
タンパク質生産のための遺伝子組み換えアロエ植物と関連方法

【課題】遺伝子組み換えアロエ植物と、アロエ植物を形質転換するための組み換えコンストラクトを提供する。
【解決手段】組み換えコンストラクトは、哺乳類のタンパク質をコードする一つ以上のDNA配列と、アロエ植物で組み換えタンパク質の発現を誘導することができる少なくとも一つのプロモーターを含む。遺伝子組み換えアロエ植物を構築し、再生する。
【解決手段】組み換えコンストラクトは、哺乳類のタンパク質をコードする一つ以上のDNA配列と、アロエ植物で組み換えタンパク質の発現を誘導することができる少なくとも一つのプロモーターを含む。遺伝子組み換えアロエ植物を構築し、再生する。
【発明の詳細な説明】
【技術分野】
【0001】
<関連出願への相互参照>
本特許出願は、2005年9月26日出願の米国特許仮出願第60/720,540号、表題“TRANSGENIC ALOE PLANTS FOR PRODUCTION OF PROTEINS AND RELATED METHODS”の優先権と利益を主張し、この出願はここにその全容が引用により本明細書に組み込まれる。
【背景技術】
【0002】
<発明の概要>
本発明は、遺伝子組み換え単子葉植物に関し、より詳細には、遺伝子組み換えアロエ植物と、遺伝子組み換えアロエ植物生産のための方法と組成、および遺伝子組み換えアロエ植物からタンパク質を抽出する方法に関する。
【0003】
<関連技術の説明>
その多くが治療目的で用いられる生物活性タンパク質の市場は拡大し続けている。現在、160以上のタンパク質から作られる医薬品が利用可能である。さらに50くらいが今後2〜3年のうちに認可される見通しである。治療タンパク質生産に対する現行需要は、既に生産能力を上回っている。業界はこの隘路を打開するために、生産能力を4倍から5倍増加させる必要があると予測されている。しかしながら、治療タンパク質の生産設備は高価で、一般的に建設に時間がかかる。従って、より安価で、かつ生産を増加させるために必要な時間を短縮し得る生産方法が必要とされる。
【0004】
いくつかの動物性タンパク質生産法が用いられる。しかしながら、これらは頻繁に病気から健康上のリスクを引き起こす。そのようなリスクは、動物と、ヒトの患者などの最終使用者の両方に影響し得る病気の二次汚染から生じ得る。従って、産生生物と最終使用者の間での二次汚染の可能性を排除するような生産方法が必要とされる。
【0005】
加えて、多くの現在の生産方法は、治療タンパク質を生産した動物もしくは宿主生物から、治療タンパク質を抽出するために、および、複合物を患者によって利用され得る状態にするために、大規模な処理を必要とする。精製後、タンパク質は、タンパク質を安定化し、患者が利用できるようにするために、アジュバントもしくは他の担体物質と混合されてもよい。しかしながら、抽出、精製、再懸濁の処理過程は、治療タンパク質の処理に関するものの中でもとりわけ複雑で煩雑であり、一般的に治療法を必要とする発展途上国での使用につながらない可能性がある。従って、治療タンパク質の抽出後処理を排除する、もしくは削減し得る簡単な生産方法が必要とされる。
【発明の開示】
【0006】
本発明に従う組成物と方法は、上述の必要性と欠点の多くを解決し、また本発明の開示を考察することで当業者に理解される、さらなる改良と利点を提供し得る。
【0007】
一態様では、本発明は目的遺伝子を安定的に組み込んだ遺伝子組み換えアロエ植物を提供し得る。他の態様では、アロエ植物のゲノムに目的のDNA配列を組み込むために、新規ベクターとコンストラクトが提供され得る。目的の配列は生物活性タンパク質をコードしていてもよい。生物活性タンパク質は、インターフェロン、免疫グロブリン、リンフォカイン、成長因子、ホルモン、血液因子、組織適合抗原、酵素、化粧品タンパク質と他の哺乳類タンパク質、もしくは他の目的タンパク質であってもよい。いくつかの態様では、目的タンパク質はヒトのタンパク質である。本発明に従うアロエ植物は、目的遺伝子を目的タンパク質に転写して翻訳し得る。一態様では、目的タンパク質の少なくともいくつかは、アロエ葉の中心部に移動する。他の態様では、目的タンパク質はアロエ葉の中心部への転移を促進するようなシグナル配列を含んでもよい。他の態様では、アロエ植物から個別細胞を分離するための新規の方法が提供され得る。さらに他の態様では、本発明は、アロエ植物にベクターを組み込み、またそのような遺伝子組み換えアロエ植物を再生産するための新規の方法を提供し得る。
【0008】
本発明に従う哺乳類タンパク質を生産する遺伝子組み換えアロエ植物は、例えば様々な生物活性タンパク質と化粧品タンパク質などの目的タンパク質の生産のために、より採算の合う代替案を提供し得る。一態様では、目的タンパク質はアロエ葉のゲル内に局在してもよいし、および/または濃縮されてもよい。この目的タンパク質の局在化は、植物からのタンパク質の除去を単純化し得る。この態様では、目的タンパク質はアロエ葉の中心部内の天然ゲルの抽出と共に共抽出(co-extract)されてもよい。従って本発明は、その遺伝子組み換え植物由来の目的タンパク質が、例えばタバコととうもろこしなどのほとんどの遺伝子組み換え植物由来のものよりも、一般的により容易に利用しやすい遺伝子組み換え植物を提供し得る。さらに、本発明はタンパク質分離のための効率的な方法を提供し得る。さらに他の態様では、目的タンパク質はアロエ植物内に特に局在していなくてもよい。しかしながら、アロエ植物の組織と生理学は、本発明の開示を考察することで当業者に理解される、目的タンパク質の生産のためのいくらかのさらなる利点をも提供し得る。
【0009】
アロエ植物は、バクテリアと酵母の中で目的タンパク質を生産するための、従来の方法に勝る様々な利点を提供することができる。アロエ植物10の利点は、単純単細胞のバクテリアと酵母は適合性が低く、かつ所望の形でタンパク質を生産するために必要とされるやり方で、タンパク質を処理する能力を含み得る。この処理は、グリコシル化などによる化学修飾と、例えばいくつかのタンパク質の折り畳みを含むことができる。さらに、動物細胞に基づく他のタンパク質生産方法に比べて、アロエ植物生産は著しい費用便益と拡張性の利点を提供し、またヒトに有害となり得る汚染のリスクを軽減し得る。
【0010】
本発明に従う遺伝子組み換えアロエ植物のアロエ葉の中心部からのタンパク質修飾ゲルは、アロエ植物から直接使用され得る。これにより、比較的複雑で高価な天然植物材料からの目的タンパク質の抽出を行う必要がなくなる可能性がある。いくつかの態様では、葉から抽出されたゲルは、タンパク質抽出もしくは処理の必要なしに直接使用され得る。ゲルは、遺伝子組み換えアロエ植物の折れた葉の切り口(open end)から機械的に抽出される髄の形であってもよい。
【0011】
本発明は、生物活性のある、および/または化粧品に応用される、ヒトおよび他の哺乳類のタンパク質の生産のための、採算の合う代替案を提供し得る。
【0012】
本発明の開示を考察することにより、当業者は本発明のさらなる改良と利点を理解するだろう。
【図面の簡単な説明】
【0013】
【図1】アロエ葉の断面組織の例をあらわす。
【図2】カルス組織を生成するための方法例をあらわす。
【図3】遺伝子組み換えアロエ植物を生成するための様々なステップを図式的に概説する。
【図4】配列情報を含むプラスミドベクターの構成要素を図式的にあらわす。
【図5】トウモロコシ由来のユビキチンプロモーターの配列情報を列挙し、ユビキチンプロモーターの領域を強調している。
【図6A】本発明の態様に従うプラスミドベクター系のコンストラクションをあらわす。
【図6B】本発明の態様に従うプラスミドベクター系のコンストラクションをあらわす。
【図7A】本発明の態様に従う別のプラスミドベクター系のコンストラクションをあらわす。
【図7B】本発明の態様に従う別のプラスミドベクター系のコンストラクションをあらわす。
【図7C】本発明の態様に従う別のプラスミドベクター系のコンストラクションをあらわす。
【図8】本発明の態様に従うさらなるプラスミドベクター系のコンストラクションをあらわす。
【図9】本発明の態様に従う組み込み系のコンストラクションをあらわす。
【0014】
全図面は、本発明の基本的教示の説明を簡略化するために示されるに過ぎない。すなわち、数、位置、配列、様々な実施形態の関係と組成に関する図面の拡張は、以下の記述を考察した後で、説明されるかもしくは当該技術分野内に含まれることになるだろう。さらに、開示された発明を実施するための様々なプロトコル、ツール、および組成物が、以下の記述を考察した後で当該技術分野内に含まれることになるだろう。
【発明を実施するための最良の形態】
【0015】
明細書で使用する“a”もしくは“an”は、一つもしくはそれ以上を意味してもよい。請求項(群)で使用するように、“含む(comprising)”という用語と併用する際、用語“a”もしくは“an”は一つもしくは一つ以上を意味してもよい。本明細書で使用する“別の(another)”は、少なくとも二つ目もしくはそれ以上を意味してもよい。
【0016】
本明細書で使用する“形質転換アロエ細胞”は、例えばAgrobacterium媒介形質転換、もしくは組み換えDNAで被覆された微小粒子を用いる打ち込み、もしくは他の手段によって、安定的に組み込まれた、非天然の組み換えDNAで形質転換された植物細胞を意味する。本発明の形質転換アロエ細胞は、オリジナルに形質転換された植物細胞であってもよく、これは微小生物として、もしくは後代の植物細胞として生存し、分化した組織に再生される。例えば、安定に導入された非天然の組み換えDNAを持つ遺伝子組み換えアロエ植物10、あるいは後代の遺伝子組み換えアロエ植物10由来の種子、もしくは花粉に再生される。
【0017】
本明細書で使用する“遺伝子組み換えアロエ植物”は、組み換えDNAの安定的な組み込みによってゲノムを改変したアロエ植物を意味する。遺伝子組み換えアロエ植物10は、オリジナルに形質転換されたアロエ細胞から再生したアロエ植物と、後世代もしくは形質転換アロエ植物10の交雑由来の後代の遺伝子組み換えアロエ植物を含む。
【0018】
本明細書で使用する“組み換えDNA”は、細胞の外側で構築され遺伝子操作されたDNAを意味し、天然DNA、cDNA、合成DNA、および/または他のDNAを包含するDNAを含む。
【0019】
本明細書で使用する“プロモーター”は、転写開始のための調節DNAを意味する。“植物プロモーター”は、その起源がアロエ細胞であるかどうかに関わらず、アロエ細胞の転写を開始することができるプロモーターであり、例えばAgrobacteriumプロモーターはアロエ細胞内で機能することがよく知られている。従って植物プロモーターは、植物、植物ウィルス、およびAgrobacteriumと根粒菌(Bradyrhizobium)バクテリアなどのバクテリアから得られるプロモーターDNAを含む。発生制御下にあるプロモーターの例としては、葉、根、もしくは種子などの特定の組織において選択的に転写を開始するプロモーターを含む。そのようなプロモーターは、“組織選択的”であると言われる。特定の組織のみで転写を開始するプロモーターは、“組織特異的”であると言われる。“細胞型”特異的プロモーターは、一つ以上の器官において特定の細胞型における発現を主に促進する(例えば、根もしくは葉における道管細胞)。“誘導”プロモーター、もしくは“抑制”プロモーターは、環境制御下にあるプロモーターである。誘導プロモーターによって転写を誘起し得る環境条件の例としては、嫌気条件、もしくは特定の化学物質、もしくは光の存在を含む。組織特異的プロモーター、組織選択的プロモーター、細胞型特異的プロモーター、および誘導プロモーターは、“非構成的”プロモーターの分類を構成する。“構成的”プロモーターは、ほとんどの条件下で活性化されているプロモーターである。そうしたプロモーターは、エンハンサー、あるいは、転写開始、転写開始部位、転写のレベル、転写終結部位、もしくは得られるリボ核酸の任意の後処理に作用する他の要素を含んでもよい。
【0020】
本明細書で使用する“遺伝子コンストラクト(genetic construct)”という用語は、遺伝子組み換えアロエ植物の作製を目的として、少なくとも二つのDNAセグメントの合成配列から成るDNA配列と定義される。特定の実施形態では、一つのセグメントは調節配列で、もう一方のセグメントは遺伝子産物をコードする。
【0021】
本明細書で使用する“連結(operably linked)”とは、DNAコンストラクトにおける二つ以上のDNAフラグメントの結合を意味し、一方のフラグメントの機能(例えばタンパク質をコードするDNA)が、もう一方のフラグメント(例えばプロモーター、終結配列など)によって制御されるようになっている。
【0022】
本明細書で使用する“転写”という用語は、DNA鋳型からRNA分子を作製することと定義される。
【0023】
本明細書で使用する“翻訳”という用語は、RNA鋳型からポリペプチドを作製することと定義される。
【0024】
本明細書で使用する“発現した”とは、“生成した(produced)”を意味し、例えばタンパク質と同起源のDNAがmRNAに転写され、それがタンパク質に翻訳された時、そのタンパク質は植物細胞内で発現していることになる。
【0025】
本発明は、様々な目的タンパク質を発現する新規の遺伝子組み換えアロエ植物10を提供してもよいし、遺伝子組み換えアロエ植物10を生産するための方法と組成を提供してもよいし、遺伝子組み換えアロエ植物10からタンパク質を抽出するための方法を提供してもよいし、また、目的タンパク質と遺伝子組み換えアロエ植物10に由来する構成要素の新規の組成を提供してもよい。本発明に従って生成される目的タンパク質は、髄18もしくはアロエ葉12の髄でのタンパク質の蓄積を促進させるような分泌シグナルを含んでもよい。一態様では、遺伝子組み換えアロエ植物10の葉のmusalegenous細胞内で、このタンパク質の蓄積の少なくとも一部分が起こってもよい。
【0026】
髄(pith)18もしくはアロエ葉12の髄(pulp)は、アロエ葉12から抽出された際に、少なくとも部分的には一般に“ゲル”と呼ばれる葉の構成要素を含む。本明細書で使用する“ゲル”は、抽出された髄18、および、次の処理の程度に関わらず、アロエ葉12から抽出される際に髄18に付随する他の付随物質を意味する。本発明の一つ以上の態様に従って、アロエ葉12から抽出したゲルは、変性した組織を持っていてもよい。特定の一態様では、ゲルの組織は少なくとも一つの外来タンパク質成分を含むように変性されてもよく、これは“タンパク質変性ゲル”と呼ばれる。付随する目的タンパク質(群)を伴うタンパク質変性ゲルの抽出により、タンパク質を容易に入手でき、および/または効率的なタンパク質分離方法が提供され得る。本発明に従って生成されるおよび/または抽出されるタンパク質変性ゲルは、タンパク質抽出の必要無しに直接使用され得る。
【0027】
本発明の態様は、例示目的として図1から図8に大まかに示される。本発明は、遺伝子組み換え植物と組成と方法を提供し、これらは概してユリ科アロエ属に応用できる。典型的には、本発明は例示目的として、Aloe vera(barbadensis miller)、Aloe ferox、およびAloe arborescenceの群から選択される種への応用に関して記載される。アロエ属は一般的に茎のない大きなロゼット型多肉性単子葉植物を含む。これらの植物を、本発明の開示を目的として概してアロエ植物10と呼ぶこととする。
【0028】
アロエ植物から生成される様々な化合物が医薬目的に使用されてきた。これらの化合物は、タンパク質変性ゲルに存在する場合、本発明に従う遺伝子組み換えアロエ植物10の生物活性タンパク質の薬理作用を補足し得る。図1は遺伝子組み換えアロエ植物10の葉12の断面図を示す。遺伝子組み換えアロエ植物10の葉は、容易に抽出可能な、タンパク質、糖質、および水のゲル状混合物をゲル内に含み、ゲルは主に髄18から得られる。このゲル状混合物は、主にアロエ葉12の中心部に位置する。ゲル内の様々な化合物は、数多くの薬理作用と医薬用途を有することがわかっている。一態様では、ゲルおよび/または髄18は、遺伝子組み換えアロエ植物10によって生成され、ゲルおよび/または髄18に局在する遺伝子組み換えタンパク質を安定化させる。
【0029】
図1は特に、表皮14、皮層もしくは葉肉16、および髄(pithもしくはpulp)18を示す。表皮14は葉の細胞の外層を形成する。本発明の一態様では、遺伝子組み換えアロエ植物は、これらの組織の内の一つ以上において、アロエ細胞によって発現される遺伝子コンストラクトを、一時的にもしくは安定的に組み込むアロエ細胞を含んでもよい。皮層16は、維管束、木質部、および師部と同様に、葉緑体の豊富な細胞を含む。髄18は、ほぼ大きな皮層細胞のみで構成される海綿状組織であり、少なくとも部分的にゲルをあらわし、そこに一つ以上の目的の遺伝子組み換えタンパク質を組み込むか、もしくは蓄積させる上で好都合となり得る。
【0030】
表皮14は典型的には細胞の単外層から成る。表皮14のすぐ下は、維管束の網状組織を含む皮層16である。維管束の外側の支えは、通常は鞘細胞によってもたらされる。維管束は三つの大まかな種類の管構造から構成される。すなわち、木質部、師部、および付随する大きな周辺環状小管(pericyclic tubules)である。木質部は水と無機物を植物の根から葉に輸送する。師部はデンプンと他の合成物質を植物の至る所に輸送する。周辺環状小管は、緩下性のあるアントラキノンを非常に多く含む(特にアロイン)、乳液もしくは樹液を含む。アントラキノンは太陽の紫外線を吸収し、一般的にアロエ植物の水貯蔵器官として機能するアロエ葉12の中心部の過熱を防ぐ。維管束の周辺環状部分は表皮14に付着しており、一方維管束の残りの部分は髄18の中に突き出ている。アロエ葉12の最深部の主要部分は、少なくとも部分的にゲルを構成する髄18である。ゲルの抽出を目的として、表皮14および皮層16は、ゲルを含む葉鞘を含むと一般的に見なされ得る。抽出されたゲルは、実質的に髄18から成るが、典型的には粘度の高い粘液性の物質であり、昔から、例えば火傷や創傷の治療などの医薬目的として、皮膚に局所塗付されてきた。
【0031】
通常、ゲルはアロエ植物のための物質貯蔵庫としてはたらく。一般的に、皮層16はアロエ植物に必要な糖質と糖タンパク質の多くを合成する。過剰に合成された糖質は、通常は髄18に輸送され、水といくつかの無機物と共に貯蔵される。糖質は師部道管によって、髄18の皮層16細胞内の大きな液胞に輸送される。その後、水が浸透圧によって糖質に引き付けられ、髄18がアロエ植物の水貯蔵器官として機能できるようになっている。
【0032】
ゲルは、アロエ葉12を縦軸に沿って折り、葉を押しつぶしてゲルを割れ目を通して周りの鞘から押し出すことにより、比較的容易に抽出される。付加的な物質がゲルに伴って葉の組織から抽出されることもある。
【0033】
<処理の概説>
本発明の一つ以上に従う遺伝子組み換えアロエ植物10は、概して、一つ以上の目的タンパク質を発現するように植物に安定的に組み込まれた一つ以上のDNAコンストラクトを含む。本発明の一つ以上に従う遺伝子組み換えアロエ植物10を作製することは、種々の新規の組成と方法を含んでもよい。典型的には、一つ以上のDNAコンストラクトは、遺伝子組み換えアロエ植物10に一つ以上のタンパク質を発現するように改良される。一態様では、一つのコンストラクトは一つのmRNAを発現してもよい。他の態様では、一つのコンストラクトは複数のmRNAを発現してもよい。なおも他の態様では、複数のコンストラクトが複数のmRNAを産出してもよい。各mRNAは一つ以上のポリペプチドを産出してもよい。
【0034】
コンストラクトをアロエ植物に導入するには、図2に大まかに示すように、アロエ細胞もしくは未分化カルス組織が典型的に使用される。アロエ細胞とカルス組織は、典型的にはアロエ植物の根もしくは葉の分裂組織、もしくは種子に由来する。DNAコンストラクトは、図3に図式的に示すように、様々な技術および/またはコンストラクトを用いてアロエ細胞に導入される。いくつかの技術は、本発明の開示を考察することで当業者に理解されるだろう。所望のコンストラクトを組み込んだアロエ細胞もしくはカルス組織は、様々な技術を使用するために選択される。典型的には、DNAコンストラクトは選択可能なマーカーを含む。いったん所望のコンストラクトが安定的にアロエ細胞もしくはカルス組織に導入されると、通常はアロエ細胞もしくはカルス組織からアロエ小植物体が発生する。DNAコンストラクトを含む遺伝子組み変え生存アロエ植物10に成長するアロエ小植物体は、所望の目的タンパク質(群)を構成的に産出するか、あるいは誘導的に産出する可能性がある。目的タンパク質は、産出される特定のタンパク質、および/またはタンパク質に付随するシグナル配列の有無に応じて、遺伝子組み換えアロエ植物10内で局在してもよいし、あるいは、遺伝子組み換えアロエ植物10の全体に概して存在してもよい。その後、特定のコンストラクトおよび/または目的の関連タンパク質に応じて、遺伝子組み換えアロエ植物10は、栄養繁殖(無性生殖)もしくは有性繁殖(有性生殖)され得る。タンパク質は、幅広い技術を用いて遺伝子組み換えアロエ植物10から分離され得る。その後タンパク質はさらに分離されるか、および/またはさらに処理されてもよい。そのような処理は、本発明の開示を考察することで当業者に理解される他の処理の中でも特に、酵素修飾、化学修飾、適切なアジュバントへの混入を含んでもよい。一態様では、特定のタンパク質の特異的用途のために、タンパク質は不活性体(前駆体)から活性体に処理されてもよい。
【0035】
<細胞の分離>
図2に図示した配列例に示すように、アロエ細胞もしくはアロエ細胞群は、典型的には所望のコンストラクトの組み込みの前にアロエ植物から分離される。DNAコンストラクトの組み込みに一般的に選ばれるアロエ細胞の種類は、その再生能に基づいて一般的に選択される。アロエ植物体からの茎分裂組織もしくは根分裂組織由来の分裂組織細胞、もしくはアロエ種子由来のアロエ胚細胞を使用してもよい。しかしながら、アロエ植物の多くの部分は、再生する能力、もしくはカルス組織を形成する能力を保持しており、これらを利用してもよい。細胞は典型的には、本発明の開示を考察することで当業者に理解される様々な技術を用いて分離される。典型的には、細胞は外科用メスを用いてアロエ植物から機械的に分離される。あるいは、本発明の開示を考察することで当業者に理解されるように、必要な細胞を分離するために、他の機械的技術もしくはその他の技術を用いてもよい。適切なアロエ細胞もしくはアロエ細胞群が分離されると、通常、アロエ細胞はカルス組織を形成するために培養下で育成される。或る技術は、アロエ細胞が最初にカルス組織に成長することを必要としなくてもよいが、カルス組織は、遺伝子組み換えアロエ植物10を作り出す能力を保持する未分化のアロエ細胞セットの源を提供する。カルス組織は典型的には固形培地上で育成される。しかしながら、カルス組織は液体培地に置くこともできるし、浮遊状態で成長させることもできる。
【0036】
分裂組織細胞は、アロエ植物の根の先端もしくは葉から分離してもよい。分裂組織アロエ細胞は頂端分裂組織から分離してもよい。典型的には、組織が分離されるアロエ植物は、若い健康なアロエ植物である。通常、アロエ植物は8インチ未満で六つ以下の初期芽を持つものが選択される。切り傷は通常、アロエ植物を断片に切ることによって、アロエ植物の芽の上に形成される。傷をつけた表面は、アロエ細胞が成長するのを促進し得る。通常、最初に傷の表面でカルス組織が成長し始める。アロエ葉12からアロエ細胞を分離するために、断片は通常、若い成長芽の遠位末端から切り取られる。その後断片部分は成長培地上に置かれる。芽の頂端分裂組織は、若いアロエ植物の基部から1インチを切って得られる。アロエ葉12は切片から除去され、断片がプレートに置かれる。分裂組織アロエ細胞は“切片”内に見られる。これらは、培養プレート上での成長維持能力(カルス組織の形成もしくは芽の再生)によって“選別”されるかもしくは“分離”される。分裂組織アロエ細胞は分離されると、その後適切な培地上で、カルス組織形成を促進する条件下で培養される。
【0037】
種子からのアロエ胚細胞の分離は、一般的に、胚を露出させるために種皮を除去することと、所望のアロエ細胞を胚から機械的に除去することを含む。アロエ種子は、典型的には滅菌され、種子の外皮が除去される。アロエ植物もしくはアロエ胚のいずれかからアロエ細胞を機械的に除去するために、外科用メスが通常用いられる。アロエ胚細胞は種皮から分離されると、その後適切な培地上で、カルス組織形成を促進する条件下で培養される。
【0038】
カルス組織が必要な際は、分離されたアロエ細胞は通常、本発明の開示を考察することで当業者に理解されるように、カルス組織の形成に好都合な条件下で育成される。典型的には、分離されたアロエ細胞は、形質転換の前に直径約1cmのサイズを有するカルス組織を形成するために、適切な固体栄養培地に播種されて成長する。アロエ細胞は典型的にはセ氏23度〜26度の間で成長する。適切な培地は固体もしくは液体の基盤をふくみ、通常、これらは、補助無機栄養素(マクロ元素(窒素、硫黄、リン、カルシウム、マグネシウム、およびカリウム)と微量元素(鉄、ホウ素、コバルト、銅、ヨウ素、マンガン、モリブデン、および亜鉛)の両方)、ならびに、糖(スクロースもしくはマルトース)およびビタミンおよび捕因子(とりわけチアミン、ナイアシン、ビオチン、ピリドキシン、ミオイノシトール)を含む有機栄養素、ならびに、アミノ酸(プロリンおよびカゼイン塩加水分解物)、さらに、主にオーキシンの源(通常(NAA)、1-ナフタレン酢酸、(IAA)、インドール-3-酢酸、もしくは(2,4-D)、2,4-ジクロロフェノキシ酢酸)およびサイトカイニンの源(通常(BAP)6-ベンジルアミノプリン)である成長調節因子を含む。これらの培地とビタミンは、通常、様々な濃度の無機栄養素とビタミンで前処方された組成で市販されており、とりわけ、MS培地(Murashige-skoog)、Gamborg培地、もしくはChu N6培地として知られている。さらに、ペトリ皿上で培養される細胞は、通常、成長培地に添加されるアガーなどの固体マトリクス支持物質を必要とする。
【0039】
<DNAコンストラクトの作製>
適切なDNAコンストラクトは、典型的には、得られる遺伝子組み換えアロエ植物10によって目的タンパク質(群)の生成が可能になるように、分離されたアロエ細胞に由来するカルス組織に導入される。本発明の開示を考察することで当業者に理解されるように、DNAコンストラクトは、分離されたアロエ細胞もしくは成熟アロエ植物に導入されてもよい。DNAコンストラクトは、典型的には、コンストラクトの増殖およびアロエ細胞への導入のために、プラスミドもしくはウィルスなどのベクターに組み込まれる。プラスミドベクターの例としては、例示目的として図4に図式的に示してあるInvitrogen(Carlsbad, California)から商標pZErOで市販されているプラスミドを含んでもよく、また、例えばこのプラスミドの機能単位のうちの一つ以上を含んでもよい。
【0040】
本発明に従うDNAコンストラクトは、アロエ細胞内で機能できるプロモーター配列、目的タンパク質をコードする配列、終結配列、および転写開始部位と転写終結部位(これら全てがアロエ細胞内で機能できる)を含むことができる。これらの構成要素は、適切に構成され結合された際には、DNAの転写とmRNAの翻訳、およびこれらそれぞれの終結をアロエ細胞内で開始することができる。DNAコンストラクトは分泌シグナルを含んでもよい。
あるいは、例えばインターフェロンなどのいくつかの目的タンパク質は、インターフェロンを分泌の標的とする、天然もしくは合成のドメインを含んでもよい。分泌シグナルは、通常、タンパク質が細胞を離れる際に切断されるか、あるいはいくつかの場合には、実質的にタンパク質の機能に影響を与えることなく、タンパク質によって保持されることもある。分泌シグナルは転移のために分泌シグナルを必要とし得る様々なタンパク質に加えられてもよい。これらの分泌シグナルは様々な植物分泌配列に由来してもよい。DNAコンストラクトもしくは関連ベクターは、少なくとも一つの選択可能なマーカーも含んでもよい。一態様では、DNAコンストラクトは、バクテリアでのベクターの増殖のための第一選択可能マーカーと、アロエ細胞での成長のための選択可能マーカーの両方を含んでもよい。加えて、DNAコンストラクトは、アロエ植物10の特定のアロエ細胞での発現のための、他の調節因子を含んでもよい。他のコンストラクトの構成要素は、転写促進のための5’リーダーとイントロン、3’非翻訳領域(ポリアデニル化シグナルおよびポリアデニル化部位など)、輸送ペプチドのためのDNAなど、付加的な調節因子を含んでもよい。所望の治療タンパク質をコードする配列などの、所望のDNA配列の増幅は、ポリメラーゼ連鎖反応によって行われ得る(U.S. Pat. No. 4,683,202および5,928,906を参照。それぞれ引用によって本明細書に組み込まれる。)。本発明の開示を考察することで、当業者は、プロモーター配列、調節配列、安定化配列、ターゲティング配列、および/または終結配列を含むDNAコンストラクトが、当業者に周知の方法を用いてその機能に影響を及ぼすように改変されてもよいことがわかるだろう。
【0041】
上述の通り、アロエ細胞で機能するプロモーターが通常は利用される。プロモーターは、典型的には、制御タンパク質と制御分子が結合し得る遺伝子因子を含む。これらのタンパク質は、典型的にはRNAポリメラーゼと他の転写因子を含む。プロモーターは、機能上の位置および/または方向で核酸配列に機能的に結合して、その配列の転写開始および/または発現を制御する。プロモーターは、核酸配列の転写活性化に関係するシス作用調節配列を意味する“エンハンサー”と併用されてもされなくてもよい。
【0042】
当然ながら、その中で発現することが望まれるアロエ植物10の、少なくとも一つの細胞型において、DNAセグメントの発現を効果的に誘導するようなプロモーターおよび/またはエンハンサーを採用することが重要となる。分子生物学分野の当業者は、一般的にタンパク質発現のためのプロモーター、エンハンサー、および細胞型の組み合わせの使用法を知っており、例えば、引用により本明細書に組み込まれるSambrook et al.(1989)を参照のこと。採用されるプロモーターは、構成的、組織特異的、誘導的、および/または、適切な条件下で導入されたDNAセグメントの高レベルの発現を誘導するのに有用であるものでもよく、例えば、組み換えタンパク質および/またはペプチドのラージスケールでの産出において有利である。プロモーターは異種であっても内因性であってもよい。DNAコンストラクトは、通常はアロエ細胞内で機能できる転写開始・終結制御シグナルと翻訳開始・終結制御シグナルを必要とする。幅広い種類の転写開始調節配列が使用され得る。転写開始を制御するDNA配列は、Agrobacterium、ウィルス、もしくは植物に由来し得る。カリフラワーモザイクウィルス(35S-CaMV)由来の35Sウィルス転写開始領域がアロエ植物10に使用され得る。アロエ植物10で使用され得る植物プロモーターは、種々の単子葉植物もしくは双子葉植物由来のリブロース-1,5-ビスリン酸カルボキシラーゼ(RUBISCO)小サブユニットプロモーター、またはトウモロコシ由来のユビキチンプロモーターも含んでもよい。他の適切なプロモーターが使用されてもよく、本発明の開示を考察することで当業者に理解され得る。もし誘導調節が望まれる場合は、ドメインは異なる起源から得られてもよく、一方の起源由来の調節領域が、もう一方の起源由来のRNAポリメラーゼ結合ドメインに結合されるようになっている。発現制御は、根、葉、種子、花、樹液などの遺伝子組み換えアロエ植物10の特定の部分における、遺伝子組み換えアロエ植物10の成長の特定の段階に対してなされてもよいし、あるいは、植物の部分と成長段階の組み合わせに対してなされてもよい。特定の成長段階もしくは組織に対する発現制御は、本発明の開示を考察することで当業者に理解される、さらなるDNA要素を必要としてもよい。
【0043】
植物細胞で活性のある数多くの他のプロモーターが文献に記載され、アロエ細胞で有効である可能性がある。これらは、植物ゲノムに存在するプロモーターだけでなく、他の起源由来のプロモーターも含む。すなわち、Agrobacterium tumefaciensの腫瘍誘導プラスミド上にあるノパリンシンターゼ(NOS)プロモーターおよびオクトピンシンターゼ(OCS)プロモーター、ならびにカリフラワーモザイクウィルスなどのcaulimovirusプロモーターを含む。さらに、種々の他のプロモーターも様々な参考文献で同定されており、カリフラワーモザイクウィルス(CaMV35S)由来の構成的プロモーターの種類を開示しているU.S. Pat. No. 5,858,742および5,322,938、イネアクチンプロモーターを開示しているU.S. Pat. No. 5,641,876、効果的な植物発現ベクターの設計に有用な5',3'およびイントロンエレメントを開示しているU.S. Patent Application Publication 2002/0192813A1、トウモロコシ葉緑体アルドラーゼプロモーターを開示しているU.S. patent application Ser. No. 09/757,089、イネグルテリンプロモーターを開示しているU.S. patent application Ser. No. 08/706,946、トウモロコシアルドラーゼ(FDA)プロモーターを開示しているU.S. patent application Ser. No. 09/757,089、トウモロコシニコチアナミンシンターゼプロモーターを開示しているU.S. patent application Ser. No. 60/310,370を含むが、これらに限定はされず、これら全ては引用により本明細書に組み込まれる。植物細胞内で機能する、これらの、また数多くの他のプロモーターは、所望の治療タンパク質の発現のために遺伝子組み換えアロエ植物内で機能し得る。
【0044】
具体的な一実施例として、トウモロコシ由来のユビキチンプロモーター(SEQ. ID. NO.44)が使用され得る。トウモロコシ由来のユビキチンプロモーターは大きなエレメントであり、長さが約2kbで、少なくとも三つの基本領域(general region)から成る。この配列は、特に関連性のある特徴にラベルを付けて図5に記載されている。ユビキチンプロモーターの第一区域は、最も5’末端側に位置しマトリックス結合領域(MAR)を含む。このMARは、ヒストンと他の核タンパク質と相互作用するエレメントで、フランキング配列をループアウト(loop out)させるように働き、それらがより容易に細胞の転写機構に接近できるようにする。また、それらはお互いに転写単位を隔離させるのを助けるが、これは、ある場所で開始された転写が、第二の配列に“読み過ごし(reading through)”されてしまうのを防ぐ上で重要である。これはアンチセンスメッセージを作り出すリスクを減らす助けとなり得る。
【0045】
第二区域は、エンハンサーエレメントと実プロモーターを含む。エンハンサーエレメントは、転写機構pol II複合体をプロモーターに結合させて転写を開始させる役目を持つ、転写因子と結合する。“プロモーター”とは、コア転写開始機構と相互作用するのに必要な必須DNAエレメントを特に意味するが、プロモーターという用語は、より一般的には転写開始に関わるDNAエレメントの全てを包含するように用いられ、またこの場合は、“ユビキチンプロモーター”とは、5’から目的のクローニング遺伝子までの2kb領域の全て、もしくはそれらの変異物を意味するように大まかに用いられる。
【0046】
第三区域は、約1kbのイントロンを含む。イントロンは、スプライシングの過程を通して、最終翻訳メッセージであるmRNAから除去された転写遺伝子の領域である。イントロンは、代わりのエンハンサーエレメントを供給することによって、さらに、スプライシングに一部関連する過程である、核から細胞質へのRNAメッセージの転移の促進により、タンパク質翻訳を増加させることによって、直接的に遺伝子発現に影響を与えることもわかっている。しかしながら、遺伝子組み変え系にイントロンが存在しても、必ずしもRNAの発現や翻訳タンパク質のレベルの増加につながるとは限らない。或る態様では、ユビキチンイントロンは、ベクターの全体のサイズを縮小するため、また、アロエでの遺伝子組み換え遺伝子発現への影響を評価するために、修飾されてもよい。
【0047】
本発明の他の態様では、アロエ植物の緑色組織で選択発現することが望まれ得る。こうした利用のための目的プロモーターは、Arabidopsis thalianaリブロース-1,5-ビスリン酸カルボキシラーゼ(Rubisco)小サブユニット(Fischhoff et al. (1992) Plant Mol Biol. 20:81-93)、アルドラーゼおよびピルビン酸オルトリン酸ジキナーゼ(PPDK)(Taniguchi et al. (2000) Plant Cell Physiol. 41(1):42-48)などの遺伝子由来のものを含んでもよい。
【0048】
上述の通り、プロモーターは、遺伝子発現を高める一助となる、複数の“エンハンサー配列”を含むように改変されてもよい。そのようなエンハンサーは当該技術分野で周知である。エンハンサー配列をコンストラクトに含めることにより、選択されたタンパク質の発現が促進され得る。これらのエンハンサーは、しばしば真核細胞で機能するプロモーターの5’から転写開始点に見つかるが、しばしば上流(5’)もしくは下流(3’)からコード配列に挿入されてもよい。いくつかの場合には、これらの5’増強エレメントはイントロンである。特に有用なエンハンサーは、イネアクチン1の5’イントロン(U.S. Pat. No. 5,641,876を参照)およびイネアクチン2遺伝子、トウモロコシアルコールデヒドロゲナーゼ遺伝子イントロン、トウモロコシ熱ショックタンパク質70遺伝子イントロン(U.S. Pat. No. 5,593,874)およびトウモロコシshrunken 1遺伝子である。
【0049】
本発明の態様に従うコンストラクトは、典型的にポリアデニル化シグナルとポリアデニル化部位を含む3’エレメントを含んでもよい。よく知られている3’ エレメントは、例えばU.S. Pat. No. 6,090,627(引用により本明細書に組み込まれる)に開示されている、nos 3’、tml 3’、tmr 3’、tms 3’、ocs 3’、tr7 3’などのAgrobacterium tumefaciens遺伝子由来のもの、小麦(Triticum aesevitum)熱ショックタンパク質17(Hsp17 3’)、小麦ユビキチン遺伝子、小麦フルクトース-1,6-ビフォスファターゼ遺伝子、イネグルテリン遺伝子、イネ乳酸デヒドロゲナーゼ遺伝子、およびイネベータチューブリン遺伝子などの、植物遺伝子由来の3’エレメント(これらは全て、引用により本明細書に組み込まれるU.S. published patent application 2002/0192813 A1に開示されている)、ならびに、エンドウマメ(Pisum sativum)リブロースビフォスフェートカルボキシラーゼ遺伝子(rbs 3’)、および宿主植物内の遺伝子由来の3’エレメントを含む。
【0050】
目的タンパク質はDNAコンストラクトに組み込まれた構造遺伝子によってコードされ得る。構造遺伝子は、哺乳類の遺伝子であってもよいし、もしくは哺乳類の遺伝子の一部であってもよい。目的の構造遺伝子は、インターフェロン、免疫グロブリン、リンフォカイン、成長因子、ホルモン、血液因子、組織適合性抗原、酵素、もしくは他のタンパク質をコードしてもよい。インターフェロンアルファ2の配列は例示目的としてSEQ. ID. NO. 33と記載される。構造遺伝子は、クラゲ由来の緑色蛍光タンパク質(GFP)などのマーカータンパク質もコードしてもよい。構造遺伝子のDNA配列は、アロエ植物10での高レベルの発現が可能になるように修飾されてもよい。アロエ植物10のコドンバイアス(codon bias)は、構造遺伝子が分離される原種のコドンバイアスと異なっていてもよい。天然の構造遺伝子は、アロエ植物10でコードされたタンパク質の産出を最適化するように設計されてもよい。さらに、構造遺伝子のDNA配列は、タンパク質の構造に干渉しない適切なグリコシル化を、アロエ植物10にもたらすように設計されてもよい。
【0051】
転写と翻訳の終結は、アロエ植物10で機能できる種々の転写終結配列と翻訳終結配列によってもたらされ得る。一態様では、終結配列はAgrobacterium由来のノパリン合成酵素(NOS)遺伝子由来の配列を含んでもよい。他の態様では、終結配列は天然アロエ遺伝子とタンパク質の終結配列由来であってもよい。
【0052】
マーカー遺伝子は、しばしばDNAコンストラクトに組み込まれる。マーカー遺伝子は、目的の構造遺伝子を含む細胞が、マーカー遺伝子を含まない全細胞集団から選択されることを可能にする。マーカー遺伝子は、とりわけ、カナマイシン、クロラムフェニコール、G418およびゲンタマイシンなどに対する抵抗性を与える酵素もしくは他のタンパク質を含んでもよい。例えばAgrobacteriumがベクターとして用いられる場合、さらに他の特定のDNA配列が必要になることもある。もしDNAが特定の細胞型を標的とする場合は、他の領域が存在してもよい。
【0053】
上記の通り、DNAコンストラクトは分泌シグナルも含んでもよい。コンストラクトは、治療タンパク質を、そのタンパク質が形成された細胞から除去する標的とし得るようなシグナルペプチドをコードする転移配列も含んでもよい。植物で機能し、特にアロエ植物などの単子葉植物で機能する種々のシグナル配列が文献で同定されている。これらは、コンストラクトもしくは目的遺伝子の中に機能的に組み込まれてもよい。一態様では、イネ(Oryza sativa)由来のアルファアミラーゼ分泌配列(SEQ. ID. NO. 29)が用いられてもよい。このシグナル配列は、“The alpha-amylase genes in Oryza sativa”Mol Gen Genet., 1990 April;221(2):235-44で述べられており、この開示はその全容が引用により本明細書に組み込まれる。あるいは、例えばインターフェロンなどのいくつかの目的遺伝子は、インターフェロンを分泌の標的とする、天然もしくは合成のドメインを含んでもよい。分泌シグナルは、典型的にはタンパク質が細胞を離れる際に切断されるか、あるいはいくつかの場合には、実質的にタンパク質の機能に影響を与えることなく、タンパク質に保持されてもよい。分泌シグナルは、転移に分泌シグナルを必要とし得る種々のタンパク質に加えられてもよい。これらの分泌配列は種々の植物分泌配列に由来してもよい。
【0054】
図6Aから図6Bは、アロエ植物から目的タンパク質を産出するためのコンストラクトの例示的なセットをあらわす。プラスミドコンストラクションは、pzUI(TGC9)とラベルされる。TGC9はヒトインターフェロンアルファ2の発現を促進するユビキチンプロモーターの全長を含み、pzUEK(TGC12)(SEQ. ID. NO. 38)は、eGFP(強化緑色蛍光タンパク質)(SEQ. ID. NO. 31)およびカナマイシン抵抗性タンパク質を含む融合タンパク質の発現を促進するユビキチンプロモーターの全長を含む。融合タンパク質は、両方のタンパク質のスクリーニング特性(選択と視覚化)を一つに兼ね備える。
【0055】
図示したように、プラスミドpzI(TGC8)は、それぞれ二つの異なる制限酵素部位を含むPCRプライマーを用いたヒトインターフェロンアルファ2遺伝子のPCR増幅によって作製される(プライマーの配列は表1を参照)。増幅されたインターフェロン(IFN)遺伝子は、5’フランキング領域にPstIおよびBamHI部位を持ち、3’フランキング領域にSacIおよびXhoI部位を持つ。PCRで作製したIFNはその後、コンストラクトpzI(TGC8)を作製し、次のクローニングのためのBamHIおよびSacI部位を導入するために、PstI/XhoIフラグメントとしてpZErOベクターにクローニングされる。このコンストラクトは、IFNアルファ遺伝子の全長と、細胞からのIFNタンパク質の分泌の誘導に関与する自身のシグナル配列を含む。これはスライド3に描かれている。これらのPCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0056】
図示したように、プラスミドベクターpzUI(TGC9)は、全長ユビキチンプロモーターの下流にIFNアルファ2遺伝子をクローニングすることによって作製される。IFNのPstI/XhoIフラグメントはpzI(TGC8)から切り離され、コンストラクトpzUI(TGC9)を作製するためにpzU(TGC1)にライゲーションされる。これはスライド3に描かれている。
【0057】
図示したように、BamHI(5’末端)およびXhoI(3’末端)制限部位を含むプライマーのセットを用いてeGFP遺伝子がPCR増幅される(プライマー配列は表1を参照)。このPCR作製eGFP遺伝子は停止コドンを含まずに作製される(最終的にeGFP-kan融合タンパク質の発現を可能にするため)。従って翻訳はeGFP配列の末端では終結しない。PCR増幅フラグメントは、制限酵素BamHIとXhoIで消化され、pZErOのBamHIおよびXhoI部位にライゲーションされ、pzEnostop(TGC10)を作製する。これはスライド3に描かれている。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0058】
図示したように、pzEK(TGC11)はeGFP遺伝子とカナマイシン抵抗性遺伝子の間で融合コンストラクトを作製する。eGFPはBamHI/XhoIフラグメントとしてpzEnostop(TGC10)から切り離され、コンストラクトpzEK(TGC11)を作製するためにpzK(TGC4)にライゲーションされる。eGFP遺伝子はイン・フレームに(in frame)、5’からkan/nosにクローニングされる(SEQ. ID. NO. 35)。
【0059】
図示したように、eGFP/kan/nosカセットを含むpzEK(TGC11)由来のBamHI/XbaIフラグメントは、eGFP/kan融合遺伝子の発現を可能にするために、pzUI(TGC9)のBamHIおよびXbaI部位にクローニングされる。このようにしてIFN配列は除去される。この新しいコンストラクトであるpzUEK(TGC12)は、全長ユビキチンプロモーターからeGFP/kan融合タンパク質を発現する(スライド4に描かれている)。この融合タンパク質は、個々のタンパク質それぞれの特徴を保持し、従って、視覚マーカーと選択剤の両方の機能を一つに兼ね備えるという明白な利点を有する。
【0060】
図7Aから図7Cは、アロエ植物から目的タンパク質を産出するためのコンストラクトの別の例示的なセットをあらわす。図示されたベクターは、二つの遺伝子が一つの転写産物として発現することを可能にするが、この二つの遺伝子はそれでもなお二つの個別のタンパク質に翻訳される。これを実現するために、IRESエレメント(内部リボソーム侵入配列)(SEQ. ID. NO. 34)を介在させることによって、二つの遺伝子は相互に分離される。IRESエレメントは、リボソームの結合と翻訳のための、第二の内部部位をもたらす。この第二部位は、3’側からIRESエレメントにクローニングされた転写産物が別々に翻訳されることを可能にする。これらのベクター、すなわちpzUSEIrK(TGC18)(SEQ. ID. NO. 43)およびpzUIIrK(TGC19)(SEQ. ID. NO. 39)は段階的に構築され、最終的に、カナマイシン抵抗性マーカーと共にeGFPもしくはIFN(それぞれ)の発現を可能にする。
【0061】
図示したように、pzEnostart(TGC13)ベクターは、制限酵素部位BamHI(5’末端)およびSacI(3’末端)を含むプライマーを用いるPCR(プライマー配列は表1)によって、eGFPを最初に増幅することによって作製される。このPCR産物は、BamHI/SacIフラグメントとしてpZErOにクローニングされる。このタンパク質はアルファアミラーゼシグナル配列との融合として発現されるので、このコンストラクトのeGFP遺伝子はATG翻訳開始部位を持たない(pzSE、TGC14の構築を参照)。これはスライド5に描かれている。これらのPCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0062】
図示したように、アルファアミラーゼ遺伝子から一本鎖のシグナル配列が合成され、その後PCRを用いる補充(fill in)によって二本鎖にされる(プライマー配列は表1)。このシグナル配列は、その後制限酵素PstIおよびBamHIで消化され、pzEnostart(TGC13)にクローニングされる。これはスライド5に描かれている。このシグナル配列はイン・フレームに、5’からeGFPにクローニングされ、シグナル配列とeGFPの融合に翻訳開始部位を提供する。得られるpzSE(TGC14)クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0063】
図示したように、ベクターpzUSE(TGC15)は、アルファアミラーゼシグナル配列(ss)-eGFP融合とユビキチンプロモーターを結合させるために構築される。pzSE(TGC14)由来のss-eGFPを含むこの遺伝子カセットは、PstI/SacIフラグメントとしてpzUI(TGC9)にクローニングされる。これは、(制限酵素PstIおよびSacIで消化することによってpzUI(TGC9)から切り離される)IFN遺伝子カセットを置換し、pzUSE(TGC15)を作製する。これはスライド5に描かれている。
【0064】
図示したように、IRESエレメントは、PCRによってpIRES2-EGFP(Clontech, BD Biosciences)から制限酵素部位SacI(5’末端)およびXhoI(3’末端)を含むプライマーで増幅される(プライマー配列は表1)。増幅されたIRESエレメントは、pzIr(TGC16)を作製するためにSacI/XhoIフラグメントとしてpZErOにクローニングされる。これはスライド6に描かれている。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0065】
図示したように、pzIr(TGC16)由来のIRESエレメントは、その後pzIrK(TGC17)を作製するためにSacI/XhoIフラグメントとしてpzK(TGC4)にクローニングされる。このようにしてIRESをkan/nos遺伝子カセットの上流に配置する。
【0066】
図示したように、IRES-kan/nos遺伝子カセットは、pzUSEIrK(TGC18)を作製するために、SacI/XbaIフラグメントとしてpzUSE(TGC15)にクローニングされる。これはスライド6に描かれている。このコンストラクトは、アルファアミラーゼシグナル配列との融合としてeGFPを発現し、eGFPを細胞からの分泌の標的とする。これにより、遺伝子組み換え植物内でのタンパク質の輸送と蓄積を視覚的に観察することが可能になる。このベクターは、カナマイシン抵抗性タンパク質(内部IRESエレメントから翻訳される)も発現し、遺伝子組み換え植物の選択を可能にする。
【0067】
図示したように、また、IRES-kan/nos遺伝子カセットは、pzUIIrK(TGC19)を作製するために、SacI/XbaIフラグメントとしてpzUI(TGC9)にクローニングされる。これはスライド7に描かれている。このコンストラクトは、分泌のためのシグナル配列と共にIFNアルファ2を発現する。このベクターは、カナマイシン抵抗性タンパク質(内部IRESエレメントから翻訳される)も発現し、遺伝子組み換え植物の選択を可能にする。TGC18とTGC19の両方とも、二つの遺伝子を一つの転写産物として発現し、第二プロモーターの必要性がなく、従って各ベクターの全体のサイズを縮小する。導入コンストラクトの全体のサイズを縮小することは、これらのエレメントが核に転移できる効率を増加し、安定的な組み込みと遺伝子組み換え植物の選択につながるので、このことは重要である。単一のプロモーター系は、かく乱フランキング遺伝子の発現のリスクや、最終的に導入遺伝子の発現に影響し得る他の状況も軽減する。
【0068】
上記のプラスミドは、他の目的コード配列をアルファインターフェロンと置換してもよい。当業者に理解されるように、これらはプラスミドの同じ遺伝子座もしくは他の位置に置換されてもよい。
【0069】
図8に図式的にあらわされるように、プロトロンビンコード配列(SEQ. ID. NO. 36)も、本発明の態様に従ってプラスミドベクターに組み込まれてもよい。プロトロンビンはトロンビンの前駆体タンパク質である。これは、可溶性フィブリノゲンを不溶性フィブリンストランドに変換する凝固タンパク質である、活性化トロンビンを切り離すために活性化因子Xによって2箇所で切断される。プロトロンビン遺伝子カセット(1874塩基対)はPCRプライマーを用いて増幅される(プライマー配列は表1)。eGFP遺伝子カセットは、制限酵素BamHIによってpzUSEIrK(TGC18)ベクターから切り離される。得られる5’突出は、その後平滑末端を作製するためにMung Beanヌクレアーゼで除去される。この平滑末端は、セ氏15度でプロトロンビン遺伝子カセットと一晩ライゲーションする前に、セ氏50度で2時間インキュベートすることによって仔牛小腸ホスファターゼ(CIP)を用いて脱リン酸化される。こうしてpzUSTIrK(TGC20)(SEQ, ID. NO. 42)が作製される。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0070】
また、図8に図式的にあらわされるように、ダームシジン(DCD)コード配列(SEQ. ID. NO. 30)も、本発明の態様に従ってプラスミドベクターに組み込まれてもよい。ダームシジンは、近年報告された広域抗菌性ペプチドで、汗腺に構成的に発現することがわかっている。このタンパク質は汗の中に分泌され、表皮表面に輸送される。PCR増幅DCD遺伝子カセットは、フランキングBamHI制限酵素部位を持つ(プライマー配列は表1)。BamHI処理したDCD遺伝子カセットは、eGFPによって占められていたpzUSEIrK(TGC18)ベクターのBamHI部位にライゲーションされ、pzUSDIrK(TGC21)ベクター(SEQ. ID. NO. 40)を作製する。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0071】
また、図8に図式的にあらわされるように、ヒト成長ホルモン(hGH)コード配列(SEQ. ID. NO. 32)も、本発明の態様に従ってプラスミドベクターに組み込まれてもよい。hGH遺伝子カセットは、フランキングBamHI制限酵素部位とともにPCR増幅によって作製される(プライマー配列は表1)。その後BamHI消化したhGH遺伝子カセットは、eGFPによって占められていたpzUSEIrK(TGC18)のBamHI部位にライゲーションされ、pzUSHIrK(TGC22)ベクター(SEQ. ID. NO. 41)を作製する。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0072】
また、図8に図式的にあらわされるように、ヒトインターフェロンガンマ(hIFNg)コード配列も、本発明の態様に従ってプラスミドベクターに組み込まれてもよい。hIFNg遺伝子カセットは、フランキングBamHI制限酵素部位とともにPCR増幅によって作製される(プライマー配列は表1)。その後BamHI消化したhIFNg遺伝子カセットは、eGFPによって占められていたpzUSEIrK(TGC18)のBamHI部位にライゲーションされ、pzUSIfgIrK(TGC23)ベクター(SEQ. ID. NO. 37)を作製する。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0073】
【表1】

【0074】
これらの配列はその独自のシグナル配列を含んでもよい。アロエ植物で機能することが示されたアルファインターフェロンのシグナル配列と同様に、プロトロンビン、ダームシジン(DCD)、ヒト成長ホルモン(hGH)、およびヒトインターフェロンガンマ(hIFNg)の天然シグナル配列も、アロエ植物で機能することが示され得る。とにかく、これらのタンパク質コード配列は、図示された図8の実施例のようにアルファアミラーゼシグナル配列を含むベクターにクローニングされ得る。
【0075】
上記のコンストラクトの作製において、形質転換に用いられる全ベクターは、250ml LB50μg/Lゼオシンで培養され、CsCl法を用いて分離される。その後ベクターは、各コンストラクトからの適切な発現を確実にするために、一過性トランスフェクション実験に用いられる。遺伝子銃を用いるトランスフェクションは、トウモロコシ、タバコ、およびアロエで実施され、eGFPの発現を視覚的に観察するか、あるいはIFNもしくはkanのいずれかの発現に対するrt-PCRによって、発現を観察した。
【0076】
<形質転換>
ベクターに組み込まれた、目的遺伝子と機能的な種々の機能単位を含むDNAコンストラクトを作製した後、DNAコンストラクトは、本発明の開示を考察することで当業者に理解される数多くの技術を用いて、アロエ細胞に導入される。これらのDNAコンストラクトは、概して安定的に形質転換されたアロエ植物10の形成を促進するために設計される。組み換えDNAで植物細胞を形質転換する数多くの方法が当該技術分野で周知であり、本発明において使用され得る。安定的なアロエ形質転換体を作製するために、ベクターに含まれるDNAコンストラクトをアロエ細胞もしくは組織に組み込むいくつかの方法は、A. tumefaciensもしくはA. rhizogenesの感染、複製欠損性ウィルスの感染、バイオリスティック形質転換、プロトプラスト形質転換、花粉への遺伝子導入、生殖器官への導入、未熟胚への導入、もしくは同様の方法を含むことができる。現在、よく用いられている植物形質転換方法のうちの二つは、Agrobacterium媒介形質転換とバイオリスティック形質転換である。
形質転換アロエ細胞もしくはカルス組織は、形質転換細胞を選択するために、典型的には適切な栄養培地で培養される。しばしば、選択培地は、非形質転換細胞を殺す毒素もしくは他の選択因子を含む。本発明の開示を考察することで当業者に理解されるように、様々な培地の交換により、目的遺伝子を含むアロエ植物10の産出が可能になる。
【0077】
形質転換の実践においては、典型的には、任意の一つの形質転換実験において、わずかな比率の標的植物細胞にのみDNAが導入される。マーカー遺伝子は、遺伝子組み換えDNAコンストラクトをゲノムに受け取って組み込むことにより、安定的に形質転換された細胞を特定するための、効率的な系を提供するために用いられる。好ましいマーカー遺伝子は、抗生物質もしくは除草剤などの選択剤に対する抵抗性を与える選択マーカーをもたらす。本発明の植物が抵抗性を示し得る除草剤のいずれも、選択マーカーとして有効な薬剤である。形質転換された可能性のある細胞は選択剤に晒される。生存細胞の集団においては、概して、抵抗性付与遺伝子が組み込まれ、細胞が生存できる十分なレベルで発現している細胞が存在する。細胞はさらに、外来DNAの安定的な組み込みを確認するためにテストされ得る。よく用いられる選択マーカー遺伝子は、カナマイシンとパロモマイシン(nptII)、ハイグロマイシンB(aph IV)およびゲンタマイシン(aac3およびaacC4)などの抗生物質に対する抵抗性を与えるもの、あるいは、グルホシネート(bar orpat)とグリホセート(aroAもしくはEPSPS)などの除草剤に対する抵抗性を与えるものを含む。そのような選択可能マーカーの例は、U.S. Pat. No. 5,550,318、5,633,435、5,780,708および6,118,047に記載され、これらは全て引用により本明細書に組み込まれる。形質転換体を視覚的に特定する能力を与える選択可能マーカーも利用することができ、例えば、ルシフェラーゼもしくは緑色蛍光タンパク質(GFP)などの、有色タンパク質もしくは蛍光タンパク質を発現する遺伝子、あるいは、様々な発色基質が知られている、ベータグルクロニダーゼもしくはuidA遺伝子(GUS)を発現する遺伝子などである。
【0078】
一般的に、標的植物の系統のゲノムに、ランダムに、すなわち非特異的な位置に、組み換えDNAを導入することが有益である。特別な場合には、例えばゲノムの既存遺伝子を置換したり、植物ゲノムの既存プロモーターを使用したり、あるいは遺伝子発現に活性があるとわかっている所定部位に組み換えポリヌクレオチドを挿入するために、部位特異的組み込みを実現するために組み換えDNA挿入を標的とすることが有益となり得る。U.S. Pat.No. 4,959,317に開示されているcre-lox、およびU.S. Pat. No. 5,527,695に開示されているFLP-FRTを含む、埋め込み物として機能することが知られているいくつかの部位特異的な組み換え系が存在する(両特許は引用により本明細書に組み込まれ、下記でより詳細に述べられる)。
【0079】
Agrobacterium tumefaciensによる植物形質転換系として、形質転換コンストラクト上に存在する付加的なエレメントは、植物ゲノムへの組み換えポリヌクレオチドの組み込みを促進する、T-DNA左右境界配列を含む。agrobacterium(A. tumefaciens)感染では、アロエカルス組織は、DNAコンストラクトを組み込むベクターを含むagrobacteriumの一晩培養物と共に室温で1時間インキュベートされる。アグロバクテリウムは適切な選択培地で培養される。選択培地は典型的にはストレプトマイシンとカナマイシンを含む。選択培地はアセトシリンゴンも含んでもよい。50μMから250μMの濃度のアセトシリンゴンは、典型的には単子葉植物の伝染性の効率を増加させる。選択培地上での培養の後、感染アロエ組織はセ氏25度で暗所で二日間、選別されずにペトリ皿の培地に移される。適切な培地の例は、アセトシリンゴンを含むMS培地である。その後、agrobacteriumを殺すために二日間にわたってセフォタキシムが加えられ得る。その後細胞は、形質転換アロエ細胞を選択するための50 mg/Lのカナマイシンと、発芽誘導成長因子(0.2 mg/L NAA、2 mg/L BAP)を含む培地上に移され、16時間の光照射で4週間にわたって培養される。その後再生芽は、土壌に移される前に根を発達させるために(約4〜6週間後)、発根培地(1/2 MS with 0.2 mg/L NAA)に移され得る。
【0080】
バイオリスティック形質転換では、アロエ細胞もしくはカルス組織は、ベクターコンストラクトを直接的に打ち込まれる。バイオリスティック形質転換の様々な実施形態と態様は、U.S. Pat. No. 5,015,580(大豆)、5,550,318(トウモロコシ)、5,538,880(トウモロコシ)、5,914,451(大豆)、6,160,208(トウモロコシ)、6,399,861(トウモロコシ)、および6,153,812(小麦)に開示され、Agrobacterium媒介形質転換はU.S. Pat. No. 5,159,135(綿)、5,824,877(大豆)、5,591,616(トウモロコシ)、および6,384,301(大豆)に開示され、これら全ては引用により本明細書に組み込まれる。さらに、例えばBio-Rad, Inc.のBiolistic PDS-1000/He particle delivery systemなど、様々なバイオリスティック形質転換装置が使用され得る。この方法では、ベクターと関連DNAコンストラクトは、金もしくはその他の粒子に結合され、アロエ細胞に直接打ち込まれる。打ち込みされたアロエ細胞は、選択培地に移される前に、暗所で4日間選択されずに培養される。この場合も、選択培地は50 mg/Lのカナマイシンを含んでもよい。形質転換体は、典型的には8〜12週間の内に形成し始め、発芽・発根培地にさらに8〜12週間移すことができる。一旦根が形成し始めれば、小植物体は土壌に移すことができる。
【0081】
形質転換は、他の様々な技術を用いて実現することもできる。そのような技術は、複製欠陥ウィルスベクター系を用いるウィルス感染、特に液体培養で培養したカルス細胞もしくはプロトプラストの電気穿孔法、および、プロトプラストへのPEGもしくは脂質媒介トランスファーを含む。その後、形質転換体の選択は、バイオリスティック形質転換で概説したものと同じ基本ステップに従うことができる。
【0082】
一旦アロエカルス組織に導入されれば、図3に大まかに示されるように、コンストラクトは植物の遺伝物質に組み込まれ得る。他の態様では、コンストラクトはアロエ植物10の遺伝物質の外側の細胞に安定的に組み込まれ得る。コンストラクトによって、目的タンパク質はいくつかの形式の転写誘導を伴わずに発現してもよいし、しなくてもよい。一態様では、一旦アロエ細胞に導入されたコンストラクトは、タンパク質合成もしくはアロエ植物10の特定の組織への輸送を誘導し得る。典型的には、これは標的シグナル配列が目的タンパク質上に含まれる際に起こる。
【0083】
<部位特異的組み込み(Cre-Lox)>
一態様では、本発明はアロエ植物に導入されたDNAコンストラクトの部位特異的組み込みもしくは切除を利用してもよい。部位特異的組み込みもしくは切除の利点は、形質転換コンストラクトが通常は宿主ゲノムに複数コピーでランダムに組み込まれるという、従来の形質転換技術に伴う問題を克服するために使用できるということである。この導入DNAの宿主細胞ゲノムへのランダム挿入は、もし外来DNAが必須遺伝子に挿入される場合、致命的になることがある。加えて、導入遺伝子の発現は、周囲のゲノムDNAに起因する“位置効果”によって影響され得る。さらに、遺伝子抑制、組み換え、および予測不可能な遺伝を含む、植物が保有する導入遺伝子の複製コピーに関連する問題のために、一般的には遺伝子組み換えアロエ植物10のゲノムに挿入されたDNAコンストラクトのコピー数を制御することが好ましく、多くの場合単一コピーのDNAコンストラクトの挿入のみが望ましい。
【0084】
導入遺伝子もしくは導入遺伝子の一部の、部位特異的組み込みもしくは切除は、相同組み換えを用いて植物で実現できる(例えば、特にその全容が引用により本明細書に組み込まれる、U.S. Pat. No. 5,527,695を参照)。相同組み換えは、相似ヌクレオチド配列を持つDNA配列の任意の対の間の反応であり、ここで二つの配列は新たな組み換えDNA種を形成するために相互作用(組み換え)する。相同組み換えの頻度は、共通ヌクレオチドDNA配列の長さが増えるにつれて増加し、また環状プラスミド分子よりも線状化プラスミド分子の方が高頻度で起こる。相同組み換えは、完全に同一とは言えない二つのDNA配列間でも起こり得るが、組み換え頻度はニ配列間の相違が増すにつれて減少する。
【0085】
導入されるDNA配列は、目的DNA分子を、宿主細胞の内在性配列と相同性を有する配列に結合することで、相同組み換えによって標的となることができる。一旦DNAがアロエ細胞に入れば、導入されるDNAを相同ゲノムDNA配列が位置する部位に挿入するために、二つの相同配列が相互作用することができる。従って、導入されるDNA上に含まれる相同配列の選択が、導入されるDNAが相同組み換えによって組み込まれる場所を決定する。例えば、もし目的DNA配列が宿主アロエ細胞の単一コピー遺伝子と相同性を持つDNA配列に結合される場合、目的DNA配列はその単一の特定部位のみに相同組み換えによって挿入される。しかしながら、もし目的DNA配列が宿主真核細胞のマルチコピー遺伝子と相同性を持つDNA配列に結合される場合、目的DNA配列は、遺伝子のコピーが位置する特定部位のそれぞれに、相同組み換えによって挿入され得る。
【0086】
DNAは、単一相互組み換え(導入DNAの全長の挿入をもたらす)もしくは二重相互組み換え(二つの組み換え現象間に位置するDNAのみの挿入をもたらす)のいずれかを含む相同組み換え反応によって、宿主ゲノムに挿入され得る。例えば、もし選択された遺伝子が位置する遺伝子位置に外来遺伝子を挿入したい場合、導入されるDNAは選択された遺伝子と相同性を持つ配列を含んでいなければならない。単一相同組み換え現象では、その後、導入されるDNA配列全体が選択された遺伝子に挿入されることになる。もう一つの方法として、二重組み換え現象は、目的DNA配列(ゲノムに挿入される配列)の各末端を、選択された遺伝子と相同性を持つDNA配列とフランキングすることによって実現できる。各相同フランキング領域を含む相同組み換え現象は、外来DNAの挿入をもたらす。従って、ゲノム相同性を持つ二つの領域間に位置するDNA配列のみが、ゲノムに組み込まれることになる。
【0087】
導入されるDNA配列は、相同組み換えによって特定のゲノム位置への挿入の標的となることができるが、高等真核生物では、相同組み換えはランダム挿入現象に比べて比較的まれな現象である。アロエ細胞では、外来DNA分子はアロエ植物ゲノムで相同配列を見つけ、約0.5〜4.2 x 10-4の頻度で組み換えを起こす。従って、相同組み換えによって組み込まれた導入DNA配列を含む任意の形質転換細胞も、ランダムに組み込まれた導入DNA配列の多数コピーを含む可能性がある。これらのランダム挿入を防ぐ一つの方法は、部位特異的リコンビナーゼ系を利用することである。概して、部位特異的リコンビナーゼ系は三つの要素から構成される。すなわち、二対のDNA配列(部位特異的組み換え配列)と特異的酵素(部位特異的リコンビナーゼ)である。部位特異的リコンビナーゼは、二つの部位特異的組み換え配列間でのみ組み換え反応を触媒する。
【0088】
バクテリオファージP1のCre/lox系(特にその全容が引用により本明細書に組み込まれるU.S. Pat. No. 5,658,772)、酵母のFLP/FRT系(Golic and Lindquist, 1989)、ファージMuのGinリコンビナーゼ(Maeser and Kahmann, 1991)、E. coliのPinリコンビナーゼ(Enomoto et al., 1983)、およびpSR1プラスミドのR/RS系(Araki et al., 1992)を含むが、これに限定されない数々の異なる部位特異的リコンビナーゼ系を本発明に従って利用できる。バクテリオファージP1 Cre/lox系および酵母FLP/FRT系は、導入遺伝子の部位特異的組み込みもしくは切除に特に有用な二つの系から構成される。これらの系では、リコンビナーゼ(CreもしくはFLP)は、介在配列を反転または切除するために、それぞれの部位特異的組み換え配列(それぞれloxもしくはFRT)と特異的に相互作用する。これら二つの系のそれぞれの配列は比較的短く(loxは34 bp、FRTは47 bp)、従って形質転換ベクターと一緒に用いるのに都合がよい。
【0089】
図8はFlp/FrtおよびCre/Loxpリコンビナーゼ系を用いる特定のプロトコル例をあらわす。こうした系では、バクテリオファージと酵母由来の部位特異的リコンビナーゼが、試験管と生体の両方でDNAを扱うツールとして使用され得る。リコンビナーゼ/組み換え部位の組み合わせは、Cre-Lox、FLP-FRT、およびR-RSを含み、ここでCre、FLP、およびRはリコンビナーゼで、Lox、FRT、およびRSは組み換え部位である。この系を用いるために、組み換えに特異的な部位を含む遺伝子組み換え植物が作製される。親となる遺伝子組み換えアロエ植物10は、選択可能マーカーを発現する、FrtおよびLox部位をタンデムに組み込んだベクターを形質転換することによって作製される。FrtおよびLox部位の両方とも、三つの要素から構成され、それぞれ13ヌクレオチドからなる2つの逆方向反復間に、8ヌクレオチドからなるスペーサー配列が存在する。逆方向反復は特定のリコンビナーゼに対するDNA結合ドメインとしてはたらき、一方スペーサーエレメントは、変更できるが相同組み換えに必須である。LoxPもしくはFRT部位のいずれかのスペーサーエレメントを改変することにより、複数の別個の組み換え部位を作ることができる。FrtおよびLox部位を改変することにより、多重部位に誘導される組み込みを可能にする系を作ることができる(図8に概説する通り)。この系は数多くの利点を持ち得る。最初の親株の世代の後、内在性遺伝子の破壊が最小限に抑えられる。形質転換効率は、部位特異的リコンビナーゼの発現によって増加する。loxおよびfrt部位をタンデムに用いることで、目的遺伝子を保持しながら選択マーカーの選択的除去が可能になる。また、一旦親株が作られれば、細胞増殖と再生能は増強される。しかしながら親株は第一段階であり、標準的な形質転換法(バイオリスティックもしくはAgrobacterium)に従って作製される。後代の遺伝子組み換え植物は、LoxPおよびFrt部位の両方、さらに目的遺伝子と選択可能マーカーを含むベクターを、Creリコンビナーゼを発現するベクターと一緒に共形質転換することによって作製される。Creリコンビナーゼの発現は、ベクターと親株のLoxP部位を通して相同組み換えを誘導する。形質転換体は選択可能マーカーの発現によって選択され、再生するように誘導される。その後のFlpリコンビナーゼ発現ベクターとの形質転換は、選択可能マーカーを除去し、次の他のLoxp部位への組み込みを可能にすることができる。
【0090】
<生存株の産出>
アロエ細胞もしくはアロエ組織の形質転換に続いて、形質転換アロエ細胞もしくはアロエ組織は、小植物体に成長され得る。選択剤に晒されても生き残ったアロエ細胞、もしくはスクリーニングアッセイで陽性となったアロエ細胞は、再生培地で培養され得、遺伝子組み換えアロエ植物10に成長させられ得る。形質転換アロエ細胞から再生した成長中のアロエ小植物体は、植物成長ミックスに移され、温室もしくは成熟のための栽培室に移す前に、例えば約85%の相対湿度、600 ppmのCO2、25〜250マイクロアインシュタインm-2s-1の光で環境制御されたチャンバーで、寒さに慣れさせることができる。形質転換アロエ植物10は、形質転換体が特定された後、最初の組織に応じて約6週間から10ヶ月で再生される。再生された形質転換アロエ植物10もしくはその子孫となる種子もしくは植物体は、通常は組み換えDNAの発現を調べられる。
【0091】
形質転換未分化カルス組織もしくは分裂組織から遺伝子組み換えアロエ植物10を再生することは、主として成長因子オーキシンとキネチンを操作することを含む。第一段階は芽の成長促進である。発芽誘導培地は、典型的には低濃度のオーキシン(0.2 mg/L NAA)と比較的高濃度のサイトカイニン(2 mg/L BAP)を含む。2%スクロースにまで減らした糖濃度(浸透圧に影響する)も、植物再生に寄与し得る。発芽培地上の細胞は、12hr/dayの光周期で25℃のインキュベーターに置かれる。形成し始めた芽は、約2 cmの長さに達した後、初期カルスから切除され、発根培地に置かれる。これらの芽は、さらに芽の発達を促進するために発芽培地に保持することもできる。発根培地は根の発達を促進し、典型的には1/2 MS培地と比較的低濃度のオーキシン(0.2 mg/L NAA)を含む。根が成長し始めた後(1〜2 cm)、小植物体は土壌に移され、徐々に相対湿度を下げることで適応させる。一旦生成されれば、遺伝子組み換えアロエ植物10は、種子、クローン増殖法、もしくは本発明の開示を考察することで当業者に理解される別の方法で、繁殖され得る。
【0092】
<植物もしくは植物細胞からのタンパク質の抽出と精製>
ベクターを介して植物細胞と植物に導入された構造タンパク質もしくは酵素は、根、塊茎、葉、種子、花、樹液を含むアロエ植物10の様々な組織において、あるいは植物の部分と発育段階の組み合わせにおいて見られる。一態様では、一つもしくは複数の目的タンパク質は、葉の中心部のゲルマトリックスに濃縮される。別の態様では、一つもしくは複数の目的タンパク質は生物活性があり、アロエ葉12から抽出されてある程度の薬効を提供する。別の態様では、一つもしくは複数の目的タンパク質、および少なくとも一つの天然アロエタンパク質、糖質および/またはゲルマトリックスに見られるその他の化合物は、患者に所望の治療を施すように相乗的に作用する。タンパク質はバイオマス全体もしくは特定の組織から抽出できる。植物細胞からのタンパク質抽出は、物理的方法と化学的方法を含んでもよい。葉もしくは樹液からのタンパク質抽出は、ろ過、超遠心分離法、化学抽出、およびアフィニティークロマトグラフィーを含んでもよい。
【0093】
一態様では、目的タンパク質は形質転換アロエ植物10の葉のゲル内に蓄積し得る。この領域は主に水であり(約99%)、有害なプロテアーゼを含まない。ゲルの抽出は、当業者に理解される従来技術である。タンパク質のいくつか、特に化粧品タンパク質は、アロエゲルを直接補うために用いられてもよく、またゲルから個別に分離される必要が全くなくてもよい。
【実施例】
【0094】
<植物細胞の分離方法>
第一の実施例では、直接使用のため、あるいはカルス発生のための芽分裂組織が、アロエ(Aloe vera、Aloe ferox、もしくはAloe arborescenceのいずれか)の茎から分離される。葉の部分は除去され、茎はアロエ葉の縦軸を通って横方向に切られる。組織はTween 20(0.05%)と次亜塩素酸塩(5%)の混合物で10分間滅菌され、その後30秒エタノールで処理し、滅菌水で三回洗浄した。切片はその後、様々な濃度の成長因子(オーキシンとサイトカイニン)と2〜3%のスクロースを含むアガー(0.8〜1%)を含むMS培地の入ったプレートに、露出面を横倒しにして置かれる。生育条件は所望の処理過程に応じて異なる。芽の発達を伴わないカルス培養は、典型的にはこれらの切片を、BAPを含まないNAA(2〜5 mg/L)か、もしくは低濃度のBAP(0.2 mg/L)を含むNAA(2〜5 mg/L)で生育させることによって開始される。未分化細胞は切られた茎の範囲に沿って成長し始め、別の皿上で継代培養されて保持され得る。発芽誘導は、こうした切片を発芽培地(0.2 mg/L NAA(もしくはIAA 0.2 mg/L)と2 mg/L BAPを含むMS培地)に置くことによって直接的に開始させることができる。NAAとIAAは両方ともオーキシンの供給源としてはたらき、また、効果のあるオーキシンとサイトカイニン両方の濃度の範囲が存在する。未熟胚由来のカルス細胞は、市販の種子から育成される。種子は最初に滅菌される(エタノールで30秒、5%次亜塩素酸塩と0.05% Tween 20で15分間、ならびに滅菌水で3回洗浄)。滅菌された種子は、その後セ氏4度で水中に一晩放置される。種皮を除去することによって未熟胚が分離される。三角形の種子の一角に小さな切れ目を作り、胚を搾り出す。その後これは、カルスを誘導するためにオーキシンのみ(NAA 2〜5 mg/L)を含むMS培地か、あるいはカルスと芽の増殖を誘導するためにオーキシンとサイトカイニン(NAA 0.2 mg/L、BAP 2 mg/L)を含むMS培地上に置かれる。糖濃度は通常2〜3%である。
【0095】
茎頂分裂組織由来のカルス細胞は、葉を全て除去した若いアロエ植物10の基部から切り取った1〜2cmの切片から育成される。切片は表面を滅菌し、MS培地とオーキシンとサイトカイニン(2 mg/L NAA 0.2 mg/L BAP)を含む1%アガーに移される。
【0096】
カルス細胞誘導は1〜2週間以内で開始する。細胞はその後これらの培養から分離され、形質転換の出発物質として使用することができる。
【0097】
[DNAコンストラクト]
1.pBI121ユビキチンインターフェロンベクター(pBI-UI)
pBI-UIの作製の骨格として、pBI121 Agrobacteriumバイナリベクターが使用される。シグナル配列を持つヒトインターフェロンアルファ2遺伝子は、5’PstIおよび3’SacI/XhoI制限部位を含む遺伝子特異プライマーを用いてヒト293細胞から増幅される。587 bpのインターフェロンPCR産物はpZErOにクローニングされ、シーケンスにかけられる。トウモロコシ由来の1962 bpユビキチン(Ubi)プロモーターエレメントも、ベクターpUBI-GFP由来の領域を増幅し、5’HindIIIおよび3’PstI制限部位にクローニングすることによって、pZErOにクローニングされる。Ubiフラグメントは、その後pZeroインターフェロンベクターにHindIII/PstI(pZErO UI)を用いてサブクローニングされる。無傷のUbiインターフェロンカセットは、その後HindIII/SacIを用いてpBI121にサブクローニングされ、CaMV 35SプロモーターとGUS遺伝子を除去して、pBI UIを得る。このベクターは、agrobacterium媒介感染および形質転換に必要な左右のTエレメント境界領域を保持し、Nosプロモーターの制御下にカナマイシン抵抗性遺伝子(NPTII)を発現する。
【0098】
2.pZErOユビキチンインターフェロンIRESカナマイシン(pZ UIIK)
このベクターは、UbiプロモーターをpZErOクローニングベクターにクローニングすることによって作製される。このフラグメントの後ろに、IRES(内部リボソーム侵入配列)とカナマイシン抵抗性遺伝子と一緒に、ヒトインターフェロンアルファ2を含むカセットがクローニングされる。この単位は単一の転写産物として発現されるが、二つの別個のタンパク質として翻訳される(IRESによってもたらされる第二転写開始部位のため)。これにより、選択可能マーカーとインターフェロン両方がUbiプロモーターの制御下で発現することが可能になる。
【0099】
3.CreおよびFlpリコンビナーゼを利用する二重プラスミド系
標的組み換えのためのcreおよびflp部位を持つ選択可能マーカーを発現する親株が最初に作製される。目的遺伝子を持つ第二トランスフェクション導入ベクターがcre部位に向けられる。その後不要な遺伝物質がflpを用いて除去される。
【0100】
[分離植物細胞へのDNAコンストラクトの導入]
1.Agrobacterium媒介遺伝子導入
Agrobacterium strain LB4404(Invitrogen, Carlsbad, CA)は、pBI UIベクターと、50μg/mlカナマイシンと100μg/mlストレプトマイシンを含むLBアガープレート上で選択された陽性クローンと共に電気穿孔された。個々のクローンは250μMアセトシリンゴンを含むLB培地でセ氏30度で一晩培養された。感染は、植物切片もしくはカルスを一晩培養に沈水させ、滅菌ろ紙上にブロットドライ(blotting dry)し、暗所でセ氏25度で250μMアセトシリンゴンを含むMSアガープレート上に播種することで行われた。感染から二日後、Agrobacteriumを殺すために、組織は200μMセフォタキシムを含むMSアガープレートに移された。その後、形質転換体を選択し芽再生を誘導するために、組織は50 mg/Lカナマイシンと0.2 mg/L NAAと2 mg/L BAPを含むMSアガープレートに移される。組織は12時間の光照射でセ氏25度で育成される。不定芽は約2 cmの長さに達するまで育成され、その後切除され、選択を続けながら発根培地上に植え替えられる。目的遺伝子を発現する小植物体は、発現レベルを分析し、土壌に移される。
【0101】
2.バイオリスティック遺伝子導入
金粒子(0.6〜1μm)がDNAコンストラクトを含むベクターで被覆される。金粒子は、浸漬させてボルテックスにかけることで70%エタノールで15 分間洗浄され、その後滅菌水で3回洗浄される。その後、金粒子は60 mg/mlの濃度まで50%グリセロールに再懸濁される。洗浄された粒子上にDNAベクターを被覆させるために、3 mgの粒子が1.5 mlマイクロフュージチューブに加えられる。これに5μlのDNAを1μg/μlの濃度で加え、50μlの2.5M CaCl2、および20μlの0.1Mスペルミジンを加え、2〜3分間ボルテックスする。これを安定させ、1〜2秒間遠心し、液体を除去する。これに140μlの70%エタノールを加え、遠心し、液体を捨てる。これに140μlの100%エタノールを加え、遠心して液体を捨てる。沈殿物に48μlの100%エタノールを加え、穏やかに再懸濁させる。遺伝子銃装置(PDS 1000/He, Bio-Rad Laboratories, Inc., Hercules, CA)は70%エタノールを用いて滅菌される。遺伝子銃装置は、マクロキャリアホルダーと停止スクリーン間の最短ギャップ距離を用いて組み立てられた(推奨設定)。被覆金粒子(8μl)はマクロキャリアの中心上にピペットされる。600〜1100 psiの破裂板を用いて、標的距離は標的組織に応じて設定された(組織は6 cm、カルスは9 cm)。
【0102】
第一の実施例では、打ち込み後に細胞は2 mg/L NAAを含むMSアガープレート上に、選択されずにセ氏25度で暗所に一週間置かれた。その後細胞は、選択のための50 mg/Lカナマイシンと、オーキシン(2 mg/L NAA)およびサイトカイニン(0.2 mg/L, 6-ベンジルアミノプリン)を含むMSアガープレートに4〜5週間以上移された。細胞はセ氏25度で12時間の光照射で培養される。
【0103】
第二の実施例では、マイクロプロジェクタイル打ち込み(microprojectile bombardment)が、MSプレート(4 g/L MS塩、1 mg/L (1000X)MS ビタミンストック、2 mg/L NAA、100 mg/L ミオイノシトール、2.76 g/L プロリン、30 g/L スクロース、100 mg/L カゼイン加水分解物、36.4 g/L ソルビトール、36.4 g/L マンニトール、2.5 g/L ゲルライト、pH 5.8)上で行われた。打ち込みされたカルスは、打ち込み後1時間にわたってMSOSM上に残され、その後MS初期培地(4 g/L MS塩、1 mg/L (1000X)MS ビタミンストック、2 mg/L NAA、100 mg/L ミオイノシトール、2 g/L プロリン、30 g/L スクロース、100 mg/L カゼイン加水分解物、2.5 g/L ゲルライト、pH 5.8)に移された。MS上で7〜10日後、打ち込みされたカルスは、形質転換細胞の選択のための50 mg/L カナマイシンを含むMSS選択培地(4 g/L MS塩、1 mg/L (1000X)MS ビタミンストック、2 mg/L NAA、100 mg/L ミオイノシトール、30 g/L スクロース、2.5 g/L ゲルライト、pH 5.8)に移された。3週間後、個々のカルス片は新鮮なMSS培地に移された。打ち込みから6〜8週間の間に、カナマイシン抵抗性クローンが選択されたカルス片から現れた。
【0104】
3.プロトプラスト
プロトプラストは、外側の細胞壁がない植物細胞である。このような細胞を作製する利点は、トランスフェクション効率を高め、こうした細胞が融合して体細胞雑種を形成できることである。体細胞雑種は異なる植物由来の遺伝子を混ぜ合わせ、全く新しいものをもたらす可能性がある。
【0105】
本発明に従ってプロトプラストを用いる技術は、細胞を液体培養で培養することを含んでもよい(対数増殖期の2〜3日間)。その後細胞を遠沈させる(1000 xg 5分間)。細胞は、セルラーゼ、マセロザイム、ペクトリアーゼを含む溶液に再懸濁される。これをセ氏25度で暗所で一晩振盪させる。遠心分離機にかけ(600 xg 5分間)、上清を除去する。プロトプラストを培地に再懸濁し、残りの酵素を除去するためにもう一度遠心する。プロトプラストは保存も増殖もできず、比較的迅速にトランスフェクションや体細胞融合に使用しなければならない。
【0106】
[DNAコンストラクトを含む生存植物の産生]
第一の実施例では、成功した形質転換体は、最初に細胞毒素(例えばカナマイシン)に対する抵抗性で選択される。安定的にトランスフェクションされた細胞は、抵抗性マーカーと目的遺伝子(例えばインターフェロンをコードする遺伝子など)の両方を発現しているはずであり、さらに正常な植物を再生できるはずである。選択を開始するために、50 mg/LのカナマイシンがMSアガープレートに加えられる。選択の経過は、細胞が複製する速度に一部依存するが、6〜10週間で起こり得る。この間に、0.2 mg/L NAAと2 mg/L BAPをMSアガープレートに加え、12 hr/dayの光周期(淘汰圧を続けながら)で芽発達を促進することにより、形質転換細胞の再生が誘導される。不定芽が発達し始めると、これらは約2 cmの長さで遺伝子発現を分析され得る。所望の遺伝子産物を発現する芽(RT-PCR、Western blot、および生物学的検定法によって分析する)は、続いて根形成を誘導するためにMSアガープレートに移される(1/2 MSと0.2 mg/L NAA)。
【0107】
第二の実施例では、遺伝子組み換えカルスの再生は、約15の小切片(約4 mm)を、ろ過滅菌したカナマイシン(50 mg/L)を加えた再生培地I(4.3 g/L MS塩、1 ml/L (1000X)MSビタミンストック、100 mg/L ミオイノシトール、60 g/L スクロース、3 g/L ゲルライト、pH 5.8)に移し、セ氏25度で暗所で2〜3週間インキュベートすることによって行われる。成熟体細胞胚は、ろ過滅菌したカナマイシン(50 mg/L)を加えた、発芽のための再生培地II(4.3 mg/L MS塩、1 ml/L (1000X)MSビタミンストック、100 mg/L ミオイノシトール、30 g/L スクロース、3 g/L ゲルライト、pH 5.8)上で、光下に移される。
【技術分野】
【0001】
<関連出願への相互参照>
本特許出願は、2005年9月26日出願の米国特許仮出願第60/720,540号、表題“TRANSGENIC ALOE PLANTS FOR PRODUCTION OF PROTEINS AND RELATED METHODS”の優先権と利益を主張し、この出願はここにその全容が引用により本明細書に組み込まれる。
【背景技術】
【0002】
<発明の概要>
本発明は、遺伝子組み換え単子葉植物に関し、より詳細には、遺伝子組み換えアロエ植物と、遺伝子組み換えアロエ植物生産のための方法と組成、および遺伝子組み換えアロエ植物からタンパク質を抽出する方法に関する。
【0003】
<関連技術の説明>
その多くが治療目的で用いられる生物活性タンパク質の市場は拡大し続けている。現在、160以上のタンパク質から作られる医薬品が利用可能である。さらに50くらいが今後2〜3年のうちに認可される見通しである。治療タンパク質生産に対する現行需要は、既に生産能力を上回っている。業界はこの隘路を打開するために、生産能力を4倍から5倍増加させる必要があると予測されている。しかしながら、治療タンパク質の生産設備は高価で、一般的に建設に時間がかかる。従って、より安価で、かつ生産を増加させるために必要な時間を短縮し得る生産方法が必要とされる。
【0004】
いくつかの動物性タンパク質生産法が用いられる。しかしながら、これらは頻繁に病気から健康上のリスクを引き起こす。そのようなリスクは、動物と、ヒトの患者などの最終使用者の両方に影響し得る病気の二次汚染から生じ得る。従って、産生生物と最終使用者の間での二次汚染の可能性を排除するような生産方法が必要とされる。
【0005】
加えて、多くの現在の生産方法は、治療タンパク質を生産した動物もしくは宿主生物から、治療タンパク質を抽出するために、および、複合物を患者によって利用され得る状態にするために、大規模な処理を必要とする。精製後、タンパク質は、タンパク質を安定化し、患者が利用できるようにするために、アジュバントもしくは他の担体物質と混合されてもよい。しかしながら、抽出、精製、再懸濁の処理過程は、治療タンパク質の処理に関するものの中でもとりわけ複雑で煩雑であり、一般的に治療法を必要とする発展途上国での使用につながらない可能性がある。従って、治療タンパク質の抽出後処理を排除する、もしくは削減し得る簡単な生産方法が必要とされる。
【発明の開示】
【0006】
本発明に従う組成物と方法は、上述の必要性と欠点の多くを解決し、また本発明の開示を考察することで当業者に理解される、さらなる改良と利点を提供し得る。
【0007】
一態様では、本発明は目的遺伝子を安定的に組み込んだ遺伝子組み換えアロエ植物を提供し得る。他の態様では、アロエ植物のゲノムに目的のDNA配列を組み込むために、新規ベクターとコンストラクトが提供され得る。目的の配列は生物活性タンパク質をコードしていてもよい。生物活性タンパク質は、インターフェロン、免疫グロブリン、リンフォカイン、成長因子、ホルモン、血液因子、組織適合抗原、酵素、化粧品タンパク質と他の哺乳類タンパク質、もしくは他の目的タンパク質であってもよい。いくつかの態様では、目的タンパク質はヒトのタンパク質である。本発明に従うアロエ植物は、目的遺伝子を目的タンパク質に転写して翻訳し得る。一態様では、目的タンパク質の少なくともいくつかは、アロエ葉の中心部に移動する。他の態様では、目的タンパク質はアロエ葉の中心部への転移を促進するようなシグナル配列を含んでもよい。他の態様では、アロエ植物から個別細胞を分離するための新規の方法が提供され得る。さらに他の態様では、本発明は、アロエ植物にベクターを組み込み、またそのような遺伝子組み換えアロエ植物を再生産するための新規の方法を提供し得る。
【0008】
本発明に従う哺乳類タンパク質を生産する遺伝子組み換えアロエ植物は、例えば様々な生物活性タンパク質と化粧品タンパク質などの目的タンパク質の生産のために、より採算の合う代替案を提供し得る。一態様では、目的タンパク質はアロエ葉のゲル内に局在してもよいし、および/または濃縮されてもよい。この目的タンパク質の局在化は、植物からのタンパク質の除去を単純化し得る。この態様では、目的タンパク質はアロエ葉の中心部内の天然ゲルの抽出と共に共抽出(co-extract)されてもよい。従って本発明は、その遺伝子組み換え植物由来の目的タンパク質が、例えばタバコととうもろこしなどのほとんどの遺伝子組み換え植物由来のものよりも、一般的により容易に利用しやすい遺伝子組み換え植物を提供し得る。さらに、本発明はタンパク質分離のための効率的な方法を提供し得る。さらに他の態様では、目的タンパク質はアロエ植物内に特に局在していなくてもよい。しかしながら、アロエ植物の組織と生理学は、本発明の開示を考察することで当業者に理解される、目的タンパク質の生産のためのいくらかのさらなる利点をも提供し得る。
【0009】
アロエ植物は、バクテリアと酵母の中で目的タンパク質を生産するための、従来の方法に勝る様々な利点を提供することができる。アロエ植物10の利点は、単純単細胞のバクテリアと酵母は適合性が低く、かつ所望の形でタンパク質を生産するために必要とされるやり方で、タンパク質を処理する能力を含み得る。この処理は、グリコシル化などによる化学修飾と、例えばいくつかのタンパク質の折り畳みを含むことができる。さらに、動物細胞に基づく他のタンパク質生産方法に比べて、アロエ植物生産は著しい費用便益と拡張性の利点を提供し、またヒトに有害となり得る汚染のリスクを軽減し得る。
【0010】
本発明に従う遺伝子組み換えアロエ植物のアロエ葉の中心部からのタンパク質修飾ゲルは、アロエ植物から直接使用され得る。これにより、比較的複雑で高価な天然植物材料からの目的タンパク質の抽出を行う必要がなくなる可能性がある。いくつかの態様では、葉から抽出されたゲルは、タンパク質抽出もしくは処理の必要なしに直接使用され得る。ゲルは、遺伝子組み換えアロエ植物の折れた葉の切り口(open end)から機械的に抽出される髄の形であってもよい。
【0011】
本発明は、生物活性のある、および/または化粧品に応用される、ヒトおよび他の哺乳類のタンパク質の生産のための、採算の合う代替案を提供し得る。
【0012】
本発明の開示を考察することにより、当業者は本発明のさらなる改良と利点を理解するだろう。
【図面の簡単な説明】
【0013】
【図1】アロエ葉の断面組織の例をあらわす。
【図2】カルス組織を生成するための方法例をあらわす。
【図3】遺伝子組み換えアロエ植物を生成するための様々なステップを図式的に概説する。
【図4】配列情報を含むプラスミドベクターの構成要素を図式的にあらわす。
【図5】トウモロコシ由来のユビキチンプロモーターの配列情報を列挙し、ユビキチンプロモーターの領域を強調している。
【図6A】本発明の態様に従うプラスミドベクター系のコンストラクションをあらわす。
【図6B】本発明の態様に従うプラスミドベクター系のコンストラクションをあらわす。
【図7A】本発明の態様に従う別のプラスミドベクター系のコンストラクションをあらわす。
【図7B】本発明の態様に従う別のプラスミドベクター系のコンストラクションをあらわす。
【図7C】本発明の態様に従う別のプラスミドベクター系のコンストラクションをあらわす。
【図8】本発明の態様に従うさらなるプラスミドベクター系のコンストラクションをあらわす。
【図9】本発明の態様に従う組み込み系のコンストラクションをあらわす。
【0014】
全図面は、本発明の基本的教示の説明を簡略化するために示されるに過ぎない。すなわち、数、位置、配列、様々な実施形態の関係と組成に関する図面の拡張は、以下の記述を考察した後で、説明されるかもしくは当該技術分野内に含まれることになるだろう。さらに、開示された発明を実施するための様々なプロトコル、ツール、および組成物が、以下の記述を考察した後で当該技術分野内に含まれることになるだろう。
【発明を実施するための最良の形態】
【0015】
明細書で使用する“a”もしくは“an”は、一つもしくはそれ以上を意味してもよい。請求項(群)で使用するように、“含む(comprising)”という用語と併用する際、用語“a”もしくは“an”は一つもしくは一つ以上を意味してもよい。本明細書で使用する“別の(another)”は、少なくとも二つ目もしくはそれ以上を意味してもよい。
【0016】
本明細書で使用する“形質転換アロエ細胞”は、例えばAgrobacterium媒介形質転換、もしくは組み換えDNAで被覆された微小粒子を用いる打ち込み、もしくは他の手段によって、安定的に組み込まれた、非天然の組み換えDNAで形質転換された植物細胞を意味する。本発明の形質転換アロエ細胞は、オリジナルに形質転換された植物細胞であってもよく、これは微小生物として、もしくは後代の植物細胞として生存し、分化した組織に再生される。例えば、安定に導入された非天然の組み換えDNAを持つ遺伝子組み換えアロエ植物10、あるいは後代の遺伝子組み換えアロエ植物10由来の種子、もしくは花粉に再生される。
【0017】
本明細書で使用する“遺伝子組み換えアロエ植物”は、組み換えDNAの安定的な組み込みによってゲノムを改変したアロエ植物を意味する。遺伝子組み換えアロエ植物10は、オリジナルに形質転換されたアロエ細胞から再生したアロエ植物と、後世代もしくは形質転換アロエ植物10の交雑由来の後代の遺伝子組み換えアロエ植物を含む。
【0018】
本明細書で使用する“組み換えDNA”は、細胞の外側で構築され遺伝子操作されたDNAを意味し、天然DNA、cDNA、合成DNA、および/または他のDNAを包含するDNAを含む。
【0019】
本明細書で使用する“プロモーター”は、転写開始のための調節DNAを意味する。“植物プロモーター”は、その起源がアロエ細胞であるかどうかに関わらず、アロエ細胞の転写を開始することができるプロモーターであり、例えばAgrobacteriumプロモーターはアロエ細胞内で機能することがよく知られている。従って植物プロモーターは、植物、植物ウィルス、およびAgrobacteriumと根粒菌(Bradyrhizobium)バクテリアなどのバクテリアから得られるプロモーターDNAを含む。発生制御下にあるプロモーターの例としては、葉、根、もしくは種子などの特定の組織において選択的に転写を開始するプロモーターを含む。そのようなプロモーターは、“組織選択的”であると言われる。特定の組織のみで転写を開始するプロモーターは、“組織特異的”であると言われる。“細胞型”特異的プロモーターは、一つ以上の器官において特定の細胞型における発現を主に促進する(例えば、根もしくは葉における道管細胞)。“誘導”プロモーター、もしくは“抑制”プロモーターは、環境制御下にあるプロモーターである。誘導プロモーターによって転写を誘起し得る環境条件の例としては、嫌気条件、もしくは特定の化学物質、もしくは光の存在を含む。組織特異的プロモーター、組織選択的プロモーター、細胞型特異的プロモーター、および誘導プロモーターは、“非構成的”プロモーターの分類を構成する。“構成的”プロモーターは、ほとんどの条件下で活性化されているプロモーターである。そうしたプロモーターは、エンハンサー、あるいは、転写開始、転写開始部位、転写のレベル、転写終結部位、もしくは得られるリボ核酸の任意の後処理に作用する他の要素を含んでもよい。
【0020】
本明細書で使用する“遺伝子コンストラクト(genetic construct)”という用語は、遺伝子組み換えアロエ植物の作製を目的として、少なくとも二つのDNAセグメントの合成配列から成るDNA配列と定義される。特定の実施形態では、一つのセグメントは調節配列で、もう一方のセグメントは遺伝子産物をコードする。
【0021】
本明細書で使用する“連結(operably linked)”とは、DNAコンストラクトにおける二つ以上のDNAフラグメントの結合を意味し、一方のフラグメントの機能(例えばタンパク質をコードするDNA)が、もう一方のフラグメント(例えばプロモーター、終結配列など)によって制御されるようになっている。
【0022】
本明細書で使用する“転写”という用語は、DNA鋳型からRNA分子を作製することと定義される。
【0023】
本明細書で使用する“翻訳”という用語は、RNA鋳型からポリペプチドを作製することと定義される。
【0024】
本明細書で使用する“発現した”とは、“生成した(produced)”を意味し、例えばタンパク質と同起源のDNAがmRNAに転写され、それがタンパク質に翻訳された時、そのタンパク質は植物細胞内で発現していることになる。
【0025】
本発明は、様々な目的タンパク質を発現する新規の遺伝子組み換えアロエ植物10を提供してもよいし、遺伝子組み換えアロエ植物10を生産するための方法と組成を提供してもよいし、遺伝子組み換えアロエ植物10からタンパク質を抽出するための方法を提供してもよいし、また、目的タンパク質と遺伝子組み換えアロエ植物10に由来する構成要素の新規の組成を提供してもよい。本発明に従って生成される目的タンパク質は、髄18もしくはアロエ葉12の髄でのタンパク質の蓄積を促進させるような分泌シグナルを含んでもよい。一態様では、遺伝子組み換えアロエ植物10の葉のmusalegenous細胞内で、このタンパク質の蓄積の少なくとも一部分が起こってもよい。
【0026】
髄(pith)18もしくはアロエ葉12の髄(pulp)は、アロエ葉12から抽出された際に、少なくとも部分的には一般に“ゲル”と呼ばれる葉の構成要素を含む。本明細書で使用する“ゲル”は、抽出された髄18、および、次の処理の程度に関わらず、アロエ葉12から抽出される際に髄18に付随する他の付随物質を意味する。本発明の一つ以上の態様に従って、アロエ葉12から抽出したゲルは、変性した組織を持っていてもよい。特定の一態様では、ゲルの組織は少なくとも一つの外来タンパク質成分を含むように変性されてもよく、これは“タンパク質変性ゲル”と呼ばれる。付随する目的タンパク質(群)を伴うタンパク質変性ゲルの抽出により、タンパク質を容易に入手でき、および/または効率的なタンパク質分離方法が提供され得る。本発明に従って生成されるおよび/または抽出されるタンパク質変性ゲルは、タンパク質抽出の必要無しに直接使用され得る。
【0027】
本発明の態様は、例示目的として図1から図8に大まかに示される。本発明は、遺伝子組み換え植物と組成と方法を提供し、これらは概してユリ科アロエ属に応用できる。典型的には、本発明は例示目的として、Aloe vera(barbadensis miller)、Aloe ferox、およびAloe arborescenceの群から選択される種への応用に関して記載される。アロエ属は一般的に茎のない大きなロゼット型多肉性単子葉植物を含む。これらの植物を、本発明の開示を目的として概してアロエ植物10と呼ぶこととする。
【0028】
アロエ植物から生成される様々な化合物が医薬目的に使用されてきた。これらの化合物は、タンパク質変性ゲルに存在する場合、本発明に従う遺伝子組み換えアロエ植物10の生物活性タンパク質の薬理作用を補足し得る。図1は遺伝子組み換えアロエ植物10の葉12の断面図を示す。遺伝子組み換えアロエ植物10の葉は、容易に抽出可能な、タンパク質、糖質、および水のゲル状混合物をゲル内に含み、ゲルは主に髄18から得られる。このゲル状混合物は、主にアロエ葉12の中心部に位置する。ゲル内の様々な化合物は、数多くの薬理作用と医薬用途を有することがわかっている。一態様では、ゲルおよび/または髄18は、遺伝子組み換えアロエ植物10によって生成され、ゲルおよび/または髄18に局在する遺伝子組み換えタンパク質を安定化させる。
【0029】
図1は特に、表皮14、皮層もしくは葉肉16、および髄(pithもしくはpulp)18を示す。表皮14は葉の細胞の外層を形成する。本発明の一態様では、遺伝子組み換えアロエ植物は、これらの組織の内の一つ以上において、アロエ細胞によって発現される遺伝子コンストラクトを、一時的にもしくは安定的に組み込むアロエ細胞を含んでもよい。皮層16は、維管束、木質部、および師部と同様に、葉緑体の豊富な細胞を含む。髄18は、ほぼ大きな皮層細胞のみで構成される海綿状組織であり、少なくとも部分的にゲルをあらわし、そこに一つ以上の目的の遺伝子組み換えタンパク質を組み込むか、もしくは蓄積させる上で好都合となり得る。
【0030】
表皮14は典型的には細胞の単外層から成る。表皮14のすぐ下は、維管束の網状組織を含む皮層16である。維管束の外側の支えは、通常は鞘細胞によってもたらされる。維管束は三つの大まかな種類の管構造から構成される。すなわち、木質部、師部、および付随する大きな周辺環状小管(pericyclic tubules)である。木質部は水と無機物を植物の根から葉に輸送する。師部はデンプンと他の合成物質を植物の至る所に輸送する。周辺環状小管は、緩下性のあるアントラキノンを非常に多く含む(特にアロイン)、乳液もしくは樹液を含む。アントラキノンは太陽の紫外線を吸収し、一般的にアロエ植物の水貯蔵器官として機能するアロエ葉12の中心部の過熱を防ぐ。維管束の周辺環状部分は表皮14に付着しており、一方維管束の残りの部分は髄18の中に突き出ている。アロエ葉12の最深部の主要部分は、少なくとも部分的にゲルを構成する髄18である。ゲルの抽出を目的として、表皮14および皮層16は、ゲルを含む葉鞘を含むと一般的に見なされ得る。抽出されたゲルは、実質的に髄18から成るが、典型的には粘度の高い粘液性の物質であり、昔から、例えば火傷や創傷の治療などの医薬目的として、皮膚に局所塗付されてきた。
【0031】
通常、ゲルはアロエ植物のための物質貯蔵庫としてはたらく。一般的に、皮層16はアロエ植物に必要な糖質と糖タンパク質の多くを合成する。過剰に合成された糖質は、通常は髄18に輸送され、水といくつかの無機物と共に貯蔵される。糖質は師部道管によって、髄18の皮層16細胞内の大きな液胞に輸送される。その後、水が浸透圧によって糖質に引き付けられ、髄18がアロエ植物の水貯蔵器官として機能できるようになっている。
【0032】
ゲルは、アロエ葉12を縦軸に沿って折り、葉を押しつぶしてゲルを割れ目を通して周りの鞘から押し出すことにより、比較的容易に抽出される。付加的な物質がゲルに伴って葉の組織から抽出されることもある。
【0033】
<処理の概説>
本発明の一つ以上に従う遺伝子組み換えアロエ植物10は、概して、一つ以上の目的タンパク質を発現するように植物に安定的に組み込まれた一つ以上のDNAコンストラクトを含む。本発明の一つ以上に従う遺伝子組み換えアロエ植物10を作製することは、種々の新規の組成と方法を含んでもよい。典型的には、一つ以上のDNAコンストラクトは、遺伝子組み換えアロエ植物10に一つ以上のタンパク質を発現するように改良される。一態様では、一つのコンストラクトは一つのmRNAを発現してもよい。他の態様では、一つのコンストラクトは複数のmRNAを発現してもよい。なおも他の態様では、複数のコンストラクトが複数のmRNAを産出してもよい。各mRNAは一つ以上のポリペプチドを産出してもよい。
【0034】
コンストラクトをアロエ植物に導入するには、図2に大まかに示すように、アロエ細胞もしくは未分化カルス組織が典型的に使用される。アロエ細胞とカルス組織は、典型的にはアロエ植物の根もしくは葉の分裂組織、もしくは種子に由来する。DNAコンストラクトは、図3に図式的に示すように、様々な技術および/またはコンストラクトを用いてアロエ細胞に導入される。いくつかの技術は、本発明の開示を考察することで当業者に理解されるだろう。所望のコンストラクトを組み込んだアロエ細胞もしくはカルス組織は、様々な技術を使用するために選択される。典型的には、DNAコンストラクトは選択可能なマーカーを含む。いったん所望のコンストラクトが安定的にアロエ細胞もしくはカルス組織に導入されると、通常はアロエ細胞もしくはカルス組織からアロエ小植物体が発生する。DNAコンストラクトを含む遺伝子組み変え生存アロエ植物10に成長するアロエ小植物体は、所望の目的タンパク質(群)を構成的に産出するか、あるいは誘導的に産出する可能性がある。目的タンパク質は、産出される特定のタンパク質、および/またはタンパク質に付随するシグナル配列の有無に応じて、遺伝子組み換えアロエ植物10内で局在してもよいし、あるいは、遺伝子組み換えアロエ植物10の全体に概して存在してもよい。その後、特定のコンストラクトおよび/または目的の関連タンパク質に応じて、遺伝子組み換えアロエ植物10は、栄養繁殖(無性生殖)もしくは有性繁殖(有性生殖)され得る。タンパク質は、幅広い技術を用いて遺伝子組み換えアロエ植物10から分離され得る。その後タンパク質はさらに分離されるか、および/またはさらに処理されてもよい。そのような処理は、本発明の開示を考察することで当業者に理解される他の処理の中でも特に、酵素修飾、化学修飾、適切なアジュバントへの混入を含んでもよい。一態様では、特定のタンパク質の特異的用途のために、タンパク質は不活性体(前駆体)から活性体に処理されてもよい。
【0035】
<細胞の分離>
図2に図示した配列例に示すように、アロエ細胞もしくはアロエ細胞群は、典型的には所望のコンストラクトの組み込みの前にアロエ植物から分離される。DNAコンストラクトの組み込みに一般的に選ばれるアロエ細胞の種類は、その再生能に基づいて一般的に選択される。アロエ植物体からの茎分裂組織もしくは根分裂組織由来の分裂組織細胞、もしくはアロエ種子由来のアロエ胚細胞を使用してもよい。しかしながら、アロエ植物の多くの部分は、再生する能力、もしくはカルス組織を形成する能力を保持しており、これらを利用してもよい。細胞は典型的には、本発明の開示を考察することで当業者に理解される様々な技術を用いて分離される。典型的には、細胞は外科用メスを用いてアロエ植物から機械的に分離される。あるいは、本発明の開示を考察することで当業者に理解されるように、必要な細胞を分離するために、他の機械的技術もしくはその他の技術を用いてもよい。適切なアロエ細胞もしくはアロエ細胞群が分離されると、通常、アロエ細胞はカルス組織を形成するために培養下で育成される。或る技術は、アロエ細胞が最初にカルス組織に成長することを必要としなくてもよいが、カルス組織は、遺伝子組み換えアロエ植物10を作り出す能力を保持する未分化のアロエ細胞セットの源を提供する。カルス組織は典型的には固形培地上で育成される。しかしながら、カルス組織は液体培地に置くこともできるし、浮遊状態で成長させることもできる。
【0036】
分裂組織細胞は、アロエ植物の根の先端もしくは葉から分離してもよい。分裂組織アロエ細胞は頂端分裂組織から分離してもよい。典型的には、組織が分離されるアロエ植物は、若い健康なアロエ植物である。通常、アロエ植物は8インチ未満で六つ以下の初期芽を持つものが選択される。切り傷は通常、アロエ植物を断片に切ることによって、アロエ植物の芽の上に形成される。傷をつけた表面は、アロエ細胞が成長するのを促進し得る。通常、最初に傷の表面でカルス組織が成長し始める。アロエ葉12からアロエ細胞を分離するために、断片は通常、若い成長芽の遠位末端から切り取られる。その後断片部分は成長培地上に置かれる。芽の頂端分裂組織は、若いアロエ植物の基部から1インチを切って得られる。アロエ葉12は切片から除去され、断片がプレートに置かれる。分裂組織アロエ細胞は“切片”内に見られる。これらは、培養プレート上での成長維持能力(カルス組織の形成もしくは芽の再生)によって“選別”されるかもしくは“分離”される。分裂組織アロエ細胞は分離されると、その後適切な培地上で、カルス組織形成を促進する条件下で培養される。
【0037】
種子からのアロエ胚細胞の分離は、一般的に、胚を露出させるために種皮を除去することと、所望のアロエ細胞を胚から機械的に除去することを含む。アロエ種子は、典型的には滅菌され、種子の外皮が除去される。アロエ植物もしくはアロエ胚のいずれかからアロエ細胞を機械的に除去するために、外科用メスが通常用いられる。アロエ胚細胞は種皮から分離されると、その後適切な培地上で、カルス組織形成を促進する条件下で培養される。
【0038】
カルス組織が必要な際は、分離されたアロエ細胞は通常、本発明の開示を考察することで当業者に理解されるように、カルス組織の形成に好都合な条件下で育成される。典型的には、分離されたアロエ細胞は、形質転換の前に直径約1cmのサイズを有するカルス組織を形成するために、適切な固体栄養培地に播種されて成長する。アロエ細胞は典型的にはセ氏23度〜26度の間で成長する。適切な培地は固体もしくは液体の基盤をふくみ、通常、これらは、補助無機栄養素(マクロ元素(窒素、硫黄、リン、カルシウム、マグネシウム、およびカリウム)と微量元素(鉄、ホウ素、コバルト、銅、ヨウ素、マンガン、モリブデン、および亜鉛)の両方)、ならびに、糖(スクロースもしくはマルトース)およびビタミンおよび捕因子(とりわけチアミン、ナイアシン、ビオチン、ピリドキシン、ミオイノシトール)を含む有機栄養素、ならびに、アミノ酸(プロリンおよびカゼイン塩加水分解物)、さらに、主にオーキシンの源(通常(NAA)、1-ナフタレン酢酸、(IAA)、インドール-3-酢酸、もしくは(2,4-D)、2,4-ジクロロフェノキシ酢酸)およびサイトカイニンの源(通常(BAP)6-ベンジルアミノプリン)である成長調節因子を含む。これらの培地とビタミンは、通常、様々な濃度の無機栄養素とビタミンで前処方された組成で市販されており、とりわけ、MS培地(Murashige-skoog)、Gamborg培地、もしくはChu N6培地として知られている。さらに、ペトリ皿上で培養される細胞は、通常、成長培地に添加されるアガーなどの固体マトリクス支持物質を必要とする。
【0039】
<DNAコンストラクトの作製>
適切なDNAコンストラクトは、典型的には、得られる遺伝子組み換えアロエ植物10によって目的タンパク質(群)の生成が可能になるように、分離されたアロエ細胞に由来するカルス組織に導入される。本発明の開示を考察することで当業者に理解されるように、DNAコンストラクトは、分離されたアロエ細胞もしくは成熟アロエ植物に導入されてもよい。DNAコンストラクトは、典型的には、コンストラクトの増殖およびアロエ細胞への導入のために、プラスミドもしくはウィルスなどのベクターに組み込まれる。プラスミドベクターの例としては、例示目的として図4に図式的に示してあるInvitrogen(Carlsbad, California)から商標pZErOで市販されているプラスミドを含んでもよく、また、例えばこのプラスミドの機能単位のうちの一つ以上を含んでもよい。
【0040】
本発明に従うDNAコンストラクトは、アロエ細胞内で機能できるプロモーター配列、目的タンパク質をコードする配列、終結配列、および転写開始部位と転写終結部位(これら全てがアロエ細胞内で機能できる)を含むことができる。これらの構成要素は、適切に構成され結合された際には、DNAの転写とmRNAの翻訳、およびこれらそれぞれの終結をアロエ細胞内で開始することができる。DNAコンストラクトは分泌シグナルを含んでもよい。
あるいは、例えばインターフェロンなどのいくつかの目的タンパク質は、インターフェロンを分泌の標的とする、天然もしくは合成のドメインを含んでもよい。分泌シグナルは、通常、タンパク質が細胞を離れる際に切断されるか、あるいはいくつかの場合には、実質的にタンパク質の機能に影響を与えることなく、タンパク質によって保持されることもある。分泌シグナルは転移のために分泌シグナルを必要とし得る様々なタンパク質に加えられてもよい。これらの分泌シグナルは様々な植物分泌配列に由来してもよい。DNAコンストラクトもしくは関連ベクターは、少なくとも一つの選択可能なマーカーも含んでもよい。一態様では、DNAコンストラクトは、バクテリアでのベクターの増殖のための第一選択可能マーカーと、アロエ細胞での成長のための選択可能マーカーの両方を含んでもよい。加えて、DNAコンストラクトは、アロエ植物10の特定のアロエ細胞での発現のための、他の調節因子を含んでもよい。他のコンストラクトの構成要素は、転写促進のための5’リーダーとイントロン、3’非翻訳領域(ポリアデニル化シグナルおよびポリアデニル化部位など)、輸送ペプチドのためのDNAなど、付加的な調節因子を含んでもよい。所望の治療タンパク質をコードする配列などの、所望のDNA配列の増幅は、ポリメラーゼ連鎖反応によって行われ得る(U.S. Pat. No. 4,683,202および5,928,906を参照。それぞれ引用によって本明細書に組み込まれる。)。本発明の開示を考察することで、当業者は、プロモーター配列、調節配列、安定化配列、ターゲティング配列、および/または終結配列を含むDNAコンストラクトが、当業者に周知の方法を用いてその機能に影響を及ぼすように改変されてもよいことがわかるだろう。
【0041】
上述の通り、アロエ細胞で機能するプロモーターが通常は利用される。プロモーターは、典型的には、制御タンパク質と制御分子が結合し得る遺伝子因子を含む。これらのタンパク質は、典型的にはRNAポリメラーゼと他の転写因子を含む。プロモーターは、機能上の位置および/または方向で核酸配列に機能的に結合して、その配列の転写開始および/または発現を制御する。プロモーターは、核酸配列の転写活性化に関係するシス作用調節配列を意味する“エンハンサー”と併用されてもされなくてもよい。
【0042】
当然ながら、その中で発現することが望まれるアロエ植物10の、少なくとも一つの細胞型において、DNAセグメントの発現を効果的に誘導するようなプロモーターおよび/またはエンハンサーを採用することが重要となる。分子生物学分野の当業者は、一般的にタンパク質発現のためのプロモーター、エンハンサー、および細胞型の組み合わせの使用法を知っており、例えば、引用により本明細書に組み込まれるSambrook et al.(1989)を参照のこと。採用されるプロモーターは、構成的、組織特異的、誘導的、および/または、適切な条件下で導入されたDNAセグメントの高レベルの発現を誘導するのに有用であるものでもよく、例えば、組み換えタンパク質および/またはペプチドのラージスケールでの産出において有利である。プロモーターは異種であっても内因性であってもよい。DNAコンストラクトは、通常はアロエ細胞内で機能できる転写開始・終結制御シグナルと翻訳開始・終結制御シグナルを必要とする。幅広い種類の転写開始調節配列が使用され得る。転写開始を制御するDNA配列は、Agrobacterium、ウィルス、もしくは植物に由来し得る。カリフラワーモザイクウィルス(35S-CaMV)由来の35Sウィルス転写開始領域がアロエ植物10に使用され得る。アロエ植物10で使用され得る植物プロモーターは、種々の単子葉植物もしくは双子葉植物由来のリブロース-1,5-ビスリン酸カルボキシラーゼ(RUBISCO)小サブユニットプロモーター、またはトウモロコシ由来のユビキチンプロモーターも含んでもよい。他の適切なプロモーターが使用されてもよく、本発明の開示を考察することで当業者に理解され得る。もし誘導調節が望まれる場合は、ドメインは異なる起源から得られてもよく、一方の起源由来の調節領域が、もう一方の起源由来のRNAポリメラーゼ結合ドメインに結合されるようになっている。発現制御は、根、葉、種子、花、樹液などの遺伝子組み換えアロエ植物10の特定の部分における、遺伝子組み換えアロエ植物10の成長の特定の段階に対してなされてもよいし、あるいは、植物の部分と成長段階の組み合わせに対してなされてもよい。特定の成長段階もしくは組織に対する発現制御は、本発明の開示を考察することで当業者に理解される、さらなるDNA要素を必要としてもよい。
【0043】
植物細胞で活性のある数多くの他のプロモーターが文献に記載され、アロエ細胞で有効である可能性がある。これらは、植物ゲノムに存在するプロモーターだけでなく、他の起源由来のプロモーターも含む。すなわち、Agrobacterium tumefaciensの腫瘍誘導プラスミド上にあるノパリンシンターゼ(NOS)プロモーターおよびオクトピンシンターゼ(OCS)プロモーター、ならびにカリフラワーモザイクウィルスなどのcaulimovirusプロモーターを含む。さらに、種々の他のプロモーターも様々な参考文献で同定されており、カリフラワーモザイクウィルス(CaMV35S)由来の構成的プロモーターの種類を開示しているU.S. Pat. No. 5,858,742および5,322,938、イネアクチンプロモーターを開示しているU.S. Pat. No. 5,641,876、効果的な植物発現ベクターの設計に有用な5',3'およびイントロンエレメントを開示しているU.S. Patent Application Publication 2002/0192813A1、トウモロコシ葉緑体アルドラーゼプロモーターを開示しているU.S. patent application Ser. No. 09/757,089、イネグルテリンプロモーターを開示しているU.S. patent application Ser. No. 08/706,946、トウモロコシアルドラーゼ(FDA)プロモーターを開示しているU.S. patent application Ser. No. 09/757,089、トウモロコシニコチアナミンシンターゼプロモーターを開示しているU.S. patent application Ser. No. 60/310,370を含むが、これらに限定はされず、これら全ては引用により本明細書に組み込まれる。植物細胞内で機能する、これらの、また数多くの他のプロモーターは、所望の治療タンパク質の発現のために遺伝子組み換えアロエ植物内で機能し得る。
【0044】
具体的な一実施例として、トウモロコシ由来のユビキチンプロモーター(SEQ. ID. NO.44)が使用され得る。トウモロコシ由来のユビキチンプロモーターは大きなエレメントであり、長さが約2kbで、少なくとも三つの基本領域(general region)から成る。この配列は、特に関連性のある特徴にラベルを付けて図5に記載されている。ユビキチンプロモーターの第一区域は、最も5’末端側に位置しマトリックス結合領域(MAR)を含む。このMARは、ヒストンと他の核タンパク質と相互作用するエレメントで、フランキング配列をループアウト(loop out)させるように働き、それらがより容易に細胞の転写機構に接近できるようにする。また、それらはお互いに転写単位を隔離させるのを助けるが、これは、ある場所で開始された転写が、第二の配列に“読み過ごし(reading through)”されてしまうのを防ぐ上で重要である。これはアンチセンスメッセージを作り出すリスクを減らす助けとなり得る。
【0045】
第二区域は、エンハンサーエレメントと実プロモーターを含む。エンハンサーエレメントは、転写機構pol II複合体をプロモーターに結合させて転写を開始させる役目を持つ、転写因子と結合する。“プロモーター”とは、コア転写開始機構と相互作用するのに必要な必須DNAエレメントを特に意味するが、プロモーターという用語は、より一般的には転写開始に関わるDNAエレメントの全てを包含するように用いられ、またこの場合は、“ユビキチンプロモーター”とは、5’から目的のクローニング遺伝子までの2kb領域の全て、もしくはそれらの変異物を意味するように大まかに用いられる。
【0046】
第三区域は、約1kbのイントロンを含む。イントロンは、スプライシングの過程を通して、最終翻訳メッセージであるmRNAから除去された転写遺伝子の領域である。イントロンは、代わりのエンハンサーエレメントを供給することによって、さらに、スプライシングに一部関連する過程である、核から細胞質へのRNAメッセージの転移の促進により、タンパク質翻訳を増加させることによって、直接的に遺伝子発現に影響を与えることもわかっている。しかしながら、遺伝子組み変え系にイントロンが存在しても、必ずしもRNAの発現や翻訳タンパク質のレベルの増加につながるとは限らない。或る態様では、ユビキチンイントロンは、ベクターの全体のサイズを縮小するため、また、アロエでの遺伝子組み換え遺伝子発現への影響を評価するために、修飾されてもよい。
【0047】
本発明の他の態様では、アロエ植物の緑色組織で選択発現することが望まれ得る。こうした利用のための目的プロモーターは、Arabidopsis thalianaリブロース-1,5-ビスリン酸カルボキシラーゼ(Rubisco)小サブユニット(Fischhoff et al. (1992) Plant Mol Biol. 20:81-93)、アルドラーゼおよびピルビン酸オルトリン酸ジキナーゼ(PPDK)(Taniguchi et al. (2000) Plant Cell Physiol. 41(1):42-48)などの遺伝子由来のものを含んでもよい。
【0048】
上述の通り、プロモーターは、遺伝子発現を高める一助となる、複数の“エンハンサー配列”を含むように改変されてもよい。そのようなエンハンサーは当該技術分野で周知である。エンハンサー配列をコンストラクトに含めることにより、選択されたタンパク質の発現が促進され得る。これらのエンハンサーは、しばしば真核細胞で機能するプロモーターの5’から転写開始点に見つかるが、しばしば上流(5’)もしくは下流(3’)からコード配列に挿入されてもよい。いくつかの場合には、これらの5’増強エレメントはイントロンである。特に有用なエンハンサーは、イネアクチン1の5’イントロン(U.S. Pat. No. 5,641,876を参照)およびイネアクチン2遺伝子、トウモロコシアルコールデヒドロゲナーゼ遺伝子イントロン、トウモロコシ熱ショックタンパク質70遺伝子イントロン(U.S. Pat. No. 5,593,874)およびトウモロコシshrunken 1遺伝子である。
【0049】
本発明の態様に従うコンストラクトは、典型的にポリアデニル化シグナルとポリアデニル化部位を含む3’エレメントを含んでもよい。よく知られている3’ エレメントは、例えばU.S. Pat. No. 6,090,627(引用により本明細書に組み込まれる)に開示されている、nos 3’、tml 3’、tmr 3’、tms 3’、ocs 3’、tr7 3’などのAgrobacterium tumefaciens遺伝子由来のもの、小麦(Triticum aesevitum)熱ショックタンパク質17(Hsp17 3’)、小麦ユビキチン遺伝子、小麦フルクトース-1,6-ビフォスファターゼ遺伝子、イネグルテリン遺伝子、イネ乳酸デヒドロゲナーゼ遺伝子、およびイネベータチューブリン遺伝子などの、植物遺伝子由来の3’エレメント(これらは全て、引用により本明細書に組み込まれるU.S. published patent application 2002/0192813 A1に開示されている)、ならびに、エンドウマメ(Pisum sativum)リブロースビフォスフェートカルボキシラーゼ遺伝子(rbs 3’)、および宿主植物内の遺伝子由来の3’エレメントを含む。
【0050】
目的タンパク質はDNAコンストラクトに組み込まれた構造遺伝子によってコードされ得る。構造遺伝子は、哺乳類の遺伝子であってもよいし、もしくは哺乳類の遺伝子の一部であってもよい。目的の構造遺伝子は、インターフェロン、免疫グロブリン、リンフォカイン、成長因子、ホルモン、血液因子、組織適合性抗原、酵素、もしくは他のタンパク質をコードしてもよい。インターフェロンアルファ2の配列は例示目的としてSEQ. ID. NO. 33と記載される。構造遺伝子は、クラゲ由来の緑色蛍光タンパク質(GFP)などのマーカータンパク質もコードしてもよい。構造遺伝子のDNA配列は、アロエ植物10での高レベルの発現が可能になるように修飾されてもよい。アロエ植物10のコドンバイアス(codon bias)は、構造遺伝子が分離される原種のコドンバイアスと異なっていてもよい。天然の構造遺伝子は、アロエ植物10でコードされたタンパク質の産出を最適化するように設計されてもよい。さらに、構造遺伝子のDNA配列は、タンパク質の構造に干渉しない適切なグリコシル化を、アロエ植物10にもたらすように設計されてもよい。
【0051】
転写と翻訳の終結は、アロエ植物10で機能できる種々の転写終結配列と翻訳終結配列によってもたらされ得る。一態様では、終結配列はAgrobacterium由来のノパリン合成酵素(NOS)遺伝子由来の配列を含んでもよい。他の態様では、終結配列は天然アロエ遺伝子とタンパク質の終結配列由来であってもよい。
【0052】
マーカー遺伝子は、しばしばDNAコンストラクトに組み込まれる。マーカー遺伝子は、目的の構造遺伝子を含む細胞が、マーカー遺伝子を含まない全細胞集団から選択されることを可能にする。マーカー遺伝子は、とりわけ、カナマイシン、クロラムフェニコール、G418およびゲンタマイシンなどに対する抵抗性を与える酵素もしくは他のタンパク質を含んでもよい。例えばAgrobacteriumがベクターとして用いられる場合、さらに他の特定のDNA配列が必要になることもある。もしDNAが特定の細胞型を標的とする場合は、他の領域が存在してもよい。
【0053】
上記の通り、DNAコンストラクトは分泌シグナルも含んでもよい。コンストラクトは、治療タンパク質を、そのタンパク質が形成された細胞から除去する標的とし得るようなシグナルペプチドをコードする転移配列も含んでもよい。植物で機能し、特にアロエ植物などの単子葉植物で機能する種々のシグナル配列が文献で同定されている。これらは、コンストラクトもしくは目的遺伝子の中に機能的に組み込まれてもよい。一態様では、イネ(Oryza sativa)由来のアルファアミラーゼ分泌配列(SEQ. ID. NO. 29)が用いられてもよい。このシグナル配列は、“The alpha-amylase genes in Oryza sativa”Mol Gen Genet., 1990 April;221(2):235-44で述べられており、この開示はその全容が引用により本明細書に組み込まれる。あるいは、例えばインターフェロンなどのいくつかの目的遺伝子は、インターフェロンを分泌の標的とする、天然もしくは合成のドメインを含んでもよい。分泌シグナルは、典型的にはタンパク質が細胞を離れる際に切断されるか、あるいはいくつかの場合には、実質的にタンパク質の機能に影響を与えることなく、タンパク質に保持されてもよい。分泌シグナルは、転移に分泌シグナルを必要とし得る種々のタンパク質に加えられてもよい。これらの分泌配列は種々の植物分泌配列に由来してもよい。
【0054】
図6Aから図6Bは、アロエ植物から目的タンパク質を産出するためのコンストラクトの例示的なセットをあらわす。プラスミドコンストラクションは、pzUI(TGC9)とラベルされる。TGC9はヒトインターフェロンアルファ2の発現を促進するユビキチンプロモーターの全長を含み、pzUEK(TGC12)(SEQ. ID. NO. 38)は、eGFP(強化緑色蛍光タンパク質)(SEQ. ID. NO. 31)およびカナマイシン抵抗性タンパク質を含む融合タンパク質の発現を促進するユビキチンプロモーターの全長を含む。融合タンパク質は、両方のタンパク質のスクリーニング特性(選択と視覚化)を一つに兼ね備える。
【0055】
図示したように、プラスミドpzI(TGC8)は、それぞれ二つの異なる制限酵素部位を含むPCRプライマーを用いたヒトインターフェロンアルファ2遺伝子のPCR増幅によって作製される(プライマーの配列は表1を参照)。増幅されたインターフェロン(IFN)遺伝子は、5’フランキング領域にPstIおよびBamHI部位を持ち、3’フランキング領域にSacIおよびXhoI部位を持つ。PCRで作製したIFNはその後、コンストラクトpzI(TGC8)を作製し、次のクローニングのためのBamHIおよびSacI部位を導入するために、PstI/XhoIフラグメントとしてpZErOベクターにクローニングされる。このコンストラクトは、IFNアルファ遺伝子の全長と、細胞からのIFNタンパク質の分泌の誘導に関与する自身のシグナル配列を含む。これはスライド3に描かれている。これらのPCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0056】
図示したように、プラスミドベクターpzUI(TGC9)は、全長ユビキチンプロモーターの下流にIFNアルファ2遺伝子をクローニングすることによって作製される。IFNのPstI/XhoIフラグメントはpzI(TGC8)から切り離され、コンストラクトpzUI(TGC9)を作製するためにpzU(TGC1)にライゲーションされる。これはスライド3に描かれている。
【0057】
図示したように、BamHI(5’末端)およびXhoI(3’末端)制限部位を含むプライマーのセットを用いてeGFP遺伝子がPCR増幅される(プライマー配列は表1を参照)。このPCR作製eGFP遺伝子は停止コドンを含まずに作製される(最終的にeGFP-kan融合タンパク質の発現を可能にするため)。従って翻訳はeGFP配列の末端では終結しない。PCR増幅フラグメントは、制限酵素BamHIとXhoIで消化され、pZErOのBamHIおよびXhoI部位にライゲーションされ、pzEnostop(TGC10)を作製する。これはスライド3に描かれている。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0058】
図示したように、pzEK(TGC11)はeGFP遺伝子とカナマイシン抵抗性遺伝子の間で融合コンストラクトを作製する。eGFPはBamHI/XhoIフラグメントとしてpzEnostop(TGC10)から切り離され、コンストラクトpzEK(TGC11)を作製するためにpzK(TGC4)にライゲーションされる。eGFP遺伝子はイン・フレームに(in frame)、5’からkan/nosにクローニングされる(SEQ. ID. NO. 35)。
【0059】
図示したように、eGFP/kan/nosカセットを含むpzEK(TGC11)由来のBamHI/XbaIフラグメントは、eGFP/kan融合遺伝子の発現を可能にするために、pzUI(TGC9)のBamHIおよびXbaI部位にクローニングされる。このようにしてIFN配列は除去される。この新しいコンストラクトであるpzUEK(TGC12)は、全長ユビキチンプロモーターからeGFP/kan融合タンパク質を発現する(スライド4に描かれている)。この融合タンパク質は、個々のタンパク質それぞれの特徴を保持し、従って、視覚マーカーと選択剤の両方の機能を一つに兼ね備えるという明白な利点を有する。
【0060】
図7Aから図7Cは、アロエ植物から目的タンパク質を産出するためのコンストラクトの別の例示的なセットをあらわす。図示されたベクターは、二つの遺伝子が一つの転写産物として発現することを可能にするが、この二つの遺伝子はそれでもなお二つの個別のタンパク質に翻訳される。これを実現するために、IRESエレメント(内部リボソーム侵入配列)(SEQ. ID. NO. 34)を介在させることによって、二つの遺伝子は相互に分離される。IRESエレメントは、リボソームの結合と翻訳のための、第二の内部部位をもたらす。この第二部位は、3’側からIRESエレメントにクローニングされた転写産物が別々に翻訳されることを可能にする。これらのベクター、すなわちpzUSEIrK(TGC18)(SEQ. ID. NO. 43)およびpzUIIrK(TGC19)(SEQ. ID. NO. 39)は段階的に構築され、最終的に、カナマイシン抵抗性マーカーと共にeGFPもしくはIFN(それぞれ)の発現を可能にする。
【0061】
図示したように、pzEnostart(TGC13)ベクターは、制限酵素部位BamHI(5’末端)およびSacI(3’末端)を含むプライマーを用いるPCR(プライマー配列は表1)によって、eGFPを最初に増幅することによって作製される。このPCR産物は、BamHI/SacIフラグメントとしてpZErOにクローニングされる。このタンパク質はアルファアミラーゼシグナル配列との融合として発現されるので、このコンストラクトのeGFP遺伝子はATG翻訳開始部位を持たない(pzSE、TGC14の構築を参照)。これはスライド5に描かれている。これらのPCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0062】
図示したように、アルファアミラーゼ遺伝子から一本鎖のシグナル配列が合成され、その後PCRを用いる補充(fill in)によって二本鎖にされる(プライマー配列は表1)。このシグナル配列は、その後制限酵素PstIおよびBamHIで消化され、pzEnostart(TGC13)にクローニングされる。これはスライド5に描かれている。このシグナル配列はイン・フレームに、5’からeGFPにクローニングされ、シグナル配列とeGFPの融合に翻訳開始部位を提供する。得られるpzSE(TGC14)クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0063】
図示したように、ベクターpzUSE(TGC15)は、アルファアミラーゼシグナル配列(ss)-eGFP融合とユビキチンプロモーターを結合させるために構築される。pzSE(TGC14)由来のss-eGFPを含むこの遺伝子カセットは、PstI/SacIフラグメントとしてpzUI(TGC9)にクローニングされる。これは、(制限酵素PstIおよびSacIで消化することによってpzUI(TGC9)から切り離される)IFN遺伝子カセットを置換し、pzUSE(TGC15)を作製する。これはスライド5に描かれている。
【0064】
図示したように、IRESエレメントは、PCRによってpIRES2-EGFP(Clontech, BD Biosciences)から制限酵素部位SacI(5’末端)およびXhoI(3’末端)を含むプライマーで増幅される(プライマー配列は表1)。増幅されたIRESエレメントは、pzIr(TGC16)を作製するためにSacI/XhoIフラグメントとしてpZErOにクローニングされる。これはスライド6に描かれている。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0065】
図示したように、pzIr(TGC16)由来のIRESエレメントは、その後pzIrK(TGC17)を作製するためにSacI/XhoIフラグメントとしてpzK(TGC4)にクローニングされる。このようにしてIRESをkan/nos遺伝子カセットの上流に配置する。
【0066】
図示したように、IRES-kan/nos遺伝子カセットは、pzUSEIrK(TGC18)を作製するために、SacI/XbaIフラグメントとしてpzUSE(TGC15)にクローニングされる。これはスライド6に描かれている。このコンストラクトは、アルファアミラーゼシグナル配列との融合としてeGFPを発現し、eGFPを細胞からの分泌の標的とする。これにより、遺伝子組み換え植物内でのタンパク質の輸送と蓄積を視覚的に観察することが可能になる。このベクターは、カナマイシン抵抗性タンパク質(内部IRESエレメントから翻訳される)も発現し、遺伝子組み換え植物の選択を可能にする。
【0067】
図示したように、また、IRES-kan/nos遺伝子カセットは、pzUIIrK(TGC19)を作製するために、SacI/XbaIフラグメントとしてpzUI(TGC9)にクローニングされる。これはスライド7に描かれている。このコンストラクトは、分泌のためのシグナル配列と共にIFNアルファ2を発現する。このベクターは、カナマイシン抵抗性タンパク質(内部IRESエレメントから翻訳される)も発現し、遺伝子組み換え植物の選択を可能にする。TGC18とTGC19の両方とも、二つの遺伝子を一つの転写産物として発現し、第二プロモーターの必要性がなく、従って各ベクターの全体のサイズを縮小する。導入コンストラクトの全体のサイズを縮小することは、これらのエレメントが核に転移できる効率を増加し、安定的な組み込みと遺伝子組み換え植物の選択につながるので、このことは重要である。単一のプロモーター系は、かく乱フランキング遺伝子の発現のリスクや、最終的に導入遺伝子の発現に影響し得る他の状況も軽減する。
【0068】
上記のプラスミドは、他の目的コード配列をアルファインターフェロンと置換してもよい。当業者に理解されるように、これらはプラスミドの同じ遺伝子座もしくは他の位置に置換されてもよい。
【0069】
図8に図式的にあらわされるように、プロトロンビンコード配列(SEQ. ID. NO. 36)も、本発明の態様に従ってプラスミドベクターに組み込まれてもよい。プロトロンビンはトロンビンの前駆体タンパク質である。これは、可溶性フィブリノゲンを不溶性フィブリンストランドに変換する凝固タンパク質である、活性化トロンビンを切り離すために活性化因子Xによって2箇所で切断される。プロトロンビン遺伝子カセット(1874塩基対)はPCRプライマーを用いて増幅される(プライマー配列は表1)。eGFP遺伝子カセットは、制限酵素BamHIによってpzUSEIrK(TGC18)ベクターから切り離される。得られる5’突出は、その後平滑末端を作製するためにMung Beanヌクレアーゼで除去される。この平滑末端は、セ氏15度でプロトロンビン遺伝子カセットと一晩ライゲーションする前に、セ氏50度で2時間インキュベートすることによって仔牛小腸ホスファターゼ(CIP)を用いて脱リン酸化される。こうしてpzUSTIrK(TGC20)(SEQ, ID. NO. 42)が作製される。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0070】
また、図8に図式的にあらわされるように、ダームシジン(DCD)コード配列(SEQ. ID. NO. 30)も、本発明の態様に従ってプラスミドベクターに組み込まれてもよい。ダームシジンは、近年報告された広域抗菌性ペプチドで、汗腺に構成的に発現することがわかっている。このタンパク質は汗の中に分泌され、表皮表面に輸送される。PCR増幅DCD遺伝子カセットは、フランキングBamHI制限酵素部位を持つ(プライマー配列は表1)。BamHI処理したDCD遺伝子カセットは、eGFPによって占められていたpzUSEIrK(TGC18)ベクターのBamHI部位にライゲーションされ、pzUSDIrK(TGC21)ベクター(SEQ. ID. NO. 40)を作製する。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0071】
また、図8に図式的にあらわされるように、ヒト成長ホルモン(hGH)コード配列(SEQ. ID. NO. 32)も、本発明の態様に従ってプラスミドベクターに組み込まれてもよい。hGH遺伝子カセットは、フランキングBamHI制限酵素部位とともにPCR増幅によって作製される(プライマー配列は表1)。その後BamHI消化したhGH遺伝子カセットは、eGFPによって占められていたpzUSEIrK(TGC18)のBamHI部位にライゲーションされ、pzUSHIrK(TGC22)ベクター(SEQ. ID. NO. 41)を作製する。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0072】
また、図8に図式的にあらわされるように、ヒトインターフェロンガンマ(hIFNg)コード配列も、本発明の態様に従ってプラスミドベクターに組み込まれてもよい。hIFNg遺伝子カセットは、フランキングBamHI制限酵素部位とともにPCR増幅によって作製される(プライマー配列は表1)。その後BamHI消化したhIFNg遺伝子カセットは、eGFPによって占められていたpzUSEIrK(TGC18)のBamHI部位にライゲーションされ、pzUSIfgIrK(TGC23)ベクター(SEQ. ID. NO. 37)を作製する。PCR作製クローンは、DNA変異が一つも無いことを確認するためにシーケンスにかけられる。
【0073】
【表1】
【0074】
これらの配列はその独自のシグナル配列を含んでもよい。アロエ植物で機能することが示されたアルファインターフェロンのシグナル配列と同様に、プロトロンビン、ダームシジン(DCD)、ヒト成長ホルモン(hGH)、およびヒトインターフェロンガンマ(hIFNg)の天然シグナル配列も、アロエ植物で機能することが示され得る。とにかく、これらのタンパク質コード配列は、図示された図8の実施例のようにアルファアミラーゼシグナル配列を含むベクターにクローニングされ得る。
【0075】
上記のコンストラクトの作製において、形質転換に用いられる全ベクターは、250ml LB50μg/Lゼオシンで培養され、CsCl法を用いて分離される。その後ベクターは、各コンストラクトからの適切な発現を確実にするために、一過性トランスフェクション実験に用いられる。遺伝子銃を用いるトランスフェクションは、トウモロコシ、タバコ、およびアロエで実施され、eGFPの発現を視覚的に観察するか、あるいはIFNもしくはkanのいずれかの発現に対するrt-PCRによって、発現を観察した。
【0076】
<形質転換>
ベクターに組み込まれた、目的遺伝子と機能的な種々の機能単位を含むDNAコンストラクトを作製した後、DNAコンストラクトは、本発明の開示を考察することで当業者に理解される数多くの技術を用いて、アロエ細胞に導入される。これらのDNAコンストラクトは、概して安定的に形質転換されたアロエ植物10の形成を促進するために設計される。組み換えDNAで植物細胞を形質転換する数多くの方法が当該技術分野で周知であり、本発明において使用され得る。安定的なアロエ形質転換体を作製するために、ベクターに含まれるDNAコンストラクトをアロエ細胞もしくは組織に組み込むいくつかの方法は、A. tumefaciensもしくはA. rhizogenesの感染、複製欠損性ウィルスの感染、バイオリスティック形質転換、プロトプラスト形質転換、花粉への遺伝子導入、生殖器官への導入、未熟胚への導入、もしくは同様の方法を含むことができる。現在、よく用いられている植物形質転換方法のうちの二つは、Agrobacterium媒介形質転換とバイオリスティック形質転換である。
形質転換アロエ細胞もしくはカルス組織は、形質転換細胞を選択するために、典型的には適切な栄養培地で培養される。しばしば、選択培地は、非形質転換細胞を殺す毒素もしくは他の選択因子を含む。本発明の開示を考察することで当業者に理解されるように、様々な培地の交換により、目的遺伝子を含むアロエ植物10の産出が可能になる。
【0077】
形質転換の実践においては、典型的には、任意の一つの形質転換実験において、わずかな比率の標的植物細胞にのみDNAが導入される。マーカー遺伝子は、遺伝子組み換えDNAコンストラクトをゲノムに受け取って組み込むことにより、安定的に形質転換された細胞を特定するための、効率的な系を提供するために用いられる。好ましいマーカー遺伝子は、抗生物質もしくは除草剤などの選択剤に対する抵抗性を与える選択マーカーをもたらす。本発明の植物が抵抗性を示し得る除草剤のいずれも、選択マーカーとして有効な薬剤である。形質転換された可能性のある細胞は選択剤に晒される。生存細胞の集団においては、概して、抵抗性付与遺伝子が組み込まれ、細胞が生存できる十分なレベルで発現している細胞が存在する。細胞はさらに、外来DNAの安定的な組み込みを確認するためにテストされ得る。よく用いられる選択マーカー遺伝子は、カナマイシンとパロモマイシン(nptII)、ハイグロマイシンB(aph IV)およびゲンタマイシン(aac3およびaacC4)などの抗生物質に対する抵抗性を与えるもの、あるいは、グルホシネート(bar orpat)とグリホセート(aroAもしくはEPSPS)などの除草剤に対する抵抗性を与えるものを含む。そのような選択可能マーカーの例は、U.S. Pat. No. 5,550,318、5,633,435、5,780,708および6,118,047に記載され、これらは全て引用により本明細書に組み込まれる。形質転換体を視覚的に特定する能力を与える選択可能マーカーも利用することができ、例えば、ルシフェラーゼもしくは緑色蛍光タンパク質(GFP)などの、有色タンパク質もしくは蛍光タンパク質を発現する遺伝子、あるいは、様々な発色基質が知られている、ベータグルクロニダーゼもしくはuidA遺伝子(GUS)を発現する遺伝子などである。
【0078】
一般的に、標的植物の系統のゲノムに、ランダムに、すなわち非特異的な位置に、組み換えDNAを導入することが有益である。特別な場合には、例えばゲノムの既存遺伝子を置換したり、植物ゲノムの既存プロモーターを使用したり、あるいは遺伝子発現に活性があるとわかっている所定部位に組み換えポリヌクレオチドを挿入するために、部位特異的組み込みを実現するために組み換えDNA挿入を標的とすることが有益となり得る。U.S. Pat.No. 4,959,317に開示されているcre-lox、およびU.S. Pat. No. 5,527,695に開示されているFLP-FRTを含む、埋め込み物として機能することが知られているいくつかの部位特異的な組み換え系が存在する(両特許は引用により本明細書に組み込まれ、下記でより詳細に述べられる)。
【0079】
Agrobacterium tumefaciensによる植物形質転換系として、形質転換コンストラクト上に存在する付加的なエレメントは、植物ゲノムへの組み換えポリヌクレオチドの組み込みを促進する、T-DNA左右境界配列を含む。agrobacterium(A. tumefaciens)感染では、アロエカルス組織は、DNAコンストラクトを組み込むベクターを含むagrobacteriumの一晩培養物と共に室温で1時間インキュベートされる。アグロバクテリウムは適切な選択培地で培養される。選択培地は典型的にはストレプトマイシンとカナマイシンを含む。選択培地はアセトシリンゴンも含んでもよい。50μMから250μMの濃度のアセトシリンゴンは、典型的には単子葉植物の伝染性の効率を増加させる。選択培地上での培養の後、感染アロエ組織はセ氏25度で暗所で二日間、選別されずにペトリ皿の培地に移される。適切な培地の例は、アセトシリンゴンを含むMS培地である。その後、agrobacteriumを殺すために二日間にわたってセフォタキシムが加えられ得る。その後細胞は、形質転換アロエ細胞を選択するための50 mg/Lのカナマイシンと、発芽誘導成長因子(0.2 mg/L NAA、2 mg/L BAP)を含む培地上に移され、16時間の光照射で4週間にわたって培養される。その後再生芽は、土壌に移される前に根を発達させるために(約4〜6週間後)、発根培地(1/2 MS with 0.2 mg/L NAA)に移され得る。
【0080】
バイオリスティック形質転換では、アロエ細胞もしくはカルス組織は、ベクターコンストラクトを直接的に打ち込まれる。バイオリスティック形質転換の様々な実施形態と態様は、U.S. Pat. No. 5,015,580(大豆)、5,550,318(トウモロコシ)、5,538,880(トウモロコシ)、5,914,451(大豆)、6,160,208(トウモロコシ)、6,399,861(トウモロコシ)、および6,153,812(小麦)に開示され、Agrobacterium媒介形質転換はU.S. Pat. No. 5,159,135(綿)、5,824,877(大豆)、5,591,616(トウモロコシ)、および6,384,301(大豆)に開示され、これら全ては引用により本明細書に組み込まれる。さらに、例えばBio-Rad, Inc.のBiolistic PDS-1000/He particle delivery systemなど、様々なバイオリスティック形質転換装置が使用され得る。この方法では、ベクターと関連DNAコンストラクトは、金もしくはその他の粒子に結合され、アロエ細胞に直接打ち込まれる。打ち込みされたアロエ細胞は、選択培地に移される前に、暗所で4日間選択されずに培養される。この場合も、選択培地は50 mg/Lのカナマイシンを含んでもよい。形質転換体は、典型的には8〜12週間の内に形成し始め、発芽・発根培地にさらに8〜12週間移すことができる。一旦根が形成し始めれば、小植物体は土壌に移すことができる。
【0081】
形質転換は、他の様々な技術を用いて実現することもできる。そのような技術は、複製欠陥ウィルスベクター系を用いるウィルス感染、特に液体培養で培養したカルス細胞もしくはプロトプラストの電気穿孔法、および、プロトプラストへのPEGもしくは脂質媒介トランスファーを含む。その後、形質転換体の選択は、バイオリスティック形質転換で概説したものと同じ基本ステップに従うことができる。
【0082】
一旦アロエカルス組織に導入されれば、図3に大まかに示されるように、コンストラクトは植物の遺伝物質に組み込まれ得る。他の態様では、コンストラクトはアロエ植物10の遺伝物質の外側の細胞に安定的に組み込まれ得る。コンストラクトによって、目的タンパク質はいくつかの形式の転写誘導を伴わずに発現してもよいし、しなくてもよい。一態様では、一旦アロエ細胞に導入されたコンストラクトは、タンパク質合成もしくはアロエ植物10の特定の組織への輸送を誘導し得る。典型的には、これは標的シグナル配列が目的タンパク質上に含まれる際に起こる。
【0083】
<部位特異的組み込み(Cre-Lox)>
一態様では、本発明はアロエ植物に導入されたDNAコンストラクトの部位特異的組み込みもしくは切除を利用してもよい。部位特異的組み込みもしくは切除の利点は、形質転換コンストラクトが通常は宿主ゲノムに複数コピーでランダムに組み込まれるという、従来の形質転換技術に伴う問題を克服するために使用できるということである。この導入DNAの宿主細胞ゲノムへのランダム挿入は、もし外来DNAが必須遺伝子に挿入される場合、致命的になることがある。加えて、導入遺伝子の発現は、周囲のゲノムDNAに起因する“位置効果”によって影響され得る。さらに、遺伝子抑制、組み換え、および予測不可能な遺伝を含む、植物が保有する導入遺伝子の複製コピーに関連する問題のために、一般的には遺伝子組み換えアロエ植物10のゲノムに挿入されたDNAコンストラクトのコピー数を制御することが好ましく、多くの場合単一コピーのDNAコンストラクトの挿入のみが望ましい。
【0084】
導入遺伝子もしくは導入遺伝子の一部の、部位特異的組み込みもしくは切除は、相同組み換えを用いて植物で実現できる(例えば、特にその全容が引用により本明細書に組み込まれる、U.S. Pat. No. 5,527,695を参照)。相同組み換えは、相似ヌクレオチド配列を持つDNA配列の任意の対の間の反応であり、ここで二つの配列は新たな組み換えDNA種を形成するために相互作用(組み換え)する。相同組み換えの頻度は、共通ヌクレオチドDNA配列の長さが増えるにつれて増加し、また環状プラスミド分子よりも線状化プラスミド分子の方が高頻度で起こる。相同組み換えは、完全に同一とは言えない二つのDNA配列間でも起こり得るが、組み換え頻度はニ配列間の相違が増すにつれて減少する。
【0085】
導入されるDNA配列は、目的DNA分子を、宿主細胞の内在性配列と相同性を有する配列に結合することで、相同組み換えによって標的となることができる。一旦DNAがアロエ細胞に入れば、導入されるDNAを相同ゲノムDNA配列が位置する部位に挿入するために、二つの相同配列が相互作用することができる。従って、導入されるDNA上に含まれる相同配列の選択が、導入されるDNAが相同組み換えによって組み込まれる場所を決定する。例えば、もし目的DNA配列が宿主アロエ細胞の単一コピー遺伝子と相同性を持つDNA配列に結合される場合、目的DNA配列はその単一の特定部位のみに相同組み換えによって挿入される。しかしながら、もし目的DNA配列が宿主真核細胞のマルチコピー遺伝子と相同性を持つDNA配列に結合される場合、目的DNA配列は、遺伝子のコピーが位置する特定部位のそれぞれに、相同組み換えによって挿入され得る。
【0086】
DNAは、単一相互組み換え(導入DNAの全長の挿入をもたらす)もしくは二重相互組み換え(二つの組み換え現象間に位置するDNAのみの挿入をもたらす)のいずれかを含む相同組み換え反応によって、宿主ゲノムに挿入され得る。例えば、もし選択された遺伝子が位置する遺伝子位置に外来遺伝子を挿入したい場合、導入されるDNAは選択された遺伝子と相同性を持つ配列を含んでいなければならない。単一相同組み換え現象では、その後、導入されるDNA配列全体が選択された遺伝子に挿入されることになる。もう一つの方法として、二重組み換え現象は、目的DNA配列(ゲノムに挿入される配列)の各末端を、選択された遺伝子と相同性を持つDNA配列とフランキングすることによって実現できる。各相同フランキング領域を含む相同組み換え現象は、外来DNAの挿入をもたらす。従って、ゲノム相同性を持つ二つの領域間に位置するDNA配列のみが、ゲノムに組み込まれることになる。
【0087】
導入されるDNA配列は、相同組み換えによって特定のゲノム位置への挿入の標的となることができるが、高等真核生物では、相同組み換えはランダム挿入現象に比べて比較的まれな現象である。アロエ細胞では、外来DNA分子はアロエ植物ゲノムで相同配列を見つけ、約0.5〜4.2 x 10-4の頻度で組み換えを起こす。従って、相同組み換えによって組み込まれた導入DNA配列を含む任意の形質転換細胞も、ランダムに組み込まれた導入DNA配列の多数コピーを含む可能性がある。これらのランダム挿入を防ぐ一つの方法は、部位特異的リコンビナーゼ系を利用することである。概して、部位特異的リコンビナーゼ系は三つの要素から構成される。すなわち、二対のDNA配列(部位特異的組み換え配列)と特異的酵素(部位特異的リコンビナーゼ)である。部位特異的リコンビナーゼは、二つの部位特異的組み換え配列間でのみ組み換え反応を触媒する。
【0088】
バクテリオファージP1のCre/lox系(特にその全容が引用により本明細書に組み込まれるU.S. Pat. No. 5,658,772)、酵母のFLP/FRT系(Golic and Lindquist, 1989)、ファージMuのGinリコンビナーゼ(Maeser and Kahmann, 1991)、E. coliのPinリコンビナーゼ(Enomoto et al., 1983)、およびpSR1プラスミドのR/RS系(Araki et al., 1992)を含むが、これに限定されない数々の異なる部位特異的リコンビナーゼ系を本発明に従って利用できる。バクテリオファージP1 Cre/lox系および酵母FLP/FRT系は、導入遺伝子の部位特異的組み込みもしくは切除に特に有用な二つの系から構成される。これらの系では、リコンビナーゼ(CreもしくはFLP)は、介在配列を反転または切除するために、それぞれの部位特異的組み換え配列(それぞれloxもしくはFRT)と特異的に相互作用する。これら二つの系のそれぞれの配列は比較的短く(loxは34 bp、FRTは47 bp)、従って形質転換ベクターと一緒に用いるのに都合がよい。
【0089】
図8はFlp/FrtおよびCre/Loxpリコンビナーゼ系を用いる特定のプロトコル例をあらわす。こうした系では、バクテリオファージと酵母由来の部位特異的リコンビナーゼが、試験管と生体の両方でDNAを扱うツールとして使用され得る。リコンビナーゼ/組み換え部位の組み合わせは、Cre-Lox、FLP-FRT、およびR-RSを含み、ここでCre、FLP、およびRはリコンビナーゼで、Lox、FRT、およびRSは組み換え部位である。この系を用いるために、組み換えに特異的な部位を含む遺伝子組み換え植物が作製される。親となる遺伝子組み換えアロエ植物10は、選択可能マーカーを発現する、FrtおよびLox部位をタンデムに組み込んだベクターを形質転換することによって作製される。FrtおよびLox部位の両方とも、三つの要素から構成され、それぞれ13ヌクレオチドからなる2つの逆方向反復間に、8ヌクレオチドからなるスペーサー配列が存在する。逆方向反復は特定のリコンビナーゼに対するDNA結合ドメインとしてはたらき、一方スペーサーエレメントは、変更できるが相同組み換えに必須である。LoxPもしくはFRT部位のいずれかのスペーサーエレメントを改変することにより、複数の別個の組み換え部位を作ることができる。FrtおよびLox部位を改変することにより、多重部位に誘導される組み込みを可能にする系を作ることができる(図8に概説する通り)。この系は数多くの利点を持ち得る。最初の親株の世代の後、内在性遺伝子の破壊が最小限に抑えられる。形質転換効率は、部位特異的リコンビナーゼの発現によって増加する。loxおよびfrt部位をタンデムに用いることで、目的遺伝子を保持しながら選択マーカーの選択的除去が可能になる。また、一旦親株が作られれば、細胞増殖と再生能は増強される。しかしながら親株は第一段階であり、標準的な形質転換法(バイオリスティックもしくはAgrobacterium)に従って作製される。後代の遺伝子組み換え植物は、LoxPおよびFrt部位の両方、さらに目的遺伝子と選択可能マーカーを含むベクターを、Creリコンビナーゼを発現するベクターと一緒に共形質転換することによって作製される。Creリコンビナーゼの発現は、ベクターと親株のLoxP部位を通して相同組み換えを誘導する。形質転換体は選択可能マーカーの発現によって選択され、再生するように誘導される。その後のFlpリコンビナーゼ発現ベクターとの形質転換は、選択可能マーカーを除去し、次の他のLoxp部位への組み込みを可能にすることができる。
【0090】
<生存株の産出>
アロエ細胞もしくはアロエ組織の形質転換に続いて、形質転換アロエ細胞もしくはアロエ組織は、小植物体に成長され得る。選択剤に晒されても生き残ったアロエ細胞、もしくはスクリーニングアッセイで陽性となったアロエ細胞は、再生培地で培養され得、遺伝子組み換えアロエ植物10に成長させられ得る。形質転換アロエ細胞から再生した成長中のアロエ小植物体は、植物成長ミックスに移され、温室もしくは成熟のための栽培室に移す前に、例えば約85%の相対湿度、600 ppmのCO2、25〜250マイクロアインシュタインm-2s-1の光で環境制御されたチャンバーで、寒さに慣れさせることができる。形質転換アロエ植物10は、形質転換体が特定された後、最初の組織に応じて約6週間から10ヶ月で再生される。再生された形質転換アロエ植物10もしくはその子孫となる種子もしくは植物体は、通常は組み換えDNAの発現を調べられる。
【0091】
形質転換未分化カルス組織もしくは分裂組織から遺伝子組み換えアロエ植物10を再生することは、主として成長因子オーキシンとキネチンを操作することを含む。第一段階は芽の成長促進である。発芽誘導培地は、典型的には低濃度のオーキシン(0.2 mg/L NAA)と比較的高濃度のサイトカイニン(2 mg/L BAP)を含む。2%スクロースにまで減らした糖濃度(浸透圧に影響する)も、植物再生に寄与し得る。発芽培地上の細胞は、12hr/dayの光周期で25℃のインキュベーターに置かれる。形成し始めた芽は、約2 cmの長さに達した後、初期カルスから切除され、発根培地に置かれる。これらの芽は、さらに芽の発達を促進するために発芽培地に保持することもできる。発根培地は根の発達を促進し、典型的には1/2 MS培地と比較的低濃度のオーキシン(0.2 mg/L NAA)を含む。根が成長し始めた後(1〜2 cm)、小植物体は土壌に移され、徐々に相対湿度を下げることで適応させる。一旦生成されれば、遺伝子組み換えアロエ植物10は、種子、クローン増殖法、もしくは本発明の開示を考察することで当業者に理解される別の方法で、繁殖され得る。
【0092】
<植物もしくは植物細胞からのタンパク質の抽出と精製>
ベクターを介して植物細胞と植物に導入された構造タンパク質もしくは酵素は、根、塊茎、葉、種子、花、樹液を含むアロエ植物10の様々な組織において、あるいは植物の部分と発育段階の組み合わせにおいて見られる。一態様では、一つもしくは複数の目的タンパク質は、葉の中心部のゲルマトリックスに濃縮される。別の態様では、一つもしくは複数の目的タンパク質は生物活性があり、アロエ葉12から抽出されてある程度の薬効を提供する。別の態様では、一つもしくは複数の目的タンパク質、および少なくとも一つの天然アロエタンパク質、糖質および/またはゲルマトリックスに見られるその他の化合物は、患者に所望の治療を施すように相乗的に作用する。タンパク質はバイオマス全体もしくは特定の組織から抽出できる。植物細胞からのタンパク質抽出は、物理的方法と化学的方法を含んでもよい。葉もしくは樹液からのタンパク質抽出は、ろ過、超遠心分離法、化学抽出、およびアフィニティークロマトグラフィーを含んでもよい。
【0093】
一態様では、目的タンパク質は形質転換アロエ植物10の葉のゲル内に蓄積し得る。この領域は主に水であり(約99%)、有害なプロテアーゼを含まない。ゲルの抽出は、当業者に理解される従来技術である。タンパク質のいくつか、特に化粧品タンパク質は、アロエゲルを直接補うために用いられてもよく、またゲルから個別に分離される必要が全くなくてもよい。
【実施例】
【0094】
<植物細胞の分離方法>
第一の実施例では、直接使用のため、あるいはカルス発生のための芽分裂組織が、アロエ(Aloe vera、Aloe ferox、もしくはAloe arborescenceのいずれか)の茎から分離される。葉の部分は除去され、茎はアロエ葉の縦軸を通って横方向に切られる。組織はTween 20(0.05%)と次亜塩素酸塩(5%)の混合物で10分間滅菌され、その後30秒エタノールで処理し、滅菌水で三回洗浄した。切片はその後、様々な濃度の成長因子(オーキシンとサイトカイニン)と2〜3%のスクロースを含むアガー(0.8〜1%)を含むMS培地の入ったプレートに、露出面を横倒しにして置かれる。生育条件は所望の処理過程に応じて異なる。芽の発達を伴わないカルス培養は、典型的にはこれらの切片を、BAPを含まないNAA(2〜5 mg/L)か、もしくは低濃度のBAP(0.2 mg/L)を含むNAA(2〜5 mg/L)で生育させることによって開始される。未分化細胞は切られた茎の範囲に沿って成長し始め、別の皿上で継代培養されて保持され得る。発芽誘導は、こうした切片を発芽培地(0.2 mg/L NAA(もしくはIAA 0.2 mg/L)と2 mg/L BAPを含むMS培地)に置くことによって直接的に開始させることができる。NAAとIAAは両方ともオーキシンの供給源としてはたらき、また、効果のあるオーキシンとサイトカイニン両方の濃度の範囲が存在する。未熟胚由来のカルス細胞は、市販の種子から育成される。種子は最初に滅菌される(エタノールで30秒、5%次亜塩素酸塩と0.05% Tween 20で15分間、ならびに滅菌水で3回洗浄)。滅菌された種子は、その後セ氏4度で水中に一晩放置される。種皮を除去することによって未熟胚が分離される。三角形の種子の一角に小さな切れ目を作り、胚を搾り出す。その後これは、カルスを誘導するためにオーキシンのみ(NAA 2〜5 mg/L)を含むMS培地か、あるいはカルスと芽の増殖を誘導するためにオーキシンとサイトカイニン(NAA 0.2 mg/L、BAP 2 mg/L)を含むMS培地上に置かれる。糖濃度は通常2〜3%である。
【0095】
茎頂分裂組織由来のカルス細胞は、葉を全て除去した若いアロエ植物10の基部から切り取った1〜2cmの切片から育成される。切片は表面を滅菌し、MS培地とオーキシンとサイトカイニン(2 mg/L NAA 0.2 mg/L BAP)を含む1%アガーに移される。
【0096】
カルス細胞誘導は1〜2週間以内で開始する。細胞はその後これらの培養から分離され、形質転換の出発物質として使用することができる。
【0097】
[DNAコンストラクト]
1.pBI121ユビキチンインターフェロンベクター(pBI-UI)
pBI-UIの作製の骨格として、pBI121 Agrobacteriumバイナリベクターが使用される。シグナル配列を持つヒトインターフェロンアルファ2遺伝子は、5’PstIおよび3’SacI/XhoI制限部位を含む遺伝子特異プライマーを用いてヒト293細胞から増幅される。587 bpのインターフェロンPCR産物はpZErOにクローニングされ、シーケンスにかけられる。トウモロコシ由来の1962 bpユビキチン(Ubi)プロモーターエレメントも、ベクターpUBI-GFP由来の領域を増幅し、5’HindIIIおよび3’PstI制限部位にクローニングすることによって、pZErOにクローニングされる。Ubiフラグメントは、その後pZeroインターフェロンベクターにHindIII/PstI(pZErO UI)を用いてサブクローニングされる。無傷のUbiインターフェロンカセットは、その後HindIII/SacIを用いてpBI121にサブクローニングされ、CaMV 35SプロモーターとGUS遺伝子を除去して、pBI UIを得る。このベクターは、agrobacterium媒介感染および形質転換に必要な左右のTエレメント境界領域を保持し、Nosプロモーターの制御下にカナマイシン抵抗性遺伝子(NPTII)を発現する。
【0098】
2.pZErOユビキチンインターフェロンIRESカナマイシン(pZ UIIK)
このベクターは、UbiプロモーターをpZErOクローニングベクターにクローニングすることによって作製される。このフラグメントの後ろに、IRES(内部リボソーム侵入配列)とカナマイシン抵抗性遺伝子と一緒に、ヒトインターフェロンアルファ2を含むカセットがクローニングされる。この単位は単一の転写産物として発現されるが、二つの別個のタンパク質として翻訳される(IRESによってもたらされる第二転写開始部位のため)。これにより、選択可能マーカーとインターフェロン両方がUbiプロモーターの制御下で発現することが可能になる。
【0099】
3.CreおよびFlpリコンビナーゼを利用する二重プラスミド系
標的組み換えのためのcreおよびflp部位を持つ選択可能マーカーを発現する親株が最初に作製される。目的遺伝子を持つ第二トランスフェクション導入ベクターがcre部位に向けられる。その後不要な遺伝物質がflpを用いて除去される。
【0100】
[分離植物細胞へのDNAコンストラクトの導入]
1.Agrobacterium媒介遺伝子導入
Agrobacterium strain LB4404(Invitrogen, Carlsbad, CA)は、pBI UIベクターと、50μg/mlカナマイシンと100μg/mlストレプトマイシンを含むLBアガープレート上で選択された陽性クローンと共に電気穿孔された。個々のクローンは250μMアセトシリンゴンを含むLB培地でセ氏30度で一晩培養された。感染は、植物切片もしくはカルスを一晩培養に沈水させ、滅菌ろ紙上にブロットドライ(blotting dry)し、暗所でセ氏25度で250μMアセトシリンゴンを含むMSアガープレート上に播種することで行われた。感染から二日後、Agrobacteriumを殺すために、組織は200μMセフォタキシムを含むMSアガープレートに移された。その後、形質転換体を選択し芽再生を誘導するために、組織は50 mg/Lカナマイシンと0.2 mg/L NAAと2 mg/L BAPを含むMSアガープレートに移される。組織は12時間の光照射でセ氏25度で育成される。不定芽は約2 cmの長さに達するまで育成され、その後切除され、選択を続けながら発根培地上に植え替えられる。目的遺伝子を発現する小植物体は、発現レベルを分析し、土壌に移される。
【0101】
2.バイオリスティック遺伝子導入
金粒子(0.6〜1μm)がDNAコンストラクトを含むベクターで被覆される。金粒子は、浸漬させてボルテックスにかけることで70%エタノールで15 分間洗浄され、その後滅菌水で3回洗浄される。その後、金粒子は60 mg/mlの濃度まで50%グリセロールに再懸濁される。洗浄された粒子上にDNAベクターを被覆させるために、3 mgの粒子が1.5 mlマイクロフュージチューブに加えられる。これに5μlのDNAを1μg/μlの濃度で加え、50μlの2.5M CaCl2、および20μlの0.1Mスペルミジンを加え、2〜3分間ボルテックスする。これを安定させ、1〜2秒間遠心し、液体を除去する。これに140μlの70%エタノールを加え、遠心し、液体を捨てる。これに140μlの100%エタノールを加え、遠心して液体を捨てる。沈殿物に48μlの100%エタノールを加え、穏やかに再懸濁させる。遺伝子銃装置(PDS 1000/He, Bio-Rad Laboratories, Inc., Hercules, CA)は70%エタノールを用いて滅菌される。遺伝子銃装置は、マクロキャリアホルダーと停止スクリーン間の最短ギャップ距離を用いて組み立てられた(推奨設定)。被覆金粒子(8μl)はマクロキャリアの中心上にピペットされる。600〜1100 psiの破裂板を用いて、標的距離は標的組織に応じて設定された(組織は6 cm、カルスは9 cm)。
【0102】
第一の実施例では、打ち込み後に細胞は2 mg/L NAAを含むMSアガープレート上に、選択されずにセ氏25度で暗所に一週間置かれた。その後細胞は、選択のための50 mg/Lカナマイシンと、オーキシン(2 mg/L NAA)およびサイトカイニン(0.2 mg/L, 6-ベンジルアミノプリン)を含むMSアガープレートに4〜5週間以上移された。細胞はセ氏25度で12時間の光照射で培養される。
【0103】
第二の実施例では、マイクロプロジェクタイル打ち込み(microprojectile bombardment)が、MSプレート(4 g/L MS塩、1 mg/L (1000X)MS ビタミンストック、2 mg/L NAA、100 mg/L ミオイノシトール、2.76 g/L プロリン、30 g/L スクロース、100 mg/L カゼイン加水分解物、36.4 g/L ソルビトール、36.4 g/L マンニトール、2.5 g/L ゲルライト、pH 5.8)上で行われた。打ち込みされたカルスは、打ち込み後1時間にわたってMSOSM上に残され、その後MS初期培地(4 g/L MS塩、1 mg/L (1000X)MS ビタミンストック、2 mg/L NAA、100 mg/L ミオイノシトール、2 g/L プロリン、30 g/L スクロース、100 mg/L カゼイン加水分解物、2.5 g/L ゲルライト、pH 5.8)に移された。MS上で7〜10日後、打ち込みされたカルスは、形質転換細胞の選択のための50 mg/L カナマイシンを含むMSS選択培地(4 g/L MS塩、1 mg/L (1000X)MS ビタミンストック、2 mg/L NAA、100 mg/L ミオイノシトール、30 g/L スクロース、2.5 g/L ゲルライト、pH 5.8)に移された。3週間後、個々のカルス片は新鮮なMSS培地に移された。打ち込みから6〜8週間の間に、カナマイシン抵抗性クローンが選択されたカルス片から現れた。
【0104】
3.プロトプラスト
プロトプラストは、外側の細胞壁がない植物細胞である。このような細胞を作製する利点は、トランスフェクション効率を高め、こうした細胞が融合して体細胞雑種を形成できることである。体細胞雑種は異なる植物由来の遺伝子を混ぜ合わせ、全く新しいものをもたらす可能性がある。
【0105】
本発明に従ってプロトプラストを用いる技術は、細胞を液体培養で培養することを含んでもよい(対数増殖期の2〜3日間)。その後細胞を遠沈させる(1000 xg 5分間)。細胞は、セルラーゼ、マセロザイム、ペクトリアーゼを含む溶液に再懸濁される。これをセ氏25度で暗所で一晩振盪させる。遠心分離機にかけ(600 xg 5分間)、上清を除去する。プロトプラストを培地に再懸濁し、残りの酵素を除去するためにもう一度遠心する。プロトプラストは保存も増殖もできず、比較的迅速にトランスフェクションや体細胞融合に使用しなければならない。
【0106】
[DNAコンストラクトを含む生存植物の産生]
第一の実施例では、成功した形質転換体は、最初に細胞毒素(例えばカナマイシン)に対する抵抗性で選択される。安定的にトランスフェクションされた細胞は、抵抗性マーカーと目的遺伝子(例えばインターフェロンをコードする遺伝子など)の両方を発現しているはずであり、さらに正常な植物を再生できるはずである。選択を開始するために、50 mg/LのカナマイシンがMSアガープレートに加えられる。選択の経過は、細胞が複製する速度に一部依存するが、6〜10週間で起こり得る。この間に、0.2 mg/L NAAと2 mg/L BAPをMSアガープレートに加え、12 hr/dayの光周期(淘汰圧を続けながら)で芽発達を促進することにより、形質転換細胞の再生が誘導される。不定芽が発達し始めると、これらは約2 cmの長さで遺伝子発現を分析され得る。所望の遺伝子産物を発現する芽(RT-PCR、Western blot、および生物学的検定法によって分析する)は、続いて根形成を誘導するためにMSアガープレートに移される(1/2 MSと0.2 mg/L NAA)。
【0107】
第二の実施例では、遺伝子組み換えカルスの再生は、約15の小切片(約4 mm)を、ろ過滅菌したカナマイシン(50 mg/L)を加えた再生培地I(4.3 g/L MS塩、1 ml/L (1000X)MSビタミンストック、100 mg/L ミオイノシトール、60 g/L スクロース、3 g/L ゲルライト、pH 5.8)に移し、セ氏25度で暗所で2〜3週間インキュベートすることによって行われる。成熟体細胞胚は、ろ過滅菌したカナマイシン(50 mg/L)を加えた、発芽のための再生培地II(4.3 mg/L MS塩、1 ml/L (1000X)MSビタミンストック、100 mg/L ミオイノシトール、30 g/L スクロース、3 g/L ゲルライト、pH 5.8)上で、光下に移される。
【特許請求の範囲】
【請求項1】
植物内で、前記植物に対して外来性の生物活性タンパク質を生成するために、遺伝子組み換えアロエ植物を生成する方法であり、
アロエ細胞の未分化カルスを、1-ナフタレン酢酸(NAA)もしくはインドール3-酢酸(IAA)および6-ベンジルアミノプリン(BAP)を含む栄養培地で培養するステップと、
前記カルスに、プロモーター、前記外来タンパク質をコードする配列、終結配列、および分泌シグナルペプチドをコードする転移配列を含む組み換えDNAコンストラクトを導入するステップと、
前記カルスを、0.2 mg/l NAAもしくはIAAおよび2 mg/ml BAPを含む発芽・選択培地で培養するステップと、
前記発芽・選択培地で再生された芽を、0.2 mg/l NAAを含む発根培地で培養するステップと、
一旦根が形成し始めたら、小植物体を土壌に移すステップと、
を含み、
前記外来タンパク質は、インターフェロン、免疫グロブリン、リンフォカイン、哺乳類成長因子、哺乳類ホルモン、血液因子、組織適合抗原、SEQ. ID. NO. 36によってコードされるプロトロンビン、およびSEQ. ID. NO. 30によってコードされるダームシジンから選択される哺乳類タンパク質である、
ことを特徴とする、方法。
【請求項2】
前記カルスはアロエ種子由来である、請求項1に記載の方法。
【請求項3】
前記カルス組織は前記組み換えDNAコンストラクトを直接打ち込まれる、請求項1もしくは2に記載の方法。
【請求項1】
植物内で、前記植物に対して外来性の生物活性タンパク質を生成するために、遺伝子組み換えアロエ植物を生成する方法であり、
アロエ細胞の未分化カルスを、1-ナフタレン酢酸(NAA)もしくはインドール3-酢酸(IAA)および6-ベンジルアミノプリン(BAP)を含む栄養培地で培養するステップと、
前記カルスに、プロモーター、前記外来タンパク質をコードする配列、終結配列、および分泌シグナルペプチドをコードする転移配列を含む組み換えDNAコンストラクトを導入するステップと、
前記カルスを、0.2 mg/l NAAもしくはIAAおよび2 mg/ml BAPを含む発芽・選択培地で培養するステップと、
前記発芽・選択培地で再生された芽を、0.2 mg/l NAAを含む発根培地で培養するステップと、
一旦根が形成し始めたら、小植物体を土壌に移すステップと、
を含み、
前記外来タンパク質は、インターフェロン、免疫グロブリン、リンフォカイン、哺乳類成長因子、哺乳類ホルモン、血液因子、組織適合抗原、SEQ. ID. NO. 36によってコードされるプロトロンビン、およびSEQ. ID. NO. 30によってコードされるダームシジンから選択される哺乳類タンパク質である、
ことを特徴とする、方法。
【請求項2】
前記カルスはアロエ種子由来である、請求項1に記載の方法。
【請求項3】
前記カルス組織は前記組み換えDNAコンストラクトを直接打ち込まれる、請求項1もしくは2に記載の方法。
【図1】

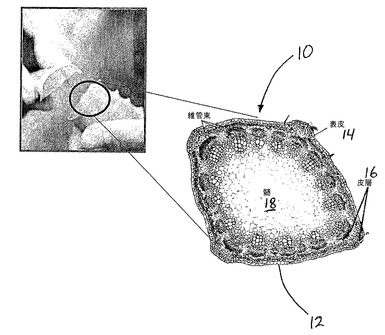
【図2】
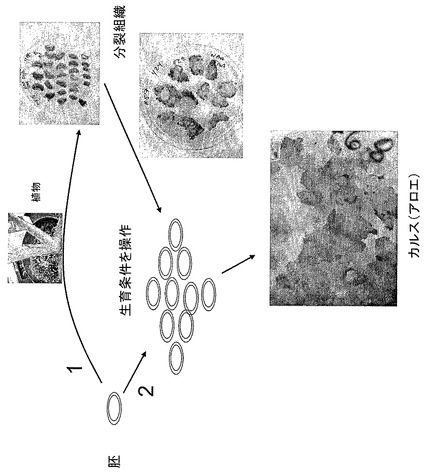
【図3】
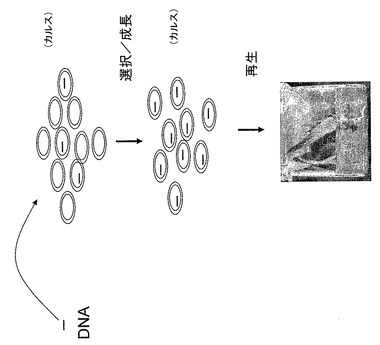
【図4】

【図5】

【図6A】
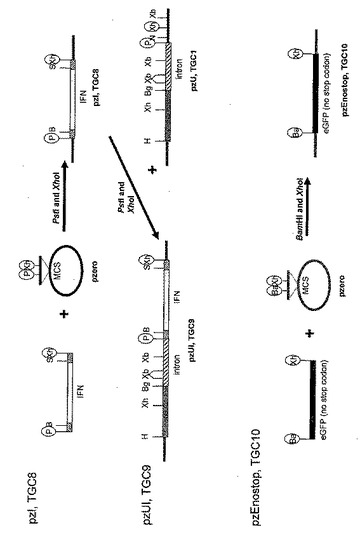
【図6B】
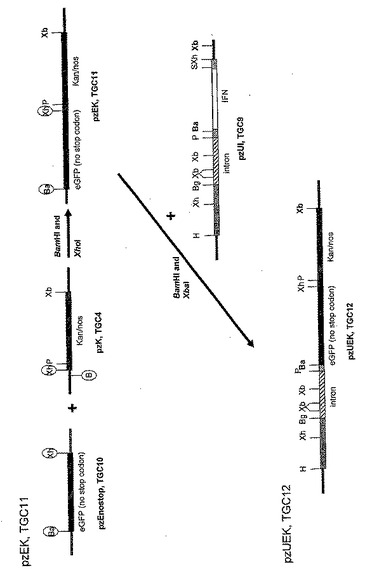
【図7A】
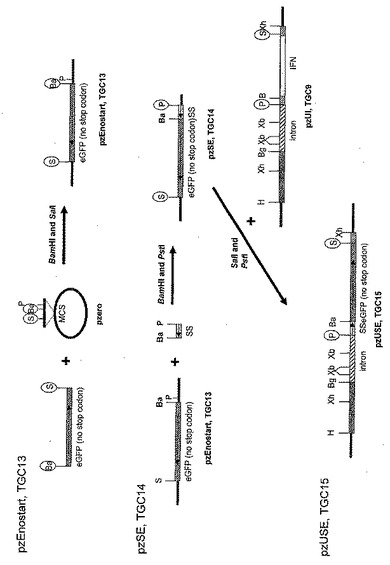
【図7B】
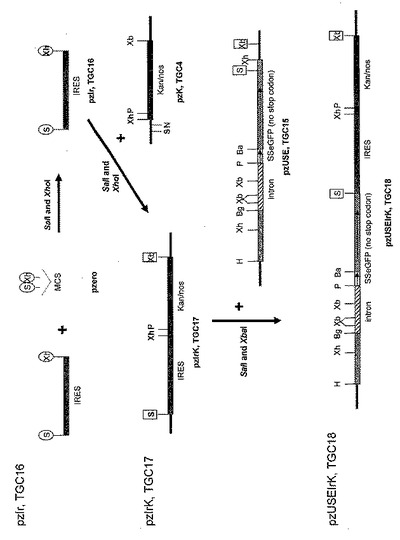
【図7C】

【図8】
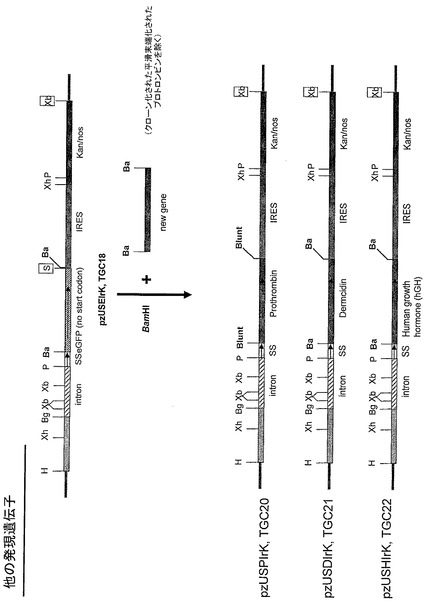
【図9】
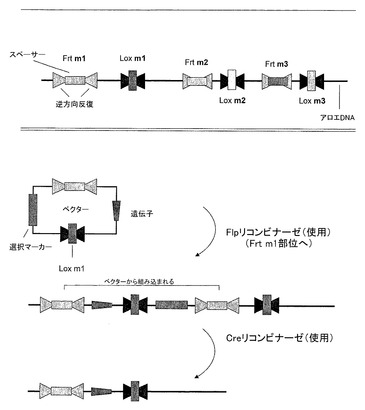
【図2】
【図3】
【図4】
【図5】
【図6A】
【図6B】
【図7A】
【図7B】
【図7C】
【図8】
【図9】
【公開番号】特開2012−196221(P2012−196221A)
【公開日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願番号】特願2012−126986(P2012−126986)
【出願日】平成24年6月4日(2012.6.4)
【分割の表示】特願2008−532506(P2008−532506)の分割
【原出願日】平成18年9月26日(2006.9.26)
【出願人】(508088649)ザグリーンセル,インク. (2)
【氏名又は名称原語表記】THEGREENCELL,INC.
【住所又は居所原語表記】7909 Tilbury Street,Bethesda,MD 20814,UNITED STATES OF AMERICA
【Fターム(参考)】
【公開日】平成24年10月18日(2012.10.18)
【国際特許分類】
【出願日】平成24年6月4日(2012.6.4)
【分割の表示】特願2008−532506(P2008−532506)の分割
【原出願日】平成18年9月26日(2006.9.26)
【出願人】(508088649)ザグリーンセル,インク. (2)
【氏名又は名称原語表記】THEGREENCELL,INC.
【住所又は居所原語表記】7909 Tilbury Street,Bethesda,MD 20814,UNITED STATES OF AMERICA
【Fターム(参考)】
[ Back to top ]