
セルロース結合ドメインが不活性化されたセロビオヒドロラーゼ活性を有するタンパク質
【課題】より比活性に優れたP.chrysosporium由来のセロビオヒドロラーゼの提供。
【解決手段】特定のアミノ酸配列において、セルラーゼ結合領域が不活性化ないし欠失しており、セロビオヒドロラーゼ活性を有するタンパク質。該タンパク質をコードするDNA。該発現用DNAコンストラクトにより形質転換された細胞。
【解決手段】特定のアミノ酸配列において、セルラーゼ結合領域が不活性化ないし欠失しており、セロビオヒドロラーゼ活性を有するタンパク質。該タンパク質をコードするDNA。該発現用DNAコンストラクトにより形質転換された細胞。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、バイオマスに含まれるセルロースを有効利用するためのセロビオヒドロラーゼ及びその利用に関する。
【背景技術】
【0002】
近年、有限である石油資源を代替するものとして、植物の光合成作用に由来するバイオマスへの期待が高まってきており、バイオマスをエネルギーや各種材料に利用するための各種の試みがなされている。なかでも、セルロースの利用が期待されている。セルロースは、糖であるグルコースがβ−1,4グリコシド結合によって縮合した高分子化合物であり、分子間水素結合により強固な結晶構造を構成している。セルロースを分解し、さらに、グルコースに糖化し発酵原料として用いるには、セルロースを単糖まで分解(糖化)するには、エンドグルカナーゼ、セロビオヒドロラーゼ及びβ−グルコシダーゼという、少なくとも3種類のセルラーゼ(セルロース分解酵素)セルラーゼの相乗効果が必要である。
【0003】
自然界においては、クロストリジウム・サーモセラム(Clostridium thermocellum)やトリコデルマ・リーゼイ(Trichoderma reesei)などある種のカビ等が強力なセルラーゼを分泌することが知られている。セルラーゼは、通常、セルロース結合ドメインという、基質であるセルロースに結合するためのドメインを有していることが多い。セルロース結合ドメインを有することにより、水不溶性のセルロースや水溶性のオリゴマーに結合し、セルロースを分解しやすくなると考えられている。しかしながら、こうしたセルロース結合ドメインを有しないセルラーゼも天然に存在しており、例えば、T. reesei由来のエンドグルカナーゼ3やシロアリ腸内細菌由来のセルラーゼなどが挙げられる。
【0004】
一方、本来的に存在するセルロース結合ドメインを人工的に欠失させたエンドグルカナーゼが開示されている(特許文献1)。この文献には、この接合菌由来のエンドグルカナーゼにおいてセルロース結合ドメインを欠失させることでエンドグルカナーゼ活性が向上したことが記載されている。
【0005】
効率的にあるいは低コストにセルロースを利用するには、強力なセルラーゼあるいは相乗効果の高いセルラーゼを提供することが必要である。ここで、セルラーゼの一種であるセロビオヒドロラーゼとして、ファネロケーテ・クリソスポリウム(Phanerochaete chrysosporium)由来のセロビオヒドロラーゼが知られている(非特許文献1)。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開第WO2002/042474号
【非特許文献】
【0007】
【非特許文献1】Uzcatsguiら、J.Biotechnol.19(2-3):271-85, 1991
【発明の概要】
【発明が解決しようとする課題】
【0008】
P. chrysosporium由来のセロビオヒドロラーゼは高い相乗効果を奏するため、さらなる比活性の向上が求められている。
【0009】
また、特許文献1に開示される特定の接合菌由来のエンドグルカナーゼについては、セルロース結合ドメインの欠失がエンドグルカナーゼ活性向上に寄与していた。セルロース結合ドメインは、セルロースに結合する部位ではあるため、通常は、当該部位がセルロース分解活性に寄与するもの考えられ、当該部位の欠失は酵素活性の低下を招くことが予想される。また、セルロース結合ドメインと触媒部位とがセルロースに対して一定の位置関係が、セルロースの分解に寄与することも想定されており、かかる場合には、セルロース結合ドメインの欠失は、著しく酵素活性を低下させることがある。ましてや、セルロース結合ドメインの欠失がセルラーゼ活性の向上に寄与するセルラーゼをアミノ酸配列等から識別することもできなかった。加えて、こうした酵素活性の変化は、基質であるセルロースの状態により変動することもあった。したがって、本来的にセルロース結合ドメインを有するセルラーゼからセルロース結合活性を不活性化させることが、そのセルラーゼの酵素活性の向上に寄与するかどうかは全く予測することはできなかった。
【0010】
そこで、本明細書の開示は、より優れた比活性を備えるセロビオヒドロラーゼを提供し、さらにその利用を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、P. chrysosporium由来のセロビオヒドロラーゼIIの活性向上を目的として、種々検討しているなか、基質過剰条件下において、見かけの酵素活性が低下する傾向があるという知見を得た。人工的な環境下でのセルロースの分解・糖化反応を考慮すると、反応効率を向上させるためには、基質濃度は著しく高い環境下での反応を行うことが求められる。したがって、このセロビオヒドロラーゼをセルロースの糖化に用いるには不利であると考えられた。そこで、本発明者らは、過剰のセルロースの存在下においては、セルロース結合ドメインが存在することにより、セロビオヒドロラーゼがセルロースから離れにくくなり、引き続き生じうる酵素反応を阻害する可能性があるとの推論に至った。そして、このセロビオヒドロラーゼからセルロース結合ドメインを欠失させてセルロース結合活性を低下させたところ、セロビオヒドロラーゼ活性が向上するという知見を得た。本明細書の開示は、これらの知見に基づいて提供される。
【0012】
本明細書の開示によれば、以下の(a)〜(d)のいずれかのアミノ酸配列において、セルロース結合ドメインが不活性化しており、セロビオヒドロラーゼ活性を有するタンパク質。
(a)配列番号2で表されるアミノ酸配列
(b)配列番号2で表されるアミノ酸配列において1又は複数個のアミノ酸の置換、欠失及び/又は付加を有するアミノ酸配列
(c)配列番号2で表されるアミノ酸配列と75%以上の同一性を有するアミノ酸配列
(d)配列番号1で表される塩基配列と相補的な塩基配列からなるDNA又はその一部とストリンジェントな条件下でハイブリダイズするDNAによってコードされるアミノ酸配列
(e)配列番号1で表される塩基配列と75%以上の同一性を有する塩基配列によってコードされるアミノ酸配列
【0013】
本明細書の開示によれば、また、上記タンパク質をコードする、DNA、当該DNAを含む発現用コンストラクト、当該発現用DNAコンストラクトにより形質転換された細胞、上記タンパク質を細胞外に分泌又は細胞表層に提示する細胞も提供される。
【0014】
本明細書の開示によれば、セルロースの分解産物の生産方法であって、上記タンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解する工程、を備える、セルロースの分解産物の生産方法が提供される。
【0015】
本明細書の開示によれば、微生物の発酵により有用物質を生産する方法であって、上記タンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解してセルロース分解産物を取得する工程と、微生物により前記セルロース分解産物を含む炭素源を発酵して、前記有用物質を生産する工程と、を備える、方法が提供される。また、微生物の発酵により有用物質を生産する方法であって、前記微生物の細胞外に分泌又は細胞表層に提示される上記タンパク質を含む、1種又は2種以上のセルラーゼの存在下、前記微生物を用いて、セルロースを少なくとも含む炭素源を発酵して前記有用物質を生産する工程、を備える、方法も提供される。
【0016】
さらに、本明細書の開示によれば、上記タンパク質を含むセルラーゼ組成物及び上記タンパク質を含む、タンパク質複合体も提供される。
【図面の簡単な説明】
【0017】
【図1】CBD部位の有無によるP. chrysosporium由来のセロビオヒドロラーゼIIの試験管内合成量の比較を示す図である。
【図2】CBD部位の有無(保有状態vs欠失状態)によるP. chrysosporium由来のセロビオヒドロラーゼIIのPSCを基質とした場合の相対比活性を示す図である。
【図3】CBD部位の有無によるP. chrysosporium由来のセロビオヒドロラーゼIIのAvicelを基質とした場合の相対比活性を示す図である。
【図4】CBD部位の有無によるT. reesei由来のエンドグルカナーゼIIのPSC、Avicel及びCMCを基質とした場合の相対比活性を示す図である。
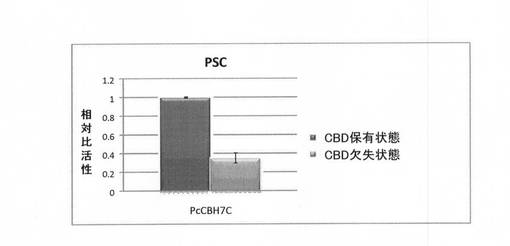
【図5】CBD部位の有無によるP. chrysosporium由来のセロビオヒドロラーゼ7CのPSCを基質とした場合の相対比活性を示す図である。
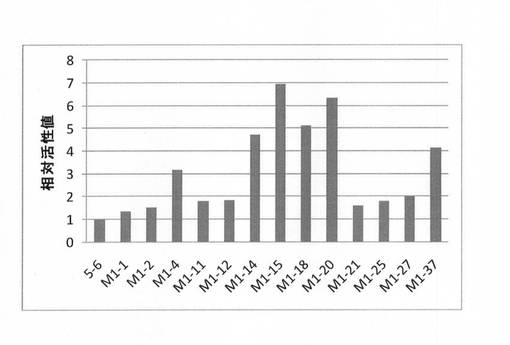
【図6】P. chrysosporium由来のセロビオヒドロラーゼIIの耐熱変異体についての52℃環境下での有利変異を持つ変異体の残存活性値を示す図である。
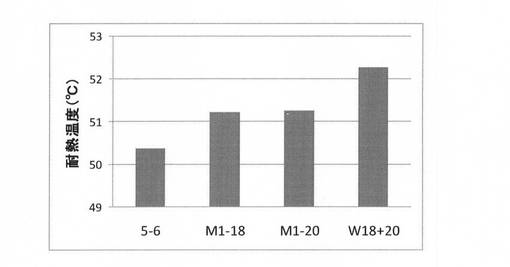
【図7】第2世代の有利変異の蓄積実験結果を示す図である。
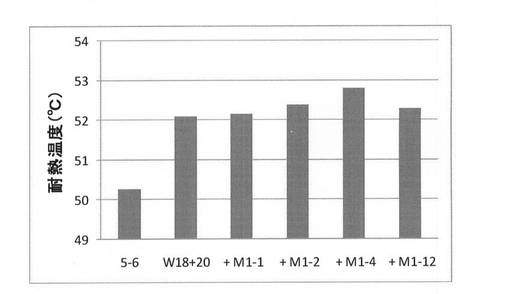
【図8】第3世代の有利変異の蓄積実験結果を示す図である。
【図9】第4世代の有利変異の蓄積実験結果を示す図である。
【図10】第5世代の有利変異の蓄積実験結果を示す図である。
【図11】第6世代の有利変異の蓄積実験結果を示す図である。
【図12】スクリーニング実験の耐熱試験結果を示す図である。
【図13】スクリーニングで得られた変異体の耐熱性評価結果を示す図である。
【図14】各世代の耐熱化変異体の耐熱試験結果を示す図である。
【図15】耐熱化変異体のCBD欠失体の活性評価結果を示す図である。
【図16】CBD欠失変異体のセルロースへの結合力の評価結果を示す図である。
【発明を実施するための形態】
【0018】
本明細書の開示は、バイオマスに含まれるセルロースを有効利用するための活性の高いPhanerochaete chrysosporium由来のセロビオヒドロラーゼ及びその利用に関する。本明細書に開示されるタンパク質は、基質であるセルロースに対する結合活性が不活性化されることで、セルロースに対する分解活性(比活性)が、当該不活性化前よりも高められている。なお、セルロース結合活性の不活性化による比活性に対する効果は、各種セルラーゼにおいて異なり、不活性化の効果を当業者といえども予測することはできなかった。また、本タンパク質におけるセロビオヒドロラーゼ活性の向上程度は、通常の変異導入による改変による活性向上と同等あるいはそれ以上であり、当業者の予測を超えるものであった。
【0019】
Phanerochaete chrysosporium由来のセロビオヒドロラーゼは、本来的に、他の1種又は2種以上のセルラーゼと組み合わせることにより高い相乗効果を発揮してセルロースを分解できるため、本明細書に開示されるタンパク質によれば、さらに活性の高いセルロース分解能を有する組み合わせを提供でき、セルロースの分解、セルロースからの有用物質の生産を効率的に行うことができるようになる。
【0020】
セルロース結合活性の不活性化による、セロビオヒドロラーゼ活性(比活性)の向上には、本来的に有していたセルロース結合ドメインによる、セルロースの過剰存在下における、非生産的なセロビオヒドロラーゼのセルロースへの吸着を抑制するものと推測されるが、推論であって、本発明を拘束するものではない。
【0021】
以下、本明細書の開示に含まれる種々の実施形態について詳細に説明する。なお、本明細書において用いる「GHF(Glycoside Hydrolase Family)」とは、CAZy(Carbohydrate active Enzymes)のホームページ(http://www.cazy.org/fam/acc_GH.html)において提供される、グリコシド加水分解酵素の分類である。
【0022】
(セルロース結合ドメインが不活性化されたPhanerochaete chrysosporium由来のセロビオヒドロラーゼ)
配列番号2で表されるアミノ酸配列は、Phanerochaete chrysosporiumのセロビオヒドロラーゼIIのアミノ酸配列をコードしている(Appl. Environ.Microbaial. 60(12),4387−4393(1994))。Phanerochaete chrysosporiumのセロビオヒドロラーゼIIは、配列番号2で表されるアミノ酸配列において、N末端側にセルロース結合ドメインを有し、リンカー領域を介して活性ドメインを有している。本タンパク質は、Phanerochaete chrysosporiumから取得したセロビオヒドロラーゼ IIのアミノ酸配列(配列番号2)において、セルロース結合活性が不活性化されているが、セロビオヒドロラーゼ活性を有するタンパク質である。このような本タンパク質は、配列番号2で表されるアミノ酸配列において以下に説明するような各種態様でセルロース結合ドメインが不活性化され、しかも、セロビオヒドロラーゼ活性を維持するアミノ酸配列を有するタンパク質である。
【0023】
(セルロース結合ドメインの不活性化の態様)
本タンパク質における、セルロース結合ドメインの不活性化の態様としては、セルロース結合ドメインの部分的又は全体の欠失が挙げられる。セルロース結合ドメインは、配列番号2で表されるアミノ酸配列の2番目から37番目までのアミノ酸36残基の領域とされており、不活性化の態様としては、このアミノ酸配列において1個以上、典型的には、複数個から全体(100%)に及ぶアミノ酸の欠失が挙げられる。好ましくは、2以上の連続したアミノ酸の欠失である。たとえば、セルロース結合ドメインを除去したアミノ酸配列を配列番号3に示す。
【0024】
アミノ酸が欠失されている領域やアミノ酸の数は、セルロース結合活性を評価することにより決定することができる。セルロース結合ドメインが部分的に欠失されている場合、セルロース結合ドメインをコードするアミノ酸配列の20%以上、好ましくは30%以上、より好ましくは40%以上、さらに好ましくは50%以上、一層好ましくは60%以上、より一層好ましくは70%以上、さらに一層好ましくは80%以上、さらに好ましくは90%以上、欠失されている。典型的には、セルロース結合ドメインのN末端側又はC末端側から連続してアミノ酸が欠失されているが、好ましくは、N末端側から連続して複数個のアミノ酸が欠失されている。
【0025】
セルロース結合ドメインの不活性化は、タンパク質においてセルロース結合ドメインの一部又は全体が欠失している態様が好ましい。アミノ酸の欠失により、ペプチド鎖が短くなり、異種タンパク質としての合成量を増大させることができる。特に、本タンパク質を、試験管内におけるタンパク質生産や遺伝子工学的に異種タンパク質として微生物宿主等において生産させる場合には、好適である。
【0026】
あるタンパク質がセルロース結合ドメイン活性(セルロース結合活性)を有しているか否か又はその程度は、例えば、タンパク質とアビセルとの結合力を測定することで評価することができる。具体的には、酢酸緩衝液pH5.0でけん濁したアビセルにタンパク質を添加し攪拌そして氷上にて一定時間静置後の遠心上清のタンパク質量を測定することで評価できる。その遠心上清にタンパク質がないかあるいは少ない時にはセルロース結合ドメイン活性を有すると言える。
【0027】
セルロース結合活性の不活性化の他の態様としては、セルロース結合ドメインにおける1以上のアミノ酸の置換及び/又は挿入が挙げられる。置換又は挿入されているアミノ酸の部位や個数、置換又は挿入されて新たにセルロース結合ドメイン中に配置されるアミノ酸の種類は、得られた変異体タンパク質につき、セルロース結合活性を評価することで適宜決定することができる。
【0028】
セルロース結合ドメインの不活性化は、配列番号2で表されるアミノ酸配列からなるセロビオヒドロラーゼIIのセルロース結合活性と比較したとき、当該結合活性より低くなっていればよく、その程度は問わないが、好ましくは、配列番号2で表されるアミノ酸配列からなるセロビオヒドロラーゼIIのセルロース結合活性の60%以下、より好ましくは50%以下、さらに好ましくは40%以下、一層好ましくは30%以下、より一層好ましくは20%以下、さらに一層好ましくは10%以下、最も好ましくは5%以下である。
【0029】
本タンパク質は、セロビオヒドロラーゼ活性を有している。セロビオヒドロラーゼ活性は、例えば、50mM酢酸緩衝液pH5.0で100倍希釈後、1%リン酸膨潤セルロース(PSC)又は2.5%Avicelを同量添加し(0.5%PSC又は1.25%Avicel)、40℃で16時間反応させ、反応液を、TZアッセイ法(Journal of Biochemical and Biophysical Methods, 11 (1985) 109-115)にて遠心上精の還元糖量を測定することにより、測定できる。セロビオヒドロラーゼ活性の程度は、特に問わないが、セルロース結合活性の不活性化により、本タンパク質のセロビオヒドロラーゼ活性が向上することがわかっている。本タンパク質のセロビオヒドロラーゼ活性は、配列番号2で表されるアミノ酸配列からなるセロビオヒドロラーゼIIのセロビオヒドロラーゼ活性と比較したとき、好ましくは、当該活性の同等又はそれ以上であり、より好ましくは110%以上、さらに好ましくは120%以上、一層好ましくは130%以上、より一層好ましくは140%以上、さらに一層好ましくは150%以上、最も好ましくは160%以上である。
【0030】
本タンパク質は、また、配列番号2で表されるアミノ酸配列と一定の関係を有するアミノ酸配列においてセルロース結合ドメインが不活性化されつつ、セロビオヒドロラーゼ活性を有するものであってもよい。すなわち、例えば、配列番号2で表されるアミノ酸配列を有するタンパク質の変異体であって、セルロース結合活性やセロビオヒドロラーゼ活性を有する改変体において、セルロース結合ドメインが不活性化された態様が挙げられる。典型的には、セロビオヒドロラーゼ活性ドメインにおいては有効な変異を有し、かつセルロース結合ドメインには、セルロース結合活性が不活性化されるような変異を備えるタンパク質が挙げられる。このような本タンパク質においては、配列番号2で表されるアミノ酸配列と一定関係のあるアミノ酸配列において各種態様でセルロース結合ドメインが不活性化されており、しかも、セロビオヒドロラーゼ活性を維持するアミノ酸配列を有している。
【0031】
こうした本タンパク質の他の一態様としては、配列番号2で表されるアミノ酸配列に対して1又は数個のアミノ酸が欠失、置換及び/又は付加されたアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質が挙げられる。配列番号2で表されるアミノ酸配列に対するアミノ酸の変異は、欠失、置換及び/又は付加は、いずれか1種類であってもよいし、2種類以上が組み合わされていてもよい。また、これらの変異の総数は、特に限定されないが、好ましくは、1個以上25個以下程度である。より好ましくは、1個以上20個以下である。さらに好ましくは1個以上15個以下であり、一層このましくは10個以下であり、より一層好ましくは5個以下である。アミノ酸置換の例としては、保存的置換が好ましく、具体的には以下のグループ内での置換が挙げられる。(グリシン、アラニン)(バリン、イソロイシン、ロイシン)(アスパラギン酸、グルタミン酸)(アスパラギン、グルタミン)(セリン、トレオニン)(リジン、アルギニン)(フェニルアラニン、チロシン)。このような態様としては、たとえば、セロビオヒドロラーゼの活性ドメインやセルロース結合ドメインに隣接するリンカードメインの一部又は全部の欠失などが挙げられる。また、セロビオヒドロラーゼ活性向上のためのアミノ酸置換の導入等が挙げられる。
【0032】
さらに他の一態様としては、配列番号2で表されるアミノ酸配列に対して70%以上の同一性を有するアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質が挙げられる。同一性は好ましくは80%以上であり、より好ましくは85%以上であり、さらに好ましくは、90%以上であり、一層好ましくは95%以上である。最も好ましくは、98%以上である。
【0033】
本明細書において同一性又は類似性とは、当該技術分野で知られているとおり、配列を比較することにより決定される、2以上のタンパク質あるいは2以上のポリヌクレオチドの間の関係である。当該技術で“同一性 ”とは、タンパク質またはポリヌクレオチド配列の間のアラインメントによって、あるいは場合によっては、一続きのそのような配列間のアラインメントによって決定されるような、タンパク質またはポリヌクレオチド配列の間の配列不変性の程度を意味する。また、類似性とは、タンパク質またはポリヌクレオチド配列の間のアラインメントによって、あるいは場合によっては、一続きの部分的な配列間のアラインメントによって決定されるような、タンパク質またはポリヌクレオチド配列の間の相関性の程度を意味する。より具体的には、配列の同一性と保存性(配列中の特定アミノ酸又は配列における物理化学特性を維持する置換)によって決定される。なお、類似性は、後述するBLASTの配列相同性検索結果においてSimilarity と称される。同一性及び類似性を決定する方法は、対比する配列間で最も長くアラインメントするように設計される方法であることが好ましい。同一性及び類似性を決定するための方法は、公衆に利用可能なプログラムとして提供されている。例えば、AltschulらによるBLAST (Basic Local Alignment Search Tool) プログラム(たとえば、Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ., J. Mol. Biol., 215: p403-410 (1990), Altschyl SF, Madden TL, Schaffer AA, Zhang J, Miller W, Lipman DJ., Nucleic Acids Res. 25: p3389-3402 (1997))を利用し決定することができる。BLASTのようなソフトウェアを用いる場合の条件は、特に限定するものではないが、デフォルト値を用いるのが好ましい。
【0034】
さらに他の一態様として、配列番号2で表されるアミノ酸配列をコードする塩基配列からなるDNAと相補的な塩基配列からなるDNA又はその一部とストリンジェントな条件でハイブリダイズするDNAによってコードされるアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質が挙げられる。ストリンジェントな条件とは、たとえば、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、塩基配列の同一性が高い核酸、すなわち、配列番号2で表されるアミノ酸配列をコードする塩基配列と80%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましく95%以上、最も好ましくは97%以上の同一性を有する塩基配列からなるDNAの相補鎖がハイブリダイズし、それより相同性が低い核酸の相補鎖がハイブリダイズしない条件が挙げられる。より具体的には、ナトリウム塩濃度が15〜750mM、好ましくは50〜750mM、より好ましくは300〜750mM、温度が25〜70℃、好ましくは50〜70℃、より好ましくは55〜65℃、ホルムアミド濃度が0〜50%、好ましくは20〜50%、より好ましくは35〜45%での条件をいう。さらに、ストリンジェントな条件では、ハイブリダイゼーション後のフィルターの洗浄条件が、通常はナトリウム塩濃度が15〜600mM、好ましくは50〜600mM、より好ましくは300〜600mM、温度が50〜70℃、好ましくは55〜70℃、より好ましくは60〜65℃である。なお、以上のことから、さらなる他の一態様として、配列番号2で表されるアミノ酸配列をコードする塩基配列と80%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましく95%以上、最も好ましくは97%以上の同一性を有する塩基配列を有するDNAによってコードされるアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質が挙げられる。
【0035】
なお、Phanerochaete chrysosporium由来のセロビオヒドロラーゼIIの活性ドメインにおける改変体のアミノ酸配列としては、特開2010-41996号公報に開示される各種の改変体のアミノ酸配列が挙げられる。また、配列番号2で表されるアミノ酸配列において10個のアミノ酸置換(T130I/S134Q/S183T/A241E/M259I/Q307R/A309S/T342H/Q381S/F382L)を有するアミノ酸配列及び同アミノ酸配列において15個のアミノ酸置換(T112S/N125K/T130I/S134Q/S183T/Q204K/F206Y/A241E/M259I/Q307R/A309S/T342H/I365V/Q381S/F382L)を含むアミノ酸配列が挙げられる。
【0036】
本タンパク質は、P. chrysosporiumから取得したセロビオヒドロラーゼ IIのアミノ酸配列(配列番号2)において、以下の表1に示される1又は2以上のアミノ酸置換に相当するアミノ酸置換を備えることができる。「アミノ酸配列において1又は2以上のアミノ酸置換に相当するアミノ酸置換を備える」とは、配列番号2で表されるアミノ酸配列に表1から選択される1又は2以上のアミノ酸置換を備えるほか、配列番号2で表されるアミノ酸配列と一定の関係を有するアミノ酸配列において、配列番号2で表されるアミノ酸配列と対比したときに、表1から選択される1又は2以上のアミノ酸置換に相当するアミノ酸置換を備えていることをいう。
【0037】
なお、アミノ酸置換を備えるとは、当該位置において、置換前のアミノ酸と置換後のアミノ酸とをそれぞれ特定して意味するほか、結果として「アミノ酸置換」によって特定されるアミノ酸を備えていればよいことを意味する。すなわち、「M259L」のアミノ酸置換を備えるとは、配列番号2において259位に相当する部位にL(ロイシン)を備えていることを意味する。以下、本タンパク質が備えることのできる変異であるアミノ酸置換について説明する。
【0038】
【表1】

【0039】
本タンパク質は、表2に示す各欄に示す1又は2以上のアミノ酸置換を備えるものであってもよい。なかでも、本タンパク質は、好ましくは、少なくとも、M259L、M259I、T342H、Q381S+F382Lから選択される1種又は2種以上に相当するアミノ酸置換を備えている。これらの各変異に組み合わせて、好ましくは、T130I及びS134の双方又はいずれかを含んでいることが好ましい。さらに、追加して、好ましくは、M259Iの変異を含んでいる。さらにまた、好ましくは、A241Eの変異を含んでいることが好ましい。さらに追加して、好ましくは、Q307R+A309Sの双方又はいずれかの変異を含んでいる。さらにまた、加えて、S183Tの変異を含んでいることが好ましい。さらに追加して、T112S、N125K、Q204K、I365V及びF206Yからなる群から選択される1又は2以上の変異を含んでいることが好ましく、より好ましくは3種以上であり、さらに好ましくは4種以上であり、一層好ましくは5種である。
【0040】
【表2】
【0041】
以上のことから、好ましい変異体としては、たとえば、少なくとも以下の表3に示す組み合わせのアミノ酸置換を備えるものが挙げられる。なかでも、第3世代以降が好ましく、より好ましくは第4世代以降、さらに好ましくは第5世代以降、一層好ましくは第6世代以降、最も好ましくは第7世代以降である。表3中、Sept25の組み合わせのアミノ酸置換を備え、さらに、T112S、N125K、Q204K、I365V及びF206Yからなる群から選択される1又は2以上のアミノ酸置換を備えるタンパク質も好ましい。
【表3】
【0042】
配列番号2で表されるアミノ酸配列において部分的な欠失を生じさせたり、アミノ酸の置換、付加を導入するには、公知の各種の手法を採用できる。例えば、配列番号2で表されるアミノ酸配列や当該配列と一定の関係を有するアミノ酸配列をコードするDNA等の遺伝子情報を改変する方法を用いることができる。例えば、アミノ酸配列において欠失部位を形成するには、DNAの一部をPCR等により増幅したり、あるいはこうして部分的に増幅したDNA断片を適宜結合することで任意の部位に欠失を導入することができる。また、例えば、アミノ酸配列に置換及び/又は付加などの変異を導入して遺伝子情報を改変して本発明のタンパク質を得るには、Kunkel法、Gapped duplex法等の公知の手法又はこれに準ずる方法を採用することができる。例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutan−K(TAKARA社製)やMutan−G(TAKARA社製))などを用いて変異の導入が行われる。また、エラー導入PCRやDNAシャッフリング等の手法により、遺伝子の変異導入やキメラ遺伝子を構築することもできる。エラー導入PCR及びDNAシャッフリング手法は、当技術分野で公知の手法であり、例えばエラー導入PCRについてはChen K, and Arnold FH. 1993, Proc. Natl. Acad. Sci. U. S. A., 90: 5618-5622を、またDNAシャフリングやカセットPCR等の分子進化工学的手法は、例えば、Kurtzman,A.L.,Govindarajan, S., Vahle, K., Jones, J. T., Heinrichs, V., Patten P. A.,Advances in directed protein evolution by recursive genetic recombination: applications to therapeutic proteins. Curr. Opinion Biotechnol.,12, 361-370, 2001、及び、Okuta, A., Ohnishi, A. and Harayama, S., PCR isolation of catechol 2,3-dioxygenase gene fragments from environmental samples and their assembly into functional genes. Gene, 212, 221-228, 1998を参照することができる。なかでも、エラープローンPCR等によりランダム変異を導入する分子進化的手法を利用する無細胞タンパク質合成系を採用することが好ましい。エラープローンPCRに適用する無細胞タンパク質合成系としては、公知のあるいは本出願人が出願した特開2006−61080号公報及び特開2003−116590号公報に記載のタンパク質合成系を用いることができる。本出願人によるこれらの無細胞タンパク質合成系を用いることで活性型の酵素を容易に得ることができる。このため、これらのタンパク質合成系が適用されたエラープローンPCRは、本発明のタンパク質を取得する手法として好ましく用いることができる。
【0043】
なお、本タンパク質は、決定されたアミノ酸配列や塩基配列に基づいて、化学的な合成方法のほか、遺伝子工学的な方法によって製造され、必要に応じて精製される。
【0044】
本タンパク質は、セルロース結合ドメインが不活性化されているために、配列番号2で表されるアミノ酸配列からなるPhanerochaete chrysosporium由来のセロビオヒドロラーゼIIやその改変体よりも、基質であるセルロースへの結合能が低減又は喪失している。一方、本タンパク質は、セロビオヒドロラーゼ活性及びエンドグルカナーゼやβ−グルコシダーゼとの相乗効果においても、配列番号2で表されるアミノ酸配列からなるタンパク質と同等以上の活性を有している。
【0045】
(セルラーゼ組成物)
本明細書の開示によれば、本タンパク質と、さらに、他のセルラーゼ、たとえば、他のセロビオヒドロラーゼ、エンドグルカナーゼ及びβ−グルコシダーゼからなる群から選択される1種又は2種以上を組み合わせの組成物が提供される。本組成物によれば、本タンパク質の優れたセロビオヒドロラーゼ活性により高いセルロース分解活性を呈することができる。本組成物に含まれる本タンパク質以外のセルラーゼとしては、特に限定しないで、公知のセルラーゼから適宜選択される。例えば、Phanerochaete chrysosporium由来でない他起源のエンドグルカナーゼが挙げられる。他起源由来のエンドグルカナーゼとしては、公知の各種エンドグルカナーゼが挙げられ、これらを単独であるいは2種類以上を適宜組み合わせて用いることができる。たとえば、GHF5に属するエンドグルカナーゼが挙げられる。GHF5に分類されるエンドグルカナーゼのなかでも、好ましくは、Trichoderma reesei由来のエンドグルカナーゼ、Aspergillus oryzae由来のエンドグルカナーゼ及びAspergillus niger由来のエンドグルカナーゼを好ましく用いることができる。より好ましくはTrichoderma reesei由来のエンドグルカナーゼAspergillus niger由来のエンドグルカナーゼである。GHF5に分類されるエンドグルカナーゼは、こうしたエンドグルカナーゼから選択される1種又は2種以上を適宜組み合わせて用いることができる。
【0046】
エンドグルカナーゼとしては、GHF12に属するエンドグルカナーゼが挙げられる。なかでも、Trichoderma reesei由来のエンドグルカナーゼ、Aspergillus niger由来のエンドグルカナーゼ及びAspergillus oryzae由来のエンドグルカナーゼが挙げられる。また、Phanerochaete chrysosporium由来のエンドグルカナーゼであってもよい。こうしたエンドグルカナーゼから選択される1種又は2種以上を適宜組み合わせて用いることができる。エンドグルカナーゼとしては、GHF7及びGHF45に属するエンドグルカナーゼであってもよい。なかでも、Trichoderma reesei由来のエンドグルカナーゼを好ましく用いることができる。
【0047】
本セルラーゼ組成物は、Phanerochaete chrysosporium由来のエンドグルカナーゼを含んでいてもよい。Phanerochaete chrysosporium由来のエンドグルカナーゼは、セロビオヒドロラーゼII及びその改変体をPhanerochaete chrysosporiumの培養物等から取得した場合においてセロビオヒドロラーゼII及びその改変体とともに容易に取得できる。
【0048】
本セルラーゼ組成物は、エンドグルカナーゼ以外の他のセルラーゼを含むことができる。例えば、GHF7に属するセロビオヒドロラーゼIを含有することができる。セロビオヒドロラーゼIは、セロビオヒドロラーゼIが、セロビオヒドロラーゼII等と協動することにより、一層セルロース分解の相乗効果が発揮される。セロビオヒドロラーゼIは、Phanerochaete chrysosporium由来であってもよいし、他起源であってもよい。
【0049】
本セルラーゼ組成物は、Phanerochaete chrysosporium以外の他起源のセルラーゼ生産菌の培養物(培養上清であってもよい)から取得された2種類以上のセルラーゼを含有していてもよい。セルラーゼ生産菌としては、特に限定されないで、適宜選択できるが、エンドグルカナーゼの起源としてTrichoderma reesei、Aspergillus aculeatus、Aspergillus niger、Aspergillus oryzae等が好ましく挙げられる。より好ましくは、Trichoderma reeseiである。
【0050】
本セルラーゼ組成物は、化学的あるいは遺伝子工学的に本タンパク質及び必要に応じて他のセルラーゼを生産し、組み合わせることによって得ることができる。本セルラーゼ組成物を構成する全てのセルラーゼを同一の宿主細胞で遺伝子工学的に生産し、その培養上清や培養菌体を回収することによって、本セルラーゼ組成物を得るようにしてもよい。
【0051】
本セルラーゼ組成物は、本タンパク質及び他のセルラーゼを、それぞれ精製したものとして含有していてもよいし、未精製タンパク質として他タンパク質やその他の成分を含んでいてもよい。また、その製剤形態は、特に限定されず、固形製剤(粉末(凍結乾燥体等)、タブレット等、顆粒等)であってもよいし、溶液(流通時においては凍結体であることが好ましい。)であってもよい。
【0052】
(タンパク質複合体)
本明細書に開示されるタンパク質複合体は、本タンパク質と、本タンパク質をコヘシン−ドックリン結合によりコヘシンタンパク質上に備えている。ある種の細菌が、複数種類のセルラーゼを保持するタンパク質構造体を自己細胞表層に構築することが知られており、当該タンパク質構造体がセルロソームとして知られている。セルロソームは、セルラーゼが、スキャホールディンタンパク質に保持されて構成されており、セルロソームとスキャホールディンタンパク質とは、それぞれが備えるドックリンドメインとコヘシンドメインとの間の結合、すなわち、コヘシン−ドッケリン結合により結合されている。公知のセルロソームにおけるドックリンドメイン及びコヘシンドメインのアミノ酸配列は既にいくつか開示されている。コヘシン−ドックリン結合は、非共有結合性であって、そのアミノ酸配列に依存した水素結合等に基づくと考えられる。したがって、こうした開示に従い、ドックリンドメインを付加した本タンパク質と1又は2以上のコヘシンドメインを備えるスキャホールディンタンパク質とにより、人工的なセルロソーム、すなわち、タンパク質複合体を構築することができる。
【0053】
本タンパク質複合体においては、本タンパク質には、ドックリンドメインが付加されている。ドックリンドメインは、コヘシン−ドックリン結合により後述するコヘシンタンパク質に本タンパク質を結合させる部位である。ドックリンドメインは、例えば、公知のセルロソーム生産微生物のセルロソームを構成するセルラーゼの一部に備えられている。本セルラーゼ複合体に用いるドックリンドメインとしては、公知の各種のセルロソーム生産微生物のセルラーゼのドックリンドメインから選択される。例えば、C. thermocellumのエンドグルカナーゼのドックリンドメインを含むアミノ酸配列が挙げられる。ドックリンリンドメインは、活性ドメインのN末端側及びC末端側のいずれの側にあってもよいが、好ましくは、C末端側に配置される。
【0054】
本タンパク質複合体は、本セルラーゼ組成物が含みうる他のセルラーゼがコヘシンタンパク質に保持されていてもよい。他のセルラーゼに関しても、本タンパク質と同様に、ドックリンドメインが付加されている。他のセルラーゼに付加されるドックリンドメインは、本タンパク質に付加されるドックリンドメインと同一であってもよいし異なっていてもよい。
【0055】
コヘシンタンパク質は、コヘシンタンパク質は、本タンパク質が備えるドックリンドメインを結合する1又は2以上のコヘシンドメインを有している。これにより、コヘシンタンパク質は、本タンパク質をコヘシン−ドックリン結合で保持でき、本タンパク質複合体の骨格として機能する。また、本タンパク質複合体は、他のセルラーゼを保持するための異なるコヘシンドメインの組み合わせあるいは配列を有するコヘシンタンパク質を備えていてもよい。
【0056】
(コヘシンドメイン)
コヘシンタンパク質が備える、1又は2以上のコヘシンドメインは、セルロソームのスキャホールディンタンパク質が備えるコヘシンドメインに由来している。コヘシンドメインは、セルロソーム生産微生物の形成するセルロソームにおけるタイプI〜III骨格タンパク質に備えられる触媒活性のあるセルラーゼ等を非共有結合で結合するドメインとして知られている(粟冠ら、蛋白質核酸酵素、Vol.44、No.10(1999)、p41-p50、Demain, A. L., et al., Microbiol Mol. Biol Rev., 69(1), 124-54(2005), Doi, R. H., et al., J. Bacterol., 185(20), 5907-5914(2003)等)。すなわち、コヘシンドメインとしては、セルロソームのタイプI骨格タンパク質上のタイプIコヘシンドメイン、同タイプII骨格タンパク質上のタイプIIコヘシンドメイン及びタイプIII骨格タンパク質上のタイプIIIコヘシンドメインが挙げられる。こうした各種タイプのコヘシンドメインとしては、各種セルロソーム生産微生物において多数その配列が決定されている。これらの各種のタイプのコヘシンのアミノ酸配列及びDNA配列は、NCBIのHP(http://www.ncbi.nlm.nih.gov/)等を介してアクセス可能な各種のタンパク質データベースやDNA配列のデータベースにより容易に取得することができる。
【0057】
コヘシンドメインは、こうしたコヘシンドメインに由来するドメインであって、キメラタンパク質のドックリンドメインと結合することができる。セルラーゼの活性ドメインへのドックリンドメインのキメラ化は活性ドメインの活性が意図せずに低下することなどにより困難な場合があることから、キメラタンパク質のドックリンドメインに応じてコヘシンドメインを選択することができる。また、コヘシン−ドックリン結合の強度等も考慮してコヘシンドメインを選択することができる。
【0058】
本タンパク質複合体にあっては、例えば、キメラタンパク質のドックリンドメインが例えば、C. thermocellumのcelAのドックリンドメインに対して、C. thermocellumのスキャホールディンタンパク質のコヘシンドメインが挙げられる。
【0059】
本タンパク質複合体は、タンパク質の複合体として、それ自体独立した形態であってもよいし、適当なキャリアに固定化ないし保持されていてもよい。また、後述するように、酵母等の微生物の表層に提示された状態であってもよい。本セルラーゼ複合体は、例えば、それぞれのタンパク質を公知のタンパク質製造方法により取得し、これらのタンパク質を接触させる条件下において、コヘシンタンパク質に対してキメラタンパク質を自己集合させることにより、本セルラーゼ複合体を取得できる。
【0060】
また、本タンパク質複合体は、その構成タンパク質のうち1又は2以上を分泌発現する微生物の培養上清又はそのタンパク質精製物を混合してすべての構成タンパク質を接触させて自己集合させることによって取得できる。なお、構成タンパク質のすべてが微生物によって分泌発現されなくてもよく、必要に応じ微生物によって生産されない構成タンパク質を別途製造して混合してもよい。また、構成タンパク質のすべてを分泌発現する微生物の場合、この微生物の培養上清に、これらタンパク質が自己集合可能な状態で含まれるため、その培養上清に、本タンパク質複合体を取得できる。
【0061】
(本タンパク質をコードするDNA、当該DNAを含む発現用コンストラクト)
本明細書に開示されるDNAは、本タンパク質をコードしている。かかるDNAは、本タンパク質をコードしている限り、異なるコドンを用いてコードされるDNAも含まれる。本明細書に開示される発現用コンストラクトは、本タンパク質をコードするDNAを含んでいる。本タンパク質をコードするDNAは、発現用コンストラクトを導入しようとする宿主細胞において本タンパク質を発現可能に保持されていればよい。例えば、宿主細胞で作動可能なプロモーターの制御下に連結されるとともに適切なターミネーターをその下流に有した状態で保持されている。プロモーターは、構成的プロモーターであっても誘導的プロモーターであってもよい。本発現用コンストラクトにおいて、このような状態のDNAは、宿主染色体内に組み込み可能に備えられていてもよいし、染色体外に保持可能に備えられていてもよい。本発現用コンストラクトには、こうした外来DNAの導入に伴って、宿主において利用可能な選択マーカー遺伝子も同時に保持されていることが好ましい。本発現用コンストラクトは、特に真核微生物、好ましくは、酵母での本タンパク質の発現用であることが好ましい。
【0062】
本発現用コンストラクトは、本タンパク質を既に説明したタンパク質複合体を構成可能な形態で発現可能に構築されていてもよい。すなわち、本タンパク質に対してドックリンドメインを付加した融合タンパク質を発現可能に形成されていてもよい。さらに、タンパク質複合体を構成可能な発現用コンストラクトとして、コヘシンタンパク質をコードするDNAを含む、コヘシンタンパク質の発現用コンストラクトを構築してもよい。こうした発現コンストラクトにおいて、融合タンパク質やコヘシンタンパク質のコード化DNAは、本タンパク質をコードするDNAと同様、各種形態でコンストラクトに備えられる。
【0063】
(発現用DNAコンストラクトにより形質転換された細胞、上記タンパク質を細胞外に分泌又は細胞表層に提示する真核微生物)
本明細書に開示される形質転換細胞は、本発現用DNAコンストラクトによって形質転換された細胞である。本形質転換細胞は、本発現用DNAコンストラクトによって形質転換されることにより、本タンパク質を発現可能となっている。
【0064】
本形質転換細胞は、本タンパク質又は本タンパク質複合体を発現可能な細胞であればよく、その種類を限定しないが、本タンパク質の生産及び回収を考慮すると、大腸菌などの細菌等が挙げられるが、好ましくは、本タンパク質によるセルロースの分解及び資化のための真核微生物である。真核微生物は、特に限定しないが、例えば、公知の各種酵母を利用できる。後述するエタノール発酵等を考慮すると、サッカロマイセス・セレビジエ(Saccharomyces cerevisiae)等のサッカロマイセス属の酵母、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)等のシゾサッカロマイセス属の酵母、キャンディダ・シェハーテ(Candida shehatae)等のキャンディダ属の酵母、ピヒア・スティピティス(Pichia stipitis)等のピヒア属の酵母、ハンセヌラ(Hansenula)属の酵母、トリコスポロン(Trichosporon)属の酵母、ブレタノマイセス(Brettanomyces)属の酵母、パチソレン(Pachysolen)属の酵母、ヤマダジマ(Yamadazyma)属の酵母、クルイベロマイセス・マーキシアヌス(Kluyveromyces marxianus)、クルイベロマイセス・ラクティス(Kluveromyces lactis)等のクルイベロマイセス属の酵母が挙げられる。なかでも、工業的利用性等の観点からサッカロマイセス属酵母が好ましい。なかでも、サッカロマイセス・セレビジエが好ましい。また、真核微生物は、アスペルギルス・オリゼ(Aspergillus oryzae)、アスペルギルス・アキュリータス(Aspergillus aculeatus)、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・ニジュランス(Aspergillus nidulans)、アスペルギルス・ソーヤ(Aspergillus soya)、アスペルギルス・カワチ(Aspergillus kawachii)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・サイトイ(Aspergillus saitoi)等の麹菌であってもよい。異種タンパク質を大量に発現させるには、セルラーゼ非生産菌がより好ましい。また、Trichoderma reeseiなどのセルラーゼ生産菌で生産させることも好ましい。本タンパク質は、セルロース結合ドメインが不活性化されているため、セルラーゼ生産菌が生産するセルロース結合ドメインを有するセルラーゼとは異なる場所で働くことができる。したがって、セルラーゼ生産菌において本タンパク質を生産させることで、全体としてセルロース分解活性の高いセルラーゼの組み合わせ(セルラーゼ製剤)を提供できることになる。
【0065】
本形質転換細胞は、本形質転換細胞のセルロースの分解への利用を考慮すると、本タンパク質を細胞外に分泌又は細胞表層に提示するものであることが好ましい。本タンパク質に細胞外分泌性や細胞表層提示性を付与するには、公知の分泌シグナルや表層提示用のシステムを用いることができる。例えば、分泌シグナルや凝集性タンパク質又はその一部のアミノ酸配列が付与される。分泌シグナルとしては、例えば、Rhizopus oryzaeやC. albicansのグルコアミラーゼ遺伝子の分泌シグナル、酵母インベルターゼリーダー、α因子リーダーなどが挙げられる。また、凝集性タンパク質としては、α−アグルチニンをコードするSAG1遺伝子の5’領域の320アミノ酸残基からなるペプチドが挙げられる。また、所望のタンパク質を細胞表層に提示するためのポリペプチドや手法は、WO01/79483号公報や、特開2003−235579号公報、WO2002/042483号パンフレット、WO2003/016525号パンフレット、特開2006−136223号公報、藤田らの文献(藤田ら,2004. Appl Environ Microbiol 70:1207-1212および藤田ら, 2002. Appl Environ Microbiol 68:5136-5141.)、村井ら, 1998. Appl Environ Microbiol 64:4857-4861.に開示されている。
【0066】
本形質転換細胞は、本タンパク質複合体を細胞外に分泌又は細胞表層に提示するものであることも好ましい。本形質転換細胞は、好ましくは、コヘシンタンパク質を細胞表層提示し、本タンパク質を細胞外に分泌して、コヘシンタンパク質に本タンパク質をコヘシン−ドッケリン結合を介して保持させて、タンパク質複合体を細胞表層に提示する。
【0067】
本タンパク質、本タンパク質複合体及びこれらを細胞外に分泌又は細胞表層に提示する細胞、好ましく酵母などの真核微生物は、向上されたセロビオヒドロラーゼ活性及び増強された相乗効果により、セルロースの分解、糖化、及び糖化と発酵とを同時進行させるCBP(連結バイオプロセス(糖化発酵同時進行))において、セルロースの分解効率を高めて、セルロースの効率的利用を具現化することができる。
【0068】
以上説明した本明細書に開示される形質転換細胞は、いずれも、モレキュラークローニング第3版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載されている方法に準じて作製することができる。真核微生物などの宿主細胞の形質転換のためのベクター及びその構築方法は、当業者において周知であって、モレキュラークローニング第3版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に開示されている。また、コヘシンタンパク質やドックリンドメインを有するタンパク質を真核微生物において発現させるためのベクター及びその構築方法も、同様に、当業者において周知である。また、形質転換にあたり、従来公知の各種方法、例えば、トランスフォーメーション法や、トランスフェクション法、接合法、プロトプラスト法、エレクトロポレーション法、リポフェクション法、酢酸リチウム法等を用いることも、同様に当業者において周知である。
【0069】
なお、本明細書の開示によれば、本セルラーゼ複合体を構成する第1〜第3のキメラタンパク質及び当該キメラタンパク質をコードするDNAも提供される。
【0070】
(セルロースの分解産物の生産方法)
本明細書に開示されるセルロースの分解産物の生産方法は、本明細書の開示によれば、本タンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解する工程、を備えることができる。本方法によれば、セロビオヒドロラーゼ活性が向上した本タンパク質を用いることにより、効率的にセルロースを分解できる。分解工程で用いる1種又は2種以上のセルラーゼとしては、本セルラーゼ組成物に含まれうる他のセルラーゼから適宜選択して用いることができる。分解工程においては、セルラーゼは、本セルラーゼ組成物として供給されてもよいし、本タンパク質複合体として提供されていてもよい。分解工程における処理温度や時間等の条件は、用いるセルラーゼの種類及びセルロースの供給形態によって適宜設定することができる。
【0071】
本明細書において、セルロースとは、バイオマスから他の成分から分離工程を経たセルロースとして供給されてもよいし、リグニン、ヘミセルロース及び/又はペクチンが共存する未処理あるいは部分的な前処理を施したバイオマスなどセルロースを含有する材料として供給されてもよい。こうしたバイオマス資源としては、稲ワラ、麦ワラ、バガス、枯れ草等の廃棄資源のほか、未利用資源であってもよい。また、本明細書において、セルロース分解産物としては、セルロースの低分子化物であればよく、グルコース、そのオリゴマー、セロビオース等が挙げられる。
【0072】
(有用物質の生産方法)
本明細書の開示によれば、微生物の発酵により有用物質を生産する方法であって、本タンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解してセルロース分解産物を取得する工程と、微生物によりセルロース分解産物を含む炭素源を発酵して、前記有用物質を生産する工程と、を備える、方法が提供される。本生産方法によれば、本タンパク質を用い、向上したセロビオヒドロラーゼ活性がセルロースの分解に寄与するため、効率的にセルロースを分解することができる。本生産方法では、セルロースの分解工程と、当該分解工程で得られたセルロース分解産物を含む炭素源を微生物により発酵する工程と、をそれぞれ独立して実施することができる。したがって、セルロースの分解工程は、本セルラーゼ組成物や本タンパク質複合体を、酵素製剤として用いてセルロースを分解し、その後、このセルロース分解物を発酵工程に供給して発酵してもよい。
【0073】
本明細書の開示によれば、発酵のための微生物の細胞外に分泌又は細胞表層に提示される本タンパク質を含む、1種又は2種以上のセルラーゼの存在下、前記微生物を用いて、セルロースを少なくとも含む炭素源を発酵して前記有用物質を生産する工程、を備えることができる。このような生産方法によれば、セロビオヒドロラーゼ活性の向上したタンパク質を細胞外に発現する発酵用微生物を用いることで、いわゆる糖化・発酵同時プロセス(CBP)を効率的に行うことができる。1種又は2種以上のセルラーゼは、全て発酵用微生物の細胞表層に提示されていてもよいし、部分的に細胞外から添加されていてもよい。こうしたセルラーゼは、発酵用微生物の表層において本タンパク質複合体を構築していてもよい。
【0074】
発酵用の微生物は、酵母などのエタノール生産微生物や麹菌等の真核微生物を好ましく用いることができる。微生物は、人工的に取得された微生物であってもよい。例えば、グルコースからの代謝系の1種又は2種以上の酵素を遺伝子組換えにより置換、追加等して得られる本来の代謝物でない化合物を産生可能に遺伝子工学的に改変したものであってもよい。このような微生物を用いることで、例えば、イソプレノド合成経路の追加によるファインケミカル(コエンザイムQ10、ビタミン及びその原料等)、解糖系の改変によるグリセリンの生産、プラスチック・化成品原料を生産するなどのバイオリファイナリー技術に適用できる。有用物質としては特に限定しないが、グルコースを利用して微生物が生成可能なものが好ましく、上記したように、バイオリファイナリー技術全般にわたる物質を対象とすることができる。
【実施例】
【0075】
以下、本発明を、実施例を挙げて具体的に説明するが、本発明はこれらの実施例に限定されるものではない。なお、以下に述べる遺伝子組換え操作は既出のMolecular Cloningに従い行った。
【実施例1】
【0076】
(セルロース結合ドメインを有しないPhanerochaete chrysosporium由来のセロビオヒドロラーゼIIの合成)
ファネロケーテ由来セロビオヒドラーゼII(PcCBH2)はセルロース結合ドメイン(CBD)と触媒ドメインがリンカーでつながった構造を持っている。今回CBD機能を喪失させるため、PcCBH2のCBD部位を取り除き、「リンカー−触媒ドメイン」からなるペプチドを合成した。
【0077】
(CBDを欠失したPhanerochaete chrysosporium由来のセロビオヒドロラーゼIIの試験内合成のためのDNA断片の調製)
pET23bのNdeI/HindIIIにファネロケーテ由来セロビオヒドラーゼ遺伝子(配列番号1)を挿入したプラスミドpET23b_PcCBH2wtをテンプレートに、2種類のオリゴマー配列(5’-GGATCTGCGGTCACGACCACCTCCGTT -3’(配列番号4), 5’- CATATGTATATCTCCTTCTTAAAGT-3’(配列番号5))をプライマーとしてインバースPCRを行った。取得したPCR産物につき、アガロースゲル電気泳動、ゲル染色にて目的バンドを取り出し、ゲル中のDNAをキット(Wizard(R) SV Gel and PCR Clean-up System、Promega)を用いて抽出した。抽出したDNA断片をポリヌクレオチドキナーゼ(New England BioLabs)で両5’- 末端をリン酸化後、ライゲーションキット(DNA Ligation kit、Takara)を用いてライゲーション反応をさせた。ライゲーション産物を大腸菌DH5α株に導入し、その形質転換体を培養後、プラスミド抽出精製キット(QIAprep Spin Miniprep Kit、Qiagen)で目的のDNA断片を含むプラスミドを取り出した。CBD部位の欠失は、シーケンスにより確認した。
【0078】
(CBDを欠失したPhanerochaete chrysosporium由来のセロビオヒドロラーゼII改変体の試験内合成のためのDNA断片の調製)
次いで、CBH2改変体として、Sept25:PcCBH2に10個のアミノ酸置換(T130I/S134Q/S183T/A241E/M259I/Q307R/A309S/T342H/Q381S/F382L)を行った改変体及びMall4:PcCBH2に15個のアミノ酸置換(T112S/N125K/T130I/S134Q/S183T/Q204K/F206Y/A241E/M259I/Q307R/A309S/T342H/I365V/Q381S/F382L)を行った改変体につき、それぞれCBDを欠失させたDNA断片を、PcCBH2についてのDNA断片を含むプラスミド構築に準じて調製した。
【0079】
(セルラーゼの試験管内合成方法)
試験管内合成の鋳型として、作製したプラスミドに対して2本のプライマー(5’- ATCTCGATCCCGCGAAATTAATAC-3’(配列番号6)、 5’- TCCGGATATAGTTCCTCCTTTCAG-3’(配列番号7))で増幅、PCR purification kit (Qiagen)で精製したPCR産物を使用した。なおこのPCR産物には、T7プロモーター配列、開始コドン、遺伝子配列、終始コドン、ターミネーター配列が含まれている。
【0080】
試験管内合成は、表4に示す組成の反応液で、26℃、3時間、in vitroでの転写翻訳共役反応を行なった。無細胞蛋白質合成反応の促進剤として、ATPの枯渇を補うためにクレアチンキナーゼをATP生成酵素(ADP+ホスホクレアチン→ATP)として、リファンピシンはRNA合成開始阻害剤、フォリン酸はRNA転写阻害剤として添加した。また、タンパク質のフォールディングを助ける因子として、各種シャペロンを高発現した大腸菌A19株から抽出したS30菌体抽出液(−DTT)を用いた。更に、S−S結合形成促進因子であるカビ由来プロテインジスルフィドイソメラーゼも添加した。なお合成されたセルラーゼ量を評価するため、蛍光標識されたリジン(FluoroTect GreenLys in vitro Translation Labeling System:プロメガ)を混ぜて合成反応を行った。
【0081】
【表4】
【実施例2】
【0082】
(試験管内合成反応におけるセルラーゼ合成量の評価)
CBD欠失状態(リンカー−触媒ドメイン)とCBD保有状態(CBD−リンカー−触媒ドメイン)とのそれぞれにつき、野生型PcCBH2及び2種類のPcCBH2改変体の試験管内合成量及び比活性を評価した。
【0083】
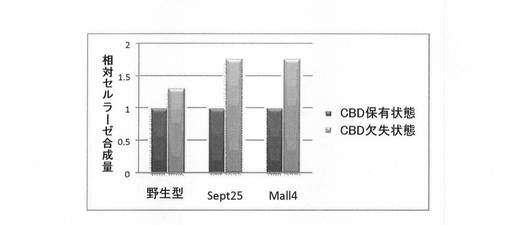
合成量の評価は、無細胞合成時に蛍光標識されたリジン(FluoroTect GreenLys in vitro Translation Labeling System:プロメガ)を取り込ませた合成産物をSDS-PAGE後、蛍光イメージアナライザー(FLA9000:富士フィルム株式会社)で検出、画像解析ソフトであるMulti Gauge(富士フィルム株式会社)での解析により行った。なお今回使用したPcCBH2野生型および改変体はそれぞれ、CBD保有状態とCBD欠失状態で同数のリジンを持っているため、SDS−PAGEでの目的バンドの蛍光量でその合成量が比較評価できる。結果を図1に示す。図1には、セルラーゼの試験管内合成量を、野生型及び改変体につき、CBD保有状態(黒)及びCBD欠失状態(灰色)として、CBD保有状態の合成量に対する相対量として示す。
【0084】
図1に示すように、野生型PcCBH2ではCBD欠失状態の方がCBD有状態よりも合成量が約1.3倍向上していた(図1)。また評価した2つのPcCBH2変異体では共に、CBD欠失状態の方がCBD保有状態よりも合成量が約1.7倍向上していた。
【実施例3】
【0085】
(セルラーゼ(セロビオヒドロラーゼ)比活性の評価)
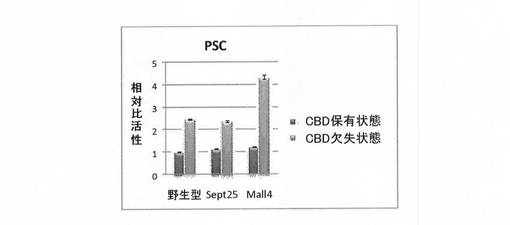
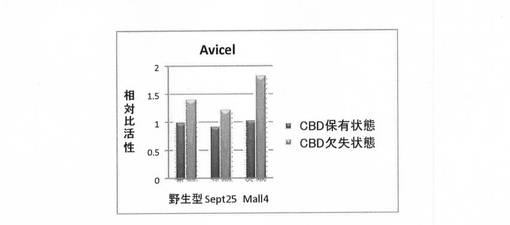
試験管内合成されたセルラーゼは、野生型及び各PcCBH2改変体でCBD保有状態及びCBD欠失状態で同じセルラーゼ量となるように試験管内合成試薬で希釈し、その活性を評価した。具体的には、50mM酢酸緩衝液pH5.0で100倍希釈後、1%リン酸膨潤セルロース(PSC)又は2.5%Avicelを同量添加し(0.5%PSC又は1.25%Avicel)、40℃で16時間反応させた。反応液を、TZアッセイ法(Journal of Biochemical and Biophysical Methods, 11 (1985) 109-115)にて遠心上精の還元糖量を測定し、セルラーゼ比活性とした。図2及び図3には、セルラーゼの比活性を、野生型及び改変体につき、CBD保有状態(黒)及びCBD欠失状態(灰色)として、CBD保有状態の活性に対する相対比活性として示す。
【0086】
(1)比活性の比較(PSCが基質の場合)
野生型PcCBH2、改変体Sept25およびMall4を用いて、PSCを基質とした条件でのCBD有欠失状態の比活性を評価した。結果は、図2に示すように、評価した野生型及び各改変体でCBD欠失状態はCBD保有状態よりも比活性が向上していた。すなわち、野生型及びSept25では2倍以上、Mall4では、4倍以上であった。
【0087】
(2)比活性の比較(Avicelが基質の場合)
野生型PcCBH2、改変体Sept25およびMall4を用いて、Avicelを基質とした条件でのCBD欠失/保有状態の比活性を評価した。結果は、図3に示すように、評価した野生型及び各改変体でCBD欠失状態はCBD保有状態よりも比活性が向上していた。すなわち、野生型では1.4倍、Sept25では1.2倍、Mall4では、1.7倍であった。
【実施例4】
【0088】
(トリコデルマ由来のエンドグルカナーゼ2のCBD欠失/保有状態での比活性比較)
本実施例では、トリコデルマ由来のエンドグルカナーゼ2(TrEG2、GHF5)を用いて、そのCBD欠失変異体を合成し、実施例3に準じて、CBD欠失/保有状態での比活性測定を行った。CBD欠失変異体のDNA断片を含むプラスミド作製は、上記エンドグルカナーゼIIの遺伝子(GenBankアクセッション番号P07982(配列番号12)に対して、プライマー配列(5’- GGGGTCCGATTTGCCGGCGTTAACAT-3’(配列番号8))及び5’- CATATGTATATCTCCTTCTTAAAGT-3’ (配列番号9))を用いる以外は、実施例1に準じてプラスミドを構築し、各ペプチドを試験管内合成した。
【0089】
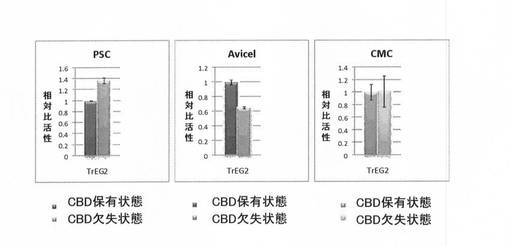
実施例3と同様に、PSC(0.5%)及びAvicel(1.25%)を基質としてCBD欠失/保有状態での比活性を評価した。さらに、基質としてカルボキシメチルセルロース(CMC、0.2%)を使用して同様にして比活性を評価した。結果を図4に示す。図4には、セルラーゼの比活性を、CBD保有状態(黒)及びCBD欠失状態(灰色)として、CBD保有状態の活性に対する相対比活性として示す。
【0090】
図4に示すように、トリコデルマ由来のエンドグルカナーゼIIについては、PSCが基質の場合には、CBD欠失状態の方がCBD保有状態よりも比活性が高いことが示された。また、Avicelが基質の場合には、CBD欠失状態の方がCBD保有状態よりも比活性が低いことが示された。さらに、CMCが基質の場合には、CBD保有状態とCBD欠失状態はほぼ同じ比活性を示した。以上のことから、GHF45とは異なるGHF2のエンドグルカナーゼでのCBD削除の効果は、基質によって様々で、一概にCBD削除による比活性の向上を示すことはできなかった。
【実施例5】
【0091】
(ファネロケーテ由来のセロビオヒドラーゼ7CのCBD欠失/保有状態での比活性比較)
PcCBH2とは異なるファネロケーテ由来のセロビオヒドラーゼ7C(PcCBH7C、GHF7)を用いて、そのCBD欠失変異体を合成し、実施例4と同様にしてCBD欠失/保有状態での比活性測定を行った。PcCBH7CのCBD欠失変異体のDNA断片を含むプラスミドの作製は、PcCBH7C遺伝子(GenBankアクセッション番号M22220H(配列番号13))に対して、プライマー配列(5’- TTAGCCGGTGTACGTGGTGTTGAGGT-3’ (配列番号10)及び5’-AAGCTTGCGGCCGCACTCGAGCA -3’ (配列番号11))を用いる以外は、実施例1に準じて行い、各ペプチドを試験管内合成した。実施例3と同様に、PSC(0.5%)を基質としてCBD欠失/保有状態での比活性を評価した。結果を図5に示す。図5には、セルラーゼの比活性を、CBD保有状態(黒)及びCBD欠失状態(灰色)として、CBD保有状態の活性に対する相対比活性として示す。
【0092】
図5に示すように、実施例3におけるPcCBH2の結果とは異なり、CBD保有状態の方がCBD欠失状態よりも比活性が高かった(図5)。以上の結果から、PcCBH2と同じファネロケーテ由来のセロビオヒドラーゼでも、CBDの欠失が比活性を向上させるわけではないことがわかる。
【実施例6】
【0093】
(耐熱化変異体)
(1)耐熱性候補変異の選定
P. chrysosporium由来のセロビオヒドロラーゼII(PcCBH2)は、触媒ドメインとセルロース結合ドメイン(CBD)がリンカーでつながった構造を持つ。以下に、触媒ドメイン内のアミノ酸置換による耐熱化変異体の取得について説明する。
【0094】
(2)耐熱性候補変異を含む変異体の構築
耐熱化への初期配列としてPcCBH2変異体5−6(配列番号14、配列番号15)を用いた。変異体5−6は、配列番号2で表されるアミノ酸配列において、そのCBDのみに、アミノ酸置換(S22P置換(塩基配列上の変異:T64C))及び同義置換(塩基配列上の変異:G9A)をそれぞれ各一つ備えており、試験管内合成において野生型PcCBH2よりもタンパク質合成量が高い変異体であることがわかっている。耐熱化変異体の取得にあたり、変異体5−6に対して、以下の表5に示す変異(アミノ酸置換)を有する変異体作成した。すなわち、これらの変異体に相当するアミノ酸配列をそれぞれコードするオリゴマーDNA及びその直近の相補鎖オリゴマーDNAをプライマーとして用いてインバースPCR法でプラスミド鎖を増幅し、5’−末端をリン酸化後、ライゲーションによるプラスミドの環状化、大腸菌への形質転換を経て取得した。各変異体の変異導入は、シーケンスを行い確認した。
【0095】
【表5】
【0096】
43種の変異体を試験管内で合成した。試験管内合成の鋳型として、(1)で取得したDNAに対してプライマー(5’- ATCTCGATCCCGCGAAATTAATAC-3’(配列番号16)、 5’- TCCGGATATAGTTCCTCCTTTCAG-3’(配列番号17))を用いて増幅させたPCR産物を、PCR purification kit (Qiagen)で精製したものを用いた。なお、このPCR産物には、T7プロモーター配列、開始コドン、遺伝子配列、終始コドン、ターミネーター配列が含まれている。
【0097】
変異体の試験管内合成は、表6に示す組成の反応液で、26℃、3時間、in vitroでの転写翻訳共役反応として行った。無細胞蛋白質合成反応の促進剤として、ATPの枯渇を補うためにクレアチンキナーゼをATP生成酵素(ADP+ホスホクレアチン→ATP)として、リファンピシンはRNA合成開始阻害剤、フォリン酸はRNA転写阻害剤として添加した。また、タンパク質のフォールディングを助ける因子として、各種シャペロンを高発現した大腸菌A19株から抽出したS30菌体抽出液(−DTT)を用いた。更に、S−S結合形成促進因子である、カビ由来プロテインジスルフィドイソメラーゼも添加した。
【0098】
【表6】
【0099】
(3)変異体の耐熱試験方法
合成した変異体を、50mM酢酸緩衝液(pH5.0)で100倍に希釈、49,50,51,52,53及び54℃の各温度で2時間保温する熱処理後、その残存活性をリン酸膨潤セルロース(PSC)分解(反応条件:40℃、16時間)後の遠心上清の還元糖量をTZアッセイ法(Journal of Biochemical and Biophysical Methods, 11 (1985) 109-115)を用いて評価した。残存活性は熱処理前の活性との比で表した。またセルラーゼの耐熱温度は上記残存活性が50%となるときの温度と定義した。
【0100】
(4)有利変異の選定
以上の耐熱性評価の結果、13個の耐熱性が向上した変異体が得られた。その耐熱性を向上させた変異を有利変異と呼ぶこととした。今回に実験で得られた有利変異を表7及び図6に示す。図6に示すように、これらの有利変異は、いずれも、52℃環境下で2時間保温後において、氷冷保持条件と比較して1.3〜7倍残存活性を向上させるものであった。
【表7】
【0101】
(5)第1世代変異体の選択
有利変異を一つずつ親配列に加算したDNAを合成し、当該DNAを鋳型として、各変異体を実施例6の(2)と同様にして試験管内合成して、(3)に記載の方法に準じて耐熱性評価を行ったところ、最も耐熱性が高かった有利変異M−18による変異体第1世代とした。
【0102】
(6)第2世代の選択
実施例2で得た変異体T342H(M1−18)に有利変異Q381S/F382L(M1−20)を導入した変異体(W1820)を実施例1と同様にして合成し、(3)に記載の方法に準じて耐熱性を評価した。結果を図7に示す。図7に示すように、有利変異を蓄積したW18+20の耐熱性はM1−18及びM1−20と比較して向上していた。この変異体を第2世代として選択した。
【0103】
(7)第3世代の選択
実施例3で得られた変異体W18+20に有利変異(M1−1,−2,−4,−12)をそれぞれ導入し、その変異体を上記(2)と同様にして合成し、上記(3)と同様にして耐熱性を評価した。結果を図8に示す。図8に示すように、有利変異T130I/S134Q(M1−4)を導入した変異体(Tri4)の耐熱性が最も向上していた。この変異体を第3世代として選択した。
【0104】
(8)第4世代の選択
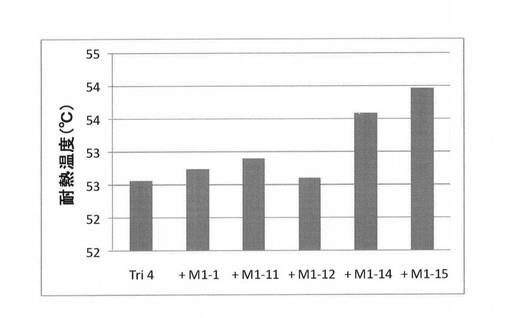
実施例4で得られた有利変異が3個導入された変異体Tri4に4つの有利変異(M1−1,−11,−12,−14,−15)をそれぞれ導入した変異体を、上記(2)と同様にして合成し、上記(3)と同様にして耐熱性を評価した。結果を図9に示す。図9に示すように、有利変異M259I(M1−15)を導入した変異体(Quard15)の耐熱性が最も向上していた。この変異体を第4世代として選択した。
【0105】
(9)第5世代の選択
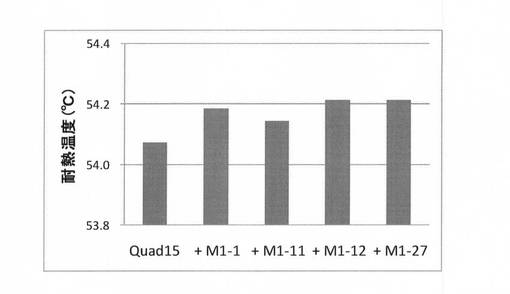
実施例5で得られた有利変異が4個導入された変異体Quard15に4つの有利変異(M1−1,−11,−12,−27)をそれぞれ導入し、その変異体を、上記(2)と同様にして合成し、上記(3)と同様にして耐熱性を評価した。結果を図10に示す。図10に示すように、耐熱評価した。その結果、有利変異A241E(M1−12)を導入した変異体(Quint12)の耐熱性が最も向上していた。この変異体を第5世代として選択した。
【0106】
(10)第6世代の選択
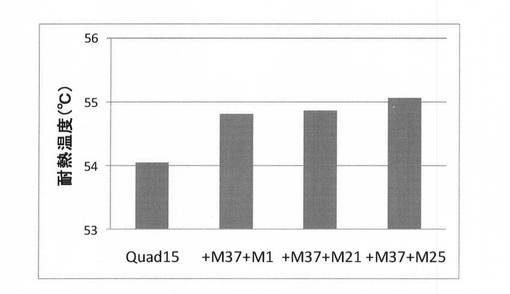
実施例6で得られた有利変異が5個導入された変異体Quint12に耐熱性への寄与度が高かった有利変異Q307R/A309S(M1−37)とその他の3つの有利変異(M1−1,−21,−25)をそれぞれ導入した変異体を、上記(2)と同様にして合成し、上記(3)と同様にして耐熱性を評価した。結果を図11に示す。図11に示すように、Quint12にQ307R/A309S及び有利変異S183T(M1−25)を導入した変異体(Sept25)の耐熱性が最も向上していた。この変異体を第6世代として選択した。
【0107】
(11)スクリーニングによる耐熱化変異体の取得
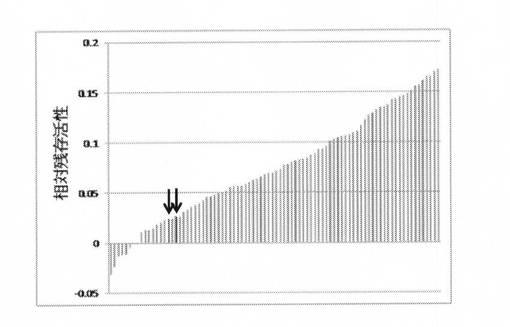
加算していない有利変異(M1−1,−2,−11,−21,−27)を変異体Sept25にばらつきを持って導入後、スクリーニングにより最も高い耐熱性を有する変異体の取得を試みた。具体的には、QuickChange Multi Site-Directed Mutagenesis Kit(Stratagene)を用いて変異を導入し、その変異体92個をクローニングした。上記(2)と同様にして試験管内合成した変異体を56℃にて2時間の熱処理後、熱処理前に対する残存活性を指標にスクリーニングを行った。結果を図12に示す。図12に示すように、熱処理に対する残存活性がSept25と比較して約5倍向上した変異体が得られた。
【0108】
(12)スクリーニングで得られた変異体の耐熱性評価
上記スクリーニングで、残存活性が高かった上位3個の変異体について、上記(3)と同様に耐熱性を評価した。結果を図13に示す。図13に示すように、評価した3変異体は共にSept25と比較して耐熱温度が1℃向上していた。
【0109】
(13)スクリーニングで得られた耐熱化変異体のアミノ酸配列の決定
上記で得られた耐熱化変異体のアミノ酸配列を塩基配列解析行うことで解析した。その結果、残りの有利変異(M1−1,−2,−11,−21,−27)がすべて入っていることが分かった。この得られたクローンを耐熱化実験での最終変異体(Mall4と呼ぶ)こととした。
【0110】
各世代の変異体における変異の組み合わせを以下の表に示す。
【表8】
【実施例7】
【0111】
(14)各世代の耐熱化変異体を耐熱性評価
これまでに得られた各世代の耐熱変異体につき、上記(3)と同様に耐熱性を評価した。結果を図14に示す。図14に示すように、その結果、有利変異を加えるに従い耐熱温度が向上していることが確認できた。初期変異体5−6の耐熱温度が49.9℃であるのに対し、耐熱化変異体Sept25は4.4℃向上した54.3℃、さらに最終変異体Mall4では5.4℃向上した54.3℃であった。
【実施例8】
【0112】
(CBD欠失PcCBH2変異体の耐熱性評価)
さらに、耐熱化変異体Mall4のCBD部位を削除した変異体を作製した。当該変異体は、実施例1に準じて試験管内合成した。Mall4及びそのCBD欠失変異体の耐熱性を評価した。また、変異体5−6の評価も併せて行った。結果を図15に示す。図15に示すように、耐熱化変異体Mall4からCBDを欠失させた変異体は、Mall4CBD保有体と同様、変異体5−6よりも、高い耐熱性を有していた。
【実施例9】
【0113】
(CBD欠失変異体のセルロースへの結合力の評価)
配列番号2で表されるアミノ酸配列の野生型PcCBH2(CBD保有)、上記の耐熱性変異体Mall4及びそれぞれのCBD欠失変異体を用いて、各セルラーゼのセルロースへの結合力をアビセルからの遊離率で評価した。具体的には、酵母で分泌発現そしてアビセルカラムで精製したそれぞれのセルラーゼ100ngを、50mM酢酸緩衝液pH5.0でけん濁したアビセル100ul(終濃度2%(w/v))に添加、攪拌後、氷上で2時間静置させた。なお、コントロール実験として、アビセル無条件でも行った。それぞれの遠心上清のセルラーゼ活性を測定し、セルラーゼのアビセルからの遊離率を算出した。結果を図16に示す。
【0114】
図16に示すように、CBD保有セルラーゼの遊離率は約0.1であったのに対しCBD失活変異体では0.3〜0.5の値を示した。以上の結果より、CBD欠失セルラーゼは、保有セルラーゼと比較して、アビセルに対する結合性が低下していた。
【配列表フリーテキスト】
【0115】
配列番号4〜13,16、17:プライマー
配列番号14,15:Phanerochaete chrysosporium由来のセロビオヒドロラーゼの変異体
【技術分野】
【0001】
本発明は、バイオマスに含まれるセルロースを有効利用するためのセロビオヒドロラーゼ及びその利用に関する。
【背景技術】
【0002】
近年、有限である石油資源を代替するものとして、植物の光合成作用に由来するバイオマスへの期待が高まってきており、バイオマスをエネルギーや各種材料に利用するための各種の試みがなされている。なかでも、セルロースの利用が期待されている。セルロースは、糖であるグルコースがβ−1,4グリコシド結合によって縮合した高分子化合物であり、分子間水素結合により強固な結晶構造を構成している。セルロースを分解し、さらに、グルコースに糖化し発酵原料として用いるには、セルロースを単糖まで分解(糖化)するには、エンドグルカナーゼ、セロビオヒドロラーゼ及びβ−グルコシダーゼという、少なくとも3種類のセルラーゼ(セルロース分解酵素)セルラーゼの相乗効果が必要である。
【0003】
自然界においては、クロストリジウム・サーモセラム(Clostridium thermocellum)やトリコデルマ・リーゼイ(Trichoderma reesei)などある種のカビ等が強力なセルラーゼを分泌することが知られている。セルラーゼは、通常、セルロース結合ドメインという、基質であるセルロースに結合するためのドメインを有していることが多い。セルロース結合ドメインを有することにより、水不溶性のセルロースや水溶性のオリゴマーに結合し、セルロースを分解しやすくなると考えられている。しかしながら、こうしたセルロース結合ドメインを有しないセルラーゼも天然に存在しており、例えば、T. reesei由来のエンドグルカナーゼ3やシロアリ腸内細菌由来のセルラーゼなどが挙げられる。
【0004】
一方、本来的に存在するセルロース結合ドメインを人工的に欠失させたエンドグルカナーゼが開示されている(特許文献1)。この文献には、この接合菌由来のエンドグルカナーゼにおいてセルロース結合ドメインを欠失させることでエンドグルカナーゼ活性が向上したことが記載されている。
【0005】
効率的にあるいは低コストにセルロースを利用するには、強力なセルラーゼあるいは相乗効果の高いセルラーゼを提供することが必要である。ここで、セルラーゼの一種であるセロビオヒドロラーゼとして、ファネロケーテ・クリソスポリウム(Phanerochaete chrysosporium)由来のセロビオヒドロラーゼが知られている(非特許文献1)。
【先行技術文献】
【特許文献】
【0006】
【特許文献1】国際公開第WO2002/042474号
【非特許文献】
【0007】
【非特許文献1】Uzcatsguiら、J.Biotechnol.19(2-3):271-85, 1991
【発明の概要】
【発明が解決しようとする課題】
【0008】
P. chrysosporium由来のセロビオヒドロラーゼは高い相乗効果を奏するため、さらなる比活性の向上が求められている。
【0009】
また、特許文献1に開示される特定の接合菌由来のエンドグルカナーゼについては、セルロース結合ドメインの欠失がエンドグルカナーゼ活性向上に寄与していた。セルロース結合ドメインは、セルロースに結合する部位ではあるため、通常は、当該部位がセルロース分解活性に寄与するもの考えられ、当該部位の欠失は酵素活性の低下を招くことが予想される。また、セルロース結合ドメインと触媒部位とがセルロースに対して一定の位置関係が、セルロースの分解に寄与することも想定されており、かかる場合には、セルロース結合ドメインの欠失は、著しく酵素活性を低下させることがある。ましてや、セルロース結合ドメインの欠失がセルラーゼ活性の向上に寄与するセルラーゼをアミノ酸配列等から識別することもできなかった。加えて、こうした酵素活性の変化は、基質であるセルロースの状態により変動することもあった。したがって、本来的にセルロース結合ドメインを有するセルラーゼからセルロース結合活性を不活性化させることが、そのセルラーゼの酵素活性の向上に寄与するかどうかは全く予測することはできなかった。
【0010】
そこで、本明細書の開示は、より優れた比活性を備えるセロビオヒドロラーゼを提供し、さらにその利用を提供することを目的とする。
【課題を解決するための手段】
【0011】
本発明者らは、P. chrysosporium由来のセロビオヒドロラーゼIIの活性向上を目的として、種々検討しているなか、基質過剰条件下において、見かけの酵素活性が低下する傾向があるという知見を得た。人工的な環境下でのセルロースの分解・糖化反応を考慮すると、反応効率を向上させるためには、基質濃度は著しく高い環境下での反応を行うことが求められる。したがって、このセロビオヒドロラーゼをセルロースの糖化に用いるには不利であると考えられた。そこで、本発明者らは、過剰のセルロースの存在下においては、セルロース結合ドメインが存在することにより、セロビオヒドロラーゼがセルロースから離れにくくなり、引き続き生じうる酵素反応を阻害する可能性があるとの推論に至った。そして、このセロビオヒドロラーゼからセルロース結合ドメインを欠失させてセルロース結合活性を低下させたところ、セロビオヒドロラーゼ活性が向上するという知見を得た。本明細書の開示は、これらの知見に基づいて提供される。
【0012】
本明細書の開示によれば、以下の(a)〜(d)のいずれかのアミノ酸配列において、セルロース結合ドメインが不活性化しており、セロビオヒドロラーゼ活性を有するタンパク質。
(a)配列番号2で表されるアミノ酸配列
(b)配列番号2で表されるアミノ酸配列において1又は複数個のアミノ酸の置換、欠失及び/又は付加を有するアミノ酸配列
(c)配列番号2で表されるアミノ酸配列と75%以上の同一性を有するアミノ酸配列
(d)配列番号1で表される塩基配列と相補的な塩基配列からなるDNA又はその一部とストリンジェントな条件下でハイブリダイズするDNAによってコードされるアミノ酸配列
(e)配列番号1で表される塩基配列と75%以上の同一性を有する塩基配列によってコードされるアミノ酸配列
【0013】
本明細書の開示によれば、また、上記タンパク質をコードする、DNA、当該DNAを含む発現用コンストラクト、当該発現用DNAコンストラクトにより形質転換された細胞、上記タンパク質を細胞外に分泌又は細胞表層に提示する細胞も提供される。
【0014】
本明細書の開示によれば、セルロースの分解産物の生産方法であって、上記タンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解する工程、を備える、セルロースの分解産物の生産方法が提供される。
【0015】
本明細書の開示によれば、微生物の発酵により有用物質を生産する方法であって、上記タンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解してセルロース分解産物を取得する工程と、微生物により前記セルロース分解産物を含む炭素源を発酵して、前記有用物質を生産する工程と、を備える、方法が提供される。また、微生物の発酵により有用物質を生産する方法であって、前記微生物の細胞外に分泌又は細胞表層に提示される上記タンパク質を含む、1種又は2種以上のセルラーゼの存在下、前記微生物を用いて、セルロースを少なくとも含む炭素源を発酵して前記有用物質を生産する工程、を備える、方法も提供される。
【0016】
さらに、本明細書の開示によれば、上記タンパク質を含むセルラーゼ組成物及び上記タンパク質を含む、タンパク質複合体も提供される。
【図面の簡単な説明】
【0017】
【図1】CBD部位の有無によるP. chrysosporium由来のセロビオヒドロラーゼIIの試験管内合成量の比較を示す図である。
【図2】CBD部位の有無(保有状態vs欠失状態)によるP. chrysosporium由来のセロビオヒドロラーゼIIのPSCを基質とした場合の相対比活性を示す図である。
【図3】CBD部位の有無によるP. chrysosporium由来のセロビオヒドロラーゼIIのAvicelを基質とした場合の相対比活性を示す図である。
【図4】CBD部位の有無によるT. reesei由来のエンドグルカナーゼIIのPSC、Avicel及びCMCを基質とした場合の相対比活性を示す図である。
【図5】CBD部位の有無によるP. chrysosporium由来のセロビオヒドロラーゼ7CのPSCを基質とした場合の相対比活性を示す図である。
【図6】P. chrysosporium由来のセロビオヒドロラーゼIIの耐熱変異体についての52℃環境下での有利変異を持つ変異体の残存活性値を示す図である。
【図7】第2世代の有利変異の蓄積実験結果を示す図である。
【図8】第3世代の有利変異の蓄積実験結果を示す図である。
【図9】第4世代の有利変異の蓄積実験結果を示す図である。
【図10】第5世代の有利変異の蓄積実験結果を示す図である。
【図11】第6世代の有利変異の蓄積実験結果を示す図である。
【図12】スクリーニング実験の耐熱試験結果を示す図である。
【図13】スクリーニングで得られた変異体の耐熱性評価結果を示す図である。
【図14】各世代の耐熱化変異体の耐熱試験結果を示す図である。
【図15】耐熱化変異体のCBD欠失体の活性評価結果を示す図である。
【図16】CBD欠失変異体のセルロースへの結合力の評価結果を示す図である。
【発明を実施するための形態】
【0018】
本明細書の開示は、バイオマスに含まれるセルロースを有効利用するための活性の高いPhanerochaete chrysosporium由来のセロビオヒドロラーゼ及びその利用に関する。本明細書に開示されるタンパク質は、基質であるセルロースに対する結合活性が不活性化されることで、セルロースに対する分解活性(比活性)が、当該不活性化前よりも高められている。なお、セルロース結合活性の不活性化による比活性に対する効果は、各種セルラーゼにおいて異なり、不活性化の効果を当業者といえども予測することはできなかった。また、本タンパク質におけるセロビオヒドロラーゼ活性の向上程度は、通常の変異導入による改変による活性向上と同等あるいはそれ以上であり、当業者の予測を超えるものであった。
【0019】
Phanerochaete chrysosporium由来のセロビオヒドロラーゼは、本来的に、他の1種又は2種以上のセルラーゼと組み合わせることにより高い相乗効果を発揮してセルロースを分解できるため、本明細書に開示されるタンパク質によれば、さらに活性の高いセルロース分解能を有する組み合わせを提供でき、セルロースの分解、セルロースからの有用物質の生産を効率的に行うことができるようになる。
【0020】
セルロース結合活性の不活性化による、セロビオヒドロラーゼ活性(比活性)の向上には、本来的に有していたセルロース結合ドメインによる、セルロースの過剰存在下における、非生産的なセロビオヒドロラーゼのセルロースへの吸着を抑制するものと推測されるが、推論であって、本発明を拘束するものではない。
【0021】
以下、本明細書の開示に含まれる種々の実施形態について詳細に説明する。なお、本明細書において用いる「GHF(Glycoside Hydrolase Family)」とは、CAZy(Carbohydrate active Enzymes)のホームページ(http://www.cazy.org/fam/acc_GH.html)において提供される、グリコシド加水分解酵素の分類である。
【0022】
(セルロース結合ドメインが不活性化されたPhanerochaete chrysosporium由来のセロビオヒドロラーゼ)
配列番号2で表されるアミノ酸配列は、Phanerochaete chrysosporiumのセロビオヒドロラーゼIIのアミノ酸配列をコードしている(Appl. Environ.Microbaial. 60(12),4387−4393(1994))。Phanerochaete chrysosporiumのセロビオヒドロラーゼIIは、配列番号2で表されるアミノ酸配列において、N末端側にセルロース結合ドメインを有し、リンカー領域を介して活性ドメインを有している。本タンパク質は、Phanerochaete chrysosporiumから取得したセロビオヒドロラーゼ IIのアミノ酸配列(配列番号2)において、セルロース結合活性が不活性化されているが、セロビオヒドロラーゼ活性を有するタンパク質である。このような本タンパク質は、配列番号2で表されるアミノ酸配列において以下に説明するような各種態様でセルロース結合ドメインが不活性化され、しかも、セロビオヒドロラーゼ活性を維持するアミノ酸配列を有するタンパク質である。
【0023】
(セルロース結合ドメインの不活性化の態様)
本タンパク質における、セルロース結合ドメインの不活性化の態様としては、セルロース結合ドメインの部分的又は全体の欠失が挙げられる。セルロース結合ドメインは、配列番号2で表されるアミノ酸配列の2番目から37番目までのアミノ酸36残基の領域とされており、不活性化の態様としては、このアミノ酸配列において1個以上、典型的には、複数個から全体(100%)に及ぶアミノ酸の欠失が挙げられる。好ましくは、2以上の連続したアミノ酸の欠失である。たとえば、セルロース結合ドメインを除去したアミノ酸配列を配列番号3に示す。
【0024】
アミノ酸が欠失されている領域やアミノ酸の数は、セルロース結合活性を評価することにより決定することができる。セルロース結合ドメインが部分的に欠失されている場合、セルロース結合ドメインをコードするアミノ酸配列の20%以上、好ましくは30%以上、より好ましくは40%以上、さらに好ましくは50%以上、一層好ましくは60%以上、より一層好ましくは70%以上、さらに一層好ましくは80%以上、さらに好ましくは90%以上、欠失されている。典型的には、セルロース結合ドメインのN末端側又はC末端側から連続してアミノ酸が欠失されているが、好ましくは、N末端側から連続して複数個のアミノ酸が欠失されている。
【0025】
セルロース結合ドメインの不活性化は、タンパク質においてセルロース結合ドメインの一部又は全体が欠失している態様が好ましい。アミノ酸の欠失により、ペプチド鎖が短くなり、異種タンパク質としての合成量を増大させることができる。特に、本タンパク質を、試験管内におけるタンパク質生産や遺伝子工学的に異種タンパク質として微生物宿主等において生産させる場合には、好適である。
【0026】
あるタンパク質がセルロース結合ドメイン活性(セルロース結合活性)を有しているか否か又はその程度は、例えば、タンパク質とアビセルとの結合力を測定することで評価することができる。具体的には、酢酸緩衝液pH5.0でけん濁したアビセルにタンパク質を添加し攪拌そして氷上にて一定時間静置後の遠心上清のタンパク質量を測定することで評価できる。その遠心上清にタンパク質がないかあるいは少ない時にはセルロース結合ドメイン活性を有すると言える。
【0027】
セルロース結合活性の不活性化の他の態様としては、セルロース結合ドメインにおける1以上のアミノ酸の置換及び/又は挿入が挙げられる。置換又は挿入されているアミノ酸の部位や個数、置換又は挿入されて新たにセルロース結合ドメイン中に配置されるアミノ酸の種類は、得られた変異体タンパク質につき、セルロース結合活性を評価することで適宜決定することができる。
【0028】
セルロース結合ドメインの不活性化は、配列番号2で表されるアミノ酸配列からなるセロビオヒドロラーゼIIのセルロース結合活性と比較したとき、当該結合活性より低くなっていればよく、その程度は問わないが、好ましくは、配列番号2で表されるアミノ酸配列からなるセロビオヒドロラーゼIIのセルロース結合活性の60%以下、より好ましくは50%以下、さらに好ましくは40%以下、一層好ましくは30%以下、より一層好ましくは20%以下、さらに一層好ましくは10%以下、最も好ましくは5%以下である。
【0029】
本タンパク質は、セロビオヒドロラーゼ活性を有している。セロビオヒドロラーゼ活性は、例えば、50mM酢酸緩衝液pH5.0で100倍希釈後、1%リン酸膨潤セルロース(PSC)又は2.5%Avicelを同量添加し(0.5%PSC又は1.25%Avicel)、40℃で16時間反応させ、反応液を、TZアッセイ法(Journal of Biochemical and Biophysical Methods, 11 (1985) 109-115)にて遠心上精の還元糖量を測定することにより、測定できる。セロビオヒドロラーゼ活性の程度は、特に問わないが、セルロース結合活性の不活性化により、本タンパク質のセロビオヒドロラーゼ活性が向上することがわかっている。本タンパク質のセロビオヒドロラーゼ活性は、配列番号2で表されるアミノ酸配列からなるセロビオヒドロラーゼIIのセロビオヒドロラーゼ活性と比較したとき、好ましくは、当該活性の同等又はそれ以上であり、より好ましくは110%以上、さらに好ましくは120%以上、一層好ましくは130%以上、より一層好ましくは140%以上、さらに一層好ましくは150%以上、最も好ましくは160%以上である。
【0030】
本タンパク質は、また、配列番号2で表されるアミノ酸配列と一定の関係を有するアミノ酸配列においてセルロース結合ドメインが不活性化されつつ、セロビオヒドロラーゼ活性を有するものであってもよい。すなわち、例えば、配列番号2で表されるアミノ酸配列を有するタンパク質の変異体であって、セルロース結合活性やセロビオヒドロラーゼ活性を有する改変体において、セルロース結合ドメインが不活性化された態様が挙げられる。典型的には、セロビオヒドロラーゼ活性ドメインにおいては有効な変異を有し、かつセルロース結合ドメインには、セルロース結合活性が不活性化されるような変異を備えるタンパク質が挙げられる。このような本タンパク質においては、配列番号2で表されるアミノ酸配列と一定関係のあるアミノ酸配列において各種態様でセルロース結合ドメインが不活性化されており、しかも、セロビオヒドロラーゼ活性を維持するアミノ酸配列を有している。
【0031】
こうした本タンパク質の他の一態様としては、配列番号2で表されるアミノ酸配列に対して1又は数個のアミノ酸が欠失、置換及び/又は付加されたアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質が挙げられる。配列番号2で表されるアミノ酸配列に対するアミノ酸の変異は、欠失、置換及び/又は付加は、いずれか1種類であってもよいし、2種類以上が組み合わされていてもよい。また、これらの変異の総数は、特に限定されないが、好ましくは、1個以上25個以下程度である。より好ましくは、1個以上20個以下である。さらに好ましくは1個以上15個以下であり、一層このましくは10個以下であり、より一層好ましくは5個以下である。アミノ酸置換の例としては、保存的置換が好ましく、具体的には以下のグループ内での置換が挙げられる。(グリシン、アラニン)(バリン、イソロイシン、ロイシン)(アスパラギン酸、グルタミン酸)(アスパラギン、グルタミン)(セリン、トレオニン)(リジン、アルギニン)(フェニルアラニン、チロシン)。このような態様としては、たとえば、セロビオヒドロラーゼの活性ドメインやセルロース結合ドメインに隣接するリンカードメインの一部又は全部の欠失などが挙げられる。また、セロビオヒドロラーゼ活性向上のためのアミノ酸置換の導入等が挙げられる。
【0032】
さらに他の一態様としては、配列番号2で表されるアミノ酸配列に対して70%以上の同一性を有するアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質が挙げられる。同一性は好ましくは80%以上であり、より好ましくは85%以上であり、さらに好ましくは、90%以上であり、一層好ましくは95%以上である。最も好ましくは、98%以上である。
【0033】
本明細書において同一性又は類似性とは、当該技術分野で知られているとおり、配列を比較することにより決定される、2以上のタンパク質あるいは2以上のポリヌクレオチドの間の関係である。当該技術で“同一性 ”とは、タンパク質またはポリヌクレオチド配列の間のアラインメントによって、あるいは場合によっては、一続きのそのような配列間のアラインメントによって決定されるような、タンパク質またはポリヌクレオチド配列の間の配列不変性の程度を意味する。また、類似性とは、タンパク質またはポリヌクレオチド配列の間のアラインメントによって、あるいは場合によっては、一続きの部分的な配列間のアラインメントによって決定されるような、タンパク質またはポリヌクレオチド配列の間の相関性の程度を意味する。より具体的には、配列の同一性と保存性(配列中の特定アミノ酸又は配列における物理化学特性を維持する置換)によって決定される。なお、類似性は、後述するBLASTの配列相同性検索結果においてSimilarity と称される。同一性及び類似性を決定する方法は、対比する配列間で最も長くアラインメントするように設計される方法であることが好ましい。同一性及び類似性を決定するための方法は、公衆に利用可能なプログラムとして提供されている。例えば、AltschulらによるBLAST (Basic Local Alignment Search Tool) プログラム(たとえば、Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ., J. Mol. Biol., 215: p403-410 (1990), Altschyl SF, Madden TL, Schaffer AA, Zhang J, Miller W, Lipman DJ., Nucleic Acids Res. 25: p3389-3402 (1997))を利用し決定することができる。BLASTのようなソフトウェアを用いる場合の条件は、特に限定するものではないが、デフォルト値を用いるのが好ましい。
【0034】
さらに他の一態様として、配列番号2で表されるアミノ酸配列をコードする塩基配列からなるDNAと相補的な塩基配列からなるDNA又はその一部とストリンジェントな条件でハイブリダイズするDNAによってコードされるアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質が挙げられる。ストリンジェントな条件とは、たとえば、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。例えば、塩基配列の同一性が高い核酸、すなわち、配列番号2で表されるアミノ酸配列をコードする塩基配列と80%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましく95%以上、最も好ましくは97%以上の同一性を有する塩基配列からなるDNAの相補鎖がハイブリダイズし、それより相同性が低い核酸の相補鎖がハイブリダイズしない条件が挙げられる。より具体的には、ナトリウム塩濃度が15〜750mM、好ましくは50〜750mM、より好ましくは300〜750mM、温度が25〜70℃、好ましくは50〜70℃、より好ましくは55〜65℃、ホルムアミド濃度が0〜50%、好ましくは20〜50%、より好ましくは35〜45%での条件をいう。さらに、ストリンジェントな条件では、ハイブリダイゼーション後のフィルターの洗浄条件が、通常はナトリウム塩濃度が15〜600mM、好ましくは50〜600mM、より好ましくは300〜600mM、温度が50〜70℃、好ましくは55〜70℃、より好ましくは60〜65℃である。なお、以上のことから、さらなる他の一態様として、配列番号2で表されるアミノ酸配列をコードする塩基配列と80%以上、好ましくは85%以上、より好ましくは90%以上、さらに好ましく95%以上、最も好ましくは97%以上の同一性を有する塩基配列を有するDNAによってコードされるアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質が挙げられる。
【0035】
なお、Phanerochaete chrysosporium由来のセロビオヒドロラーゼIIの活性ドメインにおける改変体のアミノ酸配列としては、特開2010-41996号公報に開示される各種の改変体のアミノ酸配列が挙げられる。また、配列番号2で表されるアミノ酸配列において10個のアミノ酸置換(T130I/S134Q/S183T/A241E/M259I/Q307R/A309S/T342H/Q381S/F382L)を有するアミノ酸配列及び同アミノ酸配列において15個のアミノ酸置換(T112S/N125K/T130I/S134Q/S183T/Q204K/F206Y/A241E/M259I/Q307R/A309S/T342H/I365V/Q381S/F382L)を含むアミノ酸配列が挙げられる。
【0036】
本タンパク質は、P. chrysosporiumから取得したセロビオヒドロラーゼ IIのアミノ酸配列(配列番号2)において、以下の表1に示される1又は2以上のアミノ酸置換に相当するアミノ酸置換を備えることができる。「アミノ酸配列において1又は2以上のアミノ酸置換に相当するアミノ酸置換を備える」とは、配列番号2で表されるアミノ酸配列に表1から選択される1又は2以上のアミノ酸置換を備えるほか、配列番号2で表されるアミノ酸配列と一定の関係を有するアミノ酸配列において、配列番号2で表されるアミノ酸配列と対比したときに、表1から選択される1又は2以上のアミノ酸置換に相当するアミノ酸置換を備えていることをいう。
【0037】
なお、アミノ酸置換を備えるとは、当該位置において、置換前のアミノ酸と置換後のアミノ酸とをそれぞれ特定して意味するほか、結果として「アミノ酸置換」によって特定されるアミノ酸を備えていればよいことを意味する。すなわち、「M259L」のアミノ酸置換を備えるとは、配列番号2において259位に相当する部位にL(ロイシン)を備えていることを意味する。以下、本タンパク質が備えることのできる変異であるアミノ酸置換について説明する。
【0038】
【表1】
【0039】
本タンパク質は、表2に示す各欄に示す1又は2以上のアミノ酸置換を備えるものであってもよい。なかでも、本タンパク質は、好ましくは、少なくとも、M259L、M259I、T342H、Q381S+F382Lから選択される1種又は2種以上に相当するアミノ酸置換を備えている。これらの各変異に組み合わせて、好ましくは、T130I及びS134の双方又はいずれかを含んでいることが好ましい。さらに、追加して、好ましくは、M259Iの変異を含んでいる。さらにまた、好ましくは、A241Eの変異を含んでいることが好ましい。さらに追加して、好ましくは、Q307R+A309Sの双方又はいずれかの変異を含んでいる。さらにまた、加えて、S183Tの変異を含んでいることが好ましい。さらに追加して、T112S、N125K、Q204K、I365V及びF206Yからなる群から選択される1又は2以上の変異を含んでいることが好ましく、より好ましくは3種以上であり、さらに好ましくは4種以上であり、一層好ましくは5種である。
【0040】
【表2】
【0041】
以上のことから、好ましい変異体としては、たとえば、少なくとも以下の表3に示す組み合わせのアミノ酸置換を備えるものが挙げられる。なかでも、第3世代以降が好ましく、より好ましくは第4世代以降、さらに好ましくは第5世代以降、一層好ましくは第6世代以降、最も好ましくは第7世代以降である。表3中、Sept25の組み合わせのアミノ酸置換を備え、さらに、T112S、N125K、Q204K、I365V及びF206Yからなる群から選択される1又は2以上のアミノ酸置換を備えるタンパク質も好ましい。
【表3】
【0042】
配列番号2で表されるアミノ酸配列において部分的な欠失を生じさせたり、アミノ酸の置換、付加を導入するには、公知の各種の手法を採用できる。例えば、配列番号2で表されるアミノ酸配列や当該配列と一定の関係を有するアミノ酸配列をコードするDNA等の遺伝子情報を改変する方法を用いることができる。例えば、アミノ酸配列において欠失部位を形成するには、DNAの一部をPCR等により増幅したり、あるいはこうして部分的に増幅したDNA断片を適宜結合することで任意の部位に欠失を導入することができる。また、例えば、アミノ酸配列に置換及び/又は付加などの変異を導入して遺伝子情報を改変して本発明のタンパク質を得るには、Kunkel法、Gapped duplex法等の公知の手法又はこれに準ずる方法を採用することができる。例えば部位特異的突然変異誘発法を利用した変異導入用キット(例えばMutan−K(TAKARA社製)やMutan−G(TAKARA社製))などを用いて変異の導入が行われる。また、エラー導入PCRやDNAシャッフリング等の手法により、遺伝子の変異導入やキメラ遺伝子を構築することもできる。エラー導入PCR及びDNAシャッフリング手法は、当技術分野で公知の手法であり、例えばエラー導入PCRについてはChen K, and Arnold FH. 1993, Proc. Natl. Acad. Sci. U. S. A., 90: 5618-5622を、またDNAシャフリングやカセットPCR等の分子進化工学的手法は、例えば、Kurtzman,A.L.,Govindarajan, S., Vahle, K., Jones, J. T., Heinrichs, V., Patten P. A.,Advances in directed protein evolution by recursive genetic recombination: applications to therapeutic proteins. Curr. Opinion Biotechnol.,12, 361-370, 2001、及び、Okuta, A., Ohnishi, A. and Harayama, S., PCR isolation of catechol 2,3-dioxygenase gene fragments from environmental samples and their assembly into functional genes. Gene, 212, 221-228, 1998を参照することができる。なかでも、エラープローンPCR等によりランダム変異を導入する分子進化的手法を利用する無細胞タンパク質合成系を採用することが好ましい。エラープローンPCRに適用する無細胞タンパク質合成系としては、公知のあるいは本出願人が出願した特開2006−61080号公報及び特開2003−116590号公報に記載のタンパク質合成系を用いることができる。本出願人によるこれらの無細胞タンパク質合成系を用いることで活性型の酵素を容易に得ることができる。このため、これらのタンパク質合成系が適用されたエラープローンPCRは、本発明のタンパク質を取得する手法として好ましく用いることができる。
【0043】
なお、本タンパク質は、決定されたアミノ酸配列や塩基配列に基づいて、化学的な合成方法のほか、遺伝子工学的な方法によって製造され、必要に応じて精製される。
【0044】
本タンパク質は、セルロース結合ドメインが不活性化されているために、配列番号2で表されるアミノ酸配列からなるPhanerochaete chrysosporium由来のセロビオヒドロラーゼIIやその改変体よりも、基質であるセルロースへの結合能が低減又は喪失している。一方、本タンパク質は、セロビオヒドロラーゼ活性及びエンドグルカナーゼやβ−グルコシダーゼとの相乗効果においても、配列番号2で表されるアミノ酸配列からなるタンパク質と同等以上の活性を有している。
【0045】
(セルラーゼ組成物)
本明細書の開示によれば、本タンパク質と、さらに、他のセルラーゼ、たとえば、他のセロビオヒドロラーゼ、エンドグルカナーゼ及びβ−グルコシダーゼからなる群から選択される1種又は2種以上を組み合わせの組成物が提供される。本組成物によれば、本タンパク質の優れたセロビオヒドロラーゼ活性により高いセルロース分解活性を呈することができる。本組成物に含まれる本タンパク質以外のセルラーゼとしては、特に限定しないで、公知のセルラーゼから適宜選択される。例えば、Phanerochaete chrysosporium由来でない他起源のエンドグルカナーゼが挙げられる。他起源由来のエンドグルカナーゼとしては、公知の各種エンドグルカナーゼが挙げられ、これらを単独であるいは2種類以上を適宜組み合わせて用いることができる。たとえば、GHF5に属するエンドグルカナーゼが挙げられる。GHF5に分類されるエンドグルカナーゼのなかでも、好ましくは、Trichoderma reesei由来のエンドグルカナーゼ、Aspergillus oryzae由来のエンドグルカナーゼ及びAspergillus niger由来のエンドグルカナーゼを好ましく用いることができる。より好ましくはTrichoderma reesei由来のエンドグルカナーゼAspergillus niger由来のエンドグルカナーゼである。GHF5に分類されるエンドグルカナーゼは、こうしたエンドグルカナーゼから選択される1種又は2種以上を適宜組み合わせて用いることができる。
【0046】
エンドグルカナーゼとしては、GHF12に属するエンドグルカナーゼが挙げられる。なかでも、Trichoderma reesei由来のエンドグルカナーゼ、Aspergillus niger由来のエンドグルカナーゼ及びAspergillus oryzae由来のエンドグルカナーゼが挙げられる。また、Phanerochaete chrysosporium由来のエンドグルカナーゼであってもよい。こうしたエンドグルカナーゼから選択される1種又は2種以上を適宜組み合わせて用いることができる。エンドグルカナーゼとしては、GHF7及びGHF45に属するエンドグルカナーゼであってもよい。なかでも、Trichoderma reesei由来のエンドグルカナーゼを好ましく用いることができる。
【0047】
本セルラーゼ組成物は、Phanerochaete chrysosporium由来のエンドグルカナーゼを含んでいてもよい。Phanerochaete chrysosporium由来のエンドグルカナーゼは、セロビオヒドロラーゼII及びその改変体をPhanerochaete chrysosporiumの培養物等から取得した場合においてセロビオヒドロラーゼII及びその改変体とともに容易に取得できる。
【0048】
本セルラーゼ組成物は、エンドグルカナーゼ以外の他のセルラーゼを含むことができる。例えば、GHF7に属するセロビオヒドロラーゼIを含有することができる。セロビオヒドロラーゼIは、セロビオヒドロラーゼIが、セロビオヒドロラーゼII等と協動することにより、一層セルロース分解の相乗効果が発揮される。セロビオヒドロラーゼIは、Phanerochaete chrysosporium由来であってもよいし、他起源であってもよい。
【0049】
本セルラーゼ組成物は、Phanerochaete chrysosporium以外の他起源のセルラーゼ生産菌の培養物(培養上清であってもよい)から取得された2種類以上のセルラーゼを含有していてもよい。セルラーゼ生産菌としては、特に限定されないで、適宜選択できるが、エンドグルカナーゼの起源としてTrichoderma reesei、Aspergillus aculeatus、Aspergillus niger、Aspergillus oryzae等が好ましく挙げられる。より好ましくは、Trichoderma reeseiである。
【0050】
本セルラーゼ組成物は、化学的あるいは遺伝子工学的に本タンパク質及び必要に応じて他のセルラーゼを生産し、組み合わせることによって得ることができる。本セルラーゼ組成物を構成する全てのセルラーゼを同一の宿主細胞で遺伝子工学的に生産し、その培養上清や培養菌体を回収することによって、本セルラーゼ組成物を得るようにしてもよい。
【0051】
本セルラーゼ組成物は、本タンパク質及び他のセルラーゼを、それぞれ精製したものとして含有していてもよいし、未精製タンパク質として他タンパク質やその他の成分を含んでいてもよい。また、その製剤形態は、特に限定されず、固形製剤(粉末(凍結乾燥体等)、タブレット等、顆粒等)であってもよいし、溶液(流通時においては凍結体であることが好ましい。)であってもよい。
【0052】
(タンパク質複合体)
本明細書に開示されるタンパク質複合体は、本タンパク質と、本タンパク質をコヘシン−ドックリン結合によりコヘシンタンパク質上に備えている。ある種の細菌が、複数種類のセルラーゼを保持するタンパク質構造体を自己細胞表層に構築することが知られており、当該タンパク質構造体がセルロソームとして知られている。セルロソームは、セルラーゼが、スキャホールディンタンパク質に保持されて構成されており、セルロソームとスキャホールディンタンパク質とは、それぞれが備えるドックリンドメインとコヘシンドメインとの間の結合、すなわち、コヘシン−ドッケリン結合により結合されている。公知のセルロソームにおけるドックリンドメイン及びコヘシンドメインのアミノ酸配列は既にいくつか開示されている。コヘシン−ドックリン結合は、非共有結合性であって、そのアミノ酸配列に依存した水素結合等に基づくと考えられる。したがって、こうした開示に従い、ドックリンドメインを付加した本タンパク質と1又は2以上のコヘシンドメインを備えるスキャホールディンタンパク質とにより、人工的なセルロソーム、すなわち、タンパク質複合体を構築することができる。
【0053】
本タンパク質複合体においては、本タンパク質には、ドックリンドメインが付加されている。ドックリンドメインは、コヘシン−ドックリン結合により後述するコヘシンタンパク質に本タンパク質を結合させる部位である。ドックリンドメインは、例えば、公知のセルロソーム生産微生物のセルロソームを構成するセルラーゼの一部に備えられている。本セルラーゼ複合体に用いるドックリンドメインとしては、公知の各種のセルロソーム生産微生物のセルラーゼのドックリンドメインから選択される。例えば、C. thermocellumのエンドグルカナーゼのドックリンドメインを含むアミノ酸配列が挙げられる。ドックリンリンドメインは、活性ドメインのN末端側及びC末端側のいずれの側にあってもよいが、好ましくは、C末端側に配置される。
【0054】
本タンパク質複合体は、本セルラーゼ組成物が含みうる他のセルラーゼがコヘシンタンパク質に保持されていてもよい。他のセルラーゼに関しても、本タンパク質と同様に、ドックリンドメインが付加されている。他のセルラーゼに付加されるドックリンドメインは、本タンパク質に付加されるドックリンドメインと同一であってもよいし異なっていてもよい。
【0055】
コヘシンタンパク質は、コヘシンタンパク質は、本タンパク質が備えるドックリンドメインを結合する1又は2以上のコヘシンドメインを有している。これにより、コヘシンタンパク質は、本タンパク質をコヘシン−ドックリン結合で保持でき、本タンパク質複合体の骨格として機能する。また、本タンパク質複合体は、他のセルラーゼを保持するための異なるコヘシンドメインの組み合わせあるいは配列を有するコヘシンタンパク質を備えていてもよい。
【0056】
(コヘシンドメイン)
コヘシンタンパク質が備える、1又は2以上のコヘシンドメインは、セルロソームのスキャホールディンタンパク質が備えるコヘシンドメインに由来している。コヘシンドメインは、セルロソーム生産微生物の形成するセルロソームにおけるタイプI〜III骨格タンパク質に備えられる触媒活性のあるセルラーゼ等を非共有結合で結合するドメインとして知られている(粟冠ら、蛋白質核酸酵素、Vol.44、No.10(1999)、p41-p50、Demain, A. L., et al., Microbiol Mol. Biol Rev., 69(1), 124-54(2005), Doi, R. H., et al., J. Bacterol., 185(20), 5907-5914(2003)等)。すなわち、コヘシンドメインとしては、セルロソームのタイプI骨格タンパク質上のタイプIコヘシンドメイン、同タイプII骨格タンパク質上のタイプIIコヘシンドメイン及びタイプIII骨格タンパク質上のタイプIIIコヘシンドメインが挙げられる。こうした各種タイプのコヘシンドメインとしては、各種セルロソーム生産微生物において多数その配列が決定されている。これらの各種のタイプのコヘシンのアミノ酸配列及びDNA配列は、NCBIのHP(http://www.ncbi.nlm.nih.gov/)等を介してアクセス可能な各種のタンパク質データベースやDNA配列のデータベースにより容易に取得することができる。
【0057】
コヘシンドメインは、こうしたコヘシンドメインに由来するドメインであって、キメラタンパク質のドックリンドメインと結合することができる。セルラーゼの活性ドメインへのドックリンドメインのキメラ化は活性ドメインの活性が意図せずに低下することなどにより困難な場合があることから、キメラタンパク質のドックリンドメインに応じてコヘシンドメインを選択することができる。また、コヘシン−ドックリン結合の強度等も考慮してコヘシンドメインを選択することができる。
【0058】
本タンパク質複合体にあっては、例えば、キメラタンパク質のドックリンドメインが例えば、C. thermocellumのcelAのドックリンドメインに対して、C. thermocellumのスキャホールディンタンパク質のコヘシンドメインが挙げられる。
【0059】
本タンパク質複合体は、タンパク質の複合体として、それ自体独立した形態であってもよいし、適当なキャリアに固定化ないし保持されていてもよい。また、後述するように、酵母等の微生物の表層に提示された状態であってもよい。本セルラーゼ複合体は、例えば、それぞれのタンパク質を公知のタンパク質製造方法により取得し、これらのタンパク質を接触させる条件下において、コヘシンタンパク質に対してキメラタンパク質を自己集合させることにより、本セルラーゼ複合体を取得できる。
【0060】
また、本タンパク質複合体は、その構成タンパク質のうち1又は2以上を分泌発現する微生物の培養上清又はそのタンパク質精製物を混合してすべての構成タンパク質を接触させて自己集合させることによって取得できる。なお、構成タンパク質のすべてが微生物によって分泌発現されなくてもよく、必要に応じ微生物によって生産されない構成タンパク質を別途製造して混合してもよい。また、構成タンパク質のすべてを分泌発現する微生物の場合、この微生物の培養上清に、これらタンパク質が自己集合可能な状態で含まれるため、その培養上清に、本タンパク質複合体を取得できる。
【0061】
(本タンパク質をコードするDNA、当該DNAを含む発現用コンストラクト)
本明細書に開示されるDNAは、本タンパク質をコードしている。かかるDNAは、本タンパク質をコードしている限り、異なるコドンを用いてコードされるDNAも含まれる。本明細書に開示される発現用コンストラクトは、本タンパク質をコードするDNAを含んでいる。本タンパク質をコードするDNAは、発現用コンストラクトを導入しようとする宿主細胞において本タンパク質を発現可能に保持されていればよい。例えば、宿主細胞で作動可能なプロモーターの制御下に連結されるとともに適切なターミネーターをその下流に有した状態で保持されている。プロモーターは、構成的プロモーターであっても誘導的プロモーターであってもよい。本発現用コンストラクトにおいて、このような状態のDNAは、宿主染色体内に組み込み可能に備えられていてもよいし、染色体外に保持可能に備えられていてもよい。本発現用コンストラクトには、こうした外来DNAの導入に伴って、宿主において利用可能な選択マーカー遺伝子も同時に保持されていることが好ましい。本発現用コンストラクトは、特に真核微生物、好ましくは、酵母での本タンパク質の発現用であることが好ましい。
【0062】
本発現用コンストラクトは、本タンパク質を既に説明したタンパク質複合体を構成可能な形態で発現可能に構築されていてもよい。すなわち、本タンパク質に対してドックリンドメインを付加した融合タンパク質を発現可能に形成されていてもよい。さらに、タンパク質複合体を構成可能な発現用コンストラクトとして、コヘシンタンパク質をコードするDNAを含む、コヘシンタンパク質の発現用コンストラクトを構築してもよい。こうした発現コンストラクトにおいて、融合タンパク質やコヘシンタンパク質のコード化DNAは、本タンパク質をコードするDNAと同様、各種形態でコンストラクトに備えられる。
【0063】
(発現用DNAコンストラクトにより形質転換された細胞、上記タンパク質を細胞外に分泌又は細胞表層に提示する真核微生物)
本明細書に開示される形質転換細胞は、本発現用DNAコンストラクトによって形質転換された細胞である。本形質転換細胞は、本発現用DNAコンストラクトによって形質転換されることにより、本タンパク質を発現可能となっている。
【0064】
本形質転換細胞は、本タンパク質又は本タンパク質複合体を発現可能な細胞であればよく、その種類を限定しないが、本タンパク質の生産及び回収を考慮すると、大腸菌などの細菌等が挙げられるが、好ましくは、本タンパク質によるセルロースの分解及び資化のための真核微生物である。真核微生物は、特に限定しないが、例えば、公知の各種酵母を利用できる。後述するエタノール発酵等を考慮すると、サッカロマイセス・セレビジエ(Saccharomyces cerevisiae)等のサッカロマイセス属の酵母、シゾサッカロマイセス・ポンベ(Schizosaccharomyces pombe)等のシゾサッカロマイセス属の酵母、キャンディダ・シェハーテ(Candida shehatae)等のキャンディダ属の酵母、ピヒア・スティピティス(Pichia stipitis)等のピヒア属の酵母、ハンセヌラ(Hansenula)属の酵母、トリコスポロン(Trichosporon)属の酵母、ブレタノマイセス(Brettanomyces)属の酵母、パチソレン(Pachysolen)属の酵母、ヤマダジマ(Yamadazyma)属の酵母、クルイベロマイセス・マーキシアヌス(Kluyveromyces marxianus)、クルイベロマイセス・ラクティス(Kluveromyces lactis)等のクルイベロマイセス属の酵母が挙げられる。なかでも、工業的利用性等の観点からサッカロマイセス属酵母が好ましい。なかでも、サッカロマイセス・セレビジエが好ましい。また、真核微生物は、アスペルギルス・オリゼ(Aspergillus oryzae)、アスペルギルス・アキュリータス(Aspergillus aculeatus)、アスペルギルス・ニガー(Aspergillus niger)、アスペルギルス・ニジュランス(Aspergillus nidulans)、アスペルギルス・ソーヤ(Aspergillus soya)、アスペルギルス・カワチ(Aspergillus kawachii)、アスペルギルス・アワモリ(Aspergillus awamori)、アスペルギルス・サイトイ(Aspergillus saitoi)等の麹菌であってもよい。異種タンパク質を大量に発現させるには、セルラーゼ非生産菌がより好ましい。また、Trichoderma reeseiなどのセルラーゼ生産菌で生産させることも好ましい。本タンパク質は、セルロース結合ドメインが不活性化されているため、セルラーゼ生産菌が生産するセルロース結合ドメインを有するセルラーゼとは異なる場所で働くことができる。したがって、セルラーゼ生産菌において本タンパク質を生産させることで、全体としてセルロース分解活性の高いセルラーゼの組み合わせ(セルラーゼ製剤)を提供できることになる。
【0065】
本形質転換細胞は、本形質転換細胞のセルロースの分解への利用を考慮すると、本タンパク質を細胞外に分泌又は細胞表層に提示するものであることが好ましい。本タンパク質に細胞外分泌性や細胞表層提示性を付与するには、公知の分泌シグナルや表層提示用のシステムを用いることができる。例えば、分泌シグナルや凝集性タンパク質又はその一部のアミノ酸配列が付与される。分泌シグナルとしては、例えば、Rhizopus oryzaeやC. albicansのグルコアミラーゼ遺伝子の分泌シグナル、酵母インベルターゼリーダー、α因子リーダーなどが挙げられる。また、凝集性タンパク質としては、α−アグルチニンをコードするSAG1遺伝子の5’領域の320アミノ酸残基からなるペプチドが挙げられる。また、所望のタンパク質を細胞表層に提示するためのポリペプチドや手法は、WO01/79483号公報や、特開2003−235579号公報、WO2002/042483号パンフレット、WO2003/016525号パンフレット、特開2006−136223号公報、藤田らの文献(藤田ら,2004. Appl Environ Microbiol 70:1207-1212および藤田ら, 2002. Appl Environ Microbiol 68:5136-5141.)、村井ら, 1998. Appl Environ Microbiol 64:4857-4861.に開示されている。
【0066】
本形質転換細胞は、本タンパク質複合体を細胞外に分泌又は細胞表層に提示するものであることも好ましい。本形質転換細胞は、好ましくは、コヘシンタンパク質を細胞表層提示し、本タンパク質を細胞外に分泌して、コヘシンタンパク質に本タンパク質をコヘシン−ドッケリン結合を介して保持させて、タンパク質複合体を細胞表層に提示する。
【0067】
本タンパク質、本タンパク質複合体及びこれらを細胞外に分泌又は細胞表層に提示する細胞、好ましく酵母などの真核微生物は、向上されたセロビオヒドロラーゼ活性及び増強された相乗効果により、セルロースの分解、糖化、及び糖化と発酵とを同時進行させるCBP(連結バイオプロセス(糖化発酵同時進行))において、セルロースの分解効率を高めて、セルロースの効率的利用を具現化することができる。
【0068】
以上説明した本明細書に開示される形質転換細胞は、いずれも、モレキュラークローニング第3版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に記載されている方法に準じて作製することができる。真核微生物などの宿主細胞の形質転換のためのベクター及びその構築方法は、当業者において周知であって、モレキュラークローニング第3版、カレント・プロトコールズ・イン・モレキュラー・バイオロジー等に開示されている。また、コヘシンタンパク質やドックリンドメインを有するタンパク質を真核微生物において発現させるためのベクター及びその構築方法も、同様に、当業者において周知である。また、形質転換にあたり、従来公知の各種方法、例えば、トランスフォーメーション法や、トランスフェクション法、接合法、プロトプラスト法、エレクトロポレーション法、リポフェクション法、酢酸リチウム法等を用いることも、同様に当業者において周知である。
【0069】
なお、本明細書の開示によれば、本セルラーゼ複合体を構成する第1〜第3のキメラタンパク質及び当該キメラタンパク質をコードするDNAも提供される。
【0070】
(セルロースの分解産物の生産方法)
本明細書に開示されるセルロースの分解産物の生産方法は、本明細書の開示によれば、本タンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解する工程、を備えることができる。本方法によれば、セロビオヒドロラーゼ活性が向上した本タンパク質を用いることにより、効率的にセルロースを分解できる。分解工程で用いる1種又は2種以上のセルラーゼとしては、本セルラーゼ組成物に含まれうる他のセルラーゼから適宜選択して用いることができる。分解工程においては、セルラーゼは、本セルラーゼ組成物として供給されてもよいし、本タンパク質複合体として提供されていてもよい。分解工程における処理温度や時間等の条件は、用いるセルラーゼの種類及びセルロースの供給形態によって適宜設定することができる。
【0071】
本明細書において、セルロースとは、バイオマスから他の成分から分離工程を経たセルロースとして供給されてもよいし、リグニン、ヘミセルロース及び/又はペクチンが共存する未処理あるいは部分的な前処理を施したバイオマスなどセルロースを含有する材料として供給されてもよい。こうしたバイオマス資源としては、稲ワラ、麦ワラ、バガス、枯れ草等の廃棄資源のほか、未利用資源であってもよい。また、本明細書において、セルロース分解産物としては、セルロースの低分子化物であればよく、グルコース、そのオリゴマー、セロビオース等が挙げられる。
【0072】
(有用物質の生産方法)
本明細書の開示によれば、微生物の発酵により有用物質を生産する方法であって、本タンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解してセルロース分解産物を取得する工程と、微生物によりセルロース分解産物を含む炭素源を発酵して、前記有用物質を生産する工程と、を備える、方法が提供される。本生産方法によれば、本タンパク質を用い、向上したセロビオヒドロラーゼ活性がセルロースの分解に寄与するため、効率的にセルロースを分解することができる。本生産方法では、セルロースの分解工程と、当該分解工程で得られたセルロース分解産物を含む炭素源を微生物により発酵する工程と、をそれぞれ独立して実施することができる。したがって、セルロースの分解工程は、本セルラーゼ組成物や本タンパク質複合体を、酵素製剤として用いてセルロースを分解し、その後、このセルロース分解物を発酵工程に供給して発酵してもよい。
【0073】
本明細書の開示によれば、発酵のための微生物の細胞外に分泌又は細胞表層に提示される本タンパク質を含む、1種又は2種以上のセルラーゼの存在下、前記微生物を用いて、セルロースを少なくとも含む炭素源を発酵して前記有用物質を生産する工程、を備えることができる。このような生産方法によれば、セロビオヒドロラーゼ活性の向上したタンパク質を細胞外に発現する発酵用微生物を用いることで、いわゆる糖化・発酵同時プロセス(CBP)を効率的に行うことができる。1種又は2種以上のセルラーゼは、全て発酵用微生物の細胞表層に提示されていてもよいし、部分的に細胞外から添加されていてもよい。こうしたセルラーゼは、発酵用微生物の表層において本タンパク質複合体を構築していてもよい。
【0074】
発酵用の微生物は、酵母などのエタノール生産微生物や麹菌等の真核微生物を好ましく用いることができる。微生物は、人工的に取得された微生物であってもよい。例えば、グルコースからの代謝系の1種又は2種以上の酵素を遺伝子組換えにより置換、追加等して得られる本来の代謝物でない化合物を産生可能に遺伝子工学的に改変したものであってもよい。このような微生物を用いることで、例えば、イソプレノド合成経路の追加によるファインケミカル(コエンザイムQ10、ビタミン及びその原料等)、解糖系の改変によるグリセリンの生産、プラスチック・化成品原料を生産するなどのバイオリファイナリー技術に適用できる。有用物質としては特に限定しないが、グルコースを利用して微生物が生成可能なものが好ましく、上記したように、バイオリファイナリー技術全般にわたる物質を対象とすることができる。
【実施例】
【0075】
以下、本発明を、実施例を挙げて具体的に説明するが、本発明はこれらの実施例に限定されるものではない。なお、以下に述べる遺伝子組換え操作は既出のMolecular Cloningに従い行った。
【実施例1】
【0076】
(セルロース結合ドメインを有しないPhanerochaete chrysosporium由来のセロビオヒドロラーゼIIの合成)
ファネロケーテ由来セロビオヒドラーゼII(PcCBH2)はセルロース結合ドメイン(CBD)と触媒ドメインがリンカーでつながった構造を持っている。今回CBD機能を喪失させるため、PcCBH2のCBD部位を取り除き、「リンカー−触媒ドメイン」からなるペプチドを合成した。
【0077】
(CBDを欠失したPhanerochaete chrysosporium由来のセロビオヒドロラーゼIIの試験内合成のためのDNA断片の調製)
pET23bのNdeI/HindIIIにファネロケーテ由来セロビオヒドラーゼ遺伝子(配列番号1)を挿入したプラスミドpET23b_PcCBH2wtをテンプレートに、2種類のオリゴマー配列(5’-GGATCTGCGGTCACGACCACCTCCGTT -3’(配列番号4), 5’- CATATGTATATCTCCTTCTTAAAGT-3’(配列番号5))をプライマーとしてインバースPCRを行った。取得したPCR産物につき、アガロースゲル電気泳動、ゲル染色にて目的バンドを取り出し、ゲル中のDNAをキット(Wizard(R) SV Gel and PCR Clean-up System、Promega)を用いて抽出した。抽出したDNA断片をポリヌクレオチドキナーゼ(New England BioLabs)で両5’- 末端をリン酸化後、ライゲーションキット(DNA Ligation kit、Takara)を用いてライゲーション反応をさせた。ライゲーション産物を大腸菌DH5α株に導入し、その形質転換体を培養後、プラスミド抽出精製キット(QIAprep Spin Miniprep Kit、Qiagen)で目的のDNA断片を含むプラスミドを取り出した。CBD部位の欠失は、シーケンスにより確認した。
【0078】
(CBDを欠失したPhanerochaete chrysosporium由来のセロビオヒドロラーゼII改変体の試験内合成のためのDNA断片の調製)
次いで、CBH2改変体として、Sept25:PcCBH2に10個のアミノ酸置換(T130I/S134Q/S183T/A241E/M259I/Q307R/A309S/T342H/Q381S/F382L)を行った改変体及びMall4:PcCBH2に15個のアミノ酸置換(T112S/N125K/T130I/S134Q/S183T/Q204K/F206Y/A241E/M259I/Q307R/A309S/T342H/I365V/Q381S/F382L)を行った改変体につき、それぞれCBDを欠失させたDNA断片を、PcCBH2についてのDNA断片を含むプラスミド構築に準じて調製した。
【0079】
(セルラーゼの試験管内合成方法)
試験管内合成の鋳型として、作製したプラスミドに対して2本のプライマー(5’- ATCTCGATCCCGCGAAATTAATAC-3’(配列番号6)、 5’- TCCGGATATAGTTCCTCCTTTCAG-3’(配列番号7))で増幅、PCR purification kit (Qiagen)で精製したPCR産物を使用した。なおこのPCR産物には、T7プロモーター配列、開始コドン、遺伝子配列、終始コドン、ターミネーター配列が含まれている。
【0080】
試験管内合成は、表4に示す組成の反応液で、26℃、3時間、in vitroでの転写翻訳共役反応を行なった。無細胞蛋白質合成反応の促進剤として、ATPの枯渇を補うためにクレアチンキナーゼをATP生成酵素(ADP+ホスホクレアチン→ATP)として、リファンピシンはRNA合成開始阻害剤、フォリン酸はRNA転写阻害剤として添加した。また、タンパク質のフォールディングを助ける因子として、各種シャペロンを高発現した大腸菌A19株から抽出したS30菌体抽出液(−DTT)を用いた。更に、S−S結合形成促進因子であるカビ由来プロテインジスルフィドイソメラーゼも添加した。なお合成されたセルラーゼ量を評価するため、蛍光標識されたリジン(FluoroTect GreenLys in vitro Translation Labeling System:プロメガ)を混ぜて合成反応を行った。
【0081】
【表4】
【実施例2】
【0082】
(試験管内合成反応におけるセルラーゼ合成量の評価)
CBD欠失状態(リンカー−触媒ドメイン)とCBD保有状態(CBD−リンカー−触媒ドメイン)とのそれぞれにつき、野生型PcCBH2及び2種類のPcCBH2改変体の試験管内合成量及び比活性を評価した。
【0083】
合成量の評価は、無細胞合成時に蛍光標識されたリジン(FluoroTect GreenLys in vitro Translation Labeling System:プロメガ)を取り込ませた合成産物をSDS-PAGE後、蛍光イメージアナライザー(FLA9000:富士フィルム株式会社)で検出、画像解析ソフトであるMulti Gauge(富士フィルム株式会社)での解析により行った。なお今回使用したPcCBH2野生型および改変体はそれぞれ、CBD保有状態とCBD欠失状態で同数のリジンを持っているため、SDS−PAGEでの目的バンドの蛍光量でその合成量が比較評価できる。結果を図1に示す。図1には、セルラーゼの試験管内合成量を、野生型及び改変体につき、CBD保有状態(黒)及びCBD欠失状態(灰色)として、CBD保有状態の合成量に対する相対量として示す。
【0084】
図1に示すように、野生型PcCBH2ではCBD欠失状態の方がCBD有状態よりも合成量が約1.3倍向上していた(図1)。また評価した2つのPcCBH2変異体では共に、CBD欠失状態の方がCBD保有状態よりも合成量が約1.7倍向上していた。
【実施例3】
【0085】
(セルラーゼ(セロビオヒドロラーゼ)比活性の評価)
試験管内合成されたセルラーゼは、野生型及び各PcCBH2改変体でCBD保有状態及びCBD欠失状態で同じセルラーゼ量となるように試験管内合成試薬で希釈し、その活性を評価した。具体的には、50mM酢酸緩衝液pH5.0で100倍希釈後、1%リン酸膨潤セルロース(PSC)又は2.5%Avicelを同量添加し(0.5%PSC又は1.25%Avicel)、40℃で16時間反応させた。反応液を、TZアッセイ法(Journal of Biochemical and Biophysical Methods, 11 (1985) 109-115)にて遠心上精の還元糖量を測定し、セルラーゼ比活性とした。図2及び図3には、セルラーゼの比活性を、野生型及び改変体につき、CBD保有状態(黒)及びCBD欠失状態(灰色)として、CBD保有状態の活性に対する相対比活性として示す。
【0086】
(1)比活性の比較(PSCが基質の場合)
野生型PcCBH2、改変体Sept25およびMall4を用いて、PSCを基質とした条件でのCBD有欠失状態の比活性を評価した。結果は、図2に示すように、評価した野生型及び各改変体でCBD欠失状態はCBD保有状態よりも比活性が向上していた。すなわち、野生型及びSept25では2倍以上、Mall4では、4倍以上であった。
【0087】
(2)比活性の比較(Avicelが基質の場合)
野生型PcCBH2、改変体Sept25およびMall4を用いて、Avicelを基質とした条件でのCBD欠失/保有状態の比活性を評価した。結果は、図3に示すように、評価した野生型及び各改変体でCBD欠失状態はCBD保有状態よりも比活性が向上していた。すなわち、野生型では1.4倍、Sept25では1.2倍、Mall4では、1.7倍であった。
【実施例4】
【0088】
(トリコデルマ由来のエンドグルカナーゼ2のCBD欠失/保有状態での比活性比較)
本実施例では、トリコデルマ由来のエンドグルカナーゼ2(TrEG2、GHF5)を用いて、そのCBD欠失変異体を合成し、実施例3に準じて、CBD欠失/保有状態での比活性測定を行った。CBD欠失変異体のDNA断片を含むプラスミド作製は、上記エンドグルカナーゼIIの遺伝子(GenBankアクセッション番号P07982(配列番号12)に対して、プライマー配列(5’- GGGGTCCGATTTGCCGGCGTTAACAT-3’(配列番号8))及び5’- CATATGTATATCTCCTTCTTAAAGT-3’ (配列番号9))を用いる以外は、実施例1に準じてプラスミドを構築し、各ペプチドを試験管内合成した。
【0089】
実施例3と同様に、PSC(0.5%)及びAvicel(1.25%)を基質としてCBD欠失/保有状態での比活性を評価した。さらに、基質としてカルボキシメチルセルロース(CMC、0.2%)を使用して同様にして比活性を評価した。結果を図4に示す。図4には、セルラーゼの比活性を、CBD保有状態(黒)及びCBD欠失状態(灰色)として、CBD保有状態の活性に対する相対比活性として示す。
【0090】
図4に示すように、トリコデルマ由来のエンドグルカナーゼIIについては、PSCが基質の場合には、CBD欠失状態の方がCBD保有状態よりも比活性が高いことが示された。また、Avicelが基質の場合には、CBD欠失状態の方がCBD保有状態よりも比活性が低いことが示された。さらに、CMCが基質の場合には、CBD保有状態とCBD欠失状態はほぼ同じ比活性を示した。以上のことから、GHF45とは異なるGHF2のエンドグルカナーゼでのCBD削除の効果は、基質によって様々で、一概にCBD削除による比活性の向上を示すことはできなかった。
【実施例5】
【0091】
(ファネロケーテ由来のセロビオヒドラーゼ7CのCBD欠失/保有状態での比活性比較)
PcCBH2とは異なるファネロケーテ由来のセロビオヒドラーゼ7C(PcCBH7C、GHF7)を用いて、そのCBD欠失変異体を合成し、実施例4と同様にしてCBD欠失/保有状態での比活性測定を行った。PcCBH7CのCBD欠失変異体のDNA断片を含むプラスミドの作製は、PcCBH7C遺伝子(GenBankアクセッション番号M22220H(配列番号13))に対して、プライマー配列(5’- TTAGCCGGTGTACGTGGTGTTGAGGT-3’ (配列番号10)及び5’-AAGCTTGCGGCCGCACTCGAGCA -3’ (配列番号11))を用いる以外は、実施例1に準じて行い、各ペプチドを試験管内合成した。実施例3と同様に、PSC(0.5%)を基質としてCBD欠失/保有状態での比活性を評価した。結果を図5に示す。図5には、セルラーゼの比活性を、CBD保有状態(黒)及びCBD欠失状態(灰色)として、CBD保有状態の活性に対する相対比活性として示す。
【0092】
図5に示すように、実施例3におけるPcCBH2の結果とは異なり、CBD保有状態の方がCBD欠失状態よりも比活性が高かった(図5)。以上の結果から、PcCBH2と同じファネロケーテ由来のセロビオヒドラーゼでも、CBDの欠失が比活性を向上させるわけではないことがわかる。
【実施例6】
【0093】
(耐熱化変異体)
(1)耐熱性候補変異の選定
P. chrysosporium由来のセロビオヒドロラーゼII(PcCBH2)は、触媒ドメインとセルロース結合ドメイン(CBD)がリンカーでつながった構造を持つ。以下に、触媒ドメイン内のアミノ酸置換による耐熱化変異体の取得について説明する。
【0094】
(2)耐熱性候補変異を含む変異体の構築
耐熱化への初期配列としてPcCBH2変異体5−6(配列番号14、配列番号15)を用いた。変異体5−6は、配列番号2で表されるアミノ酸配列において、そのCBDのみに、アミノ酸置換(S22P置換(塩基配列上の変異:T64C))及び同義置換(塩基配列上の変異:G9A)をそれぞれ各一つ備えており、試験管内合成において野生型PcCBH2よりもタンパク質合成量が高い変異体であることがわかっている。耐熱化変異体の取得にあたり、変異体5−6に対して、以下の表5に示す変異(アミノ酸置換)を有する変異体作成した。すなわち、これらの変異体に相当するアミノ酸配列をそれぞれコードするオリゴマーDNA及びその直近の相補鎖オリゴマーDNAをプライマーとして用いてインバースPCR法でプラスミド鎖を増幅し、5’−末端をリン酸化後、ライゲーションによるプラスミドの環状化、大腸菌への形質転換を経て取得した。各変異体の変異導入は、シーケンスを行い確認した。
【0095】
【表5】
【0096】
43種の変異体を試験管内で合成した。試験管内合成の鋳型として、(1)で取得したDNAに対してプライマー(5’- ATCTCGATCCCGCGAAATTAATAC-3’(配列番号16)、 5’- TCCGGATATAGTTCCTCCTTTCAG-3’(配列番号17))を用いて増幅させたPCR産物を、PCR purification kit (Qiagen)で精製したものを用いた。なお、このPCR産物には、T7プロモーター配列、開始コドン、遺伝子配列、終始コドン、ターミネーター配列が含まれている。
【0097】
変異体の試験管内合成は、表6に示す組成の反応液で、26℃、3時間、in vitroでの転写翻訳共役反応として行った。無細胞蛋白質合成反応の促進剤として、ATPの枯渇を補うためにクレアチンキナーゼをATP生成酵素(ADP+ホスホクレアチン→ATP)として、リファンピシンはRNA合成開始阻害剤、フォリン酸はRNA転写阻害剤として添加した。また、タンパク質のフォールディングを助ける因子として、各種シャペロンを高発現した大腸菌A19株から抽出したS30菌体抽出液(−DTT)を用いた。更に、S−S結合形成促進因子である、カビ由来プロテインジスルフィドイソメラーゼも添加した。
【0098】
【表6】
【0099】
(3)変異体の耐熱試験方法
合成した変異体を、50mM酢酸緩衝液(pH5.0)で100倍に希釈、49,50,51,52,53及び54℃の各温度で2時間保温する熱処理後、その残存活性をリン酸膨潤セルロース(PSC)分解(反応条件:40℃、16時間)後の遠心上清の還元糖量をTZアッセイ法(Journal of Biochemical and Biophysical Methods, 11 (1985) 109-115)を用いて評価した。残存活性は熱処理前の活性との比で表した。またセルラーゼの耐熱温度は上記残存活性が50%となるときの温度と定義した。
【0100】
(4)有利変異の選定
以上の耐熱性評価の結果、13個の耐熱性が向上した変異体が得られた。その耐熱性を向上させた変異を有利変異と呼ぶこととした。今回に実験で得られた有利変異を表7及び図6に示す。図6に示すように、これらの有利変異は、いずれも、52℃環境下で2時間保温後において、氷冷保持条件と比較して1.3〜7倍残存活性を向上させるものであった。
【表7】
【0101】
(5)第1世代変異体の選択
有利変異を一つずつ親配列に加算したDNAを合成し、当該DNAを鋳型として、各変異体を実施例6の(2)と同様にして試験管内合成して、(3)に記載の方法に準じて耐熱性評価を行ったところ、最も耐熱性が高かった有利変異M−18による変異体第1世代とした。
【0102】
(6)第2世代の選択
実施例2で得た変異体T342H(M1−18)に有利変異Q381S/F382L(M1−20)を導入した変異体(W1820)を実施例1と同様にして合成し、(3)に記載の方法に準じて耐熱性を評価した。結果を図7に示す。図7に示すように、有利変異を蓄積したW18+20の耐熱性はM1−18及びM1−20と比較して向上していた。この変異体を第2世代として選択した。
【0103】
(7)第3世代の選択
実施例3で得られた変異体W18+20に有利変異(M1−1,−2,−4,−12)をそれぞれ導入し、その変異体を上記(2)と同様にして合成し、上記(3)と同様にして耐熱性を評価した。結果を図8に示す。図8に示すように、有利変異T130I/S134Q(M1−4)を導入した変異体(Tri4)の耐熱性が最も向上していた。この変異体を第3世代として選択した。
【0104】
(8)第4世代の選択
実施例4で得られた有利変異が3個導入された変異体Tri4に4つの有利変異(M1−1,−11,−12,−14,−15)をそれぞれ導入した変異体を、上記(2)と同様にして合成し、上記(3)と同様にして耐熱性を評価した。結果を図9に示す。図9に示すように、有利変異M259I(M1−15)を導入した変異体(Quard15)の耐熱性が最も向上していた。この変異体を第4世代として選択した。
【0105】
(9)第5世代の選択
実施例5で得られた有利変異が4個導入された変異体Quard15に4つの有利変異(M1−1,−11,−12,−27)をそれぞれ導入し、その変異体を、上記(2)と同様にして合成し、上記(3)と同様にして耐熱性を評価した。結果を図10に示す。図10に示すように、耐熱評価した。その結果、有利変異A241E(M1−12)を導入した変異体(Quint12)の耐熱性が最も向上していた。この変異体を第5世代として選択した。
【0106】
(10)第6世代の選択
実施例6で得られた有利変異が5個導入された変異体Quint12に耐熱性への寄与度が高かった有利変異Q307R/A309S(M1−37)とその他の3つの有利変異(M1−1,−21,−25)をそれぞれ導入した変異体を、上記(2)と同様にして合成し、上記(3)と同様にして耐熱性を評価した。結果を図11に示す。図11に示すように、Quint12にQ307R/A309S及び有利変異S183T(M1−25)を導入した変異体(Sept25)の耐熱性が最も向上していた。この変異体を第6世代として選択した。
【0107】
(11)スクリーニングによる耐熱化変異体の取得
加算していない有利変異(M1−1,−2,−11,−21,−27)を変異体Sept25にばらつきを持って導入後、スクリーニングにより最も高い耐熱性を有する変異体の取得を試みた。具体的には、QuickChange Multi Site-Directed Mutagenesis Kit(Stratagene)を用いて変異を導入し、その変異体92個をクローニングした。上記(2)と同様にして試験管内合成した変異体を56℃にて2時間の熱処理後、熱処理前に対する残存活性を指標にスクリーニングを行った。結果を図12に示す。図12に示すように、熱処理に対する残存活性がSept25と比較して約5倍向上した変異体が得られた。
【0108】
(12)スクリーニングで得られた変異体の耐熱性評価
上記スクリーニングで、残存活性が高かった上位3個の変異体について、上記(3)と同様に耐熱性を評価した。結果を図13に示す。図13に示すように、評価した3変異体は共にSept25と比較して耐熱温度が1℃向上していた。
【0109】
(13)スクリーニングで得られた耐熱化変異体のアミノ酸配列の決定
上記で得られた耐熱化変異体のアミノ酸配列を塩基配列解析行うことで解析した。その結果、残りの有利変異(M1−1,−2,−11,−21,−27)がすべて入っていることが分かった。この得られたクローンを耐熱化実験での最終変異体(Mall4と呼ぶ)こととした。
【0110】
各世代の変異体における変異の組み合わせを以下の表に示す。
【表8】
【実施例7】
【0111】
(14)各世代の耐熱化変異体を耐熱性評価
これまでに得られた各世代の耐熱変異体につき、上記(3)と同様に耐熱性を評価した。結果を図14に示す。図14に示すように、その結果、有利変異を加えるに従い耐熱温度が向上していることが確認できた。初期変異体5−6の耐熱温度が49.9℃であるのに対し、耐熱化変異体Sept25は4.4℃向上した54.3℃、さらに最終変異体Mall4では5.4℃向上した54.3℃であった。
【実施例8】
【0112】
(CBD欠失PcCBH2変異体の耐熱性評価)
さらに、耐熱化変異体Mall4のCBD部位を削除した変異体を作製した。当該変異体は、実施例1に準じて試験管内合成した。Mall4及びそのCBD欠失変異体の耐熱性を評価した。また、変異体5−6の評価も併せて行った。結果を図15に示す。図15に示すように、耐熱化変異体Mall4からCBDを欠失させた変異体は、Mall4CBD保有体と同様、変異体5−6よりも、高い耐熱性を有していた。
【実施例9】
【0113】
(CBD欠失変異体のセルロースへの結合力の評価)
配列番号2で表されるアミノ酸配列の野生型PcCBH2(CBD保有)、上記の耐熱性変異体Mall4及びそれぞれのCBD欠失変異体を用いて、各セルラーゼのセルロースへの結合力をアビセルからの遊離率で評価した。具体的には、酵母で分泌発現そしてアビセルカラムで精製したそれぞれのセルラーゼ100ngを、50mM酢酸緩衝液pH5.0でけん濁したアビセル100ul(終濃度2%(w/v))に添加、攪拌後、氷上で2時間静置させた。なお、コントロール実験として、アビセル無条件でも行った。それぞれの遠心上清のセルラーゼ活性を測定し、セルラーゼのアビセルからの遊離率を算出した。結果を図16に示す。
【0114】
図16に示すように、CBD保有セルラーゼの遊離率は約0.1であったのに対しCBD失活変異体では0.3〜0.5の値を示した。以上の結果より、CBD欠失セルラーゼは、保有セルラーゼと比較して、アビセルに対する結合性が低下していた。
【配列表フリーテキスト】
【0115】
配列番号4〜13,16、17:プライマー
配列番号14,15:Phanerochaete chrysosporium由来のセロビオヒドロラーゼの変異体
【特許請求の範囲】
【請求項1】
以下の(a)〜(d)のいずれかのアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質。
(a)配列番号2で表されるアミノ酸配列
(b)配列番号2で表されるアミノ酸配列において1又は複数個のアミノ酸の置換、欠失及び/又は付加を有するアミノ酸配列
(c)配列番号2で表されるアミノ酸配列と75%以上の同一性を有するアミノ酸配列
(d)配列番号1で表される塩基配列と相補的な塩基配列からなるDNA又はその一部とストリンジェントな条件下でハイブリダイズするDNAによってコードされるアミノ酸配列
(e)配列番号1で表される塩基配列と75%以上の同一性を有する塩基配列によってコードされるアミノ酸配列
【請求項2】
前記(b)のアミノ酸配列は、配列番号2で表されるアミノ酸配列において、アミノ酸置換(T130I/S134Q/S183T/A241E/M259I/Q307R/A309S/T342H/Q381S/F382L)を有するアミノ酸配列である、請求項1に記載のタンパク質。
【請求項3】
前記(b)のアミノ酸配列は、配列番号2で表されるアミノ酸配列において、アミノ酸置換(T112S/N125K/T130I/S134Q/S183T/Q204K/F206Y/A241E/M259I/Q307R/A309S/T342H/I365V/Q381S/F382L)を有するアミノ酸配列である、請求項1に記載のタンパク質。
【請求項4】
請求項1〜3のいずれかに記載のタンパク質をコードするDNA。
【請求項5】
請求項4に記載のDNAを含む、発現用DNAコンストラクト。
【請求項6】
請求項5に記載の発現用DNAコンストラクトにより形質転換された細胞。
【請求項7】
真核微生物である、請求項6に記載の細胞。
【請求項8】
請求項1〜3のいずれかに記載のタンパク質を細胞外に分泌又は細胞表層に提示する、請求項7に記載の細胞。
【請求項9】
前記真核微生物は、非セルラーゼ生産微生物である、請求項7又は8に記載の細胞。
【請求項10】
前記真核微生物は、セルラーゼ生産微生物である、請求項7又は8に記載の細胞。
【請求項11】
セルロースの分解産物の生産方法であって、
請求項1〜3のいずれかに記載のタンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解する工程、を備える、セルロースの分解産物の生産方法。
【請求項12】
微生物の発酵により有用物質を生産する方法であって、
請求項1〜3のいずれかに記載のタンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解してセルロース分解産物を取得する工程と、
微生物により前記セルロース分解産物を含む炭素源を発酵して、前記有用物質を生産する工程と、
を備える、方法。
【請求項13】
微生物の発酵により有用物質を生産する方法であって、
前記微生物の細胞外に分泌又は細胞表層に提示される請求項1に記載のタンパク質を含む1種又は2種以上のセルラーゼの存在下、前記微生物を用いて、セルロースを少なくとも含む炭素源を発酵して前記有用物質を生産する工程、を備える、方法。
【請求項14】
請求項1〜3のいずれかに記載のタンパク質を含むセルラーゼ組成物。
【請求項15】
請求項1〜3のいずれかに記載のタンパク質を含む、タンパク質複合体。
【請求項1】
以下の(a)〜(d)のいずれかのアミノ酸配列において、セルロース結合ドメインが不活性化されており、セロビオヒドロラーゼ活性を有するタンパク質。
(a)配列番号2で表されるアミノ酸配列
(b)配列番号2で表されるアミノ酸配列において1又は複数個のアミノ酸の置換、欠失及び/又は付加を有するアミノ酸配列
(c)配列番号2で表されるアミノ酸配列と75%以上の同一性を有するアミノ酸配列
(d)配列番号1で表される塩基配列と相補的な塩基配列からなるDNA又はその一部とストリンジェントな条件下でハイブリダイズするDNAによってコードされるアミノ酸配列
(e)配列番号1で表される塩基配列と75%以上の同一性を有する塩基配列によってコードされるアミノ酸配列
【請求項2】
前記(b)のアミノ酸配列は、配列番号2で表されるアミノ酸配列において、アミノ酸置換(T130I/S134Q/S183T/A241E/M259I/Q307R/A309S/T342H/Q381S/F382L)を有するアミノ酸配列である、請求項1に記載のタンパク質。
【請求項3】
前記(b)のアミノ酸配列は、配列番号2で表されるアミノ酸配列において、アミノ酸置換(T112S/N125K/T130I/S134Q/S183T/Q204K/F206Y/A241E/M259I/Q307R/A309S/T342H/I365V/Q381S/F382L)を有するアミノ酸配列である、請求項1に記載のタンパク質。
【請求項4】
請求項1〜3のいずれかに記載のタンパク質をコードするDNA。
【請求項5】
請求項4に記載のDNAを含む、発現用DNAコンストラクト。
【請求項6】
請求項5に記載の発現用DNAコンストラクトにより形質転換された細胞。
【請求項7】
真核微生物である、請求項6に記載の細胞。
【請求項8】
請求項1〜3のいずれかに記載のタンパク質を細胞外に分泌又は細胞表層に提示する、請求項7に記載の細胞。
【請求項9】
前記真核微生物は、非セルラーゼ生産微生物である、請求項7又は8に記載の細胞。
【請求項10】
前記真核微生物は、セルラーゼ生産微生物である、請求項7又は8に記載の細胞。
【請求項11】
セルロースの分解産物の生産方法であって、
請求項1〜3のいずれかに記載のタンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解する工程、を備える、セルロースの分解産物の生産方法。
【請求項12】
微生物の発酵により有用物質を生産する方法であって、
請求項1〜3のいずれかに記載のタンパク質を含む1種又は2種以上のセルラーゼを用いて、セルロースを分解してセルロース分解産物を取得する工程と、
微生物により前記セルロース分解産物を含む炭素源を発酵して、前記有用物質を生産する工程と、
を備える、方法。
【請求項13】
微生物の発酵により有用物質を生産する方法であって、
前記微生物の細胞外に分泌又は細胞表層に提示される請求項1に記載のタンパク質を含む1種又は2種以上のセルラーゼの存在下、前記微生物を用いて、セルロースを少なくとも含む炭素源を発酵して前記有用物質を生産する工程、を備える、方法。
【請求項14】
請求項1〜3のいずれかに記載のタンパク質を含むセルラーゼ組成物。
【請求項15】
請求項1〜3のいずれかに記載のタンパク質を含む、タンパク質複合体。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
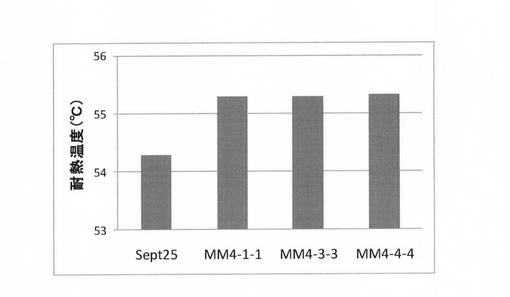
【図14】
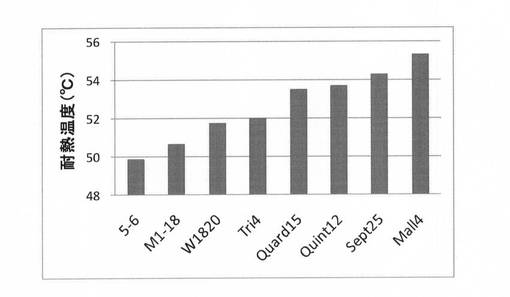
【図15】
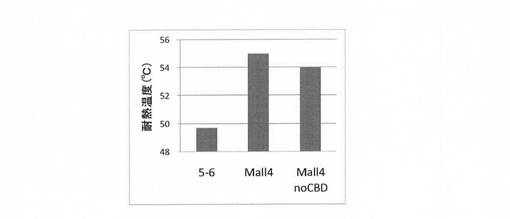
【図16】
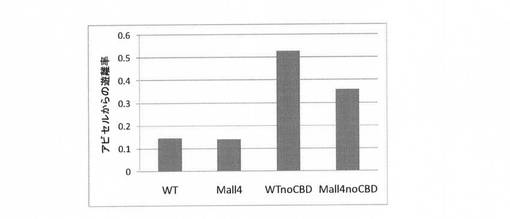
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【公開番号】特開2012−39966(P2012−39966A)
【公開日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願番号】特願2010−185614(P2010−185614)
【出願日】平成22年8月20日(2010.8.20)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度および22年度独立行政法人新エネルギー・産業技術総合開発機構「新エネルギー技術研究開発/バイオマスエネルギー等高効率転換技術開発(先導技術開発)/セルロースエタノール高効率製造のための環境調和型統合プロセス開発」の委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【Fターム(参考)】
【公開日】平成24年3月1日(2012.3.1)
【国際特許分類】
【出願日】平成22年8月20日(2010.8.20)
【国等の委託研究の成果に係る記載事項】(出願人による申告)平成21年度および22年度独立行政法人新エネルギー・産業技術総合開発機構「新エネルギー技術研究開発/バイオマスエネルギー等高効率転換技術開発(先導技術開発)/セルロースエタノール高効率製造のための環境調和型統合プロセス開発」の委託研究、産業技術力強化法第19条の適用を受ける特許出願
【出願人】(000003609)株式会社豊田中央研究所 (4,200)
【Fターム(参考)】
[ Back to top ]