
セカンドメッセンジャーを光制御するためのセルライン、システム、および方法
光活性化型分子のための種々の方法、デバイス、および組成物が提供される。そのような方法の1つは、細胞中でセカンドメッセンジャーを生成するために実施される。キメラ光応答性膜タンパク質(例えばロドプシン)を発現するためのヌクレオチド配列を1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}で改変する。光に応答してセカンドメッセンジャーを産生させるために光応答性膜タンパク質を細胞中で発現させる。
【発明の詳細な説明】
【技術分野】
【0001】
関連特許文献
本願は、「セカンドメッセンジャーを光制御するためのセルライン、システム、および方法」という名称の2008年5月29日付で提出された米国仮特許出願第61/057,108号の米国特許法第119条(e)に基づく利益を主張する。この基礎となる仮出願は全体が参照により本明細書に取り込まれる。
【0002】
電子的に提出された資料の参照による組込み
2009年4月29日に作成された「STFD195PCT_ST25.txt」というファイル名の、12,342バイトのASCII(テキスト)ファイルである、本明細書と同時に提出されたコンピューターで読取り可能なヌクレオチド配列/アミノ酸配列リストは全体が参照により取り込まれる。
【0003】
本発明は、一般的に、光刺激に応答してセカンドメッセンジャーを生成するシステムおよび手法に関し、より具体的には、それぞれ光に応答したセカンドメッセンジャー産生に関する、セルライン、ヌクレオチド配列、キメラタンパク質、およびその使用に関する。
【背景技術】
【0004】
グアニンヌクレオチド結合タンパク質(Gタンパク質)は、不活性のグアノシン二リン酸(GDP)状態と活性のグアノシン三リン酸(GTP)結合状態の間を行ったり来たりすると考えられている。これら2つの状態は細胞内でのセカンドメッセンジャーの放出と関連付けられている。放出されたセカンドメッセンジャーは、下流の細胞プロセスを制御するよう機能することができる。
【0005】
セカンドメッセンジャーには、速やかに生成/放出されるシグナル伝達分子が含まれる。これらの分子は細胞内のエフェクタータンパク質を活性化することで細胞応答を発生させる。細胞シグナル伝達系の例としては、ホスホイノシトール系、環状アデノシン一リン酸(cAMP)系、およびアラキドン酸系が含まれる。
【0006】
Gタンパク質の前記異なる状態間の変化は、Gタンパク質共役受容体(GPCR)、Gタンパク質結合型受容体(GPLR)、7回膜貫通ドメイン受容体(7TM受容体)、またはヘプタヘリカル受容体と呼ばれるタンパク質により引き起こすことができる。このタンパク質ファミリーは、種々の膜貫通型受容体を含む。これらの受容体は、細胞内へのシグナル伝達経路を活性化するにより外部刺激(例えば、光、神経伝達物質、匂い、またはホルモン)に応答する。具体的には、リガンドが結合し、伝達経路を活性化し、それによってGタンパク質をもう一方の状態に変化させる。GPCR関連活性は多くの疾患に関連するので、GPCRは多くの医薬品および治療の標的である。
【発明の概要】
【発明が解決しようとする課題】
【0007】
市場の全薬物の30%超がGタンパク質共役受容体(GPCR)を標的としており、それらの薬物の多くがセカンドメッセンジャーcAMPの産生または阻害に関連すると考えられる。cAMPが直接関わる病理学的プロセスは数多く、例えば神経生理学的疾患、内分泌学的疾患、心疾患、代謝疾患、および免疫疾患が含まれる。複雑な哺乳動物行動の研究では、技術的限界が、細胞内シグナル伝達プロセスの時空間的に正確な制御の妨げとなっていた。化学物質に基づいてcAMPレベル等のセカンドメッセンジャーレベルを調節する現在の方法は、比較的ゆっくりと機能し、神経組織や心臓組織等の特定の組織に関連して身体が用いる速いタイムスケールの活性を研究するには問題である。これらの化学的方法はしばしば、(例えば、新規治療薬のスクリーニング中における)こうした速いタイムスケールを探索するための速度を有しない。
【課題を解決するための手段】
【0008】
本発明は、セカンドメッセンジャーの生成に関連した上記の課題等の克服ならびに関連するイメージングデバイスおよびその実施に関する。本発明は数多くの実施形態および応用により例示される。その一部を以下に要約する。
【0009】
本発明の一実施形態によれば、細胞中でセカンドメッセンジャーを生成する方法が提供される。キメラ光応答性膜タンパク質(例えばロドプシン)を発現するためのヌクレオチド配列を、1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}で改変する。光に応答してセカンドメッセンジャーを産生させるために、この光応答性膜タンパク質を細胞中で発現させる。
【0010】
本発明の一実施形態によれば、細胞内メッセンジャーに関連する推定治療レジメン(例えば、薬物または電気刺激、またはこれらのセカンドメッセンジャーを介して作用する任意のもの)の有効性を評価する方法が実施される。キメラ光応答性膜タンパク質(ロドプシン)を発現するためのヌクレオチド配列を1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}で改変する。光に応答してセカンドメッセンジャーを産生させるために、光応答性膜タンパク質を細胞中で発現させる。タンパク質を光にさらす。治療効果を評価する。
【0011】
本発明の一実施形態は、1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}を有するキメラ光応答性膜タンパク質(ロドプシン)を発現する細胞に関する。
【0012】
本発明の一実施形態は、1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}を有するキメラ光応答性膜タンパク質(ロドプシン)を発現するためのヌクレオチド配列に関する。
【0013】
上記の本発明の概要は、説明される本発明の全ての実施形態または実施例を記載することを意図したものではない。添付の図面および以下の詳細な説明によりこれらの実施形態をより具体的に例示する。
【0014】
本発明は、以下の本発明の種々の実施形態の詳細な説明を添付の図面と合わせて考慮することでより完全に理解されるであろう。
【図面の簡単な説明】
【0015】
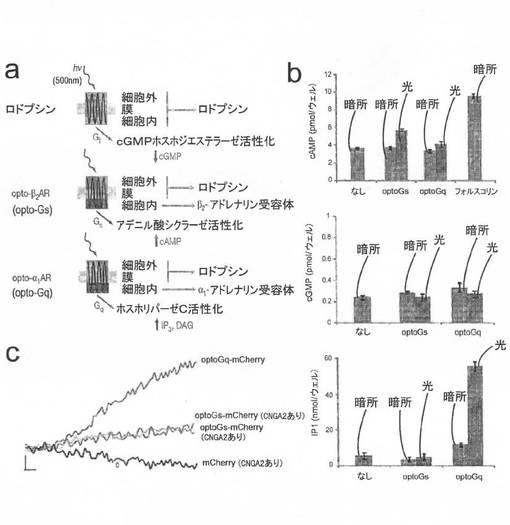
【図1】図1Aは、本発明の例示的実施形態による、optoGsおよびoptoGqを示す模式図である。 図1Bは、本発明の例示的実施形態による、トランスフェクトされていない細胞またはoptoGsもしくはoptoGqでトランスフェクトされた細胞のcAMP、cGMP、およびIP1の酵素結合免疫吸着検定法(ELISA)を示す。 図1Cは、本発明の例示的実施形態による、optoGsおよびoptoGqのmCherry融合タンパク質でトランスフェクトされた細胞のCaイメージングを示す。
【図2】本発明の例示的実施形態による、optoGsおよびoptoGqのmCherry融合タンパク質でトランスフェクトされた細胞のCaイメージングを示す。
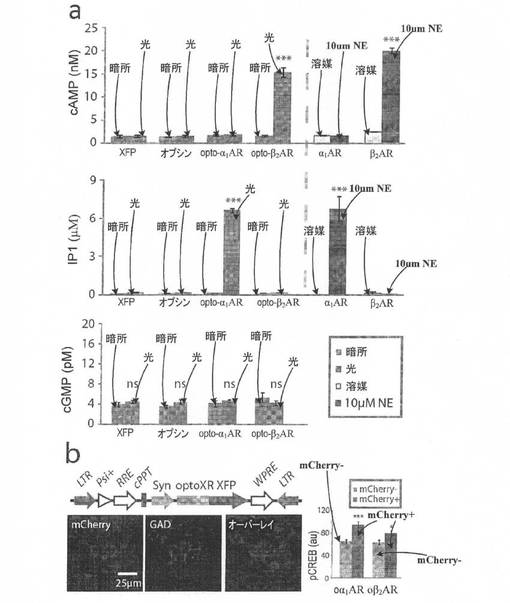
【図3】図3Aは、本発明の例示的実施形態による、種々のコンストラクトを発現するHEK細胞のcAMP、IP1、およびIP3レベルを示す。 図3Bは、本発明の例示的実施形態による、レンチウイルス発現ベクター、opto−α1AR発現細胞のGAD免疫染色、および10分間の光刺激後にoptoXR発現細胞(mCherry+)中で観察されたpCREBの活性化を示す。
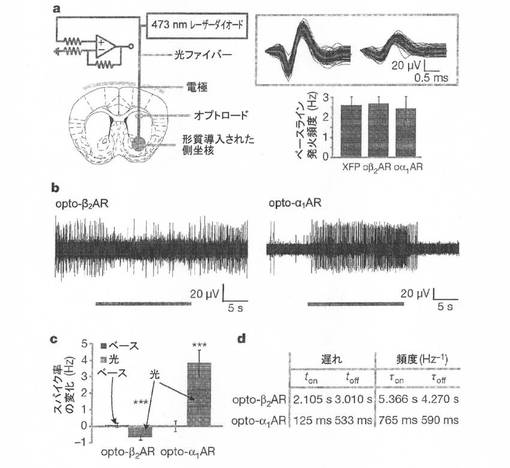
【図4】図4Aは、本発明の例示的実施形態による、記載されているコンストラクトのための、形質導入された側坐核のオプトロードターゲティング、スパイク波形、およびベースライン発火率を示す。 図4Bは、本発明の例示的実施形態による、光刺激によるインビボオプトロード記録を示す。 図4Cは、本発明の例示的実施形態による、光によるスパイク誘発頻度のベースラインに対する変化を示す。 図4Dは、本発明の例示的実施形態による、発火率変化カイネティクスを示す。
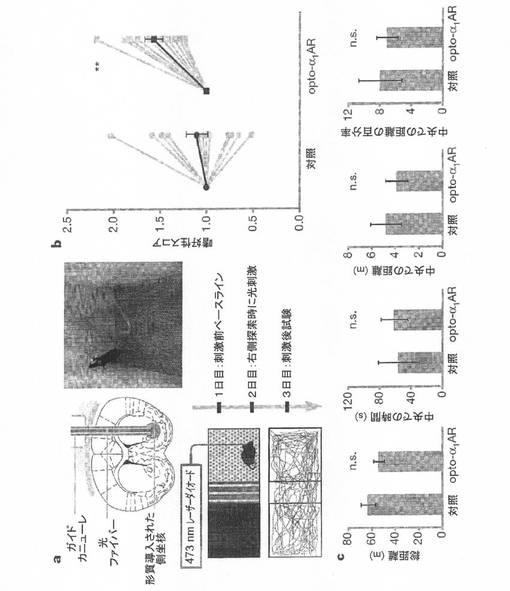
【図5】図5Aは、本発明の例示的実施形態による、形質導入された領域の定位ターゲティング、光ファイバーが植え込まれた自由行動下のマウス、場所嗜好性装置の模式図、ならびに自由に探索しているマウスの試験および軌跡を示す。 図5Bは、本発明の例示的実施形態による、対照およびopto−α1ARの嗜好性を示す。 図5Cは、本発明の例示的実施形態による、種々のオープンフィールド試験における総距離の結果を示す。
【発明を実施するための形態】
【0016】
本発明には種々の修正例および変更形態が可能であるが、その詳細は図面において例示により示され、以下に詳細に記載する。しかし、記載された特定の実施形態に本発明を限定することを意図するものではないことが理解されるべきである。逆に、本発明の精神および範囲内の全ての修正例、均等物、および代替例が含まれることが意図される。
【0017】
本発明は、種々の光学的システムおよび方法の現実的な応用を可能にする上で有用であると考えられ、本発明は、細胞内セカンドメッセンジャーレベルの光制御を扱うシステムおよび方法における使用に特に適していることが見出された。本発明はそのような応用に必ずしも限定されないが、この文脈を用いた種々の例の説明により本発明の種々の態様が理解されよう。
【0018】
本発明のある実施形態は、細胞内でセカンドメッセンジャーを放出させることにより光刺激に応答するキメラ膜タンパク質に関する。特定の例では、キメラタンパク質は異種受容体サブユニットと光に反応して光異性化によりコンホメーションを受けるタンパク質とを組み合わせたものであり、したがって、光によって活性化される。ロドプシンまたはレチニリデンタンパク質は、異種受容体サブユニットを含むように改変され得る光応答性タンパク質の例示的グループを与える。
【0019】
本発明の一実施形態によれば、7回膜貫通αヘリックスドメインを含むと考えられるタンパク質が、セカンドメッセンジャーに関連する異種受容体サブユニットを含むように改変される。細胞膜中で発現されると、このタンパク質はコンホーマル変化をすることにより光に応答する。コンホーマル変化は、セカンドメッセンジャーの放出/産生の引き金となる。
【0020】
本発明のある実施形態は、細胞内でセカンドメッセンジャーの放出を引き起こすことにより光刺激に応答するキメラ膜タンパク質、をコードするヌクレオチド配列に関する。
【0021】
本発明のある実施形態は、異種性キメラ膜タンパク質を発現する細胞に関する。このキメラ膜タンパク質は、細胞内でセカンドメッセンジャーの放出を引き起こすことにより光刺激に応答する。特定の実施形態では、キメラ膜タンパク質の発現はインビボで起こる。別の実施形態では、キメラ膜タンパク質の発現はインビトロで起こる。
【0022】
適切な受容体サブユニットを含むようにグアニンヌクレオチド結合タンパク質共役受容体タンパク質(GPCR)を改変することで任意の好適なセカンドメッセンジャーを産生するよう、本発明の実施形態を実施することができる。
【0023】
本発明の実施形態においては、種々の波長および強度の光に応答するタンパク質を使用することができる。
【0024】
本発明のある実施形態は、本明細書に開示するように、目的のセカンドメッセンジャー活性の任意の下流効果を決定するためのキメラGPCRタンパク質の使用に関する。
【0025】
本発明の実施形態は、限定されるものではないが、哺乳動物細胞、幹細胞、植物細胞、および酵母、大腸菌等の単細胞生物を含む種々の細胞型中におけるキメラGPCRタンパク質の発現に関する。
【0026】
本発明の特定の実施形態は、可視化を容易にするために蛍光タンパク質を連結したキメラタンパク質の最適化された発現、および光により誘導されるセカンドメッセンジャー活性の下流効果を研究するための様式の最適化された使用に関する。
【0027】
本発明の一実施形態は、本明細書に開示するように、特定の細胞集団中で発現させるためにその細胞集団へとキメラGPCRタンパク質を遺伝的にターゲティングすることに関する。標的細胞型中で選択的に発現される細胞型特異的プロモーターが存在する(例えば、ニューロンターゲティング用のシナプシン1;心組織用のトロポニンバリアント)。発現ベクター中でこれらのプロモーターをキメラGPCRタンパク質の上流に配置することで、目的の細胞型にタンパク質の発現をターゲティングすることができる。これには、Tet反応、ER反応、およびCre/Loxシステム等の誘導可能、反転可能、またはその他の形で制御可能なプロモーターシステムが含まれる。
【0028】
本発明の例示的一実施形態によれば、目的の細胞型中で発現されると光に応答して環状アデノシン一リン酸(cAMP)が産生される、遺伝的にコード可能なタンパク質が開発される。これは、例えば、限定されるものではないが、医薬品のスクリーニング等の細胞生理に対する下流効果を可視化するのに有用であり得る。その他の実施形態では、光に応答してセカンドメッセンジャーを放出させる異種性キメラGPCRを用いる。例示的なセカンドメッセンジャーには、cAMP、環状グアノシン一リン酸(cGMP)、イノシトール三リン酸/イノシトール1,4,5−三リン酸/トリホスホイノシトール(IP3)、およびアラキドン酸が含まれる。
【0029】
本発明の一実施形態によれば、細胞内メッセンジャーに関連する推定治療レジメン(例えば、薬物または電気刺激、またはこれらのセカンドメッセンジャーを介して作用する任意のもの)の効力を評価する方法が実施される。キメラ光応答性膜タンパク質(例えばロドプシン)を発現するためのヌクレオチド配列を、1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}で改変する。光に応答してセカンドメッセンジャーを産生させるために、光応答性膜タンパク質を細胞中で発現させる。このタンパク質を光にさらし、治療効果を評価する。
【0030】
光は、所望の刺激特性に従って与えることができる。一実施形態では、発現された膜タンパク質は数十ミリ秒以内に光に応答する。したがって、刺激プロファイルは、速く連続している一連の光パルスを含んでもよく、得られる効果は、例えばCa2+感受性色素を用いてモニターすることができる。
【0031】
一例では、細胞は最初に治療なしで刺激され得る。治療が施された後、細胞は再度刺激され得る。各試験の結果を比較することで治療の有効性を評価することができる。
【0032】
治療には、例えば、限定されるものではないが、医薬品、細胞の修飾(遺伝的修飾その他)、細胞の物理的パラメータ(例えば温度変化または電気刺激)、または生体に適用される治療レジメンなどの、広範な異なる形での実施が含まれる。
【0033】
一実施形態では、治療は、発現された膜タンパク質の光刺激である。そのような例においては、有効性は、例えば、治療されるべき障害に関連する症状をモニターすることで測定することができる。
【0034】
別の実施形態では、治療レジメンは疾患または障害のモデル化の一部として実施される。例えば、疾患モデル(細胞または動物)を用いることができ、タンパク質が発現される前にバックグラウンド/ベースライン状態を評価し、治療レジメンを評価することができる。
【0035】
実験結果は、cAMP誘導因子およびcAMP標的陽イオンチャネルの両方で細胞をトランスフェクトし、得られた活性をCa2+感受性色素を用いて可視化することで、光で惹起されるcAMPによる標的イオンチャネルの制御を可視化できることを示している。この、セカンドメッセンジャー活性の、遺伝的にコード可能な光活性化型調節物質一式は、新規治療薬のスクリーニングに有用であり得、また、ADHDや心臓のチャネル病等の多くの疾患状態にcAMPが関与するので、それ自体が治療法である。このタンパク質は、種々の他のセカンドメッセンジャー(例えばIP3)と共に使用されるように設計されてもよく、レチナール結合部位を設計するか吸収/作用スペクトルの異なるキメラロドプシンまたは錐体オプシンをキメラに選択することで光活性化のためのその他の色と共に使用されるように設計されてもよく、また、カルシウムシグナル伝達および/またはキナーゼ活性等のセカンドメッセンジャーの他の下流効果と共に使用されるように設計されてもよい。
【0036】
図1A、1B、および1Cは、開発されたセカンドメッセンジャーシグナル伝達の光活性化型誘導因子(「optoXR」)の2つの例であるoptoGsおよびoptoGqから得られた実験データを示す。これらの光活性化型誘導因子はロドプシン/GPCRキメラ体である。optoGqは、Gqシグナル伝達の光応答性制御を可能にし、optoGsは、Gsシグナル伝達の光応答性制御を可能にする。
【0037】
optoGsおよびoptoGqのどちらにおいても、暗所におけるベースラインcAMPレベルおよびIP3レベルには無視できる差しかなく、cGMP等の他のセカンドメッセンジャーの経路とのクロスオーバーはないことが示されている。optoGqの光刺激で観察されたcAMPレベルの増加は、IP3産生の予想される下流効果である。
【0038】
図1Aは、本発明の例示的実施形態による、optoGsおよびoptoGqの模式図である。各タンパク質について、ロドプシンの細胞内ループが、通常Gs(ベータ2)またはGq(アルファ1)と共役しているアドレナリン作動性タンパク質のもので置換される。この遺伝コード配列は、ヒトおよびマウスの細胞での発現用に最適化されている。得られる配列の例には、optoGs:配列番号1および配列番号2;ならびにoptoGq:配列番号3および配列番号4が含まれる。
【0039】
当業者に理解されるように、これらのタンパク質のアミノ酸配列は、実施形態を支持する非限定的な例として提示するものであり、塩基配列以外の点では同様の結果、互換可能な結果、または同等な結果をもたらす遺伝配列のバリエーション(例えば点突然変異体)も含まれる。
【0040】
図1Bは、本発明の例示的な実施形態による、トランスフェクトなしまたはoptoGsもしくはoptoGqでトランスフェクトされた細胞のcAMP(上)、cGMP(中)、およびIP1(下:IP3の分解産物)の酵素結合免疫吸着検定法(ELISA)を示す。図1Bの結果は、示されているように、504nm光(20nmバンド幅)で1ヶ所につき1分間刺激するか暗所に維持した細胞から得た。
【0041】
刺激は、環境調節倒立培養顕微鏡(inverted culture microscope;ライカ社製DMI6000B)を用いて行った。cAMPアッセイでは、アッセイの飽和したポジティブコントロールとして、一部の細胞を10μMのフォルスコリンで30分間処理した。optoGsは、光に応答してcAMPレベルを有意に増加させた。cAMPのベースラインの有意な増加やoptoGsによるcGMPレベルもしくはIP3レベルの逸脱は見られなかった。OptoGqは、光に応答して、cGMPレベルは有意に変化させず、IP3レベルを有意に増加させた。IP3産生を伴うcAMPレベルの増加は、細胞内Ca2+放出の結果であると考えられる。
【0042】
図1Cは、本発明の例示的実施形態による、optoGsおよびoptoGqのmCherry融合タンパク質でトランスフェクトした細胞のCaイメージングを示す。cAMPを検出するために、環状ヌクレオチド依存性Ca2+チャネルCNGA2のcAMP選択的変異体をoptoGsに対し過剰量トランスフェクトした。IP3は、細胞内Ca2+ストアの放出を活性化し、信頼性の高いGq活性化シグナルを与える。対照集団も、mCherry単独及び過剰量の前記変異体CNGA2でトランスフェクトした。細胞にfura−2を負荷し(20〜25分間インキュベート)、2秒毎に340nmおよび380nmを2ms照射した。optoGsおよびoptoGqのそれぞれでは、Caシグナルを生じさせるのにこの照射だけで十分であったが、対照集団では有意なシグナルは検出されなかった。
【0043】
図1は特定の実験設定で得られたデータを示しているが、本発明はこれに限定されるものではない。例えば、トランスフェクション以外の種々の送達技術が想定され、該送達技術としては、ウイルス形質導入、弾道的遺伝子導入(ballistic gene delivery;遺伝子銃)、および自発的な核酸の取込みが含まれるが、これらに限定されるものではない。
【0044】
基となるロドプシンは、アルファ2−アドレナリン受容体もしくはドーパミンD2受容体もしくはセロトニン5HT2A受容体等のGi共役受容体、またはドーパミンD1A受容体もしくは代謝型グルタミン酸受容体等のその他のGsまたはGq共役受容体等の、任意の好適な異種受容体サブユニットと共に使用されるように改変することができる。
【0045】
例示的な一実施形態によれば、基となるロドプシンはウシBos taurusに由来するタンパク質である。
【0046】
一実施形態によれば、前述した基となるロドプシン以外の基となるタンパク質を用いることもでき、前述した基となるロドプシン以外の基となるタンパク質には、錐体オプシン(赤色、緑色、または青色)、他の種のロドプシン、およびドーパミン受容体またはセロトニン受容体等のリガンド依存性受容体等の種々の7回膜貫通型タンパク質が含まれる。
【0047】
種々の実施の態様は、哺乳動物におけるインビボでの適用に関する。これらの実施の態様は、限定されるものではないが、神経回路および疾患モデルの試験および確認を含む。
【0048】
図3Aおよび3Bは、セカンドメッセンジャーシグナル伝達の光活性化型誘導因子の2つの例であるoptoGs(opto−β2AR)およびoptoGq(opto−α1AR)のインビボでの適用から得られた実験データを示す。本発明の態様は、Gタンパク質共役受容体(GPCR)に共通する構造−機能関係に影響を与えることで、受容体によって開始される生化学的シグナル伝達経路を高い時空間精度で動員および制御する、遺伝的にコードされた光学的ツール(「optoXR」)の多様なファミリーの使用および開発に関する。
【0049】
図3Aおよび3Bに示す結果は、ターゲティングされた別個のシグナル伝達経路を光に応答して選択的に動員する2つの特定のoptoXRに関するものである。これら2つのoptoXRは、インビボで側坐核中のスパイク発火に反対の影響を与え、側坐核中での正確に時間制御されたoptoXRの光刺激だけで、自由行動をしているマウスに条件付け場所嗜好性を誘導するのに十分であった。このoptoXRを用いた手法は、行動している哺乳動物における生化学的シグナル伝達の因果的影響に関する仮説を、ターゲティング可能で時間的に正確な様式で試験することを可能にする。
【0050】
GPCR間で共有されている構造−機能関係を用いて、シグナルをエフェクターに共役させる新規な伝達ロジックを有する複数の異なるオプシン/GPCR2キメラを開発し、インビボで発現させ、哺乳動物中で細胞内シグナル伝達の光制御を実施した。種々の実施の態様に従い、1または複数のキメラオプシン受容体タンパク質が、インビボにおいて哺乳動物内で機能的であり、特定の細胞にターゲティング可能であり、正確に時間制御された光パルスに応答性であるように設計される。そのような手法により、高速な光刺激(およびタンパク質応答)を用いて、正確に決められ且つ行動上適切であるタイミングで細胞内の生化学的事象について試験し特徴解析することが可能となる。いくつかの非限定的で例示的な実施の態様には、規定された細胞型における、ある範囲の時間スケールにわたる、拍動性対強直性の調節、異なる調節系間の同調性、ならびにその他の基本的な生理学的および病理学的プロセスが含まれる。
【0051】
哺乳動物での実施は成功した。例示的な一実施例では、最初にGq共役ヒトα1aアドレナリン受容体(α1AR)およびGs共役ハムスターβ2アドレナリン受容体(β2AR)の保存された残基をGt共役ウシロドプシンと整列させることで、ロドプシンの細胞内ループを特定のアドレナリン受容体の細胞内ループで置換した(図1A)。構造モデルに基づいて、Gタンパク質との共役をGtから移すように細胞内領域の交換(カルボキシ末端ドメインを含む)を各受容体について設計し、各受容体を哺乳動物中でのインビボ発現用に最適化した。種々のリガンドにより活性化すると、天然の受容体は複数の集合(ensemble)状態を取ることができ、リガンドバイアスシグナル伝達現象においてカノニカル経路および非カノニカル経路を動員する。optoXRは光を感知すると、生物学的環境に応じた様式で、1つの活性集合的状態を選択する可能性が高い。
【0052】
キメラ(opto−α1ARおよびoptoβ2AR)をコードする遺伝子を蛍光タンパク質に融合させた。機能的optoXR発現の検証は、opto−α1AR単独(Gqを介して[Ca2+]iを動員すると予想される)、またはopto−β2AR(Gsを介して環状AMPを動員すると予想される)とcAMP依存性Ca2+チャネルCNGA2−C460W/E583Mの両方でトランスフェクトしたHEK細胞中における[Ca2+]i(細胞内カルシウム濃度)のイメージングにより達成された。[Ca2+]iのレシオメトリックイメージングにより、60sの緑色光刺激(504±6nm、7mWmm−2)はいずれのoptoXRの下流でも顕著な[Ca2+]iシグナルを誘導するのに十分であるが、対照条件では誘導しないことが示され(図2)、機能的に発現していることが明らかになった。各optoXRに制御されるシグナル伝達の特異性を試験するために、形質導入されたHEK細胞に3mWmm−2、504±6nmの光を60s照射し、その後、細胞を溶解し、イムノアッセイによりcGMP、cAMP、およびIP1(IP3の分解産物)のレベルを解析した。カノニカルパターンはopto−β2ARでその分子的デザインに対応して予想されるとおりといえた。何故なら、光刺激によりopto−β2AR発現細胞中でcAMPが有意に産生され(図3A、上)、これは野生型β2ARの薬物刺激で得られるものに匹敵するとともに、IP3(図3A、中段)、[Ca2+]i(図2)や暗所での実質的な活性を動員しなかったからである。一方、光刺激は、opto−α1AR発現細胞でIP3シグナル伝達の有意なアップレギュレーションを生じさせ(図3A、中段)、これは野生型α1ARの薬物刺激により誘導されるレベルに匹敵するものであった。[Ca2+]iの上昇(図2)と共に、これらのデータは、Gqの動員について予想されるパターンを示しており、このパターンはopto−β2AR発現細胞中では見られない(図3A、上)。いずれのコンストラクトを発現している細胞の光刺激も、cGMPレベルを調節することができず(図3A、下)、このことは更に、キメラタンパク質のシグナル伝達特異性を示している。同様なアッセイから、optoXRが、天然ロドプシンの作用スペクトルに近い作用スペクトルを維持しており、生物学的に好適な光束の範囲に渡りシグナルを統合することができ、p42/p44−MAPKシグナル伝達に関して野生型受容体と同様の程度に非カノニカル経路を活性化できることが示された。
【0053】
レチナール補因子の補充が必要かどうかも含め、インタクトな神経組織中におけるoptoXRの能力を調べた。そのような試験の1つでは、(生化学的調節の標的をグリアおよび内皮細胞を初めとする他の潜在的Gs/Gq応答性細胞組織要素ではなく局所ニューロンにするため)シナプシンIプロモーターの制御下にあるoptoXR融合遺伝子を有するレンチウイルスベクター(図3B、左上)を成獣マウスの側坐核中に定位的に注入した。この戦略により、側坐核中に細胞体樹状突起区画を有するニューロン(約95%がGABA作動性中型有棘ニューロンであり、更なるサブタイプ特異性を有しない;図3B、左)が生化学的調節の標的とされ、またこれらのレンチウイルスは軸索を介して細胞に形質導入することはないため、通過繊維(fibers of passage)または求心性シナプス前終末は除外される。形質導入の2週間後、人工脳脊髄液中で側坐核の急性冠状スライスを調製し、10分間光刺激し、すぐに固定しcAMPおよびCa2+共役シグナル伝達カスケードの両方の生化学的統合因子(integrator)であるSer133−リン酸化CREB(pCREB)を染色した。外因性のレチノイドを補充しなくても、optoXR発現集団において有意に上昇したpCREBが観察されたが(図3B、右)、非照射組織では観察されなかった。
【0054】
形質導入した側坐核にターゲティングされたオプトロードを用いて、マルチユニットインビボニューロン発火を記録することで、側坐核の居所的電気活性に対するoptoXR活性化の機能面での影響を決定した(図4A)。どちらのコンストラクトにおいても暗所ではベースライン発火率に有意な差は見られなかった(図4A、右下)。光刺激は、opto−β2AR発現側坐核においてネットワーク発火を減少させた(図4Bの左のトレースは影響のカイネティクスを示す。概要データが図4Cおよび図4Dにそれぞれ示される)。これは、Gsを標的にした過去の薬理学的実験と一致していた。光刺激は、opto−α1AR発現側坐核(図4B、右;図4C、4D)において発火を増加させた。スパイク頻度のヒストグラムから、発火率へのoptoXRの影響のカイネティクスは、シグナルの電気的開始よりも生化学的開始の方に一致することが示された(図4D)。これらの電気生理学的データと前出の生化学的検証とを組み合わせれば、細胞内カスケードの差動的光活性化型制御を可能にし且つネットワークの生理を調節するようにoptoXRをインビボで機能的に発現させることができることが支持される。
【0055】
一実施例では、光遺伝学を用いて、正確にタイミングを制御されたoptoXR刺激が自由行動中のマウスの行動を調節する能力を評価した。オペラント行動に用いられる時間的に正確な様式で側坐核ニューロン内の細胞内シグナル伝達を光制御するため、携帯型ソリッドステート光送達をoptoXRのトランスジェニック発現と組み合わせた(図5A)。共焦点分析により、発現が局所的な側坐核ニューロンに限定されていることが示され、特に、求心性繊維、側坐核に突き出た遠位領域、グリア、または周辺領域中で標識は全く観察されなかった。3日間のオペラント条件付け場所嗜好性アッセイの一部として、形質導入した側坐核に光刺激をターゲティングした(図5A)。試験中毎日、動物に自由に場所嗜好性装置を探索させた(図5A、下)。1日目は、動物は光刺激を受けずに自由に装置内を探索した。2日目に、指定した条件付けされた部屋に動物が自由に進入する毎に、形質導入した領域に位置を合わせたレーザーダイオード接続光ファイバーで光パルスを10Hzで送達して、強力な報酬の間のモノアミン作動性入力の適切な強度を模倣した。経路のトレースから、柔軟な光ファイバーを用いた手法により、全ての部屋を完全且つ妨害のなしに探索できることが示された(図5A、下)。3日目に、動物は再度、光刺激を受けずに自由に装置内を探索し、条件付けた部屋で過ごした時間が2人の別の盲検評価者により定量された。特に、opto−α1ARを発現している動物は、光刺激後に、装置の条件付けされた側への嗜好性が大きく上昇したことが示された(図5B)。この時間的に正確な生化学的調節の影響は、opto−α1AR動物の別個の2つのコホートで再現可能であった(n=5〜6、P<0.05、条件付けされた部屋中での時間の各コホートについてのスチューデントのt検定;全集団では、n=11、P<0.01)。一方、その他のオプシン遺伝子opto−β2ARおよびChR2は、嗜好性の誘導についてさほど効果的ではないようであった。側坐核ニューロン中のopto−α1AR刺激の影響は報酬関連行動に特異的であり、オープンフィールド試験で同じ動物のコホートに送達された同じ光刺激が移動距離または壁の近接に対する嗜好性には有意な影響を与えなかった(図5C)ことから、不安関連行動または自発運動活性の直接的調節にまでは及ばなかった。
【0056】
上記実験に従った具体的かつ非限定的な実施例をここで記載する。記録電極(1MVタングステン、A−Mシステムズ社製)に接続した直径200mmのマルチモード光ファイバー(ソーラボ社製)からなり電極/ファイバーの先端間の距離が200〜400mmであるオプトロードを、定位フレーム(デビッド・コフ社(David Kopf Instruments)製)に固定されイソフルランで麻酔したマウスの形質導入された側坐核中に下ろして用い(電極の先端はブレグマの下4.8〜5.2mm)、インビボ記録および解析を行った。473nmダイオードレーザー(クリスタレーザー社製)の光をファイバーを介して送達した。電気シグナルをバンドパスフィルターに通し、増幅し(0.3〜1kHz、1800 Microelectrode AC Amplifier、A−Mシステムズ社製)、pClamp10.0(モレキュラーデバイス社(Molecular Devices)製)を用いて解析した。閾値によりスパイクを検出し、個々に点検して確認した。
【0057】
光ファイバー(直径200mm、ソーラボ社製)を473nmの青色ダイオードレーザー(クリスタレーザー社製)に接続し、側坐核(先端から0〜100mm)をターゲティングするカニューラに合わせ、この光ファイバーを介して光刺激を与え、行動分析を行った。ファンクションジェネレータ(Agilent 33220A)を用いてoptoXR用に50msパルス幅の光を送達した。場所嗜好性は、自由に探索できるように部屋の間の壁を除去した標準装置(SDインスツルメンツ社製)中で実施した。各部屋で過ごした時間量について、MATLAB(マス・ワークス社(Mathworks)社製)で特注の集計スクリプトを実行して、2人の独立した盲検評価者がビデオからデータを解析した。オープンフィールド試験では、40340cmの正方形のオープンフィールド中に動物を置き、場所嗜好性実験と同じパラメータを用いて光刺激を送達した。自動化されたソフトウェア(Viewpoint)を用いて、その外側の環(フィールドの残りの部分)に対する、中央の15315平方cmでの合計時間および距離についてビデオを解析した。
【0058】
表示されている場合、統計解析は、両側検定によるスチューデントのt検定(マイクロソフトエクセルで計算)または一元配置の分散分析とチューキーの事後検定(GraphPad Prism)を用いて行った。まとめた棒グラフは全て、平均値±s.e.mで表し、有意性は以下のように表した:*P<0.05、**P<0.01、***P<0.001。
【0059】
本発明の種々の実施形態の驚くべき結果および有効性を支持する更なる詳細は、参照により全体を本明細書に援用するTemporally precise in vivo control of intracellular signalling, Raag D. Airan, et al., Nature 458, 1025-1029 (23 April 2009)中に見出すことができる。
【0060】
以下の記載は、本発明の実施形態による具体的且つ非限定的な方法の詳細を提供する。この方法の多くのバリエーションが想定され、本発明の範囲内に含まれる。
【0061】
ベクター構築
opto−α1ARおよびopto−β2ARの哺乳動物コドン最適化配列(図1A中のアミノ酸配列)を合成し、pcDNA3.1にクローニングし、NotI部位を用いてmCherryまたはYFP(スタートコドンを除去)のN末端に融合させた。optoXRとmCherry/YFPのリンカーは5’GCGGCCGCC3’である。pLenti SynapsinI hChR2 mCherry WPREベクターのAgeIおよびEcoRI部位に各optoXR mCherryの導入遺伝子をクローニングすることで、SynapsinI optoXR mCherryを含むレンチウイルスベクターを構築した。
【0062】
レンチウイルスの作製
高タイターレンチウイルスを作製した。簡潔に述べると、10%FBSを含むDMEMで培養されている4層のセルファクトリー(Nunc社製)中に90%のコンフルエンスでHEK293FT細胞をプレーティングした。690μgの前述したレンチウイルスベクターおよび2種類のヘルパープラスミド(690μgのpΔCMVR8.74および460μgのpMD2.G)で細胞をコトランスフェクトした。トランスフェクションの15時間後に培地を交換した。トランスフェクションの24時間後に、5mMの酪酸ナトリウムを含む無血清UltraCULTURE(キャンブレックス社(Cambrex)製)200〜220mLで培地を交換した。トランスフェクション40時間後、今やウイルスを含む培養液上清を1000rpmで5分間スピンして細胞残渣を除去した後、0.45μmの低タンパク質結合フィルターフラスコを用いて濾過した。次いで、この清澄な上清を、SW28ローター(ベックマン社製)を用いて55,000gで2時間超遠心し、ウイルスを沈殿させた。遠心分離後、上清を捨て、得られたウイルスペレットを合計100μLの冷(4℃)PBSに溶解した。再懸濁したウイルスを7000rpmで5分間遠心分離して残りの細胞残渣およびウイルス残渣を除去した。アリコートを再使用時まで−80℃で凍結した。
【0063】
動物の外科手術および行動
スタンフォード大学の実験室用脊椎動物プロトコールに従い、10〜12週齢の雌のC57BL/6マウスを収容し、取り扱った。ウイルス溶液を以下のように右側坐核に導入した。動物をイソフルランで麻酔し、頭頂部の毛を刈り取った。イソフルラン麻酔下にいる間に動物の頭を定位フレーム(デビッド・コフ社)に固定した。頭皮を正中切開し、ブレグマの前側1.10mm、側方1.45mmに直径約1mmの開頭切開部を穿孔した。次いで、ウイルスを予め充填した33ゲージのベベル針(NanoFil、ワールド・プレシジョン・インスツルメンツ社(World Precision Instruments)製)を側坐核中に下ろし(針の先端がブレグマの腹側4.70〜4.80mm)、自動シリンジポンプ(NanoFil、ワールド・プレシジョン・インスツルメンツ社製)を用いて1.0μLのウイルスを100nL/分で注入した。注入後、3〜5分間組織を弛緩させて流体を拡散させた後、針を引き抜いた。急性スライスまたはインビボ記録実験用の動物では、開頭部にデンタルセメント(ラングデンタル社(Lang Dental)製)を充填し、VetBond(3M社製)を用いて切開部を閉じた。行動分析用の動物では、カニューレ(C316G、ペデスタルの4.5mm下で切断;プラスチクスワン社(Plastics・One)製)をペデスタルが頭蓋に面一になるよう配置した。Metabond(パーケル社(Parkell)製)およびデンタルセメント(ラングデンタル社製)を用いてカニューレを固定した。VetBondまたはセメントの乾燥後、動物をフレームから外し、その後の操作の前に少なくとも1週間回復させた。行動実験の対照動物にも実験動物と同じ操作(外科手術、カニューレの植込み、光刺激)を行い、ウイルスの代わりに溶媒(PBS)のみを注入した。場所嗜好性実験には、部屋のどちらかの側(>70%または<10%)または中央の部屋(>40%)に対するベースライン嗜好性を示さなかった動物を実験に用いた。全動物の90%超が、バイアスがかけられていないバランスのとれた場所嗜好性デザインのためのこれらの基準を満たした。
【0064】
急性スライスの調製
動物をイソフルランで麻酔し、外科手術用のハサミ(ファイン・サイエンス・ツールズ社(Fine Science Tools)製)を用いて断頭した。側坐核を含む275μm厚の冠状スライスを切り出し、64mMのNaCl、2.5mMのKCl、1.25mMのNaH2PO4、25mMのNaHCO3、10mMのグルコース、120mMのスクロース、0.5mMのCaCl2、および7mMのMgCl2を含む切断溶液(95%O2/5%CO2で平衡化)中に保存した。スライス後、スライスを切断溶液中にて32〜35℃で30分間インキュベートし、その後実験まで室温でインキュベートした。エクスビボでのoptoXR刺激では、スライスを正立顕微鏡(BX51W、オリンパス社製)のステージに乗せ、124mMのNaCl、3mMのKCl、1.25mMのNaH2PO4、26mMのNaHCO3、10mMのグルコース、2.4mMのCaCl2、および1.3mMのMgCl2を含む人工脳脊髄液(95%O2/5%CO2で平衡化)で灌流した。300WのLambda DG−4(サッター社(Sutter)製)の光を473nm±20nmのバンドパスフィルター(セムロック社(Semrock)製)に通し、4倍の対物レンズ(0.28NA)を用いて10分間スライスに当て、その後すぐに、後の解析のために固定した。
【0065】
シグナル伝達検証アッセイ
リポフェクタミン2000(インビトロジェン社製)を用いて24ウェルプレート中でHEK293FT細胞(インビトロジェン社製)をトランスフェクトし、トランスフェクションの4〜6時間後に無血清培地に交換した。Ca2+イメージングには、マトリゲルでコートされたカバーガラス上にプレーティングされた細胞に、1μMのATRを含むタイロード中、5μg/mlのfura−2 AMを含むF−127プルロニック/DMSO(プローブ社(Probes)製)を37℃、5%大気CO2にて20〜25分間負荷した。負荷後、300WのLambda DG−4(サッター社製)を制御するMetafluor(アクソン・インスツルメンツ社(Axon Instruments)製)を用いて、オリンパス社製BX51W上で340nm/380nmでカバーガラスをイメージ化した。イムノアッセイでは、トランスフェクションの18〜24時間後に、1μMのATRおよび50mMのLiCl(IP1の分解を防ぐため)を添加し、プレートを環境調節された顕微鏡(ライカ社製DMI6000;37℃、5%大気CO2)に移した。5領域/ウェルをそれぞれ1分間光刺激した(サッター社製300WのLambda DG−4;セムロック社製504/12nmバンドパスフィルター;10×0.30NA対物レンズ);3ウェル/条件。インキュベート後(cAMP/cGMP:20分間;IP1:1時間)、細胞を溶解し、HTRF(シスバイオ社(CisBio)製)およびBiotek Synergy4リーダーで解析した。
【0066】
免疫組織化学および共焦点分析
インビボ刺激後、刺激終了90分後に、氷冷した4%パラホルムアルデヒド(PFA)のPBS溶液(pH7.4)をマウスの心臓に灌流した。脳を取り出し、4%PFA中で一晩固定し、その後、30%スクロースのPBS溶液中で平衡化した。40μm厚の冠状切片を凍結ミクロトーム上で切り出し、免疫組織化学用に処理するまで抗凍結剤中で4℃にて保存した。フリーフローティング切片をPBS中で洗浄した後、0.3%のTx100および3%の正常ロバ血清(NDS)中で30分間インキュベートした。急性スライス実験では、刺激直後に、275μm厚のスライスを、氷冷した4%PFA中で1時間固定し、0.5%のTx100および3%のNDSと共にインキュベートした。MAPKアッセイでは、HEK293細胞の刺激直後に、カバーガラスを15分間固定し、0.6%のH2O2と共にインキュベートし、その後、0.1%のTx100を含む3%NDSで透過処理した。マウス抗GAD67 1:500、マサチューセッツ州ビルリカのミリポア社製;ウサギ抗cfos 1:500、カリフォルニア州サンディエゴのカルバイオケム社(Calbiochem)製;ウサギ抗リン酸化CREB Ser133 1:500、ミリポア社製について、一次抗体のインキュベートを0.01%のTx100および3%のNDS中で一晩行った。切片を洗浄し、FITCまたはCy5(ペンシルベニア州ウェストグローブのジャクソン・ラボラトリー(Jackson Laboratories)製)にコンジュゲートした二次抗体(1:1000)と共に室温で3時間インキュベートした。DAPI(1:50,000)と20分間インキュベートした後、切片を洗浄し、PVD−DABCOと一緒に顕微鏡用スライドにマウントした。残りの一次抗体オーバーナイトインキュベート物(ウサギ抗リン酸化Erk1/2;抗リン酸化MAPK p38 1:500、ウィスコンシン州マディソンのプロメガ社製;マウスモノクローナル抗ドーパミンD1受容体 1:50、ケミコン社(Chemicon)製;ウサギポリクローナル抗ドーパミンD2受容体 1:50、ミリポア社製;ヤギポリクローナル抗コリンアセチルトランスフェラーゼ 1:200、ミリポア社製)はその後、ビオチン化二次抗体(1:500、ジャクソン・ラボラトリー製)とのインキュベート、アビジン−ビオチン−西洋ワサビペルオキシダーゼ処理(ABCキット、カリフォルニア州バーリンゲームのベクター・ラボ社(Vector Labs)製)、およびTSA検出(コネティカット州シェルトンのパーキンエルマー社製)を、メーカーの取扱説明書に従い行った。
【0067】
20×/0.70NAまたは40×/1.25NAの油浸対物レンズを用いてライカ社製TCS SP5走査型レーザー顕微鏡上で共焦点蛍光画像を得た。1つの条件につき、カニューレ管の下500μmの領域内で4つの連続スタック画像を得た。DAPI染色で核を可視化し、Volocity(インプロビジョン社(Improvision)製)ソフトウェアを用いてcfosまたはpCREBの免疫反応性の平均ピクセル強度を求めた。陽性またはpCREB活性の細胞を強度閾値に基づき同定し、実験条件を隠して(blind)画像の取得および解析を行った。
【0068】
【表1】

【0069】
【表2】
【0070】
【表3】
【0071】
したがって、本発明の実施形態は、細胞内シグナル伝達の光遺伝学的制御に関し、行動中の哺乳動物内でインビボで作用し且つ非常に低い暗所活性を有しつつ、時間精度を得ることおよび天然受容体下流の複雑に絡み合った複数のシグナル伝達分子を動員するのに有用であり、したがって他の手法の個々のプラス面の多くを1つの技術中で統合するのに有用である。同様な実施形態では、無数の神経伝達物質および内分泌性ホルモン等のその他の調節物質により引き起こされる7回膜貫通依存性シグナル伝達経路の因果上の意義を直接探索する。別の実施形態では、興奮性の細胞を超えてoptoXR手法が用いられ、光ファイバーの深さのターゲティングと光遺伝学的にターゲティングされる光感受性とを多用途に統合することの利点を活かす。そのような実施形態の1つは、多様な非興奮性組織における時間的に正確な生化学的シグナル伝達の因果上の意義を探索することに関する。
【0072】
本発明の実施形態は、種々のリガンドが集合(ensemble)受容体コンホメーション状態を安定化させることで代替的な伝達カスケードと共役した受容体の細胞内作用にバイアスをかけることができる、リガンドバイアスシグナル伝達現象の考察に関する。optoXRは、薬理学的操作と同様なレベルで代替的なカスケードを誘導するように用いられる(例えばopto−β2ARは、野生型β2ARへの天然リガンドの作用と同様な変化をMAPK活性化に誘導することができる)。しかし個々のoptoXRは、リガンドバイアスシグナル伝達に寄与するコンホメーション状態の全ての制御を常に可能にするとは限らない。レチナールを基にしたツールは、哺乳動物組織中に内因性発色団が存在し、暗所での活性が非常に低いため、特に有用であり得る。光遺伝学は、速い単一成分レチナール結合モジュールに連結された多様なエフェクターの形態を取ることができ、光学の有する時間的精度を利用できる。
【0073】
本発明の実施形態は、微生物オプシンによる戦略を補完するoptoXRによる方法を利用し、行動下の哺乳動物中で作動可能で、速く且つターゲティング可能な別次元の細胞制御を提供する。
【0074】
本発明の別の実施形態によれば、異なる作用スペクトルを有する公知のオプシン遺伝子に基づいて波長をシフトさせたタイプのoptoXRが使用される。そのようなoptoXRは、生化学的および電気的制御の分離可能なチャネルを提供するのに特に有用であり得る。
【0075】
本明細書に記載した特定のタンパク質配列のバリアントも本発明の実施形態に沿っている。一部のバリアントはこれらのタンパク質配列に約75%超の相同性を有し、別のものは約80%、85%、または90%超の相同性を有する。いくつかの実施形態では、相同性は約93〜約95、または約98%と高い。本発明の組成物には、本明細書に記載したタンパク質配列および核酸配列が含まれ、記載した配列に約50%を超える相同性を有するバリアントから100%の相同性を有するバリアントまで含まれる。
【0076】
本明細書に記載した種々の実施形態は、正常な作動および疾患状態の両方における神経回路の更に洗練されたインタロゲーションおよびリバースエンジニアリングのために速い回路読取り技術と統合され得る。
【0077】
前述した種々の実施形態は単に例として記載したものであり、本発明を限定するものと解釈されるべきではない。前述の説明および例示に基づき、本明細書に例示および記載された例示的な実施形態および応用例に厳密に従わなくとも本発明に種々の修正および変更を加えることができることが当業者には容易に理解されよう。例えば、そのような変更には、産生されるセカンドメッセンジャーのバリエーションが含まれ得る。そのような修正および変更は、添付の特許請求の範囲に記載されている本発明の真の精神および範囲から逸脱しない。
【技術分野】
【0001】
関連特許文献
本願は、「セカンドメッセンジャーを光制御するためのセルライン、システム、および方法」という名称の2008年5月29日付で提出された米国仮特許出願第61/057,108号の米国特許法第119条(e)に基づく利益を主張する。この基礎となる仮出願は全体が参照により本明細書に取り込まれる。
【0002】
電子的に提出された資料の参照による組込み
2009年4月29日に作成された「STFD195PCT_ST25.txt」というファイル名の、12,342バイトのASCII(テキスト)ファイルである、本明細書と同時に提出されたコンピューターで読取り可能なヌクレオチド配列/アミノ酸配列リストは全体が参照により取り込まれる。
【0003】
本発明は、一般的に、光刺激に応答してセカンドメッセンジャーを生成するシステムおよび手法に関し、より具体的には、それぞれ光に応答したセカンドメッセンジャー産生に関する、セルライン、ヌクレオチド配列、キメラタンパク質、およびその使用に関する。
【背景技術】
【0004】
グアニンヌクレオチド結合タンパク質(Gタンパク質)は、不活性のグアノシン二リン酸(GDP)状態と活性のグアノシン三リン酸(GTP)結合状態の間を行ったり来たりすると考えられている。これら2つの状態は細胞内でのセカンドメッセンジャーの放出と関連付けられている。放出されたセカンドメッセンジャーは、下流の細胞プロセスを制御するよう機能することができる。
【0005】
セカンドメッセンジャーには、速やかに生成/放出されるシグナル伝達分子が含まれる。これらの分子は細胞内のエフェクタータンパク質を活性化することで細胞応答を発生させる。細胞シグナル伝達系の例としては、ホスホイノシトール系、環状アデノシン一リン酸(cAMP)系、およびアラキドン酸系が含まれる。
【0006】
Gタンパク質の前記異なる状態間の変化は、Gタンパク質共役受容体(GPCR)、Gタンパク質結合型受容体(GPLR)、7回膜貫通ドメイン受容体(7TM受容体)、またはヘプタヘリカル受容体と呼ばれるタンパク質により引き起こすことができる。このタンパク質ファミリーは、種々の膜貫通型受容体を含む。これらの受容体は、細胞内へのシグナル伝達経路を活性化するにより外部刺激(例えば、光、神経伝達物質、匂い、またはホルモン)に応答する。具体的には、リガンドが結合し、伝達経路を活性化し、それによってGタンパク質をもう一方の状態に変化させる。GPCR関連活性は多くの疾患に関連するので、GPCRは多くの医薬品および治療の標的である。
【発明の概要】
【発明が解決しようとする課題】
【0007】
市場の全薬物の30%超がGタンパク質共役受容体(GPCR)を標的としており、それらの薬物の多くがセカンドメッセンジャーcAMPの産生または阻害に関連すると考えられる。cAMPが直接関わる病理学的プロセスは数多く、例えば神経生理学的疾患、内分泌学的疾患、心疾患、代謝疾患、および免疫疾患が含まれる。複雑な哺乳動物行動の研究では、技術的限界が、細胞内シグナル伝達プロセスの時空間的に正確な制御の妨げとなっていた。化学物質に基づいてcAMPレベル等のセカンドメッセンジャーレベルを調節する現在の方法は、比較的ゆっくりと機能し、神経組織や心臓組織等の特定の組織に関連して身体が用いる速いタイムスケールの活性を研究するには問題である。これらの化学的方法はしばしば、(例えば、新規治療薬のスクリーニング中における)こうした速いタイムスケールを探索するための速度を有しない。
【課題を解決するための手段】
【0008】
本発明は、セカンドメッセンジャーの生成に関連した上記の課題等の克服ならびに関連するイメージングデバイスおよびその実施に関する。本発明は数多くの実施形態および応用により例示される。その一部を以下に要約する。
【0009】
本発明の一実施形態によれば、細胞中でセカンドメッセンジャーを生成する方法が提供される。キメラ光応答性膜タンパク質(例えばロドプシン)を発現するためのヌクレオチド配列を、1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}で改変する。光に応答してセカンドメッセンジャーを産生させるために、この光応答性膜タンパク質を細胞中で発現させる。
【0010】
本発明の一実施形態によれば、細胞内メッセンジャーに関連する推定治療レジメン(例えば、薬物または電気刺激、またはこれらのセカンドメッセンジャーを介して作用する任意のもの)の有効性を評価する方法が実施される。キメラ光応答性膜タンパク質(ロドプシン)を発現するためのヌクレオチド配列を1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}で改変する。光に応答してセカンドメッセンジャーを産生させるために、光応答性膜タンパク質を細胞中で発現させる。タンパク質を光にさらす。治療効果を評価する。
【0011】
本発明の一実施形態は、1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}を有するキメラ光応答性膜タンパク質(ロドプシン)を発現する細胞に関する。
【0012】
本発明の一実施形態は、1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}を有するキメラ光応答性膜タンパク質(ロドプシン)を発現するためのヌクレオチド配列に関する。
【0013】
上記の本発明の概要は、説明される本発明の全ての実施形態または実施例を記載することを意図したものではない。添付の図面および以下の詳細な説明によりこれらの実施形態をより具体的に例示する。
【0014】
本発明は、以下の本発明の種々の実施形態の詳細な説明を添付の図面と合わせて考慮することでより完全に理解されるであろう。
【図面の簡単な説明】
【0015】
【図1】図1Aは、本発明の例示的実施形態による、optoGsおよびoptoGqを示す模式図である。 図1Bは、本発明の例示的実施形態による、トランスフェクトされていない細胞またはoptoGsもしくはoptoGqでトランスフェクトされた細胞のcAMP、cGMP、およびIP1の酵素結合免疫吸着検定法(ELISA)を示す。 図1Cは、本発明の例示的実施形態による、optoGsおよびoptoGqのmCherry融合タンパク質でトランスフェクトされた細胞のCaイメージングを示す。
【図2】本発明の例示的実施形態による、optoGsおよびoptoGqのmCherry融合タンパク質でトランスフェクトされた細胞のCaイメージングを示す。
【図3】図3Aは、本発明の例示的実施形態による、種々のコンストラクトを発現するHEK細胞のcAMP、IP1、およびIP3レベルを示す。 図3Bは、本発明の例示的実施形態による、レンチウイルス発現ベクター、opto−α1AR発現細胞のGAD免疫染色、および10分間の光刺激後にoptoXR発現細胞(mCherry+)中で観察されたpCREBの活性化を示す。
【図4】図4Aは、本発明の例示的実施形態による、記載されているコンストラクトのための、形質導入された側坐核のオプトロードターゲティング、スパイク波形、およびベースライン発火率を示す。 図4Bは、本発明の例示的実施形態による、光刺激によるインビボオプトロード記録を示す。 図4Cは、本発明の例示的実施形態による、光によるスパイク誘発頻度のベースラインに対する変化を示す。 図4Dは、本発明の例示的実施形態による、発火率変化カイネティクスを示す。
【図5】図5Aは、本発明の例示的実施形態による、形質導入された領域の定位ターゲティング、光ファイバーが植え込まれた自由行動下のマウス、場所嗜好性装置の模式図、ならびに自由に探索しているマウスの試験および軌跡を示す。 図5Bは、本発明の例示的実施形態による、対照およびopto−α1ARの嗜好性を示す。 図5Cは、本発明の例示的実施形態による、種々のオープンフィールド試験における総距離の結果を示す。
【発明を実施するための形態】
【0016】
本発明には種々の修正例および変更形態が可能であるが、その詳細は図面において例示により示され、以下に詳細に記載する。しかし、記載された特定の実施形態に本発明を限定することを意図するものではないことが理解されるべきである。逆に、本発明の精神および範囲内の全ての修正例、均等物、および代替例が含まれることが意図される。
【0017】
本発明は、種々の光学的システムおよび方法の現実的な応用を可能にする上で有用であると考えられ、本発明は、細胞内セカンドメッセンジャーレベルの光制御を扱うシステムおよび方法における使用に特に適していることが見出された。本発明はそのような応用に必ずしも限定されないが、この文脈を用いた種々の例の説明により本発明の種々の態様が理解されよう。
【0018】
本発明のある実施形態は、細胞内でセカンドメッセンジャーを放出させることにより光刺激に応答するキメラ膜タンパク質に関する。特定の例では、キメラタンパク質は異種受容体サブユニットと光に反応して光異性化によりコンホメーションを受けるタンパク質とを組み合わせたものであり、したがって、光によって活性化される。ロドプシンまたはレチニリデンタンパク質は、異種受容体サブユニットを含むように改変され得る光応答性タンパク質の例示的グループを与える。
【0019】
本発明の一実施形態によれば、7回膜貫通αヘリックスドメインを含むと考えられるタンパク質が、セカンドメッセンジャーに関連する異種受容体サブユニットを含むように改変される。細胞膜中で発現されると、このタンパク質はコンホーマル変化をすることにより光に応答する。コンホーマル変化は、セカンドメッセンジャーの放出/産生の引き金となる。
【0020】
本発明のある実施形態は、細胞内でセカンドメッセンジャーの放出を引き起こすことにより光刺激に応答するキメラ膜タンパク質、をコードするヌクレオチド配列に関する。
【0021】
本発明のある実施形態は、異種性キメラ膜タンパク質を発現する細胞に関する。このキメラ膜タンパク質は、細胞内でセカンドメッセンジャーの放出を引き起こすことにより光刺激に応答する。特定の実施形態では、キメラ膜タンパク質の発現はインビボで起こる。別の実施形態では、キメラ膜タンパク質の発現はインビトロで起こる。
【0022】
適切な受容体サブユニットを含むようにグアニンヌクレオチド結合タンパク質共役受容体タンパク質(GPCR)を改変することで任意の好適なセカンドメッセンジャーを産生するよう、本発明の実施形態を実施することができる。
【0023】
本発明の実施形態においては、種々の波長および強度の光に応答するタンパク質を使用することができる。
【0024】
本発明のある実施形態は、本明細書に開示するように、目的のセカンドメッセンジャー活性の任意の下流効果を決定するためのキメラGPCRタンパク質の使用に関する。
【0025】
本発明の実施形態は、限定されるものではないが、哺乳動物細胞、幹細胞、植物細胞、および酵母、大腸菌等の単細胞生物を含む種々の細胞型中におけるキメラGPCRタンパク質の発現に関する。
【0026】
本発明の特定の実施形態は、可視化を容易にするために蛍光タンパク質を連結したキメラタンパク質の最適化された発現、および光により誘導されるセカンドメッセンジャー活性の下流効果を研究するための様式の最適化された使用に関する。
【0027】
本発明の一実施形態は、本明細書に開示するように、特定の細胞集団中で発現させるためにその細胞集団へとキメラGPCRタンパク質を遺伝的にターゲティングすることに関する。標的細胞型中で選択的に発現される細胞型特異的プロモーターが存在する(例えば、ニューロンターゲティング用のシナプシン1;心組織用のトロポニンバリアント)。発現ベクター中でこれらのプロモーターをキメラGPCRタンパク質の上流に配置することで、目的の細胞型にタンパク質の発現をターゲティングすることができる。これには、Tet反応、ER反応、およびCre/Loxシステム等の誘導可能、反転可能、またはその他の形で制御可能なプロモーターシステムが含まれる。
【0028】
本発明の例示的一実施形態によれば、目的の細胞型中で発現されると光に応答して環状アデノシン一リン酸(cAMP)が産生される、遺伝的にコード可能なタンパク質が開発される。これは、例えば、限定されるものではないが、医薬品のスクリーニング等の細胞生理に対する下流効果を可視化するのに有用であり得る。その他の実施形態では、光に応答してセカンドメッセンジャーを放出させる異種性キメラGPCRを用いる。例示的なセカンドメッセンジャーには、cAMP、環状グアノシン一リン酸(cGMP)、イノシトール三リン酸/イノシトール1,4,5−三リン酸/トリホスホイノシトール(IP3)、およびアラキドン酸が含まれる。
【0029】
本発明の一実施形態によれば、細胞内メッセンジャーに関連する推定治療レジメン(例えば、薬物または電気刺激、またはこれらのセカンドメッセンジャーを介して作用する任意のもの)の効力を評価する方法が実施される。キメラ光応答性膜タンパク質(例えばロドプシン)を発現するためのヌクレオチド配列を、1または複数の異種受容体サブユニット{例えばアドレナリン受容体(アルファ1、ベータ2)}で改変する。光に応答してセカンドメッセンジャーを産生させるために、光応答性膜タンパク質を細胞中で発現させる。このタンパク質を光にさらし、治療効果を評価する。
【0030】
光は、所望の刺激特性に従って与えることができる。一実施形態では、発現された膜タンパク質は数十ミリ秒以内に光に応答する。したがって、刺激プロファイルは、速く連続している一連の光パルスを含んでもよく、得られる効果は、例えばCa2+感受性色素を用いてモニターすることができる。
【0031】
一例では、細胞は最初に治療なしで刺激され得る。治療が施された後、細胞は再度刺激され得る。各試験の結果を比較することで治療の有効性を評価することができる。
【0032】
治療には、例えば、限定されるものではないが、医薬品、細胞の修飾(遺伝的修飾その他)、細胞の物理的パラメータ(例えば温度変化または電気刺激)、または生体に適用される治療レジメンなどの、広範な異なる形での実施が含まれる。
【0033】
一実施形態では、治療は、発現された膜タンパク質の光刺激である。そのような例においては、有効性は、例えば、治療されるべき障害に関連する症状をモニターすることで測定することができる。
【0034】
別の実施形態では、治療レジメンは疾患または障害のモデル化の一部として実施される。例えば、疾患モデル(細胞または動物)を用いることができ、タンパク質が発現される前にバックグラウンド/ベースライン状態を評価し、治療レジメンを評価することができる。
【0035】
実験結果は、cAMP誘導因子およびcAMP標的陽イオンチャネルの両方で細胞をトランスフェクトし、得られた活性をCa2+感受性色素を用いて可視化することで、光で惹起されるcAMPによる標的イオンチャネルの制御を可視化できることを示している。この、セカンドメッセンジャー活性の、遺伝的にコード可能な光活性化型調節物質一式は、新規治療薬のスクリーニングに有用であり得、また、ADHDや心臓のチャネル病等の多くの疾患状態にcAMPが関与するので、それ自体が治療法である。このタンパク質は、種々の他のセカンドメッセンジャー(例えばIP3)と共に使用されるように設計されてもよく、レチナール結合部位を設計するか吸収/作用スペクトルの異なるキメラロドプシンまたは錐体オプシンをキメラに選択することで光活性化のためのその他の色と共に使用されるように設計されてもよく、また、カルシウムシグナル伝達および/またはキナーゼ活性等のセカンドメッセンジャーの他の下流効果と共に使用されるように設計されてもよい。
【0036】
図1A、1B、および1Cは、開発されたセカンドメッセンジャーシグナル伝達の光活性化型誘導因子(「optoXR」)の2つの例であるoptoGsおよびoptoGqから得られた実験データを示す。これらの光活性化型誘導因子はロドプシン/GPCRキメラ体である。optoGqは、Gqシグナル伝達の光応答性制御を可能にし、optoGsは、Gsシグナル伝達の光応答性制御を可能にする。
【0037】
optoGsおよびoptoGqのどちらにおいても、暗所におけるベースラインcAMPレベルおよびIP3レベルには無視できる差しかなく、cGMP等の他のセカンドメッセンジャーの経路とのクロスオーバーはないことが示されている。optoGqの光刺激で観察されたcAMPレベルの増加は、IP3産生の予想される下流効果である。
【0038】
図1Aは、本発明の例示的実施形態による、optoGsおよびoptoGqの模式図である。各タンパク質について、ロドプシンの細胞内ループが、通常Gs(ベータ2)またはGq(アルファ1)と共役しているアドレナリン作動性タンパク質のもので置換される。この遺伝コード配列は、ヒトおよびマウスの細胞での発現用に最適化されている。得られる配列の例には、optoGs:配列番号1および配列番号2;ならびにoptoGq:配列番号3および配列番号4が含まれる。
【0039】
当業者に理解されるように、これらのタンパク質のアミノ酸配列は、実施形態を支持する非限定的な例として提示するものであり、塩基配列以外の点では同様の結果、互換可能な結果、または同等な結果をもたらす遺伝配列のバリエーション(例えば点突然変異体)も含まれる。
【0040】
図1Bは、本発明の例示的な実施形態による、トランスフェクトなしまたはoptoGsもしくはoptoGqでトランスフェクトされた細胞のcAMP(上)、cGMP(中)、およびIP1(下:IP3の分解産物)の酵素結合免疫吸着検定法(ELISA)を示す。図1Bの結果は、示されているように、504nm光(20nmバンド幅)で1ヶ所につき1分間刺激するか暗所に維持した細胞から得た。
【0041】
刺激は、環境調節倒立培養顕微鏡(inverted culture microscope;ライカ社製DMI6000B)を用いて行った。cAMPアッセイでは、アッセイの飽和したポジティブコントロールとして、一部の細胞を10μMのフォルスコリンで30分間処理した。optoGsは、光に応答してcAMPレベルを有意に増加させた。cAMPのベースラインの有意な増加やoptoGsによるcGMPレベルもしくはIP3レベルの逸脱は見られなかった。OptoGqは、光に応答して、cGMPレベルは有意に変化させず、IP3レベルを有意に増加させた。IP3産生を伴うcAMPレベルの増加は、細胞内Ca2+放出の結果であると考えられる。
【0042】
図1Cは、本発明の例示的実施形態による、optoGsおよびoptoGqのmCherry融合タンパク質でトランスフェクトした細胞のCaイメージングを示す。cAMPを検出するために、環状ヌクレオチド依存性Ca2+チャネルCNGA2のcAMP選択的変異体をoptoGsに対し過剰量トランスフェクトした。IP3は、細胞内Ca2+ストアの放出を活性化し、信頼性の高いGq活性化シグナルを与える。対照集団も、mCherry単独及び過剰量の前記変異体CNGA2でトランスフェクトした。細胞にfura−2を負荷し(20〜25分間インキュベート)、2秒毎に340nmおよび380nmを2ms照射した。optoGsおよびoptoGqのそれぞれでは、Caシグナルを生じさせるのにこの照射だけで十分であったが、対照集団では有意なシグナルは検出されなかった。
【0043】
図1は特定の実験設定で得られたデータを示しているが、本発明はこれに限定されるものではない。例えば、トランスフェクション以外の種々の送達技術が想定され、該送達技術としては、ウイルス形質導入、弾道的遺伝子導入(ballistic gene delivery;遺伝子銃)、および自発的な核酸の取込みが含まれるが、これらに限定されるものではない。
【0044】
基となるロドプシンは、アルファ2−アドレナリン受容体もしくはドーパミンD2受容体もしくはセロトニン5HT2A受容体等のGi共役受容体、またはドーパミンD1A受容体もしくは代謝型グルタミン酸受容体等のその他のGsまたはGq共役受容体等の、任意の好適な異種受容体サブユニットと共に使用されるように改変することができる。
【0045】
例示的な一実施形態によれば、基となるロドプシンはウシBos taurusに由来するタンパク質である。
【0046】
一実施形態によれば、前述した基となるロドプシン以外の基となるタンパク質を用いることもでき、前述した基となるロドプシン以外の基となるタンパク質には、錐体オプシン(赤色、緑色、または青色)、他の種のロドプシン、およびドーパミン受容体またはセロトニン受容体等のリガンド依存性受容体等の種々の7回膜貫通型タンパク質が含まれる。
【0047】
種々の実施の態様は、哺乳動物におけるインビボでの適用に関する。これらの実施の態様は、限定されるものではないが、神経回路および疾患モデルの試験および確認を含む。
【0048】
図3Aおよび3Bは、セカンドメッセンジャーシグナル伝達の光活性化型誘導因子の2つの例であるoptoGs(opto−β2AR)およびoptoGq(opto−α1AR)のインビボでの適用から得られた実験データを示す。本発明の態様は、Gタンパク質共役受容体(GPCR)に共通する構造−機能関係に影響を与えることで、受容体によって開始される生化学的シグナル伝達経路を高い時空間精度で動員および制御する、遺伝的にコードされた光学的ツール(「optoXR」)の多様なファミリーの使用および開発に関する。
【0049】
図3Aおよび3Bに示す結果は、ターゲティングされた別個のシグナル伝達経路を光に応答して選択的に動員する2つの特定のoptoXRに関するものである。これら2つのoptoXRは、インビボで側坐核中のスパイク発火に反対の影響を与え、側坐核中での正確に時間制御されたoptoXRの光刺激だけで、自由行動をしているマウスに条件付け場所嗜好性を誘導するのに十分であった。このoptoXRを用いた手法は、行動している哺乳動物における生化学的シグナル伝達の因果的影響に関する仮説を、ターゲティング可能で時間的に正確な様式で試験することを可能にする。
【0050】
GPCR間で共有されている構造−機能関係を用いて、シグナルをエフェクターに共役させる新規な伝達ロジックを有する複数の異なるオプシン/GPCR2キメラを開発し、インビボで発現させ、哺乳動物中で細胞内シグナル伝達の光制御を実施した。種々の実施の態様に従い、1または複数のキメラオプシン受容体タンパク質が、インビボにおいて哺乳動物内で機能的であり、特定の細胞にターゲティング可能であり、正確に時間制御された光パルスに応答性であるように設計される。そのような手法により、高速な光刺激(およびタンパク質応答)を用いて、正確に決められ且つ行動上適切であるタイミングで細胞内の生化学的事象について試験し特徴解析することが可能となる。いくつかの非限定的で例示的な実施の態様には、規定された細胞型における、ある範囲の時間スケールにわたる、拍動性対強直性の調節、異なる調節系間の同調性、ならびにその他の基本的な生理学的および病理学的プロセスが含まれる。
【0051】
哺乳動物での実施は成功した。例示的な一実施例では、最初にGq共役ヒトα1aアドレナリン受容体(α1AR)およびGs共役ハムスターβ2アドレナリン受容体(β2AR)の保存された残基をGt共役ウシロドプシンと整列させることで、ロドプシンの細胞内ループを特定のアドレナリン受容体の細胞内ループで置換した(図1A)。構造モデルに基づいて、Gタンパク質との共役をGtから移すように細胞内領域の交換(カルボキシ末端ドメインを含む)を各受容体について設計し、各受容体を哺乳動物中でのインビボ発現用に最適化した。種々のリガンドにより活性化すると、天然の受容体は複数の集合(ensemble)状態を取ることができ、リガンドバイアスシグナル伝達現象においてカノニカル経路および非カノニカル経路を動員する。optoXRは光を感知すると、生物学的環境に応じた様式で、1つの活性集合的状態を選択する可能性が高い。
【0052】
キメラ(opto−α1ARおよびoptoβ2AR)をコードする遺伝子を蛍光タンパク質に融合させた。機能的optoXR発現の検証は、opto−α1AR単独(Gqを介して[Ca2+]iを動員すると予想される)、またはopto−β2AR(Gsを介して環状AMPを動員すると予想される)とcAMP依存性Ca2+チャネルCNGA2−C460W/E583Mの両方でトランスフェクトしたHEK細胞中における[Ca2+]i(細胞内カルシウム濃度)のイメージングにより達成された。[Ca2+]iのレシオメトリックイメージングにより、60sの緑色光刺激(504±6nm、7mWmm−2)はいずれのoptoXRの下流でも顕著な[Ca2+]iシグナルを誘導するのに十分であるが、対照条件では誘導しないことが示され(図2)、機能的に発現していることが明らかになった。各optoXRに制御されるシグナル伝達の特異性を試験するために、形質導入されたHEK細胞に3mWmm−2、504±6nmの光を60s照射し、その後、細胞を溶解し、イムノアッセイによりcGMP、cAMP、およびIP1(IP3の分解産物)のレベルを解析した。カノニカルパターンはopto−β2ARでその分子的デザインに対応して予想されるとおりといえた。何故なら、光刺激によりopto−β2AR発現細胞中でcAMPが有意に産生され(図3A、上)、これは野生型β2ARの薬物刺激で得られるものに匹敵するとともに、IP3(図3A、中段)、[Ca2+]i(図2)や暗所での実質的な活性を動員しなかったからである。一方、光刺激は、opto−α1AR発現細胞でIP3シグナル伝達の有意なアップレギュレーションを生じさせ(図3A、中段)、これは野生型α1ARの薬物刺激により誘導されるレベルに匹敵するものであった。[Ca2+]iの上昇(図2)と共に、これらのデータは、Gqの動員について予想されるパターンを示しており、このパターンはopto−β2AR発現細胞中では見られない(図3A、上)。いずれのコンストラクトを発現している細胞の光刺激も、cGMPレベルを調節することができず(図3A、下)、このことは更に、キメラタンパク質のシグナル伝達特異性を示している。同様なアッセイから、optoXRが、天然ロドプシンの作用スペクトルに近い作用スペクトルを維持しており、生物学的に好適な光束の範囲に渡りシグナルを統合することができ、p42/p44−MAPKシグナル伝達に関して野生型受容体と同様の程度に非カノニカル経路を活性化できることが示された。
【0053】
レチナール補因子の補充が必要かどうかも含め、インタクトな神経組織中におけるoptoXRの能力を調べた。そのような試験の1つでは、(生化学的調節の標的をグリアおよび内皮細胞を初めとする他の潜在的Gs/Gq応答性細胞組織要素ではなく局所ニューロンにするため)シナプシンIプロモーターの制御下にあるoptoXR融合遺伝子を有するレンチウイルスベクター(図3B、左上)を成獣マウスの側坐核中に定位的に注入した。この戦略により、側坐核中に細胞体樹状突起区画を有するニューロン(約95%がGABA作動性中型有棘ニューロンであり、更なるサブタイプ特異性を有しない;図3B、左)が生化学的調節の標的とされ、またこれらのレンチウイルスは軸索を介して細胞に形質導入することはないため、通過繊維(fibers of passage)または求心性シナプス前終末は除外される。形質導入の2週間後、人工脳脊髄液中で側坐核の急性冠状スライスを調製し、10分間光刺激し、すぐに固定しcAMPおよびCa2+共役シグナル伝達カスケードの両方の生化学的統合因子(integrator)であるSer133−リン酸化CREB(pCREB)を染色した。外因性のレチノイドを補充しなくても、optoXR発現集団において有意に上昇したpCREBが観察されたが(図3B、右)、非照射組織では観察されなかった。
【0054】
形質導入した側坐核にターゲティングされたオプトロードを用いて、マルチユニットインビボニューロン発火を記録することで、側坐核の居所的電気活性に対するoptoXR活性化の機能面での影響を決定した(図4A)。どちらのコンストラクトにおいても暗所ではベースライン発火率に有意な差は見られなかった(図4A、右下)。光刺激は、opto−β2AR発現側坐核においてネットワーク発火を減少させた(図4Bの左のトレースは影響のカイネティクスを示す。概要データが図4Cおよび図4Dにそれぞれ示される)。これは、Gsを標的にした過去の薬理学的実験と一致していた。光刺激は、opto−α1AR発現側坐核(図4B、右;図4C、4D)において発火を増加させた。スパイク頻度のヒストグラムから、発火率へのoptoXRの影響のカイネティクスは、シグナルの電気的開始よりも生化学的開始の方に一致することが示された(図4D)。これらの電気生理学的データと前出の生化学的検証とを組み合わせれば、細胞内カスケードの差動的光活性化型制御を可能にし且つネットワークの生理を調節するようにoptoXRをインビボで機能的に発現させることができることが支持される。
【0055】
一実施例では、光遺伝学を用いて、正確にタイミングを制御されたoptoXR刺激が自由行動中のマウスの行動を調節する能力を評価した。オペラント行動に用いられる時間的に正確な様式で側坐核ニューロン内の細胞内シグナル伝達を光制御するため、携帯型ソリッドステート光送達をoptoXRのトランスジェニック発現と組み合わせた(図5A)。共焦点分析により、発現が局所的な側坐核ニューロンに限定されていることが示され、特に、求心性繊維、側坐核に突き出た遠位領域、グリア、または周辺領域中で標識は全く観察されなかった。3日間のオペラント条件付け場所嗜好性アッセイの一部として、形質導入した側坐核に光刺激をターゲティングした(図5A)。試験中毎日、動物に自由に場所嗜好性装置を探索させた(図5A、下)。1日目は、動物は光刺激を受けずに自由に装置内を探索した。2日目に、指定した条件付けされた部屋に動物が自由に進入する毎に、形質導入した領域に位置を合わせたレーザーダイオード接続光ファイバーで光パルスを10Hzで送達して、強力な報酬の間のモノアミン作動性入力の適切な強度を模倣した。経路のトレースから、柔軟な光ファイバーを用いた手法により、全ての部屋を完全且つ妨害のなしに探索できることが示された(図5A、下)。3日目に、動物は再度、光刺激を受けずに自由に装置内を探索し、条件付けた部屋で過ごした時間が2人の別の盲検評価者により定量された。特に、opto−α1ARを発現している動物は、光刺激後に、装置の条件付けされた側への嗜好性が大きく上昇したことが示された(図5B)。この時間的に正確な生化学的調節の影響は、opto−α1AR動物の別個の2つのコホートで再現可能であった(n=5〜6、P<0.05、条件付けされた部屋中での時間の各コホートについてのスチューデントのt検定;全集団では、n=11、P<0.01)。一方、その他のオプシン遺伝子opto−β2ARおよびChR2は、嗜好性の誘導についてさほど効果的ではないようであった。側坐核ニューロン中のopto−α1AR刺激の影響は報酬関連行動に特異的であり、オープンフィールド試験で同じ動物のコホートに送達された同じ光刺激が移動距離または壁の近接に対する嗜好性には有意な影響を与えなかった(図5C)ことから、不安関連行動または自発運動活性の直接的調節にまでは及ばなかった。
【0056】
上記実験に従った具体的かつ非限定的な実施例をここで記載する。記録電極(1MVタングステン、A−Mシステムズ社製)に接続した直径200mmのマルチモード光ファイバー(ソーラボ社製)からなり電極/ファイバーの先端間の距離が200〜400mmであるオプトロードを、定位フレーム(デビッド・コフ社(David Kopf Instruments)製)に固定されイソフルランで麻酔したマウスの形質導入された側坐核中に下ろして用い(電極の先端はブレグマの下4.8〜5.2mm)、インビボ記録および解析を行った。473nmダイオードレーザー(クリスタレーザー社製)の光をファイバーを介して送達した。電気シグナルをバンドパスフィルターに通し、増幅し(0.3〜1kHz、1800 Microelectrode AC Amplifier、A−Mシステムズ社製)、pClamp10.0(モレキュラーデバイス社(Molecular Devices)製)を用いて解析した。閾値によりスパイクを検出し、個々に点検して確認した。
【0057】
光ファイバー(直径200mm、ソーラボ社製)を473nmの青色ダイオードレーザー(クリスタレーザー社製)に接続し、側坐核(先端から0〜100mm)をターゲティングするカニューラに合わせ、この光ファイバーを介して光刺激を与え、行動分析を行った。ファンクションジェネレータ(Agilent 33220A)を用いてoptoXR用に50msパルス幅の光を送達した。場所嗜好性は、自由に探索できるように部屋の間の壁を除去した標準装置(SDインスツルメンツ社製)中で実施した。各部屋で過ごした時間量について、MATLAB(マス・ワークス社(Mathworks)社製)で特注の集計スクリプトを実行して、2人の独立した盲検評価者がビデオからデータを解析した。オープンフィールド試験では、40340cmの正方形のオープンフィールド中に動物を置き、場所嗜好性実験と同じパラメータを用いて光刺激を送達した。自動化されたソフトウェア(Viewpoint)を用いて、その外側の環(フィールドの残りの部分)に対する、中央の15315平方cmでの合計時間および距離についてビデオを解析した。
【0058】
表示されている場合、統計解析は、両側検定によるスチューデントのt検定(マイクロソフトエクセルで計算)または一元配置の分散分析とチューキーの事後検定(GraphPad Prism)を用いて行った。まとめた棒グラフは全て、平均値±s.e.mで表し、有意性は以下のように表した:*P<0.05、**P<0.01、***P<0.001。
【0059】
本発明の種々の実施形態の驚くべき結果および有効性を支持する更なる詳細は、参照により全体を本明細書に援用するTemporally precise in vivo control of intracellular signalling, Raag D. Airan, et al., Nature 458, 1025-1029 (23 April 2009)中に見出すことができる。
【0060】
以下の記載は、本発明の実施形態による具体的且つ非限定的な方法の詳細を提供する。この方法の多くのバリエーションが想定され、本発明の範囲内に含まれる。
【0061】
ベクター構築
opto−α1ARおよびopto−β2ARの哺乳動物コドン最適化配列(図1A中のアミノ酸配列)を合成し、pcDNA3.1にクローニングし、NotI部位を用いてmCherryまたはYFP(スタートコドンを除去)のN末端に融合させた。optoXRとmCherry/YFPのリンカーは5’GCGGCCGCC3’である。pLenti SynapsinI hChR2 mCherry WPREベクターのAgeIおよびEcoRI部位に各optoXR mCherryの導入遺伝子をクローニングすることで、SynapsinI optoXR mCherryを含むレンチウイルスベクターを構築した。
【0062】
レンチウイルスの作製
高タイターレンチウイルスを作製した。簡潔に述べると、10%FBSを含むDMEMで培養されている4層のセルファクトリー(Nunc社製)中に90%のコンフルエンスでHEK293FT細胞をプレーティングした。690μgの前述したレンチウイルスベクターおよび2種類のヘルパープラスミド(690μgのpΔCMVR8.74および460μgのpMD2.G)で細胞をコトランスフェクトした。トランスフェクションの15時間後に培地を交換した。トランスフェクションの24時間後に、5mMの酪酸ナトリウムを含む無血清UltraCULTURE(キャンブレックス社(Cambrex)製)200〜220mLで培地を交換した。トランスフェクション40時間後、今やウイルスを含む培養液上清を1000rpmで5分間スピンして細胞残渣を除去した後、0.45μmの低タンパク質結合フィルターフラスコを用いて濾過した。次いで、この清澄な上清を、SW28ローター(ベックマン社製)を用いて55,000gで2時間超遠心し、ウイルスを沈殿させた。遠心分離後、上清を捨て、得られたウイルスペレットを合計100μLの冷(4℃)PBSに溶解した。再懸濁したウイルスを7000rpmで5分間遠心分離して残りの細胞残渣およびウイルス残渣を除去した。アリコートを再使用時まで−80℃で凍結した。
【0063】
動物の外科手術および行動
スタンフォード大学の実験室用脊椎動物プロトコールに従い、10〜12週齢の雌のC57BL/6マウスを収容し、取り扱った。ウイルス溶液を以下のように右側坐核に導入した。動物をイソフルランで麻酔し、頭頂部の毛を刈り取った。イソフルラン麻酔下にいる間に動物の頭を定位フレーム(デビッド・コフ社)に固定した。頭皮を正中切開し、ブレグマの前側1.10mm、側方1.45mmに直径約1mmの開頭切開部を穿孔した。次いで、ウイルスを予め充填した33ゲージのベベル針(NanoFil、ワールド・プレシジョン・インスツルメンツ社(World Precision Instruments)製)を側坐核中に下ろし(針の先端がブレグマの腹側4.70〜4.80mm)、自動シリンジポンプ(NanoFil、ワールド・プレシジョン・インスツルメンツ社製)を用いて1.0μLのウイルスを100nL/分で注入した。注入後、3〜5分間組織を弛緩させて流体を拡散させた後、針を引き抜いた。急性スライスまたはインビボ記録実験用の動物では、開頭部にデンタルセメント(ラングデンタル社(Lang Dental)製)を充填し、VetBond(3M社製)を用いて切開部を閉じた。行動分析用の動物では、カニューレ(C316G、ペデスタルの4.5mm下で切断;プラスチクスワン社(Plastics・One)製)をペデスタルが頭蓋に面一になるよう配置した。Metabond(パーケル社(Parkell)製)およびデンタルセメント(ラングデンタル社製)を用いてカニューレを固定した。VetBondまたはセメントの乾燥後、動物をフレームから外し、その後の操作の前に少なくとも1週間回復させた。行動実験の対照動物にも実験動物と同じ操作(外科手術、カニューレの植込み、光刺激)を行い、ウイルスの代わりに溶媒(PBS)のみを注入した。場所嗜好性実験には、部屋のどちらかの側(>70%または<10%)または中央の部屋(>40%)に対するベースライン嗜好性を示さなかった動物を実験に用いた。全動物の90%超が、バイアスがかけられていないバランスのとれた場所嗜好性デザインのためのこれらの基準を満たした。
【0064】
急性スライスの調製
動物をイソフルランで麻酔し、外科手術用のハサミ(ファイン・サイエンス・ツールズ社(Fine Science Tools)製)を用いて断頭した。側坐核を含む275μm厚の冠状スライスを切り出し、64mMのNaCl、2.5mMのKCl、1.25mMのNaH2PO4、25mMのNaHCO3、10mMのグルコース、120mMのスクロース、0.5mMのCaCl2、および7mMのMgCl2を含む切断溶液(95%O2/5%CO2で平衡化)中に保存した。スライス後、スライスを切断溶液中にて32〜35℃で30分間インキュベートし、その後実験まで室温でインキュベートした。エクスビボでのoptoXR刺激では、スライスを正立顕微鏡(BX51W、オリンパス社製)のステージに乗せ、124mMのNaCl、3mMのKCl、1.25mMのNaH2PO4、26mMのNaHCO3、10mMのグルコース、2.4mMのCaCl2、および1.3mMのMgCl2を含む人工脳脊髄液(95%O2/5%CO2で平衡化)で灌流した。300WのLambda DG−4(サッター社(Sutter)製)の光を473nm±20nmのバンドパスフィルター(セムロック社(Semrock)製)に通し、4倍の対物レンズ(0.28NA)を用いて10分間スライスに当て、その後すぐに、後の解析のために固定した。
【0065】
シグナル伝達検証アッセイ
リポフェクタミン2000(インビトロジェン社製)を用いて24ウェルプレート中でHEK293FT細胞(インビトロジェン社製)をトランスフェクトし、トランスフェクションの4〜6時間後に無血清培地に交換した。Ca2+イメージングには、マトリゲルでコートされたカバーガラス上にプレーティングされた細胞に、1μMのATRを含むタイロード中、5μg/mlのfura−2 AMを含むF−127プルロニック/DMSO(プローブ社(Probes)製)を37℃、5%大気CO2にて20〜25分間負荷した。負荷後、300WのLambda DG−4(サッター社製)を制御するMetafluor(アクソン・インスツルメンツ社(Axon Instruments)製)を用いて、オリンパス社製BX51W上で340nm/380nmでカバーガラスをイメージ化した。イムノアッセイでは、トランスフェクションの18〜24時間後に、1μMのATRおよび50mMのLiCl(IP1の分解を防ぐため)を添加し、プレートを環境調節された顕微鏡(ライカ社製DMI6000;37℃、5%大気CO2)に移した。5領域/ウェルをそれぞれ1分間光刺激した(サッター社製300WのLambda DG−4;セムロック社製504/12nmバンドパスフィルター;10×0.30NA対物レンズ);3ウェル/条件。インキュベート後(cAMP/cGMP:20分間;IP1:1時間)、細胞を溶解し、HTRF(シスバイオ社(CisBio)製)およびBiotek Synergy4リーダーで解析した。
【0066】
免疫組織化学および共焦点分析
インビボ刺激後、刺激終了90分後に、氷冷した4%パラホルムアルデヒド(PFA)のPBS溶液(pH7.4)をマウスの心臓に灌流した。脳を取り出し、4%PFA中で一晩固定し、その後、30%スクロースのPBS溶液中で平衡化した。40μm厚の冠状切片を凍結ミクロトーム上で切り出し、免疫組織化学用に処理するまで抗凍結剤中で4℃にて保存した。フリーフローティング切片をPBS中で洗浄した後、0.3%のTx100および3%の正常ロバ血清(NDS)中で30分間インキュベートした。急性スライス実験では、刺激直後に、275μm厚のスライスを、氷冷した4%PFA中で1時間固定し、0.5%のTx100および3%のNDSと共にインキュベートした。MAPKアッセイでは、HEK293細胞の刺激直後に、カバーガラスを15分間固定し、0.6%のH2O2と共にインキュベートし、その後、0.1%のTx100を含む3%NDSで透過処理した。マウス抗GAD67 1:500、マサチューセッツ州ビルリカのミリポア社製;ウサギ抗cfos 1:500、カリフォルニア州サンディエゴのカルバイオケム社(Calbiochem)製;ウサギ抗リン酸化CREB Ser133 1:500、ミリポア社製について、一次抗体のインキュベートを0.01%のTx100および3%のNDS中で一晩行った。切片を洗浄し、FITCまたはCy5(ペンシルベニア州ウェストグローブのジャクソン・ラボラトリー(Jackson Laboratories)製)にコンジュゲートした二次抗体(1:1000)と共に室温で3時間インキュベートした。DAPI(1:50,000)と20分間インキュベートした後、切片を洗浄し、PVD−DABCOと一緒に顕微鏡用スライドにマウントした。残りの一次抗体オーバーナイトインキュベート物(ウサギ抗リン酸化Erk1/2;抗リン酸化MAPK p38 1:500、ウィスコンシン州マディソンのプロメガ社製;マウスモノクローナル抗ドーパミンD1受容体 1:50、ケミコン社(Chemicon)製;ウサギポリクローナル抗ドーパミンD2受容体 1:50、ミリポア社製;ヤギポリクローナル抗コリンアセチルトランスフェラーゼ 1:200、ミリポア社製)はその後、ビオチン化二次抗体(1:500、ジャクソン・ラボラトリー製)とのインキュベート、アビジン−ビオチン−西洋ワサビペルオキシダーゼ処理(ABCキット、カリフォルニア州バーリンゲームのベクター・ラボ社(Vector Labs)製)、およびTSA検出(コネティカット州シェルトンのパーキンエルマー社製)を、メーカーの取扱説明書に従い行った。
【0067】
20×/0.70NAまたは40×/1.25NAの油浸対物レンズを用いてライカ社製TCS SP5走査型レーザー顕微鏡上で共焦点蛍光画像を得た。1つの条件につき、カニューレ管の下500μmの領域内で4つの連続スタック画像を得た。DAPI染色で核を可視化し、Volocity(インプロビジョン社(Improvision)製)ソフトウェアを用いてcfosまたはpCREBの免疫反応性の平均ピクセル強度を求めた。陽性またはpCREB活性の細胞を強度閾値に基づき同定し、実験条件を隠して(blind)画像の取得および解析を行った。
【0068】
【表1】
【0069】
【表2】
【0070】
【表3】
【0071】
したがって、本発明の実施形態は、細胞内シグナル伝達の光遺伝学的制御に関し、行動中の哺乳動物内でインビボで作用し且つ非常に低い暗所活性を有しつつ、時間精度を得ることおよび天然受容体下流の複雑に絡み合った複数のシグナル伝達分子を動員するのに有用であり、したがって他の手法の個々のプラス面の多くを1つの技術中で統合するのに有用である。同様な実施形態では、無数の神経伝達物質および内分泌性ホルモン等のその他の調節物質により引き起こされる7回膜貫通依存性シグナル伝達経路の因果上の意義を直接探索する。別の実施形態では、興奮性の細胞を超えてoptoXR手法が用いられ、光ファイバーの深さのターゲティングと光遺伝学的にターゲティングされる光感受性とを多用途に統合することの利点を活かす。そのような実施形態の1つは、多様な非興奮性組織における時間的に正確な生化学的シグナル伝達の因果上の意義を探索することに関する。
【0072】
本発明の実施形態は、種々のリガンドが集合(ensemble)受容体コンホメーション状態を安定化させることで代替的な伝達カスケードと共役した受容体の細胞内作用にバイアスをかけることができる、リガンドバイアスシグナル伝達現象の考察に関する。optoXRは、薬理学的操作と同様なレベルで代替的なカスケードを誘導するように用いられる(例えばopto−β2ARは、野生型β2ARへの天然リガンドの作用と同様な変化をMAPK活性化に誘導することができる)。しかし個々のoptoXRは、リガンドバイアスシグナル伝達に寄与するコンホメーション状態の全ての制御を常に可能にするとは限らない。レチナールを基にしたツールは、哺乳動物組織中に内因性発色団が存在し、暗所での活性が非常に低いため、特に有用であり得る。光遺伝学は、速い単一成分レチナール結合モジュールに連結された多様なエフェクターの形態を取ることができ、光学の有する時間的精度を利用できる。
【0073】
本発明の実施形態は、微生物オプシンによる戦略を補完するoptoXRによる方法を利用し、行動下の哺乳動物中で作動可能で、速く且つターゲティング可能な別次元の細胞制御を提供する。
【0074】
本発明の別の実施形態によれば、異なる作用スペクトルを有する公知のオプシン遺伝子に基づいて波長をシフトさせたタイプのoptoXRが使用される。そのようなoptoXRは、生化学的および電気的制御の分離可能なチャネルを提供するのに特に有用であり得る。
【0075】
本明細書に記載した特定のタンパク質配列のバリアントも本発明の実施形態に沿っている。一部のバリアントはこれらのタンパク質配列に約75%超の相同性を有し、別のものは約80%、85%、または90%超の相同性を有する。いくつかの実施形態では、相同性は約93〜約95、または約98%と高い。本発明の組成物には、本明細書に記載したタンパク質配列および核酸配列が含まれ、記載した配列に約50%を超える相同性を有するバリアントから100%の相同性を有するバリアントまで含まれる。
【0076】
本明細書に記載した種々の実施形態は、正常な作動および疾患状態の両方における神経回路の更に洗練されたインタロゲーションおよびリバースエンジニアリングのために速い回路読取り技術と統合され得る。
【0077】
前述した種々の実施形態は単に例として記載したものであり、本発明を限定するものと解釈されるべきではない。前述の説明および例示に基づき、本明細書に例示および記載された例示的な実施形態および応用例に厳密に従わなくとも本発明に種々の修正および変更を加えることができることが当業者には容易に理解されよう。例えば、そのような変更には、産生されるセカンドメッセンジャーのバリエーションが含まれ得る。そのような修正および変更は、添付の特許請求の範囲に記載されている本発明の真の精神および範囲から逸脱しない。
【特許請求の範囲】
【請求項1】
細胞中でセカンドメッセンジャーを生成する方法であって、
キメラ光応答性ロドプシン系膜タンパク質を発現するためのヌクレオチド配列を1または複数の異種受容体サブユニットで改変すること;および
光に応答してセカンドメッセンジャーを産生させるために、前記光応答性膜タンパク質を細胞中で発現させること、
を含む、方法。
【請求項2】
前記1または複数の異種受容体サブユニットがアドレナリン受容体を含む、請求項1に記載の方法。
【請求項3】
前記キメラ光応答性ロドプシン系膜タンパク質が7回膜貫通型タンパク質である、請求項1に記載の方法。
【請求項4】
前記セカンドメッセンジャーが、cAMP、環状グアノシン一リン酸(cGMP)、イノシトール三リン酸/イノシトール1,4,5−三リン酸/トリホスホイノシトール(IP3)、およびアラキドン酸のいずれかである、請求項1に記載の方法。
【請求項5】
前記発現工程がインビボでなされる、請求項1に記載の方法。
【請求項6】
前記発現工程がインビトロでなされる、請求項1に記載の方法。
【請求項7】
前記発現した光応答性膜タンパク質を光刺激する工程を更に含む、請求項1に記載の方法。
【請求項8】
細胞内メッセンジャーに関連する推定治療レジメンの効力を評価する方法であって、
キメラ光応答性ロドプシン系膜タンパク質を発現するためのヌクレオチド配列を1または複数の異種受容体サブユニットで改変すること;
光に応答してセカンドメッセンジャーを産生させるために、前記光応答性膜タンパク質を細胞中で発現させること;
前記タンパク質を光にさらすこと;および
治療効果を評価すること、
を含む、方法。
【請求項9】
1または複数の異種受容体サブユニットを有するキメラ光応答性ロドプシン系膜タンパク質を発現する細胞。
【請求項10】
前記1または複数の異種受容体サブユニットが、アルファ1およびベータ2の少なくとも一方のアドレナリン受容体である、請求項9に記載の細胞。
【請求項11】
1または複数の異種受容体サブユニットを有するキメラ光応答性膜ロドプシン系タンパク質を発現するためのヌクレオチド配列。
【請求項12】
配列番号1または配列番号3に示すヌクレオチド配列およびその点突然変異体。
【請求項13】
配列番号2または配列番号4に示すアミノ酸およびその点突然変異体を発現する細胞。
【請求項1】
細胞中でセカンドメッセンジャーを生成する方法であって、
キメラ光応答性ロドプシン系膜タンパク質を発現するためのヌクレオチド配列を1または複数の異種受容体サブユニットで改変すること;および
光に応答してセカンドメッセンジャーを産生させるために、前記光応答性膜タンパク質を細胞中で発現させること、
を含む、方法。
【請求項2】
前記1または複数の異種受容体サブユニットがアドレナリン受容体を含む、請求項1に記載の方法。
【請求項3】
前記キメラ光応答性ロドプシン系膜タンパク質が7回膜貫通型タンパク質である、請求項1に記載の方法。
【請求項4】
前記セカンドメッセンジャーが、cAMP、環状グアノシン一リン酸(cGMP)、イノシトール三リン酸/イノシトール1,4,5−三リン酸/トリホスホイノシトール(IP3)、およびアラキドン酸のいずれかである、請求項1に記載の方法。
【請求項5】
前記発現工程がインビボでなされる、請求項1に記載の方法。
【請求項6】
前記発現工程がインビトロでなされる、請求項1に記載の方法。
【請求項7】
前記発現した光応答性膜タンパク質を光刺激する工程を更に含む、請求項1に記載の方法。
【請求項8】
細胞内メッセンジャーに関連する推定治療レジメンの効力を評価する方法であって、
キメラ光応答性ロドプシン系膜タンパク質を発現するためのヌクレオチド配列を1または複数の異種受容体サブユニットで改変すること;
光に応答してセカンドメッセンジャーを産生させるために、前記光応答性膜タンパク質を細胞中で発現させること;
前記タンパク質を光にさらすこと;および
治療効果を評価すること、
を含む、方法。
【請求項9】
1または複数の異種受容体サブユニットを有するキメラ光応答性ロドプシン系膜タンパク質を発現する細胞。
【請求項10】
前記1または複数の異種受容体サブユニットが、アルファ1およびベータ2の少なくとも一方のアドレナリン受容体である、請求項9に記載の細胞。
【請求項11】
1または複数の異種受容体サブユニットを有するキメラ光応答性膜ロドプシン系タンパク質を発現するためのヌクレオチド配列。
【請求項12】
配列番号1または配列番号3に示すヌクレオチド配列およびその点突然変異体。
【請求項13】
配列番号2または配列番号4に示すアミノ酸およびその点突然変異体を発現する細胞。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2011−523851(P2011−523851A)
【公表日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願番号】特願2011−511844(P2011−511844)
【出願日】平成21年5月29日(2009.5.29)
【国際出願番号】PCT/US2009/045611
【国際公開番号】WO2009/148946
【国際公開日】平成21年12月10日(2009.12.10)
【出願人】(503115205)ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティ (69)
【Fターム(参考)】
【公表日】平成23年8月25日(2011.8.25)
【国際特許分類】
【出願日】平成21年5月29日(2009.5.29)
【国際出願番号】PCT/US2009/045611
【国際公開番号】WO2009/148946
【国際公開日】平成21年12月10日(2009.12.10)
【出願人】(503115205)ボード オブ トラスティーズ オブ ザ レランド スタンフォード ジュニア ユニバーシティ (69)
【Fターム(参考)】
[ Back to top ]