
タンパク質生産速度の変更方法
タンパク質、例えば抗体の細胞分泌速度を改変する方法およびこの方法によって作成される改変細胞が開示される。この方法および改変細胞は、治療、診断または研究目的のための高レベルのタンパク質を生産するために有用である。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は、タンパク質の細胞分泌速度の変更方法に関する。
【背景技術】
【0002】
タンパク質、例えば抗体の大規模生産は、典型的には、培養される生産細胞系からのタンパク質の分泌に依存する。培養細胞によって生産される分泌タンパク質は、周囲の細胞培養培地から容易に回収かつ精製することができる。
【0003】
タンパク質の細胞分泌速度は、バイオリアクターまたは他のシステムからの分泌タンパク質の生産および精製に影響を与える重要なファクターである。一般に、より高い精製タンパク質の収量は、細胞分泌速度が比較的高い場合に達成することができる。反対に、細胞分泌速度が低すぎる場合は、タンパク質精製は実行不可能であろう。
【0004】
低分泌性細胞の問題を回避する1つのアプローチは、低分泌性細胞の集団から高分泌性のサブクローン細胞を単離することであった。典型的には、これは、高分泌性細胞系の連続希釈、スクリーニングおよび選択を限定するという数回の、かつ労力集中的なラウンドを必要とする。あるいはまた、新しい細胞系が高分泌系であろうという希望において、問題のタンパク質を生産する完全に新しい細胞系が生成される。
【0005】
高分泌性細胞系を生成する従来の各アプローチは限界を有する。例えば、低分泌性細胞の集団からサブクローニングによって高分泌性細胞系を同定することは、集団における高分泌性細胞が相対的に希少であること、ならびにいずれかの高分泌性細胞の同定に要求される広大な量の時間と労力によって制約される。
【0006】
さらに、問題の抗体またはタンパク質を生産する新しい細胞系の生成は、新しい細胞系が高分泌性ではない可能性、ならびに抗体産生細胞を再生し、そして高分泌性細胞を同定するために要求される多大な量の努力によって制約される。若干の例では、低下した分泌速度をもたらす細胞の小胞体(ER)内でのタンパク質のミスフォールディング(misfolding)により、低分泌性細胞系のみが得られることもある。
【0007】
かくして、タンパク質の細胞分泌速度を変化させる有効な方法への要求が存在する。
【発明の開示】
【0008】
本発明の1つの態様は、細胞において少なくとも1種のUPR経路構成要素の活性を調節し;そして細胞を培養する段階を含んでなる、タンパク質の細胞分泌速度の変更方法である。
【0009】
本発明のその他の態様は、細胞において少なくとも1種のアンフォールド型(unfolded)タンパク質応答(UPR)経路構成要素の活性を調節し;そして細胞を培養する段階を含んでなる、タンパク質の細胞分泌速度を改変することによって生成される、変化した細胞分泌速度をもつ形質細胞である。
【0010】
本発明のその他の態様は、細胞において少なくとも1種のUPR経路構成要素の活性を調節し、そして細胞を培養することによって生成される、変化した細胞分泌速度をもつ形質細胞を含有するトランスジェニック動物である。
【0011】
本発明のその他の態様は、配列番号:15または配列番号:16において示される配列
を有する単離された核酸である。
【0012】
限定されるものではないが、本明細書において引用される特許および特許出願を含む、すべての公表物は、完全な記述をとおして引用によって本明細書に組み入れられている。
【0013】
本明細書で使用されるところの用語「抗体」は、広範な意味において意味され、そしてポリクローナル抗体、マウス、ヒト、ヒト化およびキメラモノクローナル抗体を含むモノクローナル抗体および抗体フラグメントまたは変異体を含む、免疫グロブリンまたは抗体分子を含む。抗体は形質細胞によって構成的に発現され、そして分泌される分泌タンパク質である。また抗体は、標準的方法、例えばハイブリドーマ生成によるか、または不死化されたB細胞、例えば骨髄腫細胞または他の細胞種、例えばチャイニーズハムスター卵巣(CHO)細胞、植物細胞および昆虫細胞への抗体重鎖および/または軽鎖遺伝子のトランスフェクションによって、不死化された形質細胞を用いて生産することができる。
【0014】
抗体フラグメントまたは変異体は、ミメティボディ(mimetibody)、Fabフラグメント、F(ab’)2フラグメント、Fcフラグメント、重鎖フラグメント、軽鎖フラグメント、および少なくとも1種の抗体ペプチド鎖の一部を含有する分子を含む。そのような部分は、抗体の可変、ヒンジもしくは定常部ペプチド鎖に対応してもよい。
【0015】
本明細書で使用されるような用語「ミメティボディ」は、一般式(I):
(V1(n)−Pep(n)−Flex(n)−V2(n)−pHinge(n)−CH2(n)−CH3(n))(m) (I)
[式中、V1は免疫グロブリン可変部のN末端の少なくとも1部分であり、Pepはエピトープに結合する少なくとも1種の生物活性ペプチドであり、Flexは、ミメティボディをもう一方の方向づけと結合特性をもつようにさせることによって構造的柔軟性を与えるポリペプチドであり、V2は免疫グロブリン可変部のC末端の少なくとも1部分であり、pHingeは免疫グロブリンヒンジ部の少なくとも1部分であり、CH2は免疫グロブリンCH2定常部の少なくとも1部分であり、そしてCH3は免疫グロブリンCH3定常部の少なくとも1部分であり、ここでnおよびmは整数1〜10であってもよい]
を有するタンパク質を意味する。ミメティボディは、構築物に存在する重鎖定常ドメインアミノ酸配列に応じて、IgG1、IgG2、IgG3、IgG4、IgA、IgM、IgDおよびIgEのような免疫グロブリン分子の異なるタイプの特性および機能を模倣できる。
【0016】
本明細書で使用されるところの用語「モノクローナル抗体」(mAb)は、実質的に均一な抗体の集団から得られる抗体(または抗体フラグメント)を意味する。モノクローナル抗体は高度に特異性であり、典型的には単一な抗原決定基に対向される。修飾語句「モノクローナル」は、抗体の実質的に均一な特徴を示し、いずれか特定の方法による抗体の生産を必要としない。例えば、マウスmAbはKohler et al.,Nature 256:495(1975)のハイブリドーマ法によって作成することができる。アクセプター抗体(典型的には、ヒトのようなその他の哺乳動物種)由来の軽鎖および重鎖定常部に関連してドナー抗体(典型的には、マウス)由来の軽鎖および重鎖可変部を含有するキメラmAbは、米国特許第4,816,567号に開示された方法によって調製されてもよい。非ヒト・ドナー免疫グロブリン(典型的にはマウス)由来のCDRおよび場合によっては結合親和力を維持するように改変されたフレームワーク・サポート残基を有する1種以上の免疫グロブリンに由来する分子の残りの免疫グロブリン由来の部分を有する、ヒト化mAbは、Queen et al.,Proc.Natl Acad Sci(USA),86:10029−10032,(1989)およびHodgson et al.,Bio/Technology,9:421,(1991)において開示された技術によって得ることができる。
【0017】
いずれの非ヒト配列も欠く完全にヒトのmAbは、例えば、Lonberg et al.,Nature 368:856−859(1994);Fishwild et al.,Nature Biotechnology 14:845−851,(1996)およびMendez et al.,Nature Genetics 15:146−156,(1997)において引用される技術によってヒト免疫グロブリントランスジェニックマウスから調製されてもよい。ヒトmAbはまた、例えば、Knappik et al.,J.Mol.Biol.296:57−86,(2000)およびKrebs et al.,J,Immunol.Meth.254:67−84,(2001)において引用される技術によってファージ・ディスプレイライブラリーから調製され、そして最適化されてもよい。
【0018】
本明細書で使用されるところの用語「細胞分泌速度」は、細胞が特定のタンパク質を分泌する速度を意味する。そのような速度は、時間の変化当たりの培養培地中に存在するタンパク質の量における変化として記述されてもよく、または細胞数に標準化され、そして単位「pg/細胞/日」で表されてもよい。
【0019】
本明細書で使用されるところの用語「短い干渉性RNA(short interfering RNA)」は、標的遺伝子転写物の切断を媒介する核酸配列を意味する。短い干渉性RNA(siRNA)は、二本鎖でもまたは短いヘアピン型であってもよい。二本鎖siRNAは、2本の個々の、逆平行の、アニールされたRNA鎖または両RNAおよびDNAを含有するアニールされた核酸鎖(例えば、5’−TTTTUUUU−3’にアニールされた5’−TTTTUUUU−3’または5’−UUUU−3’にアニールされた5’−TTTT−3’)からなってもよい。短いヘアピン型のsiRNAは、siRNAとして有効なステム・ループ構造または他の二次構造を形成することができる単一RNA鎖または単一RNA:DNAハイブリッド鎖からなってもよい。当業者は、siRNAが、他の改変体、例えばヌクレオシド類似体、バックボーン改変体、および改変siRNA核酸に標的遺伝子転写物の切断をなお媒介させる他の改変体を含んでもよいことを認識している。
【0020】
本明細書で使用されるところの用語「低分子(small molecule)」は、アミノ酸残基または核酸残基のみからはならない24,000g/mol未満の分子量をもつ化合物を意味する。
【0021】
本明細書で使用されるところの用語「転写制御配列」は、遺伝子の転写に必要である核酸配列または遺伝子の転写を増大または減少させる核酸配列を意味する。
【0022】
本明細書で使用されるところの用語「UPR経路構成要素」は、UPR経路を介するシグナル伝達を媒介するか、またはUPRを活性化する、ペプチド鎖または核酸配列、例えば転写制御配列を意味する。
【0023】
本発明は、細胞によるタンパク質の細胞分泌速度を改変するために有用な方法を提供する。本発明の方法の代表的使用は、治療、診断または研究目的に有用であるタンパク質、例えば抗体のための分泌速度の増進である。
【0024】
細胞系における低いタンパク質分泌速度は、UPRを介して分泌過程を減速または停止する、細胞ERにおけるミスフォールド(misfolded)タンパク質の蓄積によって惹起できる。ER膜におけるストレス感受性タンパク質は、アンフォールド型タンパク質の過剰を検出し、そしてUPRの引き金を引く。続いて、複雑なシグナル伝達カスケードを介して、シャペロン(chaperone)タンパク質Bipおよび転写因子XBP
−1およびCHOPが上方調節される。
【0025】
シャペロンタンパク質はアンフォールド型タンパク質に結合し、そして正確な折り畳みを助ける。転写因子CHOPは、一般に細胞増殖、分化および生存のネガティブレギュレーターとされる。CHOPの上方調節は細胞周期の停止を惹起し、その結果、UPR誘導に関与する好ましくない条件に対抗する時間を細胞に与えることが観察された。
【0026】
転写因子XBP−1は、形質細胞、大量の免疫グロブリンを分泌する分化したBリンパ球を生成するために要求される。形質細胞は変化されたUPRを現し、ここでは免疫グロブリン合成の前に若干のUPR遺伝子の上方調節が起きる。これらは、XBP−1、Bip、Grp94およびp50ATF6αを含み、そしてそれらの上方調節は形質細胞の分化と適当な分泌のために必須である。(Gass,J.N.,et al.,J.Biol.Chem.227,49047−49054(2002))。これに対して、CHOPはこの推移において上方調節されず、このことは異なる種類のUPRを示唆している。これらの分子の上方調節は、細胞タンパク質分泌速度における増大またはアポトーシスのいずれかを惹起する有利または有害な効果を有する。参照、Kaufman,R.J.,Genes Dev.13,1211−1233(1999)およびCudna,R.E.,et al.,Biotechnol.Bioeng.81,56−65(2003)。 本発明の方法では、タンパク質の細胞分泌速度は、細胞における少なくとも1個のUPR経路構成要素の活性を調節し、そして細胞を培養することによって改変される。本発明の方法は、抗体のようなタンパク質の細胞分泌速度の増大または減少に備える。
【0027】
本発明の実施態様では、タンパク質の細胞分泌速度は、UPR経路構成要素をコードしている核酸により細胞を安定してトラスフェクトすることによって増大される。UPR経路構成要素は、ポリペプチドまたは核酸配列、例えば転写制御配列であってもよく、これらはUPR経路をとおしてシグナル伝達を媒介するか、またはUPRを活性化する。UPR経路構成要素の例は、Bip、XBPおよびCHOPならびに類似の活性を有する変異体を含む。UPR経路構成要素の他の例は、IRE1、PERK、ATF4、eIF2α、GRP78、GRP94、calreticulin、シャペロン、および類似の活性を有する変異体を含む(参照、例えば、Cudna and Dickson,Biotechnol.Bioeng.81,56−65(2002))。転写制御配列の例は、シス作用UPR要素(UPRE)および種々のUPR遺伝子のプロモーター中に同定されたERSEである。当業者は他のUPR経路構成要素および転写制御配列も認識できるであろう。UPR経路構成要素は、配列番号:2(マウスBip)、配列番号:4(マウスXBP−1、スプライスされた形態)、配列番号:6(マウスXBP−1、スプライスされてない形態)、配列番号:8(マウスCHOP−10)、配列番号:10(ヒトBip)、配列番号:12(ヒトXBP−1)または配列番号:14(ヒトCHOP−10)において示されるアミノ酸配列を有してもよい。UPR経路構成要素核酸は、配列番号:1(マウスBip mRNA)、配列番号:3(マウスXBP−1、スプライスされた形態 mRNA)、配列番号:5(マウスXBP−1、スプライスされてない形態,mRNA)、配列番号:7(マウスCHOP−10 mRNA)、配列番号:9(ヒトBip mRNA)、配列番号:11(ヒトXBP−1 mRNA)、配列番号:13(ヒトCHOP−10 mRNA)において示される配列を有してもよい。
【0028】
親分子に類似の活性を有するこれらの配列のバリアントもまた、本発明の方法において有用であろう。例えば、親分子に対して少なくとも80%の同一性を有する変異分子は類似の活性を有することが期待できる。2つのタンパク質配列間の同一性パーセントは、フィルタリングをターンオフ(turn off)し、そしてすべて他のデフォルト(default)セッティングを変化しないBRASTPアルゴリズムを用いて決定することができる。ポリペプチドの種々のイソ型、ポリペプチドの優性なネガティブバージョン、またはポリペプチドの共有的に改変された形態物は、親分子のバリアントの若干の例である。
【0029】
本発明のその他の態様では、タンパク質の細胞分泌速度はUPR経路構成要素の発現を減少させることによって減少できる。UPR経路構成要素、例えばBipまたはCHOPのような遺伝子の発現は、短い干渉性RNA(siRNA)分子またはアンチセンス分子による細胞の によって減少できる。
【0030】
本発明のその他の態様では、タンパク質の細胞分泌速度は、低分子の投与をとおしてUPR経路構成要素を調節することによって増大される。低分子の例は、タプシガルギン(thapsigargin)、UPR誘導剤である(Litton,J.,J.Biol.Chem.26,17067−17071(1991))。そのような低分子の他の例は、ツニカマイシン(tunicamysin)およびリポ多糖を含む。
【0031】
本発明のその他の態様では、タンパク質の細胞分泌速度は、細胞を静止増殖期に置くことによって増大される。細胞は栄養利用度を制限するか、細胞の老廃物を蓄積させるか、または細胞培養培地のpHを変化させることによって静止増殖期に置かれてもよい。当業者はまた、静止増殖期に細胞を置く他の方法を認識できる。
【0032】
本発明の方法において、代表的な細胞は、形質細胞、すなわち、抗体を分泌する分化したB細胞である。形質細胞はマウス、ヒトまたは他の動物起源から単離されてもよい。典型的には、形質細胞は、Epstein−Barrウイルスによるウイルス感染のような標準技術か、または例えば、放射線学的もしくは化学的変異誘発のような他の方法によって不死化された。不死化される形質細胞はまたがん性であってもよく、そして鉱油またはその他の化合物を動物の腹腔内に注射することによって得られてもよい。
【0033】
本発明の1つの態様では、不死化される融合パートナーは、「骨髄腫細胞」として当該技術分野において既知であるものである。骨髄腫は、一般に、多発性骨髄腫、骨髄がんを罹患している生物体から得られる不死化融合パートナーと脾臓細胞の融合により形成される。生物体は鳥類、魚類、爬虫類、哺乳類および他の動物であってもよい。骨髄腫細胞系の例は、SP2/0(American Type Culture Collection(ATCC)、Manasas,VA,CRL−1581)、NSO(European Collection of Cell Cultures(ECACC,Salisbury,Wiltshire,UK,ECACC No.85110503)、FO(ATCC CRL−1646)、およびAg653(ATCC CRL−1580)細胞系を含み、これらはマウスから得られた。ヒトから得られる骨髄腫細胞系の例は、U266細胞系(ATCC CRL−TIB−196)である。当業者は他の骨髄腫細胞系も認識することができる。
【0034】
本発明の1つの態様では、骨髄腫細胞はDNA分子により安定してトランスフェクトされる。安定してトランスフェクトされる骨髄腫細胞は、当業者には周知のトランスフェクション、スクリーニングおよび選択の方法によって作成されてもよい。細胞を安定してトランスフェクトするために使用されるDNAは、骨髄腫細胞中に任意に組み込まれてもよく、また部位特異的方式で組み込まれてもよい。そのようなDNA配列はUPR経路構成要素をコードしていてもよい。あるいはまた、安定してトランスフェクトされた核酸配列は、挿入により不活性化してもよく、またUPR構成要素、例えばUPR遺伝子または転写制御配列を欠失してもよい。
【0035】
本発明のその他の態様では、タンパク質の細胞分泌速度は、UPR経路構成要素をコードしている核酸配列の転写物を標的とするsiRNAをコードしている核酸配列により細
胞を安定してトランスフェクトすることによって増大される。そのようなsiRNAは、CHOPタンパク質をコードしている核酸配列の転写物を標的としてもよい。代表的なCHOPタンパク質は、配列番号:8または配列番号:14において示されるアミノ酸配列を有するものである。マウスCHOP−10をコードしている核酸配列の転写物を標的とするsiRNAをコードしている核酸配列による安定なトランスフェクションが有用である。代表的なマウスCHOP−10遺伝子転写物特異的siRNAは、配列番号:15または配列番号:16において示されるヌクレオチド配列を含有する。当業者は、UPR遺伝子転写物を標的とするsiRNAをコードしている他の核酸を認識できる。
【0036】
本発明の方法において有用な他の細胞は、チャイニーズ・ハムスター卵巣(CHO)細胞、昆虫細胞および植物細胞を含む。
【0037】
本発明のその他の態様では、非トランスジェニック動物に較べて上昇したレベルにおいてUPRタンパク質を構成的または誘導的に発現するトランスジェニック動物が作成できる。トランスジェニック動物を作成する技術は当該技術分野において既知である。
【0038】
本発明の方法では、細胞が培養される。細胞は懸濁液において、または付着培養物として培養されてもよい。細胞は、例えば、バイオリアクター、細胞バッグ(bag)、培養プレート、フラスコおよび当業者には周知の他の容器を含む、種々の容器において培養されてもよい。細胞は化学的に定義された培地調合物を含むすべての適当な培地において培養されてもよい。細胞培養に適当な周囲条件、例えば温度および雰囲気組成は、また当業者には周知である。細胞の培養方法もまた当業者には周知である。
【0039】
また本発明は、本発明の方法によって生成される変化した細胞分泌速度をもつ形質細胞を提供する。形質細胞は、短い干渉RNAをコードしている安定してトランスフェクトされた核酸により、UPR経路構成要素の活性を調節することによって生成されてもよい。siRNAは、CHOPタンパク質のようなUPR経路構成要素をコードしている核酸配列の転写物を標的としてもよい。提供される形質細胞は、例えば、C2−8またはC2−18細胞のようなSP2/0由来の細胞であってもよい。そのような形質細胞は精製されるべき抗体または他のポリペプチドを分泌するであろう。
【0040】
本発明は、次に示す特定の、限定されない実施例に関連してここに記述できる。
【実施例1】
【0041】
高分泌性細胞系におけるUPR遺伝子転写物レベル
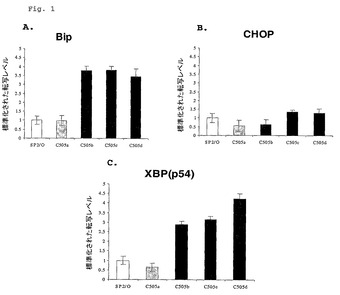
高い速度でmAbを分泌する細胞系が、低い速度で同一mAbを分泌する細胞系と比較されてUPR遺伝子転写物のレベルについて解析された。試験された細胞系は高分泌系C505B、C505CおよびC505Dを含んだ。試験された低分泌系はC505A系およびSP2/0、C505A,B,CおよびDの親の骨髄腫細胞系であった。これらの系統のすべてが、αv−インテグリンに対して特異的なヒトIgG1 mAbの重鎖および軽鎖をコードしているDNAによりトランスフェクトされた。C505B、C505CおよびC505D細胞系は、mAb分泌の高い速度に関して、連続してサブクローニング、スクリーニングおよび選択によって同定された。
【0042】
抗体分泌速度は、細胞培養培地中に24時間の間に分泌されたmAbの量を測定し、そして生存細胞数をカウントして、「pg/生存細胞/日」の単位をもつ細胞分泌速度を生成することによって決定された。C505A細胞系は、5〜7pg/生存細胞/日の低速度で抗体を分泌し、これは約5〜10μg/mL/7日の濃度に相当する。C505B系は、約15pg/生存細胞/日の速度で抗体を生産し、C505Cは、約13pg/生存細胞/日の速度で生産し、そしてC505Dは、約15pg/生存細胞/日の速度で生産
する。
【0043】
全mAb分泌性細胞系および親骨髄腫細胞系は、5%CO2の雰囲気中で、5%FBS、2mMグルタミンおよび2mMピルビン酸塩を含有するIMDM培地中37℃における懸濁液において培養された。ミコフェノール酸0.5mg/L、ヒポキサンチン2.5mg/Lおよびキサンチン50mg/Lを含有する1XMHX選択培地もまた培地には存在した。
【0044】
高発現細胞系と低発現細胞系との間のUPR遺伝子転写物レベルにおける差異は、定量PCR(Q−PCR)により調査された。細胞は対数増殖期まで増殖され、そして総RNAがRNEasyTMシステム(Qiagen Inc.,Valencia,CA)を用いて5x106細胞から単離された。Q−PCRは標準の2段階反応を用いて実施された。cDNA合成段階は、SuperscriptTM II逆転写酵素(Invitorogen Inc.,Carlsbad,CA)およびランダムヘキサマープライマーを用い、製造者によって特定された反応条件を用いて実施された。次いで、TaqmanTMQ−PCR(Applied Biosystems,Foster City,CA)が、製造者による特定されるように、ABI PRISMTM 7000HTまたは7900HT機器(Applied Biosystems,Foster City,CA)によって実施された。RNA 5,000pgが各Q−PCR反応において使用された。Q−PCRのために使用されるBip,CHOPおよびXBP特異的プライマーおよびプローブ組み合わせ物は、Primer ExpressTMソフトウエア(Applied Biosystems,Foster City,CA)を使用して設計された。cDNAレベルは、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(GAPDH)ハウスキーピング遺伝子の転写物レベルに対して標準化され、次いで、適当な親細胞系のcDNA/GAPDH転写物値に対して標準化された。PCRの初期対数期におけるデータの収集および転写物の定量は、ABI PRISM 7000HTまたは7900HT機器および関連するソフトウエアを用いて実施された。
【0045】
結果は図1において示され、そしてBipおよびXBP−1 UPR遺伝子転写物レベルは、低分泌性細胞におけるよりも高分泌性細胞系において約3〜4倍大きかったことを表している。CHOPレベルにおける相違はほとんど断定されなかった。
【実施例2】
【0046】
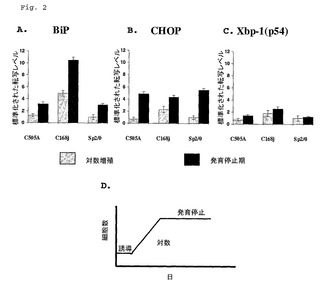
静止増殖期の細胞におけるUPR遺伝子転写物レベルおよび抗体分泌速度
静止増殖期の細胞が、対数増殖期の細胞に比較して、UPR遺伝子転写物レベルおよびmAb分泌速度について解析された。C168J細胞系は、SP2/0親骨髄腫細胞系由来のトランスフェクトーマであり、そして25〜30pg/細胞/日の速度でIgG1 mAbを分泌する。C505Aは実施例1において既に記述されたとおりである。
【0047】
C505A、C168JおよびSP2/0細胞系のIgG1 mAb分泌速度は、実施例1で記述されたように、T−75またはT−150フラスコにおいて5x106細胞を接種、培養することによって調査された。3日後、細胞は対数増殖期にあり、そして6日目に細胞は静止増殖期にあった。総細胞数および生存細胞数が両増殖期の細胞について決定された。両増殖期の細胞からの培養培地が、標準の酵素結合イムノソルベントアッセイ(ELISA)によってヒトIgGについてアッセイされた。Bip、CHOPおよびXBP−1に特異的なQ−PCRが、先の実施例1に記述されたように、対数および静止増殖期細胞について実施された。
【0048】
表1に示される結果は、C505AおよびC168J系がIgG1 mAb分泌速度を増大したことを例証している(表1)。
【0049】
【表1】
【0050】
さらに、図2に示される結果は、対数期増殖に比較して、発育停止期増殖のC505A、C168JおよびSP2/0において増大したUPR遺伝子転写物レベルを例証している。
【実施例3】
【0051】
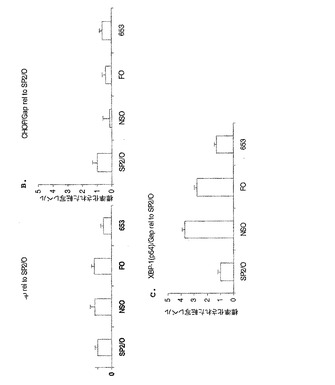
親骨髄腫細胞系におけるUPR遺伝子転写物レベル
SP2/0、NSO、FOおよびAg653親骨髄腫細胞系におけるUPR遺伝子転写物レベルが試験されて、高分泌性細胞系が、より高いレベルのUPR遺伝子を含有するか否か決定された。SP2/0、NSO、FOおよびAg653細胞は、先の実施例1で記述されたように、対数増殖期まで増殖され、総RNAが5x106細胞から単離された。Q−PCRおよび解析もまた実施例1に記述されたように実施された。図3に示される結果は、試験された全細胞において匹敵するBipおよびCHOP転写物レベルを示している。しかしながら、XBP−1転写物レベルは、試験された他の細胞タイプに比較してFOおよびNSO細胞において上昇していた。
【実施例4】
【0052】
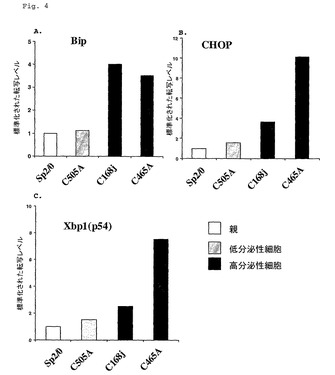
高分泌性細胞系および親骨髄腫細胞系におけるUPR遺伝子転写物レベル
高い速度でmAbを分泌する細胞系が、それらの親の細胞系と比較されてUPR遺伝子転写物のレベルについて解析された。図4〜6に示される細胞系が、先の実施例1で記述されたように、対数増殖期まで増殖され、そして総RNAが5x106細胞から単離された。Q−PCRおよび解析もまた実施例1に記述されたように実施された。
【0053】
図4〜6に示される結果は、高い速度でmAbを分泌する細胞系が、それらのSP2/0、FOおよびAg653親骨髄腫細胞系に較べて、より高いUPR遺伝子転写物レベルを有することを示している。これらの結果は、増大されたUPR遺伝子発現が、使用された親骨髄腫細胞系の同一性とは無関係に、増大された抗体分泌速度と共役されて、高分泌性細胞を生成することを示唆している。
【実施例5】
【0054】
UPR誘導後のUPRタンパク質の発現
低分泌性細胞および親骨髄腫細胞系に較べて高分泌性細胞におけるUPRタンパク質の発現が、薬理学的作用物タプシガルギンによるUPR誘導後に解析された。タプシガルギンは、筋小胞体Ca2+ATPアーゼ(SERCA)ポンプを遮断し、そしてER内腔におけるカルシウム枯渇をもたらすATPアーゼ阻害剤である。
【0055】
細胞はタプシガルギンの100nMで処理され、そしてXBP−1およびCHOPに特異的なウエスタン・ブロットが調製された。細胞は、放射線免疫沈降(RIPA)溶解バ
ッファー(1xPBS,1%N−P40、0.5%デオキシコール酸ナトリウム、0.1%ドデシル硫酸ナトリウム、1mM PMSF、およびRoche製プロテアーゼ阻害剤(Catalog No.1836153))において定められた時点で溶解され、そして清澄溶解液におけるタンパク質濃度が、標準のビシンコニニック・アシド(bicinchoninic acid)(BCA)タンパク質アッセイ(Sigma Catalog No.B9643)を用いて定量された。ウエスタン・ブロットでは、各溶解物20μgが標準のSDS−PAGEゲル上で展開され、フッ化ポリビニリデン(PVDF)メンブランに転移され、そしてXBP−1またはCHOPに特異的な抗体(Santa Cruz Biotechnology Inc.,Santa Cruz,CA)によりプローブされた。
【0056】
結果(未掲示)は、高分泌性細胞におけるUPRタンパク質が、低分泌性細胞および親骨髄腫細胞系に較べて高レベルで発現されることを示した。XBP−1に特異的なウエスタン・ブロットは、XBP−1タンパク質発現が、タプシガルギン処理された高分泌性C168j細胞において強く誘導されることを示している。このXBP−1発現の誘導は、UPRの薬理学的誘導7時間後にもっとも明確であり、そして処置後2時間くらいの初期に見ることができる。CHOPに特異的なウエスタン・ブロットは、CHOPタンパク質発現が、タプシガルギン処理された高分泌性C168j細胞において強く誘導されることを示している。CHOPは2時間目に最初に出現し、4〜7時間に最大に発現され、そして22時間では非常に低下される。
【実施例6】
【0057】
UPRタンパク質発現レベルの増大
細胞は、BipおよびXBP−1発現ベクターを用い、これらのUPRタンパク質の細胞発現レベルを高めることを試みて一過性にトランスフェクトされた。HEK293T/17(ATCC CRL−11268)細胞が、5%CO2の雰囲気中37℃で付着培養物として増殖され、5%FBS、2mMグルタミンおよび2mMピルビン酸塩を含有するIscoveの改変Dubelccoの最小必須培地(IMDM)培地において培養された。XBP−1イソ型1(NCBI Accession AF027963)、XBP−1イソ型2(NCBI Accession AF443192)またはBipをコードしているcDNAが、標準的方法を用いて一過性トランスフェクション実験のために有用なベクター中にサブクローン化された。ベクターはCMVプロモーターの制御下でZsGreen1タンパク質を発現し、かつCMVプロモーターの制御下でさらなるコーディング領域の導入のための多数のクローニング部位を有する。カナマイシン耐性遺伝子は細菌の選択のために使用された。次いで、HEK293T/17細胞が、いずれかトランスフェクトされずに残されるか、あるいは空のベクターのみ、XBP−1イソ型1発現ベクター、XBP−1イソ型2発現ベクターまたはBip発現ベクターにより一過性にトランスフェクトされた。トランスフェクションは、製造者によって指示されるようにLipofectamineTM2000試薬(Invitrogen Inc.,Carlsbad,CA)を用いて実施された。トランスフェクション後48時間に、細胞はRIPA溶解バッファーにおいて溶解され、そして清澄溶解液の等しい細胞当量がSDS−PAGEゲル上に負荷された。次いで、ウエスタンブロットが調製され、先の実施例5に記述されたようにプローブされた。次いでブロットが除去され、アクチン特異抗体で再プローブされて各列に負荷された同じタンパク質を確認した。
【0058】
結果(未掲示)は、XBP−1イソ型1(「XBP−1」を標識された列)およびXBP−1イソ型2(「X54}を標識された列)発現レベルが、これらのXBP−1イソ型をコードしている発現ベクターにより一過性にトランスフェクトされた細胞において上昇されたことを例証した。さらに、データ(未掲示)は、Bip発現レベルが、Bipをコードしている発現ベクターにより一過性にトランスフェクトされた細胞において上昇され、そしてトランスフェクトされずに残されたか、または空のベクターのみでトランスフェクトされた細胞では、Bip発現が検出されなかったことを示した。
【実施例7】
【0059】
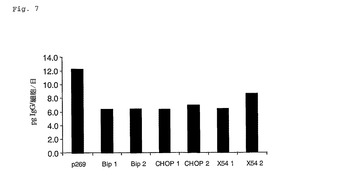
抗体分泌速度に及ぼすUPR遺伝子転写物レベルの影響
抗体分泌速度に及ぼすUPR遺伝子転写物レベルの影響が解析された。Bip、CHOPおよびXBP−1遺伝子転写物を標的とする二本鎖siRNA分子は、Ambionのインターネットに基づくsiRNA Target Finder Tool(www.ambion.com/techlib/misc/siRNA finder.html)を用いて設計され、そしてSilencerTMsiRNA Constructionキット(Ambion Inc.,Woodward,TX,Catalog No.11620)を用いて合成された。2種のsiRNAが各標的転写物のために設計された。標準条件下で培養された3x106C168J細胞が、図10に示されるようなこれらの各二本鎖siRNAの3μgを用いてエレクトロポレーションによりトランスフェクトされた。エレクトロポレートされた細胞は、先の実施例1に記述されたようにIMDM培地2mlに懸濁され、次いで、T−12プレートにおいて培養された。生存細胞濃度は8日後に測定された。またIgG1mAb濃度も、標準比濁分析技術を用いて8日後に測定された。
【0060】
図7における結果は、抗体分泌速度が、抗体分泌速度に及ぼすUPR遺伝子転写物レベルを改変することによって調節できることを示している。RNA転写物からUPR遺伝子Bip、CHOPおよびXBP−1イソ型の発現を妨げることが可能な短い干渉RNA(siRNA)による高分泌性C168J細胞のトランスフェクションは、C168J細胞のIgG1分泌速度を減少させた。
【実施例8】
【0061】
UPR遺伝子転写物および発現レベルを増大することによる抗体分泌速度の増大
UPRタンパク質の過剰発現が、細胞によるmAbのようなタンパク質の分泌速度を増大できる。タンパク質分泌性細胞系、例えばmAb分泌性細胞系は、Bip、CHOP、XBP−1および他のUPR関連のタンパク質をコードしている発現ベクターによりトランスフェクトされて、これらのタンパク質の過剰発現に影響を与えられた。細胞が、単独でも、また組み合わせてもこれらの発現ベクターを用いてトランスフェクトされる。適当なタンパク質分泌速度、例えば抗体分泌速度は、標準技術を用いてトランスフェクション後2、4および6日目に決定された。トランスフェクトされた細胞におけるタンパク質分泌速度が、トランスフェクトされない対照細胞の分泌速度に比較された。タンパク質分泌速度は、1種以上のUPRタンパク質を過剰発現する細胞において、より高いことが期待される。BipまたはXBP−1の構成的な過剰発現が、最終的にこれらの細胞においてアポトーシスを誘導する事象では、これらの遺伝子が、誘導プロモーターの下流に置かれ、そして必要な場合にのみ活性化されてもよい。
【実施例9】
【0062】
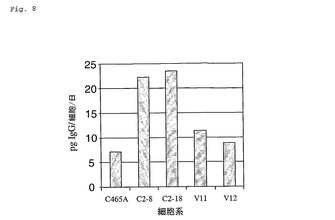
CHOP−10エンコ−ディングsiRNAにより安定してトランスフェクトされた骨髄腫細胞における抗体分泌速度の増大
CHOP−10に特異的なsiRNAを生産するDNA構築物によるC465A骨髄腫細胞の安定なトランスフェクションは、抗体分泌速度を増大した(図8)。CHOP−10遺伝子転写物を標的とする2種の異なるsiRNA分子が、Ambionのインターネットに基づくsiRNA Target Finder Tool(www.ambion.com/techlib/misc/siRNA finder.html)を用いて設計された。設計されたsiRNA分子は短いヘアピン型であった。これらのsiRNAをコードしている核酸(配列番号:15および配列番号:16)が合成され、pSILENCER4.1−neoベクター(Ambion Inc.,Woodward,TX,)のBamHIとHindIII部位中に標準法を用いてクローン化された。pSILENCER4.1−neo中への配列番号:15において示される核酸配列のクローニングはpCHOP1を生成し、一方、このベクター中への配列番号:16において示される配列のクローニングはpCHOP2を生成した。pCHOP1、pCHOP2およびpSILENCER4.1−neoプラスミドDNAが、次に、各々別々にエレクトロポレーションによってC465A骨髄腫細胞中にトランスフェクトされた。これらのベクターを含有する安定してトランスフェクトされた骨髄腫細胞が、次に、10%FBSおよびG418 300μg/mlを含有するSFM8培地において5%CO2の雰囲気中37℃での培養によって選択された。また、この選択培地は、ミコフェノール酸0.5mg/L、ヒポキサンチン2.5mg/Lおよびキサンチン50mg/Lを含有する1XMHX選択培地を含有して、C465A細胞による安定な抗体発現を維持した。
【0063】
図8に示される各細胞系は、10%FBS、1X MHXおよびG418 300μg/mlを含有するSFM8培地において6日間懸濁液において増殖された。生存細胞濃度および抗体タイターはこの6日間の間毎日決定された。生存細胞濃度は、標準の色素排除アッセイによってアッセイされ、そして培養培地中の抗体タイターは標準の比濁分析技術によってアッセイされた。C465A骨髄腫細胞系はSP2/0骨髄腫細胞系から得られ、IgG1κイソ型のヒトTNP−α特異的マウスmAbを安定して発現する。C2−8骨髄腫細胞系は、pCHOP1により安定してトランスフェクトされたC465A由来の細胞系である。C2−18骨髄腫細胞系は、pCHOP2により安定してトランスフェクトされたC465A由来の細胞系である。V11およびV12細胞系は、pSILENCER4.1−neoのみにより安定してトランスフェクトされたC465A由来の細胞系であり;両系統は独立して生成された。
【0064】
得られたデータは、pCHOP1およびpCHOP2によってコードされるCHOP−10特異的siRNAは、対照C465A、V11およびV12細胞系に較べて抗体特異的生産性を増大した(図8)。
【0065】
本発明はここに完全に記述され、付随する請求項の精神および範囲から逸脱することなく、多くの変更および修飾がそれに対して作成できることは当業者には明白であろう。
【図面の簡単な説明】
【0066】
【図1】図1は、高い速度でmAbを分泌する細胞におけるUPR遺伝子転写物レベルを示す。
【図2】図2は、静止増殖期の細胞におけるUPR遺伝子転写物レベルを示す。
【図3】図3は、親骨髄腫細胞系におけるUPR遺伝子転写物レベルの比較を示す。
【図4】図4は、Sp2/0親骨髄腫細胞系に比較して高分泌性細胞系におけるUPR遺伝子転写物レベルを示す。
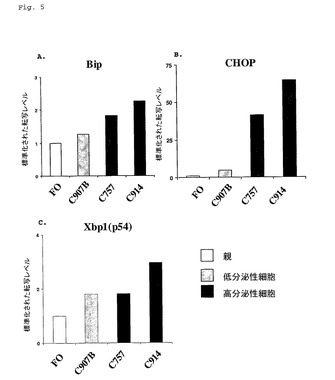
【図5】図5は、FO親骨髄腫細胞系に比較して高分泌性細胞系におけるUPR遺伝子転写物レベルを示す。
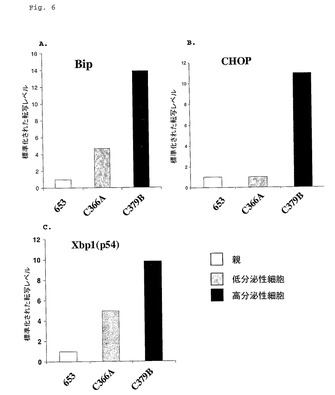
【図6】図6は、Ag−653親骨髄腫細胞系に比較して高分泌性細胞系におけるUPR遺伝子転写物レベルを示す。
【図7】図7は、UPR遺伝子転写物レベル改変の関数としての抗体分泌速度における変化を示す。
【図8】図8は、CHOP−10特異的siRNAをコードしている核酸により安定してトランスフェクトされた骨髄腫細胞における増大された抗体分泌速度を示す。
【技術分野】
【0001】
本発明は、タンパク質の細胞分泌速度の変更方法に関する。
【背景技術】
【0002】
タンパク質、例えば抗体の大規模生産は、典型的には、培養される生産細胞系からのタンパク質の分泌に依存する。培養細胞によって生産される分泌タンパク質は、周囲の細胞培養培地から容易に回収かつ精製することができる。
【0003】
タンパク質の細胞分泌速度は、バイオリアクターまたは他のシステムからの分泌タンパク質の生産および精製に影響を与える重要なファクターである。一般に、より高い精製タンパク質の収量は、細胞分泌速度が比較的高い場合に達成することができる。反対に、細胞分泌速度が低すぎる場合は、タンパク質精製は実行不可能であろう。
【0004】
低分泌性細胞の問題を回避する1つのアプローチは、低分泌性細胞の集団から高分泌性のサブクローン細胞を単離することであった。典型的には、これは、高分泌性細胞系の連続希釈、スクリーニングおよび選択を限定するという数回の、かつ労力集中的なラウンドを必要とする。あるいはまた、新しい細胞系が高分泌系であろうという希望において、問題のタンパク質を生産する完全に新しい細胞系が生成される。
【0005】
高分泌性細胞系を生成する従来の各アプローチは限界を有する。例えば、低分泌性細胞の集団からサブクローニングによって高分泌性細胞系を同定することは、集団における高分泌性細胞が相対的に希少であること、ならびにいずれかの高分泌性細胞の同定に要求される広大な量の時間と労力によって制約される。
【0006】
さらに、問題の抗体またはタンパク質を生産する新しい細胞系の生成は、新しい細胞系が高分泌性ではない可能性、ならびに抗体産生細胞を再生し、そして高分泌性細胞を同定するために要求される多大な量の努力によって制約される。若干の例では、低下した分泌速度をもたらす細胞の小胞体(ER)内でのタンパク質のミスフォールディング(misfolding)により、低分泌性細胞系のみが得られることもある。
【0007】
かくして、タンパク質の細胞分泌速度を変化させる有効な方法への要求が存在する。
【発明の開示】
【0008】
本発明の1つの態様は、細胞において少なくとも1種のUPR経路構成要素の活性を調節し;そして細胞を培養する段階を含んでなる、タンパク質の細胞分泌速度の変更方法である。
【0009】
本発明のその他の態様は、細胞において少なくとも1種のアンフォールド型(unfolded)タンパク質応答(UPR)経路構成要素の活性を調節し;そして細胞を培養する段階を含んでなる、タンパク質の細胞分泌速度を改変することによって生成される、変化した細胞分泌速度をもつ形質細胞である。
【0010】
本発明のその他の態様は、細胞において少なくとも1種のUPR経路構成要素の活性を調節し、そして細胞を培養することによって生成される、変化した細胞分泌速度をもつ形質細胞を含有するトランスジェニック動物である。
【0011】
本発明のその他の態様は、配列番号:15または配列番号:16において示される配列
を有する単離された核酸である。
【0012】
限定されるものではないが、本明細書において引用される特許および特許出願を含む、すべての公表物は、完全な記述をとおして引用によって本明細書に組み入れられている。
【0013】
本明細書で使用されるところの用語「抗体」は、広範な意味において意味され、そしてポリクローナル抗体、マウス、ヒト、ヒト化およびキメラモノクローナル抗体を含むモノクローナル抗体および抗体フラグメントまたは変異体を含む、免疫グロブリンまたは抗体分子を含む。抗体は形質細胞によって構成的に発現され、そして分泌される分泌タンパク質である。また抗体は、標準的方法、例えばハイブリドーマ生成によるか、または不死化されたB細胞、例えば骨髄腫細胞または他の細胞種、例えばチャイニーズハムスター卵巣(CHO)細胞、植物細胞および昆虫細胞への抗体重鎖および/または軽鎖遺伝子のトランスフェクションによって、不死化された形質細胞を用いて生産することができる。
【0014】
抗体フラグメントまたは変異体は、ミメティボディ(mimetibody)、Fabフラグメント、F(ab’)2フラグメント、Fcフラグメント、重鎖フラグメント、軽鎖フラグメント、および少なくとも1種の抗体ペプチド鎖の一部を含有する分子を含む。そのような部分は、抗体の可変、ヒンジもしくは定常部ペプチド鎖に対応してもよい。
【0015】
本明細書で使用されるような用語「ミメティボディ」は、一般式(I):
(V1(n)−Pep(n)−Flex(n)−V2(n)−pHinge(n)−CH2(n)−CH3(n))(m) (I)
[式中、V1は免疫グロブリン可変部のN末端の少なくとも1部分であり、Pepはエピトープに結合する少なくとも1種の生物活性ペプチドであり、Flexは、ミメティボディをもう一方の方向づけと結合特性をもつようにさせることによって構造的柔軟性を与えるポリペプチドであり、V2は免疫グロブリン可変部のC末端の少なくとも1部分であり、pHingeは免疫グロブリンヒンジ部の少なくとも1部分であり、CH2は免疫グロブリンCH2定常部の少なくとも1部分であり、そしてCH3は免疫グロブリンCH3定常部の少なくとも1部分であり、ここでnおよびmは整数1〜10であってもよい]
を有するタンパク質を意味する。ミメティボディは、構築物に存在する重鎖定常ドメインアミノ酸配列に応じて、IgG1、IgG2、IgG3、IgG4、IgA、IgM、IgDおよびIgEのような免疫グロブリン分子の異なるタイプの特性および機能を模倣できる。
【0016】
本明細書で使用されるところの用語「モノクローナル抗体」(mAb)は、実質的に均一な抗体の集団から得られる抗体(または抗体フラグメント)を意味する。モノクローナル抗体は高度に特異性であり、典型的には単一な抗原決定基に対向される。修飾語句「モノクローナル」は、抗体の実質的に均一な特徴を示し、いずれか特定の方法による抗体の生産を必要としない。例えば、マウスmAbはKohler et al.,Nature 256:495(1975)のハイブリドーマ法によって作成することができる。アクセプター抗体(典型的には、ヒトのようなその他の哺乳動物種)由来の軽鎖および重鎖定常部に関連してドナー抗体(典型的には、マウス)由来の軽鎖および重鎖可変部を含有するキメラmAbは、米国特許第4,816,567号に開示された方法によって調製されてもよい。非ヒト・ドナー免疫グロブリン(典型的にはマウス)由来のCDRおよび場合によっては結合親和力を維持するように改変されたフレームワーク・サポート残基を有する1種以上の免疫グロブリンに由来する分子の残りの免疫グロブリン由来の部分を有する、ヒト化mAbは、Queen et al.,Proc.Natl Acad Sci(USA),86:10029−10032,(1989)およびHodgson et al.,Bio/Technology,9:421,(1991)において開示された技術によって得ることができる。
【0017】
いずれの非ヒト配列も欠く完全にヒトのmAbは、例えば、Lonberg et al.,Nature 368:856−859(1994);Fishwild et al.,Nature Biotechnology 14:845−851,(1996)およびMendez et al.,Nature Genetics 15:146−156,(1997)において引用される技術によってヒト免疫グロブリントランスジェニックマウスから調製されてもよい。ヒトmAbはまた、例えば、Knappik et al.,J.Mol.Biol.296:57−86,(2000)およびKrebs et al.,J,Immunol.Meth.254:67−84,(2001)において引用される技術によってファージ・ディスプレイライブラリーから調製され、そして最適化されてもよい。
【0018】
本明細書で使用されるところの用語「細胞分泌速度」は、細胞が特定のタンパク質を分泌する速度を意味する。そのような速度は、時間の変化当たりの培養培地中に存在するタンパク質の量における変化として記述されてもよく、または細胞数に標準化され、そして単位「pg/細胞/日」で表されてもよい。
【0019】
本明細書で使用されるところの用語「短い干渉性RNA(short interfering RNA)」は、標的遺伝子転写物の切断を媒介する核酸配列を意味する。短い干渉性RNA(siRNA)は、二本鎖でもまたは短いヘアピン型であってもよい。二本鎖siRNAは、2本の個々の、逆平行の、アニールされたRNA鎖または両RNAおよびDNAを含有するアニールされた核酸鎖(例えば、5’−TTTTUUUU−3’にアニールされた5’−TTTTUUUU−3’または5’−UUUU−3’にアニールされた5’−TTTT−3’)からなってもよい。短いヘアピン型のsiRNAは、siRNAとして有効なステム・ループ構造または他の二次構造を形成することができる単一RNA鎖または単一RNA:DNAハイブリッド鎖からなってもよい。当業者は、siRNAが、他の改変体、例えばヌクレオシド類似体、バックボーン改変体、および改変siRNA核酸に標的遺伝子転写物の切断をなお媒介させる他の改変体を含んでもよいことを認識している。
【0020】
本明細書で使用されるところの用語「低分子(small molecule)」は、アミノ酸残基または核酸残基のみからはならない24,000g/mol未満の分子量をもつ化合物を意味する。
【0021】
本明細書で使用されるところの用語「転写制御配列」は、遺伝子の転写に必要である核酸配列または遺伝子の転写を増大または減少させる核酸配列を意味する。
【0022】
本明細書で使用されるところの用語「UPR経路構成要素」は、UPR経路を介するシグナル伝達を媒介するか、またはUPRを活性化する、ペプチド鎖または核酸配列、例えば転写制御配列を意味する。
【0023】
本発明は、細胞によるタンパク質の細胞分泌速度を改変するために有用な方法を提供する。本発明の方法の代表的使用は、治療、診断または研究目的に有用であるタンパク質、例えば抗体のための分泌速度の増進である。
【0024】
細胞系における低いタンパク質分泌速度は、UPRを介して分泌過程を減速または停止する、細胞ERにおけるミスフォールド(misfolded)タンパク質の蓄積によって惹起できる。ER膜におけるストレス感受性タンパク質は、アンフォールド型タンパク質の過剰を検出し、そしてUPRの引き金を引く。続いて、複雑なシグナル伝達カスケードを介して、シャペロン(chaperone)タンパク質Bipおよび転写因子XBP
−1およびCHOPが上方調節される。
【0025】
シャペロンタンパク質はアンフォールド型タンパク質に結合し、そして正確な折り畳みを助ける。転写因子CHOPは、一般に細胞増殖、分化および生存のネガティブレギュレーターとされる。CHOPの上方調節は細胞周期の停止を惹起し、その結果、UPR誘導に関与する好ましくない条件に対抗する時間を細胞に与えることが観察された。
【0026】
転写因子XBP−1は、形質細胞、大量の免疫グロブリンを分泌する分化したBリンパ球を生成するために要求される。形質細胞は変化されたUPRを現し、ここでは免疫グロブリン合成の前に若干のUPR遺伝子の上方調節が起きる。これらは、XBP−1、Bip、Grp94およびp50ATF6αを含み、そしてそれらの上方調節は形質細胞の分化と適当な分泌のために必須である。(Gass,J.N.,et al.,J.Biol.Chem.227,49047−49054(2002))。これに対して、CHOPはこの推移において上方調節されず、このことは異なる種類のUPRを示唆している。これらの分子の上方調節は、細胞タンパク質分泌速度における増大またはアポトーシスのいずれかを惹起する有利または有害な効果を有する。参照、Kaufman,R.J.,Genes Dev.13,1211−1233(1999)およびCudna,R.E.,et al.,Biotechnol.Bioeng.81,56−65(2003)。 本発明の方法では、タンパク質の細胞分泌速度は、細胞における少なくとも1個のUPR経路構成要素の活性を調節し、そして細胞を培養することによって改変される。本発明の方法は、抗体のようなタンパク質の細胞分泌速度の増大または減少に備える。
【0027】
本発明の実施態様では、タンパク質の細胞分泌速度は、UPR経路構成要素をコードしている核酸により細胞を安定してトラスフェクトすることによって増大される。UPR経路構成要素は、ポリペプチドまたは核酸配列、例えば転写制御配列であってもよく、これらはUPR経路をとおしてシグナル伝達を媒介するか、またはUPRを活性化する。UPR経路構成要素の例は、Bip、XBPおよびCHOPならびに類似の活性を有する変異体を含む。UPR経路構成要素の他の例は、IRE1、PERK、ATF4、eIF2α、GRP78、GRP94、calreticulin、シャペロン、および類似の活性を有する変異体を含む(参照、例えば、Cudna and Dickson,Biotechnol.Bioeng.81,56−65(2002))。転写制御配列の例は、シス作用UPR要素(UPRE)および種々のUPR遺伝子のプロモーター中に同定されたERSEである。当業者は他のUPR経路構成要素および転写制御配列も認識できるであろう。UPR経路構成要素は、配列番号:2(マウスBip)、配列番号:4(マウスXBP−1、スプライスされた形態)、配列番号:6(マウスXBP−1、スプライスされてない形態)、配列番号:8(マウスCHOP−10)、配列番号:10(ヒトBip)、配列番号:12(ヒトXBP−1)または配列番号:14(ヒトCHOP−10)において示されるアミノ酸配列を有してもよい。UPR経路構成要素核酸は、配列番号:1(マウスBip mRNA)、配列番号:3(マウスXBP−1、スプライスされた形態 mRNA)、配列番号:5(マウスXBP−1、スプライスされてない形態,mRNA)、配列番号:7(マウスCHOP−10 mRNA)、配列番号:9(ヒトBip mRNA)、配列番号:11(ヒトXBP−1 mRNA)、配列番号:13(ヒトCHOP−10 mRNA)において示される配列を有してもよい。
【0028】
親分子に類似の活性を有するこれらの配列のバリアントもまた、本発明の方法において有用であろう。例えば、親分子に対して少なくとも80%の同一性を有する変異分子は類似の活性を有することが期待できる。2つのタンパク質配列間の同一性パーセントは、フィルタリングをターンオフ(turn off)し、そしてすべて他のデフォルト(default)セッティングを変化しないBRASTPアルゴリズムを用いて決定することができる。ポリペプチドの種々のイソ型、ポリペプチドの優性なネガティブバージョン、またはポリペプチドの共有的に改変された形態物は、親分子のバリアントの若干の例である。
【0029】
本発明のその他の態様では、タンパク質の細胞分泌速度はUPR経路構成要素の発現を減少させることによって減少できる。UPR経路構成要素、例えばBipまたはCHOPのような遺伝子の発現は、短い干渉性RNA(siRNA)分子またはアンチセンス分子による細胞の によって減少できる。
【0030】
本発明のその他の態様では、タンパク質の細胞分泌速度は、低分子の投与をとおしてUPR経路構成要素を調節することによって増大される。低分子の例は、タプシガルギン(thapsigargin)、UPR誘導剤である(Litton,J.,J.Biol.Chem.26,17067−17071(1991))。そのような低分子の他の例は、ツニカマイシン(tunicamysin)およびリポ多糖を含む。
【0031】
本発明のその他の態様では、タンパク質の細胞分泌速度は、細胞を静止増殖期に置くことによって増大される。細胞は栄養利用度を制限するか、細胞の老廃物を蓄積させるか、または細胞培養培地のpHを変化させることによって静止増殖期に置かれてもよい。当業者はまた、静止増殖期に細胞を置く他の方法を認識できる。
【0032】
本発明の方法において、代表的な細胞は、形質細胞、すなわち、抗体を分泌する分化したB細胞である。形質細胞はマウス、ヒトまたは他の動物起源から単離されてもよい。典型的には、形質細胞は、Epstein−Barrウイルスによるウイルス感染のような標準技術か、または例えば、放射線学的もしくは化学的変異誘発のような他の方法によって不死化された。不死化される形質細胞はまたがん性であってもよく、そして鉱油またはその他の化合物を動物の腹腔内に注射することによって得られてもよい。
【0033】
本発明の1つの態様では、不死化される融合パートナーは、「骨髄腫細胞」として当該技術分野において既知であるものである。骨髄腫は、一般に、多発性骨髄腫、骨髄がんを罹患している生物体から得られる不死化融合パートナーと脾臓細胞の融合により形成される。生物体は鳥類、魚類、爬虫類、哺乳類および他の動物であってもよい。骨髄腫細胞系の例は、SP2/0(American Type Culture Collection(ATCC)、Manasas,VA,CRL−1581)、NSO(European Collection of Cell Cultures(ECACC,Salisbury,Wiltshire,UK,ECACC No.85110503)、FO(ATCC CRL−1646)、およびAg653(ATCC CRL−1580)細胞系を含み、これらはマウスから得られた。ヒトから得られる骨髄腫細胞系の例は、U266細胞系(ATCC CRL−TIB−196)である。当業者は他の骨髄腫細胞系も認識することができる。
【0034】
本発明の1つの態様では、骨髄腫細胞はDNA分子により安定してトランスフェクトされる。安定してトランスフェクトされる骨髄腫細胞は、当業者には周知のトランスフェクション、スクリーニングおよび選択の方法によって作成されてもよい。細胞を安定してトランスフェクトするために使用されるDNAは、骨髄腫細胞中に任意に組み込まれてもよく、また部位特異的方式で組み込まれてもよい。そのようなDNA配列はUPR経路構成要素をコードしていてもよい。あるいはまた、安定してトランスフェクトされた核酸配列は、挿入により不活性化してもよく、またUPR構成要素、例えばUPR遺伝子または転写制御配列を欠失してもよい。
【0035】
本発明のその他の態様では、タンパク質の細胞分泌速度は、UPR経路構成要素をコードしている核酸配列の転写物を標的とするsiRNAをコードしている核酸配列により細
胞を安定してトランスフェクトすることによって増大される。そのようなsiRNAは、CHOPタンパク質をコードしている核酸配列の転写物を標的としてもよい。代表的なCHOPタンパク質は、配列番号:8または配列番号:14において示されるアミノ酸配列を有するものである。マウスCHOP−10をコードしている核酸配列の転写物を標的とするsiRNAをコードしている核酸配列による安定なトランスフェクションが有用である。代表的なマウスCHOP−10遺伝子転写物特異的siRNAは、配列番号:15または配列番号:16において示されるヌクレオチド配列を含有する。当業者は、UPR遺伝子転写物を標的とするsiRNAをコードしている他の核酸を認識できる。
【0036】
本発明の方法において有用な他の細胞は、チャイニーズ・ハムスター卵巣(CHO)細胞、昆虫細胞および植物細胞を含む。
【0037】
本発明のその他の態様では、非トランスジェニック動物に較べて上昇したレベルにおいてUPRタンパク質を構成的または誘導的に発現するトランスジェニック動物が作成できる。トランスジェニック動物を作成する技術は当該技術分野において既知である。
【0038】
本発明の方法では、細胞が培養される。細胞は懸濁液において、または付着培養物として培養されてもよい。細胞は、例えば、バイオリアクター、細胞バッグ(bag)、培養プレート、フラスコおよび当業者には周知の他の容器を含む、種々の容器において培養されてもよい。細胞は化学的に定義された培地調合物を含むすべての適当な培地において培養されてもよい。細胞培養に適当な周囲条件、例えば温度および雰囲気組成は、また当業者には周知である。細胞の培養方法もまた当業者には周知である。
【0039】
また本発明は、本発明の方法によって生成される変化した細胞分泌速度をもつ形質細胞を提供する。形質細胞は、短い干渉RNAをコードしている安定してトランスフェクトされた核酸により、UPR経路構成要素の活性を調節することによって生成されてもよい。siRNAは、CHOPタンパク質のようなUPR経路構成要素をコードしている核酸配列の転写物を標的としてもよい。提供される形質細胞は、例えば、C2−8またはC2−18細胞のようなSP2/0由来の細胞であってもよい。そのような形質細胞は精製されるべき抗体または他のポリペプチドを分泌するであろう。
【0040】
本発明は、次に示す特定の、限定されない実施例に関連してここに記述できる。
【実施例1】
【0041】
高分泌性細胞系におけるUPR遺伝子転写物レベル
高い速度でmAbを分泌する細胞系が、低い速度で同一mAbを分泌する細胞系と比較されてUPR遺伝子転写物のレベルについて解析された。試験された細胞系は高分泌系C505B、C505CおよびC505Dを含んだ。試験された低分泌系はC505A系およびSP2/0、C505A,B,CおよびDの親の骨髄腫細胞系であった。これらの系統のすべてが、αv−インテグリンに対して特異的なヒトIgG1 mAbの重鎖および軽鎖をコードしているDNAによりトランスフェクトされた。C505B、C505CおよびC505D細胞系は、mAb分泌の高い速度に関して、連続してサブクローニング、スクリーニングおよび選択によって同定された。
【0042】
抗体分泌速度は、細胞培養培地中に24時間の間に分泌されたmAbの量を測定し、そして生存細胞数をカウントして、「pg/生存細胞/日」の単位をもつ細胞分泌速度を生成することによって決定された。C505A細胞系は、5〜7pg/生存細胞/日の低速度で抗体を分泌し、これは約5〜10μg/mL/7日の濃度に相当する。C505B系は、約15pg/生存細胞/日の速度で抗体を生産し、C505Cは、約13pg/生存細胞/日の速度で生産し、そしてC505Dは、約15pg/生存細胞/日の速度で生産
する。
【0043】
全mAb分泌性細胞系および親骨髄腫細胞系は、5%CO2の雰囲気中で、5%FBS、2mMグルタミンおよび2mMピルビン酸塩を含有するIMDM培地中37℃における懸濁液において培養された。ミコフェノール酸0.5mg/L、ヒポキサンチン2.5mg/Lおよびキサンチン50mg/Lを含有する1XMHX選択培地もまた培地には存在した。
【0044】
高発現細胞系と低発現細胞系との間のUPR遺伝子転写物レベルにおける差異は、定量PCR(Q−PCR)により調査された。細胞は対数増殖期まで増殖され、そして総RNAがRNEasyTMシステム(Qiagen Inc.,Valencia,CA)を用いて5x106細胞から単離された。Q−PCRは標準の2段階反応を用いて実施された。cDNA合成段階は、SuperscriptTM II逆転写酵素(Invitorogen Inc.,Carlsbad,CA)およびランダムヘキサマープライマーを用い、製造者によって特定された反応条件を用いて実施された。次いで、TaqmanTMQ−PCR(Applied Biosystems,Foster City,CA)が、製造者による特定されるように、ABI PRISMTM 7000HTまたは7900HT機器(Applied Biosystems,Foster City,CA)によって実施された。RNA 5,000pgが各Q−PCR反応において使用された。Q−PCRのために使用されるBip,CHOPおよびXBP特異的プライマーおよびプローブ組み合わせ物は、Primer ExpressTMソフトウエア(Applied Biosystems,Foster City,CA)を使用して設計された。cDNAレベルは、グリセルアルデヒド−3−リン酸デヒドロゲナーゼ(GAPDH)ハウスキーピング遺伝子の転写物レベルに対して標準化され、次いで、適当な親細胞系のcDNA/GAPDH転写物値に対して標準化された。PCRの初期対数期におけるデータの収集および転写物の定量は、ABI PRISM 7000HTまたは7900HT機器および関連するソフトウエアを用いて実施された。
【0045】
結果は図1において示され、そしてBipおよびXBP−1 UPR遺伝子転写物レベルは、低分泌性細胞におけるよりも高分泌性細胞系において約3〜4倍大きかったことを表している。CHOPレベルにおける相違はほとんど断定されなかった。
【実施例2】
【0046】
静止増殖期の細胞におけるUPR遺伝子転写物レベルおよび抗体分泌速度
静止増殖期の細胞が、対数増殖期の細胞に比較して、UPR遺伝子転写物レベルおよびmAb分泌速度について解析された。C168J細胞系は、SP2/0親骨髄腫細胞系由来のトランスフェクトーマであり、そして25〜30pg/細胞/日の速度でIgG1 mAbを分泌する。C505Aは実施例1において既に記述されたとおりである。
【0047】
C505A、C168JおよびSP2/0細胞系のIgG1 mAb分泌速度は、実施例1で記述されたように、T−75またはT−150フラスコにおいて5x106細胞を接種、培養することによって調査された。3日後、細胞は対数増殖期にあり、そして6日目に細胞は静止増殖期にあった。総細胞数および生存細胞数が両増殖期の細胞について決定された。両増殖期の細胞からの培養培地が、標準の酵素結合イムノソルベントアッセイ(ELISA)によってヒトIgGについてアッセイされた。Bip、CHOPおよびXBP−1に特異的なQ−PCRが、先の実施例1に記述されたように、対数および静止増殖期細胞について実施された。
【0048】
表1に示される結果は、C505AおよびC168J系がIgG1 mAb分泌速度を増大したことを例証している(表1)。
【0049】
【表1】
【0050】
さらに、図2に示される結果は、対数期増殖に比較して、発育停止期増殖のC505A、C168JおよびSP2/0において増大したUPR遺伝子転写物レベルを例証している。
【実施例3】
【0051】
親骨髄腫細胞系におけるUPR遺伝子転写物レベル
SP2/0、NSO、FOおよびAg653親骨髄腫細胞系におけるUPR遺伝子転写物レベルが試験されて、高分泌性細胞系が、より高いレベルのUPR遺伝子を含有するか否か決定された。SP2/0、NSO、FOおよびAg653細胞は、先の実施例1で記述されたように、対数増殖期まで増殖され、総RNAが5x106細胞から単離された。Q−PCRおよび解析もまた実施例1に記述されたように実施された。図3に示される結果は、試験された全細胞において匹敵するBipおよびCHOP転写物レベルを示している。しかしながら、XBP−1転写物レベルは、試験された他の細胞タイプに比較してFOおよびNSO細胞において上昇していた。
【実施例4】
【0052】
高分泌性細胞系および親骨髄腫細胞系におけるUPR遺伝子転写物レベル
高い速度でmAbを分泌する細胞系が、それらの親の細胞系と比較されてUPR遺伝子転写物のレベルについて解析された。図4〜6に示される細胞系が、先の実施例1で記述されたように、対数増殖期まで増殖され、そして総RNAが5x106細胞から単離された。Q−PCRおよび解析もまた実施例1に記述されたように実施された。
【0053】
図4〜6に示される結果は、高い速度でmAbを分泌する細胞系が、それらのSP2/0、FOおよびAg653親骨髄腫細胞系に較べて、より高いUPR遺伝子転写物レベルを有することを示している。これらの結果は、増大されたUPR遺伝子発現が、使用された親骨髄腫細胞系の同一性とは無関係に、増大された抗体分泌速度と共役されて、高分泌性細胞を生成することを示唆している。
【実施例5】
【0054】
UPR誘導後のUPRタンパク質の発現
低分泌性細胞および親骨髄腫細胞系に較べて高分泌性細胞におけるUPRタンパク質の発現が、薬理学的作用物タプシガルギンによるUPR誘導後に解析された。タプシガルギンは、筋小胞体Ca2+ATPアーゼ(SERCA)ポンプを遮断し、そしてER内腔におけるカルシウム枯渇をもたらすATPアーゼ阻害剤である。
【0055】
細胞はタプシガルギンの100nMで処理され、そしてXBP−1およびCHOPに特異的なウエスタン・ブロットが調製された。細胞は、放射線免疫沈降(RIPA)溶解バ
ッファー(1xPBS,1%N−P40、0.5%デオキシコール酸ナトリウム、0.1%ドデシル硫酸ナトリウム、1mM PMSF、およびRoche製プロテアーゼ阻害剤(Catalog No.1836153))において定められた時点で溶解され、そして清澄溶解液におけるタンパク質濃度が、標準のビシンコニニック・アシド(bicinchoninic acid)(BCA)タンパク質アッセイ(Sigma Catalog No.B9643)を用いて定量された。ウエスタン・ブロットでは、各溶解物20μgが標準のSDS−PAGEゲル上で展開され、フッ化ポリビニリデン(PVDF)メンブランに転移され、そしてXBP−1またはCHOPに特異的な抗体(Santa Cruz Biotechnology Inc.,Santa Cruz,CA)によりプローブされた。
【0056】
結果(未掲示)は、高分泌性細胞におけるUPRタンパク質が、低分泌性細胞および親骨髄腫細胞系に較べて高レベルで発現されることを示した。XBP−1に特異的なウエスタン・ブロットは、XBP−1タンパク質発現が、タプシガルギン処理された高分泌性C168j細胞において強く誘導されることを示している。このXBP−1発現の誘導は、UPRの薬理学的誘導7時間後にもっとも明確であり、そして処置後2時間くらいの初期に見ることができる。CHOPに特異的なウエスタン・ブロットは、CHOPタンパク質発現が、タプシガルギン処理された高分泌性C168j細胞において強く誘導されることを示している。CHOPは2時間目に最初に出現し、4〜7時間に最大に発現され、そして22時間では非常に低下される。
【実施例6】
【0057】
UPRタンパク質発現レベルの増大
細胞は、BipおよびXBP−1発現ベクターを用い、これらのUPRタンパク質の細胞発現レベルを高めることを試みて一過性にトランスフェクトされた。HEK293T/17(ATCC CRL−11268)細胞が、5%CO2の雰囲気中37℃で付着培養物として増殖され、5%FBS、2mMグルタミンおよび2mMピルビン酸塩を含有するIscoveの改変Dubelccoの最小必須培地(IMDM)培地において培養された。XBP−1イソ型1(NCBI Accession AF027963)、XBP−1イソ型2(NCBI Accession AF443192)またはBipをコードしているcDNAが、標準的方法を用いて一過性トランスフェクション実験のために有用なベクター中にサブクローン化された。ベクターはCMVプロモーターの制御下でZsGreen1タンパク質を発現し、かつCMVプロモーターの制御下でさらなるコーディング領域の導入のための多数のクローニング部位を有する。カナマイシン耐性遺伝子は細菌の選択のために使用された。次いで、HEK293T/17細胞が、いずれかトランスフェクトされずに残されるか、あるいは空のベクターのみ、XBP−1イソ型1発現ベクター、XBP−1イソ型2発現ベクターまたはBip発現ベクターにより一過性にトランスフェクトされた。トランスフェクションは、製造者によって指示されるようにLipofectamineTM2000試薬(Invitrogen Inc.,Carlsbad,CA)を用いて実施された。トランスフェクション後48時間に、細胞はRIPA溶解バッファーにおいて溶解され、そして清澄溶解液の等しい細胞当量がSDS−PAGEゲル上に負荷された。次いで、ウエスタンブロットが調製され、先の実施例5に記述されたようにプローブされた。次いでブロットが除去され、アクチン特異抗体で再プローブされて各列に負荷された同じタンパク質を確認した。
【0058】
結果(未掲示)は、XBP−1イソ型1(「XBP−1」を標識された列)およびXBP−1イソ型2(「X54}を標識された列)発現レベルが、これらのXBP−1イソ型をコードしている発現ベクターにより一過性にトランスフェクトされた細胞において上昇されたことを例証した。さらに、データ(未掲示)は、Bip発現レベルが、Bipをコードしている発現ベクターにより一過性にトランスフェクトされた細胞において上昇され、そしてトランスフェクトされずに残されたか、または空のベクターのみでトランスフェクトされた細胞では、Bip発現が検出されなかったことを示した。
【実施例7】
【0059】
抗体分泌速度に及ぼすUPR遺伝子転写物レベルの影響
抗体分泌速度に及ぼすUPR遺伝子転写物レベルの影響が解析された。Bip、CHOPおよびXBP−1遺伝子転写物を標的とする二本鎖siRNA分子は、Ambionのインターネットに基づくsiRNA Target Finder Tool(www.ambion.com/techlib/misc/siRNA finder.html)を用いて設計され、そしてSilencerTMsiRNA Constructionキット(Ambion Inc.,Woodward,TX,Catalog No.11620)を用いて合成された。2種のsiRNAが各標的転写物のために設計された。標準条件下で培養された3x106C168J細胞が、図10に示されるようなこれらの各二本鎖siRNAの3μgを用いてエレクトロポレーションによりトランスフェクトされた。エレクトロポレートされた細胞は、先の実施例1に記述されたようにIMDM培地2mlに懸濁され、次いで、T−12プレートにおいて培養された。生存細胞濃度は8日後に測定された。またIgG1mAb濃度も、標準比濁分析技術を用いて8日後に測定された。
【0060】
図7における結果は、抗体分泌速度が、抗体分泌速度に及ぼすUPR遺伝子転写物レベルを改変することによって調節できることを示している。RNA転写物からUPR遺伝子Bip、CHOPおよびXBP−1イソ型の発現を妨げることが可能な短い干渉RNA(siRNA)による高分泌性C168J細胞のトランスフェクションは、C168J細胞のIgG1分泌速度を減少させた。
【実施例8】
【0061】
UPR遺伝子転写物および発現レベルを増大することによる抗体分泌速度の増大
UPRタンパク質の過剰発現が、細胞によるmAbのようなタンパク質の分泌速度を増大できる。タンパク質分泌性細胞系、例えばmAb分泌性細胞系は、Bip、CHOP、XBP−1および他のUPR関連のタンパク質をコードしている発現ベクターによりトランスフェクトされて、これらのタンパク質の過剰発現に影響を与えられた。細胞が、単独でも、また組み合わせてもこれらの発現ベクターを用いてトランスフェクトされる。適当なタンパク質分泌速度、例えば抗体分泌速度は、標準技術を用いてトランスフェクション後2、4および6日目に決定された。トランスフェクトされた細胞におけるタンパク質分泌速度が、トランスフェクトされない対照細胞の分泌速度に比較された。タンパク質分泌速度は、1種以上のUPRタンパク質を過剰発現する細胞において、より高いことが期待される。BipまたはXBP−1の構成的な過剰発現が、最終的にこれらの細胞においてアポトーシスを誘導する事象では、これらの遺伝子が、誘導プロモーターの下流に置かれ、そして必要な場合にのみ活性化されてもよい。
【実施例9】
【0062】
CHOP−10エンコ−ディングsiRNAにより安定してトランスフェクトされた骨髄腫細胞における抗体分泌速度の増大
CHOP−10に特異的なsiRNAを生産するDNA構築物によるC465A骨髄腫細胞の安定なトランスフェクションは、抗体分泌速度を増大した(図8)。CHOP−10遺伝子転写物を標的とする2種の異なるsiRNA分子が、Ambionのインターネットに基づくsiRNA Target Finder Tool(www.ambion.com/techlib/misc/siRNA finder.html)を用いて設計された。設計されたsiRNA分子は短いヘアピン型であった。これらのsiRNAをコードしている核酸(配列番号:15および配列番号:16)が合成され、pSILENCER4.1−neoベクター(Ambion Inc.,Woodward,TX,)のBamHIとHindIII部位中に標準法を用いてクローン化された。pSILENCER4.1−neo中への配列番号:15において示される核酸配列のクローニングはpCHOP1を生成し、一方、このベクター中への配列番号:16において示される配列のクローニングはpCHOP2を生成した。pCHOP1、pCHOP2およびpSILENCER4.1−neoプラスミドDNAが、次に、各々別々にエレクトロポレーションによってC465A骨髄腫細胞中にトランスフェクトされた。これらのベクターを含有する安定してトランスフェクトされた骨髄腫細胞が、次に、10%FBSおよびG418 300μg/mlを含有するSFM8培地において5%CO2の雰囲気中37℃での培養によって選択された。また、この選択培地は、ミコフェノール酸0.5mg/L、ヒポキサンチン2.5mg/Lおよびキサンチン50mg/Lを含有する1XMHX選択培地を含有して、C465A細胞による安定な抗体発現を維持した。
【0063】
図8に示される各細胞系は、10%FBS、1X MHXおよびG418 300μg/mlを含有するSFM8培地において6日間懸濁液において増殖された。生存細胞濃度および抗体タイターはこの6日間の間毎日決定された。生存細胞濃度は、標準の色素排除アッセイによってアッセイされ、そして培養培地中の抗体タイターは標準の比濁分析技術によってアッセイされた。C465A骨髄腫細胞系はSP2/0骨髄腫細胞系から得られ、IgG1κイソ型のヒトTNP−α特異的マウスmAbを安定して発現する。C2−8骨髄腫細胞系は、pCHOP1により安定してトランスフェクトされたC465A由来の細胞系である。C2−18骨髄腫細胞系は、pCHOP2により安定してトランスフェクトされたC465A由来の細胞系である。V11およびV12細胞系は、pSILENCER4.1−neoのみにより安定してトランスフェクトされたC465A由来の細胞系であり;両系統は独立して生成された。
【0064】
得られたデータは、pCHOP1およびpCHOP2によってコードされるCHOP−10特異的siRNAは、対照C465A、V11およびV12細胞系に較べて抗体特異的生産性を増大した(図8)。
【0065】
本発明はここに完全に記述され、付随する請求項の精神および範囲から逸脱することなく、多くの変更および修飾がそれに対して作成できることは当業者には明白であろう。
【図面の簡単な説明】
【0066】
【図1】図1は、高い速度でmAbを分泌する細胞におけるUPR遺伝子転写物レベルを示す。
【図2】図2は、静止増殖期の細胞におけるUPR遺伝子転写物レベルを示す。
【図3】図3は、親骨髄腫細胞系におけるUPR遺伝子転写物レベルの比較を示す。
【図4】図4は、Sp2/0親骨髄腫細胞系に比較して高分泌性細胞系におけるUPR遺伝子転写物レベルを示す。
【図5】図5は、FO親骨髄腫細胞系に比較して高分泌性細胞系におけるUPR遺伝子転写物レベルを示す。
【図6】図6は、Ag−653親骨髄腫細胞系に比較して高分泌性細胞系におけるUPR遺伝子転写物レベルを示す。
【図7】図7は、UPR遺伝子転写物レベル改変の関数としての抗体分泌速度における変化を示す。
【図8】図8は、CHOP−10特異的siRNAをコードしている核酸により安定してトランスフェクトされた骨髄腫細胞における増大された抗体分泌速度を示す。
【特許請求の範囲】
【請求項1】
a)細胞において少なくとも1種のアンフォールド型(unfolded)タンパク質応答(UPR)経路構成要素の活性を調節する工程;および
b)細胞を培養する工程、
を含んでなるタンパク質の細胞分泌速度の変更方法。
【請求項2】
細胞分泌速度が増大される、請求項1の方法。
【請求項3】
UPR経路構成要素が、UPR経路構成要素をコードしている核酸により細胞を安定してトランスフェクトすることによって調節される、請求項2の方法。
【請求項4】
UPR経路構成要素の核酸が、配列番号:1、3、5、7、9、11または13において示されるアミノ酸配列を有するタンパク質をコードしている、請求項3の方法。
【請求項5】
UPR経路構成要素の核酸が、配列番号:2、4、6、8、10、12または14において示される配列を有する、請求項4の方法。
【請求項6】
UPR経路構成要素が転写制御配列である、請求項3の方法。
【請求項7】
UPR経路構成要素が低分子を投与することによって調節される、請求項2の方法。
【請求項8】
低分子がタプシガルギン(thapsigargin)、ツニカマイシン(tunicamysin)またはリポ多糖である、請求項7の方法。
【請求項9】
UPR経路構成要素が、Bip、XBP−またはCHOPである、請求項3の方法。
【請求項10】
UPR経路構成要素が、発育停止増殖期に細胞を置くことによって調節される、請求項2の方法。
【請求項11】
細胞が骨髄腫細胞である、請求項1の方法。
【請求項12】
骨髄腫細胞がSp2/0、NSO、FOまたはAg653である、請求項11の方法。
【請求項13】
骨髄腫細胞がSp2/0、NSO、FOまたはAg653のサブクローンである、請求項11の方法。
【請求項14】
タンパク質が抗体である、請求項1の方法。
【請求項15】
細胞分泌速度が減少される、請求項1の方法。
【請求項16】
UPR経路構成要素が、UPR経路構成要素をコードしている核酸の転写物を標的とする短い干渉性RNA(siRNA)をコードしている核酸により細胞を安定してトランスフェクトすることによって調節される、請求項15の方法。
【請求項17】
UPR経路構成要素が、UPR経路構成要素をコードしている核酸配列の転写物を標的とするsiRNAをコードしている核酸により細胞を安定してトランスフェクトすることによって調節される、請求項2の方法。
【請求項18】
siRNAがCHOPタンパク質をコードしている核酸配列の転写物を標的とする、請
求項17の方法。
【請求項19】
CHOPタンパク質が配列番号:8または配列番号:14において示されるアミノ酸配列を有する、請求項18の方法。
【請求項20】
核酸配列が配列番号:15または配列番号:16において示される核酸配列を含有する、請求項18の方法。
【請求項21】
a)細胞において少なくとも1種のUPR経路構成要素の活性を調節する工程;およびb)細胞を培養する工程、
によって生成される、変更された細胞分泌速度を有する形質細胞。
【請求項22】
UPR経路構成要素をコードしている核酸配列の転写物を標的とするsiRNAをコードしている核酸により細胞を安定してトランスフェクトすることによって、UPR経路構成要素の活性を調節することによって生成される、請求項21の形質細胞。
【請求項23】
CHOPタンパク質をコードしている核酸配列の転写物を標的とするsiRNAをコードしている、安定してトランスフェクトされた核酸を含有するSP2/0由来の細胞である、請求項22の形質細胞。
【請求項24】
抗体を分泌する、請求項23の形質細胞。
【請求項25】
C2−8またはC2−18細胞である、請求項23の形質細胞。
【請求項26】
請求項21の形質細胞を含有する、トランスジェニック動物。
【請求項27】
配列番号:15において示される核酸配列を含有する単離された核酸配列。
【請求項28】
配列番号:16において示される核酸配列を含有する単離された核酸配列。
【請求項1】
a)細胞において少なくとも1種のアンフォールド型(unfolded)タンパク質応答(UPR)経路構成要素の活性を調節する工程;および
b)細胞を培養する工程、
を含んでなるタンパク質の細胞分泌速度の変更方法。
【請求項2】
細胞分泌速度が増大される、請求項1の方法。
【請求項3】
UPR経路構成要素が、UPR経路構成要素をコードしている核酸により細胞を安定してトランスフェクトすることによって調節される、請求項2の方法。
【請求項4】
UPR経路構成要素の核酸が、配列番号:1、3、5、7、9、11または13において示されるアミノ酸配列を有するタンパク質をコードしている、請求項3の方法。
【請求項5】
UPR経路構成要素の核酸が、配列番号:2、4、6、8、10、12または14において示される配列を有する、請求項4の方法。
【請求項6】
UPR経路構成要素が転写制御配列である、請求項3の方法。
【請求項7】
UPR経路構成要素が低分子を投与することによって調節される、請求項2の方法。
【請求項8】
低分子がタプシガルギン(thapsigargin)、ツニカマイシン(tunicamysin)またはリポ多糖である、請求項7の方法。
【請求項9】
UPR経路構成要素が、Bip、XBP−またはCHOPである、請求項3の方法。
【請求項10】
UPR経路構成要素が、発育停止増殖期に細胞を置くことによって調節される、請求項2の方法。
【請求項11】
細胞が骨髄腫細胞である、請求項1の方法。
【請求項12】
骨髄腫細胞がSp2/0、NSO、FOまたはAg653である、請求項11の方法。
【請求項13】
骨髄腫細胞がSp2/0、NSO、FOまたはAg653のサブクローンである、請求項11の方法。
【請求項14】
タンパク質が抗体である、請求項1の方法。
【請求項15】
細胞分泌速度が減少される、請求項1の方法。
【請求項16】
UPR経路構成要素が、UPR経路構成要素をコードしている核酸の転写物を標的とする短い干渉性RNA(siRNA)をコードしている核酸により細胞を安定してトランスフェクトすることによって調節される、請求項15の方法。
【請求項17】
UPR経路構成要素が、UPR経路構成要素をコードしている核酸配列の転写物を標的とするsiRNAをコードしている核酸により細胞を安定してトランスフェクトすることによって調節される、請求項2の方法。
【請求項18】
siRNAがCHOPタンパク質をコードしている核酸配列の転写物を標的とする、請
求項17の方法。
【請求項19】
CHOPタンパク質が配列番号:8または配列番号:14において示されるアミノ酸配列を有する、請求項18の方法。
【請求項20】
核酸配列が配列番号:15または配列番号:16において示される核酸配列を含有する、請求項18の方法。
【請求項21】
a)細胞において少なくとも1種のUPR経路構成要素の活性を調節する工程;およびb)細胞を培養する工程、
によって生成される、変更された細胞分泌速度を有する形質細胞。
【請求項22】
UPR経路構成要素をコードしている核酸配列の転写物を標的とするsiRNAをコードしている核酸により細胞を安定してトランスフェクトすることによって、UPR経路構成要素の活性を調節することによって生成される、請求項21の形質細胞。
【請求項23】
CHOPタンパク質をコードしている核酸配列の転写物を標的とするsiRNAをコードしている、安定してトランスフェクトされた核酸を含有するSP2/0由来の細胞である、請求項22の形質細胞。
【請求項24】
抗体を分泌する、請求項23の形質細胞。
【請求項25】
C2−8またはC2−18細胞である、請求項23の形質細胞。
【請求項26】
請求項21の形質細胞を含有する、トランスジェニック動物。
【請求項27】
配列番号:15において示される核酸配列を含有する単離された核酸配列。
【請求項28】
配列番号:16において示される核酸配列を含有する単離された核酸配列。
【図1】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【公表番号】特表2007−537728(P2007−537728A)
【公表日】平成19年12月27日(2007.12.27)
【国際特許分類】
【出願番号】特願2007−506492(P2007−506492)
【出願日】平成17年3月31日(2005.3.31)
【国際出願番号】PCT/US2005/010605
【国際公開番号】WO2005/094355
【国際公開日】平成17年10月13日(2005.10.13)
【出願人】(503054122)セントカー・インコーポレーテツド (74)
【Fターム(参考)】
【公表日】平成19年12月27日(2007.12.27)
【国際特許分類】
【出願日】平成17年3月31日(2005.3.31)
【国際出願番号】PCT/US2005/010605
【国際公開番号】WO2005/094355
【国際公開日】平成17年10月13日(2005.10.13)
【出願人】(503054122)セントカー・インコーポレーテツド (74)
【Fターム(参考)】
[ Back to top ]