は全て、必ずしも全ての変異体についてそれぞれ結晶化して構造を解読する必要なく、本明細書に開示する情報を用いてモデリングできる。例えば、当業者は、いくつかの特殊化コンピュータプログラムの1つを使用して、変異型活性部位を有するシンターゼの設計プロセスを援助しうる。このようなプログラムの例として以下のものが挙げられる。すなわち、GRID(Goodford, 1985, J.Med.Chem., 28:849-857);MCSS(MirankerおよびKarplus, 1991, Proteins: Structure, Function and Genetics, 11:29-34);AUTODOCK(GoodsellおよびOlsen, 1990, Proteins: Structure, Function, and Genetics, 8:195-202);ならびにDOCK(Kuntzら, 1982, J.Mol.Biol., 161:269-288)である。さらに、特異的基質-活性部位相互作用、ならびにそれにより生じる変形エネルギーおよび静電相互作用を評価するための特殊なコンピュータプログラムも入手可能である。MODELLERは、タンパク質の三次元構造の相同性または比較モデリングについてよく使用されるコンピュータプログラムである。A. SaliおよびT.L.Blundell. J.Mol.Biol. 234, 779-815, 1993。モデリングされるべき予め選択されたポリペプチド配列を、結晶構造が既知である1つ以上のテルペンシンターゼとアライメントし、MODELLERプログラムを使用して空間的制約の最適条件(satisfaction)に基づいて完全原子モデルを計算する。このような制約には、とりわけ、相同的構造、蛍光分光学、NMR実験、または平均力の原子間ポテンシャル(atom-atom potentia
l of mean force)が含まれる。
【0186】
本発明は、シンターゼ変異体を作成し、その結晶構造を解読することを可能にする。さらに、本発明により、活性部位の位置、およびそれと基質との界面から、突然変異誘発のために望ましいR基の同定が可能となる。本発明の特定の実施形態は、実施例にさらに例示されている。しかし、当業者は、記載する特定の実験があくまで本発明の例示であり、本発明が請求の範囲にさらに完全に記載されていることを容易に理解するであろう。
【実施例】
【0187】
実施例1
変異型TEAS遺伝子の生成
構築物の生成および発現
全ての変異型酵素を、QuickChange方法(Stratagene)により構築した。記載したもの以外は製造元の説明書に従った。突然変異は、DNA配列決定により確認し、所望の突然変異を含むプラスミドを使用してBL-21(DE3)発現細胞を形質転換した。タンパク質を発現させ、精製し、-80℃にて保存した。
【0188】
TEAS W273S.以下のプライマーを用いて、TEAS-pET28b(+)鋳型からTEAS W273S変異体を生成した:GTTGAATGCTACTTTTCGGCATTAGGAGTTTAT(センス)(配列番号13)およびATAAACTCCTAATGCCGAAAAGTAGCATTCAAC(アンチセンス)(配列番号14)。センス鎖およびアンチセンス鎖を別々の反応で生成したこと以外は、製造元の説明書に従って突然変異誘発を行った。それぞれについて、30サイクルの1分間のプラスミドコピー、55℃でのアニーリング、および68℃での16分間の伸長を行った。二つの反応混合液を合わせ、95℃まで2.5分間加熱し、室温まで冷却してから、DpnI処理に供した。
【0189】
TEAS C440W.以下のプライマーを用いて、TEAS-pET28b(+)鋳型からTEAS C440W変異体を生成した:GCTAGTGTAATTATATGGCGAGTTATCGATGAC(センス)(配列番号15)およびGTCATCGATAACTCGCCATATAATTACACTAGC(アンチセンス)(配列番号16)。
【0190】
TEAS W273S/C440W.TEAS C440Wの生成のために記載したプライマーを用いて、TEAS W273S-pET28b(+)鋳型からTEAS C440W/W273S変異体を構築した。
【0191】
TEAS 406/407ランダムライブラリー.406位および407位にランダムなアミノ酸を有するTEAS変異体のライブラリーを生成するために、二つの50マイクロリットルQuickChange反応を、TEAS-pET28b(+)鋳型および以下のプライマーを用いて行った:GCACTAGCAACTACCACATATTACNNSNNSGCGACAACATCGTATTTGGGCATG(センス)(配列番号17)、およびCATGCCCAAATACGATGTTGTCGCSNNSNNGTAATATGTGGTAGTTGCTAGTGC(アンチセンス)(配列番号18)。ここで、NはA、C、GまたはTを指し、SはCまたはGを指す。このヌクレオチド選択により、この反応は、406位および407位において可能性のあるアミノ酸の組合せの全てをコードするプライマーを含んでいた。アミノ酸間でコドンの数を変えるための調節は行わなかった。効率的な反応を確実にするために、かつ鋳型に対する野生型プライマーのハイブリダイゼーションの優先性を最小限にするために、プライマーを、上述した突然変異を生じるために使用されるプライマーよりも長くなるように設計した。さらに、それらを使用前にHPLC精製した。18サイクルのプラスミドコピーの後、反応液を2時間DpnIと共にインキュベートし、エタノール沈殿させ、5マイクロリットルの水に再溶解させた。大腸菌NovaBlue(Novagen)細胞の4つの40マイクロリットルアリコートのそれぞれに、1.5マイクロリットルの再溶解DNAをエレクトロポレートした。回収時間の後、細胞を、カナマイシン-LB-寒天プレート上にプレートした。
新たに構築したプラスミドを発現細胞に移すために、4つのプレート全てからコロニーを掻き取り、液体LB培地中で37℃にて8時間増殖させる8mL培養を開始するのに使用した。この培養物から精製されたプラスミドを、20マイクロリットルのコンピテントBL-21(DE3)細胞を形質転換するのに使用した。
【0192】
構築物の保存のために、個々のコロニーを用いて、96-ウェル培養プレート中のカナマイシン(50マイクログラム/mL)を含有する100マイクロリットルのLB培地に播種した。A600が約1に達するまで細胞を37℃で増殖させ、100マイクロリットルの30%グリセロールのLB溶液を添加し、プレートを-80℃にて冷凍した。ランダムに選択したコロニーを、いくつかのコロニーの個々のグリセロールストックから増殖させ、プラスミドを配列決定のために抽出した。約30パーセントのコロニーが野生型であることが分かった。TEAS 406/407変異型遺伝子およびタンパク質のヌクレオチドおよびアミノ酸配列は配列番号11および12に示す。
【0193】
TEAS Y520F.配列番号2の520位にあるチロシン残基を、上述したものと同様に、プライマーを用いた部位特異的突然変異誘発によりフェニルアラニン残基に変えた。Y520Fのために、TATコドンをTTCに変えた。変異型遺伝子のヌクレオチド配列を配列番号5に示す。
【0194】
TEAS Y527F.配列番号2の527位にあるチロシン残基を、上述したものと同様に、プライマーを用いた部位特異的突然変異誘発によりフェニルアラニン残基に変えた。Y527Fのために、TEASアミノ酸配列の527位にあるTACコドンをTTCに変えた。変異型TEAS Y527F遺伝子のヌクレオチド配列を配列番号7に示す。
【0195】
TEAS W273E.配列番号2の273位にあるトリプトファン残基を、上述したものと同様に、プライマーを用いた部位特異的突然変異誘発によりフェニルアラニン残基に変えた。W273Eのために、TEASアミノ酸配列の273位にあるTGGコドンをGAGに変えた。変異型遺伝子のヌクレオチド配列を配列番号3に示す。
【0196】
実施例2
シンターゼポリペプチドの発現および単離
特に記載しない限り、変異型および非変異型TEASタンパク質は、大腸菌において発現させ、金属キレート化、陰イオン交換およびゲル濾過クロマトグラフィーにより精製した。
【0197】
ベクターpET-28b(+)(Novagen)中のTEASおよび変異型TEASタンパク質の構築物を、大腸菌細胞で発現させた。これらの酵素の典型的なタンパク質調製のために、プラスミド構築物を含む大腸菌株BL21(DE3)細胞を、4×1L TB培地において37℃にてA600=1.0まで増殖した。温度を22℃にまで下げ、IPTGを最終濃度が0.1mMになるまで添加して、タンパク質発現を誘導した。15〜20時間後、遠心分離により細胞を回収し、1gの細胞(湿重量)当たり5mLの緩衝液A(20 mM Tris、500 mM NaCl、20mMイミダゾール、pH7.9)に再懸濁し、4℃にて0.5時間攪拌した。次いで、超音波により細胞を溶解し、得られた溶解物を82,000×gにて0.7時間遠心分離した。タンパク質を含む上清を、緩衝液A中で平衡させた2〜3mL Ni2+キレート化ヒスチジンアフィニティーカラム(Qiagen)にかけ、溶出液のA280が基線に戻るまでカラムを追加の緩衝液Aで洗浄した。次いで、タンパク質を20〜200mMイミダゾール勾配の緩衝液Aで溶出した。タンパク質含有画分をプールし、緩衝液B(50mM HEPES、5 mM MgCl2、1mM DTT)に対して透析し、8mL MonoQ陽イオン交換カラム(Pharmacia)にかけた。カラムを20カラム容量の緩衝液Bで洗浄し、0〜500mM NaCl勾配の緩衝液Bでタンパク質を溶出した。得られたタンパク質を、50 mM Tris、100 mM NaCl、5 mM MgCl2、1 mM DTT、pH8.0中で、Superdex-200カラム(Pharmacia)上でのゲル濾過によりさらに精製した。次いで、精製タンパク質を、5 mM Tris、5 mM NaCl、1 mM DTT、pH8.0に対して透析し、18〜22 mg/mLまで濃縮し、必要になるまで100μLアリコートで-80℃にて保存した。
実施例3
シンターゼポリペプチドの結晶化および構造解析
結晶成長およびマイクロ播種(microseeding):全ての結晶化の試みを、懸滴気化拡散(hanging-drop vapor diffusion)方法により行った。濃縮タンパク質を、プラスチックカバースリップ上で、等容量(それぞれ2〜5μL)の貯蔵溶液と混合した。次に、0.5〜1.0 mLの貯蔵溶液を含むプラスチックの24ウェル組織培養プレートのウェル上でカバースリップを反転させ、ウェルとカバースリップとの間を真空グリースの層で密封した。プレートを4℃でインキュベートし、その間、気化拡散により懸滴中のタンパク質濃度を徐々に高めた。様々な沈殿剤および添加塩を含むpHが4.5〜9の範囲の約300の異なる貯蔵溶液を、TEAS(配列番号12)の結晶化についてアッセイした。TEASは、15%PEG 8000、100 mM MOPSO(3-[N-モルホリノ]-2-ヒドロキシプロパンスルホン酸)、200 mM酢酸マグネシウム、1mM DTT、pH6.9〜7.3)の貯蔵溶液で結晶化した。マイクロ播種のために、既存の結晶を数μLの沈殿剤溶液中で破砕し、50マイクロリットルにまで希釈した。大きい粒子を除去するための初期遠心分離の後、懸濁液を追加の沈殿剤溶液で連続希釈し、少容量の希釈種ストックをそれぞれの新しい結晶化小滴に添加した。マクロ播種(macroseeding)のために、既に早い速度で成長していない結晶(通常、小滴をセットしてから2週間後)を、2〜3滴の貯蔵溶液に連続的に通すことにより「濯いだ」。次に、結晶を、タンパク質および貯蔵溶液を含む新しい小滴に移し、初期成長時のように貯蔵溶液に対して平衡させた。個々の結晶は、内部順序(internal order)の程度が異なった。場合によっては、いくつかの結晶をスクリーニングして、低いモザイク度(mosaicity)でよく回折した結晶を同定した。
【0198】
データ回収:データを回収する前に、結晶を、結晶に浸透させる化合物を含む貯蔵溶液の小滴、または貯蔵溶液に移した。次いで、少容量の凍結保護物質溶液(15% PEG8000、100 mM MOPSO、200 mM 酢酸マグネシウム、20%エチレングリコール、1mM DTT、pH 7)を小滴に添加した。短い平衡時間(1〜5分)の後、結晶を、浸透させる化合物を添加した凍結保護物質の小滴、または凍結保護物質に移した。さらに追加の短い平衡時間の後、結晶をナイロンループ上に取り上げ、冷窒素ガス(90〜110K)の流れの中でデータ回収を速やかに開始した。
【0199】
TEAS結晶は、正方晶空間グループP41212(単位セル次元(unit cell dimensions
)は結晶により数オングストローム変動するが、平均してa=126Å、c=122Å
である)に属していた。非複合化TEAS構造を、2 mM FHPの存在下で成長させた結晶から回収したデータ(表10)に対して、初め2.8Åにまで精製した(表11)。活性部位における電子密度により、FHP、A-CおよびJ-Kループ、ならびにNH2末端にある9つの追加の残基の明確なモデリングが可能であった。精製したTEAS-FHPモデルは、残基17〜548、3つのMg2+イオン、150の水分子、ならびに1つのFHP分子から構成されていた。結合基質の存在下におけるTEASの三次元座標を表10に示す。FHPの不在下でのTEASの三次元座標を表11に示す。
【0200】
トリフルオロ-ファルネシルジホスフェート(F3-FPP)により複合化されたTEASの結晶も調製した。これらの結晶において、上手く秩序をなしたジホスフェート結合ポケットも観察した。A-CループおよびNH2末端セグメントは、はっきりとした電子密度を示した。A-Cループを活性部位に向かって中継した(translate)ところ、F3-FPPのジホスフェート部分について強力な電子密度が存在した。しかし、疎水性ポケットは、フレキシブルなままであった。J-KループおよびF3-FPPのファルネシル部分は無秩序であった。
【0201】
SwissModelプログラム(Peitsch MC(1996), Biochem. Soc. Trans., 24:274-279、ならびにGuex N.およびPeitsch MC, 1997, Electrophoresis., 18:2714-2723)のSwiss PDBビューアーインターフェースを用いて、相同性モデルを作成し、エネルギーを最小化した。VOIDOO(Kleywegt, G.J.およびJones, T.A., CCP4/ESF-EACBM Newsletter on Protein Crystallography., 29, 26-28, 1993)により活性部位容量を計算した。閉鎖した活性部位空洞を作るために、モデル化されたTEASシクラーゼ反応から得た、エネルギーを最小化されたジホスフェート部分を、TEAS D301に等価な残基に追加した。
【0202】
TEAS W273S結晶構造.FHPの存在下で2つのTEAS W273S構造を、異なる結晶から決定した。両方の結晶とも、タンパク質全体の残基(頻繁に動きやすいヘリックスD1、D2およびEを含む)についてはっきりとした主鎖および側鎖密度が明確であることから、上手く秩序をなしているように思われた。両方の結晶から得た、初期差電子密度地図から、W273S突然変異がすぐに分かった。2つの結晶をW273S-1およびW273S-2と命名した。
【0203】
各構造において、活性部位を囲むループを秩序付け、閉鎖活性部位ポケットを得た。各構造におけるA/Cループを活性部位に向かって中継し、野生型TEAS/FHP複合体で見られるような外枠の一部を形成した。しかし、W273S-1のJ/Kループが、野生型TEAS/FHP複合体で観察されるものと同じコンホメーションをとるのに対して、W273S-2中の同じループは異なるコンホメーションをとっていた。J/Kループのこのコンホメーションでは、Tyr527が、残基273の側鎖から離れていた。さらに、Tyr520およびAsp525は、Asp444の側鎖と離れて位置していた。J/Kループ、Arg266、およびN末端ループの間で先に観察された水素結合も、W273S-2構造において欠けていた。
【0204】
W273S-2コンホメーションは、W273S突然変異の結果とは思われなかった。なぜなら、これは、エピ-アリストロケン擬態デオキシカプシジオール(epi-aristolochene mimic deoxycapsidiol)が浸透した野生型TEAS結晶においても、この構造におけるデオキシカプシジオールについて電子密度が容易に明確でなかったという事実にも関わらず、観察されたからである。さらに、TEAS活性部位ループは、結晶接触から離れており、それらのコンホメーションは、結晶パッケージングの人為結果であるとは思われなかった。TEAS反応の種々の段階において、酵素のJ/Kループが異なる明確なコンホメーション中に存在し、これらの結晶構造のそれぞれが異なるコンホメーション像を獲得することが可能である。両方のW273S構造において、Arg266以外の残基、およびJ/Kループ上の残基は、野生型TEASで見られるコンホメーションから有意な再配列を経ていなかった。
【0205】
各W273S結晶構造において、活性部位における電子密度から、基質擬態FHPが複数のコンホメーションで結合することが示唆された。この密度のいくつかの領域は、変異型活性部位において結合した水分子を表している可能性がある。変異型活性部位における水分子の存在は、TEAS W273Sが複数の水酸化テルペノイド反応産物を生じるという観察と一致する。
【0206】
各W273結晶構造におけるFHP電子密度は、FHPが、野生型TEAS/FHP複合体におけるより緊密に折り畳まれたFHPコンホメーションと比べて、W273S構造においてより広がったコンホメーションで存在することを示唆するのに十分であった。W273Sの活性部位が、FHPの複数のコンホメーションと結合するという観察は、W273SがFPPを複数のテルペノイド炭化水素産物に変換するという事実と一致する。
【0207】
TEAS C440W/W273S:TEAS C440W/W273Sを、野生型TEASと同じ条件下で結晶化した。0.3 mmの結晶を、ファルネシルヒドロキシホスフェート(FHP)で飽和させた貯蔵溶液に20分間浸せきした。野生型TEASについて記載した凍結保護およびフラッシュ冷凍(flash freezing)の後、Cu-Kα放射(MacScience Corp., 日本)を用いて実験ソースからデータを回収した。非複合化TEASの出発モデル(表11)(Brookhaven Protein Database Code 5EAT(PDB 5EAT))を、水およびマグネシウムを除去し、ソフトウェアプログラムX-PLOR(A.T.Brunge, X-PLOR Version 3.1-A System for X-Ray Crystallography and NMR Yale University Press, New Haven, 1992, pp.187-207)の剛体モジュールを有する変異体データに対して配置した。バルク溶剤モデリングでの配置的および制約的b因子精製の繰返しもX-PLORで行った。この際、ソフトウェアプログラムO(Jones, TA, Zou, JY, Cowan, SW、およびKjeldgaard, M., Acta Cryst. D., 49:148-157, 1993)で手動モデル成形および調節を行った。CNSプログラムを使用したさらなる精製および地図計算の繰返しにより、有意に改善された地図を得た。この改善はバルク溶剤モデリングが改善されたためだと思われた。
【0208】
TEAS C440W:結晶の核形成がより遅く、全体的により小さかったこと以外は、野生型TEASと同じ条件下でTEAS C440Wを結晶化した。変異型結晶を、フラッシュ冷凍、およびSSRLビーム線7-1(Stanford Synchrotoon Radiation Laboratory, Menlo Park, California)でのデータ回収の前に、FHPに飽和させた貯蔵溶液中に6時間浸せきした。TEAS-FHPの出発モデル(表10)は、水分子、リガンド、および配列番号2の残基523-532を除去され、X-PLORの剛体モジュールを有するデータに対して配置した。バルク溶剤での配置的および制約的b因子精製、ならびに全体的異方性温度因子モデリングの繰返しもX-PLORで行い、ソフトウェアプログラムOで手動モデル成形および調節を行った。二重変異体の場合、電子密度地図は、CNSでの精製および地図計算の後では有意に改善されていた。
【0209】
実施例4
テルペンシンターゼ酵素アッセイ
VogeliおよびChappell, Plant Physiol.94: 1860(1990)ならびにVogeliら, Plant Physiol. 93:182(1990)に記載されたアッセイに基づき、シンターゼ活性のアッセイを行った。一般に、放射性標識(3Hまたは14C)基質を、マグネシウム塩の緩衝化溶液(特に他のものが記載されない限り200mM Tris, pH8, 50mM塩化マグネシウム、1mM DTT)中で室温にて酵素と共にインキュベートした後、炭化水素生成物をヘキサン等の有機溶媒中に選択的に抽出した。このヘキサン抽出物を一般にシリカゲルで処理して、基質の非酵素的加水分解により生成されたプレニルアルコールおよび他の含酸素化合物(これらはヘキサン中に非効率的に分配される(partition))を除去した。このヘキサン相の中に存在する炭化水素生成物をシンチレーションカウンターにより定量した。
【0210】
次により極性の高い有機溶媒(酢酸エチル等)により時折抽出を行った。含酸素化合物は酢酸エチルタイプの溶媒中により効率的に分配される。この酢酸エチル相中に存在する化合物もシンチレーションカウンターにより定量した。
【0211】
典型的な基質濃度の範囲は0.1ナノモル〜100μモルとした。幾つかのアッセイでは、基質の放射性標識を行わなかった。反応は一般に各基質濃度毎に3回行った。タンパク質濃度は、Bradford法により測定した。静止状態の動力学パラメータを測定するために、酵素濃度は、反応過程を通じて経時的生成物生成量が直線となるように選択した。
【0212】
ジテルペンシンターゼのアッセイは、典型的には、3Hゲラニルゲラニルジホスフェート(GGPP)および酵素を含む250mM Tris, 10mM 塩化マグネシウム, 1mM DTT, pH8.0を用いて行った。セスキテルペンシンターゼアッセイは、典型的には、14Cまたは 3H FPPおよび酵素を含む100mM Tris, 30mM 塩化マグネシウム, 1mM
DTT, pH8.0を用いて行った。モノテルペンシンターゼアッセイは、典型的には
、3H GPPおよび酵素を用いて行った。タンパク質によるGPPの非特異的結合についての対照として、酵素ではなくBSAを用いて全く同じ反応を設定した。
【0213】
Ag-TLCによる野生型および突然変異型のTEAS酵素の生成物分析:テルペノイド炭化水素生成物は、通常のプレートもしくは逆相プレート上での薄層クロマトグラフィーによっては容易に分離されないが、はじめにシリカプレートを硝酸銀で処理する銀染色TLC(Ag-TLC)によってある程度は分離することができる。本明細書中に記載されるAg-TLCは、Backら, Arch. Biochem. Biophys. 315: 527(1994)によって記載された手法に概ね従って行った。シリカTLCプレートを15%硝酸銀(水溶液)に浸した後、110℃で3〜5時間乾燥した。トリチウム酵素生成物(溶媒抽出物)をスポッティングした後、プレートをベンゼン:ヘキサン:酢酸エチル(50:50:1体積比)中で発色させ、En3Hance(NEN)蛍光間接撮影用スプレーを噴きつけ、フィルム上に載せて、数日〜数週間の間露光した。TLCプレートの硝酸銀処理により蛍光間接撮影用試薬の蛍光が抑制されるようであったので、一般に長時間露光する必要があった。或いは、蛍光間接撮影用スプレーを使用せずに14Cで標識した生成物を1〜2日後に検出した。
【0214】
実施例5
TEAS W273Sの活性
TEAS W273Sのジテルペンシンターゼ活性:TEAS W273S酵素および放射性標識GGPPを上記のようにインキュベートし、ヘキサンで炭化水素生成物を抽出した。次に含酸素生成物を酢酸エチルで抽出した。野生型TEASを用いた反応で定量した結果は、緩衝液のみを用いた反応のものよりも低かった。一方、TEAS W273Sの定量結果は、ヘキサン抽出および酢酸エチル抽出の両方において非常に高かった。W273SによりGGPPから形成された炭化水素生成物は、酸により触媒されたGGPPからのジホスフェートの減少によって生成された生成物とは異なるものであった。図3を参照されたい。
【0215】
TEAS W273Sのセスキテルペンシンターゼ活性:精製されたTEAS W273S突然変異体によるFPP代謝回転の生成物を、銀染色薄層クロマトグラフィー(Ag-TLC)によって分析した。主な反応生成物の1つはAg-TLCによるRfが0.7であり、これは、5-エピ-アリストロケン(Rf=0.78)およびベティスピラジエン(vetispiradiene)(Rf=0.63)のいずれとも異なるものであった。図4を参照されたい。予備GC/MSデータは、TEAS W273SによるFPP代謝回転からのヘキサン抽出物が少なくとも4つのテルペン炭化水素を含み、質量スペクトルが5-エピ-アリストロケンまたはベティスピラジエンと異なることを示した。これらの生成物のうちの1つは、ゲルマクレン(germacrene) Aと似た質量スペクトルを有していた。
【0216】
実施例6
TEAS C440W/W273Sの活性
TEAS C440W/W273Sのジテルペンシンターゼ活性:突然変異型TEAS C440W/W273S
タンパク質は、440位にトリプトファン残基および273位にセリン残基を含む。0.5μモル3H GGPP, 様々な濃度の非標識GGPP(Echelon)および酵素を用い、GGPPを用いてアッセイを行った。反応物を室温にて60分間インキュベートした。TEAS C440/W273S突然変異型タンパク質は、GGPPをヘキサン-抽出可能生成物に変換したが、野生型酵素はこの変換を行わなかった。これらの結果は、その生成物のプロフィールが野生型TEASに比べて変化したことを示した。二重突然変異体によるGGPP代謝回転のヘキサン抽出可能生成物を、Ag-TLCにより分析した。この生成物は、加水分解生成物であるゲラニルゲラニオール(Rf=0.0)とは異なる2種(Rf=0.11および0.28)を含んでいた。TEAS C440W/W273SによりGGPPから生成された生成物が加水分解生成物ゲラニルゲラニオールではないことを証明するために、サンプルをAg-TLCにより分析した。100μlの緩衝液中に3H GGPP(5μm)および酵素(40μm)を含む反応を、室温にて一晩インキュベートした。対照として、3H GGPPを反応緩衝液のみの中、およびpH1.5に調節した反応緩衝液の中で、インキュベートした。酵素的反応物および対照反応物を両方ともヘキサンで抽出し、銀染色TLCプレート上にスポッティングし、上記のように現像および露光を行った。図3に示す結果は、TEAS C440W/W273Sにより形成された生成物が、ゲラニルゲラニルジホスフェートの非酵素的分解により生成されたものと異なっていたことを示すものであった。
【0217】
TEAS C440W/W273Sのセスキテルペンシンターゼ活性
14C FPP(9μm)および酵素(160μm)を含む反応緩衝液(20μl)を用い、基
質としてFPPを用いて反応を行った。室温にて30分間インキュベートした後、TEAS C440W/W273Sによって作製された生成物をAg-TLCにより分析した。この二重突然変異体の生成物のプロフィールは、TEAS W273Sのものと似ており、それに更にRfが0.57である主な生成物が加わった。この新しい生成物は、5-エピ-アリストロケンおよびベティスピラジエンの何れとも異なるものであった。他の幾つかの生成物も形成され、そのうちの多くは銀染色TLCの際にゆっくり移動した。図4を参照されたい。
【0218】
実施例7
TEAS C440Wの活性
TEAS C440Wのジテルペンシンターゼ活性:TEAS C440Wを用いた酵素アッセイを、実施例6に記載したように実施した。図3に示すように、160μmの酵素および9μmの放射性標識GGPP(体積20μl)と共に室温にて一晩インキュベートした後、Ag-TLCによってヘキサン-抽出可能生成物は検出されなかった。
【0219】
TEAS C440Wのセスキテルペンシンターゼ活性:精製したTEAS C440Wによる放射性標識から作製された生成物のAg-TLC分析により、少なくとも1つの主なテルペノイド炭化水素生成物(Rf=0.63)の形成が検出され、これは5-エピ-アリストロケン(Rf=0.78)およびベティスピラジエンとは異なるものであった。Ag-TLCでの反応生成物のプロフィールを図4に示す。少量の低速移動生成物(Rf=0〜0.09)も形成された。
【0220】
TEAS C440Wテルペノイド炭化水素反応生成物のヘキサン抽出物のGC/MS分析により、この突然変異体が、単一の主なセスキテルペン炭化水素生成物および少数のマイナーなヒドロキシル化生成物を形成したことが確認された。この主な生成物の質量スペクトルは、スピロ環状化合物ヒネセン(hinesene)の公表されている質量スペクトルに非常に近かった。ヒネセンは、C3メチル基の立体化学構造がベティスピラジエンと異なる。
【0221】
実施例8
TEAS W273Eの活性
TEAS W273Eのセスキテルペンシンターゼ活性:基質としてFPPを用いてTEAS W273Eにより作製された生成物を測定する反応を、放射性標識したFPPを用いて本質的には上記のように行った。これらの結果は、5-エピ-アリストロケン以外に少なくとも1つの生成物が形成されたことを示した。また該結果は、FPPによるTEASのアルキル化が起こったことも示した。このアルキル化は、反応混合物中のMgCl2の存在に依存するものであった。対照実験において、煮沸したW273E-TEAS、ならびに野生型TEASおよびBSAは、アルキル化されなかった。これらの結果は、アルキル化が273位で起こったこと、および273位のアミノ酸残基はこの活性部位の一部であることを示す。
【0222】
実施例9
TEAS Y520Fの活性
TEAS Y520Fのセスキテルペンシンターゼ活性:放射性標識したFPPおよびTEAS Y520F酵素を用いた反応を、本質的には上記のように行った。反応生成物をAg-TLCおよびGC/MSにより分析した。TEAS Y520F反応の主な生成物は、基準ゲルマクレンA(authentic germacrene A)と同じGC保持時間および質量スペクトルを有していた。この生成物のGC保持時間および質量スペクトルは、5-エピ-アリストロケンとは異なっていた。
【0223】
実施例10
TEAS Y527Fの活性
TEAS Y527Fの酵素活性:大腸菌細胞中における発現を誘導し、該細胞を音波で破砕して、TEAS Y527F酵素の粗抽出物を作成した。この音波破砕物を透明化し、上清を酵素アッセイに使用した。基質としてGPPを用いたアッセイでは、生成物は観察されなかった。このことは、TEAS Y527Fが、モノテルペンシンターゼ活性を持たないことを示す。基質としてFPPを用いて反応生成物を得た。これらの生成物のAg-TLCによる分析は、5-エピ-アリストロケン以外の生成物がTEAS Y527F酵素により生成されたことを示した。
【0224】
実施例11
テルペンシンターゼ配列のアラインメント
BLOSUM62スコアリングマトリックス、ギャップ開始値=11、ギャップ延長値=1、X_ドロップオフ値=50、期待値=10、文字列の長さ=3を用い、およびコンプレキシティーの低い配列のフィルタリングを行わずに、BLASTpプログラム(NCBI)を用いて、TEAS一次アミノ酸配列(配列番号2)の残基265〜535を、リモネンシンターゼの全長アミノ酸配列(配列番号22)とアライメントさせた。表12に示すアラインメントプログラムの出力は、TEAS配列の残基527と残基528(アラインメント出力において263および264の番号を付してある)との間にギャップを含んでいた。残基321、324、345、348、349、427、452、453、454、455、458、492、496、569、572、573、577、579および580は、19 TEAS残基と最も好適なアラインメントを有するものとして選択した。TEASに見られる構造的アスペクトの空間的な配向(即ち図1および表10に示すα-ヘリックス、β-シートおよびループなど)を維持するために、リモネンシクラーゼの残基583ではなく残基580を、TEASの残基528とアライメントするものとして選択した。
【0225】
BLOSUM 62スコアリングマトリックス、ギャップ開始値=11、ギャップ延長値
=1、X_ドロップオフ値=50、期待値=10、文字列の長さ=3を用い、およびコンプレキシティーの低い配列のフィルタリングを行わずに、BLASTpプログラム(NCBI)を用いて、配列番号44のタキサジエン(taxadiene)一次アミノ酸配列の残基579〜847を含む領域を、ボルニルジホスフェートシンターゼの全長アミノ酸配列(配列番号26)とアライメントさせた。表13に示すアラインメントプログラムの出力は、ボルニルジホスフェートシンターゼ配列の残基453と残基454との間にギャップを含んでいた。ボルニルジホスフェートシンターゼの残基321、324、344、347、348、426、451、452、453、454、457、492、496、568、571、572、576、578および579は、配列番号44の検索領域配列の残基584、587、606、609、610、688、713、714、715、716、719、753、757、831、834、835、839、841および842と最も好適なアラインメントを有するものとして選択した。タキサジエンシンターゼ中に存在すると思われる構造的アスペクトの空間的な配向(即ち図1および表10に示すα-ヘリックス、β-シートおよびループなど)を維持するために、ボルニルジホスフェートシンターゼの残基453および454を、タキサジエンシンターゼの残基715および716とアライメントするものとして選択した。
【0226】
BLOSUM 50スコアリングマトリックス、ギャップ開始値=13、ギャップ延長値
=2、X_ドロップオフ値=50、期待値=10、文字列の長さ=3を用い、およびコンプレキシティーの低い配列のフィルタリングを行わずに、BLASTpプログラム(NCBI)を用いて、TEAS一次アミノ酸配列(配列番号2)の残基265〜535を、δ-セリネンシンターゼの全長アミノ酸配列(配列番号48)とアライメントさせた。アラインメントプログラムの出力を表14に示す。配列番号48の残基300、303、324、327、328、406、431、432、433、434、437、471、475、548、551、552、556、558および559を、配列番号2の残基270、273、294、297、298、376、401、402、403、404、407、440、444、516、519、520、525、527および528と最も好適なアラインメントを有するものとして選択した。
【0227】
BLOSUM 62スコアリングマトリックス、ギャップ開始値=11、ギャップ延長値=1、X_ドロップオフ値=50、期待値=10、文字列の長さ=3を用い、およびコンプレキシティーの低い配列のフィルタリングを行わずに、BLASTpプログラム(NCBI)を用いて、γ-フムレンシンターゼの一次アミノ酸配列(配列番号50)の残基307〜593を、アビエタジエン(abietadiene)シンターゼの全長アミノ酸配列(配列番号56)とアライメントさせた。アラインメントプログラムの出力を表15に示す。ジテルペンシンターゼ(配列番号56)の残基590、593、614、617、618、696、721、722、723、724、727、761、765、837、840、841、845、847および848を、セスキテルペンシンターゼ検索配列(配列番号50)の残基312、315、336、339、340、419、444、445、446、447、450、484、488、562、565、566、570、572および573と最も好適なアラインメントを有するものとして選択した。
【0228】
実施例12
新規なモノテルペンシンターゼ遺伝子の作成
ピネンシンターゼをコードするDNA配列(配列番号20)を用いて、突然変異型ピネンシンターゼ遺伝子のライブラリーを構築する。以下の9個のアミノ酸残基:L、C、C、G、H、S、L、GおよびY(配列番号20の351位、372位、480位、481位、482位、485位、519位、600位および601位に対応する)のうちの1個以上をコードするヌクレオチドにランダム突然変異を導入する。
【0229】
幾つかの場合では、ピネンシンターゼのコード配列を、配列番号20のY、I、T、F、D、YおよびSの位置に対応するアミノ酸残基348、375、376、597、605、607および608のうちの1個以上をコードするヌクレオチドにランダム突然変異を導入する。ピネンシンターゼのコード配列に時々、以下のアミノ酸残基Y、SおよびG(配列番号20の454位、479位および523位に対応する)のうちの1個以上をコードするヌクレオチドで突然変異させる。幾つかの場合では、上記9個の残基をコードするヌクレオチドにおける突然変異に加えて、さらにこれらの10個の位置における突然変異を起こす。他の場合では、上記9個の残基に突然変異を導入せずに、これらの10個の位置で突然変異を起こす。
【0230】
ピネンシンターゼのコード配列DNAをpET28b(+)ベクター中に挿入し、QuickChange(登録商標)法を用い、TEAS406/407ランダムライブラリについて実施例1で記載したものと同様のプロトコールに従って、突然変異誘発(mutagenize)する。突然変異を生じさせるために使用するプライマーを、ランダム突然変異体を作製するために、NまたはSを所望のコドン中のヌクレオチドとして用いて、実施例1に記載したように合成する。
【0231】
上記19個のピネンシンターゼアミノ酸残基のうちの1個以上における特異的突然変異は、TEASについて実施例1で記載したものと同様のプロトコールを用いて部位特異的突然変異誘発法により行う。所望の突然変異体を作成するために、突然変異させるべきコドン中に特定のA、T、CまたはG置換を有するプライマーを作製する。
【0232】
他のモノテルペンシンターゼ(例えばリモネンシンターゼ(配列番号22または58)、ミルセンシンターゼ(配列番号30)、+サビネンシンターゼ(配列番号54)、1,8 シネオール(cineole)シンターゼ(配列番号24)、および+ボルニルジホスフェートシンターゼ(配列番号26))のアミノ酸残基であって、その残基のα-炭素が表1〜6に記載した原子間距離および構造座標を有するアミノ酸残基を改変するために、上記方法と同じ方法で、ランダムおよび/または特異的突然変異を調製する。
【0233】
実施例13
新規なセスキテルペンシンターゼ遺伝子の作成
カジネンシンターゼをコードするDNA配列(配列番号33)を用いて突然変異型カジネンシンターゼのライブラリーを構築する。以下の9個のアミノ酸残基:W、I、S、G、Y、L、C、LおよびY(配列番号33によりコードされるアミノ酸残基280、301、409、410、411、414、448、527および528に対応する)のうちの1個以上をコードするヌクレオチドにランダム突然変異を導入する。
【0234】
幾つかの場合では、カジネンシンターゼのコード配列を、配列番号33によりコードされるアミノ酸残基277、304、305、524、532、534および535に対応するアミノ酸残基G、A、S、M、D、YおよびTのうちの1個以上をコードするヌクレオチドで突然変異させる。さらに、カジネンシンターゼのコード配列を時々、アミノ酸残基383、408および452(配列番号33によりコードされるアミノ酸Y、TおよびDに対応する)のうちの1個以上をコードするヌクレオチドで突然変異させる。幾つかの場合では、上記9個の残基における突然変異に加えてさらにこれら10個の突然変異を起こす。他の場合では、上記9個の残基に突然変異を導入せずに、これら10個の残基における突然変異を作製する。
【0235】
カジネンシンターゼのコード配列を、pET28b(+)ベクター中でQuickChange(登録商標)法を用いて、TEAS406/407ランダムライブラリについて実施例1で記載したものと同様のプロトコールに従って、突然変異させる。突然変異を生じさせるために使用するプライマーを、実施例11に記載したように合成する。
【0236】
上記カジネンシンターゼアミノ酸残基のうちの1個以上における特異的突然変異は、TEASについて実施例1で記載したものと同様のプロトコールを用いて部位特異的突然変異誘発法により行う。
【0237】
他のセスキテルペンシンターゼ(例えばベティスピラジエンシンターゼ(配列番号32)、ゲルマクレン Cシンターゼ(配列番号52)、E-α-ビスアボレン(bisabolene)シンターゼ(配列番号46)、γ-フムレンシンターゼ(配列番号50)、δ-セリネンシンターゼ(配列番号48)、e-b-ファルネセンシンターゼ(配列番号28)中のアミノ酸残基であって、その残基のα-炭素が表1〜6に記載した原子間距離および構造座標を有するアミノ酸残基を改変するために、上記方法と同じ方法で、ランダムおよび/または特異的突然変異を調製する。
【0238】
実施例14
新規なジテルペンシンターゼ遺伝子の作製
アビエタジエンシンターゼをコードするDNA配列(配列番号56)を用いて突然変異型アビエタジエンシンターゼのライブラリーを構築する。以下の9個のアミノ酸残基:S、S、I、A、L、V、G、FおよびY(配列番号56の593位、614位、722位、723位、724位、727位、761位、840位および841位に対応する)のうちの1個以上をコードするヌクレオチドにランダム突然変異を導入する。
【0239】
幾つかの場合では、アビエタジエンシンターゼのコード配列を、配列番号56の590位、617位、618位、837位、845位、847位および848位に対応するアミノ酸残基I、S、T、M、D、LおよびTのうちの1個以上をコードするヌクレオチドで突然変異させる。アビエタジエンシンターゼのコード配列を時々、アミノ酸残基Y、SおよびN(配列番号56の696位、721位および765位に対応する)のうちの1個以上をコードするヌクレオチドで突然変異させる。幾つかの場合では、上記9個の残基における突然変異に加えてさらにこれらの突然変異を起こす。他の場合では、上記9つの残基に突然変異を導入せずに、これらの10個の残基で突然変異を起こす。
【0240】
アビエタジエンシンターゼのコード配列をpET28b(+)ベクター中でQuickChange(登録商標)法を用いて、TEAS406/407ランダムライブラリについて実施例1で記載したものと同様のプロトコールに従って、突然変異させる。突然変異を生じさせるために使用するプライマーは、実施例11に記載したように合成する。
【0241】
上記アビエタジエンシンターゼアミノ酸残基のうちの1個以上における特異的突然変異は、TEASについて実施例1で記載したものと同様のプロトコールを用いて部位特異的突然変異誘発法により行う。
【0242】
他のジテルペンシンターゼ(例えばカスベン(casbene)シンターゼ(配列番号42)およびタキサジエンシンターゼ(配列番号44)等)中のアミノ酸残基であって、その残基のα-炭素が表1〜6に記載した原子間距離および構造座標を有するアミノ酸残基を改変するために、上記方法と同じ方法で、ランダムおよび/または特異的突然変異を調製する。
【0243】
実施例15
昆虫、哺乳動物および細菌細胞中での突然変異型シンターゼの発現
バキュロウイルス発現ベクターを用いて、実施例12、13および/または14の突然変異型シンターゼをコードする核酸を含む構築物を、昆虫スポドプテラ・フルギペルダ(Spoodoptera frugiperda)の培養細胞中に導入する。該遺伝子の発現後、この突然変異型酵素を各クローンから単離および精製する。
【0244】
SV40プロモーターを有する発現ベクターを用いて、実施例12、13および/または14の突然変異型シンターゼをコードする核酸を含む構築物を、培養したHeLa細胞中に導入する。該遺伝子の発現後、この突然変異型酵素を各クローンから単離および精製する。
【0245】
実施例1で記載したようにプラスミドベクターを用いて、実施例12、13および/または14の突然変異型シンターゼをコードする核酸を含む構築物を、大腸菌BL-21中に導入する。この突然変異型遺伝子の発現後、実施例2に記載されたようにこの突然変異型酵素を各クローンから単離および精製する。
【0246】
他の実施形態
特に記載していないが、当業者であれば、本明細書中に記載および例示した様々な特定の実施形態がいずれも、これらの特定の実施形態以外の特徴を持つようにさらに修正することができることが分かるであろう。
【0247】
本発明は、その詳細な説明に関連して記載したが、前記記載は、特許請求の範囲に定義された本発明の範囲を例示するためのものであって、これを限定するものではない。他の態様、利点および変更は、本発明の特許請求の範囲に含まれる。
【0248】
【表1】

【0249】
【表2】
【0250】
【表3】
【0251】
【表4】
【0252】
【表5】
【0253】
【表6】
【0254】
【表7】
【0255】
【表8】
【0256】
【表9】
【0257】
【表10】
【0258】
【表11】
【0259】
【表12】
【0260】
【表13】
【0261】
【表14】
【0262】
【表15】
【技術分野】
【0001】
本発明は、シンターゼタンパク質の活性部位やそこに位置するα-炭素原子やそれらの空間的関係などの情報を用いて新しい合成能を有する新規シンターゼを開発し生産する方法に関する。
【0002】
関連出願の相互参照
本出願は、1998年9月18日付けの仮出願第60/100,993号、1999年4月22日付けの仮出願第60/130,628号、および1999年8月23日付けの仮出願第60/150,262号の優先権の利益を主張するものである。
【0003】
本研究は、一部には、連邦政府機関からの基金によって支援されたものである。したがって、米国連邦政府は本発明に対して一定の権利を有する。
【背景技術】
【0004】
発明の背景
イソプレノイド化合物はさまざまな生物(例えば、植物、細菌、真菌など)によって産生される有機分子である。これまでに23,000種を超えるイソプレノイド分子が特徴づけられており、毎年何十、何百もの新しい構造が同定されている。これらの分子は種々の役割を果たすことができる。例えば、モノテルペン類は芳香剤や香味料として使用される。セスキテルペン類とジテルペン類はフェロモン、防御用薬剤、視覚色素、抗腫瘍薬、シグナル伝達経路の成分として作用しうる。トリテルペン類は膜の構成成分として、さらにステロイドホルモンと胆汁酸の前駆物質として重要な機能を果たしている。ポリプレノール類は光感受性物質およびコファクター側鎖として機能し、また天然ポリマーとしても存在しうる。
【0005】
イソプレノイド経路により産生される種々の分子化合物はモノ不飽和イソプレン単位の二リン酸エステルから作られる。イソプレンはプレニルトランスフェラーゼによって2、3、または4の倍数で一緒に付加されて、それぞれC10、C15、およびC20単位を作る。C10、C15、およびC20分子はそれぞれゲラニル二リン酸(GPP)、ファルネシル二リン酸(FPP)、およびゲラニルゲラニル二リン酸(GGPP)と呼ばれており、テルペンシンターゼの基質として作用する。
【0006】
テルペンシンターゼは化学または生物学の分野で知られている最も複雑な反応のうちの一つを介してイソプレノイド化合物の生成を触媒している。一般的に、テルペンシンターゼは約40〜100kDの分子量をもつ中程度の大きさの酵素である。酵素として、テルペンシンターゼは、絶妙な反応特異性およびキラリティーの保存と結びついた低度から中程度の代謝回転率をもつとして分類することができる。
【0007】
代謝回転は、基質の酵素への結合、基質コンホメーションの確立、基質の生成物への変換、および生成物の放出から成っている。反応は水性溶媒中でin vitroで行なうことができ、典型的にはコファクターとしてマグネシウムイオンを必要とし、得られる生成物はしばしば非常に疎水性であって、有機溶媒に分配することで回収できる。
【0008】
テルペンシンターゼ遺伝子は細菌、真菌、植物などの様々な生物中に見出される。異なるテルペンシンターゼ間でエキソンに近似する領域を交換することによって、最終的な酵素段階に関与する機能性ドメインが同定された。例えば、Nicotiana tabacum(タバコ)由来の5-エピ-アリストロケンシンターゼ(TEAS)およびヒヨス(henbane)由来のHyoscyamus muticusベチスピラジエンシンターゼ(HVS)について行なった研究により、エキソン4とエキソン6はそれぞれが反応生成物特異性に関与していることが明らかにされた。機能性ドメインを組み合わせることにより、新しい反応生成物を合成する能力がある新規の酵素が得られている(米国特許第5,824,774号)。
【0009】
多くの研究により、イソプレノイドを生成するための反応機構が提案されている。例えば、Caneら, 1985, Bioorg. Chem., 13:246-265; Wheeler and Croteau, 1987, Proc. Natl. Acad. Sci. USA, 84:4856-4859;およびPyunら, 1994, Arch. Biochem. Biophys., 308:488-496を参照されたい。これらの研究では、基質類似体および自己不活化阻害剤(Croteau, 1994, Arch. Biochem. Biophys., 251:777-782; Caneら, 1995, Biochemistry, 34:2471-2479;およびCroteau, 1993, Arch. Biochem. Biophys., 307:397-404)、ならびに触媒作用に不可欠なアミノ酸を同定しようとして化学修飾剤と部位特異的突然変異誘発(Caneら, 1995, Biochemistry, 34:2480-2488; Rajaonarivonyら, 1992, Arch. Biochem. Biophys., 296:49-57;およびRajaonarivonyら, 1992, Arch. Biochem. Biophys., 299:77-82)が採用された。しかしながら、これらの研究は、こうした技法に伴う固有の限界のため、活性部位を規定するうえで限られた成功を収めたにすぎなかった。
【発明の概要】
【0010】
本発明は、テルペンシンターゼの活性部位に存在するα-炭素原子を同定する方法、ならびに三次元空間におけるこれらの原子およびそれらの間の空間的関係を開示する。本発明はまた、そのようなα-炭素と関連したR基、および新しい反応産物を生成することができる新規テルペンシンターゼを作るためにこれらのR基を変更する方法を開示する。
【発明が解決しようとする課題】
【0011】
本発明が本出願において教示されるまで、シンターゼタンパク質の活性部位、そこに位置するアミノ酸、触媒作用に関与するアミノ酸残基、および活性部位内のα-炭素とR基の立体配置は知られていなかった。本発明は今回、シンターゼの構造を教示するのみならず、それから得られる情報を用いて新しい合成能を有する新規シンターゼを開発し生産する手段を提供する。ここに記載の方法を用いて得られたデータは、種々のイソプレノイド基質を使用して種々のイソプレノイド産物を生成することができるシンターゼ変異体の創製および生産に有用である。
【課題を解決するための手段】
【0012】
一実施形態において、本発明は、配列番号2の残基265〜535に対して約20%以上の配列同一性を有する単離されたテルペンシンターゼを特徴とする。該シンターゼは9個のα-炭素を含み、該9個のα-炭素は表6にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有する。該α-炭素のそれぞれの中心点は半径2.3Åの球内に位置する。該球のそれぞれの中心点は表5に示した構造座標を有する。該α-炭素のそれぞれは関連するR基を有しており、該シンターゼは表9に示したR基の順序配列とは異なるR基の順序配列を有する。該シンターゼは配列番号2の残基265〜535に対して約25%以上の配列同一性、または配列番号2の残基265〜535に対して約35%以上の配列同一性を有する。この種のシンターゼはモノテルペン基質、セスキテルペン基質、またはジテルペン基質からのテルペノイド産物の生成を触媒することができる。該産物は環式テルペノイド炭化水素または非環式テルペノイド炭化水素でありうる。どちらのタイプの産物もヒドロキシル化されていてもよい。α-炭素1と関連したR基は、次の群:Cys、SerおよびThrからなる群、Phe、TyrおよびTrpからなる群、Pro、GlyおよびAlaからなる群、GluおよびAspからなる群、Met、Ile、ValおよびLeuからなる群、ArgおよびLysからなる群、ならびにGln、AsnおよびHisからなる群のうちの一つより選択することができる。α-炭素2〜9と関連したR基は、表9の順序配列を有するものを除いた任意のアミノ酸でありうる。同様に、α-炭素2〜9のそれぞれと関連したR基は、Cys、SerおよびThrからなる群、Phe、TyrおよびTrpからなる群、Pro、GlyおよびAlaからなる群、GluおよびAspからなる群、Met、Ile、ValおよびLeuからなる群、ArgおよびLysからなる群、ならびにGln、AsnおよびHisからなる群から独立に選択することができる。これらの実施形態において、残りの8個のα-炭素と関連したR基は、表9の順序配列を有するものを除いた任意のアミノ酸でありうる。
【0013】
いくつかの実施形態において、α-炭素1〜9と関連したR基の順序配列は、それぞれTrp、Ile、Thr、Thr、Tyr、Leu、Cys、ThrおよびPhe、それぞれSer、Ile、Thr、Thr、Tyr、Leu、Cys、ThrおよびTyr、それぞれTrp、Ile、Thr、Thr、Tyr、Leu、Trp、ThrおよびTyr、それぞれSer、Ile、Thr、Thr、Tyr、Leu、Trp、ThrおよびTyr、またはそれぞれGlu、Ile、Thr、Thr、Tyr、Leu、Cys、ThrおよびTyrである。
【0014】
本発明はまた、所定のテルペンシンターゼポリペプチドの一次アミノ酸配列を、配列番号2の残基265〜535のアミノ酸配列にアライメントし、該所定のポリペプチドをコードする核酸に、配列番号2の残基265〜535に対して20%以上の配列同一性を有する該ポリペプチドの一次アミノ酸配列の領域中の9個のアミノ酸残基のうち1個以上のコドンで突然変異を起こさせ、ただし、該ポリペプチド中の該9個の残基は配列番号2の残基273、294、402、403、404、407、440、519および520とアライメントするものであり、そして変異型のテルペンシンターゼが産生されるように突然変異させた核酸を発現させることにより得られる、テルペンシンターゼを特徴とする。
【0015】
本発明はまた、配列番号2の残基265〜535に対して20%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、該シンターゼが16個のα-炭素を含み、該16個のα-炭素が表4にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有する、該テルペンシンターゼを特徴とする。該α-炭素のそれぞれの中心点は半径2.3Åの球内に位置する。該球のそれぞれの中心点は表3に示した構造座標を有する。該α-炭素のそれぞれは関連するR基を有しており、該シンターゼは表8に示したR基の順序配列とは異なるR基の順序配列を有する。該シンターゼは、配列番号2の残基265〜535に対して約25%以上の配列同一性、または配列番号2の残基265〜535に対して約35%以上の配列同一性を有しうる。該シンターゼはモノテルペン基質、セスキテルペン基質、またはジテルペン基質からのテルペノイド産物の生成を触媒することができる。前記産物は例えば環式テルペノイド炭化水素でありうる。α-炭素1〜16と関連した前記シンターゼ中のR基の順序配列は、それぞれCys、Trp、Ile、Ile、Ser、Thr、Thr、Tyr、Leu、Cys、Val、Thr、Tyr、Asp、PheおよびThrでありうる。
【0016】
本発明はまた、配列番号2の残基265〜535に対して20%以上の配列同一性を有する単離されたテルペンシンターゼであって、該シンターゼが19個のα-炭素を含み、該19個のα-炭素が表2にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有する、該テルペンシンターゼを特徴とする。該α-炭素のそれぞれの中心点は半径2.3Åの球内に位置する。該球のそれぞれの中心点は表1に示した構造座標を有する。該α-炭素のそれぞれは関連するR基を有しており、該シンターゼは表7に示したR基の順序配列とは異なるR基の順序配列を有する。該シンターゼは、配列番号2の残基265〜535に対して約25%以上の配列同一性、または配列番号2の残基265〜535に対して約35%以上の配列同一性を有しうる。該シンターゼはモノテルペン基質、セスキテルペン基質、またはジテルペン基質からのテルペノイド産物の生成を触媒することができる。前記産物は例えば環式テルペノイド炭化水素でありうる。
【0017】
本発明はまた、アミノ末端とカルボキシル末端とを有する第1のドメインを含む単離されたタンパク質を特徴とする。該第1のドメインは三次元空間においてグリコシルヒドロラーゼの触媒コアと構造的にアライメントするアミノ酸を含み、ここで、該グリコシルヒドロラーゼの触媒コアは、Aspergillus awamoriのグルコアミラーゼタンパク質データバンク(PDB)コード3GLYのアミノ酸36〜230、およびエンドグルカナーゼCelD PDBコード1CLCのアミノ酸36〜230からなる群より選択されるものである。該単離されたタンパク質はさらに、アミノ末端とカルボキシル末端とを有する第2のドメインを含む。該第2のドメインは三次元空間においてトリFPPシンターゼと構造的にアライメントするアミノ酸を含む。第1のドメインのカルボキシル末端は第2のドメインのアミノ末端に連結されている。第2のドメインの領域は配列番号2の残基265〜535に対して約20%以上の配列同一性を有し、かつ9個のα-炭素を含み、該9個のα-炭素は表6にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有する。該α-炭素のそれぞれの中心点は半径2.3Åの球内に位置し、該球のそれぞれの中心点は表5に示した構造座標を有する。該α-炭素のそれぞれは関連するR基を有しており、該シンターゼは表9に示したR基の順序配列とは異なるR基の順序配列を有する。前記タンパク質は配列番号2に対して約25%以上の配列同一性、または配列番号2に対して約35%以上の配列同一性を有する。前記シンターゼはモノテルペン基質、セスキテルペン基質、またはジテルペン基質からのテルペノイド産物の生成を触媒することができる。前記産物は例えば環式テルペノイド炭化水素でありうる。
【0018】
本発明はまた、配列番号20の残基343〜606に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号20の348、351、372、375、376、454、479、480、481、482、485、519、523、597、600、601、605、607および608位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸Y、L、C、I、T、Y、S、C、G、H、S、L、G、F、G、Y、D、YおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0019】
本発明はまた、配列番号22の残基316〜586に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号22の321、324、345、348、349、427、452、453、454、455、458、492、496、569、572、573、577、579および580位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、N、I、T、Y、S、I、S、G、M、L、D、A、M、Y、D、HおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0020】
本発明はまた、配列番号58の残基352〜622に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号58の357、360、381、384、385、463、487、488、489、490、493、528、532、606、609、610、614、616および617位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸Y、M、C、V、T、F、V、S、S、G、I、L、G、F、V、Y、D、YおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0021】
本発明はまた、配列番号33によりコードされるアミノ酸残基272〜540に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号33によりコードされる277、280、301、304、305、383、408、409、410、411、414、448、452、524、527、528、532、534および535位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸G、W、I、A、S、Y、T、S、G、Y、L、C、D、M、L、Y、D、YおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0022】
本発明はまた、配列番号42の残基319〜571に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号42の324、327、348、351、352、430、455、456、457、458、461、495、499、571、574、575、579、581および582位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸I、W、V、I、S、Y、T、T、G、L、V、I、N、T、S、Y、D、YおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0023】
本発明はまた、配列番号44の残基579〜847に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号44の584、587、606、609、610、688、713、714、715、716、719、753、757、831、834、835、839、841および842位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸V、S、G、Q、V、Y、S、V、G、L、C、W、N、V、F、Y、D、YおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0024】
本発明はまた、配列番号46の残基495〜767に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号46の500、503、524、527、528、606、631、632、633、634、637、674、678、751、754、755、759、761および762位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸F、L、A、Q、T、Y、S、I、G、Q、L、S、D、T、I、F、D、FおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0025】
本発明はまた、配列番号48の残基295〜564に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号48の300、303、324、327、328、406、431、432、433、434、437、471、475、548、551、552、556、558および559位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸Y、W、A、C、T、Y、S、S、G、M、L、G、D、L、I、Y、D、LおよびYとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0026】
本発明はまた、配列番号50の残基307〜578に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号50の312、315、336、339、340、419、444、445、446、447、450、484、488、562、565、566、570、572および573位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸F、W、A、M、T、Y、N、T、G、M、L、S、D、I、M、Y、D、FおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0027】
本発明はまた、配列番号52の残基264〜533に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号52の269、272、293、296、297、375、401、402、403、404、407、441、445、517、520、521、525、527および528位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、L、T、S、Y、S、A、G、Y、I、A、N、A、L、Y、D、YおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0028】
本発明はまた、配列番号56の残基585〜853に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号56の590、593、614、617、618、696、721、722、723、724、727、761、765、837、840、841、845、847および848位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸I、S、S、T、V、Y、S、I、A、L、V、G、N、M、F、Y、D、LおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0029】
本発明はまた、配列番号54の残基307〜574に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号54の312、315、336、339、340、418、443、444、445、446、449、483、487、560、563、564、566、568および569位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、I、I、T、Y、S、I、S、A、I、L、D、A、I、Y、D、DおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0030】
本発明はまた、配列番号24の残基309〜577に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号24の314、317、338、341、342、420、446、447、448、449、452、485、489、560、563、564、569、571および572位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、N、V、T、Y、I、G、G、I、L、L、D、A、I、Y、D、FおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0031】
本発明はまた、配列番号26の残基315〜584に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号26の320、323、344、347、348、426、451、452、453、454、457、492、496、568、571、572、576、578および579位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸S、W、I、A、T、Y、S、V、A、S、I、L、D、A、I、Y、D、FおよびGとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0032】
本発明はまた、配列番号28の残基265〜536に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号28の270、273、294、297、298、376、401、402、403、404、407、440、444、518、521、522、528、530および531位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸A、W、V、C、G、F、T、S、C、I、M、G、N、C、S、Y、D、YおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0033】
本発明はまた、配列番号30の残基342〜612に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号30の347、350、371、374、375、453、478、479、480、481、483、518、522、596、599、600、604、606および607位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸F、L、C、V、T、Y、S、S、A、Y、V、L、G、L、L、Y、D、FおよびSとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0034】
本発明はまた、配列番号32の残基273〜541に対して約40%以上の配列同一性を有する領域をもつ単離されたシンターゼであって、配列番号32の278、281、302、305、306、384、409、410、411、412、415、448、452、524、527、528、533、535および536位のアミノ酸残基とアライメントする該シンターゼのアミノ酸残基1個以上が、それぞれアミノ酸C、W、I、I、S、Y、T、S、T、Y、L、C、D、I、T、Y、D、YおよびTとは異なる残基である、該シンターゼを特徴とする。いくつかの実施形態において、配列同一性は約20%以上、約25%以上、または約35%以上でありうる。いくつかの実施形態において、そのようなシンターゼ中には表7に示した残基の順序配列の1以上が見出されない。
【0035】
本発明はまた、配列番号2の残基265〜535に対して20%以上の配列同一性を有する領域をもつ所定のポリペプチドにおいて、9個のアミノ酸残基を同定すること、ただし該9個のアミノ酸残基のα-炭素は表6にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有し、該α-炭素のそれぞれの中心点が半径2.3Åの球内に位置し、該球のそれぞれの中心点が表5に示した構造座標を有するものであること;および所定のポリペプチドから改変されているポリペプチドを合成すること、ただし、該改変型ポリペプチドは、所定のポリペプチド中の該α-炭素と関連したR基とは異なる該9個のα-炭素と関連したR基を1個以上有すること;を含んでなる、テルペンシンターゼの製造方法を特徴とする。前記合成工程は、該9個のα-炭素に対応するアミノ酸1個以上のコード配列が所定のポリペプチド中に存在するアミノ酸とは異なるアミノ酸のコード配列で置換されている、所定のポリペプチドをコードする核酸を作製することを含む。所定のポリペプチドは例えば配列番号2、4、6,8、10、12、20、22,24、26、28、30、32,34-40、42、44、46、48、50、52、54、56または58に示したポリペプチドのいずれか1つでありうる。
【0036】
本発明はまた、配列番号2の残基265〜535に対して20%以上の配列同一性を有する領域をもつ所定のポリペプチドにおいて、配列番号2の273、294、402、403、404、407、440、519および520位のアミノ酸残基とアライメントする9つの位置のアミノ酸残基を同定すること;および所定のポリペプチドから改変されているポリペプチドを合成すること;を含んでなる、テルペンシンターゼの使用方法を特徴とする。該新規のポリペプチドは、該9つの位置のうち1つ以上に、所定のポリペプチド中に存在するアミノ酸残基とは異なるアミノ酸残基を有することにより改変される。いくつかの実施形態において、前記同定工程は、配列番号2の270、273、294、297、298、402、403、404、407、440、516、519、520、525、527および528位のアミノ酸残基とアライメントする所定のポリペプチド中の16個のアミノ酸残基を同定することを含み、前記合成工程は、所定のポリペプチドから改変されているポリペプチドを合成することを含み、該改変型ポリペプチドが、該16の位置のうち1つ以上に、所定のポリペプチド中に存在するアミノ酸残基とは異なるアミノ酸残基を有する。いくつかの実施形態において、前記同定工程は、配列番号2の270、273、294、297、298、376、401、402、403、404、407、440、444、516、519、520、525、527および528位のアミノ酸残基とアライメントする所定のポリペプチド中の19個のアミノ酸残基を同定することを含み、前記合成工程は、所定のポリペプチドから改変されているポリペプチドを合成することを含み、該改変型ポリペプチドが、該19の位置のうち1つ以上に、所定のポリペプチド中に存在するアミノ酸残基とは異なるアミノ酸残基を有する。前記合成工程は、同定されたアミノ酸残基のうち1個以上をコードする核酸中のコード配列が所定のポリペプチド中に存在するアミノ酸とは異なるアミノ酸をコードするコード配列で置換されている、所定のポリペプチドをコードする核酸を作製することを含む。所定のポリペプチドは例えば配列番号2、4、6,8、10、12、20、22,24、26、28、30、32,34-40、42、44、46、48、50、52、54、56または58に示したポリペプ
チドのいずれか1つでありうる。前記方法はさらに、該改変型ポリペプチドとイソプレノイド基質とを、該基質が該ポリペプチドに結合するのに有効な条件下で接触させること;および該改変型ポリペプチドの、該イソプレノイド基質からの反応産物の生成を触媒する能力を測定すること;を含む。イソプレノイド基質はモノテルペン、セスキテルペン、またはジデルペンでありうる。
【0037】
本発明はまた、ポリペプチド類をコードする核酸分子の集団を作製することを含んでなる、テルペンシンターゼの製造方法を特徴とする。前記集団は、配列番号2の残基265〜535に対して約20%以上の配列同一性を有する所定のテルペンシンターゼのアミノ酸を特定する9つのコドンのうち1つ以上において相互に異なっているメンバーを含み、該9個のアミノ酸のα-炭素は表6にÅで表した原子間距離の±2.3Åであるα-炭素間の原子間距離を有し、該α-炭素のそれぞれの中心点が半径2.3Åの球内に位置し、該球のそれぞれの中心点が表5に示した構造座標を有するものである。いくつかの実施形態において、前記コドンは所定のテルペンシンターゼの表1〜2または3〜4に記載のアミノ酸を特定する。該核酸集団の一部または全部を発現させてポリペプチドの集団を作製するようにする。該ポリペプチド集団の少なくとも1つのメンバーは変異型テルペンシンターゼである。前記発現工程は該核酸集団のin vitro転写およびin vitro翻訳を含むことができる。いくつかの実施形態において、前記発現工程は、該核酸集団のメンバーを発現ベクターにクローニングすること;該発現ベクターを宿主細胞に導入し、該宿主細胞においてクローニングした核酸集団メンバーを発現させて該ポリペプチドの集団を作製すること;を含みうる。所定のテルペンシンターゼポリペプチドはモノテルペンシンターゼ、セスキテルペンシンターゼ、またはジテルペンシンターゼでありうる。宿主細胞は原核細胞または真核細胞であり、例えば、細菌細胞、真菌細胞および動物細胞(例:哺乳動物細胞または昆虫細胞)が含まれる。また、宿主細胞は植物細胞であってもよく、例えば、Graminaceae(イネ科)植物由来の細胞、Legumineae(マメ科)植物由来の細胞、Solanaceae(ナス科)植物由来の細胞、Brassicaeae(アブラナ科)植物由来の細胞、またはConifereae(針葉樹類)植物由来の細胞がある。
【0038】
本発明はまた、本明細書に記載のシンターゼをコードする核酸、および該核酸を含有する宿主細胞を特徴とする。本発明はまた、そのような核酸を含むトランスジェニック植物、またはそのような核酸を含むトランスジェニック動物細胞培養物を特徴とする。
【0039】
いくつかの実施形態において、本発明のシンターゼポリペプチドは、表5の座標を有する9個のα-炭素原子(該α-炭素間の原子間距離は表6に示した距離の±2.3Åである)を含む活性部位を有するドメインを含んでなる。これらのα-炭素原子は、三次元空間において、結合した基質または基質類似体の存在下または不在下でトリFPPシンターゼと構造的にアライメントする。別の実施形態において、本発明のシンターゼは、(i) 三次元空間において(溶解状態または結晶形態で、かつ基質が結合したまたは結合していない状態で)、(a) Aspergillus awamoriのグリコシルヒドロラーゼ(PDBコード3GLY)のアミノ酸36〜230、および(b) エンドグルカナーゼCellB(PDBコード1CLC)のアミノ酸36〜230からなる群より選択されるグリコシルヒドロラーゼ触媒コアとアライメントするアミノ酸残基を含む第1のドメイン、および(ii) 三次元空間において、基質または基質類似体が活性部位に結合したまたは結合していない状態で、トリFPPシンターゼと構造的にアライメントするアミノ酸残基を含む第2のドメイン、を含むものである。第2のドメインは表1〜2、3〜4、または5〜6の原子間距離および構造座標を有する9個、16個または19個のα-炭素原子を含む活性部位を含んでなる。これらのα-炭素原子にはイソプレノイド基質と直接または間接に相互作用しうるR基が結合されている。
【0040】
本発明はまた、触媒活性を有する変異型テルペンシンターゼの作製方法を特徴とする。この方法は、(a) 活性部位を有する触媒的に活性なテルペンシンターゼの結晶学的モデルを提供すること、および(b) 該モデルを用いて、所定のシンターゼに対して活性部位中の少なくとも1個のR基が改変されたテルペンシンターゼを設計すること、を含んでなる。本発明はまた、基質特異性が改変されているテルペンシンターゼ、該テルペンシンターゼの製造方法、およびその三次元構造の作製方法を特徴とする。
【0041】
本明細書に記載したものと同様のまたは等価の方法および材料を用いて本発明を実施することができるけれども、適当な方法および材料を以下に記載する。本明細書中で挙げた刊行物、特許出願、特許および他の文献はいずれもその全体を参照により本明細書中に組み入れるものとする。
【0042】
本発明の他の態様、実施形態、効果および特徴は本明細書から明らかになるであろう。
【図面の簡単な説明】
【0043】
【図1】図1は、Carson, M.およびBugg, C., J. Mol. Graphics 4:121(1986)のRIBBONSソフトウェアプログラムを用いて作成した、ファルネシルヒドロキシホスホネート(FHP)が結合した状態のタバコの5-エピ-アリストロケンシンターゼ(TEAS)の模式図である。
【図2A】図2Aは、α-炭素と関連するR基を示す20種の天然アミノ酸の構造である。
【図2B】図2Bは、α-炭素と関連するR基を示す20種の天然アミノ酸の構造である。
【図3】図3は、GGPPを基質として用いてTEASおよび変異体TEAS酵素により産生されたテルペノイド炭化水素産物の銀薄層クロマトグラムのオートラジオグラムである。DM:W273S/C440W変異体TEAS酵素。
【図4】図4は、FPPを基質として用いてTEASおよび変異体TEAS酵素により産生されたテルペノイド炭化水素産物の銀薄層クロマトグラムのオートラジオグラムである。
【0044】
表の説明
表1は、テルペンシンターゼの活性部位において見出された19個のα-炭素についてのX線結晶構造座標である。
【0045】
表2は、Åで表した表1の各α-炭素間の原子間距離である。各α-炭素は2.3Åの半径を有する球として空間を占有する。原子間距離は各球の中心から計算した。
【0046】
表3は、テルペンシンターゼの活性部位において見出された16個のα-炭素についてのX線結晶構造座標である。
【0047】
表4は、Åで表した表3の各α-炭素間の原子間距離である。各α-炭素は2.3Åの半径を有する球として空間を占有する。原子間距離は各球の中心から計算した。
【0048】
表5は、テルペンシンターゼの活性部位において見出された9個のα-炭素に
ついてのX線結晶構造座標である。
【0049】
表6は、Åで表した表5の各α-炭素間の原子間距離である。各α-炭素は2.3Åの半径を有する球として空間を占有する。原子間距離は各球の中心から計算した。
【0050】
表7は、表1のα-炭素に関連して見出されなかったR基の順序配列である。
【0051】
表8は、表3のα-炭素に関連して見出されなかったR基の順序配列である。
【0052】
表9は、表5のα-炭素に関連して見出されなかったR基の順序配列である。
【0053】
表10は、活性部位に基質類似体FHPが結合したTEASについてのX線構造座標である。
【0054】
表11は、基質が存在しない状態でのTEASについてのX線構造座標である。
【0055】
表12は、BLASTpアライメントプログラムを用いた、リモネンシンターゼ(配
列番号22)とTEASの残基265-535とのアライメントである。
【0056】
表13は、BLASTpアライメントプログラムを用いた、配列番号44の残基579-
847と配列番号26とのアライメントである。
【0057】
表14は、BLASTpアライメントプログラムを用いた、TEASの残基265-535と配列番号48とのアライメントである。
【0058】
表15は、BLASTpアライメントプログラムを用いた、配列番号50の残基307-593と配列番号56とのアライメントである。
【0059】
配列表の簡単な説明
配列番号1は、タバコの5-エピ-アリストロケンシンターゼ(TEAS)タンパク質のDNAコード配列である。Genbank受託番号:Q40577。
【0060】
配列番号2は、配列番号1のDNAによりコードされるタンパク質のアミノ酸配列である。
【0061】
配列番号3は、Trp273のコドンがGluのコドンに変わっているTEASタンパク質のDNAコード配列である。
【0062】
配列番号4は、配列番号3のTEAS DNAによりコードされるW273Eタンパク質のアミノ酸配列である。
【0063】
配列番号5は、Tyr520のコドンがPheのコドンに変わっているTEASタンパク質のDNAコード配列である。
【0064】
配列番号6は、配列番号5のTEAS DNAによりコードされるY520Fタンパク質のアミノ酸配列である。
【0065】
配列番号7は、Tyr527のコドンがPheのコドンに変わっているTEASタンパク質のDNAコード配列である。
【0066】
配列番号8は、配列番号7のTEAS DNAによりコードされるY527Fタンパク質のアミノ酸配列である。
【0067】
配列番号9は、Trp273のコドンがSerのコドンに変わっており、かつCys440のコドンがTrpのコドンに変わっているTEASタンパク質のDNAコード配列である。
【0068】
配列番号10は、配列番号9のTEAS DNAによりコードされるW273S/C440Wタンパク質のアミノ酸配列である。
【0069】
配列番号11は、Tyr406およびLeu407のコドンがそれぞれヌクレオチドNNSに変わっているTEASタンパク質のDNAコード配列である。
【0070】
配列番号12は、配列番号11のTEAS DNAによりコードされるY406X/L407Xタンパク質の集団のアミノ酸配列であり、但し、Xは任意の天然に存在するアミノ酸である。
【0071】
配列番号13は、DNAプライマー配列である。
【0072】
配列番号14は、DNAプライマー配列である。
【0073】
配列番号15は、DNAプライマー配列である。
【0074】
配列番号16は、DNAプライマー配列である。
【0075】
配列番号17は、DNAプライマー配列である。
【0076】
配列番号18は、DNAプライマー配列である。
【0077】
配列番号19は、アメリカオオモミ(grand fir)のピネンシンターゼのDNAコード配列である。Genbank受託番号:U87909。
【0078】
配列番号20は、配列番号19のDNAによりコードされるタンパク質のアミノ酸配列である。
【0079】
配列番号21は、オランダハッカ(spearmint)のリモネンシンターゼのDNAコード配列である。Genbank受託番号:L13459。
【0080】
配列番号22は、配列番号21のDNAによりコードされるタンパク質のアミノ酸配列である。
【0081】
配列番号23は、ヤクヨウサルビア(sage)の1,8シネオールシンターゼのDNAコード配列である。Genbank受託番号:AF051899。
【0082】
配列番号24は、配列番号23のDNAによりコードされるタンパク質のアミノ酸配列である。
【0083】
配列番号25は、ヤクヨウサルビアのボルニルジホスフェートシンターゼのDNA
コード配列である。Genbank受託番号:AF051900。
【0084】
配列番号26は、配列番号25のDNAによりコードされるタンパク質のアミノ酸配列である。
【0085】
配列番号27は、ハッカ(mint)のE-b-ファメセン(E-b-famesene)シンターゼのDNAコード配列である。Genbank受託番号:AF024615。
【0086】
配列番号28は、配列番号27のDNAによりコードされるタンパク質のアミノ酸配列である。
【0087】
配列番号29は、アメリカオオモミのミルセンシンターゼのDNAコード配列である。Genbank受託番号:U87908。
【0088】
配列番号30は、配列番号29のDNAによりコードされるタンパク質のアミノ酸配列である。
【0089】
配列番号31は、馬鈴薯のベタスピラジエン(vetaspiradiene)シンターゼのDNAコード配列である。Genbank受託番号:AB022598。
【0090】
配列番号32は、配列番号31のDNAによりコードされるタンパク質のアミノ酸配列である。
【0091】
配列番号33は、綿(cotton)のデルタ-カジネンシンターゼのゲノムDNAコード配列である。Genbank受託番号:Y18484。
【0092】
配列番号34〜40は、配列番号33のDNAによりコードされるエキソンのアミノ酸配列である。
【0093】
配列番号41は、トウゴマ(castor bean)のカスベン(casbene)シンターゼのDNAコード配列である。Genbank受託番号:L32134。
【0094】
配列番号42は、配列番号41のDNAによりコードされるタンパク質のアミノ酸配列である。
【0095】
配列番号43は、イチイ(yew)のタキサジエンシンターゼのDNAコード配列である。Genbank受託番号:U48796。
【0096】
配列番号44は、配列番号43のDNAによりコードされるタンパク質のアミノ酸配列である。
【0097】
配列番号45は、アメリカオオモミのE-アルファ-ビザボレンシンターゼのDNAコード配列である。Genbank受託番号:AF006194。
【0098】
配列番号46は、配列番号45のDNAによりコードされるタンパク質のアミノ酸配列である。
【0099】
配列番号47は、アメリカオオモミのデルタ-セリネン(delta-silenene)シンターゼのDNAコード配列である。Genbank受託番号:U92266。
【0100】
配列番号48は、配列番号47のDNAによりコードされるタンパク質のアミノ酸配列である。
【0101】
配列番号49は、アメリカオオモミのガンマ-フムレンシンターゼのDNAコード配列である。Genbank受託番号:U92267。
【0102】
配列番号50は、配列番号49のDNAによりコードされるタンパク質のアミノ酸配列である。
【0103】
配列番号51は、トマトのゲルマクレンCシンターゼのDNAコード配列である。Genbank受託番号:AF035631。
【0104】
配列番号52は、配列番号51のDNAによりコードされるタンパク質のアミノ酸配列である。
【0105】
配列番号53は、ヤクヨウサルビアの+サビネンシンターゼのDNAコード配列である。Genbank受託番号:AF051901。
【0106】
配列番号54は、配列番号53のDNAによりコードされるタンパク質のアミノ酸配列である。
【0107】
配列番号55は、アメリカオオモミのアビエタジエン(abietadiene)シンターゼのDNAコード配列である。Genbank受託番号:U50768。
【0108】
配列番号56は、配列番号55のDNAによりコードされるタンパク質のアミノ酸配列である。
【0109】
配列番号57は、アメリカオオモミのリモネンシンターゼのDNAコード配列である。Genbank受託番号:AF006193。
【0110】
配列番号58は、配列番号57のDNAによりコードされるタンパク質のアミノ酸配列である。
【0111】
詳細な説明
本明細書中では以下の用語が用いられる。
【0112】
「α-炭素」とは、アミノ酸残基中で見られるキラル炭素原子をいう。α-炭素には、アミノ基、カルボキシル基、水素原子およびR基を含む4つの置換基が共有結合している。
【0113】
「R基」とは、タンパク質内でのペプチド結合の形成に関与しないアミノ酸残基のα-炭素に結合している置換基をいう。R基は、アミノ酸の全体的な化学的特性の重要な決定要素(determinant)である。タンパク質中に見られる20の天然に存在するアミノ酸および各アミノ酸のα-炭素に結合するR基を図2A、Bに掲載する。本明細書中では、特定のアミノ酸に結合するR基について言及するために、天然に存在するアミノ酸についての3文字または1文字の略語を用いることがある。
【0114】
「天然に存在するアミノ酸」としては、タンパク質において天然に存在する20種のアミノ酸のL-異性体が含まれる。天然に存在するアミノ酸は、グリシン、
アラニン、バリン、ロイシン、イソロイシン、セリン、メチオニン、トレオニン、フェニルアラニン、チロシン、トリプトファン、システイン、プロリン、ヒスチジン、アスパラギン酸、アスパラギン、グルタミン酸、グルタミン、アルギニン、およびリシンである。特に指示しない限り、本出願中で言及する全てのアミノ酸はL形である。本明細書中では、天然に存在するアミノ酸について言及するために、3文字または1文字の略語を用いることがある。これらの略語は当業界で公知である。
【0115】
「非天然のアミノ酸」としては、タンパク質において天然には見られないアミノ酸が含まれる。本明細書中での非天然のアミノ酸の例としては、セレノシステインおよびセレノメチオニンのラセミ混合物が挙げられる。さらに、非天然のアミノ酸には、ノルロイシン、パラ-ニトロフェニルアラニン、ホモフェニルアラニン、パラ-フルオロフェニルアラニン、3-アミノ-2-ベンジルプロピオン酸、ホモアルギニン、D-フェニルアラニンなどのD形またはL形が含まれる。
【0116】
「正に荷電したアミノ酸」としては、通常の生理学的条件下で正電荷を保有するR基を有する任意の天然に存在するアミノ酸または非天然のアミノ酸が含まれる。正に荷電した天然に存在するアミノ酸の例としては、アルギニンおよびリシンが挙げられる。
【0117】
「負に荷電したアミノ酸」としては、通常の生理学的条件下で負電荷を保有するR基を有する任意の天然に存在するアミノ酸または非天然のアミノ酸が含まれる。負に荷電した天然に存在するアミノ酸の例としては、アスパラギン酸およびグルタミン酸が挙げられる。
【0118】
「疎水性のアミノ酸」としては、通常の生理学的条件下で荷電していない非極性の側鎖を有する任意の天然に存在するアミノ酸または非天然のアミノ酸が含まれる。天然に存在する疎水性のアミノ酸の例としては、ロイシン、イソロイシン、バリンおよびメチオニンが挙げられる。
【0119】
「親水性のアミノ酸」としては、荷電している極性の側鎖を有する任意の天然に存在するアミノ酸または非天然のアミノ酸が含まれる。天然に存在する親水性のアミノ酸の例としては、セリン、トレオニンおよびシステインが挙げられる。
【0120】
「突然変異テルペンシンターゼ」または「突然変異型テルペンシンターゼ」とは、一次アミノ酸配列を有するシンターゼポリペプチドをいう。該ポリペプチドの9つの残基のα-炭素の中心点は、半径が2.3Åの球内に位置する。その9つの球の中心点は、表5の構造座標、または表5の座標に合せて回転および/もしくは変換され得る座標を有する。該9つのα-炭素の間の相対的な原子間距離は、表6に示す原子間距離の±2.3Åである。各α-炭素は結合しているR基を有する。突然変異シンターゼは、非突然変異シンターゼとは、該9つのα-炭素に結合しているR基の順序配列が異なる。突然変異シンターゼは、該9つのα-炭素上に、表9に示すR基の順序配列以外のR基の順序配列を有する。該シンターゼの一次アミノ酸配列の他のα-炭素に結合しているR基は、非突然変異型シンターゼにおけるものと同じものであってもよいし、同じものでなくてもよい。
【0121】
幾つかの実施形態において、突然変異シンターゼとは、該ポリペプチドの16の残基のα-炭素の中心点が半径2.3Åの球内に位置するシンターゼをいう。該16の球の中心点は、表3の構造座標、または表3の座標に合せて回転および/もしくは変換され得る座標を有する。該9つのα-炭素の間の相対的な原子間距離は、表4に示す原子間距離の±2.3Åである。各α-炭素は結合しているR基を有する。突然変異シンターゼは、非突然変異シンターゼとは、その16のα-炭素に結合しているR基の順序配列が異なる。突然変異シンターゼは、該16のα-炭素上に、表8に示すR基の順序配列以外のR基の順序配列を有する。該シンターゼの一次アミノ酸配列の他のα-炭素に結合しているR基は、非突然変異型シンターゼにおけるものと同じものであってもよいし、同じものでなくてもよい。
【0122】
幾つかの実施形態において、突然変異シンターゼとは、該ポリペプチドの19の残基のα-炭素の中心点が半径2.3Åの球内に位置するシンターゼをいう。該19の球の中心点は、表1の三次元座標、または表1の座標に合せて回転および/もしくは変換され得る座標を有する。該19のα-炭素の間の相対的な原子間距離は、表2に示す原子間距離の±2.3Åである。各α-炭素は結合しているR基を有する。突然変異シンターゼは、非突然変異シンターゼとは、その19のα-炭素に結合しているR基の順序配列が異なる。突然変異シンターゼは、該19のα-炭素上に、表7に示すR基の順序配列以外のR基の順序配列を有する。該シンターゼの一次アミノ酸配列の他のα-炭素に結合しているR基は、非突然変異型シンターゼにおけるものと同じものであってもよいし、同じものでなくてもよい。
【0123】
「非突然変異型シンターゼ」または「非突然変異シンターゼ」としては、9、16または19のアミノ酸残基を含む一次アミノ酸配列を有するシンターゼが含まれる。これらの残基の各α-炭素の中心点は、半径2.3Åの球内に位置する。該球の中心点は、それぞれ表5、3もしくは1の三次元座標、または表5、3もしくは1の座標に合せて回転および/もしくは変換され得る座標を有する。該9、16または19のα-炭素の間の相対的な原子間距離は、それぞれ表6、4または2に示す原子間距離の±2.3Åである。各α-炭素は結合しているR基を有する。非突然変異シンターゼは、該9、16または19のα-炭素上に、それぞれ表9、8または7に示すようなR基の順序配列を有する。
【0124】
「その縮重変異体」とは、それにより、遺伝暗号の縮重のために異なるヌクレオチド配列により同じポリペプチドがコードされる遺伝子コード配列の変異体をいう。例えば、本発明のシンターゼは、一次アミノ酸配列を有する。縮重シンターゼ変異体は、異なる核酸コード配列であるが、それにもかかわらず、遺伝暗号の縮重のために同じ一次アミノ酸配列をコードする。
【0125】
「発現」とは、遺伝子または核酸分子の転写および該核酸のポリペプチドへの翻訳をいう。遺伝子の発現とはまた、真核系におけるRNAのmRNAへのプロセシングを含む。その遺伝子は、必ずしも、発現を達成するために細胞のゲノムに組み込まれる必要はない。この定義は、特定の系または特定の細胞型における発現に限られるものではなく、(限定しようとするものではないが)安定な、一過性の、in vitroでの、およびin vivoでの発現を含む。
【0126】
「プロモーター」および「プロモーター調節エレメント」とは、遺伝子の発現の調節に関与する核酸をいう。遺伝子発現を促進するために、種々の供給源に由来するプロモーター調節エレメントなどを効率的に用いることができる。プロモーター調節エレメントとしては、構成的、組織特異的、発生特異的(developmental-specific)、誘導的、サブゲノムプロモーターなどが含まれる。プロモーター調節エレメントはまた、転写効率を向上または調節する特定のエンハンサーエレメントまたはサイレンシングエレメントも含み得る。
【0127】
「活性部位」とは、テルペン基質であるGPP、FPPおよび/またはGGPPの疎水性部分を結合させるテルペンシンターゼ内の部位をいう。この活性部位は、特定の条件下で、1つ以上の反応産物が生成されるようにする生合成反応を触媒できる。
【0128】
「改変された酵素特異性」としては、対応する非突然変異型シンターゼの基質特異性およびそれにより作られる反応産物と比較した場合の、突然変異シンターゼの、特定のテルペン基質を使う能力の改変、または突然変異シンターゼからの反応産物のプロフィールの変化が含まれる。改変された特異性としては、シンターゼの、非突然変異型シンターゼと比較して異なる酵素パラメーター(Km、Vmax、など)を示す能力、および/または対応する非突然変異シンターゼにより生成されるものとは異なる産物を生成する能力を含み得る。
【0129】
「構造座標(structure coordinatesまたはstructural coordinates)」とは、結晶形態におけるシンターゼ分子の原子(散乱中心)によるX線の単色光線の回折により得られたパターンから求めた場合のフーリエ合成に関する数学的式から誘導された直交座標(x、yおよびz位置)をいう。回折データを用いて、該結晶(単位胞)中の繰り返しタンパク質単位の電子密度マップを算出する。電子密度マップを用いて、結晶の単位胞内での個々の原子の位置を確証する。本明細書中で示す構造座標の絶対値は、原子間の相対的な空間的関係を意味する。何故ならば、構造座標に帰せられる絶対値は、同じ該原子間の相対的な空間的関係を保ちながら、x、yおよび/またはz軸に沿って一緒に、または別々に回転運動および/または変換運動させることにより変えることができるからである。したがって、構造座標のセットの絶対値を表1、3または5に示す特定の値に合せて回転または変換により調節できるテルペンシンターゼは、表1、3または5のものと同じ構造座標を有すると考えられる。回転および/または変換させた後で本明細書中で示す絶対値と一致する構造座標の例は表11の座標である。
【0130】
「重原子誘導体化(heavy atom derivatization)」とは、シンターゼ結晶の化学的に改変された形態を作製する方法をいう。実際には、結晶を、該結晶中に拡散して該タンパク質の表面に結合できる重原子塩または有機金属化合物(例えば、塩化鉛、チオリンゴ酸金、チメロサール、酢酸ウラニルなど)を含む溶液に浸漬する。結合した重原子の位置は、浸漬した結晶のX線回折分析により求めることができる。次に、その情報を用いて位相の情報を構築し、次にBlundel, T.L.,およびJohnson, N.L., Protein Crystallography, Academic Press (1976)に記載のようにして、該情報を用いて該酵素の三次元構造を構築する。
【0131】
「単位胞」とは、基本的な平行六面体形のブロックをいう。そのようなブロックの規則的な集合体が結晶の全体積を構築している場合もある。各単位胞は、単位パターンの完全な表現(representation)を含んでおり、その繰返しが該結晶を構築している。
【0132】
「突然変異誘発」とは、タンパク質の一次アミノ酸配列内の特定の位置において異なるアミノ酸残基により置換して、それによりその位置に存在するR基を改変することをいう。突然変異誘発は、該タンパク質をコードする核酸のコード配列を変化させて、該核酸中の該コード配列がその位置に元々存在する残基とは異なるアミノ酸残基を特定するようにすることによって最も容易に行うことができる。
【0133】
「空間群」とは、結晶内における対称要素の配置をいう。
【0134】
「分子置換」とは、構造座標がわかっていないシンターゼの予備モデル(preliminary model)を、構造座標がわかっている分子を該未知の結晶の単位胞内で、該未知の結晶の観察される回折パターンを最もよく説明するように、配向および配置させることにより作製することをいう。次に、このモデルから位相を算出し、観察される振幅と組合せて、座標がわかっていない構造のおよそのフーリエ合成を得ることができる。次に、これを、幾つかの形態の精密化のいずれかに供して、該未知の結晶の最終的な正確な構造を得る(Lattman, E., 1985, Method in Enzymology中, 115:55-77; Rossmann, MG.,編, “The Molecular Replacement Method” 1972, Int. Sci. Rev. Ser., No. 13, Gorden & Breach, New York)。構造座標および原子間距離のマトリックスを用いれば、分子置換は、テルペンシンターゼの結晶性の突然変異体、相同体または異なる結晶形態の構造座標を求めるのに用いることができる。
【0135】
「組換えタンパク質」としては、化学的に合成された、または単離された遺伝子から生合成により誘導されたタンパク質が含まれる。
【0136】
「遺伝子」としては、ポリペプチドを作製するのに必要な情報を含む、自然に誘導された、または遺伝子操作された核酸が含まれる。
【0137】
「核酸」としては、ヌクレオチドであるグアニン、アデニン、チミン、シトシン、ウラシル、イノシンなどからなる任意の遺伝子物質が含まれる。核酸は一本鎖、二本鎖または三本鎖であってもよい。核酸は、デオキシリボ核酸であってもリボ核酸であってもよい。
【0138】
「遺伝子操作された」とは、所定の核酸中に存在するものとは異なるヌクレオチド配列を含むように改変されている遺伝子を含む。遺伝子は、合成により、または従来のクローニング、PCR、化学的遺伝子合成、直接もしくはランダムな突然変異誘発、および遺伝子シャッフリングにより操作できる。遺伝子操作された、とは、所定のタンパク質をコードする核酸の縮重変異体である遺伝子を作製するプロセスも含む。
【0139】
「第1のドメイン」とは、第1および第2の末端を有するポリペプチドを含み、そこにおいて、第1の末端は遊離アミノ基を有するアミノ末端アミノ酸を有してもよく、ペプチド結合により第2のアミノ酸に連結させてもよい。該第1の末端はまた、アセチル化などにより修飾されてもよい。該第1のドメインの第2の末端は遊離カルボキシル末端基を有してもよいし、有していなくてもよい。
【0140】
「第2のドメイン」とは、第1および第2の末端を有するポリペプチドを含み、そこにおいて、第1の末端はアミノ末端アミノ酸を有してもよく、ペプチド結合により第2のアミノ酸に連結されていてもよい。該第2のドメインの第2の末端はカルボキシル末端基を有してもよいし、有していなくてもよい。典型的には、該第2のドメインの第1の末端は、ペプチド結合を介して該第1のドメインの第2の末端に連結されている。
【0141】
「イソプレノイド基質」とは、それぞれゲラニルジホスフェート(GPP)、ファルネシルジホスフェート(FPP)およびゲラニルゲラニルジホスフェート(GGPP)と名付けられているC10、C15およびC20の分子をいう。
【0142】
「配列同一性」または「配列同一性の割合」とは、2つのタンパク質配列またはヌクレオチド配列[つまり、参照配列(query sequence)と対象配列(subject sequence)]のアライメントをとった場合の、同じ相対する位置を占めるアミノ酸またはヌクレオチドの割合(%)をいう。対象配列と参照配列との間で同一であるアミノ酸またはヌクレオチドの残基の数を数え、参照配列中の残基の数で割り、100をかける。最も高い割合の配列同一性をもたらすアライメントが見つかるまで、このプロセスを繰り返す。配列同一性の割合は、目視による確認により、および/またはMegAlign(DNASTAR, Inc., Madison, Wisconsin)またはNational Center for Biotechnology Information(NCBI)からワールドワイドウェブ上で入手可能なBLASTプログラムのような種々のコンピュータープログラムを用いて求めてもよい。配列アライメントを最大にするために、1つ以上の残基のギャップを参照配列の構造上保存されたドメイン(すなわち、α-ヘリックス、β-シートおよびループ)に挿入してもよい場合がある。
【0143】
「モノテルペン産物」とは、基質GPPからつくられる直鎖状、環状および/またはヒドロシキル化された反応産物をいう。「セスキテルペン産物」とは、基質FPPからつくられる直鎖状、環状および/またはヒドロシキル化された反応産物をいう。「ジテルペン産物」とは、基質GGPPからつくられる直鎖状、環状および/またはヒドロシキル化された反応産物をいう。
【0144】
本発明は、活性部位を含む、特定のα-炭素およびそれに結合しているR基の位置が、三次元空間において決定できるテルペンシンターゼおよびその突然変異体に関する。本発明はまた、該シンターゼの構造座標、シンターゼ相同体、突然変異体などに関する構造的情報を創り出すための該構造座標の使用、ならびにそのようなシンターゼの結晶形態に関する。さらに、本発明は、所定のテルペンシンターゼの活性部位を含む原子のα-炭素の構造座標を用いて、活性部位α-炭素原子に結合しているR基が該所定のテルペンシンターゼ中で見られるR基とは異なるシンターゼを開発する方法を提供する。さらに、本発明は、本明細書中で提供される構造的情報に基づく新規テルペンシンターゼの製造、および種々のイソプレノイド化合物を製造するためのそのようなシンターゼの使用を提供する。
【0145】
本発明はさらに、初めて、タバコの5-エピ-アリストロケンシンターゼ(TEAS)に例示されるような、基質または基質類似体の存在下または不在下で成長して、それに関連する構造座標の規定を可能にするシンターゼの結晶を提供する。該構造座標は、活性部位を含むα-炭素原子およびそれに結合しているR基の決定を可能にする。本発明の結晶は、正方晶系の空間群P41212に属し、単位胞の大きさは結晶間で数Åだけ変動するが、平均してa=126Å、c=122Å、a=b、α=90°、β=90°、およびγ=90°である。
【0146】
構造座標は、好ましくは、結合した基質または基質類似体の存在下および不在下でシンターゼについて約2.2〜約2.8Åの解像度(resolution)で得られる。活性部位に結合した基質類似体を有する場合のシンターゼの座標を表10に示す。活性部位に結合した基質類似体がない場合のシンターゼの座標は表11に示す。当業者であれば、X線結晶学により求めた構造座標のセットには標準誤差が含まれないことを理解する。したがって、本発明の目的のためには、シンターゼ、シンターゼ相同体またはその突然変異体の活性部位α-炭素が表1、3または5に示す構造座標を用いて重ねあわせた場合に±2.3Å未満の二乗平均平方根偏差を有する任意のセットの構造座標は同一であると考えられる。
【0147】
RIBBONS(CarsonおよびBugg, 1986, J. Mol. Graphics, 4:121)により作製されたシンターゼの三次元的形状の概略図を図1に示す。図1に示すシンターゼは、全体がα-ヘリックスならびに短い連結ループおよびターンからなり、第1および第2の構造ドメインに分かれて構成されている。
【0148】
1つの実施形態では、本発明の単離されたシンターゼは、表3の構造座標および表4に示す距離の±2.3Åの相対的距離を有する16の活性部位α-炭素を含む。表3の活性部位α-炭素は通常、種々の活性部位α-炭素間に介在するアミノ酸残基のために、全てが隣接しているわけではない(すなわち、シンターゼの一次アミノ酸配列において互いに隣接していない)。一方、幾つかの例においては、該活性部位α-炭素の幾つかは互いに隣接していてもよいと理解すべきである。例えば、TEAS Y527Fタンパク質(配列番号8)に示す実施形態では、一次アミノ酸配列において、それぞれ402位、403位および404位にて、活性部位α-炭素は互いに隣接しているが、残基273および294における活性部位α-炭素は分断されており、したがって隣接していない。したがって、表1、2、3、4、5または6に示す活性部位α-炭素の番号付けは単に便宜上のものにすぎず、そのようなα-炭素は、表1、3または5に示す構造座標および表2、4または6に示す±2.3Åの相対的原子間距離を達成する一次アミノ酸配列中の任意の位置に存在し得る。
【0149】
活性部位α-炭素に結合しているR基を適切に組み合わせれば、1種以上の目的の反応産物の生成を容易にし得る。本発明のテルペンシンターゼにおいて用いるために選ばれるR基の組合せは、表7、8または9に示すR基および対応する活性部位α-炭素の順序配列以外のどのような組合せであってもよい。R基およびα-炭素の適切な配列の代表例は、表3の活性部位α-炭素1〜16にそれぞれ結合しているCys、Trp、Ile、Ile、Ser、Thr、Thr、Tyr、Leu、Cys、Val、Thr、Phe、Asp、TyrおよびThrである。R基およびα-炭素の適切な配列の別の例は、表3の活性部位α-炭素1〜16のそれぞれにおけるCys、Trp、Ile、Ile、Ser、Thr、Thr、Tyr、Leu、Cys、Val、Thr、Tyr、Asp、PheおよびThrである。幾つかの実施形態では、本発明のシンターゼは、配列番号4、配列番号6、配列番号8および配列番号10に示す一次アミノ酸配列、それらをコードするDNA分子(それぞれ、配列番号3、配列番号5、配列番号7および配列番号9に示される)、ならびにそれらの縮重変異体を有する。典型的には、活性部位α-炭素で見られるR基は、天然に存在するアミノ酸において見られるものである。例えば図2A、Bを参照されたい。しかし、幾つかの実施形態では、天然に存在するアミノ酸以外のR基を用い得る。
【0150】
R基および活性部位α-炭素の配列には、反応産物を生成する突然変異テルペンシンターゼを生じさせるものもある。そのような酵素的に活性なシンターゼおよびそれらの対応する遺伝子は、例えばモノテルペン(例えばピネン)、セスキテルペン(例えばデルタ-カジネン)およびジテルペン(例えばアビエタジエン)のような既知のテルペノイド炭化水素の製造に有用である。他の酵素的に活性なシンターゼは、新規テルペノイド産物の製造に有用であり得る。
【0151】
R基および活性部位α-炭素の配列には、反応産物を所望の速度で生成しない突然変異テルペンシンターゼを生じさせるものもある。そのようなシンターゼおよびそれらの遺伝子は、酵素的に活性な突然変異シンターゼによる産物生成の分析における対照(コントロール)として有用である。そのようなシンターゼおよびそれらの遺伝子はまた、酵素的に活性な突然変異シンターゼ遺伝子の翻訳の分析においても、あるいは栄養補助剤としても有用である。そのようなシンターゼは、Sepharoseビーズに付着させて、粗調製物からのイソプレノイド化合物のアフィニティ精製に用いることができる。さらに、そのようなシンターゼおよびそれらの遺伝子はまた、種々の目的の試薬(例えばテルペンシンターゼタンパク質の発現をモニターするための免疫学的試薬)または植物育種プログラムにおけるテルペンシンターゼ遺伝子の遺伝をモニターするための核酸プローブもしくはプライマーの開発にも有用である。
【0152】
幾つかの実施形態では、シンターゼの第1のドメインのα-炭素骨格は、Aspergillus awamori由来のグルコアミラーゼ(Brookhaven Protein Database (PDB)コード3GLY)(Aleshinら, 1994, J. Mol. Biol. 238:575)およびClostridium thermocelum由来のエンドグルカナーゼCelD(PDBコードICLC)(Juyら, 1992, Nature, 357:89)により例示されるようはグリコシルヒドロラーゼの触媒コアと構造的に整列(align)し、シンターゼの第2のドメインのα-炭素骨格(活性部位を含む)は、鳥類ファルネシルジホスフェートシンターゼ(FPS)と構造的に整列し、そこにおいて、活性部位は、表1、3または5に示すα-炭素の構造座標および表2、4または6に記載の原子間距離を有する9、16または19のアミノ酸残基からなる。そのようなα-炭素は、非突然変異型シンターゼにおいて見られるものとは異なるR基の順序配列を有する。
【0153】
本発明において、第1のドメインは、水和された場合、きちんと並んだ水分子で満たされた表面の窪みを囲む8つの短い(10〜15アミノ酸残基の)ヘリックスからなる捻じれたα-バレルを形成する。第2のドメインは、疎水性で芳香族に富む活性部位ポケットを囲むα-ヘリックスの二層のバレルからなる。典型的には、この第2のドメインは基質結合部位を含む。図1に例示するように、ヘリックスHは、プロリンのようなアミノ酸によりセグメントH1およびH2の間で分断されているが、ヘリックスGによるそのヘリックス間パッキングは、ヘリックスG中でG1とG2との間で対応するキンク(kink)により収容されている。このキンク内では、トレオニンで見られるような水酸基と他のアミノ酸のカルボニル酸素との間の水素結合がヘリックスGの主鎖ヘリックス内水素結合を分断し、したがって所定の構造をとるのを助ける。
【0154】
TEASにより例示されるように、本発明のテルペンシンターゼは、ヘリックスA、BおよびC(A−Cループ)を含む第1のドメインセグメントと、ヘリックスJおよびK(J−Kループ)を含む第2のドメインとを有し得る(図1)。基質の結合の際にこれらのループが規則的に配列(ordering)することにより、閉じた、溶媒に接触不可能な活性部位ポケットが生じる。J−Kループが規則的に配列されると、基質の存在下で活性部位の入り口を締めつける蓋形の構造が形成され、活性部位ポケットに深く入り込んでいる伸びた芳香族パッチが形成される。A−Cループが規則的に配列されると、それは活性部位に向って内側に平行移動(translate)し、特定のR基をこのループ内で活性部位に、または活性部位付近に配置させる。このように、活性部位への基質の結合は、タンパク質のコンフォメーションの変化をもたらす。
【0155】
突然変異テルペンシンターゼを同定または作製するために、配列アライメントを行って、表1−2、3−4または5−6の構造座標および原子間距離を有する所定のポリペプチド中で特定の残基およびα-炭素の位置を特定し得る。この所定のポリペプチドは、該アライメントにおける対象配列として、例えば全長一次アミノ酸配列、長さ190残基の領域、長さ220残基の領域または長さ300残基の領域として用いる。このアライメントには、TEAS(配列番号2)の残基265〜535を用いることができ、これは該所定のポリペプチドとアライメントをとる参照配列として表1、3または5のα-炭素を含む。該所定のポリペプチドおよび該参照配列は、BLOSUM62スコアリングマトリックス、期待値(expect value)が10、ギャップオープン値が11、xドロップオフ値が50、ギャップイクステンション値が1、ワードサイズが3および低複雑性(low complexity)配列のフィルタリングなしを用いるBLASTp2.0.9コンピュータープログラムを用いてアライメントをとることができる。別法として、BLASTp2.0.9プログラムは、BLOSUM50スコアリングマトリックス、期待値が10、ギャップオープン値が13、xドロップオフ値が50、ギャップイクステンション値が2、ワードサイズが3および低複雑性配列のフィルタリングなしと共に用いることができる。他のパラメーター値(例えば0〜4のギャップイクステンション値)も用い得る。Altschulら, Nucl. Acids Res. 25:3389-3402を参照されたい。
【0156】
TEASの残基265〜535に対して有意な配列同一性、例えば20%以上の配列同一性、25%以上の配列同一性、35%以上の配列同一性、40%以上の配列同一性、50%以上の配列同一性、60%以上の配列同一性、70%以上の配列同一性、または80%以上の配列同一性、を有する所定のポリペプチドの領域を、表1、3または5に示すものに対応するTEASの残基と整列する特定の残基について調べる。幾つかの場合、コンピュータープログラムアライメントの出力は、アライメントプログラムによりギャップが導入されている、またはされていない表1−2、3−4または5−6の構造座標および原子間距離を有する参照配列中の該9、16または19の残基の各々に対して、所定のポリペプチド中の特定の残基を同定する。他の場合、参照配列または対象配列のいずれかにアライメントプログラムによりギャップが導入されて、関心のある参照配列中の該9、16または19の残基の1以上の間に直接的なアライメントまたはミスアライメントが起こらないようにする。いずれの場合も、出力は目視で確認でき、参照配列中のα-ヘリックスおよびβ-シートの領域が維持され、かつ対象配列中のギャップまたは挿入が参照配列のループ領域と整列するようにアライメントを調整した後で、対象配列中で特定の残基を選択できる。
【0157】
配列アライメントから、他のテルペンシンターゼがTEASの残基265〜535に対して20%以上の配列同一性を有する領域を有することが示唆される。したがって、配列番号2の残基265〜535に対して有意な配列同一性を有する配列番号4、6、8、10、12、20、22、24、26、28、30、32、34〜40、42、44、46、48、50、52、54、56または58に示すテルペンシンターゼの領域のような、TEAS以外のテルペンシンターゼの領域を参照配列として用いることができる。例えば、TEASと比較して、テキサジエンシンターゼ(配列番号44)においては大きな配列挿入物はアミノ末端に存在するか、あるいはアミノ末端ドメイン中の溶媒に暴露されるループ内に存在する。したがって、配列番号2に対して20%より高い配列同一性を有するテキサジエンシンターゼの領域は、カルボキシ末端により近接しており、例えば配列番号44の残基579〜残基847である。
【0158】
参照配列として用い得る他のテルペンシンターゼの有用な領域としては、配列番号20の残基343〜606、配列番号22の316〜586、配列番号58の残基352〜622、配列番号33の残基272〜540、配列番号42の残基319〜571、 配列番号44の残基579〜847、配列番号46の残基495〜767、配列番号48の残基295〜564、配列番号50の残基307〜578、配列番号52の残基264〜533、配列番号56の残基585〜853、配列番号54の残基307〜574、配列番号24の残基309〜577、配列番号26の残基315〜584、配列番号28の残基265〜536、配列番号30の残基342〜612、および配列番号32の残基273〜541が挙げられるが、それらに限定されない。
【0159】
参照配列中の残基と整列する対象配列中の特定の残基のうちの1つ以上は、所定のポリペプチド中では、例えば該ポリペプチドをコードする核酸中に突然変異を起こすことにより、突然変異されている。こうして作製された突然変異テルペンシンターゼは、次いで宿主細胞内で発現され、所望によりそのタンパク質を酵素活性について評価することができる。
【0160】
本発明の突然変異タンパク質は、オリゴヌクレオチド指定突然変異誘発、欠失、化学的突然変異誘発など(しかし、それらに限定されない)のような幾つかの方法により調製できる。テルペンシンターゼにおける活性部位α-炭素原子に結合している1つ以上のR基は、対応する遺伝子のヌクレオチド配列を改変することにより変えられる。例えば、突然変異は、配列番号1(TEASのヌクレオチド配列)に、次の16のα-炭素の1つ以上をコードするコドンにおいて導入することができる:α-炭素1=Cys270;α-炭素2=Trp273;α-炭素3=Ile294;α-炭素4=Ile297;α-炭素5=Ser298;α-炭素6=Thr402;α-炭素7=Thr403;α-炭素8=Tyr404;α-炭素9=Leu407;α-炭素10=Cys440;α-炭素11=Val516;α-炭素12=Thr519;α-炭素13=Tyr520;α-炭素14=Asp525;α-炭素15=Tyr527;α-炭素16=Thr528。次いで、該遺伝子を例えば細菌または植物発現系内で発現することにより、該突然変異遺伝子によりコードされるタンパク質が生産される。あるいはまた、シンターゼ突然変異体は、特定のアミノ酸を天然には存在しないアミノ酸で部位指定置換することにより作製してもよい。そのようにして、シンターゼ突然変異体は、アミノ酸残基または特定のシステインもしくはメチオニン残基をセレノシステインまたはセレノメチオニンで置換することにより作製できる。これは、野生型または突然変異型のポリペプチドを発現できる宿主生物を、天然のシステインもしくはメチオニンまたはそれらの双方が欠乏している増殖培地上で増殖させ、セレノシステイン、セレノメチオニンまたはそれらの両者のいずれかに富む増殖培地上で増殖させることにより達成できる。これらおよび類似の技法は、Sambrookら、[Molecular Cloning, A Laboratory Manual, 第2版,(1989) Cold Spring Harbor Laboratory Press]に記載されている。
【0161】
本発明の突然変異シンターゼを作製する別の適切な方法は、NoelおよびTsal(1989) J. Cell. Biochem., 40:309-320に記載の手順に基づくものである。その場合、該シンターゼをコードする核酸は、重複領域を有するオリゴヌクレオチドを用いて合成により作製でき、該オリゴヌクレオチドは、突然変異が誘導されるように特定の塩基において縮重している。
【0162】
本発明によれば、突然変異型シンターゼをコードする核酸配列は、本明細書中で記載の方法または当業者に利用可能な任意の別の方法により作製できる。関心のある核酸配列(遺伝子)の設計において、特定の発現系における発現を向上させるために、該遺伝子を再操作することが望ましいものであり得る。例えば、多くの細菌由来の遺伝子は植物系では十分に発現されないことが示されている。幾つかの場合、植物由来の遺伝子は細菌内では十分に発現しない。この現象は、用いる発現系に関して該遺伝子のG+C含量またはA+T含量が最適でないことによるものと思われる。例えば、多くの細菌遺伝子の非常に低いG+C含量により、高度にA+Tに富む植物遺伝子調節配列を模倣または複製する配列が作製されるようになる。植物に導入された遺伝子内でのA+Tに富む配列(例えば、一般に遺伝子プロモーター中で見られるTATAボックス領域)の存在は、該遺伝子の異常な転写を引き起こす可能性がある。さらに、転写されたmRNA中に存在する他の調節配列[例えば、ポリアデニル化シグナル配列(AAUAAA)またはプレmRNAスプライシングに関与する低分子の核RNAに相補的な配列]の存在は、RNAの不安定性につながり得る。したがって、遺伝子の設計における1つの目標は、特定の発現系のためのmRNAの安定性および転写精度を賦与するG+C含量を有する核酸配列を作製することである。
【0163】
遺伝暗号の重複性によりもたらされる適応性(plasticity)(すなわち、幾つかのアミノ酸は1以上のコドンにより特定されること)のために、異なる生物または生物クラスのゲノムの進化は、重複コドンの特異的な使用法をもたらしている。この「コドンバイアス」は、タンパク質コード領域の平均塩基組成において反映される。例えば、比較的低いG+C含量を有する生物は重複コドンの3番目の位置にAまたはTを有するコドンを利用し、一方高いG+C含量を有する生物は3番目の位置にGまたはCを有するコドンを利用する。したがって、発現のための遺伝子の再操作において、遺伝子を発現させようとする生物のコドンバイアスを測定したいと思うかもしれない。GenBankに寄託された特定の生物の遺伝子について調べたコドン利用を参照すれば、この情報が得られる。そのバイアスを調べた後、新規な遺伝子配列は、制限酵素部位、ならびにエキソン:イントロン接合部、ポリA付加シグナルまたはRNAポリメラーゼ終結シグナルのような転写に影響を及ぼし得る他の部位について分析することができる。
【0164】
シンターゼをコードする遺伝子は、当業者の関心に応じて適当なベクター内に配置することができ、適切な発現系を用いて発現させることができる。当業界では周知であるように、発現ベクターは、典型的には、宿主細胞内で該ベクターの複製を可能にするエレメントを含んでおり、該遺伝子を含む細胞の選択のための1つ以上の表現型マーカーを含んでもよい。発現ベクターは、典型的には、プロモーター配列、リボゾーム結合部位ならびに翻訳開始および終結配列などの発現を調節する配列を含む。発現ベクターはまた、サブゲノムプロモーター、リプレッサー遺伝子または種々のアクチベーター遺伝子のようなエレメントを含んでもよい。当業者であれば、遺伝子産物の分泌、植物色素体のような特定の細胞小器官への該産物の移動をもたらす核酸配列(米国特許第4,762,785号、同第5,451,513号および同第5,545,817号を参照)、またはアフィニティータグのようなペプチド精製をより容易にする他の配列を含ませることも選ぶであろう。
【0165】
多種多様な発現調節配列が、機能し得るように連結されれば突然変異型シンターゼの発現に有用である。そのような発現調節配列としては、例えば、動物細胞のためのSV40の初期および後期プロモーター、lac系、trp系、ファージλの主要なオペレーターおよびプロモーター系、ならびにコートタンパク質の調節領域(特に植物中のRNAウイルス由来のもの)が挙げられる。大腸菌において、有用な転写調節配列はT7 RNAポリメラーゼ結合プロモーターであり、これは、Studierら(1990)Methods Enzymology, 185:60-89に記載のようにしてpETベクターに組み込むことができる。
【0166】
発現のためには、所望の遺伝子は該発現調節配列に機能し得るように連結し、適切な読み取り枠を維持して、所望のシンターゼの産生を可能にしなければならない。多種多様な周知の発現ベクターのいずれもが、本発明では有用である。これらとしては、例えば、SV40に由来するもののような染色体DNA配列、非染色体DNA配列および合成DNA配列のセグメントからなるベクター、細菌プラスミド(大腸菌に由来するもの、例えばcol E1、pCR1、pBR322およびそれらの誘導体、pMB9)、より広い宿主範囲のプラスミド(例えばRP4)、ファージDNA(例えばファージλ、NM989、M13)、ならびにSambrookら[Molecular Cloning, A Laboratory Manual,第2版, (1989) Cold Spring Harbor Laboratory Press]により記載されているような他のそのような系が挙げられる。
【0167】
多種多様な宿主細胞が、本発明のシンターゼ突然変異体の発現に利用可能である。そのような宿主細胞としては、(限定しようとするものではないが)大腸菌、バシルス(Bacillus)およびストレプトマイセスのような細菌、真菌、酵母、動物細胞、植物細胞、昆虫細胞などが挙げられる。本発明の好ましい実施形態は、大腸菌または植物細胞において発現されるテルペンシンターゼ突然変異体を含む。該植物細胞は、懸濁液培養物または寒天ベースの培地のような固体支持体上での培養物のいずれかであり得る。
【0168】
本発明のシンターゼをコードする遺伝子はまた、トランスジェニック植物細胞内で発現させてもよい。トランスジェニック植物を作製するためには、突然変異テルペンシンターゼをコードする核酸構築物を含むベクターを植物ゲノムに挿入する。好ましくは、これらの組換えベクターは、植物ゲノムへ安定に組み込まれることができる。トランスジェニック植物の作製における1つの変化し得る要素は、選択マーカー遺伝子の選択である。選択マーカー遺伝子は、非形質転換細胞の高いバックグラウンドから形質転換細胞を同定するのに用いられる。そのような選択マーカー遺伝子としては、抗生物質カナマイシン、ネオマイシンおよびG418に対する耐性をコードするトランスポゾンTn5のアミノグルコシドホスホトランスフェラーゼ遺伝子(Aph II)、ならびにグリホセート(glyphosate)、ハイグロマイシン、メトトレキサート、ホスフィノトリシン(phosphinothricin)、イミダゾリノン、スルホニル尿素、およびトリアゾロピリミジン殺草剤(例えばクロロスルフロン、ブロモキシニル、ダラポンなど)に対する耐性または寛容性をコードする遺伝子が挙げられるが、それらに限定されない。選択マーカー遺伝子に加えて、レポーター遺伝子を用いることが好ましい場合がある。幾つかの例において、レポーター遺伝子は選択マーカーと共に用いることができる。レポーター遺伝子は、形質転換細胞の検出を可能にし、当業者の随意で用いることができる。これらのレポーター遺伝子の一覧は、K. Weisingら, 1988, Ann. Rev. Genetics, 22:421に提供されている。
【0169】
該遺伝子は、あらゆる組織においてあらゆる時期に発現するプロモーター(構成的プロモーター)、特定の組織において発現するプロモーター(組織特異的プロモーター)、特定の発生段階で発現するプロモーター(発生的プロモーター)、および/または刺激に応答して発現するプロモーター(誘導的プロモーター)により発現される。これらの選択は、当業者であれば随意に決定される。
【0170】
外来遺伝子を植物細胞に導入するため、および導入された遺伝子を安定に維持し発現する植物を得るための幾つかの技法が存在する。そのような技法としては、被覆された遺伝物質を細胞に直接的に促進すること(米国特許第4,945,050号)が挙げられる。植物はまた、アグロバクテリウム(Agrobacterium)法(米国特許第5,177,010号、同第5,104,310号、同第5,149,645号、同第5,469,976号、同第5,464,763号、同第4,940,838号、同第4,693,976号、同第5,591,616号、同第5,231,019号、同第5,463,174号、同第4,762,785号、同第5,004,863号および同第5,159,135号;欧州特許出願第116718号、同第290799号、同第320500号、同第604662号、同第627752号、同第0267159号および同第0292435号)を用いて形質転換してもよい。他の形質転換法としてはウィスカー(whiskers)法が挙げられる(米国特許第5,302,523号および同第5,464,765号を参照)。エレクトロポレーション法もまた、植物を形質転換するのに用いられている(WO87/06614、WO92/09696およびWO93/21335ならびに米国特許第5,472,869号および同第5,384,253号を参照)。米国特許第5,316,931号、同第5,589,367号、同第5,811,653号および同第5,866,785号に記載のもののようなウイルスベクター発現系も用い得る。
【0171】
植物を形質転換するための多くの技法に加えて、関心のある遺伝子と接触させる組織のタイプもまた様々なものであり得る。適切な組織としては、胚形成組織、カルス組織、胚軸組織、分裂組織などが挙げられるが、それらに限定されない。ほぼ全ての植物組織が、本明細書中に記載の適切な技法を用いて分化の間に形質転換できる。
【0172】
変異型シンターゼをコードする遺伝子は、使用する形質転換系とは関係なく、発現制御配列(植物プロモーター調節エレメント)をベクターに含ませることによって植物細胞中で該遺伝子を発現するように適合された遺伝子転移ベクターに導入されることが好ましい。植物プロモーター調節エレメントに加えて、様々な由来源から得たプロモーター調節エレメントも、植物細胞において外来遺伝子を発現するために有効に使用できる。例えば、オクトピンシンターゼプロモーター、ノパリンシンターゼプロモーター、およびマンノピン(mannopine)シンターゼプロモーター等の細菌由来のプロモーター調節エレメントを使用してもよい。カリフラワーモザイクウイルス(35Sおよび19S)等のウイルス由来のプロモーターも望ましい。植物プロモーター調節エレメントとしては、リブロース-1,6-ビスホスフェートカルボキシラーゼ小サブユニットプロモーター、β-コングリシニン(conglycinin)プロモーター、ファゼオリン(phaseolin)プロモーター、ADHプロモーター、熱ショックプロモーターおよび組織特異的プロモーター等も挙げられるがこれらに限定されない。多数のプロモーターが、入手可能であり、当業者の裁量
下で使用できる。
【0173】
全ての発現ベクターおよび発現系が、本発明の変異型遺伝子配列を発現するために同様に機能するわけではないことが理解されるべきである。宿主細胞も全てが同じ発現系に対して同等に上手く機能するわけではない。しかし、当業者は、これらのベクター、発現制御配列、および宿主を、過度の実験を要することなく、また本発明の範囲から逸脱することなく選択できるであろう。
【0174】
本発明のシンターゼを発現させた後は、それから得たタンパク質を精製して、本明細書に例示するように、構造分析、モデリング、および/または生化学的分析を行うことができる。得られるタンパク質の性質は、使用する発現系に依存しうる。例えば、遺伝子は、哺乳動物またはその他の真核生物細胞で発現された場合には、機能を変化するかもしれないしまたは変化しないかもしれないグリコシル化、リン酸化、またはその他の翻訳後修飾を生じうる潜在的シグナル配列を含み得る。タンパク質は、発現後、タンパク質生化学分野の当業者には常識的な技法を使用して、またはColliganら(1997), Current Protocols in Protein Science, Chanda, V.B., Ed., John Wiley & Sons, Inc.に記載されているように、容易に単離および精製できる。このような技法には、陽イオン交換または陰イオン交換クロマトグラフィー、ゲル濾過クロマトグラフィー等の使用が含まれることが多い。一般的に使用されうる別の技法は、アフィニティークロマトグラフィーである。アフィニティークロマトグラフィーは、抗体、基質類似体またはヒス
チジン残基(His-タグ技法)の使用を含みうる。
【0175】
本発明の変異体は、精製後、いくつかの異なる特性のいずれかによって特徴決定してもよい。例えば、このような変異体では、1つ以上の電荷ユニットの活性部位表面電荷が変化しているかもしれない。さらに、変異体は、非変異型シンターゼと比べて、基質特異性または反応産物のスペクトルが変化しているかもしれない。
【0176】
本発明は、結晶化およびその後のX線回折により変異型テルペンシンターゼの特徴決定を可能にする。ポリペプチド結晶化は、ポリペプチド濃度がその最大溶解度を超える溶液において(すなわち、ポリペプチド溶液が過飽和の状態で)生じる。このような溶液は、好ましくはポリペプチド結晶の沈殿によりポリペプチド濃度を低くすることによって、平衡状態に戻すことができる。ポリペプチド表面電荷を変化させるか、ポリペプチドとバルク水との相互作用を摂動させて結晶化をもたらす会合を促進する作用物質を添加することにより、過飽和溶液から、ポリペプチドを誘導して結晶化させることが多い。
【0177】
「沈殿剤」として知られる化合物は、ポリペプチド分子の周りにエネルギー学的に好ましくない沈殿する減損層(depleted layer)を形成することにより、濃縮溶液中のポリペプチドの溶解度を低くするためにしばしば用いられる(Weber, 1991, Advances in Protein Chemistry, 41:1-36)。沈殿剤に加えて、他の物質をポリペプチド結晶化溶液に添加する場合もある。これらには、溶液のpHを調節するための緩衝剤、およびポリペプチドの溶解度を下げるための塩が挙げられる。様々な沈殿剤が当該分野において公知であり、以下のものが挙げられる。すなわち、エタノール、3-エチル-2-4-ペンタンジオール、ならびにポリエチレングリコールなどの多くのポリグリコールである。
【0178】
よく使用されるポリペプチド結晶化方法としては、以下の技法が挙げられる。すなわち、バッチ法、懸滴法、結晶種開始法(seed initiation)および透析法である。これらの各方法では、過飽和溶液を維持することにより、核形成後の持続的な結晶化を促進することが重要である。バッチ法では、ポリペプチドを沈殿剤と混合して過飽和状態を得、容器を密封し、結晶が現れるまで置いておく。透析法では、ポリペプチドを密封した透析膜内に保持し、沈殿剤を含有する溶液に入れる。膜を介した平衡により、ポリペプチドおよび沈殿剤の濃度が高くなり、ポリペプチドを過飽和レベルに到達させる。
【0179】
好ましい懸滴技法(McPherson, 1976, J.Biol.Chem., 6300-6306)では、沈殿剤を濃縮ポリペプチド溶液に添加して、初期ポリペプチド混合液を作る。ポリペプチドおよび沈殿剤の濃度は、この初期状態においてポリペプチドが結晶化しない程度のものである。この混合液の小滴を、スライドガラスに滴下し、第2の溶液の貯蔵器上で反転させ、懸濁させる。次に、この系を密封する。典型的には、第2の溶液は、より高い濃度の沈殿剤またはその他の脱水剤を含む。沈殿剤濃度の差により、溶液よりも高い蒸気圧を有するタンパク質溶液が生じる。2つの溶液を含む系は密封されているため、平衡が確立され、ポリペプチド混合液由来の水は第2の溶液に移る。この平衡により、ポリペプチド溶液中のポリペプチドおよび沈殿剤の濃度が高くなる。ポリペプチドおよび沈殿剤の臨界濃度において、ポリペプチドの結晶が形成されうる。
【0180】
別の結晶化方法では、核形成部位を濃縮ポリペプチド溶液に導入する。通常、濃縮ポリペプチド溶液を用意し、ポリペプチドの結晶種をこの溶液に導入する。ポリペプチドおよび任意の沈殿剤の濃度が正しければ、結晶種は核形成部位を提供し、その周りにより大きい結晶が形成される。好ましい実施形態では、本発明の結晶は、沈殿剤として15%PEG 8000;200mM酢酸マグネシウムまたは塩化マグネシウム、100mM 3-(N-モルホリノ)-2-ヒドロキシプロパンスルホン酸(pH 7.0)、1mMジチオスレイトールを用いた懸滴で形成される。
【0181】
一部のタンパク質は、結晶化が困難であるかもしれない。しかし、結晶化を誘導するためにいくつかの技法を当業者は利用できる。タンパク質のアミノ末端またはカルボキシル末端におけるポリペプチドセグメントの除去により、結晶タンパク質サンプルの生成が容易になる。このようなセグメントの除去は、トリプシン、キモトリプシン、サブチリシン等のプロテアーゼを用いた、タンパク質の分子生物学的技法または処理を使用して行うことができる。このような処置は、結晶化に対して負に影響を及ぼしうるフレキシブルなポリペプチドセグメントを除去しうる。
【0182】
このように形成された結晶の用途は広範囲にわたる。例えば、高品質結晶は、変異型シンターゼの三次元構造を決定し、さらに別の変異体を設計するためのX線または中性子回折分析に適している。さらに、結晶化は、さらなる精製方法として作用しうる。いくつかの例では、ポリペプチドまたはタンパク質は、不均質混合物から結晶に結晶化する。濾過、遠心分離などによりこのような結晶を単離し、その後ポリペプチドを再溶解することにより、回折研究に必要な高品質結晶を成長させるのに適した精製溶液が得られる。この高品質結晶を、水に溶解した後に、処方して、所望に応じた他の使用用途を有する水性溶液を得ても良い。
【0183】
シンターゼは、1つ以上の結晶形態で結晶化しうるため、本発明により得られる、シンターゼまたはその一部から決定した活性部位のα-炭素の構造座標は、シンターゼの他の結晶形態の構造を解読するのに特に有用である。本明細書で得られる構造座標は、同一であるかまたは同一でないR基を有する野生型と同様に、活性部位内にα-炭素位置を有するシンターゼの構造を解読するためにも使用できる。さらに、本明細書に開示する構造座標を使用して、シンターゼの任意の機能ドメインに対して有意なアミノ酸相同性または構造的相同性を有する他のタンパク質の結晶形態の構造を決定できる。このような目的のために採用できる方法の1つとして分子置換(molecular replacement)がある。この方法では、未知の結晶構造は、それがシンターゼの別の結晶形態であるのか、変異活性部位を有するシンターゼなのか、またはシンターゼと有意な配列同一性および/もしくは構造相同性を有するなんらかの他のタンパク質の結晶なのかを、表10および/または表11に示す座標を用いて決定できる。この方法は、未知の結晶についての十分な構造形態を、最初からこのような情報を決定しようとする場合よりも効率的に提供する。さらに、本方法を用いて、問題となる所与のシンターゼが本発明の範囲に含まれるか否かを決定できる。
【0184】
本明細書にさらに開示するように、シンターゼおよびその変異体は、基質および基質類似体の存在下または不在下において結晶化できる。その後、一連の複合体の結晶構造を、分子置換により解読し、野生型のものと比較して、活性部位内のR基の適切な置換を決定するのを助けて、本発明のシンターゼ変異体を作成する。
【0185】
本発明の変異体は全て、必ずしも全ての変異体についてそれぞれ結晶化して構造を解読する必要なく、本明細書に開示する情報を用いてモデリングできる。例えば、当業者は、いくつかの特殊化コンピュータプログラムの1つを使用して、変異型活性部位を有するシンターゼの設計プロセスを援助しうる。このようなプログラムの例として以下のものが挙げられる。すなわち、GRID(Goodford, 1985, J.Med.Chem., 28:849-857);MCSS(MirankerおよびKarplus, 1991, Proteins: Structure, Function and Genetics, 11:29-34);AUTODOCK(GoodsellおよびOlsen, 1990, Proteins: Structure, Function, and Genetics, 8:195-202);ならびにDOCK(Kuntzら, 1982, J.Mol.Biol., 161:269-288)である。さらに、特異的基質-活性部位相互作用、ならびにそれにより生じる変形エネルギーおよび静電相互作用を評価するための特殊なコンピュータプログラムも入手可能である。MODELLERは、タンパク質の三次元構造の相同性または比較モデリングについてよく使用されるコンピュータプログラムである。A. SaliおよびT.L.Blundell. J.Mol.Biol. 234, 779-815, 1993。モデリングされるべき予め選択されたポリペプチド配列を、結晶構造が既知である1つ以上のテルペンシンターゼとアライメントし、MODELLERプログラムを使用して空間的制約の最適条件(satisfaction)に基づいて完全原子モデルを計算する。このような制約には、とりわけ、相同的構造、蛍光分光学、NMR実験、または平均力の原子間ポテンシャル(atom-atom potentia
l of mean force)が含まれる。
【0186】
本発明は、シンターゼ変異体を作成し、その結晶構造を解読することを可能にする。さらに、本発明により、活性部位の位置、およびそれと基質との界面から、突然変異誘発のために望ましいR基の同定が可能となる。本発明の特定の実施形態は、実施例にさらに例示されている。しかし、当業者は、記載する特定の実験があくまで本発明の例示であり、本発明が請求の範囲にさらに完全に記載されていることを容易に理解するであろう。
【実施例】
【0187】
実施例1
変異型TEAS遺伝子の生成
構築物の生成および発現
全ての変異型酵素を、QuickChange方法(Stratagene)により構築した。記載したもの以外は製造元の説明書に従った。突然変異は、DNA配列決定により確認し、所望の突然変異を含むプラスミドを使用してBL-21(DE3)発現細胞を形質転換した。タンパク質を発現させ、精製し、-80℃にて保存した。
【0188】
TEAS W273S.以下のプライマーを用いて、TEAS-pET28b(+)鋳型からTEAS W273S変異体を生成した:GTTGAATGCTACTTTTCGGCATTAGGAGTTTAT(センス)(配列番号13)およびATAAACTCCTAATGCCGAAAAGTAGCATTCAAC(アンチセンス)(配列番号14)。センス鎖およびアンチセンス鎖を別々の反応で生成したこと以外は、製造元の説明書に従って突然変異誘発を行った。それぞれについて、30サイクルの1分間のプラスミドコピー、55℃でのアニーリング、および68℃での16分間の伸長を行った。二つの反応混合液を合わせ、95℃まで2.5分間加熱し、室温まで冷却してから、DpnI処理に供した。
【0189】
TEAS C440W.以下のプライマーを用いて、TEAS-pET28b(+)鋳型からTEAS C440W変異体を生成した:GCTAGTGTAATTATATGGCGAGTTATCGATGAC(センス)(配列番号15)およびGTCATCGATAACTCGCCATATAATTACACTAGC(アンチセンス)(配列番号16)。
【0190】
TEAS W273S/C440W.TEAS C440Wの生成のために記載したプライマーを用いて、TEAS W273S-pET28b(+)鋳型からTEAS C440W/W273S変異体を構築した。
【0191】
TEAS 406/407ランダムライブラリー.406位および407位にランダムなアミノ酸を有するTEAS変異体のライブラリーを生成するために、二つの50マイクロリットルQuickChange反応を、TEAS-pET28b(+)鋳型および以下のプライマーを用いて行った:GCACTAGCAACTACCACATATTACNNSNNSGCGACAACATCGTATTTGGGCATG(センス)(配列番号17)、およびCATGCCCAAATACGATGTTGTCGCSNNSNNGTAATATGTGGTAGTTGCTAGTGC(アンチセンス)(配列番号18)。ここで、NはA、C、GまたはTを指し、SはCまたはGを指す。このヌクレオチド選択により、この反応は、406位および407位において可能性のあるアミノ酸の組合せの全てをコードするプライマーを含んでいた。アミノ酸間でコドンの数を変えるための調節は行わなかった。効率的な反応を確実にするために、かつ鋳型に対する野生型プライマーのハイブリダイゼーションの優先性を最小限にするために、プライマーを、上述した突然変異を生じるために使用されるプライマーよりも長くなるように設計した。さらに、それらを使用前にHPLC精製した。18サイクルのプラスミドコピーの後、反応液を2時間DpnIと共にインキュベートし、エタノール沈殿させ、5マイクロリットルの水に再溶解させた。大腸菌NovaBlue(Novagen)細胞の4つの40マイクロリットルアリコートのそれぞれに、1.5マイクロリットルの再溶解DNAをエレクトロポレートした。回収時間の後、細胞を、カナマイシン-LB-寒天プレート上にプレートした。
新たに構築したプラスミドを発現細胞に移すために、4つのプレート全てからコロニーを掻き取り、液体LB培地中で37℃にて8時間増殖させる8mL培養を開始するのに使用した。この培養物から精製されたプラスミドを、20マイクロリットルのコンピテントBL-21(DE3)細胞を形質転換するのに使用した。
【0192】
構築物の保存のために、個々のコロニーを用いて、96-ウェル培養プレート中のカナマイシン(50マイクログラム/mL)を含有する100マイクロリットルのLB培地に播種した。A600が約1に達するまで細胞を37℃で増殖させ、100マイクロリットルの30%グリセロールのLB溶液を添加し、プレートを-80℃にて冷凍した。ランダムに選択したコロニーを、いくつかのコロニーの個々のグリセロールストックから増殖させ、プラスミドを配列決定のために抽出した。約30パーセントのコロニーが野生型であることが分かった。TEAS 406/407変異型遺伝子およびタンパク質のヌクレオチドおよびアミノ酸配列は配列番号11および12に示す。
【0193】
TEAS Y520F.配列番号2の520位にあるチロシン残基を、上述したものと同様に、プライマーを用いた部位特異的突然変異誘発によりフェニルアラニン残基に変えた。Y520Fのために、TATコドンをTTCに変えた。変異型遺伝子のヌクレオチド配列を配列番号5に示す。
【0194】
TEAS Y527F.配列番号2の527位にあるチロシン残基を、上述したものと同様に、プライマーを用いた部位特異的突然変異誘発によりフェニルアラニン残基に変えた。Y527Fのために、TEASアミノ酸配列の527位にあるTACコドンをTTCに変えた。変異型TEAS Y527F遺伝子のヌクレオチド配列を配列番号7に示す。
【0195】
TEAS W273E.配列番号2の273位にあるトリプトファン残基を、上述したものと同様に、プライマーを用いた部位特異的突然変異誘発によりフェニルアラニン残基に変えた。W273Eのために、TEASアミノ酸配列の273位にあるTGGコドンをGAGに変えた。変異型遺伝子のヌクレオチド配列を配列番号3に示す。
【0196】
実施例2
シンターゼポリペプチドの発現および単離
特に記載しない限り、変異型および非変異型TEASタンパク質は、大腸菌において発現させ、金属キレート化、陰イオン交換およびゲル濾過クロマトグラフィーにより精製した。
【0197】
ベクターpET-28b(+)(Novagen)中のTEASおよび変異型TEASタンパク質の構築物を、大腸菌細胞で発現させた。これらの酵素の典型的なタンパク質調製のために、プラスミド構築物を含む大腸菌株BL21(DE3)細胞を、4×1L TB培地において37℃にてA600=1.0まで増殖した。温度を22℃にまで下げ、IPTGを最終濃度が0.1mMになるまで添加して、タンパク質発現を誘導した。15〜20時間後、遠心分離により細胞を回収し、1gの細胞(湿重量)当たり5mLの緩衝液A(20 mM Tris、500 mM NaCl、20mMイミダゾール、pH7.9)に再懸濁し、4℃にて0.5時間攪拌した。次いで、超音波により細胞を溶解し、得られた溶解物を82,000×gにて0.7時間遠心分離した。タンパク質を含む上清を、緩衝液A中で平衡させた2〜3mL Ni2+キレート化ヒスチジンアフィニティーカラム(Qiagen)にかけ、溶出液のA280が基線に戻るまでカラムを追加の緩衝液Aで洗浄した。次いで、タンパク質を20〜200mMイミダゾール勾配の緩衝液Aで溶出した。タンパク質含有画分をプールし、緩衝液B(50mM HEPES、5 mM MgCl2、1mM DTT)に対して透析し、8mL MonoQ陽イオン交換カラム(Pharmacia)にかけた。カラムを20カラム容量の緩衝液Bで洗浄し、0〜500mM NaCl勾配の緩衝液Bでタンパク質を溶出した。得られたタンパク質を、50 mM Tris、100 mM NaCl、5 mM MgCl2、1 mM DTT、pH8.0中で、Superdex-200カラム(Pharmacia)上でのゲル濾過によりさらに精製した。次いで、精製タンパク質を、5 mM Tris、5 mM NaCl、1 mM DTT、pH8.0に対して透析し、18〜22 mg/mLまで濃縮し、必要になるまで100μLアリコートで-80℃にて保存した。
実施例3
シンターゼポリペプチドの結晶化および構造解析
結晶成長およびマイクロ播種(microseeding):全ての結晶化の試みを、懸滴気化拡散(hanging-drop vapor diffusion)方法により行った。濃縮タンパク質を、プラスチックカバースリップ上で、等容量(それぞれ2〜5μL)の貯蔵溶液と混合した。次に、0.5〜1.0 mLの貯蔵溶液を含むプラスチックの24ウェル組織培養プレートのウェル上でカバースリップを反転させ、ウェルとカバースリップとの間を真空グリースの層で密封した。プレートを4℃でインキュベートし、その間、気化拡散により懸滴中のタンパク質濃度を徐々に高めた。様々な沈殿剤および添加塩を含むpHが4.5〜9の範囲の約300の異なる貯蔵溶液を、TEAS(配列番号12)の結晶化についてアッセイした。TEASは、15%PEG 8000、100 mM MOPSO(3-[N-モルホリノ]-2-ヒドロキシプロパンスルホン酸)、200 mM酢酸マグネシウム、1mM DTT、pH6.9〜7.3)の貯蔵溶液で結晶化した。マイクロ播種のために、既存の結晶を数μLの沈殿剤溶液中で破砕し、50マイクロリットルにまで希釈した。大きい粒子を除去するための初期遠心分離の後、懸濁液を追加の沈殿剤溶液で連続希釈し、少容量の希釈種ストックをそれぞれの新しい結晶化小滴に添加した。マクロ播種(macroseeding)のために、既に早い速度で成長していない結晶(通常、小滴をセットしてから2週間後)を、2〜3滴の貯蔵溶液に連続的に通すことにより「濯いだ」。次に、結晶を、タンパク質および貯蔵溶液を含む新しい小滴に移し、初期成長時のように貯蔵溶液に対して平衡させた。個々の結晶は、内部順序(internal order)の程度が異なった。場合によっては、いくつかの結晶をスクリーニングして、低いモザイク度(mosaicity)でよく回折した結晶を同定した。
【0198】
データ回収:データを回収する前に、結晶を、結晶に浸透させる化合物を含む貯蔵溶液の小滴、または貯蔵溶液に移した。次いで、少容量の凍結保護物質溶液(15% PEG8000、100 mM MOPSO、200 mM 酢酸マグネシウム、20%エチレングリコール、1mM DTT、pH 7)を小滴に添加した。短い平衡時間(1〜5分)の後、結晶を、浸透させる化合物を添加した凍結保護物質の小滴、または凍結保護物質に移した。さらに追加の短い平衡時間の後、結晶をナイロンループ上に取り上げ、冷窒素ガス(90〜110K)の流れの中でデータ回収を速やかに開始した。
【0199】
TEAS結晶は、正方晶空間グループP41212(単位セル次元(unit cell dimensions
)は結晶により数オングストローム変動するが、平均してa=126Å、c=122Å
である)に属していた。非複合化TEAS構造を、2 mM FHPの存在下で成長させた結晶から回収したデータ(表10)に対して、初め2.8Åにまで精製した(表11)。活性部位における電子密度により、FHP、A-CおよびJ-Kループ、ならびにNH2末端にある9つの追加の残基の明確なモデリングが可能であった。精製したTEAS-FHPモデルは、残基17〜548、3つのMg2+イオン、150の水分子、ならびに1つのFHP分子から構成されていた。結合基質の存在下におけるTEASの三次元座標を表10に示す。FHPの不在下でのTEASの三次元座標を表11に示す。
【0200】
トリフルオロ-ファルネシルジホスフェート(F3-FPP)により複合化されたTEASの結晶も調製した。これらの結晶において、上手く秩序をなしたジホスフェート結合ポケットも観察した。A-CループおよびNH2末端セグメントは、はっきりとした電子密度を示した。A-Cループを活性部位に向かって中継した(translate)ところ、F3-FPPのジホスフェート部分について強力な電子密度が存在した。しかし、疎水性ポケットは、フレキシブルなままであった。J-KループおよびF3-FPPのファルネシル部分は無秩序であった。
【0201】
SwissModelプログラム(Peitsch MC(1996), Biochem. Soc. Trans., 24:274-279、ならびにGuex N.およびPeitsch MC, 1997, Electrophoresis., 18:2714-2723)のSwiss PDBビューアーインターフェースを用いて、相同性モデルを作成し、エネルギーを最小化した。VOIDOO(Kleywegt, G.J.およびJones, T.A., CCP4/ESF-EACBM Newsletter on Protein Crystallography., 29, 26-28, 1993)により活性部位容量を計算した。閉鎖した活性部位空洞を作るために、モデル化されたTEASシクラーゼ反応から得た、エネルギーを最小化されたジホスフェート部分を、TEAS D301に等価な残基に追加した。
【0202】
TEAS W273S結晶構造.FHPの存在下で2つのTEAS W273S構造を、異なる結晶から決定した。両方の結晶とも、タンパク質全体の残基(頻繁に動きやすいヘリックスD1、D2およびEを含む)についてはっきりとした主鎖および側鎖密度が明確であることから、上手く秩序をなしているように思われた。両方の結晶から得た、初期差電子密度地図から、W273S突然変異がすぐに分かった。2つの結晶をW273S-1およびW273S-2と命名した。
【0203】
各構造において、活性部位を囲むループを秩序付け、閉鎖活性部位ポケットを得た。各構造におけるA/Cループを活性部位に向かって中継し、野生型TEAS/FHP複合体で見られるような外枠の一部を形成した。しかし、W273S-1のJ/Kループが、野生型TEAS/FHP複合体で観察されるものと同じコンホメーションをとるのに対して、W273S-2中の同じループは異なるコンホメーションをとっていた。J/Kループのこのコンホメーションでは、Tyr527が、残基273の側鎖から離れていた。さらに、Tyr520およびAsp525は、Asp444の側鎖と離れて位置していた。J/Kループ、Arg266、およびN末端ループの間で先に観察された水素結合も、W273S-2構造において欠けていた。
【0204】
W273S-2コンホメーションは、W273S突然変異の結果とは思われなかった。なぜなら、これは、エピ-アリストロケン擬態デオキシカプシジオール(epi-aristolochene mimic deoxycapsidiol)が浸透した野生型TEAS結晶においても、この構造におけるデオキシカプシジオールについて電子密度が容易に明確でなかったという事実にも関わらず、観察されたからである。さらに、TEAS活性部位ループは、結晶接触から離れており、それらのコンホメーションは、結晶パッケージングの人為結果であるとは思われなかった。TEAS反応の種々の段階において、酵素のJ/Kループが異なる明確なコンホメーション中に存在し、これらの結晶構造のそれぞれが異なるコンホメーション像を獲得することが可能である。両方のW273S構造において、Arg266以外の残基、およびJ/Kループ上の残基は、野生型TEASで見られるコンホメーションから有意な再配列を経ていなかった。
【0205】
各W273S結晶構造において、活性部位における電子密度から、基質擬態FHPが複数のコンホメーションで結合することが示唆された。この密度のいくつかの領域は、変異型活性部位において結合した水分子を表している可能性がある。変異型活性部位における水分子の存在は、TEAS W273Sが複数の水酸化テルペノイド反応産物を生じるという観察と一致する。
【0206】
各W273結晶構造におけるFHP電子密度は、FHPが、野生型TEAS/FHP複合体におけるより緊密に折り畳まれたFHPコンホメーションと比べて、W273S構造においてより広がったコンホメーションで存在することを示唆するのに十分であった。W273Sの活性部位が、FHPの複数のコンホメーションと結合するという観察は、W273SがFPPを複数のテルペノイド炭化水素産物に変換するという事実と一致する。
【0207】
TEAS C440W/W273S:TEAS C440W/W273Sを、野生型TEASと同じ条件下で結晶化した。0.3 mmの結晶を、ファルネシルヒドロキシホスフェート(FHP)で飽和させた貯蔵溶液に20分間浸せきした。野生型TEASについて記載した凍結保護およびフラッシュ冷凍(flash freezing)の後、Cu-Kα放射(MacScience Corp., 日本)を用いて実験ソースからデータを回収した。非複合化TEASの出発モデル(表11)(Brookhaven Protein Database Code 5EAT(PDB 5EAT))を、水およびマグネシウムを除去し、ソフトウェアプログラムX-PLOR(A.T.Brunge, X-PLOR Version 3.1-A System for X-Ray Crystallography and NMR Yale University Press, New Haven, 1992, pp.187-207)の剛体モジュールを有する変異体データに対して配置した。バルク溶剤モデリングでの配置的および制約的b因子精製の繰返しもX-PLORで行った。この際、ソフトウェアプログラムO(Jones, TA, Zou, JY, Cowan, SW、およびKjeldgaard, M., Acta Cryst. D., 49:148-157, 1993)で手動モデル成形および調節を行った。CNSプログラムを使用したさらなる精製および地図計算の繰返しにより、有意に改善された地図を得た。この改善はバルク溶剤モデリングが改善されたためだと思われた。
【0208】
TEAS C440W:結晶の核形成がより遅く、全体的により小さかったこと以外は、野生型TEASと同じ条件下でTEAS C440Wを結晶化した。変異型結晶を、フラッシュ冷凍、およびSSRLビーム線7-1(Stanford Synchrotoon Radiation Laboratory, Menlo Park, California)でのデータ回収の前に、FHPに飽和させた貯蔵溶液中に6時間浸せきした。TEAS-FHPの出発モデル(表10)は、水分子、リガンド、および配列番号2の残基523-532を除去され、X-PLORの剛体モジュールを有するデータに対して配置した。バルク溶剤での配置的および制約的b因子精製、ならびに全体的異方性温度因子モデリングの繰返しもX-PLORで行い、ソフトウェアプログラムOで手動モデル成形および調節を行った。二重変異体の場合、電子密度地図は、CNSでの精製および地図計算の後では有意に改善されていた。
【0209】
実施例4
テルペンシンターゼ酵素アッセイ
VogeliおよびChappell, Plant Physiol.94: 1860(1990)ならびにVogeliら, Plant Physiol. 93:182(1990)に記載されたアッセイに基づき、シンターゼ活性のアッセイを行った。一般に、放射性標識(3Hまたは14C)基質を、マグネシウム塩の緩衝化溶液(特に他のものが記載されない限り200mM Tris, pH8, 50mM塩化マグネシウム、1mM DTT)中で室温にて酵素と共にインキュベートした後、炭化水素生成物をヘキサン等の有機溶媒中に選択的に抽出した。このヘキサン抽出物を一般にシリカゲルで処理して、基質の非酵素的加水分解により生成されたプレニルアルコールおよび他の含酸素化合物(これらはヘキサン中に非効率的に分配される(partition))を除去した。このヘキサン相の中に存在する炭化水素生成物をシンチレーションカウンターにより定量した。
【0210】
次により極性の高い有機溶媒(酢酸エチル等)により時折抽出を行った。含酸素化合物は酢酸エチルタイプの溶媒中により効率的に分配される。この酢酸エチル相中に存在する化合物もシンチレーションカウンターにより定量した。
【0211】
典型的な基質濃度の範囲は0.1ナノモル〜100μモルとした。幾つかのアッセイでは、基質の放射性標識を行わなかった。反応は一般に各基質濃度毎に3回行った。タンパク質濃度は、Bradford法により測定した。静止状態の動力学パラメータを測定するために、酵素濃度は、反応過程を通じて経時的生成物生成量が直線となるように選択した。
【0212】
ジテルペンシンターゼのアッセイは、典型的には、3Hゲラニルゲラニルジホスフェート(GGPP)および酵素を含む250mM Tris, 10mM 塩化マグネシウム, 1mM DTT, pH8.0を用いて行った。セスキテルペンシンターゼアッセイは、典型的には、14Cまたは 3H FPPおよび酵素を含む100mM Tris, 30mM 塩化マグネシウム, 1mM
DTT, pH8.0を用いて行った。モノテルペンシンターゼアッセイは、典型的には
、3H GPPおよび酵素を用いて行った。タンパク質によるGPPの非特異的結合についての対照として、酵素ではなくBSAを用いて全く同じ反応を設定した。
【0213】
Ag-TLCによる野生型および突然変異型のTEAS酵素の生成物分析:テルペノイド炭化水素生成物は、通常のプレートもしくは逆相プレート上での薄層クロマトグラフィーによっては容易に分離されないが、はじめにシリカプレートを硝酸銀で処理する銀染色TLC(Ag-TLC)によってある程度は分離することができる。本明細書中に記載されるAg-TLCは、Backら, Arch. Biochem. Biophys. 315: 527(1994)によって記載された手法に概ね従って行った。シリカTLCプレートを15%硝酸銀(水溶液)に浸した後、110℃で3〜5時間乾燥した。トリチウム酵素生成物(溶媒抽出物)をスポッティングした後、プレートをベンゼン:ヘキサン:酢酸エチル(50:50:1体積比)中で発色させ、En3Hance(NEN)蛍光間接撮影用スプレーを噴きつけ、フィルム上に載せて、数日〜数週間の間露光した。TLCプレートの硝酸銀処理により蛍光間接撮影用試薬の蛍光が抑制されるようであったので、一般に長時間露光する必要があった。或いは、蛍光間接撮影用スプレーを使用せずに14Cで標識した生成物を1〜2日後に検出した。
【0214】
実施例5
TEAS W273Sの活性
TEAS W273Sのジテルペンシンターゼ活性:TEAS W273S酵素および放射性標識GGPPを上記のようにインキュベートし、ヘキサンで炭化水素生成物を抽出した。次に含酸素生成物を酢酸エチルで抽出した。野生型TEASを用いた反応で定量した結果は、緩衝液のみを用いた反応のものよりも低かった。一方、TEAS W273Sの定量結果は、ヘキサン抽出および酢酸エチル抽出の両方において非常に高かった。W273SによりGGPPから形成された炭化水素生成物は、酸により触媒されたGGPPからのジホスフェートの減少によって生成された生成物とは異なるものであった。図3を参照されたい。
【0215】
TEAS W273Sのセスキテルペンシンターゼ活性:精製されたTEAS W273S突然変異体によるFPP代謝回転の生成物を、銀染色薄層クロマトグラフィー(Ag-TLC)によって分析した。主な反応生成物の1つはAg-TLCによるRfが0.7であり、これは、5-エピ-アリストロケン(Rf=0.78)およびベティスピラジエン(vetispiradiene)(Rf=0.63)のいずれとも異なるものであった。図4を参照されたい。予備GC/MSデータは、TEAS W273SによるFPP代謝回転からのヘキサン抽出物が少なくとも4つのテルペン炭化水素を含み、質量スペクトルが5-エピ-アリストロケンまたはベティスピラジエンと異なることを示した。これらの生成物のうちの1つは、ゲルマクレン(germacrene) Aと似た質量スペクトルを有していた。
【0216】
実施例6
TEAS C440W/W273Sの活性
TEAS C440W/W273Sのジテルペンシンターゼ活性:突然変異型TEAS C440W/W273S
タンパク質は、440位にトリプトファン残基および273位にセリン残基を含む。0.5μモル3H GGPP, 様々な濃度の非標識GGPP(Echelon)および酵素を用い、GGPPを用いてアッセイを行った。反応物を室温にて60分間インキュベートした。TEAS C440/W273S突然変異型タンパク質は、GGPPをヘキサン-抽出可能生成物に変換したが、野生型酵素はこの変換を行わなかった。これらの結果は、その生成物のプロフィールが野生型TEASに比べて変化したことを示した。二重突然変異体によるGGPP代謝回転のヘキサン抽出可能生成物を、Ag-TLCにより分析した。この生成物は、加水分解生成物であるゲラニルゲラニオール(Rf=0.0)とは異なる2種(Rf=0.11および0.28)を含んでいた。TEAS C440W/W273SによりGGPPから生成された生成物が加水分解生成物ゲラニルゲラニオールではないことを証明するために、サンプルをAg-TLCにより分析した。100μlの緩衝液中に3H GGPP(5μm)および酵素(40μm)を含む反応を、室温にて一晩インキュベートした。対照として、3H GGPPを反応緩衝液のみの中、およびpH1.5に調節した反応緩衝液の中で、インキュベートした。酵素的反応物および対照反応物を両方ともヘキサンで抽出し、銀染色TLCプレート上にスポッティングし、上記のように現像および露光を行った。図3に示す結果は、TEAS C440W/W273Sにより形成された生成物が、ゲラニルゲラニルジホスフェートの非酵素的分解により生成されたものと異なっていたことを示すものであった。
【0217】
TEAS C440W/W273Sのセスキテルペンシンターゼ活性
14C FPP(9μm)および酵素(160μm)を含む反応緩衝液(20μl)を用い、基
質としてFPPを用いて反応を行った。室温にて30分間インキュベートした後、TEAS C440W/W273Sによって作製された生成物をAg-TLCにより分析した。この二重突然変異体の生成物のプロフィールは、TEAS W273Sのものと似ており、それに更にRfが0.57である主な生成物が加わった。この新しい生成物は、5-エピ-アリストロケンおよびベティスピラジエンの何れとも異なるものであった。他の幾つかの生成物も形成され、そのうちの多くは銀染色TLCの際にゆっくり移動した。図4を参照されたい。
【0218】
実施例7
TEAS C440Wの活性
TEAS C440Wのジテルペンシンターゼ活性:TEAS C440Wを用いた酵素アッセイを、実施例6に記載したように実施した。図3に示すように、160μmの酵素および9μmの放射性標識GGPP(体積20μl)と共に室温にて一晩インキュベートした後、Ag-TLCによってヘキサン-抽出可能生成物は検出されなかった。
【0219】
TEAS C440Wのセスキテルペンシンターゼ活性:精製したTEAS C440Wによる放射性標識から作製された生成物のAg-TLC分析により、少なくとも1つの主なテルペノイド炭化水素生成物(Rf=0.63)の形成が検出され、これは5-エピ-アリストロケン(Rf=0.78)およびベティスピラジエンとは異なるものであった。Ag-TLCでの反応生成物のプロフィールを図4に示す。少量の低速移動生成物(Rf=0〜0.09)も形成された。
【0220】
TEAS C440Wテルペノイド炭化水素反応生成物のヘキサン抽出物のGC/MS分析により、この突然変異体が、単一の主なセスキテルペン炭化水素生成物および少数のマイナーなヒドロキシル化生成物を形成したことが確認された。この主な生成物の質量スペクトルは、スピロ環状化合物ヒネセン(hinesene)の公表されている質量スペクトルに非常に近かった。ヒネセンは、C3メチル基の立体化学構造がベティスピラジエンと異なる。
【0221】
実施例8
TEAS W273Eの活性
TEAS W273Eのセスキテルペンシンターゼ活性:基質としてFPPを用いてTEAS W273Eにより作製された生成物を測定する反応を、放射性標識したFPPを用いて本質的には上記のように行った。これらの結果は、5-エピ-アリストロケン以外に少なくとも1つの生成物が形成されたことを示した。また該結果は、FPPによるTEASのアルキル化が起こったことも示した。このアルキル化は、反応混合物中のMgCl2の存在に依存するものであった。対照実験において、煮沸したW273E-TEAS、ならびに野生型TEASおよびBSAは、アルキル化されなかった。これらの結果は、アルキル化が273位で起こったこと、および273位のアミノ酸残基はこの活性部位の一部であることを示す。
【0222】
実施例9
TEAS Y520Fの活性
TEAS Y520Fのセスキテルペンシンターゼ活性:放射性標識したFPPおよびTEAS Y520F酵素を用いた反応を、本質的には上記のように行った。反応生成物をAg-TLCおよびGC/MSにより分析した。TEAS Y520F反応の主な生成物は、基準ゲルマクレンA(authentic germacrene A)と同じGC保持時間および質量スペクトルを有していた。この生成物のGC保持時間および質量スペクトルは、5-エピ-アリストロケンとは異なっていた。
【0223】
実施例10
TEAS Y527Fの活性
TEAS Y527Fの酵素活性:大腸菌細胞中における発現を誘導し、該細胞を音波で破砕して、TEAS Y527F酵素の粗抽出物を作成した。この音波破砕物を透明化し、上清を酵素アッセイに使用した。基質としてGPPを用いたアッセイでは、生成物は観察されなかった。このことは、TEAS Y527Fが、モノテルペンシンターゼ活性を持たないことを示す。基質としてFPPを用いて反応生成物を得た。これらの生成物のAg-TLCによる分析は、5-エピ-アリストロケン以外の生成物がTEAS Y527F酵素により生成されたことを示した。
【0224】
実施例11
テルペンシンターゼ配列のアラインメント
BLOSUM62スコアリングマトリックス、ギャップ開始値=11、ギャップ延長値=1、X_ドロップオフ値=50、期待値=10、文字列の長さ=3を用い、およびコンプレキシティーの低い配列のフィルタリングを行わずに、BLASTpプログラム(NCBI)を用いて、TEAS一次アミノ酸配列(配列番号2)の残基265〜535を、リモネンシンターゼの全長アミノ酸配列(配列番号22)とアライメントさせた。表12に示すアラインメントプログラムの出力は、TEAS配列の残基527と残基528(アラインメント出力において263および264の番号を付してある)との間にギャップを含んでいた。残基321、324、345、348、349、427、452、453、454、455、458、492、496、569、572、573、577、579および580は、19 TEAS残基と最も好適なアラインメントを有するものとして選択した。TEASに見られる構造的アスペクトの空間的な配向(即ち図1および表10に示すα-ヘリックス、β-シートおよびループなど)を維持するために、リモネンシクラーゼの残基583ではなく残基580を、TEASの残基528とアライメントするものとして選択した。
【0225】
BLOSUM 62スコアリングマトリックス、ギャップ開始値=11、ギャップ延長値
=1、X_ドロップオフ値=50、期待値=10、文字列の長さ=3を用い、およびコンプレキシティーの低い配列のフィルタリングを行わずに、BLASTpプログラム(NCBI)を用いて、配列番号44のタキサジエン(taxadiene)一次アミノ酸配列の残基579〜847を含む領域を、ボルニルジホスフェートシンターゼの全長アミノ酸配列(配列番号26)とアライメントさせた。表13に示すアラインメントプログラムの出力は、ボルニルジホスフェートシンターゼ配列の残基453と残基454との間にギャップを含んでいた。ボルニルジホスフェートシンターゼの残基321、324、344、347、348、426、451、452、453、454、457、492、496、568、571、572、576、578および579は、配列番号44の検索領域配列の残基584、587、606、609、610、688、713、714、715、716、719、753、757、831、834、835、839、841および842と最も好適なアラインメントを有するものとして選択した。タキサジエンシンターゼ中に存在すると思われる構造的アスペクトの空間的な配向(即ち図1および表10に示すα-ヘリックス、β-シートおよびループなど)を維持するために、ボルニルジホスフェートシンターゼの残基453および454を、タキサジエンシンターゼの残基715および716とアライメントするものとして選択した。
【0226】
BLOSUM 50スコアリングマトリックス、ギャップ開始値=13、ギャップ延長値
=2、X_ドロップオフ値=50、期待値=10、文字列の長さ=3を用い、およびコンプレキシティーの低い配列のフィルタリングを行わずに、BLASTpプログラム(NCBI)を用いて、TEAS一次アミノ酸配列(配列番号2)の残基265〜535を、δ-セリネンシンターゼの全長アミノ酸配列(配列番号48)とアライメントさせた。アラインメントプログラムの出力を表14に示す。配列番号48の残基300、303、324、327、328、406、431、432、433、434、437、471、475、548、551、552、556、558および559を、配列番号2の残基270、273、294、297、298、376、401、402、403、404、407、440、444、516、519、520、525、527および528と最も好適なアラインメントを有するものとして選択した。
【0227】
BLOSUM 62スコアリングマトリックス、ギャップ開始値=11、ギャップ延長値=1、X_ドロップオフ値=50、期待値=10、文字列の長さ=3を用い、およびコンプレキシティーの低い配列のフィルタリングを行わずに、BLASTpプログラム(NCBI)を用いて、γ-フムレンシンターゼの一次アミノ酸配列(配列番号50)の残基307〜593を、アビエタジエン(abietadiene)シンターゼの全長アミノ酸配列(配列番号56)とアライメントさせた。アラインメントプログラムの出力を表15に示す。ジテルペンシンターゼ(配列番号56)の残基590、593、614、617、618、696、721、722、723、724、727、761、765、837、840、841、845、847および848を、セスキテルペンシンターゼ検索配列(配列番号50)の残基312、315、336、339、340、419、444、445、446、447、450、484、488、562、565、566、570、572および573と最も好適なアラインメントを有するものとして選択した。
【0228】
実施例12
新規なモノテルペンシンターゼ遺伝子の作成
ピネンシンターゼをコードするDNA配列(配列番号20)を用いて、突然変異型ピネンシンターゼ遺伝子のライブラリーを構築する。以下の9個のアミノ酸残基:L、C、C、G、H、S、L、GおよびY(配列番号20の351位、372位、480位、481位、482位、485位、519位、600位および601位に対応する)のうちの1個以上をコードするヌクレオチドにランダム突然変異を導入する。
【0229】
幾つかの場合では、ピネンシンターゼのコード配列を、配列番号20のY、I、T、F、D、YおよびSの位置に対応するアミノ酸残基348、375、376、597、605、607および608のうちの1個以上をコードするヌクレオチドにランダム突然変異を導入する。ピネンシンターゼのコード配列に時々、以下のアミノ酸残基Y、SおよびG(配列番号20の454位、479位および523位に対応する)のうちの1個以上をコードするヌクレオチドで突然変異させる。幾つかの場合では、上記9個の残基をコードするヌクレオチドにおける突然変異に加えて、さらにこれらの10個の位置における突然変異を起こす。他の場合では、上記9個の残基に突然変異を導入せずに、これらの10個の位置で突然変異を起こす。
【0230】
ピネンシンターゼのコード配列DNAをpET28b(+)ベクター中に挿入し、QuickChange(登録商標)法を用い、TEAS406/407ランダムライブラリについて実施例1で記載したものと同様のプロトコールに従って、突然変異誘発(mutagenize)する。突然変異を生じさせるために使用するプライマーを、ランダム突然変異体を作製するために、NまたはSを所望のコドン中のヌクレオチドとして用いて、実施例1に記載したように合成する。
【0231】
上記19個のピネンシンターゼアミノ酸残基のうちの1個以上における特異的突然変異は、TEASについて実施例1で記載したものと同様のプロトコールを用いて部位特異的突然変異誘発法により行う。所望の突然変異体を作成するために、突然変異させるべきコドン中に特定のA、T、CまたはG置換を有するプライマーを作製する。
【0232】
他のモノテルペンシンターゼ(例えばリモネンシンターゼ(配列番号22または58)、ミルセンシンターゼ(配列番号30)、+サビネンシンターゼ(配列番号54)、1,8 シネオール(cineole)シンターゼ(配列番号24)、および+ボルニルジホスフェートシンターゼ(配列番号26))のアミノ酸残基であって、その残基のα-炭素が表1〜6に記載した原子間距離および構造座標を有するアミノ酸残基を改変するために、上記方法と同じ方法で、ランダムおよび/または特異的突然変異を調製する。
【0233】
実施例13
新規なセスキテルペンシンターゼ遺伝子の作成
カジネンシンターゼをコードするDNA配列(配列番号33)を用いて突然変異型カジネンシンターゼのライブラリーを構築する。以下の9個のアミノ酸残基:W、I、S、G、Y、L、C、LおよびY(配列番号33によりコードされるアミノ酸残基280、301、409、410、411、414、448、527および528に対応する)のうちの1個以上をコードするヌクレオチドにランダム突然変異を導入する。
【0234】
幾つかの場合では、カジネンシンターゼのコード配列を、配列番号33によりコードされるアミノ酸残基277、304、305、524、532、534および535に対応するアミノ酸残基G、A、S、M、D、YおよびTのうちの1個以上をコードするヌクレオチドで突然変異させる。さらに、カジネンシンターゼのコード配列を時々、アミノ酸残基383、408および452(配列番号33によりコードされるアミノ酸Y、TおよびDに対応する)のうちの1個以上をコードするヌクレオチドで突然変異させる。幾つかの場合では、上記9個の残基における突然変異に加えてさらにこれら10個の突然変異を起こす。他の場合では、上記9個の残基に突然変異を導入せずに、これら10個の残基における突然変異を作製する。
【0235】
カジネンシンターゼのコード配列を、pET28b(+)ベクター中でQuickChange(登録商標)法を用いて、TEAS406/407ランダムライブラリについて実施例1で記載したものと同様のプロトコールに従って、突然変異させる。突然変異を生じさせるために使用するプライマーを、実施例11に記載したように合成する。
【0236】
上記カジネンシンターゼアミノ酸残基のうちの1個以上における特異的突然変異は、TEASについて実施例1で記載したものと同様のプロトコールを用いて部位特異的突然変異誘発法により行う。
【0237】
他のセスキテルペンシンターゼ(例えばベティスピラジエンシンターゼ(配列番号32)、ゲルマクレン Cシンターゼ(配列番号52)、E-α-ビスアボレン(bisabolene)シンターゼ(配列番号46)、γ-フムレンシンターゼ(配列番号50)、δ-セリネンシンターゼ(配列番号48)、e-b-ファルネセンシンターゼ(配列番号28)中のアミノ酸残基であって、その残基のα-炭素が表1〜6に記載した原子間距離および構造座標を有するアミノ酸残基を改変するために、上記方法と同じ方法で、ランダムおよび/または特異的突然変異を調製する。
【0238】
実施例14
新規なジテルペンシンターゼ遺伝子の作製
アビエタジエンシンターゼをコードするDNA配列(配列番号56)を用いて突然変異型アビエタジエンシンターゼのライブラリーを構築する。以下の9個のアミノ酸残基:S、S、I、A、L、V、G、FおよびY(配列番号56の593位、614位、722位、723位、724位、727位、761位、840位および841位に対応する)のうちの1個以上をコードするヌクレオチドにランダム突然変異を導入する。
【0239】
幾つかの場合では、アビエタジエンシンターゼのコード配列を、配列番号56の590位、617位、618位、837位、845位、847位および848位に対応するアミノ酸残基I、S、T、M、D、LおよびTのうちの1個以上をコードするヌクレオチドで突然変異させる。アビエタジエンシンターゼのコード配列を時々、アミノ酸残基Y、SおよびN(配列番号56の696位、721位および765位に対応する)のうちの1個以上をコードするヌクレオチドで突然変異させる。幾つかの場合では、上記9個の残基における突然変異に加えてさらにこれらの突然変異を起こす。他の場合では、上記9つの残基に突然変異を導入せずに、これらの10個の残基で突然変異を起こす。
【0240】
アビエタジエンシンターゼのコード配列をpET28b(+)ベクター中でQuickChange(登録商標)法を用いて、TEAS406/407ランダムライブラリについて実施例1で記載したものと同様のプロトコールに従って、突然変異させる。突然変異を生じさせるために使用するプライマーは、実施例11に記載したように合成する。
【0241】
上記アビエタジエンシンターゼアミノ酸残基のうちの1個以上における特異的突然変異は、TEASについて実施例1で記載したものと同様のプロトコールを用いて部位特異的突然変異誘発法により行う。
【0242】
他のジテルペンシンターゼ(例えばカスベン(casbene)シンターゼ(配列番号42)およびタキサジエンシンターゼ(配列番号44)等)中のアミノ酸残基であって、その残基のα-炭素が表1〜6に記載した原子間距離および構造座標を有するアミノ酸残基を改変するために、上記方法と同じ方法で、ランダムおよび/または特異的突然変異を調製する。
【0243】
実施例15
昆虫、哺乳動物および細菌細胞中での突然変異型シンターゼの発現
バキュロウイルス発現ベクターを用いて、実施例12、13および/または14の突然変異型シンターゼをコードする核酸を含む構築物を、昆虫スポドプテラ・フルギペルダ(Spoodoptera frugiperda)の培養細胞中に導入する。該遺伝子の発現後、この突然変異型酵素を各クローンから単離および精製する。
【0244】
SV40プロモーターを有する発現ベクターを用いて、実施例12、13および/または14の突然変異型シンターゼをコードする核酸を含む構築物を、培養したHeLa細胞中に導入する。該遺伝子の発現後、この突然変異型酵素を各クローンから単離および精製する。
【0245】
実施例1で記載したようにプラスミドベクターを用いて、実施例12、13および/または14の突然変異型シンターゼをコードする核酸を含む構築物を、大腸菌BL-21中に導入する。この突然変異型遺伝子の発現後、実施例2に記載されたようにこの突然変異型酵素を各クローンから単離および精製する。
【0246】
他の実施形態
特に記載していないが、当業者であれば、本明細書中に記載および例示した様々な特定の実施形態がいずれも、これらの特定の実施形態以外の特徴を持つようにさらに修正することができることが分かるであろう。
【0247】
本発明は、その詳細な説明に関連して記載したが、前記記載は、特許請求の範囲に定義された本発明の範囲を例示するためのものであって、これを限定するものではない。他の態様、利点および変更は、本発明の特許請求の範囲に含まれる。
【0248】
【表1】
【0249】
【表2】
【0250】
【表3】
【0251】
【表4】
【0252】
【表5】
【0253】
【表6】
【0254】
【表7】
【0255】
【表8】
【0256】
【表9】
【0257】
【表10】
【0258】
【表11】
【0259】
【表12】
【0260】
【表13】
【0261】
【表14】
【0262】
【表15】
【特許請求の範囲】
【請求項1】
テルペノイド産物の生成を触媒し、かつ、配列番号2の残基265〜535に対して35%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、該シンターゼが19個のα-炭素を含み、該19個のα-炭素は下記のÅで表した原子間距離:
の±2.3Åであるα-炭素間の原子間距離を有し、該α-炭素のそれぞれの中心点が半径2.3Åの球内に位置し、該球のそれぞれの中心点が下記の構造座標:
を有し、該α-炭素のそれぞれが関連するR基を有しており(ここで、該19個のα-炭素の位置は、配列番号2のアミノ酸位置270, 273, 294, 297, 298, 376, 401, 402, 403, 404, 407, 440, 444, 516, 519, 520, 525, 527 および528とアラインメントするものである)、該シンターゼが下記のR基の順序配列:
とは異なるR基の順序配列を有することを特徴とする、上記シンターゼ。
【請求項2】
配列番号2の残基265〜535に対して40%以上の配列同一性を有する、請求項1に記載のシンターゼ。
【請求項3】
テルペノイド産物の生成を触媒し、かつ、配列番号2の残基265〜535に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号2のアミノ酸位置270, 273, 294, 297, 298, 376, 401, 402, 403, 404, 407, 440, 444, 516, 519, 520, 525, 527 および528のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記のR基の順序配列:
とは異なるR基の順序配列を有することを特徴とする、上記シンターゼ。
【請求項4】
配列番号2の残基265〜535に対して50%以上の配列同一性を有する、請求項3に記載のシンターゼ。
【請求項5】
配列番号2の残基265〜535に対して60%以上の配列同一性を有する、請求項3に記載のシンターゼ。
【請求項6】
配列番号2の残基265〜535に対して70%以上の配列同一性を有する、請求項3に記載のシンターゼ。
【請求項7】
配列番号2の残基265〜535に対して80%以上の配列同一性を有する、請求項3に記載のシンターゼ。
【請求項8】
テルペノイド産物の生成を触媒し、かつ、配列番号33によってコードされるアミノ酸残基272〜540に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号33によってコードされるアミノ酸位置277、280、301、304、305、383、408、409、410、411、414、448、452、524、527、528、532、534および535のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項9】
テルペノイド産物の生成を触媒し、かつ、配列番号42のアミノ酸残基319〜571に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号42のアミノ酸位置324、327、348、351、352、430、455、456、457、458、461、495、499、571、574、575、579、581および582のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項10】
テルペノイド産物の生成を触媒し、かつ、配列番号52のアミノ酸残基264〜533に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号52のアミノ酸位置269、272、293、296、297、375、401、402、403、404、407、441、445、517、520、521、525、527および528のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項11】
テルペノイド産物の生成を触媒し、かつ、配列番号56のアミノ酸残基585〜853に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号56のアミノ酸位置590、593、614、617、618、696、721、722、723、724、727、761、765、837、840、841、845、847および848のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項12】
テルペノイド産物の生成を触媒し、かつ、配列番号28のアミノ酸残基265〜536に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号28のアミノ酸位置270、273、294、297、298、376、401、402、403、404、407、440、444、518、521、522、528、530および531のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項13】
テルペノイド産物の生成を触媒し、かつ、配列番号32のアミノ酸残基307〜541に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号32のアミノ酸位置278、281、302、305、306、384、409、410、411、412、415、448、452、524、527、528、533、535および536のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項14】
モノテルペン基質からのテルペノイド産物の生成を触媒する、請求項1〜13のいずれかに記載のシンターゼ。
【請求項15】
セスキテルペン基質からのテルペノイド産物の生成を触媒する、請求項1〜13のいずれかに記載のシンターゼ。
【請求項16】
ジテルペン基質からのテルペノイド産物の生成を触媒する、請求項1〜13のいずれかに記載のシンターゼ。
【請求項17】
前記産物が環式テルペノイド炭化水素である、請求項14に記載のシンターゼ。
【請求項18】
前記産物が環式テルペノイド炭化水素である、請求項15に記載のシンターゼ。
【請求項19】
前記産物が環式テルペノイド炭化水素である、請求項16に記載のシンターゼ。
【請求項20】
請求項1〜19のいずれかに記載のシンターゼをコードする単離された核酸。
【請求項21】
請求項20記載の核酸を含む宿主細胞。
【請求項22】
原核細胞、真核細胞、昆虫細胞、および植物細胞から選択される、請求項21記載の宿主細胞。
【請求項23】
請求項1〜19のいずれかに記載のシンターゼを基質と接触させ、それによってイソプレノイド産物の形成を触媒することを含む、イソプレノイド産物の製造方法。
【請求項24】
前記基質がモノテルペン基質、セスキテルペン基質、またはジデルペン基質から選択される、請求項20記載の方法。
【請求項25】
前記テルペノイド産物が、環式テルペノイド炭化水素、非環式テルペノイド炭化水素、環式ヒドロキシル化テルペノイド炭化水素、または非環式ヒドロキシル化テルペノイド炭化水素から選択される、請求項20記載の方法。
【請求項1】
テルペノイド産物の生成を触媒し、かつ、配列番号2の残基265〜535に対して35%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、該シンターゼが19個のα-炭素を含み、該19個のα-炭素は下記のÅで表した原子間距離:
の±2.3Åであるα-炭素間の原子間距離を有し、該α-炭素のそれぞれの中心点が半径2.3Åの球内に位置し、該球のそれぞれの中心点が下記の構造座標:
を有し、該α-炭素のそれぞれが関連するR基を有しており(ここで、該19個のα-炭素の位置は、配列番号2のアミノ酸位置270, 273, 294, 297, 298, 376, 401, 402, 403, 404, 407, 440, 444, 516, 519, 520, 525, 527 および528とアラインメントするものである)、該シンターゼが下記のR基の順序配列:
とは異なるR基の順序配列を有することを特徴とする、上記シンターゼ。
【請求項2】
配列番号2の残基265〜535に対して40%以上の配列同一性を有する、請求項1に記載のシンターゼ。
【請求項3】
テルペノイド産物の生成を触媒し、かつ、配列番号2の残基265〜535に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号2のアミノ酸位置270, 273, 294, 297, 298, 376, 401, 402, 403, 404, 407, 440, 444, 516, 519, 520, 525, 527 および528のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記のR基の順序配列:
とは異なるR基の順序配列を有することを特徴とする、上記シンターゼ。
【請求項4】
配列番号2の残基265〜535に対して50%以上の配列同一性を有する、請求項3に記載のシンターゼ。
【請求項5】
配列番号2の残基265〜535に対して60%以上の配列同一性を有する、請求項3に記載のシンターゼ。
【請求項6】
配列番号2の残基265〜535に対して70%以上の配列同一性を有する、請求項3に記載のシンターゼ。
【請求項7】
配列番号2の残基265〜535に対して80%以上の配列同一性を有する、請求項3に記載のシンターゼ。
【請求項8】
テルペノイド産物の生成を触媒し、かつ、配列番号33によってコードされるアミノ酸残基272〜540に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号33によってコードされるアミノ酸位置277、280、301、304、305、383、408、409、410、411、414、448、452、524、527、528、532、534および535のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項9】
テルペノイド産物の生成を触媒し、かつ、配列番号42のアミノ酸残基319〜571に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号42のアミノ酸位置324、327、348、351、352、430、455、456、457、458、461、495、499、571、574、575、579、581および582のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項10】
テルペノイド産物の生成を触媒し、かつ、配列番号52のアミノ酸残基264〜533に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号52のアミノ酸位置269、272、293、296、297、375、401、402、403、404、407、441、445、517、520、521、525、527および528のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項11】
テルペノイド産物の生成を触媒し、かつ、配列番号56のアミノ酸残基585〜853に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号56のアミノ酸位置590、593、614、617、618、696、721、722、723、724、727、761、765、837、840、841、845、847および848のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項12】
テルペノイド産物の生成を触媒し、かつ、配列番号28のアミノ酸残基265〜536に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号28のアミノ酸位置270、273、294、297、298、376、401、402、403、404、407、440、444、518、521、522、528、530および531のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項13】
テルペノイド産物の生成を触媒し、かつ、配列番号32のアミノ酸残基307〜541に対して40%以上の配列同一性を有する領域をもつ単離されたテルペンシンターゼであって、配列番号32のアミノ酸位置278、281、302、305、306、384、409、410、411、412、415、448、452、524、527、528、533、535および536のアミノ酸残基とアラインメントする該シンターゼの一つ以上のアミノ酸残基が、下記の残基の順序配列:
とは異なる残基であることを特徴とする、上記シンターゼ。
【請求項14】
モノテルペン基質からのテルペノイド産物の生成を触媒する、請求項1〜13のいずれかに記載のシンターゼ。
【請求項15】
セスキテルペン基質からのテルペノイド産物の生成を触媒する、請求項1〜13のいずれかに記載のシンターゼ。
【請求項16】
ジテルペン基質からのテルペノイド産物の生成を触媒する、請求項1〜13のいずれかに記載のシンターゼ。
【請求項17】
前記産物が環式テルペノイド炭化水素である、請求項14に記載のシンターゼ。
【請求項18】
前記産物が環式テルペノイド炭化水素である、請求項15に記載のシンターゼ。
【請求項19】
前記産物が環式テルペノイド炭化水素である、請求項16に記載のシンターゼ。
【請求項20】
請求項1〜19のいずれかに記載のシンターゼをコードする単離された核酸。
【請求項21】
請求項20記載の核酸を含む宿主細胞。
【請求項22】
原核細胞、真核細胞、昆虫細胞、および植物細胞から選択される、請求項21記載の宿主細胞。
【請求項23】
請求項1〜19のいずれかに記載のシンターゼを基質と接触させ、それによってイソプレノイド産物の形成を触媒することを含む、イソプレノイド産物の製造方法。
【請求項24】
前記基質がモノテルペン基質、セスキテルペン基質、またはジデルペン基質から選択される、請求項20記載の方法。
【請求項25】
前記テルペノイド産物が、環式テルペノイド炭化水素、非環式テルペノイド炭化水素、環式ヒドロキシル化テルペノイド炭化水素、または非環式ヒドロキシル化テルペノイド炭化水素から選択される、請求項20記載の方法。
【図1】

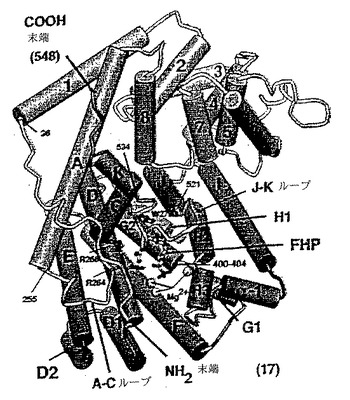
【図2A】
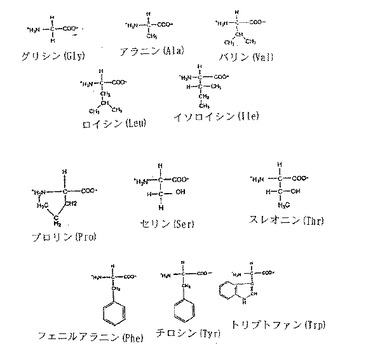
【図2B】
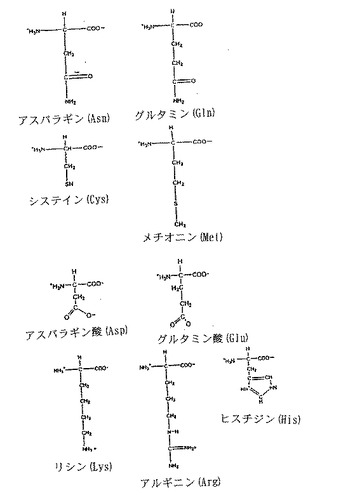
【図3】
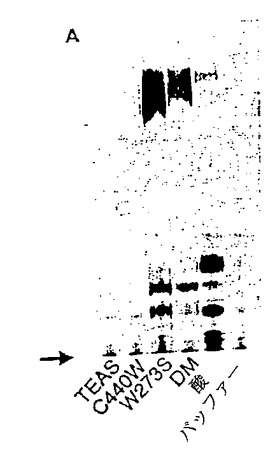
【図4】
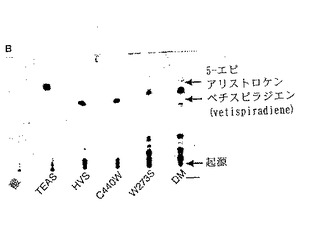
【図2A】
【図2B】
【図3】
【図4】
【公開番号】特開2011−172576(P2011−172576A)
【公開日】平成23年9月8日(2011.9.8)
【国際特許分類】
【出願番号】特願2011−65750(P2011−65750)
【出願日】平成23年3月24日(2011.3.24)
【分割の表示】特願2000−574228(P2000−574228)の分割
【原出願日】平成11年9月17日(1999.9.17)
【出願人】(501109013)ユニバーシティー オブ ケンタッキー リサーチ デパートメント (1)
【出願人】(596086505)
【氏名又は名称原語表記】The Salk Institute For Biological Studies
【住所又は居所原語表記】10010 North Torrey Pines Road,La Jolla,California 92037,United States of America
【Fターム(参考)】
【公開日】平成23年9月8日(2011.9.8)
【国際特許分類】
【出願日】平成23年3月24日(2011.3.24)
【分割の表示】特願2000−574228(P2000−574228)の分割
【原出願日】平成11年9月17日(1999.9.17)
【出願人】(501109013)ユニバーシティー オブ ケンタッキー リサーチ デパートメント (1)
【出願人】(596086505)
【氏名又は名称原語表記】The Salk Institute For Biological Studies
【住所又は居所原語表記】10010 North Torrey Pines Road,La Jolla,California 92037,United States of America
【Fターム(参考)】
[ Back to top ]