
ストレプトコッカス属細菌由来ComAタンパク質のペプチダーゼドメインの結晶
【課題】ComAタンパク質のPEPの結晶を提供する。
【解決手段】ComAタンパク質のペプチダーゼドメイン(PEP)を含むポリペプチド;又は
PEPの部分ポリペプチドであってComC切断活性を有するポリペプチド
の結晶であって、空間群P43212に属する結晶。
【解決手段】ComAタンパク質のペプチダーゼドメイン(PEP)を含むポリペプチド;又は
PEPの部分ポリペプチドであってComC切断活性を有するポリペプチド
の結晶であって、空間群P43212に属する結晶。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ストレプトコッカス属細菌由来ComAタンパク質のペプチダーゼドメイン(PEP)の結晶に関する。
【背景技術】
【0002】
口腔内常在菌としてのストレプトコッカス属細菌は歯垢内の主要な細菌群であるが、歯のう蝕に関与するだけでなく、歯周ポケットから血流に入り込むと心臓弁膜に付着してバイオフィルムを形成し心内膜炎や人工弁感染症をひきおこす。バイオフィルムとは、細菌が自ら合成・分泌する多糖類などから成るマトリックス内に共同体を形成し、固体表面に付着して生育・増殖したものである。細菌が生体内で一旦バイオフィルムを形成すると抗生物質や白血球の浸透・拡散に対して耐性を示すため、除去がきわめて困難で外科的手術が必要となる。また、ブラッシングによる口腔バイオフィルムの完全除去は不可能である。バイオフィルムの形成には細菌に広く存在する細胞間情報伝達系であるクオラムセンシングのメカニズムが重要である。したがってストレプトコッカス属細菌のクオラムセンシングのメカニズムを解明し、阻害剤を開発することは上記のStreptococcus感染症に対する有効な予防・治療法の開発に役立つと考えられる。
Streptococcusのクオラムセンシングを阻害する薬剤を開発する上で、その初発段階で機能するComAは重要なタンパク質であると考えられる。ComAはシグナルペプチド前駆体であるComCを切断すると同時に菌体外へと分泌するATP-binding cassette(ABC)トランスポーターである。しかしながら、その重要性にも関わらずComAをはじめとする、グラム陽性菌のクオラムセンシングのシグナル産生タンパク質は膜タンパク質で大量発現が困難なため構造学的解析が皆無であった。
以上のような背景からComAの立体構造の解明が渇望されている。しかしこれまで、ComAの三次元構造を解析し原子座標を明らかにし、その活性中心や基質結合部位の位置および構造を決定したという報告はない。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】J. Biol. Chem. 281, 4726-4731
【発明の概要】
【発明が解決しようとする課題】
【0004】
ComAタンパク質のペプチダーゼドメイン(PEP)の結晶を提供することを課題とする。
【課題を解決するための手段】
【0005】
一般にタンパク質の結晶化は容易ではなく、多くのタンパク質の場合、数多くの条件検討を重ねてはじめてX線結晶構造解析に供することのできる結晶が得られる。また、いかなる条件でも結晶化しないタンパク質も少なくなく、タンパク質の結晶が得られなければX線結晶構造解析を行うことができず、そのタンパク質の三次元構造を解明することはできない。
本発明者らはストレプトコッカス属細菌のComAタンパク質の三次元構造を解析するべく日夜検討を重ねた。はじめにクオラムセンシングのメカニズムが最も詳細に解析されているS. pneumoniaeのComA全長について大量発現の検討を行った。しかしながら、従来の方法をそのまま適用しても大量発現できないことが判明した。ここで本発明者らは、ComA全長の大量発現は困難であると判断し、代わりにペプチダーゼドメインのみからなるポリペプチドの大量発現を試みた。N末の150残基からなるペプチダーゼドメイン(PEP)がシグナルペプチド前駆体であるComCを切断する活性をもつことは確認済みである。ペプチダーゼドメインを含む種々の長さのポリペプチドについて大量発現を検討し、最も効率良く可溶性画分にタンパク質が発現したポリペプチドを精製し、結晶化の検討を行った。初期スクリーニングとして計480条件について検討したが、ついに結晶を得ることはできなかった。
【0006】
そこで、本発明者らは他のStreptococcus由来のPEPを加えた計6種類のPEPについても同様に大量発現、精製、及び結晶化の検討を行った。その結果、従来の方法をそのまま適用したとしてもいずれのPEPについても結晶化することはできなかった。そこで、本発明者らは試行錯誤を重ね、当初はPEPタンパク質を安定化することを目的として加えていた硫酸アンモニウムを系内から除いて結晶化を行うことによって、かつPEPとしてミュータンス菌(S. mutans)由来のPEPを採用することによって、はじめてPEPの結晶を得ることに成功した。そして、その結晶にX線を当てることにより分解能約1.9 ÅのX線回折強度が得られることを確認した。
【0007】
さらに、本発明者らは前記PEPの結晶についてX線結晶構造解析を行うことにより、その三次元構造を明らかにすることが可能であることを実証し、本発明を完成させた。
【0008】
すなわち、本発明は次の通りである:
項1.ComAタンパク質のペプチダーゼドメイン(PEP)を含むポリペプチド;又は
PEPの部分ポリペプチドであってComC切断活性を有するポリペプチド
の結晶であって、空間群P43212に属する結晶。
項2.単位格子定数がa=b=58.6±0.35 nm、及びc=112.1±0.5 nmである、項1に記載の結晶。
項3.前記ポリペプチドが、
(A) 配列番号1のアミノ酸配列からなるポリペプチド;又は
(B-1) (A)のポリペプチドのアミノ酸配列において、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチド
である、項1又は2に記載の結晶。
項4.前記ポリペプチドが、
(A) 配列番号1のアミノ酸配列からなるポリペプチド;又は
(B-2) (A)のポリペプチドのアミノ酸配列において、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されることにより、(A)のポリペプチドのアミノ酸配列に対して90%以上の相同性の範囲内で修飾されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチド
である、項1又は2に記載の結晶。
項5.項1〜4のいずれか一項に記載の結晶の製造方法であって、
硫酸アンモニウムを実質的に含まない溶液中に前記ポリペプチドを含有させた状態で結晶化することを特徴とする方法。
項6.前記溶液が、70 mMリン酸ナトリウム、及び2 mMβ−メルカプトエタノールを含有するpH 7.0の溶液に、16%ポリエチレングリコール3350、及び0.16 Mクエン酸水素二アンモニウムを含有する等量の沈殿剤を混合したものである、項5に記載の方法。
項7.さらにSitting-drop蒸気拡散法で結晶化することを特徴とする、項5又は6に記載の方法。
【発明の効果】
【0009】
本発明は、PEPの結晶化条件を初めて見出したものであり、これによりX線結晶構造解析に適した結晶の調製が可能となる。解明されたPEPの三次元構造を用いれば、活性中心および基質結合部位のアミノ酸残基とそれらの立体配置が明らかとなる。したがって、得られた座標データを用いてコンピュータースクリーニング法による阻害剤を探索することが可能となる。さらにPEPの結晶を用いて候補化合物との複合体結晶を作製し、その構造決定を行うことでより詳細な解析を行うこともできる。
【0010】
また、本発明の結晶を利用して得られた座標データは、未知の構造のPEPホモログの探索、変異体の構造のホモロジーモデリングによる推定又は分子置換法による決定のために使用できる。
【図面の簡単な説明】
【0011】
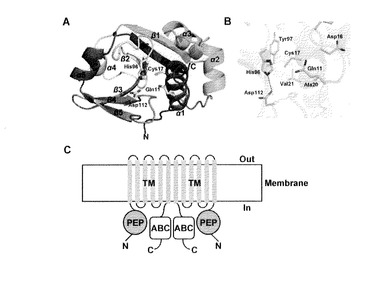
【図1】(A) MuPEP1全体構造のCartoon 表示。二次構造の各成分はレインボーカラーで示してある(N末端が青。C末端が赤。)。Catalytic triadを形成するCys17、His96、Asp112およびオキシアニオンホールを形成するGln11の側鎖はスティックで表す。(B)MuPEP1の活性中心。主鎖の原子は半透明のCartoon(緑)で示す。活性中心近傍の残基の側鎖はスティックで表す。酸素、窒素、硫黄の各原子はそれぞれ赤、青、黄で表す。点線は水素結合を示す。(C) ComAのドメイン構成の推定図。PEP, ペプチダーゼドメイン。TM, 膜貫通ドメイン。ABC, ATP-結合ドメイン。バクテリオシンのABCトランスポーターは二量体として機能すると考えられている。
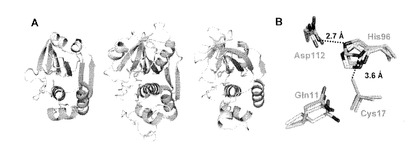
【図2】(A)MuPEP1(左)、papain(PDB ID code 9PAP)(中央)、staphopain A (PDB ID code 1CV8)(右)。それぞれの構造は活性中心の残基Gln11、Cys17、His96、Asp112のCα原子を重ね合わせ、同方向から見たもの。これらの残基はスティックモデルで示す。構造的に相同のα-ヘリックス、 α1-4(MuPEP1)およびβ-ストランド、β1-6(MuPEP1)はそれぞれシアンと緑で示す。(B)MuPEP1と他のパパイン様システインプロテアーゼの活性部位の重ね合わせ。MuPEP1のGln11、Cys17、His96、Asp112はスティックモデルで示した(緑)。これらの残基に相当するpapain(シアン)およびstaphopain A(グレー)はCα原子で重ね合わせ、スティックモデルで示した。酸素、窒素、硫黄の各原子はそれぞれ赤、青、黄で表す。
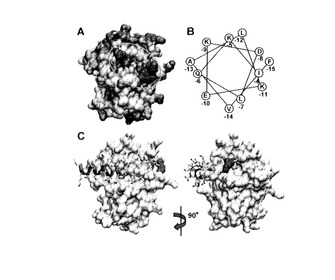
【図3】(A) HotPatchプログラム (23) の結果(表面図)。疎水性領域を赤、親水性領域を青で示した。活性部位近傍の疎水性領域をオレンジ色の線で囲った。(B) PComCのN末領域のα-ヘリカル ホイール図。PPEPとの相互作用に重要であると考えられた高度に保存されたアミノ酸(黄色)(20)。(C)MuPEP1-MuComCアシル中間体モデル。MuPEP1は表面図で示す。MuComCの側鎖はスティックモデルで示す。酸素、窒素、硫黄の各原子は赤、青、黄で示す。MuComCの4つの高度に保存された残基、Phe-15、Leu-12、Leu-7、Ile-4は球モデル(黄)で示す。2次構造の各成分はランダムコイル(緑)、ターン(シアン)、α-ヘリックス(紫)。Cys17はオレンジで、Arg93(詳細は本文)は、マゼンタで示す。右側の構造は左側の構造をY軸に対して90° 回転させたもの。
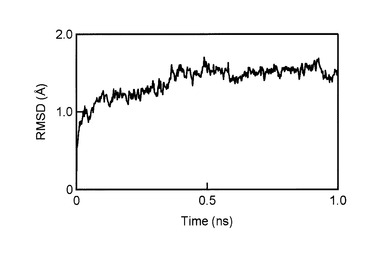
【図4】MDシミュレーション中の各Cα原子の初期構造の対応するCα原子に対するrmsdの時間経過。初期構造からのMuPEP1-MuComC複合体のrmsdを2 psごとに計算した。
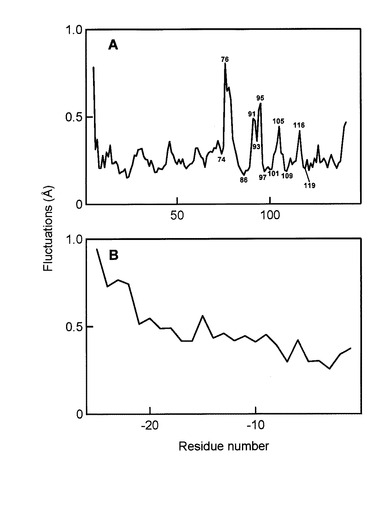
【図5】MuPEP1-MuComC複合体の各Cα原子のゆらぎ。(A)0.4ナノ秒における対応原子からの0.4ナノ秒〜1.0ナノ秒の間のMDシミュレーションでのMuPEP1(5から141の残基)の各Cα原子のRMS位置変動。主要ピークに対応する残基番号を示す。(B)0.4ナノ秒における対応原子からの0.4ナノ秒〜1.0ナノ秒の間のMDシミュレーションでのMuComC(-25から-1の残基)の各Cα原子のRMS位置変動。複合体のMuPEP1成分に対する剛体整合の後に、MuComCのRMSDを計算した。
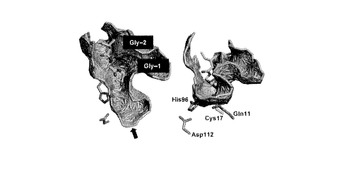
【図6】MuPEP1-MuComCアシル中間体モデル(緑)をMuPEP1(グレー)の結晶構造に重ね合わせた図。重ね合わせはGln11、Cys17、His96、Asp112のCα原子で行った。MuPEP1のクレフトの表面はPDBsum (28)によって解析し、バスケットで表わした。MuComCはGly-1、Gly-2、Ile-3のみ表示した。酸素、窒素、硫黄の各原子はそれぞれ赤、青、黄で示した。右側の構造は、左の構造の矢印で示した方向から見た図。
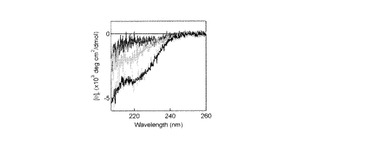
【図7】CComCの遠紫外部(207−260 nm)のCDスペクトルを変異型PPEP共存下または非共存下で測定した。波長207 nm以下のスペクトルはタンパク質の紫外線吸収によるノイズが大きいため測定できなかった。単位は平均残基楕円率で示す。CD測定は20 μM CComCおよび20 μM CComCと20 μMのCAPEPまたはA51W CAPEPまたはA67W CAPEPを用いて行った。バッファー溶液は30 mM リン酸ナトリウム、150 mM 硫酸アンモニウム、15 mM フッ化ナトリウム、0.025 mM ジチオスレイトール、pH 7.0。スペクトルはスキャン速度50 nm/分で30回の積算を行った。CComC(0% TFE中)のスペクトル(青)。CComC(35% TFE中)のスペクトル(黒)。各変異型PPEPのスペクトルをCComCと各変異型PPEPの混合液のスペクトルから差し引いた、差スペクトルは、それぞれ(CComC + CAPEP) - CAPEPは黄、 (CComC + A51W CAPEP) - A51W CAPEPは赤、(CComC + A67W CAPEP) - A67W CAPEPは緑の各色で示した。
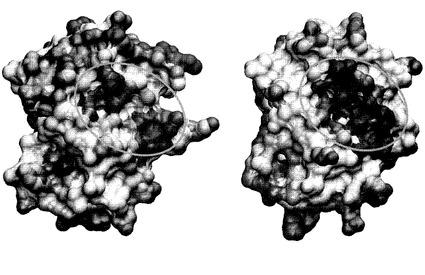
【図8】MuPEP1(左)およびPPEP(右)の推定される疎水性のくぼんだ領域。HotPatchプログラムの結果を表面図で示したもの。親水性領域を赤、疎水性領域を青で示した。オレンジの楕円形の線に囲まれた領域は、活性部位クレフトに隣接する疎水性のくぼんだ領域を示す。Cα原子の最小二乗重合に基づいて、2つの構造は、Cα原子で重ね合わせた。MuPEP1の疎水性のくぼんだ領域は、Thr50、Ala51、Leu52、Val55、His87、Leu94、Gln95、Thr132、Leu134、およびIle136から、PPEPの疎水性のくぼんだ領域は、Thr50、Ala51、Leu52、Val55、His87、Val88、Leu89、Leu94、Leu95、Thr132、Val134、およびLeu136から構成されている。
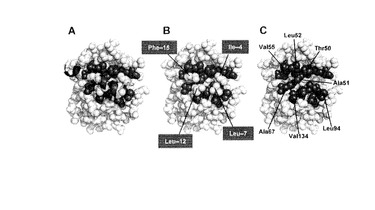
【図9】(A)PPEP-PComCアシル中間体モデル PPEPとPComCの複合体を球モデルで示した。PComCの二次構造の各成分は、それぞれランダムコイルは緑、ターンはシアン、α-ヘリックスはグレーの各色で示した。PComCの4つの保存された残基の側鎖は黄色で表した。PComCのこれら4つの残基と相互作用が予想されたPPEP側の残基を青で表した。(B)PComCの主鎖を(A)から省いた図。PComCのPhe-15はPPEPのAla51、Leu52、Val55、Ala67と、Leu-12はPPEPのAla67およびVal134、Leu-7はPPEPのLeu94と、Ile-4はPPEPのThr50とそれぞれ相互作用している。(C)PComCすべてを(A)から省き、PPEPの疎水性のくぼみを示した。
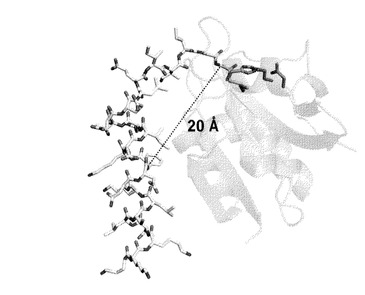
【図10】Phe-15のCα原子とGly-1のCα原子との間の距離。MuPEP1の全体構造はグレーのcartoon(半透明)によって示した。MuPEP1のGln11、Cys17、His96、およびAsp112の活性部位残基をstickによって示した。MuComCは白いstickによって示した。MuComCの高度に保存された残基の側鎖を黄色で示した。酸素、窒素、および硫黄原子を、それぞれ、赤、青、および黄色で示した。鎖線はPhe-15のCα原子とGly-1のCα原子との間の距離を示す。
【発明を実施するための形態】
【0012】
1.ComAタンパク質
本発明において使用するComAタンパク質は、特に限定されないが、ストレプトコッカス属細菌由来ComAタンパク質、又はストレプトコッカス属細菌由来ComAタンパク質の改変体であれば好ましい。さらに、ミュータンス菌(Streptococcus mutans)由来ComAタンパク質、又はミュータンス菌由来ComAタンパク質の改変体であればより好ましい。
【0013】
なお、ここでいう改変体は、天然において生じたものであってもよい。天然において生じた改変体としては、例えば突然変異や翻訳後の修飾などにより生じた改変体を挙げることができる。また、改変体は、天然に存在するComAタンパク質の遺伝子配列又はアミノ酸配列を利用して人為的に生じさせたものであってもよい。改変体には、アレル体、ホモログ、天然の変異体などであって、少なくとも90%、好ましくは95%、より好ましくは98%、さらにより好ましくは99%が、変異前のポリペプチドと相同するものが含まれる。ポリペプチドの相同性は、既知の配列分析ソフトウェア、例えばFASTAプログラムを使用した測定(Clustal,V., MethodsMol.Biol., 25, 307-318 (1994))によって解析することができる。また、SWISSPLOTSで解析することもできる。
【0014】
ミュータンス菌(Streptococcus mutans)とは、主に口腔内に存在し、う蝕の原因菌の一つであるとされるグラム陽性通性嫌気性連鎖球菌の一種である。
【0015】
ミュータンス菌由来ComAタンパク質としては、配列番号1のアミノ酸配列からなるポリペプチドを挙げることができる。ミュータンス菌由来ComAタンパク質をコードする遺伝子としては、配列番号2の遺伝子配列からなるポリヌクレオチドを挙げることができる。
【0016】
ComAタンパク質は、クオラムセンシングにおいてシグナルペプチド前駆体ComCからシグナルペプチドを切り出すと同時に菌体外へ分泌するという生理的作用を有する。この生理的作用においてPEPはComCを切断するという作用を担っている。
2.PEP含有ポリペプチド又は活性PEP部分ポリペプチド
本発明において「PEPを含むポリペプチド(PEP含有ポリペプチド)、又はPEPの部分ポリペプチドであってComC切断活性を有するポリペプチド(活性PEP部分ポリペプチド)」は、好ましくは、
(A) 配列番号1のアミノ酸配列からなるポリペプチド;又は
(B-1) (A)のポリペプチドのアミノ酸配列において、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチドである。
【0017】
上記ポリペプチド(A)は、天然に存在するミュータンス菌に由来するComAタンパク質のN末端から45番目のアミノ酸から194番目のアミノ酸までの部分ポリペプチドである。
【0018】
上記ポリペプチド(B-1)は、上記ポリペプチド(A)の改変体である。上記ポリペプチド(B-1)は、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されることにより、(A)のポリペプチドのアミノ酸配列に対して90%以上の相同性の範囲内で修飾されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチドであれば好ましい。上記相同性は、95%以上であればより好ましく、98%以上であればさらに好ましく、99%以上であればさらにより好ましい。ポリペプチドの相同性は、先述したのと同様に解析することができる。
【0019】
ComC切断活性の有無については、次のような方法で確認することができる。
PEPの活性は、50 mM Tris-HCl、150 mM硫酸アンモニウム、各濃度の基質ComCを含む、100μL(pH 7.0)の反応液中で測定した。反応開始は酵素添加によって行い、25℃下で基質消費が全体の10%以下になるような時間で測定を行った。反応は、反応溶液の入ったチューブを液体窒素で急冷することで停止した。酵素反応による生成ペプチドはBeckman System Gold HPLCシステム(Beckman-Coulter)に接続したWaters μBondasphere C8逆相カラム(Waters)により分析した。生成ペプチドを室温において流速1 mL/min、10分間の10−55%(v/v)アセトニトリル(0.1%のトリフルオロ酢酸を含む)のグラジエントにかけて分離し、210 nmの波長で検出した。PEPはミカエリス−メンテン型の酵素であるため、生成物の産生速度を算出し基質濃度に対してプロットしたs-v曲線から代謝回転数kcatおよび基質親和性Kmが求まる。
【0020】
また、簡便な切断活性の測定法としては上記の反応液中で切断したペプチドをゲル濃度16%のSDS-PAGEによって分離し、クマーシーブリリアントブルーR-250溶液により染色し目視により確認することが可能である。
【0021】
本発明において「PEP含有ポリペプチド又は活性PEP部分ポリペプチド」は、ミュータンス菌由来ComAタンパク質の60〜180番目のアミノ酸を含むポリペプチドであってもよく、同タンパク質の50〜190番目のアミノ酸を含むポリペプチドであれば好ましく、同タンパク質の45〜194番目のアミノ酸を含むポリペプチドであればより好ましく、同タンパク質の45〜194番目のアミノ酸からなるポリペプチドであればさらに好ましい。
【0022】
PEP含有ポリペプチド又は活性PEP部分ポリペプチドの取得方法としては、特に限定されないが、例えばPEP含有ポリペプチド又は活性PEP部分ポリペプチドをコードするDNAを組み込んだ発現プラスミドを微生物に導入し、この微生物にPEP含有ポリペプチド又は活性PEP部分ポリペプチドを産生させることによって取得することができる。微生物としては、特に限定されないが、例えば大腸菌等を用いることができる。ポリペプチドを効率的に産生させるために、例えばPEP含有ポリペプチド又は活性PEP部分ポリペプチドをコードするDNAに加えて薬剤耐性遺伝子も組み込んだ発現プラスミドを微生物に導入し、当該薬剤の存在下でこの微生物を培養することによって、PEP含有ポリペプチド又は活性PEP部分ポリペプチドをコードするDNAが導入された微生物を選抜することができる。薬剤としてはアンピシリン等を使用することができる。また、所定のオペロンを利用することにより発現誘導を行ってもよい。オペロンとしては、例えばラクトースオペロン等を利用することができる。この場合、イソプロピル-β-D-ガラクトピラノシドを培地中に添加することにより発現誘導を行うことができる。
【0023】
PEP含有ポリペプチド又は活性PEP部分ポリペプチドをコードするDNAを導入した微生物を培養した後にPEP含有ポリペプチド又は活性PEP部分ポリペプチドを回収することができる。なお、効率的に回収するために、PEP含有ポリペプチド又は活性PEP部分ポリペプチドを予め精製用タグを付加した形で発現させてもよい。精製用タグとしては、例えば、ヒスチジンタグ等を用いることができる。この場合、ヒスチジン結合樹脂等を用いることにより効率的に精製を行うことができる。
3.PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶
PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶は、空間群P43212に属する。
【0024】
また、好ましくは、PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶は、単位格子定数がa=b=58.6±0.7nm、及びc=112.1±1.0nmである。さらに、より好ましくは、PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶は、単位格子定数がa=b=58.6±0.35nm、及びc=112.1±0.5nmである。
【0025】
空間群・単位格子定数は、実施例に記載の方法にしたがって決定する。
【0026】
PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶は、所定条件の下、Sitting-drop蒸気拡散法(Sitting drop vapor diffusion法)によって取得することができる。
【0027】
PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶化方法においては、予めポリペプチドを分散させておく溶液中に従来タンパク質の安定化のために加えていた硫酸アンモニウムを実質的に含有させないことを特徴とする。硫酸アンモニウムを実質的に含有させないことによって、溶液中のポリペプチドの濃度を結晶化に適する程度(約5.5 mg/ml)にまで濃縮することができる。この際に使用する溶液としては、70mMリン酸ナトリウム、及び2mMβ−メルカプトエタノールを含有するpH 7.0の溶液に、さらに等量の沈殿剤(16%ポリエチレングリコール3350、及び0.16Mクエン酸水素二アンモニウム含有)を混合したものを挙げることができる。
【0028】
その他の詳しい条件は実施例に記載の通りである。
【実施例】
【0029】
以下に実施例により本発明をさらに詳細に説明するが、本発明は以下の例にのみ限定されるものではない。
1.PEPの全体構造、及びPEP-ComCアシル中間体モデル
[実験方法1]タンパク質発現および精製
結晶構造の解析方法としてMAD法を用いた。MAD法ではNativeタンパク質とセレノメチオニン(Se-Met)タンパク質を用いて解析を行う必要がある。MuPEP1のNativeおよびSeMetタンパクの結晶化には、MuPEP1の発現プラスミドpSMuP1(Biochemistry 47:2531-2538)を保持するメチオニン栄養要求性大腸菌株B834 (DE3)を使用した。この大腸菌をまず50 μg/mLのアンピシリンを含むLB培地にて37°Cで振とう培養した。菌体密度が1×108cell/mLに達したのち、この培養液の10 mLを37°Cで1時間、前培養した10 gのラクトースおよび50μg/mLのアンピシリンを含む1 LのLeMaster培地(EMBO J 9:1665-1672)に移した。この1 Lの培地には、Native MuPEP1、SeMet MuPEP1に対してそれぞれ25 mgのL-メチオニンまたはセレノ-L-メチオニンを添加した。これらの大腸菌を、37°Cにて23時間振とう培養した後に遠心分離によって回収し、20 mlの20 mM Tris-HCl、500 mM NaCl、5mMイミダゾール、pH 7.9の緩衝液に懸濁した。この細胞懸濁液を冷凍し、-80°Cで保存した。
【0030】
また、生化学的研究のためにPEPおよびComCを前述のように発現、精製した(Biochemistry 47:2531-2538)。要約すると、各発現プラスミドを保持する大腸菌株BL21(DE3)pLysSにおいて、Nativeおよび変異型の各PEPを発現させた。培養した大腸菌に0.2 mMのイソプロピル-β-D-チオガラクトピラノシド(IPTG)を加え、MuPEP1の発現は37°Cで2時間、もしくはPPEPは30°Cで5時間、誘導を行った。PEPはHis・Bind Resin(Novagen)で精製を行い、その後20 mM Tris-HCl、200 mM硫酸アンモニウム、2 mM β-メルカプトエタノール、pH 7.0 の緩衝液に対して、4°Cで透析を行い、精製標品を得た。
【0031】
ComCの発現には、各発現プラスミドを保持する大腸菌株JM109(DE3)pLysSを用い、PComCの発現は37°Cで1時間、CComCは37°Cで2時間、0.2mM IPTGによって誘導した。ComCをHis・Bind Resinによって精製し、その後AKTA高速タンパク質液体クロマトグラフィーシステム(GE Healthcare)に接続したMonoQカラムでクロマトグラフィーを行い、精製標品を得た。
【0032】
PEPおよびComCの濃度測定は、含有する芳香族アミノ酸の残基数に基づいて、分光光度法で行った(J Biol Chem 281:4726-4731)。
[実験方法2]タンパク質の結晶化およびデータ収集
Native MuPEP1の結晶を、20°CでSitting-drop蒸気拡散法によって得た(24ウェルLinbroプレートを使用)。70 mMリン酸ナトリウム、2 mM β-メルカプトエタノールに溶解した5.5 mg/mLタンパク質溶液2 μL と等量の16% ポリエチレングリコール3350、0.16 Mクエン酸2アンモニウムを含む沈殿剤と混合した。その結果、5日目に結晶が観察された。得られた結晶を24.5% ポリエチレングリコール3350、0.12 Mクエン酸 2-アンモニウムを含む沈殿剤にすばやく浸漬し、その後、液体窒素流(-173°C)中で急速冷凍した。 また、Native MuPEP1と同様の条件下で4.9 mg/mLのSeMet MuPEP1タンパク溶液から結晶を得ることができた。RIKEN Structural Genomics Beamline II BL26B2, SPring-8(兵庫県)
(J Struct Funct Genomics 7:15-22)のMarCCD 225検出器により、SeMet結晶の多波長異常分散(MAD)データセットおよびNative結晶の単一波長データセットを測定した。その後、データセットをプログラムパッケージHKL2000(Methods Enzymol 276:307-326)で処理した。
[実験方法3]構造決定および解析
測定したMADデータセットから、SHELX C/D/E 用のHKL2MAPインターフェース(J Appl Cryst 37:843-844)を使用して初期位相を計算した。結晶の空間群は、P43212であった。得られた位相をプログラムPHENIX(Acta Crystallogr D Biol Crystallogr 58:1948-1954)で改善し、続いて、プログラムARP/wARP(Nat Struct Biol 6:458-463)による自動モデル構築を行った。プログラムXtalView/X-fit(J Struct Biol 125:156-165)およびCNS(Acta Crystallogr D Biol Crystallogr 54:905-921)で、モデルの修正および精密化を行った。その後、1.9-Åの解像度でNativeのデータセットに対してモデルを修正した。CCP4パッケージソフトのプログラムPROCHECK(Acta Crystallogr D Biol Crystallogr 50:760-763)を使用して、最終モデルを作成した。結晶学的データおよび回折強度データと精密化の統計値を表S1に示す。図の描画はPyMOL(DeLano Scientific, Palo Alto, CA, USA. Delano Scientific)およびVMD(J Mol Graph 14:33-38)を使用した。
[実験方法4]分子モデリング
Xfit(J Struct Biol 125:156-165)を使用して、分子動力学シミュレーション用の初期モデルを構築した。PROPKA(Proteins 61:704-721)、PDB2PQR(Nucleic Acids Res 35:W522-W525)を使用して、これらのモデルを静電的にチェックした。タンパク質を水分子で満たされたbox中に配置し、VMD(J Mol Graph 14:33-38)を使用して電気的中性を維持するように、ナトリウムイオンをこの系に無作為に添加した。この結果、MuPEP1-MuComCモデルでは総原子数24,433個、容積67×60×63Å3、PPEPモデルは総原子数20,480個、容積57×61×64 Å3、PPEP-PComCモデルは総原子数30,511個、容積69×67×68Å3となった。
【0033】
分子動力学シミュレーションは、NAMD2.6(J Comput Chem 26:1781-1802)プログラムと、タンパク質およびリガンド用のCHARMM22フォースフィールド(J Phys Chem B 102:3586-3616)と水のTIP3Pモデル(J Chem Phys 79:926-935)を用いて実行した。ファン・デル・ワールス相互作用および実空間の静電相互作用に対する、カットオフ値(10Åから始まるスイッチング関数)は、12Åを想定した。また、周期的境界条件を使用した。長距離静電的相互作用を計算するために、particle-mesh Ewald法(J Chem Phys 98:10089-10092)を使用した。水素原子と重原子との間の全ての結合を制限するために、SHAKEアルゴリズムを使用した(J Compu Phys 23:327-341)。ステップは2 fsにした。これによりMultiple time steppingアルゴリズム(J Compu Phys 151:9-48)を使用した。共有結合および短距離の非結合相互作用を時間ステップごとに計算し、長距離静電的相互作用を2つの時間ステップごとに計算した。全てのシミュレーションで300Kの一定温度を維持するために、Langevin dynamicsを利用した。Nose-Hoover Langebin piston法(J Chem Phys 103:10252-10266) (decay period: 100 fs、damping time scale: 50 fs)を使用して、1気圧の定圧計算を行った。500回の初期エネルギー最小化の後、1 ns間、計算を行った。VMDによって、計算された軌道の解析を行った。
[考察1]PEPの全体構造
最初に、クオラムセンシングのメカニズムについて最も詳細に研究されているS. pneumoniae由来ComAのPEP(PPEP)の結晶化を行うことを検討した。
【0034】
結晶化のためにはミリグラムオーダーのPPEPのポリペプチドが必要となる。このため、PPEPのポリペプチドをできる限り高い効率で大腸菌内において発現させるため、S. pneumoniae由来ComA のうち、PEPに相当する領域を含む種々の長さのポリペプチドについて、それぞれ発現効率を比較検討することにした。(i) 1〜143番目のアミノ酸からなるポリペプチド、(ii) 1〜150番目のアミノ酸からなるポリペプチド、(iii) 1〜160番目のアミノ酸からなるポリペプチド、及び(iv) 1〜167番目のアミノ酸からなるポリペプチドについてそれぞれ検討した。なお、発現条件の最適化のため、発現誘導時に添加するIPTGの濃度や培養温度、及び培養時間等についても詳細に検討を行った。その結果、最も効率良く可溶性画分にタンパクが発現したのは、(ii) 1〜150番目のアミノ酸からなるポリペプチドであることが分かった。そのため、以降の検討ではこのポリペプチドを用いた。
【0035】
続いてPPEPの結晶化を試行したが良質な結晶を得ることができなかった。そこで、異なる菌体から6種のPEPを新たに得て、それぞれについて改めて結晶化の検討を行うことにした。
【0036】
PPEPの結晶化において種々の条件について試行錯誤を重ねるうち、系中にタンパク質安定化の目的で硫酸アンモニウムを含有させていたが、本発明者らは、この硫酸アンモニウムが結晶化の支障となっているのではないかと着想した。しかしながら、実際にこの硫酸アンモニウムを系中から除去すると、全てのPEPは溶液中で不安定となり、以降の結晶化作業を進めることができないことが判明した。ところが、さらに検討を進める中で、リン酸バッファーの濃度を70 mMにまで上げるという工夫をさらに加えることによって、唯一ミュータンス菌由来のPEP(MuPEP1)だけは、硫酸アンモニウムを系中から除去したとしても安定なままであることが分かった。しかも、MuPEP1は一般に結晶化サンプルとして必要とされる5 mg/ml以上にまで濃縮することが可能となった。これは、他に検討した種々の条件の中では到底実現することができなかったほどの高い濃度であった。
【0037】
MuPEP1の結晶化の検討においては、初期スクリーニングとして480条件についての検討を行った。その結果、1条件でのみ結晶化が可能であることが判明した。さらにその後に二次スクリーニングを行い、ポリエチレングリコール3350、及びクエン酸2アンモニウムの濃度検討を行って最適条件を決定した。
【0038】
MuPEP1の構造は1.9 Åの分解能で決定しRwork、Rfreeはそれぞれ21.2%、23.4%となった (表 1)。この構造はMuPEP1の5番目から141番目のアミノ酸残基と95個の水分子を含んでいる。N末の4残基およびC末のヒスチジンタグを含む15残基の電子密度は見えなかった。MuPEP1 はNativeタンパク質のN末44残基の疎水性領域をtruncateしたものであり、アミノ酸残基のナンバリングもtruncated formにしたがった。
【0039】
【表1】

【0040】
MuPEP1は6本のβ-ストランドと5本のヘリックスからなるα/βフォールドをとっていた(図1A)。中央に位置する6本のストランドからなる逆平行β-シートを挟むようにα-ヘリックスが配置されていて2つのサブドメインを形成している。N末側のサブドメイン(5-62番目の残基)は3本のα-ヘリックス、α1、α2およびα3 で形成されている。一方、C末側のサブドメイン(63-141番目の残基)は6本のβ-ストランドと2本のα-ヘリックスから形成されており、これらの2次構造コンポーネントはβ1、 α4、 β2、 β3、β4、 β5、α5、 β6 の順に配置されている。これら2つのサブドメインの会合面に狭いクレフトがあり、ここにCys17、His96、Asp112が形成するcatalytic triadが存在する。Cys17とHis96は、それぞれヘリックスα1 およびβストランドβ3のN末端に位置している。またAsp112はβ4 とβ5の間にあるループ上に位置している(図1A)。Cys17のSγ 原子とHis96のNδ1原子の距離は3.6 Åである。Asp112のOδ1原子とHis96のNε2 原子の距離は2.7 Åで水素結合を形成している(図1B)。
一次構造上の類似性は認められないにも関わらず、MuPEP1の結晶構造は他のパパイン様システインプロテアーゼと類似したフォールド構造を持っている(図2A)。N末側のサブドメインの3本のα-ヘリックスおよびC末側サブドメインのβ-シートとヘリックスα4 といったMuPEP1の二次構造の各成分は他のパパイン様システインプロテーゼのものと類似していた(図2A)。FATCATサーバーを用いてMuPEP1と他のパパイン様システインプロテアーゼの構造をCα 原子において重ね合わせたところpapainに対するrmsd が3.0 Å 、staphopain Aに対するrmsdが 2.7 Å となった。とりわけMuPEP1のcatalytic triadを形成する各残基とオキシアニオンホールを形成するGln11の空間配置はよく保存されている。MuPEP1のこれらの残基とpapainまたはstaphopain Aの相当する残基のCα 原子を重ね合わせたところrmsdはいずれも0.59 Åであった(図2B)。
[考察2]PEP-ComCアシル中間体モデル
PEPの基質認識メカニズムを調べるために、本発明者らはまずミュータンス菌のComC(MuComC)の大腸菌内での発現を試みたが、可溶性のタンパク質を得ることができなかった。次にMuComCを化学合成したが、できたペプチドは、ほとんどバッファーに不溶であった。また、MuPEP1のC17A変異型酵素とMuComCをグリシン6個からなるリンカーでつないだフュージョンタンパク質をデザインし、大腸菌内で発現を行ったところ可溶性のタンパク質が得られたが良質の結晶は得られなかった。本発明者らはさらに共結晶化およびソーキング法により、MuPEP1のC17A、C17S、H96A変異型酵素のいずれかと他のストレプトコッカス属細菌のComCのいずれかとの複合体結晶の作製を試みた。この方法ではいくつかの組み合わせで結晶が得られたが、ComCと複合体を形成している結晶は得られなかった。
【0041】
以上のような背景から本発明者らはMuPEP1-MuComC複合体のコンピューターモデルを作成した。まずComCのN末ヘリックスの疎水面と相互作用できそうな疎水性領域をMuPEP1のタンパク質表面から探した。この目的のため HotPatch program(J Mol Biol 369:863-879)によるタンパク質間の相互作用部位の検索をタンパク質表面の疎水性度に基づいたモードで行った。その結果、6つの疎水性領域が見つかった。しかしながら他のパパイン様プロテアーゼにおける基質配向の一般則を考慮に入れると、これらの疎水性領域のうち活性中心の近傍に位置する浅いくぼんだ領域のみがComCのN末ヘリックスを結合するサイトの候補と考えられた(図3A)。このくぼんだ領域も、MuPEP1の2つのサブドメインの会合面に位置しておりAla51、Leu52、Val55、Leu94、Leu134といった疎水性残基から形成されている。
【0042】
次に、本発明者らはMuPEP1とMuComCの間で形成されるアシル中間体を構築した。このモデルではMuPEP1の求核性のCys17がMuComCのGly-1のカルボニル炭素によってアシル化されている。また、MuComCの切断されたC末領域はこのモデルでは省略した。MuComCの主鎖Ile-3からGly-1は、ubiquitin C-terminal hydrolaseとそのインヒビターubiquitin aldehydeの複合体の構造(PDB ID code 1CMX)を参考にMuPEP1の活性部位クレフトにあてはめた。Ubiquitin aldehydeも切断部位にGly-Glyモチーフを有しており、C末端のGlyのカルボニル炭素と求核性のCys残基のSγ残基が共有結合している。このフィッティングによってIle-4の位置は制限される。MuComCのヘリックスの長さは未知なので、形成しうる最も長いIle-5からMet-25までの領域でヘリックスを形成した。そして、MuComCの保存された疎水性残基(図3B)のPhe-15、Ile-12、 Leu-7がMuPEP1の疎水性領域と相互作用するという仮定に合致するようおおよその位置にMuComCを結合させた。MuComCのこれら2つの領域をつなぎ合わせるために、Leu-7からIle-4までの主鎖をマニュアルで距離、ねじれ角が適切なコンフォーメーションをとるように調整した。最後に分子動力学(MD)シミュレーションを300 Kの温度下で行った。MDシミュレーションの間、MuPEP1-MuComC複合体のrmsdは最初の0.4 ナノ秒間でおおよそ1.5 Åまで増加し、その後1.0ナノ秒までその値を維持した(図4)。したがって1.0ナノ秒後の構造でエネルギー最小化を行った(図3C)。MDシミュレーションの0.4−1.0 ナノ秒間のMuPEP1の各Cα残基のゆらぎは4つの顕著なシグナルを示した(図5A)。これらのシグナルのピークはMuPEP1のループ構造に相当しており、それぞれβ1とβ2の間にある大きなループ構造中のα4のC末端の76番目の残基、β2とβ3の間にあるループの91番目から95番目の残基、β3とβ4の間にあるターン構造の105番目の残基とβ4とβ5の間にあるループの116番目の残基の位置に相当する。これらの位置は溶液側に露出しているため、MDシミュレーション中にMuPEP1が異常な挙動を示さなかったことを意味する。MuPEP1に対するMuComCのゆらぎはThr-22からMet-25の間で次第に増加していた(図5B)。このことより、MuComCのN末端の残基がMuPEP1から離れていることが示唆された。また、MuComCのすべての残基がラマチャンドランプロットで、適した位置に存在していた。
【0043】
図3Cのモデル中のMuComCの-25から-10までの領域がα-ヘリックス構造を保持していた。このヘリックス中の保存されているPhe-15、Ile-12とLeu-7とつづくターン構造中のLeu-7の側鎖がMuPEP1の疎水性表面に安定に結合していた。MuComCの主鎖は活性部位の入り口に位置しているLeu-7からIle-4の間で折れ曲がっている。Ile-4の側鎖はGln47とThr50の側鎖によってしっかり固定されている。一方、MuComCの親水性の残基、Lys-14、Glu-13、Lys-11、Thr-10の側鎖は溶液側に露出している。
【0044】
このモデルではMuComCのGly-GlyモチーフがMuPEP1の狭いクレフトにはまっている(図6)。このクレフトは上述の2つのサブドメインの会合面に位置している。クレフトの左側の壁(図6)は主にβ2とβ3の間にあるループ内のLys92からGln95までの残基によって形成されている。また、右側の壁はIle12からArg15までのN末鎖とα2とα3の間にあるループ内のLys46からGly48までの残基によって形成されている。活性部位の残基はクレフトの底面の一部分を形成している。Arg93の側鎖は、MDシミュレーションの後では活性部位にふたをしている(図3C マゼンタ)。このArg93のNη1と Nη2 原子はそれぞれ異なる2つの水分子を介して、Lys46の主鎖のカルボニル酸素と水素結合をしている。Arg93の側鎖のaliphaticな部分がMuComCのGly-2とファンデルワールス結合している。これらの相互作用は部分的にβ2とβ3の間にあるループのゆらぎを抑えていた(図5Aの2番目のピーク)。
2.モデルの実験による検証
[実験方法1]部位特異的変異
部位特異的変異はQuikChange II Site-Directed Mutagenesisキット(Stratagene)を使用して行った。PPEPのsingle mutationは全てpSPP1に導入した(J Biol Chem 281:4726-4731)。二重変異型PPEP、L52A/V55A PPEPは、V55AをL52A PPEPに導入した。A51W CAPEPおよびA67W CAPEPについては、C17A PPEPに各々変異を導入した(J Biol Chem 281:4726-4731)。Mutagenesisに使用したプライマーを表S2に記載する。作製した発現プラスミドについては、全てコーディング領域のヌクレオチド配列を確認した。
[実験方法2]速度論的パラメーターの決定
PPEP活性は、50 mM Tris-HCl、150 mM硫酸アンモニウム、各濃度の基質PComCを含む、100μL(pH 7.0)の反応液中で測定した。前述のように、高速液体クロマトグラフィー(HPLC)分析を行った(J Biol Chem 281:4726-4731)。要約すると、反応液をBeckman System Gold HPLCシステム(Beckman-Coulter)に接続したWaters μBondasphere C8逆相カラム(Waters)にアプライし、室温において流速1 mL/min、10分間の10−55%(v/v)アセトニトリル(0.1%のトリフルオロ酢酸を含む)のグラジエントにかけて、産生したペプチドを分析した。酵素反応で消費した基質の量は、全体の10%以下である。
[実験方法3]円二色性(CD)測定
CDスペクトルは、温度コントローラが付属したJasco分光偏光計・モデルJ-720WI(Jasco)を使用して測定した。セルは0.1cm光路長のものを使用した。緩衝液およびタンパク質の濃度は図7の凡例に記載した。
[考察]
本発明者らのモデルを検証するためにPPEPの基質結合サイトと予想される領域のアミノ酸残基に変異を導入した。この実験には唯一発現した天然のペアであるPPEP-PComCを用いた。
【0045】
上述の目的のため、新たにPPEP-PComCアシル中間体モデルを作製した。まずMuPEP1の構造をテンプレートにしてPPEPのモデルを構築した。MuPEP1の137残基を相当するPPEPのアミノ酸残基に置き換え(両タンパク質の相同性は57%)続いて分子動力学シミュレーション、エネルギー最小化を行った。得られたPPEPモデルにもMuPEP1においてHotPatchプログラムで予想されたものと同様の疎水性のくぼみが存在していた(図8)。PPEP-PComCアシル中間体モデルはMuPEP1-MuComCのモデリングと同様の手法を使って構築した。最終的に得られたモデルはMuPEP1-MuComCモデルとほぼ同様であったが、分子動力学シミュレーションの間にPComCのヘリックス構造がLeu-18とGlu-17の間で部分的にほどけていた(図9A)。
【0046】
このモデルからPComCの保存された疎水性残基との相互作用が予想されるThr50、Ala51、Leu52、Val55、Ala67、Leu94、Val134に変異を導入して活性測定を行うことにした(図9)。これらの残基の側鎖は溶液側に露出していると予想されるため変異を導入してもPPEPの全体構造にはほとんど影響ないものと考えられた。表2はこれら変異型PPEPのPComCに対する速度論的パラメーターを示す。T50S、L52A、V55A、L94A、V134Aでは触媒効率(kcat/Km)が1.8−16倍低下していた。ダブル変異を導入したL52A/V55A PPEPではPPEPのPComCに対する親和性が著しく減少していた(Kmが16倍上昇)。また、この変異型PPEPでは代謝回転数(kcat)はわずかしか低下していなかった。一方、PPEPの基質認識部位と推定される領域にTrp残基を導入すると(A51WとA67W)その立体障害によってPComCに対する親和性および代謝回転数がともに著しく減少し触媒効率がそれぞれ1/1400、1/110と著しく低下した。
【0047】
【表2】
【0048】
本発明者らは、CD測定によってComCがPEPと相互作用することで構造変化を起こすことを示してきた。A51WとA67Wの変異がひき起こす立体障害がComCのヘリックス形成にどのような影響を及ぼすのかを調べるためにA51WまたはA67W PPEP共存下、非共存下におけるComCのCDスペクトルをそれぞれ測定した。これらの変異は活性を持たないC17APPEP(CAPEP)に導入した。また、これまで調べた中でPPEPに対するKmが最も低いCComCを測定に用いた。これまでに示したようにバッファー中の20 μM CComC(図7、青)と35%トリフルオロエタノール(TFE)中(黒)の20 μM CComCはそれぞれランダムコイルとα-ヘリックス構造をとっている。CAPEP共存下でのCComCの構造変化は、CAPEP(20 μM)とCComC(20 μM)の混合液のCDとCAPEP(20 μM)のCDの差スペクトルを求めることにより調べた。CAPEP共存下でのCComCの差スペクトル(黄)は35%中で測定したCComCのスペクトルとほぼ同じであった。一方、A51W CAPEP共存下でのCComCの差スペクトル(赤)は、バッファー中で測定したCComCのスペクトルと同じであった。また、A67W CAPEP共存下でのCComCの差スペクトル(緑)のCAPEP共存下の差スペクトルに対する変化は、A51W CAPEP共存下での差スペクトルの変化と比べると中間であった。
3.考察のまとめ
(1)PEPの構造
1次構造比較から予想されていた、バクテリオシンのABCトランスポーターファミリーのペプチダーゼドメインがパパイン様システインプロテアーゼであるという仮説は、MuPEP1の3次元結晶構造によって検証することができた(図2)。C17S、C17A、H96A PPEPs とC17A MuPEP1が完全に活性を失っていた事から、PEPの触媒活性においてシステイン残基が重要であることが示された。Cys17の Sγ原子はHis96のNδ1原子から3.6 Å離れている。この距離は理想的な水素結合の距離、2.6−3.5 Åより少し長い。同様に、papainやstaphopain Aにおいても相当するCysとHis間の距離はそれぞれ、3.7 Å、4.0 Åである(図2B)。また、PROPKAプログラム(Proteins 61:704-721)から予想されるCys17、His96、Asp112のpKaは、それぞれ3.5、12、0.7であった。したがって他のパパイン様システインプロテアーゼで言われているように、Cys17とHis96は生理的条件下でthiolate-imidazoliumイオン対を形成しているものと考えられる。His96と水素結合を形成する距離にあるAsp112は、His96の空間的配置およびプロトン化を安定化しているのであろう。このようにMuPEP1はシステインプロテアーゼの触媒メカニズムに必要な全ての構造的要素を備えている。
(2)PEPの基質認識機構
PEPは、多様なタンパク質を分解するプロテアーゼと全体構造が類似しているにもかかわらず(図2A)、極めて基質特異性が高くComCと他のタンパク質を厳密に区別している。本発明者らのPEP-ComC複合体モデルは、ストレプトコッカス属細菌のPEPの基質認識の構造学的基盤を提供する。本発明者らの予想と合致して、MuPEP1の分子表面にComCと相互作用する2つの領域が見つかった。その1つは活性部位のクレフトである。このクレフトはストレプトコッカス属細菌のComCに保存されたGly-Glyモチーフを結合する(図6)。本発明者らのモデル上でMuComCのGly-1とGly-2をそれぞれAlaに改変すると、側鎖のメチル基がクレフトの側壁と立体障害を起こす。例えば、Ala-1は、PEPのThr14の主鎖と、Ala-2はGln95の主鎖とぶつかる。これまでの研究から、PComCのこれら2つのGlyをそれぞれAlaに改変したものを基質に用いてPPEPの活性を測定すると触媒効率が1/800に低下した。さらに、PPEPはシステインプロテアーゼを広く阻害するアンチパイン、ロイペプチン、E-64などによって触媒反応が阻害されない。これらの結果は、PEPのS1およびS2サイトがGly-Glyのみを認識する限定されたスペースをもつことを支持する結果である。
【0049】
もうひとつは、ComCの保存された疎水性残基(-15、-12、-7、-4番目のアミノ酸残基)と相互作用する活性部位クレフトの近傍にある疎水性の領域である(図3A、図8)。最初はComCのN末リーダー配列の全領域がα-ヘリックス構造を形成すると考えられていたが(図3B)、今回のPEP-ComC複合体モデルでは、ComCの-7番目から-4番目の領域がランダムコイルもしくはターン構造をとっている(図3C、図9A)。この疎水性表面は少しくぼんでおり、この領域を形成しているThr50、Ala51、Leu52、Val55、His87、Leu94、Thr132、Leu/Val134、Ile/Leu136はストレプトコッカス属細菌のPEP間で高度に保存されている。この領域のアミノ酸残基に変異を導入し活性測定を行ったところ、その重要性が確認された(表2)。PPEPの50、52、55、94、134番目の残基の変異ではいずれも触媒効率が1/1.8−1/16倍とある程度低下していたが、PPEPの変異型PComC、Phe-15A、Leu-12Ala、Leu-7Ala、Ile-4Alaに対する触媒効率の低下(1/31−1/180)と比較すると小さい。この事は、ComCの4つの保存性残基がPEP側の複数の残基と相互作用しているためであると考えられた。例えば、PComCのPhe-15の側鎖は、PPEPのAla51、Leu52、Val55、Ala67と相互作用している(図9B)。そして、PComCのPhe-15をAlaに変異するとこれらの相互作用が失われる一方、PEPのLeu52をAlaに変異した場合は、これらの相互作用の一部が失われるのみである。実際に、PPEPの52番目と55番目に二重変異を導入すると、PComCに対する親和性が加算的に低下した。PPEPのPhe-15Ala PComCに対するkcat/Kmは1/110倍に低下していた。また、L52A/V55A PPEPのPComCに対するkcat/Kmは1/27倍に低下していた。故に、PPEPのLeu52およびVal55の遷移状態の安定化に対する寄与はPComCのPhe-15とPPEP側の残基の相互作用の総和の68%(RTln27/RTln110)である。最小数の変異で活性を低下させ、この疎水性領域が基質結合部位であることをより明確に示すため本発明者らはこの領域に立体障害を導入した。予想された通り、A51WおよびA67W PPEPの触媒効率は著しく低下した。また、この基質結合部位と予想される領域の中央に位置するAla51に変異を導入したときの方が、辺縁部に位置するAla67に変異を導入したときよりも触媒効率の低下は大きかった。さらにA51Wの変異をMuPEP1にも導入してみたところ完全に失活していた。この結果は、この疎水性領域がすべてのストレプトコッカス属細菌のPEPにおいて重要であることを意味する。
【0050】
これら変異型PEPの活性測定の結果は、A51W CAPEPおよびA67W CAPEP共存下における、ComCのヘリックス構造変化ともよい相関を示した(図7)。これらの知見は、ComCのN末領域がPEPの疎水性領域に結合することで、ランダムコイルからヘリックスへと構造変化し、このことがPEP-ComC複合体がproductive formをとるために必要なステップであるという考えをさらに強化する。A51WおよびA67Wの変異は、kcat値を大きく下げるだけでなく、Km値も上げる。PEPはGly-Gly配列だけをもつタンパク質を切断しないことを考え合わせると、基質のGly-Gly配列周辺の主鎖は、基底状態(酵素・基質複合体)において切断部位の近傍でねじれていると考えられる。このねじれは、遷移状態で解消される。その結果、活性化エネルギーを低下させkcatを増加させる。おそらく、上述のPEPの疎水性領域とComCの間の大きな結合エネルギーはこのねじれを生じるために必要なのであろう。しかしながら、この相互作用だけでは不十分で、ComCのC末領域もPEPにしっかり結合することが必要であるし、もしくはComAの他の領域が切断部位周辺のねじれをつくるのに必要であるかもしれない。以上を明らかにするためには、基底状態や四面体型中間体、遷移状態に近い不安定な構造のモデルが必要となってくる。ここで示したアシル中間体は、基底状態および四面体型中間体いずれもと異なっている。実際に、アシル基の酸素はモデルのGln11が形成するオキシアニオンホール側を向いていない。さらに、切り出されたComCのC末領域(シグナル分子として機能するペプチド)は、細胞質側へもれ出ることなくまく貫通領域へ移行しなければならない。このように、PEPにおけるComCのC末領域の結合サイトを調べることもComAの機能を探る上で大変重要である。
本発明者らはまた、PEPのように小さなプロテアーゼがいかにして特異的に切断部位から15残基も離れた部位を認識するのかを説明するモデルを提唱する。MuPEP1-MuComCモデルでは、ComCのPhe-15のCα原子はGly-1のCα原子からおおよそ20 Åも離れている(図10)。本発明者らの知る限りでは、PEPは基質の最も広い領域を認識するプロテアーゼである。PEPのくぼんだ疎水性領域の残基のうち55番目の残基を除く全ての残基がバクテリオシンのABCトランスポーターのファミリー間で保存されている。このことから、これらのトランスポーターはComAと共通の基質認識機構を持っているものと考えられる。よって、この研究の結果は、バクテリオシンのABCトランスポーターの他のメンバーのペプチダーゼドメインを研究する上でのプロトタイプモデルを提供するものと考えられる。バクテリオシンのABCトランスポーターは、これまで原核生物にしか見つかっていないことから、ComAのペプチダーゼドメイン、とりわけMuPEP1の三次元結晶構造はストレプトコッカス属細菌のバイオフィルム形成を阻害する薬剤の開発に役立つものと期待される。
【技術分野】
【0001】
本発明は、ストレプトコッカス属細菌由来ComAタンパク質のペプチダーゼドメイン(PEP)の結晶に関する。
【背景技術】
【0002】
口腔内常在菌としてのストレプトコッカス属細菌は歯垢内の主要な細菌群であるが、歯のう蝕に関与するだけでなく、歯周ポケットから血流に入り込むと心臓弁膜に付着してバイオフィルムを形成し心内膜炎や人工弁感染症をひきおこす。バイオフィルムとは、細菌が自ら合成・分泌する多糖類などから成るマトリックス内に共同体を形成し、固体表面に付着して生育・増殖したものである。細菌が生体内で一旦バイオフィルムを形成すると抗生物質や白血球の浸透・拡散に対して耐性を示すため、除去がきわめて困難で外科的手術が必要となる。また、ブラッシングによる口腔バイオフィルムの完全除去は不可能である。バイオフィルムの形成には細菌に広く存在する細胞間情報伝達系であるクオラムセンシングのメカニズムが重要である。したがってストレプトコッカス属細菌のクオラムセンシングのメカニズムを解明し、阻害剤を開発することは上記のStreptococcus感染症に対する有効な予防・治療法の開発に役立つと考えられる。
Streptococcusのクオラムセンシングを阻害する薬剤を開発する上で、その初発段階で機能するComAは重要なタンパク質であると考えられる。ComAはシグナルペプチド前駆体であるComCを切断すると同時に菌体外へと分泌するATP-binding cassette(ABC)トランスポーターである。しかしながら、その重要性にも関わらずComAをはじめとする、グラム陽性菌のクオラムセンシングのシグナル産生タンパク質は膜タンパク質で大量発現が困難なため構造学的解析が皆無であった。
以上のような背景からComAの立体構造の解明が渇望されている。しかしこれまで、ComAの三次元構造を解析し原子座標を明らかにし、その活性中心や基質結合部位の位置および構造を決定したという報告はない。
【先行技術文献】
【非特許文献】
【0003】
【非特許文献1】J. Biol. Chem. 281, 4726-4731
【発明の概要】
【発明が解決しようとする課題】
【0004】
ComAタンパク質のペプチダーゼドメイン(PEP)の結晶を提供することを課題とする。
【課題を解決するための手段】
【0005】
一般にタンパク質の結晶化は容易ではなく、多くのタンパク質の場合、数多くの条件検討を重ねてはじめてX線結晶構造解析に供することのできる結晶が得られる。また、いかなる条件でも結晶化しないタンパク質も少なくなく、タンパク質の結晶が得られなければX線結晶構造解析を行うことができず、そのタンパク質の三次元構造を解明することはできない。
本発明者らはストレプトコッカス属細菌のComAタンパク質の三次元構造を解析するべく日夜検討を重ねた。はじめにクオラムセンシングのメカニズムが最も詳細に解析されているS. pneumoniaeのComA全長について大量発現の検討を行った。しかしながら、従来の方法をそのまま適用しても大量発現できないことが判明した。ここで本発明者らは、ComA全長の大量発現は困難であると判断し、代わりにペプチダーゼドメインのみからなるポリペプチドの大量発現を試みた。N末の150残基からなるペプチダーゼドメイン(PEP)がシグナルペプチド前駆体であるComCを切断する活性をもつことは確認済みである。ペプチダーゼドメインを含む種々の長さのポリペプチドについて大量発現を検討し、最も効率良く可溶性画分にタンパク質が発現したポリペプチドを精製し、結晶化の検討を行った。初期スクリーニングとして計480条件について検討したが、ついに結晶を得ることはできなかった。
【0006】
そこで、本発明者らは他のStreptococcus由来のPEPを加えた計6種類のPEPについても同様に大量発現、精製、及び結晶化の検討を行った。その結果、従来の方法をそのまま適用したとしてもいずれのPEPについても結晶化することはできなかった。そこで、本発明者らは試行錯誤を重ね、当初はPEPタンパク質を安定化することを目的として加えていた硫酸アンモニウムを系内から除いて結晶化を行うことによって、かつPEPとしてミュータンス菌(S. mutans)由来のPEPを採用することによって、はじめてPEPの結晶を得ることに成功した。そして、その結晶にX線を当てることにより分解能約1.9 ÅのX線回折強度が得られることを確認した。
【0007】
さらに、本発明者らは前記PEPの結晶についてX線結晶構造解析を行うことにより、その三次元構造を明らかにすることが可能であることを実証し、本発明を完成させた。
【0008】
すなわち、本発明は次の通りである:
項1.ComAタンパク質のペプチダーゼドメイン(PEP)を含むポリペプチド;又は
PEPの部分ポリペプチドであってComC切断活性を有するポリペプチド
の結晶であって、空間群P43212に属する結晶。
項2.単位格子定数がa=b=58.6±0.35 nm、及びc=112.1±0.5 nmである、項1に記載の結晶。
項3.前記ポリペプチドが、
(A) 配列番号1のアミノ酸配列からなるポリペプチド;又は
(B-1) (A)のポリペプチドのアミノ酸配列において、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチド
である、項1又は2に記載の結晶。
項4.前記ポリペプチドが、
(A) 配列番号1のアミノ酸配列からなるポリペプチド;又は
(B-2) (A)のポリペプチドのアミノ酸配列において、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されることにより、(A)のポリペプチドのアミノ酸配列に対して90%以上の相同性の範囲内で修飾されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチド
である、項1又は2に記載の結晶。
項5.項1〜4のいずれか一項に記載の結晶の製造方法であって、
硫酸アンモニウムを実質的に含まない溶液中に前記ポリペプチドを含有させた状態で結晶化することを特徴とする方法。
項6.前記溶液が、70 mMリン酸ナトリウム、及び2 mMβ−メルカプトエタノールを含有するpH 7.0の溶液に、16%ポリエチレングリコール3350、及び0.16 Mクエン酸水素二アンモニウムを含有する等量の沈殿剤を混合したものである、項5に記載の方法。
項7.さらにSitting-drop蒸気拡散法で結晶化することを特徴とする、項5又は6に記載の方法。
【発明の効果】
【0009】
本発明は、PEPの結晶化条件を初めて見出したものであり、これによりX線結晶構造解析に適した結晶の調製が可能となる。解明されたPEPの三次元構造を用いれば、活性中心および基質結合部位のアミノ酸残基とそれらの立体配置が明らかとなる。したがって、得られた座標データを用いてコンピュータースクリーニング法による阻害剤を探索することが可能となる。さらにPEPの結晶を用いて候補化合物との複合体結晶を作製し、その構造決定を行うことでより詳細な解析を行うこともできる。
【0010】
また、本発明の結晶を利用して得られた座標データは、未知の構造のPEPホモログの探索、変異体の構造のホモロジーモデリングによる推定又は分子置換法による決定のために使用できる。
【図面の簡単な説明】
【0011】
【図1】(A) MuPEP1全体構造のCartoon 表示。二次構造の各成分はレインボーカラーで示してある(N末端が青。C末端が赤。)。Catalytic triadを形成するCys17、His96、Asp112およびオキシアニオンホールを形成するGln11の側鎖はスティックで表す。(B)MuPEP1の活性中心。主鎖の原子は半透明のCartoon(緑)で示す。活性中心近傍の残基の側鎖はスティックで表す。酸素、窒素、硫黄の各原子はそれぞれ赤、青、黄で表す。点線は水素結合を示す。(C) ComAのドメイン構成の推定図。PEP, ペプチダーゼドメイン。TM, 膜貫通ドメイン。ABC, ATP-結合ドメイン。バクテリオシンのABCトランスポーターは二量体として機能すると考えられている。
【図2】(A)MuPEP1(左)、papain(PDB ID code 9PAP)(中央)、staphopain A (PDB ID code 1CV8)(右)。それぞれの構造は活性中心の残基Gln11、Cys17、His96、Asp112のCα原子を重ね合わせ、同方向から見たもの。これらの残基はスティックモデルで示す。構造的に相同のα-ヘリックス、 α1-4(MuPEP1)およびβ-ストランド、β1-6(MuPEP1)はそれぞれシアンと緑で示す。(B)MuPEP1と他のパパイン様システインプロテアーゼの活性部位の重ね合わせ。MuPEP1のGln11、Cys17、His96、Asp112はスティックモデルで示した(緑)。これらの残基に相当するpapain(シアン)およびstaphopain A(グレー)はCα原子で重ね合わせ、スティックモデルで示した。酸素、窒素、硫黄の各原子はそれぞれ赤、青、黄で表す。
【図3】(A) HotPatchプログラム (23) の結果(表面図)。疎水性領域を赤、親水性領域を青で示した。活性部位近傍の疎水性領域をオレンジ色の線で囲った。(B) PComCのN末領域のα-ヘリカル ホイール図。PPEPとの相互作用に重要であると考えられた高度に保存されたアミノ酸(黄色)(20)。(C)MuPEP1-MuComCアシル中間体モデル。MuPEP1は表面図で示す。MuComCの側鎖はスティックモデルで示す。酸素、窒素、硫黄の各原子は赤、青、黄で示す。MuComCの4つの高度に保存された残基、Phe-15、Leu-12、Leu-7、Ile-4は球モデル(黄)で示す。2次構造の各成分はランダムコイル(緑)、ターン(シアン)、α-ヘリックス(紫)。Cys17はオレンジで、Arg93(詳細は本文)は、マゼンタで示す。右側の構造は左側の構造をY軸に対して90° 回転させたもの。
【図4】MDシミュレーション中の各Cα原子の初期構造の対応するCα原子に対するrmsdの時間経過。初期構造からのMuPEP1-MuComC複合体のrmsdを2 psごとに計算した。
【図5】MuPEP1-MuComC複合体の各Cα原子のゆらぎ。(A)0.4ナノ秒における対応原子からの0.4ナノ秒〜1.0ナノ秒の間のMDシミュレーションでのMuPEP1(5から141の残基)の各Cα原子のRMS位置変動。主要ピークに対応する残基番号を示す。(B)0.4ナノ秒における対応原子からの0.4ナノ秒〜1.0ナノ秒の間のMDシミュレーションでのMuComC(-25から-1の残基)の各Cα原子のRMS位置変動。複合体のMuPEP1成分に対する剛体整合の後に、MuComCのRMSDを計算した。
【図6】MuPEP1-MuComCアシル中間体モデル(緑)をMuPEP1(グレー)の結晶構造に重ね合わせた図。重ね合わせはGln11、Cys17、His96、Asp112のCα原子で行った。MuPEP1のクレフトの表面はPDBsum (28)によって解析し、バスケットで表わした。MuComCはGly-1、Gly-2、Ile-3のみ表示した。酸素、窒素、硫黄の各原子はそれぞれ赤、青、黄で示した。右側の構造は、左の構造の矢印で示した方向から見た図。
【図7】CComCの遠紫外部(207−260 nm)のCDスペクトルを変異型PPEP共存下または非共存下で測定した。波長207 nm以下のスペクトルはタンパク質の紫外線吸収によるノイズが大きいため測定できなかった。単位は平均残基楕円率で示す。CD測定は20 μM CComCおよび20 μM CComCと20 μMのCAPEPまたはA51W CAPEPまたはA67W CAPEPを用いて行った。バッファー溶液は30 mM リン酸ナトリウム、150 mM 硫酸アンモニウム、15 mM フッ化ナトリウム、0.025 mM ジチオスレイトール、pH 7.0。スペクトルはスキャン速度50 nm/分で30回の積算を行った。CComC(0% TFE中)のスペクトル(青)。CComC(35% TFE中)のスペクトル(黒)。各変異型PPEPのスペクトルをCComCと各変異型PPEPの混合液のスペクトルから差し引いた、差スペクトルは、それぞれ(CComC + CAPEP) - CAPEPは黄、 (CComC + A51W CAPEP) - A51W CAPEPは赤、(CComC + A67W CAPEP) - A67W CAPEPは緑の各色で示した。
【図8】MuPEP1(左)およびPPEP(右)の推定される疎水性のくぼんだ領域。HotPatchプログラムの結果を表面図で示したもの。親水性領域を赤、疎水性領域を青で示した。オレンジの楕円形の線に囲まれた領域は、活性部位クレフトに隣接する疎水性のくぼんだ領域を示す。Cα原子の最小二乗重合に基づいて、2つの構造は、Cα原子で重ね合わせた。MuPEP1の疎水性のくぼんだ領域は、Thr50、Ala51、Leu52、Val55、His87、Leu94、Gln95、Thr132、Leu134、およびIle136から、PPEPの疎水性のくぼんだ領域は、Thr50、Ala51、Leu52、Val55、His87、Val88、Leu89、Leu94、Leu95、Thr132、Val134、およびLeu136から構成されている。
【図9】(A)PPEP-PComCアシル中間体モデル PPEPとPComCの複合体を球モデルで示した。PComCの二次構造の各成分は、それぞれランダムコイルは緑、ターンはシアン、α-ヘリックスはグレーの各色で示した。PComCの4つの保存された残基の側鎖は黄色で表した。PComCのこれら4つの残基と相互作用が予想されたPPEP側の残基を青で表した。(B)PComCの主鎖を(A)から省いた図。PComCのPhe-15はPPEPのAla51、Leu52、Val55、Ala67と、Leu-12はPPEPのAla67およびVal134、Leu-7はPPEPのLeu94と、Ile-4はPPEPのThr50とそれぞれ相互作用している。(C)PComCすべてを(A)から省き、PPEPの疎水性のくぼみを示した。
【図10】Phe-15のCα原子とGly-1のCα原子との間の距離。MuPEP1の全体構造はグレーのcartoon(半透明)によって示した。MuPEP1のGln11、Cys17、His96、およびAsp112の活性部位残基をstickによって示した。MuComCは白いstickによって示した。MuComCの高度に保存された残基の側鎖を黄色で示した。酸素、窒素、および硫黄原子を、それぞれ、赤、青、および黄色で示した。鎖線はPhe-15のCα原子とGly-1のCα原子との間の距離を示す。
【発明を実施するための形態】
【0012】
1.ComAタンパク質
本発明において使用するComAタンパク質は、特に限定されないが、ストレプトコッカス属細菌由来ComAタンパク質、又はストレプトコッカス属細菌由来ComAタンパク質の改変体であれば好ましい。さらに、ミュータンス菌(Streptococcus mutans)由来ComAタンパク質、又はミュータンス菌由来ComAタンパク質の改変体であればより好ましい。
【0013】
なお、ここでいう改変体は、天然において生じたものであってもよい。天然において生じた改変体としては、例えば突然変異や翻訳後の修飾などにより生じた改変体を挙げることができる。また、改変体は、天然に存在するComAタンパク質の遺伝子配列又はアミノ酸配列を利用して人為的に生じさせたものであってもよい。改変体には、アレル体、ホモログ、天然の変異体などであって、少なくとも90%、好ましくは95%、より好ましくは98%、さらにより好ましくは99%が、変異前のポリペプチドと相同するものが含まれる。ポリペプチドの相同性は、既知の配列分析ソフトウェア、例えばFASTAプログラムを使用した測定(Clustal,V., MethodsMol.Biol., 25, 307-318 (1994))によって解析することができる。また、SWISSPLOTSで解析することもできる。
【0014】
ミュータンス菌(Streptococcus mutans)とは、主に口腔内に存在し、う蝕の原因菌の一つであるとされるグラム陽性通性嫌気性連鎖球菌の一種である。
【0015】
ミュータンス菌由来ComAタンパク質としては、配列番号1のアミノ酸配列からなるポリペプチドを挙げることができる。ミュータンス菌由来ComAタンパク質をコードする遺伝子としては、配列番号2の遺伝子配列からなるポリヌクレオチドを挙げることができる。
【0016】
ComAタンパク質は、クオラムセンシングにおいてシグナルペプチド前駆体ComCからシグナルペプチドを切り出すと同時に菌体外へ分泌するという生理的作用を有する。この生理的作用においてPEPはComCを切断するという作用を担っている。
2.PEP含有ポリペプチド又は活性PEP部分ポリペプチド
本発明において「PEPを含むポリペプチド(PEP含有ポリペプチド)、又はPEPの部分ポリペプチドであってComC切断活性を有するポリペプチド(活性PEP部分ポリペプチド)」は、好ましくは、
(A) 配列番号1のアミノ酸配列からなるポリペプチド;又は
(B-1) (A)のポリペプチドのアミノ酸配列において、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチドである。
【0017】
上記ポリペプチド(A)は、天然に存在するミュータンス菌に由来するComAタンパク質のN末端から45番目のアミノ酸から194番目のアミノ酸までの部分ポリペプチドである。
【0018】
上記ポリペプチド(B-1)は、上記ポリペプチド(A)の改変体である。上記ポリペプチド(B-1)は、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されることにより、(A)のポリペプチドのアミノ酸配列に対して90%以上の相同性の範囲内で修飾されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチドであれば好ましい。上記相同性は、95%以上であればより好ましく、98%以上であればさらに好ましく、99%以上であればさらにより好ましい。ポリペプチドの相同性は、先述したのと同様に解析することができる。
【0019】
ComC切断活性の有無については、次のような方法で確認することができる。
PEPの活性は、50 mM Tris-HCl、150 mM硫酸アンモニウム、各濃度の基質ComCを含む、100μL(pH 7.0)の反応液中で測定した。反応開始は酵素添加によって行い、25℃下で基質消費が全体の10%以下になるような時間で測定を行った。反応は、反応溶液の入ったチューブを液体窒素で急冷することで停止した。酵素反応による生成ペプチドはBeckman System Gold HPLCシステム(Beckman-Coulter)に接続したWaters μBondasphere C8逆相カラム(Waters)により分析した。生成ペプチドを室温において流速1 mL/min、10分間の10−55%(v/v)アセトニトリル(0.1%のトリフルオロ酢酸を含む)のグラジエントにかけて分離し、210 nmの波長で検出した。PEPはミカエリス−メンテン型の酵素であるため、生成物の産生速度を算出し基質濃度に対してプロットしたs-v曲線から代謝回転数kcatおよび基質親和性Kmが求まる。
【0020】
また、簡便な切断活性の測定法としては上記の反応液中で切断したペプチドをゲル濃度16%のSDS-PAGEによって分離し、クマーシーブリリアントブルーR-250溶液により染色し目視により確認することが可能である。
【0021】
本発明において「PEP含有ポリペプチド又は活性PEP部分ポリペプチド」は、ミュータンス菌由来ComAタンパク質の60〜180番目のアミノ酸を含むポリペプチドであってもよく、同タンパク質の50〜190番目のアミノ酸を含むポリペプチドであれば好ましく、同タンパク質の45〜194番目のアミノ酸を含むポリペプチドであればより好ましく、同タンパク質の45〜194番目のアミノ酸からなるポリペプチドであればさらに好ましい。
【0022】
PEP含有ポリペプチド又は活性PEP部分ポリペプチドの取得方法としては、特に限定されないが、例えばPEP含有ポリペプチド又は活性PEP部分ポリペプチドをコードするDNAを組み込んだ発現プラスミドを微生物に導入し、この微生物にPEP含有ポリペプチド又は活性PEP部分ポリペプチドを産生させることによって取得することができる。微生物としては、特に限定されないが、例えば大腸菌等を用いることができる。ポリペプチドを効率的に産生させるために、例えばPEP含有ポリペプチド又は活性PEP部分ポリペプチドをコードするDNAに加えて薬剤耐性遺伝子も組み込んだ発現プラスミドを微生物に導入し、当該薬剤の存在下でこの微生物を培養することによって、PEP含有ポリペプチド又は活性PEP部分ポリペプチドをコードするDNAが導入された微生物を選抜することができる。薬剤としてはアンピシリン等を使用することができる。また、所定のオペロンを利用することにより発現誘導を行ってもよい。オペロンとしては、例えばラクトースオペロン等を利用することができる。この場合、イソプロピル-β-D-ガラクトピラノシドを培地中に添加することにより発現誘導を行うことができる。
【0023】
PEP含有ポリペプチド又は活性PEP部分ポリペプチドをコードするDNAを導入した微生物を培養した後にPEP含有ポリペプチド又は活性PEP部分ポリペプチドを回収することができる。なお、効率的に回収するために、PEP含有ポリペプチド又は活性PEP部分ポリペプチドを予め精製用タグを付加した形で発現させてもよい。精製用タグとしては、例えば、ヒスチジンタグ等を用いることができる。この場合、ヒスチジン結合樹脂等を用いることにより効率的に精製を行うことができる。
3.PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶
PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶は、空間群P43212に属する。
【0024】
また、好ましくは、PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶は、単位格子定数がa=b=58.6±0.7nm、及びc=112.1±1.0nmである。さらに、より好ましくは、PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶は、単位格子定数がa=b=58.6±0.35nm、及びc=112.1±0.5nmである。
【0025】
空間群・単位格子定数は、実施例に記載の方法にしたがって決定する。
【0026】
PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶は、所定条件の下、Sitting-drop蒸気拡散法(Sitting drop vapor diffusion法)によって取得することができる。
【0027】
PEP含有ポリペプチド又は活性PEP部分ポリペプチドの結晶化方法においては、予めポリペプチドを分散させておく溶液中に従来タンパク質の安定化のために加えていた硫酸アンモニウムを実質的に含有させないことを特徴とする。硫酸アンモニウムを実質的に含有させないことによって、溶液中のポリペプチドの濃度を結晶化に適する程度(約5.5 mg/ml)にまで濃縮することができる。この際に使用する溶液としては、70mMリン酸ナトリウム、及び2mMβ−メルカプトエタノールを含有するpH 7.0の溶液に、さらに等量の沈殿剤(16%ポリエチレングリコール3350、及び0.16Mクエン酸水素二アンモニウム含有)を混合したものを挙げることができる。
【0028】
その他の詳しい条件は実施例に記載の通りである。
【実施例】
【0029】
以下に実施例により本発明をさらに詳細に説明するが、本発明は以下の例にのみ限定されるものではない。
1.PEPの全体構造、及びPEP-ComCアシル中間体モデル
[実験方法1]タンパク質発現および精製
結晶構造の解析方法としてMAD法を用いた。MAD法ではNativeタンパク質とセレノメチオニン(Se-Met)タンパク質を用いて解析を行う必要がある。MuPEP1のNativeおよびSeMetタンパクの結晶化には、MuPEP1の発現プラスミドpSMuP1(Biochemistry 47:2531-2538)を保持するメチオニン栄養要求性大腸菌株B834 (DE3)を使用した。この大腸菌をまず50 μg/mLのアンピシリンを含むLB培地にて37°Cで振とう培養した。菌体密度が1×108cell/mLに達したのち、この培養液の10 mLを37°Cで1時間、前培養した10 gのラクトースおよび50μg/mLのアンピシリンを含む1 LのLeMaster培地(EMBO J 9:1665-1672)に移した。この1 Lの培地には、Native MuPEP1、SeMet MuPEP1に対してそれぞれ25 mgのL-メチオニンまたはセレノ-L-メチオニンを添加した。これらの大腸菌を、37°Cにて23時間振とう培養した後に遠心分離によって回収し、20 mlの20 mM Tris-HCl、500 mM NaCl、5mMイミダゾール、pH 7.9の緩衝液に懸濁した。この細胞懸濁液を冷凍し、-80°Cで保存した。
【0030】
また、生化学的研究のためにPEPおよびComCを前述のように発現、精製した(Biochemistry 47:2531-2538)。要約すると、各発現プラスミドを保持する大腸菌株BL21(DE3)pLysSにおいて、Nativeおよび変異型の各PEPを発現させた。培養した大腸菌に0.2 mMのイソプロピル-β-D-チオガラクトピラノシド(IPTG)を加え、MuPEP1の発現は37°Cで2時間、もしくはPPEPは30°Cで5時間、誘導を行った。PEPはHis・Bind Resin(Novagen)で精製を行い、その後20 mM Tris-HCl、200 mM硫酸アンモニウム、2 mM β-メルカプトエタノール、pH 7.0 の緩衝液に対して、4°Cで透析を行い、精製標品を得た。
【0031】
ComCの発現には、各発現プラスミドを保持する大腸菌株JM109(DE3)pLysSを用い、PComCの発現は37°Cで1時間、CComCは37°Cで2時間、0.2mM IPTGによって誘導した。ComCをHis・Bind Resinによって精製し、その後AKTA高速タンパク質液体クロマトグラフィーシステム(GE Healthcare)に接続したMonoQカラムでクロマトグラフィーを行い、精製標品を得た。
【0032】
PEPおよびComCの濃度測定は、含有する芳香族アミノ酸の残基数に基づいて、分光光度法で行った(J Biol Chem 281:4726-4731)。
[実験方法2]タンパク質の結晶化およびデータ収集
Native MuPEP1の結晶を、20°CでSitting-drop蒸気拡散法によって得た(24ウェルLinbroプレートを使用)。70 mMリン酸ナトリウム、2 mM β-メルカプトエタノールに溶解した5.5 mg/mLタンパク質溶液2 μL と等量の16% ポリエチレングリコール3350、0.16 Mクエン酸2アンモニウムを含む沈殿剤と混合した。その結果、5日目に結晶が観察された。得られた結晶を24.5% ポリエチレングリコール3350、0.12 Mクエン酸 2-アンモニウムを含む沈殿剤にすばやく浸漬し、その後、液体窒素流(-173°C)中で急速冷凍した。 また、Native MuPEP1と同様の条件下で4.9 mg/mLのSeMet MuPEP1タンパク溶液から結晶を得ることができた。RIKEN Structural Genomics Beamline II BL26B2, SPring-8(兵庫県)
(J Struct Funct Genomics 7:15-22)のMarCCD 225検出器により、SeMet結晶の多波長異常分散(MAD)データセットおよびNative結晶の単一波長データセットを測定した。その後、データセットをプログラムパッケージHKL2000(Methods Enzymol 276:307-326)で処理した。
[実験方法3]構造決定および解析
測定したMADデータセットから、SHELX C/D/E 用のHKL2MAPインターフェース(J Appl Cryst 37:843-844)を使用して初期位相を計算した。結晶の空間群は、P43212であった。得られた位相をプログラムPHENIX(Acta Crystallogr D Biol Crystallogr 58:1948-1954)で改善し、続いて、プログラムARP/wARP(Nat Struct Biol 6:458-463)による自動モデル構築を行った。プログラムXtalView/X-fit(J Struct Biol 125:156-165)およびCNS(Acta Crystallogr D Biol Crystallogr 54:905-921)で、モデルの修正および精密化を行った。その後、1.9-Åの解像度でNativeのデータセットに対してモデルを修正した。CCP4パッケージソフトのプログラムPROCHECK(Acta Crystallogr D Biol Crystallogr 50:760-763)を使用して、最終モデルを作成した。結晶学的データおよび回折強度データと精密化の統計値を表S1に示す。図の描画はPyMOL(DeLano Scientific, Palo Alto, CA, USA. Delano Scientific)およびVMD(J Mol Graph 14:33-38)を使用した。
[実験方法4]分子モデリング
Xfit(J Struct Biol 125:156-165)を使用して、分子動力学シミュレーション用の初期モデルを構築した。PROPKA(Proteins 61:704-721)、PDB2PQR(Nucleic Acids Res 35:W522-W525)を使用して、これらのモデルを静電的にチェックした。タンパク質を水分子で満たされたbox中に配置し、VMD(J Mol Graph 14:33-38)を使用して電気的中性を維持するように、ナトリウムイオンをこの系に無作為に添加した。この結果、MuPEP1-MuComCモデルでは総原子数24,433個、容積67×60×63Å3、PPEPモデルは総原子数20,480個、容積57×61×64 Å3、PPEP-PComCモデルは総原子数30,511個、容積69×67×68Å3となった。
【0033】
分子動力学シミュレーションは、NAMD2.6(J Comput Chem 26:1781-1802)プログラムと、タンパク質およびリガンド用のCHARMM22フォースフィールド(J Phys Chem B 102:3586-3616)と水のTIP3Pモデル(J Chem Phys 79:926-935)を用いて実行した。ファン・デル・ワールス相互作用および実空間の静電相互作用に対する、カットオフ値(10Åから始まるスイッチング関数)は、12Åを想定した。また、周期的境界条件を使用した。長距離静電的相互作用を計算するために、particle-mesh Ewald法(J Chem Phys 98:10089-10092)を使用した。水素原子と重原子との間の全ての結合を制限するために、SHAKEアルゴリズムを使用した(J Compu Phys 23:327-341)。ステップは2 fsにした。これによりMultiple time steppingアルゴリズム(J Compu Phys 151:9-48)を使用した。共有結合および短距離の非結合相互作用を時間ステップごとに計算し、長距離静電的相互作用を2つの時間ステップごとに計算した。全てのシミュレーションで300Kの一定温度を維持するために、Langevin dynamicsを利用した。Nose-Hoover Langebin piston法(J Chem Phys 103:10252-10266) (decay period: 100 fs、damping time scale: 50 fs)を使用して、1気圧の定圧計算を行った。500回の初期エネルギー最小化の後、1 ns間、計算を行った。VMDによって、計算された軌道の解析を行った。
[考察1]PEPの全体構造
最初に、クオラムセンシングのメカニズムについて最も詳細に研究されているS. pneumoniae由来ComAのPEP(PPEP)の結晶化を行うことを検討した。
【0034】
結晶化のためにはミリグラムオーダーのPPEPのポリペプチドが必要となる。このため、PPEPのポリペプチドをできる限り高い効率で大腸菌内において発現させるため、S. pneumoniae由来ComA のうち、PEPに相当する領域を含む種々の長さのポリペプチドについて、それぞれ発現効率を比較検討することにした。(i) 1〜143番目のアミノ酸からなるポリペプチド、(ii) 1〜150番目のアミノ酸からなるポリペプチド、(iii) 1〜160番目のアミノ酸からなるポリペプチド、及び(iv) 1〜167番目のアミノ酸からなるポリペプチドについてそれぞれ検討した。なお、発現条件の最適化のため、発現誘導時に添加するIPTGの濃度や培養温度、及び培養時間等についても詳細に検討を行った。その結果、最も効率良く可溶性画分にタンパクが発現したのは、(ii) 1〜150番目のアミノ酸からなるポリペプチドであることが分かった。そのため、以降の検討ではこのポリペプチドを用いた。
【0035】
続いてPPEPの結晶化を試行したが良質な結晶を得ることができなかった。そこで、異なる菌体から6種のPEPを新たに得て、それぞれについて改めて結晶化の検討を行うことにした。
【0036】
PPEPの結晶化において種々の条件について試行錯誤を重ねるうち、系中にタンパク質安定化の目的で硫酸アンモニウムを含有させていたが、本発明者らは、この硫酸アンモニウムが結晶化の支障となっているのではないかと着想した。しかしながら、実際にこの硫酸アンモニウムを系中から除去すると、全てのPEPは溶液中で不安定となり、以降の結晶化作業を進めることができないことが判明した。ところが、さらに検討を進める中で、リン酸バッファーの濃度を70 mMにまで上げるという工夫をさらに加えることによって、唯一ミュータンス菌由来のPEP(MuPEP1)だけは、硫酸アンモニウムを系中から除去したとしても安定なままであることが分かった。しかも、MuPEP1は一般に結晶化サンプルとして必要とされる5 mg/ml以上にまで濃縮することが可能となった。これは、他に検討した種々の条件の中では到底実現することができなかったほどの高い濃度であった。
【0037】
MuPEP1の結晶化の検討においては、初期スクリーニングとして480条件についての検討を行った。その結果、1条件でのみ結晶化が可能であることが判明した。さらにその後に二次スクリーニングを行い、ポリエチレングリコール3350、及びクエン酸2アンモニウムの濃度検討を行って最適条件を決定した。
【0038】
MuPEP1の構造は1.9 Åの分解能で決定しRwork、Rfreeはそれぞれ21.2%、23.4%となった (表 1)。この構造はMuPEP1の5番目から141番目のアミノ酸残基と95個の水分子を含んでいる。N末の4残基およびC末のヒスチジンタグを含む15残基の電子密度は見えなかった。MuPEP1 はNativeタンパク質のN末44残基の疎水性領域をtruncateしたものであり、アミノ酸残基のナンバリングもtruncated formにしたがった。
【0039】
【表1】
【0040】
MuPEP1は6本のβ-ストランドと5本のヘリックスからなるα/βフォールドをとっていた(図1A)。中央に位置する6本のストランドからなる逆平行β-シートを挟むようにα-ヘリックスが配置されていて2つのサブドメインを形成している。N末側のサブドメイン(5-62番目の残基)は3本のα-ヘリックス、α1、α2およびα3 で形成されている。一方、C末側のサブドメイン(63-141番目の残基)は6本のβ-ストランドと2本のα-ヘリックスから形成されており、これらの2次構造コンポーネントはβ1、 α4、 β2、 β3、β4、 β5、α5、 β6 の順に配置されている。これら2つのサブドメインの会合面に狭いクレフトがあり、ここにCys17、His96、Asp112が形成するcatalytic triadが存在する。Cys17とHis96は、それぞれヘリックスα1 およびβストランドβ3のN末端に位置している。またAsp112はβ4 とβ5の間にあるループ上に位置している(図1A)。Cys17のSγ 原子とHis96のNδ1原子の距離は3.6 Åである。Asp112のOδ1原子とHis96のNε2 原子の距離は2.7 Åで水素結合を形成している(図1B)。
一次構造上の類似性は認められないにも関わらず、MuPEP1の結晶構造は他のパパイン様システインプロテアーゼと類似したフォールド構造を持っている(図2A)。N末側のサブドメインの3本のα-ヘリックスおよびC末側サブドメインのβ-シートとヘリックスα4 といったMuPEP1の二次構造の各成分は他のパパイン様システインプロテーゼのものと類似していた(図2A)。FATCATサーバーを用いてMuPEP1と他のパパイン様システインプロテアーゼの構造をCα 原子において重ね合わせたところpapainに対するrmsd が3.0 Å 、staphopain Aに対するrmsdが 2.7 Å となった。とりわけMuPEP1のcatalytic triadを形成する各残基とオキシアニオンホールを形成するGln11の空間配置はよく保存されている。MuPEP1のこれらの残基とpapainまたはstaphopain Aの相当する残基のCα 原子を重ね合わせたところrmsdはいずれも0.59 Åであった(図2B)。
[考察2]PEP-ComCアシル中間体モデル
PEPの基質認識メカニズムを調べるために、本発明者らはまずミュータンス菌のComC(MuComC)の大腸菌内での発現を試みたが、可溶性のタンパク質を得ることができなかった。次にMuComCを化学合成したが、できたペプチドは、ほとんどバッファーに不溶であった。また、MuPEP1のC17A変異型酵素とMuComCをグリシン6個からなるリンカーでつないだフュージョンタンパク質をデザインし、大腸菌内で発現を行ったところ可溶性のタンパク質が得られたが良質の結晶は得られなかった。本発明者らはさらに共結晶化およびソーキング法により、MuPEP1のC17A、C17S、H96A変異型酵素のいずれかと他のストレプトコッカス属細菌のComCのいずれかとの複合体結晶の作製を試みた。この方法ではいくつかの組み合わせで結晶が得られたが、ComCと複合体を形成している結晶は得られなかった。
【0041】
以上のような背景から本発明者らはMuPEP1-MuComC複合体のコンピューターモデルを作成した。まずComCのN末ヘリックスの疎水面と相互作用できそうな疎水性領域をMuPEP1のタンパク質表面から探した。この目的のため HotPatch program(J Mol Biol 369:863-879)によるタンパク質間の相互作用部位の検索をタンパク質表面の疎水性度に基づいたモードで行った。その結果、6つの疎水性領域が見つかった。しかしながら他のパパイン様プロテアーゼにおける基質配向の一般則を考慮に入れると、これらの疎水性領域のうち活性中心の近傍に位置する浅いくぼんだ領域のみがComCのN末ヘリックスを結合するサイトの候補と考えられた(図3A)。このくぼんだ領域も、MuPEP1の2つのサブドメインの会合面に位置しておりAla51、Leu52、Val55、Leu94、Leu134といった疎水性残基から形成されている。
【0042】
次に、本発明者らはMuPEP1とMuComCの間で形成されるアシル中間体を構築した。このモデルではMuPEP1の求核性のCys17がMuComCのGly-1のカルボニル炭素によってアシル化されている。また、MuComCの切断されたC末領域はこのモデルでは省略した。MuComCの主鎖Ile-3からGly-1は、ubiquitin C-terminal hydrolaseとそのインヒビターubiquitin aldehydeの複合体の構造(PDB ID code 1CMX)を参考にMuPEP1の活性部位クレフトにあてはめた。Ubiquitin aldehydeも切断部位にGly-Glyモチーフを有しており、C末端のGlyのカルボニル炭素と求核性のCys残基のSγ残基が共有結合している。このフィッティングによってIle-4の位置は制限される。MuComCのヘリックスの長さは未知なので、形成しうる最も長いIle-5からMet-25までの領域でヘリックスを形成した。そして、MuComCの保存された疎水性残基(図3B)のPhe-15、Ile-12、 Leu-7がMuPEP1の疎水性領域と相互作用するという仮定に合致するようおおよその位置にMuComCを結合させた。MuComCのこれら2つの領域をつなぎ合わせるために、Leu-7からIle-4までの主鎖をマニュアルで距離、ねじれ角が適切なコンフォーメーションをとるように調整した。最後に分子動力学(MD)シミュレーションを300 Kの温度下で行った。MDシミュレーションの間、MuPEP1-MuComC複合体のrmsdは最初の0.4 ナノ秒間でおおよそ1.5 Åまで増加し、その後1.0ナノ秒までその値を維持した(図4)。したがって1.0ナノ秒後の構造でエネルギー最小化を行った(図3C)。MDシミュレーションの0.4−1.0 ナノ秒間のMuPEP1の各Cα残基のゆらぎは4つの顕著なシグナルを示した(図5A)。これらのシグナルのピークはMuPEP1のループ構造に相当しており、それぞれβ1とβ2の間にある大きなループ構造中のα4のC末端の76番目の残基、β2とβ3の間にあるループの91番目から95番目の残基、β3とβ4の間にあるターン構造の105番目の残基とβ4とβ5の間にあるループの116番目の残基の位置に相当する。これらの位置は溶液側に露出しているため、MDシミュレーション中にMuPEP1が異常な挙動を示さなかったことを意味する。MuPEP1に対するMuComCのゆらぎはThr-22からMet-25の間で次第に増加していた(図5B)。このことより、MuComCのN末端の残基がMuPEP1から離れていることが示唆された。また、MuComCのすべての残基がラマチャンドランプロットで、適した位置に存在していた。
【0043】
図3Cのモデル中のMuComCの-25から-10までの領域がα-ヘリックス構造を保持していた。このヘリックス中の保存されているPhe-15、Ile-12とLeu-7とつづくターン構造中のLeu-7の側鎖がMuPEP1の疎水性表面に安定に結合していた。MuComCの主鎖は活性部位の入り口に位置しているLeu-7からIle-4の間で折れ曲がっている。Ile-4の側鎖はGln47とThr50の側鎖によってしっかり固定されている。一方、MuComCの親水性の残基、Lys-14、Glu-13、Lys-11、Thr-10の側鎖は溶液側に露出している。
【0044】
このモデルではMuComCのGly-GlyモチーフがMuPEP1の狭いクレフトにはまっている(図6)。このクレフトは上述の2つのサブドメインの会合面に位置している。クレフトの左側の壁(図6)は主にβ2とβ3の間にあるループ内のLys92からGln95までの残基によって形成されている。また、右側の壁はIle12からArg15までのN末鎖とα2とα3の間にあるループ内のLys46からGly48までの残基によって形成されている。活性部位の残基はクレフトの底面の一部分を形成している。Arg93の側鎖は、MDシミュレーションの後では活性部位にふたをしている(図3C マゼンタ)。このArg93のNη1と Nη2 原子はそれぞれ異なる2つの水分子を介して、Lys46の主鎖のカルボニル酸素と水素結合をしている。Arg93の側鎖のaliphaticな部分がMuComCのGly-2とファンデルワールス結合している。これらの相互作用は部分的にβ2とβ3の間にあるループのゆらぎを抑えていた(図5Aの2番目のピーク)。
2.モデルの実験による検証
[実験方法1]部位特異的変異
部位特異的変異はQuikChange II Site-Directed Mutagenesisキット(Stratagene)を使用して行った。PPEPのsingle mutationは全てpSPP1に導入した(J Biol Chem 281:4726-4731)。二重変異型PPEP、L52A/V55A PPEPは、V55AをL52A PPEPに導入した。A51W CAPEPおよびA67W CAPEPについては、C17A PPEPに各々変異を導入した(J Biol Chem 281:4726-4731)。Mutagenesisに使用したプライマーを表S2に記載する。作製した発現プラスミドについては、全てコーディング領域のヌクレオチド配列を確認した。
[実験方法2]速度論的パラメーターの決定
PPEP活性は、50 mM Tris-HCl、150 mM硫酸アンモニウム、各濃度の基質PComCを含む、100μL(pH 7.0)の反応液中で測定した。前述のように、高速液体クロマトグラフィー(HPLC)分析を行った(J Biol Chem 281:4726-4731)。要約すると、反応液をBeckman System Gold HPLCシステム(Beckman-Coulter)に接続したWaters μBondasphere C8逆相カラム(Waters)にアプライし、室温において流速1 mL/min、10分間の10−55%(v/v)アセトニトリル(0.1%のトリフルオロ酢酸を含む)のグラジエントにかけて、産生したペプチドを分析した。酵素反応で消費した基質の量は、全体の10%以下である。
[実験方法3]円二色性(CD)測定
CDスペクトルは、温度コントローラが付属したJasco分光偏光計・モデルJ-720WI(Jasco)を使用して測定した。セルは0.1cm光路長のものを使用した。緩衝液およびタンパク質の濃度は図7の凡例に記載した。
[考察]
本発明者らのモデルを検証するためにPPEPの基質結合サイトと予想される領域のアミノ酸残基に変異を導入した。この実験には唯一発現した天然のペアであるPPEP-PComCを用いた。
【0045】
上述の目的のため、新たにPPEP-PComCアシル中間体モデルを作製した。まずMuPEP1の構造をテンプレートにしてPPEPのモデルを構築した。MuPEP1の137残基を相当するPPEPのアミノ酸残基に置き換え(両タンパク質の相同性は57%)続いて分子動力学シミュレーション、エネルギー最小化を行った。得られたPPEPモデルにもMuPEP1においてHotPatchプログラムで予想されたものと同様の疎水性のくぼみが存在していた(図8)。PPEP-PComCアシル中間体モデルはMuPEP1-MuComCのモデリングと同様の手法を使って構築した。最終的に得られたモデルはMuPEP1-MuComCモデルとほぼ同様であったが、分子動力学シミュレーションの間にPComCのヘリックス構造がLeu-18とGlu-17の間で部分的にほどけていた(図9A)。
【0046】
このモデルからPComCの保存された疎水性残基との相互作用が予想されるThr50、Ala51、Leu52、Val55、Ala67、Leu94、Val134に変異を導入して活性測定を行うことにした(図9)。これらの残基の側鎖は溶液側に露出していると予想されるため変異を導入してもPPEPの全体構造にはほとんど影響ないものと考えられた。表2はこれら変異型PPEPのPComCに対する速度論的パラメーターを示す。T50S、L52A、V55A、L94A、V134Aでは触媒効率(kcat/Km)が1.8−16倍低下していた。ダブル変異を導入したL52A/V55A PPEPではPPEPのPComCに対する親和性が著しく減少していた(Kmが16倍上昇)。また、この変異型PPEPでは代謝回転数(kcat)はわずかしか低下していなかった。一方、PPEPの基質認識部位と推定される領域にTrp残基を導入すると(A51WとA67W)その立体障害によってPComCに対する親和性および代謝回転数がともに著しく減少し触媒効率がそれぞれ1/1400、1/110と著しく低下した。
【0047】
【表2】
【0048】
本発明者らは、CD測定によってComCがPEPと相互作用することで構造変化を起こすことを示してきた。A51WとA67Wの変異がひき起こす立体障害がComCのヘリックス形成にどのような影響を及ぼすのかを調べるためにA51WまたはA67W PPEP共存下、非共存下におけるComCのCDスペクトルをそれぞれ測定した。これらの変異は活性を持たないC17APPEP(CAPEP)に導入した。また、これまで調べた中でPPEPに対するKmが最も低いCComCを測定に用いた。これまでに示したようにバッファー中の20 μM CComC(図7、青)と35%トリフルオロエタノール(TFE)中(黒)の20 μM CComCはそれぞれランダムコイルとα-ヘリックス構造をとっている。CAPEP共存下でのCComCの構造変化は、CAPEP(20 μM)とCComC(20 μM)の混合液のCDとCAPEP(20 μM)のCDの差スペクトルを求めることにより調べた。CAPEP共存下でのCComCの差スペクトル(黄)は35%中で測定したCComCのスペクトルとほぼ同じであった。一方、A51W CAPEP共存下でのCComCの差スペクトル(赤)は、バッファー中で測定したCComCのスペクトルと同じであった。また、A67W CAPEP共存下でのCComCの差スペクトル(緑)のCAPEP共存下の差スペクトルに対する変化は、A51W CAPEP共存下での差スペクトルの変化と比べると中間であった。
3.考察のまとめ
(1)PEPの構造
1次構造比較から予想されていた、バクテリオシンのABCトランスポーターファミリーのペプチダーゼドメインがパパイン様システインプロテアーゼであるという仮説は、MuPEP1の3次元結晶構造によって検証することができた(図2)。C17S、C17A、H96A PPEPs とC17A MuPEP1が完全に活性を失っていた事から、PEPの触媒活性においてシステイン残基が重要であることが示された。Cys17の Sγ原子はHis96のNδ1原子から3.6 Å離れている。この距離は理想的な水素結合の距離、2.6−3.5 Åより少し長い。同様に、papainやstaphopain Aにおいても相当するCysとHis間の距離はそれぞれ、3.7 Å、4.0 Åである(図2B)。また、PROPKAプログラム(Proteins 61:704-721)から予想されるCys17、His96、Asp112のpKaは、それぞれ3.5、12、0.7であった。したがって他のパパイン様システインプロテアーゼで言われているように、Cys17とHis96は生理的条件下でthiolate-imidazoliumイオン対を形成しているものと考えられる。His96と水素結合を形成する距離にあるAsp112は、His96の空間的配置およびプロトン化を安定化しているのであろう。このようにMuPEP1はシステインプロテアーゼの触媒メカニズムに必要な全ての構造的要素を備えている。
(2)PEPの基質認識機構
PEPは、多様なタンパク質を分解するプロテアーゼと全体構造が類似しているにもかかわらず(図2A)、極めて基質特異性が高くComCと他のタンパク質を厳密に区別している。本発明者らのPEP-ComC複合体モデルは、ストレプトコッカス属細菌のPEPの基質認識の構造学的基盤を提供する。本発明者らの予想と合致して、MuPEP1の分子表面にComCと相互作用する2つの領域が見つかった。その1つは活性部位のクレフトである。このクレフトはストレプトコッカス属細菌のComCに保存されたGly-Glyモチーフを結合する(図6)。本発明者らのモデル上でMuComCのGly-1とGly-2をそれぞれAlaに改変すると、側鎖のメチル基がクレフトの側壁と立体障害を起こす。例えば、Ala-1は、PEPのThr14の主鎖と、Ala-2はGln95の主鎖とぶつかる。これまでの研究から、PComCのこれら2つのGlyをそれぞれAlaに改変したものを基質に用いてPPEPの活性を測定すると触媒効率が1/800に低下した。さらに、PPEPはシステインプロテアーゼを広く阻害するアンチパイン、ロイペプチン、E-64などによって触媒反応が阻害されない。これらの結果は、PEPのS1およびS2サイトがGly-Glyのみを認識する限定されたスペースをもつことを支持する結果である。
【0049】
もうひとつは、ComCの保存された疎水性残基(-15、-12、-7、-4番目のアミノ酸残基)と相互作用する活性部位クレフトの近傍にある疎水性の領域である(図3A、図8)。最初はComCのN末リーダー配列の全領域がα-ヘリックス構造を形成すると考えられていたが(図3B)、今回のPEP-ComC複合体モデルでは、ComCの-7番目から-4番目の領域がランダムコイルもしくはターン構造をとっている(図3C、図9A)。この疎水性表面は少しくぼんでおり、この領域を形成しているThr50、Ala51、Leu52、Val55、His87、Leu94、Thr132、Leu/Val134、Ile/Leu136はストレプトコッカス属細菌のPEP間で高度に保存されている。この領域のアミノ酸残基に変異を導入し活性測定を行ったところ、その重要性が確認された(表2)。PPEPの50、52、55、94、134番目の残基の変異ではいずれも触媒効率が1/1.8−1/16倍とある程度低下していたが、PPEPの変異型PComC、Phe-15A、Leu-12Ala、Leu-7Ala、Ile-4Alaに対する触媒効率の低下(1/31−1/180)と比較すると小さい。この事は、ComCの4つの保存性残基がPEP側の複数の残基と相互作用しているためであると考えられた。例えば、PComCのPhe-15の側鎖は、PPEPのAla51、Leu52、Val55、Ala67と相互作用している(図9B)。そして、PComCのPhe-15をAlaに変異するとこれらの相互作用が失われる一方、PEPのLeu52をAlaに変異した場合は、これらの相互作用の一部が失われるのみである。実際に、PPEPの52番目と55番目に二重変異を導入すると、PComCに対する親和性が加算的に低下した。PPEPのPhe-15Ala PComCに対するkcat/Kmは1/110倍に低下していた。また、L52A/V55A PPEPのPComCに対するkcat/Kmは1/27倍に低下していた。故に、PPEPのLeu52およびVal55の遷移状態の安定化に対する寄与はPComCのPhe-15とPPEP側の残基の相互作用の総和の68%(RTln27/RTln110)である。最小数の変異で活性を低下させ、この疎水性領域が基質結合部位であることをより明確に示すため本発明者らはこの領域に立体障害を導入した。予想された通り、A51WおよびA67W PPEPの触媒効率は著しく低下した。また、この基質結合部位と予想される領域の中央に位置するAla51に変異を導入したときの方が、辺縁部に位置するAla67に変異を導入したときよりも触媒効率の低下は大きかった。さらにA51Wの変異をMuPEP1にも導入してみたところ完全に失活していた。この結果は、この疎水性領域がすべてのストレプトコッカス属細菌のPEPにおいて重要であることを意味する。
【0050】
これら変異型PEPの活性測定の結果は、A51W CAPEPおよびA67W CAPEP共存下における、ComCのヘリックス構造変化ともよい相関を示した(図7)。これらの知見は、ComCのN末領域がPEPの疎水性領域に結合することで、ランダムコイルからヘリックスへと構造変化し、このことがPEP-ComC複合体がproductive formをとるために必要なステップであるという考えをさらに強化する。A51WおよびA67Wの変異は、kcat値を大きく下げるだけでなく、Km値も上げる。PEPはGly-Gly配列だけをもつタンパク質を切断しないことを考え合わせると、基質のGly-Gly配列周辺の主鎖は、基底状態(酵素・基質複合体)において切断部位の近傍でねじれていると考えられる。このねじれは、遷移状態で解消される。その結果、活性化エネルギーを低下させkcatを増加させる。おそらく、上述のPEPの疎水性領域とComCの間の大きな結合エネルギーはこのねじれを生じるために必要なのであろう。しかしながら、この相互作用だけでは不十分で、ComCのC末領域もPEPにしっかり結合することが必要であるし、もしくはComAの他の領域が切断部位周辺のねじれをつくるのに必要であるかもしれない。以上を明らかにするためには、基底状態や四面体型中間体、遷移状態に近い不安定な構造のモデルが必要となってくる。ここで示したアシル中間体は、基底状態および四面体型中間体いずれもと異なっている。実際に、アシル基の酸素はモデルのGln11が形成するオキシアニオンホール側を向いていない。さらに、切り出されたComCのC末領域(シグナル分子として機能するペプチド)は、細胞質側へもれ出ることなくまく貫通領域へ移行しなければならない。このように、PEPにおけるComCのC末領域の結合サイトを調べることもComAの機能を探る上で大変重要である。
本発明者らはまた、PEPのように小さなプロテアーゼがいかにして特異的に切断部位から15残基も離れた部位を認識するのかを説明するモデルを提唱する。MuPEP1-MuComCモデルでは、ComCのPhe-15のCα原子はGly-1のCα原子からおおよそ20 Åも離れている(図10)。本発明者らの知る限りでは、PEPは基質の最も広い領域を認識するプロテアーゼである。PEPのくぼんだ疎水性領域の残基のうち55番目の残基を除く全ての残基がバクテリオシンのABCトランスポーターのファミリー間で保存されている。このことから、これらのトランスポーターはComAと共通の基質認識機構を持っているものと考えられる。よって、この研究の結果は、バクテリオシンのABCトランスポーターの他のメンバーのペプチダーゼドメインを研究する上でのプロトタイプモデルを提供するものと考えられる。バクテリオシンのABCトランスポーターは、これまで原核生物にしか見つかっていないことから、ComAのペプチダーゼドメイン、とりわけMuPEP1の三次元結晶構造はストレプトコッカス属細菌のバイオフィルム形成を阻害する薬剤の開発に役立つものと期待される。
【特許請求の範囲】
【請求項1】
ComAタンパク質のペプチダーゼドメイン(PEP)を含むポリペプチド;又は
PEPの部分ポリペプチドであってComC切断活性を有するポリペプチド
の結晶であって、空間群P43212に属する結晶。
【請求項2】
単位格子定数がa=b=58.6±0.35 nm、及びc=112.1±0.5 nmである、請求項1に記載の結晶。
【請求項3】
前記ポリペプチドが、
(A) 配列番号1のアミノ酸配列からなるポリペプチド;又は
(B-1) (A)のポリペプチドのアミノ酸配列において、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチド
である、請求項1又は2に記載の結晶。
【請求項1】
ComAタンパク質のペプチダーゼドメイン(PEP)を含むポリペプチド;又は
PEPの部分ポリペプチドであってComC切断活性を有するポリペプチド
の結晶であって、空間群P43212に属する結晶。
【請求項2】
単位格子定数がa=b=58.6±0.35 nm、及びc=112.1±0.5 nmである、請求項1に記載の結晶。
【請求項3】
前記ポリペプチドが、
(A) 配列番号1のアミノ酸配列からなるポリペプチド;又は
(B-1) (A)のポリペプチドのアミノ酸配列において、1若しくは複数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなり、かつComC切断活性を有するポリペプチド
である、請求項1又は2に記載の結晶。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【公開番号】特開2012−23995(P2012−23995A)
【公開日】平成24年2月9日(2012.2.9)
【国際特許分類】
【出願番号】特願2010−164123(P2010−164123)
【出願日】平成22年7月21日(2010.7.21)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り ▲1▼掲載年月日 2010年1月25日 掲載アドレス http://www.jbc.org/content/285/14/10777.abstract?sid=0aca73c4−3535−47b1−ac1a−6bc36711f190 ▲2▼掲載年月日 2010年2月23日 掲載アドレス http://www.pdb.org/pdb/explore/explore.do?structureId=3K8U
【出願人】(502437894)学校法人大阪医科大学 (8)
【出願人】(000125369)学校法人東海大学 (352)
【Fターム(参考)】
【公開日】平成24年2月9日(2012.2.9)
【国際特許分類】
【出願日】平成22年7月21日(2010.7.21)
【新規性喪失の例外の表示】特許法第30条第1項適用申請有り ▲1▼掲載年月日 2010年1月25日 掲載アドレス http://www.jbc.org/content/285/14/10777.abstract?sid=0aca73c4−3535−47b1−ac1a−6bc36711f190 ▲2▼掲載年月日 2010年2月23日 掲載アドレス http://www.pdb.org/pdb/explore/explore.do?structureId=3K8U
【出願人】(502437894)学校法人大阪医科大学 (8)
【出願人】(000125369)学校法人東海大学 (352)
【Fターム(参考)】
[ Back to top ]