
スタヒロコッカス・アウレウスORF0657Nを標的とする抗原結合タンパク質
本発明は、ORF0657n標的領域(配列番号1)に結合する抗原結合タンパク質に関する。ORF0657nはスタヒロコッカス・アウレウス(S.aureus)タンパク質である。ORF0657n標的領域はmAb lG3.BD4、mAb 2H2.BE11、mAb 13C7.BClおよびmAb 13G11.BF3結合部位により与えられる。致死モデルチャレンジにおいて、mAb 2H2.BE11およびmAb 13C7.BC1はスタヒロコッカス・アウレウス(S.aureus)感染に対する生存性の上昇をもたらした。また、mAb 2H2のIgG1またはIgG2b形態を使用するエクスビボモデルにおいて、およびmAb 2H2.BE11を使用する受動免疫マウス留置カテーテルモデルにおいて、防御が実証された。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、スタヒロコッカス・アウレウス(S.aureus)ORF0657nを標的とする抗原結合タンパク質に関する。
【背景技術】
【0002】
本出願の全体にわたって引用されている参考文献は、特許請求されている本発明の先行技術であると自認するものではない。
【0003】
スタヒロコッカス・アウレウス(Staphylococcus aureus)は、多種多様な疾患および状態を引き起こす病原体である。スタヒロコッカス・アウレウス(S.aureus)により引き起こされる疾患および状態の具体例には、菌血症、感染性心内膜炎、毛包炎、フルンケル、カルブンケル、膿痂疹、水疱性膿痂疹、蜂窩織炎、ボトリオミセス症、中毒性ショック症候群、熱傷様皮膚症候群、中枢神経系感染症、感染性および炎症性眼疾患、骨髄炎ならびに関節および骨の他の感染症、ならびに気道感染症が含まれる(The Staphylococci in Human Disease,Crossley and Archer(編),Churchill Livingstone Inc.1997)。
【0004】
スタヒロコッカス・アウレウス(S.aureus)感染およびスタヒロコッカス・アウレウス(S.aureus)の広がりを抑制するためには、免疫学に基づく方法が用いられうる。免疫学に基づく方法には受動免疫化および能動免疫化が含まれる。受動免疫化は、スタヒロコッカス・アウレウス(S.aureus)を標的とする免疫グロブリンを使用する。能動免疫化はスタヒロコッカス・アウレウス(S.aureus)に対する免疫応答を誘導する。
【発明の開示】
【0005】
発明の概要
本発明は、ORF0657n標的領域(配列番号1)に結合する抗原結合タンパク質に関する。ORF0657nはスタヒロコッカス・アウレウス(S.aureus)タンパク質である。ORF0657n標的領域はmAb lG3.BD4、mAb 2H2.BE11、mAb 13C7.BClおよびmAb 13G11.BF3結合部位により与えられる。致死モデルチャレンジにおいて、mAb 2H2.BE11およびmAb 13C7.BC1はスタヒロコッカス・アウレウス(S.aureus)感染に対する生存性の上昇をもたらした。また、mAb 2H2のIgG1またはIgG2b形態を使用するエクスビボモデルにおいて、およびmAb 2H2.BE11を使用する受動免疫マウス留置カテーテルモデルにおいても、防御が実証された。
【0006】
mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1およびmAb 13G11.BF3を産生するマウスハイブリドーマ細胞系は、ブダペスト条約に従い、2005年9月30日付けでAmerican Type Culture Collection, 10801 University Boulevard,Manassas,VA 20110−2209に寄託された。該細胞系にはATCC番号PTA−7124(mAb 2H2.BE11を産生する)、ATCC番号PTA−7125(mAb 13C7.BC1を産生する)、ATCC番号PTA−7126(mAb 1G3.BD4を産生する)およびATCC番号PTA−7127(mAb 13G11.BF3を産生する)が付与された。
【0007】
したがって、本発明の第1の態様は、第1可変領域および第2可変領域を含んでなる単離された抗原結合タンパク質に関する。第1および第2可変領域は、mAb 1G3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1標的領域およびmAb 13G11.BF3標的領域よりなる群から選ばれる1以上の標的領域に結合する。
【0008】
「単離(された)」に対する言及は、天然で見出されるものとは異なる形態を示す。そのような異なる形態は、例えば、天然で見出されるものとは異なる純度および/または天然で見出されない構造体でありうる。天然で見出されない構造体には、異なる領域が一緒になった組換え構造体、例えば、1以上のマウス相補性決定領域がヒトフレームワークスカフォールド上に挿入された又はヒト抗体の表面残基を模倣するためにマウス抗体の表面が修飾されたヒト化抗体、抗原結合タンパク質由来の1以上の相補性決定領域が別のフレームワークスカフォールド内に挿入されたハイブリッド抗体、ならびに軽可変ドメインおよび重可変ドメインをコードする遺伝子が互いにランダムに組合わされた天然ヒト配列に由来する抗体が含まれる。
【0009】
単離されたタンパク質は、好ましくは、血清タンパク質を実質的に含有しない。血清タンパク質を実質的に含有しないタンパク質は、ほとんど又は全ての血清タンパク質を欠く環境中に存在する。
【0010】
「可変領域」は重鎖または軽鎖からの抗体可変領域の構造を有する。抗体重鎖および軽鎖の可変領域は、フレームワーク上に間隔をあけて存在する3つの相補性決定領域を含有する。該相補性決定領域は主として、特定のエピトープの認識をもたらす。
【0011】
標的領域は、mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1またはmAb 13G11.BF3が結合するORF0657n領域(配列番号1)に関して定義される。例えば、mAb IG3.BD4標的領域は、mAb IG3.BD4が結合するORF0657n領域である。
【0012】
特定された標的領域に結合するタンパク質は該標的領域への結合に関してmAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1またはmAb 13G11.BF3と競合する。例えば、ORF0657nに結合するmAb 1G3.BD4と競合するタンパク質は、mAb 1G3.BD4標的領域に結合する。
【0013】
モノクローナル抗体mAb 1G3.B3、mAb 2H2.B8、mAb 13C7.D12またはmAb 13G11.C11のいずれかと競合するタンパク質は、過剰量および等量の該競合タンパク質およびモノクローナル抗体を使用した場合、ORF0657nへの該モノクローナル抗体の結合を少なくとも約20%、好ましくは少なくとも約50%軽減する。
【0014】
「タンパク質」に対する言及は連続的アミノ酸配列を示し、最小または最大サイズ限界を与えるものではない。該タンパク質中に存在する1以上のアミノ酸はグリコシル化またはジスルフィド結合形成のような翻訳後修飾を含有しうる。
【0015】
好ましい抗原結合タンパク質はモノクローナル抗体である。「モノクローナル抗体」に対する言及は、同じ又は実質的に同じ相補性決定領域および結合特異性を有する一群の抗体を示す。該抗体における変異は、該抗体が同一構築物から製造された場合に生じるものである。
【0016】
モノクローナル抗体は、例えば、特定のハイブリドーマから、および該抗体をコードする1以上の組換え遺伝子を含有する組換え細胞から製造されうる。該抗体は2以上の組換え遺伝子によりコードされることが可能であり、この場合、例えば、1つの遺伝子は重鎖をコードし、1つの遺伝子は軽鎖をコードする。
【0017】
本発明のもう1つの態様は、抗体可変領域をコードするヌクレオチド配列を含む組換え遺伝子を含有する核酸を記載する。該抗体可変領域は、mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合しうる。
【0018】
組換え遺伝子は、適切な転写およびプロセシングのための調節要素(これは翻訳および翻訳後要素を含みうる)と共にタンパク質をコードする組換え核酸を含有する。該組換え核酸は、その配列および/または形態の点で、天然では見出されない。組換え核酸の具体例には、精製された核酸、天然で見出されるものとは異なる核酸を与える一緒になった2以上の核酸領域、互いに天然で付随している1以上の核酸領域(例えば、上流または下流領域)の非存在が含まれる。
【0019】
本発明のもう1つの態様は、mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合する抗体可変領域をコードする1以上の組換え遺伝子を含んでなる組換え細胞を記載する。複数の組換え遺伝子が有用であり、例えば、この場合、1つの遺伝子は抗体重鎖またはそのフラグメント(Vh領域を含有するもの)をコードし、もう1つの核酸は抗体軽鎖またはそのフラグメント(Vl領域を含有するもの)をコードする。
【0020】
本発明のもう1つの態様は、抗体可変領域を含むタンパク質の製造方法を含む。該方法は、(a)該タンパク質が発現される条件下、タンパク質をコードする組換え核酸を含む組換え細胞を増殖させ、(b)該タンパク質を精製する段階を含む。
【0021】
本発明のもう1つの態様は医薬組成物を記載する。該組成物は治療的有効量の抗原結合タンパク質および医薬上許容される担体を含有する。
【0022】
治療的有効量は、有用な治療効果または予防効果をもたらすのに十分な量である。スタヒロコッカス・アウレウス(S.aureus)に感染した患者の場合、有効量は、以下の効果の1以上を達成するのに十分な量である:該患者におけるスタヒロコッカス・アウレウス(S.aureus)の増殖能の軽減、または該患者におけるスタヒロコッカス・アウレウス(S.aureus)の量の減少。スタヒロコッカス・アウレウス(S.aureus)に感染していない患者の場合、有効量は、以下の1以上を達成するのに十分な量である:スタヒロコッカス・アウレウス(S.aureus)感染に対する感受性の軽減、または慢性疾患を招く持続感染を該感染細菌が確立する能力の軽減。
【0023】
本発明のもう1つの態様は、溶液中または細胞上のOFR0657n抗原の存在の検出方法を記載する。該方法は、本明細書に記載の結合タンパク質を該溶液または細胞に加え、該溶液または細胞中の該抗原への該結合タンパク質の結合能を測定することを含む。測定は定量的または定性的でありうる。
【0024】
ORF0657n抗原に対する言及は、完全長ORF0657n、またはmAb 1G3.B3、mAb 2H2.B8、mAb 13C7.D12またはmAb 13G11.C11により認識されるエピトープを有するその誘導体を含む。誘導体の具体例には、トランケート化形態;および以下のアミノ酸改変の1以上を含有するORF0657nの完全長またはトランケート化形態が含まれる:1以上の付加、1以上の置換および1以上の欠失。
【0025】
本発明のもう1つの態様は、スタヒロコッカス・アウレウス(S.aureus)感染に対する患者の治療方法に関する。該方法は、本明細書に記載の抗原結合タンパク質の有効量を患者に投与する段階を含む。治療される患者はスタヒロコッカス・アウレウス(S.aureus)に感染していても感染していなくてもよい。好ましくは、患者はヒトである。
【0026】
本発明のもう1つの態様は、mAb 1G3.B3、mAb 2H2.B8、mAb 13C7.D12もしくはmAb 13G11.C11のいずれかである又はORF0657nへの結合に関してmAb IG3.B3、mAb 2H2.B8、mAb 13C7.D12もしくはmAb 13G11.C11のいずれかと競合するタンパク質を産生する細胞系を記載する。好ましい細胞系は、該タンパク質をコードする組換え核酸を含有する組換え細胞系およびハイブリドーマである。
【0027】
「含む(含んでなる)」のような非限定的用語に対する言及は追加的な要素または段階を許容する。場合によっては、「1以上」のような表現は、追加的な要素または段階の可能性を強調するために非限定的用語と共に又はそれを伴わずに用いられる。
【0028】
明示的に示されていない限り、単数表現は単数を示すと限定されるものではない。例えば、「細胞」は複数の細胞を除外するものではない。場合によっては、可能な複数の存在を強調するために、1以上のような表現が用いられる。
【0029】
本発明の他の特徴および利点は、種々の実施例(具体例)を含む本明細書に記載の更なる説明から明らかである。記載されている実施例(具体例)は、本発明の実施に有用な種々の成分および方法を例示する。該実施例(具体例)は、特許請求されている本発明を限定するものではない。本開示に基づき、当業者は、本発明の実施に有用な他の成分および方法を特定し使用することが可能である。
【0030】
発明の詳細な説明
ORF0657nは、スタヒロコッカス・アウレウス(S.aureus)の外膜に位置するスタヒロコッカス・アウレウス(S.aureus)タンパク質である。ORF0657nはスタヒロコッカス・アウレウス(S.aureus)の種々の株において良く保存されていることが判明している(Andersonら,国際公開番号WO 2005/009379、国際公開日2005年2月3日)。スタヒロコッカス・アウレウス(S.aureus)感染に対する防御免疫応答を引き起こすために、種々のORF0657n誘導体が使用されうる(Andersonら,国際公開番号WO 2005/009379、国際公開日2005年2月3日)。
【0031】
本明細書に記載の抗原結合タンパク質は、ORF0657nを認識するそれらの能力により、例えば、ORF0657nに基づく抗原の製造、特徴づけまたは研究における手段として使用されうる。適当なORF0657nエピトープを認識する抗原結合タンパク質は、スタヒロコッカス・アウレウス(S.aureus)感染を治療するための物質としても使用されうる。
【0032】
I.抗原結合タンパク質
抗原結合タンパク質は、エピトープへの特異的結合をもたらす抗体可変領域を含有する。該抗体可変領域は、例えば、完全抗体、抗体フラグメント、および抗体または抗体フラグメントの組換え誘導体において存在しうる。
【0033】
抗体は、そのクラスによって異なる構造を有する。IgGに関して、種々の抗体領域が例示されうる(図1)。IgG分子は4つのアミノ酸鎖(2つの、より長い重鎖、および2つの、より短い軽鎖)を含有する。重鎖および軽鎖はそれぞれ、定常領域および可変領域を含有する。該可変領域内には、抗原特異性をもたらす3つの超可変領域が存在する(例えば、Breitlingら,Recombinant Antibodies,John Wiley & Sons,Inc.およびSpektrum Akademischer Verlag,1999;およびLewin,Genes IV,Oxford University Press and Cell Press,1990を参照されたい)。
【0034】
該超可変領域(相補性決定領域とも称される)は、より保存されたフランキング領域(フレームワーク領域とも称される)の間に介在している。フレームワーク領域および相補性決定領域に関連したアミノ酸は、Kabatら,Sequences of Proteins of Immunological Interest,U.S.Department of Health and Human Services,1991に記載されているとおりに番号付けされ、整列(アライン)されうる。
【0035】
2つの重鎖カルボキシル領域は、Fc領域を形成するようジスルフィド結合により連結された定常領域である。Fc領域は、補体およびマクロファージの活性化のような抗体の生物活性をもたらすのに重要である。Fc領域を構成する2つの重鎖のそれぞれはヒンジ領域を介して異なるFab領域へと伸長している。
【0036】
高等脊椎動物においては、2つのクラスの軽鎖および5つのクラスの重鎖が存在する。軽鎖はκまたはλである。重鎖は抗体クラスを定め、α、δ、ε、γまたはμである。例えば、IgGはγ重鎖を有する。種々のタイプの重鎖に関しては、ヒトγ1、γ2、γ3およびγ4のようなサブクラスも存在する。重鎖は、ヒンジおよびテイル(尾)領域に、特徴的なコンホメーションをもたらす(Lewin,Genes IV,Oxford University Press and Cell Press,1990)。
【0037】
抗体可変領域を含有する抗体フラグメントには、Fv、FabおよびFab2領域が含まれる。各Fab領域は、可変領域および定常領域から構成される軽鎖と、可変領域および定常領域を含有する重鎖領域とを含有する。軽鎖は定常領域を介してジスルフィド結合により重鎖に連結される。Fab領域の軽鎖および重鎖可変領域は、抗原結合に関与するFv領域を与える。
【0038】
抗体可変領域は組換え誘導体中に存在しうる。組換え誘導体の具体例には、一本鎖抗体、ジアボディ(diabody)、トリアボディ(triabody)、テトラボディ(tetrabody)および小型抗体(miniantibody)が含まれる(Kipriyanovら,Molecular Biotechnology 26:39−60,2004)。
【0039】
該抗原結合タンパク質は、同じ又は異なるエピトープを認識する1以上の可変領域を含有しうる(Kipriyanovら,Molecular Biotechnology 26:39−60,2004)。
【0040】
II.特定された標的領域に対する抗原結合タンパク質の作製
mAb 1G3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1標的領域またはmAb 13G11.BF3標的領域に対する種々の抗原結合タンパク質が、それぞれのモノクローナル抗体から作製されうる。あるいは、結合タンパク質により認識されるエピトープを使用して、追加的な結合タンパク質を選択することが可能である。
【0041】
mAb 2H2.BE11標的領域はORF0657nのアミノ酸約76から357に位置するらしい。抗体を選択するための標的抗原として、ORF0657nのアミノ酸76から357を含有するポリペプチドまたは完全長ORF0657nが使用されうる。作製される抗体の標的領域は決定可能である。
【0042】
抗原を認識するタンパク質を選択するためには、種々の技術が利用可能である。そのような技術の具体例には、ファージ提示技術およびハイブリドーマ産生の利用が含まれる。ヒト抗体は、ゼノマウス(XenoMouse)またはトランスクロモ(Trans−Chromo)マウスのようなキメラマウスを使用して製造されうる(Azzazyら,Clinical Biochemistry 35:425−445,2002,Bergerら,Am.J.Med.Sci.324(1):14−40,2002)。
【0043】
モノクローナル抗体mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1およびmAb 13G11.BF3は、ORF0675nを認識する可変領域を含有する。抗体可変領域に基づき、ORF0657nを認識する追加的な結合タンパク質が製造されうる。追加的な結合タンパク質は、例えば、既存のモノクローナル抗体を修飾することにより、および可変領域配列情報を用いることにより製造されうる。タンパク質の構築および配列の操作は、組換え核酸技術を用いて行われうる。
【0044】
モノクローナル抗体mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1およびmAb 13G11.BF3はマウス抗体である。ヒトの治療用には、そのようなmAbに基づく好ましい結合タンパク質は、該マウス領域を認識するヒト抗マウス抗体の潜在的生成を軽減するよう設計される。
【0045】
ヒト抗マウス抗体の潜在的生成は、マウス抗体のヒト化、脱免疫化(de−immunization)およびキメラ抗体産生のような技術を用いて軽減されうる(例えば、O’Brienら,Humanization of Monoclonal Antibodies by CDR Grafting,p81−100,From Methods in Molecular Biology Vol.207:Recombinant antibodies for Cancer Therapy:Methods and Protocols(WelschofおよびKrauss編)Humana Press,Totowa,New Jersey,2003;Kipriyanovら,Molecular Biotechnology 26:39−60,2004;Gonzalesら,Tumor Biol.26:31−43,2005,Presta,Advanced Drug Delivery Reviews 58:640−656,2006,Tsurushitaら,Methods 36:69−83,2005,Roqueら,Biotechnol.Prog.20:639−654,2004を参照されたい)。
【0046】
マウス抗体は、フレームワーク領域内への相補性決定領域のグラフティングまたはリサーフェシング(resurfacing)のような技術を用いてヒト化されうる。リサーフェシング(ベニアリング(veneering;上張り)としても公知である)は、表面露出領域がヒト化されるよう可変領域を修飾することを含む。
【0047】
相補性決定領域のグラフティングは、そのような領域またはそのような領域の一部を例えばマウス由来物から取り出し該領域をヒト可変領域フレームワーク内に挿入することを含む。グラフティングに使用するヒトフレームワークは、該領域が入手された可変領域(例えば、マウスのもの)に対する配列相同性に基づいて選択されうる。グラフティングされる相補性決定領域に関連する必須フレームワーク残基も新たなフレームワーク内に供給されるべきである。
【0048】
脱免疫化(de−immunization)は、該抗体内に存在する潜在的な直鎖状T細胞エピトープを改変することを含む。該エピトープは、既知のヒトHLAクラスIおよび/またはクラスIIエピトープのバイオインフォマティクススキャンに基づいて特定されうる(Presta,Advanced Drug Delivery Reviews 58:640−656,2006)。
【0049】
キメラ抗体は、ヒト定常領域を、マウスのような異なる生物に由来する可変領域と共に含有する。該ヒト定常領域はFc領域を供与する。
【0050】
改変の追加的な例は、例えば一本鎖抗体、ジアボディ(diabody)、トリアボディ(triabody)、テトラボディ(tetrabody)および小型抗体(miniantibody)における可変領域を提供することを含む(Kipriyanovら,Molecular Biotechnology 26:39−60,2004)。該抗原結合タンパク質は、同じ又は異なるエピトープを認識する1以上の可変領域を含有しうる(同誌)。本発明の追加的な実施形態は、mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1またはmAb 13G11.BF3結合部位に対する一本鎖抗体、ジアボディ(diabody)、トリアボディ(triabody)、テトラボディ(tetrabody)および小型抗体(miniantibody)に関する。
【0051】
III.mAb 2H2.BE11標的領域に対する結合タンパク質
後記の実施例に記載されているとおり、mAb 2H2.BE11標的領域を更に詳細に特徴づけし、該可変領域のアミノ酸配列を決定した。特定されている標的領域および配列情報は、mAb 2H2.BE11標的領域に対する種々の結合タンパク質の入手を促進する。
【0052】
本発明の1つの実施形態においては、該結合タンパク質は、配列番号1のアミノ酸76から357よりなるポリペプチドに結合する。好ましくは、該結合タンパク質は、ヒト抗体、ヒト化抗体、脱免疫化(de−immunized)抗体またはキメラ抗体である。好ましい抗体は、単離された抗体およびモノクローナル抗体である。
【0053】
mAb 2H2.BE11可変領域のアミノ酸配列を配列番号20(Vh)および配列番号21(Vl)に示す。Vh内の相補性決定領域(CDR)はアミノ酸36から45,50から65および98から107において特定された。Vl内のCDRは配列番号21のアミノ酸24から33、49から55および88から96において特定された。
【0054】
Vh領域に関する他の実施形態においては、該結合タンパク質はmAb 2H2.BE11標的領域に結合し、そして、配列番号20のアミノ酸36から45またはアミノ酸36から45とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第1 Vh CDR;配列番号20のアミノ酸50から65またはアミノ酸50から65とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第2 Vh CDR;および配列番号20のアミノ酸98から107またはアミノ酸98から107とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第3 Vh CDRを含む又はそれよりなる又はそれより実質的になる。
【0055】
Vl領域に関する他の実施形態においては、該結合タンパク質はmAb 2H2.BE11標的領域に結合し、そして、配列番号21のアミノ酸24から33またはアミノ酸24から33とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第1 VlCDR;配列番号21のアミノ酸49から55またはアミノ酸49から55とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第2 Vl CDR;および配列番号21のアミノ酸88から96またはアミノ酸88から96とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第3 Vl CDRを含む又はそれよりなる又はそれより実質的になる。
【0056】
可変領域、CDR領域または抗体配列に関する「より実質的になる」なる表現は、標的への結合を有意に低下させない1以上の追加的なアミノ酸の、可能な存在を示す。
【0057】
アミノ酸の相違はアミノ酸の欠失、挿入または置換でありうる。活性を維持するようアミノ酸を置換する場合、該置換アミノ酸は、ほぼ同じ電荷、サイズ、極性および/または疎水性のような1以上の類似特性を有するべきである。
【0058】
好ましくは、アミノ酸の置換は保存的置換である。保存的置換は、あるアミノ酸を、類似特性を有する別のアミノ酸で置換するものである。表1はアミノ酸のグループの一覧を示し、ここで、該グループの1つのアミノ酸は別のアミノ酸で保存的置換される。
【0059】
【表1】
【0060】
他の実施形態においては、該Vh領域は配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20であり、および/または該Vl領域は配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21である。
【0061】
抗体に関する他の実施形態においては、該抗体は、(a)この第III節において記載されているVh領域、ならびにヒトヒンジ、CH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4からのもの)を含む重鎖、ならびに(b)この第III節において前記で記載されているVl領域と、ヒトカッパCLまたはヒトラムダCLとを含む軽鎖を含む又はそれによりなる又はそれより実質的になる。他の実施形態においては、該抗体は、(a)この第III節において記載されているVh領域、ならびにヒトヒンジ、CH1、CH2およびCH3領域(IgG1またはIgG2からのもの)を含む重鎖、ならびに(b)この第III節において前記で記載されているVl領域とヒトカッパCLとを含む軽鎖;ならびに配列番号22のアミノ酸配列より実質的になる重鎖および/または配列番号23のアミノ酸配列より実質的になる軽鎖を含む又はそれよりなる又はそれより実質的になる。
【0062】
他の実施形態においては、本明細書に記載の抗原結合タンパク質は、標的抗原に対する少なくとも約100nM、好ましくは少なくとも約30nMのアフィニティーKDをもたらすVhおよびVl領域を有する。標的抗原への結合は、アミノ酸42から486からのORF0657n断片を使用して、実施例11に記載されているとおりに測定されうる。
【0063】
他の実施形態のための好ましい結合タンパク質は抗体である。より好ましくは、該抗体は単離されている、またはモノクローナル抗体である。
【0064】
IV.タンパク質の製造
抗原結合タンパク質は、好ましくは、組換え核酸技術またはハイブリドーマの使用により製造される。組換え核酸技術は、タンパク質合成のための核酸鋳型を構築することを含む。ハイブリドーマ技術は、該抗原結合タンパク質を製造するために不死化細胞系を使用することを含む。適当な組換え核酸およびハイブリドーマ技術は当技術分野でよく知られている(例えば、Ausubel,Current Protocols in Molecular Biology,John Wiley,2005,Harlowら,Antibodies,A Laboratory Manual,Cold Spring Harbor Laboratory,1988を参照されたい)。
【0065】
抗原結合タンパク質をコードする組換え核酸は、実際に該コード化タンパク質の生産工場として働く宿主細胞内で発現されうる。該組換え核酸は、宿主細胞ゲノムから自律して又は宿主細胞ゲノムの一部として存在する該抗原結合タンパク質をコードする組換え遺伝子を与えうる。
【0066】
組換え遺伝子は、タンパク質の発現のための調節要素と共に、タンパク質をコードする核酸を含有する。一般には、組換え遺伝子内に存在する調節要素には、転写プロモーター、リボソーム結合部位、ターミネーター、および場合によって存在するオペレーターが含まれる。真核細胞内でのプロセシングのための好ましい要素はポリアデニル化シグナルである。抗体に関連するイントロンも存在しうる。抗体または抗体フラグメントの製造のための発現カセットの具体例は当技術分野でよく知られている(例えば、Persicら,Gene 187:9−18,1997,Boelら,J.Immunol.Methods 239:153−166,2000,Liangら,J.Immunol.Methods 247:119−130,2001,Tsurushitaら,Methods 36:69−83,2005)。
【0067】
遺伝暗号の縮重のため、特定のタンパク質をコードするために多数の異なるコード化核酸配列が用いられうる。ほとんど全てのアミノ酸は、異なる組合せのヌクレオチドトリプレット、すなわち「コドン」によりコードされているため、遺伝暗号の縮重が生じる。アミノ酸は、以下のとおりに、コドンによりコードされる。
A=Ala=アラニン:コドンGCA、GCC、GCG、GCU。
C=Cys=システイン:コドンUGC、UGU。
D=Asp=アスパラギン酸:コドンGAC、GAU。
E=Glu=グルタミン酸:コドンGAA、GAG。
F=Phe=フェニルアラニン:コドンUUC、UUU。
G=Gly=グリシン:コドンGGA、GGC、GGG、GGU。
H=His=ヒスチジン:コドンCAC、CAU。
I=Ile=イソロイシン:コドンAUA、AUC、AUU。
K=Lys=リシン:コドンAAA、AAG。
L=Leu=ロイシン:コドンUUA、UUG、CUA、CUC、CUG、CUU。
M=Met=メチオニン:コドンAUG。
N=Asn=アスパラギン:コドンAAC、AAU。
P=Pro=プロリン:コドンCCA、CCC、CCG、CCU。
Q=Gln=グルタミン:コドンCAA、CAG。
R=Arg=アルギニン:コドンAGA、AGG、CGA、CGC、CGG、CGU。
S=Ser=セリン:コドンAGC、AGU、UCA、UCC、UCG、UCU。T=Thr=トレオニン:コドンACA、ACC、ACG、ACU。
V=Val=バリン:コドンGUA、GUC、GUG、GUU。
W=Trp=トリプトファン:コドンUGG。
Y=Tyr=チロシン:コドンUAC、UAU。
【0068】
細胞内の組換え遺伝子の発現は、発現ベクターを使用して促進される。好ましくは、該発現ベクターは、組換え遺伝子に加えて、宿主細胞内での自律的複製のための複製起点、選択マーカー、限られた数の有用な制限酵素部位および潜在的な高コピー数をも含有する。抗体および抗体フラグメントの製造のための発現ベクターの具体例は当技術分野でよく知られている(例えば、Persicら,Gene 187:9−18,1997,Boelら,J.Immunol.Methods 239:153−166,2000,Liangら,J.Immunol Methods 247:119−130,2001,Tsurushitaら,Methods 36:69−83,2005)。
【0069】
所望により、当技術分野でよく知られた技術を用いて(例えば、Ausubel,Current Protocols in Molecular Biology,John Wiley,2005,Marksら,国際出願番号WO95/17516(国際公開日1995年6月29日)を参照されたい)、抗体をコードする核酸を宿主染色体内に組込むことが可能である。
【0070】
組換え抗原結合タンパク質の発現には、原核生物(例えば、大腸菌(E.coli)、バシラス属種(Bacillus sp)およびストレプトマイセス属種(Streptomyces sp.)(または放線菌属))および真核生物(例えば、酵母、バキュロウイルスおよび哺乳類)に由来する細胞系を含む多種多様な細胞系が使用されうる(Breitlingら,Recombinant Antibodies,John Wiley & Sons,Inc.and Spektrum Akademischer Verlag,1999,Kipriyanovら,Molecular Biotechnology 26:39−60,2004,Tsurushitaら,Methods 36:69−83,2005)
【0071】
組換え抗原結合タンパク質の発現のための好ましい宿主は哺乳類翻訳後修飾をもたらす。翻訳後修飾には、化学的修飾、例えばグリコシル化およびジスルフィド結合形成が含まれる。もう1つのタイプの翻訳後修飾はシグナルペプチドの切断である。
【0072】
適切なグリコシル化は抗体機能に重要でありうる(Yooら,Journal of Immunological Methods 261:1−20,2002,Liら,Nature Biotechnology 24(2):210−215,2006)。天然に存在する抗体は、重鎖に結合した少なくとも1つのN−結合炭水化物を含有する(Yooら,Journal of Immunological Methods 261:1−20,2002)。他のN−結合炭水化物およびO−結合炭水化物が存在することが可能であり、抗体機能に重要でありうる(同誌)。
【0073】
効率的な翻訳後修飾を得るために、哺乳類宿主細胞および非哺乳類細胞を含む種々のタイプの宿主細胞が使用されうる。哺乳類宿主細胞の具体例には、チャイニーズハムスター卵巣(CHO)、HeLa、C6、PC12、ヒト胎児腎(HEK293)および骨髄腫細胞が含まれるが、これらに限定されるものではない(Yooら,Journal of Immunological Methods 261:1−20,2002,Persicら,Gene 187:9−18,1997)。非哺乳類細胞は、ヒトグリコシル化を模倣するよう改変されうる(Liら,Nature Biotechnology 24(2):210−215,2006)。糖操作(gIycoenginnered)されたピチア・パストリス(Pichia pastoris)は、そのような改変された非哺乳類細胞の一例である(Liら,Nature Biotechnology 24(2):210−215,2006)。
【0074】
好ましい組換え遺伝子は、mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合する抗体可変領域をコードするヌクレオチド配列を含む。特定の組換え遺伝子は、一方の可変領域またはVhおよびVlの両方の領域を含有するタンパク質をコードしうる。該組換え遺伝子は抗体定常領域およびヒンジ領域をもコードしうる。所望により、1つの遺伝子が軽鎖をコードし第2の遺伝子が重鎖をコードする、組換え遺伝子の組合せを用いて、抗体が製造されうる。
【0075】
他の実施形態は、前記第II節または第III節に記載のタンパク質をコードする核酸により提供される。そのような実施形態の具体例を以下に記載する。
【0076】
Vhコード領域に関する1つの実施形態においては、該ヌクレオチド配列は、配列番号20のアミノ酸36から45またはアミノ酸36から45とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第1 Vh CDR;配列番号20のアミノ酸50から65またはアミノ酸50から65とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第2 Vh CDR;および配列番号20のアミノ酸98から107またはアミノ酸98から107とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第3 Vh CDRを含む又はそれよりなる又はそれより実質的になる可変領域をコードする。
【0077】
Vlコード領域に関する1つの実施形態においては、該ヌクレオチド配列は、配列番号21のアミノ酸24から33またはアミノ酸24から33とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第1 VlCDR;配列番号21のアミノ酸49から55またはアミノ酸49から55とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第2 Vl CDR;および配列番号21のアミノ酸88から96またはアミノ酸88から96とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第3 Vl CDRを含む又はそれよりなる又はそれより実質的になる可変領域をコードする。
【0078】
他の実施形態においては、該Vh領域は配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20であり、および該Vl領域は配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21である。
【0079】
抗体重鎖および/または軽鎖に関する他の実施形態においては、該組換え遺伝子は、(a)前記第III節に記載されているVh領域、ヒトヒンジ、CH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4サブタイプからのもの)を含む重鎖、または(b)前記第III節に記載されているVl領域と、ヒトカッパCLまたはラムダCLとを含む軽鎖を含む又はそれによりなる又はそれより実質的になるタンパク質の一方または両方をコードする。もう1つの実施形態においては、該重鎖は配列番号22のアミノ酸配列より実質的になり、該軽鎖は配列番号23のアミノ酸配列より実質的になる。
【0080】
V.抗原結合タンパク質の用途
あるORF0657n領域を含有する抗原は、スタヒロコッカス・アウレウス(S.aureus)感染に対する防御免疫応答をもたらすために使用されうる(Andersonら,国際公開番号WO2005/009379(国際公開日2005年2月3日))。ORF0657n標的領域を認識する抗原結合タンパク質は、ORF0657n抗原およびワクチンの製造、特徴づけ又は研究を促進するために使用されうる。また、適当なエピトープを認識する抗原結合タンパク質は治療用途を有しうる。
【0081】
ORF0657n関連抗原およびワクチンの製造、特徴づけ又は研究における種々の用途の具体例には以下のものが含まれる。
【0082】
1)例えばウエスタンブロットによる、ORF0657n抗原の存在の特定。
【0083】
2)例えばフローサイトメトリーによる、細胞表面上のORF0657n抗原の存在の特定。これは、例えば、スタヒロコッカス・アウレウス(S.aureus)の複数の株に関する発現の測定、およびノックアウト突然変異体の確認において有用である。
【0084】
3)受動防御実験。該抗体は、ORF0657nタンパク質の特定の領域が防御をもたらすのかどうかを判定するために致死モデルにおいて使用されうる。
【0085】
4)イムノアッセイ。該アッセイは、抗原の質、産物の生産性および安定性をモニターするために用いられうる。
【0086】
5)抗原ワクチン産物の免疫原性をモニターするためのマウス効力アッセイにおける対照として。
【0087】
6)血清学的アッセイにおいて、ORF0657n由来抗原でワクチン接種された患者に対する免疫応答を特定するために、モノクローナル抗体が競合様態で使用されうる。
【0088】
標的タンパク質の製造、特徴づけ又は研究においてモノクローナル抗体のような抗原結合タンパク質を使用するための技術は当技術分野でよく知られている(例えば、Ausubel,Current Protocols in Molecular Biology,John Wiley,2005,Harlowら,Antibodies,A Laboratory Manual,Cold Spring Harbor Laboratory,1988,Harlowら,Using Antibodies,Cold Spring Harbor Laboratory,Cold Spring Harbor,N.Y.,Cold Spring Harbor Laboratory Press,1999,Lipmanら,ILAR Journal 46:258−268,2005を参照されたい)。
【0089】
本発明の1つの実施形態においては、抗原結合タンパク質を使用して、溶液中の又はミクロスフェアに結合した又は細胞上のORF0657n抗原の存在を確認する。溶液または細胞に存在するタンパク質への該結合タンパク質の結合能は、ウエスタンブロット、酵素結合イムノソルベントアッセイ(ELISA)、フローサイトメトリーおよびルミネックス(Luminex)イムノアッセイのような種々の技術を用いて測定されうる。
【0090】
VI.治療
適当な標的領域に結合する抗原結合タンパク質を使用して、治療的および予防的治療が患者に対して行われうる。治療的治療は、スタヒロコッカス・アウレウス(S.aureus)に感染した者に対して行われる。予防的治療は一般集団または一般集団の小集団に対して行われうる。一般集団の好ましい小集団としては、スタヒロコッカス・アウレウス(S.aureus)感染のリスクの高い者が挙げられる。
【0091】
「患者」は、スタヒロコッカス・アウレウス(S.aureus)に感染しうる哺乳動物を意味する。好ましくは、患者はヒトである。しかし、他のタイプの哺乳動物、例えばウシ、ブタ、ヒツジ、ヤギ、ウサギ、ウマ、イヌ、ネコ、サル、ラットおよびマウスがスタヒロコッカス・アウレウス(S.aureus)に感染しうる。非ヒト患者の治療は、ペットおよび家畜を防御するのに、ならびに特定の治療の効力を評価するのに有用である。
【0092】
スタヒロコッカス・アウレウス(S.aureus)感染のリスクの高い者には、医療従事者、入院患者、低下した免疫系を有する患者、手術を受けている患者、カテーテルまたは血管装置のような異物インプラントを受け入れている患者、免疫低下を招く療法を受けている患者、および火傷または創傷のリスクの高い職業の者が含まれる(The Staphylococci in Human Disease,Crossley and Archer(編),Churchill Livingstone Inc.1997)。
【0093】
1つの実施形態においては、手術または異物インプラントと共に、抗原結合タンパク質を患者に投与する。「手術または異物インプラント」に対する言及は、異物インプラントの付与を伴う又は伴わない手術、および手術を伴う又は伴わない異物インプラントの付与を含む。投与の時機は、予防的治療および/または治療的治療が達成されうよう計画されうる。投与は、好ましくは、手術または移植とほぼ同時に開始されうる。
【0094】
医薬投与全般に関する指針は、例えばRemington’s Pharmaceutical Sciences 20th Edition,Gennaro編,Mack Publishing,2000;ならびにModern Pharmaceutics 2nd Edition,BankerおよびRhodes編,Marcel Dekker,Inc.,1990に記載されている。
【0095】
医薬上許容される担体は抗原結合タンパク質の保存および投与を促進する。タンパク質溶液製剤を安定化するために使用される物質には、炭水化物、アミノ酸および緩衝化塩が含まれる(Middaughら,Handbook of Experimental Pharmacology 137:33−58,1999)。
【0096】
抗原結合タンパク質は静脈内、皮下、筋肉内または粘膜のような種々の経路により投与されうる。皮下および筋肉内投与は、例えば針または噴射式注射器を使用して行われうる。鼻腔内運搬のような粘膜運搬は、吸着部位における、より長い保持時間を得るために、増強物質または粘膜付着物質を使用することを含みうる(Middaughら,Handbook of Experimental Pharmacology 137:33−58,1999)。
【0097】
適当な投与計画は、好ましくは、患者の年齢、体重、性別および医学的状態;投与経路;所望の効果;ならびに使用する個々の化合物を含む、当技術分野でよく知られた要因を考慮して決定される。有効用量範囲は約0.1mg/kg〜20mg/kg、または0.5mg/kg〜5mg/kgとなると予想される。投与頻度は該化合物の有効性および安定性によって異なりうる。投与頻度の具体例には、隔週、毎週、毎月および隔月が含まれる。
【0098】
VII.実施例
以下に記載の実施例は本発明の種々の特徴を更に詳しく例示するものである。該実施例は本発明の実施のための有用な方法をも例示する。これらの実施例は、特許請求されている本発明を限定するものではない。
【0099】
(実施例1)
ORF0657nに対するモノクローナル抗体の作製
ORF0657n−C/e(配列番号2)またはORF0657n−H/y(配列番号3)を抗原として使用して、ORF0657n(配列番号1)に対するモノクローナル抗体を作製した。ELISAおよびフローサイトメトリーにより該抗体を特定し、特徴づけした。
【0100】
マウスおよび免疫化:4〜5週齢の雌BALB/cマウスをTaconic(Germantown,N.Y.)から購入した。アルミニウムヒドロキシホスファートアジュバント上で製剤化された、大腸菌(E.coli)により産生されたORF0657n−C/e抗原または酵母により発現されたORF0657n−H/y抗原の20μgで、第0、7および21日にマウスの筋肉内に免疫化した(Andersonら,国際公開番号WO 2005/009379(国際公開日2005年2月3日))。リン酸緩衝食塩水(PBS)中の20μgのタンパク質の最終静脈内注射(i.v.)を、融合の3日前にマウスに実施した。マウスを犠死させ、細胞融合のために脾臓を摘出した。
【0101】
MAbの製造:脾臓から調製したリンパ球をマウス骨髄腫対応体SP2/0−Agl4(ATCC 1581)に、3:1の比のポリエチレングリコール1500(Boehringer Mannheim)により融合させた。20% ウシ胎児血清、ヒポキサンチン(10−4M)、チミジン(10−5M)を含有する高グルコース、ピルビン酸のダルベッコ変法イーグル培地(DMEM)を含む96ウェル平底マイクロタイタープレート内に該融合体をプレーティングし、24時間後にアミノプテリン(4×10−7M)を加えた。増殖(成長)中のハイブリドーマからの上清を、後記のとおり、ORF0657nに対する反応性に関してELISAによりスクリーニングした。陽性ウェルを限界希釈によりクローニングし、ELISA反応性に関して再試験した。モノクローナル抗体を抗体イソタイプ決定キット(Roche Diagnostics Corporation,Indianapolis,IN)で分類した。
【0102】
ELISA:コスター(Costar)培地結合性マイクロタイタープレートを、大腸菌(E.coli)により発現された、PBS中の配列番号2の50ナノグラム/ウェルで、2〜8℃で一晩コーティングした。該プレートをPBS、0.05% Tween20で3回洗浄し、1% ウシ血清アルブミン、PBS、0.05% Tween20(アッセイ希釈剤)で少なくとも1時間ブロッキングした。該プレートを前記のとおりに洗浄し、該融合ウェルまたはクローン化ハイブリドーマからの上清を加え、室温で2時間インキュベートした。該プレートを前記のとおりに洗浄し、ヤギ抗マウスIgG(H+L)−HRPコンジュゲート(Zymed)(アッセイ希釈剤中で1:8000)を加え、室温で1時間インキュベートした。アッセイプレートをTMB基質で現像し、反応を2.0N H2SO4で停止させ、OD450nmでプレートリーダーで読み取った。450nmで1.0を超える光学濃度を有するウェルを陽性とみなした。
【0103】
フローサイトメトリー:鉄枯渇条件(RPMI中)下で継代されたスタヒロコッカス・アウレウス(S.aureus)の調製されたグリセロールストックを使用して、ORF0657n結合に関してmAbを評価した。凍結グリセロールストック細胞を解凍し、PBS;1% ウシ血清アルブミン;0.1% アジ化ナトリウム、0.2% ブタIgG(Sigma)(PAAG)に、5×107 CFU/50μlの濃度まで再懸濁させた。該細胞の50μlアリコートを反応ごとに1.5mlエッペンドルフチューブ内に配置した。50マイクロリットルの該ハイブリドーマ培養物を各反応チューブに加え、室温で1時間インキュベートした。1mLのリン酸緩衝食塩水、1% ウシ血清アルブミン、0.1% アジ化ナトリウム(PAA)を該チューブに加えることにより、該細胞を洗浄した。該細胞を遠心分離(5500rpm、5分間)によりペレット化した。上清を取り出し、該細胞を100μLの二次抗体(PAAG中で1:100希釈されたFITC標識ヤギ抗マウスIg(BD Pharmingen))と混合した。インキュベーションは暗所内で室温で1時間行った。インキュベーション後、1mLのPAAを該反応混合物に加え、該細胞をペレット化し(5500rpm、5分間)、上清を取り出した。該ペレットを1mLのPBSに再懸濁させ、FAC分析のために12×75mmチューブに移した。
【0104】
チューブを、細菌細胞に関してゲート化され該細胞関連FITCの量を測定するBD−FACSCaliburフローサイトメーター装置上で測定に付した。スタヒロコッカス・アウレウス(S.aureus)の表面に結合することが判明している標準的な抗体を、各アッセイにおいて測定に付した。陰性対照を、細胞および該二次コンジュゲートの単独体として測定に付した。幾何平均値が30より大きい場合に、ハイブリドーマウェルは陽性であるとみなした。
【0105】
2つの別々の融合体は12個のモノクローナル抗体(mAb)のパネルを与えた。該mAbの全てはELISAにおいて反応性であった(表2)。これらの12個のmAbのうちの10個が細菌の表面に結合したことが、フローサイトメトリーにより実証された。該mAbの全ては、野生型タンパク質を使用するウエスタンブロット分析で陽性であった。
【0106】
【表2】
【0107】
(実施例2)
mAbのクラススイッチ
天然抗原に結合した単離されたmAbの全てはIgG1イソタイプのものであった。シフト変異体の選択により、これらの抗体はIgG2bイソタイプにクラススイッチされた(Spiraら,J.of Immunogical Methods,74:307−315,1985)。IgG2bを使用して適当なイムノアッセイを開発し、該細胞系を高密度でプレーティングした。体細胞突然変異を選択し、富化させ、ついでクローニングした。スイッチしたmAbの結合部位は依然として元のmAbと同じであったが、IgG2bサブタイプへのスイッチは、受動防御研究における、より好ましいイソタイプ(補体カスケードを開始させる)を与えた。
【0108】
【表3】
【0109】
(実施例3)
天然抗原を使用する結合阻害研究
mAb標識キット(Molecular Probes)を該製造業者の説明に従い使用して、精製抗体をアレクサフルオル(Alexafluor)−488で標識した。RPMI増殖細菌細胞の表面をちょうど飽和させるmAbの量を、標識mAbおよび未標識mAbの両方に関して測定した。表3(第1列)のmAbのそれぞれを、標識体および未標識体として使用した。
【0110】
まず、5×107個の細胞を、該細胞の表面を飽和させる濃度の該未標識mAbと共にインキュベートすることにより、該阻害アッセイを行った。この反応を室温で1時間インキュベートした。このインキュベーションの後、該反応を1mlのPAAで洗浄し、ミクロ遠心機(Hermle)において6,000RPMで5分間遠心した。上清を除去して〜50μlに減少させ、該細胞を、該細胞の表面をちょうど飽和させる量の直接標識mAbを含有する100μlのPAAGに再懸濁させた。このインキュベーションの後、該反応を1mlのPAAで洗浄し、ミクロ遠心機(Hermle)において6,000RPMで5分間遠心した。上清を除去して〜50μlに減少させ、該細胞を1mlのPBSに再懸濁させ、FAC分析のために12×75mmチューブに移した。対照として、該未標識mAbを含有する別の反応を二次アレクサフルオル(Alexafluor)−488コンジュゲート化ヤギ抗マウスIgG(H+L)(Molecularプローブ;PAAG中、1:400)で測定して、このmAbが該表面に結合することを確認した。該細胞と共に該標識mAbのみを含有する陽性対照についても実験を行った。該未標識mAbが、該標識mAbと同じエピトープに結合すれば、該細胞に関連した蛍光活性は全く無いか又は低いことになる。該未標識mAbが、該標識mAbとは異なるエピトープに結合すれば、該表面に関連した反応性のレベルは、該標識mAbのみの対照細胞と同等ということになる。
【0111】
阻害研究により、モノクローナル抗体のパネルは4つの反応性グループに分類された。
【0112】
【表4】
【0113】
(実施例4)
変性抗原および改変抗原での結合研究
結合を更に特徴づけるために、ORF0657n改変タンパク質を使用した。まず、ORF0657nをコードする核酸を発現ベクターpET−28a(Novagen)内にクローニングし、C末端6×hisタグと共に大腸菌(E.coli)内で発現させた(配列番号2)。StratageneのQuikChange XL Site−Directed Mutagenesis Kitを該製造業者の説明に従い使用して、該クローン化遺伝子を含有する発現ベクターを突然変異誘発に付した。該遺伝子は、特異的な連続的アミノ酸変化を伴って突然変異した。得られたプラスミドを、StratageneのXL10−Goldコンピテント細胞内に、該製造業者のプロトコールに従い形質転換した。QiagenのQIAprep Spin Miniprep Kitを使用して、プラスミドを形質転換体から単離した。ABIの310 DNA Sequencerを使用して配列決定することにより、形質転換体をスクリーニングした。最大数の塩基変化を示す形質転換体からのプラスミドを発現宿主HMS 174(DE3)(Novagen)内に形質転換した。Novagenの説明に従い、形質転換体を発現させた。
【0114】
異なるORF0657n改変タンパク質を使用して、ORF0657n mAb(配列番号4〜19)の多様性を確認した。これらのタンパク質を、標準的な方法を用いるドットブロットにおいて10個の異なるmAbでスクリーニングした。標準的な方法を用いるウエスタンブロットにより、陽性/陰性を確認した。このアプローチにより、抗体を、それらの結合プロファイルに従い分類した。該抗体のうちの7個は3つのグループに分類され、残りの3個の抗体(2H2.B8、8A8.E11.H3および13G11.C11)は、互いに類似しているが同一ではないプロファイルを有していた(表5)。
【0115】
【表5】
【0116】
(実施例5)
BAIcore研究
BIAcore研究において、精製ORF0657n−H/yを抗原として使用する「フットプリント分析」によりmAbを検査した。BIACORE(登録商標)によるリアルタイム生体分子相互作用分析を用いて、ペア様態結合実験を行った。BIACORE(商標登録)は、カルボキシルメチルデキストランでコーティングされた(CM5)センサーチップの表面に直接照射される偏光の屈折率の変化をモニターすることにより質量変化を検出するために、ミクロフルイディクス技術および表面プラズモン共鳴(SPR)を組合せたものである。応答単位(Response Unit)で測定される応答の変化は結合アナライト(すなわち、抗原または抗体)の量に相関されうる。
【0117】
抗スタヒロコッカス抗体(mAb 13C7.D12)を該CM5センサーチップの表面上に共有結合(固定化)させた。該固定化mAbを、まず、ORF0657nタンパク質にさらし、ついでマトリックス形態で抗体のペアにさらした。ORF0657nタンパク質+抗体ペアの各サイクルの後、20mM HClを使用して、該センサーチップの表面を固定化mAb 13C7.D12へと再生させた。各抗体ペアの全ての組合せが分析されうるよう、マトリックス形態でORF0657nタンパク質に対して8個の抗体を試験した。この実験で使用したmAbペアに関するマトリックス設計を表6に要約する。
【0118】
【表6】
【0119】
各実施において最初に結合した(捕捉された)抗原の量を正規化するために、各試験抗体/抗原複合体に関する以下の比を計算する。
【0120】
【数1】
【0121】
各抗体に残存している利用可能なエピトープの割合(%)は、以下のとおりに、マッピングペアに関して計算されうる。
【0122】
【数2】
【0123】
図2は、ペア様態の結合研究における該モノクローナル抗体の反応性の概要を示すマトリックスを例示する。モノクローナル抗体のパネルは、BIACORE(登録商標)法により、3つの反応性領域に分類された(表7を参照されたい)。
【0124】
【表7】
【0125】
(実施例6)
マウス敗血症モデルにおける受動免疫化での防御研究
モノクローナル抗体mAb 2H2.BE11およびmAb 13C7.BC1を、スタヒロコッカス・アウレウス(S.aureus)感染に対する防御をもたらすそれらの能力に関して試験した。これらの抗体はORF0657nタンパク質上の異なるエピトープを認識する。対照には、イソタイプが一致したmAbおよびPBS単体が含まれた。
【0126】
細菌チャレンジの20時間前に該mAbまたはPBSを腹腔内(i.p.)投与した。ついでマウスをLD80−90用量のスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)でチャレンジ(i.v.)し、生存をモニターした。各実験を10または20匹のマウスの群で3回反復し、10日間モニターした。未感染BALB/cマウスにおける該モノクローナル抗体に関する半減期は約8日間である。0.5mgの用量が最適であることが判明した。それらの2つのモノクローナル抗体での実験の結果を図3A〜C、4A、4Bおよび5A〜Cに示す。
【0127】
1つの実験においては、mAb 13C7.BC1は対照と比べて第10日における生存を有意に改善したが、その他の2つの反復実験においては、全体的な生存率は対照の場合に類似していた(図3A〜3C)。しかし、対照と比べて、mAb 13C7.BC1で治療されたマウスの、この10日間以内の死亡までの時間における遅延が認められた。また、3つの実験のうちの2つにおいて、mAb 2H2.BE11で治療されたマウスの死亡までの時間の遅延における類似した傾向が認められた(図5A〜5C)。
【0128】
また、最近のスタヒロコッカス・アウレウス(S.aureus)臨床分離株であるUK58を使用して、mAb 13C7.BC1の効果を調べた(図4Aおよび4B)。この株は患者の膿瘍部位から最低限度で継代された。2つの独立した実験において、結果はUK58チャレンジによる死亡までの時間における遅延を示している。
【0129】
抗体持続性研究は、該LD80−90モデルにおいては、死亡速度が速いため評価できない。したがって、亜致死チャレンジモデルを用いた。該亜致死モデルにおいては、使用したチャレンジ用量は、該LD80−90モデルで使用した用量の10%である。該亜致死チャレンジモデルを4日間にわたってモニターした。22匹のマウスの群に0.5mgの用量のmAb 13C7.BC1またはイソタイプ対照mAb(6G6)を、5×107 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)でのi.v.細菌チャレンジの20時間前に投与した。チャレンジの時点の血清中のmAbレベルを測定するために、チャレンジの直前(T=0)に、各群の2匹の動物を犠死させた。チャレンジの2、24、48、72および96時間後、各群の4匹のマウスを犠死させ、血清mAbレベルを測定した。
【0130】
この亜致死チャレンジ実験から、スタヒロコッカス・アウレウス(S.aureus)感染マウスにおけるmAb 13C7.BC1の半減期は約1日であると推定された。これとは対照的に、イソタイプ対照mAbの半減期は4日を超えると推定された(データ非表示)。これらのデータは、スタヒロコッカス・アウレウス(S.aureus)でチャレンジされたマウスにおけるmAb 13C7.BC1の特異的減少を示しており、これは、該致死モデルにおいてモニターされた10日間のかなり前に枯渇すると思われる。
【0131】
図3A〜C、4A、4Bおよび5A〜Cに例示されている8つの実験のうちの6つにおいて、該mAbを投与した群に関して約3日までの生存の改善が認められた。これらの結果は、そのようなmAbがスタヒロコッカス・アウレウス(S.aureus)チャレンジマウスの生存率に対して正の効果を及ぼすことを示している。
【0132】
(実施例7)
マウス留置カテーテルモデルにおける受動免疫化での防御研究
マウス留置カテーテルモデルをmAb 2H2.BE11と共に使用した。このモデルにおいて使用したスタヒロコッカス・アウレウス(S.aureus)株は臨床分離株MCL8538であった。この株を選択したのは、マウス敗血症モデルにおいて使用した株であるスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)と比べて、カテーテルの再現可能なコロニー形成を尚も達成しつつ、より少量の接種物の投与が可能だからである。
【0133】
ICRマウスは、頚静脈内に外科的に移植され縫合により適所に保持され該マウスの背側中央線上の出口から出るカテーテル(PE50シリコーンゴム)を有していた。術後9〜11日間、マウスを休ませた。チャレンジの24時間前に、600mcgのマウスモノクローナル抗体2H2.BE11の1回のi.p.注射投与によりマウスを受動免疫化した。第0日に、スタヒロコッカス・アウレウス(S.aureus)MCL8538のi.v.投与によりマウスをチャレンジした。接種物の用量は100μlの体積中、2〜8×105 CFUであった(実験1〜3)。この低用量は4日後に該カテーテルから自然に消失することが判明した。したがって、チャレンジの24時間後に細菌に関してカテーテルを評価した。その時点で、マウスを犠死させ、カテーテルを回収した。TSA上でカテーテル全体を培養することにより、カテーテル上の細菌の存在を評価した。該プレート上で増殖の徴候が認められたら、該カテーテルは培養陽性であると評価した。
【0134】
最初の3つの実験の2つにおいて、培養陰性カテーテルの数は、抗体2H2.BE11で受動免疫化されたマウスにおいては、イソタイプ対照抗体の場合より有意に低かった。より大きな接種物用量を用いて、4番目の実験を行った。このより過酷なチャレンジにおいては、該用量は、カテーテルの100%が再現可能に感染し対照マウスにおいてこの感染が自然には消失しない(7日間にわたるモニター)用量であると定められた。そのようなより大きな接種物サイズを用いる実験4においてもまた、2H2.BE11に対する抗体を注射したマウスにおいては、イソタイプ対照の場合と比べて、有意に少数のカテーテルが感染したことが判明した。4つの実験の結果を表8に要約する。
【0135】
【表8】
【0136】
(実施例8)
抗ORF0657nモノクローナル抗体2H2.B8(IgG1)、2H2.BE11(IgG2b)もしくは13C7.IgG2bまたはイソタイプ一致対照mAbを使用する、細菌のエクスビボ前オプソニン化
ORF0657nのモノクローナル抗体がオプソニン性であるかどうかを試験するために、致死用量のスタヒロコッカス・アウレウス(S.aureus)をモノクローナル抗体2H2.B8、2H2.BE11もしくは13C7.IgG2bまたはイソタイプ一致対照モノクローナル抗体で前オプソニン化させる受動防御実験を行った。ついで前オプソニン化細菌をマウスにi.p.投与した。これらの実験で使用した細菌はスタヒロコッカス・アウレウス(S.aureus)RN4220(野生型)またはRN4220.0657nであった。RN4220.0657n細菌を、FURボックスによる制御の非存在化でORF0657nを発現するよう操作した。したがって、それは鉄の存在下で増殖可能であり、その表面上でORF0657n抗原を尚も発現することが可能であった。あるいは、RN4220(野生型)を、該細菌表面上での0657nの発現を誘導するために、低鉄培地RPMI内で2回継代した。
【0137】
6Balb/cマウスに十分な細菌の量(6×LD100)を、穏やかに振とうしながら、800μgのIgGと共に4℃で1時間インキュベートした。ついで細菌をペレット化し、未結合mAbを除去した。抗体オプソニン化細菌を2.4mLのPBSに再懸濁させ、0.4mL(1×LD100)を5匹のマウスのそれぞれに注射した。マウスの全群に同等のCFUがもたらされること及び該mAbが該細菌を凝集させていないことが保証されるよう、チャレンジ後、TSA上のプレーティングにより各接種物を定量した。チャレンジ後3日間、生存をモニターした。この手法が有効となるためには該標的抗原が該細菌の表面上に存在しなければならないため、オプソニン化前に該細菌上で0657nが発現されることが保証されるよう配慮した。mAb 2H2.B8を使用するフローサイトメトリーにより、ORF0657nの発現をモニターした。各マウスに注射したオプソニン化細菌の用量は2〜4×109 CFU RN4220.0657n/マウスまたは1〜2×109 CFU RN4220(2×RPMI)/マウスであった。
【0138】
マウスは、イソタイプ一致対照mAbではなく2H2.B8または2H2.BE11のいずれかで前オプソニン化された場合に、致死用量のRN4220.0657nスタヒロコッカスによる死亡から防御された。該実験をIgG1イソタイプに関して2回、およびIgG2bイソタイプに関して3回反復したところ、同様の結果が得られた(表9A)。
【0139】
【表9】
【0140】
マウスは、イソタイプ一致対照mAbではなくmAb 2H2.B8で前オプソニン化された場合に、致死用量のRN4220(2×RPMI)スタヒロコッカスによる死亡から防御された。該実験を6回反復したところ、同様の結果が得られた(表9B)。
【0141】
【表10】
【0142】
マウス抗0657n 2H2は、この致死モデルにおける死亡の予防に非常に有効であった。13C7 mAbは、(図3〜6に例示されている既に記載されているモデルの場合とは対照的に)このモデルでは有効でなかった。該抗0657n mAbの全て(2H2.BE11、2H2.B8および13C7.IgG2b)がRN4220に結合し(フローサイトメトリーを用いて実証されているとおり)、全てがインビトロOPAアッセイにおいてオプソニン活性を有する。このモデルは、該マウスの腹膜における、生存を向上させるためのエピトープ特異性に関する追加的な要件を表している。
【0143】
(実施例8)
2H2 mAbで行ったエピトープマッピング研究
この実施例に記載する実験は、モノクローナル抗体2H2.BE11がORF0657n内のコンホメーションエピトープを認識するという証拠を提供する。該実験は、2H2 mAbにより認識される三次元構造におけるコンホメーションエピトープを提示するのに要求されるORF0657n内の最小配列を位置決定した。また、該実験は、2H2 mAbがORF0657nに結合すると小分子との反応から保護される特徴的なリシン残基をORF0657nの最小配列内に特定した。
【0144】
ORF0657nの配列内の典型的には9から14アミノ酸長の直鎖状エピトープを2H2 mAbが認識する潜在能力を、エピトープ抽出を用いて、配列番号1のアミノ酸42からアミノ酸486のORF0657n断片(「ORF0657t」)から調べた。詳細に説明すると、該エピトープ抽出実験のそれぞれについて、10mgの臭化シアン活性化セファロース(Amersham cat.No.17 043001)への化学的架橋により30μgの2H2 mAbを固定化した。ORF0657tのタンパク質分解消化物をGluC(Roche Applied Science cat.No.11 420 3997 001)、Asp−N(Roche Applied Science cat.No.11 054 589 001)またはキモトリプシン(Roche Applied Science cat.No.11 418 467 001)で生成させ、リニアイオントラップ(LTQ−Thermo Electron Inc)において1D/LC−MS/MSにより特徴づけた。3つの独立した実験において、いずれかのプロテアーゼに由来する特徴づけられたタンパク質分解消化物8.4μgを該固定化抗体と反応させた。未結合ペプチドを該抗体架橋ビーズから洗い落とした。潜在的結合ペプチドを低pHで溶出し、1D/LC−MS/MSにより特徴づけた。生じたタンパク質分解ペプチドはいずれも、2H2 mAbにより、高い効率および特異性では認識されなかった。このことは、2H2 mAbが直鎖状エピトープを認識しなかったことを明確に示している。
【0145】
2H2 mAbがORF0657nの直鎖状配列を認識しなかったという知見は、限られた化学切断実験により裏付けられた。ORF0657tをCNBrで化学的に2時間にわたって切断した。得られた切断産物をSDS−PAGEにより分析した。SDS−PAGE分析は、約42kDa、35kDa、25kDa、15kDaおよび10kDaの分子量を有する5つの主要バンドを示した。2H2 mAbでのウエスタンブロット分析は、該42kDaバンドのみが2H2により認識されることを明らかに示した。すべてのバンドを該SDS−PAGEから切り出し、ゲル内消化を行い、タンデム質量分析により特定された得られたペプチドをORF0657t内の対応配列と合致させた。該主要バンドの分析の結果を表10に示す。
【0146】
【表11】
【0147】
〜42kDaの分子量を有する断片の重要性をエピトープ切断により確認した。詳細に説明すると、該エピトープ切断実験のそれぞれについて、50mgの臭化シアン活性化セファロース(Amersham cat.No.17 0430 01)への化学的架橋により、210μgの2H2 mAbを固定化した。ついで50μgの無傷ORF0657nを該固定化抗体に結合させ、未結合ORF0657nをリン酸緩衝食塩水での十分な洗浄により洗い落とした。3つの独立した実験において、プロテアーゼGlu−C、トリプシン、ならびにGluC、AspN、トリプシン、キモトリプシンおよびカルボキシ−ペプチダーゼYの連続的組合せを、5時間または該連続的組合せのプロテアーゼ当たり1時間、加えた。該インキュベーション中に該プロテアーゼにより切り出されたペプチドを十分に洗い落とし、2H2 mAbに特異的に結合したORF0657n断片をSDSローディングバッファーで遊離させた。
【0148】
2H2 mAbに特異的に結合した断片をSDS−PAGEにより分析した。該エピトープ切断実験の3つ全てが、該SDS−PAGE分析において、40〜42kDaの分子量を有する唯一のバンドを示した。2H2 mAbに結合するバンドがウエスタンブロット分析により確認された。該エピトープ切断実験をGlu−Cプロテアーゼに関して反復した。今回は、2H2 mAbに特異的に結合したORF0657tの断片を酸性条件で遊離させ、リニアイオントラップ(LTQ,Thermo Electron)において1D/LC−MS/MSにより分析した。溶出したサンプルは、82〜87分(40%〜45%アセトニトリル)に、予想強度を有するシグナル(全イオン係数)を示し、42.628kDaと推定される複数の電荷状態([M+67 H]67+〜([M+30 H]30+)を示した。この特定の質量に対応するORF0657tの考えられうる断片は、42.6kDaの分子量を有するORF0657tの配列[012から382]である。
【0149】
ORF0657tのどのリシン残基が2H2 mAbの結合に際して化学反応から保護されるのかを決定するために、2H2mAbの存在下または非存在下の3つの異なる実験条件を用いて、スルホ−NHS−アセタート(Pierce Cat.No.26777)を使用する化学的標識実験を行った。表11を参照されたい。
【0150】
【表12】
【0151】
各実験について、0または3モル過剰の2H2 mAbで生成した反応産物を3つのうちの1つのプロテアーゼと共にインキュベートして、2×9個の反応混合物を得た。実験1では、GluC、AspNおよびトリプシンを使用した。実験2および3では、GluC、AspNおよびキモトリプシンを使用した。ついで該タンパク質分解ペプチドを1D/LC−MS/MSにより分析した。該反応のそれぞれについて、アセチル化および未アセチル化リシン残基の比率を、個々のペプチドの全イオン計数(TIC)の曲線下面積に基づいて算出した。ついで、同一反応条件に関して、得られた比率を該ペア間(2H2 mAbの存在下または非存在下)で比較した。2H2 mAbへのORF0657tの結合に際して最大の遮蔽を受ける、ORF0657t内のリシン残基を特定するために、全3個の反応条件で全体的な分析を行った。前記の化学的標識実験は、2H2 mAbの結合に際して最も保護されたものとして、K76、K257および潜在的にK443を特定した。化学的標識に対する保護は直接結合によるものでありうる。しかし、そのような保護は、保護部位の近傍への結合、またはORF0657nI内の長い領域にわたる構造変化によるものである可能性がある。
【0152】
要約すると、前記実験は、2H2 mAbにより認識される、ORF0657t内のエピトープが、コンホメーション性(conformational)であるという、明らかな証拠を示している。2H2 mAbにより認識されるORF0657tの断片は、ORF0675tのアミノ酸1から115に位置するN末端、およびORF0657tのアミノ酸323から357に位置するカルボキシル末端を有する。2H2 mAbの結合の際の化学的標識からの保護が、長い領域にわたる構造変化により影響されることは除外できないとしても、リシン76およびリシン275の近傍の領域が直接的な抗体相互作用に関与している可能性が高い。
【0153】
(実施例9)
2H2 mAb配列特定
一本鎖可変フラグメント(scFv)構築のための縮重プライマーPCR/重複伸長クローニング法(Krebberら,JIM 201(1):35−55,1997)を、ヒトカッパ軽鎖定常ドメインまたはscAb物質に融合された可溶性scFvのBiacoreによるハイスループットスクリーニングと組合せることにより、ハイブリドーマにより発現された2H2 IgGの可変軽(Vl)配列および可変重(Vh)配列の特定が達成された。これは、該縮重プライマー法により生じたVlフレームワーク1、4およびVhフレームワーク1、4における突然変異の精密な識別を可能にした。
【0154】
簡潔に説明すると、Total RNA Kit(商標)(Ambion Inc.)の標準的な方法を用いて、RNA物質を該ハイブリドーマ細胞系から精製した。ついで、この物質をcDNAに逆転写し、それを、該可変領域を増幅するためのPCRにおける鋳型として使用した。VlおよびVh鎖のPCR増幅のための条件は、Krebberら,JIM201(l):35−55,1997に記載されているプロトコールに基づくものであった。(Gly4Ser)4リンカー(配列番号32)が付加されて、VhおよびVlが重なり合ってVl−(Gly4Ser)4−Vh scFvを与える、第3PCR反応のためのドメインが得られるよう、プライマーを設計する。
【0155】
該可変鎖を個々に増幅するための第1セットのPCR反応を、5μlのcDNA反応物、それぞれ2μMのVlおよびVhの増幅のためのフォワードおよびリバースプライマーセットならびに高忠実度PCRマスター混合物を含有する100μlの容量中で行った。該反応を94℃で4分間変性させ、ついで94℃で30秒間、50℃で30秒間、72℃で1分間の30サイクルに付し、72℃で5分間の最終サイクルに付した。完全長PCR産物をゲル精製した。
【0156】
増幅されたVhおよびVl物質をscFvへと合体させて完全長産物を構築するために、第3PCR反応を行った。100μlの容量中で、それぞれ約20ngのVhおよびVl DNAならびに高忠実度PCRマスター混合物を94℃で5分間変性させ、ついで94℃で30秒間、60℃で30秒間および72℃で30秒間の3サイクル(プライマーの非存在下)に付した。修飾されたPCRプライマー配列番号33および配列番号34を1μMの最終濃度で加え、94℃で30秒間、60℃で1分間および72℃で1分間の30サイクルを行い、ついで72℃で7分間の加熱を行った。予想される完全長scFv PCR産物をゲル精製した。
【0157】
増幅されたscFv物質を、scAbの産生および配列分析のためにMP16可溶性発現ベクター内にクローニングした(Hayhurstら,JIM 276(1−2):185−196,2003)。MP16ベクター内への定向性(directional)クローニングのために、Sfi1での1回の制限酵素消化を行った。ついで、存在する見掛け上完全長の可変重鎖および可変軽鎖を含有するクローンをXL1−Blue細胞内でscAbとして発現させ、標準的な浸透圧ショック法を用いてペリプラズムから回収した。簡潔に説明すると、96ウェル形態において、2% グルコースおよび100μg/ml アンピシリンを含有する増殖培地内でクローンを37℃で一晩増殖させた。20μlの該一晩培養物を、0.1% グルコースおよび100μg/ml アンピシリンを含有する新たな培地に移し、0.6のOD600に達するまで増殖させた。IPTGを0.5mMの最終濃度で加えることにより、scAbの発現のために該細胞を誘導し、150rpmで振とうしながら室温で一晩インキュベートした。Qiagen Ni−NTAスーパーフロー(superflow)自動化法を用いて、該細胞からscAbを精製した。
【0158】
ORF0657tに対する結合活性に関して各scAbペリプラズム調製物を分析するために、Biacore3000表面プラズモン共鳴(SPR)装置(Upsala,Sweden)を使用した。標準的なEDC/NHSカップリングを用いて、約250共鳴単位の0657t抗原をCM5センサーチップの実験用フローセル表面に直接的に共有結合させた。タンパク質のカップリングを行うことなく、参照フローセル表面を活性化し不活性化した。ついで各調製物を該表面に通過させ、抗原へのscAbの会合および解離を測定した。各実施ごとに、10mM HClを流速20μl/分で20秒間にわたり1回注入し、ついで2分間の安定化時間を設けることにより、該表面を再生させた。全てのサンプルを2通りに実施し、バッファーのみの実施を対照として用いた。95個のクローンをスクリーニングした後、1個のクローンを、その結合活性に基づいて選択した。選択した最終的な2H2クローンは、元のハイブリドーマにより調製されたIgG物質に類似した、ORF0657tに対するそのアフィニティー、および比較配列分析に基づくものであった。
【0159】
2H2 Vh(配列番号20)およびVl(配列番号21)のアミノ酸配列は以下のとおりであった。
【0160】
【化1】
【0161】
下線部はCDRである。CDRは、カバット(Kabat)定義に基づいて特定された。コードしている核酸配列は配列番号24(Vh)および配列番号25(Vl)に示されている。
【0162】
(実施例10)
2H2 IgGキメラの発現
2H2 mAbの可変領域を、実施例9に記載されているとおりにマウスハイブリドーマからクローニングした。該可変領域の配列をPCR増幅し、重鎖可変領域をコードするDNAを、IgG1定常領域をコードするDNAにインフレームで融合させ、一方、軽鎖可変領域をコードするDNAを、カッパ定常領域をコードするDNAにインフレームで融合させた。得られた抗体発現ベクターに関するクローニング方法を以下に説明する。
【0163】
該可変領域をPCR増幅した。PCR反応は、高忠実度PCRマスター混合物、鋳型容量1μlならびにフォワードおよびリバースプライマー(それぞれ1μl)を含有する25μlの容量中で行った。PCR条件は94℃で2分間の1サイクル、94℃で1.5分間、60℃で1.5分間、72℃で1.5分間の25サイクルおよび72℃で7分間であり、反応産物を、取り出すまで4℃にし、ついでIn−Fusion法を用いて5’末端においてリーダー配列に及び3’末端において定常領域にインフレームでクローニングした。以下のプライマーを使用した。軽鎖フォワード,5’−ACAGATGCCAGATGCGATATTGTGATGACCCAGTCT(配列番号28);軽鎖リバース,5’−TGCAGCCACCGTACGTTTTATTTCCAGCTTGGTCCC(配列番号29);重鎖フォワード,5’−ACAGGTGTCCACTCGGATGTGCACCTGGTGGAGTCA(配列番号30);および重鎖リバース,5’−GCCCTTGGTGGATGCCGAGGAGACTGTGAGAGTGGT(配列番号31)。該クローンの全てのDNA配列は配列決定により確認された。
【0164】
DNA配列から推定されるアミノ酸配列を以下に示す。
【0165】
【化2】
【0166】
可変領域は下線で示されている。
【0167】
該抗体を293EBNA単層細胞内で発現させた。PE1に基づくトランスフェクション試薬を使用して、該プラスミドをトランスフェクトした。トランスフェクトされた細胞をOpti−MEM無血清培地内でインキュベートし、分泌した抗体を、プロテインA/Gアフィニティークロマトグラフィーを用いて培地から精製した。精製抗体の濃度をOD280nmにより決定し、純度をLabChip(商標)キャピラリー電気泳動により測定した。
【0168】
軽鎖および重鎖の両方の発現はヒトCMVプロモーターおよびウシ成長ホルモンポリアデニル化シグナルにより駆動された(Shiverら,Ann.N.Y.Acad.Sci.,772:198−208,1995)。前方のリーダー配列が培養培地内への抗体の分泌をもたらした。重鎖に関するリーダー配列はMEWSWVFLFFLSVTTGVHS(配列番号26)であり、軽鎖に関するリーダー配列はMSVPTQVLGLLLLWLTDARC(配列番号27)であった。該発現ベクターは、293EBNA細胞内での持続的発現のためのEBVウイルスゲノム由来のoriP、ならびに大腸菌(E.coli)における複製起点およびカナマイシン選択マーカーのための細菌配列を含有する。
【0169】
該抗体を293EBNA単層細胞内で発現させた。PE1に基づくトランスフェクション試薬を使用して、該プラスミドをトランスフェクトした。トランスフェクトされた細胞をOpti−MEM無血清培地内でインキュベートし、分泌した抗体を、プロテインA/Gアフィニティークロマトグラフィーを用いて培地から精製した。精製抗体の濃度をOD280nmにより決定し、純度をLabChipキャピラリー電気泳動により測定した。
【0170】
(実施例11)
アフィニティー測定
ハイブリドーマ物質としての2H2 mAb、scAbおよびキメラ抗体に関する比較分析を行った。2h2 mAb VhおよびVl領域をクローニングし、IgGキメラとして発現させた(実施例10に記載されているとおり)。scAbを、ヒトカッパ鎖タグに融合したscfvを産生するMP16ベクター(実施例9)内にクローニングした。以下に更に詳細に説明するとおり、該抗原アフィニティーは該構築物間で有意には異ならなかった。
【0171】
結合ドメインと抗原との間の1:1相互作用を測定するために、Biacoreの実験条件を、抗体フラグメントを測定するのか又は完全長IgGを測定するのかに応じて改変した。IgGの測定の場合には、IgGをリガンドとして表面上に捕捉し、ORF0657tをアナライトとして通過させた。抗体フラグメントの分析の場合には、ORF0657tを表面に結合させ、抗体フラグメントをアナライトとして通過させた。これは、ORF0657t抗原に対する元の2H2 mAbハイブリドーマ物質のアフィニティーが組換えクローニング後に有意な変化を何ら示さないことを示した(表12)。Biacore 3000での表面プラズモン共鳴によりデータを収集し、各アナライトを複数の濃度(1濃度当たり二重の重複実験)で通過させた。一連の全濃度を同時フィットさせるBIAevaluation(Biacore,Inc.)でデータを分析した。フィットパラメーターを表12に示す。
【0172】
【表13】
【0173】
(実施例12)
ORF0657nに基づく配列
配列番号(SEQ ID NO)4から19において強調して示されている(太字および下線で示されている)アミノ酸は、ORF0657nから変化したアミノ酸を示す。
【0174】
【化3】
【0175】
他の実施形態も特許請求の範囲内に含まれる。本明細書中には幾つかの実施形態が示され記載されているが、本発明の精神および範囲から逸脱することなく種々の改変が施されうる。
【図面の簡単な説明】
【0176】
【図1】図1はIgG分子の構造を例示する。「VL」は軽鎖可変領域を意味する。「VH」は重鎖可変領域を意味する。「CL」は軽鎖定常領域を意味する。「CH1」、「CH2」および「CH3」は重鎖定常領域である。点線はジスルフィド結合を示す。
【図2】図2は、ペア様態の結合研究における種々のモノクローナル抗体の反応性の概要を示すマトリックスを例示する。モノクローナル抗体のパネルは、BIACORE(登録商標)法により、3つの反応性領域に分類された。
【図3A】図3A〜C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1;□,mAb 6G6.A8(イソタイプ対照);または○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図3A−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3B−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.6×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3C−0.50mgのmAb13C7.BC1;0.45mgのmAb 6G6;および9.9×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図3B】図3A〜C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1;□,mAb 6G6.A8(イソタイプ対照);または○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図3A−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3B−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.6×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3C−0.50mgのmAb13C7.BC1;0.45mgのmAb 6G6;および9.9×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図3C】図3A〜C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1;□,mAb 6G6.A8(イソタイプ対照);または○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図3A−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3B−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.6×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3C−0.50mgのmAb13C7.BC1;0.45mgのmAb 6G6;および9.9×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図4A】図4Aおよび4B:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1(0.5mg);□,mAb 6G6.A8(イソタイプ対照)(0.5mg);または○,PBS(0.5ml)のi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図4Aは、2.09×108 CFUのスタヒロコッカス・アウレウス(S.aureus)UK58での結果を例示している。図4Bは、2.15×108 スタヒロコッカス・アウレウス(S.aureus)UK58での結果を例示している。
【図4B】図4Aおよび4B:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1(0.5mg);□,mAb 6G6.A8(イソタイプ対照)(0.5mg);または○,PBS(0.5ml)のi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図4Aは、2.09×108 CFUのスタヒロコッカス・アウレウス(S.aureus)UK58での結果を例示している。図4Bは、2.15×108 スタヒロコッカス・アウレウス(S.aureus)UK58での結果を例示している。
【図5A】図5A〜5C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 2H2.BE11;□,mAb 6G6.A8(イソタイプ対照);○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図5A−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5B−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および8.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5C−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図5B】図5A〜5C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 2H2.BE11;□,mAb 6G6.A8(イソタイプ対照);○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図5A−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5B−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および8.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5C−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図5C】図5A〜5C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 2H2.BE11;□,mAb 6G6.A8(イソタイプ対照);○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図5A−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5B−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および8.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5C−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【技術分野】
【0001】
本発明は、スタヒロコッカス・アウレウス(S.aureus)ORF0657nを標的とする抗原結合タンパク質に関する。
【背景技術】
【0002】
本出願の全体にわたって引用されている参考文献は、特許請求されている本発明の先行技術であると自認するものではない。
【0003】
スタヒロコッカス・アウレウス(Staphylococcus aureus)は、多種多様な疾患および状態を引き起こす病原体である。スタヒロコッカス・アウレウス(S.aureus)により引き起こされる疾患および状態の具体例には、菌血症、感染性心内膜炎、毛包炎、フルンケル、カルブンケル、膿痂疹、水疱性膿痂疹、蜂窩織炎、ボトリオミセス症、中毒性ショック症候群、熱傷様皮膚症候群、中枢神経系感染症、感染性および炎症性眼疾患、骨髄炎ならびに関節および骨の他の感染症、ならびに気道感染症が含まれる(The Staphylococci in Human Disease,Crossley and Archer(編),Churchill Livingstone Inc.1997)。
【0004】
スタヒロコッカス・アウレウス(S.aureus)感染およびスタヒロコッカス・アウレウス(S.aureus)の広がりを抑制するためには、免疫学に基づく方法が用いられうる。免疫学に基づく方法には受動免疫化および能動免疫化が含まれる。受動免疫化は、スタヒロコッカス・アウレウス(S.aureus)を標的とする免疫グロブリンを使用する。能動免疫化はスタヒロコッカス・アウレウス(S.aureus)に対する免疫応答を誘導する。
【発明の開示】
【0005】
発明の概要
本発明は、ORF0657n標的領域(配列番号1)に結合する抗原結合タンパク質に関する。ORF0657nはスタヒロコッカス・アウレウス(S.aureus)タンパク質である。ORF0657n標的領域はmAb lG3.BD4、mAb 2H2.BE11、mAb 13C7.BClおよびmAb 13G11.BF3結合部位により与えられる。致死モデルチャレンジにおいて、mAb 2H2.BE11およびmAb 13C7.BC1はスタヒロコッカス・アウレウス(S.aureus)感染に対する生存性の上昇をもたらした。また、mAb 2H2のIgG1またはIgG2b形態を使用するエクスビボモデルにおいて、およびmAb 2H2.BE11を使用する受動免疫マウス留置カテーテルモデルにおいても、防御が実証された。
【0006】
mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1およびmAb 13G11.BF3を産生するマウスハイブリドーマ細胞系は、ブダペスト条約に従い、2005年9月30日付けでAmerican Type Culture Collection, 10801 University Boulevard,Manassas,VA 20110−2209に寄託された。該細胞系にはATCC番号PTA−7124(mAb 2H2.BE11を産生する)、ATCC番号PTA−7125(mAb 13C7.BC1を産生する)、ATCC番号PTA−7126(mAb 1G3.BD4を産生する)およびATCC番号PTA−7127(mAb 13G11.BF3を産生する)が付与された。
【0007】
したがって、本発明の第1の態様は、第1可変領域および第2可変領域を含んでなる単離された抗原結合タンパク質に関する。第1および第2可変領域は、mAb 1G3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1標的領域およびmAb 13G11.BF3標的領域よりなる群から選ばれる1以上の標的領域に結合する。
【0008】
「単離(された)」に対する言及は、天然で見出されるものとは異なる形態を示す。そのような異なる形態は、例えば、天然で見出されるものとは異なる純度および/または天然で見出されない構造体でありうる。天然で見出されない構造体には、異なる領域が一緒になった組換え構造体、例えば、1以上のマウス相補性決定領域がヒトフレームワークスカフォールド上に挿入された又はヒト抗体の表面残基を模倣するためにマウス抗体の表面が修飾されたヒト化抗体、抗原結合タンパク質由来の1以上の相補性決定領域が別のフレームワークスカフォールド内に挿入されたハイブリッド抗体、ならびに軽可変ドメインおよび重可変ドメインをコードする遺伝子が互いにランダムに組合わされた天然ヒト配列に由来する抗体が含まれる。
【0009】
単離されたタンパク質は、好ましくは、血清タンパク質を実質的に含有しない。血清タンパク質を実質的に含有しないタンパク質は、ほとんど又は全ての血清タンパク質を欠く環境中に存在する。
【0010】
「可変領域」は重鎖または軽鎖からの抗体可変領域の構造を有する。抗体重鎖および軽鎖の可変領域は、フレームワーク上に間隔をあけて存在する3つの相補性決定領域を含有する。該相補性決定領域は主として、特定のエピトープの認識をもたらす。
【0011】
標的領域は、mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1またはmAb 13G11.BF3が結合するORF0657n領域(配列番号1)に関して定義される。例えば、mAb IG3.BD4標的領域は、mAb IG3.BD4が結合するORF0657n領域である。
【0012】
特定された標的領域に結合するタンパク質は該標的領域への結合に関してmAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1またはmAb 13G11.BF3と競合する。例えば、ORF0657nに結合するmAb 1G3.BD4と競合するタンパク質は、mAb 1G3.BD4標的領域に結合する。
【0013】
モノクローナル抗体mAb 1G3.B3、mAb 2H2.B8、mAb 13C7.D12またはmAb 13G11.C11のいずれかと競合するタンパク質は、過剰量および等量の該競合タンパク質およびモノクローナル抗体を使用した場合、ORF0657nへの該モノクローナル抗体の結合を少なくとも約20%、好ましくは少なくとも約50%軽減する。
【0014】
「タンパク質」に対する言及は連続的アミノ酸配列を示し、最小または最大サイズ限界を与えるものではない。該タンパク質中に存在する1以上のアミノ酸はグリコシル化またはジスルフィド結合形成のような翻訳後修飾を含有しうる。
【0015】
好ましい抗原結合タンパク質はモノクローナル抗体である。「モノクローナル抗体」に対する言及は、同じ又は実質的に同じ相補性決定領域および結合特異性を有する一群の抗体を示す。該抗体における変異は、該抗体が同一構築物から製造された場合に生じるものである。
【0016】
モノクローナル抗体は、例えば、特定のハイブリドーマから、および該抗体をコードする1以上の組換え遺伝子を含有する組換え細胞から製造されうる。該抗体は2以上の組換え遺伝子によりコードされることが可能であり、この場合、例えば、1つの遺伝子は重鎖をコードし、1つの遺伝子は軽鎖をコードする。
【0017】
本発明のもう1つの態様は、抗体可変領域をコードするヌクレオチド配列を含む組換え遺伝子を含有する核酸を記載する。該抗体可変領域は、mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合しうる。
【0018】
組換え遺伝子は、適切な転写およびプロセシングのための調節要素(これは翻訳および翻訳後要素を含みうる)と共にタンパク質をコードする組換え核酸を含有する。該組換え核酸は、その配列および/または形態の点で、天然では見出されない。組換え核酸の具体例には、精製された核酸、天然で見出されるものとは異なる核酸を与える一緒になった2以上の核酸領域、互いに天然で付随している1以上の核酸領域(例えば、上流または下流領域)の非存在が含まれる。
【0019】
本発明のもう1つの態様は、mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合する抗体可変領域をコードする1以上の組換え遺伝子を含んでなる組換え細胞を記載する。複数の組換え遺伝子が有用であり、例えば、この場合、1つの遺伝子は抗体重鎖またはそのフラグメント(Vh領域を含有するもの)をコードし、もう1つの核酸は抗体軽鎖またはそのフラグメント(Vl領域を含有するもの)をコードする。
【0020】
本発明のもう1つの態様は、抗体可変領域を含むタンパク質の製造方法を含む。該方法は、(a)該タンパク質が発現される条件下、タンパク質をコードする組換え核酸を含む組換え細胞を増殖させ、(b)該タンパク質を精製する段階を含む。
【0021】
本発明のもう1つの態様は医薬組成物を記載する。該組成物は治療的有効量の抗原結合タンパク質および医薬上許容される担体を含有する。
【0022】
治療的有効量は、有用な治療効果または予防効果をもたらすのに十分な量である。スタヒロコッカス・アウレウス(S.aureus)に感染した患者の場合、有効量は、以下の効果の1以上を達成するのに十分な量である:該患者におけるスタヒロコッカス・アウレウス(S.aureus)の増殖能の軽減、または該患者におけるスタヒロコッカス・アウレウス(S.aureus)の量の減少。スタヒロコッカス・アウレウス(S.aureus)に感染していない患者の場合、有効量は、以下の1以上を達成するのに十分な量である:スタヒロコッカス・アウレウス(S.aureus)感染に対する感受性の軽減、または慢性疾患を招く持続感染を該感染細菌が確立する能力の軽減。
【0023】
本発明のもう1つの態様は、溶液中または細胞上のOFR0657n抗原の存在の検出方法を記載する。該方法は、本明細書に記載の結合タンパク質を該溶液または細胞に加え、該溶液または細胞中の該抗原への該結合タンパク質の結合能を測定することを含む。測定は定量的または定性的でありうる。
【0024】
ORF0657n抗原に対する言及は、完全長ORF0657n、またはmAb 1G3.B3、mAb 2H2.B8、mAb 13C7.D12またはmAb 13G11.C11により認識されるエピトープを有するその誘導体を含む。誘導体の具体例には、トランケート化形態;および以下のアミノ酸改変の1以上を含有するORF0657nの完全長またはトランケート化形態が含まれる:1以上の付加、1以上の置換および1以上の欠失。
【0025】
本発明のもう1つの態様は、スタヒロコッカス・アウレウス(S.aureus)感染に対する患者の治療方法に関する。該方法は、本明細書に記載の抗原結合タンパク質の有効量を患者に投与する段階を含む。治療される患者はスタヒロコッカス・アウレウス(S.aureus)に感染していても感染していなくてもよい。好ましくは、患者はヒトである。
【0026】
本発明のもう1つの態様は、mAb 1G3.B3、mAb 2H2.B8、mAb 13C7.D12もしくはmAb 13G11.C11のいずれかである又はORF0657nへの結合に関してmAb IG3.B3、mAb 2H2.B8、mAb 13C7.D12もしくはmAb 13G11.C11のいずれかと競合するタンパク質を産生する細胞系を記載する。好ましい細胞系は、該タンパク質をコードする組換え核酸を含有する組換え細胞系およびハイブリドーマである。
【0027】
「含む(含んでなる)」のような非限定的用語に対する言及は追加的な要素または段階を許容する。場合によっては、「1以上」のような表現は、追加的な要素または段階の可能性を強調するために非限定的用語と共に又はそれを伴わずに用いられる。
【0028】
明示的に示されていない限り、単数表現は単数を示すと限定されるものではない。例えば、「細胞」は複数の細胞を除外するものではない。場合によっては、可能な複数の存在を強調するために、1以上のような表現が用いられる。
【0029】
本発明の他の特徴および利点は、種々の実施例(具体例)を含む本明細書に記載の更なる説明から明らかである。記載されている実施例(具体例)は、本発明の実施に有用な種々の成分および方法を例示する。該実施例(具体例)は、特許請求されている本発明を限定するものではない。本開示に基づき、当業者は、本発明の実施に有用な他の成分および方法を特定し使用することが可能である。
【0030】
発明の詳細な説明
ORF0657nは、スタヒロコッカス・アウレウス(S.aureus)の外膜に位置するスタヒロコッカス・アウレウス(S.aureus)タンパク質である。ORF0657nはスタヒロコッカス・アウレウス(S.aureus)の種々の株において良く保存されていることが判明している(Andersonら,国際公開番号WO 2005/009379、国際公開日2005年2月3日)。スタヒロコッカス・アウレウス(S.aureus)感染に対する防御免疫応答を引き起こすために、種々のORF0657n誘導体が使用されうる(Andersonら,国際公開番号WO 2005/009379、国際公開日2005年2月3日)。
【0031】
本明細書に記載の抗原結合タンパク質は、ORF0657nを認識するそれらの能力により、例えば、ORF0657nに基づく抗原の製造、特徴づけまたは研究における手段として使用されうる。適当なORF0657nエピトープを認識する抗原結合タンパク質は、スタヒロコッカス・アウレウス(S.aureus)感染を治療するための物質としても使用されうる。
【0032】
I.抗原結合タンパク質
抗原結合タンパク質は、エピトープへの特異的結合をもたらす抗体可変領域を含有する。該抗体可変領域は、例えば、完全抗体、抗体フラグメント、および抗体または抗体フラグメントの組換え誘導体において存在しうる。
【0033】
抗体は、そのクラスによって異なる構造を有する。IgGに関して、種々の抗体領域が例示されうる(図1)。IgG分子は4つのアミノ酸鎖(2つの、より長い重鎖、および2つの、より短い軽鎖)を含有する。重鎖および軽鎖はそれぞれ、定常領域および可変領域を含有する。該可変領域内には、抗原特異性をもたらす3つの超可変領域が存在する(例えば、Breitlingら,Recombinant Antibodies,John Wiley & Sons,Inc.およびSpektrum Akademischer Verlag,1999;およびLewin,Genes IV,Oxford University Press and Cell Press,1990を参照されたい)。
【0034】
該超可変領域(相補性決定領域とも称される)は、より保存されたフランキング領域(フレームワーク領域とも称される)の間に介在している。フレームワーク領域および相補性決定領域に関連したアミノ酸は、Kabatら,Sequences of Proteins of Immunological Interest,U.S.Department of Health and Human Services,1991に記載されているとおりに番号付けされ、整列(アライン)されうる。
【0035】
2つの重鎖カルボキシル領域は、Fc領域を形成するようジスルフィド結合により連結された定常領域である。Fc領域は、補体およびマクロファージの活性化のような抗体の生物活性をもたらすのに重要である。Fc領域を構成する2つの重鎖のそれぞれはヒンジ領域を介して異なるFab領域へと伸長している。
【0036】
高等脊椎動物においては、2つのクラスの軽鎖および5つのクラスの重鎖が存在する。軽鎖はκまたはλである。重鎖は抗体クラスを定め、α、δ、ε、γまたはμである。例えば、IgGはγ重鎖を有する。種々のタイプの重鎖に関しては、ヒトγ1、γ2、γ3およびγ4のようなサブクラスも存在する。重鎖は、ヒンジおよびテイル(尾)領域に、特徴的なコンホメーションをもたらす(Lewin,Genes IV,Oxford University Press and Cell Press,1990)。
【0037】
抗体可変領域を含有する抗体フラグメントには、Fv、FabおよびFab2領域が含まれる。各Fab領域は、可変領域および定常領域から構成される軽鎖と、可変領域および定常領域を含有する重鎖領域とを含有する。軽鎖は定常領域を介してジスルフィド結合により重鎖に連結される。Fab領域の軽鎖および重鎖可変領域は、抗原結合に関与するFv領域を与える。
【0038】
抗体可変領域は組換え誘導体中に存在しうる。組換え誘導体の具体例には、一本鎖抗体、ジアボディ(diabody)、トリアボディ(triabody)、テトラボディ(tetrabody)および小型抗体(miniantibody)が含まれる(Kipriyanovら,Molecular Biotechnology 26:39−60,2004)。
【0039】
該抗原結合タンパク質は、同じ又は異なるエピトープを認識する1以上の可変領域を含有しうる(Kipriyanovら,Molecular Biotechnology 26:39−60,2004)。
【0040】
II.特定された標的領域に対する抗原結合タンパク質の作製
mAb 1G3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1標的領域またはmAb 13G11.BF3標的領域に対する種々の抗原結合タンパク質が、それぞれのモノクローナル抗体から作製されうる。あるいは、結合タンパク質により認識されるエピトープを使用して、追加的な結合タンパク質を選択することが可能である。
【0041】
mAb 2H2.BE11標的領域はORF0657nのアミノ酸約76から357に位置するらしい。抗体を選択するための標的抗原として、ORF0657nのアミノ酸76から357を含有するポリペプチドまたは完全長ORF0657nが使用されうる。作製される抗体の標的領域は決定可能である。
【0042】
抗原を認識するタンパク質を選択するためには、種々の技術が利用可能である。そのような技術の具体例には、ファージ提示技術およびハイブリドーマ産生の利用が含まれる。ヒト抗体は、ゼノマウス(XenoMouse)またはトランスクロモ(Trans−Chromo)マウスのようなキメラマウスを使用して製造されうる(Azzazyら,Clinical Biochemistry 35:425−445,2002,Bergerら,Am.J.Med.Sci.324(1):14−40,2002)。
【0043】
モノクローナル抗体mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1およびmAb 13G11.BF3は、ORF0675nを認識する可変領域を含有する。抗体可変領域に基づき、ORF0657nを認識する追加的な結合タンパク質が製造されうる。追加的な結合タンパク質は、例えば、既存のモノクローナル抗体を修飾することにより、および可変領域配列情報を用いることにより製造されうる。タンパク質の構築および配列の操作は、組換え核酸技術を用いて行われうる。
【0044】
モノクローナル抗体mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1およびmAb 13G11.BF3はマウス抗体である。ヒトの治療用には、そのようなmAbに基づく好ましい結合タンパク質は、該マウス領域を認識するヒト抗マウス抗体の潜在的生成を軽減するよう設計される。
【0045】
ヒト抗マウス抗体の潜在的生成は、マウス抗体のヒト化、脱免疫化(de−immunization)およびキメラ抗体産生のような技術を用いて軽減されうる(例えば、O’Brienら,Humanization of Monoclonal Antibodies by CDR Grafting,p81−100,From Methods in Molecular Biology Vol.207:Recombinant antibodies for Cancer Therapy:Methods and Protocols(WelschofおよびKrauss編)Humana Press,Totowa,New Jersey,2003;Kipriyanovら,Molecular Biotechnology 26:39−60,2004;Gonzalesら,Tumor Biol.26:31−43,2005,Presta,Advanced Drug Delivery Reviews 58:640−656,2006,Tsurushitaら,Methods 36:69−83,2005,Roqueら,Biotechnol.Prog.20:639−654,2004を参照されたい)。
【0046】
マウス抗体は、フレームワーク領域内への相補性決定領域のグラフティングまたはリサーフェシング(resurfacing)のような技術を用いてヒト化されうる。リサーフェシング(ベニアリング(veneering;上張り)としても公知である)は、表面露出領域がヒト化されるよう可変領域を修飾することを含む。
【0047】
相補性決定領域のグラフティングは、そのような領域またはそのような領域の一部を例えばマウス由来物から取り出し該領域をヒト可変領域フレームワーク内に挿入することを含む。グラフティングに使用するヒトフレームワークは、該領域が入手された可変領域(例えば、マウスのもの)に対する配列相同性に基づいて選択されうる。グラフティングされる相補性決定領域に関連する必須フレームワーク残基も新たなフレームワーク内に供給されるべきである。
【0048】
脱免疫化(de−immunization)は、該抗体内に存在する潜在的な直鎖状T細胞エピトープを改変することを含む。該エピトープは、既知のヒトHLAクラスIおよび/またはクラスIIエピトープのバイオインフォマティクススキャンに基づいて特定されうる(Presta,Advanced Drug Delivery Reviews 58:640−656,2006)。
【0049】
キメラ抗体は、ヒト定常領域を、マウスのような異なる生物に由来する可変領域と共に含有する。該ヒト定常領域はFc領域を供与する。
【0050】
改変の追加的な例は、例えば一本鎖抗体、ジアボディ(diabody)、トリアボディ(triabody)、テトラボディ(tetrabody)および小型抗体(miniantibody)における可変領域を提供することを含む(Kipriyanovら,Molecular Biotechnology 26:39−60,2004)。該抗原結合タンパク質は、同じ又は異なるエピトープを認識する1以上の可変領域を含有しうる(同誌)。本発明の追加的な実施形態は、mAb 1G3.BD4、mAb 2H2.BE11、mAb 13C7.BC1またはmAb 13G11.BF3結合部位に対する一本鎖抗体、ジアボディ(diabody)、トリアボディ(triabody)、テトラボディ(tetrabody)および小型抗体(miniantibody)に関する。
【0051】
III.mAb 2H2.BE11標的領域に対する結合タンパク質
後記の実施例に記載されているとおり、mAb 2H2.BE11標的領域を更に詳細に特徴づけし、該可変領域のアミノ酸配列を決定した。特定されている標的領域および配列情報は、mAb 2H2.BE11標的領域に対する種々の結合タンパク質の入手を促進する。
【0052】
本発明の1つの実施形態においては、該結合タンパク質は、配列番号1のアミノ酸76から357よりなるポリペプチドに結合する。好ましくは、該結合タンパク質は、ヒト抗体、ヒト化抗体、脱免疫化(de−immunized)抗体またはキメラ抗体である。好ましい抗体は、単離された抗体およびモノクローナル抗体である。
【0053】
mAb 2H2.BE11可変領域のアミノ酸配列を配列番号20(Vh)および配列番号21(Vl)に示す。Vh内の相補性決定領域(CDR)はアミノ酸36から45,50から65および98から107において特定された。Vl内のCDRは配列番号21のアミノ酸24から33、49から55および88から96において特定された。
【0054】
Vh領域に関する他の実施形態においては、該結合タンパク質はmAb 2H2.BE11標的領域に結合し、そして、配列番号20のアミノ酸36から45またはアミノ酸36から45とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第1 Vh CDR;配列番号20のアミノ酸50から65またはアミノ酸50から65とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第2 Vh CDR;および配列番号20のアミノ酸98から107またはアミノ酸98から107とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第3 Vh CDRを含む又はそれよりなる又はそれより実質的になる。
【0055】
Vl領域に関する他の実施形態においては、該結合タンパク質はmAb 2H2.BE11標的領域に結合し、そして、配列番号21のアミノ酸24から33またはアミノ酸24から33とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第1 VlCDR;配列番号21のアミノ酸49から55またはアミノ酸49から55とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第2 Vl CDR;および配列番号21のアミノ酸88から96またはアミノ酸88から96とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第3 Vl CDRを含む又はそれよりなる又はそれより実質的になる。
【0056】
可変領域、CDR領域または抗体配列に関する「より実質的になる」なる表現は、標的への結合を有意に低下させない1以上の追加的なアミノ酸の、可能な存在を示す。
【0057】
アミノ酸の相違はアミノ酸の欠失、挿入または置換でありうる。活性を維持するようアミノ酸を置換する場合、該置換アミノ酸は、ほぼ同じ電荷、サイズ、極性および/または疎水性のような1以上の類似特性を有するべきである。
【0058】
好ましくは、アミノ酸の置換は保存的置換である。保存的置換は、あるアミノ酸を、類似特性を有する別のアミノ酸で置換するものである。表1はアミノ酸のグループの一覧を示し、ここで、該グループの1つのアミノ酸は別のアミノ酸で保存的置換される。
【0059】
【表1】
【0060】
他の実施形態においては、該Vh領域は配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20であり、および/または該Vl領域は配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21である。
【0061】
抗体に関する他の実施形態においては、該抗体は、(a)この第III節において記載されているVh領域、ならびにヒトヒンジ、CH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4からのもの)を含む重鎖、ならびに(b)この第III節において前記で記載されているVl領域と、ヒトカッパCLまたはヒトラムダCLとを含む軽鎖を含む又はそれによりなる又はそれより実質的になる。他の実施形態においては、該抗体は、(a)この第III節において記載されているVh領域、ならびにヒトヒンジ、CH1、CH2およびCH3領域(IgG1またはIgG2からのもの)を含む重鎖、ならびに(b)この第III節において前記で記載されているVl領域とヒトカッパCLとを含む軽鎖;ならびに配列番号22のアミノ酸配列より実質的になる重鎖および/または配列番号23のアミノ酸配列より実質的になる軽鎖を含む又はそれよりなる又はそれより実質的になる。
【0062】
他の実施形態においては、本明細書に記載の抗原結合タンパク質は、標的抗原に対する少なくとも約100nM、好ましくは少なくとも約30nMのアフィニティーKDをもたらすVhおよびVl領域を有する。標的抗原への結合は、アミノ酸42から486からのORF0657n断片を使用して、実施例11に記載されているとおりに測定されうる。
【0063】
他の実施形態のための好ましい結合タンパク質は抗体である。より好ましくは、該抗体は単離されている、またはモノクローナル抗体である。
【0064】
IV.タンパク質の製造
抗原結合タンパク質は、好ましくは、組換え核酸技術またはハイブリドーマの使用により製造される。組換え核酸技術は、タンパク質合成のための核酸鋳型を構築することを含む。ハイブリドーマ技術は、該抗原結合タンパク質を製造するために不死化細胞系を使用することを含む。適当な組換え核酸およびハイブリドーマ技術は当技術分野でよく知られている(例えば、Ausubel,Current Protocols in Molecular Biology,John Wiley,2005,Harlowら,Antibodies,A Laboratory Manual,Cold Spring Harbor Laboratory,1988を参照されたい)。
【0065】
抗原結合タンパク質をコードする組換え核酸は、実際に該コード化タンパク質の生産工場として働く宿主細胞内で発現されうる。該組換え核酸は、宿主細胞ゲノムから自律して又は宿主細胞ゲノムの一部として存在する該抗原結合タンパク質をコードする組換え遺伝子を与えうる。
【0066】
組換え遺伝子は、タンパク質の発現のための調節要素と共に、タンパク質をコードする核酸を含有する。一般には、組換え遺伝子内に存在する調節要素には、転写プロモーター、リボソーム結合部位、ターミネーター、および場合によって存在するオペレーターが含まれる。真核細胞内でのプロセシングのための好ましい要素はポリアデニル化シグナルである。抗体に関連するイントロンも存在しうる。抗体または抗体フラグメントの製造のための発現カセットの具体例は当技術分野でよく知られている(例えば、Persicら,Gene 187:9−18,1997,Boelら,J.Immunol.Methods 239:153−166,2000,Liangら,J.Immunol.Methods 247:119−130,2001,Tsurushitaら,Methods 36:69−83,2005)。
【0067】
遺伝暗号の縮重のため、特定のタンパク質をコードするために多数の異なるコード化核酸配列が用いられうる。ほとんど全てのアミノ酸は、異なる組合せのヌクレオチドトリプレット、すなわち「コドン」によりコードされているため、遺伝暗号の縮重が生じる。アミノ酸は、以下のとおりに、コドンによりコードされる。
A=Ala=アラニン:コドンGCA、GCC、GCG、GCU。
C=Cys=システイン:コドンUGC、UGU。
D=Asp=アスパラギン酸:コドンGAC、GAU。
E=Glu=グルタミン酸:コドンGAA、GAG。
F=Phe=フェニルアラニン:コドンUUC、UUU。
G=Gly=グリシン:コドンGGA、GGC、GGG、GGU。
H=His=ヒスチジン:コドンCAC、CAU。
I=Ile=イソロイシン:コドンAUA、AUC、AUU。
K=Lys=リシン:コドンAAA、AAG。
L=Leu=ロイシン:コドンUUA、UUG、CUA、CUC、CUG、CUU。
M=Met=メチオニン:コドンAUG。
N=Asn=アスパラギン:コドンAAC、AAU。
P=Pro=プロリン:コドンCCA、CCC、CCG、CCU。
Q=Gln=グルタミン:コドンCAA、CAG。
R=Arg=アルギニン:コドンAGA、AGG、CGA、CGC、CGG、CGU。
S=Ser=セリン:コドンAGC、AGU、UCA、UCC、UCG、UCU。T=Thr=トレオニン:コドンACA、ACC、ACG、ACU。
V=Val=バリン:コドンGUA、GUC、GUG、GUU。
W=Trp=トリプトファン:コドンUGG。
Y=Tyr=チロシン:コドンUAC、UAU。
【0068】
細胞内の組換え遺伝子の発現は、発現ベクターを使用して促進される。好ましくは、該発現ベクターは、組換え遺伝子に加えて、宿主細胞内での自律的複製のための複製起点、選択マーカー、限られた数の有用な制限酵素部位および潜在的な高コピー数をも含有する。抗体および抗体フラグメントの製造のための発現ベクターの具体例は当技術分野でよく知られている(例えば、Persicら,Gene 187:9−18,1997,Boelら,J.Immunol.Methods 239:153−166,2000,Liangら,J.Immunol Methods 247:119−130,2001,Tsurushitaら,Methods 36:69−83,2005)。
【0069】
所望により、当技術分野でよく知られた技術を用いて(例えば、Ausubel,Current Protocols in Molecular Biology,John Wiley,2005,Marksら,国際出願番号WO95/17516(国際公開日1995年6月29日)を参照されたい)、抗体をコードする核酸を宿主染色体内に組込むことが可能である。
【0070】
組換え抗原結合タンパク質の発現には、原核生物(例えば、大腸菌(E.coli)、バシラス属種(Bacillus sp)およびストレプトマイセス属種(Streptomyces sp.)(または放線菌属))および真核生物(例えば、酵母、バキュロウイルスおよび哺乳類)に由来する細胞系を含む多種多様な細胞系が使用されうる(Breitlingら,Recombinant Antibodies,John Wiley & Sons,Inc.and Spektrum Akademischer Verlag,1999,Kipriyanovら,Molecular Biotechnology 26:39−60,2004,Tsurushitaら,Methods 36:69−83,2005)
【0071】
組換え抗原結合タンパク質の発現のための好ましい宿主は哺乳類翻訳後修飾をもたらす。翻訳後修飾には、化学的修飾、例えばグリコシル化およびジスルフィド結合形成が含まれる。もう1つのタイプの翻訳後修飾はシグナルペプチドの切断である。
【0072】
適切なグリコシル化は抗体機能に重要でありうる(Yooら,Journal of Immunological Methods 261:1−20,2002,Liら,Nature Biotechnology 24(2):210−215,2006)。天然に存在する抗体は、重鎖に結合した少なくとも1つのN−結合炭水化物を含有する(Yooら,Journal of Immunological Methods 261:1−20,2002)。他のN−結合炭水化物およびO−結合炭水化物が存在することが可能であり、抗体機能に重要でありうる(同誌)。
【0073】
効率的な翻訳後修飾を得るために、哺乳類宿主細胞および非哺乳類細胞を含む種々のタイプの宿主細胞が使用されうる。哺乳類宿主細胞の具体例には、チャイニーズハムスター卵巣(CHO)、HeLa、C6、PC12、ヒト胎児腎(HEK293)および骨髄腫細胞が含まれるが、これらに限定されるものではない(Yooら,Journal of Immunological Methods 261:1−20,2002,Persicら,Gene 187:9−18,1997)。非哺乳類細胞は、ヒトグリコシル化を模倣するよう改変されうる(Liら,Nature Biotechnology 24(2):210−215,2006)。糖操作(gIycoenginnered)されたピチア・パストリス(Pichia pastoris)は、そのような改変された非哺乳類細胞の一例である(Liら,Nature Biotechnology 24(2):210−215,2006)。
【0074】
好ましい組換え遺伝子は、mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合する抗体可変領域をコードするヌクレオチド配列を含む。特定の組換え遺伝子は、一方の可変領域またはVhおよびVlの両方の領域を含有するタンパク質をコードしうる。該組換え遺伝子は抗体定常領域およびヒンジ領域をもコードしうる。所望により、1つの遺伝子が軽鎖をコードし第2の遺伝子が重鎖をコードする、組換え遺伝子の組合せを用いて、抗体が製造されうる。
【0075】
他の実施形態は、前記第II節または第III節に記載のタンパク質をコードする核酸により提供される。そのような実施形態の具体例を以下に記載する。
【0076】
Vhコード領域に関する1つの実施形態においては、該ヌクレオチド配列は、配列番号20のアミノ酸36から45またはアミノ酸36から45とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第1 Vh CDR;配列番号20のアミノ酸50から65またはアミノ酸50から65とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第2 Vh CDR;および配列番号20のアミノ酸98から107またはアミノ酸98から107とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第3 Vh CDRを含む又はそれよりなる又はそれより実質的になる可変領域をコードする。
【0077】
Vlコード領域に関する1つの実施形態においては、該ヌクレオチド配列は、配列番号21のアミノ酸24から33またはアミノ酸24から33とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第1 VlCDR;配列番号21のアミノ酸49から55またはアミノ酸49から55とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第2 Vl CDR;および配列番号21のアミノ酸88から96またはアミノ酸88から96とは1つのアミノ酸が異なる配列を含む又はそれよりなる又はそれより実質的になる第3 Vl CDRを含む又はそれよりなる又はそれより実質的になる可変領域をコードする。
【0078】
他の実施形態においては、該Vh領域は配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20であり、および該Vl領域は配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21である。
【0079】
抗体重鎖および/または軽鎖に関する他の実施形態においては、該組換え遺伝子は、(a)前記第III節に記載されているVh領域、ヒトヒンジ、CH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4サブタイプからのもの)を含む重鎖、または(b)前記第III節に記載されているVl領域と、ヒトカッパCLまたはラムダCLとを含む軽鎖を含む又はそれによりなる又はそれより実質的になるタンパク質の一方または両方をコードする。もう1つの実施形態においては、該重鎖は配列番号22のアミノ酸配列より実質的になり、該軽鎖は配列番号23のアミノ酸配列より実質的になる。
【0080】
V.抗原結合タンパク質の用途
あるORF0657n領域を含有する抗原は、スタヒロコッカス・アウレウス(S.aureus)感染に対する防御免疫応答をもたらすために使用されうる(Andersonら,国際公開番号WO2005/009379(国際公開日2005年2月3日))。ORF0657n標的領域を認識する抗原結合タンパク質は、ORF0657n抗原およびワクチンの製造、特徴づけ又は研究を促進するために使用されうる。また、適当なエピトープを認識する抗原結合タンパク質は治療用途を有しうる。
【0081】
ORF0657n関連抗原およびワクチンの製造、特徴づけ又は研究における種々の用途の具体例には以下のものが含まれる。
【0082】
1)例えばウエスタンブロットによる、ORF0657n抗原の存在の特定。
【0083】
2)例えばフローサイトメトリーによる、細胞表面上のORF0657n抗原の存在の特定。これは、例えば、スタヒロコッカス・アウレウス(S.aureus)の複数の株に関する発現の測定、およびノックアウト突然変異体の確認において有用である。
【0084】
3)受動防御実験。該抗体は、ORF0657nタンパク質の特定の領域が防御をもたらすのかどうかを判定するために致死モデルにおいて使用されうる。
【0085】
4)イムノアッセイ。該アッセイは、抗原の質、産物の生産性および安定性をモニターするために用いられうる。
【0086】
5)抗原ワクチン産物の免疫原性をモニターするためのマウス効力アッセイにおける対照として。
【0087】
6)血清学的アッセイにおいて、ORF0657n由来抗原でワクチン接種された患者に対する免疫応答を特定するために、モノクローナル抗体が競合様態で使用されうる。
【0088】
標的タンパク質の製造、特徴づけ又は研究においてモノクローナル抗体のような抗原結合タンパク質を使用するための技術は当技術分野でよく知られている(例えば、Ausubel,Current Protocols in Molecular Biology,John Wiley,2005,Harlowら,Antibodies,A Laboratory Manual,Cold Spring Harbor Laboratory,1988,Harlowら,Using Antibodies,Cold Spring Harbor Laboratory,Cold Spring Harbor,N.Y.,Cold Spring Harbor Laboratory Press,1999,Lipmanら,ILAR Journal 46:258−268,2005を参照されたい)。
【0089】
本発明の1つの実施形態においては、抗原結合タンパク質を使用して、溶液中の又はミクロスフェアに結合した又は細胞上のORF0657n抗原の存在を確認する。溶液または細胞に存在するタンパク質への該結合タンパク質の結合能は、ウエスタンブロット、酵素結合イムノソルベントアッセイ(ELISA)、フローサイトメトリーおよびルミネックス(Luminex)イムノアッセイのような種々の技術を用いて測定されうる。
【0090】
VI.治療
適当な標的領域に結合する抗原結合タンパク質を使用して、治療的および予防的治療が患者に対して行われうる。治療的治療は、スタヒロコッカス・アウレウス(S.aureus)に感染した者に対して行われる。予防的治療は一般集団または一般集団の小集団に対して行われうる。一般集団の好ましい小集団としては、スタヒロコッカス・アウレウス(S.aureus)感染のリスクの高い者が挙げられる。
【0091】
「患者」は、スタヒロコッカス・アウレウス(S.aureus)に感染しうる哺乳動物を意味する。好ましくは、患者はヒトである。しかし、他のタイプの哺乳動物、例えばウシ、ブタ、ヒツジ、ヤギ、ウサギ、ウマ、イヌ、ネコ、サル、ラットおよびマウスがスタヒロコッカス・アウレウス(S.aureus)に感染しうる。非ヒト患者の治療は、ペットおよび家畜を防御するのに、ならびに特定の治療の効力を評価するのに有用である。
【0092】
スタヒロコッカス・アウレウス(S.aureus)感染のリスクの高い者には、医療従事者、入院患者、低下した免疫系を有する患者、手術を受けている患者、カテーテルまたは血管装置のような異物インプラントを受け入れている患者、免疫低下を招く療法を受けている患者、および火傷または創傷のリスクの高い職業の者が含まれる(The Staphylococci in Human Disease,Crossley and Archer(編),Churchill Livingstone Inc.1997)。
【0093】
1つの実施形態においては、手術または異物インプラントと共に、抗原結合タンパク質を患者に投与する。「手術または異物インプラント」に対する言及は、異物インプラントの付与を伴う又は伴わない手術、および手術を伴う又は伴わない異物インプラントの付与を含む。投与の時機は、予防的治療および/または治療的治療が達成されうよう計画されうる。投与は、好ましくは、手術または移植とほぼ同時に開始されうる。
【0094】
医薬投与全般に関する指針は、例えばRemington’s Pharmaceutical Sciences 20th Edition,Gennaro編,Mack Publishing,2000;ならびにModern Pharmaceutics 2nd Edition,BankerおよびRhodes編,Marcel Dekker,Inc.,1990に記載されている。
【0095】
医薬上許容される担体は抗原結合タンパク質の保存および投与を促進する。タンパク質溶液製剤を安定化するために使用される物質には、炭水化物、アミノ酸および緩衝化塩が含まれる(Middaughら,Handbook of Experimental Pharmacology 137:33−58,1999)。
【0096】
抗原結合タンパク質は静脈内、皮下、筋肉内または粘膜のような種々の経路により投与されうる。皮下および筋肉内投与は、例えば針または噴射式注射器を使用して行われうる。鼻腔内運搬のような粘膜運搬は、吸着部位における、より長い保持時間を得るために、増強物質または粘膜付着物質を使用することを含みうる(Middaughら,Handbook of Experimental Pharmacology 137:33−58,1999)。
【0097】
適当な投与計画は、好ましくは、患者の年齢、体重、性別および医学的状態;投与経路;所望の効果;ならびに使用する個々の化合物を含む、当技術分野でよく知られた要因を考慮して決定される。有効用量範囲は約0.1mg/kg〜20mg/kg、または0.5mg/kg〜5mg/kgとなると予想される。投与頻度は該化合物の有効性および安定性によって異なりうる。投与頻度の具体例には、隔週、毎週、毎月および隔月が含まれる。
【0098】
VII.実施例
以下に記載の実施例は本発明の種々の特徴を更に詳しく例示するものである。該実施例は本発明の実施のための有用な方法をも例示する。これらの実施例は、特許請求されている本発明を限定するものではない。
【0099】
(実施例1)
ORF0657nに対するモノクローナル抗体の作製
ORF0657n−C/e(配列番号2)またはORF0657n−H/y(配列番号3)を抗原として使用して、ORF0657n(配列番号1)に対するモノクローナル抗体を作製した。ELISAおよびフローサイトメトリーにより該抗体を特定し、特徴づけした。
【0100】
マウスおよび免疫化:4〜5週齢の雌BALB/cマウスをTaconic(Germantown,N.Y.)から購入した。アルミニウムヒドロキシホスファートアジュバント上で製剤化された、大腸菌(E.coli)により産生されたORF0657n−C/e抗原または酵母により発現されたORF0657n−H/y抗原の20μgで、第0、7および21日にマウスの筋肉内に免疫化した(Andersonら,国際公開番号WO 2005/009379(国際公開日2005年2月3日))。リン酸緩衝食塩水(PBS)中の20μgのタンパク質の最終静脈内注射(i.v.)を、融合の3日前にマウスに実施した。マウスを犠死させ、細胞融合のために脾臓を摘出した。
【0101】
MAbの製造:脾臓から調製したリンパ球をマウス骨髄腫対応体SP2/0−Agl4(ATCC 1581)に、3:1の比のポリエチレングリコール1500(Boehringer Mannheim)により融合させた。20% ウシ胎児血清、ヒポキサンチン(10−4M)、チミジン(10−5M)を含有する高グルコース、ピルビン酸のダルベッコ変法イーグル培地(DMEM)を含む96ウェル平底マイクロタイタープレート内に該融合体をプレーティングし、24時間後にアミノプテリン(4×10−7M)を加えた。増殖(成長)中のハイブリドーマからの上清を、後記のとおり、ORF0657nに対する反応性に関してELISAによりスクリーニングした。陽性ウェルを限界希釈によりクローニングし、ELISA反応性に関して再試験した。モノクローナル抗体を抗体イソタイプ決定キット(Roche Diagnostics Corporation,Indianapolis,IN)で分類した。
【0102】
ELISA:コスター(Costar)培地結合性マイクロタイタープレートを、大腸菌(E.coli)により発現された、PBS中の配列番号2の50ナノグラム/ウェルで、2〜8℃で一晩コーティングした。該プレートをPBS、0.05% Tween20で3回洗浄し、1% ウシ血清アルブミン、PBS、0.05% Tween20(アッセイ希釈剤)で少なくとも1時間ブロッキングした。該プレートを前記のとおりに洗浄し、該融合ウェルまたはクローン化ハイブリドーマからの上清を加え、室温で2時間インキュベートした。該プレートを前記のとおりに洗浄し、ヤギ抗マウスIgG(H+L)−HRPコンジュゲート(Zymed)(アッセイ希釈剤中で1:8000)を加え、室温で1時間インキュベートした。アッセイプレートをTMB基質で現像し、反応を2.0N H2SO4で停止させ、OD450nmでプレートリーダーで読み取った。450nmで1.0を超える光学濃度を有するウェルを陽性とみなした。
【0103】
フローサイトメトリー:鉄枯渇条件(RPMI中)下で継代されたスタヒロコッカス・アウレウス(S.aureus)の調製されたグリセロールストックを使用して、ORF0657n結合に関してmAbを評価した。凍結グリセロールストック細胞を解凍し、PBS;1% ウシ血清アルブミン;0.1% アジ化ナトリウム、0.2% ブタIgG(Sigma)(PAAG)に、5×107 CFU/50μlの濃度まで再懸濁させた。該細胞の50μlアリコートを反応ごとに1.5mlエッペンドルフチューブ内に配置した。50マイクロリットルの該ハイブリドーマ培養物を各反応チューブに加え、室温で1時間インキュベートした。1mLのリン酸緩衝食塩水、1% ウシ血清アルブミン、0.1% アジ化ナトリウム(PAA)を該チューブに加えることにより、該細胞を洗浄した。該細胞を遠心分離(5500rpm、5分間)によりペレット化した。上清を取り出し、該細胞を100μLの二次抗体(PAAG中で1:100希釈されたFITC標識ヤギ抗マウスIg(BD Pharmingen))と混合した。インキュベーションは暗所内で室温で1時間行った。インキュベーション後、1mLのPAAを該反応混合物に加え、該細胞をペレット化し(5500rpm、5分間)、上清を取り出した。該ペレットを1mLのPBSに再懸濁させ、FAC分析のために12×75mmチューブに移した。
【0104】
チューブを、細菌細胞に関してゲート化され該細胞関連FITCの量を測定するBD−FACSCaliburフローサイトメーター装置上で測定に付した。スタヒロコッカス・アウレウス(S.aureus)の表面に結合することが判明している標準的な抗体を、各アッセイにおいて測定に付した。陰性対照を、細胞および該二次コンジュゲートの単独体として測定に付した。幾何平均値が30より大きい場合に、ハイブリドーマウェルは陽性であるとみなした。
【0105】
2つの別々の融合体は12個のモノクローナル抗体(mAb)のパネルを与えた。該mAbの全てはELISAにおいて反応性であった(表2)。これらの12個のmAbのうちの10個が細菌の表面に結合したことが、フローサイトメトリーにより実証された。該mAbの全ては、野生型タンパク質を使用するウエスタンブロット分析で陽性であった。
【0106】
【表2】
【0107】
(実施例2)
mAbのクラススイッチ
天然抗原に結合した単離されたmAbの全てはIgG1イソタイプのものであった。シフト変異体の選択により、これらの抗体はIgG2bイソタイプにクラススイッチされた(Spiraら,J.of Immunogical Methods,74:307−315,1985)。IgG2bを使用して適当なイムノアッセイを開発し、該細胞系を高密度でプレーティングした。体細胞突然変異を選択し、富化させ、ついでクローニングした。スイッチしたmAbの結合部位は依然として元のmAbと同じであったが、IgG2bサブタイプへのスイッチは、受動防御研究における、より好ましいイソタイプ(補体カスケードを開始させる)を与えた。
【0108】
【表3】
【0109】
(実施例3)
天然抗原を使用する結合阻害研究
mAb標識キット(Molecular Probes)を該製造業者の説明に従い使用して、精製抗体をアレクサフルオル(Alexafluor)−488で標識した。RPMI増殖細菌細胞の表面をちょうど飽和させるmAbの量を、標識mAbおよび未標識mAbの両方に関して測定した。表3(第1列)のmAbのそれぞれを、標識体および未標識体として使用した。
【0110】
まず、5×107個の細胞を、該細胞の表面を飽和させる濃度の該未標識mAbと共にインキュベートすることにより、該阻害アッセイを行った。この反応を室温で1時間インキュベートした。このインキュベーションの後、該反応を1mlのPAAで洗浄し、ミクロ遠心機(Hermle)において6,000RPMで5分間遠心した。上清を除去して〜50μlに減少させ、該細胞を、該細胞の表面をちょうど飽和させる量の直接標識mAbを含有する100μlのPAAGに再懸濁させた。このインキュベーションの後、該反応を1mlのPAAで洗浄し、ミクロ遠心機(Hermle)において6,000RPMで5分間遠心した。上清を除去して〜50μlに減少させ、該細胞を1mlのPBSに再懸濁させ、FAC分析のために12×75mmチューブに移した。対照として、該未標識mAbを含有する別の反応を二次アレクサフルオル(Alexafluor)−488コンジュゲート化ヤギ抗マウスIgG(H+L)(Molecularプローブ;PAAG中、1:400)で測定して、このmAbが該表面に結合することを確認した。該細胞と共に該標識mAbのみを含有する陽性対照についても実験を行った。該未標識mAbが、該標識mAbと同じエピトープに結合すれば、該細胞に関連した蛍光活性は全く無いか又は低いことになる。該未標識mAbが、該標識mAbとは異なるエピトープに結合すれば、該表面に関連した反応性のレベルは、該標識mAbのみの対照細胞と同等ということになる。
【0111】
阻害研究により、モノクローナル抗体のパネルは4つの反応性グループに分類された。
【0112】
【表4】
【0113】
(実施例4)
変性抗原および改変抗原での結合研究
結合を更に特徴づけるために、ORF0657n改変タンパク質を使用した。まず、ORF0657nをコードする核酸を発現ベクターpET−28a(Novagen)内にクローニングし、C末端6×hisタグと共に大腸菌(E.coli)内で発現させた(配列番号2)。StratageneのQuikChange XL Site−Directed Mutagenesis Kitを該製造業者の説明に従い使用して、該クローン化遺伝子を含有する発現ベクターを突然変異誘発に付した。該遺伝子は、特異的な連続的アミノ酸変化を伴って突然変異した。得られたプラスミドを、StratageneのXL10−Goldコンピテント細胞内に、該製造業者のプロトコールに従い形質転換した。QiagenのQIAprep Spin Miniprep Kitを使用して、プラスミドを形質転換体から単離した。ABIの310 DNA Sequencerを使用して配列決定することにより、形質転換体をスクリーニングした。最大数の塩基変化を示す形質転換体からのプラスミドを発現宿主HMS 174(DE3)(Novagen)内に形質転換した。Novagenの説明に従い、形質転換体を発現させた。
【0114】
異なるORF0657n改変タンパク質を使用して、ORF0657n mAb(配列番号4〜19)の多様性を確認した。これらのタンパク質を、標準的な方法を用いるドットブロットにおいて10個の異なるmAbでスクリーニングした。標準的な方法を用いるウエスタンブロットにより、陽性/陰性を確認した。このアプローチにより、抗体を、それらの結合プロファイルに従い分類した。該抗体のうちの7個は3つのグループに分類され、残りの3個の抗体(2H2.B8、8A8.E11.H3および13G11.C11)は、互いに類似しているが同一ではないプロファイルを有していた(表5)。
【0115】
【表5】
【0116】
(実施例5)
BAIcore研究
BIAcore研究において、精製ORF0657n−H/yを抗原として使用する「フットプリント分析」によりmAbを検査した。BIACORE(登録商標)によるリアルタイム生体分子相互作用分析を用いて、ペア様態結合実験を行った。BIACORE(商標登録)は、カルボキシルメチルデキストランでコーティングされた(CM5)センサーチップの表面に直接照射される偏光の屈折率の変化をモニターすることにより質量変化を検出するために、ミクロフルイディクス技術および表面プラズモン共鳴(SPR)を組合せたものである。応答単位(Response Unit)で測定される応答の変化は結合アナライト(すなわち、抗原または抗体)の量に相関されうる。
【0117】
抗スタヒロコッカス抗体(mAb 13C7.D12)を該CM5センサーチップの表面上に共有結合(固定化)させた。該固定化mAbを、まず、ORF0657nタンパク質にさらし、ついでマトリックス形態で抗体のペアにさらした。ORF0657nタンパク質+抗体ペアの各サイクルの後、20mM HClを使用して、該センサーチップの表面を固定化mAb 13C7.D12へと再生させた。各抗体ペアの全ての組合せが分析されうるよう、マトリックス形態でORF0657nタンパク質に対して8個の抗体を試験した。この実験で使用したmAbペアに関するマトリックス設計を表6に要約する。
【0118】
【表6】
【0119】
各実施において最初に結合した(捕捉された)抗原の量を正規化するために、各試験抗体/抗原複合体に関する以下の比を計算する。
【0120】
【数1】
【0121】
各抗体に残存している利用可能なエピトープの割合(%)は、以下のとおりに、マッピングペアに関して計算されうる。
【0122】
【数2】
【0123】
図2は、ペア様態の結合研究における該モノクローナル抗体の反応性の概要を示すマトリックスを例示する。モノクローナル抗体のパネルは、BIACORE(登録商標)法により、3つの反応性領域に分類された(表7を参照されたい)。
【0124】
【表7】
【0125】
(実施例6)
マウス敗血症モデルにおける受動免疫化での防御研究
モノクローナル抗体mAb 2H2.BE11およびmAb 13C7.BC1を、スタヒロコッカス・アウレウス(S.aureus)感染に対する防御をもたらすそれらの能力に関して試験した。これらの抗体はORF0657nタンパク質上の異なるエピトープを認識する。対照には、イソタイプが一致したmAbおよびPBS単体が含まれた。
【0126】
細菌チャレンジの20時間前に該mAbまたはPBSを腹腔内(i.p.)投与した。ついでマウスをLD80−90用量のスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)でチャレンジ(i.v.)し、生存をモニターした。各実験を10または20匹のマウスの群で3回反復し、10日間モニターした。未感染BALB/cマウスにおける該モノクローナル抗体に関する半減期は約8日間である。0.5mgの用量が最適であることが判明した。それらの2つのモノクローナル抗体での実験の結果を図3A〜C、4A、4Bおよび5A〜Cに示す。
【0127】
1つの実験においては、mAb 13C7.BC1は対照と比べて第10日における生存を有意に改善したが、その他の2つの反復実験においては、全体的な生存率は対照の場合に類似していた(図3A〜3C)。しかし、対照と比べて、mAb 13C7.BC1で治療されたマウスの、この10日間以内の死亡までの時間における遅延が認められた。また、3つの実験のうちの2つにおいて、mAb 2H2.BE11で治療されたマウスの死亡までの時間の遅延における類似した傾向が認められた(図5A〜5C)。
【0128】
また、最近のスタヒロコッカス・アウレウス(S.aureus)臨床分離株であるUK58を使用して、mAb 13C7.BC1の効果を調べた(図4Aおよび4B)。この株は患者の膿瘍部位から最低限度で継代された。2つの独立した実験において、結果はUK58チャレンジによる死亡までの時間における遅延を示している。
【0129】
抗体持続性研究は、該LD80−90モデルにおいては、死亡速度が速いため評価できない。したがって、亜致死チャレンジモデルを用いた。該亜致死モデルにおいては、使用したチャレンジ用量は、該LD80−90モデルで使用した用量の10%である。該亜致死チャレンジモデルを4日間にわたってモニターした。22匹のマウスの群に0.5mgの用量のmAb 13C7.BC1またはイソタイプ対照mAb(6G6)を、5×107 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)でのi.v.細菌チャレンジの20時間前に投与した。チャレンジの時点の血清中のmAbレベルを測定するために、チャレンジの直前(T=0)に、各群の2匹の動物を犠死させた。チャレンジの2、24、48、72および96時間後、各群の4匹のマウスを犠死させ、血清mAbレベルを測定した。
【0130】
この亜致死チャレンジ実験から、スタヒロコッカス・アウレウス(S.aureus)感染マウスにおけるmAb 13C7.BC1の半減期は約1日であると推定された。これとは対照的に、イソタイプ対照mAbの半減期は4日を超えると推定された(データ非表示)。これらのデータは、スタヒロコッカス・アウレウス(S.aureus)でチャレンジされたマウスにおけるmAb 13C7.BC1の特異的減少を示しており、これは、該致死モデルにおいてモニターされた10日間のかなり前に枯渇すると思われる。
【0131】
図3A〜C、4A、4Bおよび5A〜Cに例示されている8つの実験のうちの6つにおいて、該mAbを投与した群に関して約3日までの生存の改善が認められた。これらの結果は、そのようなmAbがスタヒロコッカス・アウレウス(S.aureus)チャレンジマウスの生存率に対して正の効果を及ぼすことを示している。
【0132】
(実施例7)
マウス留置カテーテルモデルにおける受動免疫化での防御研究
マウス留置カテーテルモデルをmAb 2H2.BE11と共に使用した。このモデルにおいて使用したスタヒロコッカス・アウレウス(S.aureus)株は臨床分離株MCL8538であった。この株を選択したのは、マウス敗血症モデルにおいて使用した株であるスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)と比べて、カテーテルの再現可能なコロニー形成を尚も達成しつつ、より少量の接種物の投与が可能だからである。
【0133】
ICRマウスは、頚静脈内に外科的に移植され縫合により適所に保持され該マウスの背側中央線上の出口から出るカテーテル(PE50シリコーンゴム)を有していた。術後9〜11日間、マウスを休ませた。チャレンジの24時間前に、600mcgのマウスモノクローナル抗体2H2.BE11の1回のi.p.注射投与によりマウスを受動免疫化した。第0日に、スタヒロコッカス・アウレウス(S.aureus)MCL8538のi.v.投与によりマウスをチャレンジした。接種物の用量は100μlの体積中、2〜8×105 CFUであった(実験1〜3)。この低用量は4日後に該カテーテルから自然に消失することが判明した。したがって、チャレンジの24時間後に細菌に関してカテーテルを評価した。その時点で、マウスを犠死させ、カテーテルを回収した。TSA上でカテーテル全体を培養することにより、カテーテル上の細菌の存在を評価した。該プレート上で増殖の徴候が認められたら、該カテーテルは培養陽性であると評価した。
【0134】
最初の3つの実験の2つにおいて、培養陰性カテーテルの数は、抗体2H2.BE11で受動免疫化されたマウスにおいては、イソタイプ対照抗体の場合より有意に低かった。より大きな接種物用量を用いて、4番目の実験を行った。このより過酷なチャレンジにおいては、該用量は、カテーテルの100%が再現可能に感染し対照マウスにおいてこの感染が自然には消失しない(7日間にわたるモニター)用量であると定められた。そのようなより大きな接種物サイズを用いる実験4においてもまた、2H2.BE11に対する抗体を注射したマウスにおいては、イソタイプ対照の場合と比べて、有意に少数のカテーテルが感染したことが判明した。4つの実験の結果を表8に要約する。
【0135】
【表8】
【0136】
(実施例8)
抗ORF0657nモノクローナル抗体2H2.B8(IgG1)、2H2.BE11(IgG2b)もしくは13C7.IgG2bまたはイソタイプ一致対照mAbを使用する、細菌のエクスビボ前オプソニン化
ORF0657nのモノクローナル抗体がオプソニン性であるかどうかを試験するために、致死用量のスタヒロコッカス・アウレウス(S.aureus)をモノクローナル抗体2H2.B8、2H2.BE11もしくは13C7.IgG2bまたはイソタイプ一致対照モノクローナル抗体で前オプソニン化させる受動防御実験を行った。ついで前オプソニン化細菌をマウスにi.p.投与した。これらの実験で使用した細菌はスタヒロコッカス・アウレウス(S.aureus)RN4220(野生型)またはRN4220.0657nであった。RN4220.0657n細菌を、FURボックスによる制御の非存在化でORF0657nを発現するよう操作した。したがって、それは鉄の存在下で増殖可能であり、その表面上でORF0657n抗原を尚も発現することが可能であった。あるいは、RN4220(野生型)を、該細菌表面上での0657nの発現を誘導するために、低鉄培地RPMI内で2回継代した。
【0137】
6Balb/cマウスに十分な細菌の量(6×LD100)を、穏やかに振とうしながら、800μgのIgGと共に4℃で1時間インキュベートした。ついで細菌をペレット化し、未結合mAbを除去した。抗体オプソニン化細菌を2.4mLのPBSに再懸濁させ、0.4mL(1×LD100)を5匹のマウスのそれぞれに注射した。マウスの全群に同等のCFUがもたらされること及び該mAbが該細菌を凝集させていないことが保証されるよう、チャレンジ後、TSA上のプレーティングにより各接種物を定量した。チャレンジ後3日間、生存をモニターした。この手法が有効となるためには該標的抗原が該細菌の表面上に存在しなければならないため、オプソニン化前に該細菌上で0657nが発現されることが保証されるよう配慮した。mAb 2H2.B8を使用するフローサイトメトリーにより、ORF0657nの発現をモニターした。各マウスに注射したオプソニン化細菌の用量は2〜4×109 CFU RN4220.0657n/マウスまたは1〜2×109 CFU RN4220(2×RPMI)/マウスであった。
【0138】
マウスは、イソタイプ一致対照mAbではなく2H2.B8または2H2.BE11のいずれかで前オプソニン化された場合に、致死用量のRN4220.0657nスタヒロコッカスによる死亡から防御された。該実験をIgG1イソタイプに関して2回、およびIgG2bイソタイプに関して3回反復したところ、同様の結果が得られた(表9A)。
【0139】
【表9】
【0140】
マウスは、イソタイプ一致対照mAbではなくmAb 2H2.B8で前オプソニン化された場合に、致死用量のRN4220(2×RPMI)スタヒロコッカスによる死亡から防御された。該実験を6回反復したところ、同様の結果が得られた(表9B)。
【0141】
【表10】
【0142】
マウス抗0657n 2H2は、この致死モデルにおける死亡の予防に非常に有効であった。13C7 mAbは、(図3〜6に例示されている既に記載されているモデルの場合とは対照的に)このモデルでは有効でなかった。該抗0657n mAbの全て(2H2.BE11、2H2.B8および13C7.IgG2b)がRN4220に結合し(フローサイトメトリーを用いて実証されているとおり)、全てがインビトロOPAアッセイにおいてオプソニン活性を有する。このモデルは、該マウスの腹膜における、生存を向上させるためのエピトープ特異性に関する追加的な要件を表している。
【0143】
(実施例8)
2H2 mAbで行ったエピトープマッピング研究
この実施例に記載する実験は、モノクローナル抗体2H2.BE11がORF0657n内のコンホメーションエピトープを認識するという証拠を提供する。該実験は、2H2 mAbにより認識される三次元構造におけるコンホメーションエピトープを提示するのに要求されるORF0657n内の最小配列を位置決定した。また、該実験は、2H2 mAbがORF0657nに結合すると小分子との反応から保護される特徴的なリシン残基をORF0657nの最小配列内に特定した。
【0144】
ORF0657nの配列内の典型的には9から14アミノ酸長の直鎖状エピトープを2H2 mAbが認識する潜在能力を、エピトープ抽出を用いて、配列番号1のアミノ酸42からアミノ酸486のORF0657n断片(「ORF0657t」)から調べた。詳細に説明すると、該エピトープ抽出実験のそれぞれについて、10mgの臭化シアン活性化セファロース(Amersham cat.No.17 043001)への化学的架橋により30μgの2H2 mAbを固定化した。ORF0657tのタンパク質分解消化物をGluC(Roche Applied Science cat.No.11 420 3997 001)、Asp−N(Roche Applied Science cat.No.11 054 589 001)またはキモトリプシン(Roche Applied Science cat.No.11 418 467 001)で生成させ、リニアイオントラップ(LTQ−Thermo Electron Inc)において1D/LC−MS/MSにより特徴づけた。3つの独立した実験において、いずれかのプロテアーゼに由来する特徴づけられたタンパク質分解消化物8.4μgを該固定化抗体と反応させた。未結合ペプチドを該抗体架橋ビーズから洗い落とした。潜在的結合ペプチドを低pHで溶出し、1D/LC−MS/MSにより特徴づけた。生じたタンパク質分解ペプチドはいずれも、2H2 mAbにより、高い効率および特異性では認識されなかった。このことは、2H2 mAbが直鎖状エピトープを認識しなかったことを明確に示している。
【0145】
2H2 mAbがORF0657nの直鎖状配列を認識しなかったという知見は、限られた化学切断実験により裏付けられた。ORF0657tをCNBrで化学的に2時間にわたって切断した。得られた切断産物をSDS−PAGEにより分析した。SDS−PAGE分析は、約42kDa、35kDa、25kDa、15kDaおよび10kDaの分子量を有する5つの主要バンドを示した。2H2 mAbでのウエスタンブロット分析は、該42kDaバンドのみが2H2により認識されることを明らかに示した。すべてのバンドを該SDS−PAGEから切り出し、ゲル内消化を行い、タンデム質量分析により特定された得られたペプチドをORF0657t内の対応配列と合致させた。該主要バンドの分析の結果を表10に示す。
【0146】
【表11】
【0147】
〜42kDaの分子量を有する断片の重要性をエピトープ切断により確認した。詳細に説明すると、該エピトープ切断実験のそれぞれについて、50mgの臭化シアン活性化セファロース(Amersham cat.No.17 0430 01)への化学的架橋により、210μgの2H2 mAbを固定化した。ついで50μgの無傷ORF0657nを該固定化抗体に結合させ、未結合ORF0657nをリン酸緩衝食塩水での十分な洗浄により洗い落とした。3つの独立した実験において、プロテアーゼGlu−C、トリプシン、ならびにGluC、AspN、トリプシン、キモトリプシンおよびカルボキシ−ペプチダーゼYの連続的組合せを、5時間または該連続的組合せのプロテアーゼ当たり1時間、加えた。該インキュベーション中に該プロテアーゼにより切り出されたペプチドを十分に洗い落とし、2H2 mAbに特異的に結合したORF0657n断片をSDSローディングバッファーで遊離させた。
【0148】
2H2 mAbに特異的に結合した断片をSDS−PAGEにより分析した。該エピトープ切断実験の3つ全てが、該SDS−PAGE分析において、40〜42kDaの分子量を有する唯一のバンドを示した。2H2 mAbに結合するバンドがウエスタンブロット分析により確認された。該エピトープ切断実験をGlu−Cプロテアーゼに関して反復した。今回は、2H2 mAbに特異的に結合したORF0657tの断片を酸性条件で遊離させ、リニアイオントラップ(LTQ,Thermo Electron)において1D/LC−MS/MSにより分析した。溶出したサンプルは、82〜87分(40%〜45%アセトニトリル)に、予想強度を有するシグナル(全イオン係数)を示し、42.628kDaと推定される複数の電荷状態([M+67 H]67+〜([M+30 H]30+)を示した。この特定の質量に対応するORF0657tの考えられうる断片は、42.6kDaの分子量を有するORF0657tの配列[012から382]である。
【0149】
ORF0657tのどのリシン残基が2H2 mAbの結合に際して化学反応から保護されるのかを決定するために、2H2mAbの存在下または非存在下の3つの異なる実験条件を用いて、スルホ−NHS−アセタート(Pierce Cat.No.26777)を使用する化学的標識実験を行った。表11を参照されたい。
【0150】
【表12】
【0151】
各実験について、0または3モル過剰の2H2 mAbで生成した反応産物を3つのうちの1つのプロテアーゼと共にインキュベートして、2×9個の反応混合物を得た。実験1では、GluC、AspNおよびトリプシンを使用した。実験2および3では、GluC、AspNおよびキモトリプシンを使用した。ついで該タンパク質分解ペプチドを1D/LC−MS/MSにより分析した。該反応のそれぞれについて、アセチル化および未アセチル化リシン残基の比率を、個々のペプチドの全イオン計数(TIC)の曲線下面積に基づいて算出した。ついで、同一反応条件に関して、得られた比率を該ペア間(2H2 mAbの存在下または非存在下)で比較した。2H2 mAbへのORF0657tの結合に際して最大の遮蔽を受ける、ORF0657t内のリシン残基を特定するために、全3個の反応条件で全体的な分析を行った。前記の化学的標識実験は、2H2 mAbの結合に際して最も保護されたものとして、K76、K257および潜在的にK443を特定した。化学的標識に対する保護は直接結合によるものでありうる。しかし、そのような保護は、保護部位の近傍への結合、またはORF0657nI内の長い領域にわたる構造変化によるものである可能性がある。
【0152】
要約すると、前記実験は、2H2 mAbにより認識される、ORF0657t内のエピトープが、コンホメーション性(conformational)であるという、明らかな証拠を示している。2H2 mAbにより認識されるORF0657tの断片は、ORF0675tのアミノ酸1から115に位置するN末端、およびORF0657tのアミノ酸323から357に位置するカルボキシル末端を有する。2H2 mAbの結合の際の化学的標識からの保護が、長い領域にわたる構造変化により影響されることは除外できないとしても、リシン76およびリシン275の近傍の領域が直接的な抗体相互作用に関与している可能性が高い。
【0153】
(実施例9)
2H2 mAb配列特定
一本鎖可変フラグメント(scFv)構築のための縮重プライマーPCR/重複伸長クローニング法(Krebberら,JIM 201(1):35−55,1997)を、ヒトカッパ軽鎖定常ドメインまたはscAb物質に融合された可溶性scFvのBiacoreによるハイスループットスクリーニングと組合せることにより、ハイブリドーマにより発現された2H2 IgGの可変軽(Vl)配列および可変重(Vh)配列の特定が達成された。これは、該縮重プライマー法により生じたVlフレームワーク1、4およびVhフレームワーク1、4における突然変異の精密な識別を可能にした。
【0154】
簡潔に説明すると、Total RNA Kit(商標)(Ambion Inc.)の標準的な方法を用いて、RNA物質を該ハイブリドーマ細胞系から精製した。ついで、この物質をcDNAに逆転写し、それを、該可変領域を増幅するためのPCRにおける鋳型として使用した。VlおよびVh鎖のPCR増幅のための条件は、Krebberら,JIM201(l):35−55,1997に記載されているプロトコールに基づくものであった。(Gly4Ser)4リンカー(配列番号32)が付加されて、VhおよびVlが重なり合ってVl−(Gly4Ser)4−Vh scFvを与える、第3PCR反応のためのドメインが得られるよう、プライマーを設計する。
【0155】
該可変鎖を個々に増幅するための第1セットのPCR反応を、5μlのcDNA反応物、それぞれ2μMのVlおよびVhの増幅のためのフォワードおよびリバースプライマーセットならびに高忠実度PCRマスター混合物を含有する100μlの容量中で行った。該反応を94℃で4分間変性させ、ついで94℃で30秒間、50℃で30秒間、72℃で1分間の30サイクルに付し、72℃で5分間の最終サイクルに付した。完全長PCR産物をゲル精製した。
【0156】
増幅されたVhおよびVl物質をscFvへと合体させて完全長産物を構築するために、第3PCR反応を行った。100μlの容量中で、それぞれ約20ngのVhおよびVl DNAならびに高忠実度PCRマスター混合物を94℃で5分間変性させ、ついで94℃で30秒間、60℃で30秒間および72℃で30秒間の3サイクル(プライマーの非存在下)に付した。修飾されたPCRプライマー配列番号33および配列番号34を1μMの最終濃度で加え、94℃で30秒間、60℃で1分間および72℃で1分間の30サイクルを行い、ついで72℃で7分間の加熱を行った。予想される完全長scFv PCR産物をゲル精製した。
【0157】
増幅されたscFv物質を、scAbの産生および配列分析のためにMP16可溶性発現ベクター内にクローニングした(Hayhurstら,JIM 276(1−2):185−196,2003)。MP16ベクター内への定向性(directional)クローニングのために、Sfi1での1回の制限酵素消化を行った。ついで、存在する見掛け上完全長の可変重鎖および可変軽鎖を含有するクローンをXL1−Blue細胞内でscAbとして発現させ、標準的な浸透圧ショック法を用いてペリプラズムから回収した。簡潔に説明すると、96ウェル形態において、2% グルコースおよび100μg/ml アンピシリンを含有する増殖培地内でクローンを37℃で一晩増殖させた。20μlの該一晩培養物を、0.1% グルコースおよび100μg/ml アンピシリンを含有する新たな培地に移し、0.6のOD600に達するまで増殖させた。IPTGを0.5mMの最終濃度で加えることにより、scAbの発現のために該細胞を誘導し、150rpmで振とうしながら室温で一晩インキュベートした。Qiagen Ni−NTAスーパーフロー(superflow)自動化法を用いて、該細胞からscAbを精製した。
【0158】
ORF0657tに対する結合活性に関して各scAbペリプラズム調製物を分析するために、Biacore3000表面プラズモン共鳴(SPR)装置(Upsala,Sweden)を使用した。標準的なEDC/NHSカップリングを用いて、約250共鳴単位の0657t抗原をCM5センサーチップの実験用フローセル表面に直接的に共有結合させた。タンパク質のカップリングを行うことなく、参照フローセル表面を活性化し不活性化した。ついで各調製物を該表面に通過させ、抗原へのscAbの会合および解離を測定した。各実施ごとに、10mM HClを流速20μl/分で20秒間にわたり1回注入し、ついで2分間の安定化時間を設けることにより、該表面を再生させた。全てのサンプルを2通りに実施し、バッファーのみの実施を対照として用いた。95個のクローンをスクリーニングした後、1個のクローンを、その結合活性に基づいて選択した。選択した最終的な2H2クローンは、元のハイブリドーマにより調製されたIgG物質に類似した、ORF0657tに対するそのアフィニティー、および比較配列分析に基づくものであった。
【0159】
2H2 Vh(配列番号20)およびVl(配列番号21)のアミノ酸配列は以下のとおりであった。
【0160】
【化1】
【0161】
下線部はCDRである。CDRは、カバット(Kabat)定義に基づいて特定された。コードしている核酸配列は配列番号24(Vh)および配列番号25(Vl)に示されている。
【0162】
(実施例10)
2H2 IgGキメラの発現
2H2 mAbの可変領域を、実施例9に記載されているとおりにマウスハイブリドーマからクローニングした。該可変領域の配列をPCR増幅し、重鎖可変領域をコードするDNAを、IgG1定常領域をコードするDNAにインフレームで融合させ、一方、軽鎖可変領域をコードするDNAを、カッパ定常領域をコードするDNAにインフレームで融合させた。得られた抗体発現ベクターに関するクローニング方法を以下に説明する。
【0163】
該可変領域をPCR増幅した。PCR反応は、高忠実度PCRマスター混合物、鋳型容量1μlならびにフォワードおよびリバースプライマー(それぞれ1μl)を含有する25μlの容量中で行った。PCR条件は94℃で2分間の1サイクル、94℃で1.5分間、60℃で1.5分間、72℃で1.5分間の25サイクルおよび72℃で7分間であり、反応産物を、取り出すまで4℃にし、ついでIn−Fusion法を用いて5’末端においてリーダー配列に及び3’末端において定常領域にインフレームでクローニングした。以下のプライマーを使用した。軽鎖フォワード,5’−ACAGATGCCAGATGCGATATTGTGATGACCCAGTCT(配列番号28);軽鎖リバース,5’−TGCAGCCACCGTACGTTTTATTTCCAGCTTGGTCCC(配列番号29);重鎖フォワード,5’−ACAGGTGTCCACTCGGATGTGCACCTGGTGGAGTCA(配列番号30);および重鎖リバース,5’−GCCCTTGGTGGATGCCGAGGAGACTGTGAGAGTGGT(配列番号31)。該クローンの全てのDNA配列は配列決定により確認された。
【0164】
DNA配列から推定されるアミノ酸配列を以下に示す。
【0165】
【化2】
【0166】
可変領域は下線で示されている。
【0167】
該抗体を293EBNA単層細胞内で発現させた。PE1に基づくトランスフェクション試薬を使用して、該プラスミドをトランスフェクトした。トランスフェクトされた細胞をOpti−MEM無血清培地内でインキュベートし、分泌した抗体を、プロテインA/Gアフィニティークロマトグラフィーを用いて培地から精製した。精製抗体の濃度をOD280nmにより決定し、純度をLabChip(商標)キャピラリー電気泳動により測定した。
【0168】
軽鎖および重鎖の両方の発現はヒトCMVプロモーターおよびウシ成長ホルモンポリアデニル化シグナルにより駆動された(Shiverら,Ann.N.Y.Acad.Sci.,772:198−208,1995)。前方のリーダー配列が培養培地内への抗体の分泌をもたらした。重鎖に関するリーダー配列はMEWSWVFLFFLSVTTGVHS(配列番号26)であり、軽鎖に関するリーダー配列はMSVPTQVLGLLLLWLTDARC(配列番号27)であった。該発現ベクターは、293EBNA細胞内での持続的発現のためのEBVウイルスゲノム由来のoriP、ならびに大腸菌(E.coli)における複製起点およびカナマイシン選択マーカーのための細菌配列を含有する。
【0169】
該抗体を293EBNA単層細胞内で発現させた。PE1に基づくトランスフェクション試薬を使用して、該プラスミドをトランスフェクトした。トランスフェクトされた細胞をOpti−MEM無血清培地内でインキュベートし、分泌した抗体を、プロテインA/Gアフィニティークロマトグラフィーを用いて培地から精製した。精製抗体の濃度をOD280nmにより決定し、純度をLabChipキャピラリー電気泳動により測定した。
【0170】
(実施例11)
アフィニティー測定
ハイブリドーマ物質としての2H2 mAb、scAbおよびキメラ抗体に関する比較分析を行った。2h2 mAb VhおよびVl領域をクローニングし、IgGキメラとして発現させた(実施例10に記載されているとおり)。scAbを、ヒトカッパ鎖タグに融合したscfvを産生するMP16ベクター(実施例9)内にクローニングした。以下に更に詳細に説明するとおり、該抗原アフィニティーは該構築物間で有意には異ならなかった。
【0171】
結合ドメインと抗原との間の1:1相互作用を測定するために、Biacoreの実験条件を、抗体フラグメントを測定するのか又は完全長IgGを測定するのかに応じて改変した。IgGの測定の場合には、IgGをリガンドとして表面上に捕捉し、ORF0657tをアナライトとして通過させた。抗体フラグメントの分析の場合には、ORF0657tを表面に結合させ、抗体フラグメントをアナライトとして通過させた。これは、ORF0657t抗原に対する元の2H2 mAbハイブリドーマ物質のアフィニティーが組換えクローニング後に有意な変化を何ら示さないことを示した(表12)。Biacore 3000での表面プラズモン共鳴によりデータを収集し、各アナライトを複数の濃度(1濃度当たり二重の重複実験)で通過させた。一連の全濃度を同時フィットさせるBIAevaluation(Biacore,Inc.)でデータを分析した。フィットパラメーターを表12に示す。
【0172】
【表13】
【0173】
(実施例12)
ORF0657nに基づく配列
配列番号(SEQ ID NO)4から19において強調して示されている(太字および下線で示されている)アミノ酸は、ORF0657nから変化したアミノ酸を示す。
【0174】
【化3】
【0175】
他の実施形態も特許請求の範囲内に含まれる。本明細書中には幾つかの実施形態が示され記載されているが、本発明の精神および範囲から逸脱することなく種々の改変が施されうる。
【図面の簡単な説明】
【0176】
【図1】図1はIgG分子の構造を例示する。「VL」は軽鎖可変領域を意味する。「VH」は重鎖可変領域を意味する。「CL」は軽鎖定常領域を意味する。「CH1」、「CH2」および「CH3」は重鎖定常領域である。点線はジスルフィド結合を示す。
【図2】図2は、ペア様態の結合研究における種々のモノクローナル抗体の反応性の概要を示すマトリックスを例示する。モノクローナル抗体のパネルは、BIACORE(登録商標)法により、3つの反応性領域に分類された。
【図3A】図3A〜C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1;□,mAb 6G6.A8(イソタイプ対照);または○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図3A−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3B−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.6×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3C−0.50mgのmAb13C7.BC1;0.45mgのmAb 6G6;および9.9×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図3B】図3A〜C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1;□,mAb 6G6.A8(イソタイプ対照);または○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図3A−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3B−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.6×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3C−0.50mgのmAb13C7.BC1;0.45mgのmAb 6G6;および9.9×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図3C】図3A〜C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1;□,mAb 6G6.A8(イソタイプ対照);または○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図3A−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3B−0.49mgのmAb 13C7.BC1;0.45mgのmAb 6G6.A8;および9.6×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図3C−0.50mgのmAb13C7.BC1;0.45mgのmAb 6G6;および9.9×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図4A】図4Aおよび4B:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1(0.5mg);□,mAb 6G6.A8(イソタイプ対照)(0.5mg);または○,PBS(0.5ml)のi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図4Aは、2.09×108 CFUのスタヒロコッカス・アウレウス(S.aureus)UK58での結果を例示している。図4Bは、2.15×108 スタヒロコッカス・アウレウス(S.aureus)UK58での結果を例示している。
【図4B】図4Aおよび4B:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 13C7.BC1(0.5mg);□,mAb 6G6.A8(イソタイプ対照)(0.5mg);または○,PBS(0.5ml)のi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図4Aは、2.09×108 CFUのスタヒロコッカス・アウレウス(S.aureus)UK58での結果を例示している。図4Bは、2.15×108 スタヒロコッカス・アウレウス(S.aureus)UK58での結果を例示している。
【図5A】図5A〜5C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 2H2.BE11;□,mAb 6G6.A8(イソタイプ対照);○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図5A−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5B−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および8.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5C−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図5B】図5A〜5C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 2H2.BE11;□,mAb 6G6.A8(イソタイプ対照);○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図5A−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5B−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および8.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5C−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【図5C】図5A〜5C:BALB/cマウス(n=20)の群を、細菌チャレンジの20時間前に、■,mAb 2H2.BE11;□,mAb 6G6.A8(イソタイプ対照);○,PBSのi.p.注射で治療した。マウスをi.v.注射によりスタヒロコッカス・アウレウス(S.aureus)でチャレンジし、生存をモニターした。図5A−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.8×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5B−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および8.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。図5C−0.43mgのmAb 2H2.BE11;0.5mgのmAb 6G6.A8;および9.3×108 CFUのスタヒロコッカス・アウレウス(S.aureus)ベッカー(Becker)。
【特許請求の範囲】
【請求項1】
mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合する、第1可変領域および第2可変領域を含んでなる単離された抗原結合タンパク質。
【請求項2】
該標的領域が、mAb 2H2.BE11標的領域であり、第1可変領域が、
配列番号20のアミノ酸36から45またはアミノ酸36から45とは1つのアミノ酸が異なる配列を含む第1 Vh CDR、
配列番号20のアミノ酸50から65またはアミノ酸50から65とは1つのアミノ酸が異なる配列を含む第2 Vh CDR、および
配列番号20のアミノ酸98から107またはアミノ酸98から107とは1つのアミノ酸が異なる配列を含む第3 Vh CDR
を含むVh領域である、請求項1記載の結合タンパク質。
【請求項3】
第2可変領域が、
配列番号21のアミノ酸24から33またはアミノ酸24から33とは1つのアミノ酸が異なる配列を含む第1 Vl CDR、
配列番号21のアミノ酸49から55またはアミノ酸49から55とは1つのアミノ酸が異なる配列を含む第2 Vl CDR、および
配列番号21のアミノ酸88から96またはアミノ酸88から96とは1つのアミノ酸が異なる配列を含む第3 Vl CDR
を含むVl領域である、請求項2記載の結合タンパク質。
【請求項4】
該結合タンパク質が抗体である、請求項3記載の結合タンパク質。
【請求項5】
該抗体がモノクローナル抗体である、請求項4記載の結合タンパク質。
【請求項6】
該Vh領域が、配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20のいずれかであり、該Vl領域が、配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21のいずれかである、請求項4記載の結合タンパク質。
【請求項7】
該結合タンパク質が、(a)該Vh領域、ならびにヒトヒンジ、CH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4サブタイプからのもの)を含む重鎖と、(b)該Vl領域、およびヒトカッパCLまたはヒトラムダCLのいずれかを含む軽鎖とを含む抗体である、請求項6記載の結合タンパク質。
【請求項8】
該Vh領域が、配列番号20のアミノ酸36から45よりなる第1 Vh CDR、配列番号20のアミノ酸50から65よりなる第2 Vh CDR、および配列番号20のアミノ酸98から107よりなる第3 Vh CDRを含み、
該Vl領域が、配列番号21のアミノ酸24から33よりなる第1 Vl CDR、配列番号21のアミノ酸49から55よりなる第2 Vl CDR、および配列番号21のアミノ酸88から96よりなる第3 Vl CDRを含む、請求項3記載の結合タンパク質。
【請求項9】
該結合タンパク質が抗体である、請求項8記載の結合タンパク質。
【請求項10】
該抗体がモノクローナル抗体である、請求項9記載の結合タンパク質。
【請求項11】
該Vh領域が、配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20のいずれかであり、該Vl領域が、配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21のいずれかである、請求項9記載の結合タンパク質。
【請求項12】
該結合タンパク質が、(a)該Vh領域、ならびにヒトヒンジ、CH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4サブタイプからのもの)を含む重鎖と、(b)該Vl領域、およびヒトカッパCLまたはヒトラムダCLのいずれかを含む軽鎖とを含む抗体である、請求項8記載の結合タンパク質。
【請求項13】
該重鎖が配列番号22のアミノ酸配列より実質的になり、該軽鎖が配列番号23のアミノ酸配列より実質的になる、請求項12記載の結合タンパク質。
【請求項14】
mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合する抗体可変領域をコードするヌクレオチド配列を含む組換え遺伝子を含んでなる核酸。
【請求項15】
該標的領域が、mAb 2H2.BE11標的領域であり、該可変領域が、
配列番号20のアミノ酸36から45またはアミノ酸36から45とは1つのアミノ酸が異なる配列を含む第1 Vh CDR、
配列番号20のアミノ酸50から65またはアミノ酸50から65とは1つのアミノ酸が異なる配列を含む第2 Vh CDR、および
配列番号20のアミノ酸98から107またはアミノ酸98から107とは1つのアミノ酸が異なる配列を含む第3 Vh CDR
を含むVh領域である、請求項14記載の核酸。
【請求項16】
該Vh領域が、配列番号20のアミノ酸36から45よりなる第1 Vh CDR、配列番号20のアミノ酸50から65よりなる第2 Vh CDR、および配列番号20のアミノ酸98から107よりなる第3 Vh CDRを含む、請求項15記載の核酸。
【請求項17】
該可変領域が、配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20のいずれかである、請求項14記載の核酸。
【請求項18】
該組換え遺伝子が、該Vh領域、ヒトヒンジ、ならびにCH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4サブタイプからのもの)を含む抗体重鎖をコードする、請求項17記載の核酸。
【請求項19】
該可変領域が、
配列番号21のアミノ酸24から33またはアミノ酸24から33とは1つのアミノ酸が異なる配列を含む第1 Vl CDR、
配列番号21のアミノ酸49から55またはアミノ酸49から55とは1つのアミノ酸が異なる配列を含む第2 Vl CDR、および
配列番号21のアミノ酸88から96またはアミノ酸88から96とは1つのアミノ酸が異なる配列を含む第3 Vl CDR
を含むVl領域である、請求項14記載の核酸。
【請求項20】
該第1 Vl領域が、配列番号21のアミノ酸24から33よりなる第1 Vl CDR、配列番号21のアミノ酸49から55よりなる第2 Vl CDR、および配列番号21のアミノ酸88から96よりなる第3 Vl CDRを含む、請求項19記載の核酸。
【請求項21】
該可変領域が、配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21のいずれかである、請求項14記載の核酸。
【請求項22】
該組換え遺伝子が、該可変領域およびヒトカッパまたはラムダCLを含む抗体軽鎖をコードする、請求項17記載の核酸。
【請求項23】
請求項14〜22のいずれか1項記載の1以上の核酸を含んでなる組換え細胞。
【請求項24】
該細胞が請求項18記載の核酸および請求項22記載の核酸の両方を含む、請求項23記載の組換え細胞。
【請求項25】
抗体可変領域を含むタンパク質の製造方法であって、
(a)該タンパク質が発現される条件下、請求項23記載の組換え細胞を増殖させ、
(b)該タンパク質を精製する段階を含んでなる製造方法。
【請求項26】
抗体可変領域を含むタンパク質の製造方法であって、
(a)該タンパク質が発現される条件下、請求項24記載の組換え細胞を増殖させ、
(b)該タンパク質を精製する段階を含んでなる製造方法。
【請求項27】
請求項1〜13のいずれか1項記載の結合タンパク質と医薬上許容される担体とを含んでなる医薬組成物。
【請求項28】
溶液中または細胞上のOFR0657n抗原の存在の検出方法であって、(a)請求項1〜13のいずれか1項記載の結合タンパク質を該溶液または該細胞に加え、(b)該溶液中に存在する該抗原への又は該細胞への該結合タンパク質の結合能を測定する段階を含んでなる検出方法。
【請求項29】
請求項1〜13のいずれか1項記載の結合タンパク質の有効量を患者に投与する段階を含んでなる、患者におけるスタヒロコッカス・アウレウス(S.aureus)感染に対する治療方法。
【請求項30】
該抗原結合タンパク質を手術または異物インプラントと共に投与する、請求項29記載の方法。
【請求項31】
mAb 1G3.B3、mAb 2H2.B8、mAb 13C7.D12またはmAb 13G11.C11のいずれかであるタンパク質、またはORF0657nへの結合に関してmAb IG3.B3、mAb 2H2.B8、mAb 13C7.D12またはmAb 13G11.C11のいずれかと競合するタンパク質を産生する細胞系。
【請求項32】
該系がATCC番号PTA−7124、ATCC番号PTA−7125、ATCC番号PTA−7126またはATCC番号PTA−7127のいずれかである、請求項31記載の細胞系。
【請求項1】
mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合する、第1可変領域および第2可変領域を含んでなる単離された抗原結合タンパク質。
【請求項2】
該標的領域が、mAb 2H2.BE11標的領域であり、第1可変領域が、
配列番号20のアミノ酸36から45またはアミノ酸36から45とは1つのアミノ酸が異なる配列を含む第1 Vh CDR、
配列番号20のアミノ酸50から65またはアミノ酸50から65とは1つのアミノ酸が異なる配列を含む第2 Vh CDR、および
配列番号20のアミノ酸98から107またはアミノ酸98から107とは1つのアミノ酸が異なる配列を含む第3 Vh CDR
を含むVh領域である、請求項1記載の結合タンパク質。
【請求項3】
第2可変領域が、
配列番号21のアミノ酸24から33またはアミノ酸24から33とは1つのアミノ酸が異なる配列を含む第1 Vl CDR、
配列番号21のアミノ酸49から55またはアミノ酸49から55とは1つのアミノ酸が異なる配列を含む第2 Vl CDR、および
配列番号21のアミノ酸88から96またはアミノ酸88から96とは1つのアミノ酸が異なる配列を含む第3 Vl CDR
を含むVl領域である、請求項2記載の結合タンパク質。
【請求項4】
該結合タンパク質が抗体である、請求項3記載の結合タンパク質。
【請求項5】
該抗体がモノクローナル抗体である、請求項4記載の結合タンパク質。
【請求項6】
該Vh領域が、配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20のいずれかであり、該Vl領域が、配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21のいずれかである、請求項4記載の結合タンパク質。
【請求項7】
該結合タンパク質が、(a)該Vh領域、ならびにヒトヒンジ、CH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4サブタイプからのもの)を含む重鎖と、(b)該Vl領域、およびヒトカッパCLまたはヒトラムダCLのいずれかを含む軽鎖とを含む抗体である、請求項6記載の結合タンパク質。
【請求項8】
該Vh領域が、配列番号20のアミノ酸36から45よりなる第1 Vh CDR、配列番号20のアミノ酸50から65よりなる第2 Vh CDR、および配列番号20のアミノ酸98から107よりなる第3 Vh CDRを含み、
該Vl領域が、配列番号21のアミノ酸24から33よりなる第1 Vl CDR、配列番号21のアミノ酸49から55よりなる第2 Vl CDR、および配列番号21のアミノ酸88から96よりなる第3 Vl CDRを含む、請求項3記載の結合タンパク質。
【請求項9】
該結合タンパク質が抗体である、請求項8記載の結合タンパク質。
【請求項10】
該抗体がモノクローナル抗体である、請求項9記載の結合タンパク質。
【請求項11】
該Vh領域が、配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20のいずれかであり、該Vl領域が、配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21のいずれかである、請求項9記載の結合タンパク質。
【請求項12】
該結合タンパク質が、(a)該Vh領域、ならびにヒトヒンジ、CH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4サブタイプからのもの)を含む重鎖と、(b)該Vl領域、およびヒトカッパCLまたはヒトラムダCLのいずれかを含む軽鎖とを含む抗体である、請求項8記載の結合タンパク質。
【請求項13】
該重鎖が配列番号22のアミノ酸配列より実質的になり、該軽鎖が配列番号23のアミノ酸配列より実質的になる、請求項12記載の結合タンパク質。
【請求項14】
mAb IG3.BD4標的領域、mAb 2H2.BE11標的領域、mAb 13C7.BC1およびmAb 13G11.BF3標的領域よりなる群から選ばれる標的領域に結合する抗体可変領域をコードするヌクレオチド配列を含む組換え遺伝子を含んでなる核酸。
【請求項15】
該標的領域が、mAb 2H2.BE11標的領域であり、該可変領域が、
配列番号20のアミノ酸36から45またはアミノ酸36から45とは1つのアミノ酸が異なる配列を含む第1 Vh CDR、
配列番号20のアミノ酸50から65またはアミノ酸50から65とは1つのアミノ酸が異なる配列を含む第2 Vh CDR、および
配列番号20のアミノ酸98から107またはアミノ酸98から107とは1つのアミノ酸が異なる配列を含む第3 Vh CDR
を含むVh領域である、請求項14記載の核酸。
【請求項16】
該Vh領域が、配列番号20のアミノ酸36から45よりなる第1 Vh CDR、配列番号20のアミノ酸50から65よりなる第2 Vh CDR、および配列番号20のアミノ酸98から107よりなる第3 Vh CDRを含む、請求項15記載の核酸。
【請求項17】
該可変領域が、配列番号20、ヒト化された配列番号20または脱免疫化(de−immunized)された配列番号20のいずれかである、請求項14記載の核酸。
【請求項18】
該組換え遺伝子が、該Vh領域、ヒトヒンジ、ならびにCH1、CH2およびCH3領域(IgG1、IgG2、IgG3またはIgG4サブタイプからのもの)を含む抗体重鎖をコードする、請求項17記載の核酸。
【請求項19】
該可変領域が、
配列番号21のアミノ酸24から33またはアミノ酸24から33とは1つのアミノ酸が異なる配列を含む第1 Vl CDR、
配列番号21のアミノ酸49から55またはアミノ酸49から55とは1つのアミノ酸が異なる配列を含む第2 Vl CDR、および
配列番号21のアミノ酸88から96またはアミノ酸88から96とは1つのアミノ酸が異なる配列を含む第3 Vl CDR
を含むVl領域である、請求項14記載の核酸。
【請求項20】
該第1 Vl領域が、配列番号21のアミノ酸24から33よりなる第1 Vl CDR、配列番号21のアミノ酸49から55よりなる第2 Vl CDR、および配列番号21のアミノ酸88から96よりなる第3 Vl CDRを含む、請求項19記載の核酸。
【請求項21】
該可変領域が、配列番号21、ヒト化された配列番号21または脱免疫化(de−immunized)された配列番号21のいずれかである、請求項14記載の核酸。
【請求項22】
該組換え遺伝子が、該可変領域およびヒトカッパまたはラムダCLを含む抗体軽鎖をコードする、請求項17記載の核酸。
【請求項23】
請求項14〜22のいずれか1項記載の1以上の核酸を含んでなる組換え細胞。
【請求項24】
該細胞が請求項18記載の核酸および請求項22記載の核酸の両方を含む、請求項23記載の組換え細胞。
【請求項25】
抗体可変領域を含むタンパク質の製造方法であって、
(a)該タンパク質が発現される条件下、請求項23記載の組換え細胞を増殖させ、
(b)該タンパク質を精製する段階を含んでなる製造方法。
【請求項26】
抗体可変領域を含むタンパク質の製造方法であって、
(a)該タンパク質が発現される条件下、請求項24記載の組換え細胞を増殖させ、
(b)該タンパク質を精製する段階を含んでなる製造方法。
【請求項27】
請求項1〜13のいずれか1項記載の結合タンパク質と医薬上許容される担体とを含んでなる医薬組成物。
【請求項28】
溶液中または細胞上のOFR0657n抗原の存在の検出方法であって、(a)請求項1〜13のいずれか1項記載の結合タンパク質を該溶液または該細胞に加え、(b)該溶液中に存在する該抗原への又は該細胞への該結合タンパク質の結合能を測定する段階を含んでなる検出方法。
【請求項29】
請求項1〜13のいずれか1項記載の結合タンパク質の有効量を患者に投与する段階を含んでなる、患者におけるスタヒロコッカス・アウレウス(S.aureus)感染に対する治療方法。
【請求項30】
該抗原結合タンパク質を手術または異物インプラントと共に投与する、請求項29記載の方法。
【請求項31】
mAb 1G3.B3、mAb 2H2.B8、mAb 13C7.D12またはmAb 13G11.C11のいずれかであるタンパク質、またはORF0657nへの結合に関してmAb IG3.B3、mAb 2H2.B8、mAb 13C7.D12またはmAb 13G11.C11のいずれかと競合するタンパク質を産生する細胞系。
【請求項32】
該系がATCC番号PTA−7124、ATCC番号PTA−7125、ATCC番号PTA−7126またはATCC番号PTA−7127のいずれかである、請求項31記載の細胞系。
【図1】

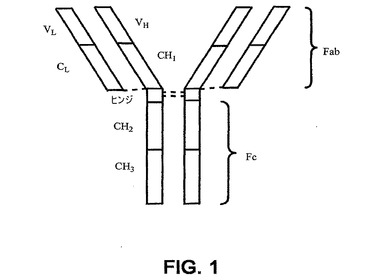
【図2】
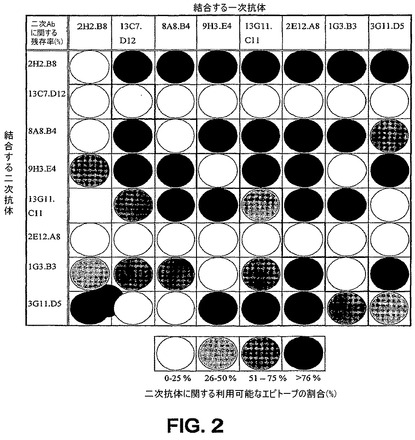
【図3A】
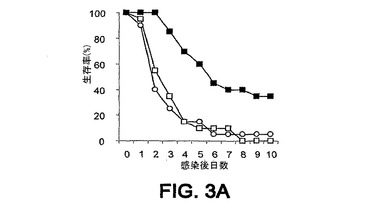
【図3B】
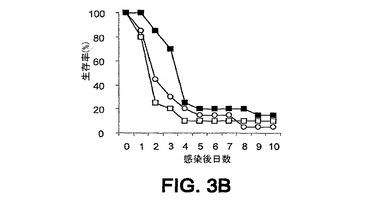
【図3C】
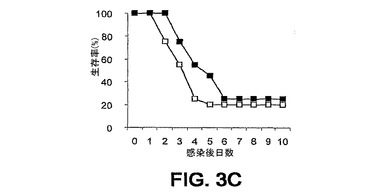
【図4A】
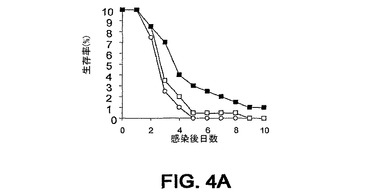
【図4B】
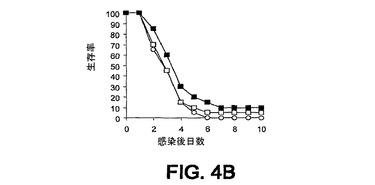
【図5A】
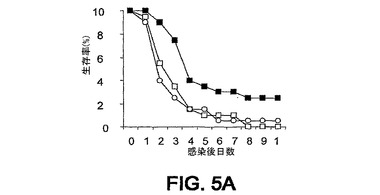
【図5B】
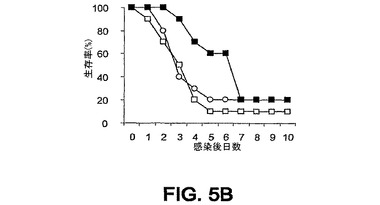
【図5C】
【図2】
【図3A】
【図3B】
【図3C】
【図4A】
【図4B】
【図5A】
【図5B】
【図5C】
【公表番号】特表2009−524428(P2009−524428A)
【公表日】平成21年7月2日(2009.7.2)
【国際特許分類】
【出願番号】特願2008−552351(P2008−552351)
【出願日】平成19年1月23日(2007.1.23)
【国際出願番号】PCT/US2007/001687
【国際公開番号】WO2007/089470
【国際公開日】平成19年8月9日(2007.8.9)
【出願人】(390023526)メルク エンド カムパニー インコーポレーテッド (924)
【氏名又は名称原語表記】MERCK & COMPANY INCOPORATED
【Fターム(参考)】
【公表日】平成21年7月2日(2009.7.2)
【国際特許分類】
【出願日】平成19年1月23日(2007.1.23)
【国際出願番号】PCT/US2007/001687
【国際公開番号】WO2007/089470
【国際公開日】平成19年8月9日(2007.8.9)
【出願人】(390023526)メルク エンド カムパニー インコーポレーテッド (924)
【氏名又は名称原語表記】MERCK & COMPANY INCOPORATED
【Fターム(参考)】
[ Back to top ]