
セーフハーバー遺伝子座への導入遺伝子の挿入のためのエンドヌクレアーゼの使用
本発明は、「セーフハーバー遺伝子座」、すなわち導入遺伝子の安全な発現を可能にする遺伝子座に位置する標的配列を切断することが可能なエンドヌクレアーゼに関する。本発明は、細胞、組織又は個体への導入遺伝子の挿入のためのかかるエンドヌクレアーゼの使用に更に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、「セーフハーバー遺伝子座」、すなわち導入遺伝子の安全な発現を可能にする遺伝子座に位置する標的配列を切断することが可能なエンドヌクレアーゼに関する。本発明は、細胞、組織又は生物への導入遺伝子の挿入のためのかかるエンドヌクレアーゼの使用に更に関する。
【背景技術】
【0002】
メガヌクレアーゼ
ホーミングエンドヌクレアーゼとも称されるメガヌクレアーゼは、生細胞における二本鎖切断及び組換えを誘導するために使用された最初のエンドヌクレアーゼであった(非特許文献1、非特許文献2、非特許文献3、非特許文献4)。しかしながら、それらの使用は、それらの狭い特異性によって長年制限されていた。数百個の天然メガヌクレアーゼが過去数年にわたって同定されたが、この多様性は依然としてゲノムの複雑性に対応するにはほとんどが不十分であり、関心の遺伝子内のメガヌクレアーゼ切断部位を発見する確率は、依然として極めて低い。これらの知見から、選ばれた配列を天然エンドヌクレアーゼと同じ選択性で切断する、個々に適合した特異性を有する人工エンドヌクレアーゼの必要性が問題となっている。
【0003】
メガヌクレアーゼは、所望の標的配列を切断するゲノム工学ツールを得るのに最適な足場として登場した(非特許文献5)。変更された特異性を有するメガヌクレアーゼの改変を可能にするコンビナトリアルアセンブリプロセスは、非特許文献6、非特許文献7、非特許文献8、非特許文献9によって記載されている。簡潔に述べると、これらのプロセスは、野生型メガヌクレアーゼの基質特異性と数ヌクレオチドだけ異なる基質特異性を有する局所改変変異体の同定に依存する。次いで、かかるタンパク質において同定された最大で4組の突然変異を、完全に再設計された結合面を有する新たなメガヌクレアーゼを生成するために、新たなタンパク質に組み立てることができる。
【0004】
これらのプロセスは2つの工程を必要とする。初めに、異なる組の突然変異を、パリンドローム標的を切断するホモ二量体変異体に組み立てる。次いで、2つのホモ二量体を、選ばれた非パリンドローム標的を切断するヘテロ二量体メガヌクレアーゼを生成するために共発現させることができる。このプロセスの第1の工程は、依然として最も難しいものであり、所与の遺伝子座を切断するメガヌクレアーゼを絶対確実に得ることができるか否かを事前に知ることはできない。実際に、全ての配列が改変メガヌクレアーゼによって同等に切断される可能性があるわけではなく、場合によっては、メガヌクレアーゼの改変は困難であることが判明し得る(非特許文献10)。
【0005】
部位特異的ゲノム修飾に好適な他の酵素
インテグラーゼ、リコンビナーゼ、トランスポザーゼ及びエンドヌクレアーゼのような特殊化された酵素が、部位特異的ゲノム修飾に提案されている。何年もの間、これらの酵素の使用は、所望の標的部位に対するそれらの天然の特異性の再標的化の難しさのために制限されたままである。実際に、これらのタンパク質の標的部位又は十分な程度の配列同一性を有する配列は、訂正される突然変異に近接する配列中又は不活性化される遺伝子内に存在しなくてはならないが、予め改変された配列の場合を除いて、通常はそのようなことはない。遺伝子療法におけるこれらのDNA修飾酵素の使用を可能にすることの主な課題は、それらのDNA結合特性を再設計する可能性に左右される。個々に適合した基質特異性を有する人工タンパク質を得ることを目的として、多くの戦略が開発されている。
【0006】
ストレプトミセスファージPhiC31に由来するインテグラーゼは、内因性遺伝子座における標的化遺伝子導入に早くから使用されていた。この酵素は、ファージ付着部位(attP)と細菌付着部位(attB)との間の部位特異的反応による細菌染色体へのファージゲノムの組換えを媒介する(非特許文献11、非特許文献12)。これはattB部位を保有するプラスミドから、偽attP部位(attP’)と呼ばれるattPとの部分同一性を有する天然のゲノム配列へと起こり得る。PhiC31インテグラーゼは、ヒトゲノムにおいてhFIXを含む幾つかの導入遺伝子を導入するために使用されている(非特許文献13、非特許文献14、非特許文献15、非特許文献16、非特許文献17)。ここでの欠点は、組込みが起こり得る部位を選ぶことができず(非特許文献16)、正確な組込みについてヒトゲノム遺伝子座内の偽attP部位に依存せざるを得ないということである。主要な組込み部位は染色体19に見られるが、数百個の他の組込み遺伝子座が同定されている(非特許文献16)。最近の研究では、attP’部位での組込みの効率及び特異性を高め、選ばれた部位を標的とする改変インテグラーゼの開発の道を開くために、PhiC31インテグラーゼを突然変異させた(非特許文献18)。しかしながら、改変インテグラーゼの開発は、標的化リコンビナーゼ系及び標的化エンドヌクレアーゼ系に焦点を合わせた同様の試みに遅れを取っている。
【0007】
バクテリオファージP1に由来するCreリコンビナーゼ又はサッカロミセス・セレビシエ(Saccharomyces cerevisiae)に由来するFlpタンパク質等の部位特異的リコンビナーゼは、それらの同族部位を含有する予め改変された配列間の組換えを誘導するために使用されている。Creリコンビナーゼは、loxPとして知られる2つの同一の34bp部位を認識し、それらの間の組換えを媒介する(非特許文献19)。長年にわたって、Cre由来リコンビナーゼの制限は、反復loxP部位又は偽loxP部位が、これら2つの部位間のDNA組込みを可能にするために存在しなければならないということであった。しかしながら、この分子のDNA結合面の指向進化を使用して、新たな特異性を有するリコンビナーゼが作り出されている(非特許文献20、非特許文献21)。Creリコンビナーゼ系は、ウイルス配列の切除を誘導するDNA標的化酵素の使用のためのフレームワークの提供にも有用であった。実際に、レトロウイルスのモロニーマウス白血病ウイルスのベクター系を用いた研究から、loxP部位を組込みレトロウイルスベクターのLTRに導入する場合、Creの発現が2つのloxP部位間の全ての配列の欠失をもたらし得ることが示されている(非特許文献22)。ごく最近では、改変Creリコンビナーゼ変異体が、細胞からHIV1型プロウイルスを切除するために使用されている(非特許文献23)。リコンビナーゼはプロウイルスLTRを標的とするように再設計され、全ての介在配列の切除を誘導するために使用されている。FRT(Flp組換え標的)配列を標的とするFlpリコンビナーゼを用いた改変も試みられており(非特許文献24)、非天然Flp組換え標的を認識する変異体が得られている(非特許文献25)。しかしながら、今日、かかる酵素による予め改変されていない遺伝子座における標的化挿入の例はない。
【0008】
Piggy Back及びSleeping Beauty等のトランスポゾンは、脊椎動物細胞への配列の挿入に効率的なツールを提供することができ、長期的発現を達成するウイルス媒介遺伝子送達の代替として提案されている(非特許文献26、非特許文献27、非特許文献28)。トランスポゾンは、DNA分子中に存在するDNA配列がトランスポザーゼの作用によって別の位置に挿入される、遺伝子送達の天然の手段である。SB100Xと呼ばれる改変SBトランスポザーゼが、このプロセスの効率を高めることが近年示されている(非特許文献28)。転位はゲノムレベルでランダムであり(例えば、SBはTAジヌクレオチドに組み込まれる)(非特許文献29)、したがって標的化アプローチのためのツールとみなすべきではない。しかしながら、更なる研究から、トランスポザーゼ触媒ドメインを特異的なDNA結合ドメインに融合させることにより、ヒト細胞において改変トランスポザーゼによって媒介される染色体転位の可能性が示されており(非特許文献30)、新たなカテゴリーの標的化ツールの開発の道が開かれている。
【0009】
遺伝子療法
10年近く前の遺伝子療法による数人のX−SCID患者の治療の成功は、遺伝子療法の分野における最も重要な節目の1つであった。この素晴らしい業績の後、別の形態のSCID、表皮水疱症及びレーバー黒内障等を含む種々の疾患に対処する他の臨床試験における大きな成功が続いた。しかしながら、これらの当初の成功は、一連の深刻な有害事象、すなわちX−SCID治療患者における白血病の発生のために長い間目立たないものであった(非特許文献31、非特許文献32、非特許文献33)。1例を除く白血病の全ての症例を化学療法によって最終的に治療することができたため、このアプローチは全体的にみて成功のように思われるが、これらの深刻な有害事象は、現行の遺伝子療法アプローチの重大なリスクを浮き彫りにしている。
【0010】
このため、当該技術分野においては、遺伝子を被験体のゲノムに挿入する安全な方法が必要とされている。
【0011】
遺伝性疾患の治療のために現在開発されている遺伝子療法プロトコルの大半は、疾患原因遺伝子の付加的かつ機能的なコピーによる変異型対立遺伝子の補完に基づいている。網膜等の非分裂組織では、このコピーの送達は、例えばアデノ随伴ウイルス(AAV)に由来する非組込みベクターを用いて達成することができる。しかしながら、増殖する運命である造血幹細胞(HSC)等の幹細胞を標的とする場合、持続的発現が問題になり、組込みベクターが必要とされる。ゲノム内に組み込まれ、宿主の染色体を用いて複製するレトロウイルスベクターが、この目的に有効であることが証明されたが、それらの挿入のランダム性は、全て遺伝子発現に関係する様々な懸念を生じている。X−SCID試験において観察された白血病の症例は、組込み部位の近傍のがん原遺伝子の活性化に明らかに関連していた。加えて、導入遺伝子の不適切な発現は代謝的問題又は免疫学的問題をもたらす可能性がある。最後に、挿入は内因性遺伝子のノックアウトをもたらす可能性がある。
【0012】
部位特異的組込みは、挿入突然変異誘発のリスクを緩和し得るため、ウイルスベクターのランダム組込みの有望な代替策であり得る(非特許文献34、非特許文献35、非特許文献5)。しかしながら、標的化組換えのツールを設計するには比較的時間がかかる。加えて、各々のツールは活性及び特異性に関してその固有の特性を有する。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】Rouet et al. PNAS 1994 91:6064-6068
【非特許文献2】Rouet et al. Mol Cell Biol. 1994 14:8096-8106
【非特許文献3】Choulika et al. Mol Cell Biol. 1995 15:1968-1973
【非特許文献4】Puchta et al. PNAS 1996 93:5055-5060
【非特許文献5】Paques et al. Curr Gen Ther. 2007 7:49-66
【非特許文献6】Arnould et al. J Mol Biol. 2006 355:443-458
【非特許文献7】Arnould et al. J Mol Biol. 2007 371:49-65
【非特許文献8】Smith et al. NAR 2006 34:e149
【非特許文献9】Grizot et al. NAR 2009 37:5405
【非特許文献10】Galetto et al. Expert Opin Biol Ther. 2009 9:1289-303
【非特許文献11】Kuhstoss et al. J Mol Biol 1991 222:897-908
【非特許文献12】Rausch et al. NAR 1991 19:5187-5189
【非特許文献13】Olivares et al. Nat Biotech 2002 20:1124-1128
【非特許文献14】Ginsburg et al. Adv Genet 2005 54:179-187
【非特許文献15】Calos Curr Gene Ther 2006 6:633-645
【非特許文献16】Chalberg et al. J Mol Biol 2006 357:28-48
【非特許文献17】Aneja et al. J Gene Med 2007 9:967-975
【非特許文献18】Keravala et al. Mol Ther 2009 17:112-120
【非特許文献19】Abremski et al. Cell 1983 32:1301-1311
【非特許文献20】Buchholz et al. Nat Biotech 2001 19:1047-1052
【非特許文献21】Santoro et al. PNAS 2002 99:4185-4190
【非特許文献22】Choulika et al. J Virol 1996 70:1792-1798
【非特許文献23】Sarkar et al. Science 2007 316:1912-1915
【非特許文献24】Buchholzt et al. Nat Biotech 1998 16:657-662
【非特許文献25】Voziyanov et al. J Mol Biol 2003 326:65-76
【非特許文献26】Izsvak et al. Mol ther 2004 9:147-156
【非特許文献27】Ivics et al. Curr Gene Ther 2006 6:593-607
【非特許文献28】Mates et al. Nat Genet 2009 41:753-761
【非特許文献29】Vigdal et al. J Mol Biol 2002 323:441-452
【非特許文献30】Ivics, et al. Mol Ther 2007 15:1137-1144
【非特許文献31】Hacein-Bey-Abina et al. Science 2003 302:415-419
【非特許文献32】Hacein-Bey-Abina et al. J Clin Invest. 2008 118:3132-3142
【非特許文献33】Howe et al. J Clin Invest. 2008 118:3143-3150
【非特許文献34】Kolb et al. Trends Biotechnol. 2005 23:399-406
【非特許文献35】Porteus et al. Nat Biotechnol. 2005 23:967-973
【発明の概要】
【発明が解決しようとする課題】
【0014】
したがって、当該技術分野では、遺伝子付加の「セーフハーバー」であるとみなされ得るゲノムの遺伝子座への導入遺伝子の標的化挿入を可能にするツールが必要とされている。加えて、このツールを、それらの配列に関わりなく導入遺伝子の挿入に使用し、それにより同じツールを用いた遺伝子療法による多数の疾患の治療を可能にすることができれば極めて有利であり得る。さらに、このツールが高い有効性でゲノムに導入遺伝子を挿入し、高いレベルで導入遺伝子の安定な発現をもたらすことができれば極めて有利であり得る。
【課題を解決するための手段】
【0015】
本発明はとりわけ、以下の実施の形態に関する(drawn to):
実施の形態1:導入遺伝子を個体のゲノムに挿入する際に使用される標的配列を切断することが可能な変異型エンドヌクレアーゼであって、
i.上記ゲノムが上記標的配列を含む遺伝子座を含み、
ii.上記標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連しない、変異型エンドヌクレアーゼ。
実施の形態2:上記導入遺伝子の挿入が、上記標的配列の近傍に位置する遺伝子の発現を実質的に変更しない、実施の形態1に記載のエンドヌクレアーゼ。
実施の形態3:上記標的配列が、最近接遺伝子から少なくとも100kb離れて位置する、実施の形態1又は2に記載のエンドヌクレアーゼ。
実施の形態4:上記RISが、幹細胞の形質導入による遺伝子療法によって治療される患者に由来する細胞において同定されたものである、実施の形態1〜3のいずれか1つに記載のエンドヌクレアーゼ。
実施の形態5:上記RISが、造血幹細胞の形質導入による遺伝子療法によって治療される患者に由来する細胞において同定されたものである、実施の形態1〜3のいずれか1つに記載のエンドヌクレアーゼ。
実施の形態6:ホーミングエンドヌクレアーゼである、実施の形態1〜5のいずれか1つに記載のエンドヌクレアーゼ。
実施の形態7:上記ホーミングエンドヌクレアーゼが、LAGLIDADGエンドヌクレアーゼファミリーの成員である、実施の形態6に記載のエンドヌクレアーゼ。
実施の形態8:上記LAGLIDADGエンドヌクレアーゼファミリーの成員がI−CreIである、実施の形態7に記載のエンドヌクレアーゼ。
実施の形態9:上記遺伝子座が、ヒト染色体6p25.1上のSH3遺伝子座、ヒト染色体7q31.2上のSH4遺伝子座、ヒト染色体21q21.1上のSH6遺伝子座、ヒト染色体13q34上のSH12遺伝子座、ヒト染色体3p12.2上のSH13遺伝子座、ヒト染色体22上のSH19遺伝子座、ヒト染色体12q21.2上のSH20遺伝子座、ヒト染色体3p24.1上のSH21遺伝子座、ヒト染色体6p12.2上のSH33遺伝子座、ヒト染色体2p16.1上のSH7遺伝子座及びヒト染色体5上のSH8遺伝子座から選択される、実施の形態1〜8のいずれか1つに記載のエンドヌクレアーゼ。
実施の形態10:導入遺伝子を細胞又は組織のゲノムに挿入するための実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼのin vitro又はex vivoでの使用。
実施の形態11:配列番号1又は配列番号42と少なくとも80%同一な配列を含む2つの単量体を含む変異型二量体I−CreIタンパク質であって、
i.上記二量体I−CreIタンパク質が、個体の遺伝子座内に位置する標的配列を切断することが可能であり、該標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連せず、
ii.上記標的配列が配列番号4の配列を含まない、変異型二量体I−CreIタンパク質。
実施の形態12:ヒト染色体6p25.1上のSH3遺伝子座内に位置する標的配列を切断することが可能である、実施の形態11に記載の二量体I−CreIタンパク質。
実施の形態13:上記標的配列が配列番号2の配列を含む、実施の形態12に記載の二量体I−CreIタンパク質。
実施の形態14:
a)配列番号1の30位、38位、70位及び75位にアミノ酸置換を含む第1の単量体と、
b)配列番号1の44位、54位、70位及び75位にアミノ酸置換を含む第2の単量体と、
を含む、実施の形態12又は13に記載の二量体I−CreIタンパク質。
実施の形態15:上記ポリペプチドが、
a)30G 38R 70D 75N 86D突然変異を含む第1の単量体、
b)
i.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
ii.44A 54L 70Q 75Y 92R 158R 162A突然変異を含む単量体、
iii.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iv.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
v.44A 54L 70Q 75N突然変異を含む単量体、
vi.44A 54L 57E 70Q 75N 158R 162A突然変異を含む単量体、及び
vii.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含む、実施の形態14に記載の二量体I−CreIタンパク質。
実施の形態16:上記ポリペプチドが、
a)30G 38R 70D 75N 81T 154G突然変異を含む第1の単量体、
b)
i.44A 54L 70Q 75N 105A 158R 162A突然変異を含む単量体、
ii.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iii.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iv.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
v.44A 54L 70Q 75N突然変異を含む単量体、及び
vi.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含む、実施の形態14に記載の二量体I−CreIタンパク質。
実施の形態17:上記ポリペプチドが、
a)30G 38R 50R 70D 75N 142R突然変異を含む第1の単量体、
b)
i.44A 54L 70Q 75N 105A 158R 162A突然変異を含む単量体、
ii.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iii.44A 54L 70Q 75Y 92R 158R 162A突然変異を含む単量体、
iv.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
v.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
vi.44A 54L 66C 70Q 71R 75N 151A 158R 162A突然変異を含む単量体、
vii.44A 54L 70Q 75N突然変異を含む単量体、
viii.44A 54L 57E 70Q 75N 158R 162A突然変異を含む単量体、及び
ix.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含む、実施の形態14に記載の二量体I−CreIタンパク質。
実施の形態18:ヒト染色体7q31.2上のSH4遺伝子座内に位置する標的配列を切断することが可能である、実施の形態11に記載の二量体I−CreIタンパク質。
実施の形態19:上記標的配列が配列番号3の配列を含む、実施の形態18に記載の二量体I−CreIタンパク質。
実施の形態20:
a)配列番号1の24位、70位、75位及び77位にアミノ酸置換を含む第1の単量体と、
b)配列番号1の24位、44位及び70位にアミノ酸置換を含む第2の単量体と、
を含む、実施の形態18又は19に記載の二量体I−CreIタンパク質。
実施の形態21:上記ポリペプチドが、
a)
i.24V 44R 68Y 70S 75Y 77N突然変異を含む単量体、
ii.24V 68A 70S 75N 77R突然変異を含む単量体、及び
iii.24V 70D 75N 77R突然変異を含む単量体からなる群から選択される第1の単量体、
b)
i.24V 44Y 70S突然変異を含む単量体、及び
ii.24V 44Y 70S 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含む、実施の形態20に記載の二量体I−CreIタンパク質。
実施の形態22:ヒト染色体21q21.1上のSH6遺伝子座内に位置する標的配列を切断することが可能である、実施の形態11に記載の二量体I−CreIタンパク質。
実施の形態23:上記標的配列が配列番号59の配列を含む、実施の形態22に記載の二量体I−CreIタンパク質。
実施の形態24:
a)配列番号1の44位、並びに任意に70位及び/又は75位にアミノ酸置換を含む第1の単量体と、
b)配列番号1の28位、40位、44位、70位及び75位にアミノ酸置換を含む第2の単量体と、
を含む、実施の形態22又は23に記載の二量体I−CreIタンパク質。
実施の形態25:上記ポリペプチドが、
a)44K 68T 70G 75N突然変異を含む第1の単量体と、
b)
i.28Q 40R 44A 70L 75N 96R 111H 144S突然変異を含む単量体、
ii.7R 28Q 40R 44A 70L 75N 85R 103T突然変異を含む単量体、
iii.28Q 40R 44A 70L 75N 103S突然変異を含む単量体、
iv.24F 27V 28Q 40R 44A 70L 75N 99R突然変異を含む単量体、
v.7R 28Q 40R 44A 70L 75N 81T突然変異を含む単量体、
vi.7R 28Q 40R 44A 70L 75N 77V突然変異を含む単量体、
vii.7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R突然変異を含む単量体、
viii.28Q 40R 44A 70L 75N突然変異を含む単量体、
ix.7R 28Q 40R 44A 70L 75N 103T突然変異を含む単量体、及び
x.28Q 34R 40R 44A 70L 75N 81V 103T 108V 160E突然変異を含む単量体からなる群から選択される第2の単量体と、
を含む、実施の形態24に記載の二量体I−CreIタンパク質。
実施の形態26:上記ポリペプチドが、
a)44K突然変異、並びに任意に70S及び/又は75N突然変異を含む第1の単量体と、
b)
i.28Q 40R 44A 70L 75N 96R 111H 144S突然変異を含む単量体、
ii.7R 28Q 40R 44A 70L 75N 85R 103T突然変異を含む単量体、
iii.28Q 40R 44A 70L 75N 103S突然変異を含む単量体、
iv.24F 27V 28Q 40R 44A 70L 75N 99R突然変異を含む単量体、
v.7R 28Q 40R 44A 70L 75N 81T突然変異を含む単量体、
vi.7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R突然変異を含む単量体、
vii.7R 28Q 40R 44A 70L 75N 103T突然変異を含む単量体、及び
viii.28Q 34R 40R 44A 70L 75N 81V 103T 108V 160E突然変異を含む単量体からなる群から選択される第2の単量体と、
を含む、実施の形態24に記載の二量体I−CreIタンパク質。
実施の形態27:実施の形態11〜26のいずれか1つに記載の二量体I−CreIタンパク質の単量体を含む融合タンパク質。
実施の形態28:上記単量体が、配列番号43の配列を含むペプチドリンカーによって連結される、実施の形態27に記載の融合タンパク質。
実施の形態29:C末端単量体がK7E突然変異及びK96E突然変異を更に含み、N末端単量体がE8K突然変異、E61R突然変異及びG19S突然変異を更に含む、実施の形態27又は28に記載の融合タンパク質。
実施の形態30:配列番号25〜配列番号40及び配列番号76〜配列番号96からなる群から選択される配列を含む、実施の形態27〜29のいずれか1つに記載の融合タンパク質。
実施の形態31:実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼ又は実施の形態11〜30のいずれか1つに記載のタンパク質をコードする核酸。
実施の形態32:実施の形態31に記載の核酸を含む発現ベクター。
実施の形態33:導入遺伝子を含む標的化構築物と、実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼ又は実施の形態11〜30のいずれか1つに記載のタンパク質によって認識される標的配列に隣接するゲノム配列に相同な2つの配列とを更に含む、実施の形態32に記載の発現ベクター。
実施の形態34:上記導入遺伝子が治療用ポリペプチドをコードする、実施の形態33に記載の発現ベクター。
実施の形態35:遺伝子療法に使用される、実施の形態32〜34のいずれか1つに記載の発現ベクター。
実施の形態36:実施の形態32に記載の発現ベクターと、
導入遺伝子を含む標的化構築物と、実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼ又は実施の形態11〜30のいずれか1つに記載のタンパク質によって認識される標的配列のゲノム配列に相同な2つの配列とを含むベクターと、
の組合せ。
実施の形態37:実施の形態32〜34のいずれか1つに記載の発現ベクター又は実施の形態36に記載の組合せと、薬学的に活性な担体とを含む医薬組成物。
実施の形態38:遺伝子療法によって個体を治療する方法であって、有効量の実施の形態32〜34のいずれか1つに記載の発現ベクター又は実施の形態36に記載の組合せを、それを必要とする個体に投与することを含む、遺伝子療法によって個体を治療する方法。
実施の形態39:個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法であって、
a)上記個体のゲノム内で、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)を選択する工程と、
b)上記RISの200kb上流及び200kb下流に伸びるゲノム領域を規定する工程と、
c)上記ゲノム領域内に位置する標的配列を切断することが可能な野生型エンドヌクレアーゼを同定するか、又は変異型エンドヌクレアーゼを構築する工程と、
を含む、個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法。
実施の形態40:導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入するための実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼ又は実施の形態11〜30のいずれか1つに記載のタンパク質又は実施の形態31に記載の核酸又は実施の形態32〜34のいずれか1つに記載の発現ベクター又は実施の形態36に記載の組合せの使用であって、治療目的でない、使用。
実施の形態41:遺伝性障害の非ヒト動物モデルを作製するための、実施の形態40に記載の使用。
実施の形態42:組換えタンパク質を製造するための、実施の形態40に記載の使用。
実施の形態43:実施の形態31に記載の核酸又は実施の形態32〜34のいずれか1つに記載の発現ベクター又は実施の形態36に記載の組合せを、そのゲノム内に含む非ヒトトランスジェニック動物。
【図面の簡単な説明】
【0016】
【図1】実施例1に記載のメガヌクレアーゼの標的配列を示す図である。図中の「SEQ ID NO:」の記載は「配列番号」を意味する。以下の図も同様である。
【図2】SCOH SH3メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図3】SCOH SH3メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図4】実施例2に記載のメガヌクレアーゼの標的配列を示す図である。
【図5】SCOH SH4メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図6】SCOH SH4メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図7】非相同末端結合(NHEJ)による二本鎖切断の修復時に小さな欠失及び挿入(InDel)の生成をもたらす機構のスキームを示す図である。
【図8】SH3メガヌクレアーゼ又はSH4メガヌクレアーゼによる切断の際の挿入部位を示す図である。
【図9】実施例5に記載のメガヌクレアーゼの標的配列を示す図である。
【図10】SCOH SH6メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図11】配列番号1のI−CreI単量体と配列番号42のI−CreI単量体との間の配列アラインメントを示す図である。
【図12】配列番号1のI−CreI単量体と、44A 54L 64A 70Q 75N 158R及び162Aの突然変異を含む2つのI−CreI単量体との間の配列アラインメントを示す図である。第1の単量体(配列番号57)は配列番号1に直接由来し、第2の単量体(配列番号58)は配列番号42に直接由来する。
【図13】実施例6〜実施例9を説明する図である。
【図14】実施例6〜実施例9を説明する図である。
【図15】実施例7を説明する図である。
【図16】実施例7を説明する図である。
【図17】実施例8を説明する図である。
【発明を実施するための形態】
【0017】
本発明者らは、標的化挿入による導入遺伝子の安全な発現を可能にするゲノム内の「セーフハーバー」遺伝子座を同定したが、ここで、(i)上記遺伝子座は、遺伝子療法によって治療される患者に由来する細胞において同定されたレトロウイルス挿入部位の近くにあり、及び(ii)上記レトロウイルス挿入は、がん又は異常細胞増殖と関連しない。以下の説明及び実施例からすぐに分かるように、本発明によるセーフハーバー遺伝子座は遺伝子のイントロン内に位置していても、又は遺伝子間領域内に位置していてもよい。
【0018】
特に、本発明者らは、エンドヌクレアーゼを、上記セーフハーバーを遺伝子付加の標的とするように改変することができることを見出した。
【0019】
より具体的には、本発明者らは、種々のセーフハーバー遺伝子座、例えば本明細書中で更に説明するSH6遺伝子座、SH3遺伝子座、SH4遺伝子座、SH12遺伝子座、SH13遺伝子座、SH19遺伝子座、SH20遺伝子座、SH21遺伝子座、SH33遺伝子座、SH7遺伝子座、SH8遺伝子座、SH18遺伝子座、SH31遺伝子座、SH38遺伝子座、SH39遺伝子座、SH41遺伝子座、SH42遺伝子座、SH43遺伝子座、SH44遺伝子座、SH45遺伝子座、SH46遺伝子座、SH47遺伝子座、SH48遺伝子座、SH49遺伝子座、SH50遺伝子座、SH51遺伝子座、SH52遺伝子座、SH70遺伝子座、SH71遺伝子座、SH72遺伝子座、SH73遺伝子座、SH74遺伝子座、SH75遺伝子座、SH101遺伝子座、SH106遺伝子座、SH107遺伝子座、SH102遺伝子座、SH105遺伝子座、SH103遺伝子座、SH104遺伝子座、SH113遺伝子座、SH109遺伝子座、SH112遺伝子座、SH108遺伝子座、SH110遺伝子座、SH114遺伝子座、SH116遺伝子座、SH111遺伝子座、SH115遺伝子座、SH121遺伝子座、SH120遺伝子座、SH122遺伝子座、SH117遺伝子座、SH118遺伝子座、SH119遺伝子座、SH123遺伝子座、SH126遺伝子座、SH128遺伝子座、SH129遺伝子座、SH124遺伝子座、SH131遺伝子座、SH125遺伝子座、SH127遺伝子座、SH130遺伝子座、SH11遺伝子座、SH17遺伝子座、SH23遺伝子座、SH34遺伝子座、SH40遺伝子座、SH53遺伝子座、SH54遺伝子座、SH55遺伝子座、SH56遺伝子座、SH57遺伝子座、SH58遺伝子座、SH59遺伝子座、SH60遺伝子座、SH61遺伝子座、SH62遺伝子座、SH65遺伝子座、SH67遺伝子座、SH68遺伝子座及びSH69遺伝子座内に位置する標的配列を認識し切断することが可能な幾つかのI−CreIメガヌクレアーゼを設計した。
【0020】
これらのメガヌクレアーゼが、それらの標的配列を効率的に切断することができることが更に示された。
【0021】
したがって、これらのメガヌクレアーゼ、並びにインテグラーゼ、リコンビナーゼ及びトランスポザーゼのような他の酵素(enzymes)を、導入遺伝子をセーフハーバーに挿入するためのツールとして使用し、それにより遺伝子療法における白血病等の有害事象の発生を回避することができる。加えて、これらのメガヌクレアーゼ、並びにインテグラーゼ、リコンビナーゼ及びトランスポザーゼのような他の酵素は、導入遺伝子の配列に関わりなく単一の標的化構築物から出発して任意の導入遺伝子をセーフハーバーに挿入するために使用することができる。
【0022】
本発明によるエンドヌクレアーゼ及びその使用
本発明はしたがって、以下のものに関する:
導入遺伝子を個体のゲノムに挿入する際に使用される標的配列を切断することが可能なエンドヌクレアーゼであって、(i)上記ゲノムが上記標的配列を含む遺伝子座を含み、(ii)上記標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連しない、エンドヌクレアーゼ;
導入遺伝子を細胞又は組織のゲノムに挿入するための標的配列を切断することが可能なエンドヌクレアーゼのin vitro又はex vivoでの使用であって、(i)上記ゲノムが上記標的配列を含む遺伝子座を含み、(ii)上記標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連しない、in vitro又はex vivoでの使用;
導入遺伝子を個体のゲノムに挿入する方法であって、(i)標的配列を切断することが可能なエンドヌクレアーゼを準備する工程であって、上記ゲノムが上記標的配列を含む遺伝子座を含み、該標的配列が、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)から最大で200kb離れて位置する、工程と、(ii)個体を導入遺伝子及び上記エンドヌクレアーゼと接触させ、それにより上記導入遺伝子を個体のゲノムの上記遺伝子座に挿入する、工程とを含む、導入遺伝子を個体のゲノムに挿入する方法。
【0023】
本明細書中で使用される場合、「エンドヌクレアーゼ」という用語は、DNA分子又はRNA分子、好ましくはDNA分子内の核酸間の結合の加水分解(切断)を触媒することが可能な任意の野生型又は変異型の酵素を指す。本発明によるエンドヌクレアーゼは、DNA分子又はRNA分子を、その配列に関係なく切断することはなく、「標的配列」又は「標的部位」と更に称される特異的なポリヌクレオチド配列でDNA分子又はRNA分子を認識し切断する。本発明によるエンドヌクレアーゼによって認識され切断される標的配列は、本発明による標的配列と称される。
【0024】
本発明によるエンドヌクレアーゼは例えば、ホーミングエンドヌクレアーゼ(非特許文献5)、改変ジンクフィンガードメインとFokI等の制限酵素の触媒ドメインとの融合から得られるキメラジンクフィンガーヌクレアーゼ(ZFN)(非特許文献35)又は化学的エンドヌクレアーゼ(Arimondo et al. Mol Cell Biol. 2006 26:324-333、Simon et al. NAR 2008 36:3531-3538、Eisenschmidt et al. NAR 2005 33:7039-7047、Cannata et al. PNAS 2008 105:9576-9581)であり得る。化学的エンドヌクレアーゼでは、化学的切断剤(cleaver)又はペプチド切断剤を核酸のポリマー又は特異的な標的配列を認識する別のDNAのいずれかと結合させ、それにより特異的な配列に対する切断活性を標的とする。
【0025】
本発明によるエンドヌクレアーゼは好ましくは、メガヌクレアーゼの名称でも知られるホーミングエンドヌクレアーゼである。かかるホーミングエンドヌクレアーゼは、当該技術分野で既知である(例えば、Stoddard, Quarterly Reviews of Biophysics, 2006, 38:49-95を参照されたい)。ホーミングエンドヌクレアーゼはDNA標的配列を認識し、一本鎖又は二本鎖の切断を生じる。ホーミングエンドヌクレアーゼは高度に特異的であり、12塩基対(bp)〜45塩基対の範囲の長さ、通常は14bp〜40bpの範囲の長さのDNA標的部位を認識する。本発明によるホーミングエンドヌクレアーゼは、例えばLAGLIDADGエンドヌクレアーゼ、HNHエンドヌクレアーゼ又はGIY−YIGエンドヌクレアーゼに相当し得る。かかるエンドヌクレアーゼの例としては、I−Sce I、I−Chu I、I−Cre I、I−Csm I、PI−Sce I、PI−Tli I、PI−Mtu I、I−Ceu I、I−Sce II、I−Sce III、HO、PI−Civ I、PI−Ctr I、PI−Aae I、PI−Bsu I、PI−Dha I、PI−Dra I、PI−Mav I、PI−Mch I、PI−Mfu I、PI−Mfl I、PI−Mga I、PI−Mgo I、PI−Min I、PI−Mka I、PI−Mle I、PI−Mma I、PI−Msh I、PI−Msm I、PI−Mth I、PI−Mtu I、PI−Mxe I、PI−Npu I、PI−Pfu I、PI−Rma I、PI−Spb I、PI−Ssp I、PI−Fac I、PI−Mja I、PI−Pho I、PI−Tag I、PI−Thy I、PI−Tko I、PI−Tsp I、I−MsoIが挙げられる。
【0026】
好ましい実施形態では、本発明によるホーミングエンドヌクレアーゼは、I−SceI、I−CreI、I−CeuI、I−MsoI及びI−DmoI等のLAGLIDADGエンドヌクレアーゼである。
【0027】
最も好ましい実施形態では、上記LAGLIDADGエンドヌクレアーゼはI−CreIである。野生型I−CreIは、22bp〜24bpの二本鎖標的配列を切断することが可能なホモ二量体ホーミングエンドヌクレアーゼである。I−CreIの野生型単量体の配列は、配列番号1として示される配列(pdbアクセッション番号1g9yのI−CreI配列に相当する)及びSwissProtアクセッション番号P05725で示される配列(特にバージョン73で示される配列、最終更新日2009年11月3日)を含む。
【0028】
本特許出願において、I−CreI変異体は、野生型I−CreI配列の最初のメチオニンの後に更なるアラニン及びC末端(C-terminal extremity)に3つの更なるアミノ酸残基を含み得る(配列番号42の配列及び図11を参照されたい)。これら3つの更なるアミノ酸残基は、野生型I−CreI配列の最後のプロリンの後の2つの更なるアラニン残基及び1つのアスパラギン酸残基からなる。これらの更なる残基は、酵素の特性に影響を与えない。明確化のために、これらの更なる残基はI−CreI又はその変異体における残基のナンバリングに影響を与えないものとする。より具体的には、本明細書中で使用されるナンバリングは、配列番号1の野生型I−CreI酵素における残基の位置のみを指す。例えば、野生型I−CreIの2番目の残基は、この変異体が最初のメチオニンの後に更なるアラニンを含むため、実際には配列番号42の変異体の3番目の残基である。
【0029】
本出願において、I−CreI変異体は、ホモ二量体(2つの同一の単量体を含むメガヌクレアーゼ)、ヘテロ二量体(2つの同一でない単量体を含むメガヌクレアーゼ)及び一本鎖であり得る。
【0030】
本発明は、野生型(天然)エンドヌクレアーゼ及び変異型エンドヌクレアーゼの両方を包含する。好ましい実施形態では、本発明によるエンドヌクレアーゼは、「変異型」エンドヌクレアーゼ、すなわち自然界に天然に存在せず、遺伝子操作又はランダム突然変異誘発によって得られるエンドヌクレアーゼである。本発明による変異型エンドヌクレアーゼは例えば、野生型、天然のエンドヌクレアーゼのアミノ酸配列の少なくとも1つの残基を異なるアミノ酸で置換することによって得ることができる。上記置換(複数も可)は、例えば部位特異的突然変異誘発及び/又はランダム突然変異誘発によって導入することができる。本発明の枠組みにおいて、かかる変異型エンドヌクレアーゼは機能的なままである、すなわち標的配列を認識し、特異的に切断する能力を保持する。
【0031】
本発明による変異型エンドヌクレアーゼは、対応する野生型エンドヌクレアーゼの標的配列とは異なる標的配列を切断する。例えば、変異型I−CreIエンドヌクレアーゼの標的配列は、配列番号4の配列とは異なる。新規の特異性を有するかかる変異型エンドヌクレアーゼを得る方法は、当該技術分野で既知である。
【0032】
本発明は、新規の特異性を有するかかる変異型エンドヌクレアーゼを、細胞、組織又は個体のゲノムの「セーフハーバー」遺伝子座に遺伝子を挿入するために使用することができるという発見に基づく。
【0033】
本明細書中で使用される場合、「遺伝子座」という用語は、染色体上のDNA配列(例えば遺伝子)の特定の物理的な位置である。本明細書中で使用される場合、「遺伝子座」という用語は、通常は染色体上のエンドヌクレアーゼの標的配列の特定の物理的な位置を指す。本発明によるエンドヌクレアーゼによって認識され切断される標的配列を含むかかる遺伝子座は、「本発明による遺伝子座」と称される。
【0034】
理想的には、セーフハーバー遺伝子座への挿入は、他の遺伝子の発現に対して影響を有しないものとする。これらの特性の試験は多工程プロセスであり、最初に候補セーフハーバー遺伝子座のプレスクリーニングをバイオインフォマティクス手段により行うことが望ましい。このため、標的化挿入が挿入突然変異誘発をもたらす可能性が低い遺伝子座を最初に同定することができる。
【0035】
本発明による遺伝子座の主要な特徴の1つは、(i)遺伝子療法の臨床試験において患者に由来する細胞でレトロウイルス挿入が観察された領域中に位置すること、及び(ii)上記レトロウイルス挿入が、がん又は異常細胞増殖と関連しなかったことである。
【0036】
実際には、本発明によるセーフハーバー遺伝子座を同定する一方法は、先の遺伝子療法試験によって生成したデータを使用することである。X−SCID試験では、LMO2遺伝子及びCCND2遺伝子の隣のレトロウイルスベクター媒介導入遺伝子の挿入が、白血病と関連することが示されている。患者におけるベクター挿入の追跡調査から、この挿入を保有する細胞の数が数年の経過後に他の修飾細胞の数を上回ることが明らかに実証されている(非特許文献31、Deichmann et al. J. of Clin. Invest. 2007 117:2225-32、Cavazzana-Calvo et al. Blood 2007 109:4575-4581)。別の臨床試験では、幾つかの遺伝子座における挿入が、2人の患者において高い増殖速度を引き起こすことが見出された(Ott et al. Nat Med 2006 12:401-9)。これらの症例においては、増殖はMDS1−EVI1遺伝子、PRDM16遺伝子又はSETBP1遺伝子の挿入活性化の結果のようであった。悪性腫瘍は当初観察されなかったが、EVII活性化は最終的に両方の患者において骨髄異形成をもたらした(Stein et al., Nat. Med 2010 16: 198-205)。より一般的には、非発がん性であっても、挿入断片の近くにある遺伝子の活性化から生じる細胞増殖が悪性腫瘍に向かう第一段階を示し、したがって安全性に関する潜在的な問題を招く可能性がある。ウイルスベクター組込みのパターン及び形質転換細胞の運命に対するその潜在的な結果をよりよく理解するために、遺伝子療法試験の患者においてレトロウイルス挿入部位(RIS)の大規模研究が幾つか行われている(Mavilio et al., Nat Med 2006:1397-1402、Recchia et al. PNAS 2006:1457-62、Aiuti, et al. J Clin Invest 2007:2233-40、Schwarzwaelder et al. J Clin Invest 2007:2241-9、Deichmann et al. J Clin Invest 2007:2225-32)。白血病又は異常細胞増殖と関連しないRISはセーフハーバーとみなすことができる。したがって、本発明による遺伝子座は、臨床試験において同定されたRISと重複するか又はその近くにあり、それにも関わらず、がん又は異常細胞増殖と関連しないことが好ましい。
【0037】
より具体的には、本発明による遺伝子座は、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)から最大で200kb、180kb、150kb、100kb又は50kb離れて位置する標的配列を含む遺伝子座と定義される。かかる遺伝子座は、本発明による「セーフハーバー」遺伝子座(又は本発明による遺伝子座)、すなわち導入遺伝子の挿入に安全な遺伝子座と称される。
【0038】
「レトロウイルス挿入部位」(RIS)とは、レトロウイルスベクターを用いた遺伝子療法によって治療される患者に由来する細胞において上記レトロウイルスベクターの挿入部位として同定されたゲノム部位を意味する。かかるRISは当該技術分野で既知である。RISとしては、Schwarzwaelder et al.(J. Clin. Invest. 2007 117:2241)、Deichmann et al.(J. of Clin. Invest. 2007 117:2225)、Aiuti et al.(J. Clin. Invest. 2007 117:2233)、Recchia et al.(PNAS 2006 103:1457)及びMavilio et al.(Nature Medicine 12:1397, 2006)に記載されるものが挙げられるが、これらに限定されない。
【0039】
「レトロウイルスベクター」とは、レトロウイルス科のウイルスに由来する任意のベクターを意味する。
【0040】
本発明によるRISは、がんとも異常細胞増殖とも関連しない。白血病又は異常細胞増殖と関連することが知られるRISは当該技術分野で既知であり、当業者であれば容易に除外することができる。白血病又は異常細胞増殖と関連することが知られるかかるRISとしては、例えばLMO2遺伝子、CCND2遺伝子、MDS1−EVI1遺伝子、PRDM16遺伝子及びSETBP1遺伝子の隣の挿入部位が挙げられる。
【0041】
本発明によるより好ましい実施形態では、セーフハーバー遺伝子座を規定するために使用されるRISは、形質導入細胞が幹細胞である臨床試験において同定されている。RISはこのように、幹細胞の形質導入による遺伝子療法によって治療される患者に由来する細胞において同定されている可能性がある。
【0042】
本発明による別の最も好ましい実施形態では、セーフハーバー遺伝子座を規定するために使用するRISは、形質導入細胞が造血幹細胞(HSC)であるSCID患者の臨床試験において同定されている。RISはこのように、造血幹細胞の形質導入による遺伝子療法によって治療される患者に由来する細胞において同定されている可能性がある。
【0043】
さらに、本発明によるRISを既定するためのより厳しい基準を使用することができる。
【0044】
RISの中でも、共通組込み部位(CIS)は、RISの統計的過剰出現が挿入時の細胞の高い増殖速度の結果であると解釈され得る遺伝子座である(Mikkers et al., 2003, Nat. Genet. 32:153、Lund et al., 2002, Nat. Genet 32 :160、Hemati et al. 2004, PLOS Biol. 2:e423、Suzuki et al., 2002, Nat. Genet. 32:166-174、Deichman et al. J. of Clin. Invest. 2007 117:2225-32)。例えば、Deichman et al.(J. of Clin. Invest. 2007 117:2225-32)は、遺伝子療法によって治療した9人のX−SCID患者に由来するRISに関する調査を行い、ヒトゲノムに明白にマップすることができる572個の独自のRISを見出した。それらの中でも、Deichman et al.は2次、3次、4次、5次及びより高次のCISを規定した。2次のCISは、30kb距離内での2個のレトロウイルス挿入の発生によって規定され、3次、4次及び5次のCISは、それぞれ50kb、100kb又は200kb内での3つ、4つ又は5つの挿入の発生によって規定された。RISのランダム分布で期待される値の33倍である、122個のRISが47個の異なるCIS遺伝子座において発見された。11個のCISが、ZNF217、VAV−3、CCND2、LMO2、MDS1、BCL2L1、NOTCH2、SOCS2、RUNX1、RUNX3及びSEPT6を含むがん原遺伝子の隣に局在することが見出された。
【0045】
最大の安全性を確保するために、CIS内に位置するRISを回避することが好ましい場合がある。したがって、本発明による好ましい実施形態では、本発明による標的配列はCISに位置しない。加えて、上記標的配列又は遺伝子座は、共通組込み部位(CIS)の一部であるRISから少なくとも50kb、100kb又は200kb離れて位置するのが好ましい。
【0046】
「共通組込み部位」(CIS)とは、臨床試験において同定されたRISが大きな比率を占める(挿入のランダム分布を仮定する)30kb、50kb、100kb又は200kbのゲノム領域を意味する。かかるCISは当該技術分野で既知であり、Schwarzwaelder et al.(J. Clin. Invest. 2007 117:2241)、Deichmann et al.(J. of Clin. Invest. 2007 117:2225)、Aiuti et al.(J. Clin. Invest. 2007 117:2233)、Recchia et al.(PNAS 2006 103:1457)、Mavilio et al.(Nature Medicine 12:1397, 2006)及びGabriel et al.(Nat. Med. 2009 15(12):143)に記載されている。
【0047】
RISの近くであることに加えて、本発明による遺伝子座への標的化組込みは、標的化細胞における必須機能の破壊をもたらさないものとする。
【0048】
したがって、本発明による具体的な実施形態では、本発明による遺伝子座への挿入は、好ましくは標的配列の近傍に位置する遺伝子、例えば最近接遺伝子の発現を実質的に変更しない。
【0049】
加えて、別の具体的な実施形態では、上記遺伝子座への遺伝要素の挿入は、好ましくは上記細胞、組織又は個体の表現型(遺伝要素の発現による表現型を除く)を実質的に変更しない。「表現型」とは、細胞、組織又は個体の目に見える形質を意味する。表現型としては、例えば生存能力、細胞増殖及び/又は成長速度が挙げられる。当業者であれば、例えば隣接遺伝子の発現パターンを分析すること、トランスクリプトームのマイクロアレイ研究を行うこと、並びに/又は増殖及び/若しくは(もしあれば)分化異常を特性評価することによって、遺伝子座が本発明によるセーフハーバー遺伝子座であるかを容易に検証することができる。
【0050】
また別の具体的な実施形態では、本発明による遺伝子座は任意の遺伝子を含まない。任意の遺伝子を含まない遺伝子座とは、任意の参照された又は既知の遺伝子を含まない遺伝子座を指す。言い換えると、かかる遺伝子座は、米国国立バイオテクノロジー情報センター(NCBI)のウェブサイト上で入手可能なもののような配列データベースによる任意の既知の遺伝子を含まない。したがって、本発明による標的配列及び/又は本発明による遺伝子座は、最近接遺伝子から少なくとも1kb、5kb、10kb、25kb、50kb、100kb、180kb、200kb、250kb、300kb、400kb又は500kb離れて有利に位置することができる。
【0051】
「遺伝子」とは、特定のタンパク質又はタンパク質の一部をコードする、染色体に沿って線状に配列したDNAの一部からなる遺伝の基本単位を意味する。遺伝子は通常、プロモーター、5’非翻訳領域、1つ又は複数のコード配列(エクソン)、任意にイントロン、3’非翻訳領域を含む。遺伝子はターミネーター、エンハンサー及び/又はサイレンサーを更に含み得る。
【0052】
「最近接遺伝子」とは、それぞれ標的配列の最も近くに、標的配列に対してセントロメリックに(centromeric)及びテロメリックに(telomeric)位置する1つ、2つ又は3つの遺伝子を意味する。
【0053】
好ましい実施形態では、本発明による遺伝子座は、導入遺伝子の安定な発現を更に可能にする。
【0054】
別の好ましい実施形態では、本発明による標的配列は、上記細胞、組織又は個体のゲノム内に1回しか存在しない。
【0055】
本発明によるかかるセーフハーバー遺伝子座を選択した後、(i)例えば実施例1、実施例2及び実施例5に記載のように、上記遺伝子座内に位置する標的配列を特異的に認識し切断する変異型エンドヌクレアーゼを構築するか、又は(ii)既知の野生型エンドヌクレアーゼが、上記遺伝子座内に位置する標的配列を切断することが可能か否かを決定することができる。代替的には、本発明によるセーフハーバー遺伝子座を選択した後、当業者は、既知の野生型エンドヌクレアーゼ又は変異型エンドヌクレアーゼによって認識され切断される標的配列をそれに挿入することができる。
【0056】
したがって、本発明は、個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法であって、
a)上記個体のゲノム内で、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)を選択及び/又は同定する工程と、
b)上記RISの200kb上流及び200kb下流に伸びるゲノム領域を規定する工程と、
c)上記ゲノム領域内に位置する標的配列を切断することが可能な野生型エンドヌクレアーゼを同定するか、又は変異型エンドヌクレアーゼを構築する工程と、
を含む、個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法に関する。
【0057】
かかるエンドヌクレアーゼは、例えば(i)最近接遺伝子の発現、並びに/又は(ii)細胞、組織若しくは個体の細胞増殖及び/若しくは成長速度を実質的に変更することなく、細胞、組織又は個体のゲノムに導入遺伝子を安全に挿入することを可能にする。
【0058】
本発明による遺伝子座に関連して上で提示する全ての基準は、当然ながら上記の方法を行う場合に適用することができる。例えば、CISの一部であるRISを除外してもよく、並びに/又は工程(b)で規定されるゲノム領域が上記RISの50kb上流及び50kb下流にしか伸びなくてもよく、並びに/又は標的配列を含む遺伝子座が任意の遺伝子を含まなくてもよい。
【0059】
本発明による遺伝子座は例えば、下記表A〜表Cに記載のSH3遺伝子座、SH4遺伝子座、SH6遺伝子座、SH12遺伝子座、SH13遺伝子座、SH19遺伝子座、SH20遺伝子座、SH21遺伝子座、SH33遺伝子座、SH7遺伝子座又はSH8遺伝子座のいずれか1つに相当し得る。
【0060】
表Aは、ヒトゲノム内の遺伝子座の位置、遺伝子座内に含まれる標的配列、最も近いRISの位置及びRISを記載する刊行物への参照、及び遺伝子座を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0061】
表Bは、本発明による遺伝子座のすぐ上流(5’側)及び下流(3’側)に位置する最近接遺伝子に関する情報を提示する。距離とは、標的配列と遺伝子の最も近いコード配列との距離を示す。
【0062】
表C及び表Dは、表Bと同様であるが、第2の最近接遺伝子及び第3の最近接遺伝子についての情報をそれぞれ提示する。
【0063】
表A’、表B’、表C’及び表D’は、一部の遺伝子座、すなわちSH3、SH4、SH6、SH8及びSH19、並びに関連するこれらの遺伝子座内の標的配列の例についての表A、表B、表C及び表Dと同様の最新情報をそれぞれ提示する。最新の局在情報は、ヒトゲノムアセンブリのGRCh37/hg19バージョンを参照することによって与えられる。
【0064】
本発明による遺伝子座は、下記表A’’〜表D’’に記載のSH18、SH31、SH38、SH39、SH41、SH42、SH43、SH44、SH45、SH46、SH47、SH48、SH49、SH50、SH51、SH52、SH70、SH71、SH72、SH73、SH74及びSH75のいずれか1つにも相当し得る。
【0065】
表A’’は、ヒトゲノム内の遺伝子座の位置、遺伝子座内に含まれる標的配列、最も近いRISの位置及びRISを記載する刊行物への参照、上記標的と最も近いRISとの距離、及び遺伝子座を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0066】
表B’’は、本発明による遺伝子座のすぐ上流(5’側)及び下流(3’側)に位置する最近接遺伝子に関する情報を提示する。距離とは、標的配列と遺伝子の最も近いコード配列との距離を示す。
【0067】
表C’’及び表D’’は、表B’’と同様であるが、第2の最近接遺伝子及び第3の最近接遺伝子についての情報をそれぞれ提示する。
【0068】
遺伝子座の位置、この遺伝子座における標的及び遺伝子は、ヒトゲノムアセンブリのGRCh37/hg19バージョンに従って与えられる。
【0069】
【表1】
【0070】
【表2】
【0071】
【表3−1】
【表3−2】
【0072】
【表4】
【0073】
【表5】
【0074】
【表6】
【0075】
【表7】
【0076】
【表8】
【0077】
【表9】
【0078】
【表10】
【0079】
【表11】
【0080】
【表12】
【0081】
本発明による遺伝子座は、下記表E及び表Fに記載のSH101、SH106、SH107、SH102、SH105、SH103、SH104、SH113、SH109、SH112、SH108、SH110、SH114、SH116、SH111、SH115、SH121、SH120、SH122、SH117、SH118、SH119、SH123、SH126、SH128、SH129、SH124、SH131、SH125、SH127及びSH130のいずれか1つにも相当し得る。
【0082】
表Eは、ヒトゲノム内の遺伝子座の位置、遺伝子座内に含まれる標的配列、最も近いRISの位置及びRISを記載する刊行物への参照、上記標的と最も近いRISとの距離、及び遺伝子座を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0083】
表Fは、本発明による遺伝子座のすぐ上流(5’側)及び下流(3’側)に位置する最近接遺伝子に関する情報を提示する。距離とは、標的配列と遺伝子の最も近いコード配列との距離を示す。
【0084】
表E及び表Fにおいて、遺伝子座の位置、この遺伝子座における標的及び遺伝子は、ヒトゲノムアセンブリのGRCh36.3/hg19バージョンに従って与えられる。
【0085】
【表13−1】
【表13−2】
【0086】
【表14】
【0087】
本発明による遺伝子座は、下記表Gに記載のSH125、SH127、SH130、SH102、SH105、SH103、SH104、SH117、SH118、SH119及びSH123のいずれか1つにも相当し得る。
【0088】
表Gは、言及した遺伝子のイントロン内に位置する標的配列の例及び該イントロン遺伝子座を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0089】
【表15】
【0090】
本発明による遺伝子座は、下記表Hに記載のSH11、SH12、SH13、SH17、SH19、SH20、SH21、SH23、SH33、SH34、SH40、SH53、SH54、SH55、SH56、SH57、SH58、SH59、SH60、SH61、SH62、SH65、SH67、SH68及びSH69のいずれか1つも含有し得る。
【0091】
表Hは、これらの遺伝子座内に含まれる標的配列及びこれらの標的配列を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0092】
【表16】
【0093】
具体的な実施形態では、本発明による遺伝子座はSH3遺伝子座である。「SH3遺伝子座」という用語は、リンパ球抗原86をコードする遺伝子に対して約120kbセントロメリックに位置するヒト染色体6の領域(例えば、染色体6の6430K〜7230K領域を示す、ワールドワイドウェブサイトncbi.nlm.nih.gov/projects/mapview/maps.cgi?TAXID=9606&CHR=6&MAPS=ideogr%2Ccntg-r%2CugHs%2Cgenes&BEG=6432845&END=7232845&thmb=onを参照されたい)及び他の種における相同領域を指す。より正確には、SH3遺伝子座は、NC_000006.11に示される配列の6850510位から6853677位に伸びる。これは配列番号54の配列を含む。
【0094】
別の具体的な実施形態では、本発明による遺伝子座はSH4遺伝子座である。SH4遺伝子座は、本明細書ではMyoDファミリー阻害剤ドメイン含有遺伝子座(MDFIC)に対して約320kbテロメリックに位置するヒト染色体7の領域として、又は別の種における相同領域として規定される(例えば、染色体7の114660K〜115660K領域を示す、ワールドワイドウェブサイトncbi.nlm.nih.gov/projects/mapview/maps.cgi?TAXID=9606&CHR=7&MAPS=ideogr,cntg-r,ugHs,genes[113908811.00%3A114908811.00]&CMD=DNを参照されたい)。より正確には、SH4遺伝子座は、NC_000007.13に示される配列の114972751位から114976380位に伸びる。これは配列番号55の配列を含む。
【0095】
本明細書中で使用される場合、「導入遺伝子」という用語は、ポリペプチドをコードする配列を指す。好ましくは、導入遺伝子によってコードされるポリペプチドは、導入遺伝子が挿入される細胞、組織又は個体において発現されないか、又は発現されるが生物学的に活性ではない。最も好ましくは、導入遺伝子は、個体の治療に有用な治療用ポリペプチドをコードする。
【0096】
本発明の枠組みにおいて、個体はヒト又は非ヒト動物であり得る。個体は好ましくはヒトである。代替的に、個体は非ヒト動物、好ましくは脊椎動物、及び/又は例えばマウス、ラット、ウサギ、チャイニーズハムスター、モルモット若しくはサル等の哺乳動物であってもよい。本発明による細胞及び組織は好ましくは、かかるヒト又は非ヒト動物に由来する。
【0097】
I−CreIに由来する本発明によるエンドヌクレアーゼ
本発明による変異型エンドヌクレアーゼは、例えば、
各々が配列番号1の配列若しくはSwissProtアクセッション番号P05725に示される配列を含む2つの単量体を含むホモ二量体タンパク質である野生型I−CreIメガヌクレアーゼ、又は
各々が配列番号42の配列を含む2つの単量体を含むI−CreIメガヌクレアーゼ
のいずれかに由来し得る。野生型標的配列を認識するかかるI−CreIメガヌクレアーゼは、新規の特異性を有するエンドヌクレアーゼの改変に好適であることが示されている。
【0098】
したがって、本発明は、セーフハーバー遺伝子座内に位置する標的配列を切断することが可能である、各々が配列番号1又は配列番号42と少なくとも80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%又は99%同一な配列を含むか又はそれらからなる2つの単量体を含むか又はそれらからなる二量体I−CreIタンパク質に関する。
【0099】
好ましくは、標的配列は配列番号4の配列を含みも、それらからなりもしない。
【0100】
最も好ましくは、本発明による二量体I−CreIタンパク質はヘテロ二量体タンパク質である。
【0101】
本発明のクエリー配列と少なくとも例えば95%「同一な」配列を有するタンパク質とは、タンパク質の配列が、クエリー配列の100個のアミノ酸当たり最大で5個のヌクレオチド突然変異を含み得る以外はクエリー配列と同一であることを意図する。言い換えると、クエリー配列と少なくとも95%同一な配列を有するタンパク質を得るには、最大で配列の5%(100個のうち5個)のアミノ酸が挿入されているか、欠失しているか、又は別のヌクレオチドで置き換えられていてもよい。2つの配列の最適アラインメント(ギャップを含む)をそれらの全長を考慮して見出すために、Needleman−Wunschグローバルアライメントアルゴリズム(Needleman and Wunsch, 1970 J. Mol. Biol. 48:443-453)を使用する「needle」プログラムを例えば使用することができる。needleプログラムは、例えばebi.ac.ukワールドワイドウェブサイト上で入手可能である。本発明に従う同一性のパーセンテージはこのように、「Gap Open」パラメータが10.0に等しく、「Gap Extend」パラメータが0.5に等しいEMBOSS::needle(グローバル)プログラム及びBlosum62マトリックスを用いて算出することができる。
【0102】
本発明による二量体I−CreIタンパク質の各々の単量体は例えば、野生型単量体の配列(配列番号1)又は配列番号42の単量体の配列と比較して少なくとも、多くとも又は約2個、5個、8個、10個、12個、15個、18個、20個又は25個の突然変異を含み得る。言い換えると、本発明による単量体は、配列番号1又は配列番号42と少なくとも、多くとも又は約2個、5個、8個、10個、12個、15個、18個、20個、25個又は30個の突然変異によって異なる配列を含む。
【0103】
本発明の枠組みにおいて、突然変異は、好ましくは1つのアミノ酸の別のアミノ酸による置換に相当する。したがって、本発明による好ましい実施形態は、配列番号1又は配列番号42とアミノ酸置換の存在によってのみ異なる、配列番号1又は配列番号42と少なくとも80%同一な配列を含む2つの単量体を含むか又はそれらからなる二量体I−CreIタンパク質に関する。
【0104】
本発明による二量体I−CreIタンパク質の単量体は、好ましくは配列番号42の配列を含むか又はそれからなる単量体に由来する。
【0105】
突然変異は、好ましくは標的配列の認識に関与するI−CreI配列の位置に位置する。実際に、かかる突然変異の導入は、新規の特異性を有するメガヌクレアーゼの設計を可能にする。
【0106】
かかる突然変異に加えて、単量体は以下のものに相当する突然変異も有し得る:
標的部位に対するタンパク質の結合特性及び/又は切断特性を改善する突然変異、例えばG19S、G19A、F54L、S79G、E80K、F87L、V105A及び/又はI132V等(例えば国際公開第2008/152524号を参照されたい);及び/又は
偏性ヘテロ二量体(例えば、国際公開第2008/093249号及びFajardo-Sanchez et al., Nucleic Acids Res. 2008 36:2163-73を参照されたい)の獲得をもたらす突然変異;及び/又は
融合タンパク質の生成に好適な突然変異、例えば融合タンパク質のC末端単量体における配列番号1の5個の主要なN末端アミノ酸残基の欠失等;及び/又は
配列番号42の単量体の場合と同様に、配列番号1の1番目及び2番目の残基の間のアラニンの挿入からなる突然変異。
【0107】
配列番号1又は配列番号42に相同な配列に加えて、本発明によるタンパク質の単量体は、配列のNH2末端及び/又はCOOH末端で付加された1つ又は複数のアミノ酸、例えばタンパク質、プロペプチド及び/又は核局在化シグナルの精製に有用なタグを含み得る。特に、本発明によるタンパク質の単量体は、配列番号42の単量体の場合と同様、配列番号1の配列のCOOH末端で付加されたAADアミノ酸を含み得る。
【0108】
本明細書において、突然変異は、配列番号1上の位置に続く、配列番号1のその位置に位置するアミノ酸を置き換えるアミノ酸の性質で示される。例えば、44A突然変異を含む単量体は、配列番号1の44位のアミノ酸(すなわちグルタミン、Q)がアラニン(A)で置き換えられたI−CreI単量体を指す。このため、この単量体は、配列番号1の野生型I−CreI単量体と、少なくともアミノ酸置換Q44Aによって異なる。上で説明したように、配列番号42のI−CreI単量体は、配列番号1のI−CreI単量体と比較して幾つかの更なるアミノ酸残基を含む(図11を参照されたい)。したがって、配列番号42上では、44A突然変異はアラニンによる配列番号42の45位のグルタミンの置き換えに相当する。
【0109】
説明のために、44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体は例えば、配列番号57の配列(この単量体が配列番号1のI−CreI単量体に直接由来する場合)又は配列番号58の配列(この単量体が配列番号42のI−CreI単量体に直接由来する場合)を有し得る。図12は2つのかかる単量体間のアラインメントを示し、これらの単量体上の44A 54L 64A 70Q 75N 158R及び162A突然変異の位置を示す。
【0110】
SH3遺伝子座、SH4遺伝子座又はSH6遺伝子座に位置する標的配列を切断することが可能な本発明による二量体I−CreIタンパク質の例を、下記に更に記載する。
【0111】
SH3遺伝子座を切断することが可能な本発明による二量体I−CreIタンパク質
好ましい実施形態では、標的配列はSH3遺伝子座(上で規定される)内に位置する。SH3内に位置する標的配列は例えば、配列番号2又は配列番号2のヌクレオチド2〜23を含むか又はそれらからなっていてもよい。実施例1は、かかる標的配列を切断することが可能な本発明によるヘテロ二量体I−CreIタンパク質の幾つかの例を開示する。加えて、他のかかるタンパク質を構築する方法は当該技術分野で既知であり、例えばPCT出願の国際公開第2006/097784号、国際公開第2006/097853号及び国際公開第2009019614号、並びに非特許文献6に記載されているものが挙げられる。
【0112】
かかる二量体タンパク質の単量体は好ましくは、配列番号1の4位、24位、26位、28位、30位、32位、33位、38位、44位、50位、54位、57位、64位、66位、70位、71位、75位、77位、81位、86位、92位、105位、142位、151位、154位、158位及び162位、好ましくは配列番号1の4位、30位、38位、44位、50位、54位、57位、64位、66位、70位、71位、75位、77位、81位、86位、92位、105位、142位、151位、154位、158位及び162位からなる群から選択される位置に位置する少なくとも1つ、好ましくは少なくとも3つ、4つ、5つ又は6つのアミノ酸置換を含む。該置換は、例えば以下の置換から選択され得る:4E、30G、38R、44A、50R、54L、57E、64A、66C、70Q、70D、71R、75N、75Y、77V、81T、86D、92R、105A、142R、151A、154G、158R、158W及び162A。二量体タンパク質は任意に1位に突然変異を含んでいてもよいが、かかる突然変異は切断活性又は切断特異性に影響を有しない。
【0113】
かかる二量体I−CreIタンパク質は、例えば以下のものを含むか又はそれらからなっていてもよい:
配列番号1と比較して、配列番号1の30位、38位、50位、70位、75位、81位、86位、142位及び154位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第1の単量体。好ましくは、該第1の単量体は、配列番号1の30位、38位、70位及び75位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:30G、38R、50R、70D、75N、81T、86D、142R及び154G。かかる単量体は例えば、配列番号1と比較して少なくとも4個、5個若しくは6個の突然変異、及び/又は配列番号1と比較して最大で4個、5個、6個、8個、10個、12個若しくは15個のアミノ酸突然変異を含み得る;及び
配列番号1と比較して、配列番号1の4位、44位、54位、57位、64位、66位、70位、71位、75位、77位、92位、105位、151位、158位及び162位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第2の単量体。好ましくは、該第2の単量体は、配列番号1の44位、54位、70位及び75位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:4E、44A、54L、57E、64A、66C、70Q、71R、75N、75Y、77V、92R、105A、151A 158R、158W及び162A。かかる単量体は例えば、配列番号1と比較して少なくとも4個、5個若しくは6個の突然変異、及び/又は配列番号1と比較して最大で4個、6個、8個、10個、12個又は15個のアミノ酸突然変異を含み得る。
【0114】
具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)30G 38R 70D 75N 86D突然変異を含む第1の単量体、
b)
i.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
ii.44A 54L 70Q 75Y 92R 158R 162A突然変異を含む単量体、
iii.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iv.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
v.44A 54L 70Q 75N突然変異を含む単量体、
vi.44A 54L 57E 70Q 75N 158R 162A突然変異を含む単量体、及び
vii.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含むか又はそれらからなる。
【0115】
別の具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)30G 38R 70D 75N 81T 154G突然変異を含む第1の単量体、
b)
i.44A 54L 70Q 75N 105A 158R 162A突然変異を含む単量体、
ii.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iii.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iv.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
v.44A 54L 70Q 75N突然変異を含む単量体、及び
vi.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含むか又はそれらからなる。
【0116】
更に別の具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)30G 38R 50R 70D 75N 142R突然変異を含む第1の単量体、
b)
i.44A 54L 70Q 75N 105A 158R 162A突然変異を含む単量体、
ii.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iii.44A 54L 70Q 75Y 92R 158R 162A突然変異を含む単量体、
iv.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
v.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
vi.44A 54L 66C 70Q 71R 75N 151A 158R 162A突然変異を含む単量体、
vii.44A 54L 70Q 75N突然変異を含む単量体、
viii.44A 54L 57E 70Q 75N 158R 162A突然変異を含む単量体、及び
ix.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含むか又はそれらからなる。
【0117】
二量体I−CreIタンパク質の単量体は、例えば偏性ヘテロ二量体の獲得を可能にする更なる突然変異も含み得る。かかる突然変異は当業者に既知であり、Fajardo-Sanchez et al.(Nucleic Acids Res. 2008 36:2163-73)に記載されるものが挙げられる。
【0118】
具体的な実施形態では、上記の単量体は配列番号42の単量体に直接由来し、配列番号42の配列と指定の突然変異の存在によってのみ異なる。
【0119】
SH4遺伝子座を切断することが可能な本発明による二量体I−CreIタンパク質
好ましい実施形態では、標的配列はSH4遺伝子座(上で規定される)内に位置する。SH4内に位置する標的配列は例えば、配列番号3又は配列番号3のヌクレオチド2〜23を含むか又はそれらからなっていてもよい。実施例2は、かかる標的配列を切断することが可能な本発明による二量体I−CreIタンパク質の幾つかの例を開示する。
【0120】
かかる二量体タンパク質の単量体は好ましくは、配列番号1の24位、44位、68位、70位、75位及び77位からなる群から選択される位置に位置する少なくとも1つ、好ましくは少なくとも3つ、4つ、5つ又は6つのアミノ酸置換を含む。該置換は、例えば以下の置換から選択され得る:24V、44R、44Y、68Y、68A、70S、70D、75Y、75N、77R、77N及び77V。
【0121】
かかる二量体I−CreIタンパク質は、例えば以下のものを含むか又はそれらからなっていてもよい:
配列番号1と比較して、配列番号1の24位、44位、68位、70位、75位及び77位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第1の単量体。好ましくは、第1の単量体は、配列番号1の24位、70位、75位及び77位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:24V、44R、68Y、68A、70D、70S、75Y、75N、77N及び77R。かかる単量体は例えば、配列番号1と比較して少なくとも4個、5個若しくは6個の突然変異、及び/又は配列番号1と比較して最大で4個、5個、6個、8個、10個、12個若しくは15個のアミノ酸突然変異を含み得る;及び
配列番号1と比較して、配列番号1の24位、44位、70位及び77位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第2の単量体。好ましくは、第2の単量体は配列番号1の24位、44位及び70位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:24V、44Y、70S及び77V。かかる単量体は例えば、配列番号1と比較して少なくとも3個若しくは4個の突然変異、及び/又は配列番号1と比較して最大で3個、4個、6個、8個、10個、12個若しくは15個のアミノ酸突然変異を含み得る。
【0122】
具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)
i.24V 44R 68Y 70S 75Y 77N突然変異を含む単量体、
ii.24V 68A 70S 75N 77R突然変異を含む単量体、及び
iii.24V 70D 75N 77R突然変異を含む単量体からなる群から選択される第1の単量体、
b)
i.24V 44Y 70S突然変異を含む単量体、及び
ii.24V 44Y 70S 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含むか又はそれらからなる。
【0123】
二量体I−CreIタンパク質の単量体は、例えば偏性ヘテロ二量体の獲得を可能にする更なる突然変異も含み得る。かかる突然変異は当業者に既知であり、Fajardo-Sanchez et al.(Nucleic Acids Res. 2008 36:2163-73)に記載されるものが挙げられる。
【0124】
具体的な実施形態では、上記の単量体は配列番号42の単量体に直接由来し、配列番号42の配列と指定の突然変異の存在によってのみ異なる。
【0125】
SH6遺伝子座を切断することが可能な本発明による二量体I−CreIタンパク質
好ましい実施形態では、標的配列はSH6遺伝子座(上で規定される)内に位置する。SH6内に位置する標的配列は例えば、配列番号59又は配列番号59のヌクレオチド2〜23を含むか又はそれらからなっていてもよい。実施例5は、かかる標的配列を切断することが可能な本発明による二量体I−CreIタンパク質の幾つかの例を開示する。
【0126】
かかる二量体タンパク質の単量体は好ましくは、配列番号1の7位、24位、27位、28位、34位、40位、44位、68位、70位、75位、77位、81位、85位、96位、99位、103位、108位、111位、121位、132位、144位及び160位からなる群から選択される位置に位置する少なくとも1つ、好ましくは少なくとも3つ、4つ、5つ又は6つのアミノ酸置換を含む。該置換は、例えば以下の置換から選択され得る:7R、24F、27V、28Q、34R、40R、44A、44K、68T、70L、70G、70S、75N、77V、81T、81V、85R、96R、99R、103T、103S、108V、111H、121E、132V、144S、160R及び160E。
【0127】
かかる二量体I−CreIタンパク質は、例えば以下のものを含むか又はそれらからなっていてもよい:
配列番号1と比較して、配列番号1の7位、24位、27位、28位、34位、40位、44位、70位、75位、77位、81位、85位、96位、99位、103位、108位、111位、121位、132位、144位及び160位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第1の単量体。好ましくは、第1の単量体は、配列番号1の28位、40位、44位、70位及び75位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:7R、24F、27V、28Q、34R、40R、44A、70L、75N、77V、81T、81V、85R、96R、99R、103T、103S、108V、111H、121E、132V、144S、160R及び(and)160E。かかる単量体は例えば、配列番号1と比較して少なくとも5個若しくは6個の突然変異、及び/又は配列番号1と比較して最大で5個、6個、8個、10個、12個、15個若しくは20個のアミノ酸突然変異を含み得る;及び
配列番号1と比較して、配列番号1の44位、68位、70位及び75位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第2の単量体。好ましくは、第2の単量体は配列番号1の44位、70位及び75位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:44K、68T、70G、70S及び75N。かかる単量体は例えば、配列番号1と比較して少なくとも3個若しくは4個の突然変異、及び/又は配列番号1と比較して最大で3個、4個、6個、8個、10個、12個若しくは15個のアミノ酸突然変異を含み得る。
【0128】
具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)44K 68T 70G 75N突然変異を含む第1の単量体と、
b)
i.28Q 40R 44A 70L 75N 96R 111H 144S突然変異を含む単量体、
ii.7R 28Q 40R 44A 70L 75N 85R 103T突然変異を含む単量体、
iii.28Q 40R 44A 70L 75N 103S突然変異を含む単量体、
iv.24F 27V 28Q 40R 44A 70L 75N 99R突然変異を含む単量体、
v.7R 28Q 40R 44A 70L 75N 81T突然変異を含む単量体、
vi.7R 28Q 40R 44A 70L 75N 77V突然変異を含む単量体、
vii.7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R突然変異を含む単量体、
viii.28Q 40R 44A 70L 75N突然変異を含む単量体、
ix.7R 28Q 40R 44A 70L 75N 103T突然変異を含む単量体、及び
x.28Q 34R 40R 44A 70L 75N 81V 103T 108V 160E突然変異を含む単量体からなる群から選択される第2の単量体と、
を含むか又はそれらからなる。
【0129】
別の具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)44K 70S 75N突然変異を含む第1の単量体と、
b)
i.28Q 40R 44A 70L 75N 96R 111H 144S突然変異を含む単量体、
ii.7R 28Q 40R 44A 70L 75N 85R 103T突然変異を含む単量体、
iii.28Q 40R 44A 70L 75N 103S突然変異を含む単量体、
iv.24F 27V 28Q 40R 44A 70L 75N 99R突然変異を含む単量体、
v.7R 28Q 40R 44A 70L 75N 81T突然変異を含む単量体、
vi.7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R突然変異を含む単量体、
vii.7R 28Q 40R 44A 70L 75N 103T突然変異を含む単量体、及び
viii.28Q 34R 40R 44A 70L 75N 81V 103T 108V 160E突然変異を含む単量体からなる群から選択される第2の単量体と、
を含むか又はそれらからなる。
【0130】
二量体I−CreIタンパク質の単量体は、例えば偏性ヘテロ二量体の獲得を可能にする更なる突然変異も含み得る。かかる突然変異は当業者に既知であり、Fajardo-Sanchez et al.(Nucleic Acids Res. 2008 36:2163-73)に記載されるものが挙げられる。
【0131】
具体的な実施形態では、上記の単量体は配列番号42の単量体に直接由来し、配列番号42の配列と指定の突然変異の存在によってのみ異なる。
【0132】
本発明による融合タンパク質
融合した二量体I−CreIタンパク質の2つの単量体を含み、親二量体I−CreIタンパク質の生物学的活性を保持する融合タンパク質を構築することができる(非特許文献9、Li et al. Nucleic Acids Res. 2009 37:1650-62、Epinat et al. Nucleic Acids Res. 2003 31:2952-62)。かかる融合タンパク質は、一般に「一本鎖メガヌクレアーゼ」と称される。
【0133】
したがって、本発明は、上で規定される二量体I−CreIタンパク質の2つの単量体又はかかる単量体の生物学的に活性な断片を含む融合タンパク質に更に関する。かかる融合タンパク質では、上で規定される二量体I−CreIタンパク質の第1の単量体及び第2の単量体は融合し、任意にペプチドリンカー等のリンカーによって互いに連結される。リンカーは例えば、配列番号43又は配列番号326を含むか又はそれらからなっていてもよい。
【0134】
本発明の範囲において、本発明に従うかかる融合タンパク質は、本発明による標的配列を切断することが可能であること、すなわち、それが由来する二量体I−CreIタンパク質と同じ標的配列を切断することが可能であることが理解される。本発明の一本鎖メガヌクレアーゼは、一本鎖偏性ヘテロ二量体メガヌクレアーゼ変異体が得られるように、上に記載のような偏性ヘテロ二量体突然変異を更に含む。
【0135】
最初のバージョンのI−CreI一本鎖(Epinat et al. NAR 2003 3:2952-2962、国際公開第03/078619号)では、一本鎖メガヌクレアーゼのN末端単量体は本質的にI−CreIアミノ酸配列の1位〜93位からなっていたが、C末端(I−CreIアミノ酸配列の8位〜163位)は、ほぼ完全なI−CreI単量体であった。ごく最近では、I−CreIホモ二量体メガヌクレアーゼに由来する一本鎖分子を設計する新たな方法は、2つのほぼ完全なC末端I−CreI単量体及びN末端I−CreI単量体であった(例えば国際公開第2009/095793号を参照されたい)。この設計はオフサイト切断及び毒性を大幅に減少させ、一方で有効性を向上させる。この一本鎖分子の構造及び安定性は二量体変異体のものとほぼ同じであり、この分子は溶液中で単量体状であるようである。あらゆる点で、この一本鎖分子は、特異性に関して至適基準(gold standard)であると考えられるI−SceIと同様に振る舞う。これらの特性から、この新たなメガヌクレアーゼの生成はゲノム手術戦略に利用可能な最良な分子バサミの一つとなり、単一遺伝子疾患、例えば重症複合免疫不全(SCID)の遺伝子修正療法を容易にする一方で、これまでの遺伝子療法アプローチの有害作用を回避する可能性がある。
【0136】
上に記載の突然変異に加えて、更なる突然変異を融合タンパク質の2つの単量体の各々の配列に導入してもよい。例えば、C末端単量体はK7E突然変異及びK96E突然変異を含んでいてもよく、N末端単量体はE8K突然変異、E61R突然変異及びG19S突然変異を含んでいてもよい。
【0137】
実施例1、実施例2及び実施例5は、本発明によるかかる融合タンパク質の幾つかの例を開示する。
【0138】
具体的な実施形態では、本発明による融合タンパク質は、配列番号25〜配列番号40及び配列番号76〜配列番号96のいずれか1つと少なくとも80%、85%、90%、95%、96%、97%、98%、99%若しくは100%同一な配列又はその少なくとも50アミノ酸、100アミノ酸、150アミノ酸若しくは200アミノ酸の断片を含むか又はそれらからなる。
【0139】
本発明による核酸、ベクター及び組合せ
導入遺伝子を細胞、組織又は動物のゲノムに挿入する場合、本発明によるエンドヌクレアーゼを上記細胞、組織又は動物にタンパク質としてではなく核酸分子として導入するのが好ましい。
【0140】
したがって、本発明は、本発明によるエンドヌクレアーゼをコードする、例えば上に記載の二量体I−CreIタンパク質又は融合タンパク質をコードする核酸に関する。エンドヌクレアーゼが二量体I−CreIタンパク質である場合、上記核酸は、各々の単量体につき1つの少なくとも2つのコード配列を含む。エンドヌクレアーゼが融合タンパク質である場合、上記核酸は少なくとも1つのコード配列を含む。エンドヌクレアーゼタンパク質は、組換えタンパク質をもたらす様々な細胞透過性ペプチドと組み合わせることができ、そのような組み合わせた分子は、エンドヌクレアーゼ単独よりもはるかに高いレベルの効率で標的細胞に入ることが可能である。これらの細胞透過性ペプチドは、Diatos S.A.によって開発されている(国際公開第01/64738号、国際公開第05/016960号、国際公開第03/018636号、国際公開第05/018650号、国際公開第07/069068号)。本出願人はこれまで、エンドヌクレアーゼ細胞透過性ペプチドの組合せが標的細胞に効率的に入ることができること及び取り込まれたエンドヌクレアーゼが、DSBを生じ、次に相同組換え事象を刺激するように標的細胞ゲノムに作用することができることを示している。本出願人は、エンドヌクレアーゼの複雑な三次元構造が細胞透過性ペプチドの存在によって影響されないこと、またエンドヌクレアーゼの全ての重要な特異性も影響を受けないままであることを示している(データは示さない)。
【0141】
本発明の別の態様は、本発明によるかかる核酸を含むベクターである。「ベクター」とは、それが連結された別の核酸を輸送することが可能な核酸分子を意味する。
【0142】
本発明において使用することができるベクターとしては、染色体性核酸、非染色体性核酸、半合成核酸又は合成核酸からなり得るウイルスベクター、プラスミド及びYACが挙げられるが、これらに限定されない。好ましいベクターは、自己複製(エピソームベクター)及び/又はそれが連結された核酸の発現(発現ベクター)が可能なものである。多数の好適なベクターが当業者に既知であり、市販されている。
【0143】
好ましい実施形態では、ベクターはウイルスベクター、例えばレトロウイルス、アデノウイルス、パルボウイルス(例えばアデノ随伴ウイルス)、コロナウイルス、マイナス鎖RNAウイルス(例えば、インフルエンザウイルス等のオルトミクソウイルス、狂犬病ウイルス及び水疱性口内炎ウイルス等のラブドウイルス、麻疹ウイルス及びセンダイウイルス等のパラミクソウイルス)、ピコルナウイルス及びアルファウイルス等のプラス鎖RNAウイルス又はアデノウイルス、ヘルペスウイルス(例えば、単純ヘルペスウイルス1型及び2型、エプスタイン・バーウイルス、サイトメガロウイルス)及びポックスウイルス(例えばワクシニアウイルス、鶏痘ウイルス及びカナリア痘ウイルス)等の二本鎖DNAウイルス等に由来するウイルスベクターである。好ましいベクターとしては、レンチウイルスベクター、特に自己不活性化レンチウイルスベクターが挙げられる。
【0144】
本発明によるエンドヌクレアーゼをコードする配列に加えて、ベクターは以下のもののような要素も含み得る:
転写制御要素及び翻訳制御要素、例えばプロモーター、エンハンサー、ポリアデニル化部位、終結シグナル、イントロン等;
多重クローニング部位;
複製起点;
選択マーカー;
導入遺伝子;及び/又は
本明細書中で規定されるようなゲノム標的部位の周囲の領域と相同性を有する配列を含む標的化構築物。
【0145】
好ましい実施形態では、上記ベクターは「発現ベクター」、すなわち少なくとも1つのコード配列が転写制御要素及び翻訳制御要素に動作可能に連結したベクターである。この実施形態の範囲において、本発明によるエンドヌクレアーゼをコードする(例えば、上に記載の二量体I−CreIタンパク質又は融合タンパク質をコードする)核酸は、転写制御要素及び翻訳制御要素に動作可能に連結する。
【0146】
好ましい実施形態では、本発明によるベクターは、導入遺伝子を含む標的化構築物と、本明細書中で規定されるような標的配列(例えば配列番号2又は配列番号3の標的配列)に隣接するゲノム配列に相同な2つの配列とを含む。標的配列に隣接するゲノム配列は、好ましくは標的部位に直接隣接する。
【0147】
かかる標的化構築物は当業者に既知である。導入遺伝子の挿入については、かかる構築物は通常、導入遺伝子が挿入される標的配列に隣接する上流(5’)ゲノム配列に相同な第1の配列と、標的配列に隣接する下流(3’)ゲノム配列に相同な第2の断片とを含む。
【0148】
「相同な」とは、別の配列に対して配列間で相同組換えをもたらすのに十分な同一性を有する、より具体的には互いに少なくとも95%の同一性、好ましくは97%の同一性、より好ましくは99%の同一性を有する配列を意図する。
【0149】
好ましくは、少なくとも50bp、好ましくは100bp超、より好ましくは200bp超の相同配列が使用される。したがって、標的化DNA構築物は、好ましくは200pb〜6000pb、より好ましくは1000pb〜2000pbである。実際に、共通DNA相同性は切断部位の上流及び下流に隣接する領域に位置し、導入されるDNA配列は2つのアーム間に位置するものとする。
【0150】
標的化構築物は、2つのホモロジーアーム間に陽性選択マーカー、最終的には第1のホモロジーアームの上流又は第2のホモロジーアームの下流に陰性選択マーカーも含み得る。マーカー(複数も可)により、相同組換えによって標的部位で関心の配列が挿入された細胞の選択が可能になる。
【0151】
導入遺伝子をSH3遺伝子座又はSH4遺伝子座に挿入するのに好適な標的化構築物を構築する方法は、実施例4に与えられる。
【0152】
本発明によるエンドヌクレアーゼ及び標的化構築物をコードする核酸は、2つの別個のベクター上に位置することもできる。したがって、本発明は2つのベクター、すなわち:
本発明による発現ベクター;及び
導入遺伝子を含む標的化構築物と、本発明による標的配列のゲノム配列に相同な2つの配列とを含むベクター
の組合せにも関する。
【0153】
本発明による薬学的使用
上に記載のベクター及び組合せは、例えば薬剤として使用することができる。特に、これらのベクター及び組合せは遺伝子療法において使用することができる。
【0154】
したがって、本発明は、薬剤として使用される本発明によるベクター又は組合せに関する。かかるベクター及び組合せでは、導入遺伝子は治療用ポリペプチドをコードする。
【0155】
特に、本発明によるベクター及び組合せを用いた遺伝子療法によって治療され得る疾患としては、X−SCID、SCID、表皮水疱症、レーバー黒内障、血友病、サラセミア、ファンコニ貧血及び筋ジストロフィーが挙げられるが、これらに限定されない。
【0156】
これらの疾患では、導入遺伝子はそれぞれ以下の治療用ポリペプチドをコードする:IL2RG、Gl7A1、Rp 65、血液第VIII因子及び血液第IX因子、ヘモグロビンA及びヘモグロビンB、Fanc−A、Fanc−C(又は他のファンコニ貧血関連遺伝子)、ジストロフィン。
【0157】
本発明は、本発明によるベクター及び組合せと、薬学的に活性な担体とを含む医薬組成物に更に関する。
【0158】
本発明は、遺伝子療法によって個体を治療する方法であって、それを必要とする個体に有効量の本発明によるベクター又は組合せを投与することを含む、遺伝子療法によって個体を治療する方法にも関する。
【0159】
「有効量」とは、治療対象の個体のゲノムへの導入遺伝子の挿入を達成するのに十分な量を意味する。当業者はかかる濃度を日常的に決定することができる。
【0160】
「それを必要とする被験体」とは、導入遺伝子の挿入によって治療又は予防することができる遺伝的疾患を患っているか又はそれを患いやすい個体を意味する。本発明の範囲における治療対象の個体は、好ましくはヒトである。
【0161】
本発明による非薬学的使用
上に記載のベクター及び組合せは、遺伝子療法だけでなく、例えば動物モデルの作製及び関心のタンパク質を発現する組換え細胞株の作製等の非薬学的使用にも利用される。
【0162】
したがって、本発明は、以下のものに関する:
治療目的でない、導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入するための本発明によるエンドヌクレアーゼ、核酸、発現ベクター又は組合せの使用;
導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入する方法であって、該細胞、組織又は非ヒト動物を、本発明によるエンドヌクレアーゼ、核酸、発現ベクター又は組合せと接触させ、それにより上記導入遺伝子を上記ゲノムに挿入することを含む、導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入する方法。
【0163】
好ましい実施形態では、上記の使用又は方法は、タンパク質産生のための組換え細胞株を得るために関心のタンパク質をコードする導入遺伝子を細胞のゲノムに挿入することを目的とする。タンパク質産生のための組換え細胞株を構築するのに好適な細胞としては、ヒト(例えばPER.C6又はHEK)、チャイニーズハムスター卵巣(CHO)細胞及びマウス(NSE0)細胞が挙げられるが、これらに限定されない。
【0164】
別の好ましい実施形態では、上記の使用は、遺伝性障害の非ヒト動物モデルを作製することを目的とする。
【0165】
本発明は、そのゲノム内に本発明による核酸、発現ベクター又は組合せを含む非ヒトトランスジェニック動物にも関する。
【0166】
学術論文又は抄録、公開特許出願、交付済み特許又は任意の他の参考文献を含む本明細書中で引用される全ての参考文献は、引用される参考文献に提示される全てのデータ、表、図面及び本文を含めて参照により本明細書中に完全に援用される。
【0167】
本発明は以下の実施例及び添付の図面に鑑みて更に評価される。
【0168】
配列の簡単な説明
配列番号1は、野生型I−CreI単量体のアミノ酸配列を示す。
【0169】
配列番号2は、SH3遺伝子座内に位置する本発明による標的配列の配列を示す。
【0170】
配列番号3は、SH4遺伝子座内に位置する本発明による標的配列の配列を示す。
【0171】
配列番号4は、野生型I−CreIホモ二量体タンパク質の標的配列の配列を示す。
【0172】
配列番号5〜配列番号10は、図1に示される配列を表す。
【0173】
配列番号11〜配列番号15は、実施例1において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0174】
配列番号16〜配列番号19は、図4に示される配列を表す。
【0175】
配列番号20〜配列番号24は、実施例2において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0176】
配列番号25〜配列番号32は、それぞれSCOH−SH3−b56−A、SCOH−SH3−b56−B、SCOH−SH3−b56−C、SCOH−SH3−b56−D、SCOH−SH3−b1−A、SCOH−SH3−b1−B、SCOH−SH3−b1−C及びSCOH−SH3−b1−Dと称される、実施例1において構築される一本鎖メガヌクレアーゼを表す。
【0177】
配列番号33〜配列番号40は、それぞれSCOH−SH4−b56−A、SCOH−SH4−b56−B、SCOH−SH4−b56−C、SCOH−SH4−b56−D、SCOH−SH4−b1−A、SCOH−SH4−b1−B、SCOH−SH4−b1−C及びSCOH−SH4−b1−Dと称される、実施例2において構築される一本鎖メガヌクレアーゼを表す。
【0178】
配列番号41は、陽性対照であるSCOH−RAGを表す。
【0179】
配列番号42は、2位に更なるアラニン及び最後のプロリンの後に3つの更なる残基を有するI−CreI単量体のアミノ酸配列を示す。
【0180】
配列番号43は、RM2リンカーのアミノ酸配列を示す。
【0181】
配列番号44〜配列番号49は、実施例3において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0182】
配列番号50〜配列番号53は、実施例4において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0183】
配列番号54〜配列番号55は、それぞれSH3遺伝子座、SH4遺伝子座及びSH6遺伝子座に含まれる配列を示す。
【0184】
配列番号57は、44A 54L 64A 70Q 75N 158R 162A突然変異を含む配列番号1の単量体に由来する単量体を示す。
【0185】
配列番号58は、44A 54L 64A 70Q 75N 158R 162A突然変異を含む配列番号42の単量体に由来する単量体を示す。
【0186】
配列番号59は、SH6遺伝子座内に位置する本発明による標的配列の配列を示す。
【0187】
配列番号60〜配列番号64は、図9に示される配列を表す。
【0188】
配列番号65〜配列番号75は、実施例5において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0189】
配列番号76〜配列番号85は、それぞれSCOH−SH6−b1−B、SCOH−SH6−b1−C、SCOH−SH6−b1−C、QCSH61−A01、QCSH61−E01、QCSH61−H0、QCSH62−A02、QCSH61−H01b、QCSH61−H01c及びQCSH61−H01dと称される、実施例5において構築される一本鎖メガヌクレアーゼを表す。
【0190】
配列番号86〜配列番号96は、SH7遺伝子座(配列番号86及び配列番号87)、SH8遺伝子座(配列番号88)、SH12遺伝子座(配列番号89)、SH13遺伝子座(配列番号90)、SH19遺伝子座(配列番号91)、SH20遺伝子座(配列番号92)、SH21遺伝子座(配列番号93〜配列番号95)及びSH33遺伝子座(配列番号96)を切断することが可能な一本鎖メガヌクレアーゼを表す。
【0191】
配列番号97〜配列番号104は、それぞれSH12遺伝子座、SH13遺伝子座、SH19遺伝子座、SH20遺伝子座、SH21遺伝子座、SH33遺伝子座、SH7遺伝子座及びSH8遺伝子座内に含まれる配列を表す。
【0192】
配列番号105〜配列番号325は、実施例6〜実施例9並びに/又は表A’、表A’’、表E、表G及び表Hのいずれか1つに開示される配列を表す。
【0193】
配列番号326は、BQYリンカーのアミノ酸配列を示す。
【実施例】
【0194】
以下の実施例において、全てのI−CreI変異体は、配列番号42のI−CreI単量体の遺伝子操作によって構築した。
【0195】
実施例1: SH3遺伝子座を標的とするメガヌクレアーゼの改変
SH3は、染色体6上に存在する24bpの非パリンドローム標的(配列番号2)を含む遺伝子座である。表Aに示されるように、SH3は、Deichmann et al.(J. of Clin. Invest. 2007 117:2225)に開示されるRISの近傍に位置する。SH3配列は、Deichmann et alに記載されるCISのいずれにも含まれない。
【0196】
配列番号2の標的配列を切断することが可能なI−CreIヘテロ二量体を、Chames et al.(Nucleic Acids Res., 2005, 33, e178)、非特許文献6、非特許文献8、非特許文献7に記載されるものに基づく方法を用いて同定した。次いで、これらのヘテロ二量体の一部を、CHO細胞におけるSH3切断を判定するための哺乳動物発現ベクターにクローニングした。次いで、これらの結果を利用して、配列番号2の標的配列に指向性を有する一本鎖メガヌクレアーゼを設計した。これらの一本鎖メガヌクレアーゼを哺乳動物発現ベクターにクローニングし、CHO細胞におけるSH3切断を試験した。SH3標的の強い切断活性を、これらの一本鎖分子について哺乳動物細胞において観察することができた。
【0197】
実施例1.1. SH3を切断するメガヌクレアーゼの同定
SH3標的配列を切断する可能性があるヘテロ二量体形態のI−CreI変異体を、遺伝子操作によって構築した。次いで、かかる変異体の対を酵母において共発現させた。共発現時に3つの分子種、すなわち2つのホモ二量体及び1つのヘテロ二量体が得られる。次いで、このヘテロ二量体が配列番号2のSH3標的配列を切断することが可能であるか否かを決定した。
【0198】
a)SH3標的配列に由来するパリンドローム配列を切断するI−CreIメガヌクレアーゼの変異体の構築
SH3配列は部分的に、図1に示される10AAT_P(配列番号5)、5AAG_P(配列番号6)、10AGG_P(配列番号7)及び5TTT_P(配列番号8)標的配列の組合せである。これらの配列は、国際PCT出願の国際公開第2006/097784号及び国際公開第2006/097853号、非特許文献6、並びに非特許文献8に記載のように得られるメガヌクレアーゼによって切断される。このため、SH3は、これらのこれまでに同定されたメガヌクレアーゼから生じるコンビナトリアル変異体によって切断されるはずである。
【0199】
2つのパリンドローム標的SH3.3及びSH3.4は、SH3に由来するものであった(図1)。SH3.3及びSH3.4はパリンドローム性であるため、ホモ二量体タンパク質によって切断されるはずである。したがって、配列番号9のSH3.3パリンドローム標的配列又は配列番号10のSH3.4パリンドローム標的配列のいずれかを切断するホモ二量体I−CreI変異体は、Chames et al.(Nucleic Acids Res., 2005, 33, e178)、非特許文献6、非特許文献8及び非特許文献7に記載されるものに基づく方法を用いて構築した。
【0200】
b)標的ベクターの構築
ゲートウェイクローニング配列に隣接するSH3標的配列に相当する配列番号11のオリゴヌクレオチドを、PROLIGOにオーダーした。このオリゴヌクレオチドは以下の配列を有する:TGGCATACAAGTTTCCAATACAAGGTACAAAGTCCTGACAATCGTCTGTCA)。一本鎖オリゴヌクレオチドのPCR増幅によって生成した二本鎖標的DNAを、pCLS1055酵母レポーターベクターにゲートウェイプロトコル(INVITROGEN)を用いてクローニングした。
【0201】
酵母レポーターベクターを、以下の遺伝子型を有するFYBL2−7Bサッカロミセス・セレビシエ株に形質転換した:MAT a、ura3Δ851、trp1Δ63、leu2Δ1、lys2Δ202。得られた株はレポーター株(MilleGen)に相当する。
【0202】
c)変異体の共発現
SH3.4配列又はSH3.3配列を切断する変異体をコードするオープンリーディングフレームを、それぞれpCLS542発現ベクター及びpCLS1107発現ベクターにクローニングした。これらの変異体に由来する酵母DNAを標準プロトコルを用いて抽出し、大腸菌を形質転換するために使用した。次いで、得られたプラスミドを使用して、酵母を同時形質転換した。形質転換体を、ロイシンを欠き、G418を含有する合成培地上で選択した。
【0203】
d)メガヌクレアーゼを共発現するクローンの接合及び酵母におけるスクリーニング
接合はコロニーグリッダー(QpixII、Genetix)を用いて行った。変異体をYPDプレートを覆うナイロンフィルター上に、低グリッド密度(4〜6スポット/cm2)を用いてグリッドした。第2のグリッドプロセスを同じフィルター上で行い、各々の標的について異なるレポーターを有する酵母株からなる第2の層をスポットした。メンブレンを固形寒天YPD富栄養培地上に置き、30℃で一晩インキュベートして接合させた。次に、フィルターを、ロイシン及びトリプトファンを欠き、ガラクトース(2%)を炭素源として含むG418を添加した合成培地に移し、37℃で5日間インキュベートして、発現ベクター及び標的ベクターを保有する2倍体を選択した。5日後、フィルターを、0.5Mリン酸ナトリウムバッファー(pH7.0)中の0.02%X−Gal、0.1%SDS、6%ジメチルホルムアミド(DMF)、7mM β−メルカプトエタノール、1%アガロースを含む固形アガロース培地上に置き、37℃でインキュベートして、β−ガラクトシダーゼ活性をモニタリングした。結果をスキャニングによって分析し、適切なソフトウェアを用いて定量化を行った。
【0204】
e)結果
異なる変異体の共発現は、58個の試験した組合せにおいてSH3標的の切断をもたらした。機能的組合せを下の表Iにまとめる。この表において、「+」はSH3標的配列に対する機能的な組合せ、すなわちヘテロ二量体がSH3標的配列を切断することが可能であることを示す。
【0205】
【表17】
【0206】
結論として、酵母においてSH3標的配列を切断することが可能な幾つかのヘテロ二量体I−CreI変異体が同定された。
【0207】
実施例1.2. CHO細胞における染色体外モデルでのSH3標的切断の検証
ヘテロ二量体を形成した場合に酵母においてSH3標的を効率的に切断することが可能なI−CreI変異体は、上の実施例1.1で記載している。CHO細胞においてSH3標的に対する最大切断活性を示すヘテロ二量体を同定するために、これらの変異体の一部の効率を、CHO細胞における染色体外アッセイを用いて比較した。CHO細胞におけるスクリーニングは、メガヌクレアーゼによる標的の切断がLagoZレポーター遺伝子(細菌lacZ遺伝子の誘導体)の相同組換え及び発現を誘導する一本鎖アニーリング(SSA)ベースのアッセイである。
【0208】
a)CHOスクリーニングのためのベクターにおけるSH3標的のクローニング
ゲートウェイクローニング配列に隣接するSH3標的配列に相当するオリゴヌクレオチドを、PROLIGOにオーダーした(配列番号12、TGGCATACAAGTTTCCAATACAAGGTACAAAGTCCTGACAATCGTCTGTCA)。一本鎖オリゴヌクレオチドのPCR増幅によって生成した二本鎖標的DNAを、ゲートウェイプロトコル(INVITROGEN)を用いてpCLS1058 CHOレポーターベクターにクローニングした。クローニングした標的をシークエンシング(MILLEGEN)によって検証した。
【0209】
b)メガヌクレアーゼの再クローニング
上の表Iの同定されたこれらの変異体をコードするオープンリーディングフレームを、pCLS2437発現ベクターにサブクローニングした。ORFを、酵母DNA上で配列番号13及び配列番号14(5’−AAAAAGCAGGCTGGCGCGCCTACACAGCGGCCTTGCCACCATG−3’及び5’−AGAAAGCTGGGTGCTAGCGCTCGAGTTATCAGTCGG−3’)のプライマーを用いてPCRによって増幅した。PCR産物をCHO発現ベクターpCLS2437に、内部断片置き換えのためのAscI制限酵素及びXhoI制限酵素を用いてクローニングした。ライゲーション工程及び大腸菌形質転換工程から得られる選択されたクローンを、シークエンシング(MILLEGEN)によって検証した。
【0210】
c)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を、Polyfect(商標)トランスフェクション試薬を用い、供給業者のプロトコル(QIAGEN)に従ってトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl(通常は、100mlの溶解バッファー(10mM Tris−HCl(pH7.5)、150mM NaCl、0.1%Triton X100、0.1mg/ml BSA、プロテアーゼ阻害剤)、10mlのMg 100Xバッファー(100mM MgCl2、35%β−メルカプトエタノール)、110mlのONPG(8mg/ml)及び780mlの0.1Mリン酸ナトリウム(pH7.5)を含有するバッファーを1リットル)添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。
【0211】
1回のアッセイにつき、150ngの標的ベクターを、両方の変異体それぞれ12.5ngと共トランスフェクトした。
【0212】
d)結果
表Iに記載の以下の4つの変異体を、pCLS2437に再クローニングした:
44A 54L 70Q 75Y 92R 158R 162A(SH3.3−MAと称される);
1V 44A 54L 64A 70Q 75N 158W 162A(SH3.3−MBと称される);
30G 38R 70D 75N 86D(SH3.4−M1と称される);及び
30G 38R 70D 75N 81T 154G(SH3.4−M2と称される)。
【0213】
これらのI−CreI変異体を、CHO染色体外アッセイにおいてSH3標的に対するヘテロ二量体としてともにアッセイした。
【0214】
表IIは、9つのヘテロ二量体から得られた機能的な組合せを示す。
【0215】
【表18】
【0216】
SH3配列の切断及び組換えの効率の分析から、I−CreI変異体の試験した4つの組合せが全て、更なる突然変異なしにそれらの切断活性を酵母からCHO細胞へと移すことが可能であったことが実証される。
【0217】
実施例1.3. 一本鎖としての共有結合アセンブリ及びSH3を切断するメガヌクレアーゼの改良
実施例1.1.において同定された変異体の共発現は、酵母におけるSH3標的の高い切断活性をもたらす。ヘテロ二量体の一部は哺乳動物発現系におけるSH3切断が検証されている(実施例1.2.)。表IIIに示される、そのうちの1つを更なる最適化のために選択した。
【0218】
【表19】
【0219】
MA x M1 SH3ヘテロ二量体は酵母における高い切断活性をもたらす。SH3.3−MAは、I−CreI野生型配列と比較して以下の突然変異を有するSH3.3カッターである:44A 54L 70Q 75Y 92R 158R 162A。SH3.4−M1は、I−CreI野生型配列と比較して以下の突然変異を有するSH3.4カッターである:30G 38R 70D 75N 86D。
【0220】
一本鎖構築物を配列番号15(AAGGSDKYNQALSKYNQALSKYNQALSGGGGS)のリンカーRM2を用いて改変し、それにより一本鎖分子:MA−リンカーRM2−M1を作製した。この設計工程において、G19S突然変異をC末端M1変異体に導入した。加えて、突然変異K7E、K96EをMA変異体に、突然変異E8K、E61RをM1変異体に導入して、更にSCOH−SH3−b1足場と呼ばれる一本鎖分子:MA(K7E K96E)−リンカーRM2−M1(E8K E61R G19S)を作り出した。一部の更なるアミノ酸置換は、これまでの研究においてI−CreI誘導体の活性を増強することが見出されている。イソロイシン132のバリンでの置き換え(I132V)がそのうちの1つである。I132V突然変異を、N末端タンパク質断片及びC末端タンパク質断片のコード配列のいずれか1つ若しくは両方に導入するか、又はいずれにも導入しなかった。
【0221】
同じ戦略を、SH3.3(44A 54L 70Q 75Y 92R 158R 162A)及びSH3.4(30G 38R 50R 70D 75N 142R)をそれぞれホモ二量体として切断する最良な変異体に基づく、SCOH−SH3−b56足場と称される第2の足場に適用した。
【0222】
得られたタンパク質を下記表IVに示す。全ての一本鎖分子を、SH3標的の切断についてCHOにおいてアッセイした。
【0223】
a)一本鎖分子のクローニング
一連の合成遺伝子アセンブリをMWG-EUROFINSにオーダーした。SH3を標的とする異なる一本鎖変異体をコードする合成遺伝子を、AscI制限部位及びXhoI制限部位を用いてpCLS1853にクローニングした。
【0224】
b)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を実施例1.2に記載のようにトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。1回のアッセイにつき、150ngの標的ベクターを、3.12ngから25ngまでの漸増量の変異体DNA(12.5ng+12.5ngのヘテロ二量体DNAに相当する25ngの一本鎖DNA)と共トランスフェクトした。最後に、トランスフェクトしたDNA変異体のDNA量は3.12ng、6.25ng、12.5ng及び25ngであった。トランスフェクトしたDNAの総量は、空ベクター(pCLS0002)を用いて175ng(標的DNA、変異体DNA、担体DNA)とした。
【0225】
d)結果
SH3標的に対する一本鎖分子の活性を、これまでに記載されたCHOアッセイを本内部対照SCOH−RAG及びI−Sce Iメガヌクレアーゼとともに用いてモニタリングした。全ての比較は、3.12ng、6.25ng、12.5ng及び25ngのトランスフェクトした変異体DNAで行った(図2及び図3)。全ての一本鎖分子が、表IVに挙げられるようにCHOアッセイにおいてSH3標的切断活性を示した。
【0226】
【表20】
【0227】
変異体は、それらが有する突然変異プロファイルに応じて、アッセイした用量で特異的な挙動を共有していた(図2及び図3)。例えば、SCOH−SH3−b1−Cは、同様のプロファイルを有し、更に活性が高い。その活性は、低い量から高い量にかけてトランスフェクトした最も低いDNA量で最大に達する。SCOH−SH3−b1−Cと比較して、分子SCOH−SH3−b56−Aは、より高いDNA用量で最大活性を有するが、SCOH−SH3−b1−C及び本内部標準と同等のレベルの活性に達する。
【0228】
記載の全ての変異体が活性であり、導入遺伝子をSH3遺伝子座へ挿入するのに使用することができる。
【0229】
実施例2: SH4遺伝子座を標的とするメガヌクレアーゼの改変
SH4は染色体7上に存在する遺伝子座である。SH4遺伝子座は配列番号3の24bpの非パリンドローム配列を含む。表Aに示されるように、SH4は、Schwarzwaelder et al.(J. Clin. Invest. 2007 117:2241)に開示されるRISの周辺に位置する。SH4配列は、Deichmann et alに記載されるCISのいずれにも含まれない。
【0230】
上の実施例1に記載のものと同様の実験を行い、配列番号3の標的配列を切断することが可能なI−CreIヘテロ二量体及び一本鎖メガヌクレアーゼを同定した。
【0231】
実施例2.1. SH4を切断するメガヌクレアーゼの同定
SH4標的配列を切断する可能性があるヘテロ二量体形態のI−CreI変異体を、遺伝子操作によって構築した。次いで、かかる変異体の対を酵母において共発現させた。共発現時に3つの分子種、すなわち2つのホモ二量体及び1つのヘテロ二量体が得られる。次いで、このヘテロ二量体が配列番号3のSH4標的配列を切断することが可能であるか否かを決定した。
【0232】
a)SH4標的配列に由来するパリンドローム配列を切断するI−CreIメガヌクレアーゼの変異体の構築
SH4配列は部分的に、図4に示される10AAA_P(配列番号4)、5ACT_P(配列番号16)、10AAA_P(配列番号4)、5GGT_P(配列番号17)標的の組合せである。これらの配列は、国際PCT出願の国際公開第2006/097784号及び国際公開第2006/097853号、非特許文献6、非特許文献8に記載のように得られる、これまでに同定されたメガヌクレアーゼによって切断される。このため、SH4は、これらのこれまでに同定されたメガヌクレアーゼから生じるコンビナトリアル変異体によって切断されるはずである。
【0233】
スクリーニング手順を、Chames et al.(Nucleic Acids Res., 2005, 33, e178)、非特許文献6、非特許文献8及び非特許文献7に記載されるものに基づく方法を用いて、以下の2つのパリンドローム配列に対して行った:配列番号18のSH4.3配列及び配列番号19のSH4.4配列。
【0234】
b)標的ベクターの構築
実験手順は、配列番号20(5’−TGGCATACAAGTTTTTAAAACACTGTACACCATTTTGACAATCGTCTGTCA−3’)のSH4標的配列に相当するオリゴヌクレオチドを使用した以外は実施例1.1に記載されるものである。
【0235】
c)変異体の共発現
酵母DNAを、pCLS542発現ベクター及びpCLS1107発現ベクターにおいてSH4.3標的及びSH4.4標的を切断する変異体から標準プロトコルを用いて抽出し、大腸菌を形質転換するために使用した。次いで、得られたプラスミドDNAを使用して、酵母株を同時形質転換した。形質転換体を、ロイシンを欠き、G418を含有する合成培地上で選択した。
【0236】
d)メガヌクレアーゼを共発現するクローンの接合及び酵母におけるスクリーニング
接合はコロニーグリッダー(QpixII、genetix)を用いて行った。変異体をYPDプレートを覆うナイロンフィルター上に、低グリッド密度(4〜6スポット/cm2)を用いてグリッドした。第2のグリッドプロセスを同じフィルター上で行い、各々の標的について異なるレポーターを有する酵母株からなる第2の層をスポットした。膜を固形寒天YPD富栄養培地上に置き、30℃で一晩インキュベートして接合させた。次に、フィルターを、ロイシン及びトリプトファンを欠き、ガラクトース(2%)を炭素源として含むG418を添加した合成培地に移し、37℃で5日間インキュベートして、発現ベクター及び標的ベクターを保有する2倍体を選択した。5日後、フィルターを、0.5Mリン酸ナトリウムバッファー(pH7.0)中の0.02%X−Gal、0.1%SDS、6%ジメチルホルムアミド(DMF)、7mM β−メルカプトエタノール、1%アガロースを含む固形アガロース培地上に置き、37℃でインキュベートして、β−ガラクトシダーゼ活性をモニタリングした。結果をスキャニングによって分析し、適切なソフトウェアを用いて定量化を行った。
【0237】
e)結果
SH4.3標的を切断する変異体及びSH4.4標的を切断する変異体の共発現は、6つの事例においてSH4標的の切断をもたらした。機能的組合せを表Vにまとめる。
【0238】
【表21】
【0239】
実施例2.2. CHO細胞における染色体外モデルでのSH4標的切断の検証
CHO細胞においてSH4標的に対して最大切断活性を示すヘテロ二量体を同定するために、SH4標的を切断する変異体の幾つかの組合せの効率を、CHO細胞における染色体外アッセイを用いて判断した。CHO細胞におけるスクリーニングは、メガヌクレアーゼによる標的の切断がLagoZレポーター遺伝子(細菌lacZ遺伝子の誘導体)の相同組換え及び発現を誘導する一本鎖アニーリング(SSA)ベースのアッセイである。
【0240】
a)CHOスクリーニングのためのベクターにおけるSH4標的のクローニング
標的を以下のようにクローニングした。ゲートウェイクローニング配列に隣接するSH4標的配列に相当する配列番号21のオリゴヌクレオチドを、PROLIGOにオーダーした(5’−TGGCATACAAGTTTTTAAAACACTGTACACCATTTTGACAATCGTCTGTCA−3’)。一本鎖オリゴヌクレオチドのPCR増幅によって生成した二本鎖標的DNAを、ゲートウェイプロトコル(INVITROGEN)を用いてCHOレポーターベクター(pCLS1058)にクローニングした。クローニングした断片をシークエンシング(MILLEGEN)によって検証した。
【0241】
b)メガヌクレアーゼの再クローニング
上で得られたSH4.5標的及びSH4.6標的を切断するI−CreI変異体のORFをpCLS2437にサブクローニングした。ORFを、酵母DNA上で配列番号22及び配列番号23(5’−AAAAAGCAGGCTGGCGCGCCTACACAGCGGCCTTGCCACCATG−3’及び5’−AGAAAGCTGGGTGCTAGCGCTCGAGTTATCAGTCGG−3’)のプライマーを用いてPCRによって増幅した。PCR産物をCHO発現ベクターpCLS2437に、内部断片置き換えのためのAscI制限部位及びNheI制限部位を用いてクローニングした。ライゲーション工程及び大腸菌形質転換工程から得られる選択されたクローンを、シークエンシング(MILLEGEN)によって検証した。
【0242】
c)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を、Polyfect(商標)トランスフェクション試薬を用い、供給業者のプロトコル(QIAGEN)に従ってトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl(通常は、100mlの溶解バッファー(10mM Tris−HCl(pH7.5)、150mM NaCl、0.1%Triton X100、0.1mg/ml BSA、プロテアーゼ阻害剤)、10mlのMg 100Xバッファー(100mM MgCl2、35%β−メルカプトエタノール)、110mlのONPG(8mg/ml)及び780mlの0.1Mリン酸ナトリウム(pH7.5)を含有するバッファーを1リットル)添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。1回のアッセイにつき、150ngの標的ベクターを、両方の変異体それぞれ12.5ng(パリンドロームSH4.3標的を切断する変異体12.5ng及びパリンドロームSH4.4標的を切断する変異体12.5ng)と共トランスフェクトした。
【0243】
d)結果
表VIに示され、上の実施例2.1に記載の4つの変異体を更なる分析に選択した。
【0244】
【表22】
【0245】
これらの変異体をpCLS2437にクローニングした。次いで、SH4.3標的又はSH4.4標的を切断するI−CreI変異体を、CHO染色体外アッセイにおいてSH4標的に対するヘテロ二量体としてともにアッセイした。SH4配列の切断及び組換えの効率の分析から、I−CreI変異体の試験した全ての組合せが、更なる突然変異なしにそれらの切断活性を酵母からCHO細胞へと移すことが可能であったことが実証される(表VII)。
【0246】
【表23】
【0247】
実施例2.3. 一本鎖としての共有結合アセンブリ及び部位特異的突然変異誘発によるSH4を切断するメガヌクレアーゼの改良
実施例2.1.に記載の変異体の共発現は、酵母におけるSH4標的の高い切断活性をもたらす。加えて、それらの一部は哺乳動物発現系におけるSH4切断が検証されている(実施例2.2.)。
【0248】
MA x M2 SH4ヘテロ二量体は酵母における高い切断活性をもたらす。SH4.3−MAは、I−CreI野生型配列と比較して以下の突然変異を有するSH4.3カッターである:24V 44R 68Y 70S 75Y 77N。SH4.4−M2は、I−CreI野生型配列と比較して以下の突然変異を有するSH4.4カッターである:24V 44Y 70S 77V。
【0249】
実施例1.3に記載のように、リンカーRM2を用いて一本鎖構築物を改変し、それによりMA−リンカーRM2−M2と称される一本鎖分子を作製した。この設計工程において、G19S突然変異をC末端M2突然変異体に導入した。加えて、更にSCOH−SH4−b1足場と呼ばれる、MA(K7E K96E)−リンカーRM2−M2(E8K E61R G19S)と称される一本鎖分子を作り出すために、K7E突然変異及びK96E突然変異をMA突然変異体に、E8K突然変異及びE61R突然変異をM2突然変異体に導入した。
【0250】
イソロイシン132のバリンへの(I132V)突然変異を、N末端タンパク質断片及びC末端タンパク質断片のコード配列のいずれか1つ若しくは両方に導入するか、又はいずれにも導入しなかった。
【0251】
同じ戦略を、SH4.3(44R 68Y 70S 75Y 77N)及びSH4.4(24V 44Y 70S 77V)に対する良好なカッターに基づく第2の足場に適用した。この足場は更にSCOH−SH4−b56足場と称される。
【0252】
得られた一本鎖構築物の設計を表VIIIに示す。一本鎖構築物を、SH4標的の切断を誘導するそれらの能力についてCHOにおいて試験した。
【0253】
a)一本鎖分子のクローニング
一連の合成遺伝子アセンブリをMWG-EUROFINSに依頼した。SH4を標的とする異なる一本鎖変異体をコードする合成遺伝子を、AscI制限部位及びXhoI制限部位を用いてpCLS1853にクローニングした。
【0254】
b)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を上に記載のようにトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。1回のアッセイにつき、150ngの標的ベクターを、3.12ngから25ngまでの漸増量の変異体DNA(12.5ng+12.5ngのヘテロ二量体DNAに相当する25ngの一本鎖DNA)と共トランスフェクトした。最後に、トランスフェクトしたDNA変異体のDNA量は3.12ng、6.25ng、12.5ng及び25ngであった。トランスフェクトしたDNAの総量は、空ベクター(pCLS0002)を用いて175ng(標的DNA、変異体DNA、担体DNA)とした。
【0255】
c)結果
表VIIIに記載の一本鎖分子を、SH4標的に対するそれらの活性について、これまでに記載されたCHOアッセイを用いて、本内部対照SCOH−RAG及びI−Sce Iメガヌクレアーゼと比較することによってモニタリングした。全ての活性評価は、3.12ng、6.25ng、12.5ng及び25ngのDNAトランスフェクト用量で行った。全ての一本鎖分子が、表VIIIで報告されるようにSH4標的に対する活性を示していた。
【0256】
【表24】
【0257】
変異体は、それらが有する突然変異プロファイルに応じて、アッセイした用量で特異的な挙動を共有していた(図5及び図6)。例えば、SCOH−SH4−b1Cは、内部標準SCOH−RAGと同じ範囲内の活性レベルを示す(。その活性は、低い量から高い量にかけて増大する。アッセイしたDNAトランスフェクト用量で、その活性はSCOH−SH4−B56Aよりも優れている。
【0258】
これらの変異体は全て、異なるレベルの強度で活性であるため、SH4ゲノム標的化に使用することができる。
【0259】
実施例3: ヒト細胞株におけるSH遺伝子座での切断活性の検出
酵母及び哺乳動物細胞(CHO K1細胞)においてSH3標的及びSH4標的を効率的に切断することが可能なI−CreI変異体が、実施例1及び実施例2において同定されている。それらの内因性DNA標的配列を切断するSH3メガヌクレアーゼ及びSH4メガヌクレアーゼの効率を次に試験した。本実施例では、SH3標的配列及びSH4標的配列を切断するように改変されたメガヌクレアーゼが、ヒト細胞においてそれらの同族内因性部位を切断することが実証される。
【0260】
非相同末端結合(NHEJ)による二本鎖切断の修復は、小さな欠失及び挿入(InDel)を生成し得る(図7)。本来は、このエラーを起こしやすい機構は細胞の生存に有害であり得るが、内因性遺伝子座でのメガヌクレアーゼ活性の迅速な指標をもたらす。
【0261】
実施例3.1: 内因性部位で誘導された突然変異誘発の検出
特異性が変化した変異体のスクリーニングに使用される、哺乳動物細胞又は酵母細胞における切断誘導組換えに基づくアッセイが、国際PCT出願の国際公開第2004/067736号、Epinat et al., Nucleic Acids Res., 2003, 31:2952-2962、Chames et al., Nucleic Acids Res., 2005, 33:e178及び非特許文献6に記載されている。これらのアッセイは、標準方法によってモニタリングすることができる機能的なLacZレポーター遺伝子をもたらす。
【0262】
pCLS1853プラスミドにクローニングしたSH3及びSH4に対する一本鎖I−CreI変異体を、この実験に使用した。実験の前日に、ヒト胎児腎臓細胞株293−H(Invitrogen)に由来する細胞を、10cmディッシュに1.2×106細胞/ディッシュの密度で播種した。翌日、細胞に3μgの空プラスミド又はメガヌクレアーゼ発現プラスミドを、lipofectamine(Invitrogen)を用いてトランスフェクトした。トランスフェクションの72時間後、細胞を回収し、新鮮培養培地で希釈した(希釈率20倍)。7日間の培養後、細胞を回収し、ゲノムDNAを抽出した。
【0263】
200ngのゲノムDNAを使用して、メガヌクレアーゼ切断部位の周囲の内因性遺伝子座をPCR増幅によって増幅した。SH3遺伝子座に相当する377bpの断片は、特異的なPCRプライマーA(配列番号44;5’−tgggggtcttactctgtttccc−3’)及びPCRプライマーB(配列番号45;5’−aggagagtccttctttggcc−3’)を用いて増幅した。SH4遺伝子座に相当する396bpの断片は、PCRプライマーC(配列番号46;5’−gagtgatagcataatgaaaacc−3’)及びPCRプライマーD(配列番号47;5’−ctcaccataagtcaactgtctc−3’)を用いて増幅した。PCR増幅を454シークエンシングシステム(454 Life Sciences)上で行い、シークエンシングサービスを提供する会社(GATC Biotech AG,Germany)によって供給される特異的なアダプター配列(配列番号48;5’−CCATCTCATCCCTGCGTGTCTCCGACTCAG−3’及び配列番号49;5’−CCTATCCCCTGTGTGCCTTGGCAGTCTCAG−3’)に隣接する断片を得た。平均して18000個の配列が2つの単位複製配列のプール(各々500ng)から得られた。シークエンシング後、種々のサンプルを、上記のアダプターの最初に導入したバーコード配列に基づき同定した。
【0264】
次いで、配列をそれぞれSH3又はSH4の切断部位における挿入又は欠失の存在について分析した。
【0265】
実施例3.2: 結果
表IXに得られた結果をまとめる。
【0266】
【表25】
【0267】
SH3遺伝子座を標的とするメガヌクレアーゼをトランスフェクトした細胞から抽出したゲノムDNAの分析によって、12841個の分析した配列のうち56個(0.44%)がSH3の認識部位内にInDel事象を有することが示された。同様に、SH4遺伝子座を標的とするメガヌクレアーゼのトランスフェクション後、8259個の分析した配列のうち18個(0.22%)がSH4の認識部位内にInDel事象を有していた。
【0268】
小さな欠失又は挿入はPCR又はシークエンシングのアーチファクト(artefacts)に関連し得るため、同じ遺伝子座を、メガヌクレアーゼを発現しないプラスミドのトランスフェクション後に分析した。SH3遺伝子座及びSH4遺伝子座の分析から、InDel事象を実質的に全く検出することができないことが明らかとなった。実際に、分析した配列の0.05%(1/2153)及び0.02%(3/12811)しか突然変異を含有しなかった。
【0269】
さらに、DNA挿入配列又はDNA欠失配列のサイズの分析(図8)から、小さな挿入(5bp未満)及び小さな欠失(10bp未満)の優勢な同様のタイプの事象が明らかとなった。
【0270】
これらのデータから、それぞれSH3遺伝子座又はSH4遺伝子座を標的とするように改変されたメガヌクレアーゼが、ヒト細胞において活性であり、それらの同族内因性配列を切断することができることが実証される。さらに、メガヌクレアーゼが、遺伝子ORFを破壊し、それにより対応する遺伝子発現産物を不活性化し得る配列内に小さなInDel事象を発生させる能力があることが示される。
【0271】
実施例4: ヒト細胞における内因性SH3及びSH4遺伝子座での遺伝子標的化
改変された一本鎖SH3メガヌクレアーゼ及び一本鎖SH4メガヌクレアーゼの切断活性を検証するために、内因性ヒトSH3遺伝子座及び内因性ヒトSH4遺伝子座で相同組換えを刺激するそれらの能力を次に評価した。細胞に、一本鎖分子SCOH−SH3−b1−C又はSCOH−SH4−b1−Cに対する哺乳動物発現プラスミド及び標的化構築物を含むベクターをトランスフェクトした。標的化構築物を含むベクター(「ドナー修復プラスミド」とも称される)は、ヒトSH3遺伝子座又はヒトSH4遺伝子座に相同な2つの配列に隣接する外因性DNA配列からなる2.8kbの配列を含有するpCLS3777プラスミド又はpCLS3778プラスミドであった。ヒトSH3遺伝子座又はヒトSH4遺伝子座に相同な配列は1.5kbの長さを有していた。メガヌクレアーゼによる天然のSH3遺伝子座又はSH4遺伝子座の切断は、ドナー修復プラスミドを修復マトリックスとして使用し得る相同組換えのための基質を生じる。このため、SH3遺伝子座又はSH4遺伝子座で標的化組込みが起こる頻度は、ゲノムSH3標的部位又はゲノムSH4標的部位の切断効率を示す。
【0272】
実施例4.1: 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼは、哺乳動物発現ベクターにクローニングされ、それぞれプラスミドpCLS2697及びpCLS2705を生じるSCOH−SH3−b1−C及びSCOH−SH4−b1−Cである。
【0273】
b)ドナー修復プラスミド
SH3遺伝子標的化実験については、ドナープラスミドは以下のものを含有していた:
左ホモロジーアームとして、SH3遺伝子座のPCR生成断片(染色体6上の6850510位〜6852051位、NC_000006.11)。この断片は1540bpの長さを有する;
右ホモロジーアームとして、SH3遺伝子座の断片(染色体6上の6852107位〜6853677位、NC_000006.11)。この断片は1571bpの長さを有する。
【0274】
SH4遺伝子標的化実験については、ドナープラスミドは以下のものを含有していた:
左ホモロジーアームとして、SH4遺伝子座のPCR生成断片(染色体7上の114972751位〜114974269位、NC_000007.13)。この断片は1519bpの長さを有する;及び
右ホモロジーアームとして、SH4遺伝子座の断片(染色体7上の114974316位〜114976380位、NC_000007.13)。この断片は2065bpの長さを有する。
【0275】
SH3及びSH4の両方について、左ホモロジーアーム及び右ホモロジーアームを、2つのCMVプロモーター及びネオマイシン耐性遺伝子を含有する2.8kbの外因性DNA断片のそれぞれ上流(AscI部位を用いる)及び下流(SbfI部位を用いる)に挿入した。得られたプラスミドはpCLS3777(SH3に対する)及びpCLS3778(SH4に対する)と称される。
【0276】
c)Sh3遺伝子及びSh4遺伝子の標的化実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞をトランスフェクトした。簡潔に述べると、2μgのドナープラスミドを3μgの一本鎖メガヌクレアーゼ発現ベクターと共トランスフェクトした。37℃で72時間のインキュベーション後、細胞をトリプシン処理し、96ウェルプレートにおいて1ウェル当たり10細胞又は100細胞で、完全培地中で平板培養した。細胞が80%〜100%コンフルエントになった時点で、ZR−96ゲノムDNAキット(Zymo research)を用い、供給業者のプロトコルに従ってゲノムDNA抽出を行った。
【0277】
d)遺伝子標的化事象のPCR分析
遺伝子標的化頻度を、以下のプライマーを用いたゲノムDNAに対するPCRによって決定した:SH3については5’−CTGTGTGCTATGATCTTGCC−3’(SH3GHGF4;配列番号50)及び5’−CCTGTCTCTTGATCAGATCC−3’(NeoR2;配列番号51)、並びにSH4については5’−GTGGCCTCTCAGTCTGTTTA−3’(SH4GHGF2;配列番号52)及び5’−AGTCATAGCCGAATAGCCTC−3’(NeoR5;配列番号53)。PCRによって、2500bp(SH3)又は2268bp(SH4)の遺伝子標的化特異的PCR産物が得られる。SH3GHGF4プライマー及びSH4GHGF2プライマーは、ドナー修復プラスミドの左ホモロジーアームの上流に位置するフォワードプライマーである。NeoRプライマーは、ドナー修復プラスミドの2つのホモロジーアームの間に挿入される外因性DNAに位置するリバースプライマーである。
【0278】
実施例4.2: 結果
ヒト胎児腎臓293H細胞に、2つの一本鎖SH3メガヌクレアーゼ又は一本鎖SH4メガヌクレアーゼの一方を発現するプラスミド及びドナー修復プラスミドpCLS3777又はpCLS3778を共トランスフェクトした。自発的組換えの対照として、293H細胞をドナー修復プラスミド単独でもトランスフェクトした。次いで、細胞を96ウェルマイクロプレートにおいて1ウェル当たり10細胞又は100細胞で平板培養した。これらの細胞に由来するゲノムDNAを遺伝子標的化について、材料及び方法の項に記載されるようにPCRによって分析した。
【0279】
メガヌクレアーゼの非存在下(修復プラスミド単独)では、10個又は100個の細胞のプールにおいて分析した22560個及び18800個の細胞(それぞれSH3及びSH4に対する)の中でPCR陽性シグナルは検出されなかった。
【0280】
これに対し、SH3メガヌクレアーゼの存在下では、100個の細胞のプールにおいて分析した18800個の細胞の中で12個の陽性クローンが検出され、それにより組換えの頻度が0.064%であることが示された。SH4メガヌクレアーゼの存在下では、10個の細胞のプールにおいて分析した3760個の細胞の中で11個の陽性クローンが検出され、組換えの頻度が0.29%であることが示された。結果を下記表Xに提示する。ここで示される組換え頻度は、平板培養した全ての細胞が再び分裂を開始したわけではないため、低く見積もったものである。平板培養時の生存率はこのため、約33%であると推定することができる。したがって、組換えの頻度はおそらく3倍低く見積もられている。
【0281】
【表26】
【0282】
これらの結果から、2つの一本鎖分子SCOH−SH3−b1−C及びSCOH−SH4−b1−Cが、それぞれ内因性SH3遺伝子座及び内因性SH4遺伝子座で高レベルの遺伝子標的化を誘導することが可能であることが実証される。
【0283】
実施例5: SH6遺伝子座を標的とするメガヌクレアーゼの改変
SH6は、染色体21上に存在する24bpの非パリンドローム標的(TTAATACCCCGTACCTAATATTGC、配列番号59)を含む遺伝子座である。SH6は、Schwarzwaelder et al.(J Clin Invest 2007:2241-9)に開示されるRISの近傍に位置する。SH6配列は、Deichmann et alに記載されるCISのいずれにも含まれない。
【0284】
実施例5.1. SH6を切断するメガヌクレアーゼの同定
SH6標的配列を切断する可能性があるヘテロ二量体形態のI−CreI変異体を、遺伝子操作によって構築した。次いで、かかる変異体の対を酵母において共発現させた。共発現時に3つの分子種、すなわち2つのホモ二量体及び1つのヘテロ二量体が得られる。次いで、このヘテロ二量体が配列番号59のSH6標的配列を切断することが可能であるか否かを決定した。
【0285】
a)SH6標的配列に由来するパリンドローム配列を切断するI−CreIメガヌクレアーゼの変異体の構築
SH6配列は部分的に、図9に示される10AAT_P(配列番号60)、5CCC_P(配列番号61)、10AAT_P(配列番号60)、5TAG_P(配列番号62)標的配列の組合せである。これらの配列は、国際PCT出願の国際公開第2006/097784号及び国際公開第2006/097853号、非特許文献6、並びに非特許文献8に記載のように得られるメガヌクレアーゼによって切断される。このため、SH6は、これらのこれまでに同定されたメガヌクレアーゼから生じるコンビナトリアル変異体によって切断されるはずである。
【0286】
2つのパリンドローム標的SH6.3及びSH6.4は、SH6に由来するものであった(図9)。SH6.3及びSH6.4はパリンドローム性であるため、ホモ二量体タンパク質によって切断されるはずである。したがって、配列番号63のSH6.3パリンドローム標的配列又は配列番号64のSH6.4パリンドローム標的配列のいずれかを切断するホモ二量体I−CreI変異体は、Chames et al.(Nucleic Acids Res., 2005, 33, e178)、非特許文献6、非特許文献8及び非特許文献7に記載されるものに基づく方法を用いて構築した。
【0287】
b)標的ベクターの構築
実験手順は、SH6標的配列(5’−TGGCATACAAGTTTTTAATACCCCGTACCTAATATTGCCAATCGTCTGTCA−3’(配列番号65))に相当するオリゴヌクレオチドを使用した以外は実施例1.1に記載されるものである。
【0288】
c)変異体の共発現
pCLS542発現ベクター及びpCLS1107発現ベクターにおいてSH6.3標的及びSH6.4標的を切断する変異体に由来する酵母DNAを、標準プロトコルを用いて抽出し、大腸菌を形質転換するために使用した。形質転換体を、ロイシンを欠き、G418を含有する合成培地上で選択した。
【0289】
d)メガヌクレアーゼを共発現するクローンの接合及び酵母におけるスクリーニング
接合はコロニーグリッダー(QpixII、Genetix)を用いて行った。変異体をYPDプレートを覆うナイロンフィルター上に、低グリッド密度(4〜6スポット/cm2)を用いてグリッドした。第2のグリッドプロセスを同じフィルター上で行い、各々の標的について異なるレポーターを有する酵母株からなる第2の層をスポットした。膜を固形寒天YPD富栄養培地上に置き、30℃で一晩インキュベートして接合させた。次に、フィルターを、ロイシン及びトリプトファンを欠き、ガラクトース(2%)を炭素源として含むG418を添加した合成培地に移し、37℃で5日間インキュベートして、発現ベクター及び標的ベクターを保有する2倍体を選択した。5日後、フィルターを、0.5Mリン酸ナトリウムバッファー(pH7.0)中の0.02%X−Gal、0.1%SDS、6%ジメチルホルムアミド(DMF)、7mM β−メルカプトエタノール、1%アガロースを含む固形アガロース培地上に置き、37℃でインキュベートして、β−ガラクトシダーゼ活性をモニタリングした。結果をスキャニングによって分析し、適切なソフトウェアを用いて定量化を行った。
【0290】
e)結果
SH6.4標的を切断する10個の変異体及びSH6.3標的を切断する2個の変異体の共発現は、2つを除く全ての事例においてSH6.1標的の切断をもたらした。これらの2つの事例は、二重形質転換体が得られなかった場合に相当する。機能的組合せを表XIにまとめる。
【0291】
【表27】
【0292】
実施例5.2. CHO細胞における染色体外モデルでのSH6標的切断の検証
ヘテロ二量体を形成した場合に酵母においてSH6標的を効率的に切断することが可能なI−CreI変異体は、上の実施例5.1で記載している。CHO細胞においてSH3標的に対して最大切断活性を示すヘテロ二量体を同定するために、これらの変異体の幾つかの効率を、CHO細胞における染色体外アッセイを用いて比較した。CHO細胞におけるスクリーニングは、メガヌクレアーゼによる標的の切断がLagoZレポーター遺伝子(細菌lacZ遺伝子の誘導体)の相同組換え及び発現を誘導する一本鎖アニーリング(SSA)ベースのアッセイである。
【0293】
a)CHOスクリーニングのためのベクターにおけるSH6標的のクローニング
標的を以下のようにクローニングした。ゲートウェイクローニング配列に隣接するSH6標的配列に相当するオリゴヌクレオチドを、PROLIGOにオーダーした(5’−TGGCATACAAGTTTTTAATACCCCGTACCTAATATTGCCAATCGTCTGTCA−3’(配列番号65))。一本鎖オリゴヌクレオチドのPCR増幅によって生成した二本鎖標的DNAを、ゲートウェイプロトコル(INVITROGEN)を用いてCHOレポーターベクター(pCLS1058)にクローニングした。クローニングした標的をシークエンシング(MILLEGEN)によって検証した。
【0294】
b)メガヌクレアーゼの再クローニング
実施例5.1で同定されたSH6.3標的及びSH6.4標的を切断するI−CreI変異体のORFをpCLS2437にサブクローニングした。ORFを、酵母DNA上で以下のプライマー:5’−AAAAAGCAGGCTGGCGCGCCTACACAGCGGCCTTGCCACCATG−3’(配列番号66)及び5’−AGAAAGCTGGGTGCTAGCGCTCGAGTTATCAGTCGG−3’(配列番号67)のプライマーを用いてPCRによって増幅した。PCR産物をCHO発現ベクターpCLS2437に、内部断片置き換えのためのAscI及びXhoIを用いてクローニングした。ライゲーション工程及び大腸菌形質転換工程から得られる選択されたクローンを、シークエンシング(MILLEGEN)によって検証した。
【0295】
c)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を、Polyfect(商標)トランスフェクション試薬を用い、供給業者のプロトコル(QIAGEN)に従ってトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl(通常は、100mlの溶解バッファー(10mM Tris−HCl(pH7.5)、150mM NaCl、0.1%Triton X100、0.1mg/ml BSA、プロテアーゼ阻害剤)、10mlのMg 100Xバッファー(100mM MgCl2、35%β−メルカプトエタノール)、110mlのONPG(8mg/ml)及び780mlの0.1Mリン酸ナトリウム(pH7.5)を含有するバッファーを1リットル)添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。1回のアッセイにつき、150ngの標的ベクターを、両方の変異体それぞれ12.5ng(パリンドロームSH6.3標的を切断する変異体12.5ng及びパリンドロームSH6.4標的を切断する変異体12.5ng)と共トランスフェクトした。
【0296】
d)結果
酵母においてSH6を切断することが可能なヘテロ二量体エンドヌクレアーゼを形成する変異体の1つの組を、CHOにおいて一時的トランスフェクションでの染色体外アッセイを用いて確認するために選んだ。
【0297】
SH6.3を切断することが可能な単量体は、以下の突然変異:44K 70S 75Nを含み(SH6−3−M1−44K 70S 75Nと称される)、SH6.4を切断することが可能な単量体は、以下の突然変異:28Q 40R 44A 70L 75N 96R 111H 144Sを含んでいた(SH6−4−MB−28Q 40R 44A 70L 75N 96R 111H 144Sと称される)。
【0298】
SH6配列の切断及び組換えの効率の分析から、I−CreI変異体の試験した組合せが、更なる突然変異なしにその切断活性を酵母からCHO細胞へと移すことが可能であったことが実証される。
【0299】
実施例5.3. 一本鎖としての共有結合アセンブリ及びSH6を切断するメガヌクレアーゼの改良
実施例5.1に記載のカッターの共発現は、酵母におけるSH6標的の高い切断活性をもたらす。それらのうちの1つは哺乳動物発現系におけるSH6切断が検証されている(実施例5.2)。
【0300】
M1 x MA SH6ヘテロ二量体は酵母における高い切断活性をもたらす。M1は、I−CreI野生型配列と比較して以下の突然変異を有するSH6.3カッターである:44K 70S 75N。MAは、I−CreI野生型配列と比較して以下の突然変異を有するSH6.4カッターである:7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R。
【0301】
リンカーRM2(AAGGSDKYNQALSKYNQALSKYNQALSGGGGS;配列番号15)を用いて一本鎖構築物を改変し、それにより一本鎖分子:MA−RM2−M1を作製した。この設計工程において、G19S突然変異をC末端M1突然変異体に導入した。加えて、更にSCOH−SH6 b1足場と呼ばれる一本鎖分子:MA(K96E)−RM2−MA(E8K E61R)を作り出すために、突然変異K96EをMA突然変異体に、突然変異E8K、E61RをM1突然変異体に導入した。
【0302】
4つの更なるアミノ酸置換が、これまでの研究においてI−CreI誘導体の活性を増強することが見出されている。これらの突然変異は、フェニルアラニン54のロイシンでの置き換え(F54L)、グルタミン酸80のリジンでの置き換え(E80K)、バリン105のアラニンでの置き換え(V105A)、及びイソロイシン132のバリンでの置き換え(I132V)に相当する。幾つかの組合せをN末端タンパク質断片及びC末端タンパク質断片のコード配列に導入し、得られたタンパク質の最初のバッチを、SH6標的の切断を誘導するそれらの能力についてアッセイした。
【0303】
a)SC−OH一本鎖構築物への更なる突然変異の導入
更なる突然変異を、Stratagene/Agilent technologies Inc製のQuikChange Multi Site−Directed Mutagenesis Kitを用い、製造業者の使用説明書に従って導入した。オリゴヌクレオチドの第1のセットを使用して、第1の単量体に相当する一本鎖分子の一部に突然変異を導入した。オリゴヌクレオチドの第2のセットを、同じ突然変異を表XIIに示される第2の単量体に相当する一本鎖分子の第2の部分に特異的に導入するように設計した。
【0304】
【表28】
【0305】
このプロセスの間に得られた単離クローンをシークエンシングして、得られる特異的な突然変異プロファイルを確認した。次いで、関心のプロファイルをCHO SSAアッセイにおいて記載の最初の構築物と比較して試験した。
【0306】
b)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を上に記載されるようにトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。
【0307】
1回のアッセイにつき、150ngの標的ベクターを、3.12ngから25ngまでの漸増量の変異体DNA(12.5ng+12.5ngのヘテロ二量体DNAに相当する25ngの一本鎖DNA)と共トランスフェクトした。最後に、トランスフェクトしたDNA変異体のDNA量は3.12ng、6.25ng、12.5ng及び25ngであった。トランスフェクトしたDNAの総量は、空ベクター(pCLS0001)を用いて175ng(標的DNA、変異体DNA、担体DNA)とした。
【0308】
c)結果
SH6標的に対するSCOH−SH6−b1−C(pCLS2796)及びSCOH−SH6−b1−B(pCLS2928)一本鎖分子(表XIIIを参照されたい)の活性を、これまでに記載されたCHOアッセイを用い、SH6.3−M1 x SH6.4−MBを形成するヘテロ二量体及び本内部対照SCOH−RAG及びI−Sce Iメガヌクレアーゼと比較することによってモニタリングした。全ての比較は、3.12ng、6.25ng、12.5ng及び25ngのトランスフェクトした変異体DNAで行った(図10)。2つの一本鎖メガヌクレアーゼは、開始(starting)ヘテロ二量体よりも効率的にSH6標的を切断することが可能であった。最良の分子SCOH−SH6−b1−Cの活性を、メガヌクレアーゼの新たなバッチにおいて上に記載されたものに更なる突然変異を導入することによって更に改善した。
【0309】
【表29】
【0310】
更なる突然変異を材料及び方法の項に従って一本鎖足場に更に導入した。得られて試験した分子を表XIVに挙げる。
【0311】
【表30】
【0312】
全ての変異体は記載の条件で活性であり、それらが有する突然変異プロファイルに応じて、アッセイした用量で特異的な挙動を共有していた(図10)。例えば、QCSH61−H01a、QCSH61−H01b、QCSH61−H01c、QCSH61−H01dは、本内部標準SCOH−RAGと同様のプロファイルを有する。これらは低用量であっても非常に活性な分子である。これらの変異体は全てSH6ゲノム標的化に使用することができた。
【0313】
実施例6: ヒト細胞における内因性SH6遺伝子座での遺伝子標的化
改変された一本鎖SH6メガヌクレアーゼの切断活性を検証するために、内因性ヒトSH6遺伝子座で相同組換えを刺激するそれらの能力を評価した。細胞に、一本鎖分子SCOH−QCSH6−H01(配列番号81;pCLS3690)又はSCOH−QC−SH6−H01−V2−7E−70R75D(配列番号85;pCLS4373)に対する哺乳動物発現プラスミド、及びヒトSH6遺伝子座に相同な2つの配列(どちらも1.5kb長)に隣接する2.8kbの外因性DNA配列を含有するドナー修復プラスミドpCLS3779(図13;配列番号279)をトランスフェクトした。メガヌクレアーゼによる天然のSH6遺伝子座の切断は、ホモロジーアームに隣接する2.8kbの外因性DNAを含有するドナー修復プラスミドを修復マトリックスとして使用し得る相同組換えのための基質を生じる。このため、SH6遺伝子座で標的化組込みが起こる頻度は、ゲノムSH6標的部位の切断効率を示す。
【0314】
実施例6.1. 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼは、哺乳動物発現ベクターにクローニングされ、それぞれプラスミドpCLS3690(図13)及びpCLS4373を生じるSCOH−QCSH6−H01(配列番号81)又はSCOH−QC−SH6−H01−V2−7E−70R75D(配列番号85)である。
【0315】
b)ドナー修復プラスミド
ドナープラスミドは、左ホモロジーアームとしてSH6遺伝子座のPCR生成1517bp断片(染色体21上の18437771位〜18439287位、NC_000021.8)、及び右ホモロジーアームとして、SH6遺伝子座の1571bp断片(染色体21上の18439343位〜18440846位、NC_000021.8)を含有する。左ホモロジーアーム及び右ホモロジーアームを、2つのCMVプロモーター及びネオマイシン耐性遺伝子を含有する2.8kbの外因性DNA断片のそれぞれ上流(AscI部位を用いる)及び下流(SbfI部位を用いる)に挿入した。得られたプラスミドはpCLS3779(図13;配列番号279)である。
【0316】
c)Sh6遺伝子標的化実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞をトランスフェクトした。簡潔に述べると、2μgのドナープラスミドを3μgの一本鎖メガヌクレアーゼ発現ベクターと共トランスフェクトした。37℃で72時間のインキュベーション後、細胞をトリプシン処理し、96ウェルプレートにおいて1ウェル当たり10細胞又は100細胞で、完全培地中で平板培養した。代替的に、37℃で72時間のインキュベーション後、細胞をトリプシン処理し、10cmディッシュ1枚当たり300細胞で、完全培地中で平板培養した。37℃で2週間のインキュベーション後、個々のクローン細胞コロニーを採取し、96ウェルプレートにおいて完全培地中で平板培養した。細胞が80%〜100%コンフルエントになった時点で、ZR−96ゲノムDNAキット(Zymo research)を用い、供給業者のプロトコルに従ってゲノムDNA抽出を行った。
【0317】
d)遺伝子標的化事象のPCR分析
遺伝子標的化の頻度を、プライマーSH6GHGF3:5’−CAATGGAGTTTTGGAGCCAC−3’(配列番号280)及びNeoR9:5’−ATCAGAGCAGCCGATTGTCT−3’(配列番号281)を用いたゲノムDNAに対するPCRによって決定した。PCRによって、2300bpの遺伝子標的化特異的PCR産物が得られる(図14)。SH6GHGF3プライマーは、ドナー修復プラスミドの左ホモロジーアームの上流に位置するフォワードプライマーである。NeoR9プライマーは、ドナー修復プラスミドの2つのホモロジーアームの間に挿入される外因性DNAに位置するリバースプライマーである。
【0318】
実施例6.2. 結果
ヒト胎児腎臓293H細胞に、2つのベクター:2つの一本鎖SH6メガヌクレアーゼの一方を発現するプラスミド及びドナー修復プラスミドpCLS3779(図13;配列番号279)を共トランスフェクトした。自発的組換えの対照として、293H細胞をドナー修復プラスミド単独でもトランスフェクトした。次いで、細胞を96ウェルマイクロプレートにおいて1ウェル当たり10細胞又は100細胞で、又は10cmディッシュ1枚当たり300細胞で平板培養し、2週間後にクローンコロニーを単離し、96ウェルマイクロプレートにおいて平板培養した。これらの細胞に由来するゲノムDNAを遺伝子標的化について、材料及び方法の項に記載されるようにPCRによって分析した。メガヌクレアーゼの非存在下(修復プラスミド単独)では、10個又は100個の細胞のプールにおいて分析した67680個の細胞の中で5個のPCR陽性シグナルが検出され、0.007%の自発的組換えの頻度が示された。対照的に、SCOH−QCSH6−H01メガヌクレアーゼ(配列番号81;pCLS3690)又はSCOH−QC−SH6−H01−V2−7E−70R75Dメガヌクレアーゼ(配列番号85;pCLS4773)の存在下では、10個又は100個の細胞のプールにおいて分析した73320個及び18800個の細胞の中で177個及び35個の陽性シグナルが検出され、それぞれ0.24%及び0.19%の組換えの頻度が示された。結果を表XVに提示する。これらの結果から、2つの一本鎖分子SCOH−QCSH6−H01(配列番号81;pCLS3690)及びSCOH−QC−SH6−H01−V2−7E−70R75D(配列番号85;pCLS4773)が、内因性sh6遺伝子座で高レベルの遺伝子標的化を誘導することが可能なことが実証される。
【0319】
【表31】
【0320】
実施例7: ヒト細胞における内因性sh6遺伝子座での遺伝子標的化後の導入遺伝子発現
sh6遺伝子座での導入遺伝子発現を支持するsh6遺伝子座の能力、改変された一本鎖SH6メガヌクレアーゼの切断活性を検証するために、ネオマイシン耐性遺伝子発現カセットを含有する修復プラスミドを用いて遺伝子標的化実験を行い、ネオマイシン含有培地中で成長する修飾細胞の能力を測定した。ネオマイシンの存在下での細胞の生存及び成長はネオマイシン耐性遺伝子の発現に依存し、したがって標的化組込み後のSH6遺伝子座での導入遺伝子の発現を示す。
【0321】
実施例7.1. 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼは、哺乳動物発現ベクターにクローニングされ、プラスミドpCLS3690を生じるSCOH−QCSH6−H01(配列番号81)である。
【0322】
b)ドナー修復プラスミド
ドナープラスミドは、左ホモロジーアームとしてSH6遺伝子座のPCR生成1517bp断片(染色体21上の18437771位〜18439287位、NC_000021.8)、及び右ホモロジーアームとして、SH6遺伝子座の1571bp断片(染色体21上の18439343位〜18440846位、NC_000021.8)を含有する。左ホモロジーアーム及び右ホモロジーアームを、2つのCMVプロモーター及びネオマイシン耐性遺伝子を含有する2.8kbの外因性DNA断片のそれぞれ上流(AscI部位を用いる)及び下流(SbfI部位を用いる)に挿入した。得られたプラスミドはpCLS3779(図13;配列番号279)である。
【0323】
c)Sh6遺伝子標的化実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞をトランスフェクトした。簡潔に述べると、2μgのドナープラスミドを3μgの一本鎖メガヌクレアーゼ発現ベクターと共トランスフェクトした。37℃で72時間のインキュベーション後、細胞をトリプシン処理し、10cmディッシュ1枚当たり300細胞で、完全培地中で平板培養した。37℃で2週間のインキュベーション後、個々のクローン細胞コロニーを採取し、96ウェルプレートにおいて完全培地中で平板培養した。37℃で1週間のインキュベーション後、細胞をトリプシン処理し、2つの96ウェル複製プレートに平板培養して、37℃でインキュベートした。細胞が80%〜100%コンフルエントになった時点で、ZR−96ゲノムDNAキット(Zymo research)を用い複製プレートの1つで、供給業者のプロトコルに従ってゲノムDNA抽出を行った。他の複製プレートを使用して遺伝子標的化クローンを単離し、それらを増殖させた。
【0324】
d)遺伝子標的化クローンのPCR同定
遺伝子標的化を、プライマーSH6GHGF3:5’−CAATGGAGTTTTGGAGCCAC−3’(配列番号280)及びNeoR9:5’−ATCAGAGCAGCCGATTGTCT−3’(配列番号281)を用いたゲノムDNAに対するPCRによって決定した。PCRによって、2300bpの遺伝子標的化特異的PCR産物が得られる(図14)。SH6GHGF3プライマーは、ドナー修復プラスミドの左ホモロジーアームの上流に位置するフォワードプライマーである。NeoR9プライマーは、ドナー修復プラスミドの2つのホモロジーアームの間に挿入される外因性DNAに位置するリバースプライマーである。
【0325】
e)サザンブロット法による標的化組込みの検証
細胞クローンに由来するゲノムDNAを、StuI制限酵素又はHindIII制限酵素(New England Biolabs)で消化し、0.8%アガロースゲル(gel)上での電気泳動によって分離し、ニトロセルロース膜に転写した。DNAプローブを、ネオマイシン耐性遺伝子に相同な25ngのDNA断片から、32P放射標識dCTP及びRediprime IIランダムプライムラベリングシステム(GE Healthcare)を用い、供給業者のプロトコルに従って調製し、ハイブリダイゼーションバッファー(20mM NaPi、7%SDS、1mM EDTA)中でプレインキュベートしたニトロセルロース膜に添加した。65℃で一晩のインキュベーションの後、膜を洗浄し、放射線写真用フィルムに感光させた。期待されるバンドの放射線写真上でのサイズは、StuI消化については5.3kb、HindIII消化については6.8kbである(図15)。
【0326】
f)ネオマイシン耐性試験:
SH6遺伝子座で標的化されるPCRによって同定された細胞クローンを、96ウェルマイクロプレートにおいて1ウェル当たり300細胞で、G418抗生物質(PAA laboratories)の存在下で平板培養した。37℃で10日間のインキュベーションの後、Vialightバイオアッセイキット(Lonza)及びVictorルミネッセンスリーダー(Perkin Elmer)を用い、供給業者のプロトコルに従って生存能力を測定した。
【0327】
実施例7.2. 結果
ヒト胎児腎臓293H細胞に、2つのベクター:2つの一本鎖SH6メガヌクレアーゼの一方を発現するプラスミド及びドナー修復プラスミドpCLS3779を共トランスフェクトした。次いで、細胞を10cmディッシュ1枚当たり300細胞で平板培養し、2週間後にクローンコロニーを単離し、96ウェルマイクロプレートにおいて平板培養した。これらの細胞に由来するゲノムDNAを遺伝子標的化について、材料及び方法の項に記載されるようにPCRによって分析した。次いで、ゲノムDNAを使用して、サザンブロット分析により標的化組込みを検証した。クローンの#7及び#8は期待されるサイズのバンドを示したが、陰性対照クローンの#5及び#6は示さなかった(図16)。これらの細胞クローンを、G418(PAA laboratories)の存在下で生存するそれらの能力について試験した。標的化組込みを有するクローン(#7及び#8)のみが、0.4mg/mlを上回る濃度のG418に対する耐性を示した(図16)。このことは、sh6遺伝子座での標的化組込みが機能的な導入遺伝子の発現を支持し得ることを示している。
【0328】
実施例8. ヒト細胞における内因性sh6遺伝子座での遺伝子標的化後の近接遺伝子の発現
近接遺伝子の発現を妨げることなく導入遺伝子組込みを支持するsh6遺伝子座の能力を検証するために、2.8kbの外因性DNA断片を含有する修復プラスミドを用いて遺伝子標的化実験を行い、標的化組込みを有する細胞クローンを同定した。sh6組込み部位の上流及び下流の遺伝子の発現を測定し、標的化組込みが起こっていない細胞クローンの発現と比較した。
【0329】
実施例8.1. 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼは、哺乳動物発現ベクターにクローニングされ、プラスミドpCLS3690を生じるSCOH−QCSH6−H01(配列番号81)である。
【0330】
b)ドナー修復プラスミド
ドナープラスミドは、左ホモロジーアームとしてSH6遺伝子座のPCR生成1517bp断片(染色体21上の18437771位〜18439287位、NC_000021.8)、及び右ホモロジーアームとして、SH6遺伝子座の1571bp断片(染色体21上の18439343位〜18440846位、NC_000021.8)を含有する。左ホモロジーアーム及び右ホモロジーアームを、2つのCMVプロモーター及びネオマイシン耐性遺伝子を含有する2.8kbの外因性DNA断片のそれぞれ上流(AscI部位を用いる)及び下流(SbfI部位を用いる)に挿入した。得られたプラスミドはpCLS3779(図13;配列番号279)である。
【0331】
c)Sh6遺伝子標的化実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞をトランスフェクトした。簡潔に述べると、2μgのドナープラスミドを3μgの一本鎖メガヌクレアーゼ発現ベクターと共トランスフェクトした。37℃で72時間のインキュベーション後、細胞をトリプシン処理し、10cmディッシュ1枚当たり300細胞で、完全培地中で平板培養した。37℃で2週間のインキュベーション後、個々のクローン細胞コロニーを採取し、96ウェルプレートにおいて完全培地中で平板培養した。37℃で1週間のインキュベーション後、細胞をトリプシン処理し、2つの96ウェル複製プレートに平板培養し、37℃でインキュベートした。細胞が80%〜100%コンフルエントになった時点で、ZR−96ゲノムDNAキット(Zymo research)を用い複製プレートの1つで、供給業者のプロトコルに従ってゲノムDNA抽出を行った。他の複製プレートを使用して遺伝子標的化クローンを単離し、それらを増殖させた。
【0332】
d)遺伝子標的化クローンのPCR同定
遺伝子標的化を、プライマーSH6GHGF3:5’−CAATGGAGTTTTGGAGCCAC−3’(配列番号280)及びNeoR9:5’−ATCAGAGCAGCCGATTGTCT−3’(配列番号281)を用いたゲノムDNAに対するPCRによって決定した。PCRによって、2300bpの遺伝子標的化特異的PCR産物が得られる(図XX)。SH6GHGF3プライマー(配列番号280)は、ドナー修復プラスミドの左ホモロジーアームの上流に位置するフォワードプライマーである。NeoR9プライマー(配列番号281)は、ドナー修復プラスミドの2つのホモロジーアームの間に挿入される外因性DNAに位置するリバースプライマーである。
【0333】
e)sh6遺伝子座の上流及び下流の遺伝子の発現
遺伝子発現を定量RT−PCRによって測定した。RNAをサブコンフルエントな細胞クローンからRNeasy RNA単離キット(Qiagen)を用いて、製造業者のプロトコルに従って単離した。3μgのRNAを使用し、Superscript III First−strandキット(Invitrogen)を用いてcDNAを生成した。定量PCRを、2連の(duplicate)サンプルにおいて、SYBR(商標)Premix Ex Taq(商標)DNA Polymerase(Lonza)を用い、Stratagene MPX3000機器上で12μlの反応につき10ngのcDNAに対して行った。各々の遺伝子について、使用したプライマーを以下の表に挙げる。
【0334】
【表32】
【0335】
サイクル閾値(threshold cycles)(Ct)を、Stratageneソフトウェアを用い、ROXリファレンス色素による正規化の後に蛍光(dRn)に対して決定した。遺伝子発現の強度を、ハウスキーピング遺伝子HPRTの発現を内部正規化因子として使用して、式2Ct(HPRT)-Ct(遺伝子)を用いて算出した。
【0336】
実施例8.2. 結果
ヒト胎児腎臓293H細胞に、2つのベクター:3つの一本鎖SH6メガヌクレアーゼのうち1つを発現するプラスミド及びドナー修復プラスミドpCLS3779を共トランスフェクトした。次いで、細胞を10cmディッシュ1枚当たり300細胞で平板培養し、2週間後にクローンコロニーを単離し、96ウェルマイクロプレートにおいて平板培養した。これらの細胞に由来するゲノムDNAを遺伝子標的化について、材料及び方法の項に記載されるようにPCRによって分析した。標的化組込みを示すクローン及び陰性対照からRNAを単離した。定量RT−PCRを行い、標的化組込みの遺伝子座の周囲の遺伝子の発現を測定した。データを図17に提示する。図17では、ハウスキーピング遺伝子HPRTでの正規化後の3つの個々の標的化クローン(KI)及び3つの個々の非標的化クローン(WT)について2連のサンプルの平均強度を示す。測定した5つの遺伝子の各々について有意差は観察されず、sh6遺伝子座での標的化組込みが近接遺伝子の発現に影響をもたらさないことが示される。
【0337】
実施例9: ヒト細胞における内因性セーフハーバー遺伝子座での突然変異誘発
改変された一本鎖セーフハーバーメガヌクレアーゼの切断活性を検証するために、内因性ヒトセーフハーバー遺伝子座での突然変異誘発を刺激するそれらの能力を評価した。細胞に、一本鎖分子に対する哺乳動物発現プラスミドをトランスフェクトした。メガヌクレアーゼによる天然のセーフハーバー遺伝子座の切断は、エラーを起こしやすいプロセスである非相同末端結合の基質を生じ、メガヌクレアーゼ標的部位で小さな挿入又は欠失をもたらし得る。このため、内因性セーフハーバー遺伝子座で突然変異が起こる頻度は、メガヌクレアーゼによるゲノム標的部位の切断効率を示す。
【0338】
実施例9.1. 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼに対するコード配列は、哺乳動物発現ベクターにクローニングされ、表XVIに挙げるプラスミドを生じた。
【0339】
【表33】
【0340】
b)セーフハーバー遺伝子座の突然変異誘発実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞に3μgの一本鎖メガヌクレアーゼ発現ベクターをトランスフェクトした。37℃で2日間〜6日間のインキュベーション後、細胞をトリプシン処理し、DNeasy血液及び組織キット(Qiagen)を用い、供給業者のプロトコルに従ってゲノムDNA抽出を行った。
【0341】
c)突然変異誘発事象のディープシークエンシング分析
突然変異誘発の頻度をディープシークエンシング分析によって決定した。オリゴヌクレオチドを各々のセーフハーバー標的の周囲のDNA断片のPCR増幅のために設計した。それらを表XVIIに挙げる。
【0342】
【表34】
【0343】
ヌクレオチドを添加し、454シークエンシングシステム(454 Life Sciences)上で、シークエンシングサービスを提供する会社(GATC Biotech AG,Germany)によって供給される特異的なアダプター配列(5’−CCATCTCATCCCTGCGTGTCTCCGACTCAG−3’;配列番号324)及び(5’−CCTATCCCCTGTGTGCCTTGGCAGTCTCAG−3’;配列番号325)に隣接する断片を得た。平均して18000個の配列が2つ又は3つの単位複製配列のプール(各々500ng)から得られた。シークエンシング後、種々のサンプルを、上記のアダプターの最初に導入したバーコード配列に基づき同定した。
【0344】
実施例9.2. 結果
ヒト胎児腎臓293H細胞に、一本鎖セーフハーバーメガヌクレアーゼを発現するプラスミドをトランスフェクトした。37℃で2日間〜6日間のインキュベーション後、ゲノムDNAを単離し、PCRを用いてメガヌクレアーゼ標的部位の周囲のゲノム配列を増幅した。次いで、各々のセーフハーバー標的の切断部位における挿入又は欠失の事象(InDel)の存在について配列を分析した。結果を表XVIIIにまとめる。
【0345】
【表35】
【0346】
実施例10: 結論
結論として、実施例1、実施例2、実施例3及び実施例5から、SH3遺伝子座、SH4遺伝子座及びSH6遺伝子座を切断することが可能なI−CreIヘテロ二量体タンパク質及び一本鎖メガヌクレアーゼの両方を得ることができることが実証される。さらに、これらのエンドヌクレアーゼは、これらの遺伝子座を強い切断活性で切断することが可能である。
【0347】
実施例4から、SH3遺伝子座及びSH4遺伝子座を切断することが可能な一本鎖メガヌクレアーゼが、ヒト細胞の標的部位への導入遺伝子の効率的な挿入を可能にすることが実証される。
【0348】
このため、これらのエンドヌクレアーゼは、個体のSH3遺伝子座、SH4遺伝子座又はSH6遺伝子座への導入遺伝子の挿入に有利に使用することができる。
【0349】
実施例6から、本発明による少なくとも2つの一本鎖分子が、内因性sh6遺伝子座で高レベルの遺伝子標的化を誘導することが可能であることが実証される。
【0350】
実施例7から、遺伝子座での標的化組込みが機能的な導入遺伝子の発現を支持し得ることが実証される。
【0351】
実施例8から、遺伝子座での標的化組込みが、標的配列の近傍に位置する5個の遺伝子の発現を実質的に変更しないことが実証される。
【0352】
実施例9から、セーフハーバー配列を標的とする種々のメガヌクレアーゼの突然変異誘発頻度(該メガヌクレアーゼによるゲノム標的部位の切断効率を示す)が実証される。
【技術分野】
【0001】
本発明は、「セーフハーバー遺伝子座」、すなわち導入遺伝子の安全な発現を可能にする遺伝子座に位置する標的配列を切断することが可能なエンドヌクレアーゼに関する。本発明は、細胞、組織又は生物への導入遺伝子の挿入のためのかかるエンドヌクレアーゼの使用に更に関する。
【背景技術】
【0002】
メガヌクレアーゼ
ホーミングエンドヌクレアーゼとも称されるメガヌクレアーゼは、生細胞における二本鎖切断及び組換えを誘導するために使用された最初のエンドヌクレアーゼであった(非特許文献1、非特許文献2、非特許文献3、非特許文献4)。しかしながら、それらの使用は、それらの狭い特異性によって長年制限されていた。数百個の天然メガヌクレアーゼが過去数年にわたって同定されたが、この多様性は依然としてゲノムの複雑性に対応するにはほとんどが不十分であり、関心の遺伝子内のメガヌクレアーゼ切断部位を発見する確率は、依然として極めて低い。これらの知見から、選ばれた配列を天然エンドヌクレアーゼと同じ選択性で切断する、個々に適合した特異性を有する人工エンドヌクレアーゼの必要性が問題となっている。
【0003】
メガヌクレアーゼは、所望の標的配列を切断するゲノム工学ツールを得るのに最適な足場として登場した(非特許文献5)。変更された特異性を有するメガヌクレアーゼの改変を可能にするコンビナトリアルアセンブリプロセスは、非特許文献6、非特許文献7、非特許文献8、非特許文献9によって記載されている。簡潔に述べると、これらのプロセスは、野生型メガヌクレアーゼの基質特異性と数ヌクレオチドだけ異なる基質特異性を有する局所改変変異体の同定に依存する。次いで、かかるタンパク質において同定された最大で4組の突然変異を、完全に再設計された結合面を有する新たなメガヌクレアーゼを生成するために、新たなタンパク質に組み立てることができる。
【0004】
これらのプロセスは2つの工程を必要とする。初めに、異なる組の突然変異を、パリンドローム標的を切断するホモ二量体変異体に組み立てる。次いで、2つのホモ二量体を、選ばれた非パリンドローム標的を切断するヘテロ二量体メガヌクレアーゼを生成するために共発現させることができる。このプロセスの第1の工程は、依然として最も難しいものであり、所与の遺伝子座を切断するメガヌクレアーゼを絶対確実に得ることができるか否かを事前に知ることはできない。実際に、全ての配列が改変メガヌクレアーゼによって同等に切断される可能性があるわけではなく、場合によっては、メガヌクレアーゼの改変は困難であることが判明し得る(非特許文献10)。
【0005】
部位特異的ゲノム修飾に好適な他の酵素
インテグラーゼ、リコンビナーゼ、トランスポザーゼ及びエンドヌクレアーゼのような特殊化された酵素が、部位特異的ゲノム修飾に提案されている。何年もの間、これらの酵素の使用は、所望の標的部位に対するそれらの天然の特異性の再標的化の難しさのために制限されたままである。実際に、これらのタンパク質の標的部位又は十分な程度の配列同一性を有する配列は、訂正される突然変異に近接する配列中又は不活性化される遺伝子内に存在しなくてはならないが、予め改変された配列の場合を除いて、通常はそのようなことはない。遺伝子療法におけるこれらのDNA修飾酵素の使用を可能にすることの主な課題は、それらのDNA結合特性を再設計する可能性に左右される。個々に適合した基質特異性を有する人工タンパク質を得ることを目的として、多くの戦略が開発されている。
【0006】
ストレプトミセスファージPhiC31に由来するインテグラーゼは、内因性遺伝子座における標的化遺伝子導入に早くから使用されていた。この酵素は、ファージ付着部位(attP)と細菌付着部位(attB)との間の部位特異的反応による細菌染色体へのファージゲノムの組換えを媒介する(非特許文献11、非特許文献12)。これはattB部位を保有するプラスミドから、偽attP部位(attP’)と呼ばれるattPとの部分同一性を有する天然のゲノム配列へと起こり得る。PhiC31インテグラーゼは、ヒトゲノムにおいてhFIXを含む幾つかの導入遺伝子を導入するために使用されている(非特許文献13、非特許文献14、非特許文献15、非特許文献16、非特許文献17)。ここでの欠点は、組込みが起こり得る部位を選ぶことができず(非特許文献16)、正確な組込みについてヒトゲノム遺伝子座内の偽attP部位に依存せざるを得ないということである。主要な組込み部位は染色体19に見られるが、数百個の他の組込み遺伝子座が同定されている(非特許文献16)。最近の研究では、attP’部位での組込みの効率及び特異性を高め、選ばれた部位を標的とする改変インテグラーゼの開発の道を開くために、PhiC31インテグラーゼを突然変異させた(非特許文献18)。しかしながら、改変インテグラーゼの開発は、標的化リコンビナーゼ系及び標的化エンドヌクレアーゼ系に焦点を合わせた同様の試みに遅れを取っている。
【0007】
バクテリオファージP1に由来するCreリコンビナーゼ又はサッカロミセス・セレビシエ(Saccharomyces cerevisiae)に由来するFlpタンパク質等の部位特異的リコンビナーゼは、それらの同族部位を含有する予め改変された配列間の組換えを誘導するために使用されている。Creリコンビナーゼは、loxPとして知られる2つの同一の34bp部位を認識し、それらの間の組換えを媒介する(非特許文献19)。長年にわたって、Cre由来リコンビナーゼの制限は、反復loxP部位又は偽loxP部位が、これら2つの部位間のDNA組込みを可能にするために存在しなければならないということであった。しかしながら、この分子のDNA結合面の指向進化を使用して、新たな特異性を有するリコンビナーゼが作り出されている(非特許文献20、非特許文献21)。Creリコンビナーゼ系は、ウイルス配列の切除を誘導するDNA標的化酵素の使用のためのフレームワークの提供にも有用であった。実際に、レトロウイルスのモロニーマウス白血病ウイルスのベクター系を用いた研究から、loxP部位を組込みレトロウイルスベクターのLTRに導入する場合、Creの発現が2つのloxP部位間の全ての配列の欠失をもたらし得ることが示されている(非特許文献22)。ごく最近では、改変Creリコンビナーゼ変異体が、細胞からHIV1型プロウイルスを切除するために使用されている(非特許文献23)。リコンビナーゼはプロウイルスLTRを標的とするように再設計され、全ての介在配列の切除を誘導するために使用されている。FRT(Flp組換え標的)配列を標的とするFlpリコンビナーゼを用いた改変も試みられており(非特許文献24)、非天然Flp組換え標的を認識する変異体が得られている(非特許文献25)。しかしながら、今日、かかる酵素による予め改変されていない遺伝子座における標的化挿入の例はない。
【0008】
Piggy Back及びSleeping Beauty等のトランスポゾンは、脊椎動物細胞への配列の挿入に効率的なツールを提供することができ、長期的発現を達成するウイルス媒介遺伝子送達の代替として提案されている(非特許文献26、非特許文献27、非特許文献28)。トランスポゾンは、DNA分子中に存在するDNA配列がトランスポザーゼの作用によって別の位置に挿入される、遺伝子送達の天然の手段である。SB100Xと呼ばれる改変SBトランスポザーゼが、このプロセスの効率を高めることが近年示されている(非特許文献28)。転位はゲノムレベルでランダムであり(例えば、SBはTAジヌクレオチドに組み込まれる)(非特許文献29)、したがって標的化アプローチのためのツールとみなすべきではない。しかしながら、更なる研究から、トランスポザーゼ触媒ドメインを特異的なDNA結合ドメインに融合させることにより、ヒト細胞において改変トランスポザーゼによって媒介される染色体転位の可能性が示されており(非特許文献30)、新たなカテゴリーの標的化ツールの開発の道が開かれている。
【0009】
遺伝子療法
10年近く前の遺伝子療法による数人のX−SCID患者の治療の成功は、遺伝子療法の分野における最も重要な節目の1つであった。この素晴らしい業績の後、別の形態のSCID、表皮水疱症及びレーバー黒内障等を含む種々の疾患に対処する他の臨床試験における大きな成功が続いた。しかしながら、これらの当初の成功は、一連の深刻な有害事象、すなわちX−SCID治療患者における白血病の発生のために長い間目立たないものであった(非特許文献31、非特許文献32、非特許文献33)。1例を除く白血病の全ての症例を化学療法によって最終的に治療することができたため、このアプローチは全体的にみて成功のように思われるが、これらの深刻な有害事象は、現行の遺伝子療法アプローチの重大なリスクを浮き彫りにしている。
【0010】
このため、当該技術分野においては、遺伝子を被験体のゲノムに挿入する安全な方法が必要とされている。
【0011】
遺伝性疾患の治療のために現在開発されている遺伝子療法プロトコルの大半は、疾患原因遺伝子の付加的かつ機能的なコピーによる変異型対立遺伝子の補完に基づいている。網膜等の非分裂組織では、このコピーの送達は、例えばアデノ随伴ウイルス(AAV)に由来する非組込みベクターを用いて達成することができる。しかしながら、増殖する運命である造血幹細胞(HSC)等の幹細胞を標的とする場合、持続的発現が問題になり、組込みベクターが必要とされる。ゲノム内に組み込まれ、宿主の染色体を用いて複製するレトロウイルスベクターが、この目的に有効であることが証明されたが、それらの挿入のランダム性は、全て遺伝子発現に関係する様々な懸念を生じている。X−SCID試験において観察された白血病の症例は、組込み部位の近傍のがん原遺伝子の活性化に明らかに関連していた。加えて、導入遺伝子の不適切な発現は代謝的問題又は免疫学的問題をもたらす可能性がある。最後に、挿入は内因性遺伝子のノックアウトをもたらす可能性がある。
【0012】
部位特異的組込みは、挿入突然変異誘発のリスクを緩和し得るため、ウイルスベクターのランダム組込みの有望な代替策であり得る(非特許文献34、非特許文献35、非特許文献5)。しかしながら、標的化組換えのツールを設計するには比較的時間がかかる。加えて、各々のツールは活性及び特異性に関してその固有の特性を有する。
【先行技術文献】
【非特許文献】
【0013】
【非特許文献1】Rouet et al. PNAS 1994 91:6064-6068
【非特許文献2】Rouet et al. Mol Cell Biol. 1994 14:8096-8106
【非特許文献3】Choulika et al. Mol Cell Biol. 1995 15:1968-1973
【非特許文献4】Puchta et al. PNAS 1996 93:5055-5060
【非特許文献5】Paques et al. Curr Gen Ther. 2007 7:49-66
【非特許文献6】Arnould et al. J Mol Biol. 2006 355:443-458
【非特許文献7】Arnould et al. J Mol Biol. 2007 371:49-65
【非特許文献8】Smith et al. NAR 2006 34:e149
【非特許文献9】Grizot et al. NAR 2009 37:5405
【非特許文献10】Galetto et al. Expert Opin Biol Ther. 2009 9:1289-303
【非特許文献11】Kuhstoss et al. J Mol Biol 1991 222:897-908
【非特許文献12】Rausch et al. NAR 1991 19:5187-5189
【非特許文献13】Olivares et al. Nat Biotech 2002 20:1124-1128
【非特許文献14】Ginsburg et al. Adv Genet 2005 54:179-187
【非特許文献15】Calos Curr Gene Ther 2006 6:633-645
【非特許文献16】Chalberg et al. J Mol Biol 2006 357:28-48
【非特許文献17】Aneja et al. J Gene Med 2007 9:967-975
【非特許文献18】Keravala et al. Mol Ther 2009 17:112-120
【非特許文献19】Abremski et al. Cell 1983 32:1301-1311
【非特許文献20】Buchholz et al. Nat Biotech 2001 19:1047-1052
【非特許文献21】Santoro et al. PNAS 2002 99:4185-4190
【非特許文献22】Choulika et al. J Virol 1996 70:1792-1798
【非特許文献23】Sarkar et al. Science 2007 316:1912-1915
【非特許文献24】Buchholzt et al. Nat Biotech 1998 16:657-662
【非特許文献25】Voziyanov et al. J Mol Biol 2003 326:65-76
【非特許文献26】Izsvak et al. Mol ther 2004 9:147-156
【非特許文献27】Ivics et al. Curr Gene Ther 2006 6:593-607
【非特許文献28】Mates et al. Nat Genet 2009 41:753-761
【非特許文献29】Vigdal et al. J Mol Biol 2002 323:441-452
【非特許文献30】Ivics, et al. Mol Ther 2007 15:1137-1144
【非特許文献31】Hacein-Bey-Abina et al. Science 2003 302:415-419
【非特許文献32】Hacein-Bey-Abina et al. J Clin Invest. 2008 118:3132-3142
【非特許文献33】Howe et al. J Clin Invest. 2008 118:3143-3150
【非特許文献34】Kolb et al. Trends Biotechnol. 2005 23:399-406
【非特許文献35】Porteus et al. Nat Biotechnol. 2005 23:967-973
【発明の概要】
【発明が解決しようとする課題】
【0014】
したがって、当該技術分野では、遺伝子付加の「セーフハーバー」であるとみなされ得るゲノムの遺伝子座への導入遺伝子の標的化挿入を可能にするツールが必要とされている。加えて、このツールを、それらの配列に関わりなく導入遺伝子の挿入に使用し、それにより同じツールを用いた遺伝子療法による多数の疾患の治療を可能にすることができれば極めて有利であり得る。さらに、このツールが高い有効性でゲノムに導入遺伝子を挿入し、高いレベルで導入遺伝子の安定な発現をもたらすことができれば極めて有利であり得る。
【課題を解決するための手段】
【0015】
本発明はとりわけ、以下の実施の形態に関する(drawn to):
実施の形態1:導入遺伝子を個体のゲノムに挿入する際に使用される標的配列を切断することが可能な変異型エンドヌクレアーゼであって、
i.上記ゲノムが上記標的配列を含む遺伝子座を含み、
ii.上記標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連しない、変異型エンドヌクレアーゼ。
実施の形態2:上記導入遺伝子の挿入が、上記標的配列の近傍に位置する遺伝子の発現を実質的に変更しない、実施の形態1に記載のエンドヌクレアーゼ。
実施の形態3:上記標的配列が、最近接遺伝子から少なくとも100kb離れて位置する、実施の形態1又は2に記載のエンドヌクレアーゼ。
実施の形態4:上記RISが、幹細胞の形質導入による遺伝子療法によって治療される患者に由来する細胞において同定されたものである、実施の形態1〜3のいずれか1つに記載のエンドヌクレアーゼ。
実施の形態5:上記RISが、造血幹細胞の形質導入による遺伝子療法によって治療される患者に由来する細胞において同定されたものである、実施の形態1〜3のいずれか1つに記載のエンドヌクレアーゼ。
実施の形態6:ホーミングエンドヌクレアーゼである、実施の形態1〜5のいずれか1つに記載のエンドヌクレアーゼ。
実施の形態7:上記ホーミングエンドヌクレアーゼが、LAGLIDADGエンドヌクレアーゼファミリーの成員である、実施の形態6に記載のエンドヌクレアーゼ。
実施の形態8:上記LAGLIDADGエンドヌクレアーゼファミリーの成員がI−CreIである、実施の形態7に記載のエンドヌクレアーゼ。
実施の形態9:上記遺伝子座が、ヒト染色体6p25.1上のSH3遺伝子座、ヒト染色体7q31.2上のSH4遺伝子座、ヒト染色体21q21.1上のSH6遺伝子座、ヒト染色体13q34上のSH12遺伝子座、ヒト染色体3p12.2上のSH13遺伝子座、ヒト染色体22上のSH19遺伝子座、ヒト染色体12q21.2上のSH20遺伝子座、ヒト染色体3p24.1上のSH21遺伝子座、ヒト染色体6p12.2上のSH33遺伝子座、ヒト染色体2p16.1上のSH7遺伝子座及びヒト染色体5上のSH8遺伝子座から選択される、実施の形態1〜8のいずれか1つに記載のエンドヌクレアーゼ。
実施の形態10:導入遺伝子を細胞又は組織のゲノムに挿入するための実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼのin vitro又はex vivoでの使用。
実施の形態11:配列番号1又は配列番号42と少なくとも80%同一な配列を含む2つの単量体を含む変異型二量体I−CreIタンパク質であって、
i.上記二量体I−CreIタンパク質が、個体の遺伝子座内に位置する標的配列を切断することが可能であり、該標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連せず、
ii.上記標的配列が配列番号4の配列を含まない、変異型二量体I−CreIタンパク質。
実施の形態12:ヒト染色体6p25.1上のSH3遺伝子座内に位置する標的配列を切断することが可能である、実施の形態11に記載の二量体I−CreIタンパク質。
実施の形態13:上記標的配列が配列番号2の配列を含む、実施の形態12に記載の二量体I−CreIタンパク質。
実施の形態14:
a)配列番号1の30位、38位、70位及び75位にアミノ酸置換を含む第1の単量体と、
b)配列番号1の44位、54位、70位及び75位にアミノ酸置換を含む第2の単量体と、
を含む、実施の形態12又は13に記載の二量体I−CreIタンパク質。
実施の形態15:上記ポリペプチドが、
a)30G 38R 70D 75N 86D突然変異を含む第1の単量体、
b)
i.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
ii.44A 54L 70Q 75Y 92R 158R 162A突然変異を含む単量体、
iii.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iv.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
v.44A 54L 70Q 75N突然変異を含む単量体、
vi.44A 54L 57E 70Q 75N 158R 162A突然変異を含む単量体、及び
vii.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含む、実施の形態14に記載の二量体I−CreIタンパク質。
実施の形態16:上記ポリペプチドが、
a)30G 38R 70D 75N 81T 154G突然変異を含む第1の単量体、
b)
i.44A 54L 70Q 75N 105A 158R 162A突然変異を含む単量体、
ii.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iii.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iv.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
v.44A 54L 70Q 75N突然変異を含む単量体、及び
vi.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含む、実施の形態14に記載の二量体I−CreIタンパク質。
実施の形態17:上記ポリペプチドが、
a)30G 38R 50R 70D 75N 142R突然変異を含む第1の単量体、
b)
i.44A 54L 70Q 75N 105A 158R 162A突然変異を含む単量体、
ii.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iii.44A 54L 70Q 75Y 92R 158R 162A突然変異を含む単量体、
iv.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
v.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
vi.44A 54L 66C 70Q 71R 75N 151A 158R 162A突然変異を含む単量体、
vii.44A 54L 70Q 75N突然変異を含む単量体、
viii.44A 54L 57E 70Q 75N 158R 162A突然変異を含む単量体、及び
ix.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含む、実施の形態14に記載の二量体I−CreIタンパク質。
実施の形態18:ヒト染色体7q31.2上のSH4遺伝子座内に位置する標的配列を切断することが可能である、実施の形態11に記載の二量体I−CreIタンパク質。
実施の形態19:上記標的配列が配列番号3の配列を含む、実施の形態18に記載の二量体I−CreIタンパク質。
実施の形態20:
a)配列番号1の24位、70位、75位及び77位にアミノ酸置換を含む第1の単量体と、
b)配列番号1の24位、44位及び70位にアミノ酸置換を含む第2の単量体と、
を含む、実施の形態18又は19に記載の二量体I−CreIタンパク質。
実施の形態21:上記ポリペプチドが、
a)
i.24V 44R 68Y 70S 75Y 77N突然変異を含む単量体、
ii.24V 68A 70S 75N 77R突然変異を含む単量体、及び
iii.24V 70D 75N 77R突然変異を含む単量体からなる群から選択される第1の単量体、
b)
i.24V 44Y 70S突然変異を含む単量体、及び
ii.24V 44Y 70S 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含む、実施の形態20に記載の二量体I−CreIタンパク質。
実施の形態22:ヒト染色体21q21.1上のSH6遺伝子座内に位置する標的配列を切断することが可能である、実施の形態11に記載の二量体I−CreIタンパク質。
実施の形態23:上記標的配列が配列番号59の配列を含む、実施の形態22に記載の二量体I−CreIタンパク質。
実施の形態24:
a)配列番号1の44位、並びに任意に70位及び/又は75位にアミノ酸置換を含む第1の単量体と、
b)配列番号1の28位、40位、44位、70位及び75位にアミノ酸置換を含む第2の単量体と、
を含む、実施の形態22又は23に記載の二量体I−CreIタンパク質。
実施の形態25:上記ポリペプチドが、
a)44K 68T 70G 75N突然変異を含む第1の単量体と、
b)
i.28Q 40R 44A 70L 75N 96R 111H 144S突然変異を含む単量体、
ii.7R 28Q 40R 44A 70L 75N 85R 103T突然変異を含む単量体、
iii.28Q 40R 44A 70L 75N 103S突然変異を含む単量体、
iv.24F 27V 28Q 40R 44A 70L 75N 99R突然変異を含む単量体、
v.7R 28Q 40R 44A 70L 75N 81T突然変異を含む単量体、
vi.7R 28Q 40R 44A 70L 75N 77V突然変異を含む単量体、
vii.7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R突然変異を含む単量体、
viii.28Q 40R 44A 70L 75N突然変異を含む単量体、
ix.7R 28Q 40R 44A 70L 75N 103T突然変異を含む単量体、及び
x.28Q 34R 40R 44A 70L 75N 81V 103T 108V 160E突然変異を含む単量体からなる群から選択される第2の単量体と、
を含む、実施の形態24に記載の二量体I−CreIタンパク質。
実施の形態26:上記ポリペプチドが、
a)44K突然変異、並びに任意に70S及び/又は75N突然変異を含む第1の単量体と、
b)
i.28Q 40R 44A 70L 75N 96R 111H 144S突然変異を含む単量体、
ii.7R 28Q 40R 44A 70L 75N 85R 103T突然変異を含む単量体、
iii.28Q 40R 44A 70L 75N 103S突然変異を含む単量体、
iv.24F 27V 28Q 40R 44A 70L 75N 99R突然変異を含む単量体、
v.7R 28Q 40R 44A 70L 75N 81T突然変異を含む単量体、
vi.7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R突然変異を含む単量体、
vii.7R 28Q 40R 44A 70L 75N 103T突然変異を含む単量体、及び
viii.28Q 34R 40R 44A 70L 75N 81V 103T 108V 160E突然変異を含む単量体からなる群から選択される第2の単量体と、
を含む、実施の形態24に記載の二量体I−CreIタンパク質。
実施の形態27:実施の形態11〜26のいずれか1つに記載の二量体I−CreIタンパク質の単量体を含む融合タンパク質。
実施の形態28:上記単量体が、配列番号43の配列を含むペプチドリンカーによって連結される、実施の形態27に記載の融合タンパク質。
実施の形態29:C末端単量体がK7E突然変異及びK96E突然変異を更に含み、N末端単量体がE8K突然変異、E61R突然変異及びG19S突然変異を更に含む、実施の形態27又は28に記載の融合タンパク質。
実施の形態30:配列番号25〜配列番号40及び配列番号76〜配列番号96からなる群から選択される配列を含む、実施の形態27〜29のいずれか1つに記載の融合タンパク質。
実施の形態31:実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼ又は実施の形態11〜30のいずれか1つに記載のタンパク質をコードする核酸。
実施の形態32:実施の形態31に記載の核酸を含む発現ベクター。
実施の形態33:導入遺伝子を含む標的化構築物と、実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼ又は実施の形態11〜30のいずれか1つに記載のタンパク質によって認識される標的配列に隣接するゲノム配列に相同な2つの配列とを更に含む、実施の形態32に記載の発現ベクター。
実施の形態34:上記導入遺伝子が治療用ポリペプチドをコードする、実施の形態33に記載の発現ベクター。
実施の形態35:遺伝子療法に使用される、実施の形態32〜34のいずれか1つに記載の発現ベクター。
実施の形態36:実施の形態32に記載の発現ベクターと、
導入遺伝子を含む標的化構築物と、実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼ又は実施の形態11〜30のいずれか1つに記載のタンパク質によって認識される標的配列のゲノム配列に相同な2つの配列とを含むベクターと、
の組合せ。
実施の形態37:実施の形態32〜34のいずれか1つに記載の発現ベクター又は実施の形態36に記載の組合せと、薬学的に活性な担体とを含む医薬組成物。
実施の形態38:遺伝子療法によって個体を治療する方法であって、有効量の実施の形態32〜34のいずれか1つに記載の発現ベクター又は実施の形態36に記載の組合せを、それを必要とする個体に投与することを含む、遺伝子療法によって個体を治療する方法。
実施の形態39:個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法であって、
a)上記個体のゲノム内で、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)を選択する工程と、
b)上記RISの200kb上流及び200kb下流に伸びるゲノム領域を規定する工程と、
c)上記ゲノム領域内に位置する標的配列を切断することが可能な野生型エンドヌクレアーゼを同定するか、又は変異型エンドヌクレアーゼを構築する工程と、
を含む、個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法。
実施の形態40:導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入するための実施の形態1〜9のいずれか1つに記載のエンドヌクレアーゼ又は実施の形態11〜30のいずれか1つに記載のタンパク質又は実施の形態31に記載の核酸又は実施の形態32〜34のいずれか1つに記載の発現ベクター又は実施の形態36に記載の組合せの使用であって、治療目的でない、使用。
実施の形態41:遺伝性障害の非ヒト動物モデルを作製するための、実施の形態40に記載の使用。
実施の形態42:組換えタンパク質を製造するための、実施の形態40に記載の使用。
実施の形態43:実施の形態31に記載の核酸又は実施の形態32〜34のいずれか1つに記載の発現ベクター又は実施の形態36に記載の組合せを、そのゲノム内に含む非ヒトトランスジェニック動物。
【図面の簡単な説明】
【0016】
【図1】実施例1に記載のメガヌクレアーゼの標的配列を示す図である。図中の「SEQ ID NO:」の記載は「配列番号」を意味する。以下の図も同様である。
【図2】SCOH SH3メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図3】SCOH SH3メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図4】実施例2に記載のメガヌクレアーゼの標的配列を示す図である。
【図5】SCOH SH4メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図6】SCOH SH4メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図7】非相同末端結合(NHEJ)による二本鎖切断の修復時に小さな欠失及び挿入(InDel)の生成をもたらす機構のスキームを示す図である。
【図8】SH3メガヌクレアーゼ又はSH4メガヌクレアーゼによる切断の際の挿入部位を示す図である。
【図9】実施例5に記載のメガヌクレアーゼの標的配列を示す図である。
【図10】SCOH SH6メガヌクレアーゼ対I−SceI及びSCOH−RAG DNAのCHOにおける用量応答を示す図である。
【図11】配列番号1のI−CreI単量体と配列番号42のI−CreI単量体との間の配列アラインメントを示す図である。
【図12】配列番号1のI−CreI単量体と、44A 54L 64A 70Q 75N 158R及び162Aの突然変異を含む2つのI−CreI単量体との間の配列アラインメントを示す図である。第1の単量体(配列番号57)は配列番号1に直接由来し、第2の単量体(配列番号58)は配列番号42に直接由来する。
【図13】実施例6〜実施例9を説明する図である。
【図14】実施例6〜実施例9を説明する図である。
【図15】実施例7を説明する図である。
【図16】実施例7を説明する図である。
【図17】実施例8を説明する図である。
【発明を実施するための形態】
【0017】
本発明者らは、標的化挿入による導入遺伝子の安全な発現を可能にするゲノム内の「セーフハーバー」遺伝子座を同定したが、ここで、(i)上記遺伝子座は、遺伝子療法によって治療される患者に由来する細胞において同定されたレトロウイルス挿入部位の近くにあり、及び(ii)上記レトロウイルス挿入は、がん又は異常細胞増殖と関連しない。以下の説明及び実施例からすぐに分かるように、本発明によるセーフハーバー遺伝子座は遺伝子のイントロン内に位置していても、又は遺伝子間領域内に位置していてもよい。
【0018】
特に、本発明者らは、エンドヌクレアーゼを、上記セーフハーバーを遺伝子付加の標的とするように改変することができることを見出した。
【0019】
より具体的には、本発明者らは、種々のセーフハーバー遺伝子座、例えば本明細書中で更に説明するSH6遺伝子座、SH3遺伝子座、SH4遺伝子座、SH12遺伝子座、SH13遺伝子座、SH19遺伝子座、SH20遺伝子座、SH21遺伝子座、SH33遺伝子座、SH7遺伝子座、SH8遺伝子座、SH18遺伝子座、SH31遺伝子座、SH38遺伝子座、SH39遺伝子座、SH41遺伝子座、SH42遺伝子座、SH43遺伝子座、SH44遺伝子座、SH45遺伝子座、SH46遺伝子座、SH47遺伝子座、SH48遺伝子座、SH49遺伝子座、SH50遺伝子座、SH51遺伝子座、SH52遺伝子座、SH70遺伝子座、SH71遺伝子座、SH72遺伝子座、SH73遺伝子座、SH74遺伝子座、SH75遺伝子座、SH101遺伝子座、SH106遺伝子座、SH107遺伝子座、SH102遺伝子座、SH105遺伝子座、SH103遺伝子座、SH104遺伝子座、SH113遺伝子座、SH109遺伝子座、SH112遺伝子座、SH108遺伝子座、SH110遺伝子座、SH114遺伝子座、SH116遺伝子座、SH111遺伝子座、SH115遺伝子座、SH121遺伝子座、SH120遺伝子座、SH122遺伝子座、SH117遺伝子座、SH118遺伝子座、SH119遺伝子座、SH123遺伝子座、SH126遺伝子座、SH128遺伝子座、SH129遺伝子座、SH124遺伝子座、SH131遺伝子座、SH125遺伝子座、SH127遺伝子座、SH130遺伝子座、SH11遺伝子座、SH17遺伝子座、SH23遺伝子座、SH34遺伝子座、SH40遺伝子座、SH53遺伝子座、SH54遺伝子座、SH55遺伝子座、SH56遺伝子座、SH57遺伝子座、SH58遺伝子座、SH59遺伝子座、SH60遺伝子座、SH61遺伝子座、SH62遺伝子座、SH65遺伝子座、SH67遺伝子座、SH68遺伝子座及びSH69遺伝子座内に位置する標的配列を認識し切断することが可能な幾つかのI−CreIメガヌクレアーゼを設計した。
【0020】
これらのメガヌクレアーゼが、それらの標的配列を効率的に切断することができることが更に示された。
【0021】
したがって、これらのメガヌクレアーゼ、並びにインテグラーゼ、リコンビナーゼ及びトランスポザーゼのような他の酵素(enzymes)を、導入遺伝子をセーフハーバーに挿入するためのツールとして使用し、それにより遺伝子療法における白血病等の有害事象の発生を回避することができる。加えて、これらのメガヌクレアーゼ、並びにインテグラーゼ、リコンビナーゼ及びトランスポザーゼのような他の酵素は、導入遺伝子の配列に関わりなく単一の標的化構築物から出発して任意の導入遺伝子をセーフハーバーに挿入するために使用することができる。
【0022】
本発明によるエンドヌクレアーゼ及びその使用
本発明はしたがって、以下のものに関する:
導入遺伝子を個体のゲノムに挿入する際に使用される標的配列を切断することが可能なエンドヌクレアーゼであって、(i)上記ゲノムが上記標的配列を含む遺伝子座を含み、(ii)上記標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連しない、エンドヌクレアーゼ;
導入遺伝子を細胞又は組織のゲノムに挿入するための標的配列を切断することが可能なエンドヌクレアーゼのin vitro又はex vivoでの使用であって、(i)上記ゲノムが上記標的配列を含む遺伝子座を含み、(ii)上記標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連しない、in vitro又はex vivoでの使用;
導入遺伝子を個体のゲノムに挿入する方法であって、(i)標的配列を切断することが可能なエンドヌクレアーゼを準備する工程であって、上記ゲノムが上記標的配列を含む遺伝子座を含み、該標的配列が、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)から最大で200kb離れて位置する、工程と、(ii)個体を導入遺伝子及び上記エンドヌクレアーゼと接触させ、それにより上記導入遺伝子を個体のゲノムの上記遺伝子座に挿入する、工程とを含む、導入遺伝子を個体のゲノムに挿入する方法。
【0023】
本明細書中で使用される場合、「エンドヌクレアーゼ」という用語は、DNA分子又はRNA分子、好ましくはDNA分子内の核酸間の結合の加水分解(切断)を触媒することが可能な任意の野生型又は変異型の酵素を指す。本発明によるエンドヌクレアーゼは、DNA分子又はRNA分子を、その配列に関係なく切断することはなく、「標的配列」又は「標的部位」と更に称される特異的なポリヌクレオチド配列でDNA分子又はRNA分子を認識し切断する。本発明によるエンドヌクレアーゼによって認識され切断される標的配列は、本発明による標的配列と称される。
【0024】
本発明によるエンドヌクレアーゼは例えば、ホーミングエンドヌクレアーゼ(非特許文献5)、改変ジンクフィンガードメインとFokI等の制限酵素の触媒ドメインとの融合から得られるキメラジンクフィンガーヌクレアーゼ(ZFN)(非特許文献35)又は化学的エンドヌクレアーゼ(Arimondo et al. Mol Cell Biol. 2006 26:324-333、Simon et al. NAR 2008 36:3531-3538、Eisenschmidt et al. NAR 2005 33:7039-7047、Cannata et al. PNAS 2008 105:9576-9581)であり得る。化学的エンドヌクレアーゼでは、化学的切断剤(cleaver)又はペプチド切断剤を核酸のポリマー又は特異的な標的配列を認識する別のDNAのいずれかと結合させ、それにより特異的な配列に対する切断活性を標的とする。
【0025】
本発明によるエンドヌクレアーゼは好ましくは、メガヌクレアーゼの名称でも知られるホーミングエンドヌクレアーゼである。かかるホーミングエンドヌクレアーゼは、当該技術分野で既知である(例えば、Stoddard, Quarterly Reviews of Biophysics, 2006, 38:49-95を参照されたい)。ホーミングエンドヌクレアーゼはDNA標的配列を認識し、一本鎖又は二本鎖の切断を生じる。ホーミングエンドヌクレアーゼは高度に特異的であり、12塩基対(bp)〜45塩基対の範囲の長さ、通常は14bp〜40bpの範囲の長さのDNA標的部位を認識する。本発明によるホーミングエンドヌクレアーゼは、例えばLAGLIDADGエンドヌクレアーゼ、HNHエンドヌクレアーゼ又はGIY−YIGエンドヌクレアーゼに相当し得る。かかるエンドヌクレアーゼの例としては、I−Sce I、I−Chu I、I−Cre I、I−Csm I、PI−Sce I、PI−Tli I、PI−Mtu I、I−Ceu I、I−Sce II、I−Sce III、HO、PI−Civ I、PI−Ctr I、PI−Aae I、PI−Bsu I、PI−Dha I、PI−Dra I、PI−Mav I、PI−Mch I、PI−Mfu I、PI−Mfl I、PI−Mga I、PI−Mgo I、PI−Min I、PI−Mka I、PI−Mle I、PI−Mma I、PI−Msh I、PI−Msm I、PI−Mth I、PI−Mtu I、PI−Mxe I、PI−Npu I、PI−Pfu I、PI−Rma I、PI−Spb I、PI−Ssp I、PI−Fac I、PI−Mja I、PI−Pho I、PI−Tag I、PI−Thy I、PI−Tko I、PI−Tsp I、I−MsoIが挙げられる。
【0026】
好ましい実施形態では、本発明によるホーミングエンドヌクレアーゼは、I−SceI、I−CreI、I−CeuI、I−MsoI及びI−DmoI等のLAGLIDADGエンドヌクレアーゼである。
【0027】
最も好ましい実施形態では、上記LAGLIDADGエンドヌクレアーゼはI−CreIである。野生型I−CreIは、22bp〜24bpの二本鎖標的配列を切断することが可能なホモ二量体ホーミングエンドヌクレアーゼである。I−CreIの野生型単量体の配列は、配列番号1として示される配列(pdbアクセッション番号1g9yのI−CreI配列に相当する)及びSwissProtアクセッション番号P05725で示される配列(特にバージョン73で示される配列、最終更新日2009年11月3日)を含む。
【0028】
本特許出願において、I−CreI変異体は、野生型I−CreI配列の最初のメチオニンの後に更なるアラニン及びC末端(C-terminal extremity)に3つの更なるアミノ酸残基を含み得る(配列番号42の配列及び図11を参照されたい)。これら3つの更なるアミノ酸残基は、野生型I−CreI配列の最後のプロリンの後の2つの更なるアラニン残基及び1つのアスパラギン酸残基からなる。これらの更なる残基は、酵素の特性に影響を与えない。明確化のために、これらの更なる残基はI−CreI又はその変異体における残基のナンバリングに影響を与えないものとする。より具体的には、本明細書中で使用されるナンバリングは、配列番号1の野生型I−CreI酵素における残基の位置のみを指す。例えば、野生型I−CreIの2番目の残基は、この変異体が最初のメチオニンの後に更なるアラニンを含むため、実際には配列番号42の変異体の3番目の残基である。
【0029】
本出願において、I−CreI変異体は、ホモ二量体(2つの同一の単量体を含むメガヌクレアーゼ)、ヘテロ二量体(2つの同一でない単量体を含むメガヌクレアーゼ)及び一本鎖であり得る。
【0030】
本発明は、野生型(天然)エンドヌクレアーゼ及び変異型エンドヌクレアーゼの両方を包含する。好ましい実施形態では、本発明によるエンドヌクレアーゼは、「変異型」エンドヌクレアーゼ、すなわち自然界に天然に存在せず、遺伝子操作又はランダム突然変異誘発によって得られるエンドヌクレアーゼである。本発明による変異型エンドヌクレアーゼは例えば、野生型、天然のエンドヌクレアーゼのアミノ酸配列の少なくとも1つの残基を異なるアミノ酸で置換することによって得ることができる。上記置換(複数も可)は、例えば部位特異的突然変異誘発及び/又はランダム突然変異誘発によって導入することができる。本発明の枠組みにおいて、かかる変異型エンドヌクレアーゼは機能的なままである、すなわち標的配列を認識し、特異的に切断する能力を保持する。
【0031】
本発明による変異型エンドヌクレアーゼは、対応する野生型エンドヌクレアーゼの標的配列とは異なる標的配列を切断する。例えば、変異型I−CreIエンドヌクレアーゼの標的配列は、配列番号4の配列とは異なる。新規の特異性を有するかかる変異型エンドヌクレアーゼを得る方法は、当該技術分野で既知である。
【0032】
本発明は、新規の特異性を有するかかる変異型エンドヌクレアーゼを、細胞、組織又は個体のゲノムの「セーフハーバー」遺伝子座に遺伝子を挿入するために使用することができるという発見に基づく。
【0033】
本明細書中で使用される場合、「遺伝子座」という用語は、染色体上のDNA配列(例えば遺伝子)の特定の物理的な位置である。本明細書中で使用される場合、「遺伝子座」という用語は、通常は染色体上のエンドヌクレアーゼの標的配列の特定の物理的な位置を指す。本発明によるエンドヌクレアーゼによって認識され切断される標的配列を含むかかる遺伝子座は、「本発明による遺伝子座」と称される。
【0034】
理想的には、セーフハーバー遺伝子座への挿入は、他の遺伝子の発現に対して影響を有しないものとする。これらの特性の試験は多工程プロセスであり、最初に候補セーフハーバー遺伝子座のプレスクリーニングをバイオインフォマティクス手段により行うことが望ましい。このため、標的化挿入が挿入突然変異誘発をもたらす可能性が低い遺伝子座を最初に同定することができる。
【0035】
本発明による遺伝子座の主要な特徴の1つは、(i)遺伝子療法の臨床試験において患者に由来する細胞でレトロウイルス挿入が観察された領域中に位置すること、及び(ii)上記レトロウイルス挿入が、がん又は異常細胞増殖と関連しなかったことである。
【0036】
実際には、本発明によるセーフハーバー遺伝子座を同定する一方法は、先の遺伝子療法試験によって生成したデータを使用することである。X−SCID試験では、LMO2遺伝子及びCCND2遺伝子の隣のレトロウイルスベクター媒介導入遺伝子の挿入が、白血病と関連することが示されている。患者におけるベクター挿入の追跡調査から、この挿入を保有する細胞の数が数年の経過後に他の修飾細胞の数を上回ることが明らかに実証されている(非特許文献31、Deichmann et al. J. of Clin. Invest. 2007 117:2225-32、Cavazzana-Calvo et al. Blood 2007 109:4575-4581)。別の臨床試験では、幾つかの遺伝子座における挿入が、2人の患者において高い増殖速度を引き起こすことが見出された(Ott et al. Nat Med 2006 12:401-9)。これらの症例においては、増殖はMDS1−EVI1遺伝子、PRDM16遺伝子又はSETBP1遺伝子の挿入活性化の結果のようであった。悪性腫瘍は当初観察されなかったが、EVII活性化は最終的に両方の患者において骨髄異形成をもたらした(Stein et al., Nat. Med 2010 16: 198-205)。より一般的には、非発がん性であっても、挿入断片の近くにある遺伝子の活性化から生じる細胞増殖が悪性腫瘍に向かう第一段階を示し、したがって安全性に関する潜在的な問題を招く可能性がある。ウイルスベクター組込みのパターン及び形質転換細胞の運命に対するその潜在的な結果をよりよく理解するために、遺伝子療法試験の患者においてレトロウイルス挿入部位(RIS)の大規模研究が幾つか行われている(Mavilio et al., Nat Med 2006:1397-1402、Recchia et al. PNAS 2006:1457-62、Aiuti, et al. J Clin Invest 2007:2233-40、Schwarzwaelder et al. J Clin Invest 2007:2241-9、Deichmann et al. J Clin Invest 2007:2225-32)。白血病又は異常細胞増殖と関連しないRISはセーフハーバーとみなすことができる。したがって、本発明による遺伝子座は、臨床試験において同定されたRISと重複するか又はその近くにあり、それにも関わらず、がん又は異常細胞増殖と関連しないことが好ましい。
【0037】
より具体的には、本発明による遺伝子座は、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)から最大で200kb、180kb、150kb、100kb又は50kb離れて位置する標的配列を含む遺伝子座と定義される。かかる遺伝子座は、本発明による「セーフハーバー」遺伝子座(又は本発明による遺伝子座)、すなわち導入遺伝子の挿入に安全な遺伝子座と称される。
【0038】
「レトロウイルス挿入部位」(RIS)とは、レトロウイルスベクターを用いた遺伝子療法によって治療される患者に由来する細胞において上記レトロウイルスベクターの挿入部位として同定されたゲノム部位を意味する。かかるRISは当該技術分野で既知である。RISとしては、Schwarzwaelder et al.(J. Clin. Invest. 2007 117:2241)、Deichmann et al.(J. of Clin. Invest. 2007 117:2225)、Aiuti et al.(J. Clin. Invest. 2007 117:2233)、Recchia et al.(PNAS 2006 103:1457)及びMavilio et al.(Nature Medicine 12:1397, 2006)に記載されるものが挙げられるが、これらに限定されない。
【0039】
「レトロウイルスベクター」とは、レトロウイルス科のウイルスに由来する任意のベクターを意味する。
【0040】
本発明によるRISは、がんとも異常細胞増殖とも関連しない。白血病又は異常細胞増殖と関連することが知られるRISは当該技術分野で既知であり、当業者であれば容易に除外することができる。白血病又は異常細胞増殖と関連することが知られるかかるRISとしては、例えばLMO2遺伝子、CCND2遺伝子、MDS1−EVI1遺伝子、PRDM16遺伝子及びSETBP1遺伝子の隣の挿入部位が挙げられる。
【0041】
本発明によるより好ましい実施形態では、セーフハーバー遺伝子座を規定するために使用されるRISは、形質導入細胞が幹細胞である臨床試験において同定されている。RISはこのように、幹細胞の形質導入による遺伝子療法によって治療される患者に由来する細胞において同定されている可能性がある。
【0042】
本発明による別の最も好ましい実施形態では、セーフハーバー遺伝子座を規定するために使用するRISは、形質導入細胞が造血幹細胞(HSC)であるSCID患者の臨床試験において同定されている。RISはこのように、造血幹細胞の形質導入による遺伝子療法によって治療される患者に由来する細胞において同定されている可能性がある。
【0043】
さらに、本発明によるRISを既定するためのより厳しい基準を使用することができる。
【0044】
RISの中でも、共通組込み部位(CIS)は、RISの統計的過剰出現が挿入時の細胞の高い増殖速度の結果であると解釈され得る遺伝子座である(Mikkers et al., 2003, Nat. Genet. 32:153、Lund et al., 2002, Nat. Genet 32 :160、Hemati et al. 2004, PLOS Biol. 2:e423、Suzuki et al., 2002, Nat. Genet. 32:166-174、Deichman et al. J. of Clin. Invest. 2007 117:2225-32)。例えば、Deichman et al.(J. of Clin. Invest. 2007 117:2225-32)は、遺伝子療法によって治療した9人のX−SCID患者に由来するRISに関する調査を行い、ヒトゲノムに明白にマップすることができる572個の独自のRISを見出した。それらの中でも、Deichman et al.は2次、3次、4次、5次及びより高次のCISを規定した。2次のCISは、30kb距離内での2個のレトロウイルス挿入の発生によって規定され、3次、4次及び5次のCISは、それぞれ50kb、100kb又は200kb内での3つ、4つ又は5つの挿入の発生によって規定された。RISのランダム分布で期待される値の33倍である、122個のRISが47個の異なるCIS遺伝子座において発見された。11個のCISが、ZNF217、VAV−3、CCND2、LMO2、MDS1、BCL2L1、NOTCH2、SOCS2、RUNX1、RUNX3及びSEPT6を含むがん原遺伝子の隣に局在することが見出された。
【0045】
最大の安全性を確保するために、CIS内に位置するRISを回避することが好ましい場合がある。したがって、本発明による好ましい実施形態では、本発明による標的配列はCISに位置しない。加えて、上記標的配列又は遺伝子座は、共通組込み部位(CIS)の一部であるRISから少なくとも50kb、100kb又は200kb離れて位置するのが好ましい。
【0046】
「共通組込み部位」(CIS)とは、臨床試験において同定されたRISが大きな比率を占める(挿入のランダム分布を仮定する)30kb、50kb、100kb又は200kbのゲノム領域を意味する。かかるCISは当該技術分野で既知であり、Schwarzwaelder et al.(J. Clin. Invest. 2007 117:2241)、Deichmann et al.(J. of Clin. Invest. 2007 117:2225)、Aiuti et al.(J. Clin. Invest. 2007 117:2233)、Recchia et al.(PNAS 2006 103:1457)、Mavilio et al.(Nature Medicine 12:1397, 2006)及びGabriel et al.(Nat. Med. 2009 15(12):143)に記載されている。
【0047】
RISの近くであることに加えて、本発明による遺伝子座への標的化組込みは、標的化細胞における必須機能の破壊をもたらさないものとする。
【0048】
したがって、本発明による具体的な実施形態では、本発明による遺伝子座への挿入は、好ましくは標的配列の近傍に位置する遺伝子、例えば最近接遺伝子の発現を実質的に変更しない。
【0049】
加えて、別の具体的な実施形態では、上記遺伝子座への遺伝要素の挿入は、好ましくは上記細胞、組織又は個体の表現型(遺伝要素の発現による表現型を除く)を実質的に変更しない。「表現型」とは、細胞、組織又は個体の目に見える形質を意味する。表現型としては、例えば生存能力、細胞増殖及び/又は成長速度が挙げられる。当業者であれば、例えば隣接遺伝子の発現パターンを分析すること、トランスクリプトームのマイクロアレイ研究を行うこと、並びに/又は増殖及び/若しくは(もしあれば)分化異常を特性評価することによって、遺伝子座が本発明によるセーフハーバー遺伝子座であるかを容易に検証することができる。
【0050】
また別の具体的な実施形態では、本発明による遺伝子座は任意の遺伝子を含まない。任意の遺伝子を含まない遺伝子座とは、任意の参照された又は既知の遺伝子を含まない遺伝子座を指す。言い換えると、かかる遺伝子座は、米国国立バイオテクノロジー情報センター(NCBI)のウェブサイト上で入手可能なもののような配列データベースによる任意の既知の遺伝子を含まない。したがって、本発明による標的配列及び/又は本発明による遺伝子座は、最近接遺伝子から少なくとも1kb、5kb、10kb、25kb、50kb、100kb、180kb、200kb、250kb、300kb、400kb又は500kb離れて有利に位置することができる。
【0051】
「遺伝子」とは、特定のタンパク質又はタンパク質の一部をコードする、染色体に沿って線状に配列したDNAの一部からなる遺伝の基本単位を意味する。遺伝子は通常、プロモーター、5’非翻訳領域、1つ又は複数のコード配列(エクソン)、任意にイントロン、3’非翻訳領域を含む。遺伝子はターミネーター、エンハンサー及び/又はサイレンサーを更に含み得る。
【0052】
「最近接遺伝子」とは、それぞれ標的配列の最も近くに、標的配列に対してセントロメリックに(centromeric)及びテロメリックに(telomeric)位置する1つ、2つ又は3つの遺伝子を意味する。
【0053】
好ましい実施形態では、本発明による遺伝子座は、導入遺伝子の安定な発現を更に可能にする。
【0054】
別の好ましい実施形態では、本発明による標的配列は、上記細胞、組織又は個体のゲノム内に1回しか存在しない。
【0055】
本発明によるかかるセーフハーバー遺伝子座を選択した後、(i)例えば実施例1、実施例2及び実施例5に記載のように、上記遺伝子座内に位置する標的配列を特異的に認識し切断する変異型エンドヌクレアーゼを構築するか、又は(ii)既知の野生型エンドヌクレアーゼが、上記遺伝子座内に位置する標的配列を切断することが可能か否かを決定することができる。代替的には、本発明によるセーフハーバー遺伝子座を選択した後、当業者は、既知の野生型エンドヌクレアーゼ又は変異型エンドヌクレアーゼによって認識され切断される標的配列をそれに挿入することができる。
【0056】
したがって、本発明は、個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法であって、
a)上記個体のゲノム内で、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)を選択及び/又は同定する工程と、
b)上記RISの200kb上流及び200kb下流に伸びるゲノム領域を規定する工程と、
c)上記ゲノム領域内に位置する標的配列を切断することが可能な野生型エンドヌクレアーゼを同定するか、又は変異型エンドヌクレアーゼを構築する工程と、
を含む、個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法に関する。
【0057】
かかるエンドヌクレアーゼは、例えば(i)最近接遺伝子の発現、並びに/又は(ii)細胞、組織若しくは個体の細胞増殖及び/若しくは成長速度を実質的に変更することなく、細胞、組織又は個体のゲノムに導入遺伝子を安全に挿入することを可能にする。
【0058】
本発明による遺伝子座に関連して上で提示する全ての基準は、当然ながら上記の方法を行う場合に適用することができる。例えば、CISの一部であるRISを除外してもよく、並びに/又は工程(b)で規定されるゲノム領域が上記RISの50kb上流及び50kb下流にしか伸びなくてもよく、並びに/又は標的配列を含む遺伝子座が任意の遺伝子を含まなくてもよい。
【0059】
本発明による遺伝子座は例えば、下記表A〜表Cに記載のSH3遺伝子座、SH4遺伝子座、SH6遺伝子座、SH12遺伝子座、SH13遺伝子座、SH19遺伝子座、SH20遺伝子座、SH21遺伝子座、SH33遺伝子座、SH7遺伝子座又はSH8遺伝子座のいずれか1つに相当し得る。
【0060】
表Aは、ヒトゲノム内の遺伝子座の位置、遺伝子座内に含まれる標的配列、最も近いRISの位置及びRISを記載する刊行物への参照、及び遺伝子座を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0061】
表Bは、本発明による遺伝子座のすぐ上流(5’側)及び下流(3’側)に位置する最近接遺伝子に関する情報を提示する。距離とは、標的配列と遺伝子の最も近いコード配列との距離を示す。
【0062】
表C及び表Dは、表Bと同様であるが、第2の最近接遺伝子及び第3の最近接遺伝子についての情報をそれぞれ提示する。
【0063】
表A’、表B’、表C’及び表D’は、一部の遺伝子座、すなわちSH3、SH4、SH6、SH8及びSH19、並びに関連するこれらの遺伝子座内の標的配列の例についての表A、表B、表C及び表Dと同様の最新情報をそれぞれ提示する。最新の局在情報は、ヒトゲノムアセンブリのGRCh37/hg19バージョンを参照することによって与えられる。
【0064】
本発明による遺伝子座は、下記表A’’〜表D’’に記載のSH18、SH31、SH38、SH39、SH41、SH42、SH43、SH44、SH45、SH46、SH47、SH48、SH49、SH50、SH51、SH52、SH70、SH71、SH72、SH73、SH74及びSH75のいずれか1つにも相当し得る。
【0065】
表A’’は、ヒトゲノム内の遺伝子座の位置、遺伝子座内に含まれる標的配列、最も近いRISの位置及びRISを記載する刊行物への参照、上記標的と最も近いRISとの距離、及び遺伝子座を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0066】
表B’’は、本発明による遺伝子座のすぐ上流(5’側)及び下流(3’側)に位置する最近接遺伝子に関する情報を提示する。距離とは、標的配列と遺伝子の最も近いコード配列との距離を示す。
【0067】
表C’’及び表D’’は、表B’’と同様であるが、第2の最近接遺伝子及び第3の最近接遺伝子についての情報をそれぞれ提示する。
【0068】
遺伝子座の位置、この遺伝子座における標的及び遺伝子は、ヒトゲノムアセンブリのGRCh37/hg19バージョンに従って与えられる。
【0069】
【表1】
【0070】
【表2】
【0071】
【表3−1】
【表3−2】
【0072】
【表4】
【0073】
【表5】
【0074】
【表6】
【0075】
【表7】
【0076】
【表8】
【0077】
【表9】
【0078】
【表10】
【0079】
【表11】
【0080】
【表12】
【0081】
本発明による遺伝子座は、下記表E及び表Fに記載のSH101、SH106、SH107、SH102、SH105、SH103、SH104、SH113、SH109、SH112、SH108、SH110、SH114、SH116、SH111、SH115、SH121、SH120、SH122、SH117、SH118、SH119、SH123、SH126、SH128、SH129、SH124、SH131、SH125、SH127及びSH130のいずれか1つにも相当し得る。
【0082】
表Eは、ヒトゲノム内の遺伝子座の位置、遺伝子座内に含まれる標的配列、最も近いRISの位置及びRISを記載する刊行物への参照、上記標的と最も近いRISとの距離、及び遺伝子座を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0083】
表Fは、本発明による遺伝子座のすぐ上流(5’側)及び下流(3’側)に位置する最近接遺伝子に関する情報を提示する。距離とは、標的配列と遺伝子の最も近いコード配列との距離を示す。
【0084】
表E及び表Fにおいて、遺伝子座の位置、この遺伝子座における標的及び遺伝子は、ヒトゲノムアセンブリのGRCh36.3/hg19バージョンに従って与えられる。
【0085】
【表13−1】
【表13−2】
【0086】
【表14】
【0087】
本発明による遺伝子座は、下記表Gに記載のSH125、SH127、SH130、SH102、SH105、SH103、SH104、SH117、SH118、SH119及びSH123のいずれか1つにも相当し得る。
【0088】
表Gは、言及した遺伝子のイントロン内に位置する標的配列の例及び該イントロン遺伝子座を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0089】
【表15】
【0090】
本発明による遺伝子座は、下記表Hに記載のSH11、SH12、SH13、SH17、SH19、SH20、SH21、SH23、SH33、SH34、SH40、SH53、SH54、SH55、SH56、SH57、SH58、SH59、SH60、SH61、SH62、SH65、SH67、SH68及びSH69のいずれか1つも含有し得る。
【0091】
表Hは、これらの遺伝子座内に含まれる標的配列及びこれらの標的配列を切断する本発明によるエンドヌクレアーゼの例を提示する。
【0092】
【表16】
【0093】
具体的な実施形態では、本発明による遺伝子座はSH3遺伝子座である。「SH3遺伝子座」という用語は、リンパ球抗原86をコードする遺伝子に対して約120kbセントロメリックに位置するヒト染色体6の領域(例えば、染色体6の6430K〜7230K領域を示す、ワールドワイドウェブサイトncbi.nlm.nih.gov/projects/mapview/maps.cgi?TAXID=9606&CHR=6&MAPS=ideogr%2Ccntg-r%2CugHs%2Cgenes&BEG=6432845&END=7232845&thmb=onを参照されたい)及び他の種における相同領域を指す。より正確には、SH3遺伝子座は、NC_000006.11に示される配列の6850510位から6853677位に伸びる。これは配列番号54の配列を含む。
【0094】
別の具体的な実施形態では、本発明による遺伝子座はSH4遺伝子座である。SH4遺伝子座は、本明細書ではMyoDファミリー阻害剤ドメイン含有遺伝子座(MDFIC)に対して約320kbテロメリックに位置するヒト染色体7の領域として、又は別の種における相同領域として規定される(例えば、染色体7の114660K〜115660K領域を示す、ワールドワイドウェブサイトncbi.nlm.nih.gov/projects/mapview/maps.cgi?TAXID=9606&CHR=7&MAPS=ideogr,cntg-r,ugHs,genes[113908811.00%3A114908811.00]&CMD=DNを参照されたい)。より正確には、SH4遺伝子座は、NC_000007.13に示される配列の114972751位から114976380位に伸びる。これは配列番号55の配列を含む。
【0095】
本明細書中で使用される場合、「導入遺伝子」という用語は、ポリペプチドをコードする配列を指す。好ましくは、導入遺伝子によってコードされるポリペプチドは、導入遺伝子が挿入される細胞、組織又は個体において発現されないか、又は発現されるが生物学的に活性ではない。最も好ましくは、導入遺伝子は、個体の治療に有用な治療用ポリペプチドをコードする。
【0096】
本発明の枠組みにおいて、個体はヒト又は非ヒト動物であり得る。個体は好ましくはヒトである。代替的に、個体は非ヒト動物、好ましくは脊椎動物、及び/又は例えばマウス、ラット、ウサギ、チャイニーズハムスター、モルモット若しくはサル等の哺乳動物であってもよい。本発明による細胞及び組織は好ましくは、かかるヒト又は非ヒト動物に由来する。
【0097】
I−CreIに由来する本発明によるエンドヌクレアーゼ
本発明による変異型エンドヌクレアーゼは、例えば、
各々が配列番号1の配列若しくはSwissProtアクセッション番号P05725に示される配列を含む2つの単量体を含むホモ二量体タンパク質である野生型I−CreIメガヌクレアーゼ、又は
各々が配列番号42の配列を含む2つの単量体を含むI−CreIメガヌクレアーゼ
のいずれかに由来し得る。野生型標的配列を認識するかかるI−CreIメガヌクレアーゼは、新規の特異性を有するエンドヌクレアーゼの改変に好適であることが示されている。
【0098】
したがって、本発明は、セーフハーバー遺伝子座内に位置する標的配列を切断することが可能である、各々が配列番号1又は配列番号42と少なくとも80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%又は99%同一な配列を含むか又はそれらからなる2つの単量体を含むか又はそれらからなる二量体I−CreIタンパク質に関する。
【0099】
好ましくは、標的配列は配列番号4の配列を含みも、それらからなりもしない。
【0100】
最も好ましくは、本発明による二量体I−CreIタンパク質はヘテロ二量体タンパク質である。
【0101】
本発明のクエリー配列と少なくとも例えば95%「同一な」配列を有するタンパク質とは、タンパク質の配列が、クエリー配列の100個のアミノ酸当たり最大で5個のヌクレオチド突然変異を含み得る以外はクエリー配列と同一であることを意図する。言い換えると、クエリー配列と少なくとも95%同一な配列を有するタンパク質を得るには、最大で配列の5%(100個のうち5個)のアミノ酸が挿入されているか、欠失しているか、又は別のヌクレオチドで置き換えられていてもよい。2つの配列の最適アラインメント(ギャップを含む)をそれらの全長を考慮して見出すために、Needleman−Wunschグローバルアライメントアルゴリズム(Needleman and Wunsch, 1970 J. Mol. Biol. 48:443-453)を使用する「needle」プログラムを例えば使用することができる。needleプログラムは、例えばebi.ac.ukワールドワイドウェブサイト上で入手可能である。本発明に従う同一性のパーセンテージはこのように、「Gap Open」パラメータが10.0に等しく、「Gap Extend」パラメータが0.5に等しいEMBOSS::needle(グローバル)プログラム及びBlosum62マトリックスを用いて算出することができる。
【0102】
本発明による二量体I−CreIタンパク質の各々の単量体は例えば、野生型単量体の配列(配列番号1)又は配列番号42の単量体の配列と比較して少なくとも、多くとも又は約2個、5個、8個、10個、12個、15個、18個、20個又は25個の突然変異を含み得る。言い換えると、本発明による単量体は、配列番号1又は配列番号42と少なくとも、多くとも又は約2個、5個、8個、10個、12個、15個、18個、20個、25個又は30個の突然変異によって異なる配列を含む。
【0103】
本発明の枠組みにおいて、突然変異は、好ましくは1つのアミノ酸の別のアミノ酸による置換に相当する。したがって、本発明による好ましい実施形態は、配列番号1又は配列番号42とアミノ酸置換の存在によってのみ異なる、配列番号1又は配列番号42と少なくとも80%同一な配列を含む2つの単量体を含むか又はそれらからなる二量体I−CreIタンパク質に関する。
【0104】
本発明による二量体I−CreIタンパク質の単量体は、好ましくは配列番号42の配列を含むか又はそれからなる単量体に由来する。
【0105】
突然変異は、好ましくは標的配列の認識に関与するI−CreI配列の位置に位置する。実際に、かかる突然変異の導入は、新規の特異性を有するメガヌクレアーゼの設計を可能にする。
【0106】
かかる突然変異に加えて、単量体は以下のものに相当する突然変異も有し得る:
標的部位に対するタンパク質の結合特性及び/又は切断特性を改善する突然変異、例えばG19S、G19A、F54L、S79G、E80K、F87L、V105A及び/又はI132V等(例えば国際公開第2008/152524号を参照されたい);及び/又は
偏性ヘテロ二量体(例えば、国際公開第2008/093249号及びFajardo-Sanchez et al., Nucleic Acids Res. 2008 36:2163-73を参照されたい)の獲得をもたらす突然変異;及び/又は
融合タンパク質の生成に好適な突然変異、例えば融合タンパク質のC末端単量体における配列番号1の5個の主要なN末端アミノ酸残基の欠失等;及び/又は
配列番号42の単量体の場合と同様に、配列番号1の1番目及び2番目の残基の間のアラニンの挿入からなる突然変異。
【0107】
配列番号1又は配列番号42に相同な配列に加えて、本発明によるタンパク質の単量体は、配列のNH2末端及び/又はCOOH末端で付加された1つ又は複数のアミノ酸、例えばタンパク質、プロペプチド及び/又は核局在化シグナルの精製に有用なタグを含み得る。特に、本発明によるタンパク質の単量体は、配列番号42の単量体の場合と同様、配列番号1の配列のCOOH末端で付加されたAADアミノ酸を含み得る。
【0108】
本明細書において、突然変異は、配列番号1上の位置に続く、配列番号1のその位置に位置するアミノ酸を置き換えるアミノ酸の性質で示される。例えば、44A突然変異を含む単量体は、配列番号1の44位のアミノ酸(すなわちグルタミン、Q)がアラニン(A)で置き換えられたI−CreI単量体を指す。このため、この単量体は、配列番号1の野生型I−CreI単量体と、少なくともアミノ酸置換Q44Aによって異なる。上で説明したように、配列番号42のI−CreI単量体は、配列番号1のI−CreI単量体と比較して幾つかの更なるアミノ酸残基を含む(図11を参照されたい)。したがって、配列番号42上では、44A突然変異はアラニンによる配列番号42の45位のグルタミンの置き換えに相当する。
【0109】
説明のために、44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体は例えば、配列番号57の配列(この単量体が配列番号1のI−CreI単量体に直接由来する場合)又は配列番号58の配列(この単量体が配列番号42のI−CreI単量体に直接由来する場合)を有し得る。図12は2つのかかる単量体間のアラインメントを示し、これらの単量体上の44A 54L 64A 70Q 75N 158R及び162A突然変異の位置を示す。
【0110】
SH3遺伝子座、SH4遺伝子座又はSH6遺伝子座に位置する標的配列を切断することが可能な本発明による二量体I−CreIタンパク質の例を、下記に更に記載する。
【0111】
SH3遺伝子座を切断することが可能な本発明による二量体I−CreIタンパク質
好ましい実施形態では、標的配列はSH3遺伝子座(上で規定される)内に位置する。SH3内に位置する標的配列は例えば、配列番号2又は配列番号2のヌクレオチド2〜23を含むか又はそれらからなっていてもよい。実施例1は、かかる標的配列を切断することが可能な本発明によるヘテロ二量体I−CreIタンパク質の幾つかの例を開示する。加えて、他のかかるタンパク質を構築する方法は当該技術分野で既知であり、例えばPCT出願の国際公開第2006/097784号、国際公開第2006/097853号及び国際公開第2009019614号、並びに非特許文献6に記載されているものが挙げられる。
【0112】
かかる二量体タンパク質の単量体は好ましくは、配列番号1の4位、24位、26位、28位、30位、32位、33位、38位、44位、50位、54位、57位、64位、66位、70位、71位、75位、77位、81位、86位、92位、105位、142位、151位、154位、158位及び162位、好ましくは配列番号1の4位、30位、38位、44位、50位、54位、57位、64位、66位、70位、71位、75位、77位、81位、86位、92位、105位、142位、151位、154位、158位及び162位からなる群から選択される位置に位置する少なくとも1つ、好ましくは少なくとも3つ、4つ、5つ又は6つのアミノ酸置換を含む。該置換は、例えば以下の置換から選択され得る:4E、30G、38R、44A、50R、54L、57E、64A、66C、70Q、70D、71R、75N、75Y、77V、81T、86D、92R、105A、142R、151A、154G、158R、158W及び162A。二量体タンパク質は任意に1位に突然変異を含んでいてもよいが、かかる突然変異は切断活性又は切断特異性に影響を有しない。
【0113】
かかる二量体I−CreIタンパク質は、例えば以下のものを含むか又はそれらからなっていてもよい:
配列番号1と比較して、配列番号1の30位、38位、50位、70位、75位、81位、86位、142位及び154位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第1の単量体。好ましくは、該第1の単量体は、配列番号1の30位、38位、70位及び75位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:30G、38R、50R、70D、75N、81T、86D、142R及び154G。かかる単量体は例えば、配列番号1と比較して少なくとも4個、5個若しくは6個の突然変異、及び/又は配列番号1と比較して最大で4個、5個、6個、8個、10個、12個若しくは15個のアミノ酸突然変異を含み得る;及び
配列番号1と比較して、配列番号1の4位、44位、54位、57位、64位、66位、70位、71位、75位、77位、92位、105位、151位、158位及び162位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第2の単量体。好ましくは、該第2の単量体は、配列番号1の44位、54位、70位及び75位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:4E、44A、54L、57E、64A、66C、70Q、71R、75N、75Y、77V、92R、105A、151A 158R、158W及び162A。かかる単量体は例えば、配列番号1と比較して少なくとも4個、5個若しくは6個の突然変異、及び/又は配列番号1と比較して最大で4個、6個、8個、10個、12個又は15個のアミノ酸突然変異を含み得る。
【0114】
具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)30G 38R 70D 75N 86D突然変異を含む第1の単量体、
b)
i.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
ii.44A 54L 70Q 75Y 92R 158R 162A突然変異を含む単量体、
iii.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iv.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
v.44A 54L 70Q 75N突然変異を含む単量体、
vi.44A 54L 57E 70Q 75N 158R 162A突然変異を含む単量体、及び
vii.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含むか又はそれらからなる。
【0115】
別の具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)30G 38R 70D 75N 81T 154G突然変異を含む第1の単量体、
b)
i.44A 54L 70Q 75N 105A 158R 162A突然変異を含む単量体、
ii.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iii.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iv.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
v.44A 54L 70Q 75N突然変異を含む単量体、及び
vi.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含むか又はそれらからなる。
【0116】
更に別の具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)30G 38R 50R 70D 75N 142R突然変異を含む第1の単量体、
b)
i.44A 54L 70Q 75N 105A 158R 162A突然変異を含む単量体、
ii.44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
iii.44A 54L 70Q 75Y 92R 158R 162A突然変異を含む単量体、
iv.4E 44A 54L 64A 70Q 75N 158R 162A突然変異を含む単量体、
v.44A 54L 64A 70Q 75N 158W 162A突然変異を含む単量体、
vi.44A 54L 66C 70Q 71R 75N 151A 158R 162A突然変異を含む単量体、
vii.44A 54L 70Q 75N突然変異を含む単量体、
viii.44A 54L 57E 70Q 75N 158R 162A突然変異を含む単量体、及び
ix.44V 54L 70Q 75N 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含むか又はそれらからなる。
【0117】
二量体I−CreIタンパク質の単量体は、例えば偏性ヘテロ二量体の獲得を可能にする更なる突然変異も含み得る。かかる突然変異は当業者に既知であり、Fajardo-Sanchez et al.(Nucleic Acids Res. 2008 36:2163-73)に記載されるものが挙げられる。
【0118】
具体的な実施形態では、上記の単量体は配列番号42の単量体に直接由来し、配列番号42の配列と指定の突然変異の存在によってのみ異なる。
【0119】
SH4遺伝子座を切断することが可能な本発明による二量体I−CreIタンパク質
好ましい実施形態では、標的配列はSH4遺伝子座(上で規定される)内に位置する。SH4内に位置する標的配列は例えば、配列番号3又は配列番号3のヌクレオチド2〜23を含むか又はそれらからなっていてもよい。実施例2は、かかる標的配列を切断することが可能な本発明による二量体I−CreIタンパク質の幾つかの例を開示する。
【0120】
かかる二量体タンパク質の単量体は好ましくは、配列番号1の24位、44位、68位、70位、75位及び77位からなる群から選択される位置に位置する少なくとも1つ、好ましくは少なくとも3つ、4つ、5つ又は6つのアミノ酸置換を含む。該置換は、例えば以下の置換から選択され得る:24V、44R、44Y、68Y、68A、70S、70D、75Y、75N、77R、77N及び77V。
【0121】
かかる二量体I−CreIタンパク質は、例えば以下のものを含むか又はそれらからなっていてもよい:
配列番号1と比較して、配列番号1の24位、44位、68位、70位、75位及び77位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第1の単量体。好ましくは、第1の単量体は、配列番号1の24位、70位、75位及び77位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:24V、44R、68Y、68A、70D、70S、75Y、75N、77N及び77R。かかる単量体は例えば、配列番号1と比較して少なくとも4個、5個若しくは6個の突然変異、及び/又は配列番号1と比較して最大で4個、5個、6個、8個、10個、12個若しくは15個のアミノ酸突然変異を含み得る;及び
配列番号1と比較して、配列番号1の24位、44位、70位及び77位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第2の単量体。好ましくは、第2の単量体は配列番号1の24位、44位及び70位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:24V、44Y、70S及び77V。かかる単量体は例えば、配列番号1と比較して少なくとも3個若しくは4個の突然変異、及び/又は配列番号1と比較して最大で3個、4個、6個、8個、10個、12個若しくは15個のアミノ酸突然変異を含み得る。
【0122】
具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)
i.24V 44R 68Y 70S 75Y 77N突然変異を含む単量体、
ii.24V 68A 70S 75N 77R突然変異を含む単量体、及び
iii.24V 70D 75N 77R突然変異を含む単量体からなる群から選択される第1の単量体、
b)
i.24V 44Y 70S突然変異を含む単量体、及び
ii.24V 44Y 70S 77V突然変異を含む単量体からなる群から選択される第2の単量体、
を含むか又はそれらからなる。
【0123】
二量体I−CreIタンパク質の単量体は、例えば偏性ヘテロ二量体の獲得を可能にする更なる突然変異も含み得る。かかる突然変異は当業者に既知であり、Fajardo-Sanchez et al.(Nucleic Acids Res. 2008 36:2163-73)に記載されるものが挙げられる。
【0124】
具体的な実施形態では、上記の単量体は配列番号42の単量体に直接由来し、配列番号42の配列と指定の突然変異の存在によってのみ異なる。
【0125】
SH6遺伝子座を切断することが可能な本発明による二量体I−CreIタンパク質
好ましい実施形態では、標的配列はSH6遺伝子座(上で規定される)内に位置する。SH6内に位置する標的配列は例えば、配列番号59又は配列番号59のヌクレオチド2〜23を含むか又はそれらからなっていてもよい。実施例5は、かかる標的配列を切断することが可能な本発明による二量体I−CreIタンパク質の幾つかの例を開示する。
【0126】
かかる二量体タンパク質の単量体は好ましくは、配列番号1の7位、24位、27位、28位、34位、40位、44位、68位、70位、75位、77位、81位、85位、96位、99位、103位、108位、111位、121位、132位、144位及び160位からなる群から選択される位置に位置する少なくとも1つ、好ましくは少なくとも3つ、4つ、5つ又は6つのアミノ酸置換を含む。該置換は、例えば以下の置換から選択され得る:7R、24F、27V、28Q、34R、40R、44A、44K、68T、70L、70G、70S、75N、77V、81T、81V、85R、96R、99R、103T、103S、108V、111H、121E、132V、144S、160R及び160E。
【0127】
かかる二量体I−CreIタンパク質は、例えば以下のものを含むか又はそれらからなっていてもよい:
配列番号1と比較して、配列番号1の7位、24位、27位、28位、34位、40位、44位、70位、75位、77位、81位、85位、96位、99位、103位、108位、111位、121位、132位、144位及び160位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第1の単量体。好ましくは、第1の単量体は、配列番号1の28位、40位、44位、70位及び75位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:7R、24F、27V、28Q、34R、40R、44A、70L、75N、77V、81T、81V、85R、96R、99R、103T、103S、108V、111H、121E、132V、144S、160R及び(and)160E。かかる単量体は例えば、配列番号1と比較して少なくとも5個若しくは6個の突然変異、及び/又は配列番号1と比較して最大で5個、6個、8個、10個、12個、15個若しくは20個のアミノ酸突然変異を含み得る;及び
配列番号1と比較して、配列番号1の44位、68位、70位及び75位からなる群から選択される位置に位置する少なくとも1つのアミノ酸置換を含む第2の単量体。好ましくは、第2の単量体は配列番号1の44位、70位及び75位に置換を含む。最も好ましくは、該置換は以下の置換から選択される:44K、68T、70G、70S及び75N。かかる単量体は例えば、配列番号1と比較して少なくとも3個若しくは4個の突然変異、及び/又は配列番号1と比較して最大で3個、4個、6個、8個、10個、12個若しくは15個のアミノ酸突然変異を含み得る。
【0128】
具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)44K 68T 70G 75N突然変異を含む第1の単量体と、
b)
i.28Q 40R 44A 70L 75N 96R 111H 144S突然変異を含む単量体、
ii.7R 28Q 40R 44A 70L 75N 85R 103T突然変異を含む単量体、
iii.28Q 40R 44A 70L 75N 103S突然変異を含む単量体、
iv.24F 27V 28Q 40R 44A 70L 75N 99R突然変異を含む単量体、
v.7R 28Q 40R 44A 70L 75N 81T突然変異を含む単量体、
vi.7R 28Q 40R 44A 70L 75N 77V突然変異を含む単量体、
vii.7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R突然変異を含む単量体、
viii.28Q 40R 44A 70L 75N突然変異を含む単量体、
ix.7R 28Q 40R 44A 70L 75N 103T突然変異を含む単量体、及び
x.28Q 34R 40R 44A 70L 75N 81V 103T 108V 160E突然変異を含む単量体からなる群から選択される第2の単量体と、
を含むか又はそれらからなる。
【0129】
別の具体的な実施形態では、本発明による二量体I−CreIタンパク質は、
a)44K 70S 75N突然変異を含む第1の単量体と、
b)
i.28Q 40R 44A 70L 75N 96R 111H 144S突然変異を含む単量体、
ii.7R 28Q 40R 44A 70L 75N 85R 103T突然変異を含む単量体、
iii.28Q 40R 44A 70L 75N 103S突然変異を含む単量体、
iv.24F 27V 28Q 40R 44A 70L 75N 99R突然変異を含む単量体、
v.7R 28Q 40R 44A 70L 75N 81T突然変異を含む単量体、
vi.7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R突然変異を含む単量体、
vii.7R 28Q 40R 44A 70L 75N 103T突然変異を含む単量体、及び
viii.28Q 34R 40R 44A 70L 75N 81V 103T 108V 160E突然変異を含む単量体からなる群から選択される第2の単量体と、
を含むか又はそれらからなる。
【0130】
二量体I−CreIタンパク質の単量体は、例えば偏性ヘテロ二量体の獲得を可能にする更なる突然変異も含み得る。かかる突然変異は当業者に既知であり、Fajardo-Sanchez et al.(Nucleic Acids Res. 2008 36:2163-73)に記載されるものが挙げられる。
【0131】
具体的な実施形態では、上記の単量体は配列番号42の単量体に直接由来し、配列番号42の配列と指定の突然変異の存在によってのみ異なる。
【0132】
本発明による融合タンパク質
融合した二量体I−CreIタンパク質の2つの単量体を含み、親二量体I−CreIタンパク質の生物学的活性を保持する融合タンパク質を構築することができる(非特許文献9、Li et al. Nucleic Acids Res. 2009 37:1650-62、Epinat et al. Nucleic Acids Res. 2003 31:2952-62)。かかる融合タンパク質は、一般に「一本鎖メガヌクレアーゼ」と称される。
【0133】
したがって、本発明は、上で規定される二量体I−CreIタンパク質の2つの単量体又はかかる単量体の生物学的に活性な断片を含む融合タンパク質に更に関する。かかる融合タンパク質では、上で規定される二量体I−CreIタンパク質の第1の単量体及び第2の単量体は融合し、任意にペプチドリンカー等のリンカーによって互いに連結される。リンカーは例えば、配列番号43又は配列番号326を含むか又はそれらからなっていてもよい。
【0134】
本発明の範囲において、本発明に従うかかる融合タンパク質は、本発明による標的配列を切断することが可能であること、すなわち、それが由来する二量体I−CreIタンパク質と同じ標的配列を切断することが可能であることが理解される。本発明の一本鎖メガヌクレアーゼは、一本鎖偏性ヘテロ二量体メガヌクレアーゼ変異体が得られるように、上に記載のような偏性ヘテロ二量体突然変異を更に含む。
【0135】
最初のバージョンのI−CreI一本鎖(Epinat et al. NAR 2003 3:2952-2962、国際公開第03/078619号)では、一本鎖メガヌクレアーゼのN末端単量体は本質的にI−CreIアミノ酸配列の1位〜93位からなっていたが、C末端(I−CreIアミノ酸配列の8位〜163位)は、ほぼ完全なI−CreI単量体であった。ごく最近では、I−CreIホモ二量体メガヌクレアーゼに由来する一本鎖分子を設計する新たな方法は、2つのほぼ完全なC末端I−CreI単量体及びN末端I−CreI単量体であった(例えば国際公開第2009/095793号を参照されたい)。この設計はオフサイト切断及び毒性を大幅に減少させ、一方で有効性を向上させる。この一本鎖分子の構造及び安定性は二量体変異体のものとほぼ同じであり、この分子は溶液中で単量体状であるようである。あらゆる点で、この一本鎖分子は、特異性に関して至適基準(gold standard)であると考えられるI−SceIと同様に振る舞う。これらの特性から、この新たなメガヌクレアーゼの生成はゲノム手術戦略に利用可能な最良な分子バサミの一つとなり、単一遺伝子疾患、例えば重症複合免疫不全(SCID)の遺伝子修正療法を容易にする一方で、これまでの遺伝子療法アプローチの有害作用を回避する可能性がある。
【0136】
上に記載の突然変異に加えて、更なる突然変異を融合タンパク質の2つの単量体の各々の配列に導入してもよい。例えば、C末端単量体はK7E突然変異及びK96E突然変異を含んでいてもよく、N末端単量体はE8K突然変異、E61R突然変異及びG19S突然変異を含んでいてもよい。
【0137】
実施例1、実施例2及び実施例5は、本発明によるかかる融合タンパク質の幾つかの例を開示する。
【0138】
具体的な実施形態では、本発明による融合タンパク質は、配列番号25〜配列番号40及び配列番号76〜配列番号96のいずれか1つと少なくとも80%、85%、90%、95%、96%、97%、98%、99%若しくは100%同一な配列又はその少なくとも50アミノ酸、100アミノ酸、150アミノ酸若しくは200アミノ酸の断片を含むか又はそれらからなる。
【0139】
本発明による核酸、ベクター及び組合せ
導入遺伝子を細胞、組織又は動物のゲノムに挿入する場合、本発明によるエンドヌクレアーゼを上記細胞、組織又は動物にタンパク質としてではなく核酸分子として導入するのが好ましい。
【0140】
したがって、本発明は、本発明によるエンドヌクレアーゼをコードする、例えば上に記載の二量体I−CreIタンパク質又は融合タンパク質をコードする核酸に関する。エンドヌクレアーゼが二量体I−CreIタンパク質である場合、上記核酸は、各々の単量体につき1つの少なくとも2つのコード配列を含む。エンドヌクレアーゼが融合タンパク質である場合、上記核酸は少なくとも1つのコード配列を含む。エンドヌクレアーゼタンパク質は、組換えタンパク質をもたらす様々な細胞透過性ペプチドと組み合わせることができ、そのような組み合わせた分子は、エンドヌクレアーゼ単独よりもはるかに高いレベルの効率で標的細胞に入ることが可能である。これらの細胞透過性ペプチドは、Diatos S.A.によって開発されている(国際公開第01/64738号、国際公開第05/016960号、国際公開第03/018636号、国際公開第05/018650号、国際公開第07/069068号)。本出願人はこれまで、エンドヌクレアーゼ細胞透過性ペプチドの組合せが標的細胞に効率的に入ることができること及び取り込まれたエンドヌクレアーゼが、DSBを生じ、次に相同組換え事象を刺激するように標的細胞ゲノムに作用することができることを示している。本出願人は、エンドヌクレアーゼの複雑な三次元構造が細胞透過性ペプチドの存在によって影響されないこと、またエンドヌクレアーゼの全ての重要な特異性も影響を受けないままであることを示している(データは示さない)。
【0141】
本発明の別の態様は、本発明によるかかる核酸を含むベクターである。「ベクター」とは、それが連結された別の核酸を輸送することが可能な核酸分子を意味する。
【0142】
本発明において使用することができるベクターとしては、染色体性核酸、非染色体性核酸、半合成核酸又は合成核酸からなり得るウイルスベクター、プラスミド及びYACが挙げられるが、これらに限定されない。好ましいベクターは、自己複製(エピソームベクター)及び/又はそれが連結された核酸の発現(発現ベクター)が可能なものである。多数の好適なベクターが当業者に既知であり、市販されている。
【0143】
好ましい実施形態では、ベクターはウイルスベクター、例えばレトロウイルス、アデノウイルス、パルボウイルス(例えばアデノ随伴ウイルス)、コロナウイルス、マイナス鎖RNAウイルス(例えば、インフルエンザウイルス等のオルトミクソウイルス、狂犬病ウイルス及び水疱性口内炎ウイルス等のラブドウイルス、麻疹ウイルス及びセンダイウイルス等のパラミクソウイルス)、ピコルナウイルス及びアルファウイルス等のプラス鎖RNAウイルス又はアデノウイルス、ヘルペスウイルス(例えば、単純ヘルペスウイルス1型及び2型、エプスタイン・バーウイルス、サイトメガロウイルス)及びポックスウイルス(例えばワクシニアウイルス、鶏痘ウイルス及びカナリア痘ウイルス)等の二本鎖DNAウイルス等に由来するウイルスベクターである。好ましいベクターとしては、レンチウイルスベクター、特に自己不活性化レンチウイルスベクターが挙げられる。
【0144】
本発明によるエンドヌクレアーゼをコードする配列に加えて、ベクターは以下のもののような要素も含み得る:
転写制御要素及び翻訳制御要素、例えばプロモーター、エンハンサー、ポリアデニル化部位、終結シグナル、イントロン等;
多重クローニング部位;
複製起点;
選択マーカー;
導入遺伝子;及び/又は
本明細書中で規定されるようなゲノム標的部位の周囲の領域と相同性を有する配列を含む標的化構築物。
【0145】
好ましい実施形態では、上記ベクターは「発現ベクター」、すなわち少なくとも1つのコード配列が転写制御要素及び翻訳制御要素に動作可能に連結したベクターである。この実施形態の範囲において、本発明によるエンドヌクレアーゼをコードする(例えば、上に記載の二量体I−CreIタンパク質又は融合タンパク質をコードする)核酸は、転写制御要素及び翻訳制御要素に動作可能に連結する。
【0146】
好ましい実施形態では、本発明によるベクターは、導入遺伝子を含む標的化構築物と、本明細書中で規定されるような標的配列(例えば配列番号2又は配列番号3の標的配列)に隣接するゲノム配列に相同な2つの配列とを含む。標的配列に隣接するゲノム配列は、好ましくは標的部位に直接隣接する。
【0147】
かかる標的化構築物は当業者に既知である。導入遺伝子の挿入については、かかる構築物は通常、導入遺伝子が挿入される標的配列に隣接する上流(5’)ゲノム配列に相同な第1の配列と、標的配列に隣接する下流(3’)ゲノム配列に相同な第2の断片とを含む。
【0148】
「相同な」とは、別の配列に対して配列間で相同組換えをもたらすのに十分な同一性を有する、より具体的には互いに少なくとも95%の同一性、好ましくは97%の同一性、より好ましくは99%の同一性を有する配列を意図する。
【0149】
好ましくは、少なくとも50bp、好ましくは100bp超、より好ましくは200bp超の相同配列が使用される。したがって、標的化DNA構築物は、好ましくは200pb〜6000pb、より好ましくは1000pb〜2000pbである。実際に、共通DNA相同性は切断部位の上流及び下流に隣接する領域に位置し、導入されるDNA配列は2つのアーム間に位置するものとする。
【0150】
標的化構築物は、2つのホモロジーアーム間に陽性選択マーカー、最終的には第1のホモロジーアームの上流又は第2のホモロジーアームの下流に陰性選択マーカーも含み得る。マーカー(複数も可)により、相同組換えによって標的部位で関心の配列が挿入された細胞の選択が可能になる。
【0151】
導入遺伝子をSH3遺伝子座又はSH4遺伝子座に挿入するのに好適な標的化構築物を構築する方法は、実施例4に与えられる。
【0152】
本発明によるエンドヌクレアーゼ及び標的化構築物をコードする核酸は、2つの別個のベクター上に位置することもできる。したがって、本発明は2つのベクター、すなわち:
本発明による発現ベクター;及び
導入遺伝子を含む標的化構築物と、本発明による標的配列のゲノム配列に相同な2つの配列とを含むベクター
の組合せにも関する。
【0153】
本発明による薬学的使用
上に記載のベクター及び組合せは、例えば薬剤として使用することができる。特に、これらのベクター及び組合せは遺伝子療法において使用することができる。
【0154】
したがって、本発明は、薬剤として使用される本発明によるベクター又は組合せに関する。かかるベクター及び組合せでは、導入遺伝子は治療用ポリペプチドをコードする。
【0155】
特に、本発明によるベクター及び組合せを用いた遺伝子療法によって治療され得る疾患としては、X−SCID、SCID、表皮水疱症、レーバー黒内障、血友病、サラセミア、ファンコニ貧血及び筋ジストロフィーが挙げられるが、これらに限定されない。
【0156】
これらの疾患では、導入遺伝子はそれぞれ以下の治療用ポリペプチドをコードする:IL2RG、Gl7A1、Rp 65、血液第VIII因子及び血液第IX因子、ヘモグロビンA及びヘモグロビンB、Fanc−A、Fanc−C(又は他のファンコニ貧血関連遺伝子)、ジストロフィン。
【0157】
本発明は、本発明によるベクター及び組合せと、薬学的に活性な担体とを含む医薬組成物に更に関する。
【0158】
本発明は、遺伝子療法によって個体を治療する方法であって、それを必要とする個体に有効量の本発明によるベクター又は組合せを投与することを含む、遺伝子療法によって個体を治療する方法にも関する。
【0159】
「有効量」とは、治療対象の個体のゲノムへの導入遺伝子の挿入を達成するのに十分な量を意味する。当業者はかかる濃度を日常的に決定することができる。
【0160】
「それを必要とする被験体」とは、導入遺伝子の挿入によって治療又は予防することができる遺伝的疾患を患っているか又はそれを患いやすい個体を意味する。本発明の範囲における治療対象の個体は、好ましくはヒトである。
【0161】
本発明による非薬学的使用
上に記載のベクター及び組合せは、遺伝子療法だけでなく、例えば動物モデルの作製及び関心のタンパク質を発現する組換え細胞株の作製等の非薬学的使用にも利用される。
【0162】
したがって、本発明は、以下のものに関する:
治療目的でない、導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入するための本発明によるエンドヌクレアーゼ、核酸、発現ベクター又は組合せの使用;
導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入する方法であって、該細胞、組織又は非ヒト動物を、本発明によるエンドヌクレアーゼ、核酸、発現ベクター又は組合せと接触させ、それにより上記導入遺伝子を上記ゲノムに挿入することを含む、導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入する方法。
【0163】
好ましい実施形態では、上記の使用又は方法は、タンパク質産生のための組換え細胞株を得るために関心のタンパク質をコードする導入遺伝子を細胞のゲノムに挿入することを目的とする。タンパク質産生のための組換え細胞株を構築するのに好適な細胞としては、ヒト(例えばPER.C6又はHEK)、チャイニーズハムスター卵巣(CHO)細胞及びマウス(NSE0)細胞が挙げられるが、これらに限定されない。
【0164】
別の好ましい実施形態では、上記の使用は、遺伝性障害の非ヒト動物モデルを作製することを目的とする。
【0165】
本発明は、そのゲノム内に本発明による核酸、発現ベクター又は組合せを含む非ヒトトランスジェニック動物にも関する。
【0166】
学術論文又は抄録、公開特許出願、交付済み特許又は任意の他の参考文献を含む本明細書中で引用される全ての参考文献は、引用される参考文献に提示される全てのデータ、表、図面及び本文を含めて参照により本明細書中に完全に援用される。
【0167】
本発明は以下の実施例及び添付の図面に鑑みて更に評価される。
【0168】
配列の簡単な説明
配列番号1は、野生型I−CreI単量体のアミノ酸配列を示す。
【0169】
配列番号2は、SH3遺伝子座内に位置する本発明による標的配列の配列を示す。
【0170】
配列番号3は、SH4遺伝子座内に位置する本発明による標的配列の配列を示す。
【0171】
配列番号4は、野生型I−CreIホモ二量体タンパク質の標的配列の配列を示す。
【0172】
配列番号5〜配列番号10は、図1に示される配列を表す。
【0173】
配列番号11〜配列番号15は、実施例1において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0174】
配列番号16〜配列番号19は、図4に示される配列を表す。
【0175】
配列番号20〜配列番号24は、実施例2において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0176】
配列番号25〜配列番号32は、それぞれSCOH−SH3−b56−A、SCOH−SH3−b56−B、SCOH−SH3−b56−C、SCOH−SH3−b56−D、SCOH−SH3−b1−A、SCOH−SH3−b1−B、SCOH−SH3−b1−C及びSCOH−SH3−b1−Dと称される、実施例1において構築される一本鎖メガヌクレアーゼを表す。
【0177】
配列番号33〜配列番号40は、それぞれSCOH−SH4−b56−A、SCOH−SH4−b56−B、SCOH−SH4−b56−C、SCOH−SH4−b56−D、SCOH−SH4−b1−A、SCOH−SH4−b1−B、SCOH−SH4−b1−C及びSCOH−SH4−b1−Dと称される、実施例2において構築される一本鎖メガヌクレアーゼを表す。
【0178】
配列番号41は、陽性対照であるSCOH−RAGを表す。
【0179】
配列番号42は、2位に更なるアラニン及び最後のプロリンの後に3つの更なる残基を有するI−CreI単量体のアミノ酸配列を示す。
【0180】
配列番号43は、RM2リンカーのアミノ酸配列を示す。
【0181】
配列番号44〜配列番号49は、実施例3において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0182】
配列番号50〜配列番号53は、実施例4において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0183】
配列番号54〜配列番号55は、それぞれSH3遺伝子座、SH4遺伝子座及びSH6遺伝子座に含まれる配列を示す。
【0184】
配列番号57は、44A 54L 64A 70Q 75N 158R 162A突然変異を含む配列番号1の単量体に由来する単量体を示す。
【0185】
配列番号58は、44A 54L 64A 70Q 75N 158R 162A突然変異を含む配列番号42の単量体に由来する単量体を示す。
【0186】
配列番号59は、SH6遺伝子座内に位置する本発明による標的配列の配列を示す。
【0187】
配列番号60〜配列番号64は、図9に示される配列を表す。
【0188】
配列番号65〜配列番号75は、実施例5において使用されるオリゴヌクレオチド、プライマー及びリンカーを表す。
【0189】
配列番号76〜配列番号85は、それぞれSCOH−SH6−b1−B、SCOH−SH6−b1−C、SCOH−SH6−b1−C、QCSH61−A01、QCSH61−E01、QCSH61−H0、QCSH62−A02、QCSH61−H01b、QCSH61−H01c及びQCSH61−H01dと称される、実施例5において構築される一本鎖メガヌクレアーゼを表す。
【0190】
配列番号86〜配列番号96は、SH7遺伝子座(配列番号86及び配列番号87)、SH8遺伝子座(配列番号88)、SH12遺伝子座(配列番号89)、SH13遺伝子座(配列番号90)、SH19遺伝子座(配列番号91)、SH20遺伝子座(配列番号92)、SH21遺伝子座(配列番号93〜配列番号95)及びSH33遺伝子座(配列番号96)を切断することが可能な一本鎖メガヌクレアーゼを表す。
【0191】
配列番号97〜配列番号104は、それぞれSH12遺伝子座、SH13遺伝子座、SH19遺伝子座、SH20遺伝子座、SH21遺伝子座、SH33遺伝子座、SH7遺伝子座及びSH8遺伝子座内に含まれる配列を表す。
【0192】
配列番号105〜配列番号325は、実施例6〜実施例9並びに/又は表A’、表A’’、表E、表G及び表Hのいずれか1つに開示される配列を表す。
【0193】
配列番号326は、BQYリンカーのアミノ酸配列を示す。
【実施例】
【0194】
以下の実施例において、全てのI−CreI変異体は、配列番号42のI−CreI単量体の遺伝子操作によって構築した。
【0195】
実施例1: SH3遺伝子座を標的とするメガヌクレアーゼの改変
SH3は、染色体6上に存在する24bpの非パリンドローム標的(配列番号2)を含む遺伝子座である。表Aに示されるように、SH3は、Deichmann et al.(J. of Clin. Invest. 2007 117:2225)に開示されるRISの近傍に位置する。SH3配列は、Deichmann et alに記載されるCISのいずれにも含まれない。
【0196】
配列番号2の標的配列を切断することが可能なI−CreIヘテロ二量体を、Chames et al.(Nucleic Acids Res., 2005, 33, e178)、非特許文献6、非特許文献8、非特許文献7に記載されるものに基づく方法を用いて同定した。次いで、これらのヘテロ二量体の一部を、CHO細胞におけるSH3切断を判定するための哺乳動物発現ベクターにクローニングした。次いで、これらの結果を利用して、配列番号2の標的配列に指向性を有する一本鎖メガヌクレアーゼを設計した。これらの一本鎖メガヌクレアーゼを哺乳動物発現ベクターにクローニングし、CHO細胞におけるSH3切断を試験した。SH3標的の強い切断活性を、これらの一本鎖分子について哺乳動物細胞において観察することができた。
【0197】
実施例1.1. SH3を切断するメガヌクレアーゼの同定
SH3標的配列を切断する可能性があるヘテロ二量体形態のI−CreI変異体を、遺伝子操作によって構築した。次いで、かかる変異体の対を酵母において共発現させた。共発現時に3つの分子種、すなわち2つのホモ二量体及び1つのヘテロ二量体が得られる。次いで、このヘテロ二量体が配列番号2のSH3標的配列を切断することが可能であるか否かを決定した。
【0198】
a)SH3標的配列に由来するパリンドローム配列を切断するI−CreIメガヌクレアーゼの変異体の構築
SH3配列は部分的に、図1に示される10AAT_P(配列番号5)、5AAG_P(配列番号6)、10AGG_P(配列番号7)及び5TTT_P(配列番号8)標的配列の組合せである。これらの配列は、国際PCT出願の国際公開第2006/097784号及び国際公開第2006/097853号、非特許文献6、並びに非特許文献8に記載のように得られるメガヌクレアーゼによって切断される。このため、SH3は、これらのこれまでに同定されたメガヌクレアーゼから生じるコンビナトリアル変異体によって切断されるはずである。
【0199】
2つのパリンドローム標的SH3.3及びSH3.4は、SH3に由来するものであった(図1)。SH3.3及びSH3.4はパリンドローム性であるため、ホモ二量体タンパク質によって切断されるはずである。したがって、配列番号9のSH3.3パリンドローム標的配列又は配列番号10のSH3.4パリンドローム標的配列のいずれかを切断するホモ二量体I−CreI変異体は、Chames et al.(Nucleic Acids Res., 2005, 33, e178)、非特許文献6、非特許文献8及び非特許文献7に記載されるものに基づく方法を用いて構築した。
【0200】
b)標的ベクターの構築
ゲートウェイクローニング配列に隣接するSH3標的配列に相当する配列番号11のオリゴヌクレオチドを、PROLIGOにオーダーした。このオリゴヌクレオチドは以下の配列を有する:TGGCATACAAGTTTCCAATACAAGGTACAAAGTCCTGACAATCGTCTGTCA)。一本鎖オリゴヌクレオチドのPCR増幅によって生成した二本鎖標的DNAを、pCLS1055酵母レポーターベクターにゲートウェイプロトコル(INVITROGEN)を用いてクローニングした。
【0201】
酵母レポーターベクターを、以下の遺伝子型を有するFYBL2−7Bサッカロミセス・セレビシエ株に形質転換した:MAT a、ura3Δ851、trp1Δ63、leu2Δ1、lys2Δ202。得られた株はレポーター株(MilleGen)に相当する。
【0202】
c)変異体の共発現
SH3.4配列又はSH3.3配列を切断する変異体をコードするオープンリーディングフレームを、それぞれpCLS542発現ベクター及びpCLS1107発現ベクターにクローニングした。これらの変異体に由来する酵母DNAを標準プロトコルを用いて抽出し、大腸菌を形質転換するために使用した。次いで、得られたプラスミドを使用して、酵母を同時形質転換した。形質転換体を、ロイシンを欠き、G418を含有する合成培地上で選択した。
【0203】
d)メガヌクレアーゼを共発現するクローンの接合及び酵母におけるスクリーニング
接合はコロニーグリッダー(QpixII、Genetix)を用いて行った。変異体をYPDプレートを覆うナイロンフィルター上に、低グリッド密度(4〜6スポット/cm2)を用いてグリッドした。第2のグリッドプロセスを同じフィルター上で行い、各々の標的について異なるレポーターを有する酵母株からなる第2の層をスポットした。メンブレンを固形寒天YPD富栄養培地上に置き、30℃で一晩インキュベートして接合させた。次に、フィルターを、ロイシン及びトリプトファンを欠き、ガラクトース(2%)を炭素源として含むG418を添加した合成培地に移し、37℃で5日間インキュベートして、発現ベクター及び標的ベクターを保有する2倍体を選択した。5日後、フィルターを、0.5Mリン酸ナトリウムバッファー(pH7.0)中の0.02%X−Gal、0.1%SDS、6%ジメチルホルムアミド(DMF)、7mM β−メルカプトエタノール、1%アガロースを含む固形アガロース培地上に置き、37℃でインキュベートして、β−ガラクトシダーゼ活性をモニタリングした。結果をスキャニングによって分析し、適切なソフトウェアを用いて定量化を行った。
【0204】
e)結果
異なる変異体の共発現は、58個の試験した組合せにおいてSH3標的の切断をもたらした。機能的組合せを下の表Iにまとめる。この表において、「+」はSH3標的配列に対する機能的な組合せ、すなわちヘテロ二量体がSH3標的配列を切断することが可能であることを示す。
【0205】
【表17】
【0206】
結論として、酵母においてSH3標的配列を切断することが可能な幾つかのヘテロ二量体I−CreI変異体が同定された。
【0207】
実施例1.2. CHO細胞における染色体外モデルでのSH3標的切断の検証
ヘテロ二量体を形成した場合に酵母においてSH3標的を効率的に切断することが可能なI−CreI変異体は、上の実施例1.1で記載している。CHO細胞においてSH3標的に対する最大切断活性を示すヘテロ二量体を同定するために、これらの変異体の一部の効率を、CHO細胞における染色体外アッセイを用いて比較した。CHO細胞におけるスクリーニングは、メガヌクレアーゼによる標的の切断がLagoZレポーター遺伝子(細菌lacZ遺伝子の誘導体)の相同組換え及び発現を誘導する一本鎖アニーリング(SSA)ベースのアッセイである。
【0208】
a)CHOスクリーニングのためのベクターにおけるSH3標的のクローニング
ゲートウェイクローニング配列に隣接するSH3標的配列に相当するオリゴヌクレオチドを、PROLIGOにオーダーした(配列番号12、TGGCATACAAGTTTCCAATACAAGGTACAAAGTCCTGACAATCGTCTGTCA)。一本鎖オリゴヌクレオチドのPCR増幅によって生成した二本鎖標的DNAを、ゲートウェイプロトコル(INVITROGEN)を用いてpCLS1058 CHOレポーターベクターにクローニングした。クローニングした標的をシークエンシング(MILLEGEN)によって検証した。
【0209】
b)メガヌクレアーゼの再クローニング
上の表Iの同定されたこれらの変異体をコードするオープンリーディングフレームを、pCLS2437発現ベクターにサブクローニングした。ORFを、酵母DNA上で配列番号13及び配列番号14(5’−AAAAAGCAGGCTGGCGCGCCTACACAGCGGCCTTGCCACCATG−3’及び5’−AGAAAGCTGGGTGCTAGCGCTCGAGTTATCAGTCGG−3’)のプライマーを用いてPCRによって増幅した。PCR産物をCHO発現ベクターpCLS2437に、内部断片置き換えのためのAscI制限酵素及びXhoI制限酵素を用いてクローニングした。ライゲーション工程及び大腸菌形質転換工程から得られる選択されたクローンを、シークエンシング(MILLEGEN)によって検証した。
【0210】
c)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を、Polyfect(商標)トランスフェクション試薬を用い、供給業者のプロトコル(QIAGEN)に従ってトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl(通常は、100mlの溶解バッファー(10mM Tris−HCl(pH7.5)、150mM NaCl、0.1%Triton X100、0.1mg/ml BSA、プロテアーゼ阻害剤)、10mlのMg 100Xバッファー(100mM MgCl2、35%β−メルカプトエタノール)、110mlのONPG(8mg/ml)及び780mlの0.1Mリン酸ナトリウム(pH7.5)を含有するバッファーを1リットル)添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。
【0211】
1回のアッセイにつき、150ngの標的ベクターを、両方の変異体それぞれ12.5ngと共トランスフェクトした。
【0212】
d)結果
表Iに記載の以下の4つの変異体を、pCLS2437に再クローニングした:
44A 54L 70Q 75Y 92R 158R 162A(SH3.3−MAと称される);
1V 44A 54L 64A 70Q 75N 158W 162A(SH3.3−MBと称される);
30G 38R 70D 75N 86D(SH3.4−M1と称される);及び
30G 38R 70D 75N 81T 154G(SH3.4−M2と称される)。
【0213】
これらのI−CreI変異体を、CHO染色体外アッセイにおいてSH3標的に対するヘテロ二量体としてともにアッセイした。
【0214】
表IIは、9つのヘテロ二量体から得られた機能的な組合せを示す。
【0215】
【表18】
【0216】
SH3配列の切断及び組換えの効率の分析から、I−CreI変異体の試験した4つの組合せが全て、更なる突然変異なしにそれらの切断活性を酵母からCHO細胞へと移すことが可能であったことが実証される。
【0217】
実施例1.3. 一本鎖としての共有結合アセンブリ及びSH3を切断するメガヌクレアーゼの改良
実施例1.1.において同定された変異体の共発現は、酵母におけるSH3標的の高い切断活性をもたらす。ヘテロ二量体の一部は哺乳動物発現系におけるSH3切断が検証されている(実施例1.2.)。表IIIに示される、そのうちの1つを更なる最適化のために選択した。
【0218】
【表19】
【0219】
MA x M1 SH3ヘテロ二量体は酵母における高い切断活性をもたらす。SH3.3−MAは、I−CreI野生型配列と比較して以下の突然変異を有するSH3.3カッターである:44A 54L 70Q 75Y 92R 158R 162A。SH3.4−M1は、I−CreI野生型配列と比較して以下の突然変異を有するSH3.4カッターである:30G 38R 70D 75N 86D。
【0220】
一本鎖構築物を配列番号15(AAGGSDKYNQALSKYNQALSKYNQALSGGGGS)のリンカーRM2を用いて改変し、それにより一本鎖分子:MA−リンカーRM2−M1を作製した。この設計工程において、G19S突然変異をC末端M1変異体に導入した。加えて、突然変異K7E、K96EをMA変異体に、突然変異E8K、E61RをM1変異体に導入して、更にSCOH−SH3−b1足場と呼ばれる一本鎖分子:MA(K7E K96E)−リンカーRM2−M1(E8K E61R G19S)を作り出した。一部の更なるアミノ酸置換は、これまでの研究においてI−CreI誘導体の活性を増強することが見出されている。イソロイシン132のバリンでの置き換え(I132V)がそのうちの1つである。I132V突然変異を、N末端タンパク質断片及びC末端タンパク質断片のコード配列のいずれか1つ若しくは両方に導入するか、又はいずれにも導入しなかった。
【0221】
同じ戦略を、SH3.3(44A 54L 70Q 75Y 92R 158R 162A)及びSH3.4(30G 38R 50R 70D 75N 142R)をそれぞれホモ二量体として切断する最良な変異体に基づく、SCOH−SH3−b56足場と称される第2の足場に適用した。
【0222】
得られたタンパク質を下記表IVに示す。全ての一本鎖分子を、SH3標的の切断についてCHOにおいてアッセイした。
【0223】
a)一本鎖分子のクローニング
一連の合成遺伝子アセンブリをMWG-EUROFINSにオーダーした。SH3を標的とする異なる一本鎖変異体をコードする合成遺伝子を、AscI制限部位及びXhoI制限部位を用いてpCLS1853にクローニングした。
【0224】
b)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を実施例1.2に記載のようにトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。1回のアッセイにつき、150ngの標的ベクターを、3.12ngから25ngまでの漸増量の変異体DNA(12.5ng+12.5ngのヘテロ二量体DNAに相当する25ngの一本鎖DNA)と共トランスフェクトした。最後に、トランスフェクトしたDNA変異体のDNA量は3.12ng、6.25ng、12.5ng及び25ngであった。トランスフェクトしたDNAの総量は、空ベクター(pCLS0002)を用いて175ng(標的DNA、変異体DNA、担体DNA)とした。
【0225】
d)結果
SH3標的に対する一本鎖分子の活性を、これまでに記載されたCHOアッセイを本内部対照SCOH−RAG及びI−Sce Iメガヌクレアーゼとともに用いてモニタリングした。全ての比較は、3.12ng、6.25ng、12.5ng及び25ngのトランスフェクトした変異体DNAで行った(図2及び図3)。全ての一本鎖分子が、表IVに挙げられるようにCHOアッセイにおいてSH3標的切断活性を示した。
【0226】
【表20】
【0227】
変異体は、それらが有する突然変異プロファイルに応じて、アッセイした用量で特異的な挙動を共有していた(図2及び図3)。例えば、SCOH−SH3−b1−Cは、同様のプロファイルを有し、更に活性が高い。その活性は、低い量から高い量にかけてトランスフェクトした最も低いDNA量で最大に達する。SCOH−SH3−b1−Cと比較して、分子SCOH−SH3−b56−Aは、より高いDNA用量で最大活性を有するが、SCOH−SH3−b1−C及び本内部標準と同等のレベルの活性に達する。
【0228】
記載の全ての変異体が活性であり、導入遺伝子をSH3遺伝子座へ挿入するのに使用することができる。
【0229】
実施例2: SH4遺伝子座を標的とするメガヌクレアーゼの改変
SH4は染色体7上に存在する遺伝子座である。SH4遺伝子座は配列番号3の24bpの非パリンドローム配列を含む。表Aに示されるように、SH4は、Schwarzwaelder et al.(J. Clin. Invest. 2007 117:2241)に開示されるRISの周辺に位置する。SH4配列は、Deichmann et alに記載されるCISのいずれにも含まれない。
【0230】
上の実施例1に記載のものと同様の実験を行い、配列番号3の標的配列を切断することが可能なI−CreIヘテロ二量体及び一本鎖メガヌクレアーゼを同定した。
【0231】
実施例2.1. SH4を切断するメガヌクレアーゼの同定
SH4標的配列を切断する可能性があるヘテロ二量体形態のI−CreI変異体を、遺伝子操作によって構築した。次いで、かかる変異体の対を酵母において共発現させた。共発現時に3つの分子種、すなわち2つのホモ二量体及び1つのヘテロ二量体が得られる。次いで、このヘテロ二量体が配列番号3のSH4標的配列を切断することが可能であるか否かを決定した。
【0232】
a)SH4標的配列に由来するパリンドローム配列を切断するI−CreIメガヌクレアーゼの変異体の構築
SH4配列は部分的に、図4に示される10AAA_P(配列番号4)、5ACT_P(配列番号16)、10AAA_P(配列番号4)、5GGT_P(配列番号17)標的の組合せである。これらの配列は、国際PCT出願の国際公開第2006/097784号及び国際公開第2006/097853号、非特許文献6、非特許文献8に記載のように得られる、これまでに同定されたメガヌクレアーゼによって切断される。このため、SH4は、これらのこれまでに同定されたメガヌクレアーゼから生じるコンビナトリアル変異体によって切断されるはずである。
【0233】
スクリーニング手順を、Chames et al.(Nucleic Acids Res., 2005, 33, e178)、非特許文献6、非特許文献8及び非特許文献7に記載されるものに基づく方法を用いて、以下の2つのパリンドローム配列に対して行った:配列番号18のSH4.3配列及び配列番号19のSH4.4配列。
【0234】
b)標的ベクターの構築
実験手順は、配列番号20(5’−TGGCATACAAGTTTTTAAAACACTGTACACCATTTTGACAATCGTCTGTCA−3’)のSH4標的配列に相当するオリゴヌクレオチドを使用した以外は実施例1.1に記載されるものである。
【0235】
c)変異体の共発現
酵母DNAを、pCLS542発現ベクター及びpCLS1107発現ベクターにおいてSH4.3標的及びSH4.4標的を切断する変異体から標準プロトコルを用いて抽出し、大腸菌を形質転換するために使用した。次いで、得られたプラスミドDNAを使用して、酵母株を同時形質転換した。形質転換体を、ロイシンを欠き、G418を含有する合成培地上で選択した。
【0236】
d)メガヌクレアーゼを共発現するクローンの接合及び酵母におけるスクリーニング
接合はコロニーグリッダー(QpixII、genetix)を用いて行った。変異体をYPDプレートを覆うナイロンフィルター上に、低グリッド密度(4〜6スポット/cm2)を用いてグリッドした。第2のグリッドプロセスを同じフィルター上で行い、各々の標的について異なるレポーターを有する酵母株からなる第2の層をスポットした。膜を固形寒天YPD富栄養培地上に置き、30℃で一晩インキュベートして接合させた。次に、フィルターを、ロイシン及びトリプトファンを欠き、ガラクトース(2%)を炭素源として含むG418を添加した合成培地に移し、37℃で5日間インキュベートして、発現ベクター及び標的ベクターを保有する2倍体を選択した。5日後、フィルターを、0.5Mリン酸ナトリウムバッファー(pH7.0)中の0.02%X−Gal、0.1%SDS、6%ジメチルホルムアミド(DMF)、7mM β−メルカプトエタノール、1%アガロースを含む固形アガロース培地上に置き、37℃でインキュベートして、β−ガラクトシダーゼ活性をモニタリングした。結果をスキャニングによって分析し、適切なソフトウェアを用いて定量化を行った。
【0237】
e)結果
SH4.3標的を切断する変異体及びSH4.4標的を切断する変異体の共発現は、6つの事例においてSH4標的の切断をもたらした。機能的組合せを表Vにまとめる。
【0238】
【表21】
【0239】
実施例2.2. CHO細胞における染色体外モデルでのSH4標的切断の検証
CHO細胞においてSH4標的に対して最大切断活性を示すヘテロ二量体を同定するために、SH4標的を切断する変異体の幾つかの組合せの効率を、CHO細胞における染色体外アッセイを用いて判断した。CHO細胞におけるスクリーニングは、メガヌクレアーゼによる標的の切断がLagoZレポーター遺伝子(細菌lacZ遺伝子の誘導体)の相同組換え及び発現を誘導する一本鎖アニーリング(SSA)ベースのアッセイである。
【0240】
a)CHOスクリーニングのためのベクターにおけるSH4標的のクローニング
標的を以下のようにクローニングした。ゲートウェイクローニング配列に隣接するSH4標的配列に相当する配列番号21のオリゴヌクレオチドを、PROLIGOにオーダーした(5’−TGGCATACAAGTTTTTAAAACACTGTACACCATTTTGACAATCGTCTGTCA−3’)。一本鎖オリゴヌクレオチドのPCR増幅によって生成した二本鎖標的DNAを、ゲートウェイプロトコル(INVITROGEN)を用いてCHOレポーターベクター(pCLS1058)にクローニングした。クローニングした断片をシークエンシング(MILLEGEN)によって検証した。
【0241】
b)メガヌクレアーゼの再クローニング
上で得られたSH4.5標的及びSH4.6標的を切断するI−CreI変異体のORFをpCLS2437にサブクローニングした。ORFを、酵母DNA上で配列番号22及び配列番号23(5’−AAAAAGCAGGCTGGCGCGCCTACACAGCGGCCTTGCCACCATG−3’及び5’−AGAAAGCTGGGTGCTAGCGCTCGAGTTATCAGTCGG−3’)のプライマーを用いてPCRによって増幅した。PCR産物をCHO発現ベクターpCLS2437に、内部断片置き換えのためのAscI制限部位及びNheI制限部位を用いてクローニングした。ライゲーション工程及び大腸菌形質転換工程から得られる選択されたクローンを、シークエンシング(MILLEGEN)によって検証した。
【0242】
c)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を、Polyfect(商標)トランスフェクション試薬を用い、供給業者のプロトコル(QIAGEN)に従ってトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl(通常は、100mlの溶解バッファー(10mM Tris−HCl(pH7.5)、150mM NaCl、0.1%Triton X100、0.1mg/ml BSA、プロテアーゼ阻害剤)、10mlのMg 100Xバッファー(100mM MgCl2、35%β−メルカプトエタノール)、110mlのONPG(8mg/ml)及び780mlの0.1Mリン酸ナトリウム(pH7.5)を含有するバッファーを1リットル)添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。1回のアッセイにつき、150ngの標的ベクターを、両方の変異体それぞれ12.5ng(パリンドロームSH4.3標的を切断する変異体12.5ng及びパリンドロームSH4.4標的を切断する変異体12.5ng)と共トランスフェクトした。
【0243】
d)結果
表VIに示され、上の実施例2.1に記載の4つの変異体を更なる分析に選択した。
【0244】
【表22】
【0245】
これらの変異体をpCLS2437にクローニングした。次いで、SH4.3標的又はSH4.4標的を切断するI−CreI変異体を、CHO染色体外アッセイにおいてSH4標的に対するヘテロ二量体としてともにアッセイした。SH4配列の切断及び組換えの効率の分析から、I−CreI変異体の試験した全ての組合せが、更なる突然変異なしにそれらの切断活性を酵母からCHO細胞へと移すことが可能であったことが実証される(表VII)。
【0246】
【表23】
【0247】
実施例2.3. 一本鎖としての共有結合アセンブリ及び部位特異的突然変異誘発によるSH4を切断するメガヌクレアーゼの改良
実施例2.1.に記載の変異体の共発現は、酵母におけるSH4標的の高い切断活性をもたらす。加えて、それらの一部は哺乳動物発現系におけるSH4切断が検証されている(実施例2.2.)。
【0248】
MA x M2 SH4ヘテロ二量体は酵母における高い切断活性をもたらす。SH4.3−MAは、I−CreI野生型配列と比較して以下の突然変異を有するSH4.3カッターである:24V 44R 68Y 70S 75Y 77N。SH4.4−M2は、I−CreI野生型配列と比較して以下の突然変異を有するSH4.4カッターである:24V 44Y 70S 77V。
【0249】
実施例1.3に記載のように、リンカーRM2を用いて一本鎖構築物を改変し、それによりMA−リンカーRM2−M2と称される一本鎖分子を作製した。この設計工程において、G19S突然変異をC末端M2突然変異体に導入した。加えて、更にSCOH−SH4−b1足場と呼ばれる、MA(K7E K96E)−リンカーRM2−M2(E8K E61R G19S)と称される一本鎖分子を作り出すために、K7E突然変異及びK96E突然変異をMA突然変異体に、E8K突然変異及びE61R突然変異をM2突然変異体に導入した。
【0250】
イソロイシン132のバリンへの(I132V)突然変異を、N末端タンパク質断片及びC末端タンパク質断片のコード配列のいずれか1つ若しくは両方に導入するか、又はいずれにも導入しなかった。
【0251】
同じ戦略を、SH4.3(44R 68Y 70S 75Y 77N)及びSH4.4(24V 44Y 70S 77V)に対する良好なカッターに基づく第2の足場に適用した。この足場は更にSCOH−SH4−b56足場と称される。
【0252】
得られた一本鎖構築物の設計を表VIIIに示す。一本鎖構築物を、SH4標的の切断を誘導するそれらの能力についてCHOにおいて試験した。
【0253】
a)一本鎖分子のクローニング
一連の合成遺伝子アセンブリをMWG-EUROFINSに依頼した。SH4を標的とする異なる一本鎖変異体をコードする合成遺伝子を、AscI制限部位及びXhoI制限部位を用いてpCLS1853にクローニングした。
【0254】
b)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を上に記載のようにトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。1回のアッセイにつき、150ngの標的ベクターを、3.12ngから25ngまでの漸増量の変異体DNA(12.5ng+12.5ngのヘテロ二量体DNAに相当する25ngの一本鎖DNA)と共トランスフェクトした。最後に、トランスフェクトしたDNA変異体のDNA量は3.12ng、6.25ng、12.5ng及び25ngであった。トランスフェクトしたDNAの総量は、空ベクター(pCLS0002)を用いて175ng(標的DNA、変異体DNA、担体DNA)とした。
【0255】
c)結果
表VIIIに記載の一本鎖分子を、SH4標的に対するそれらの活性について、これまでに記載されたCHOアッセイを用いて、本内部対照SCOH−RAG及びI−Sce Iメガヌクレアーゼと比較することによってモニタリングした。全ての活性評価は、3.12ng、6.25ng、12.5ng及び25ngのDNAトランスフェクト用量で行った。全ての一本鎖分子が、表VIIIで報告されるようにSH4標的に対する活性を示していた。
【0256】
【表24】
【0257】
変異体は、それらが有する突然変異プロファイルに応じて、アッセイした用量で特異的な挙動を共有していた(図5及び図6)。例えば、SCOH−SH4−b1Cは、内部標準SCOH−RAGと同じ範囲内の活性レベルを示す(。その活性は、低い量から高い量にかけて増大する。アッセイしたDNAトランスフェクト用量で、その活性はSCOH−SH4−B56Aよりも優れている。
【0258】
これらの変異体は全て、異なるレベルの強度で活性であるため、SH4ゲノム標的化に使用することができる。
【0259】
実施例3: ヒト細胞株におけるSH遺伝子座での切断活性の検出
酵母及び哺乳動物細胞(CHO K1細胞)においてSH3標的及びSH4標的を効率的に切断することが可能なI−CreI変異体が、実施例1及び実施例2において同定されている。それらの内因性DNA標的配列を切断するSH3メガヌクレアーゼ及びSH4メガヌクレアーゼの効率を次に試験した。本実施例では、SH3標的配列及びSH4標的配列を切断するように改変されたメガヌクレアーゼが、ヒト細胞においてそれらの同族内因性部位を切断することが実証される。
【0260】
非相同末端結合(NHEJ)による二本鎖切断の修復は、小さな欠失及び挿入(InDel)を生成し得る(図7)。本来は、このエラーを起こしやすい機構は細胞の生存に有害であり得るが、内因性遺伝子座でのメガヌクレアーゼ活性の迅速な指標をもたらす。
【0261】
実施例3.1: 内因性部位で誘導された突然変異誘発の検出
特異性が変化した変異体のスクリーニングに使用される、哺乳動物細胞又は酵母細胞における切断誘導組換えに基づくアッセイが、国際PCT出願の国際公開第2004/067736号、Epinat et al., Nucleic Acids Res., 2003, 31:2952-2962、Chames et al., Nucleic Acids Res., 2005, 33:e178及び非特許文献6に記載されている。これらのアッセイは、標準方法によってモニタリングすることができる機能的なLacZレポーター遺伝子をもたらす。
【0262】
pCLS1853プラスミドにクローニングしたSH3及びSH4に対する一本鎖I−CreI変異体を、この実験に使用した。実験の前日に、ヒト胎児腎臓細胞株293−H(Invitrogen)に由来する細胞を、10cmディッシュに1.2×106細胞/ディッシュの密度で播種した。翌日、細胞に3μgの空プラスミド又はメガヌクレアーゼ発現プラスミドを、lipofectamine(Invitrogen)を用いてトランスフェクトした。トランスフェクションの72時間後、細胞を回収し、新鮮培養培地で希釈した(希釈率20倍)。7日間の培養後、細胞を回収し、ゲノムDNAを抽出した。
【0263】
200ngのゲノムDNAを使用して、メガヌクレアーゼ切断部位の周囲の内因性遺伝子座をPCR増幅によって増幅した。SH3遺伝子座に相当する377bpの断片は、特異的なPCRプライマーA(配列番号44;5’−tgggggtcttactctgtttccc−3’)及びPCRプライマーB(配列番号45;5’−aggagagtccttctttggcc−3’)を用いて増幅した。SH4遺伝子座に相当する396bpの断片は、PCRプライマーC(配列番号46;5’−gagtgatagcataatgaaaacc−3’)及びPCRプライマーD(配列番号47;5’−ctcaccataagtcaactgtctc−3’)を用いて増幅した。PCR増幅を454シークエンシングシステム(454 Life Sciences)上で行い、シークエンシングサービスを提供する会社(GATC Biotech AG,Germany)によって供給される特異的なアダプター配列(配列番号48;5’−CCATCTCATCCCTGCGTGTCTCCGACTCAG−3’及び配列番号49;5’−CCTATCCCCTGTGTGCCTTGGCAGTCTCAG−3’)に隣接する断片を得た。平均して18000個の配列が2つの単位複製配列のプール(各々500ng)から得られた。シークエンシング後、種々のサンプルを、上記のアダプターの最初に導入したバーコード配列に基づき同定した。
【0264】
次いで、配列をそれぞれSH3又はSH4の切断部位における挿入又は欠失の存在について分析した。
【0265】
実施例3.2: 結果
表IXに得られた結果をまとめる。
【0266】
【表25】
【0267】
SH3遺伝子座を標的とするメガヌクレアーゼをトランスフェクトした細胞から抽出したゲノムDNAの分析によって、12841個の分析した配列のうち56個(0.44%)がSH3の認識部位内にInDel事象を有することが示された。同様に、SH4遺伝子座を標的とするメガヌクレアーゼのトランスフェクション後、8259個の分析した配列のうち18個(0.22%)がSH4の認識部位内にInDel事象を有していた。
【0268】
小さな欠失又は挿入はPCR又はシークエンシングのアーチファクト(artefacts)に関連し得るため、同じ遺伝子座を、メガヌクレアーゼを発現しないプラスミドのトランスフェクション後に分析した。SH3遺伝子座及びSH4遺伝子座の分析から、InDel事象を実質的に全く検出することができないことが明らかとなった。実際に、分析した配列の0.05%(1/2153)及び0.02%(3/12811)しか突然変異を含有しなかった。
【0269】
さらに、DNA挿入配列又はDNA欠失配列のサイズの分析(図8)から、小さな挿入(5bp未満)及び小さな欠失(10bp未満)の優勢な同様のタイプの事象が明らかとなった。
【0270】
これらのデータから、それぞれSH3遺伝子座又はSH4遺伝子座を標的とするように改変されたメガヌクレアーゼが、ヒト細胞において活性であり、それらの同族内因性配列を切断することができることが実証される。さらに、メガヌクレアーゼが、遺伝子ORFを破壊し、それにより対応する遺伝子発現産物を不活性化し得る配列内に小さなInDel事象を発生させる能力があることが示される。
【0271】
実施例4: ヒト細胞における内因性SH3及びSH4遺伝子座での遺伝子標的化
改変された一本鎖SH3メガヌクレアーゼ及び一本鎖SH4メガヌクレアーゼの切断活性を検証するために、内因性ヒトSH3遺伝子座及び内因性ヒトSH4遺伝子座で相同組換えを刺激するそれらの能力を次に評価した。細胞に、一本鎖分子SCOH−SH3−b1−C又はSCOH−SH4−b1−Cに対する哺乳動物発現プラスミド及び標的化構築物を含むベクターをトランスフェクトした。標的化構築物を含むベクター(「ドナー修復プラスミド」とも称される)は、ヒトSH3遺伝子座又はヒトSH4遺伝子座に相同な2つの配列に隣接する外因性DNA配列からなる2.8kbの配列を含有するpCLS3777プラスミド又はpCLS3778プラスミドであった。ヒトSH3遺伝子座又はヒトSH4遺伝子座に相同な配列は1.5kbの長さを有していた。メガヌクレアーゼによる天然のSH3遺伝子座又はSH4遺伝子座の切断は、ドナー修復プラスミドを修復マトリックスとして使用し得る相同組換えのための基質を生じる。このため、SH3遺伝子座又はSH4遺伝子座で標的化組込みが起こる頻度は、ゲノムSH3標的部位又はゲノムSH4標的部位の切断効率を示す。
【0272】
実施例4.1: 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼは、哺乳動物発現ベクターにクローニングされ、それぞれプラスミドpCLS2697及びpCLS2705を生じるSCOH−SH3−b1−C及びSCOH−SH4−b1−Cである。
【0273】
b)ドナー修復プラスミド
SH3遺伝子標的化実験については、ドナープラスミドは以下のものを含有していた:
左ホモロジーアームとして、SH3遺伝子座のPCR生成断片(染色体6上の6850510位〜6852051位、NC_000006.11)。この断片は1540bpの長さを有する;
右ホモロジーアームとして、SH3遺伝子座の断片(染色体6上の6852107位〜6853677位、NC_000006.11)。この断片は1571bpの長さを有する。
【0274】
SH4遺伝子標的化実験については、ドナープラスミドは以下のものを含有していた:
左ホモロジーアームとして、SH4遺伝子座のPCR生成断片(染色体7上の114972751位〜114974269位、NC_000007.13)。この断片は1519bpの長さを有する;及び
右ホモロジーアームとして、SH4遺伝子座の断片(染色体7上の114974316位〜114976380位、NC_000007.13)。この断片は2065bpの長さを有する。
【0275】
SH3及びSH4の両方について、左ホモロジーアーム及び右ホモロジーアームを、2つのCMVプロモーター及びネオマイシン耐性遺伝子を含有する2.8kbの外因性DNA断片のそれぞれ上流(AscI部位を用いる)及び下流(SbfI部位を用いる)に挿入した。得られたプラスミドはpCLS3777(SH3に対する)及びpCLS3778(SH4に対する)と称される。
【0276】
c)Sh3遺伝子及びSh4遺伝子の標的化実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞をトランスフェクトした。簡潔に述べると、2μgのドナープラスミドを3μgの一本鎖メガヌクレアーゼ発現ベクターと共トランスフェクトした。37℃で72時間のインキュベーション後、細胞をトリプシン処理し、96ウェルプレートにおいて1ウェル当たり10細胞又は100細胞で、完全培地中で平板培養した。細胞が80%〜100%コンフルエントになった時点で、ZR−96ゲノムDNAキット(Zymo research)を用い、供給業者のプロトコルに従ってゲノムDNA抽出を行った。
【0277】
d)遺伝子標的化事象のPCR分析
遺伝子標的化頻度を、以下のプライマーを用いたゲノムDNAに対するPCRによって決定した:SH3については5’−CTGTGTGCTATGATCTTGCC−3’(SH3GHGF4;配列番号50)及び5’−CCTGTCTCTTGATCAGATCC−3’(NeoR2;配列番号51)、並びにSH4については5’−GTGGCCTCTCAGTCTGTTTA−3’(SH4GHGF2;配列番号52)及び5’−AGTCATAGCCGAATAGCCTC−3’(NeoR5;配列番号53)。PCRによって、2500bp(SH3)又は2268bp(SH4)の遺伝子標的化特異的PCR産物が得られる。SH3GHGF4プライマー及びSH4GHGF2プライマーは、ドナー修復プラスミドの左ホモロジーアームの上流に位置するフォワードプライマーである。NeoRプライマーは、ドナー修復プラスミドの2つのホモロジーアームの間に挿入される外因性DNAに位置するリバースプライマーである。
【0278】
実施例4.2: 結果
ヒト胎児腎臓293H細胞に、2つの一本鎖SH3メガヌクレアーゼ又は一本鎖SH4メガヌクレアーゼの一方を発現するプラスミド及びドナー修復プラスミドpCLS3777又はpCLS3778を共トランスフェクトした。自発的組換えの対照として、293H細胞をドナー修復プラスミド単独でもトランスフェクトした。次いで、細胞を96ウェルマイクロプレートにおいて1ウェル当たり10細胞又は100細胞で平板培養した。これらの細胞に由来するゲノムDNAを遺伝子標的化について、材料及び方法の項に記載されるようにPCRによって分析した。
【0279】
メガヌクレアーゼの非存在下(修復プラスミド単独)では、10個又は100個の細胞のプールにおいて分析した22560個及び18800個の細胞(それぞれSH3及びSH4に対する)の中でPCR陽性シグナルは検出されなかった。
【0280】
これに対し、SH3メガヌクレアーゼの存在下では、100個の細胞のプールにおいて分析した18800個の細胞の中で12個の陽性クローンが検出され、それにより組換えの頻度が0.064%であることが示された。SH4メガヌクレアーゼの存在下では、10個の細胞のプールにおいて分析した3760個の細胞の中で11個の陽性クローンが検出され、組換えの頻度が0.29%であることが示された。結果を下記表Xに提示する。ここで示される組換え頻度は、平板培養した全ての細胞が再び分裂を開始したわけではないため、低く見積もったものである。平板培養時の生存率はこのため、約33%であると推定することができる。したがって、組換えの頻度はおそらく3倍低く見積もられている。
【0281】
【表26】
【0282】
これらの結果から、2つの一本鎖分子SCOH−SH3−b1−C及びSCOH−SH4−b1−Cが、それぞれ内因性SH3遺伝子座及び内因性SH4遺伝子座で高レベルの遺伝子標的化を誘導することが可能であることが実証される。
【0283】
実施例5: SH6遺伝子座を標的とするメガヌクレアーゼの改変
SH6は、染色体21上に存在する24bpの非パリンドローム標的(TTAATACCCCGTACCTAATATTGC、配列番号59)を含む遺伝子座である。SH6は、Schwarzwaelder et al.(J Clin Invest 2007:2241-9)に開示されるRISの近傍に位置する。SH6配列は、Deichmann et alに記載されるCISのいずれにも含まれない。
【0284】
実施例5.1. SH6を切断するメガヌクレアーゼの同定
SH6標的配列を切断する可能性があるヘテロ二量体形態のI−CreI変異体を、遺伝子操作によって構築した。次いで、かかる変異体の対を酵母において共発現させた。共発現時に3つの分子種、すなわち2つのホモ二量体及び1つのヘテロ二量体が得られる。次いで、このヘテロ二量体が配列番号59のSH6標的配列を切断することが可能であるか否かを決定した。
【0285】
a)SH6標的配列に由来するパリンドローム配列を切断するI−CreIメガヌクレアーゼの変異体の構築
SH6配列は部分的に、図9に示される10AAT_P(配列番号60)、5CCC_P(配列番号61)、10AAT_P(配列番号60)、5TAG_P(配列番号62)標的配列の組合せである。これらの配列は、国際PCT出願の国際公開第2006/097784号及び国際公開第2006/097853号、非特許文献6、並びに非特許文献8に記載のように得られるメガヌクレアーゼによって切断される。このため、SH6は、これらのこれまでに同定されたメガヌクレアーゼから生じるコンビナトリアル変異体によって切断されるはずである。
【0286】
2つのパリンドローム標的SH6.3及びSH6.4は、SH6に由来するものであった(図9)。SH6.3及びSH6.4はパリンドローム性であるため、ホモ二量体タンパク質によって切断されるはずである。したがって、配列番号63のSH6.3パリンドローム標的配列又は配列番号64のSH6.4パリンドローム標的配列のいずれかを切断するホモ二量体I−CreI変異体は、Chames et al.(Nucleic Acids Res., 2005, 33, e178)、非特許文献6、非特許文献8及び非特許文献7に記載されるものに基づく方法を用いて構築した。
【0287】
b)標的ベクターの構築
実験手順は、SH6標的配列(5’−TGGCATACAAGTTTTTAATACCCCGTACCTAATATTGCCAATCGTCTGTCA−3’(配列番号65))に相当するオリゴヌクレオチドを使用した以外は実施例1.1に記載されるものである。
【0288】
c)変異体の共発現
pCLS542発現ベクター及びpCLS1107発現ベクターにおいてSH6.3標的及びSH6.4標的を切断する変異体に由来する酵母DNAを、標準プロトコルを用いて抽出し、大腸菌を形質転換するために使用した。形質転換体を、ロイシンを欠き、G418を含有する合成培地上で選択した。
【0289】
d)メガヌクレアーゼを共発現するクローンの接合及び酵母におけるスクリーニング
接合はコロニーグリッダー(QpixII、Genetix)を用いて行った。変異体をYPDプレートを覆うナイロンフィルター上に、低グリッド密度(4〜6スポット/cm2)を用いてグリッドした。第2のグリッドプロセスを同じフィルター上で行い、各々の標的について異なるレポーターを有する酵母株からなる第2の層をスポットした。膜を固形寒天YPD富栄養培地上に置き、30℃で一晩インキュベートして接合させた。次に、フィルターを、ロイシン及びトリプトファンを欠き、ガラクトース(2%)を炭素源として含むG418を添加した合成培地に移し、37℃で5日間インキュベートして、発現ベクター及び標的ベクターを保有する2倍体を選択した。5日後、フィルターを、0.5Mリン酸ナトリウムバッファー(pH7.0)中の0.02%X−Gal、0.1%SDS、6%ジメチルホルムアミド(DMF)、7mM β−メルカプトエタノール、1%アガロースを含む固形アガロース培地上に置き、37℃でインキュベートして、β−ガラクトシダーゼ活性をモニタリングした。結果をスキャニングによって分析し、適切なソフトウェアを用いて定量化を行った。
【0290】
e)結果
SH6.4標的を切断する10個の変異体及びSH6.3標的を切断する2個の変異体の共発現は、2つを除く全ての事例においてSH6.1標的の切断をもたらした。これらの2つの事例は、二重形質転換体が得られなかった場合に相当する。機能的組合せを表XIにまとめる。
【0291】
【表27】
【0292】
実施例5.2. CHO細胞における染色体外モデルでのSH6標的切断の検証
ヘテロ二量体を形成した場合に酵母においてSH6標的を効率的に切断することが可能なI−CreI変異体は、上の実施例5.1で記載している。CHO細胞においてSH3標的に対して最大切断活性を示すヘテロ二量体を同定するために、これらの変異体の幾つかの効率を、CHO細胞における染色体外アッセイを用いて比較した。CHO細胞におけるスクリーニングは、メガヌクレアーゼによる標的の切断がLagoZレポーター遺伝子(細菌lacZ遺伝子の誘導体)の相同組換え及び発現を誘導する一本鎖アニーリング(SSA)ベースのアッセイである。
【0293】
a)CHOスクリーニングのためのベクターにおけるSH6標的のクローニング
標的を以下のようにクローニングした。ゲートウェイクローニング配列に隣接するSH6標的配列に相当するオリゴヌクレオチドを、PROLIGOにオーダーした(5’−TGGCATACAAGTTTTTAATACCCCGTACCTAATATTGCCAATCGTCTGTCA−3’(配列番号65))。一本鎖オリゴヌクレオチドのPCR増幅によって生成した二本鎖標的DNAを、ゲートウェイプロトコル(INVITROGEN)を用いてCHOレポーターベクター(pCLS1058)にクローニングした。クローニングした標的をシークエンシング(MILLEGEN)によって検証した。
【0294】
b)メガヌクレアーゼの再クローニング
実施例5.1で同定されたSH6.3標的及びSH6.4標的を切断するI−CreI変異体のORFをpCLS2437にサブクローニングした。ORFを、酵母DNA上で以下のプライマー:5’−AAAAAGCAGGCTGGCGCGCCTACACAGCGGCCTTGCCACCATG−3’(配列番号66)及び5’−AGAAAGCTGGGTGCTAGCGCTCGAGTTATCAGTCGG−3’(配列番号67)のプライマーを用いてPCRによって増幅した。PCR産物をCHO発現ベクターpCLS2437に、内部断片置き換えのためのAscI及びXhoIを用いてクローニングした。ライゲーション工程及び大腸菌形質転換工程から得られる選択されたクローンを、シークエンシング(MILLEGEN)によって検証した。
【0295】
c)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を、Polyfect(商標)トランスフェクション試薬を用い、供給業者のプロトコル(QIAGEN)に従ってトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl(通常は、100mlの溶解バッファー(10mM Tris−HCl(pH7.5)、150mM NaCl、0.1%Triton X100、0.1mg/ml BSA、プロテアーゼ阻害剤)、10mlのMg 100Xバッファー(100mM MgCl2、35%β−メルカプトエタノール)、110mlのONPG(8mg/ml)及び780mlの0.1Mリン酸ナトリウム(pH7.5)を含有するバッファーを1リットル)添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。1回のアッセイにつき、150ngの標的ベクターを、両方の変異体それぞれ12.5ng(パリンドロームSH6.3標的を切断する変異体12.5ng及びパリンドロームSH6.4標的を切断する変異体12.5ng)と共トランスフェクトした。
【0296】
d)結果
酵母においてSH6を切断することが可能なヘテロ二量体エンドヌクレアーゼを形成する変異体の1つの組を、CHOにおいて一時的トランスフェクションでの染色体外アッセイを用いて確認するために選んだ。
【0297】
SH6.3を切断することが可能な単量体は、以下の突然変異:44K 70S 75Nを含み(SH6−3−M1−44K 70S 75Nと称される)、SH6.4を切断することが可能な単量体は、以下の突然変異:28Q 40R 44A 70L 75N 96R 111H 144Sを含んでいた(SH6−4−MB−28Q 40R 44A 70L 75N 96R 111H 144Sと称される)。
【0298】
SH6配列の切断及び組換えの効率の分析から、I−CreI変異体の試験した組合せが、更なる突然変異なしにその切断活性を酵母からCHO細胞へと移すことが可能であったことが実証される。
【0299】
実施例5.3. 一本鎖としての共有結合アセンブリ及びSH6を切断するメガヌクレアーゼの改良
実施例5.1に記載のカッターの共発現は、酵母におけるSH6標的の高い切断活性をもたらす。それらのうちの1つは哺乳動物発現系におけるSH6切断が検証されている(実施例5.2)。
【0300】
M1 x MA SH6ヘテロ二量体は酵母における高い切断活性をもたらす。M1は、I−CreI野生型配列と比較して以下の突然変異を有するSH6.3カッターである:44K 70S 75N。MAは、I−CreI野生型配列と比較して以下の突然変異を有するSH6.4カッターである:7R 28Q 40R 44A 70L 75N 103T 121E 132V 160R。
【0301】
リンカーRM2(AAGGSDKYNQALSKYNQALSKYNQALSGGGGS;配列番号15)を用いて一本鎖構築物を改変し、それにより一本鎖分子:MA−RM2−M1を作製した。この設計工程において、G19S突然変異をC末端M1突然変異体に導入した。加えて、更にSCOH−SH6 b1足場と呼ばれる一本鎖分子:MA(K96E)−RM2−MA(E8K E61R)を作り出すために、突然変異K96EをMA突然変異体に、突然変異E8K、E61RをM1突然変異体に導入した。
【0302】
4つの更なるアミノ酸置換が、これまでの研究においてI−CreI誘導体の活性を増強することが見出されている。これらの突然変異は、フェニルアラニン54のロイシンでの置き換え(F54L)、グルタミン酸80のリジンでの置き換え(E80K)、バリン105のアラニンでの置き換え(V105A)、及びイソロイシン132のバリンでの置き換え(I132V)に相当する。幾つかの組合せをN末端タンパク質断片及びC末端タンパク質断片のコード配列に導入し、得られたタンパク質の最初のバッチを、SH6標的の切断を誘導するそれらの能力についてアッセイした。
【0303】
a)SC−OH一本鎖構築物への更なる突然変異の導入
更なる突然変異を、Stratagene/Agilent technologies Inc製のQuikChange Multi Site−Directed Mutagenesis Kitを用い、製造業者の使用説明書に従って導入した。オリゴヌクレオチドの第1のセットを使用して、第1の単量体に相当する一本鎖分子の一部に突然変異を導入した。オリゴヌクレオチドの第2のセットを、同じ突然変異を表XIIに示される第2の単量体に相当する一本鎖分子の第2の部分に特異的に導入するように設計した。
【0304】
【表28】
【0305】
このプロセスの間に得られた単離クローンをシークエンシングして、得られる特異的な突然変異プロファイルを確認した。次いで、関心のプロファイルをCHO SSAアッセイにおいて記載の最初の構築物と比較して試験した。
【0306】
b)哺乳動物細胞における染色体外アッセイ
CHO K1細胞を上に記載されるようにトランスフェクトした。トランスフェクションの72時間後に、培養培地を除去し、β−ガラクトシダーゼ液体アッセイのための溶解/曝露バッファーを150μl添加した。37℃でのインキュベーション後、ODを420nmで測定した。全プロセスを自動Velocity11 BioCelプラットフォーム上で行った。
【0307】
1回のアッセイにつき、150ngの標的ベクターを、3.12ngから25ngまでの漸増量の変異体DNA(12.5ng+12.5ngのヘテロ二量体DNAに相当する25ngの一本鎖DNA)と共トランスフェクトした。最後に、トランスフェクトしたDNA変異体のDNA量は3.12ng、6.25ng、12.5ng及び25ngであった。トランスフェクトしたDNAの総量は、空ベクター(pCLS0001)を用いて175ng(標的DNA、変異体DNA、担体DNA)とした。
【0308】
c)結果
SH6標的に対するSCOH−SH6−b1−C(pCLS2796)及びSCOH−SH6−b1−B(pCLS2928)一本鎖分子(表XIIIを参照されたい)の活性を、これまでに記載されたCHOアッセイを用い、SH6.3−M1 x SH6.4−MBを形成するヘテロ二量体及び本内部対照SCOH−RAG及びI−Sce Iメガヌクレアーゼと比較することによってモニタリングした。全ての比較は、3.12ng、6.25ng、12.5ng及び25ngのトランスフェクトした変異体DNAで行った(図10)。2つの一本鎖メガヌクレアーゼは、開始(starting)ヘテロ二量体よりも効率的にSH6標的を切断することが可能であった。最良の分子SCOH−SH6−b1−Cの活性を、メガヌクレアーゼの新たなバッチにおいて上に記載されたものに更なる突然変異を導入することによって更に改善した。
【0309】
【表29】
【0310】
更なる突然変異を材料及び方法の項に従って一本鎖足場に更に導入した。得られて試験した分子を表XIVに挙げる。
【0311】
【表30】
【0312】
全ての変異体は記載の条件で活性であり、それらが有する突然変異プロファイルに応じて、アッセイした用量で特異的な挙動を共有していた(図10)。例えば、QCSH61−H01a、QCSH61−H01b、QCSH61−H01c、QCSH61−H01dは、本内部標準SCOH−RAGと同様のプロファイルを有する。これらは低用量であっても非常に活性な分子である。これらの変異体は全てSH6ゲノム標的化に使用することができた。
【0313】
実施例6: ヒト細胞における内因性SH6遺伝子座での遺伝子標的化
改変された一本鎖SH6メガヌクレアーゼの切断活性を検証するために、内因性ヒトSH6遺伝子座で相同組換えを刺激するそれらの能力を評価した。細胞に、一本鎖分子SCOH−QCSH6−H01(配列番号81;pCLS3690)又はSCOH−QC−SH6−H01−V2−7E−70R75D(配列番号85;pCLS4373)に対する哺乳動物発現プラスミド、及びヒトSH6遺伝子座に相同な2つの配列(どちらも1.5kb長)に隣接する2.8kbの外因性DNA配列を含有するドナー修復プラスミドpCLS3779(図13;配列番号279)をトランスフェクトした。メガヌクレアーゼによる天然のSH6遺伝子座の切断は、ホモロジーアームに隣接する2.8kbの外因性DNAを含有するドナー修復プラスミドを修復マトリックスとして使用し得る相同組換えのための基質を生じる。このため、SH6遺伝子座で標的化組込みが起こる頻度は、ゲノムSH6標的部位の切断効率を示す。
【0314】
実施例6.1. 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼは、哺乳動物発現ベクターにクローニングされ、それぞれプラスミドpCLS3690(図13)及びpCLS4373を生じるSCOH−QCSH6−H01(配列番号81)又はSCOH−QC−SH6−H01−V2−7E−70R75D(配列番号85)である。
【0315】
b)ドナー修復プラスミド
ドナープラスミドは、左ホモロジーアームとしてSH6遺伝子座のPCR生成1517bp断片(染色体21上の18437771位〜18439287位、NC_000021.8)、及び右ホモロジーアームとして、SH6遺伝子座の1571bp断片(染色体21上の18439343位〜18440846位、NC_000021.8)を含有する。左ホモロジーアーム及び右ホモロジーアームを、2つのCMVプロモーター及びネオマイシン耐性遺伝子を含有する2.8kbの外因性DNA断片のそれぞれ上流(AscI部位を用いる)及び下流(SbfI部位を用いる)に挿入した。得られたプラスミドはpCLS3779(図13;配列番号279)である。
【0316】
c)Sh6遺伝子標的化実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞をトランスフェクトした。簡潔に述べると、2μgのドナープラスミドを3μgの一本鎖メガヌクレアーゼ発現ベクターと共トランスフェクトした。37℃で72時間のインキュベーション後、細胞をトリプシン処理し、96ウェルプレートにおいて1ウェル当たり10細胞又は100細胞で、完全培地中で平板培養した。代替的に、37℃で72時間のインキュベーション後、細胞をトリプシン処理し、10cmディッシュ1枚当たり300細胞で、完全培地中で平板培養した。37℃で2週間のインキュベーション後、個々のクローン細胞コロニーを採取し、96ウェルプレートにおいて完全培地中で平板培養した。細胞が80%〜100%コンフルエントになった時点で、ZR−96ゲノムDNAキット(Zymo research)を用い、供給業者のプロトコルに従ってゲノムDNA抽出を行った。
【0317】
d)遺伝子標的化事象のPCR分析
遺伝子標的化の頻度を、プライマーSH6GHGF3:5’−CAATGGAGTTTTGGAGCCAC−3’(配列番号280)及びNeoR9:5’−ATCAGAGCAGCCGATTGTCT−3’(配列番号281)を用いたゲノムDNAに対するPCRによって決定した。PCRによって、2300bpの遺伝子標的化特異的PCR産物が得られる(図14)。SH6GHGF3プライマーは、ドナー修復プラスミドの左ホモロジーアームの上流に位置するフォワードプライマーである。NeoR9プライマーは、ドナー修復プラスミドの2つのホモロジーアームの間に挿入される外因性DNAに位置するリバースプライマーである。
【0318】
実施例6.2. 結果
ヒト胎児腎臓293H細胞に、2つのベクター:2つの一本鎖SH6メガヌクレアーゼの一方を発現するプラスミド及びドナー修復プラスミドpCLS3779(図13;配列番号279)を共トランスフェクトした。自発的組換えの対照として、293H細胞をドナー修復プラスミド単独でもトランスフェクトした。次いで、細胞を96ウェルマイクロプレートにおいて1ウェル当たり10細胞又は100細胞で、又は10cmディッシュ1枚当たり300細胞で平板培養し、2週間後にクローンコロニーを単離し、96ウェルマイクロプレートにおいて平板培養した。これらの細胞に由来するゲノムDNAを遺伝子標的化について、材料及び方法の項に記載されるようにPCRによって分析した。メガヌクレアーゼの非存在下(修復プラスミド単独)では、10個又は100個の細胞のプールにおいて分析した67680個の細胞の中で5個のPCR陽性シグナルが検出され、0.007%の自発的組換えの頻度が示された。対照的に、SCOH−QCSH6−H01メガヌクレアーゼ(配列番号81;pCLS3690)又はSCOH−QC−SH6−H01−V2−7E−70R75Dメガヌクレアーゼ(配列番号85;pCLS4773)の存在下では、10個又は100個の細胞のプールにおいて分析した73320個及び18800個の細胞の中で177個及び35個の陽性シグナルが検出され、それぞれ0.24%及び0.19%の組換えの頻度が示された。結果を表XVに提示する。これらの結果から、2つの一本鎖分子SCOH−QCSH6−H01(配列番号81;pCLS3690)及びSCOH−QC−SH6−H01−V2−7E−70R75D(配列番号85;pCLS4773)が、内因性sh6遺伝子座で高レベルの遺伝子標的化を誘導することが可能なことが実証される。
【0319】
【表31】
【0320】
実施例7: ヒト細胞における内因性sh6遺伝子座での遺伝子標的化後の導入遺伝子発現
sh6遺伝子座での導入遺伝子発現を支持するsh6遺伝子座の能力、改変された一本鎖SH6メガヌクレアーゼの切断活性を検証するために、ネオマイシン耐性遺伝子発現カセットを含有する修復プラスミドを用いて遺伝子標的化実験を行い、ネオマイシン含有培地中で成長する修飾細胞の能力を測定した。ネオマイシンの存在下での細胞の生存及び成長はネオマイシン耐性遺伝子の発現に依存し、したがって標的化組込み後のSH6遺伝子座での導入遺伝子の発現を示す。
【0321】
実施例7.1. 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼは、哺乳動物発現ベクターにクローニングされ、プラスミドpCLS3690を生じるSCOH−QCSH6−H01(配列番号81)である。
【0322】
b)ドナー修復プラスミド
ドナープラスミドは、左ホモロジーアームとしてSH6遺伝子座のPCR生成1517bp断片(染色体21上の18437771位〜18439287位、NC_000021.8)、及び右ホモロジーアームとして、SH6遺伝子座の1571bp断片(染色体21上の18439343位〜18440846位、NC_000021.8)を含有する。左ホモロジーアーム及び右ホモロジーアームを、2つのCMVプロモーター及びネオマイシン耐性遺伝子を含有する2.8kbの外因性DNA断片のそれぞれ上流(AscI部位を用いる)及び下流(SbfI部位を用いる)に挿入した。得られたプラスミドはpCLS3779(図13;配列番号279)である。
【0323】
c)Sh6遺伝子標的化実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞をトランスフェクトした。簡潔に述べると、2μgのドナープラスミドを3μgの一本鎖メガヌクレアーゼ発現ベクターと共トランスフェクトした。37℃で72時間のインキュベーション後、細胞をトリプシン処理し、10cmディッシュ1枚当たり300細胞で、完全培地中で平板培養した。37℃で2週間のインキュベーション後、個々のクローン細胞コロニーを採取し、96ウェルプレートにおいて完全培地中で平板培養した。37℃で1週間のインキュベーション後、細胞をトリプシン処理し、2つの96ウェル複製プレートに平板培養して、37℃でインキュベートした。細胞が80%〜100%コンフルエントになった時点で、ZR−96ゲノムDNAキット(Zymo research)を用い複製プレートの1つで、供給業者のプロトコルに従ってゲノムDNA抽出を行った。他の複製プレートを使用して遺伝子標的化クローンを単離し、それらを増殖させた。
【0324】
d)遺伝子標的化クローンのPCR同定
遺伝子標的化を、プライマーSH6GHGF3:5’−CAATGGAGTTTTGGAGCCAC−3’(配列番号280)及びNeoR9:5’−ATCAGAGCAGCCGATTGTCT−3’(配列番号281)を用いたゲノムDNAに対するPCRによって決定した。PCRによって、2300bpの遺伝子標的化特異的PCR産物が得られる(図14)。SH6GHGF3プライマーは、ドナー修復プラスミドの左ホモロジーアームの上流に位置するフォワードプライマーである。NeoR9プライマーは、ドナー修復プラスミドの2つのホモロジーアームの間に挿入される外因性DNAに位置するリバースプライマーである。
【0325】
e)サザンブロット法による標的化組込みの検証
細胞クローンに由来するゲノムDNAを、StuI制限酵素又はHindIII制限酵素(New England Biolabs)で消化し、0.8%アガロースゲル(gel)上での電気泳動によって分離し、ニトロセルロース膜に転写した。DNAプローブを、ネオマイシン耐性遺伝子に相同な25ngのDNA断片から、32P放射標識dCTP及びRediprime IIランダムプライムラベリングシステム(GE Healthcare)を用い、供給業者のプロトコルに従って調製し、ハイブリダイゼーションバッファー(20mM NaPi、7%SDS、1mM EDTA)中でプレインキュベートしたニトロセルロース膜に添加した。65℃で一晩のインキュベーションの後、膜を洗浄し、放射線写真用フィルムに感光させた。期待されるバンドの放射線写真上でのサイズは、StuI消化については5.3kb、HindIII消化については6.8kbである(図15)。
【0326】
f)ネオマイシン耐性試験:
SH6遺伝子座で標的化されるPCRによって同定された細胞クローンを、96ウェルマイクロプレートにおいて1ウェル当たり300細胞で、G418抗生物質(PAA laboratories)の存在下で平板培養した。37℃で10日間のインキュベーションの後、Vialightバイオアッセイキット(Lonza)及びVictorルミネッセンスリーダー(Perkin Elmer)を用い、供給業者のプロトコルに従って生存能力を測定した。
【0327】
実施例7.2. 結果
ヒト胎児腎臓293H細胞に、2つのベクター:2つの一本鎖SH6メガヌクレアーゼの一方を発現するプラスミド及びドナー修復プラスミドpCLS3779を共トランスフェクトした。次いで、細胞を10cmディッシュ1枚当たり300細胞で平板培養し、2週間後にクローンコロニーを単離し、96ウェルマイクロプレートにおいて平板培養した。これらの細胞に由来するゲノムDNAを遺伝子標的化について、材料及び方法の項に記載されるようにPCRによって分析した。次いで、ゲノムDNAを使用して、サザンブロット分析により標的化組込みを検証した。クローンの#7及び#8は期待されるサイズのバンドを示したが、陰性対照クローンの#5及び#6は示さなかった(図16)。これらの細胞クローンを、G418(PAA laboratories)の存在下で生存するそれらの能力について試験した。標的化組込みを有するクローン(#7及び#8)のみが、0.4mg/mlを上回る濃度のG418に対する耐性を示した(図16)。このことは、sh6遺伝子座での標的化組込みが機能的な導入遺伝子の発現を支持し得ることを示している。
【0328】
実施例8. ヒト細胞における内因性sh6遺伝子座での遺伝子標的化後の近接遺伝子の発現
近接遺伝子の発現を妨げることなく導入遺伝子組込みを支持するsh6遺伝子座の能力を検証するために、2.8kbの外因性DNA断片を含有する修復プラスミドを用いて遺伝子標的化実験を行い、標的化組込みを有する細胞クローンを同定した。sh6組込み部位の上流及び下流の遺伝子の発現を測定し、標的化組込みが起こっていない細胞クローンの発現と比較した。
【0329】
実施例8.1. 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼは、哺乳動物発現ベクターにクローニングされ、プラスミドpCLS3690を生じるSCOH−QCSH6−H01(配列番号81)である。
【0330】
b)ドナー修復プラスミド
ドナープラスミドは、左ホモロジーアームとしてSH6遺伝子座のPCR生成1517bp断片(染色体21上の18437771位〜18439287位、NC_000021.8)、及び右ホモロジーアームとして、SH6遺伝子座の1571bp断片(染色体21上の18439343位〜18440846位、NC_000021.8)を含有する。左ホモロジーアーム及び右ホモロジーアームを、2つのCMVプロモーター及びネオマイシン耐性遺伝子を含有する2.8kbの外因性DNA断片のそれぞれ上流(AscI部位を用いる)及び下流(SbfI部位を用いる)に挿入した。得られたプラスミドはpCLS3779(図13;配列番号279)である。
【0331】
c)Sh6遺伝子標的化実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞をトランスフェクトした。簡潔に述べると、2μgのドナープラスミドを3μgの一本鎖メガヌクレアーゼ発現ベクターと共トランスフェクトした。37℃で72時間のインキュベーション後、細胞をトリプシン処理し、10cmディッシュ1枚当たり300細胞で、完全培地中で平板培養した。37℃で2週間のインキュベーション後、個々のクローン細胞コロニーを採取し、96ウェルプレートにおいて完全培地中で平板培養した。37℃で1週間のインキュベーション後、細胞をトリプシン処理し、2つの96ウェル複製プレートに平板培養し、37℃でインキュベートした。細胞が80%〜100%コンフルエントになった時点で、ZR−96ゲノムDNAキット(Zymo research)を用い複製プレートの1つで、供給業者のプロトコルに従ってゲノムDNA抽出を行った。他の複製プレートを使用して遺伝子標的化クローンを単離し、それらを増殖させた。
【0332】
d)遺伝子標的化クローンのPCR同定
遺伝子標的化を、プライマーSH6GHGF3:5’−CAATGGAGTTTTGGAGCCAC−3’(配列番号280)及びNeoR9:5’−ATCAGAGCAGCCGATTGTCT−3’(配列番号281)を用いたゲノムDNAに対するPCRによって決定した。PCRによって、2300bpの遺伝子標的化特異的PCR産物が得られる(図XX)。SH6GHGF3プライマー(配列番号280)は、ドナー修復プラスミドの左ホモロジーアームの上流に位置するフォワードプライマーである。NeoR9プライマー(配列番号281)は、ドナー修復プラスミドの2つのホモロジーアームの間に挿入される外因性DNAに位置するリバースプライマーである。
【0333】
e)sh6遺伝子座の上流及び下流の遺伝子の発現
遺伝子発現を定量RT−PCRによって測定した。RNAをサブコンフルエントな細胞クローンからRNeasy RNA単離キット(Qiagen)を用いて、製造業者のプロトコルに従って単離した。3μgのRNAを使用し、Superscript III First−strandキット(Invitrogen)を用いてcDNAを生成した。定量PCRを、2連の(duplicate)サンプルにおいて、SYBR(商標)Premix Ex Taq(商標)DNA Polymerase(Lonza)を用い、Stratagene MPX3000機器上で12μlの反応につき10ngのcDNAに対して行った。各々の遺伝子について、使用したプライマーを以下の表に挙げる。
【0334】
【表32】
【0335】
サイクル閾値(threshold cycles)(Ct)を、Stratageneソフトウェアを用い、ROXリファレンス色素による正規化の後に蛍光(dRn)に対して決定した。遺伝子発現の強度を、ハウスキーピング遺伝子HPRTの発現を内部正規化因子として使用して、式2Ct(HPRT)-Ct(遺伝子)を用いて算出した。
【0336】
実施例8.2. 結果
ヒト胎児腎臓293H細胞に、2つのベクター:3つの一本鎖SH6メガヌクレアーゼのうち1つを発現するプラスミド及びドナー修復プラスミドpCLS3779を共トランスフェクトした。次いで、細胞を10cmディッシュ1枚当たり300細胞で平板培養し、2週間後にクローンコロニーを単離し、96ウェルマイクロプレートにおいて平板培養した。これらの細胞に由来するゲノムDNAを遺伝子標的化について、材料及び方法の項に記載されるようにPCRによって分析した。標的化組込みを示すクローン及び陰性対照からRNAを単離した。定量RT−PCRを行い、標的化組込みの遺伝子座の周囲の遺伝子の発現を測定した。データを図17に提示する。図17では、ハウスキーピング遺伝子HPRTでの正規化後の3つの個々の標的化クローン(KI)及び3つの個々の非標的化クローン(WT)について2連のサンプルの平均強度を示す。測定した5つの遺伝子の各々について有意差は観察されず、sh6遺伝子座での標的化組込みが近接遺伝子の発現に影響をもたらさないことが示される。
【0337】
実施例9: ヒト細胞における内因性セーフハーバー遺伝子座での突然変異誘発
改変された一本鎖セーフハーバーメガヌクレアーゼの切断活性を検証するために、内因性ヒトセーフハーバー遺伝子座での突然変異誘発を刺激するそれらの能力を評価した。細胞に、一本鎖分子に対する哺乳動物発現プラスミドをトランスフェクトした。メガヌクレアーゼによる天然のセーフハーバー遺伝子座の切断は、エラーを起こしやすいプロセスである非相同末端結合の基質を生じ、メガヌクレアーゼ標的部位で小さな挿入又は欠失をもたらし得る。このため、内因性セーフハーバー遺伝子座で突然変異が起こる頻度は、メガヌクレアーゼによるゲノム標的部位の切断効率を示す。
【0338】
実施例9.1. 材料及び方法
a)メガヌクレアーゼ発現プラスミド
本実施例で使用されるメガヌクレアーゼに対するコード配列は、哺乳動物発現ベクターにクローニングされ、表XVIに挙げるプラスミドを生じた。
【0339】
【表33】
【0340】
b)セーフハーバー遺伝子座の突然変異誘発実験
ヒト胎児腎臓293H細胞(Invitrogen)を、10cmディッシュ1枚当たり1×106細胞の密度で、完全培地(2mM L−グルタミン、ペニシリン(100UI/ml)、ストレプトマイシン(100μg/ml)、アムホテリシンB(Fongizone)(0.25μg/ml)(Invitrogen-Life Science)及び10%FBSを添加したDMEM)中で平板培養した。翌日、Lipofectamine 2000トランスフェクション試薬(Invitrogen)を用い、供給業者のプロトコルに従って細胞に3μgの一本鎖メガヌクレアーゼ発現ベクターをトランスフェクトした。37℃で2日間〜6日間のインキュベーション後、細胞をトリプシン処理し、DNeasy血液及び組織キット(Qiagen)を用い、供給業者のプロトコルに従ってゲノムDNA抽出を行った。
【0341】
c)突然変異誘発事象のディープシークエンシング分析
突然変異誘発の頻度をディープシークエンシング分析によって決定した。オリゴヌクレオチドを各々のセーフハーバー標的の周囲のDNA断片のPCR増幅のために設計した。それらを表XVIIに挙げる。
【0342】
【表34】
【0343】
ヌクレオチドを添加し、454シークエンシングシステム(454 Life Sciences)上で、シークエンシングサービスを提供する会社(GATC Biotech AG,Germany)によって供給される特異的なアダプター配列(5’−CCATCTCATCCCTGCGTGTCTCCGACTCAG−3’;配列番号324)及び(5’−CCTATCCCCTGTGTGCCTTGGCAGTCTCAG−3’;配列番号325)に隣接する断片を得た。平均して18000個の配列が2つ又は3つの単位複製配列のプール(各々500ng)から得られた。シークエンシング後、種々のサンプルを、上記のアダプターの最初に導入したバーコード配列に基づき同定した。
【0344】
実施例9.2. 結果
ヒト胎児腎臓293H細胞に、一本鎖セーフハーバーメガヌクレアーゼを発現するプラスミドをトランスフェクトした。37℃で2日間〜6日間のインキュベーション後、ゲノムDNAを単離し、PCRを用いてメガヌクレアーゼ標的部位の周囲のゲノム配列を増幅した。次いで、各々のセーフハーバー標的の切断部位における挿入又は欠失の事象(InDel)の存在について配列を分析した。結果を表XVIIIにまとめる。
【0345】
【表35】
【0346】
実施例10: 結論
結論として、実施例1、実施例2、実施例3及び実施例5から、SH3遺伝子座、SH4遺伝子座及びSH6遺伝子座を切断することが可能なI−CreIヘテロ二量体タンパク質及び一本鎖メガヌクレアーゼの両方を得ることができることが実証される。さらに、これらのエンドヌクレアーゼは、これらの遺伝子座を強い切断活性で切断することが可能である。
【0347】
実施例4から、SH3遺伝子座及びSH4遺伝子座を切断することが可能な一本鎖メガヌクレアーゼが、ヒト細胞の標的部位への導入遺伝子の効率的な挿入を可能にすることが実証される。
【0348】
このため、これらのエンドヌクレアーゼは、個体のSH3遺伝子座、SH4遺伝子座又はSH6遺伝子座への導入遺伝子の挿入に有利に使用することができる。
【0349】
実施例6から、本発明による少なくとも2つの一本鎖分子が、内因性sh6遺伝子座で高レベルの遺伝子標的化を誘導することが可能であることが実証される。
【0350】
実施例7から、遺伝子座での標的化組込みが機能的な導入遺伝子の発現を支持し得ることが実証される。
【0351】
実施例8から、遺伝子座での標的化組込みが、標的配列の近傍に位置する5個の遺伝子の発現を実質的に変更しないことが実証される。
【0352】
実施例9から、セーフハーバー配列を標的とする種々のメガヌクレアーゼの突然変異誘発頻度(該メガヌクレアーゼによるゲノム標的部位の切断効率を示す)が実証される。
【特許請求の範囲】
【請求項1】
導入遺伝子を個体のゲノムに挿入する際に使用される標的配列を切断することが可能な変異型エンドヌクレアーゼであって、
i.前記ゲノムが前記標的配列を含む遺伝子座を含み、
ii.前記標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連しない、変異型エンドヌクレアーゼ。
【請求項2】
前記導入遺伝子の挿入が、前記標的配列の近傍に位置する遺伝子の発現を実質的に変更しない、請求項1に記載のエンドヌクレアーゼ。
【請求項3】
前記標的配列が、最近接遺伝子から少なくとも100kb離れて位置する、請求項1又は2に記載のエンドヌクレアーゼ。
【請求項4】
ホーミングエンドヌクレアーゼである、請求項1〜3のいずれか一項に記載のエンドヌクレアーゼ。
【請求項5】
ヒト染色体21q21.1上のSH6遺伝子座、ヒト染色体6p25.1上のSH3遺伝子座、ヒト染色体7q31.2上のSH4遺伝子座、ヒト染色体13q34上のSH12遺伝子座、ヒト染色体3p12.2上のSH13遺伝子座、ヒト染色体22上のSH19遺伝子座、ヒト染色体12q21.2上のSH20遺伝子座、ヒト染色体3p24.1上のSH21遺伝子座、ヒト染色体6p12.2上のSH33遺伝子座、ヒト染色体2p16.1上のSH7遺伝子座、ヒト染色体5上のSH8遺伝子座、SH18遺伝子座、SH31遺伝子座、SH38遺伝子座、SH39遺伝子座、SH41遺伝子座、SH42遺伝子座、SH43遺伝子座、SH44遺伝子座、SH45遺伝子座、SH46遺伝子座、SH47遺伝子座、SH48遺伝子座、SH49遺伝子座、SH50遺伝子座、SH51遺伝子座、SH52遺伝子座、SH70遺伝子座、SH71遺伝子座、SH72遺伝子座、SH73遺伝子座、SH74遺伝子座、SH75遺伝子座、SH101遺伝子座、SH106遺伝子座、SH107遺伝子座、SH102遺伝子座、SH105遺伝子座、SH103遺伝子座、SH104遺伝子座、SH113遺伝子座、SH109遺伝子座、SH112遺伝子座、SH108遺伝子座、SH110遺伝子座、SH114遺伝子座、SH116遺伝子座、SH111遺伝子座、SH115遺伝子座、SH121遺伝子座、SH120遺伝子座、SH122遺伝子座、SH117遺伝子座、SH118遺伝子座、SH119遺伝子座、SH123遺伝子座、SH126遺伝子座、SH128遺伝子座、SH129遺伝子座、SH124遺伝子座、SH131遺伝子座、SH125遺伝子座、SH127遺伝子座、SH130遺伝子座、SH11遺伝子座、SH17遺伝子座、SH23遺伝子座、SH34遺伝子座、SH40遺伝子座、SH53遺伝子座、SH54遺伝子座、SH55遺伝子座、SH56遺伝子座、SH57遺伝子座、SH58遺伝子座、SH59遺伝子座、SH60遺伝子座、SH61遺伝子座、SH62遺伝子座、SH65遺伝子座、SH67遺伝子座、SH68遺伝子座及びSH69遺伝子座からなる群から選択される遺伝子座内に位置する標的配列を切断することが可能である、請求項1〜4のいずれか一項に記載のエンドヌクレアーゼ。
【請求項6】
各々が配列番号1又は配列番号42と少なくとも80%同一な配列を含む2つの単量体を含む変異型二量体I−CreIタンパク質であって、
i.前記二量体I−CreIタンパク質が、個体の遺伝子座内に位置する標的配列を切断することが可能であり、該標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連せず、
ii.前記標的配列が配列番号4の配列を含まない、変異型二量体I−CreIタンパク質。
【請求項7】
ヒト染色体21q21.1上のSH6遺伝子座、ヒト染色体6p25.1上のSH3遺伝子座、ヒト染色体7q31.2上のSH4遺伝子座、ヒト染色体13q34上のSH12遺伝子座、ヒト染色体3p12.2上のSH13遺伝子座、ヒト染色体22上のSH19遺伝子座、ヒト染色体12q21.2上のSH20遺伝子座、ヒト染色体3p24.1上のSH21遺伝子座、ヒト染色体6p12.2上のSH33遺伝子座、ヒト染色体2p16.1上のSH7遺伝子座、ヒト染色体5上のSH8遺伝子座、SH18遺伝子座、SH31遺伝子座、SH38遺伝子座、SH39遺伝子座、SH41遺伝子座、SH42遺伝子座、SH43遺伝子座、SH44遺伝子座、SH45遺伝子座、SH46遺伝子座、SH47遺伝子座、SH48遺伝子座、SH49遺伝子座、SH50遺伝子座、SH51遺伝子座、SH52遺伝子座、SH70遺伝子座、SH71遺伝子座、SH72遺伝子座、SH73遺伝子座、SH74遺伝子座、SH75遺伝子座、SH101遺伝子座、SH106遺伝子座、SH107遺伝子座、SH102遺伝子座、SH105遺伝子座、SH103遺伝子座、SH104遺伝子座、SH113遺伝子座、SH109遺伝子座、SH112遺伝子座、SH108遺伝子座、SH110遺伝子座、SH114遺伝子座、SH116遺伝子座、SH111遺伝子座、SH115遺伝子座、SH121遺伝子座、SH120遺伝子座、SH122遺伝子座、SH117遺伝子座、SH118遺伝子座、SH119遺伝子座、SH123遺伝子座、SH126遺伝子座、SH128遺伝子座、SH129遺伝子座、SH124遺伝子座、SH131遺伝子座、SH125遺伝子座、SH127遺伝子座、SH130遺伝子座、SH11遺伝子座、SH17遺伝子座、SH23遺伝子座、SH34遺伝子座、SH40遺伝子座、SH53遺伝子座、SH54遺伝子座、SH55遺伝子座、SH56遺伝子座、SH57遺伝子座、SH58遺伝子座、SH59遺伝子座、SH60遺伝子座、SH61遺伝子座、SH62遺伝子座、SH65遺伝子座、SH67遺伝子座、SH68遺伝子座及びSH69遺伝子座からなる群から選択される遺伝子座内に位置する標的配列を切断することが可能である、請求項6に記載の二量体I−CreIタンパク質。
【請求項8】
ヒト染色体21q21.1上のSH6遺伝子座内に位置する標的配列を切断することが可能である、請求項6又は7に記載の二量体I−CreIタンパク質。
【請求項9】
前記標的配列が配列番号59の配列を含む、請求項8に記載の二量体I−CreIタンパク質。
【請求項10】
請求項6〜9のいずれか一項に記載の二量体I−CreIタンパク質の単量体を含む融合タンパク質。
【請求項11】
配列番号81、配列番号82〜配列番号85、配列番号294、配列番号295、配列番号76〜配列番号80、配列番号25〜配列番号40、配列番号86〜配列番号96、配列番号127〜配列番号150、配列番号182〜配列番号213、配列番号235〜配列番号270及び配列番号275〜配列番号278からなる群から選択される配列を含む、請求項10に記載の融合タンパク質。
【請求項12】
請求項1〜5のいずれか一項に記載のエンドヌクレアーゼ又は請求項6〜11のいずれか一項に記載のタンパク質をコードする核酸。
【請求項13】
請求項12に記載の核酸を含む発現ベクター。
【請求項14】
導入遺伝子を含む標的化構築物と、請求項1〜5のいずれか一項に記載のエンドヌクレアーゼ又は請求項6〜11のいずれか一項に記載のタンパク質によって認識される標的配列に隣接するゲノム配列に相同な2つの配列とを更に含む、請求項13に記載の発現ベクター。
【請求項15】
請求項13に記載の発現ベクター並びに、
導入遺伝子を含む標的化構築物と、請求項1〜5のいずれか一項に記載のエンドヌクレアーゼ又は請求項6〜11のいずれか一項に記載のタンパク質によって認識される標的配列のゲノム配列に相同な2つの配列とを含むベクターと、
の組合せ。
【請求項16】
請求項14に記載の発現ベクター又は請求項15に記載の組合せと、薬学的に許容可能な担体とを含む医薬組成物。
【請求項17】
導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入するための請求項1〜5のいずれか一項に記載のエンドヌクレアーゼ又は請求項6〜11のいずれか一項に記載のタンパク質又は請求項12に記載の核酸又は請求項13若しくは14に記載の発現ベクター又は請求項15に記載の組合せの使用であって、該使用は治療目的でない、使用。
【請求項18】
遺伝性障害の非ヒト動物モデルを作製するための、請求項17に記載の使用。
【請求項19】
組換えタンパク質を製造するための、請求項17に記載の使用。
【請求項20】
個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法であって、
a)前記個体のゲノム内で、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)を選択する工程と、
b)前記RISの200kb上流及び200kb下流に伸びるゲノム領域を規定する工程と、
c)前記ゲノム領域内に位置する標的配列を切断することが可能な野生型エンドヌクレアーゼを同定するか、又は変異型エンドヌクレアーゼを構築する工程と、
を含む、個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法。
【請求項1】
導入遺伝子を個体のゲノムに挿入する際に使用される標的配列を切断することが可能な変異型エンドヌクレアーゼであって、
i.前記ゲノムが前記標的配列を含む遺伝子座を含み、
ii.前記標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連しない、変異型エンドヌクレアーゼ。
【請求項2】
前記導入遺伝子の挿入が、前記標的配列の近傍に位置する遺伝子の発現を実質的に変更しない、請求項1に記載のエンドヌクレアーゼ。
【請求項3】
前記標的配列が、最近接遺伝子から少なくとも100kb離れて位置する、請求項1又は2に記載のエンドヌクレアーゼ。
【請求項4】
ホーミングエンドヌクレアーゼである、請求項1〜3のいずれか一項に記載のエンドヌクレアーゼ。
【請求項5】
ヒト染色体21q21.1上のSH6遺伝子座、ヒト染色体6p25.1上のSH3遺伝子座、ヒト染色体7q31.2上のSH4遺伝子座、ヒト染色体13q34上のSH12遺伝子座、ヒト染色体3p12.2上のSH13遺伝子座、ヒト染色体22上のSH19遺伝子座、ヒト染色体12q21.2上のSH20遺伝子座、ヒト染色体3p24.1上のSH21遺伝子座、ヒト染色体6p12.2上のSH33遺伝子座、ヒト染色体2p16.1上のSH7遺伝子座、ヒト染色体5上のSH8遺伝子座、SH18遺伝子座、SH31遺伝子座、SH38遺伝子座、SH39遺伝子座、SH41遺伝子座、SH42遺伝子座、SH43遺伝子座、SH44遺伝子座、SH45遺伝子座、SH46遺伝子座、SH47遺伝子座、SH48遺伝子座、SH49遺伝子座、SH50遺伝子座、SH51遺伝子座、SH52遺伝子座、SH70遺伝子座、SH71遺伝子座、SH72遺伝子座、SH73遺伝子座、SH74遺伝子座、SH75遺伝子座、SH101遺伝子座、SH106遺伝子座、SH107遺伝子座、SH102遺伝子座、SH105遺伝子座、SH103遺伝子座、SH104遺伝子座、SH113遺伝子座、SH109遺伝子座、SH112遺伝子座、SH108遺伝子座、SH110遺伝子座、SH114遺伝子座、SH116遺伝子座、SH111遺伝子座、SH115遺伝子座、SH121遺伝子座、SH120遺伝子座、SH122遺伝子座、SH117遺伝子座、SH118遺伝子座、SH119遺伝子座、SH123遺伝子座、SH126遺伝子座、SH128遺伝子座、SH129遺伝子座、SH124遺伝子座、SH131遺伝子座、SH125遺伝子座、SH127遺伝子座、SH130遺伝子座、SH11遺伝子座、SH17遺伝子座、SH23遺伝子座、SH34遺伝子座、SH40遺伝子座、SH53遺伝子座、SH54遺伝子座、SH55遺伝子座、SH56遺伝子座、SH57遺伝子座、SH58遺伝子座、SH59遺伝子座、SH60遺伝子座、SH61遺伝子座、SH62遺伝子座、SH65遺伝子座、SH67遺伝子座、SH68遺伝子座及びSH69遺伝子座からなる群から選択される遺伝子座内に位置する標的配列を切断することが可能である、請求項1〜4のいずれか一項に記載のエンドヌクレアーゼ。
【請求項6】
各々が配列番号1又は配列番号42と少なくとも80%同一な配列を含む2つの単量体を含む変異型二量体I−CreIタンパク質であって、
i.前記二量体I−CreIタンパク質が、個体の遺伝子座内に位置する標的配列を切断することが可能であり、該標的配列がレトロウイルス挿入部位(RIS)から最大で200kb離れて位置し、該RISが、がんとも異常細胞増殖とも関連せず、
ii.前記標的配列が配列番号4の配列を含まない、変異型二量体I−CreIタンパク質。
【請求項7】
ヒト染色体21q21.1上のSH6遺伝子座、ヒト染色体6p25.1上のSH3遺伝子座、ヒト染色体7q31.2上のSH4遺伝子座、ヒト染色体13q34上のSH12遺伝子座、ヒト染色体3p12.2上のSH13遺伝子座、ヒト染色体22上のSH19遺伝子座、ヒト染色体12q21.2上のSH20遺伝子座、ヒト染色体3p24.1上のSH21遺伝子座、ヒト染色体6p12.2上のSH33遺伝子座、ヒト染色体2p16.1上のSH7遺伝子座、ヒト染色体5上のSH8遺伝子座、SH18遺伝子座、SH31遺伝子座、SH38遺伝子座、SH39遺伝子座、SH41遺伝子座、SH42遺伝子座、SH43遺伝子座、SH44遺伝子座、SH45遺伝子座、SH46遺伝子座、SH47遺伝子座、SH48遺伝子座、SH49遺伝子座、SH50遺伝子座、SH51遺伝子座、SH52遺伝子座、SH70遺伝子座、SH71遺伝子座、SH72遺伝子座、SH73遺伝子座、SH74遺伝子座、SH75遺伝子座、SH101遺伝子座、SH106遺伝子座、SH107遺伝子座、SH102遺伝子座、SH105遺伝子座、SH103遺伝子座、SH104遺伝子座、SH113遺伝子座、SH109遺伝子座、SH112遺伝子座、SH108遺伝子座、SH110遺伝子座、SH114遺伝子座、SH116遺伝子座、SH111遺伝子座、SH115遺伝子座、SH121遺伝子座、SH120遺伝子座、SH122遺伝子座、SH117遺伝子座、SH118遺伝子座、SH119遺伝子座、SH123遺伝子座、SH126遺伝子座、SH128遺伝子座、SH129遺伝子座、SH124遺伝子座、SH131遺伝子座、SH125遺伝子座、SH127遺伝子座、SH130遺伝子座、SH11遺伝子座、SH17遺伝子座、SH23遺伝子座、SH34遺伝子座、SH40遺伝子座、SH53遺伝子座、SH54遺伝子座、SH55遺伝子座、SH56遺伝子座、SH57遺伝子座、SH58遺伝子座、SH59遺伝子座、SH60遺伝子座、SH61遺伝子座、SH62遺伝子座、SH65遺伝子座、SH67遺伝子座、SH68遺伝子座及びSH69遺伝子座からなる群から選択される遺伝子座内に位置する標的配列を切断することが可能である、請求項6に記載の二量体I−CreIタンパク質。
【請求項8】
ヒト染色体21q21.1上のSH6遺伝子座内に位置する標的配列を切断することが可能である、請求項6又は7に記載の二量体I−CreIタンパク質。
【請求項9】
前記標的配列が配列番号59の配列を含む、請求項8に記載の二量体I−CreIタンパク質。
【請求項10】
請求項6〜9のいずれか一項に記載の二量体I−CreIタンパク質の単量体を含む融合タンパク質。
【請求項11】
配列番号81、配列番号82〜配列番号85、配列番号294、配列番号295、配列番号76〜配列番号80、配列番号25〜配列番号40、配列番号86〜配列番号96、配列番号127〜配列番号150、配列番号182〜配列番号213、配列番号235〜配列番号270及び配列番号275〜配列番号278からなる群から選択される配列を含む、請求項10に記載の融合タンパク質。
【請求項12】
請求項1〜5のいずれか一項に記載のエンドヌクレアーゼ又は請求項6〜11のいずれか一項に記載のタンパク質をコードする核酸。
【請求項13】
請求項12に記載の核酸を含む発現ベクター。
【請求項14】
導入遺伝子を含む標的化構築物と、請求項1〜5のいずれか一項に記載のエンドヌクレアーゼ又は請求項6〜11のいずれか一項に記載のタンパク質によって認識される標的配列に隣接するゲノム配列に相同な2つの配列とを更に含む、請求項13に記載の発現ベクター。
【請求項15】
請求項13に記載の発現ベクター並びに、
導入遺伝子を含む標的化構築物と、請求項1〜5のいずれか一項に記載のエンドヌクレアーゼ又は請求項6〜11のいずれか一項に記載のタンパク質によって認識される標的配列のゲノム配列に相同な2つの配列とを含むベクターと、
の組合せ。
【請求項16】
請求項14に記載の発現ベクター又は請求項15に記載の組合せと、薬学的に許容可能な担体とを含む医薬組成物。
【請求項17】
導入遺伝子を細胞、組織又は非ヒト動物のゲノムに挿入するための請求項1〜5のいずれか一項に記載のエンドヌクレアーゼ又は請求項6〜11のいずれか一項に記載のタンパク質又は請求項12に記載の核酸又は請求項13若しくは14に記載の発現ベクター又は請求項15に記載の組合せの使用であって、該使用は治療目的でない、使用。
【請求項18】
遺伝性障害の非ヒト動物モデルを作製するための、請求項17に記載の使用。
【請求項19】
組換えタンパク質を製造するための、請求項17に記載の使用。
【請求項20】
個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法であって、
a)前記個体のゲノム内で、がんとも異常細胞増殖とも関連しないレトロウイルス挿入部位(RIS)を選択する工程と、
b)前記RISの200kb上流及び200kb下流に伸びるゲノム領域を規定する工程と、
c)前記ゲノム領域内に位置する標的配列を切断することが可能な野生型エンドヌクレアーゼを同定するか、又は変異型エンドヌクレアーゼを構築する工程と、
を含む、個体のゲノムへの導入遺伝子の挿入に好適なエンドヌクレアーゼを得る方法。
【図1】


【図2】
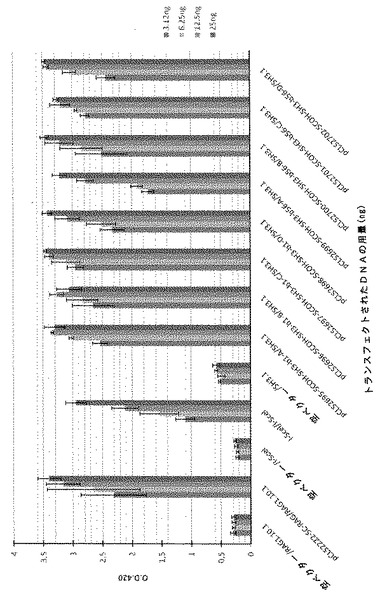
【図3】
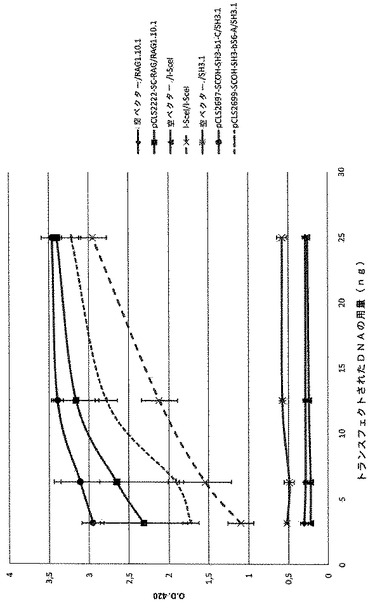
【図4】

【図5】

【図6】
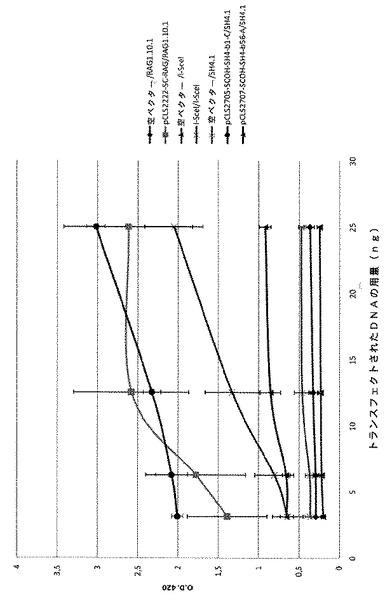
【図7】
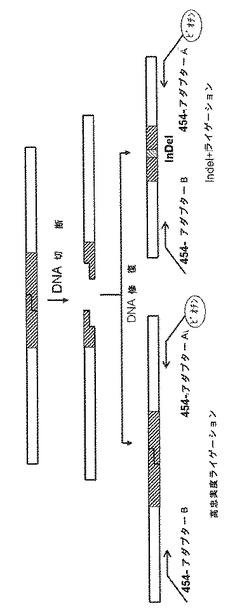
【図8】
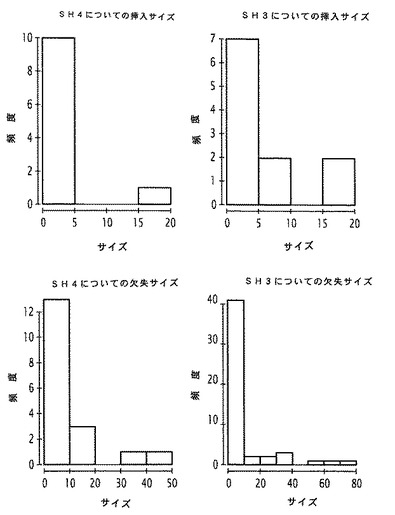
【図9】


【図10】
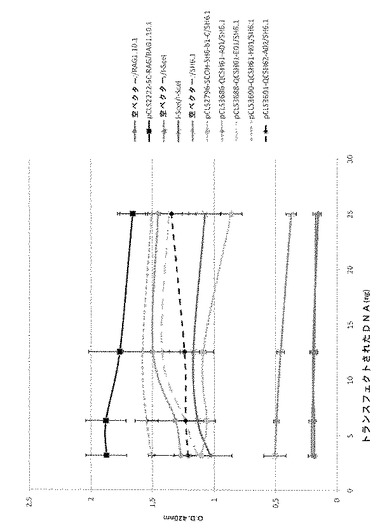
【図11】

【図12】

【図13】

【図14】

【図15】
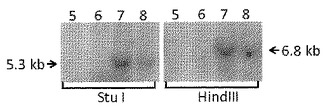
【図16】
【図17】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【公表番号】特表2013−520190(P2013−520190A)
【公表日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願番号】特願2012−554368(P2012−554368)
【出願日】平成23年2月28日(2011.2.28)
【国際出願番号】PCT/EP2011/052916
【国際公開番号】WO2011/104382
【国際公開日】平成23年9月1日(2011.9.1)
【出願人】(508245231)
【氏名又は名称原語表記】CELLECTIS
【住所又は居所原語表記】102 avenue Gaston Roussel,F−93235 Romainville Cedex,France
【Fターム(参考)】
【公表日】平成25年6月6日(2013.6.6)
【国際特許分類】
【出願日】平成23年2月28日(2011.2.28)
【国際出願番号】PCT/EP2011/052916
【国際公開番号】WO2011/104382
【国際公開日】平成23年9月1日(2011.9.1)
【出願人】(508245231)
【氏名又は名称原語表記】CELLECTIS
【住所又は居所原語表記】102 avenue Gaston Roussel,F−93235 Romainville Cedex,France
【Fターム(参考)】
[ Back to top ]