
ステロイド骨格の24位を還元する酵素をコードする遺伝子
【課題】ステロイド骨格の24位を還元する酵素をコードする遺伝子、およびステロイド類やビタミンD3類の製造方法を提供する。
【解決手段】無細胞系で、あるいは細胞または植物を用いて、外因性のステロイド骨格の24位の二重結合を還元する酵素活性を有するタンパク質の存在下またはそれをコードするDNAの発現下で、ステロイド骨格の24位の二重結合を還元する酵素的変換方法、ならびにこの方法を利用したステロイド骨格の24位の二重結合を還元した化合物の製造方法。
【解決手段】無細胞系で、あるいは細胞または植物を用いて、外因性のステロイド骨格の24位の二重結合を還元する酵素活性を有するタンパク質の存在下またはそれをコードするDNAの発現下で、ステロイド骨格の24位の二重結合を還元する酵素的変換方法、ならびにこの方法を利用したステロイド骨格の24位の二重結合を還元した化合物の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ステロイド骨格の24位を還元する酵素をコードする遺伝子に関する。本発明は、ステロイド類やビタミンD3類に関する。本発明はまた、上記化合物の生産用の細胞およびトランスジェニック酵母に関する。
【背景技術】
【0002】
コレステロールは最も重要な動物性ステロイドである。コレステロールは細胞膜の基本成分であり、細胞が生きていくための必須の成分である。物質としてのコレステロールは薬品産業でも重要であり、ウルソデオキシコール酸などコレステロールから誘導・合成される胆汁酸関連物質は医薬品としておおきな需要がある。コレステロールから誘導・合成されるビタミンD3類は、医薬品としてはもちろん、食物の栄養強化として食品や飼料に添加されている。現在、市場で市販されているコレステロールの大部分は、動物組織(ウシの脊髄や羊毛)からの抽出によっている。しかし、動物組織から直接抽出した物質を原料とすることは、BSE(牛海綿状脳症)の原因物質の持込の可能性、さらには未知の病原性物質の混入など、医薬品等の生産にはリスクがあると考えられている。このことから、動物組織の利用は、さまざまな物質生産から排除される方向にある。コレステロールは、一部においては化学合成がされているが、すべてを化学合成で行うことは価格が非常に高くなり産業として成り立たない。そこで、植物や酵母などの微生物を用いたより大量の安価な生産方法が求められてきた。
【0003】
しかしながら、植物には24位にメチル基が導入される経路以降に生産される植物ステロール類が主として生産されることから、コレステロールの供給源とはなりえなかった。また酵母を代表とする真菌類でも同じくエルゴステロール類のみを蓄積しているため、生産に直接利用することは極めて困難であった。先行例としては酵母にて24位にメチル基を導入する酵素(ステロール24-C-メチル基転移酵素)遺伝子ERG6を欠損し、7位の2重結合を還元する酵素(7-デヒドロコレステロール還元酵素)遺伝子DWF5を導入した酵母に、ヒトのステロイド骨格の24位の2重結合(Δ24、C24とC25の間の2重結合)を還元する酵素(3β-ヒドロキシコレステロールΔ24-還元酵素)遺伝子DHCR24を導入し酵母でコレステロールを生産したものが報告されている(特許文献1、特許文献2)。一般にヒトなど哺乳類の遺伝子は酵母など真菌類に導入した際の相性が悪いことが多く発現が弱いことが知られている。ヒトと相同性の高い哺乳類の遺伝子を利用した改良では、より高発現の真菌生物を作成する際に遺伝子情報が限られており、発現の改善が問題になる。また、ヒトの遺伝子を産業利用することについて倫理的な問題などがあった。
【0004】
植物においては、主に24位にメチル基が導入される経路以降に生産される植物ステロールが生産される。この経路の生合成遺伝子としてシロイヌナズナのDWF1遺伝子(非特許文献1と2)が知られており、酵素反応はステロイドのΔ24(28)(C24とC28の間の2重結合)を還元する、つまり24位のメチレン等を還元する反応を触媒する。DWF1遺伝子とDHCR24遺伝子の間には弱い相同性があることが知られている。一方、一部の植物では植物ステロールとは別にコレステロールの生産が認められる(非特許文献3)。しかし、その生合成経路は不明であり、コレステロール含量を増加させる手段は全く、不明であった。さらに、植物においてビタミンD3を生産しうる植物が知られている(非特許文献4及び5)。これらは7−デヒドロコレステロールが中間産物になっていると推定されるが、同様にビタミンD3を増加させる遺伝子レベルでの研究は行われてこなかった。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特表2007−535960号公報
【特許文献2】米国特許第7608421号明細書
【非特許文献】
【0006】
【非特許文献1】Choeら, Plant Physiol., 1999, 119:897-907
【非特許文献2】Klahreら, Plant Cell, 1998, 10:1677-1690
【非特許文献3】Eckart Eich, ‘Solanaceae and ConVolvulaceae: Secondary Metabolite,’ 2008, Springer, Heidelberg, Germany, p.368-370
【非特許文献4】Eckart Eich, ‘Solanaceae and ConVolvulaceae: Secondary Metabolite,’ 2008, Springer, Heidelberg, Germany, p.461-464
【非特許文献5】Bolandら, Plant Science 2003, 164: 357-369
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の目的は、ステロイド骨格の24位を還元する酵素をコードする遺伝子を提供することである。
【課題を解決するための手段】
【0008】
本発明者らは、上記目的を達成すべく鋭意研究を重ねた。本発明者らは植物ステロールの中でコレステロールの含量は低いが、ステロイドサポニンやステロイドアルカロイドを生成することから、コレステロールを中間産物として生成しうるナス科植物、特にSolanum属の植物に着目した(Eckart Eich, ‘Solanaceae and ConVolvulaceae: Secondary Metabolite,’ 2008, Springer, Heidelberg, Germany, p.368-370)。これら植物からはDWF1遺伝子と相同性を持つ2つの遺伝子をin silicoで探しだした。この候補遺伝子を、適正な基質を生産する酵母に、それぞれを導入することで、一方は従来から同定されている植物ステロールの合成に関わるDWF1と同じ機能を持つ遺伝子(StDWF1)であることを明らかにし、さらにもう一方は、それとは異なるステロイド骨格の24位を還元する酵素をコードする遺伝子(StDWF1H)であることを同定することができた。また、本遺伝子を発現させた酵母はコレステロールを著量生産することを示した。さらに本遺伝子は、すでに報告のあるヒトの24位を還元する酵素をコードする遺伝子よりも活性が高いことを見出し、本発明を完成させるに至った。StDWF1とStDWF1Hの相同性はきわめて高く、かつ、それぞれと、既知のヒトの24位を還元する酵素をコードする遺伝子との相同性は大きく離れており、今までの単純な遺伝子配列の相同性からは予想できない結果であり、想定外の結果といわざるを得ない。
【0009】
したがって、本発明は、以下のとおりである。
[1] コレステロールを生産する植物に由来する、ステロイド骨格の24位を還元する活性を有するタンパク質。
[2] ナス科、ゴマノハグサ科、ヤマノイモ科、またはシソ科に属する植物に由来する、[1]のタンパク質。
[3] ジャガイモ、トマトに由来する、[1]のタンパク質。
[4] 以下の(a)〜(c)からなる群から選択される[1]のタンパク質。
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質
(b) 配列番号1または配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含むタンパク質
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含むタンパク質
[5] ステロイド骨格がコレスタン誘導体のステロイド骨格である、[1]〜[4]のいずれかのタンパク質。
[6] 膜貫通領域を持ちFAD結合型配列を持つ還元酵素に属する、[1]〜[5]のいずれかのタンパク質。
[7] 以下の(a)〜(g)からなる群から選択されるDNAを含有する組換え体DNA。
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質をコードするDNA
(b) 配列番号1または配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(d) 配列番号2または配列番号4に示す塩基配列を含むDNA
(e) 配列番号2または配列番号4に示す塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(f) 配列番号2または配列番号4に示す塩基配列と90%以上の配列同一性を有する塩基配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(g) 配列番号2または配列番号4に示す塩基配列において縮重配列を含むDNA
[8] [7]の組換え体DNAを含むベクター。
[9] [7]の組換え体DNAを含む形質転換体。
[10] 形質転換体が微生物または植物を宿主として得られる形質転換体である、[9]の形質転換体。
[11] 微生物が酵母である、[10]の形質転換体。
[12] 24位が還元されたステロイドまたはその誘導体を生成する能力を有する、[9]〜[11]のいずれかの形質転換体。
[13] [9]〜[12]のいずれかの形質転換体を培地に培養し、培養物中に[4]記載のタンパク質を生成、蓄積させ、該培養物より該蛋白質を採取する[4]のタンパク質の製造法。
[14] [9]〜[12]のいずれかの形質転換体であって、24位に2重結合を持つステロイドまたはその誘導体を細胞内に生成する形質転換体を培養し、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収する、24位が還元されたステロイドまたはその誘導体の製造法。
[15] [9]〜[12]のいずれかの形質転換体を24位に2重結合を持つステロイドまたはその誘導体の存在下で培養し、該形質転換体中に24位に2重結合を持つステロイドまたはその誘導体を取り込ませ、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収する、24位が還元されたステロイドまたはその誘導体の製造法。
[16] [9]〜[12]のいずれかの形質転換体の培養物または該培養物の処理物を酵素源に用い、該酵素源、および24位に2重結合を持つステロイドまたはその誘導体を水性媒体中に存在せしめ、該媒体中に24位が還元されたステロイドまたはその誘導体を生成、蓄積させ、該媒体から24位が還元されたステロイドまたはその誘導体を採取する、24位が還元されたステロイドまたはその誘導体の製造法。
[17] 前記培養物が、培養物を遠心分離して得られる菌体もしくは培養液上清、菌体および/または培養液上清の濃縮物、乾燥物、凍結乾燥物、界面活性剤処理物、超音波処理物、機械的摩砕処理物、溶媒処理物、酵素処理物、蛋白質分画物、固定化物あるいは菌体および/または培養液上清から抽出して得られる酵素標品であることを特徴とする、[14]〜[16]のいずれかの製造法。
[18] 24位に2重結合を持つステロイドが、デスモステロール、または7−デヒドロデスモステロールであり、24位が還元されたステロイドが、コレステロール、または7−デヒドロコレステロールである、[14]〜[16]のいずれかの製造法。
[19] [18]の製造法において、24位に2重結合を持つステロイドとして7−デヒドロデスモステロールを用い、7−デヒドロコレステロールを製造し、さらに、得られた7−デヒドロコレステロールに光を当てることを含む、ビタミンD3の製造法。
[20] 前記形質転換体に蓄積された7−デヒドロコレステロールに対して光を当てるものである、[19]の製造法。
[21] (i) ゲノムDNAまたはRNAである核酸を植物から単離する工程、
(ii) (i)の核酸がRNAである場合に逆転写しcDNAを合成する工程、
(iii) (i)または(ii)の工程で得られたDNAから配列番号2、配列番号4または配列番号5に示す塩基配列を含有する遺伝子断片を増幅する工程、ならびに
(iv) DNA中に突然変異および/または多型の存在を決定する工程、
とを含む、植物におけるステロイド骨格の24位を還元する酵素をコードする遺伝子の突然変異および/または多型の存在を検出する方法。
[22] 植物がナス科植物である[21]の方法。
[23] [21]または[22]の方法によってステロイド骨格の24位を還元する酵素をコードする遺伝子の突然変異および/または多型を検出し、突然変異および/または多型を有する植物体を選抜する方法。
[24] [23]の方法により選抜された、ステロイド骨格の24位を還元する酵素をコードする遺伝子に突然変異および/または多型を有する植物体。
[25] ナス科植物である[24]の植物体。
[26] ステロイド骨格の24位を還元する酵素をコードする遺伝子の発現能またはコードするステロイド骨格の24位を還元する酵素の活性が、既存品種に対して変化している植物を選抜する、[23]の植物体を選抜する方法。
[27] [26]の方法によって選抜された、ステロイド骨格の24位を還元する酵素をコードする遺伝子の発現能が既存品種に対して変化しているか、またはステロイド骨格の24位を還元する酵素の活性が既存品種に対して変化している植物体。
[28] ナス科植物である[27]の植物体。
【発明の効果】
【0010】
本発明のステロイド骨格の24位を還元する酵素活性を有するタンパク質により、有用な生理活性を示す、コレステロール類やビタミンD3類、とりわけウルソデオキシコール酸やビタミンD3、およびそれらの置換誘導体を大量かつ安価に生産できる。本発明によれば、ステロイド骨格の24位を還元する酵素タンパク質とそれをコードするDNAの活性発現の調整を行うことができ、該遺伝子の活性が増強された植物および微生物を作製する方法、コレステロールなどのステロイド骨格の24位を還元されたコレステロール類の生産能が変更された植物が提供される。本発明により上記コレステロール類、ビタミンD3類、およびそれらの置換誘導体の過剰発現に特徴がある植物の作出が可能である。
【図面の簡単な説明】
【0011】
【図1】StDWF1HとStDWF1のアミノ酸配列について相同性をGENETYX(ゼネティックス社)で解析を行った結果を示す図である。562アミノ酸に渡って80.0%の同一性がある。
【図2】DWF1に相同なアミノ酸配列を持つ遺伝子の分子進化系統樹解析の結果を示す図である。数値はブーストラップ検定1000に対する数値である。
【図3】酵母発現用ベクターpTochigi101の構造を示す図であり、機能領域(2μ複製領域:2u ori、pUC複製領域:pUC ori、HIS3遺伝子:HIS3、アンピシリン耐性遺伝子:Amp、TDH3プロモーター、PGKターミナーター)と制限酵素部位を示す。
【図4】DWF1H発現酵母(T33, T34)のGCチャート(TIC)とマススペクトル(MS)を示す図である。
【図5】作成した形質転換酵母とプラスミドの組み合わせを示す図である。
【図6】erg6酵母とerg4 erg5酵母にDWF1H、DWF1、LKB又はDHCR24遺伝子を導入した酵母のGCチャート(TIC)とマススペクトル(MS)を示す図である。
【図7】erg4 erg5酵母でのDWF1H、DWF1、LKB又はDHCR24遺伝子を発現する酵母のGCチャート(TIC)とマススペクトル(MS)を示す図である。
【図8】ジャガイモ品種「サッシー」の塊茎の曝光前後での24位還元酵素遺伝子の発現を示す電気泳動図である。
【図9】StDWF1H遺伝子の過剰発現用ベクターの構造を示す図であり、導入する遺伝子部分のT-DNAのライトボーダー(RB)、レフトボーダー(LB)の内部の構造、制限酵素部位を示す。
【発明を実施するための形態】
【0012】
以下、本発明を詳細に説明する。
1.ステロイド骨格の24位を還元する酵素タンパク質(24位還元酵素)
本発明の酵素タンパク質は、ステロイド骨格の24位の2重結合(Δ24、C24とC25の間の2重結合)を還元する酵素活性を有するタンパク質であり、デスモステロール(コレスタ-5,24-ジエノール)、7−デヒドロデスモステロール(コレスタ-5,7,24-トリエノール)、ラノステロール(ラノスタ-8,24-ジエノール)、シクロアルテノール(シクロアルタ‐24‐エノール)などのステロイド骨格の24位に2重結合をもった化合物の24位を還元する能力を有するタンパク質である。本発明の酵素タンパク質により24位が還元されるステロイド骨格を有する化合物はコレスタン誘導体である。本明細書では、そのような変換能を、「24位還元酵素活性」、そのような変換能を有する酵素タンパク質を「24位還元酵素」と言う。また、本発明の酵素タンパク質は、膜貫通領域を持ちFAD結合型配列を持つ還元酵素に属する。
【0013】
上記酵素を含む植物には、コレステロールを生成する植物、例えばナス科のジャガイモ(Solanum tuberosum)、トマト(Solanum lycopersicum)、ルリヤナギ(Solanum glaucophyllum)、ゴマノハグサ科のジギタリス(Digitalis purpurea)、ヤマノイモ科のヤムイモ(Dioscorea sp.)やシソ科Ajuga reptans等の植物種が含まれる。
【0014】
本発明の酵素により得られる24位が還元された化合物には、植物によって合成されるコレステロールまたはその置換誘導体が含まれ、例えばコレステロール(コレスタ-5-エノール)、7−デヒドロコレステロール(コレスタ-5,7-ジエノール)、それらの塩、それらの置換誘導体などが挙げられる。
【0015】
塩としては、ナトリウム塩、カリウム塩などのアルカリ金属塩、アンモニアや、脂肪族アミン、芳香族アミン、飽和アミン、不飽和アミンなどの有機アミンとのアンモニウム塩、カルボン酸塩などが含まれる。これらの塩は、本発明方法による目的トリテルペン化合物の生成の間に形成されてもよいし、あるいは、該化合物の生成後に中和処理により塩形成を行ってもよい。
【0016】
置換誘導体は、ステロイド化合物の、例えば1位、2位、4位、6位、7位、11位、12位、14位、16位、17位、18位、19位、21位、22位、23位、26位、27位などの、24位還元酵素活性に影響の少ないと思われる位置の水素原子が、別の置換基、例えば低級アルキル基(メチル、エチル、プロピル、ブチルなど)、ハロゲン(フッ素、塩素、臭素、ヨウ素)、水酸基、エステル基(アセトキシ、プロパノイルオキシなど)、アシル基(ホルミル、アセチル、プロピオニルなど)、アルコキシ基(メトキシ、エトキシ、プロポキシなど)、アミノ基、モノ-もしくはジ-低級アルキルアミノ基(メチルアミノ、ジメチルアミノ、エチルアミノなど)、アミド基、低級アルキルアミド基(アセタミドなど)、オキソ基、シアノ基、ニトロ基、低級アルキルチオ基(メチルチオ、エチルチオなど)、スルフォニル基(メシル、エチルスルホニルなど)などの官能基で置換された化合物を含む。本発明において、コレステロールおよびコレステロールの誘導体を含めて、コレステロール類と呼ぶ。
【0017】
本発明の24位還元酵素の基質となる好ましいステロイド化合物としては、デスモステロール、7−デヒドロデスモステロール、これらの置換誘導体などのステロイド化合物が挙げられる。置換誘導体の置換位置および置換基は、上記例示と同様である。本発明の方法により、24位還元酵素の作用によって基質となる化合物から24位の二重結合が還元されたステロイド化合物が得られる。一例として、図5上にデスモステロールの24位の二重結合を還元する反応を示す。コレステロールは、デスモステロールのステロイド骨格の24位の二重結合を還元することにより生成し、7−デヒドロコレステロールは、7−デヒドロデスモステロールのステロイド骨格の24位の二重結合を還元することにより生成する。さらに、7−デヒドロコレステロールに紫外線等の光を当てることによりビタミンD3が生成する。本発明において、ビタミンD3およびビタミンD3誘導体を含めてビタミンD3類と呼ぶ。ビタミンD3の誘導体は、上記のステロイド化合物の誘導体と同様の置換基を有する誘導体をいう。
【0018】
24位還元酵素の二重結合を持つ基質は2,3-オキシドスクアレンから、ラノステロール合成酵素やシクロアルテノール合成酵素の作用によってラノステロールやシクロアルテノールが合成され、ステロイドの基本骨格が合成されることにより合成される(P.M.Dewick, Medicinal Natural Product, 3rd ed., John Wiley & Sons, 2009)。これらの合成酵素に関する配列情報およびクローニングについては、種々の動物、植物、菌類のcDNAライブラリーからクローニングされて配列決定されており、すでに公知である。ラノステロール合成酵素についてはBakerら (1995) Biochem. Biophys. Res. Commun. 213: 154-160、Sungら (1995) Bio. Pharm. Bull. 18: 1459-1461、Coreyら (1994) Proc. Natl Acad. Sci. USA 91: 2211-2215、Shiら(1994) Proc. Natl Acad. Sci. USA 91: 7370-7374.などに記載されている。シクロアルテノール合成酵素については、Coreyら(1993) Proc. Natl Acad. Sci. USA 90: 11628-11632などに記載されている。これらの合成酵素は、例えばヒト、マウス、ラット、酵母、麹菌、シロイヌナズナ、イネ、ポプラ、ブドウ、オオムギ、トマト、ダイズ、などの動物、植物、菌類に存在することが知られているので、必要に応じて、これらの生物から上記文献記載のクローニング手法を用いて上記のラノステロール合成酵素やシクロアルテノール合成酵素をコードするDNAを取得し、周知のDNA組換え技術、PCR法などを使用して該DNAを微生物細胞、菌類細胞または植物細胞に導入して該合成酵素を発現するようにすることも可能である。このようにして得られたラノステロール合成酵素やシクロアルテノール合成酵素を発現し得る形質転換菌類細胞または植物細胞から再生されたトランスジェニック菌類や植物に、さらに24位還元酵素をコードするDNAを発現可能に組み込むことによって、これらの細胞、菌類、植物を用いてコレステロールや7−デヒドロコレステロールなどの、24位が還元されたステロイド化合物を生成することが可能になる。
【0019】
本発明で使用可能な24位還元酵素は、以下のものに限定されないが、例えばジャガイモやトマト由来のものであり、それぞれ、例えば配列番号1や配列番号3に示されるアミノ酸配列を含む酵素を包含する。さらに、本発明で使用可能な該酵素は、配列番号1や配列番号3に示されるアミノ酸配列に部分的に変異を有するアミノ酸配列を含み、かつ、24位還元酵素活性を有するタンパク質を包含する。ジャガイモ由来の上記24位還元酵素をStDWF1H、トマト由来の上記24位還元酵素をSlDWF1Hと呼ぶ。
【0020】
ここで、「部分的に変異を有するアミノ酸配列」としては、配列番号1や配列番号3に示されるアミノ酸配列において1もしくは複数、好ましくは1もしくは数個、例えば、1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個、さらに好ましくは1〜3個、さらに好ましくは1個もしくは2個、のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列、または該アミノ酸配列と、BLAST、FASTAなどの相同性検索のための公知のアルゴリズム(例えば、デフォルトすなわち初期設定のパラメータを使用する。)を用いて計算したときに、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、好ましくは少なくとも90%、さらに好ましくは少なくとも95%、特に好ましくは少なくとも97%、98%もしくは99%の配列同一性を有しているアミノ酸配列が挙げられる。因みに、上記配列番号1または配列番号3に示されるアミノ酸配列を含む酵素タンパク質間の配列同一性は約70〜約80%である。
【0021】
本明細書で使用する「配列同一性」は、例えば2つのアミノ酸配列または塩基(ヌクレオチド)配列をアラインメントしたとき(ただしギャップを導入してもよいしギャップを導入しなくてもよいが、好ましくはギャップを導入する。)、ギャップを含むアミノ酸または塩基の総数に対する同一アミノ酸または塩基の数の割合(%)を指す。
【0022】
本発明の24位還元酵素は、植物体から単離された天然の24位還元酵素および遺伝子工学的手法により製造された組換え24位還元酵素を含む。
【0023】
2.24位還元酵素をコードするDNA
本明細書中で使用する「DNA」という用語は、ゲノムDNA、遺伝子、cDNAおよび化学修飾DNAを包含するものとする。
【0024】
本発明で使用する24位還元酵素をコードするDNAは、ステロイド骨格の24位を還元する活性を有する酵素をコードするDNAである。
【0025】
上記の24位還元酵素をコードするDNAは、例えば上記配列番号1や配列番号3に示されるアミノ酸配列をそれぞれコードする塩基配列を含むものであり、具体的には配列番号2や配列番号4に示される塩基配列を含むものである。
【0026】
本発明で使用可能な24位還元酵素をコードするDNAは、また、配列番号2や配列番号4に示される各塩基配列に相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズするDNA、あるいは、配列番号2や配列番号4に示される塩基配列と、BLAST、FASTAなどの相同性検索のための公知のアルゴリズム(例えば、デフォルトすなわち初期設定のパラメータを使用する。)を用いて計算したときに、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、好ましくは少なくとも90%、さらに好ましくは少なくとも95%、特に好ましくは少なくとも97%、98%もしくは99%の配列同一性を有するDNA、あるいは、これらのDNAによりコードされるタンパク質のアミノ酸配列に対して1もしくは複数、好ましくは1もしくは数個、例えば、1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個、さらに好ましくは1〜3個、さらに好ましくは1個もしくは2個、のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列を含むタンパク質をコードするDNAである。ただし該DNAは24位還元酵素活性を有するタンパク質をコードするDNAを包含する。
【0027】
これらのDNAは、配列番号2や配列番号4に示される塩基配列を含むDNAのホモログ(相同体)、アナログ(類似体)または変異体である。このようなDNAは、コレステロールを生成する植物、例えばナス科のジャガイモ(Solanum tuberosum)、トマト(Solanum lycopersicum)、ルリヤナギ(Solanum glaucophyllum)、ゴマノハグサ科のジギタリス(Digitalis purpurea)やシソ科Ajuga reptans等の植物の葉、根、種子などからハイブリダイゼーション、PCR増幅などによって得ることが可能である。
【0028】
本明細書中で使用する「ストリンジェントな条件」は、配列同一性の高いDNAがハイブリダイズする条件であり、そのような条件は当業者ならば適宜決定することができるが、例えば、「1×SSC、0.1% SDS、37℃」程度の条件であり、より厳しい(中ストリンジェントな)条件としては「0.5×SSC、0.1% SDS、42℃」程度の条件であり、さらに厳しい(高ストリンジェントな)条件としては「0.1〜0.2×SSC、0.1% SDS、65℃」程度の条件である。ハイブリダイゼーションの後でさらに、例えば0.1×SSC、0.1% SDS、55〜68℃で洗浄を行う操作を含んでもよく、この操作によってストリンジェンシーを高めることができる。ここで、1×SSCバッファーは、150 mM塩化ナトリウム、15 mMクエン酸ナトリウム、pH7.0である。
【0029】
ハイブリダイゼーション条件やPCR反応の手順については、例えばF.M. Ausbel et al., Short Protocols in Molecular Biology, 3rd ed., John Wiley & Sons, 1995などに記載されている。
【0030】
さらに、本発明で使用可能な24位還元酵素をコードするDNAは、配列番号2や配列番号4に示す塩基配列において遺伝暗号の縮重に基づく配列(縮重配列)を含むDNAも包含する。
【0031】
本発明のDNAは、上記のとおり、24位還元酵素活性を有するタンパク質、すなわち以下の(a)〜(c):
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質;
(b) 配列番号1または配列番号3に示すに示すアミノ酸配列において1または数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含み、かつ、24位還元酵素活性を有するタンパク質;
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含み、かつ、24位還元酵素活性を有するタンパク質;
のいずれかからなる群から選択されるタンパク質をコードする。
【0032】
さらに具体的には、上記DNAは、以下の(d)〜(g):
(d) 配列番号2または配列番号4に示す塩基配列を含むDNA;
(e) 配列番号2または配列番号4に示す塩基配列を含むDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、24位還元酵素活性を有するタンパク質をコードするDNA;
(f) 配列番号2または配列番号4に示す塩基配列と90%以上の配列同一性を有する塩基配列を含み、かつ、24位還元酵素活性を有するタンパク質をコードするDNA;
(g) 配列番号2または配列番号4に示す塩基配列において縮重配列を含むDNA;
のいずれかからなる群から選択される。
【0033】
3.組換えベクター
本発明のDNAは、それを発現可能にするために、制御配列を含む適切なベクターに挿入される。このようにして得られた組換え体DNAが組換えベクターである。
【0034】
ベクターとしては、原核または真核生物の細胞で使用可能なあらゆるベクターを意図し、例えば細菌(エシェリシア属、シュードモナス属、バチルス属、ロドコッカス属など)、糸状菌(アスペルギルス属、ニューロスポラ属、フザリウム属、トリコデルマ属、ペニシリウム属など)、担子菌(白色腐朽菌など)、酵母(サッカロマイセス属、ピチア属、カンジダ属など)等の微生物用ベクター、植物細胞用ベクター、昆虫細胞用ベクターなどを使用できる。
【0035】
例えば、細菌用ベクターとしては、pBR、pUC、pET、pBluescriptシリーズのベクター類などが挙げられ、酵母用ベクターとしては、非限定的にpDR196、pYES-DEST 52、YIp5、YRp17、YEp24などが挙げられ、植物細胞用ベクターとしては、非限定的にpGWB vector、pBiEl2-GUS、pIG121-Hm、pBI121、pBiHyg-HSE、pB119、pBI101、pGV3850、pABH-Hm1などが挙げられ、昆虫細胞用ベクターとしては、非限定的にpBM030、pBM034、pBK283などが挙げられる。
【0036】
本発明において使用されるベクターには、プロモーター、ターミネーター、エンハンサー、シャインダルガルノ配列、リボソーム結合配列、シグナル配列等の遺伝子の発現、調節、分泌に関する構成要素が組込まれ、必要に応じて、選択マーカー(例えば、薬剤耐性遺伝子、レポーター遺伝子)を含有する。
【0037】
プロモーターには、lacプロモーター、trpプロモーター、recAプロモーター、tacプロモーター、λPLプロモーター、T7プロモーター、CaMV35Sプロモーター、ADH1プロモーター、GALプロモーター、PHO5プロモーター、PGKプロモーター、GAPDHプロモーターなどが非限定的に含まれる。
【0038】
薬剤耐性遺伝子には、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、ハイグロマイシン耐性遺伝子などが含まれる。レポーター遺伝子には、lacZ遺伝子、GFP遺伝子、GUS遺伝子、ルシフェラーゼ遺伝子などが含まれる。その他の選択マーカーには、例えばNPTII遺伝子、ジヒドロ葉酸レダクターゼ遺伝子などが含まれる。
【0039】
遺伝子の発現、調節、分泌に関する構成要素は、その性質に応じて、それぞれが機能し得る形で組換えベクターに組み込まれることが好ましい。そのような操作は、当業者であれば適切に行うことができる。
【0040】
4.形質転換体
本発明の形質転換体は、本発明の組換えベクターを一時的もしくは恒常的に保持する形質転換体である。形質転換体は、24位還元酵素をコードするDNAを挿入した組換えベクターを、目的DNAが発現し得るように宿主細胞中に導入することにより得ることができる。該DNA断片を適当な発現ベクターのプロモーターの下流に挿入することにより、組換え体DNAを作製する。該組換え体DNAを、該発現ベクターに適合した宿主細胞に導入することにより、本発明のタンパク質を生産する形質転換体を得ることができる。
【0041】
宿主細胞としては、細菌、酵母など菌類、動物細胞、昆虫細胞、植物細胞等、目的とする遺伝子を発現できるものであればいずれも用いることができる。好ましくは、酵母など菌類、昆虫細胞または植物細胞である。
【0042】
上記の宿主細胞が、ステロイド化合物の生合成系を有している細胞、あるいは、該生合成系を有していない細胞であっても外因的に(または外来的に)ラノステロール合成酵素、シクロアルテノール合成酵素などをコードするDNAが発現可能に組み込まれた細胞のいずれかである場合には、上記の形質転換によってさらにラノステロール合成酵素、シクロアルテノール合成酵素をコードするDNAが発現可能に含まれるため、これらの細胞を適当な基質を含む培地で培養することによってラノステロールやシクロアルテノールなどステロイド類を生産することができる。
【0043】
ラノステロールやシクロアルテノールなどステロイド類の生合成系を有している細胞には、植物細胞、酵母細胞、菌類細胞、動物細胞のように本来該生合成系を備えている細胞が含まれる。また、上記の生合成系を有していない細胞には、該生合成系に関わる酵素類のゲノム領域を外因的に含む細胞などが包含される。外因的ゲノム領域は、例えばプラスミド、ファージミド、BAC、PAC、YAC、ウイルスなどのベクターに挿入されて宿主細胞内に移入されうる。いずれにしても、上記例示の細胞の形質転換によって、該細胞は、場合により誘導的に、24位還元酵素を過剰発現する。
【0044】
細菌等の原核生物を宿主細胞として用いる場合は、本発明のDNAまたは本発明の製造法に用いられるDNAを有する組換え体DNAは、原核生物中で自立複製可能であると同時に、プロモーター、リボソーム結合配列、本発明のDNAまたは本発明の製造法に用いられるDNA、転写終結配列より構成された組換え体DNAであることが好ましい。プロモーターを制御する遺伝子が含まれていてもよい。
【0045】
発現ベクターとしては、pBTrp2、pBTac1、pBTac2(いずれもベーリンガーマンハイム社)、pHelix1(ロシュ・ダイアグノスティクス社)、pKK233-2(アマシャム・ファルマシア・バイオテク社)、pSE280(インビトロジェン社)、pGEMEX-1(プロメガ社)、pQE-8(キアゲン社)、pET-3(ノバジェン社)、pKYP10(特開昭58-110600)、pKYP200[Agric.Biol.Chem.,48,669(1984)]、pLSA1[Agric.Biol.Chem.,53,277(1989)]、pGEL1[Proc.Natl.Acad.Sci.,USA,82,4306(1985)]、pBluescriptII SK(+)、pBluescript II KS(-)(ストラタジーン社)、pTrS30[エシェリヒア・コリJM109/pTrS30(FERM BP-5407)より調製]、pTrS32[エシェリヒア・コリJM109/pTrS32(FERM BP-5408)より調製]、pPAC31(WO98/12343)、pUC19[Gene,33,103(1985)]、pSTV28(タカラバイオ社)、pUC118(タカラバイオ社)、pPA1(特開昭63-233798)等を例示することができる。
【0046】
プロモーターとしては、エシェリヒア・コリ等の宿主細胞中で機能するものであればいかなるものでもよい。例えば、trpプロモーター(Ptrp)、lacプロモーター(Plac)、PLプロモーター、PRプロモーター、PSEプロモーター等の、大腸菌やファージ等に由来するプロモーター、SPO1プロモーター、SPO2プロモーター、penPプロモーター等をあげることができる。またPtrpを2つ直列させたプロモーター、tacプロモーター、lacT7プロモーター、let Iプロモーターのように人為的に設計改変されたプロモーター等も用いることができる。
【0047】
リボソーム結合配列であるシャイン−ダルガノ(Shine-Dalgarno)配列と開始コドンとの間を適当な距離(例えば6〜18塩基)に調節したプラスミドを用いることが好ましい。
【0048】
本発明のDNAまたは本発明の製造法に用いられるDNAを発現ベクターに結合させた組換え体DNAにおいては、転写終結配列は必ずしも必要ではないが、構造遺伝子の直下に転写終結配列を配置することが好ましい。
【0049】
原核生物としては、エシェリヒア属、セラチア(Serratia)属、バチルス属、ブレビバクテリウム(Brevibacterium)属、コリネバクテリウム(Corynebacterium)属、ミクロバクテリウム(Microbacterium)属、シュードモナス(Pseudomonas)属に属する微生物等をあげることができる。
【0050】
組換え体DNAの導入方法としては、上記宿主細胞へDNAを導入する方法であればいずれも用いることができ、例えば、カルシウムイオンを用いる方法[Proc.Natl.Acad.Sci.,USA,69,2110(1972)]、プロトプラスト法(特開昭63-248394)、エレクトロポレーション法[Nucleic Acids Res.,16,6127(1988)]等をあげることができる。
【0051】
酵母菌株を宿主細胞として用いる場合には、発現ベクターとして、例えば、YEp13(ATCC37115)、YEp24(ATCC37051)、YCp50(ATCC37419)、pHS19、pHS15等を用いることができる。
【0052】
プロモーターとしては、酵母菌株中で機能するものであればいずれのものを用いてもよく、例えば、PHO5プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーター、gal 1プロモーター、gal 10プロモーター、ヒートショックポリペプチドプロモーター、MFα1 プロモーター、CUP 1プロモーター等のプロモーターをあげることができる。
【0053】
宿主細胞としては、サッカロマイセス(Saccharomyces)属、シゾサッカロマイセス(Schizosaccharomyces)属、クルイベロマイセス(Kluyveromyces)属、トリコスポロン(Trichosporon)属、シワニオマイミセス(Schwanniomyces)属、ピチア(Pichia)属、またはキャンディダ(Candida)属等に属する酵母菌株をあげることができ、具体的には、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クルイベロマイセス・ラクティス(Kluyveromyces lactis)、トリコスポロン・プルランス(Trichosporon pullulans)、シワニオマイセス・アルビウス(Schwanniomyces alluvius)、ピチア・パストリス(Pichia pastoris)、キャンディダ・ウティリス(Candida utilis)等をあげることができる。
【0054】
組換え体DNAの導入方法としては、酵母にDNAを導入する方法であればいずれも用いることができ、例えば、エレクトロポレーション法[Methods Enzymol.,194,182(1990)]、スフェロプラスト法[Proc.Natl.Acad.Sci.,USA,81,4889(1984)]、酢酸リチウム法[J.Bacteriol.,153,163(1983)]等をあげることができる。
【0055】
動物細胞を宿主として用いる場合には、発現ベクターとして、例えば、pcDNAI、pcDM8(フナコシ社より市販)、pAGE107(特開平3-22979号公報)、pAS3-3(特開平2-227075号公報)、pCDM8[Nature,329,840(1987)]、pcDNAI/Amp(インビトロジェン社)、pREP4(インビトロジェン社)、pAGE103[J.Biochem,101,1307(1987)]、pAGE210、pAMo、pAMoA等を用いることができる。
【0056】
プロモーターとしては、動物細胞中で機能するものであればいずれも用いることができ、例えば、サイトメガロウイルス(CMV)のIE(immediate early)遺伝子のプロモーター、SV40の初期プロモーターあるいはメタロチオネインのプロモーター、レトロウイルスのプロモーター、ヒートショックプロモーター、SRαプロモーター等をあげることができる。また、ヒトCMVのIE遺伝子のエンハンサーをプロモーターと共に用いてもよい。
【0057】
宿主細胞としては、マウス・ミエローマ細胞、ラット・ミエローマ細胞、マウス・ハイブリドーマ細胞、ヒトの細胞であるナマルバ(Namalwa)細胞またはナマルバKJM-1細胞、ヒト胎児腎臓細胞、ヒト白血病細胞、アフリカミドリザル腎臓細胞、チャイニーズ・ハムスターの細胞であるCHO細胞、HBT5637(特開昭63-299号公報)等をあげることができる。
【0058】
マウス・ミエローマ細胞としては、SP2/0、NSO等、ラット・ミエローマ細胞としてはYB2/0等、ヒト胎児腎臓細胞としてはHEK293(ATCC CRL-1573)、ヒト白血病細胞としてはBALL-1等、アフリカミドリザル腎臓細胞としてはCOS-1、COS-7等をあげることができる。
【0059】
組換え体DNAの導入方法としては、動物細胞にDNAを導入する方法であればいずれも用いることができ、例えば、エレクトロポレーション法[Cytotechnology,3,133(1990)]、リン酸カルシウム法(特開平2-227075号公報)、リポフェクション法[Proc.Natl.Acad.Sci.,USA,84,7413(1987)]、Virology,52,456(1973)に記載の方法等をあげることができる。
【0060】
昆虫細胞を宿主として用いる場合には、例えばBaculovirus Expression Vectors,A Laboratory Manual,W.H.Freeman and Company,New York(1992)、カレント・プロトコールズ・イン・モレキュラー・バイオロジー、Molecular Biology,A Laboratory Manual、Bio/Technology,6,47(1988)等に記載された方法によって、タンパク質を生産することができる。
【0061】
即ち、組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、タンパク質を生産させることができる。
【0062】
該方法において用いられる遺伝子導入ベクターとしては、例えば、pVL1392、pVL1393、pBlueBacIII(いずれもインビトロジェン社)等をあげることができる。
【0063】
バキュロウイルスとしては、例えば、夜盗蛾科昆虫に感染するウイルスであるアウトグラファ・カリフォルニカ・ヌクレアー・ポリヘドロシス・ウイルス(Autographa californica nuclear polyhedrosis virus)等を用いることができる。
【0064】
昆虫細胞としては、スポドプテラ・フルギペルダ(Spodoptera frugiperda)の卵巣細胞、トリコプルシア・ニ(Trichoplusia ni)の卵巣細胞、カイコ卵巣由来の培養細胞等を用いることができる。
【0065】
スポドプテラ・フルギペルダの卵巣細胞としてはSf9、Sf21(バキュロウイルス・イクスプレッション・ベクターズ ア・ラボラトリー・マニュアル)等、トリコプルシア・ニの卵巣細胞としてはHigh5、BTI-TN-5B1-4(インビトロジェン社)等、カイコ卵巣由来の培養細胞としてはボンビクス・モリ(Bombyxmori)N4等をあげることができる。
【0066】
組換えウイルスを調製するための、昆虫細胞への上記組換え遺伝子導入ベクターと上記バキュロウイルスの共導入方法としては、例えば、リン酸カルシウム法(特開平2-227075)、リポフェクション法[Proc.Natl.Acad.Sci.,USA,84,7413(1987)]等をあげることができる。
【0067】
植物細胞を宿主細胞として用いる場合には、ウイルスベクター、アグロバクテリウムのTiプラスミド、Riプラスミド等をベクターとして用いるアグロバクテリウム法が好適に使用できる。タバコモザイクウイルスベクター等、植物ウイルスを利用したものをあげることができる。プロモーターとしては、植物細胞中で機能するものであればいずれのものを用いてもよく、例えば、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター、イネアクチン1プロモーター等をあげることができる。宿主細胞としては、タバコ、ジャガイモ、トマト、ニンジン、ダイズ、アブラナ、アルファルファ、イネ、コムギ、オオムギ等の植物細胞等をあげることができる。
【0068】
組換えベクターの導入方法としては、植物細胞にDNAを導入する方法であればいずれも用いることができ、例えば、アグロバクテリウム(Agrobacterium)を用いる方法(特開昭59-140885号公報、特開昭60-70080号公報、WO94/00977公報)、エレクトロポレーション法(特開昭60-251887号公報)、パーティクルガン(遺伝子銃)を用いる方法(特許第2606856号公報、特許第2517813号公報)等をあげることができる。
【0069】
酵母、動物細胞、昆虫細胞または植物細胞により発現させた場合には、糖あるいは糖鎖が付加されたタンパク質を得ることができる。さらにまた、形質転換植物体(「トランスジェニック植物」とも称する。)を作出する方法は、24位還元酵素をコードするDNAを含むベクターで形質転換された植物細胞から植物体を再生させることにより得ることができる。植物細胞からの植物体の再生は公知の方法、例えばカルス培養等により行うことができる。
【0070】
5.本発明のタンパク質の製造法
本発明のタンパク質は、本発明のタンパク質をコードするDNAを宿主細胞中で発現させる一時的もしくは恒常的な4.形質転換体であげられた形質転換体を得て、製造することができる。本発明のタンパク質をコードする部分の塩基配列を、宿主の発現に最適なコドンとなるように、塩基を置換することにより、該タンパク質の生産率を向上させることができる。
【0071】
得られる形質転換体を培地に培養し、もしくは形質転換体植物の場合は栽培し、培養物中に本発明のタンパク質を生成蓄積させ、該培養物から採取することにより、該タンパク質を製造することができる。
【0072】
本発明のタンパク質を製造するための上記形質転換体の宿主としては、細菌、酵母など菌類、動物細胞、昆虫細胞、植物細胞等、目的とする遺伝子を発現できるものであればいずれも用いることができる。好ましくは、酵母など菌類、昆虫細胞または植物細胞である。
上記形質転換体を培地に培養する方法は、宿主の培養に用いられる通常の方法に従って行うことができる。
【0073】
エシェリヒア・コリ等の原核生物あるいは酵母等の真核生物を宿主として得られた形質転換体を培養する培地としては、該生物が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行える培地であれば天然培地、合成培地のいずれを用いてもよい。
【0074】
炭素源としては、該生物が資化し得るものであればよく、グルコース、フラクトース、スクロース、これらを含有する糖蜜、デンプンあるいはデンプン加水分解物等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類等を用いることができる。
【0075】
窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸もしくは有機酸のアンモニウム塩、その他の含窒素化合物、並びに、ペプトン、肉エキス、酵母エキス、コーンスチープリカー、カゼイン加水分解物、大豆粕および大豆粕加水分解物、各種発酵菌体、およびその消化物等を用いることができる。
【0076】
無機塩としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等を用いることができる。
【0077】
培養は、通常振盪培養または深部通気攪拌培養等の好気的条件下で行う。培養温度は15〜40℃がよく、培養時間は、通常5時間〜7日間である。培養中pHは3.0〜9.0に保持する。pHの調整は、無機または有機の酸、アルカリ溶液、尿素、炭酸カルシウム、アンモニア等を用いて行う。
また、培養中必要に応じて、アンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。
【0078】
プロモーターとして誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養するときには、必要に応じてインデューサーを培地に添加してもよい。例えば、lacプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはイソプロピル−β−D−チオガラクトピラノシド等を、trpプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはインドールアクリル酸等を培地に添加してもよい。
【0079】
動物細胞を宿主として得られた形質転換体を培養する培地としては、一般に使用されているRPMI1640培地[J.Am.Med.Assoc.,199,519(1967)]、イーグル(Eagle)のMEM培地[Science,122,501(1952)]、DMEM培地[Virology,8,396(1959)]、199培地[Proc.Soc.Biol.Med.,73,1(1950)]またはこれら培地に牛胎児血清等を添加した培地等を用いることができる。
【0080】
培養は、通常pH6〜8、25〜40℃、5%CO2存在下等の条件下で1〜7日間行う。
また、培養中必要に応じて、カナマイシン、ペニシリン、ストレプトマイシン等の抗生物質を培地に添加してもよい。
【0081】
昆虫細胞を宿主として得られた形質転換体を培養する培地としては、一般に使用されているTNM-FH培地(ファーミンジェン社)、Sf-900 II SFM培地(ライフ・テクノロジーズ社)、ExCell400、ExCell405[いずれもJRHバイオサイエンシーズ社]、Grace’s Insect Medium[Nature,195,788(1962)]等を用いることができる。
【0082】
培養は、通常pH6〜7、25〜30℃等の条件下で1〜5日間行う。
また、培養中必要に応じて、ゲンタマイシン等の抗生物質を培地に添加してもよい。
【0083】
植物細胞を宿主として得られた形質転換体は、細胞として、または植物の細胞や器官に分化させて培養することができる。該形質転換体を培養する培地としては、一般に使用されているムラシゲ・アンド・スクーグ(MS)培地、ホワイト(White)培地、またはこれら培地にオーキシン、サイトカイニン等、植物ホルモンを添加した培地等を用いることができる。
【0084】
培養は、通常pH5〜9、20〜40℃の条件下で3〜60日間行う。
また、培養中必要に応じて、カナマイシン、ハイグロマイシン等の抗生物質を培地に添加してもよい。
【0085】
上記のとおり、本発明のタンパク質をコードするDNAを発現ベクターに連結した組換え体DNAを保有する微生物、昆虫細胞、動物細胞、あるいは植物細胞由来の形質転換体を、通常の培養方法に従って培養し、本発明のタンパク質を生成蓄積させ、該培養物または培養物の処理物より該タンパク質を採取することにより、該タンパク質を製造することができる。ここで、培養物の処理物とは、培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の溶媒処理物、該菌体の酵素処理物、該菌体の蛋白質分画物、該菌体の固定化物あるいは該菌体より抽出して得られる酵素標品等を含む。
【0086】
本発明のタンパク質の生産方法としては、宿主細胞内に生産させる方法、宿主細胞外に分泌させる方法、あるいは宿主細胞外膜上に生産させる方法があり、選択した方法に応じて、生産させるタンパク質の構造を変えることができる。
【0087】
本発明のタンパク質が宿主細胞内あるいは宿主細胞外膜上に生産される場合、ポールソンらの方法[J.Biol.Chem.,264,17619(1989)]、ロウらの方法[Proc.Natl.Acad.Sci.,USA,86,8227(1989)、Genes Develop.,4,1288(1990)]、または特開平05-336963号公報、WO94/23021号公報等に記載の方法を準用することにより、該タンパク質を宿主細胞外に積極的に分泌させることができる。
【0088】
すなわち、遺伝子組換えの手法を用いて、本発明のタンパク質の活性部位を含むタンパク質の手前にシグナルペプチドを付加した形で生産させることにより、該タンパク質を宿主細胞外に積極的に分泌させることができる。
【0089】
また、特開平2-227075号公報に記載されている方法に準じて、ジヒドロ葉酸還元酵素遺伝子等を用いた遺伝子増幅系を利用して生産量を上昇させることもできる。
【0090】
さらに、遺伝子導入した動物または植物の細胞を再分化させることにより、遺伝子が導入された動物個体(トランスジェニック非ヒト動物)または植物個体(トランスジェニック植物)を造成し、これらの個体を用いて本発明のタンパク質を製造することもできる。
【0091】
本発明のタンパク質を生産する形質転換体が動物個体または植物個体の場合は、通常の方法に従って、飼育または栽培し、該タンパク質を生成蓄積させ、該動物個体または植物個体より該タンパク質を採取することにより、該タンパク質を製造することができる。
【0092】
動物個体を用いて本発明のタンパク質を製造する方法としては、例えば公知の方法[Am.J.Clin.Nutr.,63,639S(1996)、Am.J.Clin.Nutr.,63,627S(1996)、Bio/Technology,9,830(1991)]に準じて遺伝子を導入して造成した動物中に本発明のタンパク質を生産する方法があげられる。
【0093】
動物個体の場合は、例えば、本発明のDNAまたは本発明の製造法に用いられるDNAを導入したトランスジェニック非ヒト動物を飼育し、本発明のタンパク質を該動物中に生成、蓄積させ、該動物中より該タンパク質を採取することにより、該タンパク質を製造することができる。該動物中の該タンパク質を生成、蓄積させる場所としては、例えば、該動物のミルク(特開昭63-309192号公報)、卵等をあげることができる。この際に用いられるプロモーターとしては、動物で機能するものであればいずれも用いることができるが、例えば、乳腺細胞特異的なプロモーターであるαカゼインプロモーター、βカゼインプロモーター、βラクトグロブリンプロモーター、ホエー酸性プロテインプロモーター等が好適に用いられる。
【0094】
植物個体を用いて本発明のタンパク質を製造する方法としては、例えば本発明のタンパク質をコードするDNAを導入したトランスジェニック植物を公知の方法[組織培養,20(1994)、組織培養,21(1995)、Trends Biotechnol.,15,45(1997)]に準じて栽培し、該タンパク質を該植物中に生成、蓄積させ、該植物中より該タンパク質を採取することにより、該タンパク質を生産する方法があげられる。
【0095】
本発明のタンパク質を生産する形質転換体を用いて製造された本発明のタンパク質を単離・精製する方法としては、通常の酵素の単離、精製法を用いることができる。
【0096】
例えば、本発明のタンパク質が、細胞内に溶解状態で生産された場合には、培養終了後、細胞を遠心分離により回収し、水系緩衝液にけん濁後、超音波破砕機、フレンチプレス、マントンガウリンホモゲナイザー、ダイノミル等により細胞を破砕し、無細胞抽出液を得る。
【0097】
該無細胞抽出液を遠心分離することにより得られる上清から、通常の酵素の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)−セファロース、DIAION HPA-75(三菱化成社製)等レジンを用いた陰イオン交換クロマトグラフィー法、S-Sepharose FF(ファルマシア社)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィー法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、精製標品を得ることができる。
【0098】
また、該タンパク質が細胞内に不溶体を形成して生産された場合は、同様に細胞を回収後破砕し、遠心分離を行うことにより得られた沈殿画分より、通常の方法により該タンパク質を回収後、該タンパク質の不溶体をタンパク質変性剤で可溶化する。
【0099】
該可溶化液を、タンパク質変性剤を含まないあるいはタンパク質変性剤の濃度がタンパク質が変性しない程度に希薄な溶液に希釈、あるいは透析し、該タンパク質を正常な立体構造に構成させた後、上記と同様の単離精製法により精製標品を得ることができる。
【0100】
本発明のタンパク質またはその糖修飾体等の誘導体が細胞外に分泌された場合には、培養上清に該タンパク質またはその糖付加体等の誘導体を回収することができる。
【0101】
即ち、該培養物を上記と同様の遠心分離等の手法により処理することにより可溶性画分を取得し、該可溶性画分から、上記と同様の単離精製法を用いることにより、精製標品を得ることができる。
【0102】
また、本発明のタンパク質を他のタンパク質との融合タンパク質として生産し、融合したタンパク質に親和性をもつ物質を用いたアフィニティークロマトグラフィーを利用して精製することもできる。例えば、ロウらの方法[Proc.Natl.Acad.Sci.,USA,86,8227(1989)、Genes Develop.,4,1288(1990)]、特開平5-336963号公報、WO94/23021号公報に記載の方法に準じて、本発明のタンパク質をプロテインAとの融合タンパク質として生産し、イムノグロブリンGを用いるアフィニティークロマトグラフィーにより精製することができる。
【0103】
また、本発明のタンパク質をFlagペプチドとの融合タンパク質として生産し、抗Flag抗体を用いるアフィニティークロマトグラフィーにより精製することができる[Proc.Natl.Acad.Sci.,USA,86,8227(1989)、Genes Develop.,4,1288(1990)]。更に、該タンパク質自身に対する抗体を用いたアフィニティークロマトグラフィーで精製することもできる。
【0104】
上記で取得されたタンパク質のアミノ酸配列情報を基に、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法により、本発明のタンパク質を製造することができる。また、Advanced ChemTech社、パーキン・エルマー社、Pharmacia社、Protein Technology Instrument社、Synthecell-Vega社、PerSeptive社、島津製作所等のペプチド合成機を利用して化学合成することもできる。
【0105】
6.24位が還元されたステロイド化合物の製造方法
本発明は、無細胞系で、あるいは細胞又は植物を用いて、外因性の24位還元酵素活性を有するタンパク質存在下または24位還元酵素をコードするDNAの発現下で、デスモステロール、7−デヒドロデスモステロール、ラノステロール、シクロアルテノールなどのステロイド骨格の24位に2重結合をもった化合物の24位を還元する方法を提供する。本発明の方法により、24位の二重結合を還元する上記化合物の中でもコレステロールの材料となるデスモステロールおよびビタミンD3の材料となる7−デヒドロデスモステロールのステロイド骨格の24位を還元する方法が重要である。
【0106】
本明細書で使用される「外因性」とは、細胞または植物が本来もつ酵素またはその遺伝子自体ではなく、該細胞または植物に、あるいは上記のステロイド化合物原料の反応系に、人為的に該酵素またはそれをコードするDNAを導入することを意味する。
【0107】
本発明はまた、上記の変換方法によってステロイド骨格の24位に2重結合をもった化合物の24位を還元されたステロイド化合物を生成し、該化合物を回収することを含む、24位を還元されたステロイド化合物の製造方法を提供する。
【0108】
本発明では、24位還元酵素は、粗製、半精製もしくは精製された、あるいは多糖類、多孔性ポリマー、多孔性無機物(例えばガラス、鉱物、セラミックス等)などの支持体などに共有結合もしくは非共有結合によって固定化された、酵素として反応系で使用されてもよいし、該酵素を産生する細胞または植物体自体が反応系で使用されてもよい。
【0109】
本発明で使用可能な24位還元酵素は膜結合型でNADPHを補酵素とする還元酵素の一種であり、上述の製造法で得ることができる。
【0110】
24位を還元されたステロイド化合物の製造は、24位還元酵素をコードするDNAが導入された形質転換体を用いて行うことができ、以下の(a)〜(c)の方法が挙げられる。
(a) 24位に二重結合を持つステロイドを細胞内に生成する形質転換体を培養して、菌体内で24位還元活性を発現させて還元体を製造する方法。該方法においては、形質転換体であって、24位に2重結合を持つステロイドまたはその誘導体を細胞内に生成する形質転換体を培養し、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収すればよい。
(b) 24位に二重結合を持つステロイドを培養の過程で形質転換体に取り込ませて、菌体内で24位還元活性を発現させて還元体を製造する方法。該方法においては、形質転換体を24位に2重結合を持つステロイドまたはその誘導体の存在下で培養し、該形質転換体中に24位に2重結合を持つステロイドまたはその誘導体を取り込ませ、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収すればよい。
(c) 培養物を酵素源として、24位に二重結合を持つステロイドを酵素源に接触させて還元体を製造する方法。該方法においては、質転換体の培養物または該培養物の処理物を酵素源に用い、該酵素源、および24位に2重結合を持つステロイドまたはその誘導体を水性媒体中に存在せしめ、該媒体中に24位が還元されたステロイドまたはその誘導体を生成、蓄積させ、該媒体から24位が還元されたステロイドまたはその誘導体を採取すればよい。
【0111】
DNA組換え技術によって、高い酵素活性を持ったタンパク質の発現が可能であるため、4.形質転換体であげられた形質転換体(例えば、酵母細胞、菌類細胞、昆虫細胞、植物細胞)の培養液に上記24位還元酵素の基質、例えばデスモステロール、7−デヒドロデスモステロール、ラノステロール、シクロアルテノール、またはそれらの置換誘導体、を添加することにより、24位を還元されたステロイド化合物を生産することができる。例えば、形質転換微生物の培養液にデスモステロール等のステロイド類を基質として投与することにより、24位を還元されたされたコレステロールを効率的に大量に生産することが可能である。酵母細胞、菌類細胞、植物細胞などは24位還元酵素の基質は内在性のものも利用できる。また、内在性の基質の含量をあらかじめ増加させるようにした細胞も利用できる。
【0112】
さらに、7−デヒドロデスモステロールの24位を還元して生成した7−デヒドロコレステロールに紫外線等の光を当てることによりビタミンD3(コレカルシフェノール)が生成する。プロビタミンD3である7−デヒドロコレステロールに紫外線等の光を当てるとプレビタミンD3((6z)-タカルシオール)が生成し、その後自然にビタミンD3へ異性化する。この際、本発明の24位還元酵素を含む形質転換体に7−デヒドロコレステロールを産生蓄積させ、該形質転換体に紫外線を当ててもよい。
【0113】
本発明は、本発明の24位還元酵素を用いて、コレステロールを製造する方法及びビタミンD3を製造する方法をも包含する。
【0114】
(1)酵素的製造法
無細胞系では、上記形質転換体の培養液から24位還元酵素を含有する無細胞抽出液を調製し、その一部を、デスモステロールまたはそれらの置換誘導体などのステロイド化合物を含有するバッファーに加えて変換反応を行うことによって、24位を還元されたステロイド化合物を製造することができる。
【0115】
収穫した細胞の懸濁液、あるいは適量のバッファー中の形質転換植物を、ホモジナイザー、超音波破砕機あるいはフレンチプレス等により破砕後、遠心分離して無細胞抽出液を得る。バッファーには、ポリペプチドの失活を防ぐため、抗酸化剤、酵素の安定化剤、ポリフェノール吸着剤、金属配位子などを添加することができる。さらに比活性を高めるにはポリペプチドを精製することが有効であり、超遠心機による遠心分離法、硫安等による塩析方、ゲル濾過法、イオン交換クロマトグラフィー法、アフィニティクロマトグラフィー法、電気泳動法などの手法を単独で、あるいは組み合わせて用いることができる。得られたポリペプチドを含むバッファーに、基質となる上記のステロイドおよび補酵素を添加してインキュベートする。補酵素としてはNADPHが利用でき、グルコース-6-リン酸とグルコース-6-リン酸デヒドロゲナーゼを用いたNADPH再構成系も併用できる。
【0116】
上記製造法において、本発明のタンパク質は、基質として用いるデスモステロール1mgあたり0.01〜100mg、好ましくは0.1mg〜10mg添加する。
【0117】
上記製造法において、基質として用いるデスモステロールは、0.1〜500g/L、好ましくは0.2〜200g/Lの濃度になるように反応水性媒体に初発または反応途中に添加する。上記製造法で用いられる水性媒体としては、24位を還元されたステロイド化合物の生成反応を阻害しない限り、いかなる成分、組成の水性媒体であってもよく、例えば、水、りん酸塩、炭酸塩、酢酸塩、ほう酸塩、クエン酸塩、トリスなどの緩衝液などをあげることができる。また、メタノール、エタノールなどのアルコール類、酢酸エチルなどのエステル類、アセトンなどのケトン類、アセトアミドなどのアミド類を含有していてもよい。
【0118】
24位を還元されたステロイド化合物の生成反応は水性媒体中、pH5〜11、好ましくはpH6〜10、20〜50℃、好ましくは25〜45℃の条件で2〜150時間、好ましくは6〜120時間行う。
【0119】
(2)形質転換体または微生物の培養物もしくは培養物の処理物を酵素源として用いる製造法
形質転換体または微生物の培養物もしくは培養物の処理物を酵素源として用いる24位を還元されたステロイド化合物の製造法としては、本発明のタンパク質を生産する能力を有する形質転換体に原料となる24位に2重結合を持つステロイドまたはその誘導体を取り込ませ、形質転換体内で酵素反応を行わせ、形質転換体中に24位を還元されたステロイド化合物を生成、蓄積させ、該形質転換体から24位を還元されたステロイド化合物を抽出する製造法を挙げることができる。原料となる24位に2重結合を持つステロイドまたはその誘導体の形質転換体への取り込みは、培養液中に24位に2重結合を持つステロイドまたはその誘導体を添加することにより、24位に2重結合を持つステロイドまたはその誘導体の存在下で形質転換体を培養することにより行うことができる。さらに、本発明のタンパク質を生産する能力を有する形質転換体の培養物もしくは培養物の処理物を酵素源に用い、該酵素源、およびステロイドを水性媒体中に存在せしめ、該媒体中に24位を還元されたステロイド化合物を生成、蓄積させ、該媒体から24位を還元されたステロイド化合物を採取する製造法をあげることができる。
【0120】
上記製造法に用いられる形質転換体としては、上記4.や5.の方法により製造することができる、本発明のタンパク質を生産する形質転換体をあげることができる。さらに宿主が産生するNADPH以外のNADPHを外部から加えることも可能である。
【0121】
培養物の処理物としては、培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の溶媒処理物、該菌体の酵素処理物、該菌体のタンパク質分画物、該菌体の固定化物あるいは該菌体より抽出して得られる酵素標品などをあげることができる。
【0122】
上記製造法において、基質として用いるステロイド類の種類、使用濃度および添加時期、並びに生産される24位を還元されたステロイド化合物は、上記(1)の酵素的製造法のものと同様である。
【0123】
また、微生物の培養物または該培養物の処理物を酵素源とした製造法において用いられる水性媒体としては、上記(1)の酵素的製造法に用いられる水性媒体に加え、酵素源として用いた形質転換体または微生物の培養液も水性媒体として用いることができる。
【0124】
また必要に応じて、水性媒体中に界面活性剤あるいは有機溶媒を添加してもよい。界面活性剤としては、ポリオキシエチレン・オクタデシルアミン(例えばナイミーンS-215、日本油脂社)などの非イオン界面活性剤、セチルトリメチルアンモニウム・ブロマイドやアルキルジメチル・ベンジルアンモニウムクロライド(例えばカチオンF2-40E、日本油脂社)などのカチオン系界面活性剤、ラウロイル・ザルコシネートなどのアニオン系界面活性剤、アルキルジメチルアミン(例えば三級アミンFB、日本油脂社)などの三級アミン類など、ジペプチドの生成を促進するものであればいずれでもよく、1種または数種を混合して使用することもできる。界面活性剤は、界面活性剤は、通常0.1〜50 g/lの濃度で用いられる。有機溶剤としては、キシレン、トルエン、脂肪族アルコール、アセトン、酢酸エチルなどが挙げられ、通常0.1〜50 ml/lの濃度で用いられる。
【0125】
培養物または該培養物の処理物を酵素源として用いる場合、該酵素源の量は、当該酵素源の比活性等により異なるが、例えば、基質として用いるステロイド化合物1mgあたり湿菌体重量として5〜1000mg、好ましくは10〜400mg添加する。
【0126】
24位を還元されたステロイド化合物の生成反応は水性媒体中、pH5〜11、好ましくはpH6〜10、20〜50℃、好ましくは25〜45℃の条件で2〜150時間、好ましくは6〜120時間行う。
【0127】
上記(1)または(2)の製造法において、水性媒体中に生成、蓄積した24位を還元されたステロイド化合物の採取は、活性炭やイオン交換樹脂などを用いる通常の方法あるいは、有機溶媒による抽出、結晶化、薄層クロマトグラフィー、高速液体クロマトグラフィー等により行うことができる。
【0128】
7.24位還元酵素遺伝子の変異、多型または遺伝子発現変異の選抜
本発明による24位を還元されたステロイド化合物の製造方法においては、植物において24位還元酵素遺伝子の突然変異、一塩基多型(SNP)等の多型、または遺伝子発現変異の存在を検出し、そのような変異や多型をもつ該遺伝子のなかから、24位還元酵素活性が高く24位を還元されたステロイド化合物の生産性を高める変異型遺伝子を選抜することができる。そのような変異は、24位還元酵素遺伝子を有する、かつ、24位を還元されたステロイド化合物生産能をもつ植物において、放射線、UV照射、変異原等の化学処理などの人工的突然変異処理、あるいは自然突然変異、によって誘導することができる。
【0129】
上記の変異型24位還元酵素遺伝子の選抜のための方法には、ゲノムDNAやmRNAを変異個体、様々な品種や育成個体の植物から単離し、mRNAの場合には逆転写しcDNAを合成したのち、DNA増幅技術の使用によりゲノムDNAまたはcDNAから24位還元酵素遺伝子を含有する遺伝子断片を増幅する工程と、このDNA中に突然変異の存在を決定する工程が含まれる。DNAやRNAを抽出する方法には市販のキット(例えばDNeasyやRNeasy(キアゲン社)など)が使用できる。cDNAを合成する方法も市販キット(例えばスーパースクリプト ファーストストランド システム(インビトロジェン社)など)を使うことができる。
【0130】
DNA増幅技術の使用により遺伝子断片を増幅する方法としては、いわゆるPCR法やLAMP法などの技術を用いることができる。これらは継続的なポリメラーゼ反応により特異的なDNA配列の増幅(つまり、コピー数を増やすこと)を達成するためにポリメラーゼを使用することを基にした、一群の技術である。この反応は、クローニングの代わりに使用することができるが、必要であるのは、核酸配列に関する情報のみである。DNAの増幅を行うために、増幅しようとするDNAの配列に相補的なプライマーを設計する。次にそのプライマーを自動DNA合成により作製する。DNA増幅方法は、当技術分野で周知であり、本明細書中で与えられる教示及び指示に基づき、当業者であれば容易に行うことができる。いくつかのPCR法(ならびに関連技術)は、例えば、米国特許第4,683,195号、同第4,683,202号、同第4,800,159号、同第4,965,188号、およびInnisら編、PCR Protocols:A guide to Method and Applicationsで述べられている。
【0131】
上記DNA中に突然変異や多型の存在を決定する工程では、塩基配列の決定(アプライドバイオシステムズ社)や、ミスマッチペアの片側を切断する酵素を用いて突然変異体を検出するTILLING法(Till et al., 2003, Genome Res 13:524-530)など変異遺伝子と正常遺伝子の相同性を利用し検出する方法を用いればよい。これらは該技術から得られた配列データを遺伝子部分に関する24位還元酵素をコードする遺伝子の塩基配列、例えば配列番号10〜16に示されるような塩基配列と比較することによって行うことができる。
【0132】
mRNA量の違いを決定する工程では、上記cDNAに対し、配列既知の24位還元酵素をコードする遺伝子の塩基配列に基づいて作製したプライマーを利用してリアルタイムPCR法(ロシュ・ダイアグノスティックス社のライトサイクラーなど)等の定量的PCRを採用すればよい。その後、例えば、ジャガイモ品種「サッシー」から得られたcDNAの量と比較することでmRNA量の違いを決定することができる。
【0133】
特に好ましい実施形態において、上述したような24位還元酵素遺伝子変異の存在の決定方法を、ジャガイモから得られた材料に適用する。
【0134】
上記の突然変異および/または多型を決定する方法により、24位還元酵素をコードする遺伝子の突然変異や多型を塩基レベルで同定することができ、さらに24位還元酵素をコードする遺伝子に突然変異および/または多型を有する植物体を選抜し、変異型24位還元酵素をコードする遺伝子を得ることができる。
【0135】
また、突然変異や多型の決定やmRNA量の違いの決定により、24位還元酵素をコードする遺伝子の発現能または24位還元酵素の活性が変化している植物を選抜することが可能になる。ここで、24位還元酵素をコードする遺伝子の発現能または24位還元酵素の活性の変化とは、人為的突然変異等の突然変異による遺伝子の発現能または24位還元酵素の活性の改変および多型による遺伝子の発現能または24位還元酵素の活性が異なっていることを意味する。
【0136】
ある植物の24位還元酵素活性の突然変異による改変は、その植物の種に含まれる既存品種に対する改変をいい、既存品種には野生型も含まれる。既存の品種は、24位還元酵素をコードする遺伝子が改変された植物を得るためのすべての品種をいい、交配、遺伝子操作等の人為的操作により作出された品種を含む。また、活性の改変において、すべての既存品種に対して、活性が変化している必要はなく、特定の既存品種に対して改変されていれば、「24位還元酵素の活性が改変された植物」に含まれる。「24位還元酵素の活性が改変された植物」は、人為的操作を受けず自然状態で突然変異により活性が改変された植物も含み、上記の選抜方法により、自然状態で活性が変化した植物を選抜することができ、新たな品種として確立することもできる。また、ある既存品種に変異誘発処理を行い、24位還元酵素の活性が改変された植物を作出した場合、比較対象は変異誘発処理を行った既存品種でもよいし、それ以外の他の既存品種でもよい。また、自然状態で選抜された活性が変化した植物又は変異誘発処理により活性が変化した植物を交配することにより、24位還元酵素をコードする遺伝子の変異が固定された新品種として得ることもできる。
【0137】
例えば、植物がジャガイモの場合、既存品種として、「男爵薯」、「メークイン」、「さやか」、「シンシア」等がある。ここで、24位還元酵素をコードする遺伝子の発現能または24位還元酵素の活性が既存品種に対して改変された植物とは、既存品種に対して24位還元酵素をコードする遺伝子の発現能が増強あるいは減少した植物を含み、さらに、24位還元酵素の活性が既存品種に対して上昇あるいは低下した植物を含む。本発明は、このような24位還元酵素をコードする遺伝子の発現能または24位還元酵素の活性が既存品種に対して改変された植物体も包含する。
【0138】
このようにして得られた24位還元酵素をコードする遺伝子に突然変異や多型を有する植物は、本発明の方法によって、24位を還元されたステロイド化合物の製造のために使用することができる。
【実施例】
【0139】
以下、本発明を、実施例を示してより詳しく説明するが、本発明の範囲はこれらに限定されるものではない。
【0140】
実施例1
ジャガイモの萌芽cDNAシークエンスから24位還元酵素遺伝子の抽出
ジャガイモの萌芽は極めて大量のグリコアルカロイドを蓄積することが知られている。グリコアルカロイドの生合成経路は不明な点が多いが、コレステロールを初発物質としていると考えられている。このことからジャガイモの萌芽は植物でコレステロールを合成できる器官であり、コレステロール生合成の遺伝子源として優れていると考えた。
【0141】
収穫後3ヶ月以上貯蔵し休眠の開けたジャガイモを20℃暗所におき、約2週間、萌芽をさせた。この萌芽を液体窒素にて破砕し、RNeasy(キアゲン社)で行い全RNAを抽出した。2μgのRNAからT7プロモーター配列を付加したプライマーを用いて逆転写反応でcDNAを合成した。DNA polymeraseとT7 RNA polymeraseを用いて全体をRNAとして増幅後、再度、逆転写反応でcDNAを合成した。これにGS FLX解析用のアダプターを用いてcDNA2本鎖を合成した。物理的に数百bpに切断後、別のGS FLX解析用のアダプターを連結し1本鎖DNAを回収した。これを鋳型としてGenome Sequencer FLX System(タカラバイオ社委託)にて約15万の発現している遺伝子配列を解析した。その結果、シロイヌナズナのコレステロールの24メチレンを還元する酵素DWF1の遺伝子と相同性の高い遺伝子断片(StDWF1、実施例2で取得した全長配列に対しアミノ酸配列の同一性が564 アミノ酸に渡って78.0% )が54個、それよりも相同性の低い遺伝子断片(StDWF1ホモローグ=StDWF1H、実施例2で取得した全長配列に対しアミノ酸配列の同一性が556 アミノ酸に渡って75.5% )が80個、存在することを見出した。
【0142】
実施例2
StDWF1H遺伝子の同定
上記遺伝子断片を使って、StDWF1Hについては、ナス科ゲノムネットワーク(http://solgenomics.net/index.pl)の配列データSGN-U268686を基に全長配列を取得するためのプライマーU931(caccATGTCGGATGCTAAGGCCC(配列番号7): caccはベクターにクローニングするための付加配列)とU932(TCAATTCGCAGGTTCATCAG)(配列番号8)を合成した。StDWF1についてはDFCI Potato Gene Index (http://compbio.dfci.harvard.edu/tgi/plant.html) の配列データTC214162を基に全長配列を取得するためのプライマーU957(caccATGACAGATGTTCAGGCTCC(配列番号9): caccはベクターにクローニングするための付加配列)とU958(TCAATCTTCAGGCTCATCAACT)(配列番号10)を合成した。実施例1の全RNAから全cDNAの合成をスーパースクリプト ファーストストランド システム(インビトロジェン社)を用いて行った。このcDNAを鋳型に用い、アニール温度55℃でPCR(30サイクル、タカラバイオ社 PrimeSTAR HS DNA Polymeraseを使用)によって遺伝子を増幅した。これをpENTR(商標)/D-TOPOエントリーベクター(インビトロジェン社)へクローニングした。得られた8個の独立クローンについてポリヌクレオチド配列を決定した。これにより得られた配列は、配列番号2と配列番号6であり、それから推定されるポリペプチド配列は配列番号1と配列番号5である。これらベクターをpTOPO-PSStDWF1HとpTOPO-PSStDWF1とした。図1はStDWF1HとStDWF1の相同性解析を解析した結果のアラインメントを示す図であり、アミノ酸配列の同一性が562アミノ酸に渡って80.0%と極めて高かった。24位還元酵素として既知であるヒトの遺伝子(DHCR24, Am J Hum Genet. 2001, 69, 685-94)との相同性は、StDWF1Hはアミノ酸配列の同一性が533アミノ酸に渡って40.3%、StDWF1は533アミノ酸に渡って39.0%であった。なおDHCR24とDWF1は522アミノ酸に渡って39.8%であった。いずれも40%程度を示す弱い相同性であり、この時点の遺伝子相同性の結果だけで機能を推定することは極めて困難であった。
【0143】
さらに、それぞれに相当するトマトの全長遺伝子配列をナス科ゲノムネットワーク(http://solgenomics.net/index.pl)の配列データからSGN-U578468であることを突き止め取得した。配列番号4であり、それから推定されるポリペプチド配列は配列番号3である。
【0144】
これら配列と、特表2007-535960号公報「コレステロール産生酵母株およびその利用」でヒト由来24位還元酵素遺伝子と相同的であると記載されているワタ(Genbank accession AAM47602.1)、イネ(同AAP53615)、エンドウLKB遺伝子(同AAK15493)さらにシロイヌナズナの24メチレンを還元する酵素(DWF1)のアミノ酸配列を比較しCLUSTALX 2.0(Larkinら(2007) Clustal W and Clustal X version 2.0. Bioinformatics, 23:2947-2948)のブーストラップ法を用いて分子系統樹を作成した(図2)。その結果、StDWF1HとSlDWF1Hは、コレステロールの24メチレンを還元する酵素(DWF1)とは、異なるクラスターに分類され、かつヒトの24位還元酵素遺伝子DHCR24とも進化の面からも類縁関係は遠いことがわかった。
【0145】
実施例3
ジャガイモStDWF5遺伝子の単離と酵母発現ベクターpTochigi101-StDWF5の構築
ナス科ゲノムネットワーク(http://solgenomics.net/index.pl)の配列データからシロイヌナズナDWF5遺伝子(ChoeらPlant J. (2000)21:431-43)と相同性の高い配列SGN-U269317を見出した。SGN-U269317配列の予想されるポリペプチド(479アミノ酸)のN末端とC末端に相当する箇所のプライマーU929(caccAATGGCGGAGTCTCAGTTG(配列番号11): caccはベクターにクローニングするための付加配列)とU930(CTAATAAATTCCTGGTATGACCC)(配列番号12)を合成した。実施例2のcDNAを鋳型として、アニール温度55℃でPCR(30サイクル、タカラバイオ社 PrimeSTAR HS DNA Polymeraseを使用)によって遺伝子を増幅した。これをpENTR(商標)/D-TOPOエントリーベクター(インビトロジェン社)へクローニングした。得られた8個の独立クローンについてポリヌクレオチド配列を決定しpTOPO-PSStDWF5を得た。pTochigi101(pYO323(ナショナルバイオリソースプロジェクトNBRP_ID = BYP563):HIS3の酵母選択マーカーを持った多コピー型プラスミド)のマルチクローニング部位にXhoI部位とSacI部位の間にTDH3プロモーター、NotI部位とAscI部位を含む合成リンカー、PGK1ターミネーターを含む、図3)のNotI部位とAscI部位の間を、pTOPO-PSStDWF5の制限酵素NotIとAscIで切り出されるStDWF5遺伝子と入れ替えることで、pTochigi101-StDWF5を作成した。このベクターを導入した酵母は恒常的にStDWF5遺伝子産物を発現し、ステロイドの7位の還元酵素反応をすることが期待できる。
【0146】
実施例4
ジャガイモStDWF1H遺伝子の酵母発現ベクターの構築
実施例2で作製した、pTOPO-PSStDWF1Hプラスミド(エントリークローン)とpYES-DEST52(インビトロジェン社)とを混合し、Gateway LR Clonase II Enzyme Mix(インビトロジェン社)を用いて塩基配列特異的な組み換え反応(attL x attR反応)によって、StDWF1H遺伝子の発現ベクターpYES-DEST52-StDWF1Hを得た。
【0147】
実施例5
erg6酵母(pTochigi101-StDWF5、pYES-DEST52-StDWF1H)における生成物の確認
実験室酵母erg6はdelta(24)-sterol C-methyltransferaseを欠損しているため7−デヒドロデスモステロールが蓄積していることが知られている。(Gaberら (1989) Mol Cell Biol 9(8):3447-56)。そこでYeast Knock Out Strain Collection(オープンバイオシステムズ社)のBY4742 erg6にpTochigi101-StDWF5とpYES-DEST52-StDWF1Hを電気穿孔法(「酵母分子遺伝学実験法」大嶋泰治編著、学会出版センター発行, 1996)を用いて形質転換を行い、独立のクローンを2つ(T33、T34)得た。
【0148】
BY4742 erg6の2コロニー(N1とN2)と、T33酵母、T34酵母を5mlのSC(0.67% Yeast Nitrogen base without amino acids、2% グルコース、100 mg/lロイシン、40 mg/lリジン、20 mg/lヒスチジン、20 mg/lウラシル)-Ura/His培地30℃、135rpm、1日間培養した。培養した酵母を3000g、10分間遠心することにより集菌し、ガラクトース(20mg/ml)を添加した10mlのSC-Ura/His-グルコース培地に懸濁し、30℃、135rpm、2日間培養した。培養酵母を3000g、10分間遠心することにより集菌し5mlの酢酸エチルを加え、20分間超音波処理した後、3分間遠心し酢酸エチル抽出物を回収した。この操作を3回繰り返した。酢酸エチル抽出物は溶媒を乾燥除去した後、N−メチル−N−トリメチルシリルトリフルオロアセトアミドを加え、80℃で30分間加熱し、トリメチルシリルエーテル体に誘導体化しGC-MS分析の試料とした。変換物の同定は標品をGCの保持時間ならびにMSスペクトルを比較することで決定した。
【0149】
BY4742 erg6の酵母(N1とN2)は主に7−デヒドロデスモステロールを蓄積していた。一方、pTochigi101-StDWF5及びpYES-DEST52-StDWF1Hの2つのベクターを保持する酵母T33とT34の抽出物(図4、T33、T34と示したGCチャート)からは、新たなピークが2つ検出され、標品との移動度、マススペクトルによって、デスモステロール(点線矢印)とコレステロールのピーク(図4の実線矢印)であることが同定された。これらはこのことからStDWF1Hは24位還元酵素をコードする遺伝子であることが明らかになった。また、定量を行ったところ、10 mlの培養物あたり7μgのコレステロールを生産していることがあきらかになった。しかし、本数値については、酵母での発現形態や培養条件を特に最適化しておらず、あくまで参考の数値として捉えるべきものである。
【0150】
実施例6
ジャガイモStDWF1遺伝子、ヒトDHCR24遺伝子、エンドウLKBの酵母発現ベクターの構築
ジャガイモStDWF1遺伝子が24位還元酵素活性を持っているかどうかを確認するため、酵母発現ベクターを構築した。実施例2で作製した、pTOPO-PSStDWF1プラスミド(エントリークローン)とpYES-DEST52とを混合し、Gateway LR Clonase II Enzyme Mix(インビトロジェン社)を用いて塩基配列特異的な組み換え反応(attL x attR反応)によって、StDWF1遺伝子の発現ベクターpYES-DEST52-StDWF1を得た。
【0151】
図2で示された系統樹解析ならびに、特表2007-535960号公報「コレステロール産生酵母株およびその利用」で24位還元酵素活性にて示されたワタ、イネ、エンドウの3つの植物遺伝子のうち、最もDHCR24と相同性の高かったエンドウのLKBについて24位還元酵素活性を持っているかどうかを確認するため、遺伝子を取得し酵母発現ベクターを構築した。食料品店で購入した生食用のエンドウの苗である豆苗を液体窒素にて破砕した。RNAの抽出はRNeasyで行い、全cDNAの合成はスーパースクリプト ファーストストランド システムを用いて行った。プライマーとしてはU959(caccATGTCTGATCTTGAGGCTCCG(配列番号13): caccはベクターにクローニングするための付加配列)とU960(TCAGTCTGGTGGTTGATCAACTTC)(配列番号14)を用いた。このcDNAを鋳型に用い、アニール温度55℃でPCR(30サイクル、タカラバイオ社 PrimeSTAR HS DNA Polymeraseを使用)によって遺伝子を増幅した。これをpENTR(商標)/D-TOPOエントリーベクター(インビトロジェン社)へクローニングした。塩基配列解析し既報の配列と同じであることを確認したpTOPO-LKBプラスミド(エントリークローン)を得た。これとpYES-DEST52とを混合し、Gateway LR Clonase II Enzyme Mix(インビトロジェン社)を用いて塩基配列特異的な組み換え反応(attL x attR反応)によって、LKB遺伝子の発現ベクターpYES-DEST52-LKBを得た。
【0152】
StDWF1Hと既報のヒトDHCR24の遺伝子産物を比較するためDHCR24遺伝子を取得し酵母発現ベクターを構築した。スーパースクリプト ファーストストランド システムに添付されていたヒトHela細胞の全RNAを用いて全cDNAの合成をスーパースクリプト ファーストストランド システムを用いて行った。プライマーとしてはU945(caccATGGAGCCCGCCGTGT(配列番号15): caccはベクターにクローニングするための付加配列)とU946(TCAGTGCCTGGCGGCCTTGC)(配列番号16)を用いた。このcDNAを鋳型に用い、アニール温度55℃でPCR(30サイクル、タカラバイオ社 PrimeSTAR HS DNA Polymeraseを使用)によって遺伝子を増幅した。これをpENTR(商標)/D-TOPOエントリーベクター(インビトロジェン社)へクローニングした。塩基配列解析し既報の配列と同じであることを確認したpTOPO-PSDHCR24プラスミド(エントリークローン)を得た。これとpYES-DEST52とを混合し、Gateway LR Clonase II Enzyme Mix(インビトロジェン社)を用いて塩基配列特異的な組み換え反応(attL x attR反応)によって、DHCR24遺伝子の発現ベクターpYES-DEST52-DHCR24を得た。
【0153】
BY4742 erg6にpTochigi101-StDWF5のみを導入したもの、pTochigi101-StDWF5とpYES-DEST52-StDWF1、pTochigi101-StDWF5とpYES-DEST52-LKB、 pTochigi101-StDWF5とpYES-DEST52-DHCR24を、電気穿孔法を用いて形質転換を行い独立のクローンを各2つずつ(T31、T32)(T35、T36)(T37、T38)(T39、T40)得た(図5)。
【0154】
実施例7
erg6酵母でStDWF5とDWF1相同遺伝子を発現した酵母における生成物の確認
T31、T32、T33、T34、T35、T36、T37、T38、T39、T40各酵母を5mlのSC-His培地、もしくはSC-Ura/His培地30℃、135rpm、1日間培養した。培養した酵母を3000g、10分間遠心することにより集菌し、ガラクトース(20mg/ml)を添加した10mlのSC-Ura/His-グルコース培地に懸濁し、30℃、135rpm、2日間培養した。培養酵母を3000g、10分間遠心することにより集菌し5mlの酢酸エチルを加え、20分間超音波処理した後、3分間遠心し酢酸エチル抽出物を回収した。この操作を3回繰り返した。酢酸エチル抽出物は溶媒を乾燥除去した後、N−メチル−N−トリメチルシリルトリフルオロアセトアミドを加え、80℃で30分間加熱し、トリメチルシリルエーテル体に誘導体化しGC-MS分析の試料とした。変換物の同定は標品をGCの保持時間ならびにMSスペクトルを比較することで決定した。
【0155】
pTochigi101-StDWF5のみを保持する酵母T31とT32は、デスモステロールを生産していた(図6)。pTochigi101-StDWF5及びpYES-DEST52-StDWF1Hの2つのベクターを保持する酵母T33とT34の抽出物は図3と同様の結果を示した。pTochigi101-StDWF5及びpYES-DEST52-StDWF1の2つのベクターを保持する酵母T35とT36の抽出物にはデスモステロールは存在するが、コレステロールは確認できなかった。pTochigi101-StDWF5及びpYES-DEST52-LKBの2つのベクターを保持する酵母T37とT38の抽出物にはデスモステロールは存在するが、コレステロールは確認できなかった。このことはStDWF1とLKB遺伝子産物は24位還元酵素活性を持たないことが明らかになった。pTochigi101-StDWF5及びpYES-DEST52-DHCR24の2つのベクターを保持する酵母T39とT40の抽出物にはコレステロールが確認できたが、その生成量はpTochigi101-StDWF5及びpYES-DEST52-StDWF1Hの2つのベクターを保持する酵母T33とT34よりも少なく、StDWF1H遺伝子導入の効果は、既報のDHCR24より優れていることが明らかとなった。
【0156】
比較例1
erg4 erg5酵母でStDWF5とDWF1相同遺伝子を発現した酵母における生成物の確認
実験室酵母erg4は24メチレンの還元酵素を欠損しているため24メチレンコレステロールが蓄積していることが知られている。(Zweytickら (2000) FEBS Lett 470(1):83-7)。ERG5遺伝子は酵母のC-22 ステロイドの不飽和化酵素をコードしていることが知られている(Skaggs ら (1996) Gene 169(1):105-9)。この酵素が存在すると、最終生産物に影響を及ぼす可能性があるが考えられた。そこでYeast Knock Out Strain Collection(オープンバイオシステムズ社)のBY4742 erg4に以下のPhleomycin耐性カセット(を用いた形質転換を行い、BY4742 erg4 erg5二重変異酵母を作成した。
【0157】
これをもとに通常のLi法(「酵母分子遺伝学実験法」大嶋泰治編著、学会出版センター発行, 1996)を用いて形質転換を行い、pTochigi101-StDWF5のみを導入したもの、BY4742 erg4 erg5にpTochigi101-StDWF5とpYES-DEST52-StDWF1H、pTochigi101-StDWF5とpYES-DEST52-StDWF1、pTochigi101-StDWF5とpYES-DEST52-LKB、 pTochigi101-StDWF5とpYES-DEST52-DHCR24を、電気穿孔法を用いて形質転換を行い独立のクローンを各2つずつ(T21、T22)(T23、T24)(T25、T26)(T27、T28)(T29、T30)を得た(図5)。
【0158】
T21、T22、T23、T24、T25、T26、T27、T28、T29、T30各酵母を5mlのSC-His培地、もしくはSC-Ura/His培地30℃、135rpm、1日間培養した。培養した酵母を3000g、10分間遠心することにより集菌し、ガラクトース(20mg/ml)を添加した10mlのSC-Ura/His-グルコース培地に懸濁し、30℃、135rpm、2日間培養した。培養酵母を3000g、10分間遠心することにより集菌し5mlの酢酸エチルを加え、20分間超音波処理した後、3分間遠心し酢酸エチル抽出物を回収した。この操作を3回繰り返した。酢酸エチル抽出物は溶媒を乾燥除去した後、N−メチル−N−トリメチルシリルトリフルオロアセトアミドを加え、80℃で30分間加熱し、トリメチルシリルエーテル体に誘導体化しGC-MS分析の試料とした。変換物の同定は標品をGCの保持時間ならびにMSスペクトルを比較することで決定した。
【0159】
BY4742 erg4 erg5にpTochigi101-StDWF5のみを導入した酵母(T21とT22)は24メチレンコレステロールを蓄積していた(図7)。pTochigi101-StDWF5及びpYES-DEST52-StDWF1Hの2つのベクターを保持する酵母T23とT24の抽出物とpTochigi101-StDWF5とpYES-DEST52-DHCR24の2つのベクターを保持する酵母T29とT30は対照のT21とT22と同じで24メチレンの還元酵素活性を持たないことが明らかになった。pTochigi101-StDWF5及びpYES-DEST52-StDWF1の2つのベクターを保持する酵母T25とT26の抽出物とpTochigi101-StDWF5及びpYES-DEST52-LKBの2つのベクターを保持する酵母T27とT28の抽出物にはカンペステロールを確認することができた。このことはStDWF1とLKB遺伝子産物は24メチレンの還元酵素活性を持ち、本ベクターならびに酵母での遺伝子発現が正常に行われていることが確認できた。
【0160】
実施例8
ジャガイモ組織での遺伝子発現解析
ジャガイモ塊茎を曝光させることで顕著にコレステロールが増加することでグリコアルカロイドが増加することが知られている。品種「サッシー」の塊茎を太陽光で2日間曝光した。曝光する前の塊茎果皮と曝光後の塊茎果皮から、RNAの抽出はRNeasyで行い、全cDNAの合成はスーパースクリプト ファーストストランド システムを用いて行った。StDWF1H遺伝子である配列番号3に基づいてプライマーU931(CACCATGTCGGATGCTAAGGCCC)(配列番号7)とU953(GGATCCATATGCGTTTCTCATTCCAACAACA)(配列番号17)を作製しRT-PCR(条件:95℃ 5分、(95℃ 30秒、55℃ 30秒、72℃ 1分)を25回、72℃ 5分)を行った。図8に示すように曝光する前に対して曝光したあとではStDWF1H遺伝子の発現が高いことがわかった。このようにステロイド含量の高い組織の同定が遺伝子の発現解析で可能であり、ステロイド化合物と組成との相関を解析することで、組織含量の増加のみならず、品種評価、QTLなどの遺伝子解析が可能になり、遺伝子発現マーカーを作製することができることが示された。
【0161】
実施例9
酵母でのビタミンD3の生産
実施例5および実施例7で明らかなようにerg6変異を持つ酵母は7−デヒドロデスモステロールを蓄積している。Yeast Knock Out Strain CollectionのBY4742 erg6にpYES-DEST52-StDWF1Hを、電気穿孔法を用いて形質転換を行い、独立のクローンを2つ(T19、T20)得た。この酵母はガラクトースを含む培地でStDWF1H遺伝子を発現させることができ7−デヒドロコレステロールを蓄積することができた。この酵母に紫外線を当てることによってビタミンD3を製造することが可能となる。酵母は元来、エルゴステロールを蓄積するが、これに紫外線をあてることでビタミンD2ができることが知られている。この際のエルゴステロールを生産している酵母への効果的な紫外線照射方法は特許3009606号公報に記載されており、あらたに作成された7−デヒドロコレステロールを蓄積する酵母への照射方法の改良の手がかりとなりうる。
【0162】
酵母からのビタミンD3の抽出、分析については、コレステロールと同様に以下の通り行うことができる、各酵母を5mlのSC-Ura/His培地30℃、135rpm、1日間培養する。培養した酵母を3000g、10分間遠心することにより集菌し、ガラクトース(20mg/ml)を添加した10mlのSC-Ura/His-グルコース培地に懸濁し、30℃、135rpm、2日間培養する。培養酵母を3000g、10分間遠心することにより集菌し5mlの酢酸エチルを加え、20分間超音波処理した後、3分間遠心し酢酸エチル抽出物を回収する。この操作を3回繰り返す。酢酸エチル抽出物は溶媒を乾燥除去した後、N−メチル−N−トリメチルシリルトリフルオロアセトアミドを加え、80℃で30分間加熱し、トリメチルシリルエーテル体に誘導体化しGC-MS分析の試料とする。標品をGCの保持時間ならびにMSスペクトルを比較することで分析、定量することができる。
【0163】
今回、使用した酵母BY4742はパン酵母由来の実験室酵母である。物質生産として優れているキャンディダ酵母も利用できる。より具体的な形質転換体の取得についてはキャンディダ・ユティリス(特開2010−75171公報)、キャンディダ・ボイディニ(特開2009−268407公報)を参照することによって、実験室酵母と同様にビタミンD3を製造する酵母を作成することが可能である。
【0164】
また、ジャーファーメンターなどを用いた攪拌培養において、7−デヒドロデスモステロールあるいはその前駆体を培地中に添加することにより、これらの物質を酵母細胞に取り込ませることができる。当該酵母細胞において、StDWF1H遺伝子を発現させることにより、7−デヒドロコレステロールを蓄積することができる。7−デヒドロコレステロールを蓄積した酵母細胞に光(好ましくは紫外線)を照射することによって、ビタミンD3を製造することができる。また、添加する物質をデスモステロールあるいはその前駆体とすれば、当該酵母細胞においてコレステロールを製造することができる。
【0165】
実施例10
植物でのコレステロールとビタミンD3の生産
ジャガイモやトマトなど一部の植物ではコレステロールを生産しているが、通常の植物ではコレステロールを生産していない。そこで、コレステロールを生産していないシロイヌナズナや、一部生産しているジャガイモのコレステロール生産量を増強する目的で植物のStDWF1H遺伝子の過剰発現形質転換体を得る実験を実施した。バイナリーベクターpKT11(特開2001-161373号公報)を基本として、カルフラワーモザイクウイルスの35S RNAプロモーター、StDWF1H遺伝子を順方向、35S RNAのターミネーターの順に連結を行い、植物形質転換用ベクターpKT254を作成した(図9)。作製したベクターをエレクトロポレーション法(GelvinとSchilperoor編, Plant Molecular Biology Manual, C2, 1-32 (1994), Kluwer Academic Publishers)により、アグロバクテリウム・ツメファシエンスGV3110株に導入した。
【0166】
シロイヌナズナの形質転換は(島本・岡田監修「モデル植物の実験プロトコール」(1996)秀潤社)に従い実施できる。ジャガイモの形質転換は(門馬(1990)植物組織培養7:57-63)に従い実施できる。以下、ジャガイモの形質転換方法について詳しく記述する。ジャガイモ品種「サッシー」(ジャパンアグリバイオ社)から得られたマイクロチューバーを2〜3mmにスライスし、アグロバクテリウム感染用の材料とする。これを上記のアグロバクテリウムの菌液に浸した後、滅菌済みの濾紙上に置いて過剰のアグロバクテリウムを除く。シャーレ内のMS培地(Zeatin 1ppm, IAA 0.1ppm, アセトシリンゴン100μM、及び寒天0.8%を含む)上に置き、培養は3日間25℃、16時間照明(光量子束密度32μE/m2s)/8時間無照明の条件下で行う。ついで、アセトシリンゴンの代わりにカルベニシリン250ppmを含んだ培地で1週間培養する。その後、さらにカナマイシン50 ppmを含む培地上に移し、2週間ごとに継代する。この間に不定芽が形成し、シュートを生じる。伸張したシュートをカルベニシン250 ppm及びカナマイシン100 ppmを含み、植物生長調節物質を含まないMS培地に置床する。発根したシュートをカナマイシン耐性の生長した植物体の中から外来遺伝子としてカナマイシン耐性遺伝子を含有する個体を得る。カナマイシン耐性遺伝子のプライマーを用いてPCRを行うことで検出し、植物体が形質転換植物体であることを確認することができる。これらの形質転換体ではコレステロールを多く蓄積していることが期待できる。
【0167】
StDWF1H遺伝子の過剰発現形質転換体でも、シロイヌナズナはDWF5遺伝子、ジャガイモは実施例3にあるStDWF5遺伝子が機能しているため、7−デヒドロコレステロールが多く蓄積することは期待できない。しかし、これらの機能が欠損している変異体(dwf5)やRNAi法などによる遺伝子発現抑制形質転換体では7−デヒドロコレステロールが蓄積することが期待できる。これに実施例9で行われる紫外線照射の方法を組み合わせることでビタミンD3を生産できる植物が得られるものと期待できる。
【産業上の利用可能性】
【0168】
本発明の24位還元酵素及びそれをコードする遺伝子を用いる、薬理活性のあるステロイド化合物の生産方法は、天然からの抽出法に比べて大量生産を可能にするため有用である。
【配列表フリーテキスト】
【0169】
配列番号7〜17: プライマー
【技術分野】
【0001】
本発明は、ステロイド骨格の24位を還元する酵素をコードする遺伝子に関する。本発明は、ステロイド類やビタミンD3類に関する。本発明はまた、上記化合物の生産用の細胞およびトランスジェニック酵母に関する。
【背景技術】
【0002】
コレステロールは最も重要な動物性ステロイドである。コレステロールは細胞膜の基本成分であり、細胞が生きていくための必須の成分である。物質としてのコレステロールは薬品産業でも重要であり、ウルソデオキシコール酸などコレステロールから誘導・合成される胆汁酸関連物質は医薬品としておおきな需要がある。コレステロールから誘導・合成されるビタミンD3類は、医薬品としてはもちろん、食物の栄養強化として食品や飼料に添加されている。現在、市場で市販されているコレステロールの大部分は、動物組織(ウシの脊髄や羊毛)からの抽出によっている。しかし、動物組織から直接抽出した物質を原料とすることは、BSE(牛海綿状脳症)の原因物質の持込の可能性、さらには未知の病原性物質の混入など、医薬品等の生産にはリスクがあると考えられている。このことから、動物組織の利用は、さまざまな物質生産から排除される方向にある。コレステロールは、一部においては化学合成がされているが、すべてを化学合成で行うことは価格が非常に高くなり産業として成り立たない。そこで、植物や酵母などの微生物を用いたより大量の安価な生産方法が求められてきた。
【0003】
しかしながら、植物には24位にメチル基が導入される経路以降に生産される植物ステロール類が主として生産されることから、コレステロールの供給源とはなりえなかった。また酵母を代表とする真菌類でも同じくエルゴステロール類のみを蓄積しているため、生産に直接利用することは極めて困難であった。先行例としては酵母にて24位にメチル基を導入する酵素(ステロール24-C-メチル基転移酵素)遺伝子ERG6を欠損し、7位の2重結合を還元する酵素(7-デヒドロコレステロール還元酵素)遺伝子DWF5を導入した酵母に、ヒトのステロイド骨格の24位の2重結合(Δ24、C24とC25の間の2重結合)を還元する酵素(3β-ヒドロキシコレステロールΔ24-還元酵素)遺伝子DHCR24を導入し酵母でコレステロールを生産したものが報告されている(特許文献1、特許文献2)。一般にヒトなど哺乳類の遺伝子は酵母など真菌類に導入した際の相性が悪いことが多く発現が弱いことが知られている。ヒトと相同性の高い哺乳類の遺伝子を利用した改良では、より高発現の真菌生物を作成する際に遺伝子情報が限られており、発現の改善が問題になる。また、ヒトの遺伝子を産業利用することについて倫理的な問題などがあった。
【0004】
植物においては、主に24位にメチル基が導入される経路以降に生産される植物ステロールが生産される。この経路の生合成遺伝子としてシロイヌナズナのDWF1遺伝子(非特許文献1と2)が知られており、酵素反応はステロイドのΔ24(28)(C24とC28の間の2重結合)を還元する、つまり24位のメチレン等を還元する反応を触媒する。DWF1遺伝子とDHCR24遺伝子の間には弱い相同性があることが知られている。一方、一部の植物では植物ステロールとは別にコレステロールの生産が認められる(非特許文献3)。しかし、その生合成経路は不明であり、コレステロール含量を増加させる手段は全く、不明であった。さらに、植物においてビタミンD3を生産しうる植物が知られている(非特許文献4及び5)。これらは7−デヒドロコレステロールが中間産物になっていると推定されるが、同様にビタミンD3を増加させる遺伝子レベルでの研究は行われてこなかった。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特表2007−535960号公報
【特許文献2】米国特許第7608421号明細書
【非特許文献】
【0006】
【非特許文献1】Choeら, Plant Physiol., 1999, 119:897-907
【非特許文献2】Klahreら, Plant Cell, 1998, 10:1677-1690
【非特許文献3】Eckart Eich, ‘Solanaceae and ConVolvulaceae: Secondary Metabolite,’ 2008, Springer, Heidelberg, Germany, p.368-370
【非特許文献4】Eckart Eich, ‘Solanaceae and ConVolvulaceae: Secondary Metabolite,’ 2008, Springer, Heidelberg, Germany, p.461-464
【非特許文献5】Bolandら, Plant Science 2003, 164: 357-369
【発明の概要】
【発明が解決しようとする課題】
【0007】
本発明の目的は、ステロイド骨格の24位を還元する酵素をコードする遺伝子を提供することである。
【課題を解決するための手段】
【0008】
本発明者らは、上記目的を達成すべく鋭意研究を重ねた。本発明者らは植物ステロールの中でコレステロールの含量は低いが、ステロイドサポニンやステロイドアルカロイドを生成することから、コレステロールを中間産物として生成しうるナス科植物、特にSolanum属の植物に着目した(Eckart Eich, ‘Solanaceae and ConVolvulaceae: Secondary Metabolite,’ 2008, Springer, Heidelberg, Germany, p.368-370)。これら植物からはDWF1遺伝子と相同性を持つ2つの遺伝子をin silicoで探しだした。この候補遺伝子を、適正な基質を生産する酵母に、それぞれを導入することで、一方は従来から同定されている植物ステロールの合成に関わるDWF1と同じ機能を持つ遺伝子(StDWF1)であることを明らかにし、さらにもう一方は、それとは異なるステロイド骨格の24位を還元する酵素をコードする遺伝子(StDWF1H)であることを同定することができた。また、本遺伝子を発現させた酵母はコレステロールを著量生産することを示した。さらに本遺伝子は、すでに報告のあるヒトの24位を還元する酵素をコードする遺伝子よりも活性が高いことを見出し、本発明を完成させるに至った。StDWF1とStDWF1Hの相同性はきわめて高く、かつ、それぞれと、既知のヒトの24位を還元する酵素をコードする遺伝子との相同性は大きく離れており、今までの単純な遺伝子配列の相同性からは予想できない結果であり、想定外の結果といわざるを得ない。
【0009】
したがって、本発明は、以下のとおりである。
[1] コレステロールを生産する植物に由来する、ステロイド骨格の24位を還元する活性を有するタンパク質。
[2] ナス科、ゴマノハグサ科、ヤマノイモ科、またはシソ科に属する植物に由来する、[1]のタンパク質。
[3] ジャガイモ、トマトに由来する、[1]のタンパク質。
[4] 以下の(a)〜(c)からなる群から選択される[1]のタンパク質。
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質
(b) 配列番号1または配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含むタンパク質
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含むタンパク質
[5] ステロイド骨格がコレスタン誘導体のステロイド骨格である、[1]〜[4]のいずれかのタンパク質。
[6] 膜貫通領域を持ちFAD結合型配列を持つ還元酵素に属する、[1]〜[5]のいずれかのタンパク質。
[7] 以下の(a)〜(g)からなる群から選択されるDNAを含有する組換え体DNA。
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質をコードするDNA
(b) 配列番号1または配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(d) 配列番号2または配列番号4に示す塩基配列を含むDNA
(e) 配列番号2または配列番号4に示す塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(f) 配列番号2または配列番号4に示す塩基配列と90%以上の配列同一性を有する塩基配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(g) 配列番号2または配列番号4に示す塩基配列において縮重配列を含むDNA
[8] [7]の組換え体DNAを含むベクター。
[9] [7]の組換え体DNAを含む形質転換体。
[10] 形質転換体が微生物または植物を宿主として得られる形質転換体である、[9]の形質転換体。
[11] 微生物が酵母である、[10]の形質転換体。
[12] 24位が還元されたステロイドまたはその誘導体を生成する能力を有する、[9]〜[11]のいずれかの形質転換体。
[13] [9]〜[12]のいずれかの形質転換体を培地に培養し、培養物中に[4]記載のタンパク質を生成、蓄積させ、該培養物より該蛋白質を採取する[4]のタンパク質の製造法。
[14] [9]〜[12]のいずれかの形質転換体であって、24位に2重結合を持つステロイドまたはその誘導体を細胞内に生成する形質転換体を培養し、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収する、24位が還元されたステロイドまたはその誘導体の製造法。
[15] [9]〜[12]のいずれかの形質転換体を24位に2重結合を持つステロイドまたはその誘導体の存在下で培養し、該形質転換体中に24位に2重結合を持つステロイドまたはその誘導体を取り込ませ、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収する、24位が還元されたステロイドまたはその誘導体の製造法。
[16] [9]〜[12]のいずれかの形質転換体の培養物または該培養物の処理物を酵素源に用い、該酵素源、および24位に2重結合を持つステロイドまたはその誘導体を水性媒体中に存在せしめ、該媒体中に24位が還元されたステロイドまたはその誘導体を生成、蓄積させ、該媒体から24位が還元されたステロイドまたはその誘導体を採取する、24位が還元されたステロイドまたはその誘導体の製造法。
[17] 前記培養物が、培養物を遠心分離して得られる菌体もしくは培養液上清、菌体および/または培養液上清の濃縮物、乾燥物、凍結乾燥物、界面活性剤処理物、超音波処理物、機械的摩砕処理物、溶媒処理物、酵素処理物、蛋白質分画物、固定化物あるいは菌体および/または培養液上清から抽出して得られる酵素標品であることを特徴とする、[14]〜[16]のいずれかの製造法。
[18] 24位に2重結合を持つステロイドが、デスモステロール、または7−デヒドロデスモステロールであり、24位が還元されたステロイドが、コレステロール、または7−デヒドロコレステロールである、[14]〜[16]のいずれかの製造法。
[19] [18]の製造法において、24位に2重結合を持つステロイドとして7−デヒドロデスモステロールを用い、7−デヒドロコレステロールを製造し、さらに、得られた7−デヒドロコレステロールに光を当てることを含む、ビタミンD3の製造法。
[20] 前記形質転換体に蓄積された7−デヒドロコレステロールに対して光を当てるものである、[19]の製造法。
[21] (i) ゲノムDNAまたはRNAである核酸を植物から単離する工程、
(ii) (i)の核酸がRNAである場合に逆転写しcDNAを合成する工程、
(iii) (i)または(ii)の工程で得られたDNAから配列番号2、配列番号4または配列番号5に示す塩基配列を含有する遺伝子断片を増幅する工程、ならびに
(iv) DNA中に突然変異および/または多型の存在を決定する工程、
とを含む、植物におけるステロイド骨格の24位を還元する酵素をコードする遺伝子の突然変異および/または多型の存在を検出する方法。
[22] 植物がナス科植物である[21]の方法。
[23] [21]または[22]の方法によってステロイド骨格の24位を還元する酵素をコードする遺伝子の突然変異および/または多型を検出し、突然変異および/または多型を有する植物体を選抜する方法。
[24] [23]の方法により選抜された、ステロイド骨格の24位を還元する酵素をコードする遺伝子に突然変異および/または多型を有する植物体。
[25] ナス科植物である[24]の植物体。
[26] ステロイド骨格の24位を還元する酵素をコードする遺伝子の発現能またはコードするステロイド骨格の24位を還元する酵素の活性が、既存品種に対して変化している植物を選抜する、[23]の植物体を選抜する方法。
[27] [26]の方法によって選抜された、ステロイド骨格の24位を還元する酵素をコードする遺伝子の発現能が既存品種に対して変化しているか、またはステロイド骨格の24位を還元する酵素の活性が既存品種に対して変化している植物体。
[28] ナス科植物である[27]の植物体。
【発明の効果】
【0010】
本発明のステロイド骨格の24位を還元する酵素活性を有するタンパク質により、有用な生理活性を示す、コレステロール類やビタミンD3類、とりわけウルソデオキシコール酸やビタミンD3、およびそれらの置換誘導体を大量かつ安価に生産できる。本発明によれば、ステロイド骨格の24位を還元する酵素タンパク質とそれをコードするDNAの活性発現の調整を行うことができ、該遺伝子の活性が増強された植物および微生物を作製する方法、コレステロールなどのステロイド骨格の24位を還元されたコレステロール類の生産能が変更された植物が提供される。本発明により上記コレステロール類、ビタミンD3類、およびそれらの置換誘導体の過剰発現に特徴がある植物の作出が可能である。
【図面の簡単な説明】
【0011】
【図1】StDWF1HとStDWF1のアミノ酸配列について相同性をGENETYX(ゼネティックス社)で解析を行った結果を示す図である。562アミノ酸に渡って80.0%の同一性がある。
【図2】DWF1に相同なアミノ酸配列を持つ遺伝子の分子進化系統樹解析の結果を示す図である。数値はブーストラップ検定1000に対する数値である。
【図3】酵母発現用ベクターpTochigi101の構造を示す図であり、機能領域(2μ複製領域:2u ori、pUC複製領域:pUC ori、HIS3遺伝子:HIS3、アンピシリン耐性遺伝子:Amp、TDH3プロモーター、PGKターミナーター)と制限酵素部位を示す。
【図4】DWF1H発現酵母(T33, T34)のGCチャート(TIC)とマススペクトル(MS)を示す図である。
【図5】作成した形質転換酵母とプラスミドの組み合わせを示す図である。
【図6】erg6酵母とerg4 erg5酵母にDWF1H、DWF1、LKB又はDHCR24遺伝子を導入した酵母のGCチャート(TIC)とマススペクトル(MS)を示す図である。
【図7】erg4 erg5酵母でのDWF1H、DWF1、LKB又はDHCR24遺伝子を発現する酵母のGCチャート(TIC)とマススペクトル(MS)を示す図である。
【図8】ジャガイモ品種「サッシー」の塊茎の曝光前後での24位還元酵素遺伝子の発現を示す電気泳動図である。
【図9】StDWF1H遺伝子の過剰発現用ベクターの構造を示す図であり、導入する遺伝子部分のT-DNAのライトボーダー(RB)、レフトボーダー(LB)の内部の構造、制限酵素部位を示す。
【発明を実施するための形態】
【0012】
以下、本発明を詳細に説明する。
1.ステロイド骨格の24位を還元する酵素タンパク質(24位還元酵素)
本発明の酵素タンパク質は、ステロイド骨格の24位の2重結合(Δ24、C24とC25の間の2重結合)を還元する酵素活性を有するタンパク質であり、デスモステロール(コレスタ-5,24-ジエノール)、7−デヒドロデスモステロール(コレスタ-5,7,24-トリエノール)、ラノステロール(ラノスタ-8,24-ジエノール)、シクロアルテノール(シクロアルタ‐24‐エノール)などのステロイド骨格の24位に2重結合をもった化合物の24位を還元する能力を有するタンパク質である。本発明の酵素タンパク質により24位が還元されるステロイド骨格を有する化合物はコレスタン誘導体である。本明細書では、そのような変換能を、「24位還元酵素活性」、そのような変換能を有する酵素タンパク質を「24位還元酵素」と言う。また、本発明の酵素タンパク質は、膜貫通領域を持ちFAD結合型配列を持つ還元酵素に属する。
【0013】
上記酵素を含む植物には、コレステロールを生成する植物、例えばナス科のジャガイモ(Solanum tuberosum)、トマト(Solanum lycopersicum)、ルリヤナギ(Solanum glaucophyllum)、ゴマノハグサ科のジギタリス(Digitalis purpurea)、ヤマノイモ科のヤムイモ(Dioscorea sp.)やシソ科Ajuga reptans等の植物種が含まれる。
【0014】
本発明の酵素により得られる24位が還元された化合物には、植物によって合成されるコレステロールまたはその置換誘導体が含まれ、例えばコレステロール(コレスタ-5-エノール)、7−デヒドロコレステロール(コレスタ-5,7-ジエノール)、それらの塩、それらの置換誘導体などが挙げられる。
【0015】
塩としては、ナトリウム塩、カリウム塩などのアルカリ金属塩、アンモニアや、脂肪族アミン、芳香族アミン、飽和アミン、不飽和アミンなどの有機アミンとのアンモニウム塩、カルボン酸塩などが含まれる。これらの塩は、本発明方法による目的トリテルペン化合物の生成の間に形成されてもよいし、あるいは、該化合物の生成後に中和処理により塩形成を行ってもよい。
【0016】
置換誘導体は、ステロイド化合物の、例えば1位、2位、4位、6位、7位、11位、12位、14位、16位、17位、18位、19位、21位、22位、23位、26位、27位などの、24位還元酵素活性に影響の少ないと思われる位置の水素原子が、別の置換基、例えば低級アルキル基(メチル、エチル、プロピル、ブチルなど)、ハロゲン(フッ素、塩素、臭素、ヨウ素)、水酸基、エステル基(アセトキシ、プロパノイルオキシなど)、アシル基(ホルミル、アセチル、プロピオニルなど)、アルコキシ基(メトキシ、エトキシ、プロポキシなど)、アミノ基、モノ-もしくはジ-低級アルキルアミノ基(メチルアミノ、ジメチルアミノ、エチルアミノなど)、アミド基、低級アルキルアミド基(アセタミドなど)、オキソ基、シアノ基、ニトロ基、低級アルキルチオ基(メチルチオ、エチルチオなど)、スルフォニル基(メシル、エチルスルホニルなど)などの官能基で置換された化合物を含む。本発明において、コレステロールおよびコレステロールの誘導体を含めて、コレステロール類と呼ぶ。
【0017】
本発明の24位還元酵素の基質となる好ましいステロイド化合物としては、デスモステロール、7−デヒドロデスモステロール、これらの置換誘導体などのステロイド化合物が挙げられる。置換誘導体の置換位置および置換基は、上記例示と同様である。本発明の方法により、24位還元酵素の作用によって基質となる化合物から24位の二重結合が還元されたステロイド化合物が得られる。一例として、図5上にデスモステロールの24位の二重結合を還元する反応を示す。コレステロールは、デスモステロールのステロイド骨格の24位の二重結合を還元することにより生成し、7−デヒドロコレステロールは、7−デヒドロデスモステロールのステロイド骨格の24位の二重結合を還元することにより生成する。さらに、7−デヒドロコレステロールに紫外線等の光を当てることによりビタミンD3が生成する。本発明において、ビタミンD3およびビタミンD3誘導体を含めてビタミンD3類と呼ぶ。ビタミンD3の誘導体は、上記のステロイド化合物の誘導体と同様の置換基を有する誘導体をいう。
【0018】
24位還元酵素の二重結合を持つ基質は2,3-オキシドスクアレンから、ラノステロール合成酵素やシクロアルテノール合成酵素の作用によってラノステロールやシクロアルテノールが合成され、ステロイドの基本骨格が合成されることにより合成される(P.M.Dewick, Medicinal Natural Product, 3rd ed., John Wiley & Sons, 2009)。これらの合成酵素に関する配列情報およびクローニングについては、種々の動物、植物、菌類のcDNAライブラリーからクローニングされて配列決定されており、すでに公知である。ラノステロール合成酵素についてはBakerら (1995) Biochem. Biophys. Res. Commun. 213: 154-160、Sungら (1995) Bio. Pharm. Bull. 18: 1459-1461、Coreyら (1994) Proc. Natl Acad. Sci. USA 91: 2211-2215、Shiら(1994) Proc. Natl Acad. Sci. USA 91: 7370-7374.などに記載されている。シクロアルテノール合成酵素については、Coreyら(1993) Proc. Natl Acad. Sci. USA 90: 11628-11632などに記載されている。これらの合成酵素は、例えばヒト、マウス、ラット、酵母、麹菌、シロイヌナズナ、イネ、ポプラ、ブドウ、オオムギ、トマト、ダイズ、などの動物、植物、菌類に存在することが知られているので、必要に応じて、これらの生物から上記文献記載のクローニング手法を用いて上記のラノステロール合成酵素やシクロアルテノール合成酵素をコードするDNAを取得し、周知のDNA組換え技術、PCR法などを使用して該DNAを微生物細胞、菌類細胞または植物細胞に導入して該合成酵素を発現するようにすることも可能である。このようにして得られたラノステロール合成酵素やシクロアルテノール合成酵素を発現し得る形質転換菌類細胞または植物細胞から再生されたトランスジェニック菌類や植物に、さらに24位還元酵素をコードするDNAを発現可能に組み込むことによって、これらの細胞、菌類、植物を用いてコレステロールや7−デヒドロコレステロールなどの、24位が還元されたステロイド化合物を生成することが可能になる。
【0019】
本発明で使用可能な24位還元酵素は、以下のものに限定されないが、例えばジャガイモやトマト由来のものであり、それぞれ、例えば配列番号1や配列番号3に示されるアミノ酸配列を含む酵素を包含する。さらに、本発明で使用可能な該酵素は、配列番号1や配列番号3に示されるアミノ酸配列に部分的に変異を有するアミノ酸配列を含み、かつ、24位還元酵素活性を有するタンパク質を包含する。ジャガイモ由来の上記24位還元酵素をStDWF1H、トマト由来の上記24位還元酵素をSlDWF1Hと呼ぶ。
【0020】
ここで、「部分的に変異を有するアミノ酸配列」としては、配列番号1や配列番号3に示されるアミノ酸配列において1もしくは複数、好ましくは1もしくは数個、例えば、1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個、さらに好ましくは1〜3個、さらに好ましくは1個もしくは2個、のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列、または該アミノ酸配列と、BLAST、FASTAなどの相同性検索のための公知のアルゴリズム(例えば、デフォルトすなわち初期設定のパラメータを使用する。)を用いて計算したときに、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、好ましくは少なくとも90%、さらに好ましくは少なくとも95%、特に好ましくは少なくとも97%、98%もしくは99%の配列同一性を有しているアミノ酸配列が挙げられる。因みに、上記配列番号1または配列番号3に示されるアミノ酸配列を含む酵素タンパク質間の配列同一性は約70〜約80%である。
【0021】
本明細書で使用する「配列同一性」は、例えば2つのアミノ酸配列または塩基(ヌクレオチド)配列をアラインメントしたとき(ただしギャップを導入してもよいしギャップを導入しなくてもよいが、好ましくはギャップを導入する。)、ギャップを含むアミノ酸または塩基の総数に対する同一アミノ酸または塩基の数の割合(%)を指す。
【0022】
本発明の24位還元酵素は、植物体から単離された天然の24位還元酵素および遺伝子工学的手法により製造された組換え24位還元酵素を含む。
【0023】
2.24位還元酵素をコードするDNA
本明細書中で使用する「DNA」という用語は、ゲノムDNA、遺伝子、cDNAおよび化学修飾DNAを包含するものとする。
【0024】
本発明で使用する24位還元酵素をコードするDNAは、ステロイド骨格の24位を還元する活性を有する酵素をコードするDNAである。
【0025】
上記の24位還元酵素をコードするDNAは、例えば上記配列番号1や配列番号3に示されるアミノ酸配列をそれぞれコードする塩基配列を含むものであり、具体的には配列番号2や配列番号4に示される塩基配列を含むものである。
【0026】
本発明で使用可能な24位還元酵素をコードするDNAは、また、配列番号2や配列番号4に示される各塩基配列に相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズするDNA、あるいは、配列番号2や配列番号4に示される塩基配列と、BLAST、FASTAなどの相同性検索のための公知のアルゴリズム(例えば、デフォルトすなわち初期設定のパラメータを使用する。)を用いて計算したときに、少なくとも60%、少なくとも70%、少なくとも80%、少なくとも85%、好ましくは少なくとも90%、さらに好ましくは少なくとも95%、特に好ましくは少なくとも97%、98%もしくは99%の配列同一性を有するDNA、あるいは、これらのDNAによりコードされるタンパク質のアミノ酸配列に対して1もしくは複数、好ましくは1もしくは数個、例えば、1〜10個、好ましくは1〜7個、さらに好ましくは1〜5個、さらに好ましくは1〜3個、さらに好ましくは1個もしくは2個、のアミノ酸が欠失、置換、挿入および/または付加されたアミノ酸配列を含むタンパク質をコードするDNAである。ただし該DNAは24位還元酵素活性を有するタンパク質をコードするDNAを包含する。
【0027】
これらのDNAは、配列番号2や配列番号4に示される塩基配列を含むDNAのホモログ(相同体)、アナログ(類似体)または変異体である。このようなDNAは、コレステロールを生成する植物、例えばナス科のジャガイモ(Solanum tuberosum)、トマト(Solanum lycopersicum)、ルリヤナギ(Solanum glaucophyllum)、ゴマノハグサ科のジギタリス(Digitalis purpurea)やシソ科Ajuga reptans等の植物の葉、根、種子などからハイブリダイゼーション、PCR増幅などによって得ることが可能である。
【0028】
本明細書中で使用する「ストリンジェントな条件」は、配列同一性の高いDNAがハイブリダイズする条件であり、そのような条件は当業者ならば適宜決定することができるが、例えば、「1×SSC、0.1% SDS、37℃」程度の条件であり、より厳しい(中ストリンジェントな)条件としては「0.5×SSC、0.1% SDS、42℃」程度の条件であり、さらに厳しい(高ストリンジェントな)条件としては「0.1〜0.2×SSC、0.1% SDS、65℃」程度の条件である。ハイブリダイゼーションの後でさらに、例えば0.1×SSC、0.1% SDS、55〜68℃で洗浄を行う操作を含んでもよく、この操作によってストリンジェンシーを高めることができる。ここで、1×SSCバッファーは、150 mM塩化ナトリウム、15 mMクエン酸ナトリウム、pH7.0である。
【0029】
ハイブリダイゼーション条件やPCR反応の手順については、例えばF.M. Ausbel et al., Short Protocols in Molecular Biology, 3rd ed., John Wiley & Sons, 1995などに記載されている。
【0030】
さらに、本発明で使用可能な24位還元酵素をコードするDNAは、配列番号2や配列番号4に示す塩基配列において遺伝暗号の縮重に基づく配列(縮重配列)を含むDNAも包含する。
【0031】
本発明のDNAは、上記のとおり、24位還元酵素活性を有するタンパク質、すなわち以下の(a)〜(c):
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質;
(b) 配列番号1または配列番号3に示すに示すアミノ酸配列において1または数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含み、かつ、24位還元酵素活性を有するタンパク質;
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含み、かつ、24位還元酵素活性を有するタンパク質;
のいずれかからなる群から選択されるタンパク質をコードする。
【0032】
さらに具体的には、上記DNAは、以下の(d)〜(g):
(d) 配列番号2または配列番号4に示す塩基配列を含むDNA;
(e) 配列番号2または配列番号4に示す塩基配列を含むDNAに相補的な塩基配列からなるDNAとストリンジェントな条件下でハイブリダイズし、かつ、24位還元酵素活性を有するタンパク質をコードするDNA;
(f) 配列番号2または配列番号4に示す塩基配列と90%以上の配列同一性を有する塩基配列を含み、かつ、24位還元酵素活性を有するタンパク質をコードするDNA;
(g) 配列番号2または配列番号4に示す塩基配列において縮重配列を含むDNA;
のいずれかからなる群から選択される。
【0033】
3.組換えベクター
本発明のDNAは、それを発現可能にするために、制御配列を含む適切なベクターに挿入される。このようにして得られた組換え体DNAが組換えベクターである。
【0034】
ベクターとしては、原核または真核生物の細胞で使用可能なあらゆるベクターを意図し、例えば細菌(エシェリシア属、シュードモナス属、バチルス属、ロドコッカス属など)、糸状菌(アスペルギルス属、ニューロスポラ属、フザリウム属、トリコデルマ属、ペニシリウム属など)、担子菌(白色腐朽菌など)、酵母(サッカロマイセス属、ピチア属、カンジダ属など)等の微生物用ベクター、植物細胞用ベクター、昆虫細胞用ベクターなどを使用できる。
【0035】
例えば、細菌用ベクターとしては、pBR、pUC、pET、pBluescriptシリーズのベクター類などが挙げられ、酵母用ベクターとしては、非限定的にpDR196、pYES-DEST 52、YIp5、YRp17、YEp24などが挙げられ、植物細胞用ベクターとしては、非限定的にpGWB vector、pBiEl2-GUS、pIG121-Hm、pBI121、pBiHyg-HSE、pB119、pBI101、pGV3850、pABH-Hm1などが挙げられ、昆虫細胞用ベクターとしては、非限定的にpBM030、pBM034、pBK283などが挙げられる。
【0036】
本発明において使用されるベクターには、プロモーター、ターミネーター、エンハンサー、シャインダルガルノ配列、リボソーム結合配列、シグナル配列等の遺伝子の発現、調節、分泌に関する構成要素が組込まれ、必要に応じて、選択マーカー(例えば、薬剤耐性遺伝子、レポーター遺伝子)を含有する。
【0037】
プロモーターには、lacプロモーター、trpプロモーター、recAプロモーター、tacプロモーター、λPLプロモーター、T7プロモーター、CaMV35Sプロモーター、ADH1プロモーター、GALプロモーター、PHO5プロモーター、PGKプロモーター、GAPDHプロモーターなどが非限定的に含まれる。
【0038】
薬剤耐性遺伝子には、カナマイシン耐性遺伝子、アンピシリン耐性遺伝子、ハイグロマイシン耐性遺伝子などが含まれる。レポーター遺伝子には、lacZ遺伝子、GFP遺伝子、GUS遺伝子、ルシフェラーゼ遺伝子などが含まれる。その他の選択マーカーには、例えばNPTII遺伝子、ジヒドロ葉酸レダクターゼ遺伝子などが含まれる。
【0039】
遺伝子の発現、調節、分泌に関する構成要素は、その性質に応じて、それぞれが機能し得る形で組換えベクターに組み込まれることが好ましい。そのような操作は、当業者であれば適切に行うことができる。
【0040】
4.形質転換体
本発明の形質転換体は、本発明の組換えベクターを一時的もしくは恒常的に保持する形質転換体である。形質転換体は、24位還元酵素をコードするDNAを挿入した組換えベクターを、目的DNAが発現し得るように宿主細胞中に導入することにより得ることができる。該DNA断片を適当な発現ベクターのプロモーターの下流に挿入することにより、組換え体DNAを作製する。該組換え体DNAを、該発現ベクターに適合した宿主細胞に導入することにより、本発明のタンパク質を生産する形質転換体を得ることができる。
【0041】
宿主細胞としては、細菌、酵母など菌類、動物細胞、昆虫細胞、植物細胞等、目的とする遺伝子を発現できるものであればいずれも用いることができる。好ましくは、酵母など菌類、昆虫細胞または植物細胞である。
【0042】
上記の宿主細胞が、ステロイド化合物の生合成系を有している細胞、あるいは、該生合成系を有していない細胞であっても外因的に(または外来的に)ラノステロール合成酵素、シクロアルテノール合成酵素などをコードするDNAが発現可能に組み込まれた細胞のいずれかである場合には、上記の形質転換によってさらにラノステロール合成酵素、シクロアルテノール合成酵素をコードするDNAが発現可能に含まれるため、これらの細胞を適当な基質を含む培地で培養することによってラノステロールやシクロアルテノールなどステロイド類を生産することができる。
【0043】
ラノステロールやシクロアルテノールなどステロイド類の生合成系を有している細胞には、植物細胞、酵母細胞、菌類細胞、動物細胞のように本来該生合成系を備えている細胞が含まれる。また、上記の生合成系を有していない細胞には、該生合成系に関わる酵素類のゲノム領域を外因的に含む細胞などが包含される。外因的ゲノム領域は、例えばプラスミド、ファージミド、BAC、PAC、YAC、ウイルスなどのベクターに挿入されて宿主細胞内に移入されうる。いずれにしても、上記例示の細胞の形質転換によって、該細胞は、場合により誘導的に、24位還元酵素を過剰発現する。
【0044】
細菌等の原核生物を宿主細胞として用いる場合は、本発明のDNAまたは本発明の製造法に用いられるDNAを有する組換え体DNAは、原核生物中で自立複製可能であると同時に、プロモーター、リボソーム結合配列、本発明のDNAまたは本発明の製造法に用いられるDNA、転写終結配列より構成された組換え体DNAであることが好ましい。プロモーターを制御する遺伝子が含まれていてもよい。
【0045】
発現ベクターとしては、pBTrp2、pBTac1、pBTac2(いずれもベーリンガーマンハイム社)、pHelix1(ロシュ・ダイアグノスティクス社)、pKK233-2(アマシャム・ファルマシア・バイオテク社)、pSE280(インビトロジェン社)、pGEMEX-1(プロメガ社)、pQE-8(キアゲン社)、pET-3(ノバジェン社)、pKYP10(特開昭58-110600)、pKYP200[Agric.Biol.Chem.,48,669(1984)]、pLSA1[Agric.Biol.Chem.,53,277(1989)]、pGEL1[Proc.Natl.Acad.Sci.,USA,82,4306(1985)]、pBluescriptII SK(+)、pBluescript II KS(-)(ストラタジーン社)、pTrS30[エシェリヒア・コリJM109/pTrS30(FERM BP-5407)より調製]、pTrS32[エシェリヒア・コリJM109/pTrS32(FERM BP-5408)より調製]、pPAC31(WO98/12343)、pUC19[Gene,33,103(1985)]、pSTV28(タカラバイオ社)、pUC118(タカラバイオ社)、pPA1(特開昭63-233798)等を例示することができる。
【0046】
プロモーターとしては、エシェリヒア・コリ等の宿主細胞中で機能するものであればいかなるものでもよい。例えば、trpプロモーター(Ptrp)、lacプロモーター(Plac)、PLプロモーター、PRプロモーター、PSEプロモーター等の、大腸菌やファージ等に由来するプロモーター、SPO1プロモーター、SPO2プロモーター、penPプロモーター等をあげることができる。またPtrpを2つ直列させたプロモーター、tacプロモーター、lacT7プロモーター、let Iプロモーターのように人為的に設計改変されたプロモーター等も用いることができる。
【0047】
リボソーム結合配列であるシャイン−ダルガノ(Shine-Dalgarno)配列と開始コドンとの間を適当な距離(例えば6〜18塩基)に調節したプラスミドを用いることが好ましい。
【0048】
本発明のDNAまたは本発明の製造法に用いられるDNAを発現ベクターに結合させた組換え体DNAにおいては、転写終結配列は必ずしも必要ではないが、構造遺伝子の直下に転写終結配列を配置することが好ましい。
【0049】
原核生物としては、エシェリヒア属、セラチア(Serratia)属、バチルス属、ブレビバクテリウム(Brevibacterium)属、コリネバクテリウム(Corynebacterium)属、ミクロバクテリウム(Microbacterium)属、シュードモナス(Pseudomonas)属に属する微生物等をあげることができる。
【0050】
組換え体DNAの導入方法としては、上記宿主細胞へDNAを導入する方法であればいずれも用いることができ、例えば、カルシウムイオンを用いる方法[Proc.Natl.Acad.Sci.,USA,69,2110(1972)]、プロトプラスト法(特開昭63-248394)、エレクトロポレーション法[Nucleic Acids Res.,16,6127(1988)]等をあげることができる。
【0051】
酵母菌株を宿主細胞として用いる場合には、発現ベクターとして、例えば、YEp13(ATCC37115)、YEp24(ATCC37051)、YCp50(ATCC37419)、pHS19、pHS15等を用いることができる。
【0052】
プロモーターとしては、酵母菌株中で機能するものであればいずれのものを用いてもよく、例えば、PHO5プロモーター、PGKプロモーター、GAPプロモーター、ADHプロモーター、gal 1プロモーター、gal 10プロモーター、ヒートショックポリペプチドプロモーター、MFα1 プロモーター、CUP 1プロモーター等のプロモーターをあげることができる。
【0053】
宿主細胞としては、サッカロマイセス(Saccharomyces)属、シゾサッカロマイセス(Schizosaccharomyces)属、クルイベロマイセス(Kluyveromyces)属、トリコスポロン(Trichosporon)属、シワニオマイミセス(Schwanniomyces)属、ピチア(Pichia)属、またはキャンディダ(Candida)属等に属する酵母菌株をあげることができ、具体的には、サッカロマイセス・セレビシエ(Saccharomyces cerevisiae)、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)、クルイベロマイセス・ラクティス(Kluyveromyces lactis)、トリコスポロン・プルランス(Trichosporon pullulans)、シワニオマイセス・アルビウス(Schwanniomyces alluvius)、ピチア・パストリス(Pichia pastoris)、キャンディダ・ウティリス(Candida utilis)等をあげることができる。
【0054】
組換え体DNAの導入方法としては、酵母にDNAを導入する方法であればいずれも用いることができ、例えば、エレクトロポレーション法[Methods Enzymol.,194,182(1990)]、スフェロプラスト法[Proc.Natl.Acad.Sci.,USA,81,4889(1984)]、酢酸リチウム法[J.Bacteriol.,153,163(1983)]等をあげることができる。
【0055】
動物細胞を宿主として用いる場合には、発現ベクターとして、例えば、pcDNAI、pcDM8(フナコシ社より市販)、pAGE107(特開平3-22979号公報)、pAS3-3(特開平2-227075号公報)、pCDM8[Nature,329,840(1987)]、pcDNAI/Amp(インビトロジェン社)、pREP4(インビトロジェン社)、pAGE103[J.Biochem,101,1307(1987)]、pAGE210、pAMo、pAMoA等を用いることができる。
【0056】
プロモーターとしては、動物細胞中で機能するものであればいずれも用いることができ、例えば、サイトメガロウイルス(CMV)のIE(immediate early)遺伝子のプロモーター、SV40の初期プロモーターあるいはメタロチオネインのプロモーター、レトロウイルスのプロモーター、ヒートショックプロモーター、SRαプロモーター等をあげることができる。また、ヒトCMVのIE遺伝子のエンハンサーをプロモーターと共に用いてもよい。
【0057】
宿主細胞としては、マウス・ミエローマ細胞、ラット・ミエローマ細胞、マウス・ハイブリドーマ細胞、ヒトの細胞であるナマルバ(Namalwa)細胞またはナマルバKJM-1細胞、ヒト胎児腎臓細胞、ヒト白血病細胞、アフリカミドリザル腎臓細胞、チャイニーズ・ハムスターの細胞であるCHO細胞、HBT5637(特開昭63-299号公報)等をあげることができる。
【0058】
マウス・ミエローマ細胞としては、SP2/0、NSO等、ラット・ミエローマ細胞としてはYB2/0等、ヒト胎児腎臓細胞としてはHEK293(ATCC CRL-1573)、ヒト白血病細胞としてはBALL-1等、アフリカミドリザル腎臓細胞としてはCOS-1、COS-7等をあげることができる。
【0059】
組換え体DNAの導入方法としては、動物細胞にDNAを導入する方法であればいずれも用いることができ、例えば、エレクトロポレーション法[Cytotechnology,3,133(1990)]、リン酸カルシウム法(特開平2-227075号公報)、リポフェクション法[Proc.Natl.Acad.Sci.,USA,84,7413(1987)]、Virology,52,456(1973)に記載の方法等をあげることができる。
【0060】
昆虫細胞を宿主として用いる場合には、例えばBaculovirus Expression Vectors,A Laboratory Manual,W.H.Freeman and Company,New York(1992)、カレント・プロトコールズ・イン・モレキュラー・バイオロジー、Molecular Biology,A Laboratory Manual、Bio/Technology,6,47(1988)等に記載された方法によって、タンパク質を生産することができる。
【0061】
即ち、組換え遺伝子導入ベクターおよびバキュロウイルスを昆虫細胞に共導入して昆虫細胞培養上清中に組換えウイルスを得た後、さらに組換えウイルスを昆虫細胞に感染させ、タンパク質を生産させることができる。
【0062】
該方法において用いられる遺伝子導入ベクターとしては、例えば、pVL1392、pVL1393、pBlueBacIII(いずれもインビトロジェン社)等をあげることができる。
【0063】
バキュロウイルスとしては、例えば、夜盗蛾科昆虫に感染するウイルスであるアウトグラファ・カリフォルニカ・ヌクレアー・ポリヘドロシス・ウイルス(Autographa californica nuclear polyhedrosis virus)等を用いることができる。
【0064】
昆虫細胞としては、スポドプテラ・フルギペルダ(Spodoptera frugiperda)の卵巣細胞、トリコプルシア・ニ(Trichoplusia ni)の卵巣細胞、カイコ卵巣由来の培養細胞等を用いることができる。
【0065】
スポドプテラ・フルギペルダの卵巣細胞としてはSf9、Sf21(バキュロウイルス・イクスプレッション・ベクターズ ア・ラボラトリー・マニュアル)等、トリコプルシア・ニの卵巣細胞としてはHigh5、BTI-TN-5B1-4(インビトロジェン社)等、カイコ卵巣由来の培養細胞としてはボンビクス・モリ(Bombyxmori)N4等をあげることができる。
【0066】
組換えウイルスを調製するための、昆虫細胞への上記組換え遺伝子導入ベクターと上記バキュロウイルスの共導入方法としては、例えば、リン酸カルシウム法(特開平2-227075)、リポフェクション法[Proc.Natl.Acad.Sci.,USA,84,7413(1987)]等をあげることができる。
【0067】
植物細胞を宿主細胞として用いる場合には、ウイルスベクター、アグロバクテリウムのTiプラスミド、Riプラスミド等をベクターとして用いるアグロバクテリウム法が好適に使用できる。タバコモザイクウイルスベクター等、植物ウイルスを利用したものをあげることができる。プロモーターとしては、植物細胞中で機能するものであればいずれのものを用いてもよく、例えば、カリフラワーモザイクウイルス(CaMV)の35Sプロモーター、イネアクチン1プロモーター等をあげることができる。宿主細胞としては、タバコ、ジャガイモ、トマト、ニンジン、ダイズ、アブラナ、アルファルファ、イネ、コムギ、オオムギ等の植物細胞等をあげることができる。
【0068】
組換えベクターの導入方法としては、植物細胞にDNAを導入する方法であればいずれも用いることができ、例えば、アグロバクテリウム(Agrobacterium)を用いる方法(特開昭59-140885号公報、特開昭60-70080号公報、WO94/00977公報)、エレクトロポレーション法(特開昭60-251887号公報)、パーティクルガン(遺伝子銃)を用いる方法(特許第2606856号公報、特許第2517813号公報)等をあげることができる。
【0069】
酵母、動物細胞、昆虫細胞または植物細胞により発現させた場合には、糖あるいは糖鎖が付加されたタンパク質を得ることができる。さらにまた、形質転換植物体(「トランスジェニック植物」とも称する。)を作出する方法は、24位還元酵素をコードするDNAを含むベクターで形質転換された植物細胞から植物体を再生させることにより得ることができる。植物細胞からの植物体の再生は公知の方法、例えばカルス培養等により行うことができる。
【0070】
5.本発明のタンパク質の製造法
本発明のタンパク質は、本発明のタンパク質をコードするDNAを宿主細胞中で発現させる一時的もしくは恒常的な4.形質転換体であげられた形質転換体を得て、製造することができる。本発明のタンパク質をコードする部分の塩基配列を、宿主の発現に最適なコドンとなるように、塩基を置換することにより、該タンパク質の生産率を向上させることができる。
【0071】
得られる形質転換体を培地に培養し、もしくは形質転換体植物の場合は栽培し、培養物中に本発明のタンパク質を生成蓄積させ、該培養物から採取することにより、該タンパク質を製造することができる。
【0072】
本発明のタンパク質を製造するための上記形質転換体の宿主としては、細菌、酵母など菌類、動物細胞、昆虫細胞、植物細胞等、目的とする遺伝子を発現できるものであればいずれも用いることができる。好ましくは、酵母など菌類、昆虫細胞または植物細胞である。
上記形質転換体を培地に培養する方法は、宿主の培養に用いられる通常の方法に従って行うことができる。
【0073】
エシェリヒア・コリ等の原核生物あるいは酵母等の真核生物を宿主として得られた形質転換体を培養する培地としては、該生物が資化し得る炭素源、窒素源、無機塩類等を含有し、形質転換体の培養を効率的に行える培地であれば天然培地、合成培地のいずれを用いてもよい。
【0074】
炭素源としては、該生物が資化し得るものであればよく、グルコース、フラクトース、スクロース、これらを含有する糖蜜、デンプンあるいはデンプン加水分解物等の炭水化物、酢酸、プロピオン酸等の有機酸、エタノール、プロパノール等のアルコール類等を用いることができる。
【0075】
窒素源としては、アンモニア、塩化アンモニウム、硫酸アンモニウム、酢酸アンモニウム、リン酸アンモニウム等の無機酸もしくは有機酸のアンモニウム塩、その他の含窒素化合物、並びに、ペプトン、肉エキス、酵母エキス、コーンスチープリカー、カゼイン加水分解物、大豆粕および大豆粕加水分解物、各種発酵菌体、およびその消化物等を用いることができる。
【0076】
無機塩としては、リン酸第一カリウム、リン酸第二カリウム、リン酸マグネシウム、硫酸マグネシウム、塩化ナトリウム、硫酸第一鉄、硫酸マンガン、硫酸銅、炭酸カルシウム等を用いることができる。
【0077】
培養は、通常振盪培養または深部通気攪拌培養等の好気的条件下で行う。培養温度は15〜40℃がよく、培養時間は、通常5時間〜7日間である。培養中pHは3.0〜9.0に保持する。pHの調整は、無機または有機の酸、アルカリ溶液、尿素、炭酸カルシウム、アンモニア等を用いて行う。
また、培養中必要に応じて、アンピシリンやテトラサイクリン等の抗生物質を培地に添加してもよい。
【0078】
プロモーターとして誘導性のプロモーターを用いた発現ベクターで形質転換した微生物を培養するときには、必要に応じてインデューサーを培地に添加してもよい。例えば、lacプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはイソプロピル−β−D−チオガラクトピラノシド等を、trpプロモーターを用いた発現ベクターで形質転換した微生物を培養するときにはインドールアクリル酸等を培地に添加してもよい。
【0079】
動物細胞を宿主として得られた形質転換体を培養する培地としては、一般に使用されているRPMI1640培地[J.Am.Med.Assoc.,199,519(1967)]、イーグル(Eagle)のMEM培地[Science,122,501(1952)]、DMEM培地[Virology,8,396(1959)]、199培地[Proc.Soc.Biol.Med.,73,1(1950)]またはこれら培地に牛胎児血清等を添加した培地等を用いることができる。
【0080】
培養は、通常pH6〜8、25〜40℃、5%CO2存在下等の条件下で1〜7日間行う。
また、培養中必要に応じて、カナマイシン、ペニシリン、ストレプトマイシン等の抗生物質を培地に添加してもよい。
【0081】
昆虫細胞を宿主として得られた形質転換体を培養する培地としては、一般に使用されているTNM-FH培地(ファーミンジェン社)、Sf-900 II SFM培地(ライフ・テクノロジーズ社)、ExCell400、ExCell405[いずれもJRHバイオサイエンシーズ社]、Grace’s Insect Medium[Nature,195,788(1962)]等を用いることができる。
【0082】
培養は、通常pH6〜7、25〜30℃等の条件下で1〜5日間行う。
また、培養中必要に応じて、ゲンタマイシン等の抗生物質を培地に添加してもよい。
【0083】
植物細胞を宿主として得られた形質転換体は、細胞として、または植物の細胞や器官に分化させて培養することができる。該形質転換体を培養する培地としては、一般に使用されているムラシゲ・アンド・スクーグ(MS)培地、ホワイト(White)培地、またはこれら培地にオーキシン、サイトカイニン等、植物ホルモンを添加した培地等を用いることができる。
【0084】
培養は、通常pH5〜9、20〜40℃の条件下で3〜60日間行う。
また、培養中必要に応じて、カナマイシン、ハイグロマイシン等の抗生物質を培地に添加してもよい。
【0085】
上記のとおり、本発明のタンパク質をコードするDNAを発現ベクターに連結した組換え体DNAを保有する微生物、昆虫細胞、動物細胞、あるいは植物細胞由来の形質転換体を、通常の培養方法に従って培養し、本発明のタンパク質を生成蓄積させ、該培養物または培養物の処理物より該タンパク質を採取することにより、該タンパク質を製造することができる。ここで、培養物の処理物とは、培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の溶媒処理物、該菌体の酵素処理物、該菌体の蛋白質分画物、該菌体の固定化物あるいは該菌体より抽出して得られる酵素標品等を含む。
【0086】
本発明のタンパク質の生産方法としては、宿主細胞内に生産させる方法、宿主細胞外に分泌させる方法、あるいは宿主細胞外膜上に生産させる方法があり、選択した方法に応じて、生産させるタンパク質の構造を変えることができる。
【0087】
本発明のタンパク質が宿主細胞内あるいは宿主細胞外膜上に生産される場合、ポールソンらの方法[J.Biol.Chem.,264,17619(1989)]、ロウらの方法[Proc.Natl.Acad.Sci.,USA,86,8227(1989)、Genes Develop.,4,1288(1990)]、または特開平05-336963号公報、WO94/23021号公報等に記載の方法を準用することにより、該タンパク質を宿主細胞外に積極的に分泌させることができる。
【0088】
すなわち、遺伝子組換えの手法を用いて、本発明のタンパク質の活性部位を含むタンパク質の手前にシグナルペプチドを付加した形で生産させることにより、該タンパク質を宿主細胞外に積極的に分泌させることができる。
【0089】
また、特開平2-227075号公報に記載されている方法に準じて、ジヒドロ葉酸還元酵素遺伝子等を用いた遺伝子増幅系を利用して生産量を上昇させることもできる。
【0090】
さらに、遺伝子導入した動物または植物の細胞を再分化させることにより、遺伝子が導入された動物個体(トランスジェニック非ヒト動物)または植物個体(トランスジェニック植物)を造成し、これらの個体を用いて本発明のタンパク質を製造することもできる。
【0091】
本発明のタンパク質を生産する形質転換体が動物個体または植物個体の場合は、通常の方法に従って、飼育または栽培し、該タンパク質を生成蓄積させ、該動物個体または植物個体より該タンパク質を採取することにより、該タンパク質を製造することができる。
【0092】
動物個体を用いて本発明のタンパク質を製造する方法としては、例えば公知の方法[Am.J.Clin.Nutr.,63,639S(1996)、Am.J.Clin.Nutr.,63,627S(1996)、Bio/Technology,9,830(1991)]に準じて遺伝子を導入して造成した動物中に本発明のタンパク質を生産する方法があげられる。
【0093】
動物個体の場合は、例えば、本発明のDNAまたは本発明の製造法に用いられるDNAを導入したトランスジェニック非ヒト動物を飼育し、本発明のタンパク質を該動物中に生成、蓄積させ、該動物中より該タンパク質を採取することにより、該タンパク質を製造することができる。該動物中の該タンパク質を生成、蓄積させる場所としては、例えば、該動物のミルク(特開昭63-309192号公報)、卵等をあげることができる。この際に用いられるプロモーターとしては、動物で機能するものであればいずれも用いることができるが、例えば、乳腺細胞特異的なプロモーターであるαカゼインプロモーター、βカゼインプロモーター、βラクトグロブリンプロモーター、ホエー酸性プロテインプロモーター等が好適に用いられる。
【0094】
植物個体を用いて本発明のタンパク質を製造する方法としては、例えば本発明のタンパク質をコードするDNAを導入したトランスジェニック植物を公知の方法[組織培養,20(1994)、組織培養,21(1995)、Trends Biotechnol.,15,45(1997)]に準じて栽培し、該タンパク質を該植物中に生成、蓄積させ、該植物中より該タンパク質を採取することにより、該タンパク質を生産する方法があげられる。
【0095】
本発明のタンパク質を生産する形質転換体を用いて製造された本発明のタンパク質を単離・精製する方法としては、通常の酵素の単離、精製法を用いることができる。
【0096】
例えば、本発明のタンパク質が、細胞内に溶解状態で生産された場合には、培養終了後、細胞を遠心分離により回収し、水系緩衝液にけん濁後、超音波破砕機、フレンチプレス、マントンガウリンホモゲナイザー、ダイノミル等により細胞を破砕し、無細胞抽出液を得る。
【0097】
該無細胞抽出液を遠心分離することにより得られる上清から、通常の酵素の単離精製法、即ち、溶媒抽出法、硫安等による塩析法、脱塩法、有機溶媒による沈殿法、ジエチルアミノエチル(DEAE)−セファロース、DIAION HPA-75(三菱化成社製)等レジンを用いた陰イオン交換クロマトグラフィー法、S-Sepharose FF(ファルマシア社)等のレジンを用いた陽イオン交換クロマトグラフィー法、ブチルセファロース、フェニルセファロース等のレジンを用いた疎水性クロマトグラフィー法、分子篩を用いたゲルろ過法、アフィニティークロマトグラフィー法、クロマトフォーカシング法、等電点電気泳動等の電気泳動法等の手法を単独あるいは組み合わせて用い、精製標品を得ることができる。
【0098】
また、該タンパク質が細胞内に不溶体を形成して生産された場合は、同様に細胞を回収後破砕し、遠心分離を行うことにより得られた沈殿画分より、通常の方法により該タンパク質を回収後、該タンパク質の不溶体をタンパク質変性剤で可溶化する。
【0099】
該可溶化液を、タンパク質変性剤を含まないあるいはタンパク質変性剤の濃度がタンパク質が変性しない程度に希薄な溶液に希釈、あるいは透析し、該タンパク質を正常な立体構造に構成させた後、上記と同様の単離精製法により精製標品を得ることができる。
【0100】
本発明のタンパク質またはその糖修飾体等の誘導体が細胞外に分泌された場合には、培養上清に該タンパク質またはその糖付加体等の誘導体を回収することができる。
【0101】
即ち、該培養物を上記と同様の遠心分離等の手法により処理することにより可溶性画分を取得し、該可溶性画分から、上記と同様の単離精製法を用いることにより、精製標品を得ることができる。
【0102】
また、本発明のタンパク質を他のタンパク質との融合タンパク質として生産し、融合したタンパク質に親和性をもつ物質を用いたアフィニティークロマトグラフィーを利用して精製することもできる。例えば、ロウらの方法[Proc.Natl.Acad.Sci.,USA,86,8227(1989)、Genes Develop.,4,1288(1990)]、特開平5-336963号公報、WO94/23021号公報に記載の方法に準じて、本発明のタンパク質をプロテインAとの融合タンパク質として生産し、イムノグロブリンGを用いるアフィニティークロマトグラフィーにより精製することができる。
【0103】
また、本発明のタンパク質をFlagペプチドとの融合タンパク質として生産し、抗Flag抗体を用いるアフィニティークロマトグラフィーにより精製することができる[Proc.Natl.Acad.Sci.,USA,86,8227(1989)、Genes Develop.,4,1288(1990)]。更に、該タンパク質自身に対する抗体を用いたアフィニティークロマトグラフィーで精製することもできる。
【0104】
上記で取得されたタンパク質のアミノ酸配列情報を基に、Fmoc法(フルオレニルメチルオキシカルボニル法)、tBoc法(t−ブチルオキシカルボニル法)等の化学合成法により、本発明のタンパク質を製造することができる。また、Advanced ChemTech社、パーキン・エルマー社、Pharmacia社、Protein Technology Instrument社、Synthecell-Vega社、PerSeptive社、島津製作所等のペプチド合成機を利用して化学合成することもできる。
【0105】
6.24位が還元されたステロイド化合物の製造方法
本発明は、無細胞系で、あるいは細胞又は植物を用いて、外因性の24位還元酵素活性を有するタンパク質存在下または24位還元酵素をコードするDNAの発現下で、デスモステロール、7−デヒドロデスモステロール、ラノステロール、シクロアルテノールなどのステロイド骨格の24位に2重結合をもった化合物の24位を還元する方法を提供する。本発明の方法により、24位の二重結合を還元する上記化合物の中でもコレステロールの材料となるデスモステロールおよびビタミンD3の材料となる7−デヒドロデスモステロールのステロイド骨格の24位を還元する方法が重要である。
【0106】
本明細書で使用される「外因性」とは、細胞または植物が本来もつ酵素またはその遺伝子自体ではなく、該細胞または植物に、あるいは上記のステロイド化合物原料の反応系に、人為的に該酵素またはそれをコードするDNAを導入することを意味する。
【0107】
本発明はまた、上記の変換方法によってステロイド骨格の24位に2重結合をもった化合物の24位を還元されたステロイド化合物を生成し、該化合物を回収することを含む、24位を還元されたステロイド化合物の製造方法を提供する。
【0108】
本発明では、24位還元酵素は、粗製、半精製もしくは精製された、あるいは多糖類、多孔性ポリマー、多孔性無機物(例えばガラス、鉱物、セラミックス等)などの支持体などに共有結合もしくは非共有結合によって固定化された、酵素として反応系で使用されてもよいし、該酵素を産生する細胞または植物体自体が反応系で使用されてもよい。
【0109】
本発明で使用可能な24位還元酵素は膜結合型でNADPHを補酵素とする還元酵素の一種であり、上述の製造法で得ることができる。
【0110】
24位を還元されたステロイド化合物の製造は、24位還元酵素をコードするDNAが導入された形質転換体を用いて行うことができ、以下の(a)〜(c)の方法が挙げられる。
(a) 24位に二重結合を持つステロイドを細胞内に生成する形質転換体を培養して、菌体内で24位還元活性を発現させて還元体を製造する方法。該方法においては、形質転換体であって、24位に2重結合を持つステロイドまたはその誘導体を細胞内に生成する形質転換体を培養し、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収すればよい。
(b) 24位に二重結合を持つステロイドを培養の過程で形質転換体に取り込ませて、菌体内で24位還元活性を発現させて還元体を製造する方法。該方法においては、形質転換体を24位に2重結合を持つステロイドまたはその誘導体の存在下で培養し、該形質転換体中に24位に2重結合を持つステロイドまたはその誘導体を取り込ませ、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収すればよい。
(c) 培養物を酵素源として、24位に二重結合を持つステロイドを酵素源に接触させて還元体を製造する方法。該方法においては、質転換体の培養物または該培養物の処理物を酵素源に用い、該酵素源、および24位に2重結合を持つステロイドまたはその誘導体を水性媒体中に存在せしめ、該媒体中に24位が還元されたステロイドまたはその誘導体を生成、蓄積させ、該媒体から24位が還元されたステロイドまたはその誘導体を採取すればよい。
【0111】
DNA組換え技術によって、高い酵素活性を持ったタンパク質の発現が可能であるため、4.形質転換体であげられた形質転換体(例えば、酵母細胞、菌類細胞、昆虫細胞、植物細胞)の培養液に上記24位還元酵素の基質、例えばデスモステロール、7−デヒドロデスモステロール、ラノステロール、シクロアルテノール、またはそれらの置換誘導体、を添加することにより、24位を還元されたステロイド化合物を生産することができる。例えば、形質転換微生物の培養液にデスモステロール等のステロイド類を基質として投与することにより、24位を還元されたされたコレステロールを効率的に大量に生産することが可能である。酵母細胞、菌類細胞、植物細胞などは24位還元酵素の基質は内在性のものも利用できる。また、内在性の基質の含量をあらかじめ増加させるようにした細胞も利用できる。
【0112】
さらに、7−デヒドロデスモステロールの24位を還元して生成した7−デヒドロコレステロールに紫外線等の光を当てることによりビタミンD3(コレカルシフェノール)が生成する。プロビタミンD3である7−デヒドロコレステロールに紫外線等の光を当てるとプレビタミンD3((6z)-タカルシオール)が生成し、その後自然にビタミンD3へ異性化する。この際、本発明の24位還元酵素を含む形質転換体に7−デヒドロコレステロールを産生蓄積させ、該形質転換体に紫外線を当ててもよい。
【0113】
本発明は、本発明の24位還元酵素を用いて、コレステロールを製造する方法及びビタミンD3を製造する方法をも包含する。
【0114】
(1)酵素的製造法
無細胞系では、上記形質転換体の培養液から24位還元酵素を含有する無細胞抽出液を調製し、その一部を、デスモステロールまたはそれらの置換誘導体などのステロイド化合物を含有するバッファーに加えて変換反応を行うことによって、24位を還元されたステロイド化合物を製造することができる。
【0115】
収穫した細胞の懸濁液、あるいは適量のバッファー中の形質転換植物を、ホモジナイザー、超音波破砕機あるいはフレンチプレス等により破砕後、遠心分離して無細胞抽出液を得る。バッファーには、ポリペプチドの失活を防ぐため、抗酸化剤、酵素の安定化剤、ポリフェノール吸着剤、金属配位子などを添加することができる。さらに比活性を高めるにはポリペプチドを精製することが有効であり、超遠心機による遠心分離法、硫安等による塩析方、ゲル濾過法、イオン交換クロマトグラフィー法、アフィニティクロマトグラフィー法、電気泳動法などの手法を単独で、あるいは組み合わせて用いることができる。得られたポリペプチドを含むバッファーに、基質となる上記のステロイドおよび補酵素を添加してインキュベートする。補酵素としてはNADPHが利用でき、グルコース-6-リン酸とグルコース-6-リン酸デヒドロゲナーゼを用いたNADPH再構成系も併用できる。
【0116】
上記製造法において、本発明のタンパク質は、基質として用いるデスモステロール1mgあたり0.01〜100mg、好ましくは0.1mg〜10mg添加する。
【0117】
上記製造法において、基質として用いるデスモステロールは、0.1〜500g/L、好ましくは0.2〜200g/Lの濃度になるように反応水性媒体に初発または反応途中に添加する。上記製造法で用いられる水性媒体としては、24位を還元されたステロイド化合物の生成反応を阻害しない限り、いかなる成分、組成の水性媒体であってもよく、例えば、水、りん酸塩、炭酸塩、酢酸塩、ほう酸塩、クエン酸塩、トリスなどの緩衝液などをあげることができる。また、メタノール、エタノールなどのアルコール類、酢酸エチルなどのエステル類、アセトンなどのケトン類、アセトアミドなどのアミド類を含有していてもよい。
【0118】
24位を還元されたステロイド化合物の生成反応は水性媒体中、pH5〜11、好ましくはpH6〜10、20〜50℃、好ましくは25〜45℃の条件で2〜150時間、好ましくは6〜120時間行う。
【0119】
(2)形質転換体または微生物の培養物もしくは培養物の処理物を酵素源として用いる製造法
形質転換体または微生物の培養物もしくは培養物の処理物を酵素源として用いる24位を還元されたステロイド化合物の製造法としては、本発明のタンパク質を生産する能力を有する形質転換体に原料となる24位に2重結合を持つステロイドまたはその誘導体を取り込ませ、形質転換体内で酵素反応を行わせ、形質転換体中に24位を還元されたステロイド化合物を生成、蓄積させ、該形質転換体から24位を還元されたステロイド化合物を抽出する製造法を挙げることができる。原料となる24位に2重結合を持つステロイドまたはその誘導体の形質転換体への取り込みは、培養液中に24位に2重結合を持つステロイドまたはその誘導体を添加することにより、24位に2重結合を持つステロイドまたはその誘導体の存在下で形質転換体を培養することにより行うことができる。さらに、本発明のタンパク質を生産する能力を有する形質転換体の培養物もしくは培養物の処理物を酵素源に用い、該酵素源、およびステロイドを水性媒体中に存在せしめ、該媒体中に24位を還元されたステロイド化合物を生成、蓄積させ、該媒体から24位を還元されたステロイド化合物を採取する製造法をあげることができる。
【0120】
上記製造法に用いられる形質転換体としては、上記4.や5.の方法により製造することができる、本発明のタンパク質を生産する形質転換体をあげることができる。さらに宿主が産生するNADPH以外のNADPHを外部から加えることも可能である。
【0121】
培養物の処理物としては、培養物の濃縮物、培養物の乾燥物、培養物を遠心分離して得られる菌体、該菌体の乾燥物、該菌体の凍結乾燥物、該菌体の界面活性剤処理物、該菌体の超音波処理物、該菌体の機械的摩砕処理物、該菌体の溶媒処理物、該菌体の酵素処理物、該菌体のタンパク質分画物、該菌体の固定化物あるいは該菌体より抽出して得られる酵素標品などをあげることができる。
【0122】
上記製造法において、基質として用いるステロイド類の種類、使用濃度および添加時期、並びに生産される24位を還元されたステロイド化合物は、上記(1)の酵素的製造法のものと同様である。
【0123】
また、微生物の培養物または該培養物の処理物を酵素源とした製造法において用いられる水性媒体としては、上記(1)の酵素的製造法に用いられる水性媒体に加え、酵素源として用いた形質転換体または微生物の培養液も水性媒体として用いることができる。
【0124】
また必要に応じて、水性媒体中に界面活性剤あるいは有機溶媒を添加してもよい。界面活性剤としては、ポリオキシエチレン・オクタデシルアミン(例えばナイミーンS-215、日本油脂社)などの非イオン界面活性剤、セチルトリメチルアンモニウム・ブロマイドやアルキルジメチル・ベンジルアンモニウムクロライド(例えばカチオンF2-40E、日本油脂社)などのカチオン系界面活性剤、ラウロイル・ザルコシネートなどのアニオン系界面活性剤、アルキルジメチルアミン(例えば三級アミンFB、日本油脂社)などの三級アミン類など、ジペプチドの生成を促進するものであればいずれでもよく、1種または数種を混合して使用することもできる。界面活性剤は、界面活性剤は、通常0.1〜50 g/lの濃度で用いられる。有機溶剤としては、キシレン、トルエン、脂肪族アルコール、アセトン、酢酸エチルなどが挙げられ、通常0.1〜50 ml/lの濃度で用いられる。
【0125】
培養物または該培養物の処理物を酵素源として用いる場合、該酵素源の量は、当該酵素源の比活性等により異なるが、例えば、基質として用いるステロイド化合物1mgあたり湿菌体重量として5〜1000mg、好ましくは10〜400mg添加する。
【0126】
24位を還元されたステロイド化合物の生成反応は水性媒体中、pH5〜11、好ましくはpH6〜10、20〜50℃、好ましくは25〜45℃の条件で2〜150時間、好ましくは6〜120時間行う。
【0127】
上記(1)または(2)の製造法において、水性媒体中に生成、蓄積した24位を還元されたステロイド化合物の採取は、活性炭やイオン交換樹脂などを用いる通常の方法あるいは、有機溶媒による抽出、結晶化、薄層クロマトグラフィー、高速液体クロマトグラフィー等により行うことができる。
【0128】
7.24位還元酵素遺伝子の変異、多型または遺伝子発現変異の選抜
本発明による24位を還元されたステロイド化合物の製造方法においては、植物において24位還元酵素遺伝子の突然変異、一塩基多型(SNP)等の多型、または遺伝子発現変異の存在を検出し、そのような変異や多型をもつ該遺伝子のなかから、24位還元酵素活性が高く24位を還元されたステロイド化合物の生産性を高める変異型遺伝子を選抜することができる。そのような変異は、24位還元酵素遺伝子を有する、かつ、24位を還元されたステロイド化合物生産能をもつ植物において、放射線、UV照射、変異原等の化学処理などの人工的突然変異処理、あるいは自然突然変異、によって誘導することができる。
【0129】
上記の変異型24位還元酵素遺伝子の選抜のための方法には、ゲノムDNAやmRNAを変異個体、様々な品種や育成個体の植物から単離し、mRNAの場合には逆転写しcDNAを合成したのち、DNA増幅技術の使用によりゲノムDNAまたはcDNAから24位還元酵素遺伝子を含有する遺伝子断片を増幅する工程と、このDNA中に突然変異の存在を決定する工程が含まれる。DNAやRNAを抽出する方法には市販のキット(例えばDNeasyやRNeasy(キアゲン社)など)が使用できる。cDNAを合成する方法も市販キット(例えばスーパースクリプト ファーストストランド システム(インビトロジェン社)など)を使うことができる。
【0130】
DNA増幅技術の使用により遺伝子断片を増幅する方法としては、いわゆるPCR法やLAMP法などの技術を用いることができる。これらは継続的なポリメラーゼ反応により特異的なDNA配列の増幅(つまり、コピー数を増やすこと)を達成するためにポリメラーゼを使用することを基にした、一群の技術である。この反応は、クローニングの代わりに使用することができるが、必要であるのは、核酸配列に関する情報のみである。DNAの増幅を行うために、増幅しようとするDNAの配列に相補的なプライマーを設計する。次にそのプライマーを自動DNA合成により作製する。DNA増幅方法は、当技術分野で周知であり、本明細書中で与えられる教示及び指示に基づき、当業者であれば容易に行うことができる。いくつかのPCR法(ならびに関連技術)は、例えば、米国特許第4,683,195号、同第4,683,202号、同第4,800,159号、同第4,965,188号、およびInnisら編、PCR Protocols:A guide to Method and Applicationsで述べられている。
【0131】
上記DNA中に突然変異や多型の存在を決定する工程では、塩基配列の決定(アプライドバイオシステムズ社)や、ミスマッチペアの片側を切断する酵素を用いて突然変異体を検出するTILLING法(Till et al., 2003, Genome Res 13:524-530)など変異遺伝子と正常遺伝子の相同性を利用し検出する方法を用いればよい。これらは該技術から得られた配列データを遺伝子部分に関する24位還元酵素をコードする遺伝子の塩基配列、例えば配列番号10〜16に示されるような塩基配列と比較することによって行うことができる。
【0132】
mRNA量の違いを決定する工程では、上記cDNAに対し、配列既知の24位還元酵素をコードする遺伝子の塩基配列に基づいて作製したプライマーを利用してリアルタイムPCR法(ロシュ・ダイアグノスティックス社のライトサイクラーなど)等の定量的PCRを採用すればよい。その後、例えば、ジャガイモ品種「サッシー」から得られたcDNAの量と比較することでmRNA量の違いを決定することができる。
【0133】
特に好ましい実施形態において、上述したような24位還元酵素遺伝子変異の存在の決定方法を、ジャガイモから得られた材料に適用する。
【0134】
上記の突然変異および/または多型を決定する方法により、24位還元酵素をコードする遺伝子の突然変異や多型を塩基レベルで同定することができ、さらに24位還元酵素をコードする遺伝子に突然変異および/または多型を有する植物体を選抜し、変異型24位還元酵素をコードする遺伝子を得ることができる。
【0135】
また、突然変異や多型の決定やmRNA量の違いの決定により、24位還元酵素をコードする遺伝子の発現能または24位還元酵素の活性が変化している植物を選抜することが可能になる。ここで、24位還元酵素をコードする遺伝子の発現能または24位還元酵素の活性の変化とは、人為的突然変異等の突然変異による遺伝子の発現能または24位還元酵素の活性の改変および多型による遺伝子の発現能または24位還元酵素の活性が異なっていることを意味する。
【0136】
ある植物の24位還元酵素活性の突然変異による改変は、その植物の種に含まれる既存品種に対する改変をいい、既存品種には野生型も含まれる。既存の品種は、24位還元酵素をコードする遺伝子が改変された植物を得るためのすべての品種をいい、交配、遺伝子操作等の人為的操作により作出された品種を含む。また、活性の改変において、すべての既存品種に対して、活性が変化している必要はなく、特定の既存品種に対して改変されていれば、「24位還元酵素の活性が改変された植物」に含まれる。「24位還元酵素の活性が改変された植物」は、人為的操作を受けず自然状態で突然変異により活性が改変された植物も含み、上記の選抜方法により、自然状態で活性が変化した植物を選抜することができ、新たな品種として確立することもできる。また、ある既存品種に変異誘発処理を行い、24位還元酵素の活性が改変された植物を作出した場合、比較対象は変異誘発処理を行った既存品種でもよいし、それ以外の他の既存品種でもよい。また、自然状態で選抜された活性が変化した植物又は変異誘発処理により活性が変化した植物を交配することにより、24位還元酵素をコードする遺伝子の変異が固定された新品種として得ることもできる。
【0137】
例えば、植物がジャガイモの場合、既存品種として、「男爵薯」、「メークイン」、「さやか」、「シンシア」等がある。ここで、24位還元酵素をコードする遺伝子の発現能または24位還元酵素の活性が既存品種に対して改変された植物とは、既存品種に対して24位還元酵素をコードする遺伝子の発現能が増強あるいは減少した植物を含み、さらに、24位還元酵素の活性が既存品種に対して上昇あるいは低下した植物を含む。本発明は、このような24位還元酵素をコードする遺伝子の発現能または24位還元酵素の活性が既存品種に対して改変された植物体も包含する。
【0138】
このようにして得られた24位還元酵素をコードする遺伝子に突然変異や多型を有する植物は、本発明の方法によって、24位を還元されたステロイド化合物の製造のために使用することができる。
【実施例】
【0139】
以下、本発明を、実施例を示してより詳しく説明するが、本発明の範囲はこれらに限定されるものではない。
【0140】
実施例1
ジャガイモの萌芽cDNAシークエンスから24位還元酵素遺伝子の抽出
ジャガイモの萌芽は極めて大量のグリコアルカロイドを蓄積することが知られている。グリコアルカロイドの生合成経路は不明な点が多いが、コレステロールを初発物質としていると考えられている。このことからジャガイモの萌芽は植物でコレステロールを合成できる器官であり、コレステロール生合成の遺伝子源として優れていると考えた。
【0141】
収穫後3ヶ月以上貯蔵し休眠の開けたジャガイモを20℃暗所におき、約2週間、萌芽をさせた。この萌芽を液体窒素にて破砕し、RNeasy(キアゲン社)で行い全RNAを抽出した。2μgのRNAからT7プロモーター配列を付加したプライマーを用いて逆転写反応でcDNAを合成した。DNA polymeraseとT7 RNA polymeraseを用いて全体をRNAとして増幅後、再度、逆転写反応でcDNAを合成した。これにGS FLX解析用のアダプターを用いてcDNA2本鎖を合成した。物理的に数百bpに切断後、別のGS FLX解析用のアダプターを連結し1本鎖DNAを回収した。これを鋳型としてGenome Sequencer FLX System(タカラバイオ社委託)にて約15万の発現している遺伝子配列を解析した。その結果、シロイヌナズナのコレステロールの24メチレンを還元する酵素DWF1の遺伝子と相同性の高い遺伝子断片(StDWF1、実施例2で取得した全長配列に対しアミノ酸配列の同一性が564 アミノ酸に渡って78.0% )が54個、それよりも相同性の低い遺伝子断片(StDWF1ホモローグ=StDWF1H、実施例2で取得した全長配列に対しアミノ酸配列の同一性が556 アミノ酸に渡って75.5% )が80個、存在することを見出した。
【0142】
実施例2
StDWF1H遺伝子の同定
上記遺伝子断片を使って、StDWF1Hについては、ナス科ゲノムネットワーク(http://solgenomics.net/index.pl)の配列データSGN-U268686を基に全長配列を取得するためのプライマーU931(caccATGTCGGATGCTAAGGCCC(配列番号7): caccはベクターにクローニングするための付加配列)とU932(TCAATTCGCAGGTTCATCAG)(配列番号8)を合成した。StDWF1についてはDFCI Potato Gene Index (http://compbio.dfci.harvard.edu/tgi/plant.html) の配列データTC214162を基に全長配列を取得するためのプライマーU957(caccATGACAGATGTTCAGGCTCC(配列番号9): caccはベクターにクローニングするための付加配列)とU958(TCAATCTTCAGGCTCATCAACT)(配列番号10)を合成した。実施例1の全RNAから全cDNAの合成をスーパースクリプト ファーストストランド システム(インビトロジェン社)を用いて行った。このcDNAを鋳型に用い、アニール温度55℃でPCR(30サイクル、タカラバイオ社 PrimeSTAR HS DNA Polymeraseを使用)によって遺伝子を増幅した。これをpENTR(商標)/D-TOPOエントリーベクター(インビトロジェン社)へクローニングした。得られた8個の独立クローンについてポリヌクレオチド配列を決定した。これにより得られた配列は、配列番号2と配列番号6であり、それから推定されるポリペプチド配列は配列番号1と配列番号5である。これらベクターをpTOPO-PSStDWF1HとpTOPO-PSStDWF1とした。図1はStDWF1HとStDWF1の相同性解析を解析した結果のアラインメントを示す図であり、アミノ酸配列の同一性が562アミノ酸に渡って80.0%と極めて高かった。24位還元酵素として既知であるヒトの遺伝子(DHCR24, Am J Hum Genet. 2001, 69, 685-94)との相同性は、StDWF1Hはアミノ酸配列の同一性が533アミノ酸に渡って40.3%、StDWF1は533アミノ酸に渡って39.0%であった。なおDHCR24とDWF1は522アミノ酸に渡って39.8%であった。いずれも40%程度を示す弱い相同性であり、この時点の遺伝子相同性の結果だけで機能を推定することは極めて困難であった。
【0143】
さらに、それぞれに相当するトマトの全長遺伝子配列をナス科ゲノムネットワーク(http://solgenomics.net/index.pl)の配列データからSGN-U578468であることを突き止め取得した。配列番号4であり、それから推定されるポリペプチド配列は配列番号3である。
【0144】
これら配列と、特表2007-535960号公報「コレステロール産生酵母株およびその利用」でヒト由来24位還元酵素遺伝子と相同的であると記載されているワタ(Genbank accession AAM47602.1)、イネ(同AAP53615)、エンドウLKB遺伝子(同AAK15493)さらにシロイヌナズナの24メチレンを還元する酵素(DWF1)のアミノ酸配列を比較しCLUSTALX 2.0(Larkinら(2007) Clustal W and Clustal X version 2.0. Bioinformatics, 23:2947-2948)のブーストラップ法を用いて分子系統樹を作成した(図2)。その結果、StDWF1HとSlDWF1Hは、コレステロールの24メチレンを還元する酵素(DWF1)とは、異なるクラスターに分類され、かつヒトの24位還元酵素遺伝子DHCR24とも進化の面からも類縁関係は遠いことがわかった。
【0145】
実施例3
ジャガイモStDWF5遺伝子の単離と酵母発現ベクターpTochigi101-StDWF5の構築
ナス科ゲノムネットワーク(http://solgenomics.net/index.pl)の配列データからシロイヌナズナDWF5遺伝子(ChoeらPlant J. (2000)21:431-43)と相同性の高い配列SGN-U269317を見出した。SGN-U269317配列の予想されるポリペプチド(479アミノ酸)のN末端とC末端に相当する箇所のプライマーU929(caccAATGGCGGAGTCTCAGTTG(配列番号11): caccはベクターにクローニングするための付加配列)とU930(CTAATAAATTCCTGGTATGACCC)(配列番号12)を合成した。実施例2のcDNAを鋳型として、アニール温度55℃でPCR(30サイクル、タカラバイオ社 PrimeSTAR HS DNA Polymeraseを使用)によって遺伝子を増幅した。これをpENTR(商標)/D-TOPOエントリーベクター(インビトロジェン社)へクローニングした。得られた8個の独立クローンについてポリヌクレオチド配列を決定しpTOPO-PSStDWF5を得た。pTochigi101(pYO323(ナショナルバイオリソースプロジェクトNBRP_ID = BYP563):HIS3の酵母選択マーカーを持った多コピー型プラスミド)のマルチクローニング部位にXhoI部位とSacI部位の間にTDH3プロモーター、NotI部位とAscI部位を含む合成リンカー、PGK1ターミネーターを含む、図3)のNotI部位とAscI部位の間を、pTOPO-PSStDWF5の制限酵素NotIとAscIで切り出されるStDWF5遺伝子と入れ替えることで、pTochigi101-StDWF5を作成した。このベクターを導入した酵母は恒常的にStDWF5遺伝子産物を発現し、ステロイドの7位の還元酵素反応をすることが期待できる。
【0146】
実施例4
ジャガイモStDWF1H遺伝子の酵母発現ベクターの構築
実施例2で作製した、pTOPO-PSStDWF1Hプラスミド(エントリークローン)とpYES-DEST52(インビトロジェン社)とを混合し、Gateway LR Clonase II Enzyme Mix(インビトロジェン社)を用いて塩基配列特異的な組み換え反応(attL x attR反応)によって、StDWF1H遺伝子の発現ベクターpYES-DEST52-StDWF1Hを得た。
【0147】
実施例5
erg6酵母(pTochigi101-StDWF5、pYES-DEST52-StDWF1H)における生成物の確認
実験室酵母erg6はdelta(24)-sterol C-methyltransferaseを欠損しているため7−デヒドロデスモステロールが蓄積していることが知られている。(Gaberら (1989) Mol Cell Biol 9(8):3447-56)。そこでYeast Knock Out Strain Collection(オープンバイオシステムズ社)のBY4742 erg6にpTochigi101-StDWF5とpYES-DEST52-StDWF1Hを電気穿孔法(「酵母分子遺伝学実験法」大嶋泰治編著、学会出版センター発行, 1996)を用いて形質転換を行い、独立のクローンを2つ(T33、T34)得た。
【0148】
BY4742 erg6の2コロニー(N1とN2)と、T33酵母、T34酵母を5mlのSC(0.67% Yeast Nitrogen base without amino acids、2% グルコース、100 mg/lロイシン、40 mg/lリジン、20 mg/lヒスチジン、20 mg/lウラシル)-Ura/His培地30℃、135rpm、1日間培養した。培養した酵母を3000g、10分間遠心することにより集菌し、ガラクトース(20mg/ml)を添加した10mlのSC-Ura/His-グルコース培地に懸濁し、30℃、135rpm、2日間培養した。培養酵母を3000g、10分間遠心することにより集菌し5mlの酢酸エチルを加え、20分間超音波処理した後、3分間遠心し酢酸エチル抽出物を回収した。この操作を3回繰り返した。酢酸エチル抽出物は溶媒を乾燥除去した後、N−メチル−N−トリメチルシリルトリフルオロアセトアミドを加え、80℃で30分間加熱し、トリメチルシリルエーテル体に誘導体化しGC-MS分析の試料とした。変換物の同定は標品をGCの保持時間ならびにMSスペクトルを比較することで決定した。
【0149】
BY4742 erg6の酵母(N1とN2)は主に7−デヒドロデスモステロールを蓄積していた。一方、pTochigi101-StDWF5及びpYES-DEST52-StDWF1Hの2つのベクターを保持する酵母T33とT34の抽出物(図4、T33、T34と示したGCチャート)からは、新たなピークが2つ検出され、標品との移動度、マススペクトルによって、デスモステロール(点線矢印)とコレステロールのピーク(図4の実線矢印)であることが同定された。これらはこのことからStDWF1Hは24位還元酵素をコードする遺伝子であることが明らかになった。また、定量を行ったところ、10 mlの培養物あたり7μgのコレステロールを生産していることがあきらかになった。しかし、本数値については、酵母での発現形態や培養条件を特に最適化しておらず、あくまで参考の数値として捉えるべきものである。
【0150】
実施例6
ジャガイモStDWF1遺伝子、ヒトDHCR24遺伝子、エンドウLKBの酵母発現ベクターの構築
ジャガイモStDWF1遺伝子が24位還元酵素活性を持っているかどうかを確認するため、酵母発現ベクターを構築した。実施例2で作製した、pTOPO-PSStDWF1プラスミド(エントリークローン)とpYES-DEST52とを混合し、Gateway LR Clonase II Enzyme Mix(インビトロジェン社)を用いて塩基配列特異的な組み換え反応(attL x attR反応)によって、StDWF1遺伝子の発現ベクターpYES-DEST52-StDWF1を得た。
【0151】
図2で示された系統樹解析ならびに、特表2007-535960号公報「コレステロール産生酵母株およびその利用」で24位還元酵素活性にて示されたワタ、イネ、エンドウの3つの植物遺伝子のうち、最もDHCR24と相同性の高かったエンドウのLKBについて24位還元酵素活性を持っているかどうかを確認するため、遺伝子を取得し酵母発現ベクターを構築した。食料品店で購入した生食用のエンドウの苗である豆苗を液体窒素にて破砕した。RNAの抽出はRNeasyで行い、全cDNAの合成はスーパースクリプト ファーストストランド システムを用いて行った。プライマーとしてはU959(caccATGTCTGATCTTGAGGCTCCG(配列番号13): caccはベクターにクローニングするための付加配列)とU960(TCAGTCTGGTGGTTGATCAACTTC)(配列番号14)を用いた。このcDNAを鋳型に用い、アニール温度55℃でPCR(30サイクル、タカラバイオ社 PrimeSTAR HS DNA Polymeraseを使用)によって遺伝子を増幅した。これをpENTR(商標)/D-TOPOエントリーベクター(インビトロジェン社)へクローニングした。塩基配列解析し既報の配列と同じであることを確認したpTOPO-LKBプラスミド(エントリークローン)を得た。これとpYES-DEST52とを混合し、Gateway LR Clonase II Enzyme Mix(インビトロジェン社)を用いて塩基配列特異的な組み換え反応(attL x attR反応)によって、LKB遺伝子の発現ベクターpYES-DEST52-LKBを得た。
【0152】
StDWF1Hと既報のヒトDHCR24の遺伝子産物を比較するためDHCR24遺伝子を取得し酵母発現ベクターを構築した。スーパースクリプト ファーストストランド システムに添付されていたヒトHela細胞の全RNAを用いて全cDNAの合成をスーパースクリプト ファーストストランド システムを用いて行った。プライマーとしてはU945(caccATGGAGCCCGCCGTGT(配列番号15): caccはベクターにクローニングするための付加配列)とU946(TCAGTGCCTGGCGGCCTTGC)(配列番号16)を用いた。このcDNAを鋳型に用い、アニール温度55℃でPCR(30サイクル、タカラバイオ社 PrimeSTAR HS DNA Polymeraseを使用)によって遺伝子を増幅した。これをpENTR(商標)/D-TOPOエントリーベクター(インビトロジェン社)へクローニングした。塩基配列解析し既報の配列と同じであることを確認したpTOPO-PSDHCR24プラスミド(エントリークローン)を得た。これとpYES-DEST52とを混合し、Gateway LR Clonase II Enzyme Mix(インビトロジェン社)を用いて塩基配列特異的な組み換え反応(attL x attR反応)によって、DHCR24遺伝子の発現ベクターpYES-DEST52-DHCR24を得た。
【0153】
BY4742 erg6にpTochigi101-StDWF5のみを導入したもの、pTochigi101-StDWF5とpYES-DEST52-StDWF1、pTochigi101-StDWF5とpYES-DEST52-LKB、 pTochigi101-StDWF5とpYES-DEST52-DHCR24を、電気穿孔法を用いて形質転換を行い独立のクローンを各2つずつ(T31、T32)(T35、T36)(T37、T38)(T39、T40)得た(図5)。
【0154】
実施例7
erg6酵母でStDWF5とDWF1相同遺伝子を発現した酵母における生成物の確認
T31、T32、T33、T34、T35、T36、T37、T38、T39、T40各酵母を5mlのSC-His培地、もしくはSC-Ura/His培地30℃、135rpm、1日間培養した。培養した酵母を3000g、10分間遠心することにより集菌し、ガラクトース(20mg/ml)を添加した10mlのSC-Ura/His-グルコース培地に懸濁し、30℃、135rpm、2日間培養した。培養酵母を3000g、10分間遠心することにより集菌し5mlの酢酸エチルを加え、20分間超音波処理した後、3分間遠心し酢酸エチル抽出物を回収した。この操作を3回繰り返した。酢酸エチル抽出物は溶媒を乾燥除去した後、N−メチル−N−トリメチルシリルトリフルオロアセトアミドを加え、80℃で30分間加熱し、トリメチルシリルエーテル体に誘導体化しGC-MS分析の試料とした。変換物の同定は標品をGCの保持時間ならびにMSスペクトルを比較することで決定した。
【0155】
pTochigi101-StDWF5のみを保持する酵母T31とT32は、デスモステロールを生産していた(図6)。pTochigi101-StDWF5及びpYES-DEST52-StDWF1Hの2つのベクターを保持する酵母T33とT34の抽出物は図3と同様の結果を示した。pTochigi101-StDWF5及びpYES-DEST52-StDWF1の2つのベクターを保持する酵母T35とT36の抽出物にはデスモステロールは存在するが、コレステロールは確認できなかった。pTochigi101-StDWF5及びpYES-DEST52-LKBの2つのベクターを保持する酵母T37とT38の抽出物にはデスモステロールは存在するが、コレステロールは確認できなかった。このことはStDWF1とLKB遺伝子産物は24位還元酵素活性を持たないことが明らかになった。pTochigi101-StDWF5及びpYES-DEST52-DHCR24の2つのベクターを保持する酵母T39とT40の抽出物にはコレステロールが確認できたが、その生成量はpTochigi101-StDWF5及びpYES-DEST52-StDWF1Hの2つのベクターを保持する酵母T33とT34よりも少なく、StDWF1H遺伝子導入の効果は、既報のDHCR24より優れていることが明らかとなった。
【0156】
比較例1
erg4 erg5酵母でStDWF5とDWF1相同遺伝子を発現した酵母における生成物の確認
実験室酵母erg4は24メチレンの還元酵素を欠損しているため24メチレンコレステロールが蓄積していることが知られている。(Zweytickら (2000) FEBS Lett 470(1):83-7)。ERG5遺伝子は酵母のC-22 ステロイドの不飽和化酵素をコードしていることが知られている(Skaggs ら (1996) Gene 169(1):105-9)。この酵素が存在すると、最終生産物に影響を及ぼす可能性があるが考えられた。そこでYeast Knock Out Strain Collection(オープンバイオシステムズ社)のBY4742 erg4に以下のPhleomycin耐性カセット(を用いた形質転換を行い、BY4742 erg4 erg5二重変異酵母を作成した。
【0157】
これをもとに通常のLi法(「酵母分子遺伝学実験法」大嶋泰治編著、学会出版センター発行, 1996)を用いて形質転換を行い、pTochigi101-StDWF5のみを導入したもの、BY4742 erg4 erg5にpTochigi101-StDWF5とpYES-DEST52-StDWF1H、pTochigi101-StDWF5とpYES-DEST52-StDWF1、pTochigi101-StDWF5とpYES-DEST52-LKB、 pTochigi101-StDWF5とpYES-DEST52-DHCR24を、電気穿孔法を用いて形質転換を行い独立のクローンを各2つずつ(T21、T22)(T23、T24)(T25、T26)(T27、T28)(T29、T30)を得た(図5)。
【0158】
T21、T22、T23、T24、T25、T26、T27、T28、T29、T30各酵母を5mlのSC-His培地、もしくはSC-Ura/His培地30℃、135rpm、1日間培養した。培養した酵母を3000g、10分間遠心することにより集菌し、ガラクトース(20mg/ml)を添加した10mlのSC-Ura/His-グルコース培地に懸濁し、30℃、135rpm、2日間培養した。培養酵母を3000g、10分間遠心することにより集菌し5mlの酢酸エチルを加え、20分間超音波処理した後、3分間遠心し酢酸エチル抽出物を回収した。この操作を3回繰り返した。酢酸エチル抽出物は溶媒を乾燥除去した後、N−メチル−N−トリメチルシリルトリフルオロアセトアミドを加え、80℃で30分間加熱し、トリメチルシリルエーテル体に誘導体化しGC-MS分析の試料とした。変換物の同定は標品をGCの保持時間ならびにMSスペクトルを比較することで決定した。
【0159】
BY4742 erg4 erg5にpTochigi101-StDWF5のみを導入した酵母(T21とT22)は24メチレンコレステロールを蓄積していた(図7)。pTochigi101-StDWF5及びpYES-DEST52-StDWF1Hの2つのベクターを保持する酵母T23とT24の抽出物とpTochigi101-StDWF5とpYES-DEST52-DHCR24の2つのベクターを保持する酵母T29とT30は対照のT21とT22と同じで24メチレンの還元酵素活性を持たないことが明らかになった。pTochigi101-StDWF5及びpYES-DEST52-StDWF1の2つのベクターを保持する酵母T25とT26の抽出物とpTochigi101-StDWF5及びpYES-DEST52-LKBの2つのベクターを保持する酵母T27とT28の抽出物にはカンペステロールを確認することができた。このことはStDWF1とLKB遺伝子産物は24メチレンの還元酵素活性を持ち、本ベクターならびに酵母での遺伝子発現が正常に行われていることが確認できた。
【0160】
実施例8
ジャガイモ組織での遺伝子発現解析
ジャガイモ塊茎を曝光させることで顕著にコレステロールが増加することでグリコアルカロイドが増加することが知られている。品種「サッシー」の塊茎を太陽光で2日間曝光した。曝光する前の塊茎果皮と曝光後の塊茎果皮から、RNAの抽出はRNeasyで行い、全cDNAの合成はスーパースクリプト ファーストストランド システムを用いて行った。StDWF1H遺伝子である配列番号3に基づいてプライマーU931(CACCATGTCGGATGCTAAGGCCC)(配列番号7)とU953(GGATCCATATGCGTTTCTCATTCCAACAACA)(配列番号17)を作製しRT-PCR(条件:95℃ 5分、(95℃ 30秒、55℃ 30秒、72℃ 1分)を25回、72℃ 5分)を行った。図8に示すように曝光する前に対して曝光したあとではStDWF1H遺伝子の発現が高いことがわかった。このようにステロイド含量の高い組織の同定が遺伝子の発現解析で可能であり、ステロイド化合物と組成との相関を解析することで、組織含量の増加のみならず、品種評価、QTLなどの遺伝子解析が可能になり、遺伝子発現マーカーを作製することができることが示された。
【0161】
実施例9
酵母でのビタミンD3の生産
実施例5および実施例7で明らかなようにerg6変異を持つ酵母は7−デヒドロデスモステロールを蓄積している。Yeast Knock Out Strain CollectionのBY4742 erg6にpYES-DEST52-StDWF1Hを、電気穿孔法を用いて形質転換を行い、独立のクローンを2つ(T19、T20)得た。この酵母はガラクトースを含む培地でStDWF1H遺伝子を発現させることができ7−デヒドロコレステロールを蓄積することができた。この酵母に紫外線を当てることによってビタミンD3を製造することが可能となる。酵母は元来、エルゴステロールを蓄積するが、これに紫外線をあてることでビタミンD2ができることが知られている。この際のエルゴステロールを生産している酵母への効果的な紫外線照射方法は特許3009606号公報に記載されており、あらたに作成された7−デヒドロコレステロールを蓄積する酵母への照射方法の改良の手がかりとなりうる。
【0162】
酵母からのビタミンD3の抽出、分析については、コレステロールと同様に以下の通り行うことができる、各酵母を5mlのSC-Ura/His培地30℃、135rpm、1日間培養する。培養した酵母を3000g、10分間遠心することにより集菌し、ガラクトース(20mg/ml)を添加した10mlのSC-Ura/His-グルコース培地に懸濁し、30℃、135rpm、2日間培養する。培養酵母を3000g、10分間遠心することにより集菌し5mlの酢酸エチルを加え、20分間超音波処理した後、3分間遠心し酢酸エチル抽出物を回収する。この操作を3回繰り返す。酢酸エチル抽出物は溶媒を乾燥除去した後、N−メチル−N−トリメチルシリルトリフルオロアセトアミドを加え、80℃で30分間加熱し、トリメチルシリルエーテル体に誘導体化しGC-MS分析の試料とする。標品をGCの保持時間ならびにMSスペクトルを比較することで分析、定量することができる。
【0163】
今回、使用した酵母BY4742はパン酵母由来の実験室酵母である。物質生産として優れているキャンディダ酵母も利用できる。より具体的な形質転換体の取得についてはキャンディダ・ユティリス(特開2010−75171公報)、キャンディダ・ボイディニ(特開2009−268407公報)を参照することによって、実験室酵母と同様にビタミンD3を製造する酵母を作成することが可能である。
【0164】
また、ジャーファーメンターなどを用いた攪拌培養において、7−デヒドロデスモステロールあるいはその前駆体を培地中に添加することにより、これらの物質を酵母細胞に取り込ませることができる。当該酵母細胞において、StDWF1H遺伝子を発現させることにより、7−デヒドロコレステロールを蓄積することができる。7−デヒドロコレステロールを蓄積した酵母細胞に光(好ましくは紫外線)を照射することによって、ビタミンD3を製造することができる。また、添加する物質をデスモステロールあるいはその前駆体とすれば、当該酵母細胞においてコレステロールを製造することができる。
【0165】
実施例10
植物でのコレステロールとビタミンD3の生産
ジャガイモやトマトなど一部の植物ではコレステロールを生産しているが、通常の植物ではコレステロールを生産していない。そこで、コレステロールを生産していないシロイヌナズナや、一部生産しているジャガイモのコレステロール生産量を増強する目的で植物のStDWF1H遺伝子の過剰発現形質転換体を得る実験を実施した。バイナリーベクターpKT11(特開2001-161373号公報)を基本として、カルフラワーモザイクウイルスの35S RNAプロモーター、StDWF1H遺伝子を順方向、35S RNAのターミネーターの順に連結を行い、植物形質転換用ベクターpKT254を作成した(図9)。作製したベクターをエレクトロポレーション法(GelvinとSchilperoor編, Plant Molecular Biology Manual, C2, 1-32 (1994), Kluwer Academic Publishers)により、アグロバクテリウム・ツメファシエンスGV3110株に導入した。
【0166】
シロイヌナズナの形質転換は(島本・岡田監修「モデル植物の実験プロトコール」(1996)秀潤社)に従い実施できる。ジャガイモの形質転換は(門馬(1990)植物組織培養7:57-63)に従い実施できる。以下、ジャガイモの形質転換方法について詳しく記述する。ジャガイモ品種「サッシー」(ジャパンアグリバイオ社)から得られたマイクロチューバーを2〜3mmにスライスし、アグロバクテリウム感染用の材料とする。これを上記のアグロバクテリウムの菌液に浸した後、滅菌済みの濾紙上に置いて過剰のアグロバクテリウムを除く。シャーレ内のMS培地(Zeatin 1ppm, IAA 0.1ppm, アセトシリンゴン100μM、及び寒天0.8%を含む)上に置き、培養は3日間25℃、16時間照明(光量子束密度32μE/m2s)/8時間無照明の条件下で行う。ついで、アセトシリンゴンの代わりにカルベニシリン250ppmを含んだ培地で1週間培養する。その後、さらにカナマイシン50 ppmを含む培地上に移し、2週間ごとに継代する。この間に不定芽が形成し、シュートを生じる。伸張したシュートをカルベニシン250 ppm及びカナマイシン100 ppmを含み、植物生長調節物質を含まないMS培地に置床する。発根したシュートをカナマイシン耐性の生長した植物体の中から外来遺伝子としてカナマイシン耐性遺伝子を含有する個体を得る。カナマイシン耐性遺伝子のプライマーを用いてPCRを行うことで検出し、植物体が形質転換植物体であることを確認することができる。これらの形質転換体ではコレステロールを多く蓄積していることが期待できる。
【0167】
StDWF1H遺伝子の過剰発現形質転換体でも、シロイヌナズナはDWF5遺伝子、ジャガイモは実施例3にあるStDWF5遺伝子が機能しているため、7−デヒドロコレステロールが多く蓄積することは期待できない。しかし、これらの機能が欠損している変異体(dwf5)やRNAi法などによる遺伝子発現抑制形質転換体では7−デヒドロコレステロールが蓄積することが期待できる。これに実施例9で行われる紫外線照射の方法を組み合わせることでビタミンD3を生産できる植物が得られるものと期待できる。
【産業上の利用可能性】
【0168】
本発明の24位還元酵素及びそれをコードする遺伝子を用いる、薬理活性のあるステロイド化合物の生産方法は、天然からの抽出法に比べて大量生産を可能にするため有用である。
【配列表フリーテキスト】
【0169】
配列番号7〜17: プライマー
【特許請求の範囲】
【請求項1】
コレステロールを生産する植物に由来する、ステロイド骨格の24位を還元する活性を有するタンパク質。
【請求項2】
ナス科、ゴマノハグサ科、ヤマノイモ科、またはシソ科に属する植物に由来する、請求項1記載のタンパク質。
【請求項3】
ジャガイモ、トマトに由来する、請求項1記載のタンパク質。
【請求項4】
以下の(a)〜(c)からなる群から選択される請求項1記載のタンパク質。
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質
(b) 配列番号1または配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含むタンパク質
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含むタンパク質
【請求項5】
ステロイド骨格がコレスタン誘導体のステロイド骨格である、請求項1〜4のいずれか1項に記載のタンパク質。
【請求項6】
膜貫通領域を持ちFAD結合型配列を持つ還元酵素に属する、請求項1〜5のいずれか1項に記載のタンパク質。
【請求項7】
以下の(a)〜(g)からなる群から選択されるDNAを含有する組換え体DNA。
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質をコードするDNA
(b) 配列番号1または配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(d) 配列番号2または配列番号4に示す塩基配列を含むDNA
(e) 配列番号2または配列番号4に示す塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(f) 配列番号2または配列番号4に示す塩基配列と90%以上の配列同一性を有する塩基配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(g) 配列番号2または配列番号4に示す塩基配列において縮重配列を含むDNA
【請求項8】
請求項7記載の組換え体DNAを含むベクター。
【請求項9】
請求項7記載の組換え体DNAを含む形質転換体。
【請求項10】
形質転換体が微生物または植物を宿主として得られる形質転換体である、請求項9記載の形質転換体。
【請求項11】
微生物が酵母である、請求項10記載の形質転換体。
【請求項12】
24位が還元されたステロイドまたはその誘導体を生成する能力を有する、請求項9〜11のいずれか1項に記載の形質転換体。
【請求項13】
請求項9〜12のいずれか1項に記載の形質転換体を培地に培養し、培養物中に請求項4記載のタンパク質を生成、蓄積させ、該培養物より該蛋白質を採取する請求項4記載のタンパク質の製造法。
【請求項14】
請求項9〜12のいずれか1項に記載の形質転換体であって、24位に2重結合を持つステロイドまたはその誘導体を細胞内に生成する形質転換体を培養し、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収する、24位が還元されたステロイドまたはその誘導体の製造法。
【請求項15】
請求項9〜12のいずれか1項に記載の形質転換体を24位に2重結合を持つステロイドまたはその誘導体の存在下で培養し、該形質転換体中に24位に2重結合を持つステロイドまたはその誘導体を取り込ませ、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収する、24位が還元されたステロイドまたはその誘導体の製造法。
【請求項16】
請求項9〜12のいずれか1項に記載の形質転換体の培養物または該培養物の処理物を酵素源に用い、該酵素源、および24位に2重結合を持つステロイドまたはその誘導体を水性媒体中に存在せしめ、該媒体中に24位が還元されたステロイドまたはその誘導体を生成、蓄積させ、該媒体から24位が還元されたステロイドまたはその誘導体を採取する、24位が還元されたステロイドまたはその誘導体の製造法。
【請求項17】
前記培養物が、培養物を遠心分離して得られる菌体もしくは培養液上清、菌体および/または培養液上清の濃縮物、乾燥物、凍結乾燥物、界面活性剤処理物、超音波処理物、機械的摩砕処理物、溶媒処理物、酵素処理物、蛋白質分画物、固定化物あるいは菌体および/または培養液上清から抽出して得られる酵素標品であることを特徴とする、請求項14〜16のいずれか1項に記載の製造法。
【請求項18】
24位に2重結合を持つステロイドが、デスモステロール、または7−デヒドロデスモステロールであり、24位が還元されたステロイドが、コレステロール、または7−デヒドロコレステロールである、請求項14〜16のいずれか1項に記載の製造法。
【請求項19】
請求項18記載の製造法において、24位に2重結合を持つステロイドとして7−デヒドロデスモステロールを用い、7−デヒドロコレステロールを製造し、さらに、得られた7−デヒドロコレステロールに光を当てることを含む、ビタミンD3の製造法。
【請求項20】
前記形質転換体に蓄積された7−デヒドロコレステロールに対して光を当てるものである、請求項19記載の製造法。
【請求項1】
コレステロールを生産する植物に由来する、ステロイド骨格の24位を還元する活性を有するタンパク質。
【請求項2】
ナス科、ゴマノハグサ科、ヤマノイモ科、またはシソ科に属する植物に由来する、請求項1記載のタンパク質。
【請求項3】
ジャガイモ、トマトに由来する、請求項1記載のタンパク質。
【請求項4】
以下の(a)〜(c)からなる群から選択される請求項1記載のタンパク質。
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質
(b) 配列番号1または配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含むタンパク質
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含むタンパク質
【請求項5】
ステロイド骨格がコレスタン誘導体のステロイド骨格である、請求項1〜4のいずれか1項に記載のタンパク質。
【請求項6】
膜貫通領域を持ちFAD結合型配列を持つ還元酵素に属する、請求項1〜5のいずれか1項に記載のタンパク質。
【請求項7】
以下の(a)〜(g)からなる群から選択されるDNAを含有する組換え体DNA。
(a) 配列番号1または配列番号3に示すアミノ酸配列を含むタンパク質をコードするDNA
(b) 配列番号1または配列番号3に示すアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換、挿入又は付加されたアミノ酸配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(c) 配列番号1または配列番号3に示すアミノ酸配列と90%以上の配列同一性を有するアミノ酸配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(d) 配列番号2または配列番号4に示す塩基配列を含むDNA
(e) 配列番号2または配列番号4に示す塩基配列と相補的な塩基配列を有するDNAとストリンジェントな条件下でハイブリダイズし、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(f) 配列番号2または配列番号4に示す塩基配列と90%以上の配列同一性を有する塩基配列を含み、かつ、ステロイド骨格の24位を還元する活性を有するタンパク質をコードするDNA
(g) 配列番号2または配列番号4に示す塩基配列において縮重配列を含むDNA
【請求項8】
請求項7記載の組換え体DNAを含むベクター。
【請求項9】
請求項7記載の組換え体DNAを含む形質転換体。
【請求項10】
形質転換体が微生物または植物を宿主として得られる形質転換体である、請求項9記載の形質転換体。
【請求項11】
微生物が酵母である、請求項10記載の形質転換体。
【請求項12】
24位が還元されたステロイドまたはその誘導体を生成する能力を有する、請求項9〜11のいずれか1項に記載の形質転換体。
【請求項13】
請求項9〜12のいずれか1項に記載の形質転換体を培地に培養し、培養物中に請求項4記載のタンパク質を生成、蓄積させ、該培養物より該蛋白質を採取する請求項4記載のタンパク質の製造法。
【請求項14】
請求項9〜12のいずれか1項に記載の形質転換体であって、24位に2重結合を持つステロイドまたはその誘導体を細胞内に生成する形質転換体を培養し、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収する、24位が還元されたステロイドまたはその誘導体の製造法。
【請求項15】
請求項9〜12のいずれか1項に記載の形質転換体を24位に2重結合を持つステロイドまたはその誘導体の存在下で培養し、該形質転換体中に24位に2重結合を持つステロイドまたはその誘導体を取り込ませ、前記形質転換体中において、24位に2重結合を持つステロイドまたはその誘導体の24位の2重結合を還元させ、形質転換体中に24位が還元されたステロイドまたはその誘導体を生成させ、該培養物から24位が還元されたステロイドまたはその誘導体を回収する、24位が還元されたステロイドまたはその誘導体の製造法。
【請求項16】
請求項9〜12のいずれか1項に記載の形質転換体の培養物または該培養物の処理物を酵素源に用い、該酵素源、および24位に2重結合を持つステロイドまたはその誘導体を水性媒体中に存在せしめ、該媒体中に24位が還元されたステロイドまたはその誘導体を生成、蓄積させ、該媒体から24位が還元されたステロイドまたはその誘導体を採取する、24位が還元されたステロイドまたはその誘導体の製造法。
【請求項17】
前記培養物が、培養物を遠心分離して得られる菌体もしくは培養液上清、菌体および/または培養液上清の濃縮物、乾燥物、凍結乾燥物、界面活性剤処理物、超音波処理物、機械的摩砕処理物、溶媒処理物、酵素処理物、蛋白質分画物、固定化物あるいは菌体および/または培養液上清から抽出して得られる酵素標品であることを特徴とする、請求項14〜16のいずれか1項に記載の製造法。
【請求項18】
24位に2重結合を持つステロイドが、デスモステロール、または7−デヒドロデスモステロールであり、24位が還元されたステロイドが、コレステロール、または7−デヒドロコレステロールである、請求項14〜16のいずれか1項に記載の製造法。
【請求項19】
請求項18記載の製造法において、24位に2重結合を持つステロイドとして7−デヒドロデスモステロールを用い、7−デヒドロコレステロールを製造し、さらに、得られた7−デヒドロコレステロールに光を当てることを含む、ビタミンD3の製造法。
【請求項20】
前記形質転換体に蓄積された7−デヒドロコレステロールに対して光を当てるものである、請求項19記載の製造法。
【図1】



【図2】
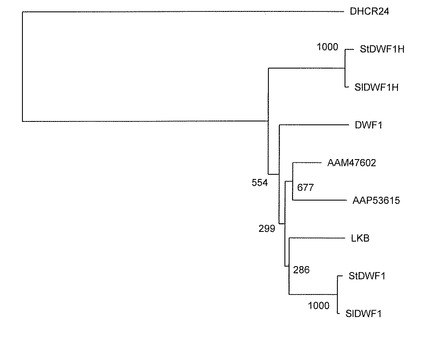
【図3】
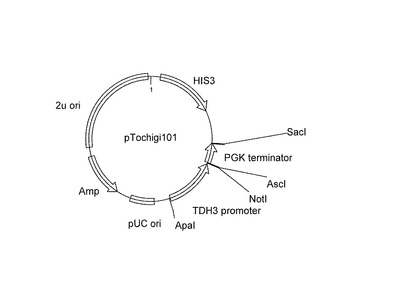
【図4】
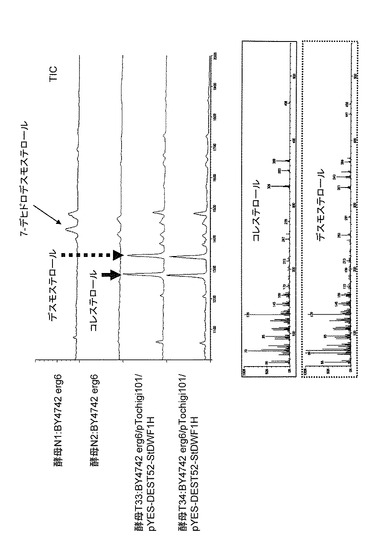
【図5】
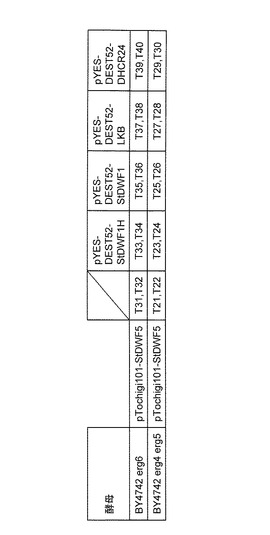
【図6】
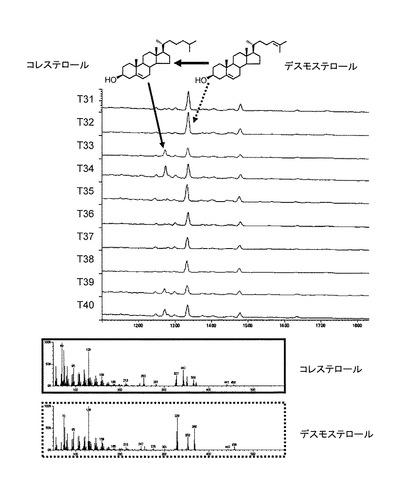
【図7】
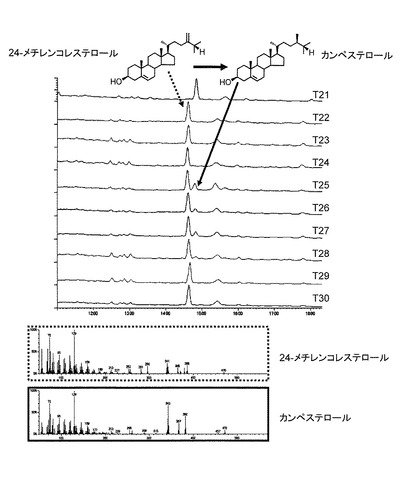
【図8】
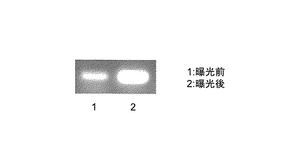
【図9】
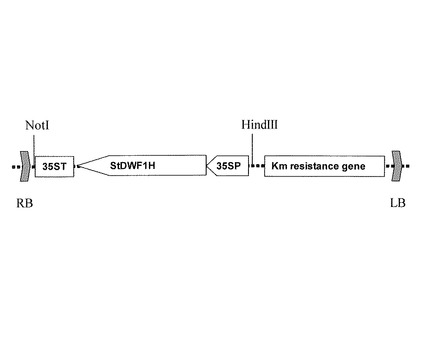
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【公開番号】特開2012−239412(P2012−239412A)
【公開日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願番号】特願2011−111888(P2011−111888)
【出願日】平成23年5月18日(2011.5.18)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度採択、独立行政法人農業・食品産業技術総合研究機構 生物系特定産業技術研究支援センター、「イノベーション創出基礎的研究推進事業」委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
【公開日】平成24年12月10日(2012.12.10)
【国際特許分類】
【出願日】平成23年5月18日(2011.5.18)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成22年度採択、独立行政法人農業・食品産業技術総合研究機構 生物系特定産業技術研究支援センター、「イノベーション創出基礎的研究推進事業」委託事業、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
[ Back to top ]