
タンパク質における繰り返しモチーフの識別方法
【課題】本発明の目的は、明確にかつ再現性よくタンパク質における繰り返しモチーフを識別することができる方法及びそのための試薬を提供することである。
【解決手段】特定配列(D)内に3’末端を有するプライマー(D1)と、 プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)と、プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C1)を用いて核酸増幅反応を行いタンパク質における繰り返しモチーフを識別する方法を提供する。
【解決手段】特定配列(D)内に3’末端を有するプライマー(D1)と、 プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)と、プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C1)を用いて核酸増幅反応を行いタンパク質における繰り返しモチーフを識別する方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、繰り返しモチーフを持つタンパク質の識別方法に関するものである。本発明は、細菌の産生する毒素タンパク質の分類、それにもとづいた該細菌の分類または病原性の推測などに際して特に有用である。
【背景技術】
【0002】
タンパク質の中には、ある一定の機能を持つと期待される特徴的なアミノ酸配列を含むモチーフ(motif)と呼ばれる領域があり、その有無によってタンパク質の機能推定を行うことができる。モチーフとしては、ロイシン−不特定6アミノ酸の繰り返しからなるDNA結合(ロイシンジッパー)モチーフ、セリン−グリシン−不特定1アミノ酸−グリシンからなるグリコサミノグリカン結合モチーフ、アルギニン−グリシン−アスパラギン酸からなる細胞付着(RGD)モチーフなど数個のアミノ酸で構成される小さなものから、遺伝子調節タンパク質に多く見られるホメオドメイン、ジンクフィンガー、ヘリックス−ループ−ヘリックス、ヘアピンβシートなど数十個のアミノ酸で構成される大きなものまで多数が知られている。
【0003】
モチーフの繰り返しにより、その機能が増強される例も報告されている。例えばヘリコバクター・ピロリ菌の病原因子の一つである細胞空胞化毒素関連タンパク質(CagA)のカルボキシル末端(C末端)領域には、C配列、D配列と名付けられたリン酸化を受けるモチーフが存在する。C配列は主に欧米人に多く感染するヘリコバクター・ピロリ菌に存在し、D配列は主に東アジア人に多く感染するヘリコバクター・ピロリ菌に存在する、それぞれ類似した配列(バリアント)である。CagAタンパク質は、胃の上皮細胞に感染したヘリコバクター・ピロリ菌によって細胞内に送り込まれるとSrcによってリン酸化を受け、Src homology phosphatase-2(SHP-2)と結合することで、Ras、ERKを介した異常な細胞増殖と細胞接着に関連するfocal adhesion kinase(FAK)を脱リン酸化することで細胞の運動能向上を引き起こす。SHP-2はCagAタンパク質のC配列またはD配列に結合することが分かっており、また、CagAタンパク質がC配列またはD配列を複数持つ場合にSHP-2との結合力が強くなり、毒性が強くなり発ガンしやすくなることが知られている(例えば、非特許文献1〜4参照)。
【0004】
このように、ある特定のモチーフを解析することにより、タンパク質の機能を推定することができる。上記の一例では、ヘリコバクター・ピロリ菌の産生するCagAタンパク質のC配列またはD配列の種類および/または繰り返し数を解析することにより、該菌の分類または病原性の推測が可能となる。
【0005】
タンパク質のモチーフを解析するための従来技術としては、例えば該タンパク質をコードする核酸の配列を決定するシークエンシング法がある。シークエンシング法では、核酸配列を解析することによりアミノ酸配列の解析に必要な情報を得ることができるが、判定可能な核酸配列の長さが実質的に500〜700塩基程度であるため、おおよそ200アミノ酸より長いモチーフ、特に繰り返しモチーフの解析は困難である。また、鋳型核酸の調製、DNAポリメラーゼ反応、ポリアクリルアミドゲル電気泳動、核酸配列の解析などを行うため多大な労力と時間が必要であり、近年の自動シークエンサーを用いることで省力化を行うことはできるが、高価な装置が必要であるという問題がある。その他の従来技術として、該タンパク質を構成するアミノ酸の配列を直接解析することも可能であるが、判定可能な長さが10アミノ酸程度と短いため、極めて小さなモチーフしか解析できない欠点を有する。
【非特許文献1】Asahiら、J. Exp. Med. 第191巻、第593〜602頁、(2000年)
【非特許文献2】Higashiら、Proc. Natl. Acad. Sci. USA 第99巻、第14428〜14433頁、(2002年)
【非特許文献3】Azumaら、J. Infect. Dis. 第189巻、第820〜827頁、(2004年)
【非特許文献4】Azumaら、J. Clin. Microbiol. 第42巻、第2508〜2517頁、(2004年)
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、上記のような問題点を解決して、明確にかつ再現性よくタンパク質のモチーフを識別することができる方法を提供することである。
【課題を解決するための手段】
【0007】
本発明者らは、上記事情に鑑み、鋭意研究の結果、本発明を完成させるに至った。
【0008】
すなわち、本発明は以下のような構成からなる。
1.タンパク質における繰り返しモチーフを識別する方法であって、下記の(a)〜(b)の工程を含み、該タンパク質をコードする核酸配列中の特定配列の繰り返し数を判定することを特徴とする方法。
(a)次の(i)、(ii)及び(iii)のプライマーを用いて、核酸増幅を行う工程:
(i)特定配列(D)内に3’末端を有するプライマー(D1)
(ii) プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)
(iii) プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C1)
(b)工程(a)で得られた1つまたはそれ以上のプライマー(D1)から得られる増幅核酸断片の種類と、プライマー(C1)から得られる増幅核酸断片の種類を解析する工程
2.タンパク質における繰り返しモチーフを識別する方法であって、下記の(a)〜(b)の工程を含み、該タンパク質をコードする核酸配列中の特定配列の繰り返し数を判定することを特徴とする方法。
(a)次の(i)、(ii)及び(iii)のプライマーを用いて、核酸増幅を行う工程:
(i)特定配列(D)内に3’末端を有するプライマー(D1)
(ii) プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)
(iii)プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)
(iv) プライマー(P2)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C2)
(b)工程(a)で得られた1つまたはそれ以上のプライマー(D1)から得られる増幅核酸断片の種類と、プライマー(C2)から得られる増幅核酸断片の種類と、プライマー(P1)および(P2)から得られる増幅核酸断片の有無を解析する工程
3.プライマー(D1)の3’末端から2番目以降の少なくとも一つのヌクレオチドがリガンドにより標識されていることを特徴とする1または2の識別方法。
4.プライマー(D1)及びプライマー(C1またはC2)の3’末端から2番目以降の少なくとも一つのヌクレオチドが、それぞれ異なるリガンドにより標識されていることを特徴とする1〜3のいずれかの識別方法。
5.リガンドが抗原、抗体、蛍光物質、発光団および酵素からなる群より選ばれた少なくとも1種以上であることを特徴とする3または4の識別方法。
6.特定配列内に3’末端を有するプライマーの3’末端から2番目以降の少なくとも一つのヌクレオチドに結合したリガンドを検出することにより、該特定配列の繰り返し数を判定することを特徴とする3〜5のいずれかの識別方法。
7.繰り返しモチーフが、ヘリコバクター・ピロリ菌のCagAタンパク質のC末端領域に存在するC配列またはD配列であることを特徴とする1〜6のいずれかの識別方法。
【発明の効果】
【0009】
本発明により、タンパク質が保有するモチーフを明確にまた簡便に識別できる方法が提供される。また本発明の方法によれば、タンパク質が繰り返しモチーフを持つ場合に、その繰り返し数も判定することができ、より詳細にモチーフやタンパク質を識別することが可能である。
【発明を実施するための最良の形態】
【0010】
以下、本発明を詳細に説明する。
本発明において、プライマーとは、識別の対象となるタンパク質をコードする核酸または該核酸を含む染色体、あるいはその断片に相補的な配列を有するオリゴヌクレオチドであって、核酸増幅のために一般的に利用される特性を有していれば良く、必要に応じて修飾されていても良い。プライマー鎖長は、9〜35塩基であればよく、好ましくは、11〜30塩基である。これらのプライマーを用いた増幅方法は特に限定はされない。既知の増幅方法を用いればよい。既知の増幅方法としては、例えば、PCR、NASBA、LCR、SDA、RCA、TMA、LAMP、ICANおよびUCAN法などがある。
【0011】
本発明において核酸の増幅とは、試料中の核酸から増幅したい核酸(標的核酸)をその固有の配列の相補性を利用して増幅することを指す。本発明におけるタンパク質の繰り返しモチーフの識別方法に適用可能な核酸増幅の方法としては、基本的には、従来の方法を用いて行うことができ、通常、一本鎖に変性させた、識別の対象となるタンパク質をコードする核酸または該核酸を含む染色体、あるいはその断片に、4種類のデオキシヌクレオシド三リン酸(dNTP)およびDNAポリメラーゼおよび2種類以上のオリゴヌクレオチドすなわちプライマーを作用させることで、標的核酸を鋳型として用いたプライマー間の配列が増幅される。また、標的核酸が検出するのに十分な量が含まれていない場合、あらかじめ前記識別の対象となるタンパク質をコードする核酸または該核酸を含む染色体、あるいはその断片を以下に示す増幅反応によって、増幅しておくことも可能である。
【0012】
核酸増幅方法としては、PCR(特公平4−67960号公報、特公平4−67957号公報)、NASBA(Nucleic acid sequence-basedamplification method;Nature 第350巻、第91頁(1991年))、LCR(国際公開89/12696号公報、特開平2−2934号公報)、SDA(Strand Displacement Amplification:Nucleic Acids Res. 第20巻、第1691頁(1992年))、RCA(国際公開90/1069号公報)、TMA(Transcription mediated amplification method;J. Clin. Microbiol. 第31巻、第3270頁(1993年))、LAMP(loop-mediated isothermal amplification method:J. Clin Microbiol. 第42巻:第1,956頁(2004年))、ICAN(isothermal and chimeric primer-initiated amplification of nucleic acids:Kekkaku. 第78巻、第533頁(2003年))、UCAN(日本癌学会(H13.9.26〜H13.9.28、横浜にて開催)または日本鑑識科学技術学会(H13.11.8〜H13.11.9、東京にて開催)参照)などが挙げられる。
【0013】
なかでもPCR法は、試料核酸、4種類のデオキシヌクレオシド三リン酸、一対のプライマー及び耐熱性DNAポリメラーゼの存在下で、変性、アニーリング、伸長の3工程からなるサイクルを繰り返すことにより、上記一対のプライマーで挟まれる試料核酸の領域を指数関数的に増幅させる方法である。すなわち、変性工程で試料の核酸を変性し、続くアニーリング工程において各プライマーと、それぞれに相補的な一本鎖試料核酸上の領域とをハイブリダイズさせ、続く伸長工程で、各プライマーを起点としてDNAポリメラーゼの働きにより鋳型となる各一本鎖試料核酸に相補的なDNA鎖を伸長させ、二本鎖DNAとする。この1サイクルにより、1本の二本鎖DNAが2本の二本鎖DNAに増幅される。従って、このサイクルをn回繰り返せば、理論上、上記一対のプライマーで挟まれた試料DNAの領域は2のn乗倍に増幅される。増幅されたDNA領域は大量に存在するので、電気泳動等の方法により容易に検出できる。よって、遺伝子増幅法を用いれば、従来では検出不可能であった、極めて微量(1分子でも可)の試料核酸をも検出することが可能であり、最近非常に広く用いられている技術である。
【0014】
増幅に使用する核酸を含有する試料は、例えば、バクテリア、動物または植物組織、個体細胞由来の溶解物などのあらゆる材料から調製することができる。該試料の調製法は特に限定されないが、例えば、患者の血液、組織から、既知の方法により調製してもよい。代表的なものとして、フェノール/クロロホルム抽出法(Biochimica et Biophysica acta 第72巻、第619〜629頁、1963年)、アルカリSDS法(Nucleic Acids Res. 第7巻、第1513〜1523頁、1979年)などの液相で行う方法がある。また、核酸の単離に核酸結合用担体を用いる系としては、ガラス粒子とヨウ化ナトリウム溶液を使用する方法(Proc. Natl. Acad. Sci. USA 第76−2巻、第615〜619頁、1979年)、ハイドロキシアパタイトを用いる方法(特開昭63−263093号公報)等がある。その他の方法としてはシリカ粒子とカオトロピックイオンを用いた方法(J.Clin.Microbiol.第28−3巻、第495〜503頁、1990年、特開平2−289596号公報)が挙げられる。また、核酸は試料中に溶解させてもよいし、固相に固定させてもよい。
【0015】
本発明において、特定配列(D)とは、タンパク質が保有するモチーフまたはモチーフに含まれる繰り返し単位をコードする核酸配列または該配列に相補的な核酸配列である。また、特定配列(C)とは、特定配列(D)に類似した核酸配列(バリアント)であり、タンパク質が保有するモチーフまたはモチーフに含まれる繰り返し単位をコードする核酸配列または該配列に相補的な核酸配列である。
【0016】
本発明のタンパク質における繰り返しモチーフの識別方法に適用可能な増幅核酸断片の解析法としては従来公知の核酸解析法を使用することができ、特に限定されるものではないが、簡便性の観点からアガロースゲル電気泳動が好ましい。アガロースゲル電気泳動を使用する場合は、一例として、特定配列(D)内に3’末端を有するプライマー(D1)と該プライマー(D1)とともに核酸増幅反応における一対のプライマーとなり得る核酸配列を含有するプライマーにより生成される増幅核酸断片と、特定配列(C)内に3’末端を有するプライマー(C1またはC2)と該プライマー(C1またはC2)とともに核酸増幅反応における一対のプライマーとなり得る核酸配列を含有するプライマーにより生成される増幅核酸断片の鎖長が異なるように各プライマーを設計する。これらのプライマーを混合して核酸を増幅することにより、特定配列(D)および/または(C)に対応するモチーフの有無および繰り返し数について、増幅された核酸断片のパターンから迅速に判定できる。また、該プライマー(D1)および該プライマー(C1またはC2)をそれぞれ異なるリガンドで標識しておけば、該プライマー(D1)により生成される増幅核酸断片と該プライマー(C1またはC2)により生成される増幅核酸断片の鎖長が同じであっても、アガロースゲル電気泳動の後で各リガンドを検出することにより増幅核酸断片が特定配列(D)または(C)のどちらに由来するかを判定することができる。
【0017】
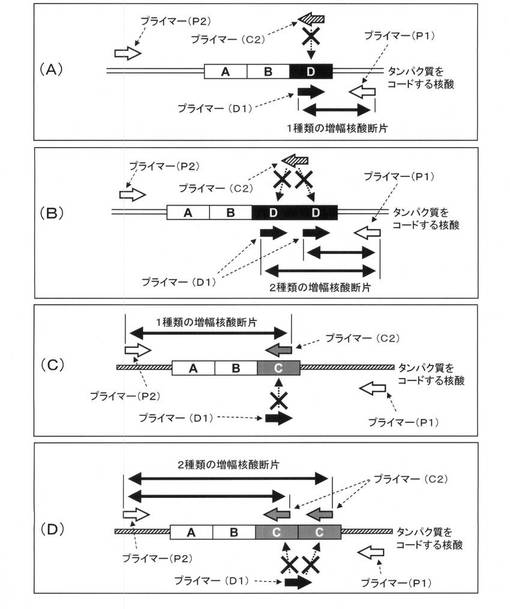
本発明の重要な開示の一つは、特定配列(D)内に3’末端を有するプライマー(D1)と、特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)と、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C2)と、特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)とを、一例として図1に示すように設定し、核酸増幅反応を行い、対象とするタンパク質において特定配列(D)および/または(C)に対応するモチーフの有無および繰り返し数について明確に判定できるような顕著な効果を見出したことにある。
【0018】
例えば、タンパク質をコードする核酸に特定配列(D)が1個存在する場合は、図1(A)に示すように、特定配列(D)内に3’末端を有するプライマー(D1)と特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)により1種類の増幅核酸断片が生成する。また、タンパク質をコードする核酸に特定配列(D)が2個存在する場合は、図1(B)に示すように、特定配列(D)内に3’末端を有するプライマー(D1)と特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)により鎖長の異なる2種類の増幅核酸断片が生成する。さらに、タンパク質をコードする核酸に特定配列(D)のバリアントである特定配列(C)が1個存在する場合は、図1(C)に示すように、特定配列(C)内に3’末端を有するプライマー(C2)と特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)により1種類の増幅核酸断片が生成する。また、タンパク質をコードする核酸に特定配列(C)が2個存在する場合は、図1(D)に示すように、特定配列(C)内に3’末端を有するプライマー(C2)と特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)により鎖長の異なる2種類の増幅核酸断片が生成する。
【0019】
本発明に用いるプライマー(D1,C1,C2,P1,P2)は、核酸増幅のために一般的に利用される特性を有していれば良く、必要に応じて修飾されていても良い。リガンドは核酸の増幅反応を阻害しない位置に結合させることが好ましい。該リガンドを結合させる位置としては、該プライマー(D1,C1,C2,P1,P2)の3’末端から2番目以降の少なくとも一つのヌクレオチドにリガンドを結合させることが好ましく、該プライマー((D1,C1,C2,P1,P2)の5’末端にリガンドを結合させることがより好ましい。
【0020】
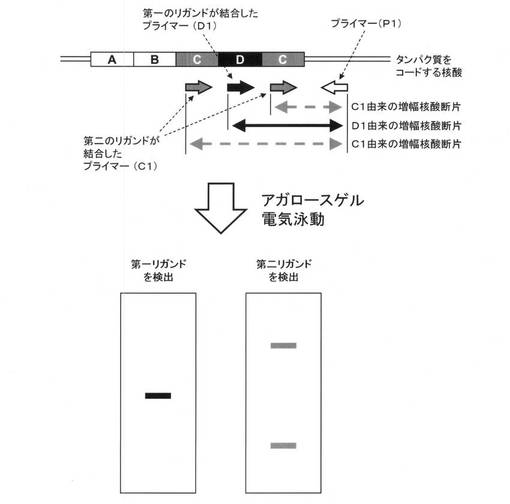
本発明の別の重要な開示の一つは、第一のリガンドが結合しており特定配列(D)内に3’末端を有するプライマー(D1)と、第二のリガンドが結合しており特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C1)と、特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)とを、一例として図2に示すように設定し、核酸増幅反応を行い、第一のリガンドおよび第二のリガンドをそれぞれ検出することにより、対象とするタンパク質において特定配列(D)および/または(C)に対応するモチーフの有無および繰り返し数について明確に判定できるような顕著な効果を見出したことにある。
【0021】
例えば、タンパク質をコードする核酸に、図2に示すように特定配列(D)および該特定配列(D)のバリアントである特定配列(C)が並んでいる場合は、図2に示すように、第一のリガンドが結合しており特定配列(D)内に3’末端を有するプライマー(D1)と特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)により1種類の増幅核酸断片が生成し、第二のリガンドが結合しており特定配列(C)内に3’末端を有するプライマー(C1)と特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)により2種類の増幅核酸断片が生成する。アガロースゲル電気泳動を行い、各リガンドを検出することにより、増幅核酸断片が特定配列(D)または(C)のどちらに由来するかを判定することができる。
【0022】
リガンドは核酸の増幅を妨げるものでないのであれば、特に限定されるものではないが、好適には抗体、オリゴヌクレオチド、レセプター、核酸特異的結合物質、アビジン、ビオチン、ジゴケシゲニン、蛍光化学物質、発光団、酵素、蛍光蛋白質、発光蛋白質、磁性体、導電性物質よりなる群より選ぶことができる。好ましくはアビジン、ビオチン、ジゴケシゲニン、蛍光化学物質、蛍光蛋白質、発光団、酵素、導電性物質がよく、さらに好ましくはアビジン、ビオチン、ジゴケシゲニン、蛍光化学物質がよく、特に好ましくは蛍光化学物質がよい。
【0023】
リガンドの検出は既知の方法であれば特に限定されるものではないが、リガンドが蛍光化学物質または蛍光蛋白質の場合、特定波長の光を照射し蛍光化学物質または蛍光蛋白質を励起させ、基底状態に変換される際に生じる特定波長の蛍光量を測定することが可能である。これらに用いられる蛍光化学物質としては、FITC、FAM、TAMRA、TexasRed、VIC、Cy3、Cy5、HEX等が挙げられ、蛍光蛋白質としては、GFP、YFP、RFP等を挙げることができる。標識が酵素である場合、酵素の基質を添加することによって生成される反応生成物を検出することによって測定が可能である。これらに用いられる酵素と基質の組み合わせとしては、アルカリフォスファターゼとパラニトロフェニルリン酸、CDP−star、AMPPD、DDAOphospate、BCIP−NBT等の組み合わせ、パーオキシダーゼとTMB、Lumi−Light(ロッシュ・ダイアグノスティックス)、SAT−1(同仁化学)等の組み合わせ、ジアホラーゼとNTB等の組み合わせ、各種オキシダーゼと基質、各種デヒドロゲナーゼと基質の組み合わせ等、反応性生物が検出されるものであれば、これらに限定されることはなく用いることが可能である。標識が磁性体の場合、磁気を検出することによって測定が可能である。標識が導電性物質、電流値を検出することによって測定が可能である。
【0024】
本発明のタンパク質における繰り返しモチーフの識別方法は、例えばヘリコバクター・ピロリ菌による発ガンの危険性を予測するために用いられることができる。上記ガンとしては、胃ガンなどを挙げることができる。上述したように、本発明の識別方法を用いれば、対象とするタンパク質の繰り返しモチーフにおいて特定配列の有無および繰り返し数について明確に判定できることを利用して、CagAタンパク質のC末端領域の多型を検出することができる。更に詳細には、本発明の識別方法を用いれば、発ガンに関係が深いC配列および/またはD配列の有無を検出することができるとともに、C配列および/またはD配列の数をも検出することができる。そして、C配列および/またはD配列の有無によって、発ガンの可能性の有無を予測することができるとともに、上記C配列および/またはD配列の数によって、発ガンの可能性の大小(C配列および/またはD配列が多いほど、発ガンの危険度が大きい)を予測することができる。一例として、欧米人に多く感染するヘリコバクター・ピロリ菌について、発ガンの可能性の有無をC配列の有無によって予測することができるとともに、発ガンの可能性の大小(C配列が多いほど、発ガンの危険度が大きい)を上記C配列の数によって予測することができる。また、東アジア人に多く感染するヘリコバクター・ピロリ菌について、発ガンの可能性の有無をD配列の有無によって予測することができるとともに、発ガンの可能性の大小(D配列が多いほど、発ガンの危険度が大きい)を上記D配列の数によって予測することができる。
【実施例】
【0025】
以下、実施例に基づき本発明をより具体的に説明する。もっとも、本発明は下記実施例に限定されるものではない。
【0026】
実施例1
ヘリコバクター・ピロリ菌のCagAタンパク質における繰り返しモチーフの識別
(1)オリゴヌクレオチドの合成
パーキンエルマー社製DNAシンセサイザー392型を用いて、ホスホアミダイト法にて、配列番号1〜3に示される塩基配列を有するオリゴヌクレオチド(以下、オリゴ1〜3と示す)を合成した。合成はマニュアルに従い、各種オリゴヌクレオチドの脱保護はアンモニア水で55℃、一夜実施した。オリゴヌクレオチドの精製はパーキンエルマー社OPCカラムにて実施した。もしくはDNA合成受託会社((株)日本バイオサービス、シグマアルドリッチジャパン(株)、オペロンバイオテクノロジー(株)等)に依頼した。
オリゴ1がセンス鎖であり、CagAタンパク質のD配列をコードする核酸配列すなわち特定配列(D)内に3’末端を有するプライマー(D1)であり、アンチセンス鎖のオリゴ3と組み合わせて増幅反応のオリゴヌクレオチドとして使用される。オリゴ2はCagAタンパク質のC配列をコードする核酸配列すなわち特定配列(C)内に3’末端を有するプライマー(C1)であり、アンチセンス鎖のオリゴ3と組み合わせて増幅反応のオリゴヌクレオチドとして使用される。オリゴ3はCagAタンパク質のファミリーをコードする核酸配列に共通のプライマーすなわちプライマー(D1)または(C1)とともに核酸増幅反応における一対のプライマーとなり得る核酸配列を含有するプライマー(P1)である。オリゴ1は5’末端をFITCにより標識され、オリゴ2は5’末端をTAMRAにより標識されている。
【0027】
(2)PCR法によるヘリコバクター・ピロリ菌cagA遺伝子の解析
表1におけるサンプル番号1〜6のヘリコバクター・ピロリ菌からそれぞれフェノール・クロロホルム法により抽出したDNA溶液をサンプルとして使用して、下記試薬を添加して、下記条件によりPCR法によるヘリコバクター・ピロリ菌のcagA遺伝子を解析した。
【0028】
【表1】

【0029】
試薬
以下の試薬を含む25μl溶液を調製した。
KOD DNAポリメラーゼ反応液
オリゴ1(5’末端をFITCにより標識) 10pmol、
オリゴ2(5’末端をTAMRAにより標識) 10pmol、
オリゴ3 10pmol、
×10緩衝液 2.5μl、
2mM dNTP 2.5μl、
25mM MgCl2 1μl、
KOD DNAポリメラーゼ 0.5U、
抽出DNA溶液 100ng
【0030】
増幅条件
94℃・5分
94℃・15秒、
60℃・30秒、
68℃・30秒(35サイクル)
25℃・15分。
【0031】
(3)アガロースゲル電気泳動を用いた検出
増幅反応液5μlを3%アガロースゲルにて電気泳動し、蛍光撮影用LED光源装置ビジレイズAE−6935(ATTO(株)製)にて検出した。オリゴ1の標識であるFITCをブルー光源下、オリゴ2の標識であるTAMRAグリーン(ロング)光源下で観察し、増幅バンドの蛍光を目視によって確認した。表2に、試料No.1〜6で示すサンプルについて、明らかに確認できた増幅バンドの数を示す。
【0032】
【表2】
【0033】
(4)結果
表2(および表1)のサンプル番号1〜6より明らかなように、本発明のタンパク質における繰り返しモチーフの識別方法によれば、ヘリコバクター・ピロリ菌のCagAタンパク質における繰り返しモチーフの種類と繰り返し数を判定することが可能である。これらの結果は、核酸配列を直接解析することにより判別に必要な情報を得ることができるが多大な労力と時間が必要であるなどの欠点を有するシークエンシング法を用いた該6株の識別結果(表1)と一致する。
【0034】
すなわち、本発明のタンパク質における繰り返しモチーフの識別方法では従来の方法と同様の識別能力を実現しながら、解析に必要な作業工程数を大幅に短縮し、容易にかつ迅速にタンパク質におけるモチーフの有無および該モチーフの繰り返し数を明確に判定することが可能である。
【産業上の利用可能性】
【0035】
本発明により、タンパク質におけるモチーフの有無および該モチーフの繰り返し数を明確に判定することが可能となり、これまでの方法のように煩雑な操作を必要とせず、迅速で容易に再現性の良い結果が得られることからも、産業界に大きく寄与することが期待される。
【図面の簡単な説明】
【0036】
【図1】各プライマーの設定方法を示す一例
【図2】各プライマーの設定方法を示す一例と、第一のリガンドおよび第二のリガンドをによる検出例
【技術分野】
【0001】
本発明は、繰り返しモチーフを持つタンパク質の識別方法に関するものである。本発明は、細菌の産生する毒素タンパク質の分類、それにもとづいた該細菌の分類または病原性の推測などに際して特に有用である。
【背景技術】
【0002】
タンパク質の中には、ある一定の機能を持つと期待される特徴的なアミノ酸配列を含むモチーフ(motif)と呼ばれる領域があり、その有無によってタンパク質の機能推定を行うことができる。モチーフとしては、ロイシン−不特定6アミノ酸の繰り返しからなるDNA結合(ロイシンジッパー)モチーフ、セリン−グリシン−不特定1アミノ酸−グリシンからなるグリコサミノグリカン結合モチーフ、アルギニン−グリシン−アスパラギン酸からなる細胞付着(RGD)モチーフなど数個のアミノ酸で構成される小さなものから、遺伝子調節タンパク質に多く見られるホメオドメイン、ジンクフィンガー、ヘリックス−ループ−ヘリックス、ヘアピンβシートなど数十個のアミノ酸で構成される大きなものまで多数が知られている。
【0003】
モチーフの繰り返しにより、その機能が増強される例も報告されている。例えばヘリコバクター・ピロリ菌の病原因子の一つである細胞空胞化毒素関連タンパク質(CagA)のカルボキシル末端(C末端)領域には、C配列、D配列と名付けられたリン酸化を受けるモチーフが存在する。C配列は主に欧米人に多く感染するヘリコバクター・ピロリ菌に存在し、D配列は主に東アジア人に多く感染するヘリコバクター・ピロリ菌に存在する、それぞれ類似した配列(バリアント)である。CagAタンパク質は、胃の上皮細胞に感染したヘリコバクター・ピロリ菌によって細胞内に送り込まれるとSrcによってリン酸化を受け、Src homology phosphatase-2(SHP-2)と結合することで、Ras、ERKを介した異常な細胞増殖と細胞接着に関連するfocal adhesion kinase(FAK)を脱リン酸化することで細胞の運動能向上を引き起こす。SHP-2はCagAタンパク質のC配列またはD配列に結合することが分かっており、また、CagAタンパク質がC配列またはD配列を複数持つ場合にSHP-2との結合力が強くなり、毒性が強くなり発ガンしやすくなることが知られている(例えば、非特許文献1〜4参照)。
【0004】
このように、ある特定のモチーフを解析することにより、タンパク質の機能を推定することができる。上記の一例では、ヘリコバクター・ピロリ菌の産生するCagAタンパク質のC配列またはD配列の種類および/または繰り返し数を解析することにより、該菌の分類または病原性の推測が可能となる。
【0005】
タンパク質のモチーフを解析するための従来技術としては、例えば該タンパク質をコードする核酸の配列を決定するシークエンシング法がある。シークエンシング法では、核酸配列を解析することによりアミノ酸配列の解析に必要な情報を得ることができるが、判定可能な核酸配列の長さが実質的に500〜700塩基程度であるため、おおよそ200アミノ酸より長いモチーフ、特に繰り返しモチーフの解析は困難である。また、鋳型核酸の調製、DNAポリメラーゼ反応、ポリアクリルアミドゲル電気泳動、核酸配列の解析などを行うため多大な労力と時間が必要であり、近年の自動シークエンサーを用いることで省力化を行うことはできるが、高価な装置が必要であるという問題がある。その他の従来技術として、該タンパク質を構成するアミノ酸の配列を直接解析することも可能であるが、判定可能な長さが10アミノ酸程度と短いため、極めて小さなモチーフしか解析できない欠点を有する。
【非特許文献1】Asahiら、J. Exp. Med. 第191巻、第593〜602頁、(2000年)
【非特許文献2】Higashiら、Proc. Natl. Acad. Sci. USA 第99巻、第14428〜14433頁、(2002年)
【非特許文献3】Azumaら、J. Infect. Dis. 第189巻、第820〜827頁、(2004年)
【非特許文献4】Azumaら、J. Clin. Microbiol. 第42巻、第2508〜2517頁、(2004年)
【発明の開示】
【発明が解決しようとする課題】
【0006】
本発明の目的は、上記のような問題点を解決して、明確にかつ再現性よくタンパク質のモチーフを識別することができる方法を提供することである。
【課題を解決するための手段】
【0007】
本発明者らは、上記事情に鑑み、鋭意研究の結果、本発明を完成させるに至った。
【0008】
すなわち、本発明は以下のような構成からなる。
1.タンパク質における繰り返しモチーフを識別する方法であって、下記の(a)〜(b)の工程を含み、該タンパク質をコードする核酸配列中の特定配列の繰り返し数を判定することを特徴とする方法。
(a)次の(i)、(ii)及び(iii)のプライマーを用いて、核酸増幅を行う工程:
(i)特定配列(D)内に3’末端を有するプライマー(D1)
(ii) プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)
(iii) プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C1)
(b)工程(a)で得られた1つまたはそれ以上のプライマー(D1)から得られる増幅核酸断片の種類と、プライマー(C1)から得られる増幅核酸断片の種類を解析する工程
2.タンパク質における繰り返しモチーフを識別する方法であって、下記の(a)〜(b)の工程を含み、該タンパク質をコードする核酸配列中の特定配列の繰り返し数を判定することを特徴とする方法。
(a)次の(i)、(ii)及び(iii)のプライマーを用いて、核酸増幅を行う工程:
(i)特定配列(D)内に3’末端を有するプライマー(D1)
(ii) プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)
(iii)プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)
(iv) プライマー(P2)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C2)
(b)工程(a)で得られた1つまたはそれ以上のプライマー(D1)から得られる増幅核酸断片の種類と、プライマー(C2)から得られる増幅核酸断片の種類と、プライマー(P1)および(P2)から得られる増幅核酸断片の有無を解析する工程
3.プライマー(D1)の3’末端から2番目以降の少なくとも一つのヌクレオチドがリガンドにより標識されていることを特徴とする1または2の識別方法。
4.プライマー(D1)及びプライマー(C1またはC2)の3’末端から2番目以降の少なくとも一つのヌクレオチドが、それぞれ異なるリガンドにより標識されていることを特徴とする1〜3のいずれかの識別方法。
5.リガンドが抗原、抗体、蛍光物質、発光団および酵素からなる群より選ばれた少なくとも1種以上であることを特徴とする3または4の識別方法。
6.特定配列内に3’末端を有するプライマーの3’末端から2番目以降の少なくとも一つのヌクレオチドに結合したリガンドを検出することにより、該特定配列の繰り返し数を判定することを特徴とする3〜5のいずれかの識別方法。
7.繰り返しモチーフが、ヘリコバクター・ピロリ菌のCagAタンパク質のC末端領域に存在するC配列またはD配列であることを特徴とする1〜6のいずれかの識別方法。
【発明の効果】
【0009】
本発明により、タンパク質が保有するモチーフを明確にまた簡便に識別できる方法が提供される。また本発明の方法によれば、タンパク質が繰り返しモチーフを持つ場合に、その繰り返し数も判定することができ、より詳細にモチーフやタンパク質を識別することが可能である。
【発明を実施するための最良の形態】
【0010】
以下、本発明を詳細に説明する。
本発明において、プライマーとは、識別の対象となるタンパク質をコードする核酸または該核酸を含む染色体、あるいはその断片に相補的な配列を有するオリゴヌクレオチドであって、核酸増幅のために一般的に利用される特性を有していれば良く、必要に応じて修飾されていても良い。プライマー鎖長は、9〜35塩基であればよく、好ましくは、11〜30塩基である。これらのプライマーを用いた増幅方法は特に限定はされない。既知の増幅方法を用いればよい。既知の増幅方法としては、例えば、PCR、NASBA、LCR、SDA、RCA、TMA、LAMP、ICANおよびUCAN法などがある。
【0011】
本発明において核酸の増幅とは、試料中の核酸から増幅したい核酸(標的核酸)をその固有の配列の相補性を利用して増幅することを指す。本発明におけるタンパク質の繰り返しモチーフの識別方法に適用可能な核酸増幅の方法としては、基本的には、従来の方法を用いて行うことができ、通常、一本鎖に変性させた、識別の対象となるタンパク質をコードする核酸または該核酸を含む染色体、あるいはその断片に、4種類のデオキシヌクレオシド三リン酸(dNTP)およびDNAポリメラーゼおよび2種類以上のオリゴヌクレオチドすなわちプライマーを作用させることで、標的核酸を鋳型として用いたプライマー間の配列が増幅される。また、標的核酸が検出するのに十分な量が含まれていない場合、あらかじめ前記識別の対象となるタンパク質をコードする核酸または該核酸を含む染色体、あるいはその断片を以下に示す増幅反応によって、増幅しておくことも可能である。
【0012】
核酸増幅方法としては、PCR(特公平4−67960号公報、特公平4−67957号公報)、NASBA(Nucleic acid sequence-basedamplification method;Nature 第350巻、第91頁(1991年))、LCR(国際公開89/12696号公報、特開平2−2934号公報)、SDA(Strand Displacement Amplification:Nucleic Acids Res. 第20巻、第1691頁(1992年))、RCA(国際公開90/1069号公報)、TMA(Transcription mediated amplification method;J. Clin. Microbiol. 第31巻、第3270頁(1993年))、LAMP(loop-mediated isothermal amplification method:J. Clin Microbiol. 第42巻:第1,956頁(2004年))、ICAN(isothermal and chimeric primer-initiated amplification of nucleic acids:Kekkaku. 第78巻、第533頁(2003年))、UCAN(日本癌学会(H13.9.26〜H13.9.28、横浜にて開催)または日本鑑識科学技術学会(H13.11.8〜H13.11.9、東京にて開催)参照)などが挙げられる。
【0013】
なかでもPCR法は、試料核酸、4種類のデオキシヌクレオシド三リン酸、一対のプライマー及び耐熱性DNAポリメラーゼの存在下で、変性、アニーリング、伸長の3工程からなるサイクルを繰り返すことにより、上記一対のプライマーで挟まれる試料核酸の領域を指数関数的に増幅させる方法である。すなわち、変性工程で試料の核酸を変性し、続くアニーリング工程において各プライマーと、それぞれに相補的な一本鎖試料核酸上の領域とをハイブリダイズさせ、続く伸長工程で、各プライマーを起点としてDNAポリメラーゼの働きにより鋳型となる各一本鎖試料核酸に相補的なDNA鎖を伸長させ、二本鎖DNAとする。この1サイクルにより、1本の二本鎖DNAが2本の二本鎖DNAに増幅される。従って、このサイクルをn回繰り返せば、理論上、上記一対のプライマーで挟まれた試料DNAの領域は2のn乗倍に増幅される。増幅されたDNA領域は大量に存在するので、電気泳動等の方法により容易に検出できる。よって、遺伝子増幅法を用いれば、従来では検出不可能であった、極めて微量(1分子でも可)の試料核酸をも検出することが可能であり、最近非常に広く用いられている技術である。
【0014】
増幅に使用する核酸を含有する試料は、例えば、バクテリア、動物または植物組織、個体細胞由来の溶解物などのあらゆる材料から調製することができる。該試料の調製法は特に限定されないが、例えば、患者の血液、組織から、既知の方法により調製してもよい。代表的なものとして、フェノール/クロロホルム抽出法(Biochimica et Biophysica acta 第72巻、第619〜629頁、1963年)、アルカリSDS法(Nucleic Acids Res. 第7巻、第1513〜1523頁、1979年)などの液相で行う方法がある。また、核酸の単離に核酸結合用担体を用いる系としては、ガラス粒子とヨウ化ナトリウム溶液を使用する方法(Proc. Natl. Acad. Sci. USA 第76−2巻、第615〜619頁、1979年)、ハイドロキシアパタイトを用いる方法(特開昭63−263093号公報)等がある。その他の方法としてはシリカ粒子とカオトロピックイオンを用いた方法(J.Clin.Microbiol.第28−3巻、第495〜503頁、1990年、特開平2−289596号公報)が挙げられる。また、核酸は試料中に溶解させてもよいし、固相に固定させてもよい。
【0015】
本発明において、特定配列(D)とは、タンパク質が保有するモチーフまたはモチーフに含まれる繰り返し単位をコードする核酸配列または該配列に相補的な核酸配列である。また、特定配列(C)とは、特定配列(D)に類似した核酸配列(バリアント)であり、タンパク質が保有するモチーフまたはモチーフに含まれる繰り返し単位をコードする核酸配列または該配列に相補的な核酸配列である。
【0016】
本発明のタンパク質における繰り返しモチーフの識別方法に適用可能な増幅核酸断片の解析法としては従来公知の核酸解析法を使用することができ、特に限定されるものではないが、簡便性の観点からアガロースゲル電気泳動が好ましい。アガロースゲル電気泳動を使用する場合は、一例として、特定配列(D)内に3’末端を有するプライマー(D1)と該プライマー(D1)とともに核酸増幅反応における一対のプライマーとなり得る核酸配列を含有するプライマーにより生成される増幅核酸断片と、特定配列(C)内に3’末端を有するプライマー(C1またはC2)と該プライマー(C1またはC2)とともに核酸増幅反応における一対のプライマーとなり得る核酸配列を含有するプライマーにより生成される増幅核酸断片の鎖長が異なるように各プライマーを設計する。これらのプライマーを混合して核酸を増幅することにより、特定配列(D)および/または(C)に対応するモチーフの有無および繰り返し数について、増幅された核酸断片のパターンから迅速に判定できる。また、該プライマー(D1)および該プライマー(C1またはC2)をそれぞれ異なるリガンドで標識しておけば、該プライマー(D1)により生成される増幅核酸断片と該プライマー(C1またはC2)により生成される増幅核酸断片の鎖長が同じであっても、アガロースゲル電気泳動の後で各リガンドを検出することにより増幅核酸断片が特定配列(D)または(C)のどちらに由来するかを判定することができる。
【0017】
本発明の重要な開示の一つは、特定配列(D)内に3’末端を有するプライマー(D1)と、特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)と、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C2)と、特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)とを、一例として図1に示すように設定し、核酸増幅反応を行い、対象とするタンパク質において特定配列(D)および/または(C)に対応するモチーフの有無および繰り返し数について明確に判定できるような顕著な効果を見出したことにある。
【0018】
例えば、タンパク質をコードする核酸に特定配列(D)が1個存在する場合は、図1(A)に示すように、特定配列(D)内に3’末端を有するプライマー(D1)と特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)により1種類の増幅核酸断片が生成する。また、タンパク質をコードする核酸に特定配列(D)が2個存在する場合は、図1(B)に示すように、特定配列(D)内に3’末端を有するプライマー(D1)と特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)により鎖長の異なる2種類の増幅核酸断片が生成する。さらに、タンパク質をコードする核酸に特定配列(D)のバリアントである特定配列(C)が1個存在する場合は、図1(C)に示すように、特定配列(C)内に3’末端を有するプライマー(C2)と特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)により1種類の増幅核酸断片が生成する。また、タンパク質をコードする核酸に特定配列(C)が2個存在する場合は、図1(D)に示すように、特定配列(C)内に3’末端を有するプライマー(C2)と特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)により鎖長の異なる2種類の増幅核酸断片が生成する。
【0019】
本発明に用いるプライマー(D1,C1,C2,P1,P2)は、核酸増幅のために一般的に利用される特性を有していれば良く、必要に応じて修飾されていても良い。リガンドは核酸の増幅反応を阻害しない位置に結合させることが好ましい。該リガンドを結合させる位置としては、該プライマー(D1,C1,C2,P1,P2)の3’末端から2番目以降の少なくとも一つのヌクレオチドにリガンドを結合させることが好ましく、該プライマー((D1,C1,C2,P1,P2)の5’末端にリガンドを結合させることがより好ましい。
【0020】
本発明の別の重要な開示の一つは、第一のリガンドが結合しており特定配列(D)内に3’末端を有するプライマー(D1)と、第二のリガンドが結合しており特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C1)と、特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)とを、一例として図2に示すように設定し、核酸増幅反応を行い、第一のリガンドおよび第二のリガンドをそれぞれ検出することにより、対象とするタンパク質において特定配列(D)および/または(C)に対応するモチーフの有無および繰り返し数について明確に判定できるような顕著な効果を見出したことにある。
【0021】
例えば、タンパク質をコードする核酸に、図2に示すように特定配列(D)および該特定配列(D)のバリアントである特定配列(C)が並んでいる場合は、図2に示すように、第一のリガンドが結合しており特定配列(D)内に3’末端を有するプライマー(D1)と特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)により1種類の増幅核酸断片が生成し、第二のリガンドが結合しており特定配列(C)内に3’末端を有するプライマー(C1)と特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)により2種類の増幅核酸断片が生成する。アガロースゲル電気泳動を行い、各リガンドを検出することにより、増幅核酸断片が特定配列(D)または(C)のどちらに由来するかを判定することができる。
【0022】
リガンドは核酸の増幅を妨げるものでないのであれば、特に限定されるものではないが、好適には抗体、オリゴヌクレオチド、レセプター、核酸特異的結合物質、アビジン、ビオチン、ジゴケシゲニン、蛍光化学物質、発光団、酵素、蛍光蛋白質、発光蛋白質、磁性体、導電性物質よりなる群より選ぶことができる。好ましくはアビジン、ビオチン、ジゴケシゲニン、蛍光化学物質、蛍光蛋白質、発光団、酵素、導電性物質がよく、さらに好ましくはアビジン、ビオチン、ジゴケシゲニン、蛍光化学物質がよく、特に好ましくは蛍光化学物質がよい。
【0023】
リガンドの検出は既知の方法であれば特に限定されるものではないが、リガンドが蛍光化学物質または蛍光蛋白質の場合、特定波長の光を照射し蛍光化学物質または蛍光蛋白質を励起させ、基底状態に変換される際に生じる特定波長の蛍光量を測定することが可能である。これらに用いられる蛍光化学物質としては、FITC、FAM、TAMRA、TexasRed、VIC、Cy3、Cy5、HEX等が挙げられ、蛍光蛋白質としては、GFP、YFP、RFP等を挙げることができる。標識が酵素である場合、酵素の基質を添加することによって生成される反応生成物を検出することによって測定が可能である。これらに用いられる酵素と基質の組み合わせとしては、アルカリフォスファターゼとパラニトロフェニルリン酸、CDP−star、AMPPD、DDAOphospate、BCIP−NBT等の組み合わせ、パーオキシダーゼとTMB、Lumi−Light(ロッシュ・ダイアグノスティックス)、SAT−1(同仁化学)等の組み合わせ、ジアホラーゼとNTB等の組み合わせ、各種オキシダーゼと基質、各種デヒドロゲナーゼと基質の組み合わせ等、反応性生物が検出されるものであれば、これらに限定されることはなく用いることが可能である。標識が磁性体の場合、磁気を検出することによって測定が可能である。標識が導電性物質、電流値を検出することによって測定が可能である。
【0024】
本発明のタンパク質における繰り返しモチーフの識別方法は、例えばヘリコバクター・ピロリ菌による発ガンの危険性を予測するために用いられることができる。上記ガンとしては、胃ガンなどを挙げることができる。上述したように、本発明の識別方法を用いれば、対象とするタンパク質の繰り返しモチーフにおいて特定配列の有無および繰り返し数について明確に判定できることを利用して、CagAタンパク質のC末端領域の多型を検出することができる。更に詳細には、本発明の識別方法を用いれば、発ガンに関係が深いC配列および/またはD配列の有無を検出することができるとともに、C配列および/またはD配列の数をも検出することができる。そして、C配列および/またはD配列の有無によって、発ガンの可能性の有無を予測することができるとともに、上記C配列および/またはD配列の数によって、発ガンの可能性の大小(C配列および/またはD配列が多いほど、発ガンの危険度が大きい)を予測することができる。一例として、欧米人に多く感染するヘリコバクター・ピロリ菌について、発ガンの可能性の有無をC配列の有無によって予測することができるとともに、発ガンの可能性の大小(C配列が多いほど、発ガンの危険度が大きい)を上記C配列の数によって予測することができる。また、東アジア人に多く感染するヘリコバクター・ピロリ菌について、発ガンの可能性の有無をD配列の有無によって予測することができるとともに、発ガンの可能性の大小(D配列が多いほど、発ガンの危険度が大きい)を上記D配列の数によって予測することができる。
【実施例】
【0025】
以下、実施例に基づき本発明をより具体的に説明する。もっとも、本発明は下記実施例に限定されるものではない。
【0026】
実施例1
ヘリコバクター・ピロリ菌のCagAタンパク質における繰り返しモチーフの識別
(1)オリゴヌクレオチドの合成
パーキンエルマー社製DNAシンセサイザー392型を用いて、ホスホアミダイト法にて、配列番号1〜3に示される塩基配列を有するオリゴヌクレオチド(以下、オリゴ1〜3と示す)を合成した。合成はマニュアルに従い、各種オリゴヌクレオチドの脱保護はアンモニア水で55℃、一夜実施した。オリゴヌクレオチドの精製はパーキンエルマー社OPCカラムにて実施した。もしくはDNA合成受託会社((株)日本バイオサービス、シグマアルドリッチジャパン(株)、オペロンバイオテクノロジー(株)等)に依頼した。
オリゴ1がセンス鎖であり、CagAタンパク質のD配列をコードする核酸配列すなわち特定配列(D)内に3’末端を有するプライマー(D1)であり、アンチセンス鎖のオリゴ3と組み合わせて増幅反応のオリゴヌクレオチドとして使用される。オリゴ2はCagAタンパク質のC配列をコードする核酸配列すなわち特定配列(C)内に3’末端を有するプライマー(C1)であり、アンチセンス鎖のオリゴ3と組み合わせて増幅反応のオリゴヌクレオチドとして使用される。オリゴ3はCagAタンパク質のファミリーをコードする核酸配列に共通のプライマーすなわちプライマー(D1)または(C1)とともに核酸増幅反応における一対のプライマーとなり得る核酸配列を含有するプライマー(P1)である。オリゴ1は5’末端をFITCにより標識され、オリゴ2は5’末端をTAMRAにより標識されている。
【0027】
(2)PCR法によるヘリコバクター・ピロリ菌cagA遺伝子の解析
表1におけるサンプル番号1〜6のヘリコバクター・ピロリ菌からそれぞれフェノール・クロロホルム法により抽出したDNA溶液をサンプルとして使用して、下記試薬を添加して、下記条件によりPCR法によるヘリコバクター・ピロリ菌のcagA遺伝子を解析した。
【0028】
【表1】
【0029】
試薬
以下の試薬を含む25μl溶液を調製した。
KOD DNAポリメラーゼ反応液
オリゴ1(5’末端をFITCにより標識) 10pmol、
オリゴ2(5’末端をTAMRAにより標識) 10pmol、
オリゴ3 10pmol、
×10緩衝液 2.5μl、
2mM dNTP 2.5μl、
25mM MgCl2 1μl、
KOD DNAポリメラーゼ 0.5U、
抽出DNA溶液 100ng
【0030】
増幅条件
94℃・5分
94℃・15秒、
60℃・30秒、
68℃・30秒(35サイクル)
25℃・15分。
【0031】
(3)アガロースゲル電気泳動を用いた検出
増幅反応液5μlを3%アガロースゲルにて電気泳動し、蛍光撮影用LED光源装置ビジレイズAE−6935(ATTO(株)製)にて検出した。オリゴ1の標識であるFITCをブルー光源下、オリゴ2の標識であるTAMRAグリーン(ロング)光源下で観察し、増幅バンドの蛍光を目視によって確認した。表2に、試料No.1〜6で示すサンプルについて、明らかに確認できた増幅バンドの数を示す。
【0032】
【表2】
【0033】
(4)結果
表2(および表1)のサンプル番号1〜6より明らかなように、本発明のタンパク質における繰り返しモチーフの識別方法によれば、ヘリコバクター・ピロリ菌のCagAタンパク質における繰り返しモチーフの種類と繰り返し数を判定することが可能である。これらの結果は、核酸配列を直接解析することにより判別に必要な情報を得ることができるが多大な労力と時間が必要であるなどの欠点を有するシークエンシング法を用いた該6株の識別結果(表1)と一致する。
【0034】
すなわち、本発明のタンパク質における繰り返しモチーフの識別方法では従来の方法と同様の識別能力を実現しながら、解析に必要な作業工程数を大幅に短縮し、容易にかつ迅速にタンパク質におけるモチーフの有無および該モチーフの繰り返し数を明確に判定することが可能である。
【産業上の利用可能性】
【0035】
本発明により、タンパク質におけるモチーフの有無および該モチーフの繰り返し数を明確に判定することが可能となり、これまでの方法のように煩雑な操作を必要とせず、迅速で容易に再現性の良い結果が得られることからも、産業界に大きく寄与することが期待される。
【図面の簡単な説明】
【0036】
【図1】各プライマーの設定方法を示す一例
【図2】各プライマーの設定方法を示す一例と、第一のリガンドおよび第二のリガンドをによる検出例
【特許請求の範囲】
【請求項1】
タンパク質における繰り返しモチーフを識別する方法であって、下記の(a)〜(b)の工程を含み、該タンパク質をコードする核酸配列中の特定配列の繰り返し数を判定することを特徴とする方法。
(a)次の(i)、(ii)及び(iii)のプライマーを用いて、核酸増幅を行う工程:
(i)特定配列(D)内に3’末端を有するプライマー(D1)
(ii) プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)
(iii) プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C1)
(b)工程(a)で得られた1つまたはそれ以上のプライマー(D1)から得られる増幅核酸断片の種類と、プライマー(C1)から得られる増幅核酸断片の種類を解析する工程
【請求項2】
タンパク質における繰り返しモチーフを識別する方法であって、下記の(a)〜(b)の工程を含み、該タンパク質をコードする核酸配列中の特定配列の繰り返し数を判定することを特徴とする方法。
(a)次の(i)、(ii)及び(iii)のプライマーを用いて、核酸増幅を行う工程:
(i)特定配列(D)内に3’末端を有するプライマー(D1)
(ii) プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)
(iii)プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)
(iv) プライマー(P2)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C2)
(b)工程(a)で得られた1つまたはそれ以上のプライマー(D1)から得られる増幅核酸断片の種類と、プライマー(C2)から得られる増幅核酸断片の種類と、プライマー(P1)および(P2)から得られる増幅核酸断片の有無を解析する工程
【請求項3】
プライマー(D1)の3’末端から2番目以降の少なくとも一つのヌクレオチドがリガンドにより標識されていることを特徴とする請求項1または2に記載の識別方法。
【請求項4】
プライマー(D1)及びプライマー(C1またはC2)の3’末端から2番目以降の少なくとも一つのヌクレオチドが、それぞれ異なるリガンドにより標識されていることを特徴とする請求項1〜3のいずれかに記載の識別方法。
【請求項5】
リガンドが抗原、抗体、蛍光物質、発光団および酵素からなる群より選ばれた少なくとも1種以上であることを特徴とする請求項3または4に記載の識別方法。
【請求項6】
特定配列内に3’末端を有するプライマーの3’末端から2番目以降の少なくとも一つのヌクレオチドに結合したリガンドを検出することにより、該特定配列の繰り返し数を判定することを特徴とする請求項3〜5のいずれかに記載の識別方法。
【請求項7】
繰り返しモチーフが、ヘリコバクター・ピロリ菌のCagAタンパク質のC末端領域に存在するC配列またはD配列であることを特徴とする請求項1〜6のいずれかに記載の識別方法。
【請求項1】
タンパク質における繰り返しモチーフを識別する方法であって、下記の(a)〜(b)の工程を含み、該タンパク質をコードする核酸配列中の特定配列の繰り返し数を判定することを特徴とする方法。
(a)次の(i)、(ii)及び(iii)のプライマーを用いて、核酸増幅を行う工程:
(i)特定配列(D)内に3’末端を有するプライマー(D1)
(ii) プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)
(iii) プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C1)
(b)工程(a)で得られた1つまたはそれ以上のプライマー(D1)から得られる増幅核酸断片の種類と、プライマー(C1)から得られる増幅核酸断片の種類を解析する工程
【請求項2】
タンパク質における繰り返しモチーフを識別する方法であって、下記の(a)〜(b)の工程を含み、該タンパク質をコードする核酸配列中の特定配列の繰り返し数を判定することを特徴とする方法。
(a)次の(i)、(ii)及び(iii)のプライマーを用いて、核酸増幅を行う工程:
(i)特定配列(D)内に3’末端を有するプライマー(D1)
(ii) プライマー(D1)と互いの伸長産物が相互の鋳型となる性質を有し、 特定配列外の2つの領域のうちいずれかの領域に3’末端を有し、該タンパク質のファミリーをコードする核酸配列に共通のプライマー(P1)
(iii)プライマー(P1)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列外の2つの領域のうち、プライマー(P1)を設定した領域とは異なる他方の領域に3’末端を有し、該タンパク質のファミリーに共通のプライマー(P2)
(iv) プライマー(P2)と互いの伸長産物が相互の鋳型となる性質を有し、特定配列(D)のバリアントである特定配列(C)内に3’末端を有するプライマー(C2)
(b)工程(a)で得られた1つまたはそれ以上のプライマー(D1)から得られる増幅核酸断片の種類と、プライマー(C2)から得られる増幅核酸断片の種類と、プライマー(P1)および(P2)から得られる増幅核酸断片の有無を解析する工程
【請求項3】
プライマー(D1)の3’末端から2番目以降の少なくとも一つのヌクレオチドがリガンドにより標識されていることを特徴とする請求項1または2に記載の識別方法。
【請求項4】
プライマー(D1)及びプライマー(C1またはC2)の3’末端から2番目以降の少なくとも一つのヌクレオチドが、それぞれ異なるリガンドにより標識されていることを特徴とする請求項1〜3のいずれかに記載の識別方法。
【請求項5】
リガンドが抗原、抗体、蛍光物質、発光団および酵素からなる群より選ばれた少なくとも1種以上であることを特徴とする請求項3または4に記載の識別方法。
【請求項6】
特定配列内に3’末端を有するプライマーの3’末端から2番目以降の少なくとも一つのヌクレオチドに結合したリガンドを検出することにより、該特定配列の繰り返し数を判定することを特徴とする請求項3〜5のいずれかに記載の識別方法。
【請求項7】
繰り返しモチーフが、ヘリコバクター・ピロリ菌のCagAタンパク質のC末端領域に存在するC配列またはD配列であることを特徴とする請求項1〜6のいずれかに記載の識別方法。
【図1】

【図2】
【図2】
【公開番号】特開2008−161111(P2008−161111A)
【公開日】平成20年7月17日(2008.7.17)
【国際特許分類】
【出願番号】特願2006−354165(P2006−354165)
【出願日】平成18年12月28日(2006.12.28)
【出願人】(000003160)東洋紡績株式会社 (3,622)
【Fターム(参考)】
【公開日】平成20年7月17日(2008.7.17)
【国際特許分類】
【出願日】平成18年12月28日(2006.12.28)
【出願人】(000003160)東洋紡績株式会社 (3,622)
【Fターム(参考)】
[ Back to top ]