
ジペプチジルアミノペプチダーゼ活性を欠くピチア・パストリスにおける治療用タンパク質の製造方法
本発明は、ジペプチジルアミノペプチダーゼ(DAP)活性を欠く酵母細胞系、特にピチア・パストリス(Pichia pastoris)において治療用タンパク質を製造するための方法および組成物に関する。STE13およびDAP2が欠失するようにピチア・パストリス(Pichia pastoris)細胞系を遺伝的に修飾することにより、DAP活性は遺伝的に排除されている。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ヒトまたは動物治療用物質として有用である糖タンパク質の、ジペプチジルアミノペプチダーゼ(DAP)活性を有さない酵母細胞系における製造のための方法および組成物に関する。
【背景技術】
【0002】
酵母は組換えタンパク質の製造のための重要な生産媒体である。酵母は真核生物であるため、それは、分泌経路で生じる翻訳後修飾の多くを含め、より高等な真核生物のものと共通の進化的過程を共有している。糖操作における最近の進歩は、ヒトにおける糖タンパク質の過程によく似た一連の酵素反応を可能にする遺伝的に修飾されたグリコシル化経路を有する酵母株ピチア・パストリス(Pichia pastoris)の細胞系を生み出している。例えば、ヒト対応物と実質的に同一である組換え糖タンパク質の、下等真核宿主細胞における製造方法を記載している米国特許第7,029,872号および第7,326,681号を参照されたい。前記方法からピチア・パストリス(Pichia pastoris)において産生されるようなヒト様シアル酸化二分岐複合体N−結合グリカンは治療用糖タンパク質の製造のための有用性を実証している。
【0003】
高等真核生物と同様に、酵母も多数のプロテアーゼを発現し、それらのうちの多くは分泌経路に局在化し、あるいはそれを通過してそれらの最終目的地へと向かう。その結果、幾つかの組換えタンパク質の望ましくないタンパク質分解が生じる可能性があり、該特異的切断は、関与するプロテアーゼのクラスに左右される。ジペプチジルアミノペプチダーゼ(DAP)は、タンパク質のN末端から2つのアミノ酸ペプチドを除去するタンパク質分解酵素のクラスである。サッカロミセス・セレビシエ(Saccharomyces cerevisiae)においては、酵素STE13およびDAP2の遺伝子がDAP活性を有するものとして特定されている。Juliusら,Cell,32:839−852,1983;Renduelesら,J.Bacteriology,169:4041−4048,1987を参照されたい。本出願者は、完全長治療用タンパク質の製造を可能にする、ピチア・パストリス(Pichia pastoris)におけるDAP活性の排除のための方法を開発した。
【発明の概要】
【0004】
発明の概括
1つの実施形態においては、本発明は、ジペプチジルアミノペプチダーゼ(DAP)活性を欠く酵母細胞系における治療用タンパク質の製造方法である。この実施形態は、DAP活性が排除されている遺伝的に修飾されたピチア・パストリス(Pichia pastoris)細胞系を、該治療用タンパク質をコードするポリヌクレオチドベクターで形質転換し、該形質転換細胞を培養して該治療用タンパク質を得ることを含む。DAP活性は、STE13、DAP2およびDPPIIIが欠失または破壊されるようにピチア・パストリス(Pichia pastoris)細胞系を修飾することにより排除されうる。もう1つの実施形態においては、DAP活性は、STE13およびDAP2が欠失または破壊されるようにピチア・パストリス(Pichia pastoris)細胞系を修飾することにより排除される。
【0005】
1つの実施形態においては、本発明は、STE13およびDAP2の欠失により組換え的に修飾されたピチア・パストリス(Pichia pastoris)細胞系を含む、治療用タンパク質の製造に使用されうる、DAP活性を欠く遺伝的に修飾された酵母細胞系である。
【0006】
更に他の実施形態においては、本発明は、TNFRII−Fc、すなわち、IgG1 Fcドメインに融合した腫瘍壊死因子受容体2(TNFRII)のエクトドメインを含む組換え融合タンパク質(TNFRII−Fc)、または顆粒球コロニー刺激因子(GCSF)の分泌血漿形態を含む組換えGCSFポリペプチドのような、ピチア・パストリス(Pichia pastoris)からの治療用タンパク質の製造方法である。
【0007】
発明の詳細な説明
定義
本明細書中で用いる「ジペプチジルアミノペプチダーゼ活性」または「DAP活性」なる語は、STE13、DAP2またはDPPIIIと称される遺伝子により産生されるポリペプチドの酵素的切断を意味する。
【0008】
本明細書中で用いる「ジペプチジルアミノペプチダーゼ活性の排除」または「DAP活性の排除」なる語は、STE13、DAP2またはDPPIIIと称される遺伝子により生じる酵素活性の非存在を意味する。
【0009】
本明細書中で用いる「治療用タンパク質」なる語は、動物またはヒトにおける疾患または状態を治療するための治療用物質として使用されうる完全長(すなわち、非トランケート化形態)の生物学的に活性なポリペプチドを意味する。本明細書中で用いられるこの用語の具体例としては、腫瘍壊死因子受容体2(TNFRII)とIgG1のFc領域(Fc)とを含む融合タンパク質TNFRI−Fc、および組換え顆粒球コロニー刺激因子(GCSF)タンパク質が挙げられる。
【0010】
本明細書中で用いる「N末端認識部位」なる語は、モチーフX−ProまたはX−Ala(ここで、Xは任意のアミノ酸であり、N末端に対して2番目の位置はプロリンまたはアラニンである)のN末端配列を有するポリペプチドを意味する。
【0011】
本明細書中で用いる「糖操作(された)ピチア・パストリス(Pichia pastoris)株」なる語は、ヒト糖タンパク質を発現するように操作されたピチア・パストリス(Pichia pastoris)株を意味する。代表的な株には、YJN201(Choiら,PNAS,100(9):5022−5027,2003);YSH44(Hamiltonら,Science,301(5637):1244−1246,2003);RDP36−1(Davidsonら,Glycobiology,14(4):1−9,2004);PBP6−5(Bobrowiczら,Glycobiology,14(9):757−766,2004);YSH597(Hamiltonら,Science,313(5792):1441−1443,2006)が含まれる。
【0012】
本明細書中で用いる「野生型株」なる語は、遺伝子STE13、DAP2またはDPPIIIがゲノムから改変、破壊または欠失されていない糖操作ピチア・パストリス(Pichia pastoris)株を意味する。
【0013】
本明細書中で用いる「欠失株」なる語は、機能性DAP活性を排除するために1つ、2つ又は全てのDAP遺伝子(すなわち、STE13およびDAP2およびDPPIII)のピチア・パストリス(Pichia pastoris)ホモログがゲノムレベルで修飾されている糖操作ピチア・パストリス(Pichia pastoris)株を意味する。これは、該遺伝子(プロモーター、オープンリーディングフレームおよびターミネーターを含む)の完全または部分的な欠失、該遺伝子またはコード化mRNAのそれぞれ転写または翻訳を改変する1以上の突然変異の導入、および該タンパク質活性を不活性化する1以上の突然変異の導入を含むが、これらに限定されるものではない。そのような欠失株の一例はYGLY8084である。
【0014】
本明細書中で用いる「欠失または破壊された」および「欠失または破壊」なる語は、酵母細胞ゲノムから産生される酵素の活性の抑制を意味し、該酵素活性の抑制は、基質タンパク質が無傷N末端を保有するまでのものに限られる。酵母宿主細胞において該酵素活性が阻害または破壊されうる具体例としては以下のものが含まれる(それらに限定されるものではない)、1)該遺伝子の発現を制御する上流または下流調節配列の欠失または破壊(これらに限定されるものではない);2)該遺伝子を非機能性にする、該酵素活性をコードする遺伝子の突然変異(ここで、「突然変異」は、該遺伝子に関する酵素活性を不能にする該遺伝子内への欠失、置換、挿入または付加を含む);3)化学的インヒビター、ペプチドインヒビターまたはタンパク質インヒビターによる該酵素活性の阻害または破壊;4)核酸に基づく発現インヒビター(例えば、アンチセンスDNAおよびsiRNA)による該酵素活性の阻害または破壊;5)転写インヒビター、または該酵素活性をコードする遺伝子の発現を制御もしくは調節する調節因子の発現もしくは活性のインヒビターによる該酵素活性の阻害または破壊;および6)得られる産物が、たとえ発現されたとしても、該分泌タンパク質と同一ではない、および機能が低下している、いずれかの手段。
【0015】
略語
以下の略語が本明細書に全体において使用される。
URA5:オロタートホスホリボシルトランスフェラーゼ(OPRTアーゼ)アイソザイム
ScSUC2:サッカロミセス・セレビシエ(S.cerevisiae)インベルターゼ
OCH1:アルファ−1,6−マンノシルトランスフェラーゼ
KlGlcNAcTr:ケイ・ラクチス(K.lactis)UDP−GlcNAc輸送体
BMT2:ベータ−マンノース−転移(ベータ−マンノース排除)
MNN4B:MNN4A様遺伝子(電荷排除)
MmGlcNAcTr:UDP−GlcNAc輸送体のマウスホモログ
PNO1:N−グリカンのホスホマンノシル化(電荷排除)
MNN4A:マンノシルトランスフェラーゼ(電荷排除)
ADE1:N−スクシニル−5−アミノイミダゾール−4−カルボキサミドリボチドシンテターゼ
MNS1:ScSEC12リーダーに融合したマウスマンノシダーゼIA触媒ドメイン
GnTI:PpSEC12リーダーに融合したヒトGlcNAcトランスフェラーゼI触媒ドメイン
HIS1:ATPホスホリボシルトランスフェラーゼ
GalTI:ScKRE2リーダーに融合したトランケート化ヒトガラクトシルトランスフェラーゼ1触媒ドメイン
GalE:サッカロミセス・セレビシエ(S.cerevisiae)UDP−グルコース4−エピメラーゼ
UDP−GalTr:UDP−ガラクトース輸送体
ARG1:アルギノスクシナートシンテターゼ
MNSII:ScMNN2リーダーに融合したドロソフィラ(Drosophila)マンノシダーゼII触媒ドメイン
GnTII:ScMNN2リーダーに融合したラットGlcNAcトランスフェラーゼII触媒ドメイン
PRO1:ガンマ−グルタミルキナーゼ
TrMNS1:ScαMATに融合した分泌性ティー・レーセイ(T.reseei)マンノシダーゼI触媒ドメイン
AOX1:アルコールオキシダーゼI
TNFRII−Fc:IgG1のFcドメインに融合したヒト腫瘍壊死因子受容体II
Zeo:ゼオシン耐性マーカー
STE13:ジペプチジルアミノペプチダーゼ
DAP2:ジペプチジルアミノペプチダーゼ
DPPIII:ジペプチジルアミノペプチダーゼ
Nat:ナーセオスリシン(nourseothricin)耐性マーカー。
【0016】
酵母における治療用タンパク質の製造
ヒトまたは他の動物から単離されるタンパク質の大部分はグリコシル化されている。治療に使用されるタンパク質のうちの約70%はグリコシル化されている。治療用タンパク質が酵母のような微生物宿主において産生され、内在性経路を用いてグリコシル化される場合、典型的には、その治療効力は著しく低下する。それにもかかわらず、そのような糖タンパク質はヒトにおいて免疫原性であり、投与後のインビボにおける半減期の減少を示す(Takeuchi,Trends in Glycoscience and Glycotechnology,9:S29−S35,1997)。
【0017】
ヒトおよび動物における特定の受容体は末端マンノース残基を認識し、血流からの該タンパク質の迅速なクリアランスを促進しうる。他の有害作用には、タンパク質フォールディング、溶解度、プロテアーゼに対する感受性、トラフィッキング、輸送、区画化、分泌、他のタンパク質もしくは因子による認識またはアレルゲン性の変化が含まれうる。したがって、グリコシル化のパターンが、ヒトまたは意図される被投与種において生じるものと同じ又は少なくとも類似した動物宿主系において治療用糖タンパク質を産生させることが必要である。ほとんどの場合、哺乳類宿主系、例えば哺乳類細胞培養が使用される。使用される系には、チャイニーズハムスター卵巣細胞(CHO)、トランスジェニック動物、植物または昆虫細胞が含まれる。そのような系において発現される組換えヒトタンパク質には更に、非ヒトグリコフォーム(glycoform)が含まれうる(Rajuら,Annals Biochem.,283(2):123−132,2000)。これらの系の全ては重大な欠点を有する。それらの欠点には、ある治療用タンパク質だけが動物または植物系における発現に適していること、該細胞培養系は典型的には非常に遅いこと、タンパク質収率が微生物発酵法に比べて低く、複雑で高価な栄養または添加物を要しうること、またはそのような系は汚染されやすいことが含まれる(これらに限定されるものではない)。
【0018】
したがって、適当な発現の欠如は、あらゆる治療用糖タンパク質の製造に対する障害となっている。微生物の発酵による糖タンパク質の製造は、高濃度のタンパク質の迅速な製造、無菌の十分に制御された製造条件または化学的に規定された増殖培地が使用可能であること、多種多様なタンパク質の発現が可能であること、および該治療用タンパク質の容易な回収(これらに限定されるものではない)を含む、既存の系に比べた場合の利点をもたらすであろう。しかし、前記のとおり、細菌系は真核生物のようにはグリコシル化治療用タンパク質を産生しない。その目的のために、ヒト糖タンパク質を産生する遺伝的に修飾されたグリコシル化経路を有する酵母細胞系、特にピチア・パストリス(Pichia pastoris)が開発されている。例えば、対応ヒト対応物と実質的に同一である組換え糖タンパク質の、下等真核宿主細胞における製造方法を記載している米国特許第7,029,872号および第7,326,681号ならびにUS 2006−0286637を参照されたい。前記方法によりピチア・パストリス(Pichia pastoris)において産生されるようなヒト様シアル酸化二分岐複合N−結合グリカンは治療用糖タンパク質の製造のための有用性を示している。
【0019】
前記ピチア・パストリス(Pichia pastoris)細胞系は、タンパク質を治療用物質としての使用に適したものにするヒト様グリコシル化パターンを有するタンパク質を産生しうるが、ピチア・パストリス(Pichia pastoris)における治療用タンパク質の発現は必ずしも、完全長ポリペプチドの産生をもたらさない。酵母発現系を効率的に使用するためには、治療用タンパク質をトランケート化するタンパク質分解のような望ましくない酵素活性が排除される必要がある。
【0020】
ジペプチジルアミノペプチダーゼ活性を欠くピチアにおける治療用タンパク質の製造方法
酵母細胞系ピチア・パストリス(Pichia pastoris)から分泌される組換え融合タンパク質TNFRII−Fcのペプチド配列(その配列は図3(配列番号3)に示されている)の分析は、産生されるペプチドの全てがN末端において2アミノ酸だけトランケート化されていたことを示した。本出願人は、ピチア・パストリス(Pichia pastoris)細胞系における2つの遺伝子STE13およびDAP2が全てのDAP活性を排除し、完全長TNFRII−Fcの産生をもたらすことを確認した。出願人はまた、ピチア・パストリス(Pichia pastoris)が第3のジペプチジルアミノペプチダーゼDPPIIIを有することを確認している。したがって、1つの実施形態においては、本発明は、DAP活性を欠く酵母細胞系における治療用タンパク質の製造方法である。この実施形態は、DAP活性が排除された遺伝的に修飾されたピチア・パストリス(Pichia pastoris)細胞系を、治療用タンパク質をコードするポリヌクレオチドベクターで形質転換し、該形質転換宿主細胞を培養して、該治療用タンパク質を産生させることを含む。DAP活性は、STE13およびDAP2および/またはDPPIIIが欠失または破壊されるようにピチア・パストリス(Pichia pastoris)細胞系を修飾することにより排除される。本発明の特定の実施形態においては、DAP活性は、STE13およびDAP2が欠失または破壊されるようにピチア・パストリス(Pichia pastoris)細胞系を修飾することにより排除された。
【0021】
もう1つの実施形態においては、本発明は、STE13およびDAP2遺伝子が欠失または破壊され全ての生じるDAP活性が排除されるように組換え的に修飾されたピチア・パストリス(Pichia pastoris)細胞系を含む、治療用タンパク質の製造に使用される酵母細胞系である。
【0022】
更に他の実施形態においては、本発明は、ポリペプチドのN末端に対して2番目の位置にアミノ酸プロリンまたはアラニンを有する治療用タンパク質、例えば、本明細書に記載されているTNFRII−Fc融合タンパク質またはGCSFタンパク質の、ピチア・パストリス(Pichia pastoris)からの製造方法である。
【0023】
ジペプチジルアミノペプチダーゼ活性
ポリペプチドのアミノ末端から2つのアミノ酸をトランケート化することが知られているプロテアーゼのサブクラスであるジペプチジルアミノペプチダーゼ(DAP)の活性に関連した遺伝子が酵母において特定されている。サッカロミセス・セレビシエ(S.cerevisiae)の非接合性アルファ−細胞突然変異体は、アルファ接合因子フェロモンの不完全なプロセッシングによるものであると考えられている(Juliusら,Cell,32(3),839−52,1983)。更に、Ste13p活性を欠損したサッカロミセス・セレビシエ(S.cerevisiae)突然変異株をスクリーニングすることにより、第2のジペプチジルペプチダーゼDap2pが特定された(Suarez RenduelesおよびWolf,Journal of Bacteriology,169(9),4041−48,1987)。サッカロミセス・セレビシエ(S.cerevisiae)STE13のピチア(Pichia)ホモログのノックアウトが、N末端にアミノ酸HG(His−Gly)を有するタンパク質のインビボでのタンパク質切断を妨げ、完全長インスリン指向性(insulinotropic)ペプチドの産生を可能にしたことも報告されている(Melarkodeら,WO 2007/148345;Prabhaら,Protein Expression and Purification,64,155−161,2009)。逆に、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)のDAP2ピチア(Pichia)ホモログの破壊はN末端タンパク質分解切断を妨げなかった(Melarkodeら,WO 2007/148345;Prabhaら,Protein Expression and Purification,64,155−161,2009)。
【0024】
Ste13pおよびDap2pは、モチーフX−P/A(ここで、Xは任意のアミノ酸であり、2番目の位置はプロリン(P)またはアラニン(A)である)を有するN末端ペプチドを切断することが別々に示されている(MisumiおよびIkehara,Handbook of Proteolytic Enzvmes,2nd edition,pp.1910−1911,Elsevier,London,2004)。本発明を例示するために本明細書中で用いるTNFRII−FcおよびGCSFタンパク質は、保存されたX−Pro Ste13pおよびDap2pモチーフに合致しており、このことは、ピチア・パストリス(Pichia pastoris)における治療用タンパク質の製造のためには、Ste13pまたはDap2pのDAP活性が排除される必要があることを示唆している。これとは対照的に、Melarkodeら,WO 2007/148345のインスリン指向性ペプチドは、Ste13pのみにより認識されるHis−Glyモチーフを有する新規非保存性N末端を有する。
【0025】
本出願人により本明細書中で示されているとおり、ピチア・パストリス(Pichia pastoris)Ste13p活性の排除はTNFRII−Fcタンパク質に対するDAP活性の部分的低下をもたらしたに過ぎず、産生されたTNFRII−FCタンパク質の半分より幾らか多くが完全長配列を有していた。驚くべきことに、本出願人は、DAP活性の完全な排除、すなわち、完全長TNFRII−Fcタンパク質の100%の産生のためには、Ste13pおよびDap2pの両方の活性がピチア・パストリス(Pichia pastoris)細胞系から排除される必要があることを見出した。同様に、両方のDAP遺伝子がピチア・パストリス(Pichia pastoris)から排除された場合にのみ、本出願人は組換えGCSFタンパク質の完全な産生を観察した。したがって、N末端X−P/Aモチーフを有する完全長タンパク質の産生のためにはピチア・パストリス(Pichia pastoris)におけるSte13pおよびDap2p活性の両方の排除を要する点で本発明は先行技術とは異なる、と当業者は認識し理解するであろう。
【0026】
公知DAP遺伝子における相同性に基づいて、当業者はPCRプライマー(それらの具体例を表1に示す)を設計し、あるいは標的生物のDNAライブラリーにおけるホモログを特定するためのプローブとして遺伝子または遺伝子断片を使用することが可能である。DAP活性を有するホモログを特定するためのピチア・パストリス(Pichia pastoris)ゲノムの本出願人の分析は、それぞれPpSTE13およびPpDAP2(図8Aおよび8B、配列番号37および38、ならびに図9Aおよび9B、配列番号39および40)と称される、STE13およびDAP2のホモログの特定をもたらした。Melarkodeら,WO 2007/148345は、ピチア・パストリス(Pichia pastoris)STE13ホモログのDNA配列を記載しており、一方、Prabhaら,Protein Expression and Purification,64:155−161,2009は、ピチア・パストリス(Pichia pastoris)Ste13pおよびDap2pホモログのタンパク質配列を記載している。出願人により本発明において作製されたSte13pホモログは、Melarkodeらにおいて既に報告されているものに合致しているようである。本出願人により本発明において作製されたDap2pタンパク質配列の大部分は、Prabhaらにより報告されているものに合致しているが、本明細書中に特定されているDap2pホモログはC末端において異なる。本出願人のホモログは以下のC末端配列を含有する:GLENYDIHVFPDSDHSIRYHNGNVIVYDKLFHWIRRAFKAGK(配列番号46)。一方、Prabhaらのホモログは以下のC末端配列を有する:GLENYDIHVFPDTIPLD(配列番号47)。前記C末端配列の下線部分までの及びそれを含むN末端配列は両方のホモログにおいて保存されている。
【0027】
表1は、ピチア・パストリス(Pichia pastoris)におけるノックアウトベクターの作製に使用される代表的プライマーの配列を示す。該プライマー配列における下線領域は、遺伝子ノックアウト断片の作製を促進するために導入された制限酵素認識部位を表す。導入された特異的制限酵素認識部位が隣のプライマー説明の欄に挙げられている。表2は、表1におけるプライマーを使用して作製されたノックアウトベクターでの形質転換の後のゲノムからのピチア(Pichia)STE13およびDAP2のノックアウトを確認するために使用されるプライマーを示す。該5’および3’プライマーセットが所望のサイズのPCR産物を与え、該ノックアウトプライマーセットが産物を与えない場合に、欠失の成功が確認される。
【0028】
【表1】
【0029】
【表2】
TNFRII−Fcで観察されるN末端トランケート化に関するこれらの遺伝子のそれぞれの役割を評価するために、ゲノムのDAP2またはSTE13遺伝子座の破壊に該ベクターを標的化するピチア・パストリス(Pichia pastoris)DAP2またはSTE13遺伝子の5’および3’領域を有する、URA5遺伝子を含有するノックアウトベクターを設計した(図1Aおよび1B)。これらのノックアウトベクターを、組換え融合タンパク質TNFRII−Fcを発現する同じuraマイナス親株(YGLY7406)内に形質転換した。該形質転換体のPCRスクリーニングの後、Ppste13およびPpdap2ノックアウト株(それぞれYGLY8084およびYGLY8090)を特定した。二重ノックアウト株、すなわち、同じ株におけるPpSTE13およびPpDAP2の両方のノックアウト体を作製するために、Ppste13欠失株YGLY8084をPpDAP2ナーセオスリシン(nourseothricin)ドミナントマーカーノックアウトベクターpGLY5019(図1D)で形質転換した。該形質転換体のPCRスクリーニングの後、Ppste13::URA5バックグラウンド親株において幾つかのPpdap2ノックアウト体を得た。代表的なPpste13/Ppdap2二重ノックアウト株をYGY8096と命名した。
【0030】
これらの株のそれぞれから分泌されたTNFRII−Fcのペプチド分析は、このポリペプチドのタンパク質分解においてSte13pは重要な役割を果たしているが、DAP活性を排除するためには両方のDAP遺伝子の欠失が必要であることを示している。該野生型株では、産生されるタンパク質の100%がN末端において2アミノ酸だけトランケート化された。STE13の単一欠失は、無傷N末端を有するペプチド集団を59%を与え、一方、DAP2の欠失は、無傷TNFRII−Fcの集団を与えたが、それは遥かに低い度合であり、該分泌ポリペプチドの15%しか完全長配列を有していなかった。二重ノックアウト株YGLY8096から分泌されたTNFRII−Fcの分析は、該ペプチド集団の100%が無傷N末端を有することを示した。表3は、単離された完全長ペプチドに対するトランケート化ペプチドのモル%を示すこの分析から得られたデータの要約である。データは各株に関する3つの重複サンプルの代表例である。uraマイナス株は組換えタンパク質の不良分泌体であるため、PpSTE13およびPpDAP2遺伝子がどちらも妨げられていない株から物質を産生させるためにYGLY7406の原栄養性親株YGLY6646を使用した。この株の遺伝子型は、無傷PpSTE13およびPpDAP2遺伝子座の両方に関して野生型として表3に記載されている。
【0031】
【表3】
この分析に基づいて、本出願人は、PpSte13pおよびppDap2pが、ピチア・パストリス(Pichia pastoris)において発現される組換えTNFRII−FcのN末端タンパク質分解に関与する唯一のプロテアーゼであり、DAP活性を排除するためには両方の欠失が必要であると判定した。同様に、野生型顆粒球コロニー刺激因子(GCSF)ポリペプチド(図5におけるタンパク質配列;配列番号6)がピチア・パストリス(Pichia pastoris)において発現された場合、そのN末端はトランケート化されていた。この物質のN末端配列決定は、それがN末端の2アミノ酸を欠くことを示した(図4Aおよび4B、それぞれ配列番号4および5)。また、GCSFのこのトランケート化形態の産生は、二重ΔPpste13/Ppdap2ノックアウトバックグラウンドにおいてGCSFを産生させることにより完全に排除された(図6)。どちらの場合にも、これらの組換えタンパク質(TNFRII−FcおよびCSF)の切断は、該ポリペプチドの(N末端に対して)2番目の位置のプロリン残基のC末端において生じる。本明細書に示されているとおり、ピチア・パストリス(Pichia pastoris)における両方のDAP関連遺伝子の欠失は、ポリペプチドのN末端から2番目の位置にプロリンを有する治療用タンパク質のトランケート化を妨げ、DAP活性の排除のためにはppSTE13およびPpDAP2のその欠失が必要であった。
【0032】
前記のどちらの治療用タンパク質の例もN末端モチーフXaa−Pro(ここで、Xは任意のアミノ酸でありうる)を有する。前記のとおり、Ste13pおよびDap2pは、モチーフXaa−Ala(ここで、Xaaは任意のアミノ酸であり、アラニン(Ala)は2番目のアミノ酸である)を同様に有するN末端ペプチドを切断することが別々に示されている(MisumiおよびIkehara,Handbook of Proteolytic Enzymes,2nd edition,pp.1910−1911,Elsevier,London,2004)。Xaa−Alaモチーフを有するタンパク質のトランケート化を防ぐためにはPpSTE13およびPpDAP2の両方のノックアウトが必要であることを実証するために、当業者は、TNFR−Fcの分泌形態における2番目のアミノ酸をプロリンからアラニンへと突然変異させることが可能である。本明細書における結果に基づけば、PpSTE13またはPpDAP2の欠失が、観察されるN末端切断の比率を減少させるらしい。DAP活性の完全な排除および生じる100%の無傷タンパク質は二重ΔPpste13/Ppdap2ノックアウト宿主株(すなわち、STE13およびDAP2の両方をノックアウトするための突然変異)のみから観察されるであろう。
【0033】
PpSte13pおよびPpDap2pの組合せ排除は、無傷N末端を有する治療用タンパク質の産生を示しているが、本出願人は、ピチア・パストリス(Pichia pastoris)が、本明細書中でPpDPPIIIと称される第3のジペプチジルアミノペプチダーゼを有することを見出した。この酵素はピチア・パストリス(Pichia pastoris)ゲノムのアノテーションにより特定され、ヒト酵素ジペプチジルペプチダーゼIII(DppIIIp)に対して43%の配列同一性を有する。ピチア・パストリス(Pichia pastoris)DppIIIpホモログは未だ特徴づけられていないが、PpSte13およびPpDap2pとは配列レベルで異なり、それぞれ10%および12%の配列同一性を有するに過ぎない。このDAP遺伝子は完全には特徴づけられていないが、PpDppIIIpが、適切な条件下で発現されると、本明細書において評価されている条件下で活性が観察されないにもかかわらず、トランケート化タンパク質を与える、と本出願人は考えている。他の増殖条件下または他のレポータータンパク質との発現により、PpDppIIIp活性が証明されうる、と当業者は認識し理解するであろう。したがって、他の適当な条件またはレポーターにおいて、ピチア・パストリス(Pichia pastoris)におけるDAP活性を完全に排除するためには、PpDppIIIが単独で或いはppSte13pおよび/またはPpDap2p活性と共に欠失または排除される必要がありうる。当業者であれば、本明細書の実施例に記載されている方法、特に実施例5の方法により、ppDPPIIIを欠失または破壊させることが可能であろう。
【0034】
DAP活性の排除のためのピチア株
商業的に入手可能な任意のピチア・パストリス(Pichia pastoris)株、例えばNRRL−Y11430(American Type Culture Collection(ATCC),Manassas,VA,カタログ番号76273)が本発明に使用可能であるが、好ましい実施形態においては、本発明に使用される株は、糖操作されたピチア・パストリス(Pichia pastoris)株、例えば、実施例3に記載されているGS5.0株、または後記の糖操作された株(これは、発現に際してヒト様グリコシル化プロファイルを与える修飾を含む)であろう。例えば、実施例3のGS5.0株の場合、そのような修飾には、Δoch1、Δpno1、Δmnn4B、Δbmt2およびΔura5の欠失、またはケイ・ラクチス(K.lactis)およびエム・ムスクルス(M.musculus)UDP−GlcNAc輸送体、エム・ムスクルス(M.musculus)α−1,2−MsnI、ホモ・サピエンス(H.sapiens)β−1,2−GlcNAcトランスフェラーゼ1活性、アール・ノルベギクス(R.norvegicus)β−1,2−GlcNAcトランスフェラーゼII活性、キイロショウジョウバエ(D.melanogaster)MnsII活性、エス・ポンベ(S.pombe)Galエピメラーゼ、キイロショウジョウバエ(D.melanogaster)UDP−Gal輸送体およびホモ・サピエンス(H.sapiens)β−1,4−ガラクトシルトランスフェラーゼ活性の挿入のために行われるものが含まれる。GS5.0株は、ガラクトースを末端とするN−グリカン、例えばGalGlcNAc2MAN3GlcNAc2、Gal2GlcNAc2Man3GlcNAc2またはそれらの混合物を有する糖タンパク質を産生しうる。他の代表的な糖操作株には、YJN201(Choiら,PNAS,100(9):5022−5027,2003)、YSH44(Hamiltonら,Science,301(5637):1244−1246,2003)、RDP36−1(Davidsonら,Glycobiology,14(4):1−9,2004)、PBP6−5(Bobrowiczら,Glyoobiology,14(9):757−766,2004)、YSH597(Hamiltonら,Science,313(5792):1441−1443,2006)が含まれる。
【0035】
ピチア・パストリス(Pichia pastoris)に加えて、本発明において有用な宿主細胞には、ピチア・パストリス(Pichia pastoris)DAP2およびSTE13遺伝子のホモログを発現する酵母が含まれる。そのような酵母は以下のものから選択されうる:ピチア・フィンランディカ(Pichia finlandica)、ピチア・トレハロフィラ(Pichia trehalophila)、ピチア・コクラメ(Pichia koclamae)、ピチア・メンブラネファシエンス(Pichia membranaefaciens)、ピチア・ミヌタ(Pichia minuta)(オガタエア・ミヌタ(Ogataea minuta)、ピチア・リンドネリ(Pichia lindneri))、ピチア・オプンチエ(Pichia opuntiae)、ピチア・テルモトレランス(Pichia thermotolerans)、ピチア・サリクタリア(Pichia salictaria)、ピチア・グエルクウム(Pichia guercuum)、ピチア・ピエペリ(Pichia pijperi)、ピチア・スチプティス(Pichia stiptis)、ピチア・メタノリカ(Pichia methanolica)、ピチア属種(Pichia sp.)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス属種(Saccharomyces sp.)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、クライベロミセス属種(Kluyveromyces sp.)、クライベロミセス・ラクチス(Kluyveromyces lactis)、カンジダ・アルビカンス(Candida albicans)。種々の酵母、例えばクライベロミセス・ラクチス(K.lactis)、ピチア・パストリス(Pichia pastoris)、ピチア・メタノリカ(Pichia methanolica)およびハンゼヌラ・ポリモルファ(Hansenula polymorpha)が細胞培養に特に適している。なぜなら、それらは高い細胞密度まで増殖可能であり、大量の組換えタンパク質を分泌しうるからである。
【0036】
酵母は、グリコシル化パターンがヒト様である又はヒト化されている糖タンパク質を発現するように、遺伝的に修飾されうる。このようにして、特定の所望のグリコフォームが組成物において優勢である糖タンパク質組成物が製造されうる。選択された内在性グリコシル化酵素を除去し、および/または該宿主細胞を遺伝的に操作し、および/または外在性酵素を供与して、哺乳類グリコシル化経路の全部または一部を模擬することにより(US 2004/0018590に記載されているとおり)、それは達成されうる。所望により、該グリコシル化の追加的遺伝的操作を行って、該糖タンパク質が、コアフコシル化を伴って又は伴わないで産生されうるようにすることが可能である。下等真核宿主細胞の使用は更に有利である。なぜなら、これらの細胞は糖タンパク質の高度に均一な組成物を産生することが可能であり、該糖タンパク質の主要グリコフォームが該組成物中の糖タンパク質の30モル%超(すなわち、30モル%を超えて)で存在することが可能となるからである。特定の態様においては、該主要グリコフォームは、該組成物中に存在する糖タンパク質の40モル%超、50モル%超、60モル%超、70モル%超、最も好ましくは80モル%超で存在しうる。
【0037】
酵母は、該グリコシル化パターンがヒト様である又はヒト化されている糖タンパク質をそれらが発現するように、遺伝的に修飾されうる。ヒト様またはヒト化N−グリカンには、ハイブリッドおよび複合N−グリカンの両方が含まれる。Gerngrossら,米国公開出願第2004/0018590に記載されているとおり、選択された内因性グリコシル化酵素を排除し、および/または外因性酵素を供給することにより、それは達成されうる。例えば、宿主細胞は、糖タンパク質上のN−グリカン上にマンノース残基を付加する1,6−マンノシルトランスフェラーゼ活性が喪失するよう選択され又は操作されうる。選択された内在性グリコシル化酵素を除去し、および/または外在性酵素を供与することにより(Gerngrossら,US 20040018590に記載されているとおり)、それは達成されうる。例えば、宿主細胞は、糖タンパク質上のN−グリカン上にマンノース残基を付加する1,6−マンノシルトランスフェラーゼ活性を欠くように選択または操作されうる。
【0038】
1つの実施形態においては、該宿主細胞は更に、α1,2−マンノシダーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、α1,2−マンノシダーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る組換え糖タンパク質の通過は、Man5GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、Man5GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生する。例えば、米国特許第7,029,872号ならびに米国公開特許出願第2004/0018590号および第2005/0170452号は、Man5GlcNAc2グリコフォームを含む糖タンパク質を産生しうる下等真核宿主細胞を開示している。
【0039】
更に詳細な実施形態においては、直前の宿主細胞は更に、GlcNAcトランスフェラーゼI(GnT I)触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、GlcNAcトランスフェラーゼI活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、GlcNAcMan5GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、ハイブリッドGlcNAcMan5GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生する。例えば、米国特許第7,029,872号ならびに米国公開特許出願第2004/0018590号および第2005/0170452号は、GlcNAcMan5GlcNAc2グリコフォームを含む糖タンパク質を産生しうる下等真核宿主細胞を開示している。前記細胞において産生された糖タンパク質をヘキソサミニダーゼでインビトロで処理して、Man5GlcNAc2グリコフォームを含む組換え糖タンパク質を産生させることが可能である。
【0040】
更に詳細な実施形態においては、直前の宿主細胞は更に、マンノシダーゼII触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、マンノシダーゼII活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、GlcNAcMan3GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、複合GlcNAcMan3GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生する。米国特許第7,029,872号および米国公開特許出願第2004/0230042号は、マンノシダーゼII酵素を発現し、GlcNAc2Man3GlcNAc2グリコフォームを主に有する糖タンパク質を産生しうる下等真核宿主細胞を開示している。前記細胞において産生された糖タンパク質をヘキソサミニダーゼでインビトロで処理して、Man3GlcNAc2グリコフォームを含む組換え糖タンパク質を産生させることが可能である。
【0041】
更に詳細な実施形態においては、直前の宿主細胞は更に、GlcNAcトランスフェラーゼII(GnT II)触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、GlcNAcトランスフェラーゼII活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、GlcNAc2Man3GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、複合GlcNAc2Man3GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生する。米国特許第7,029,872号ならびに米国公開特許出願第2004/0018590号および第2005/0170452号は、GlcNAc2Man3GlcNAc2グリコフォームを含む糖タンパク質を産生しうる下等真核宿主細胞を開示している。前記細胞において産生された糖タンパク質をヘキソサミニダーゼでインビトロで処理して、Man3GlcNAc2グリコフォームを含む組換え糖タンパク質を産生させることが可能である。
【0042】
更に詳細な実施形態においては、直前の宿主細胞は更に、ガラクトシルトランスフェラーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、ガラクトシルトランスフェラーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、複合GalGlcNAc2Man3GlcNAc2(G1)もしくは複合Gal2GlcNAc2Man3GlcNAc2(G2)グリコフォームまたはそれらの混合物を含む組換え糖タンパク質、例えば、GalGlcNAc2Man3GlcNAc2グリコフォームもしくはGal2GlcNAc2Man3GlcNAc2グリコフォームまたはそれらの混合物を主に含む組換え糖タンパク質組成物を産生する。米国特許第7,029,872号および米国公開特許出願第2006/0040353号は、Gal2GlcNAc2Man3GlcNAc2グリコフォームを含む糖タンパク質を産生しうる下等真核宿主細胞を開示している。前記細胞において産生された糖タンパク質をガラクトシダーゼでインビトロで処理して、GlcNAc2Man3GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、GlcNAc2Man3GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生させることが可能である。
【0043】
更に詳細な実施形態においては、直前の宿主細胞は更に、シアリルトランスフェラーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、シアリルトランスフェラーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、複合NANA2Gal2GlcNAc2Man3GlcNAc2グリコフォームもしくは複合NANAGal2GlcNAc2Man3GlcNAc2グリコフォームまたはそれらの混合物を主に含む組換え糖タンパク質を産生する。該宿主細胞は、N−グリカンへの転移のためのCMP−シアル酸を供与するための手段を更に含むことが有用である。米国公開特許出願第2005/0260729号は、CMP−シアル酸合成経路を有するように下等真核生物を遺伝的に操作するための方法を開示しており、米国公開特許出願第2006/0286637号は、シアル酸化糖タンパク質を産生するように真核生物を遺伝的に操作するための方法を開示している。前記細胞において産生された糖タンパク質をノイラミニダーゼでインビトロで処理して、Gal2GlcNAc2Man3GlcNAc2グリコフォームを主に含む組換え糖タンパク質を産生させることが可能である。
【0044】
前記宿主細胞はいずれも、米国公開特許出願第2004/074458号および第2007/0037248号に開示されているような二分岐(bisected)(GnT III)および/または多分岐(GnT IV、V、VIおよびIX)N−グリカン構造を有する糖タンパク質を産生させるためのGnT III、GnT IV、GnT V、GnT VIおよびGnT IXからなる群から選択される1以上のGlcNAcトランスフェラーゼを更に含みうる。
【0045】
更に詳細な実施形態においては、ハイブリッドGlcNAcMan5GlcNAc2 N−グリカンを主に有する糖タンパク質を産生する宿主細胞は更に、ガラクトシルトランスフェラーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、ガラクトシルトランスフェラーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、ハイブリッドGalGlcNAcMan5GlcNAc2グリコフォームを主に含む組換え糖タンパク質を産生する。
【0046】
更に詳細な実施形態においては、GalGlcNAcMan5GlcNAc2 N−グリカンを主に有する糖タンパク質を産生した直前の宿主細胞は更に、シアリルトランスフェラーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、シアリルトランスフェラーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、ハイブリッドNANAGalGlcNAcMan5GlcNAc2グリコフォームを含む組換え糖タンパク質を産生する。
【0047】
種々の前記宿主細胞は更に、1以上の糖輸送体、例えばUDP−GlcNAc輸送体[例えば、クライベロミセス・ラクチス(Kluyveromyces lactis)およびムス・ムスクルス(Mus musculus)UDP−GlcNAc輸送体]、UDP−ガラクトース輸送体[例えば、キイロショウジョウバエ(Drosophila melanogaster)UDP−ガラクトース輸送体]およびCMP−シアル酸輸送体(例えば、ヒトシアル酸輸送体)を含む。下等真核宿主細胞、例えば酵母および糸状菌は前記輸送体を欠くため、下等真核宿主細胞、例えば酵母および糸状菌は、前記輸送体を含むように遺伝的に操作されることが好ましい。
【0048】
宿主細胞は更に、β−マンノシルトランスフェラーゼ遺伝子(例えば、BMT1、BMT2、BMT3およびBMT4)の1以上を欠失させ又は破壊することにより、α−マンノシダーゼ耐性N−グリカンを有する糖タンパク質を排除するように米国公開特許出願第2006/0211085号を参照されたい)、ならびにホスホマンノシルトランスフェラーゼ遺伝子PNO1およびMNN4Bの一方または両方を欠失させ又は破壊することにより、ホスホマンノース残基を有する糖タンパク質を排除するように(米国特許第7,198,921号および第7,259,007号を参照されたい)、遺伝的に操作された下等真核細胞[例えば、酵母、例えば、ピチア・パストリス(Pichia pastoris)]が含まれ、これは、更に詳細な実施形態においては、MNN4A遺伝子の欠失または破壊をも含みうる。破壊は、特定の酵素をコードするオープンリーディングフレームの破壊、あるいは該オープンリーディングフレームの発現の破壊、あるいは干渉性RNA、アンチセンスRNAなどを使用する、β−マンノシルトランスフェラーゼおよび/またはホスホマンノシルトランスフェラーゼの1以上をコードするRNAの翻訳の阻害を含む。該宿主細胞は更に、特定のN−グリカン構造を産生するように修飾された前記宿主細胞のいずれかを含みうる。
【0049】
宿主細胞は更に、タンパク質O−マンノシルトランスフェラーゼ[Dol−P−Man:タンパク質(Ser/Thr)マンノシルトランスフェラーゼ遺伝子](PMT)(米国特許第5,714,377号を参照されたい)の1以上を欠失させ又は破壊することにより糖タンパク質のO−グリコシル化を制御するように遺伝的に修飾された、あるいは公開国際出願番号WO2007061631に開示されているとおりにPmtpインヒビターおよび/またはアルファ−マンノシダーゼの存在下で増殖された、あるいはそれらの両方に付された下等真核細胞[例えば酵母、例えばピチア・パストリス(Pichia pastoris)]を含む。破壊は、Pmtpをコードするオープンリーディングフレームの破壊、あるいは該オープンリーディングフレームの発現の破壊、あるいは干渉性RNA、アンチセンスRNAなどを使用する、Pmtpの1以上をコードするRNAの翻訳の阻害を含む。該宿主細胞は更に、特定のN−グリカン構造を産生するように修飾された前記宿主細胞のいずれかを含みうる。
【0050】
Pmtpインヒビターには、ベンジリデンチアゾリジンジオンが含まれるが、これに限定されるものではない。使用されうるベンジリデンチアゾリジンジオンの具体例としては、5−[[3,4−ビス(フェニルメトキシ)フェニル]メチレン]−4−オキソ−2−チオキソ−3−チアゾリジン酢酸、5−[[3−(1−フェニルエトキシ)−4−(2−フェニルエトキシ)]フェニル]メチレン]−4−オキソ−2−チオキソ−3−チアゾリジン酢酸、および5−[[3−(1−フェニル−2−ヒドロキシ)エトキシ)−4−(2−フェニルエトキシ)]フェニル]メチレン]−4−オキソ−2−チオキソ−3−チアゾリジン酢酸が挙げられる。
【0051】
特定の実施形態においては、少なくとも1つの内在性PMT遺伝子の機能または発現が低減、破壊または欠失される。例えば、特定の実施形態においては、PMT1、PMT2、PMT3およびPMT4遺伝子からなる群から選択される少なくとも1つの内在性PMT遺伝子の機能または発現が低減、破壊または欠失され、あるいは該宿主細胞は1以上のPMTインヒビターの存在下で培養される。更に詳細な実施形態においては、該宿主細胞は1以上のPMT遺伝子の欠失または破壊を含み、該宿主細胞は1以上のPmtpインヒビターの存在下で培養される。これらの実施形態の特定の態様においては、該宿主細胞は分泌性アルファ−1,2−マンノシダーゼをも発現する。
【0052】
PMT欠失もしくは破壊および/またはPmtpインヒビターは、O−グリコシル化の占有を低減することにより、すなわち、グリコシル化される糖タンパク質上のO−グリコシル化部位の総数を減少させることにより、O−グリコシル化を制御する。該細胞により分泌されるアルファ−1,2−マンノシダーゼの更なる添加は、該糖タンパク質上に存在するO−グリカンのマンノース鎖長を減少させることにより、O−グリコシル化を制御する。したがって、PMT欠失もしくは破壊および/またはPmtpインヒビターを分泌性アルファ−1,2−マンノシダーゼの発現と組合せることは、占有および鎖長を減少させることによりO−グリコシル化を制御する。個々の状況においては、個々の異種糖タンパク質(例えば、抗体)は種々の度合の効率で発現されゴルジ装置から輸送される可能性があり、したがって、PMT欠失または破壊、Pmtpインヒビターおよびアルファ−1,2−マンノシダーゼの特定の組合せを要しうるため、PMT欠失または破壊、Pmtpインヒビターおよびアルファ−1,2−マンノシダーゼの個々の組合せは実験的に決定される。もう1つの態様においては、1以上の内在性マンノシルトランスフェラーゼ酵素をコードする遺伝子が欠失される。この欠失は分泌性アルファ−1,2−マンノシダーゼおよび/またはPMTインヒビターの供与と組合されることが可能であり、あるいは分泌性アルファ−1,2−マンノシダーゼおよび/またはPMTインヒビターの供与の代わりに行われうる。
【0053】
したがって、該宿主細胞は、糖タンパク質を産生するように遺伝的に修飾された任意の宿主細胞であることが可能であり、この場合、該主要N−グリカンは、複合N−グリカン、ハイブリッドN−グリカンおよび高マンノースN−グリカンからなる群から選択され、ここで、複合N−グリカンは、Man3GlcNAc2、GlcNAC(1−4)Man3GlcNAc2、Gal(1−4)GlcNAc(1−4)Man3GlcNAc2およびNANA(1−4)Gal(1−4)Man3GlcNAc2からなる群から選択され、ハイブリッドN−グリカンは、Man5GlcNAc2、GlcNAcMan5GlcNAc2、GalGlcNAcMan5GlcNAc2およびNANAGalGlcMan5GlcNAc2からなる群から選択され、高マンノースN−グリカンは、Man6GlcNAc2、Man7GlcNAc2、Man8GlcNAc2およびMan9GlcNAc2からなる群から選択される。N−グリカン構造の具体例には以下のものが含まれるが、それらに限定されるものではない:Man5GlcNAc2、GlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc2Man3GlcNAc2、GlcNAc3Man3GlcNAc2、GlcNAc4Man3GlcNAc2、GalGlcNAc2Man3GlcNAc2、Gal2GlcNAc2Man3GlcNAc2、Gal2GlcNAc3Man3GlcNAc2、Gal2GlcNAc4Man3GlcNAc2、Gal3GlcNAc3Man3GlcNAc2、Gal3GlcNAc4Man3GlcNAc2、Gal4GlcNAc4Man3GlcNAc2、NANAGal2GlcNAc2Man3GlcNAc2、NANA2Gal2GlcNAc2Man3GlcNAc2、NANA3Gal3GlcNAc3Man3GlcNAc2およびNANA4Gal4GlcNAc4Man3GlcNAc2。
【0054】
以下の実施例においては、糖操作されたピチア・パストリス(Pichia pastoris)株を、Ste13pおよびDap2pジペプチジルアミノペプチダーゼ活性を排除するために修飾した。一般に、Ste13pおよび/またはDap2p活性は、本明細書に記載されている方法を用いて、任意のピチア・パストリス(Pichia pastoris)株から排除されうる。ただし、選択に使用されるマーカーが該株と共に使用されうる。あるいは、選択を可能にする別の栄養要求性またはドミナントマーカーが代用されうる。例えば、ベクターpGLY5018(実施例2C)および選択マーカーとしてのナーセオスリシン(nourseothricin)を使用して、商業的に入手可能な株NRRL−Y11430からSTE13を欠失させることが可能である。後記のDAP2ノックアウトベクター(実施例2B)のいずれのマーカーもNRRL−Y11430に和合性でないため、pAG32から1654bpのHygマーカーを増幅するためにプライマーSH806およびSH807(表1)の斜字表示断片に対応するプライマー(それぞれ配列番号48および配列番号49)を使用してpAG32(Goldsteinら,Yeast,15(6):507−511,1999;Erratum:Yeast,15(12):1297,1999)からヒグロマイシン標識ベクターを作製することが可能である。ついで、この後者の断片を、pGLY5019の作製に関する実施例2Cに記載されているとおりに、DAP2 5’およびDAP2 3’フランキング領域に融合させることが可能である。SfiI(New England BioLabs,Ipswich,MA)での消化の後、該ベクターをNRRL−Y11430またはそのste13ノックアウト誘導体内に形質転換し、ヒグロマイシン含有プレート上で選択して、それぞれ単一dap2Δまたは二重Δste13/dap2ノックアウト株を得ることが可能である。本明細書に記載されている方法を用いてDppIIIpジペプチジルアミノペプチダーゼ活性を排除するために修飾された糖操作ピチア・パストリス(Pichia pastoris)株を当業者がどうのようにして使用しうるのかに関する一例も示す。
【図面の簡単な説明】
【0055】
【図1A】図1は、本発明における方法において使用されるベクターの図示である。Ura5標識PpSTE13およびPpDAP2ノックアウトベクターが、それぞれ図1Aおよび1BにpGLY4520およびpGLY4521として示されている。PpSTE13およびPpDAP2ナーセオスリシン(nourseothricin)標識ベクターが、それぞれ図1Cおよび1DにpGLY5018およびpGLY5019として示されている。形質転換体におけるノックアウト断片を切り出すために用いられるSfiI制限部位が下線で示されている。PpSTE13およびPpDAP2フランキング領域が黒で太く表示されている。
【図1B】図1は、本発明における方法において使用されるベクターの図示である。Ura5標識PpSTE13およびPpDAP2ノックアウトベクターが、それぞれ図1Aおよび1BにpGLY4520およびpGLY4521として示されている。PpSTE13およびPpDAP2ナーセオスリシン(nourseothricin)標識ベクターが、それぞれ図1Cおよび1DにpGLY5018およびpGLY5019として示されている。形質転換体におけるノックアウト断片を切り出すために用いられるSfiI制限部位が下線で示されている。PpSTE13およびPpDAP2フランキング領域が黒で太く表示されている。
【図1C】図1は、本発明における方法において使用されるベクターの図示である。Ura5標識PpSTE13およびPpDAP2ノックアウトベクターが、それぞれ図1Aおよび1BにpGLY4520およびpGLY4521として示されている。PpSTE13およびPpDAP2ナーセオスリシン(nourseothricin)標識ベクターが、それぞれ図1Cおよび1DにpGLY5018およびpGLY5019として示されている。形質転換体におけるノックアウト断片を切り出すために用いられるSfiI制限部位が下線で示されている。PpSTE13およびPpDAP2フランキング領域が黒で太く表示されている。
【図1D】図1は、本発明における方法において使用されるベクターの図示である。Ura5標識PpSTE13およびPpDAP2ノックアウトベクターが、それぞれ図1Aおよび1BにpGLY4520およびpGLY4521として示されている。PpSTE13およびPpDAP2ナーセオスリシン(nourseothricin)標識ベクターが、それぞれ図1Cおよび1DにpGLY5018およびpGLY5019として示されている。形質転換体におけるノックアウト断片を切り出すために用いられるSfiI制限部位が下線で示されている。PpSTE13およびPpDAP2フランキング領域が黒で太く表示されている。
【図2A】図2は、酵母におけるDAP活性に関連したTNFRII−FcのN末端切断の図示である。図2Aは無傷分泌性TNFRII−Fc(配列番号3)の7個のN末端アミノ酸(配列番号1)を示し、図2Bは該トランケート化産物の5個のN末端アミノ酸(配列番号2)を示す。矢印は、ジペプチジルアミノペプチダーゼDap2pおよびSte13pの両方により認識される切断部位を示す。
【図2B】図2は、酵母におけるDAP活性に関連したTNFRII−FcのN末端切断の図示である。図2Aは無傷分泌性TNFRII−Fc(配列番号3)の7個のN末端アミノ酸(配列番号1)を示し、図2Bは該トランケート化産物の5個のN末端アミノ酸(配列番号2)を示す。矢印は、ジペプチジルアミノペプチダーゼDap2pおよびSte13pの両方により認識される切断部位を示す。
【図3】図3は、腫瘍壊死因子受容体2(TNFRII)とIgG1 Fc領域(Fc)とを含む融合タンパク質TNFRII−Fcのアミノ酸配列(配列番号3)である。
【図4A】図4は、酵母におけるDAP活性に関連したGCSFのN末端切断の図示である。図4Aは無傷分泌性GCSF(配列番号6)の7個のN末端アミノ酸(配列番号4)を示し、図4Bは該トランケート化産物の5個のN末端アミノ酸(配列番号5)を示す。矢印は、ジペプチジルアミノペプチダーゼDap2pおよびSte13pの両方により認識される切断部位を示す。
【図4B】図4は、酵母におけるDAP活性に関連したGCSFのN末端切断の図示である。図4Aは無傷分泌性GCSF(配列番号6)の7個のN末端アミノ酸(配列番号4)を示し、図4Bは該トランケート化産物の5個のN末端アミノ酸(配列番号5)を示す。矢印は、ジペプチジルアミノペプチダーゼDap2pおよびSte13pの両方により認識される切断部位を示す。
【図5】図5は組換え顆粒球コロニー刺激因子(GSCF)タンパク質のアミノ酸配列(配列番号6)である。
【図6】図6は、STE13遺伝子もDAP2遺伝子も欠失も破壊もされていないピチア・パストリス(Pichia pastoris)株において産生されたGCSF、すなわち、DAP切断による低分子量GCSF(レーン27〜29)、ならびにそれと比較されている、Ste13pおよびDap2p活性が排除されたピチア・パストリス(Pichia pastoris)株において産生された無傷高分子量GCSF産物(レーン32〜34)のウエスタンブロットの表示である。
【図7A】図7A〜7Cは、STE13およびDAP2のノックアウト体の作製(実施例3)に使用される糖操作株YGLY7406に関する流れ図を表す。図7Cは、単一ノックアウト糖操作株ste13(YGLY8084)およびdap2(YGLY8090)ならびに後続の二重ノックアウト株(YGLY8096)に関する流れ図を表す。
【図7B】図7A〜7Cは、STE13およびDAP2のノックアウト体の作製(実施例3)に使用される糖操作株YGLY7406に関する流れ図を表す。図7Cは、単一ノックアウト糖操作株ste13(YGLY8084)およびdap2(YGLY8090)ならびに後続の二重ノックアウト株(YGLY8096)に関する流れ図を表す。
【図7C】図7A〜7Cは、STE13およびDAP2のノックアウト体の作製(実施例3)に使用される糖操作株YGLY7406に関する流れ図を表す。図7Cは、単一ノックアウト糖操作株ste13(YGLY8084)およびdap2(YGLY8090)ならびに後続の二重ノックアウト株(YGLY8096)に関する流れ図を表す。
【図8A−1】図8Aおよび8Bはピチア・パストリス(Pichia pastoris)DAP2のcDNA配列(配列番号37)およびアミノ酸配列(配列番号38)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図8A−2】図8Aおよび8Bはピチア・パストリス(Pichia pastoris)DAP2のcDNA配列(配列番号37)およびアミノ酸配列(配列番号38)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図8A−3】図8Aおよび8Bはピチア・パストリス(Pichia pastoris)DAP2のcDNA配列(配列番号37)およびアミノ酸配列(配列番号38)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図8B】図8Aおよび8Bはピチア・パストリス(Pichia pastoris)DAP2のcDNA配列(配列番号37)およびアミノ酸配列(配列番号38)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図9A−1】図9Aおよび9Bはピチア・パストリス(Pichia pastoris)STE13のcDNA配列(配列番号39)およびアミノ酸配列(配列番号0)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図9A−2】図9Aおよび9Bはピチア・パストリス(Pichia pastoris)STE13のcDNA配列(配列番号39)およびアミノ酸配列(配列番号0)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図9A−3】図9Aおよび9Bはピチア・パストリス(Pichia pastoris)STE13のcDNA配列(配列番号39)およびアミノ酸配列(配列番号0)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図9B】図9Aおよび9Bはピチア・パストリス(Pichia pastoris)STE13のcDNA配列(配列番号39)およびアミノ酸配列(配列番号0)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図10A】図10Aおよび10Bは、それぞれpGLY4511およびpGLY4512を作製するために増幅されるPpSTE13に関する5’(配列番号41)および3’(配列番号42)DNAフランキング領域である。該フランキング領域(下線部)自体は、PpSTE13 5’フランキング領域ではEcoRI制限部位を構成するヌクレオチドに隣接しており、PpSTE13 3’フランキング領域ではHindIII制限部位を構成するヌクレオチドに隣接している。
【図10B】図10Aおよび10Bは、それぞれpGLY4511およびpGLY4512を作製するために増幅されるPpSTE13に関する5’(配列番号41)および3’(配列番号42)DNAフランキング領域である。該フランキング領域(下線部)自体は、PpSTE13 5’フランキング領域ではEcoRI制限部位を構成するヌクレオチドに隣接しており、PpSTE13 3’フランキング領域ではHindIII制限部位を構成するヌクレオチドに隣接している。
【図11A】図11Aおよび11Bは、それぞれpGLY4513およびpGLY4514を作製するために増幅されるPpDAP2に関する5’(配列番号43)および3’(配列番号44)DNAフランキング領域である。該フランキング領域(下線部)自体は、PpDAP2 5’フランキング領域ではEcoRI制限部位を構成するヌクレオチドに隣接しており、PpDAP2 3’フランキング領域ではHindIII制限部位を構成するヌクレオチドに隣接している。
【図11B】図11Aおよび11Bは、それぞれpGLY4513およびpGLY4514を作製するために増幅されるPpDAP2に関する5’(配列番号43)および3’(配列番号44)DNAフランキング領域である。該フランキング領域(下線部)自体は、PpDAP2 5’フランキング領域ではEcoRI制限部位を構成するヌクレオチドに隣接しており、PpDAP2 3’フランキング領域ではHindIII制限部位を構成するヌクレオチドに隣接している。
【図12】図12は、pAG25から増幅されるナーセオスリシン(nourseothricin)マーカーカセットのcDNA配列(配列番号45)であり、該ORFが太字で示されている。
【図13−1】図13はピチア・パストリス(Pichia pastoris)DPPIIIのcDNA(配列番号54)配列である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図13−2】図13はピチア・パストリス(Pichia pastoris)DPPIIIのcDNA(配列番号54)配列である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【実施例1】
【0056】
株、培養条件および試薬
大腸菌(Escherichia coli)株TOP10(Invitrogen,Carlsbad,CA)またはXL10−Gold(Stratagene,Santa Clara,CA)を組換えDNA実験に使用した。制限および修飾酵素はNew England BioLabs,Ipswich,MAから入手し、製造業者の指示どおりに使用した。オリゴヌクレオチドはIntegrated DNA Technologies,Coralville,IAから入手した。塩および緩衝剤はSigma,St.Louis,MOから得た。ここで使用した最少培地は1.4% 酵母窒素ベース、2% デキストロース、1.5% 寒天および4×10−5% ビオチンおよびアミノ酸(適宜補足されるもの)を含むものであった。YMDに富む培地は1% 酵母エキス、2% マルトン(martone)、2% デキストロースおよび1.5% 寒天(プレート用)である。ナーセオスリシン(nourseothricin)はUS Biologicals,Swampscott,MA(カタログ番号N5375−74)から得られ、YMDに富む培地に100μg/mlの最終濃度で加えられる。
【実施例2】
【0057】
ノックアウトベクターの作製
A.ste13::URA5ノックアウトベクターの作製
PfuUltra(商標)DNAポリメラーゼ(Stratagene,Santa Clara,CA)および鋳型としてのピチア・パストリス(Pichia pastoris)株NRRL−Y11430由来のゲノムDNAを使用して、STE13オープンリーディングフレームに対応する5’および3’フランキング領域に対応するDNA断片(配列番号41および42)を増幅した。表1に示されているプライマーペアSH774(配列番号13)およびSH775(配列番号14)ならびにSH776(配列番号15)およびSH777(配列番号16)を使用して、それぞれSTE13の5’および3’に関する771bpおよび949bpの断片を増幅した。ExTaq(商標)(TaKaRa,Bio.Inc.,Japan)の存在下の72℃で10分間のインキュベーションの後、該増幅断片をpCR2.1(Invitrogen,Carlsbad,CA)内にクローニングし、TOP10コンピテント細胞内に形質転換した。DNA配列決定は、STE13 5’およびSTE13 3’フランキング領域が正確であることを証明した。得られたベクターをそれぞれpGLY4511およびpGLY4512と命名した。
【0058】
763bpのSTE13 5’フランキング領域断片(図10A、配列番号50の下線領域により表される)を、EcoRIを使用してpGLY4511から消化し、同じ制限酵素で予め消化され子ウシ腸アルカリホスファターゼ(CIAP)で処理された、pJN396(NettおよびGerngross,Yeast,20,1279−1290,2003)に類似したピチア・パストリス(P.pastoris)URA5−ブラスターベクターpGLY13b内にサブクローニングした。URA5−ブラスターベクターは、5−フルオロオロト酸の存在下の対抗選択によるURA5マーカーの回収を促進するためのLacZ直接反復に隣接したピチア(Pichia)URA5遺伝子の機能性遺伝子断片を含有するpUC19ベクターバックボーンを有する。その新たに連結されたベクターをXL10 Goldコンピテント細胞内に形質転換し、制限分析により確認した後、得られたベクターをpGLY4518と命名した。ベクターpGLY4512を、STE13 3’フランキング領域をコードする940bp(図10B、配列番号51の下線領域により表される)断片を遊離させるためにHindIIIで消化し、同じ酵素で予め消化されCIAP処理されたpGY4518内にサブクローニングした。該連結産物をXL10 Goldコンピテント細胞内に形質転換し、制限分析後にpGLY4520と命名した。この最終的なste13::Ura5ノックアウトベクターを図1Aに図示する。
【0059】
B.dap2::URA5ノックアウトベクターの作製
表1に示されているプライマーセットSH782(配列番号21)およびSH783(配列番号22)ならびにSH784(配列番号23)およびSH785(配列番号24)を使用して、DAP2 5’および3’フランキング領域(配列番号43および44)を、前記のとおりにピチア・パストリス(Pichia pastoris)ゲノムDNAから増幅して、それぞれ1003bpおよび1142bpの断片を得た。pCR2.1内へのクローニングおよび配列決定の後、該ベクターをpGLY4513およびpGLY4514(それぞれDAP2 5’およびDAP2 3’領域をコードする)と命名した。前記実施例2Aに類似したアプローチに従い、995bpのDAP2 5’領域(図11A、配列番号52の下線領域により表される)をUra5−ブラスターベクターpGLY13b内のEcoRI部位内にサブクローニングして、中間構築物pGLY4519を得た。ついで1133bpのDAP2 3’領域(図11B、配列番号53の下線領域により表される)をpGLY4519のHindIII部位内にサブクローニングして、dap2::URA5ノックアウトベクターpGLY4521(図1Bに図示されている)を得た。
【0060】
C.ドミナントマーカーSTE13およびDAP2ノックアウトベクターの作製
PCR融合体を使用して、STE13およびDAP2の両方のノックアウトベクターを作製した。表1に示されているプライマーセットSH774(配列番号13)およびSH801(配列番号29)ならびにSH804(配列番号32)およびSH777(配列番号16)を使用し、PfuUltra(商標)DNAポリメラーゼを使用して、STE13 5’およびSTE13 3’断片をpGLY4520から増幅した。アシビア・ゴシッピィ(Ashbya gossypii)転写伸長因子(TEF)プロモーターおよびターミネーター(図12の通常文字により表される)(それぞれ、太字のオープンリーディングフレームの5’および3’側に位置する)の発現制御下でストレプトマイセス・ノウルセイ(Streptomyces noursei)からのナーセオスリシン(nourseothricin)アセチルトランスフェラーゼのオープンリーディングフレーム(図12、配列番号54の太字で表されている)を含有するナーセオスリシン(NATR)マーカーカセット(配列番号45)を、プライマーSH802(配列番号30)およびSH803(配列番号31)を使用して、pAG25(GoldsteinおよびMcCusker,Yeast,15,1541−1553,1999)から増幅した。該PCR反応をDNAアガロースゲル上で行い、それぞれSTE13 5’、STE13 3’およびNatマーカーに対応する779bp、958bpおよび1249bpの断片を単離した。ついでそれぞれ20ngを合わせ、PfuUltra(商標)DNAポリメラーゼならびにプライマーペアSH774(配列番号13)およびSH777(配列番号16)を使用して互いに融合させた。ExTaq(商標)DNAポリメラーゼ(TaKaRa,Bio.Inc.,Japan)の存在下の72℃で10分間のインキュベーションの後、該増幅(2896bp)断片をpCR2.1内にクローニングし、TOP10コンピテント細胞内に形質転換した。DNA配列決定は、該ste13::NATR融合体が正確であることを証明した。得られたベクターをそれぞれpGLY5018と命名した。このベクターを図1Cに図示する。
【0061】
同様にして、表1に示されているプライマーセットSH782(配列番号21)およびSH805(配列番号33)、SH808(配列番号36)およびSH785(配列番号24)、ならびにSH806(配列番号34)およびSH807(配列番号35)を使用して、DAP2 5’、DAP2 3’およびNATRマーカーに対応する1011bp、1151bpおよび1248bpの断片をpGLY4521およびpAG25から増幅した。単離後、各断片の20ngをプライマーペアSH782(配列番号21)およびSH785(配列番号24)と共に使用して3321bpの断片を得、これをpCR2.1内にクローニングし、配列決定し、pGLY5019と命名した。このベクターを図1Dに図示する。
【実施例3】
【0062】
STE13およびDAP2ノックアウト株の作製
GS5.0グリカンを発現するピチア・パストリス(Pichia pastoris)栄養要求性糖操作細胞系YGLY7406[Δoch1、Δpno1、Δmnn4B、Δbmt2、Δura5、K.lactisおよびM.musculus UDP−GlcNAc輸送体、M.musculus α−1,2−MnsI、H.sapiens β−1,2−GlcNAcトランスフェラーゼI、R.norvegicus β−1,2−GlcNAcトランスフェラーゼII、D.melanogaster MnsII、S.pombe Galエピメラーゼ、D.melanogaster UDP−Gal輸送体およびH.sapiens β−1,4−ガラクトシルトランスフェラーゼ](例えば、Bobrowiczら,Glycobiology,14(9):757−766,2004;Hamiltonら,Science,313(5792):14411−1443,2006);米国公開出願第20060040353号を参照されたい)を全ての操作の出発株として使用した。この株および後続の株をどのようにして作製したかに関する流れ図は、図7(A〜C)を参照されたい。GS5.0株は、一方または両方の末端の非還元性末端上のβ−1,4−ガラクトース残基を末端とする二分岐アフコシル化N−結合グリカンを有する糖タンパク質を産生しうる(Bobrowiczら,Glycobiology,14(9):757−766,2004;Hamiltonら,Science,313(5792):14411−1443,2006)。株YGLY7406は、ジペプチジルペプチダーエ活性に関するレポータータンパク質として使用される、IgG1のFcドメインに融合した完全長ヒト腫瘍壊死因子受容体II(TNFRII−Fc)を発現する。
【0063】
形質転換のために、20μgのベクターpGLY4520およびpGLY4521を制限酵素SfiI(New England Biolabs,Ipswich,MA)で消化して、4091bpのSTE13::Ura5ノックアウト断片または4516bpのDAP2::URA5ノックアウト断片を得、これらをYGLY7406内にエレクトロポレーションにより形質転換して、それぞれSTE13またはDAP2を欠失させ、該形質転換体をuraマイナス最少プレート上で選択した。各遺伝子のノックアウトの成功を、表2に示されている5’、3’およびノックアウトプライマーセットを使用して確認した。ste13Δおよびdap2Δノックアウト株を、それぞれYGLY8084およびYGLY8090と命名した。ついで、ste13Δノックアウト株YGLY8084を、3290bpのDAP2::Natノックアウト断片を遊離させるためにSfiIで予め消化されたpGLY5019で形質転換することにより、二重Δste13/dap2ノックアウト株を作製した。形質転換体を100μg/ml Nat YMDプレート上でプレーティングし、二重ノックアウトの成功を、表2に示されている5’、3’およびノックアウトプライマーセットを使用して確認した。代表的な二重ノックアウト株をYGLY8096と命名した。最初にSTE13をノックアウトし、ついでDAP2をノックアウトすることによりこの株を作製したが、本発明者らは、最初にDAP2遺伝子をノックアウトし、ついでSTE13遺伝子をノックアウトした場合に、同じ所望の表現型を有する株が得られうることを実験的に実証している。そのような株を得るために、pGLY5018をSfiIで消化し、生じた2865bpのste13::NATRノックアウト断片を、DAP2遺伝子が既にノックアウトされているYGLY8090のような株内に形質転換することが可能である。
【実施例4】
【0064】
レポータータンパク質の作製、単離および分析
1% 酵母エキス、2% ペプトン、100mM リン酸カリウムバッファー(pH6.0)、1.34% 酵母窒素ベース、0.00004% ビオチンおよび1% グリセロールからなる増殖培地としての200mlの緩衝化グリセロール−複合培地(BMGY)内でタンパク質発現を26℃で72時間行った。BMGYにおけるグリセロールの代わりに1.5% メタノールからなる20mlの緩衝化メタノール−複合培地(BMMY)内で誘導を48時間行った。
【0065】
発現後、2000rpmで15分間の遠心分離により該細胞を除去した。GE Healthcare(Chalfont St.Giles,UK,Cat #17−1281−01)のStreamliner Protein A樹脂を使用して、TNFRII−Fc融合タンパク質を上清からアフィニティークロマトグラフィーにより捕捉した。該無細胞上清培地をStreamliner Protein Aカラム(XK 16/20 1.6cm×10.0cm)上にローディングし、3カラム体積の20mM Tris−HCl(pH7.0)で5.0ml/分の流量で前平衡化した。該カラムを3カラム体積の同じバッファーで洗浄し、TNFRII−Fc融合タンパク質を7カラム体積の40mM クエン酸ナトリウム(pH3.5)で溶出した。溶出した融合タンパク質を直ちに1M Tris−HCl(pH8.0)で中和した。
【0066】
CHT(登録商標)ヒドロキシアパタイトI型40μm樹脂(Bio−Rad Laboratories,Hercules,CA,Cat #157−0040)を第2精製工程として使用した。該ヒドロキシアパタイトカラムを3カラム体積の5mM リン酸ナトリウム(pH6.5)で平衡化し、Streamliner Protein A精製TNFRII−Fc融合タンパク質を該平衡化バッファー内にバッファー交換し、該カラムに適用した。ローディング後、該カラムを3カラム体積の該平衡化バッファーで洗浄し、溶出を、0〜1000mM 塩化ナトリウムの範囲の20カラム体積にわたる勾配を展開することにより行った。TNFRII−Fc融合タンパク質は約550−650mM 塩化ナトリウムで溶出する。プール化したTNFRII−Fc融合タンパク質を、0.2μm ポリエーテルスルホン(PES)膜フィルターを使用して滅菌濾過し、4℃で保存した。
【0067】
SDS−PAGE(4〜20% Tris−HClゲル,Bio−Rad Laboratories,Hercules,CA,Cat.#161−1123)を、それらの2つのカラム精製TNFRII−Fc融合タンパク質画分に対して行い、ポリビニリデンジフルオリド(PVDF)膜(ProBlott(商標) Membranes,Applied Biosystems,Foster City,CA,Cat #400994)上に55ボルトで30分にわたってトランスファーし、ProBlott染料(Applied Biosystems,Foster City,CA)で染色した。TNFRII−Fc融合タンパク質に対応するバンドをPVDF膜から切り出し、N末端配列決定のためにTufts Core Facility,Boston,MAに送った。
【0068】
同様にして、完全長組換え顆粒球コロニー刺激因子(GCSF)タンパク質の発現を行った。しかし、TNFRII−Fcを発現させるために使用した前記のGS5.0宿主細胞の代わりに、GCSFを発現させるために使用した宿主細胞は、実施例3に記載の方法によりSTE13およびDAP2遺伝子が破壊されているピチア・パストリス(Pichia pastoris)GS2.0宿主細胞であった。GS2.0宿主細胞は、Man5GlcNAc2 N−グリカンを含む糖タンパク質を産生するように遺伝的に操作された宿主細胞である。これらの株は、ガラクトースを末端とするN−グリカンを有する糖タンパク質を産生し得ない。そのような株の具体例はNettおよびGerngross,Yeast 20:1279(2003);Choiら,Proc.Natl.Acad.Sci.USA 100:5022(2003);ならびにHamiltonら,Science 301:1244(2003)に開示されている。図6は、TNFRII−Fcを発現するGS5.0に関して前記で示されているとおりにSTE13およびDAP2遺伝子が破壊された場合、これらの宿主細胞が無傷GCSFを産生しうることを示している(STE13/DAP2株において産生されたGCSFを示すレーン27〜29を、ste13/dap2ノックアウト株において産生されたGCSFを示すレーン32〜34と比較されたい)。
【実施例5】
【0069】
PpDppIIIpの欠失
同様に、PpSte13pおよびPpDap2pを排除するための前記実施例に記載されている方法を、PpDppIIIpを排除するために用いることが可能である。1つのそのような方法は、実施例2でPpSTE13およびPpDAP2に関して示されているのに類似した方法でノックアウトベクターを設計することであろう。プライマーは、PpDPPIII遺伝子(図13、配列番号54)の5’および3’フランキング領域を増幅するように設計されるであろう。なぜなら、ゲノム配列の必須領域、例えば、機能的PpDppIIIp活性に必要な領域は省かれるからである。PpDppIIIp活性は、これらのフランキング領域を選択マーカーと組合せることにより排除されうる。適合性選択マーカーとの組合せにより、PpDppIIIp活性は、個々に、あるいはPpSte13pおよび/またはPpDap2活性の排除と共に排除されうる。PpSte13p、PpDap2pおよびPpDppIIIp活性が排除されている株は、いずれかの潜在的ジペプチジルアミノペプチダーゼ活性を欠く株を与えるであろう。
【0070】
本発明は、本発明の少数の態様の例示として意図される実施例に開示されている具体的な実施形態によっては限定されず、機能的に同等なあらゆる実施形態が本発明の範囲内である。実際、本明細書に示され記載されているものに加えて本発明の種々の修飾が当業者に明らかとなり、添付の特許請求の範囲の範囲内に含まれると意図される。
【技術分野】
【0001】
本発明は、ヒトまたは動物治療用物質として有用である糖タンパク質の、ジペプチジルアミノペプチダーゼ(DAP)活性を有さない酵母細胞系における製造のための方法および組成物に関する。
【背景技術】
【0002】
酵母は組換えタンパク質の製造のための重要な生産媒体である。酵母は真核生物であるため、それは、分泌経路で生じる翻訳後修飾の多くを含め、より高等な真核生物のものと共通の進化的過程を共有している。糖操作における最近の進歩は、ヒトにおける糖タンパク質の過程によく似た一連の酵素反応を可能にする遺伝的に修飾されたグリコシル化経路を有する酵母株ピチア・パストリス(Pichia pastoris)の細胞系を生み出している。例えば、ヒト対応物と実質的に同一である組換え糖タンパク質の、下等真核宿主細胞における製造方法を記載している米国特許第7,029,872号および第7,326,681号を参照されたい。前記方法からピチア・パストリス(Pichia pastoris)において産生されるようなヒト様シアル酸化二分岐複合体N−結合グリカンは治療用糖タンパク質の製造のための有用性を実証している。
【0003】
高等真核生物と同様に、酵母も多数のプロテアーゼを発現し、それらのうちの多くは分泌経路に局在化し、あるいはそれを通過してそれらの最終目的地へと向かう。その結果、幾つかの組換えタンパク質の望ましくないタンパク質分解が生じる可能性があり、該特異的切断は、関与するプロテアーゼのクラスに左右される。ジペプチジルアミノペプチダーゼ(DAP)は、タンパク質のN末端から2つのアミノ酸ペプチドを除去するタンパク質分解酵素のクラスである。サッカロミセス・セレビシエ(Saccharomyces cerevisiae)においては、酵素STE13およびDAP2の遺伝子がDAP活性を有するものとして特定されている。Juliusら,Cell,32:839−852,1983;Renduelesら,J.Bacteriology,169:4041−4048,1987を参照されたい。本出願者は、完全長治療用タンパク質の製造を可能にする、ピチア・パストリス(Pichia pastoris)におけるDAP活性の排除のための方法を開発した。
【発明の概要】
【0004】
発明の概括
1つの実施形態においては、本発明は、ジペプチジルアミノペプチダーゼ(DAP)活性を欠く酵母細胞系における治療用タンパク質の製造方法である。この実施形態は、DAP活性が排除されている遺伝的に修飾されたピチア・パストリス(Pichia pastoris)細胞系を、該治療用タンパク質をコードするポリヌクレオチドベクターで形質転換し、該形質転換細胞を培養して該治療用タンパク質を得ることを含む。DAP活性は、STE13、DAP2およびDPPIIIが欠失または破壊されるようにピチア・パストリス(Pichia pastoris)細胞系を修飾することにより排除されうる。もう1つの実施形態においては、DAP活性は、STE13およびDAP2が欠失または破壊されるようにピチア・パストリス(Pichia pastoris)細胞系を修飾することにより排除される。
【0005】
1つの実施形態においては、本発明は、STE13およびDAP2の欠失により組換え的に修飾されたピチア・パストリス(Pichia pastoris)細胞系を含む、治療用タンパク質の製造に使用されうる、DAP活性を欠く遺伝的に修飾された酵母細胞系である。
【0006】
更に他の実施形態においては、本発明は、TNFRII−Fc、すなわち、IgG1 Fcドメインに融合した腫瘍壊死因子受容体2(TNFRII)のエクトドメインを含む組換え融合タンパク質(TNFRII−Fc)、または顆粒球コロニー刺激因子(GCSF)の分泌血漿形態を含む組換えGCSFポリペプチドのような、ピチア・パストリス(Pichia pastoris)からの治療用タンパク質の製造方法である。
【0007】
発明の詳細な説明
定義
本明細書中で用いる「ジペプチジルアミノペプチダーゼ活性」または「DAP活性」なる語は、STE13、DAP2またはDPPIIIと称される遺伝子により産生されるポリペプチドの酵素的切断を意味する。
【0008】
本明細書中で用いる「ジペプチジルアミノペプチダーゼ活性の排除」または「DAP活性の排除」なる語は、STE13、DAP2またはDPPIIIと称される遺伝子により生じる酵素活性の非存在を意味する。
【0009】
本明細書中で用いる「治療用タンパク質」なる語は、動物またはヒトにおける疾患または状態を治療するための治療用物質として使用されうる完全長(すなわち、非トランケート化形態)の生物学的に活性なポリペプチドを意味する。本明細書中で用いられるこの用語の具体例としては、腫瘍壊死因子受容体2(TNFRII)とIgG1のFc領域(Fc)とを含む融合タンパク質TNFRI−Fc、および組換え顆粒球コロニー刺激因子(GCSF)タンパク質が挙げられる。
【0010】
本明細書中で用いる「N末端認識部位」なる語は、モチーフX−ProまたはX−Ala(ここで、Xは任意のアミノ酸であり、N末端に対して2番目の位置はプロリンまたはアラニンである)のN末端配列を有するポリペプチドを意味する。
【0011】
本明細書中で用いる「糖操作(された)ピチア・パストリス(Pichia pastoris)株」なる語は、ヒト糖タンパク質を発現するように操作されたピチア・パストリス(Pichia pastoris)株を意味する。代表的な株には、YJN201(Choiら,PNAS,100(9):5022−5027,2003);YSH44(Hamiltonら,Science,301(5637):1244−1246,2003);RDP36−1(Davidsonら,Glycobiology,14(4):1−9,2004);PBP6−5(Bobrowiczら,Glycobiology,14(9):757−766,2004);YSH597(Hamiltonら,Science,313(5792):1441−1443,2006)が含まれる。
【0012】
本明細書中で用いる「野生型株」なる語は、遺伝子STE13、DAP2またはDPPIIIがゲノムから改変、破壊または欠失されていない糖操作ピチア・パストリス(Pichia pastoris)株を意味する。
【0013】
本明細書中で用いる「欠失株」なる語は、機能性DAP活性を排除するために1つ、2つ又は全てのDAP遺伝子(すなわち、STE13およびDAP2およびDPPIII)のピチア・パストリス(Pichia pastoris)ホモログがゲノムレベルで修飾されている糖操作ピチア・パストリス(Pichia pastoris)株を意味する。これは、該遺伝子(プロモーター、オープンリーディングフレームおよびターミネーターを含む)の完全または部分的な欠失、該遺伝子またはコード化mRNAのそれぞれ転写または翻訳を改変する1以上の突然変異の導入、および該タンパク質活性を不活性化する1以上の突然変異の導入を含むが、これらに限定されるものではない。そのような欠失株の一例はYGLY8084である。
【0014】
本明細書中で用いる「欠失または破壊された」および「欠失または破壊」なる語は、酵母細胞ゲノムから産生される酵素の活性の抑制を意味し、該酵素活性の抑制は、基質タンパク質が無傷N末端を保有するまでのものに限られる。酵母宿主細胞において該酵素活性が阻害または破壊されうる具体例としては以下のものが含まれる(それらに限定されるものではない)、1)該遺伝子の発現を制御する上流または下流調節配列の欠失または破壊(これらに限定されるものではない);2)該遺伝子を非機能性にする、該酵素活性をコードする遺伝子の突然変異(ここで、「突然変異」は、該遺伝子に関する酵素活性を不能にする該遺伝子内への欠失、置換、挿入または付加を含む);3)化学的インヒビター、ペプチドインヒビターまたはタンパク質インヒビターによる該酵素活性の阻害または破壊;4)核酸に基づく発現インヒビター(例えば、アンチセンスDNAおよびsiRNA)による該酵素活性の阻害または破壊;5)転写インヒビター、または該酵素活性をコードする遺伝子の発現を制御もしくは調節する調節因子の発現もしくは活性のインヒビターによる該酵素活性の阻害または破壊;および6)得られる産物が、たとえ発現されたとしても、該分泌タンパク質と同一ではない、および機能が低下している、いずれかの手段。
【0015】
略語
以下の略語が本明細書に全体において使用される。
URA5:オロタートホスホリボシルトランスフェラーゼ(OPRTアーゼ)アイソザイム
ScSUC2:サッカロミセス・セレビシエ(S.cerevisiae)インベルターゼ
OCH1:アルファ−1,6−マンノシルトランスフェラーゼ
KlGlcNAcTr:ケイ・ラクチス(K.lactis)UDP−GlcNAc輸送体
BMT2:ベータ−マンノース−転移(ベータ−マンノース排除)
MNN4B:MNN4A様遺伝子(電荷排除)
MmGlcNAcTr:UDP−GlcNAc輸送体のマウスホモログ
PNO1:N−グリカンのホスホマンノシル化(電荷排除)
MNN4A:マンノシルトランスフェラーゼ(電荷排除)
ADE1:N−スクシニル−5−アミノイミダゾール−4−カルボキサミドリボチドシンテターゼ
MNS1:ScSEC12リーダーに融合したマウスマンノシダーゼIA触媒ドメイン
GnTI:PpSEC12リーダーに融合したヒトGlcNAcトランスフェラーゼI触媒ドメイン
HIS1:ATPホスホリボシルトランスフェラーゼ
GalTI:ScKRE2リーダーに融合したトランケート化ヒトガラクトシルトランスフェラーゼ1触媒ドメイン
GalE:サッカロミセス・セレビシエ(S.cerevisiae)UDP−グルコース4−エピメラーゼ
UDP−GalTr:UDP−ガラクトース輸送体
ARG1:アルギノスクシナートシンテターゼ
MNSII:ScMNN2リーダーに融合したドロソフィラ(Drosophila)マンノシダーゼII触媒ドメイン
GnTII:ScMNN2リーダーに融合したラットGlcNAcトランスフェラーゼII触媒ドメイン
PRO1:ガンマ−グルタミルキナーゼ
TrMNS1:ScαMATに融合した分泌性ティー・レーセイ(T.reseei)マンノシダーゼI触媒ドメイン
AOX1:アルコールオキシダーゼI
TNFRII−Fc:IgG1のFcドメインに融合したヒト腫瘍壊死因子受容体II
Zeo:ゼオシン耐性マーカー
STE13:ジペプチジルアミノペプチダーゼ
DAP2:ジペプチジルアミノペプチダーゼ
DPPIII:ジペプチジルアミノペプチダーゼ
Nat:ナーセオスリシン(nourseothricin)耐性マーカー。
【0016】
酵母における治療用タンパク質の製造
ヒトまたは他の動物から単離されるタンパク質の大部分はグリコシル化されている。治療に使用されるタンパク質のうちの約70%はグリコシル化されている。治療用タンパク質が酵母のような微生物宿主において産生され、内在性経路を用いてグリコシル化される場合、典型的には、その治療効力は著しく低下する。それにもかかわらず、そのような糖タンパク質はヒトにおいて免疫原性であり、投与後のインビボにおける半減期の減少を示す(Takeuchi,Trends in Glycoscience and Glycotechnology,9:S29−S35,1997)。
【0017】
ヒトおよび動物における特定の受容体は末端マンノース残基を認識し、血流からの該タンパク質の迅速なクリアランスを促進しうる。他の有害作用には、タンパク質フォールディング、溶解度、プロテアーゼに対する感受性、トラフィッキング、輸送、区画化、分泌、他のタンパク質もしくは因子による認識またはアレルゲン性の変化が含まれうる。したがって、グリコシル化のパターンが、ヒトまたは意図される被投与種において生じるものと同じ又は少なくとも類似した動物宿主系において治療用糖タンパク質を産生させることが必要である。ほとんどの場合、哺乳類宿主系、例えば哺乳類細胞培養が使用される。使用される系には、チャイニーズハムスター卵巣細胞(CHO)、トランスジェニック動物、植物または昆虫細胞が含まれる。そのような系において発現される組換えヒトタンパク質には更に、非ヒトグリコフォーム(glycoform)が含まれうる(Rajuら,Annals Biochem.,283(2):123−132,2000)。これらの系の全ては重大な欠点を有する。それらの欠点には、ある治療用タンパク質だけが動物または植物系における発現に適していること、該細胞培養系は典型的には非常に遅いこと、タンパク質収率が微生物発酵法に比べて低く、複雑で高価な栄養または添加物を要しうること、またはそのような系は汚染されやすいことが含まれる(これらに限定されるものではない)。
【0018】
したがって、適当な発現の欠如は、あらゆる治療用糖タンパク質の製造に対する障害となっている。微生物の発酵による糖タンパク質の製造は、高濃度のタンパク質の迅速な製造、無菌の十分に制御された製造条件または化学的に規定された増殖培地が使用可能であること、多種多様なタンパク質の発現が可能であること、および該治療用タンパク質の容易な回収(これらに限定されるものではない)を含む、既存の系に比べた場合の利点をもたらすであろう。しかし、前記のとおり、細菌系は真核生物のようにはグリコシル化治療用タンパク質を産生しない。その目的のために、ヒト糖タンパク質を産生する遺伝的に修飾されたグリコシル化経路を有する酵母細胞系、特にピチア・パストリス(Pichia pastoris)が開発されている。例えば、対応ヒト対応物と実質的に同一である組換え糖タンパク質の、下等真核宿主細胞における製造方法を記載している米国特許第7,029,872号および第7,326,681号ならびにUS 2006−0286637を参照されたい。前記方法によりピチア・パストリス(Pichia pastoris)において産生されるようなヒト様シアル酸化二分岐複合N−結合グリカンは治療用糖タンパク質の製造のための有用性を示している。
【0019】
前記ピチア・パストリス(Pichia pastoris)細胞系は、タンパク質を治療用物質としての使用に適したものにするヒト様グリコシル化パターンを有するタンパク質を産生しうるが、ピチア・パストリス(Pichia pastoris)における治療用タンパク質の発現は必ずしも、完全長ポリペプチドの産生をもたらさない。酵母発現系を効率的に使用するためには、治療用タンパク質をトランケート化するタンパク質分解のような望ましくない酵素活性が排除される必要がある。
【0020】
ジペプチジルアミノペプチダーゼ活性を欠くピチアにおける治療用タンパク質の製造方法
酵母細胞系ピチア・パストリス(Pichia pastoris)から分泌される組換え融合タンパク質TNFRII−Fcのペプチド配列(その配列は図3(配列番号3)に示されている)の分析は、産生されるペプチドの全てがN末端において2アミノ酸だけトランケート化されていたことを示した。本出願人は、ピチア・パストリス(Pichia pastoris)細胞系における2つの遺伝子STE13およびDAP2が全てのDAP活性を排除し、完全長TNFRII−Fcの産生をもたらすことを確認した。出願人はまた、ピチア・パストリス(Pichia pastoris)が第3のジペプチジルアミノペプチダーゼDPPIIIを有することを確認している。したがって、1つの実施形態においては、本発明は、DAP活性を欠く酵母細胞系における治療用タンパク質の製造方法である。この実施形態は、DAP活性が排除された遺伝的に修飾されたピチア・パストリス(Pichia pastoris)細胞系を、治療用タンパク質をコードするポリヌクレオチドベクターで形質転換し、該形質転換宿主細胞を培養して、該治療用タンパク質を産生させることを含む。DAP活性は、STE13およびDAP2および/またはDPPIIIが欠失または破壊されるようにピチア・パストリス(Pichia pastoris)細胞系を修飾することにより排除される。本発明の特定の実施形態においては、DAP活性は、STE13およびDAP2が欠失または破壊されるようにピチア・パストリス(Pichia pastoris)細胞系を修飾することにより排除された。
【0021】
もう1つの実施形態においては、本発明は、STE13およびDAP2遺伝子が欠失または破壊され全ての生じるDAP活性が排除されるように組換え的に修飾されたピチア・パストリス(Pichia pastoris)細胞系を含む、治療用タンパク質の製造に使用される酵母細胞系である。
【0022】
更に他の実施形態においては、本発明は、ポリペプチドのN末端に対して2番目の位置にアミノ酸プロリンまたはアラニンを有する治療用タンパク質、例えば、本明細書に記載されているTNFRII−Fc融合タンパク質またはGCSFタンパク質の、ピチア・パストリス(Pichia pastoris)からの製造方法である。
【0023】
ジペプチジルアミノペプチダーゼ活性
ポリペプチドのアミノ末端から2つのアミノ酸をトランケート化することが知られているプロテアーゼのサブクラスであるジペプチジルアミノペプチダーゼ(DAP)の活性に関連した遺伝子が酵母において特定されている。サッカロミセス・セレビシエ(S.cerevisiae)の非接合性アルファ−細胞突然変異体は、アルファ接合因子フェロモンの不完全なプロセッシングによるものであると考えられている(Juliusら,Cell,32(3),839−52,1983)。更に、Ste13p活性を欠損したサッカロミセス・セレビシエ(S.cerevisiae)突然変異株をスクリーニングすることにより、第2のジペプチジルペプチダーゼDap2pが特定された(Suarez RenduelesおよびWolf,Journal of Bacteriology,169(9),4041−48,1987)。サッカロミセス・セレビシエ(S.cerevisiae)STE13のピチア(Pichia)ホモログのノックアウトが、N末端にアミノ酸HG(His−Gly)を有するタンパク質のインビボでのタンパク質切断を妨げ、完全長インスリン指向性(insulinotropic)ペプチドの産生を可能にしたことも報告されている(Melarkodeら,WO 2007/148345;Prabhaら,Protein Expression and Purification,64,155−161,2009)。逆に、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)のDAP2ピチア(Pichia)ホモログの破壊はN末端タンパク質分解切断を妨げなかった(Melarkodeら,WO 2007/148345;Prabhaら,Protein Expression and Purification,64,155−161,2009)。
【0024】
Ste13pおよびDap2pは、モチーフX−P/A(ここで、Xは任意のアミノ酸であり、2番目の位置はプロリン(P)またはアラニン(A)である)を有するN末端ペプチドを切断することが別々に示されている(MisumiおよびIkehara,Handbook of Proteolytic Enzvmes,2nd edition,pp.1910−1911,Elsevier,London,2004)。本発明を例示するために本明細書中で用いるTNFRII−FcおよびGCSFタンパク質は、保存されたX−Pro Ste13pおよびDap2pモチーフに合致しており、このことは、ピチア・パストリス(Pichia pastoris)における治療用タンパク質の製造のためには、Ste13pまたはDap2pのDAP活性が排除される必要があることを示唆している。これとは対照的に、Melarkodeら,WO 2007/148345のインスリン指向性ペプチドは、Ste13pのみにより認識されるHis−Glyモチーフを有する新規非保存性N末端を有する。
【0025】
本出願人により本明細書中で示されているとおり、ピチア・パストリス(Pichia pastoris)Ste13p活性の排除はTNFRII−Fcタンパク質に対するDAP活性の部分的低下をもたらしたに過ぎず、産生されたTNFRII−FCタンパク質の半分より幾らか多くが完全長配列を有していた。驚くべきことに、本出願人は、DAP活性の完全な排除、すなわち、完全長TNFRII−Fcタンパク質の100%の産生のためには、Ste13pおよびDap2pの両方の活性がピチア・パストリス(Pichia pastoris)細胞系から排除される必要があることを見出した。同様に、両方のDAP遺伝子がピチア・パストリス(Pichia pastoris)から排除された場合にのみ、本出願人は組換えGCSFタンパク質の完全な産生を観察した。したがって、N末端X−P/Aモチーフを有する完全長タンパク質の産生のためにはピチア・パストリス(Pichia pastoris)におけるSte13pおよびDap2p活性の両方の排除を要する点で本発明は先行技術とは異なる、と当業者は認識し理解するであろう。
【0026】
公知DAP遺伝子における相同性に基づいて、当業者はPCRプライマー(それらの具体例を表1に示す)を設計し、あるいは標的生物のDNAライブラリーにおけるホモログを特定するためのプローブとして遺伝子または遺伝子断片を使用することが可能である。DAP活性を有するホモログを特定するためのピチア・パストリス(Pichia pastoris)ゲノムの本出願人の分析は、それぞれPpSTE13およびPpDAP2(図8Aおよび8B、配列番号37および38、ならびに図9Aおよび9B、配列番号39および40)と称される、STE13およびDAP2のホモログの特定をもたらした。Melarkodeら,WO 2007/148345は、ピチア・パストリス(Pichia pastoris)STE13ホモログのDNA配列を記載しており、一方、Prabhaら,Protein Expression and Purification,64:155−161,2009は、ピチア・パストリス(Pichia pastoris)Ste13pおよびDap2pホモログのタンパク質配列を記載している。出願人により本発明において作製されたSte13pホモログは、Melarkodeらにおいて既に報告されているものに合致しているようである。本出願人により本発明において作製されたDap2pタンパク質配列の大部分は、Prabhaらにより報告されているものに合致しているが、本明細書中に特定されているDap2pホモログはC末端において異なる。本出願人のホモログは以下のC末端配列を含有する:GLENYDIHVFPDSDHSIRYHNGNVIVYDKLFHWIRRAFKAGK(配列番号46)。一方、Prabhaらのホモログは以下のC末端配列を有する:GLENYDIHVFPDTIPLD(配列番号47)。前記C末端配列の下線部分までの及びそれを含むN末端配列は両方のホモログにおいて保存されている。
【0027】
表1は、ピチア・パストリス(Pichia pastoris)におけるノックアウトベクターの作製に使用される代表的プライマーの配列を示す。該プライマー配列における下線領域は、遺伝子ノックアウト断片の作製を促進するために導入された制限酵素認識部位を表す。導入された特異的制限酵素認識部位が隣のプライマー説明の欄に挙げられている。表2は、表1におけるプライマーを使用して作製されたノックアウトベクターでの形質転換の後のゲノムからのピチア(Pichia)STE13およびDAP2のノックアウトを確認するために使用されるプライマーを示す。該5’および3’プライマーセットが所望のサイズのPCR産物を与え、該ノックアウトプライマーセットが産物を与えない場合に、欠失の成功が確認される。
【0028】
【表1】
【0029】
【表2】
TNFRII−Fcで観察されるN末端トランケート化に関するこれらの遺伝子のそれぞれの役割を評価するために、ゲノムのDAP2またはSTE13遺伝子座の破壊に該ベクターを標的化するピチア・パストリス(Pichia pastoris)DAP2またはSTE13遺伝子の5’および3’領域を有する、URA5遺伝子を含有するノックアウトベクターを設計した(図1Aおよび1B)。これらのノックアウトベクターを、組換え融合タンパク質TNFRII−Fcを発現する同じuraマイナス親株(YGLY7406)内に形質転換した。該形質転換体のPCRスクリーニングの後、Ppste13およびPpdap2ノックアウト株(それぞれYGLY8084およびYGLY8090)を特定した。二重ノックアウト株、すなわち、同じ株におけるPpSTE13およびPpDAP2の両方のノックアウト体を作製するために、Ppste13欠失株YGLY8084をPpDAP2ナーセオスリシン(nourseothricin)ドミナントマーカーノックアウトベクターpGLY5019(図1D)で形質転換した。該形質転換体のPCRスクリーニングの後、Ppste13::URA5バックグラウンド親株において幾つかのPpdap2ノックアウト体を得た。代表的なPpste13/Ppdap2二重ノックアウト株をYGY8096と命名した。
【0030】
これらの株のそれぞれから分泌されたTNFRII−Fcのペプチド分析は、このポリペプチドのタンパク質分解においてSte13pは重要な役割を果たしているが、DAP活性を排除するためには両方のDAP遺伝子の欠失が必要であることを示している。該野生型株では、産生されるタンパク質の100%がN末端において2アミノ酸だけトランケート化された。STE13の単一欠失は、無傷N末端を有するペプチド集団を59%を与え、一方、DAP2の欠失は、無傷TNFRII−Fcの集団を与えたが、それは遥かに低い度合であり、該分泌ポリペプチドの15%しか完全長配列を有していなかった。二重ノックアウト株YGLY8096から分泌されたTNFRII−Fcの分析は、該ペプチド集団の100%が無傷N末端を有することを示した。表3は、単離された完全長ペプチドに対するトランケート化ペプチドのモル%を示すこの分析から得られたデータの要約である。データは各株に関する3つの重複サンプルの代表例である。uraマイナス株は組換えタンパク質の不良分泌体であるため、PpSTE13およびPpDAP2遺伝子がどちらも妨げられていない株から物質を産生させるためにYGLY7406の原栄養性親株YGLY6646を使用した。この株の遺伝子型は、無傷PpSTE13およびPpDAP2遺伝子座の両方に関して野生型として表3に記載されている。
【0031】
【表3】
この分析に基づいて、本出願人は、PpSte13pおよびppDap2pが、ピチア・パストリス(Pichia pastoris)において発現される組換えTNFRII−FcのN末端タンパク質分解に関与する唯一のプロテアーゼであり、DAP活性を排除するためには両方の欠失が必要であると判定した。同様に、野生型顆粒球コロニー刺激因子(GCSF)ポリペプチド(図5におけるタンパク質配列;配列番号6)がピチア・パストリス(Pichia pastoris)において発現された場合、そのN末端はトランケート化されていた。この物質のN末端配列決定は、それがN末端の2アミノ酸を欠くことを示した(図4Aおよび4B、それぞれ配列番号4および5)。また、GCSFのこのトランケート化形態の産生は、二重ΔPpste13/Ppdap2ノックアウトバックグラウンドにおいてGCSFを産生させることにより完全に排除された(図6)。どちらの場合にも、これらの組換えタンパク質(TNFRII−FcおよびCSF)の切断は、該ポリペプチドの(N末端に対して)2番目の位置のプロリン残基のC末端において生じる。本明細書に示されているとおり、ピチア・パストリス(Pichia pastoris)における両方のDAP関連遺伝子の欠失は、ポリペプチドのN末端から2番目の位置にプロリンを有する治療用タンパク質のトランケート化を妨げ、DAP活性の排除のためにはppSTE13およびPpDAP2のその欠失が必要であった。
【0032】
前記のどちらの治療用タンパク質の例もN末端モチーフXaa−Pro(ここで、Xは任意のアミノ酸でありうる)を有する。前記のとおり、Ste13pおよびDap2pは、モチーフXaa−Ala(ここで、Xaaは任意のアミノ酸であり、アラニン(Ala)は2番目のアミノ酸である)を同様に有するN末端ペプチドを切断することが別々に示されている(MisumiおよびIkehara,Handbook of Proteolytic Enzymes,2nd edition,pp.1910−1911,Elsevier,London,2004)。Xaa−Alaモチーフを有するタンパク質のトランケート化を防ぐためにはPpSTE13およびPpDAP2の両方のノックアウトが必要であることを実証するために、当業者は、TNFR−Fcの分泌形態における2番目のアミノ酸をプロリンからアラニンへと突然変異させることが可能である。本明細書における結果に基づけば、PpSTE13またはPpDAP2の欠失が、観察されるN末端切断の比率を減少させるらしい。DAP活性の完全な排除および生じる100%の無傷タンパク質は二重ΔPpste13/Ppdap2ノックアウト宿主株(すなわち、STE13およびDAP2の両方をノックアウトするための突然変異)のみから観察されるであろう。
【0033】
PpSte13pおよびPpDap2pの組合せ排除は、無傷N末端を有する治療用タンパク質の産生を示しているが、本出願人は、ピチア・パストリス(Pichia pastoris)が、本明細書中でPpDPPIIIと称される第3のジペプチジルアミノペプチダーゼを有することを見出した。この酵素はピチア・パストリス(Pichia pastoris)ゲノムのアノテーションにより特定され、ヒト酵素ジペプチジルペプチダーゼIII(DppIIIp)に対して43%の配列同一性を有する。ピチア・パストリス(Pichia pastoris)DppIIIpホモログは未だ特徴づけられていないが、PpSte13およびPpDap2pとは配列レベルで異なり、それぞれ10%および12%の配列同一性を有するに過ぎない。このDAP遺伝子は完全には特徴づけられていないが、PpDppIIIpが、適切な条件下で発現されると、本明細書において評価されている条件下で活性が観察されないにもかかわらず、トランケート化タンパク質を与える、と本出願人は考えている。他の増殖条件下または他のレポータータンパク質との発現により、PpDppIIIp活性が証明されうる、と当業者は認識し理解するであろう。したがって、他の適当な条件またはレポーターにおいて、ピチア・パストリス(Pichia pastoris)におけるDAP活性を完全に排除するためには、PpDppIIIが単独で或いはppSte13pおよび/またはPpDap2p活性と共に欠失または排除される必要がありうる。当業者であれば、本明細書の実施例に記載されている方法、特に実施例5の方法により、ppDPPIIIを欠失または破壊させることが可能であろう。
【0034】
DAP活性の排除のためのピチア株
商業的に入手可能な任意のピチア・パストリス(Pichia pastoris)株、例えばNRRL−Y11430(American Type Culture Collection(ATCC),Manassas,VA,カタログ番号76273)が本発明に使用可能であるが、好ましい実施形態においては、本発明に使用される株は、糖操作されたピチア・パストリス(Pichia pastoris)株、例えば、実施例3に記載されているGS5.0株、または後記の糖操作された株(これは、発現に際してヒト様グリコシル化プロファイルを与える修飾を含む)であろう。例えば、実施例3のGS5.0株の場合、そのような修飾には、Δoch1、Δpno1、Δmnn4B、Δbmt2およびΔura5の欠失、またはケイ・ラクチス(K.lactis)およびエム・ムスクルス(M.musculus)UDP−GlcNAc輸送体、エム・ムスクルス(M.musculus)α−1,2−MsnI、ホモ・サピエンス(H.sapiens)β−1,2−GlcNAcトランスフェラーゼ1活性、アール・ノルベギクス(R.norvegicus)β−1,2−GlcNAcトランスフェラーゼII活性、キイロショウジョウバエ(D.melanogaster)MnsII活性、エス・ポンベ(S.pombe)Galエピメラーゼ、キイロショウジョウバエ(D.melanogaster)UDP−Gal輸送体およびホモ・サピエンス(H.sapiens)β−1,4−ガラクトシルトランスフェラーゼ活性の挿入のために行われるものが含まれる。GS5.0株は、ガラクトースを末端とするN−グリカン、例えばGalGlcNAc2MAN3GlcNAc2、Gal2GlcNAc2Man3GlcNAc2またはそれらの混合物を有する糖タンパク質を産生しうる。他の代表的な糖操作株には、YJN201(Choiら,PNAS,100(9):5022−5027,2003)、YSH44(Hamiltonら,Science,301(5637):1244−1246,2003)、RDP36−1(Davidsonら,Glycobiology,14(4):1−9,2004)、PBP6−5(Bobrowiczら,Glyoobiology,14(9):757−766,2004)、YSH597(Hamiltonら,Science,313(5792):1441−1443,2006)が含まれる。
【0035】
ピチア・パストリス(Pichia pastoris)に加えて、本発明において有用な宿主細胞には、ピチア・パストリス(Pichia pastoris)DAP2およびSTE13遺伝子のホモログを発現する酵母が含まれる。そのような酵母は以下のものから選択されうる:ピチア・フィンランディカ(Pichia finlandica)、ピチア・トレハロフィラ(Pichia trehalophila)、ピチア・コクラメ(Pichia koclamae)、ピチア・メンブラネファシエンス(Pichia membranaefaciens)、ピチア・ミヌタ(Pichia minuta)(オガタエア・ミヌタ(Ogataea minuta)、ピチア・リンドネリ(Pichia lindneri))、ピチア・オプンチエ(Pichia opuntiae)、ピチア・テルモトレランス(Pichia thermotolerans)、ピチア・サリクタリア(Pichia salictaria)、ピチア・グエルクウム(Pichia guercuum)、ピチア・ピエペリ(Pichia pijperi)、ピチア・スチプティス(Pichia stiptis)、ピチア・メタノリカ(Pichia methanolica)、ピチア属種(Pichia sp.)、サッカロミセス・セレビシエ(Saccharomyces cerevisiae)、サッカロミセス属種(Saccharomyces sp.)、ハンゼヌラ・ポリモルファ(Hansenula polymorpha)、クライベロミセス属種(Kluyveromyces sp.)、クライベロミセス・ラクチス(Kluyveromyces lactis)、カンジダ・アルビカンス(Candida albicans)。種々の酵母、例えばクライベロミセス・ラクチス(K.lactis)、ピチア・パストリス(Pichia pastoris)、ピチア・メタノリカ(Pichia methanolica)およびハンゼヌラ・ポリモルファ(Hansenula polymorpha)が細胞培養に特に適している。なぜなら、それらは高い細胞密度まで増殖可能であり、大量の組換えタンパク質を分泌しうるからである。
【0036】
酵母は、グリコシル化パターンがヒト様である又はヒト化されている糖タンパク質を発現するように、遺伝的に修飾されうる。このようにして、特定の所望のグリコフォームが組成物において優勢である糖タンパク質組成物が製造されうる。選択された内在性グリコシル化酵素を除去し、および/または該宿主細胞を遺伝的に操作し、および/または外在性酵素を供与して、哺乳類グリコシル化経路の全部または一部を模擬することにより(US 2004/0018590に記載されているとおり)、それは達成されうる。所望により、該グリコシル化の追加的遺伝的操作を行って、該糖タンパク質が、コアフコシル化を伴って又は伴わないで産生されうるようにすることが可能である。下等真核宿主細胞の使用は更に有利である。なぜなら、これらの細胞は糖タンパク質の高度に均一な組成物を産生することが可能であり、該糖タンパク質の主要グリコフォームが該組成物中の糖タンパク質の30モル%超(すなわち、30モル%を超えて)で存在することが可能となるからである。特定の態様においては、該主要グリコフォームは、該組成物中に存在する糖タンパク質の40モル%超、50モル%超、60モル%超、70モル%超、最も好ましくは80モル%超で存在しうる。
【0037】
酵母は、該グリコシル化パターンがヒト様である又はヒト化されている糖タンパク質をそれらが発現するように、遺伝的に修飾されうる。ヒト様またはヒト化N−グリカンには、ハイブリッドおよび複合N−グリカンの両方が含まれる。Gerngrossら,米国公開出願第2004/0018590に記載されているとおり、選択された内因性グリコシル化酵素を排除し、および/または外因性酵素を供給することにより、それは達成されうる。例えば、宿主細胞は、糖タンパク質上のN−グリカン上にマンノース残基を付加する1,6−マンノシルトランスフェラーゼ活性が喪失するよう選択され又は操作されうる。選択された内在性グリコシル化酵素を除去し、および/または外在性酵素を供与することにより(Gerngrossら,US 20040018590に記載されているとおり)、それは達成されうる。例えば、宿主細胞は、糖タンパク質上のN−グリカン上にマンノース残基を付加する1,6−マンノシルトランスフェラーゼ活性を欠くように選択または操作されうる。
【0038】
1つの実施形態においては、該宿主細胞は更に、α1,2−マンノシダーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、α1,2−マンノシダーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る組換え糖タンパク質の通過は、Man5GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、Man5GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生する。例えば、米国特許第7,029,872号ならびに米国公開特許出願第2004/0018590号および第2005/0170452号は、Man5GlcNAc2グリコフォームを含む糖タンパク質を産生しうる下等真核宿主細胞を開示している。
【0039】
更に詳細な実施形態においては、直前の宿主細胞は更に、GlcNAcトランスフェラーゼI(GnT I)触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、GlcNAcトランスフェラーゼI活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、GlcNAcMan5GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、ハイブリッドGlcNAcMan5GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生する。例えば、米国特許第7,029,872号ならびに米国公開特許出願第2004/0018590号および第2005/0170452号は、GlcNAcMan5GlcNAc2グリコフォームを含む糖タンパク質を産生しうる下等真核宿主細胞を開示している。前記細胞において産生された糖タンパク質をヘキソサミニダーゼでインビトロで処理して、Man5GlcNAc2グリコフォームを含む組換え糖タンパク質を産生させることが可能である。
【0040】
更に詳細な実施形態においては、直前の宿主細胞は更に、マンノシダーゼII触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、マンノシダーゼII活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、GlcNAcMan3GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、複合GlcNAcMan3GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生する。米国特許第7,029,872号および米国公開特許出願第2004/0230042号は、マンノシダーゼII酵素を発現し、GlcNAc2Man3GlcNAc2グリコフォームを主に有する糖タンパク質を産生しうる下等真核宿主細胞を開示している。前記細胞において産生された糖タンパク質をヘキソサミニダーゼでインビトロで処理して、Man3GlcNAc2グリコフォームを含む組換え糖タンパク質を産生させることが可能である。
【0041】
更に詳細な実施形態においては、直前の宿主細胞は更に、GlcNAcトランスフェラーゼII(GnT II)触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、GlcNAcトランスフェラーゼII活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、GlcNAc2Man3GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、複合GlcNAc2Man3GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生する。米国特許第7,029,872号ならびに米国公開特許出願第2004/0018590号および第2005/0170452号は、GlcNAc2Man3GlcNAc2グリコフォームを含む糖タンパク質を産生しうる下等真核宿主細胞を開示している。前記細胞において産生された糖タンパク質をヘキソサミニダーゼでインビトロで処理して、Man3GlcNAc2グリコフォームを含む組換え糖タンパク質を産生させることが可能である。
【0042】
更に詳細な実施形態においては、直前の宿主細胞は更に、ガラクトシルトランスフェラーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、ガラクトシルトランスフェラーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、複合GalGlcNAc2Man3GlcNAc2(G1)もしくは複合Gal2GlcNAc2Man3GlcNAc2(G2)グリコフォームまたはそれらの混合物を含む組換え糖タンパク質、例えば、GalGlcNAc2Man3GlcNAc2グリコフォームもしくはGal2GlcNAc2Man3GlcNAc2グリコフォームまたはそれらの混合物を主に含む組換え糖タンパク質組成物を産生する。米国特許第7,029,872号および米国公開特許出願第2006/0040353号は、Gal2GlcNAc2Man3GlcNAc2グリコフォームを含む糖タンパク質を産生しうる下等真核宿主細胞を開示している。前記細胞において産生された糖タンパク質をガラクトシダーゼでインビトロで処理して、GlcNAc2Man3GlcNAc2グリコフォームを含む組換え糖タンパク質、例えば、GlcNAc2Man3GlcNAc2グリコフォームを主に含む組換え糖タンパク質組成物を産生させることが可能である。
【0043】
更に詳細な実施形態においては、直前の宿主細胞は更に、シアリルトランスフェラーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、シアリルトランスフェラーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、複合NANA2Gal2GlcNAc2Man3GlcNAc2グリコフォームもしくは複合NANAGal2GlcNAc2Man3GlcNAc2グリコフォームまたはそれらの混合物を主に含む組換え糖タンパク質を産生する。該宿主細胞は、N−グリカンへの転移のためのCMP−シアル酸を供与するための手段を更に含むことが有用である。米国公開特許出願第2005/0260729号は、CMP−シアル酸合成経路を有するように下等真核生物を遺伝的に操作するための方法を開示しており、米国公開特許出願第2006/0286637号は、シアル酸化糖タンパク質を産生するように真核生物を遺伝的に操作するための方法を開示している。前記細胞において産生された糖タンパク質をノイラミニダーゼでインビトロで処理して、Gal2GlcNAc2Man3GlcNAc2グリコフォームを主に含む組換え糖タンパク質を産生させることが可能である。
【0044】
前記宿主細胞はいずれも、米国公開特許出願第2004/074458号および第2007/0037248号に開示されているような二分岐(bisected)(GnT III)および/または多分岐(GnT IV、V、VIおよびIX)N−グリカン構造を有する糖タンパク質を産生させるためのGnT III、GnT IV、GnT V、GnT VIおよびGnT IXからなる群から選択される1以上のGlcNAcトランスフェラーゼを更に含みうる。
【0045】
更に詳細な実施形態においては、ハイブリッドGlcNAcMan5GlcNAc2 N−グリカンを主に有する糖タンパク質を産生する宿主細胞は更に、ガラクトシルトランスフェラーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、ガラクトシルトランスフェラーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、ハイブリッドGalGlcNAcMan5GlcNAc2グリコフォームを主に含む組換え糖タンパク質を産生する。
【0046】
更に詳細な実施形態においては、GalGlcNAcMan5GlcNAc2 N−グリカンを主に有する糖タンパク質を産生した直前の宿主細胞は更に、シアリルトランスフェラーゼ触媒ドメインを含み、該触媒ドメインは、該触媒ドメインに通常は結合していない細胞標的化シグナルペプチドに融合しており、該シグナルペプチドは、シアリルトランスフェラーゼ活性を該宿主細胞のERまたはゴルジ装置に標的化するように選択される。該宿主細胞のERまたはゴルジ装置を通る該組換え糖タンパク質の通過は、ハイブリッドNANAGalGlcNAcMan5GlcNAc2グリコフォームを含む組換え糖タンパク質を産生する。
【0047】
種々の前記宿主細胞は更に、1以上の糖輸送体、例えばUDP−GlcNAc輸送体[例えば、クライベロミセス・ラクチス(Kluyveromyces lactis)およびムス・ムスクルス(Mus musculus)UDP−GlcNAc輸送体]、UDP−ガラクトース輸送体[例えば、キイロショウジョウバエ(Drosophila melanogaster)UDP−ガラクトース輸送体]およびCMP−シアル酸輸送体(例えば、ヒトシアル酸輸送体)を含む。下等真核宿主細胞、例えば酵母および糸状菌は前記輸送体を欠くため、下等真核宿主細胞、例えば酵母および糸状菌は、前記輸送体を含むように遺伝的に操作されることが好ましい。
【0048】
宿主細胞は更に、β−マンノシルトランスフェラーゼ遺伝子(例えば、BMT1、BMT2、BMT3およびBMT4)の1以上を欠失させ又は破壊することにより、α−マンノシダーゼ耐性N−グリカンを有する糖タンパク質を排除するように米国公開特許出願第2006/0211085号を参照されたい)、ならびにホスホマンノシルトランスフェラーゼ遺伝子PNO1およびMNN4Bの一方または両方を欠失させ又は破壊することにより、ホスホマンノース残基を有する糖タンパク質を排除するように(米国特許第7,198,921号および第7,259,007号を参照されたい)、遺伝的に操作された下等真核細胞[例えば、酵母、例えば、ピチア・パストリス(Pichia pastoris)]が含まれ、これは、更に詳細な実施形態においては、MNN4A遺伝子の欠失または破壊をも含みうる。破壊は、特定の酵素をコードするオープンリーディングフレームの破壊、あるいは該オープンリーディングフレームの発現の破壊、あるいは干渉性RNA、アンチセンスRNAなどを使用する、β−マンノシルトランスフェラーゼおよび/またはホスホマンノシルトランスフェラーゼの1以上をコードするRNAの翻訳の阻害を含む。該宿主細胞は更に、特定のN−グリカン構造を産生するように修飾された前記宿主細胞のいずれかを含みうる。
【0049】
宿主細胞は更に、タンパク質O−マンノシルトランスフェラーゼ[Dol−P−Man:タンパク質(Ser/Thr)マンノシルトランスフェラーゼ遺伝子](PMT)(米国特許第5,714,377号を参照されたい)の1以上を欠失させ又は破壊することにより糖タンパク質のO−グリコシル化を制御するように遺伝的に修飾された、あるいは公開国際出願番号WO2007061631に開示されているとおりにPmtpインヒビターおよび/またはアルファ−マンノシダーゼの存在下で増殖された、あるいはそれらの両方に付された下等真核細胞[例えば酵母、例えばピチア・パストリス(Pichia pastoris)]を含む。破壊は、Pmtpをコードするオープンリーディングフレームの破壊、あるいは該オープンリーディングフレームの発現の破壊、あるいは干渉性RNA、アンチセンスRNAなどを使用する、Pmtpの1以上をコードするRNAの翻訳の阻害を含む。該宿主細胞は更に、特定のN−グリカン構造を産生するように修飾された前記宿主細胞のいずれかを含みうる。
【0050】
Pmtpインヒビターには、ベンジリデンチアゾリジンジオンが含まれるが、これに限定されるものではない。使用されうるベンジリデンチアゾリジンジオンの具体例としては、5−[[3,4−ビス(フェニルメトキシ)フェニル]メチレン]−4−オキソ−2−チオキソ−3−チアゾリジン酢酸、5−[[3−(1−フェニルエトキシ)−4−(2−フェニルエトキシ)]フェニル]メチレン]−4−オキソ−2−チオキソ−3−チアゾリジン酢酸、および5−[[3−(1−フェニル−2−ヒドロキシ)エトキシ)−4−(2−フェニルエトキシ)]フェニル]メチレン]−4−オキソ−2−チオキソ−3−チアゾリジン酢酸が挙げられる。
【0051】
特定の実施形態においては、少なくとも1つの内在性PMT遺伝子の機能または発現が低減、破壊または欠失される。例えば、特定の実施形態においては、PMT1、PMT2、PMT3およびPMT4遺伝子からなる群から選択される少なくとも1つの内在性PMT遺伝子の機能または発現が低減、破壊または欠失され、あるいは該宿主細胞は1以上のPMTインヒビターの存在下で培養される。更に詳細な実施形態においては、該宿主細胞は1以上のPMT遺伝子の欠失または破壊を含み、該宿主細胞は1以上のPmtpインヒビターの存在下で培養される。これらの実施形態の特定の態様においては、該宿主細胞は分泌性アルファ−1,2−マンノシダーゼをも発現する。
【0052】
PMT欠失もしくは破壊および/またはPmtpインヒビターは、O−グリコシル化の占有を低減することにより、すなわち、グリコシル化される糖タンパク質上のO−グリコシル化部位の総数を減少させることにより、O−グリコシル化を制御する。該細胞により分泌されるアルファ−1,2−マンノシダーゼの更なる添加は、該糖タンパク質上に存在するO−グリカンのマンノース鎖長を減少させることにより、O−グリコシル化を制御する。したがって、PMT欠失もしくは破壊および/またはPmtpインヒビターを分泌性アルファ−1,2−マンノシダーゼの発現と組合せることは、占有および鎖長を減少させることによりO−グリコシル化を制御する。個々の状況においては、個々の異種糖タンパク質(例えば、抗体)は種々の度合の効率で発現されゴルジ装置から輸送される可能性があり、したがって、PMT欠失または破壊、Pmtpインヒビターおよびアルファ−1,2−マンノシダーゼの特定の組合せを要しうるため、PMT欠失または破壊、Pmtpインヒビターおよびアルファ−1,2−マンノシダーゼの個々の組合せは実験的に決定される。もう1つの態様においては、1以上の内在性マンノシルトランスフェラーゼ酵素をコードする遺伝子が欠失される。この欠失は分泌性アルファ−1,2−マンノシダーゼおよび/またはPMTインヒビターの供与と組合されることが可能であり、あるいは分泌性アルファ−1,2−マンノシダーゼおよび/またはPMTインヒビターの供与の代わりに行われうる。
【0053】
したがって、該宿主細胞は、糖タンパク質を産生するように遺伝的に修飾された任意の宿主細胞であることが可能であり、この場合、該主要N−グリカンは、複合N−グリカン、ハイブリッドN−グリカンおよび高マンノースN−グリカンからなる群から選択され、ここで、複合N−グリカンは、Man3GlcNAc2、GlcNAC(1−4)Man3GlcNAc2、Gal(1−4)GlcNAc(1−4)Man3GlcNAc2およびNANA(1−4)Gal(1−4)Man3GlcNAc2からなる群から選択され、ハイブリッドN−グリカンは、Man5GlcNAc2、GlcNAcMan5GlcNAc2、GalGlcNAcMan5GlcNAc2およびNANAGalGlcMan5GlcNAc2からなる群から選択され、高マンノースN−グリカンは、Man6GlcNAc2、Man7GlcNAc2、Man8GlcNAc2およびMan9GlcNAc2からなる群から選択される。N−グリカン構造の具体例には以下のものが含まれるが、それらに限定されるものではない:Man5GlcNAc2、GlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc2Man3GlcNAc2、GlcNAc3Man3GlcNAc2、GlcNAc4Man3GlcNAc2、GalGlcNAc2Man3GlcNAc2、Gal2GlcNAc2Man3GlcNAc2、Gal2GlcNAc3Man3GlcNAc2、Gal2GlcNAc4Man3GlcNAc2、Gal3GlcNAc3Man3GlcNAc2、Gal3GlcNAc4Man3GlcNAc2、Gal4GlcNAc4Man3GlcNAc2、NANAGal2GlcNAc2Man3GlcNAc2、NANA2Gal2GlcNAc2Man3GlcNAc2、NANA3Gal3GlcNAc3Man3GlcNAc2およびNANA4Gal4GlcNAc4Man3GlcNAc2。
【0054】
以下の実施例においては、糖操作されたピチア・パストリス(Pichia pastoris)株を、Ste13pおよびDap2pジペプチジルアミノペプチダーゼ活性を排除するために修飾した。一般に、Ste13pおよび/またはDap2p活性は、本明細書に記載されている方法を用いて、任意のピチア・パストリス(Pichia pastoris)株から排除されうる。ただし、選択に使用されるマーカーが該株と共に使用されうる。あるいは、選択を可能にする別の栄養要求性またはドミナントマーカーが代用されうる。例えば、ベクターpGLY5018(実施例2C)および選択マーカーとしてのナーセオスリシン(nourseothricin)を使用して、商業的に入手可能な株NRRL−Y11430からSTE13を欠失させることが可能である。後記のDAP2ノックアウトベクター(実施例2B)のいずれのマーカーもNRRL−Y11430に和合性でないため、pAG32から1654bpのHygマーカーを増幅するためにプライマーSH806およびSH807(表1)の斜字表示断片に対応するプライマー(それぞれ配列番号48および配列番号49)を使用してpAG32(Goldsteinら,Yeast,15(6):507−511,1999;Erratum:Yeast,15(12):1297,1999)からヒグロマイシン標識ベクターを作製することが可能である。ついで、この後者の断片を、pGLY5019の作製に関する実施例2Cに記載されているとおりに、DAP2 5’およびDAP2 3’フランキング領域に融合させることが可能である。SfiI(New England BioLabs,Ipswich,MA)での消化の後、該ベクターをNRRL−Y11430またはそのste13ノックアウト誘導体内に形質転換し、ヒグロマイシン含有プレート上で選択して、それぞれ単一dap2Δまたは二重Δste13/dap2ノックアウト株を得ることが可能である。本明細書に記載されている方法を用いてDppIIIpジペプチジルアミノペプチダーゼ活性を排除するために修飾された糖操作ピチア・パストリス(Pichia pastoris)株を当業者がどうのようにして使用しうるのかに関する一例も示す。
【図面の簡単な説明】
【0055】
【図1A】図1は、本発明における方法において使用されるベクターの図示である。Ura5標識PpSTE13およびPpDAP2ノックアウトベクターが、それぞれ図1Aおよび1BにpGLY4520およびpGLY4521として示されている。PpSTE13およびPpDAP2ナーセオスリシン(nourseothricin)標識ベクターが、それぞれ図1Cおよび1DにpGLY5018およびpGLY5019として示されている。形質転換体におけるノックアウト断片を切り出すために用いられるSfiI制限部位が下線で示されている。PpSTE13およびPpDAP2フランキング領域が黒で太く表示されている。
【図1B】図1は、本発明における方法において使用されるベクターの図示である。Ura5標識PpSTE13およびPpDAP2ノックアウトベクターが、それぞれ図1Aおよび1BにpGLY4520およびpGLY4521として示されている。PpSTE13およびPpDAP2ナーセオスリシン(nourseothricin)標識ベクターが、それぞれ図1Cおよび1DにpGLY5018およびpGLY5019として示されている。形質転換体におけるノックアウト断片を切り出すために用いられるSfiI制限部位が下線で示されている。PpSTE13およびPpDAP2フランキング領域が黒で太く表示されている。
【図1C】図1は、本発明における方法において使用されるベクターの図示である。Ura5標識PpSTE13およびPpDAP2ノックアウトベクターが、それぞれ図1Aおよび1BにpGLY4520およびpGLY4521として示されている。PpSTE13およびPpDAP2ナーセオスリシン(nourseothricin)標識ベクターが、それぞれ図1Cおよび1DにpGLY5018およびpGLY5019として示されている。形質転換体におけるノックアウト断片を切り出すために用いられるSfiI制限部位が下線で示されている。PpSTE13およびPpDAP2フランキング領域が黒で太く表示されている。
【図1D】図1は、本発明における方法において使用されるベクターの図示である。Ura5標識PpSTE13およびPpDAP2ノックアウトベクターが、それぞれ図1Aおよび1BにpGLY4520およびpGLY4521として示されている。PpSTE13およびPpDAP2ナーセオスリシン(nourseothricin)標識ベクターが、それぞれ図1Cおよび1DにpGLY5018およびpGLY5019として示されている。形質転換体におけるノックアウト断片を切り出すために用いられるSfiI制限部位が下線で示されている。PpSTE13およびPpDAP2フランキング領域が黒で太く表示されている。
【図2A】図2は、酵母におけるDAP活性に関連したTNFRII−FcのN末端切断の図示である。図2Aは無傷分泌性TNFRII−Fc(配列番号3)の7個のN末端アミノ酸(配列番号1)を示し、図2Bは該トランケート化産物の5個のN末端アミノ酸(配列番号2)を示す。矢印は、ジペプチジルアミノペプチダーゼDap2pおよびSte13pの両方により認識される切断部位を示す。
【図2B】図2は、酵母におけるDAP活性に関連したTNFRII−FcのN末端切断の図示である。図2Aは無傷分泌性TNFRII−Fc(配列番号3)の7個のN末端アミノ酸(配列番号1)を示し、図2Bは該トランケート化産物の5個のN末端アミノ酸(配列番号2)を示す。矢印は、ジペプチジルアミノペプチダーゼDap2pおよびSte13pの両方により認識される切断部位を示す。
【図3】図3は、腫瘍壊死因子受容体2(TNFRII)とIgG1 Fc領域(Fc)とを含む融合タンパク質TNFRII−Fcのアミノ酸配列(配列番号3)である。
【図4A】図4は、酵母におけるDAP活性に関連したGCSFのN末端切断の図示である。図4Aは無傷分泌性GCSF(配列番号6)の7個のN末端アミノ酸(配列番号4)を示し、図4Bは該トランケート化産物の5個のN末端アミノ酸(配列番号5)を示す。矢印は、ジペプチジルアミノペプチダーゼDap2pおよびSte13pの両方により認識される切断部位を示す。
【図4B】図4は、酵母におけるDAP活性に関連したGCSFのN末端切断の図示である。図4Aは無傷分泌性GCSF(配列番号6)の7個のN末端アミノ酸(配列番号4)を示し、図4Bは該トランケート化産物の5個のN末端アミノ酸(配列番号5)を示す。矢印は、ジペプチジルアミノペプチダーゼDap2pおよびSte13pの両方により認識される切断部位を示す。
【図5】図5は組換え顆粒球コロニー刺激因子(GSCF)タンパク質のアミノ酸配列(配列番号6)である。
【図6】図6は、STE13遺伝子もDAP2遺伝子も欠失も破壊もされていないピチア・パストリス(Pichia pastoris)株において産生されたGCSF、すなわち、DAP切断による低分子量GCSF(レーン27〜29)、ならびにそれと比較されている、Ste13pおよびDap2p活性が排除されたピチア・パストリス(Pichia pastoris)株において産生された無傷高分子量GCSF産物(レーン32〜34)のウエスタンブロットの表示である。
【図7A】図7A〜7Cは、STE13およびDAP2のノックアウト体の作製(実施例3)に使用される糖操作株YGLY7406に関する流れ図を表す。図7Cは、単一ノックアウト糖操作株ste13(YGLY8084)およびdap2(YGLY8090)ならびに後続の二重ノックアウト株(YGLY8096)に関する流れ図を表す。
【図7B】図7A〜7Cは、STE13およびDAP2のノックアウト体の作製(実施例3)に使用される糖操作株YGLY7406に関する流れ図を表す。図7Cは、単一ノックアウト糖操作株ste13(YGLY8084)およびdap2(YGLY8090)ならびに後続の二重ノックアウト株(YGLY8096)に関する流れ図を表す。
【図7C】図7A〜7Cは、STE13およびDAP2のノックアウト体の作製(実施例3)に使用される糖操作株YGLY7406に関する流れ図を表す。図7Cは、単一ノックアウト糖操作株ste13(YGLY8084)およびdap2(YGLY8090)ならびに後続の二重ノックアウト株(YGLY8096)に関する流れ図を表す。
【図8A−1】図8Aおよび8Bはピチア・パストリス(Pichia pastoris)DAP2のcDNA配列(配列番号37)およびアミノ酸配列(配列番号38)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図8A−2】図8Aおよび8Bはピチア・パストリス(Pichia pastoris)DAP2のcDNA配列(配列番号37)およびアミノ酸配列(配列番号38)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図8A−3】図8Aおよび8Bはピチア・パストリス(Pichia pastoris)DAP2のcDNA配列(配列番号37)およびアミノ酸配列(配列番号38)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図8B】図8Aおよび8Bはピチア・パストリス(Pichia pastoris)DAP2のcDNA配列(配列番号37)およびアミノ酸配列(配列番号38)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図9A−1】図9Aおよび9Bはピチア・パストリス(Pichia pastoris)STE13のcDNA配列(配列番号39)およびアミノ酸配列(配列番号0)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図9A−2】図9Aおよび9Bはピチア・パストリス(Pichia pastoris)STE13のcDNA配列(配列番号39)およびアミノ酸配列(配列番号0)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図9A−3】図9Aおよび9Bはピチア・パストリス(Pichia pastoris)STE13のcDNA配列(配列番号39)およびアミノ酸配列(配列番号0)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図9B】図9Aおよび9Bはピチア・パストリス(Pichia pastoris)STE13のcDNA配列(配列番号39)およびアミノ酸配列(配列番号0)である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図10A】図10Aおよび10Bは、それぞれpGLY4511およびpGLY4512を作製するために増幅されるPpSTE13に関する5’(配列番号41)および3’(配列番号42)DNAフランキング領域である。該フランキング領域(下線部)自体は、PpSTE13 5’フランキング領域ではEcoRI制限部位を構成するヌクレオチドに隣接しており、PpSTE13 3’フランキング領域ではHindIII制限部位を構成するヌクレオチドに隣接している。
【図10B】図10Aおよび10Bは、それぞれpGLY4511およびpGLY4512を作製するために増幅されるPpSTE13に関する5’(配列番号41)および3’(配列番号42)DNAフランキング領域である。該フランキング領域(下線部)自体は、PpSTE13 5’フランキング領域ではEcoRI制限部位を構成するヌクレオチドに隣接しており、PpSTE13 3’フランキング領域ではHindIII制限部位を構成するヌクレオチドに隣接している。
【図11A】図11Aおよび11Bは、それぞれpGLY4513およびpGLY4514を作製するために増幅されるPpDAP2に関する5’(配列番号43)および3’(配列番号44)DNAフランキング領域である。該フランキング領域(下線部)自体は、PpDAP2 5’フランキング領域ではEcoRI制限部位を構成するヌクレオチドに隣接しており、PpDAP2 3’フランキング領域ではHindIII制限部位を構成するヌクレオチドに隣接している。
【図11B】図11Aおよび11Bは、それぞれpGLY4513およびpGLY4514を作製するために増幅されるPpDAP2に関する5’(配列番号43)および3’(配列番号44)DNAフランキング領域である。該フランキング領域(下線部)自体は、PpDAP2 5’フランキング領域ではEcoRI制限部位を構成するヌクレオチドに隣接しており、PpDAP2 3’フランキング領域ではHindIII制限部位を構成するヌクレオチドに隣接している。
【図12】図12は、pAG25から増幅されるナーセオスリシン(nourseothricin)マーカーカセットのcDNA配列(配列番号45)であり、該ORFが太字で示されている。
【図13−1】図13はピチア・パストリス(Pichia pastoris)DPPIIIのcDNA(配列番号54)配列である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【図13−2】図13はピチア・パストリス(Pichia pastoris)DPPIIIのcDNA(配列番号54)配列である。ORFが太字で示されている(約1kbのフランキング配列を伴う又は伴わない)。
【実施例1】
【0056】
株、培養条件および試薬
大腸菌(Escherichia coli)株TOP10(Invitrogen,Carlsbad,CA)またはXL10−Gold(Stratagene,Santa Clara,CA)を組換えDNA実験に使用した。制限および修飾酵素はNew England BioLabs,Ipswich,MAから入手し、製造業者の指示どおりに使用した。オリゴヌクレオチドはIntegrated DNA Technologies,Coralville,IAから入手した。塩および緩衝剤はSigma,St.Louis,MOから得た。ここで使用した最少培地は1.4% 酵母窒素ベース、2% デキストロース、1.5% 寒天および4×10−5% ビオチンおよびアミノ酸(適宜補足されるもの)を含むものであった。YMDに富む培地は1% 酵母エキス、2% マルトン(martone)、2% デキストロースおよび1.5% 寒天(プレート用)である。ナーセオスリシン(nourseothricin)はUS Biologicals,Swampscott,MA(カタログ番号N5375−74)から得られ、YMDに富む培地に100μg/mlの最終濃度で加えられる。
【実施例2】
【0057】
ノックアウトベクターの作製
A.ste13::URA5ノックアウトベクターの作製
PfuUltra(商標)DNAポリメラーゼ(Stratagene,Santa Clara,CA)および鋳型としてのピチア・パストリス(Pichia pastoris)株NRRL−Y11430由来のゲノムDNAを使用して、STE13オープンリーディングフレームに対応する5’および3’フランキング領域に対応するDNA断片(配列番号41および42)を増幅した。表1に示されているプライマーペアSH774(配列番号13)およびSH775(配列番号14)ならびにSH776(配列番号15)およびSH777(配列番号16)を使用して、それぞれSTE13の5’および3’に関する771bpおよび949bpの断片を増幅した。ExTaq(商標)(TaKaRa,Bio.Inc.,Japan)の存在下の72℃で10分間のインキュベーションの後、該増幅断片をpCR2.1(Invitrogen,Carlsbad,CA)内にクローニングし、TOP10コンピテント細胞内に形質転換した。DNA配列決定は、STE13 5’およびSTE13 3’フランキング領域が正確であることを証明した。得られたベクターをそれぞれpGLY4511およびpGLY4512と命名した。
【0058】
763bpのSTE13 5’フランキング領域断片(図10A、配列番号50の下線領域により表される)を、EcoRIを使用してpGLY4511から消化し、同じ制限酵素で予め消化され子ウシ腸アルカリホスファターゼ(CIAP)で処理された、pJN396(NettおよびGerngross,Yeast,20,1279−1290,2003)に類似したピチア・パストリス(P.pastoris)URA5−ブラスターベクターpGLY13b内にサブクローニングした。URA5−ブラスターベクターは、5−フルオロオロト酸の存在下の対抗選択によるURA5マーカーの回収を促進するためのLacZ直接反復に隣接したピチア(Pichia)URA5遺伝子の機能性遺伝子断片を含有するpUC19ベクターバックボーンを有する。その新たに連結されたベクターをXL10 Goldコンピテント細胞内に形質転換し、制限分析により確認した後、得られたベクターをpGLY4518と命名した。ベクターpGLY4512を、STE13 3’フランキング領域をコードする940bp(図10B、配列番号51の下線領域により表される)断片を遊離させるためにHindIIIで消化し、同じ酵素で予め消化されCIAP処理されたpGY4518内にサブクローニングした。該連結産物をXL10 Goldコンピテント細胞内に形質転換し、制限分析後にpGLY4520と命名した。この最終的なste13::Ura5ノックアウトベクターを図1Aに図示する。
【0059】
B.dap2::URA5ノックアウトベクターの作製
表1に示されているプライマーセットSH782(配列番号21)およびSH783(配列番号22)ならびにSH784(配列番号23)およびSH785(配列番号24)を使用して、DAP2 5’および3’フランキング領域(配列番号43および44)を、前記のとおりにピチア・パストリス(Pichia pastoris)ゲノムDNAから増幅して、それぞれ1003bpおよび1142bpの断片を得た。pCR2.1内へのクローニングおよび配列決定の後、該ベクターをpGLY4513およびpGLY4514(それぞれDAP2 5’およびDAP2 3’領域をコードする)と命名した。前記実施例2Aに類似したアプローチに従い、995bpのDAP2 5’領域(図11A、配列番号52の下線領域により表される)をUra5−ブラスターベクターpGLY13b内のEcoRI部位内にサブクローニングして、中間構築物pGLY4519を得た。ついで1133bpのDAP2 3’領域(図11B、配列番号53の下線領域により表される)をpGLY4519のHindIII部位内にサブクローニングして、dap2::URA5ノックアウトベクターpGLY4521(図1Bに図示されている)を得た。
【0060】
C.ドミナントマーカーSTE13およびDAP2ノックアウトベクターの作製
PCR融合体を使用して、STE13およびDAP2の両方のノックアウトベクターを作製した。表1に示されているプライマーセットSH774(配列番号13)およびSH801(配列番号29)ならびにSH804(配列番号32)およびSH777(配列番号16)を使用し、PfuUltra(商標)DNAポリメラーゼを使用して、STE13 5’およびSTE13 3’断片をpGLY4520から増幅した。アシビア・ゴシッピィ(Ashbya gossypii)転写伸長因子(TEF)プロモーターおよびターミネーター(図12の通常文字により表される)(それぞれ、太字のオープンリーディングフレームの5’および3’側に位置する)の発現制御下でストレプトマイセス・ノウルセイ(Streptomyces noursei)からのナーセオスリシン(nourseothricin)アセチルトランスフェラーゼのオープンリーディングフレーム(図12、配列番号54の太字で表されている)を含有するナーセオスリシン(NATR)マーカーカセット(配列番号45)を、プライマーSH802(配列番号30)およびSH803(配列番号31)を使用して、pAG25(GoldsteinおよびMcCusker,Yeast,15,1541−1553,1999)から増幅した。該PCR反応をDNAアガロースゲル上で行い、それぞれSTE13 5’、STE13 3’およびNatマーカーに対応する779bp、958bpおよび1249bpの断片を単離した。ついでそれぞれ20ngを合わせ、PfuUltra(商標)DNAポリメラーゼならびにプライマーペアSH774(配列番号13)およびSH777(配列番号16)を使用して互いに融合させた。ExTaq(商標)DNAポリメラーゼ(TaKaRa,Bio.Inc.,Japan)の存在下の72℃で10分間のインキュベーションの後、該増幅(2896bp)断片をpCR2.1内にクローニングし、TOP10コンピテント細胞内に形質転換した。DNA配列決定は、該ste13::NATR融合体が正確であることを証明した。得られたベクターをそれぞれpGLY5018と命名した。このベクターを図1Cに図示する。
【0061】
同様にして、表1に示されているプライマーセットSH782(配列番号21)およびSH805(配列番号33)、SH808(配列番号36)およびSH785(配列番号24)、ならびにSH806(配列番号34)およびSH807(配列番号35)を使用して、DAP2 5’、DAP2 3’およびNATRマーカーに対応する1011bp、1151bpおよび1248bpの断片をpGLY4521およびpAG25から増幅した。単離後、各断片の20ngをプライマーペアSH782(配列番号21)およびSH785(配列番号24)と共に使用して3321bpの断片を得、これをpCR2.1内にクローニングし、配列決定し、pGLY5019と命名した。このベクターを図1Dに図示する。
【実施例3】
【0062】
STE13およびDAP2ノックアウト株の作製
GS5.0グリカンを発現するピチア・パストリス(Pichia pastoris)栄養要求性糖操作細胞系YGLY7406[Δoch1、Δpno1、Δmnn4B、Δbmt2、Δura5、K.lactisおよびM.musculus UDP−GlcNAc輸送体、M.musculus α−1,2−MnsI、H.sapiens β−1,2−GlcNAcトランスフェラーゼI、R.norvegicus β−1,2−GlcNAcトランスフェラーゼII、D.melanogaster MnsII、S.pombe Galエピメラーゼ、D.melanogaster UDP−Gal輸送体およびH.sapiens β−1,4−ガラクトシルトランスフェラーゼ](例えば、Bobrowiczら,Glycobiology,14(9):757−766,2004;Hamiltonら,Science,313(5792):14411−1443,2006);米国公開出願第20060040353号を参照されたい)を全ての操作の出発株として使用した。この株および後続の株をどのようにして作製したかに関する流れ図は、図7(A〜C)を参照されたい。GS5.0株は、一方または両方の末端の非還元性末端上のβ−1,4−ガラクトース残基を末端とする二分岐アフコシル化N−結合グリカンを有する糖タンパク質を産生しうる(Bobrowiczら,Glycobiology,14(9):757−766,2004;Hamiltonら,Science,313(5792):14411−1443,2006)。株YGLY7406は、ジペプチジルペプチダーエ活性に関するレポータータンパク質として使用される、IgG1のFcドメインに融合した完全長ヒト腫瘍壊死因子受容体II(TNFRII−Fc)を発現する。
【0063】
形質転換のために、20μgのベクターpGLY4520およびpGLY4521を制限酵素SfiI(New England Biolabs,Ipswich,MA)で消化して、4091bpのSTE13::Ura5ノックアウト断片または4516bpのDAP2::URA5ノックアウト断片を得、これらをYGLY7406内にエレクトロポレーションにより形質転換して、それぞれSTE13またはDAP2を欠失させ、該形質転換体をuraマイナス最少プレート上で選択した。各遺伝子のノックアウトの成功を、表2に示されている5’、3’およびノックアウトプライマーセットを使用して確認した。ste13Δおよびdap2Δノックアウト株を、それぞれYGLY8084およびYGLY8090と命名した。ついで、ste13Δノックアウト株YGLY8084を、3290bpのDAP2::Natノックアウト断片を遊離させるためにSfiIで予め消化されたpGLY5019で形質転換することにより、二重Δste13/dap2ノックアウト株を作製した。形質転換体を100μg/ml Nat YMDプレート上でプレーティングし、二重ノックアウトの成功を、表2に示されている5’、3’およびノックアウトプライマーセットを使用して確認した。代表的な二重ノックアウト株をYGLY8096と命名した。最初にSTE13をノックアウトし、ついでDAP2をノックアウトすることによりこの株を作製したが、本発明者らは、最初にDAP2遺伝子をノックアウトし、ついでSTE13遺伝子をノックアウトした場合に、同じ所望の表現型を有する株が得られうることを実験的に実証している。そのような株を得るために、pGLY5018をSfiIで消化し、生じた2865bpのste13::NATRノックアウト断片を、DAP2遺伝子が既にノックアウトされているYGLY8090のような株内に形質転換することが可能である。
【実施例4】
【0064】
レポータータンパク質の作製、単離および分析
1% 酵母エキス、2% ペプトン、100mM リン酸カリウムバッファー(pH6.0)、1.34% 酵母窒素ベース、0.00004% ビオチンおよび1% グリセロールからなる増殖培地としての200mlの緩衝化グリセロール−複合培地(BMGY)内でタンパク質発現を26℃で72時間行った。BMGYにおけるグリセロールの代わりに1.5% メタノールからなる20mlの緩衝化メタノール−複合培地(BMMY)内で誘導を48時間行った。
【0065】
発現後、2000rpmで15分間の遠心分離により該細胞を除去した。GE Healthcare(Chalfont St.Giles,UK,Cat #17−1281−01)のStreamliner Protein A樹脂を使用して、TNFRII−Fc融合タンパク質を上清からアフィニティークロマトグラフィーにより捕捉した。該無細胞上清培地をStreamliner Protein Aカラム(XK 16/20 1.6cm×10.0cm)上にローディングし、3カラム体積の20mM Tris−HCl(pH7.0)で5.0ml/分の流量で前平衡化した。該カラムを3カラム体積の同じバッファーで洗浄し、TNFRII−Fc融合タンパク質を7カラム体積の40mM クエン酸ナトリウム(pH3.5)で溶出した。溶出した融合タンパク質を直ちに1M Tris−HCl(pH8.0)で中和した。
【0066】
CHT(登録商標)ヒドロキシアパタイトI型40μm樹脂(Bio−Rad Laboratories,Hercules,CA,Cat #157−0040)を第2精製工程として使用した。該ヒドロキシアパタイトカラムを3カラム体積の5mM リン酸ナトリウム(pH6.5)で平衡化し、Streamliner Protein A精製TNFRII−Fc融合タンパク質を該平衡化バッファー内にバッファー交換し、該カラムに適用した。ローディング後、該カラムを3カラム体積の該平衡化バッファーで洗浄し、溶出を、0〜1000mM 塩化ナトリウムの範囲の20カラム体積にわたる勾配を展開することにより行った。TNFRII−Fc融合タンパク質は約550−650mM 塩化ナトリウムで溶出する。プール化したTNFRII−Fc融合タンパク質を、0.2μm ポリエーテルスルホン(PES)膜フィルターを使用して滅菌濾過し、4℃で保存した。
【0067】
SDS−PAGE(4〜20% Tris−HClゲル,Bio−Rad Laboratories,Hercules,CA,Cat.#161−1123)を、それらの2つのカラム精製TNFRII−Fc融合タンパク質画分に対して行い、ポリビニリデンジフルオリド(PVDF)膜(ProBlott(商標) Membranes,Applied Biosystems,Foster City,CA,Cat #400994)上に55ボルトで30分にわたってトランスファーし、ProBlott染料(Applied Biosystems,Foster City,CA)で染色した。TNFRII−Fc融合タンパク質に対応するバンドをPVDF膜から切り出し、N末端配列決定のためにTufts Core Facility,Boston,MAに送った。
【0068】
同様にして、完全長組換え顆粒球コロニー刺激因子(GCSF)タンパク質の発現を行った。しかし、TNFRII−Fcを発現させるために使用した前記のGS5.0宿主細胞の代わりに、GCSFを発現させるために使用した宿主細胞は、実施例3に記載の方法によりSTE13およびDAP2遺伝子が破壊されているピチア・パストリス(Pichia pastoris)GS2.0宿主細胞であった。GS2.0宿主細胞は、Man5GlcNAc2 N−グリカンを含む糖タンパク質を産生するように遺伝的に操作された宿主細胞である。これらの株は、ガラクトースを末端とするN−グリカンを有する糖タンパク質を産生し得ない。そのような株の具体例はNettおよびGerngross,Yeast 20:1279(2003);Choiら,Proc.Natl.Acad.Sci.USA 100:5022(2003);ならびにHamiltonら,Science 301:1244(2003)に開示されている。図6は、TNFRII−Fcを発現するGS5.0に関して前記で示されているとおりにSTE13およびDAP2遺伝子が破壊された場合、これらの宿主細胞が無傷GCSFを産生しうることを示している(STE13/DAP2株において産生されたGCSFを示すレーン27〜29を、ste13/dap2ノックアウト株において産生されたGCSFを示すレーン32〜34と比較されたい)。
【実施例5】
【0069】
PpDppIIIpの欠失
同様に、PpSte13pおよびPpDap2pを排除するための前記実施例に記載されている方法を、PpDppIIIpを排除するために用いることが可能である。1つのそのような方法は、実施例2でPpSTE13およびPpDAP2に関して示されているのに類似した方法でノックアウトベクターを設計することであろう。プライマーは、PpDPPIII遺伝子(図13、配列番号54)の5’および3’フランキング領域を増幅するように設計されるであろう。なぜなら、ゲノム配列の必須領域、例えば、機能的PpDppIIIp活性に必要な領域は省かれるからである。PpDppIIIp活性は、これらのフランキング領域を選択マーカーと組合せることにより排除されうる。適合性選択マーカーとの組合せにより、PpDppIIIp活性は、個々に、あるいはPpSte13pおよび/またはPpDap2活性の排除と共に排除されうる。PpSte13p、PpDap2pおよびPpDppIIIp活性が排除されている株は、いずれかの潜在的ジペプチジルアミノペプチダーゼ活性を欠く株を与えるであろう。
【0070】
本発明は、本発明の少数の態様の例示として意図される実施例に開示されている具体的な実施形態によっては限定されず、機能的に同等なあらゆる実施形態が本発明の範囲内である。実際、本明細書に示され記載されているものに加えて本発明の種々の修飾が当業者に明らかとなり、添付の特許請求の範囲の範囲内に含まれると意図される。
【特許請求の範囲】
【請求項1】
ジペプチジルアミノペプチダーゼ(DAP)活性を欠く酵母宿主細胞における組換えタンパク質の製造方法であって、
a.DAP活性が排除されている遺伝的に修飾された酵母細胞を、該タンパク質をコードするポリヌクレオチドベクターで形質転換し、
b.該タンパク質の発現を誘導する条件下、該形質転換宿主細胞を培養し、
c.該形質転換宿主細胞または培地から該タンパク質を単離することを含む製造方法。
【請求項2】
該酵母細胞がピチア・パストリス(Pichia pastoris)である、請求項1記載の製造方法。
【請求項3】
酵母細胞ゲノムからのSTE13およびDAP2遺伝子の欠失または破壊によりDAP活性が排除されている、請求項1または2記載の製造方法。
【請求項4】
該宿主細胞が、ヒト様N−グリカンを含む糖タンパク質を産生するように遺伝的に操作されている、請求項1記載の酵母。
【請求項5】
該ヒト様N−グリカンが、ハイブリッドおよび複合N−グリカンからなる群から選択される、請求項4記載の酵母。
【請求項6】
Man5GlcNAc2、GlcNAcMan5GlcNAc2、GalGlcNAcMan5GlcNAc2、NANAGalGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc(1−4)Man3GlcNAc2、Gal(1−4)GlcNAc(1−4)Man3GlcNAc2およびNANA(1−4)Gal(1−4)GlcNAc(1−4)Man3GlcNAc2から選択されるN−グリカンを主として有する糖タンパク質を産生するように該宿主細胞が遺伝的に操作されている、請求項1記載の酵母。
【請求項7】
該酵母が更に、ヒト糖タンパク質を発現する、請求項1記載の製造方法。
【請求項8】
STE13およびDAP2をコードするゲノムDNAが酵母細胞ゲノムから欠失または破壊されている酵母細胞系を含む、ジペプチジルアミノペプチダーゼ(DAP)活性を欠く遺伝的に修飾された酵母細胞。
【請求項9】
該酵母細胞がピチア・パストリス(Pichia pastoris)である、請求項8記載の酵母細胞。
【請求項10】
該宿主細胞が、ヒト様N−グリカンを含む糖タンパク質を産生するように遺伝的に操作されている、請求項6記載の製造方法。
【請求項11】
該ヒト様N−グリカンが、ハイブリッドおよび複合N−グリカンからなる群から選択される、請求項10記載の製造方法。
【請求項12】
Man5GlcNAc2、GlcNAcMan5GlcNAc2、GalGlcNAcMan5GlcNAc2、NANAGalGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc(1−4)Man3GlcNAc2、Gal(1−4)GlcNAc(1−4)Man3GlcNAc2およびNANA(1−4)Gal(1−4)GlcNAc(1−4)Man3GlcNAc2から選択されるN−グリカンを主として有する糖タンパク質を産生するように該宿主細胞が遺伝的に操作されている、請求項6記載の製造方法。
【請求項13】
該タンパク質がヒト糖タンパク質である、請求項6記載の製造方法。
【請求項14】
N末端認識部位Xaa−ProまたはXaa−Ala(ここで、Xaaはいずれかのアミノ酸を表す)を有する治療用タンパク質の、酵母細胞における製造方法であって、
a.DAP活性が排除されている遺伝的に修飾された酵母細胞を、Xaa−ProおよびXaa−Alaからなる群から選択されるN末端認識部位を有する該治療用タンパク質をコードするポリヌクレオチドベクターで形質転換し、
b.該治療用タンパク質の発現を誘導する条件下、該形質転換宿主細胞を培養し、
c.該形質転換宿主細胞から該治療用タンパク質を単離することを含む製造方法。
【請求項15】
該酵母細胞がピチア・パストリス(Pichia pastoris)である、請求項8記載の製造方法。
【請求項16】
酵母細胞ゲノムからのSTE13およびDAP2の欠失または破壊によりDAP活性が排除されている、請求項14または15記載の製造方法。
【請求項17】
該宿主細胞が、ヒト様N−グリカンを含む糖タンパク質を産生するように遺伝的に操作されている、請求項14記載の製造方法。
【請求項18】
該ヒト様N−グリカンが、ハイブリッドおよび複合N−グリカンからなる群から選択される、請求項17記載の製造方法。
【請求項19】
Man5GlcNAc2、GlcNAcMan5GlcNAc2、GalGlcNAcMan5GlcNAc2、NANAGalGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc(1−4)Man3GlcNAc2、Gal(1−4)GlcNAc(1−4)Man3GlcNAc2およびNANA(1−4)Gal(1−4)GlcNAc(1−4)Man3GlcNAc2から選択されるN−グリカンを主として有する糖タンパク質を産生するように該宿主細胞が遺伝的に操作されている、請求項14記載の製造方法。
【請求項20】
該タンパク質がヒト糖タンパク質である、請求項14記載の製造方法。
【請求項1】
ジペプチジルアミノペプチダーゼ(DAP)活性を欠く酵母宿主細胞における組換えタンパク質の製造方法であって、
a.DAP活性が排除されている遺伝的に修飾された酵母細胞を、該タンパク質をコードするポリヌクレオチドベクターで形質転換し、
b.該タンパク質の発現を誘導する条件下、該形質転換宿主細胞を培養し、
c.該形質転換宿主細胞または培地から該タンパク質を単離することを含む製造方法。
【請求項2】
該酵母細胞がピチア・パストリス(Pichia pastoris)である、請求項1記載の製造方法。
【請求項3】
酵母細胞ゲノムからのSTE13およびDAP2遺伝子の欠失または破壊によりDAP活性が排除されている、請求項1または2記載の製造方法。
【請求項4】
該宿主細胞が、ヒト様N−グリカンを含む糖タンパク質を産生するように遺伝的に操作されている、請求項1記載の酵母。
【請求項5】
該ヒト様N−グリカンが、ハイブリッドおよび複合N−グリカンからなる群から選択される、請求項4記載の酵母。
【請求項6】
Man5GlcNAc2、GlcNAcMan5GlcNAc2、GalGlcNAcMan5GlcNAc2、NANAGalGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc(1−4)Man3GlcNAc2、Gal(1−4)GlcNAc(1−4)Man3GlcNAc2およびNANA(1−4)Gal(1−4)GlcNAc(1−4)Man3GlcNAc2から選択されるN−グリカンを主として有する糖タンパク質を産生するように該宿主細胞が遺伝的に操作されている、請求項1記載の酵母。
【請求項7】
該酵母が更に、ヒト糖タンパク質を発現する、請求項1記載の製造方法。
【請求項8】
STE13およびDAP2をコードするゲノムDNAが酵母細胞ゲノムから欠失または破壊されている酵母細胞系を含む、ジペプチジルアミノペプチダーゼ(DAP)活性を欠く遺伝的に修飾された酵母細胞。
【請求項9】
該酵母細胞がピチア・パストリス(Pichia pastoris)である、請求項8記載の酵母細胞。
【請求項10】
該宿主細胞が、ヒト様N−グリカンを含む糖タンパク質を産生するように遺伝的に操作されている、請求項6記載の製造方法。
【請求項11】
該ヒト様N−グリカンが、ハイブリッドおよび複合N−グリカンからなる群から選択される、請求項10記載の製造方法。
【請求項12】
Man5GlcNAc2、GlcNAcMan5GlcNAc2、GalGlcNAcMan5GlcNAc2、NANAGalGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc(1−4)Man3GlcNAc2、Gal(1−4)GlcNAc(1−4)Man3GlcNAc2およびNANA(1−4)Gal(1−4)GlcNAc(1−4)Man3GlcNAc2から選択されるN−グリカンを主として有する糖タンパク質を産生するように該宿主細胞が遺伝的に操作されている、請求項6記載の製造方法。
【請求項13】
該タンパク質がヒト糖タンパク質である、請求項6記載の製造方法。
【請求項14】
N末端認識部位Xaa−ProまたはXaa−Ala(ここで、Xaaはいずれかのアミノ酸を表す)を有する治療用タンパク質の、酵母細胞における製造方法であって、
a.DAP活性が排除されている遺伝的に修飾された酵母細胞を、Xaa−ProおよびXaa−Alaからなる群から選択されるN末端認識部位を有する該治療用タンパク質をコードするポリヌクレオチドベクターで形質転換し、
b.該治療用タンパク質の発現を誘導する条件下、該形質転換宿主細胞を培養し、
c.該形質転換宿主細胞から該治療用タンパク質を単離することを含む製造方法。
【請求項15】
該酵母細胞がピチア・パストリス(Pichia pastoris)である、請求項8記載の製造方法。
【請求項16】
酵母細胞ゲノムからのSTE13およびDAP2の欠失または破壊によりDAP活性が排除されている、請求項14または15記載の製造方法。
【請求項17】
該宿主細胞が、ヒト様N−グリカンを含む糖タンパク質を産生するように遺伝的に操作されている、請求項14記載の製造方法。
【請求項18】
該ヒト様N−グリカンが、ハイブリッドおよび複合N−グリカンからなる群から選択される、請求項17記載の製造方法。
【請求項19】
Man5GlcNAc2、GlcNAcMan5GlcNAc2、GalGlcNAcMan5GlcNAc2、NANAGalGlcNAcMan5GlcNAc2、GlcNAcMan3GlcNAc2、GlcNAc(1−4)Man3GlcNAc2、Gal(1−4)GlcNAc(1−4)Man3GlcNAc2およびNANA(1−4)Gal(1−4)GlcNAc(1−4)Man3GlcNAc2から選択されるN−グリカンを主として有する糖タンパク質を産生するように該宿主細胞が遺伝的に操作されている、請求項14記載の製造方法。
【請求項20】
該タンパク質がヒト糖タンパク質である、請求項14記載の製造方法。
【図1A】

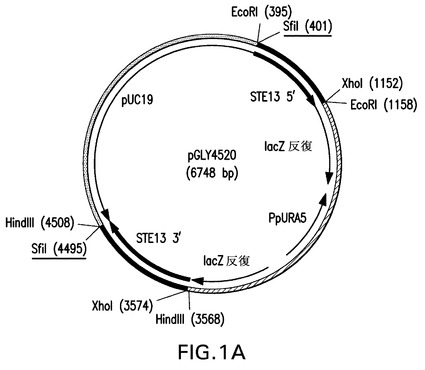
【図1B】
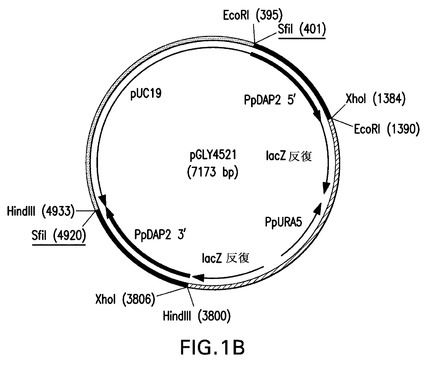
【図1C】
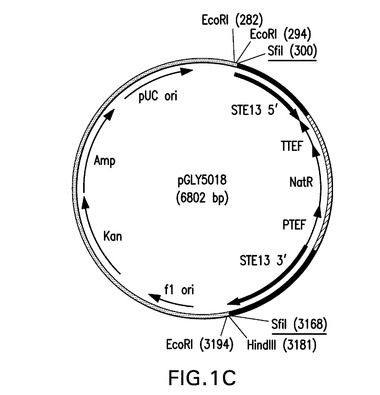
【図1D】
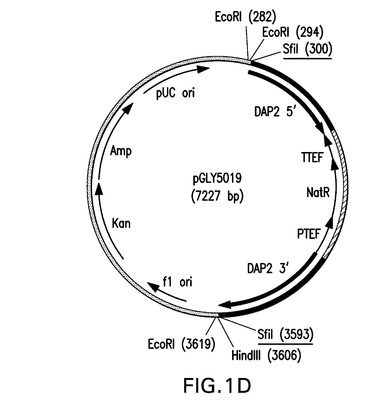
【図2A】

【図2B】

【図3】

【図4A】

【図4B】

【図5】

【図6】
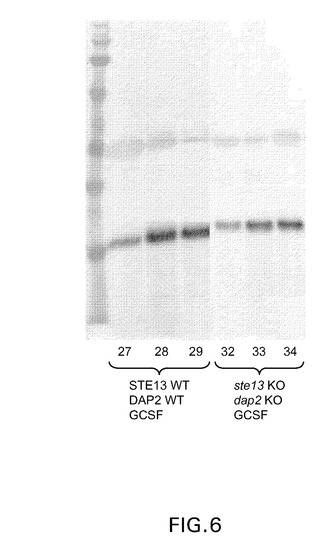
【図7A】
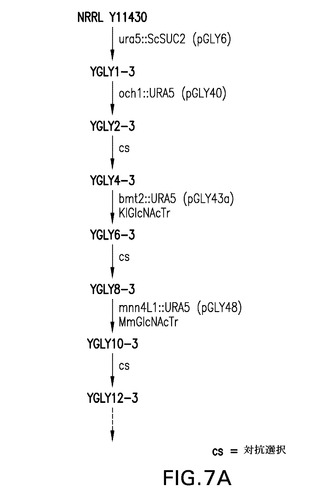
【図7B】
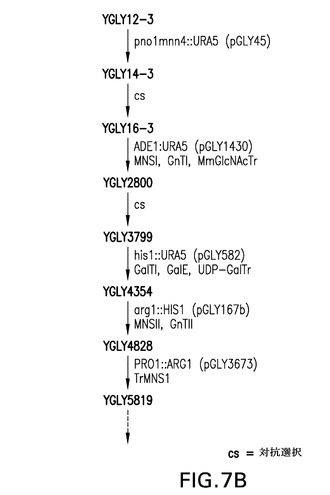
【図7C】
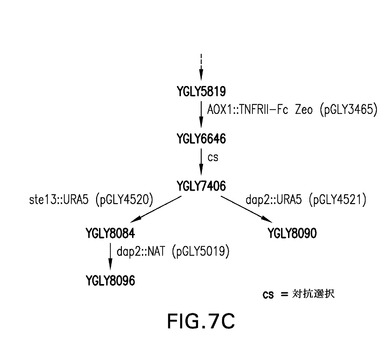
【図8A−1】

【図8A−2】

【図8A−3】

【図8B】

【図9A−1】

【図9A−2】

【図9A−3】

【図9B】

【図10A】
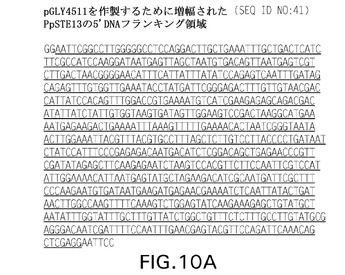
【図10B】

【図11A】

【図11B】

【図12】

【図13−1】

【図13−2】

【図1B】
【図1C】
【図1D】
【図2A】
【図2B】
【図3】
【図4A】
【図4B】
【図5】
【図6】
【図7A】
【図7B】
【図7C】
【図8A−1】
【図8A−2】
【図8A−3】
【図8B】
【図9A−1】
【図9A−2】
【図9A−3】
【図9B】
【図10A】
【図10B】
【図11A】
【図11B】
【図12】
【図13−1】
【図13−2】
【公表番号】特表2013−509181(P2013−509181A)
【公表日】平成25年3月14日(2013.3.14)
【国際特許分類】
【出願番号】特願2012−536977(P2012−536977)
【出願日】平成22年10月27日(2010.10.27)
【国際出願番号】PCT/US2010/054183
【国際公開番号】WO2011/053612
【国際公開日】平成23年5月5日(2011.5.5)
【出願人】(390023526)メルク・シャープ・エンド・ドーム・コーポレイション (924)
【Fターム(参考)】
【公表日】平成25年3月14日(2013.3.14)
【国際特許分類】
【出願日】平成22年10月27日(2010.10.27)
【国際出願番号】PCT/US2010/054183
【国際公開番号】WO2011/053612
【国際公開日】平成23年5月5日(2011.5.5)
【出願人】(390023526)メルク・シャープ・エンド・ドーム・コーポレイション (924)
【Fターム(参考)】
[ Back to top ]