
ジペプチドの製造法
本発明によれば、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質をコードするDNA、該DNAを含有する組換え体DNA、該組換え体DNAを保有する形質転換体、ジペプチドの合成活性を有する蛋白質の製造法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を用いたジペプチドの酵素的合成法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を生産する能力を有する微生物または形質転換体の培養物等を酵素源に用いたジペプチドの製造法が提供される。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ジペプチド合成活性を有する蛋白質またはジペプチド合成用蛋白質、ジペプチド合成活性を有する蛋白質の製造法、ジペプチド合成活性を有する蛋白質またはジペプチド合成用蛋白質を用いたジペプチドの製造法、ジペプチド合成活性を有する蛋白質またはジペプチド合成用蛋白質を生産する微生物または形質転換体、および該微生物または形質転換体を用いたジペプチドの製造法に関する。
【背景技術】
【0002】
ペプチドの大量合成法については、化学合成法(液相法、固相法)、酵素的合成法およびDNA組換え法を用いた生物学的合成法が知られている。現在、50残基以上の長鎖のペプチドに関しては酵素的合成法あるいは生物学的合成法が用いられ、ジペプチドに関しては化学合成法と酵素的合成法が主に用いられている。
化学合成法によるジペプチドの合成では、官能基の保護・脱保護などの操作が必要であり、またラセミ体も合成されることから、化学合成法は経済的、効率的な方法とはいえない。また、化学合成法は大量の有機溶媒等を使うため環境衛生上も好ましい方法ではない。
【0003】
酵素法によるジペプチドの合成に関しては、タンパク質分解酵素(プロテアーゼ)の逆反応を利用した方法[J.Biol.Chem.,119,707−720(1937)]、耐熱性アミノアシルt−RNA合成酵素を利用する方法(特開昭58−146539号公報、特開昭58−209991号公報、特開昭58−209992号公報、特開昭59−106298号公報)、非リボゾームペプチドシンセターゼ(以下、NRPSと称す)を利用する方法[Chem.Biol.,7,373−384(2000)、FEBS Lett.,498,42−45(2001)、米国特許第5795738号、米国特許第5652116号]が知られている。
【0004】
しかし、タンパク分解酵素の逆反応を利用した方法では、基質となるアミノ酸の官能基の保護・脱保護が必要であり、ペプチド形成反応の効率化およびペプチド分解反応の阻止が困難といった問題点がある。耐熱性アミノアシルt−RNA合成酵素を利用する方法には、酵素の発現、目的産物以外の副生反応の阻止が困難という問題点がある。NRPSを利用する方法に関しては、酵素分子が巨大なためにDNA組換え法を用いて該酵素を発現することが困難であること、補酵素である4’−ホスフォパンテテイン(4’−phosphopantetheine)の供給が必要であり、効率的な製造法とはいえない。
【0005】
一方、酵素分子量がNRPSより小さく、補酵素である4’−phosphopantetheineを必要としないγ−グルタミルシステインシンセターゼ(γ−glutamylcysteinesynthetase)、グルタチオンシンセターゼ(glutathione synthetase)、D−アラニン−D−アラニン(D−Ala−D−Ala)リガーゼ(D−Ala−D−Alaligase)、ポリ−γ−グルタミン酸シンセターゼ(poly−γ−glutamate synthetase)等の一群のペプチドシンセターゼも知られている。これらの酵素の殆どはD−アミノ酸を基質に用いる、またはγ位のカルボキシル基でのペプチド結合の形成を触媒する等の特徴を有するため、L−アミノ酸のα位カルボキシル基でペプチド結合するジペプチドの合成に用いることはできない。
【0006】
L−アミノ酸のα位カルボキシル基でのペプチド結合形成活性によるジペプチド生成が知られているのはバチルス属に属する微生物由来のジペプチド抗生物質であるバシリシン合成酵素のみである。バシリシン合成酵素は、バシリシン(L−アラニル−L−アンチカプシン、L−Ala−L−anticapsin)およびL−アラニル−L−アラニン(L−Ala−L−Ala)を合成する活性を有することは知られているが、その他のジペプチドの合成活性については知られていない[J.Ind.Microbiol.,2,201−208(1987)、Enzyme.Microbial.Technol.,29,400−406(2001)]。
【0007】
ある種の微生物は2つのアミノ酸が環状につながった環状ジペプチド(ジケトピペラジン)構造を持つ化合物を生成することも知られている[J.Nat.Prod.,59,293−296(1996)、Tetrahedron,28,2999(1972)、J.Appl.Microbiol.,86,29−53(1999)]。ジケトピペラジンの生合成については、Streptomyces acidiscabiesのThaxtomin生合成過程ではNRPSによってcyclo−(L−4−nitrotryptophyl−L−phenylalanine)構造が合成される[Mol.Microbiol.,38,794−804(2000)]こと、およびバチルス属細菌のNRPSの一部のモジュールの作用によってフェニルアラニンとプロリンからcyclo(phenylalanyl−proline)が生成すること[J.Biol.Chem.,273,22773−22781(1998)]が報告されている。
【0008】
一方、抗生物質であるアルボノルシン(albonoursin)の生産株として知られているストレプトマイセス・ノルセイ(Streptomyces noursei)ATCC11455株ではNRPS酵素とは全く類似性のない蛋白質(albC遺伝子産物)がシクロ(L−フェニルアラニル−L−ロイシン)[cyclo(L−phenylalanyl−L−leucine)]構造の合成を担っていること、albC遺伝子を導入したEscherichia coliおよびStreptomyces lividansの培養液にcyclo dipeptide oxidaseを作用させるとアルボノルシンが検出されたとの報告はある[Chemistry&Biol.,9,1355−1364(2002)]が、albC遺伝子産物が直鎖状のジペプチドを生成するとの報告はない。
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明の目的は、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質をコードするDNA、該DNAを含有する組換え体DNA、該組換え体DNAを保有する形質転換体、ジペプチドの合成活性を有する蛋白質の製造法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を用いたジペプチドの酵素的合成法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を生産する能力を有する微生物または形質転換体の培養物等を酵素源に用いたジペプチドの製造法を提供することにある。
【課題を解決するための手段】
【0010】
本発明は、以下の(1)〜(24)に関する。
(1)以下の[1]〜[3]のいずれかに記載の蛋白質(ただし、配列番号1で表されるアミノ酸配列からなる蛋白質を除く)。
[1]配列番号2で表されるアミノ酸配列を有する蛋白質
[2]配列番号1または2で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドの合成活性を有する蛋白質
[3]配列番号1または2で表されるアミノ酸配列と65%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有する蛋白質
(2)以下の[1]〜[3]のいずれかに記載のジペプチド合成用蛋白質。
[1]配列番号1で表されるアミノ酸配列を有するジペプチド合成用蛋白質
[2]配列番号1で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)
で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質
[3]配列番号1で表されるアミノ酸配列と65%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質
(3)以下の[1]〜[3]のいずれかに記載のDNA(ただし、配列番号3で表される塩基配列からなるDNAを除く)。
[1]上記(1)の蛋白質をコードするDNA
[2]配列番号4で表される塩基配列を有するDNA
[3]配列番号4で表される塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチド合成活性を有する蛋白質をコードするDNA
(4)上記(3)のDNAを含有する組換え体DNA。
【0011】
(5)上記(4)の組換え体DNAを有する形質転換体。
(6)形質転換体が、微生物を宿主として得られる形質転換体である上記(5)の形質転換体。
(7)微生物が、エシェリヒア(Escherichia)属に属する微生物である上記(6)の形質転換体。
【0012】
(8)上記(5)〜(7)のいずれか1つに記載の形質転換体を培地に培養し、培養物中に上記(1)の蛋白質を生成、蓄積させ、該培養物より該蛋白質を採取する上記(1)の蛋白質の製造法。
(9)上記(1)の蛋白質を生産する能力を有する微生物を培地に培養し、培養物中に該蛋白質を生成、蓄積させ、該培養物より該蛋白質を採取する上記(1)の蛋白質の製造法。
【0013】
(10)微生物がストレプトマイセス(Streptomyces)属に属する微生物である上記(9)の製造法。
(11)ストレプトマイセス属に属する微生物が、アルボノルシンを生産する能力を有するストレプトミセス属に属する微生物である上記(10)の製造法。
(12)アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物が、ストレプトマイセス・アルボラス(Streptomyces alborus)またはストレプトマイセス・ノルセイ(Streptomyces noursei)である上記(11)の製造法。
【0014】
(13)上記(1)の蛋白質または上記(2)のジペプチド合成用蛋白質、1種以上のアミノ酸、およびATPを水性媒体中に存在せしめ、該媒体中に式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法。
【0015】
(14)以下の[1]〜[3]から選ばれる培養物または該培養物の処理物を酵素源に用い、該酵素源、1種以上のアミノ酸を水性媒体中に存在せしめ、該媒体中に式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法。
[1]上記(5)〜(7)のいずれか1つに記載の形質転換体の培養物または該培養物の処理物
[2]上記(1)の蛋白質を生産する能力を有する微生物の培養物または該培養物の処理物
[3]上記(2)のジペプチド合成用蛋白質を生産する能力を有する微生物の培養物または該培養物の処理物
(15)上記(1)の蛋白質を生産する能力を有する微生物が、ストレプトマイセス属に属する微生物である上記(14)の製造法。
【0016】
(16)上記(2)のジペプチド合成用蛋白質を生産する能力を有する微生物が、ストレプトマイセス属に属する微生物である上記(14)の製造法。
(17)ストレプトマイセス属に属する微生物が、アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物である上記(15)または(16)の製造法。
【0017】
(18)アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物が、ストレプトマイセス・アルボラスまたはストレプトマイセス・ノルセイに属する微生物である上記(17)の製造法。
(19)上記(2)のジペプチド合成用蛋白質を生産する能力を有する微生物が、上記(2)のジペプチド合成用蛋白質をコードするDNAで形質転換された微生物である上記(14)の製造法。
(20)上記(2)のジペプチド合成用蛋白質をコードするDNAで形質転換された微生物が、エシェリヒア属に属する微生物である上記(19)の製造法。
(21)培養物の処理物が、培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の溶媒処理物、該菌体の酵素処理物、該菌体の蛋白質分画物、該菌体の固定化物あるいは該菌体より抽出して得られる酵素標品であることを特徴とする、上記(14)〜(20)のいずれか1つの製造法。
【0018】
(22)1種以上のアミノ酸が、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体である上記(13)〜(21)のいずれか1つの製造法。
(23)ジペプチドが式(II)
R3−R4 (II)
(式中、R3およびR4は同一または異なって、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体を表す)であらわされるジペプチドである上記(13)〜(22)のいずれか1つの製造法。
【0019】
(24)L体またはD体のアミノ酸が、アラニン、グルタミン、グルタミン酸、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、トリプトファン、メチオニン、セリン、スレオニン、システイン、アスパラギン、チロシン、リジン、アルギニン、ヒスチジン、アスパラギン酸、α−アミノ酪酸、アザセリン、テアニン、4−ヒドロキシプロリン、3−ヒドロキシプロリン、オルニチン、シトルリンおよび6−ジアゾ−5−オキソノルロイシンから選ばれるアミノ酸である上記(22)または(23)の製造法。
【発明の効果】
【0020】
本発明によれば、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質をコードするDNA、該DNAを含有する組換え体DNA、該組換え体DNAを保有する形質転換体、ジペプチドの合成活性を有する蛋白質の製造法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を用いたジペプチドの酵素的合成法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を生産する能力を有する微生物または形質転換体の培養物等を酵素源に用いたジペプチドの製造法を提供することができる。
【図面の簡単な説明】
【0021】
【図1】図1は、ジペプチドの合成活性を有する蛋白質の発現プラスミドベクターであるpAL−nouおよびpAL−albの構築過程を示す図である。
【符号の説明】
【0022】
Ampr:アンピシリン耐性遺伝子
lacIq:ラクトースリプレッサー遺伝子
albC:albC遺伝子またはalbC類似遺伝子
【発明を実施するための最良の形態】
【0023】
本発明の蛋白質としては、以下の[1]〜[3]記載の蛋白質(ただし、配列番号1で表されるアミノ酸配列からなる蛋白質を除く)、
[1]配列番号2で表されるアミノ酸配列を有する蛋白質、
[2]配列番号1または2で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドの合成活性を有する蛋白質、および
[3]配列番号1または2で表されるアミノ酸配列と65%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上、最も好ましくは99%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有する蛋白質、
をあげることができる。
【0024】
また、本発明のジペプチド合成用蛋白質としては、以下の[4]〜[6]記載のジペプチド合成用蛋白質、
[4]配列番号1で表されるアミノ酸配列を有するジペプチド合成用蛋白質、
[5]配列番号1で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質、
[6]配列番号1で表されるアミノ酸配列と65%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上、最も好ましくは99%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質、
をあげることができる。
【0025】
以下、上記本発明の蛋白質およびジペプチドの合成用蛋白質を合わせて本発明の蛋白質と称する場合もある。
上記において、1以上のアミノ酸が欠失、置換または付加されたアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有する蛋白質は、Molecular Cloning,A Laboratory Manual,Third Edition,Cold SpringHarbor Laboratory Press(1989)(以下、モレキュラー・クローニング第3版と略す)、Current Protocols in Molecular Biology,John Wiley & Sons(1987−1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)、NucleicAcids Research,10,6487(1982)、Proc.Natl.Acad.Sci.USA,79,6409(1982)、Gene,34,315(1985)、Nucleic Acids Research,13,4431(1985)、Proc.Natl.Acad.Sci.USA,82,488(1985)等に記載の部位特異的変異導入法を用いて、例えば配列番号1または2で表されるアミノ酸配列からなる蛋白質をコードするDNAに部位特異的変異を導入することにより、取得することができる。
【0026】
欠失、置換または付加されるアミノ酸の数は特に限定されないが、上記の部位特異的変異法等の周知の方法により欠失、置換または付加できる程度の数であり、1個から数十個、好ましくは1〜20個、より好ましくは1〜10個、さらに好ましくは1〜5個である。
配列番号1または2で表されるアミノ酸配列において1以上のアミノ酸が欠失、置換または付加されたとは、同一配列中の任意の位置において、1または複数のアミノ酸が欠失、置換または付加されていてもよい。
【0027】
アミノ酸の置換が可能なアミノ酸としては、例えば配列番号1または2で表されるアミノ酸配列を公知のアライメントソフトウェアを用いて比較したときに、すべてのアミノ酸配列において保存されていないアミノ酸をあげることができる。公知のアライメントソフトウェアとしては、例えば遺伝子解析ソフトウェアGenetyx(ソフトウェア開発株式会社)に含まれるアライメント解析ソフトをあげることができる。該解析ソフトの解析パラメータとしては、デフォルト値を用いることができる。
【0028】
また、アミノ酸の欠失または付加が可能なアミノ酸の位置としては、例えば配列番号1または2で表されるアミノ酸配列のN末端側およびC末端側をあげることができる。
欠失、置換または付加は同時に生じてもよく、置換または付加されるアミノ酸は天然型と非天然型とを問わない。天然型アミノ酸としては、L−アラニン、L−アスパラギン、L−アスパラギン酸、L−グルタミン、L−グルタミン酸、グリシン、L−ヒスチジン、L−イソロイシン、L−ロイシン、L−リジン、L−メチオニン、L−フェニルアラニン、L−プロリン、L−セリン、L−スレオニン、L−トリプトファン、L−チロシン、L−バリン、L−システインなどがあげられる。
【0029】
以下に、相互に置換可能なアミノ酸の例を示す。同一群に含まれるアミノ酸は相互に置換可能である。
A群:ロイシン、イソロイシン、ノルロイシン、バリン、ノルバリン、アラニン、2−アミノブタン酸、メチオニン、O−メチルセリン、t−ブチルグリシン、t−ブチルアラニン、シクロヘキシルアラニン
B群:アスパラギン酸、グルタミン酸、イソアスパラギン酸、イソグルタミン酸、2−アミノアジピン酸、2−アミノスベリン酸
C群:アスパラギン、グルタミン
D群:リジン、アルギニン、オルニチン、2,4−ジアミノブタン酸、2,3−ジアミノプロピオン酸
E群:プロリン、3−ヒドロキシプロリン、4−ヒドロキシプロリン
F群:セリン、スレオニン、ホモセリン
G群:フェニルアラニン、チロシン
また、本発明の蛋白質が式(I)で表されるジペプチド合成活性を有するためには、配列番号1または2で表されるアミノ酸配列との相同性が65%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上、最も好ましくは99%以上の相同性を有していることが望ましい。
【0030】
アミノ酸配列や塩基配列の相同性は、Karlin and AltschulによるアルゴリズムBLAST[Pro.Natl.Acad.Sci.USA,90,5873(1993)]やFASTA[Methods Enzymol.,183,63(1990)]を用いて決定することができる。このアルゴリズムBLASTに基づいて、BLASTNやBLASTXとよばれるプログラムが開発されている[J.Mol.Biol.,215,403(1990)]。BLASTに基づいてBLASTNによって塩基配列を解析する場合には、パラメータは例えばScore=100、wordlength=12とする。また、BLASTに基づいてBLASTXによってアミノ酸配列を解析する場合には、パラメータは例えばscore=50、wordlength=3とする。BLASTとGappedBLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov.)。
【0031】
本発明の蛋白質が、式(I)で表されるジペプチド合成活性を有する蛋白質であることを確認する手段としては、例えばDNA組換え法を用いて本発明の蛋白質を発現する形質転換体を作製し、該形質転換体を用いて本発明の蛋白質を製造した後、本発明の蛋白質、1種以上のアミノ酸、およびATPを水性媒体中に存在せしめ、該水性媒体中に上記式(I)で表されるジペプチドが生成、蓄積するか否かをHPLC等により分析する方法をあげることができる。
【0032】
本発明のDNAとしては、以下の[1]〜[3]記載のDNA(ただし、配列番号3で表される塩基配列からなるDNAを除く)、
[1]配列番号2で表されるアミノ酸配列を有する蛋白質をコードするDNA、
[2]配列番号4で表される塩基配列を有するDNA、および
[3]配列番号4で表される塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ式(I)で表されるジペプチド合成活性を有する蛋白質をコードするDNA、
をあげることができる。
【0033】
また、本発明の式(I)で表されるジペプチドの製造法に用いることができるDNAとしては、上記[1]〜[3]記載のDNAに加え、以下の[4]〜[6]記載のDNA、
[4]配列番号1で表されるアミノ酸配列を有する蛋白質をコードするDNA、
[5]配列番号3で表される塩基配列を有するDNA、および
[6]配列番号3で表される塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ式(I)で表されるジペプチド合成活性を有する蛋白質をコードするDNA、
をあげることができる。
【0034】
ここでいう「ハイブリダイズする」とは、特定の塩基配列を有するDNAまたは該DNAの一部にDNAがハイブリダイズすることである。したがって、該特定の塩基配列を有するDNAまたは該DNAの一部の塩基配列は、ノーザンまたはサザンブロット解析のプローブとして有用であるか、またはPCR解析のオリゴヌクレオチドプライマーとして使用できる長さのDNAであってもよい。プローブとして用いるDNAとしては、少なくとも100塩基以上、好ましくは200塩基以上、より好ましくは500塩基以上のDNAをあげることができるが、少なくとも10塩基以上、好ましくは15塩基以上のDNAであってもよい。
【0035】
DNAのハイブリダイゼーション実験の方法はよく知られており、例えば当業者であれば本願明細書に従い、ハイブリダイゼーションの条件を決定することができる。該ハイブリダイゼーションの条件は、モレキュラー・クローニング第2版、第3版(2001年)、Methods for General and Molecular Bacteriolgy,ASM Press(1994)、Immunologymethods manual,Academic press(Molecular)に記載の他、多数の他の標準的な教科書に従っておこなうことができる。
【0036】
上記のストリンジェントな条件とは、例えばDNAを固定化したフィルターとプローブDNAとを50%ホルムアミド、5×SSC(750mMの塩化ナトリウム、75mMのクエン酸ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハルト溶液、10%の硫酸デキストラン、および20μg/lの変性させたサケ精子DNAを含む溶液中で42℃で一晩、インキュベートした後、例えば約65℃の0.2×SSC溶液中で該フィルターを洗浄する条件が好ましいが、より低いストリンジェント条件を用いることもできる。ストリンジェンな条件の変更は、ホルムアミドの濃度調整(ホルムアミドの濃度を下げるほど低ストリンジェントになる)、塩濃度および温度条件の変更により可能である。低ストリンジェント条件としては、例えば6×SSCE(20×SSCEは、3mol/lの塩化ナトリウム、0.2mol/lのリン酸二水素ナトリウム、0.02mol/lのEDTA、pH7.4)、0.5%のSDS、30%のホルムアミド、100μg/lの変性させたサケ精子DNAを含む溶液中で、37℃で一晩インキュベートした後、50℃の1×SSC、0.1%SDS溶液を用いて洗浄する条件をあげることができる。また、さらに低いストリンジェントな条件としては、上記した低ストリンジェント条件において、高塩濃度(例えば5×SSC)の溶液を用いてハイブリダイゼーションを行った後、洗浄する条件をあげることができる。
【0037】
上記した様々な条件は、ハイブリダイゼーション実験のバックグラウンドを抑えるために用いるブロッキング試薬を添加、または変更することにより設定することもできる。上記したブロッキング試薬の添加は、条件を適合させるために、ハイブリダイゼーション条件の変更を伴ってもよい。
上記したストリンジェントな条件下でハイブリダイズ可能なDNAとしては、例えば上記したBLASTやFASTA等を用いて上記したパラメータ等に基づいて計算したときに、配列番号3または4で表される塩基配列と少なくとも90%以上、好ましくは95%以上、より好ましくは98%以上、さらに好ましくは99%以上の相同性を有するDNAをあげることができる。
【0038】
配列番号3または4で表される塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズするDNAが、式(I)で表されるジペプチドの合成活性を有する蛋白質をコードするDNAであることは、例えば上記したように、組換えDNA法を用いて該DNAにコードされる蛋白質を製造し、該蛋白質の活性を測定することにより確認することができる。
(i)本発明のDNA、および本発明の蛋白質またはジペプチドの製造法に用いられるDNAの調製
本発明のDNA、および本発明の蛋白質またはジペプチドの製造法(以下、単に本発明の製造法ともいう)に用いられるDNAは、例えば、配列番号3または4で表される塩基配列に基づき設計することができるプローブを用いた、ストレプトマイセス属に属する微生物の染色体DNAライブラリーに対するサザンハイブリダイゼーション、または配列番号3または4で表される塩基配列に基づき設計することができるプライマーDNAを用いた、ストレプトマイセス属に属する微生物の染色体DNAを鋳型としたPCR[PCR Protocols,Academic Press(1990)]により取得することができる。
【0039】
また、各種の遺伝子配列データベースに対して配列番号1または2で表されるアミノ酸配列をコードするDNAの塩基配列と75%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上、最も好ましくは99%以上の相同性を有する配列を検索し、該検索によって得られた塩基配列に基づき、該塩基配列を有する生物の染色体DNA、cDNAライブラリー等から上記した方法により本発明のDNA、または本発明の製造法に用いられるDNAを取得することもできる。
【0040】
取得したDNAをそのまま、あるいは適当な制限酵素などで切断し、常法によりベクターに組み込み、得られた組換え体DNAを宿主細胞に導入した後、通常用いられる塩基配列解析方法、例えばジデオキシ法[Proc.Natl.Acad.Sci.,USA,74,5463(1977)]あるいは373A・DNAシークエンサー(パーキン・エルマー社製)等の塩基配列分析装置を用いて分析することにより、該DNAの塩基配列を決定することができる。
【0041】
塩基配列を決定した結果、取得されたDNAが部分長であった場合は、該部分長DNAをプローブに用いた、染色体DNAライブラリーに対するサザンハイブリダイゼーション法等により、全長DNAを取得することができる。
更に、決定されたDNAの塩基配列に基づいて、パーセプティブ・バイオシステムズ社製8905型DNA合成装置等を用いて化学合成することにより目的とするDNAを調製することもできる。
【0042】
上記のようにして取得されるDNAとして、例えば、配列番号3または4で表される塩基配列を有するDNAをあげることができる。
本発明のDNAおよび本発明の製造法に用いられるDNAを組み込むベクターとしては、pBluescriptII KS(+)(ストラタジーン社製)、pDIRECT[Nucleic Acids Res.,18,6069(1990)]、pCR−Script Amp SK(+)(ストラタジーン社製)、pT7Blue(ノバジェン社製)、pCRII(インビトロジェン社製)およびpCR−TRAP(ジーンハンター社製)などをあげることができる。
【0043】
宿主細胞としては、エシェリヒア属に属する微生物などをあげることができる。エシェリヒア属に属する微生物としては、例えば、エシェリヒア・コリ(Escherichia coli)XL1−Blue、エシェリヒア・コリXL2−Blue、エシェリヒア・コリDH1、エシェリヒア・コリMC1000、エシェリヒア・コリKY3276、エシェリヒア・コリW1485、エシェリヒア・コリJM109、エシェリヒア・コリHB101、エシェリヒア・コリNo.49、エシェリヒア・コリW3110、エシェリヒア・コリNY49、エシェリヒア・コリMP347、エシェリヒア・コリNM522、エシェリヒア・コリME8415等をあげることができる。
【0044】
組換え体DNAの導入方法としては、上記宿主細胞へDNAを導入する方法であればいずれも用いることができ、例えば、カルシウムイオンを用いる方法「Proc.Natl.Acad.Sci.,USA,69,2110(1972)]、プロトプラスト法(特開昭63−248394)、エレクトロポレーション法[NucleicAcids Res.,16,6127(1988)]等をあげることができる。
上記方法によって得られる本発明の製造法に用いられるDNAを保有する微生物としては、例えば配列番号3で表される配列を有するDNAを含有する組換え体DNAを保有する微生物であるエシェリヒア・コリNM522/pAL−nouをあげることができる。
(ii)本発明の蛋白質および本発明のジペプチド合成用蛋白質の製造法
本発明の蛋白質および本発明のジペプチド合成用蛋白質は、モレキュラー・クローニング第3版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載された方法等を用い、例えば以下の方法により、上記(i)の方法により取得した本発明のDNAおよび本発明の製造法に用いられるDNAを宿主細胞中で発現させて、製造することができる。
【0045】
本発明のDNAまたは本発明の製造法に用いられるDNAをもとにして、必要に応じて、本発明の蛋白質または本発明のジペプチド合成用蛋白質をコードする部分を含む適当な長さのDNA断片を調製する。また、該蛋白質をコードする部分の塩基配列を、宿主の発現に最適なコドンとなるように、塩基を置換することにより、該蛋白質の生産率を向上させることができる。
【0046】
該DNA断片を適当な発現ベクターのプロモーターの下流に挿入することにより、組換え体DNAを作製する。
該組換え体DNAを、該発現ベクターに適合した宿主細胞に導入することにより、本発明の蛋白質を生産する形質転換体を得ることができる。
宿主細胞としては、細菌、酵母、動物細胞、昆虫細胞等、植物細胞等、目的とする遺伝子を発現できるものであればいずれも用いることができ、好ましくは細菌、より好ましくはエシェリヒア属、バチルス属またはコリネバクテリウム属に属する微生物、さらに好ましくはエシェリヒア属に属する微生物をあげることができる。
【0047】
発現ベクターとしては、上記宿主細胞において自立複製可能ないしは染色体中への組込が可能で、本発明のDNAまたは本発明の製造法に用いられるDNAを転写できる位置にプロモーターを含有しているものが用いられる。
細菌等の原核生物を宿主細胞として用いる場合は、本発明のDNAまたは本発明の製造法に用いられるDNAを有する組換え体DNAは、原核生物中で自立複製可能であると同時に、プロモーター、リボソーム結合配列、本発明のDNAまたは本発明の製造法に用いられるDNA、転写終結配列より構成された組換え体DNAであることが好ましい。プロモーターを制御する遺伝子が含まれていてもよい。
【0048】
発現ベクターとしては、pBTrp2、pBTac1、pBTac2(いずれもベーリンガーマンハイム社製)、pHelix1(ロシュ・ダイアグノスティクス社製)、pKK233−2(アマシャム・ファルマシア・バイオテク社製)、pSE280(インビトロジェン社製)、pGEMEX−1(プロメガ社製)、pQE8(キアゲン社製)、pQE60(キアゲン社製)、pET−3(ノバジェン社製)、pKYP10(特開昭58−110600)、pKYP200[Agric.Biol.Chem.,48,669(1984)]、pLSA1[Agric.Biol.Chem.,53,277(1989)]、pGEL1[Proc.Natl.Acad.Sci.,USA,82,4306(1985)]、pBluescriptIISK(+)、pBluescript II KS(−)(ストラタジーン社製)、pTrS30[エシェリヒア・コリJM109/pTrS30(FERMBP−5407)より調製]、pTrS32[エシェリヒア・コリJM109/pTrS32(FERM BP−5408)より調製]、pPAC31(WO98/12343)、pUC19[Gene,33,103(1985)]、pSTV28(宝酒造社製)、pUC118(宝酒造社製)、pPA1(特開昭63−233798)等を例示することができる。
【0049】
プロモーターとしては、エシェリヒア・コリ等の宿主細胞中で機能するものであればいかなるものでもよい。例えば、trpプロモーター(Ptrp)、lacプロモーター(Plac)、PLプロモーター、PRプロモーター、PSEプロモーター等の、大腸菌やファージ等に由来するプロモーター、SPO1プロモーター、SPO2プロモーター、penPプロモーター等をあげることができる。またPtrpを2つ直列させたプロモーター、tacプロモーター、lacT7プロモーター、let Iプロモーターのように人為的に設計改変されたプロモーター等も用いることができる。
【0050】
さらにバチルス属に属する微生物中で発現させるためのxylAプロモーター[Appl.Microbiol.Biotechnol.,35,594−599(1991)]やコリネバクテリウム(Corynebacterium)属に属する微生物中で発現させるためのP54−6プロモーター[Appl.Microbiol.Biotechnol.,53,674−679(2000)]、ストレプトマイセス属に属する微生物中で発現させるためのxylAプロモーター(GeneticManipulation of Streptomyces:a Laboratory Manual:John Innes Foundation)なども用いることができる。
【0051】
リボソーム結合配列であるシャイン−ダルガノ(Shine−Dalgarno)配列と開始コドンとの間を適当な距離(例えば6〜18塩基)に調節したプラスミドを用いることが好ましい。
本発明のDNAまたは本発明の製造法に用いられるDNAを発現ベクターに結合させた組換え体DNAにおいては、転写終結配列は必ずしも必要ではないが、構造遺伝子の直下に転写終結配列を配置することが好ましい。
【0052】
このような組換え体DNAとしては、例えばpAL−nuoおよびpAL−albをあげることができる。
原核生物としては、エシェリヒア属、セラチア(Serratia)属、バチルス属、ブレビバクテリウム(Brevibacterium)属、コリネバクテリウム(Corynebacterium)属、ミクロバクテリウム属(Microbacterium)、シュードモナス(Pseudomonas)属、アグロバクテリウム(Agrobacterium)属、アリシクロバチルス属(Alicyclobacillus)、アナベナ(Anabena)属、アナシスティス(Anacystis)属、アスロバクター(Arthrobacter)属、アゾバクター(Azotobacter)属、クロマチウム(Chromatium)属、エルビニア(Erwinia)属、メチロバクテリウム(Methylobacterium)属、フォルミディウム(Phormidium)属、ロドバクター(Rhodobacter)属、ロドシュードモナス(Rhodopseudomonas)属、ロドスピリウム(Rhodospirillum)属、セネデスムス(Scenedesmus)属、ストレプトマイセス(Streptomyces)属、シネコッカス(Synechoccus)属、ザイモモナス(Zymomonas)属等に属する微生物、例えば、エシェリヒア・コリXL1−Blue、エシェリヒア・コリXL2−Blue、エシェリヒア・コリDH1、エシェリヒア・コリDH5α、エシェリヒア・コリMC1000、エシェリヒア・コリKY3276、エシェリヒア・コリW1485、エシェリヒア・コリJM109、エシェリヒア・コリHB101、エシェリヒア・コリNo.49、エシェリヒア・コリW3110、エシェリヒア・コリNY49、エシェリヒア・コリMP347、エシェリヒア・コリNM522、バチルス・サチリスATCC33712、バチルス・メガテリウム、バチルス・エスピー(Bacillus sp.)FERM BP−6030、バチルス・アミロリケファスエンス、バチルス・コアギュランス、バチルス・リケニフォルミス、バチルス・プミルス、ブレビバクテリウム・アンモニアゲネス(Brevibacterium ammoniagenes)、ブレビバクテリウム・イマリオフィルム(Brevibacterium immariophilum)ATCC14068、ブレビバクテリウム・サッカロリティカム(Brevibacterium saccharolyticum)ATCC14066、ブレビバクテリウム・フラバム(Brevibacterium flavum)ATCC14067、ブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)ATCC13869、コリネバクテリウム・グルタミカム(Corynebacterium glutamicum)ATCC13032、コリネバクテリウム・グルタミカムATCC14297、コリネバクテリウム・アセトアシドフィルム(Corynebacterium acetoacidophilum)ATCC13870、ミクロバクテリウム・アンモニアフィルム(Microbacterium ammoniaphilum)ATCC15354、セラチア・フィカリア(Serratia ficaria)、セラチア・フォンチコラ(Serratia fonticola)、セラチア・リケファシエンス(Serratia liquefaciens)、セラチア・マルセッセンス(Serratia marcescens)、シュードモナス・エスピー(Pseudomonas sp.)D−0110、アグロバクテリウム・ラジオバクタ(Agrobacterium radiobacter)、アグロバクテリウム・リゾジーンズ(Agrobacterium rhizogenes)、アグロバクテリウム・ルビ(Agrobacterium rubi)、アナベナ・シリンドリカ(Anabaena cylindrica)、アナベナ・ドリオルム(Anabaena doliolum)、アナベナ・フロスアクア(Anabaena flos−aquae)、アースロバクター・オーレッセンス(Arthrobacter aurescens)、アースロバクター・シトレウス(Arthrobacter citreus)、アースロバクター・グロブフォルミス(Arthrobacter globformis)、アースロバクター・ヒドロカーボグルタミカス(Arthrobacter hydrocarboglutamicus)、アースロバクター・ミソレンス(Arthrobacter mysorens)、アースロバクター・ニコチアナ(Arthrobacter nicotianae)、アースロバクター・パラフィネウス(Arthrobacter paraffineus)、アースロバクター・プロトフォルミエ(Arthrobacter protophormiae)、アースロバクター・ロセオパラフィナス(Arthrobacter roseoparaffinus)、アースロバクター・スルフレウス(Arthrobacter sulfureus)、アースロバクター・ウレアファシエン(Arthrobacter ureafaciens)、クロマチウム・ブデリ(Chromatium buderi)、クロマチウム・テピダム(Chromatium tepidum)、クロマチウム・ビノサム(Chromatium vinosum)、クロマチウム・ワーミンギ(Chromatium warmingii)、クロマチウム・フルビアタティレ(Chromatium fluviatile)、エルビニア・ウレドバラ(Erwinia uredovora)、エルビニア・カロトバラ(Erwinia carotovora)、エルビニア・アナス(Erwinia ananas)、エルビニア・ヘリコラ(Erwinia herbicola)、エルビニア・パンクタタ(Erwinia punctata)、エルビニア・テレウス(Erwinia terreus)、メチロバクテリウム・ロデシアナム(Methylobacterium rhodesianum)、メチロバクテリウム・エクソトルクエンス(Methylobacterium extorquens)、フォルミディウム・エスピー(Phormidiumsp.)ATCC29409、ロドバクター・カプスラタス(Rhodobacter capsulatus)、ロドバクター・スフェロイデス(Rhodobacter sphaeroides)、ロドシュードモナス・ブラスチカ(Rhodopseudomonas blastica)、ロドシュードモナス・マリナ(Rhodopseudomonas marina)、ロドシュードモナス・パルストリス(Rhodopseudomonas palustris)、ロドスピリウム・リブラム(Rhodospirillumrubrum)、ロドスピリウム・サレキシゲンス(Rhodospirillum salexigens)、ロドスピリウム・サリナラム(Rhodospirillum salinarum)、ストレプトマイセス・アンボファシエンス(Streptomyces ambofaciens)、ストレプトマイセス・オーレオファシエンス(Streptomyces aureofaciens)、ストレプトマイセス・アウレウス(Streptomyces aureus)、ストレプトマイセス・フンジシディカ(Streptomyces fungicidicus)、ストレプトマイセス・グリセオクロモゲナス(Streptomyces griseochromogenes)、ストレプトマイセス・グリセウス(Streptomycesgriseus)、ストレプトマイセス・リビダンス(Streptomyces lividans、ストレプトマイセス・オリボグリセウス(Streptomyces olivogriseus)、ストレプトマイセス・ラメウス(Streptomyces rameus)、ストレプトマイセス・タナシエンシス(Streptomyces tanashiensis)、ストレプトマイセス・ビナセウス(Streptomyces vinaceus)、ザイモモナス・モビリス(Zymomonas mobilis)等をあげることができ、好ましくはエシェリヒア属、バチルス属、ストレプトマオセス属またはコリネバクテリウム属に属する微生物をあげることができる。
【0053】
組換え体DNAの導入方法としては、上記宿主細胞へDNAを導入する方法であればいずれも用いることができ、例えば、カルシウムイオンを用いる方法[Proc.Natl.Acad.Sci.,USA,69,2110(1972)]、プロトプラスト法(特開昭63−248394)、エレクトロポレーション法[NucleicAcids Res.,16,6127(1988)]等をあげることができる。
酵母菌株を宿主細胞として用いる場合には、発現ベクターとして、例えば、YEp13(ATCC37115)、YEp24(ATCC37051)、YCp50(ATCC37419)、pHS19、pHS15等を用いることができる。
【0054】
プロモーターとしては、酵母菌株中で機能するものであればいずれのものを用いてもよく、例えば、PHO5プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーター、gal 1プロモーター、gal 10プロモーター、ヒートショックポリペプチドプロモーター、MFα1プロモーター、CUP1プロモーター等のプロモーターをあげることができる。
宿主細胞としては、サッカロマイセス(Saccharomyces)属、シゾサッカロマイセス(Schizosaccharomyces)属、クルイベロマイセス(Kluyveromyces)属、トリコスポロン(Trichosporon)属、シワニオマイミセス(Schwanniomyces)属、ピチア(Pichia)属、またはキャンディダ(Candida)属等に属する酵母菌株をあげることができ、具体的には、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クルイベロマイセス・ラクティス(Kluyveromyces lactis)、トリコスポロン・プルランス(Trichosporon pullulans)、シワニオマイセス・アルビウス(Schwanniomyces alluvius)、ピチア・パストリス(Pichia pastoris)、キャンディダ・ウティリス(Candida utilis)等をあげることができる。
【0055】
組換え体DNAの導入方法としては、酵母にDNAを導入する方法であればいずれも用いることができ、例えば、エレクトロポレーション法[Methods Enzymol.,194,182(1990)]、スフェロプラスト法[Proc.Natl.Acad.Sci.,USA,81,4889(1984)]、酢酸リチウム法[J.Bacteriol.,153,163(1983)]等をあげることができる。
動物細胞を宿主として用いる場合には、発現ベクターとして、例えば、pcDNAI、pcDM8(フナコシ社より市販)、pAGE107(特開平3−22979)、pAS3−3(特開平2−227075)、pCDM8[Nature,329,840(1987)]、pcDNAI/Amp(インビトロジェン社製)、pREP4(インビトロジェン社製)、pAGE103[J.Biochem,101,1307(1987)]、pAGE210、pAMo、pAMoA等を用いることができる。
【0056】
プロモーターとしては、動物細胞中で機能するものであればいずれも用いることができ、例えば、サイトメガロウイルス(CMV)のIE(immediate early)遺伝子のプロモーター、SV40の初期プロモーターあるいはメタロチオネインのプロモーター、レトロウイルスのプロモーター、ヒートショックプロモーター、SRαプロモーター等をあげることができる。また、ヒトCMVのIE遺伝子のエンハンサーをプロモーターと共に用いてもよい。
【0057】
宿主細胞としては、マウス・ミエローマ細胞、ラット・ミエローマ細胞、マウス・ハイブリドーマ細胞、ヒトの細胞であるナマルバ(Namalwa)細胞またはナマルバKJM−1細胞、ヒト胎児腎臓細胞、ヒト白血病細胞、アフリカミドリザル腎臓細胞、チャイニーズ・ハムスターの細胞であるCHO細胞、HBT5637(特開昭63−299)等をあげることができる。
マウス・ミエローマ細胞としては、SP2/0、NSO等、ラット・ミエローマ細胞としてはYB2/0等、ヒト胎児腎臓細胞としてはHEK293(ATCC CRL−1573)、ヒト白血病細胞としてはBALL−1等、アフリカミドリザル腎臓細胞としてはCOS−1、COS−7等をあげることができる。
【0058】
組換え体DNAの導入方法としては、動物細胞にDNAを導入する方法であればいずれも用いることができ、例えば、エレクトロポレーション法[Cytotechnology,3,133(1990)]、リン酸カルシウム法(特開平2−227075)、リポフェクション法[Proc.Natl.Acad.Sci.,USA,84,7413(1987)]、Virology,52,456(1973)に記載の方法等をあげることができる。
【0059】
昆虫細胞を宿主として用いる場合には、例えばBaculovirus Expression Vectors,A Laboratory Manual,W.H.Freemanand Company,New York(1992)、カレント・プロトコールズ・イン・モレキュラー・バイオロジー、MolecularBiology,A Laboratory Manual、Bio/Technology,6,47(1988)等に記載された方法によって、蛋白質を生産することができる。
【0060】
即ち、組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、蛋白質を生産させることができる。
該方法において用いられる遺伝子導入ベクターとしては、例えば、pVL1392、pVL1393、pBlueBacIII(いずれもインビトロジェン社製)等をあげることができる。
【0061】
バキュロウイルスとしては、例えば、夜盗蛾科昆虫に感染するウイルスであるアウトグラファ・カリフォルニカ・ヌクレアー・ポリヘドロシス・ウイルス(Autographa californica nuclear polyhedrosis virus)等を用いることができる。
昆虫細胞としては、スポドプテラ・フルギペルダ(Spodoptera frugiperda)の卵巣細胞、トリコプルシア・ニ(Trichoplusia ni)の卵巣細胞、カイコ卵巣由来の培養細胞等を用いることができる。
【0062】
スポドプテラ・フルギペルダの卵巣細胞としてはSf9、Sf21(バキュロウイルス・イクスプレッション・ベクターズ ア・ラボラトリー・マニュアル)等、トリコプルシア・ニの卵巣細胞としてはHigh5、BTI−TN−5B1−4(インビトロジェン社製)等、カイコ卵巣由来の培養細胞としてはボンビクス・モリ(Bombyx mori)N4等をあげることができる。
組換えウイルスを調製するための、昆虫細胞への上記組換え遺伝子導入ベクターと上記バキュロウイルスの共導入方法としては、例えば、リン酸カルシウム法(特開平2−227075)、リポフェクション法[Proc.Natl.Acad.Sci.,USA,84,7413(1987)]等をあげることができる。
【0063】
植物細胞を宿主細胞として用いる場合には、発現ベクターとして、例えば、Tiプラスミド、タバコモザイクウイルスベクター等をあげることができる。
プロモーターとしては、植物細胞中で機能するものであればいずれのものを用いてもよく、例えば、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター、イネアクチン1プロモーター等をあげることができる。
【0064】
宿主細胞としては、タバコ、ジャガイモ、トマト、ニンジン、ダイズ、アブラナ、アルファルファ、イネ、コムギ、オオムギ等の植物細胞等をあげることができる。
組換えベクターの導入方法としては、植物細胞にDNAを導入する方法であればいずれも用いることができ、例えば、アグロバクテリウム(Agrobacterium)を用いる方法(特開昭59−140885、特開昭60−70080、WO94/00977)、エレクトロポレーション法(特開昭60−251887)、パーティクルガン(遺伝子銃)を用いる方法(特許第2606856、特許第2517813)等をあげることができる。
【0065】
酵母、動物細胞、昆虫細胞または植物細胞により発現させた場合には、糖あるいは糖鎖が付加された蛋白質を得ることができる。
以上のようにして得られる形質転換体を培地に培養し、培養物中に本発明の蛋白質を生成蓄積させ、該培養物から採取することにより、該蛋白質を製造することができる。
本発明の蛋白質を製造するための上記形質転換体の宿主としては、細菌、酵母、動物細胞、昆虫細胞等、植物細胞等いずれであってもよいが、好ましくは細菌、より好ましくはエシェリヒア属に属する微生物、さらに好ましくはエシェリヒア・コリに属する微生物をあげることができる。
【0066】
上記形質転換体を培地に培養する方法は、宿主の培養に用いられる通常の方法に従って行うことができる。
エシェリヒア・コリ等の原核生物あるいは酵母等の真核生物を宿主として得られた形質転換体を培養する培地としては、該生物が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行える培地であれば天然培地、合成培地のいずれを用いてもよい。
【0067】
炭素源としては、該生物が資化し得るものであればよく、グルコース、フラクトース、スクロース、これらを含有する糖蜜、デンプンあるいはデンプン加水分解物等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類等を用いることができる。
窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸もしくは有機酸のアンモニウム塩、その他の含窒素化合物、並びに、ペプトン、肉エキス、酵母エキス、コーンスチープリカー、カゼイン加水分解物、大豆粕および大豆粕加水分解物、各種発酵菌体、およびその消化物等を用いることができる。
【0068】
無機塩としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等を用いることができる。
培養は、通常振盪培養または深部通気攪拌培養等の好気的条件下で行う。培養温度は15〜40℃がよく、培養時間は、通常5時間〜7日間である。培養中pHは3.0〜9.0に保持する。pHの調整は、無機または有機の酸、アルカリ溶液、尿素、炭酸カルシウム、アンモニア等を用いて行う。
【0069】
また、培養中必要に応じて、アンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。
プロモーターとして誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養するときには、必要に応じてインデューサーを培地に添加してもよい。例えば、lacプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはイソプロピル−β−D−チオガラクトピラノシド等を、trpプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはインドールアクリル酸等を培地に添加してもよい。
【0070】
動物細胞を宿主として得られた形質転換体を培養する培地としては、一般に使用されているRPMI1640培地[J.Am.Med.Assoc.,199,519(1967)]、イーグル(Eagle)のMEM培地[Science,122,501(1952)]、DMEM培地[Virology,8,396(1959)]、199培地[Proc.Soc.Biol.Med.,73,1(1950)]またはこれら培地に牛胎児血清等を添加した培地等を用いることができる。
【0071】
培養は、通常pH6〜8、25〜40℃、5%CO2存在下等の条件下で1〜7日間行う。
また、培養中必要に応じて、カナマイシン、ペニシリン、ストレプトマイシン等の抗生物質を培地に添加してもよい。
昆虫細胞を宿主として得られた形質転換体を培養する培地としては、一般に使用されているTNM−FH培地(ファーミンジェン社製)、Sf−900 II SFM培地(ライフ・テクノロジーズ社製)、ExCell400、ExCell405[いずれもJRHバイオサイエンシーズ社製]、Grace’sInsect Medium[Nature,195,788(1962)]等を用いることができる。
【0072】
培養は、通常pH6〜7、25〜30℃等の条件下で1〜5日間行う。
また、培養中必要に応じて、ゲンタマイシン等の抗生物質を培地に添加してもよい。
植物細胞を宿主として得られた形質転換体は、細胞として、または植物の細胞や器官に分化させて培養することができる。該形質転換体を培養する培地としては、一般に使用されているムラシゲ・アンド・スクーグ(MS)培地、ホワイト(White)培地、またはこれら培地にオーキシン、サイトカイニン等、植物ホルモンを添加した培地等を用いることができる。
【0073】
培養は、通常pH5〜9、20〜40℃の条件下で3〜60日間行う。
また、培養中必要に応じて、カナマイシン、ハイグロマイシン等の抗生物質を培地に添加してもよい。
上記のとおり、本発明のDNAまたは本発明の製造法に用いられるDNAを発現ベクターに連結した組換え体DNAを保有する微生物、昆虫細胞、動物細胞、あるいは植物細胞由来の形質転換体を、通常の培養方法に従って培養し、本発明の蛋白質を生成蓄積させ、該培養物より該蛋白質を採取することにより、該蛋白質を製造することができる。
【0074】
本発明の蛋白質の生産方法としては、宿主細胞内に生産させる方法、宿主細胞外に分泌させる方法、あるいは宿主細胞外膜上に生産させる方法があり、選択した方法に応じて、使用する宿主細胞を選択し、生産させる蛋白質の構造を変更すればよい。
本発明の蛋白質が宿主細胞内あるいは宿主細胞外膜上に生産される場合、ポールソンらの方法[J.Biol.Chem.,264,17619(1989)]、ロウらの方法[Proc.Natl.Acad.Sci.,USA,86,8227(1989)、Genes Develop.,4,1288(1990)]、または特開平05−336963、WO94/23021等に記載の方法を準用することにより、該蛋白質を宿主細胞外に積極的に分泌させることができる。
【0075】
すなわち、遺伝子組換えの手法を用いて、本発明の蛋白質の活性部位を含む蛋白質の手前にシグナルペプチドを付加した形で生産させることにより、該蛋白質を宿主細胞外に積極的に分泌させることができる。
また、特開平2−227075に記載されている方法に準じて、ジヒドロ葉酸還元酵素遺伝子等を用いた遺伝子増幅系を利用して生産量を上昇させることもできる。
【0076】
さらに、遺伝子導入した動物または植物の細胞を再分化させることにより、遺伝子が導入された動物個体(トランスジェニック非ヒト動物)または植物個体(トランスジェニック植物)を造成し、これらの個体を用いて本発明の蛋白質を製造することもできる。
本発明の蛋白質を生産する形質転換体が動物個体または植物個体の場合は、通常の方法に従って、飼育または栽培し、該蛋白質を生成蓄積させ、該動物個体または植物個体より該蛋白質を採取することにより、該蛋白質を製造することができる。
【0077】
動物個体を用いて本発明の蛋白質を製造する方法としては、例えば公知の方法[Am.J.Clin.Nutr.,63,639S(1996)、Am.J.Clin.Nutr.,63,627S(1996)、Bio/Technology,9,830(1991)]に準じて遺伝子を導入して造成した動物中に本発明の蛋白質を生産する方法があげられる。
動物個体の場合は、例えば、本発明のDNAまたは本発明の製造法に用いられるDNAを導入したトランスジェニック非ヒト動物を飼育し、本発明の蛋白質を該動物中に生成、蓄積させ、該動物中より該蛋白質を採取することにより、該蛋白質を製造することができる。該動物中の生成、蓄積させる場所としては、例えば、該動物のミルク(特開昭63−309192)、卵等をあげることができる。この際に用いられるプロモーターとしては、動物で機能するものであればいずれも用いることができるが、例えば、乳腺細胞特異的なプロモーターであるαカゼインプロモーター、βカゼインプロモーター、βラクトグロブリンプロモーター、ホエー酸性プロテインプロモーター等が好適に用いられる。
【0078】
植物個体を用いて本発明の蛋白質を製造する方法としては、例えば本発明の蛋白質をコードするDNAを導入したトランスジェニック植物を公知の方法[組織培養,20(1994)、組織培養,21(1995)、Trends Biotechnol.,15,45(1997)]に準じて栽培し、該蛋白質を該植物中に生成、蓄積させ、該植物中より該蛋白質を採取することにより、該蛋白質を生産する方法があげられる。
【0079】
本発明の蛋白質を生産する形質転換体を用いて製造された本発明の蛋白質を単離・精製する方法としては、通常の酵素の単離、精製法を用いることができる。
例えば、本発明の蛋白質が、細胞内に溶解状態で生産された場合には、培養終了後、細胞を遠心分離により回収し、水系緩衝液にけん濁後、超音波破砕機、フレンチプレス、マントンガウリンホモゲナイザー、ダイノミル等により細胞を破砕し、無細胞抽出液を得る。
【0080】
該無細胞抽出液を遠心分離することにより得られる上清から、通常の酵素の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)−セファロース、DIAION HPA−75(三菱化成社製)等レジンを用いた陰イオン交換クロマトグラフィー法、S−SepharoseFF(ファルマシア社製)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィー法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、精製標品を得ることができる。
【0081】
また、該蛋白質が細胞内に不溶体を形成して生産された場合は、同様に細胞を回収後破砕し、遠心分離を行うことにより得られた沈殿画分より、通常の方法により該蛋白質を回収後、該蛋白質の不溶体を蛋白質変性剤で可溶化する。
該可溶化液を、蛋白質変性剤を含まないあるいは蛋白質変性剤の濃度が蛋白質が変性しない程度に希薄な溶液に希釈、あるいは透析し、該蛋白質を正常な立体構造に構成させた後、上記と同様の単離精製法により精製標品を得ることができる。
【0082】
本発明の蛋白質またはその糖修飾体等の誘導体が細胞外に分泌された場合には、培養上清に該蛋白質またはその糖付加体等の誘導体を回収することができる。
即ち、該培養物を上記と同様の遠心分離等の手法により処理することにより可溶性画分を取得し、該可溶性画分から、上記と同様の単離精製法を用いることにより、精製標品を得ることができる。
【0083】
このようにして取得される蛋白質として、例えば、配列番号1または2で表されるアミノ酸配列からなる蛋白質をあげることができる。
また、本発明の蛋白質を他の蛋白質との融合蛋白質として生産し、融合した蛋白質に親和性をもつ物質を用いたアフィニティークロマトグラフィーを利用して精製することもできる。
【0084】
融合させる蛋白質としては、β−ガラクトシダーゼ、プロテインA、プロテインAのIgG結合領域、クロラムフェニコール・アセチルトランスフェラーゼ、ポリ(Arg)、ポリ(Glu)、プロテインG、マルトース結合タンパク質、グルタチオンS−トランスフェラーゼ、ポリヒスチジン鎖(His−tag)、Sペプチド、DNA結合タンパク質ドメイン、Tac抗原、チオレドキシン、グリーン・フルオレッセント・プロテイン、FLAGペプチド、および任意の抗体のエピトープなどがあげられる〔山川彰夫,実験医学,13,469−474(1995)〕。
【0085】
上記した融合させる蛋白質に親和性をもつ物質としては、β−ガラクトシダーゼ、プロテインA、プロテインAのイムノグロブリンG結合領域、クロラムフェニコール・アセチルトランスフェラーゼ、ポリ(Arg)、ポリ(Glu)、プロテインG、マルトース結合タンパク質、グルタチオンS−トランスフェラーゼ、ポリヒスチジン鎖(His−tag)、Sペプチド、DNA結合タンパク質ドメイン、Tac抗原、チオレドキシン、グリーン・フルオレッセント・プロテイン、FLAGペプチドまたは任意の抗体のエピトープを認識する抗体、例えばイムノグロブリンGなどをあげることができる。
【0086】
また、上記で取得された蛋白質のアミノ酸情報を基に、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法により、本発明の蛋白質を製造することができる。また、Advanced ChemTech社、パーキン・エルマー社、Pharmacia社、Protein TechnologyInstrument社、Synthecell−Vega社、PerSeptive社、島津製作所等のペプチド合成機を利用して化学合成することもできる。
(iii)本発明のジペプチドの製造法
(1)酵素的製造法
ジペプチドの酵素的製造法としては、本発明の蛋白質またはジペプチド合成用蛋白質、1種以上のアミノ酸、およびATPを水性媒体中に存在せしめ、該媒体中に式(I)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する方法をあげることができる。
【0087】
上記製造法において、基質に用いられる1種以上のアミノ酸、好ましくは1または2種のアミノ酸としては、L体またはD体のアミノ酸、グリシン(Gly)、β−アラニン(βAla)およびそれらの誘導体から選ばれるアミノ酸、好ましくはL−アミノ酸、GlyおよびβAla並びにそれらの誘導体からなる群より選ばれるアミノ酸をあげることができ、該アミノ酸であれば、いずれのアミノ酸をいずれの組み合わせで用いてもよい。L体またはD体のアミノ酸としては、例えばアラニン(Ala)、グルタミン(Gln)、グルタミン酸(Glu)、バリン(Val)、ロイシン(Leu)、イソロイシン(Ile)、プロリン(Pro)、フェニルアラニン(Phe)、トリプトファン(Trp)、メチオニン(Met)、セリン(Ser)、スレオニン(Thr)、システイン(Cys)、アスパラギン(Asn)、チロシン(Tyr)、リジン(Lys)、アルギニン(Arg)、ヒスチジン(His)、アスパラギン酸(Asp)、α−アミノ酪酸(α−AB)、アザセリン(Azaserine)、テアニン(theanine)、4−ヒドロキシプロリン(4−HYP)、3−ヒドロキシプロリン(3−HYP)、オルニチン(Orn)、シトルリン(Cit)および6−ジアゾ−5−オキソノルロイシン(6−diazo−5−oxo−norleucine)などをあげることができ、L体のアミノ酸としては、L−Ala、L−Gln、L−Glu、Gly、L−Val、L−Leu、L−Ile、L−Pro、L−Phe、L−Trp、L−Met、L−Ser、L−Thr、L−Cys、L−Asn、L−Tyr、L−Lys、L−Arg、L−His、L−Asp、L−α−AB、L−Azaserine、L−theanine、L−4−HYP、L−3−HYP、L−Orn、L−CitおよびL−6−diazo−5−oxo−norleucineなどをあげることができる。
【0088】
アミノ酸の誘導体としては、例えばヒドロキシアミノ酸(β−ヒドロキシグルタミン、β−ヒドロキシグルタミン酸、γ−ヒドロキシグルタミン酸、α−ヒドロキシグリシン、β−ヒドロキシバリン、γ−ヒドロキシバリン、β−ヒドロキシロイシン、γ−ヒドロキシロイシン、δ−ヒドロキシロイシン、β−ヒドロキシイソロイシン、γ−ヒドロキシイソロイシン、3−ヒドロキシプロリン、4−ヒドロキシプロリン、β−ヒドロキシフェニルアラニン、3,4−ジヒドロキシフェニルアラニン、2,4,5−トリヒドロキシフェニルアラニン、β−ヒドロキシトリプトファン、5−ヒドロキシトリプトファン、α−ヒドロキシメチオニン、β−ヒドロキシセリン、γ−ヒドロキシスレオニン、S−ヒドロキシシステイン、β−ヒドロキシアスパラギン、β−ヒドロキシチロシン、β−ヒドロキシリジン、γ−ヒドロキシリジン、δ−ヒドロキシリジン、N−ヒドロキシリジン、β−ヒドロキシアルギニン、δ−ヒドロキシアルギニン、N−ヒドロキシアルギニン、β−ヒドロキシヒスチジン、β−ヒドロキシアスパラギン酸、β−ヒドロキシオルニチン、γ−ヒドロキシオルニチン、N−ヒドロキシオルニチン)、N−メチルアミノ酸(N−メチル−アラニン、N−メチル−グルタミン、N−メチル−グルタミン酸、N−メチル−グリシン、N−メチル−バリン、N−メチル−ロイシン、N−メチル−イソロイシン、N−メチル−プロリン、N−メチル−フェニルアラニン、N−メチル−トリプトファン、N−メチル−メチオニン、N−メチル−セリン、N−メチル−スレオニン、N−メチル−システイン、N−メチル−アスパラギン、N−メチル−チロシン、N−メチル−リジン、N−メチル−アルギニン、N−メチル−ヒスチジン、N−メチル−アスパラギン酸、N−メチル−オルニチン)などをあげることができる。
【0089】
上記製造法に用いられる、より好ましい1または2種のアミノ酸としては、L−Ala、L−LeuおよびL−Pheから選ばれる1または2種のアミノ酸、さらに好ましくは、L−Ala、L−LeuおよびL−Pheから選ばれる1種のアミノ酸、またはL−LeuとL−Pheとからなる2種のアミノ酸をあげることができる。
上記製造法において、本発明の蛋白質またはジペプチド合成用蛋白質は、基質として用いるアミノ酸1mgあたり0.01〜100mg、好ましくは0.1mg〜10mg添加する。
【0090】
上記製造法において、基質として用いるアミノ酸は、全量として0.1〜500g/L、好ましくは0.2〜200g/Lの濃度になるように水性媒体中に初発または反応途中に添加する。
上記製造法において、エネルギー源として用いるATPは、0.5mmol/L〜10mol/Lの濃度で用いる。
上記製造法で用いられる水性媒体としては、ジペプチドの生成反応を阻害しない限り、いかなる成分、組成の水性媒体であってもよく、例えば、水、りん酸塩、炭酸塩、酢酸塩、ほう酸塩、クエン酸塩、トリスなどの緩衝液などをあげることができる。また、メタノール、エタノールなどのアルコール類、酢酸エチルなどのエステル類、アセトンなどのケトン類、アセトアミドなどのアミド類を含有していてもよい。
【0091】
ジペプチドの生成反応は水性媒体中、pH5〜11、好ましくはpH6〜10、20〜50℃、好ましくは25〜45℃の条件で2〜150時間、好ましくは6〜120時間行う。
上記方法で製造されるジペプチドとしては、式(I)で表されるジペプチドをあげることができ、好ましくは、式(I)においてR1およびR2が同時にまたは異なって、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体であるジペプチド、より好ましくはL体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体であるジペプチドをあげることができる。
【0092】
L体のアミノ酸およびアミノ酸の誘導体としては、上記した物質をあげることができる。
上記方法で製造される、さらに好ましいジペプチドとしては、L−ロイシル−L−フェニルアラニン、L−フェニルアラニル−L−ロイシン、L−アラニル−L−アラニン、L−ロイシル−L−ロイシンおよびL−フェニルアラニル−L−フェニルアラニンなどをあげることができる。
(2)形質転換体または微生物の培養物もしくは培養物の処理物を酵素源として用いる製造法
形質転換体または微生物の培養物または培養物の処理物を酵素源として用いるジペプチドの製造法としては、本発明の蛋白質もしくはジペプチド合成用蛋白質を生産する能力を有する形質転換体、または本発明の蛋白質もしくはジペプチド合成用蛋白質を生産する能力を有する微生物の培養物もしくは培養物の処理物を酵素源に用い、該酵素源、および1種以上のアミノ酸を水性媒体中に存在せしめ、該媒体中に式(I)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法をあげることができる。
【0093】
上記製造法に用いられる形質転換体としては、上記(ii)の方法により製造することができる、本発明の蛋白質もしくはジペプチド合成用蛋白質を生産する形質転換体をあげることができる。該形質転換体の宿主としては、細菌、酵母、動物細胞、昆虫細胞等、植物細胞等をあげることができ、好ましく細菌、より好ましくはエシェリヒア属、バチルス属、ストレプトマイセス属またはコリネバクテリウム属に属する微生物をあげることができる。
【0094】
エシェリヒア属に属する微生物としては、好ましくはエシェリヒア・コリに属する微生物等をあげることができ、バチルス属に属する微生物としては好ましくはバチルス・サチリスまたはバチルス・メガテリウム等に属する微生物をあげることができ、ストレプトマイセス属に属する微生物としては、好ましくはストレプトマイセス・リビダンスをあげることができ、コリネバクテリウム属に属する微生物としては好ましくはコリネバクテリウム・グルタミカムまたはコリネバクテリウム・アンモニアゲネス等に属する微生物をあげることができる。
【0095】
上記製造法に用いられる微生物は、本発明の蛋白質もしくはジペプチド合成用蛋白質を生産する能力を有する微生物であれば、いずれの微生物であってもよく、好ましくはストレプトマイセス属に属する微生物、より好ましくはアルボノルシン合成活性を有するストレプトマイセス属に属する微生物、さらに好ましくはストレプトマイセス・アルボラスまたはストレプトマイセス・ノウルセイより選ばれる種に属する微生物、最も好ましくは、ストレプトマイセス・アルボラスIFO14147株、ストレプトマイセス・ノウルセイATCC11455株またはIFO15452株をあげることができる。
【0096】
培養物の処理物としては、培養物と同じ活性を維持している限り、公知のいかなる処理を施された培養物の処理物であってもよく、該処理物としては、例えば培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の溶媒処理物、該菌体の酵素処理物および該菌体の固定化物等の生細胞からなる処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の蛋白質分画物および該菌体より抽出して得られる酵素標品などをあげることができる。
【0097】
上記製造法において、基質として用いるアミノ酸の種類、使用濃度および添加時期、並びに生産されるジペプチドは、上記(iii)の(1)の酵素的製造法のものと同様である。
また、微生物の培養物または該培養物の処理物を酵素源とした製造法において用いられる水性媒体としては、上記(iii)の(1)の酵素的製造法に用いられる水性媒体に加え、酵素源として用いた形質転換体または微生物の培養液も水性媒体として用いることができる。
【0098】
また上記製造法においては、必要に応じて、ATPの供給源として、ATPまたは形質転換体もしくは微生物が代謝してATPを生産し得る化合物、例えばグルコースのような糖類、エタノールのようなアルコール類、酢酸のような有機酸類などを水性媒体中に加えることができる。
さらに必要に応じて、水性媒体中に界面活性剤あるいは有機溶媒を添加してもよい。界面活性剤としては、ポリオキシエチレン・オクタデシルアミン(例えばナイミーンS−215、日本油脂社製)などの非イオン界面活性剤、セチルトリメチルアンモニウム・ブロマイドやアルキルジメチル・ベンジルアンモニウムクロライド(例えばカチオンF2−40E、日本油脂社製)などのカチオン系界面活性剤、ラウロイル・ザルコシネートなどのアニオン系界面活性剤、アルキルジメチルアミン(例えば三級アミンFB、日本油脂社製)などの三級アミン類など、ジペプチドの生成を促進するものであればいずれでもよく、1種または数種を混合して使用することもできる。界面活性剤は、通常0.1〜50g/lの濃度で用いられる。有機溶剤としては、キシレン、トルエン、脂肪族アルコール、アセトン、酢酸エチルなどがあげられ、通常0.1〜50ml/lの濃度で用いられる。
【0099】
培養物または該培養物の処理物を酵素源として用いる場合、該酵素源の量は、当該酵素源の比活性等により異なるが、例えば、基質として用いるアミノ酸1mgあたり湿菌体重量として5〜1000mg、好ましくは10〜400mg添加する。
ジペプチドの生成反応は水性媒体中、pH5〜11、好ましくはpH6〜10、20〜65℃、好ましくは25〜55℃、より好ましくは30〜45℃の条件で通常1分間〜150時間、好ましくは3分間〜120時間、より好ましくは30分間〜100時間行う。
【0100】
上記(iii)の(1)または(2)の製造法において、水性媒体中に生成、蓄積したジペプチドの採取は、活性炭やイオン交換樹脂などを用いる通常の方法あるいは、有機溶媒による抽出、結晶化、薄層クロマトグラフィー、高速液体クロマトグラフィー等により行うことができる。
以下に、実施例を示すが、本発明は下記実施例に限定されるものではない。
【実施例1】
【0101】
albC遺伝子およびその類縁遺伝子の取得
ストレプトマイセス・ノウルセイのalbC遺伝子の塩基配列[Chemistry & Biol.,9,1355(2002)]に基づき、ストレプトマイセス・ノウルセイおよびストレプトマイセス・アルボラスよりalbC遺伝子およびその類縁遺伝子を、以下の方法で取得した。
まず、ストレプトマイセス・ノウルセイIFO15452株とストレプトマイセス・アルボラスIFO14147株を、それぞれ、1%グリシン添加したKM73培地[2g/lイーストエキス(ディフコ社製)、10g/l可溶性デンプン(和光純薬工業社製)]、KP培地[15g/lグルコース、10g/lグリセロール、10g/lポリペプトン(日本製薬株式会社製)、10g/l肉エキス(極東製薬工業株式会社製)、4g/l炭酸カルシウム]に植菌し28℃で一晩振とう培養した。なお、ストレプトマイセス・ノウルセイIFO15452株およびストレプトマイセス・アルボラスIFO14147株は、独立行政法人 製品評価技術基盤機構 生物資源部門[National Institute of Technology andEvaluation(NITE)Biological Resource Center(BRC)](〒292−0818 千葉県木更津市かずさ鎌足2−5−8)より分譲を受けた。
【0102】
培養後、Genetic Manipulation of Streptomyces:a Laboratory Manual:John InnesFoundationに記載の方法に従って該微生物の染色体DNAを単離精製した。
albC遺伝子の塩基配列に基づき、パーセプティブ・バイオシステムズ(Perseptive Biosystems)社製8905型DNA合成機を用いて、配列番号5および6で表される塩基配列を有するDNA(以下、それぞれプライマーA、プライマーBと呼ぶ)を合成した。プライマーAは、ストレプトマイセス・ノウルセイの染色体DNAのalbC遺伝子の開始コドンを含む領域の5’末端にNcoI認識配列を含む塩基配列を付加したものである。プライマーBは、albC遺伝子の終止コドンを含む配列と相補的な塩基配列の5’末端にBglII認識配列を含む塩基配列を付加したものである。
【0103】
上記のプライマーAおよびプライマーBをプライマーセットに、鋳型としてストトレプトマイセス・ノウルセイまたはストレプトマイセス・アルボラスの染色体DNAを用いてPCRを行った。PCRは、鋳型として0.1μgの染色体DNA、0.5μmol/Lの各プライマー、2.5unitsのEx Taq DNAポリメラーゼ(タカラバイオ社製)、5μLのExTaqDNAポリメラーゼ用×10緩衝液(タカラバイオ社製)、各200μmol/LのdNTP(dATP、dGTP、dCTPおよびdTTP)、5μLのジメチルスルホキシドを含む反応液50μLを調製し、94℃で1分間、55℃で30秒間、72℃で1分間の工程を30回繰り返すことにより行った。
【0104】
該反応液の1/10量をアガロースゲル電気泳動し、約0.7kbのDNA断片が増幅していることを確認した後、残りの反応液と等量のTE[10mmol/L Tris−HCl(pH8.0)、1mmol/L EDTA]飽和フェノール/クロロホルム(1vol/1vol)溶液を添加し、混合した。該溶液を遠心分離して得られた上層に、2倍容量の冷エタノールを加えて混合し、−80℃に30分間放置した。該溶液を遠心分離してDNAを沈殿させた後、該DNAをそれぞれ20μLのTEに溶解した。
【0105】
該溶解液それぞれ5μLを用い、増幅したDNAを制限酵素NcoIおよびBglIIで切断し、アガロースゲル電気泳動によりDNA断片を分離した後、ジーンクリーンIIキット(GENECLEANII kit、BIO 101社製)を用いて、700bpのDNA断片をそれぞれ回収した。
ファージT5プロモーターを含む発現ベクターpQE60(キアゲン社製)0.2μgを制限酵素NcoIおよびBglIIで切断後、アガロースゲル電気泳動によりDNA断片を分離し、上記と同様の方法により3.4kbのDNA断片を回収した。
【0106】
上記で得られた各放線菌由来の0.7kb断片およびpQE60由来の3.4kbの断片をライゲーションキット(タカラバイオ社製)を用いて、16℃で16時間反応させ連結した。
該反応液を用いてエシェリヒア・コリNM522株(ストラタジーン社製)を、カルシウムイオンを用いる方法[Proc.Natl.Acad.Sci.,USA,69,2110(1972)]によって形質転換し、該形質転換体を50μg/mlのアンピシリンを含むLB寒天培地[10g/lバクトトリプトン(ディフコ社製)、5g/lイーストエキス(ディフコ社製)、5g/l塩化ナトリウム、寒天20g/l]に塗布した後、30℃で一晩培養した。
【0107】
生育してきた形質転換体のコロニーより公知の方法に従ってプラスミドを抽出し、制限酵素を用いてその構造を解析することにより、ファージT5プロモーター下流にストレプトマイセス・ノウルセイ由来のDNAが連結された発現ベクターであるpAL−nou、およびストレプトマイセス・アルブラス由来のDNAが連結された発現ベクターであるpAL−albが取得されていることを確認した(図1)。
【0108】
それぞれのプラスミドに挿入された放線菌由来のDNA部分の塩基配列を塩基配列決定装置373A・DNAシークエンサーを使って決定したところ、pAL−albには配列番号1で表されるアミノ酸配列を有する蛋白質をコードするDNA、すなわち配列番号3で表される塩基配列を有するDNAが含有され、pAL−nouには配列番号2で表されるアミノ酸配列を有する蛋白質をコードするDNA、すなわち配列番号4で表される塩基配列を有するDNAが含有されていることを確認した。
【実施例2】
【0109】
菌体を酵素源に用いたジペプチドの製造
実施例1で得られたpAL−nouまたはpAL−albを保有するエシェリヒア・コリNM522(エシェリヒア・コリNM522/pAL−nou株またはNM522/pAL−alb株)およびプラスミドを保有しないNM522株を50μg/mlのアンピシリンを含む10mlのLB培地[10g/lバクトトリプトン(ディフコ社製)、5g/lイーストエキス(ディフコ社製)、5g/l塩化ナトリウム]が入った試験管(プラスミドを持たない株の場合にはアンピシリンは無添加、以下同様)に接種し、30℃で17時間培養した。この培養液0.5mlを50mlのLB培地が入った250ml容の三角フラスコにそれぞれ植菌し、30℃で1時間振とう培養した後、イソプロピル−β−D−チオガラクトピラノシド(IPTG)を終濃度1mmol/lになるように添加し、さらに4時間培養を継続した。該培養液を遠心分離し、湿菌体を取得した。
【0110】
終濃度100mg/mlの該湿菌体、60mmol/Lのリン酸カリウムバッファー(pH7.2)、10mmol/Lの塩化マグネシウム、10mmol/LのATP、1g/LのL−Leu、1g/LのL−Pheからなる3.0mlの反応液を調製し、30℃で反応を行った。反応1時間後にサンプリングし、アセトニトリルを20%(v/v)になるように加えた後、反応生成物を以下のHPLC分析により分析した。
分離カラム:ODS−HAカラム(YMC社製)
溶離液:30%(v/v)アセトニトリル
流動速度:0.6ml/min
検出:紫外吸収215nm
その結果、エシェリヒア・コリNM522/pAL−nuo株の反応液中には36.7mg/lのcyclo(L−Leu−L−Phe)の蓄積が確認されたが、エシェリヒア・コリNM522株の反応液中にはcyclo(L−Leu−L−Phe)はまったく検出されなかった。同じ反応液を以下の条件でHPLCによって分析し、直鎖状のジペプチドであるL−ロイシル−L−フェニルアラニン(L−Leu−L−Phe)およびL−フェニルアラニル−L−ロイシン(L−Phe−L−Leu)を測定した。
【0111】
直鎖状のジペプチドはF−moc化法で誘導体化した後にHPLC分析により分析した。
分離カラム:野村化学社製のODS−HG−5カラム
溶離液:
A液−6ml/L酢酸、20%(v/v)アセトニトリル(トリエチルアミンでpH4.8に調整)
B液−6ml/L酢酸、70%(v/v)アセトニトリル(トリエチルアミンでpH4.8に調整)
流動速度:0.6ml/min
検出:励起波長254nm、蛍光波長630nm
溶離液混合比:表1のとおり
【0112】
【表1】

その結果、エシェリヒア・コリNM522/pAL−nuo株の反応液中には21.9mg/lのL−Leu−L−Pheと12.0mg/lのL−Phe−L−Leuが蓄積していることが確認された。また対照株として用いたエシェリヒア・コリNM522株の反応液中にはいずれの直鎖ジペプチドも検出されなかった。このことから、本発明で取得されたシクロジペプチド合成酵素は直鎖状のジペプチドを合成する能力をもつことが明らかになった。
【実施例3】
【0113】
精製酵素を用いたジペプチドの製造(1)
実施例2と同様にエシェリヒア・コリNM522/pAL−nuo株を培養した。培養終了後、遠心分離によって湿菌体を取得し、60mmol/lのリン酸カリウムバッファー(pH7.2)で洗浄後、10mmol/lイミダゾール含有20mmol/lリン酸カリウムバッファーに懸濁した。この懸濁を4℃で超音波処理して菌体破砕液を取得した。この菌体破砕液(10ml、蛋白質0.863mgを含む)をアマシャム社製Hisタグ精製カラムに通塔し、10mmol/lイミダゾール含有20mmol/lリン酸カリウムバッファー15mlを通塔することによる洗浄を行い、HisタグつきalbC蛋白質をカラム内にて精製した。次にこのHisタグ付きのalbC蛋白質を保持するカラムに、実施例2と同組成の反応液(反応液組成:60mmol/Lのリン酸カリウムバッファー(pH7.2)、10mmol/Lの塩化マグネシウム、10mmol/LのATP、1g/LのL−Leu、1g/LのL−Pheからなる反応液)2mlを通塔し、カラム内に基質を保持した状態で、30℃にて、インキュベートした。24時間後、同組成の反応液3mlでカラム内の反応液を溶出し、実施例2と同様の方法により反応液中のシクロジペプチドおよびジペプチドを定量した。
【0114】
その結果、cyclo(L−Leu−L−Phe)6.8mg/L、L−Leu−L−Phe 28.7mg/LおよびL−Phe−L−Leu 18.5mg/Lが生成していることが分かった。ATPを含まない反応液で同様にインキュベートした場合は、シクロジペプチド、ジペプチドともに検出されなかった。
【実施例4】
【0115】
精製酵素を用いたジペプチドの製造(2)
基質のアミノ酸を他のアミノ酸に換える以外は実施例3と同様の方法で酵素反応を行い、生成物を分析した。反応液は、基質のアミノ酸を、1g/LのL−Ala、L−LeuまたはL−Pheに置き換えた以外は実施例2と同じ組成の溶液を用いた。
その結果、反応開始後24時間で、それぞれ9.41mg/LのL−Ala−L−Ala、7.85mg/LのL−Leu−L−Leu、または5.20mg/LのL−Phe−L−Pheが生成していることが分かった。
【配列表フリーテキスト】
【0116】
配列番号5−人工配列の説明:合成DNA
配列番号6−人工配列の説明:合成DNA
【技術分野】
【0001】
本発明は、ジペプチド合成活性を有する蛋白質またはジペプチド合成用蛋白質、ジペプチド合成活性を有する蛋白質の製造法、ジペプチド合成活性を有する蛋白質またはジペプチド合成用蛋白質を用いたジペプチドの製造法、ジペプチド合成活性を有する蛋白質またはジペプチド合成用蛋白質を生産する微生物または形質転換体、および該微生物または形質転換体を用いたジペプチドの製造法に関する。
【背景技術】
【0002】
ペプチドの大量合成法については、化学合成法(液相法、固相法)、酵素的合成法およびDNA組換え法を用いた生物学的合成法が知られている。現在、50残基以上の長鎖のペプチドに関しては酵素的合成法あるいは生物学的合成法が用いられ、ジペプチドに関しては化学合成法と酵素的合成法が主に用いられている。
化学合成法によるジペプチドの合成では、官能基の保護・脱保護などの操作が必要であり、またラセミ体も合成されることから、化学合成法は経済的、効率的な方法とはいえない。また、化学合成法は大量の有機溶媒等を使うため環境衛生上も好ましい方法ではない。
【0003】
酵素法によるジペプチドの合成に関しては、タンパク質分解酵素(プロテアーゼ)の逆反応を利用した方法[J.Biol.Chem.,119,707−720(1937)]、耐熱性アミノアシルt−RNA合成酵素を利用する方法(特開昭58−146539号公報、特開昭58−209991号公報、特開昭58−209992号公報、特開昭59−106298号公報)、非リボゾームペプチドシンセターゼ(以下、NRPSと称す)を利用する方法[Chem.Biol.,7,373−384(2000)、FEBS Lett.,498,42−45(2001)、米国特許第5795738号、米国特許第5652116号]が知られている。
【0004】
しかし、タンパク分解酵素の逆反応を利用した方法では、基質となるアミノ酸の官能基の保護・脱保護が必要であり、ペプチド形成反応の効率化およびペプチド分解反応の阻止が困難といった問題点がある。耐熱性アミノアシルt−RNA合成酵素を利用する方法には、酵素の発現、目的産物以外の副生反応の阻止が困難という問題点がある。NRPSを利用する方法に関しては、酵素分子が巨大なためにDNA組換え法を用いて該酵素を発現することが困難であること、補酵素である4’−ホスフォパンテテイン(4’−phosphopantetheine)の供給が必要であり、効率的な製造法とはいえない。
【0005】
一方、酵素分子量がNRPSより小さく、補酵素である4’−phosphopantetheineを必要としないγ−グルタミルシステインシンセターゼ(γ−glutamylcysteinesynthetase)、グルタチオンシンセターゼ(glutathione synthetase)、D−アラニン−D−アラニン(D−Ala−D−Ala)リガーゼ(D−Ala−D−Alaligase)、ポリ−γ−グルタミン酸シンセターゼ(poly−γ−glutamate synthetase)等の一群のペプチドシンセターゼも知られている。これらの酵素の殆どはD−アミノ酸を基質に用いる、またはγ位のカルボキシル基でのペプチド結合の形成を触媒する等の特徴を有するため、L−アミノ酸のα位カルボキシル基でペプチド結合するジペプチドの合成に用いることはできない。
【0006】
L−アミノ酸のα位カルボキシル基でのペプチド結合形成活性によるジペプチド生成が知られているのはバチルス属に属する微生物由来のジペプチド抗生物質であるバシリシン合成酵素のみである。バシリシン合成酵素は、バシリシン(L−アラニル−L−アンチカプシン、L−Ala−L−anticapsin)およびL−アラニル−L−アラニン(L−Ala−L−Ala)を合成する活性を有することは知られているが、その他のジペプチドの合成活性については知られていない[J.Ind.Microbiol.,2,201−208(1987)、Enzyme.Microbial.Technol.,29,400−406(2001)]。
【0007】
ある種の微生物は2つのアミノ酸が環状につながった環状ジペプチド(ジケトピペラジン)構造を持つ化合物を生成することも知られている[J.Nat.Prod.,59,293−296(1996)、Tetrahedron,28,2999(1972)、J.Appl.Microbiol.,86,29−53(1999)]。ジケトピペラジンの生合成については、Streptomyces acidiscabiesのThaxtomin生合成過程ではNRPSによってcyclo−(L−4−nitrotryptophyl−L−phenylalanine)構造が合成される[Mol.Microbiol.,38,794−804(2000)]こと、およびバチルス属細菌のNRPSの一部のモジュールの作用によってフェニルアラニンとプロリンからcyclo(phenylalanyl−proline)が生成すること[J.Biol.Chem.,273,22773−22781(1998)]が報告されている。
【0008】
一方、抗生物質であるアルボノルシン(albonoursin)の生産株として知られているストレプトマイセス・ノルセイ(Streptomyces noursei)ATCC11455株ではNRPS酵素とは全く類似性のない蛋白質(albC遺伝子産物)がシクロ(L−フェニルアラニル−L−ロイシン)[cyclo(L−phenylalanyl−L−leucine)]構造の合成を担っていること、albC遺伝子を導入したEscherichia coliおよびStreptomyces lividansの培養液にcyclo dipeptide oxidaseを作用させるとアルボノルシンが検出されたとの報告はある[Chemistry&Biol.,9,1355−1364(2002)]が、albC遺伝子産物が直鎖状のジペプチドを生成するとの報告はない。
【発明の開示】
【発明が解決しようとする課題】
【0009】
本発明の目的は、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質をコードするDNA、該DNAを含有する組換え体DNA、該組換え体DNAを保有する形質転換体、ジペプチドの合成活性を有する蛋白質の製造法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を用いたジペプチドの酵素的合成法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を生産する能力を有する微生物または形質転換体の培養物等を酵素源に用いたジペプチドの製造法を提供することにある。
【課題を解決するための手段】
【0010】
本発明は、以下の(1)〜(24)に関する。
(1)以下の[1]〜[3]のいずれかに記載の蛋白質(ただし、配列番号1で表されるアミノ酸配列からなる蛋白質を除く)。
[1]配列番号2で表されるアミノ酸配列を有する蛋白質
[2]配列番号1または2で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドの合成活性を有する蛋白質
[3]配列番号1または2で表されるアミノ酸配列と65%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有する蛋白質
(2)以下の[1]〜[3]のいずれかに記載のジペプチド合成用蛋白質。
[1]配列番号1で表されるアミノ酸配列を有するジペプチド合成用蛋白質
[2]配列番号1で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)
で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質
[3]配列番号1で表されるアミノ酸配列と65%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質
(3)以下の[1]〜[3]のいずれかに記載のDNA(ただし、配列番号3で表される塩基配列からなるDNAを除く)。
[1]上記(1)の蛋白質をコードするDNA
[2]配列番号4で表される塩基配列を有するDNA
[3]配列番号4で表される塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチド合成活性を有する蛋白質をコードするDNA
(4)上記(3)のDNAを含有する組換え体DNA。
【0011】
(5)上記(4)の組換え体DNAを有する形質転換体。
(6)形質転換体が、微生物を宿主として得られる形質転換体である上記(5)の形質転換体。
(7)微生物が、エシェリヒア(Escherichia)属に属する微生物である上記(6)の形質転換体。
【0012】
(8)上記(5)〜(7)のいずれか1つに記載の形質転換体を培地に培養し、培養物中に上記(1)の蛋白質を生成、蓄積させ、該培養物より該蛋白質を採取する上記(1)の蛋白質の製造法。
(9)上記(1)の蛋白質を生産する能力を有する微生物を培地に培養し、培養物中に該蛋白質を生成、蓄積させ、該培養物より該蛋白質を採取する上記(1)の蛋白質の製造法。
【0013】
(10)微生物がストレプトマイセス(Streptomyces)属に属する微生物である上記(9)の製造法。
(11)ストレプトマイセス属に属する微生物が、アルボノルシンを生産する能力を有するストレプトミセス属に属する微生物である上記(10)の製造法。
(12)アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物が、ストレプトマイセス・アルボラス(Streptomyces alborus)またはストレプトマイセス・ノルセイ(Streptomyces noursei)である上記(11)の製造法。
【0014】
(13)上記(1)の蛋白質または上記(2)のジペプチド合成用蛋白質、1種以上のアミノ酸、およびATPを水性媒体中に存在せしめ、該媒体中に式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法。
【0015】
(14)以下の[1]〜[3]から選ばれる培養物または該培養物の処理物を酵素源に用い、該酵素源、1種以上のアミノ酸を水性媒体中に存在せしめ、該媒体中に式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法。
[1]上記(5)〜(7)のいずれか1つに記載の形質転換体の培養物または該培養物の処理物
[2]上記(1)の蛋白質を生産する能力を有する微生物の培養物または該培養物の処理物
[3]上記(2)のジペプチド合成用蛋白質を生産する能力を有する微生物の培養物または該培養物の処理物
(15)上記(1)の蛋白質を生産する能力を有する微生物が、ストレプトマイセス属に属する微生物である上記(14)の製造法。
【0016】
(16)上記(2)のジペプチド合成用蛋白質を生産する能力を有する微生物が、ストレプトマイセス属に属する微生物である上記(14)の製造法。
(17)ストレプトマイセス属に属する微生物が、アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物である上記(15)または(16)の製造法。
【0017】
(18)アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物が、ストレプトマイセス・アルボラスまたはストレプトマイセス・ノルセイに属する微生物である上記(17)の製造法。
(19)上記(2)のジペプチド合成用蛋白質を生産する能力を有する微生物が、上記(2)のジペプチド合成用蛋白質をコードするDNAで形質転換された微生物である上記(14)の製造法。
(20)上記(2)のジペプチド合成用蛋白質をコードするDNAで形質転換された微生物が、エシェリヒア属に属する微生物である上記(19)の製造法。
(21)培養物の処理物が、培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の溶媒処理物、該菌体の酵素処理物、該菌体の蛋白質分画物、該菌体の固定化物あるいは該菌体より抽出して得られる酵素標品であることを特徴とする、上記(14)〜(20)のいずれか1つの製造法。
【0018】
(22)1種以上のアミノ酸が、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体である上記(13)〜(21)のいずれか1つの製造法。
(23)ジペプチドが式(II)
R3−R4 (II)
(式中、R3およびR4は同一または異なって、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体を表す)であらわされるジペプチドである上記(13)〜(22)のいずれか1つの製造法。
【0019】
(24)L体またはD体のアミノ酸が、アラニン、グルタミン、グルタミン酸、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、トリプトファン、メチオニン、セリン、スレオニン、システイン、アスパラギン、チロシン、リジン、アルギニン、ヒスチジン、アスパラギン酸、α−アミノ酪酸、アザセリン、テアニン、4−ヒドロキシプロリン、3−ヒドロキシプロリン、オルニチン、シトルリンおよび6−ジアゾ−5−オキソノルロイシンから選ばれるアミノ酸である上記(22)または(23)の製造法。
【発明の効果】
【0020】
本発明によれば、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質をコードするDNA、該DNAを含有する組換え体DNA、該組換え体DNAを保有する形質転換体、ジペプチドの合成活性を有する蛋白質の製造法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を用いたジペプチドの酵素的合成法、ジペプチドの合成活性を有する蛋白質またはジペプチド合成用蛋白質を生産する能力を有する微生物または形質転換体の培養物等を酵素源に用いたジペプチドの製造法を提供することができる。
【図面の簡単な説明】
【0021】
【図1】図1は、ジペプチドの合成活性を有する蛋白質の発現プラスミドベクターであるpAL−nouおよびpAL−albの構築過程を示す図である。
【符号の説明】
【0022】
Ampr:アンピシリン耐性遺伝子
lacIq:ラクトースリプレッサー遺伝子
albC:albC遺伝子またはalbC類似遺伝子
【発明を実施するための最良の形態】
【0023】
本発明の蛋白質としては、以下の[1]〜[3]記載の蛋白質(ただし、配列番号1で表されるアミノ酸配列からなる蛋白質を除く)、
[1]配列番号2で表されるアミノ酸配列を有する蛋白質、
[2]配列番号1または2で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドの合成活性を有する蛋白質、および
[3]配列番号1または2で表されるアミノ酸配列と65%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上、最も好ましくは99%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有する蛋白質、
をあげることができる。
【0024】
また、本発明のジペプチド合成用蛋白質としては、以下の[4]〜[6]記載のジペプチド合成用蛋白質、
[4]配列番号1で表されるアミノ酸配列を有するジペプチド合成用蛋白質、
[5]配列番号1で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質、
[6]配列番号1で表されるアミノ酸配列と65%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上、最も好ましくは99%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質、
をあげることができる。
【0025】
以下、上記本発明の蛋白質およびジペプチドの合成用蛋白質を合わせて本発明の蛋白質と称する場合もある。
上記において、1以上のアミノ酸が欠失、置換または付加されたアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有する蛋白質は、Molecular Cloning,A Laboratory Manual,Third Edition,Cold SpringHarbor Laboratory Press(1989)(以下、モレキュラー・クローニング第3版と略す)、Current Protocols in Molecular Biology,John Wiley & Sons(1987−1997)(以下、カレント・プロトコールズ・イン・モレキュラー・バイオロジーと略す)、NucleicAcids Research,10,6487(1982)、Proc.Natl.Acad.Sci.USA,79,6409(1982)、Gene,34,315(1985)、Nucleic Acids Research,13,4431(1985)、Proc.Natl.Acad.Sci.USA,82,488(1985)等に記載の部位特異的変異導入法を用いて、例えば配列番号1または2で表されるアミノ酸配列からなる蛋白質をコードするDNAに部位特異的変異を導入することにより、取得することができる。
【0026】
欠失、置換または付加されるアミノ酸の数は特に限定されないが、上記の部位特異的変異法等の周知の方法により欠失、置換または付加できる程度の数であり、1個から数十個、好ましくは1〜20個、より好ましくは1〜10個、さらに好ましくは1〜5個である。
配列番号1または2で表されるアミノ酸配列において1以上のアミノ酸が欠失、置換または付加されたとは、同一配列中の任意の位置において、1または複数のアミノ酸が欠失、置換または付加されていてもよい。
【0027】
アミノ酸の置換が可能なアミノ酸としては、例えば配列番号1または2で表されるアミノ酸配列を公知のアライメントソフトウェアを用いて比較したときに、すべてのアミノ酸配列において保存されていないアミノ酸をあげることができる。公知のアライメントソフトウェアとしては、例えば遺伝子解析ソフトウェアGenetyx(ソフトウェア開発株式会社)に含まれるアライメント解析ソフトをあげることができる。該解析ソフトの解析パラメータとしては、デフォルト値を用いることができる。
【0028】
また、アミノ酸の欠失または付加が可能なアミノ酸の位置としては、例えば配列番号1または2で表されるアミノ酸配列のN末端側およびC末端側をあげることができる。
欠失、置換または付加は同時に生じてもよく、置換または付加されるアミノ酸は天然型と非天然型とを問わない。天然型アミノ酸としては、L−アラニン、L−アスパラギン、L−アスパラギン酸、L−グルタミン、L−グルタミン酸、グリシン、L−ヒスチジン、L−イソロイシン、L−ロイシン、L−リジン、L−メチオニン、L−フェニルアラニン、L−プロリン、L−セリン、L−スレオニン、L−トリプトファン、L−チロシン、L−バリン、L−システインなどがあげられる。
【0029】
以下に、相互に置換可能なアミノ酸の例を示す。同一群に含まれるアミノ酸は相互に置換可能である。
A群:ロイシン、イソロイシン、ノルロイシン、バリン、ノルバリン、アラニン、2−アミノブタン酸、メチオニン、O−メチルセリン、t−ブチルグリシン、t−ブチルアラニン、シクロヘキシルアラニン
B群:アスパラギン酸、グルタミン酸、イソアスパラギン酸、イソグルタミン酸、2−アミノアジピン酸、2−アミノスベリン酸
C群:アスパラギン、グルタミン
D群:リジン、アルギニン、オルニチン、2,4−ジアミノブタン酸、2,3−ジアミノプロピオン酸
E群:プロリン、3−ヒドロキシプロリン、4−ヒドロキシプロリン
F群:セリン、スレオニン、ホモセリン
G群:フェニルアラニン、チロシン
また、本発明の蛋白質が式(I)で表されるジペプチド合成活性を有するためには、配列番号1または2で表されるアミノ酸配列との相同性が65%以上、好ましくは80%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上、最も好ましくは99%以上の相同性を有していることが望ましい。
【0030】
アミノ酸配列や塩基配列の相同性は、Karlin and AltschulによるアルゴリズムBLAST[Pro.Natl.Acad.Sci.USA,90,5873(1993)]やFASTA[Methods Enzymol.,183,63(1990)]を用いて決定することができる。このアルゴリズムBLASTに基づいて、BLASTNやBLASTXとよばれるプログラムが開発されている[J.Mol.Biol.,215,403(1990)]。BLASTに基づいてBLASTNによって塩基配列を解析する場合には、パラメータは例えばScore=100、wordlength=12とする。また、BLASTに基づいてBLASTXによってアミノ酸配列を解析する場合には、パラメータは例えばscore=50、wordlength=3とする。BLASTとGappedBLASTプログラムを用いる場合には、各プログラムのデフォルトパラメーターを用いる。これらの解析方法の具体的な手法は公知である(http://www.ncbi.nlm.nih.gov.)。
【0031】
本発明の蛋白質が、式(I)で表されるジペプチド合成活性を有する蛋白質であることを確認する手段としては、例えばDNA組換え法を用いて本発明の蛋白質を発現する形質転換体を作製し、該形質転換体を用いて本発明の蛋白質を製造した後、本発明の蛋白質、1種以上のアミノ酸、およびATPを水性媒体中に存在せしめ、該水性媒体中に上記式(I)で表されるジペプチドが生成、蓄積するか否かをHPLC等により分析する方法をあげることができる。
【0032】
本発明のDNAとしては、以下の[1]〜[3]記載のDNA(ただし、配列番号3で表される塩基配列からなるDNAを除く)、
[1]配列番号2で表されるアミノ酸配列を有する蛋白質をコードするDNA、
[2]配列番号4で表される塩基配列を有するDNA、および
[3]配列番号4で表される塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ式(I)で表されるジペプチド合成活性を有する蛋白質をコードするDNA、
をあげることができる。
【0033】
また、本発明の式(I)で表されるジペプチドの製造法に用いることができるDNAとしては、上記[1]〜[3]記載のDNAに加え、以下の[4]〜[6]記載のDNA、
[4]配列番号1で表されるアミノ酸配列を有する蛋白質をコードするDNA、
[5]配列番号3で表される塩基配列を有するDNA、および
[6]配列番号3で表される塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ式(I)で表されるジペプチド合成活性を有する蛋白質をコードするDNA、
をあげることができる。
【0034】
ここでいう「ハイブリダイズする」とは、特定の塩基配列を有するDNAまたは該DNAの一部にDNAがハイブリダイズすることである。したがって、該特定の塩基配列を有するDNAまたは該DNAの一部の塩基配列は、ノーザンまたはサザンブロット解析のプローブとして有用であるか、またはPCR解析のオリゴヌクレオチドプライマーとして使用できる長さのDNAであってもよい。プローブとして用いるDNAとしては、少なくとも100塩基以上、好ましくは200塩基以上、より好ましくは500塩基以上のDNAをあげることができるが、少なくとも10塩基以上、好ましくは15塩基以上のDNAであってもよい。
【0035】
DNAのハイブリダイゼーション実験の方法はよく知られており、例えば当業者であれば本願明細書に従い、ハイブリダイゼーションの条件を決定することができる。該ハイブリダイゼーションの条件は、モレキュラー・クローニング第2版、第3版(2001年)、Methods for General and Molecular Bacteriolgy,ASM Press(1994)、Immunologymethods manual,Academic press(Molecular)に記載の他、多数の他の標準的な教科書に従っておこなうことができる。
【0036】
上記のストリンジェントな条件とは、例えばDNAを固定化したフィルターとプローブDNAとを50%ホルムアミド、5×SSC(750mMの塩化ナトリウム、75mMのクエン酸ナトリウム)、50mMのリン酸ナトリウム(pH7.6)、5×デンハルト溶液、10%の硫酸デキストラン、および20μg/lの変性させたサケ精子DNAを含む溶液中で42℃で一晩、インキュベートした後、例えば約65℃の0.2×SSC溶液中で該フィルターを洗浄する条件が好ましいが、より低いストリンジェント条件を用いることもできる。ストリンジェンな条件の変更は、ホルムアミドの濃度調整(ホルムアミドの濃度を下げるほど低ストリンジェントになる)、塩濃度および温度条件の変更により可能である。低ストリンジェント条件としては、例えば6×SSCE(20×SSCEは、3mol/lの塩化ナトリウム、0.2mol/lのリン酸二水素ナトリウム、0.02mol/lのEDTA、pH7.4)、0.5%のSDS、30%のホルムアミド、100μg/lの変性させたサケ精子DNAを含む溶液中で、37℃で一晩インキュベートした後、50℃の1×SSC、0.1%SDS溶液を用いて洗浄する条件をあげることができる。また、さらに低いストリンジェントな条件としては、上記した低ストリンジェント条件において、高塩濃度(例えば5×SSC)の溶液を用いてハイブリダイゼーションを行った後、洗浄する条件をあげることができる。
【0037】
上記した様々な条件は、ハイブリダイゼーション実験のバックグラウンドを抑えるために用いるブロッキング試薬を添加、または変更することにより設定することもできる。上記したブロッキング試薬の添加は、条件を適合させるために、ハイブリダイゼーション条件の変更を伴ってもよい。
上記したストリンジェントな条件下でハイブリダイズ可能なDNAとしては、例えば上記したBLASTやFASTA等を用いて上記したパラメータ等に基づいて計算したときに、配列番号3または4で表される塩基配列と少なくとも90%以上、好ましくは95%以上、より好ましくは98%以上、さらに好ましくは99%以上の相同性を有するDNAをあげることができる。
【0038】
配列番号3または4で表される塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズするDNAが、式(I)で表されるジペプチドの合成活性を有する蛋白質をコードするDNAであることは、例えば上記したように、組換えDNA法を用いて該DNAにコードされる蛋白質を製造し、該蛋白質の活性を測定することにより確認することができる。
(i)本発明のDNA、および本発明の蛋白質またはジペプチドの製造法に用いられるDNAの調製
本発明のDNA、および本発明の蛋白質またはジペプチドの製造法(以下、単に本発明の製造法ともいう)に用いられるDNAは、例えば、配列番号3または4で表される塩基配列に基づき設計することができるプローブを用いた、ストレプトマイセス属に属する微生物の染色体DNAライブラリーに対するサザンハイブリダイゼーション、または配列番号3または4で表される塩基配列に基づき設計することができるプライマーDNAを用いた、ストレプトマイセス属に属する微生物の染色体DNAを鋳型としたPCR[PCR Protocols,Academic Press(1990)]により取得することができる。
【0039】
また、各種の遺伝子配列データベースに対して配列番号1または2で表されるアミノ酸配列をコードするDNAの塩基配列と75%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましくは95%以上、特に好ましくは98%以上、最も好ましくは99%以上の相同性を有する配列を検索し、該検索によって得られた塩基配列に基づき、該塩基配列を有する生物の染色体DNA、cDNAライブラリー等から上記した方法により本発明のDNA、または本発明の製造法に用いられるDNAを取得することもできる。
【0040】
取得したDNAをそのまま、あるいは適当な制限酵素などで切断し、常法によりベクターに組み込み、得られた組換え体DNAを宿主細胞に導入した後、通常用いられる塩基配列解析方法、例えばジデオキシ法[Proc.Natl.Acad.Sci.,USA,74,5463(1977)]あるいは373A・DNAシークエンサー(パーキン・エルマー社製)等の塩基配列分析装置を用いて分析することにより、該DNAの塩基配列を決定することができる。
【0041】
塩基配列を決定した結果、取得されたDNAが部分長であった場合は、該部分長DNAをプローブに用いた、染色体DNAライブラリーに対するサザンハイブリダイゼーション法等により、全長DNAを取得することができる。
更に、決定されたDNAの塩基配列に基づいて、パーセプティブ・バイオシステムズ社製8905型DNA合成装置等を用いて化学合成することにより目的とするDNAを調製することもできる。
【0042】
上記のようにして取得されるDNAとして、例えば、配列番号3または4で表される塩基配列を有するDNAをあげることができる。
本発明のDNAおよび本発明の製造法に用いられるDNAを組み込むベクターとしては、pBluescriptII KS(+)(ストラタジーン社製)、pDIRECT[Nucleic Acids Res.,18,6069(1990)]、pCR−Script Amp SK(+)(ストラタジーン社製)、pT7Blue(ノバジェン社製)、pCRII(インビトロジェン社製)およびpCR−TRAP(ジーンハンター社製)などをあげることができる。
【0043】
宿主細胞としては、エシェリヒア属に属する微生物などをあげることができる。エシェリヒア属に属する微生物としては、例えば、エシェリヒア・コリ(Escherichia coli)XL1−Blue、エシェリヒア・コリXL2−Blue、エシェリヒア・コリDH1、エシェリヒア・コリMC1000、エシェリヒア・コリKY3276、エシェリヒア・コリW1485、エシェリヒア・コリJM109、エシェリヒア・コリHB101、エシェリヒア・コリNo.49、エシェリヒア・コリW3110、エシェリヒア・コリNY49、エシェリヒア・コリMP347、エシェリヒア・コリNM522、エシェリヒア・コリME8415等をあげることができる。
【0044】
組換え体DNAの導入方法としては、上記宿主細胞へDNAを導入する方法であればいずれも用いることができ、例えば、カルシウムイオンを用いる方法「Proc.Natl.Acad.Sci.,USA,69,2110(1972)]、プロトプラスト法(特開昭63−248394)、エレクトロポレーション法[NucleicAcids Res.,16,6127(1988)]等をあげることができる。
上記方法によって得られる本発明の製造法に用いられるDNAを保有する微生物としては、例えば配列番号3で表される配列を有するDNAを含有する組換え体DNAを保有する微生物であるエシェリヒア・コリNM522/pAL−nouをあげることができる。
(ii)本発明の蛋白質および本発明のジペプチド合成用蛋白質の製造法
本発明の蛋白質および本発明のジペプチド合成用蛋白質は、モレキュラー・クローニング第3版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載された方法等を用い、例えば以下の方法により、上記(i)の方法により取得した本発明のDNAおよび本発明の製造法に用いられるDNAを宿主細胞中で発現させて、製造することができる。
【0045】
本発明のDNAまたは本発明の製造法に用いられるDNAをもとにして、必要に応じて、本発明の蛋白質または本発明のジペプチド合成用蛋白質をコードする部分を含む適当な長さのDNA断片を調製する。また、該蛋白質をコードする部分の塩基配列を、宿主の発現に最適なコドンとなるように、塩基を置換することにより、該蛋白質の生産率を向上させることができる。
【0046】
該DNA断片を適当な発現ベクターのプロモーターの下流に挿入することにより、組換え体DNAを作製する。
該組換え体DNAを、該発現ベクターに適合した宿主細胞に導入することにより、本発明の蛋白質を生産する形質転換体を得ることができる。
宿主細胞としては、細菌、酵母、動物細胞、昆虫細胞等、植物細胞等、目的とする遺伝子を発現できるものであればいずれも用いることができ、好ましくは細菌、より好ましくはエシェリヒア属、バチルス属またはコリネバクテリウム属に属する微生物、さらに好ましくはエシェリヒア属に属する微生物をあげることができる。
【0047】
発現ベクターとしては、上記宿主細胞において自立複製可能ないしは染色体中への組込が可能で、本発明のDNAまたは本発明の製造法に用いられるDNAを転写できる位置にプロモーターを含有しているものが用いられる。
細菌等の原核生物を宿主細胞として用いる場合は、本発明のDNAまたは本発明の製造法に用いられるDNAを有する組換え体DNAは、原核生物中で自立複製可能であると同時に、プロモーター、リボソーム結合配列、本発明のDNAまたは本発明の製造法に用いられるDNA、転写終結配列より構成された組換え体DNAであることが好ましい。プロモーターを制御する遺伝子が含まれていてもよい。
【0048】
発現ベクターとしては、pBTrp2、pBTac1、pBTac2(いずれもベーリンガーマンハイム社製)、pHelix1(ロシュ・ダイアグノスティクス社製)、pKK233−2(アマシャム・ファルマシア・バイオテク社製)、pSE280(インビトロジェン社製)、pGEMEX−1(プロメガ社製)、pQE8(キアゲン社製)、pQE60(キアゲン社製)、pET−3(ノバジェン社製)、pKYP10(特開昭58−110600)、pKYP200[Agric.Biol.Chem.,48,669(1984)]、pLSA1[Agric.Biol.Chem.,53,277(1989)]、pGEL1[Proc.Natl.Acad.Sci.,USA,82,4306(1985)]、pBluescriptIISK(+)、pBluescript II KS(−)(ストラタジーン社製)、pTrS30[エシェリヒア・コリJM109/pTrS30(FERMBP−5407)より調製]、pTrS32[エシェリヒア・コリJM109/pTrS32(FERM BP−5408)より調製]、pPAC31(WO98/12343)、pUC19[Gene,33,103(1985)]、pSTV28(宝酒造社製)、pUC118(宝酒造社製)、pPA1(特開昭63−233798)等を例示することができる。
【0049】
プロモーターとしては、エシェリヒア・コリ等の宿主細胞中で機能するものであればいかなるものでもよい。例えば、trpプロモーター(Ptrp)、lacプロモーター(Plac)、PLプロモーター、PRプロモーター、PSEプロモーター等の、大腸菌やファージ等に由来するプロモーター、SPO1プロモーター、SPO2プロモーター、penPプロモーター等をあげることができる。またPtrpを2つ直列させたプロモーター、tacプロモーター、lacT7プロモーター、let Iプロモーターのように人為的に設計改変されたプロモーター等も用いることができる。
【0050】
さらにバチルス属に属する微生物中で発現させるためのxylAプロモーター[Appl.Microbiol.Biotechnol.,35,594−599(1991)]やコリネバクテリウム(Corynebacterium)属に属する微生物中で発現させるためのP54−6プロモーター[Appl.Microbiol.Biotechnol.,53,674−679(2000)]、ストレプトマイセス属に属する微生物中で発現させるためのxylAプロモーター(GeneticManipulation of Streptomyces:a Laboratory Manual:John Innes Foundation)なども用いることができる。
【0051】
リボソーム結合配列であるシャイン−ダルガノ(Shine−Dalgarno)配列と開始コドンとの間を適当な距離(例えば6〜18塩基)に調節したプラスミドを用いることが好ましい。
本発明のDNAまたは本発明の製造法に用いられるDNAを発現ベクターに結合させた組換え体DNAにおいては、転写終結配列は必ずしも必要ではないが、構造遺伝子の直下に転写終結配列を配置することが好ましい。
【0052】
このような組換え体DNAとしては、例えばpAL−nuoおよびpAL−albをあげることができる。
原核生物としては、エシェリヒア属、セラチア(Serratia)属、バチルス属、ブレビバクテリウム(Brevibacterium)属、コリネバクテリウム(Corynebacterium)属、ミクロバクテリウム属(Microbacterium)、シュードモナス(Pseudomonas)属、アグロバクテリウム(Agrobacterium)属、アリシクロバチルス属(Alicyclobacillus)、アナベナ(Anabena)属、アナシスティス(Anacystis)属、アスロバクター(Arthrobacter)属、アゾバクター(Azotobacter)属、クロマチウム(Chromatium)属、エルビニア(Erwinia)属、メチロバクテリウム(Methylobacterium)属、フォルミディウム(Phormidium)属、ロドバクター(Rhodobacter)属、ロドシュードモナス(Rhodopseudomonas)属、ロドスピリウム(Rhodospirillum)属、セネデスムス(Scenedesmus)属、ストレプトマイセス(Streptomyces)属、シネコッカス(Synechoccus)属、ザイモモナス(Zymomonas)属等に属する微生物、例えば、エシェリヒア・コリXL1−Blue、エシェリヒア・コリXL2−Blue、エシェリヒア・コリDH1、エシェリヒア・コリDH5α、エシェリヒア・コリMC1000、エシェリヒア・コリKY3276、エシェリヒア・コリW1485、エシェリヒア・コリJM109、エシェリヒア・コリHB101、エシェリヒア・コリNo.49、エシェリヒア・コリW3110、エシェリヒア・コリNY49、エシェリヒア・コリMP347、エシェリヒア・コリNM522、バチルス・サチリスATCC33712、バチルス・メガテリウム、バチルス・エスピー(Bacillus sp.)FERM BP−6030、バチルス・アミロリケファスエンス、バチルス・コアギュランス、バチルス・リケニフォルミス、バチルス・プミルス、ブレビバクテリウム・アンモニアゲネス(Brevibacterium ammoniagenes)、ブレビバクテリウム・イマリオフィルム(Brevibacterium immariophilum)ATCC14068、ブレビバクテリウム・サッカロリティカム(Brevibacterium saccharolyticum)ATCC14066、ブレビバクテリウム・フラバム(Brevibacterium flavum)ATCC14067、ブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)ATCC13869、コリネバクテリウム・グルタミカム(Corynebacterium glutamicum)ATCC13032、コリネバクテリウム・グルタミカムATCC14297、コリネバクテリウム・アセトアシドフィルム(Corynebacterium acetoacidophilum)ATCC13870、ミクロバクテリウム・アンモニアフィルム(Microbacterium ammoniaphilum)ATCC15354、セラチア・フィカリア(Serratia ficaria)、セラチア・フォンチコラ(Serratia fonticola)、セラチア・リケファシエンス(Serratia liquefaciens)、セラチア・マルセッセンス(Serratia marcescens)、シュードモナス・エスピー(Pseudomonas sp.)D−0110、アグロバクテリウム・ラジオバクタ(Agrobacterium radiobacter)、アグロバクテリウム・リゾジーンズ(Agrobacterium rhizogenes)、アグロバクテリウム・ルビ(Agrobacterium rubi)、アナベナ・シリンドリカ(Anabaena cylindrica)、アナベナ・ドリオルム(Anabaena doliolum)、アナベナ・フロスアクア(Anabaena flos−aquae)、アースロバクター・オーレッセンス(Arthrobacter aurescens)、アースロバクター・シトレウス(Arthrobacter citreus)、アースロバクター・グロブフォルミス(Arthrobacter globformis)、アースロバクター・ヒドロカーボグルタミカス(Arthrobacter hydrocarboglutamicus)、アースロバクター・ミソレンス(Arthrobacter mysorens)、アースロバクター・ニコチアナ(Arthrobacter nicotianae)、アースロバクター・パラフィネウス(Arthrobacter paraffineus)、アースロバクター・プロトフォルミエ(Arthrobacter protophormiae)、アースロバクター・ロセオパラフィナス(Arthrobacter roseoparaffinus)、アースロバクター・スルフレウス(Arthrobacter sulfureus)、アースロバクター・ウレアファシエン(Arthrobacter ureafaciens)、クロマチウム・ブデリ(Chromatium buderi)、クロマチウム・テピダム(Chromatium tepidum)、クロマチウム・ビノサム(Chromatium vinosum)、クロマチウム・ワーミンギ(Chromatium warmingii)、クロマチウム・フルビアタティレ(Chromatium fluviatile)、エルビニア・ウレドバラ(Erwinia uredovora)、エルビニア・カロトバラ(Erwinia carotovora)、エルビニア・アナス(Erwinia ananas)、エルビニア・ヘリコラ(Erwinia herbicola)、エルビニア・パンクタタ(Erwinia punctata)、エルビニア・テレウス(Erwinia terreus)、メチロバクテリウム・ロデシアナム(Methylobacterium rhodesianum)、メチロバクテリウム・エクソトルクエンス(Methylobacterium extorquens)、フォルミディウム・エスピー(Phormidiumsp.)ATCC29409、ロドバクター・カプスラタス(Rhodobacter capsulatus)、ロドバクター・スフェロイデス(Rhodobacter sphaeroides)、ロドシュードモナス・ブラスチカ(Rhodopseudomonas blastica)、ロドシュードモナス・マリナ(Rhodopseudomonas marina)、ロドシュードモナス・パルストリス(Rhodopseudomonas palustris)、ロドスピリウム・リブラム(Rhodospirillumrubrum)、ロドスピリウム・サレキシゲンス(Rhodospirillum salexigens)、ロドスピリウム・サリナラム(Rhodospirillum salinarum)、ストレプトマイセス・アンボファシエンス(Streptomyces ambofaciens)、ストレプトマイセス・オーレオファシエンス(Streptomyces aureofaciens)、ストレプトマイセス・アウレウス(Streptomyces aureus)、ストレプトマイセス・フンジシディカ(Streptomyces fungicidicus)、ストレプトマイセス・グリセオクロモゲナス(Streptomyces griseochromogenes)、ストレプトマイセス・グリセウス(Streptomycesgriseus)、ストレプトマイセス・リビダンス(Streptomyces lividans、ストレプトマイセス・オリボグリセウス(Streptomyces olivogriseus)、ストレプトマイセス・ラメウス(Streptomyces rameus)、ストレプトマイセス・タナシエンシス(Streptomyces tanashiensis)、ストレプトマイセス・ビナセウス(Streptomyces vinaceus)、ザイモモナス・モビリス(Zymomonas mobilis)等をあげることができ、好ましくはエシェリヒア属、バチルス属、ストレプトマオセス属またはコリネバクテリウム属に属する微生物をあげることができる。
【0053】
組換え体DNAの導入方法としては、上記宿主細胞へDNAを導入する方法であればいずれも用いることができ、例えば、カルシウムイオンを用いる方法[Proc.Natl.Acad.Sci.,USA,69,2110(1972)]、プロトプラスト法(特開昭63−248394)、エレクトロポレーション法[NucleicAcids Res.,16,6127(1988)]等をあげることができる。
酵母菌株を宿主細胞として用いる場合には、発現ベクターとして、例えば、YEp13(ATCC37115)、YEp24(ATCC37051)、YCp50(ATCC37419)、pHS19、pHS15等を用いることができる。
【0054】
プロモーターとしては、酵母菌株中で機能するものであればいずれのものを用いてもよく、例えば、PHO5プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーター、gal 1プロモーター、gal 10プロモーター、ヒートショックポリペプチドプロモーター、MFα1プロモーター、CUP1プロモーター等のプロモーターをあげることができる。
宿主細胞としては、サッカロマイセス(Saccharomyces)属、シゾサッカロマイセス(Schizosaccharomyces)属、クルイベロマイセス(Kluyveromyces)属、トリコスポロン(Trichosporon)属、シワニオマイミセス(Schwanniomyces)属、ピチア(Pichia)属、またはキャンディダ(Candida)属等に属する酵母菌株をあげることができ、具体的には、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クルイベロマイセス・ラクティス(Kluyveromyces lactis)、トリコスポロン・プルランス(Trichosporon pullulans)、シワニオマイセス・アルビウス(Schwanniomyces alluvius)、ピチア・パストリス(Pichia pastoris)、キャンディダ・ウティリス(Candida utilis)等をあげることができる。
【0055】
組換え体DNAの導入方法としては、酵母にDNAを導入する方法であればいずれも用いることができ、例えば、エレクトロポレーション法[Methods Enzymol.,194,182(1990)]、スフェロプラスト法[Proc.Natl.Acad.Sci.,USA,81,4889(1984)]、酢酸リチウム法[J.Bacteriol.,153,163(1983)]等をあげることができる。
動物細胞を宿主として用いる場合には、発現ベクターとして、例えば、pcDNAI、pcDM8(フナコシ社より市販)、pAGE107(特開平3−22979)、pAS3−3(特開平2−227075)、pCDM8[Nature,329,840(1987)]、pcDNAI/Amp(インビトロジェン社製)、pREP4(インビトロジェン社製)、pAGE103[J.Biochem,101,1307(1987)]、pAGE210、pAMo、pAMoA等を用いることができる。
【0056】
プロモーターとしては、動物細胞中で機能するものであればいずれも用いることができ、例えば、サイトメガロウイルス(CMV)のIE(immediate early)遺伝子のプロモーター、SV40の初期プロモーターあるいはメタロチオネインのプロモーター、レトロウイルスのプロモーター、ヒートショックプロモーター、SRαプロモーター等をあげることができる。また、ヒトCMVのIE遺伝子のエンハンサーをプロモーターと共に用いてもよい。
【0057】
宿主細胞としては、マウス・ミエローマ細胞、ラット・ミエローマ細胞、マウス・ハイブリドーマ細胞、ヒトの細胞であるナマルバ(Namalwa)細胞またはナマルバKJM−1細胞、ヒト胎児腎臓細胞、ヒト白血病細胞、アフリカミドリザル腎臓細胞、チャイニーズ・ハムスターの細胞であるCHO細胞、HBT5637(特開昭63−299)等をあげることができる。
マウス・ミエローマ細胞としては、SP2/0、NSO等、ラット・ミエローマ細胞としてはYB2/0等、ヒト胎児腎臓細胞としてはHEK293(ATCC CRL−1573)、ヒト白血病細胞としてはBALL−1等、アフリカミドリザル腎臓細胞としてはCOS−1、COS−7等をあげることができる。
【0058】
組換え体DNAの導入方法としては、動物細胞にDNAを導入する方法であればいずれも用いることができ、例えば、エレクトロポレーション法[Cytotechnology,3,133(1990)]、リン酸カルシウム法(特開平2−227075)、リポフェクション法[Proc.Natl.Acad.Sci.,USA,84,7413(1987)]、Virology,52,456(1973)に記載の方法等をあげることができる。
【0059】
昆虫細胞を宿主として用いる場合には、例えばBaculovirus Expression Vectors,A Laboratory Manual,W.H.Freemanand Company,New York(1992)、カレント・プロトコールズ・イン・モレキュラー・バイオロジー、MolecularBiology,A Laboratory Manual、Bio/Technology,6,47(1988)等に記載された方法によって、蛋白質を生産することができる。
【0060】
即ち、組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、蛋白質を生産させることができる。
該方法において用いられる遺伝子導入ベクターとしては、例えば、pVL1392、pVL1393、pBlueBacIII(いずれもインビトロジェン社製)等をあげることができる。
【0061】
バキュロウイルスとしては、例えば、夜盗蛾科昆虫に感染するウイルスであるアウトグラファ・カリフォルニカ・ヌクレアー・ポリヘドロシス・ウイルス(Autographa californica nuclear polyhedrosis virus)等を用いることができる。
昆虫細胞としては、スポドプテラ・フルギペルダ(Spodoptera frugiperda)の卵巣細胞、トリコプルシア・ニ(Trichoplusia ni)の卵巣細胞、カイコ卵巣由来の培養細胞等を用いることができる。
【0062】
スポドプテラ・フルギペルダの卵巣細胞としてはSf9、Sf21(バキュロウイルス・イクスプレッション・ベクターズ ア・ラボラトリー・マニュアル)等、トリコプルシア・ニの卵巣細胞としてはHigh5、BTI−TN−5B1−4(インビトロジェン社製)等、カイコ卵巣由来の培養細胞としてはボンビクス・モリ(Bombyx mori)N4等をあげることができる。
組換えウイルスを調製するための、昆虫細胞への上記組換え遺伝子導入ベクターと上記バキュロウイルスの共導入方法としては、例えば、リン酸カルシウム法(特開平2−227075)、リポフェクション法[Proc.Natl.Acad.Sci.,USA,84,7413(1987)]等をあげることができる。
【0063】
植物細胞を宿主細胞として用いる場合には、発現ベクターとして、例えば、Tiプラスミド、タバコモザイクウイルスベクター等をあげることができる。
プロモーターとしては、植物細胞中で機能するものであればいずれのものを用いてもよく、例えば、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター、イネアクチン1プロモーター等をあげることができる。
【0064】
宿主細胞としては、タバコ、ジャガイモ、トマト、ニンジン、ダイズ、アブラナ、アルファルファ、イネ、コムギ、オオムギ等の植物細胞等をあげることができる。
組換えベクターの導入方法としては、植物細胞にDNAを導入する方法であればいずれも用いることができ、例えば、アグロバクテリウム(Agrobacterium)を用いる方法(特開昭59−140885、特開昭60−70080、WO94/00977)、エレクトロポレーション法(特開昭60−251887)、パーティクルガン(遺伝子銃)を用いる方法(特許第2606856、特許第2517813)等をあげることができる。
【0065】
酵母、動物細胞、昆虫細胞または植物細胞により発現させた場合には、糖あるいは糖鎖が付加された蛋白質を得ることができる。
以上のようにして得られる形質転換体を培地に培養し、培養物中に本発明の蛋白質を生成蓄積させ、該培養物から採取することにより、該蛋白質を製造することができる。
本発明の蛋白質を製造するための上記形質転換体の宿主としては、細菌、酵母、動物細胞、昆虫細胞等、植物細胞等いずれであってもよいが、好ましくは細菌、より好ましくはエシェリヒア属に属する微生物、さらに好ましくはエシェリヒア・コリに属する微生物をあげることができる。
【0066】
上記形質転換体を培地に培養する方法は、宿主の培養に用いられる通常の方法に従って行うことができる。
エシェリヒア・コリ等の原核生物あるいは酵母等の真核生物を宿主として得られた形質転換体を培養する培地としては、該生物が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行える培地であれば天然培地、合成培地のいずれを用いてもよい。
【0067】
炭素源としては、該生物が資化し得るものであればよく、グルコース、フラクトース、スクロース、これらを含有する糖蜜、デンプンあるいはデンプン加水分解物等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類等を用いることができる。
窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸もしくは有機酸のアンモニウム塩、その他の含窒素化合物、並びに、ペプトン、肉エキス、酵母エキス、コーンスチープリカー、カゼイン加水分解物、大豆粕および大豆粕加水分解物、各種発酵菌体、およびその消化物等を用いることができる。
【0068】
無機塩としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等を用いることができる。
培養は、通常振盪培養または深部通気攪拌培養等の好気的条件下で行う。培養温度は15〜40℃がよく、培養時間は、通常5時間〜7日間である。培養中pHは3.0〜9.0に保持する。pHの調整は、無機または有機の酸、アルカリ溶液、尿素、炭酸カルシウム、アンモニア等を用いて行う。
【0069】
また、培養中必要に応じて、アンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。
プロモーターとして誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養するときには、必要に応じてインデューサーを培地に添加してもよい。例えば、lacプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはイソプロピル−β−D−チオガラクトピラノシド等を、trpプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはインドールアクリル酸等を培地に添加してもよい。
【0070】
動物細胞を宿主として得られた形質転換体を培養する培地としては、一般に使用されているRPMI1640培地[J.Am.Med.Assoc.,199,519(1967)]、イーグル(Eagle)のMEM培地[Science,122,501(1952)]、DMEM培地[Virology,8,396(1959)]、199培地[Proc.Soc.Biol.Med.,73,1(1950)]またはこれら培地に牛胎児血清等を添加した培地等を用いることができる。
【0071】
培養は、通常pH6〜8、25〜40℃、5%CO2存在下等の条件下で1〜7日間行う。
また、培養中必要に応じて、カナマイシン、ペニシリン、ストレプトマイシン等の抗生物質を培地に添加してもよい。
昆虫細胞を宿主として得られた形質転換体を培養する培地としては、一般に使用されているTNM−FH培地(ファーミンジェン社製)、Sf−900 II SFM培地(ライフ・テクノロジーズ社製)、ExCell400、ExCell405[いずれもJRHバイオサイエンシーズ社製]、Grace’sInsect Medium[Nature,195,788(1962)]等を用いることができる。
【0072】
培養は、通常pH6〜7、25〜30℃等の条件下で1〜5日間行う。
また、培養中必要に応じて、ゲンタマイシン等の抗生物質を培地に添加してもよい。
植物細胞を宿主として得られた形質転換体は、細胞として、または植物の細胞や器官に分化させて培養することができる。該形質転換体を培養する培地としては、一般に使用されているムラシゲ・アンド・スクーグ(MS)培地、ホワイト(White)培地、またはこれら培地にオーキシン、サイトカイニン等、植物ホルモンを添加した培地等を用いることができる。
【0073】
培養は、通常pH5〜9、20〜40℃の条件下で3〜60日間行う。
また、培養中必要に応じて、カナマイシン、ハイグロマイシン等の抗生物質を培地に添加してもよい。
上記のとおり、本発明のDNAまたは本発明の製造法に用いられるDNAを発現ベクターに連結した組換え体DNAを保有する微生物、昆虫細胞、動物細胞、あるいは植物細胞由来の形質転換体を、通常の培養方法に従って培養し、本発明の蛋白質を生成蓄積させ、該培養物より該蛋白質を採取することにより、該蛋白質を製造することができる。
【0074】
本発明の蛋白質の生産方法としては、宿主細胞内に生産させる方法、宿主細胞外に分泌させる方法、あるいは宿主細胞外膜上に生産させる方法があり、選択した方法に応じて、使用する宿主細胞を選択し、生産させる蛋白質の構造を変更すればよい。
本発明の蛋白質が宿主細胞内あるいは宿主細胞外膜上に生産される場合、ポールソンらの方法[J.Biol.Chem.,264,17619(1989)]、ロウらの方法[Proc.Natl.Acad.Sci.,USA,86,8227(1989)、Genes Develop.,4,1288(1990)]、または特開平05−336963、WO94/23021等に記載の方法を準用することにより、該蛋白質を宿主細胞外に積極的に分泌させることができる。
【0075】
すなわち、遺伝子組換えの手法を用いて、本発明の蛋白質の活性部位を含む蛋白質の手前にシグナルペプチドを付加した形で生産させることにより、該蛋白質を宿主細胞外に積極的に分泌させることができる。
また、特開平2−227075に記載されている方法に準じて、ジヒドロ葉酸還元酵素遺伝子等を用いた遺伝子増幅系を利用して生産量を上昇させることもできる。
【0076】
さらに、遺伝子導入した動物または植物の細胞を再分化させることにより、遺伝子が導入された動物個体(トランスジェニック非ヒト動物)または植物個体(トランスジェニック植物)を造成し、これらの個体を用いて本発明の蛋白質を製造することもできる。
本発明の蛋白質を生産する形質転換体が動物個体または植物個体の場合は、通常の方法に従って、飼育または栽培し、該蛋白質を生成蓄積させ、該動物個体または植物個体より該蛋白質を採取することにより、該蛋白質を製造することができる。
【0077】
動物個体を用いて本発明の蛋白質を製造する方法としては、例えば公知の方法[Am.J.Clin.Nutr.,63,639S(1996)、Am.J.Clin.Nutr.,63,627S(1996)、Bio/Technology,9,830(1991)]に準じて遺伝子を導入して造成した動物中に本発明の蛋白質を生産する方法があげられる。
動物個体の場合は、例えば、本発明のDNAまたは本発明の製造法に用いられるDNAを導入したトランスジェニック非ヒト動物を飼育し、本発明の蛋白質を該動物中に生成、蓄積させ、該動物中より該蛋白質を採取することにより、該蛋白質を製造することができる。該動物中の生成、蓄積させる場所としては、例えば、該動物のミルク(特開昭63−309192)、卵等をあげることができる。この際に用いられるプロモーターとしては、動物で機能するものであればいずれも用いることができるが、例えば、乳腺細胞特異的なプロモーターであるαカゼインプロモーター、βカゼインプロモーター、βラクトグロブリンプロモーター、ホエー酸性プロテインプロモーター等が好適に用いられる。
【0078】
植物個体を用いて本発明の蛋白質を製造する方法としては、例えば本発明の蛋白質をコードするDNAを導入したトランスジェニック植物を公知の方法[組織培養,20(1994)、組織培養,21(1995)、Trends Biotechnol.,15,45(1997)]に準じて栽培し、該蛋白質を該植物中に生成、蓄積させ、該植物中より該蛋白質を採取することにより、該蛋白質を生産する方法があげられる。
【0079】
本発明の蛋白質を生産する形質転換体を用いて製造された本発明の蛋白質を単離・精製する方法としては、通常の酵素の単離、精製法を用いることができる。
例えば、本発明の蛋白質が、細胞内に溶解状態で生産された場合には、培養終了後、細胞を遠心分離により回収し、水系緩衝液にけん濁後、超音波破砕機、フレンチプレス、マントンガウリンホモゲナイザー、ダイノミル等により細胞を破砕し、無細胞抽出液を得る。
【0080】
該無細胞抽出液を遠心分離することにより得られる上清から、通常の酵素の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)−セファロース、DIAION HPA−75(三菱化成社製)等レジンを用いた陰イオン交換クロマトグラフィー法、S−SepharoseFF(ファルマシア社製)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィー法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、精製標品を得ることができる。
【0081】
また、該蛋白質が細胞内に不溶体を形成して生産された場合は、同様に細胞を回収後破砕し、遠心分離を行うことにより得られた沈殿画分より、通常の方法により該蛋白質を回収後、該蛋白質の不溶体を蛋白質変性剤で可溶化する。
該可溶化液を、蛋白質変性剤を含まないあるいは蛋白質変性剤の濃度が蛋白質が変性しない程度に希薄な溶液に希釈、あるいは透析し、該蛋白質を正常な立体構造に構成させた後、上記と同様の単離精製法により精製標品を得ることができる。
【0082】
本発明の蛋白質またはその糖修飾体等の誘導体が細胞外に分泌された場合には、培養上清に該蛋白質またはその糖付加体等の誘導体を回収することができる。
即ち、該培養物を上記と同様の遠心分離等の手法により処理することにより可溶性画分を取得し、該可溶性画分から、上記と同様の単離精製法を用いることにより、精製標品を得ることができる。
【0083】
このようにして取得される蛋白質として、例えば、配列番号1または2で表されるアミノ酸配列からなる蛋白質をあげることができる。
また、本発明の蛋白質を他の蛋白質との融合蛋白質として生産し、融合した蛋白質に親和性をもつ物質を用いたアフィニティークロマトグラフィーを利用して精製することもできる。
【0084】
融合させる蛋白質としては、β−ガラクトシダーゼ、プロテインA、プロテインAのIgG結合領域、クロラムフェニコール・アセチルトランスフェラーゼ、ポリ(Arg)、ポリ(Glu)、プロテインG、マルトース結合タンパク質、グルタチオンS−トランスフェラーゼ、ポリヒスチジン鎖(His−tag)、Sペプチド、DNA結合タンパク質ドメイン、Tac抗原、チオレドキシン、グリーン・フルオレッセント・プロテイン、FLAGペプチド、および任意の抗体のエピトープなどがあげられる〔山川彰夫,実験医学,13,469−474(1995)〕。
【0085】
上記した融合させる蛋白質に親和性をもつ物質としては、β−ガラクトシダーゼ、プロテインA、プロテインAのイムノグロブリンG結合領域、クロラムフェニコール・アセチルトランスフェラーゼ、ポリ(Arg)、ポリ(Glu)、プロテインG、マルトース結合タンパク質、グルタチオンS−トランスフェラーゼ、ポリヒスチジン鎖(His−tag)、Sペプチド、DNA結合タンパク質ドメイン、Tac抗原、チオレドキシン、グリーン・フルオレッセント・プロテイン、FLAGペプチドまたは任意の抗体のエピトープを認識する抗体、例えばイムノグロブリンGなどをあげることができる。
【0086】
また、上記で取得された蛋白質のアミノ酸情報を基に、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法により、本発明の蛋白質を製造することができる。また、Advanced ChemTech社、パーキン・エルマー社、Pharmacia社、Protein TechnologyInstrument社、Synthecell−Vega社、PerSeptive社、島津製作所等のペプチド合成機を利用して化学合成することもできる。
(iii)本発明のジペプチドの製造法
(1)酵素的製造法
ジペプチドの酵素的製造法としては、本発明の蛋白質またはジペプチド合成用蛋白質、1種以上のアミノ酸、およびATPを水性媒体中に存在せしめ、該媒体中に式(I)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する方法をあげることができる。
【0087】
上記製造法において、基質に用いられる1種以上のアミノ酸、好ましくは1または2種のアミノ酸としては、L体またはD体のアミノ酸、グリシン(Gly)、β−アラニン(βAla)およびそれらの誘導体から選ばれるアミノ酸、好ましくはL−アミノ酸、GlyおよびβAla並びにそれらの誘導体からなる群より選ばれるアミノ酸をあげることができ、該アミノ酸であれば、いずれのアミノ酸をいずれの組み合わせで用いてもよい。L体またはD体のアミノ酸としては、例えばアラニン(Ala)、グルタミン(Gln)、グルタミン酸(Glu)、バリン(Val)、ロイシン(Leu)、イソロイシン(Ile)、プロリン(Pro)、フェニルアラニン(Phe)、トリプトファン(Trp)、メチオニン(Met)、セリン(Ser)、スレオニン(Thr)、システイン(Cys)、アスパラギン(Asn)、チロシン(Tyr)、リジン(Lys)、アルギニン(Arg)、ヒスチジン(His)、アスパラギン酸(Asp)、α−アミノ酪酸(α−AB)、アザセリン(Azaserine)、テアニン(theanine)、4−ヒドロキシプロリン(4−HYP)、3−ヒドロキシプロリン(3−HYP)、オルニチン(Orn)、シトルリン(Cit)および6−ジアゾ−5−オキソノルロイシン(6−diazo−5−oxo−norleucine)などをあげることができ、L体のアミノ酸としては、L−Ala、L−Gln、L−Glu、Gly、L−Val、L−Leu、L−Ile、L−Pro、L−Phe、L−Trp、L−Met、L−Ser、L−Thr、L−Cys、L−Asn、L−Tyr、L−Lys、L−Arg、L−His、L−Asp、L−α−AB、L−Azaserine、L−theanine、L−4−HYP、L−3−HYP、L−Orn、L−CitおよびL−6−diazo−5−oxo−norleucineなどをあげることができる。
【0088】
アミノ酸の誘導体としては、例えばヒドロキシアミノ酸(β−ヒドロキシグルタミン、β−ヒドロキシグルタミン酸、γ−ヒドロキシグルタミン酸、α−ヒドロキシグリシン、β−ヒドロキシバリン、γ−ヒドロキシバリン、β−ヒドロキシロイシン、γ−ヒドロキシロイシン、δ−ヒドロキシロイシン、β−ヒドロキシイソロイシン、γ−ヒドロキシイソロイシン、3−ヒドロキシプロリン、4−ヒドロキシプロリン、β−ヒドロキシフェニルアラニン、3,4−ジヒドロキシフェニルアラニン、2,4,5−トリヒドロキシフェニルアラニン、β−ヒドロキシトリプトファン、5−ヒドロキシトリプトファン、α−ヒドロキシメチオニン、β−ヒドロキシセリン、γ−ヒドロキシスレオニン、S−ヒドロキシシステイン、β−ヒドロキシアスパラギン、β−ヒドロキシチロシン、β−ヒドロキシリジン、γ−ヒドロキシリジン、δ−ヒドロキシリジン、N−ヒドロキシリジン、β−ヒドロキシアルギニン、δ−ヒドロキシアルギニン、N−ヒドロキシアルギニン、β−ヒドロキシヒスチジン、β−ヒドロキシアスパラギン酸、β−ヒドロキシオルニチン、γ−ヒドロキシオルニチン、N−ヒドロキシオルニチン)、N−メチルアミノ酸(N−メチル−アラニン、N−メチル−グルタミン、N−メチル−グルタミン酸、N−メチル−グリシン、N−メチル−バリン、N−メチル−ロイシン、N−メチル−イソロイシン、N−メチル−プロリン、N−メチル−フェニルアラニン、N−メチル−トリプトファン、N−メチル−メチオニン、N−メチル−セリン、N−メチル−スレオニン、N−メチル−システイン、N−メチル−アスパラギン、N−メチル−チロシン、N−メチル−リジン、N−メチル−アルギニン、N−メチル−ヒスチジン、N−メチル−アスパラギン酸、N−メチル−オルニチン)などをあげることができる。
【0089】
上記製造法に用いられる、より好ましい1または2種のアミノ酸としては、L−Ala、L−LeuおよびL−Pheから選ばれる1または2種のアミノ酸、さらに好ましくは、L−Ala、L−LeuおよびL−Pheから選ばれる1種のアミノ酸、またはL−LeuとL−Pheとからなる2種のアミノ酸をあげることができる。
上記製造法において、本発明の蛋白質またはジペプチド合成用蛋白質は、基質として用いるアミノ酸1mgあたり0.01〜100mg、好ましくは0.1mg〜10mg添加する。
【0090】
上記製造法において、基質として用いるアミノ酸は、全量として0.1〜500g/L、好ましくは0.2〜200g/Lの濃度になるように水性媒体中に初発または反応途中に添加する。
上記製造法において、エネルギー源として用いるATPは、0.5mmol/L〜10mol/Lの濃度で用いる。
上記製造法で用いられる水性媒体としては、ジペプチドの生成反応を阻害しない限り、いかなる成分、組成の水性媒体であってもよく、例えば、水、りん酸塩、炭酸塩、酢酸塩、ほう酸塩、クエン酸塩、トリスなどの緩衝液などをあげることができる。また、メタノール、エタノールなどのアルコール類、酢酸エチルなどのエステル類、アセトンなどのケトン類、アセトアミドなどのアミド類を含有していてもよい。
【0091】
ジペプチドの生成反応は水性媒体中、pH5〜11、好ましくはpH6〜10、20〜50℃、好ましくは25〜45℃の条件で2〜150時間、好ましくは6〜120時間行う。
上記方法で製造されるジペプチドとしては、式(I)で表されるジペプチドをあげることができ、好ましくは、式(I)においてR1およびR2が同時にまたは異なって、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体であるジペプチド、より好ましくはL体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体であるジペプチドをあげることができる。
【0092】
L体のアミノ酸およびアミノ酸の誘導体としては、上記した物質をあげることができる。
上記方法で製造される、さらに好ましいジペプチドとしては、L−ロイシル−L−フェニルアラニン、L−フェニルアラニル−L−ロイシン、L−アラニル−L−アラニン、L−ロイシル−L−ロイシンおよびL−フェニルアラニル−L−フェニルアラニンなどをあげることができる。
(2)形質転換体または微生物の培養物もしくは培養物の処理物を酵素源として用いる製造法
形質転換体または微生物の培養物または培養物の処理物を酵素源として用いるジペプチドの製造法としては、本発明の蛋白質もしくはジペプチド合成用蛋白質を生産する能力を有する形質転換体、または本発明の蛋白質もしくはジペプチド合成用蛋白質を生産する能力を有する微生物の培養物もしくは培養物の処理物を酵素源に用い、該酵素源、および1種以上のアミノ酸を水性媒体中に存在せしめ、該媒体中に式(I)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法をあげることができる。
【0093】
上記製造法に用いられる形質転換体としては、上記(ii)の方法により製造することができる、本発明の蛋白質もしくはジペプチド合成用蛋白質を生産する形質転換体をあげることができる。該形質転換体の宿主としては、細菌、酵母、動物細胞、昆虫細胞等、植物細胞等をあげることができ、好ましく細菌、より好ましくはエシェリヒア属、バチルス属、ストレプトマイセス属またはコリネバクテリウム属に属する微生物をあげることができる。
【0094】
エシェリヒア属に属する微生物としては、好ましくはエシェリヒア・コリに属する微生物等をあげることができ、バチルス属に属する微生物としては好ましくはバチルス・サチリスまたはバチルス・メガテリウム等に属する微生物をあげることができ、ストレプトマイセス属に属する微生物としては、好ましくはストレプトマイセス・リビダンスをあげることができ、コリネバクテリウム属に属する微生物としては好ましくはコリネバクテリウム・グルタミカムまたはコリネバクテリウム・アンモニアゲネス等に属する微生物をあげることができる。
【0095】
上記製造法に用いられる微生物は、本発明の蛋白質もしくはジペプチド合成用蛋白質を生産する能力を有する微生物であれば、いずれの微生物であってもよく、好ましくはストレプトマイセス属に属する微生物、より好ましくはアルボノルシン合成活性を有するストレプトマイセス属に属する微生物、さらに好ましくはストレプトマイセス・アルボラスまたはストレプトマイセス・ノウルセイより選ばれる種に属する微生物、最も好ましくは、ストレプトマイセス・アルボラスIFO14147株、ストレプトマイセス・ノウルセイATCC11455株またはIFO15452株をあげることができる。
【0096】
培養物の処理物としては、培養物と同じ活性を維持している限り、公知のいかなる処理を施された培養物の処理物であってもよく、該処理物としては、例えば培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の溶媒処理物、該菌体の酵素処理物および該菌体の固定化物等の生細胞からなる処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の蛋白質分画物および該菌体より抽出して得られる酵素標品などをあげることができる。
【0097】
上記製造法において、基質として用いるアミノ酸の種類、使用濃度および添加時期、並びに生産されるジペプチドは、上記(iii)の(1)の酵素的製造法のものと同様である。
また、微生物の培養物または該培養物の処理物を酵素源とした製造法において用いられる水性媒体としては、上記(iii)の(1)の酵素的製造法に用いられる水性媒体に加え、酵素源として用いた形質転換体または微生物の培養液も水性媒体として用いることができる。
【0098】
また上記製造法においては、必要に応じて、ATPの供給源として、ATPまたは形質転換体もしくは微生物が代謝してATPを生産し得る化合物、例えばグルコースのような糖類、エタノールのようなアルコール類、酢酸のような有機酸類などを水性媒体中に加えることができる。
さらに必要に応じて、水性媒体中に界面活性剤あるいは有機溶媒を添加してもよい。界面活性剤としては、ポリオキシエチレン・オクタデシルアミン(例えばナイミーンS−215、日本油脂社製)などの非イオン界面活性剤、セチルトリメチルアンモニウム・ブロマイドやアルキルジメチル・ベンジルアンモニウムクロライド(例えばカチオンF2−40E、日本油脂社製)などのカチオン系界面活性剤、ラウロイル・ザルコシネートなどのアニオン系界面活性剤、アルキルジメチルアミン(例えば三級アミンFB、日本油脂社製)などの三級アミン類など、ジペプチドの生成を促進するものであればいずれでもよく、1種または数種を混合して使用することもできる。界面活性剤は、通常0.1〜50g/lの濃度で用いられる。有機溶剤としては、キシレン、トルエン、脂肪族アルコール、アセトン、酢酸エチルなどがあげられ、通常0.1〜50ml/lの濃度で用いられる。
【0099】
培養物または該培養物の処理物を酵素源として用いる場合、該酵素源の量は、当該酵素源の比活性等により異なるが、例えば、基質として用いるアミノ酸1mgあたり湿菌体重量として5〜1000mg、好ましくは10〜400mg添加する。
ジペプチドの生成反応は水性媒体中、pH5〜11、好ましくはpH6〜10、20〜65℃、好ましくは25〜55℃、より好ましくは30〜45℃の条件で通常1分間〜150時間、好ましくは3分間〜120時間、より好ましくは30分間〜100時間行う。
【0100】
上記(iii)の(1)または(2)の製造法において、水性媒体中に生成、蓄積したジペプチドの採取は、活性炭やイオン交換樹脂などを用いる通常の方法あるいは、有機溶媒による抽出、結晶化、薄層クロマトグラフィー、高速液体クロマトグラフィー等により行うことができる。
以下に、実施例を示すが、本発明は下記実施例に限定されるものではない。
【実施例1】
【0101】
albC遺伝子およびその類縁遺伝子の取得
ストレプトマイセス・ノウルセイのalbC遺伝子の塩基配列[Chemistry & Biol.,9,1355(2002)]に基づき、ストレプトマイセス・ノウルセイおよびストレプトマイセス・アルボラスよりalbC遺伝子およびその類縁遺伝子を、以下の方法で取得した。
まず、ストレプトマイセス・ノウルセイIFO15452株とストレプトマイセス・アルボラスIFO14147株を、それぞれ、1%グリシン添加したKM73培地[2g/lイーストエキス(ディフコ社製)、10g/l可溶性デンプン(和光純薬工業社製)]、KP培地[15g/lグルコース、10g/lグリセロール、10g/lポリペプトン(日本製薬株式会社製)、10g/l肉エキス(極東製薬工業株式会社製)、4g/l炭酸カルシウム]に植菌し28℃で一晩振とう培養した。なお、ストレプトマイセス・ノウルセイIFO15452株およびストレプトマイセス・アルボラスIFO14147株は、独立行政法人 製品評価技術基盤機構 生物資源部門[National Institute of Technology andEvaluation(NITE)Biological Resource Center(BRC)](〒292−0818 千葉県木更津市かずさ鎌足2−5−8)より分譲を受けた。
【0102】
培養後、Genetic Manipulation of Streptomyces:a Laboratory Manual:John InnesFoundationに記載の方法に従って該微生物の染色体DNAを単離精製した。
albC遺伝子の塩基配列に基づき、パーセプティブ・バイオシステムズ(Perseptive Biosystems)社製8905型DNA合成機を用いて、配列番号5および6で表される塩基配列を有するDNA(以下、それぞれプライマーA、プライマーBと呼ぶ)を合成した。プライマーAは、ストレプトマイセス・ノウルセイの染色体DNAのalbC遺伝子の開始コドンを含む領域の5’末端にNcoI認識配列を含む塩基配列を付加したものである。プライマーBは、albC遺伝子の終止コドンを含む配列と相補的な塩基配列の5’末端にBglII認識配列を含む塩基配列を付加したものである。
【0103】
上記のプライマーAおよびプライマーBをプライマーセットに、鋳型としてストトレプトマイセス・ノウルセイまたはストレプトマイセス・アルボラスの染色体DNAを用いてPCRを行った。PCRは、鋳型として0.1μgの染色体DNA、0.5μmol/Lの各プライマー、2.5unitsのEx Taq DNAポリメラーゼ(タカラバイオ社製)、5μLのExTaqDNAポリメラーゼ用×10緩衝液(タカラバイオ社製)、各200μmol/LのdNTP(dATP、dGTP、dCTPおよびdTTP)、5μLのジメチルスルホキシドを含む反応液50μLを調製し、94℃で1分間、55℃で30秒間、72℃で1分間の工程を30回繰り返すことにより行った。
【0104】
該反応液の1/10量をアガロースゲル電気泳動し、約0.7kbのDNA断片が増幅していることを確認した後、残りの反応液と等量のTE[10mmol/L Tris−HCl(pH8.0)、1mmol/L EDTA]飽和フェノール/クロロホルム(1vol/1vol)溶液を添加し、混合した。該溶液を遠心分離して得られた上層に、2倍容量の冷エタノールを加えて混合し、−80℃に30分間放置した。該溶液を遠心分離してDNAを沈殿させた後、該DNAをそれぞれ20μLのTEに溶解した。
【0105】
該溶解液それぞれ5μLを用い、増幅したDNAを制限酵素NcoIおよびBglIIで切断し、アガロースゲル電気泳動によりDNA断片を分離した後、ジーンクリーンIIキット(GENECLEANII kit、BIO 101社製)を用いて、700bpのDNA断片をそれぞれ回収した。
ファージT5プロモーターを含む発現ベクターpQE60(キアゲン社製)0.2μgを制限酵素NcoIおよびBglIIで切断後、アガロースゲル電気泳動によりDNA断片を分離し、上記と同様の方法により3.4kbのDNA断片を回収した。
【0106】
上記で得られた各放線菌由来の0.7kb断片およびpQE60由来の3.4kbの断片をライゲーションキット(タカラバイオ社製)を用いて、16℃で16時間反応させ連結した。
該反応液を用いてエシェリヒア・コリNM522株(ストラタジーン社製)を、カルシウムイオンを用いる方法[Proc.Natl.Acad.Sci.,USA,69,2110(1972)]によって形質転換し、該形質転換体を50μg/mlのアンピシリンを含むLB寒天培地[10g/lバクトトリプトン(ディフコ社製)、5g/lイーストエキス(ディフコ社製)、5g/l塩化ナトリウム、寒天20g/l]に塗布した後、30℃で一晩培養した。
【0107】
生育してきた形質転換体のコロニーより公知の方法に従ってプラスミドを抽出し、制限酵素を用いてその構造を解析することにより、ファージT5プロモーター下流にストレプトマイセス・ノウルセイ由来のDNAが連結された発現ベクターであるpAL−nou、およびストレプトマイセス・アルブラス由来のDNAが連結された発現ベクターであるpAL−albが取得されていることを確認した(図1)。
【0108】
それぞれのプラスミドに挿入された放線菌由来のDNA部分の塩基配列を塩基配列決定装置373A・DNAシークエンサーを使って決定したところ、pAL−albには配列番号1で表されるアミノ酸配列を有する蛋白質をコードするDNA、すなわち配列番号3で表される塩基配列を有するDNAが含有され、pAL−nouには配列番号2で表されるアミノ酸配列を有する蛋白質をコードするDNA、すなわち配列番号4で表される塩基配列を有するDNAが含有されていることを確認した。
【実施例2】
【0109】
菌体を酵素源に用いたジペプチドの製造
実施例1で得られたpAL−nouまたはpAL−albを保有するエシェリヒア・コリNM522(エシェリヒア・コリNM522/pAL−nou株またはNM522/pAL−alb株)およびプラスミドを保有しないNM522株を50μg/mlのアンピシリンを含む10mlのLB培地[10g/lバクトトリプトン(ディフコ社製)、5g/lイーストエキス(ディフコ社製)、5g/l塩化ナトリウム]が入った試験管(プラスミドを持たない株の場合にはアンピシリンは無添加、以下同様)に接種し、30℃で17時間培養した。この培養液0.5mlを50mlのLB培地が入った250ml容の三角フラスコにそれぞれ植菌し、30℃で1時間振とう培養した後、イソプロピル−β−D−チオガラクトピラノシド(IPTG)を終濃度1mmol/lになるように添加し、さらに4時間培養を継続した。該培養液を遠心分離し、湿菌体を取得した。
【0110】
終濃度100mg/mlの該湿菌体、60mmol/Lのリン酸カリウムバッファー(pH7.2)、10mmol/Lの塩化マグネシウム、10mmol/LのATP、1g/LのL−Leu、1g/LのL−Pheからなる3.0mlの反応液を調製し、30℃で反応を行った。反応1時間後にサンプリングし、アセトニトリルを20%(v/v)になるように加えた後、反応生成物を以下のHPLC分析により分析した。
分離カラム:ODS−HAカラム(YMC社製)
溶離液:30%(v/v)アセトニトリル
流動速度:0.6ml/min
検出:紫外吸収215nm
その結果、エシェリヒア・コリNM522/pAL−nuo株の反応液中には36.7mg/lのcyclo(L−Leu−L−Phe)の蓄積が確認されたが、エシェリヒア・コリNM522株の反応液中にはcyclo(L−Leu−L−Phe)はまったく検出されなかった。同じ反応液を以下の条件でHPLCによって分析し、直鎖状のジペプチドであるL−ロイシル−L−フェニルアラニン(L−Leu−L−Phe)およびL−フェニルアラニル−L−ロイシン(L−Phe−L−Leu)を測定した。
【0111】
直鎖状のジペプチドはF−moc化法で誘導体化した後にHPLC分析により分析した。
分離カラム:野村化学社製のODS−HG−5カラム
溶離液:
A液−6ml/L酢酸、20%(v/v)アセトニトリル(トリエチルアミンでpH4.8に調整)
B液−6ml/L酢酸、70%(v/v)アセトニトリル(トリエチルアミンでpH4.8に調整)
流動速度:0.6ml/min
検出:励起波長254nm、蛍光波長630nm
溶離液混合比:表1のとおり
【0112】
【表1】
その結果、エシェリヒア・コリNM522/pAL−nuo株の反応液中には21.9mg/lのL−Leu−L−Pheと12.0mg/lのL−Phe−L−Leuが蓄積していることが確認された。また対照株として用いたエシェリヒア・コリNM522株の反応液中にはいずれの直鎖ジペプチドも検出されなかった。このことから、本発明で取得されたシクロジペプチド合成酵素は直鎖状のジペプチドを合成する能力をもつことが明らかになった。
【実施例3】
【0113】
精製酵素を用いたジペプチドの製造(1)
実施例2と同様にエシェリヒア・コリNM522/pAL−nuo株を培養した。培養終了後、遠心分離によって湿菌体を取得し、60mmol/lのリン酸カリウムバッファー(pH7.2)で洗浄後、10mmol/lイミダゾール含有20mmol/lリン酸カリウムバッファーに懸濁した。この懸濁を4℃で超音波処理して菌体破砕液を取得した。この菌体破砕液(10ml、蛋白質0.863mgを含む)をアマシャム社製Hisタグ精製カラムに通塔し、10mmol/lイミダゾール含有20mmol/lリン酸カリウムバッファー15mlを通塔することによる洗浄を行い、HisタグつきalbC蛋白質をカラム内にて精製した。次にこのHisタグ付きのalbC蛋白質を保持するカラムに、実施例2と同組成の反応液(反応液組成:60mmol/Lのリン酸カリウムバッファー(pH7.2)、10mmol/Lの塩化マグネシウム、10mmol/LのATP、1g/LのL−Leu、1g/LのL−Pheからなる反応液)2mlを通塔し、カラム内に基質を保持した状態で、30℃にて、インキュベートした。24時間後、同組成の反応液3mlでカラム内の反応液を溶出し、実施例2と同様の方法により反応液中のシクロジペプチドおよびジペプチドを定量した。
【0114】
その結果、cyclo(L−Leu−L−Phe)6.8mg/L、L−Leu−L−Phe 28.7mg/LおよびL−Phe−L−Leu 18.5mg/Lが生成していることが分かった。ATPを含まない反応液で同様にインキュベートした場合は、シクロジペプチド、ジペプチドともに検出されなかった。
【実施例4】
【0115】
精製酵素を用いたジペプチドの製造(2)
基質のアミノ酸を他のアミノ酸に換える以外は実施例3と同様の方法で酵素反応を行い、生成物を分析した。反応液は、基質のアミノ酸を、1g/LのL−Ala、L−LeuまたはL−Pheに置き換えた以外は実施例2と同じ組成の溶液を用いた。
その結果、反応開始後24時間で、それぞれ9.41mg/LのL−Ala−L−Ala、7.85mg/LのL−Leu−L−Leu、または5.20mg/LのL−Phe−L−Pheが生成していることが分かった。
【配列表フリーテキスト】
【0116】
配列番号5−人工配列の説明:合成DNA
配列番号6−人工配列の説明:合成DNA
【特許請求の範囲】
【請求項1】
以下の[1]〜[3]のいずれかに記載の蛋白質(ただし、配列番号1で表されるアミノ酸配列からなる蛋白質を除く)。
[1]配列番号2で表されるアミノ酸配列を有する蛋白質
[2]配列番号1または2で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドの合成活性を有する蛋白質
[3]配列番号1または2で表されるアミノ酸配列と65%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有する蛋白質
【請求項2】
以下の[1]〜[3]のいずれかに記載のジペプチド合成用蛋白質。
[1]配列番号1で表されるアミノ酸配列を有するジペプチド合成用蛋白質
[2]配列番号1で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)
で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質
[3]配列番号1で表されるアミノ酸配列と65%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質
【請求項3】
以下の[1]〜[3]のいずれかに記載のDNA(ただし、配列番号3で表される塩基配列からなるDNAを除く)。
[1]請求項1記載の蛋白質をコードするDNA
[2]配列番号4で表される塩基配列を有するDNA
[3]配列番号4で表される塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチド合成活性を有する蛋白質をコードするDNA
【請求項4】
請求項3記載のDNAを含有する組換え体DNA。
【請求項5】
請求項4記載の組換え体DNAを有する形質転換体。
【請求項6】
形質転換体が、微生物を宿主として得られる形質転換体である請求項5記載の形質転換体。
【請求項7】
微生物が、エシェリヒア(Escherichia)属に属する微生物である請求項6記載の形質転換体。
【請求項8】
請求項5〜7のいずれか1項に記載の形質転換体を培地に培養し、培養物中に請求項1記載の蛋白質を生成、蓄積させ、該培養物より該蛋白質を採取する請求項1記載の蛋白質の製造法。
【請求項9】
請求項1記載の蛋白質を生産する能力を有する微生物を培地に培養し、培養物中に該蛋白質を生成、蓄積させ、該培養物より該蛋白質を採取する請求項1記載の蛋白質の製造法。
【請求項10】
微生物がストレプトマイセス(Streptomyces)属に属する微生物である請求項9記載の製造法。
【請求項11】
ストレプトマイセス属に属する微生物が、アルボノルシンを生産する能力を有するストレプトミセス属に属する微生物である請求項10記載の製造法。
【請求項12】
アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物が、ストレプトマイセス・アルボラス(Streptomyces alborus)またはストレプトマイセス・ノウルセイ(Streptomyces noursei)である請求項11記載の製造法。
【請求項13】
請求項1記載の蛋白質または請求項2記載のジペプチド合成用蛋白質、1種以上のアミノ酸、およびATPを水性媒体中に存在せしめ、該媒体中に式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法。
【請求項14】
以下の[1]〜[3]から選ばれる培養物または該培養物の処理物を酵素源に用い、該酵素源、1種以上のアミノ酸を水性媒体中に存在せしめ、該媒体中に式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表される直鎖ジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法。
[1]請求項5〜7のいずれか1項に記載の形質転換体の培養物または該培養物の処理物
[2]請求項1記載の蛋白質を生産する能力を有する微生物の培養物または該培養物の処理物
[3]請求項2記載のジペプチド合成用蛋白質を生産する能力を有する微生物の培養物または該培養物の処理物
【請求項15】
請求項1記載の蛋白質を生産する能力を有する微生物が、ストレプトマイセス属に属する微生物である請求項14記載の製造法。
【請求項16】
請求項2記載のジペプチド合成用蛋白質を生産する能力を有する微生物が、ストレプトマイセス属に属する微生物である請求項14記載の製造法。
【請求項17】
ストレプトマイセス属に属する微生物が、アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物である請求項15または16記載の製造法。
【請求項18】
アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物が、ストレプトマイセス・アルボラス(Streptomyces alborus)またはストレプトマイセス・ノウルセイ(Streptomyces noursei)に属する微生物である請求項17記載の製造法。
【請求項19】
請求項2記載のジペプチド合成用蛋白質を生産する能力を有する微生物が、請求項2記載のジペプチド合成用蛋白質をコードするDNAで形質転換された微生物である請求項14記載の製造法。
【請求項20】
請求項2記載のジペプチド合成用蛋白質をコードするDNAで形質転換された微生物が、エシェリヒア属に属する微生物である請求項19記載の製造法。
【請求項21】
培養物の処理物が、培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の溶媒処理物、該菌体の酵素処理物、該菌体の蛋白質分画物、該菌体の固定化物あるいは該菌体より抽出して得られる酵素標品であることを特徴とする、請求項14〜20のいずれか1項に記載の製造法。
【請求項22】
1種以上のアミノ酸が、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体である請求項13〜21のいずれか1項に記載の製造法。
【請求項23】
ジペプチドが式(II)
R3−R4 (II)
(式中、R3およびR4は同一または異なって、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体を表す)で表されるジペプチドである請求項13〜22のいずれか1項に記載の製造法。
【請求項24】
L体またはD体のアミノ酸が、アラニン、グルタミン、グルタミン酸、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、トリプトファン、メチオニン、セリン、スレオニン、システイン、アスパラギン、チロシン、リジン、アルギニン、ヒスチジン、アスパラギン酸、α−アミノ酪酸、アザセリン、テアニン、4−ヒドロキシプロリン、3−ヒドロキシプロリン、オルニチン、シトルリンおよび6−ジアゾ−5−オキソノルロイシンから選ばれるアミノ酸である請求項22または23記載の製造法。
【請求項1】
以下の[1]〜[3]のいずれかに記載の蛋白質(ただし、配列番号1で表されるアミノ酸配列からなる蛋白質を除く)。
[1]配列番号2で表されるアミノ酸配列を有する蛋白質
[2]配列番号1または2で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドの合成活性を有する蛋白質
[3]配列番号1または2で表されるアミノ酸配列と65%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有する蛋白質
【請求項2】
以下の[1]〜[3]のいずれかに記載のジペプチド合成用蛋白質。
[1]配列番号1で表されるアミノ酸配列を有するジペプチド合成用蛋白質
[2]配列番号1で表されるアミノ酸配列において、1以上のアミノ酸が欠失、置換または付加したアミノ酸配列からなり、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)
で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質
[3]配列番号1で表されるアミノ酸配列と65%以上の相同性を有するアミノ酸配列からなり、かつ式(I)で表されるジペプチドの合成活性を有するジペプチド合成用蛋白質
【請求項3】
以下の[1]〜[3]のいずれかに記載のDNA(ただし、配列番号3で表される塩基配列からなるDNAを除く)。
[1]請求項1記載の蛋白質をコードするDNA
[2]配列番号4で表される塩基配列を有するDNA
[3]配列番号4で表される塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチド合成活性を有する蛋白質をコードするDNA
【請求項4】
請求項3記載のDNAを含有する組換え体DNA。
【請求項5】
請求項4記載の組換え体DNAを有する形質転換体。
【請求項6】
形質転換体が、微生物を宿主として得られる形質転換体である請求項5記載の形質転換体。
【請求項7】
微生物が、エシェリヒア(Escherichia)属に属する微生物である請求項6記載の形質転換体。
【請求項8】
請求項5〜7のいずれか1項に記載の形質転換体を培地に培養し、培養物中に請求項1記載の蛋白質を生成、蓄積させ、該培養物より該蛋白質を採取する請求項1記載の蛋白質の製造法。
【請求項9】
請求項1記載の蛋白質を生産する能力を有する微生物を培地に培養し、培養物中に該蛋白質を生成、蓄積させ、該培養物より該蛋白質を採取する請求項1記載の蛋白質の製造法。
【請求項10】
微生物がストレプトマイセス(Streptomyces)属に属する微生物である請求項9記載の製造法。
【請求項11】
ストレプトマイセス属に属する微生物が、アルボノルシンを生産する能力を有するストレプトミセス属に属する微生物である請求項10記載の製造法。
【請求項12】
アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物が、ストレプトマイセス・アルボラス(Streptomyces alborus)またはストレプトマイセス・ノウルセイ(Streptomyces noursei)である請求項11記載の製造法。
【請求項13】
請求項1記載の蛋白質または請求項2記載のジペプチド合成用蛋白質、1種以上のアミノ酸、およびATPを水性媒体中に存在せしめ、該媒体中に式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表されるジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法。
【請求項14】
以下の[1]〜[3]から選ばれる培養物または該培養物の処理物を酵素源に用い、該酵素源、1種以上のアミノ酸を水性媒体中に存在せしめ、該媒体中に式(I)
R1−R2 (I)
(式中、R1およびR2は、同一または異なってアミノ酸を表す)で表される直鎖ジペプチドを生成、蓄積させ、該媒体から該ジペプチドを採取する該ジペプチドの製造法。
[1]請求項5〜7のいずれか1項に記載の形質転換体の培養物または該培養物の処理物
[2]請求項1記載の蛋白質を生産する能力を有する微生物の培養物または該培養物の処理物
[3]請求項2記載のジペプチド合成用蛋白質を生産する能力を有する微生物の培養物または該培養物の処理物
【請求項15】
請求項1記載の蛋白質を生産する能力を有する微生物が、ストレプトマイセス属に属する微生物である請求項14記載の製造法。
【請求項16】
請求項2記載のジペプチド合成用蛋白質を生産する能力を有する微生物が、ストレプトマイセス属に属する微生物である請求項14記載の製造法。
【請求項17】
ストレプトマイセス属に属する微生物が、アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物である請求項15または16記載の製造法。
【請求項18】
アルボノルシンを生産する能力を有するストレプトマイセス属に属する微生物が、ストレプトマイセス・アルボラス(Streptomyces alborus)またはストレプトマイセス・ノウルセイ(Streptomyces noursei)に属する微生物である請求項17記載の製造法。
【請求項19】
請求項2記載のジペプチド合成用蛋白質を生産する能力を有する微生物が、請求項2記載のジペプチド合成用蛋白質をコードするDNAで形質転換された微生物である請求項14記載の製造法。
【請求項20】
請求項2記載のジペプチド合成用蛋白質をコードするDNAで形質転換された微生物が、エシェリヒア属に属する微生物である請求項19記載の製造法。
【請求項21】
培養物の処理物が、培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の溶媒処理物、該菌体の酵素処理物、該菌体の蛋白質分画物、該菌体の固定化物あるいは該菌体より抽出して得られる酵素標品であることを特徴とする、請求項14〜20のいずれか1項に記載の製造法。
【請求項22】
1種以上のアミノ酸が、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体である請求項13〜21のいずれか1項に記載の製造法。
【請求項23】
ジペプチドが式(II)
R3−R4 (II)
(式中、R3およびR4は同一または異なって、L体もしくはD体のアミノ酸、グリシン、β−アラニンまたはそれらの誘導体を表す)で表されるジペプチドである請求項13〜22のいずれか1項に記載の製造法。
【請求項24】
L体またはD体のアミノ酸が、アラニン、グルタミン、グルタミン酸、バリン、ロイシン、イソロイシン、プロリン、フェニルアラニン、トリプトファン、メチオニン、セリン、スレオニン、システイン、アスパラギン、チロシン、リジン、アルギニン、ヒスチジン、アスパラギン酸、α−アミノ酪酸、アザセリン、テアニン、4−ヒドロキシプロリン、3−ヒドロキシプロリン、オルニチン、シトルリンおよび6−ジアゾ−5−オキソノルロイシンから選ばれるアミノ酸である請求項22または23記載の製造法。
【図1】

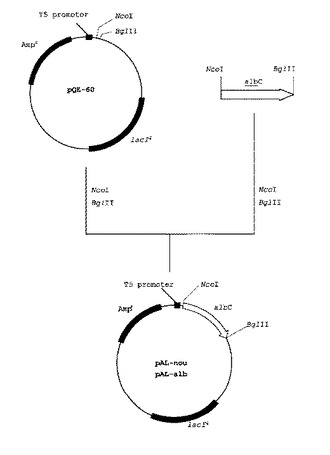
【国際公開番号】WO2005/103260
【国際公開日】平成17年11月3日(2005.11.3)
【発行日】平成19年8月16日(2007.8.16)
【国際特許分類】
【出願番号】特願2006−512590(P2006−512590)
【国際出願番号】PCT/JP2005/007626
【国際出願日】平成17年4月21日(2005.4.21)
【出願人】(000001029)協和醗酵工業株式会社 (276)
【Fターム(参考)】
【国際公開日】平成17年11月3日(2005.11.3)
【発行日】平成19年8月16日(2007.8.16)
【国際特許分類】
【国際出願番号】PCT/JP2005/007626
【国際出願日】平成17年4月21日(2005.4.21)
【出願人】(000001029)協和醗酵工業株式会社 (276)
【Fターム(参考)】
[ Back to top ]