
チンパンジーアデノウイルスワクチン担体
本発明は、チンパンジーアデノウイルス由来の組み換え複製欠損アデノウイルスベクター及びヒトE1発現細胞株において組み換えアデノウイルスを生成させるための方法を提供する。本発明はまた、増強免疫反応が望まれる免疫原をコードする導入遺伝子の送達及び発現のための使用に適切な組成物及び方法を提供する。本発明はさらに、ヒトにおける使用に適切な臨床グレードのベクターストックを生成させるための方法を提供する。特定の実施形態において、本発明は、癌の予防及び治療のための、ワクチン及び医薬組成物における腫瘍関連抗原をコードする導入遺伝子を含有するベクターの使用をもくろむ。

【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本願は、参照により本明細書中に組み込む、2004年1月23日提出の、米国仮出願第60/538,799の優先権を主張する。
【0002】
本発明は、組み換えベクターの分野、とりわけ、哺乳動物宿主において免疫反応を誘発するための組み換え複製欠損チンパンジーアデノウイルスベクターの産生及び使用に関する。
【背景技術】
【0003】
アデノウイルス(Ad)は、エンベロープのない20面体のキャプシド構造を有する、両性類、鳥類及び哺乳動物において見出される2本鎖DNAウイルスの大きなファミリーを含む(Straus,Adenovirus infections in humans.Adenoviruses.451−498,1984;Hierholzerら、J.Infect.Dis.,158:804−813,1998;Schnurr及びDondero,Intervirology.,36:79−83,1993;Jongら、J.Clin Microbiol.,37:3940−3945:1999)。レトロウイルスとは異なり、アデノウイルスは、宿主細胞のゲノムに組み込まれることなく、分裂細胞及び非分裂細胞の両方を含む、いくつかの哺乳動物種の多数の細胞タイプに形質導入することができる。
【0004】
一般に、アデノウイルスDNAは、通常、非常に安定で、形質転換又は腫瘍形成が起こらない限り、エピソーム性のままである(例えば染色体外性)。さらに、アデノウイルスベクターは、臨床グレードの組成物の製薬規模の産生に容易に持って行くことができる、よく分かっている産生系において高収量に増殖させることができる。これらの特性及びそれらのよく調べられている分子遺伝学により、組み換えアデノウイルスベクターは、ワクチン担体としての使用のための優れた候補となっている。通常、組み換えアデノウイルスベクターの産生は、非機能的となるように欠失されている又は加工されているアデノウイルス遺伝子産物の機能を補完することができるパッケージング細胞株の使用に依存している。
【0005】
現在、性質がよく調べられている2種類のヒトサブグループCアデノウイルス血清型(つまりhAd2及びhAd5)が、遺伝子治療のために使用される殆どのアデノウイルスベクターに対するウイルス骨格のソースとして広く使用されている。複製欠損ヒトアデノウイルスベクターに対してまた、様々な感染物質(例えば、ウイルス、寄生生物又は細菌病原体)及び、腫瘍関連抗原(TAA)を含む腫瘍細胞由来の様々な免疫原送達のためのワクチン担体としての試験が行われている。
【0006】
実験動物(例えば、げっ歯類、サル及び非ヒト哺乳動物)において行われた研究から、ヒトパピローマウイルス(HPV−16)のE6及びE7癌タンパク質(He、Zら、(2001)Virology,270:3583−3590)、狂犬病ウイルス糖タンパク質(Xiang,Zら、(1996)Virology,219:220−227)、Plasmodium falciparum(熱帯熱マラリア原虫)のスポロゾイト周囲タンパク質(Rodriguez,Eら、(1997)J.Immunol.158:1268−1274)ならびにその他の異種抗原由来の免疫原をコードする導入遺伝子を担う組み換え複製欠損ヒトアデノウイルスベクターが、その導入遺伝子産物に対する液性及び細胞性免疫反応の両方を誘導することが示されている。一般には、研究者らは、免疫反応を誘発すると予想される組み換えアデノウイルスベクターの高用量を用いる免疫付与プロトコールを用いて、又は、追加免疫付与として、異なる血清型由来であるが同じ導入遺伝子産物を担うアデノウイルスベクターの連続投与を利用する免疫付与プロトコールを用いることのいずれかにより、非ヒト実験系においてワクチン担体としてヒトアデノウイルスベクターを使用して、成功を報告している(Mastrangeliら、Human Gene Therapy,7:79−87(1996))。
【0007】
しかし、2型及び5型などの、遍在性ヒト血清型由来のワクチン担体は、ヒト集団において既存の液性及び細胞性免疫に直面すると予想されている。したがって、複製欠損組み換えヒトアデノウイルスが、成功裡にワクチン担体としてげっ歯類、サル及び非ヒト霊長類宿主を用いた実験系で利用されているにもかかわらず、ヒトの先天的及び適応的免疫により、ワクチン担体としてのこれらの血清型の利用が著しく制限されると予想される。この予想は、2型及び5型を含むサブグループCのアデノウイルス感染が、ヒト集団において固有であるという事実に基づくものである。その結果、大部分のヒトで、自然感染の結果として、生まれてから最初の5年間のうちに血清転換が起こっている。したがって、ヒトにおいて自然に感染し複製するウイルス由来のベクターは、ワクチン担体としての使用に対して最適な候補ではない可能性がある。
【0008】
ヒトアデノウイルス由来ベクターの使用に関する別の問題は、組み換えウイルスを増殖させるために使用される産生方法により、複製可能なアデノウイルス(RCA)が混入しているベクターストックを生じるというリスクである。これは、ヒト293細胞などのE1補完ヘルパー細胞株において存在する、組み換えベクターとアデノウイルス遺伝子との重複配列間の相同組み換えにより起こる(Graham,F.L.ら(1977)J.Gen.Virol.36:59−72)。臨床試験での使用のために調製されたベクターストックにおけるRCAの存在は、複製欠損ウイルスの移行及び拡大を促進する可能性があるため、安全性リスクの一要素である。欠損ウイルスの拡大により、宿主免疫反応が拡大され、他の不都合な免疫病理学的結果を生じる可能性がある(Fallux,F.J.ら、Human Gene Therapy 9:1909−1917(1998))。したがって、Food and Drug Administration(食品医薬品局)(FDA)及びその他の規制機関は、臨床での使用を意図されるベクター調製物中に存在し得るRCAのレベルにおける制限を確立するガイドラインを広めてきた。RCA制限を課す意図は、臨床試験で使用される組成物中での複製するアデノウイルスへの患者の曝露を確実に制限することにある。
【発明の開示】
【発明が解決しようとする課題】
【0009】
したがって、引き続き、操作しやすく、製薬スケールに持って行きやすく、長期保存でき、ヒトの補完的細胞株で高レベルで複製することができ、免疫原性が高く、ヒトアデノウイルスの共通血清型と交差反応する中和B細胞エピトープを欠いており、規制当局により広められている安全性RCA基準に合致し、ヒトでの使用に適切な初回刺激/追加免疫に使用できる、ヒト宿主での使用に適したアデノウイルスワクチン担体の開発が必要とされている。
【課題を解決するための手段】
【0010】
本発明は、チンパンジーアデノウイルス由来の組み換え複製欠損アデノウイルスベクター及びヒトE1発現細胞株においてチンパンジーアデノウイルスベクターを生成させるための方法に関する。本発明はまた、ヒトにおける使用に適切な臨床グレードのベクターストックを生成させるための方法及び、防御的及び/又は治療的免疫反応を誘発するために、開示するベクターをワクチン担体として使用するための手段を提供する。本発明はさらに、免疫原をコードする導入遺伝子を送達し、発現を支配するように設計されたワクチン組成物を調製するための、本発明の組み換えアデノウイルスを使用するための方法を提供する。ある実施形態において、本発明は、感染物質由来の免疫原をコードする導入遺伝子を含有するワクチン投与のための、ワクチン担体としての、開示するベクターの使用をもくろむ。第二の実施形態において、本発明は、癌ワクチンを調製及び投与するための、開示するベクターの使用をもくろむ。特定の実施形態において、本発明は、TAAをコードする導入遺伝子を含有する癌ワクチンの調製及び投与をもくろむ。
【0011】
ある局面において、本発明は、本明細書中で、ChAd3(配列番号1)(図5A−5K)、ChAd6(配列番号2)(図6A−6K、CV32(配列番号3)(図7A−7K)、CV33(配列番号4)(図8A−8K)及びCV23(配列番号5)(図9A−9J)と呼ばれる、5種類のチンパンジーアデノウイルス(ChAd)の完全ゲノム配列を開示する。
【0012】
ChAd3及びChAd6は、本明細書中で開示する方法に従い単離された新規アデノウイルスを表す。ChAd3及びChAd6のゲノムは、それぞれ37741及び36648塩基対の長さである。このChAd3ヘキソン遺伝子(配列番号41)は、配列番号1のヌクレオチド(nt)19086から21965(停止コドンを除く。)を含有し、ChAd3繊維遺伝子(配列番号42)は、配列番号1のnt32805から34487(停止コドンを除く。)を含有する。ChAd6ヘキソン遺伝子は、配列番号2のnt18266−21124(配列番号43)(停止コドンを除く。)を含有し、その繊維遺伝子(配列番号44)は、配列番号2のnt32218から33552(停止コドンを除く。)を含有する。全長ヘキソンペプチドの多配列アラインメントから推定される配列相同性に基づき、ChAd3は、ヒトサブグループCに分類され、ChAd6は、ヒトサブグループEに分類される。
【0013】
CV32、CV33及びCV23アデノウイルスのゲノムは、それぞれ、36,606、36,535及び32,020塩基対の長さである。CV32(Pan6)(ATCC N.VR−592)、CV33(Pan7)(ATCC N.VR−593)及びCV23(Pan5)(Esoterix Inc.,)は、全て、ヒトAd4(hAd4)に関連することが分かっている(サブグループE)(Wigand,Rら、Intervirology 1989,30:1−9)。しかし、ヘキソン配列アラインメントに基づいて、CV32は、その後、hAd4よりも、ヒトサブグループDメンバーに密接に類似するものとして特徴付けられた。
【0014】
第二の局面において、本発明は、本明細書中で開示する方法に従い単離された21種類のさらなるチンパンジーアデノウイルス(ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82)の繊維及びヘキソン遺伝子に対するヌクレオチド配列を提供する。
【0015】
ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19に対する繊維遺伝子ヌクレオチド配列は、それぞれ図10から図19に記載されており、本明細書中で、配列番号6から15(配列番号6、ChAd20);(配列番号7、ChAd4);(配列番号8、ChAd5);(配列番号9、ChAd7);(配列番号10、ChAd9);(配列番号11、ChAd10);(配列番号12、ChAd11);(配列番号13、ChAd16)(配列番号14、ChAd17)及び(配列番号15、ChAd19)と呼ばれる。
【0016】
ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82に対する繊維遺伝子ヌクレオチド配列は、本明細書中で(配列番号58、ChAd8)、(配列番号60、ChAd22)、(配列番号62、ChAd24)、(配列番号64、ChAd26)、(配列番号66、ChAd30)、(配列番号68、ChAd31)、(配列番号70、ChAd37)、(配列番号72、ChAd38)、(配列番号74、ChAd44)、(配列番号76、ChAd63)及び(配列番号78、ChAd82)と呼び、配列リストに記載する。
【0017】
ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19に対するヘキソン遺伝子ヌクレオチド配列は、図21−30にそれぞれ記載され、本明細書中で、配列番号16から25:(配列番号16、ChAd20);(配列番号17、ChAd4);(配列番号18、ChAd5);(配列番号19、ChAd7);(配列番号20、ChAd9);(配列番号21、ChAd10);(配列番号22、ChAd11);(配列番号23、ChAd16);(配列番号24、ChAd17)及び(配列番号25、ChAd19)と呼ぶ。
【0018】
ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82に対するヘキソン遺伝子ヌクレオチド配列は、本明細書中で、(配列番号97、−ChAd8)、(配列番号99、ChAd22)、(配列番号101、ChAd24)、(配列番号103、ChAd26)、(配列番号105、ChAd30)、(配列番号107、ChAd31)、(配列番号109、ChAd37)、(配列番号111、ChAd38)、(配列番号113、ChAd44)、(配列番号115、ChAd63)及び(配列番号117、ChAd82)と呼び、配列リストに記載する。
【0019】
第三の局面において、本発明は、本明細書中で開示する方法に従い単離された、21種類のさらなるチンパンジーアデノウイルス(ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82)の繊維及びヘキソンタンパク質に対するアミノ酸配列を提供する。
【0020】
本明細書中で開示され主張される繊維タンパク質は、(配列番号83、ChAd3)、(配列番号84、ChAd6)、(配列番号48、ChAd20)、(配列番号49、ChAd4)、(配列番号50、ChAd5)、(配列番号51、ChAd7)、(配列番号52、ChAd9)、(配列番号53、ChAd10)、(配列番号54、ChAd11)、(配列番号55、ChAd16)、(配列番号56、ChAd17)、(配列番号57、ChAd19)、(配列番号59、ChAd8)、(配列番号61、ChAd22)、(配列番号63、ChAd24)、(配列番号65、ChAd26)、(配列番号67、ChAd30)、(配列番号69、ChAd31)、(配列番号71、ChAd37)、(配列番号73、ChAd38)、(配列番号75、ChAd44)、(配列番号77、ChAd63)及び(配列番号79、ChAd82)と呼ばれる。図20Aから20Gは、本明細書中で開示され主張される繊維タンパク質のアミノ酸配列を、C1(配列番号85)、CV68(配列番号86)、Pan5(CV23とも呼ばれる。)(配列番号80)、Pan6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)の繊維タンパク質のアミノ酸配列と比較するアラインメントを提供する。
【0021】
本明細書中で開示され主張されるヘキソンタンパク質は、(配列番号122、ChAd3)、(配列番号123、ChAd6)、(配列番号87、ChAd20)、(配列番号88、ChAd4)、(配列番号89、ChAd5)、(配列番号90、ChAd7)、(配列番号91、ChAd9)、(配列番号92、ChAd10)、(配列番号93、ChAd11)、(配列番号94、ChAd16)、(配列番号95、ChAd17)、(配列番号96、ChAd19)、(配列番号98、ChAd8)、(配列番号100、ChAd22)、(配列番号102、ChAd24)、(配列番号104、ChAd26)、(配列番号106、ChAd30)、(配列番号108、ChAd31)、(配列番号110、ChAd37)、(配列番号112、ChAd38)、(配列番号114、ChAd44)、(配列番号116、ChAd63)及び(配列番号118、ChAd82)と呼ばれる。図31Aから31Jは、本明細書中で開示され主張されるヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、Pan5(CV23とも呼ばれる。)(配列番号119)、Pan6(CV32とも呼ばれる。)、(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)のヘキソンタンパク質のアミノ酸配列との比較を与える。ヘキソンタンパク質の多配列アラインメントにより、熟練者は、ヒトアデノウイルス血清型の提案される分類に従う系統発生解析を行うことが可能となる(Rux,J.J.ら、(2003)J.Virol.77:9553−9566)。
【0022】
代替的局面において、本発明はさらに、21種類のさらなるチンパンジーアデノウイルス分離株を提供する。ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17及びChAd19を含有する試料を、Budapest Treatyのもと、オリジナル委託物として、European Collection of Cell Cultures(ECACC、Prton Down,Salisbury,Wiltshire,SP4 0JG,United Kingdom)により、2003年12月12日に委託した。この委託物に対して、次の受託番号が割り当てられた:03121201(ChAd4)、03121202(ChAd5)、03121203(ChAd7)、03121204(ChAd9)、03121205(ChAd10)、03121206(ChAd11)、03121207(ChAd16)、03121208(ChAd17)、03121209(ChAd19)及び03121210(ChAd20)。
【0023】
ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82を含有する試料を、Budapest Treatyのもと、オリジナル委託物として、ECACC(Prton Down,Salisbury,Wiltshire,SP4 0JG,United Kingdom)により、2005年1月12日に委託した。これらの委託物に対して、次の受託番号が割り当てられた:05011201(ChAd8)、05011202(ChAd22)、05011203(ChAd24)、05011204(ChAd26)、05011205(ChAd30)、05011206(ChAd31)、05011207(ChAd37)、05011208(ChAd38)、05011209(ChAd44)、05011210(ChAd63)及び05011211(ChAd82)。
【0024】
これらの委託物は、International Recognition of the Deposit of Microorganisms of the Purposes of Patent ProcedureにおけるBudapest Treatyの条項に従い維持される。これらの委託物は、当業者に対する便宜としてのみ、なされたものであり、委託物が25U.S.C.§112のもとで要求されることの承認ではない。特許の付与において、37 C.F.R.§1.808(b)において規定される要件を除き、委託した物質の公共に対する利用可能性における制限は全て、変更不能に取り除かれる。
【0025】
さらなる局面において、本発明はまた、哺乳動物細胞、好ましくはヒト細胞に感染することができ、コードされる導入遺伝子産物の発現を支配することができる、複製欠損組み換えアデノウイルスベクターを提供する。本明細書中で示されるように、開示するベクターは、免疫反応が望まれる免疫原を含有する導入遺伝子の送達に対する使用のためのワクチン担体としての使用に適する。特定の実施形態ににおいて、本発明は、ヒトE1発現(つまりパッケージング)細胞株において高レベルで複製することができる、組み換え複製欠損チンパンジーアデノウイルスベクターを提供する。ある実施形態において、本発明は、PER.C6TM細胞において複製できる組み換えアデノウイルスを提供する。
【0026】
一般に、本発明に包含される組み換えベクターは、ヒト集団において一般に直面させられるアデノウイルス血清型に対して既存の免疫を回避するワクチン担体を提供する。とりわけ、本発明の組み換えベクターは、本明細書中で、ヒトアデノウイルス由来ベクターの共通血清型と交差反応する中和Bエピトープを欠いていることが示される、ベクター骨格配列を含有する。
【0027】
本発明はさらに、アデノウイルス部分及びプラスミド部分を含み、該アデノウイルス部分が通常、a)ウイルス左端(ITR及びパッケージングシグナル)、pIX遺伝子の一部及びウイルスゲノム右端と;b)遺伝子発現カセットと、を含む、群特異的シャトルベクターを提供する。この群特異的シャトルベクターは、同じ血清型サブグループ(つまり、サブグループA、B、C、D又はE)に割り当てられ、本明細書中で開示されるヌクレオチド配列を操作するために、及び/又は、E1領域を欠くチンパンジーアデノウイルスゲノムを含有するアデノウイルスプレプラスミドを生じる、同じサブグループに属する他のチンパンジーアデノウイルスをクローニングするために、使用することができるアデノウイルスの間で観察される、ヌクレオチド配列相同性を活用するように設計されている。
【0028】
本発明の他の局面には、アデノウイルスワクチンベクター及び/又はアデノウイルスプレプラスミドベクターを含有する宿主細胞が含まれ、該ベクターの生成方法には、アデノウイルスワクチンベクターを、アデノウイルスE1タンパク質を発現する宿主細胞に導入することと、得られたアデノウイルスワクチンベクターを回収することと、が含まれる。特定の実施形態において、本発明は、アデノウイルスE1発現ヒト細胞に、開示するアデノウイルスベクターの1つを導入することと、得られた組み換えアデノウイルスを回収することと、を含む、複製欠損チンパンジーアデノウイルスベクターを生成する方法を提供する。
【0029】
本発明の別の局面はまた、本発明のアデノウイルスベクターを含有するワクチン組成物を提供する。組み換えチンパンジーアデノウイルスベクターを含有する組成物は、単独で、又はその他のウイルス性もしくは非ウイルス性ベースのDNA/タンパク質ワクチンと組み合わせて、投与し得る。これらはまた、幅広い治療計画の一部として投与し得る。これらの組成物は、予防的又は治療的設定のいずれかで、哺乳動物宿主、好ましくはヒト宿主に投与することができる。本明細書中で示されるように、初回刺激追加免疫計画又は血清学的に区別されるAdベクターの複数回注射など、単独又は組合せ方式での開示するワクチン組成物の投与の結果、その導入遺伝子によりコードされる免疫原を特異的に認識することができる、哺乳動物における免疫反応が誘導される。
【0030】
本明細書中で開示され主張される方法の1つは、少なくとも、その野生型E1遺伝子の機能的欠失を含有し、少なくとも1個の標的抗原をコードするヌクレオチド配列の発現を支配することができるプロモーターを含む配列を担う、組み換えチンパンジーアデノウイルスベクターの十分量を、哺乳動物(未処置又は標的抗原に対して免疫活性となるように初回刺激されているもののいずれか)に投与することを含み、本組み換えベクターの投与が抗原特異的免疫反応を誘発(又は刺激)する。
【0031】
ある実施形態において、本発明は、感染物質に対して免疫反応(予防的又は治療的)を誘導するように設計された方法を提供する(例えば、ウイルス性もしくは細菌病原体又は哺乳動物寄生生物)。第二の実施形態において、本発明は、TAAなどの、自己抗原に対する寛容性を打破する、哺乳動物において免疫反応を誘導するように設計された方法を提供する。本発明のこの局面は、癌ワクチンの調製及び投与のための、ワクチン担体としての、開示するベクターの使用をもくろむ。
【0032】
本発明のさらに他の実施形態及び長所は、次の本発明の詳細な説明から容易に明らかとなろう。
【0033】
本発明の詳細な説明
本明細書及び添付の請求項の範囲を通して使用される場合、次の定義及び略語が適用される。
【0034】
「カセット」という用語は、その転写及び翻訳調節配列に従って発現されるべき、少なくとも1個の核酸配列を含有する核酸分子を指す。カセットを変化させることにより、様々な配列及び配列の組合せの発現を支配するために組み込まれるベクターが生じる。本発明との関連において、本カセットに存在する核酸配列は通常、免疫原をコードする。5’及び3’末端に存在するように操作された制限部位のために、本カセットを容易に挿入、除去又は別のカセットで置換することができる。
【0035】
「cis作用エレメント」という用語は、遺伝子を制御するヌクレオチド配列に連結されるヌクレオチド配列を指す。DNAに存在するcis作用エレメントは、転写を制御し、mRNAに転写されたものは、RNAプロセシング、ターンオーバー及びタンパク質合成を制御することができる。
【0036】
「ベクター」という用語は、宿主生物又は宿主組織にDNA断片を導入することができる、ある手段を指す。プラスミド、ウイルス(アデノウイルスを含む。)、バクテリオファージ及びコスミドを含む、様々なタイプのベクターがある。
【0037】
「プロモーター」という用語は、RNAポリメラーゼが結合する、DNA鎖上の認識部位を指す。プロモーターは、転写活性を開始し、駆動させるために、RNAポリメラーゼと開始複合体を形成する。本複合体は、エンハンサーなどの活性化配列又はサイレンサーなどの抑制配列により修飾され得る。
【0038】
「医薬的有効量」という用語は、宿主細胞に形質導入し、免疫反応を誘発するための導入遺伝子発現の十分なレベルを与える、特定の投与経路において効果的な、組み換えアデノウイルスの量を指す。
【0039】
「複製能のある」組み換えアデノウイルス(AdV)という用語は、原型を保つ、又は機能的な必須初期遺伝子(つまり、E1A、E1B、E2A、E2B及びE4)を有するアデノウイルスを指す。野生型アデノウイルスは、複製能がある。
【0040】
「複製欠損」組み換えAdVという用語は、ウイルス複製に必須の遺伝子産物を、少なくとも機能的に欠損する、又は完全除去されるように加工されているため、複製ができないようにされているアデノウイルスを指す。本発明の組み換えチンパンジーアデノウイルスベクターは、複製欠損である。
【0041】
「哺乳動物」という用語は、ヒトを含む、あらゆる哺乳動物を指す。
【0042】
核酸配列との関連における、「%配列同一性」又は「同一」という用語は、最大一致に対して並べた場合に同じである、2つの配列における残基を指す。配列同一性比較の長さは、そのゲノムの全長(例えば約36kbp)、遺伝子のオープンリーディングフレームの全長、タンパク質、サブユニット又は酵素[例えば、アデノウイルスコード領域を与える表を参照のこと。]又は少なくとも約500から5000ヌクレオチドの断片にわたり得、それが望ましい。しかし、より小さい断片(例えば、少なくとも約9ヌクレオチド、通常は少なくとも約20から24ヌクレオチド、少なくとも28から32ヌクレオチド、少なくとも36以上のヌクレオチド)の間での同一性もまた望ましい。同様に、「%配列同一性」は、タンパク質の全長にわたる、又はその断片にわたるアミノ酸配列に対して容易に決定し得る。適切であるものとして、断片は、少なくとも約8アミノ酸の長さであり、約700アミノ酸以下であり得る。適切な断片の例を本明細書中で述べる。
【0043】
初期設定で本明細書中で定義されるようなアルゴリズム及びコンピュータープログラムを用いて、同一性が容易に決定される。好ましくは、このような同一性は、タンパク質、酵素、サブユニットの全長にわたるか、又は少なくとも約8アミノ酸の長さの断片にわたる。しかし、同一性は、同一遺伝子産物が置かれている使用に適した、より短い領域に基づくものであり得る。
【0044】
一般に、アデノウイルスコンストラクト、遺伝子コンストラクトは、本明細書中に含有される遺伝子を参照して名付ける。例えば、「pChAd3 ΔE1gag」は、E1領域が欠失している、ChAd3 チンパンジーアデノウイルスゲノムを含有するプラスミドコンストラクトを指す。このプラスミドにおいて、E1領域は、ヒトCMVプロモーターと、次いでウシ成長ホルモンポリアデニル化シグナルの調節下でHIVgag遺伝子を含有する免疫原発現カセットにより置換される。同様に、pCV33DE1−E3 NSmutは、E1及びE3領域が欠失しており、ヒトCMVプロモーターと、次いでウシ成長ホルモンポリアデニル化シグナルの調節下において、HCV非構造遺伝子を含む免疫原発現カセットで置換されている、CV33 チンパンジーアデノウイルスゲノムを含有する、本明細書中で開示される第二のプラスミドコンストラクトを指す。
【0045】
略語「Ag」は抗原を指す。
【0046】
本明細書及び添付の請求項の範囲を通して使用される場合、単数体「a」、「an」及び「the」は、文脈において明確に示されない限り、複数形の意味を含む。
【0047】
アデノウイルス(Ad)は、いくつかの鳥類及び哺乳動物宿主において同定されている、エンベロープがない20面体のウイルスである。ヒトAd(hAd)は、全ての既知のヒト及び動物の多くのAd系を含む(例えば、ウシ、ブタ、イヌ、マウス、ウマ、サル及びヒツジ)、マストアデノウイルス属に属する。ヒトアデノウイルスは、ラット及びアカゲザル赤血球の赤血球凝集特性、DNA相同性、制限酵素切断パターン、G+C含量の割合及び発癌性を含む、多くの、生物学的、化学的、免疫学的及び構造的基準に基づいて、6個のサブグループ(AからF)に分けられる(Straus,1984,Adenobiruses、H.Ginsberg編、pps.451−498,New York:Plenus Press及びHorwitz,1990 Virology、B.N.Fields及びD.M.Knipe編、pps.1679−1721)。今日までに、51種類の異なる血清型が認識され、これらの赤血球凝集特性及び生物物理的及び生化学的基準に基づき、サブグループにグループ分けされいてる。
【0048】
アデノウイルスビリオンは、20面体で対称であり、血清型によって、60から90nmの直径である。この20面体キャプシドは、3つの主要なタンパク質、ヘキソン(II)、ペントンベース(III)及びノブ付き繊維(IV)ならびに、多くの少量のタンパク質(つまり、VI、VIII、IX、IIIa及びIVa2)からなる(W.C.Russel,J.Gen.Virol.,81:2573−2604(2000))。ヒトにおいて観察される、既存の免疫のある局面は、液性免疫であり、この結果、ウイルスタンパク質に対して特異的な抗体が産生され持続され得る。アデノウイルスにより誘発される液性反応は主に、主要な構造タンパク質:ヘキソン、ペントン及び繊維に対するものである。
【0049】
公表されている報告から、複数の血清型に対する抗体を含有するタイターが共通であること(Dambrosio,E.(1982)J.Hyg.(London)89:209-219)及び既存のタイターの大部分が、中和活性を有することが、分かっている。アデノウイルスに対する中和免疫は、タイプ特異的であり、アデノウイルスの特定の血清型による感染により、その血清型のみに対する免疫が与えられる。いくつかの報告から、ヘキソンに対して向けられる抗体が、最強であり、最も中和能力が高いことが示唆されている(Toogood,C.I.A.,Crompton,J.及びHay R.T.(1992)J.Gen.Virol.73,1429−1435)。したがって、タイプ特異的中和に関与するエピトープが、様々な血清型由来のヘキソン配列のアラインメントにより同定される7個の超可変領域に位置すると仮定することは、理にかなっている(Crawford−Miksza,L及びD.P.Schnurr(1996)J.Virol.70:1836−1844)。
【0050】
タイプ特異的中和抗体の存在と、同じ血清型に基づくベクターにより免疫反応を誘発できないこととの間の正相関は、未処置動物への処置動物からの免疫血清の受動転移を含む、様々な方法により分かっている。一般に、特異的なウイルス血清型に対して既存の液性免疫により、ベクター投与の治療効率が低下する。さらに、特異的ウイルス血清型に基づくベクターの投与により、同じ血清型の再投与を防御する、ベクターに対する免疫反応が誘発される。
【0051】
特定の実施形態において、本発明は、hAdの共通血清型に対する既存の免疫の結果に付随する不都合な影響を回避する方法を提供する。とりわけ、本発明は、ヒトにおいて回避しない血清型を特徴とするチンパンジーアデノウイルスベクターの使用をもくろむ。したがって、本発明は、ワクチン担体として、共通ヒト血清型と交差反応する中和B細胞エピトープを欠く、アデノウイルス(ChAd)ベクターを提供する。
【0052】
アデノウイルス特異的細胞性免疫(CMI)が、交差反応性であり得ることが報告されているが、複数血清型の繰り返し注射に基づくワクチン接種実験が、1個のベクターを用いた免疫付与スケジュールよりも高い効率であることが示されている。これらの実験はさらに、ワクチン目的のベクター投与の主な制限が、そのベクターに対する液性の既存の免疫であることを示す。ワクチン担体としてのヒトアデノウイルスの使用に付随する問題を解決することができる可能性のある方法には、既存の免疫反応に直面することが予想される、アデノウイルスのさらに高用量の投与(例えば、サブグループC血清型)及び稀少なヒト血清型に基づくベクターの使用が含まれる。しかし、ワクチンの用量を高くして使用することにより、ワクチンのコストが上昇し、望ましくない副作用のリスクが増加し、前臨床試験の結果から、ヒトの代替的血清型は、hAd5及びhAd6よりも免疫原性が低いことが示唆されている。
【0053】
本ベクターのアデノウイルス骨格エレメントに対する宿主の液性及び細胞性免疫反応の問題を回避する、及び複製能のあるアデノウイルス(RCA)が混入し得るヒトアデノウイルス由来ベクターストックの使用のリスクを最小化する試みにおいて、いくつかの非ヒトアデノウイルスの特徴が調べられ、ワクチン担体として開発されている(Soudois,Cら(2000)J.Virology,74:10639-10649;Farina,S.Fら(2001)J.Virology,75:11603-11613;Cohen,C.J.ら(2002)J.Gne.Virology,83:151-155)。既存の免疫に関連する問題を回避するための非ヒトアデノウイルス配列の使用に内在する前提は、共通ヒトアデノウイルス血清型に対する中和抗体が非ヒトウイルスを交差中和しないと思われる観察に基づくものである。しかし、ウイルス及び細胞性の要因の不適合により、非ヒト細胞株で増殖させなければならないという欠点を特徴とする、代替的ベクター系(ウシ、ヒツジ、イヌ)の大部分に対して、実用上の限界がある。
【0054】
Wilsonらは、チンパンジーの腸間膜リンパ節からもともと単離された、チンパンジーアデノウイルス タイプ68(CV68)C68に基づく複製欠損ベクターの特性を述べる報告を公表し(Basnight,Mら(1971)Am.J.Epidmiol.94:166−171)、CV68は、完全に配列決定され、ヒトアデノウイルスと全体的な構造が類似していることが分かった(Farina,S.Fら、J.Virol.75(23):11603−11613(2001))。このウイルスのゲノムは、36,521塩基対の長さであり、ヒトアデノウイルスのサブグループEと、最も類似しているものとして述べられており、配列決定されたヒトAd4オープンリーディングフレームの殆どと90%の同一性がある。CV68 ITRは、130塩基対の長さであり、主要なアデノウイルスの初期及び後期遺伝子の全てが存在する。CV68は、ヒトにおいて循環せず、共通ヒト血清型のものと交差反応する中和B細胞エピトープを欠く血清型を特徴とする。チンパンジーアデノウイルスがヒトアデノウイルスと類似しているにもかかわらず、チンパンジー血清型に対する交差反応性中和免疫は、ヒトにおいて立証されていない(Farina,S.F.ら、J.Virol.(2001)75(23):11603−13)。
【0055】
CV68由来の組み換えベクターは、コクサッキーウイルス及びアデノウイルス受容体を発現する細胞の形質導入を支持するのに十分にヒト血清型と類似であるものとして述べられている(Cohen,Cら、J.Gen.Virol.83:151−155(2002))。注目に値すべきは、CV68が、ヒトアデノウイルス5型からのE1を含有するその複製293細胞を支持するのに十分なレベルでヒトアデノウイルスと類似していることを特徴とすることである(Farina,S.F.ら、J.Virol.75(23):11603−11613(2001))。さらに、ヒト血清型5E1の隣接配列が、CV68由来ベクター配列のそれと非相同的であるという観察から、相同組み換えは起こらないと予想される。したがって、CV68由来ワクチンストックにRCAが混入する可能性は低いと推測される。
【0056】
同じ研究者グループは、続いて、マウスにおける、狂犬病ウイルス糖タンパク質に対する抗体の誘導のためのワクチン担体としての、CV68由来アデノウイルス配列の使用を報告した。CV68の複製欠損体は、E1A及びE1B遺伝子をミニジーンカセットで置換することにより作製された。狂犬病ウイルス糖タンパク質をコードする導入遺伝子産物を含有するE1欠失含有アデノウイルス組み換え体(AdC68rab.gp)で免疫付与したマウスは、狂犬病ウイルスに対する防御免疫を生じ、別の狂犬病ウイルスの致死用量を用いた病原体付与に対して耐性を保持した(Xiang,Zら、J.Virol.76(5):2667−2675(2002))。HIV−1のgagのコドン最適化した短縮型を発現する、第二のCV68コンストラクトは、最近、マウスにおいて活発なgag−特異的CD8+T細胞反応を誘導することが報告された。ワクチン誘導反応は、ワクシニアgag組み換えウイルスを用いた病原体投与に対して防御を与えることが示された(Fitzgerald,J.C.ら、J.Immunol.170:1416−1422(2003))。CV68gag又はAd5gagベクターを用いて、ヒトアデノウイルス血清型5に対して前もって免疫付与したマウスの実験的ワクチン接種から、CV68ワクチンによるものよりも、Ad5によって誘発されるウイルス病原体付与に対して、gag特異的T細胞及び防御が顕著に低下することが示された。C68gagワクチンの効率の低下は、Ad5特異的CD8+T細胞(Id)の交差反応性により生じた。
【0057】
このデータと合わせて考えると、サル由来複製欠損アデノウイルスベクターが、共通ヒト血清型に基づくベクターよりも、ヒトワクチン担体としての使用により適している可能性があることが示唆される。本明細書中で示されるように、ヒトAd5(図36)に対して強く免疫付与されたマウスを、ChAd3−gagアデノウイルスベクターで免疫付与できるという実験結果から、既存の抗ヒトAd5免疫が、ChAdベクターにより誘発されるgag特異的CMI反応を低下させなかったことが示される。これらの結果は、ヒトAd5交差反応性B及びT細胞エピトープが、ChAd3又はChAd6ベクターにおいて存在しないという結論と合致する。
【0058】
一般に、アデノウイルスゲノムは、よく調べられており、いくつかの異なる血清型が存在するにもかかわらず、特異的機能が同じように位置しつつアデノウイルスゲノムの全体的構成において、いくつかの共通の保存がある。Wilsonらにより開示されたチンパンジーアデノウイルス C1及びCV68のヌクレオチド配列及び各ウイルスのE1A、E1B、E2A、E2B、E3、E4、L1、L2、L3、L4及びL5遺伝子の位置は、米国特許第6,083,716号(Chimpanzee Adenovirus Vectors)及びPCT公開出願WO03/000851(Methods for Rapid Screening of Bacterial Transformants and Novel Simion Adenoviral proteins.)により与えられる(これらの教示は、参照により本明細書中に組み込む。)。
【0059】
アデノウイルスゲノムの端部には、逆方向末端反復(ITR)として知られている配列が含まれ、これは、ウイルス複製に必要である。ウイルスはまた、ウイルスコードプロテアーゼを含有し、これは、感染性ビリオンを生成するのに必要とされる構造タンパク質の一部のプロセシングに必要である。アデノウイルスゲノムの構造は、宿主細胞の形質導入後、ウイルス遺伝子が発現される順番で述べられる。より具体的には、ウイルス遺伝子は、転写がDNA複製開始の前に起こるか、後に起こるかに従い、初期(E)又は後期(L)遺伝子と呼ばれる。形質導入の初期において、アデノウイルスのE1、E2、E3及びE4遺伝子が、ウイルス複製に対して宿主細胞を準備するために発現される。このウイルスは、ウイルスゲノムの必須の初期領域1(E1)を欠失させることにより、複製欠損にされ得る。Brodyら、1994 Ann NY Acad Sci.,716:90−101。後期の間、ウイルス粒子の構造要素をコードする、後期遺伝子L1−L5の発現のスイッチが入る。後期遺伝子は全て、1個のプロモーターの調節下となり、ペントン(L2)、ヘキソン(L3)、100kDa 足場タンパク質(L4)及び繊維タンパク質(L5)を含むタンパク質をコードし、これらは、アデノウイルスDNAがキャプシド化されるようになる、新しいウイルスを形成する。ウイルスの血清型に依存して、1個の宿主細胞において、10,000から100,000の子孫アデノウイルス粒子が生じ得る。最終的に、アデノウイルス複製プロセスにより、細胞溶解が起こる。
【0060】
本明細書中で開示される複製欠損アデノウイルスベクターは、開示されているチンパンジー核酸配列からの特定のヌクレオチド配列の欠失により、及び導入遺伝子の挿入、発現又はその他の遺伝子操作に有用な他のDNA配列由来の配列の挿入により構築される。したがって、本明細書中で述べる組み換えチンパンジーアデノウイルスは、1又は複数のチンパンジーアデノウイルス由来のアデノウイルス配列又はチンパンジーアデノウイルスもしくはヒトアデノウイルス由来の配列を含有し得る。適切なポリヌクレオチド配列は、組み換えにより、合成により、又は天然ソースから単離して、作成され得る。本発明の特定の局面における使用に適切なアデノウイルス配列には、共通ヒト血清型と交差反応する中和B細胞エピトープを欠失している配列が含まれる。
【0061】
最低限、本発明の組み換えチンパンジーアデノウイルス(例えばベクター)は、少なくとも1個の免疫原発現カセットと組み合わせて、複製及びビリオンのキャプシド化に必要なチンパンジーアデノウイルス cis作用エレメントを含有する。通常、このcis作用エレメントは、少なくとも1個の抗原をコードする導入遺伝子を含む発現カセットに隣接する。より具体的に、本発明のベクターは、アデノウイルスの、不可欠なcis作用5’逆方向末端反復(ITR)配列(これは、複製の開始点として機能する。)、3’ITR配列、パッケージング/エンハンサードメイン及び異種分子をコードするヌクレオチド配列を含有する。本発明のベクターは、組み換えベクターが、最少のアデノウイルス配列のみを含有するか、特定遺伝子における機能的欠失(例えばE1及び/又はE3又はE4領域)のみがある全体のアデノウイルスゲノムを含有するかに関わらず、チンパンジーアデノウイルスキャプシドを含有する。
【0062】
一般に、本明細書中で開示するアデノウイルスベクターは、複製欠損アデノウイルスゲノム(このアデノウイルスゲノムは、機能的E1遺伝子を持たない。)と;a)免疫反応が望まれる少なくとも1個の免疫原をコードする核酸と、b)該免疫原をコードする核酸配列と機能可能に連結された異種(つまり、アデノウイルス配列に関して)プロモーターとを含有する免疫原発現カセットと;転写終結因子と、を含有する。
【0063】
より具体的には、本発明は、E1、E2、E3及びE4からなる群から選択される、少なくとも1個の初期遺伝子を欠く、組み換えアデノウイルスゲノムからなる複製欠損ベクターを提供する。ある実施形態において、この複製欠損ベクターは、本明細書中で開示するアデノウイルス分離株の1つ(例えば、ChAd3、ChAd6、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63又はChAd82)のE1遺伝子を、免疫原発現カセットで置換又は破壊することにより調製される。例えば、ChAd3(配列番号1)又はChA6(配列番号2)のE1遺伝子を欠失させる/破壊することにより、ベクターを調製することができる。あるいは、ChAd3、ChAd6、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82又はChAd20を含む、本明細書中で開示する他のアデノウイルス分離株のいずれか1つから、複製欠損ベクターを調製することができる。他の実施形態において、本発明の複製欠損ベクターは、機能的E3遺伝子を欠失するように、場合によっては加工されている、本明細書中で開示するChAdのうち1個に由来するアデノウイルスゲノムを含む。初期遺伝子を完全に除去するか、又はその遺伝子を非操作可能又は非機能的にすることによって、本明細書中で開示するチンパンジーアデノウイルス配列は、複製欠損になされうると理解されたい。
【0064】
本発明は、1又は複数の選択された遺伝子産物を発現するアデノウイルスの能力を破壊する「機能的欠失」などの改変があることを特徴とするベクターを包含すると理解されたい。本明細書中で使用される場合、「機能的欠失」という用語は、広く、特定の遺伝子産物を非機能的にする効果を有する改変を包含する。一般に、機能的欠失は、アデノウイルス遺伝子の部分的又は全体的欠失の形をとる。しかし、これらに限定されないがフレームシフト突然変異を導入する改変を行うことを含む、その他の操作でもまた機能的欠失を達成することを当業者は容易に認めるであろう。例えば、本発明の組み換えチンパンジーアデノウイルスベクターは、本ウイルスのアデノウイルスE1A及び/又はE1B発現能力を妨害する、又は機能的に欠失させるよう設計されている改変を導入することにより、複製欠損になされ得る。
【0065】
E2遺伝子の群からの1又は複数の遺伝子の発現を妨害する、又は機能的に欠失させるよう設計されている改変を導入することにより、複製欠損アデノウイルスベクターを得ることができることは周知である。より詳しく述べると、ポリメラーゼ遺伝子又は、末端直前のタンパク質遺伝子もしくはDNA結合タンパク質遺伝子を不活性化することにより、複製欠損ベクターを構築することができる。さらに、E4領域により発現される遺伝子の欠失又は不活性化は、複製欠損 chimp Adベクターを構築するための代替的方法である。初期遺伝子欠失又は不活性化を、さらに短縮されたベクターを作製するために、組み合わせることができる。あるいは、複製欠損ChAdベクターはまた、後期遺伝子L1からL5などの他のウイルス遺伝子において、さらなる改変を含み得る。さらに、異なる血清型から得られたヘキソン及び繊維遺伝子を組み合わせることにより、新規アデノウイルスワクチン担体を生成させることができる。ヘキソン及び繊維遺伝子シャッフリング法の利用によってもまた、研究者らは、ChAdの生物学的特性を変化させ、様々な指向性を有する、又は新しい血清学的特性を有する、ベクターの作製を促進することができる。
【0066】
本発明は、単独で、又は何らかの組合せで、その改変遺伝子の生物学的活性を効果的に破壊する、遺伝子全体又はその一部の欠失を含む組み換えアデノウイルスベクターを包含すると理解されたい。本明細書中で示されるように、E1遺伝子の機能的欠失と組み合わせて、異種のAd5 E4配列をベクターに導入することは、望ましいものであり得るが、例えば、E4領域により発現される遺伝子の機能的欠失を有する組み換えサルアデノウイルスを構築することができる。あるいは、アデノウイルス 遅初期(delayed early)E3遺伝子の機能を削除し得るが、E3の機能は、組み換えアデノウイルス粒子の産生に必要ではないため、本発明で有用なウイルスをパッケージングすることが可能な組み換え体を作製するために、この遺伝子産物を置換する必要はない。
【0067】
本発明のある実施形態において、使用される複製欠損アデノウイルスベクターは、E1及び場合によってはE3における欠失を含有する、チンパンジーサブグループCアデノウイルスである。例えば、ChAd3の場合、適切なE1欠失/破壊は、bp460からbp3542の領域に導入することができる(配列番号1を参照。)。ChAd6の場合、適切なE1欠失/破壊は、bp457からbp3425の領域に導入することができる(配列番号2を参照。)。CV32の場合、E1欠失は、好ましくは、bp456からbp3416である(配列番号3を参照。);CV33の場合、E1欠失は、好ましくは、bp456からbp3425(配列番号4を参照。)であり、CV23の場合、E1欠失は、好ましくは、bp456からbp3415(配列番号5を参照。)である。CV32及びCV33に対するE3欠失は、好ましくは、それぞれ、bp27446からbp31911(配列番号3を参照。)であり、bp27146からbp31609(配列番号4を参照。)である。当業者は、配列相同性及び多配列アラインメントに基づき、他のチンパンジー分離株に対する同等な配列を容易に決定することができる。
【0068】
E1欠失アデノウイルスベクターを構築するために、そのベクター骨格の構造及び導入遺伝子を含有する核酸配列の組成に関して多くのことを決定しなければならないことを、当業者は、容易に認めるであろう。例えば、研究者らは、E1欠失の大きさが導入遺伝子の大きさに適合するか否かを調べなければならない。適合しない場合、そのベクターの骨格にさらなる欠失を導入しなければならない。
【0069】
導入遺伝子を有する核酸配列は、遺伝子又は遺伝子の機能的部分であり得、通常は、発現カセットの形態で存在する。通常、遺伝子発現カセットには、(a)タンパク質又は関心のある抗原をコードする核酸と;(b)そのタンパク質をコードする該核酸に機能可能に連結された異種のプロモーターと;(c)転写終結シグナルと、が含まれる。本核酸は、DNA及び/又はRNAであり得、2本鎖又は1本鎖であり得る。この酸は、所望する宿主における発現のために、コドン最適化され得る(例えば、哺乳動物宿主)。
【0070】
導入遺伝子が導入される骨格内の部位及びその導入遺伝子の方向に関しても決定を行わなければならない。より具体的に、E1パラレル(5’から3’に転写)又は反パラレル(そのベクター骨格に対して、3’から5’方向に転写)方向で、導入遺伝子を挿入することができる。さらに、そのベクターが、同定されその導入遺伝子に機能可能に連結される必要があるワクチン担体としての使用のために調製されている、哺乳動物宿主細胞においてその導入遺伝子の発現を支配することが可能な適切な転写調節エレメントである。「操作可能に連結された」配列には、それらが調節する核酸配列に隣接する発現調節配列及び、トランスで作用する、又は調節される核酸配列を調節するのに離れて作用する調節配列の両方が含まれる。
【0071】
調節配列には、転写開始、終結、エンハンサー及びプロモーター配列などの適切な発現調節配列;スプライシング及びポリアデニル化シグナルなどの効率的RNAプロセシングシグナル;翻訳効率を促進する配列(例えば、コザックコンセンサス配列);タンパク質安定性を促進する配列及び場合によっては、タンパク質分泌を促進する配列が含まれる。これら及び他の共通のベクターエレメントの選択は、従来のものであり、多くの適切な配列が当業者にとり周知である(例えば、Sambrookら、及びそこで引用する文献、例えば、3.18−3.26ページ及び16.17-16.27、及びAusubelら、Current Protocols in Molecular Biology,Jhon Wiley&Sons,New York,1989)。
【0072】
具体的な実施形態において、本プロモーターは、真核RNAポリメラーゼにより認識される異種のプロモーター(つまり、アデノウイルス配列に対して。)である。好ましい実施形態において、このプロモーターは、「強力」又は「効率的」プロモーターである。強力なプロモーターの例は、前初期ヒトサイトメガロウイルスプロモーターである(Chapmanら、1991 Nucl.Acids Res 19:3979−3986(参照により組み込む。))。(CMV)なしで、又はイントロンA配列(CMV−intA)とともに、ヒトCMVプロモーターを使用することができるが、EF1αプロモーター、マウスCMVプロモーター、ラウス肉腫ウイルス(RSV)プロモーター、SV40初期/後期プロモーター及びβアクチンプロモーターを含む、強力な免疫グロブリン又はその他の真核遺伝子プロモーターなどのその他の既知のプロモーターの多くのいずれかを使用し得ることを当業者は、認識するであろう。
【0073】
本発明において使用できるプロモーターのさらなる例は、強力な免疫グロブリンプロモーター、EF1αプロモーター、マウスCMVプロモーター、ラウス肉腫ウイルスプロモーター、SV40初期/後期プロモーター及びβアクチンプロモーターであるが、本発明の方法に従い、意図する宿主において発現を実行することができるあらゆるプロモーターを使用することができることを、当業者は認め得る。このプロモーターは、Tetオペレーター配列などの調節可能配列を含み得る。転写及び発現の調節の可能性を与えるこれらのような配列は、遺伝子転写の抑制が求められる場合に有用である。
【0074】
適切な遺伝子発現カセットはまた、転写終結配列も含む。好ましい転写終結因子は、ウシ成長ホルモン終結因子である。CMV intA−BGH終結因子の、プロモーター/転写終結の組合せは、特に好ましいが、その他のプロモーター/終結因子の組合せもまた使用し得る。本明細書中で示されるように、ウシ成長ホルモン終結/ポリアデニル化シグナル(bGHpA)又は次のように定義される50ヌクレオチド長の短い合成polyAシグナル(SPA):即ちAATAAAAGATCTTTATTTTCATTAGATCTGTGTGTTGGTTTTTTGTGTG(配列番号26)である。一般に、適切な終結配列を例示する。導入遺伝子を含有する核酸配列の後であって、E3’アデノウイルスITR配列の前に、polyAシグナルを挿入する。
【0075】
本明細書中で述べられる組み換えアデノウイルスベクターには、アデノウイルスの1又は複数の株由来のアデノウイルス配列が含まれる。適切な配列は、天然のソースから得ることができ、組み換えにより、合成により、又は他の遺伝子操作もしくは化学的方法により作製することができる。特定の実施形態において、この組み換えチンパンジーアデノウイルスは、非チンパンジーアデノウイルスポリヌクレオチド配列を含有するキメラ組み換え体である。適切な非チンパンジーアデノウイルス配列は、ヒトアデノウイルス株から得ることができる。例えば、hAd5 E4(Ad5 nt32816からnt35619)又はAd5E4 orf6(Ad5 nt33193からnt34077)(Ad5 GenBank Accession No:M73260)により、ネイティブのE4領域を置換することができる。
【0076】
一般に、本発明の組み換えアデノウイルスベクターにより送達される免疫原(抗原性分子)は、コードされる産物の転写及び/又は翻訳を宿主細胞において指示するのに必要な調節配列を与えるヌクレオチド配列と組み合わせて、導入遺伝子によりコードされるポリペプチド、タンパク質又は酵素産物を含有する。この導入遺伝子の組成は、ベクターの意図する使用に依存する。例えば、哺乳動物宿主において、感染物質に対して特異的な、抗体反応又は細胞性免疫反応を誘発するために免疫原性組成物が設計されている場合、病原体特異的免疫を受容者に与えると予想される少なくとも1個の免疫原性産物をコードする核酸配列を利用することが適切である。あるいは、癌ワクチンとしての使用のためにその組成物が調製される場合、適切な導入遺伝子は、TAAなどの自己抗原の免疫原性部分を含有し得、それは、特定のTAAに対する宿主寛容性を打破するため、及び癌の発症を防ぐか、又は腫瘍成長を防ぐのに十分な長期(例えば記憶)反応を誘発するために十分な効力の防御的免疫反応を誘発する目的に関して選択される。したがって、哺乳動物宿主に感染する幅広い病原性物質(これらに限定されないが、ウイルス、寄生生物、細菌及び真菌など。)又は癌もしくは腫瘍細胞から、適切な免疫原性遺伝子産物を得ることができる。試験免疫原の特定の設定で本明細書中において本発明を説明しているが、本発明は、本明細書中で例示される抗原の使用に限定されないことを理解されたい。より具体的には、本発明は、これらに限定されないがTAAなどの、免疫原としての、異種及び自己抗原の両方の使用をもくろむ。
【0077】
ある実施形態において、本発明は、感染物質により発現される抗原(つまり免疫原)に対する免疫反応を誘導する、免疫原性組成物(例えばワクチン)を提供する。例えば、ヒト及び/又は非ヒト動物種に感染するウイルスに対する免疫反応を誘発することが望ましい。予防的及び/又は治療的免疫反応が望まれるウイルスファミリーの例には、アフソウイルス、カルディオウイルス、エンテロウイルス、へパトウイルス、パレコウイルス、リノウイルスなどの6種類の異なる属を含むピコルナウイルス科(Picornaviridae)ファミリーが含まれる。免疫反応が望まれるピコルナウイルスの例は、口蹄疫ウイルス、脳心筋炎ウイルス、ポリオウイルス、コクサッキーウイルス、ヒトA型肝炎ウイルス、ヒトパレコウイルス、リノウイルスである。カリシウイルス科(Caliciviridae)には、ウイルスのノーウォーク群により生じるヒトにおける流行性胃腸炎及び、ウサギ出血性疾患ウイルスに関するウサギにおける出血性疾患及びネコカリシウイルスにより生じる呼吸器疾患のような、動物におけるその他の疾患を伴う、様々な属が含まれる。
【0078】
免疫反応を誘発することが望まれ得るウイルスの別のファミリーは、ヒトならびに多くの様々な動物種から単離された、ウイルスを含むアストロウイルス科(Astroviridae)である。ヒトアストロウイルスは、胃腸炎及び低年齢児の下痢に関連する。あるいは、2つの属:アルファウイルス及びルビウイルスを含む、ウイルスのトガウイルス科(Togaviridae)のメンバーに対する免疫を哺乳動物宿主に付与することが望まれ得る。アルファウイルスは、関節炎(つまり、チックングニアウイルス、シンドビスウイルス)又は脳炎(つまり、東部ウマ脳炎ウイルス、西部ウマ脳炎ウイルス)などの、ヒト及び獣医学の疾患に関与する。
【0079】
風疹ウイルスは、代替的なウイルス標的を与え、ルビウイルス属のメンバーのみが、発熱及びリンパ節炎を伴う軽度発疹疾患の流行に関与する。風疹ウイルス感染はまた、妊娠初期に母親が感染した際の、胎児の異常にも関与する。フラビウイルス科(Flaviviridae)は、重要なヒトならびに動物病原体を含む、3種類の属:フラビウイルス属、ペスチウイルス属及びヘパチウイルス属からなる別のウイルスファミリーである。フラビウイルス属メンバーの多くは、発熱、脳炎及び出血熱を含む、様々な疾患を引き起こす、節足動物媒介性ヒト病原体である。デング熱ウイルス、黄熱病ウイルス、日本脳炎ウイルス、西ナイル熱ウイルス、ダニ媒介性脳炎ウイルスは、世界中が関心を持っている、又はある地域(風土病的)で関心が持たれている主要な病原体である。ペスチウイルス属には、ウシウイルス性下痢ウイルス、ブタコレラウイルス、ボーダー病ウイルスなどの、経済的に非常に重要な動物病原体が含まれる。C型肝炎ウイルスは、急性及び慢性肝炎に関与する、ヘパチウイルス(Hepaci virus)属の唯一のメンバーである。組み換えアデノウイルスにより発現されるHCVタンパク質は、世界中で1億7千万人を侵すウイルス感染の結果を制限する、防御的ならびに治療的免疫反応を誘発することができる。
【0080】
あるいは、感染に対する防御を獲得するために、組み換えアデノウイルスベクターにより、コロナウイルス科(Coronaviridae)のメンバー由来の抗原を発現させることができる。重症急性呼吸器症候群コロナウイルス(SARS−Coウイルス)に対する防御は、これらに限定されないが、ヌクレオキャプシド(N)タンパク質、ポリメラーゼ(P)タンパク質、膜(M)糖タンパク質、スパイク(S)糖タンパク質、小型エンベロープ(E)タンパク質又はそのウイルスにより発現されるその他の何らかのポリペプチドを含む、1又は複数のSARS Covタンパク質を発現する、ChAd3、4、5、6、7、9、10、11、16、17、19、20、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82を含む群から選択される1又は複数のチンパンジーアデノウイルスで免疫付与することにより獲得することができる。狂犬病ウイルスを含むラブドウイルス科(Rhabdoviridae)メンバーは、ウイルスタンパク質を発現する組み換えワクチンの標的であり得る。
【0081】
その他の可能な標的には、重症出血熱の流行に関与する、エボラ様ウイルス及びマルブルグ様ウイルス属が含まれるフィロウイルス科(Filoviridae);麻疹、呼吸器合胞体ウイルス、パラインフルエンザウイルスのような、ヒトにおいて知られる最も流行しているいくつかのウイルス及びニューカッスル病及び牛疫ウイルスなどの獣医学的に関心のあるウイルスを含むパラミクソウイルス科(Paramyxoviridae);インフルエンザA、B、Cウイルスを含むオルトミクソウイルス科(Orthomyxoviridae);リフトバレー熱、Sin Nombre(シンノンブレ)、ハンタ、プーマラウイルスなどの重要なヒト病原体を含む、節足動物から脊椎動物宿主に主に伝染するブニヤウイルス科(Bunyaviridae);リンパ球性脈絡髄膜炎、ラッサ熱、アルゼンチン出血熱、ボリビア出血熱ウイルスを含む、アレナウイルス科(Arenaviridae);主にウマ及びヒツジにおいて中枢神経疾患を引き起こすウイルスを含む、ボルナウイルス科(Bornaviridae);世界中での幼児及び低年齢児における重症下痢疾患の最も重要な原因である、ロタウイルスファミリー及び、ヒト及び他の動物両方に伝染し得るオルビウイルス(ブルータング病、伝染性出血性疾患ウイルス)を含む、レオウイルス科(Reoviridae);ヒト免疫不全ウイルス1及び2(HIV−1及びHIV−2)及びヒトT細胞白血病ウイルスタイプE1及び2(HTLV1及び2)のような、重要なヒト病原体ならびにヒツジ及びヤギに伝染するマエディ/ビスナウイルス、ウマに伝染するウマ感染性貧血ウイルス、ウシに伝染するウシ免疫不全ウイルス、ネコに伝染するネコ免疫不全ウイルスなどの非ヒトレンチウイルスを含むウイルスの大きなグループである、レトロウイルス科(Retroviridae);ポリオーマウイルス科(Polyomaviridae)グループ小型DNA癌原性ウイルス、典型ウイルスは、それぞれマウス及びアカゲザルに感染するポリオーマ及びSV40、(SV40に非常に関連しているBK及びJCウイルスは、ヒト患者から単離された。);パピローマウイルス科(Papillomaviridae)は、いぼ及びコンジローマを発生させる、ヒトを含む高等脊椎動物に感染するDNAウイルスの群からなる。パピローマウイルス感染は、ヒト及び動物両方での癌の発生に関連する。ヒトパピローマウイルスは、子宮頸癌、膣癌及び皮膚癌に関与する。ヘルペスウイルス科(Herpesviridae)は、ヒト及びその他の哺乳動物に対するいくつかの重要な病原体が分類される亜科を含む。抗原の適切なソースは、これらに限定されないが、単純ヘルペスウイルス1及び2、水疱瘡ウイルス、エプスタインバーウイルス、サイトメガロウイルス、ヒトヘルペスウイルス6A、6B及び7、カポジ肉腫関連ヘルペスウイルスである。抗原のさらに適切なソースは、サル痘ウイルス、伝染性軟属腫(Molluscum contagisum)ウイルス、天然痘ウイルスのようなポックスウイルス科(Poxviridae)のメンバー;へパドナウイルス科(Hepadnaviridae)の典型メンバーである、B型肝炎ウイルスならびに、D型肝炎(hepatitis delta)ウイルス、E型肝炎ウイルスのような、急性及び/又は慢性肝炎を引き起こすその他のウイルスである。
【0082】
本発明のアデノウイルスベクターはまた、細菌、真菌、寄生生物病原体を含む非ウイルス性病原体により発現されるタンパク質に対して、ヒト又は動物において免疫反応を刺激するように設計される免疫原性組成物の調製に適切である。例えば、本明細書中で開示するベクターは、これらに限定されないが、Staphylococcus aureus(黄色ブドウ球菌)、Streptococcus pyogenes(化膿レンサ球菌)、Streptococcus Ppneumoniae(肺炎レンサ球菌)、Vibrio cholerae(コレラ菌)、Clostridium tetani(破傷風菌)、Neisseria meningitis(髄膜炎菌)、Corynebacterium diphteriae(ジフテリア菌)、Mycobacteria tuberculosis(マイコバクテリウム・テュバキュローシス))、Mycobacteria leprae(ライ菌)、Listeria monocytogenes(リステリアモノサイトジェン)及びLegionella pneumofila(レジオネラニューモフィラ)に対するワクチンを調製するために使用することができる。予防的又は治療的ワクチンを調製することが望まれ得る、真菌及び哺乳動物寄生生物の例には、Candida albicans(カンジダアルビカンス)、Aspergillus fumigatus(アスペルギウス・フミガタス)、Histoplasma capsulatum(ヒストプラズム・カプスラーツス)、Plasmodium malariae(四日熱マラリア原虫)、Leishmania major(リーシュマニア・メジャー)、Trypanosome cruzi(トリパノソーマ・クルジ)及びbrucei(ブルシェイ)、Schistosoma haematobium(ビルハルツ住血吸虫)、mansoni(マンソン住血吸虫)及びjaponicum(日本住血吸虫);Entamoeba histolytica(エンタモエバ・ヒストリチカ)及びヒトフィラリアに関与することが知られているフィラリアの多数の種が含まれる。
【0083】
癌は、一般に、細胞周期の維持又は細胞増殖の調節に関与するポリペプチドをコードする遺伝子の脱制御を含む(例えば、増殖因子、癌遺伝子、受容体及び腫瘍抑制因子)。癌に関わる遺伝子の多くの産物は、様々な腫瘍細胞の表面で発現される。T及びBリンパ球により認識され得る様々な腫瘍抗原が、ヒト及び動物の癌で同定されている。腫瘍細胞において発現されるのに加えて、正常組織及び/又は胎児発生中にも発現されるため、抗癌ワクチン試験での使用に適切なヒト腫瘍関連抗原(TAA)の大部分は、「自己抗原」として説明される。TAAに対する標的集団の免疫寛容性により、多くの癌ワクチンが効果がないとされる理由が説明される。
【0084】
腫瘍抗原は、T/B細胞反応を刺激することができる、変化した細胞性タンパク質の例である、Ras、p53又はBcr−Ab1タンパク質などの、癌原遺伝子又は腫瘍抑制遺伝子で変化が起こる正常細胞遺伝子の発癌突然変異体により産生され得る。腫瘍抗原は、腫瘍細胞で過剰発現されるか(チロシナーゼ、GP100、MARTは、メラニン産生細胞において低レベルで正常に発現され、メラノーマにおいて過剰発現される。)、又は腫瘍細胞において異常に発現される(メラノーマ及び多くの癌腫において発現されるが、精巣及び胎盤において正常に発現される、MAGE、BAGE、GAGE。)。腫瘍抗原は、発癌ウイルスの産物であり得る:子宮頸部癌により発現される、パピローマウイルスE6及びE7タンパク質;EBV+リンパ腫及び上咽頭癌により産生される、EBV EBNA−1タンパク質;SV40誘導実験的腫瘍におけるSV40 T抗原。癌胎児性抗原は、癌細胞及び正常な発生過程(胎児)の組織において高レベルで発現されるが、成人組織では発現されない。癌胎児性抗原(CEA)及びα−フェトタンパク質(AFP)は、よく特性が調べられている癌胎児性抗原の例である。
【0085】
最近の証拠により、免疫反応を誘発する、したがってワクチン治療に対する免疫原に適切な抗原のクラスを生じさせることができるTAAの存在が裏付けられている。しかし、抗原のクラスとして、TAAは、周知のとおり弱い免疫原であり、TAAに非常に特異的なT細胞は、T細胞発生の過程で欠失するか、アネルギー化されている。したがって、特定のTAAに対する、腫瘍を有する宿主の免疫反応は非常に弱いと予想される。標的TAAに対する宿主の寛容性を打破するという固有の必要性があるため、実験的臨床ワクチン研究は、特に、TAA特異的T細胞反応を促進するTAA免疫付与ストラテジーの開発に焦点を当てている。一般に、効果的な癌ワクチンは、免疫寛容性を乗り越え、かつ、宿主の免疫反応を、予防的及び/又は防御的なレベルに上げなければならない。多くの実験的ワクチン研究における抗腫瘍効果は、TAAに対するT細胞反応に相関がある。
【0086】
代替的実施形態において、本発明は、腫瘍抗原に対する免疫反応を誘導するために使用することができる、免疫原性組成物(例えば、癌ワクチン)をもくろむ。適切な組成物は、腫瘍抗原をコードする核酸配列を含有する組み換えチンパンジーアデノウイルス及び生理学的に許容可能な担体を含有する。特定の実施形態において、カセットのコード配列エレメントは、癌関連抗原などの自己抗原由来の免疫原性ペプチド配列などの、1個の免疫原をコードし得る。ある実施形態において、免疫原をコードする核酸配列(つまり導入遺伝子)は、特定の哺乳動物種での発現のためにコドン最適化され得る。他の実施形態において、コード配列は、1又は複数のコドン最適化された腫瘍抗原などの、複数の免疫原をコードし得る。例えば、開示するアデノウイルスベクターを利用する癌ワクチンは、HER2/neu、CEA、Hepcam、PSA、PSMA、テロメラーゼ、gp100、Melan−A/MART−1、Muc−1、NY−ESO−1、サバイビン、ストロメリシン 3、チロシナーゼ、MAGE3、CML68、CML66、OY−TES−1、SSX−2、SART−1、SART−2、SART−3、NY−CO−58、NY−BR−62、hKLP2、VEGFなどの、自己抗原の組合せをコードし得る。
【0087】
効果的な癌ワクチンの開発には、ワクチン接種された患者において抗原特異的免疫を誘発するストラテジー及び活性のある免疫付与終了後に免疫反応の生成が持続することの検証が必要である。このストラテジーの成功は、標的抗原に対して向けられる測定可能な免疫反応が、癌発生又は再発に対する防御に相関するか否かに依存する。細胞性免疫及び液性免疫両方のエフェクターメカニズムにより、腫瘍細胞が殺滅されることが示されている。しかし、実験系からのデータにより、腫瘍細胞の除去に対して、抗原特異的T細胞が、最も強力な免疫メカニズムを与えることが示唆される。腫瘍特異的抗原(例えばTAA)のエフェクターT細胞による認識により、TAAが腫瘍拒絶抗原として機能することができるようになると予想される。公表されている実験から、CD8+及びCD4+ヘルパーT細胞反応の刺激が、最善の腫瘍クリアランスの達成に重要であることが示唆されている(Greenberg,P.D.(1991)Adv.Immunol.49:281−355;Pardoll,D.M.ら(1998)Curr.Opin.Immunol.10:588−94)。臨床反応(つまり効能)は、インターフェロンγ分泌細胞毒性T細胞の増加と関与する。本ワクチン担体の効率を明らかにするための、本明細書中で使用される、ELISPOTアッセイなどのアッセイの出現により、研究者らは、ワクチン接種計画に対するT細胞反応を測定できるようになり、それにより癌ワクチンの開発が促進される。
【0088】
癌ワクチンは、予防的又は治療的の何れかであり得る。癌ワクチンの予防的使用の基礎をなす一般的な仮定は、腫瘍を有する対象において、TAAが非常に弱い免疫原であるか機能的に非免疫原性であるということである。より具体的には、癌免疫学の分野において、癌に罹患した患者における免疫療法として、ワクチンを使用することができる。したがって、既存の腫瘍又は悪性腫瘍により発現されるTAAに向けられる免疫反応を誘発するように、癌ワクチンを設計することができる。したがって、特定の実施形態ににおいて、治療的癌ワクチンは、従来の治療法に対する耐性が進展しているか、従来の治療後に再発が起こる可能性が高い、腫瘍を有する患者における使用を意図する。
【0089】
アデノウイルスの高い免疫原性により、アデノウイルスベクターは、自己抗原に対する宿主寛容性を打破するために設計されたワクチン担体の関連における使用のための、特に優れた候補となる。自己免疫疾患において最初に記載された、エピトープ又は決定基拡大の現象は、MHCクラスI及びMHCクラスII限定反応の両方と関与し、HER2 neuタンパク質特異的T細胞免疫の発生と相関がある。エピトープ拡大は、免疫原性タンパク質の特定部分に対する免疫反応の生成を表し、次いで、同じタンパク質に存在する、他の抗原性決定基に対する免疫の自然な拡大が起こる。例えば、Disisらは、顆粒球−マクロファージコロニー刺激因子と混合した、HER−2タンパク質の強力なTヘルパーエピトープ由来のペプチドを含有するワクチンを投与した、HER−2/neu過剰発現悪性腫瘍がある患者の84%でエピトープ拡大を観察した(J.Clin.Oncol.(2002)20(11):2624-2632)。重要なことは、エピトープ拡大が、HER−2/neuタンパク質ドメイン反応の生成と相関したことであり、このことから、免疫付与が効果的に免疫寛容性を回避したことが示唆される。
【0090】
癌ワクチンに対する標的としての、開示するアデノウイルスベクター及び方法の使用に適切なTAAは、いくつかの特徴を保持すべきである。例えば、標的TAAは、好ましい発現プロファイルを有さなければならず、これは、正常組織と比較して、腫瘍又は悪性腫瘍組織において、選択的に発現されるか、過剰発現されることを意味する。さらに、腫瘍発生に関与するTAAは、癌進行の様々な段階で保持される可能性がより高いので、適切な標的TAAはまた、腫瘍進行及び転移を通じて保持されるべきである。適切な標的TAAはまた、腫瘍内で均一に発現される。第三に、適切な標的TAAは、絶対的な免疫寛容性に対する対象であってはならない。より具体的には、関心のあるTAAに対して認識し、かつ反応することができるT細胞は、宿主のT細胞レパートリーから完全に欠失されていないことのいくつかの証拠があるべきである(Berinstein,N.L.,J.Clin.Oncol.29(8):2197(2002))。
【0091】
癌胎児性抗原(CEA)は、抗癌ワクチンのための標的抗原としての使用のために魅力的なTAAとなる多くの特徴を有する。これは、有利な発現パターンを特徴とする、Igスーパーファミリーのメンバーである。これは、全ヒト癌の50%を超えるものにおいて、発現され、腫瘍発生プロセスに関わっており、このことから、その発現が、癌進行を通して、選択され、保持され得ることが示唆される。さらに、CEAに対する免疫寛容性が絶対的ではないことが分かっている。公表された研究から、ヒトT細胞が、CEAを発現する癌細胞を、認識し、活性化し、溶解させることが分かっている(Berinstein,N.L.,J.Clin.Oncol.29(8):2197(2002))。例えば、その後のペプチドべースのインビトロ刺激と組み合わせた、CEAを発現する組み換えワクシニアウイルスを用いた患者への免疫付与により、自己腫瘍を細胞溶解できる、CD8+MHC−限定CTLが生成した(Tsang,K.Y,ら、J.Natl.Cancer Inst.,(1995)87:982−990)。あるいは、手術後の結腸直腸癌患者に対する組み換えCEAを用いた免疫付与が、組み換えCEAに対する、弱い抗体及び細胞性応答を誘導したことが報告された(Samanci,Aら(1998)Cancer Immunol.Immunother.47:131−142)。さらに、結腸直腸癌と診断された患者への抗CEA 抗イディオタイプ抗体の投与により、抗CEA抗体が生じ、イディオタイプ特異的T細胞増殖が起こった(Foon,L,A.ら(1995)J.Clin.Invest.,96:334−342)。文献により、また、いくつかの異なるワクチン接種法を用いて、癌患者におけるCEAに対する寛容性を乗り越えることができることが示される(つまり、組み換えCEA又は組み換えオルトポックスもしくはアビポックスCEAウイルスを用いたワクチン接種、抗イディオタイプ抗体の投与、CEAアゴニストエピトープによる樹状細胞のパルシング)。
【0092】
CEAは、正常胎児結腸で発現される、正常結腸粘膜では非常に低い、癌胎児性糖タンパク質である。これは、多くの腺癌、特に結腸、膵臓、乳房、肺、直腸及び胃の腺癌においても過剰発現される。多くの結腸直腸癌及びある癌腫は、血清中で測定可能な高レベルのCEAを産生し、それにより、特に結腸直腸癌の患者において、最も広く使用される悪性の血清マーカーの1つとなっている。
【0093】
本発明の組成物及び方法における使用のための適切な免疫原を提供する第二のTAAは、HER2/erb−2(neuとも呼ばれる。)癌原遺伝子の産物である。CEAのように、HER2/neuは、有利な発現パターンを有し、絶対的寛容性の対象ではない。より具体的には、HER2/neu転写産物及び185kDポリペプチド産物の発現を検出したところ、皮膚及び乳房を含む様々な組織の正常成人上皮細胞及び胃腸組織、生殖管及び尿路では低レベルであり;胚発生中の対応する胎児組織では高レベルであった(Pressら、Oncogene 5:953−962(1990))。いくつかの証拠から、ヒト乳癌、肺癌、前立腺癌、卵巣癌、子宮内膜癌及び結腸直腸癌でのHER−2の増幅と悪性形質転換との間の関連が示唆される(Disis及びCheever,Adv.Cancer Research 71:343−371(1997))。一般に、HER2/neuの過剰発現は、癌患者に対して、予後が悪いこと及び再発率が高いことと相関がある(Slamonら、Science 244:707−712(1989))。したがって、HER2/neuタンパク質に特異的なワクチンは、多くの様々なヒト悪性腫瘍における疾患再発の予防において、広く適用でき、利用できると思われる。
【0094】
HER2/neuは、生来のチロシンキナーゼ活性を保持し、上皮細胞増殖因子(EGF)受容体に対する高い相同性を示す膜貫通糖タンパク質をコードする(Akiyama,Tら、(1986)Science 232:1644−1646)。癌免疫療法に対してHER2を標的として利用した最初の臨床研究の1つでは、HER−2特異的モノクローナル抗体Herceptinが乳癌の治療に対して用いられた(Goldenberg MM(1999)Clin.Ther.21:309−318)。これにより、自己抗原提示細胞を活性化するためのHER−2ドメインを含有する組み換え融合タンパク質の使用を含む、効果的な抗腫瘍反応を誘発するための、免疫系のT細胞アームに対する標的としてのHER−2の使用に焦点を当てたその後の試みが導かれた。公表されている報告により、neu発現性の、乳癌及び卵巣癌に罹患している多くの癌患者において、HER2/neu癌遺伝子のタンパク質産物に対する免疫反応が得られることが分かっている(例えば、抗原特異的抗体及びT細胞を産生する。)
【0095】
様々な中間体プラスミド及びシャトルベクターへの組み換えアデノウイルス配列、導入遺伝子及び他のベクターエレメントのアセンブリー及び、組み換えウイルス粒子を産生させるためのプラスミド及びベクターの使用は全て、当業者にとって周知の標準的教科書に記載されているような従来技術を使用することにより達成される(Sambrookら、Molecular Cloning:A Laboratory Manual,第二版、Cold Spring Harbor Press,Cold Spring Harbor,NY(1989))。そのような技術には、これらに限定されないが、従来のcDNAクローニング技術、アデノウイルスゲノム由来の重複オリゴヌクレオチド配列の使用、相同組み換え、ポリメラーゼ連鎖反応、標準的トランスフェクション技術、寒天重層におけるウイルスのプラーク生成及びその他の関連方法が含まれる。
【0096】
原核細胞におけるポリヌクレオチドの調製を促進するために、プラスミド型のアデノウイルスベクターを調製することが多い(アデノウイルスプレプラスミド)。このアデノウイルスプレプラスミドは、アデノウイルス部分及びプラスミド部分を含有する。このアデノウイルス部分は、基本的に、本発明のアデノウイルスベクター(E1、場合によってはE3領域が、非機能性であるか欠失している、アデノウイルス配列を含有する。)及び都合のよい制限部位に隣接した免疫原発現カセットを含有するアデノウイルス部分と同じである。
【0097】
アデノウイルスプレプラスミドのプラスミド部分は、抗生物質の発現が真核細胞において起こらないように、原核プロモーターの転写調節下に、抗生物質耐性マーカーを含有することが多い。アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子及びその他の医薬適合性の抗生物質を使用し得る。原核生物における発酵によるポリヌクレオチドの高レベル産生を促進するために、アデノウイルスプレプラスミドが原核性の複製起点を含有し、高コピー数であることが好都合である。多くの市販の原核クローニングベクターによりこれらの利益を得られる。必須でないDNA配列を除去することが望ましい。ベクターが真核細胞において複製できることもまた望ましい。これにより、ポリヌクレオチドワクチン配列の、受容者のゲノムへの組み込みのリスクが最小に抑えられる。特定の組織タイプにポリヌクレオチドの発現を限定することが望ましいかどうかを問わず、組織特異的プロモーター又はエンハンサーを使用し得る。
【0098】
アデノウイルス骨格DNA及び適切なシャトルベクター(特異的欠失を標的とするように、及び結果として得られるプラスミドに所望する制限部位を組み込むように設計される。)を用いた相同組み換えにより、アデノウイルスプレプラスミド(望ましい欠失及び挿入を有する複製欠損アデノウイルスのゲノムを含有するプラスミド)を生成させることができる。例えば、所望する欠失を考慮したアデノウイルス末端のPCR及び適切なクローニングプラスミドへの個別のセグメントの、続いて行われるクローニングなど、本技術分野で広く理解され認識される一般的方法を用いて、この目的における用途のシャトルベクターを生成させることができる。次に、アデノウイルスプレプラスミドを消化し、リン酸カルシウム共沈殿又はその他の適切な手段を介して補完的な細胞株にトランスフェクションすることができる。次に、ウイルス複製及び増幅が起こり、これは、顕著な細胞変性効果により明らかになる現象である。次に、ウイルス複製の完了後、感染細胞及び培地を回収する(一般に、トランスフェクションから、7日から10日後。)。
【0099】
一般に、所望するアデノウイルスプレプラスミドの構築及びアセンブリー後、アデノウイルスE1を発現しているヒト細胞株に導入することにより、アデノウイルスプレプラスミドをウイルスとする。パッケージング細胞株とベクターのウイルス遺伝子との間の相補により、ベクターのアデノウイルス−導入遺伝子配列を複製させ、ビリオンキャプシド中にパッケージングされ、その結果、組み換えアデノウイルスの産生が起こる。本発明の方法での使用のために、当業者とって公知のあらゆる様々な方法により、得られたウイルスを単離し、精製し得る。
【0100】
チンパンジーアデノウイルス遺伝子の、1又は複数の選択した欠失をウイルスベクターに導入したとき、産生細胞株中に存在する配列による産生プロセスの間に、欠失遺伝子産物の機能を補われ得ることは、当業者にとって容易に明らかである。したがって、パッケージングに必要であるがそのベクターにおいて機能しないアデノウイルス機能の一部又は全てを特徴とする、永久的に形質転換された細胞株により、操作した遺伝子の機能を与えることができる(例えば、E1A、E1B、E2A、E2B、E4のいずれか。)。あるいは、その機能を与えるのに必要な遺伝子を含有するコンストラクトを用いて、適切な細胞に感染又は一時的トランスフェクションを行うことにより、適切なパッケージング細胞株に対して必要なアデノウイルス機能を与えることができる。
【0101】
したがって、本発明はまた、E1発現ヒト細胞株においてチンパンジーアデノウイルスベクターを生成させる方法を提供する。より具体的には、公知の細胞株 293及びPER.C6TMを含むE1補完細胞株中に、開示されたベクターを増殖させることができる。これらの両細胞株は、アデノウイルスE1遺伝子産物を発現する。PER.C6TMは、1997年1月3日公開のWO97/00326に記載されている(これは、参照により本明細書中に組み込む。)。これは、複製欠損第一世代アデノウイルスの産生を補完するが、相同組み換えによる複製能のあるアデノウイルスの生成を防ぐように設計されているE1遺伝子セグメントにより形質導入されている初代ヒト網膜芽細胞株である。293細胞は、Grahamら(1977)J.Gen.Virol 36:59−72(これも、参照により本明細書中に組み込む。)において記載されている。「第一世代アデノウイルス」という用語が、E1領域が非機能的であるか欠失しているかのいずれかであり、場合によってはE3領域が非機能的であるか欠失している、複製欠損アデノウイルスを指すことを、当業者は、認識するであろう。
【0102】
臨床試験においてワクチン組成物としての使用を意図する複製欠損アデノウイルスベクターのバッチは、RCA不含であることが証明されているべきである(Fallaux,F.J.ら(1998)Human Gene Therapy,9:1909−1917)。実施に際し、これは、費用のかかるスクリーニングプログラムを確立し利用する必要がある、手間のかかるプロセスである。RCA生成が高頻度に起こることにより、生成したバッチに対する失敗率が高まるだけでなく、スケールアップが非常に制限されることを、当業者は認めるであろう。ベクターのヌクレオチド配列とヘルパー産生/パッケージング細胞株のゲノムに存在するアデノウイルス配列との間の配列相同性を削除することにより、相同組み換えにより生成するRCAが混入するベクターのバッチ産生の可能性が取り除かれると思われる。
【0103】
通常、組み換え複製欠損アデノウイルスベクターは、トランスにおいてE1遺伝子産物を与える細胞株中に増殖する。ベクターが、同じ又は非常に似た血清型由来である場合、トランスでの必須のE1遺伝子産物の供給は、非常に効果的である。例えば、E1欠失(つまりΔE1)グループC血清型(Ad2及びAd5)ベクターが、Ad5 E1領域を含有し、発現する、293又はPER.C6細胞において増殖できることは、周知である。しかし、293及びPER.C6産生細胞に存在するAd5 E1配列が、必ずしも非グループC血清型の複製を完全に補完し得るとは限らないことが観察されている。したがって、サブグループC以外のE1欠失血清型、例えば、サブグループA、B、D、E及びF由来のものの複製は、対応する野生ウイルスと比較して、より低効率であるか、又は293又はPER.C6細胞において全く複製し得ない。これは、Ad5(グループC)E1 B 55K遺伝子産物が、非グループC血清型のE4 orf6遺伝子産物と機能的に相互作用できないからである。
【0104】
Ad5 E1発現細胞における複製効率の低下は、様々なサブグループのベクターによって変化し得る。サブグループD及びE アデノウイルス由来のΔE1ベクターが、293及びPer.C6TM細胞において様々な効率でレスキュー可能であり、増殖可能である一方、サブグループBのΔE1ベクターは、増殖能が完全に損なわれている(Vogels Rら、(2003)Aug.Replication−deficient human Adenovirus type 35 vectors for gene transfer and vaccination:efficient human cell infection and bypass of preexisting adenovirus immunity.J Virol.77(15):8263-71)。
【0105】
Ad5 E1b 55kとベクター発現E4 orf6 タンパク質との間の相互作用が、同じサブグループのメンバー内で保存されているにもかからわず、このことは、非C血清型のE4 orf6 タンパク質が発現される場合、十分安定ではないであろうことを示す。E1b 55k/E4 orf6複合体の形成が不十分であるか不安定であることから、ΔE1ベクターの増殖が低下しなくなる。したがって、成功裡に、かつ効率よく、グループB血清型の組み換えアデノウイルスをレスキューするために、関心のある血清型のE1領域を発現する細胞株を生成させることが必要とされ得ることが、実験的に分かっている。293又はPER.C6TMなどの、Ad5 E1を発現する細胞において、発現をE1b 55Kタンパク質に限定することができる。あるいは、適切なAd5 E1発現細胞株を、Ad5 E1に加えて全体のAd5 E4領域(又はE4orf6のみ)を発現するように、改変することができる。Ad5 E1及びorf6の両方を発現する細胞株の生成は、代替的なアデノウイルス血清型を補完するのに有用である;例えば、Abrahamsenら、1997 J.Virol.8946−8951を参照のこと。
【0106】
Ad5補完細胞株へのE4(orf6)の組み込みは、トランスで血清型特異的E1遺伝子産物を与える血清型特異的細胞株の生成であるとして知られている。あるいは、非グループCベクター増殖効率は、ネイティブE4領域の、Ad5 orf6での置換によるウイルス骨格の改変により向上させ得る。ネイティブ orf6のみをAd5又は他のサブグループCウイルス(Ad1、Ad2、Ad6)由来のorf6と置換することによって、同様の結果を得ることができる。米国特許第5,849,561号は、Ad5−E4遺伝子の一部も発現するAd5−E1補完細胞株における、E1欠失非グループCアデノウイルスベクターの補完を開示する。
【0107】
Vigneらに発行された米国特許第6,127,175号は、アデノウイルスのE4領域の一部、好ましくは、orf6/orf6/7を発現する、安定的にトランスフェクションされた哺乳動物細胞株を開示する。このような細胞株は、E4領域を欠く組み換えAdゲノムの補完に有用である。
【0108】
ワクチン組成物を含む、開示するアデノウイルスベクターを含有する組成物は、本発明の重要な局面である。哺乳動物宿主、好ましくはヒト宿主に、予防的又は治療的設定のいずれかで、これらの組成物を投与することができる。宿主/ワクチンとして可能なものには、これらに限定されないが、霊長類、特にヒト及び非ヒト霊長類が含まれ、商業用動物又は家畜として重要なあらゆる非ヒト哺乳動物が含まれる。組み換えチンパンジーアデノウイルスベクターを含有する組成物は、単独で又はその他のウイルス性もしくは非ウイルス性ベースのDNA/タンパク質ワクチンと組み合わせて投与し得る。これらはまた、広範囲の治療計画の一部として投与し得る。
【0109】
本発明の特定の実施形態において、自己抗原又は腫瘍関連抗原に対する宿主の寛容性を打破するために設計された免疫付与プロトコールにおいて、開示するベクターを使用し得る。多くのTAAの同定により、癌治療に対する、活性のあるワクチン接種アプローチの開発が可能となった。プロセシングされ、提示される細胞表面抗原及び細胞内抗原の両方が、有用な標的を与える。一般に、自己抗原に対する宿主寛容性を打破する、開示する方法には、(a)抗原特異的免疫反応が望まれる自己抗原をコードするヌクレオチド配列を担う、第一のChAdベクター又はプラスミドベクターを含有する第一のワクチン組成物を投与することにより、自己抗原に対する抗原特異的反応を刺激することと、(b)少なくともゲノムのE1遺伝子の機能的欠失を含有し、そのE1遺伝子部位において配列が、初回刺激段階で送達された同じ自己抗原をコードするDNAの発現を支配することができるプロモーターを含有する、異なる血清型の組み換えChAdベクターを含有する第二のワクチン組成物を投与することにより、(a)の免疫反応を保持し、拡大することとが含まれ、それにより、自己抗原に対する寛容性を打破する効果を有する免疫反応を宿主が備える。
【0110】
したがって、熟練者は、宿主寛容性を打破するための使用に適切であり得るいくつかの異なる免疫付与プロトコールを設計するために、この開示を利用することができる。例えば、最初又は初回免疫付与が、TAAなどの特定の自己抗原をコードするプラスミドDNAを含み、何らかのその後の免疫付与が、ChAdベクターを含む、プロトコールを利用することができる。自己抗原をコードするヌクレオチド配列を含むプラスミドDNA配列は、1又は複数の注射において、電気刺激あり、又はなしで、筋肉内に送達され得る。例えば、エレクトロポレーションを介した、TAAをコードするプラスミドDNAの複数回(例えば、3もしくは4もしくは5)筋肉内注射と、それに続く、同じTAAをコードする導入遺伝子を含有するChAdベクターの1又は複数の筋肉内注射に基づく免疫付与プロトコールは、本明細書中で開示され主張される一般的方法に包含される。
【0111】
あるいは、寛容性を打破するための適切なプロトコールは、自己抗原をコードする導入遺伝子を含有するChAd又はhAdベクターを用いた、1又は複数の初回刺激免疫付与と、それに続く、同じもの又は、初回免疫付与に使用されるベクターと交差反応しないことが知られている、異なるChAdベクターを用いた、1又は複数の追加免疫付与を含み得る。例えば、初回刺激のためにChAd3を用い、追加免疫のためにChAd6を用いる免疫付与プロトコール、又は初回刺激のためにChAd3を用い、続いて追加免疫のためにChAd6及びChAd9を用いる免疫付与プロトコールを、宿主寛容性を打破するために使用し得る。特定の実施形態ににおいて、本発明は、HER2/neu、CEA、EpCaM、PSA、PSMA、テロメラーゼ、gap100、Melan−A/MART−1、Muc−1、NY−ESO−1、サバイビン、ストロメリシン 3、チロシナーゼ、MAGE3、CML68、CML66、OY−TES−1、SSX−2、SART−1、SART−2、SART−3、NY−CO−58、NY−BR−62、hKLP2、VEGFからなる群から選択される、少なくとも1個の腫瘍関連抗原を含有する自己抗原の使用をもくろむ。特定の実施形態において、本発明は、癌に罹患している哺乳動物に、TAAをコードする組み換えチンパンジーアデノウイルスを送達することによる、選択された悪性腫瘍に特異的な腫瘍関連抗原に対する免疫反応(例えば液性又は細胞性)を誘導するための方法を提供する。本発明のこの局面の好ましい実施形態において、誘発される免疫反応は、抗原特異的CD4+及びCD8+T細胞の産生を特徴とする免疫反応で構成される。
【0112】
本発明の免疫原性組成物は、予防的又は治療的設定のいずれかで、哺乳動物宿主、好ましくはヒト宿主に投与することができる。可能性のある宿主/ワクチンには、これらに限定されないが、霊長類、特にヒト及び非ヒト霊長類が含まれ、商業用動物又は家畜として重要なあらゆる非ヒト哺乳動物が含まれる。組み換えチンパンジーアデノウイルスベクターを含有する組成物は、単独で又はその他のウイルス性もしくは非ウイルス性ベースのDNA/タンパク質ワクチン組み合わせて、投与し得る。これらはまた、広範囲の治療計画の一部として投与し得る。本発明の方法での使用に適切な組成物は、緩衝液、生理食塩水もしくはリン酸緩衝生理食塩水、スクロース、その他の塩及びポリソルベートなどの、生理学的に許容可能な成分と組み合わせて、本発明の組み換えウイルスベクターを含有し得る。この組成物は、筋肉内注射時に組織に刺激を与えない。この組成物は、好ましくは、使用時まで凍結される。場合によっては、本発明のワクチン組成物は、これらに限定されないが、アジュバント、安定化剤、pH調整剤又は保存料などの、他の成分を含有するように処方し得る。このような成分は、当業者にとって周知である。
【0113】
本発明の組み換えチンパンジーアデノウイルスが、宿主細胞に形質導入され、受容者/ワクチンに対して治療的利益又は防御的免疫を与える免疫反応を誘発するのに十分な導入遺伝子の発現量を与えるために、選択した投与経路において効果的な組み換えウイルス量である「効果的な量」で、ヒト又は獣医学的宿主に投与されることが想定される。
【0114】
ワクチン受容者に導入されるべき本ワクチン組成物におけるウイルス粒子量は、使用される転写及び翻訳プロモーターの強さ及び発現される遺伝子産物の免疫原性に依存する。一般に、1x107から1x1012粒子(つまり、1x107、2x107、3x107、5x107、1x108、2x108、3x108、5x108又は1x109、2x109、3x109、5x109、)、好ましくは、約1x1010から1x1011粒子の免疫学的又は予防的有効量を、直接筋肉組織に投与する。皮下注射、皮内導入、皮膚を介した押印(impressio)及び腹腔内又は吸入送達などのその他の投与様式ももくろまれる。
【0115】
本発明の組み換えチンパンジーアデノウイルスベクターは、混合様式の初回刺激/追加免疫ワクチン接種計画の一部として、又は様々な血清型ベクターの複数注射に基づくワクチン接種計画において、単独で投与し得る。通常、少なくとも1個の免疫原を含有する初回刺激投与は、特定の病原体又は自己抗原に対する効果的な免疫反応を必要とする哺乳動物宿主に投与される。この投与は、効果的に、その後の抗原の同定において、宿主が免疫原に対してすぐに免疫反応を促進又は増強することができるように、免疫反応を刺激する。初回刺激及び追加免疫に対して代替的処方を利用する、混合様式ワクチン接種スキームの結果、免疫反応が促進される。初回刺激−追加免疫投与は通常、対象を少なくとも1回刺激することと(ウイルスベクター、プラスミド、タンパク質などによる。)、規定の時間の経過後、次に、追加免疫を行うこと(ウイルスベクター、プラスミド、タンパク質などによる。)を含む。複数免疫付与は、通常1回から4回であるが、さらに多く行い得る。初回刺激と追加免疫との間の時間の長さは、通常、約4ヶ月から1年の間で変化し得うるが、当業者が認める場合、他の時間枠を使用し得る。中和免疫が明らかとなる前に、各ベクターの複数回注射を約2週間の時間枠内で投与することができる。
【0116】
本発明のある実施形態において、アデノウイルスワクチンベクターの複数回の投与でワクチンを与え、このウイルスワクチンは、プラスミドワクチンの投与を伴う計画で与え得る。本明細書中で開示するベクターと組み合わせた使用に適切なプラスミドワクチンは、免疫原コード配列及び転写終結因子配列に機能可能に連結された、免疫原をコードする核酸配列の発現を支配することができる異種のプロモーターと組み合わせた、免疫反応の初回刺激又は増強が望ましい、少なくとも1個の免疫原をコードするプラスミドを含む。
【0117】
例えば、組み換え複製欠損チンパンジーアデノウイルスベクターの様々な血清型の複数注射を利用する投与計画を使用することができる。あるいは、プラスミドワクチンの最初の投与(つまり初回刺激投与)及び、該プラスミドワクチンにおいて送達された同じ免疫原のコード配列を含有する複製欠損組み換えチンパンジーアデノウイルスベクターを含有する第二の投与(つまり追加免疫投与)を個体に与え得る。あるいは、その個体に少なくとも1個の免疫原をコードするヒトアデノウイルスワクチンベクターの最初の投与を行い、次いで、本明細書中で開示する複製欠損組み換えチンパンジーアデノウイルスベクター(これは、初回刺激投与において送達された同じ免疫原のコード配列を含有する。)を含有する第二の投与を行い得る。第二の代替的実施形態において、本発明のベクターを含有するワクチン組成物を最初に投与し、次いで、プラスミドワクチン投与を行い得る。これらの実施形態のいずれかにおいて、ウイルスベクター又はプラスミド形態のいずれかで、同じ免疫原の複数回投与を個体に行い得る。これは、規定の最小時間をあけて投与を行い得る。
【0118】
目的の1個のタンパク質又は抗原が、本発明の組み換え複製欠損チンパンジーアデノウイルスベクターにより送達されるのに加えて、別個のビヒクルを介して、又は同じビヒクルを介してのいずれかで、2以上のタンパク質又は抗原を送達することができる。複数のオープンリーディングフレームを含むアデノウイルスプレプラスミドの生成のための、適切なシャトルプラスミドに、複数の遺伝子/機能的同等物を連結することができる。この複数の遺伝子/機能的同等物に対するオープンリーディングフレームを、別個のプロモーター及び転写終結配列に、機能可能に連結することができる。
【0119】
本明細書中で示されるように、適切な免疫付与計画において、様々なアデノウイルス血清型を用いることができる。このようなプロトコールの一例は、第一の血清型(例えばChAd3又はChAd6)の組み換えアデノウイルスベクターを含有する初回刺激投与と、その後の、第二の血清型の組み換えチンパンジーアデノウイルスベクターを含有する追加免疫投与である。代替的な実施形態において、この初回刺激投与は、異なるタンパク質/抗原をコードする遺伝子をそれぞれ含有する、別個のアデノウイルスビヒクルの混合物を含み得る。このような場合、この追加免疫投与は、初回免疫投与で送達された同じ又は類似の抗原セットをコードする遺伝物質を含有する組み換えウイルスベクターを投与する、という条件で、別個のタンパク質/抗原をコードする遺伝子をそれぞれ含有するベクターの混合物を含み得る。これらの複数の遺伝子/ベクター投与様式をさらに組み合わせることができる。特異的抗原からの複数であるが別個の成分を含む、併用様式計画に着手することは、さらに、本発明の範囲内にある。
【0120】
ヒトベクターのウイルスエレメントに対して向けられた既存の免疫により中和されないチンパンジーアデノウイルス由来の組み換えベクターを使用することにより、ワクチン担体としてのヒトAdベクターの使用に対する代替法が得られる。アデノウイルスが高免疫原性であるため、アデノウイルスベクターは、自己抗原に対する宿主寛容性を打破するために設計されたワクチン担体の関連での使用のための特に優れた候補である。さらに、ヒトウイルスと同等の効率での、ヒト細胞、特にPer.C6TM細胞株でのchimp ウイルスの増殖能により、調節の観点及び治療法又はワクチンの大規模産生に対する両方から、顕著な有利性が与えられる。したがって、本発明は、遺伝子ワクチン接種のためのワクチン担体として、単独で、又は組み合わせて、使用し得る、組み換えウイルス調製を可能にする、チンパンジーアデノウイルス配列、ベクター及びプラスミドのコレクションを提供する。
【0121】
本明細書中で言及した公表物は全て、本発明との関連で使用され得る方法及び物質を説明し開示する目的のために、参照により組み込む。明細書のいかなる開示内容も、本発明が先行技術の効力によってこのような開示に対して先行する権利を与えられていないことを認めるものではない。
【0122】
添付の図面を参照しながら本発明の好ましい実施形態を説明してきたが、本発明がまさにその実施形態に限定されず、添付の特許請求の範囲で定義される本発明の範囲又は精神から逸脱することなく、様々な変更及び改変が当業者によりなされ得ることを理解されたい。
【0123】
次の実施例は、本発明を説明するが、限定するものではない。
【実施例1】
【0124】
ChAdの単離、クローニング、配列決定及び特性決定
チンパンジーアデノウイルスの単離
ウイルス輸送培地(VTM; Microtest M4−R Multi−Microbe Transport Medium, Remel Inc.)において便検体を回収し、次にNIRC(New Iberia Research Center 4401 W. Admiral Doyle Drive New Iberia, LA 70560)で−70℃にて凍結又は直接凍結した。この検体を細胞培養物への接種のために処理するまで<−70℃で凍結保存した。この検体を細胞培養物へ接種する際、この検体を解凍し、次に過剰の氷冷ウイルス輸送培地中でボルテックスした。前記検体を懸濁液へ入れて解離させた後、それらを1500から1800rpmで10分間遠心分離した。上清を0.8μmシリンジフィルター及び0.2μmシリンジフィルターを通じて連続して濾過し、次にこの濾過した材料を細胞培養物へ接種した(シェルバイアルへ200から250μl、及び試験管培養物へ250から300μl)。処理した各検体を293細胞もしくはA549細胞を播種した試験管培養物及びシェルバイアル培養物へと接種した。
【0125】
対照(ポジティブ及びネガティブ)培養物を、1セットの試料を接種する各場合に調製した。設定におけるシェルバイアルに全て接種したら、それらを室温にて60±10分間、2000rpm(900xg)で遠心分離した。培養物中の熱損傷を防止するため、ローターの回転が停止した直後に前記バイアルを遠心分離機から取り出した。遠心分離後、交差汚染を防止するため、各バイアルにおいて新鮮な滅菌済みパスツールピペットを使用して、接種材料を前記シェルバイアルから吸引した。各洗浄につき1.0mlの新鮮な培地を使用して、前記培養物を3回洗浄した。3回目の洗浄の後、新鮮な培地(1.0ml)を各バイアルへピペットを介して添加し、前記シェルバイアルを35から37℃のインキュベーターの中で3日から4日間(約96時間)静置した。
【0126】
培養期間の終了時に、上清を前記培養物から吸引し、各洗浄に対して約1.0mlの緩衝液を使用して、免疫蛍光アッセイ(IFA)緩衝液で各バイアルにおける細胞層を2回洗浄した。1.0mLの冷アセトンを各バイアルへ添加する(2から8℃で10分間)ことにより、細胞を固定した。アセトン洗浄したスライドを、シェルバイアルカバースリップと関連付けた検体認識番号で標識した。このシェルバイアルカバースリップを、一次マウス抗アデノウイルス抗体[MAB8052、Chemicon]を使用してアデノウイルス感染細胞の蛍光標識用に処理した。蛍光顕微鏡を使用して、スライドを評価する。ウェル(1+から4+)にわたる免疫蛍光がカバーする範囲に留意して、10倍の対物レンズを使用して各調製物をスキャンした。40倍の対物レンズを使用して特異的免疫蛍光の有無を確認した。前記シェルバイアルについて記載したものと同じ手順で、試験管培養物に接種した(例えば、ネガティブ対照を最初に処理し、次いで、臨床検体及びポジティブ対照)。この接種材料を36から38℃にて60分から120分間吸着させた。吸着時間後、この検体/対照を試験管から吸引し、新鮮な培地と交換した。
【0127】
接種から3日から4日後、及び、その後週に1回、この培養試験管から培地を吸引し、1.5mlの新鮮な培地と交換した。培養試験管をCPEについて、接種後少なくとも21日間、少なくとも1日おきに目で見て観察した。チンパンジー検体で接種した培養物を対照と比較し、CPEの程度を観察することによって評価した。14日後に、CPEを示さない培養物を新鮮な試験管培養物へ通し、21日後にCPEに対して陰性である培養試験管を陰性とみなした。3+から4+のCPEの培養試験管を10秒間ボルテックスした。細胞を試験管壁から、滅菌済みの1.0mL血清学用ピペットを使用して回収し、培養上清中で懸濁した。5mLのスナップキャップ試験管に検体認識番号及び日付を付した後、−70℃で保存した。この細胞懸濁液 500μlを、培養試験管からスナップキャップ試験管へ移し、アデノウイルスを検出するための間接免疫蛍光抗体技術(シェルバイアルを染色するための手段と同等)を使用して処理するまで、2℃から8℃で最長1日間保存した。
【0128】
チンパンジーアデノウイルス増幅
ATCC又はEsoterix Inc.,. Austin, Texasから購入した野生型チンパンジーアデノウイルスCV32、CV33、CV23及びCV68(ATCC受託番号:CV32、VR−592;CV33、VR−593;)及びオリジナルの分離株を、ヒトE1発現細胞株PER.C6TM又は293を使用することによって以下のように増殖させた。簡潔には、10% ウシ胎児血清(FBS GibcoBRL, Life Technologies)、1% ペニシリン−ストレプトマイシン、2mM グルタミン及び10mM MgCl2 (Per.C6TM)を添加したダルベッコ改変イーグル培地(DMEM;GibcoBRL, Life Technologies)中で細胞を培養した。5% ウマ血清(GibcoBRL, Life Technologies)を添加したDMEM中でアデノウイルス感染を行った。細胞の100%がウイルスにより誘導される細胞病理学的効果(CPE)を呈したら、感染した細胞及び培地を回収し、凍結融解を3回繰り返して細胞溶解させた。
【0129】
野生型チンパンジーアデノウイルス(CV)ストックを全て、増幅の第一継代後に96穴プレートに播種した293細胞を感染させることによってクローニングした。ウイルスクローニングは、ウイルス増幅の第一継代で得られる細胞溶解液の希釈を制限することによって行った。5個の単離されたクローンを拾い、連続して増殖させた。増幅の3回から4回の連続した継代の後、5つの2層細胞ファクトリー(NUNC)上に播種した細胞において、アデノウイルスの大規模調製を行った(1つの細胞ファクトリーにつき2億個の細胞)。塩化セシウム密度勾配における超遠心段階を2回行い、細胞溶解液から、精製したウイルス粒子を得た。
【0130】
ウイルスゲノムDNAの配列決定
1%SDS−TEN中のプロテイナーゼK(0.5mg/ml)で(55℃で2時間)消化することにより、精製ウイルス調製物 3x1012ppから、ゲノムDNAを単離した。フェノール−クロロホルム抽出ならびにエタノール沈殿の後、このゲノムDNAを水中で再懸濁し、ゲノム配列決定に供した。
【0131】
全長のAdゲノム配列決定のため、精製ウイルスDNAを霧状にし、無作為にせん断された断片を得た。E.コリDNAポリメラーゼ及びポリヌクレオチドキナーゼのKlenow断片を用いて、このDNA断片を平滑末端化にした。平滑末端断片を低融点アガロースゲル電気泳動に供し、1から3kbの範囲の大きさの断片を精製し、pUC19ベクターのSmaI部位へとクローニングし、ショットガンライブラリを作製した。ライゲーションを使用してコンピテントXL1−Blue MRF’を形質転換した。X−gal及びIPTGを含有するLB寒天上での白色/青色スクリーニングにより陽性コロニーを同定した。プラスミドDNAの3個から4個の96穴ブロックをライブラリから単離し、pUC順方向プライマー及び逆方向プライマーを用いて配列決定した。配列決定の読み取りは全て、Phred−Phrapソフトウェアパッケージを使用して性質及びベクター配列についてスクリーニングした。このスクリーニングを通過した読み取りをコンティグに集めた。プライマーは、ギャップを閉鎖し、両端のDNA配列を決定するために、アデノウイルスDNAを直接配列決定するよう設計した。
【0132】
ChAd3(配列番号1)、ChAd6(配列番号2)、CV32(配列番号3)、CV33(配列番号4)、及びCV23(配列番号5)を含む選択されたウイルスについて完全なウイルスゲノム配列決定が行われた。表1は、ChAd3、ChAd6、Pan5(CV23)、Pan6(CV32)、Pan7(CV33)、C1及びC68アデノウイルスゲノムのヌクレオチド配列間の同一性の割合を要約するデータを提供する。アラインメントは、FASTAパッケージ第2版の一部としてのALIGNプログラムを使用して計算した(William R. Penson, University of Virginia; Myers & Miller, CABIOS 1989, 4:11−17)。
【0133】
【表1】
【0134】
新規のアデノウイルス分離株(例えば、ChAd20、ChAd4、ChAd5、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82)の特徴を調べるために、ヘキソン及び繊維遺伝子のヌクレオチド配列もプライマーウォーキングにより決定した。繊維遺伝子は、配列番号6から15(配列番号6、ChAd20;(配列番号7、ChAd4);配列番号8、ChAd5);配列番号9、ChAd7);配列番号10、ChAd9);配列番号11、ChAd10);配列番号12、ChAd11);配列番号13、ChAd16);配列番号14、ChAd17);配列番号15、ChAd19)及び(配列番号58、ChAd8)、(配列番号60、ChAd22)、(配列番号62、ChAd24)、(配列番号64、ChAd26)、(配列番号66、ChAd30)、(配列番号68、ChAd31)、(配列番号70、ChAd37)、(配列番号72、ChAd38)、(配列番号74、ChAd44)、(配列番号76、ChAd63)及び(配列番号78、ChAd82)である。図20Aから20Gは本明細書中で開示され主張される前記ChAd分離株の繊維タンパク質のアミノ酸配列の比較を与える。
【0135】
ヘキソン遺伝子配列は、配列番号16から25(配列番号16、ChAd20);配列番号17、ChAd4);配列番号18、ChAd5);配列番号19、ChAd7);配列番号20、ChAd9);配列番号21、ChAd10);配列番号22、ChAd11);配列番号23、ChAd16);配列番号24、ChAd17);配列番号25、ChAd19)、(配列番号97、ChAd8)、(配列番号99、ChAd22)、(配列番号101、ChAd24)、(配列番号103、ChAd26)、(配列番号105、ChAd30)、(配列番号107、ChAd31)、(配列番号109、ChAd37)、(配列番号111、ChAd38)、(配列番号113、ChAd44)、(配列番号115、ChAd63)及び(配列番号117、ChAd82)である。図31Aから31Jは、本明細書に開示され主張されるChAd分離株のヘキソンタンパク質のアミノ酸配列の比較を与える。
【0136】
チンパンジーアデノウイルスの分類
異なるチンパンジーアデノウイルス株の分類は、6個のサブグループへのヒトアデノウイルス血清型のすでに提唱されている分類(Horowitz, MS (1990) Adenoviridae and their replication.Virology B. N. Fields及びD.M.Knipe,編(Raven Press, New York)pp.1679−1740)に従い、Align Xプログラム(Informax, Inc)を使用することによるアミノ酸及びヌクレオチド配列アラインメントにより得た。
【0137】
新たな分離株の最初の分類は、異なる制限エンドヌクレアーゼによるウイルスゲノムの制限パターンを観察すること及びヘキソン遺伝子の超可変領域7(HVR7)の配列分析により得られた。このために、HVR7に隣接する高度に保存された領域に関して2つのプライマー:TGTCCTACCARCTCTTGCTTGA(配列番号45)及びGTGGAARGGCACGTAGCG(配列番号46)を設計した。精製ウイルスDNA又は未精製293細胞溶解液をテンプレートとして使用するPCRによりこのHVR7を増幅した後配列決定した。HVR7配列分析に基づいて、我々は、新たに単離されたウイルスを、ヒトAdウイルスについて提唱されるサブグループ(A〜F)へ分類した(Horowitz,MS(1990)Adenoviridae and their replication.Virology B.N.Fields and D.M.Knipe編(raven Press, New York)pp.1679−1740)。
【0138】
図35に示される系統発生樹はヒト及びチンパンジーのアデノウイルスへキソンアミノ酸配列のアラインメントにより得られた。この結果は、Align Xプログラム(Informax,Inc)を使用することによるヘキソンHVR7に限定したヌクレオチド配列アラインメントに基づく最初の分類と一致する。この系統樹は、PAUPSEARCH(Wisconsin Package Version 10.3,Accelrys Inc.)を使用する全長のヘキソンペプチド配列の多配列アラインメントから推定し、TREEVIEWにより可視化を行い、操作した。Wisconsin Packageにおいて実装されるPAUPSEARCHプログラムを使用してブートストラップ信頼分析を実行した。各アラインメントについて、検索基準として「Heuristic Search」を用い、最適性基準としてMaximum Parsimonyを用いて、前記プログラムを1000個の複製物に関して実行し、報告する信頼値は、50%多数決コンセンサス(50% majority−rule consensus)から得た。
【実施例2】
【0139】
ChAdシャトルベクター及び発現ベクターの構築及びレスキュー
ベクター構築及びレスキュー
ウイルスゲノムの左右両端由来のウイルスDNA配列を含有する適切なシャトルベクターを用いて、相同組み換えにより、ゲノムウイルスDNAを標準プラスミドベクターにクローニングした(図2)。下記でさらに詳しく述べるように、グループ特異的シャトルベクターを開発するために、同一血清型サブグループに分類されるウイルス間で観察される配列相同性を利用した。サブグループD及びEへ分類されるチンパンジーアデノウイルスのゲノムウイルスDNAは、両サブグループに属するウイルスをクローニングするための共通のシャトルベクターの構築が可能となるのに十分な相同性を生じた。
【0140】
サブグループD/Eシャトルベクターの構築
ChAd6ウイルスゲノムを完全に配列決定し(配列番号2)、得られた情報を使用して、サブグループD及びEのチンパンジーアデノウイルスの相同組み換えによるクローニングを促進するためにシャトルベクターを構築した。
【0141】
本明細書でpARS ChAd6−3と呼ばれるChAd6シャトルベクターの構築を図1に記載する。図32は、本明細書に記載されるクローニング実験で使用されるオリゴヌクレオチド配列(配列番号26から40及び配列番号45から46)のリストを提供する。簡潔に述べると、オリゴヌクレオチド、5’−ATGGAATTCGTTTAAACCATCATCAATAATATACCTC−3(配列番号27)及び5’−CGCTGGCACTCAAGAGTGGCCTC−3’(配列番号28)を使用して、PCRによりChAd6DNAの左端由来の457bpを増幅し、EcoRI及びSnaBIで消化し、EcoRI−SnaBIで切断しておいたpNEBAd35−2へクローニングし、pNEBChAd6−LIを得た。右のChAd6 ITR(bp36222からbp36648)を、PCRにより、オリゴヌクレオチド5’−ATGAAGCTTGTTTAAACCCATCATCAATAATATACCT−3’(配列番号29)及び5’−ATCTAGACAGCGTCCATAGCTTACCG−3’(配列番号30)を用いて増幅し、制限酵素HindIII及びXbaIで消化し、HindIII−XbaIで切断しておいたpNEBChAd6-LIへクローニングし、その結果、pNEBChAd6−RLIを得た。最後に、ChAd6ゲノムDNA配列のヌクレオチド3426から3813に対応するDNA断片を、オリゴヌクレオチド5’ATGCTACGTAGCGATCGCGTGAGTAGTGTTTGGGGGTGGGTGGG−3’(配列番号31)及び5’−TAGGCGCGCCGCTTCTCCTCGTTCAGGCTGGCG−3’(配列番号32)を用いて増幅し、SnaBI及びAscIで消化した後にSnaBI−AscIで消化しておいたpNEBChAd6−RLIにライゲーションし、その結果、pNEBChAd6−RLIdE1を得た。
【0142】
DH5a E.コリ株における組み換え及びプラスミド増幅効率を向上させるため、ChAd6の左右両方のITRを含有する1306bp断片及びpIX遺伝子断片を、pNEBChAd6−RLIdE1から、PmeI消化により切り出し、pMRKAd5SEAPからの、オリゴヌクレオチド5’−GATCTAGTTAGTTTAAACGAATTCGGATCTGCGACGCG−3’(配列番号33)及び5’TTCGATCATGTTTAAACGAAATTAAGAATTCGGATCC−3’(配列番号34)を使用するPCR増幅により得られた異なるプラスミドベクターへ移した。この最後のライゲーション段階により、ChAd6シャトルベクター pARSChAd6−3が生じた。
【0143】
サブグループCシャトルベクターの構築
ChAd3ウイルスゲノムを完全に配列決定し(配列番号1)、得られた情報を使用して、サブグループCチンパンジーアデノウイルスの相同組み換えによるクローニングを促進するためにシャトルベクターを構築した。
【0144】
簡潔には、本明細書でpChAd3EGFPと呼ばれるサブグループCチンパンジーアデノウイルスをクローニングするのに使用されるシャトルベクターを以下のように構築した。pIXコード化領域を含有するChAd3DNA断片(ヌクレオチド3542から4105)を、オリゴヌクレオチド5’−TATTCTGCGATCGCTGAGGTGGGTGAGTGGGCG−3’(配列番号35)及び5’−TAGGCGCGCCCTTAAACGGCATTTGTGGGAG−3’(配列番号36)を使用するPCRにより増幅し、SgfI−AscIにより消化し、その後SgfI−AscIで消化したpARSCV32−3へとクローニングし、pARS−ChAd3Dを得た。ChAd3右端(nt37320から37441)を、オリゴヌクレオチド5’−CGTCTAGAAGACCCGAGTCTTACCAGT−3’(配列番号37)及び5’−CGGGATCCGTTTAAACCATCATCAATAATATACCTTATT−3’(配列番号38)を用いたPCRにより増幅し、XbaI及びBamHIで消化した後、XbaI及びBamHIで制限酵素処理したpARS−ChAd3Dへライゲーションし、pARS−ChAd3RDを得た。ChAd3ウイルスDNA左端(nt1から460)を、オリゴヌクレオチド5’−ATGGAATTCGTTTAAACCATCATCAATAATATACCTT−3’(配列番号39)及び5’−ATGACGCGATCGCTGATATCCTATAATAATAAAACGCAGACTTTG−3’(配列番号40)を用いたPCRにより増幅し、EcoRI及びSgfIで消化した後、EcoRI及びSgfIで切断したpARS−ChAd3RDへクローニングし、その結果、pARS−ChAd3RLDを得た。このウイルスDNAカセットはまた、酵素切断によってプラスミドDNAからウイルスDNAが放出されるよう、両ITRの端にある制限酵素部位(PmeI)を含有するように設計された。
【0145】
サブグループBシャトルベクターの構築
サブグループBシャトルの構築は、サブグループC及びD/Eシャトルの構築についてすでに記載したストラテジーに従った。簡潔には、pARS−ChAd3RLDが左端、pIX領域、右端を、ChAd30の対応する断片と置換することによって改変した。さらに、ChAd30のE4領域を、ChAd30E4プロモーター調節の下でクローニングされるAd5E4 orf6と置換した。このシャトルプラスミドを、pChAd30 EGFPシャトルベクターと命名した。
【0146】
ΔE1チンパンジーアデノウイルスベクターの構築
サブグループB:サブグループBのチンパンジーアデノウイルスベクターをE.コリ株BJ5183における相同組み換えにより構築した。BJ5183細胞に対して、BstEII及びBst1107Iで消化したpChAd30EGFPシャトルベクターと、ChAd8及びChAd30の精製ウイルスDNAとを用いて、同時形質変換した。pIX遺伝子、直線化したpChAd30EGFPシャトルの両端に存在する右ITR DNA配列及びウイルスゲノムDNAの間の相同組み換えにより、プラスミドベクターにおける挿入が可能になり、EGFP発現カセットにより置換されるE1領域が同時に欠失した。ヒトサイトメガロウイルス(HCMV)プロモーター及びウシ成長ホルモンポリアデニル化シグナル(BghポリA)に基づいた発現カセットを構築して、分泌されるアルカリフォスファターゼ(SEAP)、EGFP、HIVgag、HCV NS領域(ChAd6シャトルベクターについて図3に記載されるようなもの。)、並びにヒト及びアカゲザル由来のCEA及びHER2 neuのような腫瘍関連抗原を発現させた。全ての発現カセットを相同組み換えによりChAd30ベクターへと挿入した。
【0147】
サブグループC:サブグループCのチンバンジーアデノウイルスベクターをE.コリ株BJ5183における相同組み換えにより構築した。BstEII及びBst1107Iで切断したpChAd3EGFPシャトルベクターと、ChAd3、ChAd11、ChAd19及びChAd20の精製ウイルスDNAと、を用いて、BJ5183細胞に対して同時形質変換を行った。pIX遺伝子、直線化したpChAd3EGFPシャトルの端に存在する右ITR DNA配列と、ウイルスゲノムDNAとの間の相同組み換えにより、プラスミドベクターにおける挿入が可能になり、EGFP発現カセットにより置換されるE1領域が同時に欠失した。ヒトサイトメガロウイルス(HCMV)プロモーター及びウシ成長ホルモンポリアデニル化シグナル(BghポリA)に基づいた発現カセットを構築して、分泌されるアルカリフォスファターゼ(SEAP)、EGFP、HIVgag、HCV NS領域(ChAd6シャトルベクターに対して図3に記載されるようなもの。)、並びにヒト及びアカゲザル由来のCEA及びHER2 neuのような腫瘍関連抗原を発現させた。
【0148】
サブグループD及びE:サブグループD及びEのチンパンジーアデノウイルスに基づいたΔE1ベクターを構築するため、シャトルベクターpARS ChAd6−3をAscIで消化し、E.コリ株 BJ5183に、CV32、CV33、CV68、ChAd4、ChAd5、ChAd6、ChAd7、ChAd9、ChAd10及びChAd16の精製ウイルスDNAを用いて同時形質転換した。pIX遺伝子及び直線化したpChAd6−3の端に存在する右ITR DNA配列と、ウイルスゲノムDNAとの間の相同組み換えにより、プラスミドベクターにおける挿入が可能となり、E1領域が同時に欠失した。(図2及び図4)。ヒトサイトメガロウイルス(HCMV)プロモーター及びウシ成長ホルモンポリアデニル化シグナル(BghポリA)に基づいた発現カセットを構築して、分泌されるアルカリフォスファターゼ(SEAP)、EGFP、HIVgag、HCV NS領域(図3)、並びにヒト及びアカゲザル由来のCEA及びHER2 neuのような腫瘍関連抗原を発現させた。図4に記載されるようなΔE1アデノウイルスプレプラスミドへの相同組み換えによって転移されるpARS ChAd6−3ベクターの1個のSnaBI部位へ全発現カセットを挿入した。
【0149】
ΔE1ベクターのレスキュー及び増幅
6cmの細胞培養皿に播種された5x106個のPER.C6TM細胞を、エンドヌクレアーゼ消化によりプラスミド配列から放出されるクローン化されたウイルスベクター10μgでトランスフェクトションした。Lipofectamine(Invitrogen)を使用してDNAトランスフェクションを行った。トランスフェクションした細胞及び培地を、トランスフェクションから5から10日後に回収し、凍結解凍により溶解した。次に、レスキューしたベクターを293細胞又はPER.C6TM細胞での連続継代により増幅した。総計1から2x109個の細胞において、5から10個の細胞ファクトリー(NUNC,Inc.)上に播種した細胞を感染させることによって大規模増幅を行った。塩化セシウム勾配上での2回の超遠心により、精製ベクター調製物を得て、10%グリセロールを含有するPBSに対して透析し、分注して−70℃にて保存した。
【実施例3】
【0150】
中和実験
本明細書で開示されるチンパンジーアデノウイルスに対する中和抗体のヒト血清における普及率を評価するため、中和アッセイを実行した。このアッセイにより、分泌型アルカリフォスファターゼ(SEAP)に対する遺伝子を担うチンパンジーアデノウイルスがヒト293細胞に対して形質導入する能力に及ぼす血清プレインキュベーションの効果を評価した。中和タイターは、ウイルスのみのポジティブ対照において観察されるSEAP活性の50%の低下を与える血清の希釈度として定義される。
【0151】
CV33−SEAP、CV32−SEAP及びChAd3−SEAPの2x106個から1.5x107個の自然粒子を完全培地100μlに希釈し、完全培地に希釈したヒト又はチンパンジー血清等量に添加した。各血清試料を様々な希釈率で試験した(1/18希釈から開始して、1/4希釈を5回行い、1:4608まで。)。試料を37℃で1時間プレインキュベーションし、その後96穴プレートへ播種した293細胞(1個のウェルあたり3x104個の細胞)へ添加した。インキュベーションの1時間後に接種材料を除去し、細胞に新鮮培地を再供給し、24時間後、培地50μlを取り出し、SEAP活性を化学発光アッセイにより測定した。ウイルスのみのポジティブ対照において観察されるSEAP活性の50%低下を与える血清の希釈度として、中和タイターを定義する。100検体のヒト血清のパネルをChAd中和活性について検査した。並行して同じパネルをAd5 SEAPベクターに関して検査した。
【0152】
【表2】
【0153】
表2で与える結果は、チンパンジーアデノウイルス由来のベクターに対して向けられた、中和抗体のヒト血清における普及率が非常に低いことを示す。CV32ベクターにおいて200の閾値を超えるタイターを示すものは、4検体の血清のみであるのに対し、ChAd3 SEAPベクターについては、8検体が200を超えるタイターを示した。一方、検査したチンパンジー血清のパネルは、抗チンパンジーAd免疫性の普及率が非常に高いことを示した。これらの知見から、予想したように、チンパンジーAdに基づくベクターがヒトにおいて中和される機会が非常に少ないことが確認された。それゆえこれらの知見は、Ad5などの共通のヒトAd血清型に基づいたウイルスベクターの投与を制限する、既存の抗ヒトAd免疫性の問題に対する理想的な解決を呈示する。
【実施例4】
【0154】
ChAdベクター親和性
Ad5ベクター及びChAdベクターを介した遺伝子導入の効率を、異なる組織由来のヒト初代細胞のパネルにおけるEGFP発現により評価した。ヒト軟骨細胞、骨芽細胞、ケラチン生成細胞、メラニン細胞、骨格筋細胞及びメラニン細胞を製造者の説明書に従って培養した。ヒト単球細胞、未成熟及び成熟樹状細胞(DC)を記載されているように得た(Romani,N.ら、1996,J.Immunol.Methods,196,137.)。形質導入された蛍光細胞をFACS分析により検出した。試験するヒト初代細胞のパネルには、心臓循環器疾患、関節リウマチ、組織工学(骨、皮膚及び軟骨)及びワクチン接種の分野におけるインビボ及びエクスビボでの遺伝子導入を利用した異なる治療ストラテジーに対する重要な標的細胞である細胞が含まれる。図38Aから図38Dで示す結果は、異なる細胞タイプの感染の差異に基づく効率により示されるように、CARに代わる受容体を、異なるチンパンジーアデノウイルスが認識できることが示唆される。
【0155】
マウス免疫付与実験
方法及び材料
免疫付与プロトコール及び脾臓細胞/PBMC調製
免疫付与:緩衝液 0.1mLで希釈した、選択したアデノウイルスを用いてマウスに免疫付与した。各ベクター用量を50μlずつ2つに分割し、マウスの両側の大腿四頭筋に注射した。
【0156】
脾臓細胞調製:マウスを注射3週間後に屠殺し、それらの脾臓を摘出し、R10(10%FCS、55mM 2−メルカプトエタノール、1M HEPES緩衝液、2mM L−グルタミン、RPMI培地1640中の1xペニシリン−ストレプトマイシン溶液) 10mLに移した。スチール製の網を介して脾臓を細分化し、R10 2mLでこの網を洗浄した後、脾臓細胞を50mLのファルコンチューブに移し、1200rpmで10分間室温(rt)にて遠心分離した。上清を除去し、ACK溶解緩衝液(Gibco BRL Formulation#79−0422DG)3mLを添加した。細胞を5分間rtでインキュベーションした。1xPBS 45mlを添加し、試験管を上述のように遠心分離した。R10 30mLで洗浄した後、細胞をR10 5mL中で再懸濁し、70mナイロン細胞濾過器(Falcon2350)を介して濾過した。細胞 10μLをTurk溶液(Merck 040417345)990μLで希釈し、計数した。最終的に、細胞をR10中で、107個細胞/mLに希釈した。
【0157】
末梢血単核球(PBMC)調製:マウス血液試料(150μL)を2mLのエッペンドルフチューブへ50μL PBS/2%EDTAとともに移した。1mL ACK緩衝液を各チューブへ添加した。RTで5分間穏やかに混合し、インキュベーションした。試料を微量遠心器において1500rpmで5分間遠心分離した。上清を除去し、同じ免疫付与コホート由来の白血球ペレットを合わせた。ACK緩衝液インキュベーションを繰り返した後、PBMCのペレットを、R10培地 1mLに再懸濁した。
【0158】
IFN−γELISPOTアッセイ
Millipore MAIP45プレートを、PBSで2.5μg/mLに希釈した、100μl/ウェルの精製ラット抗マウスIFN−γモノクローナル抗体(Pharmingen、cat.551216)でコーティングし、4℃で一晩(o/n)インキュベーションした。プレートを滅菌済みPBSで2回洗浄し、CO2インキュベーターの中でR10 200μl/ウェルとともに2時間インキュベーションすることにより、非特異的結合部位をブロックした。HIVgagを発現するAdベクターによる免疫付与実験において、9マーのペプチド(AMQMLKETI、Balb/CマウスにおいてマッピングされるCD8HIVGAGエピトープ)(配列番号47)をR10中で2μg/mLに希釈し、50μl/ウェルの量でウェルに添加した。HCV−NS発現ベクターを用いて行った免疫付与実験において、NS3ヘリカーゼドメインをカバーするペプチドのプールならびにヘリカーゼドメインに含まれるマッピングされたCD8エピトープを呈示する9マーのペプチドを使用した。ヒトCEA抗原を発現するChAdによる免疫化実験を、全アミノ酸配列をカバーする、重複する15マーペプチドのプールにより評価した。対照としてDMSO及びコンカナバリンAを使用した。細胞を各ウェルへ5x105及び2.5x105の量で添加した。CO2インキュベーターにおいて一晩インキュベーションした後、プレートを0.05% Tween20/PBSで洗浄し、アッセイ緩衝液(5%FBS、0.005% Tween20、PBS)で1/250に希釈したビオチン化ラット抗マウスIFN−γモノクローナル抗体(PharMingen、Cat.554410) 50μl/ウェルを添加した。プレートを4℃で一晩インキュベーションし、上述のように洗浄した。ストレプトアビジン−アルカリフォスファターゼ複合体(BD554065)をアッセイ緩衝液で1/2500に希釈し、50μL/ウェルの量で室温にて2時間添加した。洗浄後、BCIP/NBT 1ステップ溶液(PiercE34042)50μL/ウェルを添加してプレートを発色させた。ウェルを脱イオン水で洗浄することにより反応を停止させた。スポットをELISPOTリーダーにより自動計数した。
【0159】
マウス IFN−γ細胞内染色(ICS)
脾臓細胞をR10 1mL中2x106個細胞に希釈し、2μg/mLの濃度の上述と同じ抗原で刺激した。対照として、DMSO及びブドウ球菌エンテロトキシンB(SEB)を使用した。CO2インキュベーター中で一晩インキュベーションした後、細胞をFACS緩衝液(1%FCS、0.01%NaN3、PBS)で洗浄し、精製した抗マウスCD16/CD32Fcブロック(クローン2.4G2、Pharmingen、cat.553142)を1/25に希釈し、100μL/試料の量で添加し、4℃で15分間インキュベーションした。細胞をFACS緩衝液で洗浄し、FACS緩衝液で1:50に希釈したAPC標識抗マウスCD3e(クローン145−2C11、Pharmingen#553066)、PE標識抗マウスCD4(クローンL3T4、BD Pharmingen、cat.553142)及びPerCP標識抗マウスCD8a(クローン53−6.7、Pharmingen、cat.553036)を100μL/試料の量で添加した。細胞をrtで30分間インキュベーションし、洗浄し、固定し、浸透させ(Becton Dickinson、FACS Perm2)、PermWash(100μL/試料)中で1:50に希釈したFITC標識抗マウスIFN−γ(Pharmingen、cat.554411)とともにRTにて30分間インキュベーションした。洗浄後、細胞を1% ホルムアルデヒド/PBS 500μLに再懸濁し、CellQuestソフトウェア(Becton Dickinson)を用いて細胞内サイトカイン染色(ICS)をFACS−Caliburフローサイトメーターで分析した。
【実施例5】
【0160】
ChAdベクターはマウスにおいて強いCMI反応を誘発する
本明細書に開示されるChAdベクターが細胞性免疫反応(CMI)を誘発する能力を、HIVgag導入遺伝子を発現するベクターを使用してマウスにおいて評価した。簡潔に述べると、5匹のBalb/Cマウスの群に、様々なベクターを、マウス1匹あたり105から1010vpの、10倍ずつ増加する用量で注射した。
【0161】
注射から3週間後に、脾臓細胞中のgag特異的CD8+T細胞を定量することにより、免疫反応の強さを調べた。Balb/CマウスにおいてマッピングされるgagCD8+T細胞エピトープを再現するペプチドによるインビトロ刺激後に、ELISPOTアッセイによって又はIFN−γ細胞内染色及びFACS分析によって、IFN−γ分泌CD8+T細胞の数を調べた。
【0162】
表3で報告する5匹の免疫付与動物から得られた結果は、9マーのCD8+gagエピトープ又はgagペプチドプールとのインキュベーション後の、脾臓細胞100万個あたりのスポット形成細胞数として示す。gagペプチドプールは、全gag配列を包含する10−aaまで重複する20−aaペプチドからなる。ポジティブの値を太字で示す。
【0163】
表3で与えるデータから、本明細書に開示され主張されるChAdベクターの投与により、hAd5によって誘発される反応に匹敵する強い細胞性免疫反応が誘発されることが示される。ポジティブの免疫付与結果(免疫付与ブレイクポイント)を生じる最低のベクター用量で観察することによって、発明者らは、106ppベクター用量でのブレイクポイントを有するもっとも有力なサブグループCのChAd3gagである、様々なベクターの効力を評価した。免疫付与ブレイクポイントによる評価を図33に示す。
【0164】
【表3】
【実施例6】
【0165】
ChAd3ベクター及びCV33GAGベクターはGAG特異的CD8+T細胞を特徴とするCMI反応を誘発する。
【0166】
HIVgag導入遺伝子を含むChAdベクターに反応して誘発されるCMI反応の特徴を調べるために、様々な用量のベクターで免疫付与した5匹のマウスのコホートからプールされた脾臓細胞を細胞内IFN−γ染色により分析した。表3及び表4に示されるデータは個別の実験において回収された。
【0167】
脾臓細胞を R10 1mLで2x106個細胞に希釈し、2μg/mLの濃度で上述と同じ抗原で刺激した。対照として、DMSO及びSEB(ブドウ球菌エンテロトキシンB)を使用した。CO2インキュベーターで一晩インキュベーションした後、細胞をFACS緩衝液(1%FCS、0.01%NaN3、PBS)で洗浄し、精製抗マウスCD16/CD32Fcブロック(クローン2.4G2、Pharmingen、cat.553142)を1/25に希釈し、100μL/試料の量で添加し、4℃で15分間インキュベーションした。細胞をFACS緩衝液で洗浄し、FACS緩衝液中で1:50希釈した、APC標識抗マウスCD3e(クローン145−2C11、Pharmingen、cat.553066)、PE標識抗マウスCD4(クローンL3T4、BD Pharmingen、cat.553142)及びPerCP標識抗マウスCD8a(クローン53−6.7、Pharmingen、cat.553036)を100μL/試料の量で添加した。細胞を室温で30分間インキュベーションし、洗浄し、固定し、浸透させ(Becton Dickson、FACS Perm2)、PermWash(100μl/試料)で1:50に希釈したFITC標識抗マウスIFN−γ(Pharmingen、cat.554411)とともにRTで30分間インキュベーションした。洗浄後、細胞を1%ホルムアルデヒド/PBS 500μLに再懸濁し、CellQuestソフトウェア(Becton Dickinson)を用いてFACS−Caliburフローサイトメーターで分析した。
【0168】
表4は、gag特異的CD8+T細胞又はCD4+T細胞のいずれかであるgag特異的CD3+T細胞の割合を要約するデータを与える。ポジティブの結果を太字で示す。本明細書で与えるデータから、様々な血清型サブグループ(つまりサブグループC、D及びE)へと分類されるウイルス由来のChAdベクターにより誘発される免疫反応の細胞特性が同様であり、gag特異的反応は全て、CD8+T細胞を主に特徴とすることが示される。さらに、多量のベクターでgag特異的CD4+反応が全ての免疫付与実験で明らかになることを留意すること。ICSアッセイにより、ChAd3ベクターが106個のベクター用量で抗gagCD8+反応を刺激できることが確認された。
【0169】
【表4】
【実施例7】
【0170】
ChAdベクターは、HCV NS特異的T細胞反応を誘発する。
【0171】
CV32−NSmutベクター及びCV33−NSmutベクターの効力をMRKAd6NSmutの効力に対して、C57/Black6において評価した。107から109vp/マウスの、10倍ずつ増加する量のベクターを動物に注射した。単回注射の3週間後に、NS3タンパク質のヘリカーゼドメインにおいてマッピングされるCD8+T細胞エピトープを再現する9マーペプチドでT細胞を刺激して、IFN−γELISPOT及びIFN−γ細胞内染色により、CMIを分析した。表5で与えるデータは、NS3 9マーペプチド非存在下(偽)又は存在下におけるインキュベーション後の、100万個の脾臓細胞あたりのスポット形成細胞数を要約する。
【0172】
HCV−NSを発現するCV32ベクター及びCV33ベクターの両者は強いT細胞反応を刺激することが本データから示される。CV32ベクターに対する最初の陽性結果が109vp/用量を注射することにより得られたという観察に基づき、CV32DE1E3 NSmutベクターの免疫付与効力は、ヒトサブグループCのAd6DE1E3 NSmutベクターよりも、約1/100と、低いと思われる。MRKAd6NSmutを使用する並行した実験から、107vp/動物の用量が細胞性免疫性を刺激するのに十分であることが示された。それゆえ本結果から、gag発現ベクターによる実験(表3参照)においても観察されたヒトサブグループCベクター(hAd5及びhAd6など)に対して、CV32由来のベクターの免疫付与効力が低いことが確認される。
【0173】
【表5】
【実施例8】
【0174】
抗Ad5既存免疫はChAd3gagにより誘発される抗GAG CMIを抑制しない。
【0175】
高い血清普及率の(seroprevalent)Ad5に対する既存免疫性のChAd3免疫付与への影響を調べるために、1コホートにつきBalbCマウス5匹の4コホートに対して、大腿四頭筋にAd5野生型1回1010vpを第0週及び第2週の2回注射してプレ免疫付与した。対照として1コホート5匹の2コホートのマウスに緩衝液のみを同じ時点で注射した。次に、Ad5でプレ免疫付与したマウスのコホートに対して、Ad5gagベクター又はChAd3gagベクターのいずれか 106vp又は107vp/マウスで免疫付与した。対照(未処理の)マウスのコホートを、Ad5gagベクター又はChAd3gagベクター 106vp/マウスで免疫付与した。
【0176】
抗Ad5及びChAd3の中和免疫性を、Ad5ベクター及びChAd3 SEAPベクターを使用して上述の中和アッセイにより第4週に評価した。BalbCマウスにおいてマッピングされるgagエピトープを含有するgag9マーペプチドで刺激した、精製脾臓細胞においてELISPOT分析により抗gag免疫性を評価した。図36で報告する結果から、抗Ad5免疫性がChAd3gagにより誘発される抗gagCMIを抑制しないのに対し、予測されるように、抗Ad5免疫性がAd5gag免疫付与を完全に遮断することが示された。
【実施例9】
【0177】
ChAd3hCEA免疫化はヒトCEAを発現するトランスジェニックマウスにおいて強いCEA特異的免疫反応を誘発する。
【0178】
本明細書で開示し主張するChAdベクターが自己抗原に対して免疫反応を誘発し、それゆえにその寛容性を打破する能力も、ヒトCEAを発現するトランスジェニックマウスにおいて評価した(Clarke,Pら、Cancer Res.(1988)58(7):1469−77.)。
【0179】
8匹のマウスのコホートの大腿四頭筋にChAd3hCEA又はAd5hCEA 1010vpを既に述べたように注射した。aa497から端(aa703)までのヒトCEAアミノ酸配列を包含する15マーペプチドのプールで刺激したPBMCに対して、CEAに対する免疫反応を75日目まで毎週モニタリングした。CEAペプチドプールインキュベーションに反応した、CD4−CD8+T細胞分泌性インターフェロン−γを調べるICSによって、抗CEA免疫性を評価した。
【0180】
図37で報告する結果から、ChAd3hCEAベクター免疫付与が、同じ導入遺伝子を発現するAd5よりも、ヒトCEAに対するCD8+T細胞反応をより持続するよう刺激することが示される。
【0181】
霊長類免疫付与実験
方法及び材料
免疫付与プロトコール
本明細書に開示し主張するChAdベクターがアカゲザル(本明細書でサルと呼ばれる。)においてCMIを誘発する能力も評価した。サルを麻酔し(ケタミン/キシラジン)、本ワクチンをツベルクリンシリンジ(Becton−Dickinson)を使用して両側の三角筋へ0.5mlずつ筋肉内に送達した。全ての場合において、このサルの体重は、3から10kgであり、各ワクチンの用量は緩衝液1ml中で投与した。
【0182】
免疫付与手法の間、何回か回収した血液試料から、血清及び末梢血単核球(PBMC)を調製した。動物の取り扱い及び処置は全て、the Guide for Care and Use of Laboratory animals,Institute of Laboratory animal Resources,National Research Councilに記載の原則に従い、the Institutional animal Care and Use Committeeにより承認される基準に従った。
【0183】
ELISPOTアッセイ
アカゲザルについてのIFN−γELISPOTアッセイを、いくつかの改変を行い、上述のプロトコール(Allenら、2001 J.Virol.75(2):738−749)に従って実行した。gag特異的刺激について、10−aaの重複がある全HIV−1gag配列を包含する20−aaのペプチドからペプチドプールを調製した(Synpep Corp., Dublin, CA)。HCV NS特異的刺激について、10−aaの重複があるNS3からNS5bの全HCV−NS配列を包含する15−aaペプチドから、6個のペプチドプールを調製した。
【0184】
HER2 neu及びCEA特異的刺激を、10−aaの重複がある全タンパク質の配列を包含する15−aaのペプチドで実行した。
【0185】
2から4x105個の末梢血単核球(PBMC)50μlを各ウェルへ添加した。小さなサイズを80fLでカットオフして、細胞をBeckman Coulter Z2粒子アナライザーを使用して計数した。培地 50μl又はペプチドあたり8μg/mlのgagペプチドプールのいずれかをこのPBMCへ添加した。この試料を37℃、5%CO2で20時間から24時間インキュベーションした。このように、スポットを発色させ、プレートを特注の画像化装置及びImageProプラットフォーム(Silver Spring, MD)に基づいた自動計数サブルーチンを使用して処理した。計数を106個の細胞インプットに対して正規化した。
【0186】
細胞内サイトカイン染色(ICS)
2x106 PBMC/mL完全RPMI培地 1mL(17x100mmの丸底ポリプロピレンチューブ(Sarstedt,Newton,NC)に入れた。)に、抗hCD28(クローンL293、Becton−Dickinson)及び抗hCD49d(クローンL25、Becton−Dickinson)モノクローナル抗体を、終濃度1μg/mLとなるように添加した。gag特異的刺激については、ペプチドプール 10μL(ペプチド1個につき0.4mg/ml)を添加した。同様の条件をHCV NS特異的刺激について使用した。このチューブを37℃で1時間インキュベーションした後、5mg/mLのブレフェルジンA(Sigma)20μlLを添加した。細胞を37℃、5%CO2、90%湿度で16時間インキュベーションした。冷PBS/2%FBS 4mLを各チューブへ添加し、細胞を1200rpmで10分間遠心分離してペレットを得た。細胞をPBS/2%FBS中に再懸濁し、いくつかの蛍光標識mAbを使用して表面マーカーに対して染色した(30分間、4℃)(そのモノクローナル抗体は、チューブ1本あたり20μlの、抗hCD3−APC、クローンFN−18(Biosource);20μl 抗hCD8−PerCP、クローンSK1(Becton Dickinson, Franklin Lakes, NJ);及び20μL 抗hCD4−PE、クローンSK3(Becton Dickinson)であった。)。この段階から、試料を暗所で取り扱った。細胞を洗浄し、750μL 1xFACS Perm緩衝液(Becton Dickinson)中で室温にて10分間インキュベーションした。この細胞をペレットにし、PBS/2%FBS中に再懸濁し、FITC−抗hIFN−γ、クローンMD−1(Biosource)0.1μgを添加した。30分間のインキュベーションの後、細胞を洗浄し、PBS中に再懸濁した。Becton Dickinson FACSCalibur機器の4色チャネル全部を使用して試料を分析した。本データを分析するため、下部側及び前方に散乱するリンパ球集団を最初にゲーティングした。サイトカイン陽性反応についての共通の蛍光のカットオフをCD4+及びCD8+の両集団について使用し、試料の偽反応及びgagペプチド反応チューブの両者に対して使用した。
【実施例10】
【0187】
ChAdΔE1−gagベクターを使用する同種初回刺激−追加免疫法はサルにおけるgag特異的T細胞を誘発する。
【0188】
1コホート3匹の動物のコホートに以下のコンストラクトのいずれかを第0週及び第4週で筋肉内注射により与えた。すなわちCV32ΔE1−gag 1010vp又はCV33ΔE1−gag 1010vp又はMRKAd5ΔE1gag 1010vp及び108vpであった。4週間隔で回収したPBMCを、ELISPOTアッセイにより分析した。Gagペプチドプール非存在下(偽)又は存在下でのインキュベーション後の、100万個のPBMCあたりのスポット形成細胞数を示す表6で与える結果から、非ヒト霊長類においてgag特異的T細胞の有意なレベルをCV32ΔE1−gag及びCV33ΔE1gagの両者が誘導できることが確認される。興味深いことに、単回投与(第4週)後、CV32ΔE1−gag反応は、108vp/投与のMRKAd5ΔE1−gagに匹敵し、1010vp/投与のMRKAd5−gagのそれより低かったことに留意すること。1010vp/投与のCV33ΔE1−gagは、1010vp/投与のMRKAd5−gagのそれに匹敵する反応を誘導する。この結果は2回目の投与後の第8週で確認された。
【0189】
【表6】
【実施例11】
【0190】
ChAdベクターは、異種初回刺激−追加免疫法においてHCV NS特異的T細胞反応を誘発する。
【0191】
個別の実験において、2匹及び3匹のサルのグループに第0週、第4週で動物1匹につき108vp又は1010vpでMRK Ad6NSoptmutベクターを用いて免疫付与を行った。この動物を第24週で、同じ用量の同じウイルスで追加免疫し、1010個vp/動物でCV33−NSmutにより第104週に再度追加免疫を行った。この結果は、HCV NSペプチドプールの非存在下(偽)又は存在下でのインキュベーション後の100万個のPBMCあたりのスポット形成細胞数を要約する表7及び表8に示す。
【0192】
IFN−γELISPOTにより評価されるように、T細胞免疫性は1010vpを注射した動物において最初の投与後第4週で(表8)、及び108vpで注射した動物において第8週(2回目の投与後)で(表7)ピークの反応を示した。この反応は第24週での注射により促進されなかった(同種追加免疫)のに対し、強い促進効果がCV33−NSmutによる注射の後に観察された(異種追加免疫)。
【0193】
【表7】
【0194】
【表8】
【0195】
チンパンジーAdベクターによる異種追加免疫の効率を第二の実験において評価した。3匹のサルのコホートに対して、第0週及び第4週に、MRKAd5gag(1010vp/動物)、MRKAd6NSmut(1010vp/動物)又は両ベクター(各ベクターにつき1010vp/動物)の組み合わせで免疫付与し、その後第24週で同じ抗原で追加免疫した(同種追加免疫)。同種追加免疫を同じ抗原で実行し、異種追加免疫をCV33gag、CV32NSmut又は前記2つのベクターの組み合わせで実行した。表9で与える結果は、HCV NSペプチドプール非存在下(偽)又は存在下でのインキュベーション後、100万個のPBMCあたりのスポット形成細胞数を要約する。
【0196】
同じコホートを、第51週に、CV33gag(1010vp/動物)、CV32NSmut(1010vp/動物)及び前記2つのベクターの組み合わせ(各ベクターにつき1010vp/動物)で再度追加免疫した。表9で与える結果はさらに、この反応がワクチンの初回用量の注射後第4週で観察されるピークを下回るため、同種追加免疫が効果的ではなかったことを示す。異種チンパンジーベクターによる免疫付与後第54週で、IFN−γELISPOTにより強い免疫促進効果が測定された。
【0197】
【表9】
【実施例12】
【0198】
TAAを含むChAdベクターによるワクチン接種は寛容性を打破し、サルにおけるTAA特異的T細胞反応を誘発する。
【0199】
チンパンジーアデノウイルスベクターが自己抗原に対する寛容性を打破し、ヒトアデノウイルスベクターにより初回刺激された免疫反応を促進するためのチンパンジーベクターの有用性を証明するのに十分に免疫原性があるか否かを調べるために設定した実験を、1コホート4匹のサルのコホートにおいて実施した。腫瘍関連抗原CEAを含むAd5DE1 RhCEA(1011vp)の3回の注射を第0週、第2週及び第4週に行って動物に免疫付与した後、第16週、第18週及び第20週に、CV33DE1 RhCEA(1011vp)によるワクチン接種を行った。アカゲザルCEAペプチドを用いて、IFNγELISPOTにより、T細胞反応を測定した。
【0200】
アカゲザルCEA C及びDペプチドプール非存在下(DMSO)又は存在下のインキュベーション後の100万個のPBMCあたりのスポット形成細胞数を与える図34に報告される結果により、2つの異なるAd血清型によるワクチン接種に基づく免疫付与プロトコールが、非ヒト霊長類において、CEAに対してT細胞反応を持続させるに到ることが確認される。
【0201】
本発明はその特定の好ましい実施形態に関して詳細に記載されているが、改変及びバリエーションが、記載され、主張される本発明の精神及び範囲内であることは理解されよう。
【図面の簡単な説明】
【0202】
【図1】図1は、ChAd6シャトルベクター(pARS ChAd6−3)を構築するために使用されるクローニングストラテジーを要約する概略図である。
【図2】図2は、E.コリ株BJ5183での相同組み換えによりChAd6ウイルスゲノムをクローニングするために使用されるクローニングストラテジーを説明する概略図である。
【図3】図3は、pARS ChAd6−3 GAG:pARS ChAd6−3 SEAP;pARS ChAd6−3 EGFP及びpARS ChAd6−3 NS MUTを含む、様々なChAd6シャトルプラスミドのエレメントを説明する概略図である。
【図4】図4は、ChAd6ΔE1発現ベクターをクローニングするために使用される相同組み換えスキームを説明する概略図である。
【図5A】図5Aは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5B】図5Bは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5C】図5Cは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5D】図5Dは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5E】図5Eは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5F】図5Fは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5G】図5Gは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5H】図5Hは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5I】図5Iは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5J】図5Jは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5K】図5Kは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図6A】図6Aは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6B】図6Bは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6C】図6Cは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6D】図6Dは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6E】図6Eは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6F】図6Fは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6G】図6Gは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6H】図6Hは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6I】図6Iは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6J】図6Jは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6K】図6Kは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図7A】図7Aは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7B】図7Bは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7C】図7Cは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7D】図7Dは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7E】図7Eは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7F】図7Fは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7G】図7Gは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7H】図7Hは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7I】図7Iは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7J】図7Jは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7K】図7Kは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図8A】図8Aは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8B】図8Bは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8C】図8Cは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8D】図8Dは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8E】図8Eは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8F】図8Fは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8G】図8Gは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8H】図8Hは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8I】図8Iは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8J】図8Jは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8K】図8Kは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図9A】図9Aは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9B】図9Bは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9C】図9Cは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9D】図9Dは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9E】図9Eは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9F】図9Fは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9G】図9Gは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9H】図9Hは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9I】図9Iは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9J】図9Jは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図10】図10は、ChAd20(配列番号6)の繊維遺伝子のヌクレオチド配列を与える。
【図11】図11は、ChAd4(配列番号7)の繊維遺伝子のヌクレオチド配列を与える。
【図12】図12は、ChAd5(配列番号8)の繊維遺伝子のヌクレオチド配列を与える。
【図13】図13は、ChAd7(配列番号9)の繊維遺伝子のヌクレオチド配列を与える。
【図14】図14は、ChAd9(配列番号10)の繊維遺伝子のヌクレオチド配列を与える。
【図15】図15は、ChAd10(配列番号11)の繊維遺伝子のヌクレオチド配列を与える。
【図16】図16は、ChAd11(配列番号12)の繊維遺伝子のヌクレオチド配列を与える。
【図17】図17は、ChAd16(配列番号13)の繊維遺伝子のヌクレオチド配列を与える。
【図18】図18は、ChAd17(配列番号14)の繊維遺伝子のヌクレオチド配列を与える。
【図19】図19は、ChAd19(配列番号15)の繊維遺伝子のヌクレオチド配列を与える。
【図20A】図20Aは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20B】図20Bは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20C】図20Cは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20D】図20Dは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20E】図20Eは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20F】図20Fは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20G】図20Gは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図21】図21は、ChAd20(配列番号16)のヘキソン遺伝子のヌクレオチド配列を与える。
【図22】図22は、ChAd4(配列番号17)のヘキソン遺伝子のヌクレオチド配列を与える。
【図23】図23は、ChAd5(配列番号18)のヘキソン遺伝子のヌクレオチド配列を与える。
【図24】図24は、ChAd7(配列番号19)のヘキソン遺伝子のヌクレオチド配列を与える。
【図25】図25は、ChAd9(配列番号20)のヘキソン遺伝子のヌクレオチド配列を与える。
【図26】図26は、ChAd10(配列番号21)のヘキソン遺伝子のヌクレオチド配列を与える。
【図27】図27は、ChAd11(配列番号22)のヘキソン遺伝子のヌクレオチド配列を与える。
【図28】図28は、ChAd16(配列番号23)のヘキソン遺伝子のヌクレオチド配列を与える。
【図29】図29は、ChAd17(配列番号24)のヘキソン遺伝子のヌクレオチド配列を与える。
【図30】図30は、ChAd19(配列番号25)のヘキソン遺伝子のヌクレオチド配列を与える。
【図31A】図31Aは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31B】図31Bは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31C】図31Cは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31D】図31Dは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31E】図31Eは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31F】図31Fは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31G】図31Gは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31H】図31Hは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31I】図31Iは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31J】図31Jは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図32】図32は、本明細書中で開示される、オリゴマー及びプライマーを含む、人工的配列、配列番号26から40及び配列番号45及び46のリストを与える。
【図33】図33は、様々な血清型サブグループ(つまり、サブグループC、E及びD)に属するChAdベクターの免疫付与ブレイクポイントを図示したものである。gag−発現する、ChAd3、ChAd11、ChAd20、CV33、CV68、ChAd6、ChAd9、ChAd10、CV32、ChAd4、ChAd7及びChAd16ベクターで免疫付与したマウスにおいてタイター決定実験を行うことにより、測定可能な免疫反応を誘発する最低用量を調べた。
【図34】図34は、ヒトアデノウイルスベクター(MRKAd5 ThCEA)と、次いで、12週間の間隔後、チンパンジーアデノウイルスベクター(CV33 RhCEA)で連続的に免疫付与したアカゲザルにおいて誘発されるCEA特異的T細胞反応のグラフである。IFNγ−ELISPOTアッセイにより免疫反応を評価した。データは、アカゲザルCEA C及びDペプチドプールの非存在下(DMSO)及び存在下でのインキュベーション後の、末梢血単核球(PBMC)百万個あたりのスポット形成細胞(SFC)の数を示す。
【図35】図35は、PAUPSEARCH(Wisconsin Package Version 10.3,Accelrys Inc.)を用いて、TREEVIEWにより視覚化し操作した、全長ヘキソンペプチド配列の多配列アラインメントから推定される、ヒト及びチンパンジーアデノウイルスの系統樹を与える。
【図36】図36は、hAd5に前もって曝露したマウスへのChAd3及びhAd5 gagベクターの投与に対する反応において得られた免疫付与結果を表すグラフである。精製脾臓細胞を用いて、IFNγ−ELISPOTにより、免疫付与から3週間後に、細胞性免疫を評価した。
【図37】図37は、ChAd3hCEA及びAd5hCEAで免疫付与したヒトCEAトランスジェニックマウスにおいて誘発された、抗CEA CMIのキネティクスを表すグラフである。CEAペプチドプールで刺激したPBMCのICSにより、CMIを評価した。結果は、IFNγ+CD8+/総PBMCの%として表す。
【図38A】図38Aは、EGFPを発現し、様々なサブグループ(B、C、D、E)に属する様々なChAdベクターの、moi 50、250及び1250に曝露した様々なヒト初代細胞の感染効率を表すグラフである。結果は、蛍光細胞/総細胞の%として表す。
【図38B】図38Bは、EGFPを発現し、様々なサブグループ(B、C、D、E)に属する様々なChAdベクターの、moi 50、250及び1250に曝露した様々なヒト初代細胞の感染効率を表すグラフである。結果は、蛍光細胞/総細胞の%として表す。
【図38C】図38Cは、EGFPを発現し、様々なサブグループ(B、C、D、E)に属する様々なChAdベクターの、moi 50、250及び1250に曝露した様々なヒト初代細胞の感染効率を表すグラフである。結果は、蛍光細胞/総細胞の%として表す。
【図38D】図38Dは、EGFPを発現し、様々なサブグループ(B、C、D、E)に属する様々なChAdベクターの、moi 50、250及び1250に曝露した様々なヒト初代細胞の感染効率を表すグラフである。結果は、蛍光細胞/総細胞の%として表す。
【技術分野】
【0001】
関連出願の相互参照
本願は、参照により本明細書中に組み込む、2004年1月23日提出の、米国仮出願第60/538,799の優先権を主張する。
【0002】
本発明は、組み換えベクターの分野、とりわけ、哺乳動物宿主において免疫反応を誘発するための組み換え複製欠損チンパンジーアデノウイルスベクターの産生及び使用に関する。
【背景技術】
【0003】
アデノウイルス(Ad)は、エンベロープのない20面体のキャプシド構造を有する、両性類、鳥類及び哺乳動物において見出される2本鎖DNAウイルスの大きなファミリーを含む(Straus,Adenovirus infections in humans.Adenoviruses.451−498,1984;Hierholzerら、J.Infect.Dis.,158:804−813,1998;Schnurr及びDondero,Intervirology.,36:79−83,1993;Jongら、J.Clin Microbiol.,37:3940−3945:1999)。レトロウイルスとは異なり、アデノウイルスは、宿主細胞のゲノムに組み込まれることなく、分裂細胞及び非分裂細胞の両方を含む、いくつかの哺乳動物種の多数の細胞タイプに形質導入することができる。
【0004】
一般に、アデノウイルスDNAは、通常、非常に安定で、形質転換又は腫瘍形成が起こらない限り、エピソーム性のままである(例えば染色体外性)。さらに、アデノウイルスベクターは、臨床グレードの組成物の製薬規模の産生に容易に持って行くことができる、よく分かっている産生系において高収量に増殖させることができる。これらの特性及びそれらのよく調べられている分子遺伝学により、組み換えアデノウイルスベクターは、ワクチン担体としての使用のための優れた候補となっている。通常、組み換えアデノウイルスベクターの産生は、非機能的となるように欠失されている又は加工されているアデノウイルス遺伝子産物の機能を補完することができるパッケージング細胞株の使用に依存している。
【0005】
現在、性質がよく調べられている2種類のヒトサブグループCアデノウイルス血清型(つまりhAd2及びhAd5)が、遺伝子治療のために使用される殆どのアデノウイルスベクターに対するウイルス骨格のソースとして広く使用されている。複製欠損ヒトアデノウイルスベクターに対してまた、様々な感染物質(例えば、ウイルス、寄生生物又は細菌病原体)及び、腫瘍関連抗原(TAA)を含む腫瘍細胞由来の様々な免疫原送達のためのワクチン担体としての試験が行われている。
【0006】
実験動物(例えば、げっ歯類、サル及び非ヒト哺乳動物)において行われた研究から、ヒトパピローマウイルス(HPV−16)のE6及びE7癌タンパク質(He、Zら、(2001)Virology,270:3583−3590)、狂犬病ウイルス糖タンパク質(Xiang,Zら、(1996)Virology,219:220−227)、Plasmodium falciparum(熱帯熱マラリア原虫)のスポロゾイト周囲タンパク質(Rodriguez,Eら、(1997)J.Immunol.158:1268−1274)ならびにその他の異種抗原由来の免疫原をコードする導入遺伝子を担う組み換え複製欠損ヒトアデノウイルスベクターが、その導入遺伝子産物に対する液性及び細胞性免疫反応の両方を誘導することが示されている。一般には、研究者らは、免疫反応を誘発すると予想される組み換えアデノウイルスベクターの高用量を用いる免疫付与プロトコールを用いて、又は、追加免疫付与として、異なる血清型由来であるが同じ導入遺伝子産物を担うアデノウイルスベクターの連続投与を利用する免疫付与プロトコールを用いることのいずれかにより、非ヒト実験系においてワクチン担体としてヒトアデノウイルスベクターを使用して、成功を報告している(Mastrangeliら、Human Gene Therapy,7:79−87(1996))。
【0007】
しかし、2型及び5型などの、遍在性ヒト血清型由来のワクチン担体は、ヒト集団において既存の液性及び細胞性免疫に直面すると予想されている。したがって、複製欠損組み換えヒトアデノウイルスが、成功裡にワクチン担体としてげっ歯類、サル及び非ヒト霊長類宿主を用いた実験系で利用されているにもかかわらず、ヒトの先天的及び適応的免疫により、ワクチン担体としてのこれらの血清型の利用が著しく制限されると予想される。この予想は、2型及び5型を含むサブグループCのアデノウイルス感染が、ヒト集団において固有であるという事実に基づくものである。その結果、大部分のヒトで、自然感染の結果として、生まれてから最初の5年間のうちに血清転換が起こっている。したがって、ヒトにおいて自然に感染し複製するウイルス由来のベクターは、ワクチン担体としての使用に対して最適な候補ではない可能性がある。
【0008】
ヒトアデノウイルス由来ベクターの使用に関する別の問題は、組み換えウイルスを増殖させるために使用される産生方法により、複製可能なアデノウイルス(RCA)が混入しているベクターストックを生じるというリスクである。これは、ヒト293細胞などのE1補完ヘルパー細胞株において存在する、組み換えベクターとアデノウイルス遺伝子との重複配列間の相同組み換えにより起こる(Graham,F.L.ら(1977)J.Gen.Virol.36:59−72)。臨床試験での使用のために調製されたベクターストックにおけるRCAの存在は、複製欠損ウイルスの移行及び拡大を促進する可能性があるため、安全性リスクの一要素である。欠損ウイルスの拡大により、宿主免疫反応が拡大され、他の不都合な免疫病理学的結果を生じる可能性がある(Fallux,F.J.ら、Human Gene Therapy 9:1909−1917(1998))。したがって、Food and Drug Administration(食品医薬品局)(FDA)及びその他の規制機関は、臨床での使用を意図されるベクター調製物中に存在し得るRCAのレベルにおける制限を確立するガイドラインを広めてきた。RCA制限を課す意図は、臨床試験で使用される組成物中での複製するアデノウイルスへの患者の曝露を確実に制限することにある。
【発明の開示】
【発明が解決しようとする課題】
【0009】
したがって、引き続き、操作しやすく、製薬スケールに持って行きやすく、長期保存でき、ヒトの補完的細胞株で高レベルで複製することができ、免疫原性が高く、ヒトアデノウイルスの共通血清型と交差反応する中和B細胞エピトープを欠いており、規制当局により広められている安全性RCA基準に合致し、ヒトでの使用に適切な初回刺激/追加免疫に使用できる、ヒト宿主での使用に適したアデノウイルスワクチン担体の開発が必要とされている。
【課題を解決するための手段】
【0010】
本発明は、チンパンジーアデノウイルス由来の組み換え複製欠損アデノウイルスベクター及びヒトE1発現細胞株においてチンパンジーアデノウイルスベクターを生成させるための方法に関する。本発明はまた、ヒトにおける使用に適切な臨床グレードのベクターストックを生成させるための方法及び、防御的及び/又は治療的免疫反応を誘発するために、開示するベクターをワクチン担体として使用するための手段を提供する。本発明はさらに、免疫原をコードする導入遺伝子を送達し、発現を支配するように設計されたワクチン組成物を調製するための、本発明の組み換えアデノウイルスを使用するための方法を提供する。ある実施形態において、本発明は、感染物質由来の免疫原をコードする導入遺伝子を含有するワクチン投与のための、ワクチン担体としての、開示するベクターの使用をもくろむ。第二の実施形態において、本発明は、癌ワクチンを調製及び投与するための、開示するベクターの使用をもくろむ。特定の実施形態において、本発明は、TAAをコードする導入遺伝子を含有する癌ワクチンの調製及び投与をもくろむ。
【0011】
ある局面において、本発明は、本明細書中で、ChAd3(配列番号1)(図5A−5K)、ChAd6(配列番号2)(図6A−6K、CV32(配列番号3)(図7A−7K)、CV33(配列番号4)(図8A−8K)及びCV23(配列番号5)(図9A−9J)と呼ばれる、5種類のチンパンジーアデノウイルス(ChAd)の完全ゲノム配列を開示する。
【0012】
ChAd3及びChAd6は、本明細書中で開示する方法に従い単離された新規アデノウイルスを表す。ChAd3及びChAd6のゲノムは、それぞれ37741及び36648塩基対の長さである。このChAd3ヘキソン遺伝子(配列番号41)は、配列番号1のヌクレオチド(nt)19086から21965(停止コドンを除く。)を含有し、ChAd3繊維遺伝子(配列番号42)は、配列番号1のnt32805から34487(停止コドンを除く。)を含有する。ChAd6ヘキソン遺伝子は、配列番号2のnt18266−21124(配列番号43)(停止コドンを除く。)を含有し、その繊維遺伝子(配列番号44)は、配列番号2のnt32218から33552(停止コドンを除く。)を含有する。全長ヘキソンペプチドの多配列アラインメントから推定される配列相同性に基づき、ChAd3は、ヒトサブグループCに分類され、ChAd6は、ヒトサブグループEに分類される。
【0013】
CV32、CV33及びCV23アデノウイルスのゲノムは、それぞれ、36,606、36,535及び32,020塩基対の長さである。CV32(Pan6)(ATCC N.VR−592)、CV33(Pan7)(ATCC N.VR−593)及びCV23(Pan5)(Esoterix Inc.,)は、全て、ヒトAd4(hAd4)に関連することが分かっている(サブグループE)(Wigand,Rら、Intervirology 1989,30:1−9)。しかし、ヘキソン配列アラインメントに基づいて、CV32は、その後、hAd4よりも、ヒトサブグループDメンバーに密接に類似するものとして特徴付けられた。
【0014】
第二の局面において、本発明は、本明細書中で開示する方法に従い単離された21種類のさらなるチンパンジーアデノウイルス(ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82)の繊維及びヘキソン遺伝子に対するヌクレオチド配列を提供する。
【0015】
ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19に対する繊維遺伝子ヌクレオチド配列は、それぞれ図10から図19に記載されており、本明細書中で、配列番号6から15(配列番号6、ChAd20);(配列番号7、ChAd4);(配列番号8、ChAd5);(配列番号9、ChAd7);(配列番号10、ChAd9);(配列番号11、ChAd10);(配列番号12、ChAd11);(配列番号13、ChAd16)(配列番号14、ChAd17)及び(配列番号15、ChAd19)と呼ばれる。
【0016】
ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82に対する繊維遺伝子ヌクレオチド配列は、本明細書中で(配列番号58、ChAd8)、(配列番号60、ChAd22)、(配列番号62、ChAd24)、(配列番号64、ChAd26)、(配列番号66、ChAd30)、(配列番号68、ChAd31)、(配列番号70、ChAd37)、(配列番号72、ChAd38)、(配列番号74、ChAd44)、(配列番号76、ChAd63)及び(配列番号78、ChAd82)と呼び、配列リストに記載する。
【0017】
ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19に対するヘキソン遺伝子ヌクレオチド配列は、図21−30にそれぞれ記載され、本明細書中で、配列番号16から25:(配列番号16、ChAd20);(配列番号17、ChAd4);(配列番号18、ChAd5);(配列番号19、ChAd7);(配列番号20、ChAd9);(配列番号21、ChAd10);(配列番号22、ChAd11);(配列番号23、ChAd16);(配列番号24、ChAd17)及び(配列番号25、ChAd19)と呼ぶ。
【0018】
ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82に対するヘキソン遺伝子ヌクレオチド配列は、本明細書中で、(配列番号97、−ChAd8)、(配列番号99、ChAd22)、(配列番号101、ChAd24)、(配列番号103、ChAd26)、(配列番号105、ChAd30)、(配列番号107、ChAd31)、(配列番号109、ChAd37)、(配列番号111、ChAd38)、(配列番号113、ChAd44)、(配列番号115、ChAd63)及び(配列番号117、ChAd82)と呼び、配列リストに記載する。
【0019】
第三の局面において、本発明は、本明細書中で開示する方法に従い単離された、21種類のさらなるチンパンジーアデノウイルス(ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82)の繊維及びヘキソンタンパク質に対するアミノ酸配列を提供する。
【0020】
本明細書中で開示され主張される繊維タンパク質は、(配列番号83、ChAd3)、(配列番号84、ChAd6)、(配列番号48、ChAd20)、(配列番号49、ChAd4)、(配列番号50、ChAd5)、(配列番号51、ChAd7)、(配列番号52、ChAd9)、(配列番号53、ChAd10)、(配列番号54、ChAd11)、(配列番号55、ChAd16)、(配列番号56、ChAd17)、(配列番号57、ChAd19)、(配列番号59、ChAd8)、(配列番号61、ChAd22)、(配列番号63、ChAd24)、(配列番号65、ChAd26)、(配列番号67、ChAd30)、(配列番号69、ChAd31)、(配列番号71、ChAd37)、(配列番号73、ChAd38)、(配列番号75、ChAd44)、(配列番号77、ChAd63)及び(配列番号79、ChAd82)と呼ばれる。図20Aから20Gは、本明細書中で開示され主張される繊維タンパク質のアミノ酸配列を、C1(配列番号85)、CV68(配列番号86)、Pan5(CV23とも呼ばれる。)(配列番号80)、Pan6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)の繊維タンパク質のアミノ酸配列と比較するアラインメントを提供する。
【0021】
本明細書中で開示され主張されるヘキソンタンパク質は、(配列番号122、ChAd3)、(配列番号123、ChAd6)、(配列番号87、ChAd20)、(配列番号88、ChAd4)、(配列番号89、ChAd5)、(配列番号90、ChAd7)、(配列番号91、ChAd9)、(配列番号92、ChAd10)、(配列番号93、ChAd11)、(配列番号94、ChAd16)、(配列番号95、ChAd17)、(配列番号96、ChAd19)、(配列番号98、ChAd8)、(配列番号100、ChAd22)、(配列番号102、ChAd24)、(配列番号104、ChAd26)、(配列番号106、ChAd30)、(配列番号108、ChAd31)、(配列番号110、ChAd37)、(配列番号112、ChAd38)、(配列番号114、ChAd44)、(配列番号116、ChAd63)及び(配列番号118、ChAd82)と呼ばれる。図31Aから31Jは、本明細書中で開示され主張されるヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、Pan5(CV23とも呼ばれる。)(配列番号119)、Pan6(CV32とも呼ばれる。)、(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)のヘキソンタンパク質のアミノ酸配列との比較を与える。ヘキソンタンパク質の多配列アラインメントにより、熟練者は、ヒトアデノウイルス血清型の提案される分類に従う系統発生解析を行うことが可能となる(Rux,J.J.ら、(2003)J.Virol.77:9553−9566)。
【0022】
代替的局面において、本発明はさらに、21種類のさらなるチンパンジーアデノウイルス分離株を提供する。ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17及びChAd19を含有する試料を、Budapest Treatyのもと、オリジナル委託物として、European Collection of Cell Cultures(ECACC、Prton Down,Salisbury,Wiltshire,SP4 0JG,United Kingdom)により、2003年12月12日に委託した。この委託物に対して、次の受託番号が割り当てられた:03121201(ChAd4)、03121202(ChAd5)、03121203(ChAd7)、03121204(ChAd9)、03121205(ChAd10)、03121206(ChAd11)、03121207(ChAd16)、03121208(ChAd17)、03121209(ChAd19)及び03121210(ChAd20)。
【0023】
ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82を含有する試料を、Budapest Treatyのもと、オリジナル委託物として、ECACC(Prton Down,Salisbury,Wiltshire,SP4 0JG,United Kingdom)により、2005年1月12日に委託した。これらの委託物に対して、次の受託番号が割り当てられた:05011201(ChAd8)、05011202(ChAd22)、05011203(ChAd24)、05011204(ChAd26)、05011205(ChAd30)、05011206(ChAd31)、05011207(ChAd37)、05011208(ChAd38)、05011209(ChAd44)、05011210(ChAd63)及び05011211(ChAd82)。
【0024】
これらの委託物は、International Recognition of the Deposit of Microorganisms of the Purposes of Patent ProcedureにおけるBudapest Treatyの条項に従い維持される。これらの委託物は、当業者に対する便宜としてのみ、なされたものであり、委託物が25U.S.C.§112のもとで要求されることの承認ではない。特許の付与において、37 C.F.R.§1.808(b)において規定される要件を除き、委託した物質の公共に対する利用可能性における制限は全て、変更不能に取り除かれる。
【0025】
さらなる局面において、本発明はまた、哺乳動物細胞、好ましくはヒト細胞に感染することができ、コードされる導入遺伝子産物の発現を支配することができる、複製欠損組み換えアデノウイルスベクターを提供する。本明細書中で示されるように、開示するベクターは、免疫反応が望まれる免疫原を含有する導入遺伝子の送達に対する使用のためのワクチン担体としての使用に適する。特定の実施形態ににおいて、本発明は、ヒトE1発現(つまりパッケージング)細胞株において高レベルで複製することができる、組み換え複製欠損チンパンジーアデノウイルスベクターを提供する。ある実施形態において、本発明は、PER.C6TM細胞において複製できる組み換えアデノウイルスを提供する。
【0026】
一般に、本発明に包含される組み換えベクターは、ヒト集団において一般に直面させられるアデノウイルス血清型に対して既存の免疫を回避するワクチン担体を提供する。とりわけ、本発明の組み換えベクターは、本明細書中で、ヒトアデノウイルス由来ベクターの共通血清型と交差反応する中和Bエピトープを欠いていることが示される、ベクター骨格配列を含有する。
【0027】
本発明はさらに、アデノウイルス部分及びプラスミド部分を含み、該アデノウイルス部分が通常、a)ウイルス左端(ITR及びパッケージングシグナル)、pIX遺伝子の一部及びウイルスゲノム右端と;b)遺伝子発現カセットと、を含む、群特異的シャトルベクターを提供する。この群特異的シャトルベクターは、同じ血清型サブグループ(つまり、サブグループA、B、C、D又はE)に割り当てられ、本明細書中で開示されるヌクレオチド配列を操作するために、及び/又は、E1領域を欠くチンパンジーアデノウイルスゲノムを含有するアデノウイルスプレプラスミドを生じる、同じサブグループに属する他のチンパンジーアデノウイルスをクローニングするために、使用することができるアデノウイルスの間で観察される、ヌクレオチド配列相同性を活用するように設計されている。
【0028】
本発明の他の局面には、アデノウイルスワクチンベクター及び/又はアデノウイルスプレプラスミドベクターを含有する宿主細胞が含まれ、該ベクターの生成方法には、アデノウイルスワクチンベクターを、アデノウイルスE1タンパク質を発現する宿主細胞に導入することと、得られたアデノウイルスワクチンベクターを回収することと、が含まれる。特定の実施形態において、本発明は、アデノウイルスE1発現ヒト細胞に、開示するアデノウイルスベクターの1つを導入することと、得られた組み換えアデノウイルスを回収することと、を含む、複製欠損チンパンジーアデノウイルスベクターを生成する方法を提供する。
【0029】
本発明の別の局面はまた、本発明のアデノウイルスベクターを含有するワクチン組成物を提供する。組み換えチンパンジーアデノウイルスベクターを含有する組成物は、単独で、又はその他のウイルス性もしくは非ウイルス性ベースのDNA/タンパク質ワクチンと組み合わせて、投与し得る。これらはまた、幅広い治療計画の一部として投与し得る。これらの組成物は、予防的又は治療的設定のいずれかで、哺乳動物宿主、好ましくはヒト宿主に投与することができる。本明細書中で示されるように、初回刺激追加免疫計画又は血清学的に区別されるAdベクターの複数回注射など、単独又は組合せ方式での開示するワクチン組成物の投与の結果、その導入遺伝子によりコードされる免疫原を特異的に認識することができる、哺乳動物における免疫反応が誘導される。
【0030】
本明細書中で開示され主張される方法の1つは、少なくとも、その野生型E1遺伝子の機能的欠失を含有し、少なくとも1個の標的抗原をコードするヌクレオチド配列の発現を支配することができるプロモーターを含む配列を担う、組み換えチンパンジーアデノウイルスベクターの十分量を、哺乳動物(未処置又は標的抗原に対して免疫活性となるように初回刺激されているもののいずれか)に投与することを含み、本組み換えベクターの投与が抗原特異的免疫反応を誘発(又は刺激)する。
【0031】
ある実施形態において、本発明は、感染物質に対して免疫反応(予防的又は治療的)を誘導するように設計された方法を提供する(例えば、ウイルス性もしくは細菌病原体又は哺乳動物寄生生物)。第二の実施形態において、本発明は、TAAなどの、自己抗原に対する寛容性を打破する、哺乳動物において免疫反応を誘導するように設計された方法を提供する。本発明のこの局面は、癌ワクチンの調製及び投与のための、ワクチン担体としての、開示するベクターの使用をもくろむ。
【0032】
本発明のさらに他の実施形態及び長所は、次の本発明の詳細な説明から容易に明らかとなろう。
【0033】
本発明の詳細な説明
本明細書及び添付の請求項の範囲を通して使用される場合、次の定義及び略語が適用される。
【0034】
「カセット」という用語は、その転写及び翻訳調節配列に従って発現されるべき、少なくとも1個の核酸配列を含有する核酸分子を指す。カセットを変化させることにより、様々な配列及び配列の組合せの発現を支配するために組み込まれるベクターが生じる。本発明との関連において、本カセットに存在する核酸配列は通常、免疫原をコードする。5’及び3’末端に存在するように操作された制限部位のために、本カセットを容易に挿入、除去又は別のカセットで置換することができる。
【0035】
「cis作用エレメント」という用語は、遺伝子を制御するヌクレオチド配列に連結されるヌクレオチド配列を指す。DNAに存在するcis作用エレメントは、転写を制御し、mRNAに転写されたものは、RNAプロセシング、ターンオーバー及びタンパク質合成を制御することができる。
【0036】
「ベクター」という用語は、宿主生物又は宿主組織にDNA断片を導入することができる、ある手段を指す。プラスミド、ウイルス(アデノウイルスを含む。)、バクテリオファージ及びコスミドを含む、様々なタイプのベクターがある。
【0037】
「プロモーター」という用語は、RNAポリメラーゼが結合する、DNA鎖上の認識部位を指す。プロモーターは、転写活性を開始し、駆動させるために、RNAポリメラーゼと開始複合体を形成する。本複合体は、エンハンサーなどの活性化配列又はサイレンサーなどの抑制配列により修飾され得る。
【0038】
「医薬的有効量」という用語は、宿主細胞に形質導入し、免疫反応を誘発するための導入遺伝子発現の十分なレベルを与える、特定の投与経路において効果的な、組み換えアデノウイルスの量を指す。
【0039】
「複製能のある」組み換えアデノウイルス(AdV)という用語は、原型を保つ、又は機能的な必須初期遺伝子(つまり、E1A、E1B、E2A、E2B及びE4)を有するアデノウイルスを指す。野生型アデノウイルスは、複製能がある。
【0040】
「複製欠損」組み換えAdVという用語は、ウイルス複製に必須の遺伝子産物を、少なくとも機能的に欠損する、又は完全除去されるように加工されているため、複製ができないようにされているアデノウイルスを指す。本発明の組み換えチンパンジーアデノウイルスベクターは、複製欠損である。
【0041】
「哺乳動物」という用語は、ヒトを含む、あらゆる哺乳動物を指す。
【0042】
核酸配列との関連における、「%配列同一性」又は「同一」という用語は、最大一致に対して並べた場合に同じである、2つの配列における残基を指す。配列同一性比較の長さは、そのゲノムの全長(例えば約36kbp)、遺伝子のオープンリーディングフレームの全長、タンパク質、サブユニット又は酵素[例えば、アデノウイルスコード領域を与える表を参照のこと。]又は少なくとも約500から5000ヌクレオチドの断片にわたり得、それが望ましい。しかし、より小さい断片(例えば、少なくとも約9ヌクレオチド、通常は少なくとも約20から24ヌクレオチド、少なくとも28から32ヌクレオチド、少なくとも36以上のヌクレオチド)の間での同一性もまた望ましい。同様に、「%配列同一性」は、タンパク質の全長にわたる、又はその断片にわたるアミノ酸配列に対して容易に決定し得る。適切であるものとして、断片は、少なくとも約8アミノ酸の長さであり、約700アミノ酸以下であり得る。適切な断片の例を本明細書中で述べる。
【0043】
初期設定で本明細書中で定義されるようなアルゴリズム及びコンピュータープログラムを用いて、同一性が容易に決定される。好ましくは、このような同一性は、タンパク質、酵素、サブユニットの全長にわたるか、又は少なくとも約8アミノ酸の長さの断片にわたる。しかし、同一性は、同一遺伝子産物が置かれている使用に適した、より短い領域に基づくものであり得る。
【0044】
一般に、アデノウイルスコンストラクト、遺伝子コンストラクトは、本明細書中に含有される遺伝子を参照して名付ける。例えば、「pChAd3 ΔE1gag」は、E1領域が欠失している、ChAd3 チンパンジーアデノウイルスゲノムを含有するプラスミドコンストラクトを指す。このプラスミドにおいて、E1領域は、ヒトCMVプロモーターと、次いでウシ成長ホルモンポリアデニル化シグナルの調節下でHIVgag遺伝子を含有する免疫原発現カセットにより置換される。同様に、pCV33DE1−E3 NSmutは、E1及びE3領域が欠失しており、ヒトCMVプロモーターと、次いでウシ成長ホルモンポリアデニル化シグナルの調節下において、HCV非構造遺伝子を含む免疫原発現カセットで置換されている、CV33 チンパンジーアデノウイルスゲノムを含有する、本明細書中で開示される第二のプラスミドコンストラクトを指す。
【0045】
略語「Ag」は抗原を指す。
【0046】
本明細書及び添付の請求項の範囲を通して使用される場合、単数体「a」、「an」及び「the」は、文脈において明確に示されない限り、複数形の意味を含む。
【0047】
アデノウイルス(Ad)は、いくつかの鳥類及び哺乳動物宿主において同定されている、エンベロープがない20面体のウイルスである。ヒトAd(hAd)は、全ての既知のヒト及び動物の多くのAd系を含む(例えば、ウシ、ブタ、イヌ、マウス、ウマ、サル及びヒツジ)、マストアデノウイルス属に属する。ヒトアデノウイルスは、ラット及びアカゲザル赤血球の赤血球凝集特性、DNA相同性、制限酵素切断パターン、G+C含量の割合及び発癌性を含む、多くの、生物学的、化学的、免疫学的及び構造的基準に基づいて、6個のサブグループ(AからF)に分けられる(Straus,1984,Adenobiruses、H.Ginsberg編、pps.451−498,New York:Plenus Press及びHorwitz,1990 Virology、B.N.Fields及びD.M.Knipe編、pps.1679−1721)。今日までに、51種類の異なる血清型が認識され、これらの赤血球凝集特性及び生物物理的及び生化学的基準に基づき、サブグループにグループ分けされいてる。
【0048】
アデノウイルスビリオンは、20面体で対称であり、血清型によって、60から90nmの直径である。この20面体キャプシドは、3つの主要なタンパク質、ヘキソン(II)、ペントンベース(III)及びノブ付き繊維(IV)ならびに、多くの少量のタンパク質(つまり、VI、VIII、IX、IIIa及びIVa2)からなる(W.C.Russel,J.Gen.Virol.,81:2573−2604(2000))。ヒトにおいて観察される、既存の免疫のある局面は、液性免疫であり、この結果、ウイルスタンパク質に対して特異的な抗体が産生され持続され得る。アデノウイルスにより誘発される液性反応は主に、主要な構造タンパク質:ヘキソン、ペントン及び繊維に対するものである。
【0049】
公表されている報告から、複数の血清型に対する抗体を含有するタイターが共通であること(Dambrosio,E.(1982)J.Hyg.(London)89:209-219)及び既存のタイターの大部分が、中和活性を有することが、分かっている。アデノウイルスに対する中和免疫は、タイプ特異的であり、アデノウイルスの特定の血清型による感染により、その血清型のみに対する免疫が与えられる。いくつかの報告から、ヘキソンに対して向けられる抗体が、最強であり、最も中和能力が高いことが示唆されている(Toogood,C.I.A.,Crompton,J.及びHay R.T.(1992)J.Gen.Virol.73,1429−1435)。したがって、タイプ特異的中和に関与するエピトープが、様々な血清型由来のヘキソン配列のアラインメントにより同定される7個の超可変領域に位置すると仮定することは、理にかなっている(Crawford−Miksza,L及びD.P.Schnurr(1996)J.Virol.70:1836−1844)。
【0050】
タイプ特異的中和抗体の存在と、同じ血清型に基づくベクターにより免疫反応を誘発できないこととの間の正相関は、未処置動物への処置動物からの免疫血清の受動転移を含む、様々な方法により分かっている。一般に、特異的なウイルス血清型に対して既存の液性免疫により、ベクター投与の治療効率が低下する。さらに、特異的ウイルス血清型に基づくベクターの投与により、同じ血清型の再投与を防御する、ベクターに対する免疫反応が誘発される。
【0051】
特定の実施形態において、本発明は、hAdの共通血清型に対する既存の免疫の結果に付随する不都合な影響を回避する方法を提供する。とりわけ、本発明は、ヒトにおいて回避しない血清型を特徴とするチンパンジーアデノウイルスベクターの使用をもくろむ。したがって、本発明は、ワクチン担体として、共通ヒト血清型と交差反応する中和B細胞エピトープを欠く、アデノウイルス(ChAd)ベクターを提供する。
【0052】
アデノウイルス特異的細胞性免疫(CMI)が、交差反応性であり得ることが報告されているが、複数血清型の繰り返し注射に基づくワクチン接種実験が、1個のベクターを用いた免疫付与スケジュールよりも高い効率であることが示されている。これらの実験はさらに、ワクチン目的のベクター投与の主な制限が、そのベクターに対する液性の既存の免疫であることを示す。ワクチン担体としてのヒトアデノウイルスの使用に付随する問題を解決することができる可能性のある方法には、既存の免疫反応に直面することが予想される、アデノウイルスのさらに高用量の投与(例えば、サブグループC血清型)及び稀少なヒト血清型に基づくベクターの使用が含まれる。しかし、ワクチンの用量を高くして使用することにより、ワクチンのコストが上昇し、望ましくない副作用のリスクが増加し、前臨床試験の結果から、ヒトの代替的血清型は、hAd5及びhAd6よりも免疫原性が低いことが示唆されている。
【0053】
本ベクターのアデノウイルス骨格エレメントに対する宿主の液性及び細胞性免疫反応の問題を回避する、及び複製能のあるアデノウイルス(RCA)が混入し得るヒトアデノウイルス由来ベクターストックの使用のリスクを最小化する試みにおいて、いくつかの非ヒトアデノウイルスの特徴が調べられ、ワクチン担体として開発されている(Soudois,Cら(2000)J.Virology,74:10639-10649;Farina,S.Fら(2001)J.Virology,75:11603-11613;Cohen,C.J.ら(2002)J.Gne.Virology,83:151-155)。既存の免疫に関連する問題を回避するための非ヒトアデノウイルス配列の使用に内在する前提は、共通ヒトアデノウイルス血清型に対する中和抗体が非ヒトウイルスを交差中和しないと思われる観察に基づくものである。しかし、ウイルス及び細胞性の要因の不適合により、非ヒト細胞株で増殖させなければならないという欠点を特徴とする、代替的ベクター系(ウシ、ヒツジ、イヌ)の大部分に対して、実用上の限界がある。
【0054】
Wilsonらは、チンパンジーの腸間膜リンパ節からもともと単離された、チンパンジーアデノウイルス タイプ68(CV68)C68に基づく複製欠損ベクターの特性を述べる報告を公表し(Basnight,Mら(1971)Am.J.Epidmiol.94:166−171)、CV68は、完全に配列決定され、ヒトアデノウイルスと全体的な構造が類似していることが分かった(Farina,S.Fら、J.Virol.75(23):11603−11613(2001))。このウイルスのゲノムは、36,521塩基対の長さであり、ヒトアデノウイルスのサブグループEと、最も類似しているものとして述べられており、配列決定されたヒトAd4オープンリーディングフレームの殆どと90%の同一性がある。CV68 ITRは、130塩基対の長さであり、主要なアデノウイルスの初期及び後期遺伝子の全てが存在する。CV68は、ヒトにおいて循環せず、共通ヒト血清型のものと交差反応する中和B細胞エピトープを欠く血清型を特徴とする。チンパンジーアデノウイルスがヒトアデノウイルスと類似しているにもかかわらず、チンパンジー血清型に対する交差反応性中和免疫は、ヒトにおいて立証されていない(Farina,S.F.ら、J.Virol.(2001)75(23):11603−13)。
【0055】
CV68由来の組み換えベクターは、コクサッキーウイルス及びアデノウイルス受容体を発現する細胞の形質導入を支持するのに十分にヒト血清型と類似であるものとして述べられている(Cohen,Cら、J.Gen.Virol.83:151−155(2002))。注目に値すべきは、CV68が、ヒトアデノウイルス5型からのE1を含有するその複製293細胞を支持するのに十分なレベルでヒトアデノウイルスと類似していることを特徴とすることである(Farina,S.F.ら、J.Virol.75(23):11603−11613(2001))。さらに、ヒト血清型5E1の隣接配列が、CV68由来ベクター配列のそれと非相同的であるという観察から、相同組み換えは起こらないと予想される。したがって、CV68由来ワクチンストックにRCAが混入する可能性は低いと推測される。
【0056】
同じ研究者グループは、続いて、マウスにおける、狂犬病ウイルス糖タンパク質に対する抗体の誘導のためのワクチン担体としての、CV68由来アデノウイルス配列の使用を報告した。CV68の複製欠損体は、E1A及びE1B遺伝子をミニジーンカセットで置換することにより作製された。狂犬病ウイルス糖タンパク質をコードする導入遺伝子産物を含有するE1欠失含有アデノウイルス組み換え体(AdC68rab.gp)で免疫付与したマウスは、狂犬病ウイルスに対する防御免疫を生じ、別の狂犬病ウイルスの致死用量を用いた病原体付与に対して耐性を保持した(Xiang,Zら、J.Virol.76(5):2667−2675(2002))。HIV−1のgagのコドン最適化した短縮型を発現する、第二のCV68コンストラクトは、最近、マウスにおいて活発なgag−特異的CD8+T細胞反応を誘導することが報告された。ワクチン誘導反応は、ワクシニアgag組み換えウイルスを用いた病原体投与に対して防御を与えることが示された(Fitzgerald,J.C.ら、J.Immunol.170:1416−1422(2003))。CV68gag又はAd5gagベクターを用いて、ヒトアデノウイルス血清型5に対して前もって免疫付与したマウスの実験的ワクチン接種から、CV68ワクチンによるものよりも、Ad5によって誘発されるウイルス病原体付与に対して、gag特異的T細胞及び防御が顕著に低下することが示された。C68gagワクチンの効率の低下は、Ad5特異的CD8+T細胞(Id)の交差反応性により生じた。
【0057】
このデータと合わせて考えると、サル由来複製欠損アデノウイルスベクターが、共通ヒト血清型に基づくベクターよりも、ヒトワクチン担体としての使用により適している可能性があることが示唆される。本明細書中で示されるように、ヒトAd5(図36)に対して強く免疫付与されたマウスを、ChAd3−gagアデノウイルスベクターで免疫付与できるという実験結果から、既存の抗ヒトAd5免疫が、ChAdベクターにより誘発されるgag特異的CMI反応を低下させなかったことが示される。これらの結果は、ヒトAd5交差反応性B及びT細胞エピトープが、ChAd3又はChAd6ベクターにおいて存在しないという結論と合致する。
【0058】
一般に、アデノウイルスゲノムは、よく調べられており、いくつかの異なる血清型が存在するにもかかわらず、特異的機能が同じように位置しつつアデノウイルスゲノムの全体的構成において、いくつかの共通の保存がある。Wilsonらにより開示されたチンパンジーアデノウイルス C1及びCV68のヌクレオチド配列及び各ウイルスのE1A、E1B、E2A、E2B、E3、E4、L1、L2、L3、L4及びL5遺伝子の位置は、米国特許第6,083,716号(Chimpanzee Adenovirus Vectors)及びPCT公開出願WO03/000851(Methods for Rapid Screening of Bacterial Transformants and Novel Simion Adenoviral proteins.)により与えられる(これらの教示は、参照により本明細書中に組み込む。)。
【0059】
アデノウイルスゲノムの端部には、逆方向末端反復(ITR)として知られている配列が含まれ、これは、ウイルス複製に必要である。ウイルスはまた、ウイルスコードプロテアーゼを含有し、これは、感染性ビリオンを生成するのに必要とされる構造タンパク質の一部のプロセシングに必要である。アデノウイルスゲノムの構造は、宿主細胞の形質導入後、ウイルス遺伝子が発現される順番で述べられる。より具体的には、ウイルス遺伝子は、転写がDNA複製開始の前に起こるか、後に起こるかに従い、初期(E)又は後期(L)遺伝子と呼ばれる。形質導入の初期において、アデノウイルスのE1、E2、E3及びE4遺伝子が、ウイルス複製に対して宿主細胞を準備するために発現される。このウイルスは、ウイルスゲノムの必須の初期領域1(E1)を欠失させることにより、複製欠損にされ得る。Brodyら、1994 Ann NY Acad Sci.,716:90−101。後期の間、ウイルス粒子の構造要素をコードする、後期遺伝子L1−L5の発現のスイッチが入る。後期遺伝子は全て、1個のプロモーターの調節下となり、ペントン(L2)、ヘキソン(L3)、100kDa 足場タンパク質(L4)及び繊維タンパク質(L5)を含むタンパク質をコードし、これらは、アデノウイルスDNAがキャプシド化されるようになる、新しいウイルスを形成する。ウイルスの血清型に依存して、1個の宿主細胞において、10,000から100,000の子孫アデノウイルス粒子が生じ得る。最終的に、アデノウイルス複製プロセスにより、細胞溶解が起こる。
【0060】
本明細書中で開示される複製欠損アデノウイルスベクターは、開示されているチンパンジー核酸配列からの特定のヌクレオチド配列の欠失により、及び導入遺伝子の挿入、発現又はその他の遺伝子操作に有用な他のDNA配列由来の配列の挿入により構築される。したがって、本明細書中で述べる組み換えチンパンジーアデノウイルスは、1又は複数のチンパンジーアデノウイルス由来のアデノウイルス配列又はチンパンジーアデノウイルスもしくはヒトアデノウイルス由来の配列を含有し得る。適切なポリヌクレオチド配列は、組み換えにより、合成により、又は天然ソースから単離して、作成され得る。本発明の特定の局面における使用に適切なアデノウイルス配列には、共通ヒト血清型と交差反応する中和B細胞エピトープを欠失している配列が含まれる。
【0061】
最低限、本発明の組み換えチンパンジーアデノウイルス(例えばベクター)は、少なくとも1個の免疫原発現カセットと組み合わせて、複製及びビリオンのキャプシド化に必要なチンパンジーアデノウイルス cis作用エレメントを含有する。通常、このcis作用エレメントは、少なくとも1個の抗原をコードする導入遺伝子を含む発現カセットに隣接する。より具体的に、本発明のベクターは、アデノウイルスの、不可欠なcis作用5’逆方向末端反復(ITR)配列(これは、複製の開始点として機能する。)、3’ITR配列、パッケージング/エンハンサードメイン及び異種分子をコードするヌクレオチド配列を含有する。本発明のベクターは、組み換えベクターが、最少のアデノウイルス配列のみを含有するか、特定遺伝子における機能的欠失(例えばE1及び/又はE3又はE4領域)のみがある全体のアデノウイルスゲノムを含有するかに関わらず、チンパンジーアデノウイルスキャプシドを含有する。
【0062】
一般に、本明細書中で開示するアデノウイルスベクターは、複製欠損アデノウイルスゲノム(このアデノウイルスゲノムは、機能的E1遺伝子を持たない。)と;a)免疫反応が望まれる少なくとも1個の免疫原をコードする核酸と、b)該免疫原をコードする核酸配列と機能可能に連結された異種(つまり、アデノウイルス配列に関して)プロモーターとを含有する免疫原発現カセットと;転写終結因子と、を含有する。
【0063】
より具体的には、本発明は、E1、E2、E3及びE4からなる群から選択される、少なくとも1個の初期遺伝子を欠く、組み換えアデノウイルスゲノムからなる複製欠損ベクターを提供する。ある実施形態において、この複製欠損ベクターは、本明細書中で開示するアデノウイルス分離株の1つ(例えば、ChAd3、ChAd6、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63又はChAd82)のE1遺伝子を、免疫原発現カセットで置換又は破壊することにより調製される。例えば、ChAd3(配列番号1)又はChA6(配列番号2)のE1遺伝子を欠失させる/破壊することにより、ベクターを調製することができる。あるいは、ChAd3、ChAd6、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82又はChAd20を含む、本明細書中で開示する他のアデノウイルス分離株のいずれか1つから、複製欠損ベクターを調製することができる。他の実施形態において、本発明の複製欠損ベクターは、機能的E3遺伝子を欠失するように、場合によっては加工されている、本明細書中で開示するChAdのうち1個に由来するアデノウイルスゲノムを含む。初期遺伝子を完全に除去するか、又はその遺伝子を非操作可能又は非機能的にすることによって、本明細書中で開示するチンパンジーアデノウイルス配列は、複製欠損になされうると理解されたい。
【0064】
本発明は、1又は複数の選択された遺伝子産物を発現するアデノウイルスの能力を破壊する「機能的欠失」などの改変があることを特徴とするベクターを包含すると理解されたい。本明細書中で使用される場合、「機能的欠失」という用語は、広く、特定の遺伝子産物を非機能的にする効果を有する改変を包含する。一般に、機能的欠失は、アデノウイルス遺伝子の部分的又は全体的欠失の形をとる。しかし、これらに限定されないがフレームシフト突然変異を導入する改変を行うことを含む、その他の操作でもまた機能的欠失を達成することを当業者は容易に認めるであろう。例えば、本発明の組み換えチンパンジーアデノウイルスベクターは、本ウイルスのアデノウイルスE1A及び/又はE1B発現能力を妨害する、又は機能的に欠失させるよう設計されている改変を導入することにより、複製欠損になされ得る。
【0065】
E2遺伝子の群からの1又は複数の遺伝子の発現を妨害する、又は機能的に欠失させるよう設計されている改変を導入することにより、複製欠損アデノウイルスベクターを得ることができることは周知である。より詳しく述べると、ポリメラーゼ遺伝子又は、末端直前のタンパク質遺伝子もしくはDNA結合タンパク質遺伝子を不活性化することにより、複製欠損ベクターを構築することができる。さらに、E4領域により発現される遺伝子の欠失又は不活性化は、複製欠損 chimp Adベクターを構築するための代替的方法である。初期遺伝子欠失又は不活性化を、さらに短縮されたベクターを作製するために、組み合わせることができる。あるいは、複製欠損ChAdベクターはまた、後期遺伝子L1からL5などの他のウイルス遺伝子において、さらなる改変を含み得る。さらに、異なる血清型から得られたヘキソン及び繊維遺伝子を組み合わせることにより、新規アデノウイルスワクチン担体を生成させることができる。ヘキソン及び繊維遺伝子シャッフリング法の利用によってもまた、研究者らは、ChAdの生物学的特性を変化させ、様々な指向性を有する、又は新しい血清学的特性を有する、ベクターの作製を促進することができる。
【0066】
本発明は、単独で、又は何らかの組合せで、その改変遺伝子の生物学的活性を効果的に破壊する、遺伝子全体又はその一部の欠失を含む組み換えアデノウイルスベクターを包含すると理解されたい。本明細書中で示されるように、E1遺伝子の機能的欠失と組み合わせて、異種のAd5 E4配列をベクターに導入することは、望ましいものであり得るが、例えば、E4領域により発現される遺伝子の機能的欠失を有する組み換えサルアデノウイルスを構築することができる。あるいは、アデノウイルス 遅初期(delayed early)E3遺伝子の機能を削除し得るが、E3の機能は、組み換えアデノウイルス粒子の産生に必要ではないため、本発明で有用なウイルスをパッケージングすることが可能な組み換え体を作製するために、この遺伝子産物を置換する必要はない。
【0067】
本発明のある実施形態において、使用される複製欠損アデノウイルスベクターは、E1及び場合によってはE3における欠失を含有する、チンパンジーサブグループCアデノウイルスである。例えば、ChAd3の場合、適切なE1欠失/破壊は、bp460からbp3542の領域に導入することができる(配列番号1を参照。)。ChAd6の場合、適切なE1欠失/破壊は、bp457からbp3425の領域に導入することができる(配列番号2を参照。)。CV32の場合、E1欠失は、好ましくは、bp456からbp3416である(配列番号3を参照。);CV33の場合、E1欠失は、好ましくは、bp456からbp3425(配列番号4を参照。)であり、CV23の場合、E1欠失は、好ましくは、bp456からbp3415(配列番号5を参照。)である。CV32及びCV33に対するE3欠失は、好ましくは、それぞれ、bp27446からbp31911(配列番号3を参照。)であり、bp27146からbp31609(配列番号4を参照。)である。当業者は、配列相同性及び多配列アラインメントに基づき、他のチンパンジー分離株に対する同等な配列を容易に決定することができる。
【0068】
E1欠失アデノウイルスベクターを構築するために、そのベクター骨格の構造及び導入遺伝子を含有する核酸配列の組成に関して多くのことを決定しなければならないことを、当業者は、容易に認めるであろう。例えば、研究者らは、E1欠失の大きさが導入遺伝子の大きさに適合するか否かを調べなければならない。適合しない場合、そのベクターの骨格にさらなる欠失を導入しなければならない。
【0069】
導入遺伝子を有する核酸配列は、遺伝子又は遺伝子の機能的部分であり得、通常は、発現カセットの形態で存在する。通常、遺伝子発現カセットには、(a)タンパク質又は関心のある抗原をコードする核酸と;(b)そのタンパク質をコードする該核酸に機能可能に連結された異種のプロモーターと;(c)転写終結シグナルと、が含まれる。本核酸は、DNA及び/又はRNAであり得、2本鎖又は1本鎖であり得る。この酸は、所望する宿主における発現のために、コドン最適化され得る(例えば、哺乳動物宿主)。
【0070】
導入遺伝子が導入される骨格内の部位及びその導入遺伝子の方向に関しても決定を行わなければならない。より具体的に、E1パラレル(5’から3’に転写)又は反パラレル(そのベクター骨格に対して、3’から5’方向に転写)方向で、導入遺伝子を挿入することができる。さらに、そのベクターが、同定されその導入遺伝子に機能可能に連結される必要があるワクチン担体としての使用のために調製されている、哺乳動物宿主細胞においてその導入遺伝子の発現を支配することが可能な適切な転写調節エレメントである。「操作可能に連結された」配列には、それらが調節する核酸配列に隣接する発現調節配列及び、トランスで作用する、又は調節される核酸配列を調節するのに離れて作用する調節配列の両方が含まれる。
【0071】
調節配列には、転写開始、終結、エンハンサー及びプロモーター配列などの適切な発現調節配列;スプライシング及びポリアデニル化シグナルなどの効率的RNAプロセシングシグナル;翻訳効率を促進する配列(例えば、コザックコンセンサス配列);タンパク質安定性を促進する配列及び場合によっては、タンパク質分泌を促進する配列が含まれる。これら及び他の共通のベクターエレメントの選択は、従来のものであり、多くの適切な配列が当業者にとり周知である(例えば、Sambrookら、及びそこで引用する文献、例えば、3.18−3.26ページ及び16.17-16.27、及びAusubelら、Current Protocols in Molecular Biology,Jhon Wiley&Sons,New York,1989)。
【0072】
具体的な実施形態において、本プロモーターは、真核RNAポリメラーゼにより認識される異種のプロモーター(つまり、アデノウイルス配列に対して。)である。好ましい実施形態において、このプロモーターは、「強力」又は「効率的」プロモーターである。強力なプロモーターの例は、前初期ヒトサイトメガロウイルスプロモーターである(Chapmanら、1991 Nucl.Acids Res 19:3979−3986(参照により組み込む。))。(CMV)なしで、又はイントロンA配列(CMV−intA)とともに、ヒトCMVプロモーターを使用することができるが、EF1αプロモーター、マウスCMVプロモーター、ラウス肉腫ウイルス(RSV)プロモーター、SV40初期/後期プロモーター及びβアクチンプロモーターを含む、強力な免疫グロブリン又はその他の真核遺伝子プロモーターなどのその他の既知のプロモーターの多くのいずれかを使用し得ることを当業者は、認識するであろう。
【0073】
本発明において使用できるプロモーターのさらなる例は、強力な免疫グロブリンプロモーター、EF1αプロモーター、マウスCMVプロモーター、ラウス肉腫ウイルスプロモーター、SV40初期/後期プロモーター及びβアクチンプロモーターであるが、本発明の方法に従い、意図する宿主において発現を実行することができるあらゆるプロモーターを使用することができることを、当業者は認め得る。このプロモーターは、Tetオペレーター配列などの調節可能配列を含み得る。転写及び発現の調節の可能性を与えるこれらのような配列は、遺伝子転写の抑制が求められる場合に有用である。
【0074】
適切な遺伝子発現カセットはまた、転写終結配列も含む。好ましい転写終結因子は、ウシ成長ホルモン終結因子である。CMV intA−BGH終結因子の、プロモーター/転写終結の組合せは、特に好ましいが、その他のプロモーター/終結因子の組合せもまた使用し得る。本明細書中で示されるように、ウシ成長ホルモン終結/ポリアデニル化シグナル(bGHpA)又は次のように定義される50ヌクレオチド長の短い合成polyAシグナル(SPA):即ちAATAAAAGATCTTTATTTTCATTAGATCTGTGTGTTGGTTTTTTGTGTG(配列番号26)である。一般に、適切な終結配列を例示する。導入遺伝子を含有する核酸配列の後であって、E3’アデノウイルスITR配列の前に、polyAシグナルを挿入する。
【0075】
本明細書中で述べられる組み換えアデノウイルスベクターには、アデノウイルスの1又は複数の株由来のアデノウイルス配列が含まれる。適切な配列は、天然のソースから得ることができ、組み換えにより、合成により、又は他の遺伝子操作もしくは化学的方法により作製することができる。特定の実施形態において、この組み換えチンパンジーアデノウイルスは、非チンパンジーアデノウイルスポリヌクレオチド配列を含有するキメラ組み換え体である。適切な非チンパンジーアデノウイルス配列は、ヒトアデノウイルス株から得ることができる。例えば、hAd5 E4(Ad5 nt32816からnt35619)又はAd5E4 orf6(Ad5 nt33193からnt34077)(Ad5 GenBank Accession No:M73260)により、ネイティブのE4領域を置換することができる。
【0076】
一般に、本発明の組み換えアデノウイルスベクターにより送達される免疫原(抗原性分子)は、コードされる産物の転写及び/又は翻訳を宿主細胞において指示するのに必要な調節配列を与えるヌクレオチド配列と組み合わせて、導入遺伝子によりコードされるポリペプチド、タンパク質又は酵素産物を含有する。この導入遺伝子の組成は、ベクターの意図する使用に依存する。例えば、哺乳動物宿主において、感染物質に対して特異的な、抗体反応又は細胞性免疫反応を誘発するために免疫原性組成物が設計されている場合、病原体特異的免疫を受容者に与えると予想される少なくとも1個の免疫原性産物をコードする核酸配列を利用することが適切である。あるいは、癌ワクチンとしての使用のためにその組成物が調製される場合、適切な導入遺伝子は、TAAなどの自己抗原の免疫原性部分を含有し得、それは、特定のTAAに対する宿主寛容性を打破するため、及び癌の発症を防ぐか、又は腫瘍成長を防ぐのに十分な長期(例えば記憶)反応を誘発するために十分な効力の防御的免疫反応を誘発する目的に関して選択される。したがって、哺乳動物宿主に感染する幅広い病原性物質(これらに限定されないが、ウイルス、寄生生物、細菌及び真菌など。)又は癌もしくは腫瘍細胞から、適切な免疫原性遺伝子産物を得ることができる。試験免疫原の特定の設定で本明細書中において本発明を説明しているが、本発明は、本明細書中で例示される抗原の使用に限定されないことを理解されたい。より具体的には、本発明は、これらに限定されないがTAAなどの、免疫原としての、異種及び自己抗原の両方の使用をもくろむ。
【0077】
ある実施形態において、本発明は、感染物質により発現される抗原(つまり免疫原)に対する免疫反応を誘導する、免疫原性組成物(例えばワクチン)を提供する。例えば、ヒト及び/又は非ヒト動物種に感染するウイルスに対する免疫反応を誘発することが望ましい。予防的及び/又は治療的免疫反応が望まれるウイルスファミリーの例には、アフソウイルス、カルディオウイルス、エンテロウイルス、へパトウイルス、パレコウイルス、リノウイルスなどの6種類の異なる属を含むピコルナウイルス科(Picornaviridae)ファミリーが含まれる。免疫反応が望まれるピコルナウイルスの例は、口蹄疫ウイルス、脳心筋炎ウイルス、ポリオウイルス、コクサッキーウイルス、ヒトA型肝炎ウイルス、ヒトパレコウイルス、リノウイルスである。カリシウイルス科(Caliciviridae)には、ウイルスのノーウォーク群により生じるヒトにおける流行性胃腸炎及び、ウサギ出血性疾患ウイルスに関するウサギにおける出血性疾患及びネコカリシウイルスにより生じる呼吸器疾患のような、動物におけるその他の疾患を伴う、様々な属が含まれる。
【0078】
免疫反応を誘発することが望まれ得るウイルスの別のファミリーは、ヒトならびに多くの様々な動物種から単離された、ウイルスを含むアストロウイルス科(Astroviridae)である。ヒトアストロウイルスは、胃腸炎及び低年齢児の下痢に関連する。あるいは、2つの属:アルファウイルス及びルビウイルスを含む、ウイルスのトガウイルス科(Togaviridae)のメンバーに対する免疫を哺乳動物宿主に付与することが望まれ得る。アルファウイルスは、関節炎(つまり、チックングニアウイルス、シンドビスウイルス)又は脳炎(つまり、東部ウマ脳炎ウイルス、西部ウマ脳炎ウイルス)などの、ヒト及び獣医学の疾患に関与する。
【0079】
風疹ウイルスは、代替的なウイルス標的を与え、ルビウイルス属のメンバーのみが、発熱及びリンパ節炎を伴う軽度発疹疾患の流行に関与する。風疹ウイルス感染はまた、妊娠初期に母親が感染した際の、胎児の異常にも関与する。フラビウイルス科(Flaviviridae)は、重要なヒトならびに動物病原体を含む、3種類の属:フラビウイルス属、ペスチウイルス属及びヘパチウイルス属からなる別のウイルスファミリーである。フラビウイルス属メンバーの多くは、発熱、脳炎及び出血熱を含む、様々な疾患を引き起こす、節足動物媒介性ヒト病原体である。デング熱ウイルス、黄熱病ウイルス、日本脳炎ウイルス、西ナイル熱ウイルス、ダニ媒介性脳炎ウイルスは、世界中が関心を持っている、又はある地域(風土病的)で関心が持たれている主要な病原体である。ペスチウイルス属には、ウシウイルス性下痢ウイルス、ブタコレラウイルス、ボーダー病ウイルスなどの、経済的に非常に重要な動物病原体が含まれる。C型肝炎ウイルスは、急性及び慢性肝炎に関与する、ヘパチウイルス(Hepaci virus)属の唯一のメンバーである。組み換えアデノウイルスにより発現されるHCVタンパク質は、世界中で1億7千万人を侵すウイルス感染の結果を制限する、防御的ならびに治療的免疫反応を誘発することができる。
【0080】
あるいは、感染に対する防御を獲得するために、組み換えアデノウイルスベクターにより、コロナウイルス科(Coronaviridae)のメンバー由来の抗原を発現させることができる。重症急性呼吸器症候群コロナウイルス(SARS−Coウイルス)に対する防御は、これらに限定されないが、ヌクレオキャプシド(N)タンパク質、ポリメラーゼ(P)タンパク質、膜(M)糖タンパク質、スパイク(S)糖タンパク質、小型エンベロープ(E)タンパク質又はそのウイルスにより発現されるその他の何らかのポリペプチドを含む、1又は複数のSARS Covタンパク質を発現する、ChAd3、4、5、6、7、9、10、11、16、17、19、20、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82を含む群から選択される1又は複数のチンパンジーアデノウイルスで免疫付与することにより獲得することができる。狂犬病ウイルスを含むラブドウイルス科(Rhabdoviridae)メンバーは、ウイルスタンパク質を発現する組み換えワクチンの標的であり得る。
【0081】
その他の可能な標的には、重症出血熱の流行に関与する、エボラ様ウイルス及びマルブルグ様ウイルス属が含まれるフィロウイルス科(Filoviridae);麻疹、呼吸器合胞体ウイルス、パラインフルエンザウイルスのような、ヒトにおいて知られる最も流行しているいくつかのウイルス及びニューカッスル病及び牛疫ウイルスなどの獣医学的に関心のあるウイルスを含むパラミクソウイルス科(Paramyxoviridae);インフルエンザA、B、Cウイルスを含むオルトミクソウイルス科(Orthomyxoviridae);リフトバレー熱、Sin Nombre(シンノンブレ)、ハンタ、プーマラウイルスなどの重要なヒト病原体を含む、節足動物から脊椎動物宿主に主に伝染するブニヤウイルス科(Bunyaviridae);リンパ球性脈絡髄膜炎、ラッサ熱、アルゼンチン出血熱、ボリビア出血熱ウイルスを含む、アレナウイルス科(Arenaviridae);主にウマ及びヒツジにおいて中枢神経疾患を引き起こすウイルスを含む、ボルナウイルス科(Bornaviridae);世界中での幼児及び低年齢児における重症下痢疾患の最も重要な原因である、ロタウイルスファミリー及び、ヒト及び他の動物両方に伝染し得るオルビウイルス(ブルータング病、伝染性出血性疾患ウイルス)を含む、レオウイルス科(Reoviridae);ヒト免疫不全ウイルス1及び2(HIV−1及びHIV−2)及びヒトT細胞白血病ウイルスタイプE1及び2(HTLV1及び2)のような、重要なヒト病原体ならびにヒツジ及びヤギに伝染するマエディ/ビスナウイルス、ウマに伝染するウマ感染性貧血ウイルス、ウシに伝染するウシ免疫不全ウイルス、ネコに伝染するネコ免疫不全ウイルスなどの非ヒトレンチウイルスを含むウイルスの大きなグループである、レトロウイルス科(Retroviridae);ポリオーマウイルス科(Polyomaviridae)グループ小型DNA癌原性ウイルス、典型ウイルスは、それぞれマウス及びアカゲザルに感染するポリオーマ及びSV40、(SV40に非常に関連しているBK及びJCウイルスは、ヒト患者から単離された。);パピローマウイルス科(Papillomaviridae)は、いぼ及びコンジローマを発生させる、ヒトを含む高等脊椎動物に感染するDNAウイルスの群からなる。パピローマウイルス感染は、ヒト及び動物両方での癌の発生に関連する。ヒトパピローマウイルスは、子宮頸癌、膣癌及び皮膚癌に関与する。ヘルペスウイルス科(Herpesviridae)は、ヒト及びその他の哺乳動物に対するいくつかの重要な病原体が分類される亜科を含む。抗原の適切なソースは、これらに限定されないが、単純ヘルペスウイルス1及び2、水疱瘡ウイルス、エプスタインバーウイルス、サイトメガロウイルス、ヒトヘルペスウイルス6A、6B及び7、カポジ肉腫関連ヘルペスウイルスである。抗原のさらに適切なソースは、サル痘ウイルス、伝染性軟属腫(Molluscum contagisum)ウイルス、天然痘ウイルスのようなポックスウイルス科(Poxviridae)のメンバー;へパドナウイルス科(Hepadnaviridae)の典型メンバーである、B型肝炎ウイルスならびに、D型肝炎(hepatitis delta)ウイルス、E型肝炎ウイルスのような、急性及び/又は慢性肝炎を引き起こすその他のウイルスである。
【0082】
本発明のアデノウイルスベクターはまた、細菌、真菌、寄生生物病原体を含む非ウイルス性病原体により発現されるタンパク質に対して、ヒト又は動物において免疫反応を刺激するように設計される免疫原性組成物の調製に適切である。例えば、本明細書中で開示するベクターは、これらに限定されないが、Staphylococcus aureus(黄色ブドウ球菌)、Streptococcus pyogenes(化膿レンサ球菌)、Streptococcus Ppneumoniae(肺炎レンサ球菌)、Vibrio cholerae(コレラ菌)、Clostridium tetani(破傷風菌)、Neisseria meningitis(髄膜炎菌)、Corynebacterium diphteriae(ジフテリア菌)、Mycobacteria tuberculosis(マイコバクテリウム・テュバキュローシス))、Mycobacteria leprae(ライ菌)、Listeria monocytogenes(リステリアモノサイトジェン)及びLegionella pneumofila(レジオネラニューモフィラ)に対するワクチンを調製するために使用することができる。予防的又は治療的ワクチンを調製することが望まれ得る、真菌及び哺乳動物寄生生物の例には、Candida albicans(カンジダアルビカンス)、Aspergillus fumigatus(アスペルギウス・フミガタス)、Histoplasma capsulatum(ヒストプラズム・カプスラーツス)、Plasmodium malariae(四日熱マラリア原虫)、Leishmania major(リーシュマニア・メジャー)、Trypanosome cruzi(トリパノソーマ・クルジ)及びbrucei(ブルシェイ)、Schistosoma haematobium(ビルハルツ住血吸虫)、mansoni(マンソン住血吸虫)及びjaponicum(日本住血吸虫);Entamoeba histolytica(エンタモエバ・ヒストリチカ)及びヒトフィラリアに関与することが知られているフィラリアの多数の種が含まれる。
【0083】
癌は、一般に、細胞周期の維持又は細胞増殖の調節に関与するポリペプチドをコードする遺伝子の脱制御を含む(例えば、増殖因子、癌遺伝子、受容体及び腫瘍抑制因子)。癌に関わる遺伝子の多くの産物は、様々な腫瘍細胞の表面で発現される。T及びBリンパ球により認識され得る様々な腫瘍抗原が、ヒト及び動物の癌で同定されている。腫瘍細胞において発現されるのに加えて、正常組織及び/又は胎児発生中にも発現されるため、抗癌ワクチン試験での使用に適切なヒト腫瘍関連抗原(TAA)の大部分は、「自己抗原」として説明される。TAAに対する標的集団の免疫寛容性により、多くの癌ワクチンが効果がないとされる理由が説明される。
【0084】
腫瘍抗原は、T/B細胞反応を刺激することができる、変化した細胞性タンパク質の例である、Ras、p53又はBcr−Ab1タンパク質などの、癌原遺伝子又は腫瘍抑制遺伝子で変化が起こる正常細胞遺伝子の発癌突然変異体により産生され得る。腫瘍抗原は、腫瘍細胞で過剰発現されるか(チロシナーゼ、GP100、MARTは、メラニン産生細胞において低レベルで正常に発現され、メラノーマにおいて過剰発現される。)、又は腫瘍細胞において異常に発現される(メラノーマ及び多くの癌腫において発現されるが、精巣及び胎盤において正常に発現される、MAGE、BAGE、GAGE。)。腫瘍抗原は、発癌ウイルスの産物であり得る:子宮頸部癌により発現される、パピローマウイルスE6及びE7タンパク質;EBV+リンパ腫及び上咽頭癌により産生される、EBV EBNA−1タンパク質;SV40誘導実験的腫瘍におけるSV40 T抗原。癌胎児性抗原は、癌細胞及び正常な発生過程(胎児)の組織において高レベルで発現されるが、成人組織では発現されない。癌胎児性抗原(CEA)及びα−フェトタンパク質(AFP)は、よく特性が調べられている癌胎児性抗原の例である。
【0085】
最近の証拠により、免疫反応を誘発する、したがってワクチン治療に対する免疫原に適切な抗原のクラスを生じさせることができるTAAの存在が裏付けられている。しかし、抗原のクラスとして、TAAは、周知のとおり弱い免疫原であり、TAAに非常に特異的なT細胞は、T細胞発生の過程で欠失するか、アネルギー化されている。したがって、特定のTAAに対する、腫瘍を有する宿主の免疫反応は非常に弱いと予想される。標的TAAに対する宿主の寛容性を打破するという固有の必要性があるため、実験的臨床ワクチン研究は、特に、TAA特異的T細胞反応を促進するTAA免疫付与ストラテジーの開発に焦点を当てている。一般に、効果的な癌ワクチンは、免疫寛容性を乗り越え、かつ、宿主の免疫反応を、予防的及び/又は防御的なレベルに上げなければならない。多くの実験的ワクチン研究における抗腫瘍効果は、TAAに対するT細胞反応に相関がある。
【0086】
代替的実施形態において、本発明は、腫瘍抗原に対する免疫反応を誘導するために使用することができる、免疫原性組成物(例えば、癌ワクチン)をもくろむ。適切な組成物は、腫瘍抗原をコードする核酸配列を含有する組み換えチンパンジーアデノウイルス及び生理学的に許容可能な担体を含有する。特定の実施形態において、カセットのコード配列エレメントは、癌関連抗原などの自己抗原由来の免疫原性ペプチド配列などの、1個の免疫原をコードし得る。ある実施形態において、免疫原をコードする核酸配列(つまり導入遺伝子)は、特定の哺乳動物種での発現のためにコドン最適化され得る。他の実施形態において、コード配列は、1又は複数のコドン最適化された腫瘍抗原などの、複数の免疫原をコードし得る。例えば、開示するアデノウイルスベクターを利用する癌ワクチンは、HER2/neu、CEA、Hepcam、PSA、PSMA、テロメラーゼ、gp100、Melan−A/MART−1、Muc−1、NY−ESO−1、サバイビン、ストロメリシン 3、チロシナーゼ、MAGE3、CML68、CML66、OY−TES−1、SSX−2、SART−1、SART−2、SART−3、NY−CO−58、NY−BR−62、hKLP2、VEGFなどの、自己抗原の組合せをコードし得る。
【0087】
効果的な癌ワクチンの開発には、ワクチン接種された患者において抗原特異的免疫を誘発するストラテジー及び活性のある免疫付与終了後に免疫反応の生成が持続することの検証が必要である。このストラテジーの成功は、標的抗原に対して向けられる測定可能な免疫反応が、癌発生又は再発に対する防御に相関するか否かに依存する。細胞性免疫及び液性免疫両方のエフェクターメカニズムにより、腫瘍細胞が殺滅されることが示されている。しかし、実験系からのデータにより、腫瘍細胞の除去に対して、抗原特異的T細胞が、最も強力な免疫メカニズムを与えることが示唆される。腫瘍特異的抗原(例えばTAA)のエフェクターT細胞による認識により、TAAが腫瘍拒絶抗原として機能することができるようになると予想される。公表されている実験から、CD8+及びCD4+ヘルパーT細胞反応の刺激が、最善の腫瘍クリアランスの達成に重要であることが示唆されている(Greenberg,P.D.(1991)Adv.Immunol.49:281−355;Pardoll,D.M.ら(1998)Curr.Opin.Immunol.10:588−94)。臨床反応(つまり効能)は、インターフェロンγ分泌細胞毒性T細胞の増加と関与する。本ワクチン担体の効率を明らかにするための、本明細書中で使用される、ELISPOTアッセイなどのアッセイの出現により、研究者らは、ワクチン接種計画に対するT細胞反応を測定できるようになり、それにより癌ワクチンの開発が促進される。
【0088】
癌ワクチンは、予防的又は治療的の何れかであり得る。癌ワクチンの予防的使用の基礎をなす一般的な仮定は、腫瘍を有する対象において、TAAが非常に弱い免疫原であるか機能的に非免疫原性であるということである。より具体的には、癌免疫学の分野において、癌に罹患した患者における免疫療法として、ワクチンを使用することができる。したがって、既存の腫瘍又は悪性腫瘍により発現されるTAAに向けられる免疫反応を誘発するように、癌ワクチンを設計することができる。したがって、特定の実施形態ににおいて、治療的癌ワクチンは、従来の治療法に対する耐性が進展しているか、従来の治療後に再発が起こる可能性が高い、腫瘍を有する患者における使用を意図する。
【0089】
アデノウイルスの高い免疫原性により、アデノウイルスベクターは、自己抗原に対する宿主寛容性を打破するために設計されたワクチン担体の関連における使用のための、特に優れた候補となる。自己免疫疾患において最初に記載された、エピトープ又は決定基拡大の現象は、MHCクラスI及びMHCクラスII限定反応の両方と関与し、HER2 neuタンパク質特異的T細胞免疫の発生と相関がある。エピトープ拡大は、免疫原性タンパク質の特定部分に対する免疫反応の生成を表し、次いで、同じタンパク質に存在する、他の抗原性決定基に対する免疫の自然な拡大が起こる。例えば、Disisらは、顆粒球−マクロファージコロニー刺激因子と混合した、HER−2タンパク質の強力なTヘルパーエピトープ由来のペプチドを含有するワクチンを投与した、HER−2/neu過剰発現悪性腫瘍がある患者の84%でエピトープ拡大を観察した(J.Clin.Oncol.(2002)20(11):2624-2632)。重要なことは、エピトープ拡大が、HER−2/neuタンパク質ドメイン反応の生成と相関したことであり、このことから、免疫付与が効果的に免疫寛容性を回避したことが示唆される。
【0090】
癌ワクチンに対する標的としての、開示するアデノウイルスベクター及び方法の使用に適切なTAAは、いくつかの特徴を保持すべきである。例えば、標的TAAは、好ましい発現プロファイルを有さなければならず、これは、正常組織と比較して、腫瘍又は悪性腫瘍組織において、選択的に発現されるか、過剰発現されることを意味する。さらに、腫瘍発生に関与するTAAは、癌進行の様々な段階で保持される可能性がより高いので、適切な標的TAAはまた、腫瘍進行及び転移を通じて保持されるべきである。適切な標的TAAはまた、腫瘍内で均一に発現される。第三に、適切な標的TAAは、絶対的な免疫寛容性に対する対象であってはならない。より具体的には、関心のあるTAAに対して認識し、かつ反応することができるT細胞は、宿主のT細胞レパートリーから完全に欠失されていないことのいくつかの証拠があるべきである(Berinstein,N.L.,J.Clin.Oncol.29(8):2197(2002))。
【0091】
癌胎児性抗原(CEA)は、抗癌ワクチンのための標的抗原としての使用のために魅力的なTAAとなる多くの特徴を有する。これは、有利な発現パターンを特徴とする、Igスーパーファミリーのメンバーである。これは、全ヒト癌の50%を超えるものにおいて、発現され、腫瘍発生プロセスに関わっており、このことから、その発現が、癌進行を通して、選択され、保持され得ることが示唆される。さらに、CEAに対する免疫寛容性が絶対的ではないことが分かっている。公表された研究から、ヒトT細胞が、CEAを発現する癌細胞を、認識し、活性化し、溶解させることが分かっている(Berinstein,N.L.,J.Clin.Oncol.29(8):2197(2002))。例えば、その後のペプチドべースのインビトロ刺激と組み合わせた、CEAを発現する組み換えワクシニアウイルスを用いた患者への免疫付与により、自己腫瘍を細胞溶解できる、CD8+MHC−限定CTLが生成した(Tsang,K.Y,ら、J.Natl.Cancer Inst.,(1995)87:982−990)。あるいは、手術後の結腸直腸癌患者に対する組み換えCEAを用いた免疫付与が、組み換えCEAに対する、弱い抗体及び細胞性応答を誘導したことが報告された(Samanci,Aら(1998)Cancer Immunol.Immunother.47:131−142)。さらに、結腸直腸癌と診断された患者への抗CEA 抗イディオタイプ抗体の投与により、抗CEA抗体が生じ、イディオタイプ特異的T細胞増殖が起こった(Foon,L,A.ら(1995)J.Clin.Invest.,96:334−342)。文献により、また、いくつかの異なるワクチン接種法を用いて、癌患者におけるCEAに対する寛容性を乗り越えることができることが示される(つまり、組み換えCEA又は組み換えオルトポックスもしくはアビポックスCEAウイルスを用いたワクチン接種、抗イディオタイプ抗体の投与、CEAアゴニストエピトープによる樹状細胞のパルシング)。
【0092】
CEAは、正常胎児結腸で発現される、正常結腸粘膜では非常に低い、癌胎児性糖タンパク質である。これは、多くの腺癌、特に結腸、膵臓、乳房、肺、直腸及び胃の腺癌においても過剰発現される。多くの結腸直腸癌及びある癌腫は、血清中で測定可能な高レベルのCEAを産生し、それにより、特に結腸直腸癌の患者において、最も広く使用される悪性の血清マーカーの1つとなっている。
【0093】
本発明の組成物及び方法における使用のための適切な免疫原を提供する第二のTAAは、HER2/erb−2(neuとも呼ばれる。)癌原遺伝子の産物である。CEAのように、HER2/neuは、有利な発現パターンを有し、絶対的寛容性の対象ではない。より具体的には、HER2/neu転写産物及び185kDポリペプチド産物の発現を検出したところ、皮膚及び乳房を含む様々な組織の正常成人上皮細胞及び胃腸組織、生殖管及び尿路では低レベルであり;胚発生中の対応する胎児組織では高レベルであった(Pressら、Oncogene 5:953−962(1990))。いくつかの証拠から、ヒト乳癌、肺癌、前立腺癌、卵巣癌、子宮内膜癌及び結腸直腸癌でのHER−2の増幅と悪性形質転換との間の関連が示唆される(Disis及びCheever,Adv.Cancer Research 71:343−371(1997))。一般に、HER2/neuの過剰発現は、癌患者に対して、予後が悪いこと及び再発率が高いことと相関がある(Slamonら、Science 244:707−712(1989))。したがって、HER2/neuタンパク質に特異的なワクチンは、多くの様々なヒト悪性腫瘍における疾患再発の予防において、広く適用でき、利用できると思われる。
【0094】
HER2/neuは、生来のチロシンキナーゼ活性を保持し、上皮細胞増殖因子(EGF)受容体に対する高い相同性を示す膜貫通糖タンパク質をコードする(Akiyama,Tら、(1986)Science 232:1644−1646)。癌免疫療法に対してHER2を標的として利用した最初の臨床研究の1つでは、HER−2特異的モノクローナル抗体Herceptinが乳癌の治療に対して用いられた(Goldenberg MM(1999)Clin.Ther.21:309−318)。これにより、自己抗原提示細胞を活性化するためのHER−2ドメインを含有する組み換え融合タンパク質の使用を含む、効果的な抗腫瘍反応を誘発するための、免疫系のT細胞アームに対する標的としてのHER−2の使用に焦点を当てたその後の試みが導かれた。公表されている報告により、neu発現性の、乳癌及び卵巣癌に罹患している多くの癌患者において、HER2/neu癌遺伝子のタンパク質産物に対する免疫反応が得られることが分かっている(例えば、抗原特異的抗体及びT細胞を産生する。)
【0095】
様々な中間体プラスミド及びシャトルベクターへの組み換えアデノウイルス配列、導入遺伝子及び他のベクターエレメントのアセンブリー及び、組み換えウイルス粒子を産生させるためのプラスミド及びベクターの使用は全て、当業者にとって周知の標準的教科書に記載されているような従来技術を使用することにより達成される(Sambrookら、Molecular Cloning:A Laboratory Manual,第二版、Cold Spring Harbor Press,Cold Spring Harbor,NY(1989))。そのような技術には、これらに限定されないが、従来のcDNAクローニング技術、アデノウイルスゲノム由来の重複オリゴヌクレオチド配列の使用、相同組み換え、ポリメラーゼ連鎖反応、標準的トランスフェクション技術、寒天重層におけるウイルスのプラーク生成及びその他の関連方法が含まれる。
【0096】
原核細胞におけるポリヌクレオチドの調製を促進するために、プラスミド型のアデノウイルスベクターを調製することが多い(アデノウイルスプレプラスミド)。このアデノウイルスプレプラスミドは、アデノウイルス部分及びプラスミド部分を含有する。このアデノウイルス部分は、基本的に、本発明のアデノウイルスベクター(E1、場合によってはE3領域が、非機能性であるか欠失している、アデノウイルス配列を含有する。)及び都合のよい制限部位に隣接した免疫原発現カセットを含有するアデノウイルス部分と同じである。
【0097】
アデノウイルスプレプラスミドのプラスミド部分は、抗生物質の発現が真核細胞において起こらないように、原核プロモーターの転写調節下に、抗生物質耐性マーカーを含有することが多い。アンピシリン耐性遺伝子、ネオマイシン耐性遺伝子及びその他の医薬適合性の抗生物質を使用し得る。原核生物における発酵によるポリヌクレオチドの高レベル産生を促進するために、アデノウイルスプレプラスミドが原核性の複製起点を含有し、高コピー数であることが好都合である。多くの市販の原核クローニングベクターによりこれらの利益を得られる。必須でないDNA配列を除去することが望ましい。ベクターが真核細胞において複製できることもまた望ましい。これにより、ポリヌクレオチドワクチン配列の、受容者のゲノムへの組み込みのリスクが最小に抑えられる。特定の組織タイプにポリヌクレオチドの発現を限定することが望ましいかどうかを問わず、組織特異的プロモーター又はエンハンサーを使用し得る。
【0098】
アデノウイルス骨格DNA及び適切なシャトルベクター(特異的欠失を標的とするように、及び結果として得られるプラスミドに所望する制限部位を組み込むように設計される。)を用いた相同組み換えにより、アデノウイルスプレプラスミド(望ましい欠失及び挿入を有する複製欠損アデノウイルスのゲノムを含有するプラスミド)を生成させることができる。例えば、所望する欠失を考慮したアデノウイルス末端のPCR及び適切なクローニングプラスミドへの個別のセグメントの、続いて行われるクローニングなど、本技術分野で広く理解され認識される一般的方法を用いて、この目的における用途のシャトルベクターを生成させることができる。次に、アデノウイルスプレプラスミドを消化し、リン酸カルシウム共沈殿又はその他の適切な手段を介して補完的な細胞株にトランスフェクションすることができる。次に、ウイルス複製及び増幅が起こり、これは、顕著な細胞変性効果により明らかになる現象である。次に、ウイルス複製の完了後、感染細胞及び培地を回収する(一般に、トランスフェクションから、7日から10日後。)。
【0099】
一般に、所望するアデノウイルスプレプラスミドの構築及びアセンブリー後、アデノウイルスE1を発現しているヒト細胞株に導入することにより、アデノウイルスプレプラスミドをウイルスとする。パッケージング細胞株とベクターのウイルス遺伝子との間の相補により、ベクターのアデノウイルス−導入遺伝子配列を複製させ、ビリオンキャプシド中にパッケージングされ、その結果、組み換えアデノウイルスの産生が起こる。本発明の方法での使用のために、当業者とって公知のあらゆる様々な方法により、得られたウイルスを単離し、精製し得る。
【0100】
チンパンジーアデノウイルス遺伝子の、1又は複数の選択した欠失をウイルスベクターに導入したとき、産生細胞株中に存在する配列による産生プロセスの間に、欠失遺伝子産物の機能を補われ得ることは、当業者にとって容易に明らかである。したがって、パッケージングに必要であるがそのベクターにおいて機能しないアデノウイルス機能の一部又は全てを特徴とする、永久的に形質転換された細胞株により、操作した遺伝子の機能を与えることができる(例えば、E1A、E1B、E2A、E2B、E4のいずれか。)。あるいは、その機能を与えるのに必要な遺伝子を含有するコンストラクトを用いて、適切な細胞に感染又は一時的トランスフェクションを行うことにより、適切なパッケージング細胞株に対して必要なアデノウイルス機能を与えることができる。
【0101】
したがって、本発明はまた、E1発現ヒト細胞株においてチンパンジーアデノウイルスベクターを生成させる方法を提供する。より具体的には、公知の細胞株 293及びPER.C6TMを含むE1補完細胞株中に、開示されたベクターを増殖させることができる。これらの両細胞株は、アデノウイルスE1遺伝子産物を発現する。PER.C6TMは、1997年1月3日公開のWO97/00326に記載されている(これは、参照により本明細書中に組み込む。)。これは、複製欠損第一世代アデノウイルスの産生を補完するが、相同組み換えによる複製能のあるアデノウイルスの生成を防ぐように設計されているE1遺伝子セグメントにより形質導入されている初代ヒト網膜芽細胞株である。293細胞は、Grahamら(1977)J.Gen.Virol 36:59−72(これも、参照により本明細書中に組み込む。)において記載されている。「第一世代アデノウイルス」という用語が、E1領域が非機能的であるか欠失しているかのいずれかであり、場合によってはE3領域が非機能的であるか欠失している、複製欠損アデノウイルスを指すことを、当業者は、認識するであろう。
【0102】
臨床試験においてワクチン組成物としての使用を意図する複製欠損アデノウイルスベクターのバッチは、RCA不含であることが証明されているべきである(Fallaux,F.J.ら(1998)Human Gene Therapy,9:1909−1917)。実施に際し、これは、費用のかかるスクリーニングプログラムを確立し利用する必要がある、手間のかかるプロセスである。RCA生成が高頻度に起こることにより、生成したバッチに対する失敗率が高まるだけでなく、スケールアップが非常に制限されることを、当業者は認めるであろう。ベクターのヌクレオチド配列とヘルパー産生/パッケージング細胞株のゲノムに存在するアデノウイルス配列との間の配列相同性を削除することにより、相同組み換えにより生成するRCAが混入するベクターのバッチ産生の可能性が取り除かれると思われる。
【0103】
通常、組み換え複製欠損アデノウイルスベクターは、トランスにおいてE1遺伝子産物を与える細胞株中に増殖する。ベクターが、同じ又は非常に似た血清型由来である場合、トランスでの必須のE1遺伝子産物の供給は、非常に効果的である。例えば、E1欠失(つまりΔE1)グループC血清型(Ad2及びAd5)ベクターが、Ad5 E1領域を含有し、発現する、293又はPER.C6細胞において増殖できることは、周知である。しかし、293及びPER.C6産生細胞に存在するAd5 E1配列が、必ずしも非グループC血清型の複製を完全に補完し得るとは限らないことが観察されている。したがって、サブグループC以外のE1欠失血清型、例えば、サブグループA、B、D、E及びF由来のものの複製は、対応する野生ウイルスと比較して、より低効率であるか、又は293又はPER.C6細胞において全く複製し得ない。これは、Ad5(グループC)E1 B 55K遺伝子産物が、非グループC血清型のE4 orf6遺伝子産物と機能的に相互作用できないからである。
【0104】
Ad5 E1発現細胞における複製効率の低下は、様々なサブグループのベクターによって変化し得る。サブグループD及びE アデノウイルス由来のΔE1ベクターが、293及びPer.C6TM細胞において様々な効率でレスキュー可能であり、増殖可能である一方、サブグループBのΔE1ベクターは、増殖能が完全に損なわれている(Vogels Rら、(2003)Aug.Replication−deficient human Adenovirus type 35 vectors for gene transfer and vaccination:efficient human cell infection and bypass of preexisting adenovirus immunity.J Virol.77(15):8263-71)。
【0105】
Ad5 E1b 55kとベクター発現E4 orf6 タンパク質との間の相互作用が、同じサブグループのメンバー内で保存されているにもかからわず、このことは、非C血清型のE4 orf6 タンパク質が発現される場合、十分安定ではないであろうことを示す。E1b 55k/E4 orf6複合体の形成が不十分であるか不安定であることから、ΔE1ベクターの増殖が低下しなくなる。したがって、成功裡に、かつ効率よく、グループB血清型の組み換えアデノウイルスをレスキューするために、関心のある血清型のE1領域を発現する細胞株を生成させることが必要とされ得ることが、実験的に分かっている。293又はPER.C6TMなどの、Ad5 E1を発現する細胞において、発現をE1b 55Kタンパク質に限定することができる。あるいは、適切なAd5 E1発現細胞株を、Ad5 E1に加えて全体のAd5 E4領域(又はE4orf6のみ)を発現するように、改変することができる。Ad5 E1及びorf6の両方を発現する細胞株の生成は、代替的なアデノウイルス血清型を補完するのに有用である;例えば、Abrahamsenら、1997 J.Virol.8946−8951を参照のこと。
【0106】
Ad5補完細胞株へのE4(orf6)の組み込みは、トランスで血清型特異的E1遺伝子産物を与える血清型特異的細胞株の生成であるとして知られている。あるいは、非グループCベクター増殖効率は、ネイティブE4領域の、Ad5 orf6での置換によるウイルス骨格の改変により向上させ得る。ネイティブ orf6のみをAd5又は他のサブグループCウイルス(Ad1、Ad2、Ad6)由来のorf6と置換することによって、同様の結果を得ることができる。米国特許第5,849,561号は、Ad5−E4遺伝子の一部も発現するAd5−E1補完細胞株における、E1欠失非グループCアデノウイルスベクターの補完を開示する。
【0107】
Vigneらに発行された米国特許第6,127,175号は、アデノウイルスのE4領域の一部、好ましくは、orf6/orf6/7を発現する、安定的にトランスフェクションされた哺乳動物細胞株を開示する。このような細胞株は、E4領域を欠く組み換えAdゲノムの補完に有用である。
【0108】
ワクチン組成物を含む、開示するアデノウイルスベクターを含有する組成物は、本発明の重要な局面である。哺乳動物宿主、好ましくはヒト宿主に、予防的又は治療的設定のいずれかで、これらの組成物を投与することができる。宿主/ワクチンとして可能なものには、これらに限定されないが、霊長類、特にヒト及び非ヒト霊長類が含まれ、商業用動物又は家畜として重要なあらゆる非ヒト哺乳動物が含まれる。組み換えチンパンジーアデノウイルスベクターを含有する組成物は、単独で又はその他のウイルス性もしくは非ウイルス性ベースのDNA/タンパク質ワクチンと組み合わせて投与し得る。これらはまた、広範囲の治療計画の一部として投与し得る。
【0109】
本発明の特定の実施形態において、自己抗原又は腫瘍関連抗原に対する宿主の寛容性を打破するために設計された免疫付与プロトコールにおいて、開示するベクターを使用し得る。多くのTAAの同定により、癌治療に対する、活性のあるワクチン接種アプローチの開発が可能となった。プロセシングされ、提示される細胞表面抗原及び細胞内抗原の両方が、有用な標的を与える。一般に、自己抗原に対する宿主寛容性を打破する、開示する方法には、(a)抗原特異的免疫反応が望まれる自己抗原をコードするヌクレオチド配列を担う、第一のChAdベクター又はプラスミドベクターを含有する第一のワクチン組成物を投与することにより、自己抗原に対する抗原特異的反応を刺激することと、(b)少なくともゲノムのE1遺伝子の機能的欠失を含有し、そのE1遺伝子部位において配列が、初回刺激段階で送達された同じ自己抗原をコードするDNAの発現を支配することができるプロモーターを含有する、異なる血清型の組み換えChAdベクターを含有する第二のワクチン組成物を投与することにより、(a)の免疫反応を保持し、拡大することとが含まれ、それにより、自己抗原に対する寛容性を打破する効果を有する免疫反応を宿主が備える。
【0110】
したがって、熟練者は、宿主寛容性を打破するための使用に適切であり得るいくつかの異なる免疫付与プロトコールを設計するために、この開示を利用することができる。例えば、最初又は初回免疫付与が、TAAなどの特定の自己抗原をコードするプラスミドDNAを含み、何らかのその後の免疫付与が、ChAdベクターを含む、プロトコールを利用することができる。自己抗原をコードするヌクレオチド配列を含むプラスミドDNA配列は、1又は複数の注射において、電気刺激あり、又はなしで、筋肉内に送達され得る。例えば、エレクトロポレーションを介した、TAAをコードするプラスミドDNAの複数回(例えば、3もしくは4もしくは5)筋肉内注射と、それに続く、同じTAAをコードする導入遺伝子を含有するChAdベクターの1又は複数の筋肉内注射に基づく免疫付与プロトコールは、本明細書中で開示され主張される一般的方法に包含される。
【0111】
あるいは、寛容性を打破するための適切なプロトコールは、自己抗原をコードする導入遺伝子を含有するChAd又はhAdベクターを用いた、1又は複数の初回刺激免疫付与と、それに続く、同じもの又は、初回免疫付与に使用されるベクターと交差反応しないことが知られている、異なるChAdベクターを用いた、1又は複数の追加免疫付与を含み得る。例えば、初回刺激のためにChAd3を用い、追加免疫のためにChAd6を用いる免疫付与プロトコール、又は初回刺激のためにChAd3を用い、続いて追加免疫のためにChAd6及びChAd9を用いる免疫付与プロトコールを、宿主寛容性を打破するために使用し得る。特定の実施形態ににおいて、本発明は、HER2/neu、CEA、EpCaM、PSA、PSMA、テロメラーゼ、gap100、Melan−A/MART−1、Muc−1、NY−ESO−1、サバイビン、ストロメリシン 3、チロシナーゼ、MAGE3、CML68、CML66、OY−TES−1、SSX−2、SART−1、SART−2、SART−3、NY−CO−58、NY−BR−62、hKLP2、VEGFからなる群から選択される、少なくとも1個の腫瘍関連抗原を含有する自己抗原の使用をもくろむ。特定の実施形態において、本発明は、癌に罹患している哺乳動物に、TAAをコードする組み換えチンパンジーアデノウイルスを送達することによる、選択された悪性腫瘍に特異的な腫瘍関連抗原に対する免疫反応(例えば液性又は細胞性)を誘導するための方法を提供する。本発明のこの局面の好ましい実施形態において、誘発される免疫反応は、抗原特異的CD4+及びCD8+T細胞の産生を特徴とする免疫反応で構成される。
【0112】
本発明の免疫原性組成物は、予防的又は治療的設定のいずれかで、哺乳動物宿主、好ましくはヒト宿主に投与することができる。可能性のある宿主/ワクチンには、これらに限定されないが、霊長類、特にヒト及び非ヒト霊長類が含まれ、商業用動物又は家畜として重要なあらゆる非ヒト哺乳動物が含まれる。組み換えチンパンジーアデノウイルスベクターを含有する組成物は、単独で又はその他のウイルス性もしくは非ウイルス性ベースのDNA/タンパク質ワクチン組み合わせて、投与し得る。これらはまた、広範囲の治療計画の一部として投与し得る。本発明の方法での使用に適切な組成物は、緩衝液、生理食塩水もしくはリン酸緩衝生理食塩水、スクロース、その他の塩及びポリソルベートなどの、生理学的に許容可能な成分と組み合わせて、本発明の組み換えウイルスベクターを含有し得る。この組成物は、筋肉内注射時に組織に刺激を与えない。この組成物は、好ましくは、使用時まで凍結される。場合によっては、本発明のワクチン組成物は、これらに限定されないが、アジュバント、安定化剤、pH調整剤又は保存料などの、他の成分を含有するように処方し得る。このような成分は、当業者にとって周知である。
【0113】
本発明の組み換えチンパンジーアデノウイルスが、宿主細胞に形質導入され、受容者/ワクチンに対して治療的利益又は防御的免疫を与える免疫反応を誘発するのに十分な導入遺伝子の発現量を与えるために、選択した投与経路において効果的な組み換えウイルス量である「効果的な量」で、ヒト又は獣医学的宿主に投与されることが想定される。
【0114】
ワクチン受容者に導入されるべき本ワクチン組成物におけるウイルス粒子量は、使用される転写及び翻訳プロモーターの強さ及び発現される遺伝子産物の免疫原性に依存する。一般に、1x107から1x1012粒子(つまり、1x107、2x107、3x107、5x107、1x108、2x108、3x108、5x108又は1x109、2x109、3x109、5x109、)、好ましくは、約1x1010から1x1011粒子の免疫学的又は予防的有効量を、直接筋肉組織に投与する。皮下注射、皮内導入、皮膚を介した押印(impressio)及び腹腔内又は吸入送達などのその他の投与様式ももくろまれる。
【0115】
本発明の組み換えチンパンジーアデノウイルスベクターは、混合様式の初回刺激/追加免疫ワクチン接種計画の一部として、又は様々な血清型ベクターの複数注射に基づくワクチン接種計画において、単独で投与し得る。通常、少なくとも1個の免疫原を含有する初回刺激投与は、特定の病原体又は自己抗原に対する効果的な免疫反応を必要とする哺乳動物宿主に投与される。この投与は、効果的に、その後の抗原の同定において、宿主が免疫原に対してすぐに免疫反応を促進又は増強することができるように、免疫反応を刺激する。初回刺激及び追加免疫に対して代替的処方を利用する、混合様式ワクチン接種スキームの結果、免疫反応が促進される。初回刺激−追加免疫投与は通常、対象を少なくとも1回刺激することと(ウイルスベクター、プラスミド、タンパク質などによる。)、規定の時間の経過後、次に、追加免疫を行うこと(ウイルスベクター、プラスミド、タンパク質などによる。)を含む。複数免疫付与は、通常1回から4回であるが、さらに多く行い得る。初回刺激と追加免疫との間の時間の長さは、通常、約4ヶ月から1年の間で変化し得うるが、当業者が認める場合、他の時間枠を使用し得る。中和免疫が明らかとなる前に、各ベクターの複数回注射を約2週間の時間枠内で投与することができる。
【0116】
本発明のある実施形態において、アデノウイルスワクチンベクターの複数回の投与でワクチンを与え、このウイルスワクチンは、プラスミドワクチンの投与を伴う計画で与え得る。本明細書中で開示するベクターと組み合わせた使用に適切なプラスミドワクチンは、免疫原コード配列及び転写終結因子配列に機能可能に連結された、免疫原をコードする核酸配列の発現を支配することができる異種のプロモーターと組み合わせた、免疫反応の初回刺激又は増強が望ましい、少なくとも1個の免疫原をコードするプラスミドを含む。
【0117】
例えば、組み換え複製欠損チンパンジーアデノウイルスベクターの様々な血清型の複数注射を利用する投与計画を使用することができる。あるいは、プラスミドワクチンの最初の投与(つまり初回刺激投与)及び、該プラスミドワクチンにおいて送達された同じ免疫原のコード配列を含有する複製欠損組み換えチンパンジーアデノウイルスベクターを含有する第二の投与(つまり追加免疫投与)を個体に与え得る。あるいは、その個体に少なくとも1個の免疫原をコードするヒトアデノウイルスワクチンベクターの最初の投与を行い、次いで、本明細書中で開示する複製欠損組み換えチンパンジーアデノウイルスベクター(これは、初回刺激投与において送達された同じ免疫原のコード配列を含有する。)を含有する第二の投与を行い得る。第二の代替的実施形態において、本発明のベクターを含有するワクチン組成物を最初に投与し、次いで、プラスミドワクチン投与を行い得る。これらの実施形態のいずれかにおいて、ウイルスベクター又はプラスミド形態のいずれかで、同じ免疫原の複数回投与を個体に行い得る。これは、規定の最小時間をあけて投与を行い得る。
【0118】
目的の1個のタンパク質又は抗原が、本発明の組み換え複製欠損チンパンジーアデノウイルスベクターにより送達されるのに加えて、別個のビヒクルを介して、又は同じビヒクルを介してのいずれかで、2以上のタンパク質又は抗原を送達することができる。複数のオープンリーディングフレームを含むアデノウイルスプレプラスミドの生成のための、適切なシャトルプラスミドに、複数の遺伝子/機能的同等物を連結することができる。この複数の遺伝子/機能的同等物に対するオープンリーディングフレームを、別個のプロモーター及び転写終結配列に、機能可能に連結することができる。
【0119】
本明細書中で示されるように、適切な免疫付与計画において、様々なアデノウイルス血清型を用いることができる。このようなプロトコールの一例は、第一の血清型(例えばChAd3又はChAd6)の組み換えアデノウイルスベクターを含有する初回刺激投与と、その後の、第二の血清型の組み換えチンパンジーアデノウイルスベクターを含有する追加免疫投与である。代替的な実施形態において、この初回刺激投与は、異なるタンパク質/抗原をコードする遺伝子をそれぞれ含有する、別個のアデノウイルスビヒクルの混合物を含み得る。このような場合、この追加免疫投与は、初回免疫投与で送達された同じ又は類似の抗原セットをコードする遺伝物質を含有する組み換えウイルスベクターを投与する、という条件で、別個のタンパク質/抗原をコードする遺伝子をそれぞれ含有するベクターの混合物を含み得る。これらの複数の遺伝子/ベクター投与様式をさらに組み合わせることができる。特異的抗原からの複数であるが別個の成分を含む、併用様式計画に着手することは、さらに、本発明の範囲内にある。
【0120】
ヒトベクターのウイルスエレメントに対して向けられた既存の免疫により中和されないチンパンジーアデノウイルス由来の組み換えベクターを使用することにより、ワクチン担体としてのヒトAdベクターの使用に対する代替法が得られる。アデノウイルスが高免疫原性であるため、アデノウイルスベクターは、自己抗原に対する宿主寛容性を打破するために設計されたワクチン担体の関連での使用のための特に優れた候補である。さらに、ヒトウイルスと同等の効率での、ヒト細胞、特にPer.C6TM細胞株でのchimp ウイルスの増殖能により、調節の観点及び治療法又はワクチンの大規模産生に対する両方から、顕著な有利性が与えられる。したがって、本発明は、遺伝子ワクチン接種のためのワクチン担体として、単独で、又は組み合わせて、使用し得る、組み換えウイルス調製を可能にする、チンパンジーアデノウイルス配列、ベクター及びプラスミドのコレクションを提供する。
【0121】
本明細書中で言及した公表物は全て、本発明との関連で使用され得る方法及び物質を説明し開示する目的のために、参照により組み込む。明細書のいかなる開示内容も、本発明が先行技術の効力によってこのような開示に対して先行する権利を与えられていないことを認めるものではない。
【0122】
添付の図面を参照しながら本発明の好ましい実施形態を説明してきたが、本発明がまさにその実施形態に限定されず、添付の特許請求の範囲で定義される本発明の範囲又は精神から逸脱することなく、様々な変更及び改変が当業者によりなされ得ることを理解されたい。
【0123】
次の実施例は、本発明を説明するが、限定するものではない。
【実施例1】
【0124】
ChAdの単離、クローニング、配列決定及び特性決定
チンパンジーアデノウイルスの単離
ウイルス輸送培地(VTM; Microtest M4−R Multi−Microbe Transport Medium, Remel Inc.)において便検体を回収し、次にNIRC(New Iberia Research Center 4401 W. Admiral Doyle Drive New Iberia, LA 70560)で−70℃にて凍結又は直接凍結した。この検体を細胞培養物への接種のために処理するまで<−70℃で凍結保存した。この検体を細胞培養物へ接種する際、この検体を解凍し、次に過剰の氷冷ウイルス輸送培地中でボルテックスした。前記検体を懸濁液へ入れて解離させた後、それらを1500から1800rpmで10分間遠心分離した。上清を0.8μmシリンジフィルター及び0.2μmシリンジフィルターを通じて連続して濾過し、次にこの濾過した材料を細胞培養物へ接種した(シェルバイアルへ200から250μl、及び試験管培養物へ250から300μl)。処理した各検体を293細胞もしくはA549細胞を播種した試験管培養物及びシェルバイアル培養物へと接種した。
【0125】
対照(ポジティブ及びネガティブ)培養物を、1セットの試料を接種する各場合に調製した。設定におけるシェルバイアルに全て接種したら、それらを室温にて60±10分間、2000rpm(900xg)で遠心分離した。培養物中の熱損傷を防止するため、ローターの回転が停止した直後に前記バイアルを遠心分離機から取り出した。遠心分離後、交差汚染を防止するため、各バイアルにおいて新鮮な滅菌済みパスツールピペットを使用して、接種材料を前記シェルバイアルから吸引した。各洗浄につき1.0mlの新鮮な培地を使用して、前記培養物を3回洗浄した。3回目の洗浄の後、新鮮な培地(1.0ml)を各バイアルへピペットを介して添加し、前記シェルバイアルを35から37℃のインキュベーターの中で3日から4日間(約96時間)静置した。
【0126】
培養期間の終了時に、上清を前記培養物から吸引し、各洗浄に対して約1.0mlの緩衝液を使用して、免疫蛍光アッセイ(IFA)緩衝液で各バイアルにおける細胞層を2回洗浄した。1.0mLの冷アセトンを各バイアルへ添加する(2から8℃で10分間)ことにより、細胞を固定した。アセトン洗浄したスライドを、シェルバイアルカバースリップと関連付けた検体認識番号で標識した。このシェルバイアルカバースリップを、一次マウス抗アデノウイルス抗体[MAB8052、Chemicon]を使用してアデノウイルス感染細胞の蛍光標識用に処理した。蛍光顕微鏡を使用して、スライドを評価する。ウェル(1+から4+)にわたる免疫蛍光がカバーする範囲に留意して、10倍の対物レンズを使用して各調製物をスキャンした。40倍の対物レンズを使用して特異的免疫蛍光の有無を確認した。前記シェルバイアルについて記載したものと同じ手順で、試験管培養物に接種した(例えば、ネガティブ対照を最初に処理し、次いで、臨床検体及びポジティブ対照)。この接種材料を36から38℃にて60分から120分間吸着させた。吸着時間後、この検体/対照を試験管から吸引し、新鮮な培地と交換した。
【0127】
接種から3日から4日後、及び、その後週に1回、この培養試験管から培地を吸引し、1.5mlの新鮮な培地と交換した。培養試験管をCPEについて、接種後少なくとも21日間、少なくとも1日おきに目で見て観察した。チンパンジー検体で接種した培養物を対照と比較し、CPEの程度を観察することによって評価した。14日後に、CPEを示さない培養物を新鮮な試験管培養物へ通し、21日後にCPEに対して陰性である培養試験管を陰性とみなした。3+から4+のCPEの培養試験管を10秒間ボルテックスした。細胞を試験管壁から、滅菌済みの1.0mL血清学用ピペットを使用して回収し、培養上清中で懸濁した。5mLのスナップキャップ試験管に検体認識番号及び日付を付した後、−70℃で保存した。この細胞懸濁液 500μlを、培養試験管からスナップキャップ試験管へ移し、アデノウイルスを検出するための間接免疫蛍光抗体技術(シェルバイアルを染色するための手段と同等)を使用して処理するまで、2℃から8℃で最長1日間保存した。
【0128】
チンパンジーアデノウイルス増幅
ATCC又はEsoterix Inc.,. Austin, Texasから購入した野生型チンパンジーアデノウイルスCV32、CV33、CV23及びCV68(ATCC受託番号:CV32、VR−592;CV33、VR−593;)及びオリジナルの分離株を、ヒトE1発現細胞株PER.C6TM又は293を使用することによって以下のように増殖させた。簡潔には、10% ウシ胎児血清(FBS GibcoBRL, Life Technologies)、1% ペニシリン−ストレプトマイシン、2mM グルタミン及び10mM MgCl2 (Per.C6TM)を添加したダルベッコ改変イーグル培地(DMEM;GibcoBRL, Life Technologies)中で細胞を培養した。5% ウマ血清(GibcoBRL, Life Technologies)を添加したDMEM中でアデノウイルス感染を行った。細胞の100%がウイルスにより誘導される細胞病理学的効果(CPE)を呈したら、感染した細胞及び培地を回収し、凍結融解を3回繰り返して細胞溶解させた。
【0129】
野生型チンパンジーアデノウイルス(CV)ストックを全て、増幅の第一継代後に96穴プレートに播種した293細胞を感染させることによってクローニングした。ウイルスクローニングは、ウイルス増幅の第一継代で得られる細胞溶解液の希釈を制限することによって行った。5個の単離されたクローンを拾い、連続して増殖させた。増幅の3回から4回の連続した継代の後、5つの2層細胞ファクトリー(NUNC)上に播種した細胞において、アデノウイルスの大規模調製を行った(1つの細胞ファクトリーにつき2億個の細胞)。塩化セシウム密度勾配における超遠心段階を2回行い、細胞溶解液から、精製したウイルス粒子を得た。
【0130】
ウイルスゲノムDNAの配列決定
1%SDS−TEN中のプロテイナーゼK(0.5mg/ml)で(55℃で2時間)消化することにより、精製ウイルス調製物 3x1012ppから、ゲノムDNAを単離した。フェノール−クロロホルム抽出ならびにエタノール沈殿の後、このゲノムDNAを水中で再懸濁し、ゲノム配列決定に供した。
【0131】
全長のAdゲノム配列決定のため、精製ウイルスDNAを霧状にし、無作為にせん断された断片を得た。E.コリDNAポリメラーゼ及びポリヌクレオチドキナーゼのKlenow断片を用いて、このDNA断片を平滑末端化にした。平滑末端断片を低融点アガロースゲル電気泳動に供し、1から3kbの範囲の大きさの断片を精製し、pUC19ベクターのSmaI部位へとクローニングし、ショットガンライブラリを作製した。ライゲーションを使用してコンピテントXL1−Blue MRF’を形質転換した。X−gal及びIPTGを含有するLB寒天上での白色/青色スクリーニングにより陽性コロニーを同定した。プラスミドDNAの3個から4個の96穴ブロックをライブラリから単離し、pUC順方向プライマー及び逆方向プライマーを用いて配列決定した。配列決定の読み取りは全て、Phred−Phrapソフトウェアパッケージを使用して性質及びベクター配列についてスクリーニングした。このスクリーニングを通過した読み取りをコンティグに集めた。プライマーは、ギャップを閉鎖し、両端のDNA配列を決定するために、アデノウイルスDNAを直接配列決定するよう設計した。
【0132】
ChAd3(配列番号1)、ChAd6(配列番号2)、CV32(配列番号3)、CV33(配列番号4)、及びCV23(配列番号5)を含む選択されたウイルスについて完全なウイルスゲノム配列決定が行われた。表1は、ChAd3、ChAd6、Pan5(CV23)、Pan6(CV32)、Pan7(CV33)、C1及びC68アデノウイルスゲノムのヌクレオチド配列間の同一性の割合を要約するデータを提供する。アラインメントは、FASTAパッケージ第2版の一部としてのALIGNプログラムを使用して計算した(William R. Penson, University of Virginia; Myers & Miller, CABIOS 1989, 4:11−17)。
【0133】
【表1】
【0134】
新規のアデノウイルス分離株(例えば、ChAd20、ChAd4、ChAd5、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82)の特徴を調べるために、ヘキソン及び繊維遺伝子のヌクレオチド配列もプライマーウォーキングにより決定した。繊維遺伝子は、配列番号6から15(配列番号6、ChAd20;(配列番号7、ChAd4);配列番号8、ChAd5);配列番号9、ChAd7);配列番号10、ChAd9);配列番号11、ChAd10);配列番号12、ChAd11);配列番号13、ChAd16);配列番号14、ChAd17);配列番号15、ChAd19)及び(配列番号58、ChAd8)、(配列番号60、ChAd22)、(配列番号62、ChAd24)、(配列番号64、ChAd26)、(配列番号66、ChAd30)、(配列番号68、ChAd31)、(配列番号70、ChAd37)、(配列番号72、ChAd38)、(配列番号74、ChAd44)、(配列番号76、ChAd63)及び(配列番号78、ChAd82)である。図20Aから20Gは本明細書中で開示され主張される前記ChAd分離株の繊維タンパク質のアミノ酸配列の比較を与える。
【0135】
ヘキソン遺伝子配列は、配列番号16から25(配列番号16、ChAd20);配列番号17、ChAd4);配列番号18、ChAd5);配列番号19、ChAd7);配列番号20、ChAd9);配列番号21、ChAd10);配列番号22、ChAd11);配列番号23、ChAd16);配列番号24、ChAd17);配列番号25、ChAd19)、(配列番号97、ChAd8)、(配列番号99、ChAd22)、(配列番号101、ChAd24)、(配列番号103、ChAd26)、(配列番号105、ChAd30)、(配列番号107、ChAd31)、(配列番号109、ChAd37)、(配列番号111、ChAd38)、(配列番号113、ChAd44)、(配列番号115、ChAd63)及び(配列番号117、ChAd82)である。図31Aから31Jは、本明細書に開示され主張されるChAd分離株のヘキソンタンパク質のアミノ酸配列の比較を与える。
【0136】
チンパンジーアデノウイルスの分類
異なるチンパンジーアデノウイルス株の分類は、6個のサブグループへのヒトアデノウイルス血清型のすでに提唱されている分類(Horowitz, MS (1990) Adenoviridae and their replication.Virology B. N. Fields及びD.M.Knipe,編(Raven Press, New York)pp.1679−1740)に従い、Align Xプログラム(Informax, Inc)を使用することによるアミノ酸及びヌクレオチド配列アラインメントにより得た。
【0137】
新たな分離株の最初の分類は、異なる制限エンドヌクレアーゼによるウイルスゲノムの制限パターンを観察すること及びヘキソン遺伝子の超可変領域7(HVR7)の配列分析により得られた。このために、HVR7に隣接する高度に保存された領域に関して2つのプライマー:TGTCCTACCARCTCTTGCTTGA(配列番号45)及びGTGGAARGGCACGTAGCG(配列番号46)を設計した。精製ウイルスDNA又は未精製293細胞溶解液をテンプレートとして使用するPCRによりこのHVR7を増幅した後配列決定した。HVR7配列分析に基づいて、我々は、新たに単離されたウイルスを、ヒトAdウイルスについて提唱されるサブグループ(A〜F)へ分類した(Horowitz,MS(1990)Adenoviridae and their replication.Virology B.N.Fields and D.M.Knipe編(raven Press, New York)pp.1679−1740)。
【0138】
図35に示される系統発生樹はヒト及びチンパンジーのアデノウイルスへキソンアミノ酸配列のアラインメントにより得られた。この結果は、Align Xプログラム(Informax,Inc)を使用することによるヘキソンHVR7に限定したヌクレオチド配列アラインメントに基づく最初の分類と一致する。この系統樹は、PAUPSEARCH(Wisconsin Package Version 10.3,Accelrys Inc.)を使用する全長のヘキソンペプチド配列の多配列アラインメントから推定し、TREEVIEWにより可視化を行い、操作した。Wisconsin Packageにおいて実装されるPAUPSEARCHプログラムを使用してブートストラップ信頼分析を実行した。各アラインメントについて、検索基準として「Heuristic Search」を用い、最適性基準としてMaximum Parsimonyを用いて、前記プログラムを1000個の複製物に関して実行し、報告する信頼値は、50%多数決コンセンサス(50% majority−rule consensus)から得た。
【実施例2】
【0139】
ChAdシャトルベクター及び発現ベクターの構築及びレスキュー
ベクター構築及びレスキュー
ウイルスゲノムの左右両端由来のウイルスDNA配列を含有する適切なシャトルベクターを用いて、相同組み換えにより、ゲノムウイルスDNAを標準プラスミドベクターにクローニングした(図2)。下記でさらに詳しく述べるように、グループ特異的シャトルベクターを開発するために、同一血清型サブグループに分類されるウイルス間で観察される配列相同性を利用した。サブグループD及びEへ分類されるチンパンジーアデノウイルスのゲノムウイルスDNAは、両サブグループに属するウイルスをクローニングするための共通のシャトルベクターの構築が可能となるのに十分な相同性を生じた。
【0140】
サブグループD/Eシャトルベクターの構築
ChAd6ウイルスゲノムを完全に配列決定し(配列番号2)、得られた情報を使用して、サブグループD及びEのチンパンジーアデノウイルスの相同組み換えによるクローニングを促進するためにシャトルベクターを構築した。
【0141】
本明細書でpARS ChAd6−3と呼ばれるChAd6シャトルベクターの構築を図1に記載する。図32は、本明細書に記載されるクローニング実験で使用されるオリゴヌクレオチド配列(配列番号26から40及び配列番号45から46)のリストを提供する。簡潔に述べると、オリゴヌクレオチド、5’−ATGGAATTCGTTTAAACCATCATCAATAATATACCTC−3(配列番号27)及び5’−CGCTGGCACTCAAGAGTGGCCTC−3’(配列番号28)を使用して、PCRによりChAd6DNAの左端由来の457bpを増幅し、EcoRI及びSnaBIで消化し、EcoRI−SnaBIで切断しておいたpNEBAd35−2へクローニングし、pNEBChAd6−LIを得た。右のChAd6 ITR(bp36222からbp36648)を、PCRにより、オリゴヌクレオチド5’−ATGAAGCTTGTTTAAACCCATCATCAATAATATACCT−3’(配列番号29)及び5’−ATCTAGACAGCGTCCATAGCTTACCG−3’(配列番号30)を用いて増幅し、制限酵素HindIII及びXbaIで消化し、HindIII−XbaIで切断しておいたpNEBChAd6-LIへクローニングし、その結果、pNEBChAd6−RLIを得た。最後に、ChAd6ゲノムDNA配列のヌクレオチド3426から3813に対応するDNA断片を、オリゴヌクレオチド5’ATGCTACGTAGCGATCGCGTGAGTAGTGTTTGGGGGTGGGTGGG−3’(配列番号31)及び5’−TAGGCGCGCCGCTTCTCCTCGTTCAGGCTGGCG−3’(配列番号32)を用いて増幅し、SnaBI及びAscIで消化した後にSnaBI−AscIで消化しておいたpNEBChAd6−RLIにライゲーションし、その結果、pNEBChAd6−RLIdE1を得た。
【0142】
DH5a E.コリ株における組み換え及びプラスミド増幅効率を向上させるため、ChAd6の左右両方のITRを含有する1306bp断片及びpIX遺伝子断片を、pNEBChAd6−RLIdE1から、PmeI消化により切り出し、pMRKAd5SEAPからの、オリゴヌクレオチド5’−GATCTAGTTAGTTTAAACGAATTCGGATCTGCGACGCG−3’(配列番号33)及び5’TTCGATCATGTTTAAACGAAATTAAGAATTCGGATCC−3’(配列番号34)を使用するPCR増幅により得られた異なるプラスミドベクターへ移した。この最後のライゲーション段階により、ChAd6シャトルベクター pARSChAd6−3が生じた。
【0143】
サブグループCシャトルベクターの構築
ChAd3ウイルスゲノムを完全に配列決定し(配列番号1)、得られた情報を使用して、サブグループCチンパンジーアデノウイルスの相同組み換えによるクローニングを促進するためにシャトルベクターを構築した。
【0144】
簡潔には、本明細書でpChAd3EGFPと呼ばれるサブグループCチンパンジーアデノウイルスをクローニングするのに使用されるシャトルベクターを以下のように構築した。pIXコード化領域を含有するChAd3DNA断片(ヌクレオチド3542から4105)を、オリゴヌクレオチド5’−TATTCTGCGATCGCTGAGGTGGGTGAGTGGGCG−3’(配列番号35)及び5’−TAGGCGCGCCCTTAAACGGCATTTGTGGGAG−3’(配列番号36)を使用するPCRにより増幅し、SgfI−AscIにより消化し、その後SgfI−AscIで消化したpARSCV32−3へとクローニングし、pARS−ChAd3Dを得た。ChAd3右端(nt37320から37441)を、オリゴヌクレオチド5’−CGTCTAGAAGACCCGAGTCTTACCAGT−3’(配列番号37)及び5’−CGGGATCCGTTTAAACCATCATCAATAATATACCTTATT−3’(配列番号38)を用いたPCRにより増幅し、XbaI及びBamHIで消化した後、XbaI及びBamHIで制限酵素処理したpARS−ChAd3Dへライゲーションし、pARS−ChAd3RDを得た。ChAd3ウイルスDNA左端(nt1から460)を、オリゴヌクレオチド5’−ATGGAATTCGTTTAAACCATCATCAATAATATACCTT−3’(配列番号39)及び5’−ATGACGCGATCGCTGATATCCTATAATAATAAAACGCAGACTTTG−3’(配列番号40)を用いたPCRにより増幅し、EcoRI及びSgfIで消化した後、EcoRI及びSgfIで切断したpARS−ChAd3RDへクローニングし、その結果、pARS−ChAd3RLDを得た。このウイルスDNAカセットはまた、酵素切断によってプラスミドDNAからウイルスDNAが放出されるよう、両ITRの端にある制限酵素部位(PmeI)を含有するように設計された。
【0145】
サブグループBシャトルベクターの構築
サブグループBシャトルの構築は、サブグループC及びD/Eシャトルの構築についてすでに記載したストラテジーに従った。簡潔には、pARS−ChAd3RLDが左端、pIX領域、右端を、ChAd30の対応する断片と置換することによって改変した。さらに、ChAd30のE4領域を、ChAd30E4プロモーター調節の下でクローニングされるAd5E4 orf6と置換した。このシャトルプラスミドを、pChAd30 EGFPシャトルベクターと命名した。
【0146】
ΔE1チンパンジーアデノウイルスベクターの構築
サブグループB:サブグループBのチンパンジーアデノウイルスベクターをE.コリ株BJ5183における相同組み換えにより構築した。BJ5183細胞に対して、BstEII及びBst1107Iで消化したpChAd30EGFPシャトルベクターと、ChAd8及びChAd30の精製ウイルスDNAとを用いて、同時形質変換した。pIX遺伝子、直線化したpChAd30EGFPシャトルの両端に存在する右ITR DNA配列及びウイルスゲノムDNAの間の相同組み換えにより、プラスミドベクターにおける挿入が可能になり、EGFP発現カセットにより置換されるE1領域が同時に欠失した。ヒトサイトメガロウイルス(HCMV)プロモーター及びウシ成長ホルモンポリアデニル化シグナル(BghポリA)に基づいた発現カセットを構築して、分泌されるアルカリフォスファターゼ(SEAP)、EGFP、HIVgag、HCV NS領域(ChAd6シャトルベクターについて図3に記載されるようなもの。)、並びにヒト及びアカゲザル由来のCEA及びHER2 neuのような腫瘍関連抗原を発現させた。全ての発現カセットを相同組み換えによりChAd30ベクターへと挿入した。
【0147】
サブグループC:サブグループCのチンバンジーアデノウイルスベクターをE.コリ株BJ5183における相同組み換えにより構築した。BstEII及びBst1107Iで切断したpChAd3EGFPシャトルベクターと、ChAd3、ChAd11、ChAd19及びChAd20の精製ウイルスDNAと、を用いて、BJ5183細胞に対して同時形質変換を行った。pIX遺伝子、直線化したpChAd3EGFPシャトルの端に存在する右ITR DNA配列と、ウイルスゲノムDNAとの間の相同組み換えにより、プラスミドベクターにおける挿入が可能になり、EGFP発現カセットにより置換されるE1領域が同時に欠失した。ヒトサイトメガロウイルス(HCMV)プロモーター及びウシ成長ホルモンポリアデニル化シグナル(BghポリA)に基づいた発現カセットを構築して、分泌されるアルカリフォスファターゼ(SEAP)、EGFP、HIVgag、HCV NS領域(ChAd6シャトルベクターに対して図3に記載されるようなもの。)、並びにヒト及びアカゲザル由来のCEA及びHER2 neuのような腫瘍関連抗原を発現させた。
【0148】
サブグループD及びE:サブグループD及びEのチンパンジーアデノウイルスに基づいたΔE1ベクターを構築するため、シャトルベクターpARS ChAd6−3をAscIで消化し、E.コリ株 BJ5183に、CV32、CV33、CV68、ChAd4、ChAd5、ChAd6、ChAd7、ChAd9、ChAd10及びChAd16の精製ウイルスDNAを用いて同時形質転換した。pIX遺伝子及び直線化したpChAd6−3の端に存在する右ITR DNA配列と、ウイルスゲノムDNAとの間の相同組み換えにより、プラスミドベクターにおける挿入が可能となり、E1領域が同時に欠失した。(図2及び図4)。ヒトサイトメガロウイルス(HCMV)プロモーター及びウシ成長ホルモンポリアデニル化シグナル(BghポリA)に基づいた発現カセットを構築して、分泌されるアルカリフォスファターゼ(SEAP)、EGFP、HIVgag、HCV NS領域(図3)、並びにヒト及びアカゲザル由来のCEA及びHER2 neuのような腫瘍関連抗原を発現させた。図4に記載されるようなΔE1アデノウイルスプレプラスミドへの相同組み換えによって転移されるpARS ChAd6−3ベクターの1個のSnaBI部位へ全発現カセットを挿入した。
【0149】
ΔE1ベクターのレスキュー及び増幅
6cmの細胞培養皿に播種された5x106個のPER.C6TM細胞を、エンドヌクレアーゼ消化によりプラスミド配列から放出されるクローン化されたウイルスベクター10μgでトランスフェクトションした。Lipofectamine(Invitrogen)を使用してDNAトランスフェクションを行った。トランスフェクションした細胞及び培地を、トランスフェクションから5から10日後に回収し、凍結解凍により溶解した。次に、レスキューしたベクターを293細胞又はPER.C6TM細胞での連続継代により増幅した。総計1から2x109個の細胞において、5から10個の細胞ファクトリー(NUNC,Inc.)上に播種した細胞を感染させることによって大規模増幅を行った。塩化セシウム勾配上での2回の超遠心により、精製ベクター調製物を得て、10%グリセロールを含有するPBSに対して透析し、分注して−70℃にて保存した。
【実施例3】
【0150】
中和実験
本明細書で開示されるチンパンジーアデノウイルスに対する中和抗体のヒト血清における普及率を評価するため、中和アッセイを実行した。このアッセイにより、分泌型アルカリフォスファターゼ(SEAP)に対する遺伝子を担うチンパンジーアデノウイルスがヒト293細胞に対して形質導入する能力に及ぼす血清プレインキュベーションの効果を評価した。中和タイターは、ウイルスのみのポジティブ対照において観察されるSEAP活性の50%の低下を与える血清の希釈度として定義される。
【0151】
CV33−SEAP、CV32−SEAP及びChAd3−SEAPの2x106個から1.5x107個の自然粒子を完全培地100μlに希釈し、完全培地に希釈したヒト又はチンパンジー血清等量に添加した。各血清試料を様々な希釈率で試験した(1/18希釈から開始して、1/4希釈を5回行い、1:4608まで。)。試料を37℃で1時間プレインキュベーションし、その後96穴プレートへ播種した293細胞(1個のウェルあたり3x104個の細胞)へ添加した。インキュベーションの1時間後に接種材料を除去し、細胞に新鮮培地を再供給し、24時間後、培地50μlを取り出し、SEAP活性を化学発光アッセイにより測定した。ウイルスのみのポジティブ対照において観察されるSEAP活性の50%低下を与える血清の希釈度として、中和タイターを定義する。100検体のヒト血清のパネルをChAd中和活性について検査した。並行して同じパネルをAd5 SEAPベクターに関して検査した。
【0152】
【表2】
【0153】
表2で与える結果は、チンパンジーアデノウイルス由来のベクターに対して向けられた、中和抗体のヒト血清における普及率が非常に低いことを示す。CV32ベクターにおいて200の閾値を超えるタイターを示すものは、4検体の血清のみであるのに対し、ChAd3 SEAPベクターについては、8検体が200を超えるタイターを示した。一方、検査したチンパンジー血清のパネルは、抗チンパンジーAd免疫性の普及率が非常に高いことを示した。これらの知見から、予想したように、チンパンジーAdに基づくベクターがヒトにおいて中和される機会が非常に少ないことが確認された。それゆえこれらの知見は、Ad5などの共通のヒトAd血清型に基づいたウイルスベクターの投与を制限する、既存の抗ヒトAd免疫性の問題に対する理想的な解決を呈示する。
【実施例4】
【0154】
ChAdベクター親和性
Ad5ベクター及びChAdベクターを介した遺伝子導入の効率を、異なる組織由来のヒト初代細胞のパネルにおけるEGFP発現により評価した。ヒト軟骨細胞、骨芽細胞、ケラチン生成細胞、メラニン細胞、骨格筋細胞及びメラニン細胞を製造者の説明書に従って培養した。ヒト単球細胞、未成熟及び成熟樹状細胞(DC)を記載されているように得た(Romani,N.ら、1996,J.Immunol.Methods,196,137.)。形質導入された蛍光細胞をFACS分析により検出した。試験するヒト初代細胞のパネルには、心臓循環器疾患、関節リウマチ、組織工学(骨、皮膚及び軟骨)及びワクチン接種の分野におけるインビボ及びエクスビボでの遺伝子導入を利用した異なる治療ストラテジーに対する重要な標的細胞である細胞が含まれる。図38Aから図38Dで示す結果は、異なる細胞タイプの感染の差異に基づく効率により示されるように、CARに代わる受容体を、異なるチンパンジーアデノウイルスが認識できることが示唆される。
【0155】
マウス免疫付与実験
方法及び材料
免疫付与プロトコール及び脾臓細胞/PBMC調製
免疫付与:緩衝液 0.1mLで希釈した、選択したアデノウイルスを用いてマウスに免疫付与した。各ベクター用量を50μlずつ2つに分割し、マウスの両側の大腿四頭筋に注射した。
【0156】
脾臓細胞調製:マウスを注射3週間後に屠殺し、それらの脾臓を摘出し、R10(10%FCS、55mM 2−メルカプトエタノール、1M HEPES緩衝液、2mM L−グルタミン、RPMI培地1640中の1xペニシリン−ストレプトマイシン溶液) 10mLに移した。スチール製の網を介して脾臓を細分化し、R10 2mLでこの網を洗浄した後、脾臓細胞を50mLのファルコンチューブに移し、1200rpmで10分間室温(rt)にて遠心分離した。上清を除去し、ACK溶解緩衝液(Gibco BRL Formulation#79−0422DG)3mLを添加した。細胞を5分間rtでインキュベーションした。1xPBS 45mlを添加し、試験管を上述のように遠心分離した。R10 30mLで洗浄した後、細胞をR10 5mL中で再懸濁し、70mナイロン細胞濾過器(Falcon2350)を介して濾過した。細胞 10μLをTurk溶液(Merck 040417345)990μLで希釈し、計数した。最終的に、細胞をR10中で、107個細胞/mLに希釈した。
【0157】
末梢血単核球(PBMC)調製:マウス血液試料(150μL)を2mLのエッペンドルフチューブへ50μL PBS/2%EDTAとともに移した。1mL ACK緩衝液を各チューブへ添加した。RTで5分間穏やかに混合し、インキュベーションした。試料を微量遠心器において1500rpmで5分間遠心分離した。上清を除去し、同じ免疫付与コホート由来の白血球ペレットを合わせた。ACK緩衝液インキュベーションを繰り返した後、PBMCのペレットを、R10培地 1mLに再懸濁した。
【0158】
IFN−γELISPOTアッセイ
Millipore MAIP45プレートを、PBSで2.5μg/mLに希釈した、100μl/ウェルの精製ラット抗マウスIFN−γモノクローナル抗体(Pharmingen、cat.551216)でコーティングし、4℃で一晩(o/n)インキュベーションした。プレートを滅菌済みPBSで2回洗浄し、CO2インキュベーターの中でR10 200μl/ウェルとともに2時間インキュベーションすることにより、非特異的結合部位をブロックした。HIVgagを発現するAdベクターによる免疫付与実験において、9マーのペプチド(AMQMLKETI、Balb/CマウスにおいてマッピングされるCD8HIVGAGエピトープ)(配列番号47)をR10中で2μg/mLに希釈し、50μl/ウェルの量でウェルに添加した。HCV−NS発現ベクターを用いて行った免疫付与実験において、NS3ヘリカーゼドメインをカバーするペプチドのプールならびにヘリカーゼドメインに含まれるマッピングされたCD8エピトープを呈示する9マーのペプチドを使用した。ヒトCEA抗原を発現するChAdによる免疫化実験を、全アミノ酸配列をカバーする、重複する15マーペプチドのプールにより評価した。対照としてDMSO及びコンカナバリンAを使用した。細胞を各ウェルへ5x105及び2.5x105の量で添加した。CO2インキュベーターにおいて一晩インキュベーションした後、プレートを0.05% Tween20/PBSで洗浄し、アッセイ緩衝液(5%FBS、0.005% Tween20、PBS)で1/250に希釈したビオチン化ラット抗マウスIFN−γモノクローナル抗体(PharMingen、Cat.554410) 50μl/ウェルを添加した。プレートを4℃で一晩インキュベーションし、上述のように洗浄した。ストレプトアビジン−アルカリフォスファターゼ複合体(BD554065)をアッセイ緩衝液で1/2500に希釈し、50μL/ウェルの量で室温にて2時間添加した。洗浄後、BCIP/NBT 1ステップ溶液(PiercE34042)50μL/ウェルを添加してプレートを発色させた。ウェルを脱イオン水で洗浄することにより反応を停止させた。スポットをELISPOTリーダーにより自動計数した。
【0159】
マウス IFN−γ細胞内染色(ICS)
脾臓細胞をR10 1mL中2x106個細胞に希釈し、2μg/mLの濃度の上述と同じ抗原で刺激した。対照として、DMSO及びブドウ球菌エンテロトキシンB(SEB)を使用した。CO2インキュベーター中で一晩インキュベーションした後、細胞をFACS緩衝液(1%FCS、0.01%NaN3、PBS)で洗浄し、精製した抗マウスCD16/CD32Fcブロック(クローン2.4G2、Pharmingen、cat.553142)を1/25に希釈し、100μL/試料の量で添加し、4℃で15分間インキュベーションした。細胞をFACS緩衝液で洗浄し、FACS緩衝液で1:50に希釈したAPC標識抗マウスCD3e(クローン145−2C11、Pharmingen#553066)、PE標識抗マウスCD4(クローンL3T4、BD Pharmingen、cat.553142)及びPerCP標識抗マウスCD8a(クローン53−6.7、Pharmingen、cat.553036)を100μL/試料の量で添加した。細胞をrtで30分間インキュベーションし、洗浄し、固定し、浸透させ(Becton Dickinson、FACS Perm2)、PermWash(100μL/試料)中で1:50に希釈したFITC標識抗マウスIFN−γ(Pharmingen、cat.554411)とともにRTにて30分間インキュベーションした。洗浄後、細胞を1% ホルムアルデヒド/PBS 500μLに再懸濁し、CellQuestソフトウェア(Becton Dickinson)を用いて細胞内サイトカイン染色(ICS)をFACS−Caliburフローサイトメーターで分析した。
【実施例5】
【0160】
ChAdベクターはマウスにおいて強いCMI反応を誘発する
本明細書に開示されるChAdベクターが細胞性免疫反応(CMI)を誘発する能力を、HIVgag導入遺伝子を発現するベクターを使用してマウスにおいて評価した。簡潔に述べると、5匹のBalb/Cマウスの群に、様々なベクターを、マウス1匹あたり105から1010vpの、10倍ずつ増加する用量で注射した。
【0161】
注射から3週間後に、脾臓細胞中のgag特異的CD8+T細胞を定量することにより、免疫反応の強さを調べた。Balb/CマウスにおいてマッピングされるgagCD8+T細胞エピトープを再現するペプチドによるインビトロ刺激後に、ELISPOTアッセイによって又はIFN−γ細胞内染色及びFACS分析によって、IFN−γ分泌CD8+T細胞の数を調べた。
【0162】
表3で報告する5匹の免疫付与動物から得られた結果は、9マーのCD8+gagエピトープ又はgagペプチドプールとのインキュベーション後の、脾臓細胞100万個あたりのスポット形成細胞数として示す。gagペプチドプールは、全gag配列を包含する10−aaまで重複する20−aaペプチドからなる。ポジティブの値を太字で示す。
【0163】
表3で与えるデータから、本明細書に開示され主張されるChAdベクターの投与により、hAd5によって誘発される反応に匹敵する強い細胞性免疫反応が誘発されることが示される。ポジティブの免疫付与結果(免疫付与ブレイクポイント)を生じる最低のベクター用量で観察することによって、発明者らは、106ppベクター用量でのブレイクポイントを有するもっとも有力なサブグループCのChAd3gagである、様々なベクターの効力を評価した。免疫付与ブレイクポイントによる評価を図33に示す。
【0164】
【表3】
【実施例6】
【0165】
ChAd3ベクター及びCV33GAGベクターはGAG特異的CD8+T細胞を特徴とするCMI反応を誘発する。
【0166】
HIVgag導入遺伝子を含むChAdベクターに反応して誘発されるCMI反応の特徴を調べるために、様々な用量のベクターで免疫付与した5匹のマウスのコホートからプールされた脾臓細胞を細胞内IFN−γ染色により分析した。表3及び表4に示されるデータは個別の実験において回収された。
【0167】
脾臓細胞を R10 1mLで2x106個細胞に希釈し、2μg/mLの濃度で上述と同じ抗原で刺激した。対照として、DMSO及びSEB(ブドウ球菌エンテロトキシンB)を使用した。CO2インキュベーターで一晩インキュベーションした後、細胞をFACS緩衝液(1%FCS、0.01%NaN3、PBS)で洗浄し、精製抗マウスCD16/CD32Fcブロック(クローン2.4G2、Pharmingen、cat.553142)を1/25に希釈し、100μL/試料の量で添加し、4℃で15分間インキュベーションした。細胞をFACS緩衝液で洗浄し、FACS緩衝液中で1:50希釈した、APC標識抗マウスCD3e(クローン145−2C11、Pharmingen、cat.553066)、PE標識抗マウスCD4(クローンL3T4、BD Pharmingen、cat.553142)及びPerCP標識抗マウスCD8a(クローン53−6.7、Pharmingen、cat.553036)を100μL/試料の量で添加した。細胞を室温で30分間インキュベーションし、洗浄し、固定し、浸透させ(Becton Dickson、FACS Perm2)、PermWash(100μl/試料)で1:50に希釈したFITC標識抗マウスIFN−γ(Pharmingen、cat.554411)とともにRTで30分間インキュベーションした。洗浄後、細胞を1%ホルムアルデヒド/PBS 500μLに再懸濁し、CellQuestソフトウェア(Becton Dickinson)を用いてFACS−Caliburフローサイトメーターで分析した。
【0168】
表4は、gag特異的CD8+T細胞又はCD4+T細胞のいずれかであるgag特異的CD3+T細胞の割合を要約するデータを与える。ポジティブの結果を太字で示す。本明細書で与えるデータから、様々な血清型サブグループ(つまりサブグループC、D及びE)へと分類されるウイルス由来のChAdベクターにより誘発される免疫反応の細胞特性が同様であり、gag特異的反応は全て、CD8+T細胞を主に特徴とすることが示される。さらに、多量のベクターでgag特異的CD4+反応が全ての免疫付与実験で明らかになることを留意すること。ICSアッセイにより、ChAd3ベクターが106個のベクター用量で抗gagCD8+反応を刺激できることが確認された。
【0169】
【表4】
【実施例7】
【0170】
ChAdベクターは、HCV NS特異的T細胞反応を誘発する。
【0171】
CV32−NSmutベクター及びCV33−NSmutベクターの効力をMRKAd6NSmutの効力に対して、C57/Black6において評価した。107から109vp/マウスの、10倍ずつ増加する量のベクターを動物に注射した。単回注射の3週間後に、NS3タンパク質のヘリカーゼドメインにおいてマッピングされるCD8+T細胞エピトープを再現する9マーペプチドでT細胞を刺激して、IFN−γELISPOT及びIFN−γ細胞内染色により、CMIを分析した。表5で与えるデータは、NS3 9マーペプチド非存在下(偽)又は存在下におけるインキュベーション後の、100万個の脾臓細胞あたりのスポット形成細胞数を要約する。
【0172】
HCV−NSを発現するCV32ベクター及びCV33ベクターの両者は強いT細胞反応を刺激することが本データから示される。CV32ベクターに対する最初の陽性結果が109vp/用量を注射することにより得られたという観察に基づき、CV32DE1E3 NSmutベクターの免疫付与効力は、ヒトサブグループCのAd6DE1E3 NSmutベクターよりも、約1/100と、低いと思われる。MRKAd6NSmutを使用する並行した実験から、107vp/動物の用量が細胞性免疫性を刺激するのに十分であることが示された。それゆえ本結果から、gag発現ベクターによる実験(表3参照)においても観察されたヒトサブグループCベクター(hAd5及びhAd6など)に対して、CV32由来のベクターの免疫付与効力が低いことが確認される。
【0173】
【表5】
【実施例8】
【0174】
抗Ad5既存免疫はChAd3gagにより誘発される抗GAG CMIを抑制しない。
【0175】
高い血清普及率の(seroprevalent)Ad5に対する既存免疫性のChAd3免疫付与への影響を調べるために、1コホートにつきBalbCマウス5匹の4コホートに対して、大腿四頭筋にAd5野生型1回1010vpを第0週及び第2週の2回注射してプレ免疫付与した。対照として1コホート5匹の2コホートのマウスに緩衝液のみを同じ時点で注射した。次に、Ad5でプレ免疫付与したマウスのコホートに対して、Ad5gagベクター又はChAd3gagベクターのいずれか 106vp又は107vp/マウスで免疫付与した。対照(未処理の)マウスのコホートを、Ad5gagベクター又はChAd3gagベクター 106vp/マウスで免疫付与した。
【0176】
抗Ad5及びChAd3の中和免疫性を、Ad5ベクター及びChAd3 SEAPベクターを使用して上述の中和アッセイにより第4週に評価した。BalbCマウスにおいてマッピングされるgagエピトープを含有するgag9マーペプチドで刺激した、精製脾臓細胞においてELISPOT分析により抗gag免疫性を評価した。図36で報告する結果から、抗Ad5免疫性がChAd3gagにより誘発される抗gagCMIを抑制しないのに対し、予測されるように、抗Ad5免疫性がAd5gag免疫付与を完全に遮断することが示された。
【実施例9】
【0177】
ChAd3hCEA免疫化はヒトCEAを発現するトランスジェニックマウスにおいて強いCEA特異的免疫反応を誘発する。
【0178】
本明細書で開示し主張するChAdベクターが自己抗原に対して免疫反応を誘発し、それゆえにその寛容性を打破する能力も、ヒトCEAを発現するトランスジェニックマウスにおいて評価した(Clarke,Pら、Cancer Res.(1988)58(7):1469−77.)。
【0179】
8匹のマウスのコホートの大腿四頭筋にChAd3hCEA又はAd5hCEA 1010vpを既に述べたように注射した。aa497から端(aa703)までのヒトCEAアミノ酸配列を包含する15マーペプチドのプールで刺激したPBMCに対して、CEAに対する免疫反応を75日目まで毎週モニタリングした。CEAペプチドプールインキュベーションに反応した、CD4−CD8+T細胞分泌性インターフェロン−γを調べるICSによって、抗CEA免疫性を評価した。
【0180】
図37で報告する結果から、ChAd3hCEAベクター免疫付与が、同じ導入遺伝子を発現するAd5よりも、ヒトCEAに対するCD8+T細胞反応をより持続するよう刺激することが示される。
【0181】
霊長類免疫付与実験
方法及び材料
免疫付与プロトコール
本明細書に開示し主張するChAdベクターがアカゲザル(本明細書でサルと呼ばれる。)においてCMIを誘発する能力も評価した。サルを麻酔し(ケタミン/キシラジン)、本ワクチンをツベルクリンシリンジ(Becton−Dickinson)を使用して両側の三角筋へ0.5mlずつ筋肉内に送達した。全ての場合において、このサルの体重は、3から10kgであり、各ワクチンの用量は緩衝液1ml中で投与した。
【0182】
免疫付与手法の間、何回か回収した血液試料から、血清及び末梢血単核球(PBMC)を調製した。動物の取り扱い及び処置は全て、the Guide for Care and Use of Laboratory animals,Institute of Laboratory animal Resources,National Research Councilに記載の原則に従い、the Institutional animal Care and Use Committeeにより承認される基準に従った。
【0183】
ELISPOTアッセイ
アカゲザルについてのIFN−γELISPOTアッセイを、いくつかの改変を行い、上述のプロトコール(Allenら、2001 J.Virol.75(2):738−749)に従って実行した。gag特異的刺激について、10−aaの重複がある全HIV−1gag配列を包含する20−aaのペプチドからペプチドプールを調製した(Synpep Corp., Dublin, CA)。HCV NS特異的刺激について、10−aaの重複があるNS3からNS5bの全HCV−NS配列を包含する15−aaペプチドから、6個のペプチドプールを調製した。
【0184】
HER2 neu及びCEA特異的刺激を、10−aaの重複がある全タンパク質の配列を包含する15−aaのペプチドで実行した。
【0185】
2から4x105個の末梢血単核球(PBMC)50μlを各ウェルへ添加した。小さなサイズを80fLでカットオフして、細胞をBeckman Coulter Z2粒子アナライザーを使用して計数した。培地 50μl又はペプチドあたり8μg/mlのgagペプチドプールのいずれかをこのPBMCへ添加した。この試料を37℃、5%CO2で20時間から24時間インキュベーションした。このように、スポットを発色させ、プレートを特注の画像化装置及びImageProプラットフォーム(Silver Spring, MD)に基づいた自動計数サブルーチンを使用して処理した。計数を106個の細胞インプットに対して正規化した。
【0186】
細胞内サイトカイン染色(ICS)
2x106 PBMC/mL完全RPMI培地 1mL(17x100mmの丸底ポリプロピレンチューブ(Sarstedt,Newton,NC)に入れた。)に、抗hCD28(クローンL293、Becton−Dickinson)及び抗hCD49d(クローンL25、Becton−Dickinson)モノクローナル抗体を、終濃度1μg/mLとなるように添加した。gag特異的刺激については、ペプチドプール 10μL(ペプチド1個につき0.4mg/ml)を添加した。同様の条件をHCV NS特異的刺激について使用した。このチューブを37℃で1時間インキュベーションした後、5mg/mLのブレフェルジンA(Sigma)20μlLを添加した。細胞を37℃、5%CO2、90%湿度で16時間インキュベーションした。冷PBS/2%FBS 4mLを各チューブへ添加し、細胞を1200rpmで10分間遠心分離してペレットを得た。細胞をPBS/2%FBS中に再懸濁し、いくつかの蛍光標識mAbを使用して表面マーカーに対して染色した(30分間、4℃)(そのモノクローナル抗体は、チューブ1本あたり20μlの、抗hCD3−APC、クローンFN−18(Biosource);20μl 抗hCD8−PerCP、クローンSK1(Becton Dickinson, Franklin Lakes, NJ);及び20μL 抗hCD4−PE、クローンSK3(Becton Dickinson)であった。)。この段階から、試料を暗所で取り扱った。細胞を洗浄し、750μL 1xFACS Perm緩衝液(Becton Dickinson)中で室温にて10分間インキュベーションした。この細胞をペレットにし、PBS/2%FBS中に再懸濁し、FITC−抗hIFN−γ、クローンMD−1(Biosource)0.1μgを添加した。30分間のインキュベーションの後、細胞を洗浄し、PBS中に再懸濁した。Becton Dickinson FACSCalibur機器の4色チャネル全部を使用して試料を分析した。本データを分析するため、下部側及び前方に散乱するリンパ球集団を最初にゲーティングした。サイトカイン陽性反応についての共通の蛍光のカットオフをCD4+及びCD8+の両集団について使用し、試料の偽反応及びgagペプチド反応チューブの両者に対して使用した。
【実施例10】
【0187】
ChAdΔE1−gagベクターを使用する同種初回刺激−追加免疫法はサルにおけるgag特異的T細胞を誘発する。
【0188】
1コホート3匹の動物のコホートに以下のコンストラクトのいずれかを第0週及び第4週で筋肉内注射により与えた。すなわちCV32ΔE1−gag 1010vp又はCV33ΔE1−gag 1010vp又はMRKAd5ΔE1gag 1010vp及び108vpであった。4週間隔で回収したPBMCを、ELISPOTアッセイにより分析した。Gagペプチドプール非存在下(偽)又は存在下でのインキュベーション後の、100万個のPBMCあたりのスポット形成細胞数を示す表6で与える結果から、非ヒト霊長類においてgag特異的T細胞の有意なレベルをCV32ΔE1−gag及びCV33ΔE1gagの両者が誘導できることが確認される。興味深いことに、単回投与(第4週)後、CV32ΔE1−gag反応は、108vp/投与のMRKAd5ΔE1−gagに匹敵し、1010vp/投与のMRKAd5−gagのそれより低かったことに留意すること。1010vp/投与のCV33ΔE1−gagは、1010vp/投与のMRKAd5−gagのそれに匹敵する反応を誘導する。この結果は2回目の投与後の第8週で確認された。
【0189】
【表6】
【実施例11】
【0190】
ChAdベクターは、異種初回刺激−追加免疫法においてHCV NS特異的T細胞反応を誘発する。
【0191】
個別の実験において、2匹及び3匹のサルのグループに第0週、第4週で動物1匹につき108vp又は1010vpでMRK Ad6NSoptmutベクターを用いて免疫付与を行った。この動物を第24週で、同じ用量の同じウイルスで追加免疫し、1010個vp/動物でCV33−NSmutにより第104週に再度追加免疫を行った。この結果は、HCV NSペプチドプールの非存在下(偽)又は存在下でのインキュベーション後の100万個のPBMCあたりのスポット形成細胞数を要約する表7及び表8に示す。
【0192】
IFN−γELISPOTにより評価されるように、T細胞免疫性は1010vpを注射した動物において最初の投与後第4週で(表8)、及び108vpで注射した動物において第8週(2回目の投与後)で(表7)ピークの反応を示した。この反応は第24週での注射により促進されなかった(同種追加免疫)のに対し、強い促進効果がCV33−NSmutによる注射の後に観察された(異種追加免疫)。
【0193】
【表7】
【0194】
【表8】
【0195】
チンパンジーAdベクターによる異種追加免疫の効率を第二の実験において評価した。3匹のサルのコホートに対して、第0週及び第4週に、MRKAd5gag(1010vp/動物)、MRKAd6NSmut(1010vp/動物)又は両ベクター(各ベクターにつき1010vp/動物)の組み合わせで免疫付与し、その後第24週で同じ抗原で追加免疫した(同種追加免疫)。同種追加免疫を同じ抗原で実行し、異種追加免疫をCV33gag、CV32NSmut又は前記2つのベクターの組み合わせで実行した。表9で与える結果は、HCV NSペプチドプール非存在下(偽)又は存在下でのインキュベーション後、100万個のPBMCあたりのスポット形成細胞数を要約する。
【0196】
同じコホートを、第51週に、CV33gag(1010vp/動物)、CV32NSmut(1010vp/動物)及び前記2つのベクターの組み合わせ(各ベクターにつき1010vp/動物)で再度追加免疫した。表9で与える結果はさらに、この反応がワクチンの初回用量の注射後第4週で観察されるピークを下回るため、同種追加免疫が効果的ではなかったことを示す。異種チンパンジーベクターによる免疫付与後第54週で、IFN−γELISPOTにより強い免疫促進効果が測定された。
【0197】
【表9】
【実施例12】
【0198】
TAAを含むChAdベクターによるワクチン接種は寛容性を打破し、サルにおけるTAA特異的T細胞反応を誘発する。
【0199】
チンパンジーアデノウイルスベクターが自己抗原に対する寛容性を打破し、ヒトアデノウイルスベクターにより初回刺激された免疫反応を促進するためのチンパンジーベクターの有用性を証明するのに十分に免疫原性があるか否かを調べるために設定した実験を、1コホート4匹のサルのコホートにおいて実施した。腫瘍関連抗原CEAを含むAd5DE1 RhCEA(1011vp)の3回の注射を第0週、第2週及び第4週に行って動物に免疫付与した後、第16週、第18週及び第20週に、CV33DE1 RhCEA(1011vp)によるワクチン接種を行った。アカゲザルCEAペプチドを用いて、IFNγELISPOTにより、T細胞反応を測定した。
【0200】
アカゲザルCEA C及びDペプチドプール非存在下(DMSO)又は存在下のインキュベーション後の100万個のPBMCあたりのスポット形成細胞数を与える図34に報告される結果により、2つの異なるAd血清型によるワクチン接種に基づく免疫付与プロトコールが、非ヒト霊長類において、CEAに対してT細胞反応を持続させるに到ることが確認される。
【0201】
本発明はその特定の好ましい実施形態に関して詳細に記載されているが、改変及びバリエーションが、記載され、主張される本発明の精神及び範囲内であることは理解されよう。
【図面の簡単な説明】
【0202】
【図1】図1は、ChAd6シャトルベクター(pARS ChAd6−3)を構築するために使用されるクローニングストラテジーを要約する概略図である。
【図2】図2は、E.コリ株BJ5183での相同組み換えによりChAd6ウイルスゲノムをクローニングするために使用されるクローニングストラテジーを説明する概略図である。
【図3】図3は、pARS ChAd6−3 GAG:pARS ChAd6−3 SEAP;pARS ChAd6−3 EGFP及びpARS ChAd6−3 NS MUTを含む、様々なChAd6シャトルプラスミドのエレメントを説明する概略図である。
【図4】図4は、ChAd6ΔE1発現ベクターをクローニングするために使用される相同組み換えスキームを説明する概略図である。
【図5A】図5Aは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5B】図5Bは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5C】図5Cは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5D】図5Dは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5E】図5Eは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5F】図5Fは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5G】図5Gは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5H】図5Hは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5I】図5Iは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5J】図5Jは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図5K】図5Kは、ChAd3(配列番号1)のゲノムヌクレオチド配列を与える。
【図6A】図6Aは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6B】図6Bは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6C】図6Cは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6D】図6Dは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6E】図6Eは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6F】図6Fは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6G】図6Gは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6H】図6Hは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6I】図6Iは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6J】図6Jは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図6K】図6Kは、ChAd6(配列番号2)のゲノムヌクレオチド配列を与える。
【図7A】図7Aは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7B】図7Bは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7C】図7Cは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7D】図7Dは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7E】図7Eは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7F】図7Fは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7G】図7Gは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7H】図7Hは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7I】図7Iは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7J】図7Jは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図7K】図7Kは、CV32(配列番号3)のゲノムヌクレオチド配列を与える。
【図8A】図8Aは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8B】図8Bは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8C】図8Cは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8D】図8Dは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8E】図8Eは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8F】図8Fは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8G】図8Gは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8H】図8Hは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8I】図8Iは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8J】図8Jは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図8K】図8Kは、CV33(配列番号4)のゲノムヌクレオチド配列を与える。
【図9A】図9Aは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9B】図9Bは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9C】図9Cは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9D】図9Dは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9E】図9Eは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9F】図9Fは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9G】図9Gは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9H】図9Hは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9I】図9Iは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図9J】図9Jは、CV23(配列番号5)のゲノムヌクレオチド配列を与える。
【図10】図10は、ChAd20(配列番号6)の繊維遺伝子のヌクレオチド配列を与える。
【図11】図11は、ChAd4(配列番号7)の繊維遺伝子のヌクレオチド配列を与える。
【図12】図12は、ChAd5(配列番号8)の繊維遺伝子のヌクレオチド配列を与える。
【図13】図13は、ChAd7(配列番号9)の繊維遺伝子のヌクレオチド配列を与える。
【図14】図14は、ChAd9(配列番号10)の繊維遺伝子のヌクレオチド配列を与える。
【図15】図15は、ChAd10(配列番号11)の繊維遺伝子のヌクレオチド配列を与える。
【図16】図16は、ChAd11(配列番号12)の繊維遺伝子のヌクレオチド配列を与える。
【図17】図17は、ChAd16(配列番号13)の繊維遺伝子のヌクレオチド配列を与える。
【図18】図18は、ChAd17(配列番号14)の繊維遺伝子のヌクレオチド配列を与える。
【図19】図19は、ChAd19(配列番号15)の繊維遺伝子のヌクレオチド配列を与える。
【図20A】図20Aは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20B】図20Bは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20C】図20Cは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20D】図20Dは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20E】図20Eは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20F】図20Fは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図20G】図20Gは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82の繊維タンパク質のアミノ酸配列の、C1(配列番号85)、CV68(配列番号86)、PAN5(CV23とも呼ばれる。)(配列番号80)、PAN6(CV32とも呼ばれる。)(配列番号81)及びPan7(CV33とも呼ばれる。)(配列番号82)からの参照繊維タンパク質配列との比較を与える。
【図21】図21は、ChAd20(配列番号16)のヘキソン遺伝子のヌクレオチド配列を与える。
【図22】図22は、ChAd4(配列番号17)のヘキソン遺伝子のヌクレオチド配列を与える。
【図23】図23は、ChAd5(配列番号18)のヘキソン遺伝子のヌクレオチド配列を与える。
【図24】図24は、ChAd7(配列番号19)のヘキソン遺伝子のヌクレオチド配列を与える。
【図25】図25は、ChAd9(配列番号20)のヘキソン遺伝子のヌクレオチド配列を与える。
【図26】図26は、ChAd10(配列番号21)のヘキソン遺伝子のヌクレオチド配列を与える。
【図27】図27は、ChAd11(配列番号22)のヘキソン遺伝子のヌクレオチド配列を与える。
【図28】図28は、ChAd16(配列番号23)のヘキソン遺伝子のヌクレオチド配列を与える。
【図29】図29は、ChAd17(配列番号24)のヘキソン遺伝子のヌクレオチド配列を与える。
【図30】図30は、ChAd19(配列番号25)のヘキソン遺伝子のヌクレオチド配列を与える。
【図31A】図31Aは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31B】図31Bは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31C】図31Cは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31D】図31Dは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31E】図31Eは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31F】図31Fは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31G】図31Gは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31H】図31Hは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31I】図31Iは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図31J】図31Jは、ChAd3、ChAd4、ChAd5、ChAd6、ChAd7、ChAd8、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd20、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82のヘキソンタンパク質のアミノ酸配列の、C1(配列番号124)、CV68(配列番号125)、PAN5(CV23とも呼ばれる。)(配列番号119)、PAN6(CV32とも呼ばれる。)(配列番号120)及びPan7(CV33とも呼ばれる。)(配列番号121)からの参照繊維タンパク質配列との比較を与える。
【図32】図32は、本明細書中で開示される、オリゴマー及びプライマーを含む、人工的配列、配列番号26から40及び配列番号45及び46のリストを与える。
【図33】図33は、様々な血清型サブグループ(つまり、サブグループC、E及びD)に属するChAdベクターの免疫付与ブレイクポイントを図示したものである。gag−発現する、ChAd3、ChAd11、ChAd20、CV33、CV68、ChAd6、ChAd9、ChAd10、CV32、ChAd4、ChAd7及びChAd16ベクターで免疫付与したマウスにおいてタイター決定実験を行うことにより、測定可能な免疫反応を誘発する最低用量を調べた。
【図34】図34は、ヒトアデノウイルスベクター(MRKAd5 ThCEA)と、次いで、12週間の間隔後、チンパンジーアデノウイルスベクター(CV33 RhCEA)で連続的に免疫付与したアカゲザルにおいて誘発されるCEA特異的T細胞反応のグラフである。IFNγ−ELISPOTアッセイにより免疫反応を評価した。データは、アカゲザルCEA C及びDペプチドプールの非存在下(DMSO)及び存在下でのインキュベーション後の、末梢血単核球(PBMC)百万個あたりのスポット形成細胞(SFC)の数を示す。
【図35】図35は、PAUPSEARCH(Wisconsin Package Version 10.3,Accelrys Inc.)を用いて、TREEVIEWにより視覚化し操作した、全長ヘキソンペプチド配列の多配列アラインメントから推定される、ヒト及びチンパンジーアデノウイルスの系統樹を与える。
【図36】図36は、hAd5に前もって曝露したマウスへのChAd3及びhAd5 gagベクターの投与に対する反応において得られた免疫付与結果を表すグラフである。精製脾臓細胞を用いて、IFNγ−ELISPOTにより、免疫付与から3週間後に、細胞性免疫を評価した。
【図37】図37は、ChAd3hCEA及びAd5hCEAで免疫付与したヒトCEAトランスジェニックマウスにおいて誘発された、抗CEA CMIのキネティクスを表すグラフである。CEAペプチドプールで刺激したPBMCのICSにより、CMIを評価した。結果は、IFNγ+CD8+/総PBMCの%として表す。
【図38A】図38Aは、EGFPを発現し、様々なサブグループ(B、C、D、E)に属する様々なChAdベクターの、moi 50、250及び1250に曝露した様々なヒト初代細胞の感染効率を表すグラフである。結果は、蛍光細胞/総細胞の%として表す。
【図38B】図38Bは、EGFPを発現し、様々なサブグループ(B、C、D、E)に属する様々なChAdベクターの、moi 50、250及び1250に曝露した様々なヒト初代細胞の感染効率を表すグラフである。結果は、蛍光細胞/総細胞の%として表す。
【図38C】図38Cは、EGFPを発現し、様々なサブグループ(B、C、D、E)に属する様々なChAdベクターの、moi 50、250及び1250に曝露した様々なヒト初代細胞の感染効率を表すグラフである。結果は、蛍光細胞/総細胞の%として表す。
【図38D】図38Dは、EGFPを発現し、様々なサブグループ(B、C、D、E)に属する様々なChAdベクターの、moi 50、250及び1250に曝露した様々なヒト初代細胞の感染効率を表すグラフである。結果は、蛍光細胞/総細胞の%として表す。
【特許請求の範囲】
【請求項1】
a)配列番号1;
b)配列番号2;及び
c)(a)又は(b)の配列に相補的な核酸配列
からなる群から選択される、単離されたチンパンジー核酸配列。
【請求項2】
a)配列番号16から25、41、43、97、99、101、103、105、107、109、111、113、115及び117からなる群から選択される、ヘキソン遺伝子配列;並びに
b)配列番号6から15、42、44、58、60、62、64、66、68、70、72、74、76及び78からなる群から選択される、繊維遺伝子配列
の群から選択されるヘキソン及び繊維核酸配列の任意の組合せを含む、単離された組み換えチンパンジー血清型。
【請求項3】
ECACC受託番号03121201(ChAd4)、03121202(ChAd5)、03121203(ChAd7)、03121204(ChAd9)、03121205(ChAd10)、03121206(ChAd11)、03121207(ChAd16)、03121208(ChAd17)、03121209(ChAd19)及び03121210(ChAd20)からなる分離株の群から選択される、アデノウイルス由来のヘキソン及び繊維核酸配列の任意の組合せを含む、単離された組み換えチンパンジー血清型。
【請求項4】
ECACC受託番号03121201(ChAd4)、03121202(ChAd5)、03121203(ChAd7)、03121204(ChAd9)、03121205(ChAd10)、03121206(ChAd11)、03121207(ChAd16)、03121208(ChAd17)、03121209(ChAd19)及び03121210(ChAd20)からなる分離株の群から選択される、アデノウイルス由来のヌクレオチド配列と、哺乳動物細胞においてその導入遺伝子の発現を支配する調節配列に操作可能に連結された少なくとも1個の免疫原をコードする導入遺伝子と、を含み、アデノウイルスE1、E2、E3及びE4からなる群から選択される、少なくとも1個のアデノウイルス遺伝子を含むヌクレオチドを欠く、複製欠損チンパンジーアデノウイルス(ChAd)ベクター。
【請求項5】
配列番号1又は配列番号2に記載のヌクレオチド配列と、哺乳動物細胞においてその導入遺伝子の発現を支配する調節配列に操作可能に連結された少なくとも1個の免疫原をコードする導入遺伝子を含み、アデノウイルスE1、E2、E3及びE4からなる群から選択される、少なくとも1個の遺伝子を含む、配列番号1又は配列番号2のヌクレオチド配列を欠く、複製欠損チンパンジーアデノウイルス(ChAd)ベクター。
【請求項6】
配列番号1のbp460からbp3542又は配列番号2のbp457からbp3425の領域における、E1ヌクレオチド配列での欠損/破壊を含む、複製欠損ChAdベクター。
【請求項7】
前記ベクターが、HIV、HBV、HCV、HPV、HSV1、HSV2、SARS CoV、Plasmodium malariae(四日熱マラリア原虫)、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB及びMycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)からなる群から選択される、導入遺伝子を含む、請求項6に記載のChAdベクター。
【請求項8】
前記ベクターが、配列番号1のbp460からbp3542又は配列番号2のbp457からbp3425の領域におけるE1ヌクレオチド配列での欠損/破壊を含む、該ベクターが、さらに、少なくとも1個の癌関連抗原(TAA)をコードする導入遺伝子を含む、請求項6に記載のChAdベクター。
【請求項9】
少なくとも1個のTAAが、HER2 NEU、CEA、EPCAM、PSA、PSMA、テロメラーゼ、GP100、MELAN−A/MART−1、MUC−1、NY−ESO−1、サバイビン、ストロメリシン 3、チロシナーゼ、MAGE3、CML68、CML66、OY−TES−1、SSX−2、SART−1、SART−2、SART−3、NY−CO−58、NY−BR−62、HKLP2、5T4及びVEGFR2からなる群から選択される、請求項8に記載のChAdベクター。
【請求項10】
E1a、E1b、E2a、E2b、E4 orfs 1,2、3、4、5、6、6/7、pIX、IVa2、領域L1、L2、L3、L4、L5からなる群から選択される、1又は複数のアデノウイルス領域を発現し、請求項1又は請求項2に記載の核酸分子を含む、宿主細胞。
【請求項11】
請求項5に記載のアデノウイルスベクターをアデノウイルスE−1発現ヒト細胞に導入することと、得られるアデノウイルスを回収することと、を含む、複製欠損チンパンジーアデノウイルスベクターを生成させる方法。
【請求項12】
前記ヒト細胞が、293細胞又はPER.C6TM細胞である、請求項11に記載の方法。
【請求項13】
請求項4から請求項6のいずれか1項に記載の複製欠損ChAdベクターを含む、ワクチン組成物。
【請求項14】
配列番号1に記載のヌクレオチド配列を含む、アデノウイルスE1発現ヒト細胞。
【請求項15】
配列番号2に記載のヌクレオチド配列を含む、アデノウイルスE1発現ヒト細胞。
【請求項16】
少なくともそのE1遺伝子の機能的欠損と、標的抗原をコードするヌクレオチド配列と、該標的抗原をコードする該ヌクレオチド配列の発現を支配することが可能なプロモーター配列と、を含有するチンパンジーアデノウイルスゲノムを含む組み換えChAdベクターの十分量を哺乳動物に投与することを含み、該chAdベクターの投与が増強反応を誘発する、該哺乳動物における抗原特異的免疫反応を増強する方法。
【請求項17】
前記ChAdベクターが、そのE1遺伝子の完全欠失を含み、さらに、該ベクターが、そのE3遺伝子の欠失を場合によっては含む、請求項16に記載の方法。
【請求項18】
前記増強された免疫反応が、HIV、HBV、HCV、HPV、HSV1、HSV2、SARS CoV、Plasmodium malariae(四日熱マラリア原虫)、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB及びMycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)からなる群から選択される、感染物質由来の抗原に特異的である、請求項16に記載の方法。
【請求項19】
前記免疫反応が、TAAに特異的な増強免疫反応である、請求項16に記載の方法。
【請求項20】
前記増強された免疫反応が、抗原特異的CD8+T細胞の産生を含む、請求項19に記載の方法。
【請求項21】
前記増強された免疫反応が、抗原特異的CD8+T細胞の産生を含む、請求項16に記載の方法。
【請求項22】
少なくともそのE1遺伝子の機能的欠損と、標的抗原をコードするヌクレオチドと、該標的抗原をコードする該ヌクレオチド配列の発現を支配することが可能なプロモーター配列と、を含有するチンパンジーアデノウイルスゲノムを含むChAdベクターの十分量を哺乳動物に投与することを含み、該ChAdベクターの投与が一次免疫反応を誘発する、未処置哺乳動物において免疫反応を誘発する方法。
【請求項23】
前記一次免疫反応が、これらに限定されないが、HIV、HCV、HPV、HSV1、HSV2、SARS Cov、Plasmodium malariae(四日熱マラリア原虫)、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB、Mycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)などの、感染物質由来の抗原に特異的である、請求項22に記載の方法。
【請求項24】
前記免疫反応が、前記哺乳動物が寛容性を有するTAAに特異的な一次免疫反応である、請求項16に記載の方法。
【請求項25】
前記組み換えアデノウイルスが、配列番号87から96、98、100,102、104、106、108、110、112、114、116、118、122及び123からなる群から選択される、アミノ酸配列からなるヘキソンペプチドをコードするヌクレオチド配列を含む、請求項16に記載の方法。
【請求項26】
前記組み換えアデノウイルスが、配列番号48から57、59、61、63、65、67、69、71、73、75、77、79、83及び84からなる群から選択されるアミノ酸配列からなる繊維タンパク質をコードするヌクレオチド配列を含む、請求項16のいずれか1つに記載の方法。
【請求項27】
(a)抗原特異的免疫反応が望まれる感染物質抗原をコードするヌクレオチド配列を含む第一のワクチン組成物を投与することによって、感染物質抗原に反応させるために、宿主に初回刺激を行う段階、
(b)少なくともそのE1遺伝子の機能的欠失、及び該E1遺伝子欠失の部位において、該初回刺激段階で送達される同じ感染物質抗原をコードするDNAの発現を支配することが可能であるプロモーターを含む配列、を含有する、組み換えChAdベクターを含む第二のワクチン組成物を投与することによって、段階(a)の免疫反応を増強する段階、
を含み、
該追加免疫組成物の投与が、防御的免疫を付与する効果を有する免疫反応を誘発する、HIV、HCV、HPV、HSV1、HSV2、SARS、Plasmodium malariae(四日熱マラリア原虫)、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB及びMycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)からなる群から選択される、感染物質由来の抗原に対して、免疫反応を誘導する方法。
【請求項28】
前記第一のワクチン組成物が、電気刺激と組み合わせて筋肉内に投与される、プラスミドDNAを含む、請求項27に記載の方法。
【請求項29】
前記第二のワクチン組成物が、HIV、HCV、HPV、HSV1、HSV2、SARS、マラリア、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB及びMycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)からなる群から選択される感染物質由来の抗原をコードするDNAを含むChAdベクターを含む、請求項27に記載の方法。
【請求項30】
前記免疫反応が抗原特異的CD8+T細胞の産生を含む、請求項27に記載の方法。
【請求項31】
前記ChAdベクターが、ChAd3、ChAd6、ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82からなる群から選択されるアデノウイルス由来である、請求項30に記載の方法。
【請求項32】
前記ChAdベクターが、配列番号87から96、98、100、102、104、106、108、110、112、114、116、118、122及び123からなる群から選択される、アミノ酸配列からなるヘキソンペプチドをコードするヌクレオチド配列を含む、請求項30に記載の方法。
【請求項33】
前記ChAdベクターが、配列番号48から57、59、61、63、65、67、69、71、73、75、77、79、83及び84からなる群から選択される、アミノ酸配列からなる繊維タンパク質をコードするヌクレオチド配列を含む、請求項30に記載の方法。
【請求項34】
前記第一及び第二のワクチン組成物が、異なる血清型により特徴付けられるChAdベクターである、請求項30に記載の方法。
【請求項35】
(a)抗原特異的免疫反応が望まれる自己抗原をコードするヌクレオチド配列を含む第一のワクチン組成物を投与することによって、自己抗原に対して反応させるために宿主に初回刺激を行うことと(それによって、一次反応を誘発する。);
(b)少なくともそのE1遺伝子の機能的欠失を含有し、該E1遺伝子欠失の部位において、初回刺激段階で送達される同じ自己抗原をコードするDNAの発現を支配することが可能であるプロモーターを含む配列,を含有する、組み換えChAdベクターを含む第二のワクチン組成物を投与することによって、段階(a)の一次免疫反応を増強することと、を含み、
該追加免疫組成物の投与が、自己抗原に対する宿主の寛容性を打破する効果を有する免疫反応を誘発する、自己抗原に対する宿主寛容性を打破する方法。
【請求項36】
前記第一のワクチン組成物が、電気刺激と組み合わせて筋肉内に投与される、プラスミドDNAを含む、請求項35に記載の方法。
【請求項37】
前記第二のワクチン組成物が、HER2 NEU、CEA、HEPCAM、PSA、PSMA、テロメラーゼ、GP100、MELAN−A/MART−1、MUC−1、NY−ESO−1、サバイビン、ストロメリシン 3、チロシナーゼ、MAGE3、CML68、CML66、OY−TES−1、SSX−2、SART−1、SART−2、SART−3、NY−CO−58、NY−BR−62、HKLP2、5T4及びVEGFR2からなる群から選択される、自己抗原をコードするDNAを含むChAdベクターを含む、請求項35に記載の方法。
【請求項38】
前記免疫反応が、抗原特異的CD8+T細胞の産生を含む、請求項35に記載の方法。
【請求項39】
前記ChAdベクターが、ChAd3、ChAd6、ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82からなる群から選択される、アデノウイルス由来である、請求項35に記載の方法。
【請求項40】
前記ChAdベクターが、配列番号87から96、98、100、102、104、106、108、110、112、114、116、118、122及び123からなる群から選択されるアミノ酸配列からなるヘキソンペプチドをコードするヌクレオチド配列を含む、請求項35に記載の方法。
【請求項41】
前記ChAdベクターが、配列番号48から57、59、61、63、65、67、69、71、73、75、77、79、83及び84からなる群から選択されるアミノ酸配列からなる繊維ペプチドをコードするヌクレオチド配列を含む、請求項35に記載の方法。
【請求項42】
前記第一及び第二のワクチン組成物が両者とも、異なる血清型により特徴付けられるChAdベクターである、請求項35に記載の方法。
【請求項1】
a)配列番号1;
b)配列番号2;及び
c)(a)又は(b)の配列に相補的な核酸配列
からなる群から選択される、単離されたチンパンジー核酸配列。
【請求項2】
a)配列番号16から25、41、43、97、99、101、103、105、107、109、111、113、115及び117からなる群から選択される、ヘキソン遺伝子配列;並びに
b)配列番号6から15、42、44、58、60、62、64、66、68、70、72、74、76及び78からなる群から選択される、繊維遺伝子配列
の群から選択されるヘキソン及び繊維核酸配列の任意の組合せを含む、単離された組み換えチンパンジー血清型。
【請求項3】
ECACC受託番号03121201(ChAd4)、03121202(ChAd5)、03121203(ChAd7)、03121204(ChAd9)、03121205(ChAd10)、03121206(ChAd11)、03121207(ChAd16)、03121208(ChAd17)、03121209(ChAd19)及び03121210(ChAd20)からなる分離株の群から選択される、アデノウイルス由来のヘキソン及び繊維核酸配列の任意の組合せを含む、単離された組み換えチンパンジー血清型。
【請求項4】
ECACC受託番号03121201(ChAd4)、03121202(ChAd5)、03121203(ChAd7)、03121204(ChAd9)、03121205(ChAd10)、03121206(ChAd11)、03121207(ChAd16)、03121208(ChAd17)、03121209(ChAd19)及び03121210(ChAd20)からなる分離株の群から選択される、アデノウイルス由来のヌクレオチド配列と、哺乳動物細胞においてその導入遺伝子の発現を支配する調節配列に操作可能に連結された少なくとも1個の免疫原をコードする導入遺伝子と、を含み、アデノウイルスE1、E2、E3及びE4からなる群から選択される、少なくとも1個のアデノウイルス遺伝子を含むヌクレオチドを欠く、複製欠損チンパンジーアデノウイルス(ChAd)ベクター。
【請求項5】
配列番号1又は配列番号2に記載のヌクレオチド配列と、哺乳動物細胞においてその導入遺伝子の発現を支配する調節配列に操作可能に連結された少なくとも1個の免疫原をコードする導入遺伝子を含み、アデノウイルスE1、E2、E3及びE4からなる群から選択される、少なくとも1個の遺伝子を含む、配列番号1又は配列番号2のヌクレオチド配列を欠く、複製欠損チンパンジーアデノウイルス(ChAd)ベクター。
【請求項6】
配列番号1のbp460からbp3542又は配列番号2のbp457からbp3425の領域における、E1ヌクレオチド配列での欠損/破壊を含む、複製欠損ChAdベクター。
【請求項7】
前記ベクターが、HIV、HBV、HCV、HPV、HSV1、HSV2、SARS CoV、Plasmodium malariae(四日熱マラリア原虫)、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB及びMycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)からなる群から選択される、導入遺伝子を含む、請求項6に記載のChAdベクター。
【請求項8】
前記ベクターが、配列番号1のbp460からbp3542又は配列番号2のbp457からbp3425の領域におけるE1ヌクレオチド配列での欠損/破壊を含む、該ベクターが、さらに、少なくとも1個の癌関連抗原(TAA)をコードする導入遺伝子を含む、請求項6に記載のChAdベクター。
【請求項9】
少なくとも1個のTAAが、HER2 NEU、CEA、EPCAM、PSA、PSMA、テロメラーゼ、GP100、MELAN−A/MART−1、MUC−1、NY−ESO−1、サバイビン、ストロメリシン 3、チロシナーゼ、MAGE3、CML68、CML66、OY−TES−1、SSX−2、SART−1、SART−2、SART−3、NY−CO−58、NY−BR−62、HKLP2、5T4及びVEGFR2からなる群から選択される、請求項8に記載のChAdベクター。
【請求項10】
E1a、E1b、E2a、E2b、E4 orfs 1,2、3、4、5、6、6/7、pIX、IVa2、領域L1、L2、L3、L4、L5からなる群から選択される、1又は複数のアデノウイルス領域を発現し、請求項1又は請求項2に記載の核酸分子を含む、宿主細胞。
【請求項11】
請求項5に記載のアデノウイルスベクターをアデノウイルスE−1発現ヒト細胞に導入することと、得られるアデノウイルスを回収することと、を含む、複製欠損チンパンジーアデノウイルスベクターを生成させる方法。
【請求項12】
前記ヒト細胞が、293細胞又はPER.C6TM細胞である、請求項11に記載の方法。
【請求項13】
請求項4から請求項6のいずれか1項に記載の複製欠損ChAdベクターを含む、ワクチン組成物。
【請求項14】
配列番号1に記載のヌクレオチド配列を含む、アデノウイルスE1発現ヒト細胞。
【請求項15】
配列番号2に記載のヌクレオチド配列を含む、アデノウイルスE1発現ヒト細胞。
【請求項16】
少なくともそのE1遺伝子の機能的欠損と、標的抗原をコードするヌクレオチド配列と、該標的抗原をコードする該ヌクレオチド配列の発現を支配することが可能なプロモーター配列と、を含有するチンパンジーアデノウイルスゲノムを含む組み換えChAdベクターの十分量を哺乳動物に投与することを含み、該chAdベクターの投与が増強反応を誘発する、該哺乳動物における抗原特異的免疫反応を増強する方法。
【請求項17】
前記ChAdベクターが、そのE1遺伝子の完全欠失を含み、さらに、該ベクターが、そのE3遺伝子の欠失を場合によっては含む、請求項16に記載の方法。
【請求項18】
前記増強された免疫反応が、HIV、HBV、HCV、HPV、HSV1、HSV2、SARS CoV、Plasmodium malariae(四日熱マラリア原虫)、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB及びMycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)からなる群から選択される、感染物質由来の抗原に特異的である、請求項16に記載の方法。
【請求項19】
前記免疫反応が、TAAに特異的な増強免疫反応である、請求項16に記載の方法。
【請求項20】
前記増強された免疫反応が、抗原特異的CD8+T細胞の産生を含む、請求項19に記載の方法。
【請求項21】
前記増強された免疫反応が、抗原特異的CD8+T細胞の産生を含む、請求項16に記載の方法。
【請求項22】
少なくともそのE1遺伝子の機能的欠損と、標的抗原をコードするヌクレオチドと、該標的抗原をコードする該ヌクレオチド配列の発現を支配することが可能なプロモーター配列と、を含有するチンパンジーアデノウイルスゲノムを含むChAdベクターの十分量を哺乳動物に投与することを含み、該ChAdベクターの投与が一次免疫反応を誘発する、未処置哺乳動物において免疫反応を誘発する方法。
【請求項23】
前記一次免疫反応が、これらに限定されないが、HIV、HCV、HPV、HSV1、HSV2、SARS Cov、Plasmodium malariae(四日熱マラリア原虫)、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB、Mycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)などの、感染物質由来の抗原に特異的である、請求項22に記載の方法。
【請求項24】
前記免疫反応が、前記哺乳動物が寛容性を有するTAAに特異的な一次免疫反応である、請求項16に記載の方法。
【請求項25】
前記組み換えアデノウイルスが、配列番号87から96、98、100,102、104、106、108、110、112、114、116、118、122及び123からなる群から選択される、アミノ酸配列からなるヘキソンペプチドをコードするヌクレオチド配列を含む、請求項16に記載の方法。
【請求項26】
前記組み換えアデノウイルスが、配列番号48から57、59、61、63、65、67、69、71、73、75、77、79、83及び84からなる群から選択されるアミノ酸配列からなる繊維タンパク質をコードするヌクレオチド配列を含む、請求項16のいずれか1つに記載の方法。
【請求項27】
(a)抗原特異的免疫反応が望まれる感染物質抗原をコードするヌクレオチド配列を含む第一のワクチン組成物を投与することによって、感染物質抗原に反応させるために、宿主に初回刺激を行う段階、
(b)少なくともそのE1遺伝子の機能的欠失、及び該E1遺伝子欠失の部位において、該初回刺激段階で送達される同じ感染物質抗原をコードするDNAの発現を支配することが可能であるプロモーターを含む配列、を含有する、組み換えChAdベクターを含む第二のワクチン組成物を投与することによって、段階(a)の免疫反応を増強する段階、
を含み、
該追加免疫組成物の投与が、防御的免疫を付与する効果を有する免疫反応を誘発する、HIV、HCV、HPV、HSV1、HSV2、SARS、Plasmodium malariae(四日熱マラリア原虫)、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB及びMycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)からなる群から選択される、感染物質由来の抗原に対して、免疫反応を誘導する方法。
【請求項28】
前記第一のワクチン組成物が、電気刺激と組み合わせて筋肉内に投与される、プラスミドDNAを含む、請求項27に記載の方法。
【請求項29】
前記第二のワクチン組成物が、HIV、HCV、HPV、HSV1、HSV2、SARS、マラリア、エボラウイルス、ウエストナイルウイルス、デング熱ウイルス、インフルエンザA、インフルエンザB及びMycobacterium tubercolosis(マイコバクテリウム・テュバキュローシス)からなる群から選択される感染物質由来の抗原をコードするDNAを含むChAdベクターを含む、請求項27に記載の方法。
【請求項30】
前記免疫反応が抗原特異的CD8+T細胞の産生を含む、請求項27に記載の方法。
【請求項31】
前記ChAdベクターが、ChAd3、ChAd6、ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82からなる群から選択されるアデノウイルス由来である、請求項30に記載の方法。
【請求項32】
前記ChAdベクターが、配列番号87から96、98、100、102、104、106、108、110、112、114、116、118、122及び123からなる群から選択される、アミノ酸配列からなるヘキソンペプチドをコードするヌクレオチド配列を含む、請求項30に記載の方法。
【請求項33】
前記ChAdベクターが、配列番号48から57、59、61、63、65、67、69、71、73、75、77、79、83及び84からなる群から選択される、アミノ酸配列からなる繊維タンパク質をコードするヌクレオチド配列を含む、請求項30に記載の方法。
【請求項34】
前記第一及び第二のワクチン組成物が、異なる血清型により特徴付けられるChAdベクターである、請求項30に記載の方法。
【請求項35】
(a)抗原特異的免疫反応が望まれる自己抗原をコードするヌクレオチド配列を含む第一のワクチン組成物を投与することによって、自己抗原に対して反応させるために宿主に初回刺激を行うことと(それによって、一次反応を誘発する。);
(b)少なくともそのE1遺伝子の機能的欠失を含有し、該E1遺伝子欠失の部位において、初回刺激段階で送達される同じ自己抗原をコードするDNAの発現を支配することが可能であるプロモーターを含む配列,を含有する、組み換えChAdベクターを含む第二のワクチン組成物を投与することによって、段階(a)の一次免疫反応を増強することと、を含み、
該追加免疫組成物の投与が、自己抗原に対する宿主の寛容性を打破する効果を有する免疫反応を誘発する、自己抗原に対する宿主寛容性を打破する方法。
【請求項36】
前記第一のワクチン組成物が、電気刺激と組み合わせて筋肉内に投与される、プラスミドDNAを含む、請求項35に記載の方法。
【請求項37】
前記第二のワクチン組成物が、HER2 NEU、CEA、HEPCAM、PSA、PSMA、テロメラーゼ、GP100、MELAN−A/MART−1、MUC−1、NY−ESO−1、サバイビン、ストロメリシン 3、チロシナーゼ、MAGE3、CML68、CML66、OY−TES−1、SSX−2、SART−1、SART−2、SART−3、NY−CO−58、NY−BR−62、HKLP2、5T4及びVEGFR2からなる群から選択される、自己抗原をコードするDNAを含むChAdベクターを含む、請求項35に記載の方法。
【請求項38】
前記免疫反応が、抗原特異的CD8+T細胞の産生を含む、請求項35に記載の方法。
【請求項39】
前記ChAdベクターが、ChAd3、ChAd6、ChAd20、ChAd4、ChAd5、ChAd7、ChAd9、ChAd10、ChAd11、ChAd16、ChAd17、ChAd19、ChAd8、ChAd22、ChAd24、ChAd26、ChAd30、ChAd31、ChAd37、ChAd38、ChAd44、ChAd63及びChAd82からなる群から選択される、アデノウイルス由来である、請求項35に記載の方法。
【請求項40】
前記ChAdベクターが、配列番号87から96、98、100、102、104、106、108、110、112、114、116、118、122及び123からなる群から選択されるアミノ酸配列からなるヘキソンペプチドをコードするヌクレオチド配列を含む、請求項35に記載の方法。
【請求項41】
前記ChAdベクターが、配列番号48から57、59、61、63、65、67、69、71、73、75、77、79、83及び84からなる群から選択されるアミノ酸配列からなる繊維ペプチドをコードするヌクレオチド配列を含む、請求項35に記載の方法。
【請求項42】
前記第一及び第二のワクチン組成物が両者とも、異なる血清型により特徴付けられるChAdベクターである、請求項35に記載の方法。
【図1】

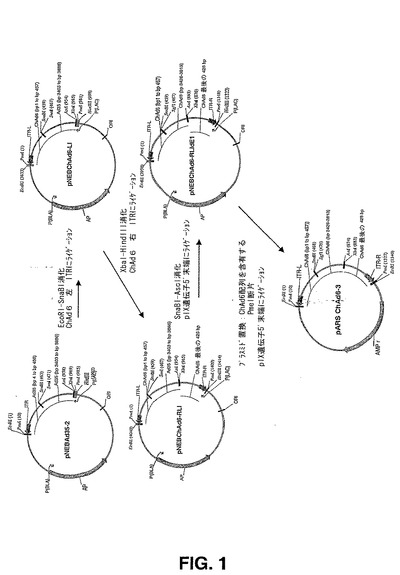
【図2】
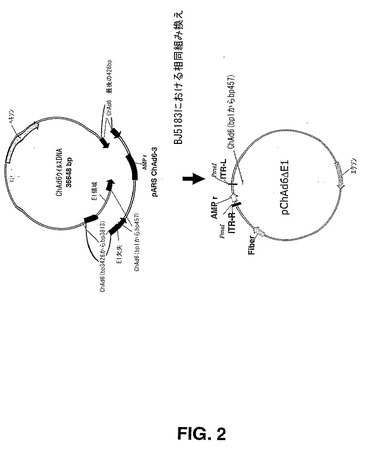
【図3】
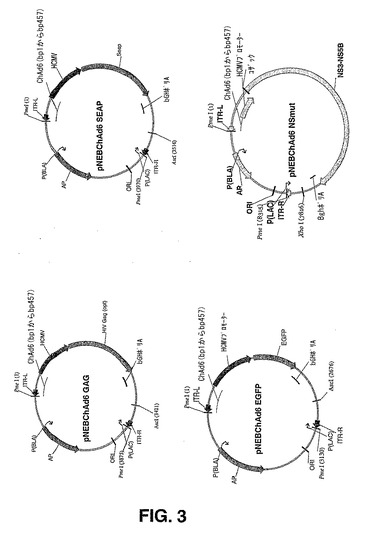
【図4】
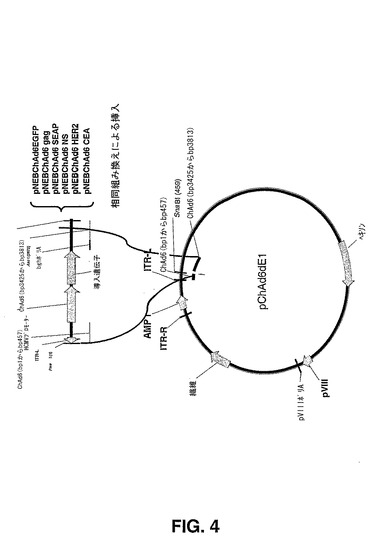
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図5I】
【図5J】
【図5K】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
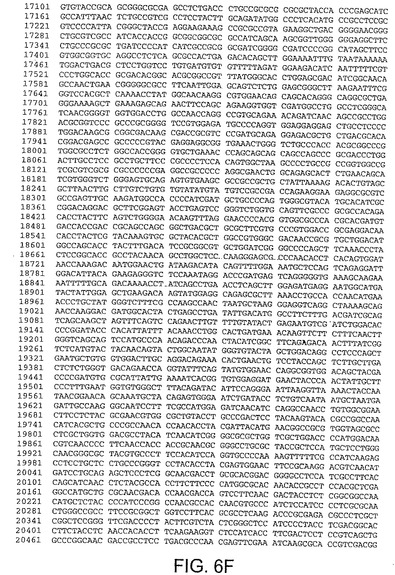
【図6G】
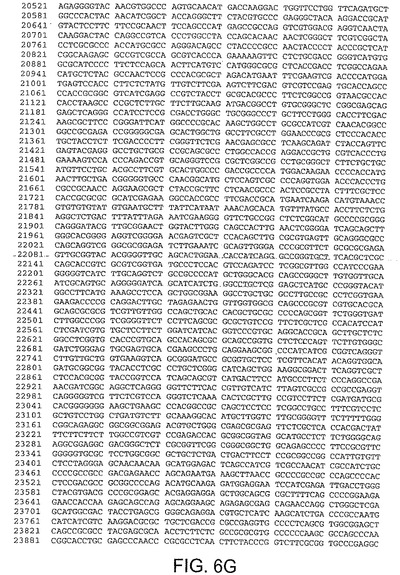
【図6H】
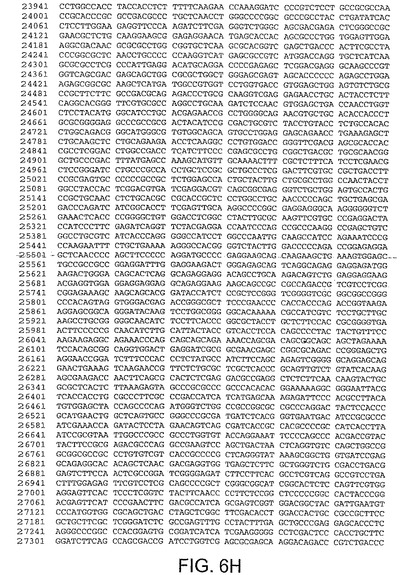
【図6I】
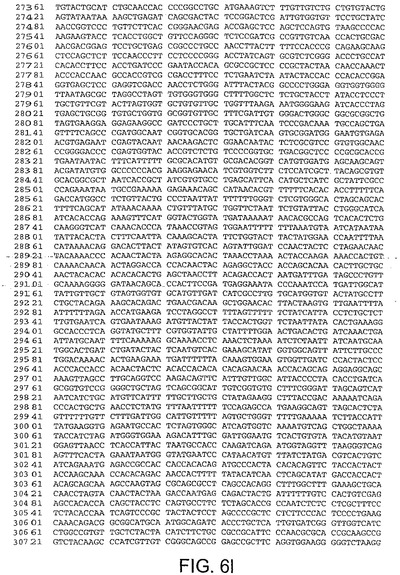
【図6J】
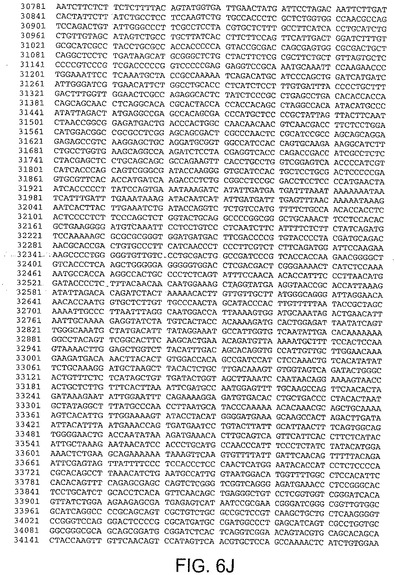
【図6K】
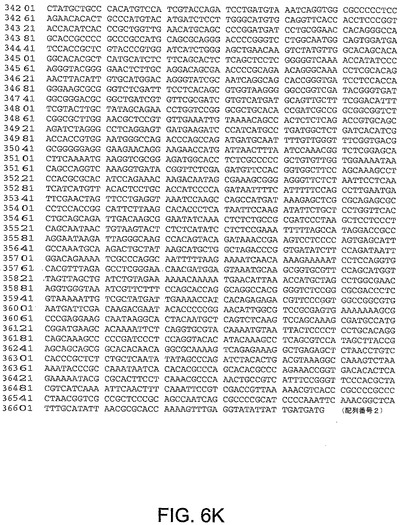
【図7A】

【図7B】
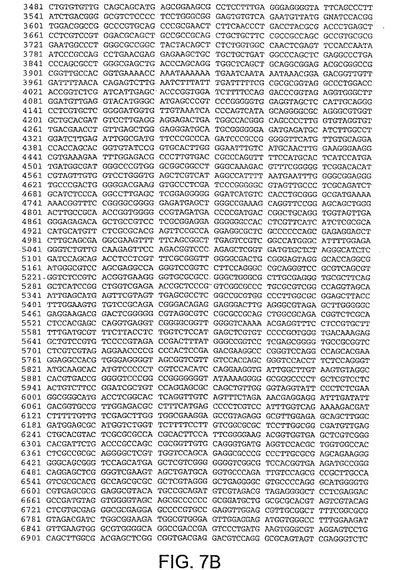
【図7C】
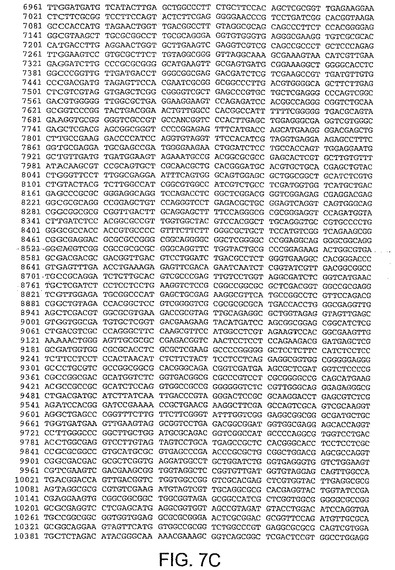
【図7D】
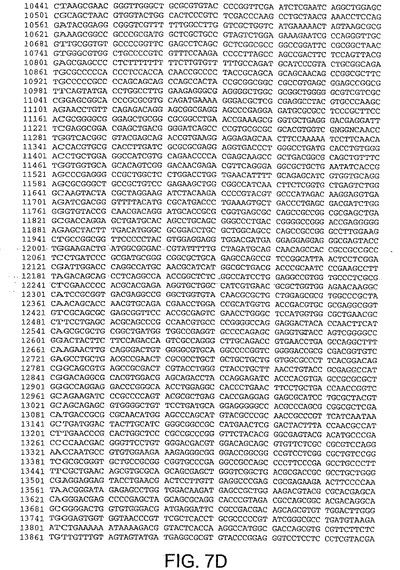
【図7E】
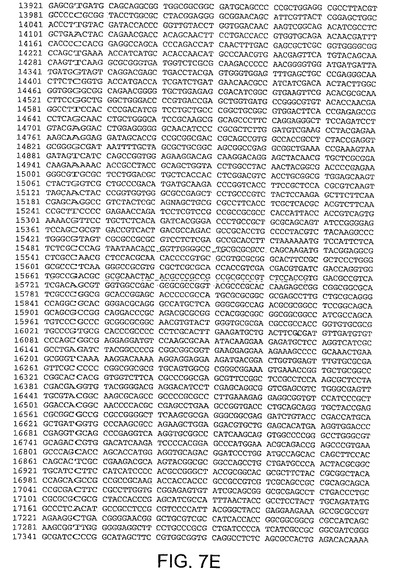
【図7F】
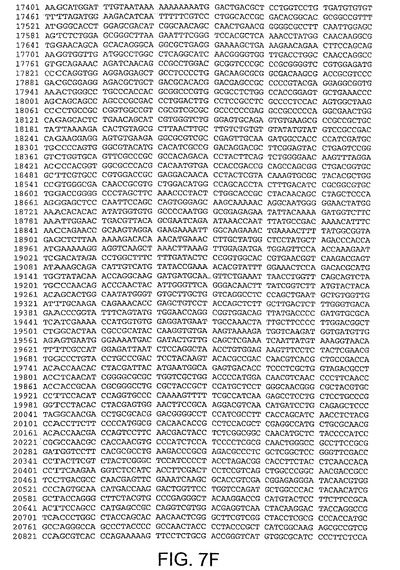
【図7G】
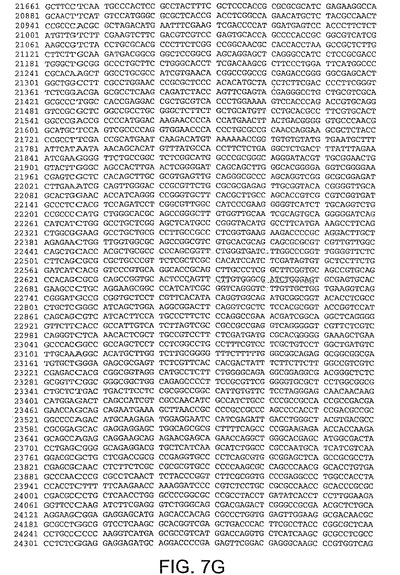
【図7H】
【図7I】

【図7J】
【図7K】

【図8A】
【図8B】
【図8C】
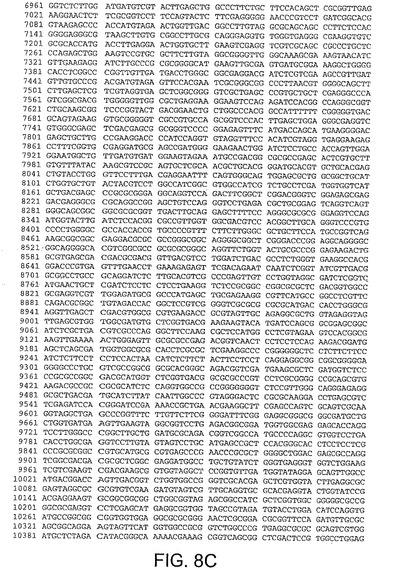
【図8D】
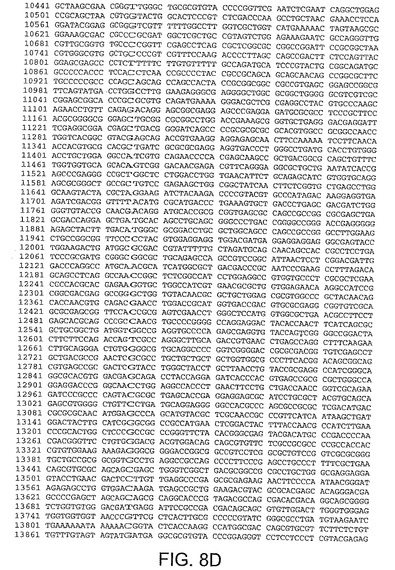
【図8E】
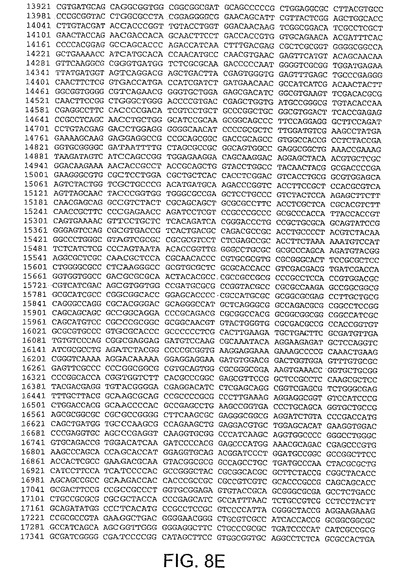
【図8F】
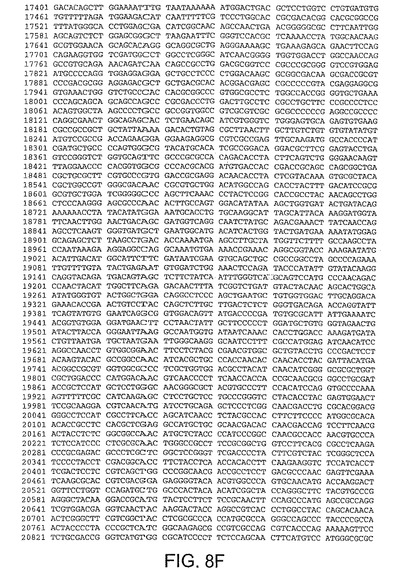
【図8G】
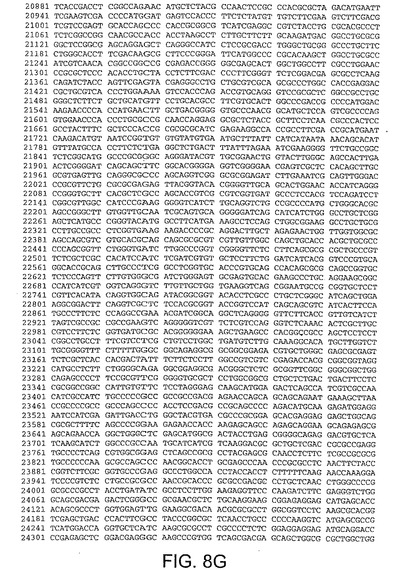
【図8H】
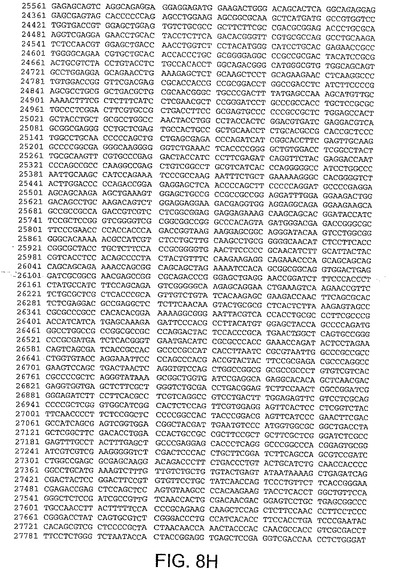
【図8I】
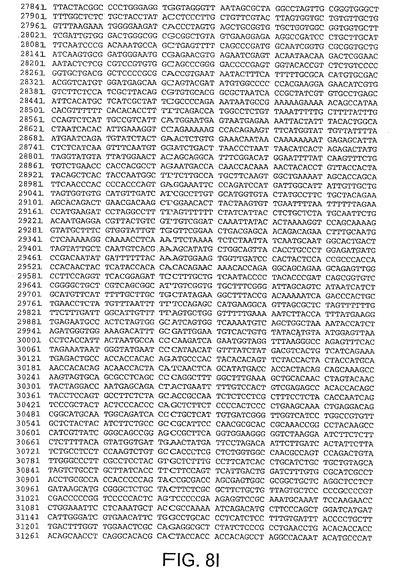
【図8J】
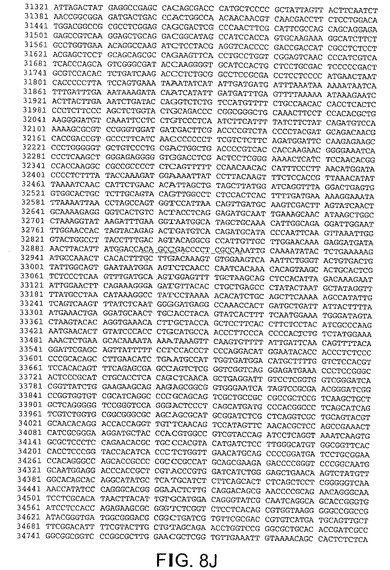
【図8K】

【図9A】

【図9B】

【図9C】

【図9D】

【図9E】

【図9F】

【図9G】

【図9H】

【図9I】

【図9J】

【図10】

【図11】
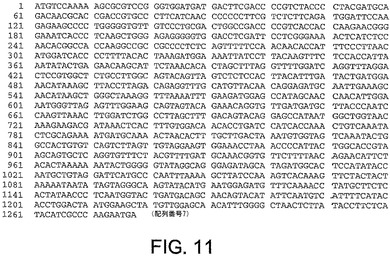
【図12】

【図13】

【図14】
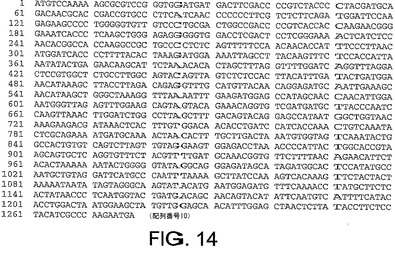
【図15】
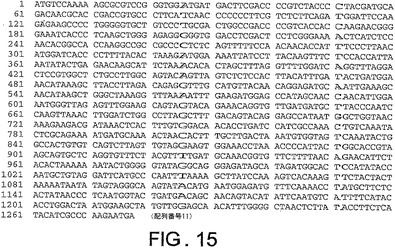
【図16】

【図17】

【図18】

【図19】

【図20A】

【図20B】

【図20C】

【図20D】

【図20E】

【図20F】

【図20G】

【図21】
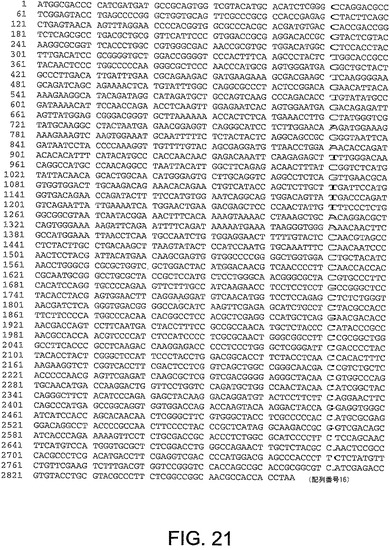
【図22】
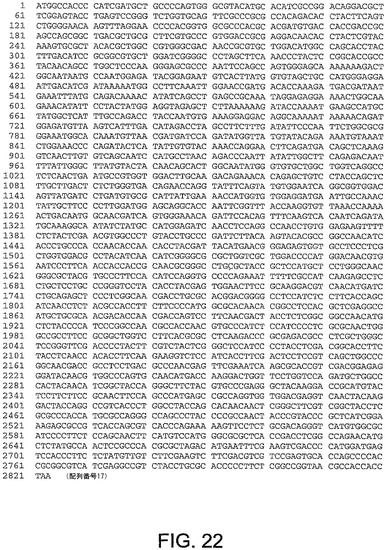
【図23】
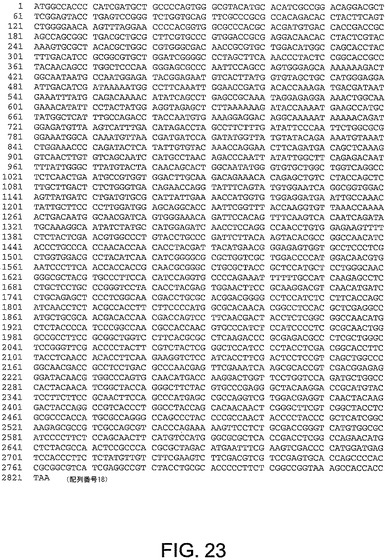
【図24】
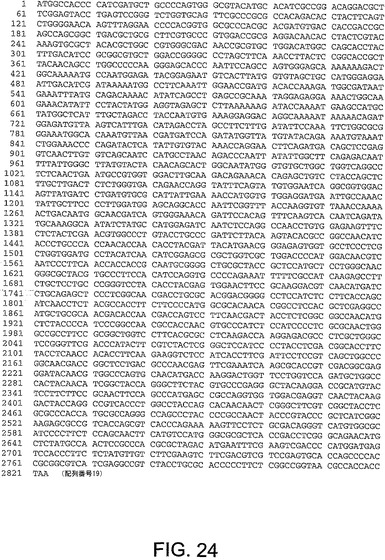
【図25】
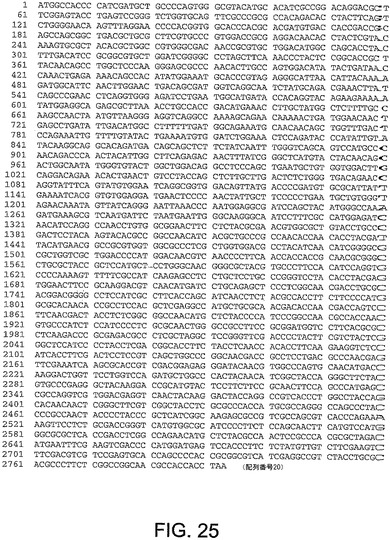
【図26】
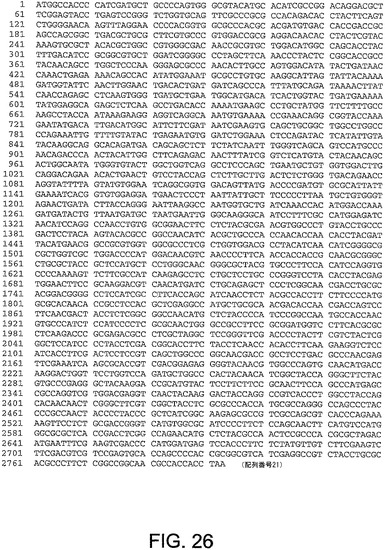
【図27】
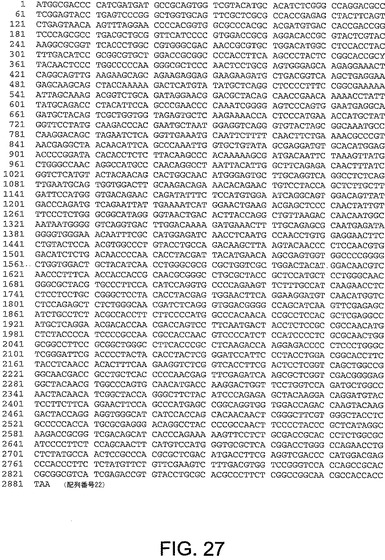
【図28】
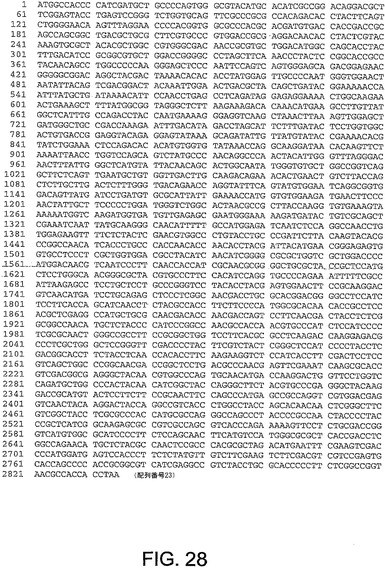
【図29】
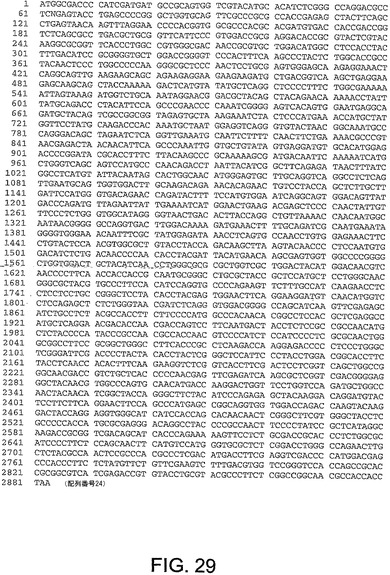
【図30】
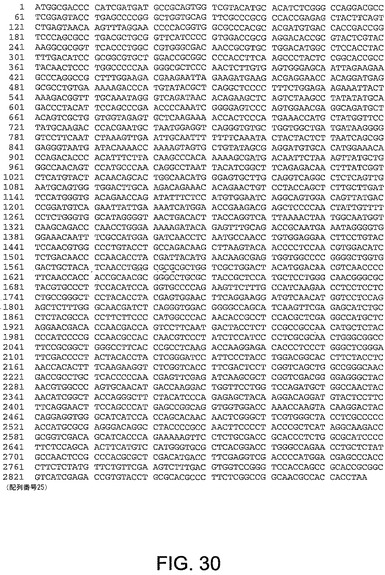
【図31A】

【図31B】

【図31C】

【図31D】

【図31E】

【図31F】

【図31G】

【図31H】

【図31I】

【図31J】

【図32】

【図33】
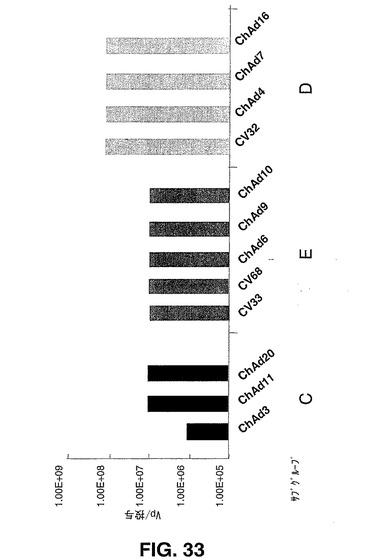
【図34】
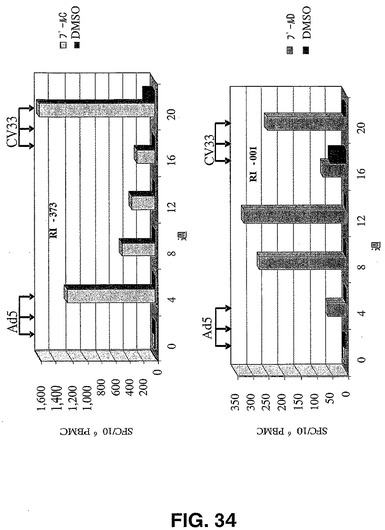
【図35】
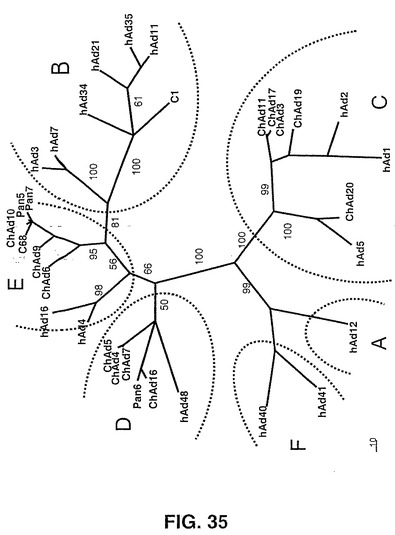
【図36】
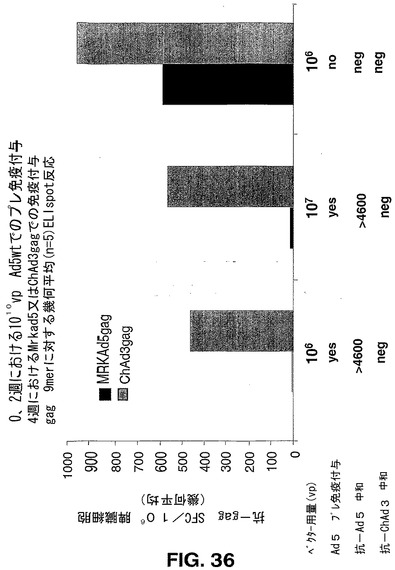
【図37】
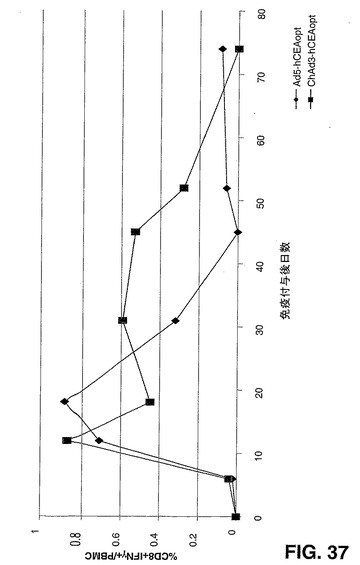
【図38A】
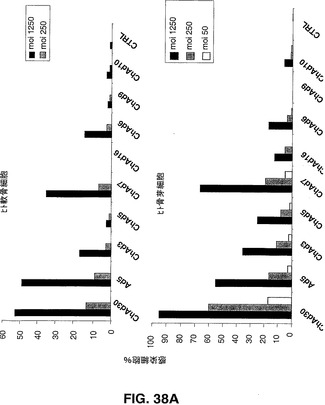
【図38B】
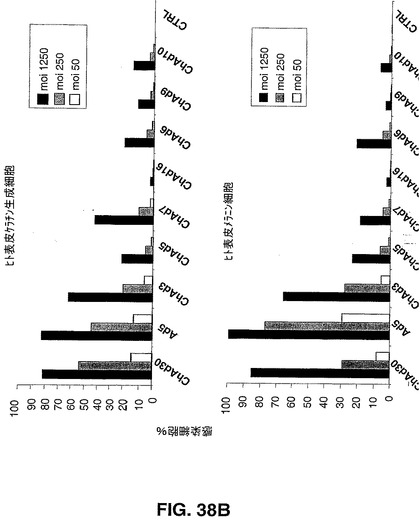
【図38C】
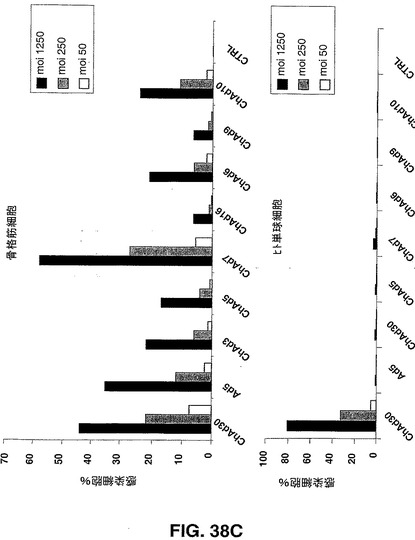
【図38D】
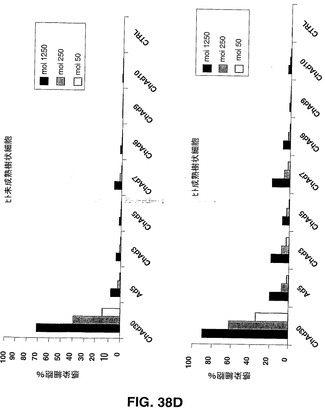
【図2】
【図3】
【図4】
【図5A】
【図5B】
【図5C】
【図5D】
【図5E】
【図5F】
【図5G】
【図5H】
【図5I】
【図5J】
【図5K】
【図6A】
【図6B】
【図6C】
【図6D】
【図6E】
【図6F】
【図6G】
【図6H】
【図6I】
【図6J】
【図6K】
【図7A】
【図7B】
【図7C】
【図7D】
【図7E】
【図7F】
【図7G】
【図7H】
【図7I】
【図7J】
【図7K】
【図8A】
【図8B】
【図8C】
【図8D】
【図8E】
【図8F】
【図8G】
【図8H】
【図8I】
【図8J】
【図8K】
【図9A】
【図9B】
【図9C】
【図9D】
【図9E】
【図9F】
【図9G】
【図9H】
【図9I】
【図9J】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20A】
【図20B】
【図20C】
【図20D】
【図20E】
【図20F】
【図20G】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28】
【図29】
【図30】
【図31A】
【図31B】
【図31C】
【図31D】
【図31E】
【図31F】
【図31G】
【図31H】
【図31I】
【図31J】
【図32】
【図33】
【図34】
【図35】
【図36】
【図37】
【図38A】
【図38B】
【図38C】
【図38D】
【公表番号】特表2007−518414(P2007−518414A)
【公表日】平成19年7月12日(2007.7.12)
【国際特許分類】
【出願番号】特願2006−550042(P2006−550042)
【出願日】平成17年1月18日(2005.1.18)
【国際出願番号】PCT/EP2005/000558
【国際公開番号】WO2005/071093
【国際公開日】平成17年8月4日(2005.8.4)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
【公表日】平成19年7月12日(2007.7.12)
【国際特許分類】
【出願日】平成17年1月18日(2005.1.18)
【国際出願番号】PCT/EP2005/000558
【国際公開番号】WO2005/071093
【国際公開日】平成17年8月4日(2005.8.4)
【出願人】(501209427)イステイチユート・デイ・リチエルケ・デイ・ビオロジア・モレコラーレ・ピ・アンジエレツテイ・エツセ・ピー・アー (90)
【Fターム(参考)】
[ Back to top ]