
ソマトスタチン類似体
本発明は、概ね、X1CX2WKX3CT(ここで、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVである)からなるアミノ酸配列を有する単離されたペプチドに関し、これらは任意の順列組み合わせであり、それぞれのアミノ酸はL配置であり、ペプチドは2つのシステイン残基の間にジスルフィド結合を含む。本発明はまた、概ね、上記ペプチドを含む融合タンパク質、上記ペプチドまたは上記融合タンパク質をコードするポリヌクレオチド、上記ペプチドまたは上記融合タンパク質の製造方法、上記ペプチドまたは上記融合タンパク質を含む医薬、および、これらの使用に関する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、概ね、天然には存在せず生物学的に合成されるペプチド類に関し、当該ペプチド類をコードするポリヌクレオチド類に関し、当該ペプチド類の製造方法に関し、当該ペプチド類の用途に関する。本発明のペプチド類は、医薬に(例えば、ホルモン障害、癌および出血性障害の治療に)用いられうる。本発明はまた、概ね、安定性が向上したペプチド類の修飾体に関し、かような修飾ペプチドの作製方法に関し、当該修飾ペプチド類の用途に関する。特に、本発明は、ポリエチレングリコールなどのポリマーを用いて化学的に修飾されうる、生物学的に合成されるペプチド類に関し、末端肥大症、消化管ホルモン産生腫瘍などの腫瘍、および消化管出血の治療における当該ペプチド類またはその修飾体の用途に関する。
【背景技術】
【0002】
末端肥大症は、頭、手および足の骨、並びに軟部組織の肥大を特徴とする、生活に支障を来すほどのホルモン障害であり、早期の死をもたらす。末端肥大症への罹患率は、1年間に人口100万人あたり3症例であり、その有病率は100万人当たり60人である。この疾患の症状は知らぬ間に進行することから、臨床症状の発現から診断がなされるまでに7〜10年遅れることもよくある。末端肥大症の原因は、通常は下垂体腺腫による下垂体からの成長ホルモン(GH)の過剰分泌である下垂体の成長ホルモン分泌細胞が増殖することで成長ホルモンが異常に高いレベルで分泌され、末端肥大症の典型的な臨床症状がもたらされる。
【0003】
成長ホルモンは191アミノ酸の、4つのヘリックスの束のタンパク質、およびより少量の176アミノ酸体として分泌される。成長ホルモンは、下垂体門脈循環を往来する視床下部放出ホルモンおよび視床下部抑制ホルモンを介した視床下部の制御下で脈動により循環系に進入する。次いで、成長ホルモン分泌細胞表面の特異的な受容体に直接作用する。成長ホルモンは、(i)末梢のインスリン様成長因子1(IGF−1)の合成を誘導し、(ii)循環(エンドクリン)並びに局所(オートクリンおよびパラクリン)IGF−1により誘導される細胞増殖を誘導し、(iii)アポトーシスを阻害する。末端肥大症の過剰な罹患率および死亡率は、GHレベルおよびIGF−1レベルの長期にわたる上昇の結果である。これら2つのホルモンのレベルを障害にわたって注意深く制御することで、患者の福利が改善され、正常な寿命が取り戻される。
【0004】
現在の末端肥大症の治療のなかでも、外科的手術は成長ホルモンを分泌する微小腺腫を除去する目的でよく行われる。放射線治療は通常、薬物療法に抵抗性の、または耐性を示す患者において、手術後に再発するか生き残っている腫瘍のために取って置かれる。
【0005】
また、現在の治療のなかには、成長ホルモンの分泌を抑制する天然のホルモンであるソマトスタチンの化学合成された類似体を投与するというものもある。かような類似体の1つは、酢酸オクトレオチド(ノバルティスファーマシューティカルズ社によりサンドスタチン(登録商標)の商品名で市販されている)である。この薬は過去20年間にわたって、末端肥大症の治療法として成功を収め、臨床上も認められてきた。酢酸オクトレオチド(体系的なIUPAC名は(4R,7S,10S,13R,16S,19R)-10-(4-アミノブチル)-19-[[(2R)-2-アミノ-3-フェニル-プロパノイル]アミノ]-16-ベンジル-N-[(2R,3R)-1,3-ジヒドロキシブタン-2-イル]-7-(1-ヒドロキシエチル)-13-(1H-インドール-3-イルメチル)-6,9,12,15,18-ペンタオキソ-1,2-ジチア-5,8,11,14,17-ペンタアザシクロイコサン-4-カルボキサミドである)は、固相化学合成法(Bauer et al., (1982) Life Sciences, Vol. 31, pp. 1133-1140)により製造されるオクタペプチドである。化学合成された酢酸オクトレオチドのアミノ酸配列は:
【0006】
【化1】

【0007】
である。
【0008】
他の化学合成されたソマトスタチン類似体は、酢酸ランレオチド(体系的なIUPAC名は(4S,7S,10S,13R,16S,19S)-10-(4-アミノブチル)-19-[[(2R)-2-アミノ-3-ナフタレン-2-イル-プロパノイル]アミノ]-N-[(1S,2R)-1-カルバモイル-2-ヒドロキシ-プロピル]-16-[(4-ヒドロキシフェニル)メチル]-13-(1H-インドール-3-イルメチル)-6,9,12,15,18-ペンタオキソ-7-プロパン-2-イル-1,2-ジチア-5,8,11,14,17-ペンタアザシクロイコサン-4-カルボキサミドである)である。この薬は、イプセンファーマシューティカルズによりソマツリンLA(登録商標)の商品名で市販されている。
【0009】
酢酸ランレオチドのアミノ酸配列は:
【0010】
【化2】
【0011】
である。
【0012】
特に、酢酸ランレオチドは1位に(D)アラニンを、4位に(D)トリプトファンを含んでいる。また、2つのシステイン残基にわたってジスルフィド結合を有してもいる。酢酸ランレオチドは構造的に酢酸オクトレオチドと非常に類似しており、酢酸オクトレオチドにとてもよく似たソマトスタチン受容体結合プロファイルを示し、酢酸オクトレオチドと同様の適応症を有している(Weckbecker, G. et al., (2003) Nature Reviews, Vol. 2, pp. 999-1017)。
【0013】
天然のホルモンであるソマトスタチンは、βターンの角にTrp残基およびLys残基が存在することに起因して逆平行に伸びたβシートが存在することで、そのコンホメーションに環状の部位を有していることが知られている。酢酸オクトレオチドおよび酢酸ランレオチドでは、D−Trp4の導入によってファーマコフォアのβターンを安定化するように同様に作用する。
【0014】
また、ジスルフィド架橋の形成によるペプチドの環化によってタンパク質やペプチド(ソマトスタチンも含む)の化学的安定性や代謝安定性が付与されることも一般的に知られている。この理由から、化学合成された酢酸オクトレオチドおよび酢酸ランレオチドには、架橋ユニットであるCys2−S−S−Cys7が化学的に導入されている。酢酸オクトレオチドの場合には、ペプチドの代謝安定性が3倍に上昇する(Bauer et al., (1982) Life Sciences, Vol. 31, pp. 1133-1140; Cai et al., (1986) Proc. Natl. Acad. Sci. (USA), Vol. 83, pp. 1896-1900)。
【0015】
これらをまとめると、2つのDアミノ酸の挿入およびジスルフィド架橋の形成によってこれらの化学合成8アミノ酸ペプチドの血清半減期は長くなる。酢酸オクトレオチドの場合には、数分間が1.5時間まで長くなる(Harris AG., (1994) Somatostatin and somatostatin analogues: pharmacokinetics and pharmacodynamic effects. Gut, Vol.35, pp. 1-4)
酢酸オクトレオチドおよび酢酸ランレオチドは、ソマトスタチン受容体サブタイプ2(SST2)およびソマトスタチン受容体サブタイプ5(SST5)に高親和性で結合する。ソマトスタチン受容体サブタイプ3に対してこれらは中程度の親和性で結合する。ソマトスタチン受容体サブタイプ1またはソマトスタチン受容体サブタイプ4に対してこれらは結合しない。また、酢酸オクトレオチドおよび酢酸ランレオチドはD2ドーパミン受容体に結合することが示されている。
【0016】
酢酸オクトレオチドの生物活性には、その構造中のPhe3−Trp4−Lys5−Thr6のみが必須であることが知られている。酢酸オクトレオチドがソマトスタチン受容体に結合すると、成長ホルモンの分泌および成長ホルモン分泌細胞の増殖を抑制するように下垂体に信号を送り、また、肝臓に作用してIGF−1の合成を阻害する。SST2およびSST5受容体に対するソマトスタチン受容体のリガンドとして、酢酸オクトレオチドはGHおよびIGF−1のレベルを低下させ、腫瘍増殖を抑制し、肝臓により媒介されるGHの受容体への結合・作用を阻害する。GH受容体のアンタゴニストとして、酢酸オクトレオチドはGH受容体のシグナル伝達を阻害し、これにより末梢血中のIGF−1レベルが低下する。
【0017】
末端肥大症の患者における酢酸オクトレオチドの投与に応答した成長ホルモン分泌の抑制は、ソマトスタチン受容体サブタイプの利用可能性に依存する。酢酸オクトレオチドおよび酢酸ランレオチドの直接的な抗腫瘍活性が、腫瘍細胞において発現するソマトスタチン受容体を介した実験腫瘍モデルで実証されている。これらの抗増殖作用は、細胞分裂の阻害およびアポトーシスの誘導の結果である。ソマトスタチン受容体に化学合成ソマトスタチン類似体が結合すると、特異的なシグナル伝達経路が開始される。このようにして、それぞれのソマトスタチン受容体サブタイプは異なる生物学的作用を媒介することができる。これらの機構を媒介する受容体サブタイプは、SST1、SST2、SST4およびSST5である。ヒト腫瘍の多くは複数のソマトスタチン受容体サブタイプを発現しているが、主なものはSST2である。ソマトスタチン類似体である酢酸オクトレオチドおよび酢酸ランレオチドは、SST2受容体に対して高い親和性を有している。
【0018】
ソマトスタチンおよびその合成類似体はまた、多くの間接的な抗腫瘍作用を示す。これらのなかには、腫瘍増殖を促進する成長因子およびホルモンの放出の阻害がある。これらのソマトスタチン類似体の間接的な効果により生じる腫瘍増殖の低下は、成長因子や、IGF−1および成長ホルモンなどのホルモン類の合成および分泌の抑制(並びに作用の減退)を含む。ソマトスタチン類似体は、中枢および末梢の双方の機構によって成長ホルモン−IGF−1系を抑制する。下垂体の成長ホルモンの放出の阻害を媒介する主要な受容体サブタイプはSST2およびSST5である。ソマトスタチン類似体はまた、STAT5bの脱リン酸化およびIGF−1遺伝子の転写の低下をもたらすSST媒介性のチロシンホスファターゼの活性化を介して、成長ホルモンにより誘導される肝臓のIGF−1の産生を阻害する(Weckbecker, G. et al., (2003) Nature Reviews, Vol. 2, pp. 999-1017; Susini C & Buscail L, (2006) Ann of Oncology Vol. 17, pp. 1733-1742)。
【0019】
一方、ソマトスタチン受容体サブタイプの利用可能性は、患者血清中の血清成長ホルモンおよびIGF−1の濃度に酢酸オクトレオチド療法が与える長期にわたる効果を予測する。血清の成長ホルモンおよびIGF−1が最も抑制されるのは、皮下で300〜600μg/日の用量である。酢酸オクトレオチドはまず、成長ホルモンのレベルを正常なレベルにまで低下させるために、数ヶ月間の間、少なくとも日に3回(半減期がたった1.5時間であるため)、皮下注射として患者に投与される。成長ホルモンの過剰産生が正常レベルにまで低下したら、サンドスタチン(登録商標)長期間作用型などの徐放性ポリマーベースの複合体デポ製剤を患者に(数週間にわたって)移植することもよく行われる。これは、殿筋の深部に筋肉内注射することにより投与され、三角筋に投与することはできない。これは、大きい殿筋と比べて三角筋ではサンドスタチンがもたらす痛みが過剰であることによる。このポリマーベースの徐放性デポから酢酸オクトレオチドが徐放されるということは、酢酸オクトレオチドは通常、痛みのない皮下注射により投与される場合の毎週と比較して、2週間おきに(痛みを伴って)投与されうることを意味する。
【0020】
デポ製剤(すなわち、長期間作用放出型の酢酸オクトレオチド(サンドスタチン(登録商標)LAR(登録商標)として市販されている))により、14日おきの注射による投与が可能となる。これらの製剤は、酢酸オクトレオチドの有効なレベルを維持することができる。報告によれば、酢酸オクトレオチドによる治療の間9年間追跡された患者の80%で、成長ホルモンのレベルが2.5μg/L未満であり、かつ、IGF−1のレベルが正常であったとされている。ユーゴナディズム(Eugonadism)は性腺機能低下症を有する末端肥大症患者の3分の2に残っている(S. Farooqi et al. (1999) Pituitary, Vol. 2, pp. 79-88; A.N. Paisley and P.J. Trainer, (2003) Current opinion in Pharmacology, Vol. 3, pp. 672-677; S. Melmed. (2006) New England Journal of Medicine, Vol. 355, pp. 2558-2573)。
【0021】
酢酸オクトレオチドの有効性の決定因子としては、治療前の成長ホルモンのレベル、腫瘍での十分なSST2およびSST5の発現の存在・不存在、薬物の用量、状態の評価に用いられる生化学的指標、並びに治療に対する患者の忠実さなどが挙げられる。腫瘍塊の縮小は患者の50%で起こるが、酢酸オクトレオチドを用いた治療を止めると回復してしまう。
【0022】
また、外科的な全部の切除を必要としない巨大腺腫の外科的減量によれば、その後の酢酸オクトレオチドによる治療の有効性が向上する。薬を服用した患者の80%超が、頭痛や末梢軟組織の膨張などの症状が改善されたと報告している。腫瘍の縮小および生化学的な制御は、必ずしも並行して起こるとは限らない。よって、酢酸オクトレオチドはGHおよびIGF−1のレベルの生化学的な制御を変化させることができなかった手術の後や、成長ホルモンのレベルが上昇したままであったことによる放射線治療の後に投与されることが多い。酢酸オクトレオチドを用いた一次的な医学的治療は有効かつ安全である。患者が手術または放射線治療を受けているか否かにかかわらず、長期にわたる薬物投与に対する同等の生化学的応答が達成されうることから、一次的な医学的治療は、(i)巨大な細胞外腫瘍を有し、中枢圧縮作用(central compressive effect)の証拠を示さない患者、(ii)薄弱すぎて手術を受けることができない患者、および(iii)手術を辞退した患者に対して提案されうる。
【0023】
酢酸オクトレオチドおよび酢酸ランレオチドはまた、消化管関連のカルシノイド腫瘍や転移性のカルシノイド腫瘍の治療にも用いられうる。胃腸管のカルシノイドの全体的な発生率は年間84人/百万人と見積もられている。胃腸管のカルシノイド腫瘍は、全てのカルシノイド腫瘍の90%を占めている。酢酸オクトレオチドを用いた治療により、カルシノイド症候群を患っている患者の50〜80%の症状および生活の質が改善される。また、酢酸オクトレオチドおよび酢酸ランレオチドは、腫瘍の発達を停止させることができる。
【0024】
酢酸オクトレオチドは、血管作動性腸管ペプチドを分泌する腫瘍(VIPoma)を有する患者における下痢の治療に治療上有効であることが示されており、他の原因による重篤な難治性の下痢の治療に用いられている。毒性学においてもまた、酢酸オクトレオチドはスルホニルウレアの過剰摂取後の長期にわたる再発性低血糖の治療に用いられており、インスリンの過剰分泌の抑制を補助する目的で、膵島細胞症を有する乳幼児に用いられている。食道静脈瘤が疑われる患者において、酢酸オクトレオチドは出欠の抑制を補助する目的で投与されうる。さらに、酢酸オクトレオチドは、慢性膵炎からくる痛みを有する患者の治療や、胸腺腫瘍の治療、および糖尿病に起因する眼疾患の治療にも用いられうる。酢酸ランレオチドも酢酸オクトレオチドと同様の適応症を有している。
【0025】
酢酸オクトレオチドや酢酸ランレオチドのような数年間にわたって恒常的に服用する必要のある小分子ペプチドベースの医薬すべてにおける主要な実用上の欠点は、全世界で用いられる医薬として製造するための費用対効果のよい大量生産法がないということである。現時点では、天然のペプチドおよび設計されたペプチドのいずれの製造にも化学的なペプチド合成が用いられているが、このアプローチは非常にコストがかかる。その結果、患者における酢酸オクトレオチドの臨床的な使用は法外に高価なものとなっている。末端肥大症の患者の場合、イギリスで1人の患者を治療するコストは、約16000ポンド/年である。悪性のカルシノイド腫瘍を有する患者に酢酸オクトレオチドを用いると、イギリスで1人の患者を治療するコストは約32000ポンド/年である。
【0026】
医薬としてのペプチド類の工業的な生産に対する主な障害は、経済的な大量生産である。酢酸オクトレオチドおよび酢酸ランレオチドの製造においては、段階的なFmoc固相化学合成とそれに続く分子内ジスルフィド結合の形成が必要とされる。D−Trpの存在下でペプチドの代謝安定性を3倍に改善するための分子内ジスルフィド結合の形成は主要な問題となりうる。また、化学合成時の側鎖の保護も必要である。これらの化学的な問題により、収率は14%と低い結果となっている。酢酸オクトレオチドの固相合成に伴う他の問題としては、(i)C末端のCys残基のラセミ化、(ii)ペプチドの集合の非効率性、(iii)有害な副作用、(iv)樹脂上での非効率なジスルフィド結合の形成、(v)ジスルフィド結合形成時のD−Trpの修飾、(vi)アミノリシスによる不完全で非効率なペプチド−樹脂の開裂、(v)断片のカップリング時のラセミ化、および、(vi)複雑な精製工程、が挙げられる。さらに、(a)L−PheをD−Pheで置換しなければならないとき、(b)L−TrpをD−Trpで置換しなければならないとき、(c)直鎖ペプチドの安定性を向上させる目的でジスルフィド結合を形成するために直鎖ペプチドを環化する必要があるときに、化学的なペプチドベースの合成のコストは著しく高騰する。酢酸ランレオチドの固相合成についても同様の問題が存在する。
【0027】
よって、代わりとなる、より経済的かつ効率的な、酢酸オクトレオチドや酢酸ランレオチドのような小ペプチドの合成方法が必要とされている。この点で、細菌や酵母での組換えDNA法がよく用いられる。また、タンパク質の生物活性においてグリコシル化が役割を担っていないことが示されているのであれば、汎用の宿主である大腸菌においてタンパク質の製造を実施することで相当のコストおよび製造上の利点が存在する。この技術により、多くの組換えタンパク質が大腸菌で製造されている。
【0028】
しかしながら、かような系では、組換えDNA技術を用いることでタンパク質の高い生産性は達成されるものの、きわめて小さい天然のペプチド類についての問題として、より大きな融合タンパク質の一部として製造される必要があるということがある。このため、天然のペプチドの遺伝子はより大きいキャリアタンパク質の遺伝子と連結され、次いで、大腸菌などの宿主細胞中で、単一の大きいタンパク質として、融合タンパク質が発現する。タンパク質の合成の後、所望のペプチドは融合相手から切断されなければならない。実用上は、このアプローチによる主な問題として、非常に小さいペプチドは、ほとんどの微生物の細胞質に見られる酵素によるタンパク質分解を非常に受けやすい(サイズが非常に小さいことで、高度に配列した三次構造を有しえなくなる)ということがある。したがって、小ペプチドは製造された宿主の細胞内で急速に、かつ細胞の他の細胞質成分から首尾よく単離されうる前に、分解されることが多い。
【0029】
また、多くの異種タンパク質(すなわち、宿主細胞において天然には産生されないタンパク質)が、宿主細胞中で大量に製造されることに起因する毒性作用によって細菌/酵母の増殖を妨げることが確認されているという別の問題もある。さらに、これらの外来ペプチド産物は宿主細胞中で不安定なことが多い。このことは、微生物中で融合タンパク質の一部としてペプチドを発現させるときには、当該ペプチドの発現を安定化させるために多くの努力を要するということを意味する。これらの問題は特に、微生物系において非常に短いペプチド鎖の発現を必要とする場合に関係があり、このことは所望のペプチド配列がより大きい融合タンパク質の一部として発現する場合でさえそうである。
【0030】
組換えDNA技術を用いた組換えペプチドの生産が大規模で達成され、しかもかような生産が経済的なものとされうるとしても、細菌/酵母溶解物、そして融合タンパク質から非常に小さいペプチドを単離し精製するコストが高いことで、費用効率の高い医薬を製造するという問題はこじれてしまう。細菌または酵母からペプチドを生産するためのこれらの下流の処理工程は、実質的な追加の生産コストに寄与することが多い。例えば、細菌または酵母の細胞からのペプチドの最初の回収には、細胞の破砕および溶解、破砕・溶解された細胞からの封入体(すなわち、凝集した、それゆえに不溶性のタンパク質)の単離、溶解性の融合タンパク質を得るための単離された封入体の溶解、並びに、融合タンパク質の切断とその後のキャリアタンパク質からのペプチドの分離などの、複数かつ独特の処理工程が必要とされうる。よって、大規模で費用効率の高い生産を現実のものとするためには、非常に小さい組換えペプチドの生産のすべての形態を改善し、最適化することが望まれる。
【0031】
ペプチドベースの医薬については、効果的な薬物の送達を達成することにも問題がある。ポリマーベースのマトリックスを用いる場合、ペプチドの制御放出性の薬物送達システムの重要でかつ十分に解決されてはいない問題の1つは、生分解性ポリマーベースのマトリックス中に配合された後のペプチドの安定性である。マイクロスフェアに一般的に用いられているポリ(乳酸)およびポリ(乳酸−co−グリコール酸)のポリマーの分解により、ペプチドを含有するマイクロスフェアの内部には酸性度の高い微小環境(すなわち、pH1.5)が形成されることが多い。このことは、これらのマトリックス中に配合されたペプチドの不安定性の主要な原因となっていることが知られている。このペプチドの不安定性は、ポリ(乳酸)およびポリ(乳酸−co−グリコール酸)のマイクロスフェアの分解と、これによって生じる乳酸およびグリコール酸のユニットでの配合されたペプチドのアシル化により共有結合修飾によるものである。アシル化により、ペプチドの筋肉内のデポ注射部位からの吸収速度は遅くなる。いまでは、多くの機関によって、分解しているポリ(乳酸)およびポリ(乳酸−co−グリコール酸)のマイクロスフェア内部でのペプチド薬剤のアシル化は、新規でかつ有効性の高い治療薬としての生物活性ペプチドベースの薬物の、安定した、持続的な、うまい送達のために依然として克服する必要のある主要な障害として捉えられている(Werle M. et al., Amino Acids, (2006) Vol. 30; pp. 351-367)。
【0032】
この問題は、濃度が異なり(42.5%、21.3%、および8.5%;wt/wt)、37℃でのpH値も異なる(2.25、1.47、および1.85)3つの乳酸溶液中での酢酸オクトレオチドのアシル化反応の調査によるある研究において説明されている(Na et al., AAPS PharmSciTech., 2003, 4(4): article 72)。これらのインキュベーションの間、酢酸オクトレオチドのアシル化産物を、時間依存的かつ濃度依存的に測定した。42.5%乳酸溶液(pH2.25)では、30日間のインキュベーション後の完全な酢酸オクトレオチドの残存量は、出発物質のたった51%であった。これは、生物学的に不活性なアシル化酢酸オクトレオチドの量が増加した結果である。徐放性製剤の利用に向けた他のアプローチとしてはPEG化が挙げられ、これはタンパク質またはペプチドの安定性および半減期を増加させるのに用いられうる。ペプチドおよびタンパク質をPEG化することの治療上有用な効果としては、物理学的および熱的な安定性の向上、循環半減期の増加、免疫原性および抗原性の低下、並びに、毒性の低下が挙げられる。ペプチドおよびタンパク質をPEG化することで、有機溶媒への曝露に対して、天然のペプチドおよびタンパク質よりもずっと高い安定性をも示す。また、PEG化されたタンパク質のマイクロスフェアは、PEG化されていないペプチドおよびタンパク質と比較すると、異なる薬物放出プロファイルをも示し、初期のペプチドおよびタンパク質の一度の放出を抑制する。しかしこれまで、これらのPEG化ベースのアプローチは、分子量(MWt)が10kDaを超えるタンパク質ベースの薬物に限定されていた。また、安定化の問題は凝集や変性などのタンパク質の物理的安定性を改善するための主なものである。
【発明の概要】
【0033】
本発明者らは、この度、天然には存在しない新規なペプチドをコードするポリヌクレオチドを製造した。このポリヌクレオチドは、上述の新規なペプチドを製造するのに用いられうる。このペプチドは、酢酸オクトレオチドにおける4つの重要なアミノ酸の三次元結合部位構造を保持している。このペプチドは、宿主細胞中で経済的に生産されうるし、工業的規模で、費用効果の高いいくつかの工程で、宿主細胞から単離されうる。このペプチドはまた、自身の化学的安定性および代謝安定性を実質的に向上させ、これにより患者における治療有効性を実質的に高める目的で、費用効果の高い追加の化学修飾を伴ってもよい。
【0034】
以下の発明の記載では、添付の図面を参照する:
【図面の簡単な説明】
【0035】
【図1】図1は、化学的に合成された酢酸オクトレオチドを示す。丸で囲んだアミノ酸(すなわち、Phe−(D)Trp−Lys−Thr)のみがソマトスタチン細胞の表面受容体に結合する。
【図2】図2は、化学的に合成された酢酸オクトレオチド(分子量1018Da)の分子モデルを示しており、2つのD−アミノ酸を含有している。標識されたアミノ酸(すなわち、Phe−(D)Trp−Lys−Thr)がソマトスタチン細胞の表面受容体に結合する。
【図3】図3は、本発明による、生物学的に合成されたオクトレオチド(L−アミノ酸Phe−Cys−Phe−Trp−Lys−Thr−Cys−Thrを含有する)の分子モデルを、化学合成された酢酸オクトレオチド(2つのD−アミノ酸を含有する)の分子モデルと重ねて示す。太い灰色線が生物学的に合成されたオクタペプチド(すべてL−アミノ酸)を示し、細い黒線が化学合成された酢酸オクトレオチド(2つのD−アミノ酸)を示す。
【図4】図4は、本発明によって生物学的に合成され、次いで3つの炭素の架橋(ここにポリエチレングリコールが共有結合している)の挿入により化学修飾されたオクトレオチドの分子モデルを示す。
【図5】図5は、本発明によって生物学的に合成され、次いで3つの炭素の架橋(ここにポリエチレングリコールが共有結合している)の挿入により化学修飾されたオクトレオチドの分子モデルを示す。PEG化されたオクタペプチドの各化学成分が示されている:(a)点線の丸は、受容体に結合するアミノ酸Phe−Trp−Lys−Thrを示す;(b)破線は残りのアミノ酸(すなわち、Phe−Cys−ジスルフィド結合−Thr)を示す;(c)連続した線はポリエチレングリコールを示す。
【図6】図6は、本発明によって生物学的に合成され、次いでオクタペプチドのジスルフィド結合を横切る3つの炭素の架橋(CCCで示す)の挿入により化学修飾されたオクトレオチドの分子モデルを示す。この3つの炭素の架橋には、ポリエチレングリコールが共有結合しうる。
【図7】図7は、本発明の生物学的に合成されたオクタペプチドの柔軟性を示す。公開されている分子モデリングのプロトコール(Zloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083)を用い、アミノ酸配列FCFWKTCTを有する生物学的に合成されたオクタペプチドが、シミュレーション中の骨格原子の標準偏差(RMSD)の値を記録することによりモニターされうる柔軟度を示すことが示された。RMSD値の範囲は1〜4.4Åである。これは、3つの炭素の架橋を介してポリエチレングリコールが結合してもオクタペプチドの柔軟性が損なわれないことを示している。よって、このジスルフィド部位特異的にPEG化されたオクタペプチドは、その生物学的ターゲットと有効に相互作用できる程度に十分に柔軟なままでいられるであろう。
【図8】図8は、ポリエチレングリコールが共有結合した3つの炭素の架橋の挿入により化学修飾された、本発明によるL−アミノ酸を含有するアミノ酸配列FCFWKTCTを有する生物学的に合成されたオクタペプチドの分子モデルを、D‐アミノ酸を含有する化学合成された酢酸オクトレオチドの分子モデルと重ねて示す。これら2つの構造はエネルギー的に最低の配座異性体(すなわち、最も安定な形態)であり、これらはZloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083で公開されたのと同一の分子モデリング法を用いて予測された。太い灰色線が生物学的に合成されたオクタペプチドを示し、細い黒線が化学合成された酢酸オクトレオチド(2つのD−アミノ酸)を示す。
【図9】図9は、実施例2に記載のオクタペプチドプラスミドのバージョン1をクローニングするのに用いたアミノ酸配列およびヌクレオチド配列を示す。オクタペプチドFCFWKTCTをコードする配列に下線を付している。EcoRIおよびXhoIの制限部位の配列を太字で示している。
【図10】図10は、本発明のプラスミドベクターのバージョン1を示し、実施例2に記載の、プラスミドオクタペプチドのpGex4.3クローン5(メチオニン残基に連結したグルタチオントランスフェラーゼとそれに続く挿入オクタペプチドのコード配列を含む)とpGex4.3ベクターとのヌクレオチド配列のアライメントを示す。オクタペプチド*=オクタペプチドFCFWKTCTおよびクローニング配列。オクタペプチドFCFWKTCTをコードする配列に下線を付している。ベクターおよびpGex系の説明はGEライフサイエンセズによるGST融合系のハンドブックに記載されている(http://www4.gelifesciences.com/aptrix/upp00919.nsf/Content/87478CFA7E09E0C7C1256EB400417E59/$file/18115758.pdf)。
【図11】図11は、実施例3に記載のオクタペプチドプラスミドのバージョン2をクローニングするのに用いたアミノ酸配列およびヌクレオチド配列を示す。これは、TEV認識部位を含んでいる。X=任意のアミノ酸(プロリンを除く)。オクタペプチドFCFWKTCTおよびそれをコードする配列に下線を付している。
【図12】図12は、本発明のプラスミドベクターのバージョン2を示す。また、図12は、実施例3に記載のプラスミドオクタペプチドのpGex4.3と、追加のTEV開裂部位の配列を有するオクタペプチドFCFWKTCTのコード配列とのヌクレオチド配列のアライメントを示す。オクタペプチドFCFWKTCTをコードする配列に下線を付している。TS8v2は、TEVプロテアーゼ認識配列に連結したグルタチオントランスフェラーゼとそれに続く挿入オクタペプチドのコード配列を含む。
【図13】図13は、実施例2に記載のSjGST−TM−オクタペプチド(バージョン1)をコードするプラスミドから発現したタンパク質のSDS−PAGEゲルを示す(レーン1:マーカー;レーン2:SjGST−TM−オクタペプチド(細菌の全溶解物として);レーン3:SjGST−TM−オクタペプチド(グルタチオンアガロースカラムの流出物);レーン4:溶出したSjGST−TM−オクタペプチドタンパク質(単一種として27kDaの分子量を有する))。
【図14】図14は、実施例3に記載のSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)をコードするプラスミドから発現したタンパク質のSDS−PAGEゲルを示す。レーン1:分子量マーカー;レーン2:誘導細胞からの全溶解物;レーン3:溶出したSjGST−TEV−オクタペプチド融合タンパク質(分子量=28.4kDa)。
【図15】図15は、実施例5に記載の化学合成された酢酸オクトレオチドのマトリクス支援レーザー脱離イオン化質量分析法(MALDI−TOF−MS)による分析を示す。化学合成された酢酸オクトレオチドの理論質量は1018Daである。化学合成された酢酸オクトレオチドの実測質量は1019Daであった。化学合成された酢酸オクトレオチドNa+の理論質量は1041Daである。化学合成された酢酸オクトレオチドNa+の実測質量は1041Daであった。
【図16】図16は、実施例6に記載のSjGST−TM−オクタペプチド融合タンパク質バージョン1の実測質量のMALDI−TOF−MSによる分析を示す。SjGSTの理論質量は25498Daである。SjGStに付加されたオクタペプチドの理論質量は2259Daである。これにより、SjGSST−TM−オクタペプチドバージョン1融合タンパク質の合計理論質量は27757Daとなる。このSjGSST−TM−オクタペプチドの実測質量は27751Daであった。質量誤差百分率は0.02%である。理論質量と実測質量との間で0.1%の質量誤差百分率は許容されうる。
【図17】図17は、実施例7に記載の、トロンビンによる開裂後のSjGSTタンパク質(バージョン1)の実測質量のMALDI−TOF−MSによる分析を示す。精製された融合タンパク質の溶液を、100mM DTTを用いて還元し、PD−10ゲル濾過カラムによりバッファーを10mMリン酸ナトリウムバッファー(2mM EDTAを含有、pH7.8)に交換した。次いで、融合タンパク質の溶液を、10mMリン酸ナトリウムバッファー(pH7.8)中、37℃にて24時間トロンビン消化に供した。この溶液に、3mM DTTを添加し、正しくないジスルフィド結合の形成を防いだ。次いでこれをMALDI−TOF−MS分析に供した。SjGSST−TM−オクタペプチド融合タンパク質バージョン1の理論質量は27757Daである。トロンビンにより開裂した後のSjGSTタンパク質の理論質量は26150Daである。トロンビンにより開裂した後のSjGSTタンパク質の実測質量は26153Daであった。質量誤差百分率は0.01%である。トロンビン開裂後のオクタペプチドの理論質量は1607Daである。トロンビン開裂後のオクタペプチドの実測質量については、この特定のMALDI−TOFスペクトルでは低分子量の分子に対する解像度が低いため、測定できなかった(図18)。
【図18】図18は、実施例8に記載の、精製されたSjGST−TM−オクタペプチド融合タンパク質バージョン1から、シアノゲンブロマイドによりメチオニン部位で化学的に開裂した後の本発明のFCFWKTCTの配列を有するオクタペプチドのMALDI−TOF−MS分析を示す。精製されたタンパク質の溶液を、100mM DTTを用いて還元し、PD−10ゲル濾過カラムによりバッファーを10mMリン酸ナトリウムバッファー(2mM EDTAを含有、pH7.8)に交換した。次いで、この溶液を、10mMリン酸ナトリウムバッファー(pH7.8)中、37℃にて24時間の、オクタペプチドに直接隣接したメチオニン残基のシアノゲンブロマイド(すなわち、化学的)媒介性の開裂に供した。得られた溶液に3mM DTTを添加し、正しくないジスルフィド結合の形成を防いだ。次いでこれをMALDI−TOF−MS分析に供した。このオクタペプチドの理論質量は1035Daである。この生物学的に合成されたオクタペプチドの、メチオニンでの化学的開裂後の実測質量は1034Daであった。質量誤差百分率は0.09%である。化学合成された酢酸オクトレオチドと、生物学的に合成されたオクタペプチドFCFWKTCTとの実測質量の差は16Daである。これは、酢酸オクトレオチドがそのC末端残基としてL−スレオニノールを有するように化学的に修飾されているためである。生物学的なオクタペプチドFCFWKTCTの場合には、天然のアミノ酸であるL−スレオニンがC末端残基として存在している。
【図19】図19は、実施例9に記載の、ラクトースを用いた大腸菌でのSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)の自己誘導、およびその封入体からの精製を示す。レーン1:分子量マーカー;レーン2:大腸菌の誘導細胞からの全溶解物;レーン3:ソニケーション後の大腸菌細胞溶解物由来の上清;レーン4:2M尿素および100mM Tris(pH12.5)を用いた処理後の細胞ペレット由来の上清;レーン5:2M尿素および100mM Tris(pH12.5)を用いた処理とそれに続く塩酸によるpH8への調整後の細胞ペレット由来の上清;レーン6:2M尿素および100mM Tris(pH12.5)で溶解され、続いて塩酸によるpH8への調整を経た大腸菌細胞由来の残存ペレット。各レーンにおいて、17kDaに見られるタンパク質のバンドは、細胞を消化する目的で添加されたリゾチームである。
【図20】図20は、実施例10に記載の、化学合成された酢酸オクトレオチドの3つの炭素の架橋PEG化を示す。比較のために、酢酸オクトレオチドのMALDI−TOF−MSスペクトルを図15に示す。図20は、化学合成された酢酸オクトレオチドが、ジスルフィド架橋を横切る3つの炭素の架橋を介して、5kDaのポリエチレングリコールに共有結合していることを示す。このPEG化された酢酸オクトレオチドの理論質量は6092Da(すなわち、ポリエチレングリコール5073Da+酢酸オクトレオチド1019Da)である。ポリエチレングリコールの実測分子量は5073Daである。PEG化された酢酸オクトレオチドの実測分子量は6097Daである。質量誤差百分率は0.08%である。
【図21】図21は、実施例11に記載のように、SDS−PAGEを用いて、ポリエチレングリコールが共有結合する3つの炭素の架橋の化学的な挿入によるSjGST−TM−オクタペプチド融合タンパク質(バージョン1)の修飾を示す。レーン1:分子量マーカー;レーン2:27757Daの理論分子量を有する、精製されたSjGST−TM−オクタペプチド(バージョン1);レーン3:4℃にて72時間のSjGST−TM−OCTとの5Daのポリエチレングリコールのジスルフィド部位特異的な架橋反応とそれに続く37℃にて24時間のトロンビンによる消化。SjGST−TM−オクタペプチドのタンパク質バンド27.7kDaは消失し、モノPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化)、ジPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化、および、GST中のジスルフィド結合のモノPEG化)、並びに、トリPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化、および、GST中の2つのジスルフィド結合のジPEG化)が出現した;レーン4:反応バッファー中における5kDaのポリエチレングリコール試薬(3等量濃度)のジスルフィド部位特異的な架橋。
【図22】図22は、実施例12に記載の、MALDI−TOF−MS分析を用いて、ポリエチレングリコールが共有結合した3つの炭素の架橋の化学的な挿入によるSjGST−TM−オクタペプチド(バージョン1)の修飾を示している。この図は、トロンビン消化され、ジスルフィド部位特異的にPEG化されたSjGST−TM−オクタペプチド(バージョン1)タンパク質溶液のMALDI−TOF−MSを示す。SjGST−TM−オクタペプチド(バージョン1)融合タンパク質の理論質量は27757Daである。トロンビンにより開裂したSjGSTの大きい断片の理論質量は26150Daである。トロンビンによる理論上の小さい開裂産物は、1607Daの断片である。このことから、オクタペプチド(1607Da)に連結されたポリエチレングリコール(5073Da)の理論質量は6680Daとなる。オクタペプチドに連結したポリエチレングリコールの実測質量は6692Daであった。質量誤差は0.1%である。
【図23】図23は、BALB/Cマウスの血清成長ホルモン(GH)レベルの低下における本発明のペプチドの3つの実施例の効果を(化学合成された酢酸オクトレオチドと比較して)示す。図23AはマウスのGHレベル(ng/ml)を示す。図23Bは、化学合成された酢酸オクトレオチドと比較したGHの抑制百分率を示す。
【発明を実施するための形態】
【0036】
第1の形態において、本発明は、X1CX2WKX3CTのアミノ酸配列からなる単離されたペプチドを提供し、この際、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVであり、これらは任意の順列組み合わせであり、それぞれのアミノ酸はL配置であり、ペプチドは2つのシステイン残基の間にジスルフィド結合を含む。よって、本発明のペプチドは、以下のアミノ酸配列のいずれかを有しうる:FCFWKTCT、FCFWKVCT、FCYWKTCT、FCYWKVCT、ACFWKTCT、ACFWKVCT、ACYWKTCTまたはACYWKVCT。好ましくは、本発明のペプチドは、FCFWKTCTまたはACYWKVCTのアミノ酸配列からなる。
【0037】
本明細書において、アミノ酸配列は、標準的な一文字または三文字のアミノ酸表記を用いて規定され、G=グリシン(Gly)、P=プロリン(Pro)、A=アラニン(Ala)、V=バリン(Val)、L=ロイシン(Leu)、I=イソロイシン(Ile)、M=メチオニン(Met)、C=システイン(Cys)、F=フェニルアラニン(Phe)、Y=チロシン(Tyr)、W=トリプトファン(Trp)、H=ヒスチジン(His)、K=リジン(Lys)、R=アルギニン(Arg)、Q=グルタミン(Gln)、N=アスパラギン(Asn)、E=グルタミン酸(Glu)、D=アスパラギン酸(Asp)、S=セリン(Ser)、およびT=スレオニン(Thr)である。
【0038】
各アミノ酸についてのDNAコドンを表1に示す:
【0039】
【表1】
【0040】
各アミノ酸が(D配置とは逆の)L配置で存在し、2つのシステイン残基間にジスルフィド結合を含む本発明の単離されたペプチドは、化学合成された酢酸オクトレオチドと同一の三次元結合部位構造を保持しており、よって、化学合成された酢酸オクトレオチドと同一の活性を保持している。酢酸オクトレオチドがD−Phe1残基およびD−Trp4残基を含んでおり、酢酸ランレオチドがD−Ala1残基およびD−Trp4残基を含んでいるのに対し、本発明のペプチドは、すべてL−エナンチオマーからなっている。
【0041】
本発明のペプチドは、末端に化学修飾を含んでもよい。かような修飾によれば、例えば、プロテアーゼによる分解に対する感受性を低下させることなどにより、ペプチドの安定性が向上しうる。
【0042】
本発明のペプチドFCFWKTCTの分子モデリングによれば、その予測される三次元化学構造は化学合成された酢酸オクトレオチドのものと有意には異なっていないことが示されている。図3に示すように、ペプチドFCFWKTCTにおけるアミノ酸の相対位置は、化学合成された酢酸オクトレオチドのアミノ酸と同様の位置に配置されている。最も重要なこととして、Phe3、Trp4、Lys5およびThr6は、本発明のペプチドにおいて、酢酸オクトレオチドにおけるのと同一の相対位置に存在している。リジン5は異なる方向に位置するように見えるが、その直鎖状の性質からすれば溶液中では非常に柔軟であることになる。このことは、リジン5について図3に示す2つの位置の間の差異が生物学的には有意でないことを意味する。図3において標識されているアミノ酸は、オクトレオチドの生物活性に必須であることが知られている;それらは酢酸オクトレオチドのその細胞表面受容体への結合に関与しているのである。このように、本発明のペプチドFCFWKTCTは、酢酸オクトレオチドと同一の三次元化学結合部位の特性を有しており、分子モデリングの研究によれば、生体系において同様に作用すると予測されている。本発明のペプチドのそれぞれの三次元配座も、分子モデリング研究により、同様に酢酸オクトレオチドまたは酢酸ランレオチドと同一の三次元化学結合部位の特性を保持すると予測されている。
【0043】
本発明のペプチドは、好ましくは当該ペプチドをコードするポリヌクレオチドの生物学的宿主における発現により作製され、これによれば、ペプチドの生産について生物有機体のタンパク質発現系に依存することになる。よって本発明のペプチドは、酢酸オクトレオチドや酢酸ランレオチドを生産するのに用いられる化学合成法とは対照的に、生物学的に合成されることが好ましい。本発明のペプチドの生物学的な合成はまた、2つのシステイン残基間で天然にジスルフィド結合を形成することになる。よって、他の形態では、本発明は、本発明のペプチドをコードするポリヌクレオチドを提供する。当該ポリヌクレオチドは、好ましくは、本発明のオクタペプチドの8アミノ酸を、天然のホルモンであるソマトスタチンの残りのアミノ酸とは切り離してコードする。よって、ソマトスタチンをコードする天然のゲノムDNAは、本発明の範囲から除外される。上記ペプチドを発現可能な任意の生物学的宿主が用いられうる。適切な宿主は本技術分野において公知であり、細菌、酵母、昆虫細胞、および動物細胞が挙げられる。
【0044】
本発明のペプチドをコードするポリヌクレオチドは、DNAまたはRNAでありうる。よって、本明細書ではDNA配列を記載するが、等価なRNA配列も用いられうるものとする。
【0045】
本発明のペプチドFCFWKTCTをコードするポリヌクレオチドは、コアとなるヌクレオチド配列5’−TTYTGYTTYTGGAARACNTGYACN−3’を含み、この際、Y=CまたはTであり、R=AまたはGであり、N=A、C、T、またはGであり、任意の順列組み合わせが可能である。細菌中での発現のために上記ポリヌクレオチドをベクターに導入する際、ポリヌクレオチドの好ましい配列は、細菌での最も一般的なコドン使用頻度に基づき、5’−TTCTGTTTTTGGAAAACCTGTACC−3’である。ペプチドFCFWKTCTをコードする16個の異なるヌクレオチド配列を表2に示し、下線を付した配列は最も一般的に用いられている細菌のコドンに対応している。本発明のポリヌクレオチドは、表2に示す配列のいずれか1つを含みうるが、これらの配列には限定されない。ペプチドFCFWKTCTをコードするポリヌクレオチドを酵母宿主細胞中で発現させるときには、好ましいコドンは酵母において最も一般的なコドン使用頻度に従って選択されうる。
【0046】
【表2】
【0047】
ACYWKVCT、FCFWKVCT、FCYWKTCT、FCYWKVCT、ACFWKTCT、ACFWKVCTまたはACYWKTCTのいずれかのペプチドをコードするポリヌクレオチドは、それぞれ下記の表3〜9に示すコア配列を有する。表3〜9に示すコア配列において、Y=CまたはTであり、R=AまたはGであり、N=A、C、TまたはGであり、任意の順列組み合わせが可能である。各ペプチドをコードする16個の異なるヌクレオチド配列の例を表3〜9に示し、下線を付した配列が、最も一般的に用いられている細菌のコドンに対応する。本発明のポリヌクレオチドは、表3〜9に示す配列のいずれかを含みうるが、これらの配列には限定されない。ポリヌクレオチドが酵母宿主細胞中で発現する場合、好ましいコドンは、酵母における最も一般的なコドン使用頻度に従って選択されうる。
【0048】
【表3】
【0049】
【表4】
【0050】
【表5】
【0051】
【表6】
【0052】
【表7】
【0053】
【表8】
【0054】
【表9】
【0055】
本発明のペプチドをコードするポリヌクレオチドは、宿主細胞への形質転換のためのヌクレオチドベクター中に導入されうる。当該ベクターは、DNAベクターまたはRNAベクターでありうる。当該ベクターは、プラスミドベクターであってもよい。好ましくは、当該ベクターは、本発明のペプチドをコードするポリヌクレオチドに作動可能なように連結されたプロモーター配列を含む。適当なヌクレオチドベクターとしては、pGex4.3(GEヘルスケア)などの市販のプラスミドベクターが挙げられ、他の代表的な例を表3に示す。Hosfield T et al., (1998) Biotechniques, Vol. 25, pp. 306-309,di Guan et al., (1988), Gene, 67; 21-30、または、Makrides (1996), Microbiol Rev., Vol. 60; 512-538に記載の任意のベクターもまた、用いられうる。
【0056】
よって、本発明は、本発明のペプチドをコードする発現ベクターを提供する。本発明において用いられうる市販の発現系の例を表10に示すが、これらの例には限定されない。
【0057】
【表10】
【0058】
好ましくは、本発明のペプチドは、宿主細胞からのその単離を単純化する目的で、融合タンパク質の一部として発現する。融合タンパク質は、本発明のペプチドおよび1または2以上のキャリアタンパク質を含む単一のポリペプチドである。このキャリアタンパク質は、融合タンパク質が生産される発現系から単離されうる任意のタンパク質でありうる。
【0059】
本発明のペプチドを含む融合タンパク質は、検出されて宿主細胞から単離されうる任意のキャリアタンパク質を含みうる。好ましくは、このキャリアタンパク質は異種タンパク質(すなわち、宿主細胞で天然にはつくられないタンパク質)である。このキャリアタンパク質は分子タグとして機能し、本発明のペプチドの単離を可能にする。例えば、融合タンパク質は、グルタチオン−S−トランスフェラーゼ(GST)(GEヘルスケア)、ポリヒスチジンタグタンパク質(クロンテック)、マルトース結合タンパク質(ニューイングランドバイオラボ)、ビオチン化融合タンパク質(プロメガ)、カルモジュリン結合ペプチド(ストラタジーン)、β−ラクタマーゼ(ジェランティス)などの任意の市販のタンパク質タグ、および本技術分野において公知の他のものを(キャリアタンパク質として)含みうる。また、イオン交換クロマトグラフィによる融合タンパク質の精製とその後の適当な酵素を用いた消化を可能にするために、融合タンパク質にイオン電荷を付与する任意のタンパク質(例えば、C末端に導入されうるポリアルギニン)(Fuchs SM & Raines RT, (2005) Protein Science Vol. 14; pp 1538-1544)。
【0060】
よって、他の形態において、本発明は、本発明のペプチドおよび1または2以上のキャリアタンパク質を含む融合タンパク質を提供する。本発明はまた、本発明のペプチドおよびキャリアタンパク質を含む融合タンパク質をコードするポリヌクレオチドをも提供する。本発明の融合タンパク質をコードするポリヌクレオチドは、本明細書に記載の任意のベクター中に導入されうる。キャリアタンパク質をコードするヌクレオチド配列は、上述したコアヌクレオチド配列(または本発明のペプチドをコードする任意の他の配列)の5’末端のすぐ上流にインフレームで位置してもよいし、当該コア配列のすぐ下流に位置してもよい。あるいは、融合タンパク質をコードするヌクレオチド配列は、ペプチドとキャリアタンパク質との間のスペーサー領域をコードしてもよい。このスペーサー領域は、キャリアタンパク質の正しい折り畳みを可能とするようにペプチドとキャリアタンパク質との間に十分な空間を提供しうる。
【0061】
さらに、このスペーサー領域は、融合タンパク質の発現後にキャリアタンパク質からのペプチドの分離を可能とすべく、1または2以上の切断部位(1または2以上の酵素的または化学的な切断部位)をコードしてもよい。本発明のペプチドを含む融合タンパク質中に導入されうる酵素的な切断部位としては、(a)TEVプロテアーゼ切断部位(Glu-Asn-Leu-Tyr-Phe-Gln* X(ここで、Xは任意のアミノ酸(プロリンを除く)であり、*は酵素切断部位を表す));(b)エンテロキナーゼ切断部位(Asp-Asp-Asp-Asp-Lys*);(c)因子Xaプロテアーゼ切断部位(Ile-Glu/Asp-Gly-Arg*);(d)Arg−Cプロテイナーゼ切断部位(X-X-R*X-X-)(ここで、Xは任意のアミノ酸(プロリンを除く)であり、存在する場合には、ペプチドのN末端の上流が、切断時にいかなる他のN末端アミノ酸もなしに-Phe-Cys-Phe-Trp-Lys-Thr-Cys-Thrの配列を生成する);(e)タバコベインモットリングウイルス(TVMV)およびインテイン由来のプロテアーゼによる切断部位などの、本技術分野において公知の他のもの、が挙げられる。化学的な切断部位の適当な例としては、(a)シアノゲンブロマイドによる切断のためのMet;ギ酸による切断のためのAsp;および、(c)本技術分野において公知の他の切断部位。
【0062】
上述した切断用プロテアーゼの認識配列のDNAコドンを表11に示す。
【0063】
【表11】
【0064】
切断部位をコードするヌクレオチド配列は、本発明のペプチドをコードするコアヌクレオチド配列のすぐ5’に位置しうる。よって、ヌクレオチド配列GARAAYTTATAYTTYCAR(TEVプロテアーゼ切断部位をコードする;Y=CまたはTであり、RはAまたはGであり、任意の順列組み合わせが可能である)は本発明のペプチドをコードするコアヌクレオチド配列のすぐ5’に位置しうる。例えば、ペプチドFCFWKTCTおよびTEVプロテアーゼ切断部位をコードする本発明のポリヌクレオチドは、コアヌクレオチド配列5’-GARAAYTTATAYTTYCARTTYTGYTTYTGGAARACNTGYACN-3’を含むことができ、Y=CまたはTであり、R=AまたはGであり、N=A、C、TまたはGであり、任意の順列組み合わせが可能である。ポリヌクレオチドが細菌での発現用のベクター中に導入される場合、細菌での最も一般的なコドン使用頻度に基づき、好ましい配列は5’ GAA AAC CTG TAT TTT CAG TTC TGT TTT TGG AAA ACC TGTACC 3’である。酵母の好ましいコドンを含む他の配列が、酵母宿主中でタンパク質を発現させる際には用いられうる。融合タンパク質のペプチド部分をコードする他のヌクレオチド配列の例を、表2〜9に示し、下線を付した配列は、最も一般的に用いられている細菌のコドンに対応する。本発明の融合タンパク質をコードするポリヌクレオチドは、表2〜9に示す配列の任意のものを含みうるが、これらの配列には限定されない。本発明の融合タンパク質を生産するのに用いられうる適当な発現系および精製系としては、市販のpETシステム(ノバジェン)、Ni−NTA精製システム(キアゲン)、pMALタンパク質融合・精製システム(ニューイングランドバイオラボ)、ピンポイント(商標)Xaタンパク質精製システム(プロメガ)、CBPカルモジュリン結合ペプチドアフィニティタグシステム(ストラタジーン)、INTEIN(商標)およびINTEIN−TWINシステム(ニューイングランドバイオラボ)、EndoproteinAceシステム(ジェランティス)、および他のもの(例えば、表10を参照)が挙げられる。
【0065】
1982年に、生物活性断片の配座が安定したソマトスタチンの類似体として酢酸オクトレオチドが初めて化学合成されて以来、他のソマトスタチン類似体についての研究は専ら、化学合成される他のペプチド断片に集中していた(Weckbecker, G. et al., (2003) Nature Reviews, Vol. 2, pp. 999-1017)。本発明のペプチドの生物学的な生産により、(1)大腸菌等の宿主細胞中でジスルフィド結合が自然に形成され、(2)酢酸オクトレオチドの三次元の化学構造を保持しており、(3)酢酸オクトレオチドの細胞ベースの毒性(D−アミノ酸(大腸菌等の有機体の増殖を阻害する)の存在による(Meister, (1965) Biochemistry of the amino acids. New York Academic Press))の問題を解決し、(4)大腸菌に対して毒性を示さない封入体中でのペプチドの生産を可能とする、そのようなペプチドの安価な製造方法が提供される。したがって、本発明によれば、上記ペプチドの高収率で低コストでの生物学的な生産が可能となる。
【0066】
特にD−アミノ酸の生きている有機体における毒性効果について述べると、いくつかのD−アミノ酸(D−トリプトファンなど)は大腸菌に対して毒性を示しうることが知られている(Soutourina et al., (2004) J. Biological Chemistry; Vol 279(41): pp 42560 - 42565)。これは、D−アミノ酸は生物界に見られるものの、リボソームのタンパク質合成酵素はD−アミノ酸が天然のポリペプチド中には導入されないように保証しているためである。天然のポリペプチドからのD−アミノ酸の排除の第1段階は、アミノアシル−tRNA合成酵素により担われている。これは、D−アミノ酸が3つの異なる機構によって細胞毒性をもたらしていることによる。上記細胞毒性の第1の機構は、D−アミノ酸がポリペプチド中に導入されると機能しないタンパク質が生成するというものである。このことは、L−アミノ酸による細胞のタンパク質翻訳機構の立体特異性にも反映している。上記細胞毒性の第2の機構は、D−アミノ酸が12アミノ酸ベースのペプチド中に導入された実験によって示されている;トリチウム標識チミジンの生細胞への導入により測定されるように、D−アミノ酸は細胞成長および細胞増殖を有意に低下させることがわかっている(Hayry et al., (1995) The FASEB journal, Vol 9; pp 1336 - 1344)。上記細胞毒性の第3の機構は、ペルオキシソームのD−アミノ酸オキシダーゼ(D−AAO)が代謝および生きている有機体からの特定のD−アミノ酸の除去に有効であるということである。これは、D−AAOがD−アミノ酸を対応するα−ケトン酸およびNH3へと代謝することによる。このプロセスでは、補酵素であるフラビンアデニンジヌクレオチドの還元型から酸化型への再生により細胞毒性のある過酸化水素が生成し、次いでこれがより毒性の高い反応性酸素種を生成する。これらのフリーラジカルは、重篤な細胞障害の原因となる(Krug et al., (2007) Am J. Physiol. Renal Physiol. Vol 293; pp F382 - F390)。
【0067】
他の形態において、本発明は、本発明のペプチドの製造方法を提供し、当該方法は、本発明のポリヌクレオチドまたはベクターを、当該ポリヌクレオチドまたは当該ベクターを発現できる宿主細胞中に導入し、当該宿主細胞から当該ペプチドを単離することを含む。本発明のペプチドをコードするポリヌクレオチドまたは本発明の融合タンパク質をコードするポリヌクレオチドが用いられうる。宿主細胞中で本発明のペプチドを生産することにより、各アミノ酸はL配座で提供される。好ましくは、上記ポリヌクレオチドまたはベクターは、上述したように、本発明のペプチドおよびキャリアタンパク質を含む融合タンパク質をコードするヌクレオチド配列を含有する。よって、本発明のペプチドの精製工程は、好ましくは、宿主細胞から融合タンパク質を単離し、当該融合タンパク質から上記ペプチドを単離することを含む。
【0068】
本発明のペプチドおよび融合タンパク質は、単純に生産され単離されうるため、低コストである。本発明のペプチドまたは融合タンパク質をコードするポリヌクレオチドまたはベクターは、当業者に公知の任意の手段によって適切な宿主細胞中に導入されうる。宿主細胞の培養条件は、ペプチドまたは融合タンパク質が最高の収率で生産されるように最適化されうる。例えば、lacプロモーターの誘導体(tac、pac、rac)は最も強力な細菌のプロモーターであり、これらのいずれもが本発明のベクターに用いられうる。これらは、大腸菌における外来遺伝子の誘導された過剰発現によく用いられる。しかしながら、工業的な醗酵プロセスにこれらを用いることは、誘導剤であるイソプロピル−β−D−チオガラクトピラノシド(IPTG)のコストが高いことにより制限される。本発明の方法での使用に適した大規模培養系では誘導剤としてラクトースを用いることができる。これは、ラクトースがIPTGと同等の効率で本発明のベクターを誘導することができるためである。大腸菌では、誘導剤としてラクトースを用いることができるし、グルコースの存在下であっても炭素/エネルギー源として用いることができる(Neubauer P. et al., (1994) FEMS Microbiol Rev. Vol. 14; pp. 99-102; Vasala A. et al., (2005) J. Biotechnol. Vol. 117; pp. 421-431)。この低コストのアプローチを、改変培地および改変増殖条件を利用した自己誘導と組み合わせてもよい(Studier FW., (2005) Protein Expression and Purification Vol. 41; pp. 207-234)。大腸菌およびラクトースを用いたSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)の高収率で低コストの自己誘導と、それに続く大腸菌中の封入体からの融合タンパク質の単離の例を、実施例9および図19に示す。
【0069】
ペプチドまたは融合タンパク質の化学的な修飾を、ペプチドの安定性を改善し、ペプチドの単離方法を単純化させる目的で、ペプチドまたは融合タンパク質の発現の後に行なってもよい。
【0070】
次いで、本技術分野において公知の手法により、ペプチドは宿主細胞の成分から単離されうる。ペプチドが融合タンパク質の一部として発現している場合には、当該ペプチドは本技術分野において公知の手法により、融合タンパク質から単離されうる。特に注目すべきは、封入体からのタンパク質の単離である。なぜならば、これによってかなりの製造上および経済的なプロセスの利点があるためである。これらの利点としては:(a)可溶性のタンパク質として大量に存在すると宿主細胞に対して毒性を示すタンパク質が高レベルで発現するが、封入体として存在することで毒性を示さないこと;(b)封入体のサイズおよび密度が細胞の混入物質とは異なることで、細胞からの封入体の単離が容易であること;(c)細胞のプロテアーゼによるタンパク質分解の攻撃に対して抵抗性であることにより、発現したタンパク質の分解が防止されること;並びに(d)混入物質がより少なく、封入体中で対象タンパク質が均一であること、が挙げられる。
【0071】
封入体を単純に洗浄すると不純物が除去され、天然タンパク質の収率はより改善される。これらの利点により、タンパク質の商業的な生産のために大腸菌中の封入体として発現する組換えタンパク質が広く開発されている。封入体からの生物活性なタンパク質を回収する際には、タンパク質の凝集体を充分に可溶化し、次いで、可溶化したタンパク質を生物活性な形態へとリフォールディングする必要がある。封入体中のタンパク質は折り畳みの過程の中間段階で存在し、かなりの量の二次構造を有していることが示されている。この天然様の二次構造を乱すことなく封入体からのタンパク質を可溶化できれば、リフォールディング時のタンパク質凝集の程度も低くなり、生物活性のタンパク質の回収率も高くなる。したがって、生物活性タンパク質の回収率の改善には、ランダムコイルのタンパク質構造を生じることなく封入体の凝集物をマイルドに可溶化させることが重要である。1つのアプローチとして、タンパク質の等電点から離れたpHショックをタンパク質凝集体に与えることである。これにより、極めて低濃度の変性剤の存在下でタンパク質は可溶性となる。封入体のタンパク質がかようなマイルドな条件下でひとたび可溶化されれば、それに続く精製タンパク質のリフォールディングも容易となり、生物活性タンパク質の回収率も高くなる。これに関連して、2M尿素ではタンパク質の構造が開かれず、タンパク質の天然の二次構造が保存されることが示されている(Rathore A.S. et al., (2003) Biotechnology Progress Vol. 19; pp. 1541-1546; Lee Y.S. et al., (2003), Biotechnology & Applied Biochemistry Vol. 38; pp. 9-13; DeNardo, S.J. et al., (2003) Clinical Cancer Research Vol. 9; pp. 3854s-3864s; Pezza J.A. et al., (2004) Chemical Communications pp. 2412-2413; Singh S.R. & Panda AK. (2005) Journal of Bioscience and Bioengineering. Vol. 99; pp. 303-310; Rajan R.S. et al., (2006) Protein Science Vol. 15; pp. 1063-1075; Wang F. et al., (2008) Nuclear Medicine and Biology Vol. 35; pp. 665-671)。よって、本発明のペプチドの製造方法は、本発明のペプチドまたは本発明の融合タンパク質を、宿主細胞中の封入体から単離することを含みうる。
【0072】
また、従来の液相でのリフォールディングプロセスの際のタンパク質凝集の問題は、膨張層吸着クロマトグラフィーと組み合わせた固相でのリフォールディング手法を用いることで回避されうる。その結果、細胞ホモジネートからの封入体中のタンパク質は、高収率で正確にリフォールディングされる(Cho T.H. et al., (2002) Bioseparation Vol. 10, pp. 189-196)。このことは、大腸菌において生産されるヒト上皮成長因子について示されている。膨張層吸着クロマトグラフィーは、カチオン交換によって同時にタンパク質を捕捉し、希釈培地から細胞バイオマスを除去する。また、ハイスループットで行うこともでき、その結果、高収率(>90%)となり、精製因子も80%超の純度で20倍となる。かようなタンパク質の精製プロセスは効率的であり、例えば本発明のペプチドの大規模かつ費用対効果の高い製造に採用されうる(Lee Y.S. et al., (2003), Biotechnology & Applied Biochemistry Vol. 38; pp. 9-13)。
【0073】
融合タンパク質および/または化学修飾された融合タンパク質を単離した後、酵素的または化学的な切断部位を介してこれを切断してもよい。例えば、TEV酵素切断部位を含む融合タンパク質の部位特異的な切断は、ヒスチジンタグ付きのTEV酵素を用いて行われうる。ニッケルアガロースカラムを通過させてヒスチジンタグ付きのTEVプロテアーゼを除去することにより、ペプチドを精製することができる。グルタチオンアガロースおよびニッケルアガロースカラムを再生させることで、これらを再利用することができる。それぞれの手法についての個別の工程に関する完全な詳細は、当業者に周知である(Structural Genomics Consortium et al., (2008) Nature Methods, Vol. 5(2), pp. 135-146)。
【0074】
上述したように、本発明のペプチドは、その安定性を改善する目的で化学的に修飾されてもよく、これによりペプチドの半減期が延びて治療効果も改善される。かような修飾により、上記ペプチドは、1年またはそれ以上の期間にわたる使用に適した長期療法として用いられうる、費用対効果に優れた治療用製品に特に適したものとなる。本発明の修飾されたペプチドの有効性が改善されることで、酢酸オクトレオチドまたは酢酸ランレオチドにより可能な現行の投薬計画と比較して、より長期間にわたって患者に投与するのに特に適したものとなる。
【0075】
例えば、下記の文献に詳細に記載されているジスルフィド部位特異的なPEG化の手法などにより、ペプチドおよび/または融合タンパク質をPEG化してもよい:Shaunak et al., (2006) Nature Chemical Biology. Vol. 2, pp. 312-313; Zloh et al., (2006) Physical Chemistry., Vol. F-3-O, pp. 347-349; Brocchini et al., (2006) Nature Protocols, Vol. 1(5), pp. 2241-2252 ; Balan et al., (2007) Bioconjugate Chemistry, Vol. 18, pp. 61-76; Godwin et al., (2007) Theoretical Chemical Accounts, Vol. 117, pp. 259-265; Zloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083 ; Brocchini et al., (2008), Advanced Drug Delivery Reviews, Vol. 60(1), pp. 3-12。ポリエチレングリコールのサイズは、5kDa〜40kDaでありうる。好ましいサイズは、20kDa〜30kDaである。
【0076】
小さいタンパク質およびペプチドは、大きいタンパク質よりも多くのジスルフィド結合を有する傾向にある。これは、前者が自信の比較的少ない疎水相互作用を補償する必要があるためである。溶媒に接近できるジスルフィド結合は通常ほとんどのタンパク質に存在することから、このジスルフィド結合を化学的に減らして2つの遊離したシステインの硫黄原子としつつ、タンパク質またはペプチドの三次構造を維持することが可能である。次いで、PEG化はこれらの2つのシステイン硫黄原子を3炭素の架橋を介して再結合させるビスアルキル化によって達成されうる。このアプローチの利点は、遊離システインを導入する(これはタンパク質またはペプチドの不溶性を上昇させる)ためのタンパク質の組換え的な改変を必要とすることなく、選択的かつ効率的なチオールの付加反応が行われうることである。より詳細には、このアプローチ手法は、すべての天然のジスルフィド結合において、双方の硫黄原子の化学的な反応性を利用するものである。
【0077】
2つのシステイン残基の2つの硫黄原子の間に3炭素の架橋を挿入する予備的な工程により、(a)安定性が向上するという特有の利点を有し、(b)上記3炭素の架橋その後のさらなる修飾の潜在性を付与する、修飾ペプチドが得られる。
【0078】
よって、本発明はまた、X1CX2WKX3CTのアミノ酸配列からなる単離されたペプチドを提供し、この際、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVであり、任意の順列組み合わせが可能であり、それぞれのアミノ酸はL配置であり、2つのシステイン残基の2つの硫黄原子は3つの炭素原子を介して連結されている。よって、当該ペプチドは、以下のアミノ酸配列のいずれかを有しうる:FCFWKTCT、FCFWKVCT、FCYWKTCT、FCYWKVCT、ACFWKTCT、ACFWKVCT、ACYWKTCTまたはACYWKVCT。好ましくは、上記ペプチドは、FCFWKTCTまたはACYWKVCTのアミノ酸配列からなる。こうして、本発明のペプチドにおける2つのシステイン残基の間には安定化相互作用が働く。例えば、ジスルフィド結合に3炭素の架橋を挿入することで修飾して、さらなる安定化相互作用としてもよい。
【0079】
本発明者らによる分子モデリング研究により、2つのシステイン残基の2つの硫黄原子の間に3炭素原子を含むFCFWKTCTペプチドは、化学合成された酢酸オクトレオチドにきわめて類似した三次元配座を保持していることが示されている(図6を参照)。したがって、かようなペプチドもまた、未修飾の、生物学的に合成された本発明のペプチドについて本明細書に記載されたのと同一の用途(特に、医療用途)に用いられうる。2つのシステイン残基の2つの硫黄原子の間に3炭素原子を含む本発明のペプチドのそれぞれの三次元配置もまた、分子モデリング研究により、酢酸オクトレオチドまたは酢酸ランレオチドと同一の三次元化学結合部位の特性を保持していることが予測されている。したがって、かようなペプチドは、酢酸オクトレオチドまたは酢酸ランレオチドと同一の活性を保持している。
【0080】
ポリエチレングリコール分子などの親水性ポリマーが、3炭素架橋に共有結合していてもよい。好ましくは、ポリエチレングリコール分子は5kDa〜50kDaの分子量を有し、より好ましくは10kDa〜40kDaの分子量を有し、最も好ましくは20kDa〜30kDaの分子量を有する。あるいは、当業者に公知のポリエチレングリコール以外の親水性ポリマーが、3炭素架橋に共有結合していてもよい。
【0081】
PEG化試薬は、好ましくは、ポリエチレングリコール試薬の末端の共役部位として、置換されたプロペニル基を有する。この共役部位は、電子吸引性基(例えば、カルボニル)、α,βー不飽和二重結合、およびスルフィン酸として脱離しやすいα,β−スルホニル基を含みうる。電子吸引性基は、チオール付加を促進し、αプロトンのpKaを低下させて、これにより脱離反応が進行しうる。このような化学的な官能性の並置により、潜在的な交差共役系が生じる。ポリエチレングリコールモノスルホン中の共役二重結合は、一連の双方向的で連続した付加−脱離反応を引き起こす。第1のチオレートが付加するとスルホン酸誘導体の脱離が生じうる。これにより、第2のチオレートの付加のためのα,β’位に位置する別の共役二重結合が生じる。これらの2つのチオールがタンパク質のジスルフィド結合由来であれば、もともとのジスルフィドのシステインの硫黄原子間に3炭素の架橋が形成される。この新たな3炭素のジスルフィド架橋は、元の生物学的に合成されたジスルフィド結合よりも、化学的なおよび代謝による分解、並びにプロテオリシス分解に対してずっと安定である。
【0082】
本発明のPEG化ペプチドの調製方法は、2〜3の工程で低コストで行われうる。本発明のペプチドを含む融合タンパク質は、生産された後に続いて上述したようにPEG化されうる。次いで、この融合タンパク質は、イオン交換カラムまたはサイズ排除カラムまたはGST−融合タンパク質用のグルタチオンアガロースアフィニティー精製カラムなどのアフィニティーカラムに溶液を通すことで、未反応のポリエチレングリコールから精製されうる。後者の場合、GST−融合タンパク質はグルタチオンアガロースカラムに結合するようになる。次いで、(例えば、50カラム体積の0.1%トライトンX−114含有PBSとそれに続く20カラム体積のPBSを用いて)カラムを洗浄することで、エンドトキシンが除去されうる。エンドトキシンは洗浄バッファー中に溶出する。好ましくは、これらすべての操作が4℃で行われる。
【0083】
本発明のペプチドのPEG化は、合成されうる化学合成酢酸オクトレオチドの異なるPEG化体の混合物と比較して、好ましくはモノPEG化ペプチドを生成するものである。PEG化タンパク質は、サイズ排除クロマトグラフィーにより、より小さい非PEG化ペプチドから分離されうる。
【0084】
従来公知の発表によると、分子モデリング研究は、化学合成の結果および生物学的アッセイの結果を高い信頼性で予測するのに、化学に基づく実験と有効に組み合わされうる;Shaunak et al., (2006) Nature Chemical Biology. Vol. 2, pp. 312-313; Zloh et al., (2006). Physical Chemistry., Vol. F-3-O, pp. 347-349; Brocchini et al., (2006) Nature Protocols, Vol. 1(5), pp. 2241-2252 ; Balan et al., (2007) Bioconjugate Chemistry, Vol. 18, pp. 61-76; Godwin et al., (2007) Theoretical Chemical Accounts, Vol. 117, pp. 259-265; Zloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083 ; Brocchini et al., (2008), Advanced Drug Delivery Reviews, Vol. 60(1), pp. 3-12。このように、モデリング研究によれば、タンパク質またはぺぷの接近可能なジスルフィド結合への3炭素の架橋の挿入の構造的な効果を高い信頼性で予測することができる。公知のプロトコールは、タンパク質データベース(例えば、タンパク質データバンク(Protein Data Bank, PDB - www.pdb.org))に対して照会を行い、タンパク質の表面に近く、化学的に修飾されうる少なくとも1つのジスルフィド結合を有するタンパク質を発見するために開発されてきた。また、上記プロトコールは、3炭素の架橋によってタンパク質の三次構造が失われてしまうか否かを計算的に予測するのにも用いられうる。このアプローチを用いて、3炭素の架橋の挿入後のタンパク質の生物活性な表面の変化を正確に決定したり、3炭素の架橋へのポリエチレングリコールの連結がタンパク質の生物活性に及ぼす効果を決定したりすることができる。本明細書に記載の分子モデリング研究は、統合された分子モデリングパッケージであるMaestro v6.5およびMacromodel v9.1を用いて行われた。
【0085】
従来、ポリエチレングリコールが溶液中に存在すると、そのアルブミンとの負の優先相互作用のために、ヒトアルブミンの天然のコンパクトな状態を安定化することが知られている。ポリエチレングリコール分子とアルブミンとの間の相互作用は熱力学的に不利であることから(Farruggia et al., (1999), Int. J. Biol. Macromol., Vol. 26, pp. 23-33)、ポリエチレングリコールはタンパク質またはペプチドの生物活性な表面を阻害しないと考えられる。このことは計算的に確認されている;ポリエチレングリコールは、本発明のペプチドとは独立して折り畳まれる。ポリエチレングリコール分子は、タンパク質の周りで自身を「包む」ことはない(Zloh et al., (2006) Proceedings of 6th European Conference on Computational Chemistry, Slovakia)。しかしながら、本発明のペプチドとポリエチレングリコールとのサイズは有意に異なるため、当業者であれば、ポリエチレングリコールがペプチドを巻き込み、それによりペプチドの生物活性を阻害するものと予測していた。本発明者らは、驚くべきことに、本発明のペプチドのPEG化はペプチドの生物活性を阻害しないことを示したのである。
【0086】
PEG化のプロセスは通常、精製されたタンパク質を用いて行われる。これには、所望のPEG化タンパク質を回収するための複数工程の精製プロセスをさらに必要である。インビトロでのタンパク質のリフォールディングおよびPEG化が統合されれば、特に封入体のタンパク質については生産物の生物学的処理に有利であることが知られている。生物活性でPEG化されたタンパク質を封入体から直接得る目的でPEG化をタンパク質の再生プロセスと統合すると、下流の処理能力が有意に改善される。このことは、PEG化リパーゼについて実証されている(Kim M.Y. et al., (2007) Journal of Biotechnology Vol. 131, pp. 177-179; Choi W.C. et al., (2005) Process Biochem. Vol. 40, pp. 1967-1972)。このリパーゼは、尿素およびDTTを用いて変性され、次いでポリエチレングリコールで修飾されていた。この共役ポリエチレングリコール分子はリパーゼのリフォールディングを邪魔しなかった。よって、PEG化およびタンパク質のリフォールディングのプロセスを単一の処理工程へと統合することができた。換言すれば、封入体からの可溶化タンパク質は、単一の処理操作でPEG化され、リフォールディングされうるのである。同様の手法が本発明にも適用されうる。よって、本発明のPEG化ペプチドの製造方法は、宿主細胞中の封入体からの本発明のペプチドまたは融合タンパク質の単離と、それに続く統合されたPEG化およびPEG化ペプチドのリフォールディングとを含みうる。本発明のペプチドまたは融合タンパク質にポリエチレングリコールが共有結合すると、ペプチドまたは融合タンパク質の溶解性がかなり向上し、それによってタンパク質の凝集体の形成が抑制されるというさらなる利点もある(Rajan RS., (2006) Protein Science Vol: 15; pp. 1063-1075)。酢酸オクトレオチドおよび酢酸ランレオチドの不溶性は、工業的な製造上の主要な課題である。
【0087】
他の形態において、本発明は、医薬に用いるための、本発明のペプチドまたはPEG化ペプチドを提供する。本発明のペプチドまたはPEG化ペプチドは、ホルモン障害、特に成長ホルモンの過剰発現に起因するホルモン障害の治療に用いられうる。本発明のペプチドまたはPEG化ペプチドを用いて治療されうるホルモン障害の例としては、末端肥大症および巨人症が挙げられる(S. Farooqi et al., (1999) Pituitary, Vol. 2, pp. 79-88; A.N. Paisley and P.J. Trainer, (2003) Current opinion in Pharmacology, Vol. 3, pp. 672-677; S. Melmed, (2006) New England Journal of Medicine, Vol. 355, pp. 2558-2573)。
【0088】
また、本発明のペプチドまたはPEG化ペプチドは、癌の治療に、特に消化管関連のカルシノイド腫瘍や転移性のカルシノイド腫瘍などの消化管ホルモン分泌癌の治療に用いられうる。
【0089】
本発明のペプチドまたはPEG化ペプチドはまた、酢酸オクトレオチドまたは酢酸ランレオチドが治療上有効であることが知られている任意の疾患または症状の治療にも用いられうる。例えば、本発明のペプチドまたはPEG化ペプチドは、血管作動性腸管ペプチドを分泌する腫瘍(VIPoma)を有する患者における下痢の治療に、および、他の原因による重篤で難治性の下痢の治療にも用いられうる。本発明のペプチドまたはPEG化ペプチドはまた、スルホニルウレアの過剰摂取後の長期にわたる再発性低血糖の治療に、膵島細胞症の乳幼児におけるインスリンの過剰分泌の抑制を補助するのに、および、食道動脈瘤が疑われる患者における出血の抑制を補助するのにも、用いられうる。本発明のペプチドまたはPEG化ペプチドはまた、慢性膵炎による疼痛を有する患者の治療や、胸腺腫瘍、および、糖尿病に起因する眼疾患の治療にも用いられうる(S. Farooqi et al., (1999) Pituitary, Vol. 2, pp. 79-88; A.N. Paisley and P.J. Trainer, (2003) Current opinion in Pharmacology, Vol. 3, pp. 672-677; S. Melmed. (2006) New England Journal of Medicine, Vol. 355, pp. 2558-2573)。
【0090】
よって、本発明は、ホルモン障害(末端肥大症および巨人症);癌(消化管関連のカルシノイド腫瘍や転移性のカルシノイド腫瘍など);血管作動性腸管ペプチドを分泌する腫瘍(VIPoma)を有する患者における下痢;重篤で難治性の下痢;スルホニルウレアの過剰摂取後の長期にわたる再発性低血糖;膵島細胞症の乳幼児におけるインスリンの過剰分泌;食道動脈瘤が疑われる患者における出血、の治療に用いるための、本発明のペプチドまたはPEG化ペプチドを提供する。
【0091】
放射性標識されたソマトスタチン類似体は、ソマトスタチン受容体シンチグラフィーの有用な診断ツールである。上記類似体は、微小なSST発現腫瘍を検出・位置決めするのに用いられ、原発性および転移性の内分泌腫瘍をピンポイントで特定するのに特に有用である。標的化放射線療法の毒性により、現在では並行した開発の場により、SST発現腫瘍を有する患者を治療するための放射性標識されたソマトスタチン類似体の使用が検討されている。内分泌腫瘍の特異的な適応症として、イットリウム90(OctreoTher(登録商標))またはルテチウムなどのエネルギー性のアイソトープを含む新たな化合物は、放射線の深部への侵入により、腫瘍を有効に治療できるものと期待されている。イットリウム標識された化合物であるDOTATOCはまた、患者の60%で疾患を安定化させ、患者の20%では他覚的反応を示すことも判明している(Weckbecker, G. et al., (2003) Nature Reviews, Vol. 2, pp. 999-1017; Susini C & Buscail L, (2006) Ann of Oncology Vol. 17, pp. 1733-1742; Asnacios A et al. (2008) J. Clin. Oncology Vol. 26, pp. 963-970)。本発明はまた、腫瘍可視化剤として用いるための、本発明のペプチドまたはPEG化ペプチドをも提供する。例えば、放射性標識などの検出可能な標識を、2つの硫黄原子間に3炭素の架橋を有する本発明のペプチドに結合させることができ、これにより当該ペプチドが受容体に結合したときに、腫瘍部位において標識ペプチドを検出することが可能となる。上記標識は、上記3炭素の架橋に直接結合してもよいし、上記3炭素の架橋に結合したポリエチレングリコール分子に結合してもよい。本技術分野において公知の放射性標識以外の検出可能な標識もまた、用いられうる。
【0092】
さらに、本発明は、ホルモン障害(末端肥大症および巨人症);癌(消化管関連のカルシノイド腫瘍や転移性のカルシノイド腫瘍など);血管作動性腸管ペプチドを分泌する腫瘍を有する患者における下痢;重篤で難治性の下痢;スルホニルウレアの過剰摂取後の長期にわたる再発性低血糖;膵島細胞症の乳幼児におけるインスリンの過剰分泌;食道動脈瘤が疑われる患者における出血;並びに、糖尿病に起因する眼疾患、の治療のための医薬の製造における、本発明のペプチドまたはPEG化ペプチドの使用を提供する。
【0093】
本発明はまた、上述した疾患のいずれかに罹患した患者の治療方法であって、本発明のペプチドまたはPEG化ペプチドを患者に投与することを含む治療方法をも提供する。
【0094】
本発明はまた、本発明のペプチドまたはPEG化ペプチドおよび製薬上許容されうる担体または希釈剤を含む医薬組成物をも提供する。本発明のペプチドまたは当該ペプチドのPEG化体は、酢酸塩まどの製薬上許容されうる塩として提供されてもよい。上記医薬組成物は、患者に対する所望の投与方法に応じて、任意の適当な形態でありうる。上記医薬組成物は、単回用量製剤であってもよいし、通常は密封容器中で提供され、キットの一部として提供されてもよい。かようなキットは通常(必ずしもそうとは限らないが)、使用説明書を含む。当該キットは、複数の上記単回用量製剤を含みうる。上記医薬組成物は、上述した障害のいずれかの治療用に製剤化されうる。推奨される治療上の用量は、300〜600μg/日または20mg/月であり、2〜200mg/月の範囲である。上記医薬組成物は、毎日、週3回、週1回、2週に1回、または月1回で投与されうる。
【0095】
上記医薬組成物は、例えば、非経口(皮下、筋肉内、静脈内もしくは皮内など)、経口(バッカルもしくは舌下など)、呼吸器経由の、直腸経由の、経鼻、または経皮の経路により、任意の適当な経路による投与に適用されうる。皮下の投与経路が好ましい。かような組成物は、例えば滅菌条件下で活性成分を担体または賦形剤と混合するといった薬学の技術分野において公知の任意の手法により調製されうる。
【0096】
非経口投与に適用される医薬組成物としては、抗酸化剤、バッファー、静菌薬および製剤を非投与者の血液または組織と実質的に等張にしうる溶質を含有してもよい水性または非水性の滅菌注射液;並びに、懸濁化剤および増粘剤を含有してもよい水性または非水性の懸濁剤が挙げられる。注射溶液に用いられうる賦形剤としては、例えば、水、アルコール、ポリオール、グリセリンおよび植物油が挙げられる。上記組成物は、単回用量または複数回用量の容器(例えば、密封されたアンプルまたはバイアル)中に提供されることができ、使用直前に、例えば注射水といった滅菌液体を添加するだけでよい凍結乾燥条件下で貯蔵されうる。即時調製用の注射液および懸濁液は、滅菌された粉末、顆粒、および錠剤から調製されうる。
【0097】
経口投与用の医薬組成物は、カプセルまたは錠剤として;粉末または顆粒として;溶液、シロップまたは懸濁液として(水性または非水性の液体中で;または食用のフォームもしくはホイップとして;またはエマルジョンとして)、提供されうる。錠剤または硬ゼラチンカプセル用の適当な賦形剤としては、ラクトース、トウモロコシでんぷんまたはその誘導体、ステアリン酸またはその塩が挙げられる。軟ゼラチンカプセル用の適当な賦形剤としては、例えば、植物油、ワックス、脂肪、半固体もしくは液状のポリオールなどが挙げられる。液剤およびシロップの調製に用いられうる賦形剤としては、例えば、水、ポリオールおよび糖が挙げられる。懸濁液の調製には、水中油または油中水型の懸濁液を提供するのに油脂(例えば、植物油)が用いられうる。
【0098】
経皮投与に適用される医薬組成物は、長期間にわたって被投与者の上皮とじかに接触したままとされることを前提とした任意のパッチとして提供されうる。例えば、活性成分は、Pharmaceutical Research, 3(6):318 (1986)に概説されるようなイオントフォレシスによりパッチから送達されうる。
【0099】
本発明の方法において用いられうる他の製剤は、経皮送達デバイス(「パッチ」)を用いる。かような経皮パッチは、本発明のペプチドまたはPEG化ペプチドを制御された量で連続的または非連続的に注入するのに用いられうる。医薬製剤の送達のための経皮パッチの構築および使用については、本技術分野において周知の事項である。かようなパッチは、連続的な、パルス状の、または必要に応じた医薬製剤の送達用に構築されうる。
【0100】
他の製剤は、本発明のペプチドまたはPEG化ペプチドの制御された徐放を可能とする生分解性のマイクロスフェアを利用するものである。かような製剤は、合成のポリマーまたはコポリマーまたはハイドロゲルのインプラントを含みうる。かような製剤は、注射、吸入、経鼻または経口の投与が可能である。医薬製剤の送達のための生分解性マイクロスフェアの構築および使用については、本技術分野において周知の事項である(例えば、米国特許第6706289号)。
【0101】
また、本発明によるジスルフィド部位特異的なPEG化オクタペプチドを生分解性マイクロスフェアに配合することで、異なる放出パターン(例えば、オクタペプチドの低減された初期の放出とそれに続く長期にわたる安定した持続的な薬物の放出)および分解するマイクロスフェア内でのオクタペプチドの不活化に対する安定性を有する非常に有効でかつ効率的な徐放性システムが得られる。
【0102】
皮下、静脈内、筋肉内など用に適した製剤のための、適当な医薬担体並びに製剤化および投与の技術は、本技術分野において周知の手法のいずれかにより準備されうる(Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Pa., 20th edition, 2000を参照)。
【0103】
直腸投与に適用される医薬組成物は、坐剤または浣腸剤として提供されうる。経鼻投与に適用される医薬組成物(担体が固形状の場合)は、鼻の近くに置かれた粉末の容器から嗅がれるように(すなわち、鼻道を介した急速吸入により)投与される例えば20〜500ミクロンの粒子径を有する崩壊性の粉末を含む。経鼻スプレーまたは経鼻ドロップとしての投与のための、担体が液状の場合の適当な組成物としては、活性成分の水溶液または油の溶液が挙げられる。吸入による投与に適用される医薬組成物としては、種々のタイプの定量圧縮エアロゾル、ネブライザーまたは吸入器により生じる微粒子のダストまたはミストが挙げられる。
【0104】
上記医薬組成物は、保存料、可溶化剤、安定化剤、湿潤化剤、乳化剤、甘味料、着色料、着香料、塩、バッファー、被覆剤または抗酸化剤を含有してもよい。上記医薬組成物はまた、本発明のペプチドに加えて、治療上の活性剤を含有してもよい。
【0105】
本発明の物質の用量は、治療される疾患または障害、治療される個体の年齢および症状などに応じて広い範囲で変動する可能性があり、最終的には医師が用いられる適当な用量を決定する。この用量は適当な回数だけ繰り返されてもよい。仮に副作用が発生した場合には、通常の臨床実務に従って用量の量および/または頻度を減らすこともできる。
【0106】
また、本発明は、上述した疾患のいずれかに罹患した患者の治療方法であって、本発明の医薬組成物を患者に投与することを含む治療方法を提供する。
【0107】
本発明のそれぞれの形態の好ましい特徴は、変更すべきところは変更した上で、他の形態のそれぞれに対しても同様に好ましい。本明細書に記載の先行技術文献は、法が許す限り最大限に引用される。
【実施例】
【0108】
以下の非限定的な実施例を参照することにより、本発明をより詳細に説明する。
【0109】
実施例1:本発明のペプチドの予測構造の分子モデリング
これらの実験には、公知(Zloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083)の分子モデリングプロトコールを用いた。PDBのファイルをMaestroにインポートし、適当なリンカーおよび10kDaのポリエチレングリコール鎖を有する3炭素の架橋の挿入により、ジスルフィド結合を修飾した。このモデルを最小化し、Macromodelソフトフェア、OPLS-2005力場、およびGB/SA暗溶媒モデルを用いて、300Kでの2ピコ秒の確率的シミュレーションに供した。構造のスナップショットについて得られた軌跡を分析した。それによれば、生物学的に合成されたオクタペプチドは10kDaのPEGによって包まれるようにはならないことが示唆された(図4および図5)。このことは、重要なことに、生物学的に合成されたオクタペプチドの生物活性な表面は依然として自身の生物学的な受容体と相互作用することが可能であることを意味する。
【0110】
生物学的に合成されたオクタペプチドは、シミュレーション中の骨格原子の標準偏差(RMSD)の値を記録することによりモニターされうる柔軟度を示す(図6)。RMSDの値の範囲は、1〜4.4Åである(図7)。これにより、3炭素の架橋を介してポリエチレングリコールが連結しても、オクタペプチドの柔軟性が損なわれないということが示される。したがって、このジスルフィド部位特異的なPEG化オクタペプチドは、依然として自身の生物学的な重合体結合部位と効果的に相互作用することができる程度に充分に柔軟なままなのである。
【0111】
生物学的に合成されたオクタペプチドの配座にPEG化が及ぼす影響を、生物学的に合成されたオクタペプチドおよびジスルフィド部位特異的なPEG化オクタペプチドについての配座探索を用いて詳細に評価した。2つの硫黄原子間の距離は2.04Å(生物学的に合成されたオクタペプチド)から3.94Å(ジスルフィド部位特異的なPEG化オクタペプチド)へと増加したとしても、これら2つのペプチドの最低の(すなわち、最も安定な)エネルギーは、当該オクタペプチドの環状部位の骨格原子と標準偏差0.58Åで類似していることが判明した。また、ジスルフィド部位特異的なPEG化オクタペプチドの受容体との相互作用に関与するアミノ酸の方向は極めて類似している(図8)。このことから、大きなポリエチレングリコール分子が連結したときに、オクタペプチド上の生物学的に関与する表面は依然として生物学的に関与する受容体への提示に用いられうるということが示唆される。
【0112】
これらのモデリング研究の結果から、生物学的に合成されたオクタペプチドのジスルフィド結合上にポリ(エチレングリコール)分子が存在しても、当該オクタペプチドの受容体結合部位との相互作用は阻害されないことが示される。
【0113】
実施例2:生物学的に合成されるオクタペプチド(バージョン1)用のDNA配列を有するpGex4.3プラスミドの作製
構築した第1のプラスミドは、N末端からC末端に向かって、以下の構成成分をコードしていた:日本住血吸虫のグルタチオン−S−トランスフェラーゼ(GST)+トロンビン切断部位(T)+メチオニン(M)+オクタペプチド、を有するベクター。
【0114】
以下の8アミノ酸に対応するヌクレオチドを含む二本鎖の63bpのDNA配列を構築した:
【0115】
【化3】
【0116】
このDNA配列はまた、EcoRIおよびXhoIの制限酵素部位の認識配列をも含んでいた。これにより、上記DNA配列は、pGex4.3融合タンパク質ベクターへのクローニングに適合するものであった。そして、プラスミドを構築した。
【0117】
上記DNA配列は、最も一般的な細菌のアミノ酸のコドン使用頻度に基づくものであった。別のDNA配列(例えば、上述したようなもの)もまた、用いられうる。また、シアノゲンブロマイドによるアミノ酸の切断を促進する目的で、上記オクタペプチドのDNA配列の5’末端にはMet配列を挿入した。当該DNA配列の3’末端には、2つのタンデム終止コドンが含まれていた。
【0118】
図9に示す63塩基対のオクタペプチドDNA配列を、2つの部分的に重複する相補的なオリゴヌクレオチド(オクタペプチドフォワード:5’ gga tcc ccg aat tcc atg ttc tgt ttt tgg aaa acc tgt acc 3’およびオクタペプチドリバース:5’gcg gcc gct cga gtc cta tta ggt aca ggt ttt cca aaa aca 3’)をハイブリダイズすることにより作製した。各オリゴヌクレオチドの1mMアリコート2μLずつをPCR増幅ミックス中でハイブリダイズさせ、20μLのPCR反応ミックス(Sigma JumpStart mix)中で5サイクルのPCR増幅により伸長させた。
【0119】
次いで、この二本鎖オリゴヌクレオチド配列を2倍体積のエタノールで沈殿させ、20μLの水に再懸濁し、EcoRIおよびXhoI制限酵素を用いて1時間消化した。次いで、切断された配列をエタノールで沈殿させ、20μLの水に再懸濁し、その2μLを、プロメガ迅速ライゲーションキットを用いて、予めEcoRIおよびXhoI制限酵素で切断しておいたpGex4.3ベクター(2μL)中にライゲーションした。ヌクレオチドベクターの配列の比較を図10に示す。ライゲーションの後、構築物を用いてJM109コンピテント細胞(プロメガ)を形質転換させた。
【0120】
形質転換コロニーを、オクタペプチドプライマー5’ gga tcc ccg aat tcc atg ttc tgt ttt tgg aaa acc tgt acc 3’およびベクターpGex4.3プライマー5’ccg gga gct gca tgt gtc aga gg 3’を用いたインサートの存在のPCR増幅により確認した。
【0121】
次いで、陽性コロニーを増殖させ、プラスミドを抽出し、DNA配列をシーケンシングにより確認して、インサートが正しいリーディングフレームにあることを確かめた。次いで、このpGex−TS8プラスミドにより、大腸菌BL−21コンピテント細胞(プロメガ)を形質転換させた。
【0122】
実施例3:オクタペプチド(バージョン2)用のDNA配列を有するpGex4.3プラスミドの作製
第2のプラスミドを構築したが、このプラスミドは、N末端からC末端に向かって、以下の構成成分をコードしていた:日本住血吸虫のグルタチオン−S−トランスフェラーゼ(GST)+トロンビン切断部位(T)+TEVプロテアーゼ切断部位+オクタペプチド、を有するベクター。
【0123】
プラスミドのバージョン2は、TTC TGT TTT TGG AAA ACC TGTACCの前にTEV切断配列を含んでいた。このTEV切断配列は、図11に示すようにMetコドンを置換したものである。78塩基対のオクタペプチドDNA配列を、2つの部分的に重複する相補的なオリゴヌクレオチド(オクタペプチドTEVフォワード:5’ GGA TCC CCGAAT TCC GAA AAC CTG TAT TTT CAGTTC TGTTTT TGG 3’およびオクタペプチドTEVリバース:5’GCG GCC GCT CGA GTC CTATTA GGT ACA GGT TTT CCA AAA ACA GAA 3’)をハイブリダイズすることにより作製した。実施例1においてプラスミドのバージョン1について説明したように、オリゴヌクレオチドの伸長、制限酵素による消化、ライゲーション、およびクローニングを行なった。
【0124】
ヌクレオチドベクターの配列の比較を図12に示す。
【0125】
プラスミドSj−GST−T−TEV−オクタペプチドをトランスフェクションしたBL21大腸菌の小規模な50mL培養物を2×TY培地中で増殖させた。この培養物を、IPTG(0.1mM)を用いて3時間誘導し、回収した。1mg/mLリゾチーム、50mMトリスpH8.5、10%グリセロール、10mM DTT、0.5%デオキシコール酸ナトリウム、およびプロテアーゼ阻害剤カクテル(シグマ)を含有する溶解バッファー中で、細胞を溶解させた。溶解物をソ二ケーティングプローブを用いて30秒間で3回ソニケーションし、16000gで10分間遠心分離し、清澄な上清をGST−アガロースカラムにアプライした。PBSでよく洗浄した後、40mMグルタチオン、50mMトリスpH8を用いて融合タンパク質を溶出させた。選択した画分を、4〜20%ポリアクリルアミドゲル上で分析した。レーン1:分子量マーカー;レーン2:誘導細胞からの全溶解物;レーン3:溶出した融合タンパク質(分子量=28.4kDa)。
【0126】
実施例4:オクタペプチド融合タンパク質の誘導およびその精製
オクタペプチド融合タンパク質(実施例1および2によるバージョン1および2)をコードするプラスミドベクターでトランスフェクションされた小規模(20〜200mL)および中規模(1〜5L)の細菌培養物を、大腸菌BL21中で発現させた。この培養物を2×TY培地中で増殖させ、0.1mM IPTGを用い、室温(25℃)にてタンパク質を一晩誘導した。大腸菌細胞合計46gを遠心分離により回収した。これを、細胞破砕機を用いて溶解させた。溶解バッファーは、PBS、10mM DTT、10mM EDTA、5%グリセロールおよび1%トリトンX−100を含んでいた。可溶性タンパク質を、遠心分離により分離した。封入体中の不溶性タンパク質を、2M尿素(pH12.5)を添加することにより可溶化させた。不溶性タンパク質が溶解したら、水を用いて5倍に希釈し、1N HClを用いて溶液のpHを7.5とした。
【0127】
上清をグルタチオンアガロースアフィニティー精製カラムに通すことにより、可溶化した融合タンパク質を分離した。この段階で、融合タンパク質はグルタチオンアガロースカラムに結合していた。次いで、カラムを4℃にて50カラム体積のPBS(0.1%トリトンX−114を含有)とそれに続く20カラム体積のPBSで洗浄することにより、エンドトキシンを除去した。このアプローチを用いることで、エンドトキシンは洗浄バッファー中に溶出した。次いで、ニッケルアガロースカラムを通すことにより、ヒスチジンタグTEV酵素を除去した。グルタチオンアガロースカラムおよびニッケルアガロースカラムを再生させることで、これら双方の試薬を再利用可能なものとした。上記手法のそれぞれの個別の工程の完全な詳細は当業者に公知である(Structural Genomics Consortium et al., (2008) Nature Methods, Vol. 5(2), pp. 135-146)。
【0128】
図13および図14に示すように、溶出画分のサンプルを5〜20%のプレキャストポリアクリルアミドゲル(インビトロジェン)で分離し、クマシーブルータンパク質染色により染色した。
【0129】
実施例5:MALDI−TOF−MSを用いた酢酸オクトレオチドの質量の決定
MALDI−TOF−MSを用いて、化学合成された酢酸オクトレオチドの質量を決定する目的で、この実験を行なった。化学合成された酢酸オクトレオチドの理論質量は1018Daである。図15に示すように、化学合成された酢酸オクトレオチドの実測質量は1019Daであった。
【0130】
化学合成された酢酸オクトレオチドNa+の理論質量は1041Daである。図15に示すように、化学合成された酢酸オクトレオチドNa+の実測質量は1041Daであった。
【0131】
実施例6:結合したグルタチオン(GSH)を有さない単離SjGST−TM−オクタペプチド融合タンパク質(バージョン1)の質量の決定
結合したグルタチオン(GSH)を有さない精製SjGST−TM−オクタペプチド融合タンパク質(バージョン1)の質量を決定する目的で、この実験を行なった。SjGST−TM−オクタペプチドの溶液は、ジチオスレイトール(DTT)(3mM)を含んでいた。
【0132】
SjGST−TM−オクタペプチドバージョン1融合タンパク質の理論質量は27757Daである。図16に示すように、SjGST−TM−オクタペプチドバージョン1の実測質量は27751Daであった。質量誤差百分率は0.02%である。理論質量と実測質量との間の質量誤差百分率が0.1%であれば許容されうる。
【0133】
この実験により、融合タンパク質の結合グルタチオン(GSH)による質量の増加はないことが確認された。SjGSTの理論質量は25498Daである。さらにアミノ酸およびオクタペプチドがSjGSTに付加すると、理論質量は2259Da増加する。還元型グルタチオン(GSH)の理論質量は307Daである。メチオニンの理論質量は131Daである。
【0134】
実施例7:精製融合タンパク質(バージョン1)におけるオクタペプチドの存在の同定
精製融合タンパク質(バージョン1)におけるオクタペプチドの存在を同定する目的で、この実験を行なった。100mM DTTを用いて精製タンパク質の溶液を還元し、PD−10ゲル濾過カラムによりバッファーを10mMリン酸ナトリウムバッファー(2mM EDTA、pH7.8を含有)に置換した。次いで、融合タンパク質の溶液を、10mMリン酸ナトリウムバッファー(pH7.8)中、37℃にて24時間、トロンビン消化に供した。この溶液に3mM DTTを添加し(ジスルフィドの不正確な精製を防ぐ目的で)、次いでMALDI−TOF−MS分析に供した。
【0135】
SjGST−TM−オクタペプチドバージョン1融合タンパク質の理論質量は27757Daである。トロンビンによる理論上の切断産物は1607Daの断片である。トロンビンで切断されたSjGSTの大きい断片の理論質量は26150Daである。
【0136】
図17に示すように、トロンビンで切断されたSjGSTの大きい断片の実測質量は26153Daであった。質量誤差百分率は0.01%である。トロンビン切断後のオクタペプチドの理論質量は1607Daである。トロンビン切断後のオクタペプチドの実測質量は1607Daである。実施例8に記載するように、トロンビン切断後のオクタペプチドの実測質量については、この特定のMALDI−TOFスペクトルでは低分子量の分子に対する解像度が低いため、測定できなかった。
【0137】
実施例8:精製SjGST−TM−オクタペプチド融合タンパク質(バージョン1)におけるオクタペプチドの存在の同定
精製SjGST−TM−オクタペプチド融合タンパク質(バージョン1)におけるオクタペプチドの存在を同定する目的で、この実験を行なった。精製されたタンパク質の溶液を、100mM DTTを用いて還元し、PD−10ゲル濾過カラムによりバッファーを10mMリン酸ナトリウムバッファー(2mM EDTAを含有、pH7.8)に交換した。次いで、この溶液を、10mMリン酸ナトリウムバッファー(pH7.8)中、37℃にて24時間の、オクタペプチドに直接隣接したメチオニン残基のシアノゲンブロマイド媒介性の切断に供した。得られた溶液に3mM DTTを添加し(正しくないジスルフィド結合の形成を防ぐ目的)、次いでこれをMALDI−TOF−MS分析に供した。
【0138】
このオクタペプチドの理論質量は1035Daである。図18に示すように、この生物学的に合成されたオクタペプチドの、メチオニンでの化学的切断後の実測質量は1034Daであった。質量誤差百分率は0.09%である。化学合成された酢酸オクトレオチドと、生物学的に合成されたオクタペプチドFCFWKTCTとの実測質量の差は16Daである。これは、酢酸オクトレオチドがそのC末端残基としてL−スレオニノールを有するように化学的に修飾されているためである。生物学的なオクタペプチドFCFWKTCTの場合には、天然のアミノ酸であるL−スレオニンがC末端残基として存在している。
【0139】
実施例9:大腸菌におけるSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)のラクトースによる自己誘導と、封入体からのその精製
大腸菌におけるSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)のラクトースによる自己誘導と、封入体からのその単純な精製を実証する目的で、この実験を行なった。1日目に、BL21DE3細胞中のバージョン2プラスミドの培養物5mLをLB培地に播種し、37℃にて8時間増殖させた。次いでこの培地を新鮮なLB培地(5mL)で1/100に希釈し、一晩増殖させた。2日目に、この培地を再度、LB培地中に1/100(5mL)に希釈し、37℃にて8時間増殖させた。次いでこの培地を25mM Na2HPO4、25mM KH2PO4、20mM NH4Cl、2mM Na2SO4、2mM MgSO4、0.5%グルコース、0.25%アスパラギン酸塩を含有する培地中に1/100に希釈し、37℃にて一晩増殖させた。3日目に、培地を200mLの1%トリプトン、0.5%酵母抽出物、0.05%グルコース、0.2%ラクトース、0.5%グリセロール、2mM MgSO4、25mM Na2HPO4、25mM KH2PO4、50mM NH4Cl、2mM Na2SO4を含有する培地中に播種した。次いで、細胞を振盪しながら28℃にて26時間増殖させ、回収した。
【0140】
次いで、50mLの培地からの細胞ペレットを処理して、封入体を得た。50mMトリス(pH8.5)、10%グリセロール、10mM DTT、および0.5%デオキシコール酸塩を含有する溶解バッファー5mLにこのペレットを溶解させた。リゾチーム(1mg/mL)500μLを添加し、溶液を室温にて10分間インキュベートした。溶解物を30秒間ずつソニケーションし、次いで16000gで10分間遠心分離した。上清を回収し、ペレットを5mLの溶解バッファー中に再懸濁させ、30秒間ソニケーションし、さらに遠心分離した。この処理を3〜4回繰り返し、それぞれのソニケーション後に上清を回収した。残留ペレットを2M尿素および10mMトリス(pH12.5)に再溶解させた。
【0141】
図19は、BL21DE3大腸菌細胞中の分子量28.4kDaのSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)のラクトースによる自己誘導を示す。レーン1:分子量マーカー;レーン2:大腸菌の誘導細胞からの全溶解物;レーン3:ソニケーション後の大腸菌細胞溶解物由来の上清;レーン4:2M尿素および100mM Tris(pH12.5)を用いた処理後の細胞ペレット由来の上清;レーン5:2M尿素および100mM Tris(pH12.5)を用いた処理とそれに続く塩酸によるpH8への調整後の細胞ペレット由来の上清;レーン6:2M尿素および100mM Tris(pH12.5)で溶解され、続いて塩酸によるpH8への調整を経た大腸菌細胞由来の残存ペレット。各レーンにおいて、17kDaに見られるタンパク質のバンドは、細胞を消化する目的で添加されたリゾチームである。
【0142】
実施例10:化学合成酢酸オクトレオチドの3炭素架橋のPEG化
10mM EDTAを含有する50mMリン酸ナトリウムバッファー(pH7.8)中で調製した化学合成酢酸オクトレオチドの溶液(0.25mg/mL、0.25μmol)に、TCEP HCl(70μg、0.25μmol、5mg/mL溶液を14μL)を室温にて1時間添加した。次いで、この還元された酢酸オクトレオチドにポリエチレングリコールビススルホン(1.35mg、0.25μmol、1当量)を添加した。反応溶液を穏やかにかき混ぜ、4℃にて一晩静置した。次いでこの溶液を、PD−10脱塩カラムを用いた脱イオン水に対する脱塩処理に供した。最大量のPEG化酢酸オクトレオチドを含有する共役画分(すなわち、第3の1mL画分)を、MALDI−TOF−MSにより分析した。
【0143】
比較のために、酢酸オクトレオチドのMALDI−TOF−MSスペクトルを図15に示す。図20は、化学合成された酢酸オクトレオチドが、ジスルフィド架橋を横切る3つの炭素の架橋を介して、5kDaのポリエチレングリコールに共有結合していることを示す。
【0144】
このPEG化された酢酸オクトレオチドの理論質量は6092Da(すなわち、ポリエチレングリコール5073Da+酢酸オクトレオチド1019Da)である。ポリエチレングリコールの実測分子量は5073Daである。PEG化された酢酸オクトレオチドの実測分子量は6097Daである。質量誤差百分率は0.08%である。
【0145】
実施例11:ポリエチレングリコールが共有結合した3炭素架橋の化学的な挿入による、SjGST−TM−オクタペプチド融合タンパク質(バージョン1)の修飾のSDS−PAGEによる確認
50mMリン酸ナトリウムバッファー(pH7.4)中のSjGST−TM−オクタペプチド(バージョン1)(0.5mg/mL、1mL)融合タンパク質の精製溶液に、100mM DTT(15.4mg)を添加した。次いで、この溶液を室温にて30分間インキュベートし、次いでセファデックスG−25PD−10カラムを用いてバッファーを50mMリン酸ナトリウムバッファー(10mM EDTAを含有、pH7.8)に置換した。この溶液に、ジスルフィド部位特異的に架橋する5kDaポリエチレングリコール(0.29mg、5μmol、3当量)を添加した。得られた溶液を4℃にて72時間インキュベートした。次いで、この混合物にウシトロンビン(シグマ−アルドリッチ、融合タンパク質に対して0.1当量)を添加し、37℃にて24時間インキュベートした。次いで、得られた溶液をSDS−PAGEおよびMALDI−TOF−MSにより分析した。
【0146】
図21において、レーン1:分子量マーカー;レーン2:27757Daの理論分子量を有する、精製されたSjGST−TM−オクタペプチド(バージョン1);レーン3:4℃にて72時間のSjGST−TM−OCTとの5Daのポリエチレングリコールのジスルフィド部位特異的な架橋反応とそれに続く37℃にて24時間のトロンビンによる消化。SjGST−TM−オクタペプチドのタンパク質バンド27.7kDaは消失し、モノPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化)、ジPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化、および、GST中のジスルフィド結合のモノPEG化)、並びに、トリPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化、および、GST中の2つのジスルフィド結合のジPEG化)が出現した;レーン4:反応バッファー中における5kDaのポリエチレングリコール試薬(3等量濃度)のジスルフィド部位特異的な架橋。
【0147】
実施例12:ポリエチレングリコールが共有結合した3炭素架橋の化学的な挿入による、SjGST−TM−オクタペプチド融合タンパク質(バージョン1)の修飾のMALDI−TOF−MSによる確認
実施例11に記載のタンパク質溶液を100mM DTTを用いて還元し、PD−10カラムを用いてバッファーを50mMリン酸ナトリウムバッファー(pH7.8、10mM EDTAを含有)に置換した。次いで、還元されたSjGST−TM−オクタペプチド(バージョン1)の溶液を、4℃にて24時間、ジスルフィド部位特異的なPEG化に供した。得られた溶液を37℃にて72時間トロンビン消化に供し、MALDI−TOF−MS分析に供した。
【0148】
図22は、トロンビン消化され、ジスルフィド部位特異的にPEG化されたSjGST−TM−オクタペプチド(バージョン1)タンパク質溶液のMALDI−TOF−MSを示す。
【0149】
SjGST−TM−オクタペプチド(バージョン1)融合タンパク質の理論質量は27757Daである。トロンビンにより切断したSjGSTの大きい断片の理論質量は26150Daである。トロンビンによる理論上の小さい切断産物は、1607Daの断片である。このことから、オクタペプチド(1607Da)に連結されたポリエチレングリコール(5073Da)の理論質量は6680Daとなる。オクタペプチドに連結したポリエチレングリコールの実測質量は6692Daであった。質量誤差は0.1%である。
【0150】
実施例13:インビボでの成長ホルモンレベルに及ぼすオクタペプチドの効果
上記の実施例で詳述した化学的処理を用い、3つのオクタペプチド(FCFWKTCTおよびACYWKVCTおよびACYWKTCT)に対して、ポリエチレングリコールが共有結合した3炭素架橋を化学的に挿入した。入手可能な最高グレードの化学物質を、臨床グレードの超純水とともに、バッファー(50mMリン酸ナトリウムバッファー(10mM EDTAを含有、pH7.8))および透析に用いた。すべての溶液を、使用前には30秒間アルゴンでパージした。凝集を防止すべく、反応混合物中でのタンパク質の濃度上昇を回避した。新鮮に調製したTCEP HCl(5mg/mL)を用い、無水条件下、室温にて1時間、タンパク質を還元した。還元タンパク質については、熱または直射日光に晒さないようにした。還元タンパク質を激しく振盪することも避けた。30kDaの3炭素架橋ポリエチレングリコールビススルホンに対するこのタンパク質の比は1:1であった。この共役反応溶液を穏やかにかき混ぜて30kDaの3炭素架橋ポリエチレングリコールビススルホンを溶解させ、次いで溶液を暗所に4℃にて12時間静置した。次いで、5〜7kDaの分子量カットオフを有するピアース透析チャンバを用いてこの溶液を透析して、30kDaのポリエチレングリコールに共有結合していないオクタペプチドを除去した。2時間後、4時間後、および6時間後に、それぞれ1Lの透析水の交換を行なった。次いで凍結乾燥することにより、水および他の揮発性物質を除去した。
【0151】
インビボの動物実験では、体重約20gの雄性BALB/Cマウスを用いた。それぞれの皮下注射は、250μLの注射水に溶解したPEG化オクタペプチドを含んでいた。各注射で投与されたオクタペプチドのみの重量は125μgであった。それぞれのマウスは合計で4回の注射を4日おきに投与された。注射は、首の後部の皮膚をつまみ、皮下に行った。最後の注射の30時間後に動物を屠殺した。市販の酵素イムノアッセイを用いた成長ホルモンの測定のため、血清を回収した。臨床上の毒性は観察されなかった。
【0152】
結果(n=4)を図23に示す。この動物実験では、血清中成長ホルモン(GH)レベルの低下について、3つのすべてのPEG化オクタペプチドが化学合成オクトレオチドよりも高い活性を示した。PEG化されたFCFWKTCTオクタペプチドは、化学合成オクトレオチドと比べると、血清成長ホルモンを70%多く低下させた。PEG化されたACYWKVCTオクタペプチドは、化学合成オクトレオチドと比べると、血清成長ホルモンを78%多く低下させた。PEG化されたACYWKTCTオクタペプチドは、化学合成オクトレオチドと比べると、血清成長ホルモンを94%多く低下させた。
【技術分野】
【0001】
本発明は、概ね、天然には存在せず生物学的に合成されるペプチド類に関し、当該ペプチド類をコードするポリヌクレオチド類に関し、当該ペプチド類の製造方法に関し、当該ペプチド類の用途に関する。本発明のペプチド類は、医薬に(例えば、ホルモン障害、癌および出血性障害の治療に)用いられうる。本発明はまた、概ね、安定性が向上したペプチド類の修飾体に関し、かような修飾ペプチドの作製方法に関し、当該修飾ペプチド類の用途に関する。特に、本発明は、ポリエチレングリコールなどのポリマーを用いて化学的に修飾されうる、生物学的に合成されるペプチド類に関し、末端肥大症、消化管ホルモン産生腫瘍などの腫瘍、および消化管出血の治療における当該ペプチド類またはその修飾体の用途に関する。
【背景技術】
【0002】
末端肥大症は、頭、手および足の骨、並びに軟部組織の肥大を特徴とする、生活に支障を来すほどのホルモン障害であり、早期の死をもたらす。末端肥大症への罹患率は、1年間に人口100万人あたり3症例であり、その有病率は100万人当たり60人である。この疾患の症状は知らぬ間に進行することから、臨床症状の発現から診断がなされるまでに7〜10年遅れることもよくある。末端肥大症の原因は、通常は下垂体腺腫による下垂体からの成長ホルモン(GH)の過剰分泌である下垂体の成長ホルモン分泌細胞が増殖することで成長ホルモンが異常に高いレベルで分泌され、末端肥大症の典型的な臨床症状がもたらされる。
【0003】
成長ホルモンは191アミノ酸の、4つのヘリックスの束のタンパク質、およびより少量の176アミノ酸体として分泌される。成長ホルモンは、下垂体門脈循環を往来する視床下部放出ホルモンおよび視床下部抑制ホルモンを介した視床下部の制御下で脈動により循環系に進入する。次いで、成長ホルモン分泌細胞表面の特異的な受容体に直接作用する。成長ホルモンは、(i)末梢のインスリン様成長因子1(IGF−1)の合成を誘導し、(ii)循環(エンドクリン)並びに局所(オートクリンおよびパラクリン)IGF−1により誘導される細胞増殖を誘導し、(iii)アポトーシスを阻害する。末端肥大症の過剰な罹患率および死亡率は、GHレベルおよびIGF−1レベルの長期にわたる上昇の結果である。これら2つのホルモンのレベルを障害にわたって注意深く制御することで、患者の福利が改善され、正常な寿命が取り戻される。
【0004】
現在の末端肥大症の治療のなかでも、外科的手術は成長ホルモンを分泌する微小腺腫を除去する目的でよく行われる。放射線治療は通常、薬物療法に抵抗性の、または耐性を示す患者において、手術後に再発するか生き残っている腫瘍のために取って置かれる。
【0005】
また、現在の治療のなかには、成長ホルモンの分泌を抑制する天然のホルモンであるソマトスタチンの化学合成された類似体を投与するというものもある。かような類似体の1つは、酢酸オクトレオチド(ノバルティスファーマシューティカルズ社によりサンドスタチン(登録商標)の商品名で市販されている)である。この薬は過去20年間にわたって、末端肥大症の治療法として成功を収め、臨床上も認められてきた。酢酸オクトレオチド(体系的なIUPAC名は(4R,7S,10S,13R,16S,19R)-10-(4-アミノブチル)-19-[[(2R)-2-アミノ-3-フェニル-プロパノイル]アミノ]-16-ベンジル-N-[(2R,3R)-1,3-ジヒドロキシブタン-2-イル]-7-(1-ヒドロキシエチル)-13-(1H-インドール-3-イルメチル)-6,9,12,15,18-ペンタオキソ-1,2-ジチア-5,8,11,14,17-ペンタアザシクロイコサン-4-カルボキサミドである)は、固相化学合成法(Bauer et al., (1982) Life Sciences, Vol. 31, pp. 1133-1140)により製造されるオクタペプチドである。化学合成された酢酸オクトレオチドのアミノ酸配列は:
【0006】
【化1】
【0007】
である。
【0008】
他の化学合成されたソマトスタチン類似体は、酢酸ランレオチド(体系的なIUPAC名は(4S,7S,10S,13R,16S,19S)-10-(4-アミノブチル)-19-[[(2R)-2-アミノ-3-ナフタレン-2-イル-プロパノイル]アミノ]-N-[(1S,2R)-1-カルバモイル-2-ヒドロキシ-プロピル]-16-[(4-ヒドロキシフェニル)メチル]-13-(1H-インドール-3-イルメチル)-6,9,12,15,18-ペンタオキソ-7-プロパン-2-イル-1,2-ジチア-5,8,11,14,17-ペンタアザシクロイコサン-4-カルボキサミドである)である。この薬は、イプセンファーマシューティカルズによりソマツリンLA(登録商標)の商品名で市販されている。
【0009】
酢酸ランレオチドのアミノ酸配列は:
【0010】
【化2】
【0011】
である。
【0012】
特に、酢酸ランレオチドは1位に(D)アラニンを、4位に(D)トリプトファンを含んでいる。また、2つのシステイン残基にわたってジスルフィド結合を有してもいる。酢酸ランレオチドは構造的に酢酸オクトレオチドと非常に類似しており、酢酸オクトレオチドにとてもよく似たソマトスタチン受容体結合プロファイルを示し、酢酸オクトレオチドと同様の適応症を有している(Weckbecker, G. et al., (2003) Nature Reviews, Vol. 2, pp. 999-1017)。
【0013】
天然のホルモンであるソマトスタチンは、βターンの角にTrp残基およびLys残基が存在することに起因して逆平行に伸びたβシートが存在することで、そのコンホメーションに環状の部位を有していることが知られている。酢酸オクトレオチドおよび酢酸ランレオチドでは、D−Trp4の導入によってファーマコフォアのβターンを安定化するように同様に作用する。
【0014】
また、ジスルフィド架橋の形成によるペプチドの環化によってタンパク質やペプチド(ソマトスタチンも含む)の化学的安定性や代謝安定性が付与されることも一般的に知られている。この理由から、化学合成された酢酸オクトレオチドおよび酢酸ランレオチドには、架橋ユニットであるCys2−S−S−Cys7が化学的に導入されている。酢酸オクトレオチドの場合には、ペプチドの代謝安定性が3倍に上昇する(Bauer et al., (1982) Life Sciences, Vol. 31, pp. 1133-1140; Cai et al., (1986) Proc. Natl. Acad. Sci. (USA), Vol. 83, pp. 1896-1900)。
【0015】
これらをまとめると、2つのDアミノ酸の挿入およびジスルフィド架橋の形成によってこれらの化学合成8アミノ酸ペプチドの血清半減期は長くなる。酢酸オクトレオチドの場合には、数分間が1.5時間まで長くなる(Harris AG., (1994) Somatostatin and somatostatin analogues: pharmacokinetics and pharmacodynamic effects. Gut, Vol.35, pp. 1-4)
酢酸オクトレオチドおよび酢酸ランレオチドは、ソマトスタチン受容体サブタイプ2(SST2)およびソマトスタチン受容体サブタイプ5(SST5)に高親和性で結合する。ソマトスタチン受容体サブタイプ3に対してこれらは中程度の親和性で結合する。ソマトスタチン受容体サブタイプ1またはソマトスタチン受容体サブタイプ4に対してこれらは結合しない。また、酢酸オクトレオチドおよび酢酸ランレオチドはD2ドーパミン受容体に結合することが示されている。
【0016】
酢酸オクトレオチドの生物活性には、その構造中のPhe3−Trp4−Lys5−Thr6のみが必須であることが知られている。酢酸オクトレオチドがソマトスタチン受容体に結合すると、成長ホルモンの分泌および成長ホルモン分泌細胞の増殖を抑制するように下垂体に信号を送り、また、肝臓に作用してIGF−1の合成を阻害する。SST2およびSST5受容体に対するソマトスタチン受容体のリガンドとして、酢酸オクトレオチドはGHおよびIGF−1のレベルを低下させ、腫瘍増殖を抑制し、肝臓により媒介されるGHの受容体への結合・作用を阻害する。GH受容体のアンタゴニストとして、酢酸オクトレオチドはGH受容体のシグナル伝達を阻害し、これにより末梢血中のIGF−1レベルが低下する。
【0017】
末端肥大症の患者における酢酸オクトレオチドの投与に応答した成長ホルモン分泌の抑制は、ソマトスタチン受容体サブタイプの利用可能性に依存する。酢酸オクトレオチドおよび酢酸ランレオチドの直接的な抗腫瘍活性が、腫瘍細胞において発現するソマトスタチン受容体を介した実験腫瘍モデルで実証されている。これらの抗増殖作用は、細胞分裂の阻害およびアポトーシスの誘導の結果である。ソマトスタチン受容体に化学合成ソマトスタチン類似体が結合すると、特異的なシグナル伝達経路が開始される。このようにして、それぞれのソマトスタチン受容体サブタイプは異なる生物学的作用を媒介することができる。これらの機構を媒介する受容体サブタイプは、SST1、SST2、SST4およびSST5である。ヒト腫瘍の多くは複数のソマトスタチン受容体サブタイプを発現しているが、主なものはSST2である。ソマトスタチン類似体である酢酸オクトレオチドおよび酢酸ランレオチドは、SST2受容体に対して高い親和性を有している。
【0018】
ソマトスタチンおよびその合成類似体はまた、多くの間接的な抗腫瘍作用を示す。これらのなかには、腫瘍増殖を促進する成長因子およびホルモンの放出の阻害がある。これらのソマトスタチン類似体の間接的な効果により生じる腫瘍増殖の低下は、成長因子や、IGF−1および成長ホルモンなどのホルモン類の合成および分泌の抑制(並びに作用の減退)を含む。ソマトスタチン類似体は、中枢および末梢の双方の機構によって成長ホルモン−IGF−1系を抑制する。下垂体の成長ホルモンの放出の阻害を媒介する主要な受容体サブタイプはSST2およびSST5である。ソマトスタチン類似体はまた、STAT5bの脱リン酸化およびIGF−1遺伝子の転写の低下をもたらすSST媒介性のチロシンホスファターゼの活性化を介して、成長ホルモンにより誘導される肝臓のIGF−1の産生を阻害する(Weckbecker, G. et al., (2003) Nature Reviews, Vol. 2, pp. 999-1017; Susini C & Buscail L, (2006) Ann of Oncology Vol. 17, pp. 1733-1742)。
【0019】
一方、ソマトスタチン受容体サブタイプの利用可能性は、患者血清中の血清成長ホルモンおよびIGF−1の濃度に酢酸オクトレオチド療法が与える長期にわたる効果を予測する。血清の成長ホルモンおよびIGF−1が最も抑制されるのは、皮下で300〜600μg/日の用量である。酢酸オクトレオチドはまず、成長ホルモンのレベルを正常なレベルにまで低下させるために、数ヶ月間の間、少なくとも日に3回(半減期がたった1.5時間であるため)、皮下注射として患者に投与される。成長ホルモンの過剰産生が正常レベルにまで低下したら、サンドスタチン(登録商標)長期間作用型などの徐放性ポリマーベースの複合体デポ製剤を患者に(数週間にわたって)移植することもよく行われる。これは、殿筋の深部に筋肉内注射することにより投与され、三角筋に投与することはできない。これは、大きい殿筋と比べて三角筋ではサンドスタチンがもたらす痛みが過剰であることによる。このポリマーベースの徐放性デポから酢酸オクトレオチドが徐放されるということは、酢酸オクトレオチドは通常、痛みのない皮下注射により投与される場合の毎週と比較して、2週間おきに(痛みを伴って)投与されうることを意味する。
【0020】
デポ製剤(すなわち、長期間作用放出型の酢酸オクトレオチド(サンドスタチン(登録商標)LAR(登録商標)として市販されている))により、14日おきの注射による投与が可能となる。これらの製剤は、酢酸オクトレオチドの有効なレベルを維持することができる。報告によれば、酢酸オクトレオチドによる治療の間9年間追跡された患者の80%で、成長ホルモンのレベルが2.5μg/L未満であり、かつ、IGF−1のレベルが正常であったとされている。ユーゴナディズム(Eugonadism)は性腺機能低下症を有する末端肥大症患者の3分の2に残っている(S. Farooqi et al. (1999) Pituitary, Vol. 2, pp. 79-88; A.N. Paisley and P.J. Trainer, (2003) Current opinion in Pharmacology, Vol. 3, pp. 672-677; S. Melmed. (2006) New England Journal of Medicine, Vol. 355, pp. 2558-2573)。
【0021】
酢酸オクトレオチドの有効性の決定因子としては、治療前の成長ホルモンのレベル、腫瘍での十分なSST2およびSST5の発現の存在・不存在、薬物の用量、状態の評価に用いられる生化学的指標、並びに治療に対する患者の忠実さなどが挙げられる。腫瘍塊の縮小は患者の50%で起こるが、酢酸オクトレオチドを用いた治療を止めると回復してしまう。
【0022】
また、外科的な全部の切除を必要としない巨大腺腫の外科的減量によれば、その後の酢酸オクトレオチドによる治療の有効性が向上する。薬を服用した患者の80%超が、頭痛や末梢軟組織の膨張などの症状が改善されたと報告している。腫瘍の縮小および生化学的な制御は、必ずしも並行して起こるとは限らない。よって、酢酸オクトレオチドはGHおよびIGF−1のレベルの生化学的な制御を変化させることができなかった手術の後や、成長ホルモンのレベルが上昇したままであったことによる放射線治療の後に投与されることが多い。酢酸オクトレオチドを用いた一次的な医学的治療は有効かつ安全である。患者が手術または放射線治療を受けているか否かにかかわらず、長期にわたる薬物投与に対する同等の生化学的応答が達成されうることから、一次的な医学的治療は、(i)巨大な細胞外腫瘍を有し、中枢圧縮作用(central compressive effect)の証拠を示さない患者、(ii)薄弱すぎて手術を受けることができない患者、および(iii)手術を辞退した患者に対して提案されうる。
【0023】
酢酸オクトレオチドおよび酢酸ランレオチドはまた、消化管関連のカルシノイド腫瘍や転移性のカルシノイド腫瘍の治療にも用いられうる。胃腸管のカルシノイドの全体的な発生率は年間84人/百万人と見積もられている。胃腸管のカルシノイド腫瘍は、全てのカルシノイド腫瘍の90%を占めている。酢酸オクトレオチドを用いた治療により、カルシノイド症候群を患っている患者の50〜80%の症状および生活の質が改善される。また、酢酸オクトレオチドおよび酢酸ランレオチドは、腫瘍の発達を停止させることができる。
【0024】
酢酸オクトレオチドは、血管作動性腸管ペプチドを分泌する腫瘍(VIPoma)を有する患者における下痢の治療に治療上有効であることが示されており、他の原因による重篤な難治性の下痢の治療に用いられている。毒性学においてもまた、酢酸オクトレオチドはスルホニルウレアの過剰摂取後の長期にわたる再発性低血糖の治療に用いられており、インスリンの過剰分泌の抑制を補助する目的で、膵島細胞症を有する乳幼児に用いられている。食道静脈瘤が疑われる患者において、酢酸オクトレオチドは出欠の抑制を補助する目的で投与されうる。さらに、酢酸オクトレオチドは、慢性膵炎からくる痛みを有する患者の治療や、胸腺腫瘍の治療、および糖尿病に起因する眼疾患の治療にも用いられうる。酢酸ランレオチドも酢酸オクトレオチドと同様の適応症を有している。
【0025】
酢酸オクトレオチドや酢酸ランレオチドのような数年間にわたって恒常的に服用する必要のある小分子ペプチドベースの医薬すべてにおける主要な実用上の欠点は、全世界で用いられる医薬として製造するための費用対効果のよい大量生産法がないということである。現時点では、天然のペプチドおよび設計されたペプチドのいずれの製造にも化学的なペプチド合成が用いられているが、このアプローチは非常にコストがかかる。その結果、患者における酢酸オクトレオチドの臨床的な使用は法外に高価なものとなっている。末端肥大症の患者の場合、イギリスで1人の患者を治療するコストは、約16000ポンド/年である。悪性のカルシノイド腫瘍を有する患者に酢酸オクトレオチドを用いると、イギリスで1人の患者を治療するコストは約32000ポンド/年である。
【0026】
医薬としてのペプチド類の工業的な生産に対する主な障害は、経済的な大量生産である。酢酸オクトレオチドおよび酢酸ランレオチドの製造においては、段階的なFmoc固相化学合成とそれに続く分子内ジスルフィド結合の形成が必要とされる。D−Trpの存在下でペプチドの代謝安定性を3倍に改善するための分子内ジスルフィド結合の形成は主要な問題となりうる。また、化学合成時の側鎖の保護も必要である。これらの化学的な問題により、収率は14%と低い結果となっている。酢酸オクトレオチドの固相合成に伴う他の問題としては、(i)C末端のCys残基のラセミ化、(ii)ペプチドの集合の非効率性、(iii)有害な副作用、(iv)樹脂上での非効率なジスルフィド結合の形成、(v)ジスルフィド結合形成時のD−Trpの修飾、(vi)アミノリシスによる不完全で非効率なペプチド−樹脂の開裂、(v)断片のカップリング時のラセミ化、および、(vi)複雑な精製工程、が挙げられる。さらに、(a)L−PheをD−Pheで置換しなければならないとき、(b)L−TrpをD−Trpで置換しなければならないとき、(c)直鎖ペプチドの安定性を向上させる目的でジスルフィド結合を形成するために直鎖ペプチドを環化する必要があるときに、化学的なペプチドベースの合成のコストは著しく高騰する。酢酸ランレオチドの固相合成についても同様の問題が存在する。
【0027】
よって、代わりとなる、より経済的かつ効率的な、酢酸オクトレオチドや酢酸ランレオチドのような小ペプチドの合成方法が必要とされている。この点で、細菌や酵母での組換えDNA法がよく用いられる。また、タンパク質の生物活性においてグリコシル化が役割を担っていないことが示されているのであれば、汎用の宿主である大腸菌においてタンパク質の製造を実施することで相当のコストおよび製造上の利点が存在する。この技術により、多くの組換えタンパク質が大腸菌で製造されている。
【0028】
しかしながら、かような系では、組換えDNA技術を用いることでタンパク質の高い生産性は達成されるものの、きわめて小さい天然のペプチド類についての問題として、より大きな融合タンパク質の一部として製造される必要があるということがある。このため、天然のペプチドの遺伝子はより大きいキャリアタンパク質の遺伝子と連結され、次いで、大腸菌などの宿主細胞中で、単一の大きいタンパク質として、融合タンパク質が発現する。タンパク質の合成の後、所望のペプチドは融合相手から切断されなければならない。実用上は、このアプローチによる主な問題として、非常に小さいペプチドは、ほとんどの微生物の細胞質に見られる酵素によるタンパク質分解を非常に受けやすい(サイズが非常に小さいことで、高度に配列した三次構造を有しえなくなる)ということがある。したがって、小ペプチドは製造された宿主の細胞内で急速に、かつ細胞の他の細胞質成分から首尾よく単離されうる前に、分解されることが多い。
【0029】
また、多くの異種タンパク質(すなわち、宿主細胞において天然には産生されないタンパク質)が、宿主細胞中で大量に製造されることに起因する毒性作用によって細菌/酵母の増殖を妨げることが確認されているという別の問題もある。さらに、これらの外来ペプチド産物は宿主細胞中で不安定なことが多い。このことは、微生物中で融合タンパク質の一部としてペプチドを発現させるときには、当該ペプチドの発現を安定化させるために多くの努力を要するということを意味する。これらの問題は特に、微生物系において非常に短いペプチド鎖の発現を必要とする場合に関係があり、このことは所望のペプチド配列がより大きい融合タンパク質の一部として発現する場合でさえそうである。
【0030】
組換えDNA技術を用いた組換えペプチドの生産が大規模で達成され、しかもかような生産が経済的なものとされうるとしても、細菌/酵母溶解物、そして融合タンパク質から非常に小さいペプチドを単離し精製するコストが高いことで、費用効率の高い医薬を製造するという問題はこじれてしまう。細菌または酵母からペプチドを生産するためのこれらの下流の処理工程は、実質的な追加の生産コストに寄与することが多い。例えば、細菌または酵母の細胞からのペプチドの最初の回収には、細胞の破砕および溶解、破砕・溶解された細胞からの封入体(すなわち、凝集した、それゆえに不溶性のタンパク質)の単離、溶解性の融合タンパク質を得るための単離された封入体の溶解、並びに、融合タンパク質の切断とその後のキャリアタンパク質からのペプチドの分離などの、複数かつ独特の処理工程が必要とされうる。よって、大規模で費用効率の高い生産を現実のものとするためには、非常に小さい組換えペプチドの生産のすべての形態を改善し、最適化することが望まれる。
【0031】
ペプチドベースの医薬については、効果的な薬物の送達を達成することにも問題がある。ポリマーベースのマトリックスを用いる場合、ペプチドの制御放出性の薬物送達システムの重要でかつ十分に解決されてはいない問題の1つは、生分解性ポリマーベースのマトリックス中に配合された後のペプチドの安定性である。マイクロスフェアに一般的に用いられているポリ(乳酸)およびポリ(乳酸−co−グリコール酸)のポリマーの分解により、ペプチドを含有するマイクロスフェアの内部には酸性度の高い微小環境(すなわち、pH1.5)が形成されることが多い。このことは、これらのマトリックス中に配合されたペプチドの不安定性の主要な原因となっていることが知られている。このペプチドの不安定性は、ポリ(乳酸)およびポリ(乳酸−co−グリコール酸)のマイクロスフェアの分解と、これによって生じる乳酸およびグリコール酸のユニットでの配合されたペプチドのアシル化により共有結合修飾によるものである。アシル化により、ペプチドの筋肉内のデポ注射部位からの吸収速度は遅くなる。いまでは、多くの機関によって、分解しているポリ(乳酸)およびポリ(乳酸−co−グリコール酸)のマイクロスフェア内部でのペプチド薬剤のアシル化は、新規でかつ有効性の高い治療薬としての生物活性ペプチドベースの薬物の、安定した、持続的な、うまい送達のために依然として克服する必要のある主要な障害として捉えられている(Werle M. et al., Amino Acids, (2006) Vol. 30; pp. 351-367)。
【0032】
この問題は、濃度が異なり(42.5%、21.3%、および8.5%;wt/wt)、37℃でのpH値も異なる(2.25、1.47、および1.85)3つの乳酸溶液中での酢酸オクトレオチドのアシル化反応の調査によるある研究において説明されている(Na et al., AAPS PharmSciTech., 2003, 4(4): article 72)。これらのインキュベーションの間、酢酸オクトレオチドのアシル化産物を、時間依存的かつ濃度依存的に測定した。42.5%乳酸溶液(pH2.25)では、30日間のインキュベーション後の完全な酢酸オクトレオチドの残存量は、出発物質のたった51%であった。これは、生物学的に不活性なアシル化酢酸オクトレオチドの量が増加した結果である。徐放性製剤の利用に向けた他のアプローチとしてはPEG化が挙げられ、これはタンパク質またはペプチドの安定性および半減期を増加させるのに用いられうる。ペプチドおよびタンパク質をPEG化することの治療上有用な効果としては、物理学的および熱的な安定性の向上、循環半減期の増加、免疫原性および抗原性の低下、並びに、毒性の低下が挙げられる。ペプチドおよびタンパク質をPEG化することで、有機溶媒への曝露に対して、天然のペプチドおよびタンパク質よりもずっと高い安定性をも示す。また、PEG化されたタンパク質のマイクロスフェアは、PEG化されていないペプチドおよびタンパク質と比較すると、異なる薬物放出プロファイルをも示し、初期のペプチドおよびタンパク質の一度の放出を抑制する。しかしこれまで、これらのPEG化ベースのアプローチは、分子量(MWt)が10kDaを超えるタンパク質ベースの薬物に限定されていた。また、安定化の問題は凝集や変性などのタンパク質の物理的安定性を改善するための主なものである。
【発明の概要】
【0033】
本発明者らは、この度、天然には存在しない新規なペプチドをコードするポリヌクレオチドを製造した。このポリヌクレオチドは、上述の新規なペプチドを製造するのに用いられうる。このペプチドは、酢酸オクトレオチドにおける4つの重要なアミノ酸の三次元結合部位構造を保持している。このペプチドは、宿主細胞中で経済的に生産されうるし、工業的規模で、費用効果の高いいくつかの工程で、宿主細胞から単離されうる。このペプチドはまた、自身の化学的安定性および代謝安定性を実質的に向上させ、これにより患者における治療有効性を実質的に高める目的で、費用効果の高い追加の化学修飾を伴ってもよい。
【0034】
以下の発明の記載では、添付の図面を参照する:
【図面の簡単な説明】
【0035】
【図1】図1は、化学的に合成された酢酸オクトレオチドを示す。丸で囲んだアミノ酸(すなわち、Phe−(D)Trp−Lys−Thr)のみがソマトスタチン細胞の表面受容体に結合する。
【図2】図2は、化学的に合成された酢酸オクトレオチド(分子量1018Da)の分子モデルを示しており、2つのD−アミノ酸を含有している。標識されたアミノ酸(すなわち、Phe−(D)Trp−Lys−Thr)がソマトスタチン細胞の表面受容体に結合する。
【図3】図3は、本発明による、生物学的に合成されたオクトレオチド(L−アミノ酸Phe−Cys−Phe−Trp−Lys−Thr−Cys−Thrを含有する)の分子モデルを、化学合成された酢酸オクトレオチド(2つのD−アミノ酸を含有する)の分子モデルと重ねて示す。太い灰色線が生物学的に合成されたオクタペプチド(すべてL−アミノ酸)を示し、細い黒線が化学合成された酢酸オクトレオチド(2つのD−アミノ酸)を示す。
【図4】図4は、本発明によって生物学的に合成され、次いで3つの炭素の架橋(ここにポリエチレングリコールが共有結合している)の挿入により化学修飾されたオクトレオチドの分子モデルを示す。
【図5】図5は、本発明によって生物学的に合成され、次いで3つの炭素の架橋(ここにポリエチレングリコールが共有結合している)の挿入により化学修飾されたオクトレオチドの分子モデルを示す。PEG化されたオクタペプチドの各化学成分が示されている:(a)点線の丸は、受容体に結合するアミノ酸Phe−Trp−Lys−Thrを示す;(b)破線は残りのアミノ酸(すなわち、Phe−Cys−ジスルフィド結合−Thr)を示す;(c)連続した線はポリエチレングリコールを示す。
【図6】図6は、本発明によって生物学的に合成され、次いでオクタペプチドのジスルフィド結合を横切る3つの炭素の架橋(CCCで示す)の挿入により化学修飾されたオクトレオチドの分子モデルを示す。この3つの炭素の架橋には、ポリエチレングリコールが共有結合しうる。
【図7】図7は、本発明の生物学的に合成されたオクタペプチドの柔軟性を示す。公開されている分子モデリングのプロトコール(Zloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083)を用い、アミノ酸配列FCFWKTCTを有する生物学的に合成されたオクタペプチドが、シミュレーション中の骨格原子の標準偏差(RMSD)の値を記録することによりモニターされうる柔軟度を示すことが示された。RMSD値の範囲は1〜4.4Åである。これは、3つの炭素の架橋を介してポリエチレングリコールが結合してもオクタペプチドの柔軟性が損なわれないことを示している。よって、このジスルフィド部位特異的にPEG化されたオクタペプチドは、その生物学的ターゲットと有効に相互作用できる程度に十分に柔軟なままでいられるであろう。
【図8】図8は、ポリエチレングリコールが共有結合した3つの炭素の架橋の挿入により化学修飾された、本発明によるL−アミノ酸を含有するアミノ酸配列FCFWKTCTを有する生物学的に合成されたオクタペプチドの分子モデルを、D‐アミノ酸を含有する化学合成された酢酸オクトレオチドの分子モデルと重ねて示す。これら2つの構造はエネルギー的に最低の配座異性体(すなわち、最も安定な形態)であり、これらはZloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083で公開されたのと同一の分子モデリング法を用いて予測された。太い灰色線が生物学的に合成されたオクタペプチドを示し、細い黒線が化学合成された酢酸オクトレオチド(2つのD−アミノ酸)を示す。
【図9】図9は、実施例2に記載のオクタペプチドプラスミドのバージョン1をクローニングするのに用いたアミノ酸配列およびヌクレオチド配列を示す。オクタペプチドFCFWKTCTをコードする配列に下線を付している。EcoRIおよびXhoIの制限部位の配列を太字で示している。
【図10】図10は、本発明のプラスミドベクターのバージョン1を示し、実施例2に記載の、プラスミドオクタペプチドのpGex4.3クローン5(メチオニン残基に連結したグルタチオントランスフェラーゼとそれに続く挿入オクタペプチドのコード配列を含む)とpGex4.3ベクターとのヌクレオチド配列のアライメントを示す。オクタペプチド*=オクタペプチドFCFWKTCTおよびクローニング配列。オクタペプチドFCFWKTCTをコードする配列に下線を付している。ベクターおよびpGex系の説明はGEライフサイエンセズによるGST融合系のハンドブックに記載されている(http://www4.gelifesciences.com/aptrix/upp00919.nsf/Content/87478CFA7E09E0C7C1256EB400417E59/$file/18115758.pdf)。
【図11】図11は、実施例3に記載のオクタペプチドプラスミドのバージョン2をクローニングするのに用いたアミノ酸配列およびヌクレオチド配列を示す。これは、TEV認識部位を含んでいる。X=任意のアミノ酸(プロリンを除く)。オクタペプチドFCFWKTCTおよびそれをコードする配列に下線を付している。
【図12】図12は、本発明のプラスミドベクターのバージョン2を示す。また、図12は、実施例3に記載のプラスミドオクタペプチドのpGex4.3と、追加のTEV開裂部位の配列を有するオクタペプチドFCFWKTCTのコード配列とのヌクレオチド配列のアライメントを示す。オクタペプチドFCFWKTCTをコードする配列に下線を付している。TS8v2は、TEVプロテアーゼ認識配列に連結したグルタチオントランスフェラーゼとそれに続く挿入オクタペプチドのコード配列を含む。
【図13】図13は、実施例2に記載のSjGST−TM−オクタペプチド(バージョン1)をコードするプラスミドから発現したタンパク質のSDS−PAGEゲルを示す(レーン1:マーカー;レーン2:SjGST−TM−オクタペプチド(細菌の全溶解物として);レーン3:SjGST−TM−オクタペプチド(グルタチオンアガロースカラムの流出物);レーン4:溶出したSjGST−TM−オクタペプチドタンパク質(単一種として27kDaの分子量を有する))。
【図14】図14は、実施例3に記載のSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)をコードするプラスミドから発現したタンパク質のSDS−PAGEゲルを示す。レーン1:分子量マーカー;レーン2:誘導細胞からの全溶解物;レーン3:溶出したSjGST−TEV−オクタペプチド融合タンパク質(分子量=28.4kDa)。
【図15】図15は、実施例5に記載の化学合成された酢酸オクトレオチドのマトリクス支援レーザー脱離イオン化質量分析法(MALDI−TOF−MS)による分析を示す。化学合成された酢酸オクトレオチドの理論質量は1018Daである。化学合成された酢酸オクトレオチドの実測質量は1019Daであった。化学合成された酢酸オクトレオチドNa+の理論質量は1041Daである。化学合成された酢酸オクトレオチドNa+の実測質量は1041Daであった。
【図16】図16は、実施例6に記載のSjGST−TM−オクタペプチド融合タンパク質バージョン1の実測質量のMALDI−TOF−MSによる分析を示す。SjGSTの理論質量は25498Daである。SjGStに付加されたオクタペプチドの理論質量は2259Daである。これにより、SjGSST−TM−オクタペプチドバージョン1融合タンパク質の合計理論質量は27757Daとなる。このSjGSST−TM−オクタペプチドの実測質量は27751Daであった。質量誤差百分率は0.02%である。理論質量と実測質量との間で0.1%の質量誤差百分率は許容されうる。
【図17】図17は、実施例7に記載の、トロンビンによる開裂後のSjGSTタンパク質(バージョン1)の実測質量のMALDI−TOF−MSによる分析を示す。精製された融合タンパク質の溶液を、100mM DTTを用いて還元し、PD−10ゲル濾過カラムによりバッファーを10mMリン酸ナトリウムバッファー(2mM EDTAを含有、pH7.8)に交換した。次いで、融合タンパク質の溶液を、10mMリン酸ナトリウムバッファー(pH7.8)中、37℃にて24時間トロンビン消化に供した。この溶液に、3mM DTTを添加し、正しくないジスルフィド結合の形成を防いだ。次いでこれをMALDI−TOF−MS分析に供した。SjGSST−TM−オクタペプチド融合タンパク質バージョン1の理論質量は27757Daである。トロンビンにより開裂した後のSjGSTタンパク質の理論質量は26150Daである。トロンビンにより開裂した後のSjGSTタンパク質の実測質量は26153Daであった。質量誤差百分率は0.01%である。トロンビン開裂後のオクタペプチドの理論質量は1607Daである。トロンビン開裂後のオクタペプチドの実測質量については、この特定のMALDI−TOFスペクトルでは低分子量の分子に対する解像度が低いため、測定できなかった(図18)。
【図18】図18は、実施例8に記載の、精製されたSjGST−TM−オクタペプチド融合タンパク質バージョン1から、シアノゲンブロマイドによりメチオニン部位で化学的に開裂した後の本発明のFCFWKTCTの配列を有するオクタペプチドのMALDI−TOF−MS分析を示す。精製されたタンパク質の溶液を、100mM DTTを用いて還元し、PD−10ゲル濾過カラムによりバッファーを10mMリン酸ナトリウムバッファー(2mM EDTAを含有、pH7.8)に交換した。次いで、この溶液を、10mMリン酸ナトリウムバッファー(pH7.8)中、37℃にて24時間の、オクタペプチドに直接隣接したメチオニン残基のシアノゲンブロマイド(すなわち、化学的)媒介性の開裂に供した。得られた溶液に3mM DTTを添加し、正しくないジスルフィド結合の形成を防いだ。次いでこれをMALDI−TOF−MS分析に供した。このオクタペプチドの理論質量は1035Daである。この生物学的に合成されたオクタペプチドの、メチオニンでの化学的開裂後の実測質量は1034Daであった。質量誤差百分率は0.09%である。化学合成された酢酸オクトレオチドと、生物学的に合成されたオクタペプチドFCFWKTCTとの実測質量の差は16Daである。これは、酢酸オクトレオチドがそのC末端残基としてL−スレオニノールを有するように化学的に修飾されているためである。生物学的なオクタペプチドFCFWKTCTの場合には、天然のアミノ酸であるL−スレオニンがC末端残基として存在している。
【図19】図19は、実施例9に記載の、ラクトースを用いた大腸菌でのSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)の自己誘導、およびその封入体からの精製を示す。レーン1:分子量マーカー;レーン2:大腸菌の誘導細胞からの全溶解物;レーン3:ソニケーション後の大腸菌細胞溶解物由来の上清;レーン4:2M尿素および100mM Tris(pH12.5)を用いた処理後の細胞ペレット由来の上清;レーン5:2M尿素および100mM Tris(pH12.5)を用いた処理とそれに続く塩酸によるpH8への調整後の細胞ペレット由来の上清;レーン6:2M尿素および100mM Tris(pH12.5)で溶解され、続いて塩酸によるpH8への調整を経た大腸菌細胞由来の残存ペレット。各レーンにおいて、17kDaに見られるタンパク質のバンドは、細胞を消化する目的で添加されたリゾチームである。
【図20】図20は、実施例10に記載の、化学合成された酢酸オクトレオチドの3つの炭素の架橋PEG化を示す。比較のために、酢酸オクトレオチドのMALDI−TOF−MSスペクトルを図15に示す。図20は、化学合成された酢酸オクトレオチドが、ジスルフィド架橋を横切る3つの炭素の架橋を介して、5kDaのポリエチレングリコールに共有結合していることを示す。このPEG化された酢酸オクトレオチドの理論質量は6092Da(すなわち、ポリエチレングリコール5073Da+酢酸オクトレオチド1019Da)である。ポリエチレングリコールの実測分子量は5073Daである。PEG化された酢酸オクトレオチドの実測分子量は6097Daである。質量誤差百分率は0.08%である。
【図21】図21は、実施例11に記載のように、SDS−PAGEを用いて、ポリエチレングリコールが共有結合する3つの炭素の架橋の化学的な挿入によるSjGST−TM−オクタペプチド融合タンパク質(バージョン1)の修飾を示す。レーン1:分子量マーカー;レーン2:27757Daの理論分子量を有する、精製されたSjGST−TM−オクタペプチド(バージョン1);レーン3:4℃にて72時間のSjGST−TM−OCTとの5Daのポリエチレングリコールのジスルフィド部位特異的な架橋反応とそれに続く37℃にて24時間のトロンビンによる消化。SjGST−TM−オクタペプチドのタンパク質バンド27.7kDaは消失し、モノPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化)、ジPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化、および、GST中のジスルフィド結合のモノPEG化)、並びに、トリPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化、および、GST中の2つのジスルフィド結合のジPEG化)が出現した;レーン4:反応バッファー中における5kDaのポリエチレングリコール試薬(3等量濃度)のジスルフィド部位特異的な架橋。
【図22】図22は、実施例12に記載の、MALDI−TOF−MS分析を用いて、ポリエチレングリコールが共有結合した3つの炭素の架橋の化学的な挿入によるSjGST−TM−オクタペプチド(バージョン1)の修飾を示している。この図は、トロンビン消化され、ジスルフィド部位特異的にPEG化されたSjGST−TM−オクタペプチド(バージョン1)タンパク質溶液のMALDI−TOF−MSを示す。SjGST−TM−オクタペプチド(バージョン1)融合タンパク質の理論質量は27757Daである。トロンビンにより開裂したSjGSTの大きい断片の理論質量は26150Daである。トロンビンによる理論上の小さい開裂産物は、1607Daの断片である。このことから、オクタペプチド(1607Da)に連結されたポリエチレングリコール(5073Da)の理論質量は6680Daとなる。オクタペプチドに連結したポリエチレングリコールの実測質量は6692Daであった。質量誤差は0.1%である。
【図23】図23は、BALB/Cマウスの血清成長ホルモン(GH)レベルの低下における本発明のペプチドの3つの実施例の効果を(化学合成された酢酸オクトレオチドと比較して)示す。図23AはマウスのGHレベル(ng/ml)を示す。図23Bは、化学合成された酢酸オクトレオチドと比較したGHの抑制百分率を示す。
【発明を実施するための形態】
【0036】
第1の形態において、本発明は、X1CX2WKX3CTのアミノ酸配列からなる単離されたペプチドを提供し、この際、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVであり、これらは任意の順列組み合わせであり、それぞれのアミノ酸はL配置であり、ペプチドは2つのシステイン残基の間にジスルフィド結合を含む。よって、本発明のペプチドは、以下のアミノ酸配列のいずれかを有しうる:FCFWKTCT、FCFWKVCT、FCYWKTCT、FCYWKVCT、ACFWKTCT、ACFWKVCT、ACYWKTCTまたはACYWKVCT。好ましくは、本発明のペプチドは、FCFWKTCTまたはACYWKVCTのアミノ酸配列からなる。
【0037】
本明細書において、アミノ酸配列は、標準的な一文字または三文字のアミノ酸表記を用いて規定され、G=グリシン(Gly)、P=プロリン(Pro)、A=アラニン(Ala)、V=バリン(Val)、L=ロイシン(Leu)、I=イソロイシン(Ile)、M=メチオニン(Met)、C=システイン(Cys)、F=フェニルアラニン(Phe)、Y=チロシン(Tyr)、W=トリプトファン(Trp)、H=ヒスチジン(His)、K=リジン(Lys)、R=アルギニン(Arg)、Q=グルタミン(Gln)、N=アスパラギン(Asn)、E=グルタミン酸(Glu)、D=アスパラギン酸(Asp)、S=セリン(Ser)、およびT=スレオニン(Thr)である。
【0038】
各アミノ酸についてのDNAコドンを表1に示す:
【0039】
【表1】
【0040】
各アミノ酸が(D配置とは逆の)L配置で存在し、2つのシステイン残基間にジスルフィド結合を含む本発明の単離されたペプチドは、化学合成された酢酸オクトレオチドと同一の三次元結合部位構造を保持しており、よって、化学合成された酢酸オクトレオチドと同一の活性を保持している。酢酸オクトレオチドがD−Phe1残基およびD−Trp4残基を含んでおり、酢酸ランレオチドがD−Ala1残基およびD−Trp4残基を含んでいるのに対し、本発明のペプチドは、すべてL−エナンチオマーからなっている。
【0041】
本発明のペプチドは、末端に化学修飾を含んでもよい。かような修飾によれば、例えば、プロテアーゼによる分解に対する感受性を低下させることなどにより、ペプチドの安定性が向上しうる。
【0042】
本発明のペプチドFCFWKTCTの分子モデリングによれば、その予測される三次元化学構造は化学合成された酢酸オクトレオチドのものと有意には異なっていないことが示されている。図3に示すように、ペプチドFCFWKTCTにおけるアミノ酸の相対位置は、化学合成された酢酸オクトレオチドのアミノ酸と同様の位置に配置されている。最も重要なこととして、Phe3、Trp4、Lys5およびThr6は、本発明のペプチドにおいて、酢酸オクトレオチドにおけるのと同一の相対位置に存在している。リジン5は異なる方向に位置するように見えるが、その直鎖状の性質からすれば溶液中では非常に柔軟であることになる。このことは、リジン5について図3に示す2つの位置の間の差異が生物学的には有意でないことを意味する。図3において標識されているアミノ酸は、オクトレオチドの生物活性に必須であることが知られている;それらは酢酸オクトレオチドのその細胞表面受容体への結合に関与しているのである。このように、本発明のペプチドFCFWKTCTは、酢酸オクトレオチドと同一の三次元化学結合部位の特性を有しており、分子モデリングの研究によれば、生体系において同様に作用すると予測されている。本発明のペプチドのそれぞれの三次元配座も、分子モデリング研究により、同様に酢酸オクトレオチドまたは酢酸ランレオチドと同一の三次元化学結合部位の特性を保持すると予測されている。
【0043】
本発明のペプチドは、好ましくは当該ペプチドをコードするポリヌクレオチドの生物学的宿主における発現により作製され、これによれば、ペプチドの生産について生物有機体のタンパク質発現系に依存することになる。よって本発明のペプチドは、酢酸オクトレオチドや酢酸ランレオチドを生産するのに用いられる化学合成法とは対照的に、生物学的に合成されることが好ましい。本発明のペプチドの生物学的な合成はまた、2つのシステイン残基間で天然にジスルフィド結合を形成することになる。よって、他の形態では、本発明は、本発明のペプチドをコードするポリヌクレオチドを提供する。当該ポリヌクレオチドは、好ましくは、本発明のオクタペプチドの8アミノ酸を、天然のホルモンであるソマトスタチンの残りのアミノ酸とは切り離してコードする。よって、ソマトスタチンをコードする天然のゲノムDNAは、本発明の範囲から除外される。上記ペプチドを発現可能な任意の生物学的宿主が用いられうる。適切な宿主は本技術分野において公知であり、細菌、酵母、昆虫細胞、および動物細胞が挙げられる。
【0044】
本発明のペプチドをコードするポリヌクレオチドは、DNAまたはRNAでありうる。よって、本明細書ではDNA配列を記載するが、等価なRNA配列も用いられうるものとする。
【0045】
本発明のペプチドFCFWKTCTをコードするポリヌクレオチドは、コアとなるヌクレオチド配列5’−TTYTGYTTYTGGAARACNTGYACN−3’を含み、この際、Y=CまたはTであり、R=AまたはGであり、N=A、C、T、またはGであり、任意の順列組み合わせが可能である。細菌中での発現のために上記ポリヌクレオチドをベクターに導入する際、ポリヌクレオチドの好ましい配列は、細菌での最も一般的なコドン使用頻度に基づき、5’−TTCTGTTTTTGGAAAACCTGTACC−3’である。ペプチドFCFWKTCTをコードする16個の異なるヌクレオチド配列を表2に示し、下線を付した配列は最も一般的に用いられている細菌のコドンに対応している。本発明のポリヌクレオチドは、表2に示す配列のいずれか1つを含みうるが、これらの配列には限定されない。ペプチドFCFWKTCTをコードするポリヌクレオチドを酵母宿主細胞中で発現させるときには、好ましいコドンは酵母において最も一般的なコドン使用頻度に従って選択されうる。
【0046】
【表2】
【0047】
ACYWKVCT、FCFWKVCT、FCYWKTCT、FCYWKVCT、ACFWKTCT、ACFWKVCTまたはACYWKTCTのいずれかのペプチドをコードするポリヌクレオチドは、それぞれ下記の表3〜9に示すコア配列を有する。表3〜9に示すコア配列において、Y=CまたはTであり、R=AまたはGであり、N=A、C、TまたはGであり、任意の順列組み合わせが可能である。各ペプチドをコードする16個の異なるヌクレオチド配列の例を表3〜9に示し、下線を付した配列が、最も一般的に用いられている細菌のコドンに対応する。本発明のポリヌクレオチドは、表3〜9に示す配列のいずれかを含みうるが、これらの配列には限定されない。ポリヌクレオチドが酵母宿主細胞中で発現する場合、好ましいコドンは、酵母における最も一般的なコドン使用頻度に従って選択されうる。
【0048】
【表3】
【0049】
【表4】
【0050】
【表5】
【0051】
【表6】
【0052】
【表7】
【0053】
【表8】
【0054】
【表9】
【0055】
本発明のペプチドをコードするポリヌクレオチドは、宿主細胞への形質転換のためのヌクレオチドベクター中に導入されうる。当該ベクターは、DNAベクターまたはRNAベクターでありうる。当該ベクターは、プラスミドベクターであってもよい。好ましくは、当該ベクターは、本発明のペプチドをコードするポリヌクレオチドに作動可能なように連結されたプロモーター配列を含む。適当なヌクレオチドベクターとしては、pGex4.3(GEヘルスケア)などの市販のプラスミドベクターが挙げられ、他の代表的な例を表3に示す。Hosfield T et al., (1998) Biotechniques, Vol. 25, pp. 306-309,di Guan et al., (1988), Gene, 67; 21-30、または、Makrides (1996), Microbiol Rev., Vol. 60; 512-538に記載の任意のベクターもまた、用いられうる。
【0056】
よって、本発明は、本発明のペプチドをコードする発現ベクターを提供する。本発明において用いられうる市販の発現系の例を表10に示すが、これらの例には限定されない。
【0057】
【表10】
【0058】
好ましくは、本発明のペプチドは、宿主細胞からのその単離を単純化する目的で、融合タンパク質の一部として発現する。融合タンパク質は、本発明のペプチドおよび1または2以上のキャリアタンパク質を含む単一のポリペプチドである。このキャリアタンパク質は、融合タンパク質が生産される発現系から単離されうる任意のタンパク質でありうる。
【0059】
本発明のペプチドを含む融合タンパク質は、検出されて宿主細胞から単離されうる任意のキャリアタンパク質を含みうる。好ましくは、このキャリアタンパク質は異種タンパク質(すなわち、宿主細胞で天然にはつくられないタンパク質)である。このキャリアタンパク質は分子タグとして機能し、本発明のペプチドの単離を可能にする。例えば、融合タンパク質は、グルタチオン−S−トランスフェラーゼ(GST)(GEヘルスケア)、ポリヒスチジンタグタンパク質(クロンテック)、マルトース結合タンパク質(ニューイングランドバイオラボ)、ビオチン化融合タンパク質(プロメガ)、カルモジュリン結合ペプチド(ストラタジーン)、β−ラクタマーゼ(ジェランティス)などの任意の市販のタンパク質タグ、および本技術分野において公知の他のものを(キャリアタンパク質として)含みうる。また、イオン交換クロマトグラフィによる融合タンパク質の精製とその後の適当な酵素を用いた消化を可能にするために、融合タンパク質にイオン電荷を付与する任意のタンパク質(例えば、C末端に導入されうるポリアルギニン)(Fuchs SM & Raines RT, (2005) Protein Science Vol. 14; pp 1538-1544)。
【0060】
よって、他の形態において、本発明は、本発明のペプチドおよび1または2以上のキャリアタンパク質を含む融合タンパク質を提供する。本発明はまた、本発明のペプチドおよびキャリアタンパク質を含む融合タンパク質をコードするポリヌクレオチドをも提供する。本発明の融合タンパク質をコードするポリヌクレオチドは、本明細書に記載の任意のベクター中に導入されうる。キャリアタンパク質をコードするヌクレオチド配列は、上述したコアヌクレオチド配列(または本発明のペプチドをコードする任意の他の配列)の5’末端のすぐ上流にインフレームで位置してもよいし、当該コア配列のすぐ下流に位置してもよい。あるいは、融合タンパク質をコードするヌクレオチド配列は、ペプチドとキャリアタンパク質との間のスペーサー領域をコードしてもよい。このスペーサー領域は、キャリアタンパク質の正しい折り畳みを可能とするようにペプチドとキャリアタンパク質との間に十分な空間を提供しうる。
【0061】
さらに、このスペーサー領域は、融合タンパク質の発現後にキャリアタンパク質からのペプチドの分離を可能とすべく、1または2以上の切断部位(1または2以上の酵素的または化学的な切断部位)をコードしてもよい。本発明のペプチドを含む融合タンパク質中に導入されうる酵素的な切断部位としては、(a)TEVプロテアーゼ切断部位(Glu-Asn-Leu-Tyr-Phe-Gln* X(ここで、Xは任意のアミノ酸(プロリンを除く)であり、*は酵素切断部位を表す));(b)エンテロキナーゼ切断部位(Asp-Asp-Asp-Asp-Lys*);(c)因子Xaプロテアーゼ切断部位(Ile-Glu/Asp-Gly-Arg*);(d)Arg−Cプロテイナーゼ切断部位(X-X-R*X-X-)(ここで、Xは任意のアミノ酸(プロリンを除く)であり、存在する場合には、ペプチドのN末端の上流が、切断時にいかなる他のN末端アミノ酸もなしに-Phe-Cys-Phe-Trp-Lys-Thr-Cys-Thrの配列を生成する);(e)タバコベインモットリングウイルス(TVMV)およびインテイン由来のプロテアーゼによる切断部位などの、本技術分野において公知の他のもの、が挙げられる。化学的な切断部位の適当な例としては、(a)シアノゲンブロマイドによる切断のためのMet;ギ酸による切断のためのAsp;および、(c)本技術分野において公知の他の切断部位。
【0062】
上述した切断用プロテアーゼの認識配列のDNAコドンを表11に示す。
【0063】
【表11】
【0064】
切断部位をコードするヌクレオチド配列は、本発明のペプチドをコードするコアヌクレオチド配列のすぐ5’に位置しうる。よって、ヌクレオチド配列GARAAYTTATAYTTYCAR(TEVプロテアーゼ切断部位をコードする;Y=CまたはTであり、RはAまたはGであり、任意の順列組み合わせが可能である)は本発明のペプチドをコードするコアヌクレオチド配列のすぐ5’に位置しうる。例えば、ペプチドFCFWKTCTおよびTEVプロテアーゼ切断部位をコードする本発明のポリヌクレオチドは、コアヌクレオチド配列5’-GARAAYTTATAYTTYCARTTYTGYTTYTGGAARACNTGYACN-3’を含むことができ、Y=CまたはTであり、R=AまたはGであり、N=A、C、TまたはGであり、任意の順列組み合わせが可能である。ポリヌクレオチドが細菌での発現用のベクター中に導入される場合、細菌での最も一般的なコドン使用頻度に基づき、好ましい配列は5’ GAA AAC CTG TAT TTT CAG TTC TGT TTT TGG AAA ACC TGTACC 3’である。酵母の好ましいコドンを含む他の配列が、酵母宿主中でタンパク質を発現させる際には用いられうる。融合タンパク質のペプチド部分をコードする他のヌクレオチド配列の例を、表2〜9に示し、下線を付した配列は、最も一般的に用いられている細菌のコドンに対応する。本発明の融合タンパク質をコードするポリヌクレオチドは、表2〜9に示す配列の任意のものを含みうるが、これらの配列には限定されない。本発明の融合タンパク質を生産するのに用いられうる適当な発現系および精製系としては、市販のpETシステム(ノバジェン)、Ni−NTA精製システム(キアゲン)、pMALタンパク質融合・精製システム(ニューイングランドバイオラボ)、ピンポイント(商標)Xaタンパク質精製システム(プロメガ)、CBPカルモジュリン結合ペプチドアフィニティタグシステム(ストラタジーン)、INTEIN(商標)およびINTEIN−TWINシステム(ニューイングランドバイオラボ)、EndoproteinAceシステム(ジェランティス)、および他のもの(例えば、表10を参照)が挙げられる。
【0065】
1982年に、生物活性断片の配座が安定したソマトスタチンの類似体として酢酸オクトレオチドが初めて化学合成されて以来、他のソマトスタチン類似体についての研究は専ら、化学合成される他のペプチド断片に集中していた(Weckbecker, G. et al., (2003) Nature Reviews, Vol. 2, pp. 999-1017)。本発明のペプチドの生物学的な生産により、(1)大腸菌等の宿主細胞中でジスルフィド結合が自然に形成され、(2)酢酸オクトレオチドの三次元の化学構造を保持しており、(3)酢酸オクトレオチドの細胞ベースの毒性(D−アミノ酸(大腸菌等の有機体の増殖を阻害する)の存在による(Meister, (1965) Biochemistry of the amino acids. New York Academic Press))の問題を解決し、(4)大腸菌に対して毒性を示さない封入体中でのペプチドの生産を可能とする、そのようなペプチドの安価な製造方法が提供される。したがって、本発明によれば、上記ペプチドの高収率で低コストでの生物学的な生産が可能となる。
【0066】
特にD−アミノ酸の生きている有機体における毒性効果について述べると、いくつかのD−アミノ酸(D−トリプトファンなど)は大腸菌に対して毒性を示しうることが知られている(Soutourina et al., (2004) J. Biological Chemistry; Vol 279(41): pp 42560 - 42565)。これは、D−アミノ酸は生物界に見られるものの、リボソームのタンパク質合成酵素はD−アミノ酸が天然のポリペプチド中には導入されないように保証しているためである。天然のポリペプチドからのD−アミノ酸の排除の第1段階は、アミノアシル−tRNA合成酵素により担われている。これは、D−アミノ酸が3つの異なる機構によって細胞毒性をもたらしていることによる。上記細胞毒性の第1の機構は、D−アミノ酸がポリペプチド中に導入されると機能しないタンパク質が生成するというものである。このことは、L−アミノ酸による細胞のタンパク質翻訳機構の立体特異性にも反映している。上記細胞毒性の第2の機構は、D−アミノ酸が12アミノ酸ベースのペプチド中に導入された実験によって示されている;トリチウム標識チミジンの生細胞への導入により測定されるように、D−アミノ酸は細胞成長および細胞増殖を有意に低下させることがわかっている(Hayry et al., (1995) The FASEB journal, Vol 9; pp 1336 - 1344)。上記細胞毒性の第3の機構は、ペルオキシソームのD−アミノ酸オキシダーゼ(D−AAO)が代謝および生きている有機体からの特定のD−アミノ酸の除去に有効であるということである。これは、D−AAOがD−アミノ酸を対応するα−ケトン酸およびNH3へと代謝することによる。このプロセスでは、補酵素であるフラビンアデニンジヌクレオチドの還元型から酸化型への再生により細胞毒性のある過酸化水素が生成し、次いでこれがより毒性の高い反応性酸素種を生成する。これらのフリーラジカルは、重篤な細胞障害の原因となる(Krug et al., (2007) Am J. Physiol. Renal Physiol. Vol 293; pp F382 - F390)。
【0067】
他の形態において、本発明は、本発明のペプチドの製造方法を提供し、当該方法は、本発明のポリヌクレオチドまたはベクターを、当該ポリヌクレオチドまたは当該ベクターを発現できる宿主細胞中に導入し、当該宿主細胞から当該ペプチドを単離することを含む。本発明のペプチドをコードするポリヌクレオチドまたは本発明の融合タンパク質をコードするポリヌクレオチドが用いられうる。宿主細胞中で本発明のペプチドを生産することにより、各アミノ酸はL配座で提供される。好ましくは、上記ポリヌクレオチドまたはベクターは、上述したように、本発明のペプチドおよびキャリアタンパク質を含む融合タンパク質をコードするヌクレオチド配列を含有する。よって、本発明のペプチドの精製工程は、好ましくは、宿主細胞から融合タンパク質を単離し、当該融合タンパク質から上記ペプチドを単離することを含む。
【0068】
本発明のペプチドおよび融合タンパク質は、単純に生産され単離されうるため、低コストである。本発明のペプチドまたは融合タンパク質をコードするポリヌクレオチドまたはベクターは、当業者に公知の任意の手段によって適切な宿主細胞中に導入されうる。宿主細胞の培養条件は、ペプチドまたは融合タンパク質が最高の収率で生産されるように最適化されうる。例えば、lacプロモーターの誘導体(tac、pac、rac)は最も強力な細菌のプロモーターであり、これらのいずれもが本発明のベクターに用いられうる。これらは、大腸菌における外来遺伝子の誘導された過剰発現によく用いられる。しかしながら、工業的な醗酵プロセスにこれらを用いることは、誘導剤であるイソプロピル−β−D−チオガラクトピラノシド(IPTG)のコストが高いことにより制限される。本発明の方法での使用に適した大規模培養系では誘導剤としてラクトースを用いることができる。これは、ラクトースがIPTGと同等の効率で本発明のベクターを誘導することができるためである。大腸菌では、誘導剤としてラクトースを用いることができるし、グルコースの存在下であっても炭素/エネルギー源として用いることができる(Neubauer P. et al., (1994) FEMS Microbiol Rev. Vol. 14; pp. 99-102; Vasala A. et al., (2005) J. Biotechnol. Vol. 117; pp. 421-431)。この低コストのアプローチを、改変培地および改変増殖条件を利用した自己誘導と組み合わせてもよい(Studier FW., (2005) Protein Expression and Purification Vol. 41; pp. 207-234)。大腸菌およびラクトースを用いたSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)の高収率で低コストの自己誘導と、それに続く大腸菌中の封入体からの融合タンパク質の単離の例を、実施例9および図19に示す。
【0069】
ペプチドまたは融合タンパク質の化学的な修飾を、ペプチドの安定性を改善し、ペプチドの単離方法を単純化させる目的で、ペプチドまたは融合タンパク質の発現の後に行なってもよい。
【0070】
次いで、本技術分野において公知の手法により、ペプチドは宿主細胞の成分から単離されうる。ペプチドが融合タンパク質の一部として発現している場合には、当該ペプチドは本技術分野において公知の手法により、融合タンパク質から単離されうる。特に注目すべきは、封入体からのタンパク質の単離である。なぜならば、これによってかなりの製造上および経済的なプロセスの利点があるためである。これらの利点としては:(a)可溶性のタンパク質として大量に存在すると宿主細胞に対して毒性を示すタンパク質が高レベルで発現するが、封入体として存在することで毒性を示さないこと;(b)封入体のサイズおよび密度が細胞の混入物質とは異なることで、細胞からの封入体の単離が容易であること;(c)細胞のプロテアーゼによるタンパク質分解の攻撃に対して抵抗性であることにより、発現したタンパク質の分解が防止されること;並びに(d)混入物質がより少なく、封入体中で対象タンパク質が均一であること、が挙げられる。
【0071】
封入体を単純に洗浄すると不純物が除去され、天然タンパク質の収率はより改善される。これらの利点により、タンパク質の商業的な生産のために大腸菌中の封入体として発現する組換えタンパク質が広く開発されている。封入体からの生物活性なタンパク質を回収する際には、タンパク質の凝集体を充分に可溶化し、次いで、可溶化したタンパク質を生物活性な形態へとリフォールディングする必要がある。封入体中のタンパク質は折り畳みの過程の中間段階で存在し、かなりの量の二次構造を有していることが示されている。この天然様の二次構造を乱すことなく封入体からのタンパク質を可溶化できれば、リフォールディング時のタンパク質凝集の程度も低くなり、生物活性のタンパク質の回収率も高くなる。したがって、生物活性タンパク質の回収率の改善には、ランダムコイルのタンパク質構造を生じることなく封入体の凝集物をマイルドに可溶化させることが重要である。1つのアプローチとして、タンパク質の等電点から離れたpHショックをタンパク質凝集体に与えることである。これにより、極めて低濃度の変性剤の存在下でタンパク質は可溶性となる。封入体のタンパク質がかようなマイルドな条件下でひとたび可溶化されれば、それに続く精製タンパク質のリフォールディングも容易となり、生物活性タンパク質の回収率も高くなる。これに関連して、2M尿素ではタンパク質の構造が開かれず、タンパク質の天然の二次構造が保存されることが示されている(Rathore A.S. et al., (2003) Biotechnology Progress Vol. 19; pp. 1541-1546; Lee Y.S. et al., (2003), Biotechnology & Applied Biochemistry Vol. 38; pp. 9-13; DeNardo, S.J. et al., (2003) Clinical Cancer Research Vol. 9; pp. 3854s-3864s; Pezza J.A. et al., (2004) Chemical Communications pp. 2412-2413; Singh S.R. & Panda AK. (2005) Journal of Bioscience and Bioengineering. Vol. 99; pp. 303-310; Rajan R.S. et al., (2006) Protein Science Vol. 15; pp. 1063-1075; Wang F. et al., (2008) Nuclear Medicine and Biology Vol. 35; pp. 665-671)。よって、本発明のペプチドの製造方法は、本発明のペプチドまたは本発明の融合タンパク質を、宿主細胞中の封入体から単離することを含みうる。
【0072】
また、従来の液相でのリフォールディングプロセスの際のタンパク質凝集の問題は、膨張層吸着クロマトグラフィーと組み合わせた固相でのリフォールディング手法を用いることで回避されうる。その結果、細胞ホモジネートからの封入体中のタンパク質は、高収率で正確にリフォールディングされる(Cho T.H. et al., (2002) Bioseparation Vol. 10, pp. 189-196)。このことは、大腸菌において生産されるヒト上皮成長因子について示されている。膨張層吸着クロマトグラフィーは、カチオン交換によって同時にタンパク質を捕捉し、希釈培地から細胞バイオマスを除去する。また、ハイスループットで行うこともでき、その結果、高収率(>90%)となり、精製因子も80%超の純度で20倍となる。かようなタンパク質の精製プロセスは効率的であり、例えば本発明のペプチドの大規模かつ費用対効果の高い製造に採用されうる(Lee Y.S. et al., (2003), Biotechnology & Applied Biochemistry Vol. 38; pp. 9-13)。
【0073】
融合タンパク質および/または化学修飾された融合タンパク質を単離した後、酵素的または化学的な切断部位を介してこれを切断してもよい。例えば、TEV酵素切断部位を含む融合タンパク質の部位特異的な切断は、ヒスチジンタグ付きのTEV酵素を用いて行われうる。ニッケルアガロースカラムを通過させてヒスチジンタグ付きのTEVプロテアーゼを除去することにより、ペプチドを精製することができる。グルタチオンアガロースおよびニッケルアガロースカラムを再生させることで、これらを再利用することができる。それぞれの手法についての個別の工程に関する完全な詳細は、当業者に周知である(Structural Genomics Consortium et al., (2008) Nature Methods, Vol. 5(2), pp. 135-146)。
【0074】
上述したように、本発明のペプチドは、その安定性を改善する目的で化学的に修飾されてもよく、これによりペプチドの半減期が延びて治療効果も改善される。かような修飾により、上記ペプチドは、1年またはそれ以上の期間にわたる使用に適した長期療法として用いられうる、費用対効果に優れた治療用製品に特に適したものとなる。本発明の修飾されたペプチドの有効性が改善されることで、酢酸オクトレオチドまたは酢酸ランレオチドにより可能な現行の投薬計画と比較して、より長期間にわたって患者に投与するのに特に適したものとなる。
【0075】
例えば、下記の文献に詳細に記載されているジスルフィド部位特異的なPEG化の手法などにより、ペプチドおよび/または融合タンパク質をPEG化してもよい:Shaunak et al., (2006) Nature Chemical Biology. Vol. 2, pp. 312-313; Zloh et al., (2006) Physical Chemistry., Vol. F-3-O, pp. 347-349; Brocchini et al., (2006) Nature Protocols, Vol. 1(5), pp. 2241-2252 ; Balan et al., (2007) Bioconjugate Chemistry, Vol. 18, pp. 61-76; Godwin et al., (2007) Theoretical Chemical Accounts, Vol. 117, pp. 259-265; Zloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083 ; Brocchini et al., (2008), Advanced Drug Delivery Reviews, Vol. 60(1), pp. 3-12。ポリエチレングリコールのサイズは、5kDa〜40kDaでありうる。好ましいサイズは、20kDa〜30kDaである。
【0076】
小さいタンパク質およびペプチドは、大きいタンパク質よりも多くのジスルフィド結合を有する傾向にある。これは、前者が自信の比較的少ない疎水相互作用を補償する必要があるためである。溶媒に接近できるジスルフィド結合は通常ほとんどのタンパク質に存在することから、このジスルフィド結合を化学的に減らして2つの遊離したシステインの硫黄原子としつつ、タンパク質またはペプチドの三次構造を維持することが可能である。次いで、PEG化はこれらの2つのシステイン硫黄原子を3炭素の架橋を介して再結合させるビスアルキル化によって達成されうる。このアプローチの利点は、遊離システインを導入する(これはタンパク質またはペプチドの不溶性を上昇させる)ためのタンパク質の組換え的な改変を必要とすることなく、選択的かつ効率的なチオールの付加反応が行われうることである。より詳細には、このアプローチ手法は、すべての天然のジスルフィド結合において、双方の硫黄原子の化学的な反応性を利用するものである。
【0077】
2つのシステイン残基の2つの硫黄原子の間に3炭素の架橋を挿入する予備的な工程により、(a)安定性が向上するという特有の利点を有し、(b)上記3炭素の架橋その後のさらなる修飾の潜在性を付与する、修飾ペプチドが得られる。
【0078】
よって、本発明はまた、X1CX2WKX3CTのアミノ酸配列からなる単離されたペプチドを提供し、この際、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVであり、任意の順列組み合わせが可能であり、それぞれのアミノ酸はL配置であり、2つのシステイン残基の2つの硫黄原子は3つの炭素原子を介して連結されている。よって、当該ペプチドは、以下のアミノ酸配列のいずれかを有しうる:FCFWKTCT、FCFWKVCT、FCYWKTCT、FCYWKVCT、ACFWKTCT、ACFWKVCT、ACYWKTCTまたはACYWKVCT。好ましくは、上記ペプチドは、FCFWKTCTまたはACYWKVCTのアミノ酸配列からなる。こうして、本発明のペプチドにおける2つのシステイン残基の間には安定化相互作用が働く。例えば、ジスルフィド結合に3炭素の架橋を挿入することで修飾して、さらなる安定化相互作用としてもよい。
【0079】
本発明者らによる分子モデリング研究により、2つのシステイン残基の2つの硫黄原子の間に3炭素原子を含むFCFWKTCTペプチドは、化学合成された酢酸オクトレオチドにきわめて類似した三次元配座を保持していることが示されている(図6を参照)。したがって、かようなペプチドもまた、未修飾の、生物学的に合成された本発明のペプチドについて本明細書に記載されたのと同一の用途(特に、医療用途)に用いられうる。2つのシステイン残基の2つの硫黄原子の間に3炭素原子を含む本発明のペプチドのそれぞれの三次元配置もまた、分子モデリング研究により、酢酸オクトレオチドまたは酢酸ランレオチドと同一の三次元化学結合部位の特性を保持していることが予測されている。したがって、かようなペプチドは、酢酸オクトレオチドまたは酢酸ランレオチドと同一の活性を保持している。
【0080】
ポリエチレングリコール分子などの親水性ポリマーが、3炭素架橋に共有結合していてもよい。好ましくは、ポリエチレングリコール分子は5kDa〜50kDaの分子量を有し、より好ましくは10kDa〜40kDaの分子量を有し、最も好ましくは20kDa〜30kDaの分子量を有する。あるいは、当業者に公知のポリエチレングリコール以外の親水性ポリマーが、3炭素架橋に共有結合していてもよい。
【0081】
PEG化試薬は、好ましくは、ポリエチレングリコール試薬の末端の共役部位として、置換されたプロペニル基を有する。この共役部位は、電子吸引性基(例えば、カルボニル)、α,βー不飽和二重結合、およびスルフィン酸として脱離しやすいα,β−スルホニル基を含みうる。電子吸引性基は、チオール付加を促進し、αプロトンのpKaを低下させて、これにより脱離反応が進行しうる。このような化学的な官能性の並置により、潜在的な交差共役系が生じる。ポリエチレングリコールモノスルホン中の共役二重結合は、一連の双方向的で連続した付加−脱離反応を引き起こす。第1のチオレートが付加するとスルホン酸誘導体の脱離が生じうる。これにより、第2のチオレートの付加のためのα,β’位に位置する別の共役二重結合が生じる。これらの2つのチオールがタンパク質のジスルフィド結合由来であれば、もともとのジスルフィドのシステインの硫黄原子間に3炭素の架橋が形成される。この新たな3炭素のジスルフィド架橋は、元の生物学的に合成されたジスルフィド結合よりも、化学的なおよび代謝による分解、並びにプロテオリシス分解に対してずっと安定である。
【0082】
本発明のPEG化ペプチドの調製方法は、2〜3の工程で低コストで行われうる。本発明のペプチドを含む融合タンパク質は、生産された後に続いて上述したようにPEG化されうる。次いで、この融合タンパク質は、イオン交換カラムまたはサイズ排除カラムまたはGST−融合タンパク質用のグルタチオンアガロースアフィニティー精製カラムなどのアフィニティーカラムに溶液を通すことで、未反応のポリエチレングリコールから精製されうる。後者の場合、GST−融合タンパク質はグルタチオンアガロースカラムに結合するようになる。次いで、(例えば、50カラム体積の0.1%トライトンX−114含有PBSとそれに続く20カラム体積のPBSを用いて)カラムを洗浄することで、エンドトキシンが除去されうる。エンドトキシンは洗浄バッファー中に溶出する。好ましくは、これらすべての操作が4℃で行われる。
【0083】
本発明のペプチドのPEG化は、合成されうる化学合成酢酸オクトレオチドの異なるPEG化体の混合物と比較して、好ましくはモノPEG化ペプチドを生成するものである。PEG化タンパク質は、サイズ排除クロマトグラフィーにより、より小さい非PEG化ペプチドから分離されうる。
【0084】
従来公知の発表によると、分子モデリング研究は、化学合成の結果および生物学的アッセイの結果を高い信頼性で予測するのに、化学に基づく実験と有効に組み合わされうる;Shaunak et al., (2006) Nature Chemical Biology. Vol. 2, pp. 312-313; Zloh et al., (2006). Physical Chemistry., Vol. F-3-O, pp. 347-349; Brocchini et al., (2006) Nature Protocols, Vol. 1(5), pp. 2241-2252 ; Balan et al., (2007) Bioconjugate Chemistry, Vol. 18, pp. 61-76; Godwin et al., (2007) Theoretical Chemical Accounts, Vol. 117, pp. 259-265; Zloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083 ; Brocchini et al., (2008), Advanced Drug Delivery Reviews, Vol. 60(1), pp. 3-12。このように、モデリング研究によれば、タンパク質またはぺぷの接近可能なジスルフィド結合への3炭素の架橋の挿入の構造的な効果を高い信頼性で予測することができる。公知のプロトコールは、タンパク質データベース(例えば、タンパク質データバンク(Protein Data Bank, PDB - www.pdb.org))に対して照会を行い、タンパク質の表面に近く、化学的に修飾されうる少なくとも1つのジスルフィド結合を有するタンパク質を発見するために開発されてきた。また、上記プロトコールは、3炭素の架橋によってタンパク質の三次構造が失われてしまうか否かを計算的に予測するのにも用いられうる。このアプローチを用いて、3炭素の架橋の挿入後のタンパク質の生物活性な表面の変化を正確に決定したり、3炭素の架橋へのポリエチレングリコールの連結がタンパク質の生物活性に及ぼす効果を決定したりすることができる。本明細書に記載の分子モデリング研究は、統合された分子モデリングパッケージであるMaestro v6.5およびMacromodel v9.1を用いて行われた。
【0085】
従来、ポリエチレングリコールが溶液中に存在すると、そのアルブミンとの負の優先相互作用のために、ヒトアルブミンの天然のコンパクトな状態を安定化することが知られている。ポリエチレングリコール分子とアルブミンとの間の相互作用は熱力学的に不利であることから(Farruggia et al., (1999), Int. J. Biol. Macromol., Vol. 26, pp. 23-33)、ポリエチレングリコールはタンパク質またはペプチドの生物活性な表面を阻害しないと考えられる。このことは計算的に確認されている;ポリエチレングリコールは、本発明のペプチドとは独立して折り畳まれる。ポリエチレングリコール分子は、タンパク質の周りで自身を「包む」ことはない(Zloh et al., (2006) Proceedings of 6th European Conference on Computational Chemistry, Slovakia)。しかしながら、本発明のペプチドとポリエチレングリコールとのサイズは有意に異なるため、当業者であれば、ポリエチレングリコールがペプチドを巻き込み、それによりペプチドの生物活性を阻害するものと予測していた。本発明者らは、驚くべきことに、本発明のペプチドのPEG化はペプチドの生物活性を阻害しないことを示したのである。
【0086】
PEG化のプロセスは通常、精製されたタンパク質を用いて行われる。これには、所望のPEG化タンパク質を回収するための複数工程の精製プロセスをさらに必要である。インビトロでのタンパク質のリフォールディングおよびPEG化が統合されれば、特に封入体のタンパク質については生産物の生物学的処理に有利であることが知られている。生物活性でPEG化されたタンパク質を封入体から直接得る目的でPEG化をタンパク質の再生プロセスと統合すると、下流の処理能力が有意に改善される。このことは、PEG化リパーゼについて実証されている(Kim M.Y. et al., (2007) Journal of Biotechnology Vol. 131, pp. 177-179; Choi W.C. et al., (2005) Process Biochem. Vol. 40, pp. 1967-1972)。このリパーゼは、尿素およびDTTを用いて変性され、次いでポリエチレングリコールで修飾されていた。この共役ポリエチレングリコール分子はリパーゼのリフォールディングを邪魔しなかった。よって、PEG化およびタンパク質のリフォールディングのプロセスを単一の処理工程へと統合することができた。換言すれば、封入体からの可溶化タンパク質は、単一の処理操作でPEG化され、リフォールディングされうるのである。同様の手法が本発明にも適用されうる。よって、本発明のPEG化ペプチドの製造方法は、宿主細胞中の封入体からの本発明のペプチドまたは融合タンパク質の単離と、それに続く統合されたPEG化およびPEG化ペプチドのリフォールディングとを含みうる。本発明のペプチドまたは融合タンパク質にポリエチレングリコールが共有結合すると、ペプチドまたは融合タンパク質の溶解性がかなり向上し、それによってタンパク質の凝集体の形成が抑制されるというさらなる利点もある(Rajan RS., (2006) Protein Science Vol: 15; pp. 1063-1075)。酢酸オクトレオチドおよび酢酸ランレオチドの不溶性は、工業的な製造上の主要な課題である。
【0087】
他の形態において、本発明は、医薬に用いるための、本発明のペプチドまたはPEG化ペプチドを提供する。本発明のペプチドまたはPEG化ペプチドは、ホルモン障害、特に成長ホルモンの過剰発現に起因するホルモン障害の治療に用いられうる。本発明のペプチドまたはPEG化ペプチドを用いて治療されうるホルモン障害の例としては、末端肥大症および巨人症が挙げられる(S. Farooqi et al., (1999) Pituitary, Vol. 2, pp. 79-88; A.N. Paisley and P.J. Trainer, (2003) Current opinion in Pharmacology, Vol. 3, pp. 672-677; S. Melmed, (2006) New England Journal of Medicine, Vol. 355, pp. 2558-2573)。
【0088】
また、本発明のペプチドまたはPEG化ペプチドは、癌の治療に、特に消化管関連のカルシノイド腫瘍や転移性のカルシノイド腫瘍などの消化管ホルモン分泌癌の治療に用いられうる。
【0089】
本発明のペプチドまたはPEG化ペプチドはまた、酢酸オクトレオチドまたは酢酸ランレオチドが治療上有効であることが知られている任意の疾患または症状の治療にも用いられうる。例えば、本発明のペプチドまたはPEG化ペプチドは、血管作動性腸管ペプチドを分泌する腫瘍(VIPoma)を有する患者における下痢の治療に、および、他の原因による重篤で難治性の下痢の治療にも用いられうる。本発明のペプチドまたはPEG化ペプチドはまた、スルホニルウレアの過剰摂取後の長期にわたる再発性低血糖の治療に、膵島細胞症の乳幼児におけるインスリンの過剰分泌の抑制を補助するのに、および、食道動脈瘤が疑われる患者における出血の抑制を補助するのにも、用いられうる。本発明のペプチドまたはPEG化ペプチドはまた、慢性膵炎による疼痛を有する患者の治療や、胸腺腫瘍、および、糖尿病に起因する眼疾患の治療にも用いられうる(S. Farooqi et al., (1999) Pituitary, Vol. 2, pp. 79-88; A.N. Paisley and P.J. Trainer, (2003) Current opinion in Pharmacology, Vol. 3, pp. 672-677; S. Melmed. (2006) New England Journal of Medicine, Vol. 355, pp. 2558-2573)。
【0090】
よって、本発明は、ホルモン障害(末端肥大症および巨人症);癌(消化管関連のカルシノイド腫瘍や転移性のカルシノイド腫瘍など);血管作動性腸管ペプチドを分泌する腫瘍(VIPoma)を有する患者における下痢;重篤で難治性の下痢;スルホニルウレアの過剰摂取後の長期にわたる再発性低血糖;膵島細胞症の乳幼児におけるインスリンの過剰分泌;食道動脈瘤が疑われる患者における出血、の治療に用いるための、本発明のペプチドまたはPEG化ペプチドを提供する。
【0091】
放射性標識されたソマトスタチン類似体は、ソマトスタチン受容体シンチグラフィーの有用な診断ツールである。上記類似体は、微小なSST発現腫瘍を検出・位置決めするのに用いられ、原発性および転移性の内分泌腫瘍をピンポイントで特定するのに特に有用である。標的化放射線療法の毒性により、現在では並行した開発の場により、SST発現腫瘍を有する患者を治療するための放射性標識されたソマトスタチン類似体の使用が検討されている。内分泌腫瘍の特異的な適応症として、イットリウム90(OctreoTher(登録商標))またはルテチウムなどのエネルギー性のアイソトープを含む新たな化合物は、放射線の深部への侵入により、腫瘍を有効に治療できるものと期待されている。イットリウム標識された化合物であるDOTATOCはまた、患者の60%で疾患を安定化させ、患者の20%では他覚的反応を示すことも判明している(Weckbecker, G. et al., (2003) Nature Reviews, Vol. 2, pp. 999-1017; Susini C & Buscail L, (2006) Ann of Oncology Vol. 17, pp. 1733-1742; Asnacios A et al. (2008) J. Clin. Oncology Vol. 26, pp. 963-970)。本発明はまた、腫瘍可視化剤として用いるための、本発明のペプチドまたはPEG化ペプチドをも提供する。例えば、放射性標識などの検出可能な標識を、2つの硫黄原子間に3炭素の架橋を有する本発明のペプチドに結合させることができ、これにより当該ペプチドが受容体に結合したときに、腫瘍部位において標識ペプチドを検出することが可能となる。上記標識は、上記3炭素の架橋に直接結合してもよいし、上記3炭素の架橋に結合したポリエチレングリコール分子に結合してもよい。本技術分野において公知の放射性標識以外の検出可能な標識もまた、用いられうる。
【0092】
さらに、本発明は、ホルモン障害(末端肥大症および巨人症);癌(消化管関連のカルシノイド腫瘍や転移性のカルシノイド腫瘍など);血管作動性腸管ペプチドを分泌する腫瘍を有する患者における下痢;重篤で難治性の下痢;スルホニルウレアの過剰摂取後の長期にわたる再発性低血糖;膵島細胞症の乳幼児におけるインスリンの過剰分泌;食道動脈瘤が疑われる患者における出血;並びに、糖尿病に起因する眼疾患、の治療のための医薬の製造における、本発明のペプチドまたはPEG化ペプチドの使用を提供する。
【0093】
本発明はまた、上述した疾患のいずれかに罹患した患者の治療方法であって、本発明のペプチドまたはPEG化ペプチドを患者に投与することを含む治療方法をも提供する。
【0094】
本発明はまた、本発明のペプチドまたはPEG化ペプチドおよび製薬上許容されうる担体または希釈剤を含む医薬組成物をも提供する。本発明のペプチドまたは当該ペプチドのPEG化体は、酢酸塩まどの製薬上許容されうる塩として提供されてもよい。上記医薬組成物は、患者に対する所望の投与方法に応じて、任意の適当な形態でありうる。上記医薬組成物は、単回用量製剤であってもよいし、通常は密封容器中で提供され、キットの一部として提供されてもよい。かようなキットは通常(必ずしもそうとは限らないが)、使用説明書を含む。当該キットは、複数の上記単回用量製剤を含みうる。上記医薬組成物は、上述した障害のいずれかの治療用に製剤化されうる。推奨される治療上の用量は、300〜600μg/日または20mg/月であり、2〜200mg/月の範囲である。上記医薬組成物は、毎日、週3回、週1回、2週に1回、または月1回で投与されうる。
【0095】
上記医薬組成物は、例えば、非経口(皮下、筋肉内、静脈内もしくは皮内など)、経口(バッカルもしくは舌下など)、呼吸器経由の、直腸経由の、経鼻、または経皮の経路により、任意の適当な経路による投与に適用されうる。皮下の投与経路が好ましい。かような組成物は、例えば滅菌条件下で活性成分を担体または賦形剤と混合するといった薬学の技術分野において公知の任意の手法により調製されうる。
【0096】
非経口投与に適用される医薬組成物としては、抗酸化剤、バッファー、静菌薬および製剤を非投与者の血液または組織と実質的に等張にしうる溶質を含有してもよい水性または非水性の滅菌注射液;並びに、懸濁化剤および増粘剤を含有してもよい水性または非水性の懸濁剤が挙げられる。注射溶液に用いられうる賦形剤としては、例えば、水、アルコール、ポリオール、グリセリンおよび植物油が挙げられる。上記組成物は、単回用量または複数回用量の容器(例えば、密封されたアンプルまたはバイアル)中に提供されることができ、使用直前に、例えば注射水といった滅菌液体を添加するだけでよい凍結乾燥条件下で貯蔵されうる。即時調製用の注射液および懸濁液は、滅菌された粉末、顆粒、および錠剤から調製されうる。
【0097】
経口投与用の医薬組成物は、カプセルまたは錠剤として;粉末または顆粒として;溶液、シロップまたは懸濁液として(水性または非水性の液体中で;または食用のフォームもしくはホイップとして;またはエマルジョンとして)、提供されうる。錠剤または硬ゼラチンカプセル用の適当な賦形剤としては、ラクトース、トウモロコシでんぷんまたはその誘導体、ステアリン酸またはその塩が挙げられる。軟ゼラチンカプセル用の適当な賦形剤としては、例えば、植物油、ワックス、脂肪、半固体もしくは液状のポリオールなどが挙げられる。液剤およびシロップの調製に用いられうる賦形剤としては、例えば、水、ポリオールおよび糖が挙げられる。懸濁液の調製には、水中油または油中水型の懸濁液を提供するのに油脂(例えば、植物油)が用いられうる。
【0098】
経皮投与に適用される医薬組成物は、長期間にわたって被投与者の上皮とじかに接触したままとされることを前提とした任意のパッチとして提供されうる。例えば、活性成分は、Pharmaceutical Research, 3(6):318 (1986)に概説されるようなイオントフォレシスによりパッチから送達されうる。
【0099】
本発明の方法において用いられうる他の製剤は、経皮送達デバイス(「パッチ」)を用いる。かような経皮パッチは、本発明のペプチドまたはPEG化ペプチドを制御された量で連続的または非連続的に注入するのに用いられうる。医薬製剤の送達のための経皮パッチの構築および使用については、本技術分野において周知の事項である。かようなパッチは、連続的な、パルス状の、または必要に応じた医薬製剤の送達用に構築されうる。
【0100】
他の製剤は、本発明のペプチドまたはPEG化ペプチドの制御された徐放を可能とする生分解性のマイクロスフェアを利用するものである。かような製剤は、合成のポリマーまたはコポリマーまたはハイドロゲルのインプラントを含みうる。かような製剤は、注射、吸入、経鼻または経口の投与が可能である。医薬製剤の送達のための生分解性マイクロスフェアの構築および使用については、本技術分野において周知の事項である(例えば、米国特許第6706289号)。
【0101】
また、本発明によるジスルフィド部位特異的なPEG化オクタペプチドを生分解性マイクロスフェアに配合することで、異なる放出パターン(例えば、オクタペプチドの低減された初期の放出とそれに続く長期にわたる安定した持続的な薬物の放出)および分解するマイクロスフェア内でのオクタペプチドの不活化に対する安定性を有する非常に有効でかつ効率的な徐放性システムが得られる。
【0102】
皮下、静脈内、筋肉内など用に適した製剤のための、適当な医薬担体並びに製剤化および投与の技術は、本技術分野において周知の手法のいずれかにより準備されうる(Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, Pa., 20th edition, 2000を参照)。
【0103】
直腸投与に適用される医薬組成物は、坐剤または浣腸剤として提供されうる。経鼻投与に適用される医薬組成物(担体が固形状の場合)は、鼻の近くに置かれた粉末の容器から嗅がれるように(すなわち、鼻道を介した急速吸入により)投与される例えば20〜500ミクロンの粒子径を有する崩壊性の粉末を含む。経鼻スプレーまたは経鼻ドロップとしての投与のための、担体が液状の場合の適当な組成物としては、活性成分の水溶液または油の溶液が挙げられる。吸入による投与に適用される医薬組成物としては、種々のタイプの定量圧縮エアロゾル、ネブライザーまたは吸入器により生じる微粒子のダストまたはミストが挙げられる。
【0104】
上記医薬組成物は、保存料、可溶化剤、安定化剤、湿潤化剤、乳化剤、甘味料、着色料、着香料、塩、バッファー、被覆剤または抗酸化剤を含有してもよい。上記医薬組成物はまた、本発明のペプチドに加えて、治療上の活性剤を含有してもよい。
【0105】
本発明の物質の用量は、治療される疾患または障害、治療される個体の年齢および症状などに応じて広い範囲で変動する可能性があり、最終的には医師が用いられる適当な用量を決定する。この用量は適当な回数だけ繰り返されてもよい。仮に副作用が発生した場合には、通常の臨床実務に従って用量の量および/または頻度を減らすこともできる。
【0106】
また、本発明は、上述した疾患のいずれかに罹患した患者の治療方法であって、本発明の医薬組成物を患者に投与することを含む治療方法を提供する。
【0107】
本発明のそれぞれの形態の好ましい特徴は、変更すべきところは変更した上で、他の形態のそれぞれに対しても同様に好ましい。本明細書に記載の先行技術文献は、法が許す限り最大限に引用される。
【実施例】
【0108】
以下の非限定的な実施例を参照することにより、本発明をより詳細に説明する。
【0109】
実施例1:本発明のペプチドの予測構造の分子モデリング
これらの実験には、公知(Zloh et al., (2007) Nature Protocols, Vol. 2, pp. 1070-1083)の分子モデリングプロトコールを用いた。PDBのファイルをMaestroにインポートし、適当なリンカーおよび10kDaのポリエチレングリコール鎖を有する3炭素の架橋の挿入により、ジスルフィド結合を修飾した。このモデルを最小化し、Macromodelソフトフェア、OPLS-2005力場、およびGB/SA暗溶媒モデルを用いて、300Kでの2ピコ秒の確率的シミュレーションに供した。構造のスナップショットについて得られた軌跡を分析した。それによれば、生物学的に合成されたオクタペプチドは10kDaのPEGによって包まれるようにはならないことが示唆された(図4および図5)。このことは、重要なことに、生物学的に合成されたオクタペプチドの生物活性な表面は依然として自身の生物学的な受容体と相互作用することが可能であることを意味する。
【0110】
生物学的に合成されたオクタペプチドは、シミュレーション中の骨格原子の標準偏差(RMSD)の値を記録することによりモニターされうる柔軟度を示す(図6)。RMSDの値の範囲は、1〜4.4Åである(図7)。これにより、3炭素の架橋を介してポリエチレングリコールが連結しても、オクタペプチドの柔軟性が損なわれないということが示される。したがって、このジスルフィド部位特異的なPEG化オクタペプチドは、依然として自身の生物学的な重合体結合部位と効果的に相互作用することができる程度に充分に柔軟なままなのである。
【0111】
生物学的に合成されたオクタペプチドの配座にPEG化が及ぼす影響を、生物学的に合成されたオクタペプチドおよびジスルフィド部位特異的なPEG化オクタペプチドについての配座探索を用いて詳細に評価した。2つの硫黄原子間の距離は2.04Å(生物学的に合成されたオクタペプチド)から3.94Å(ジスルフィド部位特異的なPEG化オクタペプチド)へと増加したとしても、これら2つのペプチドの最低の(すなわち、最も安定な)エネルギーは、当該オクタペプチドの環状部位の骨格原子と標準偏差0.58Åで類似していることが判明した。また、ジスルフィド部位特異的なPEG化オクタペプチドの受容体との相互作用に関与するアミノ酸の方向は極めて類似している(図8)。このことから、大きなポリエチレングリコール分子が連結したときに、オクタペプチド上の生物学的に関与する表面は依然として生物学的に関与する受容体への提示に用いられうるということが示唆される。
【0112】
これらのモデリング研究の結果から、生物学的に合成されたオクタペプチドのジスルフィド結合上にポリ(エチレングリコール)分子が存在しても、当該オクタペプチドの受容体結合部位との相互作用は阻害されないことが示される。
【0113】
実施例2:生物学的に合成されるオクタペプチド(バージョン1)用のDNA配列を有するpGex4.3プラスミドの作製
構築した第1のプラスミドは、N末端からC末端に向かって、以下の構成成分をコードしていた:日本住血吸虫のグルタチオン−S−トランスフェラーゼ(GST)+トロンビン切断部位(T)+メチオニン(M)+オクタペプチド、を有するベクター。
【0114】
以下の8アミノ酸に対応するヌクレオチドを含む二本鎖の63bpのDNA配列を構築した:
【0115】
【化3】
【0116】
このDNA配列はまた、EcoRIおよびXhoIの制限酵素部位の認識配列をも含んでいた。これにより、上記DNA配列は、pGex4.3融合タンパク質ベクターへのクローニングに適合するものであった。そして、プラスミドを構築した。
【0117】
上記DNA配列は、最も一般的な細菌のアミノ酸のコドン使用頻度に基づくものであった。別のDNA配列(例えば、上述したようなもの)もまた、用いられうる。また、シアノゲンブロマイドによるアミノ酸の切断を促進する目的で、上記オクタペプチドのDNA配列の5’末端にはMet配列を挿入した。当該DNA配列の3’末端には、2つのタンデム終止コドンが含まれていた。
【0118】
図9に示す63塩基対のオクタペプチドDNA配列を、2つの部分的に重複する相補的なオリゴヌクレオチド(オクタペプチドフォワード:5’ gga tcc ccg aat tcc atg ttc tgt ttt tgg aaa acc tgt acc 3’およびオクタペプチドリバース:5’gcg gcc gct cga gtc cta tta ggt aca ggt ttt cca aaa aca 3’)をハイブリダイズすることにより作製した。各オリゴヌクレオチドの1mMアリコート2μLずつをPCR増幅ミックス中でハイブリダイズさせ、20μLのPCR反応ミックス(Sigma JumpStart mix)中で5サイクルのPCR増幅により伸長させた。
【0119】
次いで、この二本鎖オリゴヌクレオチド配列を2倍体積のエタノールで沈殿させ、20μLの水に再懸濁し、EcoRIおよびXhoI制限酵素を用いて1時間消化した。次いで、切断された配列をエタノールで沈殿させ、20μLの水に再懸濁し、その2μLを、プロメガ迅速ライゲーションキットを用いて、予めEcoRIおよびXhoI制限酵素で切断しておいたpGex4.3ベクター(2μL)中にライゲーションした。ヌクレオチドベクターの配列の比較を図10に示す。ライゲーションの後、構築物を用いてJM109コンピテント細胞(プロメガ)を形質転換させた。
【0120】
形質転換コロニーを、オクタペプチドプライマー5’ gga tcc ccg aat tcc atg ttc tgt ttt tgg aaa acc tgt acc 3’およびベクターpGex4.3プライマー5’ccg gga gct gca tgt gtc aga gg 3’を用いたインサートの存在のPCR増幅により確認した。
【0121】
次いで、陽性コロニーを増殖させ、プラスミドを抽出し、DNA配列をシーケンシングにより確認して、インサートが正しいリーディングフレームにあることを確かめた。次いで、このpGex−TS8プラスミドにより、大腸菌BL−21コンピテント細胞(プロメガ)を形質転換させた。
【0122】
実施例3:オクタペプチド(バージョン2)用のDNA配列を有するpGex4.3プラスミドの作製
第2のプラスミドを構築したが、このプラスミドは、N末端からC末端に向かって、以下の構成成分をコードしていた:日本住血吸虫のグルタチオン−S−トランスフェラーゼ(GST)+トロンビン切断部位(T)+TEVプロテアーゼ切断部位+オクタペプチド、を有するベクター。
【0123】
プラスミドのバージョン2は、TTC TGT TTT TGG AAA ACC TGTACCの前にTEV切断配列を含んでいた。このTEV切断配列は、図11に示すようにMetコドンを置換したものである。78塩基対のオクタペプチドDNA配列を、2つの部分的に重複する相補的なオリゴヌクレオチド(オクタペプチドTEVフォワード:5’ GGA TCC CCGAAT TCC GAA AAC CTG TAT TTT CAGTTC TGTTTT TGG 3’およびオクタペプチドTEVリバース:5’GCG GCC GCT CGA GTC CTATTA GGT ACA GGT TTT CCA AAA ACA GAA 3’)をハイブリダイズすることにより作製した。実施例1においてプラスミドのバージョン1について説明したように、オリゴヌクレオチドの伸長、制限酵素による消化、ライゲーション、およびクローニングを行なった。
【0124】
ヌクレオチドベクターの配列の比較を図12に示す。
【0125】
プラスミドSj−GST−T−TEV−オクタペプチドをトランスフェクションしたBL21大腸菌の小規模な50mL培養物を2×TY培地中で増殖させた。この培養物を、IPTG(0.1mM)を用いて3時間誘導し、回収した。1mg/mLリゾチーム、50mMトリスpH8.5、10%グリセロール、10mM DTT、0.5%デオキシコール酸ナトリウム、およびプロテアーゼ阻害剤カクテル(シグマ)を含有する溶解バッファー中で、細胞を溶解させた。溶解物をソ二ケーティングプローブを用いて30秒間で3回ソニケーションし、16000gで10分間遠心分離し、清澄な上清をGST−アガロースカラムにアプライした。PBSでよく洗浄した後、40mMグルタチオン、50mMトリスpH8を用いて融合タンパク質を溶出させた。選択した画分を、4〜20%ポリアクリルアミドゲル上で分析した。レーン1:分子量マーカー;レーン2:誘導細胞からの全溶解物;レーン3:溶出した融合タンパク質(分子量=28.4kDa)。
【0126】
実施例4:オクタペプチド融合タンパク質の誘導およびその精製
オクタペプチド融合タンパク質(実施例1および2によるバージョン1および2)をコードするプラスミドベクターでトランスフェクションされた小規模(20〜200mL)および中規模(1〜5L)の細菌培養物を、大腸菌BL21中で発現させた。この培養物を2×TY培地中で増殖させ、0.1mM IPTGを用い、室温(25℃)にてタンパク質を一晩誘導した。大腸菌細胞合計46gを遠心分離により回収した。これを、細胞破砕機を用いて溶解させた。溶解バッファーは、PBS、10mM DTT、10mM EDTA、5%グリセロールおよび1%トリトンX−100を含んでいた。可溶性タンパク質を、遠心分離により分離した。封入体中の不溶性タンパク質を、2M尿素(pH12.5)を添加することにより可溶化させた。不溶性タンパク質が溶解したら、水を用いて5倍に希釈し、1N HClを用いて溶液のpHを7.5とした。
【0127】
上清をグルタチオンアガロースアフィニティー精製カラムに通すことにより、可溶化した融合タンパク質を分離した。この段階で、融合タンパク質はグルタチオンアガロースカラムに結合していた。次いで、カラムを4℃にて50カラム体積のPBS(0.1%トリトンX−114を含有)とそれに続く20カラム体積のPBSで洗浄することにより、エンドトキシンを除去した。このアプローチを用いることで、エンドトキシンは洗浄バッファー中に溶出した。次いで、ニッケルアガロースカラムを通すことにより、ヒスチジンタグTEV酵素を除去した。グルタチオンアガロースカラムおよびニッケルアガロースカラムを再生させることで、これら双方の試薬を再利用可能なものとした。上記手法のそれぞれの個別の工程の完全な詳細は当業者に公知である(Structural Genomics Consortium et al., (2008) Nature Methods, Vol. 5(2), pp. 135-146)。
【0128】
図13および図14に示すように、溶出画分のサンプルを5〜20%のプレキャストポリアクリルアミドゲル(インビトロジェン)で分離し、クマシーブルータンパク質染色により染色した。
【0129】
実施例5:MALDI−TOF−MSを用いた酢酸オクトレオチドの質量の決定
MALDI−TOF−MSを用いて、化学合成された酢酸オクトレオチドの質量を決定する目的で、この実験を行なった。化学合成された酢酸オクトレオチドの理論質量は1018Daである。図15に示すように、化学合成された酢酸オクトレオチドの実測質量は1019Daであった。
【0130】
化学合成された酢酸オクトレオチドNa+の理論質量は1041Daである。図15に示すように、化学合成された酢酸オクトレオチドNa+の実測質量は1041Daであった。
【0131】
実施例6:結合したグルタチオン(GSH)を有さない単離SjGST−TM−オクタペプチド融合タンパク質(バージョン1)の質量の決定
結合したグルタチオン(GSH)を有さない精製SjGST−TM−オクタペプチド融合タンパク質(バージョン1)の質量を決定する目的で、この実験を行なった。SjGST−TM−オクタペプチドの溶液は、ジチオスレイトール(DTT)(3mM)を含んでいた。
【0132】
SjGST−TM−オクタペプチドバージョン1融合タンパク質の理論質量は27757Daである。図16に示すように、SjGST−TM−オクタペプチドバージョン1の実測質量は27751Daであった。質量誤差百分率は0.02%である。理論質量と実測質量との間の質量誤差百分率が0.1%であれば許容されうる。
【0133】
この実験により、融合タンパク質の結合グルタチオン(GSH)による質量の増加はないことが確認された。SjGSTの理論質量は25498Daである。さらにアミノ酸およびオクタペプチドがSjGSTに付加すると、理論質量は2259Da増加する。還元型グルタチオン(GSH)の理論質量は307Daである。メチオニンの理論質量は131Daである。
【0134】
実施例7:精製融合タンパク質(バージョン1)におけるオクタペプチドの存在の同定
精製融合タンパク質(バージョン1)におけるオクタペプチドの存在を同定する目的で、この実験を行なった。100mM DTTを用いて精製タンパク質の溶液を還元し、PD−10ゲル濾過カラムによりバッファーを10mMリン酸ナトリウムバッファー(2mM EDTA、pH7.8を含有)に置換した。次いで、融合タンパク質の溶液を、10mMリン酸ナトリウムバッファー(pH7.8)中、37℃にて24時間、トロンビン消化に供した。この溶液に3mM DTTを添加し(ジスルフィドの不正確な精製を防ぐ目的で)、次いでMALDI−TOF−MS分析に供した。
【0135】
SjGST−TM−オクタペプチドバージョン1融合タンパク質の理論質量は27757Daである。トロンビンによる理論上の切断産物は1607Daの断片である。トロンビンで切断されたSjGSTの大きい断片の理論質量は26150Daである。
【0136】
図17に示すように、トロンビンで切断されたSjGSTの大きい断片の実測質量は26153Daであった。質量誤差百分率は0.01%である。トロンビン切断後のオクタペプチドの理論質量は1607Daである。トロンビン切断後のオクタペプチドの実測質量は1607Daである。実施例8に記載するように、トロンビン切断後のオクタペプチドの実測質量については、この特定のMALDI−TOFスペクトルでは低分子量の分子に対する解像度が低いため、測定できなかった。
【0137】
実施例8:精製SjGST−TM−オクタペプチド融合タンパク質(バージョン1)におけるオクタペプチドの存在の同定
精製SjGST−TM−オクタペプチド融合タンパク質(バージョン1)におけるオクタペプチドの存在を同定する目的で、この実験を行なった。精製されたタンパク質の溶液を、100mM DTTを用いて還元し、PD−10ゲル濾過カラムによりバッファーを10mMリン酸ナトリウムバッファー(2mM EDTAを含有、pH7.8)に交換した。次いで、この溶液を、10mMリン酸ナトリウムバッファー(pH7.8)中、37℃にて24時間の、オクタペプチドに直接隣接したメチオニン残基のシアノゲンブロマイド媒介性の切断に供した。得られた溶液に3mM DTTを添加し(正しくないジスルフィド結合の形成を防ぐ目的)、次いでこれをMALDI−TOF−MS分析に供した。
【0138】
このオクタペプチドの理論質量は1035Daである。図18に示すように、この生物学的に合成されたオクタペプチドの、メチオニンでの化学的切断後の実測質量は1034Daであった。質量誤差百分率は0.09%である。化学合成された酢酸オクトレオチドと、生物学的に合成されたオクタペプチドFCFWKTCTとの実測質量の差は16Daである。これは、酢酸オクトレオチドがそのC末端残基としてL−スレオニノールを有するように化学的に修飾されているためである。生物学的なオクタペプチドFCFWKTCTの場合には、天然のアミノ酸であるL−スレオニンがC末端残基として存在している。
【0139】
実施例9:大腸菌におけるSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)のラクトースによる自己誘導と、封入体からのその精製
大腸菌におけるSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)のラクトースによる自己誘導と、封入体からのその単純な精製を実証する目的で、この実験を行なった。1日目に、BL21DE3細胞中のバージョン2プラスミドの培養物5mLをLB培地に播種し、37℃にて8時間増殖させた。次いでこの培地を新鮮なLB培地(5mL)で1/100に希釈し、一晩増殖させた。2日目に、この培地を再度、LB培地中に1/100(5mL)に希釈し、37℃にて8時間増殖させた。次いでこの培地を25mM Na2HPO4、25mM KH2PO4、20mM NH4Cl、2mM Na2SO4、2mM MgSO4、0.5%グルコース、0.25%アスパラギン酸塩を含有する培地中に1/100に希釈し、37℃にて一晩増殖させた。3日目に、培地を200mLの1%トリプトン、0.5%酵母抽出物、0.05%グルコース、0.2%ラクトース、0.5%グリセロール、2mM MgSO4、25mM Na2HPO4、25mM KH2PO4、50mM NH4Cl、2mM Na2SO4を含有する培地中に播種した。次いで、細胞を振盪しながら28℃にて26時間増殖させ、回収した。
【0140】
次いで、50mLの培地からの細胞ペレットを処理して、封入体を得た。50mMトリス(pH8.5)、10%グリセロール、10mM DTT、および0.5%デオキシコール酸塩を含有する溶解バッファー5mLにこのペレットを溶解させた。リゾチーム(1mg/mL)500μLを添加し、溶液を室温にて10分間インキュベートした。溶解物を30秒間ずつソニケーションし、次いで16000gで10分間遠心分離した。上清を回収し、ペレットを5mLの溶解バッファー中に再懸濁させ、30秒間ソニケーションし、さらに遠心分離した。この処理を3〜4回繰り返し、それぞれのソニケーション後に上清を回収した。残留ペレットを2M尿素および10mMトリス(pH12.5)に再溶解させた。
【0141】
図19は、BL21DE3大腸菌細胞中の分子量28.4kDaのSjGST−TEV−オクタペプチド融合タンパク質(バージョン2)のラクトースによる自己誘導を示す。レーン1:分子量マーカー;レーン2:大腸菌の誘導細胞からの全溶解物;レーン3:ソニケーション後の大腸菌細胞溶解物由来の上清;レーン4:2M尿素および100mM Tris(pH12.5)を用いた処理後の細胞ペレット由来の上清;レーン5:2M尿素および100mM Tris(pH12.5)を用いた処理とそれに続く塩酸によるpH8への調整後の細胞ペレット由来の上清;レーン6:2M尿素および100mM Tris(pH12.5)で溶解され、続いて塩酸によるpH8への調整を経た大腸菌細胞由来の残存ペレット。各レーンにおいて、17kDaに見られるタンパク質のバンドは、細胞を消化する目的で添加されたリゾチームである。
【0142】
実施例10:化学合成酢酸オクトレオチドの3炭素架橋のPEG化
10mM EDTAを含有する50mMリン酸ナトリウムバッファー(pH7.8)中で調製した化学合成酢酸オクトレオチドの溶液(0.25mg/mL、0.25μmol)に、TCEP HCl(70μg、0.25μmol、5mg/mL溶液を14μL)を室温にて1時間添加した。次いで、この還元された酢酸オクトレオチドにポリエチレングリコールビススルホン(1.35mg、0.25μmol、1当量)を添加した。反応溶液を穏やかにかき混ぜ、4℃にて一晩静置した。次いでこの溶液を、PD−10脱塩カラムを用いた脱イオン水に対する脱塩処理に供した。最大量のPEG化酢酸オクトレオチドを含有する共役画分(すなわち、第3の1mL画分)を、MALDI−TOF−MSにより分析した。
【0143】
比較のために、酢酸オクトレオチドのMALDI−TOF−MSスペクトルを図15に示す。図20は、化学合成された酢酸オクトレオチドが、ジスルフィド架橋を横切る3つの炭素の架橋を介して、5kDaのポリエチレングリコールに共有結合していることを示す。
【0144】
このPEG化された酢酸オクトレオチドの理論質量は6092Da(すなわち、ポリエチレングリコール5073Da+酢酸オクトレオチド1019Da)である。ポリエチレングリコールの実測分子量は5073Daである。PEG化された酢酸オクトレオチドの実測分子量は6097Daである。質量誤差百分率は0.08%である。
【0145】
実施例11:ポリエチレングリコールが共有結合した3炭素架橋の化学的な挿入による、SjGST−TM−オクタペプチド融合タンパク質(バージョン1)の修飾のSDS−PAGEによる確認
50mMリン酸ナトリウムバッファー(pH7.4)中のSjGST−TM−オクタペプチド(バージョン1)(0.5mg/mL、1mL)融合タンパク質の精製溶液に、100mM DTT(15.4mg)を添加した。次いで、この溶液を室温にて30分間インキュベートし、次いでセファデックスG−25PD−10カラムを用いてバッファーを50mMリン酸ナトリウムバッファー(10mM EDTAを含有、pH7.8)に置換した。この溶液に、ジスルフィド部位特異的に架橋する5kDaポリエチレングリコール(0.29mg、5μmol、3当量)を添加した。得られた溶液を4℃にて72時間インキュベートした。次いで、この混合物にウシトロンビン(シグマ−アルドリッチ、融合タンパク質に対して0.1当量)を添加し、37℃にて24時間インキュベートした。次いで、得られた溶液をSDS−PAGEおよびMALDI−TOF−MSにより分析した。
【0146】
図21において、レーン1:分子量マーカー;レーン2:27757Daの理論分子量を有する、精製されたSjGST−TM−オクタペプチド(バージョン1);レーン3:4℃にて72時間のSjGST−TM−OCTとの5Daのポリエチレングリコールのジスルフィド部位特異的な架橋反応とそれに続く37℃にて24時間のトロンビンによる消化。SjGST−TM−オクタペプチドのタンパク質バンド27.7kDaは消失し、モノPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化)、ジPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化、および、GST中のジスルフィド結合のモノPEG化)、並びに、トリPEG化タンパク質のバンド(すなわち、ペプチドのジスルフィド結合のモノPEG化、および、GST中の2つのジスルフィド結合のジPEG化)が出現した;レーン4:反応バッファー中における5kDaのポリエチレングリコール試薬(3等量濃度)のジスルフィド部位特異的な架橋。
【0147】
実施例12:ポリエチレングリコールが共有結合した3炭素架橋の化学的な挿入による、SjGST−TM−オクタペプチド融合タンパク質(バージョン1)の修飾のMALDI−TOF−MSによる確認
実施例11に記載のタンパク質溶液を100mM DTTを用いて還元し、PD−10カラムを用いてバッファーを50mMリン酸ナトリウムバッファー(pH7.8、10mM EDTAを含有)に置換した。次いで、還元されたSjGST−TM−オクタペプチド(バージョン1)の溶液を、4℃にて24時間、ジスルフィド部位特異的なPEG化に供した。得られた溶液を37℃にて72時間トロンビン消化に供し、MALDI−TOF−MS分析に供した。
【0148】
図22は、トロンビン消化され、ジスルフィド部位特異的にPEG化されたSjGST−TM−オクタペプチド(バージョン1)タンパク質溶液のMALDI−TOF−MSを示す。
【0149】
SjGST−TM−オクタペプチド(バージョン1)融合タンパク質の理論質量は27757Daである。トロンビンにより切断したSjGSTの大きい断片の理論質量は26150Daである。トロンビンによる理論上の小さい切断産物は、1607Daの断片である。このことから、オクタペプチド(1607Da)に連結されたポリエチレングリコール(5073Da)の理論質量は6680Daとなる。オクタペプチドに連結したポリエチレングリコールの実測質量は6692Daであった。質量誤差は0.1%である。
【0150】
実施例13:インビボでの成長ホルモンレベルに及ぼすオクタペプチドの効果
上記の実施例で詳述した化学的処理を用い、3つのオクタペプチド(FCFWKTCTおよびACYWKVCTおよびACYWKTCT)に対して、ポリエチレングリコールが共有結合した3炭素架橋を化学的に挿入した。入手可能な最高グレードの化学物質を、臨床グレードの超純水とともに、バッファー(50mMリン酸ナトリウムバッファー(10mM EDTAを含有、pH7.8))および透析に用いた。すべての溶液を、使用前には30秒間アルゴンでパージした。凝集を防止すべく、反応混合物中でのタンパク質の濃度上昇を回避した。新鮮に調製したTCEP HCl(5mg/mL)を用い、無水条件下、室温にて1時間、タンパク質を還元した。還元タンパク質については、熱または直射日光に晒さないようにした。還元タンパク質を激しく振盪することも避けた。30kDaの3炭素架橋ポリエチレングリコールビススルホンに対するこのタンパク質の比は1:1であった。この共役反応溶液を穏やかにかき混ぜて30kDaの3炭素架橋ポリエチレングリコールビススルホンを溶解させ、次いで溶液を暗所に4℃にて12時間静置した。次いで、5〜7kDaの分子量カットオフを有するピアース透析チャンバを用いてこの溶液を透析して、30kDaのポリエチレングリコールに共有結合していないオクタペプチドを除去した。2時間後、4時間後、および6時間後に、それぞれ1Lの透析水の交換を行なった。次いで凍結乾燥することにより、水および他の揮発性物質を除去した。
【0151】
インビボの動物実験では、体重約20gの雄性BALB/Cマウスを用いた。それぞれの皮下注射は、250μLの注射水に溶解したPEG化オクタペプチドを含んでいた。各注射で投与されたオクタペプチドのみの重量は125μgであった。それぞれのマウスは合計で4回の注射を4日おきに投与された。注射は、首の後部の皮膚をつまみ、皮下に行った。最後の注射の30時間後に動物を屠殺した。市販の酵素イムノアッセイを用いた成長ホルモンの測定のため、血清を回収した。臨床上の毒性は観察されなかった。
【0152】
結果(n=4)を図23に示す。この動物実験では、血清中成長ホルモン(GH)レベルの低下について、3つのすべてのPEG化オクタペプチドが化学合成オクトレオチドよりも高い活性を示した。PEG化されたFCFWKTCTオクタペプチドは、化学合成オクトレオチドと比べると、血清成長ホルモンを70%多く低下させた。PEG化されたACYWKVCTオクタペプチドは、化学合成オクトレオチドと比べると、血清成長ホルモンを78%多く低下させた。PEG化されたACYWKTCTオクタペプチドは、化学合成オクトレオチドと比べると、血清成長ホルモンを94%多く低下させた。
【特許請求の範囲】
【請求項1】
【化1】
式中、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVであり、これらは任意の順列組み合わせであり、それぞれのアミノ酸はL配置であり、ペプチドは2つのシステイン残基の間にジスルフィド結合を含む、
のアミノ酸配列からなる単離されたペプチド。
【請求項2】
【化2】
式中、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVであり、これらは任意の順列組み合わせであり、それぞれのアミノ酸はL配置であり、2つのシステイン残基の2つの硫黄原子は3つの炭素原子を介して連結されている、
のアミノ酸配列からなる単離されたペプチド。
【請求項3】
前記3つの炭素原子の1つに共有結合している親水性ポリマーをさらに含む、請求項2に記載のペプチド。
【請求項4】
前記親水性ポリマーがポリエチレングリコールである、請求項3に記載のペプチド。
【請求項5】
請求項1〜4のいずれか1項に記載のペプチドと、1または2以上のキャリアタンパク質と、を含む、融合タンパク質。
【請求項6】
請求項1に記載のペプチドをコードする第1のヌクレオチド配列を含む、ポリヌクレオチド。
【請求項7】
前記第1のヌクレオチド配列が:
【化3】
式中、YはCまたはTであり、RはAまたはGであり、NはA、C、TまたはGであり、これらは任意の順列組み合わせである、
である、請求項6に記載のポリヌクレオチド。
【請求項8】
キャリアタンパク質をコードする第2のヌクレオチド配列をさらに含む、請求項6または7に記載のポリヌクレオチド。
【請求項9】
請求項6〜8のいずれか1項に記載のポリヌクレオチドを含むベクター。
【請求項10】
請求項6〜8のいずれか1項に記載のポリヌクレオチドに作動可能なように連結されたプロモーターをさらに含む、請求項9に記載のベクター。
【請求項11】
請求項1に記載のペプチドの製造方法であって、
請求項6〜8のいずれか1項に記載のポリヌクレオチドまたは請求項9もしくは10に記載のベクターを、前記ポリヌクレオチドまたはベクターを発現できる宿主細胞に導入し、前記宿主細胞から前記ペプチドを単離することを含む、製造方法。
【請求項12】
前記宿主細胞が、細菌細胞または酵母である、請求項11に記載の製造方法。
【請求項13】
請求項2に記載のペプチドの製造方法であって、
請求項6〜8のいずれか1項に記載のポリヌクレオチドまたは請求項9もしくは10に記載のベクターを、前記ポリヌクレオチドまたはベクターを発現できる宿主細胞に導入し、前記宿主細胞から前記ペプチドを単離し、2つのシステイン残基の2つの硫黄原子間に3炭素架橋を導入することを含む、製造方法。
【請求項14】
3つの炭素原子の1つに対して、親水性ポリマーを共有結合させる工程をさらに含む、請求項13に記載の製造方法。
【請求項15】
前記親水性ポリマーはポリエチレングリコールである、請求項14に記載の製造方法。
【請求項16】
医薬に用いるための、請求項1〜4のいずれか1項に記載のペプチド。
【請求項17】
ホルモン障害、癌、血管作動性腸管ペプチドを分泌する腫瘍の患者における下痢、重篤で難治性の下痢、長期にわたる再発性低血糖、膵島細胞症の乳幼児におけるインスリンの過剰分泌、および食道動脈瘤が疑われる患者における出血、および糖尿病に起因する眼疾患の治療に用いるための、請求項16に記載のペプチド。
【請求項18】
請求項1〜4のいずれか1項に記載のペプチドおよび製薬上許容されうる担体を含む、薬剤組成物。
【請求項1】
【化1】
式中、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVであり、これらは任意の順列組み合わせであり、それぞれのアミノ酸はL配置であり、ペプチドは2つのシステイン残基の間にジスルフィド結合を含む、
のアミノ酸配列からなる単離されたペプチド。
【請求項2】
【化2】
式中、X1はFまたはAであり、X2はFまたはYであり、X3はTまたはVであり、これらは任意の順列組み合わせであり、それぞれのアミノ酸はL配置であり、2つのシステイン残基の2つの硫黄原子は3つの炭素原子を介して連結されている、
のアミノ酸配列からなる単離されたペプチド。
【請求項3】
前記3つの炭素原子の1つに共有結合している親水性ポリマーをさらに含む、請求項2に記載のペプチド。
【請求項4】
前記親水性ポリマーがポリエチレングリコールである、請求項3に記載のペプチド。
【請求項5】
請求項1〜4のいずれか1項に記載のペプチドと、1または2以上のキャリアタンパク質と、を含む、融合タンパク質。
【請求項6】
請求項1に記載のペプチドをコードする第1のヌクレオチド配列を含む、ポリヌクレオチド。
【請求項7】
前記第1のヌクレオチド配列が:
【化3】
式中、YはCまたはTであり、RはAまたはGであり、NはA、C、TまたはGであり、これらは任意の順列組み合わせである、
である、請求項6に記載のポリヌクレオチド。
【請求項8】
キャリアタンパク質をコードする第2のヌクレオチド配列をさらに含む、請求項6または7に記載のポリヌクレオチド。
【請求項9】
請求項6〜8のいずれか1項に記載のポリヌクレオチドを含むベクター。
【請求項10】
請求項6〜8のいずれか1項に記載のポリヌクレオチドに作動可能なように連結されたプロモーターをさらに含む、請求項9に記載のベクター。
【請求項11】
請求項1に記載のペプチドの製造方法であって、
請求項6〜8のいずれか1項に記載のポリヌクレオチドまたは請求項9もしくは10に記載のベクターを、前記ポリヌクレオチドまたはベクターを発現できる宿主細胞に導入し、前記宿主細胞から前記ペプチドを単離することを含む、製造方法。
【請求項12】
前記宿主細胞が、細菌細胞または酵母である、請求項11に記載の製造方法。
【請求項13】
請求項2に記載のペプチドの製造方法であって、
請求項6〜8のいずれか1項に記載のポリヌクレオチドまたは請求項9もしくは10に記載のベクターを、前記ポリヌクレオチドまたはベクターを発現できる宿主細胞に導入し、前記宿主細胞から前記ペプチドを単離し、2つのシステイン残基の2つの硫黄原子間に3炭素架橋を導入することを含む、製造方法。
【請求項14】
3つの炭素原子の1つに対して、親水性ポリマーを共有結合させる工程をさらに含む、請求項13に記載の製造方法。
【請求項15】
前記親水性ポリマーはポリエチレングリコールである、請求項14に記載の製造方法。
【請求項16】
医薬に用いるための、請求項1〜4のいずれか1項に記載のペプチド。
【請求項17】
ホルモン障害、癌、血管作動性腸管ペプチドを分泌する腫瘍の患者における下痢、重篤で難治性の下痢、長期にわたる再発性低血糖、膵島細胞症の乳幼児におけるインスリンの過剰分泌、および食道動脈瘤が疑われる患者における出血、および糖尿病に起因する眼疾患の治療に用いるための、請求項16に記載のペプチド。
【請求項18】
請求項1〜4のいずれか1項に記載のペプチドおよび製薬上許容されうる担体を含む、薬剤組成物。
【図1】

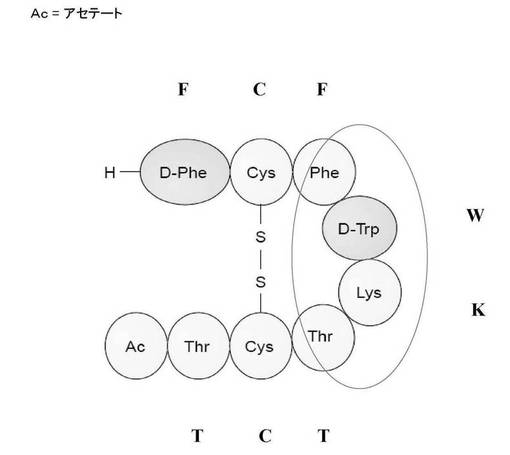
【図2】

【図3】
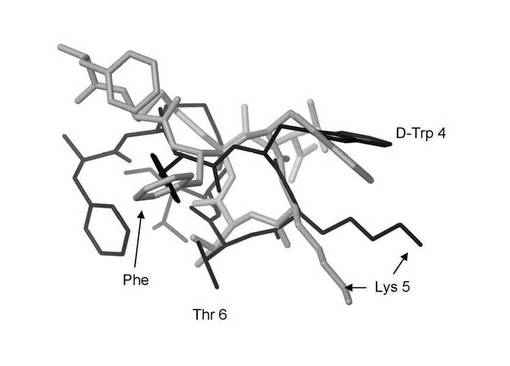
【図4】
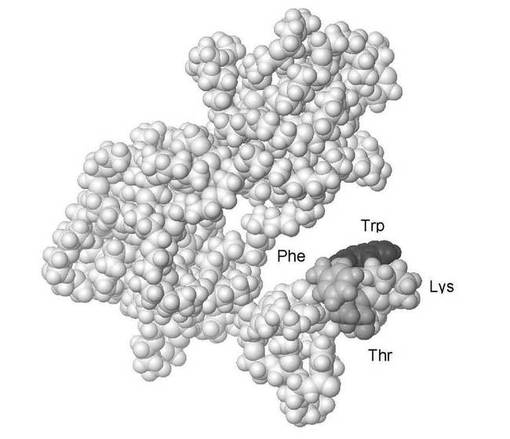
【図5】
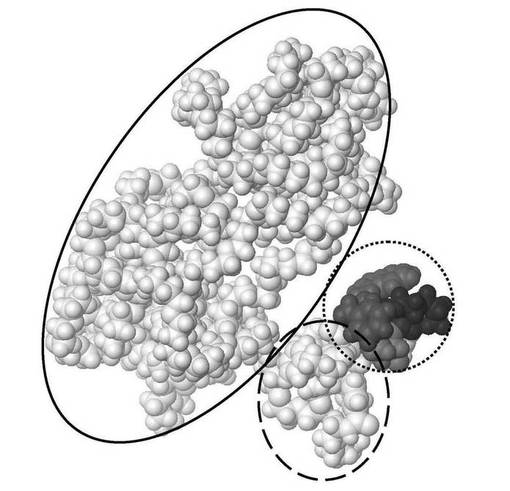
【図6】
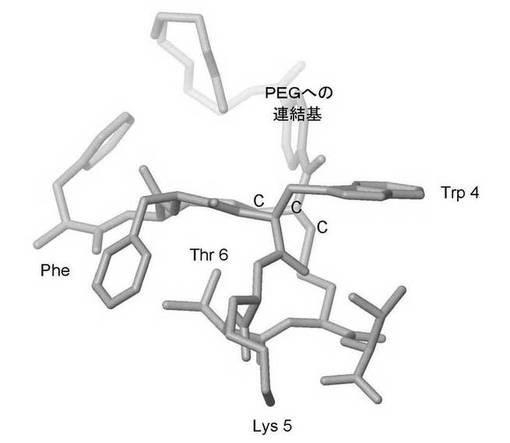
【図7】
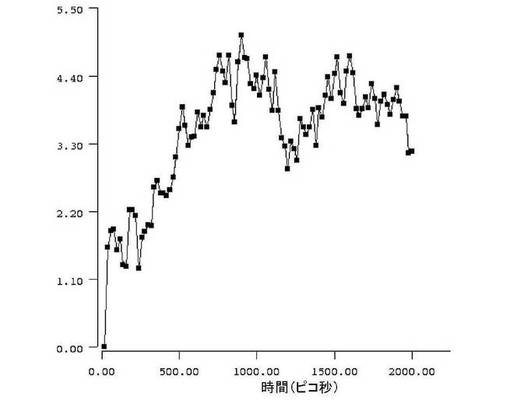
【図8】
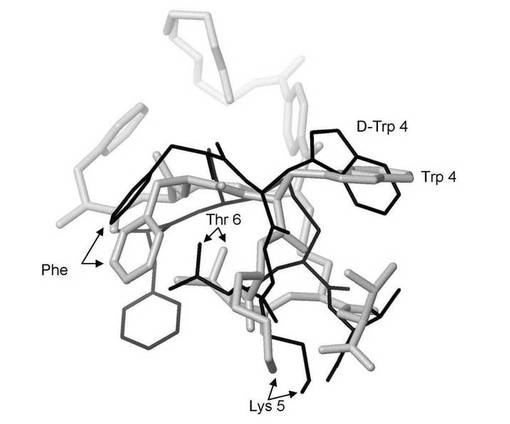
【図9】

【図10】

【図11】

【図12】

【図13】
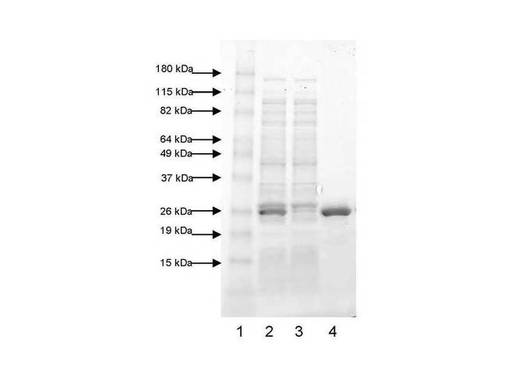
【図14】
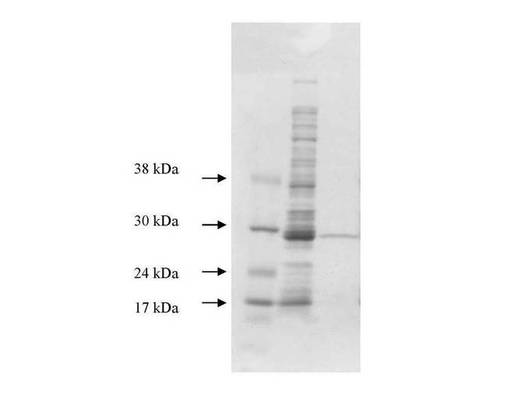
【図15】
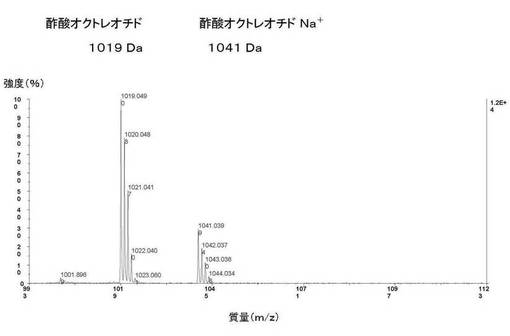
【図16】
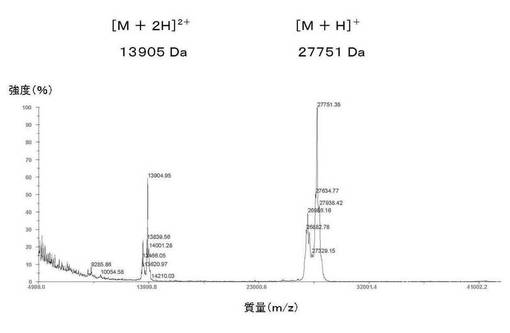
【図17】
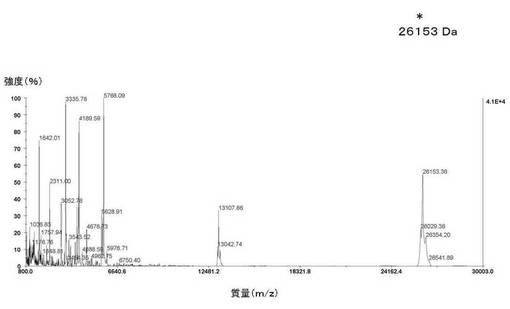
【図18】
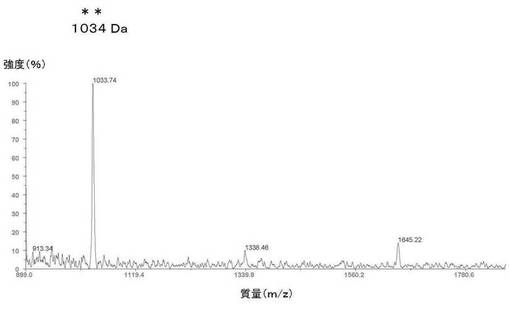
【図19】
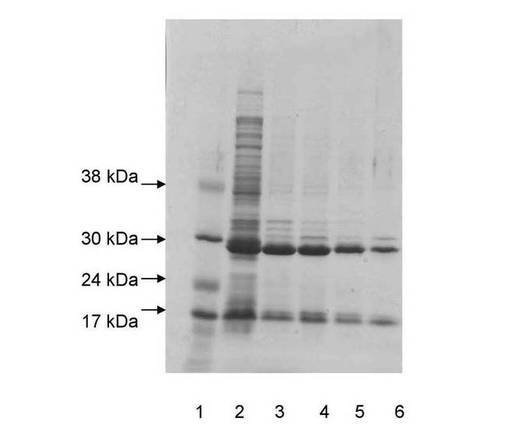
【図20】
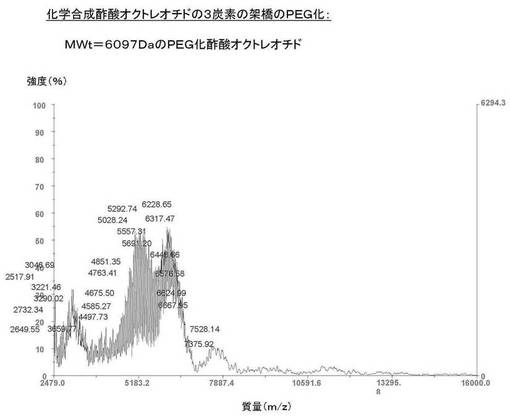
【図21】

【図22】
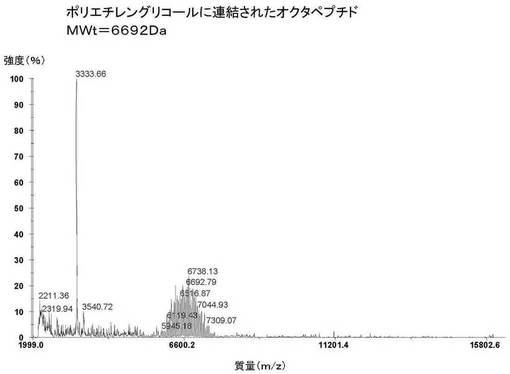
【図23】
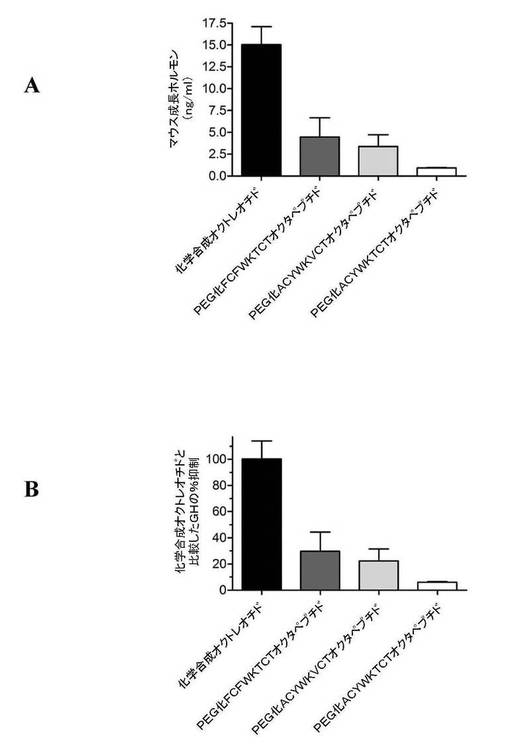
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【公表番号】特表2012−504409(P2012−504409A)
【公表日】平成24年2月23日(2012.2.23)
【国際特許分類】
【出願番号】特願2011−529614(P2011−529614)
【出願日】平成21年9月30日(2009.9.30)
【国際出願番号】PCT/GB2009/002316
【国際公開番号】WO2010/038010
【国際公開日】平成22年4月8日(2010.4.8)
【出願人】(500165809)インペリアル・イノベイションズ・リミテッド (32)
【Fターム(参考)】
【公表日】平成24年2月23日(2012.2.23)
【国際特許分類】
【出願日】平成21年9月30日(2009.9.30)
【国際出願番号】PCT/GB2009/002316
【国際公開番号】WO2010/038010
【国際公開日】平成22年4月8日(2010.4.8)
【出願人】(500165809)インペリアル・イノベイションズ・リミテッド (32)
【Fターム(参考)】
[ Back to top ]