
タンパク質表面のリモデリング

【課題】タンパク質をより安定させるために、および/または凝集を防止するために慣例的に修飾するシステムを提供する。
【解決手段】凝集は、タンパク質の不良な挙動の主な原因である。より安定した変異体を生成するためにタンパク質を修飾するシステムが提供される。前記方法は、より親水性の高い残基(例えば、荷電アミノ酸)に変異させるのに適した、タンパク質表面の非保存性疎水性アミノ酸残基を同定する工程を伴う。表面の任意の数の残基が、より可溶性が高く、凝集に対するより高い抵抗力を持ち、リフォールディング能力が高く、および/または種々の条件における安定性が高い変異体を生成するために変更される場合がある。本発明はまた、本発明の技術により生成される理論上の正味電荷が高いGFP、ストレプトアビジン、およびGST変異体も提供する。また、いずれかの対象タンパク質においてこのような修飾を実施するためのキットも提供される。
【解決手段】凝集は、タンパク質の不良な挙動の主な原因である。より安定した変異体を生成するためにタンパク質を修飾するシステムが提供される。前記方法は、より親水性の高い残基(例えば、荷電アミノ酸)に変異させるのに適した、タンパク質表面の非保存性疎水性アミノ酸残基を同定する工程を伴う。表面の任意の数の残基が、より可溶性が高く、凝集に対するより高い抵抗力を持ち、リフォールディング能力が高く、および/または種々の条件における安定性が高い変異体を生成するために変更される場合がある。本発明はまた、本発明の技術により生成される理論上の正味電荷が高いGFP、ストレプトアビジン、およびGST変異体も提供する。また、いずれかの対象タンパク質においてこのような修飾を実施するためのキットも提供される。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願
この出願は、米国特許法§119(e)の下、2006年6月2日に出願された米国仮特許出願第60/810,364号(この全体の内容は、参考として本明細書に援用される)への優先権を主張する。
【0002】
(連邦政府支援研究に関する声明)
本明細書に記載の研究は、国立衛生研究所の助成金(GM065400)の支援を一部受けている。米国政府は、本発明における一定の権利を有する。
【背景技術】
【0003】
タンパク質は細胞の働き者である。タンパク質は、化学反応物質を触媒したり、生体系においてシグナルを変換したり、細胞の構造成分や細胞外マトリックスを提供したり、メッセンジャーとして作用したりなどの働きを行う。タンパク質の誤動作の主な原因の1つに凝集がある。これは単に研究室だけの問題ではなく、アルツハイマー病などの多くの疾病における問題である。計算により設計したタンパク質においては、凝集は特に面倒な問題である。例えば、TOP7は、新規のフォールドを有する計算により設計したタンパク質である。TOP7のより長い型であるTOP7 extendedは、きわめて凝集の影響を受けやすい。TOP7exは、主に不溶性凝集体として発現される。
【発明の概要】
【発明が解決しようとする課題】
【0004】
より多くのタンパク質が、生体系を研究するためのツールとして使用するために設計または修飾されており、あるいはより多くのタンパク質(野生型または修飾)が治療薬として使用されていることから、これらのタンパク質をより安定させるために、および/または凝集を防止するために慣例的に修飾するシステムが必要とされている。
【課題を解決するための手段】
【0005】
本発明は、タンパク質をより安定させるために修飾するシステムを提供する。本発明は、タンパク質表面の疎水性領域を修飾することにより、タンパク質の超熱力学的特性を改善することができるという認識に基づいている。本発明のシステムは、対象タンパク質の可溶性の改善、タンパク質の凝集に対する抵抗力の改善、および/またはタンパク質の復元能力の改善に特に有用である。これらの特性はすべて、タンパク質の産生、タンパク質の精製、ならびに治療薬および研究ツールとしてのタンパク質の使用に特に有用である。
【0006】
一態様において、本発明は、タンパク質の凝集に対する抵抗力、可溶性、リフォールディング能力、および/または広範な条件における一般的な安定性を向上させるために、タンパク質の一次配列を変更する方法を提供する。好ましくは、修飾タンパク質の活性は、修飾されていないタンパク質とほぼまたは実質的に同じである。ある実施形態において、修飾タンパク質は、野生型タンパク質の活性の少なくとも50%、75%、90%または95%を維持する。一実施形態において、前記方法は、(a)対象タンパク質の表面残基を同定する工程と;(b)対象タンパク質に関連する他のタンパク質の中で高度に保存されていない特定の表面残基を同定する(すなわち、どのアミノ酸が前記タンパク質の活性または機能に必須でないかを決定する)手順と;(c)同定した非保存表面残基の疎水性を決定する工程と;(e)少なくとも1つ以上の同定した疎水性非保存残基を、より極性が高いかまたは生理学的なpHで荷電したアミノ酸で置換する工程と、を含む。上述の手順はそれぞれ、当該技術分野で既知のいずれかの技法、コンピュータソフトウェア、アルゴリズム、パラダイムなどを使用して行われる場合がある。生成した修飾タンパク質は、求めている活性および/または所望の特性について試験される場合がある。ある実施形態において、修飾タンパク質はより安定性が高い。ある実施形態において、修飾タンパク質は、より凝集の影響を受けにくい。通常、本発明の方法は、生理学的なpHでタンパク質の正味電荷(正電荷または負電荷)を増大させる。
【0007】
別の態様において、本発明は、タンパク質を「超荷電する」ことにより、タンパク質の凝集への抵抗、可溶性、リフォールディング能力、および/または広範な条件における一般的な安定性を向上させるために、タンパク質の一次配列を変更する方法を提供する。すなわち、修飾タンパク質の全体正味電荷は、野生型タンパク質に比べて増大する(正電荷または負電荷のいずれか)。好ましくは、修飾タンパク質の活性は、修飾されていないタンパク質とほぼまたは実質的に同じである。ある実施形態において、前記方法は、(a)対象タンパク質の表面残基を同定する工程と;(b)対象タンパク質に関する他のタンパク質の中で高度に保存されていない特定の表面残基を同定する(すなわち、どのアミノ酸が前記タンパク質の活性または機能に必須でないかを決定する)手順と;(c)同定した非保存表面残基の親水性を決定する工程と;(e)少なくとも1つ以上の同定した、荷電または極性を有する、溶媒に曝露した非保存残基を、生理学的なpHで荷電した荷電アミノ酸で置換する工程と、を含む。ある実施形態においては、負に荷電した「超荷電」タンパク質を生成するために、修飾用に同定した残基を、アスパラギン酸(Asp)またはグルタミン酸(Glu)残基のいずれかに変異させる。その他のある実施形態においては、正に荷電した「超荷電」タンパク質を生成するために、修飾用に同定した残基を、リジン(Lys)またはアルギニン(Arg)残基のいずれかに変異させる。上述の手順はそれぞれ、当該技術分野で既知のいずれかの技法、コンピュータソフトウェア、アルゴリズム、パラダイムなどを使用して行われる場合がある。生成した修飾タンパク質は、求めている活性および/または所望の特性について試験される場合がある。ある実施形態において、修飾タンパク質「(超荷電タンパク質」)はより安定している。ある実施形態において、修飾タンパク質は、より凝集の影響を受けにくい。通常、本発明の方法は、生理学的なpHでタンパク質の正味電荷(正電荷または負電荷)を増大させる。
【0008】
Protein Data Bank(PDB)に掲載されるタンパク質の80%超の理論上の正味電荷は、±10の範囲内である。本発明により生成される修飾タンパク質は通常、正味電荷が−10未満または+10超である。ある実施形態において、修飾タンパク質は、正味電荷が−20未満または+20超である。ある実施形態において、修飾タンパク質は、正味電荷が−30未満または+30超である。ある実施形態において、修飾タンパク質は、正味電荷が−40未満または+40超である。ある実施形態において、修飾タンパク質は、正味電荷が−50未満または+50超である。修飾タンパク質は正確に折り畳み、生物活性を維持することができる。
【0009】
いずれのタンパク質も本発明のシステムを使用して修飾される場合があり、本発明のシステムによって生成されるタンパク質変異体は、変異タンパク質をコードするポリヌクレオチドまたはベクター、および変異タンパク質を発現する細胞と同様に、本発明の一部であると考えられる。本発明のシステムは、いくつかの新規の緑色蛍光タンパク質(GFP)の変異体を生成するのに使用されてきた。これらの変異体は蛍光性を維持する一方で、広範な環境において現在のGFP型よりも高い安定性を有する。本発明のGFPは、長期間にわたり凝集を誘導する環境においても凝集に対して免疫を有し、煮沸により変性した後でも蛍光タンパク質にリフォールディングすることができる。本発明のシステムは、ストレプトアビジンやグルタチオン‐S‐トランスフェラーゼ(GST)の新規変異体の生成にも使用されてきた。これらの変異体は、加熱しても生物活性を維持し、可溶性を維持する。本発明は、本発明のGFP、ストレプトアビジン、およびGSTタンパク質配列をコードするポリヌクレオチド配列、これらのヌクレオチド配列のいずれかを含むベクター、ならびにこのようなポリヌクレオチド配列もしくはベクターを含むか、または本発明の変異体を発現する細胞も含む。ある実施形態において、本発明は、本発明の変異体を過剰発現する細菌または他の細胞を含む。本発明の変異体は、当該技術分野で既知の種々の生物学的アッセイに使用される場合がある。例えば、超荷電GFPは、現在GFPをレポータータンパク質として使用しているいずれかのアッセイで使用される場合がある。
【0010】
別の態様において、本発明は、本発明のシステムによって修飾された他のタンパク質を提供する。これらの修飾タンパク質は、好ましくは元の活性の大部分を維持する。ある実施形態において、修飾タンパク質は、未修飾型の活性の少なくとも99%、98%、95%または90%を維持する。修飾タンパク質は、より可溶性が高く、凝集に対するより高い抵抗力を持ち、リフォールディング能力が高く、および/または種々の条件における安定性が高い場合がある。本発明のシステムにより修飾されるタンパク質には、疎水性タンパク質、組換えタンパク質、膜タンパク質、構造タンパク質、酵素、細胞外タンパク質、治療用タンパク質(例えば、インシュリン、サイトカイン、免疫グロブリン、免疫グロブリンの断片など)、受容体、細胞シグナル伝達タンパク質、細胞質タンパク質、核タンパク質、転写因子などが含まれる。ある特定の実施形態において、前記タンパク質は、ヒト医学や獣医学で使用する治療用タンパク質である。ある実施形態において、前記タンパク質は、非天然タンパク質、例えば、計算により設計したタンパク質である。他の実施形態において、前記タンパク質は、ハイブリッドタンパク質、融合タンパク質、変質タンパク質、変異タンパク質、遺伝子合成タンパク質、または手動により変質させたその他いずれかのタンパク質である。
【0011】
また、本発明を実施するためのキットも提供される。前記キットには、凝集に対する抵抗力を高めるか、復元能力を高めるか、または全体的な安定性を高めるために対象タンパク質を修飾する上で必要となる試薬が含まれる場合がある。このようなキットには、ポリヌクレオチド、コンピュータソフトウェア、ヌクレオチド、プライマー、ベクター、細胞系、説明書、プレート、媒質、緩衝液、酵素、Eppendorf管、部位特異的突然変異誘発キットなどのすべてかまたはいくつかが含まれる場合がある。好ましくは、前記キットは、実験環境で使用するのに好都合に包装される。研究者は通常、本発明の技法を使用して、修飾するタンパク質のDNAコード配列を提供する。
【0012】
定義
「アミノ酸」:「アミノ酸」という用語は、タンパク質の基本構造サブユニットを指す。アルファアミノ酸は、アミノ基、カルボキシル基、水素原子、およびすべてが中心の炭素原子に結合している側鎖(すなわち、R基)からなる。この中心の炭素原子は、カルボキシル基に隣接していることから、アルファ炭素と呼ばれる。天然アミノ酸は、グリシン、アラニン、バリン、ロイシン、イソロイシン、フェニルアラニン、チロシン、トリプトファン、システイン、メチオニン、セリン、トレオニン、リジン、アルギニン、ヒスチジン、アスパラギン酸塩、グルタミン酸塩、アスパラギン、グルタミン酸塩、およびプロリンを含めた20個が存在する。疎水性アミノ酸には、アラニン、バリン、ロイシン、イソロイシンおよびフェニルアラニンが含まれる。芳香族アミノ酸には、フェニルアラニン、チロシン、トリプトファンおよびヒスチジンが含まれる。極性アミノ酸には、チロシン、システイン、セリン、トレオニン、リジン、アルギニン、ヒスチジン、アスパラギン酸塩、グルタミン酸塩、アスパラギン、およびグルタミンが含まれる。含硫アミノ酸には、システインおよびメチオニンが含まれる。塩基性アミノ酸には、リジン、アルギニンおよびヒスチジンが含まれる。酸性アミノ酸には、アスパラギン酸塩およびグルタミン酸塩が含まれる。非天然アミノ酸もまたタンパク質に挿入されている。ある実施形態において、「アミノ酸」という用語を使用する場合は、20個の天然アミノ酸を指している。
【0013】
「抗体」:「抗体」という用語は、天然であるか、またはすべてもしくは一部が合成により産生されているかにかかわらず、免疫グロブリンを指す。特異的な結合能力を維持するそのすべての誘導体も本用語に包含される。本用語はまた、免疫グロブリン結合ドメインに相同であるか大部分が相同である結合ドメインを有するいずれのタンパク質をも網羅する。これらのタンパク質は、天然供給源に由来する場合もあれば、または一部もしくはすべてが合成により産生される場合もある。抗体はモノクローナルである場合もあれば、ポリクローナルである場合もある。抗体は、IgG、IgM、IgA、IgDおよびIgEというヒトクラスのいずれかを含めた、いずれかの免疫グロブリンクラスの一員である場合がある。
【0014】
「保存された」:「保存された」という用語は、比較している複数の関連する配列の同じ位置で不変的に生じる、ポリヌクレオチド配列またはアミノ酸配列それぞれのヌクレオチドまたはアミノ酸残基を指す。相対的に保存されたヌクレオチドまたはアミノ酸は、配列のその他の位置に生じるヌクレオチドまたはアミノ酸よりも関連性が高い配列の中で保存されるものである。
【0015】
「相同」:本明細書で使用される「相同」という用語は、ヌクレオチドまたはアミノ酸配列のレベルで高度に関連する核酸またはタンパク質を指す、当該技術分野で理解されている用語である。互いに相同である核酸またはタンパク質は、相同体と呼ばれる。相同という用語は、2つの配列(すなわち、ヌクレオチド配列またはアミノ酸)間の配列類似性の程度を指す場合がある。本明細書で言及される相同率の数値は、2つの配列間で可能な最大の相同性、すなわち、2つの配列が多くの一致した(相同な)位置を有するように整列される場合の相同率を表す。相同性は、以下の文献に記載のような既知の方法によって容易に算出することができる:Computational Molecular Biology,Lesk,A.M,ed.,Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.,ed.,Academic Press,New York,1993;Sequence Analysis in Molecular Biology,von Heinje,G., Academic Press,1987;Computer Analysis of
Sequence Data,Part I,Griffin,A.M.,and Griffin,H.G.,eds.,Humana Press,New Jersey,1994;およびSequence Analysis Primer,Gribskov,M.and Devereux,J.,eds.,M Stockton Press,New York,1991(それぞれ参考として本明細書で援用される)。配列間の相同性を決定するのに通常使用される方法には、参考として本明細書で援用される、Carillo,H.,and Lipman,D.,SIAM J Applied Math.,48:1073(1988)に開示される方法が含まれるが、これらに限定されない。相同性を決定する技法は、公的に利用可能なコンピュータプログラムで体系化される。2つの配列間の相同性を決定するコンピュータソフトウェアの例には、GCGプログラムパッケージ(Devereux,J.,et al.,Nucleic Acids Research,12(1),387(1984))、BLASTP、BLASTN、およびFASTA(Atschul,S.F.et al.,J Molec.Biol.,215,403(1990))が含まれるが、これらに限定されない。
【0016】
「相同」という用語は必ず、少なくとも2つの配列(ヌクレオチド配列またはアミノ酸配列)間の比較を指す。本発明によれば、2つのヌクレオチド配列がコードするポリペプチドが、少なくとも20個のアミノ酸からなる少なくとも1つのストレッチについて、少なくとも約50〜60%同一であり、好ましくは約70%同一である場合に、2つのヌクレオチド配列が相同であると見なされる。好ましくは、相同なヌクレオチド配列はまた、少なくとも4〜5個の一意に定義されるアミノ酸からなるストレッチをコードする能力を特徴とする。互いに関連するこれらのアミノ酸の同一性および近似間隔の両方が、相同であると見なされるヌクレオチド配列において考えられなければならない。長さが60個未満のヌクレオチドであるヌクレオチド配列では、少なくとも4〜5個の一意に定義されるアミノ酸からなるストレッチをコードする能力によって相同性が決定される。
【0017】
「ペプチド」または「タンパク質」:本発明によれば、「ペプチド」または「タンパク質」は、ペプチド結合により結合した少なくとも3個のアミノ酸からなるストリングを含む。「タンパク質」および「ペプチド」という用語は、交換可能に使用される場合がある。本発明のペプチドは、好ましくは天然アミノ酸のみを含有するが、非天然アミノ酸(すなわち、天然には生じないがポリペプチド鎖に組み込むことができる化合物)、および/または当該技術分野で既知のアミノ酸類似体が代わりに使用される場合がある。また、本発明のペプチド中の1つ以上のアミノ酸は、例えば、炭水化物基、リン酸基、ファルネシル基、イソファルネシル基、脂肪酸基、結合用リンカー、官能化、または他の修飾(例えば、アルファアミノ化)などの化学物質を添加することにより、修飾される場合がある。好ましい実施形態において、ペプチドの修飾により、より安定した(例えば、in vivoでの半減期がより長い)ペプチドが生じる。これらの修飾には、ペプチドの環化、Dアミノ酸の混入などが含まれる場合がある。どの修飾も、ペプチドの所望の生物活性を実質的に阻害してはならない。ある実施形態においては、ペプチドの修飾により、より生物活性の高いペプチドが生じる。
【0018】
「ポリヌクレオチド」または「オリゴヌクレオチド」:ポリヌクレオチドまたはオリゴヌクレオチドは、ヌクレオチドのポリマーを指す。通常、ポリヌクレオチドは、少なくとも3個のヌクレオチドを含む。このポリマーには、天然ヌクレオシド(すなわち、アデノシン、チミジン、グアノシン、シチジン、ウリジン、デオキシアデノシン、デオキシチミジン、デオキシグアノシン、およびデオキシシチジン)、ヌクレオシド類似体(例えば、2‐アミノアデノシン、2‐チオチミジン、イノシン、ピロロ‐ピリミジン、3‐メチルアデノシン、C5‐プロピニルシチジン、C5‐プロピニルウリジン、C5‐ブロモウリジン、C5‐フルオロウリジン、C5‐ヨードウリジン、C5‐メチルシチジン、7‐デアザアデノシン、7‐デアザグアノシン、8‐オキソアデノシン、8‐オキソグアノシン、O(6)‐メチルグアニン、および2‐チオシチジン)、化学修飾塩基、生物学的修飾塩基(例えば、メチル化塩基)、介在基、修飾糖(例えば、2’‐フルオロリボース、リボース、2’‐デオキシリボース、アラビノース、およびヘキソース)、および/または修飾リン酸基(例えば、モノチオリン酸塩および5’‐N‐ホスホロアミダイトリンケージ)が含まれる場合がある。
【0019】
「小分子」:本明細書で使用される「小分子」という用語は、実験室で調製するか、または天然で認められる非ペプチド性非オリゴマー有機化合物を指す。本明細書で使用される小分子は、「天然物のような」化合物を指してもよいが、この「小分子」という用語は「天然物のような」化合物に限定されない。むしろ、小分子は通常、いくつかの炭素間結合を含有し、1500未満の分子量を有することを特徴とするが、但し、この特徴は本発明において限定することを目的としたものではない。その他のある好ましい実施形態においては、天然物のような小分子が使用される。
【0020】
「安定した」:タンパク質を言及するのに本明細書で使用される「安定した」という用語は、タンパク質の安定性のいずれかの態様を指す。元の野生型タンパク質と比べて安定した修飾タンパク質は、より可溶性が高い、凝集に対してより高い抵抗力を持つ、変性に対してより高い抵抗力を持つ、アンフォールディングに対してより高い抵抗力を持つ、不適切または不要なフォールディングに対してより高い抵抗力を持つ、復元能力が高い、熱安定性が高い、種々の環境(例えば、pH、塩濃度、洗浄剤の存在、変性剤の存在など)における安定性が高い、非水性環境における安定性が高い、といった特徴の内のいずれか1つ以上を有する。ある実施形態において、安定した修飾タンパク質は、上述の特徴の少なくとも2つを示す。ある実施形態において、安定した修飾タンパク質は、上述の特徴の少なくとも3つを示す。このような特徴により、活性タンパク質をより高いレベルで産生することが可能となる場合がある。例えば、修飾タンパク質は、タンパク質の非修飾型よりも、凝集することなくより高いレベルで過剰発現させることができる。また、このような特徴により、治療薬または研究ツールとしてタンパク質を使用することが可能となる場合がある。
本発明はまた、以下の項目を提供する。
(項目1)
対象タンパク質の安定性を改善する方法であって、
対象タンパク質に関連する他のタンパク質の中で高度に保存されていない、該対象タンパク質の表面残基を同定する工程と;
複数の非保存表面残基を、生理学的なpHで正に荷電したアミノ酸残基で置換する工程と、
を含む、方法。
(項目2)
対象タンパク質の安定性を改善する方法であって、
対象タンパク質に関連する他のタンパク質の中で高度に保存されていない、該対象タンパク質の表面残基を同定する工程と;
複数の非保存表面残基を、生理学的なpHで負に荷電したアミノ酸残基で置換する工程と、
を含む、方法。
(項目3)
上記非保存表面残基が疎水性である、項目1または2に記載の方法。
(項目4)
上記非保存表面残基が親水性であるか、または負に荷電している、項目1に記載の方法。
(項目5)
上記非保存表面残基が親水性であるか、または正に荷電している、項目2に記載の方法。
(項目6)
対象タンパク質の安定性を改善する方法であって、
対象タンパク質の表面残基を同定する工程と;
該対象タンパク質に関連する他のタンパク質の中で高度に保存されていない、該対象タンパク質の表面残基を同定する工程と;
該同定した非保存表面残基のそれぞれに疎水性値を割り当てる工程と;
少なくとも1個の表面残基を、生理学的なpHで荷電したアミノ酸残基で置換する工程であって、該置換されている残基がすべて同じ型の残基に変異し、該残基の型が正に荷電した残基であるか、または負に荷電した残基のいずれかである、工程と、
を含む、方法。
(項目7)
上記置換工程が、少なくとも1個の疎水性表面残基を置換する工程を含む、項目6に記載の方法。
(項目8)
上記置換工程が、少なくとも1個の親水性表面残基を置換する工程を含む、項目6に記載の方法。
(項目9)
上記置換工程が、少なくとも1個の荷電した表面残基を置換する工程を含む、項目6に記載の方法。
(項目10)
上記置換工程が、少なくとも1個の表面残基をリジン残基で置換する工程を含む、項目6に記載の方法。
(項目11)
上記置換工程が、少なくとも1個の表面残基をアスパラギン酸残基またはグルタミン酸残基で置換する工程を含む、項目6に記載の方法。
(項目12)
上記置換工程が、少なくとも2個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目13)
上記置換工程が、少なくとも5個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目14)
上記置換工程が、少なくとも10個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目15)
上記置換工程が、少なくとも20個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目16)
上記置換工程が、少なくとも30個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目17)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHでより大きな正味電荷を有する修飾タンパク質が生成される、項目1、2または6に記載の方法。
(項目18)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHでより負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目19)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも−5より負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目20)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも−10より負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目21)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも−15より負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目22)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも−20より負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目23)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHでより正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目24)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも+5より正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目25)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも+10より正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目26)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも+15より正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目27)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも+20より正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目28)
上記対象タンパク質が凝集しやすいタンパク質である、項目1、2または6に記載の方法。
(項目29)
上記対象タンパク質が疎水性タンパク質である、項目1、2または6に記載の方法。
(項目30)
上記対象タンパク質が膜タンパク質である、項目1、2または6に記載の方法。
(項目31)
上記対象タンパク質が、過剰発現しにくいタンパク質である、項目1、2または6に記載の方法。
(項目32)
上記対象タンパク質が、精製しにくいタンパク質である、項目1、2または6に記載の方法。
(項目33)
上記対象タンパク質が受容体である、項目1、2または6に記載の方法。
(項目34)
上記対象タンパク質が転写因子である、項目1、2または6に記載の方法。
(項目35)
上記対象タンパク質が酵素である、項目1、2または6に記載の方法。
(項目36)
上記対象タンパク質が構造タンパク質である、項目1、2または6に記載の方法。
(項目37)
上記対象タンパク質が蛍光タンパク質である、項目1、2または6に記載の方法。
(項目38)
上記対象タンパク質が緑色蛍光タンパク質(GFP)である、項目1、2または6に記載の方法。
(項目39)
上記対象タンパク質が細胞外タンパク質である、項目1、2または6に記載の方法。
(項目40)
上記対象タンパク質がストレプトアビジンである、項目1、2または6に記載の方法。
(項目41)
上記対象タンパク質がグルタチオン‐S‐トランスフェラーゼである、項目1、2または6に記載の方法。
(項目42)
上記表面残基を同定する工程が、上記タンパク質の三次元構造をコンピュータモデリングする工程を含む、項目1、2または6に記載の方法。
(項目43)
上記表面残基を同定する工程が、アルゴリズムを使用して、残基がタンパク質の表面に見出されるか否かを予測する工程を含む、項目1、2または6に記載の方法。
(項目44)
上記表面残基を同定する工程が、閾値未満のavNAPSA値を有する残基を同定する工程を含む、項目1、2または6に記載の方法。
(項目45)
上記高度に保存されていない表面残基を同定する工程が、上記タンパク質のアミノ酸配列を、同じタンパク質ファミリーに由来する少なくとも1つの他のタンパク質と整列させる工程を含む、項目1、2または6に記載の方法。
(項目46)
上記高度に保存されていない表面残基を同定する工程が、上記タンパク質のアミノ酸配列を、異なる種に由来する該タンパク質の少なくとも1つの他のアミノ酸配列と整列させる工程を含む、項目1、2または6に記載の方法。
(項目47)
上記整列が、少なくとも2つの他のタンパク質配列を使用して行われる、項目45または46に記載の方法。
(項目48)
上記整列が、少なくとも3つの他のタンパク質配列を使用して行われる、項目45または46に記載の方法。
(項目49)
上記整列が、少なくとも5つの他のタンパク質配列を使用して行われる、項目45または46に記載の方法。
(項目50)
上記疎水性値を割り当てる工程が、オクタノール/水のP値を使用する工程を含む、項目6に記載の方法。
(項目51)
上記置換工程が、上記同定した疎水性表面残基を、生理学的なpHで荷電した天然アミノ酸残基で置換するように、上記タンパク質配列に突然変異を起こさせる工程を含む、項目1、2または6に記載の方法。
(項目52)
上記天然アミノ酸残基が、リジン、グルタミン酸、アスパラギン酸、ヒスチジンおよびアルギニンからなる群から選択される、項目51に記載の方法。
(項目53)
上記天然アミノ酸残基が、リジン、ヒスチジンおよびアルギニンからなる群から選択される、項目51に記載の方法。
(項目54)
上記天然アミノ酸残基がリジンである、項目51に記載の方法。
(項目55)
上記天然アミノ酸残基が、グルタミン酸およびアスパラギン酸からなる群から選択される、項目51に記載の方法。
(項目56)
上記置換工程が、上記同定した表面残基の部位特異的突然変異誘発を含む、項目1、2または6に記載の方法。
(項目57)
上記置換工程が、上記同定した表面残基のPCR突然突然変異誘発を含む、項目1、2または6に記載の方法。
(項目58)
アミノ酸配列:
【化21】

の緑色蛍光タンパク質(+36 GFP)。
(項目59)
項目58に記載の緑色蛍光タンパク質をコードするポリヌクレオチド。
(項目60)
配列:
【化22】
の項目59に記載のポリヌクレオチド。
(項目61)
アミノ酸配列:
【化23】
の緑色蛍光タンパク質(+49 GFP)。
(項目62)
項目61に記載の緑色蛍光タンパク質をコードするポリヌクレオチド。
(項目63)
配列:
【化24】
の項目62に記載のポリヌクレオチド。
(項目64)
アミノ酸配列:
【化25】
の緑色蛍光タンパク質(‐29 GFP)。
(項目65)
項目64に記載の緑色蛍光タンパク質をコードするポリヌクレオチド。
(項目66)
配列:
【化26】
の項目65に記載のポリヌクレオチド。
(項目67)
アミノ酸配列:
【化27】
のストレプトアビジン(−40 SAV)。
(項目68)
項目67に記載のストレプトアビジンをコードするポリヌクレオチド。
(項目69)
配列:
【化28】
の項目68に記載のポリヌクレオチド。
(項目70)
アミノ酸配列:
【化29】
のストレプトアビジン(+52 SAV)。
(項目71)
項目70に記載のストレプトアビジンをコードするポリヌクレオチド。
(項目72)
配列:
【化30】
の項目71に記載のポリヌクレオチド。
(項目73)
アミノ酸配列:
【化31】
のグルタチオン‐S‐トランスフェラーゼ(−40 GST)。
(項目74)
項目73に記載のストレプトアビジンをコードするポリヌクレオチド。
(項目75)
配列:
【化32】
の項目74に記載のポリヌクレオチド。
(項目76)
項目1〜57のいずれか1項に記載の方法により修飾されるタンパク質。
(項目77)
項目76に記載のタンパク質をコードするポリヌクレオチド。
(項目78)
項目1〜57のいずれか1項に記載の方法を実行するためのキット。
【図面の簡単な説明】
【0021】
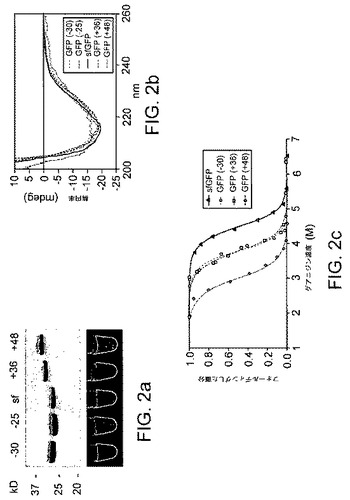
【図1】超荷電した緑色蛍光タンパク質(GFP)を示す。(a)GFP変異体のタンパク質配列であり、フルオロフォア形成残基を緑色で強調表示し、負に荷電した残基を赤色で強調表示し、正に荷電した残基を青色で強調表示した。(b)−25kT/e(赤色)から+25kT/e(青色)の範囲で着色した、sfGFP(左)、GFP(+36)(中央)、およびGFP(−30)(右)の静電表面電位である。
【図2】GFP変異体の分子内特性を示す。(a)精製したGFP変異体の染色およびUV蛍光。各レーンおよび管は0.2μgのタンパク質を含有する。(b)GFP変異体の円二色性スペクトル。(c)グアニジンにより誘導したアンフォールディングで測定したGFP変異体の熱力学的安定性。
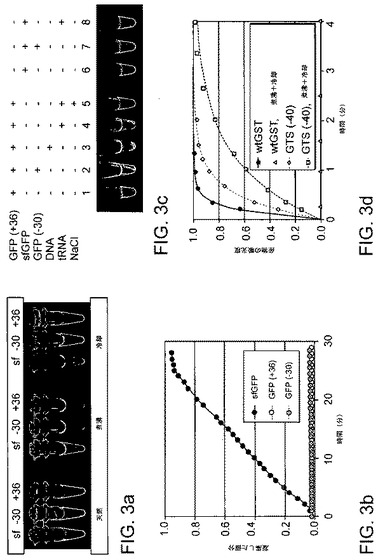
【図3】超荷電タンパク質の分子間特性を示す。(a)精製したGFP変異体のUV照射試料(「天然」)と、100℃にて1分間加熱した試料(「煮沸」)と、次いで25℃にて2時間冷却した試料(「冷却」)。(b)GFP変異体の凝集を25℃の40% TFEで誘導し、直角の光散乱によって監視した。(c)超荷電GFPが逆荷電した高分子に可逆的に付着する。試料1:30μLの25mM Tris、pH7.0および100mM NaCl中のGFP(+36)6μg。試料2:GFP(−30)6μgを試料1に添加。試料3:サケ精子DNA30μgを試料1に添加。試料4:大腸菌tRNA 20μgを試料1に添加。試料5:1MのNaClを試料4に添加。試料6〜8:GFP(+36)の代わりにsfGFPを使用した以外は、試料1、2および4それぞれと同じ。すべての試料はミクロ遠心分離機で軽く撹拌した後、紫外線で視覚化した。(d)GST変異体の酵素アッセイ。反応物は、GST変異体0.5mg/mL、クロロジニトロベンゼン20mM、グルタチオン20mM、およびリン酸カリウム100mM、pH6.5を含有した。産物形成を340nmで監視した結果、煮沸および冷却後の反応速度(kobs)は、野生型GSTが6分−1、GST(−40)が2.2分−1、GST(−40)が0.9分−1であることが認められた。
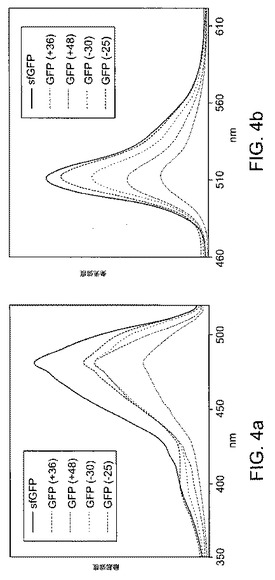
【図4】GFP変異体の(a)励起および(b)発光スペクトルを示す。各試料は、490nmの発色団吸光度で定量化により等しい量のタンパク質を含有した。
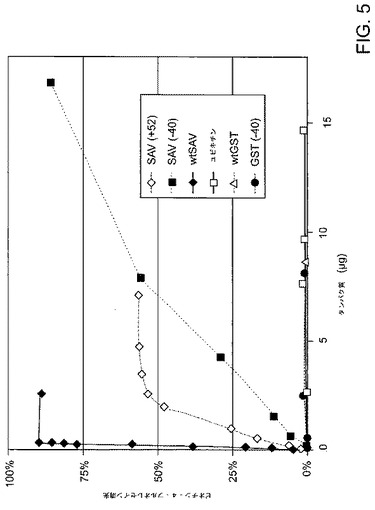
【図5】結合依存性のビオチン‐4‐フルオレスセイン(Invitrogen)を監視することによって、既述の通りに測定した(参考として本明細書で援用される、Kadaら、Rapid estimation of avidin and streptavidin by fluorescence quenching or fluorescence polarization.Biochim.Biophys.Acta 1427,44‐48(1999))、ストレプトアビジン変異体のビオチン結合活性を示す。タンパク質試料を、0.3μMのビオチン‐4‐フルオレスセイン(B4F)、100mMのNaCI、1mMのEDTA、0.1mg/mLのウシ血清アルブミン(BSA)、50mMのリン酸カリウム、pH7.5に滴定した。526nmの蛍光消光を、Perkin‐Elmer LS50B蛍光分光計で470nmの励起により測定した。測定値は、600倍の過剰な非蛍光性ビオチンを含有する対照滴定液に正規化した。凡例内の下の3つのタンパク質を陰性対照として含めた。
【発明を実施するための形態】
【0022】
本発明の特定の好ましい実施形態の詳細な説明
本発明は、タンパク質をより安定させるために修飾するシステムを提供する。前記システムは、タンパク質表面の非保存アミノ酸を、より極性の高いまたは荷電したアミノ酸残基に変更することによって機能すると考えられる。修飾するアミノ酸残基は、疎水性である場合もあれば、親水性である場合もあり、荷電している場合もあり、あるいはこれらの組み合わせである場合もある。いずれのタンパク質も、より安定した変異体を産生するために、本発明のシステムを使用して修飾される場合がある。これらの表面残基の修飾によって、タンパク質の超熱力学的特性が改善されることが明らかにされている。タンパク質が治療薬として使用されることが多くなっており、かつ研究ツールとしても依然として使用されていることから、タンパク質をより安定させるために変質させるシステムが重要であり、有用である。本発明の方法によって修飾されるタンパク質は通常、凝集に対する抵抗力を持ち、リフォールディング能力が高く、不適切なフォールディングに対する抵抗力を持ち、可溶性が高く、加熱や洗浄剤の存在といった変性条件を含めた広範な条件において一般的により高い安定性を有する。
【0023】
いずれのタンパク質も、本発明のシステムを使用してより安定した変異体を生成するために修飾される場合がある。天然だけでなく非天然のタンパク質(例えば、改変タンパク質)も修飾される場合がある。修飾される場合があるタンパク質の例には、受容体、膜結合タンパク質、膜貫通タンパク質、酵素、転写因子、細胞外タンパク質、治療用タンパク質、サイトカイン、メッセンジャータンパク質、DNA結合タンパク質、RNA結合タンパク質、シグナル変換に関与するタンパク質、構造タンパク質、細胞質タンパク質、核タンパク質、疎水性タンパク質、親水性タンパク質などが含まれる。修飾するタンパク質は、いずれの種の植物、動物または微生物からも得られる場合がある。ある実施形態において、タンパク質は哺乳類タンパク質である。ある実施形態において、タンパク質はヒトタンパク質である。ある実施形態において、タンパク質は、通常研究で使用される生体から得られる。例えば、修飾するタンパク質は、霊長類(例えば、類人猿、サル)、齧歯類(例えば、ウサギ、ハムスター、アレチネズミ)、ブタ、イヌ、ネコ、魚(例えば、ゼブラフィッシュ)、線虫(例えば、シノラブディス・エレガンス)、酵母菌(例えば、サッカロミセス・セレビジエ)、または細菌(例えば、大腸菌)から得られる場合がある。
【0024】
本発明のシステムは、凝集の影響を受けやすいか、または安定性の問題を有するタンパク質を修飾するのに特に有用である。前記システムはまた、過剰発現しているタンパク質を修飾するのに使用される場合もある。例えば、組換えにより産生されている治療用タンパク質は、本発明のシステムによる修飾が有用な場合がある。このような修飾された治療用タンパク質は、産生や精製が容易なだけでなく、タンパク質の保管や使用においてもより高い安定性を有する場合がある。
【0025】
本発明のシステムは、対象タンパク質の非保存表面残基を同定する工程と、これらの残基のいくつかを、親水性を有するか、極性を有するか、または生理学的なpHで荷電した残基で置換する工程を伴う。本発明のシステムは、タンパク質を修飾する方法だけでなく、タンパク質をより安定させるために修飾するのに有用な試薬およびキットも含む。
【0026】
修飾するタンパク質の表面残基は、当該技術分野で既知のいずれかの方法を使用して同定される。ある実施形態において、表面残基は、タンパク質のコンピュータモデリングによって同定される。ある実施形態において、タンパク質の三次元構造は、既知であり、および/または決定されており、タンパク質の構造を視覚化することによって表面残基が同定される。他の実施形態において、表面残基は、コンピュータソフトウェアを使用して予測される。ある特定の実施形態においては、Average Neighbor Atoms per Sidechain Atom(AvNAPSA)が、表面露出の予測に使用される。AvNAPSAは、コンピュータプログラムとして導入された、表面露出の自動化された測度である。付録Aを参照されたい。低いAvNAPSA値が表面露出残基を示し、高いAvNAPSA値がタンパク質内部の残基を示す。ある実施形態においては、本ソフトウェアが、タンパク質の二次構造および/または三次構造の予測に使用され、表面残基がこの予測に基づいて同定される。他の実施形態において、表面残基の予測は、残基の疎水性および親水性、ならびにタンパク質の一次配列におけるこれらのクラスタリングに基づいて行われる。In silico法の他にも、例えば、プロテアーゼ切断、表面改質などの種々の生化学的技法を使用して、タンパク質の表面残基が同定される場合もある。
【0027】
次いで、表面残基の中で、保存されている残基か、またはタンパク質の機能に重要な残基が決定される。保存残基の同定は、当該技術分野で既知のいずれかの方法を使用して決定することができる。ある実施形態において、保存残基は、対象タンパク質の一次配列を関連タンパク質と整列させることによって同定される。これらの関連するタンパク質は、同じタンパク質ファミリーに由来する場合がある。例えば、タンパク質が免疫グロブリンである場合は、他の免疫グロブリン配列が使用される場合がある。関連タンパク質はまた、異なる種に由来する同じタンパク質である場合もある。例えば、異なる種に由来する同じタンパク質の配列を整列させることによって、保存残基が同定される場合がある。他の例を挙げると、機能または生物活性が類似するタンパク質を整列させる場合がある。好ましくは、タンパク質中に保存されたアミノ酸を決定するのに、2、3、4、5、6、7、8、9または10種類の配列が使用される。ある実施形態においては、配列の50%、60%、70%、75%、80%または90%超が特定の位置に同じアミノ酸を有する場合に、残基は保存されていると見なされる。他の実施形態においては、配列の50%、60%、70%、75%、80%または90%超が特定の位置に同じかまたは類似するアミノ酸(例えば、バリン、ロイシンおよびイソロイシン;グリシンおよびアラニン;グルタミンおよびアスパラギン;またはアスパラギン酸塩およびグルタミン酸塩)を有する場合に、残基が保存されていると見なされる。本明細書に記載の通り、タンパク質配列の整列および比較に利用可能なソフトウェアパッケージは数多く存在する。当業者であれば理解するであろうが、最初に保存残基を決定する場合もあれば、あるいは最初に表面残基を決定する場合もある。順序は重要でない。ある実施形態においては、コンピュータソフトウェアパッケージで、表面残基と保存残基が同時に決定される場合がある。また、タンパク質の突然変異生成によって、タンパク質中の重要な残基が同定される場合もある。例えば、タンパク質のアラニン走査を使用して、タンパク質中の重要なアミノ酸残基を決定することができる。他の実施形態においては、部位特異的突然変異誘発が使用される場合がある。
【0028】
タンパク質の非保存表面残基が同定されれば、それぞれの残基は疎水性または親水性であると同定される。ある実施形態において、残基には疎水性スコアが割り当てられる。例えば、それぞれの非保存表面残基には、オクタノール/水のlogP値が割り当てられる場合がある。また、他の疎水性パラメータも使用される場合がある。このようなアミノ酸のスケールについては、以下で考察されている:Janin,“Surface and
Inside Volumes in Globular Proteins,”Nature 277:491‐92,1979;Wolfendenら、“Affinities of Amino Acid Side Chains for Solvent Water,”Biochemistry 20:849‐855,1981;Kyteら、“A Simple Method for Displaying the Hydropathic Character of a Protein,”J.Mol.Biol.157:105‐132,1982;Roseら、“Hydrophobicity of Amino Acid Residues in Globular
Proteins,”Science 229:834‐838,1985;Cornetteら、“Hydrophobicity Scales and Computational Techniques for Detecting Amphipathic Structures in Proteins,”J.Mol.Biol.195:659‐685,1987;Charton and Charton,“The
Structure Dependence of Amino Acid Hydrophobicity Parameters,“J.Theor.Biol.99:629‐644,1982(それぞれ参考として援用される)。これらの疎水性パラメータのいずれかが、どの非保存残基を修飾するかを決定するのに、本発明の方法で使用される場合がある。ある実施形態においては、親水性または荷電残基が修飾用に同定される。
【0029】
次いで、少なくとも1つの同定された非保存または失活表面残基が修飾用に選択される。ある実施形態においては、疎水性残基が修飾用に選択される。他の実施形態においては、親水性および/または荷電残基が修飾用に選択される。ある実施形態においては、複数の残基が修飾用に選択される。ある実施形態においては、1、2、3、4、5、6、7、8、9または10個の同定された残基が修飾用に選択される。ある実施形態においては、10個を超える、15個を超えるまたは20個を超える残基が修飾用に選択される。当業者であれば理解するであろうが、タンパク質が大きくなるほど、修飾が必要となる残基も多くなる。また、タンパク質がより疎水性であるか、または凝集もしくは沈降の影響を受けやすいほど、修飾が必要となる残基も多くなる。ある実施形態においては、それぞれ修飾が異なるタンパク質の複数の変異体が産生され、生物活性および安定性に最も優れた変異体を決定するために試験される。
【0030】
ある実施形態においては、修飾用に選択した残基を、より親水性の高い残基(荷電残基を含む)に変異させる。通常、残基は、より親水性の高い天然アミノ酸に変異させる。ある実施形態において、残基は、生理学的なpHで荷電したアミノ酸に変異させる。例えば、アルギニン、アスパラギン酸塩、グルタミン酸塩、ヒスチジンまたはリジンに残基を変化する場合がある。ある実施形態においては、修飾するすべての残基を、同じ残基に変化させる。例えば、選択したすべての残基をグルタミン酸塩の残基に変化させる。他の実施形態において、選択した残基は、異なる残基に変化させるが、最終残基はすべて、生理学的なpHで正に荷電している場合もあれば、負に荷電している場合もある。ある実施形態においては、負に荷電したタンパク質を生成するために、変異させるすべての残基を、グルタミン酸塩および/またはアスパラギン酸塩の残基に変換させる。ある実施形態において、正に荷電したタンパク質を生成するために、変異させるすべての残基をリジンの残基に変換させる。例えば、修飾用に選択したすべての残基は、アスパラギン、グルタミン、リジン、および/またはアルギニンであり、これらの残基をアスパラギン酸塩またはグルタミン酸塩の残基に変異させる。他の例を挙げると、修飾用に選択したすべての残基は、アスパラギン酸塩、グルタミン酸塩、アスパラギンおよび/またはグルタミンであり、これらの残基をリジンに変異させる。この手法により、タンパク質の正味電荷を最大限度まで修飾することが可能となる。
【0031】
他の実施形態においては、修飾タンパク質の正味電荷を非修飾タンパク質の正味電荷と同じに維持するように、タンパク質が修飾される場合がある。さらに他の実施形態においては、表面における荷電残基の総数を増加させながら、タンパク質の全体的な正味電荷を減少させるように、タンパク質が修飾される場合がある。ある実施形態においては、理論上の正味電荷を、少なくとも+1、+2、+3、+4、+5、+10、+15、+20、+25、+30または+35増加させる。ある実施形態においては、理論上の正味電荷を、少なくとも−1、−2、−3、−4、−5、−10、−15、−20、−25、−30または−35減少させる。ある実施形態においては、選択したアミノ酸を、非イオン極性残基(例えば、システイン、セリン、トレオニン、チロシン、グルタミン、アスパラギン)に変化させる。
【0032】
タンパク質におけるこれらの修飾および突然変異は、当該技術分野で既知のいずれかの技法を使用して達成される場合がある。このような変化をタンパク質配列に導入する組換えDNA技術は、当該技術分野で周知である。ある実施形態においては、タンパク質をコードするポリヌクレオチドの部位特異的突然変異誘発によって修飾が行われる。突然変異を導入する他の技法については、それぞれが参考として本明細書で援用される、Molecular Cloning:A Laboratory Manual,2nd Ed.,ed.by Sambrook,Fritsch,and Maniatis(Cold Spring Harbor Laboratory Press:1989);Methods in Enzymology(Academic Press,Inc.,N.Y.);Ausubelら、Current Protocols in Molecular Biology(John Wiley&Sons,Inc.,New
York,1999)で考察されている。修飾タンパク質は発現され、試験される。ある実施形態においては、一連の変異体が調製され、それぞれの変異体が、生物活性および安定性を決定するために試験される。後の使用のために選択する変異体は、最も安定性が高いものである場合もあれば、最も活性の高いものである場合もあり、あるいは全体として活性と安定性が最も高い組み合わせを有するものである場合もある。最初の群の変異体を調製した後、最初の群から学習した内容に基づいて、追加の群の変異体が調製される場合がある。通常作成した変異体は、当該技術分野で既知の組換え法を使用して過剰発現させる。
【0033】
本発明のシステムは、GFPの変異体の生成に使用されてきた。これらの変異体は、より安定性が高く、蛍光を維持することが明らかにされている。Aequorea victoria由来のGFPが、参考として本明細書で援用される、GenBank受託番号P42212に記載されている。この野生型GFPのアミノ酸配列は、以下の通りである:
【0034】
【化1】
野生型GFPの理論上の正味電荷は−7であるが、本発明のシステムを使用して、理論上の正味電荷が−29、−30、−25、+36、+48および+49である変異体が生成された。+36のGFPを95℃に加熱した後でも、変異タンパク質の100%が可溶性を有し、タンパク質が70%以上の蛍光を維持する。
【0035】
生成したGFP変異体のアミノ酸配列には、以下が含まれる。
【0036】
【化2】
【0037】
【化3】
当業者であれば理解するであろうが、相同タンパク質も本発明の適用範囲に含まれると見なされる。例えば、上述の配列のいずれかに対して60%、70%、80%、90%、95%または100%相同である20、30、40、50または100個のアミノ酸からなるストレッチを含むいずれのタンパク質も、本発明の一部に含まれると見なされる。さらに、付加および欠失変異体も本発明で企図される。ある実施形態においては、上述の配列のいずれかに示す変異残基を有するいずれのGFPも、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0038】
また、上述のGFP変異体をコードするいずれのDNA配列も、本発明の適用範囲に含まれる。上述の変異体それぞれをコードするDNA配列の例は、以下の通りである。
【0039】
【化4】
【0040】
【化5】
上述の配列に相同なポリヌクレオチド配列も、本発明の適用範囲に含まれる。ある実施形態において、ポリヌクレオチド配列は、上述の配列のいずれか1つに対して60%、70%、80%、90%、95%、98%、99%または100%相同である50、100または150個のヌクレオチドからなるストレッチを含む。本発明はまた、1つ以上のヌクレオチドが上述の配列の1つに挿入されるか、またはこれらの1つから欠失する配列も含む。上述の配列のいずれかに示す突然変異を有するいずれのポリヌクレオチド配列も、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0041】
本発明はまた、本明細書における本発明の配列のいずれか、または本発明のシステムを使用して修飾されるその他いずれかの配列(DNAもしくはタンパク質)を含むベクター(例えば、プラスミド、コスミド、ウイルスなど)も提供する。ある実施形態において、前記ベクターには、プロモーター、エンハンサー、リボソーム結合部位などの要素、および細胞内で本発明のGFP変異体を過剰発現するのに有用な配列が含まれる。本発明はまた、本発明の配列またはベクターを含む細胞も含む。ある実施形態において、前記細胞は、変異体GFPを過剰発現する。前記細胞は、細菌性細胞(例えば、大腸菌)、真菌細胞(例えば、ピキアパストリス)、酵母細胞(例えば、サッカロミセス・セレビシエ)、哺乳類細胞(例えば、CHO細胞)、またはヒト細胞である場合がある。
【0042】
本発明のシステムは、ストレプトアビジン変異体の生成に使用されてきた。これらの変異体は、ビオチンを結合する可溶性四量体を形成することが明らかにされている。この野生型ストレプトアビジンのアミノ酸配列は、以下の通りである。
【0043】
【化6】
野生型ストレプトアビジンの理論上の正味電荷は−4であるが、本発明のシステムを使用して、理論上の正味電荷が−40および+52である変異体が生成された。前記変異体を100℃に加熱した後でも、タンパク質は可溶性を維持した。
【0044】
生成したストレプトアビジン変異体のアミノ酸配列には、以下が含まれる。
【0045】
【化7】
当業者であれば理解するであろうが、相同タンパク質も本発明の適用範囲に含まれると見なされる。例えば、上述の配列のいずれかに対して60%、70%、80%、90%、95%または100%相同である20、30、40、50または100個のアミノ酸からなるストレッチを含むいずれのタンパク質も、本発明の一部に含まれると見なされる。さらに、付加および欠失変異体も本発明で企図される。ある実施形態においては、上述の配列のいずれかに示す変異残基を有するいずれのストレプトアビジンも、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0046】
上述のストレプトアビジン変異体をコードするいずれのDNA配列も、本発明の適用範囲に含まれる。上述の変異体それぞれをコードするDNA配列の例は、以下の通りである。
【0047】
【化8】
上述の配列に相同なポリヌクレオチド配列も、本発明の適用範囲に含まれる。ある実施形態において、ポリヌクレオチド配列は、上述の配列のいずれか1つに対して60%、70%、80%、90%、95%、98%、99%または100%相同である50、100、または150個のヌクレオチドからなるストレッチを含む。本発明はまた、1つ以上のヌクレオチドが上述の配列の1つに挿入されるか、またはこれらの1つから欠失する配列も含む。上述の配列のいずれかに示す突然変異を有するいずれのポリヌクレオチド配列も、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0048】
本発明はまた、本明細書における本発明の配列のいずれか、または本発明のシステムを使用して修飾されるその他いずれかの配列(DNAもしくはタンパク質)を含むベクター(例えば、プラスミド、コスミド、ウイルスなど)も提供する。ある実施形態において、前記ベクターには、プロモーター、エンハンサー、リボソーム結合部位などの要素、および細胞内で本発明のストレプトアビジン変異体を過剰発現するのに有用な配列が含まれる。本発明はまた、本発明の配列またはベクターを含む細胞も含む。ある実施形態において、前記細胞は、変異体ストレプトアビジンを過剰発現する。前記細胞は、細菌性細胞(例えば、大腸菌)、真菌細胞(例えば、ピキアパストリス)、酵母細胞(例えば、サッカロミセス・セレビシエ)、哺乳類細胞(例えば、CHO細胞)、またはヒト細胞である場合がある。
【0049】
本発明のシステムは、グルタチオン‐S‐トランスフェラーゼ(GST)の変異体の生成に使用されてきた。これらの変異体は、野生型GSTの触媒作用を維持することが明らかにされている。この野生型GSTのアミノ酸配列は、以下の通りである。
【0050】
【化9】
野生型GSTの理論上の正味電荷は+2であるが、本発明のシステムを使用して、理論上の正味電荷が−40である変異体が生成された。この変異体は、野生型GSTよりも比活性がわずかに2.7倍低いクロロニトロベンゼンへのグルタチオンの付加を触媒する。この変異体を100℃に加熱した後でも、タンパク質は可溶性を維持し、タンパク質は冷却時に触媒作用の40%を回復した。
【0051】
GST変異体のアミノ酸配列には、以下が含まれる。
【0052】
【化10】
当業者であれば理解するであろうが、相同タンパク質も本発明の適用範囲に含まれると見なされる。例えば、上述の配列のいずれかに対して60%、70%、80%、90%、95%または100%相同である20、30、40、50または100個のアミノ酸からなるストレッチを含むいずれのタンパク質も、本発明の一部に含まれると見なされる。さらに、付加および欠失変異体も本発明で企図される。ある実施形態においては、上述の配列のいずれかに示す変異残基を有するいずれのストレプトアビジンも、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0053】
上述のGST変異体をコードするいずれのDNA配列も、本発明の適用範囲に含まれる。上述の変異体それぞれをコードするDNA配列の例は、以下の通りである。
【0054】
【化11】
本発明はまた、本明細書における本発明の配列のいずれか、または本発明のシステムを使用して修飾されるその他いずれかの配列(DNAもしくはタンパク質)を含むベクター(例えば、プラスミド、コスミド、ウイルスなど)も提供する。ある実施形態において、前記ベクターには、プロモーター、エンハンサー、リボソーム結合部位などの要素、および細胞内で本発明のGST変異体を過剰発現するのに有用な配列が含まれる。本発明はまた、本発明の配列またはベクターを含む細胞も含む。ある実施形態において、前記細胞は、変異体GSTを過剰発現する。前記細胞は、細菌性細胞(例えば、大腸菌)、真菌細胞(例えば、ピキアパストリス)、酵母細胞(例えば、サッカロミセス・セレビシエ)、哺乳類細胞(例えば、CHO細胞)、またはヒト細胞である場合がある。
【0055】
本発明はまた、より安定したタンパク質変異体を産生するために対象タンパク質を修飾するキットも含む。これらのキットは通常、より安定したタンパク質変異体を生成するのに必要な試薬のすべてまたは大部分も含む。ある実施形態において、前記キットは、研究者が本発明の方法に基づいてより安定した変異タンパク質を設計するのを支援するコンピュータソフトウェアも含む。前記キットはまた、試薬、プライマー、オリゴヌクレオチド、ヌクレオチド、酵素、緩衝液、細胞、媒質、プレート、管、説明書、ベクターなどのすべてまたはいくつかを含む場合もある。前記キットを使用した研究により、通常は、より安定した変異体を生成するために変異させるDNA配列が提供される。内容物は通常、実験室で好都合に使用するように包装される。
【0056】
本発明のこれらおよび他の態様は、以下の実施例を考慮することによりさらに理解されるであろう。これらの実施例は、本発明のある特定の実施形態を例示することを目的としたものであり、特許請求の範囲に定義される本発明の適用範囲を限定することを目的としたものではない。
【実施例】
【0057】
(実施例1) タンパク質の超荷電により並外れた回復力がもたらされる
ヒトの疾患において周知の原因であるタンパク質凝集(それぞれ参考として本明細書で援用される、Cohen,F.E.;Kelly,J.W.,Nature 2003,426,(6968),905‐9;Chiti,F.;Dobson,C.M.,Annu Rev Biochem 2006,75,333‐66)はまた、治療薬や診断薬としてタンパク質を使用する上で直面する主要な問題である(それぞれ参考として本明細書で援用される、Frokjaer,S.;Otzen,D.E.,Nat Rev Drug Discov 2005,4,(4),298‐306;Fowler,S.B.;Poon,S.;Muff,R.;Chiti,F.;Dobson,C.M;Zurdo,J.,Proc Natl Acad Sci USA 2005,102,(29),10105‐10)。タンパク質凝集の問題の見識は、天然タンパク質に関する研究から得られたものである。タンパク質は、正味電荷がゼロとなる等電点において可溶性が最低であることがかねてから知られていた(参考として本明細書で援用される、Loeb,J.,J Gen Physiol 1921,4,547‐555)。より最近になって、正味電荷のわずかな差(±3の荷電単位)により、球状タンパク質の変異体における(参考として本明細書で援用される、Chiti,F.;Stefani,M.;Taddei,N.;Ramponi,G.;Dobson,C.M.,Nature
2003,424,(6950),805‐8)、ならびにまた本質的に無秩序なペプチドにおける(参考として本明細書で援用される、Pawar,A.P.;Dubay,K.F.;Zurdo,J.;Chiti,F.;Vendruscolo,M.;Dobson,C.M.,J Mol Biol 2005,350,(2),379‐92)凝集の傾向が予測されることが明らかにされている。タンパク質によっては正味電荷の著しい変化を許容できるものもあるという最近の証拠(例えば、表面リジンの徹底的な化学的アセチル化の後に炭酸脱水酵素が触媒作用を維持するという所見(参考として本明細書で援用される、Gudiksenら、J Am Chem Soc 2005,127,(13),4707‐14))とともに、これらの所見によって本発明者等は、正味電荷を増加させるように表面を広範に変異させるプロセス(本発明者等は本明細書において「超荷電」と呼ぶ)によって、フォールディングや機能を停止させることなく、いくつかのタンパク質の可溶性および凝集抵抗を大幅に増大させられる可能性があると結論付けた。
【0058】
まず、本発明者等は、最近報告されたばかりの、フォールディング効率や変性剤への抵抗が高度に最適化された、「スーパーフォルダーGFP」(sfGFP)と呼ばれる最新の緑色蛍光タンパク質(GFP)の変異体((参考として本明細書で援用される、Pedelacqら、Nat Biotechnol 2006,24,(1),79‐88))から始めた。スーパーフォルダーGFPの正味電荷は、野生型GFPの正味電荷とほぼ同じ−7であった。本発明者等は、単純なアルゴリズムの誘導のもとでアミノ酸の溶媒曝露を算出して(「材料および方法」を参照)、最も溶媒に曝露した残基の内の29個を正に荷電したアミノ酸に変異させることにより、理論上の正味電荷が+36であるGFPの超荷電変異体を設計した(図1)。sfGFPまたはGFP(+36)のいずれかをコードする遺伝子の発現により、緑色蛍光の細菌が著しく生じた。タンパク質の精製後、GFP(+36)の蛍光特性を測定し、sfGFPの蛍光特性と非常に類似していることが判明した。この所見に勇気付けられ、本発明者等は、正味電荷が+48、−25および−30であるさらなる超荷電GFPを設計および精製し、これらすべてもまたsfGFPと類似する蛍光を示すことが判明した(図2a)。すべての超荷電GFP変異体が、sfGFPと類似した円二色性のスペクトルを示し、これらのタンパク質が類似の二次構造含量を有することが示唆された(図2b)。前記超荷電GFP変異体の熱力学的安定性は、36個もの突然変異が存在したにもかかわらず、sfGFPの熱力学的安定性よりも若干低いだけであった(1.0〜4.1kcal/mol、図2cおよび表1)。
【0059】
sfGFPは、GFPの最適化において長い歴史を持つ産物であるが(参考として本明細書で援用される、Giepmansら、Science 2006,312,(5771),217‐24)、熱アンフォールディングや化学的アンフォールディングにより誘導される凝集の影響を依然として受けやすい。sfGFPを100℃に加熱すると、定量沈降反応と不可逆的な蛍光損失が誘導された(図3a)。対照的に、超荷電GFP(+36)およびGFP(−30)は、100℃に加熱しても可溶性を維持し、冷却時に大部分の蛍光を回復した(図3a)。重要な点として、40%の2,2,2‐トリフルオロエタノール(TFE)は25℃にて数分以内にsfGFPの完全な凝集を誘導したのに対して、超荷電GFP(+36)および(−30)の変異体は、同じ条件下でも何時間にもわたりほとんど凝集を損なわず、蛍光を損失しなかった(図3b)。
【0060】
この注目すべき凝集抵抗の他にも、超荷電GFP変異体は、対立荷電の高度に荷電した高分子に対して強力な可逆的結合力を示した(図3c)。1:1の化学量論組成で混合したGFP(+36)とGFP(−30)は、すぐに緑色蛍光性共沈物を形成し、フォールディングしたタンパク質の会合を示した。同様に、GFP(+36)は、高濃度のRNAまたはDNAとともに共沈した。NaClの付加は、これらの複合体を溶解するのに十分なものであり、このことは、形成に関する静電学的な根拠と一致していた。対照的に、sfGFPは、GFP(−30)、RNA、またはDNAの付加による影響を受けなかった(図3c)。
【0061】
次いで、本発明者等は、超荷電の原則がGFP以外のタンパク質(単量体であり、十分に保護された蛍光団を有する)にも適用できるかどうかを決定することを試みた。この目的を達成するために、本発明者等は超荷電プロセスを、GFPとは無関係の2つのタンパク質に適用した。ストレプトアビジンは、全正味電荷が−4である四量体である。溶媒曝露アルゴリズムを使用して、本発明者等は、正味電荷が−40または+52である2つの超荷電ストレプトアビジン変異体を設計した。いずれの超荷電ストレプトアビジン変異体も、親和性が減少したにもかかわらず、ビオチンを結合する可溶性四量体を形成することができた。
【0062】
全正味電荷が+2である二量体のグルタチオン‐S‐トランスフェラーゼ(GST)を超荷電し、野生型GSTの比活性度よりも比活性度がわずかに2.7倍低いクロロジニトロベンゼンへのグルタチオンの付加を触媒する、正味電荷が−40である二量体を得た(図3d)。さらに、超荷電ストレプトアビジンと超荷電GSTは、sfGFPと同様に定量的および不可逆的に沈降した野生型の相当物とは対照的に、100℃に加熱しても可溶性を維持した(表1)。さらに、GST(−40)は、冷却時に触媒作用を40%回復した(図3d)。
【0063】
要約すると、本発明者等は、種々の構造および機能の単量体および多量体タンパク質は、単にそれらの溶媒曝露残基を類似する荷電アミノ酸で置換することによって、「超荷電」することができることを証明した。超荷電することによって、タンパク質の分子間特性が大きく変化し、著しい凝集に対する抵抗力、および「分子ベルクロ」のような逆荷電した高分子とフォールディング形態で結合する能力がもたらされる。本発明者等は、これらの異常な分子間特性が、超荷電プロセスによりほとんど変化しなかった荷電アミノ酸の合計数からではなく、高正味電荷により生じることに注目している(表1)。
【0064】
これらの劇的な分子間効果とは対照的に、フォールディング、蛍光、リガンド結合、および酵素触媒反応を含めた、本明細書で考察する7つの超荷電タンパク質の分子内特性は、ほとんど損なわれていないままであった。したがって、超荷電は、凝集傾向を低減させ、かつその機能を停止することなくタンパク質の可溶性を改善するのに有用な手法であると考えられる。これらの原則は、凝集を含む予測不能なタンパク質ハンドリング特性が重要な課題のままである、de novoタンパク質設計事業において、特に有用である場合がある。超荷電天然タンパク質の上述の結果を考慮すると、設計したタンパク質の凝集抵抗は、フォールディングしたタンパク質の外部に存在すると予測された位置における類似する荷電アミノ酸の頻度を増加させるように、設計プロセスにバイアスをかけることによって、さらに改善できるであろうと推測される可能性がある。
【0065】
超荷電タンパク質は、タンパク質表面に注目すべき可塑性を示し、溶媒曝露残基の突然変異耐性から生じる機会を強調する。例えば、いくつかのタンパク質の熱力学的安定性は、合理的に操作された荷電間相互作用によって増強できることが最近明らかにされている(参考として本明細書で援用される、Stricklerら、Biochemistry
2006,45,(9),2761‐6)。タンパク質の超荷電は、タンパク質凝集に外部抵抗を与えるために、種々の方法でこの可塑性を活用できることを証明している。本発明者等の所見は、ユビキチンからのすべての荷電の除去により、そのフォールディングは損なわれなかったが、可溶性が大いに損なわれたという予備研究の結果と一致している(参考として本明細書で援用される、Loladze et al,Protein Sci 2002,11,(1),174‐7)。
【0066】
また、これらの所見によって、天然タンパク質の適度な正味電荷分布が明らかされるとも考えられる(それぞれ参考として本明細書で援用される、Knightら、ProcNatl Acad Sci USA 2004,101,(22),8390‐5;Gitlinら、Angew Chem Int Ed Engl 2006,45,(19),3022‐60)。例えば、タンパク質データバンク(PDB)ポリペプチドの84%の正味電荷は、±10の範囲内にある。本発明者等の結果は、高正味電荷がアンフォールディングを強要するのに十分な静電反発力を生成するという仮説に反論している。実際に、GFP(+48)は、現在PDBに存在するいずれかのポリペプチドより高い陽性の正味電荷を有するが、フォールディング能力および蛍光度を維持する。それに対して、本発明者等の所見は、非特異的分子間付着があまりにも多くの高度に荷電した天然タンパク質の進化を冷遇した可能性があることを示唆している。RNAに結合するリボソームタンパク質L3(+36)およびL15(+44)、またはカルシウム陽イオンに結合するカルセケストリン(−80)などの、正味電荷がきわめて高いほとんどすべての天然のタンパク質が、必須の細胞性機能の一部として逆荷電種と結合する。
【0067】
材料および方法
設計手順および超荷電タンパク質配列:溶媒曝露残基(以下灰色で示す)を、公表された構造データ(それぞれ参考として本明細書で援用される、Weber,P.C.,Ohlendorf,D.H.,Wendoloski,J.J.&Salemme,F.R.Structural origins of high‐affinity biotin binding to streptavidin.Science 243,85‐88(1989);Dirr,H.,Reinemer,P.&Huber,R.Refined crystal structure of porcine class Pi glutathione S‐transferase(pGST Pl‐1)at 2.1 A resolution.J Mol Biol 243,72‐92(1994);Pedelacq,J.D.,Cabantous,S.,Tran,T.,Terwilliger,T.C.&Waldo,G.S.Engineering and characterization of a superfolder green fluorescent protein.Nat Biotechnol 24,79‐88(2006))から、AvNAPSA値(AvNAPSAとは側鎖原子当たりの平均的な隣接原子(10Å以内)である)が150未満の構造であると同定した。荷電するかまたは高い極性を有する溶媒曝露残基(DERKNQ)を、陰性超荷電(赤色)の場合はAspもしくはGluのいずれかに、陽性超荷電(青色)の場合はLysもしくはArgに変異させた。緑色蛍光タンパク質(GFP)変異体中で変異させるためのさらなる表面露出位置を、GFP相同体内のこれらの位置における配列変化に基づいて選択した。ストレプトアビジン(SAV)とグルタチオン‐S‐トランスフェラーゼ(GST)の超荷電設計プロセスは全自動であり、まず残基は溶媒曝露によって分類された後、大部分が溶媒曝露した、荷電するかまたは高い極性を有する残基を、陽性超荷電の場合はLysに、または陰性超荷電の場合はGlu(但し、開始残基がAsnの場合はAspに)のいずれかに変異させた。
【0068】
【化12】
タンパク質の発現および精製:大腸菌コドン使用に最適化した合成遺伝子を、DNA2.0(Novagen)から購入し、pET発現ベクターにクローニングして、15℃にて5〜10時間大腸菌BL21(DE3)pLysS中に過剰発現させた。遠心分離によって細胞を採取し、超音波処理によって溶解した。Ni‐NTAアガロースクロマトグラフィー(Qiagen)によりタンパク質を精製して、100mMのNaCl、50mMのリン酸カリウム、pH7.5に緩衝液を交換し、限外ろ過(Millipore)によって濃縮した。すべてのGFP変異体を天然条件下で精製した。野生型ストレプトアビジンはPromegaから購入した。超荷電GSTの場合と同様に、超荷電ストレプトアビジン変異体を変性条件下で精製し、野生型ストレプトアビジンについて以前に報告した通りにリフォールディングした(参考として本明細書で援用される、Thompsonら、Construction and expression of a synthetic streptavidin‐encoding gene in Escherichia coli.Gene 136,243‐246(1993))。野生型GSTを天然または変性条件のいずれかで精製し、類似した活性タンパク質を得た。
【0069】
静電表面電位の算出(図1b):−30および+48の超荷電GFP変異体のモデルは、スーパーフォルダーGFPの結晶構造に基づいていた(参考として本明細書で援用される、Pedelacqら、Engineering and characterization of a superfolder green fluorescent protein.Nat Biotechnol 24,79‐88(2006))。静電電位を、APBSを使用して算出し(参考として本明細書で援用される、Bakerら、Electrostatics of nanosystems:application to microtubules and the ribosome.Proc Natl Acad Sci USA 98,10037‐10041(2001))、−25kT/e(赤色)から+25kT/e(青色)までのスケールを使用して、PyMolで表現した(参考として本明細書で援用される、Delano,W.L.,The PyMOL Molecular Graphics System,www.pymol.org(2002))。
【0070】
タンパク質染色および紫外線誘導性蛍光(図2a):10%の変性ポリアクリルアミドゲル中で電気泳動を行うことによって、それぞれのGFP変異体0.2μgを分析し、クマシーブリリアントブルー染色で染色した。100mMのNaClを有する25mMのTris、pH8.0中の同タンパク質試料0.2μgを、0.2mLのEppendorf管に入れて、紫外線下で(360nm)写真を撮影した。
【0071】
熱変性および凝集(図3a):25mMのTris、pH8.0、100mMのNaCl、および10mMのベータ‐メルカプトエタノール(BME)中で、精製したGFP変異体を2mg/mLに希釈した後、紫外線下で写真を撮影した(「天然」)。これらの試料を100℃にて1分間加熱した後、紫外線下で再度写真を撮影した(「煮沸」)。最後に、これらの試料を室温にて2時間冷却した後、紫外線下で再度写真を撮影した(「冷却」)。
【0072】
化学的に誘導された凝集(図3b):2,2,2‐トリフルオロエタノール(TFE)を添加して、1.5mg/mLのタンパク質、25mMのTris、pH7.0、10mMのBME、および40%のTFEを有する溶液を生成した。25℃の凝集を直角の光散乱によって監視した。
【0073】
サイズ排除クロマトグラフィー(表1):SAVおよびGST変異体の多量体状態を、Superdex 75ゲルろ過カラムで20〜50μgのタンパク質を分析することにより測定した。緩衝液は、100mMのNaCl、50mMのリン酸カリウム、pH7.5であった。分子量は、同一条件下で個別に分析した既知の分子量の一連の単量体タンパク質の基準と比較することによって測定した。
【0074】
(表1 算出したおよび実験により測定したタンパク質特性)
【0075】
【表1】
npos:正に荷電したアミノ酸の数(単量体当たり)
nneg:負に荷電したアミノ酸の数
ncharged:荷電アミノ酸の総数
Qnet:中性pHにおける理論上の等電点
pI:算出した等電点
n.d.:測定せず
a グアニジン変性により測定(図2c)。
b サイズ排除クロマトグラフィーにより測定。
c 100℃にて5分間の加熱、25℃への冷却、軽い遠心分離後に上清に残留するタンパク質の割合。
d 大腸菌に発現しなかったタンパク質。
【0076】
他の実施形態
以上の説明が単に本発明のある好ましい実施形態を表しているにすぎないことを、当業者であれば容易に理解するであろう。以下の特許請求の範囲で定める本発明の趣旨または適用範囲から逸脱することなく、上述の手順および組成物の種々の変更および改変を行うことが可能である。
【0077】
付録A
【0078】
【化13】
【0079】
【化14】
【0080】
【化15】
【0081】
【化16】
【0082】
【化17】
【0083】
【化18】
【0084】
【化19】
【0085】
【化20】
【技術分野】
【0001】
関連出願
この出願は、米国特許法§119(e)の下、2006年6月2日に出願された米国仮特許出願第60/810,364号(この全体の内容は、参考として本明細書に援用される)への優先権を主張する。
【0002】
(連邦政府支援研究に関する声明)
本明細書に記載の研究は、国立衛生研究所の助成金(GM065400)の支援を一部受けている。米国政府は、本発明における一定の権利を有する。
【背景技術】
【0003】
タンパク質は細胞の働き者である。タンパク質は、化学反応物質を触媒したり、生体系においてシグナルを変換したり、細胞の構造成分や細胞外マトリックスを提供したり、メッセンジャーとして作用したりなどの働きを行う。タンパク質の誤動作の主な原因の1つに凝集がある。これは単に研究室だけの問題ではなく、アルツハイマー病などの多くの疾病における問題である。計算により設計したタンパク質においては、凝集は特に面倒な問題である。例えば、TOP7は、新規のフォールドを有する計算により設計したタンパク質である。TOP7のより長い型であるTOP7 extendedは、きわめて凝集の影響を受けやすい。TOP7exは、主に不溶性凝集体として発現される。
【発明の概要】
【発明が解決しようとする課題】
【0004】
より多くのタンパク質が、生体系を研究するためのツールとして使用するために設計または修飾されており、あるいはより多くのタンパク質(野生型または修飾)が治療薬として使用されていることから、これらのタンパク質をより安定させるために、および/または凝集を防止するために慣例的に修飾するシステムが必要とされている。
【課題を解決するための手段】
【0005】
本発明は、タンパク質をより安定させるために修飾するシステムを提供する。本発明は、タンパク質表面の疎水性領域を修飾することにより、タンパク質の超熱力学的特性を改善することができるという認識に基づいている。本発明のシステムは、対象タンパク質の可溶性の改善、タンパク質の凝集に対する抵抗力の改善、および/またはタンパク質の復元能力の改善に特に有用である。これらの特性はすべて、タンパク質の産生、タンパク質の精製、ならびに治療薬および研究ツールとしてのタンパク質の使用に特に有用である。
【0006】
一態様において、本発明は、タンパク質の凝集に対する抵抗力、可溶性、リフォールディング能力、および/または広範な条件における一般的な安定性を向上させるために、タンパク質の一次配列を変更する方法を提供する。好ましくは、修飾タンパク質の活性は、修飾されていないタンパク質とほぼまたは実質的に同じである。ある実施形態において、修飾タンパク質は、野生型タンパク質の活性の少なくとも50%、75%、90%または95%を維持する。一実施形態において、前記方法は、(a)対象タンパク質の表面残基を同定する工程と;(b)対象タンパク質に関連する他のタンパク質の中で高度に保存されていない特定の表面残基を同定する(すなわち、どのアミノ酸が前記タンパク質の活性または機能に必須でないかを決定する)手順と;(c)同定した非保存表面残基の疎水性を決定する工程と;(e)少なくとも1つ以上の同定した疎水性非保存残基を、より極性が高いかまたは生理学的なpHで荷電したアミノ酸で置換する工程と、を含む。上述の手順はそれぞれ、当該技術分野で既知のいずれかの技法、コンピュータソフトウェア、アルゴリズム、パラダイムなどを使用して行われる場合がある。生成した修飾タンパク質は、求めている活性および/または所望の特性について試験される場合がある。ある実施形態において、修飾タンパク質はより安定性が高い。ある実施形態において、修飾タンパク質は、より凝集の影響を受けにくい。通常、本発明の方法は、生理学的なpHでタンパク質の正味電荷(正電荷または負電荷)を増大させる。
【0007】
別の態様において、本発明は、タンパク質を「超荷電する」ことにより、タンパク質の凝集への抵抗、可溶性、リフォールディング能力、および/または広範な条件における一般的な安定性を向上させるために、タンパク質の一次配列を変更する方法を提供する。すなわち、修飾タンパク質の全体正味電荷は、野生型タンパク質に比べて増大する(正電荷または負電荷のいずれか)。好ましくは、修飾タンパク質の活性は、修飾されていないタンパク質とほぼまたは実質的に同じである。ある実施形態において、前記方法は、(a)対象タンパク質の表面残基を同定する工程と;(b)対象タンパク質に関する他のタンパク質の中で高度に保存されていない特定の表面残基を同定する(すなわち、どのアミノ酸が前記タンパク質の活性または機能に必須でないかを決定する)手順と;(c)同定した非保存表面残基の親水性を決定する工程と;(e)少なくとも1つ以上の同定した、荷電または極性を有する、溶媒に曝露した非保存残基を、生理学的なpHで荷電した荷電アミノ酸で置換する工程と、を含む。ある実施形態においては、負に荷電した「超荷電」タンパク質を生成するために、修飾用に同定した残基を、アスパラギン酸(Asp)またはグルタミン酸(Glu)残基のいずれかに変異させる。その他のある実施形態においては、正に荷電した「超荷電」タンパク質を生成するために、修飾用に同定した残基を、リジン(Lys)またはアルギニン(Arg)残基のいずれかに変異させる。上述の手順はそれぞれ、当該技術分野で既知のいずれかの技法、コンピュータソフトウェア、アルゴリズム、パラダイムなどを使用して行われる場合がある。生成した修飾タンパク質は、求めている活性および/または所望の特性について試験される場合がある。ある実施形態において、修飾タンパク質「(超荷電タンパク質」)はより安定している。ある実施形態において、修飾タンパク質は、より凝集の影響を受けにくい。通常、本発明の方法は、生理学的なpHでタンパク質の正味電荷(正電荷または負電荷)を増大させる。
【0008】
Protein Data Bank(PDB)に掲載されるタンパク質の80%超の理論上の正味電荷は、±10の範囲内である。本発明により生成される修飾タンパク質は通常、正味電荷が−10未満または+10超である。ある実施形態において、修飾タンパク質は、正味電荷が−20未満または+20超である。ある実施形態において、修飾タンパク質は、正味電荷が−30未満または+30超である。ある実施形態において、修飾タンパク質は、正味電荷が−40未満または+40超である。ある実施形態において、修飾タンパク質は、正味電荷が−50未満または+50超である。修飾タンパク質は正確に折り畳み、生物活性を維持することができる。
【0009】
いずれのタンパク質も本発明のシステムを使用して修飾される場合があり、本発明のシステムによって生成されるタンパク質変異体は、変異タンパク質をコードするポリヌクレオチドまたはベクター、および変異タンパク質を発現する細胞と同様に、本発明の一部であると考えられる。本発明のシステムは、いくつかの新規の緑色蛍光タンパク質(GFP)の変異体を生成するのに使用されてきた。これらの変異体は蛍光性を維持する一方で、広範な環境において現在のGFP型よりも高い安定性を有する。本発明のGFPは、長期間にわたり凝集を誘導する環境においても凝集に対して免疫を有し、煮沸により変性した後でも蛍光タンパク質にリフォールディングすることができる。本発明のシステムは、ストレプトアビジンやグルタチオン‐S‐トランスフェラーゼ(GST)の新規変異体の生成にも使用されてきた。これらの変異体は、加熱しても生物活性を維持し、可溶性を維持する。本発明は、本発明のGFP、ストレプトアビジン、およびGSTタンパク質配列をコードするポリヌクレオチド配列、これらのヌクレオチド配列のいずれかを含むベクター、ならびにこのようなポリヌクレオチド配列もしくはベクターを含むか、または本発明の変異体を発現する細胞も含む。ある実施形態において、本発明は、本発明の変異体を過剰発現する細菌または他の細胞を含む。本発明の変異体は、当該技術分野で既知の種々の生物学的アッセイに使用される場合がある。例えば、超荷電GFPは、現在GFPをレポータータンパク質として使用しているいずれかのアッセイで使用される場合がある。
【0010】
別の態様において、本発明は、本発明のシステムによって修飾された他のタンパク質を提供する。これらの修飾タンパク質は、好ましくは元の活性の大部分を維持する。ある実施形態において、修飾タンパク質は、未修飾型の活性の少なくとも99%、98%、95%または90%を維持する。修飾タンパク質は、より可溶性が高く、凝集に対するより高い抵抗力を持ち、リフォールディング能力が高く、および/または種々の条件における安定性が高い場合がある。本発明のシステムにより修飾されるタンパク質には、疎水性タンパク質、組換えタンパク質、膜タンパク質、構造タンパク質、酵素、細胞外タンパク質、治療用タンパク質(例えば、インシュリン、サイトカイン、免疫グロブリン、免疫グロブリンの断片など)、受容体、細胞シグナル伝達タンパク質、細胞質タンパク質、核タンパク質、転写因子などが含まれる。ある特定の実施形態において、前記タンパク質は、ヒト医学や獣医学で使用する治療用タンパク質である。ある実施形態において、前記タンパク質は、非天然タンパク質、例えば、計算により設計したタンパク質である。他の実施形態において、前記タンパク質は、ハイブリッドタンパク質、融合タンパク質、変質タンパク質、変異タンパク質、遺伝子合成タンパク質、または手動により変質させたその他いずれかのタンパク質である。
【0011】
また、本発明を実施するためのキットも提供される。前記キットには、凝集に対する抵抗力を高めるか、復元能力を高めるか、または全体的な安定性を高めるために対象タンパク質を修飾する上で必要となる試薬が含まれる場合がある。このようなキットには、ポリヌクレオチド、コンピュータソフトウェア、ヌクレオチド、プライマー、ベクター、細胞系、説明書、プレート、媒質、緩衝液、酵素、Eppendorf管、部位特異的突然変異誘発キットなどのすべてかまたはいくつかが含まれる場合がある。好ましくは、前記キットは、実験環境で使用するのに好都合に包装される。研究者は通常、本発明の技法を使用して、修飾するタンパク質のDNAコード配列を提供する。
【0012】
定義
「アミノ酸」:「アミノ酸」という用語は、タンパク質の基本構造サブユニットを指す。アルファアミノ酸は、アミノ基、カルボキシル基、水素原子、およびすべてが中心の炭素原子に結合している側鎖(すなわち、R基)からなる。この中心の炭素原子は、カルボキシル基に隣接していることから、アルファ炭素と呼ばれる。天然アミノ酸は、グリシン、アラニン、バリン、ロイシン、イソロイシン、フェニルアラニン、チロシン、トリプトファン、システイン、メチオニン、セリン、トレオニン、リジン、アルギニン、ヒスチジン、アスパラギン酸塩、グルタミン酸塩、アスパラギン、グルタミン酸塩、およびプロリンを含めた20個が存在する。疎水性アミノ酸には、アラニン、バリン、ロイシン、イソロイシンおよびフェニルアラニンが含まれる。芳香族アミノ酸には、フェニルアラニン、チロシン、トリプトファンおよびヒスチジンが含まれる。極性アミノ酸には、チロシン、システイン、セリン、トレオニン、リジン、アルギニン、ヒスチジン、アスパラギン酸塩、グルタミン酸塩、アスパラギン、およびグルタミンが含まれる。含硫アミノ酸には、システインおよびメチオニンが含まれる。塩基性アミノ酸には、リジン、アルギニンおよびヒスチジンが含まれる。酸性アミノ酸には、アスパラギン酸塩およびグルタミン酸塩が含まれる。非天然アミノ酸もまたタンパク質に挿入されている。ある実施形態において、「アミノ酸」という用語を使用する場合は、20個の天然アミノ酸を指している。
【0013】
「抗体」:「抗体」という用語は、天然であるか、またはすべてもしくは一部が合成により産生されているかにかかわらず、免疫グロブリンを指す。特異的な結合能力を維持するそのすべての誘導体も本用語に包含される。本用語はまた、免疫グロブリン結合ドメインに相同であるか大部分が相同である結合ドメインを有するいずれのタンパク質をも網羅する。これらのタンパク質は、天然供給源に由来する場合もあれば、または一部もしくはすべてが合成により産生される場合もある。抗体はモノクローナルである場合もあれば、ポリクローナルである場合もある。抗体は、IgG、IgM、IgA、IgDおよびIgEというヒトクラスのいずれかを含めた、いずれかの免疫グロブリンクラスの一員である場合がある。
【0014】
「保存された」:「保存された」という用語は、比較している複数の関連する配列の同じ位置で不変的に生じる、ポリヌクレオチド配列またはアミノ酸配列それぞれのヌクレオチドまたはアミノ酸残基を指す。相対的に保存されたヌクレオチドまたはアミノ酸は、配列のその他の位置に生じるヌクレオチドまたはアミノ酸よりも関連性が高い配列の中で保存されるものである。
【0015】
「相同」:本明細書で使用される「相同」という用語は、ヌクレオチドまたはアミノ酸配列のレベルで高度に関連する核酸またはタンパク質を指す、当該技術分野で理解されている用語である。互いに相同である核酸またはタンパク質は、相同体と呼ばれる。相同という用語は、2つの配列(すなわち、ヌクレオチド配列またはアミノ酸)間の配列類似性の程度を指す場合がある。本明細書で言及される相同率の数値は、2つの配列間で可能な最大の相同性、すなわち、2つの配列が多くの一致した(相同な)位置を有するように整列される場合の相同率を表す。相同性は、以下の文献に記載のような既知の方法によって容易に算出することができる:Computational Molecular Biology,Lesk,A.M,ed.,Oxford University Press,New York,1988;Biocomputing:Informatics and Genome Projects,Smith,D.W.,ed.,Academic Press,New York,1993;Sequence Analysis in Molecular Biology,von Heinje,G., Academic Press,1987;Computer Analysis of
Sequence Data,Part I,Griffin,A.M.,and Griffin,H.G.,eds.,Humana Press,New Jersey,1994;およびSequence Analysis Primer,Gribskov,M.and Devereux,J.,eds.,M Stockton Press,New York,1991(それぞれ参考として本明細書で援用される)。配列間の相同性を決定するのに通常使用される方法には、参考として本明細書で援用される、Carillo,H.,and Lipman,D.,SIAM J Applied Math.,48:1073(1988)に開示される方法が含まれるが、これらに限定されない。相同性を決定する技法は、公的に利用可能なコンピュータプログラムで体系化される。2つの配列間の相同性を決定するコンピュータソフトウェアの例には、GCGプログラムパッケージ(Devereux,J.,et al.,Nucleic Acids Research,12(1),387(1984))、BLASTP、BLASTN、およびFASTA(Atschul,S.F.et al.,J Molec.Biol.,215,403(1990))が含まれるが、これらに限定されない。
【0016】
「相同」という用語は必ず、少なくとも2つの配列(ヌクレオチド配列またはアミノ酸配列)間の比較を指す。本発明によれば、2つのヌクレオチド配列がコードするポリペプチドが、少なくとも20個のアミノ酸からなる少なくとも1つのストレッチについて、少なくとも約50〜60%同一であり、好ましくは約70%同一である場合に、2つのヌクレオチド配列が相同であると見なされる。好ましくは、相同なヌクレオチド配列はまた、少なくとも4〜5個の一意に定義されるアミノ酸からなるストレッチをコードする能力を特徴とする。互いに関連するこれらのアミノ酸の同一性および近似間隔の両方が、相同であると見なされるヌクレオチド配列において考えられなければならない。長さが60個未満のヌクレオチドであるヌクレオチド配列では、少なくとも4〜5個の一意に定義されるアミノ酸からなるストレッチをコードする能力によって相同性が決定される。
【0017】
「ペプチド」または「タンパク質」:本発明によれば、「ペプチド」または「タンパク質」は、ペプチド結合により結合した少なくとも3個のアミノ酸からなるストリングを含む。「タンパク質」および「ペプチド」という用語は、交換可能に使用される場合がある。本発明のペプチドは、好ましくは天然アミノ酸のみを含有するが、非天然アミノ酸(すなわち、天然には生じないがポリペプチド鎖に組み込むことができる化合物)、および/または当該技術分野で既知のアミノ酸類似体が代わりに使用される場合がある。また、本発明のペプチド中の1つ以上のアミノ酸は、例えば、炭水化物基、リン酸基、ファルネシル基、イソファルネシル基、脂肪酸基、結合用リンカー、官能化、または他の修飾(例えば、アルファアミノ化)などの化学物質を添加することにより、修飾される場合がある。好ましい実施形態において、ペプチドの修飾により、より安定した(例えば、in vivoでの半減期がより長い)ペプチドが生じる。これらの修飾には、ペプチドの環化、Dアミノ酸の混入などが含まれる場合がある。どの修飾も、ペプチドの所望の生物活性を実質的に阻害してはならない。ある実施形態においては、ペプチドの修飾により、より生物活性の高いペプチドが生じる。
【0018】
「ポリヌクレオチド」または「オリゴヌクレオチド」:ポリヌクレオチドまたはオリゴヌクレオチドは、ヌクレオチドのポリマーを指す。通常、ポリヌクレオチドは、少なくとも3個のヌクレオチドを含む。このポリマーには、天然ヌクレオシド(すなわち、アデノシン、チミジン、グアノシン、シチジン、ウリジン、デオキシアデノシン、デオキシチミジン、デオキシグアノシン、およびデオキシシチジン)、ヌクレオシド類似体(例えば、2‐アミノアデノシン、2‐チオチミジン、イノシン、ピロロ‐ピリミジン、3‐メチルアデノシン、C5‐プロピニルシチジン、C5‐プロピニルウリジン、C5‐ブロモウリジン、C5‐フルオロウリジン、C5‐ヨードウリジン、C5‐メチルシチジン、7‐デアザアデノシン、7‐デアザグアノシン、8‐オキソアデノシン、8‐オキソグアノシン、O(6)‐メチルグアニン、および2‐チオシチジン)、化学修飾塩基、生物学的修飾塩基(例えば、メチル化塩基)、介在基、修飾糖(例えば、2’‐フルオロリボース、リボース、2’‐デオキシリボース、アラビノース、およびヘキソース)、および/または修飾リン酸基(例えば、モノチオリン酸塩および5’‐N‐ホスホロアミダイトリンケージ)が含まれる場合がある。
【0019】
「小分子」:本明細書で使用される「小分子」という用語は、実験室で調製するか、または天然で認められる非ペプチド性非オリゴマー有機化合物を指す。本明細書で使用される小分子は、「天然物のような」化合物を指してもよいが、この「小分子」という用語は「天然物のような」化合物に限定されない。むしろ、小分子は通常、いくつかの炭素間結合を含有し、1500未満の分子量を有することを特徴とするが、但し、この特徴は本発明において限定することを目的としたものではない。その他のある好ましい実施形態においては、天然物のような小分子が使用される。
【0020】
「安定した」:タンパク質を言及するのに本明細書で使用される「安定した」という用語は、タンパク質の安定性のいずれかの態様を指す。元の野生型タンパク質と比べて安定した修飾タンパク質は、より可溶性が高い、凝集に対してより高い抵抗力を持つ、変性に対してより高い抵抗力を持つ、アンフォールディングに対してより高い抵抗力を持つ、不適切または不要なフォールディングに対してより高い抵抗力を持つ、復元能力が高い、熱安定性が高い、種々の環境(例えば、pH、塩濃度、洗浄剤の存在、変性剤の存在など)における安定性が高い、非水性環境における安定性が高い、といった特徴の内のいずれか1つ以上を有する。ある実施形態において、安定した修飾タンパク質は、上述の特徴の少なくとも2つを示す。ある実施形態において、安定した修飾タンパク質は、上述の特徴の少なくとも3つを示す。このような特徴により、活性タンパク質をより高いレベルで産生することが可能となる場合がある。例えば、修飾タンパク質は、タンパク質の非修飾型よりも、凝集することなくより高いレベルで過剰発現させることができる。また、このような特徴により、治療薬または研究ツールとしてタンパク質を使用することが可能となる場合がある。
本発明はまた、以下の項目を提供する。
(項目1)
対象タンパク質の安定性を改善する方法であって、
対象タンパク質に関連する他のタンパク質の中で高度に保存されていない、該対象タンパク質の表面残基を同定する工程と;
複数の非保存表面残基を、生理学的なpHで正に荷電したアミノ酸残基で置換する工程と、
を含む、方法。
(項目2)
対象タンパク質の安定性を改善する方法であって、
対象タンパク質に関連する他のタンパク質の中で高度に保存されていない、該対象タンパク質の表面残基を同定する工程と;
複数の非保存表面残基を、生理学的なpHで負に荷電したアミノ酸残基で置換する工程と、
を含む、方法。
(項目3)
上記非保存表面残基が疎水性である、項目1または2に記載の方法。
(項目4)
上記非保存表面残基が親水性であるか、または負に荷電している、項目1に記載の方法。
(項目5)
上記非保存表面残基が親水性であるか、または正に荷電している、項目2に記載の方法。
(項目6)
対象タンパク質の安定性を改善する方法であって、
対象タンパク質の表面残基を同定する工程と;
該対象タンパク質に関連する他のタンパク質の中で高度に保存されていない、該対象タンパク質の表面残基を同定する工程と;
該同定した非保存表面残基のそれぞれに疎水性値を割り当てる工程と;
少なくとも1個の表面残基を、生理学的なpHで荷電したアミノ酸残基で置換する工程であって、該置換されている残基がすべて同じ型の残基に変異し、該残基の型が正に荷電した残基であるか、または負に荷電した残基のいずれかである、工程と、
を含む、方法。
(項目7)
上記置換工程が、少なくとも1個の疎水性表面残基を置換する工程を含む、項目6に記載の方法。
(項目8)
上記置換工程が、少なくとも1個の親水性表面残基を置換する工程を含む、項目6に記載の方法。
(項目9)
上記置換工程が、少なくとも1個の荷電した表面残基を置換する工程を含む、項目6に記載の方法。
(項目10)
上記置換工程が、少なくとも1個の表面残基をリジン残基で置換する工程を含む、項目6に記載の方法。
(項目11)
上記置換工程が、少なくとも1個の表面残基をアスパラギン酸残基またはグルタミン酸残基で置換する工程を含む、項目6に記載の方法。
(項目12)
上記置換工程が、少なくとも2個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目13)
上記置換工程が、少なくとも5個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目14)
上記置換工程が、少なくとも10個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目15)
上記置換工程が、少なくとも20個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目16)
上記置換工程が、少なくとも30個の表面残基を置換する工程を含む、項目1、2または6に記載の方法。
(項目17)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHでより大きな正味電荷を有する修飾タンパク質が生成される、項目1、2または6に記載の方法。
(項目18)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHでより負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目19)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも−5より負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目20)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも−10より負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目21)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも−15より負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目22)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも−20より負に荷電した対象修飾タンパク質が生成される、項目2または6に記載の方法。
(項目23)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHでより正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目24)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも+5より正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目25)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも+10より正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目26)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも+15より正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目27)
上記方法によって、対象となる元のタンパク質よりも、生理学的なpHで少なくとも+20より正に荷電した対象修飾タンパク質が生成される、項目1または6に記載の方法。
(項目28)
上記対象タンパク質が凝集しやすいタンパク質である、項目1、2または6に記載の方法。
(項目29)
上記対象タンパク質が疎水性タンパク質である、項目1、2または6に記載の方法。
(項目30)
上記対象タンパク質が膜タンパク質である、項目1、2または6に記載の方法。
(項目31)
上記対象タンパク質が、過剰発現しにくいタンパク質である、項目1、2または6に記載の方法。
(項目32)
上記対象タンパク質が、精製しにくいタンパク質である、項目1、2または6に記載の方法。
(項目33)
上記対象タンパク質が受容体である、項目1、2または6に記載の方法。
(項目34)
上記対象タンパク質が転写因子である、項目1、2または6に記載の方法。
(項目35)
上記対象タンパク質が酵素である、項目1、2または6に記載の方法。
(項目36)
上記対象タンパク質が構造タンパク質である、項目1、2または6に記載の方法。
(項目37)
上記対象タンパク質が蛍光タンパク質である、項目1、2または6に記載の方法。
(項目38)
上記対象タンパク質が緑色蛍光タンパク質(GFP)である、項目1、2または6に記載の方法。
(項目39)
上記対象タンパク質が細胞外タンパク質である、項目1、2または6に記載の方法。
(項目40)
上記対象タンパク質がストレプトアビジンである、項目1、2または6に記載の方法。
(項目41)
上記対象タンパク質がグルタチオン‐S‐トランスフェラーゼである、項目1、2または6に記載の方法。
(項目42)
上記表面残基を同定する工程が、上記タンパク質の三次元構造をコンピュータモデリングする工程を含む、項目1、2または6に記載の方法。
(項目43)
上記表面残基を同定する工程が、アルゴリズムを使用して、残基がタンパク質の表面に見出されるか否かを予測する工程を含む、項目1、2または6に記載の方法。
(項目44)
上記表面残基を同定する工程が、閾値未満のavNAPSA値を有する残基を同定する工程を含む、項目1、2または6に記載の方法。
(項目45)
上記高度に保存されていない表面残基を同定する工程が、上記タンパク質のアミノ酸配列を、同じタンパク質ファミリーに由来する少なくとも1つの他のタンパク質と整列させる工程を含む、項目1、2または6に記載の方法。
(項目46)
上記高度に保存されていない表面残基を同定する工程が、上記タンパク質のアミノ酸配列を、異なる種に由来する該タンパク質の少なくとも1つの他のアミノ酸配列と整列させる工程を含む、項目1、2または6に記載の方法。
(項目47)
上記整列が、少なくとも2つの他のタンパク質配列を使用して行われる、項目45または46に記載の方法。
(項目48)
上記整列が、少なくとも3つの他のタンパク質配列を使用して行われる、項目45または46に記載の方法。
(項目49)
上記整列が、少なくとも5つの他のタンパク質配列を使用して行われる、項目45または46に記載の方法。
(項目50)
上記疎水性値を割り当てる工程が、オクタノール/水のP値を使用する工程を含む、項目6に記載の方法。
(項目51)
上記置換工程が、上記同定した疎水性表面残基を、生理学的なpHで荷電した天然アミノ酸残基で置換するように、上記タンパク質配列に突然変異を起こさせる工程を含む、項目1、2または6に記載の方法。
(項目52)
上記天然アミノ酸残基が、リジン、グルタミン酸、アスパラギン酸、ヒスチジンおよびアルギニンからなる群から選択される、項目51に記載の方法。
(項目53)
上記天然アミノ酸残基が、リジン、ヒスチジンおよびアルギニンからなる群から選択される、項目51に記載の方法。
(項目54)
上記天然アミノ酸残基がリジンである、項目51に記載の方法。
(項目55)
上記天然アミノ酸残基が、グルタミン酸およびアスパラギン酸からなる群から選択される、項目51に記載の方法。
(項目56)
上記置換工程が、上記同定した表面残基の部位特異的突然変異誘発を含む、項目1、2または6に記載の方法。
(項目57)
上記置換工程が、上記同定した表面残基のPCR突然突然変異誘発を含む、項目1、2または6に記載の方法。
(項目58)
アミノ酸配列:
【化21】
の緑色蛍光タンパク質(+36 GFP)。
(項目59)
項目58に記載の緑色蛍光タンパク質をコードするポリヌクレオチド。
(項目60)
配列:
【化22】
の項目59に記載のポリヌクレオチド。
(項目61)
アミノ酸配列:
【化23】
の緑色蛍光タンパク質(+49 GFP)。
(項目62)
項目61に記載の緑色蛍光タンパク質をコードするポリヌクレオチド。
(項目63)
配列:
【化24】
の項目62に記載のポリヌクレオチド。
(項目64)
アミノ酸配列:
【化25】
の緑色蛍光タンパク質(‐29 GFP)。
(項目65)
項目64に記載の緑色蛍光タンパク質をコードするポリヌクレオチド。
(項目66)
配列:
【化26】
の項目65に記載のポリヌクレオチド。
(項目67)
アミノ酸配列:
【化27】
のストレプトアビジン(−40 SAV)。
(項目68)
項目67に記載のストレプトアビジンをコードするポリヌクレオチド。
(項目69)
配列:
【化28】
の項目68に記載のポリヌクレオチド。
(項目70)
アミノ酸配列:
【化29】
のストレプトアビジン(+52 SAV)。
(項目71)
項目70に記載のストレプトアビジンをコードするポリヌクレオチド。
(項目72)
配列:
【化30】
の項目71に記載のポリヌクレオチド。
(項目73)
アミノ酸配列:
【化31】
のグルタチオン‐S‐トランスフェラーゼ(−40 GST)。
(項目74)
項目73に記載のストレプトアビジンをコードするポリヌクレオチド。
(項目75)
配列:
【化32】
の項目74に記載のポリヌクレオチド。
(項目76)
項目1〜57のいずれか1項に記載の方法により修飾されるタンパク質。
(項目77)
項目76に記載のタンパク質をコードするポリヌクレオチド。
(項目78)
項目1〜57のいずれか1項に記載の方法を実行するためのキット。
【図面の簡単な説明】
【0021】
【図1】超荷電した緑色蛍光タンパク質(GFP)を示す。(a)GFP変異体のタンパク質配列であり、フルオロフォア形成残基を緑色で強調表示し、負に荷電した残基を赤色で強調表示し、正に荷電した残基を青色で強調表示した。(b)−25kT/e(赤色)から+25kT/e(青色)の範囲で着色した、sfGFP(左)、GFP(+36)(中央)、およびGFP(−30)(右)の静電表面電位である。
【図2】GFP変異体の分子内特性を示す。(a)精製したGFP変異体の染色およびUV蛍光。各レーンおよび管は0.2μgのタンパク質を含有する。(b)GFP変異体の円二色性スペクトル。(c)グアニジンにより誘導したアンフォールディングで測定したGFP変異体の熱力学的安定性。
【図3】超荷電タンパク質の分子間特性を示す。(a)精製したGFP変異体のUV照射試料(「天然」)と、100℃にて1分間加熱した試料(「煮沸」)と、次いで25℃にて2時間冷却した試料(「冷却」)。(b)GFP変異体の凝集を25℃の40% TFEで誘導し、直角の光散乱によって監視した。(c)超荷電GFPが逆荷電した高分子に可逆的に付着する。試料1:30μLの25mM Tris、pH7.0および100mM NaCl中のGFP(+36)6μg。試料2:GFP(−30)6μgを試料1に添加。試料3:サケ精子DNA30μgを試料1に添加。試料4:大腸菌tRNA 20μgを試料1に添加。試料5:1MのNaClを試料4に添加。試料6〜8:GFP(+36)の代わりにsfGFPを使用した以外は、試料1、2および4それぞれと同じ。すべての試料はミクロ遠心分離機で軽く撹拌した後、紫外線で視覚化した。(d)GST変異体の酵素アッセイ。反応物は、GST変異体0.5mg/mL、クロロジニトロベンゼン20mM、グルタチオン20mM、およびリン酸カリウム100mM、pH6.5を含有した。産物形成を340nmで監視した結果、煮沸および冷却後の反応速度(kobs)は、野生型GSTが6分−1、GST(−40)が2.2分−1、GST(−40)が0.9分−1であることが認められた。
【図4】GFP変異体の(a)励起および(b)発光スペクトルを示す。各試料は、490nmの発色団吸光度で定量化により等しい量のタンパク質を含有した。
【図5】結合依存性のビオチン‐4‐フルオレスセイン(Invitrogen)を監視することによって、既述の通りに測定した(参考として本明細書で援用される、Kadaら、Rapid estimation of avidin and streptavidin by fluorescence quenching or fluorescence polarization.Biochim.Biophys.Acta 1427,44‐48(1999))、ストレプトアビジン変異体のビオチン結合活性を示す。タンパク質試料を、0.3μMのビオチン‐4‐フルオレスセイン(B4F)、100mMのNaCI、1mMのEDTA、0.1mg/mLのウシ血清アルブミン(BSA)、50mMのリン酸カリウム、pH7.5に滴定した。526nmの蛍光消光を、Perkin‐Elmer LS50B蛍光分光計で470nmの励起により測定した。測定値は、600倍の過剰な非蛍光性ビオチンを含有する対照滴定液に正規化した。凡例内の下の3つのタンパク質を陰性対照として含めた。
【発明を実施するための形態】
【0022】
本発明の特定の好ましい実施形態の詳細な説明
本発明は、タンパク質をより安定させるために修飾するシステムを提供する。前記システムは、タンパク質表面の非保存アミノ酸を、より極性の高いまたは荷電したアミノ酸残基に変更することによって機能すると考えられる。修飾するアミノ酸残基は、疎水性である場合もあれば、親水性である場合もあり、荷電している場合もあり、あるいはこれらの組み合わせである場合もある。いずれのタンパク質も、より安定した変異体を産生するために、本発明のシステムを使用して修飾される場合がある。これらの表面残基の修飾によって、タンパク質の超熱力学的特性が改善されることが明らかにされている。タンパク質が治療薬として使用されることが多くなっており、かつ研究ツールとしても依然として使用されていることから、タンパク質をより安定させるために変質させるシステムが重要であり、有用である。本発明の方法によって修飾されるタンパク質は通常、凝集に対する抵抗力を持ち、リフォールディング能力が高く、不適切なフォールディングに対する抵抗力を持ち、可溶性が高く、加熱や洗浄剤の存在といった変性条件を含めた広範な条件において一般的により高い安定性を有する。
【0023】
いずれのタンパク質も、本発明のシステムを使用してより安定した変異体を生成するために修飾される場合がある。天然だけでなく非天然のタンパク質(例えば、改変タンパク質)も修飾される場合がある。修飾される場合があるタンパク質の例には、受容体、膜結合タンパク質、膜貫通タンパク質、酵素、転写因子、細胞外タンパク質、治療用タンパク質、サイトカイン、メッセンジャータンパク質、DNA結合タンパク質、RNA結合タンパク質、シグナル変換に関与するタンパク質、構造タンパク質、細胞質タンパク質、核タンパク質、疎水性タンパク質、親水性タンパク質などが含まれる。修飾するタンパク質は、いずれの種の植物、動物または微生物からも得られる場合がある。ある実施形態において、タンパク質は哺乳類タンパク質である。ある実施形態において、タンパク質はヒトタンパク質である。ある実施形態において、タンパク質は、通常研究で使用される生体から得られる。例えば、修飾するタンパク質は、霊長類(例えば、類人猿、サル)、齧歯類(例えば、ウサギ、ハムスター、アレチネズミ)、ブタ、イヌ、ネコ、魚(例えば、ゼブラフィッシュ)、線虫(例えば、シノラブディス・エレガンス)、酵母菌(例えば、サッカロミセス・セレビジエ)、または細菌(例えば、大腸菌)から得られる場合がある。
【0024】
本発明のシステムは、凝集の影響を受けやすいか、または安定性の問題を有するタンパク質を修飾するのに特に有用である。前記システムはまた、過剰発現しているタンパク質を修飾するのに使用される場合もある。例えば、組換えにより産生されている治療用タンパク質は、本発明のシステムによる修飾が有用な場合がある。このような修飾された治療用タンパク質は、産生や精製が容易なだけでなく、タンパク質の保管や使用においてもより高い安定性を有する場合がある。
【0025】
本発明のシステムは、対象タンパク質の非保存表面残基を同定する工程と、これらの残基のいくつかを、親水性を有するか、極性を有するか、または生理学的なpHで荷電した残基で置換する工程を伴う。本発明のシステムは、タンパク質を修飾する方法だけでなく、タンパク質をより安定させるために修飾するのに有用な試薬およびキットも含む。
【0026】
修飾するタンパク質の表面残基は、当該技術分野で既知のいずれかの方法を使用して同定される。ある実施形態において、表面残基は、タンパク質のコンピュータモデリングによって同定される。ある実施形態において、タンパク質の三次元構造は、既知であり、および/または決定されており、タンパク質の構造を視覚化することによって表面残基が同定される。他の実施形態において、表面残基は、コンピュータソフトウェアを使用して予測される。ある特定の実施形態においては、Average Neighbor Atoms per Sidechain Atom(AvNAPSA)が、表面露出の予測に使用される。AvNAPSAは、コンピュータプログラムとして導入された、表面露出の自動化された測度である。付録Aを参照されたい。低いAvNAPSA値が表面露出残基を示し、高いAvNAPSA値がタンパク質内部の残基を示す。ある実施形態においては、本ソフトウェアが、タンパク質の二次構造および/または三次構造の予測に使用され、表面残基がこの予測に基づいて同定される。他の実施形態において、表面残基の予測は、残基の疎水性および親水性、ならびにタンパク質の一次配列におけるこれらのクラスタリングに基づいて行われる。In silico法の他にも、例えば、プロテアーゼ切断、表面改質などの種々の生化学的技法を使用して、タンパク質の表面残基が同定される場合もある。
【0027】
次いで、表面残基の中で、保存されている残基か、またはタンパク質の機能に重要な残基が決定される。保存残基の同定は、当該技術分野で既知のいずれかの方法を使用して決定することができる。ある実施形態において、保存残基は、対象タンパク質の一次配列を関連タンパク質と整列させることによって同定される。これらの関連するタンパク質は、同じタンパク質ファミリーに由来する場合がある。例えば、タンパク質が免疫グロブリンである場合は、他の免疫グロブリン配列が使用される場合がある。関連タンパク質はまた、異なる種に由来する同じタンパク質である場合もある。例えば、異なる種に由来する同じタンパク質の配列を整列させることによって、保存残基が同定される場合がある。他の例を挙げると、機能または生物活性が類似するタンパク質を整列させる場合がある。好ましくは、タンパク質中に保存されたアミノ酸を決定するのに、2、3、4、5、6、7、8、9または10種類の配列が使用される。ある実施形態においては、配列の50%、60%、70%、75%、80%または90%超が特定の位置に同じアミノ酸を有する場合に、残基は保存されていると見なされる。他の実施形態においては、配列の50%、60%、70%、75%、80%または90%超が特定の位置に同じかまたは類似するアミノ酸(例えば、バリン、ロイシンおよびイソロイシン;グリシンおよびアラニン;グルタミンおよびアスパラギン;またはアスパラギン酸塩およびグルタミン酸塩)を有する場合に、残基が保存されていると見なされる。本明細書に記載の通り、タンパク質配列の整列および比較に利用可能なソフトウェアパッケージは数多く存在する。当業者であれば理解するであろうが、最初に保存残基を決定する場合もあれば、あるいは最初に表面残基を決定する場合もある。順序は重要でない。ある実施形態においては、コンピュータソフトウェアパッケージで、表面残基と保存残基が同時に決定される場合がある。また、タンパク質の突然変異生成によって、タンパク質中の重要な残基が同定される場合もある。例えば、タンパク質のアラニン走査を使用して、タンパク質中の重要なアミノ酸残基を決定することができる。他の実施形態においては、部位特異的突然変異誘発が使用される場合がある。
【0028】
タンパク質の非保存表面残基が同定されれば、それぞれの残基は疎水性または親水性であると同定される。ある実施形態において、残基には疎水性スコアが割り当てられる。例えば、それぞれの非保存表面残基には、オクタノール/水のlogP値が割り当てられる場合がある。また、他の疎水性パラメータも使用される場合がある。このようなアミノ酸のスケールについては、以下で考察されている:Janin,“Surface and
Inside Volumes in Globular Proteins,”Nature 277:491‐92,1979;Wolfendenら、“Affinities of Amino Acid Side Chains for Solvent Water,”Biochemistry 20:849‐855,1981;Kyteら、“A Simple Method for Displaying the Hydropathic Character of a Protein,”J.Mol.Biol.157:105‐132,1982;Roseら、“Hydrophobicity of Amino Acid Residues in Globular
Proteins,”Science 229:834‐838,1985;Cornetteら、“Hydrophobicity Scales and Computational Techniques for Detecting Amphipathic Structures in Proteins,”J.Mol.Biol.195:659‐685,1987;Charton and Charton,“The
Structure Dependence of Amino Acid Hydrophobicity Parameters,“J.Theor.Biol.99:629‐644,1982(それぞれ参考として援用される)。これらの疎水性パラメータのいずれかが、どの非保存残基を修飾するかを決定するのに、本発明の方法で使用される場合がある。ある実施形態においては、親水性または荷電残基が修飾用に同定される。
【0029】
次いで、少なくとも1つの同定された非保存または失活表面残基が修飾用に選択される。ある実施形態においては、疎水性残基が修飾用に選択される。他の実施形態においては、親水性および/または荷電残基が修飾用に選択される。ある実施形態においては、複数の残基が修飾用に選択される。ある実施形態においては、1、2、3、4、5、6、7、8、9または10個の同定された残基が修飾用に選択される。ある実施形態においては、10個を超える、15個を超えるまたは20個を超える残基が修飾用に選択される。当業者であれば理解するであろうが、タンパク質が大きくなるほど、修飾が必要となる残基も多くなる。また、タンパク質がより疎水性であるか、または凝集もしくは沈降の影響を受けやすいほど、修飾が必要となる残基も多くなる。ある実施形態においては、それぞれ修飾が異なるタンパク質の複数の変異体が産生され、生物活性および安定性に最も優れた変異体を決定するために試験される。
【0030】
ある実施形態においては、修飾用に選択した残基を、より親水性の高い残基(荷電残基を含む)に変異させる。通常、残基は、より親水性の高い天然アミノ酸に変異させる。ある実施形態において、残基は、生理学的なpHで荷電したアミノ酸に変異させる。例えば、アルギニン、アスパラギン酸塩、グルタミン酸塩、ヒスチジンまたはリジンに残基を変化する場合がある。ある実施形態においては、修飾するすべての残基を、同じ残基に変化させる。例えば、選択したすべての残基をグルタミン酸塩の残基に変化させる。他の実施形態において、選択した残基は、異なる残基に変化させるが、最終残基はすべて、生理学的なpHで正に荷電している場合もあれば、負に荷電している場合もある。ある実施形態においては、負に荷電したタンパク質を生成するために、変異させるすべての残基を、グルタミン酸塩および/またはアスパラギン酸塩の残基に変換させる。ある実施形態において、正に荷電したタンパク質を生成するために、変異させるすべての残基をリジンの残基に変換させる。例えば、修飾用に選択したすべての残基は、アスパラギン、グルタミン、リジン、および/またはアルギニンであり、これらの残基をアスパラギン酸塩またはグルタミン酸塩の残基に変異させる。他の例を挙げると、修飾用に選択したすべての残基は、アスパラギン酸塩、グルタミン酸塩、アスパラギンおよび/またはグルタミンであり、これらの残基をリジンに変異させる。この手法により、タンパク質の正味電荷を最大限度まで修飾することが可能となる。
【0031】
他の実施形態においては、修飾タンパク質の正味電荷を非修飾タンパク質の正味電荷と同じに維持するように、タンパク質が修飾される場合がある。さらに他の実施形態においては、表面における荷電残基の総数を増加させながら、タンパク質の全体的な正味電荷を減少させるように、タンパク質が修飾される場合がある。ある実施形態においては、理論上の正味電荷を、少なくとも+1、+2、+3、+4、+5、+10、+15、+20、+25、+30または+35増加させる。ある実施形態においては、理論上の正味電荷を、少なくとも−1、−2、−3、−4、−5、−10、−15、−20、−25、−30または−35減少させる。ある実施形態においては、選択したアミノ酸を、非イオン極性残基(例えば、システイン、セリン、トレオニン、チロシン、グルタミン、アスパラギン)に変化させる。
【0032】
タンパク質におけるこれらの修飾および突然変異は、当該技術分野で既知のいずれかの技法を使用して達成される場合がある。このような変化をタンパク質配列に導入する組換えDNA技術は、当該技術分野で周知である。ある実施形態においては、タンパク質をコードするポリヌクレオチドの部位特異的突然変異誘発によって修飾が行われる。突然変異を導入する他の技法については、それぞれが参考として本明細書で援用される、Molecular Cloning:A Laboratory Manual,2nd Ed.,ed.by Sambrook,Fritsch,and Maniatis(Cold Spring Harbor Laboratory Press:1989);Methods in Enzymology(Academic Press,Inc.,N.Y.);Ausubelら、Current Protocols in Molecular Biology(John Wiley&Sons,Inc.,New
York,1999)で考察されている。修飾タンパク質は発現され、試験される。ある実施形態においては、一連の変異体が調製され、それぞれの変異体が、生物活性および安定性を決定するために試験される。後の使用のために選択する変異体は、最も安定性が高いものである場合もあれば、最も活性の高いものである場合もあり、あるいは全体として活性と安定性が最も高い組み合わせを有するものである場合もある。最初の群の変異体を調製した後、最初の群から学習した内容に基づいて、追加の群の変異体が調製される場合がある。通常作成した変異体は、当該技術分野で既知の組換え法を使用して過剰発現させる。
【0033】
本発明のシステムは、GFPの変異体の生成に使用されてきた。これらの変異体は、より安定性が高く、蛍光を維持することが明らかにされている。Aequorea victoria由来のGFPが、参考として本明細書で援用される、GenBank受託番号P42212に記載されている。この野生型GFPのアミノ酸配列は、以下の通りである:
【0034】
【化1】
野生型GFPの理論上の正味電荷は−7であるが、本発明のシステムを使用して、理論上の正味電荷が−29、−30、−25、+36、+48および+49である変異体が生成された。+36のGFPを95℃に加熱した後でも、変異タンパク質の100%が可溶性を有し、タンパク質が70%以上の蛍光を維持する。
【0035】
生成したGFP変異体のアミノ酸配列には、以下が含まれる。
【0036】
【化2】
【0037】
【化3】
当業者であれば理解するであろうが、相同タンパク質も本発明の適用範囲に含まれると見なされる。例えば、上述の配列のいずれかに対して60%、70%、80%、90%、95%または100%相同である20、30、40、50または100個のアミノ酸からなるストレッチを含むいずれのタンパク質も、本発明の一部に含まれると見なされる。さらに、付加および欠失変異体も本発明で企図される。ある実施形態においては、上述の配列のいずれかに示す変異残基を有するいずれのGFPも、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0038】
また、上述のGFP変異体をコードするいずれのDNA配列も、本発明の適用範囲に含まれる。上述の変異体それぞれをコードするDNA配列の例は、以下の通りである。
【0039】
【化4】
【0040】
【化5】
上述の配列に相同なポリヌクレオチド配列も、本発明の適用範囲に含まれる。ある実施形態において、ポリヌクレオチド配列は、上述の配列のいずれか1つに対して60%、70%、80%、90%、95%、98%、99%または100%相同である50、100または150個のヌクレオチドからなるストレッチを含む。本発明はまた、1つ以上のヌクレオチドが上述の配列の1つに挿入されるか、またはこれらの1つから欠失する配列も含む。上述の配列のいずれかに示す突然変異を有するいずれのポリヌクレオチド配列も、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0041】
本発明はまた、本明細書における本発明の配列のいずれか、または本発明のシステムを使用して修飾されるその他いずれかの配列(DNAもしくはタンパク質)を含むベクター(例えば、プラスミド、コスミド、ウイルスなど)も提供する。ある実施形態において、前記ベクターには、プロモーター、エンハンサー、リボソーム結合部位などの要素、および細胞内で本発明のGFP変異体を過剰発現するのに有用な配列が含まれる。本発明はまた、本発明の配列またはベクターを含む細胞も含む。ある実施形態において、前記細胞は、変異体GFPを過剰発現する。前記細胞は、細菌性細胞(例えば、大腸菌)、真菌細胞(例えば、ピキアパストリス)、酵母細胞(例えば、サッカロミセス・セレビシエ)、哺乳類細胞(例えば、CHO細胞)、またはヒト細胞である場合がある。
【0042】
本発明のシステムは、ストレプトアビジン変異体の生成に使用されてきた。これらの変異体は、ビオチンを結合する可溶性四量体を形成することが明らかにされている。この野生型ストレプトアビジンのアミノ酸配列は、以下の通りである。
【0043】
【化6】
野生型ストレプトアビジンの理論上の正味電荷は−4であるが、本発明のシステムを使用して、理論上の正味電荷が−40および+52である変異体が生成された。前記変異体を100℃に加熱した後でも、タンパク質は可溶性を維持した。
【0044】
生成したストレプトアビジン変異体のアミノ酸配列には、以下が含まれる。
【0045】
【化7】
当業者であれば理解するであろうが、相同タンパク質も本発明の適用範囲に含まれると見なされる。例えば、上述の配列のいずれかに対して60%、70%、80%、90%、95%または100%相同である20、30、40、50または100個のアミノ酸からなるストレッチを含むいずれのタンパク質も、本発明の一部に含まれると見なされる。さらに、付加および欠失変異体も本発明で企図される。ある実施形態においては、上述の配列のいずれかに示す変異残基を有するいずれのストレプトアビジンも、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0046】
上述のストレプトアビジン変異体をコードするいずれのDNA配列も、本発明の適用範囲に含まれる。上述の変異体それぞれをコードするDNA配列の例は、以下の通りである。
【0047】
【化8】
上述の配列に相同なポリヌクレオチド配列も、本発明の適用範囲に含まれる。ある実施形態において、ポリヌクレオチド配列は、上述の配列のいずれか1つに対して60%、70%、80%、90%、95%、98%、99%または100%相同である50、100、または150個のヌクレオチドからなるストレッチを含む。本発明はまた、1つ以上のヌクレオチドが上述の配列の1つに挿入されるか、またはこれらの1つから欠失する配列も含む。上述の配列のいずれかに示す突然変異を有するいずれのポリヌクレオチド配列も、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0048】
本発明はまた、本明細書における本発明の配列のいずれか、または本発明のシステムを使用して修飾されるその他いずれかの配列(DNAもしくはタンパク質)を含むベクター(例えば、プラスミド、コスミド、ウイルスなど)も提供する。ある実施形態において、前記ベクターには、プロモーター、エンハンサー、リボソーム結合部位などの要素、および細胞内で本発明のストレプトアビジン変異体を過剰発現するのに有用な配列が含まれる。本発明はまた、本発明の配列またはベクターを含む細胞も含む。ある実施形態において、前記細胞は、変異体ストレプトアビジンを過剰発現する。前記細胞は、細菌性細胞(例えば、大腸菌)、真菌細胞(例えば、ピキアパストリス)、酵母細胞(例えば、サッカロミセス・セレビシエ)、哺乳類細胞(例えば、CHO細胞)、またはヒト細胞である場合がある。
【0049】
本発明のシステムは、グルタチオン‐S‐トランスフェラーゼ(GST)の変異体の生成に使用されてきた。これらの変異体は、野生型GSTの触媒作用を維持することが明らかにされている。この野生型GSTのアミノ酸配列は、以下の通りである。
【0050】
【化9】
野生型GSTの理論上の正味電荷は+2であるが、本発明のシステムを使用して、理論上の正味電荷が−40である変異体が生成された。この変異体は、野生型GSTよりも比活性がわずかに2.7倍低いクロロニトロベンゼンへのグルタチオンの付加を触媒する。この変異体を100℃に加熱した後でも、タンパク質は可溶性を維持し、タンパク質は冷却時に触媒作用の40%を回復した。
【0051】
GST変異体のアミノ酸配列には、以下が含まれる。
【0052】
【化10】
当業者であれば理解するであろうが、相同タンパク質も本発明の適用範囲に含まれると見なされる。例えば、上述の配列のいずれかに対して60%、70%、80%、90%、95%または100%相同である20、30、40、50または100個のアミノ酸からなるストレッチを含むいずれのタンパク質も、本発明の一部に含まれると見なされる。さらに、付加および欠失変異体も本発明で企図される。ある実施形態においては、上述の配列のいずれかに示す変異残基を有するいずれのストレプトアビジンも、本発明の一部に含まれると見なされる。ある実施形態において、前記配列は、上述の配列のいずれかに示す2、3、4、5、6、7、8、9、10個以上の突然変異を含む。
【0053】
上述のGST変異体をコードするいずれのDNA配列も、本発明の適用範囲に含まれる。上述の変異体それぞれをコードするDNA配列の例は、以下の通りである。
【0054】
【化11】
本発明はまた、本明細書における本発明の配列のいずれか、または本発明のシステムを使用して修飾されるその他いずれかの配列(DNAもしくはタンパク質)を含むベクター(例えば、プラスミド、コスミド、ウイルスなど)も提供する。ある実施形態において、前記ベクターには、プロモーター、エンハンサー、リボソーム結合部位などの要素、および細胞内で本発明のGST変異体を過剰発現するのに有用な配列が含まれる。本発明はまた、本発明の配列またはベクターを含む細胞も含む。ある実施形態において、前記細胞は、変異体GSTを過剰発現する。前記細胞は、細菌性細胞(例えば、大腸菌)、真菌細胞(例えば、ピキアパストリス)、酵母細胞(例えば、サッカロミセス・セレビシエ)、哺乳類細胞(例えば、CHO細胞)、またはヒト細胞である場合がある。
【0055】
本発明はまた、より安定したタンパク質変異体を産生するために対象タンパク質を修飾するキットも含む。これらのキットは通常、より安定したタンパク質変異体を生成するのに必要な試薬のすべてまたは大部分も含む。ある実施形態において、前記キットは、研究者が本発明の方法に基づいてより安定した変異タンパク質を設計するのを支援するコンピュータソフトウェアも含む。前記キットはまた、試薬、プライマー、オリゴヌクレオチド、ヌクレオチド、酵素、緩衝液、細胞、媒質、プレート、管、説明書、ベクターなどのすべてまたはいくつかを含む場合もある。前記キットを使用した研究により、通常は、より安定した変異体を生成するために変異させるDNA配列が提供される。内容物は通常、実験室で好都合に使用するように包装される。
【0056】
本発明のこれらおよび他の態様は、以下の実施例を考慮することによりさらに理解されるであろう。これらの実施例は、本発明のある特定の実施形態を例示することを目的としたものであり、特許請求の範囲に定義される本発明の適用範囲を限定することを目的としたものではない。
【実施例】
【0057】
(実施例1) タンパク質の超荷電により並外れた回復力がもたらされる
ヒトの疾患において周知の原因であるタンパク質凝集(それぞれ参考として本明細書で援用される、Cohen,F.E.;Kelly,J.W.,Nature 2003,426,(6968),905‐9;Chiti,F.;Dobson,C.M.,Annu Rev Biochem 2006,75,333‐66)はまた、治療薬や診断薬としてタンパク質を使用する上で直面する主要な問題である(それぞれ参考として本明細書で援用される、Frokjaer,S.;Otzen,D.E.,Nat Rev Drug Discov 2005,4,(4),298‐306;Fowler,S.B.;Poon,S.;Muff,R.;Chiti,F.;Dobson,C.M;Zurdo,J.,Proc Natl Acad Sci USA 2005,102,(29),10105‐10)。タンパク質凝集の問題の見識は、天然タンパク質に関する研究から得られたものである。タンパク質は、正味電荷がゼロとなる等電点において可溶性が最低であることがかねてから知られていた(参考として本明細書で援用される、Loeb,J.,J Gen Physiol 1921,4,547‐555)。より最近になって、正味電荷のわずかな差(±3の荷電単位)により、球状タンパク質の変異体における(参考として本明細書で援用される、Chiti,F.;Stefani,M.;Taddei,N.;Ramponi,G.;Dobson,C.M.,Nature
2003,424,(6950),805‐8)、ならびにまた本質的に無秩序なペプチドにおける(参考として本明細書で援用される、Pawar,A.P.;Dubay,K.F.;Zurdo,J.;Chiti,F.;Vendruscolo,M.;Dobson,C.M.,J Mol Biol 2005,350,(2),379‐92)凝集の傾向が予測されることが明らかにされている。タンパク質によっては正味電荷の著しい変化を許容できるものもあるという最近の証拠(例えば、表面リジンの徹底的な化学的アセチル化の後に炭酸脱水酵素が触媒作用を維持するという所見(参考として本明細書で援用される、Gudiksenら、J Am Chem Soc 2005,127,(13),4707‐14))とともに、これらの所見によって本発明者等は、正味電荷を増加させるように表面を広範に変異させるプロセス(本発明者等は本明細書において「超荷電」と呼ぶ)によって、フォールディングや機能を停止させることなく、いくつかのタンパク質の可溶性および凝集抵抗を大幅に増大させられる可能性があると結論付けた。
【0058】
まず、本発明者等は、最近報告されたばかりの、フォールディング効率や変性剤への抵抗が高度に最適化された、「スーパーフォルダーGFP」(sfGFP)と呼ばれる最新の緑色蛍光タンパク質(GFP)の変異体((参考として本明細書で援用される、Pedelacqら、Nat Biotechnol 2006,24,(1),79‐88))から始めた。スーパーフォルダーGFPの正味電荷は、野生型GFPの正味電荷とほぼ同じ−7であった。本発明者等は、単純なアルゴリズムの誘導のもとでアミノ酸の溶媒曝露を算出して(「材料および方法」を参照)、最も溶媒に曝露した残基の内の29個を正に荷電したアミノ酸に変異させることにより、理論上の正味電荷が+36であるGFPの超荷電変異体を設計した(図1)。sfGFPまたはGFP(+36)のいずれかをコードする遺伝子の発現により、緑色蛍光の細菌が著しく生じた。タンパク質の精製後、GFP(+36)の蛍光特性を測定し、sfGFPの蛍光特性と非常に類似していることが判明した。この所見に勇気付けられ、本発明者等は、正味電荷が+48、−25および−30であるさらなる超荷電GFPを設計および精製し、これらすべてもまたsfGFPと類似する蛍光を示すことが判明した(図2a)。すべての超荷電GFP変異体が、sfGFPと類似した円二色性のスペクトルを示し、これらのタンパク質が類似の二次構造含量を有することが示唆された(図2b)。前記超荷電GFP変異体の熱力学的安定性は、36個もの突然変異が存在したにもかかわらず、sfGFPの熱力学的安定性よりも若干低いだけであった(1.0〜4.1kcal/mol、図2cおよび表1)。
【0059】
sfGFPは、GFPの最適化において長い歴史を持つ産物であるが(参考として本明細書で援用される、Giepmansら、Science 2006,312,(5771),217‐24)、熱アンフォールディングや化学的アンフォールディングにより誘導される凝集の影響を依然として受けやすい。sfGFPを100℃に加熱すると、定量沈降反応と不可逆的な蛍光損失が誘導された(図3a)。対照的に、超荷電GFP(+36)およびGFP(−30)は、100℃に加熱しても可溶性を維持し、冷却時に大部分の蛍光を回復した(図3a)。重要な点として、40%の2,2,2‐トリフルオロエタノール(TFE)は25℃にて数分以内にsfGFPの完全な凝集を誘導したのに対して、超荷電GFP(+36)および(−30)の変異体は、同じ条件下でも何時間にもわたりほとんど凝集を損なわず、蛍光を損失しなかった(図3b)。
【0060】
この注目すべき凝集抵抗の他にも、超荷電GFP変異体は、対立荷電の高度に荷電した高分子に対して強力な可逆的結合力を示した(図3c)。1:1の化学量論組成で混合したGFP(+36)とGFP(−30)は、すぐに緑色蛍光性共沈物を形成し、フォールディングしたタンパク質の会合を示した。同様に、GFP(+36)は、高濃度のRNAまたはDNAとともに共沈した。NaClの付加は、これらの複合体を溶解するのに十分なものであり、このことは、形成に関する静電学的な根拠と一致していた。対照的に、sfGFPは、GFP(−30)、RNA、またはDNAの付加による影響を受けなかった(図3c)。
【0061】
次いで、本発明者等は、超荷電の原則がGFP以外のタンパク質(単量体であり、十分に保護された蛍光団を有する)にも適用できるかどうかを決定することを試みた。この目的を達成するために、本発明者等は超荷電プロセスを、GFPとは無関係の2つのタンパク質に適用した。ストレプトアビジンは、全正味電荷が−4である四量体である。溶媒曝露アルゴリズムを使用して、本発明者等は、正味電荷が−40または+52である2つの超荷電ストレプトアビジン変異体を設計した。いずれの超荷電ストレプトアビジン変異体も、親和性が減少したにもかかわらず、ビオチンを結合する可溶性四量体を形成することができた。
【0062】
全正味電荷が+2である二量体のグルタチオン‐S‐トランスフェラーゼ(GST)を超荷電し、野生型GSTの比活性度よりも比活性度がわずかに2.7倍低いクロロジニトロベンゼンへのグルタチオンの付加を触媒する、正味電荷が−40である二量体を得た(図3d)。さらに、超荷電ストレプトアビジンと超荷電GSTは、sfGFPと同様に定量的および不可逆的に沈降した野生型の相当物とは対照的に、100℃に加熱しても可溶性を維持した(表1)。さらに、GST(−40)は、冷却時に触媒作用を40%回復した(図3d)。
【0063】
要約すると、本発明者等は、種々の構造および機能の単量体および多量体タンパク質は、単にそれらの溶媒曝露残基を類似する荷電アミノ酸で置換することによって、「超荷電」することができることを証明した。超荷電することによって、タンパク質の分子間特性が大きく変化し、著しい凝集に対する抵抗力、および「分子ベルクロ」のような逆荷電した高分子とフォールディング形態で結合する能力がもたらされる。本発明者等は、これらの異常な分子間特性が、超荷電プロセスによりほとんど変化しなかった荷電アミノ酸の合計数からではなく、高正味電荷により生じることに注目している(表1)。
【0064】
これらの劇的な分子間効果とは対照的に、フォールディング、蛍光、リガンド結合、および酵素触媒反応を含めた、本明細書で考察する7つの超荷電タンパク質の分子内特性は、ほとんど損なわれていないままであった。したがって、超荷電は、凝集傾向を低減させ、かつその機能を停止することなくタンパク質の可溶性を改善するのに有用な手法であると考えられる。これらの原則は、凝集を含む予測不能なタンパク質ハンドリング特性が重要な課題のままである、de novoタンパク質設計事業において、特に有用である場合がある。超荷電天然タンパク質の上述の結果を考慮すると、設計したタンパク質の凝集抵抗は、フォールディングしたタンパク質の外部に存在すると予測された位置における類似する荷電アミノ酸の頻度を増加させるように、設計プロセスにバイアスをかけることによって、さらに改善できるであろうと推測される可能性がある。
【0065】
超荷電タンパク質は、タンパク質表面に注目すべき可塑性を示し、溶媒曝露残基の突然変異耐性から生じる機会を強調する。例えば、いくつかのタンパク質の熱力学的安定性は、合理的に操作された荷電間相互作用によって増強できることが最近明らかにされている(参考として本明細書で援用される、Stricklerら、Biochemistry
2006,45,(9),2761‐6)。タンパク質の超荷電は、タンパク質凝集に外部抵抗を与えるために、種々の方法でこの可塑性を活用できることを証明している。本発明者等の所見は、ユビキチンからのすべての荷電の除去により、そのフォールディングは損なわれなかったが、可溶性が大いに損なわれたという予備研究の結果と一致している(参考として本明細書で援用される、Loladze et al,Protein Sci 2002,11,(1),174‐7)。
【0066】
また、これらの所見によって、天然タンパク質の適度な正味電荷分布が明らかされるとも考えられる(それぞれ参考として本明細書で援用される、Knightら、ProcNatl Acad Sci USA 2004,101,(22),8390‐5;Gitlinら、Angew Chem Int Ed Engl 2006,45,(19),3022‐60)。例えば、タンパク質データバンク(PDB)ポリペプチドの84%の正味電荷は、±10の範囲内にある。本発明者等の結果は、高正味電荷がアンフォールディングを強要するのに十分な静電反発力を生成するという仮説に反論している。実際に、GFP(+48)は、現在PDBに存在するいずれかのポリペプチドより高い陽性の正味電荷を有するが、フォールディング能力および蛍光度を維持する。それに対して、本発明者等の所見は、非特異的分子間付着があまりにも多くの高度に荷電した天然タンパク質の進化を冷遇した可能性があることを示唆している。RNAに結合するリボソームタンパク質L3(+36)およびL15(+44)、またはカルシウム陽イオンに結合するカルセケストリン(−80)などの、正味電荷がきわめて高いほとんどすべての天然のタンパク質が、必須の細胞性機能の一部として逆荷電種と結合する。
【0067】
材料および方法
設計手順および超荷電タンパク質配列:溶媒曝露残基(以下灰色で示す)を、公表された構造データ(それぞれ参考として本明細書で援用される、Weber,P.C.,Ohlendorf,D.H.,Wendoloski,J.J.&Salemme,F.R.Structural origins of high‐affinity biotin binding to streptavidin.Science 243,85‐88(1989);Dirr,H.,Reinemer,P.&Huber,R.Refined crystal structure of porcine class Pi glutathione S‐transferase(pGST Pl‐1)at 2.1 A resolution.J Mol Biol 243,72‐92(1994);Pedelacq,J.D.,Cabantous,S.,Tran,T.,Terwilliger,T.C.&Waldo,G.S.Engineering and characterization of a superfolder green fluorescent protein.Nat Biotechnol 24,79‐88(2006))から、AvNAPSA値(AvNAPSAとは側鎖原子当たりの平均的な隣接原子(10Å以内)である)が150未満の構造であると同定した。荷電するかまたは高い極性を有する溶媒曝露残基(DERKNQ)を、陰性超荷電(赤色)の場合はAspもしくはGluのいずれかに、陽性超荷電(青色)の場合はLysもしくはArgに変異させた。緑色蛍光タンパク質(GFP)変異体中で変異させるためのさらなる表面露出位置を、GFP相同体内のこれらの位置における配列変化に基づいて選択した。ストレプトアビジン(SAV)とグルタチオン‐S‐トランスフェラーゼ(GST)の超荷電設計プロセスは全自動であり、まず残基は溶媒曝露によって分類された後、大部分が溶媒曝露した、荷電するかまたは高い極性を有する残基を、陽性超荷電の場合はLysに、または陰性超荷電の場合はGlu(但し、開始残基がAsnの場合はAspに)のいずれかに変異させた。
【0068】
【化12】
タンパク質の発現および精製:大腸菌コドン使用に最適化した合成遺伝子を、DNA2.0(Novagen)から購入し、pET発現ベクターにクローニングして、15℃にて5〜10時間大腸菌BL21(DE3)pLysS中に過剰発現させた。遠心分離によって細胞を採取し、超音波処理によって溶解した。Ni‐NTAアガロースクロマトグラフィー(Qiagen)によりタンパク質を精製して、100mMのNaCl、50mMのリン酸カリウム、pH7.5に緩衝液を交換し、限外ろ過(Millipore)によって濃縮した。すべてのGFP変異体を天然条件下で精製した。野生型ストレプトアビジンはPromegaから購入した。超荷電GSTの場合と同様に、超荷電ストレプトアビジン変異体を変性条件下で精製し、野生型ストレプトアビジンについて以前に報告した通りにリフォールディングした(参考として本明細書で援用される、Thompsonら、Construction and expression of a synthetic streptavidin‐encoding gene in Escherichia coli.Gene 136,243‐246(1993))。野生型GSTを天然または変性条件のいずれかで精製し、類似した活性タンパク質を得た。
【0069】
静電表面電位の算出(図1b):−30および+48の超荷電GFP変異体のモデルは、スーパーフォルダーGFPの結晶構造に基づいていた(参考として本明細書で援用される、Pedelacqら、Engineering and characterization of a superfolder green fluorescent protein.Nat Biotechnol 24,79‐88(2006))。静電電位を、APBSを使用して算出し(参考として本明細書で援用される、Bakerら、Electrostatics of nanosystems:application to microtubules and the ribosome.Proc Natl Acad Sci USA 98,10037‐10041(2001))、−25kT/e(赤色)から+25kT/e(青色)までのスケールを使用して、PyMolで表現した(参考として本明細書で援用される、Delano,W.L.,The PyMOL Molecular Graphics System,www.pymol.org(2002))。
【0070】
タンパク質染色および紫外線誘導性蛍光(図2a):10%の変性ポリアクリルアミドゲル中で電気泳動を行うことによって、それぞれのGFP変異体0.2μgを分析し、クマシーブリリアントブルー染色で染色した。100mMのNaClを有する25mMのTris、pH8.0中の同タンパク質試料0.2μgを、0.2mLのEppendorf管に入れて、紫外線下で(360nm)写真を撮影した。
【0071】
熱変性および凝集(図3a):25mMのTris、pH8.0、100mMのNaCl、および10mMのベータ‐メルカプトエタノール(BME)中で、精製したGFP変異体を2mg/mLに希釈した後、紫外線下で写真を撮影した(「天然」)。これらの試料を100℃にて1分間加熱した後、紫外線下で再度写真を撮影した(「煮沸」)。最後に、これらの試料を室温にて2時間冷却した後、紫外線下で再度写真を撮影した(「冷却」)。
【0072】
化学的に誘導された凝集(図3b):2,2,2‐トリフルオロエタノール(TFE)を添加して、1.5mg/mLのタンパク質、25mMのTris、pH7.0、10mMのBME、および40%のTFEを有する溶液を生成した。25℃の凝集を直角の光散乱によって監視した。
【0073】
サイズ排除クロマトグラフィー(表1):SAVおよびGST変異体の多量体状態を、Superdex 75ゲルろ過カラムで20〜50μgのタンパク質を分析することにより測定した。緩衝液は、100mMのNaCl、50mMのリン酸カリウム、pH7.5であった。分子量は、同一条件下で個別に分析した既知の分子量の一連の単量体タンパク質の基準と比較することによって測定した。
【0074】
(表1 算出したおよび実験により測定したタンパク質特性)
【0075】
【表1】
npos:正に荷電したアミノ酸の数(単量体当たり)
nneg:負に荷電したアミノ酸の数
ncharged:荷電アミノ酸の総数
Qnet:中性pHにおける理論上の等電点
pI:算出した等電点
n.d.:測定せず
a グアニジン変性により測定(図2c)。
b サイズ排除クロマトグラフィーにより測定。
c 100℃にて5分間の加熱、25℃への冷却、軽い遠心分離後に上清に残留するタンパク質の割合。
d 大腸菌に発現しなかったタンパク質。
【0076】
他の実施形態
以上の説明が単に本発明のある好ましい実施形態を表しているにすぎないことを、当業者であれば容易に理解するであろう。以下の特許請求の範囲で定める本発明の趣旨または適用範囲から逸脱することなく、上述の手順および組成物の種々の変更および改変を行うことが可能である。
【0077】
付録A
【0078】
【化13】
【0079】
【化14】
【0080】
【化15】
【0081】
【化16】
【0082】
【化17】
【0083】
【化18】
【0084】
【化19】
【0085】
【化20】
【特許請求の範囲】
【請求項1】
本明細書の一部に記載の発明。
【請求項1】
本明細書の一部に記載の発明。
【図1】


【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2013−63093(P2013−63093A)
【公開日】平成25年4月11日(2013.4.11)
【国際特許分類】
【外国語出願】
【出願番号】特願2013−4389(P2013−4389)
【出願日】平成25年1月15日(2013.1.15)
【分割の表示】特願2009−513478(P2009−513478)の分割
【原出願日】平成19年6月1日(2007.6.1)
【出願人】(502072134)プレジデント アンド フェロウズ オブ ハーバード カレッジ (92)
【氏名又は名称原語表記】President and Fellows of Harvard College
【Fターム(参考)】
【公開日】平成25年4月11日(2013.4.11)
【国際特許分類】
【出願番号】特願2013−4389(P2013−4389)
【出願日】平成25年1月15日(2013.1.15)
【分割の表示】特願2009−513478(P2009−513478)の分割
【原出願日】平成19年6月1日(2007.6.1)
【出願人】(502072134)プレジデント アンド フェロウズ オブ ハーバード カレッジ (92)
【氏名又は名称原語表記】President and Fellows of Harvard College
【Fターム(参考)】
[ Back to top ]