
セラミックス結合ペプチド
【課題】セラミックス表面への目的とするタンパク質や細胞の結合・解離を簡便に制御しうる方法や、当該制御方法に用いることができる、セラミックス表面に可逆的な結合・解離能を有する新規のペプチド及びその利用方法を提供すること。
【解決手段】本発明者らは、チタン酸バリウム(BT)粒子に、多様なペプチド配列をファージ粒子上に提示するファージ集団を接触させ、BT粒子に結合したファージ粒子を回収し、得られたファージ粒子を大腸菌中で増殖させ、次いで、この増殖させたファージ粒子を、再度BT粒子に接触させるパニング操作を繰り返すことにより、BT粒子に特異的に結合するファージクローンを得た。さらに、このファージクローンの提示するペプチドが他の数種類のセラミックス表面にも結合すること、また、その結合がバリウムイオン等の金属イオンの濃度を調整することにより解離することを見い出した。
【解決手段】本発明者らは、チタン酸バリウム(BT)粒子に、多様なペプチド配列をファージ粒子上に提示するファージ集団を接触させ、BT粒子に結合したファージ粒子を回収し、得られたファージ粒子を大腸菌中で増殖させ、次いで、この増殖させたファージ粒子を、再度BT粒子に接触させるパニング操作を繰り返すことにより、BT粒子に特異的に結合するファージクローンを得た。さらに、このファージクローンの提示するペプチドが他の数種類のセラミックス表面にも結合すること、また、その結合がバリウムイオン等の金属イオンの濃度を調整することにより解離することを見い出した。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、セラミックス表面への可逆的な結合・解離能を有するペプチドや、かかるペプチドを用いたセラミックス表面への目的タンパク質の結合・解離を制御するセラミックス表面の修飾方法や、表面修飾されたセラミックス等に関する。
【背景技術】
【0002】
ファージとは大腸菌などバクテリアに感染するウイルスであり、自身のDNAがタンパク質の殻で包み込まれた構造をしている。この外皮タンパク質はファージのDNAを設計図として構築されるため、このファージDNAに任意のDNA配列を挿入することで、「さや」のタンパク質に任意のアミノ酸(ペプチド)を付加することができる。このような性質を利用して、多様なペプチドがファージの外皮タンパク質に付加されたファージ集団が、ファージライブラリとして市販されている。通常のファージライブラリの多様性は109種類にもなる。
【0003】
ファージディスプレイ法は、ファージライブラリを用いて生体分子に結合するペプチド配列を同定するための手法であったが、近年、金属等の無機物質を認識するペプチド配列を特定するためにも用いられるようになってきている。これまでに、チタン(特許文献1)、ナノ黒鉛構造体(特許文献2)、GaAs(非特許文献1)等に結合するペプチドがファージディスプレイ法により同定されている。
【0004】
さらに、非特許文献2は、ファージディスプレイ法を用いたスクリーニングにより、水溶性前駆体溶液からチタン酸バリウム(BaTiO3)の結晶形成を促進するペプチドを同定している。しかし、非特許文献2では、スクリーニングによりチタン酸バリウム結晶に結合するファージクローンを取得しているに過ぎず、これらのファージクローンの提示するペプチド自体が実際にチタン酸バリウム結晶に結合するかどうかについては明らかにしていない。
【0005】
セラミックスは、歯科インプラント材料をはじめとして様々な医療用デバイスとして応用の可能性が考えられている。このような医療用デバイスでは、表面を修飾することにより、生体適合性を高めることが重要である。また、セラミックス表面を機能性成分により修飾することにより、分子認識を行うナノデバイスの開発なども可能である。しかし、セラミックス表面に対する詳細な結合特性の解析がなされたペプチドは、未だ報告されていない。
【0006】
【特許文献1】国際公開第05/010031号パンフレット
【特許文献2】特開2004−121154号公報
【非特許文献1】Whaley SR et al., Nature 405: 665-668, 2000
【非特許文献2】Ahmad G et al., J Am Chem Soc 130: 4-5, 2008
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明の課題は、セラミックス表面への目的とするタンパク質や細胞の結合(複合)・解離を簡便に制御しうる方法や、当該制御方法に用いることができる、セラミックス表面に可逆的な結合・解離能を有する新規のペプチド及びその利用方法を提供することにある。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決するため鋭意研究し、チタン酸バリウム(以下「BaTiO3」又は「BT」ということもある)粒子に、多様なペプチド配列をファージ粒子上に提示するファージ集団を接触させ、BT粒子に結合したファージ粒子を回収し、得られたファージ粒子を大腸菌中で増殖させ、次いで、この増殖させたファージ粒子を、再度BT粒子に接触させるパニング操作を繰り返すことにより、まずBT粒子に特異的に結合するファージクローンを得ることに成功した。次に、このファージクローンの提示するペプチドについて解析し、このペプチドが他の数種類のセラミックス表面にも結合すること、また、その結合がバリウムイオン等の金属イオンにより解離することを見い出し、本発明を完成するに至った。
【0009】
すなわち本発明は、(1)配列番号11又は19に示されるアミノ酸配列からなるペプチドや、(2)セラミックス表面への可逆的な結合・解離能を有するペプチドであって、配列番号11又は19に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列や、(3)配列番号11に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列が、配列番号1、14又は16に示されるアミノ酸配列であることを特徴とする上記(2)記載のペプチドや、(4)セラミックス表面への可逆的な結合・解離が、金属イオンにより制御されることを特徴とする上記(2)記載のペプチドや、(5)金属イオンが、Ba2+、Ni2+、Zn2+及びCa2+から選択される少なくとも1以上の金属イオンであることを特徴とする上記(2)記載のペプチドに関する。
【0010】
また本発明は、(6)セラミックスが、金属酸化物を主成分とするセラミックスであることを特徴とする上記(2)記載のペプチドや、(7)金属酸化物が、遷移金属酸化物であることを特徴とする上記(6)記載のペプチドや、(8)遷移金属酸化物が、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物であることを特徴とする上記(7)記載のペプチドや、(9)検出可能なマーカーで標識された上記(1)〜(8)のいずれか記載のペプチドや、(10)上記(1)〜(8)のいずれか記載のペプチドと、機能性ペプチド又は機能性タンパク質とを結合させた融合ペプチドや、(11)上記(1)〜(8)のいずれか記載のペプチドを、その粒子表面上に提示することを特徴とする、セラミックス表面への可逆的な結合・解離能を有するファージや、(12)上記(1)〜(8)のいずれか記載のペプチドを認識する抗体や、(13)上記(1)〜(8)のいずれか記載のペプチドをコードするDNAや、(14)上記(13)記載のDNAを含み、かつセラミックス表面への可逆的な結合・解離能を有するペプチドを発現することができる組換えベクターや、(15)上記(14)記載の組換えベクターが導入された形質転換体に関する。
【0011】
さらに本発明は、(16)セラミックス表面への可逆的な結合・解離能を有するペプチド、又は該ペプチドと機能性タンパク質との融合ペプチドを介しての、セラミックス表面と目的とするタンパク質との結合又は解離を、所定の金属イオンの濃度を調整することによって行うことを特徴とするセラミックス表面とタンパク質との結合・解離を制御する方法や、(17)セラミックス表面と、セラミックス表面への可逆的な結合・解離能を有するペプチドと目的とするタンパク質との融合ペプチドとの結合又は解離を、所定の金属イオンの濃度を調整することによって行うことを特徴とするセラミックス表面と融合ペプチドとの結合・解離を制御する方法や、(18)セラミックス表面への可逆的な結合・解離能を有するペプチドとして、上記(1)〜(9)のいずれか記載のペプチドを用いることを特徴とする上記(15)又は(16)記載の方法や、(19)融合ペプチドとして、上記(10)記載の融合ペプチドを用いることを特徴とする上記(15)又は(16)記載の方法や、(20)金属イオンが、Ba2+、Ni2+、Zn2+、及びCa2+から選択される少なくとも1以上の金属イオンであることを特徴とする上記(16)〜(19)記載の方法に関する。
【0012】
また本発明は、(21)セラミックスが、金属酸化物を主成分とするセラミックスであることを特徴とする上記(16)〜(20)のいずれか記載の方法や、(22)金属酸化物が、遷移金属酸化物であることを特徴とする上記(21)記載の方法や、(23)遷移金属酸化物が、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物であることを特徴とする上記(22)記載の方法や、(24)上記(16)〜(23)のいずれか記載の方法により、セラミックス表面に所定のタンパク質又は所定の細胞を脱着する方法や、(25)上記(1)〜(9)のいずれか記載のペプチド、又は上記(10)記載の融合ペプチドで表面修飾されたセラミックスや、(26)上記(1)〜(9)のいずれか記載のペプチド、又は上記(10)記載の融合ペプチドを介して、表面に所定のタンパク質又は所定の細胞が結合したセラミックスに関する。
【発明の効果】
【0013】
本発明のセラミック表面に特異的に結合するペプチドは、簡単に化学合成することができ、また、遺伝子工学的手法を用いることで他のタンパク質等と融合させた融合ペプチドを作製することができ、セラミックス表面の修飾に用いることができる。また、本発明のペプチドや融合ペプチドと、セラミックス表面との結合・解離は、金属イオンによって制御することが可能であるため、タンパク質精製など幅広い分野での応用が可能である。
【発明を実施するための最良の形態】
【0014】
本発明のペプチドとしては、配列番号11に示されるアミノ酸配列Ala−Glu−Pro−His−Pro−Ser−Ala−Ala−His−Lys−Lys−Lys−Gln−Ser−Ser−Pro−Lys−Ser−Glu−Asn−Arg−Lys(AEPHPSAAKKKQSSPKSENRK;以下、「BTBP−1」ということもある)や、配列番号19に示されるアミノ酸配列Lys−Ala−Lys−Ala−Lys−Ala−Lys−Ala−Lys−Ala−Lys−Ala(KAKAKAKAKAKA;以下「(KA)6」ということもある)からなるペプチドや、セラミックス表面への可逆的な結合・解離能を有するペプチドであって、配列番号11に示されるアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列や、配列番号19に示されるアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列のいずれかのアミノ酸配列からなるペプチドであれば特に制限されるものではなく、ここで「セラミックスへの可逆的な結合・解離能を有する」とは、セラミックスへの結合量のペプチド濃度依存性を調べたとき、結合量が平衡濃度10μM以上では飽和傾向を示し、0.1μM以下では飽和量の1/2以下に低下する程度の結合能を持っていることを意味する。上記の配列番号11に示されるアミノ酸配列において数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるペプチドであって、かつセラミックスへの可逆的な結合・解離能を有するペプチドとして、例えば、Ala−Val−Gly−Leu−Ser−Pro−Asp−Gly−Ser−Arg−Gly−Val−Gly−Gly−Gly−Ser−Ala−Glu−Pro−His−Pro−Ser−Ala−Ala−His−Lys−Lys−Lys−Gln−Ser−Ser−Pro−Lys−Ser−Glu−Asn−Arg−Lys−Val−Pro−Phe−Tyr−Ser−His−Ser−Gly−Gly−Ser−Gly−Thr−Ser−Arg−Thr−Pro−Ile−Leu−Gly(配列番号1)や、Ser−Ala−Ala−His−Lys−Lys−Lys−Gln−Ser−Ser−Pro−Lys−Ser−Glu−Asn−Arg(配列番号14)や、Ala−Gln−Pro−His−Pro−Ser−Ala−Ala−His−Lys−Lys−Lys−Gln−Ser−Ser−Pro(配列番号16)等を具体的に挙げることができる。上記セラミックスとしては、特に制限されるものではないが、好ましくは金属酸化物を主成分とするセラミックス、より好ましくは遷移金属酸化物を主成分とするセラミックスであり、なかでも、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物を主成分とするセラミックス等であることが特に好ましい。より具体的には、上記セラミックスとして、チタン酸化物系(チタン酸バリウム(BaTiO3)、チタン酸ストロンチウム(SrTiO3)、酸化チタン(TiO2)など)、ジルコニウム酸化物系(酸化ジルコニウム(ZrO2)など)、スズ酸化物系(スズ酸バリウム(BaSnO3)など)等を好適に例示することができる。
【0015】
また、本発明のペプチドのセラミックス表面への可逆的な結合・解離は、金属イオンにより制御することができる。上記金属イオンとしては、本発明のペプチドのセラミックス表面への結合・解離を制御するものであれば特に制限されるものではないが、Ba2+、Ni2+、Zn2+、Ca2+等を例として挙げることができる。本発明のペプチドのセラミックス表面への可逆的な結合・解離は、水溶液中に含まれる上記金属イオンの濃度により制御することが可能である。すなわち、セラミックスと本発明のペプチドとを解離用金属イオン非存在下又は結合用金属イオンの存在下に(金属イオンを含む溶液により)結合させ、次に、セラミックスに結合した本発明のペプチドを、解離用金属イオンの存在下に解離させることができる。また、上記解離用金属イオンを含む溶液の金属イオン濃度を低下させることにより、解離しているセラミックスと本発明のペプチドとを再結合させることもできる。上記解離用金属イオンや結合用金属イオンは、本発明のペプチドの種類やセラミックスの種類により異なる。
【0016】
アミノ配列が異なる本発明のペプチドは、それぞれ異なる種類のセラミックスに対して、可逆的な結合・解離能を有する。例えば、本発明のペプチドBTBP−1は、チタン酸バリウム(BaTiO3)、チタン酸ストロンチウム(SrTiO3)、酸化チタン(TiO2)及び酸化ジルコニウム(ZrO2)に対して優れた結合・解離能を有するが、本発明のペプチド(KA)6は、チタン酸バリウム(BaTiO3)、チタン酸ストロンチウム(SrTiO3)及びスズ酸バリウム(BaSnO3)に対して優れた結合・解離能を有する。また、本発明のペプチドとセラミックとの可逆的な結合・解離は、異なる種類の金属イオンにより制御される。例えば、BTBP−1とチタン酸バリウム(BaTiO3)はMg2+やNi2+やZn2+の存在下に結合する(結合を阻害しない)が、チタン酸バリウムに結合したBTBP−1はBa2+の存在下に解離する。一方、(KA)6とチタン酸バリウム(BaTiO3)はMg2+の存在下に結合する(結合を阻害しない)が、チタン酸バリウムに結合した(KA)6はBa2+、Ni2+、Zn2+のいずれによっても解離する。以上のように、本発明のペプチドをセラミックス表面への可逆的な結合・解離能を有するペプチドとして使用する方法も本発明の一形態として包含される。
【0017】
上記本発明ペプチドは、そのアミノ酸配列に従って、一般的な化学合成法により製造することができる。化学合成法には、通常の液相法及び固相法によるペプチド合成法が包含され、より詳しくは、アミノ酸配列情報に基づいて、各アミノ酸を1個ずつ逐次結合させ鎖を延長させていくステップワイズエロゲーション法と、アミノ酸数個からなるフラグメントを予め合成し、次いで各フラグメントをカップリング反応させるフラグメント・コンデンセーション法とが包含される。本発明のペプチドを合成するには、そのいずれの方法をも採用することができる。上記ペプチド合成に採用される縮合法も、公知の各種方法に従うことができる。その具体例としては、例えばアジド法、混合酸無水物法、DCC法、活性エステル法、酸化還元法、DPPA(ジフェニルホスホリルアジド)法、DCC+添加物(1−ヒドロキシベンゾトリアゾール、N−ヒドロキシサクシンアミド、N−ヒドロキシ−5−ノルボルネン−2,3−ジカルボキシイミド等)、ウッドワード法等を例示できる。これら各方法に利用できる溶媒もこの種ペプチド縮合反応に使用されることがよく知られている一般的なものから適宜選択することができ、例えばジメチルホルムアミド(DMF)、ジメチルスルホキシド(DMSO)、ヘキサホスホロアミド、ジオキサン、テトラヒドロフラン(THF)、酢酸エチル等及びこれらの混合溶媒等を挙げることができる。
【0018】
さらに、上記本発明のペプチドは、本発明のセラミックスへの可逆的な結合・解離能を有するペプチドをコードするDNAの塩基配列情報により、遺伝子工学的手法を用いて常法により調製することもできる。このようにして得られる本発明のセラミックスに結合能を有するペプチド、通常の方法に従って、例えばイオン交換樹脂、分配クロマトグラフィー、ゲルクロマトグラフィー、アフィニティークロマトグラフィー、高速液体クロマトグラフィー(HPLC)、向流分配法等のペプチド化学の分野で汎用されている方法に従って、適宜その精製を行うことができる。
【0019】
上記本発明のペプチドは、検出可能なマーカーで標識されていてもよく、上記検出可能なマーカーとしては、従来知られているペプチド標識用のマーカーであれば特に制限されるものではなく、例えば、3H、14C、125I等の放射性同位体や、ダンシルクロライド、テトラメチルローダミンイソチオシアネート等の蛍光物質や、ビオチン、ジゴキシゲニンのような生物学的に関連する結合構造や、生物発光化合物や、化学発光化合物や、金属キレート等を具体的に挙げることができる。なかでも、本発明のペプチドBTBP−1、(KA)6をそれぞれビオチンで標識した、ビオチン標識BTBP−1やビオチン標識(KA)6は、後述の実施例でも示されているように、セラミックスへの可逆的な結合・解離能を有しており、特に好例として挙げることができる。また、ビオチン標識BTBP−1やビオチン標識(KA)6は、ストレプトアビジン修飾抗体と併用することにより、標的となる所定の抗原タンパク質(目的タンパク質)を着脱自在に操作しうることから、例えば、抗原タンパク質の分離・精製に有利に用いることができる。
【0020】
また、本発明のペプチドは、機能性ペプチド又は機能性タンパク質に結合した融合ペプチドであってもよい。本発明の融合ペプチドとしては、本発明のペプチドと、機能性ペプチド又は機能性タンパク質とが結合しているものであればどのようなものでもよく、上記機能性タンパク質としては、例えば、抗体、抗原、プロテインA(特にプロテインAのZZドメイン)等の免疫関連タンパク質や、アルカリフォスファターゼ、HRP等の酵素、抗体のFc領域、GFP等の蛍光物質などのマーカータンパク質や、BMP、レクチン、アビジン等の結合能を有する結合性タンパク質を具体的に挙げることができ、また、上記機能性ペプチドとしては、例えば、HA、FLAG、Myc等のエピトープタグや、GST、マルトース結合タンパク質(MBP)、ビオチン化ペプチド 、オリゴヒスチジン等の親和性タグなどを具体的に例示することができる。なかでも、本発明のセラミックスへの可逆的な結合・解離能を有するペプチドとプロテインAのZZドメインとを結合させた融合ペプチドは、セラミックスへの優れた結合能を有するとともに、抗体との結合能も有していることから、生物学的解析ツールとして広く応用が可能であり、さらに、金属イオンによりセラミックス表面から解離させることが可能であるため、前記抗体が認識する標的抗原タンパク質等の精製にも非常に有用である。また、本発明のセラミックスへの可逆的な結合・解離能を有するペプチドとBMPとを結合させた融合ペプチドは、この融合ペプチドを介して、BMPと結合するBMPレセプターを表面に発現する間葉系細胞や繊維芽細胞等をセラミックス材料表面に集積するとともに骨芽細胞への分化が促進し、その結果、セラミックス材料表面への骨の形成が促進され、セラミックスと骨を良好に定着させる上で有用である。
【0021】
本発明のセラミックスへの可逆的な結合・解離能を有するファージとしては、本発明のセラミックスに結合能を有するペプチドをその粒子表面上に提示するファージであればどのようなものでもよく、かかるセラミックスに結合能を有するファージは、ファージライブラリスクリーニングの過程で、セラミックスに強く結合したペプチド提示ファージを、その他のファージ集団から分離することにより得られる他、本発明のセラミックスに結合有するペプチドをコードするDNAを、常法によりファージミドベクターに組み込んで大腸菌等の宿主細胞を形質転換し、ヘルパーファージを感染させることで得ることもできる。
【0022】
本発明のペプチドを認識する抗体としては、モノクローナル抗体、ポリクローナル抗体、キメラ抗体、一本鎖抗体、ヒト化抗体等の免疫特異的な抗体を具体的に挙げることができ、これらは上記本発明のペプチドを抗原として用いて常法により作製することができるが、なかでもモノクローナル抗体がその特異性の点でより好ましい。かかる本発明のペプチドに対する抗体は、慣用のプロトコールを用いて、動物(好ましくはヒト以外)に本発明のペプチド又はその断片を免疫することにより産生され、例えばモノクローナル抗体の調製には、連続細胞系の培養物により産生される抗体をもたらす、ハイブリドーマ法(Nature 256, 495-497, 1975)、トリオーマ法、ヒトB細胞ハイブリドーマ法(Immunology Today 4, 72, 1983)及びEBV−ハイブリドーマ法(MONOCLONAL ANTIBODIES AND CANCER THERAPY, pp.77-96, Alan R.Liss, Inc., 1985)など任意の方法を用いることができる。以下に、本発明のペプチドのひとつであるBTBP−1に対して特異的に結合するマウスのモノクローナル抗体、すなわち抗BTBP−1モノクローナル抗体の作製方法を説明する。
【0023】
上記抗BTBP−1モノクローナル抗体は、抗BTBP−1モノクローナル抗体産生ハイブリドーマをインビボ又はインビトロで常法により培養することにより生産することができる。例えば、インビボ系においては、齧歯動物、好ましくはマウス又はラットの腹腔内で培養することにより、またインビトロ系においては、動物細胞培養用培地で培養することにより得ることができる。インビトロ系でハイブリドーマを培養するための培地としては、ストレプトマイシンやペニシリン等の抗生物質を含むRPMI1640又はMEM等の細胞培養培地を例示することができる。抗BTBP−1モノクローナル抗体産生ハイブリドーマは、例えば、BTBP−1を用いてBALB/cマウスを免疫し、免疫されたマウスの脾臓細胞とマウスNS−1細胞(ATCC TIB−18)とを、常法により細胞融合させ、免疫蛍光染色パターンによりスクリーニングすることにより、抗BTBP−1モノクローナル抗体産生ハイブリドーマを作出することができる。また、かかるモノクローナル抗体の分離・精製方法としては、タンパク質の精製に一般的に用いられる方法であればどのような方法でもよく、アフィニティークロマトグラフィー等の液体クロマトグラフィーを具体的に例示することができる。
【0024】
また、上記BTBP−1に対する一本鎖抗体をつくるためには、一本鎖抗体の調製法(米国特許第4,946,778号)を適用することができる。また、ヒト化抗体を発現させるために、トランスジェニックマウス又は他の哺乳動物等を利用したり、上記抗体を用いて、そのBTBP−1を発現するクローンを単離・同定したり、アフィニティークロマトグラフィーでそのポリペプチドを精製することもできる。さらに、上記抗BTBP−1モノクローナル抗体等の抗体に、例えば、FITC(フルオレセインイソシアネート)又はテトラメチルローダミンイソシアネート等の蛍光物質や、125I、32P、14C、35S又は3H等のラジオアイソトープや、アルカリホスファターゼ、ペルオキシダーゼ、β−ガラクトシダーゼ又はフィコエリトリン等の酵素で標識したものや、グリーン蛍光タンパク質(GFP)等の蛍光発光タンパク質などを融合させた融合タンパク質を用いることによって、上記BTBP−1を検出・定量等を行うことができる。上記抗体を用いた免疫学的測定方法としては、RIA法、ELISA法、蛍光抗体法、プラーク法、スポット法、血球凝集反応法、オクタロニー法等の方法を挙げることができる。
【0025】
本発明の組換えベクターとしては、前記本発明のDNAを含み、かつセラミックスへの可逆的な結合・解離能を有するペプチドを発現することができる組換えベクターであれば特に制限されず、本発明の組換えベクターは、本発明のDNAを発現ベクター、好ましくは発現プラスミドベクターに適切に挿入することにより構築することができる。かかる発現ベクターとしては、宿主細胞において自立複製可能であるものや、あるいは宿主細胞の染色体中へ組込み可能であるものが好ましく、また、本発明のDNAが転写できる位置にプロモーター、エンハンサー、ターミネーター等の制御配列を含有しているものを好適に使用することができる。
【0026】
上記発現ベクターとして、例えば、pCMV6−XL3(OriGene Technologies Inc.社製)、EGFP-C1(Clontech社製)、pGBT−9(Clontech社製)、pcDNAI(フナコシ社製)、pcDM8(フナコシ社製)、pAGE107(Cytotechnology, 3,133, 1990)、pCDM8(Nature, 329, 840, 1987)、pcDNAI/AmP(Invitrogen社製)、pREP4(Invitrogen社製)、pAGE103(J.Blochem., 101, 1307,1987)、pAGE210等を例示することができる。また、プロモーターとしては、例えば、サイトメガロウイルス(ヒトCMV)のIE(immediate early)遺伝子のプロモーター、SV40の初期プロモーター、レトロウイルスのプロモーター、メタロチオネインプロモーター、ヒートショックプロモーター、SRαプロモーター等を挙げることができる。さらに、プロモーターの下流に蛍光蛋白質をコードする遺伝子等のレポーター遺伝子を融合することができる。蛍光蛋白質としては、緑色蛍光蛋白質(Green Fluorescence Protein(GFP))、赤色蛍光蛋白質(Cyan Fluorescence Protein(CFP))、青色蛍光蛋白質(Blue Fluorescence Protein(BFP))、黄色蛍光蛋白質(Yellow Fluorescence Protein(YFP))、ルシフェラーゼ(luciferase)を例示することができる。
【0027】
また、本発明の形質転換体としては、上記本発明の組換えベクターが宿主細胞に導入され、かつ本発明のセラミックスへの可逆的な結合・解離能を有するペプチドを発現する形質転換体であれば特に制限されるものではなく、形質転換酵母、形質転換植物(細胞、組織、個体)、形質転換細菌、形質転換動物(細胞、組織、個体)等を挙げることができるが、形質転換動物細胞が好ましい。
【0028】
本発明のセラミックス表面とタンパク質との結合・解離を制御する方法としては、セラミックス表面への可逆的な結合・解離能を有するペプチド(以下「本結合・解離ペプチド」ということがある)、又は該ペプチドと機能性タンパク質との融合ペプチドを介しての、セラミックス表面と目的とするタンパク質(又は目的とする細胞)との結合又は解離を、所定の金属イオンの濃度を調整することによって制御する方法[制御方法I]や、セラミックス表面と、本結合・解離ペプチドと目的とするタンパク質との融合ペプチドとの結合又は解離を、所定の金属イオンの濃度を調整することによって制御する方法[制御方法II]であれば特に制限されず、かかる本発明の制御方法により、セラミックス表面に所定のタンパク質(目的タンパク質)又は所定の細胞(目的細胞)を脱着、すなわち修飾することができる。上記本結合・解離ペプチドとしては、前記本発明のペプチドの他、BTBP−1と同様にファージライブラリを用いたスクリーニングにより、また(KA)6と同様に塩基性アミノ酸に富むペプチドを用いたスクリーニングにより見出すことができる。また、機能性タンパク質については前述したタンパク質を用いることができ、この機能性タンパク質が目的タンパク質になることもある。例えば、本結合・解離ペプチド−プロテインAのZZドメインとの融合ペプチドのとき、目的タンパク質はこの融合ペプチドを介してセラミックス表面に結合・解離する抗体ということになり、同様に、本結合・解離ペプチド−抗体との融合ペプチドのとき、目的タンパク質はこの融合ペプチドを介してセラミックス表面に結合・解離する抗原タンパク質ということになる。他方、目的とするタンパク質が抗体のとき、本結合・解離ペプチド−抗体との融合ペプチドにおける構成成分である抗体が目的タンパク質となる。
【0029】
本発明の制御方法Iとして、本結合・解離ペプチド又は融合ペプチドを含む溶液と、セラミックスとを解離用金属イオン非存在下又は結合用金属イオンの存在下に(あるいは、結合用金属イオン濃度を高くして)反応させることにより、セラミックス表面に本結合・解離ペプチド又は融合ペプチドを結合させ、次いで本結合・解離ペプチド又は融合ペプチドと目的タンパク質とを結合させる方法や、目的タンパク質が結合した本結合・解離ペプチド又は融合ペプチドが結合したセラミックスから、解離用金属イオンの存在下(あるいは、解離用金属イオン濃度を高くして)又は結合用金属イオンの非存在下に、目的タンパク質が結合した本結合・解離ペプチド又は融合ペプチドを解離させる方法を例示することができ、具体的には、本結合・解離ペプチドとしてビオチン標識ペプチドを使用するとき、目的タンパク質としてストレプトアビジン結合目的タンパク質を用いる方法や、融合ペプチドとしてプロテインAのZZドメインとの融合ペプチドを使用するとき、目的タンパク質として抗体を用いる方法や、融合ペプチドとしてBMP融合ペプチドを使用するとき、目的とする細胞をBMPレセプター発現間葉系細胞とする方法を挙げることができる。また、本発明の制御方法IIとして、目的タンパク質を構成成分とする融合ペプチドを含む溶液と、セラミックスとを解離用金属イオン非存在下又は結合用金属イオンの存在下(あるいは、結合用金属イオン濃度を高くして)に反応させることにより、セラミックス表面に融合ペプチドを結合させる方法や、融合ペプチドが結合したセラミックスから、解離用金属イオンの存在下(あるいは、解離用金属イオン濃度を高くして)又は結合用金属イオンの非存在下に、融合ペプチドを解離させる方法を例示することができる。本発明の制御方法や脱着(修飾)方法により、本発明のペプチド又は融合ペプチドが表面に結合したセラミックスや、目的とするタンパク質や細胞が表面に結合したセラミックス、すなわち本発明の表面修飾セラミックスを得ることができる。
【0030】
上記セラミックスとしては、特に制限されるものではないが、好ましくは金属酸化物を主成分とするセラミックス、より好ましくは遷移金属酸化物を主成分とするセラミックスであり、なかでも、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物を主成分とするセラミックス等であることが特に好ましい。より具体的には、上記セラミックスとして、チタン酸化物系(チタン酸バリウム(BaTiO3)、チタン酸ストロンチウム(SrTiO3)、酸化チタン(TiO2)など)、ジルコニウム酸化物系(酸化ジルコニウム(ZrO2)など)、スズ酸化物系(スズ酸バリウム(BaSnO3)など)等を好適に例示することができる。また、上記溶液に含まれる本発明のペプチド又は融合ペプチドの濃度としては、好ましくは0.1μM以上、より好ましくは0.5μM以上、さらに好ましくは1μM以上である。発明の修飾方法により、セラミックス表面に結合した本発明のペプチド又は融合ペプチドは、金属イオンにより解離させることができ、上記金属イオンとしては、特に制限されないが、Ba2+、Ni2+、Zn2+、Ca2+等を好例として挙げることができる。上記本発明の修飾方法により表面修飾されたセラミックスは、例えば、歯科インプラント材料などの移植用医療デバイス、分子認識を行うナノデバイスやバイオセンサ、タンパク質精製や細胞単離用の担体などに利用できる他、セラミックスの機械的な特性、電子材料としての機能(圧電性、強誘電性など)、ナノメートルオーダーの微粉末が安価に大量に合成できる等の製造的な利点を活かして、種々の用途に用いることができる。
【0031】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
【実施例1】
【0032】
[ファージディスプレイ法によるチタン酸バリウム結合ペプチドの取得]
1.使用したファージライブラリ
スクリーニングには、Ph.D-12 peptide phage display kit(New England Biolabs社製;以下D−12という)及びPh.D-C7C peptide phage display kit (BioLabs社製;以下D−C7Cという) を用いた。また、同じ製品でも製造ロットによって違う配列の集団となっている可能性があるため、D−12については異なる3つのロット(lot3、lot5、lot6)を実験に用いた。D−12は、12残基の直線状ランダムペプチド配列を提示するライブラリであり、D−C7Cは2つのシステイン間に7つのランダム配列が挿入されているライブラリである。
【0033】
2.ファージライブラリを用いたスクリーニング
スクリーニング実験や、ファージの力価(濃度)測定等は、メーカーが提示しているプロコトルに従って行った。以下にその概略を示す。
ファージを構成するタンパク質のうち、提示ペプチド以外のタンパク質が非特異的にチタン酸バリウム(BT)に結合することを防ぐため、あらかじめBT表面にウシ血清アルブミン(BSA)を吸着させ、ブロッキングを行った。2mgのBT粉末を、2%のBSAを含むTBS(50mMトリスヒドロキシメチルアミノメタン(トリス)、150mM塩化ナトリウムを含む水溶液、pH7.5)10mLに懸濁した。分散状態にある懸濁液1mLを分取し、遠心分離機によりBT粉末を沈殿させた後、上清を捨て、再び2%BSAを含むTBS 1mLで懸濁し30分間攪拌した。攪拌後、遠心分離でBT粉末を沈殿させて上清を除去し、界面活性剤tween20を0.1%含むTBS(TBST)1mLを加えて懸濁する洗浄操作を1回行った。
【0034】
ブロッキング済みのBT粉末懸濁液1mLに、ペプチドファージライブラリ10μLを投入し2時間攪拌した。攪拌後、TBSTでの洗浄操作を3回行った。遠心分離でBT粉末を沈殿させた後、今度は0.2Mグリシン塩酸水溶液(pH2.2)1mLを加え、10分間回転攪拌し、BTに結合しているファージを溶出させた。溶出液を遠心してBT粉末を沈殿させた後、上清を回収し、1Mのトリス水溶液(pH9.1)150μLで中和した。
得られたファージは大腸菌に感染させ増幅させた。ファージの溶出液を対数増殖中のER2738株を含む培地20mLに添加し、37℃で4.5時間振盪培養した。その後、培養液を遠心分離し大腸菌を沈殿させ、ファージを含む上清を回収した。増幅したファージのうち約1012pfu分を、再び、ブロッキング済みのBT粉末に投与し、2時間のインキュベートの後、TBSTで洗浄、グリシン塩酸(pH2.2)で溶出、中和するという一連の実験(パニング)を6回繰り返した。
【0035】
3.BT結合ペプチドの取得
D−12ファージライブラリのlot5を用いた実験において、4ラウンド目のファージ溶出液に含まれるファージをクローン化し、ランダムにピックアップした28種類についてDNA解析を行った。DNA解析は、ベックマンコールター社製のシーケンサーCEQ2000と、試薬DTCS Quick start kitを使用した。実験方法はメーカー提示のプロコトルに従った。この解析結果によると、28クローンのうち18が同じ配列を示しており、このクローンをφ403Nと命名した。そのほかに、2個以上同じ配列を示したクローンが2種類あり、それぞれφ404N、φ410Nと命名した。これらのクローン化ファージのBTへの結合力を調べた結果を図1に示す。結合率とは、溶出して回収されたファージ量を、投入したファージ量で除したものであり、φM13KEとは、ペプチドを提示していない(野生型の)ファージである。φ403Nは他より際立って高いBTへの結合率を示すことが明らかとなった。
一方、他のライブラリ(D−12のlot3及びlot6、D−C7C)を実験に用いたスクリーニングでは、BTBP−1と同一の配列が取得できた場合もあれば、異なる配列が取得された場合もあった。これら他のライブラリーによるスクリーニングの結果の詳細に関しては、参考例として後述する。
【実施例2】
【0036】
[BT結合クローンφ403Nのペプチド配列解析]
1.φ403NのDNA及びアミノ酸配列の決定
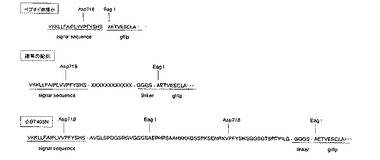
実施例1でD−12ライブラリより取得されたファージクローンφ403N、φ404N及びφ410Nについてファージの持つDNAを解析した。それぞれのその提示ペプチドのアミノ酸配列は、φ403Nは配列番号1に示される57残基のアミノ酸配列、φ404Nは配列番号2に示される11残基のアミノ酸配列、φ410Nは配列番号3に示される12残基の配列であることが明らかとなった。D−12ライブラリは12残基のペプチドを提示するよう設計されたものであるため、このφ403Nは、何らかの理由で長い配列が挿入されてできたものである。図2にこの理由についての仮説を示す。ファージライブラリは、ファージの先端のタンパク質g3p(シグナル配列の部分は、ファージが形成される際に切り離される)をコードするDNAに、ランダムなペプチドをコードするランダムなDNA配列を挿入することで形成される。図2の通常の配列では、Xで表されるランダムな12残基のペプチドがg3pタンパク質のN末端側にGGGSという4残基のアミノ酸のリンカーを介して接続されていることを示している。しかし、今回取得されてきたφ403Nのアミノ酸配列は、図2に示すように、ランダムなペプチド配列をコードするDNA断片が3つタンデムに連なって挿入されている。これはファージライブラリの製造上、ある一定の確率で起こりえる現象であると思われる。このため、DNA配列上は、シグナル配列とリンカーの間の57残基がg3pの末端に提示されていることになる。
【0037】
2.φ403N提示ペプチドのBT結合領域の決定
このDNA配列から予想される57残基のペプチドが、実際に発現しているかどうかを確認するため、ファージを構成するタンパク質をSDSアクリルアミドゲル電気泳動して、想定分子量付近のバンドを切り取り、アミノ酸配列分析した。この結果では、57残基のうちN末端のAVG〜16残基、および、AVG〜37残基が削られたタンパク質が検出された。しかし、これらのうち、どちらがBTに強く結合しているかは区別できないし、アミノ酸配列分析では検出できないほど微量のタンパク質が結合に寄与していることも否定できない。また、ペプチド配列が57残基と長いと応用上使用しにくいため、ペプチド配列のうちBTとの結合に寄与している部分の配列を同定して絞り込む必要がある。φ403Nのファージをベースに遺伝子を組み替え、図3に示すようなペプチド配列を持つファージ9種類を作製した。それぞれのファージクローンのアミノ酸配列は、以下の配列番号と対応する。
φ403N:配列番号1
φ1581:配列番号4
φ1583:配列番号5
φ1597:配列番号6
φ1599:配列番号7
φ1601:配列番号8
φ1603:配列番号9
φ1605:配列番号10
φ1611:配列番号11
φ1612:配列番号12
【0038】
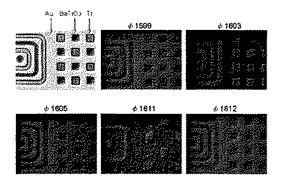
これらの遺伝子を大腸菌に導入してファージを形成させ、それぞれのBT結合能をしらべた。結合実験には、Si基板上に全面成膜されたBTスパッタ薄膜上にAuとTi薄膜がパターニングされているチップを用いた(図4、左上パネル)。φ1581、φ1583、φ1597、及びφ1601は、ファージが形成されないか、DNAに意図しない変異が入ったファージしか得られなかった。このため実験には、φ1599、φ1603、φ1605、φ1611、及びφ1612を用いた。実験は次の手順で行った。チップを2%BSAを含むTBSTでブロッキングし、約1010pfu/mLのファージを含むTBST中で30minインキュベートし、TBSTで洗浄した。さらに、anti-fd bacteriophage biotin conjugate(Sigma-Aldrich社製)を含むTBST中で15分間インキュベートした後、TBSTで洗浄した。続いて、ストレプトアビジンつき蛍光体(Qdot(登録商標)655 streptavidin conjugate、Invitrogen社製)を含むTBSTで15分間インキュベートした後、TBSTで洗浄した。このチップを水溶液に浸した状態で顕微鏡で蛍光観察した。その結果、図4に示すように、φ1611が22残基という短い配列でもBTへ特異的に結合すること分かった。このφ1611の提示ペプチドをBTBP−1(AEPHPSAAKKKQSSPKSENRK:配列番号11)と命名した。
【実施例3】
【0039】
[BTBP−1ペプチドのBT結合能の確認]
これまでの実験により、BTBP−1はファージに提示された状態で結合能力を持つことは確認されたので、次に、化学合成によって作製したBTBP−1単独でBT結合能力を持つかどうかを確認する実験を行った。完全長BTBP−1(配列番号11)の合成ペプチドを作製するとともに、BTBP−1のN末端側から18番目から22番目までのアミノ酸を削ったペプチドΔ(18−22)(配列番号13)、1番目から5番目のアミノ酸を削ったペプチドΔ(1−5)(配列番号14)、及び両末端の5残基のアミノ酸を削ったペプチドΔ(1−5、18−22) (配列番号15)の3つ変異ペプチドを作製した。実験には、実施例2(図4)と同様のチップを用い、N末端側がビオチン分子で修飾された合成ペプチドを使用した。実験手順としては、まずチップを2%BSAを含むTBSTでブロッキングし、40μMのビオチン化ペプチドを含むTBSTで15分間インキュベートし、TBSTで洗浄した後、ストレプトアビジンつき蛍光体を含むTBSTで15分間インキュベートし、TBSTで洗浄した。このチップを水溶液に浸した状態で顕微鏡で蛍光観察した。結果を図5に示す。この結果から、BTBP‐1単独の状態でもBTに結合すること、1−5番目を削った場合(Δ(1−5))でもBTに結合することが確認できた。なお、18−22番目を削った場合に大きく結合力が低下しているが、ここには塩基性アミノ酸(R、K)が2つ含まれている。このため、結合には塩基性のアミノ酸が重要であると推測できる。
【実施例4】
【0040】
[BTBP−1とプロテインAとの融合タンパク質の作製]
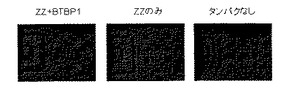
プロテインA(ProteinA)は抗体と結合する能力があるタンパク質である。このタンパクの抗体と結合する部位(Zドメイン)2つから構成されるタンパク質とBTBP‐1の融合タンパク質(ZZ+BTBP−1)を作製し、BTへの結合能力と抗体の捕捉能力を確認する実験を行った。実験手順は、実施例2と同じチップを、2%BSAを含むTBSTでブロッキングした後、融合タンパク質(ZZ+BTBP−1)を0.5μMを含むTBSTをインキュベートさせた。TBSTで洗浄した後、抗体(anti-fd bacteriophagebiotin conjugate)を含むTBSTでインキュベートし、さらにTBSTで洗浄した。最後に、ストレプトアビジンつき蛍光体を含むTBSTで15分間インキュベートし、TBSTで洗浄後、純水で洗浄し、チップを乾燥状態で観察した。結果を図6に示す。BTBP−1とZZの融合タンパク(ZZ+BTBP−1)が、BT表面に吸着し、さらに抗体を捕捉していることが確認された。一方、ZZのみ(BTBP−1を融合していないZZ)やいずれのタンパクを含まないTBST中でインキュベートした場合には、BTへの抗体の結合は融合タンパクに比べて小さかった。
【実施例5】
【0041】
[BTBP−1の配列特異性]
BTBP−1の配列の特異性を調査するため、いくつかの種類のペプチドを合成し、それらの結合能力を調査した。BTBP−1の2つのグルタミン酸(E)をグルタミン(Q)に置き換えたペプチドBTBP−1(E→Q)(配列番号16)、(DK)6(配列番号17)、(DA)6(配列番号18)、及び(KA)6(配列番号19)の4種類のペプチドを合成し、それぞれのN末端側をビオチン分子で修飾した。実施例2と同じチップを、2%BSAを含むTBSTでブロッキングし、1μMのビオチン化ペプチドを含むTBSTで15分間インキュベートした。TBSTで洗浄した後、ストレプトアビジンつき蛍光体を含むTBSTで15分間インキュベートし、さらにTBSTで洗浄した。最後に純水で洗浄し、チップを乾燥状態で観察した。結果を図7に示す。BTBP−1(E→Q)は、BTBP−1と同様に結合したことから、BTとの結合に2つの負電荷は重要でないことが分かった。また、リジン(K)とアラニン(A)が交互に6個ずつ並んだ配列(KA)6もBTに結合することが明らかとなった。このことから、塩基性アミノ酸が酸性アミノ酸よりも多く含まれることが結合に重要であることが示唆された。ただし、動的光散乱測定により(KA)6、(DA)6、(DK)6のペプチドの粒子径(分子サイズ)を計測してみたところ、100nm程度のクラスターとして存在していることが分かった(粒子系はバッファーに依存して変化する)。これにより、(KA)6については多価提示と同様な効果が発生し、結合能力が増強されている可能性もある。
【実施例6】
【0042】
[金属イオンによるBT結合ペプチドへの阻害作用]
BTBP−1のBT結合に及ぼす、水溶液中の金属イオンの影響を検討した。Mg2+、Ca2+、Ba2+、Ni2+、Zn2+の各イオンが1mMの濃度で含まれる溶液中でBTBP−1及び(KA)6ペプチドをBTと結合させた。用いたチップは実施例2と同じもので、ストレプトアビジンつきの蛍光体でラベルしたものを蛍光顕微鏡で観察した。その蛍光の強さを数値化して表した結果を図8に示す。Ba2+により、蛍光強度が落ちており、BTBP−1及び(KA)6ペプチドとBTとの結合を阻害していることが明らかとなった。また、BTBP−1と(KA)6では、Ni2+又はZn2+を投入したときの挙動が異なっており、ここにBTBP‐1の配列特異性が現れている。
【0043】
さらに、上記のような金属イオンによる結合阻害作用により、既に結合しているペプチドを、金属イオンを含む溶液によって解離できるかを調査した。図9の「解離処理あり」は、BTBP−1を結合させた後、1mMのBaCl2を含むTBSTに15分間浸漬した後、ストレプトアビジンつき蛍光体をつけたものである。「解離処理なし」は、ペプチドを結合させた後、Ba2+を含まない液に15分間浸した後、ストレプトアビジンつき蛍光体をつけたものである。この結果から、ペプチドはBa2+を含む液に浸すことによって解離を促進できることが分かった。なお、チップは実施例2のものとは異なり、赤く光っている部分がBTの領域で、その他の部分はTiである。また、図10は、同じチップにペプチドを結合させ、先にストレプトアビジン付き蛍光体を結合させて一旦蛍光顕微鏡で観察した後、再び10mMのBaCl2を含むTBST中に浸して解離するかどうかを調べた結果である。ペプチドにストレプトアビジンが結合することによって解離のスピードは低下しているが、Ba2+による解離の促進効果は明らかに確認できる。
【実施例7】
【0044】
[ペプチド濃度の結合量に及ぼす影響]
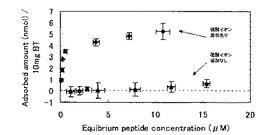
BTBP‐1とBTとの結合におけるペプチド濃度の影響を調査した。BTの粉末(粒径1μm弱)10mgを用意し、1〜16μMのBTBP−1を含む1mLのHBST溶液(50mM HEPES、150mM NaCl、0.1% tween20)と混合し、2時間回転攪拌した。その後、遠心分離で粉末を沈殿させ、上清のペプチド濃度を測定した。濃度の測定は、アミノ基と反応して蛍光を発する試薬(CBQCA、invitrogen社製)をペプチドと反応させ、蛍光分光測定計(FP-6500、日本分光社製)で測定した。BTと混合していない8μMのBTBP−1溶液も合わせて測定した。また、BTは水溶液中に置くと、最表面に存在するBa2+が溶出することが知られている。これまでのようなチップを用いた実験では表面積が小さいために溶出するBa2+の量も少なく問題はなかったが、今回の実験では溶液中のBa2+イオンの量は無視できないレベルになる。そこで、Ba2+を除去するためSO42−を加えてBaSO4として沈殿させることにした。HBSTに対して、2mMのNa2SO4を加えた実験を合わせて行った。なお、BTには若干の光触媒作用がある可能性がり、ペプチドや界面活性剤が分解される恐れがあるので、2時間の回転攪拌は遮光した状態で行っている。結果を図11に示す。まず、硫酸イオンを添加しないと、Ba2+の影響で結合がかなり阻害されていることが分かる。硫酸イオンを添加した状態では、0.1μM程度の低い値から立ち上がりが見られ、3μMからは濃度依存的な結合量の増加が確認された。
【実施例8】
【0045】
[ペプチドの多価提示]
1分子のタンパク質に複数のペプチドを提示させることで、タンパク質の無機材料(セラミックス)に対する結合力を向上させることができるかどうかについて検討した。10nMのストレプトアビジン付きの蛍光体(Qdot、Invitorogen社製)と200nMのビオチン標識BTBP−1を予め混合し、先にビオチン-アビジン複合体の形成させた。ストレプトアビジン付きの蛍光体は1粒子に対して、4量体を形成するストレプトアビジン分子が複数結合している。このため、1粒子のQdotに対して十数個〜数十個程度のペプチドが提示されていると考えられる。この複合体をチップに結合させた結果を図12に示す。図12(a)に示すように、Qdotの濃度が10nMと薄くてもチップ上での結合が確認することができた。一方(b)は、ペプチド200nMをチップに対して予め結合させておき、その後、ストレプトアビジン付きの蛍光体でラベルしたものである。この場合、(a)に比べるとペプチド濃度が同じにも関わらず結合量は(a)よりも極端に小さかった。すなわち、タンパク質の固相化にあたり、先にペプチドを無機材料に対して結合させた後、ビオチンアビジン等の結合を利用して固相化する場合は、ペプチドの濃度を0.1〜1μM程度以上にする必要があるが、タンパク質に先にペプチドを複数結合しておき(手法は化学結合でもビオチン-アビジンでもよい)それを固相化する場合には、タンパク質濃度は10nM程度でも結合させることが可能であることが分かった。どちらの方法を適用するかは、アプリケーションによって異なると考えられる。
【実施例9】
【0046】
[結合の阻害に必要な金属イオンの濃度]
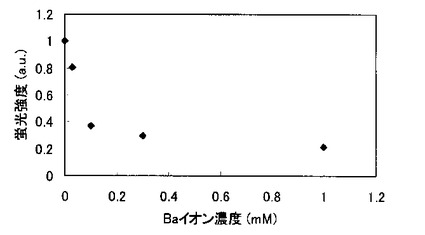
結合の阻害に必要な金属イオンの濃度を検討した。1μMのBTBP−1溶液に、BaCl2を0、0.03、0.1、0.3、1.0 mMとなるように加え、チップを浸し、ストレプトアビジン付き蛍光体でラベルし、蛍光顕微鏡で観察した。蛍光強度とバリウム濃度の関係を図13に示す。結合の阻害には30μM程度の濃度が必要であることが分かった。ただし、ペプチドの濃度、結合させたい無機材料の表面積によっては、この値は変化する可能性がある。
【実施例10】
【0047】
[他のセラミックス表面への結合]
これまでの実験結果により、BTBP−1と(KA)6は、Au等と比較してBTに優先的に結合することが示された。次に、BT以外のセラミックス材料(SrTiO3、TiO2、BaSnO3、ZrO2)に対する結合について調査した。各種セラミックスの粉末(粒径1μm弱)10mgを用意し、8μMのBTBP‐1又は(KA)6を含むHBST溶液と混合し、2時間回転攪拌した。その後、遠心分離で粉末を沈殿させ、上清のペプチド濃度を測定した。濃度の測定は、ペプチドをアミノ基と反応して蛍光を発する試薬(CBQCA、Invitrogen社製)と反応させ、蛍光分光測定計(FP-6500、日本分光社製)で測定した。セラミックスと混合していない8μM濃度のペプチド溶液も合わせて測定した。
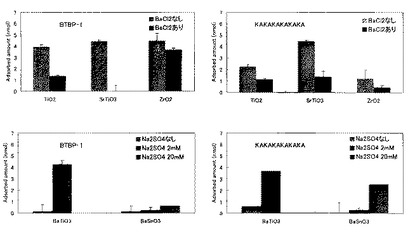
また、同時に、Ba2+の影響についても検討するため、SrTiO3(比表面積4.34m2/g)、TiO2(比表面積2.75m2/g)、ZrO2(比表面積21.2m2/g)に対しては、ペプチド溶液に1mMのBaCl2を添加し、その効果を見た。BaSnO3(比表面積2.11m2/g)については自身からBa2+が溶出し、それだけで4.2mM程度の濃度に達していたため、Na2SO4を2mMおよび10mM添加し、BaSO4の形でBa2+を除去し、その影響を見た。BaTiO3(比表面積3.43m2/g)については、既出のデータ(吸着量-平衡濃度グラフ)の再掲であるが、溶出したBa2+の濃度は0.5mM程度であり、添加したNa2SO4は2mMである。これらの実験の結果を図14に示す。BTBP−1は、(KA)6と比較して、ZrO2への結合量が4倍、TiO2への結合量が2倍ほど高かったまた、各材料により程度の差はあるがBa2+イオンの結合阻害効果は認められた。
BTBP−1のBaSnO3への結合量はNa2SO4を添加することによるBa2+イオン除去によっても大きくは増加しなかった。(KA)6のBaSnO3への結合量はNa2SO4を添加することによって増加した。このように、金属イオンの添加効果は材料によって一様ではないので、金属イオンはペプチドに直接作用する可能性のほか、材料表面の方に結合することで、ペプチドの吸着挙動に影響を与えている可能性も示唆された。
【実施例11】
【0048】
[参考例1:他のライブラリからのスクリーニング結果]
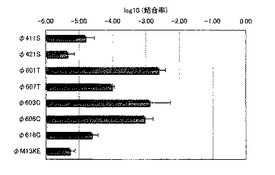
実施例1において、D−12ファージライブラリのlot5から、BT結合ファージクローンのスクリーニングに成功した。一方、その他のライブラリ(D−12のlot3及びlot6、D−C7C)からは、BTBP−1と同一の配列が取得できた場合もあれば、異なる配列が取得された場合もあった。以下にその詳細を示す。実施例1に記載の方法で、スクリーニングを行い、得られたファージをクローン化した。それぞれの配列を提示したファージの結合率を図15に示す。末尾のSはD−12ライブラリのlot6、TはD−12ライブラリのlot3、CはD−C7Cライブラリから得られたものであることを示す。Lot6では、φ403Nと同じ配列のファージも同時にスクリーニングされていたが(図15では示していない)、lot3ではφ403Nと同じものはスクリーニングされなかった。
【0049】
各アミノ酸配列を、図16及び配列番号20〜26に示す。φ411S、φ421Sは正常に12残基のペプチドを提示していたが、BTへの結合率は低かった。φ601Tはφ403Nと同様にDNA断片が3つ挿入された配列であり、塩基性のアミノ酸が比較的多いと言える。φ607Tは正常に12残基のペプチドとなっていたが、BTへの結合率は中程度であった。φ603Cは何らかの異常によって、2番目の位置に本来システインであるべきアミノ酸がセリンに変わっているが、短い配列でも比較的高い結合率を示している。φ606Cは、これもDNA断片が複数入っている配列であったが、BTへの結合力は高かった。φ616Cは、本来C7Cとなるべき配列から大きく異なっており、BTへの結合率も小さかった。
【0050】
φ603Cは短い配列でもBTへの高い結合力が観測されたので、φ603Cが提示していた配列の合成ペプチドを作製し、パターニングチップを用いてペプチド単体での結合特性を確認した。その結果、図17に示すように、ペプチドとチップのインキュベート時間が15分ではほとんど結合が見られず、10時間のインキュベートでようやく結合が確認できた。BTBP−1は15分でも十分な結合が確認できていたので、φ603Cが提示するペプチドはBTBP−1に比べると結合力が劣ることが判った。ただし、10残基という配列の短さの割には結合力が高いと言える。
【0051】
[参考例2:チタン酸バリウムの粗い粉末を用いたスクリーニング結果]
前述のように、φ403N(BTBP−1)のスクリーニングの際には、溶液1mL中に粒径約1μmのチタン酸バリウム粉末約0.2mgを分散させ用いた。これに対し、粒の粗いチタン酸バリウム粉末(チタン酸バリウムの焼結体を粗く砕いて75−300μmに粒径を揃えたもの)10mgを1mL中に分散させたものに対するパニング実験を行った。スクリーニングされたものをDNA解析すると、配列番号27〜31に示す5種類のアミノ配列が得られた。それぞれのファージクローンを作製し、結合率を調べた結果、φBT401とφBT403が高い結合率を示した(図18)。しかし、チップを用いた蛍光観察実験での結果は、図16の結果と異なっていた(図19、BaCl2なし)。φBT401は若干結合しているように見えるが、BaTiO3の領域と、Auの領域と同じように結合しており、φ403Nのような材料特異性がみられない。φBT403はペプチド非提示のφM13KEとほとんど結合量が変わらなかった。これには、スクリーニング時とチップの実験時で溶液中のバリウムイオンの濃度が異なることが影響していると考えられた。チップの場合、BaTiO3の表面積は粉末10mgに比べて5桁程度小さく、さらにチップを作製する工程で表面のバリウムはすでに溶出している可能性がある。そのため、粉末とは異なる結果になったと考えられる。そこで、バリウムイオンを添加した状態で、チップへの結合特性を調べた(図19、1mM BaCl2)。すると、BaTiO3部位への結合が回復することが確認された。興味深いことに、Ti金属部位への結合も確認され、BaTiO3への結合量よりもむしろ多かった。これは、チタン金属の表面が酸化しており、この酸化チタン表面に対してバリウムイオンが吸着し、そこを認識しているのだと思われる。よって、φBT401とφBT403の提示する配列は、バリウムなどの金属イオンが吸着したチタン酸化物表面を認識するペプチド配列である可能性があると考えられる。これらは、S、T、Y、Qなどのルイス塩基となるアミノ酸が多いのが特徴で、電荷をもつアミノ酸は比較的少ない。塩基性のアミノ酸は吸着したバリウムイオンと親和性が低く、酸性のアミノ酸は不可逆的な吸着となり得るので(K. Imamura et al., J Biosci Bioeng 1: 7-12 2007)ファージからのセレクションのときに溶出しなかった可能性がある。チタンに吸着するイオンとして、Bi、Cd、Co、Cr、Cu、Fe、Ge、In、Mn、Ni、Pb、Sb、Sn、Te、Tl、V、Zn(E. Vassileva et al., Analyst 121: 607-612, 1996)、Ba、Ca、Sr(MA. Malati and E. Smith, Power Technol 22: 279-282, 1979)が知られている。
【図面の簡単な説明】
【0052】
【図1】ファージクローンφ403N、φ404N及びφ410Nのチタン酸バリウム(BT)への結合力を調べた結果を示す図である。結合率とは、溶出して回収されたファージ量を、投入したファージ量で除したものであり、φM13KEとは、ペプチドを提示していない(野生型の)ファージである。
【図2】本発明のファージクローンφ403Nの提示ペプチドを示す図である。ランダムなペプチド配列をコードするDNA断片が3つタンデムに連なって挿入された配列となっている。
【図3】本発明のファージクローンφ403Nの提示ペプチド(配列番号1)と、9種類の変異ペプチド(配列番号4〜12)の配列を示す図である。
【図4】Si基板上に全面成膜されたBTスパッタ薄膜上にAuとTi薄膜がパターニングされているチップを用いて、φ1599、φ1603、φ1605、φ1611、及びφ1612の特異的結合を調べた結果を示す図である。
【図5】本発明のBTBP−1(配列番号11)とその3つの変異ペプチドΔ(18−22)(配列番号13)、Δ(1−5)(配列番号14)、Δ(1−5、18−22) (配列番号15)のBTへの特異的結合を調べた結果を示す図である。
【図6】本発明の融合ペプチドZZ+BTBP−1がBTに特異的に結合するとともに、抗体を補足することができることを示す図である。
【図7】BTBP−1の2つのグルタミン酸(E)をグルタミン(Q)に置き換えたペプチドBTBP−1(E→Q)(配列番号16)、(DK)6(配列番号17)、(DA)6(配列番号18)、及び(KA)6(配列番号19)の4種類のペプチドのBTへの特異的結合を示す図である。
【図8】BTBP−1及び(KA)6ペプチドのBTへの結合に及ぼす、金属イオン(Mg2+、Ca2+、Ba2+、Ni2+、Zn2+)の影響を示す図である。
【図9】既に結合しているペプチドが、金属イオンを含む溶液によって解離するかどうかを調べた結果を示す図である。「解離処理あり」は、ペプチドを結合させた後、1mMのBaCl2を含むTBSTに15分間浸漬した後に、ストレプトアビジンを反応させた結果である。また、「解離処理なし」は、ペプチドを結合させた後、Ba2+を含まない液に15分間浸した後にストレプトアビジンを反応させた結果である。
【図10】既に結合しているペプチドが、金属イオンを含む溶液によって解離するかどうかを調べた結果を示す図である。あらかじめチップに結合したペプチドにあらかじめストレプトアビジンを反応させた後、1mMのBaCl2を含むTBSTに一晩浸漬した後に結合を調べたものである。
【図11】硫酸イオンの存在下/非存在下での、BTBP−1とBTとの結合におけるペプチド濃度の影響を調査した結果である。
【図12】(a)ビオチン標識BTBP−1とQdotとを予め結合させた後、チップに吸着させた結果、及び(b)ビオチン標識BTBP−1を先にチップに吸着させた後に、Qdotを反応させた結果を示す図である。
【図13】1 μMのBTBP−1溶液に、BaCl2を0、0.03、0.1、0.3、1.0 mMとなるように加え、チップへの結合を調べた結果を示す図である。
【図14】様々なセラミックへのBTBP−1及び(KA)6の結合を調べた結果を示す図である。
【図15】ファージライブラリD−12のlot3及びlot6、ファージライブラリD−C7Cからのスクリーニングにより得られたファージ結合率を示す図である。末尾のSはD−12ライブラリのLot6、TはD−12ライブラリのLot3、CはD−C7Cライブラリから得られたものであることを示す。
【図16】図13に示されるファージクローンの提示アミノ酸配列(配列番号20〜26)を示す図である。
【図17】φ603Cの提示ペプチドを合成し、その結合特性を確認した結果を示す図である。
【図18】チタン酸バリウムの粗い粉末を用いたスクリーニングにより得られたファージクローンのBTへの結合を調べた結果を示す図である。
【図19】1mM BaCl2非存在下/存在下での、ファージクローンのBTへの結合を調べた結果を示す図である。
【技術分野】
【0001】
本発明は、セラミックス表面への可逆的な結合・解離能を有するペプチドや、かかるペプチドを用いたセラミックス表面への目的タンパク質の結合・解離を制御するセラミックス表面の修飾方法や、表面修飾されたセラミックス等に関する。
【背景技術】
【0002】
ファージとは大腸菌などバクテリアに感染するウイルスであり、自身のDNAがタンパク質の殻で包み込まれた構造をしている。この外皮タンパク質はファージのDNAを設計図として構築されるため、このファージDNAに任意のDNA配列を挿入することで、「さや」のタンパク質に任意のアミノ酸(ペプチド)を付加することができる。このような性質を利用して、多様なペプチドがファージの外皮タンパク質に付加されたファージ集団が、ファージライブラリとして市販されている。通常のファージライブラリの多様性は109種類にもなる。
【0003】
ファージディスプレイ法は、ファージライブラリを用いて生体分子に結合するペプチド配列を同定するための手法であったが、近年、金属等の無機物質を認識するペプチド配列を特定するためにも用いられるようになってきている。これまでに、チタン(特許文献1)、ナノ黒鉛構造体(特許文献2)、GaAs(非特許文献1)等に結合するペプチドがファージディスプレイ法により同定されている。
【0004】
さらに、非特許文献2は、ファージディスプレイ法を用いたスクリーニングにより、水溶性前駆体溶液からチタン酸バリウム(BaTiO3)の結晶形成を促進するペプチドを同定している。しかし、非特許文献2では、スクリーニングによりチタン酸バリウム結晶に結合するファージクローンを取得しているに過ぎず、これらのファージクローンの提示するペプチド自体が実際にチタン酸バリウム結晶に結合するかどうかについては明らかにしていない。
【0005】
セラミックスは、歯科インプラント材料をはじめとして様々な医療用デバイスとして応用の可能性が考えられている。このような医療用デバイスでは、表面を修飾することにより、生体適合性を高めることが重要である。また、セラミックス表面を機能性成分により修飾することにより、分子認識を行うナノデバイスの開発なども可能である。しかし、セラミックス表面に対する詳細な結合特性の解析がなされたペプチドは、未だ報告されていない。
【0006】
【特許文献1】国際公開第05/010031号パンフレット
【特許文献2】特開2004−121154号公報
【非特許文献1】Whaley SR et al., Nature 405: 665-668, 2000
【非特許文献2】Ahmad G et al., J Am Chem Soc 130: 4-5, 2008
【発明の開示】
【発明が解決しようとする課題】
【0007】
本発明の課題は、セラミックス表面への目的とするタンパク質や細胞の結合(複合)・解離を簡便に制御しうる方法や、当該制御方法に用いることができる、セラミックス表面に可逆的な結合・解離能を有する新規のペプチド及びその利用方法を提供することにある。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決するため鋭意研究し、チタン酸バリウム(以下「BaTiO3」又は「BT」ということもある)粒子に、多様なペプチド配列をファージ粒子上に提示するファージ集団を接触させ、BT粒子に結合したファージ粒子を回収し、得られたファージ粒子を大腸菌中で増殖させ、次いで、この増殖させたファージ粒子を、再度BT粒子に接触させるパニング操作を繰り返すことにより、まずBT粒子に特異的に結合するファージクローンを得ることに成功した。次に、このファージクローンの提示するペプチドについて解析し、このペプチドが他の数種類のセラミックス表面にも結合すること、また、その結合がバリウムイオン等の金属イオンにより解離することを見い出し、本発明を完成するに至った。
【0009】
すなわち本発明は、(1)配列番号11又は19に示されるアミノ酸配列からなるペプチドや、(2)セラミックス表面への可逆的な結合・解離能を有するペプチドであって、配列番号11又は19に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列や、(3)配列番号11に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列が、配列番号1、14又は16に示されるアミノ酸配列であることを特徴とする上記(2)記載のペプチドや、(4)セラミックス表面への可逆的な結合・解離が、金属イオンにより制御されることを特徴とする上記(2)記載のペプチドや、(5)金属イオンが、Ba2+、Ni2+、Zn2+及びCa2+から選択される少なくとも1以上の金属イオンであることを特徴とする上記(2)記載のペプチドに関する。
【0010】
また本発明は、(6)セラミックスが、金属酸化物を主成分とするセラミックスであることを特徴とする上記(2)記載のペプチドや、(7)金属酸化物が、遷移金属酸化物であることを特徴とする上記(6)記載のペプチドや、(8)遷移金属酸化物が、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物であることを特徴とする上記(7)記載のペプチドや、(9)検出可能なマーカーで標識された上記(1)〜(8)のいずれか記載のペプチドや、(10)上記(1)〜(8)のいずれか記載のペプチドと、機能性ペプチド又は機能性タンパク質とを結合させた融合ペプチドや、(11)上記(1)〜(8)のいずれか記載のペプチドを、その粒子表面上に提示することを特徴とする、セラミックス表面への可逆的な結合・解離能を有するファージや、(12)上記(1)〜(8)のいずれか記載のペプチドを認識する抗体や、(13)上記(1)〜(8)のいずれか記載のペプチドをコードするDNAや、(14)上記(13)記載のDNAを含み、かつセラミックス表面への可逆的な結合・解離能を有するペプチドを発現することができる組換えベクターや、(15)上記(14)記載の組換えベクターが導入された形質転換体に関する。
【0011】
さらに本発明は、(16)セラミックス表面への可逆的な結合・解離能を有するペプチド、又は該ペプチドと機能性タンパク質との融合ペプチドを介しての、セラミックス表面と目的とするタンパク質との結合又は解離を、所定の金属イオンの濃度を調整することによって行うことを特徴とするセラミックス表面とタンパク質との結合・解離を制御する方法や、(17)セラミックス表面と、セラミックス表面への可逆的な結合・解離能を有するペプチドと目的とするタンパク質との融合ペプチドとの結合又は解離を、所定の金属イオンの濃度を調整することによって行うことを特徴とするセラミックス表面と融合ペプチドとの結合・解離を制御する方法や、(18)セラミックス表面への可逆的な結合・解離能を有するペプチドとして、上記(1)〜(9)のいずれか記載のペプチドを用いることを特徴とする上記(15)又は(16)記載の方法や、(19)融合ペプチドとして、上記(10)記載の融合ペプチドを用いることを特徴とする上記(15)又は(16)記載の方法や、(20)金属イオンが、Ba2+、Ni2+、Zn2+、及びCa2+から選択される少なくとも1以上の金属イオンであることを特徴とする上記(16)〜(19)記載の方法に関する。
【0012】
また本発明は、(21)セラミックスが、金属酸化物を主成分とするセラミックスであることを特徴とする上記(16)〜(20)のいずれか記載の方法や、(22)金属酸化物が、遷移金属酸化物であることを特徴とする上記(21)記載の方法や、(23)遷移金属酸化物が、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物であることを特徴とする上記(22)記載の方法や、(24)上記(16)〜(23)のいずれか記載の方法により、セラミックス表面に所定のタンパク質又は所定の細胞を脱着する方法や、(25)上記(1)〜(9)のいずれか記載のペプチド、又は上記(10)記載の融合ペプチドで表面修飾されたセラミックスや、(26)上記(1)〜(9)のいずれか記載のペプチド、又は上記(10)記載の融合ペプチドを介して、表面に所定のタンパク質又は所定の細胞が結合したセラミックスに関する。
【発明の効果】
【0013】
本発明のセラミック表面に特異的に結合するペプチドは、簡単に化学合成することができ、また、遺伝子工学的手法を用いることで他のタンパク質等と融合させた融合ペプチドを作製することができ、セラミックス表面の修飾に用いることができる。また、本発明のペプチドや融合ペプチドと、セラミックス表面との結合・解離は、金属イオンによって制御することが可能であるため、タンパク質精製など幅広い分野での応用が可能である。
【発明を実施するための最良の形態】
【0014】
本発明のペプチドとしては、配列番号11に示されるアミノ酸配列Ala−Glu−Pro−His−Pro−Ser−Ala−Ala−His−Lys−Lys−Lys−Gln−Ser−Ser−Pro−Lys−Ser−Glu−Asn−Arg−Lys(AEPHPSAAKKKQSSPKSENRK;以下、「BTBP−1」ということもある)や、配列番号19に示されるアミノ酸配列Lys−Ala−Lys−Ala−Lys−Ala−Lys−Ala−Lys−Ala−Lys−Ala(KAKAKAKAKAKA;以下「(KA)6」ということもある)からなるペプチドや、セラミックス表面への可逆的な結合・解離能を有するペプチドであって、配列番号11に示されるアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列や、配列番号19に示されるアミノ酸配列において1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列のいずれかのアミノ酸配列からなるペプチドであれば特に制限されるものではなく、ここで「セラミックスへの可逆的な結合・解離能を有する」とは、セラミックスへの結合量のペプチド濃度依存性を調べたとき、結合量が平衡濃度10μM以上では飽和傾向を示し、0.1μM以下では飽和量の1/2以下に低下する程度の結合能を持っていることを意味する。上記の配列番号11に示されるアミノ酸配列において数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列からなるペプチドであって、かつセラミックスへの可逆的な結合・解離能を有するペプチドとして、例えば、Ala−Val−Gly−Leu−Ser−Pro−Asp−Gly−Ser−Arg−Gly−Val−Gly−Gly−Gly−Ser−Ala−Glu−Pro−His−Pro−Ser−Ala−Ala−His−Lys−Lys−Lys−Gln−Ser−Ser−Pro−Lys−Ser−Glu−Asn−Arg−Lys−Val−Pro−Phe−Tyr−Ser−His−Ser−Gly−Gly−Ser−Gly−Thr−Ser−Arg−Thr−Pro−Ile−Leu−Gly(配列番号1)や、Ser−Ala−Ala−His−Lys−Lys−Lys−Gln−Ser−Ser−Pro−Lys−Ser−Glu−Asn−Arg(配列番号14)や、Ala−Gln−Pro−His−Pro−Ser−Ala−Ala−His−Lys−Lys−Lys−Gln−Ser−Ser−Pro(配列番号16)等を具体的に挙げることができる。上記セラミックスとしては、特に制限されるものではないが、好ましくは金属酸化物を主成分とするセラミックス、より好ましくは遷移金属酸化物を主成分とするセラミックスであり、なかでも、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物を主成分とするセラミックス等であることが特に好ましい。より具体的には、上記セラミックスとして、チタン酸化物系(チタン酸バリウム(BaTiO3)、チタン酸ストロンチウム(SrTiO3)、酸化チタン(TiO2)など)、ジルコニウム酸化物系(酸化ジルコニウム(ZrO2)など)、スズ酸化物系(スズ酸バリウム(BaSnO3)など)等を好適に例示することができる。
【0015】
また、本発明のペプチドのセラミックス表面への可逆的な結合・解離は、金属イオンにより制御することができる。上記金属イオンとしては、本発明のペプチドのセラミックス表面への結合・解離を制御するものであれば特に制限されるものではないが、Ba2+、Ni2+、Zn2+、Ca2+等を例として挙げることができる。本発明のペプチドのセラミックス表面への可逆的な結合・解離は、水溶液中に含まれる上記金属イオンの濃度により制御することが可能である。すなわち、セラミックスと本発明のペプチドとを解離用金属イオン非存在下又は結合用金属イオンの存在下に(金属イオンを含む溶液により)結合させ、次に、セラミックスに結合した本発明のペプチドを、解離用金属イオンの存在下に解離させることができる。また、上記解離用金属イオンを含む溶液の金属イオン濃度を低下させることにより、解離しているセラミックスと本発明のペプチドとを再結合させることもできる。上記解離用金属イオンや結合用金属イオンは、本発明のペプチドの種類やセラミックスの種類により異なる。
【0016】
アミノ配列が異なる本発明のペプチドは、それぞれ異なる種類のセラミックスに対して、可逆的な結合・解離能を有する。例えば、本発明のペプチドBTBP−1は、チタン酸バリウム(BaTiO3)、チタン酸ストロンチウム(SrTiO3)、酸化チタン(TiO2)及び酸化ジルコニウム(ZrO2)に対して優れた結合・解離能を有するが、本発明のペプチド(KA)6は、チタン酸バリウム(BaTiO3)、チタン酸ストロンチウム(SrTiO3)及びスズ酸バリウム(BaSnO3)に対して優れた結合・解離能を有する。また、本発明のペプチドとセラミックとの可逆的な結合・解離は、異なる種類の金属イオンにより制御される。例えば、BTBP−1とチタン酸バリウム(BaTiO3)はMg2+やNi2+やZn2+の存在下に結合する(結合を阻害しない)が、チタン酸バリウムに結合したBTBP−1はBa2+の存在下に解離する。一方、(KA)6とチタン酸バリウム(BaTiO3)はMg2+の存在下に結合する(結合を阻害しない)が、チタン酸バリウムに結合した(KA)6はBa2+、Ni2+、Zn2+のいずれによっても解離する。以上のように、本発明のペプチドをセラミックス表面への可逆的な結合・解離能を有するペプチドとして使用する方法も本発明の一形態として包含される。
【0017】
上記本発明ペプチドは、そのアミノ酸配列に従って、一般的な化学合成法により製造することができる。化学合成法には、通常の液相法及び固相法によるペプチド合成法が包含され、より詳しくは、アミノ酸配列情報に基づいて、各アミノ酸を1個ずつ逐次結合させ鎖を延長させていくステップワイズエロゲーション法と、アミノ酸数個からなるフラグメントを予め合成し、次いで各フラグメントをカップリング反応させるフラグメント・コンデンセーション法とが包含される。本発明のペプチドを合成するには、そのいずれの方法をも採用することができる。上記ペプチド合成に採用される縮合法も、公知の各種方法に従うことができる。その具体例としては、例えばアジド法、混合酸無水物法、DCC法、活性エステル法、酸化還元法、DPPA(ジフェニルホスホリルアジド)法、DCC+添加物(1−ヒドロキシベンゾトリアゾール、N−ヒドロキシサクシンアミド、N−ヒドロキシ−5−ノルボルネン−2,3−ジカルボキシイミド等)、ウッドワード法等を例示できる。これら各方法に利用できる溶媒もこの種ペプチド縮合反応に使用されることがよく知られている一般的なものから適宜選択することができ、例えばジメチルホルムアミド(DMF)、ジメチルスルホキシド(DMSO)、ヘキサホスホロアミド、ジオキサン、テトラヒドロフラン(THF)、酢酸エチル等及びこれらの混合溶媒等を挙げることができる。
【0018】
さらに、上記本発明のペプチドは、本発明のセラミックスへの可逆的な結合・解離能を有するペプチドをコードするDNAの塩基配列情報により、遺伝子工学的手法を用いて常法により調製することもできる。このようにして得られる本発明のセラミックスに結合能を有するペプチド、通常の方法に従って、例えばイオン交換樹脂、分配クロマトグラフィー、ゲルクロマトグラフィー、アフィニティークロマトグラフィー、高速液体クロマトグラフィー(HPLC)、向流分配法等のペプチド化学の分野で汎用されている方法に従って、適宜その精製を行うことができる。
【0019】
上記本発明のペプチドは、検出可能なマーカーで標識されていてもよく、上記検出可能なマーカーとしては、従来知られているペプチド標識用のマーカーであれば特に制限されるものではなく、例えば、3H、14C、125I等の放射性同位体や、ダンシルクロライド、テトラメチルローダミンイソチオシアネート等の蛍光物質や、ビオチン、ジゴキシゲニンのような生物学的に関連する結合構造や、生物発光化合物や、化学発光化合物や、金属キレート等を具体的に挙げることができる。なかでも、本発明のペプチドBTBP−1、(KA)6をそれぞれビオチンで標識した、ビオチン標識BTBP−1やビオチン標識(KA)6は、後述の実施例でも示されているように、セラミックスへの可逆的な結合・解離能を有しており、特に好例として挙げることができる。また、ビオチン標識BTBP−1やビオチン標識(KA)6は、ストレプトアビジン修飾抗体と併用することにより、標的となる所定の抗原タンパク質(目的タンパク質)を着脱自在に操作しうることから、例えば、抗原タンパク質の分離・精製に有利に用いることができる。
【0020】
また、本発明のペプチドは、機能性ペプチド又は機能性タンパク質に結合した融合ペプチドであってもよい。本発明の融合ペプチドとしては、本発明のペプチドと、機能性ペプチド又は機能性タンパク質とが結合しているものであればどのようなものでもよく、上記機能性タンパク質としては、例えば、抗体、抗原、プロテインA(特にプロテインAのZZドメイン)等の免疫関連タンパク質や、アルカリフォスファターゼ、HRP等の酵素、抗体のFc領域、GFP等の蛍光物質などのマーカータンパク質や、BMP、レクチン、アビジン等の結合能を有する結合性タンパク質を具体的に挙げることができ、また、上記機能性ペプチドとしては、例えば、HA、FLAG、Myc等のエピトープタグや、GST、マルトース結合タンパク質(MBP)、ビオチン化ペプチド 、オリゴヒスチジン等の親和性タグなどを具体的に例示することができる。なかでも、本発明のセラミックスへの可逆的な結合・解離能を有するペプチドとプロテインAのZZドメインとを結合させた融合ペプチドは、セラミックスへの優れた結合能を有するとともに、抗体との結合能も有していることから、生物学的解析ツールとして広く応用が可能であり、さらに、金属イオンによりセラミックス表面から解離させることが可能であるため、前記抗体が認識する標的抗原タンパク質等の精製にも非常に有用である。また、本発明のセラミックスへの可逆的な結合・解離能を有するペプチドとBMPとを結合させた融合ペプチドは、この融合ペプチドを介して、BMPと結合するBMPレセプターを表面に発現する間葉系細胞や繊維芽細胞等をセラミックス材料表面に集積するとともに骨芽細胞への分化が促進し、その結果、セラミックス材料表面への骨の形成が促進され、セラミックスと骨を良好に定着させる上で有用である。
【0021】
本発明のセラミックスへの可逆的な結合・解離能を有するファージとしては、本発明のセラミックスに結合能を有するペプチドをその粒子表面上に提示するファージであればどのようなものでもよく、かかるセラミックスに結合能を有するファージは、ファージライブラリスクリーニングの過程で、セラミックスに強く結合したペプチド提示ファージを、その他のファージ集団から分離することにより得られる他、本発明のセラミックスに結合有するペプチドをコードするDNAを、常法によりファージミドベクターに組み込んで大腸菌等の宿主細胞を形質転換し、ヘルパーファージを感染させることで得ることもできる。
【0022】
本発明のペプチドを認識する抗体としては、モノクローナル抗体、ポリクローナル抗体、キメラ抗体、一本鎖抗体、ヒト化抗体等の免疫特異的な抗体を具体的に挙げることができ、これらは上記本発明のペプチドを抗原として用いて常法により作製することができるが、なかでもモノクローナル抗体がその特異性の点でより好ましい。かかる本発明のペプチドに対する抗体は、慣用のプロトコールを用いて、動物(好ましくはヒト以外)に本発明のペプチド又はその断片を免疫することにより産生され、例えばモノクローナル抗体の調製には、連続細胞系の培養物により産生される抗体をもたらす、ハイブリドーマ法(Nature 256, 495-497, 1975)、トリオーマ法、ヒトB細胞ハイブリドーマ法(Immunology Today 4, 72, 1983)及びEBV−ハイブリドーマ法(MONOCLONAL ANTIBODIES AND CANCER THERAPY, pp.77-96, Alan R.Liss, Inc., 1985)など任意の方法を用いることができる。以下に、本発明のペプチドのひとつであるBTBP−1に対して特異的に結合するマウスのモノクローナル抗体、すなわち抗BTBP−1モノクローナル抗体の作製方法を説明する。
【0023】
上記抗BTBP−1モノクローナル抗体は、抗BTBP−1モノクローナル抗体産生ハイブリドーマをインビボ又はインビトロで常法により培養することにより生産することができる。例えば、インビボ系においては、齧歯動物、好ましくはマウス又はラットの腹腔内で培養することにより、またインビトロ系においては、動物細胞培養用培地で培養することにより得ることができる。インビトロ系でハイブリドーマを培養するための培地としては、ストレプトマイシンやペニシリン等の抗生物質を含むRPMI1640又はMEM等の細胞培養培地を例示することができる。抗BTBP−1モノクローナル抗体産生ハイブリドーマは、例えば、BTBP−1を用いてBALB/cマウスを免疫し、免疫されたマウスの脾臓細胞とマウスNS−1細胞(ATCC TIB−18)とを、常法により細胞融合させ、免疫蛍光染色パターンによりスクリーニングすることにより、抗BTBP−1モノクローナル抗体産生ハイブリドーマを作出することができる。また、かかるモノクローナル抗体の分離・精製方法としては、タンパク質の精製に一般的に用いられる方法であればどのような方法でもよく、アフィニティークロマトグラフィー等の液体クロマトグラフィーを具体的に例示することができる。
【0024】
また、上記BTBP−1に対する一本鎖抗体をつくるためには、一本鎖抗体の調製法(米国特許第4,946,778号)を適用することができる。また、ヒト化抗体を発現させるために、トランスジェニックマウス又は他の哺乳動物等を利用したり、上記抗体を用いて、そのBTBP−1を発現するクローンを単離・同定したり、アフィニティークロマトグラフィーでそのポリペプチドを精製することもできる。さらに、上記抗BTBP−1モノクローナル抗体等の抗体に、例えば、FITC(フルオレセインイソシアネート)又はテトラメチルローダミンイソシアネート等の蛍光物質や、125I、32P、14C、35S又は3H等のラジオアイソトープや、アルカリホスファターゼ、ペルオキシダーゼ、β−ガラクトシダーゼ又はフィコエリトリン等の酵素で標識したものや、グリーン蛍光タンパク質(GFP)等の蛍光発光タンパク質などを融合させた融合タンパク質を用いることによって、上記BTBP−1を検出・定量等を行うことができる。上記抗体を用いた免疫学的測定方法としては、RIA法、ELISA法、蛍光抗体法、プラーク法、スポット法、血球凝集反応法、オクタロニー法等の方法を挙げることができる。
【0025】
本発明の組換えベクターとしては、前記本発明のDNAを含み、かつセラミックスへの可逆的な結合・解離能を有するペプチドを発現することができる組換えベクターであれば特に制限されず、本発明の組換えベクターは、本発明のDNAを発現ベクター、好ましくは発現プラスミドベクターに適切に挿入することにより構築することができる。かかる発現ベクターとしては、宿主細胞において自立複製可能であるものや、あるいは宿主細胞の染色体中へ組込み可能であるものが好ましく、また、本発明のDNAが転写できる位置にプロモーター、エンハンサー、ターミネーター等の制御配列を含有しているものを好適に使用することができる。
【0026】
上記発現ベクターとして、例えば、pCMV6−XL3(OriGene Technologies Inc.社製)、EGFP-C1(Clontech社製)、pGBT−9(Clontech社製)、pcDNAI(フナコシ社製)、pcDM8(フナコシ社製)、pAGE107(Cytotechnology, 3,133, 1990)、pCDM8(Nature, 329, 840, 1987)、pcDNAI/AmP(Invitrogen社製)、pREP4(Invitrogen社製)、pAGE103(J.Blochem., 101, 1307,1987)、pAGE210等を例示することができる。また、プロモーターとしては、例えば、サイトメガロウイルス(ヒトCMV)のIE(immediate early)遺伝子のプロモーター、SV40の初期プロモーター、レトロウイルスのプロモーター、メタロチオネインプロモーター、ヒートショックプロモーター、SRαプロモーター等を挙げることができる。さらに、プロモーターの下流に蛍光蛋白質をコードする遺伝子等のレポーター遺伝子を融合することができる。蛍光蛋白質としては、緑色蛍光蛋白質(Green Fluorescence Protein(GFP))、赤色蛍光蛋白質(Cyan Fluorescence Protein(CFP))、青色蛍光蛋白質(Blue Fluorescence Protein(BFP))、黄色蛍光蛋白質(Yellow Fluorescence Protein(YFP))、ルシフェラーゼ(luciferase)を例示することができる。
【0027】
また、本発明の形質転換体としては、上記本発明の組換えベクターが宿主細胞に導入され、かつ本発明のセラミックスへの可逆的な結合・解離能を有するペプチドを発現する形質転換体であれば特に制限されるものではなく、形質転換酵母、形質転換植物(細胞、組織、個体)、形質転換細菌、形質転換動物(細胞、組織、個体)等を挙げることができるが、形質転換動物細胞が好ましい。
【0028】
本発明のセラミックス表面とタンパク質との結合・解離を制御する方法としては、セラミックス表面への可逆的な結合・解離能を有するペプチド(以下「本結合・解離ペプチド」ということがある)、又は該ペプチドと機能性タンパク質との融合ペプチドを介しての、セラミックス表面と目的とするタンパク質(又は目的とする細胞)との結合又は解離を、所定の金属イオンの濃度を調整することによって制御する方法[制御方法I]や、セラミックス表面と、本結合・解離ペプチドと目的とするタンパク質との融合ペプチドとの結合又は解離を、所定の金属イオンの濃度を調整することによって制御する方法[制御方法II]であれば特に制限されず、かかる本発明の制御方法により、セラミックス表面に所定のタンパク質(目的タンパク質)又は所定の細胞(目的細胞)を脱着、すなわち修飾することができる。上記本結合・解離ペプチドとしては、前記本発明のペプチドの他、BTBP−1と同様にファージライブラリを用いたスクリーニングにより、また(KA)6と同様に塩基性アミノ酸に富むペプチドを用いたスクリーニングにより見出すことができる。また、機能性タンパク質については前述したタンパク質を用いることができ、この機能性タンパク質が目的タンパク質になることもある。例えば、本結合・解離ペプチド−プロテインAのZZドメインとの融合ペプチドのとき、目的タンパク質はこの融合ペプチドを介してセラミックス表面に結合・解離する抗体ということになり、同様に、本結合・解離ペプチド−抗体との融合ペプチドのとき、目的タンパク質はこの融合ペプチドを介してセラミックス表面に結合・解離する抗原タンパク質ということになる。他方、目的とするタンパク質が抗体のとき、本結合・解離ペプチド−抗体との融合ペプチドにおける構成成分である抗体が目的タンパク質となる。
【0029】
本発明の制御方法Iとして、本結合・解離ペプチド又は融合ペプチドを含む溶液と、セラミックスとを解離用金属イオン非存在下又は結合用金属イオンの存在下に(あるいは、結合用金属イオン濃度を高くして)反応させることにより、セラミックス表面に本結合・解離ペプチド又は融合ペプチドを結合させ、次いで本結合・解離ペプチド又は融合ペプチドと目的タンパク質とを結合させる方法や、目的タンパク質が結合した本結合・解離ペプチド又は融合ペプチドが結合したセラミックスから、解離用金属イオンの存在下(あるいは、解離用金属イオン濃度を高くして)又は結合用金属イオンの非存在下に、目的タンパク質が結合した本結合・解離ペプチド又は融合ペプチドを解離させる方法を例示することができ、具体的には、本結合・解離ペプチドとしてビオチン標識ペプチドを使用するとき、目的タンパク質としてストレプトアビジン結合目的タンパク質を用いる方法や、融合ペプチドとしてプロテインAのZZドメインとの融合ペプチドを使用するとき、目的タンパク質として抗体を用いる方法や、融合ペプチドとしてBMP融合ペプチドを使用するとき、目的とする細胞をBMPレセプター発現間葉系細胞とする方法を挙げることができる。また、本発明の制御方法IIとして、目的タンパク質を構成成分とする融合ペプチドを含む溶液と、セラミックスとを解離用金属イオン非存在下又は結合用金属イオンの存在下(あるいは、結合用金属イオン濃度を高くして)に反応させることにより、セラミックス表面に融合ペプチドを結合させる方法や、融合ペプチドが結合したセラミックスから、解離用金属イオンの存在下(あるいは、解離用金属イオン濃度を高くして)又は結合用金属イオンの非存在下に、融合ペプチドを解離させる方法を例示することができる。本発明の制御方法や脱着(修飾)方法により、本発明のペプチド又は融合ペプチドが表面に結合したセラミックスや、目的とするタンパク質や細胞が表面に結合したセラミックス、すなわち本発明の表面修飾セラミックスを得ることができる。
【0030】
上記セラミックスとしては、特に制限されるものではないが、好ましくは金属酸化物を主成分とするセラミックス、より好ましくは遷移金属酸化物を主成分とするセラミックスであり、なかでも、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物を主成分とするセラミックス等であることが特に好ましい。より具体的には、上記セラミックスとして、チタン酸化物系(チタン酸バリウム(BaTiO3)、チタン酸ストロンチウム(SrTiO3)、酸化チタン(TiO2)など)、ジルコニウム酸化物系(酸化ジルコニウム(ZrO2)など)、スズ酸化物系(スズ酸バリウム(BaSnO3)など)等を好適に例示することができる。また、上記溶液に含まれる本発明のペプチド又は融合ペプチドの濃度としては、好ましくは0.1μM以上、より好ましくは0.5μM以上、さらに好ましくは1μM以上である。発明の修飾方法により、セラミックス表面に結合した本発明のペプチド又は融合ペプチドは、金属イオンにより解離させることができ、上記金属イオンとしては、特に制限されないが、Ba2+、Ni2+、Zn2+、Ca2+等を好例として挙げることができる。上記本発明の修飾方法により表面修飾されたセラミックスは、例えば、歯科インプラント材料などの移植用医療デバイス、分子認識を行うナノデバイスやバイオセンサ、タンパク質精製や細胞単離用の担体などに利用できる他、セラミックスの機械的な特性、電子材料としての機能(圧電性、強誘電性など)、ナノメートルオーダーの微粉末が安価に大量に合成できる等の製造的な利点を活かして、種々の用途に用いることができる。
【0031】
以下、実施例により本発明をより具体的に説明するが、本発明の技術的範囲はこれらの例示に限定されるものではない。
【実施例1】
【0032】
[ファージディスプレイ法によるチタン酸バリウム結合ペプチドの取得]
1.使用したファージライブラリ
スクリーニングには、Ph.D-12 peptide phage display kit(New England Biolabs社製;以下D−12という)及びPh.D-C7C peptide phage display kit (BioLabs社製;以下D−C7Cという) を用いた。また、同じ製品でも製造ロットによって違う配列の集団となっている可能性があるため、D−12については異なる3つのロット(lot3、lot5、lot6)を実験に用いた。D−12は、12残基の直線状ランダムペプチド配列を提示するライブラリであり、D−C7Cは2つのシステイン間に7つのランダム配列が挿入されているライブラリである。
【0033】
2.ファージライブラリを用いたスクリーニング
スクリーニング実験や、ファージの力価(濃度)測定等は、メーカーが提示しているプロコトルに従って行った。以下にその概略を示す。
ファージを構成するタンパク質のうち、提示ペプチド以外のタンパク質が非特異的にチタン酸バリウム(BT)に結合することを防ぐため、あらかじめBT表面にウシ血清アルブミン(BSA)を吸着させ、ブロッキングを行った。2mgのBT粉末を、2%のBSAを含むTBS(50mMトリスヒドロキシメチルアミノメタン(トリス)、150mM塩化ナトリウムを含む水溶液、pH7.5)10mLに懸濁した。分散状態にある懸濁液1mLを分取し、遠心分離機によりBT粉末を沈殿させた後、上清を捨て、再び2%BSAを含むTBS 1mLで懸濁し30分間攪拌した。攪拌後、遠心分離でBT粉末を沈殿させて上清を除去し、界面活性剤tween20を0.1%含むTBS(TBST)1mLを加えて懸濁する洗浄操作を1回行った。
【0034】
ブロッキング済みのBT粉末懸濁液1mLに、ペプチドファージライブラリ10μLを投入し2時間攪拌した。攪拌後、TBSTでの洗浄操作を3回行った。遠心分離でBT粉末を沈殿させた後、今度は0.2Mグリシン塩酸水溶液(pH2.2)1mLを加え、10分間回転攪拌し、BTに結合しているファージを溶出させた。溶出液を遠心してBT粉末を沈殿させた後、上清を回収し、1Mのトリス水溶液(pH9.1)150μLで中和した。
得られたファージは大腸菌に感染させ増幅させた。ファージの溶出液を対数増殖中のER2738株を含む培地20mLに添加し、37℃で4.5時間振盪培養した。その後、培養液を遠心分離し大腸菌を沈殿させ、ファージを含む上清を回収した。増幅したファージのうち約1012pfu分を、再び、ブロッキング済みのBT粉末に投与し、2時間のインキュベートの後、TBSTで洗浄、グリシン塩酸(pH2.2)で溶出、中和するという一連の実験(パニング)を6回繰り返した。
【0035】
3.BT結合ペプチドの取得
D−12ファージライブラリのlot5を用いた実験において、4ラウンド目のファージ溶出液に含まれるファージをクローン化し、ランダムにピックアップした28種類についてDNA解析を行った。DNA解析は、ベックマンコールター社製のシーケンサーCEQ2000と、試薬DTCS Quick start kitを使用した。実験方法はメーカー提示のプロコトルに従った。この解析結果によると、28クローンのうち18が同じ配列を示しており、このクローンをφ403Nと命名した。そのほかに、2個以上同じ配列を示したクローンが2種類あり、それぞれφ404N、φ410Nと命名した。これらのクローン化ファージのBTへの結合力を調べた結果を図1に示す。結合率とは、溶出して回収されたファージ量を、投入したファージ量で除したものであり、φM13KEとは、ペプチドを提示していない(野生型の)ファージである。φ403Nは他より際立って高いBTへの結合率を示すことが明らかとなった。
一方、他のライブラリ(D−12のlot3及びlot6、D−C7C)を実験に用いたスクリーニングでは、BTBP−1と同一の配列が取得できた場合もあれば、異なる配列が取得された場合もあった。これら他のライブラリーによるスクリーニングの結果の詳細に関しては、参考例として後述する。
【実施例2】
【0036】
[BT結合クローンφ403Nのペプチド配列解析]
1.φ403NのDNA及びアミノ酸配列の決定
実施例1でD−12ライブラリより取得されたファージクローンφ403N、φ404N及びφ410Nについてファージの持つDNAを解析した。それぞれのその提示ペプチドのアミノ酸配列は、φ403Nは配列番号1に示される57残基のアミノ酸配列、φ404Nは配列番号2に示される11残基のアミノ酸配列、φ410Nは配列番号3に示される12残基の配列であることが明らかとなった。D−12ライブラリは12残基のペプチドを提示するよう設計されたものであるため、このφ403Nは、何らかの理由で長い配列が挿入されてできたものである。図2にこの理由についての仮説を示す。ファージライブラリは、ファージの先端のタンパク質g3p(シグナル配列の部分は、ファージが形成される際に切り離される)をコードするDNAに、ランダムなペプチドをコードするランダムなDNA配列を挿入することで形成される。図2の通常の配列では、Xで表されるランダムな12残基のペプチドがg3pタンパク質のN末端側にGGGSという4残基のアミノ酸のリンカーを介して接続されていることを示している。しかし、今回取得されてきたφ403Nのアミノ酸配列は、図2に示すように、ランダムなペプチド配列をコードするDNA断片が3つタンデムに連なって挿入されている。これはファージライブラリの製造上、ある一定の確率で起こりえる現象であると思われる。このため、DNA配列上は、シグナル配列とリンカーの間の57残基がg3pの末端に提示されていることになる。
【0037】
2.φ403N提示ペプチドのBT結合領域の決定
このDNA配列から予想される57残基のペプチドが、実際に発現しているかどうかを確認するため、ファージを構成するタンパク質をSDSアクリルアミドゲル電気泳動して、想定分子量付近のバンドを切り取り、アミノ酸配列分析した。この結果では、57残基のうちN末端のAVG〜16残基、および、AVG〜37残基が削られたタンパク質が検出された。しかし、これらのうち、どちらがBTに強く結合しているかは区別できないし、アミノ酸配列分析では検出できないほど微量のタンパク質が結合に寄与していることも否定できない。また、ペプチド配列が57残基と長いと応用上使用しにくいため、ペプチド配列のうちBTとの結合に寄与している部分の配列を同定して絞り込む必要がある。φ403Nのファージをベースに遺伝子を組み替え、図3に示すようなペプチド配列を持つファージ9種類を作製した。それぞれのファージクローンのアミノ酸配列は、以下の配列番号と対応する。
φ403N:配列番号1
φ1581:配列番号4
φ1583:配列番号5
φ1597:配列番号6
φ1599:配列番号7
φ1601:配列番号8
φ1603:配列番号9
φ1605:配列番号10
φ1611:配列番号11
φ1612:配列番号12
【0038】
これらの遺伝子を大腸菌に導入してファージを形成させ、それぞれのBT結合能をしらべた。結合実験には、Si基板上に全面成膜されたBTスパッタ薄膜上にAuとTi薄膜がパターニングされているチップを用いた(図4、左上パネル)。φ1581、φ1583、φ1597、及びφ1601は、ファージが形成されないか、DNAに意図しない変異が入ったファージしか得られなかった。このため実験には、φ1599、φ1603、φ1605、φ1611、及びφ1612を用いた。実験は次の手順で行った。チップを2%BSAを含むTBSTでブロッキングし、約1010pfu/mLのファージを含むTBST中で30minインキュベートし、TBSTで洗浄した。さらに、anti-fd bacteriophage biotin conjugate(Sigma-Aldrich社製)を含むTBST中で15分間インキュベートした後、TBSTで洗浄した。続いて、ストレプトアビジンつき蛍光体(Qdot(登録商標)655 streptavidin conjugate、Invitrogen社製)を含むTBSTで15分間インキュベートした後、TBSTで洗浄した。このチップを水溶液に浸した状態で顕微鏡で蛍光観察した。その結果、図4に示すように、φ1611が22残基という短い配列でもBTへ特異的に結合すること分かった。このφ1611の提示ペプチドをBTBP−1(AEPHPSAAKKKQSSPKSENRK:配列番号11)と命名した。
【実施例3】
【0039】
[BTBP−1ペプチドのBT結合能の確認]
これまでの実験により、BTBP−1はファージに提示された状態で結合能力を持つことは確認されたので、次に、化学合成によって作製したBTBP−1単独でBT結合能力を持つかどうかを確認する実験を行った。完全長BTBP−1(配列番号11)の合成ペプチドを作製するとともに、BTBP−1のN末端側から18番目から22番目までのアミノ酸を削ったペプチドΔ(18−22)(配列番号13)、1番目から5番目のアミノ酸を削ったペプチドΔ(1−5)(配列番号14)、及び両末端の5残基のアミノ酸を削ったペプチドΔ(1−5、18−22) (配列番号15)の3つ変異ペプチドを作製した。実験には、実施例2(図4)と同様のチップを用い、N末端側がビオチン分子で修飾された合成ペプチドを使用した。実験手順としては、まずチップを2%BSAを含むTBSTでブロッキングし、40μMのビオチン化ペプチドを含むTBSTで15分間インキュベートし、TBSTで洗浄した後、ストレプトアビジンつき蛍光体を含むTBSTで15分間インキュベートし、TBSTで洗浄した。このチップを水溶液に浸した状態で顕微鏡で蛍光観察した。結果を図5に示す。この結果から、BTBP‐1単独の状態でもBTに結合すること、1−5番目を削った場合(Δ(1−5))でもBTに結合することが確認できた。なお、18−22番目を削った場合に大きく結合力が低下しているが、ここには塩基性アミノ酸(R、K)が2つ含まれている。このため、結合には塩基性のアミノ酸が重要であると推測できる。
【実施例4】
【0040】
[BTBP−1とプロテインAとの融合タンパク質の作製]
プロテインA(ProteinA)は抗体と結合する能力があるタンパク質である。このタンパクの抗体と結合する部位(Zドメイン)2つから構成されるタンパク質とBTBP‐1の融合タンパク質(ZZ+BTBP−1)を作製し、BTへの結合能力と抗体の捕捉能力を確認する実験を行った。実験手順は、実施例2と同じチップを、2%BSAを含むTBSTでブロッキングした後、融合タンパク質(ZZ+BTBP−1)を0.5μMを含むTBSTをインキュベートさせた。TBSTで洗浄した後、抗体(anti-fd bacteriophagebiotin conjugate)を含むTBSTでインキュベートし、さらにTBSTで洗浄した。最後に、ストレプトアビジンつき蛍光体を含むTBSTで15分間インキュベートし、TBSTで洗浄後、純水で洗浄し、チップを乾燥状態で観察した。結果を図6に示す。BTBP−1とZZの融合タンパク(ZZ+BTBP−1)が、BT表面に吸着し、さらに抗体を捕捉していることが確認された。一方、ZZのみ(BTBP−1を融合していないZZ)やいずれのタンパクを含まないTBST中でインキュベートした場合には、BTへの抗体の結合は融合タンパクに比べて小さかった。
【実施例5】
【0041】
[BTBP−1の配列特異性]
BTBP−1の配列の特異性を調査するため、いくつかの種類のペプチドを合成し、それらの結合能力を調査した。BTBP−1の2つのグルタミン酸(E)をグルタミン(Q)に置き換えたペプチドBTBP−1(E→Q)(配列番号16)、(DK)6(配列番号17)、(DA)6(配列番号18)、及び(KA)6(配列番号19)の4種類のペプチドを合成し、それぞれのN末端側をビオチン分子で修飾した。実施例2と同じチップを、2%BSAを含むTBSTでブロッキングし、1μMのビオチン化ペプチドを含むTBSTで15分間インキュベートした。TBSTで洗浄した後、ストレプトアビジンつき蛍光体を含むTBSTで15分間インキュベートし、さらにTBSTで洗浄した。最後に純水で洗浄し、チップを乾燥状態で観察した。結果を図7に示す。BTBP−1(E→Q)は、BTBP−1と同様に結合したことから、BTとの結合に2つの負電荷は重要でないことが分かった。また、リジン(K)とアラニン(A)が交互に6個ずつ並んだ配列(KA)6もBTに結合することが明らかとなった。このことから、塩基性アミノ酸が酸性アミノ酸よりも多く含まれることが結合に重要であることが示唆された。ただし、動的光散乱測定により(KA)6、(DA)6、(DK)6のペプチドの粒子径(分子サイズ)を計測してみたところ、100nm程度のクラスターとして存在していることが分かった(粒子系はバッファーに依存して変化する)。これにより、(KA)6については多価提示と同様な効果が発生し、結合能力が増強されている可能性もある。
【実施例6】
【0042】
[金属イオンによるBT結合ペプチドへの阻害作用]
BTBP−1のBT結合に及ぼす、水溶液中の金属イオンの影響を検討した。Mg2+、Ca2+、Ba2+、Ni2+、Zn2+の各イオンが1mMの濃度で含まれる溶液中でBTBP−1及び(KA)6ペプチドをBTと結合させた。用いたチップは実施例2と同じもので、ストレプトアビジンつきの蛍光体でラベルしたものを蛍光顕微鏡で観察した。その蛍光の強さを数値化して表した結果を図8に示す。Ba2+により、蛍光強度が落ちており、BTBP−1及び(KA)6ペプチドとBTとの結合を阻害していることが明らかとなった。また、BTBP−1と(KA)6では、Ni2+又はZn2+を投入したときの挙動が異なっており、ここにBTBP‐1の配列特異性が現れている。
【0043】
さらに、上記のような金属イオンによる結合阻害作用により、既に結合しているペプチドを、金属イオンを含む溶液によって解離できるかを調査した。図9の「解離処理あり」は、BTBP−1を結合させた後、1mMのBaCl2を含むTBSTに15分間浸漬した後、ストレプトアビジンつき蛍光体をつけたものである。「解離処理なし」は、ペプチドを結合させた後、Ba2+を含まない液に15分間浸した後、ストレプトアビジンつき蛍光体をつけたものである。この結果から、ペプチドはBa2+を含む液に浸すことによって解離を促進できることが分かった。なお、チップは実施例2のものとは異なり、赤く光っている部分がBTの領域で、その他の部分はTiである。また、図10は、同じチップにペプチドを結合させ、先にストレプトアビジン付き蛍光体を結合させて一旦蛍光顕微鏡で観察した後、再び10mMのBaCl2を含むTBST中に浸して解離するかどうかを調べた結果である。ペプチドにストレプトアビジンが結合することによって解離のスピードは低下しているが、Ba2+による解離の促進効果は明らかに確認できる。
【実施例7】
【0044】
[ペプチド濃度の結合量に及ぼす影響]
BTBP‐1とBTとの結合におけるペプチド濃度の影響を調査した。BTの粉末(粒径1μm弱)10mgを用意し、1〜16μMのBTBP−1を含む1mLのHBST溶液(50mM HEPES、150mM NaCl、0.1% tween20)と混合し、2時間回転攪拌した。その後、遠心分離で粉末を沈殿させ、上清のペプチド濃度を測定した。濃度の測定は、アミノ基と反応して蛍光を発する試薬(CBQCA、invitrogen社製)をペプチドと反応させ、蛍光分光測定計(FP-6500、日本分光社製)で測定した。BTと混合していない8μMのBTBP−1溶液も合わせて測定した。また、BTは水溶液中に置くと、最表面に存在するBa2+が溶出することが知られている。これまでのようなチップを用いた実験では表面積が小さいために溶出するBa2+の量も少なく問題はなかったが、今回の実験では溶液中のBa2+イオンの量は無視できないレベルになる。そこで、Ba2+を除去するためSO42−を加えてBaSO4として沈殿させることにした。HBSTに対して、2mMのNa2SO4を加えた実験を合わせて行った。なお、BTには若干の光触媒作用がある可能性がり、ペプチドや界面活性剤が分解される恐れがあるので、2時間の回転攪拌は遮光した状態で行っている。結果を図11に示す。まず、硫酸イオンを添加しないと、Ba2+の影響で結合がかなり阻害されていることが分かる。硫酸イオンを添加した状態では、0.1μM程度の低い値から立ち上がりが見られ、3μMからは濃度依存的な結合量の増加が確認された。
【実施例8】
【0045】
[ペプチドの多価提示]
1分子のタンパク質に複数のペプチドを提示させることで、タンパク質の無機材料(セラミックス)に対する結合力を向上させることができるかどうかについて検討した。10nMのストレプトアビジン付きの蛍光体(Qdot、Invitorogen社製)と200nMのビオチン標識BTBP−1を予め混合し、先にビオチン-アビジン複合体の形成させた。ストレプトアビジン付きの蛍光体は1粒子に対して、4量体を形成するストレプトアビジン分子が複数結合している。このため、1粒子のQdotに対して十数個〜数十個程度のペプチドが提示されていると考えられる。この複合体をチップに結合させた結果を図12に示す。図12(a)に示すように、Qdotの濃度が10nMと薄くてもチップ上での結合が確認することができた。一方(b)は、ペプチド200nMをチップに対して予め結合させておき、その後、ストレプトアビジン付きの蛍光体でラベルしたものである。この場合、(a)に比べるとペプチド濃度が同じにも関わらず結合量は(a)よりも極端に小さかった。すなわち、タンパク質の固相化にあたり、先にペプチドを無機材料に対して結合させた後、ビオチンアビジン等の結合を利用して固相化する場合は、ペプチドの濃度を0.1〜1μM程度以上にする必要があるが、タンパク質に先にペプチドを複数結合しておき(手法は化学結合でもビオチン-アビジンでもよい)それを固相化する場合には、タンパク質濃度は10nM程度でも結合させることが可能であることが分かった。どちらの方法を適用するかは、アプリケーションによって異なると考えられる。
【実施例9】
【0046】
[結合の阻害に必要な金属イオンの濃度]
結合の阻害に必要な金属イオンの濃度を検討した。1μMのBTBP−1溶液に、BaCl2を0、0.03、0.1、0.3、1.0 mMとなるように加え、チップを浸し、ストレプトアビジン付き蛍光体でラベルし、蛍光顕微鏡で観察した。蛍光強度とバリウム濃度の関係を図13に示す。結合の阻害には30μM程度の濃度が必要であることが分かった。ただし、ペプチドの濃度、結合させたい無機材料の表面積によっては、この値は変化する可能性がある。
【実施例10】
【0047】
[他のセラミックス表面への結合]
これまでの実験結果により、BTBP−1と(KA)6は、Au等と比較してBTに優先的に結合することが示された。次に、BT以外のセラミックス材料(SrTiO3、TiO2、BaSnO3、ZrO2)に対する結合について調査した。各種セラミックスの粉末(粒径1μm弱)10mgを用意し、8μMのBTBP‐1又は(KA)6を含むHBST溶液と混合し、2時間回転攪拌した。その後、遠心分離で粉末を沈殿させ、上清のペプチド濃度を測定した。濃度の測定は、ペプチドをアミノ基と反応して蛍光を発する試薬(CBQCA、Invitrogen社製)と反応させ、蛍光分光測定計(FP-6500、日本分光社製)で測定した。セラミックスと混合していない8μM濃度のペプチド溶液も合わせて測定した。
また、同時に、Ba2+の影響についても検討するため、SrTiO3(比表面積4.34m2/g)、TiO2(比表面積2.75m2/g)、ZrO2(比表面積21.2m2/g)に対しては、ペプチド溶液に1mMのBaCl2を添加し、その効果を見た。BaSnO3(比表面積2.11m2/g)については自身からBa2+が溶出し、それだけで4.2mM程度の濃度に達していたため、Na2SO4を2mMおよび10mM添加し、BaSO4の形でBa2+を除去し、その影響を見た。BaTiO3(比表面積3.43m2/g)については、既出のデータ(吸着量-平衡濃度グラフ)の再掲であるが、溶出したBa2+の濃度は0.5mM程度であり、添加したNa2SO4は2mMである。これらの実験の結果を図14に示す。BTBP−1は、(KA)6と比較して、ZrO2への結合量が4倍、TiO2への結合量が2倍ほど高かったまた、各材料により程度の差はあるがBa2+イオンの結合阻害効果は認められた。
BTBP−1のBaSnO3への結合量はNa2SO4を添加することによるBa2+イオン除去によっても大きくは増加しなかった。(KA)6のBaSnO3への結合量はNa2SO4を添加することによって増加した。このように、金属イオンの添加効果は材料によって一様ではないので、金属イオンはペプチドに直接作用する可能性のほか、材料表面の方に結合することで、ペプチドの吸着挙動に影響を与えている可能性も示唆された。
【実施例11】
【0048】
[参考例1:他のライブラリからのスクリーニング結果]
実施例1において、D−12ファージライブラリのlot5から、BT結合ファージクローンのスクリーニングに成功した。一方、その他のライブラリ(D−12のlot3及びlot6、D−C7C)からは、BTBP−1と同一の配列が取得できた場合もあれば、異なる配列が取得された場合もあった。以下にその詳細を示す。実施例1に記載の方法で、スクリーニングを行い、得られたファージをクローン化した。それぞれの配列を提示したファージの結合率を図15に示す。末尾のSはD−12ライブラリのlot6、TはD−12ライブラリのlot3、CはD−C7Cライブラリから得られたものであることを示す。Lot6では、φ403Nと同じ配列のファージも同時にスクリーニングされていたが(図15では示していない)、lot3ではφ403Nと同じものはスクリーニングされなかった。
【0049】
各アミノ酸配列を、図16及び配列番号20〜26に示す。φ411S、φ421Sは正常に12残基のペプチドを提示していたが、BTへの結合率は低かった。φ601Tはφ403Nと同様にDNA断片が3つ挿入された配列であり、塩基性のアミノ酸が比較的多いと言える。φ607Tは正常に12残基のペプチドとなっていたが、BTへの結合率は中程度であった。φ603Cは何らかの異常によって、2番目の位置に本来システインであるべきアミノ酸がセリンに変わっているが、短い配列でも比較的高い結合率を示している。φ606Cは、これもDNA断片が複数入っている配列であったが、BTへの結合力は高かった。φ616Cは、本来C7Cとなるべき配列から大きく異なっており、BTへの結合率も小さかった。
【0050】
φ603Cは短い配列でもBTへの高い結合力が観測されたので、φ603Cが提示していた配列の合成ペプチドを作製し、パターニングチップを用いてペプチド単体での結合特性を確認した。その結果、図17に示すように、ペプチドとチップのインキュベート時間が15分ではほとんど結合が見られず、10時間のインキュベートでようやく結合が確認できた。BTBP−1は15分でも十分な結合が確認できていたので、φ603Cが提示するペプチドはBTBP−1に比べると結合力が劣ることが判った。ただし、10残基という配列の短さの割には結合力が高いと言える。
【0051】
[参考例2:チタン酸バリウムの粗い粉末を用いたスクリーニング結果]
前述のように、φ403N(BTBP−1)のスクリーニングの際には、溶液1mL中に粒径約1μmのチタン酸バリウム粉末約0.2mgを分散させ用いた。これに対し、粒の粗いチタン酸バリウム粉末(チタン酸バリウムの焼結体を粗く砕いて75−300μmに粒径を揃えたもの)10mgを1mL中に分散させたものに対するパニング実験を行った。スクリーニングされたものをDNA解析すると、配列番号27〜31に示す5種類のアミノ配列が得られた。それぞれのファージクローンを作製し、結合率を調べた結果、φBT401とφBT403が高い結合率を示した(図18)。しかし、チップを用いた蛍光観察実験での結果は、図16の結果と異なっていた(図19、BaCl2なし)。φBT401は若干結合しているように見えるが、BaTiO3の領域と、Auの領域と同じように結合しており、φ403Nのような材料特異性がみられない。φBT403はペプチド非提示のφM13KEとほとんど結合量が変わらなかった。これには、スクリーニング時とチップの実験時で溶液中のバリウムイオンの濃度が異なることが影響していると考えられた。チップの場合、BaTiO3の表面積は粉末10mgに比べて5桁程度小さく、さらにチップを作製する工程で表面のバリウムはすでに溶出している可能性がある。そのため、粉末とは異なる結果になったと考えられる。そこで、バリウムイオンを添加した状態で、チップへの結合特性を調べた(図19、1mM BaCl2)。すると、BaTiO3部位への結合が回復することが確認された。興味深いことに、Ti金属部位への結合も確認され、BaTiO3への結合量よりもむしろ多かった。これは、チタン金属の表面が酸化しており、この酸化チタン表面に対してバリウムイオンが吸着し、そこを認識しているのだと思われる。よって、φBT401とφBT403の提示する配列は、バリウムなどの金属イオンが吸着したチタン酸化物表面を認識するペプチド配列である可能性があると考えられる。これらは、S、T、Y、Qなどのルイス塩基となるアミノ酸が多いのが特徴で、電荷をもつアミノ酸は比較的少ない。塩基性のアミノ酸は吸着したバリウムイオンと親和性が低く、酸性のアミノ酸は不可逆的な吸着となり得るので(K. Imamura et al., J Biosci Bioeng 1: 7-12 2007)ファージからのセレクションのときに溶出しなかった可能性がある。チタンに吸着するイオンとして、Bi、Cd、Co、Cr、Cu、Fe、Ge、In、Mn、Ni、Pb、Sb、Sn、Te、Tl、V、Zn(E. Vassileva et al., Analyst 121: 607-612, 1996)、Ba、Ca、Sr(MA. Malati and E. Smith, Power Technol 22: 279-282, 1979)が知られている。
【図面の簡単な説明】
【0052】
【図1】ファージクローンφ403N、φ404N及びφ410Nのチタン酸バリウム(BT)への結合力を調べた結果を示す図である。結合率とは、溶出して回収されたファージ量を、投入したファージ量で除したものであり、φM13KEとは、ペプチドを提示していない(野生型の)ファージである。
【図2】本発明のファージクローンφ403Nの提示ペプチドを示す図である。ランダムなペプチド配列をコードするDNA断片が3つタンデムに連なって挿入された配列となっている。
【図3】本発明のファージクローンφ403Nの提示ペプチド(配列番号1)と、9種類の変異ペプチド(配列番号4〜12)の配列を示す図である。
【図4】Si基板上に全面成膜されたBTスパッタ薄膜上にAuとTi薄膜がパターニングされているチップを用いて、φ1599、φ1603、φ1605、φ1611、及びφ1612の特異的結合を調べた結果を示す図である。
【図5】本発明のBTBP−1(配列番号11)とその3つの変異ペプチドΔ(18−22)(配列番号13)、Δ(1−5)(配列番号14)、Δ(1−5、18−22) (配列番号15)のBTへの特異的結合を調べた結果を示す図である。
【図6】本発明の融合ペプチドZZ+BTBP−1がBTに特異的に結合するとともに、抗体を補足することができることを示す図である。
【図7】BTBP−1の2つのグルタミン酸(E)をグルタミン(Q)に置き換えたペプチドBTBP−1(E→Q)(配列番号16)、(DK)6(配列番号17)、(DA)6(配列番号18)、及び(KA)6(配列番号19)の4種類のペプチドのBTへの特異的結合を示す図である。
【図8】BTBP−1及び(KA)6ペプチドのBTへの結合に及ぼす、金属イオン(Mg2+、Ca2+、Ba2+、Ni2+、Zn2+)の影響を示す図である。
【図9】既に結合しているペプチドが、金属イオンを含む溶液によって解離するかどうかを調べた結果を示す図である。「解離処理あり」は、ペプチドを結合させた後、1mMのBaCl2を含むTBSTに15分間浸漬した後に、ストレプトアビジンを反応させた結果である。また、「解離処理なし」は、ペプチドを結合させた後、Ba2+を含まない液に15分間浸した後にストレプトアビジンを反応させた結果である。
【図10】既に結合しているペプチドが、金属イオンを含む溶液によって解離するかどうかを調べた結果を示す図である。あらかじめチップに結合したペプチドにあらかじめストレプトアビジンを反応させた後、1mMのBaCl2を含むTBSTに一晩浸漬した後に結合を調べたものである。
【図11】硫酸イオンの存在下/非存在下での、BTBP−1とBTとの結合におけるペプチド濃度の影響を調査した結果である。
【図12】(a)ビオチン標識BTBP−1とQdotとを予め結合させた後、チップに吸着させた結果、及び(b)ビオチン標識BTBP−1を先にチップに吸着させた後に、Qdotを反応させた結果を示す図である。
【図13】1 μMのBTBP−1溶液に、BaCl2を0、0.03、0.1、0.3、1.0 mMとなるように加え、チップへの結合を調べた結果を示す図である。
【図14】様々なセラミックへのBTBP−1及び(KA)6の結合を調べた結果を示す図である。
【図15】ファージライブラリD−12のlot3及びlot6、ファージライブラリD−C7Cからのスクリーニングにより得られたファージ結合率を示す図である。末尾のSはD−12ライブラリのLot6、TはD−12ライブラリのLot3、CはD−C7Cライブラリから得られたものであることを示す。
【図16】図13に示されるファージクローンの提示アミノ酸配列(配列番号20〜26)を示す図である。
【図17】φ603Cの提示ペプチドを合成し、その結合特性を確認した結果を示す図である。
【図18】チタン酸バリウムの粗い粉末を用いたスクリーニングにより得られたファージクローンのBTへの結合を調べた結果を示す図である。
【図19】1mM BaCl2非存在下/存在下での、ファージクローンのBTへの結合を調べた結果を示す図である。
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)からなるペプチド。
(a)配列番号11に示されるアミノ酸配列;
(b)配列番号19に示されるアミノ酸配列;
【請求項2】
セラミックス表面への可逆的な結合・解離能を有するペプチドであって、以下の(c)又は(d)からなるペプチド。
(c)配列番号11に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列;
(d)配列番号19に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列;
【請求項3】
(c)配列番号11に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列が、配列番号1、14又は16に示されるアミノ酸配列であることを特徴とする請求項2記載のペプチド。
【請求項4】
セラミックス表面への可逆的な結合・解離が、金属イオンにより制御されることを特徴とする請求項2記載のペプチド。
【請求項5】
金属イオンが、Ba2+、Ni2+、Zn2+及びCa2+から選択される少なくとも1以上の金属イオンであることを特徴とする請求項2記載のペプチド。
【請求項6】
セラミックスが、金属酸化物を主成分とするセラミックスであることを特徴とする請求項2記載のペプチド。
【請求項7】
金属酸化物が、遷移金属酸化物であることを特徴とする請求項6記載のペプチド。
【請求項8】
遷移金属酸化物が、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物であることを特徴とする請求項7記載のペプチド。
【請求項9】
検出可能なマーカーで標識された請求項1〜8のいずれか記載のペプチド。
【請求項10】
請求項1〜8のいずれか記載のペプチドと、機能性ペプチド又は機能性タンパク質とを結合させた融合ペプチド。
【請求項11】
請求項1〜8のいずれか記載のペプチドを、その粒子表面上に提示することを特徴とする、セラミックス表面への可逆的な結合・解離能を有するファージ。
【請求項12】
請求項1〜8のいずれか記載のペプチドを認識する抗体。
【請求項13】
請求項1〜8のいずれか記載のペプチドをコードするDNA。
【請求項14】
請求項13記載のDNAを含み、かつセラミックス表面への可逆的な結合・解離能を有するペプチドを発現することができる組換えベクター。
【請求項15】
請求項14記載の組換えベクターが導入された形質転換体。
【請求項16】
セラミックス表面への可逆的な結合・解離能を有するペプチド、又は該ペプチドと機能性タンパク質との融合ペプチドを介しての、セラミックス表面と目的とするタンパク質との結合又は解離を、所定の金属イオンの濃度を調整することによって行うことを特徴とするセラミックス表面とタンパク質との結合・解離を制御する方法。
【請求項17】
セラミックス表面と、セラミックス表面への可逆的な結合・解離能を有するペプチドと目的とするタンパク質との融合ペプチドとの結合又は解離を、所定の金属イオンの濃度を調整することによって行うことを特徴とするセラミックス表面と融合ペプチドとの結合・解離を制御する方法。
【請求項18】
セラミックス表面への可逆的な結合・解離能を有するペプチドとして、請求項1〜9のいずれか記載のペプチドを用いることを特徴とする請求項15又は16記載の方法。
【請求項19】
融合ペプチドとして、請求項10記載の融合ペプチドを用いることを特徴とする請求項15又は16記載の方法。
【請求項20】
金属イオンが、Ba2+、Ni2+、Zn2+、及びCa2+から選択される少なくとも1以上の金属イオンであることを特徴とする請求項16〜19記載の方法。
【請求項21】
セラミックスが、金属酸化物を主成分とするセラミックスであることを特徴とする請求項16〜20のいずれか記載の方法。
【請求項22】
金属酸化物が、遷移金属酸化物であることを特徴とする請求項21記載の方法。
【請求項23】
遷移金属酸化物が、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物であることを特徴とする請求項22記載の方法。
【請求項24】
請求項16〜23のいずれか記載の方法により、セラミックス表面に所定のタンパク質又は所定の細胞を脱着する方法。
【請求項25】
請求項1〜9のいずれか記載のペプチド、又は請求項10記載の融合ペプチドで表面修飾されたセラミックス。
【請求項26】
請求項1〜9のいずれか記載のペプチド、又は請求項10記載の融合ペプチドを介して、表面に所定のタンパク質又は所定の細胞が結合したセラミックス。
【請求項1】
以下の(a)又は(b)からなるペプチド。
(a)配列番号11に示されるアミノ酸配列;
(b)配列番号19に示されるアミノ酸配列;
【請求項2】
セラミックス表面への可逆的な結合・解離能を有するペプチドであって、以下の(c)又は(d)からなるペプチド。
(c)配列番号11に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列;
(d)配列番号19に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列;
【請求項3】
(c)配列番号11に示されるアミノ酸配列において、1若しくは数個のアミノ酸が欠失、置換若しくは付加されたアミノ酸配列が、配列番号1、14又は16に示されるアミノ酸配列であることを特徴とする請求項2記載のペプチド。
【請求項4】
セラミックス表面への可逆的な結合・解離が、金属イオンにより制御されることを特徴とする請求項2記載のペプチド。
【請求項5】
金属イオンが、Ba2+、Ni2+、Zn2+及びCa2+から選択される少なくとも1以上の金属イオンであることを特徴とする請求項2記載のペプチド。
【請求項6】
セラミックスが、金属酸化物を主成分とするセラミックスであることを特徴とする請求項2記載のペプチド。
【請求項7】
金属酸化物が、遷移金属酸化物であることを特徴とする請求項6記載のペプチド。
【請求項8】
遷移金属酸化物が、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物であることを特徴とする請求項7記載のペプチド。
【請求項9】
検出可能なマーカーで標識された請求項1〜8のいずれか記載のペプチド。
【請求項10】
請求項1〜8のいずれか記載のペプチドと、機能性ペプチド又は機能性タンパク質とを結合させた融合ペプチド。
【請求項11】
請求項1〜8のいずれか記載のペプチドを、その粒子表面上に提示することを特徴とする、セラミックス表面への可逆的な結合・解離能を有するファージ。
【請求項12】
請求項1〜8のいずれか記載のペプチドを認識する抗体。
【請求項13】
請求項1〜8のいずれか記載のペプチドをコードするDNA。
【請求項14】
請求項13記載のDNAを含み、かつセラミックス表面への可逆的な結合・解離能を有するペプチドを発現することができる組換えベクター。
【請求項15】
請求項14記載の組換えベクターが導入された形質転換体。
【請求項16】
セラミックス表面への可逆的な結合・解離能を有するペプチド、又は該ペプチドと機能性タンパク質との融合ペプチドを介しての、セラミックス表面と目的とするタンパク質との結合又は解離を、所定の金属イオンの濃度を調整することによって行うことを特徴とするセラミックス表面とタンパク質との結合・解離を制御する方法。
【請求項17】
セラミックス表面と、セラミックス表面への可逆的な結合・解離能を有するペプチドと目的とするタンパク質との融合ペプチドとの結合又は解離を、所定の金属イオンの濃度を調整することによって行うことを特徴とするセラミックス表面と融合ペプチドとの結合・解離を制御する方法。
【請求項18】
セラミックス表面への可逆的な結合・解離能を有するペプチドとして、請求項1〜9のいずれか記載のペプチドを用いることを特徴とする請求項15又は16記載の方法。
【請求項19】
融合ペプチドとして、請求項10記載の融合ペプチドを用いることを特徴とする請求項15又は16記載の方法。
【請求項20】
金属イオンが、Ba2+、Ni2+、Zn2+、及びCa2+から選択される少なくとも1以上の金属イオンであることを特徴とする請求項16〜19記載の方法。
【請求項21】
セラミックスが、金属酸化物を主成分とするセラミックスであることを特徴とする請求項16〜20のいずれか記載の方法。
【請求項22】
金属酸化物が、遷移金属酸化物であることを特徴とする請求項21記載の方法。
【請求項23】
遷移金属酸化物が、チタン酸化物、ジルコニウム酸化物、又はそれらの混合物であることを特徴とする請求項22記載の方法。
【請求項24】
請求項16〜23のいずれか記載の方法により、セラミックス表面に所定のタンパク質又は所定の細胞を脱着する方法。
【請求項25】
請求項1〜9のいずれか記載のペプチド、又は請求項10記載の融合ペプチドで表面修飾されたセラミックス。
【請求項26】
請求項1〜9のいずれか記載のペプチド、又は請求項10記載の融合ペプチドを介して、表面に所定のタンパク質又は所定の細胞が結合したセラミックス。
【図1】



【図2】
【図3】

【図4】
【図5】

【図6】
【図7】

【図8】

【図9】

【図10】

【図11】
【図12】

【図13】
【図14】
【図15】
【図16】

【図17】

【図18】

【図19】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【公開番号】特開2011−217608(P2011−217608A)
【公開日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願番号】特願2008−185845(P2008−185845)
【出願日】平成20年7月17日(2008.7.17)
【出願人】(000006231)株式会社村田製作所 (3,635)
【出願人】(000173588)公益財団法人がん研究会 (34)
【Fターム(参考)】
【公開日】平成23年11月4日(2011.11.4)
【国際特許分類】
【出願日】平成20年7月17日(2008.7.17)
【出願人】(000006231)株式会社村田製作所 (3,635)
【出願人】(000173588)公益財団法人がん研究会 (34)
【Fターム(参考)】
[ Back to top ]