
ゼニゴケ由来の不飽和脂肪酸合成系酵素遺伝子及びその利用
【課題】 高等植物中でアラキドン酸やエイコサペンタエン酸等の高度不飽和脂肪酸を効率よく生産すること。
【解決手段】 同一種のゼニゴケ(Marchantia polymorpha)から、Δ5脂肪酸不飽和化酵素遺伝子、Δ6脂肪酸不飽和化酵素遺伝子及びΔ6脂肪酸鎖長延長酵素遺伝子を単離する。これらの遺伝子を高等植物に導入し発現させることにより、アラキドン酸やエイコサペンタエン酸を生産し得る形質転換植物体を取得する。
【解決手段】 同一種のゼニゴケ(Marchantia polymorpha)から、Δ5脂肪酸不飽和化酵素遺伝子、Δ6脂肪酸不飽和化酵素遺伝子及びΔ6脂肪酸鎖長延長酵素遺伝子を単離する。これらの遺伝子を高等植物に導入し発現させることにより、アラキドン酸やエイコサペンタエン酸を生産し得る形質転換植物体を取得する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、ゼニゴケ(Marchantia polymorpha)由来の不飽和脂
肪酸合成系遺伝子、すなわち、Δ5脂肪酸不飽和化酵素、Δ6脂肪酸不飽和化酵素、及び
Δ6脂肪酸鎖長延長酵素の遺伝子とその利用に関するものである。
【背景技術】
【0002】
アラキドン酸、エイコサペンタエン酸(以下、適宜「EPA」と略記する)等の高度不
飽和脂肪酸(Polyunsaturated fatty acid;PUFA)は、
ヒトにおいて神経系を中心とした細胞膜脂質に多く含まれている。これらの高度不飽和脂
肪酸は、プロスタグランジンやロイコトリエンといった、生理活性物質の前駆体として作
用しており、薬理学的に非常に重要である。近年、アラキドン酸やEPAを含む健康食品
も販売されている。また、脂肪酸は、洗剤や生分解性プラスチックの原料にもなるため、
素材としても注目されている。
【0003】
高度不飽和脂肪酸は、現在、培養微生物又は魚油からの抽出により生産されている。こ
のため、生産コストが高いこと、エネルギー使用量・廃棄物量が多くなること、特に魚油
から調製する方法では、魚資源が限られていることが問題になっている。
【0004】
アラキドン酸及びEPAは、それぞれリノール酸及びα−リノレン酸を起点として、Δ
6不飽和化、脂肪酸鎖長延長及びΔ5不飽和化という3つの連続した反応により生合成さ
れると考えられている。これらの反応は、それぞれ、Δ6脂肪酸不飽和化酵素(以下、「
Δ6不飽和化酵素」と略記する)、Δ6脂肪酸鎖長延長酵素(以下、「Δ6鎖長延長酵素
」と略記する)及びΔ5脂肪酸不飽和化酵素(以下、「Δ5不飽和化酵素」と略記する)
により触媒される。
【0005】
Δ6不飽和化酵素の遺伝子は、いくつかの植物種でクローン化されている。例えば、ケ
イ藻、ヒメツリガネゴケ、ヤノウエノアカゴケ、ボリジ、ムラサキ、サクラソウ及びアネ
モネからΔ6不飽和化酵素の遺伝子がクローン化されている。また、植物以外では、糸状
菌、線虫、ラン藻、ラット及びヒトからΔ6不飽和化酵素遺伝子がクローン化されている
(非特許文献1:「Eur.J.Biochem.269,p4105,2002」、非
特許文献2:「Plant J.15,p39,1998」、非特許文献3:「Eur.
J.Biochem.,267.p3801,2000」、非特許文献4:「Proc.
Natl.Acad.Sci.USA 94,p4211,1997」、非特許文献5:
「Lipids 37,417,2002」、非特許文献6:「FEBS Lett.5
42,p100,2003」、非特許文献7:「Whitney et al.,Pla
nta Epub 2003」、非特許文献8:「Lipids 34,p649,19
99」、非特許文献9:「Gene,238,p445 1999」、非特許文献10:
「Biochem J.330,p611 1998」、非特許文献11:「Plant
Mol.Biol.,22,p293 1993」、非特許文献12:「Bioche
m.Biophys.res.Commun.255,p575,1999」、非特許文
献13:「J.Biol.Chem.274,p471,1999」参照)。また、ラン
藻のものを除いて、これらのΔ6不飽和化酵素は、いずれもN末端にシトクロムb5ドメ
インが存在する。
【0006】
Δ6鎖長延長酵素の遺伝子は、最初に糸状菌及び線虫からクローン化された(非特許文
献14:「Proc.Natl.Acad.Sci.USA 97,p8284,200
0」、非特許文献15:「Proc.Natl.Acad.Sci.USA 97,p6
421,2000」参照)。植物種では、唯一ヒメツリガネからクローン化されている(
非特許文献16:「Plant J.31,p255,2002」参照)。
【0007】
酵母(Saccharomyces cerevisiae)には、スフィンゴ脂質の
長鎖飽和アシル鎖合成に関与するELO2蛋白及びELO3蛋白が存在し(非特許文献1
7:「J.Biol.Chem.,272,p17376,1997」参照)、Δ6鎖長
延長酵素のアミノ酸配列は、これらと相同性を示す。一方、植物には、別タイプの脂肪酸
鎖長延長酵素であるβ−ケトアシルCoA合成酵素(KCS)が存在する。この酵素は、
長鎖飽和/一価不飽和脂肪酸の鎖長延長を触媒する(非特許文献15及び非特許文献18
:「Plant Cell 7,p309,1995」参照)。しかしながら、Δ6鎖長
延長酵素遺伝子及び酵母ELO2/ELO3遺伝子は、KCS遺伝子との直接的な進化上
の関係は見られない(非特許文献15及び16参照)。
【0008】
Δ5不飽和化酵素の遺伝子は、糸状菌から、初めてクローン化された(非特許文献19
:「J.Biol.Chem.273,p29360,1998」、非特許文献20:「
J.Biol.Chem.273,p19055」参照)。Δ5不飽和化酵素の構造はΔ
6不飽和化酵素と共通しており、N末端にシトクロムb5ドメインを有する。Δ5不飽和
化酵素遺伝子は、ケイ藻、線虫、ラット、ヒト、ヒメツリガネゴケなどからクローン化さ
れている(非特許文献1,非特許文献21:「FEBS Lett.439,p215,
1998」、非特許文献22:「Arch.Biochem.Biophys.391,
p8,2001」、非特許文献23:「J.Biol.Chem.274,p37335
,1999」、非特許文献24:「J.Biol.Chem.278,35115,20
03」参照)。
【0009】
陸上植物は、コケ植物(コケ植物門(Bryophyta))、シダ植物、裸子植物及
び被子植物から構成されている。コケ植物は陸上植物の中で最も古くに分岐した群であり
、セン類(セン類網(Bryosida))、タイ類(タイ類網(Hepaticops
ida))及びツノゴケ類の3つのグループから構成されている。ゼニゴケは、上記生物
のうちヒメツリガネゴケに分類上最も近いが、ヒメツリゴケはセン類に属しており、ゼニ
ゴケはタイ類網の中のゼニゴケ亞網(Marchantiidae)に属している。上記
3つのグループは、約4億3千年前には、すでに分岐していたことは確かである。したが
って、同じコケといっても、ヒメツリガネゴケとゼニゴケとは、例えば2億年前に分化し
たシロイナズナとイネとの違いどころでなく、進化上大きく異なる(非特許文献25:「
1103615143968_0.html」参照)。
【0010】
ゼニゴケ由来の高度不飽和脂肪酸合成系酵素遺伝子としては、上記のKCS様の鎖長延
長酵素遺伝子のMpFAE2及びMpFAE3が取得されている(非特許文献26:「B
iosci.Biotechnol.Biochem.67,p605,2003」、非
特許文献27:「Biosci.Biotechnol.Biochem.67,p16
67,2003」参照)。しかしながら、MpFAE2及びMpFAE3はΔ6鎖長延長
酵素遺伝子ではない。
【0011】
上述のように、多くの高度脂肪酸合成系酵素遺伝子が様々な生物種からクローン化され
ているが、アラキドン酸、EPA等の、C20以上で不飽和度4以上の高度不飽和脂肪酸
を植物中で生産させた例は少ない。このような例として、ケイ藻由来のΔ6不飽和化酵素
及びΔ5不飽和化酵素と、ヒメツリガネゴケ由来のΔ6鎖長延長酵素とをアマで発現させ
、アラキドン酸及びEPAを生産させた報告があるが、その詳細は不明である(非特許文
献24参照)。
【0012】
上述のように、アラキドン酸やEPA等の高度不飽和脂肪酸の生産は、培養微生物又は
魚油からの抽出により生産されているため、生産コストが高いこと、エネルギー使用量・
廃棄物量が多くなること、魚資源が限られていること等の問題点を有している。アラキド
ン酸やEPA等の高度不飽和脂肪酸は、分子内に二重結合を複数有するというユニークな
物性を持っているため、様々な工業用途(例えばフィルム、生分解性プラスチック、機能
性繊維、潤滑油、洗剤の素材等)にも利用可能となる。このような高度不飽和脂肪酸を遺
伝子組み換え植物により生産することにより、生産コストを低減でき、同時に、より環境
にやさしい生産プロセスを実現することができると期待される。遺伝子組み換え技術を用
いて、これらの高度不飽和脂肪酸を、油糧植物で大量生産できるようになれば、安価な多
目的原料として非常に有用である。
【0013】
一方、植物に異種生物の遺伝子を発現させる場合、その遺伝子が植物内でどの程度良好
に機能するかは、転写、翻訳、その後の修飾などの過程があるため、予想することは困難
である。特に、複数の異種生物の遺伝子を発現させる場合、上記非特許文献24のように
異なる生物種由来の複数の遺伝子を発現させるより、同一の種に由来する複数の遺伝子を
発現させるほうが植物内で良好に機能することが予想される。また、最初の陸上植物であ
るコケ類のゼニゴケは、高等植物のモデル系として注目されており、その遺伝子は植物内
で良好に機能することが期待できる。したがって、ゼニゴケ由来の高度不飽和脂肪酸合成
酵素遺伝子、すなわちΔ5不飽和化酵素遺伝子、Δ6不飽和化酵素遺伝子及びΔ6鎖長延
長酵素遺伝子を取得することができれば、これらの遺伝子を植物に導入することにより、
アラキドン酸やEPAが植物内で効率よく蓄積されることが期待できる。
【0014】
また、ゼニゴケと同じコケ植物のヒメツリガネゴケからはΔ5不飽和化酵素遺伝子、Δ
6不飽和化酵素遺伝子及びΔ6鎖長延長酵素遺伝子がクローン化されているが、ゼニゴケ
とヒメツリガネゴケは進化上大きく異なっており、ヒメツリガネゴケの遺伝子に基づいて
ゼニゴケの遺伝子を取得することは、現在の技術水準をもってしても容易にできることで
はない。
【発明の概要】
【発明が解決しようとする課題】
【0015】
本発明は、上記従来の問題点に鑑みなされたものであって、その目的は、高等植物中で
アラキドン酸やEPAを生産し得る、ゼニゴケ(Marchantia polymor
pha)由来の不飽和脂肪酸合成酵素遺伝子、すなわちΔ5不飽和化酵素遺伝子、Δ6不
飽和化酵素遺伝子及びΔ6鎖長延長酵素遺伝子、及びその利用法を提供することにある。
【課題を解決するための手段】
【0016】
本発明者らは、上記の課題を解決するために鋭意検討した結果、ゼニゴケ(March
antia polymorpha)由来のcDNAクローンから、上記Δ6不飽和化酵
素、Δ5不飽和化酵素及びΔ6鎖長延長酵素をコードする遺伝子を同定し、更にこれらの
遺伝子をメタノール資化性酵母(Pichia pastoris)に導入して発現させ
ることに成功し、これらの遺伝子を発現させたたんぱく質が、それぞれΔ6不飽和化、Δ
5不飽和化及びΔ6鎖長延長の酵素活性を有することを見出し、本発明を完成するに至っ
た。すなわち、本発明は、以下の発明を包含する。
【0017】
(1)配列番号1に示される塩基配列からなるDNAの全部又は一部、あるいは当該D
NAと相補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイ
ブリダイズし、かつΔ6脂肪酸不飽和化活性を有するたんぱく質をコードするゼニゴケ目
生物由来の遺伝子。
【0018】
(2)Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号1に示される塩基配列を有する
遺伝子。(b)配列番号1に示される塩基配列からなるDNA、又は当該DNAと相補的
な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0019】
(3)Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号1に示される塩基配列のうち、
253ないし1698番目の塩基配列を有する遺伝子。(b)配列番号1に示される塩基
配列のうち、253ないし1698番目の塩基配列からなるDNA、又は当該DNAと相
補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0020】
(4)Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号2に示されるアミノ酸配列から
なるたんぱく質をコードする遺伝子。(b)配列番号2に示されるアミノ酸配列の1個又
はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなる
たんぱく質をコードする遺伝子。
【0021】
(5)配列番号3に示される塩基配列からなるDNAの全部又は一部、あるいは当該D
NAと相補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイ
ブリダイズし、かつΔ6脂肪酸鎖長延長活性を有するたんぱく質をコードするゼニゴケ目
生物由来の遺伝子。
【0022】
(6)Δ6脂肪酸鎖長延長活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号3に示される塩基配列を有する
遺伝子。(b)配列番号3に示される塩基配列からなるDNA、又は当該DNAと相補的
な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0023】
(7)Δ6脂肪酸鎖長延長活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号1に示される塩基配列のうち、
194ないし1066番目の塩基配列を有する遺伝子。(b)配列番号1に示される塩基
配列のうち、194ないし1066番目の塩基配列からなるDNA、又は当該DNAと相
補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0024】
(8)Δ6脂肪酸鎖長延長活性を有するゼニゴケ目由来のたんぱく質をコードする、下
記(a)又は(b)に記載の遺伝子。(a)配列番号4に示されるアミノ酸配列からなる
たんぱく質、(b)配列番号4に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が
置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする
遺伝子。
【0025】
(9)配列番号5に示される塩基配列からなるDNAの全部又は一部、あるいは当該D
NAと相補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイ
ブリダイズし、かつΔ5脂肪酸不飽和化活性を有するたんぱく質をコードするゼニゴケ目
生物由来の遺伝子。
【0026】
(10)Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードす
る、下記(a)又は(b)に記載の遺伝子。(a)配列番号5に示される塩基配列を有す
る遺伝子。(b)配列番号5に示す塩基配列からなるDNA、又は当該DNAと相補的な
塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0027】
(11)Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードす
る、下記(a)又は(b)に記載の遺伝子。(a)配列番号5に示される塩基配列のうち
、375ないし1829番目の塩基配列を有する遺伝子。(b)配列番号5に示される塩
基配列のうち、375ないし1829番目の塩基配列からなるDNA、又は当該DNAと
相補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子
。
【0028】
(12)Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードす
る、下記(a)又は(b)に記載の遺伝子。(a)配列番号6に示されるアミノ酸配列か
らなるたんぱく質、(b)配列番号6に示されるアミノ酸配列の1個又はそれ以上のアミ
ノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコー
ドする遺伝子。
【0029】
(13)上記(1)〜(12)の何れかに記載の遺伝子によってコードされるたんぱく
質。
【0030】
(14)以下の(a)又は(b)記載のたんぱく質。(a)配列番号2に示されるアミ
ノ酸配列からなるたんぱく質。(b)配列番号2に示されるアミノ酸配列において、1個
又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からな
り、かつ、Δ6脂肪酸不飽和化活性を有するたんぱく質。
【0031】
(15)以下の(a)又は(b)記載のたんぱく質。(a)配列番号4に示されるアミ
ノ酸配列からなるたんぱく質。(b)配列番号4に示されるアミノ酸配列において、1個
又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からな
り、かつ、Δ6脂肪酸鎖長延長活性を有するたんぱく質。
【0032】
(16)以下の(a)又は(b)記載のたんぱく質。(a)配列番号6に示されるアミ
ノ酸配列からなるたんぱく質。(b)配列番号6に示されるアミノ酸配列において、1個
又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からな
り、かつ、Δ5脂肪酸不飽和化活性を有するたんぱく質。
【0033】
(17)上記(13)〜(16)の何れかに記載のたんぱく質を認識する抗体。
【0034】
(18)少なくとも上記(1)〜(12)の何れかに記載の遺伝子を含む組換え発現ベ
クター。
【0035】
(19)少なくとも上記(1)〜(12)の何れかに記載の遺伝子を導入してなる形質
転換体。
【0036】
(20)少なくとも上記(1)〜(12)の何れかに記載の遺伝子が発現可能に導入さ
れた植物体、もしくは当該植物体と同一の性質を有する当該植物体の子孫となる植物体、
又は当該植物体の組織。
【0037】
(21)少なくとも上記(1)〜(12)の何れかに記載の遺伝子が発現可能に導入さ
れ、脂肪酸組成が改変された植物体、もしくは当該植物体と同一の性質を有する当該植物
体の子孫となる植物体、又は当該植物体の組織。
【0038】
(22)上記(20)又は(21)に記載の植物体の繁殖材料。
【0039】
(23)上記(21)に記載の植物体又は植物体の組織を用いる脂肪酸生産方法。
【0040】
(24)上記(23)に記載の脂肪酸生産方法により得られた、γ−リノレン酸、ジホ
モ−γ−リノレン酸、アラキドン酸、ステアリドン酸、エイコサテトラエン酸、およびエ
イコサペンタエン酸から選択される少なくとも1つ含む素材。
【0041】
(25)少なくとも(1)〜(12)の何れかに記載の遺伝子を用いて脂肪酸組成を改
変する方法。
【0042】
(26)上記(1)〜(12)の何れかに記載の遺伝子における少なくとも一部の塩基
配列又はその相補配列をプローブとして用いた遺伝子検出器具。
【0043】
(27)上記(13)〜(16)の何れかに記載のたんぱく質を用いて、当該たんぱく
質を調節する遺伝子、又は当該たんぱく質を調節する物質をスクリーニングする方法。
【0044】
(28)上記(27)に記載のスクリーニング方法により得られた遺伝子又は物質。
【0045】
なお、本明細書において、特に断らない限り、A、C、GおよびTは、アデニン、シト
シン、グアニンおよびチミンの各塩基を示す。
【0046】
本発明のさらに他の目的、特徴、および優れた点は、以下に示す記載によって十分わか
るであろう。また、本発明の利益は、添付図面を参照した次の説明で明白になるであろう
。
【図面の簡単な説明】
【0047】
【図1】実施例6で用いたMpDES6遺伝子、MpELO1遺伝子及びMpDES5遺伝子の各遺伝子の発現カセットが連結されたコンストラクトの構築手順を示す説明図である。
【発明を実施するための形態】
【0048】
本発明の実施の一形態について説明すれば、以下の通りである。なお、本発明は、これ
に限定されるものではない。
【0049】
以下、アラキドン酸及びエイコサペンタエン酸(EPA)の合成経路、本発明に係る遺
伝子、本発明に係るたんぱく質、本発明に係るたんぱく質及び遺伝子の取得方法、並びに
本発明に係る遺伝子及びたんぱく質の利用方法(有用性)の順で、本発明を詳細に説明す
る。
【0050】
(1)アラキドン酸及びエイコサペンタエン酸(EPA)の合成経路
アラキドン酸及びエイコサペンタエン酸(EPA)は、それぞれリノール酸及びα−リ
ノレン酸を起点として、Δ6不飽和化、Δ6鎖長延長及びΔ5不飽和化という3つの連続
した反応により、生合成されると考えられる。これらの反応は、それぞれΔ6不飽和化酵
素、Δ6鎖長延長酵素及びΔ5不飽和化酵素により触媒され、それぞれ、n−6経路(ア
ラキドン酸合成経路)及びn−3経路(EPA合成経路)と呼ばれている。
【0051】
これまでに報告されているΔ6不飽和化酵素、Δ6鎖長延長酵素及び、Δ5不飽和化酵
素は、いずれもn−6及びn−3経路の両方に関与していることが示されている。すなわ
ち、Δ6不飽和化酵素は、n−6経路ではリノール酸(18:2D9,12、18は炭素
数を表し、2は二重結合の数を表し、9、12は二重結合の位置を表す。以下同様である
。)をg−リノレン酸(GLA;18:3D6,9,12)に変換し、n−3経路ではa
−リノレン酸(ALA;18:3D9,12,15)をステアリドン酸(STA;18:
4D6,9,12,15)に変換する。Δ6鎖長延長酵素は、n−6経路ではGLAをジ
ホモ−γ−リノレン酸(DGLA;20:3Δ8,11,14)に変換し、n−3経路で
はSTAをエイコサテトラエン酸(ETA;20:4Δ8,11,14,17)に変換す
る。Δ5不飽和化酵素は、n−6経路ではDGLAをアラキドン酸(20:4Δ5,8,
11,14)に、n−3経路ではETAをエイコサペンタエン酸(EPA;20:5Δ5
,8,11,14,17)に変換する。
【0052】
(2)本発明に係る遺伝子
〔本発明に係るΔ6不飽和化酵素遺伝子〕
本発明に係るΔ6不飽和化酵素遺伝子は、Δ6脂肪酸不飽和化活性を有するたんぱく質
をコードするゼニゴケ目生物由来の遺伝子であり、以下の条件に適合する遺伝子であれば
よい。
【0053】
1.配列番号1に示される塩基配列を有する遺伝子。
【0054】
2.配列番号1に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基配
列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0055】
3.配列番号1に示される塩基配列からなるDNAの一部、又は当該DNAと相補的な
塩基配列からなるDNAの一部とストリンジェントな条件でハイブリダイズする遺伝子。
【0056】
4.配列番号1に示される塩基配列のうち、253ないし1698番目の塩基配列を有
する遺伝子。なお、配列番号1に示される塩基配列の253ないし1698番目の塩基配
列は配列番号2に示されるアミノ酸配列からなるたんぱく質に翻訳される領域である。
【0057】
5.配列番号1に示される塩基配列のうち、253ないし1698番目の塩基配列から
なるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな条
件でハイブリダイズする遺伝子。
【0058】
6.配列番号2に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0059】
7.配列番号2に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0060】
〔本発明に係るΔ6鎖長延長酵素遺伝子〕
本発明に係るΔ6鎖長延長酵素遺伝子は、Δ6脂肪酸鎖長延長酵素活性を有するたんぱ
く質をコードするゼニゴケ目生物由来の遺伝子であり、以下の条件に適合する遺伝子であ
ればよい。
【0061】
1.配列番号3に示される塩基配列を有する遺伝子。
【0062】
2.配列番号3に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基配
列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0063】
3.配列番号3に示される塩基配列からなるDNAの一部、又は当該DNAと相補的な
塩基配列からなるDNAの一部とストリンジェントな条件でハイブリダイズする遺伝子。
【0064】
4.配列番号3に示される塩基配列のうち、194ないし1066番目の塩基配列を有
する遺伝子。なお、配列番号3に示される塩基配列の194ないし1066番目の塩基配
列は配列番号4に示されるアミノ酸配列からなるたんぱく質に翻訳される領域である。
【0065】
5.配列番号3に示される塩基配列のうち、194ないし1066番目の塩基配列から
なるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな条
件でハイブリダイズする遺伝子。
【0066】
6.配列番号4に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0067】
7.配列番号4に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0068】
〔本発明に係るΔ5不飽和化酵素遺伝子〕
本発明に係るΔ5不飽和化酵素遺伝子は、Δ5脂肪酸不飽和化活性を有するたんぱく質
をコードするゼニゴケ目生物由来の遺伝子であり、以下の条件に適合する遺伝子であれば
よい。
【0069】
1.配列番号5に示される塩基配列を有する遺伝子。
【0070】
2.配列番号5に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基配
列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0071】
3.配列番号5に示される塩基配列からなるDNAの一部、又は当該DNAと相補的な
塩基配列からなるDNAの一部とストリンジェントな条件でハイブリダイズする遺伝子。
【0072】
4.配列番号5に示される塩基配列のうち、375ないし1829番目の塩基配列を有
する遺伝子。なお、配列番号5に示される塩基配列の375ないし1829番目の塩基配
列は配列番号6に示されるアミノ酸配列からなるたんぱく質に翻訳される領域である。
【0073】
5.配列番号5に示される塩基配列のうち、375ないし1829番目の塩基配列から
なるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな条
件でハイブリダイズする遺伝子。
【0074】
6.配列番号6に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0075】
7.配列番号6に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0076】
なお、上記「ストリンジェントな条件」とは、少なくとも90%の同一性、好ましくは
少なくとも95%の同一性、最も好ましくは少なくとも97%の同一性が配列間に存在す
るときにのみハイブリダイゼーションが起こることを意味する。
【0077】
上記ハイブリダイゼーションは、J.Sambrook et al.Molecul
ar Cloning,A Laboratory Manual,2d Ed.,Co
ld Spring Harbor Laboratory(1989)に記載されてい
る方法等、従来公知の方法で行うことができる。通常、温度が高いほど、塩濃度が低いほ
どストリンジェンシーは高くなり(ハイブリダイズし難くなる)、より相同な遺伝子を取
得することができる。ハイブリダイゼーションの条件としては、従来公知の条件を好適に
用いることができ、特に限定しないが、例えば、42℃、6×SSPC、50%ホルムア
ミド、1%SDS、100μg/ml salmon sperm DNA、5×デンハ
ルト液(ただし、1×SSPE;0.18M 塩化ナトリウム、10mM リン酸ナトリ
ウム、pH7.7、1mM EDTA)が挙げられる。
【0078】
上記「ゼニゴケ目生物」とは、ゼニゴケ(Marchantia polymorph
a)に限定されるものではなく、ゼニゴケ亞網ゼニゴケ目(Marchantiales
)に属する生物が含まれる。これらのうち、Monoclea forsteri(Mo
nocleales)、Corsinia coriandrina(Marchant
iales)、Oximitra paleacea(Marchantiales)、
Ricciocarpos natans(Marchantiales)、Ricca
huebeneriana(Marchantiales)、Ricca fluit
ans(Marchantiales)、Ricca duplex(Marchant
iales)、Ricca canaliculata(Marchantiales)
、Ricca bifurca(Marchantiales)、Ricca cili
ifera(Marchantiales)、Ricca glauca(Marcha
ntiales)、Ricca sorocarpa(Marchantiales)、
Ricca warnstorfii(Marchantiales)、Ricca m
ichelii(Marchantiales)、Ricca papillosa(M
archantiales)及びRicca zachariae(Marchanti
ales)には、超長鎖長高度不飽和脂肪酸が存在することが知られている(Prog.
Lipid Res.32,p281,1993参照)。これらの生物からΔ6不飽和化
酵素、Δ6鎖長延長酵素及びΔ5不飽和化酵素の遺伝子を取得することは現在の技術水準
を持ってすれば容易である。例えば、近縁生物の同じ機能を有する酵素をコードする遺伝
子は、クロスハイブリダイゼーションすることが一般に知られている。
【0079】
本発明の遺伝子は、2本鎖DNAのみならず、それを構成するセンス鎖及びアンチセン
ス鎖といった各1本鎖DNAやRNAを包含する。アンチセンス鎖は、プローブとして又
はアンチセンス化合物として利用できる。DNAには、例えばクローニングや化学合成技
術又はそれらの組み合わせで得られるようなcDNAやゲノムDNAなどが含まれる。さ
らに、本発明の遺伝子は、非翻訳領域(UTR)の配列やベクター配列(発現ベクター配
列を含む)などの配列を含むものであってもよい。
【0080】
(3)本発明に係るたんぱく質
〔本発明に係るΔ6不飽和化酵素たんぱく質〕
本発明に係るΔ6不飽和化酵素たんぱく質は、ゼニゴケ目生物由来のたんぱく質であっ
てΔ6脂肪酸不飽和化活性を有するたんぱく質であればよい。より具体的には、以下に示
すたんぱく質であればよい。
【0081】
1.上記(2)に記載した本発明に係るΔ6不飽和化酵素遺伝子によってコードされる
たんぱく質。
【0082】
2.配列番号2に示されるアミノ酸配列からなるたんぱく質。
【0083】
3.配列番号2に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質。
【0084】
〔本発明に係るΔ6鎖長延長酵素たんぱく質〕
本発明に係るΔ6鎖長延長酵素たんぱく質は、ゼニゴケ目生物由来のたんぱく質であっ
てΔ6脂肪酸鎖長延長活性を有するたんぱく質であればよい。より具体的には、以下に示
すたんぱく質であればよい。
【0085】
1.上記(2)に記載した本発明に係るΔ6鎖長延長酵素遺伝子によってコードされる
たんぱく質。
【0086】
2.配列番号4に示されるアミノ酸配列からなるたんぱく質。
【0087】
3.配列番号4に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質。
【0088】
〔本発明に係るΔ5不飽和化酵素たんぱく質〕
本発明に係るΔ5不飽和化酵素たんぱく質は、ゼニゴケ目生物由来のたんぱく質であっ
てΔ5脂肪酸不飽和化活性を有するたんぱく質であればよい。より具体的には、以下に示
すたんぱく質であればよい。
【0089】
1.上記(2)に記載した本発明に係るΔ5不飽和化酵素遺伝子によってコードされる
たんぱく質。
【0090】
2.配列番号6に示されるアミノ酸配列からなるたんぱく質。
【0091】
3.配列番号6に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質。
【0092】
上記「Δ6脂肪酸不飽和化活性」とは、リノール酸又はα−リノレン酸に対して基質特
異性を有し、それぞれγ−リノレン酸又はステアリドン酸に変換する作用を意味する。ま
た、上記「Δ6脂肪酸鎖長延長活性」とは、γ−リノレン酸又はステアリドン酸に対して
基質特異性を有し、それぞれジホモ−γ−リノレン酸又はエイコサテトラエン酸に変換す
る作用を意味する。また、上記「Δ5脂肪酸不飽和化活性」とは、ジホモ−γ−リノレン
酸又はエイコサテトラエン酸に対して基質特異性を有し、それぞれアラキドン酸又はエイ
コサペンタエン酸(EPA)に変換する作用を意味する。
【0093】
上記「1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加された」とは
、部位特異的突然変異誘発法等の公知の変異たんぱく質作製法により置換、欠失、挿入、
及び/又は付加できる程度の数(好ましくは10個以下、より好ましくは7個以下、さら
に好ましくは5個以下)のアミノ酸が置換、欠失、挿入、及び/又は付加されることを意
味する。このような変異たんぱく質は、公知の変異たんぱく質作製法により人為的に導入
された変異を有するたんぱく質に限定されるものではなく、天然に存在する同様の変異た
んぱく質を単離精製したものであってもよい。
【0094】
なお、本発明のたんぱく質は、アミノ酸がペプチド結合してなるポリペプチドであれば
よいが、これに限定されるものではなく、ポリペプチド以外の構造を含む複合たんぱく質
であってもよい。ここでいうポリペプチド以外の構造としては、糖鎖やイソプレノイド基
等を挙げることができるが、特に限定されるものではない。
【0095】
また、本発明のたんぱく質は、付加的なポリペプチドを含むものであってもよい。この
ようなポリペプチドが付加される場合としては、例えば、HisやMyc、Flag等に
よって本発明のたんぱく質がエピトープ標識されるような場合が挙げられる。
【0096】
また、本発明のたんぱく質は、前述した本発明の遺伝子(本発明のたんぱく質をコード
する遺伝子)を宿主細胞に導入して、そのたんぱく質を細胞内発現させた状態であっても
よいし、細胞、組織などから単離精製された状態であってもよい。また、上記宿主細胞で
の発現条件によっては、本発明のたんぱく質は、他のたんぱく質とつながった融合たんぱ
く質であってもよい。さらに本発明のたんぱく質は、化学合成されたものであってもよい
。
【0097】
(4)本発明に係るたんぱく質及び遺伝子の取得方法
本発明に係るたんぱく質及び遺伝子の取得方法(生産方法)は特に限定されるものでは
ないが、代表的な方法として次に示す各方法を挙げることができる。
【0098】
〔たんぱく質の取得方法〕
本発明のたんぱく質を取得する方法(生産方法)は、上述したように特に限定されるも
のではないが、まず、本発明のたんぱく質を発現する細胞、組織などから単純精製する方
法を挙げることができる。精製方法も特に限定されるものではなく、公知の方法で細胞や
組織から細胞抽出液を調製し、この細胞抽出液を公知の方法、例えばカラム等を用いて精
製すればよい。
【0099】
また、本発明のたんぱく質を取得する方法として、遺伝子組み換え技術等を用いる方法
も挙げられる。この場合、例えば、本発明の遺伝子をベクターなどに組み込んだ後、公知
の方法により発現可能に宿主細胞に導入し、細胞内で翻訳されて得られる上記たんぱく質
を精製するという方法などを採用することができる。
【0100】
なお、このように宿主に外来遺伝子を導入する場合、外来遺伝子の発現のため宿主内で
機能するプロモーターを組み入れた発現ベクター及び宿主には様々なものが存在するので
、目的に応じたものを選択すればよい。産生されたたんぱく質を精製する方法は、用いた
宿主、たんぱく質の性質によって異なるが、タグの利用等によって比較的容易に目的のた
んぱく質を精製することが可能である。
【0101】
変異たんぱく質を作製する方法についても、特に限定されるものではない。例えば、部
位特異的突然変異誘発法(Hashimoto−Gotoh,Gene 152,271
−275(1995)他)、PCR法を利用して塩基配列に点変異を導入し変異たんぱく
質を作製する方法、あるいはトランスポゾンの挿入による突然変異株作製法などの周知の
変異たんぱく質作製法を用いることができる。変異たんぱく質の作製には市販のキットを
利用してもよい。
【0102】
本発明のたんぱく質の取得方法は上述の方法限定されることはなく、例えば、化学合成
されたものであってもよい。例えば、無細胞系のたんぱく質合成液を利用して本発明の遺
伝子から本発明のたんぱく質を合成してもよい。
【0103】
〔遺伝子の取得方法〕
本発明の遺伝子を取得する方法(生産方法)も特に限定されるものではないが、例えば
、ディファレンシャルスクリーニング(サブトラクションクローニング)を利用する方法
を挙げることができる。この方法では、公知の技術に従って、試験管内での直接的ハイブ
リダイゼーションを繰り返し、目的のcDNA(本発明の遺伝子)を濃縮すればよい。
【0104】
上記ディファレンシャルスクリーニングにおける各ステップについては、通常用いられ
る条件の下で行えばよい。これによって得られたクローンは、制限酵素地図の作成及びそ
の塩基配列決定(シークエンシング)によって、さらに詳しく解析することができる。こ
れらの解析によって、本発明の遺伝子配列を含むDNA断片を取得したか容易に確認する
ことができる。
【0105】
また、本発明の遺伝子を取得する方法として、公知の技術により、本発明の遺伝子を含
むDNA断片を単離し、クローニングする方法が挙げられる。例えば、本発明の遺伝子の
塩基配列の一部と特異的にハイブリダイズするプローブを調製し、ゲノムDNAライブラ
リーやcDNAライブラリーをスクリーニングすればよい。このようなプローブとしては
、本発明の遺伝子の塩基配列又はその相補配列の少なくとも一部に特異的にハイブリダイ
ズするプローブであれば、いずれの配列・長さのものを用いてもよい。
【0106】
また、上記プローブの配列を、上述したゼニゴケ間で良好に保存されている領域の中か
ら選択し、他のゼニゴケのゲノムDNA(又はcDNA)ライブラリーをスクリーニング
すれば、上記たんぱく質と同様の機能を有する相同分子や類縁分子をコードする遺伝子を
単離しクローニングできる。
【0107】
あるいは、本発明の遺伝子を取得する方法として、PCR等の増幅手段を用いる方法を
挙げることができる。例えば、本発明の遺伝子のcDNA配列のうち、5’側及び3’側
の配列(又はその相補配列)の中からそれぞれプライマーを調製し、これらプライマーを
用いてゲノムDNA(又はcDNA)等を鋳型にしてPCR等を行い、両プライマー間に
挟まれるDNA領域を増幅することで、本発明の遺伝子を含むDNA断片を大量に取得で
きる。
【0108】
(5)本発明に係る遺伝子及びたんぱく質の利用方法(有用性)
(5−1)組換え発現ベクター
本発明に係る組換え発現ベクターは、前記(2)に記載した本発明に係る遺伝子を含む
ものであれば、特に限定されるものではない。例えば、cDNAが挿入された組換え発現
ベクターが挙げられる。組換え発現ベクターの作製には、プラスミド、ファージ、又はコ
スミドなどを用いることができるが特に限定されるものではない。また、作製方法も公知
の方法を用いて行えばよい。
【0109】
ベクターの具体的な種類は特に限定されるものではなく、ホスト細胞中で発現可能なベ
クターを適宜選択すればよい。すなわち、ホスト細胞の種類に応じて、確実に遺伝子を発
現させるために適宜プロモーター配列を選択し、これと本発明の遺伝子を各種プラスミド
等に組み込んだものを発現ベクターとして用いればよい。
【0110】
本発明の遺伝子がホスト細胞に導入されたか否か、さらにはホスト細胞中で確実に発現
しているか否かを確認するために、各種マーカーを用いてもよい。例えば、ホスト細胞中
で欠失している遺伝子をマーカーとして用い、このマーカーと本発明の遺伝子とを含むプ
ラスミド等を発現ベクターとしてホスト細胞に導入する。これによってマーカー遺伝子の
発現から本発明の遺伝子の導入を確認することができる。あるいは、本発明のたんぱく質
を融合たんぱく質として発現させてもよく、例えば、オワンクラゲ由来の緑色蛍光たんぱ
く質GFP(Green Fluorescent Protein)をマーカーとして
用い、本発明のたんぱく質をGFP融合たんぱく質として発現させてもよい。
【0111】
上記ホスト細胞は、特に限定されるものではなく、従来公知の各種細胞を好適に用いる
ことができる。具体的には、例えば、大腸菌(Escherichia coli)等の
細菌、酵母(出芽酵母Saccharomyces cerevisiae、分裂酵母S
chizosaccharomyces pombe)、線虫(Caenorhabdi
tis elegans)、アフリカツメガエル(Xenopus laevis)の卵
母細胞等を挙げることができるが、特に限定されるものではない。
【0112】
上記発現ベクターをホスト細胞に導入する方法、すなわち形質転換方法も特に限定され
るものではなく、電気穿孔法、リン酸カルシウム法、リポソ−ム法、DEAEデキストラ
ン法等の従来公知の方法を好適に用いることができる。また、例えば、本発明のたんぱく
質を昆虫で転移発現させる場合には、バキュロウイルスを用いた発現系を採用することが
できる。
【0113】
(5−2)形質転換体
本発明に係る形質転換体は、前記(2)に記載した本発明に係る遺伝子導入された形質
転換体であれば、特に限定されるものではない。ここで「形質転換体」とは、細胞・組織
・器官のみならず、生物個体を含む意味である。
【0114】
形質転換体の作製方法(生産方法)は特に限定されるものではないが、例えば、上述し
た組換え発現ベクターをホスト細胞に導入して形質転換する方法を挙げることができる。
また、形質転換の対象となる生物も特に限定されるものではなく、上記ホスト細胞で例示
した各種微生物や動物を挙げることができる。
【0115】
本発明に係る形質転換体は、本発明に係る遺伝子が発現可能に導入された植物体、もし
くは当該植物体と同一の性質を有する当該植物体の子孫となる植物体、又は当該植物体の
組織の組織であることが好ましい。このような形質転換植物により、低コストかつ環境に
やさしい生産プロセスでアラキドン酸やEPA等の高度不飽和脂肪酸を生産することがで
きる。
【0116】
ここで「遺伝子が発現可能に導入された」とは、公知の遺伝子工学的手法(遺伝子操作
技術)により、対象細胞(宿主細胞)内に発現可能に導入されることを意味する。
【0117】
植物体の形質転換に用いられる組換え発現ベクターは、当該植物細胞内で挿入遺伝子を
発現させることが可能なものであれば特に限定しない。例えば、植物細胞内で恒常的に遺
伝子を発現させるプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモ
ーター)を有するベクターや、外的な刺激により誘導的に活性化されるプロモーターを有
するベクターを用いることができる。なお、この植物細胞には、種々の形態の植物細胞、
例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルスなどが含まれる。
【0118】
植物細胞への組み換え発現ベクターの導入には、ポリエチレングリコール法、電気穿孔
法(エレクトロポレーション法)、アグロバクテリウムを介する方法、パーティクルガン
法など、当業者に公知の種々の方法を用いることができる。形質転換細胞から植物体の再
生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。
【0119】
例えば、タバコにおいて形質転換植物体を作出する手法については、形質転換したアグ
ロバクテリウムをタバコリーフディスクに感染させる方法、ポリエチレングリコールを用
いてプロトプラストへ遺伝子を導入し植物体に再生させる方法、電気パルスによりプロト
プラストへ遺伝子導入し植物体を再生させる方法、パーティクルガン法により細胞へ遺伝
子を直接導入し植物体を再生させる方法など、いくつかの技術が既に確立されている。本
発明においては、これらの方法を好適に用いることができる。
【0120】
また、タバコは、シロイヌナズナと並んで遺伝子工学的手法を用いる植物育種のモデル
植物である。このタバコにおいて、アラキドン酸やEPAの含量が増加した形質転換体を
取得できるということは、植物全般において形質転換体を取得することができるといって
も過言ではない。なお、本明細書では、後述する実施例に示すように、タバコだけでなく
、イネの形質転換体をも取得しており、本発明によれば、あらゆる種類の形質転換植物体
を取得できることを実証している。
【0121】
例えば、イネにおいて形質転換植物体を作出する手法については、ポリエチレングリコ
ールを用いてプロトプラストへ遺伝子を導入し、植物体に再生させる方法、電気パルスに
よりプロトプラストへ遺伝子導入し、植物体を再生させる方法、パーティクルガン法によ
り細胞へ遺伝子を直接導入し、植物体を再生させる方法など、いくつかの技術が既に確立
されている。本発明においては、これらの方法を好適に用いることができる。
【0122】
上記形質転換植物体がイネである場合、イネ内のアラキドン酸やEPAの含量が増加す
るので、この形質転換植物体から得られる種子、すなわち、米を食べることで、容易にア
ラキドン酸やEPA等の高度不飽和脂肪酸を、体内に摂取することが可能になる。したが
って、イネの形質転換植物体は、食糧としての価値が高く、食品産業、農業分野に極めて
有用である。また、現在あまり利用されていない米ぬか、モミガラ、ヒコバエ等でアラキ
ドン酸やEPAを生産すれば、ここからこれら脂肪酸を抽出することにより、健康食品の
原料として有効利用できる。また、家畜の飼料としても利用できる。
【0123】
ゲノム内に本発明の遺伝子が導入された形質転換植物体がいったん得られれば、当該植
物体から有性生殖又は無性生殖により子孫を得ることができる。また、当該植物体、又は
、その子孫、あるいは、クローンから、繁殖材料(例えば、種子、果実、切穂、塊茎、塊
根、株、カルス、プロトプラストなど)を得て、それらを基に当該植物体を量産すること
も可能である。したがって、本発明には、本発明の遺伝子が発現可能に導入された植物体
、もしくは、当該植物体と同一の性質を有する当該植物体の子孫となる植物体、又は、当
該植物体の組織、あるいは、当該植物体の繁殖材料も含まれる。
【0124】
また、当該植物体、当該植物体と同一の性質を有する当該植物体の子孫となる植物体、
及び、当該植物体の組織には、栄養増殖された植物体も含まれる。栄養増殖は、栄養生殖
、クローン成長とも呼ばれ、挿し芽、挿し木などによる増殖が一般的であり、試験管内で
は、葉、茎、根などの器官からの植物体の再分化やカルスによる増殖が可能である。植物
種によっては、枝の先端が特殊な冬芽を作る、腋芽が多肉化する、花がムカゴ化する、芋
を形成するなどの場合がある。
【0125】
さらに、本発明に係る遺伝子が発現可能に導入され、脂肪酸組成が改変された植物体、
もしくは当該植物体と同一の性質を有する当該植物体の子孫となる植物体、又は当該植物
体の組織、当該植物体の繁殖材料も本発明に含まれる。「脂肪酸組成が改変された」とは
形質転換前の植物体における脂肪酸組成と形質転換後における植物体の脂肪酸組成が異な
っていることを意味する。例えば、本来脂肪酸組成にアラキドン酸やEPAが含まれてい
なかった植物を本発明に係る遺伝子で形質転換することにより、形質転換植物の脂肪酸組
成にアラキドン酸やEPAが含まれる場合等を挙げることができる。
【0126】
(5−3)脂肪酸生産方法
本発明には、本発明に係る遺伝子で形質転換され、脂肪酸組成が改変された植物体又は
植物体の組織を用いて脂肪酸を生産する方法が含まれる。
【0127】
例えば、上述のようにアラキドン酸やEPAの含量が増加した本発明に係る形質転換植
物から製造された食用油はアラキドン酸やEPAの含量が高く、価値の高いものになる。
上記形質転換植物体の種子、果実、切穂、塊茎、塊根等も、アラキドン酸やEPAを含む
食料として価値の高いものになる。
【0128】
(5−4)素材
本発明には、上述の脂肪酸生産方法により得られた物質、すなわち、γ−リノレン酸、
ジホモ−γ−リノレン酸、アラキドン酸、ステアリドン酸、エイコサテトラエン酸、エイ
コサペンタエン酸を少なくとも1つ含む素材も含まれる。この「素材」とは、上述の食料
としての種子、果実、切穂、塊茎、又は、塊根の他に、工業原料用途に利用できる素材全
般を意味する。
【0129】
上記素材としては、例えば、アラキドン酸やEPAを含む健康食品、フィルム、生分解
性プラスチック、機能性繊維、潤滑油、洗剤の素材等が挙げられる。上記の不飽和脂肪酸
は、分子内に二重結合を複数有するという、ユニークな物性を持つ。このため、例えば、
本発明の形質転換植物体により、アラキドン酸やEPAを生産させることにより、生産コ
ストを低減できる。また、本発明により、環境にやさしい生産プロセスを実現できる。
【0130】
(5−5)脂肪酸組成改変方法
本発明には、本発明に係る遺伝子を用いて脂肪酸組成を改変する方法が含まれる。例え
ば、上述のように本発明に係る遺伝子を導入した形質転換体を作製することによりホスト
細胞の脂肪酸組成を改変することが可能となる。脂肪酸組成を改変する対象は特に限定さ
れるものではなく、植物以外にも動物、細菌、酵母等、あらゆる生物を対象とすることが
可能である。
【0131】
(5−6)遺伝子検出器具
本発明に係る遺伝子検出器具は、本発明に係る遺伝子の少なくとも一部の塩基配列又は
その相補配列をプローブとして用いたものである。遺伝子検出器具は、種々の条件下にお
いて、本発明の遺伝子の発現パタ−ンの検出・測定などに利用することができる。
【0132】
本発明の遺伝子検出器具としては、例えば、本発明の遺伝子と特異的にハイブリダイズ
する上記プローブを基盤(担体)上に固定化したDNAチップが挙げられる。ここで「D
NAチップ」とは、主として、合成したオリゴヌクレオチドをプローブに用いる合成型D
NAチップを意味するが、PCR産物などのcDNAをプローブに用いる貼り付け型DN
Aマイクロアレイをも包含するものとする。
【0133】
プローブとして用いる配列は、cDNA配列の中から特徴的な配列を特定する従来公知
の方法によって決定することができる。具体的には、例えば、SAGE:Serial
Analysis of Gene Expression法(Science 276
:1268,1997;Cell 88:243,1997;Science 270:
484,1995;Nature 389:300,1997;米国特許第5,695,
937号)等を挙げることができる。
【0134】
なお、DNAチップの製造には、公知の方法を採用すればよい。例えば、オリゴヌクレ
オチドとして合成オリゴヌクレオチドを使用する場合には、フォトリソグラフィ−技術と
固相法DNA合成技術との組み合わせにより、基盤上で該オリゴヌクレオチドを合成すれ
ばよい。一方、オリゴヌクレオチドとしてcDNAを用いる場合には、アレイ機を用いて
基盤上に貼り付ければよい。
【0135】
また、一般的なDNAチップと同様、パーフェクトマッチプローブ(オリゴヌクレオチ
ド)と、該パーフェクトマッチプローブにおいて一塩基置換されたミスマッチプローブと
を配置して遺伝子の検出精度をより向上させてもよい。さらに、異なる遺伝子を並行して
検出するために、複数種のオリゴヌクレオチドを同一の基盤上に固定してDNAチップを
構成してもよい。
【0136】
本発明に係る遺伝子検出器具は、上記例示したDNAチップに限定されるものではなく
、本発明に係る遺伝子の少なくとも一部の塩基配列又はその相補配列をプローブとして用
いたものであればよい。
【0137】
(5−7)抗体
本発明に係る抗体は、本発明に係るたんぱく質、又はその部分たんぱく質・部分ペプチ
ドを抗原として、公知の方法によりポリクロ−ナル抗体又はモノクロ−ナル抗体として得
られる抗体である。公知の方法としては、例えば、文献(Harlowらの「Antib
odies:A laboratory manual(Cold Spring Ha
rbor Laboratory,New York(1988))、岩崎らの「単クロ
ーン抗体ハイブリドーマとELISA,講談社(1991)」)に記載の方法が挙げられ
る。このようにして得られる抗体は、本発明のたんぱく質の検出・測定などに利用できる
。
【0138】
(5−8)スクリーニング方法
本発明に係るスクリーニング方法は、本発明に係るたんぱく質を用いて、当該たんぱく
質を調節する遺伝子、又は当該たんぱく質を調節する物質をスクリーニングする方法であ
る。本発明のスクリーニング方法としては、物質間の結合の有無や解離の有無を調べる従
来公知の種々の方法を適用することができ、特に限定されるものではない。例えば、本発
明に係るたんぱく質の活性(Δ6不飽和化活性、Δ6鎖長延長活性及び/又はΔ5不飽和
化活性)を促進するような物質のスクリーニングを挙げることができる。
【0139】
また、本発明には、上記スクリーニング方法により得られた遺伝子又は物質も含まれる
。
【0140】
以下、実施例を示し、本発明についてさらに詳しく説明するが、もちろん、本発明は以
下の実施例に限定されるものではなく、細部については様々な態様が可能であることはい
うまでもない。さらに、本発明は上述した実施形態に限定されるものではなく、請求項に
示した範囲で種々の変更が可能であり、開示された技術的手段を適宜組み合わせて得られ
る実施形態についても本発明の技術的範囲に含まれる。
【実施例】
【0141】
本実施例において、実験手法は、特に断らない限り、Molecular Cloni
ng(Sambrook et.al.Cold Spring Harbour La
boratory Press,1989)に記載されている方法に従った。
【0142】
〔実施例1:ゼニゴケ由来のΔ6不飽和化酵素遺伝子の単離〕
これまでにクローニングされたΔ6不飽和化酵素のアミノ酸配列の比較により、アミノ
酸配列Trp−Trp−Lys−(Glu/Asp)−Lys−His−Asn(配列番
号37)及びTrp−Phe−Thr−Gly−Gly−Leu−Asn(配列番号38
)が保存されていることがわかった。そこで、ゼニゴケ由来のΔ6不飽和化酵素遺伝子を
単離するために、上記のアミノ酸配列をコードする下記の縮退プライマーを用いた。
dD6DES−F 5’−TGGTGGAA(A/G)GA(A/G/T/C)AA(A
/G)CA(T/C)AA−3’(配列番号7)
dD6DES−R 5’−(A/G)TTIA(A/G)ICCICCIGT(A/G)
AACCA−3’ (配列番号8)
(Iはイノシン、()内は複数の塩基)
【0143】
試料にはE系統ゼニゴケ(Transgenic Res.9,p179,2000参
照)の葉状体を用いた。葉状体からのpoly(A)+RNAの単離については、文献(
Biosci.Biotechnol.Biochem.67,p605,2003;B
iosci.Biotechnol.Biochem.67,p1667,2003)に
記載の方法に従った。単離したpoly(A)+RNA 1.5μlを、Ready−T
o−Go T−primed First Strand kit(Amersham社
製)を用いてcDNAに逆転写した。PCRは、上記cDNA約10ngを鋳型とし、上
記プライマー(dD6DES−F及びdD6DES−R)及び酵素(Takara Ex
Taq、Takara社製)0.5Uとを用いて、製造者の推奨する方法で行った。反
応液量は20μlとし、GeneAmp PCR system 9700(PE Ap
plied Biosystems社製)を用いて、94℃2分間保持後、94℃で1分
間、45℃で1.5分間、72℃で2分間の反応を35回繰り返し、その後4℃に冷却し
た。
【0144】
得られたPCR産物を1%(w/v)アガロ−スゲルで電気泳動し、従来のΔ6不飽和
化酵素のアミノ酸配列から予想されるサイズを有する増幅断片を、Prep−A Gen
e(Bio−rad社製)を用いてゲルより回収した。回収した増幅断片をpT7Blu
e Vector(Takara社製)に連結し、大腸菌Electro−max DH
10B cells(Invitrogen社製,Carlsbad,CA)に形質転換
した。
【0145】
BigDye Terminator Cycle Sequencing kit(
Applied Biosystems社製)及びautomated sequenc
er ABI PRISM 377(Applied Biosystems社製)を用
いて得られた全クローンの塩基配列を決定し、目的のcDNA配列をもつものを探索した
。
【0146】
さらに、全長cDNA配列を取得するために、5’−RACE及び3’−RACEを行
った。これには、5’−RACE System for Rapid Amplifi
cation of cDNA Ends Version 2.0(Invitrog
en社製)、Ready−To−Go T−primed First Strand
kit(Amersham社製)及び下記のプライマー(MpDES6−02R及びMp
DES6−01F)を用い、製造者の推奨する方法で行った。
MpDES6−02R:5’−AAGTTGCCTTCGATGTTTCTGG−3’(
配列番号9)
MpDES6−01F:5’−GCTCGCCTGGAGCAAGGAAAATC−3’
(配列番号10)
【0147】
その結果、1種類のホモログ遺伝子候補が単離され、この遺伝子をMpDES6遺伝子
とした。単離されたMpDES6遺伝子のcDNAの長さ(ポリA部分を除く)は、2,
522bpであり、そのコードする推定アミノ酸配列は481残基であった。その塩基配
列を配列番号1、アミノ酸配列を配列番号2に示した。
【0148】
MpDES6cDNAの推定アミノ酸配列をヒメツリガネゴケのΔ6不飽和化酵素のア
ミノ酸配列と比較した結果、47.5%の同一性しか示さなかった。
【0149】
〔実施例2:ゼニゴケ由来のΔ6鎖長延長酵素遺伝子の単離〕
これまでにクローニングされたΔ6鎖長延長酵素のアミノ酸配列の比較により、アミノ
酸配列Val−Glu−Phe−Met−Asp−Thr−Val(配列番号39)及び
Lys−Tyr−Leu−Phe−Trp−Gly−Arg(配列番号40)が保存され
ていることがわかった。そこで、ゼニゴケ由来のΔ6鎖長延長酵素遺伝子を単離するため
に、上記のアミノ酸配列をコードする下記の縮退プライマーを用いた。
dD6ELO−F 5’−GTIGA(A/G)TT(T/C)ATGGA(T/C)A
CIGT−3’ (配列番号11)
dD6ELO−R 5’−C(G/T)ICCCCA(A/G)AAIA(A/G)(A
/G)TA(T/C)TT−3’(配列番号12)
【0150】
上記のプライマー(dD6ELO−F及びdD6ELO−R)を用いてPCRをい、得
られたDNA断片をサブクローニングした。得られたクローンの塩基配列を決定し、目的
のcDNA配列をもつクローンについて下記のプライマー(MpELO1−02R及びM
pELO1−01F)を用いて完全長cDNAを取得した。なお、実験材料及び方法は、
実施例1と同様である。
MpELO1−02R:5’−GCGAGCTTTCTCGTTCTTTCCC−3’(
配列番号13)
MpELO1−01F:5’−TATGATTTTGAAGCGCAACACG−3’(
配列番号14)
【0151】
その結果、1種類のホモログ遺伝子候補が単離され、この遺伝子をMpELO1遺伝子
とした。MpELO1遺伝子のcDNAの長さ(ポリA部分を除く)は、1,559bp
で、推定アミノ酸配列は290残基であった。その塩基配列を配列番号3、アミノ酸配列
を配列番号4に示した。
【0152】
MpELO1cDNAの推定アミノ酸配列をヒメツリガネゴケのΔ6鎖延長酵素のアミ
ノ酸配列と比較した結果、同一性は62.7%であった。
【0153】
〔実施例3:ゼニゴケ由来のΔ5不飽和化酵素遺伝子の単離〕
他生物種のΔ5不飽和化酵素は、そのN末端にシトクロムb5ドメインを有する。この
ことから、ゼニゴケ由来のΔ5不飽和化酵素遺伝子は、Δ6不飽和化酵素遺伝子と同じく
、シトクロムb5ドメイン融合型不飽和化酵素遺伝子ファミリーに属することが予想され
た。しかしながら、ケイ藻や真菌においては、Δ5不飽和化酵素及びΔ6不飽和化酵素間
のアミノ酸配列レベルでの相同性は非常に低い。そこで、糸状菌(M.alpina)の
Δ5不飽和化酵素及びΔ6不飽和化酵素間のアミノ酸配列を比較した結果、縮退プライマ
ー設計に最低限必要な4から5残基程度の連続した保存配列が局所的に存在することを見
出した。驚くべきことに、これらの保存配列は、異種のΔ5不飽和化酵素のアミノ酸配列
間より、同種内のΔ5不飽和化酵素とΔ6不飽和化酵素との間で、より保存されているこ
とを見出した。このことから、シトクロムb5ドメイン融合型不飽和化酵素遺伝子には、
種特異的な保存配列が存在する場合があると考えられた。そこで、上述のMpDES6と
、Genetics 159,p981,2001に記載されている機能未知のMpDE
Sとで、塩基配列を鋭意比較検討した結果、2箇所のアミノ酸配列(I(E/N)(G/
D)KVYDV(配列番号41)、及び、DPDI(Q/D)(Y/T)(M/V)P(
配列番号42))のアミノ酸配列が保存されていることを見出した。それぞれのアミノ酸
配列に対応する縮退プライマーの配列を次に示す。
dD5DES−F 5’−AT(A/T/C)(A/G)AIG(A/G)IAA(A/
G)TITA(T/C)GA(T/C)GT−3’(配列番号15)
dD5DES−R 5’−GGIA(T/C)I(G/T)(A/T)IT(G/C)(
A/G/T)AT(A/G)TCIGG(A/G)TC−3’(配列番号16)
【0154】
上記プライマー(dD5DES−F及びdD5DES−R)を用いてPCRを行い、得
られたDNA断片をサブクローニングした。得られたクローンの塩基配列を決定し、目的
のcDNA配列をもつクローンについて下記のプライマー(MpDES5−02R及びM
pDES5−01F)を用いて完全長cDNAを取得した。なお、実験材料及び方法は、
実施例1と同様である。
MpDES5−02R:5’−GTGTGTACGATCCGTGGTTACC−3’(
配列番号17)
MpDES5−01F:5’−AAGGCGGGACAGGATTCAACAC−3’(
配列番号18)
【0155】
その結果、ゼニゴケ由来のΔ5不飽和化酵素の候補として、2種類の長さの異なるクロ
ーンc1及びc2を単離した(c1:2,427bp、c2:2,285bp)。c1と
c2との塩基配列を比較したところ、5’非翻訳領域で、オルタナティブ・スプライシン
グ(選択的スプライシング)が起こっていることが明らかとなった。オルタナティブ・ス
プライシングによる読み枠の変化はなく、いずれのクローンも484個のアミノ酸をコー
ドしていた(配列番号6)。以下、2,427bpの長さのクローンc1をMpDES5
遺伝子(配列番号5)とし、以下の実施例に用いた。
【0156】
MpDES5cDNAの推定アミノ酸配列を糸状菌(M.alpina)のΔ5不飽和
化酵素のアミノ酸配列と比較した結果、31.4%の同一性であった。ゼニゴケと近縁な
ヒメツリガネゴケのΔ5不飽和化酵素に関しては、配列情報が公表されていないため、こ
こでは比較を行わなかった。
【0157】
〔実施例4:メタノール資化性酵母(Pichica pastoris)を用いた機能
解析〕
MpDES6、MpELO1及びMpDES5のcDNAの機能を調べるために、まず
個々のORFを、メタノール誘導性プロモーターAOX1の下流に配置したコンストラク
トを作製した。これらのコンストラクトをメタノール資化性酵母(Pichia pas
toris)に導入し、その脂肪酸組成を解析した。MpDES6、MpELO1、及び
、MpDES5のcDNA塩基配列のORF部分を、下記のプライマーを用いて、PCR
により増幅した。
(MpDES6ORF増幅用プライマー)
MpD6−17F:5’−GGAATTCGCGATGGCCTCGTCCACCACC
AC−3’(配列番号19)
MpD6−18F:5’−GGAATTCTACTTTCGCAGCGTATGCTAC
C−3’(配列番号20)
(MpELO1ORF増幅用プライマー)
MpD6ELO1−15F:5’−GGAATTCGCGATGGAGGCGTACGA
GATGG−3’(配列番号21)
MpD6ELO1−16F:5’−GGAATTCTTCTGCCTTTTTGCTCT
TGATC−3’(配列番号22)
(MpDES5ORF増幅用プライマー)
MpD5−11F:5’−GTTGAATTCGACAGTTATGCCGCCACAC
GC−3’(配列番号23)
MpD5−12R:5’−GTTGAATTCAGGCCCAAAGCATGCTGTC
AC−3’(配列番号24)
【0158】
これらのプライマーは、下線で示したEcoRI認識配列を含んでおり、これを以下の
クローニングに利用した。また、PCRにはPyrobest DNA polymer
ase(Takara社製)0.5Uを用い、製造者の推奨する方法に従って反応液量2
0μlで行った。反応条件は、94℃で2分間保持後、94℃で1分間、57℃で1分間
、72℃で1分間の反応を25回繰り返し、その後4℃に冷却した。得られた各ORF断
片をEcoRIで消化した後、実施例1に記載の方法でゲル精製を行った。そして、メタ
ノール資化性酵母の発現ベクターpPICZA(マーカー:ゼオシン耐性遺伝子,Inv
itrogen社製)内のメタノール誘導性プロモーター5’AOX1下流のEcoRI
部位にセンス方向に連結した。
【0159】
各々の発現コンストラクト及び対照としてのpPICZAベクターを、Pichia
EasyComp kit(Invitrogen社製)を用いてメタノール資化性酵母
のPPY1系統に導入し、ゼオシン耐性をマーカーとして形質転換体を取得した。なお、
メタノール資化性酵母は、Δ6不飽和化酵素の基質であるリノール酸とα−リノレン酸を
合成することができるが、アラキドン酸やEPAのその他の前駆体を合成することはでき
ない。
【0160】
導入した遺伝子を発現させるため、EasySelect Pichia Expre
ssion Kit(Invitrogen社製)を用い、当該キットの推奨する方法に
従いて、各形質転換体を1.0%グリセロールのみを炭素源とする最小培地中でOD(6
00nm)が0.5になるまで培養した後、0.5%メタノールのみを炭素源とする最小
培地中で30℃、3日間、飽和状態になるまで培養した。その後、各々の形質転換体の脂
肪酸組成を、GC−MSを用いて公知の方法(Biosci.Biotechnol.B
iochem.67,p605,2003)により測定した。
【0161】
MpDES6遺伝子を発現させた形質転換体では、Δ6不飽和化酵素の反応産物である
g−リノレン酸及びステアリドン酸が、全脂肪酸のそれぞれ7.4%及び0.7%新たに
検出された。対照としてpPICZAベクターを導入した酵母ではこれらは検出されなか
った。以上のことから、MpDES6はΔ6不飽和化酵素をコードしていることが示され
た。
【0162】
MpELO1遺伝子を発現させた形質転換体では、g−リノレン酸を添加した場合にジ
ホモ−g−リノレン酸が全脂肪酸の14.1%、ステアリドン酸を添加した場合にエイコ
サテトラエン酸が1.5%新たに検出された。対照としてpPICZAベクターを導入し
た酵母では、これらは検出されなかった。以上のことから、MpELO1はΔ6鎖長延長
酵素をコードしていることが示された。
【0163】
MpDES5遺伝子を発現させた形質転換体では、ジホモ−g−リノレン酸を添加した
場合にアラキドン酸が全脂肪酸の1.1%、ステアリドン酸を添加した場合にエイコサペ
ンタエン酸(EPA)が0.1%検出された。対照としてpPICZAベクターを導入し
た酵母ではこれらは検出されなかった。以上から、MpDES5は、Δ5不飽和化酵素を
コードすることが示された。
【0164】
以上のようにゼニゴケから、Δ6不飽和化酵素、Δ6鎖長延長酵素及びΔ5不飽和化酵
素をコードする遺伝子として、それぞれMpDES6、MpELO1及びMpDES5を
取得することができた。
【0165】
〔実施例5:メタノール資化性酵母(P.pastoris)におけるゼニゴケ高度不飽
和脂肪酸生合成系の再構成〕
MpDES6、MpELO1及びMpDES5を共発現させるために、上記実施例4で
作製した、EcoRI消化したMpELO1及びMpDES5のORF増幅断片を、各々
別のメタノール資化性酵母発現ベクターpPIC3K(マーカー:HIS4遺伝子、In
vitrogen社製社)及びpPIC6A(マーカー:ブラスチシジン耐性遺伝子、I
nvitrogen社製社)の5’AOX1プロモーター下流のEcoRI部位にセンス
方向に連結した。また、MpDES6に関しては、実施例4で作製した発現ベクターを用
いた。以下各発現ベクターを、それぞれpPICZA−MpDES6、pPIC3K−M
pELO1及びpPIC6A−MpDES5と表記する。
【0166】
まず、pPICZA−MpDES6、又は、対照としてのpPICZAベクターのみを
、上記実施例4で用いたメタノール資化性酵母PPY1系統と同じ脂肪酸組成を持つメタ
ノール資化性酵母PPY12系統(his4,arg4)に形質転換し、ゼオシン耐性を
マーカーにして形質転換体を取得した。続いて、pPIC3K−MpELO1、又は、対
照としてのpPIC3Kベクターのみを、それぞれ、pPICZA−MpDES6、又は
pPICZAのみがゲノム中に組み込まれた形質転換体に導入し、ヒスチジン合成能をマ
ーカーにして形質転換体を取得した。最後に、pPIC6A−MpDES5、又は、対照
としてのpPIC6Aベクターのみを、pPICZA−MpDES6及びpPIC3K−
MpELO1又はpPICZA及びpPIC3Kがゲノム中に組み込まれた上記形質転換
体に導入し、ブラストジン耐性をマーカーとして形質転換体を取得した。
【0167】
得られた2種類又は3種類の遺伝子が導入された形質転換体を用いて、ゼニゴケのアラ
キドン酸/EPA生合成系の再構成実験を行った。まず、上記2種類の遺伝子(MpDE
S6及びMpELO1)を導入した。形質転換体を用いて、MpDES6たんぱく質及び
MpELO1たんぱく質をメタノール資化性酵母内で共発現させた。その結果、Δ6不飽
和化反応産物であるg−リノレン酸(全脂肪酸の2.9%)及びステアリドン酸(全脂肪
酸の0.4%)に加え、それらの鎖長延長反応産物であるジホモ−g−リノレン酸(全脂
肪酸の2.8%)及びエイコサテトラエン酸(全脂肪酸の0.2%)が生じた。対照とし
た形質転換体では、これらの脂肪酸は検出されなかった。上記形質転換体に、さらにMp
DES5を導入した形質転換体では、g−リノレン酸(全脂肪酸の2.8%)、ステアリ
ドン酸(全脂肪酸の0.5%)、ジホモ−g−リノレン酸(全脂肪酸の1.5%)及びエ
イコサテトラエン酸(全脂肪酸の0.1%)の4種の脂肪酸に加えて、アラキドン酸(全
脂肪酸の0.1%)及びエイコサペンタエン酸(EPA、全脂肪酸の0.03%)の生成
が認められた。対照とした形質転換体では、これらの脂肪酸は検出されなかった。この結
果から、ゼニゴケ由来のΔ6不飽和化酵素、Δ6鎖長延長酵素及びΔ5不飽和化酵素の遺
伝子を発現させることで、ゼニゴケ以外の生物種においても、高度不飽和脂肪酸生合成系
の再構築が可能であることが示された。
【0168】
〔実施例6:イネに導入するベクターの構築及び当該ベクターのイネへの導入〕
イネにおいて、MpDES6、MpELO1及びMpDES5遺伝子を発現させるため
に、以下の(i)〜(iv)に示す手順で発現コンストラクトを作製した。また、作製手
順を図1に示した。
【0169】
(i)pBI221(TOYOBO社製)のカリフラワ−モザイクウイルス(CaMV
)35Sプロモーター、及び、NOSターミネーター間に設計した以下のプライマーを用
いたPCRにより、β−Glucuronidase(GUS)遺伝子部分を除いた発現
ベクターp35S−NOSを作製した。
MK001(F):5’−CGGGATCCTCTCCTGGCGCACCATCGTC
−3’(配列番号25)
MK002(R):5’−GGGGTACCAACGCGCTTTCCCACCAACG
−3’(配列番号26)
【0170】
なお、プライマーMK001(F)は下線で示すBamHI認識配列を含み、GUS遺
伝子の3’末端にアニールし、プライマーMK002(R)はGUS遺伝子の5’末端に
アニールする(BamHIサイトがアニール部位の上流に存在する)。PCRにはPyr
obest DNA polymerase(Takara社製)0.5Uを用い、製造
者の推奨する方法に従って反応液量50μlで行った。反応条件は、96℃5分間保持後
、94℃で30秒間、68℃で4分間の反応を30回繰り返し、その後4℃に冷却した。
得られた各ORF断片をBamHI消化した後、実施例1に記載の方法でゲル精製を行っ
た後、自己連結させた。
【0171】
(ii)次にp35S−NOSのXbaIサイトに、MpDES6遺伝子、MpELO
1遺伝子及びMpDES5遺伝子のORFを各々連結した。ORF増幅には、下線で示す
XbaI認識配列を含む以下のプライマーを用いた。
(MpDES6 ORF増幅用プライマー)
MpD6−21F:5’−GCTCTAGAGCGATGGCCTCGTCCACCAC
C−3’(配列番号27)
MpD6−11R:5’−GCTCTAGACTATACTTTCGCAGCGTATG
C−3’(配列番号28)
(MpELO1 ORF増幅用プライマー)
MpD6ELO1−18F:5’−GCTCTAGAGCGATGGAGGCGTACG
AGATGG−3’(配列番号29)
MpD6ELO1−13R:5’−GCTCTAGATTATTCTGCCTTTTTG
CTC−3’ (配列番号30)
(MpDES5 ORF増幅用プライマー)
MpD5−22F:5’−GCTCTAGAGACAGTTATGCCGCCACACG
C−3’(配列番号31)
MpD5−23R:5’−GCTCTAGAAGGCCCAAAGCATGCTGTCA
C−3’(配列番号32)
【0172】
PCRにはPyrobest DNA polymerase(Takara社製)0
.5Uを用い、製造者の推奨する方法に従って、反応液量20μlで行った。反応条件は
、94℃で2分間保持後、94℃で1分間、57℃で1分間、72℃で1分間の反応を2
5回繰り返し、その後4℃に冷却した。得られた各ORF断片をXbaIで消化した後、
実施例1に記載の方法でゲル精製を行いクローニングに用いた。
【0173】
(iii)得られた各遺伝子の発現コンストラクト(それぞれをp35S−MpDES
6、p35S−MpELO1、p35S−MpDES5と表記する)がいずれもCaMV
35Sプロモーター5’末端にPstIサイトを、NOSターミネーター3’末端にEc
oRIサイトを持つことを利用して、上記3遺伝子の発現カセットを連結した。まず、以
下に示すプライマーを用いてp35S−MpDES5を鋳型としてPCRを行い、MpD
ES5遺伝子の発現カセット部分を増幅し、p35S−MpDES6のCaMV35Sプ
ロモーター5’末端にあるPstIサイトにクローニングした(図1参照)。
(MpDES5 遺伝子発現カセット増幅用プライマー)
M13R:5’−CAGGAAACAGCTATGACC−3’(配列番号33)
NOS−R4−PST:5’−AAACTGCAGATTCCCGATCTAGTAAC
ATAG−3’(配列番号34)
【0174】
なお、M13Rプライマーは、CaMV35Sプロモーター上流のベクター配列にアニ
ールする。また、NOS−R4−PSTプライマーは、下線で示すPstI認識配列を含
み、NOSターミネーター3’末端にアニールする。同じくNOSターミネーター3’末
端に存在するEcoRIサイトは含まない。
【0175】
PCRにはPyrobest DNA polymerase(Takara社製)0
.5Uを用い、製造者の推奨する方法に従って、反応液量20μlで行った。反応条件は
、94℃で2分間保持後、94℃で1分間、57℃で1分間、72℃で1分間の反応を2
5回繰り返し、その後4℃に冷却した。得られたDNA断片をPstIで消化した後、実
施例1に記載の方法でゲル精製を行い、MpDES6遺伝子の発現カセットを含むプラス
ミド(p35S−MpDES6)のPstIサイトにクローニングした。
【0176】
(iv)上記で得られた、MpDES5及びMpDES6遺伝子の発現カセットを連結
させたコンストラクト(p35S−MpDES5/35S−MpDES6と表記する)に
、さらにMpELO1遺伝子の発現カセットを連結した。以下のプライマーを用いてp3
5S−MpELO1を鋳型としてPCRを行い、MpELO1遺伝子の発現カセット部分
を増幅し、MpDES6遺伝子発現カセット内のNOSターミネーター3’末端にあるE
coRIサイトにクローニングした。
(MpELO1 遺伝子発現カセット増幅用プライマー)
35S−F3−EI:5’−CCGGAATTCGCATGCCTGCAGGTCCCC
AGA−3’(配列番号35)
M13F:5’−TGTAAAACGACGGCCAGT−3’ (配列番号36)
【0177】
なお、35S−F3−EIプライマーは、下線で示すEcoRI認識配列を含み、Ca
MV35Sプロモーターの5’末端にアニールする。また、M13FプライマーはNOS
ターミネーター下流のベクター配列にアニールする。
【0178】
PCRにはPyrobest DNA polymerase(Takara社製)0
.5Uを用い、製造者の推奨する方法に従って、反応液量20μlで行った。反応条件は
、94℃で2分間保持後、94℃で1分間、57℃で1分間、72℃で1分間の反応を2
5回繰り返し、その後4℃に冷却した。得られたDNA断片をEcoRIで消化した後、
実施例1に記載の方法でゲル精製を行い、MpDES5及びMpDES6遺伝子の発現カ
セットを連結させたコンストラクト(p35S−MpDES5/35S−MpDES6)
のEcoRI部位にクローニングした。
【0179】
以上の操作により、3遺伝子の発現カセットが、MpDES5,MpDES6,MpE
LO1の順で連結した発現コンストラクト(p35S−MpDES5/35S−MpDE
S6/p35S−MpELO1)を作製した。
【0180】
このようにして得られた上記コンストラクトを、公知の方法(Genes Genet
.Syst.73,p219,1998)で、ビアラフォスを選抜マーカーとして持つプ
ラスミドとともに、パーティクルガンでイネに導入し、形質転換イネを取得した。
【0181】
〔実施例7:タバコ(N.tabacum SR−1)におけるゼニゴケ高度不飽和脂肪
酸合成系の再構成〕
本実施例では、上述のゼニゴケ由来の不飽和脂肪酸合成酵素遺伝子、すなわち、MpD
ES6遺伝子、MpDES5遺伝子、およびMpELO遺伝子が、植物体中で良好に機能
することを確認した。
【0182】
具体的には、タバコにMpDES6遺伝子、MpDES5遺伝子、およびMpELO遺
伝子を導入し、このタバコ中でアラキドン酸等が生産されることを確認した。比較対照と
して、糸状菌(M.alpina)由来のΔ6不飽和化酵素遺伝子(MaDES6)、Δ
5不飽和化酵素遺伝子(MaDES5)、Δ6脂肪酸鎖長延長酵素(MaELO)の3遺
伝子を導入したタバコを作製した。
【0183】
(i)糸状菌由来遺伝子を含むベクター(pSPB1519)の構築
pE2113(Mitsuhara et al.Plant Cell Physi
ol.37,45−59 1996)は、エンハンサー配列の繰り返しを有するカリフラ
ワーモザイクウィルス35S(El235S)プロモーター、および、ノパリンシンター
ゼ(nos)ターミネーターを有する。
【0184】
pE2113をSnaBIで消化した後、XhoIリンカー(TAKARA社)と連結
することで、プラスミドを作製した。このプラスミドをSacIで消化後、平滑末端化し
、さらにBamHIリンカー(TAKARA社)と連結することによって、pUE7を作
製した。pUE7をHindIIIとEcoRIで消化することによって得られるDNA
断片のうちEl235Sプロモーターを有する断片と、HindIIIとEcoRIで消
化した植物形質転換用バイナリーベクターpBINPLUS(van Engelen
et al.Transgenic research 4,p288,1995)とを
連結することで、pSPB505を作製した。
【0185】
一方、MaDES6遺伝子を含むプラスミドであるpMLD101をXhoIで消化後
、さらにBamHIで部分消化することで、約1.6kbのDNA断片を得た。この断片
と、pSPB505をXhoIとBamHIで消化して得られるバイナリーベクター部分
のDNA断片とを連結することで、pSPB559を作製した。
【0186】
pUCAP(van Engelen et al.Transgenic rese
arch 4,p288,1995)をAscIで消化した後平滑末端化し、PacIリ
ンカーと連結することで、pUCAPPを作製した。
【0187】
pE2113をSnaBIで消化した後、BamHIリンカー(TAKARA社)と連
結することで、pUE6を作製した。pUE6をSacIで消化した後、平滑末端化し、
SalIリンカー(TAKARA社)と連結することで、pUE8を作製した。pUE8
をHindIIIおよびEcoRIで消化して得られるDNA断片のうちEl235Sプ
ロモーターを有する断片を、pUCAPPのHindIII−EcoRI間に挿入するこ
とで、プラスミドを作製した。このプラスミドをBamHIとSalIで消化して得られ
たDNA断片と、MaELO遺伝子のcDNAをBamHIおよびXhoIで消化して得
られたDNA断片とを連結し、pSPB1130を作製した。pSPB1130をPac
Iで消化し、得られる約2.3kbのDNA断片を、pBINPLUSのPacIサイト
に挿入した。MaELO遺伝子の転写方向と、pBINPLUS上のnptII遺伝子の
転写方向とが、同じ向きになっているプラスミドを選択し、そのプラスミドをpSPB1
157Pとした。
【0188】
また、上述したpSPB559をPacIで消化した後、平滑末端化し、AscIリン
カーと連結することで、pSPB559Aを作製した。そして、pSPB559AをAs
cIで消化して得られるMaDES6遺伝子を含むDNA断片を、pSPB1157Pの
AscIサイトに挿入することで、pSPB1157を作製した。
【0189】
pCGP1364(Plant Cell Physiol.36,1023,199
5)をHindIIIおよびSacIIで消化して得られる約1.3kbのDNA断片と
、pCGP1364をPstIで消化し、平滑末端化した後さらにSacIIで消化して
得られる約2.9kbのDNA断片と、pUCAPAをSacIで消化し、平滑末端化し
た後さらにHindIIIで消化して得られる約2.7kbのDNA断片とを連結するこ
とにより、pSPB184を得た。
【0190】
一方、MaDES5遺伝子がサブクローニングされたpCRIIベクターから、Xba
IとKpnIによる消化によってMaDES5遺伝子を含むDNA断片を切り出した。こ
のDNA断片と、上述のpSPB184をXbaIとKpnIで消化して得られたDNA
断片とを連結することで、pSPB1519Aを作製した。pSPB1519AをAsc
Iで消化し、得られたDNA断片をpSPB1157のAscIサイトに挿入し、pSP
B1519を作製した。このpSPB1519上で、MaDES6遺伝子、MaDES5
遺伝子、およびMaELO遺伝子は、同じ向きに転写され、同じ構成的プロモーターの制
御下にある。
【0191】
(ii)ゼニゴケ由来遺伝子のベクター(pSPB2368)の構築
pUCAP(van Engelen et al.Transgenic Rese
arch 4,288−290,1995)をAscIで消化した後SgfIリンカーと
連結し、さらに、PacIで消化した後FseIリンカーと連結することで、pUCSA
PFを作製した。また、pBINPLUSにも同様の処理を施し、pBINSAPFを作
製した。
【0192】
他にサブクローニング用のベクターとして、pUC19をHindIIIで消化した後
、PacIリンカーと連結し、さらにEcoRIで消化した後FseIリンカーと連結す
ることで、pUCPFを作製した。また、pUC19をHindIIIで消化した後Sg
fIリンカーと連結し、さらに、EcoRIで消化した後AscIリンカーと連結するこ
とで、pUCSAを作製した。pUCSAPFのHindIII−XbaI間にEl23
5Sを、SacI−EcoRI間にマノピン合成酵素(mas)遺伝子ターミネーターを
それぞれ挿入したベクターを、XbaIおよびSacIで消化した後、末端を平滑化する
ことで、pSPB2353Aを得た。pSPB2353Aの平滑末端に、p35S−Mp
DES6からXbaIで切り出して末端を平滑化したMaDES6遺伝子を含むDNA断
片を連結し、pSPB2353作製した。
【0193】
pUCSAのHindIII−XbaI間にEl235Sを、SacI−EcoRI間
にマノピン合成酵素(mas)遺伝子ターミネーターをそれぞれ挿入したベクターを、X
baI、SacIで消化することで、pSPB2355Aを作製した。
【0194】
一方、p35S−MpELO1を鋳型として、下記のプライマー、XbaMpELOf
およびSacMpELOrを用いてPCRを行った。
XbaMpELOf:5‘−AGTCTCTAGAGCGATGGAGGCGTACG−
3’(配列番号43)
SacMpELOr:5‘−CAGTGAGCTCGGTGTCTTATTCTGCC−
3’(配列番号44)
【0195】
PCRには、酵素として高精度なKOD+DNAポリメラーゼ(東洋紡)を用い、94
℃で2分間保持した後、94℃で15秒、68℃で1−3分のサイクルを25回繰り返し
た。このようにして調製されたMpELODNA断片をXbaIおよびSacIで消化し
たものを、上述のpSPB2355Aに連結し、pSPB2355を作製した。さらに、
pSPB2355をSgfI、AscIで消化して得られたDNA断片を、SgfIおよ
びAscIで消化したpSPB2353に連結し、pSPB2361を作製した。
【0196】
pUCPFのHindIII−XbaIサイトにEl235Sを、SacI−EcoR
Iサイトにマノピン合成酵素(mas)遺伝子ターミネーターをそれぞれ挿入したベクタ
ーを、XbaIおよびSacIで消化することで、pSPB2352Aを作製した。
【0197】
一方、p35S−MpDES5を鋳型に、下記のプライマー、XbaMpD5fおよび
SacMpD5rを用いてPCRを行った。PCRの反応条件は、上記のPCR条件と同
様である。
XbaMpD5f:5’−AGCTTCTAGAGCCATGCCGCCACACGCC
C−3’(配列番号45)
SacMpD5r:5’−CAGTGAGCTCTCAGCCATCCAGTCGT−3
’ (配列番号46)
【0198】
PCRによって調製されたMpD5DNA断片をXbaI、SacIで消化したものを
、pSPB2352Aに連結し、pSPB2352を作製した。
【0199】
pBINSAPFをPacIおよびFseIで消化して得られたDNA断片に、pSP
B2352よりPacIおよびFseIで切り出したMpDES5遺伝子を含むDNA断
片を連結し、pSPB2368Aを作製した。さらにpSPB2368AをSgfIおよ
びPacIで消化したものに、pSPB2361よりSgfIおよびPacIで切り出さ
れたMpDES6遺伝子およびMpELO遺伝子を含むDNA断片を連結することで、p
SPB2368を得た。このプラスミド上でMpDES6遺伝子、MpDES5遺伝子、
およびMpELO遺伝子は、同じ向きに転写され、同じ構成的プロモーターの制御下にあ
る。
【0200】
(iii)タバコへの遺伝子導入
引き続いて、公知の方法(Plant J.5,81,1994)に基づいて、pSP
B2368またはpSPB1519を用いてAgrobacterium tumefa
ciens(菌株:AglO(Lazo et al.1991,Biotechnol
ogy9:963−967))を形質転換した。このpSPB2368またはpSPB1
519を有する形質転換体アグロバクテリウムを、タバコリーフディスクに感染させた。
このようにして得られた組換えタバコの葉から、RNeasyPlantminiKit
(キアゲン)を用いてRNAを抽出し、定法に従って、RT−PCRにより、導入した遺
伝子が発現している系統を選択した。
【0201】
このようにして得られた糸状菌由来の酵素遺伝子から成るpSPB1519を導入した
タバコ(pSPB1519形質転換タバコ)、ゼニゴケ由来の酵素遺伝子から成るpSP
B2368を導入したタバコ(pSPB2368形質転換タバコ)の葉から、公知の方法
(藤野安彦編(1978)生物化学実験法9 学会出版センター、山田晃弘編(1989
)生物化学実験法24 学会出版センター)にしたがって、脂質の抽出を行った。得られ
た脂質をガスクロマトグラフィー(Hewlett Packard社 HP−6800
)で分析し、その結果を表1に示す。
【0202】
なお、コントロールとして、遺伝子が導入されていないタバコの葉を用いて、同様の分
析を行った。
【0203】
【表1】

【0204】
本実施例のガスクロマトグラフィーによる分析条件を以下に示す。
(ガスクロマトグラフィー分析条件)
カラム:Supelco SP−2330、Fused Silica Capil
lary Column、30m x 0.32mm ID、0.2μm
温度:Inj:240℃、Det:250℃、Oven:180℃ 3分、180℃
→220℃(2℃/min)
カラム流量:30cm/sec、圧力200kPa、検出器 FID
【0205】
クロマトグラム中の各ピークは標準脂肪酸のメチルエステルのリテンションタイムとG
C−MASS(Hewlett Packard社、HP−5973)分析により決定し
、またピーク面積より各脂肪酸の割合を決定した。
【0206】
表1において、Controlはコントロールを、pSPB2368はpSPB236
8形質転換タバコを、pSPB1519はpSPB1519形質転換タバコを表す。
【0207】
表1に示す結果から、糸状菌由来の酵素遺伝子から成るpSPB1519を導入したタ
バコ(pSPB1519形質転換タバコ)では、ジホモγリノレン酸の蓄積は確認できた
が、アラキドン酸の蓄積は認められなかった。一方、ゼニゴケ由来の酵素遺伝子から成る
pSPB2368を導入したタバコ(pSPB2368形質転換タバコ)ではアラキドン
酸だけでなく、エイコサテトラエン酸およびエイコサペンタエン酸の蓄積も確認できた。
このことから、高等植物においては、糸状菌由来の酵素よりもゼニゴケ由来の酵素の方が
機能的に優れており、リノール酸やαリノレン酸を基質として、アラキドン酸を始めとす
る高度不飽和脂肪酸を合成できると考えられた。
【0208】
Abbadiらは、ケイソウ(Phaeodactylum tricornutum
)のΔ6不飽和化酵素、Δ5不飽和化酵素、およびヒメツリガネゴケ(Physcomi
trella patens)の鎖長延長酵素の遺伝子を、タバコおよびアマ(Linu
m usitatissimum)に導入することで、アラキドン酸を、タバコの種子に
1.5%、アマの種子に1.0%蓄積させたことを報告している(Amine Abba
di et al.Plant Cell 16,2734−2748,2004)。
【0209】
本実施例においては、ゼニゴケ由来のΔ6不飽和化酵素、Δ5不飽和化酵素および鎖長
延長酵素をタバコに導入したことで、タバコの葉にアラキドン酸を10%以上蓄積させて
いる。この結果から、本実施例のpSPB2368形質転換タバコは、上述の報告と比較
しても、より効率的に高度不飽和脂肪酸を合成できる能力があることが示唆された。
【0210】
また、ハプト藻(Isochrysis galbana)のΔ9鎖長延長酵素、ミド
リムシ(Englena gracilis)のΔ8不飽和化酵素、糸状菌のΔ5不飽和
化酵素の3種類の遺伝子を用いて、シロイヌナズナ(Arabidopsis thal
iana)で脂質の改変を行った報告では、葉中でアラキドン酸が総脂質に対して6.6
mol%、C20以上の脂質が22.5mol%であった(Baoxiu Qi et
al.Nature Biotechnology 22,739−745,2004)
。この報告では、Δ6不飽和化酵素、Δ5不飽和化酵素、および鎖長延長酵素を用いる経
路とは別の修飾経路で高度不飽和脂肪酸を作っており、単純比較は出来ないが、ゼニゴケ
の酵素を用いたほうがより多くの高度不飽和脂肪酸を蓄積することがわかった。
【0211】
総脂質の30%以上が改変脂肪酸になっているpSPB2368形質転換タバコでは、
形態的な異常は見られなかった。さらに、稔性にも問題なく、多くの種子をつけたことか
ら、異所的な高度不飽和脂肪酸の増加が植物の生育に及ぼす影響は少ないと考えられた。
【0212】
これまでに報告されている遺伝子組換え植物におけるC20以上の高度不飽和脂肪酸の
生産は総脂質の20%前後、アラキドン酸に限ると6%前後が限界である。しかし、本実
施例のように、ゼニゴケ由来の脂肪酸合成酵素を用いることにより、この限界を打ち破り
、より多くの高度不飽和脂肪酸を植物で生産することが出来ると考えられる。
【0213】
尚、発明を実施するための最良の形態の項においてなした具体的な実施態様または実施
例は、あくまでも、本発明の技術内容を明らかにするものであって、そのような具体例に
のみ限定して狭義に解釈されるべきものではなく、本発明の精神と次に記載する特許請求
の範囲内で、いろいろと変更して実施することができるものである。
【産業上の利用可能性】
【0214】
本発明に係る遺伝子は、同一種のゼニゴケから単離されたΔ5不飽和化酵素遺伝子、Δ
6不飽和化酵素遺伝子及びΔ6鎖長延長酵素遺伝子である。したがって、これら3種類の
遺伝子を植物内で同時に発現させた場合に、異なる生物種由来の複数の遺伝子を発現させ
るより、植物内で良好に機能するという効果を奏する。さらに、ゼニゴケは、高等植物の
モデル系と考えられるため、これらの遺伝子は、植物以外の生物種由来の遺伝子より植物
内で良好に機能することができるという効果を奏する。
【0215】
また、本発明に係る形質転換体は、アラキドン酸やエイコサペンタエン酸(EPA)等
の高度不飽和脂肪酸を生産することができるという効果を奏する。特に、本発明に係る形
質転換植物は、低コストかつ環境にやさしい生産プロセスでアラキドン酸やEPA等の高
度不飽和脂肪酸を生産することができるという効果を奏する。こうして得られたアラキド
ン酸やEPAは、安価な多目的材料として活用できるという効果を奏する。さらに、上記
形質転換植物体を食料として用いた場合、アラキドン酸やEPAの含量が高い食料として
の価値が高まるという効果を奏する。
【0216】
以上にように、本発明の遺伝子・たんぱく質は、アラキドン酸やEPAの生産に有用で
ある。また、本発明の遺伝子を発現可能に導入した形質転換体は、製薬産業、食品産業、
各種素材産業等において、アラキドン酸やEPAを生産するうえで、極めて有用である。
また、特に、上記形質転換体が植物体である場合、植物体内のアラキドン酸やEPAの含
量が増加するので、農業分野等において非常に有用である。
【技術分野】
【0001】
本発明は、ゼニゴケ(Marchantia polymorpha)由来の不飽和脂
肪酸合成系遺伝子、すなわち、Δ5脂肪酸不飽和化酵素、Δ6脂肪酸不飽和化酵素、及び
Δ6脂肪酸鎖長延長酵素の遺伝子とその利用に関するものである。
【背景技術】
【0002】
アラキドン酸、エイコサペンタエン酸(以下、適宜「EPA」と略記する)等の高度不
飽和脂肪酸(Polyunsaturated fatty acid;PUFA)は、
ヒトにおいて神経系を中心とした細胞膜脂質に多く含まれている。これらの高度不飽和脂
肪酸は、プロスタグランジンやロイコトリエンといった、生理活性物質の前駆体として作
用しており、薬理学的に非常に重要である。近年、アラキドン酸やEPAを含む健康食品
も販売されている。また、脂肪酸は、洗剤や生分解性プラスチックの原料にもなるため、
素材としても注目されている。
【0003】
高度不飽和脂肪酸は、現在、培養微生物又は魚油からの抽出により生産されている。こ
のため、生産コストが高いこと、エネルギー使用量・廃棄物量が多くなること、特に魚油
から調製する方法では、魚資源が限られていることが問題になっている。
【0004】
アラキドン酸及びEPAは、それぞれリノール酸及びα−リノレン酸を起点として、Δ
6不飽和化、脂肪酸鎖長延長及びΔ5不飽和化という3つの連続した反応により生合成さ
れると考えられている。これらの反応は、それぞれ、Δ6脂肪酸不飽和化酵素(以下、「
Δ6不飽和化酵素」と略記する)、Δ6脂肪酸鎖長延長酵素(以下、「Δ6鎖長延長酵素
」と略記する)及びΔ5脂肪酸不飽和化酵素(以下、「Δ5不飽和化酵素」と略記する)
により触媒される。
【0005】
Δ6不飽和化酵素の遺伝子は、いくつかの植物種でクローン化されている。例えば、ケ
イ藻、ヒメツリガネゴケ、ヤノウエノアカゴケ、ボリジ、ムラサキ、サクラソウ及びアネ
モネからΔ6不飽和化酵素の遺伝子がクローン化されている。また、植物以外では、糸状
菌、線虫、ラン藻、ラット及びヒトからΔ6不飽和化酵素遺伝子がクローン化されている
(非特許文献1:「Eur.J.Biochem.269,p4105,2002」、非
特許文献2:「Plant J.15,p39,1998」、非特許文献3:「Eur.
J.Biochem.,267.p3801,2000」、非特許文献4:「Proc.
Natl.Acad.Sci.USA 94,p4211,1997」、非特許文献5:
「Lipids 37,417,2002」、非特許文献6:「FEBS Lett.5
42,p100,2003」、非特許文献7:「Whitney et al.,Pla
nta Epub 2003」、非特許文献8:「Lipids 34,p649,19
99」、非特許文献9:「Gene,238,p445 1999」、非特許文献10:
「Biochem J.330,p611 1998」、非特許文献11:「Plant
Mol.Biol.,22,p293 1993」、非特許文献12:「Bioche
m.Biophys.res.Commun.255,p575,1999」、非特許文
献13:「J.Biol.Chem.274,p471,1999」参照)。また、ラン
藻のものを除いて、これらのΔ6不飽和化酵素は、いずれもN末端にシトクロムb5ドメ
インが存在する。
【0006】
Δ6鎖長延長酵素の遺伝子は、最初に糸状菌及び線虫からクローン化された(非特許文
献14:「Proc.Natl.Acad.Sci.USA 97,p8284,200
0」、非特許文献15:「Proc.Natl.Acad.Sci.USA 97,p6
421,2000」参照)。植物種では、唯一ヒメツリガネからクローン化されている(
非特許文献16:「Plant J.31,p255,2002」参照)。
【0007】
酵母(Saccharomyces cerevisiae)には、スフィンゴ脂質の
長鎖飽和アシル鎖合成に関与するELO2蛋白及びELO3蛋白が存在し(非特許文献1
7:「J.Biol.Chem.,272,p17376,1997」参照)、Δ6鎖長
延長酵素のアミノ酸配列は、これらと相同性を示す。一方、植物には、別タイプの脂肪酸
鎖長延長酵素であるβ−ケトアシルCoA合成酵素(KCS)が存在する。この酵素は、
長鎖飽和/一価不飽和脂肪酸の鎖長延長を触媒する(非特許文献15及び非特許文献18
:「Plant Cell 7,p309,1995」参照)。しかしながら、Δ6鎖長
延長酵素遺伝子及び酵母ELO2/ELO3遺伝子は、KCS遺伝子との直接的な進化上
の関係は見られない(非特許文献15及び16参照)。
【0008】
Δ5不飽和化酵素の遺伝子は、糸状菌から、初めてクローン化された(非特許文献19
:「J.Biol.Chem.273,p29360,1998」、非特許文献20:「
J.Biol.Chem.273,p19055」参照)。Δ5不飽和化酵素の構造はΔ
6不飽和化酵素と共通しており、N末端にシトクロムb5ドメインを有する。Δ5不飽和
化酵素遺伝子は、ケイ藻、線虫、ラット、ヒト、ヒメツリガネゴケなどからクローン化さ
れている(非特許文献1,非特許文献21:「FEBS Lett.439,p215,
1998」、非特許文献22:「Arch.Biochem.Biophys.391,
p8,2001」、非特許文献23:「J.Biol.Chem.274,p37335
,1999」、非特許文献24:「J.Biol.Chem.278,35115,20
03」参照)。
【0009】
陸上植物は、コケ植物(コケ植物門(Bryophyta))、シダ植物、裸子植物及
び被子植物から構成されている。コケ植物は陸上植物の中で最も古くに分岐した群であり
、セン類(セン類網(Bryosida))、タイ類(タイ類網(Hepaticops
ida))及びツノゴケ類の3つのグループから構成されている。ゼニゴケは、上記生物
のうちヒメツリガネゴケに分類上最も近いが、ヒメツリゴケはセン類に属しており、ゼニ
ゴケはタイ類網の中のゼニゴケ亞網(Marchantiidae)に属している。上記
3つのグループは、約4億3千年前には、すでに分岐していたことは確かである。したが
って、同じコケといっても、ヒメツリガネゴケとゼニゴケとは、例えば2億年前に分化し
たシロイナズナとイネとの違いどころでなく、進化上大きく異なる(非特許文献25:「
1103615143968_0.html」参照)。
【0010】
ゼニゴケ由来の高度不飽和脂肪酸合成系酵素遺伝子としては、上記のKCS様の鎖長延
長酵素遺伝子のMpFAE2及びMpFAE3が取得されている(非特許文献26:「B
iosci.Biotechnol.Biochem.67,p605,2003」、非
特許文献27:「Biosci.Biotechnol.Biochem.67,p16
67,2003」参照)。しかしながら、MpFAE2及びMpFAE3はΔ6鎖長延長
酵素遺伝子ではない。
【0011】
上述のように、多くの高度脂肪酸合成系酵素遺伝子が様々な生物種からクローン化され
ているが、アラキドン酸、EPA等の、C20以上で不飽和度4以上の高度不飽和脂肪酸
を植物中で生産させた例は少ない。このような例として、ケイ藻由来のΔ6不飽和化酵素
及びΔ5不飽和化酵素と、ヒメツリガネゴケ由来のΔ6鎖長延長酵素とをアマで発現させ
、アラキドン酸及びEPAを生産させた報告があるが、その詳細は不明である(非特許文
献24参照)。
【0012】
上述のように、アラキドン酸やEPA等の高度不飽和脂肪酸の生産は、培養微生物又は
魚油からの抽出により生産されているため、生産コストが高いこと、エネルギー使用量・
廃棄物量が多くなること、魚資源が限られていること等の問題点を有している。アラキド
ン酸やEPA等の高度不飽和脂肪酸は、分子内に二重結合を複数有するというユニークな
物性を持っているため、様々な工業用途(例えばフィルム、生分解性プラスチック、機能
性繊維、潤滑油、洗剤の素材等)にも利用可能となる。このような高度不飽和脂肪酸を遺
伝子組み換え植物により生産することにより、生産コストを低減でき、同時に、より環境
にやさしい生産プロセスを実現することができると期待される。遺伝子組み換え技術を用
いて、これらの高度不飽和脂肪酸を、油糧植物で大量生産できるようになれば、安価な多
目的原料として非常に有用である。
【0013】
一方、植物に異種生物の遺伝子を発現させる場合、その遺伝子が植物内でどの程度良好
に機能するかは、転写、翻訳、その後の修飾などの過程があるため、予想することは困難
である。特に、複数の異種生物の遺伝子を発現させる場合、上記非特許文献24のように
異なる生物種由来の複数の遺伝子を発現させるより、同一の種に由来する複数の遺伝子を
発現させるほうが植物内で良好に機能することが予想される。また、最初の陸上植物であ
るコケ類のゼニゴケは、高等植物のモデル系として注目されており、その遺伝子は植物内
で良好に機能することが期待できる。したがって、ゼニゴケ由来の高度不飽和脂肪酸合成
酵素遺伝子、すなわちΔ5不飽和化酵素遺伝子、Δ6不飽和化酵素遺伝子及びΔ6鎖長延
長酵素遺伝子を取得することができれば、これらの遺伝子を植物に導入することにより、
アラキドン酸やEPAが植物内で効率よく蓄積されることが期待できる。
【0014】
また、ゼニゴケと同じコケ植物のヒメツリガネゴケからはΔ5不飽和化酵素遺伝子、Δ
6不飽和化酵素遺伝子及びΔ6鎖長延長酵素遺伝子がクローン化されているが、ゼニゴケ
とヒメツリガネゴケは進化上大きく異なっており、ヒメツリガネゴケの遺伝子に基づいて
ゼニゴケの遺伝子を取得することは、現在の技術水準をもってしても容易にできることで
はない。
【発明の概要】
【発明が解決しようとする課題】
【0015】
本発明は、上記従来の問題点に鑑みなされたものであって、その目的は、高等植物中で
アラキドン酸やEPAを生産し得る、ゼニゴケ(Marchantia polymor
pha)由来の不飽和脂肪酸合成酵素遺伝子、すなわちΔ5不飽和化酵素遺伝子、Δ6不
飽和化酵素遺伝子及びΔ6鎖長延長酵素遺伝子、及びその利用法を提供することにある。
【課題を解決するための手段】
【0016】
本発明者らは、上記の課題を解決するために鋭意検討した結果、ゼニゴケ(March
antia polymorpha)由来のcDNAクローンから、上記Δ6不飽和化酵
素、Δ5不飽和化酵素及びΔ6鎖長延長酵素をコードする遺伝子を同定し、更にこれらの
遺伝子をメタノール資化性酵母(Pichia pastoris)に導入して発現させ
ることに成功し、これらの遺伝子を発現させたたんぱく質が、それぞれΔ6不飽和化、Δ
5不飽和化及びΔ6鎖長延長の酵素活性を有することを見出し、本発明を完成するに至っ
た。すなわち、本発明は、以下の発明を包含する。
【0017】
(1)配列番号1に示される塩基配列からなるDNAの全部又は一部、あるいは当該D
NAと相補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイ
ブリダイズし、かつΔ6脂肪酸不飽和化活性を有するたんぱく質をコードするゼニゴケ目
生物由来の遺伝子。
【0018】
(2)Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号1に示される塩基配列を有する
遺伝子。(b)配列番号1に示される塩基配列からなるDNA、又は当該DNAと相補的
な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0019】
(3)Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号1に示される塩基配列のうち、
253ないし1698番目の塩基配列を有する遺伝子。(b)配列番号1に示される塩基
配列のうち、253ないし1698番目の塩基配列からなるDNA、又は当該DNAと相
補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0020】
(4)Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号2に示されるアミノ酸配列から
なるたんぱく質をコードする遺伝子。(b)配列番号2に示されるアミノ酸配列の1個又
はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなる
たんぱく質をコードする遺伝子。
【0021】
(5)配列番号3に示される塩基配列からなるDNAの全部又は一部、あるいは当該D
NAと相補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイ
ブリダイズし、かつΔ6脂肪酸鎖長延長活性を有するたんぱく質をコードするゼニゴケ目
生物由来の遺伝子。
【0022】
(6)Δ6脂肪酸鎖長延長活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号3に示される塩基配列を有する
遺伝子。(b)配列番号3に示される塩基配列からなるDNA、又は当該DNAと相補的
な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0023】
(7)Δ6脂肪酸鎖長延長活性を有するゼニゴケ目生物由来のたんぱく質をコードする
、下記(a)又は(b)に記載の遺伝子。(a)配列番号1に示される塩基配列のうち、
194ないし1066番目の塩基配列を有する遺伝子。(b)配列番号1に示される塩基
配列のうち、194ないし1066番目の塩基配列からなるDNA、又は当該DNAと相
補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0024】
(8)Δ6脂肪酸鎖長延長活性を有するゼニゴケ目由来のたんぱく質をコードする、下
記(a)又は(b)に記載の遺伝子。(a)配列番号4に示されるアミノ酸配列からなる
たんぱく質、(b)配列番号4に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が
置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする
遺伝子。
【0025】
(9)配列番号5に示される塩基配列からなるDNAの全部又は一部、あるいは当該D
NAと相補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイ
ブリダイズし、かつΔ5脂肪酸不飽和化活性を有するたんぱく質をコードするゼニゴケ目
生物由来の遺伝子。
【0026】
(10)Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードす
る、下記(a)又は(b)に記載の遺伝子。(a)配列番号5に示される塩基配列を有す
る遺伝子。(b)配列番号5に示す塩基配列からなるDNA、又は当該DNAと相補的な
塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0027】
(11)Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードす
る、下記(a)又は(b)に記載の遺伝子。(a)配列番号5に示される塩基配列のうち
、375ないし1829番目の塩基配列を有する遺伝子。(b)配列番号5に示される塩
基配列のうち、375ないし1829番目の塩基配列からなるDNA、又は当該DNAと
相補的な塩基配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子
。
【0028】
(12)Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードす
る、下記(a)又は(b)に記載の遺伝子。(a)配列番号6に示されるアミノ酸配列か
らなるたんぱく質、(b)配列番号6に示されるアミノ酸配列の1個又はそれ以上のアミ
ノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコー
ドする遺伝子。
【0029】
(13)上記(1)〜(12)の何れかに記載の遺伝子によってコードされるたんぱく
質。
【0030】
(14)以下の(a)又は(b)記載のたんぱく質。(a)配列番号2に示されるアミ
ノ酸配列からなるたんぱく質。(b)配列番号2に示されるアミノ酸配列において、1個
又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からな
り、かつ、Δ6脂肪酸不飽和化活性を有するたんぱく質。
【0031】
(15)以下の(a)又は(b)記載のたんぱく質。(a)配列番号4に示されるアミ
ノ酸配列からなるたんぱく質。(b)配列番号4に示されるアミノ酸配列において、1個
又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からな
り、かつ、Δ6脂肪酸鎖長延長活性を有するたんぱく質。
【0032】
(16)以下の(a)又は(b)記載のたんぱく質。(a)配列番号6に示されるアミ
ノ酸配列からなるたんぱく質。(b)配列番号6に示されるアミノ酸配列において、1個
又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からな
り、かつ、Δ5脂肪酸不飽和化活性を有するたんぱく質。
【0033】
(17)上記(13)〜(16)の何れかに記載のたんぱく質を認識する抗体。
【0034】
(18)少なくとも上記(1)〜(12)の何れかに記載の遺伝子を含む組換え発現ベ
クター。
【0035】
(19)少なくとも上記(1)〜(12)の何れかに記載の遺伝子を導入してなる形質
転換体。
【0036】
(20)少なくとも上記(1)〜(12)の何れかに記載の遺伝子が発現可能に導入さ
れた植物体、もしくは当該植物体と同一の性質を有する当該植物体の子孫となる植物体、
又は当該植物体の組織。
【0037】
(21)少なくとも上記(1)〜(12)の何れかに記載の遺伝子が発現可能に導入さ
れ、脂肪酸組成が改変された植物体、もしくは当該植物体と同一の性質を有する当該植物
体の子孫となる植物体、又は当該植物体の組織。
【0038】
(22)上記(20)又は(21)に記載の植物体の繁殖材料。
【0039】
(23)上記(21)に記載の植物体又は植物体の組織を用いる脂肪酸生産方法。
【0040】
(24)上記(23)に記載の脂肪酸生産方法により得られた、γ−リノレン酸、ジホ
モ−γ−リノレン酸、アラキドン酸、ステアリドン酸、エイコサテトラエン酸、およびエ
イコサペンタエン酸から選択される少なくとも1つ含む素材。
【0041】
(25)少なくとも(1)〜(12)の何れかに記載の遺伝子を用いて脂肪酸組成を改
変する方法。
【0042】
(26)上記(1)〜(12)の何れかに記載の遺伝子における少なくとも一部の塩基
配列又はその相補配列をプローブとして用いた遺伝子検出器具。
【0043】
(27)上記(13)〜(16)の何れかに記載のたんぱく質を用いて、当該たんぱく
質を調節する遺伝子、又は当該たんぱく質を調節する物質をスクリーニングする方法。
【0044】
(28)上記(27)に記載のスクリーニング方法により得られた遺伝子又は物質。
【0045】
なお、本明細書において、特に断らない限り、A、C、GおよびTは、アデニン、シト
シン、グアニンおよびチミンの各塩基を示す。
【0046】
本発明のさらに他の目的、特徴、および優れた点は、以下に示す記載によって十分わか
るであろう。また、本発明の利益は、添付図面を参照した次の説明で明白になるであろう
。
【図面の簡単な説明】
【0047】
【図1】実施例6で用いたMpDES6遺伝子、MpELO1遺伝子及びMpDES5遺伝子の各遺伝子の発現カセットが連結されたコンストラクトの構築手順を示す説明図である。
【発明を実施するための形態】
【0048】
本発明の実施の一形態について説明すれば、以下の通りである。なお、本発明は、これ
に限定されるものではない。
【0049】
以下、アラキドン酸及びエイコサペンタエン酸(EPA)の合成経路、本発明に係る遺
伝子、本発明に係るたんぱく質、本発明に係るたんぱく質及び遺伝子の取得方法、並びに
本発明に係る遺伝子及びたんぱく質の利用方法(有用性)の順で、本発明を詳細に説明す
る。
【0050】
(1)アラキドン酸及びエイコサペンタエン酸(EPA)の合成経路
アラキドン酸及びエイコサペンタエン酸(EPA)は、それぞれリノール酸及びα−リ
ノレン酸を起点として、Δ6不飽和化、Δ6鎖長延長及びΔ5不飽和化という3つの連続
した反応により、生合成されると考えられる。これらの反応は、それぞれΔ6不飽和化酵
素、Δ6鎖長延長酵素及びΔ5不飽和化酵素により触媒され、それぞれ、n−6経路(ア
ラキドン酸合成経路)及びn−3経路(EPA合成経路)と呼ばれている。
【0051】
これまでに報告されているΔ6不飽和化酵素、Δ6鎖長延長酵素及び、Δ5不飽和化酵
素は、いずれもn−6及びn−3経路の両方に関与していることが示されている。すなわ
ち、Δ6不飽和化酵素は、n−6経路ではリノール酸(18:2D9,12、18は炭素
数を表し、2は二重結合の数を表し、9、12は二重結合の位置を表す。以下同様である
。)をg−リノレン酸(GLA;18:3D6,9,12)に変換し、n−3経路ではa
−リノレン酸(ALA;18:3D9,12,15)をステアリドン酸(STA;18:
4D6,9,12,15)に変換する。Δ6鎖長延長酵素は、n−6経路ではGLAをジ
ホモ−γ−リノレン酸(DGLA;20:3Δ8,11,14)に変換し、n−3経路で
はSTAをエイコサテトラエン酸(ETA;20:4Δ8,11,14,17)に変換す
る。Δ5不飽和化酵素は、n−6経路ではDGLAをアラキドン酸(20:4Δ5,8,
11,14)に、n−3経路ではETAをエイコサペンタエン酸(EPA;20:5Δ5
,8,11,14,17)に変換する。
【0052】
(2)本発明に係る遺伝子
〔本発明に係るΔ6不飽和化酵素遺伝子〕
本発明に係るΔ6不飽和化酵素遺伝子は、Δ6脂肪酸不飽和化活性を有するたんぱく質
をコードするゼニゴケ目生物由来の遺伝子であり、以下の条件に適合する遺伝子であれば
よい。
【0053】
1.配列番号1に示される塩基配列を有する遺伝子。
【0054】
2.配列番号1に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基配
列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0055】
3.配列番号1に示される塩基配列からなるDNAの一部、又は当該DNAと相補的な
塩基配列からなるDNAの一部とストリンジェントな条件でハイブリダイズする遺伝子。
【0056】
4.配列番号1に示される塩基配列のうち、253ないし1698番目の塩基配列を有
する遺伝子。なお、配列番号1に示される塩基配列の253ないし1698番目の塩基配
列は配列番号2に示されるアミノ酸配列からなるたんぱく質に翻訳される領域である。
【0057】
5.配列番号1に示される塩基配列のうち、253ないし1698番目の塩基配列から
なるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな条
件でハイブリダイズする遺伝子。
【0058】
6.配列番号2に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0059】
7.配列番号2に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0060】
〔本発明に係るΔ6鎖長延長酵素遺伝子〕
本発明に係るΔ6鎖長延長酵素遺伝子は、Δ6脂肪酸鎖長延長酵素活性を有するたんぱ
く質をコードするゼニゴケ目生物由来の遺伝子であり、以下の条件に適合する遺伝子であ
ればよい。
【0061】
1.配列番号3に示される塩基配列を有する遺伝子。
【0062】
2.配列番号3に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基配
列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0063】
3.配列番号3に示される塩基配列からなるDNAの一部、又は当該DNAと相補的な
塩基配列からなるDNAの一部とストリンジェントな条件でハイブリダイズする遺伝子。
【0064】
4.配列番号3に示される塩基配列のうち、194ないし1066番目の塩基配列を有
する遺伝子。なお、配列番号3に示される塩基配列の194ないし1066番目の塩基配
列は配列番号4に示されるアミノ酸配列からなるたんぱく質に翻訳される領域である。
【0065】
5.配列番号3に示される塩基配列のうち、194ないし1066番目の塩基配列から
なるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな条
件でハイブリダイズする遺伝子。
【0066】
6.配列番号4に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0067】
7.配列番号4に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0068】
〔本発明に係るΔ5不飽和化酵素遺伝子〕
本発明に係るΔ5不飽和化酵素遺伝子は、Δ5脂肪酸不飽和化活性を有するたんぱく質
をコードするゼニゴケ目生物由来の遺伝子であり、以下の条件に適合する遺伝子であれば
よい。
【0069】
1.配列番号5に示される塩基配列を有する遺伝子。
【0070】
2.配列番号5に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基配
列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【0071】
3.配列番号5に示される塩基配列からなるDNAの一部、又は当該DNAと相補的な
塩基配列からなるDNAの一部とストリンジェントな条件でハイブリダイズする遺伝子。
【0072】
4.配列番号5に示される塩基配列のうち、375ないし1829番目の塩基配列を有
する遺伝子。なお、配列番号5に示される塩基配列の375ないし1829番目の塩基配
列は配列番号6に示されるアミノ酸配列からなるたんぱく質に翻訳される領域である。
【0073】
5.配列番号5に示される塩基配列のうち、375ないし1829番目の塩基配列から
なるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな条
件でハイブリダイズする遺伝子。
【0074】
6.配列番号6に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0075】
7.配列番号6に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【0076】
なお、上記「ストリンジェントな条件」とは、少なくとも90%の同一性、好ましくは
少なくとも95%の同一性、最も好ましくは少なくとも97%の同一性が配列間に存在す
るときにのみハイブリダイゼーションが起こることを意味する。
【0077】
上記ハイブリダイゼーションは、J.Sambrook et al.Molecul
ar Cloning,A Laboratory Manual,2d Ed.,Co
ld Spring Harbor Laboratory(1989)に記載されてい
る方法等、従来公知の方法で行うことができる。通常、温度が高いほど、塩濃度が低いほ
どストリンジェンシーは高くなり(ハイブリダイズし難くなる)、より相同な遺伝子を取
得することができる。ハイブリダイゼーションの条件としては、従来公知の条件を好適に
用いることができ、特に限定しないが、例えば、42℃、6×SSPC、50%ホルムア
ミド、1%SDS、100μg/ml salmon sperm DNA、5×デンハ
ルト液(ただし、1×SSPE;0.18M 塩化ナトリウム、10mM リン酸ナトリ
ウム、pH7.7、1mM EDTA)が挙げられる。
【0078】
上記「ゼニゴケ目生物」とは、ゼニゴケ(Marchantia polymorph
a)に限定されるものではなく、ゼニゴケ亞網ゼニゴケ目(Marchantiales
)に属する生物が含まれる。これらのうち、Monoclea forsteri(Mo
nocleales)、Corsinia coriandrina(Marchant
iales)、Oximitra paleacea(Marchantiales)、
Ricciocarpos natans(Marchantiales)、Ricca
huebeneriana(Marchantiales)、Ricca fluit
ans(Marchantiales)、Ricca duplex(Marchant
iales)、Ricca canaliculata(Marchantiales)
、Ricca bifurca(Marchantiales)、Ricca cili
ifera(Marchantiales)、Ricca glauca(Marcha
ntiales)、Ricca sorocarpa(Marchantiales)、
Ricca warnstorfii(Marchantiales)、Ricca m
ichelii(Marchantiales)、Ricca papillosa(M
archantiales)及びRicca zachariae(Marchanti
ales)には、超長鎖長高度不飽和脂肪酸が存在することが知られている(Prog.
Lipid Res.32,p281,1993参照)。これらの生物からΔ6不飽和化
酵素、Δ6鎖長延長酵素及びΔ5不飽和化酵素の遺伝子を取得することは現在の技術水準
を持ってすれば容易である。例えば、近縁生物の同じ機能を有する酵素をコードする遺伝
子は、クロスハイブリダイゼーションすることが一般に知られている。
【0079】
本発明の遺伝子は、2本鎖DNAのみならず、それを構成するセンス鎖及びアンチセン
ス鎖といった各1本鎖DNAやRNAを包含する。アンチセンス鎖は、プローブとして又
はアンチセンス化合物として利用できる。DNAには、例えばクローニングや化学合成技
術又はそれらの組み合わせで得られるようなcDNAやゲノムDNAなどが含まれる。さ
らに、本発明の遺伝子は、非翻訳領域(UTR)の配列やベクター配列(発現ベクター配
列を含む)などの配列を含むものであってもよい。
【0080】
(3)本発明に係るたんぱく質
〔本発明に係るΔ6不飽和化酵素たんぱく質〕
本発明に係るΔ6不飽和化酵素たんぱく質は、ゼニゴケ目生物由来のたんぱく質であっ
てΔ6脂肪酸不飽和化活性を有するたんぱく質であればよい。より具体的には、以下に示
すたんぱく質であればよい。
【0081】
1.上記(2)に記載した本発明に係るΔ6不飽和化酵素遺伝子によってコードされる
たんぱく質。
【0082】
2.配列番号2に示されるアミノ酸配列からなるたんぱく質。
【0083】
3.配列番号2に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質。
【0084】
〔本発明に係るΔ6鎖長延長酵素たんぱく質〕
本発明に係るΔ6鎖長延長酵素たんぱく質は、ゼニゴケ目生物由来のたんぱく質であっ
てΔ6脂肪酸鎖長延長活性を有するたんぱく質であればよい。より具体的には、以下に示
すたんぱく質であればよい。
【0085】
1.上記(2)に記載した本発明に係るΔ6鎖長延長酵素遺伝子によってコードされる
たんぱく質。
【0086】
2.配列番号4に示されるアミノ酸配列からなるたんぱく質。
【0087】
3.配列番号4に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質。
【0088】
〔本発明に係るΔ5不飽和化酵素たんぱく質〕
本発明に係るΔ5不飽和化酵素たんぱく質は、ゼニゴケ目生物由来のたんぱく質であっ
てΔ5脂肪酸不飽和化活性を有するたんぱく質であればよい。より具体的には、以下に示
すたんぱく質であればよい。
【0089】
1.上記(2)に記載した本発明に係るΔ5不飽和化酵素遺伝子によってコードされる
たんぱく質。
【0090】
2.配列番号6に示されるアミノ酸配列からなるたんぱく質。
【0091】
3.配列番号6に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失、
挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質。
【0092】
上記「Δ6脂肪酸不飽和化活性」とは、リノール酸又はα−リノレン酸に対して基質特
異性を有し、それぞれγ−リノレン酸又はステアリドン酸に変換する作用を意味する。ま
た、上記「Δ6脂肪酸鎖長延長活性」とは、γ−リノレン酸又はステアリドン酸に対して
基質特異性を有し、それぞれジホモ−γ−リノレン酸又はエイコサテトラエン酸に変換す
る作用を意味する。また、上記「Δ5脂肪酸不飽和化活性」とは、ジホモ−γ−リノレン
酸又はエイコサテトラエン酸に対して基質特異性を有し、それぞれアラキドン酸又はエイ
コサペンタエン酸(EPA)に変換する作用を意味する。
【0093】
上記「1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加された」とは
、部位特異的突然変異誘発法等の公知の変異たんぱく質作製法により置換、欠失、挿入、
及び/又は付加できる程度の数(好ましくは10個以下、より好ましくは7個以下、さら
に好ましくは5個以下)のアミノ酸が置換、欠失、挿入、及び/又は付加されることを意
味する。このような変異たんぱく質は、公知の変異たんぱく質作製法により人為的に導入
された変異を有するたんぱく質に限定されるものではなく、天然に存在する同様の変異た
んぱく質を単離精製したものであってもよい。
【0094】
なお、本発明のたんぱく質は、アミノ酸がペプチド結合してなるポリペプチドであれば
よいが、これに限定されるものではなく、ポリペプチド以外の構造を含む複合たんぱく質
であってもよい。ここでいうポリペプチド以外の構造としては、糖鎖やイソプレノイド基
等を挙げることができるが、特に限定されるものではない。
【0095】
また、本発明のたんぱく質は、付加的なポリペプチドを含むものであってもよい。この
ようなポリペプチドが付加される場合としては、例えば、HisやMyc、Flag等に
よって本発明のたんぱく質がエピトープ標識されるような場合が挙げられる。
【0096】
また、本発明のたんぱく質は、前述した本発明の遺伝子(本発明のたんぱく質をコード
する遺伝子)を宿主細胞に導入して、そのたんぱく質を細胞内発現させた状態であっても
よいし、細胞、組織などから単離精製された状態であってもよい。また、上記宿主細胞で
の発現条件によっては、本発明のたんぱく質は、他のたんぱく質とつながった融合たんぱ
く質であってもよい。さらに本発明のたんぱく質は、化学合成されたものであってもよい
。
【0097】
(4)本発明に係るたんぱく質及び遺伝子の取得方法
本発明に係るたんぱく質及び遺伝子の取得方法(生産方法)は特に限定されるものでは
ないが、代表的な方法として次に示す各方法を挙げることができる。
【0098】
〔たんぱく質の取得方法〕
本発明のたんぱく質を取得する方法(生産方法)は、上述したように特に限定されるも
のではないが、まず、本発明のたんぱく質を発現する細胞、組織などから単純精製する方
法を挙げることができる。精製方法も特に限定されるものではなく、公知の方法で細胞や
組織から細胞抽出液を調製し、この細胞抽出液を公知の方法、例えばカラム等を用いて精
製すればよい。
【0099】
また、本発明のたんぱく質を取得する方法として、遺伝子組み換え技術等を用いる方法
も挙げられる。この場合、例えば、本発明の遺伝子をベクターなどに組み込んだ後、公知
の方法により発現可能に宿主細胞に導入し、細胞内で翻訳されて得られる上記たんぱく質
を精製するという方法などを採用することができる。
【0100】
なお、このように宿主に外来遺伝子を導入する場合、外来遺伝子の発現のため宿主内で
機能するプロモーターを組み入れた発現ベクター及び宿主には様々なものが存在するので
、目的に応じたものを選択すればよい。産生されたたんぱく質を精製する方法は、用いた
宿主、たんぱく質の性質によって異なるが、タグの利用等によって比較的容易に目的のた
んぱく質を精製することが可能である。
【0101】
変異たんぱく質を作製する方法についても、特に限定されるものではない。例えば、部
位特異的突然変異誘発法(Hashimoto−Gotoh,Gene 152,271
−275(1995)他)、PCR法を利用して塩基配列に点変異を導入し変異たんぱく
質を作製する方法、あるいはトランスポゾンの挿入による突然変異株作製法などの周知の
変異たんぱく質作製法を用いることができる。変異たんぱく質の作製には市販のキットを
利用してもよい。
【0102】
本発明のたんぱく質の取得方法は上述の方法限定されることはなく、例えば、化学合成
されたものであってもよい。例えば、無細胞系のたんぱく質合成液を利用して本発明の遺
伝子から本発明のたんぱく質を合成してもよい。
【0103】
〔遺伝子の取得方法〕
本発明の遺伝子を取得する方法(生産方法)も特に限定されるものではないが、例えば
、ディファレンシャルスクリーニング(サブトラクションクローニング)を利用する方法
を挙げることができる。この方法では、公知の技術に従って、試験管内での直接的ハイブ
リダイゼーションを繰り返し、目的のcDNA(本発明の遺伝子)を濃縮すればよい。
【0104】
上記ディファレンシャルスクリーニングにおける各ステップについては、通常用いられ
る条件の下で行えばよい。これによって得られたクローンは、制限酵素地図の作成及びそ
の塩基配列決定(シークエンシング)によって、さらに詳しく解析することができる。こ
れらの解析によって、本発明の遺伝子配列を含むDNA断片を取得したか容易に確認する
ことができる。
【0105】
また、本発明の遺伝子を取得する方法として、公知の技術により、本発明の遺伝子を含
むDNA断片を単離し、クローニングする方法が挙げられる。例えば、本発明の遺伝子の
塩基配列の一部と特異的にハイブリダイズするプローブを調製し、ゲノムDNAライブラ
リーやcDNAライブラリーをスクリーニングすればよい。このようなプローブとしては
、本発明の遺伝子の塩基配列又はその相補配列の少なくとも一部に特異的にハイブリダイ
ズするプローブであれば、いずれの配列・長さのものを用いてもよい。
【0106】
また、上記プローブの配列を、上述したゼニゴケ間で良好に保存されている領域の中か
ら選択し、他のゼニゴケのゲノムDNA(又はcDNA)ライブラリーをスクリーニング
すれば、上記たんぱく質と同様の機能を有する相同分子や類縁分子をコードする遺伝子を
単離しクローニングできる。
【0107】
あるいは、本発明の遺伝子を取得する方法として、PCR等の増幅手段を用いる方法を
挙げることができる。例えば、本発明の遺伝子のcDNA配列のうち、5’側及び3’側
の配列(又はその相補配列)の中からそれぞれプライマーを調製し、これらプライマーを
用いてゲノムDNA(又はcDNA)等を鋳型にしてPCR等を行い、両プライマー間に
挟まれるDNA領域を増幅することで、本発明の遺伝子を含むDNA断片を大量に取得で
きる。
【0108】
(5)本発明に係る遺伝子及びたんぱく質の利用方法(有用性)
(5−1)組換え発現ベクター
本発明に係る組換え発現ベクターは、前記(2)に記載した本発明に係る遺伝子を含む
ものであれば、特に限定されるものではない。例えば、cDNAが挿入された組換え発現
ベクターが挙げられる。組換え発現ベクターの作製には、プラスミド、ファージ、又はコ
スミドなどを用いることができるが特に限定されるものではない。また、作製方法も公知
の方法を用いて行えばよい。
【0109】
ベクターの具体的な種類は特に限定されるものではなく、ホスト細胞中で発現可能なベ
クターを適宜選択すればよい。すなわち、ホスト細胞の種類に応じて、確実に遺伝子を発
現させるために適宜プロモーター配列を選択し、これと本発明の遺伝子を各種プラスミド
等に組み込んだものを発現ベクターとして用いればよい。
【0110】
本発明の遺伝子がホスト細胞に導入されたか否か、さらにはホスト細胞中で確実に発現
しているか否かを確認するために、各種マーカーを用いてもよい。例えば、ホスト細胞中
で欠失している遺伝子をマーカーとして用い、このマーカーと本発明の遺伝子とを含むプ
ラスミド等を発現ベクターとしてホスト細胞に導入する。これによってマーカー遺伝子の
発現から本発明の遺伝子の導入を確認することができる。あるいは、本発明のたんぱく質
を融合たんぱく質として発現させてもよく、例えば、オワンクラゲ由来の緑色蛍光たんぱ
く質GFP(Green Fluorescent Protein)をマーカーとして
用い、本発明のたんぱく質をGFP融合たんぱく質として発現させてもよい。
【0111】
上記ホスト細胞は、特に限定されるものではなく、従来公知の各種細胞を好適に用いる
ことができる。具体的には、例えば、大腸菌(Escherichia coli)等の
細菌、酵母(出芽酵母Saccharomyces cerevisiae、分裂酵母S
chizosaccharomyces pombe)、線虫(Caenorhabdi
tis elegans)、アフリカツメガエル(Xenopus laevis)の卵
母細胞等を挙げることができるが、特に限定されるものではない。
【0112】
上記発現ベクターをホスト細胞に導入する方法、すなわち形質転換方法も特に限定され
るものではなく、電気穿孔法、リン酸カルシウム法、リポソ−ム法、DEAEデキストラ
ン法等の従来公知の方法を好適に用いることができる。また、例えば、本発明のたんぱく
質を昆虫で転移発現させる場合には、バキュロウイルスを用いた発現系を採用することが
できる。
【0113】
(5−2)形質転換体
本発明に係る形質転換体は、前記(2)に記載した本発明に係る遺伝子導入された形質
転換体であれば、特に限定されるものではない。ここで「形質転換体」とは、細胞・組織
・器官のみならず、生物個体を含む意味である。
【0114】
形質転換体の作製方法(生産方法)は特に限定されるものではないが、例えば、上述し
た組換え発現ベクターをホスト細胞に導入して形質転換する方法を挙げることができる。
また、形質転換の対象となる生物も特に限定されるものではなく、上記ホスト細胞で例示
した各種微生物や動物を挙げることができる。
【0115】
本発明に係る形質転換体は、本発明に係る遺伝子が発現可能に導入された植物体、もし
くは当該植物体と同一の性質を有する当該植物体の子孫となる植物体、又は当該植物体の
組織の組織であることが好ましい。このような形質転換植物により、低コストかつ環境に
やさしい生産プロセスでアラキドン酸やEPA等の高度不飽和脂肪酸を生産することがで
きる。
【0116】
ここで「遺伝子が発現可能に導入された」とは、公知の遺伝子工学的手法(遺伝子操作
技術)により、対象細胞(宿主細胞)内に発現可能に導入されることを意味する。
【0117】
植物体の形質転換に用いられる組換え発現ベクターは、当該植物細胞内で挿入遺伝子を
発現させることが可能なものであれば特に限定しない。例えば、植物細胞内で恒常的に遺
伝子を発現させるプロモーター(例えば、カリフラワーモザイクウイルスの35Sプロモ
ーター)を有するベクターや、外的な刺激により誘導的に活性化されるプロモーターを有
するベクターを用いることができる。なお、この植物細胞には、種々の形態の植物細胞、
例えば、懸濁培養細胞、プロトプラスト、葉の切片、カルスなどが含まれる。
【0118】
植物細胞への組み換え発現ベクターの導入には、ポリエチレングリコール法、電気穿孔
法(エレクトロポレーション法)、アグロバクテリウムを介する方法、パーティクルガン
法など、当業者に公知の種々の方法を用いることができる。形質転換細胞から植物体の再
生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。
【0119】
例えば、タバコにおいて形質転換植物体を作出する手法については、形質転換したアグ
ロバクテリウムをタバコリーフディスクに感染させる方法、ポリエチレングリコールを用
いてプロトプラストへ遺伝子を導入し植物体に再生させる方法、電気パルスによりプロト
プラストへ遺伝子導入し植物体を再生させる方法、パーティクルガン法により細胞へ遺伝
子を直接導入し植物体を再生させる方法など、いくつかの技術が既に確立されている。本
発明においては、これらの方法を好適に用いることができる。
【0120】
また、タバコは、シロイヌナズナと並んで遺伝子工学的手法を用いる植物育種のモデル
植物である。このタバコにおいて、アラキドン酸やEPAの含量が増加した形質転換体を
取得できるということは、植物全般において形質転換体を取得することができるといって
も過言ではない。なお、本明細書では、後述する実施例に示すように、タバコだけでなく
、イネの形質転換体をも取得しており、本発明によれば、あらゆる種類の形質転換植物体
を取得できることを実証している。
【0121】
例えば、イネにおいて形質転換植物体を作出する手法については、ポリエチレングリコ
ールを用いてプロトプラストへ遺伝子を導入し、植物体に再生させる方法、電気パルスに
よりプロトプラストへ遺伝子導入し、植物体を再生させる方法、パーティクルガン法によ
り細胞へ遺伝子を直接導入し、植物体を再生させる方法など、いくつかの技術が既に確立
されている。本発明においては、これらの方法を好適に用いることができる。
【0122】
上記形質転換植物体がイネである場合、イネ内のアラキドン酸やEPAの含量が増加す
るので、この形質転換植物体から得られる種子、すなわち、米を食べることで、容易にア
ラキドン酸やEPA等の高度不飽和脂肪酸を、体内に摂取することが可能になる。したが
って、イネの形質転換植物体は、食糧としての価値が高く、食品産業、農業分野に極めて
有用である。また、現在あまり利用されていない米ぬか、モミガラ、ヒコバエ等でアラキ
ドン酸やEPAを生産すれば、ここからこれら脂肪酸を抽出することにより、健康食品の
原料として有効利用できる。また、家畜の飼料としても利用できる。
【0123】
ゲノム内に本発明の遺伝子が導入された形質転換植物体がいったん得られれば、当該植
物体から有性生殖又は無性生殖により子孫を得ることができる。また、当該植物体、又は
、その子孫、あるいは、クローンから、繁殖材料(例えば、種子、果実、切穂、塊茎、塊
根、株、カルス、プロトプラストなど)を得て、それらを基に当該植物体を量産すること
も可能である。したがって、本発明には、本発明の遺伝子が発現可能に導入された植物体
、もしくは、当該植物体と同一の性質を有する当該植物体の子孫となる植物体、又は、当
該植物体の組織、あるいは、当該植物体の繁殖材料も含まれる。
【0124】
また、当該植物体、当該植物体と同一の性質を有する当該植物体の子孫となる植物体、
及び、当該植物体の組織には、栄養増殖された植物体も含まれる。栄養増殖は、栄養生殖
、クローン成長とも呼ばれ、挿し芽、挿し木などによる増殖が一般的であり、試験管内で
は、葉、茎、根などの器官からの植物体の再分化やカルスによる増殖が可能である。植物
種によっては、枝の先端が特殊な冬芽を作る、腋芽が多肉化する、花がムカゴ化する、芋
を形成するなどの場合がある。
【0125】
さらに、本発明に係る遺伝子が発現可能に導入され、脂肪酸組成が改変された植物体、
もしくは当該植物体と同一の性質を有する当該植物体の子孫となる植物体、又は当該植物
体の組織、当該植物体の繁殖材料も本発明に含まれる。「脂肪酸組成が改変された」とは
形質転換前の植物体における脂肪酸組成と形質転換後における植物体の脂肪酸組成が異な
っていることを意味する。例えば、本来脂肪酸組成にアラキドン酸やEPAが含まれてい
なかった植物を本発明に係る遺伝子で形質転換することにより、形質転換植物の脂肪酸組
成にアラキドン酸やEPAが含まれる場合等を挙げることができる。
【0126】
(5−3)脂肪酸生産方法
本発明には、本発明に係る遺伝子で形質転換され、脂肪酸組成が改変された植物体又は
植物体の組織を用いて脂肪酸を生産する方法が含まれる。
【0127】
例えば、上述のようにアラキドン酸やEPAの含量が増加した本発明に係る形質転換植
物から製造された食用油はアラキドン酸やEPAの含量が高く、価値の高いものになる。
上記形質転換植物体の種子、果実、切穂、塊茎、塊根等も、アラキドン酸やEPAを含む
食料として価値の高いものになる。
【0128】
(5−4)素材
本発明には、上述の脂肪酸生産方法により得られた物質、すなわち、γ−リノレン酸、
ジホモ−γ−リノレン酸、アラキドン酸、ステアリドン酸、エイコサテトラエン酸、エイ
コサペンタエン酸を少なくとも1つ含む素材も含まれる。この「素材」とは、上述の食料
としての種子、果実、切穂、塊茎、又は、塊根の他に、工業原料用途に利用できる素材全
般を意味する。
【0129】
上記素材としては、例えば、アラキドン酸やEPAを含む健康食品、フィルム、生分解
性プラスチック、機能性繊維、潤滑油、洗剤の素材等が挙げられる。上記の不飽和脂肪酸
は、分子内に二重結合を複数有するという、ユニークな物性を持つ。このため、例えば、
本発明の形質転換植物体により、アラキドン酸やEPAを生産させることにより、生産コ
ストを低減できる。また、本発明により、環境にやさしい生産プロセスを実現できる。
【0130】
(5−5)脂肪酸組成改変方法
本発明には、本発明に係る遺伝子を用いて脂肪酸組成を改変する方法が含まれる。例え
ば、上述のように本発明に係る遺伝子を導入した形質転換体を作製することによりホスト
細胞の脂肪酸組成を改変することが可能となる。脂肪酸組成を改変する対象は特に限定さ
れるものではなく、植物以外にも動物、細菌、酵母等、あらゆる生物を対象とすることが
可能である。
【0131】
(5−6)遺伝子検出器具
本発明に係る遺伝子検出器具は、本発明に係る遺伝子の少なくとも一部の塩基配列又は
その相補配列をプローブとして用いたものである。遺伝子検出器具は、種々の条件下にお
いて、本発明の遺伝子の発現パタ−ンの検出・測定などに利用することができる。
【0132】
本発明の遺伝子検出器具としては、例えば、本発明の遺伝子と特異的にハイブリダイズ
する上記プローブを基盤(担体)上に固定化したDNAチップが挙げられる。ここで「D
NAチップ」とは、主として、合成したオリゴヌクレオチドをプローブに用いる合成型D
NAチップを意味するが、PCR産物などのcDNAをプローブに用いる貼り付け型DN
Aマイクロアレイをも包含するものとする。
【0133】
プローブとして用いる配列は、cDNA配列の中から特徴的な配列を特定する従来公知
の方法によって決定することができる。具体的には、例えば、SAGE:Serial
Analysis of Gene Expression法(Science 276
:1268,1997;Cell 88:243,1997;Science 270:
484,1995;Nature 389:300,1997;米国特許第5,695,
937号)等を挙げることができる。
【0134】
なお、DNAチップの製造には、公知の方法を採用すればよい。例えば、オリゴヌクレ
オチドとして合成オリゴヌクレオチドを使用する場合には、フォトリソグラフィ−技術と
固相法DNA合成技術との組み合わせにより、基盤上で該オリゴヌクレオチドを合成すれ
ばよい。一方、オリゴヌクレオチドとしてcDNAを用いる場合には、アレイ機を用いて
基盤上に貼り付ければよい。
【0135】
また、一般的なDNAチップと同様、パーフェクトマッチプローブ(オリゴヌクレオチ
ド)と、該パーフェクトマッチプローブにおいて一塩基置換されたミスマッチプローブと
を配置して遺伝子の検出精度をより向上させてもよい。さらに、異なる遺伝子を並行して
検出するために、複数種のオリゴヌクレオチドを同一の基盤上に固定してDNAチップを
構成してもよい。
【0136】
本発明に係る遺伝子検出器具は、上記例示したDNAチップに限定されるものではなく
、本発明に係る遺伝子の少なくとも一部の塩基配列又はその相補配列をプローブとして用
いたものであればよい。
【0137】
(5−7)抗体
本発明に係る抗体は、本発明に係るたんぱく質、又はその部分たんぱく質・部分ペプチ
ドを抗原として、公知の方法によりポリクロ−ナル抗体又はモノクロ−ナル抗体として得
られる抗体である。公知の方法としては、例えば、文献(Harlowらの「Antib
odies:A laboratory manual(Cold Spring Ha
rbor Laboratory,New York(1988))、岩崎らの「単クロ
ーン抗体ハイブリドーマとELISA,講談社(1991)」)に記載の方法が挙げられ
る。このようにして得られる抗体は、本発明のたんぱく質の検出・測定などに利用できる
。
【0138】
(5−8)スクリーニング方法
本発明に係るスクリーニング方法は、本発明に係るたんぱく質を用いて、当該たんぱく
質を調節する遺伝子、又は当該たんぱく質を調節する物質をスクリーニングする方法であ
る。本発明のスクリーニング方法としては、物質間の結合の有無や解離の有無を調べる従
来公知の種々の方法を適用することができ、特に限定されるものではない。例えば、本発
明に係るたんぱく質の活性(Δ6不飽和化活性、Δ6鎖長延長活性及び/又はΔ5不飽和
化活性)を促進するような物質のスクリーニングを挙げることができる。
【0139】
また、本発明には、上記スクリーニング方法により得られた遺伝子又は物質も含まれる
。
【0140】
以下、実施例を示し、本発明についてさらに詳しく説明するが、もちろん、本発明は以
下の実施例に限定されるものではなく、細部については様々な態様が可能であることはい
うまでもない。さらに、本発明は上述した実施形態に限定されるものではなく、請求項に
示した範囲で種々の変更が可能であり、開示された技術的手段を適宜組み合わせて得られ
る実施形態についても本発明の技術的範囲に含まれる。
【実施例】
【0141】
本実施例において、実験手法は、特に断らない限り、Molecular Cloni
ng(Sambrook et.al.Cold Spring Harbour La
boratory Press,1989)に記載されている方法に従った。
【0142】
〔実施例1:ゼニゴケ由来のΔ6不飽和化酵素遺伝子の単離〕
これまでにクローニングされたΔ6不飽和化酵素のアミノ酸配列の比較により、アミノ
酸配列Trp−Trp−Lys−(Glu/Asp)−Lys−His−Asn(配列番
号37)及びTrp−Phe−Thr−Gly−Gly−Leu−Asn(配列番号38
)が保存されていることがわかった。そこで、ゼニゴケ由来のΔ6不飽和化酵素遺伝子を
単離するために、上記のアミノ酸配列をコードする下記の縮退プライマーを用いた。
dD6DES−F 5’−TGGTGGAA(A/G)GA(A/G/T/C)AA(A
/G)CA(T/C)AA−3’(配列番号7)
dD6DES−R 5’−(A/G)TTIA(A/G)ICCICCIGT(A/G)
AACCA−3’ (配列番号8)
(Iはイノシン、()内は複数の塩基)
【0143】
試料にはE系統ゼニゴケ(Transgenic Res.9,p179,2000参
照)の葉状体を用いた。葉状体からのpoly(A)+RNAの単離については、文献(
Biosci.Biotechnol.Biochem.67,p605,2003;B
iosci.Biotechnol.Biochem.67,p1667,2003)に
記載の方法に従った。単離したpoly(A)+RNA 1.5μlを、Ready−T
o−Go T−primed First Strand kit(Amersham社
製)を用いてcDNAに逆転写した。PCRは、上記cDNA約10ngを鋳型とし、上
記プライマー(dD6DES−F及びdD6DES−R)及び酵素(Takara Ex
Taq、Takara社製)0.5Uとを用いて、製造者の推奨する方法で行った。反
応液量は20μlとし、GeneAmp PCR system 9700(PE Ap
plied Biosystems社製)を用いて、94℃2分間保持後、94℃で1分
間、45℃で1.5分間、72℃で2分間の反応を35回繰り返し、その後4℃に冷却し
た。
【0144】
得られたPCR産物を1%(w/v)アガロ−スゲルで電気泳動し、従来のΔ6不飽和
化酵素のアミノ酸配列から予想されるサイズを有する増幅断片を、Prep−A Gen
e(Bio−rad社製)を用いてゲルより回収した。回収した増幅断片をpT7Blu
e Vector(Takara社製)に連結し、大腸菌Electro−max DH
10B cells(Invitrogen社製,Carlsbad,CA)に形質転換
した。
【0145】
BigDye Terminator Cycle Sequencing kit(
Applied Biosystems社製)及びautomated sequenc
er ABI PRISM 377(Applied Biosystems社製)を用
いて得られた全クローンの塩基配列を決定し、目的のcDNA配列をもつものを探索した
。
【0146】
さらに、全長cDNA配列を取得するために、5’−RACE及び3’−RACEを行
った。これには、5’−RACE System for Rapid Amplifi
cation of cDNA Ends Version 2.0(Invitrog
en社製)、Ready−To−Go T−primed First Strand
kit(Amersham社製)及び下記のプライマー(MpDES6−02R及びMp
DES6−01F)を用い、製造者の推奨する方法で行った。
MpDES6−02R:5’−AAGTTGCCTTCGATGTTTCTGG−3’(
配列番号9)
MpDES6−01F:5’−GCTCGCCTGGAGCAAGGAAAATC−3’
(配列番号10)
【0147】
その結果、1種類のホモログ遺伝子候補が単離され、この遺伝子をMpDES6遺伝子
とした。単離されたMpDES6遺伝子のcDNAの長さ(ポリA部分を除く)は、2,
522bpであり、そのコードする推定アミノ酸配列は481残基であった。その塩基配
列を配列番号1、アミノ酸配列を配列番号2に示した。
【0148】
MpDES6cDNAの推定アミノ酸配列をヒメツリガネゴケのΔ6不飽和化酵素のア
ミノ酸配列と比較した結果、47.5%の同一性しか示さなかった。
【0149】
〔実施例2:ゼニゴケ由来のΔ6鎖長延長酵素遺伝子の単離〕
これまでにクローニングされたΔ6鎖長延長酵素のアミノ酸配列の比較により、アミノ
酸配列Val−Glu−Phe−Met−Asp−Thr−Val(配列番号39)及び
Lys−Tyr−Leu−Phe−Trp−Gly−Arg(配列番号40)が保存され
ていることがわかった。そこで、ゼニゴケ由来のΔ6鎖長延長酵素遺伝子を単離するため
に、上記のアミノ酸配列をコードする下記の縮退プライマーを用いた。
dD6ELO−F 5’−GTIGA(A/G)TT(T/C)ATGGA(T/C)A
CIGT−3’ (配列番号11)
dD6ELO−R 5’−C(G/T)ICCCCA(A/G)AAIA(A/G)(A
/G)TA(T/C)TT−3’(配列番号12)
【0150】
上記のプライマー(dD6ELO−F及びdD6ELO−R)を用いてPCRをい、得
られたDNA断片をサブクローニングした。得られたクローンの塩基配列を決定し、目的
のcDNA配列をもつクローンについて下記のプライマー(MpELO1−02R及びM
pELO1−01F)を用いて完全長cDNAを取得した。なお、実験材料及び方法は、
実施例1と同様である。
MpELO1−02R:5’−GCGAGCTTTCTCGTTCTTTCCC−3’(
配列番号13)
MpELO1−01F:5’−TATGATTTTGAAGCGCAACACG−3’(
配列番号14)
【0151】
その結果、1種類のホモログ遺伝子候補が単離され、この遺伝子をMpELO1遺伝子
とした。MpELO1遺伝子のcDNAの長さ(ポリA部分を除く)は、1,559bp
で、推定アミノ酸配列は290残基であった。その塩基配列を配列番号3、アミノ酸配列
を配列番号4に示した。
【0152】
MpELO1cDNAの推定アミノ酸配列をヒメツリガネゴケのΔ6鎖延長酵素のアミ
ノ酸配列と比較した結果、同一性は62.7%であった。
【0153】
〔実施例3:ゼニゴケ由来のΔ5不飽和化酵素遺伝子の単離〕
他生物種のΔ5不飽和化酵素は、そのN末端にシトクロムb5ドメインを有する。この
ことから、ゼニゴケ由来のΔ5不飽和化酵素遺伝子は、Δ6不飽和化酵素遺伝子と同じく
、シトクロムb5ドメイン融合型不飽和化酵素遺伝子ファミリーに属することが予想され
た。しかしながら、ケイ藻や真菌においては、Δ5不飽和化酵素及びΔ6不飽和化酵素間
のアミノ酸配列レベルでの相同性は非常に低い。そこで、糸状菌(M.alpina)の
Δ5不飽和化酵素及びΔ6不飽和化酵素間のアミノ酸配列を比較した結果、縮退プライマ
ー設計に最低限必要な4から5残基程度の連続した保存配列が局所的に存在することを見
出した。驚くべきことに、これらの保存配列は、異種のΔ5不飽和化酵素のアミノ酸配列
間より、同種内のΔ5不飽和化酵素とΔ6不飽和化酵素との間で、より保存されているこ
とを見出した。このことから、シトクロムb5ドメイン融合型不飽和化酵素遺伝子には、
種特異的な保存配列が存在する場合があると考えられた。そこで、上述のMpDES6と
、Genetics 159,p981,2001に記載されている機能未知のMpDE
Sとで、塩基配列を鋭意比較検討した結果、2箇所のアミノ酸配列(I(E/N)(G/
D)KVYDV(配列番号41)、及び、DPDI(Q/D)(Y/T)(M/V)P(
配列番号42))のアミノ酸配列が保存されていることを見出した。それぞれのアミノ酸
配列に対応する縮退プライマーの配列を次に示す。
dD5DES−F 5’−AT(A/T/C)(A/G)AIG(A/G)IAA(A/
G)TITA(T/C)GA(T/C)GT−3’(配列番号15)
dD5DES−R 5’−GGIA(T/C)I(G/T)(A/T)IT(G/C)(
A/G/T)AT(A/G)TCIGG(A/G)TC−3’(配列番号16)
【0154】
上記プライマー(dD5DES−F及びdD5DES−R)を用いてPCRを行い、得
られたDNA断片をサブクローニングした。得られたクローンの塩基配列を決定し、目的
のcDNA配列をもつクローンについて下記のプライマー(MpDES5−02R及びM
pDES5−01F)を用いて完全長cDNAを取得した。なお、実験材料及び方法は、
実施例1と同様である。
MpDES5−02R:5’−GTGTGTACGATCCGTGGTTACC−3’(
配列番号17)
MpDES5−01F:5’−AAGGCGGGACAGGATTCAACAC−3’(
配列番号18)
【0155】
その結果、ゼニゴケ由来のΔ5不飽和化酵素の候補として、2種類の長さの異なるクロ
ーンc1及びc2を単離した(c1:2,427bp、c2:2,285bp)。c1と
c2との塩基配列を比較したところ、5’非翻訳領域で、オルタナティブ・スプライシン
グ(選択的スプライシング)が起こっていることが明らかとなった。オルタナティブ・ス
プライシングによる読み枠の変化はなく、いずれのクローンも484個のアミノ酸をコー
ドしていた(配列番号6)。以下、2,427bpの長さのクローンc1をMpDES5
遺伝子(配列番号5)とし、以下の実施例に用いた。
【0156】
MpDES5cDNAの推定アミノ酸配列を糸状菌(M.alpina)のΔ5不飽和
化酵素のアミノ酸配列と比較した結果、31.4%の同一性であった。ゼニゴケと近縁な
ヒメツリガネゴケのΔ5不飽和化酵素に関しては、配列情報が公表されていないため、こ
こでは比較を行わなかった。
【0157】
〔実施例4:メタノール資化性酵母(Pichica pastoris)を用いた機能
解析〕
MpDES6、MpELO1及びMpDES5のcDNAの機能を調べるために、まず
個々のORFを、メタノール誘導性プロモーターAOX1の下流に配置したコンストラク
トを作製した。これらのコンストラクトをメタノール資化性酵母(Pichia pas
toris)に導入し、その脂肪酸組成を解析した。MpDES6、MpELO1、及び
、MpDES5のcDNA塩基配列のORF部分を、下記のプライマーを用いて、PCR
により増幅した。
(MpDES6ORF増幅用プライマー)
MpD6−17F:5’−GGAATTCGCGATGGCCTCGTCCACCACC
AC−3’(配列番号19)
MpD6−18F:5’−GGAATTCTACTTTCGCAGCGTATGCTAC
C−3’(配列番号20)
(MpELO1ORF増幅用プライマー)
MpD6ELO1−15F:5’−GGAATTCGCGATGGAGGCGTACGA
GATGG−3’(配列番号21)
MpD6ELO1−16F:5’−GGAATTCTTCTGCCTTTTTGCTCT
TGATC−3’(配列番号22)
(MpDES5ORF増幅用プライマー)
MpD5−11F:5’−GTTGAATTCGACAGTTATGCCGCCACAC
GC−3’(配列番号23)
MpD5−12R:5’−GTTGAATTCAGGCCCAAAGCATGCTGTC
AC−3’(配列番号24)
【0158】
これらのプライマーは、下線で示したEcoRI認識配列を含んでおり、これを以下の
クローニングに利用した。また、PCRにはPyrobest DNA polymer
ase(Takara社製)0.5Uを用い、製造者の推奨する方法に従って反応液量2
0μlで行った。反応条件は、94℃で2分間保持後、94℃で1分間、57℃で1分間
、72℃で1分間の反応を25回繰り返し、その後4℃に冷却した。得られた各ORF断
片をEcoRIで消化した後、実施例1に記載の方法でゲル精製を行った。そして、メタ
ノール資化性酵母の発現ベクターpPICZA(マーカー:ゼオシン耐性遺伝子,Inv
itrogen社製)内のメタノール誘導性プロモーター5’AOX1下流のEcoRI
部位にセンス方向に連結した。
【0159】
各々の発現コンストラクト及び対照としてのpPICZAベクターを、Pichia
EasyComp kit(Invitrogen社製)を用いてメタノール資化性酵母
のPPY1系統に導入し、ゼオシン耐性をマーカーとして形質転換体を取得した。なお、
メタノール資化性酵母は、Δ6不飽和化酵素の基質であるリノール酸とα−リノレン酸を
合成することができるが、アラキドン酸やEPAのその他の前駆体を合成することはでき
ない。
【0160】
導入した遺伝子を発現させるため、EasySelect Pichia Expre
ssion Kit(Invitrogen社製)を用い、当該キットの推奨する方法に
従いて、各形質転換体を1.0%グリセロールのみを炭素源とする最小培地中でOD(6
00nm)が0.5になるまで培養した後、0.5%メタノールのみを炭素源とする最小
培地中で30℃、3日間、飽和状態になるまで培養した。その後、各々の形質転換体の脂
肪酸組成を、GC−MSを用いて公知の方法(Biosci.Biotechnol.B
iochem.67,p605,2003)により測定した。
【0161】
MpDES6遺伝子を発現させた形質転換体では、Δ6不飽和化酵素の反応産物である
g−リノレン酸及びステアリドン酸が、全脂肪酸のそれぞれ7.4%及び0.7%新たに
検出された。対照としてpPICZAベクターを導入した酵母ではこれらは検出されなか
った。以上のことから、MpDES6はΔ6不飽和化酵素をコードしていることが示され
た。
【0162】
MpELO1遺伝子を発現させた形質転換体では、g−リノレン酸を添加した場合にジ
ホモ−g−リノレン酸が全脂肪酸の14.1%、ステアリドン酸を添加した場合にエイコ
サテトラエン酸が1.5%新たに検出された。対照としてpPICZAベクターを導入し
た酵母では、これらは検出されなかった。以上のことから、MpELO1はΔ6鎖長延長
酵素をコードしていることが示された。
【0163】
MpDES5遺伝子を発現させた形質転換体では、ジホモ−g−リノレン酸を添加した
場合にアラキドン酸が全脂肪酸の1.1%、ステアリドン酸を添加した場合にエイコサペ
ンタエン酸(EPA)が0.1%検出された。対照としてpPICZAベクターを導入し
た酵母ではこれらは検出されなかった。以上から、MpDES5は、Δ5不飽和化酵素を
コードすることが示された。
【0164】
以上のようにゼニゴケから、Δ6不飽和化酵素、Δ6鎖長延長酵素及びΔ5不飽和化酵
素をコードする遺伝子として、それぞれMpDES6、MpELO1及びMpDES5を
取得することができた。
【0165】
〔実施例5:メタノール資化性酵母(P.pastoris)におけるゼニゴケ高度不飽
和脂肪酸生合成系の再構成〕
MpDES6、MpELO1及びMpDES5を共発現させるために、上記実施例4で
作製した、EcoRI消化したMpELO1及びMpDES5のORF増幅断片を、各々
別のメタノール資化性酵母発現ベクターpPIC3K(マーカー:HIS4遺伝子、In
vitrogen社製社)及びpPIC6A(マーカー:ブラスチシジン耐性遺伝子、I
nvitrogen社製社)の5’AOX1プロモーター下流のEcoRI部位にセンス
方向に連結した。また、MpDES6に関しては、実施例4で作製した発現ベクターを用
いた。以下各発現ベクターを、それぞれpPICZA−MpDES6、pPIC3K−M
pELO1及びpPIC6A−MpDES5と表記する。
【0166】
まず、pPICZA−MpDES6、又は、対照としてのpPICZAベクターのみを
、上記実施例4で用いたメタノール資化性酵母PPY1系統と同じ脂肪酸組成を持つメタ
ノール資化性酵母PPY12系統(his4,arg4)に形質転換し、ゼオシン耐性を
マーカーにして形質転換体を取得した。続いて、pPIC3K−MpELO1、又は、対
照としてのpPIC3Kベクターのみを、それぞれ、pPICZA−MpDES6、又は
pPICZAのみがゲノム中に組み込まれた形質転換体に導入し、ヒスチジン合成能をマ
ーカーにして形質転換体を取得した。最後に、pPIC6A−MpDES5、又は、対照
としてのpPIC6Aベクターのみを、pPICZA−MpDES6及びpPIC3K−
MpELO1又はpPICZA及びpPIC3Kがゲノム中に組み込まれた上記形質転換
体に導入し、ブラストジン耐性をマーカーとして形質転換体を取得した。
【0167】
得られた2種類又は3種類の遺伝子が導入された形質転換体を用いて、ゼニゴケのアラ
キドン酸/EPA生合成系の再構成実験を行った。まず、上記2種類の遺伝子(MpDE
S6及びMpELO1)を導入した。形質転換体を用いて、MpDES6たんぱく質及び
MpELO1たんぱく質をメタノール資化性酵母内で共発現させた。その結果、Δ6不飽
和化反応産物であるg−リノレン酸(全脂肪酸の2.9%)及びステアリドン酸(全脂肪
酸の0.4%)に加え、それらの鎖長延長反応産物であるジホモ−g−リノレン酸(全脂
肪酸の2.8%)及びエイコサテトラエン酸(全脂肪酸の0.2%)が生じた。対照とし
た形質転換体では、これらの脂肪酸は検出されなかった。上記形質転換体に、さらにMp
DES5を導入した形質転換体では、g−リノレン酸(全脂肪酸の2.8%)、ステアリ
ドン酸(全脂肪酸の0.5%)、ジホモ−g−リノレン酸(全脂肪酸の1.5%)及びエ
イコサテトラエン酸(全脂肪酸の0.1%)の4種の脂肪酸に加えて、アラキドン酸(全
脂肪酸の0.1%)及びエイコサペンタエン酸(EPA、全脂肪酸の0.03%)の生成
が認められた。対照とした形質転換体では、これらの脂肪酸は検出されなかった。この結
果から、ゼニゴケ由来のΔ6不飽和化酵素、Δ6鎖長延長酵素及びΔ5不飽和化酵素の遺
伝子を発現させることで、ゼニゴケ以外の生物種においても、高度不飽和脂肪酸生合成系
の再構築が可能であることが示された。
【0168】
〔実施例6:イネに導入するベクターの構築及び当該ベクターのイネへの導入〕
イネにおいて、MpDES6、MpELO1及びMpDES5遺伝子を発現させるため
に、以下の(i)〜(iv)に示す手順で発現コンストラクトを作製した。また、作製手
順を図1に示した。
【0169】
(i)pBI221(TOYOBO社製)のカリフラワ−モザイクウイルス(CaMV
)35Sプロモーター、及び、NOSターミネーター間に設計した以下のプライマーを用
いたPCRにより、β−Glucuronidase(GUS)遺伝子部分を除いた発現
ベクターp35S−NOSを作製した。
MK001(F):5’−CGGGATCCTCTCCTGGCGCACCATCGTC
−3’(配列番号25)
MK002(R):5’−GGGGTACCAACGCGCTTTCCCACCAACG
−3’(配列番号26)
【0170】
なお、プライマーMK001(F)は下線で示すBamHI認識配列を含み、GUS遺
伝子の3’末端にアニールし、プライマーMK002(R)はGUS遺伝子の5’末端に
アニールする(BamHIサイトがアニール部位の上流に存在する)。PCRにはPyr
obest DNA polymerase(Takara社製)0.5Uを用い、製造
者の推奨する方法に従って反応液量50μlで行った。反応条件は、96℃5分間保持後
、94℃で30秒間、68℃で4分間の反応を30回繰り返し、その後4℃に冷却した。
得られた各ORF断片をBamHI消化した後、実施例1に記載の方法でゲル精製を行っ
た後、自己連結させた。
【0171】
(ii)次にp35S−NOSのXbaIサイトに、MpDES6遺伝子、MpELO
1遺伝子及びMpDES5遺伝子のORFを各々連結した。ORF増幅には、下線で示す
XbaI認識配列を含む以下のプライマーを用いた。
(MpDES6 ORF増幅用プライマー)
MpD6−21F:5’−GCTCTAGAGCGATGGCCTCGTCCACCAC
C−3’(配列番号27)
MpD6−11R:5’−GCTCTAGACTATACTTTCGCAGCGTATG
C−3’(配列番号28)
(MpELO1 ORF増幅用プライマー)
MpD6ELO1−18F:5’−GCTCTAGAGCGATGGAGGCGTACG
AGATGG−3’(配列番号29)
MpD6ELO1−13R:5’−GCTCTAGATTATTCTGCCTTTTTG
CTC−3’ (配列番号30)
(MpDES5 ORF増幅用プライマー)
MpD5−22F:5’−GCTCTAGAGACAGTTATGCCGCCACACG
C−3’(配列番号31)
MpD5−23R:5’−GCTCTAGAAGGCCCAAAGCATGCTGTCA
C−3’(配列番号32)
【0172】
PCRにはPyrobest DNA polymerase(Takara社製)0
.5Uを用い、製造者の推奨する方法に従って、反応液量20μlで行った。反応条件は
、94℃で2分間保持後、94℃で1分間、57℃で1分間、72℃で1分間の反応を2
5回繰り返し、その後4℃に冷却した。得られた各ORF断片をXbaIで消化した後、
実施例1に記載の方法でゲル精製を行いクローニングに用いた。
【0173】
(iii)得られた各遺伝子の発現コンストラクト(それぞれをp35S−MpDES
6、p35S−MpELO1、p35S−MpDES5と表記する)がいずれもCaMV
35Sプロモーター5’末端にPstIサイトを、NOSターミネーター3’末端にEc
oRIサイトを持つことを利用して、上記3遺伝子の発現カセットを連結した。まず、以
下に示すプライマーを用いてp35S−MpDES5を鋳型としてPCRを行い、MpD
ES5遺伝子の発現カセット部分を増幅し、p35S−MpDES6のCaMV35Sプ
ロモーター5’末端にあるPstIサイトにクローニングした(図1参照)。
(MpDES5 遺伝子発現カセット増幅用プライマー)
M13R:5’−CAGGAAACAGCTATGACC−3’(配列番号33)
NOS−R4−PST:5’−AAACTGCAGATTCCCGATCTAGTAAC
ATAG−3’(配列番号34)
【0174】
なお、M13Rプライマーは、CaMV35Sプロモーター上流のベクター配列にアニ
ールする。また、NOS−R4−PSTプライマーは、下線で示すPstI認識配列を含
み、NOSターミネーター3’末端にアニールする。同じくNOSターミネーター3’末
端に存在するEcoRIサイトは含まない。
【0175】
PCRにはPyrobest DNA polymerase(Takara社製)0
.5Uを用い、製造者の推奨する方法に従って、反応液量20μlで行った。反応条件は
、94℃で2分間保持後、94℃で1分間、57℃で1分間、72℃で1分間の反応を2
5回繰り返し、その後4℃に冷却した。得られたDNA断片をPstIで消化した後、実
施例1に記載の方法でゲル精製を行い、MpDES6遺伝子の発現カセットを含むプラス
ミド(p35S−MpDES6)のPstIサイトにクローニングした。
【0176】
(iv)上記で得られた、MpDES5及びMpDES6遺伝子の発現カセットを連結
させたコンストラクト(p35S−MpDES5/35S−MpDES6と表記する)に
、さらにMpELO1遺伝子の発現カセットを連結した。以下のプライマーを用いてp3
5S−MpELO1を鋳型としてPCRを行い、MpELO1遺伝子の発現カセット部分
を増幅し、MpDES6遺伝子発現カセット内のNOSターミネーター3’末端にあるE
coRIサイトにクローニングした。
(MpELO1 遺伝子発現カセット増幅用プライマー)
35S−F3−EI:5’−CCGGAATTCGCATGCCTGCAGGTCCCC
AGA−3’(配列番号35)
M13F:5’−TGTAAAACGACGGCCAGT−3’ (配列番号36)
【0177】
なお、35S−F3−EIプライマーは、下線で示すEcoRI認識配列を含み、Ca
MV35Sプロモーターの5’末端にアニールする。また、M13FプライマーはNOS
ターミネーター下流のベクター配列にアニールする。
【0178】
PCRにはPyrobest DNA polymerase(Takara社製)0
.5Uを用い、製造者の推奨する方法に従って、反応液量20μlで行った。反応条件は
、94℃で2分間保持後、94℃で1分間、57℃で1分間、72℃で1分間の反応を2
5回繰り返し、その後4℃に冷却した。得られたDNA断片をEcoRIで消化した後、
実施例1に記載の方法でゲル精製を行い、MpDES5及びMpDES6遺伝子の発現カ
セットを連結させたコンストラクト(p35S−MpDES5/35S−MpDES6)
のEcoRI部位にクローニングした。
【0179】
以上の操作により、3遺伝子の発現カセットが、MpDES5,MpDES6,MpE
LO1の順で連結した発現コンストラクト(p35S−MpDES5/35S−MpDE
S6/p35S−MpELO1)を作製した。
【0180】
このようにして得られた上記コンストラクトを、公知の方法(Genes Genet
.Syst.73,p219,1998)で、ビアラフォスを選抜マーカーとして持つプ
ラスミドとともに、パーティクルガンでイネに導入し、形質転換イネを取得した。
【0181】
〔実施例7:タバコ(N.tabacum SR−1)におけるゼニゴケ高度不飽和脂肪
酸合成系の再構成〕
本実施例では、上述のゼニゴケ由来の不飽和脂肪酸合成酵素遺伝子、すなわち、MpD
ES6遺伝子、MpDES5遺伝子、およびMpELO遺伝子が、植物体中で良好に機能
することを確認した。
【0182】
具体的には、タバコにMpDES6遺伝子、MpDES5遺伝子、およびMpELO遺
伝子を導入し、このタバコ中でアラキドン酸等が生産されることを確認した。比較対照と
して、糸状菌(M.alpina)由来のΔ6不飽和化酵素遺伝子(MaDES6)、Δ
5不飽和化酵素遺伝子(MaDES5)、Δ6脂肪酸鎖長延長酵素(MaELO)の3遺
伝子を導入したタバコを作製した。
【0183】
(i)糸状菌由来遺伝子を含むベクター(pSPB1519)の構築
pE2113(Mitsuhara et al.Plant Cell Physi
ol.37,45−59 1996)は、エンハンサー配列の繰り返しを有するカリフラ
ワーモザイクウィルス35S(El235S)プロモーター、および、ノパリンシンター
ゼ(nos)ターミネーターを有する。
【0184】
pE2113をSnaBIで消化した後、XhoIリンカー(TAKARA社)と連結
することで、プラスミドを作製した。このプラスミドをSacIで消化後、平滑末端化し
、さらにBamHIリンカー(TAKARA社)と連結することによって、pUE7を作
製した。pUE7をHindIIIとEcoRIで消化することによって得られるDNA
断片のうちEl235Sプロモーターを有する断片と、HindIIIとEcoRIで消
化した植物形質転換用バイナリーベクターpBINPLUS(van Engelen
et al.Transgenic research 4,p288,1995)とを
連結することで、pSPB505を作製した。
【0185】
一方、MaDES6遺伝子を含むプラスミドであるpMLD101をXhoIで消化後
、さらにBamHIで部分消化することで、約1.6kbのDNA断片を得た。この断片
と、pSPB505をXhoIとBamHIで消化して得られるバイナリーベクター部分
のDNA断片とを連結することで、pSPB559を作製した。
【0186】
pUCAP(van Engelen et al.Transgenic rese
arch 4,p288,1995)をAscIで消化した後平滑末端化し、PacIリ
ンカーと連結することで、pUCAPPを作製した。
【0187】
pE2113をSnaBIで消化した後、BamHIリンカー(TAKARA社)と連
結することで、pUE6を作製した。pUE6をSacIで消化した後、平滑末端化し、
SalIリンカー(TAKARA社)と連結することで、pUE8を作製した。pUE8
をHindIIIおよびEcoRIで消化して得られるDNA断片のうちEl235Sプ
ロモーターを有する断片を、pUCAPPのHindIII−EcoRI間に挿入するこ
とで、プラスミドを作製した。このプラスミドをBamHIとSalIで消化して得られ
たDNA断片と、MaELO遺伝子のcDNAをBamHIおよびXhoIで消化して得
られたDNA断片とを連結し、pSPB1130を作製した。pSPB1130をPac
Iで消化し、得られる約2.3kbのDNA断片を、pBINPLUSのPacIサイト
に挿入した。MaELO遺伝子の転写方向と、pBINPLUS上のnptII遺伝子の
転写方向とが、同じ向きになっているプラスミドを選択し、そのプラスミドをpSPB1
157Pとした。
【0188】
また、上述したpSPB559をPacIで消化した後、平滑末端化し、AscIリン
カーと連結することで、pSPB559Aを作製した。そして、pSPB559AをAs
cIで消化して得られるMaDES6遺伝子を含むDNA断片を、pSPB1157Pの
AscIサイトに挿入することで、pSPB1157を作製した。
【0189】
pCGP1364(Plant Cell Physiol.36,1023,199
5)をHindIIIおよびSacIIで消化して得られる約1.3kbのDNA断片と
、pCGP1364をPstIで消化し、平滑末端化した後さらにSacIIで消化して
得られる約2.9kbのDNA断片と、pUCAPAをSacIで消化し、平滑末端化し
た後さらにHindIIIで消化して得られる約2.7kbのDNA断片とを連結するこ
とにより、pSPB184を得た。
【0190】
一方、MaDES5遺伝子がサブクローニングされたpCRIIベクターから、Xba
IとKpnIによる消化によってMaDES5遺伝子を含むDNA断片を切り出した。こ
のDNA断片と、上述のpSPB184をXbaIとKpnIで消化して得られたDNA
断片とを連結することで、pSPB1519Aを作製した。pSPB1519AをAsc
Iで消化し、得られたDNA断片をpSPB1157のAscIサイトに挿入し、pSP
B1519を作製した。このpSPB1519上で、MaDES6遺伝子、MaDES5
遺伝子、およびMaELO遺伝子は、同じ向きに転写され、同じ構成的プロモーターの制
御下にある。
【0191】
(ii)ゼニゴケ由来遺伝子のベクター(pSPB2368)の構築
pUCAP(van Engelen et al.Transgenic Rese
arch 4,288−290,1995)をAscIで消化した後SgfIリンカーと
連結し、さらに、PacIで消化した後FseIリンカーと連結することで、pUCSA
PFを作製した。また、pBINPLUSにも同様の処理を施し、pBINSAPFを作
製した。
【0192】
他にサブクローニング用のベクターとして、pUC19をHindIIIで消化した後
、PacIリンカーと連結し、さらにEcoRIで消化した後FseIリンカーと連結す
ることで、pUCPFを作製した。また、pUC19をHindIIIで消化した後Sg
fIリンカーと連結し、さらに、EcoRIで消化した後AscIリンカーと連結するこ
とで、pUCSAを作製した。pUCSAPFのHindIII−XbaI間にEl23
5Sを、SacI−EcoRI間にマノピン合成酵素(mas)遺伝子ターミネーターを
それぞれ挿入したベクターを、XbaIおよびSacIで消化した後、末端を平滑化する
ことで、pSPB2353Aを得た。pSPB2353Aの平滑末端に、p35S−Mp
DES6からXbaIで切り出して末端を平滑化したMaDES6遺伝子を含むDNA断
片を連結し、pSPB2353作製した。
【0193】
pUCSAのHindIII−XbaI間にEl235Sを、SacI−EcoRI間
にマノピン合成酵素(mas)遺伝子ターミネーターをそれぞれ挿入したベクターを、X
baI、SacIで消化することで、pSPB2355Aを作製した。
【0194】
一方、p35S−MpELO1を鋳型として、下記のプライマー、XbaMpELOf
およびSacMpELOrを用いてPCRを行った。
XbaMpELOf:5‘−AGTCTCTAGAGCGATGGAGGCGTACG−
3’(配列番号43)
SacMpELOr:5‘−CAGTGAGCTCGGTGTCTTATTCTGCC−
3’(配列番号44)
【0195】
PCRには、酵素として高精度なKOD+DNAポリメラーゼ(東洋紡)を用い、94
℃で2分間保持した後、94℃で15秒、68℃で1−3分のサイクルを25回繰り返し
た。このようにして調製されたMpELODNA断片をXbaIおよびSacIで消化し
たものを、上述のpSPB2355Aに連結し、pSPB2355を作製した。さらに、
pSPB2355をSgfI、AscIで消化して得られたDNA断片を、SgfIおよ
びAscIで消化したpSPB2353に連結し、pSPB2361を作製した。
【0196】
pUCPFのHindIII−XbaIサイトにEl235Sを、SacI−EcoR
Iサイトにマノピン合成酵素(mas)遺伝子ターミネーターをそれぞれ挿入したベクタ
ーを、XbaIおよびSacIで消化することで、pSPB2352Aを作製した。
【0197】
一方、p35S−MpDES5を鋳型に、下記のプライマー、XbaMpD5fおよび
SacMpD5rを用いてPCRを行った。PCRの反応条件は、上記のPCR条件と同
様である。
XbaMpD5f:5’−AGCTTCTAGAGCCATGCCGCCACACGCC
C−3’(配列番号45)
SacMpD5r:5’−CAGTGAGCTCTCAGCCATCCAGTCGT−3
’ (配列番号46)
【0198】
PCRによって調製されたMpD5DNA断片をXbaI、SacIで消化したものを
、pSPB2352Aに連結し、pSPB2352を作製した。
【0199】
pBINSAPFをPacIおよびFseIで消化して得られたDNA断片に、pSP
B2352よりPacIおよびFseIで切り出したMpDES5遺伝子を含むDNA断
片を連結し、pSPB2368Aを作製した。さらにpSPB2368AをSgfIおよ
びPacIで消化したものに、pSPB2361よりSgfIおよびPacIで切り出さ
れたMpDES6遺伝子およびMpELO遺伝子を含むDNA断片を連結することで、p
SPB2368を得た。このプラスミド上でMpDES6遺伝子、MpDES5遺伝子、
およびMpELO遺伝子は、同じ向きに転写され、同じ構成的プロモーターの制御下にあ
る。
【0200】
(iii)タバコへの遺伝子導入
引き続いて、公知の方法(Plant J.5,81,1994)に基づいて、pSP
B2368またはpSPB1519を用いてAgrobacterium tumefa
ciens(菌株:AglO(Lazo et al.1991,Biotechnol
ogy9:963−967))を形質転換した。このpSPB2368またはpSPB1
519を有する形質転換体アグロバクテリウムを、タバコリーフディスクに感染させた。
このようにして得られた組換えタバコの葉から、RNeasyPlantminiKit
(キアゲン)を用いてRNAを抽出し、定法に従って、RT−PCRにより、導入した遺
伝子が発現している系統を選択した。
【0201】
このようにして得られた糸状菌由来の酵素遺伝子から成るpSPB1519を導入した
タバコ(pSPB1519形質転換タバコ)、ゼニゴケ由来の酵素遺伝子から成るpSP
B2368を導入したタバコ(pSPB2368形質転換タバコ)の葉から、公知の方法
(藤野安彦編(1978)生物化学実験法9 学会出版センター、山田晃弘編(1989
)生物化学実験法24 学会出版センター)にしたがって、脂質の抽出を行った。得られ
た脂質をガスクロマトグラフィー(Hewlett Packard社 HP−6800
)で分析し、その結果を表1に示す。
【0202】
なお、コントロールとして、遺伝子が導入されていないタバコの葉を用いて、同様の分
析を行った。
【0203】
【表1】
【0204】
本実施例のガスクロマトグラフィーによる分析条件を以下に示す。
(ガスクロマトグラフィー分析条件)
カラム:Supelco SP−2330、Fused Silica Capil
lary Column、30m x 0.32mm ID、0.2μm
温度:Inj:240℃、Det:250℃、Oven:180℃ 3分、180℃
→220℃(2℃/min)
カラム流量:30cm/sec、圧力200kPa、検出器 FID
【0205】
クロマトグラム中の各ピークは標準脂肪酸のメチルエステルのリテンションタイムとG
C−MASS(Hewlett Packard社、HP−5973)分析により決定し
、またピーク面積より各脂肪酸の割合を決定した。
【0206】
表1において、Controlはコントロールを、pSPB2368はpSPB236
8形質転換タバコを、pSPB1519はpSPB1519形質転換タバコを表す。
【0207】
表1に示す結果から、糸状菌由来の酵素遺伝子から成るpSPB1519を導入したタ
バコ(pSPB1519形質転換タバコ)では、ジホモγリノレン酸の蓄積は確認できた
が、アラキドン酸の蓄積は認められなかった。一方、ゼニゴケ由来の酵素遺伝子から成る
pSPB2368を導入したタバコ(pSPB2368形質転換タバコ)ではアラキドン
酸だけでなく、エイコサテトラエン酸およびエイコサペンタエン酸の蓄積も確認できた。
このことから、高等植物においては、糸状菌由来の酵素よりもゼニゴケ由来の酵素の方が
機能的に優れており、リノール酸やαリノレン酸を基質として、アラキドン酸を始めとす
る高度不飽和脂肪酸を合成できると考えられた。
【0208】
Abbadiらは、ケイソウ(Phaeodactylum tricornutum
)のΔ6不飽和化酵素、Δ5不飽和化酵素、およびヒメツリガネゴケ(Physcomi
trella patens)の鎖長延長酵素の遺伝子を、タバコおよびアマ(Linu
m usitatissimum)に導入することで、アラキドン酸を、タバコの種子に
1.5%、アマの種子に1.0%蓄積させたことを報告している(Amine Abba
di et al.Plant Cell 16,2734−2748,2004)。
【0209】
本実施例においては、ゼニゴケ由来のΔ6不飽和化酵素、Δ5不飽和化酵素および鎖長
延長酵素をタバコに導入したことで、タバコの葉にアラキドン酸を10%以上蓄積させて
いる。この結果から、本実施例のpSPB2368形質転換タバコは、上述の報告と比較
しても、より効率的に高度不飽和脂肪酸を合成できる能力があることが示唆された。
【0210】
また、ハプト藻(Isochrysis galbana)のΔ9鎖長延長酵素、ミド
リムシ(Englena gracilis)のΔ8不飽和化酵素、糸状菌のΔ5不飽和
化酵素の3種類の遺伝子を用いて、シロイヌナズナ(Arabidopsis thal
iana)で脂質の改変を行った報告では、葉中でアラキドン酸が総脂質に対して6.6
mol%、C20以上の脂質が22.5mol%であった(Baoxiu Qi et
al.Nature Biotechnology 22,739−745,2004)
。この報告では、Δ6不飽和化酵素、Δ5不飽和化酵素、および鎖長延長酵素を用いる経
路とは別の修飾経路で高度不飽和脂肪酸を作っており、単純比較は出来ないが、ゼニゴケ
の酵素を用いたほうがより多くの高度不飽和脂肪酸を蓄積することがわかった。
【0211】
総脂質の30%以上が改変脂肪酸になっているpSPB2368形質転換タバコでは、
形態的な異常は見られなかった。さらに、稔性にも問題なく、多くの種子をつけたことか
ら、異所的な高度不飽和脂肪酸の増加が植物の生育に及ぼす影響は少ないと考えられた。
【0212】
これまでに報告されている遺伝子組換え植物におけるC20以上の高度不飽和脂肪酸の
生産は総脂質の20%前後、アラキドン酸に限ると6%前後が限界である。しかし、本実
施例のように、ゼニゴケ由来の脂肪酸合成酵素を用いることにより、この限界を打ち破り
、より多くの高度不飽和脂肪酸を植物で生産することが出来ると考えられる。
【0213】
尚、発明を実施するための最良の形態の項においてなした具体的な実施態様または実施
例は、あくまでも、本発明の技術内容を明らかにするものであって、そのような具体例に
のみ限定して狭義に解釈されるべきものではなく、本発明の精神と次に記載する特許請求
の範囲内で、いろいろと変更して実施することができるものである。
【産業上の利用可能性】
【0214】
本発明に係る遺伝子は、同一種のゼニゴケから単離されたΔ5不飽和化酵素遺伝子、Δ
6不飽和化酵素遺伝子及びΔ6鎖長延長酵素遺伝子である。したがって、これら3種類の
遺伝子を植物内で同時に発現させた場合に、異なる生物種由来の複数の遺伝子を発現させ
るより、植物内で良好に機能するという効果を奏する。さらに、ゼニゴケは、高等植物の
モデル系と考えられるため、これらの遺伝子は、植物以外の生物種由来の遺伝子より植物
内で良好に機能することができるという効果を奏する。
【0215】
また、本発明に係る形質転換体は、アラキドン酸やエイコサペンタエン酸(EPA)等
の高度不飽和脂肪酸を生産することができるという効果を奏する。特に、本発明に係る形
質転換植物は、低コストかつ環境にやさしい生産プロセスでアラキドン酸やEPA等の高
度不飽和脂肪酸を生産することができるという効果を奏する。こうして得られたアラキド
ン酸やEPAは、安価な多目的材料として活用できるという効果を奏する。さらに、上記
形質転換植物体を食料として用いた場合、アラキドン酸やEPAの含量が高い食料として
の価値が高まるという効果を奏する。
【0216】
以上にように、本発明の遺伝子・たんぱく質は、アラキドン酸やEPAの生産に有用で
ある。また、本発明の遺伝子を発現可能に導入した形質転換体は、製薬産業、食品産業、
各種素材産業等において、アラキドン酸やEPAを生産するうえで、極めて有用である。
また、特に、上記形質転換体が植物体である場合、植物体内のアラキドン酸やEPAの含
量が増加するので、農業分野等において非常に有用である。
【特許請求の範囲】
【請求項1】
配列番号1に示される塩基配列からなるDNAの全部又は一部、あるいは当該DNAと相
補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイブリダイ
ズし、かつΔ6脂肪酸不飽和化活性を有するたんぱく質をコードするゼニゴケ目生物由来
の遺伝子。
【請求項2】
Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号1に示される塩基配列を有する遺伝子。
(b)配列番号1に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基
配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【請求項3】
Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号1に示される塩基配列のうち、253ないし1698番目の塩基配列を
有する遺伝子。
(b)配列番号1に示される塩基配列のうち、253ないし1698番目の塩基配列か
らなるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな
条件でハイブリダイズする遺伝子。
【請求項4】
Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号2に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
(b)配列番号2に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失
、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【請求項5】
配列番号3に示される塩基配列からなるDNAの全部又は一部、あるいは当該DNAと相
補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイブリダイ
ズし、かつΔ6脂肪酸鎖長延長活性を有するたんぱく質をコードするゼニゴケ目生物由来
の遺伝子。
【請求項6】
Δ6脂肪酸鎖長延長活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号3に示される塩基配列を有する遺伝子。
(b)配列番号3に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基
配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【請求項7】
Δ6脂肪酸鎖長延長活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号1に示される塩基配列のうち、194ないし1066番目の塩基配列を
有する遺伝子。
(b)配列番号1に示される塩基配列のうち、194ないし1066番目の塩基配列か
らなるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな
条件でハイブリダイズする遺伝子。
【請求項8】
Δ6脂肪酸鎖長延長活性を有するゼニゴケ目由来のたんぱく質をコードする、下記(a)
又は(b)に記載の遺伝子。
(a)配列番号4に示されるアミノ酸配列からなるたんぱく質、
(b)配列番号4に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失
、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【請求項9】
配列番号5に示される塩基配列からなるDNAの全部又は一部、あるいは当該DNAと相
補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイブリダイ
ズし、かつΔ5脂肪酸不飽和化活性を有するたんぱく質をコードするゼニゴケ目生物由来
の遺伝子。
【請求項10】
Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号5に示される塩基配列を有する遺伝子。
(b)配列番号5に示す塩基配列からなるDNA、又は当該DNAと相補的な塩基配列
からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【請求項11】
Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号5に示される塩基配列のうち、375ないし1829番目の塩基配列を
有する遺伝子。
(b)配列番号5に示される塩基配列のうち、375ないし1829番目の塩基配列か
らなるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな
条件でハイブリダイズする遺伝子。
【請求項12】
Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号6に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
(b)配列番号6に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失
、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【請求項13】
請求項1〜12の何れか1項に記載の遺伝子によってコードされるたんぱく質。
【請求項14】
以下の(a)又は(b)記載のたんぱく質。
(a)配列番号2に示されるアミノ酸配列からなるたんぱく質。
(b)配列番号2に示されるアミノ酸配列において、1個又はそれ以上のアミノ酸が置
換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ、Δ6脂肪酸不飽和
化活性を有するたんぱく質。
【請求項15】
以下の(a)又は(b)記載のたんぱく質。
(a)配列番号4に示されるアミノ酸配列からなるたんぱく質。
(b)配列番号4に示されるアミノ酸配列において、1個又はそれ以上のアミノ酸が置
換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ、Δ6脂肪酸鎖長延
長活性を有するたんぱく質。
【請求項16】
以下の(a)又は(b)記載のたんぱく質。
(a)配列番号6に示されるアミノ酸配列からなるたんぱく質。
(b)配列番号6に示されるアミノ酸配列において、1個又はそれ以上のアミノ酸が置
換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ、Δ5脂肪酸不飽和
化活性を有するたんぱく質。
【請求項17】
請求項13〜16の何れか1項に記載のたんぱく質を認識する抗体。
【請求項18】
少なくとも請求項1〜12の何れか1項に記載の遺伝子を含む組換え発現ベクター。
【請求項19】
少なくとも請求項1〜12の何れか1項に記載の遺伝子を導入してなる形質転換体。
【請求項20】
少なくとも請求項1〜12の何れか1項に記載の遺伝子が発現可能に導入された植物体、
もしくは当該植物体と同一の性質を有する当該植物体の子孫となる植物体、又は当該植物
体の組織。
【請求項21】
少なくとも請求項1〜12の何れか1項に記載の遺伝子が発現可能に導入され、脂肪酸組
成が改変された植物体、もしくは当該植物体と同一の性質を有する当該植物体の子孫とな
る植物体、又は当該植物体の組織。
【請求項22】
請求項20又は21に記載の植物体の繁殖材料。
【請求項23】
請求項21に記載の植物体又は植物体の組織を用いることを特徴とする脂肪酸生産方法。
【請求項24】
請求項23に記載の脂肪酸生産方法により得られた、γ−リノレン酸、ジホモ−γ−リノ
レン酸、アラキドン酸、ステアリドン酸、エイコサテトラエン酸、およびエイコサペンタ
エン酸から選択される少なくとも1つ含む素材。
【請求項25】
少なくとも請求項1〜12の何れか1項に記載の遺伝子を用いて脂肪酸組成を改変する方
法。
【請求項26】
請求項1〜12の何れか1項に記載の遺伝子における少なくとも一部の塩基配列又はその
相補配列をプローブとして用いた遺伝子検出器具。
【請求項27】
請求項13〜16の何れか1項に記載のたんぱく質を用いて、当該たんぱく質を調節する
遺伝子、又は当該たんぱく質を調節する物質をスクリーニングする方法。
【請求項28】
請求項27に記載のスクリーニング方法により得られた遺伝子又は物質。
【請求項1】
配列番号1に示される塩基配列からなるDNAの全部又は一部、あるいは当該DNAと相
補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイブリダイ
ズし、かつΔ6脂肪酸不飽和化活性を有するたんぱく質をコードするゼニゴケ目生物由来
の遺伝子。
【請求項2】
Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号1に示される塩基配列を有する遺伝子。
(b)配列番号1に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基
配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【請求項3】
Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号1に示される塩基配列のうち、253ないし1698番目の塩基配列を
有する遺伝子。
(b)配列番号1に示される塩基配列のうち、253ないし1698番目の塩基配列か
らなるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな
条件でハイブリダイズする遺伝子。
【請求項4】
Δ6脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号2に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
(b)配列番号2に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失
、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【請求項5】
配列番号3に示される塩基配列からなるDNAの全部又は一部、あるいは当該DNAと相
補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイブリダイ
ズし、かつΔ6脂肪酸鎖長延長活性を有するたんぱく質をコードするゼニゴケ目生物由来
の遺伝子。
【請求項6】
Δ6脂肪酸鎖長延長活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号3に示される塩基配列を有する遺伝子。
(b)配列番号3に示される塩基配列からなるDNA、又は当該DNAと相補的な塩基
配列からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【請求項7】
Δ6脂肪酸鎖長延長活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号1に示される塩基配列のうち、194ないし1066番目の塩基配列を
有する遺伝子。
(b)配列番号1に示される塩基配列のうち、194ないし1066番目の塩基配列か
らなるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな
条件でハイブリダイズする遺伝子。
【請求項8】
Δ6脂肪酸鎖長延長活性を有するゼニゴケ目由来のたんぱく質をコードする、下記(a)
又は(b)に記載の遺伝子。
(a)配列番号4に示されるアミノ酸配列からなるたんぱく質、
(b)配列番号4に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失
、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【請求項9】
配列番号5に示される塩基配列からなるDNAの全部又は一部、あるいは当該DNAと相
補的な塩基配列からなるDNAの全部又は一部とストリンジェントな条件でハイブリダイ
ズし、かつΔ5脂肪酸不飽和化活性を有するたんぱく質をコードするゼニゴケ目生物由来
の遺伝子。
【請求項10】
Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号5に示される塩基配列を有する遺伝子。
(b)配列番号5に示す塩基配列からなるDNA、又は当該DNAと相補的な塩基配列
からなるDNAとストリンジェントな条件でハイブリダイズする遺伝子。
【請求項11】
Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号5に示される塩基配列のうち、375ないし1829番目の塩基配列を
有する遺伝子。
(b)配列番号5に示される塩基配列のうち、375ないし1829番目の塩基配列か
らなるDNA、又は当該DNAと相補的な塩基配列からなるDNAとストリンジェントな
条件でハイブリダイズする遺伝子。
【請求項12】
Δ5脂肪酸不飽和化活性を有するゼニゴケ目生物由来のたんぱく質をコードする、下記(
a)又は(b)に記載の遺伝子。
(a)配列番号6に示されるアミノ酸配列からなるたんぱく質をコードする遺伝子。
(b)配列番号6に示されるアミノ酸配列の1個又はそれ以上のアミノ酸が置換、欠失
、挿入、及び/又は付加されたアミノ酸配列からなるたんぱく質をコードする遺伝子。
【請求項13】
請求項1〜12の何れか1項に記載の遺伝子によってコードされるたんぱく質。
【請求項14】
以下の(a)又は(b)記載のたんぱく質。
(a)配列番号2に示されるアミノ酸配列からなるたんぱく質。
(b)配列番号2に示されるアミノ酸配列において、1個又はそれ以上のアミノ酸が置
換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ、Δ6脂肪酸不飽和
化活性を有するたんぱく質。
【請求項15】
以下の(a)又は(b)記載のたんぱく質。
(a)配列番号4に示されるアミノ酸配列からなるたんぱく質。
(b)配列番号4に示されるアミノ酸配列において、1個又はそれ以上のアミノ酸が置
換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ、Δ6脂肪酸鎖長延
長活性を有するたんぱく質。
【請求項16】
以下の(a)又は(b)記載のたんぱく質。
(a)配列番号6に示されるアミノ酸配列からなるたんぱく質。
(b)配列番号6に示されるアミノ酸配列において、1個又はそれ以上のアミノ酸が置
換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ、Δ5脂肪酸不飽和
化活性を有するたんぱく質。
【請求項17】
請求項13〜16の何れか1項に記載のたんぱく質を認識する抗体。
【請求項18】
少なくとも請求項1〜12の何れか1項に記載の遺伝子を含む組換え発現ベクター。
【請求項19】
少なくとも請求項1〜12の何れか1項に記載の遺伝子を導入してなる形質転換体。
【請求項20】
少なくとも請求項1〜12の何れか1項に記載の遺伝子が発現可能に導入された植物体、
もしくは当該植物体と同一の性質を有する当該植物体の子孫となる植物体、又は当該植物
体の組織。
【請求項21】
少なくとも請求項1〜12の何れか1項に記載の遺伝子が発現可能に導入され、脂肪酸組
成が改変された植物体、もしくは当該植物体と同一の性質を有する当該植物体の子孫とな
る植物体、又は当該植物体の組織。
【請求項22】
請求項20又は21に記載の植物体の繁殖材料。
【請求項23】
請求項21に記載の植物体又は植物体の組織を用いることを特徴とする脂肪酸生産方法。
【請求項24】
請求項23に記載の脂肪酸生産方法により得られた、γ−リノレン酸、ジホモ−γ−リノ
レン酸、アラキドン酸、ステアリドン酸、エイコサテトラエン酸、およびエイコサペンタ
エン酸から選択される少なくとも1つ含む素材。
【請求項25】
少なくとも請求項1〜12の何れか1項に記載の遺伝子を用いて脂肪酸組成を改変する方
法。
【請求項26】
請求項1〜12の何れか1項に記載の遺伝子における少なくとも一部の塩基配列又はその
相補配列をプローブとして用いた遺伝子検出器具。
【請求項27】
請求項13〜16の何れか1項に記載のたんぱく質を用いて、当該たんぱく質を調節する
遺伝子、又は当該たんぱく質を調節する物質をスクリーニングする方法。
【請求項28】
請求項27に記載のスクリーニング方法により得られた遺伝子又は物質。
【図1】

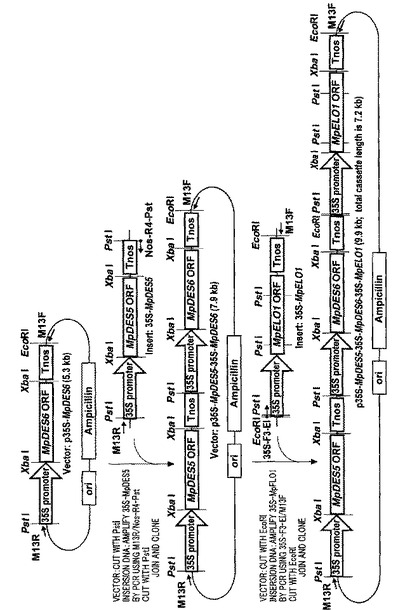
【公開番号】特開2011−123(P2011−123A)
【公開日】平成23年1月6日(2011.1.6)
【国際特許分類】
【出願番号】特願2010−183860(P2010−183860)
【出願日】平成22年8月19日(2010.8.19)
【分割の表示】特願2005−516506(P2005−516506)の分割
【原出願日】平成16年12月22日(2004.12.22)
【出願人】(309007911)サントリーホールディングス株式会社 (307)
【Fターム(参考)】
【公開日】平成23年1月6日(2011.1.6)
【国際特許分類】
【出願日】平成22年8月19日(2010.8.19)
【分割の表示】特願2005−516506(P2005−516506)の分割
【原出願日】平成16年12月22日(2004.12.22)
【出願人】(309007911)サントリーホールディングス株式会社 (307)
【Fターム(参考)】
[ Back to top ]