
スフィンゴ脂質富化麹とその製造方法
【課題】大量かつ安定的に、安全なスフィンゴ脂質を製造する方法を提供する。
【解決手段】蒸きょうした製麹原料に麹菌を植菌して培養を行う製麹において、盛り以降の工程を、湿度75%以上の条件下で、場合により製麹時間が計46時間以上となるまで、行うことを特徴とする、スフィンゴ脂質富化麹の製造方法。
【解決手段】蒸きょうした製麹原料に麹菌を植菌して培養を行う製麹において、盛り以降の工程を、湿度75%以上の条件下で、場合により製麹時間が計46時間以上となるまで、行うことを特徴とする、スフィンゴ脂質富化麹の製造方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、スフィンゴ脂質富化麹及びその製造方法に関する。
【背景技術】
【0002】
スフィンゴ脂質とは、スフィンゴイド塩基(長鎖塩基)を構造骨格としてもつ複合脂質の総称である。動物細胞のスフィンゴ脂質の骨格はスフィンゴシンであるが、植物や真菌では主にフィトスフィンゴシンである(非特許文献1)。
【0003】
スフィンゴ脂質には、スフィンゴイド塩基のアミノ基にアシル基がアミド結合したセラミドや、セラミドに糖、リン、硫黄、アミノ酸など様々な極性基が付加した複合スフィンゴ脂質などがある。なかでも糖が結合したスフィンゴ糖脂質は、真核生物やバクテリアの一部などに普遍的に存在しており、糖鎖部分の構造は様々であることが知られている。グルコースが結合したグルコシルセラミドは、セレブロシドとも呼ばれ植物、真菌、動物に共通してみられる代表的なスフィンゴ糖脂質である(非特許文献2)。
【0004】
セラミドはヒトの皮膚の角質層の細胞間脂質の主成分として約50%を占め、皮膚の保湿性や柔軟性にかかわっていると言われている。加齢により減少し、しわ、ドライスキン、肌荒れの原因となるほか、アトピー性皮膚炎にも関与していると考えられている。従来はセラミドを補充する方法として、安価な供給源である牛脳由来のスフィンゴ脂質が用いられていたが、BSE(牛海綿状脳症)発症の原因であるプリオンが脳幹部位に蓄積していることが明らかとなって以来、食品・化粧品用途での利用は不可能となっている。また、合成セラミドは安全性の観点から食品素材としては利用の制限がある。
【0005】
牛脳の代替供給源として、現在では主に農産加工副産物である大豆油さいやビール粕、米や小麦などの穀類の胚芽、コンニャク芋などが利用されているが、含有量が微量であり、食品としてそのまま利用しにくいことから、有機溶媒などによる抽出・濃縮が行なわれている。このため製造に多大なコストがかり、製品が非常に高価であるという問題があった(特許文献1、2)。
【0006】
また、酵母、キノコなどの真菌類からもセレブロシドの抽出が行われている。酵母においてはセレブロシドが存在する種は限られているため(非特許文献3)、サッカロミセス・クルイベリ(Saccharomyces kluyveri)などのセレブロシド生産菌を用いて、ビートモラセスを炭素源とし、様々な環境ストレスを与えることによりセレブロシドの生産量、蓄積量を高める製造方法も検討されている(特許文献3)。酵母は高温培養時には菌体量あたりのスフィンゴ脂質が増加し、低温培養時には減少することが分かっている(非特許文献4、5)。このように、培養条件の違いによって、菌体量あたりのスフィンゴ脂質量は変動することが知られている。しかしながら、この方法においても有機溶媒抽出や冷凍処理などが用いられており、工程が煩雑であり、多大なコストがかかるほか、環境への負荷も懸念されるなどの問題があった。
【0007】
国内で伝統的な発酵食品の製造に利用されてきた麹菌についても、液体培養時にセラミドやセレブロシドといったスフィンゴ脂質を生産するという報告がある(非特許文献6、7)。しかし、仮に麹菌の液体培養において培養条件の検討を行い、麹菌のスフィンゴ脂質含量を増加させたとしても、菌体だけをろ過回収して食するような食経験が無いため、有機溶媒抽出等の処理が必要となり、前述のような問題点が生じる。
【0008】
一方で、麹菌の固体培養物は、伝統的に「麹」として丸ごと食する食習慣があるため、食品として麹菌スフィンゴ脂質を利用するには非常に適している。そのため固体培養によってスフィンゴ脂質量を増大させる方法の確立が望まれている。
【0009】
しかしながら固体培養時、すなわち麹にした際のスフィンゴ脂質の生産能力や分子種についての報告は無く、高生産のための培養条件の検討はもちろん、麹をスフィンゴ脂質供給源として利用するための検討さえ行なわれてきていない。
【0010】
麹菌による固体培養(すなわち、製麹)について述べると、水分量や湿度、時間、温度等の培養条件に関する検討は、一般に穀物の糖質や蛋白質を分解する酵素を生産させるためや、種麹となる胞子を着生させることが目的で行われてきた。
【0011】
一般的な製麹法では、床もみ(すなわち種麹植菌)を行った後、蒸米を堆積し、布などで包む等して乾燥を防いで湿度を維持し、かつ乾燥が進む原因となる手入れをほとんど行わず、切り返しを行うのみで培養を行う。引き続いて「盛り」とは、床もみ後、一般的には18〜20時間前後になって、麹菌の生育が旺盛になりかけたものを麹蓋に入れる等、小区分に分けて品温管理をし易くする作業を指す。麹蓋に入れて棚培養する工程は、麹を薄く盛り、麹の水分蒸発を進める工程でもある。盛りまでを「床時代」、盛り以降を「棚時代」と呼ぶ。棚培養期間中に適宜麹をかき混ぜ、酸素の供給や炭酸ガスの排出とともに水分を発散させて、品温・状貌の均一化をはかる。非特許文献8によると、麹菌植菌後23〜28時間後に麹をかき混ぜる操作を仲仕事、または昼仕事といい、仲仕事後5〜7時間目に攪拌する操作を仕舞仕事または留仕事という。また、麹菌の代謝によっても水分が生じるため、製麹後半では麹室内の乾湿差を多く取り、通風製麹の場合は培養35時間頃から発酵熱により品温が上昇するため、送風を行う事で品温を下げるとともに水分の発散を促している(非特許文献8)。
【0012】
また培養水分量や湿度に関しては、伝統的にも限定吸水などによって蒸米水分を30%前後に制御し、製麹の後半(盛り以降)は麹室あるいは培養室の湿度を低下させて麹の水分含量を低下させ、酵素生産量を高める方法が採用されている。さらに、非特許文献8によると、「一般的にいって、湿気の多い麹は酵素力が弱く、粕が多く、清酒の色も濃い。また雑菌も多いので好ましくない。乾燥して、よくはぜた麹は酵素力が強く、清酒の色も淡く、味も軽い。」とある。すなわち水分が多いと酵素活性が低下してしまうので、製麹後半に麹の水分含量を低下させる方法が採用されてきた。
【0013】
製麹の後半に水分含量を低下させる事は、蒸発潜熱により品温の過昇を防ぐ以外に、麹の保存性を高める目的もある。非特許文献9によると、製麹中に品温を下降させる手段として、最も有効なのは蒸発潜熱を奪うことであり、蒸発潜熱を奪うために最も効率がよいのは、環境の相対湿度を低くすることであると記載されている。
【0014】
このため、通常の製麹法では、製麹により水分が蒸発するため、白米麹では蒸米水分が35%であれば、出麹水分は18%程度となり、醤油麹では初期水分が45%前後であれば、出麹水分は30%前後となる(非特許文献8、非特許文献10)。また製麹日数が長く、1週間となる種麹作成後の水分含量も25%程度と、仕込み時の水分含量より大きく減少している。
【0015】
製麹時間については、非特許文献8によると、「米麹の製造は麹菌分生子(種麹)を蒸米に植えつけ、30℃から40℃にわたる温度経過を経て、約40時間の培養によって行われる。」、「製麹時間の標準は酒母麹で48時間、掛麹で43時間内外である。」とある。つまり製麹時間は白米麹の場合、43〜48時間が一般的である。
【0016】
醤油麹などでは4日麹のように長時間(3日間)培養するものがある。また種麹作成のように長い特殊な場合では1週間程度製麹するものがある。これらは蛋白分解酵素(酸性カルボキシペプチダーゼ等)を高生産させること、あるいは胞子(分生子)を着生させることが目的である。これら製麹においても後半は麹室あるいは培養室の湿度を低下させて麹の水分含量を低下させる。
【0017】
このように、製麹時間については従来製麹の目的が酵素生産、あるいは胞子生産であることから、一般的には43〜48時間程度、醤油麹の場合は2〜3日程度、種麹作製のように特殊な場合では1週間とされてきた。
【0018】
以上のように、従来の産業利用上の製麹では、スフィンゴ脂質含量を高くするという観点で製麹条件(固体培養条件)の検討はなされていない。
【0019】
【特許文献1】特開平4−282317号公報
【特許文献2】特開平11−279586号公報
【特許文献3】特開2005−185126公報
【非特許文献1】Chem.Phys.Lipids,5,6−43(1970)
【非特許文献2】CMLS,Cell.Mol.Life Sci.,60,919−941(2003)
【非特許文献3】FEMS Yeast Research,2,533−538(2002)
【非特許文献4】The Journal Of BIOLOGICAL CHEMISTRY,272,48,30196−30200,1997
【非特許文献5】JOURNAL OF OLEO SCIENCE,53,3,127−133(2004)
【非特許文献6】日本農芸化学誌,第49巻,第4号,205−212(1975)
【非特許文献7】Biochimica et Biophysica Acta,486,161−171(1977)
【非特許文献8】麹学,日本醸造協会発行,村上英也 編著(1986)
【非特許文献9】新・みそ技術ハンドブック,全国味噌技術会発行(2006)
【非特許文献10】増補 醤油の科学と技術,日本醸造協会発行、栃倉辰六郎 編著
【発明の開示】
【発明が解決しようとする課題】
【0020】
スフィンゴ脂質を効率的に取得するためには、真菌類などの微生物を供給源とすることが適している。しかし、酵母やキノコ類を供給源とした場合、その形態や香味ゆえにいずれも食品としてはそのまま使用しづらく、有機溶媒等による濃縮が必須となるために、食品としては極めて高価なものになることから利用の制限があった。また麹菌は液体培養においてスフィンゴ脂質を生産することが知られるが、食用産業利用上有利な固体培養でスフィンゴ脂質を高生産するための最適な方法がなかった。
【0021】
したがって、本発明は、麹菌の固体培養によって、煩雑な工程を経ることなく、また食品組成物としてそのまま用いるために有機溶媒抽出や濃縮などを行うことなく、大量かつ安定的に、安全なスフィンゴ脂質を製造する方法を提供することを目的とする。
【課題を解決するための手段】
【0022】
本発明者らは、上記課題を解決するべく鋭意検討を重ねた結果、驚くべきことに、製麹における盛り以降の工程を、高湿度条件下で、場合により一般的な製麹時間(例えば46時間)よりも長い時間行うことにより、スフィンゴ脂質量を通常よりも顕著に増大できることを見出した。
【0023】
すなわち、本発明は以下の特徴を包含する。
(1)蒸きょうした製麹原料に麹菌を植菌して培養を行う製麹において、盛り以降の工程を、湿度75%以上の条件下で行うことを特徴とする、スフィンゴ脂質富化麹の製造方法。
(2)盛り以降の工程を湿度85%以上の条件下で行うことを特徴とする上記(1)記載の方法。
【0024】
(3)製麹時間が計46〜160時間であることを特徴とする上記(1)記載の方法。
(4)植菌後18〜32時間の時点で製麹温度を38〜42℃とすることを特徴とする、上記(3)記載の方法。
(5)麹菌はアスペルギルス・オリゼ(Aspergillus oryzae)又はアスペルギルス・ソーエ(Aspergillus sojae)であることを特徴とする上記(1)記載の方法。
【0025】
(6)製麹原料はイネ科植物であることを特徴とする上記(1)記載の方法。
(7)イネ科植物は米又は大麦であることを特徴とする上記(1)記載の方法。
(8)蒸きょうした製麹原料の水分量が30〜36重量%であることを特徴とする上記(1)記載の方法。
【0026】
(9)スフィンゴ脂質はセレブロシドであることを特徴とする上記(1)記載の方法。
(10)上記(1)〜(9)のいずれか記載の方法で製造されたスフィンゴ脂質富化麹。
(11)乾燥重量1g当たり少なくとも136μg以上のスフィンゴ脂質を含有することを特徴とする上記(10)記載のスフィンゴ脂質富化麹。
(12)上記(10)又は(11)記載のスフィンゴ脂質富化麹を含有する飲食品。
【発明の効果】
【0027】
本発明によれば、簡便かつ安価で、安全なスフィンゴ脂質富化麹を製造することができる。
【発明を実施するための最良の形態】
【0028】
本明細書で使用する「製麹」とは、蒸きょうした製麹原料に麹菌を植菌して培養する、当業者に公知の製麹方法(例えば、蓋麹法、箱麹法、床麹法、機械麹法など)を指す。これらの製麹方法は、蒸きょうした製麹原料に麹菌を植菌した後に培養する工程を含み、その間、一般的に「引き込み」、「盛り」、「仲仕事」、「仕舞仕事」といった処理を含んでいる。
【0029】
「盛り」とは、「床もみ」(すなわち種麹植菌)後、麹菌の生育が旺盛になりかけたものを小区分に分けて品温管理をしやすくする作業を指す。一般的に製麹において、盛りまでを「床時代」、盛り以降を「棚時代」と呼ぶ。床時代とは床もみを行った後、蒸した製麹原料を堆積し、布などで包むなどして湿度を維持し、乾燥を防ぐ工程であり、乾燥が進む原因となる手入れをほとんど行わず、切り返しを行うのみで培養を行う。
【0030】
本明細書において、植菌後から盛りまでの工程(すなわち床時代)を製麹前半、盛り以降の工程(すなわち棚時代)を製麹後半と称する。
【0031】
製麹前半における製麹温度は、使用する麹菌の菌体生育の至適温度に応じて、一般的には28〜42℃、例えば30〜37℃とされる。ただし、製麹前半の製麹温度をおよそ38℃以上の比較的高温とする場合、麹菌が生育阻害を受ける虞がある。なお、製麹温度とは、麹の品温を意味する。製麹温度は、麹菌の生育に伴う発酵熱を制御することによって、又は製麹における周囲温度を制御することによって調節することができる。
【0032】
製麹前半の所要時間は、一般的には18時間〜20時間である。生育が旺盛になった状態のまま長時間放置すると発酵熱を除くことができず、品温が制御できなくなるうえに、原料が密集しているため製麹の生育が抑制されてしまうため、この時点で盛りを行い、製麹後半に移行する。
【0033】
一方、製麹後半は、麹を薄く盛り、麹の水分蒸発を進める工程であり、この培養工程の中で、「仲仕事」、「仕舞仕事」と呼ばれる手入れ作業が行われる。上記非特許文献8によれば、出麹までに蒸発する総水分量の75%が仲仕事までの間に蒸発することが記載されている。
【0034】
製麹の詳細については、例えば上記非特許文献8及び発酵ハンドブック((財)バイオインダストリー協会 発酵と代謝研究会編,2001年)などを参照されたい。
【0035】
本発明のスフィンゴ脂質富化麹の製造方法(以下、本発明の方法ともいう)において、下記で特に規定されない限り、製麹を上記のような当業者に公知の製麹と同様にして行うことができること、当業者は適宜上記条件を変更できることに留意すべきである。
【0036】
具体的に、本発明の方法は、製麹において、製麹後半を湿度75%以上の条件下で行うことを特徴とする。これにより、製麹後半における麹水分の蒸散を抑制することができ、その結果、スフィンゴ脂質の生産を増加することができる。清酒、醤油麹の製造などに使用されている従来の製麹技術は、上記の通り、酵素生産量の増加及び保存性を高める観点から製麹後半において湿度を低下させることによって麹の水分含量を低下させている点で、本発明の方法と異なる。
【0037】
製麹後半は、湿度85%以上の条件下で行うことがより好ましい。湿度75%以上の条件は、従来の製麹技術と比較して製造される麹中のスフィンゴ脂質含量を増加させるのに十分であるが、下記実施例に示されるように、製麹後半における相対湿度を85%以上にすることにより、製造されるセレブロシド量をより増加させることができる。
【0038】
本発明の方法はまた、製麹において、製麹後半を、製麹時間が計46時間以上となるまで行うことを特徴とする。これにより、製造される麹に含まれるスフィンゴ脂質の量を経時的に増加させることができる。従来の製麹技術は、湿度を高く維持したまま通常の43〜48時間(醤油麹の場合は2〜3日間)の製麹時間を越えるような培養を行なっていない点で、本発明の方法と異なる。本発明の方法における製麹時間の上限は特に制限されず、スフィンゴ脂質の量が経時的に増加する限り製麹後半を継続することができる。本発明において、製麹時間は、例えば計46〜160時間、より好ましくは72〜160時間とすることができる。
【0039】
本発明の方法において、製麹前半における湿度条件は特に制限されず、従来の製麹と同様の条件で行うことができる。ただし、本発明の方法により製造される麹は、上記の通り出麹水分の減少が抑制されたものであることが好ましいところ、製麹前半においても高湿度条件、例えば湿度75%以上、好ましくは湿度85%以上の条件下で行うことが好ましい点に留意する。本発明において、製麹を通じて湿度が85%以上に維持されることが最も好ましい。
【0040】
また本発明の方法において、従来の製麹時間を越えるような製麹時間で製麹後半を行う場合には、植菌後18〜40時間、好ましくは植菌後18〜32時間の時点で製麹温度を38〜42℃に温度シフトすることが好ましい。従来の製麹時間よりも長時間で製麹後半を行う場合には、麹の味覚上好ましくない胞子の着生の問題が顕著となる。そこで、植菌後18時間〜40時間、好ましくは植菌後18〜32時間の時点で、製麹温度を38〜42℃に温度シフトすることにより、胞子の着生を抑制することができる。
【0041】
本発明の方法において、蒸きょうした製麹原料の水分量が30〜36重量%であることが好ましい。蒸きょう後の製麹原料の水分量が30%以下であるか又は36%以上であると、麹菌の立ち上がりが抑制されてしまう虞があるからである。
【0042】
本発明の方法で使用される製麹原料は、麹菌を生やして麹にできるものであれば特に制限されず、例えば穀類、擬穀類、豆類、ゴマ類、芋類等を使用することができる。具体的に、穀類としては、白米、玄米、黒米、赤米、大麦、小麦などを、擬穀類としてはソバ、アマランサス、キヌアなど、豆類としては大豆、小豆、黒豆、ヒヨコ豆などを挙げることができるが、これらに限定されるものではない。麹の原料として穀類、例えばイネ科植物を使用することが好ましく、イネ科植物として白米や、大麦などの麦類を使用することが特に好ましい。
【0043】
本発明の方法に用いることができる麹菌は、スフィンゴ脂質を生産できるものであれば特に限定されないが、例えばアスペルギルス・オリゼ(Aspergillus oryzae)や、その近縁のアスペルギルス・ソーエ(Aspergillus sojae)などを挙げることができる。A.オリゼとして具体的には市販の種麹や、登録番号RIB40、NBRC4214、JCM2228又はATCC36261で指定される特定の菌株などを用いることができる。また、A.ソーエとしては、市販の種麹や、登録番号JCM2226、NBRC4239又はNBRC5241で指定される特定の菌株などを用いることができる。A.オリゼとしてNBRC4214、JCM2228が、そしてA.ソーエとしてJCM2226が白米、大麦の両方の麹においてセレブロシド生産量が高いことから、本発明で使用するのに特に適している。また、上記麹菌以外の麹菌として、大麦麹の場合にA.オリゼ登録番号NBRC4255、ATCC11494、ATCC16868、NBRC5786、JCM2245、NBRC5240などを、米麹の場合にA.オリゼ登録番号JCM2227、JCM2173、JCM2229、NBRC4261などを使用することができる。
【0044】
上記のようにして製造される麹は、スフィンゴ脂質(特にセレブロシド)を豊富に含むものである。具体的には、従来の製麹時間、製麹湿度にて製麹された麹に比べて、スフィンゴ脂質の総量を約1.7倍〜約6.7倍まで高めることができる。また、本発明の方法により製造されるスフィンゴ脂質富化麹は、乾燥重量1g当たり少なくとも136μg以上、好ましくは248μg以上のスフィンゴ脂質を含むことができる。なお、麹中のスフィンゴ脂質の量は、薄層クロマトグラフィー、高速液体クロマトグラフィーなどを使用することによって測定することができる。
【0045】
このように、本発明の方法は、煩雑な条件でストレスをかける工程などを必要とせず、製造されるスフィンゴ脂質富化麹も、有機溶媒を用いた抽出や、濃縮をすることなく、そのまま飲食品として提供することができるため、労力とコストを低減でき、なおかつ安全性が高いといった利点を有する。また、食品用途に供することを踏まえ、長時間培養であっても、植菌後18〜40時間、好ましくは植菌後18〜32時間の時点で製麹温度を38〜42℃に温度シフトすることにより、味覚的に望ましくない胞子の着生を抑制することができるという利点も有する。
【0046】
したがって、本発明は、本発明に係るスフィンゴ脂質富化麹を含有する飲食品を包含する。
【0047】
本発明に係る飲食品は、飲食品原料の一部として用いたり、或いは飲食品の製造工程又は製造後に添加又は配合することにより得ることができる。本発明に係る飲食品は、健康食品、機能性食品、特定保健用食品、栄養補助食品などとして提供することもできる。
【0048】
本発明に係る飲食品の形態は特に制限されず、例えばこれに制限されるものではないが、ヨーグルト、ドリンクヨーグルト、ジュース、牛乳、豆乳、酒類(アルコール性飲料)、コーヒー、紅茶、煎茶、ウーロン茶、スポーツ飲料等の各種飲料や、クッキー、パン、ケーキ、煎餅などの焼き菓子、羊羹などの和菓子、プリン、ゼリー、アイスクリーム類などの冷菓、チューインガム、キャンディ等の菓子類や、クラッカー、チップス等のスナック類や、うどん、そば等の麺類や、かまぼこ、ハム、魚肉ソーセージ等の魚肉練り製品や、みそ、しょう油、ドレッシング、マヨネーズ、甘味料等の調味類や、豆腐、こんにゃく、その他佃煮、サラダ、スープ等の各種総菜などを例示することができる。
【0049】
また、飲食品中の有効成分としてのスフィンゴ脂質富化麹の摂取量は、通常、2.4〜4.8g/日、好ましくは4.4〜8.8g/日程度とすることができる。
【0050】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
【実施例】
【0051】
A.オリゼは液体培養時、グルコシルセラミドやセラミドといったスフィンゴ脂質を生産することが知られており、A.ニガー(A.niger)はその他にガラクトシルセラミドも生産するという報告がある(上記非特許文献2)。すなわち、麹菌は種によって異なる分子種を生産すると考えられる。現在、各種麹菌が上記以外に、どのような複合スフィンゴ脂質を生産しているかについては未解明であるため、以下の実施例では、代表的な麹菌としてA.オリゼを選び、スフィンゴ脂質の中でも分析例が多いグルコシルセラミド、すなわちセレブロシド量を測定することで、総スフィンゴ脂質量の評価を行った。
【0052】
[実施例1] 米麹セレブロシド標準品の精製と構造解析
米麹セレブロシド標準品の調製法を以下に示した。A.オリゼ(A.oryzae)を使用して製麹した米麹5.4kgを粉砕し、5倍量のクロロホルム−メタノール(2:1)で2回抽出、濾過して得られた抽出液を濃縮、乾固後、0.2M KOH−メタノール溶液にて37℃で2時間振盪し、グリセロ脂質を分解しアルカリ安定脂質を得た。中和後、濃縮、乾固し、クロロホルム−メタノール−0.8%NaCl(8:4:3)に再溶解した。振盪混合し、遠心分離後、下層を濃縮した。
【0053】
これをケイ酸カラムクロマトグラフィーに供し、クロロホルム-メタノールを用いて遊離脂肪酸等を除いた。再度ケイ酸カラムクロマトグラフィーに供した後、セレブロシド画分として白色の固形油脂状物407.9mgを得た。その後、わずかに混在する遊離脂肪酸を除去するため、さらにHPLC−示差屈折検出器((株)島津製作所製)を用いて精製を行なった。カラムはYMC Pack ODS−A;20×250mm、カラムオーブンは40℃、移動相にはメタノールを用いて、アイソクラティック溶出を行い、流速6ml/minとした。得られた精製品は、薄層クロマトに供したところ単一スポットとなり、また以下に示したように主要分子がグルコシルセラミドと同定されたため、これを標準品としてセレブロシドの定量を行った。
【0054】
主要分子種の構造解析を行った。すなわち、HPLC−示差屈折検出器((株)島津製作所製)を用いて、前述の条件で、精製セレブロシドに含まれる主要分子を分取した。得られた主要3分子(保持時間28.7分、32.1分、37.4分)について、以下に示すように構造解析を実施した。
【0055】
保持時間28.7分の分子をメタノールに溶解し、HRESI−MS(+)を測定した。この結果m/z 776.57456に(M+Na)+イオンが観測され、分子式はC43H79NO9(C43H79NO9Na cald 776.56525 (D+4.57mmu)と決定した。
【0056】
次にこの分子を重ピリジン(C6D5N)に溶解し、核磁気共鳴(NMR)解析を行った。1H及び13C NMRスペクトルの特徴からこの分子がセレブロシドであると同定された。まず糖部分の1H、13C化学シフト値及びJHH値から、構成糖はグルコースであり、これがβ−結合(J1”,2”=7.8Hz)で長鎖塩基と結合していること(=β−グルコシルセラミド)が判明した。次に1H−1H COSY、HMQC、HMBCスペクトルを測定し、脂肪酸及び長鎖塩基部分の構造について解析を行った。
【0057】
脂肪酸については1H−1H COSYスペクトルにおいてH−2’(δ5.10)−H−3’(δ6.10、J3’,4’=15.7Hz)−H−4’(δ6.16)というvicinal spin結合が観測されること、また1H NMRで末端メチルはδ0.85にtripletとして観測されることから、直鎖の2−ヒドロキシ、3−トランス脂肪酸であることが明らかとなった。次に脂肪酸の炭素数を明らかにするために、この分子をメタノール性0.9N HClを用いて100℃、18時間還流することで分解(メタノリシス)し、脂肪酸メチルエステルとした。ヘキサン抽出で脂肪酸メチルエステルを精製後、これをトリメチルシリル(TMS)誘導体化し、GC/MS解析を実施した。この結果、m/z 369に(M−15)+のピークが観測され、この分子を構成する脂肪酸炭素数はC18と決定した。
【0058】
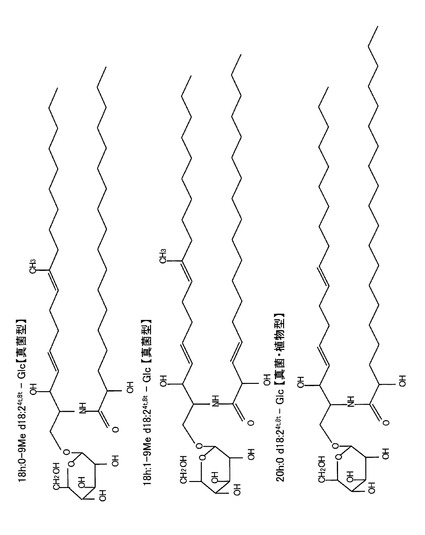
長鎖塩基については1H−1H COSYスペクトルにおいてH−1(δ4.23、δ4.69)−H−2(δ4.79)−H−3(δ4.74)−H−4(δ5.99、J4,5=15.8Hz)−H−5(δ5.92)−H−6(δ2.14)−H−7(δ2.14)−H−8(δ5.24)というvicinal spin networkが観測されること及びHMBCスペクトルで9−CH3(δ1.60)からC−8(δ123.3)、C−9(δ13.5)に1H−13C遠隔スピン結合が観測されることから、9−メチル−4トランス、8−トランス−スフィンガジエニン構造を有していることが明らかとなった。また1H NMRで末端メチルはδ0.85にtripletとして観測されるため、アルキル鎖部分は直鎖であることも明らかであった。この時点で既に脂肪酸の炭素数は明らかとなっているため、長鎖塩基を構成する炭素数は分子式から糖(C6)及び脂肪酸(C18)の炭素数を引いたC18であると決定された。その結果、この分子の構造は図1に示すように18h:1−9Me d18:24t,8t−Glcであることが一義的に決定された。また、この構造は真菌類に特徴的な構造であった(非特許文献2)。
【0059】
保持時間32.1分の分子をメタノールに溶解し、HRESI−MS(+)を測定した。この結果、m/z 778.58403に(M+Na)+イオンが観測され、分子式はC43H81NO9(C43H81NO9Na cald 778.58090 (D+3.13mmu)と決定した。
【0060】
次にこの分子を重ピリジン(C6D5N)に溶解し、核磁気共鳴(NMR)解析を行った。1H及び13C NMRスペクトルは18h:1−9Me d18:24t,8t−Glc(保持時間28.7分)によく類似していた。糖部分の1H、13C化学シフト値及びJHH値はほぼ同一であることから、構成糖はグルコースであり、これがβ−結合(J1”,2”=7.8Hz)で長鎖塩基と結合していること(=β−グルコシルセラミド)が判明した。次に1H−1H COSY、HMQC、HMBCスペクトルを測定し、脂肪酸及び長鎖塩基部分の構造について解析を行った。
【0061】
脂肪酸については1H−1H COSYスペクトルにおいてH−2’(δ4.56)−H−3’(δ2.12)というvicinal spin結合が観測されたこと、また1H NMRで末端メチルがδ0.85にtripletとして観測されることから、直鎖の2−ヒドロキシ脂肪酸であることが明らかとなった。次に脂肪酸の炭素数を明らかにするためにメタノリシスを行い、脂肪酸メチルエステルとした。ヘキサン抽出で脂肪酸メチルエステルを精製後、これをトリメチルシリル(TMS)誘導体化し、GC/MS解析を実施した。この結果、m/z 371に(M−15)+のピークが観測され、この分子を構成する脂肪酸炭素数はC18と決定した(=2−ヒドロキシステアリン酸)。
【0062】
長鎖塩基については1H−1H COSYスペクトル及びHMBCスペクトルにおいて18h:1−9Me d18:24t,8t−Glcとまったく同じ相関が認められることから、この分子の長鎖塩基も9−メチル−4トランス,8−トランス−スフィンガジエニン構造を有していることが明らかとなった。また1H NMRで末端メチルはδ0.85にtripletとして観測されるため、アルキル鎖部分も直鎖であることも明らかであった。既に脂肪酸の炭素数は明らかとなっているため、長鎖塩基を構成する炭素数は分子式から糖(C6)及び脂肪酸(C18)の炭素数を引いたC18あると決定された。その結果、この分子の構造は図1に示すように18h:0−9Me d18:24t,8t−Glcであることが一義的に決定された。また、この構造は真菌類に特徴的な構造であった(非特許文献2)。
【0063】
保持時間37.4分の分子をメタノールに溶解し、HRESI−MS(+)を測定した。この結果、m/z 792.60158に(M+Na)+イオンが観測され、F5の分子式はC44H83NO9(C44H83NO9Na cald 792.59655(Δ+0.94mmu)と決定した。
【0064】
次にこの分子を重ピリジン(C6D5N)に溶解し、核磁気共鳴(NMR解析)を行った。1H及び13C NMRスペクトルは18h:0−9Me d18:24t,8t−Glc(保持時間32.1分)によく類似していた。糖部分の1H、13C化学シフト値及びJHH値についても18h:0−9Me d18:24t,8t−Glcと同一であることから、構成糖もグルコースであり、これがβ−結合(J1”,2”=7.8Hz)で長鎖塩基と結合していること(=β−グルコシルセラミド)が判明した。次に1H−1H COSY、HMQC、HMBCスペクトルを測定し、脂肪酸及び長鎖塩基部分の構造について解析を行った。
【0065】
脂肪酸については1H−1H COSYスペクトルにおいて18h:0−9Me d18:24t,8t−Glcと同様にH−2’(δ4.56)−H−3’(δ2.12)というvicinal spin結合が観測されること、また1H NMRで末端メチルがδ0.85にtripletとして観測されることから、直鎖の2−ヒドロキシ脂肪酸であることが明らかとなった。次に脂肪酸の炭素数を明らかにするために、メタノリシスを行い、脂肪酸メチルエステルとした。精製後トリメチルシリル(TMS)誘導体化し、GC/MS解析を実施した。この結果、m/z 399に(M−15)+のピークが観測され、F5を構成する脂肪酸炭素数はC20と決定した(=2−ヒドロキシアラキジン酸)。
【0066】
長鎖塩基については1H−1H COSYスペクトルでH−1(δ4.23、δ4.69)−H−2(δ4.79)−H−3(δ4.74)−H−4(δ5.99、J4,5=15.8Hz)−H−5(δ5.92)−H−6(δ2.16)−H−7(δ2.16)−H−8(δ5.46)−H−9(δ5.46)というvicinal spin networkが観測されること及び18h:0−9Me d18:24t,8t−Glcで観測された9−CH3の信号が観測されないことから、4トランス、8−トランス−スフィンガジエニン構造を有していることが明らかとなった。また1H NMRで末端メチルはδ0.85にtripletとして観測されるため、アルキル鎖部分は直鎖であることも明らかであった。既に脂肪酸炭素数は明らかとなっているため、長鎖塩基を構成する炭素数は分子式から糖(C6)及び脂肪酸(C20)の炭素数を引いたC18であると決定され、この結果、この分子の構造は図1に示すように20h:0 d18:24t,8t−Glcであることが一義的に決定された。また、この構造は真菌類及び植物に特徴的な構造であり(非特許文献2)、米由来のセレブロシドも含まれていると考えられる。
【0067】
続いて、セレブロシドの精製品をLC/MS(LCMS−2010EV島津製作所)によって分析し、主要3分子の量比を調べた。イオン化モードはESI(−)、カラムはCadenza CD−C18 150×2mm、移動相はTHF/メタノール(1/9 v/v)を用いてアイソクラティック溶出を行い、流速は0.2ml/mlとした。その結果、それぞれの分子種の量比は、以下のとおりであることが示唆された。このことから、真菌に特徴的な分子である18h:0−9Me d18:24t,8t−Glcが、米麹中のセレブロシドの大半を占めることが示された。
【0068】
【表1】

【0069】
[実施例2] 米麹、大麦麹中のセレブロシド含量の測定法
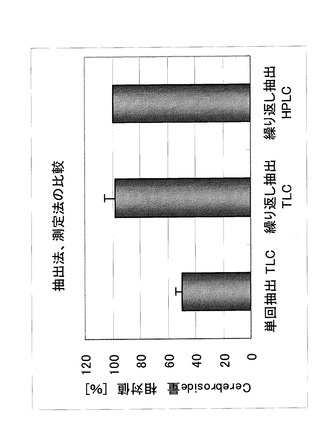
実施例3〜7におけるセレブロシドの定量は、各検体から有機溶剤を用いて抽出後、実施例1で得られたセレブロシドを標準品として用い、薄層クロマトグラフィー(TLC)又は高速液体クロマトグラフィー(HPLC)によって行った。TLCとHPLCのどちらの測定法においても、定量値がほぼ一致していることを確認した(図2)。
【0070】
また、麹からのセレブロシドの抽出は以下に示すように単回または、繰り返し抽出により行った。単回抽出は、より簡便な方法によって、各実験区におけるセレブロシド含量の比較を行うために実施し、繰り返し抽出は実際に麹に含まれるセレブロシド含量を定量する目的で実施した(実施例3〜6については単回抽出を行い、実施例7については繰り返し抽出を行った)。各抽出法で得られる定量値について、比較データの一例を図2に示した。検討の結果、麹中のセレブロシド含量によって、単回抽出の抽出効率は変動し、繰り返し抽出で得られる米麹中のセレブロシド含量は、単回抽出で得られる定量値の約1.5〜2倍程度に相当することが示された。
【0071】
単回抽出によるセレブロシド量の測定法を以下に示した。麹を凍結乾燥後、粉砕物1gに対してクロロホルム−メタノール溶液(2:1)3mlを添加し、40℃で30分間超音波処理を行った。これを遠心分離して得られた上清1mlに対し、0.8%NaCl溶液を250μl添加後、振盪混合した。これを遠心分離して得られた下層を回収し、一定量を測定に用いた。
【0072】
繰り返し抽出による総セレブロシド含量の測定法を以下に示した。米麹を凍結乾燥後、粉砕物1gに対し、クロロホルム−メタノール溶液(2:1)3mlを添加し、40℃で30分間超音波処理を行い、遠心分離後に上清を得た。計3回の抽出を行った後、上清全量に1/4量の0.8%NaCl溶液を添加し、振盪混合した。これを遠心分離して得られた下層を回収した。上清にクロロホルムを添加して、さらに2回の抽出を行った。下層全量を一定量に合わせ測定を行い、1gあたりに含まれるセレブロシド量を算出した。
【0073】
TLC−デンシトメトリー法によるセレブロシド定量法を以下に示す。抽出した試料をケイ酸薄層クロマトグラフィー(メルク社製、商品名:HPTLC silica−60 F254)にかけた。クロロホルム−メタノール(95:15)で展開後、オルシノール硫酸試薬(オルシノール:0.2% w/v、 硫酸:11.4% v/v)を噴霧し、加熱後に出現する赤紫色のセレブロシドのスポットについて、デンシトメトリーを用いて定量を行った。
【0074】
HPLC−コロナCAD検出器によるセレブロシド定量法を以下に示した。定量法は、HPLC−蒸発光散乱検出器法(J.Oleo Sci., Vol.53,No.3, p.127−133, 2004)を基に、より検出感度を上げるために検出部分を改良して行った。すなわち移動相は、A液:クロロホルム、B液:95%メタノール、を用いて、グラジエント溶出を行った。
【0075】
【表2】
【0076】
カラムは、シリカカラム:Inertsil SIL−100A (4.6×150mm、5μm)を用い、カラムオーブン:40℃、流速:1ml/minとした。また、検出器については蒸発光散乱検出器よりも感度、定量範囲、再現性が優れているとされるコロナCAD検出器を用いた。コロナCAD検出器は、ガス:窒素ガス、操作時ガス圧:35psiとした。
【0077】
[実施例3] 各種麹菌のセレブロシド生産量
蒸米および蒸大麦に対し、各種市販種麹10株及び菌株保存機関より入手した各種麹菌55株(A.oryzae、A.sojae)を0.1%の割合で接種し、品温35℃、相対湿度80%以上とし、46時間製麹した。得られた麹を凍結乾燥し、粉砕後に乾燥麹中のセレブロシド量を実施例1の単回抽出及びTLCによって測定した。
【0078】
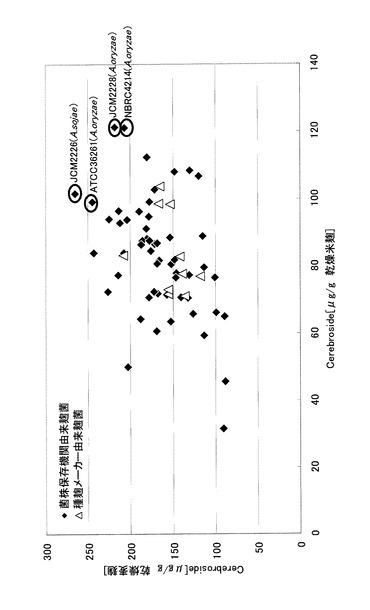
図3に、大麦麹及び米麹に含まれる各種麹菌(A.oryzae、A.sojae)のセレブロシド生産量を示した。原料が米、大麦のいずれの場合においても、セレブロシド含量が高い株について、菌株名と入手元の菌株保存機関における識別番号を記した。A.oryzaeにおいてはNBRC4214、JCM2228、ATCC36261を、A.sojaeとしてはJCM2226が、米、大麦、いずれの培地においてもセレブロシド生産量が特に高いことが示された。また、全ての菌株において原料を大麦にした場合の方が、米の場合より生産量が高く(1.1倍〜4倍程度)なった。
【0079】
[実施例4] 米麹、大麦麹での長時間培養のスフィンゴ脂質量の経時変化
蒸米に対して粉末型麹菌(A.oryzae:NBRC4214)、蒸大麦に対して粉末型麹菌(A.oryzae:市販種麹)をそれぞれ0.1%の割合で接種し、品温35℃、相対湿度85%以上とし、24〜32時間後に品温を40℃として88時間製麹した。製麹中に経時的に麹を採取、凍結乾燥し、乾燥麹中のセレブロシド量を実施例1の単回抽出及びTLCによって測定した。
【0080】
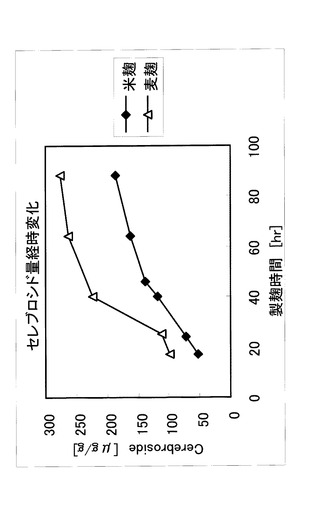
製麹中のセレブロシド量の経時変化を図4に示した。米麹、大麦麹どちらにおいても、上記培養条件によって麹中のセレブロシド量が経時的に増加することがわかった。
【0081】
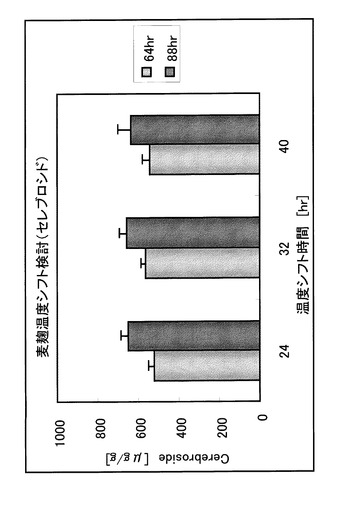
[実施例5] 温度シフト時間検討
温度シフトのタイミングの影響を評価するために、蒸大麦に対し、粉末型麹菌(A.oryzae:市販種麹)を0.1%の割合で接種し、品温35℃、相対湿度85%以上とし、24、32、40時間後に品温を40℃として88時間製麹した。調製した大麦麹のセレブロシド量は実施例1の単回抽出及びTLCによって測定した。
【0082】
出麹サンプルのセレブロシド量を図5に示した。24時間に温度シフトした場合は、64時間製麹では32〜40時間にシフトした場合よりもわずかに低かったが、88時間製麹では温度シフトのタイミングに関わらず、セレブロシド量はほぼ一定であった。
【0083】
胞子着生スコアを表3に示す。24〜32時間に温度シフトした場合と比べて、40時間にシフトした場合では胞子着生が多く、24〜32時間に温度シフトすることが好ましいと考えられた。
【0084】
【表3】
【0085】
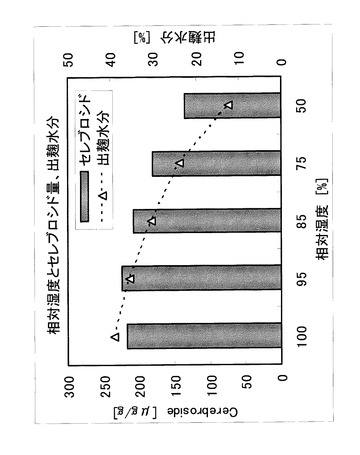
[実施例6] 米麹による湿度条件検討
温度シフト後の湿度の影響を評価するために、蒸米に対し、粉末型麹菌(A.oryzae:NBRC4214)を0.1%の割合で接種し、品温35℃、相対湿度85%以上とし、32時間後に品温を40℃とし、相対湿度を100、95、85、75、50%として88時間製麹した。相対湿度はそれぞれ、水、6.5%NaOH、15.8%NaOH、飽和NaCl、飽和硝酸カルシウム溶液を用いて調湿した。調製した米麹のセレブロシド量は、実施例1の単回抽出及びTLCによって測定した。
【0086】
製麹後半の相対湿度とセレブロシド量、出麹水分の関係を図6に示した。この結果より、温度シフト後の湿度に応じて出麹水分が低くなり、相対湿度が85%以下になるとセレブロシド量も低下することがわかった。出麹水分は相対湿度に比例して低くなっており、85%以下では麹中水分の減少によってセレブロシドの生産が抑制されたと考えられた。
【0087】
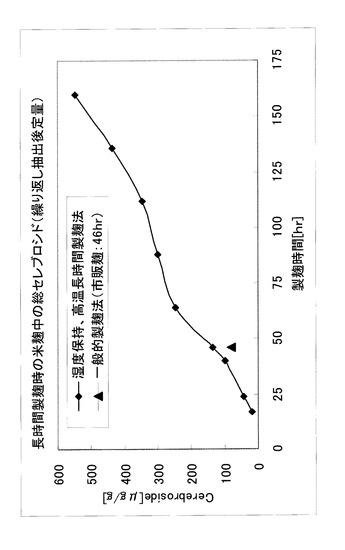
[実施例7] (米麹による長時間製麹検討)
実施例1〜6の結果を踏まえ、製麹後半の湿度保持、高温条件下における長時間製麹検討を実施した。蒸米に対し、粉末型麹菌(A.oryzae:NBRC4214)を0.1%の割合で接種し、品温35℃、相対湿度85%以上とし、32時間後に品温を40℃として160時間製麹した。調製した米麹から、総セレブロシド含量を、実施例1の繰り返し抽出及びHPLCによって測定した。また対照として、一般的な製麹時間(46時間)、湿度及び温度で製麹された市販麹についても、同様の測定法で総セレブロシド含量を測定した。
【0088】
調製した米麹中のセレブロシド量の経時変化を図7に示した。市販麹に含まれる総セレブロシド量は81μg/g乾燥麹となった。これに対し、製麹後半の湿度保持、高温条件下における長時間製麹を行った場合、製麹46時間における総セレブロシド量は136μg/g乾燥麹となり、製麹時間が同じ市販麹と比較すると約1.7倍であった。さらに160時間製麹によって、総セレブロシド量は546μg/g乾燥麹となり、市販麹の約6.7倍にまで達した。
【0089】
このことから、本発明者らは、麹菌固体培養において、製麹後半の湿度保持、高温長時間製麹によって、胞子着生をせず、一般的な46時間製麹と比較して、セレブロシド量が6.7倍増加することを見出した。
【0090】
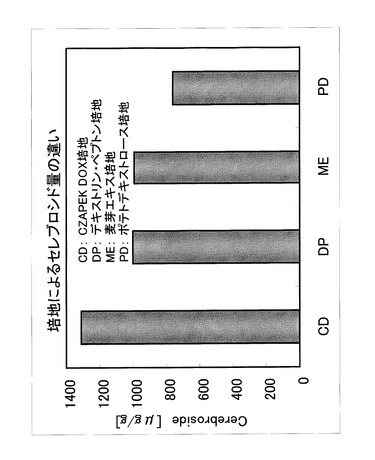
[比較例1] 各種液体培地における菌体あたりのセレブロシド量
培地条件の違いが菌体あたりのセレブロシド量に及ぼす影響を評価するために、CZAPEK DOX培地(2%Glucose、0.3%NaNO3、0.2%KCl、0.1%KH2PO4、0.05%MgSO4・7H2O)、デキストリン・ペプトン培地(2%デキストリン、1%ペプトン、0.5%KH2PO4、0.1%NaNO3、0.05%MgSO4・7H2O)、麦芽エキス培地(2%モルトエキス)およびポテトデキストロース培地(2.4%ポテトデキストロース)に対し、胞子数が105/mlとなるように粉末型麹菌(A.oryzae:NBRC4214)を接種し、30℃、120rpmで72hr培養した。培養後、菌体を回収・洗浄後、凍結乾燥し、実施例1の単回抽出及びTLCによって乾燥菌体あたりのセレブロシド量を求めた。
【0091】
各培地における乾燥菌体あたりのセレブロシド量を図8に示す。図8から分かるとおり、酵母における報告と同様に、麹菌においても培地条件の違いによって菌体あたりのセレブロシド量が変動することがわかった。これは、単純に菌体量が増えればスフィンゴ脂質も増大するものではないことを示唆している。
【産業上の利用可能性】
【0092】
本発明に係るスフィンゴ脂質富化麹の製造方法は、煩雑な工程を必要とせず、従来微生物によるスフィンゴ脂質製造において行われていたような、有機溶媒を用いた抽出や濃縮をすることも必要としないため、労力とコストを低減できるばかりでなく、食品組成物として安全性が高く、非常に有用なスフィンゴ脂質富化麹を提供することができる。
【図面の簡単な説明】
【0093】
【図1】図1は、米麹由来のグルコシルセラミド主要分子種の構造を示す。
【図2】図2は、セレブロシドの測定における薄層クロマトグラフィー(TLC)と高速液体クロマトグラフィー(HPLC)との比較を示す。
【図3】図3は、各種麹菌のセレブロシド生産量を示す。
【図4】図4は、本発明の方法による製麹におけるセレブロシド量の経時的変化を示す。
【図5】図5は、本発明の方法における温度シフト時間とセレブロシド量との関係を示す。
【図6】図6は、本発明の方法における相対湿度とセレブロシド量、出麹水分との関係を示す。
【図7】図7は、長時間湿度保持条件下での米麹中の総セレブロシド量の経時的変化を示す。
【図8】図8は、各種液体培地における乾燥菌体当たりのセレブロシド量を示す。
【技術分野】
【0001】
本発明は、スフィンゴ脂質富化麹及びその製造方法に関する。
【背景技術】
【0002】
スフィンゴ脂質とは、スフィンゴイド塩基(長鎖塩基)を構造骨格としてもつ複合脂質の総称である。動物細胞のスフィンゴ脂質の骨格はスフィンゴシンであるが、植物や真菌では主にフィトスフィンゴシンである(非特許文献1)。
【0003】
スフィンゴ脂質には、スフィンゴイド塩基のアミノ基にアシル基がアミド結合したセラミドや、セラミドに糖、リン、硫黄、アミノ酸など様々な極性基が付加した複合スフィンゴ脂質などがある。なかでも糖が結合したスフィンゴ糖脂質は、真核生物やバクテリアの一部などに普遍的に存在しており、糖鎖部分の構造は様々であることが知られている。グルコースが結合したグルコシルセラミドは、セレブロシドとも呼ばれ植物、真菌、動物に共通してみられる代表的なスフィンゴ糖脂質である(非特許文献2)。
【0004】
セラミドはヒトの皮膚の角質層の細胞間脂質の主成分として約50%を占め、皮膚の保湿性や柔軟性にかかわっていると言われている。加齢により減少し、しわ、ドライスキン、肌荒れの原因となるほか、アトピー性皮膚炎にも関与していると考えられている。従来はセラミドを補充する方法として、安価な供給源である牛脳由来のスフィンゴ脂質が用いられていたが、BSE(牛海綿状脳症)発症の原因であるプリオンが脳幹部位に蓄積していることが明らかとなって以来、食品・化粧品用途での利用は不可能となっている。また、合成セラミドは安全性の観点から食品素材としては利用の制限がある。
【0005】
牛脳の代替供給源として、現在では主に農産加工副産物である大豆油さいやビール粕、米や小麦などの穀類の胚芽、コンニャク芋などが利用されているが、含有量が微量であり、食品としてそのまま利用しにくいことから、有機溶媒などによる抽出・濃縮が行なわれている。このため製造に多大なコストがかり、製品が非常に高価であるという問題があった(特許文献1、2)。
【0006】
また、酵母、キノコなどの真菌類からもセレブロシドの抽出が行われている。酵母においてはセレブロシドが存在する種は限られているため(非特許文献3)、サッカロミセス・クルイベリ(Saccharomyces kluyveri)などのセレブロシド生産菌を用いて、ビートモラセスを炭素源とし、様々な環境ストレスを与えることによりセレブロシドの生産量、蓄積量を高める製造方法も検討されている(特許文献3)。酵母は高温培養時には菌体量あたりのスフィンゴ脂質が増加し、低温培養時には減少することが分かっている(非特許文献4、5)。このように、培養条件の違いによって、菌体量あたりのスフィンゴ脂質量は変動することが知られている。しかしながら、この方法においても有機溶媒抽出や冷凍処理などが用いられており、工程が煩雑であり、多大なコストがかかるほか、環境への負荷も懸念されるなどの問題があった。
【0007】
国内で伝統的な発酵食品の製造に利用されてきた麹菌についても、液体培養時にセラミドやセレブロシドといったスフィンゴ脂質を生産するという報告がある(非特許文献6、7)。しかし、仮に麹菌の液体培養において培養条件の検討を行い、麹菌のスフィンゴ脂質含量を増加させたとしても、菌体だけをろ過回収して食するような食経験が無いため、有機溶媒抽出等の処理が必要となり、前述のような問題点が生じる。
【0008】
一方で、麹菌の固体培養物は、伝統的に「麹」として丸ごと食する食習慣があるため、食品として麹菌スフィンゴ脂質を利用するには非常に適している。そのため固体培養によってスフィンゴ脂質量を増大させる方法の確立が望まれている。
【0009】
しかしながら固体培養時、すなわち麹にした際のスフィンゴ脂質の生産能力や分子種についての報告は無く、高生産のための培養条件の検討はもちろん、麹をスフィンゴ脂質供給源として利用するための検討さえ行なわれてきていない。
【0010】
麹菌による固体培養(すなわち、製麹)について述べると、水分量や湿度、時間、温度等の培養条件に関する検討は、一般に穀物の糖質や蛋白質を分解する酵素を生産させるためや、種麹となる胞子を着生させることが目的で行われてきた。
【0011】
一般的な製麹法では、床もみ(すなわち種麹植菌)を行った後、蒸米を堆積し、布などで包む等して乾燥を防いで湿度を維持し、かつ乾燥が進む原因となる手入れをほとんど行わず、切り返しを行うのみで培養を行う。引き続いて「盛り」とは、床もみ後、一般的には18〜20時間前後になって、麹菌の生育が旺盛になりかけたものを麹蓋に入れる等、小区分に分けて品温管理をし易くする作業を指す。麹蓋に入れて棚培養する工程は、麹を薄く盛り、麹の水分蒸発を進める工程でもある。盛りまでを「床時代」、盛り以降を「棚時代」と呼ぶ。棚培養期間中に適宜麹をかき混ぜ、酸素の供給や炭酸ガスの排出とともに水分を発散させて、品温・状貌の均一化をはかる。非特許文献8によると、麹菌植菌後23〜28時間後に麹をかき混ぜる操作を仲仕事、または昼仕事といい、仲仕事後5〜7時間目に攪拌する操作を仕舞仕事または留仕事という。また、麹菌の代謝によっても水分が生じるため、製麹後半では麹室内の乾湿差を多く取り、通風製麹の場合は培養35時間頃から発酵熱により品温が上昇するため、送風を行う事で品温を下げるとともに水分の発散を促している(非特許文献8)。
【0012】
また培養水分量や湿度に関しては、伝統的にも限定吸水などによって蒸米水分を30%前後に制御し、製麹の後半(盛り以降)は麹室あるいは培養室の湿度を低下させて麹の水分含量を低下させ、酵素生産量を高める方法が採用されている。さらに、非特許文献8によると、「一般的にいって、湿気の多い麹は酵素力が弱く、粕が多く、清酒の色も濃い。また雑菌も多いので好ましくない。乾燥して、よくはぜた麹は酵素力が強く、清酒の色も淡く、味も軽い。」とある。すなわち水分が多いと酵素活性が低下してしまうので、製麹後半に麹の水分含量を低下させる方法が採用されてきた。
【0013】
製麹の後半に水分含量を低下させる事は、蒸発潜熱により品温の過昇を防ぐ以外に、麹の保存性を高める目的もある。非特許文献9によると、製麹中に品温を下降させる手段として、最も有効なのは蒸発潜熱を奪うことであり、蒸発潜熱を奪うために最も効率がよいのは、環境の相対湿度を低くすることであると記載されている。
【0014】
このため、通常の製麹法では、製麹により水分が蒸発するため、白米麹では蒸米水分が35%であれば、出麹水分は18%程度となり、醤油麹では初期水分が45%前後であれば、出麹水分は30%前後となる(非特許文献8、非特許文献10)。また製麹日数が長く、1週間となる種麹作成後の水分含量も25%程度と、仕込み時の水分含量より大きく減少している。
【0015】
製麹時間については、非特許文献8によると、「米麹の製造は麹菌分生子(種麹)を蒸米に植えつけ、30℃から40℃にわたる温度経過を経て、約40時間の培養によって行われる。」、「製麹時間の標準は酒母麹で48時間、掛麹で43時間内外である。」とある。つまり製麹時間は白米麹の場合、43〜48時間が一般的である。
【0016】
醤油麹などでは4日麹のように長時間(3日間)培養するものがある。また種麹作成のように長い特殊な場合では1週間程度製麹するものがある。これらは蛋白分解酵素(酸性カルボキシペプチダーゼ等)を高生産させること、あるいは胞子(分生子)を着生させることが目的である。これら製麹においても後半は麹室あるいは培養室の湿度を低下させて麹の水分含量を低下させる。
【0017】
このように、製麹時間については従来製麹の目的が酵素生産、あるいは胞子生産であることから、一般的には43〜48時間程度、醤油麹の場合は2〜3日程度、種麹作製のように特殊な場合では1週間とされてきた。
【0018】
以上のように、従来の産業利用上の製麹では、スフィンゴ脂質含量を高くするという観点で製麹条件(固体培養条件)の検討はなされていない。
【0019】
【特許文献1】特開平4−282317号公報
【特許文献2】特開平11−279586号公報
【特許文献3】特開2005−185126公報
【非特許文献1】Chem.Phys.Lipids,5,6−43(1970)
【非特許文献2】CMLS,Cell.Mol.Life Sci.,60,919−941(2003)
【非特許文献3】FEMS Yeast Research,2,533−538(2002)
【非特許文献4】The Journal Of BIOLOGICAL CHEMISTRY,272,48,30196−30200,1997
【非特許文献5】JOURNAL OF OLEO SCIENCE,53,3,127−133(2004)
【非特許文献6】日本農芸化学誌,第49巻,第4号,205−212(1975)
【非特許文献7】Biochimica et Biophysica Acta,486,161−171(1977)
【非特許文献8】麹学,日本醸造協会発行,村上英也 編著(1986)
【非特許文献9】新・みそ技術ハンドブック,全国味噌技術会発行(2006)
【非特許文献10】増補 醤油の科学と技術,日本醸造協会発行、栃倉辰六郎 編著
【発明の開示】
【発明が解決しようとする課題】
【0020】
スフィンゴ脂質を効率的に取得するためには、真菌類などの微生物を供給源とすることが適している。しかし、酵母やキノコ類を供給源とした場合、その形態や香味ゆえにいずれも食品としてはそのまま使用しづらく、有機溶媒等による濃縮が必須となるために、食品としては極めて高価なものになることから利用の制限があった。また麹菌は液体培養においてスフィンゴ脂質を生産することが知られるが、食用産業利用上有利な固体培養でスフィンゴ脂質を高生産するための最適な方法がなかった。
【0021】
したがって、本発明は、麹菌の固体培養によって、煩雑な工程を経ることなく、また食品組成物としてそのまま用いるために有機溶媒抽出や濃縮などを行うことなく、大量かつ安定的に、安全なスフィンゴ脂質を製造する方法を提供することを目的とする。
【課題を解決するための手段】
【0022】
本発明者らは、上記課題を解決するべく鋭意検討を重ねた結果、驚くべきことに、製麹における盛り以降の工程を、高湿度条件下で、場合により一般的な製麹時間(例えば46時間)よりも長い時間行うことにより、スフィンゴ脂質量を通常よりも顕著に増大できることを見出した。
【0023】
すなわち、本発明は以下の特徴を包含する。
(1)蒸きょうした製麹原料に麹菌を植菌して培養を行う製麹において、盛り以降の工程を、湿度75%以上の条件下で行うことを特徴とする、スフィンゴ脂質富化麹の製造方法。
(2)盛り以降の工程を湿度85%以上の条件下で行うことを特徴とする上記(1)記載の方法。
【0024】
(3)製麹時間が計46〜160時間であることを特徴とする上記(1)記載の方法。
(4)植菌後18〜32時間の時点で製麹温度を38〜42℃とすることを特徴とする、上記(3)記載の方法。
(5)麹菌はアスペルギルス・オリゼ(Aspergillus oryzae)又はアスペルギルス・ソーエ(Aspergillus sojae)であることを特徴とする上記(1)記載の方法。
【0025】
(6)製麹原料はイネ科植物であることを特徴とする上記(1)記載の方法。
(7)イネ科植物は米又は大麦であることを特徴とする上記(1)記載の方法。
(8)蒸きょうした製麹原料の水分量が30〜36重量%であることを特徴とする上記(1)記載の方法。
【0026】
(9)スフィンゴ脂質はセレブロシドであることを特徴とする上記(1)記載の方法。
(10)上記(1)〜(9)のいずれか記載の方法で製造されたスフィンゴ脂質富化麹。
(11)乾燥重量1g当たり少なくとも136μg以上のスフィンゴ脂質を含有することを特徴とする上記(10)記載のスフィンゴ脂質富化麹。
(12)上記(10)又は(11)記載のスフィンゴ脂質富化麹を含有する飲食品。
【発明の効果】
【0027】
本発明によれば、簡便かつ安価で、安全なスフィンゴ脂質富化麹を製造することができる。
【発明を実施するための最良の形態】
【0028】
本明細書で使用する「製麹」とは、蒸きょうした製麹原料に麹菌を植菌して培養する、当業者に公知の製麹方法(例えば、蓋麹法、箱麹法、床麹法、機械麹法など)を指す。これらの製麹方法は、蒸きょうした製麹原料に麹菌を植菌した後に培養する工程を含み、その間、一般的に「引き込み」、「盛り」、「仲仕事」、「仕舞仕事」といった処理を含んでいる。
【0029】
「盛り」とは、「床もみ」(すなわち種麹植菌)後、麹菌の生育が旺盛になりかけたものを小区分に分けて品温管理をしやすくする作業を指す。一般的に製麹において、盛りまでを「床時代」、盛り以降を「棚時代」と呼ぶ。床時代とは床もみを行った後、蒸した製麹原料を堆積し、布などで包むなどして湿度を維持し、乾燥を防ぐ工程であり、乾燥が進む原因となる手入れをほとんど行わず、切り返しを行うのみで培養を行う。
【0030】
本明細書において、植菌後から盛りまでの工程(すなわち床時代)を製麹前半、盛り以降の工程(すなわち棚時代)を製麹後半と称する。
【0031】
製麹前半における製麹温度は、使用する麹菌の菌体生育の至適温度に応じて、一般的には28〜42℃、例えば30〜37℃とされる。ただし、製麹前半の製麹温度をおよそ38℃以上の比較的高温とする場合、麹菌が生育阻害を受ける虞がある。なお、製麹温度とは、麹の品温を意味する。製麹温度は、麹菌の生育に伴う発酵熱を制御することによって、又は製麹における周囲温度を制御することによって調節することができる。
【0032】
製麹前半の所要時間は、一般的には18時間〜20時間である。生育が旺盛になった状態のまま長時間放置すると発酵熱を除くことができず、品温が制御できなくなるうえに、原料が密集しているため製麹の生育が抑制されてしまうため、この時点で盛りを行い、製麹後半に移行する。
【0033】
一方、製麹後半は、麹を薄く盛り、麹の水分蒸発を進める工程であり、この培養工程の中で、「仲仕事」、「仕舞仕事」と呼ばれる手入れ作業が行われる。上記非特許文献8によれば、出麹までに蒸発する総水分量の75%が仲仕事までの間に蒸発することが記載されている。
【0034】
製麹の詳細については、例えば上記非特許文献8及び発酵ハンドブック((財)バイオインダストリー協会 発酵と代謝研究会編,2001年)などを参照されたい。
【0035】
本発明のスフィンゴ脂質富化麹の製造方法(以下、本発明の方法ともいう)において、下記で特に規定されない限り、製麹を上記のような当業者に公知の製麹と同様にして行うことができること、当業者は適宜上記条件を変更できることに留意すべきである。
【0036】
具体的に、本発明の方法は、製麹において、製麹後半を湿度75%以上の条件下で行うことを特徴とする。これにより、製麹後半における麹水分の蒸散を抑制することができ、その結果、スフィンゴ脂質の生産を増加することができる。清酒、醤油麹の製造などに使用されている従来の製麹技術は、上記の通り、酵素生産量の増加及び保存性を高める観点から製麹後半において湿度を低下させることによって麹の水分含量を低下させている点で、本発明の方法と異なる。
【0037】
製麹後半は、湿度85%以上の条件下で行うことがより好ましい。湿度75%以上の条件は、従来の製麹技術と比較して製造される麹中のスフィンゴ脂質含量を増加させるのに十分であるが、下記実施例に示されるように、製麹後半における相対湿度を85%以上にすることにより、製造されるセレブロシド量をより増加させることができる。
【0038】
本発明の方法はまた、製麹において、製麹後半を、製麹時間が計46時間以上となるまで行うことを特徴とする。これにより、製造される麹に含まれるスフィンゴ脂質の量を経時的に増加させることができる。従来の製麹技術は、湿度を高く維持したまま通常の43〜48時間(醤油麹の場合は2〜3日間)の製麹時間を越えるような培養を行なっていない点で、本発明の方法と異なる。本発明の方法における製麹時間の上限は特に制限されず、スフィンゴ脂質の量が経時的に増加する限り製麹後半を継続することができる。本発明において、製麹時間は、例えば計46〜160時間、より好ましくは72〜160時間とすることができる。
【0039】
本発明の方法において、製麹前半における湿度条件は特に制限されず、従来の製麹と同様の条件で行うことができる。ただし、本発明の方法により製造される麹は、上記の通り出麹水分の減少が抑制されたものであることが好ましいところ、製麹前半においても高湿度条件、例えば湿度75%以上、好ましくは湿度85%以上の条件下で行うことが好ましい点に留意する。本発明において、製麹を通じて湿度が85%以上に維持されることが最も好ましい。
【0040】
また本発明の方法において、従来の製麹時間を越えるような製麹時間で製麹後半を行う場合には、植菌後18〜40時間、好ましくは植菌後18〜32時間の時点で製麹温度を38〜42℃に温度シフトすることが好ましい。従来の製麹時間よりも長時間で製麹後半を行う場合には、麹の味覚上好ましくない胞子の着生の問題が顕著となる。そこで、植菌後18時間〜40時間、好ましくは植菌後18〜32時間の時点で、製麹温度を38〜42℃に温度シフトすることにより、胞子の着生を抑制することができる。
【0041】
本発明の方法において、蒸きょうした製麹原料の水分量が30〜36重量%であることが好ましい。蒸きょう後の製麹原料の水分量が30%以下であるか又は36%以上であると、麹菌の立ち上がりが抑制されてしまう虞があるからである。
【0042】
本発明の方法で使用される製麹原料は、麹菌を生やして麹にできるものであれば特に制限されず、例えば穀類、擬穀類、豆類、ゴマ類、芋類等を使用することができる。具体的に、穀類としては、白米、玄米、黒米、赤米、大麦、小麦などを、擬穀類としてはソバ、アマランサス、キヌアなど、豆類としては大豆、小豆、黒豆、ヒヨコ豆などを挙げることができるが、これらに限定されるものではない。麹の原料として穀類、例えばイネ科植物を使用することが好ましく、イネ科植物として白米や、大麦などの麦類を使用することが特に好ましい。
【0043】
本発明の方法に用いることができる麹菌は、スフィンゴ脂質を生産できるものであれば特に限定されないが、例えばアスペルギルス・オリゼ(Aspergillus oryzae)や、その近縁のアスペルギルス・ソーエ(Aspergillus sojae)などを挙げることができる。A.オリゼとして具体的には市販の種麹や、登録番号RIB40、NBRC4214、JCM2228又はATCC36261で指定される特定の菌株などを用いることができる。また、A.ソーエとしては、市販の種麹や、登録番号JCM2226、NBRC4239又はNBRC5241で指定される特定の菌株などを用いることができる。A.オリゼとしてNBRC4214、JCM2228が、そしてA.ソーエとしてJCM2226が白米、大麦の両方の麹においてセレブロシド生産量が高いことから、本発明で使用するのに特に適している。また、上記麹菌以外の麹菌として、大麦麹の場合にA.オリゼ登録番号NBRC4255、ATCC11494、ATCC16868、NBRC5786、JCM2245、NBRC5240などを、米麹の場合にA.オリゼ登録番号JCM2227、JCM2173、JCM2229、NBRC4261などを使用することができる。
【0044】
上記のようにして製造される麹は、スフィンゴ脂質(特にセレブロシド)を豊富に含むものである。具体的には、従来の製麹時間、製麹湿度にて製麹された麹に比べて、スフィンゴ脂質の総量を約1.7倍〜約6.7倍まで高めることができる。また、本発明の方法により製造されるスフィンゴ脂質富化麹は、乾燥重量1g当たり少なくとも136μg以上、好ましくは248μg以上のスフィンゴ脂質を含むことができる。なお、麹中のスフィンゴ脂質の量は、薄層クロマトグラフィー、高速液体クロマトグラフィーなどを使用することによって測定することができる。
【0045】
このように、本発明の方法は、煩雑な条件でストレスをかける工程などを必要とせず、製造されるスフィンゴ脂質富化麹も、有機溶媒を用いた抽出や、濃縮をすることなく、そのまま飲食品として提供することができるため、労力とコストを低減でき、なおかつ安全性が高いといった利点を有する。また、食品用途に供することを踏まえ、長時間培養であっても、植菌後18〜40時間、好ましくは植菌後18〜32時間の時点で製麹温度を38〜42℃に温度シフトすることにより、味覚的に望ましくない胞子の着生を抑制することができるという利点も有する。
【0046】
したがって、本発明は、本発明に係るスフィンゴ脂質富化麹を含有する飲食品を包含する。
【0047】
本発明に係る飲食品は、飲食品原料の一部として用いたり、或いは飲食品の製造工程又は製造後に添加又は配合することにより得ることができる。本発明に係る飲食品は、健康食品、機能性食品、特定保健用食品、栄養補助食品などとして提供することもできる。
【0048】
本発明に係る飲食品の形態は特に制限されず、例えばこれに制限されるものではないが、ヨーグルト、ドリンクヨーグルト、ジュース、牛乳、豆乳、酒類(アルコール性飲料)、コーヒー、紅茶、煎茶、ウーロン茶、スポーツ飲料等の各種飲料や、クッキー、パン、ケーキ、煎餅などの焼き菓子、羊羹などの和菓子、プリン、ゼリー、アイスクリーム類などの冷菓、チューインガム、キャンディ等の菓子類や、クラッカー、チップス等のスナック類や、うどん、そば等の麺類や、かまぼこ、ハム、魚肉ソーセージ等の魚肉練り製品や、みそ、しょう油、ドレッシング、マヨネーズ、甘味料等の調味類や、豆腐、こんにゃく、その他佃煮、サラダ、スープ等の各種総菜などを例示することができる。
【0049】
また、飲食品中の有効成分としてのスフィンゴ脂質富化麹の摂取量は、通常、2.4〜4.8g/日、好ましくは4.4〜8.8g/日程度とすることができる。
【0050】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
【実施例】
【0051】
A.オリゼは液体培養時、グルコシルセラミドやセラミドといったスフィンゴ脂質を生産することが知られており、A.ニガー(A.niger)はその他にガラクトシルセラミドも生産するという報告がある(上記非特許文献2)。すなわち、麹菌は種によって異なる分子種を生産すると考えられる。現在、各種麹菌が上記以外に、どのような複合スフィンゴ脂質を生産しているかについては未解明であるため、以下の実施例では、代表的な麹菌としてA.オリゼを選び、スフィンゴ脂質の中でも分析例が多いグルコシルセラミド、すなわちセレブロシド量を測定することで、総スフィンゴ脂質量の評価を行った。
【0052】
[実施例1] 米麹セレブロシド標準品の精製と構造解析
米麹セレブロシド標準品の調製法を以下に示した。A.オリゼ(A.oryzae)を使用して製麹した米麹5.4kgを粉砕し、5倍量のクロロホルム−メタノール(2:1)で2回抽出、濾過して得られた抽出液を濃縮、乾固後、0.2M KOH−メタノール溶液にて37℃で2時間振盪し、グリセロ脂質を分解しアルカリ安定脂質を得た。中和後、濃縮、乾固し、クロロホルム−メタノール−0.8%NaCl(8:4:3)に再溶解した。振盪混合し、遠心分離後、下層を濃縮した。
【0053】
これをケイ酸カラムクロマトグラフィーに供し、クロロホルム-メタノールを用いて遊離脂肪酸等を除いた。再度ケイ酸カラムクロマトグラフィーに供した後、セレブロシド画分として白色の固形油脂状物407.9mgを得た。その後、わずかに混在する遊離脂肪酸を除去するため、さらにHPLC−示差屈折検出器((株)島津製作所製)を用いて精製を行なった。カラムはYMC Pack ODS−A;20×250mm、カラムオーブンは40℃、移動相にはメタノールを用いて、アイソクラティック溶出を行い、流速6ml/minとした。得られた精製品は、薄層クロマトに供したところ単一スポットとなり、また以下に示したように主要分子がグルコシルセラミドと同定されたため、これを標準品としてセレブロシドの定量を行った。
【0054】
主要分子種の構造解析を行った。すなわち、HPLC−示差屈折検出器((株)島津製作所製)を用いて、前述の条件で、精製セレブロシドに含まれる主要分子を分取した。得られた主要3分子(保持時間28.7分、32.1分、37.4分)について、以下に示すように構造解析を実施した。
【0055】
保持時間28.7分の分子をメタノールに溶解し、HRESI−MS(+)を測定した。この結果m/z 776.57456に(M+Na)+イオンが観測され、分子式はC43H79NO9(C43H79NO9Na cald 776.56525 (D+4.57mmu)と決定した。
【0056】
次にこの分子を重ピリジン(C6D5N)に溶解し、核磁気共鳴(NMR)解析を行った。1H及び13C NMRスペクトルの特徴からこの分子がセレブロシドであると同定された。まず糖部分の1H、13C化学シフト値及びJHH値から、構成糖はグルコースであり、これがβ−結合(J1”,2”=7.8Hz)で長鎖塩基と結合していること(=β−グルコシルセラミド)が判明した。次に1H−1H COSY、HMQC、HMBCスペクトルを測定し、脂肪酸及び長鎖塩基部分の構造について解析を行った。
【0057】
脂肪酸については1H−1H COSYスペクトルにおいてH−2’(δ5.10)−H−3’(δ6.10、J3’,4’=15.7Hz)−H−4’(δ6.16)というvicinal spin結合が観測されること、また1H NMRで末端メチルはδ0.85にtripletとして観測されることから、直鎖の2−ヒドロキシ、3−トランス脂肪酸であることが明らかとなった。次に脂肪酸の炭素数を明らかにするために、この分子をメタノール性0.9N HClを用いて100℃、18時間還流することで分解(メタノリシス)し、脂肪酸メチルエステルとした。ヘキサン抽出で脂肪酸メチルエステルを精製後、これをトリメチルシリル(TMS)誘導体化し、GC/MS解析を実施した。この結果、m/z 369に(M−15)+のピークが観測され、この分子を構成する脂肪酸炭素数はC18と決定した。
【0058】
長鎖塩基については1H−1H COSYスペクトルにおいてH−1(δ4.23、δ4.69)−H−2(δ4.79)−H−3(δ4.74)−H−4(δ5.99、J4,5=15.8Hz)−H−5(δ5.92)−H−6(δ2.14)−H−7(δ2.14)−H−8(δ5.24)というvicinal spin networkが観測されること及びHMBCスペクトルで9−CH3(δ1.60)からC−8(δ123.3)、C−9(δ13.5)に1H−13C遠隔スピン結合が観測されることから、9−メチル−4トランス、8−トランス−スフィンガジエニン構造を有していることが明らかとなった。また1H NMRで末端メチルはδ0.85にtripletとして観測されるため、アルキル鎖部分は直鎖であることも明らかであった。この時点で既に脂肪酸の炭素数は明らかとなっているため、長鎖塩基を構成する炭素数は分子式から糖(C6)及び脂肪酸(C18)の炭素数を引いたC18であると決定された。その結果、この分子の構造は図1に示すように18h:1−9Me d18:24t,8t−Glcであることが一義的に決定された。また、この構造は真菌類に特徴的な構造であった(非特許文献2)。
【0059】
保持時間32.1分の分子をメタノールに溶解し、HRESI−MS(+)を測定した。この結果、m/z 778.58403に(M+Na)+イオンが観測され、分子式はC43H81NO9(C43H81NO9Na cald 778.58090 (D+3.13mmu)と決定した。
【0060】
次にこの分子を重ピリジン(C6D5N)に溶解し、核磁気共鳴(NMR)解析を行った。1H及び13C NMRスペクトルは18h:1−9Me d18:24t,8t−Glc(保持時間28.7分)によく類似していた。糖部分の1H、13C化学シフト値及びJHH値はほぼ同一であることから、構成糖はグルコースであり、これがβ−結合(J1”,2”=7.8Hz)で長鎖塩基と結合していること(=β−グルコシルセラミド)が判明した。次に1H−1H COSY、HMQC、HMBCスペクトルを測定し、脂肪酸及び長鎖塩基部分の構造について解析を行った。
【0061】
脂肪酸については1H−1H COSYスペクトルにおいてH−2’(δ4.56)−H−3’(δ2.12)というvicinal spin結合が観測されたこと、また1H NMRで末端メチルがδ0.85にtripletとして観測されることから、直鎖の2−ヒドロキシ脂肪酸であることが明らかとなった。次に脂肪酸の炭素数を明らかにするためにメタノリシスを行い、脂肪酸メチルエステルとした。ヘキサン抽出で脂肪酸メチルエステルを精製後、これをトリメチルシリル(TMS)誘導体化し、GC/MS解析を実施した。この結果、m/z 371に(M−15)+のピークが観測され、この分子を構成する脂肪酸炭素数はC18と決定した(=2−ヒドロキシステアリン酸)。
【0062】
長鎖塩基については1H−1H COSYスペクトル及びHMBCスペクトルにおいて18h:1−9Me d18:24t,8t−Glcとまったく同じ相関が認められることから、この分子の長鎖塩基も9−メチル−4トランス,8−トランス−スフィンガジエニン構造を有していることが明らかとなった。また1H NMRで末端メチルはδ0.85にtripletとして観測されるため、アルキル鎖部分も直鎖であることも明らかであった。既に脂肪酸の炭素数は明らかとなっているため、長鎖塩基を構成する炭素数は分子式から糖(C6)及び脂肪酸(C18)の炭素数を引いたC18あると決定された。その結果、この分子の構造は図1に示すように18h:0−9Me d18:24t,8t−Glcであることが一義的に決定された。また、この構造は真菌類に特徴的な構造であった(非特許文献2)。
【0063】
保持時間37.4分の分子をメタノールに溶解し、HRESI−MS(+)を測定した。この結果、m/z 792.60158に(M+Na)+イオンが観測され、F5の分子式はC44H83NO9(C44H83NO9Na cald 792.59655(Δ+0.94mmu)と決定した。
【0064】
次にこの分子を重ピリジン(C6D5N)に溶解し、核磁気共鳴(NMR解析)を行った。1H及び13C NMRスペクトルは18h:0−9Me d18:24t,8t−Glc(保持時間32.1分)によく類似していた。糖部分の1H、13C化学シフト値及びJHH値についても18h:0−9Me d18:24t,8t−Glcと同一であることから、構成糖もグルコースであり、これがβ−結合(J1”,2”=7.8Hz)で長鎖塩基と結合していること(=β−グルコシルセラミド)が判明した。次に1H−1H COSY、HMQC、HMBCスペクトルを測定し、脂肪酸及び長鎖塩基部分の構造について解析を行った。
【0065】
脂肪酸については1H−1H COSYスペクトルにおいて18h:0−9Me d18:24t,8t−Glcと同様にH−2’(δ4.56)−H−3’(δ2.12)というvicinal spin結合が観測されること、また1H NMRで末端メチルがδ0.85にtripletとして観測されることから、直鎖の2−ヒドロキシ脂肪酸であることが明らかとなった。次に脂肪酸の炭素数を明らかにするために、メタノリシスを行い、脂肪酸メチルエステルとした。精製後トリメチルシリル(TMS)誘導体化し、GC/MS解析を実施した。この結果、m/z 399に(M−15)+のピークが観測され、F5を構成する脂肪酸炭素数はC20と決定した(=2−ヒドロキシアラキジン酸)。
【0066】
長鎖塩基については1H−1H COSYスペクトルでH−1(δ4.23、δ4.69)−H−2(δ4.79)−H−3(δ4.74)−H−4(δ5.99、J4,5=15.8Hz)−H−5(δ5.92)−H−6(δ2.16)−H−7(δ2.16)−H−8(δ5.46)−H−9(δ5.46)というvicinal spin networkが観測されること及び18h:0−9Me d18:24t,8t−Glcで観測された9−CH3の信号が観測されないことから、4トランス、8−トランス−スフィンガジエニン構造を有していることが明らかとなった。また1H NMRで末端メチルはδ0.85にtripletとして観測されるため、アルキル鎖部分は直鎖であることも明らかであった。既に脂肪酸炭素数は明らかとなっているため、長鎖塩基を構成する炭素数は分子式から糖(C6)及び脂肪酸(C20)の炭素数を引いたC18であると決定され、この結果、この分子の構造は図1に示すように20h:0 d18:24t,8t−Glcであることが一義的に決定された。また、この構造は真菌類及び植物に特徴的な構造であり(非特許文献2)、米由来のセレブロシドも含まれていると考えられる。
【0067】
続いて、セレブロシドの精製品をLC/MS(LCMS−2010EV島津製作所)によって分析し、主要3分子の量比を調べた。イオン化モードはESI(−)、カラムはCadenza CD−C18 150×2mm、移動相はTHF/メタノール(1/9 v/v)を用いてアイソクラティック溶出を行い、流速は0.2ml/mlとした。その結果、それぞれの分子種の量比は、以下のとおりであることが示唆された。このことから、真菌に特徴的な分子である18h:0−9Me d18:24t,8t−Glcが、米麹中のセレブロシドの大半を占めることが示された。
【0068】
【表1】
【0069】
[実施例2] 米麹、大麦麹中のセレブロシド含量の測定法
実施例3〜7におけるセレブロシドの定量は、各検体から有機溶剤を用いて抽出後、実施例1で得られたセレブロシドを標準品として用い、薄層クロマトグラフィー(TLC)又は高速液体クロマトグラフィー(HPLC)によって行った。TLCとHPLCのどちらの測定法においても、定量値がほぼ一致していることを確認した(図2)。
【0070】
また、麹からのセレブロシドの抽出は以下に示すように単回または、繰り返し抽出により行った。単回抽出は、より簡便な方法によって、各実験区におけるセレブロシド含量の比較を行うために実施し、繰り返し抽出は実際に麹に含まれるセレブロシド含量を定量する目的で実施した(実施例3〜6については単回抽出を行い、実施例7については繰り返し抽出を行った)。各抽出法で得られる定量値について、比較データの一例を図2に示した。検討の結果、麹中のセレブロシド含量によって、単回抽出の抽出効率は変動し、繰り返し抽出で得られる米麹中のセレブロシド含量は、単回抽出で得られる定量値の約1.5〜2倍程度に相当することが示された。
【0071】
単回抽出によるセレブロシド量の測定法を以下に示した。麹を凍結乾燥後、粉砕物1gに対してクロロホルム−メタノール溶液(2:1)3mlを添加し、40℃で30分間超音波処理を行った。これを遠心分離して得られた上清1mlに対し、0.8%NaCl溶液を250μl添加後、振盪混合した。これを遠心分離して得られた下層を回収し、一定量を測定に用いた。
【0072】
繰り返し抽出による総セレブロシド含量の測定法を以下に示した。米麹を凍結乾燥後、粉砕物1gに対し、クロロホルム−メタノール溶液(2:1)3mlを添加し、40℃で30分間超音波処理を行い、遠心分離後に上清を得た。計3回の抽出を行った後、上清全量に1/4量の0.8%NaCl溶液を添加し、振盪混合した。これを遠心分離して得られた下層を回収した。上清にクロロホルムを添加して、さらに2回の抽出を行った。下層全量を一定量に合わせ測定を行い、1gあたりに含まれるセレブロシド量を算出した。
【0073】
TLC−デンシトメトリー法によるセレブロシド定量法を以下に示す。抽出した試料をケイ酸薄層クロマトグラフィー(メルク社製、商品名:HPTLC silica−60 F254)にかけた。クロロホルム−メタノール(95:15)で展開後、オルシノール硫酸試薬(オルシノール:0.2% w/v、 硫酸:11.4% v/v)を噴霧し、加熱後に出現する赤紫色のセレブロシドのスポットについて、デンシトメトリーを用いて定量を行った。
【0074】
HPLC−コロナCAD検出器によるセレブロシド定量法を以下に示した。定量法は、HPLC−蒸発光散乱検出器法(J.Oleo Sci., Vol.53,No.3, p.127−133, 2004)を基に、より検出感度を上げるために検出部分を改良して行った。すなわち移動相は、A液:クロロホルム、B液:95%メタノール、を用いて、グラジエント溶出を行った。
【0075】
【表2】
【0076】
カラムは、シリカカラム:Inertsil SIL−100A (4.6×150mm、5μm)を用い、カラムオーブン:40℃、流速:1ml/minとした。また、検出器については蒸発光散乱検出器よりも感度、定量範囲、再現性が優れているとされるコロナCAD検出器を用いた。コロナCAD検出器は、ガス:窒素ガス、操作時ガス圧:35psiとした。
【0077】
[実施例3] 各種麹菌のセレブロシド生産量
蒸米および蒸大麦に対し、各種市販種麹10株及び菌株保存機関より入手した各種麹菌55株(A.oryzae、A.sojae)を0.1%の割合で接種し、品温35℃、相対湿度80%以上とし、46時間製麹した。得られた麹を凍結乾燥し、粉砕後に乾燥麹中のセレブロシド量を実施例1の単回抽出及びTLCによって測定した。
【0078】
図3に、大麦麹及び米麹に含まれる各種麹菌(A.oryzae、A.sojae)のセレブロシド生産量を示した。原料が米、大麦のいずれの場合においても、セレブロシド含量が高い株について、菌株名と入手元の菌株保存機関における識別番号を記した。A.oryzaeにおいてはNBRC4214、JCM2228、ATCC36261を、A.sojaeとしてはJCM2226が、米、大麦、いずれの培地においてもセレブロシド生産量が特に高いことが示された。また、全ての菌株において原料を大麦にした場合の方が、米の場合より生産量が高く(1.1倍〜4倍程度)なった。
【0079】
[実施例4] 米麹、大麦麹での長時間培養のスフィンゴ脂質量の経時変化
蒸米に対して粉末型麹菌(A.oryzae:NBRC4214)、蒸大麦に対して粉末型麹菌(A.oryzae:市販種麹)をそれぞれ0.1%の割合で接種し、品温35℃、相対湿度85%以上とし、24〜32時間後に品温を40℃として88時間製麹した。製麹中に経時的に麹を採取、凍結乾燥し、乾燥麹中のセレブロシド量を実施例1の単回抽出及びTLCによって測定した。
【0080】
製麹中のセレブロシド量の経時変化を図4に示した。米麹、大麦麹どちらにおいても、上記培養条件によって麹中のセレブロシド量が経時的に増加することがわかった。
【0081】
[実施例5] 温度シフト時間検討
温度シフトのタイミングの影響を評価するために、蒸大麦に対し、粉末型麹菌(A.oryzae:市販種麹)を0.1%の割合で接種し、品温35℃、相対湿度85%以上とし、24、32、40時間後に品温を40℃として88時間製麹した。調製した大麦麹のセレブロシド量は実施例1の単回抽出及びTLCによって測定した。
【0082】
出麹サンプルのセレブロシド量を図5に示した。24時間に温度シフトした場合は、64時間製麹では32〜40時間にシフトした場合よりもわずかに低かったが、88時間製麹では温度シフトのタイミングに関わらず、セレブロシド量はほぼ一定であった。
【0083】
胞子着生スコアを表3に示す。24〜32時間に温度シフトした場合と比べて、40時間にシフトした場合では胞子着生が多く、24〜32時間に温度シフトすることが好ましいと考えられた。
【0084】
【表3】
【0085】
[実施例6] 米麹による湿度条件検討
温度シフト後の湿度の影響を評価するために、蒸米に対し、粉末型麹菌(A.oryzae:NBRC4214)を0.1%の割合で接種し、品温35℃、相対湿度85%以上とし、32時間後に品温を40℃とし、相対湿度を100、95、85、75、50%として88時間製麹した。相対湿度はそれぞれ、水、6.5%NaOH、15.8%NaOH、飽和NaCl、飽和硝酸カルシウム溶液を用いて調湿した。調製した米麹のセレブロシド量は、実施例1の単回抽出及びTLCによって測定した。
【0086】
製麹後半の相対湿度とセレブロシド量、出麹水分の関係を図6に示した。この結果より、温度シフト後の湿度に応じて出麹水分が低くなり、相対湿度が85%以下になるとセレブロシド量も低下することがわかった。出麹水分は相対湿度に比例して低くなっており、85%以下では麹中水分の減少によってセレブロシドの生産が抑制されたと考えられた。
【0087】
[実施例7] (米麹による長時間製麹検討)
実施例1〜6の結果を踏まえ、製麹後半の湿度保持、高温条件下における長時間製麹検討を実施した。蒸米に対し、粉末型麹菌(A.oryzae:NBRC4214)を0.1%の割合で接種し、品温35℃、相対湿度85%以上とし、32時間後に品温を40℃として160時間製麹した。調製した米麹から、総セレブロシド含量を、実施例1の繰り返し抽出及びHPLCによって測定した。また対照として、一般的な製麹時間(46時間)、湿度及び温度で製麹された市販麹についても、同様の測定法で総セレブロシド含量を測定した。
【0088】
調製した米麹中のセレブロシド量の経時変化を図7に示した。市販麹に含まれる総セレブロシド量は81μg/g乾燥麹となった。これに対し、製麹後半の湿度保持、高温条件下における長時間製麹を行った場合、製麹46時間における総セレブロシド量は136μg/g乾燥麹となり、製麹時間が同じ市販麹と比較すると約1.7倍であった。さらに160時間製麹によって、総セレブロシド量は546μg/g乾燥麹となり、市販麹の約6.7倍にまで達した。
【0089】
このことから、本発明者らは、麹菌固体培養において、製麹後半の湿度保持、高温長時間製麹によって、胞子着生をせず、一般的な46時間製麹と比較して、セレブロシド量が6.7倍増加することを見出した。
【0090】
[比較例1] 各種液体培地における菌体あたりのセレブロシド量
培地条件の違いが菌体あたりのセレブロシド量に及ぼす影響を評価するために、CZAPEK DOX培地(2%Glucose、0.3%NaNO3、0.2%KCl、0.1%KH2PO4、0.05%MgSO4・7H2O)、デキストリン・ペプトン培地(2%デキストリン、1%ペプトン、0.5%KH2PO4、0.1%NaNO3、0.05%MgSO4・7H2O)、麦芽エキス培地(2%モルトエキス)およびポテトデキストロース培地(2.4%ポテトデキストロース)に対し、胞子数が105/mlとなるように粉末型麹菌(A.oryzae:NBRC4214)を接種し、30℃、120rpmで72hr培養した。培養後、菌体を回収・洗浄後、凍結乾燥し、実施例1の単回抽出及びTLCによって乾燥菌体あたりのセレブロシド量を求めた。
【0091】
各培地における乾燥菌体あたりのセレブロシド量を図8に示す。図8から分かるとおり、酵母における報告と同様に、麹菌においても培地条件の違いによって菌体あたりのセレブロシド量が変動することがわかった。これは、単純に菌体量が増えればスフィンゴ脂質も増大するものではないことを示唆している。
【産業上の利用可能性】
【0092】
本発明に係るスフィンゴ脂質富化麹の製造方法は、煩雑な工程を必要とせず、従来微生物によるスフィンゴ脂質製造において行われていたような、有機溶媒を用いた抽出や濃縮をすることも必要としないため、労力とコストを低減できるばかりでなく、食品組成物として安全性が高く、非常に有用なスフィンゴ脂質富化麹を提供することができる。
【図面の簡単な説明】
【0093】
【図1】図1は、米麹由来のグルコシルセラミド主要分子種の構造を示す。
【図2】図2は、セレブロシドの測定における薄層クロマトグラフィー(TLC)と高速液体クロマトグラフィー(HPLC)との比較を示す。
【図3】図3は、各種麹菌のセレブロシド生産量を示す。
【図4】図4は、本発明の方法による製麹におけるセレブロシド量の経時的変化を示す。
【図5】図5は、本発明の方法における温度シフト時間とセレブロシド量との関係を示す。
【図6】図6は、本発明の方法における相対湿度とセレブロシド量、出麹水分との関係を示す。
【図7】図7は、長時間湿度保持条件下での米麹中の総セレブロシド量の経時的変化を示す。
【図8】図8は、各種液体培地における乾燥菌体当たりのセレブロシド量を示す。
【特許請求の範囲】
【請求項1】
蒸きょうした製麹原料に麹菌を植菌して培養を行う製麹において、盛り以降の工程を、湿度75%以上の条件下で行うことを特徴とする、スフィンゴ脂質富化麹の製造方法。
【請求項2】
盛り以降の工程を湿度85%以上の条件下で行うことを特徴とする請求項1記載の方法。
【請求項3】
製麹時間が計46〜160時間であることを特徴とする請求項1記載の方法。
【請求項4】
植菌後18〜32時間の時点で製麹温度を38〜42℃とすることを特徴とする、請求項3記載の方法。
【請求項5】
麹菌はアスペルギルス・オリゼ(Aspergillus oryzae)又はアスペルギルス・ソーエ(Aspergillus sojae)であることを特徴とする請求項1記載の方法。
【請求項6】
製麹原料はイネ科植物であることを特徴とする請求項1記載の方法。
【請求項7】
イネ科植物は米又は大麦であることを特徴とする請求項6記載の方法。
【請求項8】
蒸きょうした製麹原料の水分量が30〜36重量%であることを特徴とする請求項1記載の方法。
【請求項9】
スフィンゴ脂質はセレブロシドであることを特徴とする請求項1記載の方法。
【請求項10】
請求項1〜9のいずれか1項記載の方法で製造されたスフィンゴ脂質富化麹。
【請求項11】
乾燥重量1g当たり少なくとも136μg以上のスフィンゴ脂質を含有することを特徴とする請求項10記載のスフィンゴ脂質富化麹。
【請求項12】
請求項10又は11記載のスフィンゴ脂質富化麹を含有する飲食品。
【請求項1】
蒸きょうした製麹原料に麹菌を植菌して培養を行う製麹において、盛り以降の工程を、湿度75%以上の条件下で行うことを特徴とする、スフィンゴ脂質富化麹の製造方法。
【請求項2】
盛り以降の工程を湿度85%以上の条件下で行うことを特徴とする請求項1記載の方法。
【請求項3】
製麹時間が計46〜160時間であることを特徴とする請求項1記載の方法。
【請求項4】
植菌後18〜32時間の時点で製麹温度を38〜42℃とすることを特徴とする、請求項3記載の方法。
【請求項5】
麹菌はアスペルギルス・オリゼ(Aspergillus oryzae)又はアスペルギルス・ソーエ(Aspergillus sojae)であることを特徴とする請求項1記載の方法。
【請求項6】
製麹原料はイネ科植物であることを特徴とする請求項1記載の方法。
【請求項7】
イネ科植物は米又は大麦であることを特徴とする請求項6記載の方法。
【請求項8】
蒸きょうした製麹原料の水分量が30〜36重量%であることを特徴とする請求項1記載の方法。
【請求項9】
スフィンゴ脂質はセレブロシドであることを特徴とする請求項1記載の方法。
【請求項10】
請求項1〜9のいずれか1項記載の方法で製造されたスフィンゴ脂質富化麹。
【請求項11】
乾燥重量1g当たり少なくとも136μg以上のスフィンゴ脂質を含有することを特徴とする請求項10記載のスフィンゴ脂質富化麹。
【請求項12】
請求項10又は11記載のスフィンゴ脂質富化麹を含有する飲食品。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公開番号】特開2010−22279(P2010−22279A)
【公開日】平成22年2月4日(2010.2.4)
【国際特許分類】
【出願番号】特願2008−187747(P2008−187747)
【出願日】平成20年7月18日(2008.7.18)
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
【公開日】平成22年2月4日(2010.2.4)
【国際特許分類】
【出願日】平成20年7月18日(2008.7.18)
【出願人】(000253503)キリンホールディングス株式会社 (247)
【Fターム(参考)】
[ Back to top ]