
タキサス種の細胞培養によるタキサンの増強生産
【課題】全ての既知のTaxus種(例えばbrevifolia、canadensis、cuspidata、
baccata、globosa、floridana、wallichiana、mediaおよびchinensis)から、
タキソール、バッカチンIII、および他のタキソール様化合物、またはタキサンを非常な
高収量で生産する方法を提供すること。
【解決手段】全てのTaxus種から得られる種々のタキサンの収量を増強するための、培
養条件(即ち培地組成および操作方法)の特殊な改良点が発見された。特に好ましい増強
剤には、銀イオンまたは複合体、ジャスモン酸(特にメチルエステル)、オーキシン関連
成長調節因子、およびフェニルプロパノイド系路阻害剤(例えば3,4-メチレンジオキシ-6
-ニトロ桂皮酸)が含まれる。これらの増強剤は、単独または複数の組み合わせ、または他
の生産増強条件とともに用いられ得る。
baccata、globosa、floridana、wallichiana、mediaおよびchinensis)から、
タキソール、バッカチンIII、および他のタキソール様化合物、またはタキサンを非常な
高収量で生産する方法を提供すること。
【解決手段】全てのTaxus種から得られる種々のタキサンの収量を増強するための、培
養条件(即ち培地組成および操作方法)の特殊な改良点が発見された。特に好ましい増強
剤には、銀イオンまたは複合体、ジャスモン酸(特にメチルエステル)、オーキシン関連
成長調節因子、およびフェニルプロパノイド系路阻害剤(例えば3,4-メチレンジオキシ-6
-ニトロ桂皮酸)が含まれる。これらの増強剤は、単独または複数の組み合わせ、または他
の生産増強条件とともに用いられ得る。
【発明の詳細な説明】
【技術分野】
【0001】
本発明はタキサス(Taxus)種の細胞培養によるタキソール、バッカチンIIIおよびその
他のタキサンの増強生産および回収のための方法に関する。
(背景技術)
関連技術分野
タキサン供給課題
タキソールはセイヨウイチイ、Taxus brevifolia(Waniら、1971,J.Am.Chem.Soc.,93,2325-2327)の樹皮から単離されたジテルペノイド・アルカロイドである。タキソール
に対する興味は、国立がん研究所(NCI)が大規模な検索計画において、樹皮の粗抽出液が
抗腫瘍活性を示すことを発見したことから端を発した。それ以来、臨床的な試行により、
タキソールが難治の卵巣癌、および乳癌およびその他の癌に対して非常に効果的であるこ
とが確認されてきた。タキソールは、その細胞傷害性の基本的に異なる機構、すなわち微
小管の脱重合を阻害することによるがゆえに、化学療法における大発見とされてきた(Row
inskyら1 990,J.Natl.CancerInst.,82,1247-1259を参照)。
【0002】
タキソールにおける絶望的な要因は供給である。樹皮由来のタキソールは第一の商業薬
品源として生産中止されており、大量生産は半合成、すなわち側鎖を植物由来の前駆体で
ある10-デアセチルバッカチンIIIへ化学的に結合することにより実行されている。学術
研究室による全合成はタキソールへの生存可能である商業的な手段として信頼は薄い。それ
ゆえにコスト的に効果的であり、環境に優しく、およびタキソールの急増する需要に絶え
得る堅実な供給源の開発が急務である。
【0003】
タキソールに加え、関連タキサン分子の商業的生産のための方法を開発することが急務
である。タキソテアなどのタキソール誘導体が既に世界の市場に導入されている。さらに
、利点の大きい活性を有する新規タキサン誘導体の発見および開発に多大な研究活性が集
まっている。その利点とは、そこから得られるあらゆる誘導体が効果的に合成されるよう
な適切な最初の「骨格」分子の、増加する大量の需要を作り出すようなものである。
【0004】
そのような分子の一例は前述の前駆体である10-デアセチルバッカチンIIIであり、それ
は半合成タキソールの開始点として使用される。タキソールおよびその他の誘導体の半合
成生産のためのもう一つの望ましい開始分子はバッカチンIIIである。バッカチンIIIは通
常、植物において主要なタキサンとして蓄積せず、それゆえこの分子の容易な大量の自然
源は存在しない。しかし、この分子はタキソールに化学的に近いため、非常に望ましい半
合成のための開始点である。例えば、10-デアセチルバッカチンIIIではなくバッカチンII
Iが開始点であるならば、10-デアセチルバッカチンIIIの10位のアセチル化を必要とする
工程は省略される。
【0005】
本発明はタキソール、バッカチンIIIおよびその他のタキサンの商業的な生産のための
、植物細胞培養を基にした方法の開発に関連するものである。
植物由来の化学薬品の源としての組織培養
様々な培養計画のもとでの植物細胞の分裂、成長、および二次代謝物の生産の能力は多
くの研究グループによって十分に示されている。まずは、2つの化合物、シコニン(赤色
色素および抗炎症剤)およびジンセンゴシド(東洋医学における強壮剤)は日本において組
織培養法により生産される。他の多くの方法は報告の通り、商品化に近いものであり、バ
ニリン、ベルベリンおよびロスマリン酸(Payneら、1991,"Plant Cell and Tissue Cultur
es in LiquidSystems,"ハンザー出版、ミュンヘン、を参照)を含む。
【0006】
タキソール、バッカチンIII、およびタキサンのための植物細胞培養方法の利点は多く
、(i)細胞培養法は産物が無限であり、連続的および一定の供給を保証し、害虫、災害
、および季節の変動に左右されない、(ii)細胞培養は大きなバイオリアクターの中で培養
が可能であり、環境の条件を操作することにより、興味の対象である化合物の過剰生産を
誘発することが可能である、(iii)細胞培養は樹皮と比較して単一の範囲の化合物を生
産し、または画一化する分離および精製がかなり不要である、(iv)細胞培養法は農業を基
にした方法よりも需要の変化に迅速に順応することも可能である、(v)タキソール、バ
ッカチンIIIまたはその他の前駆体の供給の他に、細胞培養法は、有利な生物活性の側面
を示しまたはその他の生物活性を有する誘導体への変換も可能であるタキサン化合物の生
産もまた可能である。
【0007】
無菌の、大規模の植物細胞培養は本来費用がかかり、細胞培養法は、これらの費用が高
い生産性によって相殺されるときのみ商業的に妥当なものとなる。あらゆる植物種および
標的代謝産物は異なり、個別の系毎に異なるアプローチが必要である。本発明はタキソー
ル、バッカチンIIIおよびタキサンの生産のための生産性の高い植物細胞培養を得るため
の独創的で技術的なアプローチに焦点を合わせたものである。
【0008】
木本植物および針葉樹の組織培養における問題
文献上の歴史的な調査は、草本植物は比較的容易に培養操作ができるが、木本植物およ
び針葉樹の生産的培養の達成は困難を伴うことを示唆する。
【0009】
裸子植物および針葉樹の培養を生産する二次代謝産物の成長は一般的に低かった。例え
ば、BerlinおよびWhite,(1988,Phytochemistry,27,127-132)はThujaoccidentalis
の培養はそのバイオマスが18日間で約30%しか増加しないことを発見した。Van Udenら(19
90.Plant CellReports,9,257-260)はCallitris drummondiiの懸濁液が21日間で20-50
%のバイオマスの増加を報告した。Westgateら(1991,Appl.Microbiol.Biotechnol.,34
,798-803)は裸子植物Cephalotaxusharringtoniaの懸濁液につき約10日間の倍加時間を
報告した。Bornman(1983,Physiol.Plant.57,5-16)によって要約されているように、
エゾマツ懸濁液(Piceaabies)のための培地の開発に対しては大量の努力が注がれた。こ
れらの研究の集成は、裸子植物の懸濁液は実際に早く成長する能力を有すること、しかし
一般法則は適用され得ないこと、および異なる細胞系列に対する培地の設定は独立に工夫
しなければならないことを示している。
【0010】
裸子植物の培養間の二次代謝産物の生産性の調査もまた、草本植物と比較して高速生合
成を含むことの難しさを指摘している。例えば、Cephalotaxus harring toniaの培養は、
テルペンアルカロイドを親の植物に見られる量のわずか1%から3%の程度で生産した(Delfe
lおよびRothfus,1977,Phytochemistry,16,1595-1598)。成功した誘導においてさえ、
Heinstein(1985,Journalof Natural Products,48,1-9)は親の植物において生産され
た程度に接近することのみが可能であった(約0.04%乾燥質量総アルカロイド)。Van Uden
ら(1990)はポドフィロトキシンを生産するために、針葉樹Callitrisdrummondiiの懸濁培
養を誘導することが可能であったが、針によって生産された量の10分の1の程度のみであ
った。Thujaoccidentalisの有意な程度のモノテルペン(10-20mg/L)およびジテルペノイ
ド・デヒドロフェルギノール(2-8mg/L)を生産する能力はBerlinら(1988)によって説得力
をもって示されている。しかし、これらの結果は低成長速度(18日間で30%バイオマス増加
)および低細胞密度(リットルあたり5から7グラム乾燥質量)の培養とともに得られた。
【0011】
タキサン生産のための細胞培養
裸子植物懸濁液において直面する早い成長および高生産性の達成における困難は、Taxu
s細胞培養におけるタキサンの生産に対するこれまでの報告に一般的に反映されている。
【0012】
Jaziriら(1991,J.Pharm.Belg.,46,93-99)は近年Taxusbaccataのカルス培養を開
始したが、イムノソーベントアッセイを用いたタキソールの検出は不可能であった。Wick
remesinheおよびArteca(1991,PlantPhysiol.,96,(補足)p.97)はTaxus media(cv
.hicksii)のカルス培養における0.009%乾燥質量タキソールの存在を報告したが、倍加時
間、細胞密度、および報告されたタキソールの生産に要した時間についての詳細は示され
なかった。
【0013】
米国特許第5,019,504号(Christenら、1991)はTaxusbrevifoliの細胞培養によるタキサ
ンおよびタキサン様化合物の産生および回収について記載した。これらの研究者は2から4
週間の時間枠で1から3mg/Lの程度のタキソール生産を報告した。彼らはまた、「3-4週間
で5-10倍」の細胞質量の上昇を報告しており、これは約7から12日の倍加時間に相当する
。
【0014】
タキサン力価および体積上の生産性の有意な上昇は、経済的に成育可能な植物細胞培養
法によって、計画された1年あたり数百キログラムの年間需要を供給する事を可能とする
ために必要とされている。
【発明の概要】
【課題を解決するための手段】
【0015】
発明の概要
本発明の対象は、早い成長、高い細胞密度、および高い細胞生存率を促進するための特
別な環境条件の開発を含む(この研究において報告する成長の特性は、従来の結果にはる
かに勝るものである)。
【0016】
本発明の目的は細胞系列の注意深い選択、培地条件の注意深い選択および操作、増強因
子の取り込み、および方法を操作する様式の注意深い選択によって、高速でタキサンを生
産することである。
【0017】
本発明の目的は培地の組成および環境条件を変化させることによってタキサンの生産を
操作する能力を含む。特に、主要なタキサン産物としてタキソールまたはバッカチンIII
の生産を細胞に促進させること、および/または副産物セファロマニンの生産を抑制する
ことも目的であり、それによって後工程での分離および精製の問題に、みごとな生物学的
解決法を提供する。これらおよびその他の目的は本発明の一つまたはそれ以上の態様によ
って達成される。
【0018】
本発明者らは、タキソール、バッカチンIII、およびその他のタキソール様化合物、ま
たはタキサンが、既知のあらゆるタキサン種、すなわち、brevifolia,canadensis,cusp
idata,baccata,globosa,floridana,wallichiana,mediaおよびchinensisから非常に
高い収量で産生されることを発見した。さらに、本発明の方法により、タキソール、バッ
カチンIII、およびその他のタキサンを既に報告されているよりも非常に短い時間で得る
ことが可能である。特に、本発明はTaxus chinensis種は急速な成長および極めて大量の
タキソール、バッカチンIII、およびタキサンの短時間での生産が可能であることを発見
した。Taxuschinensis種によって、本発明者らはタキソール、バッカチンIII、およびタ
キサンを他のTaxus種の組織培養から得られる量をはるかに上回る量で生産する細胞を操
作することを可能にした。
【0019】
培養条件(すなわち、培地組成および操作状態)の特定の修飾は、任意のTaxus種の細胞
培養から得られる様々なタキサンの収量を増強することが発見された。特に好ましい増強
剤は、銀イオンまたは複合体、ジャスモニン酸(特にメチルエステル)、オーキシン関連成
長調節剤、および3,4-メチレンジオキシ-6-ニトロ桂皮酸などのフェニルプロパノイド経
路の阻害剤を含む。増強剤は単独でまたは互いに組み合わせて、または収量を増強するそ
の他の条件で使用することも可能である。T.chinensisの植物細胞培養からのタキサンの
収量はこれらの一つまたはそれ以上の条件の使用によって特有に増強されると同時に、全
Taxus種からのタキサンの収量がこれらの条件の使用により向上することがわかった。
【0020】
一つの態様においては、本発明は、一つまたはそれ以上の栄養培地における成長および
産物形成条件下での懸濁培養でTaxus種の細胞を培養すること、および前述の細胞または
前述の細胞培養の前述の培地、または、カルスまたは懸濁培養に由来する細胞およびフェ
ニルプロパノイド代謝の阻害剤を含む栄養培地の両方から1種またはそれ以上のタキサン
を回収することを含むTaxus種の細胞培養における高収量のタキサンを生産する方法を供
給する。フェニルプロパノイド代謝の適切な阻害剤は3,4-メチレンジオキシ-6-ニトロ桂
皮酸、3,4-メチレンジオキシ桂皮酸、3,4-メチレンジオキシフェニルプロピオン酸、3,4-
メチレンジオキシフェニル酢酸、3,4-メチレンジオキシ安息香酸、3,4-トランス-ジメト
キシ桂皮酸、4-ヒドロキシ桂皮酸、フェニルプロピオン酸、フルオロフェニルアラニン、
1-アミノベンゾトリアゾール、2-ヒドロキシ-4,6-ジメトキシ安息香酸、SKF-525A、シュ
ウ酸アンモニウム、ビニルイミダゾール、ジエチルジチオカルバミン酸、およびシナピン
酸を含む。
【0021】
より好ましい態様においては、本発明の方法において使用されている一つまたはそれ以
上の栄養培地の少なくとも一つはまた、エチレン作用の阻害剤でもあり得る別の増強剤;
ジャスモン酸またはジャスモン酸のエステル;またはオーキシン関連成長調節因子をも含
む。特定のより好ましい態様においては、その他の増強剤とは、銀含有化合物、または銀
錯体、または銀イオンであるところのエチレン作用の阻害剤である。別の特定の好ましい
態様においては、その他の増強剤はジャスモン酸またはそのアルキルエステルであり、お
よびさらに好ましくは、ジャスモン酸にエステル化されているアルキル基は1から6個の炭
素原子を有する。さらにより好ましい態様においては、増強剤はジャスモン酸またはその
アルキルエステルであり、およびその培地はまた銀含有化合物、銀錯体または銀イオンを
も含む。別の特定の好ましい態様においては、その他の増強剤はインドール酢酸、ピクロ
ラム、α-ナフタレン酢酸、インドール酪酸、2,4-ジクロロフェノキシ酢酸、3,7-ジクロ
ロ-8-キノリンカルボキシル酸、または3,6-ジクロロ-o-アニス酸などのオーキシン関連成
長調節因子である。
【0022】
別の態様においては、本発明は、一つまたはそれ以上の栄養培地における成長および産
物形成条件下での懸濁培養でTaxus種の細胞を培養し、および前述の細胞または前述の細
胞と前述の培地の両方から1種またはそれ以上のタキサンを回収することを含むT axus種
の細胞培養における高収量のタキサンを生産する方法を提供するが、その際細胞はカルス
または懸濁培養に由来し、および、栄養培地は、ジャスモン酸またはジャスモン酸のエス
テルまたはオーキシン関連成長調節因子でありうる少なくとも1種類の増強剤とともに銀
含有化合物、または銀錯体、または銀イオンの形態で900μMまたはそれ以下の濃度の銀
を含む。好ましい態様においては、増強剤はジャスモン酸またはジャスモン酸のエステル
であり、銀の増強剤に対するモル比は9.5未満である。別の好ましい態様においては、増
強剤はオーキシン関連成長調節因子であり、銀の増強剤に対するモル比は少なくとも0.01
1である。
【0023】
上記の態様のいずれにおいても、一つまたはそれ以上の栄養培地は、α-フェニルアラ
ニン、β-フェニルアラニン、またはその混合物でも有り得るタキサン前駆体をも含んで
よい。上記の態様のいずれにおいても、一つまたはそれ以上の栄養培地は、グルタミン、
グルタミン酸、アスパラギン酸またはこれらのアミノ酸の混合物をも含んでよく、または
細胞の培養に使用する一つまたはそれ以上の栄養培地は、マルトース、スクロース、グル
コースおよび/またはフルクトースを炭素源として、好ましくは一次炭素源として含んで
よい。ある態様においては、栄養培地は細胞培養成長のためのものと、タキソールおよび
タキサン産生のための物が同一である。別の態様においては、1種類またはそれ以上のタ
キサンの産生は栄養培地の組成物を変化させることによって培養中に誘導される。好まし
い態様においては、培養における培地は周期的に交換され、一般的に培地の交換は、培養
からのタキサンの周期的な除去を成し遂げる。好ましくは、前述のTaxus種の細胞は給餌-
バッチ工程(fed-batchprocess)により培養される。
【0024】
一般的に、タキソールまたはバッカチンIIIおよび/またはその他のタキサンは、前述
の細胞または前述の細胞培養の前述の培地、またはその両方から回収される。一般的に、
本発明に従ったTaxus種の培養は、タキサン産生の期間を通して平均15mg/L/日のタキサン
の平均体積生産性を提供する。タキソールの平均体積生産性は一般的に、タキソール産生
の期間の推定で少なくとも10mg/L/日である。バッカチンIIIの平均体積生産性は一般的に
、タキサン産生の期間の推定で少なくとも15mg/L/日である。
好ましくは、本発明の方法に従って培養する細胞はTaxus種の細胞であり、その種はT.
brevifolia,T.canadensis,T,chinensis,T.cuspidata,T.baccata,T.globosa,T
.floridana,T.wallichianaまたはT.mediaであり得る。好ましくは、本発明の方法に
使用したTaxus種の細胞は、増強剤を含まない培地でのカルス培養または懸濁培養におけ
るELISAにより背景値を上回るタキソールを生産する細胞である。より好ましくは、本発
明の方法に使用したTaxus種の細胞は、チオ硫酸銀、メチルジャスモン酸およびオーキシ
ンを含む培地において10mg/Lの平均体積生産性をもって懸濁培養においてタキサンを生産
する細胞である。
【図面の簡単な説明】
【0025】
【図1】培地Aにおける一般的バッチ成長周期を通してのTaxuschinensis懸濁培養系列K-1におけるバイオマスの増加。誤差線は一双のフラスコから測定される標準偏差を表す。
【図2】15日実験におけるタキソール(A)および総タキサン(B)生産性に対する第9日および第12日の培地交換の効果。各々の箱の数字は産物が生産された時間の間隔(日)を表す。細胞内ボックスの影のついた部分は実験開始時に細胞接種原に存在したタキソールまたは総タキサンを表す。すべての処理は二連で行われた。Taxus chinensis懸濁培養系列K-1は表2において述べられた培地Aとともに使用された。
【図3】実施例7.3において使用した標準Gro-Luxランプ(GTESylvania,Danvers,MA)のスペクトル特性。
【図4A】Taxuschinesis細胞懸濁物K-1におけるタキサン産生。クロマトグラムの10から40分の部分を示した。選択されたタキサンのピークのダイオード配列走査は227nmのピークを持つ特徴的なタキサンUV吸光スペクトルを示した。
【図4B】Taxuschinesis細胞懸濁物K-1におけるタキサン産生。クロマトグラムの10から40分の部分を示した。選択されたタキサンのピークのダイオード配列走査は227nmのピークを持つ特徴的なタキサンUV吸光スペクトルを示した。
【図4C】Taxuschinesis細胞懸濁物K-1におけるタキサン産生。クロマトグラムの10から40分の部分を示した。選択されたタキサンのピークのダイオード配列走査は227nmのピークを持つ特徴的なタキサンUV吸光スペクトルを示した。
【図4D】Taxuschinesis細胞懸濁物K-1におけるタキサン産生。クロマトグラムの10から40分の部分を示した。選択されたタキサンのピークのダイオード配列走査は227nmのピークを持つ特徴的なタキサンUV吸光スペクトルを示した。
【図5A】培地CにおけるTaxuschinensis細胞系列K-1による長期間培養後のタキソールおよびタキサン産生。Aは既知および未知のタキサンに対するデータを表にしたものである。
【図5B】培地CにおけるTaxuschinensis細胞系列K-1による長期間培養後のタキソールおよびタキサン産生。Bは25から42日の期間におけるタキソールおよびタキサンの生産の増加を示す。
【図6A】細胞培養の上清におけるタキソールのMS/MSによる確認。図Aは本来のタキソールのイオンスプレ-APCI質量分析を示す。
【図6B】細胞培養の上清におけるタキソールのMS/MSによる確認。図Bは親ピーク(m/z 871=タキソール+NH4+)の娘イオンスペクトルを示す。
【図6C】細胞培養の上清におけるタキソールのMS/MSによる確認。図Cは細胞培養の粗抽出液から得られたイオンスプレーAPCIスペクトルを表し、タキソールに特徴的なm/z 854および871を示す。
【図6D】細胞培養の上清におけるタキソールのMS/MSによる確認。図Dはm/z 871に対応する娘スペクトルを示し、細胞培養上清におけるタキソールの存在を示す決定的な証拠を提供する。
【発明を実施するための形態】
【0026】
発明の詳細な説明
植物は長い間、薬剤および特別な化学物質の重要な原料を供給してきた。これらの産物
は一般的に、回収した植物材料の抽出または化学的合成を通して得られていた。タキソー
ルおよびタキサンは自然産物の検索から近年発見された最も重要な分類の抗癌剤の一つと
なってきた。
【0027】
本明細書で使用されている「タキソール様化合物」または「タキサン」という用語は、
タキサン環を有するジテルペノイド化合物を記述するために互換的に使用する。タキサン
はそれ自身が抗悪性物活性を有し、または生物活性を有する化合物を産生するように修飾
されることも可能である。「総タキサン」という用語は以下の実施例5に記載されている
ような特徴的なUV吸光度を示す全てのタキサンについて言及する。
【0028】
本明細書で使用されている「カルス」という用語は、構造的に未分化であり、固体化さ
れた培地において培養される培養植物細胞の集団を記述するために使用される。本明細書
で使用されている「懸濁培養」という用語は、液体栄養培地において分散する構造的に未
分化な細胞を記載するために使用する。懸濁培養は様々な凝集の段階にある細胞を含むこ
とが理解されよう。凝集の大きさの範囲は、本発明に記載されている懸濁液において、直
径10ミクロン(単一細胞またはほぼ未凝集の細胞)から、数千の細胞から成る直径数ミリメ
ートルの凝集に至る範囲の大きさをもって計測される。
【0029】
本発明において有用な植物材料はあらゆる既知のTaxus種、たとえばbrevifolia,canad
ensis,cuspidata,bacatta,globosa,floridana,wallichiana(yunnanensisとも言及さ
れる),media,fastigiataおよびchinensis(sumatrama,celebica,およびspeciosaなど
の同義の種,およびchinensisvar.mairei亜種を含む)から得ることが可能である。特
に、本発明者らは有意な量のタキソール、バッカチンIII、およびタキサンを高い体積生
産性で産生する能力を有するものとしてTaxus chinensis種を同定した。
【0030】
本発明者らによって、特定のタキサン内容量は、植物種により、および組織源および特
定の樹木に由来する植物種内において変化することが発見されている。タキサン産生のた
めの高収量の源および培養を選択することが、医療使用のための十分な量のタキサンの供
給に向けての重要な第一歩である。
【0031】
商業的適合性の基準
商業的な魅力およびタキサン産生のための与えられた植物細胞培養を基にした工程の成
功性を判断するために数々の基準を使用することが可能である。そのような基準は、発酵
費用、後での回収の容易性、および生産量を含む、工程の鍵となる要因の効果を特徴化お
よび実証すべきである。ここに記載される基準は、ブロス力価および体積生産性である。
【0032】
ブロス力価は全体のブロスにおける産物の濃度として定義され、通常1リットルのブロ
スあたりのミリグラム数(mg/L)として表現される。定義により、全ブロス力価は産物の細
胞内および細胞外の部分は区別しない。ブロス力価は典型的にはバッチまたは給餌-バッ
チ工程の効率評価に使用される。より高いブロス力価は、与えられた反応器の体積に対し
て産物の体積がより大きいこと、およびそれに付随して単位生産費用がより低いことを意
味する。同様に、高力価産物は通常高収量で回収することが容易であり、単位生産費用の
更なる改善へと導く。
【0033】
体積生産性は、単位反応体積あたり単位時間あたりに生産された産物の量として定義さ
れ、1リットルあたり1日あたりのミリグラム数の単位で表現される。タキサン産生の目的
のために、時間の尺度は、収集および回収に先立つ、生産尺度において生産が起きる間の
時間枠として定義する。体積生産性はバッチおよび給餌-バッチ工程の基準としての力価
を補足するもので、産物が産生の間に除去される工程、例えば周期的培地交換または別の
除去方法、の評価に特に有用である。高い体積生産性は、与えられた時間を通して与えら
れた反応体積に対する産物体積が大きいこと、およびそれに付随して単位生産費用がより
低いこと、および全工程の性能の向上を意味する。
【0034】
体積生産性が生物学的工程の本来の能力を計測するために使用される場合、例えば工程
開発の初期の段階においては、生産周期の最も生産的な部分、すなわち、生合成の速度が
その最大であるときの短い期間での生産性を測定することが有用である。これは一般的に
、最大瞬間体積生産性と呼ばれる。しかし、工程の性能の計測において、より適切な基準
は、全体の生産時期を通して測定される生産性である平均体積生産性である。明らかに、
最大の平均体積生産性を達成するためには最大瞬間生産性が生産時期の大部分を通して維
持されなければならない。別に規定しないかぎり、「体積生産性」という用語は全体の生
産時期に対して決定された平均体積生産性を意味する。一般に、生産時期は、生育培地を
産生培地に置き換えること、またはタキサン産生において有意な増強を誘発する増強剤を
添加することによる栄養培地の組成物の変化によって開始する。
【0035】
Taxus細胞株の開始
Taxus植物材料はその他の大陸と同様に北アメリカ全域から集められ得る。培養は生育
に適したTaxusの組織を選択することから始められる。樹皮、形成層、針葉、幹、種子、
球果、および根を含む植物のどの部分由来の組織でもカルスを誘導するために選択され得
る。しかしながら、タキソールの収量を最良にするためには針葉および植物の分裂組織の
部分が望ましい。最も望ましいのは針葉の新芽(生え始めてから1から3ヶ月)であり、これ
は一般にそれが黄緑色をしていることで同定され得る。"新芽"という語はその年の成育期
のうちに生えた針葉を広く指している。
【0036】
培養物への汚染を防ぐために、組織は培地への導入に先だって表面を滅菌されるべきで
ある。CLOROX(CloroxCompanyの脱色剤の登録商標)処理等のどんな滅菌技術も有効である
。それに加えて、セフォキシチン、ベンレート、クロクサシリン、アンピシリン、ゲンタ
マイシン硫酸塩、およびフォスフォマイシンのような抗生物質が植物材料の表面滅菌に用
いられ得る。
【0037】
カルスの生育
培養物は典型的には生育中の形態、増殖量、生成物の性質、およびその他の性質に多様
性を示す。個々の細胞株は培地の成分に対する親和性に多様性があり、数多くの異なる増
殖培地がカルスの誘導および増殖に用いられ得る。
【0038】
適切な培地の組成は培養されている生物種によって異なる。異なる種に対する望ましい
培地は表3に挙げられる。例えば、Taxuschinensisに対する望ましい増殖栄養培地は、他
のものも用いられ得るものの、A,D,I,J,K,L,M,O,Pである。これらの培地は表2に挙げた成
分を含むことが望ましい。技術を有する技術者はその量のいくらかの変化は細胞増殖に大
きな影響を与えないことが認識し得るが、表2に示した量でその成分を含む培地を用いて
培養が行われることが望ましい。
【0039】
例えば、培地Aが用いられるならば、成長ホルモンまたは調節物質は1ppbから10ppmの間
の量で、望ましくは2ppbから1ppmの間の量で培地に含まれる。培地Dが用いられるならば
、成長ホルモンまたは調節物質は1ppbから10ppmの間の量で、望ましくは2ppbから2ppmの
間の量で培地に含まれる。その他の培地成分は表2に示された量の1/10から3倍までの濃度
で含まれ得る。
【0040】
タキサンを大量に生成することはTaxus細胞を懸濁培養で増やすことによって可能にな
る。一般に、懸濁培養はカルス培養に成功した培地を用いて開始され得る。しかしながら
、懸濁培養の必要性および特にタキサンの高い効率での生成のために、培地の修正が望ま
しい。Taxus細胞が修正され、本発明の方法によって特性を調節された培地中で培養され
た場合、培養物からの1種類あるいはそれ以上のタキサンの収量は有意に増加する。
【0041】
本明細書で用いられる"栄養培地"という語は、植物細胞のカルスの増殖および懸濁培養
に適した培地を記載するために用いられる。"栄養培地"という語は一般的な語であり、"
生育培地"および"産生培地"の両方を含む。"生育培地"と言う語は培養細胞の迅速な生育
を可能にする栄養培地について記載するために用いられる。"産生培地"と言う語は培養細
胞内でタキソール、バッカチンIIIおよびタキサンの生合成を可能にする栄養培地を指す
。産生培地内で生育が起こり得、生育培地内で生成が起こり得、また最適な生育と生成が
一つの栄養培地内で起こり得ることが理解されよう。
【0042】
懸濁増殖
Taxusの懸濁培養では迅速な増殖速度と他の植物細胞培養のように迅速で高い細胞濃度
が可能である。しかしながら最適な細胞濃度は細胞株によって異なるので任意のどんな細
胞株にたいしても迅速に最適化できるような手法が考えられなければならない。
【0043】
様々なTaxus種の培養物は多量および微量栄養塩類、炭素源、窒素源、ビタミン類、有
機酸、天然および合成植物成長調節因子を含む栄養培地に移すことで生育させられる。特
にTaxus細胞の懸濁培養用の栄養培地は典型的にはカルシウム、マグネシウム、ナトリウ
ム、カリウム、リン酸、硫酸、塩素、硝酸、およびアンモニウムと言った多量栄養分およ
び銅、鉄、マンガン、モリブデン、亜鉛、ホウ素、コバルト、ヨウ素およびニッケルとい
った微量栄養分を補給する無機塩類を含む。この培地はまた、典型的にはミオイノシトー
ル、チアミン、アスコルビン酸、ニコチン酸、葉酸、ピリドキシンおよび場合によっ.て
ビオチン、パントテン酸、ナイアシン、およびその類似物等のビタミン類をも含む。これ
らの成分は表2に列挙された濃度の1/30から30倍の範囲の濃度で存在し得、望ましくは表2
に列挙された濃度の1/20から20倍の濃度で、さらに望ましくは表2に列挙された濃度の1/3
から3倍の濃度で、および最も望ましくは表2に列挙された濃度で含まれ得る。
【0044】
栄養培地はまた1種類またはそれ以上の炭素源、および典型的には栄養培地内の全炭素
の50%以上を供給するものである1次炭素源を含む。1次炭素源は望ましくはラクトース、
ガラクトース、ラフィノース、マンノース、セロビオース、アラビソース、キシロース、
ソルビトール、または望ましくはグルコース、フルクトース、スクロース、またはマルト
ースである。1次炭素源の濃度は0.05%(w/v)から10%(w/v)の範囲で有り得、望ましくは0.1
%(w/v)から8%(w/v)の範囲で有り得る。
【0045】
栄養培地はまた、多量栄養塩類の形で加えられる任意の窒素源に加えて、望ましくは少
なくとも部分的には有機栄養源(例えばグルタミン、グルタミン酸、およびアスパラギン
酸のような1種類又はそれ以上のアミノ酸、またはタンパク質加水分解物)によって供給さ
れる窒素源をも含む。これらの有機窒素源は0.1mMから60mMの、望ましくは1から30mMの範
囲の濃度の窒素を供給し得る。この培地はまた酢酸、ピルビン酸、クエン酸、オキソグル
タル酸、コハク酸、フマル酸、マレイン酸およびそれらの類似物のような1種類またはそ
れ以上の有機酸をも含み得る。これらの成分は培地中に、0.1mMから30mMの濃度で、望ま
しくは0.5mMから20mMの濃度で含まれ得る。
【0046】
この培地はまた典型的にはピクロラム、インドール酢酸、1-ナフタレン酢酸、インドー
ル酪酸、2,4-ジクロロフェノキシ酢酸、3,7-ジクロロ-8-キノリンカルボキシル酸、3,6-
ジクロロ-o-アニス酸およびその類似物のようなオーキシン関連成長調節因子、N6-ベンジ
ルアデニン、6-[γ,γ-ジメチルアリルアミノ]プリン、キネチン、ゼアチン、N-フェニル
-N'-1,2,3-チヂアゾール-5イル尿素(チジアズロン)および関連フェニル尿素誘導体および
その類似物のようなサイトキニン関連成長調節因子、GA3、GA4、GA7およびGA誘導体のよ
うなジベレリン、アブシジン酸やその誘導体、ブラシノステロイド、およびエチレン関連
成長調節因子を含む1種類またはそれ以上の天然または合成植物成長調節因子をも含む。
さらに別の適切なオーキシン関連植物成長調節因子は以下に列挙される。栄養培地は、例
えば1種類以上のオーキシン関連調節因子、1種類以上のサイトカイニン関連調節因子のよ
うな単一の種別に属する1つ以上の成長調節因子を含み得ることに注意されたい。成長調
節因子は培地中に10-10Mから10-3Mの間の濃度で、望ましくは10-8から3x10-5Mで、さらに
望ましくは表2にあげられた濃度で含まれることが望ましい。
【0047】
他に記述がない限り、本明細書において定義される生育培地は、カルス培養培地および
産生培地の通常の技術で最適化するために適切な開始点を提供する。当該技術分野におけ
る技能を有するものにとってはある種別の成分および任意の種別の成分を添加し、修飾し
、操作することによって最適な結果を達成することは日常的技術範囲である。ある種の培
地修飾は表および下記の実施例の中に供給される。
【0048】
液体培養物は空気のような好気的な環境に曝され、振とうされるか、あるいは撹拌され
ることで培養成分の適切な混合を可能にされることが望ましい。培養物は、適切な条件お
よび/または環境下では0℃から33℃の範囲が可能であるものの、23℃から27℃の間の温度
に保たれる。pHは約3から7の間で、望ましくは4から6の間で有り得る。培養物は様々な期
間の間完全な暗黒から完全な照明(狭い周波数帯および/または広いスペクトル)下までの
範囲の照明条件下で生育される。
【0049】
倍化速度は時間依存的にバイオマスの増加を調べることとともに、単に通常の部分培養
の間の生育指数を調べることによって測定される。リットルあたり15から24グラムの最大
乾燥重量濃度が達成される。様々なTaxus種懸濁物の生育特性は実施例4に詳述される。
【0050】
タキサン生成条件
懸濁培養物における2次代謝産物の形成が生育と同時に起こる場合、その代謝は生育に
関連していると呼ばれ、1種類の培地を形成することが良好な生育と多量の生成を達成す
るのに十分で有り得る。他の多くの系においては迅速な生育と高度の生成物形成は同時に
は起こらないことが分かっている。このような場合は生育および生成相は分離され、それ
ぞれの相に対する培地が独立に開発される(Payne et al.1991,Plamt Cell and Tissue
Culture inLlquid Systems,Hanser publishers,Munichに概説されている)。Taxusにお
けるタキサン生成の場合は、生育および生成物形成は分離され得、独立な培地がそれぞれ
に対して開発された。
【0051】
本発明の望ましい様式においては細胞増殖相の間の培地組成はタキサン生成相の間の培
地組成とは異なる。例えば、炭素源、特に1次炭素源の種類と量が生育相と生成相では異
なり得る。産生培地は生育培地におけるよりも多量の糖を含むことが望ましい。産生培地
における糖の量は生育相における場合と比べて生成相において2から20倍多いことがより
望ましい。1次炭素源はラクトース、ガラクトース、ラフィノース、マンノース、セロビ
オース、アラビノース、キシロース、ソルビトールであることが望ましく、またはグルコ
ース、フルクトース、スクロース、またはマルトースであることが望ましい。1次炭素源
の濃度は005%(w/v)から10%(w/v)の間で有り得、0.1%(w/v)から8%(w/v)で有ることが望ま
しい。タキソールまたはバッカチンの生成のために特に望ましい炭素源はマルトース、ス
クロース、グルコースおよび/またはフルクトースである。特に望ましい態様においては
これらの糖は初期の栄養培地中に少なくとも3.5%の濃度で含まれる。
【0052】
ビタミン、アミノ酸のような有機窒素源を含む有機添加物の種類およびレベルは、下記
の増強剤の存在またはレベルと同様に、培地において変化し、または異なるであろう。天
然または合成の植物成長調節物質の同一性およびレベルは、培地間で異なるであろう。同
様に、多量元素および微量元素の塩のレベルおよび種類も、生育および産生培地間で異な
るであろう。好ましくは、塩含量は生育培地に比べ産生培地において減少され、所望によ
り硝酸塩および硫酸塩は不均等に減少され、かつ、より好ましくは減少度は2-20倍の係数
による減少である。しかし、この培養系に対し単一の生育/産生培地を設定することも可
能であることは理解されよう。
【0053】
本発明の産生培地は、タキサン形成のみならず、タキソールまたはバッカチンIIIとい
った特定のタキサンの生成につながる直接的な細胞の生合成をも増加させる。さらに、セ
ファロマニンのような夾雑副産物の生成は、樹皮組織に比べ最小化される。本発明の産生
培地はまた、細胞の生存能力及び生合成の増加を促進し、さらに有意なレベルの産物を細
胞外培地へ分泌させる。これらの特性は、タキサン生成の効率的な商業的スケール手法の
運用において非常に重要である。
【0054】
細胞および培地よりタキソールおよびタキサンを抽出および回収する手法は、慣用され
た技術に基づく(例えば実施例5を参照)。イムノアッセイ(ELISA)技術は、商業的に入
手可能なキットとしてHawaii Biotechnologyにより提供されるプロトコールに主として基
づいた(本明細書に参照文献として記載されたGrothausら,1995,Journal of Natural P
roducts,58,1003-1014をも参照)。抗体は、タキソールまたはバッカチンIIIといった
いずれのタキサンにも特異的であってよく、または、タキサン骨格に対し、より非特異的
であってもよい。高速液体クロマトグラフィーの手法は、実施例5に詳述された既存のプ
ロトコールより僅かに変更された。本発明において用いられた条件の下で、タキサンピー
クの明確な分離が達成され、その結果、正確な検出および定量がなされた。タキサンでな
い成分が共溶出する可能性があるため、タキサンピークのスペクトル純度は、ピーク領域
を混合する前にダイオードアレイにより慣例法で検査された。タキサン標準品の保持時間
は実施例5に示されており、かつクロマトグラムの例示は図4に含まれている。
【0055】
高等植物にとって、光は植物本体、培養細胞いずれにおいても二次代謝における有効な
因子である。光の強度および波長の双方が重要である(SeibertおよびK adkade 1980,"P
lant TissueCulture as a Source of Biochemicals"E.J.Staba編,CRC Press,Boca Ra
ton,Florida,pp123-141)。例えば、通常フラボノイドおよびアントシアニン生合成は
、強度の強い連続光により促進され、一方、他の代謝産物については、暗所で培養された
細胞が好ましい。培養細胞の緑化または光合成能力の増加は、産物生成または産物の範囲
をも増加させるであろう。本発明者らの研究は、広範囲のみならず特異的な狭い範囲の光
源の利用にも関する。実施例7.3.に示されたように、光照射はタキソールの蓄積のみなら
ず、培地への分泌の増加をも引き起こす可能性がある。タキソール生成に対する光の刺激
効果は、タキサンの生合成に関する特異な制御機構の存在を示唆する。光受容体の特性お
よび光により誘導される刺激の生化学的特性は未だ明らかでない。しかし、本発明による
と、増強剤の導入が、最適の結果に対する光の役割をより重要でないものにする。
【0056】
不揮発性の溶存栄養分に加え、気体成分、主に酸素、二酸化炭素、およびエチレン(植
物ホルモン)は、生育および産物生成において重要な役割を担っている。2つのパラメー
ターが重要である。生育およびタキソール生成を促進する溶存気体濃度は明らかに重要で
ある。なぜなら、それらは反応装置を操作するする条件を規定するからである。さらに、
最適な特定の濃度が維持されるよう、消費または生成の効率を反応装置の設計に考慮する
必要がある。
【0057】
呼吸における重要性の他に、酸素は二次代謝産物の生合成の効率にも劇的な影響を与え
る。二次生合成経路において酸素を要求する過程に対する高い飽和定数は、反応装置内に
おいて細胞が高い酸素レベルに曝されていることを要求する。高い成長率を維持するため
のCO2添加の重要性が報告されている。植物ホルモンであるエチレンは、二次代謝を含む
植物の生育および発生の全ての側面において、多面発現的な役割を担う(例えば、Payne
ら,1991を参照)。
【0058】
本発明者らは、ある気体濃度の様式が、培養細胞における生育および二次代謝を促進す
ることを見出している。例えば、酸素濃度の範囲は、1%空気飽和から200%空気飽和まで、
および好ましくは10%から100%の範囲、および最も好ましくは25%から95%の範囲において
、培養と両立する。二酸化炭素濃度の範囲は、0.03%(培地と平衡にある気相におけるv/v
)から15%(v/v)まで、および好ましくは0.3%から8%(v/v)の範囲において、培養と両立す
る。溶存気体の最適濃度は、細胞代謝により異なる可能性があり、例えば、速い成長を行
っている細胞は、典型的に高めの酸素レベルを好み、かつ、高めの二酸化炭素レベルに対
して感受性がより低いタキサン生合成中の細胞とは異なる最適値を有するであろう。最適
値はまた、培養の速度論によっても異なる可能性があり、例えば、遅滞期にある細胞は、
対数増殖期にある細胞とは異なる溶存気体濃度を好むであろう。
【0059】
溶存気体は、他の培養成分および増強剤と多様な様式により相互作用するであろう。例
えば、酸素の要求は、生合成の誘導または刺激により変化するであろう。傷害反応として
の呼吸効率の上昇は、植物培養細胞が誘導を受けた際に一般的に観察される。エリシター
または刺激物は、それらの機能をエチレンを通じ仲介し、または二次代謝の促進とは独立
にエチレン生成に影響を与えるであろう。その場合、微生物由来のエリシター調製物をエ
チレンに置換し、かつエリシター調製物中の他の微生物由来成分に関連する毒性を恐らく
抑制することが望ましいであろう。あるいは、エチレンの作用を抑制し、従ってエリシタ
ーまたは刺激物が二次代謝をより独占的、従ってより効率的な方法で促進できるようにす
ることは有利であろう。下記のように、エチレン作用に影響を与えることが知られている
成分である銀イオンは、実際タキサン生合成を有利に変化させる。
【0060】
増強剤
二次代謝産物の生成は複雑な過程であり、究極的に二次代謝産物に変換されるべき前駆
体を生成しかつ順々に修飾する、多数の異なる酵素の協調した作用を要求する。同時に、
二次代謝産物生成は、他の酵素が目的の代謝産物の前駆体を代謝し、二次代謝産物を構築
するのに必要な前駆体プールを流失させる場合には低下するであろう。
【0061】
低い生成または引き続く変換による利用可能な前駆体の量の制限、前駆体または中間産
物の下流中間産物への変換の制限、または関連酵素の活性の制限は、二次代謝産物の生成
を限定するであろう。いずれの特定の培養系においても、二次代謝産物が生成される効率
は、前駆体が二次代謝産物に変換される経路において律速段階を形成する、これらの限定
要因のうちの一つにより制御されるであろう。律速段階の原因となる限定要因の緩和は、
当該培養系の二次代謝産物生成効率を、経路の他の過程が律速となる点にまで上昇させる
であろう。生成効率全体を限定する特定の過程は、限定要因を緩和する作用と同様に、培
養系間で異なるであろう。
【0062】
タキサンは一連の多数の酵素過程を通じて生成される二次代謝産物であり、本発明者ら
は、タキサン生合成における一つまたはそれ以上の効率限定過程を緩和する、複数のクラ
スの増強剤をつきとめた。これらの増強剤のうち一つのタキサン生成細胞への添加は、タ
キサンの生成効率を増加させる。さらに本発明者らは、本明細書中に記載されている増強
剤の利用が、大部分のタキサン生成培養細胞において、少なくともいくつかの促進効果を
示すであろうことを決定しており、生成効率全体が単一の効率限定過程ではなく、複数の
限定要因の間の複雑な相互作用により決定されていることを示唆している。促進の程度は
、一旦ある種の限定要因が緩和された際にタキサン生合成における他の過程の相対的限定
効果を決定する、特定の細胞の状態に依存するであろうが、いずれか一つの限定要因の緩
和はタキサン生成を増加させるであろう。多種の限定要因間の相互作用に影響する培養条
件は、細胞の遺伝的構成、培地の組成および気体環境、温度、光照射および過程のプロト
コールを含み、かつ特定の培養系に添加される増強剤は、本明細書中に示されたように個
別の増強剤の効果を比較することにより経験的に決定されるであろう、当該培養系の限定
要因の観点より通常選択されるであろう。さらに、培養系に一つより多い増強剤が存在す
る場合に、タキサン生成の更なる増加が達成されるであろうことが発見されている。
【0063】
本発明の意図する代表的な増強剤は表1に例示されている。本発明の増強剤は、複数の
一般的なクラスに分類されるであろう。それらのクラスは、抗褐色化剤、抗老化剤、抗エ
チレン剤、オーキシン関連成長調節物質、前駆体、阻害剤、エリシター、刺激物およびジ
ャスモン酸関連化合物といった植物成長調節物質である。
【0064】
本発明の意図する増強剤の一つのクラスは、抗褐色化剤である。本明細書中で使用され
ている「抗褐色化剤」の用語は、細胞培養過程における色素の形成を阻害するため栄養培
地に添加される成分を示す。これらの色素は、細胞成長、生存能力、および産物生成に有
害な影響を及ぼすことが一般的に観察される、フェノール類および関連化合物を含む。本
発明の栄養培地において用いられる典型的な抗褐色化剤は、アスコルビン酸である。典型
的には、抗褐色化剤は培地中に10ppbから1000ppmの範囲の濃度となるよう添加されるであ
ろう。
【0065】
他の種別の増強剤は抗老化剤である。抗老化剤は細胞を老化より防護する、生物学的ま
たは非生物学的起源の物質である。この様な薬剤は例えば、老化を促進する化合物の生成
を防ぐ、老化促進物質の活性を抑える、ラジカル除去剤または抗オキシダント活性を提供
する、細胞性の膜及び小器官の無傷な状態を守護する、またはその他の機構により反応可
能である。この様な薬剤にはエチレン反応の拮抗剤、スペルミン、スペルミジン、ジアミ
ノプロパン、及びその類似物質等のポリアミン及びその代謝産物、抗褐色化剤、フェノー
ル産物の阻害剤、及びラジカル除去剤、例えば還元型グルタチオン、没食子酸プロピル、
及びβ-メルカプトエタノールアミン等のスルフヒドリル化合物を含む。
【0066】
抗エチレン剤はエチレン生成またはエチレン反応を阻害する物質として定義される。エ
チレン代謝を阻害する抗エチレン剤は、エチレン生合成拮抗剤及びエチレン反応拮抗剤へ
とさらに分類される可能性がある。エチレン生合成拮抗剤は、エチレンの生合成経路を阻
害する化合物である、この生合成経路に沿った阻害される酵素の例はACC合成酵素、ACC酸
化酵素、及びエチレン酸化酵素を含む。エチレン生合成拮抗剤の例はα-アミノイソ酪酸
、アセチルサリチル酸、メトキシビニルグリシン、アミノオキシ酢酸、及びその類似物を
含む。
【0067】
エチレン活性拮抗剤の例は、銀を含有する化合物、銀錯体、または銀イオン、二酸化炭
素、1-メチルシクロプロペン、2,5-ノルボルナジエン、トランス-シクロオクテン、シス-
ブテン、ジアゾ-シクロペンタジエン、及びその類似物を含む。適切な銀塩は硝酸銀、チ
オ硫酸銀、リン酸銀、安息香酸銀、硫酸銀、トルエン硫酸の銀塩、塩化銀、酸化銀、酢酸
銀、ペンタフルオロプロピオン酸銀、シアン酸銀、乳酸の銀塩、ヘキサフルオロリン酸銀
、亜硝酸銀、及びクエン酸の三銀塩を含む。多種の銀塩によるタキサン生合成の増強の例
は例10に示す。
【0068】
抗エチレン剤は培地に10ppbから1000ppmの程度で含有させることが可能である。培地に
銀が含まれる場合、銀は濃度が900μM以下、好ましくは500μM以下、更に好ましくは20
0μM以下になるよう添加する。培地に銀が含まれる場合、抗エチレン剤は濃度が10nM以
上、好ましくは100nM、更に好ましくは1μM、及び一般的には10μM添加する。
【0069】
本発明において想定される増強剤は、植物成長調節剤、特にオーキシン、オーキシン様
の活性を持つ化合物、及びオーキシン拮抗剤を含むオーキシン関連成長調節剤を含む。オ
ーキシン関連成長調節剤は、一般的には濃度が10-10Mと10-3Mの間で、好ましくは10-8と1
0-5Mの間で培地に含ませる。最も好まれるオーキシン関連成長調節剤の例は、1-ナフタレ
ン酢酸、2-ナフタレン酢酸、1-ナフタレンアセトアミド/ナフチルアセトアミド、N-(1-ナ
フチル)フタルアミド酸、1-ナフトキシ酢酸、2-ナフトキシ酢酸、β-ナフトキシ酢酸、1-
ナフトキシアセトアミド、3-塩化フェノキシ酢酸、4-塩化フェノキシ酢酸、4-ヨウ化フェ
ノキシ酢酸、インドールアセトアミド、インドール酢酸、インドイル酢酸、インドールア
セチルロイシン、γ-(3-インドール)酪酸、4-アミノ-3,5,6-トリクロロピコリン酸、4-ア
ミノ-3,5,6-トリクロロピクリン酸メチルエステル、3,6-ジクロロ-o-アニス酸、3,7-ジク
ロロ-8-キノリンカルボン酸、フェニル酢酸、2-ヨウ化フェニル酢酸、3-ヨウ化フェニル
酢酸、2-メトキシフェニル酢酸、クロロプロファム(イソプロピル-3-クロロカルバニレ
ート)、4-塩化インドール-3-酢酸、5-塩化インドール-3-酢酸、5-臭化-4-塩化-3-インド
イル酪酸、インドールアセチルフェニルアラニン、インドールアセチルグリシン、インド
ールアセチルアラニン、4-塩化インドール、p-塩化フェノキシイソ酪酸、1-ピレノキシル
安息香酸、リソホスファチジン酸、1-ナフチル-N-メチルカルバミン酸、及びエチル-5-塩
化-1H-インダゾール-3-イル酢酸-3-インドールブタン酸を含む。その他の好まれるオーキ
シン関連成長調節剤の例は、ナフタレン-2,6-ジカルボキシル酸、ジ無水ナフタレン-1,4,
5,8-テトラカルボキシル酸、ナフタレン-2-スルホンアミド、無水4-アミノ-3,6-ジスルホ
-1,8-ナフタリン酸、3,5-ジメチルフェノキシ酢酸、1,8-ナフタルイミド、2,4-ジクロロ
フェノキシ酢酸、2,3-ジクロロフェノキシ酢酸、2,3,5-トリクロロフェノキシ酢酸、2-メ
チル-4-塩化フェノキシ酢酸、ニトロフェノキシ酢酸、DL-α-(2,4-ジクロロフェノキシ)
プロピオン酸、D-α-(2,4-ジクロロフェノキシ)プロピオン酸、4-臭化フェノキシ酢酸、4
-フッ化フェノキシ酢酸、2-ヒドロキシフェノキシ酢酸、5-塩化インドール、6-塩化-3-イ
ンドイル酢酸、5-フッ化インドール、5-塩化インドール-2-カルボン酸、3-塩化インドー
ル-2-カルボン酸、インドール-3-ピルビン酸、5-臭化-4-塩化-3-インドイル酪酸、6-塩化
-3-インドイル酪酸、キノリン-2-チオグリコール酸、アミノフェニル酢酸、3-ニトロフェ
ニル酢酸、3-塩化-4-ヒドロキシ安息香酸、クロルフルレノール(2-クロロ-9-ヒドロキシ
フルオレン-9-カルボン酸)、6-塩化-3-インドイル酢酸、N-(6-アミノヘキシル)-5-塩化-1
-ナフタレンスルホンアミドヒドロクロライド、2-塩化-3(2,3-ジクロロフェニル)プロピ
オンニトリル、o-塩化フェノキシ酢酸、6,7-ジメトキシ-1,2-ベンゾイソキサゾール-3-酢
酸、3-オキソ-1,2-ベンゾイソチアゾリン-2-イル酢酸、マストパラン、2,3,5-トリヨード
安息香酸、2-(3-塩化フェノキシ)プロパン酸、及びメコプロップ(2-(4-クロロ-2-メチル
フェノキシ)プロパン酸)を含む。他のオーキシン関連成長調節剤に適した例は、ナフトエ
酸ヒドラジド、2,4-ジブロモフェノキシ酢酸、3-トリフルオロメチルフェノキシ酢酸、オ
キシインドール、インドール-2-カルボン酸、インドール-3-乳酸、β-(3-インドール)プ
ロピオン酸、2-臭化フェニル酢酸、3-臭化フェニル酢酸、2-塩化フェニル酢酸、3-塩化フ
ェニル酢酸、2-メチルフェニル酢酸、3-メチルフェニル酢酸、3-トリフルオロメチルフェ
ニル酢酸、3-メチルチオフェニル酢酸、フェニルプロピオン酸、4-塩化-2-メチルフェニ
ルチオ酢酸、2-塩化安息香酸、3-塩化安息香酸、2,3-ジクロロ安息香酸、3,4-ジクロロ安
息香酸、2,3,5-トリクロロ安息香酸、2,4,6-トリクロロ安息香酸、2-ベンゾチアゾールオ
キシ酢酸、2-塩化-3-(2,3-ジクロロフェニル)プロプオンニトリル、2,4-ジアミノ-s-トリ
アジン、無水ナフタル酸、ジケグラック(Dikegulac)(2,3:4,6-ビス-O-(1-メチルエチリデ
ン)-α-L-キシロ-2-ヘキスロフラノソン酸(hexulofuranosonicacid))、クロルフルレノ
ールメチルエステル、2-(p-塩化フェノキシ)-2-メチルプロピオン酸、2-塩化9-ヒドロキ
シフルオレン-9-カルボン酸、2,4,6-トリクロロフェノキシ酢酸、2-(p-クロロフェノキシ
)-2-メチルプロピオン酸、エチル4-(塩化-o-トリルオキシ)酪酸、[N-(1,3-ジメチル-1H-
ピラゾール-5-イル)-2-(3,5,6-トリクロロ-2-ピリジニル)オキシ]アセトアミド、4-塩化-
2-オキソベンゾチアゾリン-3-イル-酢酸、2-(2,4-ジクロロフェノキシ)プロパン酸、2-(2
,4,5-トリクロロフェノキシ)プロパン酸、4-フッ化フェニル酢酸、3-ヒドロキシフェニル
酢酸、オルソニル(2(β-クロロ-β-シアノエチル)-6-クロロトルエン)、3,4,5-トリメト
キシ桂皮酸、2(3,4-ジクロロフェノキシ)トリエチルアミン、インドール-3-プロピオン酸
、イオキシニルナトリウム(Sodium Ioxynil)(4-ヒドロキシ-3,5-ジヨードベンゾニトリル
)、2-ベンゾチアゾール酢酸、及び(3-フェニル-1,2,4-チアジアゾール-5-イル)チオ酢酸
を含む。
【0070】
他の種別の植物成長調節剤も増強剤として栄養培地に含ませることが可能である。これ
らは、N6-ベンジルアデニン、6-[γ,γ-ジメチルアリルアミノ]プリン、キネチン、ゼア
チン、N-フェニル-N'-1,2,3-チジアゾール-5-イル尿素(チジアズロン)、及び関連フェニ
ル尿素誘導体、及びその類似物等の、サイトカイニン関連成長調節剤、GA3、GA4、GA7、
及びGA誘導体等のジベレリン類、アブシジン酸及びその誘導体、ブラシノステロイド、及
びエチレン関連成長調節剤を含む。これらの成長調節剤は濃度が10-10Mと10-3Mの間で、
好ましくは10-8Mと10-5Mの間で培地に含ませることが可能である。
【0071】
他の種別の増強剤は前駆体または生合成前駆体である。本明細書中で使用する「前駆体
」という用語は、栄養培地に加えられ細胞により代謝され及び取り込まれたタキソール及
びタキサンになる化合物を示すために使用される。適した前駆体は、酢酸、ピルビン酸及
びその類似物、α-フェニルアラニン、β-フェニルアラニン(3-アミノ-3-フェニルプロピ
オン酸)、フェニルイソセリン、N-ベンゾイルフェニルイソセリン、安息香酸、シキミ酸
、グルタミン、桂皮酸、及びその類似物などイソプレノイド化合物の前駆体を含む。これ
らの分子の誘導体も又前駆体として適している。
【0072】
他の種の増強剤は阻害剤である。阻害剤は酵素性または他の細胞性活性を阻害する化合
物である。本明細書で使用する「代謝阻害剤」という用語は、栄養培地に加えられ、特定
の生合成経路を阻害する化合物を示すために使用される。例えば、代謝阻害剤は、初期の
生合成前駆体に関し競合する種々の経路を阻害することにより、タキソール、バッカチン
III、または他のタキサン生合成を増強するために使用することが可能である。この種の
特に効果的な増強剤は、桂皮酸またはその誘導体の合成または代謝を阻害することが可能
な化合物であるフェニルプロパノイドの代謝阻害剤を含む。これら化合物は、好ましくは
p-クマル酸、4-フッ化-DL-チロシン、4-メトキシ安息香酸、3-ジメチルアミノ安息香酸、
4-メトキシ桂皮酸、4-ニトロ桂皮酸エチルエステル、4-ニトロシンナムアルデヒド、メル
カプトエタノール、4-ヒドロキシクマリン、シンナミルフルオレン、2-シアノ-4-ヒドロ
キシ桂皮酸、シンナミルイデンマロン酸、4-ジメチルアミノ桂皮酸、N-シンナミルピペラ
ジン、N-トランス-シンナモイルイミダゾール、2-アミノインダン-2-ホスホン酸、ベンジ
ルヒドロキシルアミン、プロカイン、モネシン、N-(4-ヒドロキシフェニル)グリシン、3-
(4-ヒドロキシフェニル)プロピオン酸、3-(2-ヒドロキシフェニル)プロピオン酸、さらに
好ましくは、D-フェニルアラニン、N-(2-メルカプトプロピオニル)グリシン、及びその酢
酸塩複合体、DL-メタフルオロフェニルアラニン、p-フッ化-DL-フェニルアラニン、ジチ
オスレイトール、4-フッ化桂皮酸、トランス-3,4-ジフルオロ桂皮酸、3,4-ジフルオロ-D-
フェニルアラニン、ジエチルジチオカルバン酸、4-フッ化-(1-アミノ-2-フェニルエチル)
ホスホン酸、3,4-メチレンジオキシ安息香酸、及び最も好ましくは、3,4-メチレンジオキ
シ-6-ニトロ桂皮酸、3,4-メチレンジオキシ桂皮酸、3-[3,4-メチレンジオキシフェニル]
プロピオン酸、3,4-メチレンジオキシフェニル酢酸、4-フッ化-L-フェニルアラニン、4-
ヒドロキシフェニルピルビン酸、4-フッ化-DL-チロシン、トランス3,4-ジメトキシ桂皮酸
、フェニルプロピオル酸、L-2-ヒドロキシ-3-フェニルプロピオン酸、2-ヒドロキシ-4,6-
ジメトキシ安息香酸、SKF-525A(α-フェニル-α-プロピルベンゼン酢酸の2-(ジエチルア
ミノ)エチルエステル)、ビニルイミダゾール、オキサル酸アンモニウム、シナプ酸、及び
1-アミノベンゾトリアゾール、及び関連類似物質を含む。培地に含ませる場合、阻害剤は
濃度が10ppbと1000ppmの間で、好ましくは濃度が100ppbと100ppmの間で、更に好ましくは
濃度が1ppmと50ppmのあいだで加える。
【0073】
培養細胞中のタキソール、バッカチンIII、及び他の関連タキサンの収量を増進するた
めに、本発明者らは多くのアプローチを試みた。生産性を増強させる方法の一つは、エリ
シターと呼ばれるものの使用である。本明細書で使用する「エリシター」という用語は、
植物または培養植物細胞に添加したときに二次的代謝産物の増加を引き起こす、生物学的
及び非生物学的起源の化合物を意味するために使用する(Eilert 1987,"Cell Culture and
SomaticGenetics of Plants,"Vol.4,F.Constabel and I.K.Vasil(eds.), Academic Pre
ss,NewYork,pp.153-196;Ebel,1984,Bioregulators:Chemistry and Uses.257-271;and Da
rvill etal.,1984,Ann.Rev.Plant Physiol.,35,243-275)。多くの異なる化合物がその起
源の性質及びその細胞代謝における活性の方法に依存してエリシターとして機能すること
が可能である。当該研究分野において、本発明は二つの主要な種類のエリシターを使用し
た。1)通常は選別された菌類、細菌類、及び酵母の集団の細胞壁抽出物または濾過物、及
びその精製画分を含む生物的エリシター。2)生物起源の化合物と同様な、化学的なストレ
ス剤を含む非生物的エリシター(表1に一覧されたエリシターを参照)。加えて、重金属
イオンを含む塩及び複合体も、効果的な非生物的エリシターとみなすことが可能である。
これらは、コバルト、ニッケル、ランタン、セレニウム、バナジウム、鉛、カドミウム、
クロム、アルミニウム、ヨウ素、バリウム、ビスマス、リチウム、ルビジウム、ストロン
チウム、及び金などの例を含む。特筆すべきは、エリシター効果を伝達するある種の化合
物、例えば下記に示すジャスモン酸関連化合物もエリシターとみなすことが可能であると
いうことである。
【0074】
Christenらは(1991)Taxusbrevifoliaの懸濁によるタキソールの生産のための菌類のエ
リシター及び選別された化合物の使用について報告している、しかし、エリシター処理に
起因するタキソール蓄積程度の増加については特定していない。
【0075】
一般的には、2種類のエリシター効果(タキサンの培地中への分泌並びにタキサンの培
養細胞への蓄積)発生の範囲はエリシター間で及び種間で異なるが、両種のエリシターは
共に効果的である。最高の生産性増加はキトサングルタメート、リケナン、フェルラ酸、
及び安息香酸により達成される。キトサン及びリケナンは微生物の細胞壁由来の多糖類複
合体である。キトサンは単独で用いると培地に不溶であり、及び毒性があり、及び永久的
に細胞に損傷を与える。一方キトサングルタメートは培地に容易に溶け、及び細胞の生育
に影響を与えない。フェルラ酸及び安息香酸は生物起源の合成化学物質であり、及び一般
的には生体系では抗オキシダントとして使用される。
【0076】
エリシター及び代謝ストレス剤は、本発明によれば、タキソール、バッカチンIII及び
全タキサンを生産させ及び培養組織への分泌を最大にするために、エリシターの特異性、
濃度、タイミングおよび添加継続時間を培養年齢及び培地組成の関数として評価して使用
されるであろう。
【0077】
本発明において意図される他の種別の増強剤は刺激剤である。本明細書で使用する刺激
剤という用語は、栄養培地に加えられ、特定の生合成経路をしげきまたは活性化する(例
えば生合成を導く)化合物を示すために使用される。
【0078】
ジャスモン酸関連化合物は二次的代謝生合成を刺激することによりエリシター効果反応
に寄与する種別の化合物である。ジャスモン酸関連化合物は、ジャスモン酸、及びメチル
ジャスモン酸、エチルジャスモン酸、プロピルジャスモン酸、ブチルジャスモン酸、ペン
チルジャスモン酸、ヘキシルジャスモン酸などのそのアルキルエステル、ジヒドロジャス
モン酸、及びメチルジヒドロジャスモン酸、エチルジヒドロジャスモン酸、n-プロピルジ
ヒドロジャスモン酸、ブチルジヒドロジャスモン酸、ペンチルジヒドロジャスモン酸、ヘ
キシルジヒドロジャスモン酸などのそのアルキルエステル、エピメチルジャスモン酸、フ
ッ化メチルジャスモン酸、シス-ジャスモン、イソジャスモン、テトラヒドロジャスモン
、12-オキソフィトジエン酸、ジヒドロジャスモン、ジャスモニル酢酸、アプリトン、ア
ミルシクロペンテノン、ヘキシルシクロペンテノン、ヘキシルシクロペンタノン、及びそ
の関連誘導体及び類似物質を含む。ジャスモン酸関連化合物は濃度が10-9Mと10-3Mの間で
、好ましくは濃度が10-6と5×10-4Mの間で、更に好ましくは濃度が10-5Mと2×10-4Mの間
で培地に含ませる。特筆すべきは一種以上のジャスモン酸関連化合物を栄養培地に含ませ
ることが可能であるということである。ジャスモン酸関連化合物、オーキシン関連成長調
節剤、前駆体、及びその他の栄養物質は、これら化合物が培養中において代謝されること
により濃度が変化するということは当業者により認識されよう。特に示さない限り、本明
細書において列挙された濃度は栄養培地における初期濃度を記載する。
【0079】
以下に述べる最低二種以上の種類の増強剤からの増強剤を組み合わせると、いかなる一
種の薬剤を単独で用いたときに観察される最高の増強をも上回るTaxus細胞によるタキサ
ン生産の増強を示す。これらの種別の増強剤は、エリシター、ジャスモン酸関連化合物、
エチレン作用の阻害剤、フェニルプロパノイド代謝の阻害剤、抗老化剤、前駆体、及びオ
ーキシン関連成長調節剤である。つまり、好まれる手法において、本発明は、少なくとも
二種のこれらの薬剤群から選択された増強剤の存在下におけるTaxus種細胞の培養により
一種またはそれ以上のタキサンの生産を増強させる方法を提供する。
【0080】
好まれるタキサンの生産方法はエチレン作用の原型阻害剤である銀を、少なくとも一種
の他の増強剤と組み合わせて用いる、特に好まれる方法においては、その他の増強剤はジ
ャスモン酸メチルであるか、または3,4-メチレンジオキシニトロ桂皮酸等のフェニルプロ
パノイド代謝の阻害剤である。
【0081】
お互いに組み合わせて使用する場合、ジャスモン酸関連化合物及びエチレン作用阻害剤
は培地にお互いに特定の割合で含ませることができる。例えば、ジャスモン酸メチル及び
チオ硫酸銀を組み合わせて用いる場合、ジャスモン酸メチルのチオ硫酸銀に対するモル比
は0.0001から9.5の範囲の間で、好ましくは0.001と8の間の範囲で、更に好ましくは0.1と
7の範囲の間で、最も好ましくは1と5の範囲の間でになるであろう。
【0082】
お互いに組み合わせて使用する場合、オーキシン関連成長調節剤及びエチレン作用阻害
剤は培地にお互いに特定の割合で含ませることができる。例えば、オーキシン関連成長調
節剤及びチオ硫酸銀を組み合わせて用いる場合、オーキシン関連成長調節剤のチオ硫酸銀
に対するモル比は0.011から1000の範囲の間で、好ましくは0.015と100の間の範囲で、更
に好ましくは0.02と50の範囲の間で、最も好ましくは0.05と30の範囲の間になるであろう
。
【0083】
一般的には、Taxus細胞をタキサンの生産のために培養する場合、一種またはそれ以上
のオーキシン関連成長調節剤を培養培地に加える。オーキシン関連成長制御因子の存在は
細胞の成長を促進するが、より重要な事項は培養によるタキサンの生産を増強することで
ある。少なくとも一種の他の増強剤をオーキシン関連成長制御因子と同時に添加すること
により、さらなる増強を得ることが可能である。
【0084】
本発明において好まれる方法において、一種またはそれ以上の増強剤を十分量培地に添
加することにより、一種またはそれ以上のタキサンの生産量をを増強剤が存在しないとき
の生産量の相対比で、少なくとも3倍、好ましくは少なくとも5倍、更に好ましくは少なく
とも10倍、更に好ましくは少なくとも30倍に増強する。他の本発明において好まれる方法
において、一種またはそれ以上の増強剤を十分量培地に添加することにより、タキソール
の容積測定生産性を、少なくとも10mg/L/day、好ましくは少なくとも15mg/L/day、更に好
ましくは少なくとも22mg/L/dayに増強する。他の本発明において好まれる方法において、
一種またはそれ以上の増強剤を十分量培地に添加することにより、タキソールの全ブロス
力価を、少なくとも150mg/L、好ましくは少なくとも200mg/L、更に好ましくは少なくとも
350mg/Lに増強する。他の本発明において好まれる方法において、一種またはそれ以上の
増強剤を十分量培地に添加することにより、バッカチンIIIの容積測定生産性を、少なく
とも15mg/L/day、好ましくは少なくとも20mg/L/day)更に好ましくは少なくとも25mg/L/da
yに増強する。他の本発明において好まれる方法において、一種またはそれ以上の増強剤
を十分量培地に添加することにより、バッカチンIIIの全ブロス力価を、少なくとも100mg
/L、好ましくは少なくとも150mg/L、更に好ましくは少なくとも250mg/Lに増強する。他の
本発明において好まれる方法において、一種またはそれ以上の増強剤を十分量培地に添加
することにより、タキサンの容積測定生産性を、少なくとも15mg/L/day、好ましくは少な
くとも25mg/L/day、更に好ましくは少なくとも40mg/L/dayに増強する。他の本発明におい
て好まれる方法において、一種またはそれ以上の増強剤を十分量培地に添加することによ
り、タキサンの全ブロス力価を、少なくとも200mg/L、好ましくは少なくとも300mg/L、更
に好ましくは少なくとも400mg/Lに増強する。
【0085】
以上に増強剤として示した多くの化合物は他の植物系においても使用された。これら非
Taxus系における公式化、管理、及び適切な生理的濃度水準は当該技術者に、本発明と同
値のこれら薬剤を適用する手引きを提供する。
【0086】
細胞材料
本発明の方法において培養に適する細胞はTaxusのいかなる種由来でもあることができ
る。好ましくは、細胞は、生来タキサンを比較的高収量で生産する細胞系由来であると良
い。特に、この様な細胞は一種またはそれ以上のタキサンを標準条件下で高水準で生産す
る、または標準条件下でタキサンの高い平均容積測定生産性を示す能力を有している。適
切な細胞系は、細胞系の細胞を標準タキサン生産条件下で培養し、及び培養液における生
産された一種またはそれ以上のタキサンの水準を観察し、または以下に述べる手段により
培養液中の細胞による一種またはそれ以上のタキサンの平均容積生産性を決定することに
より単離することが可能である。
【0087】
試験手段産生培地において使用される細胞は特定の細胞系に適応した適当な培地におい
て生育する。対数増殖期の終了の後、細胞の一部をタキサンの生産試験のために培養する
。カルス培養は固形培地が使用してもよいが、生産培養は、一般的に液体培地で行われる
。生産培養においては、細胞は表2におけるN培地、スクロースを7%(w/v)マルトースで代
用するのを除いては表2におけるのと同様のN培地、または特定の細胞系の生育及び維持を
最大限にする栄養培地において培養される。生産培養においては、細胞密度は新鮮な重量
を基本として15-20パーセント(w/v)の範囲であるべきである。細胞は10-20日間、25度+/-
2度において暗条件で培養する。液体培養は適切に揺り動かし及び空気にさらすべきであ
る、例えば、旋回式振盪機で120-180rpm。
【0088】
細胞系の特徴を評価する生産培養は適した増強剤を含む。一般的には六種の選択的増強
カクテル(最高五種の増強剤の組み合わせ)を各々の細胞系に対し試験する。組み合わせ
は以下の表Aに示す。
【0089】
培養の終了時に培養における個々のタキサンの力価を、本明細書で示したELISA分析を
行うことにより測定することができる、または、培養において生産されたタキサンのプロ
フィールを、実施例5において示したようにHPLC解析により決定することができる。好ま
れる細胞系は一種またはそれ以上のタキサンを、一種またはそれ以上の増強カクテルにお
ける最低目標タキサン水準を越えて生産する。好まれる細胞系は、少なくとも一種の増強
カクテルにおいて、及び、更に好ましくは二種またはそれ以上の増強カクテルにおいて、
力価及び生産性の双方の目標水準を越える。適した細胞系の生産培養終了時の最低目標タ
キサン力価は、少なくとも100mg/Lタキサンである。あるいは、生産培養行程終了時の最
低平均容量測定生産性目標は10mg/L/dayタキサンである。さらに好まれる細胞系は、生産
培養終了時の最低タキサン力価は少なくとも100mg/Lタキソールまたは200mg/Lバッカチン
III、または生産培養行程終了時の平均容量測定生産性は10mg/L/dayタキソールまたは15m
g/L/dayバッカチンIIIに達する。
【0090】
【化1】

【0091】
様々な種に適した産生培地は表5に列挙するが、その他のものも使用できる可能性があ
る。例えば、表2の培地B、C及びNは、Taxuschinensisの産生培地として特に適している
。培地は表2に列挙した成分を含むことが望ましい。これらの培地は、主要な及び重要で
はない無機塩、有機物及び成長ホルモンまたは成長調節剤を、一般には表2に示した各培
地成分の濃度の、好ましくは1/10から始まり3倍の範囲で、量において含むことが望まし
い。培地BまたはNを使用した場合、成長調節剤を特に、量において01ppmから20ppmの間で
、好ましくは1ppmから10ppmの間で培地に含ませる。培地CまたはNを使用した場合、成長
調節剤を好ましくは0.1ppmから5ppmの範囲水準で含ませる。
【0092】
本発明における意図において、本明細書で示した培地において、他の慣例的組成物(有
機物、ビタミン、アミノ酸、前駆体、活性剤及び阻害剤など)の代用、成長調節剤を含む
様々な組成物の添加または削除、または割合の変更等の装飾を、成長とタキサン生産を表
2において示した培地において観察されるものと同等またはそれ以上にするために行う可
能性があることは、当該技術者により理解されている。
【0093】
操作工程の方法
植物細胞培養工程の操作方法は、栄養、細胞、および産物が時間について加えられ、ま
たはのぞかれる方法(Payne et al.1991)を参照する。最初に全栄養分が供給され、培養
期間の最後に細胞および産物よりなる培養内容が回収される操作方法は「1段階バッチ工
程」と呼ばれる。バッチ工程が2つの連続した期、成長期、および産生期の間に、培地が
換えられる方法では、操作法は「2段階バッチ工程」と呼ばれる。本発明の観点において
、生育培地から産生培地への変換は、急な工程変更、または連続的一連の工程による段階
的、または段階的変更により引き起こされる。ある極端な例において、段階的変更は次第
に大きく変化する組成の培地への段階的置換により成し遂げられる。また別の場合、段階
的変更は成長期培養への産生培地の一つまたはそれ以上の化合物の供給により成し遂げら
れる。これは、給餌-バッチ工程の一例である。
【0094】
「給餌-バッチ(fed-batch)」工程においては、栄養物、および/または一つまたはそれ
以上の増強剤などの特異的培地物質が培養の間、定期的または連続的に供給される。ある
成分を最初にバッチ法によって栄養培地に加え、次いで給餌-バッチ法によって加えても
よく、または専ら給餌-バッチ法によってのみ加えてもよいことに注意すべきである。
【0095】
給餌-バッチ工程の使用で、細胞を長期にわたる産生状態にすることが可能で、事実、
細胞の産生能力は増強されることが発見された。実施例15、17、および表16、および18に
示したように、給餌-バッチ工程において加える栄養物および増強剤は全タキサン生産量
および、タキソールおよびバッカチンIIIとしての特定のタキサンの合成効果において有
意な改良が見られる。さらにこの操作法はさまざまな異なる培養条件下におけるさまざま
な異なる細胞系についても適用可能であることがわかった。
【0096】
成分を給餌-バッチ式で添加することは、特定の成分の濃度が培養中で低いレベルで保
たれなければならないとき特に有利である。たとえば、基質の阻害効果を回避する場合な
どである。同様に、給餌-バッチ式の添加は、栄養培地に最初に成分を加えた時に細胞が
消極的反応を示すとき、または、化学量論的に有意量の化合物は溶解度や有毒限界量のた
めに加えることができないときに有利である。さらに、ある成分を含む給餌溶液の連続的
または連続(定期的)給餌-バッチ添加は、パルス式の添加のような急激な方法で成分が
加えられると、細胞が消極的反応を示す場合に特に好ましい。給餌-バッチ法で加えられ
ることにより好ましい反応を細胞が示す特定の化合物には、アルファーおよびベータフェ
ニルアラニンなどのタキサン前駆体や炭素源としてのマルトース、フルクトース、および
グルコース、アミノ酸としてのグルタミン、グルタミン酸、アスパラギン酸、マクロ栄養
物質としてリン酸、カルシウム、、およびマグネシウム、およびオーキシン-関連成長調
節因子およびジャスモン酸関連化合物などの増強剤などが含まれる。
【0097】
該当技術分野において通常の知識を有する技術者には、供給される成分の組成は、タキ
サン産物の増加のための産生期間の延長やより高いバイオマス濃度を得るための増殖期の
延長などの、望みの結果を得るために変化させうることは明白であろう。最適な産生活性
および効果を達成するための最適条件の選択は、本明細書の開示に基づいて通常の知識を
有する技術者には容易である。
【0098】
本明細書において、培地交換とは培養からの使用済み培地の除去、および引き続き行わ
れる新鮮培地の添加を意味する。細胞はおおむね操作の間中、培養中に維持される。本発
明の方法において、培地交換法は容量当たりのタキサンの高産生を可能にし、且つ持続さ
せ、バッチ法に比べ、優れた工程効率および全産生レベルの向上に有用である。培地交換
法を行う場合、細胞外産物はその後の回収および精製過程は、他の操作法の場合より容易
である。
【0099】
実施例14および表15に示したように、培地交換はタキサン全般および、タキソール、バ
ッカチンIIIおよび、10-脱アセチルバッカチンIIIなどの特定のタキサンの高産生活性を
維持することを成功させる。さらに、この操作法はバッチ操作法に比べ、タキサン全般お
よび、タキソールおよびバッカチンIIIなどの特定のタキサンに対し、容積当たりの産生
増加ももたらす。さらに、この操作法はさまざまな異なる培養条件下におけるさまざまな
異なる細胞系に対し、適用可能である。実施例7.3に示したように、3日毎の使用済み培地
の除去および、新鮮培地の再添加は、増殖条件でのタキサンおよびタキソールの産生の有
意な増強や、同様に細胞外産物の量の増加をもたらす。
【0100】
培地交換の刺激的効果は、細胞内産物の除去によって、フィードバック阻害や産物の分
解が回避されるためであろう。細胞内産物除去による、2次的代謝物の産生および培養液
への分泌へのこのような好ましい効果は、多くの文献中でも、Robins and Rhodes(1986,A
ppl.Microbiol.Biotechnol.,24,35-41)およびAsadaand Shuler(1989.Appl.Microbiol.B
iotechnol.,30,475-481)により報告されている。定期的な使用済み培地の除去は、上記の
利点を含み、さらに他の、非タキサン、阻害物質(フェノール化合物など)が培地から除
去されることにより、2次的生合成への抑制を取り除く作用をする。
【0101】
活発に生合成を行っている細胞への新鮮培地の再添加は、減少した必須栄養物質の供給
による産生増強をもたらすであろう。例えば、Miyasaka et al. (1986, Phitochemistry,
25,637-640)は、単なるスクロースの培地添加によりSa1via miltiorhizaの静止期の細
胞を刺激し、ジテルペン代謝産物、クリプトタンシノンおよびフェルギノールの産生をさ
せることを報告している。おそらく、静止期において炭素の限界により、生合成が止まっ
ていたのだろう。本発明において用いた定期的培地交換法は上記の要素の結果として有益
となるだろう。培地交換の量、交換頻度、および交換する培地の組成はさまざまでありう
ることは理解されよう。培地交換による生合成および分泌の活性化能は、連続的、半連続
的、または給餌-バッチ法において、効率的商業的工程の操作や構築にとって重要な意味
を持つ。
【0102】
バッチ培養の内容の全部でないが大部分が回収されて、連続的細胞増殖および産生のた
めに新鮮な培地の添加がされると、その工程は「繰り返し-汲み出し-充填」操作に似て、
「半-連続工程」と呼ばれる。新鮮培地が連続的に供給されて流れ出る培地が連続的に除
去されると、この工程は「連続的」と呼ばれる。細胞が反応装置内に保持される場合、工
程は「パーフュージョン法」と呼ばれる。細胞が連続的に流れ出る培地とともに除去され
ると、その連続的工程は「恒成分培養槽(chemostat)」と呼ばれる。
【0103】
このようなさまざまな操作工程の方法がここで記載したタキサン-合成系に適用できる
ことは理解されよう。
【実施例】
【0104】
以下の実施例は、本発明を実施するのに用いた材料と方法のさらなる記載のために提供
される。実施例は説明を意図しており、いかなる方法においても発明を限定するものでは
ない。
【0105】
実施例1 カルスの開始
Taxus植物材料は多くの野生型および培養系の植物から得られた。材料は研究室におい
て到着後すぐに処理するか、または4℃で使用時まで保存した。
【0106】
材料は最初に希釈セッケン溶液で洗浄し、水でリンスし、CLOROX溶液(1%ハイポクロラ
イト、pH7.0)で10分間、表面を滅菌した。材料は滅菌条件下で3回滅菌水でリンスした。
針は1%ポリビニルピロリドン(PVP)100mg/lアスコルビン酸溶液中で切断される。針は切断
末端を培地E(表2参照)中にして静置した。30から40の植物組織片を培地プレート当たり
に培養した。組織片を含むプレートは25±1℃の暗黒下で培養した。プレートは毎日、微
生物の汚染を調べ、汚染が見られたときは、非汚染針を取り出して新鮮な培地Eプレート
の中に置いた。実質的カルス形成を観察し、カルスを20日目に組織片から分離し、表3に
示したさまざまなカルス増殖培地へと移した。たとえばTaxus chinensisのカルスは培地D
に移した(表2参照)。この最初の手順は非常に効率がよく、低い汚染率および高い90%以
上の最初の組織片のカルス誘導率へとつながった。同様の手順をTaxus brevifolia,Taxus
canadensis,Taxuscuspidata,Taxus baccata,Taxus globosa,Taxus floridana,Taxus wa
llichiana,Taxusmedia,およびTaxus chinensisの開始培養に対しても首尾よく用いるこ
とができた。
【0107】
実施例2 カルスの増殖
組織片から取り出したカルスは、25±1℃の暗黒下で培養した。健康なカルス部分を新
鮮な培地に7から10日毎に移植したが、この移植の頻度は褐色化を防ぎ、長くカルスを維
持するために非常に重要であった。さまざまな種のカルスに対して好ましい成長および維
持培地を表3にまとめた。
【0108】
実施例3 懸濁培養の開始
1gの生重量のカルス材料を、各種に対して適当な培地(表3参照)25mlを含む125mlの3
角フラスコに無菌的に接種した。例えば培地DはTaxas chinensisに用いられた。フラスコ
にシリコンキャップ(Bello,NJ)を被せ、旋回振盪機上に120rpmで24±1℃暗黒下で置い
た。懸濁培養はおよそ3から10日で形成された。最初に、培地はミラクロスフィルター(C
albiochem)を含むブフナー漏斗を用いフラスコ内容物を吸引濾過することにより交換し
、すべての生物を新鮮な培地に再懸濁した。細胞が増殖したとき、1から2g(生重量)の細
胞を一般に新たな25mlの新鮮な培地を含む125mlフラスコに移し、その後毎週副培養(サ
ブカルチャー)した。
【0109】
実施例4 懸濁細胞の成長
典型的成長率および細胞濃度は表4にまとめた代表的種の懸濁培養において達成される
。
【0110】
ある詳細な例として、時間ごとの生物重量(生重量および乾燥重量)の増加はTaxus ch
inensis系統K-1について図1に示した。最大増殖率はもっとも急速に生物重量が増殖曲線
上で増加した点における勾配を取ることで測定した。Taxus chinensisの細胞培養は最大
倍加時間2.5日で増殖した。この増殖率はTaxus種の懸濁培養について以前報告されていた
ものより有意に高い。例えば、Christen et al.(1991)は、3から4週間の培養で生物重量
は5から10倍に増加すると報告した。これは、Taxusbrevifolia懸濁培養についての平均
倍加時間7から12日に換算される。
【0111】
高濃度での培養能力は、細胞培養過程の量的産生能力を最大にするのに重要である。Ta
xus brevifoliaの培養では、1lの培養当たり乾燥重量1g以下にしか届かない(Christen
etal.(1991)にあるデータより計算される)が、Taxus chinens isの懸濁培養では18日
の成長の後1l当たり8から20gの乾燥重量に到達することが可能である。細胞の生存率は0
.05%フルオレセインジアセテートの溶液(Widholm,1972,StainTechnol.,47,189-194)に
よりアセトン中で染色し、倒立蛍光顕微鏡下(Olympus IMT-2,Japan)で青色光の励起によ
り緑の蛍光を発した細胞を数えることにより決定した。細胞の生存率は増殖期を通じ90%
以上であった。
【0112】
急速増殖条件下での高生存率を維持しながらの細胞の高細胞濃度への培養能力は、タキ
ソール、バッカチンIII、およびタキサンの産生のための植物培養過程の経済的操作に対
しての前もって必須となる重要な要素である。
【0113】
実施例5 タキソール、バッカチンIII、および他のタキサン類の解析
5.1.ELISA法
ELISA解析(HawaiiBiotech♯TA-01)を用いて、細胞培養抽出液中のタキソールの検出を
行った。この方法は高感受性(0.1ng/mL)を提供した。しかしポリクローナル抗体が用いら
れるため、他のタキサンとの交差反応が見られた。画分回収式の調製用(解析スケール)
HPLCは、10-デアセチルタキソール、7-キシロシル-10-デアセチルタキソール、セファロ
マニン、10-デアセチル-7-エピタキソール、7エピタキソール、並びに他の未同定タキサ
ンとの交差反応を示した。そのような交差反応にも関わらずこの方法はタキサン産物の検
出に特に有用であり、多くの細胞系統が素早くスクリーニングされることが見いだされた
。優位なタキサンの産生を示す細胞抽出液は、概略を後に示すHPLC法を用い詳細に解析し
た。
【0114】
モノクローナルELISA解析(HawaiiBiotech#TA-02)も、細胞抽出液中のタキソールの
検出に用いた。この方法は高い感受性(0.1ng/mL)および有意な低い交差反応を提供する
。
【0115】
5.2 タキソール、バッカチンIII、および他のタキサンの抽出
タキサンの上清からの抽出は存在濃度に依存していくつかの方法により成し遂げられた
。十分量のタキサン(およそ1-5mg/L)が液体倍地中に存在するとき、試料はとても迅速か
つ効率的に調製された。培地(2mL)を完全に乾燥し(真空中で)、測定された量のメタノ
ール(0.5-2.0mL)を加えた。この混合物を超音波により完全に溶解するまで、または試料
の分散が達成されるまで振盪した。HPLC解析に先立ち遠心により固体を除去した。量的回
収は1mg/Lのレベルであったが、検出レベルは0.1mg/L以下であった。
【0116】
タキサンの培養上清中の濃度が非常に低いとき(1mg/L以下)は培地は3倍量のメチレ
ンクロライドとイソプロピルアルコール(IPA)との混合液(9:1用量で)により抽出した。有
機層を乾燥で減少させ、計量したメタノールの(50-250ml)中に再構成した。多くの抽出で
は90-95%のタキソール、セファロメニン、およびバッカチンIIIが0.6mg/Lのレベルで回収
された。
【0117】
上清のタキサン濃度が5mg/Lを越えるときはより迅速な試料調製を用いた。上清1に対
し(容量)3の0.1%酢酸を含むメタノールに混合する。この混合を30分間音波撹拌し、濾
過し、HPLC解析を行った。
【0118】
全ブロス試料(細胞を含む培養上清)は手順の節で記述したのと同様の方法で調製した
。培地1に対し(容量)3の0.1%酢酸を含むメタノールに混合した。この混合物は30分間
超音波撹拌し、さらに30分間静置し、濾過し、HPLC解析した。
【0119】
細胞物質は、新鮮に回収した細胞を凍結し(-5℃)および引き続き真空乾燥し、およびメ
タノールソックスレーを50サイクル繰り返し行うことにより抽出した。メタノールは回転
式真空機により濃縮し(100倍)、試料はHPLCにより解析した。一般に、70から80%のタキ
サンは10から15%の測定可能な分解とともに回収された。ソックスレーに先立つ試料の乾
燥を十分に行うと、タキソールの分解は5%以下にとどまることが後に判明した。
【0120】
固体培地およびカルスの抽出はタキサンレベルが低いときの細胞の抽出と等しい。しか
し、最終メタノール抽出でのメチレンクロライド/IPAと水の分離はいつも引き起こされる
。タキサンレベルが5mg/Lを越えるとき、全培地の抽出法は固体培地上のカルスの試料の
調製に適用される。
【0121】
5.3 高性能液体クロマトグラフィー法
高性能液体クロマトグラフィー解析(HPLC)は、CM3500/CM3200ポンプから構成されるLDC
2勾配高圧混合解析系、CM4100可変容量自動試料調製機、およびコンピューターに接続し
たSM5000光ダイオード光線検出器を用い、高-炭素注入ディフェニルカラム(Supelco,5μM
,4.6mm X25cm)において行われた。EldexCH150カラムオーブンにより、カラム温度は35℃
に制御された。タキサンのHPLC定量解析は、以下の2成分勾配溶出法を用い行った。
【0122】
【化2】
【0123】
クロマトグラフィー法は類似のいくつかの公表された方法(Witherup et al.1989,J.Li
q.Chromatog.,12,2117-2132)を用い、トリフルオロ酢酸を含むリン酸緩衝液の使用および
長勾配の適用などの改良とともに行われた。これらの改良は混合物からのタキソールおよ
び他のタキサン類の分解能を有意に向上させる。タキサン類の比保留時間は以下に示す。
タキソールは、用いたカラムとハードウェアに依存して31分から33分の間で溶出する。
【0124】
【化3】
【0125】
タキソール、セファロマニン、およびバッカチンIIIの保留時間は国立ガン協会(Nation
al CancerInstitute)より得た真性試料を用いて決定した。上にまとめた他のタキサン類
の保留時間は、HauserChemical(Boulder,CO)から提供された解析標準品と比較した。既
知のタキサン類の同定は保留時間および紫外線スペクトルの比較に基づいて行った。タキ
ソールおよびバッカチンIIIの紫外線スペクトルと類似のUVスペクトルを示すが、これら
のタキサン類の比保留時間を示さない化合物はタキサン類と考えられる。タキソール、セ
ファロマニン、およびバッカチンIIIの定量は真性物質から決定された係数に基づいて行
った。10-デアセチルバッカチンIIIの定量はバッカチンIIIに対し決定された係数を用い
行われた。必要な場合、他のタキサン類の定量はタキソールおよびバッカチンIIIの計算
に用いた係数により行われた。用語「全タキサン類」はタキソールおよびバッカチンIII
と類似の特異的UVスペクトルを示したタキサン類の合計を表す。Taxus培養において同定
された全タキサン類は、10-デアセチルバッカチンIII、9-ディヒドロバッカチンIII、7-
エピ-10-デアセチルバッカチンIII、バッカチンIII、9-ディヒドロ-13-アセチルバッカチ
ンIII、7-キシロシル-10-デアセチルセファロマニン、7-キシロシル-10-デアセチルタキ
ソール、7-エピバッカチンIII、10-デアセチルタキソール、7-キシロシルタキソール、セ
ファロマニン、7-エピ-10-デアセチルタキソール、タキソール、2-ベンゾイル-2-デアセ
チル-1-ヒドロキシバッカチンI、タキソールC、7-エピタキソール、および2-ベンゾイル-
2-デアセチルバッカチンIを含む。
【0126】
タキサン類の特異的なUV吸光度を示さないが、重量スペクトルにおいて特異的なタキサ
ン-重量-断片化特性を示す、タキサン類もTaxus細胞培養において観察された。Taxus細胞
中で産生されたそのようなタキサン類の例には、タクスウンナニンC(Taxuyunnanine C)お
よびその類似体およびその誘導体が挙げられる。
【0127】
各標準品(10μL)は典型的に注入され(最初標準品、次いで3または4試料)、各3成分
の面積を227nmクロマトグラムから計算した。各成分の係数はデータの線形-最小-2乗解析
により得られた。10μLの各試料が注入され、注入当たりの量は標準データ回帰に基づき
計算された。これらの結果は1リットル当たりの総量または乾燥重量パーセントに換算さ
れた。図4に上清試料の典型的クロマトグラムを示す。
【0128】
5.4 迅速高性能液体クロマトグラフィー法
上記の方法に加え、いくつかのHPLCの迅速な方法がより多くの試料を処理できるように
するために開発された。以下にこれらの方法のうち2つの詳細を示す。
【0129】
方法1)迅速高性能液体クロマトグラフィー(HPLC)は、環境温度において前述のハード
ウェアを用い、PhenomenexCurosil-Gカラム(5μM,4.6mm X 25cm 4.6mm X 3cmガードつ
き)行った。タキサン類の定量HPLC解析は以下に示す2成分勾配溶出法で行った。
【0130】
【化4】
【0131】
タキサン類に観察された比保留時間は以下に示す。タキソールは用いたカラムおよびハ
ードウェアに依存して約8分で溶出した。
【0132】
【化5】
【0133】
タキソール、バッカチンIII、および10-デアセチルバッカチンIIIの標準品は50mg/L,10
mg/L、および1mg/Lレベルで調製した。標準品を最初に注入し、その後9試料を注入し、各
3成分の面積を227nmクロマトグラムから算出した。各成分の係数はデータの線形最小2乗
解析により得られた。10μLの各試料を注入し、1リットル当たりの量を試料の希釈およ
び標準試料のデータ回帰に基づくピーク面積から計算した。
【0134】
方法2)迅速高性能液体クロマトグラフィー(HPLC)は前述のハードウェアを用い、環境
温度において、PhenomenexIB-SILカラム(3μM,4.6X15cm、4.6mm X 3cmのガードつき)で
行った。タキサン類の定量HPLC解析は以下の2成分勾配溶出法で行った。
【0135】
【化6】
【0136】
タキサン類に観察された比保留時間は以下に示す。タキソールは用いたカラムおよびハ
ードウェアに依存して約9.5分で溶出した。
【0137】
【化7】
【0138】
定量は前述のように行った。
上記の方法の流速および勾配間隔および時間についての修飾によって、植物細胞培養解
析にの最適なクロマトグラフィーが行われるこ々も見いだされた。
【0139】
5.4. タキソールのMS/MS確認
細胞培養上清中のタキソールの同定のため、フローインジェクションとイオンスプレー
常圧化学イオン化とを組み合わせたMS/MS法(図6に示される)を用い確かめた。図6にあ
るデータを得るのに用いた手順の詳細を以下に示す。重量スペクトル計測機:Sciex API
3の3重4極子常圧イオン化源。窒素を遮断ガスとしてして用い、アルゴンをCIDスペクトル
の衝突ガスとして用いた。 界面:イオンスプレー界面はイオン蒸発イオン化機(電気ス
プレー)により産生された。ゼロガスを噴霧ガスとして用いた。LCポンプ:5μL/分で作
動するABI140B二重シリンジポンプ装置を使用。溶剤:50/50アセトニトリル/H2O2mM NH4
OAc+0.1%ギ酸。注入量:5μL、スペクトルは全てフローインジェクション注入で求めた
。この方法により、細胞培養試料中におけるタキソールの明確な存在の確認およびHPLCの
結果によく一致する定量結果が得られた。
【0140】
実施例6 さまざまな種からのタキソールの生産
さまざまなTaxus種培養からのタキソールの生産を表5にまとめた。カルスは20日暗黒下
で各種毎に示される固体培地において培養された。細胞および培地を一緒に乾燥し、メタ
ノール抽出し、示したようにELISAまたはHPLCにより検定した。
【0141】
実施例7
7.1 増殖培地における生産
タキソールおよびタキサン類の生産は、Taxus chinensis細胞系統K1の増殖培地Aへの移
植の最初の2日間に開始した。15日において観察される最大タキソールは8.81μg/フラス
コであり、0.44mg/Lに相当する。このうち46.1%は細胞外培地に存在した。15日目に、全
タキサン濃度は72.87μg/フラスコ、または3.6mg/Lとなり、このうち58.6%が細胞外培地
に存在した。生存率は蛍光染色(実施例4)により計測され常に90%以上であった。このこ
とは細胞外タキソールおよびタキサン類は細胞の溶解によるものではなく、分泌によるも
のであることを示唆する。
【0142】
タキソール、バッカチンIIIおよび関連タキサン類の生産レベルを、タキサン生合成用
の増強剤を添加しないさまざまな異なる増殖条件(表2および他の実施例において詳しく
述べた)において、さまざまに異なる細胞系統により決定した。これらの回収データは細
胞は増殖に最適な条件で培養され、タキサン生合成条件でないとき、タキソール生合成レ
ベルは典型的には0.5mg/L以下であり、常に2mg/L未満であり、容量当たりのタキソール生
産活性は典型的には0.03mg/L/日から0.07mg/L/日の範囲にあり、常に0.3mg/L/日未満であ
った。同様にバッカチンIII生産レベルは典型的には0.5mg/L以下であり、常に1mg/L以下
であった。容量当たりのバッカチンIIIの生産活性は典型的には0.03mg/L/日以下であり、
常に0.15mg/L/日未満であった。同様に全タキサン価のレベルは典型的には5mg/L未満であ
り、常に20mg/L以下であった。容量当たりの全タキサン生産活性は、典型的には1mg/L/日
未満であり、常に3mg/L/日未満であった。
【0143】
7.2. 生産増強のための培地交換
タキソールおよび全タキサン生産活性の有意な改良は、9日目における無菌的培地Aの
吸引除去および新鮮培地の添加、および同じ処置の12日目の繰り返しによって認められた
。実験は15日間で打ち切られ、結果は図2に示される。培地交換による重要な生産活性の
増加は表6にまとめた。タキソールおよびタキサン類の総生産量は、上記処置を施さない
対照と比較して4.6倍に高まった。重要なことに、細胞外培地から回収されるタキソール
は4.9倍、全タキサン類は5.9倍に、培地交換しなかった対照に比較して高まった。
【0144】
タキソールおよび全タキサン生産活性の増加、およびさらに細胞外への産物の蓄積を引
き起こす注目すべき能力は、効率的操作、細胞の再使用を伴う連続的工程、およびその後
の精製工程の簡易化のために重要である。
【0145】
7.3. 増殖培地におけるタキサン生産に対する光の効果
光は光合成だけでなく、植物細胞培養中の2次的代謝にもさまざまな面で重要な役割を
果たす(Seibertand Kadkade 1980)。実施例4、7.1および7.2に記載した実験は暗黒条件
で行ったが、Taxuschinensis培養の光への反応をここで記述する。
【0146】
1グラムの生重量の7日目Taxus chinensis系統K1の細胞を125ml三角フラスコ中の25m
lの生育培地A(表2参照)に移植し、24±1℃において回転振盪機で120rpmで培養した。
同じフラスコを暗黒条件および標準Gro Luxランプに3フィート離しておいた。ランプのス
ペクトル特異性は図3に示される。結果は表7に示される。
【0147】
培養を光に曝すことは全タキサンのレベルおよび細胞外の蓄積に影響しなかった。しか
しタキサンの内容は有意に2つの処理で変化した。たとえば光下で培養したものはタキソ
ールを暗黒条件下のものより2.8倍も多く生産した。細胞外タキソールの割合もまた暗黒
処理のものより有意に高かった(76%対56%)。光処理の使用、特に特異的スペクトルでの使
用は、タキソールの生産のための細胞培養工程において有用でありうる。
【0148】
実施例8 エリシター
エリシターという単語は、植物細胞培養物に加えられたとき二次代謝の増加をひきおこ
す生物学上の(または生物の)、および非生物学上の(または無生物の)起源を有する化
合物を意味する。
【0149】
多くのエリシターが有効であるとわかっているが、代表例、すなわち、キトサングルタ
メートの使用が、本明細書中に詳しく記載されている。キトサンは、いくつかの植物細胞
培養物組織においてはエリシターとして以前に試されたが、褐色化および生存率の損失な
どの毒性を伴う反応は、キトサンの使用を非実用的にする(Beaumont and Knorr 1987,Bio
technol.Lett.9,377-382)。事実、そのような毒性反応は、論文において報告された多く
のエリシターの共通の欠点である。特にキトサングルタメートなどの化学的に修飾された
キトサンの使用により、毒性効果を回避しつつ、タキソールおよびタキサン生合成を誘導
することは新しいアプローチである。
【0150】
培地Dにおいて7日から8日成長させたTaxuschinensis系統K-1の懸濁液はmirachloth(
Calbiochem)フィルターを有する滅菌ブフナー漏斗の使用で無菌的に吸引濾過された。新
鮮な2グラム重量の細胞は、125mlエルレンマイヤーフラスコにおいて25mlのC
培地(表2参照)へ無菌的に移された。0.05%のキトサングルタメート溶液は、新たに調
製され、および0.22μmカートリッジフィルターを通して濾過滅菌された。1g乾燥重量
当たり165ミリグラムのエリシターの濃度となるように、この溶液の825μlを実験
の開始時にフラスコに加えた。フラスコは、暗やみにおいて110rpmで旋回撹拌器で2
4プラスマイナス1℃で保温培養された。フラスコは15日で破壊的に試料とされ、およ
び成長、細胞の色および培地の観察および細胞の生存率が記録された。試料は実施例5に
おいて記載されたようにタキサンの分析に付した。この実験の結果は表8において示され
ている。
【0151】
エリシター処理は、非処理の対照標準よりも細胞当たりの全タキサン生産量(0.53%対0.
42%乾燥重量タキサン)が多く、ささやかな改良の結果となった。エリシターの非毒性の性
質は、両方の処理において観察される高生存率(75%-80%)から明白である。実際に、対照
標準に比べてエリシター処理において増加した乾燥重量は再現性をもって観察された(14.
2g/1対10.1g/l乾燥重量)。細胞の高濃度も加味すると、エリシター処理においては全タキ
サンの1.8倍の力価、すなわち75.8mg/l対対照標準42.4mg/lの結果となった。
【0152】
細胞当たりの基準(0.098%対0.054%乾燥重量タキソール、1.8倍増加)および力価におい
ての比較(13.9mg/l対5.4mg/l、2.6倍増加)の両方で、エリシター処理はタキソール生合成
の増加の結果をもたらしたる。分泌の程度も、対照標準に比べエリシター処理のほうがよ
り高かった(85%対72%細胞外産物)。
【0153】
本明細書中に記載されているエリシター処理は、タキソール産物の増加、より有利な産
物の側面、産物分泌の増加および細胞の高生存率の保持力の結果となる。これらの生産の
特性は、タキソール生産の細胞の培養過程にかなりの改良を表す。
【0154】
実施例9 産生培地開発
実施例6において記載された濃度を越えるタキソール生産力の増加のための努力におい
ては、特別な「産生培地」を計画するために栄養濃度は処理された。培地Dにおいて成長
させたTaxus chinensis系統K-1の7日から8日たった懸濁液は、MIRACLOTH(アクリル酸
のバインダーを含むレーヨンポリエステル布地)フィルター(Calbiochem)を有する滅菌ブ
フナー漏斗の使用で無菌的に吸引濾過された。新鮮な500mg重量の細胞は、5mlの
産生培地BおよびC(表2参照)へ無菌的に移された。その容器は暗やみにおいて110
rpmで旋回撹拌器で24プラスマイナス1℃で18、25および42日のいろいろな時間の
期間で保温培養された。処理は破壊的に試料とされ、および成長、細胞の色および培地の
観察、および細胞の生存率が記録された。試料は実施例5において記載されたようにタキ
サンの分析をされた。この実験の結果は表9において示されている。
【0155】
9.1 18日培養の結果
タキサンおよびタキソールのかなりの濃度の生産によりTaxus chinensis細胞培養は培
地の組成物の変化に応答する。これらのデータは表9において要約され、および試料のク
ロマトグラムは図4において示されている。培地Bにおいては、99.8mg/lの全タキサンが
生産され、24.1mg/lのタキソールを含んだ。培地Cにおいては、110mg/lの全タキサンが
生産され、21.3mg/lのタキソールを含んだ。乾燥重量の基準では、細胞は、培地Bにおい
ては0.18%乾燥重量タキソール、培地Cにおいては0.065%乾燥重量タキソールを生産した
。
【0156】
9.2 長期培養
Taxuschinensis(K-1系統)の25および42日の長期培養後のタキソールおよびタキ
サンの生産は、培地Cにおいて研究され、その結果は図5において要約されている。以下
に重大な観察が要約されている:
(i) Tuxus培養懸濁液は、タキソールおよびほかのタキサンのかなりの濃度を生産す
ることができる。もっとも高い集積は42日で起こり、0.32%の乾燥重量タキソール、お
よび0.62%の乾燥重量タキサンを含んだ;最終の培地体積に基づくタキソールの力価153mg
/lおよび全タキサンの力価295mg/lに相当した。タンデム質量分光測定法によるこの試料
の分析は、図6において示されたようなタキソールの存在を確認した。MS/MSによる定量
は、HPLCと非常に良い一致を示した。
【0157】
(ii) 25日から42日の間のタキソール生合成の速度は、17日間においては直線
的な生産であると仮定すると、1日につき、1リットル当たり7.6mgタキソールであった
。この速度は、最初の25日においての生産速度よりも有意に高い。25日および42日
の間の全タキサンの生合成の速度は、1日につき、1リットル当たり12.3mgであった。タ
キソール、バッカチンIII、および全タキサンの生産力の体積の平均は、それぞれ、3.6、
0.5、および7.0mg/l/dayであった。
【0158】
(iii) 実施例7において記載された急速増殖条件などの(タキサン生合成は増加され
ない)急速増殖条件と比較して、産生培地の定式化は特定のタキソール含有量において4
5倍の増加の誘導も可能である。
【0159】
(iv)望ましくないタキサン産物を最小限度にし、産物スペクトルを希望する最終産物
タキソールへの生合成を漏斗を通り抜けるようにする処理も可能であった。たとえば、2
5日では、タキソールは全タキサンの28%を構成しおよび42日では、全タキサンの5
2%を構成したが、それに対して生育培地(実施例7.1参照)においては、タキソールは
全タキサンの12.2%しか構成しなかった。処理した産物の側面のこの能力は、下流部
門の精製および調節結果の純度に関わる産物に重要な影響を有するだろう。たとえば、タ
キサン副産物、セファロマニンの生産を押さえる能力は樹皮からのタキソールの精製に比
べて、後の工程での精製を非常に単純化させる可能性がある。
【0160】
(v)Taxus細胞培養物はタキソール(42日で87%)およびほかのタキサンのかな
りの量の分泌を誘導した。細胞外タキソールおよびタキサンの存在は細胞溶解液に帰する
というよりはむしろ分泌に帰するということは、いくつかの独立した観察により確証され
る:(a)連続した生合成が25日と42日の間に起こり、このことは細胞は生きていてお
よび活動したことを示唆した。独立した観察は産生培地においては18日後に70%以上
の生存率が観察されたということを示した。(b)異なるタキサンの異なる割合が分泌され
た。もし細胞が溶解するとしたなら、培地においての割合は異なるタキサンでも同じであ
るはずである。
【0161】
(vi) このTaxus細胞系統が産物濃度が非常に高い細胞外環境において生長し、および
高い割合でタキソールを生産する能力をもつことは特に注目すべき価値がある。
(vii) これらの結果が得られるTaxus細胞系統は細胞の高濃度の増殖を急速にするこ
とも可能であり、および急速増殖条件下で20世代後にも報告された生産性を明白にし、
このことはその安定性および商業上の潜在能力を証明する。
【0162】
以前に報告された結果よりも、本明細書中に記載されている条件下でのTaxus chinensi
sの細胞系統により生産されるタキソールおよびタキサンの濃度は35から150倍高い
。たとえば、Christenet al.(1991)は培養の2から4週間後Taxus brevifoliaの培養懸
濁液より1から3mg/lのタキソールの生産を報告した。WickeramesinheおよびArteca(199
1)はTaxus mediaの細胞培養においては、乾燥重量の0.009%のタキソールの生産を報告し
た。
【0163】
要約すると、我々のデータはTaxus chinensis培養の念入りな開始および選択をもって
、および特別に処理された生育培地条件をもって、細胞は高濃度の増殖の誘導も可能だろ
う。これらの細胞が産生培地条件に移されたとき、細胞は長期間に高い生存率を維持して
タキソールおよびほかのタキサンのかなりの濃度を生合成および分泌することも可能であ
る。提起的な培地の交換、光、および産生培地をもつエリシターの混合は、多くの協同性
の生産性の増加の結果となる。これらの特性は、組織培養技術の使用のタキソールおよび
タキサンの生産の十分な商業上の過程の重大な必要条件である。
【0164】
実施例10
10.1 銀の使用のタキサン生産の増加
銀、化合物を含む銀の形態において、銀混合体、あるいは銀イオンは、Taxus種の細胞
培養においてのタキソール、バッカチンIII、およびタキサン生合成の有用な増加因子で
あることが発見された。銀およびほかの増加因子の組み合わせもまたタキサン生産の高い
割合の獲得および維持においては有用であることが発見された。
【0165】
培地L(表2)において培養されたKS1A Taxus chinensis懸濁液の7日たった細胞はMI
RACLOTH(Calbiochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌的に吸引
濾過された。15%から20%(w/v)の範囲において新鮮な細胞の濃度を与えるため、約0
.75から1グラム重量の新鮮な細胞は表10において示された所定の組成物の培養培地の4
から5mlへ植えつけられた。その容器は暗やみにおいて110rpmで旋回撹拌器(1”throw)
で25プラスマイナス1℃で保温培養された。蒸発分は滅菌された蒸留水の付加により補
正された。全液の試料(すなわち細胞外および細胞内のタキサンの両方)は周期的な間隔
で得られ、および実施例5において概説された方法にしたがってHPLCにより処理およ
び分析がされた。
【0166】
表10において要約されたデータはタキソール、バッカチンIII、およびほかのタキサ
ンの生産は化合物を含むいろいろな銀によりうまく増加させられるということを指し示す
。化合物および異なる逆イオンを含む異なる銀の多様性の増加を示している表10におい
て説明したように、この増加は第一に培地にある銀の存在に帰する。これらの生産の濃度
は増加されない培養において観察された生産の濃度(その生産濃度は実施例7において詳
しく述べられている)よりも重大に高い。
【0167】
10.2 チオ硫酸銀の使用のタキサン生産の増加
毒性の考慮および調製物および保管の容易に基づくと、チオ硫酸銀は二次的な実験にお
いて使用される。チオ硫酸銀の調製のために使われる方法は以下であった:1.98gのチオ
硫酸ナトリウム(5水和物)は80mlの水に溶解された。いきおいよく全体をかき回して
20mlの0.1M硝酸銀溶液が加えられ、その結果100mlの20mMのチオ硫酸銀の貯蔵溶液と
なった。チオ硫酸カリウムは同等の効果のある結果をもつチオ硫酸ナトリウムの適所に使
用されることも可能であった。その貯蔵溶液は所定の実験の開始時に細胞培養培地のなか
へ0.22μMカートリッジフィルターを用いて滅菌濾過された。チオ硫酸銀溶液と同様なも
う一つの調製方法もまた適合される。細胞培養実験計画案は表10において記載された実
験のために記載されたものと同様であった。
【0168】
表11はTaxuschinensisの異なる多くの細胞培養の一つの増加因子として銀の使用に
より獲得されたデータを要約した。これらのデータは銀は一般的にタキサン生合成の基本
的な増加をもたらすということを示す。いかなる所定の場合において観察された特別な産
物の側面は細胞の系統および培養培地の特性を反映する。銀イオン/混合体は成長調節、
炭素源、塩、微量元素、およびそのようなものなどの生合成の好む培地においてはほかの
因子と組み合わせにおいて使用されるときタキサン生産の増加においてとくに効果的だろ
う。
【0169】
実施例11 ジャスモン酸メチルおよびジャスモン酸関連化合物の使用のタキサン生産
の増加
ジャスモン酸および関連化合物と同様に、ジャスモン酸のメチルエステル(ジャスモン
酸メチル)は、Tuxus種の細胞培養においてタキサン生合成の増加因子として有用である
ことが発見された。ジャスモン酸メチルおよびほかの増加因子の結合もまたタキサン生産
の高い割合の獲得および維持においては有用であることが発見された。
【0170】
培地M(表2)において培養されたTaxus chinensis懸濁液の7日たった細胞はMIRACLO
TH(Calbiochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌的に吸引濾過さ
れた。15%から20%(w/v)の範囲において新鮮な細胞の濃度で、細胞は表12におい
て示された所定の組成物の培養培地の4から5mlへ植えつけられた。その培養物は暗やみ
において120または180rpm(容器の大きさに依存する)で旋回撹拌器(1”throw)で
24プラスマイナス1℃で保温培養された。蒸発分は滅菌された蒸留水の付加により補正
された。全液の試料(すなわち細胞外および細胞内のタキサンの両方)は周期的な間隔で
得られ、および実施例5において概説された方法にしたがってHPLCにより処理および
分析がされた。
【0171】
表12はいくつかの代表的なTaxus chinensis細胞系統の増加因子としてジャスモン酸
およびジャスモン酸メチルエステルの使用により獲得されたデータを要約した。これらの
データはジャスモン酸およびジャスモン酸メチルエステルは一般的にタキサン生合成の基
本的な増加をもたらすということを示す。いかなる所定の場合において観察された特別な
産物の側面は細胞の系統および培養培地の特性を反映する。これらの増加因子の存在にお
いて獲得されたこれらの生産の濃度は増加されない培養において観察された生産の濃度(
その生産濃度は実施例7において詳しく述べられている)よりも重大に高い。
【0172】
ジャスモン酸、ジャスモン酸メチルエステル、および関連化合物は、ほかの増加因子、
成長調節、炭素源、塩、微量元素、およびそのようなものなどの生合成の好む培地におい
てはほかの因子と結合において使角されるときタキサン生合成の効果的増加因子である。
【0173】
実施例12 3,4-メチレンジオキシ-6-ニトロ桂皮酸の使用のタキサン生産の増加
桂皮酸類似化合物、3,4-メチレンジオキシ-6-ニトロ桂皮酸(MDNA)および関連化合物はT
uxus種の細胞培養においてタキサン生合成の有用な増加因子であることが発見された。MD
NAおよびほかの増加因子の結合もまたタキサン生産の高い割合の獲得および維持において
は有用であることが発見された。
【0174】
培地M(表2)において培養されたTaxus chinensis懸濁液培養SS122-42の7日たった
細胞はMIRACLOTH(Calbiochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌
的に吸引濾過された。細胞は15%から20%(w/v)の新鮮な細胞の濃度の培養培地条件
へ植えつけられた。その容器は暗やみにおいて180rpmで旋回撹拌器(1”throw)で24
プラスマイナス1℃で保温培養された。処理された培養物は試料になりおよびいろいろな
時間点で実施例5において記載された方法を使用して分析がされた。蒸発分は滅菌された
蒸留水の周期的な間隔の付加により補正された。全液の試料(すなわち細胞外および細胞
内のタキサンの両方)は周期的な間隔で得られ、および実施例5において概説された方法
にしたがってHPLCにより処理および分析がされた。
【0175】
表13はTaxuschinensis細胞培養においてタキサン生合成の増加因子として3,4-メ
チレンジオキシニトロ桂皮酸の使用により獲得されたデータを要約した。これらのデータ
はMDNAは一般的にタキサン生合成の基本的な増加をもたらすということを示す。培地IIに
おいて、すなわちMDNAおよび銀の存在においての培養は、タキサンの生産をより増加させ
る。いかなる所定の場合において観察された特別な産物の側面は細胞の系統および培養培
地の特性を反映する。これらの生産の濃度は増加されない培養において観察された生産の
濃度(その生産濃度は実施例7において詳しく述べられている)よりも重大に高い。
【0176】
実施例13 増加因子の組み合わせの使用のタキサン生産の増加
いろいろな増加因子は、タキサン生産においては組み合わせにおいて使用され、重大お
よび協同性の改良を与えた。
【0177】
培地P(SS64-412)、培地O(SS64-561,SS64-571)、培地I(SS124-77,SS85-26)、培地M(
SS122-29)(これらの培地の組成物は表2において記載されてある)において培養されたT
axus chinensis懸濁液培養の7日たった細胞はMIRACLOTH(Calbiochem)フィルターにあわ
せた滅菌したブフナー漏斗の使用で無菌的に吸引濾過された。細胞は20%(w/v)の新鮮
な重量の濃度の(表14において指し示されている)培養培地へ植えつけられた。その培
養物は暗やみにおいて180rpmで旋回撹拌器(1”throw)で24プラスマイナス1℃で保
温培養された。蒸発分は滅菌された蒸留水の周期的な間隔の付加により補正された。全液
の試料(すなわち細胞外および細胞内のタキサンの両方)は周期的な間隔で得られ、およ
び実施例5において概説された方法にしたがってHPLCにより処理および分析がされた
。
【0178】
表14はTaxuschinensis細胞培養においてタキソール、バッカチンIII、およびタキサ
ン生合成の増加因子のいろいろな組み合わせの使用により獲得されたデータを要約した。
そのデータは増加因子の組み合わせにより個々の因子でみられるタキサン生産の増加、お
よび増加されない条件においての生産濃度(その生産濃度は実施例7において詳しく述べ
られている)よりたくさんのタキサン生産の増加を立証する。
【0179】
実施例14 培地交換によるタキサン生産の増加
この実施例は培地において高い生産性は培地成分の補充および消費された培地の除去に
より維持されるのも可能であるということを立証する。
【0180】
細胞は最初は培地M(Paella)、培地I(SS29-3A5)、および培地I(SS45-146)において培
養された。これらの培養培地の詳しい組成物は表2において記載されている。これらの細
胞系統の7日たった細胞はMIRACLOTH(Calbiochem)フィルターにあわせた滅菌したブフナ
ー漏斗の使用で無菌的に吸引濾過された。新鮮な約1.5グラム重量の細胞は表15におい
て指し示されたそれぞれの培養培地の4.5m1へ植えつけられた。その容器は暗やみにおい
て120rpmで旋回撹拌器(1”throw)で24プラスマイナス1℃で保温培養された。蒸発
分は滅菌された蒸留水の周期的な間隔の付加により補正された。培地交換処理のために、
容器において後ろの細胞を残して、消費された産生培地は一回分の培養の10から11日
後に滅菌したピペットを使用して吸引して取り除かれた。その消費された上清は実施例5
において記載された方法を使用して細胞外タキサンの分析がされた。最初の1回分の培養
と同じ組成物の新鮮な培養培地は生産力のある細胞を含む容器に加えられた。その細胞は
上で記載された同じ環境条件下で培養された。培地交換の周期は培養のさらに10から1
1日後に繰り返された。一回分の生産の全細胞外タキサンは表15において培地交換の生
産の全細胞外タキサンと比較されている。培地交換濃縮の値は細胞懸濁液培養の体積(す
なわち、5.75ml)により分けられた細胞外培地において生産されたタキサンの全量を意味
する。
【0181】
表15は細胞は延長期間に生産力のある状態を維持されることも可能であるということ
を指し示し、および実際に、細胞の生産性は繰り返された培地交換により増加されるとい
うことを指し示す。繰り返された培地交換による増加は異なる増加条件の集まり、および
細胞培養の多様性に便利である。
【0182】
そのデータは増加されない条件においての生産濃度(その生産濃度は実施例7において
詳しく述べられている)よりたくさんのタキサン生産の増加を立証する。
実施例15 給餌バッチ操作によるタキサン生産の増加
培地I(CR-128,SS36-245)、および培地L(SS36-359)(これらの培地の組成物は表2に
おいて記載されている)において培養された細胞系統の7日たった細胞はMIRACLOTH (Cal
biochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌的に吸引濾過された。
新鮮な約1グラム重量の細胞は表16.a.において指し示された所定の組成物の培養培地
の4m1へ植えつけられた。その容器は暗やみにおいて120rpmで旋回撹拌器(1”throw)で
24プラスマイナス1℃で保温培養された。蒸発分は滅菌された蒸留水の周期的な間隔の
付加により補正された。給餌バッチ操作のため、あらかじめ決められた組成物の滅菌した
飼料溶液は、たとえば11の培養液当たり1日あたり10mlの飼料溶液のように、あらか
じめ決められた飼料を与える速度で培養溶液のなかへ連続的に飼料を与えられた。飼料溶
液の組成物および給餌の実験計画案を含む、給餌バッチ操作の詳細は表16.b.において
記載されている。処理された培養物は実施例5において記載された方法の使用で試料とさ
れおよび分析がされた。
【0183】
表16.a.は細胞は延長期間に生産力のある状態を維持されることも可能であるという
ことを指し示し、および実際に、細胞の生産性は給餌バッチ操作により増加されることも
可能であるということを指し示し、その結果バッカチンIII、タキソール、およびほかの
タキサンの高濃度の蓄積となる。特別なタキサンの関連する量は給餌実験計画案および細
胞系統および培養条件の試料の組成物の相互作用を反映する。この表もまたフェニルアラ
ニンの給餌はほかのタキサンと比例してタキソールの生産の増加の結果となるということ
を指し示す。
【0184】
そのデータは増加されない条件においての生産濃度(その生産濃度は実施例7において
詳しく述べられている)よりたくさんのタキサン生産の増加を立証する。
実施例16 増加因子の組み合わせの使用のタキサン生合成の増加
種々の増加因子が、タキソール、バッカチンIII、およびタキサン生産において組み合
わされて使用され、重大および協同性の改良を与えた。
【0185】
培地M(その培地の組成物は表2において記載されてある)において培養されたTaxus
chinensis懸濁液培養(SS122-41,cr427,SS122-30,cr857,cr452)の7日たった細胞はMIRACL
OTH(Calbiochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌的に吸引濾過
された。表17において別な方法で記載されることなしに細胞は20%(w/v)の新鮮な重
量の濃度の(表17において指し示されている)培養培地へ植えつけられた。その培養物
は暗やみにおいて180rpmで旋回撹拌器(1”throw)で24プラスマイナス1℃で保温
培養された。蒸発分は滅菌された蒸留水の必要に応じたの付加により補正された。全液の
試料(すなわち細胞外および細胞内のタキサンの両方)は周期的な間隔で得られ、および
実施例5において概説された方法にしたがってHPLCにより処理および分析がされた。
【0186】
表17はTaxuschinensis細胞培養においてタキソールおよびタキサン生合成の増加因
子のいろいろな組み合わせの使用により獲得されたデータを要約した。そのデータは増加
因子の組み合わせにより個々の因子でみられるタキサン生産の増加、および増加されない
条件(その詳細は実施例7において用意されている)よりたくさんのタキサン生産の増加
を立証する。
【0187】
実施例17 給餌バッチ操作によるタキサン生産の増加
培地M(SS122-41)(これらの培地の組成物は表2において記載されている)において培
養された細胞系統の7日たった細胞はMiracloth(Calbiochem)フィルターにあわせた滅菌
したブフナー漏斗の使用で無菌的に吸引濾過された。新鮮な約1グラム重量の細胞は表1
8.a.において指し示された所定の組成物の培養培地の4mlへ植えつけられた。その容器
は暗やみにおいて120rpmで旋回撹拌器(1”throw)で24プラスマイナス2℃で保温
培養された。蒸発分は滅菌された蒸留水の付加により補正された。給餌バッチ操作のため
、あらかじめ決められた組成物の滅菌した飼料溶液は培養溶液のなかへ連続的に飼料を与
えられた。飼料溶液の組成物および給餌の実験計画案を含む、給餌バッチ操作の詳細は表
18.b.において記載されている。処理された培養物は実施例5において記載された方法
の使用で試料とされおよび分析がされた。
【0188】
表18.a.は細胞は延長期間に生産力のある状態を維持されうるということを指し示し
、および実際に、細胞の容積測定の生産性は給餌バッチ操作により増加されうるというこ
とを指し示し、その結果バッカチンIII、タキソール、およびほかのタキサンの高濃度の
蓄積となる。特別なタキサンの関連する量は給餌実験計画案および細胞系統および培養条
件の試料の組成物の相互作用を反映する。
【0189】
そのデータは増加されない条件においての生産濃度(その生産濃度は実施例7において
詳しく述べられている)よりたくさんのタキサン生産の増加を立証する。
ほかの側面、利点および修飾は発明が関係する作品において理解されるかもしれないが
、理解の明快の目的のため特別な実施態様の組み合わせにおいて図解および例としたいく
つかの詳細において前記の発明が記載された。発明の予知に限界はないが、前記の記載お
よび例は図解される。その作品において理解された目的のような発明を実行するための上
記に記載された方法の修飾は発明の余地の中に意味され、このことは追加された請求項に
よりのみ限られる。
【0190】
この詳述において言及されたすべての刊行物および特許申請はこの発明が関係する作品
において理解されたそれらの理解の水準を示す。それぞれ独立した刊行物または特許申請
が参考文献により編入されたことを特別におよび独立に指し示されたように全ての刊行物
および特許申請は同じ程度の参考文献によりここに編入された。
【0191】
【表1−1−1】
【0192】
【表1−1−2】
【0193】
【表1−1−3】
【0194】
【表1−2】
【0195】
【表1−3−1】
【0196】
【表1−3−2】
【0197】
【表1−4】
【0198】
【表1−5−1】
【0199】
【表1−5−2】
【0200】
【表1−5−3】
【0201】
【表1−6−1】
【0202】
【表1−6−2】
【0203】
【表2−1】
【0204】
【表2−2】
【0205】
【表3】
【0206】
【表4】
【0207】
【表5】
【0208】
【表6】
【0209】
【表7】
【0210】
【表8】
【0211】
【表9】
【0212】
【表10】
【0213】
【表11】
【0214】
【表12】
【0215】
【表13】
【0216】
【表14】
【0217】
【表15】
【0218】
【表16−1】
【0219】
【表16−2】
【0220】
【表17】
【0221】
【表18−1】
【0222】
【表18−2】
【技術分野】
【0001】
本発明はタキサス(Taxus)種の細胞培養によるタキソール、バッカチンIIIおよびその
他のタキサンの増強生産および回収のための方法に関する。
(背景技術)
関連技術分野
タキサン供給課題
タキソールはセイヨウイチイ、Taxus brevifolia(Waniら、1971,J.Am.Chem.Soc.,93,2325-2327)の樹皮から単離されたジテルペノイド・アルカロイドである。タキソール
に対する興味は、国立がん研究所(NCI)が大規模な検索計画において、樹皮の粗抽出液が
抗腫瘍活性を示すことを発見したことから端を発した。それ以来、臨床的な試行により、
タキソールが難治の卵巣癌、および乳癌およびその他の癌に対して非常に効果的であるこ
とが確認されてきた。タキソールは、その細胞傷害性の基本的に異なる機構、すなわち微
小管の脱重合を阻害することによるがゆえに、化学療法における大発見とされてきた(Row
inskyら1 990,J.Natl.CancerInst.,82,1247-1259を参照)。
【0002】
タキソールにおける絶望的な要因は供給である。樹皮由来のタキソールは第一の商業薬
品源として生産中止されており、大量生産は半合成、すなわち側鎖を植物由来の前駆体で
ある10-デアセチルバッカチンIIIへ化学的に結合することにより実行されている。学術
研究室による全合成はタキソールへの生存可能である商業的な手段として信頼は薄い。それ
ゆえにコスト的に効果的であり、環境に優しく、およびタキソールの急増する需要に絶え
得る堅実な供給源の開発が急務である。
【0003】
タキソールに加え、関連タキサン分子の商業的生産のための方法を開発することが急務
である。タキソテアなどのタキソール誘導体が既に世界の市場に導入されている。さらに
、利点の大きい活性を有する新規タキサン誘導体の発見および開発に多大な研究活性が集
まっている。その利点とは、そこから得られるあらゆる誘導体が効果的に合成されるよう
な適切な最初の「骨格」分子の、増加する大量の需要を作り出すようなものである。
【0004】
そのような分子の一例は前述の前駆体である10-デアセチルバッカチンIIIであり、それ
は半合成タキソールの開始点として使用される。タキソールおよびその他の誘導体の半合
成生産のためのもう一つの望ましい開始分子はバッカチンIIIである。バッカチンIIIは通
常、植物において主要なタキサンとして蓄積せず、それゆえこの分子の容易な大量の自然
源は存在しない。しかし、この分子はタキソールに化学的に近いため、非常に望ましい半
合成のための開始点である。例えば、10-デアセチルバッカチンIIIではなくバッカチンII
Iが開始点であるならば、10-デアセチルバッカチンIIIの10位のアセチル化を必要とする
工程は省略される。
【0005】
本発明はタキソール、バッカチンIIIおよびその他のタキサンの商業的な生産のための
、植物細胞培養を基にした方法の開発に関連するものである。
植物由来の化学薬品の源としての組織培養
様々な培養計画のもとでの植物細胞の分裂、成長、および二次代謝物の生産の能力は多
くの研究グループによって十分に示されている。まずは、2つの化合物、シコニン(赤色
色素および抗炎症剤)およびジンセンゴシド(東洋医学における強壮剤)は日本において組
織培養法により生産される。他の多くの方法は報告の通り、商品化に近いものであり、バ
ニリン、ベルベリンおよびロスマリン酸(Payneら、1991,"Plant Cell and Tissue Cultur
es in LiquidSystems,"ハンザー出版、ミュンヘン、を参照)を含む。
【0006】
タキソール、バッカチンIII、およびタキサンのための植物細胞培養方法の利点は多く
、(i)細胞培養法は産物が無限であり、連続的および一定の供給を保証し、害虫、災害
、および季節の変動に左右されない、(ii)細胞培養は大きなバイオリアクターの中で培養
が可能であり、環境の条件を操作することにより、興味の対象である化合物の過剰生産を
誘発することが可能である、(iii)細胞培養は樹皮と比較して単一の範囲の化合物を生
産し、または画一化する分離および精製がかなり不要である、(iv)細胞培養法は農業を基
にした方法よりも需要の変化に迅速に順応することも可能である、(v)タキソール、バ
ッカチンIIIまたはその他の前駆体の供給の他に、細胞培養法は、有利な生物活性の側面
を示しまたはその他の生物活性を有する誘導体への変換も可能であるタキサン化合物の生
産もまた可能である。
【0007】
無菌の、大規模の植物細胞培養は本来費用がかかり、細胞培養法は、これらの費用が高
い生産性によって相殺されるときのみ商業的に妥当なものとなる。あらゆる植物種および
標的代謝産物は異なり、個別の系毎に異なるアプローチが必要である。本発明はタキソー
ル、バッカチンIIIおよびタキサンの生産のための生産性の高い植物細胞培養を得るため
の独創的で技術的なアプローチに焦点を合わせたものである。
【0008】
木本植物および針葉樹の組織培養における問題
文献上の歴史的な調査は、草本植物は比較的容易に培養操作ができるが、木本植物およ
び針葉樹の生産的培養の達成は困難を伴うことを示唆する。
【0009】
裸子植物および針葉樹の培養を生産する二次代謝産物の成長は一般的に低かった。例え
ば、BerlinおよびWhite,(1988,Phytochemistry,27,127-132)はThujaoccidentalis
の培養はそのバイオマスが18日間で約30%しか増加しないことを発見した。Van Udenら(19
90.Plant CellReports,9,257-260)はCallitris drummondiiの懸濁液が21日間で20-50
%のバイオマスの増加を報告した。Westgateら(1991,Appl.Microbiol.Biotechnol.,34
,798-803)は裸子植物Cephalotaxusharringtoniaの懸濁液につき約10日間の倍加時間を
報告した。Bornman(1983,Physiol.Plant.57,5-16)によって要約されているように、
エゾマツ懸濁液(Piceaabies)のための培地の開発に対しては大量の努力が注がれた。こ
れらの研究の集成は、裸子植物の懸濁液は実際に早く成長する能力を有すること、しかし
一般法則は適用され得ないこと、および異なる細胞系列に対する培地の設定は独立に工夫
しなければならないことを示している。
【0010】
裸子植物の培養間の二次代謝産物の生産性の調査もまた、草本植物と比較して高速生合
成を含むことの難しさを指摘している。例えば、Cephalotaxus harring toniaの培養は、
テルペンアルカロイドを親の植物に見られる量のわずか1%から3%の程度で生産した(Delfe
lおよびRothfus,1977,Phytochemistry,16,1595-1598)。成功した誘導においてさえ、
Heinstein(1985,Journalof Natural Products,48,1-9)は親の植物において生産され
た程度に接近することのみが可能であった(約0.04%乾燥質量総アルカロイド)。Van Uden
ら(1990)はポドフィロトキシンを生産するために、針葉樹Callitrisdrummondiiの懸濁培
養を誘導することが可能であったが、針によって生産された量の10分の1の程度のみであ
った。Thujaoccidentalisの有意な程度のモノテルペン(10-20mg/L)およびジテルペノイ
ド・デヒドロフェルギノール(2-8mg/L)を生産する能力はBerlinら(1988)によって説得力
をもって示されている。しかし、これらの結果は低成長速度(18日間で30%バイオマス増加
)および低細胞密度(リットルあたり5から7グラム乾燥質量)の培養とともに得られた。
【0011】
タキサン生産のための細胞培養
裸子植物懸濁液において直面する早い成長および高生産性の達成における困難は、Taxu
s細胞培養におけるタキサンの生産に対するこれまでの報告に一般的に反映されている。
【0012】
Jaziriら(1991,J.Pharm.Belg.,46,93-99)は近年Taxusbaccataのカルス培養を開
始したが、イムノソーベントアッセイを用いたタキソールの検出は不可能であった。Wick
remesinheおよびArteca(1991,PlantPhysiol.,96,(補足)p.97)はTaxus media(cv
.hicksii)のカルス培養における0.009%乾燥質量タキソールの存在を報告したが、倍加時
間、細胞密度、および報告されたタキソールの生産に要した時間についての詳細は示され
なかった。
【0013】
米国特許第5,019,504号(Christenら、1991)はTaxusbrevifoliの細胞培養によるタキサ
ンおよびタキサン様化合物の産生および回収について記載した。これらの研究者は2から4
週間の時間枠で1から3mg/Lの程度のタキソール生産を報告した。彼らはまた、「3-4週間
で5-10倍」の細胞質量の上昇を報告しており、これは約7から12日の倍加時間に相当する
。
【0014】
タキサン力価および体積上の生産性の有意な上昇は、経済的に成育可能な植物細胞培養
法によって、計画された1年あたり数百キログラムの年間需要を供給する事を可能とする
ために必要とされている。
【発明の概要】
【課題を解決するための手段】
【0015】
発明の概要
本発明の対象は、早い成長、高い細胞密度、および高い細胞生存率を促進するための特
別な環境条件の開発を含む(この研究において報告する成長の特性は、従来の結果にはる
かに勝るものである)。
【0016】
本発明の目的は細胞系列の注意深い選択、培地条件の注意深い選択および操作、増強因
子の取り込み、および方法を操作する様式の注意深い選択によって、高速でタキサンを生
産することである。
【0017】
本発明の目的は培地の組成および環境条件を変化させることによってタキサンの生産を
操作する能力を含む。特に、主要なタキサン産物としてタキソールまたはバッカチンIII
の生産を細胞に促進させること、および/または副産物セファロマニンの生産を抑制する
ことも目的であり、それによって後工程での分離および精製の問題に、みごとな生物学的
解決法を提供する。これらおよびその他の目的は本発明の一つまたはそれ以上の態様によ
って達成される。
【0018】
本発明者らは、タキソール、バッカチンIII、およびその他のタキソール様化合物、ま
たはタキサンが、既知のあらゆるタキサン種、すなわち、brevifolia,canadensis,cusp
idata,baccata,globosa,floridana,wallichiana,mediaおよびchinensisから非常に
高い収量で産生されることを発見した。さらに、本発明の方法により、タキソール、バッ
カチンIII、およびその他のタキサンを既に報告されているよりも非常に短い時間で得る
ことが可能である。特に、本発明はTaxus chinensis種は急速な成長および極めて大量の
タキソール、バッカチンIII、およびタキサンの短時間での生産が可能であることを発見
した。Taxuschinensis種によって、本発明者らはタキソール、バッカチンIII、およびタ
キサンを他のTaxus種の組織培養から得られる量をはるかに上回る量で生産する細胞を操
作することを可能にした。
【0019】
培養条件(すなわち、培地組成および操作状態)の特定の修飾は、任意のTaxus種の細胞
培養から得られる様々なタキサンの収量を増強することが発見された。特に好ましい増強
剤は、銀イオンまたは複合体、ジャスモニン酸(特にメチルエステル)、オーキシン関連成
長調節剤、および3,4-メチレンジオキシ-6-ニトロ桂皮酸などのフェニルプロパノイド経
路の阻害剤を含む。増強剤は単独でまたは互いに組み合わせて、または収量を増強するそ
の他の条件で使用することも可能である。T.chinensisの植物細胞培養からのタキサンの
収量はこれらの一つまたはそれ以上の条件の使用によって特有に増強されると同時に、全
Taxus種からのタキサンの収量がこれらの条件の使用により向上することがわかった。
【0020】
一つの態様においては、本発明は、一つまたはそれ以上の栄養培地における成長および
産物形成条件下での懸濁培養でTaxus種の細胞を培養すること、および前述の細胞または
前述の細胞培養の前述の培地、または、カルスまたは懸濁培養に由来する細胞およびフェ
ニルプロパノイド代謝の阻害剤を含む栄養培地の両方から1種またはそれ以上のタキサン
を回収することを含むTaxus種の細胞培養における高収量のタキサンを生産する方法を供
給する。フェニルプロパノイド代謝の適切な阻害剤は3,4-メチレンジオキシ-6-ニトロ桂
皮酸、3,4-メチレンジオキシ桂皮酸、3,4-メチレンジオキシフェニルプロピオン酸、3,4-
メチレンジオキシフェニル酢酸、3,4-メチレンジオキシ安息香酸、3,4-トランス-ジメト
キシ桂皮酸、4-ヒドロキシ桂皮酸、フェニルプロピオン酸、フルオロフェニルアラニン、
1-アミノベンゾトリアゾール、2-ヒドロキシ-4,6-ジメトキシ安息香酸、SKF-525A、シュ
ウ酸アンモニウム、ビニルイミダゾール、ジエチルジチオカルバミン酸、およびシナピン
酸を含む。
【0021】
より好ましい態様においては、本発明の方法において使用されている一つまたはそれ以
上の栄養培地の少なくとも一つはまた、エチレン作用の阻害剤でもあり得る別の増強剤;
ジャスモン酸またはジャスモン酸のエステル;またはオーキシン関連成長調節因子をも含
む。特定のより好ましい態様においては、その他の増強剤とは、銀含有化合物、または銀
錯体、または銀イオンであるところのエチレン作用の阻害剤である。別の特定の好ましい
態様においては、その他の増強剤はジャスモン酸またはそのアルキルエステルであり、お
よびさらに好ましくは、ジャスモン酸にエステル化されているアルキル基は1から6個の炭
素原子を有する。さらにより好ましい態様においては、増強剤はジャスモン酸またはその
アルキルエステルであり、およびその培地はまた銀含有化合物、銀錯体または銀イオンを
も含む。別の特定の好ましい態様においては、その他の増強剤はインドール酢酸、ピクロ
ラム、α-ナフタレン酢酸、インドール酪酸、2,4-ジクロロフェノキシ酢酸、3,7-ジクロ
ロ-8-キノリンカルボキシル酸、または3,6-ジクロロ-o-アニス酸などのオーキシン関連成
長調節因子である。
【0022】
別の態様においては、本発明は、一つまたはそれ以上の栄養培地における成長および産
物形成条件下での懸濁培養でTaxus種の細胞を培養し、および前述の細胞または前述の細
胞と前述の培地の両方から1種またはそれ以上のタキサンを回収することを含むT axus種
の細胞培養における高収量のタキサンを生産する方法を提供するが、その際細胞はカルス
または懸濁培養に由来し、および、栄養培地は、ジャスモン酸またはジャスモン酸のエス
テルまたはオーキシン関連成長調節因子でありうる少なくとも1種類の増強剤とともに銀
含有化合物、または銀錯体、または銀イオンの形態で900μMまたはそれ以下の濃度の銀
を含む。好ましい態様においては、増強剤はジャスモン酸またはジャスモン酸のエステル
であり、銀の増強剤に対するモル比は9.5未満である。別の好ましい態様においては、増
強剤はオーキシン関連成長調節因子であり、銀の増強剤に対するモル比は少なくとも0.01
1である。
【0023】
上記の態様のいずれにおいても、一つまたはそれ以上の栄養培地は、α-フェニルアラ
ニン、β-フェニルアラニン、またはその混合物でも有り得るタキサン前駆体をも含んで
よい。上記の態様のいずれにおいても、一つまたはそれ以上の栄養培地は、グルタミン、
グルタミン酸、アスパラギン酸またはこれらのアミノ酸の混合物をも含んでよく、または
細胞の培養に使用する一つまたはそれ以上の栄養培地は、マルトース、スクロース、グル
コースおよび/またはフルクトースを炭素源として、好ましくは一次炭素源として含んで
よい。ある態様においては、栄養培地は細胞培養成長のためのものと、タキソールおよび
タキサン産生のための物が同一である。別の態様においては、1種類またはそれ以上のタ
キサンの産生は栄養培地の組成物を変化させることによって培養中に誘導される。好まし
い態様においては、培養における培地は周期的に交換され、一般的に培地の交換は、培養
からのタキサンの周期的な除去を成し遂げる。好ましくは、前述のTaxus種の細胞は給餌-
バッチ工程(fed-batchprocess)により培養される。
【0024】
一般的に、タキソールまたはバッカチンIIIおよび/またはその他のタキサンは、前述
の細胞または前述の細胞培養の前述の培地、またはその両方から回収される。一般的に、
本発明に従ったTaxus種の培養は、タキサン産生の期間を通して平均15mg/L/日のタキサン
の平均体積生産性を提供する。タキソールの平均体積生産性は一般的に、タキソール産生
の期間の推定で少なくとも10mg/L/日である。バッカチンIIIの平均体積生産性は一般的に
、タキサン産生の期間の推定で少なくとも15mg/L/日である。
好ましくは、本発明の方法に従って培養する細胞はTaxus種の細胞であり、その種はT.
brevifolia,T.canadensis,T,chinensis,T.cuspidata,T.baccata,T.globosa,T
.floridana,T.wallichianaまたはT.mediaであり得る。好ましくは、本発明の方法に
使用したTaxus種の細胞は、増強剤を含まない培地でのカルス培養または懸濁培養におけ
るELISAにより背景値を上回るタキソールを生産する細胞である。より好ましくは、本発
明の方法に使用したTaxus種の細胞は、チオ硫酸銀、メチルジャスモン酸およびオーキシ
ンを含む培地において10mg/Lの平均体積生産性をもって懸濁培養においてタキサンを生産
する細胞である。
【図面の簡単な説明】
【0025】
【図1】培地Aにおける一般的バッチ成長周期を通してのTaxuschinensis懸濁培養系列K-1におけるバイオマスの増加。誤差線は一双のフラスコから測定される標準偏差を表す。
【図2】15日実験におけるタキソール(A)および総タキサン(B)生産性に対する第9日および第12日の培地交換の効果。各々の箱の数字は産物が生産された時間の間隔(日)を表す。細胞内ボックスの影のついた部分は実験開始時に細胞接種原に存在したタキソールまたは総タキサンを表す。すべての処理は二連で行われた。Taxus chinensis懸濁培養系列K-1は表2において述べられた培地Aとともに使用された。
【図3】実施例7.3において使用した標準Gro-Luxランプ(GTESylvania,Danvers,MA)のスペクトル特性。
【図4A】Taxuschinesis細胞懸濁物K-1におけるタキサン産生。クロマトグラムの10から40分の部分を示した。選択されたタキサンのピークのダイオード配列走査は227nmのピークを持つ特徴的なタキサンUV吸光スペクトルを示した。
【図4B】Taxuschinesis細胞懸濁物K-1におけるタキサン産生。クロマトグラムの10から40分の部分を示した。選択されたタキサンのピークのダイオード配列走査は227nmのピークを持つ特徴的なタキサンUV吸光スペクトルを示した。
【図4C】Taxuschinesis細胞懸濁物K-1におけるタキサン産生。クロマトグラムの10から40分の部分を示した。選択されたタキサンのピークのダイオード配列走査は227nmのピークを持つ特徴的なタキサンUV吸光スペクトルを示した。
【図4D】Taxuschinesis細胞懸濁物K-1におけるタキサン産生。クロマトグラムの10から40分の部分を示した。選択されたタキサンのピークのダイオード配列走査は227nmのピークを持つ特徴的なタキサンUV吸光スペクトルを示した。
【図5A】培地CにおけるTaxuschinensis細胞系列K-1による長期間培養後のタキソールおよびタキサン産生。Aは既知および未知のタキサンに対するデータを表にしたものである。
【図5B】培地CにおけるTaxuschinensis細胞系列K-1による長期間培養後のタキソールおよびタキサン産生。Bは25から42日の期間におけるタキソールおよびタキサンの生産の増加を示す。
【図6A】細胞培養の上清におけるタキソールのMS/MSによる確認。図Aは本来のタキソールのイオンスプレ-APCI質量分析を示す。
【図6B】細胞培養の上清におけるタキソールのMS/MSによる確認。図Bは親ピーク(m/z 871=タキソール+NH4+)の娘イオンスペクトルを示す。
【図6C】細胞培養の上清におけるタキソールのMS/MSによる確認。図Cは細胞培養の粗抽出液から得られたイオンスプレーAPCIスペクトルを表し、タキソールに特徴的なm/z 854および871を示す。
【図6D】細胞培養の上清におけるタキソールのMS/MSによる確認。図Dはm/z 871に対応する娘スペクトルを示し、細胞培養上清におけるタキソールの存在を示す決定的な証拠を提供する。
【発明を実施するための形態】
【0026】
発明の詳細な説明
植物は長い間、薬剤および特別な化学物質の重要な原料を供給してきた。これらの産物
は一般的に、回収した植物材料の抽出または化学的合成を通して得られていた。タキソー
ルおよびタキサンは自然産物の検索から近年発見された最も重要な分類の抗癌剤の一つと
なってきた。
【0027】
本明細書で使用されている「タキソール様化合物」または「タキサン」という用語は、
タキサン環を有するジテルペノイド化合物を記述するために互換的に使用する。タキサン
はそれ自身が抗悪性物活性を有し、または生物活性を有する化合物を産生するように修飾
されることも可能である。「総タキサン」という用語は以下の実施例5に記載されている
ような特徴的なUV吸光度を示す全てのタキサンについて言及する。
【0028】
本明細書で使用されている「カルス」という用語は、構造的に未分化であり、固体化さ
れた培地において培養される培養植物細胞の集団を記述するために使用される。本明細書
で使用されている「懸濁培養」という用語は、液体栄養培地において分散する構造的に未
分化な細胞を記載するために使用する。懸濁培養は様々な凝集の段階にある細胞を含むこ
とが理解されよう。凝集の大きさの範囲は、本発明に記載されている懸濁液において、直
径10ミクロン(単一細胞またはほぼ未凝集の細胞)から、数千の細胞から成る直径数ミリメ
ートルの凝集に至る範囲の大きさをもって計測される。
【0029】
本発明において有用な植物材料はあらゆる既知のTaxus種、たとえばbrevifolia,canad
ensis,cuspidata,bacatta,globosa,floridana,wallichiana(yunnanensisとも言及さ
れる),media,fastigiataおよびchinensis(sumatrama,celebica,およびspeciosaなど
の同義の種,およびchinensisvar.mairei亜種を含む)から得ることが可能である。特
に、本発明者らは有意な量のタキソール、バッカチンIII、およびタキサンを高い体積生
産性で産生する能力を有するものとしてTaxus chinensis種を同定した。
【0030】
本発明者らによって、特定のタキサン内容量は、植物種により、および組織源および特
定の樹木に由来する植物種内において変化することが発見されている。タキサン産生のた
めの高収量の源および培養を選択することが、医療使用のための十分な量のタキサンの供
給に向けての重要な第一歩である。
【0031】
商業的適合性の基準
商業的な魅力およびタキサン産生のための与えられた植物細胞培養を基にした工程の成
功性を判断するために数々の基準を使用することが可能である。そのような基準は、発酵
費用、後での回収の容易性、および生産量を含む、工程の鍵となる要因の効果を特徴化お
よび実証すべきである。ここに記載される基準は、ブロス力価および体積生産性である。
【0032】
ブロス力価は全体のブロスにおける産物の濃度として定義され、通常1リットルのブロ
スあたりのミリグラム数(mg/L)として表現される。定義により、全ブロス力価は産物の細
胞内および細胞外の部分は区別しない。ブロス力価は典型的にはバッチまたは給餌-バッ
チ工程の効率評価に使用される。より高いブロス力価は、与えられた反応器の体積に対し
て産物の体積がより大きいこと、およびそれに付随して単位生産費用がより低いことを意
味する。同様に、高力価産物は通常高収量で回収することが容易であり、単位生産費用の
更なる改善へと導く。
【0033】
体積生産性は、単位反応体積あたり単位時間あたりに生産された産物の量として定義さ
れ、1リットルあたり1日あたりのミリグラム数の単位で表現される。タキサン産生の目的
のために、時間の尺度は、収集および回収に先立つ、生産尺度において生産が起きる間の
時間枠として定義する。体積生産性はバッチおよび給餌-バッチ工程の基準としての力価
を補足するもので、産物が産生の間に除去される工程、例えば周期的培地交換または別の
除去方法、の評価に特に有用である。高い体積生産性は、与えられた時間を通して与えら
れた反応体積に対する産物体積が大きいこと、およびそれに付随して単位生産費用がより
低いこと、および全工程の性能の向上を意味する。
【0034】
体積生産性が生物学的工程の本来の能力を計測するために使用される場合、例えば工程
開発の初期の段階においては、生産周期の最も生産的な部分、すなわち、生合成の速度が
その最大であるときの短い期間での生産性を測定することが有用である。これは一般的に
、最大瞬間体積生産性と呼ばれる。しかし、工程の性能の計測において、より適切な基準
は、全体の生産時期を通して測定される生産性である平均体積生産性である。明らかに、
最大の平均体積生産性を達成するためには最大瞬間生産性が生産時期の大部分を通して維
持されなければならない。別に規定しないかぎり、「体積生産性」という用語は全体の生
産時期に対して決定された平均体積生産性を意味する。一般に、生産時期は、生育培地を
産生培地に置き換えること、またはタキサン産生において有意な増強を誘発する増強剤を
添加することによる栄養培地の組成物の変化によって開始する。
【0035】
Taxus細胞株の開始
Taxus植物材料はその他の大陸と同様に北アメリカ全域から集められ得る。培養は生育
に適したTaxusの組織を選択することから始められる。樹皮、形成層、針葉、幹、種子、
球果、および根を含む植物のどの部分由来の組織でもカルスを誘導するために選択され得
る。しかしながら、タキソールの収量を最良にするためには針葉および植物の分裂組織の
部分が望ましい。最も望ましいのは針葉の新芽(生え始めてから1から3ヶ月)であり、これ
は一般にそれが黄緑色をしていることで同定され得る。"新芽"という語はその年の成育期
のうちに生えた針葉を広く指している。
【0036】
培養物への汚染を防ぐために、組織は培地への導入に先だって表面を滅菌されるべきで
ある。CLOROX(CloroxCompanyの脱色剤の登録商標)処理等のどんな滅菌技術も有効である
。それに加えて、セフォキシチン、ベンレート、クロクサシリン、アンピシリン、ゲンタ
マイシン硫酸塩、およびフォスフォマイシンのような抗生物質が植物材料の表面滅菌に用
いられ得る。
【0037】
カルスの生育
培養物は典型的には生育中の形態、増殖量、生成物の性質、およびその他の性質に多様
性を示す。個々の細胞株は培地の成分に対する親和性に多様性があり、数多くの異なる増
殖培地がカルスの誘導および増殖に用いられ得る。
【0038】
適切な培地の組成は培養されている生物種によって異なる。異なる種に対する望ましい
培地は表3に挙げられる。例えば、Taxuschinensisに対する望ましい増殖栄養培地は、他
のものも用いられ得るものの、A,D,I,J,K,L,M,O,Pである。これらの培地は表2に挙げた成
分を含むことが望ましい。技術を有する技術者はその量のいくらかの変化は細胞増殖に大
きな影響を与えないことが認識し得るが、表2に示した量でその成分を含む培地を用いて
培養が行われることが望ましい。
【0039】
例えば、培地Aが用いられるならば、成長ホルモンまたは調節物質は1ppbから10ppmの間
の量で、望ましくは2ppbから1ppmの間の量で培地に含まれる。培地Dが用いられるならば
、成長ホルモンまたは調節物質は1ppbから10ppmの間の量で、望ましくは2ppbから2ppmの
間の量で培地に含まれる。その他の培地成分は表2に示された量の1/10から3倍までの濃度
で含まれ得る。
【0040】
タキサンを大量に生成することはTaxus細胞を懸濁培養で増やすことによって可能にな
る。一般に、懸濁培養はカルス培養に成功した培地を用いて開始され得る。しかしながら
、懸濁培養の必要性および特にタキサンの高い効率での生成のために、培地の修正が望ま
しい。Taxus細胞が修正され、本発明の方法によって特性を調節された培地中で培養され
た場合、培養物からの1種類あるいはそれ以上のタキサンの収量は有意に増加する。
【0041】
本明細書で用いられる"栄養培地"という語は、植物細胞のカルスの増殖および懸濁培養
に適した培地を記載するために用いられる。"栄養培地"という語は一般的な語であり、"
生育培地"および"産生培地"の両方を含む。"生育培地"と言う語は培養細胞の迅速な生育
を可能にする栄養培地について記載するために用いられる。"産生培地"と言う語は培養細
胞内でタキソール、バッカチンIIIおよびタキサンの生合成を可能にする栄養培地を指す
。産生培地内で生育が起こり得、生育培地内で生成が起こり得、また最適な生育と生成が
一つの栄養培地内で起こり得ることが理解されよう。
【0042】
懸濁増殖
Taxusの懸濁培養では迅速な増殖速度と他の植物細胞培養のように迅速で高い細胞濃度
が可能である。しかしながら最適な細胞濃度は細胞株によって異なるので任意のどんな細
胞株にたいしても迅速に最適化できるような手法が考えられなければならない。
【0043】
様々なTaxus種の培養物は多量および微量栄養塩類、炭素源、窒素源、ビタミン類、有
機酸、天然および合成植物成長調節因子を含む栄養培地に移すことで生育させられる。特
にTaxus細胞の懸濁培養用の栄養培地は典型的にはカルシウム、マグネシウム、ナトリウ
ム、カリウム、リン酸、硫酸、塩素、硝酸、およびアンモニウムと言った多量栄養分およ
び銅、鉄、マンガン、モリブデン、亜鉛、ホウ素、コバルト、ヨウ素およびニッケルとい
った微量栄養分を補給する無機塩類を含む。この培地はまた、典型的にはミオイノシトー
ル、チアミン、アスコルビン酸、ニコチン酸、葉酸、ピリドキシンおよび場合によっ.て
ビオチン、パントテン酸、ナイアシン、およびその類似物等のビタミン類をも含む。これ
らの成分は表2に列挙された濃度の1/30から30倍の範囲の濃度で存在し得、望ましくは表2
に列挙された濃度の1/20から20倍の濃度で、さらに望ましくは表2に列挙された濃度の1/3
から3倍の濃度で、および最も望ましくは表2に列挙された濃度で含まれ得る。
【0044】
栄養培地はまた1種類またはそれ以上の炭素源、および典型的には栄養培地内の全炭素
の50%以上を供給するものである1次炭素源を含む。1次炭素源は望ましくはラクトース、
ガラクトース、ラフィノース、マンノース、セロビオース、アラビソース、キシロース、
ソルビトール、または望ましくはグルコース、フルクトース、スクロース、またはマルト
ースである。1次炭素源の濃度は0.05%(w/v)から10%(w/v)の範囲で有り得、望ましくは0.1
%(w/v)から8%(w/v)の範囲で有り得る。
【0045】
栄養培地はまた、多量栄養塩類の形で加えられる任意の窒素源に加えて、望ましくは少
なくとも部分的には有機栄養源(例えばグルタミン、グルタミン酸、およびアスパラギン
酸のような1種類又はそれ以上のアミノ酸、またはタンパク質加水分解物)によって供給さ
れる窒素源をも含む。これらの有機窒素源は0.1mMから60mMの、望ましくは1から30mMの範
囲の濃度の窒素を供給し得る。この培地はまた酢酸、ピルビン酸、クエン酸、オキソグル
タル酸、コハク酸、フマル酸、マレイン酸およびそれらの類似物のような1種類またはそ
れ以上の有機酸をも含み得る。これらの成分は培地中に、0.1mMから30mMの濃度で、望ま
しくは0.5mMから20mMの濃度で含まれ得る。
【0046】
この培地はまた典型的にはピクロラム、インドール酢酸、1-ナフタレン酢酸、インドー
ル酪酸、2,4-ジクロロフェノキシ酢酸、3,7-ジクロロ-8-キノリンカルボキシル酸、3,6-
ジクロロ-o-アニス酸およびその類似物のようなオーキシン関連成長調節因子、N6-ベンジ
ルアデニン、6-[γ,γ-ジメチルアリルアミノ]プリン、キネチン、ゼアチン、N-フェニル
-N'-1,2,3-チヂアゾール-5イル尿素(チジアズロン)および関連フェニル尿素誘導体および
その類似物のようなサイトキニン関連成長調節因子、GA3、GA4、GA7およびGA誘導体のよ
うなジベレリン、アブシジン酸やその誘導体、ブラシノステロイド、およびエチレン関連
成長調節因子を含む1種類またはそれ以上の天然または合成植物成長調節因子をも含む。
さらに別の適切なオーキシン関連植物成長調節因子は以下に列挙される。栄養培地は、例
えば1種類以上のオーキシン関連調節因子、1種類以上のサイトカイニン関連調節因子のよ
うな単一の種別に属する1つ以上の成長調節因子を含み得ることに注意されたい。成長調
節因子は培地中に10-10Mから10-3Mの間の濃度で、望ましくは10-8から3x10-5Mで、さらに
望ましくは表2にあげられた濃度で含まれることが望ましい。
【0047】
他に記述がない限り、本明細書において定義される生育培地は、カルス培養培地および
産生培地の通常の技術で最適化するために適切な開始点を提供する。当該技術分野におけ
る技能を有するものにとってはある種別の成分および任意の種別の成分を添加し、修飾し
、操作することによって最適な結果を達成することは日常的技術範囲である。ある種の培
地修飾は表および下記の実施例の中に供給される。
【0048】
液体培養物は空気のような好気的な環境に曝され、振とうされるか、あるいは撹拌され
ることで培養成分の適切な混合を可能にされることが望ましい。培養物は、適切な条件お
よび/または環境下では0℃から33℃の範囲が可能であるものの、23℃から27℃の間の温度
に保たれる。pHは約3から7の間で、望ましくは4から6の間で有り得る。培養物は様々な期
間の間完全な暗黒から完全な照明(狭い周波数帯および/または広いスペクトル)下までの
範囲の照明条件下で生育される。
【0049】
倍化速度は時間依存的にバイオマスの増加を調べることとともに、単に通常の部分培養
の間の生育指数を調べることによって測定される。リットルあたり15から24グラムの最大
乾燥重量濃度が達成される。様々なTaxus種懸濁物の生育特性は実施例4に詳述される。
【0050】
タキサン生成条件
懸濁培養物における2次代謝産物の形成が生育と同時に起こる場合、その代謝は生育に
関連していると呼ばれ、1種類の培地を形成することが良好な生育と多量の生成を達成す
るのに十分で有り得る。他の多くの系においては迅速な生育と高度の生成物形成は同時に
は起こらないことが分かっている。このような場合は生育および生成相は分離され、それ
ぞれの相に対する培地が独立に開発される(Payne et al.1991,Plamt Cell and Tissue
Culture inLlquid Systems,Hanser publishers,Munichに概説されている)。Taxusにお
けるタキサン生成の場合は、生育および生成物形成は分離され得、独立な培地がそれぞれ
に対して開発された。
【0051】
本発明の望ましい様式においては細胞増殖相の間の培地組成はタキサン生成相の間の培
地組成とは異なる。例えば、炭素源、特に1次炭素源の種類と量が生育相と生成相では異
なり得る。産生培地は生育培地におけるよりも多量の糖を含むことが望ましい。産生培地
における糖の量は生育相における場合と比べて生成相において2から20倍多いことがより
望ましい。1次炭素源はラクトース、ガラクトース、ラフィノース、マンノース、セロビ
オース、アラビノース、キシロース、ソルビトールであることが望ましく、またはグルコ
ース、フルクトース、スクロース、またはマルトースであることが望ましい。1次炭素源
の濃度は005%(w/v)から10%(w/v)の間で有り得、0.1%(w/v)から8%(w/v)で有ることが望ま
しい。タキソールまたはバッカチンの生成のために特に望ましい炭素源はマルトース、ス
クロース、グルコースおよび/またはフルクトースである。特に望ましい態様においては
これらの糖は初期の栄養培地中に少なくとも3.5%の濃度で含まれる。
【0052】
ビタミン、アミノ酸のような有機窒素源を含む有機添加物の種類およびレベルは、下記
の増強剤の存在またはレベルと同様に、培地において変化し、または異なるであろう。天
然または合成の植物成長調節物質の同一性およびレベルは、培地間で異なるであろう。同
様に、多量元素および微量元素の塩のレベルおよび種類も、生育および産生培地間で異な
るであろう。好ましくは、塩含量は生育培地に比べ産生培地において減少され、所望によ
り硝酸塩および硫酸塩は不均等に減少され、かつ、より好ましくは減少度は2-20倍の係数
による減少である。しかし、この培養系に対し単一の生育/産生培地を設定することも可
能であることは理解されよう。
【0053】
本発明の産生培地は、タキサン形成のみならず、タキソールまたはバッカチンIIIとい
った特定のタキサンの生成につながる直接的な細胞の生合成をも増加させる。さらに、セ
ファロマニンのような夾雑副産物の生成は、樹皮組織に比べ最小化される。本発明の産生
培地はまた、細胞の生存能力及び生合成の増加を促進し、さらに有意なレベルの産物を細
胞外培地へ分泌させる。これらの特性は、タキサン生成の効率的な商業的スケール手法の
運用において非常に重要である。
【0054】
細胞および培地よりタキソールおよびタキサンを抽出および回収する手法は、慣用され
た技術に基づく(例えば実施例5を参照)。イムノアッセイ(ELISA)技術は、商業的に入
手可能なキットとしてHawaii Biotechnologyにより提供されるプロトコールに主として基
づいた(本明細書に参照文献として記載されたGrothausら,1995,Journal of Natural P
roducts,58,1003-1014をも参照)。抗体は、タキソールまたはバッカチンIIIといった
いずれのタキサンにも特異的であってよく、または、タキサン骨格に対し、より非特異的
であってもよい。高速液体クロマトグラフィーの手法は、実施例5に詳述された既存のプ
ロトコールより僅かに変更された。本発明において用いられた条件の下で、タキサンピー
クの明確な分離が達成され、その結果、正確な検出および定量がなされた。タキサンでな
い成分が共溶出する可能性があるため、タキサンピークのスペクトル純度は、ピーク領域
を混合する前にダイオードアレイにより慣例法で検査された。タキサン標準品の保持時間
は実施例5に示されており、かつクロマトグラムの例示は図4に含まれている。
【0055】
高等植物にとって、光は植物本体、培養細胞いずれにおいても二次代謝における有効な
因子である。光の強度および波長の双方が重要である(SeibertおよびK adkade 1980,"P
lant TissueCulture as a Source of Biochemicals"E.J.Staba編,CRC Press,Boca Ra
ton,Florida,pp123-141)。例えば、通常フラボノイドおよびアントシアニン生合成は
、強度の強い連続光により促進され、一方、他の代謝産物については、暗所で培養された
細胞が好ましい。培養細胞の緑化または光合成能力の増加は、産物生成または産物の範囲
をも増加させるであろう。本発明者らの研究は、広範囲のみならず特異的な狭い範囲の光
源の利用にも関する。実施例7.3.に示されたように、光照射はタキソールの蓄積のみなら
ず、培地への分泌の増加をも引き起こす可能性がある。タキソール生成に対する光の刺激
効果は、タキサンの生合成に関する特異な制御機構の存在を示唆する。光受容体の特性お
よび光により誘導される刺激の生化学的特性は未だ明らかでない。しかし、本発明による
と、増強剤の導入が、最適の結果に対する光の役割をより重要でないものにする。
【0056】
不揮発性の溶存栄養分に加え、気体成分、主に酸素、二酸化炭素、およびエチレン(植
物ホルモン)は、生育および産物生成において重要な役割を担っている。2つのパラメー
ターが重要である。生育およびタキソール生成を促進する溶存気体濃度は明らかに重要で
ある。なぜなら、それらは反応装置を操作するする条件を規定するからである。さらに、
最適な特定の濃度が維持されるよう、消費または生成の効率を反応装置の設計に考慮する
必要がある。
【0057】
呼吸における重要性の他に、酸素は二次代謝産物の生合成の効率にも劇的な影響を与え
る。二次生合成経路において酸素を要求する過程に対する高い飽和定数は、反応装置内に
おいて細胞が高い酸素レベルに曝されていることを要求する。高い成長率を維持するため
のCO2添加の重要性が報告されている。植物ホルモンであるエチレンは、二次代謝を含む
植物の生育および発生の全ての側面において、多面発現的な役割を担う(例えば、Payne
ら,1991を参照)。
【0058】
本発明者らは、ある気体濃度の様式が、培養細胞における生育および二次代謝を促進す
ることを見出している。例えば、酸素濃度の範囲は、1%空気飽和から200%空気飽和まで、
および好ましくは10%から100%の範囲、および最も好ましくは25%から95%の範囲において
、培養と両立する。二酸化炭素濃度の範囲は、0.03%(培地と平衡にある気相におけるv/v
)から15%(v/v)まで、および好ましくは0.3%から8%(v/v)の範囲において、培養と両立す
る。溶存気体の最適濃度は、細胞代謝により異なる可能性があり、例えば、速い成長を行
っている細胞は、典型的に高めの酸素レベルを好み、かつ、高めの二酸化炭素レベルに対
して感受性がより低いタキサン生合成中の細胞とは異なる最適値を有するであろう。最適
値はまた、培養の速度論によっても異なる可能性があり、例えば、遅滞期にある細胞は、
対数増殖期にある細胞とは異なる溶存気体濃度を好むであろう。
【0059】
溶存気体は、他の培養成分および増強剤と多様な様式により相互作用するであろう。例
えば、酸素の要求は、生合成の誘導または刺激により変化するであろう。傷害反応として
の呼吸効率の上昇は、植物培養細胞が誘導を受けた際に一般的に観察される。エリシター
または刺激物は、それらの機能をエチレンを通じ仲介し、または二次代謝の促進とは独立
にエチレン生成に影響を与えるであろう。その場合、微生物由来のエリシター調製物をエ
チレンに置換し、かつエリシター調製物中の他の微生物由来成分に関連する毒性を恐らく
抑制することが望ましいであろう。あるいは、エチレンの作用を抑制し、従ってエリシタ
ーまたは刺激物が二次代謝をより独占的、従ってより効率的な方法で促進できるようにす
ることは有利であろう。下記のように、エチレン作用に影響を与えることが知られている
成分である銀イオンは、実際タキサン生合成を有利に変化させる。
【0060】
増強剤
二次代謝産物の生成は複雑な過程であり、究極的に二次代謝産物に変換されるべき前駆
体を生成しかつ順々に修飾する、多数の異なる酵素の協調した作用を要求する。同時に、
二次代謝産物生成は、他の酵素が目的の代謝産物の前駆体を代謝し、二次代謝産物を構築
するのに必要な前駆体プールを流失させる場合には低下するであろう。
【0061】
低い生成または引き続く変換による利用可能な前駆体の量の制限、前駆体または中間産
物の下流中間産物への変換の制限、または関連酵素の活性の制限は、二次代謝産物の生成
を限定するであろう。いずれの特定の培養系においても、二次代謝産物が生成される効率
は、前駆体が二次代謝産物に変換される経路において律速段階を形成する、これらの限定
要因のうちの一つにより制御されるであろう。律速段階の原因となる限定要因の緩和は、
当該培養系の二次代謝産物生成効率を、経路の他の過程が律速となる点にまで上昇させる
であろう。生成効率全体を限定する特定の過程は、限定要因を緩和する作用と同様に、培
養系間で異なるであろう。
【0062】
タキサンは一連の多数の酵素過程を通じて生成される二次代謝産物であり、本発明者ら
は、タキサン生合成における一つまたはそれ以上の効率限定過程を緩和する、複数のクラ
スの増強剤をつきとめた。これらの増強剤のうち一つのタキサン生成細胞への添加は、タ
キサンの生成効率を増加させる。さらに本発明者らは、本明細書中に記載されている増強
剤の利用が、大部分のタキサン生成培養細胞において、少なくともいくつかの促進効果を
示すであろうことを決定しており、生成効率全体が単一の効率限定過程ではなく、複数の
限定要因の間の複雑な相互作用により決定されていることを示唆している。促進の程度は
、一旦ある種の限定要因が緩和された際にタキサン生合成における他の過程の相対的限定
効果を決定する、特定の細胞の状態に依存するであろうが、いずれか一つの限定要因の緩
和はタキサン生成を増加させるであろう。多種の限定要因間の相互作用に影響する培養条
件は、細胞の遺伝的構成、培地の組成および気体環境、温度、光照射および過程のプロト
コールを含み、かつ特定の培養系に添加される増強剤は、本明細書中に示されたように個
別の増強剤の効果を比較することにより経験的に決定されるであろう、当該培養系の限定
要因の観点より通常選択されるであろう。さらに、培養系に一つより多い増強剤が存在す
る場合に、タキサン生成の更なる増加が達成されるであろうことが発見されている。
【0063】
本発明の意図する代表的な増強剤は表1に例示されている。本発明の増強剤は、複数の
一般的なクラスに分類されるであろう。それらのクラスは、抗褐色化剤、抗老化剤、抗エ
チレン剤、オーキシン関連成長調節物質、前駆体、阻害剤、エリシター、刺激物およびジ
ャスモン酸関連化合物といった植物成長調節物質である。
【0064】
本発明の意図する増強剤の一つのクラスは、抗褐色化剤である。本明細書中で使用され
ている「抗褐色化剤」の用語は、細胞培養過程における色素の形成を阻害するため栄養培
地に添加される成分を示す。これらの色素は、細胞成長、生存能力、および産物生成に有
害な影響を及ぼすことが一般的に観察される、フェノール類および関連化合物を含む。本
発明の栄養培地において用いられる典型的な抗褐色化剤は、アスコルビン酸である。典型
的には、抗褐色化剤は培地中に10ppbから1000ppmの範囲の濃度となるよう添加されるであ
ろう。
【0065】
他の種別の増強剤は抗老化剤である。抗老化剤は細胞を老化より防護する、生物学的ま
たは非生物学的起源の物質である。この様な薬剤は例えば、老化を促進する化合物の生成
を防ぐ、老化促進物質の活性を抑える、ラジカル除去剤または抗オキシダント活性を提供
する、細胞性の膜及び小器官の無傷な状態を守護する、またはその他の機構により反応可
能である。この様な薬剤にはエチレン反応の拮抗剤、スペルミン、スペルミジン、ジアミ
ノプロパン、及びその類似物質等のポリアミン及びその代謝産物、抗褐色化剤、フェノー
ル産物の阻害剤、及びラジカル除去剤、例えば還元型グルタチオン、没食子酸プロピル、
及びβ-メルカプトエタノールアミン等のスルフヒドリル化合物を含む。
【0066】
抗エチレン剤はエチレン生成またはエチレン反応を阻害する物質として定義される。エ
チレン代謝を阻害する抗エチレン剤は、エチレン生合成拮抗剤及びエチレン反応拮抗剤へ
とさらに分類される可能性がある。エチレン生合成拮抗剤は、エチレンの生合成経路を阻
害する化合物である、この生合成経路に沿った阻害される酵素の例はACC合成酵素、ACC酸
化酵素、及びエチレン酸化酵素を含む。エチレン生合成拮抗剤の例はα-アミノイソ酪酸
、アセチルサリチル酸、メトキシビニルグリシン、アミノオキシ酢酸、及びその類似物を
含む。
【0067】
エチレン活性拮抗剤の例は、銀を含有する化合物、銀錯体、または銀イオン、二酸化炭
素、1-メチルシクロプロペン、2,5-ノルボルナジエン、トランス-シクロオクテン、シス-
ブテン、ジアゾ-シクロペンタジエン、及びその類似物を含む。適切な銀塩は硝酸銀、チ
オ硫酸銀、リン酸銀、安息香酸銀、硫酸銀、トルエン硫酸の銀塩、塩化銀、酸化銀、酢酸
銀、ペンタフルオロプロピオン酸銀、シアン酸銀、乳酸の銀塩、ヘキサフルオロリン酸銀
、亜硝酸銀、及びクエン酸の三銀塩を含む。多種の銀塩によるタキサン生合成の増強の例
は例10に示す。
【0068】
抗エチレン剤は培地に10ppbから1000ppmの程度で含有させることが可能である。培地に
銀が含まれる場合、銀は濃度が900μM以下、好ましくは500μM以下、更に好ましくは20
0μM以下になるよう添加する。培地に銀が含まれる場合、抗エチレン剤は濃度が10nM以
上、好ましくは100nM、更に好ましくは1μM、及び一般的には10μM添加する。
【0069】
本発明において想定される増強剤は、植物成長調節剤、特にオーキシン、オーキシン様
の活性を持つ化合物、及びオーキシン拮抗剤を含むオーキシン関連成長調節剤を含む。オ
ーキシン関連成長調節剤は、一般的には濃度が10-10Mと10-3Mの間で、好ましくは10-8と1
0-5Mの間で培地に含ませる。最も好まれるオーキシン関連成長調節剤の例は、1-ナフタレ
ン酢酸、2-ナフタレン酢酸、1-ナフタレンアセトアミド/ナフチルアセトアミド、N-(1-ナ
フチル)フタルアミド酸、1-ナフトキシ酢酸、2-ナフトキシ酢酸、β-ナフトキシ酢酸、1-
ナフトキシアセトアミド、3-塩化フェノキシ酢酸、4-塩化フェノキシ酢酸、4-ヨウ化フェ
ノキシ酢酸、インドールアセトアミド、インドール酢酸、インドイル酢酸、インドールア
セチルロイシン、γ-(3-インドール)酪酸、4-アミノ-3,5,6-トリクロロピコリン酸、4-ア
ミノ-3,5,6-トリクロロピクリン酸メチルエステル、3,6-ジクロロ-o-アニス酸、3,7-ジク
ロロ-8-キノリンカルボン酸、フェニル酢酸、2-ヨウ化フェニル酢酸、3-ヨウ化フェニル
酢酸、2-メトキシフェニル酢酸、クロロプロファム(イソプロピル-3-クロロカルバニレ
ート)、4-塩化インドール-3-酢酸、5-塩化インドール-3-酢酸、5-臭化-4-塩化-3-インド
イル酪酸、インドールアセチルフェニルアラニン、インドールアセチルグリシン、インド
ールアセチルアラニン、4-塩化インドール、p-塩化フェノキシイソ酪酸、1-ピレノキシル
安息香酸、リソホスファチジン酸、1-ナフチル-N-メチルカルバミン酸、及びエチル-5-塩
化-1H-インダゾール-3-イル酢酸-3-インドールブタン酸を含む。その他の好まれるオーキ
シン関連成長調節剤の例は、ナフタレン-2,6-ジカルボキシル酸、ジ無水ナフタレン-1,4,
5,8-テトラカルボキシル酸、ナフタレン-2-スルホンアミド、無水4-アミノ-3,6-ジスルホ
-1,8-ナフタリン酸、3,5-ジメチルフェノキシ酢酸、1,8-ナフタルイミド、2,4-ジクロロ
フェノキシ酢酸、2,3-ジクロロフェノキシ酢酸、2,3,5-トリクロロフェノキシ酢酸、2-メ
チル-4-塩化フェノキシ酢酸、ニトロフェノキシ酢酸、DL-α-(2,4-ジクロロフェノキシ)
プロピオン酸、D-α-(2,4-ジクロロフェノキシ)プロピオン酸、4-臭化フェノキシ酢酸、4
-フッ化フェノキシ酢酸、2-ヒドロキシフェノキシ酢酸、5-塩化インドール、6-塩化-3-イ
ンドイル酢酸、5-フッ化インドール、5-塩化インドール-2-カルボン酸、3-塩化インドー
ル-2-カルボン酸、インドール-3-ピルビン酸、5-臭化-4-塩化-3-インドイル酪酸、6-塩化
-3-インドイル酪酸、キノリン-2-チオグリコール酸、アミノフェニル酢酸、3-ニトロフェ
ニル酢酸、3-塩化-4-ヒドロキシ安息香酸、クロルフルレノール(2-クロロ-9-ヒドロキシ
フルオレン-9-カルボン酸)、6-塩化-3-インドイル酢酸、N-(6-アミノヘキシル)-5-塩化-1
-ナフタレンスルホンアミドヒドロクロライド、2-塩化-3(2,3-ジクロロフェニル)プロピ
オンニトリル、o-塩化フェノキシ酢酸、6,7-ジメトキシ-1,2-ベンゾイソキサゾール-3-酢
酸、3-オキソ-1,2-ベンゾイソチアゾリン-2-イル酢酸、マストパラン、2,3,5-トリヨード
安息香酸、2-(3-塩化フェノキシ)プロパン酸、及びメコプロップ(2-(4-クロロ-2-メチル
フェノキシ)プロパン酸)を含む。他のオーキシン関連成長調節剤に適した例は、ナフトエ
酸ヒドラジド、2,4-ジブロモフェノキシ酢酸、3-トリフルオロメチルフェノキシ酢酸、オ
キシインドール、インドール-2-カルボン酸、インドール-3-乳酸、β-(3-インドール)プ
ロピオン酸、2-臭化フェニル酢酸、3-臭化フェニル酢酸、2-塩化フェニル酢酸、3-塩化フ
ェニル酢酸、2-メチルフェニル酢酸、3-メチルフェニル酢酸、3-トリフルオロメチルフェ
ニル酢酸、3-メチルチオフェニル酢酸、フェニルプロピオン酸、4-塩化-2-メチルフェニ
ルチオ酢酸、2-塩化安息香酸、3-塩化安息香酸、2,3-ジクロロ安息香酸、3,4-ジクロロ安
息香酸、2,3,5-トリクロロ安息香酸、2,4,6-トリクロロ安息香酸、2-ベンゾチアゾールオ
キシ酢酸、2-塩化-3-(2,3-ジクロロフェニル)プロプオンニトリル、2,4-ジアミノ-s-トリ
アジン、無水ナフタル酸、ジケグラック(Dikegulac)(2,3:4,6-ビス-O-(1-メチルエチリデ
ン)-α-L-キシロ-2-ヘキスロフラノソン酸(hexulofuranosonicacid))、クロルフルレノ
ールメチルエステル、2-(p-塩化フェノキシ)-2-メチルプロピオン酸、2-塩化9-ヒドロキ
シフルオレン-9-カルボン酸、2,4,6-トリクロロフェノキシ酢酸、2-(p-クロロフェノキシ
)-2-メチルプロピオン酸、エチル4-(塩化-o-トリルオキシ)酪酸、[N-(1,3-ジメチル-1H-
ピラゾール-5-イル)-2-(3,5,6-トリクロロ-2-ピリジニル)オキシ]アセトアミド、4-塩化-
2-オキソベンゾチアゾリン-3-イル-酢酸、2-(2,4-ジクロロフェノキシ)プロパン酸、2-(2
,4,5-トリクロロフェノキシ)プロパン酸、4-フッ化フェニル酢酸、3-ヒドロキシフェニル
酢酸、オルソニル(2(β-クロロ-β-シアノエチル)-6-クロロトルエン)、3,4,5-トリメト
キシ桂皮酸、2(3,4-ジクロロフェノキシ)トリエチルアミン、インドール-3-プロピオン酸
、イオキシニルナトリウム(Sodium Ioxynil)(4-ヒドロキシ-3,5-ジヨードベンゾニトリル
)、2-ベンゾチアゾール酢酸、及び(3-フェニル-1,2,4-チアジアゾール-5-イル)チオ酢酸
を含む。
【0070】
他の種別の植物成長調節剤も増強剤として栄養培地に含ませることが可能である。これ
らは、N6-ベンジルアデニン、6-[γ,γ-ジメチルアリルアミノ]プリン、キネチン、ゼア
チン、N-フェニル-N'-1,2,3-チジアゾール-5-イル尿素(チジアズロン)、及び関連フェニ
ル尿素誘導体、及びその類似物等の、サイトカイニン関連成長調節剤、GA3、GA4、GA7、
及びGA誘導体等のジベレリン類、アブシジン酸及びその誘導体、ブラシノステロイド、及
びエチレン関連成長調節剤を含む。これらの成長調節剤は濃度が10-10Mと10-3Mの間で、
好ましくは10-8Mと10-5Mの間で培地に含ませることが可能である。
【0071】
他の種別の増強剤は前駆体または生合成前駆体である。本明細書中で使用する「前駆体
」という用語は、栄養培地に加えられ細胞により代謝され及び取り込まれたタキソール及
びタキサンになる化合物を示すために使用される。適した前駆体は、酢酸、ピルビン酸及
びその類似物、α-フェニルアラニン、β-フェニルアラニン(3-アミノ-3-フェニルプロピ
オン酸)、フェニルイソセリン、N-ベンゾイルフェニルイソセリン、安息香酸、シキミ酸
、グルタミン、桂皮酸、及びその類似物などイソプレノイド化合物の前駆体を含む。これ
らの分子の誘導体も又前駆体として適している。
【0072】
他の種の増強剤は阻害剤である。阻害剤は酵素性または他の細胞性活性を阻害する化合
物である。本明細書で使用する「代謝阻害剤」という用語は、栄養培地に加えられ、特定
の生合成経路を阻害する化合物を示すために使用される。例えば、代謝阻害剤は、初期の
生合成前駆体に関し競合する種々の経路を阻害することにより、タキソール、バッカチン
III、または他のタキサン生合成を増強するために使用することが可能である。この種の
特に効果的な増強剤は、桂皮酸またはその誘導体の合成または代謝を阻害することが可能
な化合物であるフェニルプロパノイドの代謝阻害剤を含む。これら化合物は、好ましくは
p-クマル酸、4-フッ化-DL-チロシン、4-メトキシ安息香酸、3-ジメチルアミノ安息香酸、
4-メトキシ桂皮酸、4-ニトロ桂皮酸エチルエステル、4-ニトロシンナムアルデヒド、メル
カプトエタノール、4-ヒドロキシクマリン、シンナミルフルオレン、2-シアノ-4-ヒドロ
キシ桂皮酸、シンナミルイデンマロン酸、4-ジメチルアミノ桂皮酸、N-シンナミルピペラ
ジン、N-トランス-シンナモイルイミダゾール、2-アミノインダン-2-ホスホン酸、ベンジ
ルヒドロキシルアミン、プロカイン、モネシン、N-(4-ヒドロキシフェニル)グリシン、3-
(4-ヒドロキシフェニル)プロピオン酸、3-(2-ヒドロキシフェニル)プロピオン酸、さらに
好ましくは、D-フェニルアラニン、N-(2-メルカプトプロピオニル)グリシン、及びその酢
酸塩複合体、DL-メタフルオロフェニルアラニン、p-フッ化-DL-フェニルアラニン、ジチ
オスレイトール、4-フッ化桂皮酸、トランス-3,4-ジフルオロ桂皮酸、3,4-ジフルオロ-D-
フェニルアラニン、ジエチルジチオカルバン酸、4-フッ化-(1-アミノ-2-フェニルエチル)
ホスホン酸、3,4-メチレンジオキシ安息香酸、及び最も好ましくは、3,4-メチレンジオキ
シ-6-ニトロ桂皮酸、3,4-メチレンジオキシ桂皮酸、3-[3,4-メチレンジオキシフェニル]
プロピオン酸、3,4-メチレンジオキシフェニル酢酸、4-フッ化-L-フェニルアラニン、4-
ヒドロキシフェニルピルビン酸、4-フッ化-DL-チロシン、トランス3,4-ジメトキシ桂皮酸
、フェニルプロピオル酸、L-2-ヒドロキシ-3-フェニルプロピオン酸、2-ヒドロキシ-4,6-
ジメトキシ安息香酸、SKF-525A(α-フェニル-α-プロピルベンゼン酢酸の2-(ジエチルア
ミノ)エチルエステル)、ビニルイミダゾール、オキサル酸アンモニウム、シナプ酸、及び
1-アミノベンゾトリアゾール、及び関連類似物質を含む。培地に含ませる場合、阻害剤は
濃度が10ppbと1000ppmの間で、好ましくは濃度が100ppbと100ppmの間で、更に好ましくは
濃度が1ppmと50ppmのあいだで加える。
【0073】
培養細胞中のタキソール、バッカチンIII、及び他の関連タキサンの収量を増進するた
めに、本発明者らは多くのアプローチを試みた。生産性を増強させる方法の一つは、エリ
シターと呼ばれるものの使用である。本明細書で使用する「エリシター」という用語は、
植物または培養植物細胞に添加したときに二次的代謝産物の増加を引き起こす、生物学的
及び非生物学的起源の化合物を意味するために使用する(Eilert 1987,"Cell Culture and
SomaticGenetics of Plants,"Vol.4,F.Constabel and I.K.Vasil(eds.), Academic Pre
ss,NewYork,pp.153-196;Ebel,1984,Bioregulators:Chemistry and Uses.257-271;and Da
rvill etal.,1984,Ann.Rev.Plant Physiol.,35,243-275)。多くの異なる化合物がその起
源の性質及びその細胞代謝における活性の方法に依存してエリシターとして機能すること
が可能である。当該研究分野において、本発明は二つの主要な種類のエリシターを使用し
た。1)通常は選別された菌類、細菌類、及び酵母の集団の細胞壁抽出物または濾過物、及
びその精製画分を含む生物的エリシター。2)生物起源の化合物と同様な、化学的なストレ
ス剤を含む非生物的エリシター(表1に一覧されたエリシターを参照)。加えて、重金属
イオンを含む塩及び複合体も、効果的な非生物的エリシターとみなすことが可能である。
これらは、コバルト、ニッケル、ランタン、セレニウム、バナジウム、鉛、カドミウム、
クロム、アルミニウム、ヨウ素、バリウム、ビスマス、リチウム、ルビジウム、ストロン
チウム、及び金などの例を含む。特筆すべきは、エリシター効果を伝達するある種の化合
物、例えば下記に示すジャスモン酸関連化合物もエリシターとみなすことが可能であると
いうことである。
【0074】
Christenらは(1991)Taxusbrevifoliaの懸濁によるタキソールの生産のための菌類のエ
リシター及び選別された化合物の使用について報告している、しかし、エリシター処理に
起因するタキソール蓄積程度の増加については特定していない。
【0075】
一般的には、2種類のエリシター効果(タキサンの培地中への分泌並びにタキサンの培
養細胞への蓄積)発生の範囲はエリシター間で及び種間で異なるが、両種のエリシターは
共に効果的である。最高の生産性増加はキトサングルタメート、リケナン、フェルラ酸、
及び安息香酸により達成される。キトサン及びリケナンは微生物の細胞壁由来の多糖類複
合体である。キトサンは単独で用いると培地に不溶であり、及び毒性があり、及び永久的
に細胞に損傷を与える。一方キトサングルタメートは培地に容易に溶け、及び細胞の生育
に影響を与えない。フェルラ酸及び安息香酸は生物起源の合成化学物質であり、及び一般
的には生体系では抗オキシダントとして使用される。
【0076】
エリシター及び代謝ストレス剤は、本発明によれば、タキソール、バッカチンIII及び
全タキサンを生産させ及び培養組織への分泌を最大にするために、エリシターの特異性、
濃度、タイミングおよび添加継続時間を培養年齢及び培地組成の関数として評価して使用
されるであろう。
【0077】
本発明において意図される他の種別の増強剤は刺激剤である。本明細書で使用する刺激
剤という用語は、栄養培地に加えられ、特定の生合成経路をしげきまたは活性化する(例
えば生合成を導く)化合物を示すために使用される。
【0078】
ジャスモン酸関連化合物は二次的代謝生合成を刺激することによりエリシター効果反応
に寄与する種別の化合物である。ジャスモン酸関連化合物は、ジャスモン酸、及びメチル
ジャスモン酸、エチルジャスモン酸、プロピルジャスモン酸、ブチルジャスモン酸、ペン
チルジャスモン酸、ヘキシルジャスモン酸などのそのアルキルエステル、ジヒドロジャス
モン酸、及びメチルジヒドロジャスモン酸、エチルジヒドロジャスモン酸、n-プロピルジ
ヒドロジャスモン酸、ブチルジヒドロジャスモン酸、ペンチルジヒドロジャスモン酸、ヘ
キシルジヒドロジャスモン酸などのそのアルキルエステル、エピメチルジャスモン酸、フ
ッ化メチルジャスモン酸、シス-ジャスモン、イソジャスモン、テトラヒドロジャスモン
、12-オキソフィトジエン酸、ジヒドロジャスモン、ジャスモニル酢酸、アプリトン、ア
ミルシクロペンテノン、ヘキシルシクロペンテノン、ヘキシルシクロペンタノン、及びそ
の関連誘導体及び類似物質を含む。ジャスモン酸関連化合物は濃度が10-9Mと10-3Mの間で
、好ましくは濃度が10-6と5×10-4Mの間で、更に好ましくは濃度が10-5Mと2×10-4Mの間
で培地に含ませる。特筆すべきは一種以上のジャスモン酸関連化合物を栄養培地に含ませ
ることが可能であるということである。ジャスモン酸関連化合物、オーキシン関連成長調
節剤、前駆体、及びその他の栄養物質は、これら化合物が培養中において代謝されること
により濃度が変化するということは当業者により認識されよう。特に示さない限り、本明
細書において列挙された濃度は栄養培地における初期濃度を記載する。
【0079】
以下に述べる最低二種以上の種類の増強剤からの増強剤を組み合わせると、いかなる一
種の薬剤を単独で用いたときに観察される最高の増強をも上回るTaxus細胞によるタキサ
ン生産の増強を示す。これらの種別の増強剤は、エリシター、ジャスモン酸関連化合物、
エチレン作用の阻害剤、フェニルプロパノイド代謝の阻害剤、抗老化剤、前駆体、及びオ
ーキシン関連成長調節剤である。つまり、好まれる手法において、本発明は、少なくとも
二種のこれらの薬剤群から選択された増強剤の存在下におけるTaxus種細胞の培養により
一種またはそれ以上のタキサンの生産を増強させる方法を提供する。
【0080】
好まれるタキサンの生産方法はエチレン作用の原型阻害剤である銀を、少なくとも一種
の他の増強剤と組み合わせて用いる、特に好まれる方法においては、その他の増強剤はジ
ャスモン酸メチルであるか、または3,4-メチレンジオキシニトロ桂皮酸等のフェニルプロ
パノイド代謝の阻害剤である。
【0081】
お互いに組み合わせて使用する場合、ジャスモン酸関連化合物及びエチレン作用阻害剤
は培地にお互いに特定の割合で含ませることができる。例えば、ジャスモン酸メチル及び
チオ硫酸銀を組み合わせて用いる場合、ジャスモン酸メチルのチオ硫酸銀に対するモル比
は0.0001から9.5の範囲の間で、好ましくは0.001と8の間の範囲で、更に好ましくは0.1と
7の範囲の間で、最も好ましくは1と5の範囲の間でになるであろう。
【0082】
お互いに組み合わせて使用する場合、オーキシン関連成長調節剤及びエチレン作用阻害
剤は培地にお互いに特定の割合で含ませることができる。例えば、オーキシン関連成長調
節剤及びチオ硫酸銀を組み合わせて用いる場合、オーキシン関連成長調節剤のチオ硫酸銀
に対するモル比は0.011から1000の範囲の間で、好ましくは0.015と100の間の範囲で、更
に好ましくは0.02と50の範囲の間で、最も好ましくは0.05と30の範囲の間になるであろう
。
【0083】
一般的には、Taxus細胞をタキサンの生産のために培養する場合、一種またはそれ以上
のオーキシン関連成長調節剤を培養培地に加える。オーキシン関連成長制御因子の存在は
細胞の成長を促進するが、より重要な事項は培養によるタキサンの生産を増強することで
ある。少なくとも一種の他の増強剤をオーキシン関連成長制御因子と同時に添加すること
により、さらなる増強を得ることが可能である。
【0084】
本発明において好まれる方法において、一種またはそれ以上の増強剤を十分量培地に添
加することにより、一種またはそれ以上のタキサンの生産量をを増強剤が存在しないとき
の生産量の相対比で、少なくとも3倍、好ましくは少なくとも5倍、更に好ましくは少なく
とも10倍、更に好ましくは少なくとも30倍に増強する。他の本発明において好まれる方法
において、一種またはそれ以上の増強剤を十分量培地に添加することにより、タキソール
の容積測定生産性を、少なくとも10mg/L/day、好ましくは少なくとも15mg/L/day、更に好
ましくは少なくとも22mg/L/dayに増強する。他の本発明において好まれる方法において、
一種またはそれ以上の増強剤を十分量培地に添加することにより、タキソールの全ブロス
力価を、少なくとも150mg/L、好ましくは少なくとも200mg/L、更に好ましくは少なくとも
350mg/Lに増強する。他の本発明において好まれる方法において、一種またはそれ以上の
増強剤を十分量培地に添加することにより、バッカチンIIIの容積測定生産性を、少なく
とも15mg/L/day、好ましくは少なくとも20mg/L/day)更に好ましくは少なくとも25mg/L/da
yに増強する。他の本発明において好まれる方法において、一種またはそれ以上の増強剤
を十分量培地に添加することにより、バッカチンIIIの全ブロス力価を、少なくとも100mg
/L、好ましくは少なくとも150mg/L、更に好ましくは少なくとも250mg/Lに増強する。他の
本発明において好まれる方法において、一種またはそれ以上の増強剤を十分量培地に添加
することにより、タキサンの容積測定生産性を、少なくとも15mg/L/day、好ましくは少な
くとも25mg/L/day、更に好ましくは少なくとも40mg/L/dayに増強する。他の本発明におい
て好まれる方法において、一種またはそれ以上の増強剤を十分量培地に添加することによ
り、タキサンの全ブロス力価を、少なくとも200mg/L、好ましくは少なくとも300mg/L、更
に好ましくは少なくとも400mg/Lに増強する。
【0085】
以上に増強剤として示した多くの化合物は他の植物系においても使用された。これら非
Taxus系における公式化、管理、及び適切な生理的濃度水準は当該技術者に、本発明と同
値のこれら薬剤を適用する手引きを提供する。
【0086】
細胞材料
本発明の方法において培養に適する細胞はTaxusのいかなる種由来でもあることができ
る。好ましくは、細胞は、生来タキサンを比較的高収量で生産する細胞系由来であると良
い。特に、この様な細胞は一種またはそれ以上のタキサンを標準条件下で高水準で生産す
る、または標準条件下でタキサンの高い平均容積測定生産性を示す能力を有している。適
切な細胞系は、細胞系の細胞を標準タキサン生産条件下で培養し、及び培養液における生
産された一種またはそれ以上のタキサンの水準を観察し、または以下に述べる手段により
培養液中の細胞による一種またはそれ以上のタキサンの平均容積生産性を決定することに
より単離することが可能である。
【0087】
試験手段産生培地において使用される細胞は特定の細胞系に適応した適当な培地におい
て生育する。対数増殖期の終了の後、細胞の一部をタキサンの生産試験のために培養する
。カルス培養は固形培地が使用してもよいが、生産培養は、一般的に液体培地で行われる
。生産培養においては、細胞は表2におけるN培地、スクロースを7%(w/v)マルトースで代
用するのを除いては表2におけるのと同様のN培地、または特定の細胞系の生育及び維持を
最大限にする栄養培地において培養される。生産培養においては、細胞密度は新鮮な重量
を基本として15-20パーセント(w/v)の範囲であるべきである。細胞は10-20日間、25度+/-
2度において暗条件で培養する。液体培養は適切に揺り動かし及び空気にさらすべきであ
る、例えば、旋回式振盪機で120-180rpm。
【0088】
細胞系の特徴を評価する生産培養は適した増強剤を含む。一般的には六種の選択的増強
カクテル(最高五種の増強剤の組み合わせ)を各々の細胞系に対し試験する。組み合わせ
は以下の表Aに示す。
【0089】
培養の終了時に培養における個々のタキサンの力価を、本明細書で示したELISA分析を
行うことにより測定することができる、または、培養において生産されたタキサンのプロ
フィールを、実施例5において示したようにHPLC解析により決定することができる。好ま
れる細胞系は一種またはそれ以上のタキサンを、一種またはそれ以上の増強カクテルにお
ける最低目標タキサン水準を越えて生産する。好まれる細胞系は、少なくとも一種の増強
カクテルにおいて、及び、更に好ましくは二種またはそれ以上の増強カクテルにおいて、
力価及び生産性の双方の目標水準を越える。適した細胞系の生産培養終了時の最低目標タ
キサン力価は、少なくとも100mg/Lタキサンである。あるいは、生産培養行程終了時の最
低平均容量測定生産性目標は10mg/L/dayタキサンである。さらに好まれる細胞系は、生産
培養終了時の最低タキサン力価は少なくとも100mg/Lタキソールまたは200mg/Lバッカチン
III、または生産培養行程終了時の平均容量測定生産性は10mg/L/dayタキソールまたは15m
g/L/dayバッカチンIIIに達する。
【0090】
【化1】
【0091】
様々な種に適した産生培地は表5に列挙するが、その他のものも使用できる可能性があ
る。例えば、表2の培地B、C及びNは、Taxuschinensisの産生培地として特に適している
。培地は表2に列挙した成分を含むことが望ましい。これらの培地は、主要な及び重要で
はない無機塩、有機物及び成長ホルモンまたは成長調節剤を、一般には表2に示した各培
地成分の濃度の、好ましくは1/10から始まり3倍の範囲で、量において含むことが望まし
い。培地BまたはNを使用した場合、成長調節剤を特に、量において01ppmから20ppmの間で
、好ましくは1ppmから10ppmの間で培地に含ませる。培地CまたはNを使用した場合、成長
調節剤を好ましくは0.1ppmから5ppmの範囲水準で含ませる。
【0092】
本発明における意図において、本明細書で示した培地において、他の慣例的組成物(有
機物、ビタミン、アミノ酸、前駆体、活性剤及び阻害剤など)の代用、成長調節剤を含む
様々な組成物の添加または削除、または割合の変更等の装飾を、成長とタキサン生産を表
2において示した培地において観察されるものと同等またはそれ以上にするために行う可
能性があることは、当該技術者により理解されている。
【0093】
操作工程の方法
植物細胞培養工程の操作方法は、栄養、細胞、および産物が時間について加えられ、ま
たはのぞかれる方法(Payne et al.1991)を参照する。最初に全栄養分が供給され、培養
期間の最後に細胞および産物よりなる培養内容が回収される操作方法は「1段階バッチ工
程」と呼ばれる。バッチ工程が2つの連続した期、成長期、および産生期の間に、培地が
換えられる方法では、操作法は「2段階バッチ工程」と呼ばれる。本発明の観点において
、生育培地から産生培地への変換は、急な工程変更、または連続的一連の工程による段階
的、または段階的変更により引き起こされる。ある極端な例において、段階的変更は次第
に大きく変化する組成の培地への段階的置換により成し遂げられる。また別の場合、段階
的変更は成長期培養への産生培地の一つまたはそれ以上の化合物の供給により成し遂げら
れる。これは、給餌-バッチ工程の一例である。
【0094】
「給餌-バッチ(fed-batch)」工程においては、栄養物、および/または一つまたはそれ
以上の増強剤などの特異的培地物質が培養の間、定期的または連続的に供給される。ある
成分を最初にバッチ法によって栄養培地に加え、次いで給餌-バッチ法によって加えても
よく、または専ら給餌-バッチ法によってのみ加えてもよいことに注意すべきである。
【0095】
給餌-バッチ工程の使用で、細胞を長期にわたる産生状態にすることが可能で、事実、
細胞の産生能力は増強されることが発見された。実施例15、17、および表16、および18に
示したように、給餌-バッチ工程において加える栄養物および増強剤は全タキサン生産量
および、タキソールおよびバッカチンIIIとしての特定のタキサンの合成効果において有
意な改良が見られる。さらにこの操作法はさまざまな異なる培養条件下におけるさまざま
な異なる細胞系についても適用可能であることがわかった。
【0096】
成分を給餌-バッチ式で添加することは、特定の成分の濃度が培養中で低いレベルで保
たれなければならないとき特に有利である。たとえば、基質の阻害効果を回避する場合な
どである。同様に、給餌-バッチ式の添加は、栄養培地に最初に成分を加えた時に細胞が
消極的反応を示すとき、または、化学量論的に有意量の化合物は溶解度や有毒限界量のた
めに加えることができないときに有利である。さらに、ある成分を含む給餌溶液の連続的
または連続(定期的)給餌-バッチ添加は、パルス式の添加のような急激な方法で成分が
加えられると、細胞が消極的反応を示す場合に特に好ましい。給餌-バッチ法で加えられ
ることにより好ましい反応を細胞が示す特定の化合物には、アルファーおよびベータフェ
ニルアラニンなどのタキサン前駆体や炭素源としてのマルトース、フルクトース、および
グルコース、アミノ酸としてのグルタミン、グルタミン酸、アスパラギン酸、マクロ栄養
物質としてリン酸、カルシウム、、およびマグネシウム、およびオーキシン-関連成長調
節因子およびジャスモン酸関連化合物などの増強剤などが含まれる。
【0097】
該当技術分野において通常の知識を有する技術者には、供給される成分の組成は、タキ
サン産物の増加のための産生期間の延長やより高いバイオマス濃度を得るための増殖期の
延長などの、望みの結果を得るために変化させうることは明白であろう。最適な産生活性
および効果を達成するための最適条件の選択は、本明細書の開示に基づいて通常の知識を
有する技術者には容易である。
【0098】
本明細書において、培地交換とは培養からの使用済み培地の除去、および引き続き行わ
れる新鮮培地の添加を意味する。細胞はおおむね操作の間中、培養中に維持される。本発
明の方法において、培地交換法は容量当たりのタキサンの高産生を可能にし、且つ持続さ
せ、バッチ法に比べ、優れた工程効率および全産生レベルの向上に有用である。培地交換
法を行う場合、細胞外産物はその後の回収および精製過程は、他の操作法の場合より容易
である。
【0099】
実施例14および表15に示したように、培地交換はタキサン全般および、タキソール、バ
ッカチンIIIおよび、10-脱アセチルバッカチンIIIなどの特定のタキサンの高産生活性を
維持することを成功させる。さらに、この操作法はバッチ操作法に比べ、タキサン全般お
よび、タキソールおよびバッカチンIIIなどの特定のタキサンに対し、容積当たりの産生
増加ももたらす。さらに、この操作法はさまざまな異なる培養条件下におけるさまざまな
異なる細胞系に対し、適用可能である。実施例7.3に示したように、3日毎の使用済み培地
の除去および、新鮮培地の再添加は、増殖条件でのタキサンおよびタキソールの産生の有
意な増強や、同様に細胞外産物の量の増加をもたらす。
【0100】
培地交換の刺激的効果は、細胞内産物の除去によって、フィードバック阻害や産物の分
解が回避されるためであろう。細胞内産物除去による、2次的代謝物の産生および培養液
への分泌へのこのような好ましい効果は、多くの文献中でも、Robins and Rhodes(1986,A
ppl.Microbiol.Biotechnol.,24,35-41)およびAsadaand Shuler(1989.Appl.Microbiol.B
iotechnol.,30,475-481)により報告されている。定期的な使用済み培地の除去は、上記の
利点を含み、さらに他の、非タキサン、阻害物質(フェノール化合物など)が培地から除
去されることにより、2次的生合成への抑制を取り除く作用をする。
【0101】
活発に生合成を行っている細胞への新鮮培地の再添加は、減少した必須栄養物質の供給
による産生増強をもたらすであろう。例えば、Miyasaka et al. (1986, Phitochemistry,
25,637-640)は、単なるスクロースの培地添加によりSa1via miltiorhizaの静止期の細
胞を刺激し、ジテルペン代謝産物、クリプトタンシノンおよびフェルギノールの産生をさ
せることを報告している。おそらく、静止期において炭素の限界により、生合成が止まっ
ていたのだろう。本発明において用いた定期的培地交換法は上記の要素の結果として有益
となるだろう。培地交換の量、交換頻度、および交換する培地の組成はさまざまでありう
ることは理解されよう。培地交換による生合成および分泌の活性化能は、連続的、半連続
的、または給餌-バッチ法において、効率的商業的工程の操作や構築にとって重要な意味
を持つ。
【0102】
バッチ培養の内容の全部でないが大部分が回収されて、連続的細胞増殖および産生のた
めに新鮮な培地の添加がされると、その工程は「繰り返し-汲み出し-充填」操作に似て、
「半-連続工程」と呼ばれる。新鮮培地が連続的に供給されて流れ出る培地が連続的に除
去されると、この工程は「連続的」と呼ばれる。細胞が反応装置内に保持される場合、工
程は「パーフュージョン法」と呼ばれる。細胞が連続的に流れ出る培地とともに除去され
ると、その連続的工程は「恒成分培養槽(chemostat)」と呼ばれる。
【0103】
このようなさまざまな操作工程の方法がここで記載したタキサン-合成系に適用できる
ことは理解されよう。
【実施例】
【0104】
以下の実施例は、本発明を実施するのに用いた材料と方法のさらなる記載のために提供
される。実施例は説明を意図しており、いかなる方法においても発明を限定するものでは
ない。
【0105】
実施例1 カルスの開始
Taxus植物材料は多くの野生型および培養系の植物から得られた。材料は研究室におい
て到着後すぐに処理するか、または4℃で使用時まで保存した。
【0106】
材料は最初に希釈セッケン溶液で洗浄し、水でリンスし、CLOROX溶液(1%ハイポクロラ
イト、pH7.0)で10分間、表面を滅菌した。材料は滅菌条件下で3回滅菌水でリンスした。
針は1%ポリビニルピロリドン(PVP)100mg/lアスコルビン酸溶液中で切断される。針は切断
末端を培地E(表2参照)中にして静置した。30から40の植物組織片を培地プレート当たり
に培養した。組織片を含むプレートは25±1℃の暗黒下で培養した。プレートは毎日、微
生物の汚染を調べ、汚染が見られたときは、非汚染針を取り出して新鮮な培地Eプレート
の中に置いた。実質的カルス形成を観察し、カルスを20日目に組織片から分離し、表3に
示したさまざまなカルス増殖培地へと移した。たとえばTaxus chinensisのカルスは培地D
に移した(表2参照)。この最初の手順は非常に効率がよく、低い汚染率および高い90%以
上の最初の組織片のカルス誘導率へとつながった。同様の手順をTaxus brevifolia,Taxus
canadensis,Taxuscuspidata,Taxus baccata,Taxus globosa,Taxus floridana,Taxus wa
llichiana,Taxusmedia,およびTaxus chinensisの開始培養に対しても首尾よく用いるこ
とができた。
【0107】
実施例2 カルスの増殖
組織片から取り出したカルスは、25±1℃の暗黒下で培養した。健康なカルス部分を新
鮮な培地に7から10日毎に移植したが、この移植の頻度は褐色化を防ぎ、長くカルスを維
持するために非常に重要であった。さまざまな種のカルスに対して好ましい成長および維
持培地を表3にまとめた。
【0108】
実施例3 懸濁培養の開始
1gの生重量のカルス材料を、各種に対して適当な培地(表3参照)25mlを含む125mlの3
角フラスコに無菌的に接種した。例えば培地DはTaxas chinensisに用いられた。フラスコ
にシリコンキャップ(Bello,NJ)を被せ、旋回振盪機上に120rpmで24±1℃暗黒下で置い
た。懸濁培養はおよそ3から10日で形成された。最初に、培地はミラクロスフィルター(C
albiochem)を含むブフナー漏斗を用いフラスコ内容物を吸引濾過することにより交換し
、すべての生物を新鮮な培地に再懸濁した。細胞が増殖したとき、1から2g(生重量)の細
胞を一般に新たな25mlの新鮮な培地を含む125mlフラスコに移し、その後毎週副培養(サ
ブカルチャー)した。
【0109】
実施例4 懸濁細胞の成長
典型的成長率および細胞濃度は表4にまとめた代表的種の懸濁培養において達成される
。
【0110】
ある詳細な例として、時間ごとの生物重量(生重量および乾燥重量)の増加はTaxus ch
inensis系統K-1について図1に示した。最大増殖率はもっとも急速に生物重量が増殖曲線
上で増加した点における勾配を取ることで測定した。Taxus chinensisの細胞培養は最大
倍加時間2.5日で増殖した。この増殖率はTaxus種の懸濁培養について以前報告されていた
ものより有意に高い。例えば、Christen et al.(1991)は、3から4週間の培養で生物重量
は5から10倍に増加すると報告した。これは、Taxusbrevifolia懸濁培養についての平均
倍加時間7から12日に換算される。
【0111】
高濃度での培養能力は、細胞培養過程の量的産生能力を最大にするのに重要である。Ta
xus brevifoliaの培養では、1lの培養当たり乾燥重量1g以下にしか届かない(Christen
etal.(1991)にあるデータより計算される)が、Taxus chinens isの懸濁培養では18日
の成長の後1l当たり8から20gの乾燥重量に到達することが可能である。細胞の生存率は0
.05%フルオレセインジアセテートの溶液(Widholm,1972,StainTechnol.,47,189-194)に
よりアセトン中で染色し、倒立蛍光顕微鏡下(Olympus IMT-2,Japan)で青色光の励起によ
り緑の蛍光を発した細胞を数えることにより決定した。細胞の生存率は増殖期を通じ90%
以上であった。
【0112】
急速増殖条件下での高生存率を維持しながらの細胞の高細胞濃度への培養能力は、タキ
ソール、バッカチンIII、およびタキサンの産生のための植物培養過程の経済的操作に対
しての前もって必須となる重要な要素である。
【0113】
実施例5 タキソール、バッカチンIII、および他のタキサン類の解析
5.1.ELISA法
ELISA解析(HawaiiBiotech♯TA-01)を用いて、細胞培養抽出液中のタキソールの検出を
行った。この方法は高感受性(0.1ng/mL)を提供した。しかしポリクローナル抗体が用いら
れるため、他のタキサンとの交差反応が見られた。画分回収式の調製用(解析スケール)
HPLCは、10-デアセチルタキソール、7-キシロシル-10-デアセチルタキソール、セファロ
マニン、10-デアセチル-7-エピタキソール、7エピタキソール、並びに他の未同定タキサ
ンとの交差反応を示した。そのような交差反応にも関わらずこの方法はタキサン産物の検
出に特に有用であり、多くの細胞系統が素早くスクリーニングされることが見いだされた
。優位なタキサンの産生を示す細胞抽出液は、概略を後に示すHPLC法を用い詳細に解析し
た。
【0114】
モノクローナルELISA解析(HawaiiBiotech#TA-02)も、細胞抽出液中のタキソールの
検出に用いた。この方法は高い感受性(0.1ng/mL)および有意な低い交差反応を提供する
。
【0115】
5.2 タキソール、バッカチンIII、および他のタキサンの抽出
タキサンの上清からの抽出は存在濃度に依存していくつかの方法により成し遂げられた
。十分量のタキサン(およそ1-5mg/L)が液体倍地中に存在するとき、試料はとても迅速か
つ効率的に調製された。培地(2mL)を完全に乾燥し(真空中で)、測定された量のメタノ
ール(0.5-2.0mL)を加えた。この混合物を超音波により完全に溶解するまで、または試料
の分散が達成されるまで振盪した。HPLC解析に先立ち遠心により固体を除去した。量的回
収は1mg/Lのレベルであったが、検出レベルは0.1mg/L以下であった。
【0116】
タキサンの培養上清中の濃度が非常に低いとき(1mg/L以下)は培地は3倍量のメチレ
ンクロライドとイソプロピルアルコール(IPA)との混合液(9:1用量で)により抽出した。有
機層を乾燥で減少させ、計量したメタノールの(50-250ml)中に再構成した。多くの抽出で
は90-95%のタキソール、セファロメニン、およびバッカチンIIIが0.6mg/Lのレベルで回収
された。
【0117】
上清のタキサン濃度が5mg/Lを越えるときはより迅速な試料調製を用いた。上清1に対
し(容量)3の0.1%酢酸を含むメタノールに混合する。この混合を30分間音波撹拌し、濾
過し、HPLC解析を行った。
【0118】
全ブロス試料(細胞を含む培養上清)は手順の節で記述したのと同様の方法で調製した
。培地1に対し(容量)3の0.1%酢酸を含むメタノールに混合した。この混合物は30分間
超音波撹拌し、さらに30分間静置し、濾過し、HPLC解析した。
【0119】
細胞物質は、新鮮に回収した細胞を凍結し(-5℃)および引き続き真空乾燥し、およびメ
タノールソックスレーを50サイクル繰り返し行うことにより抽出した。メタノールは回転
式真空機により濃縮し(100倍)、試料はHPLCにより解析した。一般に、70から80%のタキ
サンは10から15%の測定可能な分解とともに回収された。ソックスレーに先立つ試料の乾
燥を十分に行うと、タキソールの分解は5%以下にとどまることが後に判明した。
【0120】
固体培地およびカルスの抽出はタキサンレベルが低いときの細胞の抽出と等しい。しか
し、最終メタノール抽出でのメチレンクロライド/IPAと水の分離はいつも引き起こされる
。タキサンレベルが5mg/Lを越えるとき、全培地の抽出法は固体培地上のカルスの試料の
調製に適用される。
【0121】
5.3 高性能液体クロマトグラフィー法
高性能液体クロマトグラフィー解析(HPLC)は、CM3500/CM3200ポンプから構成されるLDC
2勾配高圧混合解析系、CM4100可変容量自動試料調製機、およびコンピューターに接続し
たSM5000光ダイオード光線検出器を用い、高-炭素注入ディフェニルカラム(Supelco,5μM
,4.6mm X25cm)において行われた。EldexCH150カラムオーブンにより、カラム温度は35℃
に制御された。タキサンのHPLC定量解析は、以下の2成分勾配溶出法を用い行った。
【0122】
【化2】
【0123】
クロマトグラフィー法は類似のいくつかの公表された方法(Witherup et al.1989,J.Li
q.Chromatog.,12,2117-2132)を用い、トリフルオロ酢酸を含むリン酸緩衝液の使用および
長勾配の適用などの改良とともに行われた。これらの改良は混合物からのタキソールおよ
び他のタキサン類の分解能を有意に向上させる。タキサン類の比保留時間は以下に示す。
タキソールは、用いたカラムとハードウェアに依存して31分から33分の間で溶出する。
【0124】
【化3】
【0125】
タキソール、セファロマニン、およびバッカチンIIIの保留時間は国立ガン協会(Nation
al CancerInstitute)より得た真性試料を用いて決定した。上にまとめた他のタキサン類
の保留時間は、HauserChemical(Boulder,CO)から提供された解析標準品と比較した。既
知のタキサン類の同定は保留時間および紫外線スペクトルの比較に基づいて行った。タキ
ソールおよびバッカチンIIIの紫外線スペクトルと類似のUVスペクトルを示すが、これら
のタキサン類の比保留時間を示さない化合物はタキサン類と考えられる。タキソール、セ
ファロマニン、およびバッカチンIIIの定量は真性物質から決定された係数に基づいて行
った。10-デアセチルバッカチンIIIの定量はバッカチンIIIに対し決定された係数を用い
行われた。必要な場合、他のタキサン類の定量はタキソールおよびバッカチンIIIの計算
に用いた係数により行われた。用語「全タキサン類」はタキソールおよびバッカチンIII
と類似の特異的UVスペクトルを示したタキサン類の合計を表す。Taxus培養において同定
された全タキサン類は、10-デアセチルバッカチンIII、9-ディヒドロバッカチンIII、7-
エピ-10-デアセチルバッカチンIII、バッカチンIII、9-ディヒドロ-13-アセチルバッカチ
ンIII、7-キシロシル-10-デアセチルセファロマニン、7-キシロシル-10-デアセチルタキ
ソール、7-エピバッカチンIII、10-デアセチルタキソール、7-キシロシルタキソール、セ
ファロマニン、7-エピ-10-デアセチルタキソール、タキソール、2-ベンゾイル-2-デアセ
チル-1-ヒドロキシバッカチンI、タキソールC、7-エピタキソール、および2-ベンゾイル-
2-デアセチルバッカチンIを含む。
【0126】
タキサン類の特異的なUV吸光度を示さないが、重量スペクトルにおいて特異的なタキサ
ン-重量-断片化特性を示す、タキサン類もTaxus細胞培養において観察された。Taxus細胞
中で産生されたそのようなタキサン類の例には、タクスウンナニンC(Taxuyunnanine C)お
よびその類似体およびその誘導体が挙げられる。
【0127】
各標準品(10μL)は典型的に注入され(最初標準品、次いで3または4試料)、各3成分
の面積を227nmクロマトグラムから計算した。各成分の係数はデータの線形-最小-2乗解析
により得られた。10μLの各試料が注入され、注入当たりの量は標準データ回帰に基づき
計算された。これらの結果は1リットル当たりの総量または乾燥重量パーセントに換算さ
れた。図4に上清試料の典型的クロマトグラムを示す。
【0128】
5.4 迅速高性能液体クロマトグラフィー法
上記の方法に加え、いくつかのHPLCの迅速な方法がより多くの試料を処理できるように
するために開発された。以下にこれらの方法のうち2つの詳細を示す。
【0129】
方法1)迅速高性能液体クロマトグラフィー(HPLC)は、環境温度において前述のハード
ウェアを用い、PhenomenexCurosil-Gカラム(5μM,4.6mm X 25cm 4.6mm X 3cmガードつ
き)行った。タキサン類の定量HPLC解析は以下に示す2成分勾配溶出法で行った。
【0130】
【化4】
【0131】
タキサン類に観察された比保留時間は以下に示す。タキソールは用いたカラムおよびハ
ードウェアに依存して約8分で溶出した。
【0132】
【化5】
【0133】
タキソール、バッカチンIII、および10-デアセチルバッカチンIIIの標準品は50mg/L,10
mg/L、および1mg/Lレベルで調製した。標準品を最初に注入し、その後9試料を注入し、各
3成分の面積を227nmクロマトグラムから算出した。各成分の係数はデータの線形最小2乗
解析により得られた。10μLの各試料を注入し、1リットル当たりの量を試料の希釈およ
び標準試料のデータ回帰に基づくピーク面積から計算した。
【0134】
方法2)迅速高性能液体クロマトグラフィー(HPLC)は前述のハードウェアを用い、環境
温度において、PhenomenexIB-SILカラム(3μM,4.6X15cm、4.6mm X 3cmのガードつき)で
行った。タキサン類の定量HPLC解析は以下の2成分勾配溶出法で行った。
【0135】
【化6】
【0136】
タキサン類に観察された比保留時間は以下に示す。タキソールは用いたカラムおよびハ
ードウェアに依存して約9.5分で溶出した。
【0137】
【化7】
【0138】
定量は前述のように行った。
上記の方法の流速および勾配間隔および時間についての修飾によって、植物細胞培養解
析にの最適なクロマトグラフィーが行われるこ々も見いだされた。
【0139】
5.4. タキソールのMS/MS確認
細胞培養上清中のタキソールの同定のため、フローインジェクションとイオンスプレー
常圧化学イオン化とを組み合わせたMS/MS法(図6に示される)を用い確かめた。図6にあ
るデータを得るのに用いた手順の詳細を以下に示す。重量スペクトル計測機:Sciex API
3の3重4極子常圧イオン化源。窒素を遮断ガスとしてして用い、アルゴンをCIDスペクトル
の衝突ガスとして用いた。 界面:イオンスプレー界面はイオン蒸発イオン化機(電気ス
プレー)により産生された。ゼロガスを噴霧ガスとして用いた。LCポンプ:5μL/分で作
動するABI140B二重シリンジポンプ装置を使用。溶剤:50/50アセトニトリル/H2O2mM NH4
OAc+0.1%ギ酸。注入量:5μL、スペクトルは全てフローインジェクション注入で求めた
。この方法により、細胞培養試料中におけるタキソールの明確な存在の確認およびHPLCの
結果によく一致する定量結果が得られた。
【0140】
実施例6 さまざまな種からのタキソールの生産
さまざまなTaxus種培養からのタキソールの生産を表5にまとめた。カルスは20日暗黒下
で各種毎に示される固体培地において培養された。細胞および培地を一緒に乾燥し、メタ
ノール抽出し、示したようにELISAまたはHPLCにより検定した。
【0141】
実施例7
7.1 増殖培地における生産
タキソールおよびタキサン類の生産は、Taxus chinensis細胞系統K1の増殖培地Aへの移
植の最初の2日間に開始した。15日において観察される最大タキソールは8.81μg/フラス
コであり、0.44mg/Lに相当する。このうち46.1%は細胞外培地に存在した。15日目に、全
タキサン濃度は72.87μg/フラスコ、または3.6mg/Lとなり、このうち58.6%が細胞外培地
に存在した。生存率は蛍光染色(実施例4)により計測され常に90%以上であった。このこ
とは細胞外タキソールおよびタキサン類は細胞の溶解によるものではなく、分泌によるも
のであることを示唆する。
【0142】
タキソール、バッカチンIIIおよび関連タキサン類の生産レベルを、タキサン生合成用
の増強剤を添加しないさまざまな異なる増殖条件(表2および他の実施例において詳しく
述べた)において、さまざまに異なる細胞系統により決定した。これらの回収データは細
胞は増殖に最適な条件で培養され、タキサン生合成条件でないとき、タキソール生合成レ
ベルは典型的には0.5mg/L以下であり、常に2mg/L未満であり、容量当たりのタキソール生
産活性は典型的には0.03mg/L/日から0.07mg/L/日の範囲にあり、常に0.3mg/L/日未満であ
った。同様にバッカチンIII生産レベルは典型的には0.5mg/L以下であり、常に1mg/L以下
であった。容量当たりのバッカチンIIIの生産活性は典型的には0.03mg/L/日以下であり、
常に0.15mg/L/日未満であった。同様に全タキサン価のレベルは典型的には5mg/L未満であ
り、常に20mg/L以下であった。容量当たりの全タキサン生産活性は、典型的には1mg/L/日
未満であり、常に3mg/L/日未満であった。
【0143】
7.2. 生産増強のための培地交換
タキソールおよび全タキサン生産活性の有意な改良は、9日目における無菌的培地Aの
吸引除去および新鮮培地の添加、および同じ処置の12日目の繰り返しによって認められた
。実験は15日間で打ち切られ、結果は図2に示される。培地交換による重要な生産活性の
増加は表6にまとめた。タキソールおよびタキサン類の総生産量は、上記処置を施さない
対照と比較して4.6倍に高まった。重要なことに、細胞外培地から回収されるタキソール
は4.9倍、全タキサン類は5.9倍に、培地交換しなかった対照に比較して高まった。
【0144】
タキソールおよび全タキサン生産活性の増加、およびさらに細胞外への産物の蓄積を引
き起こす注目すべき能力は、効率的操作、細胞の再使用を伴う連続的工程、およびその後
の精製工程の簡易化のために重要である。
【0145】
7.3. 増殖培地におけるタキサン生産に対する光の効果
光は光合成だけでなく、植物細胞培養中の2次的代謝にもさまざまな面で重要な役割を
果たす(Seibertand Kadkade 1980)。実施例4、7.1および7.2に記載した実験は暗黒条件
で行ったが、Taxuschinensis培養の光への反応をここで記述する。
【0146】
1グラムの生重量の7日目Taxus chinensis系統K1の細胞を125ml三角フラスコ中の25m
lの生育培地A(表2参照)に移植し、24±1℃において回転振盪機で120rpmで培養した。
同じフラスコを暗黒条件および標準Gro Luxランプに3フィート離しておいた。ランプのス
ペクトル特異性は図3に示される。結果は表7に示される。
【0147】
培養を光に曝すことは全タキサンのレベルおよび細胞外の蓄積に影響しなかった。しか
しタキサンの内容は有意に2つの処理で変化した。たとえば光下で培養したものはタキソ
ールを暗黒条件下のものより2.8倍も多く生産した。細胞外タキソールの割合もまた暗黒
処理のものより有意に高かった(76%対56%)。光処理の使用、特に特異的スペクトルでの使
用は、タキソールの生産のための細胞培養工程において有用でありうる。
【0148】
実施例8 エリシター
エリシターという単語は、植物細胞培養物に加えられたとき二次代謝の増加をひきおこ
す生物学上の(または生物の)、および非生物学上の(または無生物の)起源を有する化
合物を意味する。
【0149】
多くのエリシターが有効であるとわかっているが、代表例、すなわち、キトサングルタ
メートの使用が、本明細書中に詳しく記載されている。キトサンは、いくつかの植物細胞
培養物組織においてはエリシターとして以前に試されたが、褐色化および生存率の損失な
どの毒性を伴う反応は、キトサンの使用を非実用的にする(Beaumont and Knorr 1987,Bio
technol.Lett.9,377-382)。事実、そのような毒性反応は、論文において報告された多く
のエリシターの共通の欠点である。特にキトサングルタメートなどの化学的に修飾された
キトサンの使用により、毒性効果を回避しつつ、タキソールおよびタキサン生合成を誘導
することは新しいアプローチである。
【0150】
培地Dにおいて7日から8日成長させたTaxuschinensis系統K-1の懸濁液はmirachloth(
Calbiochem)フィルターを有する滅菌ブフナー漏斗の使用で無菌的に吸引濾過された。新
鮮な2グラム重量の細胞は、125mlエルレンマイヤーフラスコにおいて25mlのC
培地(表2参照)へ無菌的に移された。0.05%のキトサングルタメート溶液は、新たに調
製され、および0.22μmカートリッジフィルターを通して濾過滅菌された。1g乾燥重量
当たり165ミリグラムのエリシターの濃度となるように、この溶液の825μlを実験
の開始時にフラスコに加えた。フラスコは、暗やみにおいて110rpmで旋回撹拌器で2
4プラスマイナス1℃で保温培養された。フラスコは15日で破壊的に試料とされ、およ
び成長、細胞の色および培地の観察および細胞の生存率が記録された。試料は実施例5に
おいて記載されたようにタキサンの分析に付した。この実験の結果は表8において示され
ている。
【0151】
エリシター処理は、非処理の対照標準よりも細胞当たりの全タキサン生産量(0.53%対0.
42%乾燥重量タキサン)が多く、ささやかな改良の結果となった。エリシターの非毒性の性
質は、両方の処理において観察される高生存率(75%-80%)から明白である。実際に、対照
標準に比べてエリシター処理において増加した乾燥重量は再現性をもって観察された(14.
2g/1対10.1g/l乾燥重量)。細胞の高濃度も加味すると、エリシター処理においては全タキ
サンの1.8倍の力価、すなわち75.8mg/l対対照標準42.4mg/lの結果となった。
【0152】
細胞当たりの基準(0.098%対0.054%乾燥重量タキソール、1.8倍増加)および力価におい
ての比較(13.9mg/l対5.4mg/l、2.6倍増加)の両方で、エリシター処理はタキソール生合成
の増加の結果をもたらしたる。分泌の程度も、対照標準に比べエリシター処理のほうがよ
り高かった(85%対72%細胞外産物)。
【0153】
本明細書中に記載されているエリシター処理は、タキソール産物の増加、より有利な産
物の側面、産物分泌の増加および細胞の高生存率の保持力の結果となる。これらの生産の
特性は、タキソール生産の細胞の培養過程にかなりの改良を表す。
【0154】
実施例9 産生培地開発
実施例6において記載された濃度を越えるタキソール生産力の増加のための努力におい
ては、特別な「産生培地」を計画するために栄養濃度は処理された。培地Dにおいて成長
させたTaxus chinensis系統K-1の7日から8日たった懸濁液は、MIRACLOTH(アクリル酸
のバインダーを含むレーヨンポリエステル布地)フィルター(Calbiochem)を有する滅菌ブ
フナー漏斗の使用で無菌的に吸引濾過された。新鮮な500mg重量の細胞は、5mlの
産生培地BおよびC(表2参照)へ無菌的に移された。その容器は暗やみにおいて110
rpmで旋回撹拌器で24プラスマイナス1℃で18、25および42日のいろいろな時間の
期間で保温培養された。処理は破壊的に試料とされ、および成長、細胞の色および培地の
観察、および細胞の生存率が記録された。試料は実施例5において記載されたようにタキ
サンの分析をされた。この実験の結果は表9において示されている。
【0155】
9.1 18日培養の結果
タキサンおよびタキソールのかなりの濃度の生産によりTaxus chinensis細胞培養は培
地の組成物の変化に応答する。これらのデータは表9において要約され、および試料のク
ロマトグラムは図4において示されている。培地Bにおいては、99.8mg/lの全タキサンが
生産され、24.1mg/lのタキソールを含んだ。培地Cにおいては、110mg/lの全タキサンが
生産され、21.3mg/lのタキソールを含んだ。乾燥重量の基準では、細胞は、培地Bにおい
ては0.18%乾燥重量タキソール、培地Cにおいては0.065%乾燥重量タキソールを生産した
。
【0156】
9.2 長期培養
Taxuschinensis(K-1系統)の25および42日の長期培養後のタキソールおよびタキ
サンの生産は、培地Cにおいて研究され、その結果は図5において要約されている。以下
に重大な観察が要約されている:
(i) Tuxus培養懸濁液は、タキソールおよびほかのタキサンのかなりの濃度を生産す
ることができる。もっとも高い集積は42日で起こり、0.32%の乾燥重量タキソール、お
よび0.62%の乾燥重量タキサンを含んだ;最終の培地体積に基づくタキソールの力価153mg
/lおよび全タキサンの力価295mg/lに相当した。タンデム質量分光測定法によるこの試料
の分析は、図6において示されたようなタキソールの存在を確認した。MS/MSによる定量
は、HPLCと非常に良い一致を示した。
【0157】
(ii) 25日から42日の間のタキソール生合成の速度は、17日間においては直線
的な生産であると仮定すると、1日につき、1リットル当たり7.6mgタキソールであった
。この速度は、最初の25日においての生産速度よりも有意に高い。25日および42日
の間の全タキサンの生合成の速度は、1日につき、1リットル当たり12.3mgであった。タ
キソール、バッカチンIII、および全タキサンの生産力の体積の平均は、それぞれ、3.6、
0.5、および7.0mg/l/dayであった。
【0158】
(iii) 実施例7において記載された急速増殖条件などの(タキサン生合成は増加され
ない)急速増殖条件と比較して、産生培地の定式化は特定のタキソール含有量において4
5倍の増加の誘導も可能である。
【0159】
(iv)望ましくないタキサン産物を最小限度にし、産物スペクトルを希望する最終産物
タキソールへの生合成を漏斗を通り抜けるようにする処理も可能であった。たとえば、2
5日では、タキソールは全タキサンの28%を構成しおよび42日では、全タキサンの5
2%を構成したが、それに対して生育培地(実施例7.1参照)においては、タキソールは
全タキサンの12.2%しか構成しなかった。処理した産物の側面のこの能力は、下流部
門の精製および調節結果の純度に関わる産物に重要な影響を有するだろう。たとえば、タ
キサン副産物、セファロマニンの生産を押さえる能力は樹皮からのタキソールの精製に比
べて、後の工程での精製を非常に単純化させる可能性がある。
【0160】
(v)Taxus細胞培養物はタキソール(42日で87%)およびほかのタキサンのかな
りの量の分泌を誘導した。細胞外タキソールおよびタキサンの存在は細胞溶解液に帰する
というよりはむしろ分泌に帰するということは、いくつかの独立した観察により確証され
る:(a)連続した生合成が25日と42日の間に起こり、このことは細胞は生きていてお
よび活動したことを示唆した。独立した観察は産生培地においては18日後に70%以上
の生存率が観察されたということを示した。(b)異なるタキサンの異なる割合が分泌され
た。もし細胞が溶解するとしたなら、培地においての割合は異なるタキサンでも同じであ
るはずである。
【0161】
(vi) このTaxus細胞系統が産物濃度が非常に高い細胞外環境において生長し、および
高い割合でタキソールを生産する能力をもつことは特に注目すべき価値がある。
(vii) これらの結果が得られるTaxus細胞系統は細胞の高濃度の増殖を急速にするこ
とも可能であり、および急速増殖条件下で20世代後にも報告された生産性を明白にし、
このことはその安定性および商業上の潜在能力を証明する。
【0162】
以前に報告された結果よりも、本明細書中に記載されている条件下でのTaxus chinensi
sの細胞系統により生産されるタキソールおよびタキサンの濃度は35から150倍高い
。たとえば、Christenet al.(1991)は培養の2から4週間後Taxus brevifoliaの培養懸
濁液より1から3mg/lのタキソールの生産を報告した。WickeramesinheおよびArteca(199
1)はTaxus mediaの細胞培養においては、乾燥重量の0.009%のタキソールの生産を報告し
た。
【0163】
要約すると、我々のデータはTaxus chinensis培養の念入りな開始および選択をもって
、および特別に処理された生育培地条件をもって、細胞は高濃度の増殖の誘導も可能だろ
う。これらの細胞が産生培地条件に移されたとき、細胞は長期間に高い生存率を維持して
タキソールおよびほかのタキサンのかなりの濃度を生合成および分泌することも可能であ
る。提起的な培地の交換、光、および産生培地をもつエリシターの混合は、多くの協同性
の生産性の増加の結果となる。これらの特性は、組織培養技術の使用のタキソールおよび
タキサンの生産の十分な商業上の過程の重大な必要条件である。
【0164】
実施例10
10.1 銀の使用のタキサン生産の増加
銀、化合物を含む銀の形態において、銀混合体、あるいは銀イオンは、Taxus種の細胞
培養においてのタキソール、バッカチンIII、およびタキサン生合成の有用な増加因子で
あることが発見された。銀およびほかの増加因子の組み合わせもまたタキサン生産の高い
割合の獲得および維持においては有用であることが発見された。
【0165】
培地L(表2)において培養されたKS1A Taxus chinensis懸濁液の7日たった細胞はMI
RACLOTH(Calbiochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌的に吸引
濾過された。15%から20%(w/v)の範囲において新鮮な細胞の濃度を与えるため、約0
.75から1グラム重量の新鮮な細胞は表10において示された所定の組成物の培養培地の4
から5mlへ植えつけられた。その容器は暗やみにおいて110rpmで旋回撹拌器(1”throw)
で25プラスマイナス1℃で保温培養された。蒸発分は滅菌された蒸留水の付加により補
正された。全液の試料(すなわち細胞外および細胞内のタキサンの両方)は周期的な間隔
で得られ、および実施例5において概説された方法にしたがってHPLCにより処理およ
び分析がされた。
【0166】
表10において要約されたデータはタキソール、バッカチンIII、およびほかのタキサ
ンの生産は化合物を含むいろいろな銀によりうまく増加させられるということを指し示す
。化合物および異なる逆イオンを含む異なる銀の多様性の増加を示している表10におい
て説明したように、この増加は第一に培地にある銀の存在に帰する。これらの生産の濃度
は増加されない培養において観察された生産の濃度(その生産濃度は実施例7において詳
しく述べられている)よりも重大に高い。
【0167】
10.2 チオ硫酸銀の使用のタキサン生産の増加
毒性の考慮および調製物および保管の容易に基づくと、チオ硫酸銀は二次的な実験にお
いて使用される。チオ硫酸銀の調製のために使われる方法は以下であった:1.98gのチオ
硫酸ナトリウム(5水和物)は80mlの水に溶解された。いきおいよく全体をかき回して
20mlの0.1M硝酸銀溶液が加えられ、その結果100mlの20mMのチオ硫酸銀の貯蔵溶液と
なった。チオ硫酸カリウムは同等の効果のある結果をもつチオ硫酸ナトリウムの適所に使
用されることも可能であった。その貯蔵溶液は所定の実験の開始時に細胞培養培地のなか
へ0.22μMカートリッジフィルターを用いて滅菌濾過された。チオ硫酸銀溶液と同様なも
う一つの調製方法もまた適合される。細胞培養実験計画案は表10において記載された実
験のために記載されたものと同様であった。
【0168】
表11はTaxuschinensisの異なる多くの細胞培養の一つの増加因子として銀の使用に
より獲得されたデータを要約した。これらのデータは銀は一般的にタキサン生合成の基本
的な増加をもたらすということを示す。いかなる所定の場合において観察された特別な産
物の側面は細胞の系統および培養培地の特性を反映する。銀イオン/混合体は成長調節、
炭素源、塩、微量元素、およびそのようなものなどの生合成の好む培地においてはほかの
因子と組み合わせにおいて使用されるときタキサン生産の増加においてとくに効果的だろ
う。
【0169】
実施例11 ジャスモン酸メチルおよびジャスモン酸関連化合物の使用のタキサン生産
の増加
ジャスモン酸および関連化合物と同様に、ジャスモン酸のメチルエステル(ジャスモン
酸メチル)は、Tuxus種の細胞培養においてタキサン生合成の増加因子として有用である
ことが発見された。ジャスモン酸メチルおよびほかの増加因子の結合もまたタキサン生産
の高い割合の獲得および維持においては有用であることが発見された。
【0170】
培地M(表2)において培養されたTaxus chinensis懸濁液の7日たった細胞はMIRACLO
TH(Calbiochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌的に吸引濾過さ
れた。15%から20%(w/v)の範囲において新鮮な細胞の濃度で、細胞は表12におい
て示された所定の組成物の培養培地の4から5mlへ植えつけられた。その培養物は暗やみ
において120または180rpm(容器の大きさに依存する)で旋回撹拌器(1”throw)で
24プラスマイナス1℃で保温培養された。蒸発分は滅菌された蒸留水の付加により補正
された。全液の試料(すなわち細胞外および細胞内のタキサンの両方)は周期的な間隔で
得られ、および実施例5において概説された方法にしたがってHPLCにより処理および
分析がされた。
【0171】
表12はいくつかの代表的なTaxus chinensis細胞系統の増加因子としてジャスモン酸
およびジャスモン酸メチルエステルの使用により獲得されたデータを要約した。これらの
データはジャスモン酸およびジャスモン酸メチルエステルは一般的にタキサン生合成の基
本的な増加をもたらすということを示す。いかなる所定の場合において観察された特別な
産物の側面は細胞の系統および培養培地の特性を反映する。これらの増加因子の存在にお
いて獲得されたこれらの生産の濃度は増加されない培養において観察された生産の濃度(
その生産濃度は実施例7において詳しく述べられている)よりも重大に高い。
【0172】
ジャスモン酸、ジャスモン酸メチルエステル、および関連化合物は、ほかの増加因子、
成長調節、炭素源、塩、微量元素、およびそのようなものなどの生合成の好む培地におい
てはほかの因子と結合において使角されるときタキサン生合成の効果的増加因子である。
【0173】
実施例12 3,4-メチレンジオキシ-6-ニトロ桂皮酸の使用のタキサン生産の増加
桂皮酸類似化合物、3,4-メチレンジオキシ-6-ニトロ桂皮酸(MDNA)および関連化合物はT
uxus種の細胞培養においてタキサン生合成の有用な増加因子であることが発見された。MD
NAおよびほかの増加因子の結合もまたタキサン生産の高い割合の獲得および維持において
は有用であることが発見された。
【0174】
培地M(表2)において培養されたTaxus chinensis懸濁液培養SS122-42の7日たった
細胞はMIRACLOTH(Calbiochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌
的に吸引濾過された。細胞は15%から20%(w/v)の新鮮な細胞の濃度の培養培地条件
へ植えつけられた。その容器は暗やみにおいて180rpmで旋回撹拌器(1”throw)で24
プラスマイナス1℃で保温培養された。処理された培養物は試料になりおよびいろいろな
時間点で実施例5において記載された方法を使用して分析がされた。蒸発分は滅菌された
蒸留水の周期的な間隔の付加により補正された。全液の試料(すなわち細胞外および細胞
内のタキサンの両方)は周期的な間隔で得られ、および実施例5において概説された方法
にしたがってHPLCにより処理および分析がされた。
【0175】
表13はTaxuschinensis細胞培養においてタキサン生合成の増加因子として3,4-メ
チレンジオキシニトロ桂皮酸の使用により獲得されたデータを要約した。これらのデータ
はMDNAは一般的にタキサン生合成の基本的な増加をもたらすということを示す。培地IIに
おいて、すなわちMDNAおよび銀の存在においての培養は、タキサンの生産をより増加させ
る。いかなる所定の場合において観察された特別な産物の側面は細胞の系統および培養培
地の特性を反映する。これらの生産の濃度は増加されない培養において観察された生産の
濃度(その生産濃度は実施例7において詳しく述べられている)よりも重大に高い。
【0176】
実施例13 増加因子の組み合わせの使用のタキサン生産の増加
いろいろな増加因子は、タキサン生産においては組み合わせにおいて使用され、重大お
よび協同性の改良を与えた。
【0177】
培地P(SS64-412)、培地O(SS64-561,SS64-571)、培地I(SS124-77,SS85-26)、培地M(
SS122-29)(これらの培地の組成物は表2において記載されてある)において培養されたT
axus chinensis懸濁液培養の7日たった細胞はMIRACLOTH(Calbiochem)フィルターにあわ
せた滅菌したブフナー漏斗の使用で無菌的に吸引濾過された。細胞は20%(w/v)の新鮮
な重量の濃度の(表14において指し示されている)培養培地へ植えつけられた。その培
養物は暗やみにおいて180rpmで旋回撹拌器(1”throw)で24プラスマイナス1℃で保
温培養された。蒸発分は滅菌された蒸留水の周期的な間隔の付加により補正された。全液
の試料(すなわち細胞外および細胞内のタキサンの両方)は周期的な間隔で得られ、およ
び実施例5において概説された方法にしたがってHPLCにより処理および分析がされた
。
【0178】
表14はTaxuschinensis細胞培養においてタキソール、バッカチンIII、およびタキサ
ン生合成の増加因子のいろいろな組み合わせの使用により獲得されたデータを要約した。
そのデータは増加因子の組み合わせにより個々の因子でみられるタキサン生産の増加、お
よび増加されない条件においての生産濃度(その生産濃度は実施例7において詳しく述べ
られている)よりたくさんのタキサン生産の増加を立証する。
【0179】
実施例14 培地交換によるタキサン生産の増加
この実施例は培地において高い生産性は培地成分の補充および消費された培地の除去に
より維持されるのも可能であるということを立証する。
【0180】
細胞は最初は培地M(Paella)、培地I(SS29-3A5)、および培地I(SS45-146)において培
養された。これらの培養培地の詳しい組成物は表2において記載されている。これらの細
胞系統の7日たった細胞はMIRACLOTH(Calbiochem)フィルターにあわせた滅菌したブフナ
ー漏斗の使用で無菌的に吸引濾過された。新鮮な約1.5グラム重量の細胞は表15におい
て指し示されたそれぞれの培養培地の4.5m1へ植えつけられた。その容器は暗やみにおい
て120rpmで旋回撹拌器(1”throw)で24プラスマイナス1℃で保温培養された。蒸発
分は滅菌された蒸留水の周期的な間隔の付加により補正された。培地交換処理のために、
容器において後ろの細胞を残して、消費された産生培地は一回分の培養の10から11日
後に滅菌したピペットを使用して吸引して取り除かれた。その消費された上清は実施例5
において記載された方法を使用して細胞外タキサンの分析がされた。最初の1回分の培養
と同じ組成物の新鮮な培養培地は生産力のある細胞を含む容器に加えられた。その細胞は
上で記載された同じ環境条件下で培養された。培地交換の周期は培養のさらに10から1
1日後に繰り返された。一回分の生産の全細胞外タキサンは表15において培地交換の生
産の全細胞外タキサンと比較されている。培地交換濃縮の値は細胞懸濁液培養の体積(す
なわち、5.75ml)により分けられた細胞外培地において生産されたタキサンの全量を意味
する。
【0181】
表15は細胞は延長期間に生産力のある状態を維持されることも可能であるということ
を指し示し、および実際に、細胞の生産性は繰り返された培地交換により増加されるとい
うことを指し示す。繰り返された培地交換による増加は異なる増加条件の集まり、および
細胞培養の多様性に便利である。
【0182】
そのデータは増加されない条件においての生産濃度(その生産濃度は実施例7において
詳しく述べられている)よりたくさんのタキサン生産の増加を立証する。
実施例15 給餌バッチ操作によるタキサン生産の増加
培地I(CR-128,SS36-245)、および培地L(SS36-359)(これらの培地の組成物は表2に
おいて記載されている)において培養された細胞系統の7日たった細胞はMIRACLOTH (Cal
biochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌的に吸引濾過された。
新鮮な約1グラム重量の細胞は表16.a.において指し示された所定の組成物の培養培地
の4m1へ植えつけられた。その容器は暗やみにおいて120rpmで旋回撹拌器(1”throw)で
24プラスマイナス1℃で保温培養された。蒸発分は滅菌された蒸留水の周期的な間隔の
付加により補正された。給餌バッチ操作のため、あらかじめ決められた組成物の滅菌した
飼料溶液は、たとえば11の培養液当たり1日あたり10mlの飼料溶液のように、あらか
じめ決められた飼料を与える速度で培養溶液のなかへ連続的に飼料を与えられた。飼料溶
液の組成物および給餌の実験計画案を含む、給餌バッチ操作の詳細は表16.b.において
記載されている。処理された培養物は実施例5において記載された方法の使用で試料とさ
れおよび分析がされた。
【0183】
表16.a.は細胞は延長期間に生産力のある状態を維持されることも可能であるという
ことを指し示し、および実際に、細胞の生産性は給餌バッチ操作により増加されることも
可能であるということを指し示し、その結果バッカチンIII、タキソール、およびほかの
タキサンの高濃度の蓄積となる。特別なタキサンの関連する量は給餌実験計画案および細
胞系統および培養条件の試料の組成物の相互作用を反映する。この表もまたフェニルアラ
ニンの給餌はほかのタキサンと比例してタキソールの生産の増加の結果となるということ
を指し示す。
【0184】
そのデータは増加されない条件においての生産濃度(その生産濃度は実施例7において
詳しく述べられている)よりたくさんのタキサン生産の増加を立証する。
実施例16 増加因子の組み合わせの使用のタキサン生合成の増加
種々の増加因子が、タキソール、バッカチンIII、およびタキサン生産において組み合
わされて使用され、重大および協同性の改良を与えた。
【0185】
培地M(その培地の組成物は表2において記載されてある)において培養されたTaxus
chinensis懸濁液培養(SS122-41,cr427,SS122-30,cr857,cr452)の7日たった細胞はMIRACL
OTH(Calbiochem)フィルターにあわせた滅菌したブフナー漏斗の使用で無菌的に吸引濾過
された。表17において別な方法で記載されることなしに細胞は20%(w/v)の新鮮な重
量の濃度の(表17において指し示されている)培養培地へ植えつけられた。その培養物
は暗やみにおいて180rpmで旋回撹拌器(1”throw)で24プラスマイナス1℃で保温
培養された。蒸発分は滅菌された蒸留水の必要に応じたの付加により補正された。全液の
試料(すなわち細胞外および細胞内のタキサンの両方)は周期的な間隔で得られ、および
実施例5において概説された方法にしたがってHPLCにより処理および分析がされた。
【0186】
表17はTaxuschinensis細胞培養においてタキソールおよびタキサン生合成の増加因
子のいろいろな組み合わせの使用により獲得されたデータを要約した。そのデータは増加
因子の組み合わせにより個々の因子でみられるタキサン生産の増加、および増加されない
条件(その詳細は実施例7において用意されている)よりたくさんのタキサン生産の増加
を立証する。
【0187】
実施例17 給餌バッチ操作によるタキサン生産の増加
培地M(SS122-41)(これらの培地の組成物は表2において記載されている)において培
養された細胞系統の7日たった細胞はMiracloth(Calbiochem)フィルターにあわせた滅菌
したブフナー漏斗の使用で無菌的に吸引濾過された。新鮮な約1グラム重量の細胞は表1
8.a.において指し示された所定の組成物の培養培地の4mlへ植えつけられた。その容器
は暗やみにおいて120rpmで旋回撹拌器(1”throw)で24プラスマイナス2℃で保温
培養された。蒸発分は滅菌された蒸留水の付加により補正された。給餌バッチ操作のため
、あらかじめ決められた組成物の滅菌した飼料溶液は培養溶液のなかへ連続的に飼料を与
えられた。飼料溶液の組成物および給餌の実験計画案を含む、給餌バッチ操作の詳細は表
18.b.において記載されている。処理された培養物は実施例5において記載された方法
の使用で試料とされおよび分析がされた。
【0188】
表18.a.は細胞は延長期間に生産力のある状態を維持されうるということを指し示し
、および実際に、細胞の容積測定の生産性は給餌バッチ操作により増加されうるというこ
とを指し示し、その結果バッカチンIII、タキソール、およびほかのタキサンの高濃度の
蓄積となる。特別なタキサンの関連する量は給餌実験計画案および細胞系統および培養条
件の試料の組成物の相互作用を反映する。
【0189】
そのデータは増加されない条件においての生産濃度(その生産濃度は実施例7において
詳しく述べられている)よりたくさんのタキサン生産の増加を立証する。
ほかの側面、利点および修飾は発明が関係する作品において理解されるかもしれないが
、理解の明快の目的のため特別な実施態様の組み合わせにおいて図解および例としたいく
つかの詳細において前記の発明が記載された。発明の予知に限界はないが、前記の記載お
よび例は図解される。その作品において理解された目的のような発明を実行するための上
記に記載された方法の修飾は発明の余地の中に意味され、このことは追加された請求項に
よりのみ限られる。
【0190】
この詳述において言及されたすべての刊行物および特許申請はこの発明が関係する作品
において理解されたそれらの理解の水準を示す。それぞれ独立した刊行物または特許申請
が参考文献により編入されたことを特別におよび独立に指し示されたように全ての刊行物
および特許申請は同じ程度の参考文献によりここに編入された。
【0191】
【表1−1−1】
【0192】
【表1−1−2】
【0193】
【表1−1−3】
【0194】
【表1−2】
【0195】
【表1−3−1】
【0196】
【表1−3−2】
【0197】
【表1−4】
【0198】
【表1−5−1】
【0199】
【表1−5−2】
【0200】
【表1−5−3】
【0201】
【表1−6−1】
【0202】
【表1−6−2】
【0203】
【表2−1】
【0204】
【表2−2】
【0205】
【表3】
【0206】
【表4】
【0207】
【表5】
【0208】
【表6】
【0209】
【表7】
【0210】
【表8】
【0211】
【表9】
【0212】
【表10】
【0213】
【表11】
【0214】
【表12】
【0215】
【表13】
【0216】
【表14】
【0217】
【表15】
【0218】
【表16−1】
【0219】
【表16−2】
【0220】
【表17】
【0221】
【表18−1】
【0222】
【表18−2】
【特許請求の範囲】
【請求項1】
明細書に記載の発明。
【請求項1】
明細書に記載の発明。
【図1】

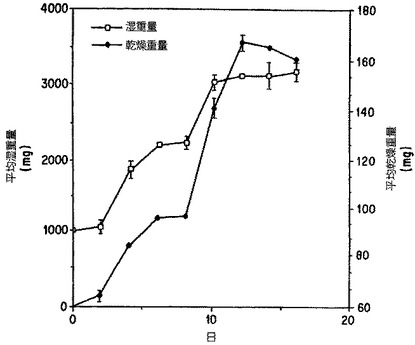
【図2】
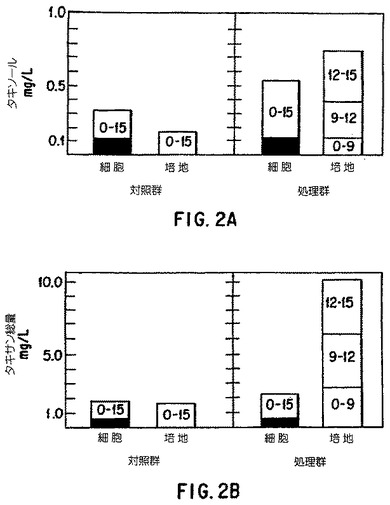
【図3】
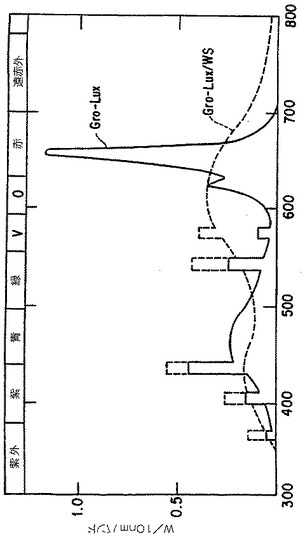
【図4A】
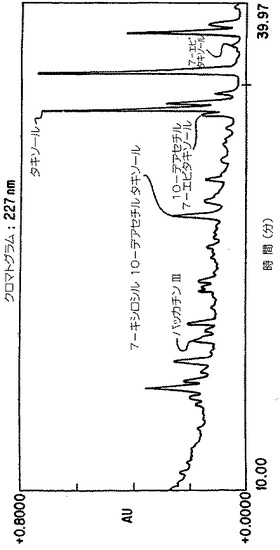
【図4B】
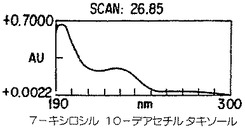
【図4C】
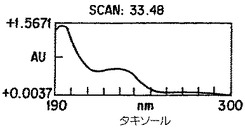
【図4D】
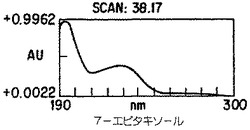
【図5A】
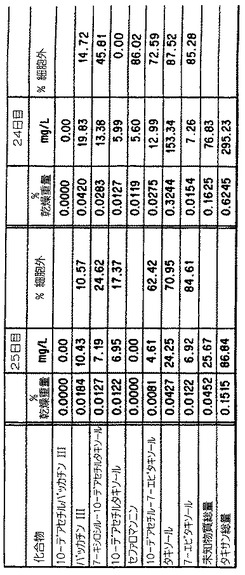
【図5B】
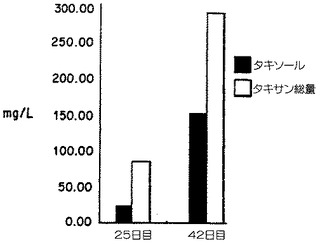
【図6A】
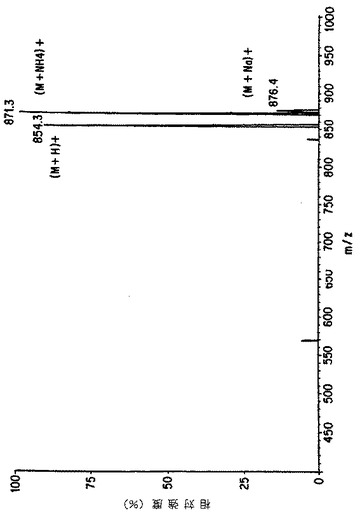
【図6B】
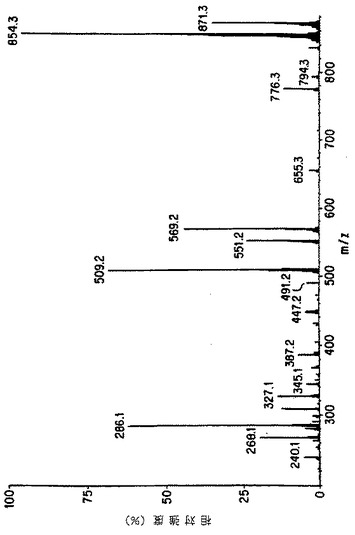
【図6C】
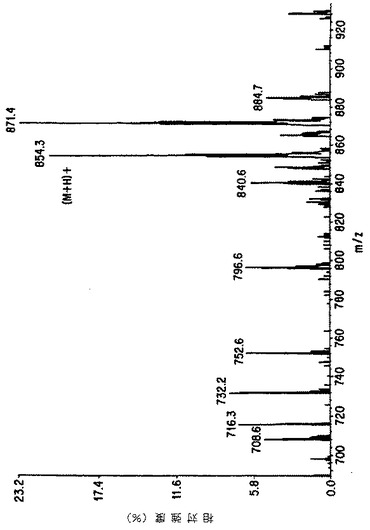
【図6D】
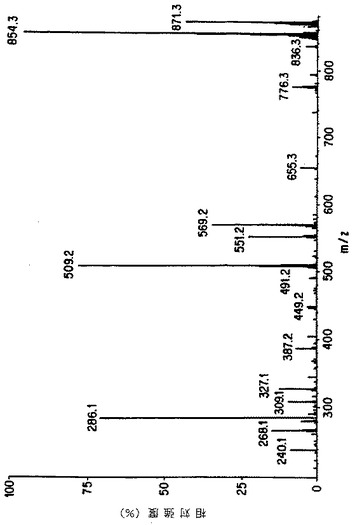
【図2】
【図3】
【図4A】
【図4B】
【図4C】
【図4D】
【図5A】
【図5B】
【図6A】
【図6B】
【図6C】
【図6D】
【公開番号】特開2011−212029(P2011−212029A)
【公開日】平成23年10月27日(2011.10.27)
【国際特許分類】
【出願番号】特願2011−170533(P2011−170533)
【出願日】平成23年8月3日(2011.8.3)
【分割の表示】特願2008−43232(P2008−43232)の分割
【原出願日】平成9年5月27日(1997.5.27)
【出願人】(507041179)ファイトン ホールディングス, インコーポレイテッド (5)
【Fターム(参考)】
【公開日】平成23年10月27日(2011.10.27)
【国際特許分類】
【出願日】平成23年8月3日(2011.8.3)
【分割の表示】特願2008−43232(P2008−43232)の分割
【原出願日】平成9年5月27日(1997.5.27)
【出願人】(507041179)ファイトン ホールディングス, インコーポレイテッド (5)
【Fターム(参考)】
[ Back to top ]