
ステロール側鎖の20位と22位の結合を切断する活性を有する酵素
【課題】産業上有用なステロイドホルモン生合成の第一段階を触媒する重要な酵素であるP450scc酵素について高活性なP450scc酵素を取得すること。
【解決手段】以下の理化学的性質を有するステロール側鎖切断酵素。
(1)作用:本明細書で定義する式(I)で表されるステロールに作用し、ステロール側鎖部分の20位と22位の炭素−炭素結合を切断する活性により該結合を切断して本明細書で定義する式(II)で表される化合物を生成する。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が10%以上であり、かつコレステロールからプレグネノロンへの変換率が10%以上である。
【解決手段】以下の理化学的性質を有するステロール側鎖切断酵素。
(1)作用:本明細書で定義する式(I)で表されるステロールに作用し、ステロール側鎖部分の20位と22位の炭素−炭素結合を切断する活性により該結合を切断して本明細書で定義する式(II)で表される化合物を生成する。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が10%以上であり、かつコレステロールからプレグネノロンへの変換率が10%以上である。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ステロール側鎖の20位と22位の結合を切断して医薬又は医薬中間体として産業上有用な化合物であるプレグネノロン、プロゲステロン又は7−デヒドロプレグネノロンに変換する活性を有する酵素、該酵素をコードするDNA、該DNAをベクターに組み込んで得られる形質転換体、並びに該形質転換体を用いるプレグネノロン、プロゲステロン、7−デヒドロプレグネノロンの製造方法、該酵素に対する抗体などに関する。
【背景技術】
【0002】
プレグネノロン、プロゲステロン、7−デヒドロプレグネノロンは、医薬および医薬中間体として産業上有用な化合物である。プレグネノロンの既存の製造法としては、コレステロール、ジオスゲニンやスティグマステロールといった天然由来のステロール化合物を原料とし、複数の有機合成反応により合成される(非特許文献1)。しかし、これらの手法は、工程数が多く、目的生産物の収率が低く、工業的生産方法としては問題点がある。一方、生化学的な手段としては、動物由来の酵素side chain cleavage cytochrome P450(P450scc)によるステロール変換方法が想定されるものの、動物由来酵素は酵素活性が低く、また、宿主となる細胞は培養に高価な培地が必要となり増殖速度が遅いなど、その生産方法は確立されていない(非特許文献2)。
さらに、ウシ由来のP450sccをサッカロミセス・セレビシエに形質転換し、電子伝達系を構成するアドレノドキシンおよびアドレノドキシンレダクターゼ存在下で、コレステロールからプレグネノロンに変換する試みも行われているが、生産性は非常に低く工業レベルの生産性は得られていない(特許文献1)。
【0003】
【非特許文献1】J. Org. Chem., 1979, 44, 1583
【非特許文献2】Proc. Natl. Acad. Sci. USA, 1988, 85, 1988
【特許文献1】特許第2963711号公報
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明が解決しようとする課題は、産業上有用なステロイドホルモン生合成の第一段階を触媒する重要な酵素であるP450scc酵素について高活性なP450scc酵素を取得すること、並びに取得したP450scc酵素を用いてステロイドホルモン合成に必要なプレグネノロン、プロゲステロン、及び7−デヒドロプレグネノロンを安価かつ簡便に生産するための方法を提供することである。
【課題を解決するための手段】
【0005】
本発明者らは、上記課題を解決するためにステロール側鎖の20位と22位の結合を切断することによるプレグネノロン等の製造方法について鋭意検討した結果、ステロール化合物側鎖の20位と22位の結合を切断する活性を有する酵素を、初めて微生物からクローニングすることに成功した。本発明者らは、該酵素をコードする遺伝子の形質転換体を作製し、組換え菌を用いて該遺伝子を発現させた該酵素形質転換体細胞、該形質転換体処理物および/または培養液を、原料となるステロール化合物に作用させることにより、高濃度で目的物であるプレグネノロン、プロゲステロン、7−デヒドロプレグネノロンを得ることができることを見出した。本発明はこれらの知見に基づいて完成したものである。
【0006】
即ち、本発明によれば、以下の発明が提供される。
1) 以下の理化学的性質を有するステロール側鎖切断酵素。
(1)作用:下記式(I)で表されるステロールに作用し、ステロール側鎖部分の20位と22位の炭素−炭素結合を切断する活性により該結合を切断して下記式(II)で表される化合物を生成する。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が10%以上であり、かつコレステロールからプレグネノロンへの変換率が10%以上である。
【化1】

[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【0007】
2) 以下の理化学的性質を有する、1)に記載のステロール側鎖切断酵素。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が50%以上であり、かつコレステロールからプレグネノロンへの変換率が50%以上である。
3) さらに以下の理化学的性質を有する、1)又は2)に記載の酵素。
(3)至適pH:7.5〜8.0
(4)作用至適温度:15〜20℃
(5)熱安定性:20℃で140時間保存後に30%以上の酵素活性が保持される。
(6)分子量:アミノ酸配列からの推定分子量が53〜54KDaであり、SDS電気泳動による測定で50〜56KDa
【0008】
4) さらに以下の理化学的性質を有する、1)から3)の何れかに記載の酵素。
(7)反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上である。
(8)基質濃度を10μMから100μMに上昇させた場合の活性の低下率が35%以下である。
【0009】
5) 微生物に由来する酵素である、1)から4)の何れかに記載の酵素。
【0010】
6) 以下の(a)、(b)または(c)のいずれかのタンパク質;
(a)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b)配列番号2に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質、
(c)配列番号2に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質。
【0011】
7) ステロール側鎖の20位と22位の結合を切断する活性について、基質濃度を10μMから100μMに上昇させた場合の活性の低下率が30%以下である、6)に記載のタンパク質。
【0012】
8) 6)又は7)に記載のタンパク質をコードするDNA。
9) 以下の(a)、(b)または(c)のいずれかのDNA;
(a)配列番号1に記載の塩基配列を有するDNA、
(b)配列番号1に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA、
(c)配列番号1に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA。
【0013】
10) 8)または9)に記載のDNAを有する組み換えベクター。
11) 8)または9)に記載のDNAあるいは10)に記載の組換えベクターを宿主細胞に導入した形質転換体。
12) 宿主細胞が大腸菌、放線菌またはシュードモナス属微生物である11)に記載の形質転換体。
【0014】
13) 6)または7)に記載のタンパク質、あるいは11)または12)に記載の形質転換体を含む、ステロール側鎖の20位と22位の結合を切断する反応を触媒するための酵素組成物。
14) 11)または12)に記載の形質転換体により産生される、6)または7)に記載のタンパク質。
【0015】
15) 1)から5)の何れかに記載の酵素あるいは6)、7)または14)に記載のタンパク質に対する抗体又はその部分フラグメント。
16) モノクローナル抗体である、15)に記載の抗体又はその部分フラグメント。
17) 1)から5)の何れかに記載の酵素あるいは6)、7)または14)に記載のタンパク質が有するステロール側鎖の20位と22位の結合を切断する活性を中和する作用を有する、15)または16)に記載の抗体又はその部分フラグメント。
【0016】
18) 11)又は12)に記載の形質転換体を培養することを特徴とする、6)又は7)に記載のタンパク質の製造方法。
【0017】
19) 式(I)で表されるステロールに、下記(i)から(vi):
(i) 1)から5)の何れかに記載の酵素、
(ii) 該酵素を産生する微生物またはその処理物、
(iii) 6)、7)または14)に記載のタンパク質、
(iv) 11)または12)に記載の形質転換体またはその処理物、
(v) 以下の(a)、(b)または(c)のいずれかのタンパク質、又は
(vi) 以下の(a)、(b)または(c)のいずれかのタンパク質をコードするDNA、以下の(d)、(e)または(f)のいずれかのDNA、もしくは上記DNAを有する組み換えベクターを宿主細胞に導入した形質転換体またはその処理物;
の何れかを反応させることを特徴とする、式(II)で表される化合物の製造方法。
(a)配列番号26に記載のアミノ酸配列からなるタンパク質、
(b)配列番号26に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質、
(c)配列番号26に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質、
(d)配列番号4に記載の塩基配列を有するDNA、
(e)配列番号4に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質をコードするDNA、
(f)配列番号4に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質をコードするDNA。
【化2】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、
母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【0018】
20) 式(I)で表されるステロールが、コレステロール、4-コレステン-3-オン、7-デヒドロコレステロール、エルゴステロール、β-シトステロール、スティグマステロール、カンペステロール、デスモステロール、 (20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、又は(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンである、19)に記載の方法。
21) 式(II)で表される化合物が、プレグネノロン、プロゲステロン又は7−デヒドロプレグネノロンである、19)又は20)に記載の方法。
【発明の効果】
【0019】
本発明により、ステロール化合物側鎖の20位と22位の結合を切断する活性を有する新規酵素が提供される。本発明の酵素を用いることによって、医薬および医薬中間体として産業上有用な化合物であるプレグネノロン、プロゲステロン、及び7−デヒドロプレグネノロンを効率的に製造することが可能になった。一般に動物由来のP450scc酵素は、微生物を宿主として発現させると不溶性タンパク質となり活性体として本タンパク質を得ることは困難であるが、本発明により初めて提供される微生物由来のP450scc酵素は、宿主として微生物を用いて発現させた場合に可溶性タンパク質となり容易に活性体として本タンパク質を得られることから、本発明の酵素の産業上の有用性は高い。
【発明を実施するための最良の形態】
【0020】
以下、本発明の実施の形態について詳細に説明する。
(1)本発明の酵素
本発明の酵素は、以下の理化学的性質を有するステロール側鎖切断酵素である。
(1)作用:下記式(I)で表されるステロールに作用し、ステロール側鎖部分の20位と22位の炭素−炭素結合を切断する活性により該結合を切断して下記式(II)で表される化合物を生成する。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは70%以上)であり、かつコレステロールからプレグネノロンへの変換率が10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは65%以上)である。
【化3】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【0021】
式(I)は、好ましくは、コレステロール、4-コレステン-3-オン、7-デヒドロコレステロール、エルゴステロール、β-シトステロール、スティグマステロール、カンペステロール、デスモステロール、 (20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンであり、より好ましくは、コレステロール、4-コレステン-3-オン、7-デヒドロコレステロールである。
【0022】
【化4】
【0023】
式(II)は、好ましくは、プレグネノロン、プロゲステロン、7-デヒドロプレグネノロンである。
【化5】
【0024】
上記の通り、本発明の酵素について、4-コレステン-3-オンからプロゲステロンへの変換反応率は10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは70%以上)であり、かつコレステロールからプレグネノロンへの変換率は10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは65%以上)である。他の基質の変換率は特に限定されるものではないが、好ましい変換率を以下に示す。
【0025】
7-デヒドロコレステロールから7−デヒドロプレグネノロンへの変換率は、10%以上(好ましくは20%以上、より好ましくは25%以上)である。
エルゴステロールから7−デヒドロプレグネノロンへの変換率は、1%以上(好ましくは2%以上)である。
β-シトステロールからプレグネノロンへの変換率は、10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは40%以上)である。
スティグマステロールからプレグネノロンへの変換率は、1%以上(好ましくは2%以上、より好ましくは5%以上)である。
カンペステロールからプレグネノロンへの変換率は、10%以上(好ましくは20%以上、より好ましくは25%以上)である。
デスモステロールからプレグネノロンへの変換率は10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは65%以上)である。
(20S)-20-ヒドロキシコレスト-4-エン-3-オンからプロゲステロンへの変換率は10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは40%以上)である。
(22R)-22-ヒドロキシコレスト-4-エン-3-オンからプロゲステロンへの変換率は10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは40%以上)である。
(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オンからプロゲステロンへの変換率は10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは35%以上)である。
(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンからプロゲステロンへの変換率は10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは40%以上)である。
ラノステロール(Lano)の変換率は0%である。
【0026】
本発明の酵素は、好ましくは、さらに以下の理化学的性質を有していてもよい。
(3)至適pH:7.5〜8.0
(4)作用至適温度:15〜20℃
(5)熱安定性:20℃で140時間保存後に30%以上の酵素活性が保持される。
(6)分子量:アミノ酸配列からの推定分子量が53〜54KDaであり、SDS電気泳動による測定で50〜56KDa
【0027】
本発明の酵素は、好ましくは、さらに以下の理化学的性質を有していてもよい。
(7)反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上、好ましくは45mmol/分/mol以上である。
(8)基質濃度を10μMから100μMに上昇させた場合の活性の低下率が35%以下である。
【0028】
VmaxおよびKmは、基質濃度を0.1〜500μMで4-コレステン-3-オンの変換を行い基質濃度と酵素活性の関係を調べることによって求めることができる。また、基質濃度を10μMから100μMに上昇させた場合の活性の低下率も、基質濃度を0.1〜500μMで4-コレステン-3-オンの変換を行い基質濃度と酵素活性の関係を調べることによって求めることができる。具体的には、反応液は1 mlで220 pmolのCYP204A1もしくは250 pmolのCYPSS、96μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、2 mM NADHと2 mM NADPHとなるように調製し、Tris-HClでpH 7.5にする。基質である4-コレステン-3-オンは終濃度が0.1, 0,5, 1, 2, 5, 10, 20, 50, 100, 500μMとなるようにDMSOに溶かしたものを20μl加える。反応はNADHとNADPHの添加により開始し、200 rpm、15℃で60分間行うことができる。
【0029】
本発明の酵素の由来は、特に限定されるものではないが、好ましくは、微生物に由来する酵素であり、例えば、Sphingomonas subterranea (例えば、Sphingomonas subterranea NBRC16086など)、又はNovosphingobium aromaticivorans (例えば、Novosphingobium aromaticivorans ATCC 700278など)などから取得することができる。
【0030】
(2)本発明の酵素の取得方法
本発明の酵素の、例えば、上記したSphingomonas subterranea又はNovosphingobium aromaticivoransなどの微生物から、通常の酵素の抽出、精製方法によって得ることができる。抽出方法として具体的には、例えばハサミ等による細片化、ホモジナイズ、音波処理、浸透ショック法、凍結融解法等の細胞破砕による抽出、界面活性剤抽出等や、これらの組み合わせ等の処理操作が挙げられる。また、精製方法として具体的には、例えば硫酸アンモニウム(硫安)や硫酸ナトリウム等による塩析、遠心分離、透析、限外濾過法、吸着クロマトグラフィー、イオン交換クロマトグラフィー、疎水性クロマトグラフィー、逆相クロマトグラフィー、ゲル濾過法、ゲル浸透クロマトグラフィー、アフィニティークロマトグラフィー、電気泳動法、ザイモグラフィー等や、これらの組合わせ等の処理操作が挙げられる。
【0031】
また、本発明の酵素の遺伝子を公知の方法に従ってクローニングし、適当な宿主に導入して、発現させることにより本発明の酵素 を得ることができる。例えば、本明細書に記載した本発明の酵素をコードする遺伝子の塩基配列の情報に基づいて本発明の酵素の遺伝子に特異的なプローブ又はプライマーを設計し、当該プローブ又はプライマーを用いてSphingomonas subterranea又はNovosphingobium aromaticivoransなどの微生物のDNAライブラリー(例えば、ゲノムDNAライブラリー、又はcDNAライブラリーなど)から本発明の酵素をコードするDNAを単離又は増幅し、これを遺伝子組換え技術によりベクターに入れ、宿主細胞に導入し、そこで発現させることにより、本発明の酵素を得ることができる。
【0032】
(3)本発明のタンパク質
本発明によれば、以下の(a)、(b)または(c)のいずれかのタンパク質が提供される。
(a)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b)配列番号2に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質、
(c)配列番号2に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質。
【0033】
好ましくは、本発明のタンパク質は、ステロール側鎖の20位と22位の結合を切断する活性について、基質濃度を10μMから100μMに上昇させた場合の活性の低下率が35%以下である。
【0034】
本発明において、「1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列」における「1もしくは数個」とは、例えば、1から20個程度を意味し、好ましくは1から10個程度を意味し、さらに好ましくは1から5個程度を意味する。
【0035】
本発明のタンパク質の取得方法としては、下記(4)に記載の本発明のDNAを適当な方法により転写/翻訳する方法が好ましく用いられる。具体的には、適当な発現用ベクター若しくは適当なベクターに適当なプロモーターとともに挿入した組換えベクターを作製し、この組換えベクターで適当な宿主微生物を形質転換したり、適当な培養細胞に導入することにより発現させ、これを精製することにより取得することができる。
【0036】
(4)本発明のDNA
本発明によれば、上記した本発明のタンパク質をコードするDNAが提供される。
本発明のDNAの具体例としては、以下の(a)、(b)または(c)のいずれかのDNAが挙げられる。
(a)配列番号1に記載の塩基配列を有するDNA、
(b)配列番号1に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA、
(c)配列番号1に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA。
【0037】
本発明の酵素をコードする遺伝子は、Sphingomonas subterranea及びNovosphingobium aromaticivoransからクローニングされたものであるが、本発明によりその塩基配列が決定されたので、この塩基配列に基づいて合成することも可能である。また、この塩基配列に基づいて作製したオリゴヌクレオチドをプローブとするハイブリダイゼーションによって、又は前記配列に基づいて作製したオリゴヌクレオチドをプライマーとするPCRによって、Sphingomonas subterranea及びNovosphingobium aromaticivoransなどの微生物のDNAライブラリー(例えば、ゲノムDNAライブラリー、又はcDNAライブラリーなど)から単離することもできる。また、本発明の酵素の遺伝子は、天然の微生物から分離されたもののみならず、通常用いられるDNA合成装置、例えばアプライド・バイオシステムズ(Applied Biosystems)社製394DNA/RNAシンセサイザーを用いて合成されたものであってもよい。
【0038】
本発明において、「1もしくは数個の塩基が欠失、置換および/または付加された塩基配列」における「1もしくは数個」とは、例えば、1から30個程度を意味し、好ましくは1から20個程度を意味し、さらに好ましくは1から10個程度を意味し、さらに好ましくは1から5個程度を意味する。
【0039】
本発明において、「ストリンジェントな条件下でハイブリダイズすることができる塩基配列」とは、配列番号1に記載の塩基配列あるいはその相補配列を有するDNAとBLAST解析で80%以上、好ましくは90%以上、さらに好ましくは95%以上の相同性を有する塩基配列を含むDNA等が挙げられる。また、ストリンジェントな条件下のハイブリダイゼーションとは、通常のハイブリダイゼーション緩衝液中で、温度が40〜70℃、好ましくは60〜65℃等で反応を行い、塩濃度が15mM〜300mM、好ましくは15mM〜60mM等の洗浄液中で洗浄を行う方法に従って行うことができる。
【0040】
酵素活性を実質的に害さないアミノ酸の欠失、置換および/または付加は、当業者であれば容易に選択することができる。また、酵素活性に実質的に影響を与えないアミノ酸の欠失、置換および/または付加を有する本発明の酵素をコードするDNAは、例えば、自然突然変異株又は変種から取得することができる。
【0041】
配列番号1に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有する遺伝子の作製は、突然変異剤を用いる方法や部位特異的変異法等の通常の変異操作によって得ることができる。これらは、例えばサイトダイレクテドミュータジェネシスキット(宝酒造)や、クイックチェンジサイトダイレクテッドミュータジェネシスキット(ストラタジーン社製)等の市販キットで容易に行うことができる。
【0042】
(5)本発明の組み換えベクター、及び形質転換体
本発明によればさらに、本発明のDNAを有する組み換えベクターが提供される。組み換えベクターを調製する際には、通常、本発明のDNAを宿主微生物に適したプロモーターとともに、このプロモーターの下流に本発明のDNAのコード領域の5'末端側が連結されるようにして、ベクターに挿入する。あるいはプロモーターを含む発現ベクターを用い、これに本発明のDNAを挿入してもよい。
【0043】
発現ベクターとしては、宿主微生物内で複製増殖可能であれば特に制限されるものではないが、プラスミドベクター、シャトルベクターおよびファージベクターが挙げられる。具体的なプラスミドベクターとしては、pBR322、pUC18、pHSG298、pUC118、pSTV28、pTWV228、pHY300PLK(以上のプラスミドベクターは、例えば宝酒造(株)から購入できる)、pKK223−3、pPL−ラムダインデューシブルエクスプレッションベクター(Lambda Inducible Expression Vector)(以上は、例えばファルマシア社から購入できる)、大腸菌−コリネ型細菌のシャトルベクターとしては、例えば、特開平3−210184号公報に記載のプラスミドpCRY30;特開平2−276575号公報に記載のプラスミドpCRY21、pCRY2KE、pCRY2KX、pCRY31、pCRY3KEおよびpCRY3KX;特開平1−191686号公報に記載のプラスミドpCRY2およびpCRY3;特開昭58−67679号公報に記載のpAM330;特開昭58−77895号公報に記載のpHM1519;特開昭58−192900号公報に記載のpAJ655、pAJ611およびpAJ1844;特開昭57−134500号公報に記載のpCG1;特開昭58−35197号公報に記載のpCG2;特開昭57−183799号公報に記載のpCG4およびpCG11等、あるいはこれらの誘導体等を挙げることができる。
また、ファージベクターとしては、(λFixIIベクタ−(Stratagene社から購入できる)等を挙げることができる。
【0044】
本発明の酵素をコードするDNAを発現させるためのプロモーターは、宿主微生物が保有するプロモーターを一般に用いることができるが、それに限られるものではなく、本発明の酵素の遺伝子の転写を開始させるための原核生物由来の塩基配列であればいかなるプロモーターであっても良い。具体的には、ラクトースオペロンのプロモーター、トリプトファンオペロンのプロモーター、λファージ由来のPLプロモーター、トリプトファンラクトース雑種(tac)プロモーター[H. A. Bose et al., Proc. Natl. Acad. Sci. U. S. A., Vol.80, p.21 (1983)]等が挙げられる。これらのプロモーターのうち、発現効率を向上させる目的で、誘導性のあるプロモーターを使用することもできる。例えば、上記ラクトースオペロンのプロモーターの場合には、ラクトースやイソプロピル−β−D−チオガラクトシド(IPTG)を添加することにより遺伝子発現を誘導することができる。
【0045】
本発明によれば、さらに本発明のDNA又は組換えベクターを宿主細胞に導入した形質転換体が提供される。本発明のDNA又は組み換えベクターを導入する宿主としては、特に限定されるものではないが、エシェリヒア(Escherichia)属細菌(例えば、大腸菌)、放線菌(Actinomycetes)属細菌、バチルス(Bacillus)属細菌、セラチア(Serratia)属細菌、シュードモナス(Pseudomonas)属細菌、コリネバクテリウム(Corynebacterium)属細菌、ブレビバクテリウム(Brevibacterium)属細菌、ロドコッカス(Rhodococcus)属細菌、ラクトバチルス(Lactobacillus)属細菌、ストレプトマイセス(Streptomyces)属細菌、サーマス(Thermus)属細菌、ストレプトコッカス(Streptococcus)属細菌等を好適に用いることができる。好ましくは、宿主細胞は大腸菌、放線菌またはシュードモナス属微生物である。
【0046】
具体的には、エシェリヒア・コリ(Escherichia coli)、バチルス・サチリス(Bacillus subtilis)、バチルス・ブレビス(Bacillus brevis)、バチルス・ステアロサーモフィラス(Bacillus stearothermophilus)、セラチア・マーセッセンス(Serratia marcescens)、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・アエルギノサ(Pseudomonas aeruginosa)、コリネバクテリウム・グルタミカム(Corynebacterium glutamicum)、ブレビバクテリウム・フラバム(Brevibacterium flavum)、ブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)、ロドコッカス・エリスロポリス(Rhodococcus erythropolis)、サーマス・サーモフィラス(Thermus thermophilus)、ストレプトコッカス・ラクティス(Streptococcus lactis)、ラクトバチルス・カゼイ(Lactobacillus casei)、ストレプトマイセス・リビダンス(Streptomyces lividans)等を用いることができる。
【0047】
上記宿主微生物への遺伝子の導入法としては、コンピテントセル法[Journal of Molecular Biology, Vol.53, p.159 (1970)]、パルス波通電法[J. Indust.Microbiol., Vol.5, p.159 (1990)]等による形質転換法、ファージを用いた形質導入法[E. Ohtsubo, Genetics, Vol.64, p.189 (1970)]、接合伝達法[J. G. C. Ottow, Ann. Rev.Microbiol., Vol.29, p.80 (1975)]、細胞融合法[M.H. Gabor, J. Bacteriol., Vol.137, p.1346 (1979)]等を用いることができる。これらの方法から、宿主微生物に適した方法を適宜選択すればよい。
【0048】
上記のような発現ベクターを用いた発現方法の他に、プロモーターを連結した本発明の酵素をコードするDNAを、宿主微生物の染色体中に直接導入する相同組換え技術あるいはトランスポゾンや挿入配列等を用いて導入する技術によっても発現させることができる。従って、本発明の形質転換体は、本発明の酵素が発現していればよく、遺伝子の導入の方法は限定されない。
【0049】
(6)本発明の形質転換体を用いた本発明の酵素の製造
本発明によれば、上記(5)のようにして得られる形質転換体を培養し、その培養物から本発明の酵素を採取することができる。
形質転換体の培養は、炭素源、窒素源、無機塩、各種ビタミン等を含む通常の栄養培地で行うことができ、炭素源としては、例えばブドウ糖、ショ糖、果糖、麦芽糖等の糖類、エタノール、メタノール等のアルコール類、クエン酸、リンゴ酸、コハク酸、マレイン酸、フマル酸等の有機酸類、廃糖蜜等が用いられる。窒素源としては、例えばアンモニア、硫酸アンモニウム、塩化アンモニウム、硝酸アンモニウム、尿素等がそれぞれ単独もしくは混合して用いられる。また、無機塩としては、例えばリン酸一水素カリウム、リン酸二水素カリウム、硫酸マグネシウム等が用いられる。この他にペプトン、肉エキス、酵母エキス、コーンステイープリカー、カザミノ酸、ビオチン等の各種ビタミン等の栄養素を培地に添加することができる。
【0050】
培養は、通常、通気撹拌、振とう等の好気条件下で行う。培養温度は、宿主微生物の生育し得る温度であれば特に制限はなく、また、培養途中のpHについても宿主微生物が生育し得るpHであれば特に制限はない。培養中のpH調整は、酸またはアルカリを添加して行うことができる。
【0051】
培養物からの酵素の採取は、酵素の活性を指標として公知の採取方法により行うことができる。酵素は必ずしも均一にまで精製される必要はなく、用途に応じた精製度まで精製すればよい。
【0052】
本発明で用いられる粗精製画分又は精製酵素としては、形質転換体を培養した培養液から分離した菌体はもちろんのこと、培養液、菌体を超音波、圧擦等の手段で破砕して得られる破砕物、該破砕物を水等で抽出して得られる、本発明の酵素 を含有する抽出物、該抽出物に更に硫安塩析、カラムクロマトグラフィー等の処理を行って得られる本発明の酵素の粗酵素標品又は精製した酵素標品を使用してもよい。さらに、上記菌体、破砕物、抽出物、粗精製画分または精製酵素を担体に固定化したものも使用することができる。
【0053】
これら菌体、菌体破砕物、抽出物または精製酵素の固定化は、それ自体既知の通常用いられている方法に従い、アクリルアミドモノマー、アルギン酸、またはカラギーナン等の適当な担体に菌体等を固定化させる方法により行うことができる。例えば、菌体を担体に固定化する場合には、培養物から回収されたまま、あるいは適当な緩衝液、例えば0.02〜0.2M程度のリン酸緩衝液(pH6〜10)等で洗浄された菌体を使用することができる。
【0054】
(7)本発明の抗体
本発明によれば、本発明の酵素又はタンパク質に対する抗体又はその部分フラグメントが提供される。本発明の抗体は、ポリクローナル抗体又はモノクローナル抗体の何れでもよく、好ましくはモノクローナル抗体である。
【0055】
本発明の抗体の調製方法としては、通常用いられる公知の方法を用いることができ、抗原として用いられるポリペプチドについても、公知の方法に従って抗原性が高くエピトープ(抗原決定基)として適した配列を選択して用いることができる。上記の抗原として用いるポリペプチドは、公知の方法に従って合成した合成ペプチドでも、また本発明の酵素又はタンパク質そのものを用いることもできる。抗原となるポリペプチドは、公知の方法に従って適当な溶液等に調製して、哺乳動物、例えばウサギ、マウス、ラット等に免疫を行えばよいが、安定的な免疫を行ったり抗体価を高めるために抗原ペプチドを適当なキャリアタンパク質とのコンジュゲートにして用いたり、アジュバント等を加えて免疫を行うのが好ましい。
【0056】
免疫に際しての抗原の投与経路は特に限定されず、例えば皮下、腹腔内、静脈内、あるいは筋肉内等のいずれの経路を用いてもよい。具体的には、例えばBALB/cマウスに抗原ポリペプチドを数日〜数週間おきに数回接種する方法等が用いられる。また、抗原の摂取量としては、抗原がポリペプチドの場合0.3〜0.5mg/1回程度が好ましいが、ポリペプチドの種類、また免疫する動物種によっては適宜調節される。
【0057】
免疫後、適宜試験的に採血を行って固相酵素免疫検定法(以下、これを「ELISA法」と称することがある)やウエスタンブロッティング等の方法で抗体価の上昇を確認し、十分に抗体価の上昇した動物から採血を行う。これに抗体の調製に用いられる適当な処理を行えばポリクローナル抗体を得ることができる。具体的には、例えば、公知の方法に従い血清から抗体成分を精製した精製抗体を取得する方法等が挙げられる。抗体成分の精製は、遠析、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等の方法を用いることができる。
【0058】
また、該動物の脾臓細胞とミエローマ細胞とを用いて公知の方法に従って融合させたハイブリドーマを用いる(Milstein,et al.,Nature,256, 495(1975))ことによりモノクローナル抗体を作製することもできる。モノクローナル抗体は、例えば以下の方法により取得することができる。
【0059】
まず、上記した抗原の免疫により抗体価の高まった動物から抗体産生細胞を取得する。抗体産生細胞は、形質細胞、及びその前駆細胞であるリンパ球であり、これは個体の何れから取得してもよいが、好ましくは脾臓、リンパ節、末梢血等から取得する。これらの細胞と融合させるミエローマとしては、一般的にはマウスから得られた株化細胞、例えば8−アザグアニン耐性マウス(BALB/c由来等)ミエローマ細胞株であるP3X63-Ag8.653(ATCC:CRL-1580)、P3-NS1/1Ag4.1(理研セルバンク:RCB0095)等が好ましく用いられる。細胞の融合は、抗体産生細胞とミエローマ細胞を適当な割合で混合し、適当な細胞融合培地、例えばRPMI1640やイスコフ改変ダルベッコ培地(IMDM)、あるいはダルベッコ改変イーグル培地(DMEM)等に、50%ポリエチレングリコール(PEG)を溶解したもの等を用いることにより行うことができる。また電気融合法(U. Zimmer- mann. et al., Naturwissenschaften,68, 577(1981))によっても行うことができる。
【0060】
ハイブリドーマは、用いたミエローマ細胞株が8−アザグアニン耐性株であることを利用して適量のヒポキサンチン・アミノプテリン・チミジン(HAT)液を含む正常培地(HAT培地)中で5%CO2、37℃で適当時間培養することにより選択することができる。この選択方法は用いるミエローマ細胞株によって適宜選択して用いることができる。選択されたハイブリドーマが産生する抗体の抗体価を上記した方法により解析し、抗体価の高い抗体を産生するハイブリドーマを限界希釈法等により分離し、分離した融合細胞を適当な培地で培養して得られる培養上清から硫安分画、アフィニティクロマトグラフィー等の適当な方法により精製してモノクローナル抗体を得ることができる。また精製には市販のモノクローナル抗体精製キットを用いることもできる。さらには、免疫した動物と同系統の動物、またはヌードマウス等の腹腔内で上記で得られた抗体産生ハイブリドーマを増殖させることにより、本発明のモノクローナル抗体を大量に含む腹水を得ることもできる。
【0061】
また、一価性抗体等の部分フラグメントは、例えばパパイン等を用いてFab部分とFc部分を切断し、アフィニティカラム等を用いてFab部分を回収することによって作製することができる。
【0062】
かくして得られる本発明の酵素又はタンパク質と特異的に結合する抗体は、本発明の酵素又はタンパク質に特異的に結合することによって、該酵素又はタンパク質が有するステロール側鎖の20位と22位の結合を切断する活性を阻害する中和抗体として用いることもできる。
【0063】
(8)本発明の酵素を用いたプレグネノロン、プロゲステロン、7−デヒドロプレグネノロンの製造方法
本発明によれば、上記した式(I)で表されるステロールに、本発明の酵素、該酵素を産生する微生物またはその処理物、本発明のタンパク質、本発明の形質転換体またはその処理物、あるいは配列番号26に記載のアミノ酸配列からなるタンパク質(CYP204A1)又はその変異体、当該タンパク質をコードするDNAもしくは上記DNAを有する組み換えベクターを宿主細胞に導入した形質転換体またはその処理物を適切な電子伝達系存在下で反応させることによって、式(II)で表される化合物を製造することができる。本願の酵素の電子伝達系とは、還元型ニコチンアミドアデニンジヌクレオチド(NADH)または還元型ニコチンアミドアデニンジヌクレオチドリン酸(NADPH)、及び2種の蛋白質により構成される。すなわち、NAD(P)H−フェレドキシン酸化還元活性を有するフェレドキシンレダクターゼ、および本願酵素への電子伝達活性を有するフェレドキシンである。従って、適切な電子伝達系とは様々なフェレドキシンレダクターゼとフェレドキシンの組み合わせによる本願酵素活性の検討の結果、反応の進行が確認されたフェレドキシンレダクターゼとフェレドキシンの組み合わせを指す。フェレドキシンレダクターゼは、例えば以下の中から選択される;ホウレン草由来のフェレドキシンレダクターゼ、シュードモナス プチダ(Pseudomonas putida)由来プチダレドキシンレダクターゼ、動物由来のアドレノドキシンレダクターゼ、スフィンゴモナス サブテラニア(Sphingomonas subterranea)由来フェレドキシンレダクターゼ、ノボスフィンゴビウム アロマティシボランス(Novosphingobium aromaticivorans)由来フェレドキシンレダクターゼ、エシェリヒア コリ(Escherichia coli)由来フラボドキシンレダクターゼやフェレドキシンレダクターゼ、またはその他のフェレドキシンレダクターゼ。フェレドキシンは、例えば以下の中から選択される;ホウレン草由来のフェレドキシン、シュードモナス プチダ(Pseudomonas putida)由来プチダレドキシン、動物由来のアドレノドキシン、スフィンゴモナス サブテラニア(Sphingomonas subterranea)由来フェレドキシン、ノボスフィンゴビウム アロマティシボランス(Novosphingobium aromaticivorans)由来フェレドキシン、エシェリヒア コリ(Escherichia coli)由来フラボドキシンやフェレドキシン、またはその他のフェレドキシン型蛋白質。
【0064】
反応の方法は特に限定されず、本発明の酵素又は該酵素を産生する微生物などを含む液体に、基質となる式(I)で表されるステロールを加え、適当な温度(例えば、10℃から40℃程度)で反応させることができる。これにより、プレグネノロン、プロゲステロン、又は7-デヒドロプレグネノロンなどの式(II)で表される化合物を製造することができる。あるいは本発明の酵素を産生する微生物が,内在的に式(I)で表されるステロールを産生する場合でも,これを反応させて式(II)で表される化合物を製造することができる。
【0065】
また、上記の方法で得られた式(II)で表される化合物に、17α−ステロイド水酸化酵素、21−ステロイド水酸化酵素、11β−ステロイド水酸化酵素を適当な温度で反応させて、ハイドロコルチゾンを製造することができる。
【0066】
さらに、本発明の酵素を産生する微生物(内在的に式(I)で表されるステロールを産生する微生物が好ましい)に、17α−ステロイド水酸化酵素、21-ステロイド水酸化酵素、11β-ステロイド水酸化酵素の遺伝子(Molecular and Cellular Endocrinology 1990, 73, 73-80)を導入してこれらの蛋白質(酵素)を発現する能力を与え、この形質転換体を用いてハイドロコルチゾンを製造することもできる。
【0067】
反応混合物から目的の式(II)で表される化合物を分画する方法は特に限定されず、当業者に公知の分離又は精製のための手法を用いることができる。例えば、溶媒抽出、晶析、樹脂吸着、カラムクロマトグラフィー等により行うことができるが、これに限定されるものではない。
【0068】
以下の実施例により本発明をさらに詳細に説明するが、本発明は実施例によって限定されるものではない。
【実施例】
【0069】
以下の実施例において一般的な実験方法はSambrookらの実験書(Sambrook, J., and Russel, D. W.: Molecular cloning: a laboratory manual. 3rd edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 2001)に従った。
【0070】
実施例1:Sphingomonas subterranea NBRC16086からのP450遺伝子のクローニング
Sphingomonas subterranea NBRC 16086をL培地(1%トリプトン、0.5%酵母エキス、0.5%NaCl、0.1%グルコース、pH7.2 )に接種して30℃で一晩培養して得られた菌体からゲノムDNAを抽出した。ゲノムDNAの抽出にはDNA抽出キットISOPLANT (和光純薬工業社製)を用いた。このゲノムDNAよりプライマーCYP-1F: 5'-CGAGATCTGATGAACTTCATGGGCCAGCGCAT-3' (配列番号8)およびプライマーCYP-2R: 5'-CGAGATCTGCAGCGGTGGACACCCCCGCCAAA-3' (配列番号9)を用いてシトクロムP450遺伝子をポリメラーゼ連鎖反応(PCR) を行って増幅した。
この時のPCRの条件は以下のとおりである。
【0071】
(反応液組成)
滅菌精製水 15μl
2倍濃縮GC buffer I (タカラバイオ社製) 25μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2.5 mM) 8μl
プライマーCYP-1F (50 p mol/μl) 0.5μl
プライマーCYP-2R (50 p mol/μl) 0.5μl
S. subterranea NBRC 16086 ゲノムDNA (100 ng/μl) 0.5μl
LA Taq ポリメラーゼ (5 units/μl,タカラバイオ社製) 0.5μl
【0072】
(温度条件)
94℃/3分、(94℃/30秒, 55℃/30秒, 72℃/90秒)30サイクル、72℃/5分
【0073】
この反応の結果、増幅した1.0 kb (キロベースペア)の遺伝子断片の塩基配列を決定した。この塩基配列情報を基にインバースPCRを行ってシトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含む1.8 kb DNA領域の全塩基配列:配列番号3)を決定した。また、配列番号3の1番目から1419番目の塩基配列を配列番号1に示し、配列番号1に記載の塩基配列によりコードされるアミノ酸配列を配列番号2に示す。
【0074】
BP215, CYP ORF, Fdx ORF
CYP ORF:1-1419
Fdx ORF: 1430-1762
【0075】
(配列番号3)
ATGGCAAGTGCAGCAGCGGGTGCAGACGGCCTTCCCTTGCTCGATGGCGGCGTGCCGCTGCTCGGGCACCTCGCCCAGTTCTTTCGCGATCCGGTTTCCGTCCTGAAGCGCGGCTATCGCTCGAAGGGCCGGCTCTTCGCGATGAACTTCATGGGCCAGCGCATGAACGTGATGCTGGGGCCGGAACACAACCGCTTCTTCTTCGAGGAAACCGACAAGCTGCTCTCGATCCGAGAGTCGATGCCGTTCTTCCTCAAGATGTTCTCGCCCGAGTTCTACTCGTTTGCCGAGATGGACGAATACCTGCGCCAGCGCGCGATCATCATGCCCCGGTTCAAGGCGGCATCGATGAAGCAGTACGTGCCGGTGATGGTCGAGGAATCGCTGAACCTCGTCGAACGGCTGGGTGAGGAAGGCGAGTTCGACCTGATCCCGACGCTGGGCCCCGTGGTAATGGACATCGCCGCGCACAGCTTCATGGGGCGCGAATTCCACGAGAAGCTTGGCCACGAATTCTTCGAGCTGTTCCGCGACTTCTCGGGCGGCATGGAGTTCGTGCTGCCGCTGTGGCTGCCGACACCAAAGATGGTGAAAAGCCAGCGCGCCAAGAAGAAGCTCCACGCGATCCTGCAATCGTGGATCGACAAGCGCCGCGCCAGTCCGCTCGATCCGCCCGACTTCTTCCAGACGATGATCGAGACGAAGTATCCCGATGGCCGCGCCGTGCCCGACGAGATCATCCGGCACCTGATCCTGCTGCTGGTCTGGGCGGGCCACGAAACCACCGCCGGGCAAGTCAGCTGGGCGCTGGCCGACCTCCTCCAGAACCCCGGCTACCAGCAGGTGCTGCGCGAAGAGGTCGCCTCGCTGCTCGGCGGCAGCGACGGGCGCGCACTGGGCTGGGAGCAGGCCGTGGCGATGGAGAAGATGGACCTCGCCCTGCGCGAGACAGAGCGGCTCCACCCCGTCGCCTACATGCTCAGCCGCAAGGCCAGTGCCGATATCGAGCGCGACGGCTATCGCATCCGCAAGGGCGAATTCGTGCTGCTCGCGCCCTCGGTCAGCCATCGCATGGAAGAAACCTTCCGCAATGCCGATGCCTATGACCCCGAGCGCTTCAACCCGCAGAACCCGGACGCGCAGATCGAGAGCAACTCGCTGATCGGCTTCGGCGGCGGGGTGCATCGCTGTGCGGGGGTAAACTTCGCGCGCATGGAAATGAAGGTGCTGGTGGCGATCCTGCTGCAGAACTTCGACATGGAGCTGATCGACGAAGTGCGCCCCATCGCCGGCGCATCGACCTACTGGCCCGCCCAGCCCTGCCGCGTGCGCTACAAGCGGCGCAAGCTGGATGGCGCGGGTGGAAGCGCGGACGTGGCGGCGCTCGCCAAGGCGGCCGGTTGCCCGGCCCATGCATGAGGCGCACTGAATGGCCAAGGTGACTTTCGTCCAGCCGGACGGCTCGGCGCGGACTTGCGTGAACTTCGAGGGCATGACCCTGATGCAGCTCGCGGTCGGCAACCTTGTCGACGGGATCGACGCGCTGTGCGGCGGCATGATGCAGTGCGCCACCTGCCATTGCTACATCGACCCGGACTGGCTTGACCGCACCGGCCCCGCCCGGCCGGAAGAACGCGAGATGCTCGAGGCCATCGATGGCGTCGAGATCCGCCCGAACAGCCGCCTGTCCTGCCAGGTCCAGCTTGGCGAGGAGTTGGACGGGCTGGTGGTGCACATTCCGGCGGAGCAACCGGGAGTTTAG
【0076】
実施例2:Sphingomonas subterranea NBRC 16086由来シトクロムP450遺伝子の大腸菌発現株BP215の作成
実施例1の塩基配列を基にしてシトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含むDNA領域をPCRで増幅した。プライマーCYP-3F: 5'-CCCCATATGGCAAGTGCAGCAGCGGGTGCAGACGG-3' (配列番号10) およびプライマーCYP-4R: 5'-GCACTAGTCTAAACTCCCGGTTGCTCCGG-3' (配列番号11)を用いた。この時のPCRの条件は以下のとおりである。
【0077】
(反応液組成)
滅菌精製水 31.5 μl
10倍濃縮 PCR buffer (東洋紡社製) 5 μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2 mM) 5 μl
25 mM MgSO4 2 μl
プライマーCYP-3F (50 p mol/μl) 1 μl
プライマーCYP-4R (50 p mol/μl) 1 μl
S. subterranea NBRC 16086 ゲノムDNA (100 ng/μl) 1 μl
KOD plus ポリメラーゼ (1 units/μl,東洋紡社製) 1 μl
DMSO 2.5 μl
【0078】
(温度条件)
95℃/5分、(98℃/30秒, 60℃/30秒, 68℃/2分)30サイクル、68℃/10分
【0079】
この反応の結果増幅した、シトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含む1.8 kb DNA断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50μl 中で制限酵素Nde IおよびSpe Iで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.8 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製した。この断片を大腸菌発現ベクターpT7NS-camAB(有澤および久米田、国際公開公報2003/087381パンフレット)のNde IおよびSpe I部位にT4 DNAリガーゼを用いて連結して、大腸菌DH5aに形質転換することで、プラスミドpCYPSS-camABを構築した。このプラスミドを大腸菌BL21(DE3)に形質転換して得られた菌株をBP215と命名した。
【0080】
実施例3:Novosphingobium aromaticivorans ATCC 700278からのシトクロムP450遺伝子(CYP204A1遺伝子)のクローニングと大腸菌発現株BP172の作成
Novosphingobium aromaticivorans ATCC 700278をL培地に接種して30℃で2日間培養して得られた菌体からゲノムDNAを抽出した。ゲノムDNAの抽出にはDNA抽出キットISOPLANT (和光純薬工業社製)を用いた。このゲノムDNAよりプライマーCYP-5F: 5'-GCCCATATGGCAAGAGCTGCGACTGCGGCC-3' (配列番号12) およびプライマーCYP-6R: 5'-GCACTAGTCTAAACTCCCGGTTGCTCCGG-3' (配列番号13)を用いてシトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含む断片を増幅した。この時のPCRの条件は以下のとおりである。
【0081】
(反応液組成)
滅菌精製水 15μl
2倍濃縮GC buffer I (タカラバイオ社製) 25μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2.5 mM) 8μl
プライマーCYP-5F (50 p mol/μl) 0.5μl
プライマーCYP-6R (50 p mol/μl) 0.5μl
N. aromaticivorans ATCC 700278ゲノムDNA (100 ng/μl) 0.5μl
LA Taq ポリメラーゼ (5 units/μl,タカラバイオ社製) 0.5μl
【0082】
(温度条件)
94℃/3分、(98℃/20秒, 63℃/30秒, 68℃/2分)30サイクル、72℃/5分
【0083】
この反応の結果増幅した、シトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含む1.8 kb DNA断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50μl 中で制限酵素Nde IおよびSpe Iで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.8 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製した。この断片を大腸菌発現ベクターpT7-camABのNde IおよびSpe I部位にT4 DNAリガーゼを用いて連結して、大腸菌DH5aに形質転換することで、プラスミドpCYP204A1-camABを構築した。このプラスミドを大腸菌BL21(DE3)に形質転換して得られた菌株をBP172と命名した。CYP204A1遺伝子によりコードされるアミノ酸配列を配列番号26に示す。
【0084】
BP172, CYP ORF, Fdx ORF
CYP ORF: 1-1422
Fdx ORF: 1433-1765
(配列番号4)
ATGGCAAGAGCTGCGACTGCGGCCGGTAATGGCCTTCCCTTGCTCGATGGAGGCGTGCCGCTCCTCGGGCATCTCGCACAGTTCTTCCGCGATCCGGTTTCGGTACTCAAGCGCGGATACCGCTCGAAGGGGCGGCTCTTCGCGATGAACTTCATGGGCCAGCGCATGAACGTGATGCTGGGTCCGGAACACAACCGCTTCTTCTTCGAGGAGACGGACAAGCTGCTCTCGATCCGGGAGTCGATGCCGTTCTTCCTCAAGATGTTCTCGCCCGAGTTCTATTCGTTCGCGGAAATGGACGAGTACCTGCGCCAGCGCTCGATCATCATGCCCCGCTTCAAGGCGGCATCGATGAAGCAGTACGTGCCGGTCATGGTCGAGGAATCGCTTAACCTGGTCGAGCGGCTGGGCGAGGAAGGCGAGTTCGACCTGATCCCGACGCTGGGCCCGGTGGTAATGGACATCGCCGCGCACAGCTTCATGGGACGCGAGTTCCACGAGAAGCTGGGGCATGAGTTCTTCGAACTCTTCCGCGATTTTTCGGGAGGCATGGAATTCGTCCTGCCGCTGTGGCTGCCGACACCCAAGATGGTCAAGTCACAGCGCGCGAAGAGGAAGCTCCACGCCATCCTGCAATCGTGGATCGACAAGCGCCGCGCCGCCCCGCTCGATCCGCCCGATTTCTTCCAGACGATGATCGAGACGAAGTATCCCGATGGCCGCCCGGTGCCCGACGAGATCATCCGCCACCTGATCCTCCTTCTCGTCTGGGCAGGGCACGAGACGACCGCCGGGCAGGTGAGCTGGGCGCTGGCGGACCTCCTTCAGAACCCGGACTACCAGAAGGTGCTGCGCGGCGAGATATCGTCGCTGCTGGGCGGCAGCGACGGGCGCGACCTTGGCTGGGAACAGGCCGTGGCGATGGAGAAGATGGACCTTGCCCTGCGCGAGACCGAGCGGCTCCATCCGGTCGCCTACATGCTCAGCCGCAAGGCGCGGGCCGATATCGAGCGCGACGGCTATGTCATCCGCAAGGGCGAGTTCGTGCTGCTTGCGCCTTCGGTCAGCCACCGCATGGAAGAGACGTTCCGCAATCCCGATGCCTATGACCCGGAACGCTTCAACCCGGCCAACCCCGATGCGCAGATCGAAAGCAATTCGTTGATCGGCTTTGGCGGGGGTGTCCACCGCTGCGCGGGCGTGAACTTCGCGCGGATGGAGATGAAGGTGCTGGTGGCGATCCTGCTCCAGAACTTCGACATGGAGCTGATGGACGAAGTGCGGCCCATCGCGGGCGCATCGACCTACTGGCCCGCCCAGCCCTGCCGGGTGCGCTATCGGCGGCGCAAGCTCGACGGGTCGGAGGCAGGTGCGGACATGGCGGCGCTGGCCCGAGCCGCCGGCTGCCCGGCGCATACGTGAGGGAGGCCTGATGGCCAAGGTGACTTTCGTCCAGCCGGACGGATCGCAGCGAACCTGCGTGAACTTCGAAGGCATGACGTTGATGCAGCTCGCAGTGGGCAATCTCGTCGACGGGATCGACGCGCTGTGCGGCGGCATGATGCAGTGCGCGACCTGCCATTGCTGGATCGACCCCGAATGGATCGGCCGCACCGGCATGGCCGGACCCGATGAGCGGGCAATGCTGGAAGCGATCGAGGGCGTCGAGATCCGTCCCGAAAGCCGCCTGTCCTGCCAGGTACAGCTTGGCGAAGAACTTGACGGGCTGGTCGTGCGCATTCCACCGGAGCAACCGGGAGTTTAG
【0085】
実施例4: CYP204A1遺伝子のBacillus megaterium での発現
pCYP204A1-camABを鋳型にしてプライマーCYP-7F: 5'-CCGCTAGCGCAAGAGCTGCGACTGCGGCC-3'(配列番号14)およびCYP-8R: 5'-GCGGATCCCTAAACTCCCGGTTGCTCCGG -3'(配列番号15)を用いてCYP204A1遺伝子およびその下流に存在するフェレドキシン遺伝子を含む断片を増幅した。この時のPCRの条件は以下のとおりである。
【0086】
滅菌精製水 33 μl
10倍濃縮 PCR buffer (東洋紡社製) 25 μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2.0 mM) 8 μl
25 mM MgSO4 2 μl
プライマーCYP-7F (50 p mol/μl) 0.5 μl
プライマーCYP-8R (50 p mol/μl) 0.5 μl
DMSO 1.5 μl
pCYP204A1-camAB DNA (100 ng/μl) 0.5 μl
KOD plus ポリメラーゼ (1 units/μl,東洋紡社製) 1.0 μl
【0087】
(温度条件)
94℃/2分、(94℃/20秒, 55℃/30秒, 68℃/3分)20サイクル、72℃/5分
【0088】
この反応の結果増幅した、CYP204A1遺伝子およびその下流に存在するフェレドキシン遺伝子を含む1.8 kb DNA断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50 μl 中で制限酵素Nhe IおよびBam HIで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.8 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製した。この断片をBacillus megaterium発現ベクターpHW1520 (MoBiTec社製)のSpe IおよびBam HI部位にT4 DNAリガーゼを用いて連結して、大腸菌DH5aに形質転換することで、プラスミドpXYL204CBを構築した。このプラスミドをBacillus megateriumコンピテント細胞(MoBiTec社製)に形質転換して菌株Bacillus megaterium(pXYL204CB)を得た。この菌株を、テトラサイクリン10mg/ml(終濃度)を添加したTB培地2 mlに植菌し、37℃、16時間、220rpmで振とう培養した。この前培養液250μlをテトラサイクリン10mg/mlを添加したTB培地2 5 mlに加え、37℃で2.5時間振とう培養後、50 % Xylose 250μlを添加し、30℃、125 rpmで6時間振とう培養した。この培養液から遠心分離により回収した菌体を変換用緩衝液(50 mM KPB、2%グリセロール[pH 7.4]) 5 mlに懸濁し、菌体懸濁液とした。この菌体懸濁液2 mlに4-Cholesten-3-oneメタノール溶液(100 mg/ml) 2μlを加えて、振とう(220rpm)しながら30℃で24時間インキュベートした。その後、この反応液に2 mlの酢酸エチルを加えて、ボルテックスした後、遠心機で4,000 rpmで10分間遠心した。得られた酢酸エチル相1.5 mlを濃縮乾固し、その後200μlのメタノールに溶解しHPLC分析に供した。結果、24.5%の変換率で4Cholestene-3-oneをProgesteroneに変換した。
【0089】
HPLC条件
Column:Inertsil ODS-3 4.6 X 50 mm
flow rate:1.2ml/min
temp.:40℃
detect:PDA(240nm)
gradient: 0/ 4/ 5/5.5(min)
40/100/100/40(%B)
HPLCチャートを図1に示す。
【0090】
実施例5:酵素の諸性質
(1)発現ベクターの構築
CYP204A1、CYPSS、CYP11A遺伝子を含むDNA断片を増幅する為に以下のプライマーを作成した。
CYP204A1-1F GCCCATATGGCAAGAGCTGCGACTGCGGCC(配列番号16)
CYP204A1-2R GCCCTCGAGTCACGTATGCGCCGGGCAGCC(配列番号17)
CYPSS-1F GCCCATATGGCAAGTGCAGCAGCGGGTGCA(配列番号18)
CYPSS-2R GCCCTCGAGTCATGCATGGGCCGGGCAACC(配列番号19)
CYP11A-1F GCCCATATGGCTTCAACACGTAGTCCTCGC(配列番号20)
CYP11A-2R GCCCTCGAGTCACTGCTGGGTTGCTTCCTG(配列番号21)
【0091】
次に、これらのプライマーとpCYP204A1-camAB、pCYPSS-camAB、pCYP11A-ARXを鋳型としてKOD plus (TOYOBO)を用いてPCR反応を行った。この時のPCR条件は以下の通りである。
【0092】
(反応液組成)
10倍濃縮PCR buffer for KOD plus 5μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP 各2 mM) 5μl
25 mM MgSO4 2μl
Primer-1F (50 pmol/μl) 0.5μl
Primer-2R (50 pmol/μl) 0.5μl
KOD plus 1μl
DMSO 3.75μl
鋳型 (50 ng/μl) 0.5μl
滅菌精製水 31.75μl
【0093】
(温度条件)
95℃/3分、(98℃/20秒、55℃/30秒、72℃/2分)25サイクル、72℃/5分
【0094】
この反応の結果、CYP204A1、CYPSS、CYP11Aにおいて約1.4 kbpの大きさのDNA断片(以下、DNA断片-A, B, Cという)が増幅された。Wizard SV Gel and PCR Clean-Up System (PROMEGA)で精製し、NdeIとXhoIにより消化した。このサンプルをアガロースゲル電気泳動にかけて分画し、約1.4 kbpの大きさのDNA断片をアガロースゲルから切り出した。Wizard SV Gel and PCR Clean-Up SystemによりNdeIとXhoIにより消化されたDNA断片-A, B, Cを精製し、NdeIとXhoIにより消化したpET22bにDNA Ligation Kit ver 2.1 (TaKaRa)により連結し、プラスミドpET22-CYP204A1、pET22-CYPSS、pET22-CYP11Aを得た。
【0095】
(2)CYP204A1、CYPSS、CYP11A誘導菌体の作成
上記(1)で調製したプラスミドpET22-CYP204A1、pET22-CYPSS、pET22-CYP11Aを用い、大腸菌BL21 (DE3)を形質転換した。こうして得られた形質転換体を50μg/mlのカルベニシリンを含む1 mlのTB培地に植菌し、37℃で14時間培養した。この1 mlの培養液を50μg/mlのカルベニシリンとOvernight Express Autoinduction system 1 (Merck)を含む100 mlのTB培地に植菌し、25℃で24時間培養および目的タンパク質の誘導を行った。遠心により菌体を回収し、20 mlのCV buffer (50 mM KPB (Potassium phosphate buffer), 10% Glycerol, 1 mM EDTA, 2 mM dithiothreitol, 1 mM D-Glucose)に懸濁して-80℃で細胞懸濁液を保存した。
【0096】
(3)無細胞抽出液の調製
上記(2)で調製した細胞懸濁液20 mlに680μlのX10 Bug Buster (Novagen)と20μlのBenzonase (Novagen)と2 mlの40 mg/ml Lysozymeを加えて、30℃で30分間攪拌して細胞を破砕し、遠心により無細胞抽出液を得た。得られた約20 mlの無細胞抽出液を1 LのBuffer R (20 mM KPB pH 7.4, 20% glycerol)で2回透析し、その間に析出した沈殿を遠心により除いた。ここで上清として得られた無細胞抽出液を以後の実験に使用した。
【0097】
(4)一酸化炭素結合スペクトル解析によるP450の発現確認
得られた無細胞抽出液を2つに分けて片方に一酸化炭素をバブリングした。一酸化炭素バブリング有無の両方のサンプルにジチオナイトを加え、差スペクトルを測定した。CYP204A1とCYPSSでは450 nm付近に極大吸収を持つピークが得られ、P450の発現が確認されたが、CYP11Aでは得られず活性型のP450がサンプル中に存在しないことが示唆された。この結果より以後の実験はCYP204A1とCYPSSで行うこととした。
【0098】
(5)至適pHの検討
上記(3)で調製した無細胞抽出液を用いて、様々なpHで4-コレステン-3-オンの変換を行った。反応液は1 ml BufferR中で215 pmol/ml CYP204A1もしくは367 pmol/ml CYPSS、64μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、50μg/ml 4-Cholesten-3-one、2 mM NADHと2 mM NADPHとなるように調製し、1 mlの反応液中に様々なpHの200 mM Bufferを562μl加えてpHを調整した。反応液作成後にpHを測定するとKPB pH 6.2, KPB pH 6.7, KPB pH 7.1, KPB pH 7.4, KPB pH 7.9, Tris-HCl pH 7.5, Tris-HCl pH 8.2, Tris-HCl pH 8.7, TES pH 6.6, TES pH 6.8, TES pH 7.3, TES pH 7.5, TES pH 8.0であった。反応はNADHとNADPHの添加により開始し、200 rpm、30℃で14時間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。各pH条件における4-コレステン-3-オンからプロゲステロンへの酵素活性を図2(及び表1)に示した。酵素活性は1時間、1 mol P450あたりのプロゲステロン生成量gで示した。この結果より、CYP204A1とCYPSSはTris-HCl pH 7.5で最も活性が高いことが示された。
【0099】
(6)至適温度の検討
上記(3)で調製した無細胞抽出液を用いて、様々なpHで4-コレステン-3-オンの変換を行った。反応液は1 mlで215 pmol/ml CYP204A1もしくは391 pmol/ml CYPSS、64μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、50μg/ml 4-コレステン-3-オン、2 mM NADHと2 mM NADPHとなるように調製し、Tris-HClでpH 7.5にした。反応はNADHとNADPHの添加により開始し、200 rpm、温度条件をそれぞれ10, 15, 20, 25, 30℃として4時間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。各温度条件における4-コレステン-3-オンからプロゲステロンへの酵素活性を図3(及び表1)に示した。この結果より、CYP204A1は15から20℃で、CYPSSは15℃で最も活性が高いことが示された。
【0100】
【表1】
【0101】
(7)pH安定性試験
上記(3)で調製した0.5 mlの無細胞抽出液と様々なpHの0.5 mlの50 mM Bufferと混合してpHを測定して6℃で140時間保存した。酵素液のpHはAcB (acetate buffer) pH 6.0, KPB pH 6.7, KPB pH 7.2, KPB pH 7.6, TES pH 7.5, Tris-HCl pH 8.1, Tris-HCl pH 8.9であった。調製した様々なpHの無細胞抽出液を用いて4-コレステン-3-オンの変換を行った。酵素液を様々なpHのBufferで混ぜた直後と140時間保存したときに反応を行い、安定性を試験した。反応液は1 mlでCYP204A1もしくはCYPSSの各pHの酵素液、64 μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、50μg/ml 4-コレステン-3-オン、2 mM NADHと2 mM NADPHとなるように調製し、200 mM Tris-HCを562μl加えてpH 7.5にした。様々なpHの酵素液を加えても反応液がpH 7.5であることを確認した。反応はNADHとNADPHの添加により開始し、200 rpm、15℃で4時間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。0時間と140時間の4-コレステン-3-オンからプロゲステロンへの酵素活性を図4と表2に示した。この結果より、CYP204A1、CYPSSともにKPB pH 7.6で最も安定であることが示された。
【0102】
【表2】
【0103】
(8)温度安定性試験
上記(3)で調製した無細胞抽出液を様々な温度で140時間保存した。保存した温度は6, 10, 20, 30, 37, 45℃とした。保存前の酵素液と140時間保存後のサンプルを用いて4-コレステン-3-オンの変換を行い、安定性を試験した。反応液は1 mlで各温度で保存したCYP204A1もしくはCYPSS酵素液、64μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、50μg/ml 4-コレステン-3-オン、2 mM NADHと2 mM NADPHとなるように調製した。反応はNADHとNADPHの添加により開始し、200 rpm、15℃で4時間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。0時間と140時間の4-コレステン-3-オンからプロゲステロンへの酵素活性を図5と表2に示した。この結果より、CYP204A1、CYPSSともに保存温度が下がるに従って安定性が上昇することが示された。特に、20℃で140時間保存した場合、CYP204A1は47%の活性が残存し、CYPSSは39%の活性が残存した。
【0104】
【表3】
【0105】
(9)CYP204A1とCYPSSのKinetics解析
上記(3)で調製した無細胞抽出液を用いて、基質濃度を0.1〜500μMで4-コレステン-3-オンの変換を行い基質濃度と酵素活性の関係を調べた。反応液は1 mlで220 pmolのCYP204A1もしくは250 pmolのCYPSS、96μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、2 mM NADHと2 mM NADPHとなるように調製し、Tris-HClでpH 7.5にした。4-コレステン-3-オンは終濃度が0.1, 0.5, 1, 2, 5, 10, 20, 50, 100, 500μMとなるようにDMSOに溶かしたものを20μl加えた。反応はNADHとNADPHの添加により開始し、200 rpm、15℃で60分間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。基質濃度と酵素活性の関係を図6 (CYP204A1)と図7 (CYPSS)に示した。また、Lineweaver-Burk plotを作成して図8(及び表4) (CYP204A1)と図9(及び表4) (CYPSS)に示した。算出されたKinetics parametersを表5に示した。この結果より、CYP204A1とCYPSSのKm値は同等の1.0と1.1μMでVmaxは47.4と78.7 mmol/min/molとCYPSSの方が約1.7倍高いことが示された。また、図6と図7の比較によりCYPSSの方が基質阻害がかかりにくいことが示された。
【0106】
【表4】
【0107】
【表5】
【0108】
上記の結果の概要を以下に示す。
至適pH: pH 7.5 〜8.0(CYP204A1, CYPSSともに)
作用至適温度:15〜20℃ (CYP204A1, CYPSSともに)
pH安定性:中性付近で安定。もっとも安定だったのはKPB pH 7.6 (CYPSS, CYP204A1ともに)
熱安定性:20℃で140時間保存後に30%以上の酵素活性が保持される。(CYPSS, CYP204A1ともに)
Kinetics解析:Vmax CYPSSはCYP204A1に対して約1.7倍高い
Km値 CYPSSとCYP204A1はほぼ同等(1μM)
基質阻害:4-コレステン-3-オンにより、基質阻害がかかることが分かったがCYPSSの方がCYP204A1に比べて基質阻害がかかりにくい。4-コレステン-3-オンの濃度が10μMのときのCYPSSまたはCYP204A1の酵素活性にくらべ、100μMのときのそれぞれの酵素活性は90%または67%であった。すなわち、基質濃度を10μMから100μMに上昇させた場合の活性の低下率は10%または33%であった。
【0109】
実施例6:BP215株による4-コレステン-3-オンの変換反応(プロゲステロンの調製)
大腸菌発現BP215株をM9混合培地(Na2HPO4: 0.68%, KH2PO4: 0.3%, NaCl: 0.05%, NH4Cl: 0.01%, Casamino acid: 1%, D-Glucose: 0.4%, CaCl2: 0.1mM, MgCl2: 1mM, Thiamine: 0.002%, FeSO4: 0.1mM, Carbenicillin: 0.005%)2mlに植菌し、30℃で17時間振とう培養し前培養液を得た。この前培養液0.25mlをOvernight ExpressTM Autoinduction System(Novagen Cat. No. 71300-4)を用いた本培養培地(Na2HPO4: 0.68%, KH2PO4: 0.3%, NaCl: 0.05%, NH4Cl: 0.01%, Casamino acid: 1%, CaCl2: 0.1mM, MgCl2: 1mM, Thiamine: 0.002%, FeSO4: 0.1mM, Carbenicillin: 0.005%, Overnight ExpressTM溶液:7%, 5-Aminolevlic acid: 0.008%)25mlに植菌し、25℃で24時間振とう培養した。この本培養液5mlを遠心分離(3500rpm, 10分間)して得た菌体にCV2バッファー(glycerol: 2%, Carbenicillin: 0.005%, Isopropyl βD-1-thiogalactopyranoside: 0.1mMを含む50mM(pH)燐酸カリウム緩衝液)1mlを加えた後、4-Cholesten-3-oneの1%メタノール溶液0.01mlおよびMethyl-β-cyclodextrinの25%水溶液0.02mlを加え28℃で5時間振とうし反応した。反応終了後、メタノール4mlを加え良く混合し、遠心分離して上層をHPLCで分析した。保持時間2.6分にプロゲステロンのピークを検出し、プロゲステロン標品と保持時間およびMSスペクトル(m/z=315 M+1)が一致した。蓄積量は78.0 mg/l(変換:78%)であった。また、4-Cholesten-3-one(保持時間8.1分)は検出されなかった。HPLCを図10に示す、MSを図11に示す。
【0110】
分析条件:Chromolith speed rod RP18e(50x4.6mm)カラムで50%→100%CH3CNグラジエント(0〜3min) 100%CH3CN(3〜8min)を移動相とし流速2ml/min、CADTM(Charged Aerosol Detection)、MSで検出した。
【0111】
実施例7:BP215株によるコレステロールの変換反応
基質をコレステロールとして実施例6と同様に反応後、処理分析した結果、保持時間2.5分にプレグネノロンのピークを検出し、プレグネノロン標品と保持時間およびMSスペクトル(m/z=317M+1)が一致した。蓄積量は68.2 mg/l(変換:68.2%)であった。また、Cholesterol(保持時間8.5分)は3.7%残留していた。HPLCを図12に示す、MSを図13に示す。
【0112】
実施例8:BP215株による7-デヒドロコレステロール、エルゴステロールの変換反応
基質を7-デヒドロコレステロールとし実施例6と同様に反応後、処理分析した結果、保持時間2.1分に7-デヒドロプレグネノロンのピーク(m/z=315M+1)を検出した。蓄積量は34.3 mg/l(変換:34.3 %)であった。また、7-Dehydrochlesterol(保持時間7.5分)は54%残留していた。HPLCを図14に示す、MSを図15に示す。
【0113】
基質をエルゴステロールとし実施例6と同様に反応後、処理分析した結果、保持時間2.1分に7-デヒドロプレグネノロンのピークを検出した。蓄積量は2.8 mg/l(変換:2.8 %)であった。
【0114】
実施例9:BP215株による各種基質の変換反応
基質をβ-シトステロール、スティグマステロール、カンペステロールおよびデスモステロールとしてそれぞれ別々に実施例6と同様に反応後、処理分析した結果、保持時間2.5分にプレグネノロンのピークを検出し、プレグネノロン標品と保持時間およびMSスペクトル(m/z=317M+1)が一致した。蓄積量はそれぞれ40.9 mg/l(変換:40.9 %)、5.8 mg/l(変換:5.8 %)、27.9 mg/l(変換:27.9 %)、71.7 mg/l(変換:71.7 %)であった。また、添加した基質(保持時間はそれぞれ10.1、9.3、9.2、7.2分)はそれぞれ55.9%,70.8%,25.5%,21.3%残留していた。なお、β-シトステロールは生成物が微量であった為、反応液を酢酸エチルで抽出し5倍に濃縮して分析した。
【0115】
実施例10:(20R,22R)-20,22-ジヒドロキシ-4-コレスト-4-エン-3-オン、及び(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンの化学合成
20,22-ジヒドロキシコレスト-4-エン-3-オンの2つの立体異性体の合成は、文献既知の合成方法を用いて20,22-ジヒドロキシコレステロールを経由したのちに、常法の酸化反応を用いて実施した。
【0116】
(A)20,22-ジヒドロキシコレステロール(10-A)の合成
Steroids(2004),69,483/ B.Watanabe et al.に、化合物(10-A)の2つの異性体[(20R,22R)体および(20R,22S)体]の化学合成が記載されており、その実験方法に従って同様に(10-A)の合成を実施した。
【0117】
概要は、プレグネノロンを出発原料として、t-ブチルジメチルシリルクロライドで3位を保護基にして、4-メチル-1-ペンチンを付加させて(20R)-3β-(t-ブチルジメチルシリロキシ)コレスタ-5-エン-22-イン-20-オルを得た。これをLindlar触媒を用いてcis-オレフィンに還元して(20R,22Z)-3β-(t-ブチルジメチルシリロキシ)コレスタ-5,22-ジエン-20-オルとし、これをVO(acac)2/TBHPにより22,23-エポキシド(異性体混合物)に変換して、これを異性体混合物のまま分離せず、LiAlH4で還元して20,22-ジヒドロキシ体に変換して、TBAFで3位の脱保護をして、(20R,22RS)-20,22-ジヒドロキシコレステロール(10-A)を得た。
【0118】
(B)20,22-ジヒドロキシコレスト-4-エン-3-オン(10-B)の合成
前出文献方法を参考に、まず(10-A)から(20R,22RS)-20,22-O-イソプロピリデンジオキシコレステロールを得た[PPTS存在下、2,2-ジメトキシプロパンを作用]。
これを常法どおりOppenauer酸化により4-エン-3-オン体に変換した。この実験手順を具体的に記す。
【0119】
十分に窒素置換したDean-Stark装置内で、(20R,22RS)-20,22-O-イソプロピリデンジオキシコレステロール(3.0g)をトルエン100mLに溶解し、1-メチル-4-ピペリドン(8.1mL,10eq)を添加したのち、反応温度を130-140℃にして、Al(OiPr)3 を(2.7g,2eq) 添加して4hrかくはんした。反応終了後、ジエチルエーテルで希釈して常法処理したのち、シリカゲルクロマトグラフィー精製[ヘキサン/酢酸エチル=8/1〜5/1]を行い、(20R,22RS)-20,22-O-イソプロピリデンジオキシコレスト-4-エン-3-オンを得た(2.9g, y97%)。
最後に(20R,22RS)-20,22-O-イソプロピリデンジオキシコレスト-4-エン-3-オンを脱アセトニド保護を行うことで(10-B)へ変換され、さらに2つの立体異性体を別々に取得することが出来た。以下に具体的な実験手順を記す。
【0120】
(20R,22RS)-20,22-O-イソプロピリデンジオキシコレスト-4-エン-3-オン(2.9g)を、THF(20mL)+メタノール(20mL)中で室温でかくはんしながら、2N塩酸(1.0mL)および過塩素酸(0.8mL)を添加して、60hr反応した。クロロホルムで希釈して常法処理したのち、シリカゲルクロマトグラフィー精製をおこない[ヘキサン/酢酸エチル=7/1〜2/1]、脱保護生成物(1.48g, y47%)と、未反応原料(1.24g)を分離取得した。
脱保護生成物からは、ヘキサン/酢酸エチルで再結晶して、白色固体(1.1g)を得た。
【0121】
1H-NMR(400MHz,CDCl3) δ: 0.90(d,J=6.6Hz,26-H3),0.90(d,J=6.8Hz,27-H3),0.91(s,18-H3), 1.19(s,19-H3),1.26(s,21-H3),3.24(dd,J=9.4Hz,5.4Hz,22-H),5.73(s,4-H)
→ (20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オン(10-B)
[非天然型異性体; 本発明による新規化合物である。]
【0122】
回収した未反応原料(590mg)を、THF(15mL)+メタノール(15mL)中で室温で攪拌しながら、過塩素酸(1.4mL)を添加して10時間反応した。クロロホルムで希釈して常法処理したのち、シリカゲルクロマトグラフィー精製[ヘキサン/酢酸エチル=7/1〜2/1]を行い、脱保護生成物(217mg, y40%)を得た。
【0123】
1H-NMR(500MHz,CDCl3) δ: 0.89(d,J=6.3Hz,26-H3),0.91(d,J=6.4Hz,27-H3),0.93(s,18-H3), 1.19(s,19-H3),1.21(s,21-H3),3.38(d,J=8.4Hz,22-H),5.73(s,4-H)
→(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン(10-B)
[天然型異性体; (引用) Helv.Chim.Acta,(2006),89,813/W.Zhang et al. ]
【0124】
実施例11:BP215株による各種基質の変換反応
(20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンを基質としてそれぞれ別々に実施例6と同様に反応し、生成物をHPLC分析した結果、プロゲステロンの生成を確認した。変換率はそれぞれ41.0%、42.3%、38.1%および69.8%であった。また、添加した基質(保持時間はそれぞれ4.9、4.4、3.8、3.5分)はそれぞれ4.8%,2.7%,0.6%,0.6%残留していた。
【0125】
また、(20S) -20-ヒドロキシ-コレステロール、(22S) -22-ヒドロキシ-コレステロール、(22R)-22-ヒドロキシ-コレステロール、(20R, 22R) -20,22-ジヒドロキシ-コレステロールを基質としてそれぞれ別々に実施例6と同様に反応後、処理分析した結果、各基質に対応する側鎖開裂体のプレグネノロンの生成を確認した。蓄積量はそれぞれ89.2mg/L(変換:89.2 %)、37.5mg/L(変換:37.5%)、80.0mg/L(変換:80 %)、53.0mg/L(変換:53.0 %)であった。また、添加した基質はそれぞれ0%、23.5%、14.8%、0%残留していた。
【0126】
実施例12:BP172株を用いた各種基質の変換反応
4-コレステン-3-オン、コレステロール、7-デヒドロコレステロール、エルゴステロール、β-シトステロール、スティグマステロール、カンペステロール、デスモステロール、(20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンを基質としてそれぞれ別々に実施例6と同様に反応し生成物をHPLC分析した結果、各基質に対応する側鎖開裂体のプロゲステロン、プレグネノロンまたは7-デヒドロプレグネノロンの生成を確認した。変換率はそれぞれ84.0%、71.2%、28.5%、2.2%、47.2%、6.5%、31.3%、66.1%、63.9%、80.9%、61.3%および41.7%であった。
【0127】
また、(20S) -20-ヒドロキシ-コレステロール、(22S) -22-ヒドロキシ-コレステロール、(22R)-22-ヒドロキシ-コレステロール、(20R, 22R)-20,22-ジヒドロキシ-コレステロールを基質としてそれぞれ別々に実施例6と同様に反応後、処理分析した結果、各基質に対応する側鎖開裂体のプレグネノロンの生成を確認した。蓄積量はそれぞれ68.2mg/l (変換:68.2%)、12.9mg/l (変換:12.9%)、69.5mg/l(変換:69.5 %)、51.6mg/l(変換:51.6 %)であった。また、添加した基質はそれぞれ25.2%、60.1%、42.8%、0%残留していた。
【0128】
参考例1:CYP11A発現大腸菌の作成
市販のヒト睾丸cDNA(Origene Technologies, Inc.)を鋳型として、プライマーCYP11A-1F: 5'-CCGCATATGGCTTCAACACGTAGTCCTCGCCCCTTCAATGAGATC-3' (配列番号22) およびプライマーCYP11A-2R: 5'-GCACTAGTTCACTGCTGGGTTGCTTCCTGGTTAAA-3' (配列番号23)を用いてCYP11A1遺伝子をポリメラーゼ連鎖反応(PCR) を行って増幅した。
この時のPCRの条件は以下のとおりである。
【0129】
(反応液組成)
滅菌精製水 34μl
10倍濃縮 PCR buffer (東洋紡社製) 5μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2 mM) 5μl
25 mM MgSO4 2μl
プライマーCYP11A-1F (50 p mol/μl) 0.5μl
プライマーCYP11A-2R (50 p mol/μl) 0.5μl
ヒト睾丸cDNA (10 ng/μl) 2μl
KOD plus ポリメラーゼ (1 units/μl,東洋紡社製) 1μl
【0130】
(温度条件)
94℃/3分、(94℃/30秒, 55℃/60秒, 72℃/90秒)30サイクル、72℃/5分
【0131】
この反応の結果、増幅した1.45 kbのCYP11A1遺伝子断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50μl 中で制限酵素Nde IおよびSpe Iで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.45 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製した。この断片を大腸菌発現ベクターpT7NS-camABのNde IおよびSpe I部位にT4 DNAリガーゼを用いて連結して、大腸菌DH5αに形質転換することで、プラスミドpCYP11A-camABを構築した。次いで、このプラスミドのSpe IおよびBam HI部位に予め調製しておいたウシ由来アドレノドキシン還元酵素−アドレノドキシン遺伝子(adr-adx)断片をT4 DNAリガーゼを用いて挿入、連結して、大腸菌DH5αに形質転換することで、プラスミドpCYP11A-ARXを構築した。このプラスミドを大腸菌BL21(DE3)に形質転換して得られた菌株をBL21CYP11と命名した。この時用いたadx-adr断片はプラスミドpKARX (Sawada et. al. Eur. J. Biochem. 265, 950-956, 1999)を鋳型として、プライマーbAdxR-1F: 5'-GCACTAGTAAGGAAACAGACCATGTCCACACAGGAGCAGACCCCC-3'(配列番号24) およびプライマーbAdx-2R: 5'-CGTGATCAGGCATCAGACACGGCATCAGG-3'(配列番号25)を用いて以下の条件でポリメラーゼ連鎖反応(PCR) を行って増幅した。
【0132】
(反応液組成)
滅菌精製水 33μl
10倍濃縮 PCR buffer (東洋紡社製) 5μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2 mM) 5μl
25 mM MgSO4 2μl
プライマーbAdxR-1F (50 p mol/μl) 1μl
プライマーbAdx-2R (50 p mol/μl) 1μl
pKARX DNA (10 ng/μl) 0.5μl
DMSO 1.5μl
KOD plus ポリメラーゼ (1 units/μl,東洋紡社製) 1μl
【0133】
(温度条件)
94℃/3分、(94℃/30秒, 60℃/30秒, 72℃/120秒)25サイクル、72℃/5分
【0134】
この反応の結果、増幅した1.85 kbのadr-adx遺伝子断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50μl 中で制限酵素Spe IおよびFba Iで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.85 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製して、上述のpCYP11A-ARX構築に使用した。
【0135】
CYP11A1のDNA配列
CYP11A1, human
Coding region; 45-1610
Mature peptide; 162-1610
(配列番号5)
GGGCGCTGAAGTGGAGCAGGTACAGTCACAGCTGTGGGGACAGCATGCTGGCCAAGGGTCTTCCCCCACGCTCAGTCCTGGTCAAAGGCTACCAGACCTTTCTGAGTGCCCCCAGGGAGGGGCTGGGGCGTCTCAGGGTGCCCACTGGCGAGGGAGCTGGCATCTCCACCCGCAGTCCTCGCCCCTTCAATGAGATCCCCTCTCCTGGTGACAATGGCTGGCTAAACCTGTACCATTTCTGGAGGGAGACGGGCACACACAAAGTCCACCTTCACCATGTCCAGAATTTCCAGAAGTATGGCCCGATTTACAGGGAGAAGCTCGGCAACGTGGAGTCGGTTTATGTCATCGACCCTGAAGATGTGGCCCTTCTCTTTAAGTCCGAGGGCCCCAACCCAGAACGATTCCTCATCCCGCCCTGGGTCGCCTATCACCAGTATTACCAGAGACCCATAGGAGTCCTGTTGAAGAAGTCGGCAGCCTGGAAGAAAGACCGGGTGGCCCTGAACCAGGAGGTGATGGCTCCAGAGGCCACCAAGAACTTTTTGCCCCTGTTGGATGCAGTGTCTCGGGACTTCGTCAGTGTCCTGCACAGGCGCATCAAGAAGGCGGGCTCCGGAAATTACTCGGGGGACATCAGTGATGACCTGTTCCGCTTTGCCTTTGAGTCCATCACTAACGTCATTTTTGGGGAGCGCCAGGGGATGCTGGAGGAAGTAGTGAACCCCGAGGCCCAGCGATTCATTGATGCCATCTACCAGATGTTCCACACCAGCGTCCCCATGCTCAACCTTCCCCCAGACCTGTTCCGTCTGTTCAGGACCAAGACCTGGAAGGACCATGTGGCTGCATGGGACGTGATTTTCAGTAAAGCTGACATATACACCCAGAACTTCTACTGGGAATTGAGACAGAAAGGAAGTGTTCACCACGATTACCGTGGCATGCTCTACAGACTCCTGGGAGACAGCAAGATGTCCTTCGAGGACATCAAGGCCAACGTCACAGAGATGCTGGCAGGAGGGGTGGACACGACGTCCATGACCCTGCAGTGGCACTTGTATGAGATGGCACGCAACCTGAAGGTGCAGGATATGCTGCGGGCAGAGGTCTTGGCTGCGCGGCACCAGGCCCAGGGAGACATGGCCACGATGCTACAGCTGGTCCCCCTCCTCAAAGCCAGCATCAAGGAGACACTAAGACTTCACCCCATCTCCGTGACCCTGCAGAGATATCTTGTAAATGACTTGGTTCTTCGAGATTACATGATTCCTGCCAAGACACTGGTGCAAGTGGCCATCTATGCTCTGGGCCGAGAGCCCACCTTCTTCTTCGACCCGGAAAATTTTGACCCAACCCGATGGCTGAGCAAAGACAAGAACATCACCTACTTCCGGAACTTGGGCTTTGGCTGGGGTGTGCGGCAGTGTCTGGGACGGCGGATCGCTGAGCTAGAGATGACCATCTTCCTCATCAATATGCTGGAGAACTTCAGAGTTGAAATCCAACACCTCAGCGATGTGGGCACCACATTCAACCTCATTCTGATGCCTGAAAAGCCCATCTCCTTCACCTTCTGGCCCTTTAACCAGGAAGCAACCCAGCAGTGATCAGAGAGGATGGCCTGCAGCCACATGGGAGGAAGGCCCAGGGGTGGGGCCCATGGGGTCTCTGCATCTTCAGTCGTCTGTCCCAAGTCCTGCTCCTTTCTGCCCAGCCTGCTCAGCAGGTTGAATGGGTTCTCAGTGGTCACCTTCCTCAGCTCAGCTGGGCCACTCCTCTTCACCCACCCCATGGAGACAATAAACAGCTGAACCATCG
【0136】
ウシadrenodoxin 遺伝子DNA配列
coding region: 165-722
mature pepyide: 339-680
(配列番号6)
TGGCGAACGCATCAGGCCCTCTAGGGAACCCCGGCCCTGAGAGTCCGCTGCGCCTCTCTTCTCTCCGCAGACCCGCGCGGCGTCAGCCGCCCGTACTCGCTGAGTCTGCCCTCGCGGCCGCTGCCGTGCGGCCTGCGTCGCCTTCCGTAGTTCCCCGACAGGCTATGGCCGCACGCCTCCTGCGCGTCGCCTCCGCCGCCCTCGGCGACACGGCCGGCCGGTGGCGGCTTCTCGTGCGACCGCGCGCGGGAGCCGGCGGCCTCCGTGGGAGCCGAGGACCGGGCCTGGGCGGCGGCGCAGTGGCGACGCGGACGCTGAGCGTATCGGGGCGAGCGCAGAGCAGCTCAGAAGATAAAATAACAGTCCACTTTATAAACCGTGATGGTGAAACATTAACAACCAAAGGAAAAATTGGTGACTCTCTGCTAGATGTTGTGGTTCAAAATAATCTAGATATTGATGGTTTTGGTGCATGTGAGGGAACCTTGGCTTGTTCTACCTGTCACCTCATCTTTGAACAGCACATATTTGAGAAATTGGAAGCAATCACTGATGAGGAGAATGACATGCTTGATCTGGCATATGGACTAACAGATAGATCGCGGTTGGGCTGCCAGATCTGTTTGACAAAGGCTATGGACAATATGACTGTTCGAGTACCTGATGCCGTGTCTGATGCCAGAGAGTCCATTGATATGGGCATGAACTCCTCAAAGATAGAATAAATAGGAATATTTTCACAACATTTTACCCTATTTTTATAATTATTATTTCTTAATGTAATTAAATGAGAACATGGATGAATGGATTTATCATTATGACTAGATTTACTAAATTCATCTGATACACTGCTGAATTTTGTAGTTCTGAAAGTATCGCCATTTTTATTTTGATTAAATTATAAAAACAAATATTAGAAGTAGTTAGTATGAAAAACCTTATATATTTTACCTGTGATTGATCAGCAACATAAATCTTACGCCTACCTACCTATAAGTAAGTAGGTTAAAAAAGGGTGTCATTATCTCTCTTGGATGTGGGTGGAGGCAGAGATGTAAGGCCTTGTCTAGGGTCATACTGCTAATTAGAAGGTCAGTACTAGAACTGTACATGTTTAGGTCGAAACTGGGGAAATTAGGGATCTCATTTAAAATCTGAGCACAGATAATAAAGCACTGTCAAAGATGTTTGGTCTCAATTCTGACTGAGGTGGAGGAGTAAACTAAAAATTAAATGTGTTTTTTTTTTTTTTAATCTTGATCAGAATGGTCTGTCTTTGTAATCTTTAAAAAGTAATGCTTATTTATAAATTTCTCCCTGTGACAGAATGGATATCTTACGGATTAGGTTTTCTCTGTCCTAAGAAGGGAACGAGGGGGCTACCATCCCCTCGGCTAGGACAGCCACGTGGTGTGCAGTGAGTATGTAGGCAGGGTCTGGAGGACGGGGGGCTTGAGCTTAAACACATCCATGCATGGCAGATATTTGTATTTTATCTTTATTGTGAATAAACTGATAAATAATAAT
【0137】
ウシadrenodoxin 還元酵素遺伝子 DNA配列
coding region: 52-1527
mature peptide: 148-1527
(配列番号7)
CGCGGGGCAGCCGGCAGAGTCTGGTCCCTGGGCTGGTTCCTCTTCCCAAGCATGGCTCCGCGCTGTTGGCGCTGGTGGCCCTGGTCGTCTTGGACTCGGACTCGGCTGCCTCCTTCCAGGAGCATCCAGAACTTCGGCCAGCACTTCTCCACACAGGAGCAGACCCCCCAGATCTGTGTGGTGGGCAGTGGCCCAGCTGGCTTTTACACGGCCCAGCACCTGCTAAAGCACCACTCCCGGGCCCACGTGGATATCTACGAGAAACAGCTGGTGCCCTTCCGCCTGGTGCGCGTTTGGCTGGCGCTGACCACCCCGAGGTCAAGAATGTTACTCAACACCTTTACCCAGACGGCCCGCTCTGACCGCTGTGCCTTCTATGGCAACGTGGAGGTGGGCAGGGATGTGACTGTGCAGGAGCTGCGGGTGTATCGCCTAACCGCCGTGGTGCTGAGCTATGGGGCAGAGGACCATCAGGCCCTGGATATCCCTGGTGAGGAGTTGCCCGGCGTGTTCTCGGCCCGGGCCTTTGTGGGCTGGTACAATGGGCTTCCTGAGAACCGGGAGCTGGCCCCGGACCTGAGCTGTGACACAGCCGTGATTCTGGGGCAGGGGAATGTGGCTCTGGACGTGGCCCGGATCCTGCTGACCCCCCCCGACCACCTGGAGAAAACGGACATCACTGAGGCCGCCCTGGGAGCCCTGAGACAGAGTCGGGTGAAGACGGTGTGGATCGTGGGCCGACGTGGACCCCTACAAGTGGCCTTCACCATAAAGGAGCTTCGGGAGATGATTCAGTTACCAGGAACTCGGCCCATGTTGGATCCTGCGGATTTCTTGGGTCTCCAGGACAGAATCAGGGAGGCCGCTCGCCCGAGGAAGCGGCTGATGGAACTGCTGCTTCGAACAGCCACGGAGAAGCCAGGGGTGGAGGAGGCTGCCCGCCGGGCATCAGCCTCCCGTGCCTGGGGCCTCCGCTTCTTCCGAAGCCCGCAGCAGGTCCTGCGCCTGCCAGATGGGCGGGCGCGACGATCCGCCTGGCAGTCACCAGAACTGGAGGGCATTGGAGAGGCACACCCGGGCAGTGCCCACTGGGGATGTGGAGGACCTCCCTGTGGGCTGGTGCTGAGCAGCATTGGGTATAAGAGCCGCCCCATCGACCCCAGTGTGCCCTTTGACCCCAAGCTCGGGGTTGTCCCCAATATGGAGGGCCGGGTTGTGGATGTGCCAGGCCTCTACTGCAGCGGCTGGGTGAAGCGGGGACCCACAGGTGTCATCACCACCACCATGACCGACAGCTTCCTCACCGGCCAGATTCTGCTACAGGACCTGAAGGCCGGGCACCTGCCGTCTGGCCCCAGGCCGGGCTCTGCATTCATCAAGGCCCTGCTGGACAGCCGAGGGGTCTGGCCCGTGTCTTTCTCGGACTGGGAGAAACTGGATGCTGAGGAGGTGTCCCGGGGCCAGGCCTCGGGGAAGCCCAGAGAGAAGCTGCTGGATCCTCAGGAGATGCTGCGGCTGCTGGGGCACTGAGCCTAGATCCCAGCCCCGCTGGTTGCAGAGAGAAGAGGAGGGTGAGCCCATGATCCCAGCCCAGCTCAGAGAAGAGAGGAGGCGCGCTGGACAGCGGACGAGAGCGTGCGGGGTCAGCCTGAGCGGGACTCTGCGACCCCAGCTGCGTCGTCTGCCCGTGCCTGGCATACACCTGCCCTGGCTGCCTCTTCTCCAGGGGCGGTGGGAGCGACTTTCTGGAGCTAGGTCACTGCTGCCAGTGTGGGTACCTTTCACAAGGAGATAACCTTAGTTAGGGATGGAGGCAGGTACAGGCTGACCTCCGTCCCTCCTGTCTCTCTCCTGCTGGACTGTGGAGGGTCCCCAGGTCAGGAATATGCTGGAAATAAAGCACCTGCCACCTAG
【0138】
参考例2:CYP11A発現大腸菌による各種基質の変換反応
4-コレステン-3-オン(4Chol)、コレステロール(Chol)、7-デヒドロコレステロール(7-DHC)、エルゴステロール(Ergo)、β-シトステロール(Sito)、スティグマステロール(Stigma)、カンペステロール(Cam)およびデスモステロール(Des)、(20S)-20-ヒドロキシコレスト-4-エン-3-オン(20-OH)、(22R)-22-ヒドロキシコレスト-4-エン-3-オン(22-OH)、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン(R-diol)、(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オン(S-diol)、ラノステロール(Lano)を基質としてそれぞれ別々に実施例6と同様に反応し、反応液を酢酸エチル1mlで分液し反応物および基質を抽出しHPLC分析した結果、各基質に対応する側鎖開裂体のプロゲステロン、プレグネノロンまたは7-デヒドロプレグネノロンの生成を確認した。変換率はそれぞれ0%、2.6%、0.7%、0%、0.8%、0%、1.0%、1.2%、0%、0.4%、4.3%および0%であり、基質の残存はそれぞれ100%、99%、99.2%、100%、99.7%、100%、99.1%、99.6%、100%、99.7%、95%および100%であった。
【0139】
また、 (20S)-20-ヒドロキシ-コレステロール、(22S)-22-ヒドロキシ-コレステロール、(22R)-22-ヒドロキシ-コレステロール、(20R, 22R)-20,22-ジヒドロキシ-コレステロールを基質としてそれぞれ別々に実施例6と同様に反応後、処理分析した結果、各基質に対応する側鎖開裂体のプレグネノロンの生成を確認した。蓄積量はそれぞれ16.3mg/l(変換:16.3%)、6.1mg/l(変換:6.1%)、28.0mg/l(変換:28.0%)、32.3mg/l(変換:32.3%)であった。また、添加した基質はそれぞれ98.8%、92.6%、78.6%、51.6%残留していた。
【0140】
各基質に対する変換反応率のまとめを以下の表6に示す。
【0141】
【表6】
【0142】
実施例13:SDS-PAGE
BL21CYP11A、BP172及びBP215を、アンピシリン50μg/ml(終濃度)を添加したTB培地2 mlに各々植菌し、30℃、16時間、220 rpmで振とう培養した。この前培養液250μlを本培養用TB培地2 5 ml(アンピシリン50μg/ml〈終濃度〉、5-アミノレブリン酸40μg/ml〈終濃度〉、誘導剤:Overnight Express Autoinduction System1〈メルク社製〉)に加え、25℃で24時間振とう培養した。この培養液から遠心分離により回収した菌体を緩衝液(50 mM KPB、2%グリセロール[pH 7.4]) 5 mlに懸濁し、菌体懸濁液とした。この菌体懸濁液2 mlに×10 Bug Buster 66.7μl (Novagen社製)、Benzonase 0.67μl (Novagen社製)、40 mg/ml Lysozymeを添加し、30℃で20分間振とうして溶菌させた。 この菌体溶菌液を全タンパク質液とした。また、菌体溶菌液を遠心分離後、遠心上清を可溶性画分タンパク質液とした。不溶性画分タンパク質は菌体溶菌液を遠心分離後、遠心上清を全て除去し、緩衝液で3回洗浄後、再度緩衝液に懸濁して調製した。これらの全タンパク質液、可溶性画分タンパク質液、不溶性画分タンパク質液はSDS-PAGE用サンプルバッファー(BIO-RAD社製)20μlと各々混合し、SDS-PAGE泳動用サンプルとした。SDS-PAGE泳動用サンプルは95℃、5 min加熱後、ポリアクリルアミドゲル(第一化学薬品社製)で電気泳動した(300V、30mA、60分)。電気泳動後、ポリアクリルアミドゲルをトレイに移し、染色−脱色液(タカラバイオ社製)を適量添加し、染色及び脱色を行った。結果を図16に示す。
【図面の簡単な説明】
【0143】
【図1】図1は、CYP204A1遺伝子を発現するBacillus megateriumの菌体を用いて4-Cholesten-3-oneをProgesteroneに変換したことを示すHPLCチャートを示す。
【図2】図2は、本発明の酵素の至適pHを検討した結果を示す。
【図3】図3は、本発明の酵素の至適温度を検討した結果を示す。
【図4】図4は、本発明の酵素のpH安定性試験の結果を示す。
【図5】図5は、本発明の酵素の温度安定性試験の結果を示す。
【図6】図6は、本発明の酵素(CYP204A1)の基質濃度と酵素活性の関係を示す。
【図7】図7は、本発明の酵素(CYPSS)の基質濃度と酵素活性の関係を示す。
【図8】図8は、本発明の酵素(CYP204A1)のLineweaver-Burk plotを示す。
【図9】図9は、本発明の酵素(CYPSS)のLineweaver-Burk plotを示す。
【図10】図10は、BP215株による4-コレステン-3-オンの変換反応のHPLCを示す。
【図11】図11は、BP215株による4-コレステン-3-オンの変換反応のMSを示す。
【図12】図12は、BP215株によるコレステロールの変換反応のHPLCを示す。
【図13】図13は、BP215株によるコレステロールの変換反応のMSを示す。
【図14】図14は、BP215株による7-デヒドロコレステロールの変換反応のHPLCを示す。
【図15】図15は、BP215株による7-デヒドロコレステロールの変換反応のMSを示す。
【図16】図16は、BL21CYP11A、BP172及びBP215を用いてSDS-PAGEを行った結果を示す。
【技術分野】
【0001】
本発明は、ステロール側鎖の20位と22位の結合を切断して医薬又は医薬中間体として産業上有用な化合物であるプレグネノロン、プロゲステロン又は7−デヒドロプレグネノロンに変換する活性を有する酵素、該酵素をコードするDNA、該DNAをベクターに組み込んで得られる形質転換体、並びに該形質転換体を用いるプレグネノロン、プロゲステロン、7−デヒドロプレグネノロンの製造方法、該酵素に対する抗体などに関する。
【背景技術】
【0002】
プレグネノロン、プロゲステロン、7−デヒドロプレグネノロンは、医薬および医薬中間体として産業上有用な化合物である。プレグネノロンの既存の製造法としては、コレステロール、ジオスゲニンやスティグマステロールといった天然由来のステロール化合物を原料とし、複数の有機合成反応により合成される(非特許文献1)。しかし、これらの手法は、工程数が多く、目的生産物の収率が低く、工業的生産方法としては問題点がある。一方、生化学的な手段としては、動物由来の酵素side chain cleavage cytochrome P450(P450scc)によるステロール変換方法が想定されるものの、動物由来酵素は酵素活性が低く、また、宿主となる細胞は培養に高価な培地が必要となり増殖速度が遅いなど、その生産方法は確立されていない(非特許文献2)。
さらに、ウシ由来のP450sccをサッカロミセス・セレビシエに形質転換し、電子伝達系を構成するアドレノドキシンおよびアドレノドキシンレダクターゼ存在下で、コレステロールからプレグネノロンに変換する試みも行われているが、生産性は非常に低く工業レベルの生産性は得られていない(特許文献1)。
【0003】
【非特許文献1】J. Org. Chem., 1979, 44, 1583
【非特許文献2】Proc. Natl. Acad. Sci. USA, 1988, 85, 1988
【特許文献1】特許第2963711号公報
【発明の開示】
【発明が解決しようとする課題】
【0004】
本発明が解決しようとする課題は、産業上有用なステロイドホルモン生合成の第一段階を触媒する重要な酵素であるP450scc酵素について高活性なP450scc酵素を取得すること、並びに取得したP450scc酵素を用いてステロイドホルモン合成に必要なプレグネノロン、プロゲステロン、及び7−デヒドロプレグネノロンを安価かつ簡便に生産するための方法を提供することである。
【課題を解決するための手段】
【0005】
本発明者らは、上記課題を解決するためにステロール側鎖の20位と22位の結合を切断することによるプレグネノロン等の製造方法について鋭意検討した結果、ステロール化合物側鎖の20位と22位の結合を切断する活性を有する酵素を、初めて微生物からクローニングすることに成功した。本発明者らは、該酵素をコードする遺伝子の形質転換体を作製し、組換え菌を用いて該遺伝子を発現させた該酵素形質転換体細胞、該形質転換体処理物および/または培養液を、原料となるステロール化合物に作用させることにより、高濃度で目的物であるプレグネノロン、プロゲステロン、7−デヒドロプレグネノロンを得ることができることを見出した。本発明はこれらの知見に基づいて完成したものである。
【0006】
即ち、本発明によれば、以下の発明が提供される。
1) 以下の理化学的性質を有するステロール側鎖切断酵素。
(1)作用:下記式(I)で表されるステロールに作用し、ステロール側鎖部分の20位と22位の炭素−炭素結合を切断する活性により該結合を切断して下記式(II)で表される化合物を生成する。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が10%以上であり、かつコレステロールからプレグネノロンへの変換率が10%以上である。
【化1】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【0007】
2) 以下の理化学的性質を有する、1)に記載のステロール側鎖切断酵素。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が50%以上であり、かつコレステロールからプレグネノロンへの変換率が50%以上である。
3) さらに以下の理化学的性質を有する、1)又は2)に記載の酵素。
(3)至適pH:7.5〜8.0
(4)作用至適温度:15〜20℃
(5)熱安定性:20℃で140時間保存後に30%以上の酵素活性が保持される。
(6)分子量:アミノ酸配列からの推定分子量が53〜54KDaであり、SDS電気泳動による測定で50〜56KDa
【0008】
4) さらに以下の理化学的性質を有する、1)から3)の何れかに記載の酵素。
(7)反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上である。
(8)基質濃度を10μMから100μMに上昇させた場合の活性の低下率が35%以下である。
【0009】
5) 微生物に由来する酵素である、1)から4)の何れかに記載の酵素。
【0010】
6) 以下の(a)、(b)または(c)のいずれかのタンパク質;
(a)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b)配列番号2に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質、
(c)配列番号2に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質。
【0011】
7) ステロール側鎖の20位と22位の結合を切断する活性について、基質濃度を10μMから100μMに上昇させた場合の活性の低下率が30%以下である、6)に記載のタンパク質。
【0012】
8) 6)又は7)に記載のタンパク質をコードするDNA。
9) 以下の(a)、(b)または(c)のいずれかのDNA;
(a)配列番号1に記載の塩基配列を有するDNA、
(b)配列番号1に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA、
(c)配列番号1に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA。
【0013】
10) 8)または9)に記載のDNAを有する組み換えベクター。
11) 8)または9)に記載のDNAあるいは10)に記載の組換えベクターを宿主細胞に導入した形質転換体。
12) 宿主細胞が大腸菌、放線菌またはシュードモナス属微生物である11)に記載の形質転換体。
【0014】
13) 6)または7)に記載のタンパク質、あるいは11)または12)に記載の形質転換体を含む、ステロール側鎖の20位と22位の結合を切断する反応を触媒するための酵素組成物。
14) 11)または12)に記載の形質転換体により産生される、6)または7)に記載のタンパク質。
【0015】
15) 1)から5)の何れかに記載の酵素あるいは6)、7)または14)に記載のタンパク質に対する抗体又はその部分フラグメント。
16) モノクローナル抗体である、15)に記載の抗体又はその部分フラグメント。
17) 1)から5)の何れかに記載の酵素あるいは6)、7)または14)に記載のタンパク質が有するステロール側鎖の20位と22位の結合を切断する活性を中和する作用を有する、15)または16)に記載の抗体又はその部分フラグメント。
【0016】
18) 11)又は12)に記載の形質転換体を培養することを特徴とする、6)又は7)に記載のタンパク質の製造方法。
【0017】
19) 式(I)で表されるステロールに、下記(i)から(vi):
(i) 1)から5)の何れかに記載の酵素、
(ii) 該酵素を産生する微生物またはその処理物、
(iii) 6)、7)または14)に記載のタンパク質、
(iv) 11)または12)に記載の形質転換体またはその処理物、
(v) 以下の(a)、(b)または(c)のいずれかのタンパク質、又は
(vi) 以下の(a)、(b)または(c)のいずれかのタンパク質をコードするDNA、以下の(d)、(e)または(f)のいずれかのDNA、もしくは上記DNAを有する組み換えベクターを宿主細胞に導入した形質転換体またはその処理物;
の何れかを反応させることを特徴とする、式(II)で表される化合物の製造方法。
(a)配列番号26に記載のアミノ酸配列からなるタンパク質、
(b)配列番号26に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質、
(c)配列番号26に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質、
(d)配列番号4に記載の塩基配列を有するDNA、
(e)配列番号4に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質をコードするDNA、
(f)配列番号4に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質をコードするDNA。
【化2】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、
母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【0018】
20) 式(I)で表されるステロールが、コレステロール、4-コレステン-3-オン、7-デヒドロコレステロール、エルゴステロール、β-シトステロール、スティグマステロール、カンペステロール、デスモステロール、 (20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、又は(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンである、19)に記載の方法。
21) 式(II)で表される化合物が、プレグネノロン、プロゲステロン又は7−デヒドロプレグネノロンである、19)又は20)に記載の方法。
【発明の効果】
【0019】
本発明により、ステロール化合物側鎖の20位と22位の結合を切断する活性を有する新規酵素が提供される。本発明の酵素を用いることによって、医薬および医薬中間体として産業上有用な化合物であるプレグネノロン、プロゲステロン、及び7−デヒドロプレグネノロンを効率的に製造することが可能になった。一般に動物由来のP450scc酵素は、微生物を宿主として発現させると不溶性タンパク質となり活性体として本タンパク質を得ることは困難であるが、本発明により初めて提供される微生物由来のP450scc酵素は、宿主として微生物を用いて発現させた場合に可溶性タンパク質となり容易に活性体として本タンパク質を得られることから、本発明の酵素の産業上の有用性は高い。
【発明を実施するための最良の形態】
【0020】
以下、本発明の実施の形態について詳細に説明する。
(1)本発明の酵素
本発明の酵素は、以下の理化学的性質を有するステロール側鎖切断酵素である。
(1)作用:下記式(I)で表されるステロールに作用し、ステロール側鎖部分の20位と22位の炭素−炭素結合を切断する活性により該結合を切断して下記式(II)で表される化合物を生成する。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは70%以上)であり、かつコレステロールからプレグネノロンへの変換率が10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは65%以上)である。
【化3】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【0021】
式(I)は、好ましくは、コレステロール、4-コレステン-3-オン、7-デヒドロコレステロール、エルゴステロール、β-シトステロール、スティグマステロール、カンペステロール、デスモステロール、 (20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンであり、より好ましくは、コレステロール、4-コレステン-3-オン、7-デヒドロコレステロールである。
【0022】
【化4】
【0023】
式(II)は、好ましくは、プレグネノロン、プロゲステロン、7-デヒドロプレグネノロンである。
【化5】
【0024】
上記の通り、本発明の酵素について、4-コレステン-3-オンからプロゲステロンへの変換反応率は10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは70%以上)であり、かつコレステロールからプレグネノロンへの変換率は10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは65%以上)である。他の基質の変換率は特に限定されるものではないが、好ましい変換率を以下に示す。
【0025】
7-デヒドロコレステロールから7−デヒドロプレグネノロンへの変換率は、10%以上(好ましくは20%以上、より好ましくは25%以上)である。
エルゴステロールから7−デヒドロプレグネノロンへの変換率は、1%以上(好ましくは2%以上)である。
β-シトステロールからプレグネノロンへの変換率は、10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは40%以上)である。
スティグマステロールからプレグネノロンへの変換率は、1%以上(好ましくは2%以上、より好ましくは5%以上)である。
カンペステロールからプレグネノロンへの変換率は、10%以上(好ましくは20%以上、より好ましくは25%以上)である。
デスモステロールからプレグネノロンへの変換率は10%以上(好ましくは30%以上、より好ましくは50%以上、さらに好ましくは60%以上、特に好ましくは65%以上)である。
(20S)-20-ヒドロキシコレスト-4-エン-3-オンからプロゲステロンへの変換率は10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは40%以上)である。
(22R)-22-ヒドロキシコレスト-4-エン-3-オンからプロゲステロンへの変換率は10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは40%以上)である。
(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オンからプロゲステロンへの変換率は10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは35%以上)である。
(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンからプロゲステロンへの変換率は10%以上(好ましくは20%以上、より好ましくは30%以上、さらに好ましくは40%以上)である。
ラノステロール(Lano)の変換率は0%である。
【0026】
本発明の酵素は、好ましくは、さらに以下の理化学的性質を有していてもよい。
(3)至適pH:7.5〜8.0
(4)作用至適温度:15〜20℃
(5)熱安定性:20℃で140時間保存後に30%以上の酵素活性が保持される。
(6)分子量:アミノ酸配列からの推定分子量が53〜54KDaであり、SDS電気泳動による測定で50〜56KDa
【0027】
本発明の酵素は、好ましくは、さらに以下の理化学的性質を有していてもよい。
(7)反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上、好ましくは45mmol/分/mol以上である。
(8)基質濃度を10μMから100μMに上昇させた場合の活性の低下率が35%以下である。
【0028】
VmaxおよびKmは、基質濃度を0.1〜500μMで4-コレステン-3-オンの変換を行い基質濃度と酵素活性の関係を調べることによって求めることができる。また、基質濃度を10μMから100μMに上昇させた場合の活性の低下率も、基質濃度を0.1〜500μMで4-コレステン-3-オンの変換を行い基質濃度と酵素活性の関係を調べることによって求めることができる。具体的には、反応液は1 mlで220 pmolのCYP204A1もしくは250 pmolのCYPSS、96μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、2 mM NADHと2 mM NADPHとなるように調製し、Tris-HClでpH 7.5にする。基質である4-コレステン-3-オンは終濃度が0.1, 0,5, 1, 2, 5, 10, 20, 50, 100, 500μMとなるようにDMSOに溶かしたものを20μl加える。反応はNADHとNADPHの添加により開始し、200 rpm、15℃で60分間行うことができる。
【0029】
本発明の酵素の由来は、特に限定されるものではないが、好ましくは、微生物に由来する酵素であり、例えば、Sphingomonas subterranea (例えば、Sphingomonas subterranea NBRC16086など)、又はNovosphingobium aromaticivorans (例えば、Novosphingobium aromaticivorans ATCC 700278など)などから取得することができる。
【0030】
(2)本発明の酵素の取得方法
本発明の酵素の、例えば、上記したSphingomonas subterranea又はNovosphingobium aromaticivoransなどの微生物から、通常の酵素の抽出、精製方法によって得ることができる。抽出方法として具体的には、例えばハサミ等による細片化、ホモジナイズ、音波処理、浸透ショック法、凍結融解法等の細胞破砕による抽出、界面活性剤抽出等や、これらの組み合わせ等の処理操作が挙げられる。また、精製方法として具体的には、例えば硫酸アンモニウム(硫安)や硫酸ナトリウム等による塩析、遠心分離、透析、限外濾過法、吸着クロマトグラフィー、イオン交換クロマトグラフィー、疎水性クロマトグラフィー、逆相クロマトグラフィー、ゲル濾過法、ゲル浸透クロマトグラフィー、アフィニティークロマトグラフィー、電気泳動法、ザイモグラフィー等や、これらの組合わせ等の処理操作が挙げられる。
【0031】
また、本発明の酵素の遺伝子を公知の方法に従ってクローニングし、適当な宿主に導入して、発現させることにより本発明の酵素 を得ることができる。例えば、本明細書に記載した本発明の酵素をコードする遺伝子の塩基配列の情報に基づいて本発明の酵素の遺伝子に特異的なプローブ又はプライマーを設計し、当該プローブ又はプライマーを用いてSphingomonas subterranea又はNovosphingobium aromaticivoransなどの微生物のDNAライブラリー(例えば、ゲノムDNAライブラリー、又はcDNAライブラリーなど)から本発明の酵素をコードするDNAを単離又は増幅し、これを遺伝子組換え技術によりベクターに入れ、宿主細胞に導入し、そこで発現させることにより、本発明の酵素を得ることができる。
【0032】
(3)本発明のタンパク質
本発明によれば、以下の(a)、(b)または(c)のいずれかのタンパク質が提供される。
(a)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b)配列番号2に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質、
(c)配列番号2に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質。
【0033】
好ましくは、本発明のタンパク質は、ステロール側鎖の20位と22位の結合を切断する活性について、基質濃度を10μMから100μMに上昇させた場合の活性の低下率が35%以下である。
【0034】
本発明において、「1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列」における「1もしくは数個」とは、例えば、1から20個程度を意味し、好ましくは1から10個程度を意味し、さらに好ましくは1から5個程度を意味する。
【0035】
本発明のタンパク質の取得方法としては、下記(4)に記載の本発明のDNAを適当な方法により転写/翻訳する方法が好ましく用いられる。具体的には、適当な発現用ベクター若しくは適当なベクターに適当なプロモーターとともに挿入した組換えベクターを作製し、この組換えベクターで適当な宿主微生物を形質転換したり、適当な培養細胞に導入することにより発現させ、これを精製することにより取得することができる。
【0036】
(4)本発明のDNA
本発明によれば、上記した本発明のタンパク質をコードするDNAが提供される。
本発明のDNAの具体例としては、以下の(a)、(b)または(c)のいずれかのDNAが挙げられる。
(a)配列番号1に記載の塩基配列を有するDNA、
(b)配列番号1に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA、
(c)配列番号1に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA。
【0037】
本発明の酵素をコードする遺伝子は、Sphingomonas subterranea及びNovosphingobium aromaticivoransからクローニングされたものであるが、本発明によりその塩基配列が決定されたので、この塩基配列に基づいて合成することも可能である。また、この塩基配列に基づいて作製したオリゴヌクレオチドをプローブとするハイブリダイゼーションによって、又は前記配列に基づいて作製したオリゴヌクレオチドをプライマーとするPCRによって、Sphingomonas subterranea及びNovosphingobium aromaticivoransなどの微生物のDNAライブラリー(例えば、ゲノムDNAライブラリー、又はcDNAライブラリーなど)から単離することもできる。また、本発明の酵素の遺伝子は、天然の微生物から分離されたもののみならず、通常用いられるDNA合成装置、例えばアプライド・バイオシステムズ(Applied Biosystems)社製394DNA/RNAシンセサイザーを用いて合成されたものであってもよい。
【0038】
本発明において、「1もしくは数個の塩基が欠失、置換および/または付加された塩基配列」における「1もしくは数個」とは、例えば、1から30個程度を意味し、好ましくは1から20個程度を意味し、さらに好ましくは1から10個程度を意味し、さらに好ましくは1から5個程度を意味する。
【0039】
本発明において、「ストリンジェントな条件下でハイブリダイズすることができる塩基配列」とは、配列番号1に記載の塩基配列あるいはその相補配列を有するDNAとBLAST解析で80%以上、好ましくは90%以上、さらに好ましくは95%以上の相同性を有する塩基配列を含むDNA等が挙げられる。また、ストリンジェントな条件下のハイブリダイゼーションとは、通常のハイブリダイゼーション緩衝液中で、温度が40〜70℃、好ましくは60〜65℃等で反応を行い、塩濃度が15mM〜300mM、好ましくは15mM〜60mM等の洗浄液中で洗浄を行う方法に従って行うことができる。
【0040】
酵素活性を実質的に害さないアミノ酸の欠失、置換および/または付加は、当業者であれば容易に選択することができる。また、酵素活性に実質的に影響を与えないアミノ酸の欠失、置換および/または付加を有する本発明の酵素をコードするDNAは、例えば、自然突然変異株又は変種から取得することができる。
【0041】
配列番号1に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有する遺伝子の作製は、突然変異剤を用いる方法や部位特異的変異法等の通常の変異操作によって得ることができる。これらは、例えばサイトダイレクテドミュータジェネシスキット(宝酒造)や、クイックチェンジサイトダイレクテッドミュータジェネシスキット(ストラタジーン社製)等の市販キットで容易に行うことができる。
【0042】
(5)本発明の組み換えベクター、及び形質転換体
本発明によればさらに、本発明のDNAを有する組み換えベクターが提供される。組み換えベクターを調製する際には、通常、本発明のDNAを宿主微生物に適したプロモーターとともに、このプロモーターの下流に本発明のDNAのコード領域の5'末端側が連結されるようにして、ベクターに挿入する。あるいはプロモーターを含む発現ベクターを用い、これに本発明のDNAを挿入してもよい。
【0043】
発現ベクターとしては、宿主微生物内で複製増殖可能であれば特に制限されるものではないが、プラスミドベクター、シャトルベクターおよびファージベクターが挙げられる。具体的なプラスミドベクターとしては、pBR322、pUC18、pHSG298、pUC118、pSTV28、pTWV228、pHY300PLK(以上のプラスミドベクターは、例えば宝酒造(株)から購入できる)、pKK223−3、pPL−ラムダインデューシブルエクスプレッションベクター(Lambda Inducible Expression Vector)(以上は、例えばファルマシア社から購入できる)、大腸菌−コリネ型細菌のシャトルベクターとしては、例えば、特開平3−210184号公報に記載のプラスミドpCRY30;特開平2−276575号公報に記載のプラスミドpCRY21、pCRY2KE、pCRY2KX、pCRY31、pCRY3KEおよびpCRY3KX;特開平1−191686号公報に記載のプラスミドpCRY2およびpCRY3;特開昭58−67679号公報に記載のpAM330;特開昭58−77895号公報に記載のpHM1519;特開昭58−192900号公報に記載のpAJ655、pAJ611およびpAJ1844;特開昭57−134500号公報に記載のpCG1;特開昭58−35197号公報に記載のpCG2;特開昭57−183799号公報に記載のpCG4およびpCG11等、あるいはこれらの誘導体等を挙げることができる。
また、ファージベクターとしては、(λFixIIベクタ−(Stratagene社から購入できる)等を挙げることができる。
【0044】
本発明の酵素をコードするDNAを発現させるためのプロモーターは、宿主微生物が保有するプロモーターを一般に用いることができるが、それに限られるものではなく、本発明の酵素の遺伝子の転写を開始させるための原核生物由来の塩基配列であればいかなるプロモーターであっても良い。具体的には、ラクトースオペロンのプロモーター、トリプトファンオペロンのプロモーター、λファージ由来のPLプロモーター、トリプトファンラクトース雑種(tac)プロモーター[H. A. Bose et al., Proc. Natl. Acad. Sci. U. S. A., Vol.80, p.21 (1983)]等が挙げられる。これらのプロモーターのうち、発現効率を向上させる目的で、誘導性のあるプロモーターを使用することもできる。例えば、上記ラクトースオペロンのプロモーターの場合には、ラクトースやイソプロピル−β−D−チオガラクトシド(IPTG)を添加することにより遺伝子発現を誘導することができる。
【0045】
本発明によれば、さらに本発明のDNA又は組換えベクターを宿主細胞に導入した形質転換体が提供される。本発明のDNA又は組み換えベクターを導入する宿主としては、特に限定されるものではないが、エシェリヒア(Escherichia)属細菌(例えば、大腸菌)、放線菌(Actinomycetes)属細菌、バチルス(Bacillus)属細菌、セラチア(Serratia)属細菌、シュードモナス(Pseudomonas)属細菌、コリネバクテリウム(Corynebacterium)属細菌、ブレビバクテリウム(Brevibacterium)属細菌、ロドコッカス(Rhodococcus)属細菌、ラクトバチルス(Lactobacillus)属細菌、ストレプトマイセス(Streptomyces)属細菌、サーマス(Thermus)属細菌、ストレプトコッカス(Streptococcus)属細菌等を好適に用いることができる。好ましくは、宿主細胞は大腸菌、放線菌またはシュードモナス属微生物である。
【0046】
具体的には、エシェリヒア・コリ(Escherichia coli)、バチルス・サチリス(Bacillus subtilis)、バチルス・ブレビス(Bacillus brevis)、バチルス・ステアロサーモフィラス(Bacillus stearothermophilus)、セラチア・マーセッセンス(Serratia marcescens)、シュードモナス・プチダ(Pseudomonas putida)、シュードモナス・アエルギノサ(Pseudomonas aeruginosa)、コリネバクテリウム・グルタミカム(Corynebacterium glutamicum)、ブレビバクテリウム・フラバム(Brevibacterium flavum)、ブレビバクテリウム・ラクトファーメンタム(Brevibacterium lactofermentum)、ロドコッカス・エリスロポリス(Rhodococcus erythropolis)、サーマス・サーモフィラス(Thermus thermophilus)、ストレプトコッカス・ラクティス(Streptococcus lactis)、ラクトバチルス・カゼイ(Lactobacillus casei)、ストレプトマイセス・リビダンス(Streptomyces lividans)等を用いることができる。
【0047】
上記宿主微生物への遺伝子の導入法としては、コンピテントセル法[Journal of Molecular Biology, Vol.53, p.159 (1970)]、パルス波通電法[J. Indust.Microbiol., Vol.5, p.159 (1990)]等による形質転換法、ファージを用いた形質導入法[E. Ohtsubo, Genetics, Vol.64, p.189 (1970)]、接合伝達法[J. G. C. Ottow, Ann. Rev.Microbiol., Vol.29, p.80 (1975)]、細胞融合法[M.H. Gabor, J. Bacteriol., Vol.137, p.1346 (1979)]等を用いることができる。これらの方法から、宿主微生物に適した方法を適宜選択すればよい。
【0048】
上記のような発現ベクターを用いた発現方法の他に、プロモーターを連結した本発明の酵素をコードするDNAを、宿主微生物の染色体中に直接導入する相同組換え技術あるいはトランスポゾンや挿入配列等を用いて導入する技術によっても発現させることができる。従って、本発明の形質転換体は、本発明の酵素が発現していればよく、遺伝子の導入の方法は限定されない。
【0049】
(6)本発明の形質転換体を用いた本発明の酵素の製造
本発明によれば、上記(5)のようにして得られる形質転換体を培養し、その培養物から本発明の酵素を採取することができる。
形質転換体の培養は、炭素源、窒素源、無機塩、各種ビタミン等を含む通常の栄養培地で行うことができ、炭素源としては、例えばブドウ糖、ショ糖、果糖、麦芽糖等の糖類、エタノール、メタノール等のアルコール類、クエン酸、リンゴ酸、コハク酸、マレイン酸、フマル酸等の有機酸類、廃糖蜜等が用いられる。窒素源としては、例えばアンモニア、硫酸アンモニウム、塩化アンモニウム、硝酸アンモニウム、尿素等がそれぞれ単独もしくは混合して用いられる。また、無機塩としては、例えばリン酸一水素カリウム、リン酸二水素カリウム、硫酸マグネシウム等が用いられる。この他にペプトン、肉エキス、酵母エキス、コーンステイープリカー、カザミノ酸、ビオチン等の各種ビタミン等の栄養素を培地に添加することができる。
【0050】
培養は、通常、通気撹拌、振とう等の好気条件下で行う。培養温度は、宿主微生物の生育し得る温度であれば特に制限はなく、また、培養途中のpHについても宿主微生物が生育し得るpHであれば特に制限はない。培養中のpH調整は、酸またはアルカリを添加して行うことができる。
【0051】
培養物からの酵素の採取は、酵素の活性を指標として公知の採取方法により行うことができる。酵素は必ずしも均一にまで精製される必要はなく、用途に応じた精製度まで精製すればよい。
【0052】
本発明で用いられる粗精製画分又は精製酵素としては、形質転換体を培養した培養液から分離した菌体はもちろんのこと、培養液、菌体を超音波、圧擦等の手段で破砕して得られる破砕物、該破砕物を水等で抽出して得られる、本発明の酵素 を含有する抽出物、該抽出物に更に硫安塩析、カラムクロマトグラフィー等の処理を行って得られる本発明の酵素の粗酵素標品又は精製した酵素標品を使用してもよい。さらに、上記菌体、破砕物、抽出物、粗精製画分または精製酵素を担体に固定化したものも使用することができる。
【0053】
これら菌体、菌体破砕物、抽出物または精製酵素の固定化は、それ自体既知の通常用いられている方法に従い、アクリルアミドモノマー、アルギン酸、またはカラギーナン等の適当な担体に菌体等を固定化させる方法により行うことができる。例えば、菌体を担体に固定化する場合には、培養物から回収されたまま、あるいは適当な緩衝液、例えば0.02〜0.2M程度のリン酸緩衝液(pH6〜10)等で洗浄された菌体を使用することができる。
【0054】
(7)本発明の抗体
本発明によれば、本発明の酵素又はタンパク質に対する抗体又はその部分フラグメントが提供される。本発明の抗体は、ポリクローナル抗体又はモノクローナル抗体の何れでもよく、好ましくはモノクローナル抗体である。
【0055】
本発明の抗体の調製方法としては、通常用いられる公知の方法を用いることができ、抗原として用いられるポリペプチドについても、公知の方法に従って抗原性が高くエピトープ(抗原決定基)として適した配列を選択して用いることができる。上記の抗原として用いるポリペプチドは、公知の方法に従って合成した合成ペプチドでも、また本発明の酵素又はタンパク質そのものを用いることもできる。抗原となるポリペプチドは、公知の方法に従って適当な溶液等に調製して、哺乳動物、例えばウサギ、マウス、ラット等に免疫を行えばよいが、安定的な免疫を行ったり抗体価を高めるために抗原ペプチドを適当なキャリアタンパク質とのコンジュゲートにして用いたり、アジュバント等を加えて免疫を行うのが好ましい。
【0056】
免疫に際しての抗原の投与経路は特に限定されず、例えば皮下、腹腔内、静脈内、あるいは筋肉内等のいずれの経路を用いてもよい。具体的には、例えばBALB/cマウスに抗原ポリペプチドを数日〜数週間おきに数回接種する方法等が用いられる。また、抗原の摂取量としては、抗原がポリペプチドの場合0.3〜0.5mg/1回程度が好ましいが、ポリペプチドの種類、また免疫する動物種によっては適宜調節される。
【0057】
免疫後、適宜試験的に採血を行って固相酵素免疫検定法(以下、これを「ELISA法」と称することがある)やウエスタンブロッティング等の方法で抗体価の上昇を確認し、十分に抗体価の上昇した動物から採血を行う。これに抗体の調製に用いられる適当な処理を行えばポリクローナル抗体を得ることができる。具体的には、例えば、公知の方法に従い血清から抗体成分を精製した精製抗体を取得する方法等が挙げられる。抗体成分の精製は、遠析、イオン交換クロマトグラフィー、アフィニティークロマトグラフィー等の方法を用いることができる。
【0058】
また、該動物の脾臓細胞とミエローマ細胞とを用いて公知の方法に従って融合させたハイブリドーマを用いる(Milstein,et al.,Nature,256, 495(1975))ことによりモノクローナル抗体を作製することもできる。モノクローナル抗体は、例えば以下の方法により取得することができる。
【0059】
まず、上記した抗原の免疫により抗体価の高まった動物から抗体産生細胞を取得する。抗体産生細胞は、形質細胞、及びその前駆細胞であるリンパ球であり、これは個体の何れから取得してもよいが、好ましくは脾臓、リンパ節、末梢血等から取得する。これらの細胞と融合させるミエローマとしては、一般的にはマウスから得られた株化細胞、例えば8−アザグアニン耐性マウス(BALB/c由来等)ミエローマ細胞株であるP3X63-Ag8.653(ATCC:CRL-1580)、P3-NS1/1Ag4.1(理研セルバンク:RCB0095)等が好ましく用いられる。細胞の融合は、抗体産生細胞とミエローマ細胞を適当な割合で混合し、適当な細胞融合培地、例えばRPMI1640やイスコフ改変ダルベッコ培地(IMDM)、あるいはダルベッコ改変イーグル培地(DMEM)等に、50%ポリエチレングリコール(PEG)を溶解したもの等を用いることにより行うことができる。また電気融合法(U. Zimmer- mann. et al., Naturwissenschaften,68, 577(1981))によっても行うことができる。
【0060】
ハイブリドーマは、用いたミエローマ細胞株が8−アザグアニン耐性株であることを利用して適量のヒポキサンチン・アミノプテリン・チミジン(HAT)液を含む正常培地(HAT培地)中で5%CO2、37℃で適当時間培養することにより選択することができる。この選択方法は用いるミエローマ細胞株によって適宜選択して用いることができる。選択されたハイブリドーマが産生する抗体の抗体価を上記した方法により解析し、抗体価の高い抗体を産生するハイブリドーマを限界希釈法等により分離し、分離した融合細胞を適当な培地で培養して得られる培養上清から硫安分画、アフィニティクロマトグラフィー等の適当な方法により精製してモノクローナル抗体を得ることができる。また精製には市販のモノクローナル抗体精製キットを用いることもできる。さらには、免疫した動物と同系統の動物、またはヌードマウス等の腹腔内で上記で得られた抗体産生ハイブリドーマを増殖させることにより、本発明のモノクローナル抗体を大量に含む腹水を得ることもできる。
【0061】
また、一価性抗体等の部分フラグメントは、例えばパパイン等を用いてFab部分とFc部分を切断し、アフィニティカラム等を用いてFab部分を回収することによって作製することができる。
【0062】
かくして得られる本発明の酵素又はタンパク質と特異的に結合する抗体は、本発明の酵素又はタンパク質に特異的に結合することによって、該酵素又はタンパク質が有するステロール側鎖の20位と22位の結合を切断する活性を阻害する中和抗体として用いることもできる。
【0063】
(8)本発明の酵素を用いたプレグネノロン、プロゲステロン、7−デヒドロプレグネノロンの製造方法
本発明によれば、上記した式(I)で表されるステロールに、本発明の酵素、該酵素を産生する微生物またはその処理物、本発明のタンパク質、本発明の形質転換体またはその処理物、あるいは配列番号26に記載のアミノ酸配列からなるタンパク質(CYP204A1)又はその変異体、当該タンパク質をコードするDNAもしくは上記DNAを有する組み換えベクターを宿主細胞に導入した形質転換体またはその処理物を適切な電子伝達系存在下で反応させることによって、式(II)で表される化合物を製造することができる。本願の酵素の電子伝達系とは、還元型ニコチンアミドアデニンジヌクレオチド(NADH)または還元型ニコチンアミドアデニンジヌクレオチドリン酸(NADPH)、及び2種の蛋白質により構成される。すなわち、NAD(P)H−フェレドキシン酸化還元活性を有するフェレドキシンレダクターゼ、および本願酵素への電子伝達活性を有するフェレドキシンである。従って、適切な電子伝達系とは様々なフェレドキシンレダクターゼとフェレドキシンの組み合わせによる本願酵素活性の検討の結果、反応の進行が確認されたフェレドキシンレダクターゼとフェレドキシンの組み合わせを指す。フェレドキシンレダクターゼは、例えば以下の中から選択される;ホウレン草由来のフェレドキシンレダクターゼ、シュードモナス プチダ(Pseudomonas putida)由来プチダレドキシンレダクターゼ、動物由来のアドレノドキシンレダクターゼ、スフィンゴモナス サブテラニア(Sphingomonas subterranea)由来フェレドキシンレダクターゼ、ノボスフィンゴビウム アロマティシボランス(Novosphingobium aromaticivorans)由来フェレドキシンレダクターゼ、エシェリヒア コリ(Escherichia coli)由来フラボドキシンレダクターゼやフェレドキシンレダクターゼ、またはその他のフェレドキシンレダクターゼ。フェレドキシンは、例えば以下の中から選択される;ホウレン草由来のフェレドキシン、シュードモナス プチダ(Pseudomonas putida)由来プチダレドキシン、動物由来のアドレノドキシン、スフィンゴモナス サブテラニア(Sphingomonas subterranea)由来フェレドキシン、ノボスフィンゴビウム アロマティシボランス(Novosphingobium aromaticivorans)由来フェレドキシン、エシェリヒア コリ(Escherichia coli)由来フラボドキシンやフェレドキシン、またはその他のフェレドキシン型蛋白質。
【0064】
反応の方法は特に限定されず、本発明の酵素又は該酵素を産生する微生物などを含む液体に、基質となる式(I)で表されるステロールを加え、適当な温度(例えば、10℃から40℃程度)で反応させることができる。これにより、プレグネノロン、プロゲステロン、又は7-デヒドロプレグネノロンなどの式(II)で表される化合物を製造することができる。あるいは本発明の酵素を産生する微生物が,内在的に式(I)で表されるステロールを産生する場合でも,これを反応させて式(II)で表される化合物を製造することができる。
【0065】
また、上記の方法で得られた式(II)で表される化合物に、17α−ステロイド水酸化酵素、21−ステロイド水酸化酵素、11β−ステロイド水酸化酵素を適当な温度で反応させて、ハイドロコルチゾンを製造することができる。
【0066】
さらに、本発明の酵素を産生する微生物(内在的に式(I)で表されるステロールを産生する微生物が好ましい)に、17α−ステロイド水酸化酵素、21-ステロイド水酸化酵素、11β-ステロイド水酸化酵素の遺伝子(Molecular and Cellular Endocrinology 1990, 73, 73-80)を導入してこれらの蛋白質(酵素)を発現する能力を与え、この形質転換体を用いてハイドロコルチゾンを製造することもできる。
【0067】
反応混合物から目的の式(II)で表される化合物を分画する方法は特に限定されず、当業者に公知の分離又は精製のための手法を用いることができる。例えば、溶媒抽出、晶析、樹脂吸着、カラムクロマトグラフィー等により行うことができるが、これに限定されるものではない。
【0068】
以下の実施例により本発明をさらに詳細に説明するが、本発明は実施例によって限定されるものではない。
【実施例】
【0069】
以下の実施例において一般的な実験方法はSambrookらの実験書(Sambrook, J., and Russel, D. W.: Molecular cloning: a laboratory manual. 3rd edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 2001)に従った。
【0070】
実施例1:Sphingomonas subterranea NBRC16086からのP450遺伝子のクローニング
Sphingomonas subterranea NBRC 16086をL培地(1%トリプトン、0.5%酵母エキス、0.5%NaCl、0.1%グルコース、pH7.2 )に接種して30℃で一晩培養して得られた菌体からゲノムDNAを抽出した。ゲノムDNAの抽出にはDNA抽出キットISOPLANT (和光純薬工業社製)を用いた。このゲノムDNAよりプライマーCYP-1F: 5'-CGAGATCTGATGAACTTCATGGGCCAGCGCAT-3' (配列番号8)およびプライマーCYP-2R: 5'-CGAGATCTGCAGCGGTGGACACCCCCGCCAAA-3' (配列番号9)を用いてシトクロムP450遺伝子をポリメラーゼ連鎖反応(PCR) を行って増幅した。
この時のPCRの条件は以下のとおりである。
【0071】
(反応液組成)
滅菌精製水 15μl
2倍濃縮GC buffer I (タカラバイオ社製) 25μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2.5 mM) 8μl
プライマーCYP-1F (50 p mol/μl) 0.5μl
プライマーCYP-2R (50 p mol/μl) 0.5μl
S. subterranea NBRC 16086 ゲノムDNA (100 ng/μl) 0.5μl
LA Taq ポリメラーゼ (5 units/μl,タカラバイオ社製) 0.5μl
【0072】
(温度条件)
94℃/3分、(94℃/30秒, 55℃/30秒, 72℃/90秒)30サイクル、72℃/5分
【0073】
この反応の結果、増幅した1.0 kb (キロベースペア)の遺伝子断片の塩基配列を決定した。この塩基配列情報を基にインバースPCRを行ってシトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含む1.8 kb DNA領域の全塩基配列:配列番号3)を決定した。また、配列番号3の1番目から1419番目の塩基配列を配列番号1に示し、配列番号1に記載の塩基配列によりコードされるアミノ酸配列を配列番号2に示す。
【0074】
BP215, CYP ORF, Fdx ORF
CYP ORF:1-1419
Fdx ORF: 1430-1762
【0075】
(配列番号3)
ATGGCAAGTGCAGCAGCGGGTGCAGACGGCCTTCCCTTGCTCGATGGCGGCGTGCCGCTGCTCGGGCACCTCGCCCAGTTCTTTCGCGATCCGGTTTCCGTCCTGAAGCGCGGCTATCGCTCGAAGGGCCGGCTCTTCGCGATGAACTTCATGGGCCAGCGCATGAACGTGATGCTGGGGCCGGAACACAACCGCTTCTTCTTCGAGGAAACCGACAAGCTGCTCTCGATCCGAGAGTCGATGCCGTTCTTCCTCAAGATGTTCTCGCCCGAGTTCTACTCGTTTGCCGAGATGGACGAATACCTGCGCCAGCGCGCGATCATCATGCCCCGGTTCAAGGCGGCATCGATGAAGCAGTACGTGCCGGTGATGGTCGAGGAATCGCTGAACCTCGTCGAACGGCTGGGTGAGGAAGGCGAGTTCGACCTGATCCCGACGCTGGGCCCCGTGGTAATGGACATCGCCGCGCACAGCTTCATGGGGCGCGAATTCCACGAGAAGCTTGGCCACGAATTCTTCGAGCTGTTCCGCGACTTCTCGGGCGGCATGGAGTTCGTGCTGCCGCTGTGGCTGCCGACACCAAAGATGGTGAAAAGCCAGCGCGCCAAGAAGAAGCTCCACGCGATCCTGCAATCGTGGATCGACAAGCGCCGCGCCAGTCCGCTCGATCCGCCCGACTTCTTCCAGACGATGATCGAGACGAAGTATCCCGATGGCCGCGCCGTGCCCGACGAGATCATCCGGCACCTGATCCTGCTGCTGGTCTGGGCGGGCCACGAAACCACCGCCGGGCAAGTCAGCTGGGCGCTGGCCGACCTCCTCCAGAACCCCGGCTACCAGCAGGTGCTGCGCGAAGAGGTCGCCTCGCTGCTCGGCGGCAGCGACGGGCGCGCACTGGGCTGGGAGCAGGCCGTGGCGATGGAGAAGATGGACCTCGCCCTGCGCGAGACAGAGCGGCTCCACCCCGTCGCCTACATGCTCAGCCGCAAGGCCAGTGCCGATATCGAGCGCGACGGCTATCGCATCCGCAAGGGCGAATTCGTGCTGCTCGCGCCCTCGGTCAGCCATCGCATGGAAGAAACCTTCCGCAATGCCGATGCCTATGACCCCGAGCGCTTCAACCCGCAGAACCCGGACGCGCAGATCGAGAGCAACTCGCTGATCGGCTTCGGCGGCGGGGTGCATCGCTGTGCGGGGGTAAACTTCGCGCGCATGGAAATGAAGGTGCTGGTGGCGATCCTGCTGCAGAACTTCGACATGGAGCTGATCGACGAAGTGCGCCCCATCGCCGGCGCATCGACCTACTGGCCCGCCCAGCCCTGCCGCGTGCGCTACAAGCGGCGCAAGCTGGATGGCGCGGGTGGAAGCGCGGACGTGGCGGCGCTCGCCAAGGCGGCCGGTTGCCCGGCCCATGCATGAGGCGCACTGAATGGCCAAGGTGACTTTCGTCCAGCCGGACGGCTCGGCGCGGACTTGCGTGAACTTCGAGGGCATGACCCTGATGCAGCTCGCGGTCGGCAACCTTGTCGACGGGATCGACGCGCTGTGCGGCGGCATGATGCAGTGCGCCACCTGCCATTGCTACATCGACCCGGACTGGCTTGACCGCACCGGCCCCGCCCGGCCGGAAGAACGCGAGATGCTCGAGGCCATCGATGGCGTCGAGATCCGCCCGAACAGCCGCCTGTCCTGCCAGGTCCAGCTTGGCGAGGAGTTGGACGGGCTGGTGGTGCACATTCCGGCGGAGCAACCGGGAGTTTAG
【0076】
実施例2:Sphingomonas subterranea NBRC 16086由来シトクロムP450遺伝子の大腸菌発現株BP215の作成
実施例1の塩基配列を基にしてシトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含むDNA領域をPCRで増幅した。プライマーCYP-3F: 5'-CCCCATATGGCAAGTGCAGCAGCGGGTGCAGACGG-3' (配列番号10) およびプライマーCYP-4R: 5'-GCACTAGTCTAAACTCCCGGTTGCTCCGG-3' (配列番号11)を用いた。この時のPCRの条件は以下のとおりである。
【0077】
(反応液組成)
滅菌精製水 31.5 μl
10倍濃縮 PCR buffer (東洋紡社製) 5 μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2 mM) 5 μl
25 mM MgSO4 2 μl
プライマーCYP-3F (50 p mol/μl) 1 μl
プライマーCYP-4R (50 p mol/μl) 1 μl
S. subterranea NBRC 16086 ゲノムDNA (100 ng/μl) 1 μl
KOD plus ポリメラーゼ (1 units/μl,東洋紡社製) 1 μl
DMSO 2.5 μl
【0078】
(温度条件)
95℃/5分、(98℃/30秒, 60℃/30秒, 68℃/2分)30サイクル、68℃/10分
【0079】
この反応の結果増幅した、シトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含む1.8 kb DNA断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50μl 中で制限酵素Nde IおよびSpe Iで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.8 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製した。この断片を大腸菌発現ベクターpT7NS-camAB(有澤および久米田、国際公開公報2003/087381パンフレット)のNde IおよびSpe I部位にT4 DNAリガーゼを用いて連結して、大腸菌DH5aに形質転換することで、プラスミドpCYPSS-camABを構築した。このプラスミドを大腸菌BL21(DE3)に形質転換して得られた菌株をBP215と命名した。
【0080】
実施例3:Novosphingobium aromaticivorans ATCC 700278からのシトクロムP450遺伝子(CYP204A1遺伝子)のクローニングと大腸菌発現株BP172の作成
Novosphingobium aromaticivorans ATCC 700278をL培地に接種して30℃で2日間培養して得られた菌体からゲノムDNAを抽出した。ゲノムDNAの抽出にはDNA抽出キットISOPLANT (和光純薬工業社製)を用いた。このゲノムDNAよりプライマーCYP-5F: 5'-GCCCATATGGCAAGAGCTGCGACTGCGGCC-3' (配列番号12) およびプライマーCYP-6R: 5'-GCACTAGTCTAAACTCCCGGTTGCTCCGG-3' (配列番号13)を用いてシトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含む断片を増幅した。この時のPCRの条件は以下のとおりである。
【0081】
(反応液組成)
滅菌精製水 15μl
2倍濃縮GC buffer I (タカラバイオ社製) 25μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2.5 mM) 8μl
プライマーCYP-5F (50 p mol/μl) 0.5μl
プライマーCYP-6R (50 p mol/μl) 0.5μl
N. aromaticivorans ATCC 700278ゲノムDNA (100 ng/μl) 0.5μl
LA Taq ポリメラーゼ (5 units/μl,タカラバイオ社製) 0.5μl
【0082】
(温度条件)
94℃/3分、(98℃/20秒, 63℃/30秒, 68℃/2分)30サイクル、72℃/5分
【0083】
この反応の結果増幅した、シトクロムP450遺伝子およびその下流に存在するフェレドキシン遺伝子を含む1.8 kb DNA断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50μl 中で制限酵素Nde IおよびSpe Iで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.8 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製した。この断片を大腸菌発現ベクターpT7-camABのNde IおよびSpe I部位にT4 DNAリガーゼを用いて連結して、大腸菌DH5aに形質転換することで、プラスミドpCYP204A1-camABを構築した。このプラスミドを大腸菌BL21(DE3)に形質転換して得られた菌株をBP172と命名した。CYP204A1遺伝子によりコードされるアミノ酸配列を配列番号26に示す。
【0084】
BP172, CYP ORF, Fdx ORF
CYP ORF: 1-1422
Fdx ORF: 1433-1765
(配列番号4)
ATGGCAAGAGCTGCGACTGCGGCCGGTAATGGCCTTCCCTTGCTCGATGGAGGCGTGCCGCTCCTCGGGCATCTCGCACAGTTCTTCCGCGATCCGGTTTCGGTACTCAAGCGCGGATACCGCTCGAAGGGGCGGCTCTTCGCGATGAACTTCATGGGCCAGCGCATGAACGTGATGCTGGGTCCGGAACACAACCGCTTCTTCTTCGAGGAGACGGACAAGCTGCTCTCGATCCGGGAGTCGATGCCGTTCTTCCTCAAGATGTTCTCGCCCGAGTTCTATTCGTTCGCGGAAATGGACGAGTACCTGCGCCAGCGCTCGATCATCATGCCCCGCTTCAAGGCGGCATCGATGAAGCAGTACGTGCCGGTCATGGTCGAGGAATCGCTTAACCTGGTCGAGCGGCTGGGCGAGGAAGGCGAGTTCGACCTGATCCCGACGCTGGGCCCGGTGGTAATGGACATCGCCGCGCACAGCTTCATGGGACGCGAGTTCCACGAGAAGCTGGGGCATGAGTTCTTCGAACTCTTCCGCGATTTTTCGGGAGGCATGGAATTCGTCCTGCCGCTGTGGCTGCCGACACCCAAGATGGTCAAGTCACAGCGCGCGAAGAGGAAGCTCCACGCCATCCTGCAATCGTGGATCGACAAGCGCCGCGCCGCCCCGCTCGATCCGCCCGATTTCTTCCAGACGATGATCGAGACGAAGTATCCCGATGGCCGCCCGGTGCCCGACGAGATCATCCGCCACCTGATCCTCCTTCTCGTCTGGGCAGGGCACGAGACGACCGCCGGGCAGGTGAGCTGGGCGCTGGCGGACCTCCTTCAGAACCCGGACTACCAGAAGGTGCTGCGCGGCGAGATATCGTCGCTGCTGGGCGGCAGCGACGGGCGCGACCTTGGCTGGGAACAGGCCGTGGCGATGGAGAAGATGGACCTTGCCCTGCGCGAGACCGAGCGGCTCCATCCGGTCGCCTACATGCTCAGCCGCAAGGCGCGGGCCGATATCGAGCGCGACGGCTATGTCATCCGCAAGGGCGAGTTCGTGCTGCTTGCGCCTTCGGTCAGCCACCGCATGGAAGAGACGTTCCGCAATCCCGATGCCTATGACCCGGAACGCTTCAACCCGGCCAACCCCGATGCGCAGATCGAAAGCAATTCGTTGATCGGCTTTGGCGGGGGTGTCCACCGCTGCGCGGGCGTGAACTTCGCGCGGATGGAGATGAAGGTGCTGGTGGCGATCCTGCTCCAGAACTTCGACATGGAGCTGATGGACGAAGTGCGGCCCATCGCGGGCGCATCGACCTACTGGCCCGCCCAGCCCTGCCGGGTGCGCTATCGGCGGCGCAAGCTCGACGGGTCGGAGGCAGGTGCGGACATGGCGGCGCTGGCCCGAGCCGCCGGCTGCCCGGCGCATACGTGAGGGAGGCCTGATGGCCAAGGTGACTTTCGTCCAGCCGGACGGATCGCAGCGAACCTGCGTGAACTTCGAAGGCATGACGTTGATGCAGCTCGCAGTGGGCAATCTCGTCGACGGGATCGACGCGCTGTGCGGCGGCATGATGCAGTGCGCGACCTGCCATTGCTGGATCGACCCCGAATGGATCGGCCGCACCGGCATGGCCGGACCCGATGAGCGGGCAATGCTGGAAGCGATCGAGGGCGTCGAGATCCGTCCCGAAAGCCGCCTGTCCTGCCAGGTACAGCTTGGCGAAGAACTTGACGGGCTGGTCGTGCGCATTCCACCGGAGCAACCGGGAGTTTAG
【0085】
実施例4: CYP204A1遺伝子のBacillus megaterium での発現
pCYP204A1-camABを鋳型にしてプライマーCYP-7F: 5'-CCGCTAGCGCAAGAGCTGCGACTGCGGCC-3'(配列番号14)およびCYP-8R: 5'-GCGGATCCCTAAACTCCCGGTTGCTCCGG -3'(配列番号15)を用いてCYP204A1遺伝子およびその下流に存在するフェレドキシン遺伝子を含む断片を増幅した。この時のPCRの条件は以下のとおりである。
【0086】
滅菌精製水 33 μl
10倍濃縮 PCR buffer (東洋紡社製) 25 μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2.0 mM) 8 μl
25 mM MgSO4 2 μl
プライマーCYP-7F (50 p mol/μl) 0.5 μl
プライマーCYP-8R (50 p mol/μl) 0.5 μl
DMSO 1.5 μl
pCYP204A1-camAB DNA (100 ng/μl) 0.5 μl
KOD plus ポリメラーゼ (1 units/μl,東洋紡社製) 1.0 μl
【0087】
(温度条件)
94℃/2分、(94℃/20秒, 55℃/30秒, 68℃/3分)20サイクル、72℃/5分
【0088】
この反応の結果増幅した、CYP204A1遺伝子およびその下流に存在するフェレドキシン遺伝子を含む1.8 kb DNA断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50 μl 中で制限酵素Nhe IおよびBam HIで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.8 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製した。この断片をBacillus megaterium発現ベクターpHW1520 (MoBiTec社製)のSpe IおよびBam HI部位にT4 DNAリガーゼを用いて連結して、大腸菌DH5aに形質転換することで、プラスミドpXYL204CBを構築した。このプラスミドをBacillus megateriumコンピテント細胞(MoBiTec社製)に形質転換して菌株Bacillus megaterium(pXYL204CB)を得た。この菌株を、テトラサイクリン10mg/ml(終濃度)を添加したTB培地2 mlに植菌し、37℃、16時間、220rpmで振とう培養した。この前培養液250μlをテトラサイクリン10mg/mlを添加したTB培地2 5 mlに加え、37℃で2.5時間振とう培養後、50 % Xylose 250μlを添加し、30℃、125 rpmで6時間振とう培養した。この培養液から遠心分離により回収した菌体を変換用緩衝液(50 mM KPB、2%グリセロール[pH 7.4]) 5 mlに懸濁し、菌体懸濁液とした。この菌体懸濁液2 mlに4-Cholesten-3-oneメタノール溶液(100 mg/ml) 2μlを加えて、振とう(220rpm)しながら30℃で24時間インキュベートした。その後、この反応液に2 mlの酢酸エチルを加えて、ボルテックスした後、遠心機で4,000 rpmで10分間遠心した。得られた酢酸エチル相1.5 mlを濃縮乾固し、その後200μlのメタノールに溶解しHPLC分析に供した。結果、24.5%の変換率で4Cholestene-3-oneをProgesteroneに変換した。
【0089】
HPLC条件
Column:Inertsil ODS-3 4.6 X 50 mm
flow rate:1.2ml/min
temp.:40℃
detect:PDA(240nm)
gradient: 0/ 4/ 5/5.5(min)
40/100/100/40(%B)
HPLCチャートを図1に示す。
【0090】
実施例5:酵素の諸性質
(1)発現ベクターの構築
CYP204A1、CYPSS、CYP11A遺伝子を含むDNA断片を増幅する為に以下のプライマーを作成した。
CYP204A1-1F GCCCATATGGCAAGAGCTGCGACTGCGGCC(配列番号16)
CYP204A1-2R GCCCTCGAGTCACGTATGCGCCGGGCAGCC(配列番号17)
CYPSS-1F GCCCATATGGCAAGTGCAGCAGCGGGTGCA(配列番号18)
CYPSS-2R GCCCTCGAGTCATGCATGGGCCGGGCAACC(配列番号19)
CYP11A-1F GCCCATATGGCTTCAACACGTAGTCCTCGC(配列番号20)
CYP11A-2R GCCCTCGAGTCACTGCTGGGTTGCTTCCTG(配列番号21)
【0091】
次に、これらのプライマーとpCYP204A1-camAB、pCYPSS-camAB、pCYP11A-ARXを鋳型としてKOD plus (TOYOBO)を用いてPCR反応を行った。この時のPCR条件は以下の通りである。
【0092】
(反応液組成)
10倍濃縮PCR buffer for KOD plus 5μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP 各2 mM) 5μl
25 mM MgSO4 2μl
Primer-1F (50 pmol/μl) 0.5μl
Primer-2R (50 pmol/μl) 0.5μl
KOD plus 1μl
DMSO 3.75μl
鋳型 (50 ng/μl) 0.5μl
滅菌精製水 31.75μl
【0093】
(温度条件)
95℃/3分、(98℃/20秒、55℃/30秒、72℃/2分)25サイクル、72℃/5分
【0094】
この反応の結果、CYP204A1、CYPSS、CYP11Aにおいて約1.4 kbpの大きさのDNA断片(以下、DNA断片-A, B, Cという)が増幅された。Wizard SV Gel and PCR Clean-Up System (PROMEGA)で精製し、NdeIとXhoIにより消化した。このサンプルをアガロースゲル電気泳動にかけて分画し、約1.4 kbpの大きさのDNA断片をアガロースゲルから切り出した。Wizard SV Gel and PCR Clean-Up SystemによりNdeIとXhoIにより消化されたDNA断片-A, B, Cを精製し、NdeIとXhoIにより消化したpET22bにDNA Ligation Kit ver 2.1 (TaKaRa)により連結し、プラスミドpET22-CYP204A1、pET22-CYPSS、pET22-CYP11Aを得た。
【0095】
(2)CYP204A1、CYPSS、CYP11A誘導菌体の作成
上記(1)で調製したプラスミドpET22-CYP204A1、pET22-CYPSS、pET22-CYP11Aを用い、大腸菌BL21 (DE3)を形質転換した。こうして得られた形質転換体を50μg/mlのカルベニシリンを含む1 mlのTB培地に植菌し、37℃で14時間培養した。この1 mlの培養液を50μg/mlのカルベニシリンとOvernight Express Autoinduction system 1 (Merck)を含む100 mlのTB培地に植菌し、25℃で24時間培養および目的タンパク質の誘導を行った。遠心により菌体を回収し、20 mlのCV buffer (50 mM KPB (Potassium phosphate buffer), 10% Glycerol, 1 mM EDTA, 2 mM dithiothreitol, 1 mM D-Glucose)に懸濁して-80℃で細胞懸濁液を保存した。
【0096】
(3)無細胞抽出液の調製
上記(2)で調製した細胞懸濁液20 mlに680μlのX10 Bug Buster (Novagen)と20μlのBenzonase (Novagen)と2 mlの40 mg/ml Lysozymeを加えて、30℃で30分間攪拌して細胞を破砕し、遠心により無細胞抽出液を得た。得られた約20 mlの無細胞抽出液を1 LのBuffer R (20 mM KPB pH 7.4, 20% glycerol)で2回透析し、その間に析出した沈殿を遠心により除いた。ここで上清として得られた無細胞抽出液を以後の実験に使用した。
【0097】
(4)一酸化炭素結合スペクトル解析によるP450の発現確認
得られた無細胞抽出液を2つに分けて片方に一酸化炭素をバブリングした。一酸化炭素バブリング有無の両方のサンプルにジチオナイトを加え、差スペクトルを測定した。CYP204A1とCYPSSでは450 nm付近に極大吸収を持つピークが得られ、P450の発現が確認されたが、CYP11Aでは得られず活性型のP450がサンプル中に存在しないことが示唆された。この結果より以後の実験はCYP204A1とCYPSSで行うこととした。
【0098】
(5)至適pHの検討
上記(3)で調製した無細胞抽出液を用いて、様々なpHで4-コレステン-3-オンの変換を行った。反応液は1 ml BufferR中で215 pmol/ml CYP204A1もしくは367 pmol/ml CYPSS、64μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、50μg/ml 4-Cholesten-3-one、2 mM NADHと2 mM NADPHとなるように調製し、1 mlの反応液中に様々なpHの200 mM Bufferを562μl加えてpHを調整した。反応液作成後にpHを測定するとKPB pH 6.2, KPB pH 6.7, KPB pH 7.1, KPB pH 7.4, KPB pH 7.9, Tris-HCl pH 7.5, Tris-HCl pH 8.2, Tris-HCl pH 8.7, TES pH 6.6, TES pH 6.8, TES pH 7.3, TES pH 7.5, TES pH 8.0であった。反応はNADHとNADPHの添加により開始し、200 rpm、30℃で14時間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。各pH条件における4-コレステン-3-オンからプロゲステロンへの酵素活性を図2(及び表1)に示した。酵素活性は1時間、1 mol P450あたりのプロゲステロン生成量gで示した。この結果より、CYP204A1とCYPSSはTris-HCl pH 7.5で最も活性が高いことが示された。
【0099】
(6)至適温度の検討
上記(3)で調製した無細胞抽出液を用いて、様々なpHで4-コレステン-3-オンの変換を行った。反応液は1 mlで215 pmol/ml CYP204A1もしくは391 pmol/ml CYPSS、64μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、50μg/ml 4-コレステン-3-オン、2 mM NADHと2 mM NADPHとなるように調製し、Tris-HClでpH 7.5にした。反応はNADHとNADPHの添加により開始し、200 rpm、温度条件をそれぞれ10, 15, 20, 25, 30℃として4時間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。各温度条件における4-コレステン-3-オンからプロゲステロンへの酵素活性を図3(及び表1)に示した。この結果より、CYP204A1は15から20℃で、CYPSSは15℃で最も活性が高いことが示された。
【0100】
【表1】
【0101】
(7)pH安定性試験
上記(3)で調製した0.5 mlの無細胞抽出液と様々なpHの0.5 mlの50 mM Bufferと混合してpHを測定して6℃で140時間保存した。酵素液のpHはAcB (acetate buffer) pH 6.0, KPB pH 6.7, KPB pH 7.2, KPB pH 7.6, TES pH 7.5, Tris-HCl pH 8.1, Tris-HCl pH 8.9であった。調製した様々なpHの無細胞抽出液を用いて4-コレステン-3-オンの変換を行った。酵素液を様々なpHのBufferで混ぜた直後と140時間保存したときに反応を行い、安定性を試験した。反応液は1 mlでCYP204A1もしくはCYPSSの各pHの酵素液、64 μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、50μg/ml 4-コレステン-3-オン、2 mM NADHと2 mM NADPHとなるように調製し、200 mM Tris-HCを562μl加えてpH 7.5にした。様々なpHの酵素液を加えても反応液がpH 7.5であることを確認した。反応はNADHとNADPHの添加により開始し、200 rpm、15℃で4時間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。0時間と140時間の4-コレステン-3-オンからプロゲステロンへの酵素活性を図4と表2に示した。この結果より、CYP204A1、CYPSSともにKPB pH 7.6で最も安定であることが示された。
【0102】
【表2】
【0103】
(8)温度安定性試験
上記(3)で調製した無細胞抽出液を様々な温度で140時間保存した。保存した温度は6, 10, 20, 30, 37, 45℃とした。保存前の酵素液と140時間保存後のサンプルを用いて4-コレステン-3-オンの変換を行い、安定性を試験した。反応液は1 mlで各温度で保存したCYP204A1もしくはCYPSS酵素液、64μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、50μg/ml 4-コレステン-3-オン、2 mM NADHと2 mM NADPHとなるように調製した。反応はNADHとNADPHの添加により開始し、200 rpm、15℃で4時間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。0時間と140時間の4-コレステン-3-オンからプロゲステロンへの酵素活性を図5と表2に示した。この結果より、CYP204A1、CYPSSともに保存温度が下がるに従って安定性が上昇することが示された。特に、20℃で140時間保存した場合、CYP204A1は47%の活性が残存し、CYPSSは39%の活性が残存した。
【0104】
【表3】
【0105】
(9)CYP204A1とCYPSSのKinetics解析
上記(3)で調製した無細胞抽出液を用いて、基質濃度を0.1〜500μMで4-コレステン-3-オンの変換を行い基質濃度と酵素活性の関係を調べた。反応液は1 mlで220 pmolのCYP204A1もしくは250 pmolのCYPSS、96μg/mlほうれん草由来フェレドキシン、0.1 U/mlほうれん草由来フェレドキシン還元酵素、3 U/mlグルコースデヒドロゲナーゼ、60 mMグルコース、2 mM NADHと2 mM NADPHとなるように調製し、Tris-HClでpH 7.5にした。4-コレステン-3-オンは終濃度が0.1, 0.5, 1, 2, 5, 10, 20, 50, 100, 500μMとなるようにDMSOに溶かしたものを20μl加えた。反応はNADHとNADPHの添加により開始し、200 rpm、15℃で60分間行った。1.5 ml酢酸エチルを添加して反応を止め、抽出を行い、再度、0.75 ml酢酸エチルで抽出を行った。得られた酢酸エチル相をエバポレーターにより乾固し、200μlメタノールで残渣を溶解した。HPLC分析によりプロゲステロンの検出を行った。基質濃度と酵素活性の関係を図6 (CYP204A1)と図7 (CYPSS)に示した。また、Lineweaver-Burk plotを作成して図8(及び表4) (CYP204A1)と図9(及び表4) (CYPSS)に示した。算出されたKinetics parametersを表5に示した。この結果より、CYP204A1とCYPSSのKm値は同等の1.0と1.1μMでVmaxは47.4と78.7 mmol/min/molとCYPSSの方が約1.7倍高いことが示された。また、図6と図7の比較によりCYPSSの方が基質阻害がかかりにくいことが示された。
【0106】
【表4】
【0107】
【表5】
【0108】
上記の結果の概要を以下に示す。
至適pH: pH 7.5 〜8.0(CYP204A1, CYPSSともに)
作用至適温度:15〜20℃ (CYP204A1, CYPSSともに)
pH安定性:中性付近で安定。もっとも安定だったのはKPB pH 7.6 (CYPSS, CYP204A1ともに)
熱安定性:20℃で140時間保存後に30%以上の酵素活性が保持される。(CYPSS, CYP204A1ともに)
Kinetics解析:Vmax CYPSSはCYP204A1に対して約1.7倍高い
Km値 CYPSSとCYP204A1はほぼ同等(1μM)
基質阻害:4-コレステン-3-オンにより、基質阻害がかかることが分かったがCYPSSの方がCYP204A1に比べて基質阻害がかかりにくい。4-コレステン-3-オンの濃度が10μMのときのCYPSSまたはCYP204A1の酵素活性にくらべ、100μMのときのそれぞれの酵素活性は90%または67%であった。すなわち、基質濃度を10μMから100μMに上昇させた場合の活性の低下率は10%または33%であった。
【0109】
実施例6:BP215株による4-コレステン-3-オンの変換反応(プロゲステロンの調製)
大腸菌発現BP215株をM9混合培地(Na2HPO4: 0.68%, KH2PO4: 0.3%, NaCl: 0.05%, NH4Cl: 0.01%, Casamino acid: 1%, D-Glucose: 0.4%, CaCl2: 0.1mM, MgCl2: 1mM, Thiamine: 0.002%, FeSO4: 0.1mM, Carbenicillin: 0.005%)2mlに植菌し、30℃で17時間振とう培養し前培養液を得た。この前培養液0.25mlをOvernight ExpressTM Autoinduction System(Novagen Cat. No. 71300-4)を用いた本培養培地(Na2HPO4: 0.68%, KH2PO4: 0.3%, NaCl: 0.05%, NH4Cl: 0.01%, Casamino acid: 1%, CaCl2: 0.1mM, MgCl2: 1mM, Thiamine: 0.002%, FeSO4: 0.1mM, Carbenicillin: 0.005%, Overnight ExpressTM溶液:7%, 5-Aminolevlic acid: 0.008%)25mlに植菌し、25℃で24時間振とう培養した。この本培養液5mlを遠心分離(3500rpm, 10分間)して得た菌体にCV2バッファー(glycerol: 2%, Carbenicillin: 0.005%, Isopropyl βD-1-thiogalactopyranoside: 0.1mMを含む50mM(pH)燐酸カリウム緩衝液)1mlを加えた後、4-Cholesten-3-oneの1%メタノール溶液0.01mlおよびMethyl-β-cyclodextrinの25%水溶液0.02mlを加え28℃で5時間振とうし反応した。反応終了後、メタノール4mlを加え良く混合し、遠心分離して上層をHPLCで分析した。保持時間2.6分にプロゲステロンのピークを検出し、プロゲステロン標品と保持時間およびMSスペクトル(m/z=315 M+1)が一致した。蓄積量は78.0 mg/l(変換:78%)であった。また、4-Cholesten-3-one(保持時間8.1分)は検出されなかった。HPLCを図10に示す、MSを図11に示す。
【0110】
分析条件:Chromolith speed rod RP18e(50x4.6mm)カラムで50%→100%CH3CNグラジエント(0〜3min) 100%CH3CN(3〜8min)を移動相とし流速2ml/min、CADTM(Charged Aerosol Detection)、MSで検出した。
【0111】
実施例7:BP215株によるコレステロールの変換反応
基質をコレステロールとして実施例6と同様に反応後、処理分析した結果、保持時間2.5分にプレグネノロンのピークを検出し、プレグネノロン標品と保持時間およびMSスペクトル(m/z=317M+1)が一致した。蓄積量は68.2 mg/l(変換:68.2%)であった。また、Cholesterol(保持時間8.5分)は3.7%残留していた。HPLCを図12に示す、MSを図13に示す。
【0112】
実施例8:BP215株による7-デヒドロコレステロール、エルゴステロールの変換反応
基質を7-デヒドロコレステロールとし実施例6と同様に反応後、処理分析した結果、保持時間2.1分に7-デヒドロプレグネノロンのピーク(m/z=315M+1)を検出した。蓄積量は34.3 mg/l(変換:34.3 %)であった。また、7-Dehydrochlesterol(保持時間7.5分)は54%残留していた。HPLCを図14に示す、MSを図15に示す。
【0113】
基質をエルゴステロールとし実施例6と同様に反応後、処理分析した結果、保持時間2.1分に7-デヒドロプレグネノロンのピークを検出した。蓄積量は2.8 mg/l(変換:2.8 %)であった。
【0114】
実施例9:BP215株による各種基質の変換反応
基質をβ-シトステロール、スティグマステロール、カンペステロールおよびデスモステロールとしてそれぞれ別々に実施例6と同様に反応後、処理分析した結果、保持時間2.5分にプレグネノロンのピークを検出し、プレグネノロン標品と保持時間およびMSスペクトル(m/z=317M+1)が一致した。蓄積量はそれぞれ40.9 mg/l(変換:40.9 %)、5.8 mg/l(変換:5.8 %)、27.9 mg/l(変換:27.9 %)、71.7 mg/l(変換:71.7 %)であった。また、添加した基質(保持時間はそれぞれ10.1、9.3、9.2、7.2分)はそれぞれ55.9%,70.8%,25.5%,21.3%残留していた。なお、β-シトステロールは生成物が微量であった為、反応液を酢酸エチルで抽出し5倍に濃縮して分析した。
【0115】
実施例10:(20R,22R)-20,22-ジヒドロキシ-4-コレスト-4-エン-3-オン、及び(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンの化学合成
20,22-ジヒドロキシコレスト-4-エン-3-オンの2つの立体異性体の合成は、文献既知の合成方法を用いて20,22-ジヒドロキシコレステロールを経由したのちに、常法の酸化反応を用いて実施した。
【0116】
(A)20,22-ジヒドロキシコレステロール(10-A)の合成
Steroids(2004),69,483/ B.Watanabe et al.に、化合物(10-A)の2つの異性体[(20R,22R)体および(20R,22S)体]の化学合成が記載されており、その実験方法に従って同様に(10-A)の合成を実施した。
【0117】
概要は、プレグネノロンを出発原料として、t-ブチルジメチルシリルクロライドで3位を保護基にして、4-メチル-1-ペンチンを付加させて(20R)-3β-(t-ブチルジメチルシリロキシ)コレスタ-5-エン-22-イン-20-オルを得た。これをLindlar触媒を用いてcis-オレフィンに還元して(20R,22Z)-3β-(t-ブチルジメチルシリロキシ)コレスタ-5,22-ジエン-20-オルとし、これをVO(acac)2/TBHPにより22,23-エポキシド(異性体混合物)に変換して、これを異性体混合物のまま分離せず、LiAlH4で還元して20,22-ジヒドロキシ体に変換して、TBAFで3位の脱保護をして、(20R,22RS)-20,22-ジヒドロキシコレステロール(10-A)を得た。
【0118】
(B)20,22-ジヒドロキシコレスト-4-エン-3-オン(10-B)の合成
前出文献方法を参考に、まず(10-A)から(20R,22RS)-20,22-O-イソプロピリデンジオキシコレステロールを得た[PPTS存在下、2,2-ジメトキシプロパンを作用]。
これを常法どおりOppenauer酸化により4-エン-3-オン体に変換した。この実験手順を具体的に記す。
【0119】
十分に窒素置換したDean-Stark装置内で、(20R,22RS)-20,22-O-イソプロピリデンジオキシコレステロール(3.0g)をトルエン100mLに溶解し、1-メチル-4-ピペリドン(8.1mL,10eq)を添加したのち、反応温度を130-140℃にして、Al(OiPr)3 を(2.7g,2eq) 添加して4hrかくはんした。反応終了後、ジエチルエーテルで希釈して常法処理したのち、シリカゲルクロマトグラフィー精製[ヘキサン/酢酸エチル=8/1〜5/1]を行い、(20R,22RS)-20,22-O-イソプロピリデンジオキシコレスト-4-エン-3-オンを得た(2.9g, y97%)。
最後に(20R,22RS)-20,22-O-イソプロピリデンジオキシコレスト-4-エン-3-オンを脱アセトニド保護を行うことで(10-B)へ変換され、さらに2つの立体異性体を別々に取得することが出来た。以下に具体的な実験手順を記す。
【0120】
(20R,22RS)-20,22-O-イソプロピリデンジオキシコレスト-4-エン-3-オン(2.9g)を、THF(20mL)+メタノール(20mL)中で室温でかくはんしながら、2N塩酸(1.0mL)および過塩素酸(0.8mL)を添加して、60hr反応した。クロロホルムで希釈して常法処理したのち、シリカゲルクロマトグラフィー精製をおこない[ヘキサン/酢酸エチル=7/1〜2/1]、脱保護生成物(1.48g, y47%)と、未反応原料(1.24g)を分離取得した。
脱保護生成物からは、ヘキサン/酢酸エチルで再結晶して、白色固体(1.1g)を得た。
【0121】
1H-NMR(400MHz,CDCl3) δ: 0.90(d,J=6.6Hz,26-H3),0.90(d,J=6.8Hz,27-H3),0.91(s,18-H3), 1.19(s,19-H3),1.26(s,21-H3),3.24(dd,J=9.4Hz,5.4Hz,22-H),5.73(s,4-H)
→ (20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オン(10-B)
[非天然型異性体; 本発明による新規化合物である。]
【0122】
回収した未反応原料(590mg)を、THF(15mL)+メタノール(15mL)中で室温で攪拌しながら、過塩素酸(1.4mL)を添加して10時間反応した。クロロホルムで希釈して常法処理したのち、シリカゲルクロマトグラフィー精製[ヘキサン/酢酸エチル=7/1〜2/1]を行い、脱保護生成物(217mg, y40%)を得た。
【0123】
1H-NMR(500MHz,CDCl3) δ: 0.89(d,J=6.3Hz,26-H3),0.91(d,J=6.4Hz,27-H3),0.93(s,18-H3), 1.19(s,19-H3),1.21(s,21-H3),3.38(d,J=8.4Hz,22-H),5.73(s,4-H)
→(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン(10-B)
[天然型異性体; (引用) Helv.Chim.Acta,(2006),89,813/W.Zhang et al. ]
【0124】
実施例11:BP215株による各種基質の変換反応
(20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンを基質としてそれぞれ別々に実施例6と同様に反応し、生成物をHPLC分析した結果、プロゲステロンの生成を確認した。変換率はそれぞれ41.0%、42.3%、38.1%および69.8%であった。また、添加した基質(保持時間はそれぞれ4.9、4.4、3.8、3.5分)はそれぞれ4.8%,2.7%,0.6%,0.6%残留していた。
【0125】
また、(20S) -20-ヒドロキシ-コレステロール、(22S) -22-ヒドロキシ-コレステロール、(22R)-22-ヒドロキシ-コレステロール、(20R, 22R) -20,22-ジヒドロキシ-コレステロールを基質としてそれぞれ別々に実施例6と同様に反応後、処理分析した結果、各基質に対応する側鎖開裂体のプレグネノロンの生成を確認した。蓄積量はそれぞれ89.2mg/L(変換:89.2 %)、37.5mg/L(変換:37.5%)、80.0mg/L(変換:80 %)、53.0mg/L(変換:53.0 %)であった。また、添加した基質はそれぞれ0%、23.5%、14.8%、0%残留していた。
【0126】
実施例12:BP172株を用いた各種基質の変換反応
4-コレステン-3-オン、コレステロール、7-デヒドロコレステロール、エルゴステロール、β-シトステロール、スティグマステロール、カンペステロール、デスモステロール、(20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンを基質としてそれぞれ別々に実施例6と同様に反応し生成物をHPLC分析した結果、各基質に対応する側鎖開裂体のプロゲステロン、プレグネノロンまたは7-デヒドロプレグネノロンの生成を確認した。変換率はそれぞれ84.0%、71.2%、28.5%、2.2%、47.2%、6.5%、31.3%、66.1%、63.9%、80.9%、61.3%および41.7%であった。
【0127】
また、(20S) -20-ヒドロキシ-コレステロール、(22S) -22-ヒドロキシ-コレステロール、(22R)-22-ヒドロキシ-コレステロール、(20R, 22R)-20,22-ジヒドロキシ-コレステロールを基質としてそれぞれ別々に実施例6と同様に反応後、処理分析した結果、各基質に対応する側鎖開裂体のプレグネノロンの生成を確認した。蓄積量はそれぞれ68.2mg/l (変換:68.2%)、12.9mg/l (変換:12.9%)、69.5mg/l(変換:69.5 %)、51.6mg/l(変換:51.6 %)であった。また、添加した基質はそれぞれ25.2%、60.1%、42.8%、0%残留していた。
【0128】
参考例1:CYP11A発現大腸菌の作成
市販のヒト睾丸cDNA(Origene Technologies, Inc.)を鋳型として、プライマーCYP11A-1F: 5'-CCGCATATGGCTTCAACACGTAGTCCTCGCCCCTTCAATGAGATC-3' (配列番号22) およびプライマーCYP11A-2R: 5'-GCACTAGTTCACTGCTGGGTTGCTTCCTGGTTAAA-3' (配列番号23)を用いてCYP11A1遺伝子をポリメラーゼ連鎖反応(PCR) を行って増幅した。
この時のPCRの条件は以下のとおりである。
【0129】
(反応液組成)
滅菌精製水 34μl
10倍濃縮 PCR buffer (東洋紡社製) 5μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2 mM) 5μl
25 mM MgSO4 2μl
プライマーCYP11A-1F (50 p mol/μl) 0.5μl
プライマーCYP11A-2R (50 p mol/μl) 0.5μl
ヒト睾丸cDNA (10 ng/μl) 2μl
KOD plus ポリメラーゼ (1 units/μl,東洋紡社製) 1μl
【0130】
(温度条件)
94℃/3分、(94℃/30秒, 55℃/60秒, 72℃/90秒)30サイクル、72℃/5分
【0131】
この反応の結果、増幅した1.45 kbのCYP11A1遺伝子断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50μl 中で制限酵素Nde IおよびSpe Iで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.45 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製した。この断片を大腸菌発現ベクターpT7NS-camABのNde IおよびSpe I部位にT4 DNAリガーゼを用いて連結して、大腸菌DH5αに形質転換することで、プラスミドpCYP11A-camABを構築した。次いで、このプラスミドのSpe IおよびBam HI部位に予め調製しておいたウシ由来アドレノドキシン還元酵素−アドレノドキシン遺伝子(adr-adx)断片をT4 DNAリガーゼを用いて挿入、連結して、大腸菌DH5αに形質転換することで、プラスミドpCYP11A-ARXを構築した。このプラスミドを大腸菌BL21(DE3)に形質転換して得られた菌株をBL21CYP11と命名した。この時用いたadx-adr断片はプラスミドpKARX (Sawada et. al. Eur. J. Biochem. 265, 950-956, 1999)を鋳型として、プライマーbAdxR-1F: 5'-GCACTAGTAAGGAAACAGACCATGTCCACACAGGAGCAGACCCCC-3'(配列番号24) およびプライマーbAdx-2R: 5'-CGTGATCAGGCATCAGACACGGCATCAGG-3'(配列番号25)を用いて以下の条件でポリメラーゼ連鎖反応(PCR) を行って増幅した。
【0132】
(反応液組成)
滅菌精製水 33μl
10倍濃縮 PCR buffer (東洋紡社製) 5μl
dNTP混合溶液 (dATP, dGTP, dTTP, dCTP各2 mM) 5μl
25 mM MgSO4 2μl
プライマーbAdxR-1F (50 p mol/μl) 1μl
プライマーbAdx-2R (50 p mol/μl) 1μl
pKARX DNA (10 ng/μl) 0.5μl
DMSO 1.5μl
KOD plus ポリメラーゼ (1 units/μl,東洋紡社製) 1μl
【0133】
(温度条件)
94℃/3分、(94℃/30秒, 60℃/30秒, 72℃/120秒)25サイクル、72℃/5分
【0134】
この反応の結果、増幅した1.85 kbのadr-adx遺伝子断片をQIAquick PCR purification Kit (キアゲン社製)を用いて精製した。このDNA溶液を50μl 中で制限酵素Spe IおよびFba Iで消化したのち、0.8%アガロースゲルにて電気泳動した。泳動後、ゲルから切り出した1.85 kb DNA断片を含むゲル切片から同断片をQIAquick Gel Extraction Kit (キアゲン社製)をもちいて回収・精製して、上述のpCYP11A-ARX構築に使用した。
【0135】
CYP11A1のDNA配列
CYP11A1, human
Coding region; 45-1610
Mature peptide; 162-1610
(配列番号5)
GGGCGCTGAAGTGGAGCAGGTACAGTCACAGCTGTGGGGACAGCATGCTGGCCAAGGGTCTTCCCCCACGCTCAGTCCTGGTCAAAGGCTACCAGACCTTTCTGAGTGCCCCCAGGGAGGGGCTGGGGCGTCTCAGGGTGCCCACTGGCGAGGGAGCTGGCATCTCCACCCGCAGTCCTCGCCCCTTCAATGAGATCCCCTCTCCTGGTGACAATGGCTGGCTAAACCTGTACCATTTCTGGAGGGAGACGGGCACACACAAAGTCCACCTTCACCATGTCCAGAATTTCCAGAAGTATGGCCCGATTTACAGGGAGAAGCTCGGCAACGTGGAGTCGGTTTATGTCATCGACCCTGAAGATGTGGCCCTTCTCTTTAAGTCCGAGGGCCCCAACCCAGAACGATTCCTCATCCCGCCCTGGGTCGCCTATCACCAGTATTACCAGAGACCCATAGGAGTCCTGTTGAAGAAGTCGGCAGCCTGGAAGAAAGACCGGGTGGCCCTGAACCAGGAGGTGATGGCTCCAGAGGCCACCAAGAACTTTTTGCCCCTGTTGGATGCAGTGTCTCGGGACTTCGTCAGTGTCCTGCACAGGCGCATCAAGAAGGCGGGCTCCGGAAATTACTCGGGGGACATCAGTGATGACCTGTTCCGCTTTGCCTTTGAGTCCATCACTAACGTCATTTTTGGGGAGCGCCAGGGGATGCTGGAGGAAGTAGTGAACCCCGAGGCCCAGCGATTCATTGATGCCATCTACCAGATGTTCCACACCAGCGTCCCCATGCTCAACCTTCCCCCAGACCTGTTCCGTCTGTTCAGGACCAAGACCTGGAAGGACCATGTGGCTGCATGGGACGTGATTTTCAGTAAAGCTGACATATACACCCAGAACTTCTACTGGGAATTGAGACAGAAAGGAAGTGTTCACCACGATTACCGTGGCATGCTCTACAGACTCCTGGGAGACAGCAAGATGTCCTTCGAGGACATCAAGGCCAACGTCACAGAGATGCTGGCAGGAGGGGTGGACACGACGTCCATGACCCTGCAGTGGCACTTGTATGAGATGGCACGCAACCTGAAGGTGCAGGATATGCTGCGGGCAGAGGTCTTGGCTGCGCGGCACCAGGCCCAGGGAGACATGGCCACGATGCTACAGCTGGTCCCCCTCCTCAAAGCCAGCATCAAGGAGACACTAAGACTTCACCCCATCTCCGTGACCCTGCAGAGATATCTTGTAAATGACTTGGTTCTTCGAGATTACATGATTCCTGCCAAGACACTGGTGCAAGTGGCCATCTATGCTCTGGGCCGAGAGCCCACCTTCTTCTTCGACCCGGAAAATTTTGACCCAACCCGATGGCTGAGCAAAGACAAGAACATCACCTACTTCCGGAACTTGGGCTTTGGCTGGGGTGTGCGGCAGTGTCTGGGACGGCGGATCGCTGAGCTAGAGATGACCATCTTCCTCATCAATATGCTGGAGAACTTCAGAGTTGAAATCCAACACCTCAGCGATGTGGGCACCACATTCAACCTCATTCTGATGCCTGAAAAGCCCATCTCCTTCACCTTCTGGCCCTTTAACCAGGAAGCAACCCAGCAGTGATCAGAGAGGATGGCCTGCAGCCACATGGGAGGAAGGCCCAGGGGTGGGGCCCATGGGGTCTCTGCATCTTCAGTCGTCTGTCCCAAGTCCTGCTCCTTTCTGCCCAGCCTGCTCAGCAGGTTGAATGGGTTCTCAGTGGTCACCTTCCTCAGCTCAGCTGGGCCACTCCTCTTCACCCACCCCATGGAGACAATAAACAGCTGAACCATCG
【0136】
ウシadrenodoxin 遺伝子DNA配列
coding region: 165-722
mature pepyide: 339-680
(配列番号6)
TGGCGAACGCATCAGGCCCTCTAGGGAACCCCGGCCCTGAGAGTCCGCTGCGCCTCTCTTCTCTCCGCAGACCCGCGCGGCGTCAGCCGCCCGTACTCGCTGAGTCTGCCCTCGCGGCCGCTGCCGTGCGGCCTGCGTCGCCTTCCGTAGTTCCCCGACAGGCTATGGCCGCACGCCTCCTGCGCGTCGCCTCCGCCGCCCTCGGCGACACGGCCGGCCGGTGGCGGCTTCTCGTGCGACCGCGCGCGGGAGCCGGCGGCCTCCGTGGGAGCCGAGGACCGGGCCTGGGCGGCGGCGCAGTGGCGACGCGGACGCTGAGCGTATCGGGGCGAGCGCAGAGCAGCTCAGAAGATAAAATAACAGTCCACTTTATAAACCGTGATGGTGAAACATTAACAACCAAAGGAAAAATTGGTGACTCTCTGCTAGATGTTGTGGTTCAAAATAATCTAGATATTGATGGTTTTGGTGCATGTGAGGGAACCTTGGCTTGTTCTACCTGTCACCTCATCTTTGAACAGCACATATTTGAGAAATTGGAAGCAATCACTGATGAGGAGAATGACATGCTTGATCTGGCATATGGACTAACAGATAGATCGCGGTTGGGCTGCCAGATCTGTTTGACAAAGGCTATGGACAATATGACTGTTCGAGTACCTGATGCCGTGTCTGATGCCAGAGAGTCCATTGATATGGGCATGAACTCCTCAAAGATAGAATAAATAGGAATATTTTCACAACATTTTACCCTATTTTTATAATTATTATTTCTTAATGTAATTAAATGAGAACATGGATGAATGGATTTATCATTATGACTAGATTTACTAAATTCATCTGATACACTGCTGAATTTTGTAGTTCTGAAAGTATCGCCATTTTTATTTTGATTAAATTATAAAAACAAATATTAGAAGTAGTTAGTATGAAAAACCTTATATATTTTACCTGTGATTGATCAGCAACATAAATCTTACGCCTACCTACCTATAAGTAAGTAGGTTAAAAAAGGGTGTCATTATCTCTCTTGGATGTGGGTGGAGGCAGAGATGTAAGGCCTTGTCTAGGGTCATACTGCTAATTAGAAGGTCAGTACTAGAACTGTACATGTTTAGGTCGAAACTGGGGAAATTAGGGATCTCATTTAAAATCTGAGCACAGATAATAAAGCACTGTCAAAGATGTTTGGTCTCAATTCTGACTGAGGTGGAGGAGTAAACTAAAAATTAAATGTGTTTTTTTTTTTTTTAATCTTGATCAGAATGGTCTGTCTTTGTAATCTTTAAAAAGTAATGCTTATTTATAAATTTCTCCCTGTGACAGAATGGATATCTTACGGATTAGGTTTTCTCTGTCCTAAGAAGGGAACGAGGGGGCTACCATCCCCTCGGCTAGGACAGCCACGTGGTGTGCAGTGAGTATGTAGGCAGGGTCTGGAGGACGGGGGGCTTGAGCTTAAACACATCCATGCATGGCAGATATTTGTATTTTATCTTTATTGTGAATAAACTGATAAATAATAAT
【0137】
ウシadrenodoxin 還元酵素遺伝子 DNA配列
coding region: 52-1527
mature peptide: 148-1527
(配列番号7)
CGCGGGGCAGCCGGCAGAGTCTGGTCCCTGGGCTGGTTCCTCTTCCCAAGCATGGCTCCGCGCTGTTGGCGCTGGTGGCCCTGGTCGTCTTGGACTCGGACTCGGCTGCCTCCTTCCAGGAGCATCCAGAACTTCGGCCAGCACTTCTCCACACAGGAGCAGACCCCCCAGATCTGTGTGGTGGGCAGTGGCCCAGCTGGCTTTTACACGGCCCAGCACCTGCTAAAGCACCACTCCCGGGCCCACGTGGATATCTACGAGAAACAGCTGGTGCCCTTCCGCCTGGTGCGCGTTTGGCTGGCGCTGACCACCCCGAGGTCAAGAATGTTACTCAACACCTTTACCCAGACGGCCCGCTCTGACCGCTGTGCCTTCTATGGCAACGTGGAGGTGGGCAGGGATGTGACTGTGCAGGAGCTGCGGGTGTATCGCCTAACCGCCGTGGTGCTGAGCTATGGGGCAGAGGACCATCAGGCCCTGGATATCCCTGGTGAGGAGTTGCCCGGCGTGTTCTCGGCCCGGGCCTTTGTGGGCTGGTACAATGGGCTTCCTGAGAACCGGGAGCTGGCCCCGGACCTGAGCTGTGACACAGCCGTGATTCTGGGGCAGGGGAATGTGGCTCTGGACGTGGCCCGGATCCTGCTGACCCCCCCCGACCACCTGGAGAAAACGGACATCACTGAGGCCGCCCTGGGAGCCCTGAGACAGAGTCGGGTGAAGACGGTGTGGATCGTGGGCCGACGTGGACCCCTACAAGTGGCCTTCACCATAAAGGAGCTTCGGGAGATGATTCAGTTACCAGGAACTCGGCCCATGTTGGATCCTGCGGATTTCTTGGGTCTCCAGGACAGAATCAGGGAGGCCGCTCGCCCGAGGAAGCGGCTGATGGAACTGCTGCTTCGAACAGCCACGGAGAAGCCAGGGGTGGAGGAGGCTGCCCGCCGGGCATCAGCCTCCCGTGCCTGGGGCCTCCGCTTCTTCCGAAGCCCGCAGCAGGTCCTGCGCCTGCCAGATGGGCGGGCGCGACGATCCGCCTGGCAGTCACCAGAACTGGAGGGCATTGGAGAGGCACACCCGGGCAGTGCCCACTGGGGATGTGGAGGACCTCCCTGTGGGCTGGTGCTGAGCAGCATTGGGTATAAGAGCCGCCCCATCGACCCCAGTGTGCCCTTTGACCCCAAGCTCGGGGTTGTCCCCAATATGGAGGGCCGGGTTGTGGATGTGCCAGGCCTCTACTGCAGCGGCTGGGTGAAGCGGGGACCCACAGGTGTCATCACCACCACCATGACCGACAGCTTCCTCACCGGCCAGATTCTGCTACAGGACCTGAAGGCCGGGCACCTGCCGTCTGGCCCCAGGCCGGGCTCTGCATTCATCAAGGCCCTGCTGGACAGCCGAGGGGTCTGGCCCGTGTCTTTCTCGGACTGGGAGAAACTGGATGCTGAGGAGGTGTCCCGGGGCCAGGCCTCGGGGAAGCCCAGAGAGAAGCTGCTGGATCCTCAGGAGATGCTGCGGCTGCTGGGGCACTGAGCCTAGATCCCAGCCCCGCTGGTTGCAGAGAGAAGAGGAGGGTGAGCCCATGATCCCAGCCCAGCTCAGAGAAGAGAGGAGGCGCGCTGGACAGCGGACGAGAGCGTGCGGGGTCAGCCTGAGCGGGACTCTGCGACCCCAGCTGCGTCGTCTGCCCGTGCCTGGCATACACCTGCCCTGGCTGCCTCTTCTCCAGGGGCGGTGGGAGCGACTTTCTGGAGCTAGGTCACTGCTGCCAGTGTGGGTACCTTTCACAAGGAGATAACCTTAGTTAGGGATGGAGGCAGGTACAGGCTGACCTCCGTCCCTCCTGTCTCTCTCCTGCTGGACTGTGGAGGGTCCCCAGGTCAGGAATATGCTGGAAATAAAGCACCTGCCACCTAG
【0138】
参考例2:CYP11A発現大腸菌による各種基質の変換反応
4-コレステン-3-オン(4Chol)、コレステロール(Chol)、7-デヒドロコレステロール(7-DHC)、エルゴステロール(Ergo)、β-シトステロール(Sito)、スティグマステロール(Stigma)、カンペステロール(Cam)およびデスモステロール(Des)、(20S)-20-ヒドロキシコレスト-4-エン-3-オン(20-OH)、(22R)-22-ヒドロキシコレスト-4-エン-3-オン(22-OH)、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン(R-diol)、(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オン(S-diol)、ラノステロール(Lano)を基質としてそれぞれ別々に実施例6と同様に反応し、反応液を酢酸エチル1mlで分液し反応物および基質を抽出しHPLC分析した結果、各基質に対応する側鎖開裂体のプロゲステロン、プレグネノロンまたは7-デヒドロプレグネノロンの生成を確認した。変換率はそれぞれ0%、2.6%、0.7%、0%、0.8%、0%、1.0%、1.2%、0%、0.4%、4.3%および0%であり、基質の残存はそれぞれ100%、99%、99.2%、100%、99.7%、100%、99.1%、99.6%、100%、99.7%、95%および100%であった。
【0139】
また、 (20S)-20-ヒドロキシ-コレステロール、(22S)-22-ヒドロキシ-コレステロール、(22R)-22-ヒドロキシ-コレステロール、(20R, 22R)-20,22-ジヒドロキシ-コレステロールを基質としてそれぞれ別々に実施例6と同様に反応後、処理分析した結果、各基質に対応する側鎖開裂体のプレグネノロンの生成を確認した。蓄積量はそれぞれ16.3mg/l(変換:16.3%)、6.1mg/l(変換:6.1%)、28.0mg/l(変換:28.0%)、32.3mg/l(変換:32.3%)であった。また、添加した基質はそれぞれ98.8%、92.6%、78.6%、51.6%残留していた。
【0140】
各基質に対する変換反応率のまとめを以下の表6に示す。
【0141】
【表6】
【0142】
実施例13:SDS-PAGE
BL21CYP11A、BP172及びBP215を、アンピシリン50μg/ml(終濃度)を添加したTB培地2 mlに各々植菌し、30℃、16時間、220 rpmで振とう培養した。この前培養液250μlを本培養用TB培地2 5 ml(アンピシリン50μg/ml〈終濃度〉、5-アミノレブリン酸40μg/ml〈終濃度〉、誘導剤:Overnight Express Autoinduction System1〈メルク社製〉)に加え、25℃で24時間振とう培養した。この培養液から遠心分離により回収した菌体を緩衝液(50 mM KPB、2%グリセロール[pH 7.4]) 5 mlに懸濁し、菌体懸濁液とした。この菌体懸濁液2 mlに×10 Bug Buster 66.7μl (Novagen社製)、Benzonase 0.67μl (Novagen社製)、40 mg/ml Lysozymeを添加し、30℃で20分間振とうして溶菌させた。 この菌体溶菌液を全タンパク質液とした。また、菌体溶菌液を遠心分離後、遠心上清を可溶性画分タンパク質液とした。不溶性画分タンパク質は菌体溶菌液を遠心分離後、遠心上清を全て除去し、緩衝液で3回洗浄後、再度緩衝液に懸濁して調製した。これらの全タンパク質液、可溶性画分タンパク質液、不溶性画分タンパク質液はSDS-PAGE用サンプルバッファー(BIO-RAD社製)20μlと各々混合し、SDS-PAGE泳動用サンプルとした。SDS-PAGE泳動用サンプルは95℃、5 min加熱後、ポリアクリルアミドゲル(第一化学薬品社製)で電気泳動した(300V、30mA、60分)。電気泳動後、ポリアクリルアミドゲルをトレイに移し、染色−脱色液(タカラバイオ社製)を適量添加し、染色及び脱色を行った。結果を図16に示す。
【図面の簡単な説明】
【0143】
【図1】図1は、CYP204A1遺伝子を発現するBacillus megateriumの菌体を用いて4-Cholesten-3-oneをProgesteroneに変換したことを示すHPLCチャートを示す。
【図2】図2は、本発明の酵素の至適pHを検討した結果を示す。
【図3】図3は、本発明の酵素の至適温度を検討した結果を示す。
【図4】図4は、本発明の酵素のpH安定性試験の結果を示す。
【図5】図5は、本発明の酵素の温度安定性試験の結果を示す。
【図6】図6は、本発明の酵素(CYP204A1)の基質濃度と酵素活性の関係を示す。
【図7】図7は、本発明の酵素(CYPSS)の基質濃度と酵素活性の関係を示す。
【図8】図8は、本発明の酵素(CYP204A1)のLineweaver-Burk plotを示す。
【図9】図9は、本発明の酵素(CYPSS)のLineweaver-Burk plotを示す。
【図10】図10は、BP215株による4-コレステン-3-オンの変換反応のHPLCを示す。
【図11】図11は、BP215株による4-コレステン-3-オンの変換反応のMSを示す。
【図12】図12は、BP215株によるコレステロールの変換反応のHPLCを示す。
【図13】図13は、BP215株によるコレステロールの変換反応のMSを示す。
【図14】図14は、BP215株による7-デヒドロコレステロールの変換反応のHPLCを示す。
【図15】図15は、BP215株による7-デヒドロコレステロールの変換反応のMSを示す。
【図16】図16は、BL21CYP11A、BP172及びBP215を用いてSDS-PAGEを行った結果を示す。
【特許請求の範囲】
【請求項1】
以下の理化学的性質を有するステロール側鎖切断酵素。
(1)作用:下記式(I)で表されるステロールに作用し、ステロール側鎖部分の20位と22位の炭素−炭素結合を切断する活性により該結合を切断して下記式(II)で表される化合物を生成する。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が10%以上であり、かつコレステロールからプレグネノロンへの変換率が10%以上である。
【化1】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【請求項2】
以下の理化学的性質を有する、請求項1に記載のステロール側鎖切断酵素。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が50%以上であり、かつコレステロールからプレグネノロンへの変換率が50%以上である。
【請求項3】
さらに以下の理化学的性質を有する、請求項1又は2に記載の酵素。
(3)至適pH:7.5〜8.0
(4)作用至適温度:15〜20℃
(5)熱安定性:20℃で140時間保存後に30%以上の酵素活性が保持される。
(6)分子量:アミノ酸配列からの推定分子量が53〜54KDaであり、SDS電気泳動による測定で50〜56KDa
【請求項4】
さらに以下の理化学的性質を有する、請求項1から3の何れかに記載の酵素。
(7)反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上である。
(8)基質濃度を10μMから100μMに上昇させた場合の活性の低下率が35%以下である。
【請求項5】
微生物に由来する酵素である、請求項1から4の何れかに記載の酵素。
【請求項6】
以下の(a)、(b)または(c)のいずれかのタンパク質;
(a)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b)配列番号2に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質、
(c)配列番号2に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質。
【請求項7】
ステロール側鎖の20位と22位の結合を切断する活性について、基質濃度を10μMから100μMに上昇させた場合の活性の低下率が30%以下である、請求項6に記載のタンパク質。
【請求項8】
請求項6又は7に記載のタンパク質をコードするDNA。
【請求項9】
以下の(a)、(b)または(c)のいずれかのDNA;
(a)配列番号1に記載の塩基配列を有するDNA、
(b)配列番号1に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA、
(c)配列番号1に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA。
【請求項10】
請求項8または9に記載のDNAを有する組み換えベクター。
【請求項11】
請求項8または9に記載のDNAあるいは請求項10に記載の組換えベクターを宿主細胞に導入した形質転換体。
【請求項12】
宿主細胞が大腸菌、放線菌またはシュードモナス属微生物である請求項11に記載の形質転換体。
【請求項13】
請求項6または7に記載のタンパク質、あるいは請求項11または12に記載の形質転換体を含む、ステロール側鎖の20位と22位の結合を切断する反応を触媒するための酵素組成物。
【請求項14】
請求項11または12に記載の形質転換体により産生される、請求項6または7に記載のタンパク質。
【請求項15】
請求項1から5の何れかに記載の酵素あるいは請求項6、7または14に記載のタンパク質に対する抗体又はその部分フラグメント。
【請求項16】
モノクローナル抗体である、請求項15に記載の抗体又はその部分フラグメント。
【請求項17】
請求項1から5の何れかに記載の酵素あるいは請求項6、7または14に記載のタンパク質が有するステロール側鎖の20位と22位の結合を切断する活性を中和する作用を有する、請求項15または16に記載の抗体又はその部分フラグメント。
【請求項18】
請求項11又は12に記載の形質転換体を培養することを特徴とする、請求項6又は7に記載のタンパク質の製造方法。
【請求項19】
式(I)で表されるステロールに、下記(i)から(vi):
(i) 請求項1から5の何れかに記載の酵素、
(ii) 該酵素を産生する微生物またはその処理物、
(iii) 請求項6、7または14に記載のタンパク質、
(iv) 請求項11または12に記載の形質転換体またはその処理物、
(v) 以下の(a)、(b)または(c)のいずれかのタンパク質、又は
(vi) 以下の(a)、(b)または(c)のいずれかのタンパク質をコードするDNA、以下の(d)、(e)または(f)のいずれかのDNA、もしくは上記DNAを有する組み換えベクターを宿主細胞に導入した形質転換体またはその処理物;
の何れかを反応させることを特徴とする、式(II)で表される化合物の製造方法。
(a)配列番号26に記載のアミノ酸配列からなるタンパク質、
(b)配列番号26に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質、
(c)配列番号26に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質、
(d)配列番号4に記載の塩基配列を有するDNA、
(e)配列番号4に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質をコードするDNA、
(f)配列番号4に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質をコードするDNA。
【化2】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、
母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【請求項20】
式(I)で表されるステロールが、コレステロール、4-コレステン-3-オン、7-デヒドロコレステロール、エルゴステロール、β-シトステロール、スティグマステロール、カンペステロール、デスモステロール、 (20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、又は(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンである、請求項19に記載の方法。
【請求項21】
式(II)で表される化合物が、プレグネノロン、プロゲステロン又は7−デヒドロプレグネノロンである、請求項19又は20に記載の方法。
【請求項1】
以下の理化学的性質を有するステロール側鎖切断酵素。
(1)作用:下記式(I)で表されるステロールに作用し、ステロール側鎖部分の20位と22位の炭素−炭素結合を切断する活性により該結合を切断して下記式(II)で表される化合物を生成する。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が10%以上であり、かつコレステロールからプレグネノロンへの変換率が10%以上である。
【化1】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【請求項2】
以下の理化学的性質を有する、請求項1に記載のステロール側鎖切断酵素。
(2)基質特異性:該酵素を産生する微生物を、4-コレステン-3-オンまたはコレステロールを100μg/ml含む水溶液に28℃で5時間反応させたときの、4-コレステン-3-オンからプロゲステロンへの変換反応率が50%以上であり、かつコレステロールからプレグネノロンへの変換率が50%以上である。
【請求項3】
さらに以下の理化学的性質を有する、請求項1又は2に記載の酵素。
(3)至適pH:7.5〜8.0
(4)作用至適温度:15〜20℃
(5)熱安定性:20℃で140時間保存後に30%以上の酵素活性が保持される。
(6)分子量:アミノ酸配列からの推定分子量が53〜54KDaであり、SDS電気泳動による測定で50〜56KDa
【請求項4】
さらに以下の理化学的性質を有する、請求項1から3の何れかに記載の酵素。
(7)反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上である。
(8)基質濃度を10μMから100μMに上昇させた場合の活性の低下率が35%以下である。
【請求項5】
微生物に由来する酵素である、請求項1から4の何れかに記載の酵素。
【請求項6】
以下の(a)、(b)または(c)のいずれかのタンパク質;
(a)配列番号2に記載のアミノ酸配列からなるタンパク質、
(b)配列番号2に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質、
(c)配列番号2に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質。
【請求項7】
ステロール側鎖の20位と22位の結合を切断する活性について、基質濃度を10μMから100μMに上昇させた場合の活性の低下率が30%以下である、請求項6に記載のタンパク質。
【請求項8】
請求項6又は7に記載のタンパク質をコードするDNA。
【請求項9】
以下の(a)、(b)または(c)のいずれかのDNA;
(a)配列番号1に記載の塩基配列を有するDNA、
(b)配列番号1に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA、
(c)配列番号1に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が50mmol/分/mol以上であるタンパク質をコードするDNA。
【請求項10】
請求項8または9に記載のDNAを有する組み換えベクター。
【請求項11】
請求項8または9に記載のDNAあるいは請求項10に記載の組換えベクターを宿主細胞に導入した形質転換体。
【請求項12】
宿主細胞が大腸菌、放線菌またはシュードモナス属微生物である請求項11に記載の形質転換体。
【請求項13】
請求項6または7に記載のタンパク質、あるいは請求項11または12に記載の形質転換体を含む、ステロール側鎖の20位と22位の結合を切断する反応を触媒するための酵素組成物。
【請求項14】
請求項11または12に記載の形質転換体により産生される、請求項6または7に記載のタンパク質。
【請求項15】
請求項1から5の何れかに記載の酵素あるいは請求項6、7または14に記載のタンパク質に対する抗体又はその部分フラグメント。
【請求項16】
モノクローナル抗体である、請求項15に記載の抗体又はその部分フラグメント。
【請求項17】
請求項1から5の何れかに記載の酵素あるいは請求項6、7または14に記載のタンパク質が有するステロール側鎖の20位と22位の結合を切断する活性を中和する作用を有する、請求項15または16に記載の抗体又はその部分フラグメント。
【請求項18】
請求項11又は12に記載の形質転換体を培養することを特徴とする、請求項6又は7に記載のタンパク質の製造方法。
【請求項19】
式(I)で表されるステロールに、下記(i)から(vi):
(i) 請求項1から5の何れかに記載の酵素、
(ii) 該酵素を産生する微生物またはその処理物、
(iii) 請求項6、7または14に記載のタンパク質、
(iv) 請求項11または12に記載の形質転換体またはその処理物、
(v) 以下の(a)、(b)または(c)のいずれかのタンパク質、又は
(vi) 以下の(a)、(b)または(c)のいずれかのタンパク質をコードするDNA、以下の(d)、(e)または(f)のいずれかのDNA、もしくは上記DNAを有する組み換えベクターを宿主細胞に導入した形質転換体またはその処理物;
の何れかを反応させることを特徴とする、式(II)で表される化合物の製造方法。
(a)配列番号26に記載のアミノ酸配列からなるタンパク質、
(b)配列番号26に記載のアミノ酸配列において1もしくは数個のアミノ酸が欠失、置換および/または付加されたアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質、
(c)配列番号26に記載のアミノ酸配列と95%以上の相同性を有するアミノ酸配列からなり、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質、
(d)配列番号4に記載の塩基配列を有するDNA、
(e)配列番号4に記載の塩基配列において1もしくは数個の塩基が欠失、置換および/または付加された塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質をコードするDNA、
(f)配列番号4に記載の塩基配列あるいはその相補配列を有するDNAとストリンジェントな条件下でハイブリダイズすることができる塩基配列を有し、かつステロール側鎖の20位と22位の結合を切断する活性を有し、上記活性の反応速度パラメーターである最大速度(Vmax)が40mmol/分/mol以上であるタンパク質をコードするDNA。
【化2】
[式(I)において、母核部分は、ステロイド類が有するA、B、C、D環からなる構造をもち、
その1位から17位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置に炭素−炭素不飽和結合を有しており、
その1位から19位(10位と13位をのぞく)のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OXで表される基
(式中、X=Hで表される水酸基、X=COR1[R1は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、X=R2[R2は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、X=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Xが糖類の1位炭素であるOグリコシル基、またはXが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。
側鎖部分は、Rは、環状部分を有していてもよい炭素数10以下の直鎖状のアルキル基、アルケニル基もしくはアルキニル基であり、または環状部分を有していてもよい炭素数10以下の分岐状のアルキル基、アルケニル基もしくはアルキニル基であり、
20位、21位およびR内のいずれかの0箇所又は1箇所以上の位置の炭素上には、それぞれ独立して、
式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、Yが糖類の1位炭素であるOグリコシル基、またはYが当該炭素の隣接炭素原子であるエポキシ基)、
または式;=Oで表されるケト基で置換されている。]
[式(II)において、
母核部分は、式(I)の母核部分と同義である。
側鎖部分において、21位の炭素上には0又は1つの、式;−OYで表される基
(式中、Y=Hで表される水酸基、Y=COR3[R3は水素原子または炭素数10以下のアルキル基、アルケニル基、アルキニル基もしくは芳香族炭化水素]で表されるアシロキシル基、Y=R4[R4は酸素原子で置換されていてもよい炭素数10以下のアルキル基、アルケニル基またはアルキニル基]で表されるOアルキル基、Y=SO3M[Mは水素原子、アルカリ金属またはアルカリ土類金属]で表される硫酸エステル類、またはYが糖類の1位炭素であるOグリコシル基)、
または式;=Oで表されるケト基で置換されている。]
【請求項20】
式(I)で表されるステロールが、コレステロール、4-コレステン-3-オン、7-デヒドロコレステロール、エルゴステロール、β-シトステロール、スティグマステロール、カンペステロール、デスモステロール、 (20S)-20-ヒドロキシコレスト-4-エン-3-オン、(22R)-22-ヒドロキシコレスト-4-エン-3-オン、(20R,22R)-20,22-ジヒドロキシコレスト-4-エン-3-オン、又は(20R,22S)-20,22-ジヒドロキシコレスト-4-エン-3-オンである、請求項19に記載の方法。
【請求項21】
式(II)で表される化合物が、プレグネノロン、プロゲステロン又は7−デヒドロプレグネノロンである、請求項19又は20に記載の方法。
【図1】

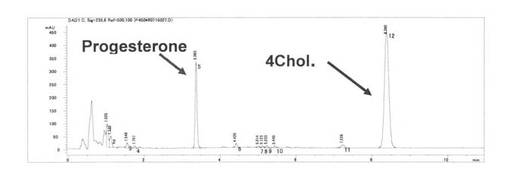
【図2】
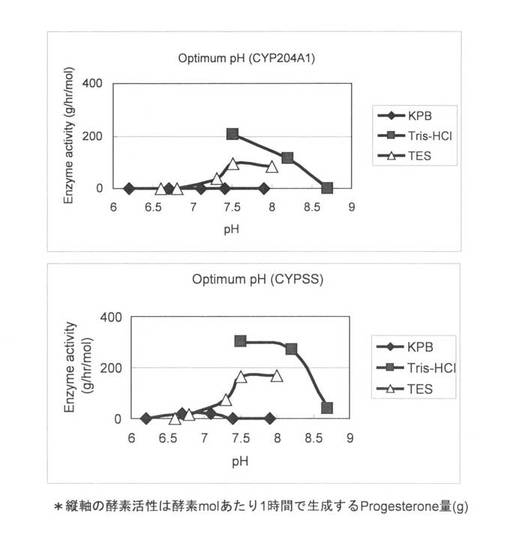
【図3】
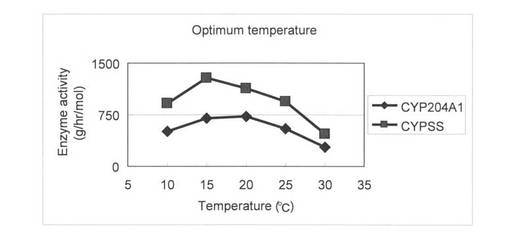
【図4】
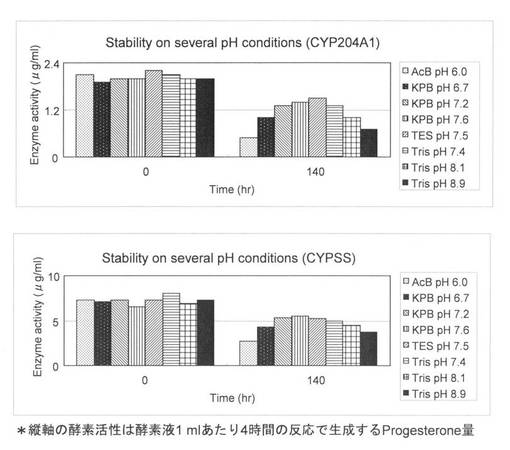
【図5】
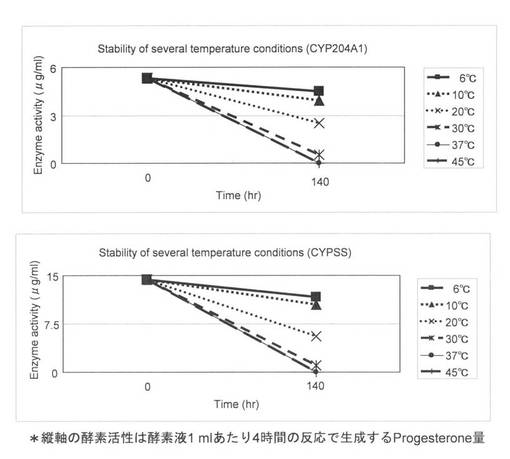
【図6】
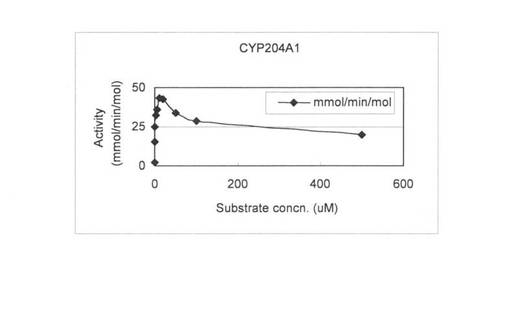
【図7】
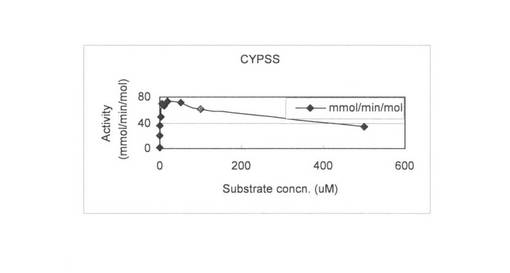
【図8】
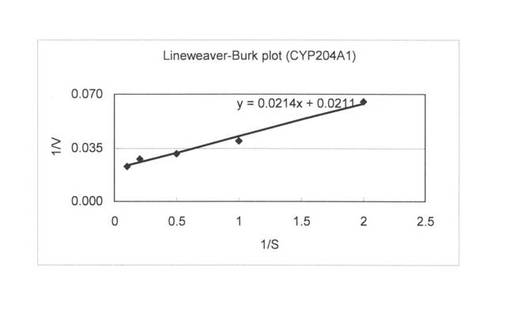
【図9】
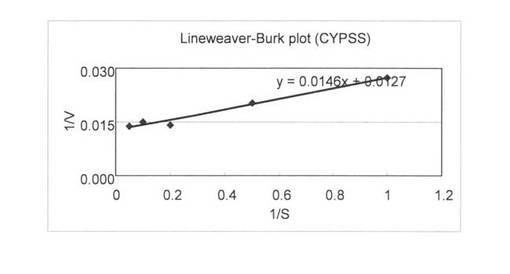
【図10】
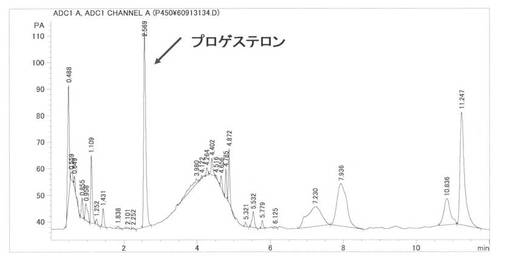
【図11】
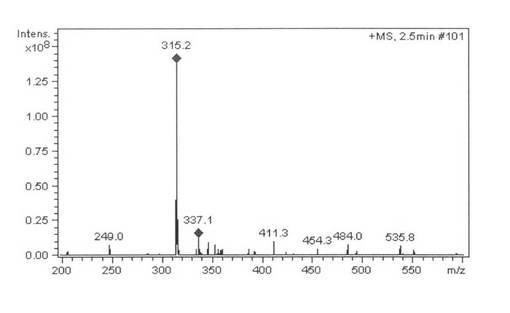
【図12】
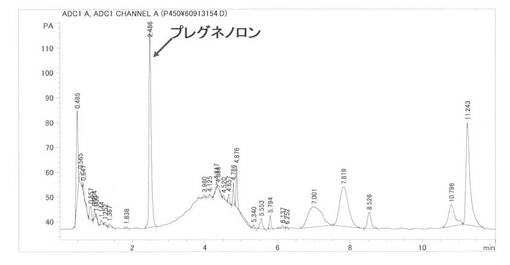
【図13】
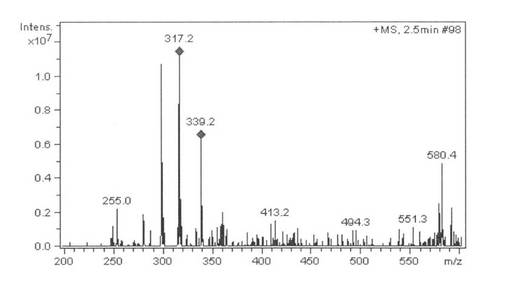
【図14】
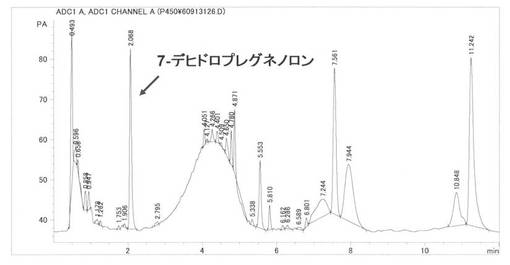
【図15】
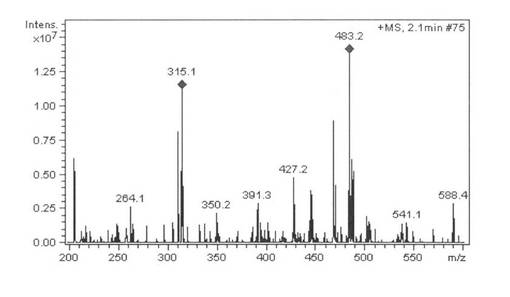
【図16】
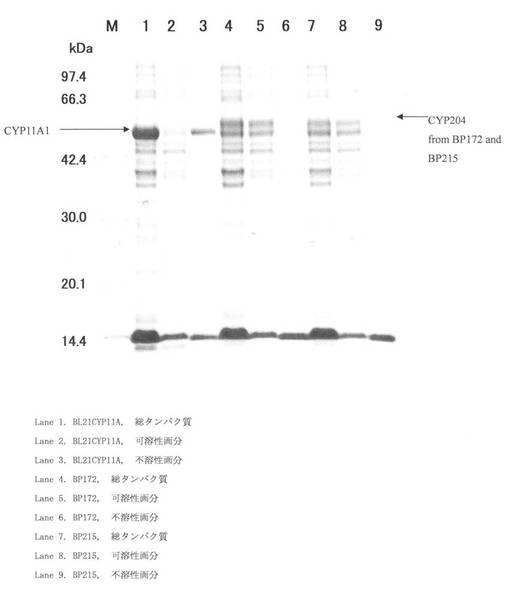
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公開番号】特開2009−103(P2009−103A)
【公開日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願番号】特願2008−134957(P2008−134957)
【出願日】平成20年5月23日(2008.5.23)
【出願人】(000005968)三菱化学株式会社 (4,356)
【Fターム(参考)】
【公開日】平成21年1月8日(2009.1.8)
【国際特許分類】
【出願日】平成20年5月23日(2008.5.23)
【出願人】(000005968)三菱化学株式会社 (4,356)
【Fターム(参考)】
[ Back to top ]