
タクロリムスの調製のための方法
ストレプトマイセス・ツクバエンシス(Streptomyces tsukubaensis)(S.ツクバエンシス)の遺伝子改変株は、これらの遺伝子改変株の培養によるタクロリムスまたはこの塩もしくは誘導体の調製のための改善された発酵方法のために使用されうる。アリルマロニルCoAの生合成を可能にする新規遺伝子は、アリルマロニル伸長単位でのポリケチド生成に使用されうる。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、免疫抑制化合物タクロリムスの調製のための方法に関する。微生物ストレプトマイセス・ツクバエンシス(Streptomyces tsukubaensis)(S.ツクバエンシス)の遺伝子改変株、およびこれらの遺伝子改変株を培養し、次いで免疫抑制化合物を単離することによるタクロリムス(FK−506)またはこの塩もしくは誘導体の調製のための改善された発酵プロセスも対象とする。
【0002】
さらに本発明は、アリルマロニルCoA生合成経路のための、詳細にはS.ツクバエンシス中でタクロリムス生合成遺伝子クラスターに位置する酵素をコードする新規遺伝子およびS.ツクバエンシスの遺伝子改変株であって、これらの遺伝子が過剰発現および/または不活性化されている新規遺伝子および遺伝子改変株に関する。
【背景技術】
【0003】
WO 2010/004304、MossらはFK−506またはFK−520を生成する株を記載している。
【0004】
FK−506(Fermentekカタログ番号506)とも称されるタクロリムスは、23員環マクロライドラクトンであり、ポリケチド群に属する。タクロリムスは、1980年代に土壌細菌ストレプトマイセス・ツクバエンシスの発酵培地から最初に単離された。抗生物質マクロライド化合物タクロリムスは、例えば1984年にKinoら(J.Antibiotics 40、1249−1255頁、1984)において報告された。その後タクロリムスは、さまざまな微生物、すなわちストレプトマイセス属種(Streptomyces sp)MA6858(US5,116,756)ATCC55098、ストレプトマイセス・ツクバエンシス NRRL18488(EP−B 0356399およびUS5,200,411)、ストレプトマイセス・クラブリゲルス(Streptomyces clavuligerus) CKD 1119(KR−B 100485877)またはストレプトマイセス・グラウセセンス(Streptomyces glaucescens)MTCC 5115(US2007 191415)などの土壌細菌を使用することにより微生物の天然生成物として調製された。
【0005】
生成物タクロリムスは、酵素ペプチジルプロピルイソメラーゼの活性を低下させるその効果およびタンパク質イムノフィリンFKBP12(FK506結合タンパク質)に結合するその効果により免疫抑制活性を示す。タクロリムスならびに構造的に類似しているポリケチド、アスコマイシンおよびラパマイシンは、生理学的に活性となるために高度に保存されたタンパク質シクロフィリンFKBP12への最初の結合を必要とする。ラパマイシン/FKBP12複合体はmTOR(ラパマイシンの哺乳類での標的)、細胞環境を感受するためおよび翻訳開始を制御するための中心制御器として作用すると考えられるセリン−スレオニンキナーゼへ結合する(例えば、Easton J.B.およびHoughton P.J.、2004、Expert Opin Ther Targets、8(6):551−64頁を参照されたい。)。しかしタクロリムス/FKBP12複合体は、シクロスポリンと同様に、さまざまな細胞標的に結合することが見出され、カルシニューリンのホスファターゼ活性を阻害する(Allison A.C.、2000、Immunopharmacology;47(2−3):63−83頁を参照されたい。)。
【0006】
タクロリムスは、例えば臓器移植後に免疫抑制のためにしばしば使用される。さらにタクロリムスおよびその誘導体は、喘息、炎症性疾患および過剰増殖性皮膚疾患など多数の疾患を治療することにおいて効果的であることが示されている。タクロリムスおよびラパマイシン、シクロスポリンなどの他の免疫抑制剤またはこれらの組合せも種々の自己免疫疾患の治療において有用である。長年にわたりカルシニューリン阻害剤(例えばシクロスポリンおよびタクロリムス)は、免疫抑制療法の中心である。これら2つの化合物は、細胞性免疫応答の強力な抑制因子であり、過去20年間における臓器移植の転帰を顕著に改善している(Allison A.C.、2000、Immunopharmacology;47(2−3):63−83頁を参照されたい。)。
【0007】
マクロライド、ラパマイシン、アスコマイシンおよびタクロリムスの遺伝子クラスターを含む、微生物由来の多数の医学的に重要な薬物の生合成経路をコードしている遺伝子クラスターは、既にクローン化され、配列決定されている。タクロリムス遺伝子クラスターのクローン化に関しては、ポリケチド合成酵素(PKS)をコードしている遺伝子を主に包含する部分配列が文献において報告された(Motamedi H.およびShafiee A.、1998、Eur J Biochem;256(3):528−34頁を参照されたい。)。一方、科学者らは、2000年にアスコマイシン遺伝子クラスターのクローン化を報告した(Wu Kら、2000、Gene;251(1):81−90頁、US6,503,737を参照されたい。)。タクロリムスは、構造的におよび生合成的起源についてアスコマイシン(FK520)およびラパマイシンに類似している(Reynoldsら、;Drugs and the Pharmaceutical Sciences,1997、82、497−520頁を参照されたい。)。これらすべては、ポリケチド(PKS)と非リボソーム性ペプチド生合成経路(NRPS)との組合せによって合成されうる(McDaniel Rら、2005、Chem Rev;105(2):543−58頁を参照されたい。)。
【0008】
タクロリムスおよびアスコマイシンは構造的に類似している。唯一の構造的差異としてタクロリムスの炭素21におけるアリル側鎖が、アスコマイシンにおいてはエチル側鎖によって置換されている。タクロリムス(FK506)およびアスコマイシン(FK520)化合物の構造は、式(Ia)および(Ib)に示されている。
【0009】
アスコマイシンおよびタクロリムスの構造は、生合成機構を考慮して4つのステップに分割されうる複合的生合成経路:
1.独特なシキミ酸由来スターターを使用する鎖の開始、
2.大部分のPKS由来化合物に共通の鎖の伸長、
3.L−ピペコリン酸の組み込みによる鎖の終結および環化ならびに
4.PKS後処理
を既に示唆している。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】国際公開第2010/004304号
【特許文献2】米国特許第5116756号明細書
【特許文献3】欧州特許第0356399号明細書
【特許文献4】米国特許第5200411号明細書
【特許文献5】韓国特許第100485877号明細書
【特許文献6】米国特許出願公開第2007/191415号明細書
【特許文献7】米国特許第6503737号明細書
【非特許文献】
【0011】
【非特許文献1】Kinoら、J.Antibiotics 40、1249−1255頁、1984
【非特許文献2】Easton J.B.およびHoughton P.J. 2004、Expert Opin Ther Targets、8(6):551−64頁
【非特許文献3】Allison A.C.、2000、Immunopharmacology;47(2−3)63−83頁
【非特許文献4】Motamedi H.およびShafiee A.、1998、Eur J Biochem;256(3)528−34頁
【非特許文献5】Wu Kら、2000、Gene;251(1)81−90頁
【非特許文献6】Reynoldsら、;Drugs and the Pharmaceutical Sciences,1997、82、497−520頁
【非特許文献7】McDaniel Rら、2005、Chem Rev;105(2):543−58頁
【発明の概要】
【発明が解決しようとする課題】
【0012】
タクロリムス発酵プロセスにおいて、望ましくないアスコマイシン(FK520)生成物も不純物として生成され、それによりタクロリムスの最終収量が低下し、タクロリムスの下流単離プロセスに著しい追加的経費を生じる。
【0013】
【化1】
【課題を解決するための手段】
【0014】
詳細には本発明は、薬剤の調製のためおよびヒトを含む哺乳類の治療のための上に定義した式(Ia)もしくは式(Ib)の化合物またはこれらの光学異性体、薬学的に許容される塩、水和物、溶媒和化合物もしくは多形体の使用に関する。本発明は、ヒトを含む動物における状態または疾患の予防および/または治療のための薬剤の調製のための化合物の使用に関する。
【0015】
エチルマロニルCoAは、アスコマイシン(FK520)生合成のための重要な基本単位として周知である。S.ハイグロスコピウス(S.hygroscopius)において見出された生合成アスコマイシンクラスターに位置し、アスコマイシン生合成に関与する以下の遺伝子は、最先端として報告されており(Liuら、Journal of Bacteriology、1999、181、6806−6813;Liuら、Metabolic Engineering、2001、3、40−48頁を参照されたい。)、関連があると考えられる:
a)クロトニルCoA還元酵素(ccr遺伝子)をコードする遺伝子、
b)B12依存性イソブチリルCoAムターゼ(icm遺伝子)をコードする遺伝子、
c)エチルマロニルCoAムターゼ(ecm遺伝子)をコードする遺伝子。
【0016】
いくつかのポリケチド由来化合物のポリケチド鎖アセンブリの特定の段階においてエチルマロニルCoA伸長単位を提供する生合成経路は、最先端として記載されている(Wuら、loc.cit;Reynoldsら、loc.citを参照されたい。)。エチルマロニルCoA伸長単位は、ブチリルCoAのカルボキシル化反応によって誘導されると考えられる。ブチリルCoAをもたらす少なくとも2つの経路がストレプトマイセスにおいて同定されている。一経路は2つの酢酸単位の縮合を含み、それによりアセト酢酸CoA活性化化合物を形成し、これはさらにクロトニルCoAに加工され、クロトニルCoA還元酵素(ccr)によって触媒されるクロトニルCoAからブチリルCoAへの還元の重要なステップを通じて処理される(Wallace K.K.ら、1995、Eur J Biochem;233:954−962頁を参照されたい。)。この遺伝子は、一次代謝遺伝子セット中に位置していることが見出されたS.コリヌス(S.collinus)において最初に同定された。後にその相同体が、エチルマロニルCoAを前駆体として必要とするいくつかの化合物をコードしている生合成クラスターにおいて同定された(Croppら、2001、J Ind Microbiol Biotechnol;27:368−377頁を参照されたい。)。
【0017】
近年、ロドバクター・スフェロイデス(Rhodobacter sphaeroides)中のccr相同体が、クロトニルCoAのエチルマロニルCoAへの還元カルボキシル化を触媒するクロトニルCoAカルボキシラーゼ/還元酵素活性を有する酵素をコードしていることが示された(Erb T.J.ら、2007、Proc Natl Acad Sci USA;25:10631−10636頁を参照されたい。)。さらにこの酵素は、イソクエン酸リアーゼ遺伝子を欠失している微生物の酢酸を唯一の炭素源とした増殖を可能にする「エチルマロニル−CoA経路」と称される新たに発見された酢酸同化経路の一部であることが示された。この経路は、いくつかのステップを含むと予測されている。先の予測のとおりこれは2つのアセチルCoA単位の縮合および続くクロトニルCoAへの加工によって開始される。次いでクロトニルCoAは、最初に(2R)−エチルマロニルCoAにエピマー化され、次にビタミンB12依存性エチルマロニルCoAムターゼ(ecm)によってメチルスクシニルCoAに転換される(2S)−エチルマロニルCoAにCcrによって直接転換される(Erb T.J.ら、2008、J Biol Chem;283:32283−32293頁を参照されたい。)。続いて、メチルスクシニルCoAは、メチルスクシニルCoA脱水素酵素によってメサコニルCoAに転換され、次のステップにおいてメサコニルCoAは、メサコニルCoA脱水酵素によってβ−メチルマリルCoAに転換される。最後に、β−メチルマリルCoAは、グリオキシル酸と、次に既知の代謝経路に入るプロピオニルCoAとに切断される。グリオキシル酸はアセチルCoAと縮合されてリンゴ酸を生じることができ、プロピオニルCoAはメチルマロニルCoAおよび次いでスクシニルCoAにカルボキシル化されうる。興味深いことにいくつかのストレプトマイセス属種は、ecm経路に関与するいくつかの遺伝子をクラスターとしてコードし、唯一の炭素源としての酢酸酪酸または脂肪酸での増殖の際にこの経路がストレプトマイセスにおいて活性であることを示唆している(Akopiants K.ら、J Ind Microbiol Biotechnol;33:141−150頁を参照されたい。)。
【0018】
バリン代謝物、イソブチリルCoAの異性化ステップを通じて進行し、ブチリルCoAを形成し、補酵素B12依存性イソブチリルCoAムターゼ(icm)によって触媒される第2経路が提案されている(Reynolds K.ら、1988、J Chem Soc Perkin Trans 1;3195−3207頁;およびZerbe−Burkhardt K.ら、1998、J Biol Chem;273:6508−6517頁を参照されたい。)。
【0019】
タクロリムス(FK506)生成生物に関してアリルマロニルCoA前駆体の代謝出発物およびその生合成に関与する遺伝子は、従来技術において同定されていない。
本発明は、アリルマロニルCoAの供給のための新規生合成経路を提供する。驚くべきことに、FK506ポリケチド合成酵素(PKS)クラスターの左腕に位置しているアリルマロニルCoA生合成に関与する遺伝子群が見出された。FK506生合成遺伝子クラスターの最も左側の新たに得られた配列は、アリルマロニルCoA生合成に関与する遺伝子をコードすることが見出され、タクロリムス遺伝子クラスターの「Allyl(All)クラスター」または「Allyl(All)サブクラスター」と称される(表1を参照されたい。)。
【0020】
本発明は、FK506ポリケチド合成酵素(PKS)遺伝子クラスターの左側に位置する、アリルマロニルCoA前駆体(基本単位)の生合成に関与する遺伝子を含有するこの新規ヌクレオチド配列に関する。
【0021】
FK506生成株の改善、具体的には収率および純度に関しての改善のためのこれらの配列の種々の使用も本発明に含まれる。本発明は、タクロリムス、アスコマイシンまたは関連化合物の選択的生成を可能にする不均一に発現された「Allylサブクラスター」を有する遺伝子改変株も提供する。
【0022】
アスコマイシンなどの副産物の形成を低減または消失させることによるおよび/またはタクロリムスの生成の収率を増加させることによるタクロリムス生成のための改善された方法を提供することは本発明の目的の1つである。
【0023】
アリルマロニルCoA対エチルマロニルCoAの伸長単位の利用能は、発酵プロセスの最後での所望の生成物タクロリムス(FK506)または副産物としてのアスコマイシン(FK520)それぞれの最終割合において重要な役割を演じることが見出された。したがって本発明は、所望の生成物と副産物との顕著に改善された割合をもたらす、前記伸長単位の割合を制御するいくつかの可能性を提供する。
【0024】
この態様において本発明は、遺伝子改変株によって、好ましくは生成物アスコマイシン(FK520)の生合成が顕著に低減されうるまたは消失されうるストレプトマイセス・ツクバエンシス(NRRL18488)によって実施されるタクロリムスの改善された発酵調製を記載する。好ましくはタクロリムスの収率は、一定のままであるまたは増加している。したがって本方法は、エチルマロニルCoA供給を低減する(もしくは消失させる)手段を使用することによっておよび/またはアリルマロニルCoA生合成を増加させることによって発酵生成の既知の方法の不利益を顕著に低減する。本方法は、アスコマイシン生成を低減させる(または消失させる)ことができ、タクロリムスの生成収率を維持または増加させうる。
【0025】
本方法の最後における一方でのタクロリムスおよび他方でのアスコマイシンの最終割合は、エチルマロニルCoAおよび/またはアリルマロニルCoAの代謝に関与するこれらの遺伝子の不活性化および/または過剰発現によって制御されうる。
さらに本出願は微生物の遺伝子改変株であって、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【図面の簡単な説明】
【0026】
【図1】ストレプトマイセス・ツクバエンシスの「All遺伝子」サブクラスター(15080bp)の遺伝子構成を示す図である。
【図2】ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891の抽出された発酵培地中のタクロリムスおよびアスコマイシンの存在をモニターするためのLC−MSクロマトグラム(MRM)を示すグラフである。アスコマイシンは、野生型株(列2)およびallA、allK、allRおよびallD遺伝子を過剰発現している変異株(列4)によって生成される。一方タクロリムスは、野生型株(列1)によって生成されず、allA、allK、allRおよびallDが過剰発現される場合(列3)にだけ生成される。
【発明を実施するための形態】
【0027】
本発明は、タクロリムスの調製のための方法であって、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養のステップを含み、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む方法を対象とする。
【0028】
本発明は、ストレプトマイセス属に属する微生物の遺伝子改変株であって、前記微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株も記載する。
詳細には本出願によるタクロリムスの調製のための方法は、以下のステップの少なくとも1つを含む:
a)微生物の遺伝子改変株の作製;
b)微生物の遺伝子改変株の栄養培地への添加および微生物の前記株を培養することを含む種培地の調製;
c)微生物の遺伝子改変株の種培地のバイオリアクター中の栄養培地への添加、微生物の前記株を培養することおよびタクロリムスの生成を含む主発酵;
d)発酵培地からの生成物の分離および精製(回収)。
【0029】
詳細には本発明によるタクロリムスの調製のための方法は、本出願に記載の微生物の遺伝子改変株の作製、種培地の調製および微生物の前記遺伝子改変株を培養することを含む主発酵を含む。好ましくは本出願によるタクロリムスの調製のための方法は、上に記載のステップa)からd)を含む。方法ステップa)からd)は下により詳細に記載される。
【0030】
好ましい実施形態において本発明は、上に記載のタクロリムスの調製のための方法であって、微生物の遺伝子改変株の培養ステップを含み、前記微生物が好ましくはストレプトマイセス属から選択される細菌である方法に関する。具体的には前記微生物は、アクチノマイセス目(Actinomycetales)から選択されうる。好ましいのは、PKS活性を有するまたは異種性に発現されるPKSもしくはこの一部を有するアクチノマイセス目に属する微生物である。より具体的には微生物は、ストレプトマイセス属から選択されうる。具体的にはストレプトマイセス・ツクバエンシス、ストレプトマイセス属種、ATCC55098またはストレプトマイセス・ハイグロスコピカス変種アスコマイセティカス(Streptomyces hygroscopicus var.ascomyceticus)ATCC14891が使用されうる。
【0031】
詳細には微生物は、ストレプトマイセス・ツクバエンシス、好ましくはストレプトマイセス・ツクバエンシス(NRRL18488)の遺伝子改変株から選択される。さらに微生物は、ストレプトマイセス・ハイグロスコピカスの遺伝子改変株から選択されうる。
【0032】
さらに本発明の方法を実施するために以下の微生物を使用することも検討される:ストレプトマイセス・ツクバエンシスNo.9993(Ferm BP−927)、ストレプトマイセス・ハイグロスコピカス亜種ハイグロスコピカス(Streptomyces hygroscopicus subsp.hygroscopicus)(DSM40822)、ストレプトマイセス属種AA6554、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6475 ATCC14891、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6678 ATCC55087、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6674、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC55276、ストレプトマイセス・ハイグロスコピカス亜種アスコマイセティカスATCC14891、ストレプトマイセス・カナマイセティカス(Streptomyces kanamyceticus)KCC S−0433、ストレプトマイセス・クラブリゲルス(Streptomyces clavuligerus)CKD1119、ストレプトマイセス・ハイグロスコピカス亜種ヤクシマエンシス(Streptomyces hygroscopicus subsp.yakushimaensis)、ストレプトマイセス属種DSM7348、ミクロモノスポラ(Micromonospora)属種(新種)A92−306401 DSM8429、ストレプトマイセス属種MA6548およびストレプトマイセス属種MA6858 ATCC55098。
【0033】
他の態様において本発明は、本出願に記載のタクロリムスの調製のための方法であって、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の条件下で実施される方法に関する。詳細にはアリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体は、上に記載の主発酵プロセス(ステップc)で発酵培地に添加される。
【0034】
さらに本出願に記載の発酵プロセスは、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくは本発明において提供されるストレプトマイセス・ツクバエンシスの遺伝子改変株を培養することおよび制御されたやり方でアリルマロニル−、エチルマロニル−および/もしくはプロピルマロニルCoAならびに/またはアリルマロニル−、エチルマロニル−および/もしくはプロピルマロニルCoAの前駆体を発酵培地に添加することを含みうる。これにより実質的に純粋なタクロリムス、アスコマイシンまたはそれぞれの関連化合物が得られる。この意味において、配列番号4に記載の不活性化された少なくともAllR遺伝子を有する、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株が使用される必要がある。
【0035】
他の実施形態において本出願に記載のタクロリムスの調製のための方法は、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養のステップであって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む培養のステップを含む。上に記載のヌクレオチド配列は、表1に記載されており別表に列挙されている。
【0036】
FK506の生合成のために必要なORF(オープンリーディングフレーム)をコードする遺伝子クラスター全体は配列決定された。PKS(ポリケチド合成酵素)遺伝子を含有するクラスターの中央部分の公表された配列(Motamedi H.およびShafiee A.1998;Eur J Biochem;256(3):528−34頁を参照されたい。)に加えて、ポリケチド合成酵素クラスターの左および右の周辺部に存在する遺伝子の配列が取得され、アノテーションされた。遺伝子クラスターの左側では、多数のORFが同定され、その大部分はアスコマイシン(FK520)生合成クラスターに存在する遺伝子にいかなる相同性も示さなかった(Wu.Kら、2000、Gene;251(1)81−90頁、US6503737を参照されたい。)。これらのORFをクエリーとして使用するBLAST検索は、表1に示すとおり同定された遺伝子の推定上の機能を明らかにした。遺伝子をコードするこのAllylクラスター(またはAllylサブクラスター)は、タクロリムスの生合成に特異的に必要なアリルマロニルCoA前駆体の生合成に関与する。さらに、遺伝子をコードするAllylサブクラスターは、エチルマロニルCoAの生合成に関与することが見出された。
【0037】
さらなる態様において、本発明は、配列番号2から配列番号10に記載のいくつかの遺伝子から構成される配列番号1に記載の配列および1つまたは複数のヌクレオチド追加、欠失、置換または逆位を含むこれらの変種を含むストレプトマイセス属に属する微生物のエチルマロニルCoAまたはアリルマロニルCoA生合成に関与するAllylサブクラスターのヌクレオチド配列に関する。Allylサブクラスターのヌクレオチド配列は表1に示されており、添付の別表に列挙されている(配列番号1から配列番号10)。
【0038】
【表1】
【0039】
変異株におけるエチルマロニルCoAおよびアリルマロニルCoAの合成を遮断することにより、これらの基本単位を供給することによってどのポリケチドが生成されるかを制御できる新規方法が可能であることが見出された。酸化還元酵素(クロトニルCoA還元酵素に類似)をコードするAllR遺伝子の不活性化は、タクロリムスの生成を完全に消失させたことが見出された。しかしアリルマロニルCoA前駆体の外部添加は、発酵培地中のアスコマイシン生成の消失を伴うタクロリムスの排他的生成を可能にする。AllR遺伝子の不活性化は、アスコマイシン(FK−520)の生成を完全に消失させたことも見出された。しかしエチルマロニルCoA前駆体の外部添加は、発酵培地中のタクロリムス生成の消失を伴うアスコマイシン(FK−520)の排他的産生を可能にする。この方法は任意のアスコマイシンまたはタクロリムス生成生物において可能である。アスコマイシン生成生物中でのエチルマロニルCoAの合成を遮断するための方法は、エチルマロニルCoA生合成に関与する遺伝子がアスコマイシン生成生物において解明されていることから当業者に周知である(Wuら、2000、Gene251、81−90頁)。ccr相同体の不活性化は、そのような生物の調製における最初のステップである。アスコマイシンおよびタクロリムス合成のためのPKSが高度に相同性であることから、アリルマロニルCoAまたはエチルマロニルCoAを取り込むモジュール7の無差別性がすべてのアスコマイシン/タクロリムス生成生物のPKSについて予測されうる。
【0040】
本発明の一実施形態において、本出願に記載のタクロリムスの調製のための方法は、ストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップであって、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される培養ステップを含む。
【0041】
さまざまなアリルマロニルCoA前駆体は使用されうる。より詳細にはアリルマロニルSNAC(アリルマロニル−(N−アセチル−システアミン)チオエステル)の添加が好ましい。本発明に関してアリルマロニルCoA前駆体はアリルマロニルCoAの類似体も含む。
【0042】
別法としてアリルマロン酸またはこの類似体は、前記方法のために使用される株の追加的遺伝子改変を考慮すると、前記方法において直接使用されうる。この場合マロニルCoA合成酵素およびジクサルボン酸キャリアタンパク質は、使用される微生物中で発現される必要がある。増殖培地中に加えられるジカルボン酸化合物の取り込みが可能である遺伝子改変微生物の一例は、Lomboら、Biotechnol.Prog.2001、17、612−617頁によって記載されている。基質としてのアリルマロン酸に対するマロニルCoA合成酵素の十分な活性は、Pohlら、J.Am.Chem.Soc.2001、123、5822−5823頁によって記載されている。
【0043】
詳細には上に記載の方法は、ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップであって、微生物の遺伝物質が
配列番号3に記載のAllK遺伝子および
配列番号4に記載のAllR遺伝子
の群から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む培養ステップを含む。
【0044】
本発明のさらなる実施形態において上に記載の方法は、ストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップであって、微生物の遺伝物質が配列番号1に記載の追加的不活性化遺伝子ccr遺伝子を含む培養ステップを含む。
【0045】
この詳細な態様において本発明は、ストレプトマイセス・ツクバエンシスの以下の株:
a)ストレプトマイセス・ツクバエンシスF499
受託番号DSM22507で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488ΔallRと記載される。
b)ストレプトマイセス・ツクバエンシスF872
受託番号DSM22509で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488ΔallKと記載される。
に関する。
【0046】
本発明のさらなる実施形態において上に記載の方法は、ストレプトマイセス属に属する微生物、好ましくはストレプトマイセス・ツクバエンシスの株の遺伝子改変株の培養ステップであって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターから選択される少なくとも1つの不活性化された遺伝子を含む培養ステップであって、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される培養ステップを含む。
【0047】
好ましい実施形態において上に記載の方法は、ストレプトマイセス属に属する微生物の遺伝子改変株の培養のステップであって、微生物の遺伝物質が配列番号3に記載のAllK遺伝子および配列番号4に記載のAllR遺伝子の群から選択される少なくとも1つの不活性化された遺伝子を含む培養ステップを含み、前記方法はアリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される。
好ましい実施形態において上に記載の方法は、ストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養のステップであって、微生物の遺伝物質が不活性化された配列番号4に記載のAllR遺伝子および不活性化された配列番号11に記載のccr遺伝子を含む培養ステップを含み、前記方法はアリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される。
【0048】
さらに本発明は、ストレプトマイセス属に属する微生物のエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する1つまたは複数の遺伝子をコードし、配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載の配列ならびに1つまたは複数のヌクレオチドの追加、欠失、置換および/または逆位を含むこれらの変種を含むヌクレオチド配列に関する。
【0049】
詳細には上に記載のヌクレオチド配列は、配列番号1に少なくとも50%のヌクレオチド同一性を有する。
【0050】
同様に本発明によって提供されるのは、上に記載のヌクレオチド配列であって、少なくとも60%、好ましくは少なくとも70%、好ましくは少なくとも80%、好ましくは少なくとも90%のヌクレオチド配列同一性を、配列番号2から配列番号10に記載のいくつかの遺伝子から構成される配列番号1に記載の配列に有するヌクレオチド配列である。ヌクレオチド配列の上に記載の変種は、ストレプトマイセス属に属する生物のエチルマロニルCoAまたはアリルマロニルCoAの生合成に関与する酵素をコードする能力を保持しているべきである。
【0051】
このように本発明は、表1に列挙し配列番号2から配列番号10に記載のすべての個々の遺伝子のヌクレオチド配列および配列番号13から配列番号21に記載のこれらのアミノ酸生成物ならびに1つまたは複数のヌクレオチド追加、欠失、置換および/または逆位を含むこれらの変種であって、少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも70%、好ましくは少なくとも80%、好ましくは少なくとも90%のヌクレオチド同一性を配列番号2から配列番号10に記載のいくつかの遺伝子から構成される配列による配列に有するヌクレオチド配列を提供する。
【0052】
提案された生合成経路は以下のとおり記載されうる。アリルマロニルCoAを得るためにC5前駆体、バレリルCoAまたは4−ペンテノイル−CoAが最初に形成されるべきである。アセト酢酸およびクロトニルCoAを通じて2つの酢酸単位から形成されるエチルマロニル−CoAと同様に、プロピルマロニルCoAまたはアリルマロニルCoAは1つの3炭素単位および1つの2炭素単位から生成できる。比較的短いポリケチド合成酵素(PKS)様酵素複合体によって単一ステップでの縮合反応を実行する、遺伝子AllA(アシル転移酵素)およびAllK(ケト合成酵素)は同定されている。さらに最終アリルマロニルCoA、C5前駆体に加工される前に5炭素中間体(3−オキソペンタン酸またはこの還元中間体)がどのようにCoAの添加によって活性化されるかまたはFK506PKS複合体に直接転移されるかは明確ではない。しかし最終合成アリル前駆体での供給実験は、伸長物を最終アリルマロニルCoA形態に組み込むFK506PKSの能力を示している。アセトアセチルCoA還元酵素に相同な推定上のAllS遺伝子は、3−ヒドロキシペンタノイルCoAを生じる最初の還元ステップを実行する。次いで追加的酸化還元酵素(AllR、クロトニルCoAカルボキシラーゼ/還元酵素、AllDおよびAllOと同様)は、この化合物をさらにプロピルマロニルCoAに還元でき、AllP、P450モノオキシゲナーゼと共にOH基および続いて二重結合も4位に導入でき、アリルマロニルCoAを生じる。次いでこれは対応するPKSモジュール4のアシル転移酵素(AT4)遺伝子によって新生ポリケチド鎖に導入される。Allylクラスターの一部として、AllMとして示されたORFが同定された。この遺伝子産物は、メチオニンγリアーゼに相同性を示し、プロピオニルCoA(最初の5炭素中間体のAllAによる生合成に必要な開始単位)に直ちに代謝される2−オキソ酪酸にメチオニンを転換すると考えられる。プロピオニルCoA前駆体は、2次代謝の間に限定されうる。したがってAllM遺伝子産物の存在は、最終産物FK−506の収率を増加させるためのプロピオニルCoAの十分な供給のために必要であると考えられる。
【0053】
提案された経路は、FK−506生成微生物の発酵培地中のエチルマロニルCoAの由来およびそれによりFK−520の由来も説明する。アリルマロニルCoAの生合成に関与するS.ツクバエンシスのFK−506生合成クラスターの≫Allylサブクラスター≪中に存在する遺伝子は、エチルマロニルCoAも生成できる。すなわちエノイルCoA還元酵素(遺伝子AllR)は、次いで上に記載の経路と同様のやり方で処理されるC5単位(3−オキソペンタン酸またはこの還元中間体)の代わりにアセト酢酸(C4)単位への限定的無差別性をほぼ確実に示す。次のステップは両前駆体について同様のやり方で進行するが、C4単位はP450調節水酸化および二重結合の形成に対する基質にはならないと考えられている。エノイルCoA還元酵素/カルボキシラーゼ(AllR)遺伝子の欠失は、この生合成経路ならびにFK−506およびFK−520の生成を完全に消失させることが見出された。ケトアシル合成酵素(AllK)遺伝子は、FK−506の産生だけを完全に消失させることが見出され、FK−520の産生は野生型S.ツクバエンシス株のレベルのままである。
【0054】
興味深いことに、FK−520/FK−506生合成のためのエチルマロニルCoAおよびアリルマロニルCoA(またはプロピルマロニルCoA)伸長単位を生じる上に記載の反応ステップは、「エチルマロニルCoA経路」の最初の反応からエチルマロニルCoA中間体が形成される点までに機構的に酷似している。この初期代謝経路は、大部分のストレプトマイセス属種の酢酸を唯一の炭素源とした増殖に最も高い可能性で関与し、クロトニルCoAカルボキシラーゼ/還元酵素(ccr)によって産生され、さらにエチルマロニルCoAムターゼ(ecm)によってメチルスクシニルCoAに転換されるエチルマロニルCoAを中間体化合物として含む。明らかに、所与の増殖条件下で活性であるならば、この経路がS.ツクバエンシス中でのエチルマロニルCoAの重要な供給源も表しうることが期待される。しかし、ccr(エチルマロニルCoAを産生する。)およびecm(これをさらに処理する。)を含む「エチルマロニルCoA経路」をコードする多数の遺伝子は、ストレプトマイセスにおいて同じオペロンに位置し、共転写されると考えられている。そのような遺伝子構造は、両遺伝子が協調して作用しなければならず、基質が最終産物への代謝経路を迅速に通過しなければならない場合には理想的であるが、単一の中間体(すなわち2次代謝のための基本単位としてのエチルマロニルCoA)の十分な量を提供するためには不十分である場合がある。これは、伸長単位としてエチルマロニルCoAを必要とするポリケチド生合成のための多数の遺伝子クラスターがccr遺伝子相同体(この内容において生成されたエチルマロニルCoAを直ちに消費するecm遺伝子は除く)の追加的コピーをも含有することの理由についての説明を提供できる。
【0055】
含まれる反応は機構的に類似しているが、S.ツクバエンシスでのFK−506およびFK−520生合成のための基本単位としてのアリルマロニルCoA(プロピルマロニルCoA)およびエチルマロニルCoAを提供する酵素と、この生物の「エチルマロニルCoA経路」の酵素とは1つの重要な観点において異なっている。「エチルマロニルCoA経路」はエチルマロニルCoA中間体を特異的に含み、ごく少量のプロピルマロニルCoAが最終的に形成される場合があるが、allサブクラスターにコードされる酵素は異なる特異性を示す。これらは、アリルマロニルCoA(プロピルマロニルCoA)を優先的に生成するが、エチルマロニルCoAは少量だけ産生される。経路のいくつかの酵素(すなわちAllR)は4炭素および5炭素中間体を同様の効率で受容できるが、他はアリルマロニルCoA(プロピルマロニルCoA)を排他的に生じる基質に対して非常に特異的である場合があることから恐らく量は比較的少ない。ストレプトマイセス酵素によるFK506の生成のための発酵プロセスとの関連で、両代謝系は(これらの発現が完全に異なる因子によってほぼ確実に誘導されるけれども)発現されうる。allサブクラスターの遺伝子はFK−506生合成クラスターの他の遺伝子と一緒に転写されると考えられるが、一度2次代謝が誘導されると「エチルマロニルCoA経路」酵素の発現および活性は増殖条件、最も重要なことには増殖培地の組成に依存すると考えられる。
【0056】
明確にはエチルマロニルCoAの濃度、すなわちFK−520の収率を可能な限り低く保つために、増殖培地の組成は「エチルマロニルCoA経路」の遺伝子が転写されないまたは最小レベルで転写されるように適合されるべきである。別法としてecm経路に加わりストレプトマイセスにおいて同じオペロン上に通常コードされる酵素の発現は、エチルマロニルCoA濃度を低く保持するために操作される必要がある。クロトニルCoAカルボキシラーゼ/還元酵素をコードするccr遺伝子は、不活性化されるべきである一方で、エチルマロニルCoAムターゼ(ecm)、メチルスクシニルCoA脱水素酵素、メサコニルCoA脱水酵素およびβ−メチルマリルCoAリアーゼをコードする他の4つの遺伝子は強力な構成的プロモーターを使用して過剰発現されるべきである。しかし、増殖条件および培地とは無関係に細胞中のエチルマロニルCoAの完全な消失が望ましい場合は、両方の経路を同じ機構的ステップで妨害する標的変異を導入する必要がある場合がある。本発明において記載のとおり、このステップは、ccr相同体、「エチルマロニルCoA経路」のccr遺伝子およびアリルマロニルCoA(プロピルマロニルCoA)の供給に主に関与するallサブクラスターのallR遺伝子の両方の不活性化でありうる。
【0057】
本発明の一目的はタクロリムスの調製のための方法であって、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップを含み、微生物の遺伝物質がアリルマロニルCoA対エチルマロニルCoAの細胞内プールの比率を改善するように改変されている方法を提供することである。
【0058】
この実施形態において本発明に記載のタクロリムスの調製のための方法は、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップであって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの過剰発現された遺伝子を含む培養ステップを含む。上に記載のヌクレオチド配列は、表1に記載されており添付の別表に列挙されている。
【0059】
本出願は、アリルマロニルCoAおよび/またはエチルマロニルCoA前駆体合成に必要なタンパク質をコードする、不活性化および/または過剰発現された表1に列挙した1つまたは複数の遺伝子を有する1つまたは複数のS.ツクバエンシス株の作製を含む。
【0060】
より詳細には本態様において本発明は、ストレプトマイセス・ツクバエンシスの遺伝子改変株の作製であって、前記株がタクロリムス(FK−506)および/またはアスコマイシン(FK−520)生合成のための重要な基本単位の供給に関与する少なくとも1つの不活性化された遺伝子を含む遺伝子改変株の作製を対象とする。これらの遺伝子改変株は、タクロリムスの改善された産生のために有用であることが見出されている。
【0061】
微生物の遺伝子改変のための方法、例えば遺伝子コピー数の増幅は当技術分野において周知である。他の例は、強力なプロモーターの制御下での前記遺伝子の追加的コピーの挿入である。さらにこの意味における他の例は「All」サブクラスターにおいて見出された転写制御因子の活性の改変である。
【0062】
この意味において本発明は、アリルマロニルCoA前駆体の細胞内プールを改善するおよびまたはエチルマロニルCoAの細胞内プールを低減することによるタクロリムスの発酵調製のためのストレプトマイセス・ツクバエンシスの改善された株を提供する。表1および配列番号1に記載の新規ヌクレオチド配列は、アリルマロニルCoAの生合成に関与する前記遺伝子の発現を制御するために使用されうる。
【0063】
本発明の一実施形態において本出願に記載のタクロリムスの調製のための方法は、微生物の遺伝子改変株の培養ステップであって、微生物の遺伝物質が配列番号11に記載のクロトニルCoA還元酵素(ccr)遺伝子および配列番号12に記載のエチルマロニルCoAムターゼ(ecm)から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む培養ステップを含む。ヌクレオチド配列は添付の別表に列挙されている。
【0064】
本出願は、ストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株であって、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0065】
詳細には本出願は、上に記載のストレプトマイセス属に属する微生物の遺伝子改変株であって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0066】
詳細には本発明は、上に記載のストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株であって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子および配列番号4に記載のAllR遺伝子の群から選択される少なくとも1つの過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0067】
詳細には本出願は、上に記載のストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株であって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子および配列番号3に記載のAllK遺伝子の群から選択される少なくとも1つの過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0068】
他に、さらに類似する実施形態においてアリルマロニルCoA前駆体の生合成に関与する遺伝子を含有する新規ヌクレオチド配列は、タクロリムスの生成のための組換え株の構築を当業者に可能にする。この意味で任意のアスコマイシン生成微生物がタクロリムスを生成するために操作されうる。この意味において適切な微生物は、これだけに限らないがストレプトマイセス・ハイグロスコピカスでありうる。これは、当技術分野において周知の方法による前記ヌクレオチド配列の前記宿主株への転移によって達成されうる。
【0069】
さらに本発明は、微生物の遺伝子改変株であって、天然のまたは操作されたポリケチド合成活性を有する微生物の遺伝子改変株を対象とする。
【0070】
本発明は、微生物の遺伝物質が
クロトニルCoA還元酵素の遠位相同体をコードする配列番号4に記載のAllR遺伝子、
チトクロムP450酵素をコードする配列番号8に記載のAllP遺伝子、
アシル転移酵素およびアシルキャリアタンパク質をコードする配列番号2に記載のAllA遺伝子、
ケトアシル合成酵素をコードする配列番号3に記載のAllK遺伝子、
アシルCoA脱水素酵素をコードする配列番号5に記載のAllD遺伝子、
メチオニンγリアーゼ酵素およびアシルキャリアタンパク質をコードする配列番号6に記載のAllM遺伝子、
転写制御因子をコードする配列番号7に記載のAllN遺伝子、
アシルCoA酸化還元酵素をコードする配列番号9に記載のAllO遺伝子および
アセトアセチルCoA還元酵素をコードする配列番号10に記載のAllS遺伝子、
の群から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含むストレプトマイセス・ツクバエンシスの遺伝子改変株に関する。
【0071】
遺伝子のヌクレオチド配列(DNA配列)および遺伝子産物のアミノ酸配列は、添付の別表のそれぞれ配列番号2から配列番号10および配列番号13から配列番号21に示されている。
【0072】
この詳細な態様において本発明は、ストレプトマイセス・ツクバエンシスの以下の株:
(a)ストレプトマイセス・ツクバエンシスF499
受託番号DSM22507で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 ΔallRと記載される。
(b)ストレプトマイセス・ツクバエンシスF872
受託番号DSM22509で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 ΔallKと記載される。
(c)ストレプトマイセス・ツクバエンシスF917
受託番号DSM22511で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 AllP+と記載される。
に関する。
【0073】
さらにアリルマロニルCoA前駆体の生合成に関与する遺伝子を含有するヌクレオチド配列は、新規ハイブリッドポリケチドを生成するために使用されうる。これは、当技術分野において周知の方法による前記ヌクレオチド配列の任意のポリケチド生成宿主株微生物への転移によって達成されうる。公知であるポリケチド合成酵素(PKS)モジュールの無差別性により、少なくとも1つの炭素位置にアリル側鎖を有する少なくとも痕跡量の新規ハイブリッドポリケチド化合物を期待できる。
【0074】
遺伝子改変(前記遺伝子の過剰発現または不活性化)のために使用されうる微生物は、細菌から選択されうる。より詳細には前記微生物は、アクチノマイセス目から選択される。好ましくはPKS(ポリケチド合成酵素)活性を有するアクチノマイセス目に属する微生物である。別法として過剰発現のための生物は、異種性に発現されたPKSまたはこの一部を有する任意の微生物であってよい。
【0075】
より詳細には微生物は、ストレプトマイセス属から選択される。具体的にはストレプトマイセス・ツクバエンシス、ストレプトマイセス属種ATCC55098またはストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891が使用されうる。
【0076】
さらに本発明の方法を実施するために以下の微生物を使用することも企図される:ストレプトマイセス・ツクバエンシスNo.9993(Ferm BP−927)、ストレプトマイセス・ハイグロスコピカス亜種ハイグロスコピカス(DSM40822)、ストレプトマイセス属種AA6554、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6475 ATCC14891、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6678 ATCC55087、ストレプトマイセス・ハグロスコピカス変種アスコマイセティカスMA6674、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC55276、ストレプトマイセス・ハイグロスコピカス亜種アスコマイセティカスATCC14891、ストレプトマイセス・カナマイセティカスKCCS−0433、ストレプトマイセス・クラブリゲルスCKD1119、ストレプトマイセス・ハイグロスコピカス亜種ヤクシマエンシス、ストレプトマイセス属種DSM7348、ミクロモノスポラ属種(新種)A92−306401 DSM8429、ストレプトマイセス属種MA6548およびストレプトマイセス属種MA6858 ATCC55098。
【0077】
一実施形態において本発明は、S.ツクバエンシス染色体上の特定のFK506遺伝子クラスターにクラスター化していない遺伝子の第2群の使用に言及する。これらの遺伝子は、ブチリルCoAおよびエチルマロニルCoA、アスコマイシン(FK520)の生合成のための前駆体の代謝に関与している。
【0078】
S.ツクバエンシスにおけるエチルマロニルCoA供給を減少させるまたは消失させるために、いくつかの戦略が行われた。この観点において本発明は、アスコマイシン生合成についての基本単位、エチルマロニルCoAの代謝に関与し、以下の酵素:
a)クロトニルCoA還元酵素(本明細書以下においてそれぞれccr、ccr遺伝子と称される。)
b)エチルマロニルCoAムターゼ(本明細書以下においてそれぞれecm、ecm遺伝子と称される。)
をコードする遺伝子のヌクレオチド配列の使用に関する。
【0079】
ヌクレオチド配列およびアミノ酸配列は、添付の別表に列挙されている(配列番号11、配列番号12を参照されたい。)。
【0080】
この態様において本出願は、タクロリムスの調製のための方法であって、微生物、好ましくはストレプトマイセス属の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップを含み、微生物の遺伝物質が配列番号11に記載のクロトニルCoA還元酵素(ccr)遺伝子および配列番号12に記載のエチルマロニルCoAムターゼ(ecm)遺伝子から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む方法を対象とする。
【0081】
詳細にはタクロリムスの調製のための方法は、微生物、好ましくはストレプトマイセス属、の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの株の培養ステップであって、微生物の遺伝物質が不活性化された配列番号11に記載のクロトニルCoA還元酵素(ccr)遺伝子を含む培養ステップを含む。
【0082】
詳細にはタクロリムスの調製のための方法は、微生物、好ましくはストレプトマイセス属、の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの株の培養ステップであって、微生物の遺伝物質が過剰発現された配列番号12に記載のエチルマロニルCoAムターゼ(ecm)遺伝子を含む培養ステップを含む。
【0083】
タクロリムスの調製のためのこの改善された方法は、アスコマイシン(FK520)の顕著に低下したまたは消失した生成を示し、これによりタクロリムス精製プロセスを著しく簡素化する。
【0084】
ccrおよびicmの遺伝子相同体は、ブチリルCoAおよび続くエチルマロニルCoAの生合成に関与していくつかの生物において同定された。補酵素B12依存性イソブチリルCoAムターゼ(icm;ブタノイルCoA:2−メチルプロパノイルCoAムターゼ、EC5.4.99.13)は、周知のメチルマロニルCoAムターゼ(mem)反応に密接に関連する反応においてイソブチリルCoAとn−ブチリルCoAとの間の可逆的転位を触媒する。クロトニルCoA還元酵素(EC.1.3.1.38、アシルCoA:NADP’−トランス−2−酸化還元酵素)は、クロトニルCoAのブチリルCoA(エチルマロニルCoA生合成のための重要な基質)への転換を触媒するまたはある環境下ではクロトニルCoAをエチルマロニルCoAに直接転換する。ecm(エチルマロニルCoAムターゼ)の遺伝子相同体は、エチルマロニルCoAをメチルスクシニルCoAに転換し、それにより混合物中でエチル置換された生成物の割合を効果的に変化させることが示されている。
【0085】
クロトニルCoA還元酵素遺伝子(ccr遺伝子)およびエチルマロニルCoAムターゼ遺伝子(ecm遺伝子)がさまざまな微生物においていくつかの代謝および生合成経路に関与することは周知である。ポリケチド生合成、詳細にはエチルマロニルCoAの生合成におけるecmおよびccrの役割は、記載されている(例えば、the role of ecm in production of monensin in S.cinnamonensis、Zhang W.ら、2001、J.Bacteriol;183:2071−2080頁)。
【0086】
したがって本発明は、エチルマロニルCoAの細胞内レベルが低下している微生物を作製する方法を記載する。これは、エチルマロニルCoAの生合成に関与する酵素を遮断することおよび/またはエチルマロニルCoAを消費する酵素を過剰発現することによって達成される。
【0087】
ストレプトマイセス・ツクバエンシスにおけるFK506/520の生合成のための重要な基本単位の生合成に特異的に関与するクロトニルCoA還元酵素(ccr)の活性が改変された。追加的に、エチルマロニルCoAのさらなる処理に関与するエチルマロニルCoAムターゼ(ecm)は、タクロリムス生成株ストレプトマイセス・ツクバエンシスにおいて過剰発現された。
【0088】
一実施形態において本出願は、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株であって微生物の遺伝物質が、
クロトニルCoA還元酵素をコードする配列番号11に記載のccr遺伝子および
エチルマロニルCoAムターゼをコードする配列番号12に記載のecm遺伝子
の群から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0089】
好ましい実施形態において本出願に記載の微生物の遺伝子改変株は、本出願に記載の2つの不活性化および/または過剰発現された遺伝子を含む二重改変微生物である。二重改変微生物は、野生型微生物および単一変異体と比較して大幅に改善された特徴を有することが見出された。
【0090】
クロトニルCoA還元酵素遺伝子相同体(ccr)の不活性化およびエチルマロニルCoAムターゼ(ecm)の過剰発現は、アスコマイシン生合成の顕著な低下によって特徴付けられるタクロリムスの生成のための改善された方法を導く。試験した増殖条件下では、ムターゼ触媒反応はブチリルCoAからイソブチリルCoAへ進行すると考えられる。エチルマロニルCoAムターゼ(ecm)遺伝子は、生成株中で過剰発現された場合にエチルマロニルCoAを効果的に消費することが見出された。
【0091】
一態様において本発明は、遺伝子改変微生物、好ましくはストレプトマイセス属の遺伝子改変微生物、より好ましくはストレプトマイセス・ツクバエンシスの株の遺伝子改変微生物であって、微生物の遺伝物質が不活性化された配列番号11に記載のクロトニルCoA還元酵素遺伝子(ccr)を含む遺伝子改変微生物を提供する。微生物中のそのような遺伝子を不活性化する方法は、例として手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり当技術分野において公知である。
【0092】
他の態様において本発明は、遺伝子改変微生物、好ましくはストレプトマイセス属の遺伝子改変微生物、より好ましくはストレプトマイセス・ツクバエンシスの株の遺伝子改変微生物であって、微生物の遺伝物質が過剰発現された配列番号12に記載のエチルマロニルCoAムターゼ遺伝子(ecm)を含む遺伝子改変微生物を提供する。微生物中のそのような遺伝子を過剰発現させる方法は、例として手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり当技術分野において公知である。
【0093】
さらなる態様において本発明は、上に記載のとおり不活性化された配列番号11に記載のクロトニルCoA還元酵素遺伝子(ccr)および過剰発現された配列番号12に記載のエチルマロニルCoAムターゼ遺伝子(ecm)を含む、遺伝子改変された微生物、好ましくはストレプトマイセス属の、より好ましくはストレプトマイセス・ツクバエンシスの株を提供する。この態様において、二重改変微生物の使用は、野生型微生物および上に記載の単一変異体と比較して大幅に改善された特徴を有する。
【0094】
具体的には本発明は、ストレプトマイセス・ツクバエンシスの以下の株:
a)ストレプトマイセス・ツクバエンシスF130
受託番号DSM22506で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 Δccr:Tsと記載される。
b)ストレプトマイセス・ツクバエンシスF879
受託番号DSM22510で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 ecm+と記載される。
に関する。
【0095】
新たに開発された株は、著しく低下したアスコマイシン含有量を示し、タクロリムス発酵生成の顕著な改善を表す。
【0096】
本発明によってさらに提供されるのは、配列番号11に記載のccr遺伝子配列、配列番号23に記載のアミノ酸配列を有するクロトニルCoA還元酵素(Ccr)をコードする配列および1つまたは複数のヌクレオチドの追加、欠失、置換または逆位を含むこれらの変種を含む、微生物ストレプトマイセス・ツクバエンシスNRRL18488のエチルマロニルCoA生合成に関与する遺伝子をコードする上に記載のヌクレオチド配列である。同様に本発明によって提供されるのは、配列番号12に記載のecm遺伝子配列、配列番号24に記載のアミノ酸配列を有するエチルマロニルCoAムターゼ(ecm)をコードする配列および1つまたは複数のヌクレオチドの追加、欠失、置換または逆位を含むこれらの変種を含む微生物ストレプトマイセス・ツクバエンシスNRRL18488のエチルマロニルCoA消費に関与する遺伝子をコードするヌクレオチド配列である。同様に本発明によって提供されるのは、上に記載のヌクレオチド配列であって、少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも70%、好ましくは少なくとも80%、好ましくは少なくとも90%の同一性を配列番号11および配列番号12に記載の配列に有するヌクレオチド配列である。
【0097】
本発明は、タクロリムスの調製のための改善された方法、具体的には産業用発酵方法であって、微生物の遺伝子改変株の培養のステップを含み、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む方法を提供する。前記方法は、上に記載の微生物の少なくとも1つの遺伝子改変株を使用することによって実施されうる。以下にこの方法をより詳細に記載する:
詳細には本発明による方法は以下のステップを含む。
【0098】
ステップa)微生物の遺伝子改変株の作製
好ましくはタクロリムスの生成のための本方法は、本発明によって記載される微生物株の遺伝子改変のステップを包含する。この意味において本発明は、具体的にはストレプトマイセス・ツクバエンシスの遺伝子改変株であって、株の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む遺伝子改変株に関する。
【0099】
本発明によるさらなる改変のためのストレプトマイセス・ツクバエンシスを作製するための手順は、当業者に周知である。さらにストレプトマイセス・ツクバエンシスの適切な株は、商業的に入手可能である。適切な株の具体的非限定的な一例は、受託番号NRRL18488を有する野生型ストレプトマイセス・ツクバエンシスである。
【0100】
微生物中の遺伝子を不活性化する方法は、例えば手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり当技術分野において公知である。
【0101】
好ましい実施形態において、微生物、好ましくはS.ツクバエンシスの株の遺伝子改変株の芽胞形成培地中での芽胞の調製、濃縮された芽胞懸濁物の調製のそれぞれが実施される。
【0102】
ステップb)種培地の調製
本発明に記載のストレプトマイセス・ツクバエンシスの改変株の培養は、当業者に周知の方法によって実施されうる。ストレプトマイセス・ツクバエンシスの培養プロセスは、例えば手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載されている。
好ましくはタクロリムス生成のための主発酵プロセスにおいて使用されうる種微生物の生成は、前記遺伝子改変微生物の芽胞形態から開始する。この点において本出願による方法は、記載された遺伝子改変微生物の、好ましくはストレプトマイセス・ツクバエンシスの芽胞貯蔵物の調製を含み、場合によって記載された遺伝子改変微生物の濃縮された芽胞懸濁物を含む。芽胞形態のこの調製は、マンガン塩を含む芽胞形成培地を使用するなどの当技術分野において周知の方法を使用して実施されうる。好ましくは微生物の遺伝子改変株のこの芽胞貯蔵物または濃縮芽胞懸濁物は、栄養培地への接種によって栄養種培地を作製するために使用される。記載の微生物の栄養形態の生成は、芽胞を含む比較的少量の種培地での接種を伴って調製されるべきである。
【0103】
種培地は、バイオリアクターに無菌的に移植されうる。
【0104】
原理上は種微生物の培養は主発酵プロセス(下のステップcに記載)と同様の条件下(例えばpHおよび温度)で実施されうる。
【0105】
ステップc)主発酵プロセス
好ましくは本出願において記載の遺伝子改変微生物を使用する主発酵プロセスは、バイオリアクター中で、詳細には撹拌および/または通気下で実施される。好ましくは本出願において記載のタクロリムスの生成のための方法は、同化炭素源、窒素源、リン酸源および鉱質源を含有する水性栄養培地(生成培地)中、液内好気性条件下で実施される。主発酵プロセスにおいて前記培地から生成されたタクロリムスの単離は、さらなる分離ステップ(ステップd)において実施されうる。
【0106】
好ましくは、主発酵プロセスは、ステップb)で得られた種微生物での生成培地の接種(詳細にはリアクターへの無菌的移植による)を含む。接種のために微生物の栄養形態を使用することは好ましい。
主発酵プロセスにおけるリアクターへの栄養培地(生成培地)の添加は、1回または複数回バッチ法でまたは継続的方法で実施されうる。栄養培地(生成培地)の添加は発酵プロセス前および/またはプロセス中に実施されうる。
【0107】
栄養培地中の好ましい炭素源は、下に例示のとおりデキストリン、ブドウ糖、可溶性デンプン、グリセロール、乳酸、麦芽糖、果糖、糖蜜およびショ糖から選択されうる。
【0108】
栄養培地中の好ましい窒素源は、酵母エキス、大豆ペプトン、大豆ミール、細菌ペプトン、カゼイン加水分解物、L−リジン、硫酸アンモニウム、コーンスティープリカーなどである。
【0109】
炭酸カルシウム、塩化ナトリウム、リン酸ナトリウムまたはリン酸カリウム、マグネシウム、マンガン、亜鉛、鉄および他の塩などの無機/鉱質塩も培地に添加されうる。
【0110】
本方法による主発酵プロセスは、約6.3から8.5のpH範囲内、20から35℃の温度範囲内で実施される。好ましくは、pHは約6.5から8.3の範囲内であり、温度は約23から31℃の範囲内である。
好ましくは、生成培養物は、80から約300時間、より好ましくは約130から280時間インキュベートされる。
【0111】
タクロリムスの生成は、生成培地の撹拌および通気を伴う好気性条件で実施されうる。培養混合物の撹拌および通気は、種々の方法で遂行されうる。生成培地の撹拌は、プロペラまたは類似の機械的装置によって提供されることができ、発酵条件および規模に応じてさまざまな程度に変更されうる。通気速度は、バイオリアクターの可動容積に対して1.0から2.5VVM(1分間あたり液体の単位容積あたりで流れる気体の容積(1分間あたり容積あたりでの容積))の範囲内で変更されうる。
【0112】
発酵プロセス用のさらなる周知の添加物は、具体的には主発酵プロセスにおいて添加されうる。培養培地の過度な起泡性を防ぐために、シリコン油、脂肪油、植物油などの消泡剤が添加されえた。具体的にはシリコンを基にする消泡剤が培養培地の過度な起泡性を防ぐために発酵プロセス中に添加されうる。
【0113】
ステップd)発酵培地からの生成物の分離および精製(回収)
発酵培地由来のタクロリムスは、生物学的に活性な物質の回収のために慣用される従来法によって分離および精製されうる。生成されたタクロリムスは、有機溶媒、すなわちアセトン、エタノールまたはメタノール中での抽出によって発酵培地から回収されうる。
【0114】
場合によって抽出物は次いで濃縮され、粗製タクロリムスを得るためにアセトン:水混合物を使用してXAD16吸着剤でカラムクロマトグラフィーされうる。粗製物質は、純粋なタクロリムスを得るために調製用HPLC上でさらに精製されうる。
【0115】
本出願による方法ステップの好ましい実施形態は、以下のとおり要約されうる:
i)固体芽胞形成アガー培地上で、S.ツクバエンシス株を28℃、10から14日間培養することによる前記S.ツクバエンシス株からのS.ツクバエンシス芽胞貯蔵物の調製。
ii)S.ツクバエンシス株の芽胞は濃縮芽胞懸濁物を作製するためおよび栄養培地600mlへの接種のために回収され、振とう機220rpmで24−32時間、栄養培地のPMV 10−20%が達成されるまでインキュベートした。回収時のpHは、6.8から7.3の範囲内である。
iii)生成培地に接種するための所望容量の10%までの種栄養接種物の増殖。栄養培地を含む10リットル発酵槽は、栄養種で接種され、24から28時間、28℃で培養される。
iv)栄養種培地の10%は生成培地100Lに接種するために使用される。発酵プロセスは、120から180時間、28℃で硫酸または水酸化ナトリウム溶液を使用してpHを7.0−7.2に維持して実施される。
【0116】
以下に非限定的な例として芽胞形成、種増殖および主発酵プロセスのための好ましい培地が記載される:
1)可溶性デンプン(10g/L)、塩化ナトリウム(1g/l)、硫酸アンモニウム(2g/L)、炭酸カルシウム(2g/L)、K2HPO4(1g/l)、MgSO4×7H2O(1g/L)、FeSO4×7H2O(0.001g/L)、MnCl2×4H2O(0.001g/L)、ZnSO4×7H2O(0.001g/L)および細菌学的アガー(20g/L)を含む固体アガー芽胞形成培地(ISP4)。pHは滅菌の前に塩基または酸の適切な添加によって7.0±0.2に調整されるべきである。
2)大豆ミール(2.5g/L)、デキストリン(10g/L)、ブドウ糖(1g/L)、酵母エキス(5g/L)、カゼイン加水分解物(7g/L)、K2HPO4(0.2g/L)、NaCl(0.5g/L)、MnCl2×4H2O(0.005g/L)、FeSO4×7H2O(0.025g/L)、ZnSO4×7H2O(0.001g/L)、MgSO4×7H2O(0.005g/L)およびCaCl2(0.02g/L)を含む種培地。pHは滅菌の前に塩基または酸の適切な添加によって7.0±0.2に調整されるべきである。
3)デキストリン(60−120g/L)、ブドウ糖(0−15g/L)、大豆ミール(5−20g/L)、大豆ペプトン(5−20g/L)、グリセロール(5−20g/L)、L−リジン(1−7.5g/L)、K2HPO4(0.5−2g/L)、CaCO3(1−5g/L)およびポリエチレングリコール(1−5g/L)を含有する生成培地。pHは、滅菌の前に塩基または酸の適切な添加によって7.0±0.2に調整されるべきである。
【0117】
本発明の他の好ましい実施形態において主発酵プロセス(ステップc)は、アリルマロニルCoAまたはアリルマロニルCoAの少なくとも1つの前駆体(類似体)の外部添加の下で実施される。本発明に関してアリルマロニルCoA前駆体は、アリルマロニルCoAの類似体も含む。
【0118】
この点において本発明は、タクロリムスの調製のための方法、具体的には産業用発酵方法であって、ストレプトマイセス属に属する微生物の遺伝子改変株の培養のステップを含み、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含み、アリルマロニルCoAおよびまたはアリルマロニルCoAの少なくとも1つの前駆体(類似体)の外部添加の下で実施される方法を提供する。
【0119】
詳細には本発明は、S.ツクバエンシスの遺伝子改変株を培養することによるタクロリムス(または関連化合物)の発酵調製であって、アリルマロニル−およびエチルマロニルCoA活性化前駆体を提供する1つまたは複数の生合成経路が、例えばAllR遺伝子の不活性化および発酵プロセス中にアリルマロニルCoAまたはエチルマロニルCoA SNAC前駆体を制御されたやり方で培養培地に添加または供給することによって、不活性化されているタクロリムスの発酵調製を対象とする。これは野生型株と比較して顕著な量でのタクロリムスまたはアスコマイシン化合物の産生を結果としてもたらす。したがって本明細書に記載の新規方法は、タクロリムスまたはアスコマイシンが排他的に産生される方法をもたらし、この方法は下流プロセスを顕著に簡素化する。
アリルマロニルCoAもしくはエチルマロニルCoAまたは本明細書に記載の少なくとも1つのこれらの前駆体(類似体)の外部添加の下での、ストレプトマイセス属に属する微生物の遺伝子改変株、具体的にはS.ツクバエンシスの遺伝子改変株を使用する純粋なタクロリムスまたは純粋なアスコマイシンの生成のための発酵プロセスは、既に記載されているものと同様に同化炭素源および窒素源を含有する水性栄養培地中において液内好気性条件下で実施されうる。
【0120】
好ましくはアリルマロニルまたはエチルマロニルエステル前駆体の標的濃度範囲は、0.1から5.0g/Lの範囲であり、より好ましくは0.5から3.0g/Lの範囲である。標的濃度は、有形データ記憶媒体に好ましくは発酵プロセスの開始前に記録されうる。供給プロセスについては、さまざまなアリルマロニルまたはエチルマロニルエステル前駆体が使用されうる。アリルマロニルまたはエチルマロニルN−アセチルシステアミン(SNAC)チオエステルは好ましい。1つのもしくは2つのチオエステルの両方またはこれらの混合物は使用されうる。
【0121】
発酵培地由来のタクロリムスは、生物学的に活性な物質の回収のために慣用される従来法によって分離および精製されうる。したがって前記方法は、下流プロセスにおいて調製用HPLCが使用される必要がある古典的方法と比較して顕著な費用優位性を有する。
【0122】
さらに本発明は、タクロリムス発酵生成のための方法における上に記載のストレプトマイセス・ツクバエンシスの遺伝子改変株の使用に関する。
【0123】
さらなる態様において本発明は、上に記載のストレプトマイセス・ツクバエンシスの遺伝子改変株の培養によって生成されたタクロリムスまたは薬学的に許容されるこの誘導体もしくは類似体を含む医薬組成物に関する。用語「類似体」または「誘導体」は、参照分子に構造的に類似しているが参照分子の1つまたは複数の特定の置換基を代替の置換基で置き換えるために標的化されおよび制御されたやり方で改変されており、それにより参照分子に構造的に類似している分子が作製されている分子を意味して、本明細書において従来の薬学的意味で使用される。追加的に当業者に周知の方法を使用して、改善された治療有効性、すなわち特定の標的受容体型でのより高い効力および/または選択性、哺乳類脳血管関門を透過する高いまたは低い能力のいずれか(例えばより高いまたはより低い脳血管関門透過率)、よりまれな副作用などを有する化合物タクロリムスの類似体および誘導体は作製されうる。
【0124】
本発明の組成物と関連して使用される句「薬学的に許容される」は、哺乳類、例えばヒトに投与された場合に生理学的に許容可能で、典型的には有害反応を生じない分子実体およびそのような組成物の他の成分を意味する。好ましくは本明細書において使用される用語「薬学的に許容される」は、連邦もしくは州政府の監督官庁によって認可されているまたは例えば米国薬局方もしくは哺乳類、より詳細にはヒトにおける使用について他の一般に認められた薬局方に列挙されていることを意味する。
【0125】
本発明のさらなる実施形態は、式(I)の少なくとも1つの化合物もしくはこの多形性形態またはこの薬学的に許容される塩および少なくとも1つのさらなる薬学的に許容可能な添加剤を含む医薬組成物である。医薬組成物は、さらなる薬物化合物も含みうる。本発明の医薬組成物において式(I)の化合物、詳細にはタクロリムスまたはこの多形性形態もしくは薬学的に許容される塩は、前記化合物またはこの薬学的に許容される塩を1日分の投与のための投与単位あたりで例えば0.1から4000mg、好ましくは1から2000mgを含有する投与単位として製剤される。本発明の全ての態様、具体的には医学的なものに関して化合物または組成物の投与は、最終的には主治医によって決定され、使用される化合物、動物種、性別、年齢、体重、症状の重症度、投与の方法、有害反応および/または他の禁忌症などの因子を考慮に入れる投与計画を有する。具体的に決定される投与量範囲は、患者の進行度および回復が完全に観察される標準的な設計の臨床試験によって決定されうる。そのような試験は、ヒトでの開始用量として動物での最大耐用量の低い百分率を使用する用量を漸増させる設計を使用する場合がある。
【0126】
本発明による生理学的に許容される化合物は、(成人患者に対して)連日投与計画で、例えば0.01mg/kg(治療される哺乳類の体重1kgあたりのmg)から100mg/kgの間、好ましくは0.1mg/kgから75mg/kgの間の経口投与量で通常投与される。
配列番号および遺伝子機能は表2に示される:
【0127】
【表2】
【実施例】
【0128】
以下の実施例は本発明を例示するためのものである。
【0129】
実験手順
以下は、「ccr」、「AllK」、「AllR」、「AllP」および「ecm」遺伝子をクローン化および分析するためならびにこれらの遺伝子相同体を使用するストレプトマイセス・ツクバエンシス変異体の作製のための実験手順の詳細な例である。同様に含まれるのは、これらの株を使用する発酵手順ならびにFK506およびFK520生成収率の判定の例である。分子生物学または微生物学の当業者に公知である標準的技術の追加的詳細および使用される具体的な酵素の名称は、例えば手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載されている。
【0130】
実施例1:
ストレプトマイセス・ツクバエンシス株の維持および芽胞調製
ストレプトマイセス・ツクバエンシス株の菌糸体を芽胞形成培地ISP4上で8−14日間、28℃でコンフルエント菌叢として増殖させた。ISP4培地は:
【0131】
【表3】
を含んでいた。
【0132】
pHは、1M NaOHで7.0に調整した。滅菌は、121±2℃、120±10kPaで20分間実施した。8−14日間の増殖後、芽胞を回収し、グリセロール(20%)中、−20℃で使用まで保存した。
【0133】
実施例2:
ストレプトマイセス・ツクバエンシスゲノムDNAの調製
ストレプトマイセス・ツクバエンシスNRRL18488(野生型)の芽胞を250mlエルレンマイヤーフラスコ中のTSB培地(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)50mlを接種するために使用し、28℃で24時間振とう(210rpm)させて維持した。培養物は、24時間28℃で増殖させた。菌糸体を遠心分離で回収し、ゲノムDNAをPureLink Genomic DNA Mini Kit(Invitrogen)をキット製造者の説明書に従って使用して調製した。DNAをTE緩衝液(SambrookおよびRussell、2000、Molecular Cloning:A Laboratory Manual、ISBN−978−087969577−4)100μlに再懸濁した。
【0134】
実施例3:
単離されたゲノムDNAをGATC AG、ドイツのRoche FLX technologyで配列決定した。FK506生合成遺伝子クラスターに属する配列連続断片が同定された場合、ORF(オープンリーディングフレーム)の位置および向きをFramePlot beta4.0 Software(Ishikawaら、FEMS Microbiol.Lett.174(1999)251−253頁)を使用して分析した。予測アミノ酸配列を使用して、相同性検索をGenBank databasesでBLASTpおよびBLASTxアルゴリズムを使用してNCBI(NCBI/Blast: http://blast.ncbi.nlm.nih.gov/Blast.cgi)で実行した。これらの結果に基づき、保存ドメイン検索によって支持された推定遺伝子機能を割り当てた。
合計15個のORFをfkbCから左に同定した。これらの内の6個はFK520クラスターに存在するORFに高い類似性を示し(fkbL−リジンシクロデアミナーゼ、fkbG、fkbH、fkbI、fkbJ、fkbK−遺伝子5個はメトキシマロニル−ACP生合成に関与する。)、9個の遺伝子は「アリルサブクラスター」を形成すると予測された。このサブクラスターは、アシル転移酵素およびアシルキャリアタンパク質ドメインを含有する遺伝子1個、ケトアシル合成酵素ドメインを含有する遺伝子1個、遠位クロトニルCoA還元酵素ドメイン1個、遠位アシルCoA脱水素酵素ドメイン含有相同体2個、メチオニンγリアーゼ含有遺伝子1個、AsnCファミリー制御遺伝子1個、P450モノオキシゲナーゼ遺伝子1個および遠位アセトアセチルCoA還元酵素相同体1個からなる。
【0135】
実施例4:
ccr遺伝子の破壊のためのベクターの構築、不活性化されたccr遺伝子を有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:ccr遺伝子の広領域の増幅のためのプライマーをストレプトマイセス・セリカラー(Streptomyces coelicolor)(タンパク質ID:NP_630556.1、遺伝子ID:1101912)由来のccr遺伝子を鋳型としたBLAST検索を使用して見出されたさまざまなストレプトマイセス属種の他の周知のccr遺伝子の保存領域および得られたBLAST結果のClustalWパイルアップ配列比較に基づいて設計した。したがってXbaI制限酵素部位を有するccr−F2(GTCTAGACCACATCATCGGCTCCGACC)およびEcoRI制限酵素部位を有するccr−R1(CGAATTCACGCCGACCTTGCCCTGGTGC)をccr遺伝子の917bp長領域を増幅するために作製した。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスのゲノムDNAをBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR反応はPfuポリメラーゼ(New England Biolabs)および製造者によって提供された緩衝液で、最終容量100μl中、200μM dNTP、10%グリセロール、0.5μMの各プライマー、鋳型S.ツクバエンシスゲノムDNAおよそ50ngおよび酵素2.5単位の存在下で30サイクル実施した。全30サイクルの温度プロファイルは、95℃、45秒間(変性ステップ)、69℃、45秒間(アニーリングステップ)および72℃、1分間(伸長ステップ)であった。PCR増幅産物をpUC19クローニングベクターにクローン化した。クローン化したPCR増幅産物の配列分析によって、それぞれの部分ccr配列を確認した。
c)相同組換えによるS.ツクバエンシスccr遺伝子の破壊において使用されるpKC1139に基づく温度感受性ベクターおよびpKC1132に基づく自殺ベクターの構築:ccr遺伝子のF2−R1増幅断片をpUC19からEcoRIおよびXbaI制限酵素部位によって切り出し、EcoRIおよびXbaI制限酵素で予め切断したベクターpKC1139およびpKC1132に連結反応によって移行させ、pKC1139−ccrおよびpKC1132−ccrを作製した。ベクターpKC1139は、E.コリ(E.coli)での複製のための正常なpUC19に基づくOriを含有しているが、34℃を超える高い温度では機能できないストレプトマイセスでの複製のための温度感受性Oriは含有していない(Biermanら、1992 Gene.116(1):43−9頁、Muthら、1989 Mol Gen Genet.219(3):341−348頁)。ベクターpKC1132はE.コリでの複製のための正常pUC19に基づくOriだけを含有するが、ストレプトマイセスでの複製を維持できず(Biermanら、1992 Gene.116(1):43−9頁)、したがって相同組換えによるゲノムへの組込みなしで失われる。次いでベクターpKC1139−ccr1およびpKC1132−ccrをccr遺伝子中で切断するBamHI制限酵素を使用して開き、917bp長断片を一方の346bpと他方の571bpとに分割し、次いでDNAポリメラーゼIラージ(Klenow)断片で末端平滑化した。次いでチオストレプトン(Ts)耐性を付与するカセットをpKC1139−ccrおよびpKC1132−ccr BamHI切断、末端平滑化ベクターに導入した。p330ベクターをXbaI制限酵素で消化し、Tsカセットを含有するおよそ961bp長断片をDNAポリメラーゼIラージ(Klenow)断片で末端平滑化し、pKC1139−ccrおよびpKC1132−ccrベクターに連結し、pKC1139−ccrTsおよびpKC1132−ccrTsベクターをそれぞれ作製した。
d)ccr遺伝子破壊のためのベクターのS.ツクバエンシスNRRL18488株への導入:プラスミド構築物pKC1139−ccrTsおよびpKC1132−ccrTsを、接合性プラスミドpUZ8002を含有するエレクトロコンピテントE.コリ株ET12567に形質転換によって導入した(Pagetら、1999 J.Bacteriol.181、204−211頁)。プラスミドpUZ8002は接合性線毛の構築のために必要な全ての遺伝子を含有しているが、転移の起点を欠いており、したがって宿主細胞に残留する(Jonesら、1997 Mol.Microbiol.23:169−178頁)。接合手順をKieserら、2000(Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり行った。ベクターpKC1139−ccrTsでの形質転換だけが接合完了体を生じた。接合完了体は、アプラマイシン50μg/mlを添加した実施例1に記載の芽胞形成培地上で、28℃で増殖させた。
e)チオストレプトン耐性付与カセットによって破壊されたccr遺伝子を有するS.ツクバエンシスの安定2次組換え株についての選択:アプラマイシン50μg/mlを添加した芽胞形成培地上で、28℃で増殖させたベクターpKC1139−ccrTsのS.ツクバエンシス接合完了体を、チオストレプトン(25μg/ml)を含む液体TSB培地に接種し、振とうフラスコ中、28℃、210rpmで、良好な種培養物を生成するためにアプラマイシンの添加をせずに増殖させた。24時間後、種培養物を、チオストレプトン(25μg/ml)を含む新鮮TSB培地を含む新たな振とうフラスコ中に継代培養し、24℃より高く温度を上昇させて210rpmで増殖させた。34℃より高い温度ではpKC1139に基づくベクターは、複製できず、S.ツクバエンシスゲノムに組み込まれ、それにより一次組換え体が生じる。培養物を、チオストレプトン(25μg/ml)を含む新鮮TSB培地を含む新たな振とうフラスコ中にさらに数回継代培養し、次いでチオストレプトンを含む芽胞形成培地に播種し、28℃で増殖させた。回収した芽胞をろ過し、系列希釈物を芽胞形成培地のプレート上に作製した。5−8日後、単一コロニーを、チオストレプトンを含む芽胞形成培地のプレート上およびチオストレプトンおよびアプラマイシンを含む芽胞形成培地のプレート上にパッチした。一次組換え体は、アプラマイシンにいまだ耐性である一方で2次組換え体はアプラマイシン耐性を欠失しており、アプラマイシンを含む芽胞形成培地上で増殖できない。選択された2次組換え株は次節に記載のサザンハイブリダイゼーションによっても確認した。
f)S.ツクバエンシスccr破壊変異体ゲノムDNAのサザンハイブリダイゼーションによる分析:DIG標識プラスミドDNAプローブの調製:プラスミドpKC1139−ccrTsの挿入部分(TsRカセットを挿入されたccr遺伝子の一部)だけをプローブとして使用し、ベクターをEcoRIおよびXbaI制限酵素で消化し、1744bp長断片をWizard SV GelおよびPCR Clean−Up System(Promega)で精製した。およそ1μgの溶出断片をDIG High Prime DNA Labelling and Detection Starter Kit I(Roche)を使用するランダムプライムド標識技術によってジゴキシゲニン−dUTPで一晩標識した。
【0136】
i)DNA移行:S.ツクバエンシス野生型のゲノムDNAおよびpKC1139−ccrTs変異体(推定1次組換え体および2次組換え体)をPureLink Genomic DNA Mini Kit(Invitrogen)をキットの製造者の説明書に従って使用して調製した。単離された各ゲノムDNAおよそ10μgをNaeI制限酵素で消化した。消化終了時にDNA断片を1%アガロースゲル中、20V、12時間の電気泳動によって分離し、正に荷電したHybond−N+nylon membrane(GE Healthcare)にアルカリキャピラリーブロッティング法(Southern,E.M.、1975、J.Mol.Biol.、98:503)を使用して6時間転写した。
【0137】
ii)ハイブリダイゼーション:ナイロン膜に固定化したDNAへのDIG標識DNAプローブのプレハイブリダイゼーションおよびハイブリダイゼーションをDIG High Prime DNA Labeling and Detection Starter Kit I製造者(Roche)によって示されたとおりに実施した。プレハイブリダイゼーションは、供給されたプレハイブリダイゼーション溶液中、1時間、50℃で実施した。ハイブリダイゼーション用に変性DNA(90℃、10分間)1μgを50℃に予め加熱した新鮮プレハイブリダイゼーション溶液に加えた。一晩のハイブリダイゼーション後に膜を以下のとおり洗浄した:2×SSC、0.1%SDS、室温、5分間で2回洗浄および0.5×SSC、0.1%SDS、55℃、15分間で2回洗浄。
【0138】
iii)免疫学的検出:ハイブリダイズしたプローブを抗ジゴキシゲニン−AP Fab断片で免疫検出し、比色分析基質NBT/BCIPでDIG High Prime DNA Labeling and Detection Starter Kit I供給者(Roche)によって示されたとおり可視化した。
【0139】
iv)予測されるバンドを分析し、pKC1139−ccrTsベクターから得られる真の2次組換え体を記載のサザンハイブリダイゼーション手順によって確認した。ccr破壊株S.ツクバエンシスF130をタクロリムスおよびアスコマイシン生成についてのさらなる検査のために選択した。
g)pKC1139−ccrTsを使用する2次相同組換えから得られたチオストレプトン耐性ccr破壊変異体の発酵タクロリムス生成。
【0140】
i)種培養プロセス:種培地は:
【0141】
【表4】
を含んでいた。
【0142】
pHを1M NaOHで7.0に調整した。ブドウ糖を含まないこの培地50mlを250mlエルレンマイヤーフラスコにろ過し、発泡栓で栓をして滅菌した。滅菌は121±2℃、120±10kPaで25分間実施した。滅菌後、滅菌したブドウ糖懸濁液を加えた。滅菌種培地50μg/mlチオストレプトンを加え、ストレプトマイセス・ツクバエンシス株の芽胞(1%v/v)を種培地に接種し、振とう機で28℃、250rpm、24−48時間、好気性条件下でインキュベートした。
【0143】
ii)主発酵プロセス:上記の種培養物のおよそ10%v/vを、50ml発酵培地を含む250mlエルレンマイヤーフラスコへの接種に使用した。発酵培地は:
【0144】
【表5】
を含んでいた。
【0145】
発酵培地のpHを滅菌前に1M NaOHで7.0に調整した。滅菌は121℃で25分間実施した。発酵は振とう機で28℃、250rpm、6−7日間実施した。
【0146】
h)pKC1139−ccrTsを使用した2次相同組換えから得られたチオストレプトン耐性ccr破壊変異体のHPLCでのタクロリムスおよびアスコマイシン生成の判定:
【0147】
タクロリムスおよびアスコマイシンの判定のための方法:タクロリムスまたはアスコマイシン生成の判定のための分析を適切なカラムおよび実行条件を使用する無勾配逆相HPLCによって実施した:カラム:Nucleosil−100 C18(150×4.0mm、粒径3μm)、流速1.5ml/分、T℃=60℃、移動相:560ml水、335mlアセトニトリル、70ml MTBEおよび0.2ml 85%H3PO4、検出210nm、試料注入20μl。
試料中のタクロリムスおよびアスコマイシン含有量の定量を、タクロリムスが12.5分で溶出され、アスコマイシンが11.5分で溶出されたタクロリムスおよびアスコマイシンの外部標準を使用して実施した。結果は試料中のタクロリムス生成と比較したアスコマイシン生成の%で表す。
【0148】
試料調製:十分に振とうした培地5mlにメタノール5mlを加え、試料を抽出するために試料を1時間振とう機に置いた。抽出後、培地のメタノール抽出物1mlを1.5mlチューブに取り、10分間、14000rpmで遠心分離した。上清0.8mlを、HPLC分析を実施するためにバイアルに移した。
【0149】
【表6】
【0150】
破壊されたccr遺伝子を有する単離された2次組換え変異体は、野生型株ストレプトマイセス・ツクバエンシスNRRL18488と比較して平均で40%のアスコマイシン生成の低下を示した。
【0151】
実施例5:
AllR遺伝子の破壊のためのベクターの構築、不活性化されたAllRを有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:実施例4に記載のとおり、454に基づく全ゲノム配列分析をS.ツクバエンシスのゲノムに使用し、それはAllR遺伝子に隣接する領域の既知のDNA配列に基づいてAllR遺伝子に隣接する領域の増幅のためのプライマーを設計することを可能にした。したがって、HindIII制限酵素部位を有するAllR−F1(CAAGCTTCACCGGTCCCGGGCTC)およびNdeI制限酵素部位を有するAllR−R1(GCATATGGTCCGGTTCGGGGGTGGG)をAllR遺伝子の上流領域を増幅するために作製し、NdeIを有するAllR−F2(GGGTCACATATGGCGAACTACCGGG)およびEcoRI制限酵素部位を有するAllR−R2(CGAATTCTGTGGGCCGACCTCACCCA)をAllR遺伝子の下流領域を増幅するために作製した。プライマーAllR−R1とAllR−F2との間、894bp(298アミノ酸)間隙を1335bp AllR遺伝子のほとんど全体の欠失のために作製した。2つの重複断片をNdeI制限酵素部位で組合せる場合、読み枠が保存され下流遺伝子への影響を最小にした標的AllR遺伝子の「インフレーム」欠失を可能にする。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAを実施例4のii節の記載と同じ条件を使用し、AllR−F1+AllR−R1およびAllR−F2+AllR−R2プライマー組合せを使用してBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR増幅産物をpUC19クローニングベクターにクローン化し、クローン化したPCR産物の配列分析によって、これらそれぞれのDNA配列を確認した。
c)相同組換えによるS.ツクバエンシスAllR遺伝子の破壊に使用するpKC1139に基づく温度感受性ベクターおよびpKC1132に基づく自殺ベクターの構築:AllR遺伝子に隣接する領域のF1−R1およびF2−R2増幅断片をそれぞれ制限酵素HindIIIとNdeIとによっておよびEcoRIとNdeIとによってpUC19から切り出した。ベクターpKC1139およびpKC1132は、予めEcoRIおよびHindIII制限酵素で切断し、2つの断片を組合せ、各標的ベクターに連結し、それぞれpKC1139−AllRおよびpKC1132−AllRを作製した。次いで実施例4のiv節に記載のとおりベクターpKC1139−AllRおよびpKC1132−AllRを、NdeI制限酵素を使用して開き、抗生物質耐性付与カセットを挿入した。AllRについてさまざまなカセットを使用した;チオストレプトン(Ts)耐性カセットは実施例4に記載のとおりp330ベクターから得た、エリスロマイシン(Er)耐性カセットはpIJ4026ベクターからベクターをEcoRIおよびXbaIで消化し、1690bp長断片を末端平滑化することによって得たならびにカナマイシン(Kn)耐性カセットはpSuperCos1(REF!)ベクターからベクターをSmaIおよびHindIIIで消化し1323bp長断片を末端平滑化することによって得た。各カセットを、開いたpKC1132−AllRおよびpKC1139−AllRベクターに連結し、それぞれpKC1132−AllRTs、pKC1132−AllREr、pKC1132−AllRKn、pKC1139−AllRTs、pKC1139−AllRErおよびpKC1139−AllRKnベクターを作製した。
d)AllR遺伝子の破壊のためのS.ツクバエンシスNRRL18488株へのベクターの導入:プラスミド構築物pKC1139−AllRTs、pKC1139−AllRErおよびpKC1139−AllRKnをS.ツクバエンシスNRRL18488株に実施例4dの記載と同じ接合手順によって導入した。
e)抗生物質耐性付与カセットによって破壊されたAllR遺伝子を有するS.ツクバエンシスの安定2次組換え株の選択:ベクターpKC1139−AllRTs、pKC1139−AllRErおよびpKC1139−AllRKn、pKC1139−ccrTsのS.ツクバエンシス接合完了体についてのチオストレプトン抗生物質付与カセットによって破壊されたAllR遺伝子を有するS.ツクバエンシスの安定2次組換え株の選択のための手順は、実施例6における記載と同様であった。継代培養を、対応する抗生物質(それに対する耐性カセットがAllR遺伝子に隣接する領域間に挿入されている。)を含む液体TSB培地で実施した。選択された2次組換え株は、もはやアプラマイシン中では増殖できないが、耐性カセットがAllR遺伝子に隣接する領域間に挿入されている第2の抗生物質中では増殖できた。
f)予め不活性化ccr遺伝子が付与されているS.ツクバエンシスF130株へのAllR遺伝子の破壊のためのベクターの導入:プラスミドpKC1139−AllRErおよびpKC1139−AllRKnを実施例4に記載のS.ツクバエンシスF130 ccr破壊株に、実施例4dの記載と同じ接合手順によって導入した。
g)チオストレプトン耐性付与カセットによって破壊されたccr遺伝子およびエリスロマイシン耐性付与カセットによって破壊されたAllR遺伝子を有するS.ツクバエンシスの安定2次組換え株の選択:破壊されたccrおよびAllR両遺伝子を有するS.ツクバエンシスF130の誘導体、S.ツクバエンシスの安定2次組換え株を選択するための手順は、実施例4eの記載と同様であった。継代培養を、対応する抗生物質(それに対する耐性カセットが、AllR遺伝子に隣接する領域間に挿入されている。)を含む液体TSB培地で実施した。選択された2次組換え株は、もはやアプラマイシン中では増殖できないが、チオストレプトン(ccr遺伝子を破壊している)、および耐性カセットが、AllR遺伝子に隣接する領域間に挿入されている、第2の抗生物質中では両方とも増殖できた。
h)pKC1139−AllRErを使用する2次相同組換えによって得たエリスロマイシン耐性AllR破壊変異体の発酵でのタクロリムス生成:エリスロマイシン耐性AllR破壊変異体についてのタクロリムス生成は、培養の開始時にエリスロマイシン50μg/mlを発酵培地に添加したことを除いて実施例4gに記載のとおり実施した。
i)ccrおよびAllR二重破壊変異体による発酵でのタクロリムス生成:エリスロマイシン耐性ccrおよびAllR破壊変異体についてのタクロリムス生成は、培養の開始時にチオストレプトンおよびエリスロマイシンを各50μg/mlの濃度で発酵培地に添加したことを除いて実施例4gに記載のとおり実施した。供給実験用にアリルマロニルCoAおよびエチルマロニルCoA前駆体、ジエチルアリルマロニルSNAC、アリルマロニルジSNACおよびエチルマロニルSNACを発酵培地に濃度5−20mMで添加した。
j)pKC1139−AllRErを使用する2次組換えによって得たエリスロマイシン耐性AllR破壊変異体のHPLCでのタクロリムスおよびアスコマイシン生成の判定:
エリスロマイシン耐性AllR破壊変異体のタクロリムスおよびアスコマイシン生成の判定のために実施例4hでの記載と同じ方法を使用した。
【0152】
【表7】
【0153】
破壊されたAllR遺伝子を有する単離された2次組換え変異体は、タクロリムス生成の完全な消失を生じた。アリルマロニルSNACまたはアリルマロニルジSNACを発酵培地に添加した場合タクロリムスの生成が再確立された。同様にエチルマロニルSNACを添加した場合アスコマイシン生成が再確立された。
k)ccrおよびAllR二重破壊変異体のHPLCでのタクロリムスおよびアスコマイシン生成の判定:
ccrおよびAllR二重破壊変異体のタクロリムスおよびアスコマイシン生成の判定のために、実施例4hに記載の方法を使用した。
【0154】
【表8】
【0155】
タクロリムスの完全な消失が不活性化されたccrおよびAllR遺伝子を有する単離された2次組換え変異体で生じた。アリルマロニルSNACまたはアリルマロニルジSNACを発酵培地に添加した場合、タクロリムスの生成が再確立された。同様にエチルマロニルSNACを添加した場合アスコマイシン生成が再確立された。
【0156】
実施例6:
allK遺伝子の破壊のためのベクターの構築、不活性化されたallKを有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:454に基づく全ゲノム配列決定をS.ツクバエンシスのゲノムに使用し、それはallK遺伝子に隣接する領域の既知のDNA配列に基づいてallK遺伝子に隣接する領域の増幅のためのプライマーを設計することを可能にした。それにより、EcoRI制限酵素部位を有するallK−F1(AGAATTCGTTACGGGGAGACGGCATCCCGG)およびallK−R1(AGGATCCGGGCGGGCTCGTCGCGGT)を、内部BamHI制限酵素部位を含むallK遺伝子の上流領域を増幅するために作製し、BamHIを有するallK−F2(TGGATCCGGCGCGTATCGCCAACCGCTAC)およびHindIII制限酵素部位を有するallK−R2(AAAGCTTCCCGGTAGTTCGCCATATGTGACCCG)をallK遺伝子の下流領域を増幅するために作製した。内部BamHI制限酵素部位とallK−F2との間、1698bp(566アミノ酸)間隙を、2388bp allK遺伝子のほとんど全体の欠失のために作製した。2つの重複断片をBamHI制限酵素部位で組合せる場合、読み枠が保存され下流遺伝子への影響を最小にしての標的allK遺伝子の欠失が可能になる。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAを実施例4bのii節での記載と同じ条件を使用し、allK−F1+allK−R1およびallK−F2+allK−R2プライマー組合せを使用してBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR増幅産物をpUC19クローニングベクターにクローン化し、クローン化したPCR産物の配列分析によって、これらそれぞれのDNA配列を確認した。
c)相同組換えによるS.ツクバエンシスallK遺伝子の破壊に使用するためのpKC1139に基づく温度感受性ベクターおよびpKC1132に基づく自殺ベクターの構築:allK遺伝子に隣接する領域のF1−R1およびF2−R2増幅断片をそれぞれ制限酵素EcoRIとBamHIとによっておよびHindIIとBamHIとによってpUC19から切り出した。ベクターpKC1139およびpKC1132は、予め制限酵素EcoRIおよびHindIIIで切断し、2つの断片を組合せ、各標的ベクターに連結し、それぞれpKC1139−allKおよびpKC1132−allKを作製した。
d)allK遺伝子のインフレーム破壊のためのベクターのS.ツクバエンシスNRRL18488株への導入:実施例4dでの記載と同じ接合手順によってプラスミド構築物pKC1139−allKをS.ツクバエンシスNRRL18488に導入した。
e)破壊されたallK遺伝子を有するS.ツクバエンシスの安定2次組換え株の選択:アプラマイシン50μg/mlを添加した芽胞形成培地上、28℃で増殖させたベクターpKC1139−allKのS.ツクバエンシス接合完了体を液体TSB培地に接種し、良好な種培養物を生成するためにアプラマイシン50μg/mlを添加して振とうフラスコ中、28℃、210rpmで増殖させた。24時間後、種培養物をアプラマイシン50μg/mlを添加した新鮮TSB培地を含む新たな振とうフラスコ中に継代培養し、34℃より高く温度を上昇させて210rpmで増殖させた。34℃より高い温度ではpKC1139に基づくベクターは複製できず、S.ツクバエンシスゲノムに組み込まれ、それにより一次組換え体が得られる。次いで培養物を、アプラマイシンを含まない新鮮TSB培地を含む新たな振とうフラスコ中に数回継代培養し、相同組換えによってpKC1139に基づくベクターが除去され、次いで芽胞形成培地に播種し、28℃で増殖させた。回収した芽胞をろ過し、系列希釈物を芽胞形成培地のプレート上に作製した。5−8日後、芽胞形成培地のプレート上およびアプラマイシンを含む芽胞形成培地のプレート上に単一コロニーをパッチした。一次組換え体は、アプラマイシンにまだ耐性である一方で2次組換え体はアプラマイシン耐性を失っており、アプラマイシンを含む芽胞形成培地上で増殖できない。allK遺伝子の所望のインフレーム欠失を有する真の2次組換え変異体を、実施例4fに記載のサザンハイブリダイゼーションを使用して野生型復帰変異体と照らし合わせて同定した。
f)pKC1139−allKを使用する2次相同組換えから得られたallK破壊変異体の発酵でのタクロリムス生成:allK破壊変異体についてタクロリムス生成を、抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
g)allK破壊変異体による発酵でのタクロリムス生成:タクロリムス生成allK破壊変異体を実施例4gに記載のとおり実施した。供給実験のためにアリルマロニルCoAおよびエチルマロニルCoA前駆体、アリルマロニルSNAC、アリルマロニルジSNACおよびエチルマロニルSNACを濃度5−20mMで発酵培地に添加した。
h)pKC1139−allKを使用する2次組換えから得られたallK破壊変異体のHPLCでのタクロリムスおよびアスコマイシン生成の判定:allK破壊変異体のタクロリムスおよびアスコマイシン生成の判定のために実施例4hの記載と同じ方法を使用した。
【0157】
【表9】
【0158】
破壊されたallK遺伝子を有する単離された2次組換え変異体は、タクロリムスおよびジヒドロタクロリムス生成の完全な消失を生じた。アリルマロニルSNACまたはアリルマロニルジSNACを発酵培地に添加した場合、タクロリムスの生成が再確立された。同様にエチルマロニルSNACを添加した場合、アスコマイシン生成が大幅に増加した。
【0159】
実施例7:
AllP遺伝子の過剰発現用のベクターの構築、過剰発現されたAllPを有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)AllP遺伝子の化学合成:454−全ゲノム配列決定に基づいてS.ツクバエンシスNRRL18488のゲノムのDNA配列を、ORFの5’末端にNdeI制限酵素部位およびORFの3’末端にXbaI制限酵素部位を付与するAllP遺伝子のDNA配列を設計および合成するために使用した。
b)AllP遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:合成したAllP遺伝子を制限酵素NdeIおよびXbaIによって切断した。Erm*プロモーター配列をpSE152ベクターのマルチクローニング部位にEcoRIおよびXbaI制限酵素部位を使用して予め導入し、NdeI部位をプロモーター配列の3’末端に作製した。このベクターをNdeIおよびXbaIで切断し、AllP遺伝子のNdeIおよびXbaI断片と連結してpSET−152−AllPプラスミドを作製した。
c)S.ツクバエンシスNRRL18488株へのAllP遺伝子の過剰発現:
プラスミド構築物pSET152−AllPをS.ツクバエンシスNRRL18488株に実施例4dに記載の接合手順によって導入した。
d)pSET152−AllPプラスミドを付与するS.ツクバエンシス変異体のタクロリムスおよびアスコマイシン生成の判定
タクロリムスおよびアスコマイシン生成の判定のためにHPLC法を実施例4hに記載のとおり使用した。
【0160】
【表10】
【0161】
AllP遺伝子が過剰発現された単離された組換え変異体は、野生型株ストレプトマイセス・ツクバエンシスNRRL18488と比較して25%増加したタクロリムス生成を示した。
【0162】
実施例8:
ecm遺伝子の過剰発現のためのベクターの構築、過剰発現されたecm遺伝子を有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)ecm遺伝子の化学合成:454−全ゲノム配列決定に基づいてS.ツクバエンシスNRRL18488のゲノムのDNA配列を、ORFの5’末端にNdel制限酵素部位およびORFの3’末端にXbaI制限酵素部位を付与するecm遺伝子のDNA配列を設計および化学合成するために使用した。
b)AllP遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:合成したecm遺伝子断片を制限酵素NdeIおよびXbaIによって切断した。Erm*プロモーター配列をpSET152ベクターのマルチクローニング部位にEcoRIおよびXbaI制限酵素部位を使用して予め導入し、NdeI部位を前記プロモーター配列の3’末端に追加した。このベクターをNdeIおよびXbaIで切断し、AllP遺伝子のNdeIおよびXbaI断片と連結してpSET−152−ecm構築物を作製した。
c)S.ツクバエンシスNRRL18488株へのecm遺伝子の過剰発現のためのベクターの導入:
プラスミド構築物pSET152−ecmをS.ツクバエンシスNRRL18488株に実施例4dに記載の接合手順によって導入した。
d)ecm過剰発現変異体におけるタクロリムスおよびアスコマイシン生成のHPLC分析
ecm過剰発現変異体のタクロリムスおよびアスコマイシン生成の判定のために実施例4hに記載のとおりの方法を使用した。
【0163】
【表11】
【0164】
過剰発現されたecm遺伝子を有する単離された組換え変異体は、野生型株ストレプトマイセス・ツクバエンシスNRRL18488と比較して20%減少したアスコマイシン生成を示した。
【0165】
実施例9:
allAおよびallK遺伝子の過剰発現のためのベクターの構築、このベクターを有するS.ツクバエンシスNRRL18488のallK不活性化株の相補性ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:実施例4に記載のとおり、454に基づく全ゲノム配列決定をS.ツクバエンシスのゲノムに使用し、それはallAからallD遺伝子のallサブクラスター領域の増幅のためのプライマーの設計を可能にした。それによりNdeI制限酵素部位を有するAT−exp−F1(ACATATGCTCGGGTCGTTCGTTACGGGGAG)およびXbaI制限酵素部位を有するAT―exp―R1(ATCTAGAACGTGGGTCATCGGCTGGTCCTTG)をこれら2つのORF(allAおよびallK)を増幅するために作製した。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAをBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR反応はPhusionポリメラーゼ(Finnzymes)および製造者によって提供された緩衝液で、最終容量50μl中の200μM dNTP、3%DMSO、0.5μMの各プライマー、鋳型S.ツクバエンシスゲノムDNAおよそ50ngおよび酵素2.5単位の存在下で30サイクル実施した。最初の5サイクルの温度プロファイルは、98℃、15秒間(変性ステップ)、65℃、30秒間(アニーリングステップ)および72℃、3分15秒間(伸長ステップ)であった。残りの25サイクルの温度プロファイルは98℃、15秒間(変性ステップ)、60℃、30秒間(アニーリングステップ)および72℃、3分15秒間(伸長ステップ)であった。PCR増幅産物をpUC19クローニングベクターにクローン化した。クローン化したPCR産物の配列分析によってallAおよびallK遺伝子の配列を確認した。
c)allAおよびallK遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:3.9kb挿入物を制限酵素NdeIおよびXbaIによってpUC19ベクターから切り出した。ErmE*プロモーター配列をpSET152ベクターのマルチクローニング部位にEcoRIおよびXbaI制限酵素部位を使用して予め導入し、NdeI部位をプロモーター配列の3’末端に作製した。このベクターをNdeIおよびXbaIで切断し、続いてallAおよびallK遺伝子を含むNdeIおよびXbaI断片と連結してpSET152−ermE*−allA−Kプラスミドを作製した。
d)実施例6において得た不活性化allK遺伝子を有するS.ツクバエンシス株へのallAおよびallK遺伝子の導入:プラスミド構築物pSET152−ermE*−allA−KをS.ツクバエンシスNRRL18488のallK不活性化株に実施例4dに記載の接合手順によって導入した。
e)pSET152−ermE*−allA−Kベクターで形質転換したallK破壊変異体の発酵でのタクロリムス生成:形質転換した変異体に関してタクロリムス生成を、抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
f)pSET152−ermE*−allA−Kプラスミドを付与されたallK不活性化S.ツクバエンシス変異体のタクロリムスおよびアスコマイシン生成の判定
【0166】
【表12】
【0167】
allK遺伝子活性の減失がallAおよびallKの過剰発現によって相補されたallK不活性化株の単離された組換え変異体は、回復したタクロリムス生成を示した。
【0168】
実施例10:
allA、allK、allR、allD遺伝子の過剰発現のためのベクターの構築、このベクターを有するS.ツクバエンシスNRRL18488のallR不活性化株の相補性ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:実施例4に記載のとおり、454に基づく全ゲノム配列決定をS.ツクバエンシスのゲノムに使用し、それはallAからallD遺伝子のallサブクラスター領域の増幅のためのプライマーの設計を可能にした。それによりNdeI制限酵素部位を有するAT−exp−F1(ACATATGCTCGGGTCGTTCGTTACGGGGAG)およびXbaI制限酵素部位を有するAT―exp―R2(ATCTAGAACATGCCCTAGGTACGTTTCGCGG)をこれら4つのORF(allA、allK、allRおよびallD)を増幅するために作製した。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAをBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR反応はPhusionポリメラーゼ(Finnzymes)および製造者によって提供された緩衝液で、最終容量50μl中の200μM dNTP、3%DMSO、0.5μMの各プライマー、鋳型S.ツクバエンシスゲノムDNAおよそ50ngおよび酵素2.5単位の存在下で30サイクル実施した。最初の5サイクルの温度プロファイルは、98℃、15秒間(変性ステップ)、65℃、30秒間(アニーリングステップ)および72℃、3分15秒間(伸長ステップ)であった。残りの25サイクルの温度プロファイルは98℃、15秒間(変性ステップ)、60℃、30秒間(アニーリングステップ)および72℃、3分15秒間(伸長ステップ)であった。PCR増幅産物をpUC19クローニングベクターにクローン化した。クローン化したPCR産物の配列分析によってallA、allK、allRおよびallD遺伝子の配列を確認した。
c)allA、allK、allRおよびallD遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:6.4kb挿入物を制限酵素NdeIおよびXbaIによってpUC19ベクターから切り出した。ErmE*プロモーター配列をpSET152ベクターのマルチクローニング部位にEcoRIおよびXbaI制限酵素部位を使用して予め導入し、NdeI部位をプロモーター配列の3’末端に作製した。このベクターをNdeIおよびXbaIで切断し、続いてallA、allK、allRおよびallD遺伝子を含むNdeIおよびXbaI断片と連結してpSET152−ermE*−allA−Dプラスミドを作製した。
d)実施例5において得た不活性化allR遺伝子を有するS.ツクバエンシス株へのallA、allK、allRおよびallD遺伝子の導入:プラスミド構築物pSET152−ermE*−allA−DをS.ツクバエンシスNRRL18488のallR不活性化株に実施例4dに記載の接合手順によって導入した。
e)pSET152−ermE*−allA−Dベクターで形質転換したallR不活性化変異体の発酵でのタクロリムス生成:形質転換した変異体に関してタクロリムス生成を、抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
f)pSET152−ermE*−allA−Dプラスミドを付与されたallR不活性化S.ツクバエンシス変異体のタクロリムスおよびアスコマイシン生成の判定
【0169】
【表13】
【0170】
allR遺伝子活性の減失がallA、allK、allRおよびallDの過剰発現によって相補されたallR不活性化株の単離された組換え変異体は、タクロリムスおよびアスコマイシンの回復した生成を示した。
【0171】
実施例11:
プラスミドpSET152−ermE*−allA−Dのストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891株への導入、この株でのallA、allK、allRおよびallD遺伝子の過剰発現ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)allA、allK、allRおよびallD遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:pSET152−ermE*−allA−Dプラスミドを実施例10に記載のとおり調製した。
b)allA、allK、allRおよびallD遺伝子の、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891への導入:プラスミド構築物pSET152−ermE*−allA−Dを、接合性プラスミドpUZ8002を含有するエレクトロコンピテントE.コリ株ET12567への形質転換によって導入した(Pagetら、1999 J.Bacteriol.181、204−211頁)。プラスミドpUZ8002は接合性線毛の構築に必要な全ての遺伝子を含有しているが、転移の起点を欠いており、したがって宿主細胞に残留する(Jonesら、1997 Mol.Microbiol.23:169−178頁)。ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891の接合手順をKieserら、2000(Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり行った。接合完了体は、アプラマイシン50μg/mlを添加した実施例1に記載の芽胞形成培地上、28℃で増殖させた。
c)pSET152−ermE*−allA−Dベクターで形質転換した株の発酵でのアスコマイシンおよびタクロリムス生成:形質転換した変異体のアスコマイシンおよびタクロリムス生成のための発酵プロセスは抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
d)allA、allK、allRおよびallD遺伝子を過剰発現しているストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891変異体のLC−MS/MSでのタクロリムスおよびアスコマイシンの生成の判定:
【0172】
試料調製:十分に振とうした培地5mlにメタノール5mlを加え、試料を抽出するために試料を1時間振とう機に置いた。抽出後、培地のメタノール抽出物1mlを1.5mlチューブに取り、10分間、14000rpmで遠心分離した。上清0.8mlをLC−MS/MS分析を実施するためにバイアルに移した。
【0173】
タクロリムスおよびアスコマイシン判定のための方法:培養培地中のFK506およびFK520の同一性をLC−MS/MS分析によって判定した。本発明者らは、Phenomenexからの逆相カラム(Gemini C18 column、5μm、150mm×2mm i.d.)を使用するWatters Micromass Quattro micro detectorと共にAgilent 1100 series LC−MS systemを使用した。分離は、流速0.250ml/分、溶媒Aとして0.5%TFA、溶媒Bとしてアセトニトリルでの勾配溶出によって実施した。勾配プログラムは:60%A、0分;60−20%A、0−17分;20−60%A、17−18分;60%A、18−30分であり、注入容量10μL、カラム温度45℃を使用した。質量選択検出器(Waters、Quattro micro API)は分析物の陽イオン化にコーン電圧20Vおよびキャピラリー電圧3.5kVを使用するエレクトロスプレーイオン化を備えていた。乾燥窒素を350℃に加熱し、乾燥ガス流量は400l/時間に、衝突エネルギーは20eVにした。ESI+ポジティブモードにおいては、FK506のナトリウムイオン([M+Na]+)の捕獲に対応するm/z=826.5のイオンが最も強く、他の調査者の結果と一致した(Yuan J.ら、2008、J.Chromatogr.、B.Anal.Technol.Biomed Life Sci.868、34−41頁を参照されたい。)。FK506同一性確認に関して、多重反応モニタリングモードを使用し、遷移FK506 m/z=826.5[M+Na]+→m/z=616.4を記録した。同様にFK520モニタリングについても遷移m/z=814.5[M+Na]+→m/z=604.4を記録した。タクロリムスはおよそ18.2分に溶出され、アスコマイシンはおよそ17.5−17.7分に溶出された。FK506とFK520との存在量は判定できず、したがって両化合物の存在または不在だけが確立された(図2)。
【0174】
【表14】
【0175】
allA、allK、allRおよびallD遺伝子が過剰発現されているストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891の単離された組換え変異体は、この株で通常生成されるアスコマイシンに加えてタクロリムスの生成も示した。
【0176】
実施例12:
allサブクラスターの大部分(allDからallS)の欠失のためのベクターの構築、allサブクラスターのこの部分を欠失しているS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:実施例4に記載のとおり、454に基づく全ゲノム配列決定をS.ツクバエンシスのゲノムに使用し、それはallD遺伝子の上流およびallS遺伝子の下流の領域の既知のDNA配列に基づいてallDとallS遺伝子との間の領域に隣接する領域の増幅のためのプライマーを設計することを可能にした。これにより、BamHI制限酵素部位を有するKS−delF2b(TGGATCCGGCGCGTATCGCCAACCGCTAC)およびXbaI制限酵素部位を有するAllRexpR1(ATCTAGAGGCGTTCGGATTCGCTCACCG)をallD遺伝子の上流領域を増幅するために作製し、XbaIを有するClusDelF2(ATCTAGAGGGCGGATACGTCACCGGCG)およびHindIII制限酵素部位を有するClusDelR2b(CCAAGCTTATGGAGATCATCGAAGGCAGC)をallS遺伝子の下流領域を増幅するために作製した。プライマーAllRexpR1とClusDelF2の間、6262bpの間隙をallD、allM、allN、allP、allOおよびallS遺伝子の欠失のために作製した。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAをBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR反応はPhusionポリメラーゼ(Finnzymes)および製造者によって提供された緩衝液で、最終容量50μl中の200μM dNTP、3%DMSO、0.5μMの各プライマー、鋳型S.ツクバエンシスゲノムDNAおよそ50ngおよび酵素2.5単位の存在下で30サイクル実施した。最初の5サイクルの温度プロファイルは、98℃、15秒間(変性ステップ)、65℃、30秒間(アニーリングステップ)および72℃、1分間(伸長ステップ)であった。残りの25サイクルの温度プロファイルは98℃、15秒間(変性ステップ)、60℃、30秒間(アニーリングステップ)および72℃、1分間(伸長ステップ)であった。PCR増幅産物をpUC19クローニングベクターにクローン化した。クローン化したPCR産物の配列分析によって増幅された上流および下流領域の配列を確認した。
c)相同組換えによるS.ツクバエンシスゲノムのallD−allS領域の破壊に使用するpKC1139に基づく温度感受性ベクターの構築:隣接領域のKS−delF2b−AllRexpR1およびClusDelF2−ClusDelR2の増幅断片をそれぞれ制限酵素BamHIとXbaIとによっておよびXbaIとHindIIIとによってpUC19から切り出した。ベクターpKC1139は、予め制限酵素BamHIおよびHindIIIで切断し、2つの断片を組合せ、各標的ベクターに連結し、pKC1139−ClusDelを作製した。
d)allサブクラスターのallD−allS領域の欠失のためのベクターの、S.ツクバエンシスNRRL18488株への導入:プラスミド構築物pKC1139−ClusDelをS.ツクバエンシスNRRL18488株に実施例4dの記載と同じ接合手順によって導入した。
e)allD−allS領域を欠失しているS.ツクバエンシスの安定2次組換え株の選択:2次組換え体の選択は、実施例6dの記載と同じ手順を使用して実施した。
f)pKC1139−ClusDelを使用する2次相同組換えによって得たallD−allS領域を欠失している変異体の発酵でのタクロリムス生成:allD−allS欠失変異体のタクロリムス生成のための発酵プロセスを、抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
g)pKC1139−ClusDelを使用する2次組換えによって得たallD−allS欠失変異体のHPLCでのタクロリムシスおよびアスコマイシン生成の判定:allD−allS欠失変異体のタクロリムスおよびアスコマイシン生成の判定のために、実施例4hにおける記載と同じ方法を使用した。
【0177】
【表15】
【0178】
破壊されたallD−allS領域を有する単離された2次組換え変異体は、タクロリムスおよびアスコマイシン生成の完全な消失を生じた。
【技術分野】
【0001】
本発明は、免疫抑制化合物タクロリムスの調製のための方法に関する。微生物ストレプトマイセス・ツクバエンシス(Streptomyces tsukubaensis)(S.ツクバエンシス)の遺伝子改変株、およびこれらの遺伝子改変株を培養し、次いで免疫抑制化合物を単離することによるタクロリムス(FK−506)またはこの塩もしくは誘導体の調製のための改善された発酵プロセスも対象とする。
【0002】
さらに本発明は、アリルマロニルCoA生合成経路のための、詳細にはS.ツクバエンシス中でタクロリムス生合成遺伝子クラスターに位置する酵素をコードする新規遺伝子およびS.ツクバエンシスの遺伝子改変株であって、これらの遺伝子が過剰発現および/または不活性化されている新規遺伝子および遺伝子改変株に関する。
【背景技術】
【0003】
WO 2010/004304、MossらはFK−506またはFK−520を生成する株を記載している。
【0004】
FK−506(Fermentekカタログ番号506)とも称されるタクロリムスは、23員環マクロライドラクトンであり、ポリケチド群に属する。タクロリムスは、1980年代に土壌細菌ストレプトマイセス・ツクバエンシスの発酵培地から最初に単離された。抗生物質マクロライド化合物タクロリムスは、例えば1984年にKinoら(J.Antibiotics 40、1249−1255頁、1984)において報告された。その後タクロリムスは、さまざまな微生物、すなわちストレプトマイセス属種(Streptomyces sp)MA6858(US5,116,756)ATCC55098、ストレプトマイセス・ツクバエンシス NRRL18488(EP−B 0356399およびUS5,200,411)、ストレプトマイセス・クラブリゲルス(Streptomyces clavuligerus) CKD 1119(KR−B 100485877)またはストレプトマイセス・グラウセセンス(Streptomyces glaucescens)MTCC 5115(US2007 191415)などの土壌細菌を使用することにより微生物の天然生成物として調製された。
【0005】
生成物タクロリムスは、酵素ペプチジルプロピルイソメラーゼの活性を低下させるその効果およびタンパク質イムノフィリンFKBP12(FK506結合タンパク質)に結合するその効果により免疫抑制活性を示す。タクロリムスならびに構造的に類似しているポリケチド、アスコマイシンおよびラパマイシンは、生理学的に活性となるために高度に保存されたタンパク質シクロフィリンFKBP12への最初の結合を必要とする。ラパマイシン/FKBP12複合体はmTOR(ラパマイシンの哺乳類での標的)、細胞環境を感受するためおよび翻訳開始を制御するための中心制御器として作用すると考えられるセリン−スレオニンキナーゼへ結合する(例えば、Easton J.B.およびHoughton P.J.、2004、Expert Opin Ther Targets、8(6):551−64頁を参照されたい。)。しかしタクロリムス/FKBP12複合体は、シクロスポリンと同様に、さまざまな細胞標的に結合することが見出され、カルシニューリンのホスファターゼ活性を阻害する(Allison A.C.、2000、Immunopharmacology;47(2−3):63−83頁を参照されたい。)。
【0006】
タクロリムスは、例えば臓器移植後に免疫抑制のためにしばしば使用される。さらにタクロリムスおよびその誘導体は、喘息、炎症性疾患および過剰増殖性皮膚疾患など多数の疾患を治療することにおいて効果的であることが示されている。タクロリムスおよびラパマイシン、シクロスポリンなどの他の免疫抑制剤またはこれらの組合せも種々の自己免疫疾患の治療において有用である。長年にわたりカルシニューリン阻害剤(例えばシクロスポリンおよびタクロリムス)は、免疫抑制療法の中心である。これら2つの化合物は、細胞性免疫応答の強力な抑制因子であり、過去20年間における臓器移植の転帰を顕著に改善している(Allison A.C.、2000、Immunopharmacology;47(2−3):63−83頁を参照されたい。)。
【0007】
マクロライド、ラパマイシン、アスコマイシンおよびタクロリムスの遺伝子クラスターを含む、微生物由来の多数の医学的に重要な薬物の生合成経路をコードしている遺伝子クラスターは、既にクローン化され、配列決定されている。タクロリムス遺伝子クラスターのクローン化に関しては、ポリケチド合成酵素(PKS)をコードしている遺伝子を主に包含する部分配列が文献において報告された(Motamedi H.およびShafiee A.、1998、Eur J Biochem;256(3):528−34頁を参照されたい。)。一方、科学者らは、2000年にアスコマイシン遺伝子クラスターのクローン化を報告した(Wu Kら、2000、Gene;251(1):81−90頁、US6,503,737を参照されたい。)。タクロリムスは、構造的におよび生合成的起源についてアスコマイシン(FK520)およびラパマイシンに類似している(Reynoldsら、;Drugs and the Pharmaceutical Sciences,1997、82、497−520頁を参照されたい。)。これらすべては、ポリケチド(PKS)と非リボソーム性ペプチド生合成経路(NRPS)との組合せによって合成されうる(McDaniel Rら、2005、Chem Rev;105(2):543−58頁を参照されたい。)。
【0008】
タクロリムスおよびアスコマイシンは構造的に類似している。唯一の構造的差異としてタクロリムスの炭素21におけるアリル側鎖が、アスコマイシンにおいてはエチル側鎖によって置換されている。タクロリムス(FK506)およびアスコマイシン(FK520)化合物の構造は、式(Ia)および(Ib)に示されている。
【0009】
アスコマイシンおよびタクロリムスの構造は、生合成機構を考慮して4つのステップに分割されうる複合的生合成経路:
1.独特なシキミ酸由来スターターを使用する鎖の開始、
2.大部分のPKS由来化合物に共通の鎖の伸長、
3.L−ピペコリン酸の組み込みによる鎖の終結および環化ならびに
4.PKS後処理
を既に示唆している。
【先行技術文献】
【特許文献】
【0010】
【特許文献1】国際公開第2010/004304号
【特許文献2】米国特許第5116756号明細書
【特許文献3】欧州特許第0356399号明細書
【特許文献4】米国特許第5200411号明細書
【特許文献5】韓国特許第100485877号明細書
【特許文献6】米国特許出願公開第2007/191415号明細書
【特許文献7】米国特許第6503737号明細書
【非特許文献】
【0011】
【非特許文献1】Kinoら、J.Antibiotics 40、1249−1255頁、1984
【非特許文献2】Easton J.B.およびHoughton P.J. 2004、Expert Opin Ther Targets、8(6):551−64頁
【非特許文献3】Allison A.C.、2000、Immunopharmacology;47(2−3)63−83頁
【非特許文献4】Motamedi H.およびShafiee A.、1998、Eur J Biochem;256(3)528−34頁
【非特許文献5】Wu Kら、2000、Gene;251(1)81−90頁
【非特許文献6】Reynoldsら、;Drugs and the Pharmaceutical Sciences,1997、82、497−520頁
【非特許文献7】McDaniel Rら、2005、Chem Rev;105(2):543−58頁
【発明の概要】
【発明が解決しようとする課題】
【0012】
タクロリムス発酵プロセスにおいて、望ましくないアスコマイシン(FK520)生成物も不純物として生成され、それによりタクロリムスの最終収量が低下し、タクロリムスの下流単離プロセスに著しい追加的経費を生じる。
【0013】
【化1】
【課題を解決するための手段】
【0014】
詳細には本発明は、薬剤の調製のためおよびヒトを含む哺乳類の治療のための上に定義した式(Ia)もしくは式(Ib)の化合物またはこれらの光学異性体、薬学的に許容される塩、水和物、溶媒和化合物もしくは多形体の使用に関する。本発明は、ヒトを含む動物における状態または疾患の予防および/または治療のための薬剤の調製のための化合物の使用に関する。
【0015】
エチルマロニルCoAは、アスコマイシン(FK520)生合成のための重要な基本単位として周知である。S.ハイグロスコピウス(S.hygroscopius)において見出された生合成アスコマイシンクラスターに位置し、アスコマイシン生合成に関与する以下の遺伝子は、最先端として報告されており(Liuら、Journal of Bacteriology、1999、181、6806−6813;Liuら、Metabolic Engineering、2001、3、40−48頁を参照されたい。)、関連があると考えられる:
a)クロトニルCoA還元酵素(ccr遺伝子)をコードする遺伝子、
b)B12依存性イソブチリルCoAムターゼ(icm遺伝子)をコードする遺伝子、
c)エチルマロニルCoAムターゼ(ecm遺伝子)をコードする遺伝子。
【0016】
いくつかのポリケチド由来化合物のポリケチド鎖アセンブリの特定の段階においてエチルマロニルCoA伸長単位を提供する生合成経路は、最先端として記載されている(Wuら、loc.cit;Reynoldsら、loc.citを参照されたい。)。エチルマロニルCoA伸長単位は、ブチリルCoAのカルボキシル化反応によって誘導されると考えられる。ブチリルCoAをもたらす少なくとも2つの経路がストレプトマイセスにおいて同定されている。一経路は2つの酢酸単位の縮合を含み、それによりアセト酢酸CoA活性化化合物を形成し、これはさらにクロトニルCoAに加工され、クロトニルCoA還元酵素(ccr)によって触媒されるクロトニルCoAからブチリルCoAへの還元の重要なステップを通じて処理される(Wallace K.K.ら、1995、Eur J Biochem;233:954−962頁を参照されたい。)。この遺伝子は、一次代謝遺伝子セット中に位置していることが見出されたS.コリヌス(S.collinus)において最初に同定された。後にその相同体が、エチルマロニルCoAを前駆体として必要とするいくつかの化合物をコードしている生合成クラスターにおいて同定された(Croppら、2001、J Ind Microbiol Biotechnol;27:368−377頁を参照されたい。)。
【0017】
近年、ロドバクター・スフェロイデス(Rhodobacter sphaeroides)中のccr相同体が、クロトニルCoAのエチルマロニルCoAへの還元カルボキシル化を触媒するクロトニルCoAカルボキシラーゼ/還元酵素活性を有する酵素をコードしていることが示された(Erb T.J.ら、2007、Proc Natl Acad Sci USA;25:10631−10636頁を参照されたい。)。さらにこの酵素は、イソクエン酸リアーゼ遺伝子を欠失している微生物の酢酸を唯一の炭素源とした増殖を可能にする「エチルマロニル−CoA経路」と称される新たに発見された酢酸同化経路の一部であることが示された。この経路は、いくつかのステップを含むと予測されている。先の予測のとおりこれは2つのアセチルCoA単位の縮合および続くクロトニルCoAへの加工によって開始される。次いでクロトニルCoAは、最初に(2R)−エチルマロニルCoAにエピマー化され、次にビタミンB12依存性エチルマロニルCoAムターゼ(ecm)によってメチルスクシニルCoAに転換される(2S)−エチルマロニルCoAにCcrによって直接転換される(Erb T.J.ら、2008、J Biol Chem;283:32283−32293頁を参照されたい。)。続いて、メチルスクシニルCoAは、メチルスクシニルCoA脱水素酵素によってメサコニルCoAに転換され、次のステップにおいてメサコニルCoAは、メサコニルCoA脱水酵素によってβ−メチルマリルCoAに転換される。最後に、β−メチルマリルCoAは、グリオキシル酸と、次に既知の代謝経路に入るプロピオニルCoAとに切断される。グリオキシル酸はアセチルCoAと縮合されてリンゴ酸を生じることができ、プロピオニルCoAはメチルマロニルCoAおよび次いでスクシニルCoAにカルボキシル化されうる。興味深いことにいくつかのストレプトマイセス属種は、ecm経路に関与するいくつかの遺伝子をクラスターとしてコードし、唯一の炭素源としての酢酸酪酸または脂肪酸での増殖の際にこの経路がストレプトマイセスにおいて活性であることを示唆している(Akopiants K.ら、J Ind Microbiol Biotechnol;33:141−150頁を参照されたい。)。
【0018】
バリン代謝物、イソブチリルCoAの異性化ステップを通じて進行し、ブチリルCoAを形成し、補酵素B12依存性イソブチリルCoAムターゼ(icm)によって触媒される第2経路が提案されている(Reynolds K.ら、1988、J Chem Soc Perkin Trans 1;3195−3207頁;およびZerbe−Burkhardt K.ら、1998、J Biol Chem;273:6508−6517頁を参照されたい。)。
【0019】
タクロリムス(FK506)生成生物に関してアリルマロニルCoA前駆体の代謝出発物およびその生合成に関与する遺伝子は、従来技術において同定されていない。
本発明は、アリルマロニルCoAの供給のための新規生合成経路を提供する。驚くべきことに、FK506ポリケチド合成酵素(PKS)クラスターの左腕に位置しているアリルマロニルCoA生合成に関与する遺伝子群が見出された。FK506生合成遺伝子クラスターの最も左側の新たに得られた配列は、アリルマロニルCoA生合成に関与する遺伝子をコードすることが見出され、タクロリムス遺伝子クラスターの「Allyl(All)クラスター」または「Allyl(All)サブクラスター」と称される(表1を参照されたい。)。
【0020】
本発明は、FK506ポリケチド合成酵素(PKS)遺伝子クラスターの左側に位置する、アリルマロニルCoA前駆体(基本単位)の生合成に関与する遺伝子を含有するこの新規ヌクレオチド配列に関する。
【0021】
FK506生成株の改善、具体的には収率および純度に関しての改善のためのこれらの配列の種々の使用も本発明に含まれる。本発明は、タクロリムス、アスコマイシンまたは関連化合物の選択的生成を可能にする不均一に発現された「Allylサブクラスター」を有する遺伝子改変株も提供する。
【0022】
アスコマイシンなどの副産物の形成を低減または消失させることによるおよび/またはタクロリムスの生成の収率を増加させることによるタクロリムス生成のための改善された方法を提供することは本発明の目的の1つである。
【0023】
アリルマロニルCoA対エチルマロニルCoAの伸長単位の利用能は、発酵プロセスの最後での所望の生成物タクロリムス(FK506)または副産物としてのアスコマイシン(FK520)それぞれの最終割合において重要な役割を演じることが見出された。したがって本発明は、所望の生成物と副産物との顕著に改善された割合をもたらす、前記伸長単位の割合を制御するいくつかの可能性を提供する。
【0024】
この態様において本発明は、遺伝子改変株によって、好ましくは生成物アスコマイシン(FK520)の生合成が顕著に低減されうるまたは消失されうるストレプトマイセス・ツクバエンシス(NRRL18488)によって実施されるタクロリムスの改善された発酵調製を記載する。好ましくはタクロリムスの収率は、一定のままであるまたは増加している。したがって本方法は、エチルマロニルCoA供給を低減する(もしくは消失させる)手段を使用することによっておよび/またはアリルマロニルCoA生合成を増加させることによって発酵生成の既知の方法の不利益を顕著に低減する。本方法は、アスコマイシン生成を低減させる(または消失させる)ことができ、タクロリムスの生成収率を維持または増加させうる。
【0025】
本方法の最後における一方でのタクロリムスおよび他方でのアスコマイシンの最終割合は、エチルマロニルCoAおよび/またはアリルマロニルCoAの代謝に関与するこれらの遺伝子の不活性化および/または過剰発現によって制御されうる。
さらに本出願は微生物の遺伝子改変株であって、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【図面の簡単な説明】
【0026】
【図1】ストレプトマイセス・ツクバエンシスの「All遺伝子」サブクラスター(15080bp)の遺伝子構成を示す図である。
【図2】ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891の抽出された発酵培地中のタクロリムスおよびアスコマイシンの存在をモニターするためのLC−MSクロマトグラム(MRM)を示すグラフである。アスコマイシンは、野生型株(列2)およびallA、allK、allRおよびallD遺伝子を過剰発現している変異株(列4)によって生成される。一方タクロリムスは、野生型株(列1)によって生成されず、allA、allK、allRおよびallDが過剰発現される場合(列3)にだけ生成される。
【発明を実施するための形態】
【0027】
本発明は、タクロリムスの調製のための方法であって、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養のステップを含み、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む方法を対象とする。
【0028】
本発明は、ストレプトマイセス属に属する微生物の遺伝子改変株であって、前記微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株も記載する。
詳細には本出願によるタクロリムスの調製のための方法は、以下のステップの少なくとも1つを含む:
a)微生物の遺伝子改変株の作製;
b)微生物の遺伝子改変株の栄養培地への添加および微生物の前記株を培養することを含む種培地の調製;
c)微生物の遺伝子改変株の種培地のバイオリアクター中の栄養培地への添加、微生物の前記株を培養することおよびタクロリムスの生成を含む主発酵;
d)発酵培地からの生成物の分離および精製(回収)。
【0029】
詳細には本発明によるタクロリムスの調製のための方法は、本出願に記載の微生物の遺伝子改変株の作製、種培地の調製および微生物の前記遺伝子改変株を培養することを含む主発酵を含む。好ましくは本出願によるタクロリムスの調製のための方法は、上に記載のステップa)からd)を含む。方法ステップa)からd)は下により詳細に記載される。
【0030】
好ましい実施形態において本発明は、上に記載のタクロリムスの調製のための方法であって、微生物の遺伝子改変株の培養ステップを含み、前記微生物が好ましくはストレプトマイセス属から選択される細菌である方法に関する。具体的には前記微生物は、アクチノマイセス目(Actinomycetales)から選択されうる。好ましいのは、PKS活性を有するまたは異種性に発現されるPKSもしくはこの一部を有するアクチノマイセス目に属する微生物である。より具体的には微生物は、ストレプトマイセス属から選択されうる。具体的にはストレプトマイセス・ツクバエンシス、ストレプトマイセス属種、ATCC55098またはストレプトマイセス・ハイグロスコピカス変種アスコマイセティカス(Streptomyces hygroscopicus var.ascomyceticus)ATCC14891が使用されうる。
【0031】
詳細には微生物は、ストレプトマイセス・ツクバエンシス、好ましくはストレプトマイセス・ツクバエンシス(NRRL18488)の遺伝子改変株から選択される。さらに微生物は、ストレプトマイセス・ハイグロスコピカスの遺伝子改変株から選択されうる。
【0032】
さらに本発明の方法を実施するために以下の微生物を使用することも検討される:ストレプトマイセス・ツクバエンシスNo.9993(Ferm BP−927)、ストレプトマイセス・ハイグロスコピカス亜種ハイグロスコピカス(Streptomyces hygroscopicus subsp.hygroscopicus)(DSM40822)、ストレプトマイセス属種AA6554、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6475 ATCC14891、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6678 ATCC55087、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6674、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC55276、ストレプトマイセス・ハイグロスコピカス亜種アスコマイセティカスATCC14891、ストレプトマイセス・カナマイセティカス(Streptomyces kanamyceticus)KCC S−0433、ストレプトマイセス・クラブリゲルス(Streptomyces clavuligerus)CKD1119、ストレプトマイセス・ハイグロスコピカス亜種ヤクシマエンシス(Streptomyces hygroscopicus subsp.yakushimaensis)、ストレプトマイセス属種DSM7348、ミクロモノスポラ(Micromonospora)属種(新種)A92−306401 DSM8429、ストレプトマイセス属種MA6548およびストレプトマイセス属種MA6858 ATCC55098。
【0033】
他の態様において本発明は、本出願に記載のタクロリムスの調製のための方法であって、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の条件下で実施される方法に関する。詳細にはアリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体は、上に記載の主発酵プロセス(ステップc)で発酵培地に添加される。
【0034】
さらに本出願に記載の発酵プロセスは、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくは本発明において提供されるストレプトマイセス・ツクバエンシスの遺伝子改変株を培養することおよび制御されたやり方でアリルマロニル−、エチルマロニル−および/もしくはプロピルマロニルCoAならびに/またはアリルマロニル−、エチルマロニル−および/もしくはプロピルマロニルCoAの前駆体を発酵培地に添加することを含みうる。これにより実質的に純粋なタクロリムス、アスコマイシンまたはそれぞれの関連化合物が得られる。この意味において、配列番号4に記載の不活性化された少なくともAllR遺伝子を有する、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株が使用される必要がある。
【0035】
他の実施形態において本出願に記載のタクロリムスの調製のための方法は、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養のステップであって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む培養のステップを含む。上に記載のヌクレオチド配列は、表1に記載されており別表に列挙されている。
【0036】
FK506の生合成のために必要なORF(オープンリーディングフレーム)をコードする遺伝子クラスター全体は配列決定された。PKS(ポリケチド合成酵素)遺伝子を含有するクラスターの中央部分の公表された配列(Motamedi H.およびShafiee A.1998;Eur J Biochem;256(3):528−34頁を参照されたい。)に加えて、ポリケチド合成酵素クラスターの左および右の周辺部に存在する遺伝子の配列が取得され、アノテーションされた。遺伝子クラスターの左側では、多数のORFが同定され、その大部分はアスコマイシン(FK520)生合成クラスターに存在する遺伝子にいかなる相同性も示さなかった(Wu.Kら、2000、Gene;251(1)81−90頁、US6503737を参照されたい。)。これらのORFをクエリーとして使用するBLAST検索は、表1に示すとおり同定された遺伝子の推定上の機能を明らかにした。遺伝子をコードするこのAllylクラスター(またはAllylサブクラスター)は、タクロリムスの生合成に特異的に必要なアリルマロニルCoA前駆体の生合成に関与する。さらに、遺伝子をコードするAllylサブクラスターは、エチルマロニルCoAの生合成に関与することが見出された。
【0037】
さらなる態様において、本発明は、配列番号2から配列番号10に記載のいくつかの遺伝子から構成される配列番号1に記載の配列および1つまたは複数のヌクレオチド追加、欠失、置換または逆位を含むこれらの変種を含むストレプトマイセス属に属する微生物のエチルマロニルCoAまたはアリルマロニルCoA生合成に関与するAllylサブクラスターのヌクレオチド配列に関する。Allylサブクラスターのヌクレオチド配列は表1に示されており、添付の別表に列挙されている(配列番号1から配列番号10)。
【0038】
【表1】
【0039】
変異株におけるエチルマロニルCoAおよびアリルマロニルCoAの合成を遮断することにより、これらの基本単位を供給することによってどのポリケチドが生成されるかを制御できる新規方法が可能であることが見出された。酸化還元酵素(クロトニルCoA還元酵素に類似)をコードするAllR遺伝子の不活性化は、タクロリムスの生成を完全に消失させたことが見出された。しかしアリルマロニルCoA前駆体の外部添加は、発酵培地中のアスコマイシン生成の消失を伴うタクロリムスの排他的生成を可能にする。AllR遺伝子の不活性化は、アスコマイシン(FK−520)の生成を完全に消失させたことも見出された。しかしエチルマロニルCoA前駆体の外部添加は、発酵培地中のタクロリムス生成の消失を伴うアスコマイシン(FK−520)の排他的産生を可能にする。この方法は任意のアスコマイシンまたはタクロリムス生成生物において可能である。アスコマイシン生成生物中でのエチルマロニルCoAの合成を遮断するための方法は、エチルマロニルCoA生合成に関与する遺伝子がアスコマイシン生成生物において解明されていることから当業者に周知である(Wuら、2000、Gene251、81−90頁)。ccr相同体の不活性化は、そのような生物の調製における最初のステップである。アスコマイシンおよびタクロリムス合成のためのPKSが高度に相同性であることから、アリルマロニルCoAまたはエチルマロニルCoAを取り込むモジュール7の無差別性がすべてのアスコマイシン/タクロリムス生成生物のPKSについて予測されうる。
【0040】
本発明の一実施形態において、本出願に記載のタクロリムスの調製のための方法は、ストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップであって、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される培養ステップを含む。
【0041】
さまざまなアリルマロニルCoA前駆体は使用されうる。より詳細にはアリルマロニルSNAC(アリルマロニル−(N−アセチル−システアミン)チオエステル)の添加が好ましい。本発明に関してアリルマロニルCoA前駆体はアリルマロニルCoAの類似体も含む。
【0042】
別法としてアリルマロン酸またはこの類似体は、前記方法のために使用される株の追加的遺伝子改変を考慮すると、前記方法において直接使用されうる。この場合マロニルCoA合成酵素およびジクサルボン酸キャリアタンパク質は、使用される微生物中で発現される必要がある。増殖培地中に加えられるジカルボン酸化合物の取り込みが可能である遺伝子改変微生物の一例は、Lomboら、Biotechnol.Prog.2001、17、612−617頁によって記載されている。基質としてのアリルマロン酸に対するマロニルCoA合成酵素の十分な活性は、Pohlら、J.Am.Chem.Soc.2001、123、5822−5823頁によって記載されている。
【0043】
詳細には上に記載の方法は、ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップであって、微生物の遺伝物質が
配列番号3に記載のAllK遺伝子および
配列番号4に記載のAllR遺伝子
の群から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む培養ステップを含む。
【0044】
本発明のさらなる実施形態において上に記載の方法は、ストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップであって、微生物の遺伝物質が配列番号1に記載の追加的不活性化遺伝子ccr遺伝子を含む培養ステップを含む。
【0045】
この詳細な態様において本発明は、ストレプトマイセス・ツクバエンシスの以下の株:
a)ストレプトマイセス・ツクバエンシスF499
受託番号DSM22507で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488ΔallRと記載される。
b)ストレプトマイセス・ツクバエンシスF872
受託番号DSM22509で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488ΔallKと記載される。
に関する。
【0046】
本発明のさらなる実施形態において上に記載の方法は、ストレプトマイセス属に属する微生物、好ましくはストレプトマイセス・ツクバエンシスの株の遺伝子改変株の培養ステップであって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターから選択される少なくとも1つの不活性化された遺伝子を含む培養ステップであって、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される培養ステップを含む。
【0047】
好ましい実施形態において上に記載の方法は、ストレプトマイセス属に属する微生物の遺伝子改変株の培養のステップであって、微生物の遺伝物質が配列番号3に記載のAllK遺伝子および配列番号4に記載のAllR遺伝子の群から選択される少なくとも1つの不活性化された遺伝子を含む培養ステップを含み、前記方法はアリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される。
好ましい実施形態において上に記載の方法は、ストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養のステップであって、微生物の遺伝物質が不活性化された配列番号4に記載のAllR遺伝子および不活性化された配列番号11に記載のccr遺伝子を含む培養ステップを含み、前記方法はアリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される。
【0048】
さらに本発明は、ストレプトマイセス属に属する微生物のエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する1つまたは複数の遺伝子をコードし、配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載の配列ならびに1つまたは複数のヌクレオチドの追加、欠失、置換および/または逆位を含むこれらの変種を含むヌクレオチド配列に関する。
【0049】
詳細には上に記載のヌクレオチド配列は、配列番号1に少なくとも50%のヌクレオチド同一性を有する。
【0050】
同様に本発明によって提供されるのは、上に記載のヌクレオチド配列であって、少なくとも60%、好ましくは少なくとも70%、好ましくは少なくとも80%、好ましくは少なくとも90%のヌクレオチド配列同一性を、配列番号2から配列番号10に記載のいくつかの遺伝子から構成される配列番号1に記載の配列に有するヌクレオチド配列である。ヌクレオチド配列の上に記載の変種は、ストレプトマイセス属に属する生物のエチルマロニルCoAまたはアリルマロニルCoAの生合成に関与する酵素をコードする能力を保持しているべきである。
【0051】
このように本発明は、表1に列挙し配列番号2から配列番号10に記載のすべての個々の遺伝子のヌクレオチド配列および配列番号13から配列番号21に記載のこれらのアミノ酸生成物ならびに1つまたは複数のヌクレオチド追加、欠失、置換および/または逆位を含むこれらの変種であって、少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも70%、好ましくは少なくとも80%、好ましくは少なくとも90%のヌクレオチド同一性を配列番号2から配列番号10に記載のいくつかの遺伝子から構成される配列による配列に有するヌクレオチド配列を提供する。
【0052】
提案された生合成経路は以下のとおり記載されうる。アリルマロニルCoAを得るためにC5前駆体、バレリルCoAまたは4−ペンテノイル−CoAが最初に形成されるべきである。アセト酢酸およびクロトニルCoAを通じて2つの酢酸単位から形成されるエチルマロニル−CoAと同様に、プロピルマロニルCoAまたはアリルマロニルCoAは1つの3炭素単位および1つの2炭素単位から生成できる。比較的短いポリケチド合成酵素(PKS)様酵素複合体によって単一ステップでの縮合反応を実行する、遺伝子AllA(アシル転移酵素)およびAllK(ケト合成酵素)は同定されている。さらに最終アリルマロニルCoA、C5前駆体に加工される前に5炭素中間体(3−オキソペンタン酸またはこの還元中間体)がどのようにCoAの添加によって活性化されるかまたはFK506PKS複合体に直接転移されるかは明確ではない。しかし最終合成アリル前駆体での供給実験は、伸長物を最終アリルマロニルCoA形態に組み込むFK506PKSの能力を示している。アセトアセチルCoA還元酵素に相同な推定上のAllS遺伝子は、3−ヒドロキシペンタノイルCoAを生じる最初の還元ステップを実行する。次いで追加的酸化還元酵素(AllR、クロトニルCoAカルボキシラーゼ/還元酵素、AllDおよびAllOと同様)は、この化合物をさらにプロピルマロニルCoAに還元でき、AllP、P450モノオキシゲナーゼと共にOH基および続いて二重結合も4位に導入でき、アリルマロニルCoAを生じる。次いでこれは対応するPKSモジュール4のアシル転移酵素(AT4)遺伝子によって新生ポリケチド鎖に導入される。Allylクラスターの一部として、AllMとして示されたORFが同定された。この遺伝子産物は、メチオニンγリアーゼに相同性を示し、プロピオニルCoA(最初の5炭素中間体のAllAによる生合成に必要な開始単位)に直ちに代謝される2−オキソ酪酸にメチオニンを転換すると考えられる。プロピオニルCoA前駆体は、2次代謝の間に限定されうる。したがってAllM遺伝子産物の存在は、最終産物FK−506の収率を増加させるためのプロピオニルCoAの十分な供給のために必要であると考えられる。
【0053】
提案された経路は、FK−506生成微生物の発酵培地中のエチルマロニルCoAの由来およびそれによりFK−520の由来も説明する。アリルマロニルCoAの生合成に関与するS.ツクバエンシスのFK−506生合成クラスターの≫Allylサブクラスター≪中に存在する遺伝子は、エチルマロニルCoAも生成できる。すなわちエノイルCoA還元酵素(遺伝子AllR)は、次いで上に記載の経路と同様のやり方で処理されるC5単位(3−オキソペンタン酸またはこの還元中間体)の代わりにアセト酢酸(C4)単位への限定的無差別性をほぼ確実に示す。次のステップは両前駆体について同様のやり方で進行するが、C4単位はP450調節水酸化および二重結合の形成に対する基質にはならないと考えられている。エノイルCoA還元酵素/カルボキシラーゼ(AllR)遺伝子の欠失は、この生合成経路ならびにFK−506およびFK−520の生成を完全に消失させることが見出された。ケトアシル合成酵素(AllK)遺伝子は、FK−506の産生だけを完全に消失させることが見出され、FK−520の産生は野生型S.ツクバエンシス株のレベルのままである。
【0054】
興味深いことに、FK−520/FK−506生合成のためのエチルマロニルCoAおよびアリルマロニルCoA(またはプロピルマロニルCoA)伸長単位を生じる上に記載の反応ステップは、「エチルマロニルCoA経路」の最初の反応からエチルマロニルCoA中間体が形成される点までに機構的に酷似している。この初期代謝経路は、大部分のストレプトマイセス属種の酢酸を唯一の炭素源とした増殖に最も高い可能性で関与し、クロトニルCoAカルボキシラーゼ/還元酵素(ccr)によって産生され、さらにエチルマロニルCoAムターゼ(ecm)によってメチルスクシニルCoAに転換されるエチルマロニルCoAを中間体化合物として含む。明らかに、所与の増殖条件下で活性であるならば、この経路がS.ツクバエンシス中でのエチルマロニルCoAの重要な供給源も表しうることが期待される。しかし、ccr(エチルマロニルCoAを産生する。)およびecm(これをさらに処理する。)を含む「エチルマロニルCoA経路」をコードする多数の遺伝子は、ストレプトマイセスにおいて同じオペロンに位置し、共転写されると考えられている。そのような遺伝子構造は、両遺伝子が協調して作用しなければならず、基質が最終産物への代謝経路を迅速に通過しなければならない場合には理想的であるが、単一の中間体(すなわち2次代謝のための基本単位としてのエチルマロニルCoA)の十分な量を提供するためには不十分である場合がある。これは、伸長単位としてエチルマロニルCoAを必要とするポリケチド生合成のための多数の遺伝子クラスターがccr遺伝子相同体(この内容において生成されたエチルマロニルCoAを直ちに消費するecm遺伝子は除く)の追加的コピーをも含有することの理由についての説明を提供できる。
【0055】
含まれる反応は機構的に類似しているが、S.ツクバエンシスでのFK−506およびFK−520生合成のための基本単位としてのアリルマロニルCoA(プロピルマロニルCoA)およびエチルマロニルCoAを提供する酵素と、この生物の「エチルマロニルCoA経路」の酵素とは1つの重要な観点において異なっている。「エチルマロニルCoA経路」はエチルマロニルCoA中間体を特異的に含み、ごく少量のプロピルマロニルCoAが最終的に形成される場合があるが、allサブクラスターにコードされる酵素は異なる特異性を示す。これらは、アリルマロニルCoA(プロピルマロニルCoA)を優先的に生成するが、エチルマロニルCoAは少量だけ産生される。経路のいくつかの酵素(すなわちAllR)は4炭素および5炭素中間体を同様の効率で受容できるが、他はアリルマロニルCoA(プロピルマロニルCoA)を排他的に生じる基質に対して非常に特異的である場合があることから恐らく量は比較的少ない。ストレプトマイセス酵素によるFK506の生成のための発酵プロセスとの関連で、両代謝系は(これらの発現が完全に異なる因子によってほぼ確実に誘導されるけれども)発現されうる。allサブクラスターの遺伝子はFK−506生合成クラスターの他の遺伝子と一緒に転写されると考えられるが、一度2次代謝が誘導されると「エチルマロニルCoA経路」酵素の発現および活性は増殖条件、最も重要なことには増殖培地の組成に依存すると考えられる。
【0056】
明確にはエチルマロニルCoAの濃度、すなわちFK−520の収率を可能な限り低く保つために、増殖培地の組成は「エチルマロニルCoA経路」の遺伝子が転写されないまたは最小レベルで転写されるように適合されるべきである。別法としてecm経路に加わりストレプトマイセスにおいて同じオペロン上に通常コードされる酵素の発現は、エチルマロニルCoA濃度を低く保持するために操作される必要がある。クロトニルCoAカルボキシラーゼ/還元酵素をコードするccr遺伝子は、不活性化されるべきである一方で、エチルマロニルCoAムターゼ(ecm)、メチルスクシニルCoA脱水素酵素、メサコニルCoA脱水酵素およびβ−メチルマリルCoAリアーゼをコードする他の4つの遺伝子は強力な構成的プロモーターを使用して過剰発現されるべきである。しかし、増殖条件および培地とは無関係に細胞中のエチルマロニルCoAの完全な消失が望ましい場合は、両方の経路を同じ機構的ステップで妨害する標的変異を導入する必要がある場合がある。本発明において記載のとおり、このステップは、ccr相同体、「エチルマロニルCoA経路」のccr遺伝子およびアリルマロニルCoA(プロピルマロニルCoA)の供給に主に関与するallサブクラスターのallR遺伝子の両方の不活性化でありうる。
【0057】
本発明の一目的はタクロリムスの調製のための方法であって、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップを含み、微生物の遺伝物質がアリルマロニルCoA対エチルマロニルCoAの細胞内プールの比率を改善するように改変されている方法を提供することである。
【0058】
この実施形態において本発明に記載のタクロリムスの調製のための方法は、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップであって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの過剰発現された遺伝子を含む培養ステップを含む。上に記載のヌクレオチド配列は、表1に記載されており添付の別表に列挙されている。
【0059】
本出願は、アリルマロニルCoAおよび/またはエチルマロニルCoA前駆体合成に必要なタンパク質をコードする、不活性化および/または過剰発現された表1に列挙した1つまたは複数の遺伝子を有する1つまたは複数のS.ツクバエンシス株の作製を含む。
【0060】
より詳細には本態様において本発明は、ストレプトマイセス・ツクバエンシスの遺伝子改変株の作製であって、前記株がタクロリムス(FK−506)および/またはアスコマイシン(FK−520)生合成のための重要な基本単位の供給に関与する少なくとも1つの不活性化された遺伝子を含む遺伝子改変株の作製を対象とする。これらの遺伝子改変株は、タクロリムスの改善された産生のために有用であることが見出されている。
【0061】
微生物の遺伝子改変のための方法、例えば遺伝子コピー数の増幅は当技術分野において周知である。他の例は、強力なプロモーターの制御下での前記遺伝子の追加的コピーの挿入である。さらにこの意味における他の例は「All」サブクラスターにおいて見出された転写制御因子の活性の改変である。
【0062】
この意味において本発明は、アリルマロニルCoA前駆体の細胞内プールを改善するおよびまたはエチルマロニルCoAの細胞内プールを低減することによるタクロリムスの発酵調製のためのストレプトマイセス・ツクバエンシスの改善された株を提供する。表1および配列番号1に記載の新規ヌクレオチド配列は、アリルマロニルCoAの生合成に関与する前記遺伝子の発現を制御するために使用されうる。
【0063】
本発明の一実施形態において本出願に記載のタクロリムスの調製のための方法は、微生物の遺伝子改変株の培養ステップであって、微生物の遺伝物質が配列番号11に記載のクロトニルCoA還元酵素(ccr)遺伝子および配列番号12に記載のエチルマロニルCoAムターゼ(ecm)から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む培養ステップを含む。ヌクレオチド配列は添付の別表に列挙されている。
【0064】
本出願は、ストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株であって、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0065】
詳細には本出願は、上に記載のストレプトマイセス属に属する微生物の遺伝子改変株であって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0066】
詳細には本発明は、上に記載のストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株であって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子および配列番号4に記載のAllR遺伝子の群から選択される少なくとも1つの過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0067】
詳細には本出願は、上に記載のストレプトマイセス属に属する微生物の遺伝子改変株、好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株であって、微生物の遺伝物質が配列番号2に記載のAllA遺伝子および配列番号3に記載のAllK遺伝子の群から選択される少なくとも1つの過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0068】
他に、さらに類似する実施形態においてアリルマロニルCoA前駆体の生合成に関与する遺伝子を含有する新規ヌクレオチド配列は、タクロリムスの生成のための組換え株の構築を当業者に可能にする。この意味で任意のアスコマイシン生成微生物がタクロリムスを生成するために操作されうる。この意味において適切な微生物は、これだけに限らないがストレプトマイセス・ハイグロスコピカスでありうる。これは、当技術分野において周知の方法による前記ヌクレオチド配列の前記宿主株への転移によって達成されうる。
【0069】
さらに本発明は、微生物の遺伝子改変株であって、天然のまたは操作されたポリケチド合成活性を有する微生物の遺伝子改変株を対象とする。
【0070】
本発明は、微生物の遺伝物質が
クロトニルCoA還元酵素の遠位相同体をコードする配列番号4に記載のAllR遺伝子、
チトクロムP450酵素をコードする配列番号8に記載のAllP遺伝子、
アシル転移酵素およびアシルキャリアタンパク質をコードする配列番号2に記載のAllA遺伝子、
ケトアシル合成酵素をコードする配列番号3に記載のAllK遺伝子、
アシルCoA脱水素酵素をコードする配列番号5に記載のAllD遺伝子、
メチオニンγリアーゼ酵素およびアシルキャリアタンパク質をコードする配列番号6に記載のAllM遺伝子、
転写制御因子をコードする配列番号7に記載のAllN遺伝子、
アシルCoA酸化還元酵素をコードする配列番号9に記載のAllO遺伝子および
アセトアセチルCoA還元酵素をコードする配列番号10に記載のAllS遺伝子、
の群から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含むストレプトマイセス・ツクバエンシスの遺伝子改変株に関する。
【0071】
遺伝子のヌクレオチド配列(DNA配列)および遺伝子産物のアミノ酸配列は、添付の別表のそれぞれ配列番号2から配列番号10および配列番号13から配列番号21に示されている。
【0072】
この詳細な態様において本発明は、ストレプトマイセス・ツクバエンシスの以下の株:
(a)ストレプトマイセス・ツクバエンシスF499
受託番号DSM22507で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 ΔallRと記載される。
(b)ストレプトマイセス・ツクバエンシスF872
受託番号DSM22509で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 ΔallKと記載される。
(c)ストレプトマイセス・ツクバエンシスF917
受託番号DSM22511で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 AllP+と記載される。
に関する。
【0073】
さらにアリルマロニルCoA前駆体の生合成に関与する遺伝子を含有するヌクレオチド配列は、新規ハイブリッドポリケチドを生成するために使用されうる。これは、当技術分野において周知の方法による前記ヌクレオチド配列の任意のポリケチド生成宿主株微生物への転移によって達成されうる。公知であるポリケチド合成酵素(PKS)モジュールの無差別性により、少なくとも1つの炭素位置にアリル側鎖を有する少なくとも痕跡量の新規ハイブリッドポリケチド化合物を期待できる。
【0074】
遺伝子改変(前記遺伝子の過剰発現または不活性化)のために使用されうる微生物は、細菌から選択されうる。より詳細には前記微生物は、アクチノマイセス目から選択される。好ましくはPKS(ポリケチド合成酵素)活性を有するアクチノマイセス目に属する微生物である。別法として過剰発現のための生物は、異種性に発現されたPKSまたはこの一部を有する任意の微生物であってよい。
【0075】
より詳細には微生物は、ストレプトマイセス属から選択される。具体的にはストレプトマイセス・ツクバエンシス、ストレプトマイセス属種ATCC55098またはストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891が使用されうる。
【0076】
さらに本発明の方法を実施するために以下の微生物を使用することも企図される:ストレプトマイセス・ツクバエンシスNo.9993(Ferm BP−927)、ストレプトマイセス・ハイグロスコピカス亜種ハイグロスコピカス(DSM40822)、ストレプトマイセス属種AA6554、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6475 ATCC14891、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスMA6678 ATCC55087、ストレプトマイセス・ハグロスコピカス変種アスコマイセティカスMA6674、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC55276、ストレプトマイセス・ハイグロスコピカス亜種アスコマイセティカスATCC14891、ストレプトマイセス・カナマイセティカスKCCS−0433、ストレプトマイセス・クラブリゲルスCKD1119、ストレプトマイセス・ハイグロスコピカス亜種ヤクシマエンシス、ストレプトマイセス属種DSM7348、ミクロモノスポラ属種(新種)A92−306401 DSM8429、ストレプトマイセス属種MA6548およびストレプトマイセス属種MA6858 ATCC55098。
【0077】
一実施形態において本発明は、S.ツクバエンシス染色体上の特定のFK506遺伝子クラスターにクラスター化していない遺伝子の第2群の使用に言及する。これらの遺伝子は、ブチリルCoAおよびエチルマロニルCoA、アスコマイシン(FK520)の生合成のための前駆体の代謝に関与している。
【0078】
S.ツクバエンシスにおけるエチルマロニルCoA供給を減少させるまたは消失させるために、いくつかの戦略が行われた。この観点において本発明は、アスコマイシン生合成についての基本単位、エチルマロニルCoAの代謝に関与し、以下の酵素:
a)クロトニルCoA還元酵素(本明細書以下においてそれぞれccr、ccr遺伝子と称される。)
b)エチルマロニルCoAムターゼ(本明細書以下においてそれぞれecm、ecm遺伝子と称される。)
をコードする遺伝子のヌクレオチド配列の使用に関する。
【0079】
ヌクレオチド配列およびアミノ酸配列は、添付の別表に列挙されている(配列番号11、配列番号12を参照されたい。)。
【0080】
この態様において本出願は、タクロリムスの調製のための方法であって、微生物、好ましくはストレプトマイセス属の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株の培養ステップを含み、微生物の遺伝物質が配列番号11に記載のクロトニルCoA還元酵素(ccr)遺伝子および配列番号12に記載のエチルマロニルCoAムターゼ(ecm)遺伝子から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む方法を対象とする。
【0081】
詳細にはタクロリムスの調製のための方法は、微生物、好ましくはストレプトマイセス属、の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの株の培養ステップであって、微生物の遺伝物質が不活性化された配列番号11に記載のクロトニルCoA還元酵素(ccr)遺伝子を含む培養ステップを含む。
【0082】
詳細にはタクロリムスの調製のための方法は、微生物、好ましくはストレプトマイセス属、の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの株の培養ステップであって、微生物の遺伝物質が過剰発現された配列番号12に記載のエチルマロニルCoAムターゼ(ecm)遺伝子を含む培養ステップを含む。
【0083】
タクロリムスの調製のためのこの改善された方法は、アスコマイシン(FK520)の顕著に低下したまたは消失した生成を示し、これによりタクロリムス精製プロセスを著しく簡素化する。
【0084】
ccrおよびicmの遺伝子相同体は、ブチリルCoAおよび続くエチルマロニルCoAの生合成に関与していくつかの生物において同定された。補酵素B12依存性イソブチリルCoAムターゼ(icm;ブタノイルCoA:2−メチルプロパノイルCoAムターゼ、EC5.4.99.13)は、周知のメチルマロニルCoAムターゼ(mem)反応に密接に関連する反応においてイソブチリルCoAとn−ブチリルCoAとの間の可逆的転位を触媒する。クロトニルCoA還元酵素(EC.1.3.1.38、アシルCoA:NADP’−トランス−2−酸化還元酵素)は、クロトニルCoAのブチリルCoA(エチルマロニルCoA生合成のための重要な基質)への転換を触媒するまたはある環境下ではクロトニルCoAをエチルマロニルCoAに直接転換する。ecm(エチルマロニルCoAムターゼ)の遺伝子相同体は、エチルマロニルCoAをメチルスクシニルCoAに転換し、それにより混合物中でエチル置換された生成物の割合を効果的に変化させることが示されている。
【0085】
クロトニルCoA還元酵素遺伝子(ccr遺伝子)およびエチルマロニルCoAムターゼ遺伝子(ecm遺伝子)がさまざまな微生物においていくつかの代謝および生合成経路に関与することは周知である。ポリケチド生合成、詳細にはエチルマロニルCoAの生合成におけるecmおよびccrの役割は、記載されている(例えば、the role of ecm in production of monensin in S.cinnamonensis、Zhang W.ら、2001、J.Bacteriol;183:2071−2080頁)。
【0086】
したがって本発明は、エチルマロニルCoAの細胞内レベルが低下している微生物を作製する方法を記載する。これは、エチルマロニルCoAの生合成に関与する酵素を遮断することおよび/またはエチルマロニルCoAを消費する酵素を過剰発現することによって達成される。
【0087】
ストレプトマイセス・ツクバエンシスにおけるFK506/520の生合成のための重要な基本単位の生合成に特異的に関与するクロトニルCoA還元酵素(ccr)の活性が改変された。追加的に、エチルマロニルCoAのさらなる処理に関与するエチルマロニルCoAムターゼ(ecm)は、タクロリムス生成株ストレプトマイセス・ツクバエンシスにおいて過剰発現された。
【0088】
一実施形態において本出願は、微生物、好ましくはストレプトマイセス属に属する微生物の遺伝子改変株、より好ましくはストレプトマイセス・ツクバエンシスの遺伝子改変株であって微生物の遺伝物質が、
クロトニルCoA還元酵素をコードする配列番号11に記載のccr遺伝子および
エチルマロニルCoAムターゼをコードする配列番号12に記載のecm遺伝子
の群から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む微生物の遺伝子改変株を対象とする。
【0089】
好ましい実施形態において本出願に記載の微生物の遺伝子改変株は、本出願に記載の2つの不活性化および/または過剰発現された遺伝子を含む二重改変微生物である。二重改変微生物は、野生型微生物および単一変異体と比較して大幅に改善された特徴を有することが見出された。
【0090】
クロトニルCoA還元酵素遺伝子相同体(ccr)の不活性化およびエチルマロニルCoAムターゼ(ecm)の過剰発現は、アスコマイシン生合成の顕著な低下によって特徴付けられるタクロリムスの生成のための改善された方法を導く。試験した増殖条件下では、ムターゼ触媒反応はブチリルCoAからイソブチリルCoAへ進行すると考えられる。エチルマロニルCoAムターゼ(ecm)遺伝子は、生成株中で過剰発現された場合にエチルマロニルCoAを効果的に消費することが見出された。
【0091】
一態様において本発明は、遺伝子改変微生物、好ましくはストレプトマイセス属の遺伝子改変微生物、より好ましくはストレプトマイセス・ツクバエンシスの株の遺伝子改変微生物であって、微生物の遺伝物質が不活性化された配列番号11に記載のクロトニルCoA還元酵素遺伝子(ccr)を含む遺伝子改変微生物を提供する。微生物中のそのような遺伝子を不活性化する方法は、例として手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり当技術分野において公知である。
【0092】
他の態様において本発明は、遺伝子改変微生物、好ましくはストレプトマイセス属の遺伝子改変微生物、より好ましくはストレプトマイセス・ツクバエンシスの株の遺伝子改変微生物であって、微生物の遺伝物質が過剰発現された配列番号12に記載のエチルマロニルCoAムターゼ遺伝子(ecm)を含む遺伝子改変微生物を提供する。微生物中のそのような遺伝子を過剰発現させる方法は、例として手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり当技術分野において公知である。
【0093】
さらなる態様において本発明は、上に記載のとおり不活性化された配列番号11に記載のクロトニルCoA還元酵素遺伝子(ccr)および過剰発現された配列番号12に記載のエチルマロニルCoAムターゼ遺伝子(ecm)を含む、遺伝子改変された微生物、好ましくはストレプトマイセス属の、より好ましくはストレプトマイセス・ツクバエンシスの株を提供する。この態様において、二重改変微生物の使用は、野生型微生物および上に記載の単一変異体と比較して大幅に改善された特徴を有する。
【0094】
具体的には本発明は、ストレプトマイセス・ツクバエンシスの以下の株:
a)ストレプトマイセス・ツクバエンシスF130
受託番号DSM22506で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 Δccr:Tsと記載される。
b)ストレプトマイセス・ツクバエンシスF879
受託番号DSM22510で2009年4月23日にBraunschweig/ドイツのDSMZ−Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH(German Collection of Microorganism and Cell Cultures)に寄託された。この株はNRRL18488 ecm+と記載される。
に関する。
【0095】
新たに開発された株は、著しく低下したアスコマイシン含有量を示し、タクロリムス発酵生成の顕著な改善を表す。
【0096】
本発明によってさらに提供されるのは、配列番号11に記載のccr遺伝子配列、配列番号23に記載のアミノ酸配列を有するクロトニルCoA還元酵素(Ccr)をコードする配列および1つまたは複数のヌクレオチドの追加、欠失、置換または逆位を含むこれらの変種を含む、微生物ストレプトマイセス・ツクバエンシスNRRL18488のエチルマロニルCoA生合成に関与する遺伝子をコードする上に記載のヌクレオチド配列である。同様に本発明によって提供されるのは、配列番号12に記載のecm遺伝子配列、配列番号24に記載のアミノ酸配列を有するエチルマロニルCoAムターゼ(ecm)をコードする配列および1つまたは複数のヌクレオチドの追加、欠失、置換または逆位を含むこれらの変種を含む微生物ストレプトマイセス・ツクバエンシスNRRL18488のエチルマロニルCoA消費に関与する遺伝子をコードするヌクレオチド配列である。同様に本発明によって提供されるのは、上に記載のヌクレオチド配列であって、少なくとも50%、好ましくは少なくとも60%、好ましくは少なくとも70%、好ましくは少なくとも80%、好ましくは少なくとも90%の同一性を配列番号11および配列番号12に記載の配列に有するヌクレオチド配列である。
【0097】
本発明は、タクロリムスの調製のための改善された方法、具体的には産業用発酵方法であって、微生物の遺伝子改変株の培養のステップを含み、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む方法を提供する。前記方法は、上に記載の微生物の少なくとも1つの遺伝子改変株を使用することによって実施されうる。以下にこの方法をより詳細に記載する:
詳細には本発明による方法は以下のステップを含む。
【0098】
ステップa)微生物の遺伝子改変株の作製
好ましくはタクロリムスの生成のための本方法は、本発明によって記載される微生物株の遺伝子改変のステップを包含する。この意味において本発明は、具体的にはストレプトマイセス・ツクバエンシスの遺伝子改変株であって、株の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む遺伝子改変株に関する。
【0099】
本発明によるさらなる改変のためのストレプトマイセス・ツクバエンシスを作製するための手順は、当業者に周知である。さらにストレプトマイセス・ツクバエンシスの適切な株は、商業的に入手可能である。適切な株の具体的非限定的な一例は、受託番号NRRL18488を有する野生型ストレプトマイセス・ツクバエンシスである。
【0100】
微生物中の遺伝子を不活性化する方法は、例えば手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり当技術分野において公知である。
【0101】
好ましい実施形態において、微生物、好ましくはS.ツクバエンシスの株の遺伝子改変株の芽胞形成培地中での芽胞の調製、濃縮された芽胞懸濁物の調製のそれぞれが実施される。
【0102】
ステップb)種培地の調製
本発明に記載のストレプトマイセス・ツクバエンシスの改変株の培養は、当業者に周知の方法によって実施されうる。ストレプトマイセス・ツクバエンシスの培養プロセスは、例えば手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載されている。
好ましくはタクロリムス生成のための主発酵プロセスにおいて使用されうる種微生物の生成は、前記遺伝子改変微生物の芽胞形態から開始する。この点において本出願による方法は、記載された遺伝子改変微生物の、好ましくはストレプトマイセス・ツクバエンシスの芽胞貯蔵物の調製を含み、場合によって記載された遺伝子改変微生物の濃縮された芽胞懸濁物を含む。芽胞形態のこの調製は、マンガン塩を含む芽胞形成培地を使用するなどの当技術分野において周知の方法を使用して実施されうる。好ましくは微生物の遺伝子改変株のこの芽胞貯蔵物または濃縮芽胞懸濁物は、栄養培地への接種によって栄養種培地を作製するために使用される。記載の微生物の栄養形態の生成は、芽胞を含む比較的少量の種培地での接種を伴って調製されるべきである。
【0103】
種培地は、バイオリアクターに無菌的に移植されうる。
【0104】
原理上は種微生物の培養は主発酵プロセス(下のステップcに記載)と同様の条件下(例えばpHおよび温度)で実施されうる。
【0105】
ステップc)主発酵プロセス
好ましくは本出願において記載の遺伝子改変微生物を使用する主発酵プロセスは、バイオリアクター中で、詳細には撹拌および/または通気下で実施される。好ましくは本出願において記載のタクロリムスの生成のための方法は、同化炭素源、窒素源、リン酸源および鉱質源を含有する水性栄養培地(生成培地)中、液内好気性条件下で実施される。主発酵プロセスにおいて前記培地から生成されたタクロリムスの単離は、さらなる分離ステップ(ステップd)において実施されうる。
【0106】
好ましくは、主発酵プロセスは、ステップb)で得られた種微生物での生成培地の接種(詳細にはリアクターへの無菌的移植による)を含む。接種のために微生物の栄養形態を使用することは好ましい。
主発酵プロセスにおけるリアクターへの栄養培地(生成培地)の添加は、1回または複数回バッチ法でまたは継続的方法で実施されうる。栄養培地(生成培地)の添加は発酵プロセス前および/またはプロセス中に実施されうる。
【0107】
栄養培地中の好ましい炭素源は、下に例示のとおりデキストリン、ブドウ糖、可溶性デンプン、グリセロール、乳酸、麦芽糖、果糖、糖蜜およびショ糖から選択されうる。
【0108】
栄養培地中の好ましい窒素源は、酵母エキス、大豆ペプトン、大豆ミール、細菌ペプトン、カゼイン加水分解物、L−リジン、硫酸アンモニウム、コーンスティープリカーなどである。
【0109】
炭酸カルシウム、塩化ナトリウム、リン酸ナトリウムまたはリン酸カリウム、マグネシウム、マンガン、亜鉛、鉄および他の塩などの無機/鉱質塩も培地に添加されうる。
【0110】
本方法による主発酵プロセスは、約6.3から8.5のpH範囲内、20から35℃の温度範囲内で実施される。好ましくは、pHは約6.5から8.3の範囲内であり、温度は約23から31℃の範囲内である。
好ましくは、生成培養物は、80から約300時間、より好ましくは約130から280時間インキュベートされる。
【0111】
タクロリムスの生成は、生成培地の撹拌および通気を伴う好気性条件で実施されうる。培養混合物の撹拌および通気は、種々の方法で遂行されうる。生成培地の撹拌は、プロペラまたは類似の機械的装置によって提供されることができ、発酵条件および規模に応じてさまざまな程度に変更されうる。通気速度は、バイオリアクターの可動容積に対して1.0から2.5VVM(1分間あたり液体の単位容積あたりで流れる気体の容積(1分間あたり容積あたりでの容積))の範囲内で変更されうる。
【0112】
発酵プロセス用のさらなる周知の添加物は、具体的には主発酵プロセスにおいて添加されうる。培養培地の過度な起泡性を防ぐために、シリコン油、脂肪油、植物油などの消泡剤が添加されえた。具体的にはシリコンを基にする消泡剤が培養培地の過度な起泡性を防ぐために発酵プロセス中に添加されうる。
【0113】
ステップd)発酵培地からの生成物の分離および精製(回収)
発酵培地由来のタクロリムスは、生物学的に活性な物質の回収のために慣用される従来法によって分離および精製されうる。生成されたタクロリムスは、有機溶媒、すなわちアセトン、エタノールまたはメタノール中での抽出によって発酵培地から回収されうる。
【0114】
場合によって抽出物は次いで濃縮され、粗製タクロリムスを得るためにアセトン:水混合物を使用してXAD16吸着剤でカラムクロマトグラフィーされうる。粗製物質は、純粋なタクロリムスを得るために調製用HPLC上でさらに精製されうる。
【0115】
本出願による方法ステップの好ましい実施形態は、以下のとおり要約されうる:
i)固体芽胞形成アガー培地上で、S.ツクバエンシス株を28℃、10から14日間培養することによる前記S.ツクバエンシス株からのS.ツクバエンシス芽胞貯蔵物の調製。
ii)S.ツクバエンシス株の芽胞は濃縮芽胞懸濁物を作製するためおよび栄養培地600mlへの接種のために回収され、振とう機220rpmで24−32時間、栄養培地のPMV 10−20%が達成されるまでインキュベートした。回収時のpHは、6.8から7.3の範囲内である。
iii)生成培地に接種するための所望容量の10%までの種栄養接種物の増殖。栄養培地を含む10リットル発酵槽は、栄養種で接種され、24から28時間、28℃で培養される。
iv)栄養種培地の10%は生成培地100Lに接種するために使用される。発酵プロセスは、120から180時間、28℃で硫酸または水酸化ナトリウム溶液を使用してpHを7.0−7.2に維持して実施される。
【0116】
以下に非限定的な例として芽胞形成、種増殖および主発酵プロセスのための好ましい培地が記載される:
1)可溶性デンプン(10g/L)、塩化ナトリウム(1g/l)、硫酸アンモニウム(2g/L)、炭酸カルシウム(2g/L)、K2HPO4(1g/l)、MgSO4×7H2O(1g/L)、FeSO4×7H2O(0.001g/L)、MnCl2×4H2O(0.001g/L)、ZnSO4×7H2O(0.001g/L)および細菌学的アガー(20g/L)を含む固体アガー芽胞形成培地(ISP4)。pHは滅菌の前に塩基または酸の適切な添加によって7.0±0.2に調整されるべきである。
2)大豆ミール(2.5g/L)、デキストリン(10g/L)、ブドウ糖(1g/L)、酵母エキス(5g/L)、カゼイン加水分解物(7g/L)、K2HPO4(0.2g/L)、NaCl(0.5g/L)、MnCl2×4H2O(0.005g/L)、FeSO4×7H2O(0.025g/L)、ZnSO4×7H2O(0.001g/L)、MgSO4×7H2O(0.005g/L)およびCaCl2(0.02g/L)を含む種培地。pHは滅菌の前に塩基または酸の適切な添加によって7.0±0.2に調整されるべきである。
3)デキストリン(60−120g/L)、ブドウ糖(0−15g/L)、大豆ミール(5−20g/L)、大豆ペプトン(5−20g/L)、グリセロール(5−20g/L)、L−リジン(1−7.5g/L)、K2HPO4(0.5−2g/L)、CaCO3(1−5g/L)およびポリエチレングリコール(1−5g/L)を含有する生成培地。pHは、滅菌の前に塩基または酸の適切な添加によって7.0±0.2に調整されるべきである。
【0117】
本発明の他の好ましい実施形態において主発酵プロセス(ステップc)は、アリルマロニルCoAまたはアリルマロニルCoAの少なくとも1つの前駆体(類似体)の外部添加の下で実施される。本発明に関してアリルマロニルCoA前駆体は、アリルマロニルCoAの類似体も含む。
【0118】
この点において本発明は、タクロリムスの調製のための方法、具体的には産業用発酵方法であって、ストレプトマイセス属に属する微生物の遺伝子改変株の培養のステップを含み、微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含み、アリルマロニルCoAおよびまたはアリルマロニルCoAの少なくとも1つの前駆体(類似体)の外部添加の下で実施される方法を提供する。
【0119】
詳細には本発明は、S.ツクバエンシスの遺伝子改変株を培養することによるタクロリムス(または関連化合物)の発酵調製であって、アリルマロニル−およびエチルマロニルCoA活性化前駆体を提供する1つまたは複数の生合成経路が、例えばAllR遺伝子の不活性化および発酵プロセス中にアリルマロニルCoAまたはエチルマロニルCoA SNAC前駆体を制御されたやり方で培養培地に添加または供給することによって、不活性化されているタクロリムスの発酵調製を対象とする。これは野生型株と比較して顕著な量でのタクロリムスまたはアスコマイシン化合物の産生を結果としてもたらす。したがって本明細書に記載の新規方法は、タクロリムスまたはアスコマイシンが排他的に産生される方法をもたらし、この方法は下流プロセスを顕著に簡素化する。
アリルマロニルCoAもしくはエチルマロニルCoAまたは本明細書に記載の少なくとも1つのこれらの前駆体(類似体)の外部添加の下での、ストレプトマイセス属に属する微生物の遺伝子改変株、具体的にはS.ツクバエンシスの遺伝子改変株を使用する純粋なタクロリムスまたは純粋なアスコマイシンの生成のための発酵プロセスは、既に記載されているものと同様に同化炭素源および窒素源を含有する水性栄養培地中において液内好気性条件下で実施されうる。
【0120】
好ましくはアリルマロニルまたはエチルマロニルエステル前駆体の標的濃度範囲は、0.1から5.0g/Lの範囲であり、より好ましくは0.5から3.0g/Lの範囲である。標的濃度は、有形データ記憶媒体に好ましくは発酵プロセスの開始前に記録されうる。供給プロセスについては、さまざまなアリルマロニルまたはエチルマロニルエステル前駆体が使用されうる。アリルマロニルまたはエチルマロニルN−アセチルシステアミン(SNAC)チオエステルは好ましい。1つのもしくは2つのチオエステルの両方またはこれらの混合物は使用されうる。
【0121】
発酵培地由来のタクロリムスは、生物学的に活性な物質の回収のために慣用される従来法によって分離および精製されうる。したがって前記方法は、下流プロセスにおいて調製用HPLCが使用される必要がある古典的方法と比較して顕著な費用優位性を有する。
【0122】
さらに本発明は、タクロリムス発酵生成のための方法における上に記載のストレプトマイセス・ツクバエンシスの遺伝子改変株の使用に関する。
【0123】
さらなる態様において本発明は、上に記載のストレプトマイセス・ツクバエンシスの遺伝子改変株の培養によって生成されたタクロリムスまたは薬学的に許容されるこの誘導体もしくは類似体を含む医薬組成物に関する。用語「類似体」または「誘導体」は、参照分子に構造的に類似しているが参照分子の1つまたは複数の特定の置換基を代替の置換基で置き換えるために標的化されおよび制御されたやり方で改変されており、それにより参照分子に構造的に類似している分子が作製されている分子を意味して、本明細書において従来の薬学的意味で使用される。追加的に当業者に周知の方法を使用して、改善された治療有効性、すなわち特定の標的受容体型でのより高い効力および/または選択性、哺乳類脳血管関門を透過する高いまたは低い能力のいずれか(例えばより高いまたはより低い脳血管関門透過率)、よりまれな副作用などを有する化合物タクロリムスの類似体および誘導体は作製されうる。
【0124】
本発明の組成物と関連して使用される句「薬学的に許容される」は、哺乳類、例えばヒトに投与された場合に生理学的に許容可能で、典型的には有害反応を生じない分子実体およびそのような組成物の他の成分を意味する。好ましくは本明細書において使用される用語「薬学的に許容される」は、連邦もしくは州政府の監督官庁によって認可されているまたは例えば米国薬局方もしくは哺乳類、より詳細にはヒトにおける使用について他の一般に認められた薬局方に列挙されていることを意味する。
【0125】
本発明のさらなる実施形態は、式(I)の少なくとも1つの化合物もしくはこの多形性形態またはこの薬学的に許容される塩および少なくとも1つのさらなる薬学的に許容可能な添加剤を含む医薬組成物である。医薬組成物は、さらなる薬物化合物も含みうる。本発明の医薬組成物において式(I)の化合物、詳細にはタクロリムスまたはこの多形性形態もしくは薬学的に許容される塩は、前記化合物またはこの薬学的に許容される塩を1日分の投与のための投与単位あたりで例えば0.1から4000mg、好ましくは1から2000mgを含有する投与単位として製剤される。本発明の全ての態様、具体的には医学的なものに関して化合物または組成物の投与は、最終的には主治医によって決定され、使用される化合物、動物種、性別、年齢、体重、症状の重症度、投与の方法、有害反応および/または他の禁忌症などの因子を考慮に入れる投与計画を有する。具体的に決定される投与量範囲は、患者の進行度および回復が完全に観察される標準的な設計の臨床試験によって決定されうる。そのような試験は、ヒトでの開始用量として動物での最大耐用量の低い百分率を使用する用量を漸増させる設計を使用する場合がある。
【0126】
本発明による生理学的に許容される化合物は、(成人患者に対して)連日投与計画で、例えば0.01mg/kg(治療される哺乳類の体重1kgあたりのmg)から100mg/kgの間、好ましくは0.1mg/kgから75mg/kgの間の経口投与量で通常投与される。
配列番号および遺伝子機能は表2に示される:
【0127】
【表2】
【実施例】
【0128】
以下の実施例は本発明を例示するためのものである。
【0129】
実験手順
以下は、「ccr」、「AllK」、「AllR」、「AllP」および「ecm」遺伝子をクローン化および分析するためならびにこれらの遺伝子相同体を使用するストレプトマイセス・ツクバエンシス変異体の作製のための実験手順の詳細な例である。同様に含まれるのは、これらの株を使用する発酵手順ならびにFK506およびFK520生成収率の判定の例である。分子生物学または微生物学の当業者に公知である標準的技術の追加的詳細および使用される具体的な酵素の名称は、例えば手引書「Practical Streptomyces genetics」(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載されている。
【0130】
実施例1:
ストレプトマイセス・ツクバエンシス株の維持および芽胞調製
ストレプトマイセス・ツクバエンシス株の菌糸体を芽胞形成培地ISP4上で8−14日間、28℃でコンフルエント菌叢として増殖させた。ISP4培地は:
【0131】
【表3】
を含んでいた。
【0132】
pHは、1M NaOHで7.0に調整した。滅菌は、121±2℃、120±10kPaで20分間実施した。8−14日間の増殖後、芽胞を回収し、グリセロール(20%)中、−20℃で使用まで保存した。
【0133】
実施例2:
ストレプトマイセス・ツクバエンシスゲノムDNAの調製
ストレプトマイセス・ツクバエンシスNRRL18488(野生型)の芽胞を250mlエルレンマイヤーフラスコ中のTSB培地(Kieserら、2000、Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)50mlを接種するために使用し、28℃で24時間振とう(210rpm)させて維持した。培養物は、24時間28℃で増殖させた。菌糸体を遠心分離で回収し、ゲノムDNAをPureLink Genomic DNA Mini Kit(Invitrogen)をキット製造者の説明書に従って使用して調製した。DNAをTE緩衝液(SambrookおよびRussell、2000、Molecular Cloning:A Laboratory Manual、ISBN−978−087969577−4)100μlに再懸濁した。
【0134】
実施例3:
単離されたゲノムDNAをGATC AG、ドイツのRoche FLX technologyで配列決定した。FK506生合成遺伝子クラスターに属する配列連続断片が同定された場合、ORF(オープンリーディングフレーム)の位置および向きをFramePlot beta4.0 Software(Ishikawaら、FEMS Microbiol.Lett.174(1999)251−253頁)を使用して分析した。予測アミノ酸配列を使用して、相同性検索をGenBank databasesでBLASTpおよびBLASTxアルゴリズムを使用してNCBI(NCBI/Blast: http://blast.ncbi.nlm.nih.gov/Blast.cgi)で実行した。これらの結果に基づき、保存ドメイン検索によって支持された推定遺伝子機能を割り当てた。
合計15個のORFをfkbCから左に同定した。これらの内の6個はFK520クラスターに存在するORFに高い類似性を示し(fkbL−リジンシクロデアミナーゼ、fkbG、fkbH、fkbI、fkbJ、fkbK−遺伝子5個はメトキシマロニル−ACP生合成に関与する。)、9個の遺伝子は「アリルサブクラスター」を形成すると予測された。このサブクラスターは、アシル転移酵素およびアシルキャリアタンパク質ドメインを含有する遺伝子1個、ケトアシル合成酵素ドメインを含有する遺伝子1個、遠位クロトニルCoA還元酵素ドメイン1個、遠位アシルCoA脱水素酵素ドメイン含有相同体2個、メチオニンγリアーゼ含有遺伝子1個、AsnCファミリー制御遺伝子1個、P450モノオキシゲナーゼ遺伝子1個および遠位アセトアセチルCoA還元酵素相同体1個からなる。
【0135】
実施例4:
ccr遺伝子の破壊のためのベクターの構築、不活性化されたccr遺伝子を有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:ccr遺伝子の広領域の増幅のためのプライマーをストレプトマイセス・セリカラー(Streptomyces coelicolor)(タンパク質ID:NP_630556.1、遺伝子ID:1101912)由来のccr遺伝子を鋳型としたBLAST検索を使用して見出されたさまざまなストレプトマイセス属種の他の周知のccr遺伝子の保存領域および得られたBLAST結果のClustalWパイルアップ配列比較に基づいて設計した。したがってXbaI制限酵素部位を有するccr−F2(GTCTAGACCACATCATCGGCTCCGACC)およびEcoRI制限酵素部位を有するccr−R1(CGAATTCACGCCGACCTTGCCCTGGTGC)をccr遺伝子の917bp長領域を増幅するために作製した。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスのゲノムDNAをBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR反応はPfuポリメラーゼ(New England Biolabs)および製造者によって提供された緩衝液で、最終容量100μl中、200μM dNTP、10%グリセロール、0.5μMの各プライマー、鋳型S.ツクバエンシスゲノムDNAおよそ50ngおよび酵素2.5単位の存在下で30サイクル実施した。全30サイクルの温度プロファイルは、95℃、45秒間(変性ステップ)、69℃、45秒間(アニーリングステップ)および72℃、1分間(伸長ステップ)であった。PCR増幅産物をpUC19クローニングベクターにクローン化した。クローン化したPCR増幅産物の配列分析によって、それぞれの部分ccr配列を確認した。
c)相同組換えによるS.ツクバエンシスccr遺伝子の破壊において使用されるpKC1139に基づく温度感受性ベクターおよびpKC1132に基づく自殺ベクターの構築:ccr遺伝子のF2−R1増幅断片をpUC19からEcoRIおよびXbaI制限酵素部位によって切り出し、EcoRIおよびXbaI制限酵素で予め切断したベクターpKC1139およびpKC1132に連結反応によって移行させ、pKC1139−ccrおよびpKC1132−ccrを作製した。ベクターpKC1139は、E.コリ(E.coli)での複製のための正常なpUC19に基づくOriを含有しているが、34℃を超える高い温度では機能できないストレプトマイセスでの複製のための温度感受性Oriは含有していない(Biermanら、1992 Gene.116(1):43−9頁、Muthら、1989 Mol Gen Genet.219(3):341−348頁)。ベクターpKC1132はE.コリでの複製のための正常pUC19に基づくOriだけを含有するが、ストレプトマイセスでの複製を維持できず(Biermanら、1992 Gene.116(1):43−9頁)、したがって相同組換えによるゲノムへの組込みなしで失われる。次いでベクターpKC1139−ccr1およびpKC1132−ccrをccr遺伝子中で切断するBamHI制限酵素を使用して開き、917bp長断片を一方の346bpと他方の571bpとに分割し、次いでDNAポリメラーゼIラージ(Klenow)断片で末端平滑化した。次いでチオストレプトン(Ts)耐性を付与するカセットをpKC1139−ccrおよびpKC1132−ccr BamHI切断、末端平滑化ベクターに導入した。p330ベクターをXbaI制限酵素で消化し、Tsカセットを含有するおよそ961bp長断片をDNAポリメラーゼIラージ(Klenow)断片で末端平滑化し、pKC1139−ccrおよびpKC1132−ccrベクターに連結し、pKC1139−ccrTsおよびpKC1132−ccrTsベクターをそれぞれ作製した。
d)ccr遺伝子破壊のためのベクターのS.ツクバエンシスNRRL18488株への導入:プラスミド構築物pKC1139−ccrTsおよびpKC1132−ccrTsを、接合性プラスミドpUZ8002を含有するエレクトロコンピテントE.コリ株ET12567に形質転換によって導入した(Pagetら、1999 J.Bacteriol.181、204−211頁)。プラスミドpUZ8002は接合性線毛の構築のために必要な全ての遺伝子を含有しているが、転移の起点を欠いており、したがって宿主細胞に残留する(Jonesら、1997 Mol.Microbiol.23:169−178頁)。接合手順をKieserら、2000(Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり行った。ベクターpKC1139−ccrTsでの形質転換だけが接合完了体を生じた。接合完了体は、アプラマイシン50μg/mlを添加した実施例1に記載の芽胞形成培地上で、28℃で増殖させた。
e)チオストレプトン耐性付与カセットによって破壊されたccr遺伝子を有するS.ツクバエンシスの安定2次組換え株についての選択:アプラマイシン50μg/mlを添加した芽胞形成培地上で、28℃で増殖させたベクターpKC1139−ccrTsのS.ツクバエンシス接合完了体を、チオストレプトン(25μg/ml)を含む液体TSB培地に接種し、振とうフラスコ中、28℃、210rpmで、良好な種培養物を生成するためにアプラマイシンの添加をせずに増殖させた。24時間後、種培養物を、チオストレプトン(25μg/ml)を含む新鮮TSB培地を含む新たな振とうフラスコ中に継代培養し、24℃より高く温度を上昇させて210rpmで増殖させた。34℃より高い温度ではpKC1139に基づくベクターは、複製できず、S.ツクバエンシスゲノムに組み込まれ、それにより一次組換え体が生じる。培養物を、チオストレプトン(25μg/ml)を含む新鮮TSB培地を含む新たな振とうフラスコ中にさらに数回継代培養し、次いでチオストレプトンを含む芽胞形成培地に播種し、28℃で増殖させた。回収した芽胞をろ過し、系列希釈物を芽胞形成培地のプレート上に作製した。5−8日後、単一コロニーを、チオストレプトンを含む芽胞形成培地のプレート上およびチオストレプトンおよびアプラマイシンを含む芽胞形成培地のプレート上にパッチした。一次組換え体は、アプラマイシンにいまだ耐性である一方で2次組換え体はアプラマイシン耐性を欠失しており、アプラマイシンを含む芽胞形成培地上で増殖できない。選択された2次組換え株は次節に記載のサザンハイブリダイゼーションによっても確認した。
f)S.ツクバエンシスccr破壊変異体ゲノムDNAのサザンハイブリダイゼーションによる分析:DIG標識プラスミドDNAプローブの調製:プラスミドpKC1139−ccrTsの挿入部分(TsRカセットを挿入されたccr遺伝子の一部)だけをプローブとして使用し、ベクターをEcoRIおよびXbaI制限酵素で消化し、1744bp長断片をWizard SV GelおよびPCR Clean−Up System(Promega)で精製した。およそ1μgの溶出断片をDIG High Prime DNA Labelling and Detection Starter Kit I(Roche)を使用するランダムプライムド標識技術によってジゴキシゲニン−dUTPで一晩標識した。
【0136】
i)DNA移行:S.ツクバエンシス野生型のゲノムDNAおよびpKC1139−ccrTs変異体(推定1次組換え体および2次組換え体)をPureLink Genomic DNA Mini Kit(Invitrogen)をキットの製造者の説明書に従って使用して調製した。単離された各ゲノムDNAおよそ10μgをNaeI制限酵素で消化した。消化終了時にDNA断片を1%アガロースゲル中、20V、12時間の電気泳動によって分離し、正に荷電したHybond−N+nylon membrane(GE Healthcare)にアルカリキャピラリーブロッティング法(Southern,E.M.、1975、J.Mol.Biol.、98:503)を使用して6時間転写した。
【0137】
ii)ハイブリダイゼーション:ナイロン膜に固定化したDNAへのDIG標識DNAプローブのプレハイブリダイゼーションおよびハイブリダイゼーションをDIG High Prime DNA Labeling and Detection Starter Kit I製造者(Roche)によって示されたとおりに実施した。プレハイブリダイゼーションは、供給されたプレハイブリダイゼーション溶液中、1時間、50℃で実施した。ハイブリダイゼーション用に変性DNA(90℃、10分間)1μgを50℃に予め加熱した新鮮プレハイブリダイゼーション溶液に加えた。一晩のハイブリダイゼーション後に膜を以下のとおり洗浄した:2×SSC、0.1%SDS、室温、5分間で2回洗浄および0.5×SSC、0.1%SDS、55℃、15分間で2回洗浄。
【0138】
iii)免疫学的検出:ハイブリダイズしたプローブを抗ジゴキシゲニン−AP Fab断片で免疫検出し、比色分析基質NBT/BCIPでDIG High Prime DNA Labeling and Detection Starter Kit I供給者(Roche)によって示されたとおり可視化した。
【0139】
iv)予測されるバンドを分析し、pKC1139−ccrTsベクターから得られる真の2次組換え体を記載のサザンハイブリダイゼーション手順によって確認した。ccr破壊株S.ツクバエンシスF130をタクロリムスおよびアスコマイシン生成についてのさらなる検査のために選択した。
g)pKC1139−ccrTsを使用する2次相同組換えから得られたチオストレプトン耐性ccr破壊変異体の発酵タクロリムス生成。
【0140】
i)種培養プロセス:種培地は:
【0141】
【表4】
を含んでいた。
【0142】
pHを1M NaOHで7.0に調整した。ブドウ糖を含まないこの培地50mlを250mlエルレンマイヤーフラスコにろ過し、発泡栓で栓をして滅菌した。滅菌は121±2℃、120±10kPaで25分間実施した。滅菌後、滅菌したブドウ糖懸濁液を加えた。滅菌種培地50μg/mlチオストレプトンを加え、ストレプトマイセス・ツクバエンシス株の芽胞(1%v/v)を種培地に接種し、振とう機で28℃、250rpm、24−48時間、好気性条件下でインキュベートした。
【0143】
ii)主発酵プロセス:上記の種培養物のおよそ10%v/vを、50ml発酵培地を含む250mlエルレンマイヤーフラスコへの接種に使用した。発酵培地は:
【0144】
【表5】
を含んでいた。
【0145】
発酵培地のpHを滅菌前に1M NaOHで7.0に調整した。滅菌は121℃で25分間実施した。発酵は振とう機で28℃、250rpm、6−7日間実施した。
【0146】
h)pKC1139−ccrTsを使用した2次相同組換えから得られたチオストレプトン耐性ccr破壊変異体のHPLCでのタクロリムスおよびアスコマイシン生成の判定:
【0147】
タクロリムスおよびアスコマイシンの判定のための方法:タクロリムスまたはアスコマイシン生成の判定のための分析を適切なカラムおよび実行条件を使用する無勾配逆相HPLCによって実施した:カラム:Nucleosil−100 C18(150×4.0mm、粒径3μm)、流速1.5ml/分、T℃=60℃、移動相:560ml水、335mlアセトニトリル、70ml MTBEおよび0.2ml 85%H3PO4、検出210nm、試料注入20μl。
試料中のタクロリムスおよびアスコマイシン含有量の定量を、タクロリムスが12.5分で溶出され、アスコマイシンが11.5分で溶出されたタクロリムスおよびアスコマイシンの外部標準を使用して実施した。結果は試料中のタクロリムス生成と比較したアスコマイシン生成の%で表す。
【0148】
試料調製:十分に振とうした培地5mlにメタノール5mlを加え、試料を抽出するために試料を1時間振とう機に置いた。抽出後、培地のメタノール抽出物1mlを1.5mlチューブに取り、10分間、14000rpmで遠心分離した。上清0.8mlを、HPLC分析を実施するためにバイアルに移した。
【0149】
【表6】
【0150】
破壊されたccr遺伝子を有する単離された2次組換え変異体は、野生型株ストレプトマイセス・ツクバエンシスNRRL18488と比較して平均で40%のアスコマイシン生成の低下を示した。
【0151】
実施例5:
AllR遺伝子の破壊のためのベクターの構築、不活性化されたAllRを有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:実施例4に記載のとおり、454に基づく全ゲノム配列分析をS.ツクバエンシスのゲノムに使用し、それはAllR遺伝子に隣接する領域の既知のDNA配列に基づいてAllR遺伝子に隣接する領域の増幅のためのプライマーを設計することを可能にした。したがって、HindIII制限酵素部位を有するAllR−F1(CAAGCTTCACCGGTCCCGGGCTC)およびNdeI制限酵素部位を有するAllR−R1(GCATATGGTCCGGTTCGGGGGTGGG)をAllR遺伝子の上流領域を増幅するために作製し、NdeIを有するAllR−F2(GGGTCACATATGGCGAACTACCGGG)およびEcoRI制限酵素部位を有するAllR−R2(CGAATTCTGTGGGCCGACCTCACCCA)をAllR遺伝子の下流領域を増幅するために作製した。プライマーAllR−R1とAllR−F2との間、894bp(298アミノ酸)間隙を1335bp AllR遺伝子のほとんど全体の欠失のために作製した。2つの重複断片をNdeI制限酵素部位で組合せる場合、読み枠が保存され下流遺伝子への影響を最小にした標的AllR遺伝子の「インフレーム」欠失を可能にする。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAを実施例4のii節の記載と同じ条件を使用し、AllR−F1+AllR−R1およびAllR−F2+AllR−R2プライマー組合せを使用してBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR増幅産物をpUC19クローニングベクターにクローン化し、クローン化したPCR産物の配列分析によって、これらそれぞれのDNA配列を確認した。
c)相同組換えによるS.ツクバエンシスAllR遺伝子の破壊に使用するpKC1139に基づく温度感受性ベクターおよびpKC1132に基づく自殺ベクターの構築:AllR遺伝子に隣接する領域のF1−R1およびF2−R2増幅断片をそれぞれ制限酵素HindIIIとNdeIとによっておよびEcoRIとNdeIとによってpUC19から切り出した。ベクターpKC1139およびpKC1132は、予めEcoRIおよびHindIII制限酵素で切断し、2つの断片を組合せ、各標的ベクターに連結し、それぞれpKC1139−AllRおよびpKC1132−AllRを作製した。次いで実施例4のiv節に記載のとおりベクターpKC1139−AllRおよびpKC1132−AllRを、NdeI制限酵素を使用して開き、抗生物質耐性付与カセットを挿入した。AllRについてさまざまなカセットを使用した;チオストレプトン(Ts)耐性カセットは実施例4に記載のとおりp330ベクターから得た、エリスロマイシン(Er)耐性カセットはpIJ4026ベクターからベクターをEcoRIおよびXbaIで消化し、1690bp長断片を末端平滑化することによって得たならびにカナマイシン(Kn)耐性カセットはpSuperCos1(REF!)ベクターからベクターをSmaIおよびHindIIIで消化し1323bp長断片を末端平滑化することによって得た。各カセットを、開いたpKC1132−AllRおよびpKC1139−AllRベクターに連結し、それぞれpKC1132−AllRTs、pKC1132−AllREr、pKC1132−AllRKn、pKC1139−AllRTs、pKC1139−AllRErおよびpKC1139−AllRKnベクターを作製した。
d)AllR遺伝子の破壊のためのS.ツクバエンシスNRRL18488株へのベクターの導入:プラスミド構築物pKC1139−AllRTs、pKC1139−AllRErおよびpKC1139−AllRKnをS.ツクバエンシスNRRL18488株に実施例4dの記載と同じ接合手順によって導入した。
e)抗生物質耐性付与カセットによって破壊されたAllR遺伝子を有するS.ツクバエンシスの安定2次組換え株の選択:ベクターpKC1139−AllRTs、pKC1139−AllRErおよびpKC1139−AllRKn、pKC1139−ccrTsのS.ツクバエンシス接合完了体についてのチオストレプトン抗生物質付与カセットによって破壊されたAllR遺伝子を有するS.ツクバエンシスの安定2次組換え株の選択のための手順は、実施例6における記載と同様であった。継代培養を、対応する抗生物質(それに対する耐性カセットがAllR遺伝子に隣接する領域間に挿入されている。)を含む液体TSB培地で実施した。選択された2次組換え株は、もはやアプラマイシン中では増殖できないが、耐性カセットがAllR遺伝子に隣接する領域間に挿入されている第2の抗生物質中では増殖できた。
f)予め不活性化ccr遺伝子が付与されているS.ツクバエンシスF130株へのAllR遺伝子の破壊のためのベクターの導入:プラスミドpKC1139−AllRErおよびpKC1139−AllRKnを実施例4に記載のS.ツクバエンシスF130 ccr破壊株に、実施例4dの記載と同じ接合手順によって導入した。
g)チオストレプトン耐性付与カセットによって破壊されたccr遺伝子およびエリスロマイシン耐性付与カセットによって破壊されたAllR遺伝子を有するS.ツクバエンシスの安定2次組換え株の選択:破壊されたccrおよびAllR両遺伝子を有するS.ツクバエンシスF130の誘導体、S.ツクバエンシスの安定2次組換え株を選択するための手順は、実施例4eの記載と同様であった。継代培養を、対応する抗生物質(それに対する耐性カセットが、AllR遺伝子に隣接する領域間に挿入されている。)を含む液体TSB培地で実施した。選択された2次組換え株は、もはやアプラマイシン中では増殖できないが、チオストレプトン(ccr遺伝子を破壊している)、および耐性カセットが、AllR遺伝子に隣接する領域間に挿入されている、第2の抗生物質中では両方とも増殖できた。
h)pKC1139−AllRErを使用する2次相同組換えによって得たエリスロマイシン耐性AllR破壊変異体の発酵でのタクロリムス生成:エリスロマイシン耐性AllR破壊変異体についてのタクロリムス生成は、培養の開始時にエリスロマイシン50μg/mlを発酵培地に添加したことを除いて実施例4gに記載のとおり実施した。
i)ccrおよびAllR二重破壊変異体による発酵でのタクロリムス生成:エリスロマイシン耐性ccrおよびAllR破壊変異体についてのタクロリムス生成は、培養の開始時にチオストレプトンおよびエリスロマイシンを各50μg/mlの濃度で発酵培地に添加したことを除いて実施例4gに記載のとおり実施した。供給実験用にアリルマロニルCoAおよびエチルマロニルCoA前駆体、ジエチルアリルマロニルSNAC、アリルマロニルジSNACおよびエチルマロニルSNACを発酵培地に濃度5−20mMで添加した。
j)pKC1139−AllRErを使用する2次組換えによって得たエリスロマイシン耐性AllR破壊変異体のHPLCでのタクロリムスおよびアスコマイシン生成の判定:
エリスロマイシン耐性AllR破壊変異体のタクロリムスおよびアスコマイシン生成の判定のために実施例4hでの記載と同じ方法を使用した。
【0152】
【表7】
【0153】
破壊されたAllR遺伝子を有する単離された2次組換え変異体は、タクロリムス生成の完全な消失を生じた。アリルマロニルSNACまたはアリルマロニルジSNACを発酵培地に添加した場合タクロリムスの生成が再確立された。同様にエチルマロニルSNACを添加した場合アスコマイシン生成が再確立された。
k)ccrおよびAllR二重破壊変異体のHPLCでのタクロリムスおよびアスコマイシン生成の判定:
ccrおよびAllR二重破壊変異体のタクロリムスおよびアスコマイシン生成の判定のために、実施例4hに記載の方法を使用した。
【0154】
【表8】
【0155】
タクロリムスの完全な消失が不活性化されたccrおよびAllR遺伝子を有する単離された2次組換え変異体で生じた。アリルマロニルSNACまたはアリルマロニルジSNACを発酵培地に添加した場合、タクロリムスの生成が再確立された。同様にエチルマロニルSNACを添加した場合アスコマイシン生成が再確立された。
【0156】
実施例6:
allK遺伝子の破壊のためのベクターの構築、不活性化されたallKを有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:454に基づく全ゲノム配列決定をS.ツクバエンシスのゲノムに使用し、それはallK遺伝子に隣接する領域の既知のDNA配列に基づいてallK遺伝子に隣接する領域の増幅のためのプライマーを設計することを可能にした。それにより、EcoRI制限酵素部位を有するallK−F1(AGAATTCGTTACGGGGAGACGGCATCCCGG)およびallK−R1(AGGATCCGGGCGGGCTCGTCGCGGT)を、内部BamHI制限酵素部位を含むallK遺伝子の上流領域を増幅するために作製し、BamHIを有するallK−F2(TGGATCCGGCGCGTATCGCCAACCGCTAC)およびHindIII制限酵素部位を有するallK−R2(AAAGCTTCCCGGTAGTTCGCCATATGTGACCCG)をallK遺伝子の下流領域を増幅するために作製した。内部BamHI制限酵素部位とallK−F2との間、1698bp(566アミノ酸)間隙を、2388bp allK遺伝子のほとんど全体の欠失のために作製した。2つの重複断片をBamHI制限酵素部位で組合せる場合、読み枠が保存され下流遺伝子への影響を最小にしての標的allK遺伝子の欠失が可能になる。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAを実施例4bのii節での記載と同じ条件を使用し、allK−F1+allK−R1およびallK−F2+allK−R2プライマー組合せを使用してBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR増幅産物をpUC19クローニングベクターにクローン化し、クローン化したPCR産物の配列分析によって、これらそれぞれのDNA配列を確認した。
c)相同組換えによるS.ツクバエンシスallK遺伝子の破壊に使用するためのpKC1139に基づく温度感受性ベクターおよびpKC1132に基づく自殺ベクターの構築:allK遺伝子に隣接する領域のF1−R1およびF2−R2増幅断片をそれぞれ制限酵素EcoRIとBamHIとによっておよびHindIIとBamHIとによってpUC19から切り出した。ベクターpKC1139およびpKC1132は、予め制限酵素EcoRIおよびHindIIIで切断し、2つの断片を組合せ、各標的ベクターに連結し、それぞれpKC1139−allKおよびpKC1132−allKを作製した。
d)allK遺伝子のインフレーム破壊のためのベクターのS.ツクバエンシスNRRL18488株への導入:実施例4dでの記載と同じ接合手順によってプラスミド構築物pKC1139−allKをS.ツクバエンシスNRRL18488に導入した。
e)破壊されたallK遺伝子を有するS.ツクバエンシスの安定2次組換え株の選択:アプラマイシン50μg/mlを添加した芽胞形成培地上、28℃で増殖させたベクターpKC1139−allKのS.ツクバエンシス接合完了体を液体TSB培地に接種し、良好な種培養物を生成するためにアプラマイシン50μg/mlを添加して振とうフラスコ中、28℃、210rpmで増殖させた。24時間後、種培養物をアプラマイシン50μg/mlを添加した新鮮TSB培地を含む新たな振とうフラスコ中に継代培養し、34℃より高く温度を上昇させて210rpmで増殖させた。34℃より高い温度ではpKC1139に基づくベクターは複製できず、S.ツクバエンシスゲノムに組み込まれ、それにより一次組換え体が得られる。次いで培養物を、アプラマイシンを含まない新鮮TSB培地を含む新たな振とうフラスコ中に数回継代培養し、相同組換えによってpKC1139に基づくベクターが除去され、次いで芽胞形成培地に播種し、28℃で増殖させた。回収した芽胞をろ過し、系列希釈物を芽胞形成培地のプレート上に作製した。5−8日後、芽胞形成培地のプレート上およびアプラマイシンを含む芽胞形成培地のプレート上に単一コロニーをパッチした。一次組換え体は、アプラマイシンにまだ耐性である一方で2次組換え体はアプラマイシン耐性を失っており、アプラマイシンを含む芽胞形成培地上で増殖できない。allK遺伝子の所望のインフレーム欠失を有する真の2次組換え変異体を、実施例4fに記載のサザンハイブリダイゼーションを使用して野生型復帰変異体と照らし合わせて同定した。
f)pKC1139−allKを使用する2次相同組換えから得られたallK破壊変異体の発酵でのタクロリムス生成:allK破壊変異体についてタクロリムス生成を、抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
g)allK破壊変異体による発酵でのタクロリムス生成:タクロリムス生成allK破壊変異体を実施例4gに記載のとおり実施した。供給実験のためにアリルマロニルCoAおよびエチルマロニルCoA前駆体、アリルマロニルSNAC、アリルマロニルジSNACおよびエチルマロニルSNACを濃度5−20mMで発酵培地に添加した。
h)pKC1139−allKを使用する2次組換えから得られたallK破壊変異体のHPLCでのタクロリムスおよびアスコマイシン生成の判定:allK破壊変異体のタクロリムスおよびアスコマイシン生成の判定のために実施例4hの記載と同じ方法を使用した。
【0157】
【表9】
【0158】
破壊されたallK遺伝子を有する単離された2次組換え変異体は、タクロリムスおよびジヒドロタクロリムス生成の完全な消失を生じた。アリルマロニルSNACまたはアリルマロニルジSNACを発酵培地に添加した場合、タクロリムスの生成が再確立された。同様にエチルマロニルSNACを添加した場合、アスコマイシン生成が大幅に増加した。
【0159】
実施例7:
AllP遺伝子の過剰発現用のベクターの構築、過剰発現されたAllPを有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)AllP遺伝子の化学合成:454−全ゲノム配列決定に基づいてS.ツクバエンシスNRRL18488のゲノムのDNA配列を、ORFの5’末端にNdeI制限酵素部位およびORFの3’末端にXbaI制限酵素部位を付与するAllP遺伝子のDNA配列を設計および合成するために使用した。
b)AllP遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:合成したAllP遺伝子を制限酵素NdeIおよびXbaIによって切断した。Erm*プロモーター配列をpSE152ベクターのマルチクローニング部位にEcoRIおよびXbaI制限酵素部位を使用して予め導入し、NdeI部位をプロモーター配列の3’末端に作製した。このベクターをNdeIおよびXbaIで切断し、AllP遺伝子のNdeIおよびXbaI断片と連結してpSET−152−AllPプラスミドを作製した。
c)S.ツクバエンシスNRRL18488株へのAllP遺伝子の過剰発現:
プラスミド構築物pSET152−AllPをS.ツクバエンシスNRRL18488株に実施例4dに記載の接合手順によって導入した。
d)pSET152−AllPプラスミドを付与するS.ツクバエンシス変異体のタクロリムスおよびアスコマイシン生成の判定
タクロリムスおよびアスコマイシン生成の判定のためにHPLC法を実施例4hに記載のとおり使用した。
【0160】
【表10】
【0161】
AllP遺伝子が過剰発現された単離された組換え変異体は、野生型株ストレプトマイセス・ツクバエンシスNRRL18488と比較して25%増加したタクロリムス生成を示した。
【0162】
実施例8:
ecm遺伝子の過剰発現のためのベクターの構築、過剰発現されたecm遺伝子を有するS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)ecm遺伝子の化学合成:454−全ゲノム配列決定に基づいてS.ツクバエンシスNRRL18488のゲノムのDNA配列を、ORFの5’末端にNdel制限酵素部位およびORFの3’末端にXbaI制限酵素部位を付与するecm遺伝子のDNA配列を設計および化学合成するために使用した。
b)AllP遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:合成したecm遺伝子断片を制限酵素NdeIおよびXbaIによって切断した。Erm*プロモーター配列をpSET152ベクターのマルチクローニング部位にEcoRIおよびXbaI制限酵素部位を使用して予め導入し、NdeI部位を前記プロモーター配列の3’末端に追加した。このベクターをNdeIおよびXbaIで切断し、AllP遺伝子のNdeIおよびXbaI断片と連結してpSET−152−ecm構築物を作製した。
c)S.ツクバエンシスNRRL18488株へのecm遺伝子の過剰発現のためのベクターの導入:
プラスミド構築物pSET152−ecmをS.ツクバエンシスNRRL18488株に実施例4dに記載の接合手順によって導入した。
d)ecm過剰発現変異体におけるタクロリムスおよびアスコマイシン生成のHPLC分析
ecm過剰発現変異体のタクロリムスおよびアスコマイシン生成の判定のために実施例4hに記載のとおりの方法を使用した。
【0163】
【表11】
【0164】
過剰発現されたecm遺伝子を有する単離された組換え変異体は、野生型株ストレプトマイセス・ツクバエンシスNRRL18488と比較して20%減少したアスコマイシン生成を示した。
【0165】
実施例9:
allAおよびallK遺伝子の過剰発現のためのベクターの構築、このベクターを有するS.ツクバエンシスNRRL18488のallK不活性化株の相補性ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:実施例4に記載のとおり、454に基づく全ゲノム配列決定をS.ツクバエンシスのゲノムに使用し、それはallAからallD遺伝子のallサブクラスター領域の増幅のためのプライマーの設計を可能にした。それによりNdeI制限酵素部位を有するAT−exp−F1(ACATATGCTCGGGTCGTTCGTTACGGGGAG)およびXbaI制限酵素部位を有するAT―exp―R1(ATCTAGAACGTGGGTCATCGGCTGGTCCTTG)をこれら2つのORF(allAおよびallK)を増幅するために作製した。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAをBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR反応はPhusionポリメラーゼ(Finnzymes)および製造者によって提供された緩衝液で、最終容量50μl中の200μM dNTP、3%DMSO、0.5μMの各プライマー、鋳型S.ツクバエンシスゲノムDNAおよそ50ngおよび酵素2.5単位の存在下で30サイクル実施した。最初の5サイクルの温度プロファイルは、98℃、15秒間(変性ステップ)、65℃、30秒間(アニーリングステップ)および72℃、3分15秒間(伸長ステップ)であった。残りの25サイクルの温度プロファイルは98℃、15秒間(変性ステップ)、60℃、30秒間(アニーリングステップ)および72℃、3分15秒間(伸長ステップ)であった。PCR増幅産物をpUC19クローニングベクターにクローン化した。クローン化したPCR産物の配列分析によってallAおよびallK遺伝子の配列を確認した。
c)allAおよびallK遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:3.9kb挿入物を制限酵素NdeIおよびXbaIによってpUC19ベクターから切り出した。ErmE*プロモーター配列をpSET152ベクターのマルチクローニング部位にEcoRIおよびXbaI制限酵素部位を使用して予め導入し、NdeI部位をプロモーター配列の3’末端に作製した。このベクターをNdeIおよびXbaIで切断し、続いてallAおよびallK遺伝子を含むNdeIおよびXbaI断片と連結してpSET152−ermE*−allA−Kプラスミドを作製した。
d)実施例6において得た不活性化allK遺伝子を有するS.ツクバエンシス株へのallAおよびallK遺伝子の導入:プラスミド構築物pSET152−ermE*−allA−KをS.ツクバエンシスNRRL18488のallK不活性化株に実施例4dに記載の接合手順によって導入した。
e)pSET152−ermE*−allA−Kベクターで形質転換したallK破壊変異体の発酵でのタクロリムス生成:形質転換した変異体に関してタクロリムス生成を、抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
f)pSET152−ermE*−allA−Kプラスミドを付与されたallK不活性化S.ツクバエンシス変異体のタクロリムスおよびアスコマイシン生成の判定
【0166】
【表12】
【0167】
allK遺伝子活性の減失がallAおよびallKの過剰発現によって相補されたallK不活性化株の単離された組換え変異体は、回復したタクロリムス生成を示した。
【0168】
実施例10:
allA、allK、allR、allD遺伝子の過剰発現のためのベクターの構築、このベクターを有するS.ツクバエンシスNRRL18488のallR不活性化株の相補性ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:実施例4に記載のとおり、454に基づく全ゲノム配列決定をS.ツクバエンシスのゲノムに使用し、それはallAからallD遺伝子のallサブクラスター領域の増幅のためのプライマーの設計を可能にした。それによりNdeI制限酵素部位を有するAT−exp−F1(ACATATGCTCGGGTCGTTCGTTACGGGGAG)およびXbaI制限酵素部位を有するAT―exp―R2(ATCTAGAACATGCCCTAGGTACGTTTCGCGG)をこれら4つのORF(allA、allK、allRおよびallD)を増幅するために作製した。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAをBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR反応はPhusionポリメラーゼ(Finnzymes)および製造者によって提供された緩衝液で、最終容量50μl中の200μM dNTP、3%DMSO、0.5μMの各プライマー、鋳型S.ツクバエンシスゲノムDNAおよそ50ngおよび酵素2.5単位の存在下で30サイクル実施した。最初の5サイクルの温度プロファイルは、98℃、15秒間(変性ステップ)、65℃、30秒間(アニーリングステップ)および72℃、3分15秒間(伸長ステップ)であった。残りの25サイクルの温度プロファイルは98℃、15秒間(変性ステップ)、60℃、30秒間(アニーリングステップ)および72℃、3分15秒間(伸長ステップ)であった。PCR増幅産物をpUC19クローニングベクターにクローン化した。クローン化したPCR産物の配列分析によってallA、allK、allRおよびallD遺伝子の配列を確認した。
c)allA、allK、allRおよびallD遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:6.4kb挿入物を制限酵素NdeIおよびXbaIによってpUC19ベクターから切り出した。ErmE*プロモーター配列をpSET152ベクターのマルチクローニング部位にEcoRIおよびXbaI制限酵素部位を使用して予め導入し、NdeI部位をプロモーター配列の3’末端に作製した。このベクターをNdeIおよびXbaIで切断し、続いてallA、allK、allRおよびallD遺伝子を含むNdeIおよびXbaI断片と連結してpSET152−ermE*−allA−Dプラスミドを作製した。
d)実施例5において得た不活性化allR遺伝子を有するS.ツクバエンシス株へのallA、allK、allRおよびallD遺伝子の導入:プラスミド構築物pSET152−ermE*−allA−DをS.ツクバエンシスNRRL18488のallR不活性化株に実施例4dに記載の接合手順によって導入した。
e)pSET152−ermE*−allA−Dベクターで形質転換したallR不活性化変異体の発酵でのタクロリムス生成:形質転換した変異体に関してタクロリムス生成を、抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
f)pSET152−ermE*−allA−Dプラスミドを付与されたallR不活性化S.ツクバエンシス変異体のタクロリムスおよびアスコマイシン生成の判定
【0169】
【表13】
【0170】
allR遺伝子活性の減失がallA、allK、allRおよびallDの過剰発現によって相補されたallR不活性化株の単離された組換え変異体は、タクロリムスおよびアスコマイシンの回復した生成を示した。
【0171】
実施例11:
プラスミドpSET152−ermE*−allA−Dのストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891株への導入、この株でのallA、allK、allRおよびallD遺伝子の過剰発現ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)allA、allK、allRおよびallD遺伝子の過剰発現のためのpSET152に基づく発現ベクターの構築:pSET152−ermE*−allA−Dプラスミドを実施例10に記載のとおり調製した。
b)allA、allK、allRおよびallD遺伝子の、ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891への導入:プラスミド構築物pSET152−ermE*−allA−Dを、接合性プラスミドpUZ8002を含有するエレクトロコンピテントE.コリ株ET12567への形質転換によって導入した(Pagetら、1999 J.Bacteriol.181、204−211頁)。プラスミドpUZ8002は接合性線毛の構築に必要な全ての遺伝子を含有しているが、転移の起点を欠いており、したがって宿主細胞に残留する(Jonesら、1997 Mol.Microbiol.23:169−178頁)。ストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891の接合手順をKieserら、2000(Practical Streptomyces genetics、A laboratory Manual.ISBN0−7084−0623−8)に記載のとおり行った。接合完了体は、アプラマイシン50μg/mlを添加した実施例1に記載の芽胞形成培地上、28℃で増殖させた。
c)pSET152−ermE*−allA−Dベクターで形質転換した株の発酵でのアスコマイシンおよびタクロリムス生成:形質転換した変異体のアスコマイシンおよびタクロリムス生成のための発酵プロセスは抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
d)allA、allK、allRおよびallD遺伝子を過剰発現しているストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891変異体のLC−MS/MSでのタクロリムスおよびアスコマイシンの生成の判定:
【0172】
試料調製:十分に振とうした培地5mlにメタノール5mlを加え、試料を抽出するために試料を1時間振とう機に置いた。抽出後、培地のメタノール抽出物1mlを1.5mlチューブに取り、10分間、14000rpmで遠心分離した。上清0.8mlをLC−MS/MS分析を実施するためにバイアルに移した。
【0173】
タクロリムスおよびアスコマイシン判定のための方法:培養培地中のFK506およびFK520の同一性をLC−MS/MS分析によって判定した。本発明者らは、Phenomenexからの逆相カラム(Gemini C18 column、5μm、150mm×2mm i.d.)を使用するWatters Micromass Quattro micro detectorと共にAgilent 1100 series LC−MS systemを使用した。分離は、流速0.250ml/分、溶媒Aとして0.5%TFA、溶媒Bとしてアセトニトリルでの勾配溶出によって実施した。勾配プログラムは:60%A、0分;60−20%A、0−17分;20−60%A、17−18分;60%A、18−30分であり、注入容量10μL、カラム温度45℃を使用した。質量選択検出器(Waters、Quattro micro API)は分析物の陽イオン化にコーン電圧20Vおよびキャピラリー電圧3.5kVを使用するエレクトロスプレーイオン化を備えていた。乾燥窒素を350℃に加熱し、乾燥ガス流量は400l/時間に、衝突エネルギーは20eVにした。ESI+ポジティブモードにおいては、FK506のナトリウムイオン([M+Na]+)の捕獲に対応するm/z=826.5のイオンが最も強く、他の調査者の結果と一致した(Yuan J.ら、2008、J.Chromatogr.、B.Anal.Technol.Biomed Life Sci.868、34−41頁を参照されたい。)。FK506同一性確認に関して、多重反応モニタリングモードを使用し、遷移FK506 m/z=826.5[M+Na]+→m/z=616.4を記録した。同様にFK520モニタリングについても遷移m/z=814.5[M+Na]+→m/z=604.4を記録した。タクロリムスはおよそ18.2分に溶出され、アスコマイシンはおよそ17.5−17.7分に溶出された。FK506とFK520との存在量は判定できず、したがって両化合物の存在または不在だけが確立された(図2)。
【0174】
【表14】
【0175】
allA、allK、allRおよびallD遺伝子が過剰発現されているストレプトマイセス・ハイグロスコピカス変種アスコマイセティカスATCC14891の単離された組換え変異体は、この株で通常生成されるアスコマイシンに加えてタクロリムスの生成も示した。
【0176】
実施例12:
allサブクラスターの大部分(allDからallS)の欠失のためのベクターの構築、allサブクラスターのこの部分を欠失しているS.ツクバエンシスNRRL18488の変異株の構築ならびに前記変異体のタクロリムスおよびアスコマイシン生成の分析
a)プライマーの設計:実施例4に記載のとおり、454に基づく全ゲノム配列決定をS.ツクバエンシスのゲノムに使用し、それはallD遺伝子の上流およびallS遺伝子の下流の領域の既知のDNA配列に基づいてallDとallS遺伝子との間の領域に隣接する領域の増幅のためのプライマーを設計することを可能にした。これにより、BamHI制限酵素部位を有するKS−delF2b(TGGATCCGGCGCGTATCGCCAACCGCTAC)およびXbaI制限酵素部位を有するAllRexpR1(ATCTAGAGGCGTTCGGATTCGCTCACCG)をallD遺伝子の上流領域を増幅するために作製し、XbaIを有するClusDelF2(ATCTAGAGGGCGGATACGTCACCGGCG)およびHindIII制限酵素部位を有するClusDelR2b(CCAAGCTTATGGAGATCATCGAAGGCAGC)をallS遺伝子の下流領域を増幅するために作製した。プライマーAllRexpR1とClusDelF2の間、6262bpの間隙をallD、allM、allN、allP、allOおよびallS遺伝子の欠失のために作製した。
b)DNA断片のPCR増幅:実施例2で得られたS.ツクバエンシスゲノムDNAをBiorad iCycler Thermal Cyclerを使用してPCR増幅した。PCR反応はPhusionポリメラーゼ(Finnzymes)および製造者によって提供された緩衝液で、最終容量50μl中の200μM dNTP、3%DMSO、0.5μMの各プライマー、鋳型S.ツクバエンシスゲノムDNAおよそ50ngおよび酵素2.5単位の存在下で30サイクル実施した。最初の5サイクルの温度プロファイルは、98℃、15秒間(変性ステップ)、65℃、30秒間(アニーリングステップ)および72℃、1分間(伸長ステップ)であった。残りの25サイクルの温度プロファイルは98℃、15秒間(変性ステップ)、60℃、30秒間(アニーリングステップ)および72℃、1分間(伸長ステップ)であった。PCR増幅産物をpUC19クローニングベクターにクローン化した。クローン化したPCR産物の配列分析によって増幅された上流および下流領域の配列を確認した。
c)相同組換えによるS.ツクバエンシスゲノムのallD−allS領域の破壊に使用するpKC1139に基づく温度感受性ベクターの構築:隣接領域のKS−delF2b−AllRexpR1およびClusDelF2−ClusDelR2の増幅断片をそれぞれ制限酵素BamHIとXbaIとによっておよびXbaIとHindIIIとによってpUC19から切り出した。ベクターpKC1139は、予め制限酵素BamHIおよびHindIIIで切断し、2つの断片を組合せ、各標的ベクターに連結し、pKC1139−ClusDelを作製した。
d)allサブクラスターのallD−allS領域の欠失のためのベクターの、S.ツクバエンシスNRRL18488株への導入:プラスミド構築物pKC1139−ClusDelをS.ツクバエンシスNRRL18488株に実施例4dの記載と同じ接合手順によって導入した。
e)allD−allS領域を欠失しているS.ツクバエンシスの安定2次組換え株の選択:2次組換え体の選択は、実施例6dの記載と同じ手順を使用して実施した。
f)pKC1139−ClusDelを使用する2次相同組換えによって得たallD−allS領域を欠失している変異体の発酵でのタクロリムス生成:allD−allS欠失変異体のタクロリムス生成のための発酵プロセスを、抗生物質を増殖培地に添加せずに実施例4gに記載のとおり実施した。
g)pKC1139−ClusDelを使用する2次組換えによって得たallD−allS欠失変異体のHPLCでのタクロリムシスおよびアスコマイシン生成の判定:allD−allS欠失変異体のタクロリムスおよびアスコマイシン生成の判定のために、実施例4hにおける記載と同じ方法を使用した。
【0177】
【表15】
【0178】
破壊されたallD−allS領域を有する単離された2次組換え変異体は、タクロリムスおよびアスコマイシン生成の完全な消失を生じた。
【特許請求の範囲】
【請求項1】
微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、タクロリムスの調製のための方法。
【請求項2】
微生物の遺伝子改変株の培養ステップを含み、前記微生物がストレプトマイセス(Streptomyces)属から選択される細菌である、請求項1に記載の方法。
【請求項3】
前記微生物がストレプトマイセス・ツクバエンシス(Streptomyces tsukubaensis)の遺伝子改変株から選択される、請求項1または2のいずれかに記載の方法。
【請求項4】
微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が配列番号11に記載のクロトニルCoA還元酵素(ccr)遺伝子および配列番号12に記載のエチルマロニルCoAムターゼ(ecm)から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、請求項1から3のいずれかに記載の方法。
【請求項5】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が、配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、請求項1から4のいずれかに記載の方法。
【請求項6】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される、請求項1から5のいずれかに記載の方法。
【請求項7】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子および配列番号8に記載のAllP遺伝子の群から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、請求項1から6のいずれかに記載の方法。
【請求項8】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が配列番号4に記載のAllR遺伝子および配列番号8に記載のAllP遺伝子の群から選択される少なくとも1つの過剰発現された遺伝子を含む、請求項1から7のいずれかに記載の方法。
【請求項9】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が配列番号3に記載のAllK遺伝子および配列番号4に記載のAllR遺伝子の群から選択される少なくとも1つの不活性化された遺伝子を含み、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される、請求項1から8のいずれかに記載の方法。
【請求項10】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が不活性化AllR遺伝子および不活性化ccr遺伝子を含み、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される、請求項1から9のいずれかに記載の方法。
【請求項11】
微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、ストレプトマイセス属に属する微生物の遺伝子改変株。
【請求項12】
微生物の遺伝物質が、配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、請求項11に記載の微生物の遺伝子改変株。
【請求項13】
天然のまたは遺伝子操作されたポリケチド合成活性を有する、請求項11または12のいずれかに記載の微生物の遺伝子改変株。
【請求項14】
配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載の配列ならびに1つまたは複数のヌクレオチド追加、欠失、置換および/または逆位を含むこれらの変種を含む、ストレプトマイセス属に属する微生物のエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する1つまたは複数の遺伝子をコードするヌクレオチド配列。
【請求項15】
配列番号1と少なくとも50%のヌクレオチド同一性を有する、請求項14に記載のヌクレオチド配列。
【請求項1】
微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、タクロリムスの調製のための方法。
【請求項2】
微生物の遺伝子改変株の培養ステップを含み、前記微生物がストレプトマイセス(Streptomyces)属から選択される細菌である、請求項1に記載の方法。
【請求項3】
前記微生物がストレプトマイセス・ツクバエンシス(Streptomyces tsukubaensis)の遺伝子改変株から選択される、請求項1または2のいずれかに記載の方法。
【請求項4】
微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が配列番号11に記載のクロトニルCoA還元酵素(ccr)遺伝子および配列番号12に記載のエチルマロニルCoAムターゼ(ecm)から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、請求項1から3のいずれかに記載の方法。
【請求項5】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が、配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、請求項1から4のいずれかに記載の方法。
【請求項6】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される、請求項1から5のいずれかに記載の方法。
【請求項7】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子および配列番号8に記載のAllP遺伝子の群から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、請求項1から6のいずれかに記載の方法。
【請求項8】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が配列番号4に記載のAllR遺伝子および配列番号8に記載のAllP遺伝子の群から選択される少なくとも1つの過剰発現された遺伝子を含む、請求項1から7のいずれかに記載の方法。
【請求項9】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が配列番号3に記載のAllK遺伝子および配列番号4に記載のAllR遺伝子の群から選択される少なくとも1つの不活性化された遺伝子を含み、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される、請求項1から8のいずれかに記載の方法。
【請求項10】
ストレプトマイセス属に属する微生物の遺伝子改変株の培養ステップを含み、前記微生物の遺伝物質が不活性化AllR遺伝子および不活性化ccr遺伝子を含み、アリルマロニルCoAおよび/またはアリルマロニルCoAの少なくとも1つの前駆体の外部添加の下で実施される、請求項1から9のいずれかに記載の方法。
【請求項11】
微生物の遺伝物質がエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、ストレプトマイセス属に属する微生物の遺伝子改変株。
【請求項12】
微生物の遺伝物質が、配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載のAllylサブクラスターの遺伝子配列から選択される少なくとも1つの不活性化および/または過剰発現された遺伝子を含む、請求項11に記載の微生物の遺伝子改変株。
【請求項13】
天然のまたは遺伝子操作されたポリケチド合成活性を有する、請求項11または12のいずれかに記載の微生物の遺伝子改変株。
【請求項14】
配列番号2に記載のAllA遺伝子、配列番号3に記載のAllK遺伝子、配列番号4に記載のAllR遺伝子、配列番号5に記載のAllD遺伝子、配列番号6に記載のAllM遺伝子、配列番号7に記載のAllN遺伝子、配列番号8に記載のAllP遺伝子、配列番号9に記載のAllO遺伝子および配列番号10に記載のAllS遺伝子から構成される配列番号1に記載の配列ならびに1つまたは複数のヌクレオチド追加、欠失、置換および/または逆位を含むこれらの変種を含む、ストレプトマイセス属に属する微生物のエチルマロニルCoAおよび/またはアリルマロニルCoAの代謝および/または生合成に関与する1つまたは複数の遺伝子をコードするヌクレオチド配列。
【請求項15】
配列番号1と少なくとも50%のヌクレオチド同一性を有する、請求項14に記載のヌクレオチド配列。
【図1】

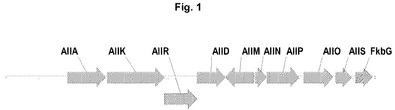
【図2】
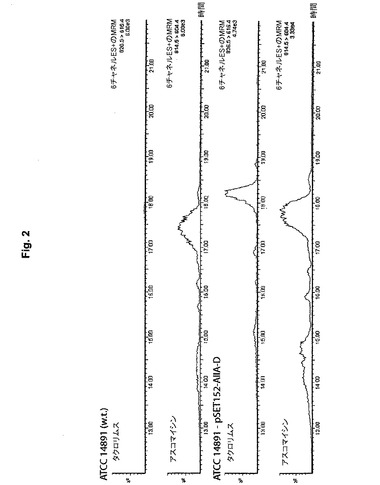
【図2】
【公表番号】特表2012−532595(P2012−532595A)
【公表日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願番号】特願2012−519011(P2012−519011)
【出願日】平成22年7月9日(2010.7.9)
【国際出願番号】PCT/EP2010/059896
【国際公開番号】WO2011/004008
【国際公開日】平成23年1月13日(2011.1.13)
【出願人】(504359293)レツク・フアーマシユーテイカルズ・デー・デー (60)
【Fターム(参考)】
【公表日】平成24年12月20日(2012.12.20)
【国際特許分類】
【出願日】平成22年7月9日(2010.7.9)
【国際出願番号】PCT/EP2010/059896
【国際公開番号】WO2011/004008
【国際公開日】平成23年1月13日(2011.1.13)
【出願人】(504359293)レツク・フアーマシユーテイカルズ・デー・デー (60)
【Fターム(参考)】
[ Back to top ]