
タンパク質と糖鎖との相互作用を分析する方法
糖鎖に相互作用を示すタンパク質の基板への固定化方法の最適条件を検討した結果、スライドガラス表面にGTMSコートを施すことにより、従来よりS/N比の高い条件で固定化されることが判明した。さらに、複数の穴を有するラバーが貼り付けられた複数の反応槽を有する基板を用いること、また、レクチンをスポットし、PBSTで洗浄することで、糖鎖−レクチン間の弱い相互作用をより感度良く検出することに成功した。また、エバネッセント励起方式のスキャナーを導入することにより、プローブ溶液の洗浄除去操作を行うことなく、レクチン−糖鎖間の相互作用を検出することが可能となった。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、タンパク質と糖鎖との相互作用を分析する方法に関する。
【背景技術】
【0002】
生体の生命機能を担う主役であるタンパク質が、細胞社会の中において秩序正しく機能を発揮するためには、糖鎖修飾をはじめとする翻訳後修飾が大変に重要な役割を担っている。生体内のほとんどのタンパク質は糖鎖による修飾をうけており、タンパク質に付加した糖鎖がウィルスの感染、原虫の寄生、感染、毒素の結合、ホルモンの結合、受精、発生分化、タンパク質の安定性、がん細胞転移、アポトーシスなど、生命現象の様々な場で重要な役割を果たしていることが近年次々と明らかになってきた。
【0003】
糖鎖機能の解析のためには、まずその糖鎖の構造解析が欠かせない。今後も糖鎖構造解析法の重要性は増すことが予想される。しかし糖鎖の構造解析は多大な時間と労力、経験を要することから、従来の手法に基づき完全な構造決定を目指すのではなく、より簡便に、高速、高感度、かつ高精度に多彩な糖鎖構造の特徴を抽出し、相互識別できるシステム開発が期待されていた。
【0004】
マイクロアレイは多種のDNA・タンパク質等の固定化試料を高密度に固相担体(ガラス・メンブレン・シリコンチップ)上にスポット状に固定化したものの総称であり、固定化した多種の試料スポットに対して特異的に結合する分子(以下プローブ)の有無を検出することが可能である。プローブ分子には一般的には蛍光標識化されたものが用いられており、プローブ溶液をアレイ表面と反応させた後に、蛍光検出スキャナーで観察することにより各試料スポットに結合したプローブ分子の定量的解析を行うことができる。米国Affimetrix社がDNAマイクロアレイを開発して以来、マイクロアレイは非常に広範な研究分野において使用され、人類に様々な新しい知見をもたらしてきた。
【0005】
第三の生命鎖と呼ばれる糖鎖の構造・機能情報を研究する上で、マイクロアレイを利用した糖鎖と糖鎖に相互作用を示すタンパク質(糖結合性タンパク質、例えば、レクチン等)の迅速かつ高感度な大規模相互作用解析が可能となれば、基礎研究から医療診断や産業応用に至るまで、広範囲に利用されうる非常に有用なツールになることが考えられる。
【0006】
糖鎖と糖鎖に相互作用を示すタンパク質間の結合は抗原抗体反応の一般的な解離定数(Kd=10−8以下)等に比べて、一般的に弱い相互作用であることが知られており、これらの解離定数(Kd)は10−6Mかそれ以上であることが多い。また糖鎖と糖鎖に相互作用を示すタンパク質間の相互作用は比較的速い解離−会合反応から成り立っていることが知られており、結果的に一般的なタンパク質間相互作用や相補的ヌクレオチド断片間の相互作用に比べ、洗浄操作などにより解離側に平衡が傾きやすい。例えば、レクチンを糖タンパク質固定化カラム等にて精製を行う際にも、レクチンの結合が弱い場合は洗浄操作中にレクチンがカラム外に流出してしまう現象がしばしば観察される。
【0007】
従来のスライドガラスを用いた一般的なマイクロアレイ技術では、プローブ溶液を固定化試料と接触させて結合反応に至らしめる過程後にプローブ溶液を洗浄・除去し、ジェットガスや遠心機を用いてスライドガラスに付着している水分を完全に除去する操作を行った後に、マイクロアレイスキャナーを用いてイメージングを行う。これは一般的なマイクロアレイリーダーが、水分の付着した状態でスライドガラス上の蛍光を観察することが出来ないことに起因している。このようにスキャニングの前段階でプローブ溶液を除去しても、相補的ヌクレオチド断片や抗原−抗体反応等、強い結合を示す相互作用においては解離速度定数が十分に小さいため、プローブ分子の解離反応は容易に進行しないと考えられる。しかし、糖鎖と糖鎖に相互作用を示すタンパク質間に一般的に見られる解離速度定数の大きい、すなわち弱い相互作用の観察に際しては、プローブ溶液の除去・洗浄操作を行う時点で糖鎖と糖鎖に相互作用を示すタンパク質間の解離反応が進行してしまい、平衡状態下での正確な相互作用情報を得ることが困難になる。このため、マイクロアレイにおいて糖鎖と糖鎖に相互作用を示すタンパク質間の平衡状態での相互作用情報を精密に解析する場合、このプローブ溶液の洗浄操作が大きな問題点となる。
【0008】
DNAマイクロアレイは現在大変広範囲に利用されている。タンパク質のマイクロアレイについてもDNAの転写産物であるタンパク質の生体内での機能を解明する等の基礎研究分野やタンパク質の量的・質的な変化に基づいた診断・判断等の応用分野においても、将来の利用が期待されており、研究面においても世界中で活発な研究が行われてきた。しかしタンパク質マイクロアレイの開発・普及はDNAマイクロアレイに比べ、圧倒的に遅れているのが現状である。この原因の一つとして、様々に異なる性質を持つタンパク質試料を、活性を保ったままの状態で、一定の割合で固定化していくという工程が技術的に大変に難しいということが、以前より多くの研究者によって指摘されている。
【0009】
タンパク質をアレイ上に固定化する方法として、最も初期に開発された方法としてPVDF膜(非特許文献1)に代表されるメンブレン上にタンパク質を物理的に吸着させる方法が挙げられる。転写因子など一部のタンパク質では、ある程度の活性を維持できると報告されているが、一般性には乏しい。またメンブレン上に固定化する場合、アレイの高密度化に限界があった。高密度化を達成するために、金属やガラスなどの固体表面に対してタンパク質を固定化していく方向で研究が進められてきたが、一般にタンパク質は金属やガラスなどの固体表面への接触によって変性しやすいという性質を持つ。このため、固体表面とタンパク質を架橋するための何らかのリンカーを使用する固定化方法が、鋭意研究・開発されてきた。
【0010】
タンパク質の変性の問題を軽減する手法の一例としては、スライドガラス上に厚さ10〜100μmのポリアクリルアミドのパッドを接合して、これにタンパク質をスポットする手法が挙げられる(非特許文献2および3)。この場合、タンパク質は3次元空間に固定化されるために、2次元表面に固定化する手法に比べて、量的に100倍以上の向上が望めるという。また、タンパク質を多孔性ポリアクリルアミドゲル内にアミノ基を介して固定化する方法もある(非特許文献4)。しかしこれらの方法は高価で特殊なスライドガラスを作製する必要があり、一般的な普及には至っていない。また固定化されたタンパク質の層に厚みがあることは、検出法によっては好ましくないこともある。
【0011】
現在最も精力的に研究されている固相へのタンパク質固定化法として、何らかのタグを付加した形でタンパク質を発現し、タンパク質のタグ部分を介した固相担体への固定化を行う方法がある。この方法ではタンパク質の有効リガンド濃度が向上するといった効果やタンパク質の向きをそろえることが原理的に可能であるといわれている。このような手法の一例として、オリゴヒスチジンタグを介して、ニッケル錯体で表面修飾した基板へ固定化する方法や(非特許文献5)、アビジン−ビオチンを介した固定化法(特許文献1)を挙げることができる。
【0012】
このような方法はタンパク質の活性を保ったまま固定化する、あるいは固定化率をそろえるという点で有効な方法であると考えられる。しかしマイクロアレイに固定化しようとするすべてのタンパク質について遺伝子レベルでタグを付加し、大腸菌、ないし無細胞系等で発現及び精製を行うことは多額のコストと莫大な労力が必要となると考えられる点で、現時点で一般的な研究者が自在かつ個々の要求に合わせる形で気軽に利用することは難しい。
【0013】
これに対し、タンパク質の官能基を固相担体との固定化に利用する方法は、天然から抽出したままのタンパク質や市販のタンパク質試料をそのまま固定化し、マイクロアレイに利用できるという特徴をもつ。タンパク質中のアミノ基を介した固相担体への固定化方法としては、固相表面に結合させた活性エステル基を介してタンパク質を固定化する方法や、固相表面に配置したエポキシ基を介してタンパク質を固定化する手法が挙げられる(非特許文献6)。タンパク質をアミノ基を介して固定化する方法はシンプルな方法であるが、市販のタンパク質や生体抽出・成分、特定のタグのない組み換えタンパク質等においても簡便に固定化することができるため、個々の使用者が自分の目的に合わせ、自在にタンパク質を選択し、迅速かつ安価に目的にあったマイクロアレイに最適化して使用することができる。タンパク質のアミノ基を介した固定化方法の欠点としては、タンパク質中のリジン残基の数が個々のタンパク質によって異なることや、固定化に用いられるリジン残基の位置によってはタンパク質の不活化が起こる可能性等が挙げられる。
【0014】
【特許文献1】特許出願2001−520104
【特許文献2】特許公開平8−201383
【特許文献3】特許公表2002−544485
【非特許文献1】L.J.Holt,K.Bussow,G.Walter,I.M.Tomlinson,Nucleic Acids Res.,15,E72,2000
【非特許文献2】D.Guschin,G.Yershov,A.Zaslavsky,A.Gemmell,V.Shick,D.Proudnikov,P.Arenkov,A.Mirzabekov,Anal.Biochem.,250,203−211,1997
【非特許文献3】A.Lueking,M.Horn,H.Eickhoff,K.Bussow,H.Lehrach,G.Walter,Anal.Biochem.,270,103−111,1999
【非特許文献4】P.Mitchell,Nat.Biotechnol.,20,225−229,2002
【非特許文献5】H.Zhu,M.Bilgin,R.Bangham,D.Hall,A.Casamayor,P.Bertone,N.Lan,R.Jansen,S.Bidlingmaier,T.Houfek,T.Mitchell,P.Miller,R.A.Dean,M.Gerstein,M.Snyder,Science,293,2101−2105,2001
【非特許文献6】H.Zhu,J.F.Klemic,S.Chang,P.Bertone,A.Casamayor,K.G.Klemic,D.Smith,M.Gerstein,M.A.Reed,M.Snyder,Nat.Genetics.26,283−289,2000
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明の目的は、より簡便に、高速、高感度、かつ高精度に糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法を提供することにある。
【課題を解決するための手段】
【0016】
本発明者らは、上記の課題を解決するために鋭意研究を行った。
タンパク質のアミノ基を介した固定化方法では、一般的にタンパク質内のランダムな位置で固相担体との架橋が起こるため、ある確率で活性が損なわれてしまう分子もありうるが、逆に、ある確率でタンパク質の活性が生き残る可能性が高い(タンパク質が全滅する危険性が小さい)。多数の性質の異なるタンパク質や性質が未知であるタンパク質を固定化していくことを想定したマイクロアレイにおいては、このような性質は実用上、非常に有用である。
【0017】
そこで、本発明者らは、レクチンアレイを作製するに当たりマイクロアレイの実用面・応用面での有用性、解析目的対象物に対する即応性、市場での競争力(コスト面も含む)等を総合的に考慮すると現時点ではタンパク質のアミノ基を利用した固定化法が最適であると判断した。本発明ではアミノ基を介した固定化法の最適条件をシグナル−ノイズ比(S/N比)とタンパク質固定化密度の向上を目指し多角的に検討した。
【0018】
その結果、スライドガラス表面に3−グリシドキシプロピルトリメトキシシラン(GTMS)コートを施すことにより、タンパク質試料は、従来製品よりS/N比の高い条件で固定化された。さらに、本発明者らは、複数の穴を有するラバーを貼り付けることで、ガラス上に複数の反応槽を形成させた基板を作成し、レクチンのスポット周囲に正確に蛍光標識化プローブ溶液を満たすことを可能にした。また、レクチンをスポットし、PBSTで洗浄することで、レクチン固定化密度の向上を図り、糖鎖−レクチン間の弱い相互作用をより感度良く検出することに成功した。
【0019】
本発明では、レクチン−糖鎖間の平衡状態での相互作用情報を精密に解析する上での大きな問題点であったプローブ溶液の洗浄除去操作が不要となり、従来法では洗浄時に洗い流されてしまうような弱い相互作用をも検出することが可能となった。
【0020】
このように、固定化方法および検出方法の改良より、レクチンアレイの実用化が可能となった。すなわち本発明は、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法に関し、以下の〔1〕〜〔22〕を提供するものである。
〔1〕以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)糖鎖に相互作用を示すタンパク質が固定化された基板に蛍光標識した被検糖鎖または被検複合糖質を接触させる工程
(b)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
〔2〕糖鎖に相互作用を示すタンパク質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に糖鎖に相互作用を示すタンパク質が固定化された基板である、〔1〕に記載の方法。
〔3〕エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、〔2〕に記載の方法。
〔4〕以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)被検複合糖質が固定化された基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程
(b)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
〔5〕被検複合糖質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に被検複合糖質が固定化された基板である、〔4〕に記載の方法。
〔6〕エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、〔5〕に記載の方法。
〔7〕以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板に被検複合糖質を接触させる工程
(b)工程(a)で得られた基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程
(c)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
〔8〕複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板である、〔7〕に記載の方法。
〔9〕エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、〔8〕に記載の方法。
〔10〕複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、〔7〕〜〔9〕のいずれかに記載の方法。
〔11〕糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、〔1〕〜〔10〕のいずれかに記載の方法。
〔12〕励起光がエバネッセント波である、〔1〕〜〔11〕のいずれかに記載の方法。
〔13〕複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、〔1〕〜〔12〕のいずれかに記載の方法。
〔14〕エポキシ基を活性基として有する化合物がコートされた基板であって、糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板。
〔15〕エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、〔14〕に記載の基板。
〔16〕複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、〔14〕または〔15〕に記載の基板。
〔17〕糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、〔14〕または〔15〕に記載の基板。
〔18〕複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、〔14〕〜〔17〕のいずれかに記載の基板。
〔19〕以下の工程を含む、基板の製造方法。
(a)基板にエポキシ基を活性基として有する化合物をコートする工程
(b)工程(a)で得られた基板に糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質を固定化する工程
〔20〕複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、〔19〕に記載の方法。
〔21〕糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、〔19〕に記載の方法。
〔22〕複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、〔19〕〜〔21〕のいずれかに記載の方法。
【図面の簡単な説明】
【0021】
【図1】本発明のシステムの構成図である。検出手段は、マイクロアレイスキャナー装置である。
【図2】本発明のシステムにおけるコンピューターの構成図である。記憶手段6には、マイクロアレイスキャナー装置を利用したシステムの処理を実行するためのプログラム61〜63が少なくとも格納されている。記憶手段(データベース)7には、糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報、および/または該相互作用情報の組み合わせパターン情報が格納されている。
【図3】ガラス表面に対するGTMSの反応過程を示す図である。GTMSのアルコキシシリル基は水で加水分解されてシラノール基となる。このシラノール基は不安定で、経時変化により部分的に縮合してオリゴマー状態になり、続いてガラス表面に水素結合的に吸着する。その後、ガラスを乾燥処理することでガラス表面のシラノール基と脱水縮合反応が起こり、強固な共有結合となる。
【図4】本実施例で用いた8つの反応槽を形成させた基板を示す図である。新規設計した8穴ラバーの厚さは1mmであり、専用のアジャスター上でスライドガラスと密着させることによってスポット周囲に正確に蛍光標識化糖プローブ溶液を満たすことが可能となる。反応槽内に満たす試料の最適量は50μLである。
【図5】2種のレクチンを固定化したアレイ上にCy3−ASF溶液を加えた、レクチンアレイの性能実験の概念図である。
【図6】固定化時のレクチン溶液濃度とスポットの蛍光強度との関係を示す図及び写真である。親和定数の高いレクチン−糖鎖間の相互作用の検出においては、スポットするレクチン試料の濃度を1mg/mL以上に高濃度化することがシグナル強度の向上に有効であることが分かった。
【図7】レクチン−糖鎖相互作用の検出と、阻害糖による相互作用への影響を示す図及び写真である。RCA120のスポットにおいて強い蛍光が、またEW29(Ch)のスポットにおいては中程度の蛍光が観察された。
【図8】阻害糖のレクチン−糖鎖相互作用への影響をグラフとして示す図である。ラクトース(競合阻害糖)共存下で実験を行った。共存するラクトース(競合阻害糖)の濃度増加に従って、スポットの蛍光強度が減少していることから、蛍光糖タンパク質プローブの結合はレクチンと糖鎖間の糖特異的結合反応であるということが確認できた。
【図9】各糖鎖関連アレイと糖鎖の相互作用の模式図である。A:レクチンアレイ、およびプローブとして糖鎖を用いた場合の模式図である。蛍光標識糖鎖の構造類推に用いることができる。なお、糖鎖は、間接的に蛍光標識することもできる。B:レクチンアレイ、およびプローブとして糖タンパク質を用いた場合の模式図である。タンパク質上の糖鎖構造の類推に用いることができる。なお、糖鎖は、間接的に蛍光標識することもできる。C:糖ペプチドアレイ、およびプローブとしてレクチンを用いた場合の模式図である。分画したペプチド上の糖鎖構造の類推に用いることができる。D:糖タンパク質アレイ、およびプローブとしてレクチンを用いた場合の模式図である。2次元分画したタンパク質上の糖鎖構造の類推に用いることができる。E:抗体アレイ、およびプローブとしてレクチンを用いた場合の模式図である。クルードな糖タンパク質上の糖鎖構造の類推に用いることができる。
【図10】レクチン−抗体ハイブリッドアレイの実施例を示す写真である。具体的には、7種のレクチンと2種の抗体を固定化したアレイに対して、各モデルタンパク質を蛍光標識化したプローブ溶液を接触させ、平行化した後にエバネッセント励起型スキャナーを用いて観察した結果を示す。
【図11】8つの反応槽を使用して、同一スライド上に異なる濃度の阻害糖を共存させて、相互作用の阻害を観察した結果を示す写真及び図である。(A)RCA120とASFの結合にラクトースを加えたケース、(B)ConAとRNase Bの結合にマンノースを加えたケースである。
【図12】マウス肝由来糖ペプチドサンプルのHPLC分画フラクションを固定化した、糖ペプチドアレイの検出実験の結果を示す図及び写真である。図中、Fucはフコース認識レクチン群、Siaはシアル酸認識レクチン群、Lacはラクトース認識レクチン群、Galはガラクトース認識レクチン群、GalNAcはN−アセチルガラクトサミン認識レクチン群、Manはマンノース認識レクチン群、Chitinはキチン認識レクチン群を意味する。
【図13】レクチンアレイの検出に糖ペプチドプローブを用いる実験の結果を示す写真及び図である。
【図14】生体由来のクルードな試料をプローブ化した際のレクチンアレイによる解析結果を示す写真である。
【発明を実施するための最良の形態】
【0022】
本発明は、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法を提供する。本発明の方法は、糖鎖に相互作用を示すタンパク質が固定化された基板に蛍光標識した被検糖鎖または被検複合糖質を接触させる工程、および、基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程を含む。
【0023】
本発明における糖鎖としては、例えば、糖タンパク質系糖鎖(N−結合型糖鎖とO−結合型糖鎖)、糖脂質系糖鎖、グリコサミノグリカン系糖鎖、または多糖類由来オリゴ糖鎖などが挙げられる。また、1)N−結合型糖鎖としては、高マンノース型・混成型・複合型からなるN−結合型糖鎖など、2)O−結合型糖鎖としては、ムチン型(O−GalNAc)・O−Fuc型・O−Man型・O−Glc型などからなるO−結合型糖鎖など、3)糖脂質系糖鎖としては、ガングリオ系列・グロボ系列ウクト・ネオラクト系列糖鎖など、4)グリコサミノグリカン系糖鎖としては、ヒアルロン酸・ケラタン硫酸・ヘパリン・ヘパラン硫酸・コンドロイチン硫酸・デルマタン硫酸など、5)多糖類由来オリゴ糖鎖としては、キチン、セルロース、カードラン、ラミナリン、デキストラン、デンプン、グリコーゲン、アラビノガラクタン、アルギン酸、フルクタン、フコイダン、キシランなどに由来するオリゴ糖鎖などが例示できる。
【0024】
その他の糖鎖としては、M3・M5A・Hybrid(monoagalacto,bisect)・NA1・NA1(α1−6Fuc)・NA2(monoagalacto)・NA2(monoagalacto,bisect)・NA2・NA2(α1−6Fuc)・A2・NA2(bisect)・NA3・NA3(α1−6Fuc)・NA4・NA4(α1−6Fuc)・NA5(pentaagalacto,bisect)・Lactose・GA2・GA1・GM3−NeuAc・GM3−NeuGc・GM1・GM2・GD1a・GD1b・GD3・Gb3・Gb4・Forssman・LNnT・LNT・Galili pentasaccharide・B−hexasaccharide・LNFP−I・LNFP−II(Lea)・LNFP−III(LeX)・LNFP−II(Leb)・A−hexasaccharide・A−heptasaccharide・B−pentasaccharide・6’Sialyl Lactose・pLNH・βGalLac・βGal2Lac・LN3・GN3・GN4・maltotriose・Sialyl LeXなどを挙げることができる。
【0025】
また、本発明において、複合糖質とは、糖鎖を持つ生体内高分子の総称である。本発明の複合糖質としては、糖タンパク質(糖ペプチドも含む)、プロテオグリカン、糖脂質が挙げられる。
【0026】
本発明の糖鎖に相互作用を示すタンパク質には、糖鎖に相互作用を示すペプチドも含まれる。本発明の糖鎖に相互作用を示すタンパク質としては、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、これらの変異体、または糖鎖に相互作用を示す抗体などが挙げられる。
【0027】
上記レクチンとしては、動・植物、真菌、細菌、ウィルスなどから得られる様々な分子家系に属するレクチン、すなわち、細菌を含むすべての生物界で見出されるリシンB鎖関連の「R型レクチン」、真核生物全般に存在し糖タンパク質のフォールディングに関与する「カルネキシン・カルレティキュリン」、多細胞動物に広く存在し、「セレクチン」、「コレクチン」等代表的なレクチンを多く含むカルシウム要求性の「C型レクチン」、動物界に広く分布しガラクトースに特異性を示す「ガレクチン」、植物豆科で大きな家系を形成する「豆科レクチン」、およびこれと構造類似性をもち動物細胞内輸送に関わる「L型レクチン」、リソソーム酵素の細胞内輸送に関わるマンノース6−リン酸結合性の「P型レクチン」、グリコサミノグリカンをはじめとする酸性糖鎖に結合する「アネキシン」、免疫グロブリン超家系に属し「シグレック」を含む「I型レクチン」などが挙げられる。
【0028】
その他のレクチンとしては、ACA(センニンコクレクチン)・BPL(ムラサキモクワンジュレクチン)・ConA(タチナタマメレクチン)・DBA(Horsegramレクチン)・DSA(ヨウシュチョウセンアサガオレクチン)・ECA(デイゴマメレクチン)・EEL(Spindle Treeレクチン)・GNA(ユキノハナレクチン)・GSL I(グリフォニアマメレクチン)・GSL II(グリフォニアマメレクチン)・HHL(アマリリスレクチン)・ジャカリン(ジャックフルーツレクチン)・LBA(リママメレクチン)・LCA(レンズマメレクチン)・LEL(トマトレクチン)・LTL(ロータスマメレクチン)・MPA(アメリカハリグワレクチン)・NPA(ラッパズイセンレクチン)・PHA−E(インゲンマメレクチン)・PHA−L(インゲンマメレクチン)・PNA(ピーナッツレクチン)・PSA(エンドウレクチン)・PTL−I(シカクマメレクチン)・PTL−II(シカクマメレクチン)・PWM(ヨウシュヤマゴボウレクチン)RCA120(ヒママメレクチン)・SBA(ダイズレクチン)・SJA(エンジュレクチン)・SNA(セイヨウニワトコレクチン)・SSA(ニホンニワトコレクチン)・STL(ジャガイモレクチン)・TJA−I(キカラスウリレクチン)・TJA−II(キカラスウリレクチン)・UDA(Common Stinging Nettleレクチン)・UEA I(ハリエニシダレクチン)・VFA(ソラマメレクチン)・VVA(ヘアリーベッチレクチン)・WFA(フジレクチン)・WGA(パンコムギレクチン)などを挙げることができる。
【0029】
上記糖結合ドメインを有する酵素タンパク質としては、各種グリコシダーゼ(キシラナーゼ、グルカナーゼ)、および糖転移酵素(UDP−GalNAc:ポリペプチドGalNAc転移酵素)などが例示できる。また、糖鎖に親和性を有するサイトカインとしては、インターロイキン−2(IL−2)、インターロイキン−12(IL−12)、腫瘍壊死因子α(TNF−α)、繊維芽細胞成長因子(FGF)などが例示できる。また、糖鎖に相互作用を示す抗体としては、糖鎖関連腫瘍マーカー(CA19−9、フォルスマン抗原、T抗原、Tn抗原、シアリルT抗原)、血液型関連糖鎖(A,B,H,Lea,Lex抗原)、分化関連抗原(Ii,SSEA−1−4)に対する抗体などが例示できる。
【0030】
本発明における基板としては、ガラス、石英ガラス、合成石英ガラスなどが例示できるが、これらに限定されるものではない。また、本発明における糖鎖に相互作用を示すタンパク質が固定化された基板は、好ましくは、エポキシ基を活性基として有する化合物がコートされた基板に糖鎖に相互作用を示すタンパク質が固定化された基板である。
【0031】
エポキシ基を活性基として有する化合物としては、好ましくは3−グリシドキシプロピルトリメトキシシラン(GTMS)が挙げられるが、これに限定されない。その他に、2−(3、4エポキシシクロヘキシル)エチルトリメトキシシラン、3−グリシドキシプロピルメチルジエトキシシラン、3−グリシドキシプロピルトリエトキシシラン、または分岐したスペーサーの先端にエポキシ基を複数持つシランカップリング化合物で、好ましくはスペーサーとしてポリエチレングリコールやタンパク質、ビオチン・アビジン等を含む化合物などが例示できる。
【0032】
本発明の、糖鎖に相互作用を示すタンパク質が固定化された基板は、下記の方法で作成することができる。
【0033】
まず、基板にエポキシ基を活性基として有する化合物をコートする。例えば、エポキシ基を活性基として有する化合物としてGTMSを用いる場合、実施例に記載の方法で行うことができる。具体的には、スライドガラスを10%KOH/MeOH溶液に浸し、容器ごと振盪させた状態で1時間放置しガラス表面を処理し、これを十分量の精製水(ミリQ水)により洗浄した後、60℃のオーブン内で乾燥させる。次にスライドガラスを2%GTMSアセトン溶液に浸し、遮光下で容器ごと振盪させながら1時間反応させる。
GTMSのアルコキシシリル基は水で加水分解されてシラノール基となっており、このシラノール基は不安定で、経時変化により部分的に結合してオリゴマー状態になり、続いてガラス表面に水素結合的に吸着する。反応後、スライドガラスを110℃のオーブン内で8時間乾燥させる。乾燥処理により、ガラス表面のシラノール基と脱水縮合反応が起こり、強力な共有結合となる。一連のGTMSコーティング方法を図3に示す。
【0034】
次に、エポキシ基を活性基として有する化合物をコーティングした基板に、糖鎖に相互作用を示すタンパク質を固定化する。糖鎖に相互作用を示すタンパク質の固定化は、共有結合によって行われる。具体的には、該基板にアミノ基を活性基として有する化合物をスポットし、反応させることで固定化することができる。スポッターとしては、日本レーザ電子社製STAMPMANなどを利用することができる。糖鎖に相互作用を示すタンパク質がレクチンである場合、スポットするレクチンの濃度は、特に限定はないが、1mg/mL以上であることが好ましい。さらに、より好ましくは、スポット処理後Tween20を含むPBS溶液(PBST)で洗浄することにより、未結合レクチンを除去することができる。
【0035】
上記糖鎖に相互作用を示すタンパク質が固定化された基板は、複数の反応槽を形成させた基板であることが好ましい。より好ましくは、複数の穴を有するラバーを貼り付けることで、複数の反応槽を形成させた基板である。一例としては、実施例に記載のように、糖鎖に相互作用を示すタンパク質を固定化したスライドガラスに対し、本発明者らが設計・開発した8穴ラバーを所定の位置に貼り付け、8つの反応槽を作製させる。この8穴ラバーには8つの長方形の穴が規則正しく空いており、スライドガラスに貼り付けたときに8つの反応槽を形成することができる。この反応槽に蛍光標識化プローブ溶液を満たすことで、糖鎖に相互作用を示すタンパク質との接触を円滑に行うことが可能になる。また、この反応槽は8穴ラバーに限定されるものではなく、例えばガラス表面の非スポット領域を撥水コートすることで反応場を形成することも可能である。より好ましくは反応場を多数化することである。本発明においては、同一基板上に複数種の糖鎖に相互作用を示すタンパク質をスポットして用いることもできる。
【0036】
本発明の方法では、上記糖鎖に相互作用を示すタンパク質が固定化された基板に蛍光標識した被検糖鎖または被検複合糖質を接触させる。
【0037】
本発明において、被検糖鎖または被検複合糖質の蛍光標識剤としては、2−アミノピリジン、Cy3、Cy3.5、Cy5、テトラメチルローダミン、フルオレセイン骨格を持つ蛍光色素各種、モレキュラープローブス社製蛍光色素Alexaシリーズ、量子ドット蛍光色素が挙げられるが、糖鎖を蛍光標識する性質を有する物質であれば、これらに限定されない。
【0038】
被検糖鎖または被検複合糖質は、直接的または間接的に蛍光標識することができる。被検糖鎖と予め蛍光標識した糖鎖に相互作用を示すタンパク質を結合させることにより、被検糖鎖を間接的に蛍光標識できる。
【0039】
また、予め蛍光標識したタンパク質であって、被検複合糖質の糖鎖以外の部分に相互作用を示すタンパク質(例えば被検複合糖質の糖鎖以外の部分に相互作用を示す抗体)と被検複合糖質を結合させることにより、被検複合糖質を間接的に蛍光標識できる。
【0040】
また、予め蛍光標識したタンパク質であって糖鎖に相互作用を示すタンパク質と被検複合糖質を結合させることによっても、被検複合糖質を間接的に蛍光標識できる(この場合、被検複合糖質が有する糖鎖のうち、糖鎖に相互作用を示すタンパク質が結合した糖鎖以外の糖鎖が基板に結合する)。
【0041】
例えば、標的複合糖質の糖鎖以外の部分に相互作用を示す予め蛍光標識した抗体を、標的複合糖質試料を含むサンプル・血液・体液・生体抽出成分・食品成分等のクルードな試料に作用させることで、クルードな試料から標的複合糖質を分離・精製することなく選択的に蛍光標識できる。また、予め蛍光標識した上記抗体を作用させたクルードな試料を、例えばレクチンが固定化された基板に接触させることで、標的複合糖質の糖鎖情報だけを選択的に観察できる。
【0042】
本発明の方法では、次いで、基板を洗浄せずに、励起光を作用させて、それぞれの糖鎖に相互作用を示すタンパク質に対する被検糖鎖または被検複合糖質の相互作用を測定する。
【0043】
糖鎖に相互作用を示すタンパク質と被検糖鎖または被検複合糖質の相互作用は一般的に良く知られているタンパク質間相互作用などに比べて弱いため、プローブ溶液の除去・洗浄操作を行うことで糖鎖に相互作用を示すタンパク質と被検糖鎖または被検複合糖質間の解離反応が進行してしまい、平衡状態下での正確な相互作用情報を得ることが出来ないケースが生じていた。
【0044】
本発明者らは上記問題を、プローブ溶液の洗浄を行うことなく、励起光を作用させて励起される蛍光の強度を測定することで解決した。より具体的には、励起光を基板の固定化されていない面から入射して、励起された蛍光を検出する測定方法である。本発明における励起光としては、特に限定はなく、白色光から切り出した光源、好ましくは単一波長からなるレーザ光、より好ましくはエバネッセント波が挙げられる。励起光の検出は、エバネッセント型励起マイクロアレイスキャナーの使用が好ましいが、その他に共焦点型マイクロアレイスキャナーを使用することもできる。
【0045】
例えば、エバネッセント励起方式では励起光をガラス内部で全反射させた際に界面からの高さ200〜300nm(励起波長の半分程度)の範囲に、エバネッセント光と呼ばれる微弱光がしみ出し、このエバネッセント光により蛍光物質を励起するために、プローブ分子を含む溶液をスライドガラス上に接触させた状態で励起光を入射して蛍光を観察する際にも、ブラウン運動をしているプローブ分子をほとんど励起することなく、結合反応に預かるプローブ分子を選択的に観察することができる。
【0046】
本発明の方法は、蛍光標識した被検糖鎖と基板に固定化された糖鎖に相互作用を示すタンパク質との相互作用を利用した糖鎖構造解析に用いることができる。すなわち本発明の方法は、糖鎖構造解析を迅速・簡便に行う糖鎖プロファイラーとしての応用が期待される。例えば、蛋白質核酸酵素2003年8月号増刊Vol.48.NO.11に記載のプロファイリング方法が応用可能である。
【0047】
すなわち、本発明は、コンピューターを用いた糖鎖構造分析システムに利用可能である。該システムを使用することで、被検糖鎖が既知構造を有している場合は被検糖鎖の構造を同定することができ、被検糖鎖が未知構造を有している場合であっても被検糖鎖中に存在する特徴的な構造(α2−3シアル酸・α2−6シアル酸・α1−3ガラクトース・α1−3フコース・α1−6フコース・バイセクトN−アセチルグルコサミン・硫酸化など)の予測、ないし既知構造糖鎖との類似性を指摘することができる。このシステムは、蛍光標識した被検糖鎖を接触させた糖鎖に相互作用を示す種々のタンパク質がそれぞれ固定化された基板をマイクロアレイスキャナー装置にセットすると、自動的に、被検糖鎖の構造が表示されるシステムである。本発明のシステムでは、糖鎖に相互作用を示す種々のタンパク質がそれぞれ固定化された基板に蛍光標識した被検糖鎖を接触させる工程を自動化することもできる。すなわち、微小流路系を基板上の反応槽に導き、流路内に送液する溶液の種類・濃度・流速をコントロールすることで、ブロッキングやブロッキング液の洗浄除去工程、蛍光標識糖鎖溶液の接触工程を一元的にコントロールすることができる。本発明のシステムには質量分析や、酵素消化を組み合わせることもでき、これらの方法を用いることでさらに信頼度の高いデータを得ることができるため、大変に有用である。
【0048】
図1に本発明のシステム構成図の一例を示す。マイクロアレイスキャナー装置を用いたシステムは以下から構成される。
(a)糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報が格納されている記憶手段(データベース)
(b)糖鎖に相互作用を示す種々のタンパク質がそれぞれ固定化された基板に、蛍光標識した被検糖鎖を接触させ、洗浄操作を行わずに、該基板に励起光を入射し、励起される蛍光の強度を検出する検出手段
(c)検出された蛍光強度の組み合わせ情報を、(a)に格納されている情報と照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出する演算手段を含むコンピューター
(d)選出結果を表示する表示手段
【0049】
データベースについては、図1のようにコンピューターの外部にある場合、図2のようにコンピューター内部にある場合、共に許容される。また、該データベースを利用することで、限定されたレクチン数であっても多種類の糖鎖を識別することが可能である。理論的には10種類のレクチンであってもその特異性が異なる場合、識別可能な糖鎖の種類は610=60,466,176となり、事実上自然界に存在する殆どの糖鎖構造の識別が可能である。
【0050】
図2に、本発明のシステムにおけるコンピューター構成図の一例を示す。入力手段1と出力手段2がバス線3に接続されている。一時記憶手段4は、入力された情報、および算出された情報などを一時的に記憶する。中央処理装置(CPU)5は、本発明のプログラムの命令を受けて各種演算を行う。記憶手段(データベース)7には、糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報、および/または該相互作用情報の組み合わせパターン情報が格納されている。相互作用情報として、本発明のマイクロアレイスキャナー装置を利用した方法やシステムによって得られた蛍光強度情報、また、これまでに確立されている様々な実験系から得られた情報が利用可能である。
【0051】
記憶手段6には、本発明の処理を実行するためのプログラムを含む各種プログラムが格納されている。本発明の処理を実行するためのプログラムには、入力された蛍光強度の組み合わせ情報を、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報と照合し、組み合わせ情報のパターンが一致する構造既知糖鎖(データベースに格納されている構造既知糖鎖情報)を1つないし複数選出するプログラム61、表示プログラム62、およびこれらを制御するためのプログラム63が少なくとも含まれる。
【0052】
相互作用情報の組み合わせ情報の照合過程においては、相互作用情報の組み合わせ情報の値同士を比較してもよい。プログラム61には、例えば、入力された蛍光強度の組み合わせ情報の値とデータベースに格納されている相互作用情報の組み合わせ情報の値を比較して、その値の近さから構造既知糖鎖を1つないし複数選出する機能が組み込まれていてもよい。
【0053】
また、相互作用情報の組み合わせ情報の照合過程においては、相互作用情報の組み合わせ情報をパターン化して、そのパターン同士を比較してもよい。このような観点から、記憶手段6には、プログラム61の代わりに(または、プログラム61とともに)、入力された蛍光強度の組み合わせ情報をパターン化するプログラム61−1、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報をパターン化するプログラム61−2、および、入力された蛍光強度の組み合わせ情報のパターンを、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報のパターンと照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出するプログラム61−3が格納されていてもよい。このパターン化に際しては、適切な内部標準を用いて相互作用情報の規格化を行うことも可能である。プログラム61−1や61−2には、例えば、任意の設定スレッショルド値が入力されることで、そのスレッショルドの範囲に相互作用情報をレベル分けし、コード化する(各レベルに対して、例えば異なる数字や異なる色を当てはめる)機能が組み込まれている。
【0054】
プログラム61−3は、プログラム61−1を実行して得られるパターンを、プログラム61−2を実行して得られるパターンと照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出するプログラムである。なお、データベースにパターン情報が格納されている場合は、プログラム61−3は、プログラム61−1を実行して得られるパターンを、データベースに格納されているパターンと照合し、パターンが一致する構造既知糖鎖を1つないし複数選出する。プログラム61−3には、例えば、構造既知糖鎖のコードと被検糖鎖のコードを比較して、被検糖鎖とコードが一致する構造既知糖鎖を選出する機能が組み込まれている。
【0055】
プログラム62は、例えば、蛍光強度情報の表示、相互作用情報の表示、選出された構造既知糖鎖情報の表示などを行う。
【0056】
本発明においては、上述のプログラムを1つのプログラムにまとめることもできる。
【0057】
本発明のシステムにより実行される処理のフローの一例としては、まず、蛍光標識した被検糖鎖を接触させた糖鎖に相互作用を示す種々のタンパク質がそれぞれ固定化された基板がマイクロアレイスキャナー装置にセットされた場合に、該基板に励起光が入射され、励起される蛍光の強度が検出される。複数の基板がマイクロアレイスキャナー装置にセットされた場合は、該複数の基板が順次自動的に検出部に固定され、スキャニングが行われる。処理のフローの一例としては、次いで、蛍光強度情報が自動的にコンピューターに入力される。入力された情報は、コンピューターの記憶手段または一時記憶手段に格納しておくことができる。また、蛍光強度情報は、データベースに格納されてもよい。蛍光強度情報を蓄積することで、今まで存在しえなかった大規模かつ実用性の高い、糖鎖に相互作用を示すタンパク質に対する糖鎖の相互作用情報データベースを構築することができる。
【0058】
本発明では、中央処理装置(CPU)などの演算手段が記憶手段中のプログラム62の指令を受け、記憶手段または一時記憶手段に格納された蛍光強度情報を読み出し、該蛍光強度情報を表示することもできる。例えば、あらかじめ性状を十分に調査した基準となる糖鎖に相互作用を示すタンパク質試料スポット(内部標準スポット)の発する蛍光強度を基準として、各スポットの輝度値を補正した値を表示することができる。内部標準スポットは複数であってもよい。
【0059】
処理のフローの一例としては、次いで、入力された蛍光強度の組み合わせ情報を、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報と照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出する。この処理工程は、中央処理装置(CPU)などの演算手段が記憶手段中のプログラム61の指令を受け、記憶手段または一時記憶手段に格納された蛍光強度の組み合わせ情報とデータベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報を読み出し、それぞれの組み合わせ情報を照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出する。選出された構造既知糖鎖情報は、コンピューターの記憶手段または一時記憶手段に格納しておくことができる。
【0060】
データベースがコンピューターの外部にある場合は、中央処理装置(CPU)などの演算手段が記憶手段中のプログラム61の指令を受け、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報をコンピューターに入力し、記憶手段または一時記憶手段に格納された蛍光強度の組み合わせ情報を読み出し、それぞれの組み合わせ情報を照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出する。
【0061】
なお、プログラム61の代わりに、プログラム61−1〜61−3を用いるときも、同様なフローで処理される。
【0062】
処理のフローの一例としては、次いで、選出結果が表示手段によって表示される。この処理工程では、中央処理装置(CPU)などの演算手段が記憶手段中のプログラム62の指令を受け、記憶手段または一時記憶手段に格納された構造既知糖鎖情報を読み出し、表示する。
【0063】
本発明は、被検複合糖質が固定化された基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程、および、基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法を提供する。
【0064】
本発明における、被検複合糖質が固定化された基板は、好ましくは、エポキシ基を活性基として有する化合物がコートされた基板に被検複合糖質が固定化された基板であり、より好ましくは、エポキシ基を活性基として有する化合物が、GTMSである基板である。各工程は、上述の方法と同様である。本発明の方法は、基板に固定化された被検複合糖質の糖鎖付加状況を蛍光標識レクチン等により調べることで、生体にあるタンパク質を分離後、各タンパク質成分それぞれについての糖鎖付加状況をタンパク質の一団について一度に調べる(グライコフォーム解析)ことができる手段として有用である。またタンパク質製剤等の糖鎖部分の品質管理チェック等に応用が期待される。
【0065】
本発明は、また、複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板に被検複合糖質を接触させる工程、該工程で得られた基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程、および、基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法を提供する。
【0066】
本発明における、被検複合糖質が固定化された基板は、好ましくは、エポキシ基を活性基として有する化合物がコートされた基板に被検複合糖質が固定化された基板であり、より好ましくは、エポキシ基を活性基として有する化合物が、GTMSである基板である。また、本発明における複合糖質の糖鎖以外の領域に相互作用を示すタンパク質は、好ましくは、複合糖質の糖鎖以外の領域に相互作用を示す抗体である。また、糖鎖に相互作用を示すタンパク質は、好ましくは、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、これらの変異体、または糖鎖に相互作用を示す抗体である。各工程は、上述の方法と基本的に同様であるが、さらに、複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板であって、被検複合糖質を接触させた基板に、蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程を含む。該工程は、例えば、複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板に対し、被検複合糖質試料を含むサンプル・血液・体液・生体抽出成分・食品成分等の原液または希釈溶液を接触させ、十分な反応時間を確保した後に、PBSないしPBSTで十分に洗浄して被検複合糖質以外の成分を除く。その後に糖鎖部分に相互作用を示す蛍光標識プローブ溶液を接触させることで行うことができる。
【0067】
本発明は、異なる複合糖質の混合物中から、目的としたタンパク質の糖鎖修飾の状況を迅速かつ簡便に調べる方法として有用であり、血液・体液・生体抽出成分・食品成分等の原液または希釈溶液等の様々な成分の混合溶液を精製することなく、目的タンパク質の糖鎖修飾の状況を観察することで、診断・治療状況の把握等を行うような分野での応用が考えられる。
【0068】
本発明は、(a)基板にエポキシ基を活性基として有する化合物をコートする工程、および、(b)工程(a)で得られた基板に糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質を固定化する工程を含む、基板の製造方法を提供する。該方法の工程は、詳しくは、上述のとおりである。本発明はまた、上記方法で製造された基板も提供する。
【0069】
すなわち、本発明は、エポキシ基を活性基として有する化合物がコートされた基板であって、糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板を提供する。本発明の基板としては、好ましくは、エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である基板である。また、好ましくは、複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である基板、あるいは、糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、これらの変異体、または糖鎖に相互作用を示す抗体である、基板である。エポキシ基を活性基として有する化合物がレクチンである場合、スポットするレクチンの濃度は、特に限定はないが、1mg/mL以上であることが好ましい。さらに、より好ましくは、PBSTで洗浄することでレクチンが固定化された基板である。本発明の基板は、複数の反応槽を形成させた基板であることが好ましい。
なお、本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0070】
以下、本発明を実施例により、さらに具体的に説明するが、本発明はこれら実施例に制限されるものではない。
〔実施例1〕レクチンアレイを利用した糖鎖とレクチンとの相互作用解析
(1)蛍光標識化糖タンパク質プローブ(Cy3−ASF)の調製
蛍光標識化糖タンパク質プローブとして、アシアロフェツイン(SIGMA社、以下ASF)を550nm付近に吸収極大波長を持つ蛍光色素であるCy3 Mono−reactive Dye(アマシャムファルマシア社、以下Cy3)を用いて蛍光標識化して調製した。ASFはN−結合型糖鎖とO−結合型糖鎖を3本ずつ分子中に持ち、かつ糖鎖中の非還元末端のシアル酸キャップが部分的に外れている糖鎖構造を持つことが知られている。ASFを0.1M炭酸緩衝液(pH9.3)に終濃度1mg/mLになるよう調製した後、1mLについて1.0mgのCy3粉末と混合させ、1時間、適時攪拌しながら暗所で反応させた。
【0071】
次に担体としてSephadex G−25を用いたゲルろ過クロマトグラフィーにより、遊離のCy3とCy3−ASFを分離回収し、精製したCy3−ASFについて吸光光度計を用いて濃度及び蛍光標識効率を測定した。タンパク質ベースの収率は35−40%、蛍光標識効率(1タンパク質分子あたりの蛍光色素数)は約3.0であった。
【0072】
(2)スライドガラスへのGTMSコーティング
エポキシ基を活性基として有する3−グリシドキシプロピルトリメトキシシラン(信越シリコーン社、以降GTMS)をコーティングしたスライドガラス(図3)を用い、レクチンをガラス表面に固定化した。GTMSコーティングは、松浪硝子工業社製のスライドガラスを用い、以下の手順で行った。スライドガラスを10%KOH/MeOH溶液に浸し、容器ごと振盪させた状態で1時間放置しガラス表面を処理した。これを十分量の精製水(ミリQ水)により洗浄した後、60℃のオーブン内で乾燥させた。次にスライドガラスを2%GTMSアセトン溶液に浸し、遮光下で容器ごと振盪させながら1時間反応させた。反応後、110℃のオーブン内で8時間乾燥させた後、十分量の精製水により洗浄し、乾燥させた。
【0073】
(3)スライドガラスへのレクチンの固定化
(2)のGTMSコーティングを施したスライドガラスにレクチンをスポットした。マイクロアレイスポッターとして日本レーザ電子社製STAMPMANを使用し、先端直径0.40mmのスタンプピンを使用してスポットを行ことで、直径約0.6−0.7mmのスポットをスライドガラス上に配置した。スポットする各レクチンを濃度1mg/mL(レクチンによっては一部0.25mg/mL)となるようにリン酸緩衝生理食塩水、pH7.4(以下PBS)に溶解した。これを96穴のPCR用マイクロタイタープレート(コーニング社)の各反応槽に10μLずつ分注したものを、マイクロアレイスポッターにセットした。
【0074】
スライドガラスへのレクチンの固定化操作に際しては以下の条件をマイクロアレイスポッター付属のコンピューターに記憶させ、スタンプピン動作プログラムを実行させた。まずスタンプピンを96穴PCR用マイクロタイタープレート内の固定化試料溶液中に1秒間浸した後に引き上げ、スライドガラス表面の所定の位置に1秒間接触させた。この動作を1スポットごとに繰り返しながら、同一試料溶液から横1列に4点スポットを行った後、スタンプピンの洗浄工程を行った。洗浄工程ではスタンプピンの針先を0.05%SDS溶液に2秒間浸し、スタンプピンをバキューム装置内で15秒間乾燥させ、さらに精製水に2秒間浸した後に、バキューム装置内で15秒間乾燥、最後にエタノールに2秒間浸してからバキューム装置内で15秒間の乾燥操作を行った。
【0075】
本実施例では様々な糖結合特異性をもったレクチン4種(RCA120、SSA、組み換え放線菌由来キシラナーゼのキシラン結合ドメイン(以下XBD)、組み換えミミズ29kDaレクチン由来のC末端側ドメイン(以下EW29(Ch))とネガティブコントロール1種(ウシ血清アルブミン(以下BSA))の計5種のタンパク質をスポットした。RCA120、BSAについてはSIGMAより購入したもの、SSAは生化学工業より購入したもの、XBD,EW29(Ch)については本発明者らの研究室で大腸菌にて発現・精製したものを用いた。
【0076】
(4)非スポット面のブロッキング
レクチン溶液をスポット処理後1時間反応させガラス表面に固定化した後、未結合レクチンを洗浄した。洗浄は0.1%Tween20を含むPBS溶液(PBST)を数回ピペットでスライドガラスに吹き付けるようにかけて洗浄した後、PBSを用いてさらに十分に洗浄した。
【0077】
このレクチン固定化後のスライドガラスに対し、本発明者らが設計・開発した8穴ラバーを所定の位置に貼り付け、8つの反応槽を作製した(図4)。この8穴ラバーは厚さ1mmからなる黒色のシリコンゴム製で、縦横9.5×7.5mmからなる8つの長方形の穴が規則正しく空いており、スライドガラスに貼り付けたときに8つの反応槽を形成することができる。この反応槽に50μL程度の試料を加えれば、内部を十分量の試料溶液で満たすことができる。
【0078】
レクチンをスポットした領域以外のガラス表面には活性基であるエポキシ基が残存しているため、非スポット面に対するブロッキング操作を行った。なお、ブロッキング剤には高純度BSA(SIGMA)を使用した。8つの反応槽内に1%BSAを含むPBS溶液を50μLずつ満たし、湿度を90%以上に保った保存容器中で4℃、1時間放置し、スライドガラス上の非スポット面へのブロッキングを行った。反応の間はガラス表面の乾燥防止に留意した。
【0079】
次にスライドガラス上のブロッキング溶液を除去し、PBSを用いて十分に洗浄した後、水分を除去した。タンパク質固定化後はガラス表面の乾燥によるタンパク質の変性や、乾燥に伴うバックグラウンドの上昇を防ぐため、可及的速やかに次の操作に移った。
【0080】
(5)プローブ溶液の添加とスキャニング
(4)で作製したレクチン固定化スライドガラス上の反応槽に相互作用を解析したい蛍光標識化糖タンパク質プローブ溶液を加えた。蛍光標識化糖タンパク質プローブは終濃度10μg/mLになるようPBSに溶解したものを調製し、それぞれの反応槽に50μLを滴下した。
【0081】
レクチン−糖鎖間の反応が平衡に達するまで静置した後に、エバネッセント励起方式マイクロアレイスキャナーであるGTMAS Scan III(日本レーザ電子社)を使用してスライドガラス端面より励起光を入射し、励起されて生じた蛍光発光を、スライドガラス下面に配置されているICCD(イメージインテンシファイアー付電荷結合素子)カメラで検出した。スライドガラスのほぼ全面に対する蛍光イメージをスキャニングし、得られたイメージ画像をTIFFファイル形式(1枚につき約100M Bite)にて保存した。スキャニング時のパラメーターはGain「5000倍」、積算回数「4回」、露光時間「33msec」で統一した。
【0082】
(6)スキャニング画像の数値化
スキャニング画像の数値化には市販のマイクロアレイ用解析ソフトであるArray−Pro Analyzer(Version 4.0 for Windows(登録商標)、Media Cybernetics社)を使用した。各スポットの輝度を上記解析ソフトを用いて算出し、非スポット領域の輝度をバックグラウンド値とした。各スポットの輝度からバックグラウンド値を差し引いたものを正味の輝度値とし、横列に4点並べた同一試料由来のスポットごとに平均値と標準偏差を算出した。
【0083】
以降は各レクチン試料に対するプローブの結合に対してはこの同一試料由来の4点の平均輝度値を用いて評価を行った。以降に示す各レクチンアレイの性能評価は操作(2)〜(6)の一連の行程を経た後に行われた。
【0084】
(7)GTMSコートスライドガラスの性能評価
上述のように作製したGTMSコートスライドガラスの性能を、既存のスライドガラス(6種)と比較評価した。すなわち表面コートされた各スライドガラスに、あらかじめCy3標識したレクチン(100μg/mL)をアレイ状に固定化し、(3)〜(6)の行程を経た後に、スポッティング領域の輝度値(S)と非スポッティング表面の輝度値(N)からS/N比を算出した。その結果、表1に示すように(2)の行程で作製したGTMSコートスライドガラスの輝度値は、最高値を示したスライドガラスAの2分の1程度にとどまったが、バックグラウンドが非常に低いため、そのS/Nは16.1となり、今回評価したスライドガラスの中で最良値を示した。
【0085】
【表1】

【0086】
(8)アレイ上の固定化レクチン濃度の検討(図5および6)
RCA120とConAはそれぞれ複合型糖鎖と高マンノース型糖鎖に対し高い親和性をもつことが知られる代表的なレクチンである。これらのレクチンを様々な濃度で調製し、同一試料について横に4点並べてアレイ状にスポットした。このアレイの各反応槽に対して10μg/mLのCy3−ASFを各50μLずつアプライして結合反応を起こさせた後に、スキャナーで蛍光を観察した。
【0087】
上述したようにASFはN−結合型糖鎖とO−結合型糖鎖を3本ずつ分子中に持ち、かつ糖鎖中の非還元末端のシアル酸キャップが外れラクトサミン構造が突出した糖鎖構造を持つことが知られている。よってRCA120とConAを固定化したレクチンアレイに対し、Cy3−ASFを添加した実験系においては、RCA120が非常に強い親和性を示し、ConAが弱い親和性を示すことが予想された。
【0088】
実験の結果、RCA120のスポットが強い蛍光を発したのに対し、ConAのスポットは同条件のRCA120のスポットに比べて1/3程度の蛍光強度を示すに止まった。ConAが弱いながらも複合型糖鎖を持つASFに結合したのは、N−結合型糖鎖のうちASFに主として存在する3本鎖型糖鎖には結合できないものの、少量存在するとされる2本鎖型糖鎖には結合できるためと考えられた。またこのデータから同一試料由来の4点についての標準偏差(SD)は約±20%程度になるということが分かった(図6)。
【0089】
次にスポット時のレクチン濃度と蛍光強度の関係をグラフ化したところ、スポット時のレクチン濃度と蛍光強度の間に正の相関関係があり、スポットするレクチン試料の濃度を1mg/mL以上に高濃度化することで、効果的にシグナル強度を向上することができることが分かった。すなわち親和定数の小さい(結合の弱い)レクチン−糖鎖間の相互作用の検出は固定化レクチン濃度を上げる事により可能となることが分かった(図6)。
【0090】
(9)レクチンアレイの性能評価
様々な糖特異性をもったレクチン4種(RCA120、SSA、XBD、EW29(Ch))、とネガティブコントロール1種(BSA)の計5種のタンパク質を同一試料について横に4点並べてアレイ状にスポットした。このアレイについて10μg/mLのCy3−ASFを各50μLずつ滴下し、スキャナーで蛍光を観察した。
【0091】
実験の結果FACでラクトサミン構造に親和性があると確認することができたRCA120、EW29(Ch)の2種のレクチンのスポットにおいて蛍光シグナルが観察された(図7)。またそれぞれの蛍光強度を比較すると、RCA120のスポットにおいて強い蛍光が、EW29(Ch)のスポットにおいては中程度の蛍光が観察され、FAC解析データと一致した。
【0092】
また同条件のアレイに対しラクトース(競合阻害糖)共存下で同様の実験を行ったところ、阻害糖の濃度上昇に伴って、スポットの蛍光強度の減少が観察された(図8)。このことから、蛍光糖タンパク質プローブとの結合はレクチンと糖鎖間の糖特異的結合反応によるものであることを確認することが出来た。
【0093】
〔実施例2〕他の糖鎖関連アレイへの応用
糖鎖関連アレイには、糖鎖をプローブに用いるレクチンアレイの他、糖タンパク質をプローブに用いるレクチンアレイ、レクチンをプローブに用いる糖ペプチドアレイ、レクチンをプローブに用いる糖タンパク質アレイ、レクチンをプローブに用いる抗体アレイの5タイプが挙げられる。図9に、各糖鎖関連アレイと糖鎖の相互作用の模式図を示す。
【0094】
それぞれのアレイを使用して得られた蛍光強度情報をコード化する。輝度値の数値は16bitで取り込まれた数値であるため、0−65355の間の値となる。そこでGain「1000倍」での(輝度値−バックグラウンド値)の値を10000刻みで6段階に分類することで0から5までのコード化を行った。Gain「1000倍」で輝度信号値が飽和している場合は、Gainを下げて観察しGain「1000倍」時の値に換算する。信号値が飽和しているかどうかの判定は、Gain値を変化させた際にGain値と信号強度の相関の直線性が保たれているかどうかで判断する。
【0095】
現在のスレッショルド値 (輝度値−バックグラウンド値)
0−10000 レベル0
10001−20000 レベル1
20001−30000 レベル2
30001−40000 レベル3
40001−50000 レベル4
50001以上 レベル5
【0096】
〔実施例3〕 レクチン−抗体を同一区画内にスポットしたアレイによる解析(図10)
1.材料と方法
(1)モデル糖タンパク質の蛍光標識化プローブの調製
本実験例ではレクチンアレイ上に固定化するレクチンとして、様々な糖結合特異性を持つレクチンとして6種(RCA120、ECA、ConA、GNA、SSA、SNA)を選択した。またネガティブコントロールとして糖鎖と結合しないタンパク質であるBSAを選択した。また今回の実験では、プローブのコアタンパク部分を認識する抗フェツイン抗体と抗RNase抗体の2種を、レクチンと同じ区画内にスポットした。GNA、SNAはVECTOR社より購入したもの、BSAについてはSIGMA社より購入したもの、RCA120、ECA、ConA、SSAは生化学工業より購入したものを用いた。
【0097】
モデル蛍光標識化糖タンパク質プローブは、ASF、FET、ウシ膵由来リボヌクレアーゼB(RNase B)、またネガティブコントロールとしてウシ膵由来リボヌクレアーゼA(RNase A)、BSAなどのタンパク質(全てSIGMA社より購入)を550nm付近に吸収極大波長を持つ蛍光色素であるCy3 Mono−reactive Dye(アマシャムファルマシア社、以下Cy3)を用いて蛍光標識化することで調製した。プローブ作製時は前述のタンパク質を0.1M炭酸緩衝液(pH9.3)に終濃度1mg/mLになるよう調製した後、1.0mgのCy3粉末と混合させ、1時間、適時攪拌しながら暗所で反応させ、担体としてSephadex G−25を用いたゲルろ過クロマトグラフィーにより、Cy3標識タンパク質を精製した。
【0098】
(2)スライドガラスへのGTMSコーティング
エポキシ基を活性基として有する3−グリシドキシプロピルトリメトキシシラン(信越シリコーン社、以降GTMS)をコーティングしたスライドガラス(図3)を用い、レクチンをガラス表面に固定化した。GTMSコーティングは、松浪硝子工業社製のスライドガラスを用い、以下の手順で行った。スライドガラスを10%KOH/MeOH溶液に浸し、容器ごと振盪させた状態で1時間放置しガラス表面を処理した。これを十分量の精製水(ミリQ水)により洗浄した後、60℃のオーブン内で乾燥させた。次にスライドガラスを2%GTMSアセトン溶液に浸し、遮光下で容器ごと振盪させながら1時間反応させた。反応後、110℃のオーブン内で8時間乾燥させた後、十分量の精製水により洗浄し、乾燥させた。
【0099】
(3)レクチンアレイの作製
(2)で作製したGTMSコートスライドガラスに対してレクチンをスポットし、レクチンアレイを作製した。マイクロアレイスポッターとして日本レーザ電子社製STAMPMANを使用し、先端直径0.40mmのスタンプピンを用い、直径約0.5mmのスポットをスライドガラス上に配置した。スライドガラスへのレクチンの固定化操作に際しては以下の条件をマイクロアレイスポッター付属のコンピューターに記憶させ、スタンプピン動作プログラムを実行させた。まずスタンプピンを96穴PCR用マイクロタイタープレート内の固定化試料溶液中に1秒間浸した後に引き上げ、スライドガラス表面の所定の位置に1秒間接触させた。この動作を1スポットごとに繰り返しながら、同一試料溶液から横1列に6点スポットを行った後、スタンプピンの洗浄を行った。洗浄工程ではスタンプピンの針先を0.05%SDS溶液に2秒間浸し、スタンプピンをバキューム装置内で15秒間乾燥させ、さらに精製水に2秒間浸した後に、バキューム装置内で15秒間乾燥、最後にエタノールに2秒間浸してからバキューム装置内で15秒間の乾燥操作を行った。
【0100】
(4)非スポット面のブロッキング
上記の操作でレクチンをスポットしたスライド上に、シリコンゴム製の8穴ラバーを接着して8つの反応槽を作製した。この反応槽に0.1% Tween20を含むPBS溶液(PBST)を満たすことで、スライド固層に結合しなかった余剰レクチンを洗浄・除去した。次に1% BSAを溶解したPBS溶液を各反応槽に200μLずつ満たした後に、湿度を90%以上に保った保存容器中で4℃、1時間放置することで非レクチンスポット領域のブロッキング操作を行った。
【0101】
(5)プローブ溶液の添加とスキャン
ブロッキングを終えたスライドに対し、約100ng/mLに調整した各蛍光標識化糖タンパク質プローブ溶液を各反応槽に50μLずつ加えることで、アレイ上にプローブ溶液を接触させた。
【0102】
レクチン−糖鎖間の反応が平衡に達するまで静置した後に、エバネッセント励起方式マイクロアレイスキャナーであるGTMAS Scan III(日本レーザ電子社)を使用してスライドガラス端面より励起光を入射し、励起されて生じた蛍光発光を、スライドガラス下面に配置されているICCD(イメージインテンシファイアー付電荷結合素子)カメラで検出した。スライドガラスのほぼ全面に対する蛍光イメージをスキャンした後、得られたイメージ画像をTIFFファイル形式にて保存した。スキャン時の各パラメーターは、Gain「4000倍」、積算回数「8回」、露光時間「110msec」で統一した。
【0103】
(6)スキャン画像の数値化
スキャン画像の数値化には市販のマイクロアレイ用解析ソフトであるArray−Pro Analyzer Ver.4.5(Media Cybernetics社)を使用した。各スポットの輝度を上記解析ソフトで算出し、非スポット領域の輝度をバックグラウンド値とした。各スポットの輝度からバックグラウンド値を差し引いたものを正味の輝度値とし、横列に5点並べた同一試料由来のスポットごとに平均値と標準偏差を算出した。
以降は各レクチン試料に対するプローブの結合に対してはこの同一試料由来の5点の平均輝度値を用いて評価を行った。
【0104】
2.結果と考察
今回の実験では多様な特異性を持つレクチンを並べたアレイを構成し、既知の糖鎖構造を持つ糖タンパク質プローブをアプライした際の各レクチンスポットの蛍光パターンから、レクチンアレイとしての性能評価を行った。また本実験では、プローブのコアタンパク部分の情報も同時に取得することを目指し、糖タンパク質プローブのコアタンパク部分を認識する抗体もレクチンと並べてスポットしたアレイを作製した。モデル糖タンパク質としては、コアタンパク部分の構造は同一だが修飾糖鎖部分の構造が異なるタンパク質として、ASFとFETの組み合わせと、RNase AとRNase Bの組み合わせを選択した。
【0105】
実験(A)・(B)では、プローブとして用いたASFとFETの糖鎖プロファイルを比較した(図10A及びB)。FETはN−結合型糖鎖とO−結合型糖鎖を3本ずつ分子中に持ち、各糖鎖構造の非還元末端が高度にシアル酸で修飾されていることが知られている。一方、ASFはFET糖鎖のうち末端シアル酸を酵素或いは酸処理によって除去したタンパク質であり、ラクトサミン構造が露出している。このためASFプローブを用いた場合、ラクトサミンを認識するRCA120やECAのスポットが観察され、FETプローブを用いた場合、シアル酸を認識するSSA、SNAのスポットが観察されると予想された。また抗体スポットに対してはFETもASFもコアタンパク部分は同一であるため、両プローブ共に結合が観察されると予想された。実験の結果、末端ラクトサミン構造を持つASFプローブではRCA120やECAの非常に明るいスポットが検出されたが(図10A)、ラクトサミン構造がシアル酸でキャッピングされているFETプローブではRCA120、ECAのスポットは非常に暗くなった(図10B)。この結果はRCA120、ECAがラクトサミン構造を強く認識し、ラクトサミン構造の末端がシアル酸でキャップされると親和性が大幅に低下するという従来の知見と一致していた。また実験(B)ではFETプローブに特徴的なシアル酸の存在に対応して、シアル酸認識レクチンであるSSAやSNAのスポットが観察された(図10B)。ConAのスポットでは実験(A)、(B)共に弱い蛍光が観察された(図10A及びB)。これはN−結合型糖鎖のうち、主として存在する3本鎖型糖鎖には親和性が低いものの、少量存在するとされる2本鎖型糖鎖には結合できるためと考えられた。共通コアタンパク部分を認識するFET抗体のスポットは、実験(A)、(B)において蛍光が観察された(図10A及びB)。
【0106】
実験(C)・(D)ではRNase AとRNase Bプローブの糖鎖プロファイルの相違を比較した(図10C及びD)。RNase Bは、ハイマンノース型のN−結合型糖鎖を分子内に1本のみ持ち、RNase Aは、コアタンパク部分はRNase Bと同一の構造を持つが糖鎖を全く持たないことが知られている。これらのプローブをレクチンアレイに接触させた場合、両プローブ共に抗RNase抗体スポットに対しては親和性を示すが、レクチンのスポットに対する反応性に相違が生じる(RNase BプローブはConAなどのマンノース認識レクチンに親和性を示すのに対し、RNase Aは親和性を示さない)ことが予想された。実験の結果、抗体に対する反応性では、両プローブにおいて共通コアタンパク部分を認識するRNase抗体との結合が観察された。一方レクチンに対する反応性では、ハイマンノース構造を持つRNase BでConAのスポットにおいて結合が観察されたが、糖鎖を持たないRNase Aではレクチンのスポットにおいて、結合による蛍光は観察されなかった。実験(E)ではネガティブコントロールとして、糖鎖を持たないタンパク質であるBSAを用いた。BSAプローブでは予想通り、抗体・レクチンスポットともに結合は観察されなかった(図10E)。
【0107】
実験(A)〜(E)を通して、微量の糖タンパク質試料から迅速に、レクチンの糖結合特異性を反映したプロファイルを得ることができた(図10A〜E)。また今回の実験では、レクチンと抗体を同一アレイ上にスポットすることで、糖タンパク質のコアタンパク部分と、修飾糖鎖部分の情報を同時並行的に一枚のスライド上で取得することができた。同一スライド上で同時並行解析を行うことで、各反応槽間の実験条件(温度、反応時間等)を揃えた形での観察が可能となるという利点が生じた。
【0108】
〔実施例4〕 レクチンアレイを用いた阻害濃度解析(図11)
1.材料と方法
先の実験で観察されたレクチンとプローブ分子間の結合が、糖鎖を介した特異的結合であることを確認する為に競合阻害糖を用いた阻害実験を行った。実験(A)ではスライドガラス上の8つの反応槽内にRCA120をスポットしてアレイを構成した上で、競合阻害糖(ラクトース)の濃度を変えた8種類のASFプローブ溶液を同時に接触させ、結合反応の阻害を観察した(図11A)。実験(B)では固定化レクチンとしてConAを用い、プローブとしてRNase B、競合阻害糖としてマンノースを用い、同様の操作で結合の阻害を観察した(図11B)。アレイ作成時に要した材料・操作は、実施例3と同じであるため記述を省略する。
【0109】
2.結果と考察
実験の結果、競合阻害糖の濃度上昇に伴ってスポットの蛍光強度の減少が観察され(図11)、阻害曲線をカーブフィッティングすることにより、阻害物質固有の半阻害濃度を算出することができた。この結果から、蛍光糖タンパク質プローブとの結合はレクチンと糖鎖間の特異的結合反応によるものであることを確認できた。またこのような阻害実験を用い、半阻害濃度を算出することで結合の強さを評価したり、結合する相手分子を探索することが可能であることが示された。
【0110】
〔実施例5〕 レクチンアレイの糖ペプチドプローブによる検出(図12)
1.材料と方法
(1)糖ペプチドプローブの調製
実施例3(1)に記述した方法でCy3−ASFを調製した後に、このCy3−ASFをトリプシン処理により断片化し、Cy3−ASFペプチドを調製した。
(2)スライドガラスへのGTMSコーティング
実施例3(2)に記述した方法で行った。
(3)レクチンアレイの作製
固定化するレクチンは、各レクチンの持つ主要な糖認識能ごとにグループ分けを行った上で、フコース認識レクチン5種、シアル酸認識レクチン6種、ラクトサミン構造認識レクチン3種、ガラクトース認識レクチン6種、ガラクトサミン認識レクチン11種、マンノース認識レクチン4種、キチン構造認識レクチン5種の計40種類のレクチンを選択し、スライドガラスに固定化したアレイを作製した。実験操作は実施例3(3)に記述した方法で行った。
(4)非スポット面のブロッキング
(5)プローブ溶液の添加とスキャン
(6)スキャン画像の数値化
上記(4)〜(6)の操作については、Cy3−ASFペプチドプローブを用い、実施例3(4)〜(6)に記述した方法と同様の操作にて、実験を行った。
【0111】
2.結果と考察
実験の結果、レクチンアレイに糖ペプチドプローブを供することで、糖鎖構造を反映した糖鎖プロファイルを得ることができた。得られた糖鎖プロファイルは酵素消化する前のASFと同等であり、レクチンアレイに糖タンパク質だけでなく糖タンパク質のペプチド消化産物をプローブとして利用できることが示された。この技術を用いれば糖ペプチドをHPLC等で分画した後にプローブとしてレクチンアレイに供することで糖ペプチド各成分の糖鎖プロファイルの観察が可能になり、有用である。
【0112】
〔実施例6〕 糖ペプチドアレイによる、糖ペプチド上の糖鎖プロファイリング(図13)
1.材料と方法
(1)レクチンプローブの調製
本実施例ではレクチンプローブとして、ラクトサミン構造を強く認識するRCA120、ネガティブコントロールとして糖結合能を持たないBSAを使用した。蛍光標識レクチンプローブは蛍光色素であるCy3を用いて蛍光標識して調製した。レクチンは0.1M炭酸緩衝液(pH9.3)中に終濃度1mg/mLになるよう溶解した後、1mL当たり1.0mgのCy3粉末と混合させ、1時間、適時攪拌しながら暗所で反応させた。反応後は限外ろ過フィルターキットを用いて未反応のCy3色素を除去した。
(2)スライドガラスへのGTMSコーティング
GTMSコートスライドは実施例5(2)に記述した方法で行った。
(3)糖ペプチドアレイの作製
(2)で作製したGTMSコートスライドガラス(図3)に対して糖ペプチドをスポットして糖ペプチドアレイを作製した。マイクロアレイスポッターとして日本レーザ電子社製STAMPMANを使用し、先端直径0.40mmのスタンプピンを使用してスポットを行ことで、直径約0.5mmのスポットをスライドガラス上に配置した。
本実験では固定化した糖ペプチドサンプルは、マウス肝臓の可溶性画分からレクチンカラムを用いて精製した糖タンパク質画分をトリプシンによりペプチド断片化後、HPLCにより分画・分取した糖ペプチドを用いた。スライドガラスへの糖ペプチドの固定化操作に際しては以下の条件をマイクロアレイスポッター付属のコンピューターに記憶させ、スタンプピン動作プログラムを実行させた。まずスタンプピンを96穴PCR用マイクロタイタープレート内の固定化試料溶液中に1秒間浸した後に引き上げ、スライドガラス表面の所定の位置に1秒間接触させた。この動作を1スポットごとに繰り返しながら、同一試料溶液から横1列に6点スポットを行った後、スタンプピンの洗浄を行った。洗浄工程ではスタンプピンの針先を0.05%SDS溶液に2秒間浸し、スタンプピンをバキューム装置内で15秒間乾燥させ、さらに精製水に2秒間浸した後に、バキューム装置内で15秒間乾燥、最後にエタノールに2秒間浸してからバキューム装置内で15秒間の乾燥操作を行った。
(4)非スポット面のブロッキング
(5)プローブ溶液の添加とスキャン
(6)スキャン画像の数値化
上記(4)〜(6)の操作については、Cy3−RCA120プローブとCy3−BSAプローブを用い、実施例3(4)〜(6)に記述した方法に従い、実験操作を行った。
【0113】
2.結果と考察
本実験の結果、糖ペプチドをアレイ化することで、糖ペプチドに付加した糖鎖の構造情報を、ハイスループットかつ容易に得られることが示された。糖ペプチドアレイの作成方法としては、1)精製糖ペプチドを固定化する、2)クルードの糖ペプチドを固定化する、3)HPLCで分離した糖ペプチドフラクションを固定化する、などが考えられる。今回の実験ではHPLCで分離した各フラクションをアレイ化したものを作製でき、有用であることを示すことができた。
【0114】
従来、HPLCで分画したフラクションのうち、どこに糖ペプチドが含まれているかという情報を得ることは、UV吸収や蛍光等でモニターできないために容易ではなかった。本実験の結果、各糖ペプチドフラクションを固定化したアレイを用いることで、どのフラクションにどのような糖鎖構造を持った糖ペプチドがあるのかを容易に知ることが可能となることが分かった。
【0115】
本法を利用することで、解析対象の糖鎖を持つ糖ペプチドがどのフラクションに含まれているのかといった情報を迅速に得ることができるため、多数のフラクションから効率的に糖鎖の含まれるフラクションのみを選出し、質量分析など他の解析へと供すること等が可能になる。また解析対象の糖ペプチドの糖鎖構造情報や、どのようなレクチンが結合するかという情報がない場合、トリプシン消化前の糖タンパク質の状態でレクチンアレイを用いた分析を行い、糖鎖プロファイリングを行うことで、当該糖タンパク質に結合するレクチンを数十種類のレクチンの中から絞り込むことが出来る。このような情報が得られることで、従来多数のレクチンを用いて総当り的に行っていたレクチンブロットなどの操作が簡略化され、大幅に時間と労力を節約できるなどの利点がある。
【0116】
〔実施例7〕 レクチンアレイに対し、クルード生体試料をプローブとして用いた実験(図14)
1.材料と方法
レクチンアレイを用いて糖タンパク質混合物、特に生体由来混合試料のプロファイリングを行うことで、生体内の糖タンパク質の糖鎖付加状態を解析する。
【0117】
マウス肝臓より抽出・精製した糖タンパク質サンプルとマウス脳より抽出・精製した糖タンパク質サンプルをCy3標識化してプローブとした後に、40種レクチンの固定化されたレクチンアレイと接触させて、マウス抽出糖タンパク質混合物全体の糖鎖プロファイルを観察した。
アレイ作製時に要した材料・操作は、実施例3と同じであるため記述を省略する。
【0118】
2.結果と考察
マウス脳由来糖タンパク質プローブ(図14A)と肝臓由来糖タンパク質プローブ(図14B)では、明らかに異なった糖鎖プロファイルが観察された。特にシアル酸認識のレクチン群で、両プローブ間に顕著な相違が見られた。脳では糖タンパク質に対するシアル酸の付加が少ないことが知られており、この事実は実験結果の傾向と一致した。このようにクルード系を用いた実験では、試料全体の糖鎖付加情報を迅速・簡便に得ることができるため、血液成分や臓器等の、個体間や病態間での糖鎖構造の差を一斉比較解析する目的での使用に適している。
【0119】
次に先の実験で用いた脳由来糖タンパク質プローブに対し、競合阻害糖としてラクトースを10mM加え、糖鎖プロファイルを観察した(図14C)。その結果、競合阻害により蛍光シグナルのパターンに変化が観察され、主にラクトサミン認識レクチンのシグナルの減弱が観察された。この実験で示したように、よりクルードな試料に対するレクチンアレイによる糖鎖プロファイリングにおいても、様々な阻害糖の添加実験を併用し、獲得される情報を絞り込んだ上で比較解析することで、より簡便かつ迅速に個体間の比較解析を行うことができる。
【産業上の利用可能性】
【0120】
本発明により、レクチン−糖鎖間の相互作用を溶液中で平衡状態のまま観察する方法が初めて実用化された。この技術は従来のレクチンブロットのような相互作用の有無(0か1か)ではなく、中間部分の情報、すなわち結合の強度情報を輝度値から得ることができる(例えば0から6段階)。このことはn種のレクチン−糖鎖間の相互作用において、従来は2n通りであった情報量が6n通りへと格段に増したことを意味する。この技術は今後さらに高密度化、高精度化していくことにより糖鎖構造解析、及び関連するその他の糖鎖工学諸分野の発展に大きな貢献をもたらすことが期待される。さらには、様々な用途に向けレクチン−糖鎖間の相互作用解析用のアレイを作製することで、血液・体液・組織抽出物等の原液または希釈溶液からの診断・判定への利用や、糖タンパク質製品の品質管理等への応用が期待される。
【技術分野】
【0001】
本発明は、タンパク質と糖鎖との相互作用を分析する方法に関する。
【背景技術】
【0002】
生体の生命機能を担う主役であるタンパク質が、細胞社会の中において秩序正しく機能を発揮するためには、糖鎖修飾をはじめとする翻訳後修飾が大変に重要な役割を担っている。生体内のほとんどのタンパク質は糖鎖による修飾をうけており、タンパク質に付加した糖鎖がウィルスの感染、原虫の寄生、感染、毒素の結合、ホルモンの結合、受精、発生分化、タンパク質の安定性、がん細胞転移、アポトーシスなど、生命現象の様々な場で重要な役割を果たしていることが近年次々と明らかになってきた。
【0003】
糖鎖機能の解析のためには、まずその糖鎖の構造解析が欠かせない。今後も糖鎖構造解析法の重要性は増すことが予想される。しかし糖鎖の構造解析は多大な時間と労力、経験を要することから、従来の手法に基づき完全な構造決定を目指すのではなく、より簡便に、高速、高感度、かつ高精度に多彩な糖鎖構造の特徴を抽出し、相互識別できるシステム開発が期待されていた。
【0004】
マイクロアレイは多種のDNA・タンパク質等の固定化試料を高密度に固相担体(ガラス・メンブレン・シリコンチップ)上にスポット状に固定化したものの総称であり、固定化した多種の試料スポットに対して特異的に結合する分子(以下プローブ)の有無を検出することが可能である。プローブ分子には一般的には蛍光標識化されたものが用いられており、プローブ溶液をアレイ表面と反応させた後に、蛍光検出スキャナーで観察することにより各試料スポットに結合したプローブ分子の定量的解析を行うことができる。米国Affimetrix社がDNAマイクロアレイを開発して以来、マイクロアレイは非常に広範な研究分野において使用され、人類に様々な新しい知見をもたらしてきた。
【0005】
第三の生命鎖と呼ばれる糖鎖の構造・機能情報を研究する上で、マイクロアレイを利用した糖鎖と糖鎖に相互作用を示すタンパク質(糖結合性タンパク質、例えば、レクチン等)の迅速かつ高感度な大規模相互作用解析が可能となれば、基礎研究から医療診断や産業応用に至るまで、広範囲に利用されうる非常に有用なツールになることが考えられる。
【0006】
糖鎖と糖鎖に相互作用を示すタンパク質間の結合は抗原抗体反応の一般的な解離定数(Kd=10−8以下)等に比べて、一般的に弱い相互作用であることが知られており、これらの解離定数(Kd)は10−6Mかそれ以上であることが多い。また糖鎖と糖鎖に相互作用を示すタンパク質間の相互作用は比較的速い解離−会合反応から成り立っていることが知られており、結果的に一般的なタンパク質間相互作用や相補的ヌクレオチド断片間の相互作用に比べ、洗浄操作などにより解離側に平衡が傾きやすい。例えば、レクチンを糖タンパク質固定化カラム等にて精製を行う際にも、レクチンの結合が弱い場合は洗浄操作中にレクチンがカラム外に流出してしまう現象がしばしば観察される。
【0007】
従来のスライドガラスを用いた一般的なマイクロアレイ技術では、プローブ溶液を固定化試料と接触させて結合反応に至らしめる過程後にプローブ溶液を洗浄・除去し、ジェットガスや遠心機を用いてスライドガラスに付着している水分を完全に除去する操作を行った後に、マイクロアレイスキャナーを用いてイメージングを行う。これは一般的なマイクロアレイリーダーが、水分の付着した状態でスライドガラス上の蛍光を観察することが出来ないことに起因している。このようにスキャニングの前段階でプローブ溶液を除去しても、相補的ヌクレオチド断片や抗原−抗体反応等、強い結合を示す相互作用においては解離速度定数が十分に小さいため、プローブ分子の解離反応は容易に進行しないと考えられる。しかし、糖鎖と糖鎖に相互作用を示すタンパク質間に一般的に見られる解離速度定数の大きい、すなわち弱い相互作用の観察に際しては、プローブ溶液の除去・洗浄操作を行う時点で糖鎖と糖鎖に相互作用を示すタンパク質間の解離反応が進行してしまい、平衡状態下での正確な相互作用情報を得ることが困難になる。このため、マイクロアレイにおいて糖鎖と糖鎖に相互作用を示すタンパク質間の平衡状態での相互作用情報を精密に解析する場合、このプローブ溶液の洗浄操作が大きな問題点となる。
【0008】
DNAマイクロアレイは現在大変広範囲に利用されている。タンパク質のマイクロアレイについてもDNAの転写産物であるタンパク質の生体内での機能を解明する等の基礎研究分野やタンパク質の量的・質的な変化に基づいた診断・判断等の応用分野においても、将来の利用が期待されており、研究面においても世界中で活発な研究が行われてきた。しかしタンパク質マイクロアレイの開発・普及はDNAマイクロアレイに比べ、圧倒的に遅れているのが現状である。この原因の一つとして、様々に異なる性質を持つタンパク質試料を、活性を保ったままの状態で、一定の割合で固定化していくという工程が技術的に大変に難しいということが、以前より多くの研究者によって指摘されている。
【0009】
タンパク質をアレイ上に固定化する方法として、最も初期に開発された方法としてPVDF膜(非特許文献1)に代表されるメンブレン上にタンパク質を物理的に吸着させる方法が挙げられる。転写因子など一部のタンパク質では、ある程度の活性を維持できると報告されているが、一般性には乏しい。またメンブレン上に固定化する場合、アレイの高密度化に限界があった。高密度化を達成するために、金属やガラスなどの固体表面に対してタンパク質を固定化していく方向で研究が進められてきたが、一般にタンパク質は金属やガラスなどの固体表面への接触によって変性しやすいという性質を持つ。このため、固体表面とタンパク質を架橋するための何らかのリンカーを使用する固定化方法が、鋭意研究・開発されてきた。
【0010】
タンパク質の変性の問題を軽減する手法の一例としては、スライドガラス上に厚さ10〜100μmのポリアクリルアミドのパッドを接合して、これにタンパク質をスポットする手法が挙げられる(非特許文献2および3)。この場合、タンパク質は3次元空間に固定化されるために、2次元表面に固定化する手法に比べて、量的に100倍以上の向上が望めるという。また、タンパク質を多孔性ポリアクリルアミドゲル内にアミノ基を介して固定化する方法もある(非特許文献4)。しかしこれらの方法は高価で特殊なスライドガラスを作製する必要があり、一般的な普及には至っていない。また固定化されたタンパク質の層に厚みがあることは、検出法によっては好ましくないこともある。
【0011】
現在最も精力的に研究されている固相へのタンパク質固定化法として、何らかのタグを付加した形でタンパク質を発現し、タンパク質のタグ部分を介した固相担体への固定化を行う方法がある。この方法ではタンパク質の有効リガンド濃度が向上するといった効果やタンパク質の向きをそろえることが原理的に可能であるといわれている。このような手法の一例として、オリゴヒスチジンタグを介して、ニッケル錯体で表面修飾した基板へ固定化する方法や(非特許文献5)、アビジン−ビオチンを介した固定化法(特許文献1)を挙げることができる。
【0012】
このような方法はタンパク質の活性を保ったまま固定化する、あるいは固定化率をそろえるという点で有効な方法であると考えられる。しかしマイクロアレイに固定化しようとするすべてのタンパク質について遺伝子レベルでタグを付加し、大腸菌、ないし無細胞系等で発現及び精製を行うことは多額のコストと莫大な労力が必要となると考えられる点で、現時点で一般的な研究者が自在かつ個々の要求に合わせる形で気軽に利用することは難しい。
【0013】
これに対し、タンパク質の官能基を固相担体との固定化に利用する方法は、天然から抽出したままのタンパク質や市販のタンパク質試料をそのまま固定化し、マイクロアレイに利用できるという特徴をもつ。タンパク質中のアミノ基を介した固相担体への固定化方法としては、固相表面に結合させた活性エステル基を介してタンパク質を固定化する方法や、固相表面に配置したエポキシ基を介してタンパク質を固定化する手法が挙げられる(非特許文献6)。タンパク質をアミノ基を介して固定化する方法はシンプルな方法であるが、市販のタンパク質や生体抽出・成分、特定のタグのない組み換えタンパク質等においても簡便に固定化することができるため、個々の使用者が自分の目的に合わせ、自在にタンパク質を選択し、迅速かつ安価に目的にあったマイクロアレイに最適化して使用することができる。タンパク質のアミノ基を介した固定化方法の欠点としては、タンパク質中のリジン残基の数が個々のタンパク質によって異なることや、固定化に用いられるリジン残基の位置によってはタンパク質の不活化が起こる可能性等が挙げられる。
【0014】
【特許文献1】特許出願2001−520104
【特許文献2】特許公開平8−201383
【特許文献3】特許公表2002−544485
【非特許文献1】L.J.Holt,K.Bussow,G.Walter,I.M.Tomlinson,Nucleic Acids Res.,15,E72,2000
【非特許文献2】D.Guschin,G.Yershov,A.Zaslavsky,A.Gemmell,V.Shick,D.Proudnikov,P.Arenkov,A.Mirzabekov,Anal.Biochem.,250,203−211,1997
【非特許文献3】A.Lueking,M.Horn,H.Eickhoff,K.Bussow,H.Lehrach,G.Walter,Anal.Biochem.,270,103−111,1999
【非特許文献4】P.Mitchell,Nat.Biotechnol.,20,225−229,2002
【非特許文献5】H.Zhu,M.Bilgin,R.Bangham,D.Hall,A.Casamayor,P.Bertone,N.Lan,R.Jansen,S.Bidlingmaier,T.Houfek,T.Mitchell,P.Miller,R.A.Dean,M.Gerstein,M.Snyder,Science,293,2101−2105,2001
【非特許文献6】H.Zhu,J.F.Klemic,S.Chang,P.Bertone,A.Casamayor,K.G.Klemic,D.Smith,M.Gerstein,M.A.Reed,M.Snyder,Nat.Genetics.26,283−289,2000
【発明の開示】
【発明が解決しようとする課題】
【0015】
本発明の目的は、より簡便に、高速、高感度、かつ高精度に糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法を提供することにある。
【課題を解決するための手段】
【0016】
本発明者らは、上記の課題を解決するために鋭意研究を行った。
タンパク質のアミノ基を介した固定化方法では、一般的にタンパク質内のランダムな位置で固相担体との架橋が起こるため、ある確率で活性が損なわれてしまう分子もありうるが、逆に、ある確率でタンパク質の活性が生き残る可能性が高い(タンパク質が全滅する危険性が小さい)。多数の性質の異なるタンパク質や性質が未知であるタンパク質を固定化していくことを想定したマイクロアレイにおいては、このような性質は実用上、非常に有用である。
【0017】
そこで、本発明者らは、レクチンアレイを作製するに当たりマイクロアレイの実用面・応用面での有用性、解析目的対象物に対する即応性、市場での競争力(コスト面も含む)等を総合的に考慮すると現時点ではタンパク質のアミノ基を利用した固定化法が最適であると判断した。本発明ではアミノ基を介した固定化法の最適条件をシグナル−ノイズ比(S/N比)とタンパク質固定化密度の向上を目指し多角的に検討した。
【0018】
その結果、スライドガラス表面に3−グリシドキシプロピルトリメトキシシラン(GTMS)コートを施すことにより、タンパク質試料は、従来製品よりS/N比の高い条件で固定化された。さらに、本発明者らは、複数の穴を有するラバーを貼り付けることで、ガラス上に複数の反応槽を形成させた基板を作成し、レクチンのスポット周囲に正確に蛍光標識化プローブ溶液を満たすことを可能にした。また、レクチンをスポットし、PBSTで洗浄することで、レクチン固定化密度の向上を図り、糖鎖−レクチン間の弱い相互作用をより感度良く検出することに成功した。
【0019】
本発明では、レクチン−糖鎖間の平衡状態での相互作用情報を精密に解析する上での大きな問題点であったプローブ溶液の洗浄除去操作が不要となり、従来法では洗浄時に洗い流されてしまうような弱い相互作用をも検出することが可能となった。
【0020】
このように、固定化方法および検出方法の改良より、レクチンアレイの実用化が可能となった。すなわち本発明は、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法に関し、以下の〔1〕〜〔22〕を提供するものである。
〔1〕以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)糖鎖に相互作用を示すタンパク質が固定化された基板に蛍光標識した被検糖鎖または被検複合糖質を接触させる工程
(b)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
〔2〕糖鎖に相互作用を示すタンパク質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に糖鎖に相互作用を示すタンパク質が固定化された基板である、〔1〕に記載の方法。
〔3〕エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、〔2〕に記載の方法。
〔4〕以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)被検複合糖質が固定化された基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程
(b)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
〔5〕被検複合糖質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に被検複合糖質が固定化された基板である、〔4〕に記載の方法。
〔6〕エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、〔5〕に記載の方法。
〔7〕以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板に被検複合糖質を接触させる工程
(b)工程(a)で得られた基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程
(c)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
〔8〕複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板である、〔7〕に記載の方法。
〔9〕エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、〔8〕に記載の方法。
〔10〕複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、〔7〕〜〔9〕のいずれかに記載の方法。
〔11〕糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、〔1〕〜〔10〕のいずれかに記載の方法。
〔12〕励起光がエバネッセント波である、〔1〕〜〔11〕のいずれかに記載の方法。
〔13〕複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、〔1〕〜〔12〕のいずれかに記載の方法。
〔14〕エポキシ基を活性基として有する化合物がコートされた基板であって、糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板。
〔15〕エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、〔14〕に記載の基板。
〔16〕複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、〔14〕または〔15〕に記載の基板。
〔17〕糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、〔14〕または〔15〕に記載の基板。
〔18〕複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、〔14〕〜〔17〕のいずれかに記載の基板。
〔19〕以下の工程を含む、基板の製造方法。
(a)基板にエポキシ基を活性基として有する化合物をコートする工程
(b)工程(a)で得られた基板に糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質を固定化する工程
〔20〕複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、〔19〕に記載の方法。
〔21〕糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、〔19〕に記載の方法。
〔22〕複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、〔19〕〜〔21〕のいずれかに記載の方法。
【図面の簡単な説明】
【0021】
【図1】本発明のシステムの構成図である。検出手段は、マイクロアレイスキャナー装置である。
【図2】本発明のシステムにおけるコンピューターの構成図である。記憶手段6には、マイクロアレイスキャナー装置を利用したシステムの処理を実行するためのプログラム61〜63が少なくとも格納されている。記憶手段(データベース)7には、糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報、および/または該相互作用情報の組み合わせパターン情報が格納されている。
【図3】ガラス表面に対するGTMSの反応過程を示す図である。GTMSのアルコキシシリル基は水で加水分解されてシラノール基となる。このシラノール基は不安定で、経時変化により部分的に縮合してオリゴマー状態になり、続いてガラス表面に水素結合的に吸着する。その後、ガラスを乾燥処理することでガラス表面のシラノール基と脱水縮合反応が起こり、強固な共有結合となる。
【図4】本実施例で用いた8つの反応槽を形成させた基板を示す図である。新規設計した8穴ラバーの厚さは1mmであり、専用のアジャスター上でスライドガラスと密着させることによってスポット周囲に正確に蛍光標識化糖プローブ溶液を満たすことが可能となる。反応槽内に満たす試料の最適量は50μLである。
【図5】2種のレクチンを固定化したアレイ上にCy3−ASF溶液を加えた、レクチンアレイの性能実験の概念図である。
【図6】固定化時のレクチン溶液濃度とスポットの蛍光強度との関係を示す図及び写真である。親和定数の高いレクチン−糖鎖間の相互作用の検出においては、スポットするレクチン試料の濃度を1mg/mL以上に高濃度化することがシグナル強度の向上に有効であることが分かった。
【図7】レクチン−糖鎖相互作用の検出と、阻害糖による相互作用への影響を示す図及び写真である。RCA120のスポットにおいて強い蛍光が、またEW29(Ch)のスポットにおいては中程度の蛍光が観察された。
【図8】阻害糖のレクチン−糖鎖相互作用への影響をグラフとして示す図である。ラクトース(競合阻害糖)共存下で実験を行った。共存するラクトース(競合阻害糖)の濃度増加に従って、スポットの蛍光強度が減少していることから、蛍光糖タンパク質プローブの結合はレクチンと糖鎖間の糖特異的結合反応であるということが確認できた。
【図9】各糖鎖関連アレイと糖鎖の相互作用の模式図である。A:レクチンアレイ、およびプローブとして糖鎖を用いた場合の模式図である。蛍光標識糖鎖の構造類推に用いることができる。なお、糖鎖は、間接的に蛍光標識することもできる。B:レクチンアレイ、およびプローブとして糖タンパク質を用いた場合の模式図である。タンパク質上の糖鎖構造の類推に用いることができる。なお、糖鎖は、間接的に蛍光標識することもできる。C:糖ペプチドアレイ、およびプローブとしてレクチンを用いた場合の模式図である。分画したペプチド上の糖鎖構造の類推に用いることができる。D:糖タンパク質アレイ、およびプローブとしてレクチンを用いた場合の模式図である。2次元分画したタンパク質上の糖鎖構造の類推に用いることができる。E:抗体アレイ、およびプローブとしてレクチンを用いた場合の模式図である。クルードな糖タンパク質上の糖鎖構造の類推に用いることができる。
【図10】レクチン−抗体ハイブリッドアレイの実施例を示す写真である。具体的には、7種のレクチンと2種の抗体を固定化したアレイに対して、各モデルタンパク質を蛍光標識化したプローブ溶液を接触させ、平行化した後にエバネッセント励起型スキャナーを用いて観察した結果を示す。
【図11】8つの反応槽を使用して、同一スライド上に異なる濃度の阻害糖を共存させて、相互作用の阻害を観察した結果を示す写真及び図である。(A)RCA120とASFの結合にラクトースを加えたケース、(B)ConAとRNase Bの結合にマンノースを加えたケースである。
【図12】マウス肝由来糖ペプチドサンプルのHPLC分画フラクションを固定化した、糖ペプチドアレイの検出実験の結果を示す図及び写真である。図中、Fucはフコース認識レクチン群、Siaはシアル酸認識レクチン群、Lacはラクトース認識レクチン群、Galはガラクトース認識レクチン群、GalNAcはN−アセチルガラクトサミン認識レクチン群、Manはマンノース認識レクチン群、Chitinはキチン認識レクチン群を意味する。
【図13】レクチンアレイの検出に糖ペプチドプローブを用いる実験の結果を示す写真及び図である。
【図14】生体由来のクルードな試料をプローブ化した際のレクチンアレイによる解析結果を示す写真である。
【発明を実施するための最良の形態】
【0022】
本発明は、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法を提供する。本発明の方法は、糖鎖に相互作用を示すタンパク質が固定化された基板に蛍光標識した被検糖鎖または被検複合糖質を接触させる工程、および、基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程を含む。
【0023】
本発明における糖鎖としては、例えば、糖タンパク質系糖鎖(N−結合型糖鎖とO−結合型糖鎖)、糖脂質系糖鎖、グリコサミノグリカン系糖鎖、または多糖類由来オリゴ糖鎖などが挙げられる。また、1)N−結合型糖鎖としては、高マンノース型・混成型・複合型からなるN−結合型糖鎖など、2)O−結合型糖鎖としては、ムチン型(O−GalNAc)・O−Fuc型・O−Man型・O−Glc型などからなるO−結合型糖鎖など、3)糖脂質系糖鎖としては、ガングリオ系列・グロボ系列ウクト・ネオラクト系列糖鎖など、4)グリコサミノグリカン系糖鎖としては、ヒアルロン酸・ケラタン硫酸・ヘパリン・ヘパラン硫酸・コンドロイチン硫酸・デルマタン硫酸など、5)多糖類由来オリゴ糖鎖としては、キチン、セルロース、カードラン、ラミナリン、デキストラン、デンプン、グリコーゲン、アラビノガラクタン、アルギン酸、フルクタン、フコイダン、キシランなどに由来するオリゴ糖鎖などが例示できる。
【0024】
その他の糖鎖としては、M3・M5A・Hybrid(monoagalacto,bisect)・NA1・NA1(α1−6Fuc)・NA2(monoagalacto)・NA2(monoagalacto,bisect)・NA2・NA2(α1−6Fuc)・A2・NA2(bisect)・NA3・NA3(α1−6Fuc)・NA4・NA4(α1−6Fuc)・NA5(pentaagalacto,bisect)・Lactose・GA2・GA1・GM3−NeuAc・GM3−NeuGc・GM1・GM2・GD1a・GD1b・GD3・Gb3・Gb4・Forssman・LNnT・LNT・Galili pentasaccharide・B−hexasaccharide・LNFP−I・LNFP−II(Lea)・LNFP−III(LeX)・LNFP−II(Leb)・A−hexasaccharide・A−heptasaccharide・B−pentasaccharide・6’Sialyl Lactose・pLNH・βGalLac・βGal2Lac・LN3・GN3・GN4・maltotriose・Sialyl LeXなどを挙げることができる。
【0025】
また、本発明において、複合糖質とは、糖鎖を持つ生体内高分子の総称である。本発明の複合糖質としては、糖タンパク質(糖ペプチドも含む)、プロテオグリカン、糖脂質が挙げられる。
【0026】
本発明の糖鎖に相互作用を示すタンパク質には、糖鎖に相互作用を示すペプチドも含まれる。本発明の糖鎖に相互作用を示すタンパク質としては、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、これらの変異体、または糖鎖に相互作用を示す抗体などが挙げられる。
【0027】
上記レクチンとしては、動・植物、真菌、細菌、ウィルスなどから得られる様々な分子家系に属するレクチン、すなわち、細菌を含むすべての生物界で見出されるリシンB鎖関連の「R型レクチン」、真核生物全般に存在し糖タンパク質のフォールディングに関与する「カルネキシン・カルレティキュリン」、多細胞動物に広く存在し、「セレクチン」、「コレクチン」等代表的なレクチンを多く含むカルシウム要求性の「C型レクチン」、動物界に広く分布しガラクトースに特異性を示す「ガレクチン」、植物豆科で大きな家系を形成する「豆科レクチン」、およびこれと構造類似性をもち動物細胞内輸送に関わる「L型レクチン」、リソソーム酵素の細胞内輸送に関わるマンノース6−リン酸結合性の「P型レクチン」、グリコサミノグリカンをはじめとする酸性糖鎖に結合する「アネキシン」、免疫グロブリン超家系に属し「シグレック」を含む「I型レクチン」などが挙げられる。
【0028】
その他のレクチンとしては、ACA(センニンコクレクチン)・BPL(ムラサキモクワンジュレクチン)・ConA(タチナタマメレクチン)・DBA(Horsegramレクチン)・DSA(ヨウシュチョウセンアサガオレクチン)・ECA(デイゴマメレクチン)・EEL(Spindle Treeレクチン)・GNA(ユキノハナレクチン)・GSL I(グリフォニアマメレクチン)・GSL II(グリフォニアマメレクチン)・HHL(アマリリスレクチン)・ジャカリン(ジャックフルーツレクチン)・LBA(リママメレクチン)・LCA(レンズマメレクチン)・LEL(トマトレクチン)・LTL(ロータスマメレクチン)・MPA(アメリカハリグワレクチン)・NPA(ラッパズイセンレクチン)・PHA−E(インゲンマメレクチン)・PHA−L(インゲンマメレクチン)・PNA(ピーナッツレクチン)・PSA(エンドウレクチン)・PTL−I(シカクマメレクチン)・PTL−II(シカクマメレクチン)・PWM(ヨウシュヤマゴボウレクチン)RCA120(ヒママメレクチン)・SBA(ダイズレクチン)・SJA(エンジュレクチン)・SNA(セイヨウニワトコレクチン)・SSA(ニホンニワトコレクチン)・STL(ジャガイモレクチン)・TJA−I(キカラスウリレクチン)・TJA−II(キカラスウリレクチン)・UDA(Common Stinging Nettleレクチン)・UEA I(ハリエニシダレクチン)・VFA(ソラマメレクチン)・VVA(ヘアリーベッチレクチン)・WFA(フジレクチン)・WGA(パンコムギレクチン)などを挙げることができる。
【0029】
上記糖結合ドメインを有する酵素タンパク質としては、各種グリコシダーゼ(キシラナーゼ、グルカナーゼ)、および糖転移酵素(UDP−GalNAc:ポリペプチドGalNAc転移酵素)などが例示できる。また、糖鎖に親和性を有するサイトカインとしては、インターロイキン−2(IL−2)、インターロイキン−12(IL−12)、腫瘍壊死因子α(TNF−α)、繊維芽細胞成長因子(FGF)などが例示できる。また、糖鎖に相互作用を示す抗体としては、糖鎖関連腫瘍マーカー(CA19−9、フォルスマン抗原、T抗原、Tn抗原、シアリルT抗原)、血液型関連糖鎖(A,B,H,Lea,Lex抗原)、分化関連抗原(Ii,SSEA−1−4)に対する抗体などが例示できる。
【0030】
本発明における基板としては、ガラス、石英ガラス、合成石英ガラスなどが例示できるが、これらに限定されるものではない。また、本発明における糖鎖に相互作用を示すタンパク質が固定化された基板は、好ましくは、エポキシ基を活性基として有する化合物がコートされた基板に糖鎖に相互作用を示すタンパク質が固定化された基板である。
【0031】
エポキシ基を活性基として有する化合物としては、好ましくは3−グリシドキシプロピルトリメトキシシラン(GTMS)が挙げられるが、これに限定されない。その他に、2−(3、4エポキシシクロヘキシル)エチルトリメトキシシラン、3−グリシドキシプロピルメチルジエトキシシラン、3−グリシドキシプロピルトリエトキシシラン、または分岐したスペーサーの先端にエポキシ基を複数持つシランカップリング化合物で、好ましくはスペーサーとしてポリエチレングリコールやタンパク質、ビオチン・アビジン等を含む化合物などが例示できる。
【0032】
本発明の、糖鎖に相互作用を示すタンパク質が固定化された基板は、下記の方法で作成することができる。
【0033】
まず、基板にエポキシ基を活性基として有する化合物をコートする。例えば、エポキシ基を活性基として有する化合物としてGTMSを用いる場合、実施例に記載の方法で行うことができる。具体的には、スライドガラスを10%KOH/MeOH溶液に浸し、容器ごと振盪させた状態で1時間放置しガラス表面を処理し、これを十分量の精製水(ミリQ水)により洗浄した後、60℃のオーブン内で乾燥させる。次にスライドガラスを2%GTMSアセトン溶液に浸し、遮光下で容器ごと振盪させながら1時間反応させる。
GTMSのアルコキシシリル基は水で加水分解されてシラノール基となっており、このシラノール基は不安定で、経時変化により部分的に結合してオリゴマー状態になり、続いてガラス表面に水素結合的に吸着する。反応後、スライドガラスを110℃のオーブン内で8時間乾燥させる。乾燥処理により、ガラス表面のシラノール基と脱水縮合反応が起こり、強力な共有結合となる。一連のGTMSコーティング方法を図3に示す。
【0034】
次に、エポキシ基を活性基として有する化合物をコーティングした基板に、糖鎖に相互作用を示すタンパク質を固定化する。糖鎖に相互作用を示すタンパク質の固定化は、共有結合によって行われる。具体的には、該基板にアミノ基を活性基として有する化合物をスポットし、反応させることで固定化することができる。スポッターとしては、日本レーザ電子社製STAMPMANなどを利用することができる。糖鎖に相互作用を示すタンパク質がレクチンである場合、スポットするレクチンの濃度は、特に限定はないが、1mg/mL以上であることが好ましい。さらに、より好ましくは、スポット処理後Tween20を含むPBS溶液(PBST)で洗浄することにより、未結合レクチンを除去することができる。
【0035】
上記糖鎖に相互作用を示すタンパク質が固定化された基板は、複数の反応槽を形成させた基板であることが好ましい。より好ましくは、複数の穴を有するラバーを貼り付けることで、複数の反応槽を形成させた基板である。一例としては、実施例に記載のように、糖鎖に相互作用を示すタンパク質を固定化したスライドガラスに対し、本発明者らが設計・開発した8穴ラバーを所定の位置に貼り付け、8つの反応槽を作製させる。この8穴ラバーには8つの長方形の穴が規則正しく空いており、スライドガラスに貼り付けたときに8つの反応槽を形成することができる。この反応槽に蛍光標識化プローブ溶液を満たすことで、糖鎖に相互作用を示すタンパク質との接触を円滑に行うことが可能になる。また、この反応槽は8穴ラバーに限定されるものではなく、例えばガラス表面の非スポット領域を撥水コートすることで反応場を形成することも可能である。より好ましくは反応場を多数化することである。本発明においては、同一基板上に複数種の糖鎖に相互作用を示すタンパク質をスポットして用いることもできる。
【0036】
本発明の方法では、上記糖鎖に相互作用を示すタンパク質が固定化された基板に蛍光標識した被検糖鎖または被検複合糖質を接触させる。
【0037】
本発明において、被検糖鎖または被検複合糖質の蛍光標識剤としては、2−アミノピリジン、Cy3、Cy3.5、Cy5、テトラメチルローダミン、フルオレセイン骨格を持つ蛍光色素各種、モレキュラープローブス社製蛍光色素Alexaシリーズ、量子ドット蛍光色素が挙げられるが、糖鎖を蛍光標識する性質を有する物質であれば、これらに限定されない。
【0038】
被検糖鎖または被検複合糖質は、直接的または間接的に蛍光標識することができる。被検糖鎖と予め蛍光標識した糖鎖に相互作用を示すタンパク質を結合させることにより、被検糖鎖を間接的に蛍光標識できる。
【0039】
また、予め蛍光標識したタンパク質であって、被検複合糖質の糖鎖以外の部分に相互作用を示すタンパク質(例えば被検複合糖質の糖鎖以外の部分に相互作用を示す抗体)と被検複合糖質を結合させることにより、被検複合糖質を間接的に蛍光標識できる。
【0040】
また、予め蛍光標識したタンパク質であって糖鎖に相互作用を示すタンパク質と被検複合糖質を結合させることによっても、被検複合糖質を間接的に蛍光標識できる(この場合、被検複合糖質が有する糖鎖のうち、糖鎖に相互作用を示すタンパク質が結合した糖鎖以外の糖鎖が基板に結合する)。
【0041】
例えば、標的複合糖質の糖鎖以外の部分に相互作用を示す予め蛍光標識した抗体を、標的複合糖質試料を含むサンプル・血液・体液・生体抽出成分・食品成分等のクルードな試料に作用させることで、クルードな試料から標的複合糖質を分離・精製することなく選択的に蛍光標識できる。また、予め蛍光標識した上記抗体を作用させたクルードな試料を、例えばレクチンが固定化された基板に接触させることで、標的複合糖質の糖鎖情報だけを選択的に観察できる。
【0042】
本発明の方法では、次いで、基板を洗浄せずに、励起光を作用させて、それぞれの糖鎖に相互作用を示すタンパク質に対する被検糖鎖または被検複合糖質の相互作用を測定する。
【0043】
糖鎖に相互作用を示すタンパク質と被検糖鎖または被検複合糖質の相互作用は一般的に良く知られているタンパク質間相互作用などに比べて弱いため、プローブ溶液の除去・洗浄操作を行うことで糖鎖に相互作用を示すタンパク質と被検糖鎖または被検複合糖質間の解離反応が進行してしまい、平衡状態下での正確な相互作用情報を得ることが出来ないケースが生じていた。
【0044】
本発明者らは上記問題を、プローブ溶液の洗浄を行うことなく、励起光を作用させて励起される蛍光の強度を測定することで解決した。より具体的には、励起光を基板の固定化されていない面から入射して、励起された蛍光を検出する測定方法である。本発明における励起光としては、特に限定はなく、白色光から切り出した光源、好ましくは単一波長からなるレーザ光、より好ましくはエバネッセント波が挙げられる。励起光の検出は、エバネッセント型励起マイクロアレイスキャナーの使用が好ましいが、その他に共焦点型マイクロアレイスキャナーを使用することもできる。
【0045】
例えば、エバネッセント励起方式では励起光をガラス内部で全反射させた際に界面からの高さ200〜300nm(励起波長の半分程度)の範囲に、エバネッセント光と呼ばれる微弱光がしみ出し、このエバネッセント光により蛍光物質を励起するために、プローブ分子を含む溶液をスライドガラス上に接触させた状態で励起光を入射して蛍光を観察する際にも、ブラウン運動をしているプローブ分子をほとんど励起することなく、結合反応に預かるプローブ分子を選択的に観察することができる。
【0046】
本発明の方法は、蛍光標識した被検糖鎖と基板に固定化された糖鎖に相互作用を示すタンパク質との相互作用を利用した糖鎖構造解析に用いることができる。すなわち本発明の方法は、糖鎖構造解析を迅速・簡便に行う糖鎖プロファイラーとしての応用が期待される。例えば、蛋白質核酸酵素2003年8月号増刊Vol.48.NO.11に記載のプロファイリング方法が応用可能である。
【0047】
すなわち、本発明は、コンピューターを用いた糖鎖構造分析システムに利用可能である。該システムを使用することで、被検糖鎖が既知構造を有している場合は被検糖鎖の構造を同定することができ、被検糖鎖が未知構造を有している場合であっても被検糖鎖中に存在する特徴的な構造(α2−3シアル酸・α2−6シアル酸・α1−3ガラクトース・α1−3フコース・α1−6フコース・バイセクトN−アセチルグルコサミン・硫酸化など)の予測、ないし既知構造糖鎖との類似性を指摘することができる。このシステムは、蛍光標識した被検糖鎖を接触させた糖鎖に相互作用を示す種々のタンパク質がそれぞれ固定化された基板をマイクロアレイスキャナー装置にセットすると、自動的に、被検糖鎖の構造が表示されるシステムである。本発明のシステムでは、糖鎖に相互作用を示す種々のタンパク質がそれぞれ固定化された基板に蛍光標識した被検糖鎖を接触させる工程を自動化することもできる。すなわち、微小流路系を基板上の反応槽に導き、流路内に送液する溶液の種類・濃度・流速をコントロールすることで、ブロッキングやブロッキング液の洗浄除去工程、蛍光標識糖鎖溶液の接触工程を一元的にコントロールすることができる。本発明のシステムには質量分析や、酵素消化を組み合わせることもでき、これらの方法を用いることでさらに信頼度の高いデータを得ることができるため、大変に有用である。
【0048】
図1に本発明のシステム構成図の一例を示す。マイクロアレイスキャナー装置を用いたシステムは以下から構成される。
(a)糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報が格納されている記憶手段(データベース)
(b)糖鎖に相互作用を示す種々のタンパク質がそれぞれ固定化された基板に、蛍光標識した被検糖鎖を接触させ、洗浄操作を行わずに、該基板に励起光を入射し、励起される蛍光の強度を検出する検出手段
(c)検出された蛍光強度の組み合わせ情報を、(a)に格納されている情報と照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出する演算手段を含むコンピューター
(d)選出結果を表示する表示手段
【0049】
データベースについては、図1のようにコンピューターの外部にある場合、図2のようにコンピューター内部にある場合、共に許容される。また、該データベースを利用することで、限定されたレクチン数であっても多種類の糖鎖を識別することが可能である。理論的には10種類のレクチンであってもその特異性が異なる場合、識別可能な糖鎖の種類は610=60,466,176となり、事実上自然界に存在する殆どの糖鎖構造の識別が可能である。
【0050】
図2に、本発明のシステムにおけるコンピューター構成図の一例を示す。入力手段1と出力手段2がバス線3に接続されている。一時記憶手段4は、入力された情報、および算出された情報などを一時的に記憶する。中央処理装置(CPU)5は、本発明のプログラムの命令を受けて各種演算を行う。記憶手段(データベース)7には、糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報、および/または該相互作用情報の組み合わせパターン情報が格納されている。相互作用情報として、本発明のマイクロアレイスキャナー装置を利用した方法やシステムによって得られた蛍光強度情報、また、これまでに確立されている様々な実験系から得られた情報が利用可能である。
【0051】
記憶手段6には、本発明の処理を実行するためのプログラムを含む各種プログラムが格納されている。本発明の処理を実行するためのプログラムには、入力された蛍光強度の組み合わせ情報を、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報と照合し、組み合わせ情報のパターンが一致する構造既知糖鎖(データベースに格納されている構造既知糖鎖情報)を1つないし複数選出するプログラム61、表示プログラム62、およびこれらを制御するためのプログラム63が少なくとも含まれる。
【0052】
相互作用情報の組み合わせ情報の照合過程においては、相互作用情報の組み合わせ情報の値同士を比較してもよい。プログラム61には、例えば、入力された蛍光強度の組み合わせ情報の値とデータベースに格納されている相互作用情報の組み合わせ情報の値を比較して、その値の近さから構造既知糖鎖を1つないし複数選出する機能が組み込まれていてもよい。
【0053】
また、相互作用情報の組み合わせ情報の照合過程においては、相互作用情報の組み合わせ情報をパターン化して、そのパターン同士を比較してもよい。このような観点から、記憶手段6には、プログラム61の代わりに(または、プログラム61とともに)、入力された蛍光強度の組み合わせ情報をパターン化するプログラム61−1、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報をパターン化するプログラム61−2、および、入力された蛍光強度の組み合わせ情報のパターンを、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報のパターンと照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出するプログラム61−3が格納されていてもよい。このパターン化に際しては、適切な内部標準を用いて相互作用情報の規格化を行うことも可能である。プログラム61−1や61−2には、例えば、任意の設定スレッショルド値が入力されることで、そのスレッショルドの範囲に相互作用情報をレベル分けし、コード化する(各レベルに対して、例えば異なる数字や異なる色を当てはめる)機能が組み込まれている。
【0054】
プログラム61−3は、プログラム61−1を実行して得られるパターンを、プログラム61−2を実行して得られるパターンと照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出するプログラムである。なお、データベースにパターン情報が格納されている場合は、プログラム61−3は、プログラム61−1を実行して得られるパターンを、データベースに格納されているパターンと照合し、パターンが一致する構造既知糖鎖を1つないし複数選出する。プログラム61−3には、例えば、構造既知糖鎖のコードと被検糖鎖のコードを比較して、被検糖鎖とコードが一致する構造既知糖鎖を選出する機能が組み込まれている。
【0055】
プログラム62は、例えば、蛍光強度情報の表示、相互作用情報の表示、選出された構造既知糖鎖情報の表示などを行う。
【0056】
本発明においては、上述のプログラムを1つのプログラムにまとめることもできる。
【0057】
本発明のシステムにより実行される処理のフローの一例としては、まず、蛍光標識した被検糖鎖を接触させた糖鎖に相互作用を示す種々のタンパク質がそれぞれ固定化された基板がマイクロアレイスキャナー装置にセットされた場合に、該基板に励起光が入射され、励起される蛍光の強度が検出される。複数の基板がマイクロアレイスキャナー装置にセットされた場合は、該複数の基板が順次自動的に検出部に固定され、スキャニングが行われる。処理のフローの一例としては、次いで、蛍光強度情報が自動的にコンピューターに入力される。入力された情報は、コンピューターの記憶手段または一時記憶手段に格納しておくことができる。また、蛍光強度情報は、データベースに格納されてもよい。蛍光強度情報を蓄積することで、今まで存在しえなかった大規模かつ実用性の高い、糖鎖に相互作用を示すタンパク質に対する糖鎖の相互作用情報データベースを構築することができる。
【0058】
本発明では、中央処理装置(CPU)などの演算手段が記憶手段中のプログラム62の指令を受け、記憶手段または一時記憶手段に格納された蛍光強度情報を読み出し、該蛍光強度情報を表示することもできる。例えば、あらかじめ性状を十分に調査した基準となる糖鎖に相互作用を示すタンパク質試料スポット(内部標準スポット)の発する蛍光強度を基準として、各スポットの輝度値を補正した値を表示することができる。内部標準スポットは複数であってもよい。
【0059】
処理のフローの一例としては、次いで、入力された蛍光強度の組み合わせ情報を、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報と照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出する。この処理工程は、中央処理装置(CPU)などの演算手段が記憶手段中のプログラム61の指令を受け、記憶手段または一時記憶手段に格納された蛍光強度の組み合わせ情報とデータベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報を読み出し、それぞれの組み合わせ情報を照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出する。選出された構造既知糖鎖情報は、コンピューターの記憶手段または一時記憶手段に格納しておくことができる。
【0060】
データベースがコンピューターの外部にある場合は、中央処理装置(CPU)などの演算手段が記憶手段中のプログラム61の指令を受け、データベースに格納されている糖鎖に相互作用を示す種々のタンパク質に対する複数の糖鎖の相互作用情報の組み合わせ情報をコンピューターに入力し、記憶手段または一時記憶手段に格納された蛍光強度の組み合わせ情報を読み出し、それぞれの組み合わせ情報を照合し、組み合わせ情報のパターンが一致する構造既知糖鎖を1つないし複数選出する。
【0061】
なお、プログラム61の代わりに、プログラム61−1〜61−3を用いるときも、同様なフローで処理される。
【0062】
処理のフローの一例としては、次いで、選出結果が表示手段によって表示される。この処理工程では、中央処理装置(CPU)などの演算手段が記憶手段中のプログラム62の指令を受け、記憶手段または一時記憶手段に格納された構造既知糖鎖情報を読み出し、表示する。
【0063】
本発明は、被検複合糖質が固定化された基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程、および、基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法を提供する。
【0064】
本発明における、被検複合糖質が固定化された基板は、好ましくは、エポキシ基を活性基として有する化合物がコートされた基板に被検複合糖質が固定化された基板であり、より好ましくは、エポキシ基を活性基として有する化合物が、GTMSである基板である。各工程は、上述の方法と同様である。本発明の方法は、基板に固定化された被検複合糖質の糖鎖付加状況を蛍光標識レクチン等により調べることで、生体にあるタンパク質を分離後、各タンパク質成分それぞれについての糖鎖付加状況をタンパク質の一団について一度に調べる(グライコフォーム解析)ことができる手段として有用である。またタンパク質製剤等の糖鎖部分の品質管理チェック等に応用が期待される。
【0065】
本発明は、また、複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板に被検複合糖質を接触させる工程、該工程で得られた基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程、および、基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法を提供する。
【0066】
本発明における、被検複合糖質が固定化された基板は、好ましくは、エポキシ基を活性基として有する化合物がコートされた基板に被検複合糖質が固定化された基板であり、より好ましくは、エポキシ基を活性基として有する化合物が、GTMSである基板である。また、本発明における複合糖質の糖鎖以外の領域に相互作用を示すタンパク質は、好ましくは、複合糖質の糖鎖以外の領域に相互作用を示す抗体である。また、糖鎖に相互作用を示すタンパク質は、好ましくは、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、これらの変異体、または糖鎖に相互作用を示す抗体である。各工程は、上述の方法と基本的に同様であるが、さらに、複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板であって、被検複合糖質を接触させた基板に、蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程を含む。該工程は、例えば、複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板に対し、被検複合糖質試料を含むサンプル・血液・体液・生体抽出成分・食品成分等の原液または希釈溶液を接触させ、十分な反応時間を確保した後に、PBSないしPBSTで十分に洗浄して被検複合糖質以外の成分を除く。その後に糖鎖部分に相互作用を示す蛍光標識プローブ溶液を接触させることで行うことができる。
【0067】
本発明は、異なる複合糖質の混合物中から、目的としたタンパク質の糖鎖修飾の状況を迅速かつ簡便に調べる方法として有用であり、血液・体液・生体抽出成分・食品成分等の原液または希釈溶液等の様々な成分の混合溶液を精製することなく、目的タンパク質の糖鎖修飾の状況を観察することで、診断・治療状況の把握等を行うような分野での応用が考えられる。
【0068】
本発明は、(a)基板にエポキシ基を活性基として有する化合物をコートする工程、および、(b)工程(a)で得られた基板に糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質を固定化する工程を含む、基板の製造方法を提供する。該方法の工程は、詳しくは、上述のとおりである。本発明はまた、上記方法で製造された基板も提供する。
【0069】
すなわち、本発明は、エポキシ基を活性基として有する化合物がコートされた基板であって、糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板を提供する。本発明の基板としては、好ましくは、エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である基板である。また、好ましくは、複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である基板、あるいは、糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、これらの変異体、または糖鎖に相互作用を示す抗体である、基板である。エポキシ基を活性基として有する化合物がレクチンである場合、スポットするレクチンの濃度は、特に限定はないが、1mg/mL以上であることが好ましい。さらに、より好ましくは、PBSTで洗浄することでレクチンが固定化された基板である。本発明の基板は、複数の反応槽を形成させた基板であることが好ましい。
なお、本明細書において引用された全ての先行技術文献は、参照として本明細書に組み入れられる。
【実施例】
【0070】
以下、本発明を実施例により、さらに具体的に説明するが、本発明はこれら実施例に制限されるものではない。
〔実施例1〕レクチンアレイを利用した糖鎖とレクチンとの相互作用解析
(1)蛍光標識化糖タンパク質プローブ(Cy3−ASF)の調製
蛍光標識化糖タンパク質プローブとして、アシアロフェツイン(SIGMA社、以下ASF)を550nm付近に吸収極大波長を持つ蛍光色素であるCy3 Mono−reactive Dye(アマシャムファルマシア社、以下Cy3)を用いて蛍光標識化して調製した。ASFはN−結合型糖鎖とO−結合型糖鎖を3本ずつ分子中に持ち、かつ糖鎖中の非還元末端のシアル酸キャップが部分的に外れている糖鎖構造を持つことが知られている。ASFを0.1M炭酸緩衝液(pH9.3)に終濃度1mg/mLになるよう調製した後、1mLについて1.0mgのCy3粉末と混合させ、1時間、適時攪拌しながら暗所で反応させた。
【0071】
次に担体としてSephadex G−25を用いたゲルろ過クロマトグラフィーにより、遊離のCy3とCy3−ASFを分離回収し、精製したCy3−ASFについて吸光光度計を用いて濃度及び蛍光標識効率を測定した。タンパク質ベースの収率は35−40%、蛍光標識効率(1タンパク質分子あたりの蛍光色素数)は約3.0であった。
【0072】
(2)スライドガラスへのGTMSコーティング
エポキシ基を活性基として有する3−グリシドキシプロピルトリメトキシシラン(信越シリコーン社、以降GTMS)をコーティングしたスライドガラス(図3)を用い、レクチンをガラス表面に固定化した。GTMSコーティングは、松浪硝子工業社製のスライドガラスを用い、以下の手順で行った。スライドガラスを10%KOH/MeOH溶液に浸し、容器ごと振盪させた状態で1時間放置しガラス表面を処理した。これを十分量の精製水(ミリQ水)により洗浄した後、60℃のオーブン内で乾燥させた。次にスライドガラスを2%GTMSアセトン溶液に浸し、遮光下で容器ごと振盪させながら1時間反応させた。反応後、110℃のオーブン内で8時間乾燥させた後、十分量の精製水により洗浄し、乾燥させた。
【0073】
(3)スライドガラスへのレクチンの固定化
(2)のGTMSコーティングを施したスライドガラスにレクチンをスポットした。マイクロアレイスポッターとして日本レーザ電子社製STAMPMANを使用し、先端直径0.40mmのスタンプピンを使用してスポットを行ことで、直径約0.6−0.7mmのスポットをスライドガラス上に配置した。スポットする各レクチンを濃度1mg/mL(レクチンによっては一部0.25mg/mL)となるようにリン酸緩衝生理食塩水、pH7.4(以下PBS)に溶解した。これを96穴のPCR用マイクロタイタープレート(コーニング社)の各反応槽に10μLずつ分注したものを、マイクロアレイスポッターにセットした。
【0074】
スライドガラスへのレクチンの固定化操作に際しては以下の条件をマイクロアレイスポッター付属のコンピューターに記憶させ、スタンプピン動作プログラムを実行させた。まずスタンプピンを96穴PCR用マイクロタイタープレート内の固定化試料溶液中に1秒間浸した後に引き上げ、スライドガラス表面の所定の位置に1秒間接触させた。この動作を1スポットごとに繰り返しながら、同一試料溶液から横1列に4点スポットを行った後、スタンプピンの洗浄工程を行った。洗浄工程ではスタンプピンの針先を0.05%SDS溶液に2秒間浸し、スタンプピンをバキューム装置内で15秒間乾燥させ、さらに精製水に2秒間浸した後に、バキューム装置内で15秒間乾燥、最後にエタノールに2秒間浸してからバキューム装置内で15秒間の乾燥操作を行った。
【0075】
本実施例では様々な糖結合特異性をもったレクチン4種(RCA120、SSA、組み換え放線菌由来キシラナーゼのキシラン結合ドメイン(以下XBD)、組み換えミミズ29kDaレクチン由来のC末端側ドメイン(以下EW29(Ch))とネガティブコントロール1種(ウシ血清アルブミン(以下BSA))の計5種のタンパク質をスポットした。RCA120、BSAについてはSIGMAより購入したもの、SSAは生化学工業より購入したもの、XBD,EW29(Ch)については本発明者らの研究室で大腸菌にて発現・精製したものを用いた。
【0076】
(4)非スポット面のブロッキング
レクチン溶液をスポット処理後1時間反応させガラス表面に固定化した後、未結合レクチンを洗浄した。洗浄は0.1%Tween20を含むPBS溶液(PBST)を数回ピペットでスライドガラスに吹き付けるようにかけて洗浄した後、PBSを用いてさらに十分に洗浄した。
【0077】
このレクチン固定化後のスライドガラスに対し、本発明者らが設計・開発した8穴ラバーを所定の位置に貼り付け、8つの反応槽を作製した(図4)。この8穴ラバーは厚さ1mmからなる黒色のシリコンゴム製で、縦横9.5×7.5mmからなる8つの長方形の穴が規則正しく空いており、スライドガラスに貼り付けたときに8つの反応槽を形成することができる。この反応槽に50μL程度の試料を加えれば、内部を十分量の試料溶液で満たすことができる。
【0078】
レクチンをスポットした領域以外のガラス表面には活性基であるエポキシ基が残存しているため、非スポット面に対するブロッキング操作を行った。なお、ブロッキング剤には高純度BSA(SIGMA)を使用した。8つの反応槽内に1%BSAを含むPBS溶液を50μLずつ満たし、湿度を90%以上に保った保存容器中で4℃、1時間放置し、スライドガラス上の非スポット面へのブロッキングを行った。反応の間はガラス表面の乾燥防止に留意した。
【0079】
次にスライドガラス上のブロッキング溶液を除去し、PBSを用いて十分に洗浄した後、水分を除去した。タンパク質固定化後はガラス表面の乾燥によるタンパク質の変性や、乾燥に伴うバックグラウンドの上昇を防ぐため、可及的速やかに次の操作に移った。
【0080】
(5)プローブ溶液の添加とスキャニング
(4)で作製したレクチン固定化スライドガラス上の反応槽に相互作用を解析したい蛍光標識化糖タンパク質プローブ溶液を加えた。蛍光標識化糖タンパク質プローブは終濃度10μg/mLになるようPBSに溶解したものを調製し、それぞれの反応槽に50μLを滴下した。
【0081】
レクチン−糖鎖間の反応が平衡に達するまで静置した後に、エバネッセント励起方式マイクロアレイスキャナーであるGTMAS Scan III(日本レーザ電子社)を使用してスライドガラス端面より励起光を入射し、励起されて生じた蛍光発光を、スライドガラス下面に配置されているICCD(イメージインテンシファイアー付電荷結合素子)カメラで検出した。スライドガラスのほぼ全面に対する蛍光イメージをスキャニングし、得られたイメージ画像をTIFFファイル形式(1枚につき約100M Bite)にて保存した。スキャニング時のパラメーターはGain「5000倍」、積算回数「4回」、露光時間「33msec」で統一した。
【0082】
(6)スキャニング画像の数値化
スキャニング画像の数値化には市販のマイクロアレイ用解析ソフトであるArray−Pro Analyzer(Version 4.0 for Windows(登録商標)、Media Cybernetics社)を使用した。各スポットの輝度を上記解析ソフトを用いて算出し、非スポット領域の輝度をバックグラウンド値とした。各スポットの輝度からバックグラウンド値を差し引いたものを正味の輝度値とし、横列に4点並べた同一試料由来のスポットごとに平均値と標準偏差を算出した。
【0083】
以降は各レクチン試料に対するプローブの結合に対してはこの同一試料由来の4点の平均輝度値を用いて評価を行った。以降に示す各レクチンアレイの性能評価は操作(2)〜(6)の一連の行程を経た後に行われた。
【0084】
(7)GTMSコートスライドガラスの性能評価
上述のように作製したGTMSコートスライドガラスの性能を、既存のスライドガラス(6種)と比較評価した。すなわち表面コートされた各スライドガラスに、あらかじめCy3標識したレクチン(100μg/mL)をアレイ状に固定化し、(3)〜(6)の行程を経た後に、スポッティング領域の輝度値(S)と非スポッティング表面の輝度値(N)からS/N比を算出した。その結果、表1に示すように(2)の行程で作製したGTMSコートスライドガラスの輝度値は、最高値を示したスライドガラスAの2分の1程度にとどまったが、バックグラウンドが非常に低いため、そのS/Nは16.1となり、今回評価したスライドガラスの中で最良値を示した。
【0085】
【表1】
【0086】
(8)アレイ上の固定化レクチン濃度の検討(図5および6)
RCA120とConAはそれぞれ複合型糖鎖と高マンノース型糖鎖に対し高い親和性をもつことが知られる代表的なレクチンである。これらのレクチンを様々な濃度で調製し、同一試料について横に4点並べてアレイ状にスポットした。このアレイの各反応槽に対して10μg/mLのCy3−ASFを各50μLずつアプライして結合反応を起こさせた後に、スキャナーで蛍光を観察した。
【0087】
上述したようにASFはN−結合型糖鎖とO−結合型糖鎖を3本ずつ分子中に持ち、かつ糖鎖中の非還元末端のシアル酸キャップが外れラクトサミン構造が突出した糖鎖構造を持つことが知られている。よってRCA120とConAを固定化したレクチンアレイに対し、Cy3−ASFを添加した実験系においては、RCA120が非常に強い親和性を示し、ConAが弱い親和性を示すことが予想された。
【0088】
実験の結果、RCA120のスポットが強い蛍光を発したのに対し、ConAのスポットは同条件のRCA120のスポットに比べて1/3程度の蛍光強度を示すに止まった。ConAが弱いながらも複合型糖鎖を持つASFに結合したのは、N−結合型糖鎖のうちASFに主として存在する3本鎖型糖鎖には結合できないものの、少量存在するとされる2本鎖型糖鎖には結合できるためと考えられた。またこのデータから同一試料由来の4点についての標準偏差(SD)は約±20%程度になるということが分かった(図6)。
【0089】
次にスポット時のレクチン濃度と蛍光強度の関係をグラフ化したところ、スポット時のレクチン濃度と蛍光強度の間に正の相関関係があり、スポットするレクチン試料の濃度を1mg/mL以上に高濃度化することで、効果的にシグナル強度を向上することができることが分かった。すなわち親和定数の小さい(結合の弱い)レクチン−糖鎖間の相互作用の検出は固定化レクチン濃度を上げる事により可能となることが分かった(図6)。
【0090】
(9)レクチンアレイの性能評価
様々な糖特異性をもったレクチン4種(RCA120、SSA、XBD、EW29(Ch))、とネガティブコントロール1種(BSA)の計5種のタンパク質を同一試料について横に4点並べてアレイ状にスポットした。このアレイについて10μg/mLのCy3−ASFを各50μLずつ滴下し、スキャナーで蛍光を観察した。
【0091】
実験の結果FACでラクトサミン構造に親和性があると確認することができたRCA120、EW29(Ch)の2種のレクチンのスポットにおいて蛍光シグナルが観察された(図7)。またそれぞれの蛍光強度を比較すると、RCA120のスポットにおいて強い蛍光が、EW29(Ch)のスポットにおいては中程度の蛍光が観察され、FAC解析データと一致した。
【0092】
また同条件のアレイに対しラクトース(競合阻害糖)共存下で同様の実験を行ったところ、阻害糖の濃度上昇に伴って、スポットの蛍光強度の減少が観察された(図8)。このことから、蛍光糖タンパク質プローブとの結合はレクチンと糖鎖間の糖特異的結合反応によるものであることを確認することが出来た。
【0093】
〔実施例2〕他の糖鎖関連アレイへの応用
糖鎖関連アレイには、糖鎖をプローブに用いるレクチンアレイの他、糖タンパク質をプローブに用いるレクチンアレイ、レクチンをプローブに用いる糖ペプチドアレイ、レクチンをプローブに用いる糖タンパク質アレイ、レクチンをプローブに用いる抗体アレイの5タイプが挙げられる。図9に、各糖鎖関連アレイと糖鎖の相互作用の模式図を示す。
【0094】
それぞれのアレイを使用して得られた蛍光強度情報をコード化する。輝度値の数値は16bitで取り込まれた数値であるため、0−65355の間の値となる。そこでGain「1000倍」での(輝度値−バックグラウンド値)の値を10000刻みで6段階に分類することで0から5までのコード化を行った。Gain「1000倍」で輝度信号値が飽和している場合は、Gainを下げて観察しGain「1000倍」時の値に換算する。信号値が飽和しているかどうかの判定は、Gain値を変化させた際にGain値と信号強度の相関の直線性が保たれているかどうかで判断する。
【0095】
現在のスレッショルド値 (輝度値−バックグラウンド値)
0−10000 レベル0
10001−20000 レベル1
20001−30000 レベル2
30001−40000 レベル3
40001−50000 レベル4
50001以上 レベル5
【0096】
〔実施例3〕 レクチン−抗体を同一区画内にスポットしたアレイによる解析(図10)
1.材料と方法
(1)モデル糖タンパク質の蛍光標識化プローブの調製
本実験例ではレクチンアレイ上に固定化するレクチンとして、様々な糖結合特異性を持つレクチンとして6種(RCA120、ECA、ConA、GNA、SSA、SNA)を選択した。またネガティブコントロールとして糖鎖と結合しないタンパク質であるBSAを選択した。また今回の実験では、プローブのコアタンパク部分を認識する抗フェツイン抗体と抗RNase抗体の2種を、レクチンと同じ区画内にスポットした。GNA、SNAはVECTOR社より購入したもの、BSAについてはSIGMA社より購入したもの、RCA120、ECA、ConA、SSAは生化学工業より購入したものを用いた。
【0097】
モデル蛍光標識化糖タンパク質プローブは、ASF、FET、ウシ膵由来リボヌクレアーゼB(RNase B)、またネガティブコントロールとしてウシ膵由来リボヌクレアーゼA(RNase A)、BSAなどのタンパク質(全てSIGMA社より購入)を550nm付近に吸収極大波長を持つ蛍光色素であるCy3 Mono−reactive Dye(アマシャムファルマシア社、以下Cy3)を用いて蛍光標識化することで調製した。プローブ作製時は前述のタンパク質を0.1M炭酸緩衝液(pH9.3)に終濃度1mg/mLになるよう調製した後、1.0mgのCy3粉末と混合させ、1時間、適時攪拌しながら暗所で反応させ、担体としてSephadex G−25を用いたゲルろ過クロマトグラフィーにより、Cy3標識タンパク質を精製した。
【0098】
(2)スライドガラスへのGTMSコーティング
エポキシ基を活性基として有する3−グリシドキシプロピルトリメトキシシラン(信越シリコーン社、以降GTMS)をコーティングしたスライドガラス(図3)を用い、レクチンをガラス表面に固定化した。GTMSコーティングは、松浪硝子工業社製のスライドガラスを用い、以下の手順で行った。スライドガラスを10%KOH/MeOH溶液に浸し、容器ごと振盪させた状態で1時間放置しガラス表面を処理した。これを十分量の精製水(ミリQ水)により洗浄した後、60℃のオーブン内で乾燥させた。次にスライドガラスを2%GTMSアセトン溶液に浸し、遮光下で容器ごと振盪させながら1時間反応させた。反応後、110℃のオーブン内で8時間乾燥させた後、十分量の精製水により洗浄し、乾燥させた。
【0099】
(3)レクチンアレイの作製
(2)で作製したGTMSコートスライドガラスに対してレクチンをスポットし、レクチンアレイを作製した。マイクロアレイスポッターとして日本レーザ電子社製STAMPMANを使用し、先端直径0.40mmのスタンプピンを用い、直径約0.5mmのスポットをスライドガラス上に配置した。スライドガラスへのレクチンの固定化操作に際しては以下の条件をマイクロアレイスポッター付属のコンピューターに記憶させ、スタンプピン動作プログラムを実行させた。まずスタンプピンを96穴PCR用マイクロタイタープレート内の固定化試料溶液中に1秒間浸した後に引き上げ、スライドガラス表面の所定の位置に1秒間接触させた。この動作を1スポットごとに繰り返しながら、同一試料溶液から横1列に6点スポットを行った後、スタンプピンの洗浄を行った。洗浄工程ではスタンプピンの針先を0.05%SDS溶液に2秒間浸し、スタンプピンをバキューム装置内で15秒間乾燥させ、さらに精製水に2秒間浸した後に、バキューム装置内で15秒間乾燥、最後にエタノールに2秒間浸してからバキューム装置内で15秒間の乾燥操作を行った。
【0100】
(4)非スポット面のブロッキング
上記の操作でレクチンをスポットしたスライド上に、シリコンゴム製の8穴ラバーを接着して8つの反応槽を作製した。この反応槽に0.1% Tween20を含むPBS溶液(PBST)を満たすことで、スライド固層に結合しなかった余剰レクチンを洗浄・除去した。次に1% BSAを溶解したPBS溶液を各反応槽に200μLずつ満たした後に、湿度を90%以上に保った保存容器中で4℃、1時間放置することで非レクチンスポット領域のブロッキング操作を行った。
【0101】
(5)プローブ溶液の添加とスキャン
ブロッキングを終えたスライドに対し、約100ng/mLに調整した各蛍光標識化糖タンパク質プローブ溶液を各反応槽に50μLずつ加えることで、アレイ上にプローブ溶液を接触させた。
【0102】
レクチン−糖鎖間の反応が平衡に達するまで静置した後に、エバネッセント励起方式マイクロアレイスキャナーであるGTMAS Scan III(日本レーザ電子社)を使用してスライドガラス端面より励起光を入射し、励起されて生じた蛍光発光を、スライドガラス下面に配置されているICCD(イメージインテンシファイアー付電荷結合素子)カメラで検出した。スライドガラスのほぼ全面に対する蛍光イメージをスキャンした後、得られたイメージ画像をTIFFファイル形式にて保存した。スキャン時の各パラメーターは、Gain「4000倍」、積算回数「8回」、露光時間「110msec」で統一した。
【0103】
(6)スキャン画像の数値化
スキャン画像の数値化には市販のマイクロアレイ用解析ソフトであるArray−Pro Analyzer Ver.4.5(Media Cybernetics社)を使用した。各スポットの輝度を上記解析ソフトで算出し、非スポット領域の輝度をバックグラウンド値とした。各スポットの輝度からバックグラウンド値を差し引いたものを正味の輝度値とし、横列に5点並べた同一試料由来のスポットごとに平均値と標準偏差を算出した。
以降は各レクチン試料に対するプローブの結合に対してはこの同一試料由来の5点の平均輝度値を用いて評価を行った。
【0104】
2.結果と考察
今回の実験では多様な特異性を持つレクチンを並べたアレイを構成し、既知の糖鎖構造を持つ糖タンパク質プローブをアプライした際の各レクチンスポットの蛍光パターンから、レクチンアレイとしての性能評価を行った。また本実験では、プローブのコアタンパク部分の情報も同時に取得することを目指し、糖タンパク質プローブのコアタンパク部分を認識する抗体もレクチンと並べてスポットしたアレイを作製した。モデル糖タンパク質としては、コアタンパク部分の構造は同一だが修飾糖鎖部分の構造が異なるタンパク質として、ASFとFETの組み合わせと、RNase AとRNase Bの組み合わせを選択した。
【0105】
実験(A)・(B)では、プローブとして用いたASFとFETの糖鎖プロファイルを比較した(図10A及びB)。FETはN−結合型糖鎖とO−結合型糖鎖を3本ずつ分子中に持ち、各糖鎖構造の非還元末端が高度にシアル酸で修飾されていることが知られている。一方、ASFはFET糖鎖のうち末端シアル酸を酵素或いは酸処理によって除去したタンパク質であり、ラクトサミン構造が露出している。このためASFプローブを用いた場合、ラクトサミンを認識するRCA120やECAのスポットが観察され、FETプローブを用いた場合、シアル酸を認識するSSA、SNAのスポットが観察されると予想された。また抗体スポットに対してはFETもASFもコアタンパク部分は同一であるため、両プローブ共に結合が観察されると予想された。実験の結果、末端ラクトサミン構造を持つASFプローブではRCA120やECAの非常に明るいスポットが検出されたが(図10A)、ラクトサミン構造がシアル酸でキャッピングされているFETプローブではRCA120、ECAのスポットは非常に暗くなった(図10B)。この結果はRCA120、ECAがラクトサミン構造を強く認識し、ラクトサミン構造の末端がシアル酸でキャップされると親和性が大幅に低下するという従来の知見と一致していた。また実験(B)ではFETプローブに特徴的なシアル酸の存在に対応して、シアル酸認識レクチンであるSSAやSNAのスポットが観察された(図10B)。ConAのスポットでは実験(A)、(B)共に弱い蛍光が観察された(図10A及びB)。これはN−結合型糖鎖のうち、主として存在する3本鎖型糖鎖には親和性が低いものの、少量存在するとされる2本鎖型糖鎖には結合できるためと考えられた。共通コアタンパク部分を認識するFET抗体のスポットは、実験(A)、(B)において蛍光が観察された(図10A及びB)。
【0106】
実験(C)・(D)ではRNase AとRNase Bプローブの糖鎖プロファイルの相違を比較した(図10C及びD)。RNase Bは、ハイマンノース型のN−結合型糖鎖を分子内に1本のみ持ち、RNase Aは、コアタンパク部分はRNase Bと同一の構造を持つが糖鎖を全く持たないことが知られている。これらのプローブをレクチンアレイに接触させた場合、両プローブ共に抗RNase抗体スポットに対しては親和性を示すが、レクチンのスポットに対する反応性に相違が生じる(RNase BプローブはConAなどのマンノース認識レクチンに親和性を示すのに対し、RNase Aは親和性を示さない)ことが予想された。実験の結果、抗体に対する反応性では、両プローブにおいて共通コアタンパク部分を認識するRNase抗体との結合が観察された。一方レクチンに対する反応性では、ハイマンノース構造を持つRNase BでConAのスポットにおいて結合が観察されたが、糖鎖を持たないRNase Aではレクチンのスポットにおいて、結合による蛍光は観察されなかった。実験(E)ではネガティブコントロールとして、糖鎖を持たないタンパク質であるBSAを用いた。BSAプローブでは予想通り、抗体・レクチンスポットともに結合は観察されなかった(図10E)。
【0107】
実験(A)〜(E)を通して、微量の糖タンパク質試料から迅速に、レクチンの糖結合特異性を反映したプロファイルを得ることができた(図10A〜E)。また今回の実験では、レクチンと抗体を同一アレイ上にスポットすることで、糖タンパク質のコアタンパク部分と、修飾糖鎖部分の情報を同時並行的に一枚のスライド上で取得することができた。同一スライド上で同時並行解析を行うことで、各反応槽間の実験条件(温度、反応時間等)を揃えた形での観察が可能となるという利点が生じた。
【0108】
〔実施例4〕 レクチンアレイを用いた阻害濃度解析(図11)
1.材料と方法
先の実験で観察されたレクチンとプローブ分子間の結合が、糖鎖を介した特異的結合であることを確認する為に競合阻害糖を用いた阻害実験を行った。実験(A)ではスライドガラス上の8つの反応槽内にRCA120をスポットしてアレイを構成した上で、競合阻害糖(ラクトース)の濃度を変えた8種類のASFプローブ溶液を同時に接触させ、結合反応の阻害を観察した(図11A)。実験(B)では固定化レクチンとしてConAを用い、プローブとしてRNase B、競合阻害糖としてマンノースを用い、同様の操作で結合の阻害を観察した(図11B)。アレイ作成時に要した材料・操作は、実施例3と同じであるため記述を省略する。
【0109】
2.結果と考察
実験の結果、競合阻害糖の濃度上昇に伴ってスポットの蛍光強度の減少が観察され(図11)、阻害曲線をカーブフィッティングすることにより、阻害物質固有の半阻害濃度を算出することができた。この結果から、蛍光糖タンパク質プローブとの結合はレクチンと糖鎖間の特異的結合反応によるものであることを確認できた。またこのような阻害実験を用い、半阻害濃度を算出することで結合の強さを評価したり、結合する相手分子を探索することが可能であることが示された。
【0110】
〔実施例5〕 レクチンアレイの糖ペプチドプローブによる検出(図12)
1.材料と方法
(1)糖ペプチドプローブの調製
実施例3(1)に記述した方法でCy3−ASFを調製した後に、このCy3−ASFをトリプシン処理により断片化し、Cy3−ASFペプチドを調製した。
(2)スライドガラスへのGTMSコーティング
実施例3(2)に記述した方法で行った。
(3)レクチンアレイの作製
固定化するレクチンは、各レクチンの持つ主要な糖認識能ごとにグループ分けを行った上で、フコース認識レクチン5種、シアル酸認識レクチン6種、ラクトサミン構造認識レクチン3種、ガラクトース認識レクチン6種、ガラクトサミン認識レクチン11種、マンノース認識レクチン4種、キチン構造認識レクチン5種の計40種類のレクチンを選択し、スライドガラスに固定化したアレイを作製した。実験操作は実施例3(3)に記述した方法で行った。
(4)非スポット面のブロッキング
(5)プローブ溶液の添加とスキャン
(6)スキャン画像の数値化
上記(4)〜(6)の操作については、Cy3−ASFペプチドプローブを用い、実施例3(4)〜(6)に記述した方法と同様の操作にて、実験を行った。
【0111】
2.結果と考察
実験の結果、レクチンアレイに糖ペプチドプローブを供することで、糖鎖構造を反映した糖鎖プロファイルを得ることができた。得られた糖鎖プロファイルは酵素消化する前のASFと同等であり、レクチンアレイに糖タンパク質だけでなく糖タンパク質のペプチド消化産物をプローブとして利用できることが示された。この技術を用いれば糖ペプチドをHPLC等で分画した後にプローブとしてレクチンアレイに供することで糖ペプチド各成分の糖鎖プロファイルの観察が可能になり、有用である。
【0112】
〔実施例6〕 糖ペプチドアレイによる、糖ペプチド上の糖鎖プロファイリング(図13)
1.材料と方法
(1)レクチンプローブの調製
本実施例ではレクチンプローブとして、ラクトサミン構造を強く認識するRCA120、ネガティブコントロールとして糖結合能を持たないBSAを使用した。蛍光標識レクチンプローブは蛍光色素であるCy3を用いて蛍光標識して調製した。レクチンは0.1M炭酸緩衝液(pH9.3)中に終濃度1mg/mLになるよう溶解した後、1mL当たり1.0mgのCy3粉末と混合させ、1時間、適時攪拌しながら暗所で反応させた。反応後は限外ろ過フィルターキットを用いて未反応のCy3色素を除去した。
(2)スライドガラスへのGTMSコーティング
GTMSコートスライドは実施例5(2)に記述した方法で行った。
(3)糖ペプチドアレイの作製
(2)で作製したGTMSコートスライドガラス(図3)に対して糖ペプチドをスポットして糖ペプチドアレイを作製した。マイクロアレイスポッターとして日本レーザ電子社製STAMPMANを使用し、先端直径0.40mmのスタンプピンを使用してスポットを行ことで、直径約0.5mmのスポットをスライドガラス上に配置した。
本実験では固定化した糖ペプチドサンプルは、マウス肝臓の可溶性画分からレクチンカラムを用いて精製した糖タンパク質画分をトリプシンによりペプチド断片化後、HPLCにより分画・分取した糖ペプチドを用いた。スライドガラスへの糖ペプチドの固定化操作に際しては以下の条件をマイクロアレイスポッター付属のコンピューターに記憶させ、スタンプピン動作プログラムを実行させた。まずスタンプピンを96穴PCR用マイクロタイタープレート内の固定化試料溶液中に1秒間浸した後に引き上げ、スライドガラス表面の所定の位置に1秒間接触させた。この動作を1スポットごとに繰り返しながら、同一試料溶液から横1列に6点スポットを行った後、スタンプピンの洗浄を行った。洗浄工程ではスタンプピンの針先を0.05%SDS溶液に2秒間浸し、スタンプピンをバキューム装置内で15秒間乾燥させ、さらに精製水に2秒間浸した後に、バキューム装置内で15秒間乾燥、最後にエタノールに2秒間浸してからバキューム装置内で15秒間の乾燥操作を行った。
(4)非スポット面のブロッキング
(5)プローブ溶液の添加とスキャン
(6)スキャン画像の数値化
上記(4)〜(6)の操作については、Cy3−RCA120プローブとCy3−BSAプローブを用い、実施例3(4)〜(6)に記述した方法に従い、実験操作を行った。
【0113】
2.結果と考察
本実験の結果、糖ペプチドをアレイ化することで、糖ペプチドに付加した糖鎖の構造情報を、ハイスループットかつ容易に得られることが示された。糖ペプチドアレイの作成方法としては、1)精製糖ペプチドを固定化する、2)クルードの糖ペプチドを固定化する、3)HPLCで分離した糖ペプチドフラクションを固定化する、などが考えられる。今回の実験ではHPLCで分離した各フラクションをアレイ化したものを作製でき、有用であることを示すことができた。
【0114】
従来、HPLCで分画したフラクションのうち、どこに糖ペプチドが含まれているかという情報を得ることは、UV吸収や蛍光等でモニターできないために容易ではなかった。本実験の結果、各糖ペプチドフラクションを固定化したアレイを用いることで、どのフラクションにどのような糖鎖構造を持った糖ペプチドがあるのかを容易に知ることが可能となることが分かった。
【0115】
本法を利用することで、解析対象の糖鎖を持つ糖ペプチドがどのフラクションに含まれているのかといった情報を迅速に得ることができるため、多数のフラクションから効率的に糖鎖の含まれるフラクションのみを選出し、質量分析など他の解析へと供すること等が可能になる。また解析対象の糖ペプチドの糖鎖構造情報や、どのようなレクチンが結合するかという情報がない場合、トリプシン消化前の糖タンパク質の状態でレクチンアレイを用いた分析を行い、糖鎖プロファイリングを行うことで、当該糖タンパク質に結合するレクチンを数十種類のレクチンの中から絞り込むことが出来る。このような情報が得られることで、従来多数のレクチンを用いて総当り的に行っていたレクチンブロットなどの操作が簡略化され、大幅に時間と労力を節約できるなどの利点がある。
【0116】
〔実施例7〕 レクチンアレイに対し、クルード生体試料をプローブとして用いた実験(図14)
1.材料と方法
レクチンアレイを用いて糖タンパク質混合物、特に生体由来混合試料のプロファイリングを行うことで、生体内の糖タンパク質の糖鎖付加状態を解析する。
【0117】
マウス肝臓より抽出・精製した糖タンパク質サンプルとマウス脳より抽出・精製した糖タンパク質サンプルをCy3標識化してプローブとした後に、40種レクチンの固定化されたレクチンアレイと接触させて、マウス抽出糖タンパク質混合物全体の糖鎖プロファイルを観察した。
アレイ作製時に要した材料・操作は、実施例3と同じであるため記述を省略する。
【0118】
2.結果と考察
マウス脳由来糖タンパク質プローブ(図14A)と肝臓由来糖タンパク質プローブ(図14B)では、明らかに異なった糖鎖プロファイルが観察された。特にシアル酸認識のレクチン群で、両プローブ間に顕著な相違が見られた。脳では糖タンパク質に対するシアル酸の付加が少ないことが知られており、この事実は実験結果の傾向と一致した。このようにクルード系を用いた実験では、試料全体の糖鎖付加情報を迅速・簡便に得ることができるため、血液成分や臓器等の、個体間や病態間での糖鎖構造の差を一斉比較解析する目的での使用に適している。
【0119】
次に先の実験で用いた脳由来糖タンパク質プローブに対し、競合阻害糖としてラクトースを10mM加え、糖鎖プロファイルを観察した(図14C)。その結果、競合阻害により蛍光シグナルのパターンに変化が観察され、主にラクトサミン認識レクチンのシグナルの減弱が観察された。この実験で示したように、よりクルードな試料に対するレクチンアレイによる糖鎖プロファイリングにおいても、様々な阻害糖の添加実験を併用し、獲得される情報を絞り込んだ上で比較解析することで、より簡便かつ迅速に個体間の比較解析を行うことができる。
【産業上の利用可能性】
【0120】
本発明により、レクチン−糖鎖間の相互作用を溶液中で平衡状態のまま観察する方法が初めて実用化された。この技術は従来のレクチンブロットのような相互作用の有無(0か1か)ではなく、中間部分の情報、すなわち結合の強度情報を輝度値から得ることができる(例えば0から6段階)。このことはn種のレクチン−糖鎖間の相互作用において、従来は2n通りであった情報量が6n通りへと格段に増したことを意味する。この技術は今後さらに高密度化、高精度化していくことにより糖鎖構造解析、及び関連するその他の糖鎖工学諸分野の発展に大きな貢献をもたらすことが期待される。さらには、様々な用途に向けレクチン−糖鎖間の相互作用解析用のアレイを作製することで、血液・体液・組織抽出物等の原液または希釈溶液からの診断・判定への利用や、糖タンパク質製品の品質管理等への応用が期待される。
【特許請求の範囲】
【請求項1】
以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)糖鎖に相互作用を示すタンパク質が固定化された基板に蛍光標識した被検糖鎖または被検複合糖質を接触させる工程
(b)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
【請求項2】
糖鎖に相互作用を示すタンパク質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に糖鎖に相互作用を示すタンパク質が固定化された基板である、請求項1に記載の方法。
【請求項3】
エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、請求項2に記載の方法。
【請求項4】
以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)被検複合糖質が固定化された基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程
(b)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
【請求項5】
被検複合糖質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に被検複合糖質が固定化された基板である、請求項4に記載の方法。
【請求項6】
エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、請求項5に記載の方法。
【請求項7】
以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板に被検複合糖質を接触させる工程
(b)工程(a)で得られた基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程
(c)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
【請求項8】
複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板である、請求項7に記載の方法。
【請求項9】
エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、請求項8に記載の方法。
【請求項10】
複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、請求項7〜9のいずれかに記載の方法。
【請求項11】
糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、請求項1〜10のいずれかに記載の方法。
【請求項12】
励起光がエバネッセント波である、請求項1〜11のいずれかに記載の方法。
【請求項13】
複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、請求項1〜12のいずれかに記載の方法。
【請求項14】
エポキシ基を活性基として有する化合物がコートされた基板であって、糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板。
【請求項15】
エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、請求項14に記載の基板。
【請求項16】
複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、請求項14または15に記載の基板。
【請求項17】
糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、請求項14または15に記載の基板。
【請求項18】
複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、請求項14〜17のいずれかに記載の基板。
【請求項19】
以下の工程を含む、基板の製造方法。
(a)基板にエポキシ基を活性基として有する化合物をコートする工程
(b)工程(a)で得られた基板に糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質を固定化する工程
【請求項20】
複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、請求項19に記載の方法。
【請求項21】
糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、請求項19に記載の方法。
【請求項22】
複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、請求項19〜21のいずれかに記載の方法。
【請求項1】
以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)糖鎖に相互作用を示すタンパク質が固定化された基板に蛍光標識した被検糖鎖または被検複合糖質を接触させる工程
(b)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
【請求項2】
糖鎖に相互作用を示すタンパク質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に糖鎖に相互作用を示すタンパク質が固定化された基板である、請求項1に記載の方法。
【請求項3】
エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、請求項2に記載の方法。
【請求項4】
以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)被検複合糖質が固定化された基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程
(b)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
【請求項5】
被検複合糖質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に被検複合糖質が固定化された基板である、請求項4に記載の方法。
【請求項6】
エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、請求項5に記載の方法。
【請求項7】
以下の工程を含む、糖鎖に相互作用を示すタンパク質と糖鎖との相互作用を分析する方法。
(a)複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板に被検複合糖質を接触させる工程
(b)工程(a)で得られた基板に蛍光標識した糖鎖に相互作用を示すタンパク質を接触させる工程
(c)基板を洗浄せずに、励起光を作用させて、励起される蛍光の強度を測定する工程
【請求項8】
複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板が、エポキシ基を活性基として有する化合物がコートされた基板に複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板である、請求項7に記載の方法。
【請求項9】
エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、請求項8に記載の方法。
【請求項10】
複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、請求項7〜9のいずれかに記載の方法。
【請求項11】
糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、請求項1〜10のいずれかに記載の方法。
【請求項12】
励起光がエバネッセント波である、請求項1〜11のいずれかに記載の方法。
【請求項13】
複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、請求項1〜12のいずれかに記載の方法。
【請求項14】
エポキシ基を活性基として有する化合物がコートされた基板であって、糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が固定化された基板。
【請求項15】
エポキシ基を活性基として有する化合物が、3−グリシドキシプロピルトリメトキシシラン(GTMS)である、請求項14に記載の基板。
【請求項16】
複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、請求項14または15に記載の基板。
【請求項17】
糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、請求項14または15に記載の基板。
【請求項18】
複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、請求項14〜17のいずれかに記載の基板。
【請求項19】
以下の工程を含む、基板の製造方法。
(a)基板にエポキシ基を活性基として有する化合物をコートする工程
(b)工程(a)で得られた基板に糖鎖に相互作用を示すタンパク質、または複合糖質の糖鎖以外の領域に相互作用を示すタンパク質を固定化する工程
【請求項20】
複合糖質の糖鎖以外の領域に相互作用を示すタンパク質が、複合糖質の糖鎖以外の領域に相互作用を示す抗体である、請求項19に記載の方法。
【請求項21】
糖鎖に相互作用を示すタンパク質が、レクチン、糖結合ドメインを有する酵素タンパク質、糖鎖に親和性を有するサイトカイン、または糖鎖に相互作用を示す抗体である、請求項19に記載の方法。
【請求項22】
複合糖質が、糖タンパク質、プロテオグリカン、または糖脂質である、請求項19〜21のいずれかに記載の方法。
【図1】

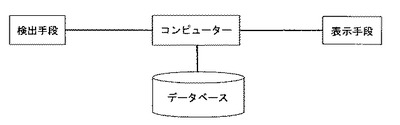
【図2】
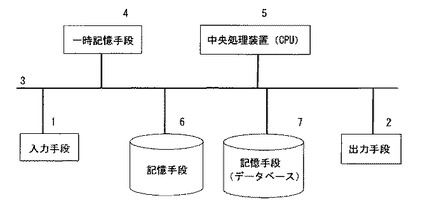
【図3】
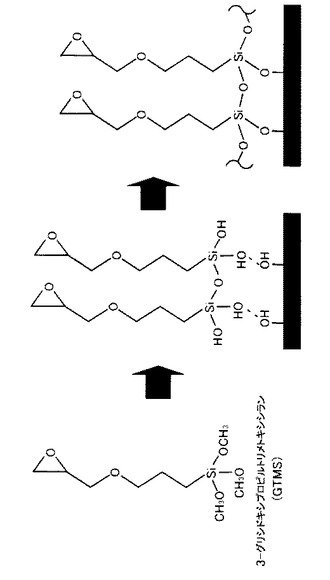
【図4】
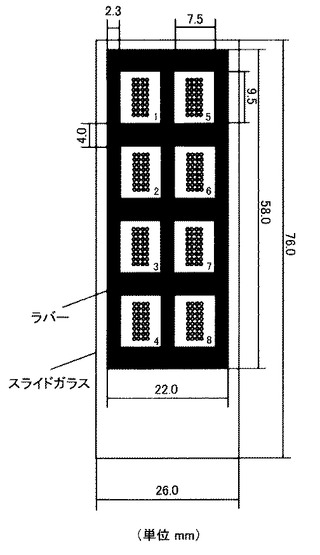
【図5】
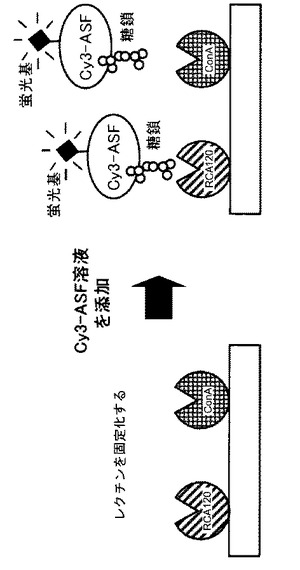
【図6】
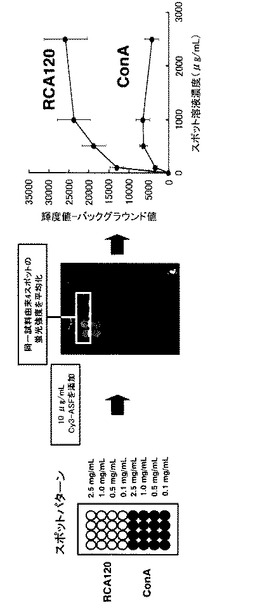
【図7】
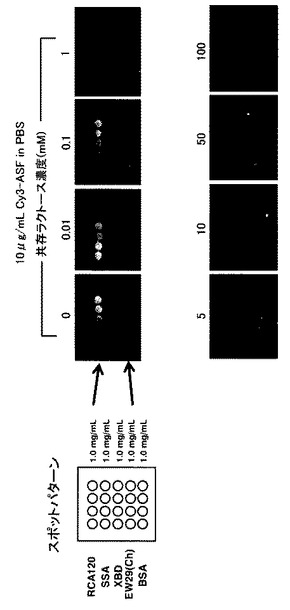
【図8】
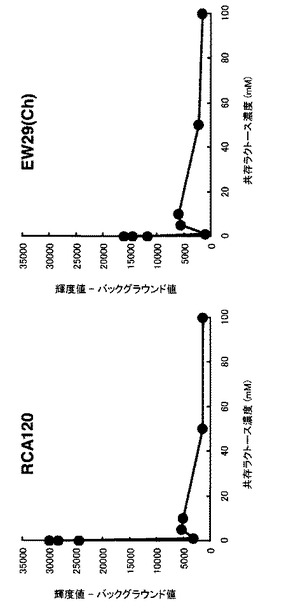
【図9】
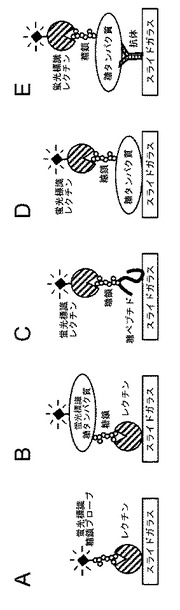
【図10】
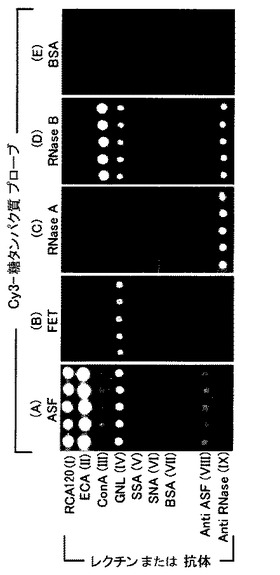
【図11】
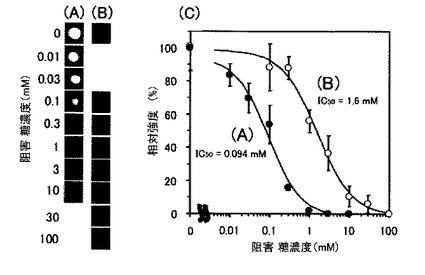
【図12】
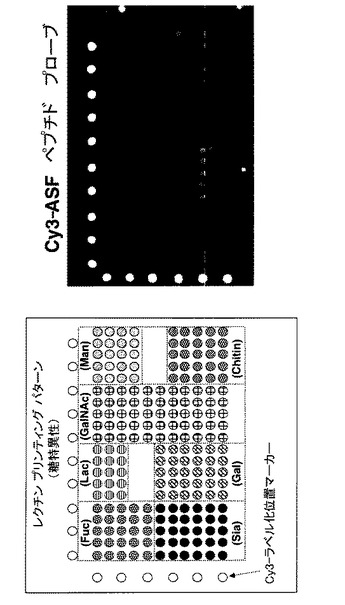
【図13】
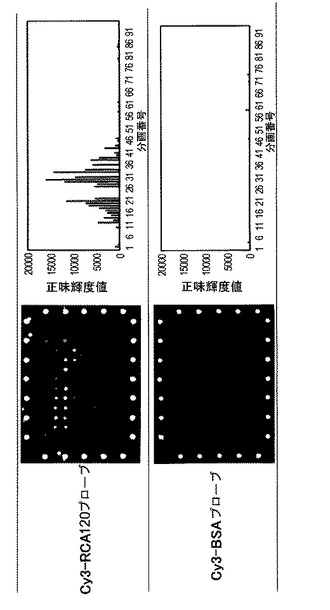
【図14】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【国際公開番号】WO2005/064333
【国際公開日】平成17年7月14日(2005.7.14)
【発行日】平成19年12月20日(2007.12.20)
【国際特許分類】
【出願番号】特願2005−516636(P2005−516636)
【国際出願番号】PCT/JP2004/019333
【国際出願日】平成16年12月24日(2004.12.24)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
【国際公開日】平成17年7月14日(2005.7.14)
【発行日】平成19年12月20日(2007.12.20)
【国際特許分類】
【国際出願番号】PCT/JP2004/019333
【国際出願日】平成16年12月24日(2004.12.24)
【出願人】(301021533)独立行政法人産業技術総合研究所 (6,529)
【Fターム(参考)】
[ Back to top ]