
テネイシンCに対する抗体
【課題】テネイシンCに対する特異的結合メンバー、特に、ヒトテネイシンCに対するヒト抗体の提供。
【解決手段】細胞外マトリックスタンパク質テネイシンCに対する特異的結合メンバー、特に、テネイシンCのドメインA1、ドメインCおよびドメインDに対するscFv抗体分子。標識、細胞毒性分子またはサイトカインに結合している抗テネイシンC特異的結合メンバー。診断および治療(特に癌の)における抗テネイシンC特異的結合メンバーの使用。
【解決手段】細胞外マトリックスタンパク質テネイシンCに対する特異的結合メンバー、特に、テネイシンCのドメインA1、ドメインCおよびドメインDに対するscFv抗体分子。標識、細胞毒性分子またはサイトカインに結合している抗テネイシンC特異的結合メンバー。診断および治療(特に癌の)における抗テネイシンC特異的結合メンバーの使用。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、テネイシンCに対する特異的結合メンバー、特に、ヒトテネイシンCに対するヒト抗体に関する。これらの特異的結合メンバーは、ある範囲の治療用途、例えば癌の診断および治療における治療用途を有する。
【背景技術】
【0002】
テネイシンCは、細胞接着をモジュレーションする、細胞外マトリックスの、大きな六量体糖タンパク質である。それは細胞増殖および細胞遊走のような過程に関与しており、形態形成および胚発生中に生じる変化で、腫瘍形成または血管新生中の組織構造の変化に関連している。
【0003】
テネイシンCのドメイン構造の概略を図1に示す。テネイシンCの幾つかのアイソフォームは、ドメインA1からドメインDにわたるこのタンパク質の中央部分における(複数の)ドメインの含有を招きうる選択的スプライシングの結果として生じうる[Borsi Lら, Int J Cancer 1992; 52:688-692, Carnemolla Bら, Eur J Biochem 1992; 205:561-567]。ドメインA1-Dは選択的スプライシングのメカニズムにより「ブロック(塊)」として挿入または除去されて「テネイシンC大(large)」および「テネイシンC小(small)」分子を与えうると以前には考えられていた[Borsi Lら, J Biol Chem 1995; 270:6243-6245]。テネイシンCの大アイソフォームの強力な過剰発現が多数の腫瘍で報告されており[Borsi 1992 前掲]、それぞれドメインA1およびDに特異的な2つのモノクローナル抗体が臨床において詳細に特徴づけられている[Riva Pら, Int J Cancer 1992; 51:7-13, Riva Pら, Cancer Res 1995; 55:5952s-5956s, Paganelli Gら, Eur J Nucl Med 1994; 21:314-321, Reardon DAら, J Clin Oncol 2002; 20:1389-1397, Bigner DDら, J Clin Oncol 1998; 16:2202-2212]。
【0004】
しかし、テネイシンCの大アイソフォーム間の分子的不均一性の増加を招く、選択スプライシングメカニズムの、より複雑な調節が生じることが、最近になって明らかになった。例えば、テネイシンCの更なる(extra)ドメインCが、免疫テネイシンCのその他の選択的スプライシングされたドメインより制限された発現パターンを示し[Carnemolla Bら, Am J Pathol 1999; 154:1345-1352]、組織化学的方法により示される顕著な血管周囲染色を伴うことが報告されている。テネイシンCのCドメインは、ほとんどの正常成体組織においては検出不能であるが、高い等級(grade)の星状細胞腫[Carnemolla Bら, Am J Pathol 1999; 154:1345-1352]および他の腫瘍型においては過剰発現される。大テネイシンCアイソフォーム間の不均一性に関する更なる裏付けが転写分析から得られており、これは、大テネイシンC転写産物が不均一な組成を特徴とすることを証明した[Katenkamp Kら, J Pathol 2004; 203:771-779]。翻訳後修飾(例えば、グリコシル化)の存在または非存在により更なるレベルの複雑性がもたらされ、これが個々のタンパク質ドメインの表面上の或るエピトープを修飾し、それらを、特異的なモノクローナル抗体に対するin vitroまたはin vivoでの特異的分子認識に利用不可能にする。
【0005】
所定の実質的に任意のタンパク質に特異的な抗体の迅速な単離が、既存の方法でin vitroで達成されうるとしても、そのような抗体が生物学的試料における又は疾患の動物モデルにおけるエピトープを認識することは明らかではない。in vivoでの結合の欠如をもたらす考えられうる理由には、エピトープの翻訳後修飾、エピトープのマスキング、および不十分な抗体特異性または安定性が含まれる。したがって、モノクローナル抗体のスクリーニングに通常用いられる酵素結合イムノソルベントアッセイ(ELISA)のような典型的な固相アッセイにおける組換え抗原(または抗原断片)とのモノクローナル抗体の反応性のみに基づいて、実際の用途へのモノクローナル抗体の適合性を評価することは困難である。
【0006】
したがって、診断用途および治療用途へのモノクローナル抗体の適合性を評価するためには、テネイシンC大アイソフォームの個々のドメインに対するモノクローナル抗体を個別に分析する必要がある。
【発明の概要】
【0007】
本発明者らは、テネイシンCの選択的スプライシングされた領域内の種々のエピトープに特異的なヒトモノクローナル抗体フラグメントを単離した。これらの抗体は、生物学的試料中の大テネイシンCアイソフォームを認識するそれらの能力により特徴づけられ、また、密接に関連した抗原間で顕著な差異を示す、ELISAアッセイにおける高特異的結合により特徴づけられる。
【0008】
本発明の1つの態様は、ヒトテネイシンC、特にテネイシンC大アイソフォームに結合する特異的結合メンバーを提供する。
【0009】
好ましい特異的結合メンバーは腫瘍特異的であり、正常組織と比べて腫瘍組織に優先的に結合する。特異的結合メンバーは、例えば、正常組織と比べて優先的に腫瘍組織のストロマならびに/または新血管構造および血管周囲構造に結合しうる。
【0010】
特異的結合メンバーは、テネイシンC小アイソフォームと比べてテネイシンC大アイソフォームに優先的に結合しうる。
【0011】
いくつかの実施形態においては、特異的結合メンバーはテネイシンCのA1ドメインに結合しうる。適当な特異的結合メンバーは、
配列番号2の4A1-F16 VHドメイン、配列番号12の3A1-D5 VHドメイン、ならびに配列番号5、配列番号6、配列番号7および配列番号13から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
配列番号4または配列番号50のVLドメインならびに配列番号8、配列番号9および配列番号10から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0012】
4A1-F16はF16とも称される。
【0013】
例えば、適当な特異的結合メンバーは、配列番号4もしくは配列番号50の4A1-F16/3A1-D5 VLドメインおよび/または配列番号2の4A1-F16 VHドメインまたは配列番号12の3A1-D5 VHドメインを含みうる。
【0014】
他の実施形態においては、特異的結合メンバーはテネイシンCのCドメインに結合しうる。適当な特異的結合メンバーは、
E10 VHドメイン(配列番号15または配列番号48)、A12 VHドメイン(配列番号25)、F4およびG11 VHドメイン(配列番号29または配列番号45)ならびに配列番号18、配列番号19、配列番号20、配列番号26および配列番号27から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
配列番号17もしくは配列番号81のVLドメイン、配列番号31もしくは配列番号83のアミノ酸配列を有するF4 VLドメイン、配列番号35もしくは配列番号47のアミノ酸配列を有するG11 VLドメインならびに配列番号21、配列番号22、配列番号23、配列番号32および配列番号33から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0015】
したがって、1つの例においては、特異的結合メンバーは、
E10 VHドメイン(配列番号15または配列番号48)、A12 VHドメイン(配列番号25)ならびに配列番号18、配列番号19、配列番号20、配列番号26および配列番号27から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
配列番号17もしくは配列番号81のVLドメインならびに配列番号21、配列番号22および配列番号23から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0016】
例えば、適当な特異的結合メンバーは、配列番号17もしくは配列番号81のE10/A12 VLドメインおよび/または配列番号15もしくは配列番号48のE10 VHドメインまたは配列番号25のA12 VHドメインを含みうる。
【0017】
もう1つの例においては、特異的結合メンバーは、
F4およびG11 VHドメイン(配列番号29または配列番号45)ならびに配列番号18、配列番号19および配列番号20から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
F4 VLドメイン(配列番号31もしくは配列番号83)、G11 VLドメイン(配列番号35または配列番号47)ならびに配列番号32、配列番号33、配列番号22および配列番号23から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0018】
例えば、適当な特異的結合メンバーは、配列番号29もしくは配列番号45のF4/G11 VHドメインおよび/または配列番号31もしくは配列番号83のF4 VLドメインまたは配列番号35もしくは配列番号47のG11 VLドメインを含みうる。
【0019】
もう1つの実施形態においては、特異的結合メンバーはテネイシンCのドメインDに結合する。好ましくは、特異的結合メンバーはヒトドメインDに結合する。それはマウスアイソフォームと交差反応しうる。本発明者らは、ドメインDに特異的な抗体分子を単離した。元のクローンはF4Sと称される。本発明者らは、D11と称される、F4Sのアフィニティ(親和性)成熟変異体をも開発した。本発明者らは更に、P12と称される、D11のアフィニティ成熟変異体を開発した。
【0020】
したがって、適当な特異的結合メンバーは、
配列番号58のF4S VHドメイン、配列番号55のD11 VHドメイン、配列番号52のP12 VHドメインならびに配列番号61、配列番号62、配列番号63、配列番号64、配列番号65および配列番号66から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
配列番号53のP12およびD11 VLドメイン、配列番号60のF4S VLドメインならびに配列番号67、配列番号68、配列番号69、配列番号70および配列番号71から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0021】
後記で詳しく説明するとおり、抗原への結合にはVHドメインのみが用いられることもあるが、一般には、VHドメインはVLドメインとペア形成して抗体抗原結合部位を与える。1つの好ましい実施形態においては、本明細書に記載のVHドメイン(すなわち、配列番号2、12、15、25、29、45、48、52、56または58)を対応VLドメイン(すなわち、配列番号4または配列番号50(4A1-F16および3A1-D5の場合)、配列番号17または配列番号81(A12およびE10の場合))、配列番号31、配列番号83、配列番号35または配列番号47(F4およびG11の場合)、配列番号4または配列番号50、配列番号54(P12およびD11の場合)あるいは配列番号60(F4Sの場合))とペア形成させて、VHおよびVLの両ドメインを含む抗体抗原結合部位(例えば、4A1-F16 VHおよびVLドメイン、3A1-D5 VHおよびVLドメイン、E10 VHおよびVLドメイン、A12 VHおよびVLドメイン、F4 VHおよびVLドメイン、G11 VHおよびVLドメイン、P12 VHおよびVLドメイン、D11 VHおよびVLドメイン、あるいはF4S VHおよびVLを含む部位)を形成させる。他の実施形態においては、VHドメインを、対応VL以外のVLドメイン、好ましくは、テネイシンCの同一ドメインに結合する特異的結合メンバーからのVLドメインとペア形成させることが可能である。軽鎖の乱交性(promiscuity)は当技術分野で十分に確認されている。
【0022】
1以上のCDRを、本明細書に開示されているVHまたはVLドメインから採用し、適当なフレームワーク内に含有させることが可能である。これについては後記で更に詳しく説明する。CDRは一般にはKabatにしたがい定義される。好ましくは、VHドメインおよび/またはVLドメインはCDR1、CDR2およびCDR3を含む。4A1-F16 VH CDR1を配列番号5に示す。3A1-D5 VH CDR1を配列番号13に示す。4A1-F16および3A1-D5 VH CDR 2および3をそれぞれ配列番号6および7に示す。4A1-F16および3A1-D5 VL CDR 1、2および3をそれぞれ配列番号8、9および10に示す。E10 VH CDR 1および2を配列番号18および19に示す。A12 VH CDR 1および2を配列番号26および27に示す。E10およびA12 VH CDR3を配列番号23に示す。E10およびA12 VL CDR 1、2および3をそれぞれ配列番号21、22および23に示す。F4およびG11 VH CDR 1、2および3をそれぞれ配列番号18、19および20に示す。F4 VL CDR 1、2および3をそれぞれ配列番号32、33および23に示す。G11 VL CDR 1、2および3をそれぞれ配列番号32、22および23を示す。P12 VH CDRをそれぞれ配列番号61、63および66に示す。P12 VL CDRをそれぞれ配列番号67、69および71に示す。D11 VH CDRをそれぞれ配列番号62、64および66に示す。D11 VL CDRをそれぞれ配列番号67、69および71に示す。F4S VH CDRをそれぞれ配列番号62、65および66に示す。F4S VL CDRをそれぞれ配列番号68、70および71に示す。
【0023】
いくつかの実施形態においては、特異的結合メンバーは、配列番号7のアミノ酸配列を有するCDR3、配列番号6のアミノ酸配列を有するCDR2および配列番号5またはより好ましくは配列番号13のアミノ酸配列を有するCDR1の1以上を含む抗体VHドメインを含みうる。
【0024】
他の実施形態においては、特異的結合メンバーは、配列番号20のアミノ酸配列を有するCDR3、配列番号27またはより好ましくは配列番号19のアミノ酸配列を有するCDR2および配列番号26またはより好ましくは配列番号18のアミノ酸配列を有するCDR1の1以上を含む抗体VHドメインを含みうる。
【0025】
特異的結合メンバーは、配列番号23のアミノ酸配列を有するCDR3、配列番号22または配列番号33のアミノ酸配列を有するCDR2および配列番号21または配列番号32のアミノ酸配列を有するCDR1の1以上を含む抗体VHドメインを含みうる。
【0026】
好ましくは、特異的結合ドメインは、本明細書中に更に詳しく説明されているscFvである。VHドメインとVLドメインとを、ペプチドリンカー、例えば配列番号37に記載のアミノ酸配列を有するリンカーを介して連結することが可能である。通常、該リンカーは、モチーフの縦列反復の1以上を含むアミノ酸配列を有する。典型的には、該モチーフは5残基配列であり、好ましくは、該残基のうちの少なくとも4個はGlyまたはSerである。該5残基のうちの4個がGlyまたはSerである場合には、その他の残基はAlaでありうる。より好ましくは、該5残基のそれぞれはGlyまたはSerである。好ましいモチーフはGGGGS、SSSSG、GSGSAおよびGGSGG(それぞれ配列番号76、77、78および79)である。好ましくは、該モチーフは該配列内で隣接しており、該反復間に介在ヌクレオチドは存在しない。該リンカー配列は、1〜5個、好ましくは3〜4個の、モチーフの反復を含む又はそれよりなることが可能である。例えば、3個の縦列反復を有するリンカーは以下のアミノ酸配列の1つを有しうる:
GGGGSGGGGSGGGGS - 配列番号39、
SSSSGSSSSGSSSSG - 配列番号41、
GSGSAGSGSAGSGSA - 配列番号42、
GGSGGGGSGGGGSGG - 配列番号43。
【0027】
本明細書に記載の配列を有しテネイシンCに対する特異的結合メンバーにおいて使用されうるVHおよびVLドメインならびにCDRの変異体は、配列の改変または突然変異およびスクリーニングの方法により得ることが可能である。そのような方法も本明細書により提供される。
【0028】
本明細書に具体的に開示されている配列を有するVHおよびVLドメインのいずれかの可変ドメインアミノ酸配列変異体を、本明細書に記載のとおり、本発明において使用することが可能である。個々の変異体は、約20個未満の改変、約15個未満の改変、約10個未満の改変、約5個未満の改変、4、3、2または1個の改変でありうる、1以上のアミノ酸配列の改変(アミノ酸残基の付加、欠失、置換および/または挿入)を含みうる。改変は1以上のフレームワーク領域および/または1以上のCDRにおいて施されうる。特に、改変はVH CDR1、VH CDR2および/またはVH CDR3、特にVH CDR3において施されうる。
【0029】
本発明の特異的結合メンバーは、特異的結合メンバー、本明細書に開示されているVHおよび/またはVLドメインあるいは本明細書に開示されているVH CDRあるいはこれらの任意の変異体を含みテネイシンC大アイソフォーム(特にそのA1またはCドメイン)に結合する任意の特異的結合メンバーと、抗原に対する結合に関して競合するものでありうる。結合メンバー間の競合はin vitroで容易にアッセイされうる。例えば、これはELISAを使用して行うことが可能であり、および/または他の非タグ付け結合メンバーの存在下で検出されうる1つの結合メンバーに特異的レポーター分子をタグ付けして、同一エピトープまたは重複エピトープに結合する特異的結合メンバーの特定を可能にすることにより行うことが可能である。
【0030】
したがって、本発明のもう1つの態様は、テネイシンCへの結合に関して4A1-F16、3A1-D5、E10、A12、F4、G11、P12、D11およびF4Sの1以上と競合するヒト抗体抗原結合部位を含む特異的結合メンバーを提供する。
【0031】
4A1-F16、3A1-D5、E10、A12、F4 G11、P12、D11またはF4Sと競合しうるテネイシンC大アイソフォームに対する抗体を得るためには、種々の方法が当技術分野において利用可能である。好ましくは、そのような抗体は、テネイシンC小アイソフォームと比べてテネイシンC大アイソフォームに優先的に結合する。
【0032】
もう1つの態様においては、本発明は、本発明の特異的結合メンバーのライブラリーと抗原とを接触させ、該抗原に結合しうる該ライブラリーの特異的結合メンバーの1以上を選択しることを含む、抗原に結合しうる1以上の特異的結合メンバーの入手方法を提供する。
【0033】
該ライブラリーはバクテリオファージ粒子の表面上に提示されることが可能であり、各粒子は、その表面上に提示される抗体VH可変ドメインをコードする核酸、および場合によっては、存在する場合には提示VLドメインをコードする核酸を含有する。
【0034】
抗原への結合能を有しバクテリオファージ粒子上に提示される特異的結合メンバーの選択の後、選択した特異的結合メンバーを提示するバクテリオファージ粒子から核酸を取り出すことが可能である。そのような核酸は、選択した特異的結合メンバーを提示するバクテリオファージ粒子から取り出した核酸の配列を有する核酸からの発現による、特異的結合メンバーまたは抗体VH可変ドメイン(場合によっては抗体VL可変ドメイン)の後続の産生において使用することが可能である。
【0035】
選択した特異的結合メンバ−の抗体VH可変ドメインのアミノ酸配列を有する抗体VH可変ドメインは、そのようなVHドメインを含む特異的結合メンバーの場合と同様に、単離された形態で提供されうる。
【0036】
テネイシンCへの結合能、また、4A1-F16、3A1-D5、E10、A12、F4、G11、P12、D11またはF4Sと競合する能力を、更に試験することが可能である。
【0037】
本発明の特異的結合メンバーは、4A1-F16、3A1-D5、E10、A12、F4、G11、P12、D11またはF4Sの1以上のアフィニティで、あるいはより大きな又はより小さなアフィニティで、テネイシンCに結合しうる。
【0038】
異なる特異的結合メンバーの結合アフィニティを、適当な条件下で比較することが可能である。
【0039】
抗体配列に加えて、本発明の特異的結合メンバーは、例えば折り畳まれた(folded)ドメインのようなペプチドまたはポリペプチドを形成する他のアミノ酸、あるいは抗原への結合能のほかに別の機能特性を該分子に付与する他のアミノ酸を含みうる。
【0040】
本発明の特異的結合メンバーは、検出可能な標識、例えば腫瘍検出を容易にする物質、例えば放射性核種または発蛍光団を含有することが可能であり、あるいは、治療方法における使用のために、殺生物事象を誘発しうる物質、例えば放射性核種、光増感剤、薬物、サイトカイン、凝血促進因子、毒素または酵素(例えば、ペプチド結合またはリンカーを介して)に結合されうる。
【0041】
他の態様においては、本発明は、本発明の特異的結合メンバーVHまたはVLドメインをコードする配列を含む単離された核酸を提供し、また、特異的結合メンバーVHドメインおよび/またはVLドメインの産生を引き起こす条件下で該核酸を発現させ、それを回収することを含む、本発明の特異的結合メンバーVHドメインおよび/またはVLドメインの製造方法を提供する。
【0042】
本明細書に記載の特異的結合メンバーは、ヒトまたは動物の身体の治療または診断方法、例えば、ヒト患者における疾患または障害、特に増殖性障害、例えば癌の(予防的治療を含みうる)治療方法において使用されることが可能であり、該方法は、特異的結合メンバーの有効量を該患者に投与することを含む。
【0043】
本発明のもう1つの態様は、本明細書に開示されている抗体VH可変ドメインおよび/またはVL可変ドメインをコードする核酸、一般には単離された核酸を提供する。
【0044】
本発明のもう1つの態様は、本明細書に開示されているVH CDRまたはVL CDR配列、特に、配列番号5、6、7、13、18、19、20、26および27から選ばれるVH CDRまたは配列番号8、9、10、21、22、23、32および33から選ばれるVL CDRをコードする核酸、一般には単離された核酸を提供する。
【0045】
もう1つの態様は、本発明の核酸で形質転換された宿主細胞を提供する。
【0046】
さらにもう1つの態様は、コード核酸からの発現を引き起こさせることを含む、抗体VH可変ドメインの製造方法を提供する。そのような方法は、該抗体VH可変ドメインの産生のための条件下で宿主細胞を培養することを含みうる。
【0047】
本発明の他の態様として、VL可変ドメインならびにVHおよび/またはVLドメインを含む特異的結合メンバーの製造のための類似方法を提供する。
【0048】
製造方法は、該産物の単離および/または精製の工程を含みうる。
【0049】
製造方法は、少なくとも1つの追加的成分、例えば製薬上許容される賦形剤を含む組成物中に該産物を製剤化することを含みうる。
【0050】
本発明のこれらの及び他の態様を後記で更に詳しく説明する。
【図面の簡単な説明】
【0051】
【図1】図1は、小(A)および大(B)テネイシンCアイソフォームの概要図である。いくつかのフィブロネクチンIII型様ドメインは選択的スプライシングに付されて、該分子内に含まれるか(B)または含まれない(A)。
【図2】図2は、U87腫瘍異種移植片担持ヌードマウスにおける4A1-F16-SIP抗体の生体分布を示す。(A)値は、注射の3時間後(上)および6時間後(下)の、3匹のマウスの群内の組織1g当たりの注射用量の平均%を表す。誤差棒線は標準偏差を表す。
【図3】図3は、U87腫瘍異種移植片担持ヌードマウスにおける4A1-F16-SIP抗体の生体分布を示す。(A)値は、注射の24時間後(上)および48時間後(下)の、3匹のマウスの群内の組織1g当たりの注射用量の平均%を表す。誤差棒線は標準偏差を表す。
【図4】図4は、図2および3に示した生体分布データの別の表示を示す。
【図5】図5はSIP(P12)の生体分布を示す。3.5μgのSIP(P12)を、皮下U87ヒト神経膠芽細胞腫を担持するU87腫瘍異種移植片担持ヌードマウスにおける4A1-F16-SIP抗体の生体分布を示す。各時点は4匹の動物により表されている。
【発明を実施するための形態】
【0052】
用語
特異的結合メンバー
これは、お互いに対する結合特異性を有する分子のペアのメンバーを意味する。特異的結合ペアのメンバーは天然由来であることが可能であり、あるいは完全または部分的に合成的に製造されうる。分子のペアの一方のメンバーは、分子のペアの他方のメンバーの個々の空間的および極性組織構造体に特異的に結合し従ってそれに相補的であるその表面上の領域または腔を有する。したがって、該ペアのメンバーは、お互いに特異的に結合する特性を有する。特異的結合ペアのタイプの例としては、抗原-抗体、ビオチン-アビジン、ホルモン-ホルモン受容体、受容体-リガンド、酵素-基質が挙げられる。本出願は抗原-抗体タイプの反応に関する。
【0053】
抗体
これは、天然由来の又は部分的もしくは完全に合成的に製造された免疫グロブリンを意味する。この用語は、抗体結合ドメインである又は抗体結合ドメインに実質的に相同である結合ドメインを有する任意のポリペプチドまたはタンパク質をも包含する。抗体の例としては、免疫グロブリンアイタイプおよびそれらのアイタイプサブクラス;抗原結合ドメインを含むフラグメント、例えばFab、scFv、Fv、dAb、Fd;ならびにダイアボディ(diabody)が挙げられる。モノクローナル抗体および他の抗体を採用することが可能であり、元の抗体の特異性を保有する他の抗体またはキメラ分子を製造するために組換えDNA技術を用いることが可能である。そのような技術は、異なる免疫グロブリンの定常領域、または定常領域 + フレームワーク領域へ、抗体の免疫グロブリン可変領域または相補性決定領域(CDR)をコードするDNAを導入することを含みうる。例えば、EP-A-184187、GB 2188638AまたはEP-A-239400を参照されたい。ハイブリドーマまたは他の抗体産生細胞を、産生される抗体の結合特異性を改変するものであっても改変しないものであってもよい遺伝的突然変異または他の変化に付すことが可能である。
【0054】
抗体は多数の方法で修飾されうるため、「抗体」なる語は、要求される特異性を有する結合ドメインを有する任意の特異的結合メンバーまたは基質を包含すると解釈されるべきである。したがって、この用語は、天然由来または完全もしくは部分的に合成由来である、免疫グロブリン結合ドメインを含む任意のポリペプチドを含む、抗体の抗体フラグメント、誘導体、機能的等価体およびホモログを包含する。したがって、別のポリペプチドに融合した免疫グロブリン結合ドメインまたは等価体を含むキメラ分子が包含される。キメラ抗体のクローニングおよび発現はEP-A-0120694およびEP-A-0125023に記載されている。
【0055】
全抗体のフラグメントは、抗原に結合する機能を果たしうることが示されている。結合性フラグメントの例としては、(i)VL、VH、CLおよびCH1ドメインよりなるFabフラグメント;(ii)VHおよびCH1ドメインよりなるFdフラグメント;(iii)単一抗体のVLおよびVHドメインよりなるFvフラグメント;(iv)VHドメインよりなるdAbフラグメント(Ward, E.S.ら, Nature 341, 544-546 (1989));(v)単離CDR領域;(vi)2つの連結されたFabフラグメントを含む二価フラグメントであるF(ab')2フラグメント;(vii)VHドメインとVLドメインとが一緒になって抗原結合部位を形成することを可能にするペプチドリンカーにより該VHドメインおよび該VLドメインが連結されている一本鎖Fv分子(scFv)(Birdら, Science, 242, 423-426, 1988; Hustonら, PNAS USA, 85, 5879-5883, 1988);(viii)二重特異性一本鎖Fv二量体(PCT/US92/09965);ならびに(ix)遺伝子融合により構築された多価または多重特異性フラグメントである「ダイアボディ(diabody)」(WO94/13804; P. Holligerら, Proc. Natl. Acad. Sci. USA 90 6444-6448, 1993)が挙げられる。Fv、scFvまたはダイアボディ分子は、VHドメインとVLドメインとを連結するジスルフィド架橋の含有により安定化されうる(Y. Reiterら, Nature Biotech 14 1239-1245 1996)。CH3ドメインに連結されたscFvを含むミニボディ(minibody)も製造されうる(S. Huら, Cancer Res. 56 3055-3061 1996)。
【0056】
ダイアボディはポリペプチドの多量体であり、各ポリペプチドは、免疫グロブリン軽鎖の結合領域を含む第1ドメインと免疫グロブリン重鎖の結合領域を含む第2ドメインとを含み、それらの2つのドメインは(例えばペプチドリンカーにより)連結されてはいるが、互いに一緒になって抗原結合部位を形成することはなく、抗原結合部位は、該多量体内の1つのポリペプチドの第1ドメインと該多量体内の別のポリペプチドの第2ドメインとの会合により形成される(WO94/13804)。
【0057】
二重特異性抗体を使用する場合には、これらは通常の二重特異性抗体であることが可能であり、それらは、種々の方法により製造することが可能であり(Holliger, P.およびWinter G. Current Opinion Biotechnol. 4, 446-449 (1993))、例えば、化学的に又はハイブリッドハイブリドーマから製造することが可能であり、あるいは前記の任意の二重特異性抗体フラグメントでありうる。ダイアボディおよびscFvは、可変ドメインのみを使用して、Fc領域を伴わずに構築することが可能であり、これは、抗イディオタイプ反応の効果を潜在的に軽減する。
【0058】
二重特異性全抗体と比べて、二重特異性ダイアボディは特に有用でありうる。なぜなら、それは大腸菌(E. coli)内で容易に構築され発現されうるからである。適当な結合特異性のダイアボディ(および例えば抗体フラグメントのような多数の他のポリペプチド)は、ファージ提示(ファージディスプレイ)(WO94/13804)を用いて、ライブラリーから容易に選択されうる。ダイアボディの一方のアームを、例えば抗原Xに対する特異性を有する定常なものにしたい場合には、他方のアームが可変性となり適当な特異性の抗体が選択されるライブラリーを作製することが可能である。二重特異性全抗体は、「ノブ・イントゥ・ホール(knobs-into-holes)」操作(J. B. B. Ridgewayら, Protein Eng. 9 616-621, 1996)により作製することが可能である。
【0059】
抗原結合ドメイン
これは、抗原の一部または全体に特異的に結合しそれに相補的である領域を含む、抗体の部分を意味する。抗原が大きい場合には、抗体は該抗原の特定の部分に結合しうるに過ぎず、その部分はエピトープと称される。抗原結合ドメインは1以上の抗体可変ドメイン(例えば、VHドメインよりなる、いわゆるFd抗体フラグメント)により提供されうる。好ましくは、抗原結合ドメインは抗体軽鎖可変領域(VL)および抗体重鎖可変領域(VH)を含む。
【0060】
特異的
これは、特異的結合ペアの一方のメンバーがその特異的結合パートナー以外の分子へのいずれの有意な結合をも示さない状況を示すために用いられうる。例えば、テネイシンCに特異的な抗体は、例えばフィブロネクチンのような細胞外マトリックスの他の成分への結合をほとんど又は全く示さないであろう。同様に、テネイシンC大アイソフォームに特異的な抗体はテネイシンC小アイソフォームへの結合をほとんど又は全く示さないであろう。この用語は、例えば、抗原結合ドメインが、多数の抗原により担持される特定のエピトープに特異的である場合にも適用可能であり、この場合、該抗原結合ドメインを担持する特異的結合メンバーは、該エピトープを担持する種々の抗原に結合しうるであろう。
【0061】
含む(含んでなる)
これは、一般には、包含するという意味で用いられ、すなわち、1以上の特徴または成分の存在を許容するものである。
【0062】
単離(された)
これは、本発明の特異的結合メンバーまたはそのような結合メンバーをコードする核酸が本発明に合致している状態を意味する。メンバーおよび核酸は、それらが天然で会合している物質(例えば、それらの天然環境中でそれらと一緒に存在する他のポリペプチドまたは核酸)、またはin vitroもしくはin vivoで実施される組換えDNA技術による製造におけるそれらの製造環境(例えば、細胞培養)中に一緒に存在する物質を含有しない又は実質的に含有しないであろう。メンバーおよび核酸は希釈剤またはアジュバントと共に製剤化されることが可能であり、その場合であっても、実際の目的においては尚も単離されている。例えば、イムノアッセイにおいて使用するためにマイクロタイタープレートをコートするために該メンバーを使用する場合には、該メンバーをゼラチンまたは他の担体と混合し、あるいは該メンバーを診断または療法において使用する場合には、該メンバーを、製薬上許容される担体または希釈剤と混合する。特異的結合メンバーは、天然で、または異種真核細胞の系(例えば、CHOまたはNS0(ECACC 85110503)細胞)によりグリコシル化されることが可能であり、あるいはそれらは(例えば、原核細胞における発現により製造される場合には)非グリコシル化形態でありうる。
【0063】
「記載されているのと実質的に同じ」は、本発明の関連CDRまたはVHもしくはVLドメインが、本明細書に記載の配列を有する特定されている領域と同一または非常に類似していることを意味する。「非常に類似」は、該CDRおよび/またはVHもしくはVLドメインにおいて、1〜5個、好ましくは1〜4個、例えば1〜3個、または1個、または2個、または3個、または4個の置換が施されうることを意味する。
【0064】
本発明のCDRを担持するための構造は、一般には、再構成免疫グロブリン遺伝子によりコードされる天然に存在するVHおよびVL抗体可変ドメインのCDRに対応する位置に該CDRが位置する、抗体重鎖もしくは軽鎖配列またはその本質的部分のものであろう。免疫グロブリン可変ドメインの構造および位置は、Kabat, E.A.ら, Sequences of Proteins of Immunological Interest. 4th Edition. US Department of Health and Human Services. 1987およびその改訂版(現在では、インターネット上(http://immuno.bme.nwu.edu)で入手可能である)を参考にして決定されうる。
【0065】
好ましくは、本明細書に記載されているのと実質的に同じCDRアミノ酸配列は、ヒト可変ドメインまたはその本質的部分におけるCDRとして担持される。
【0066】
本発明において使用する可変ドメインは、任意の生殖系列から入手可能であり、あるいはヒト可変ドメインから再構成されることが可能であり、あるいは公知ヒト可変ドメインのコンセンサス配列に基づく合成可変ドメインでありうる。本発明のCDR配列(例えば、CDR1、CDR2またはCDR3)は、組換えDNA技術を用いて、CDR(例えば、対応するCDR1、CDR2またはCDR3)を欠く可変ドメインのレパートリー内に導入されうる。
【0067】
例えば、Marksら(Bio/Technology, 1992, 10:779-783)は、CDR3を欠くVH可変ドメインのレパートリーを得るために、抗体可変ドメイン領域の5'末端に対する又は該5'末端に隣接したコンセンサスプライマーをヒトVH遺伝子の第3フレームワーク領域に対するコンセンサスプライマーと共に使用する、抗体可変ドメインのレパートリーの製造方法を記載している。Marksらは更に、このレパートリーを特定の抗体のCDR3と如何にして組合せうるかを記載している。類似した技術を用いて、本発明のCDR由来配列を、CDRを欠くVHまたはVLドメインのレパートリーとシャッフルし、そのシャッフルされた完全なVHまたはVLドメインをコグネイトVLまたはVHドメインと組合せて、本発明の特異的結合メンバーを得ることが可能である。ついで該レパートリーを、例えばWO92/01047のファージ提示系のような適当な宿主系において提示させ、適当な特異的結合メンバーを選択することが可能である。レパートリーは、〜104個以上のメンバー、例えば、106〜108または1010メンバーよりなることが可能である。
【0068】
類似したシャッフリングまたはコンビナトリアル(組合せ)技術もStemmer (Nature, 1994, 370:389-391)により開示されており、彼はβ-ラクタマーゼ遺伝子に関する技術を記載しているが、該アプローチが抗体の作製に用いられうることを見出している。
【0069】
もう1つの代替法は、可変ドメイン全体における突然変異を得るために1以上の選択されたVHおよび/またはVL遺伝子のランダム突然変異誘発を用いて、本発明のCDR由来配列を担持する新規VHまたはVL領域を作製することである。そのような技術はGramら(1992, Proc. Natl. Acad. Sci. USA, 89:3576-3580)により記載されており、彼は変異導入型PCRを用いている。
【0070】
用いうるもう1つの方法は、突然変異誘発をVHまたはVL遺伝子のCDR領域へ導くことである。そのような技術はBarbasら(1994, Proc. Natl. Acad. Sci., USA, 91:3809-3813)およびSchierら(1996, J. Mol. Biol. 263:551-567)により開示されている。
【0071】
前記技術のすべては当技術分野において自体公知であり、基本的には本発明の一部を構成するものではない。当業者であれば、当技術分野における通常の方法を用いて本発明の特異的結合メンバーを得るためにそのような技術を用いることが可能であろう。
【0072】
本発明のもう1つの態様は、本明細書に記載のVHドメインのアミノ酸配列内の1以上のアミノ酸の付加、欠失、置換または挿入により、該VHドメインのアミノ酸配列変異体であるVHドメインを得、所望により、このようにして得られたVHドメインを1以上のVLドメインと組合せ、該VHドメインまたはVH/VL組合せ体を試験して、テネイシンCに特異的な特異的結合メンバーまたは抗体抗原結合ドメインを特定することを含む、テネイシンCに特異的な抗体抗原結合ドメインの入手方法を提供する。該VLドメインは、本明細書に記載されているのと実質的に同じアミノ酸配列を有しうる。
【0073】
いくつかの実施形態においては、前記の1以上のアミノ酸はVHドメインのCDR(例えば、CDR1、CDR2および/またはCDR3)の1以上において付加、欠失、置換または挿入されうる。
【0074】
本明細書に記載のVLドメインのアミノ酸配列内の1以上のアミノ酸の付加、欠失、置換または挿入により、該VLドメインのアミノ酸配列変異体であるVLドメインを得、このようにして得られたVLドメインを1以上のVHドメインと組合せ、該VH/VL組合せ体を試験して、テネイシンCに特異的な特異的結合メンバーまたは抗体抗原結合ドメインを特定することを含む、類似方法を用いることが可能である。該VHドメインは、本明細書に記載されているのと実質的に同じアミノ酸配列を有することが可能であり、あるいは前記のとおりに得られた、本明細書に記載されているのと実質的に同じVHドメインのアミノ酸配列変異体でありうる。
【0075】
いくつかの実施形態においては、前記の1以上のアミノ酸はVLドメインのCDRの1以上において付加、欠失、置換または挿入されうる。
【0076】
本発明のもう1つの態様は、
(a)置換すべきCDRを含むか又はCDRコード領域を欠くVHドメインをコードする核酸の出発レパートリーを準備し、
(b)該レパートリーを、VH CDRに関して本明細書に記載されているのと実質的に同じアミノ酸配列をコードする供与核酸と組合せて、該供与核酸を該レパートリー内のCDR領域内に挿入して、VHドメインをコードする核酸の産物レパートリーを得、
(c)該産物レパートリーの核酸を発現させ、
(d)テネイシンC抗原に特異的な特異的結合メンバーを選択し、
(e)該特異的結合メンバーまたはそれをコードする核酸を回収することを含む、テネイシンCに特異的な特異的結合メンバーの製造方法を提供する。
【0077】
該CDRはVH CDR1、CDR2またはCDR3でありうる。
【0078】
ここでもまた、本発明のVL CDRを、置換すべきCDRを含むか又はCDRコード領域を欠くVLドメインをコードする核酸のレパートリーと組合せる、類似方法を用いることが可能である。
【0079】
該CDRはVL CDR1、CDR2またはCDR3でありうる。
【0080】
同様に、1以上または全3個のCDRをVHまたはVLドメインのレパートリー内にグラフティングし、ついでそれを、テネイシンCに特異的な、特にテネイシンC大アイソフォームに特異的な特異的結合メンバーに関してスクリーニングすることが可能である。
【0081】
免疫グロブリン可変ドメインの本質的部分は、少なくともそれらの3つのCDR領域を、それらの介在フレームワーク領域と共に含む。好ましくは、該部分はまた、第1および第4フレームワーク領域の一方または両方の少なくとも約50%を含み、該50%は第1フレームワーク領域のC末端半分および第4フレームワークのN末端半分である。該可変ドメインの本質的部分のN末端またはC末端の追加的残基は、天然に存在する可変ドメイン領域と通常は一緒になっていないものでありうる。例えば、組換えDNA技術により得られる本発明の特異的結合メンバーの構築は、クローニングまたは他の操作工程を促進するために導入されるリンカーによりコードされるNBまたはC末端残基の導入をもたらしうる。他の操作工程は、本発明の可変ドメインを、免疫グロブリン重鎖、他の可変ドメイン(例えば、ダイアボディの製造の場合)またはタンパク質標識(後記で更に詳しく説明される)を含む他のタンパク質配列へ連結するためのリンカーの導入を含む。
【0082】
本発明の好ましい態様においては、VHおよびVLドメインのペアを含む特異的結合メンバーが好ましいが、VHまたはVLのいずれかのドメイン配列に基づく単一結合ドメインも本発明の他の態様を構成する。単一免疫グロブリンドメイン、特にVHドメインは標的抗原に特異的に結合しうることが公知である。
【0083】
単一鎖特異的結合ドメインのいずれかの場合には、これらのドメインを使用して、テネイシンCに結合しうる2ドメイン特異的結合メンバーを形成しうる相補性ドメインに関してスクリーニングすることが可能である。
【0084】
これは、WO92/01047に開示されているいわゆる階層的二重組合せアプローチを用いるファージ提示スクリーニング法により達成することが可能であり、この場合、HまたはL鎖クローンのいずれかを含有する個々のコロニーを用いて、もう一方の鎖(LまたはH)をコードするクローンの完全ライブラリーに感染させ、得られた2本鎖特異的結合メンバーを、例えばその参考文献に記載されているファージ提示技術により選択する。この技術はMarksら(同誌)にも開示されている。
【0085】
本発明の特異的結合メンバーは更に、抗体定常領域またはその一部を含みうる。例えば、VLドメインをそのC末端において、ヒトCκまたはCλ鎖(好ましくはCλ鎖)を含む抗体軽鎖定常ドメインに結合させることが可能である。同様に、VHドメインに基づく特異的結合メンバーをそのC末端において、任意の抗体アイタイプ(例えば、IgG、IgA、IgEおよびIgM)および任意のアイタイプサブクラス(特にIgG1およびIgG4)に由来する免疫グロブリン重鎖の全体または一部に結合させることが可能である。
【0086】
本発明の特異的結合メンバーを、検出可能または機能的な標識で標識することが可能である。
【0087】
検出可能な標識には、放射性核種、例えばヨウ素131、イットリウム90、インジウム111およびテクネチウム99が含まれる。それらは、抗体イメージングの技術分野において公知の通常の化学を用いて、本発明の抗体に結合させることが可能である。放射性同位体で標識された特異的結合メンバーを使用して、放射線を特異的標的(例えば、腫瘍)へ選択的に運搬することが可能である。これは、後記のとおり、腫瘍をイメージングするのに又は細胞毒性量の放射線を運搬するのに有用でありうる。
【0088】
他の検出可能な標識には、ホースラディッシュペルオキシダーゼのような酵素標識、特異的コグネイト検出可能部分(例えば、標識アビジン)への結合を介して検出されうるビオチンのような化学的部分、フルオレセイン、ローダミン、フィコエリトリンおよびテキサス・レッドのような蛍光色素、ならびにCy7(Amersham Pharmacia)およびAlexa750(Molecular probes)のようなシアニン色素誘導体を含む近赤外発蛍光団が含まれうる。
【0089】
他の実施形態においては、検出可能な標識には、超音波により検出可能なマイクロバブル(microbubble)誘導体(Joseph Sら, Pharm Res. 2004 Jun;21(6):920-6)または磁気粒子(Schellenberger EAら, Bioconjug. Chem. 2004 Sep-Oct; 15(5): 1062-7)が含まれうる。
【0090】
機能的標識には、殺生物事象を誘発しうる又は抗癌作用を有する物質が含まれうる。適当な標識には、放射性核種、光増感剤、毒素ポリペプチド、毒性小分子および他の薬物、サイトカイン(例えば、IL2、IL12、TNF)、ケモカイン、凝血促進因子(例えば、組織因子)、酵素、リポソーム、および免疫応答因子(例えば、D. Neri (2004) CHIMIA “Tumor Targeting” vol. 58, p. 723-726を参照されたい)が含まれる。
【0091】
放射性核種には、ヨウ素131、イットリウム90、インジウム111およびテクネチウム99が含まれ、前記で更に詳しく説明されている。
【0092】
毒素ポリペプチドまたはペプチドは細胞毒性またはアポトーシス活性を有し、微生物、植物、動物またはヒト由来でありうる。いくつかの実施形態においては、毒素ポリペプチドは特異的結合メンバーの定常領域内に直接的に挿入されうる。毒素ポリペプチドの例には、Pseudomonas外毒素、リシンα鎖およびアンギオゲニンが含まれる。
【0093】
毒性小分子には、例えばDNA複合体形成物質または細胞周期インヒビターを含む、細胞毒性活性を有する化合物が含まれる。いくつかの実施形態においては、毒性分子は、pH感受性または酵素感受性リンカー(例えば、イミン結合を含有するリンカー)の切断により、標的細胞の近傍で遊離されうる。毒性小分子の例には、メイタンシン(maytansine)、カリケアミシン(calicheamicin)、エポチロン(epothilone)およびチューブリシン(tubulysin)ならびにそれらの誘導体が含まれる。
【0094】
免疫応答因子には、免疫エフェクター細胞に結合する特異的結合メンバーが含まれうる。特異的結合メンバーの結合は、標的細胞に対する細胞性免疫応答を惹起しうる。
【0095】
好ましい実施形態においては、本発明の特異的結合メンバーはサイトカインに結合している。該特異的結合メンバーまたはそのポリペプチド成分(例えば、抗体または多鎖抗体フラグメント(例えば、Fab)の重鎖または軽鎖)とサイトカインとを含む融合タンパク質を製造することが可能である。したがって、例えば、本発明の特異的結合メンバーのVHドメインまたはVLドメインをサイトカインに融合させることが可能である。典型的には、該特異的結合メンバーまたはその成分とサイトカインとはペプチドリンカー(例えば、約5〜25残基、例えば10〜20残基、好ましくは約15残基のペプチド)を介して連結される。ペプチドリンカーの適当な例は本明細書に記載されている。好ましくは、サイトカインはIL2、より好ましくはヒトIL2である。サイトカインは特異的結合メンバーまたはそのポリペプチド成分の上流(N末端)または下流(C末端)に融合されうる。好ましい実施形態は、本発明の特異的結合メンバー(特に抗体分子、例えばscFv分子)とIL2とを含む融合タンパク質である。そのような融合タンパク質のアミノ酸配列、およびそれらをコードするヌクレオチド配列を含む核酸は、本発明の一部を構成する。
【0096】
本発明の特異的結合メンバーは、例えば腫瘍イメージングのような診断方法、または例えば癌病態に対するヒトもしくは動物対象における治療において有用でありうる。
【0097】
したがって、本発明の他の態様は、提供される特異的結合メンバーを投与することを含む治療方法、そのような特異的結合メンバーを含む医薬組成物、および投与のための医薬の製造における、例えば、該特異的結合メンバーを製薬上許容される賦形剤と共に製剤化することを含む医薬または医薬組成物の製造方法における、そのような特異的結合メンバーの使用を提供する。
【0098】
治療方法において使用する特異的結合メンバーは、好ましくは、抗腫瘍作用を惹起する機能的標識に結合または連結される。好ましい実施形態においては、前記のとおり、特異的結合メンバーは、サイトカイン、例えばIL2に結合または連結される。
【0099】
治療上の利益を得るために本明細書に記載の特異的結合メンバーが使用されうる臨床適応症には、増殖性障害、例えば前癌性および悪性の新生物および腫瘍(例えば、組織細胞腫、神経膠腫、星状細胞腫、骨腫瘍)、癌(例えば、肺癌、小細胞肺癌、胃腸癌、大腸癌、結腸癌、乳癌、卵巣癌、前立腺癌、精巣癌、肝癌、腎癌、膀胱癌、膵癌、脳癌、肉腫、骨肉種、カポジ肉腫、黒色腫)、白血病および脈管形成性疾患が含まれる。
【0100】
前癌性または悪性状態は、限定的なものではないが肺、結腸、乳房、卵巣、前立腺、肝臓、膵臓、脳および皮膚を含む任意の細胞型において生じうる。
【0101】
本明細書に記載の治療に適した増殖性障害は、テネイシンC大アイソフォーム(特に、A1またはCドメインを含む大アイソフォーム)の発現が増強または上昇している細胞または組織の存在により特徴づけられうる。
【0102】
本発明においては、提供される組成物は個体に投与されうる。投与は、好ましくは、「治療的有効量」で行われ、これは、患者に対する利益を示すのに十分な量である。そのような利益は少なくとも1つの症状の少なくとも改善でありうる。実際の投与量ならびに投与の率および時間経過は、治療されるものの性質および重症度に左右されるであろう。治療の処方(例えば、投与量などの決定)は一般開業医および他の医師の責任でなされる。抗体の適当な用量は当技術分野でよく知られている。Ledermann J.A.ら, (1991) Int J. Cancer 47: 659-664; Bagshawe K.D.ら, (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4: 915-922を参照されたい。
【0103】
厳密な用量は、該抗体が診断用なのか治療用なのか、治療すべき領域のサイズおよび位置、該抗体の厳密な性質(例えば、全抗体、フラグメントまたはダイアボディ)ならびに任意の検出可能な標識または該抗体に結合している他の分子の性質を含む多数の要因に左右されるであろう。典型的な抗体の用量は全身用途には0.5mg〜100g、および局所用途には10μg〜1mgの範囲内となろう。典型的には、該抗体は全抗体、好ましくはIgG1またはIgG4アイタイプであろう。これは成人患者の1回の治療のための用量であり、これは小児および乳児の場合には比例的に調節されることが可能であり、他の抗体フォーマットに関しても、分子量に比例して調節されうる。治療は、医師の判断により、毎日、週2回、毎週または毎月の間隔で反復されうる。
【0104】
本発明の特異的結合メンバーは、通常、該特異的結合メンバーに加えて少なくとも1つの成分を含みうる医薬組成物の形態で投与されるであろう。
【0105】
かくして、本発明の医薬組成物、および本発明に従う使用のための医薬組成物は、有効成分に加えて、製薬上許容される賦形剤、担体、バッファー、安定剤または当業者によく知られた他の物質を含みうる。そのような物質は無毒性であるべきであり、有効成分の効力を損なうべきではない。担体または他の物質の厳密な性質は投与経路に左右され、投与経路は経口または注射、例えば静脈内注射でありうる。
【0106】
経口投与用の医薬組成物は錠剤、カプセル剤、散剤または液体形態でありうる。錠剤は、固体担体、例えばゼラチン、またはアジュバントを含みうる。液体医薬組成物は、一般には、液体担体、例えば水、石油、動物もしくは植物油、鉱油または合成油を含む。生理食塩水、デキストロースもしくは他の糖溶液、またはグリコール、例えばエチレングリコール、プロピレングリコールもしくはポリエチレングリコールを加えることが可能である。
【0107】
静脈内注射、または罹患部位への注射の場合には、有効成分は、発熱物質を含有せず適当なpH、等張性および安定性を有する非経口的に許容される水溶液の形態である。当業者であれば、例えば、食塩注射剤、リンゲル注射剤、乳酸添加リンゲル注射剤のような等張性ビヒクルを使用して、適当な溶液を製造することが十分に可能である。必要に応じて、保存剤、安定剤、バッファー、抗酸化剤および/または他の添加剤を加えることが可能である。
【0108】
組成物は、治療すべき状態に応じて、単独で、または他の治療と組合せて同時もしくは連続的に投与することが可能である。他の治療は、鎮痛薬、例えば非ステロイド抗炎症薬(例えば、アスピリン(aspirin)、パラセタモール(paracetamol)、イブプロフェン(ibuprofen)またはケトプロフェン(ketoprofen))、またはオピエート、例えばモルヒネ、または制吐薬の適当な用量の投与を含みうる。
【0109】
本発明のもう1つの態様は、本明細書に記載の抗体を個体に投与し、該個体における腫瘍細胞への該抗体の結合を検出することを含む、腫瘍細胞を検出および/またはイメージングするための方法を提供する。
【0110】
そのような方法において使用するための好ましい抗体は、検出可能な標識、例えば放射性核種または発蛍光団に結合または連結されうる。
【0111】
本発明の方法は、本発明で提供する特異的結合メンバーの、テネイシンCへの結合を引き起こさせる又は可能にすることを含みうる。記載されているとおり、そのような結合は、例えば、特異的結合メンバーまたは特異的結合メンバーをコードする核酸の投与の後に、in vivoで生じうる。
【0112】
テネイシンCへの特異的結合メンバーの結合の量は測定可能である。いくつかの実施形態においては、個体から得たサンプルへの特異的結合メンバーの結合を測定することが可能である。他の実施形態においては、例えば、個体の体内の腫瘍をイメージングまたは検出して、抗原への特異的結合メンバーの結合をin vivoで測定することが可能である。
【0113】
定量は、診断上関心が持たれうる抗原の量に関連づけられうる。
【0114】
抗体の結合は、任意の適当な手段により測定することが可能である。例えば、抗体をレポーター分子または検出可能な標識に連結または結合させ、サンプル上の該標識またはレポーターの存在、量または局在を測定することが可能である。
【0115】
in vivoでの抗体の結合を、例えば分子イメージングの方法において、放射能検出(例えば、PET、SPECT)、近赤外蛍光イメージング(例えば、拡散光学的断層撮影(diffuse optical tomography)、内視鏡検査)、超音波(例えば、標的化マイクロバブル(targeted microbubble)誘導体を使用)およびMRI(標的化磁気粒子を使用)により測定することが可能である。
【0116】
他の実施形態においては、抗体の結合は、例えばELISA、ウエスタンブロット法、免疫細胞化学法、免疫沈降法またはアフィニティクロマトグラフィーにおいてin vitroで生じうる。
【0117】
したがって、腫瘍細胞を検出および/またはイメージングするための方法は、本明細書に記載の抗体を、個体から得たサンプルと接触させ、該サンプル中の腫瘍細胞への該抗体の結合を検出することを含む。
【0118】
そのようなin vitro法において使用するための好ましい抗体はレポーター分子に結合または連結されうる。レポーター分子は放射性核種、発蛍光団、りん光体またはレーザー色素(スペクトル的に分離した吸収または放射特性を有するもの)でありうる。適当な発蛍光団には、フルオレセイン、ローダミン、フィコエリトリンおよびテキサス・レッドが含まれる。適当な発色色素にはジアミノベンジジンが含まれる。in vivoイメージングには、放射性核種または発蛍光団が好ましい。
【0119】
他のレポーターには、高分子コロイド粒子、または粒子状物質、例えば着色されたラテックスビーズ、検出可能なシグナルの視認、電子的検出またはその他の記録を直接的または間接的にもたらしうる、磁性もしくは常磁性および生物学的もしくは化学的に活性な物質が含まれる。これらの分子は、例えば、発色もしくは変色する又は電気的特性の変化をもたらす反応を触媒する酵素でありうる。それらは分子的に励起されて、エネルギー状態間の電子遷移が、特徴的なスペクトル吸収または放射をもたらしうる。それらには、バイオセンサーと共に使用される化学物質が含まれうる。ビオチン/アビジンまたはビオチン/ストレプトアビジンおよびアルカリホスファターゼ検出系が使用されうる。
【0120】
結合の測定方法は本発明の主題ではなく、当業者は、自分の好み及び全般的知識に基づいて適当な方法を選択することが可能である。
【0121】
本発明は更に、本明細書に記載されているのと実質的に同じアミノ酸を有するCDRを含むVドメインまたは本明細書に記載されているのと実質的に同じアミノ酸配列を有するVドメインを含みテネイシンCに結合する任意の特異的結合メンバー(例えば、本明細書に記載の特異的結合メンバー)とテネイシンCへの結合に関して競合する特異的結合メンバーに拡張される。結合メンバー間の競合はin vitroで容易にアッセイされうる。例えば、これは、他の非タグ付け結合メンバーの存在下で検出されうる1つの結合メンバーに特異的レポーター分子をタグ付けして、同一エピトープまたは重複エピトープに結合する特異的結合メンバーの特定を可能にすることにより行うことが可能である。競合は、例えばELISAのような標準的な技術を用いて測定することが可能である。
【0122】
競合に関する試験では、抗原のペプチド断片、特に、所定のエピトープを含むペプチドを使用することが可能である。エピトープ配列および1以上のアミノ酸(いずれかの末端に位置する)を有するペプチドを使用することが可能である。そのようなペプチドは、特定されている配列より「実質的になる」と称されうる。本発明の特異的結合メンバーは、抗原へのその結合が、与えられた配列を有する又は含むペプチドにより抑制されるものでありうる。これに関する試験では、いずれかの配列および1以上のアミノ酸を有するペプチドを使用することが可能である。適当なペプチドは、テネイシンCのD、Cおよび/またはA1ドメインの配列から得ることが可能である。
【0123】
特異的ペプチドに結合する特異的結合メンバーは、例えば、該ペプチドでのパンニングによりファージ提示ライブラリーから単離されうる。
【0124】
本発明は更に、本発明の特異的結合メンバーをコードする単離された核酸を提供する。核酸にはDNAおよびRNAが含まれる。好ましい態様においては、本発明は、前記のとおりの本発明のCDRまたはVHもしくはVLドメインをコードする核酸を提供する。
【0125】
本発明はまた、前記のとおりの少なくとも1つのポリヌクレオチドを含むプラスミド、ベクター、転写または発現カセットの形態の構築物を提供する。
【0126】
本発明はまた、前記のとおりの1以上の構築物を含む組換え宿主細胞を提供する。提供する任意のCDR、VHもしくはVLドメインまたは特異的結合メンバーをコードする核酸自体が本発明の1つの態様を構成する。また、コード産物をコードする核酸からの発現を含む、該コード産物の製造方法も同様に、本発明の1つの態様を構成する。発現は、適当な条件下、該核酸を含有する組換え宿主細胞を培養することにより簡便に達成されうる。発現による産生の後、VHもしくはVLドメインまたは特異的結合メンバーを、任意の適当な技術を用いて単離および/または精製し、適宜使用することが可能である。
【0127】
本発明の特異的結合メンバー、VHおよび/またはVLドメインならびにコード核酸分子およびベクターは、単離および/または精製された形態で提供されうる。例えば、それらは、それらの天然環境から、実質的に純粋または均一な形態で、単離および/または精製されることが可能であり、あるいは核酸の場合には、それらは、要求される機能を有するポリペプチドをコードする配列以外の核酸または遺伝子源を含有しない又は実質的に含有しない。本発明の核酸はDNAまたはRNAを含むことが可能であり、完全または部分的に合成物でありうる。本明細書に記載のヌクレオチド配列に対する言及は、特定された配列を有するDNA分子を包含し、文脈と矛盾しない限り、Tの代わりにUが用いられている特定された配列を有するRNA分子を包含する。
【0128】
多種多様な宿主細胞におけるポリペプチドのクローニングおよび発現のための系はよく知られている。適当な宿主細胞には、細菌、哺乳類細胞、酵母およびバキュロウイルス系が含まれる。異種ポリペプチドの発現のために当技術分野において利用可能な哺乳類細胞系には、チャイニーズハムスター卵巣細胞、HeLa細胞、乳児ハムスター腎細胞、NSOマウス黒色腫細胞、その他多数が含まれる。一般的な好ましい細菌宿主は大腸菌(E. coli)である。
【0129】
原核細胞、例えば大腸菌(E. coli)における抗体および抗体フラグメントの発現は当技術分野において十分に確立されている。レビューとしては、例えば、Pluckthun, A. Bio/Technology 9: 545-551 (1991)を参照されたい。培養内の真核細胞における発現も、特異的結合メンバーの製造のための選択肢として当業者に利用可能である。最近の概観のためには、例えば、M.E. (1993) Curr. Opinion Biotech. 4: 573-576; Trill J.J.ら, (1995) Curr. Opinion Biotech 6: 553-560を参照されたい。
【0130】
適宜、プロモーター配列、ターミネーター配列、ポリアデニル化配列、エンハンサー配列、マーカー遺伝子および他の配列を含む適当な調節配列を含有する適当なベクターを選択し、または構築することが可能である。ベクターは、適宜、プラスミド、ウイルス、例えば「ファージ」、またはファージミドでありうる。さらなる詳細は、例えば、Molecular Cloning: a Laboratory Manual: 2nd edition, Sambrookら, 1989, Cold Spring Harbor Laboratory Pressを参照されたい。例えば核酸構築物の調製、突然変異誘発、配列決定、細胞内へのDNAの導入および遺伝子発現における核酸の操作ならびにタンパク質の分析のための多数の公知技術およびプロトコールは、Current Protocols in Molecular Biology, Second Edition, Ausubelら編, John Wiley & Sons, 1992に詳細に記載されている。Sambrookら及びAusubelらの開示を参照により本明細書に組み入れることとする。
【0131】
したがって、本発明のもう1つの態様は、本明細書に開示されている核酸を含有する宿主細胞を提供する。さらにもう1つの態様は、そのような核酸を宿主細胞内に導入することを含む方法を提供する。該導入には、任意の利用可能な技術を用いることが可能である。真核細胞の場合には、適当な技術にはリン酸カルシウムトランスフェクション、DEAE-デキストラン、エレクトロポレーション、リポソーム媒介トランスフェクション、およびレトロウイルスまたは他のウイルス(例えば、ワクシニア、あるいは昆虫細胞の場合にはバキュロウイルス)を使用する形質導入が含まれうる。細菌細胞の場合には、適当な技術には塩化カルシウム形質転換、エレクトロポレーション、およびバクテリオファージを使用するトランスフェクションが含まれうる。
【0132】
該導入の後、例えば、該遺伝子の発現のための条件下で宿主細胞を培養することにより、該核酸からの発現を引き起こさせ又は可能にすることが可能である。
【0133】
1つの実施形態においては、本発明の核酸は宿主細胞のゲノム(例えば、染色体)内に組込まれる。組込みは、標準的な技術により、該ゲノムとの組換えを促進する配列の含有により促進されうる。
【0134】
本発明はまた、前記の特異的結合メンバーまたはポリペプチドを発現させるために発現系において前記構築物を使用することを含む方法を提供する。
【0135】
本発明の他の方法は、本明細書に記載の特異的結合メンバーを、検出可能な標識または抗癌物質に結合または連結させることを含む。
【0136】
適当な標識または物質は前記で説明されている。標識および物質は、標準的な化学的手段を用いて、特異的結合メンバーに結合されることが可能である。
【0137】
本発明の態様および実施形態は、以下の実験に関する実施例により例示される。
【0138】
本発明の種々の他の態様および実施形態は、本開示を考慮すれば当業者に明らかであろう。本明細書中に挙げられているすべての文書の全体を参照により本明細書に組み入れることとする。
【0139】
本発明は、前記の特徴の各組合せおよび副組合せ(サブコンビネーション)を含む。
【0140】
以下、本発明の或る態様および実施形態を、実施例により及び図面および後記の表を参考にして例示する。
【実施例1】
【0141】
血清の非存在下にpH 7.5で培養した正常ヒト皮膚繊維芽細胞(NHDF)から単離したmRNAから、ヒトテネイシンCのドメインA1をコードする遺伝子を、PCRオリゴTnC-A1 BamHI ba (cgggatcctccactgaacaagcccctgag)およびTnC-A1 BglII for (ggagatctttcccctgtggaggcctcagc) [16]を使用して、RT-PCRによりクローニングした。ついで該遺伝子をpQE12細菌発現ベクター(Qiagen)内にクローニングし、大腸菌(E. coli)TG-1において発現させた。ニッケルがローディングされたNi-NTAセファロース樹脂(Qiagen)を使用して、Hisタグにより精製を行った。
【0142】
精製されたドメインA1を、選択前に、スルホ-NHS-SS-ビオチン(Pierce)を使用してビオチン化した。Viti Fら, Methods Enzymol 2000; 326:480-505に記載されているとおりに、バイオパンニングを行った。簡潔に説明すると、ビオチン化タンパク質(最終濃度10-7 M)を、予めブロッキングされた600μlのETH-2ライブラリーファージと共に30分間インキュベートした。5.3×107個のストレプトアビジンでコートされた磁気ビーズ(Dynal)を加えることにより、結合ファージを捕捉した。十分な洗浄後、該ビオチンリンカー内のジスルフィド結合を還元することにより、選択されたファージを溶出した。単離されたファージをTG-1内で増幅し、ポリエチレングリコール沈殿により上清から濃縮した。3ラウンドのパンニングの後、144個の単離抗体クローンを、Viti Fら, Methods Enzymol 2000; 326:480-505に記載されているとおりに行うELISAによりスクリーニングした。ストレプトアビジンでコートされたELISAプレート(StreptaWell High Bind, Roche)をビオチン化抗原と共にインキュベートし、scFv抗体フラグメントを発現する誘導されたモノクローナル大腸菌(E. coli)TG-1培養の上清を加え、結合抗体をM2モノクローナル抗体で、ついで抗マウス免疫グロブリンG-HRPコンジュゲートで検出した。最高シグナルを有する抗体クローンを、BIAcore 3000装置を使用するリアルタイム相互作用分析により分析した。3つの最良のクローンをプロテインA-セファロースカラム上で精製し、種々の腫瘍凍結切片上で免疫組織化学により試験した。
【0143】
免疫組織化学において選択的染色パターンを示したクローンの1つであるscFv(3A1-D5)をアフィニティ成熟のために選択した。ファージミドベクターpDN322(Pini Aら, J Biol Chem 1998; 273:21769-21776)において、PCRオリゴヌクレオチドLMB3 long ba(caggaaacagctatgaccatgattac, 抗体遺伝子の5'末端の上流でプライミング)およびDP47CDR1mut for (agcctggcggacccagctcgcmnnmnnmnngctaaaggtgaatccagaggctgc)を使用して、可変重鎖(VH CDR1)の相補性決定領域1内の31-33位にランダム突然変異を挿入することにより、アフィニティ成熟ライブラリーを構築した。抗体遺伝子の3'末端を、プライマーDP47CDR1 ba(gagctgggtccgccaggctcc)およびfdseq long for(gacgttagtaaatgaattttctgtatgagg)を使用して増幅した。それらの2つの断片を、プライマーLMB3 long baおよびfdseq long forを使用するPCRにより合体させた。
【0144】
アフィニティ成熟ライブラリーのバイオパンニングをビオチン化抗原で行った。2ラウンドのパンニング(10-8Mビオチン化抗原を使用する以外は前記のとおり)の後、合計382個の抗体クローンをELISAによりスクリーニングした。ELISAにおいて陽性であった69個のクローンを、個々の抗体クローンのkoff値を評価するためにM2-ELISA(Scheuermann Jら, J Immunol Methods 2003; 276:129-134)により更に特徴づけした。簡潔に説明すると、scFv抗体を含有する上清を、抗Flag M2モノクローナル抗体(SIGMA)でコートされた表面上に加えた。ビオチン化抗原を加え、平衡になるまでインキュベートした後、過剰の非ビオチン化抗原を競合体として加えた。それぞれ0、30、60、90および120分の競合時間の後、ストレプトアビジン-HRPコンジュゲートを使用してビオチン化抗原の残存画分を検出した。
【0145】
種々のELISA陽性クローンをそれらの解離プロファイルに基づきランク付けするために、BIAcoreを使用した。抗原でコートされたセファロースカラム上での精製のために、5個の最良クローンを選択した。精製された抗体をサイズ排除クロマトグラフィーに付し、BIAcore 3000を使用してアフィニティ定数および動力学的パラメーターを決定するために、得られた単量体scFv抗体フラグメント画分を使用した。
【0146】
ヒト免疫グロブリンEのCH4ドメインをコードする遺伝子の5'末端の前にscFv(4A1-F16)配列を遺伝的に融合させることにより、最良の解離定数(KD)を有する抗体4A1-F16を二価ミニボディフォーマット内にクローニングして、4A1-F16-SIPと称される小免疫タンパク質(small immune protein)(SIP)(Borsi Lら, Int J Cancer 2002; 102:75-85)を得た。該CH4ドメインはホモ二量体化を促進して、より高いアビディテイにより該抗体の機能的アフィニティを増加させる。
【0147】
ヒト頭部-および-頚部癌組織上でテネイシンC抗体フラグメントscFv(3A1-D5)の抗A1ドメインを使用して、免疫組織化学的方法を行った。scFv抗体のC末端に付加したペプチド性FLAGタグを使用し、モノクローナル抗FLAG抗体M2(SIGMA)を次いでAPAAP系(DAKO)を使用して、一次抗体を検出した。scFv(3A1-D5)およびscFv(4A1-F16)は、種々の頭部-および-頚部の凍結切片上で腫瘍ストロマおよび新血管構造を強く染色することが観察されたが、それらは健常ドナーの正常口腔粘膜とは反応しなかった。
【0148】
4A1-F16-SIPの腫瘍標的化能を以下のとおりに評価した。
【0149】
Balb/C nu/nuマウスにおいて、マウス1匹当たり3×106個のU87ヒト神経膠芽細胞腫細胞(ATCC)の皮下注射により腫瘍を誘発させた。注射の20〜25日後、腫瘍が300〜1500 mm3のサイズに達した際に、放射能標識抗体を注射した。
【0150】
該抗体の調製は、スーパーデックス(superdex)75上のサイズ排除クロマトグラフィーによるアフィニティ精製4A1-F16-SIP抗体の更なる精製を含むものであった。ホモ二量体形態(75kDa)を表す画分を集め、ついでヨウ素源(iodogen)(Pierce)を使用してヨウ素-125(Amersham)で標識した。
【0151】
約5μgの抗体を腫瘍担持マウスの尾静脈内に静脈内注射した。それぞれ3、6、24および48時間後、3匹のマウスを犠死させ、器官を摘出し、I-125の蓄積をγカウンターで測定した。
【実施例2】
【0152】
ヒトテネイシンCのドメインCをコードする遺伝子をベクターpQE12[Carnemolla Bら, Am J Pathol 1999; 154:1345-1352, Balza Eら, FEBS Lett 1993; 332:39-43]内にクローニングし、大腸菌(E. coli)TG-1内で発現させた。ニッケルがローディングされたNi-NTAセファロース樹脂(Qiagen)を使用して、Hisタグにより精製を行った。
【0153】
精製されたドメインCを、選択前に、スルホ-NHS-SS-ビオチン(Pierce)を使用してビオチン化した。ストレプトアビジンおよびアビジンでコートされたプレート上で別のパンニング法を用いること以外はViti Fら, Methods Enzymol 2000; 326:480-505に記載されているとおりに、バイオパンニングを行った。簡潔に説明すると、ビオチン化タンパク質(最終濃度10-7 M)を、予めブロッキングされた600μlのETH-2ライブラリーファージと共に30分間インキュベートした。アビジン(第1および第3ラウンドのパンニング)またはストレプトアビジン(第2ラウンドのパンニング)でコートされたプラスチックマイクロタイタープレート上で、結合ファージを捕捉した。十分な洗浄後、該ビオチンリンカー内のジスルフィド結合を還元することにより、選択されたファージを溶出した。単離されたファージをTG-1内で増幅し、ポリエチレングリコール沈殿により上清から濃縮した。3ラウンドのパンニングの後、数十個の抗体クローンを、Viti Fら, Methods Enzymol 2000; 326:480-505に記載されているとおりに行うELISAによりスクリーニングした。
【0154】
ストレプトアビジンでコートされたELISAプレート(StreptaWell High Bind, Roche)をビオチン化抗原と共にインキュベートし、scFv抗体フラグメントを発現する誘導されたモノクローナル大腸菌(E. coli)TG-1培養の上清を加え、結合抗体を抗ペプチドタグ抗体で、ついで抗マウス免疫グロブリンG-HRPコンジュゲートで検出した。最高シグナルを有する抗体クローンを、BIAcore 3000装置を使用するリアルタイム相互作用分析により分析した。3つの最良のクローンをプロテインA-セファロースカラム上で精製し、種々の腫瘍凍結切片上で免疫組織化学により試験した。免疫組織化学において選択的染色パターンを示したクローンの1つであるscFv(A12)をアフィニティ成熟のために選択した。ファージミドベクターpHEN1[Hoogenboom HRら, Nucleic Acids Res 1991; 19:4133-4137]において、PCRオリゴヌクレオチドLMB3 long ba(caggaaacagctatgaccatgattac, 抗体遺伝子の5'末端の上流でプライミング)、DP47CDR1 for(ctggagcctggcggacccagctcatmnnmnnmnngctaaaggtgaatccagaggctgc)、DP47CDR1 ba(tgggtccgccaggctccag)、DP47CDR2 for(gcccttcacggagtctgcgtagtatgtmnnaccaccmnnmnnmnnaatagctgagacccactcc)、DP47CDR2 ba(acatactacgcagactccgtgaagggc)およびfdseq long for(gacgttagtaaatgaattttctgtatgagg)を使用して、VH CDR1内の31-33位およびVH CDR2内の52、52a、53および56位にランダム突然変異を挿入することにより、アフィニティ成熟ライブラリーを構築した。
【0155】
アフィニティ成熟ライブラリーのバイオパンニングをビオチン化抗原で行った。2ラウンドのパンニング(前記のとおり)の後、合計382個の抗体クローンをELISAによりスクリーニングした。ELISAにおいて陽性であったクローンをBIAcoreにより更に特徴づけして、種々のELISA陽性クローンをそれらの解離プロファイルに基づきランク付けした。試験したクローンのうち、scFv(E10)が最も有望な性能を示した。
【0156】
ヒト神経膠芽細胞腫多形態組織上でテネイシンC抗体フラグメントscFv(A12)の抗Cドメインを使用して、免疫組織化学的方法を行った。scFv抗体のC末端に付加したペプチド性mycタグを使用し、モノクローナル抗mycタグ抗体E10を次いでAPAAP系(DAKO)を使用して、一次抗体を検出した。これらの抗体は、神経膠芽細胞腫凍結切片上の免疫組織化学により示されるとおり、血管周囲構造体と特異的に反応することが観察された。
【実施例3】
【0157】
実施例2に記載されているものに類似したプロトコールを用いて、テネイシンCのCドメインに対する他のscFvを単離し、それぞれF4およびG11と命名した。F4およびG11は、2アミノ酸だけ異なるVLドメインを有し、同一VHを有する。VHおよびVLドメインを、それぞれ配列番号36または配列番号38によりコードされる配列番号37または配列番号39に記載のアミノ酸配列を有するペプチドリンカーを介して連結した。
【実施例4】
【0158】
テネイシンCのヒトドメインD(TnC-D)をコードする遺伝子を、ヒト黒色腫細胞系SKMel-28から単離された全RNAから、PCRオリゴTnC-D BamHI ba(cgggatccgttacagaagccgaaccggaa - 配列番号72)およびTnC-D BglII for(cgggatccgttacagaagccgaaccggaa - 配列番号73)[16]を使用するRT-PCRによりクローニングした。増幅された断片を、該タンパク質のC末端にHisタグを導入する細菌発現ベクターpQE12内にサブクローニングした。TnC-A1のマウスアイソフォームをコードする遺伝子を、浸潤性管癌組織由来の発現配列タグ(EST)クローンから、プライマーmmTnC-A1 EcoBa(agaattcattaaagaggagaaattaactatgagaggatcctccacggaagaagtgccttc - 配列番号74)およびmmTnC-A1 BglFo(tgagatcttgtccctgtggaggtctcggc - 配列番号75)を使用するPCRにより単離し、ベクターpQE12(Qiagen)内にクローニングした。
【0159】
ETHライブラリー[26]を100nM ビオチン化ヒトTnC-Dでパンニングし、結合ファージを、ストレプトアビジンでコートされた磁気ビーズで捕捉した。3ラウンドのパンニングの後、188個のクローンから得た上清をビオチン化ヒトTnC-D上およびビオチン化マウスTnC-D上のELISAによりスクリーニングした。該ヒトアイソフォームを認識したクローンのほとんどはTnC-Dのマウスアイソフォームとも反応した。
【0160】
ELISA陽性クローンの上清を第2 ELISAにおいて試験し、BIAcoreによりスクリーニングした。6個の最良のクローンを詳細に特徴づけし、scFv(F4)が最良クローンであることが確認された。scFv(F4)は、ヒトおよびマウスの両方のTnC-Dに同一アフィニティで結合した。
【0161】
scFv(F4)は専ら、ELISAにおいて、ドメインDを含有するタンパク質を認識し、ドメインDに構造的に関連しているタンパク質とは反応しなかった。しかし、それは免疫組織化学法において腫瘍構造体と非常に弱く反応したに過ぎず、バックグラウンドから識別するのが困難なシグナルを与えたに過ぎなかった。これはおそらく、scFv(F4)の低いアフィニティによるものであろう。
【0162】
scFv(F4)を、CDRループ突然変異誘発[26]によるアフィニティ成熟のための鋳型として使用した。ランダム化のために選択した残基はVL CDR1の30、31、32位ならびにVH CDR2の50、52および53位(Kabatに従う番号付け;[30])であった。該ライブラリーを、VLのC末端にmycタグを含有するファージミドベクターpHEN[31]内にクローニングした。
【0163】
アフィニティ成熟ライブラリーファージでの第1ラウンドのパンニングをビオチン化ヒトTnC-Dで行い、第2ラウンドをビオチン化マウスTnC-Dで行った。750個のクローンをビオチン化ヒトTnC-D上のELISAによりスクリーニングしたところ、約5%が陽性であった。BIAcoreおよびELISAによる該抗体の更なる特徴づけの後、scFv(D11)が最良クローンとして特定された。BIAcoreによる単量体画分でのアフィニティ測定は、scFv(D11)が、親クローンscFv(F4)に比べて2〜4倍改善されたアフィニティを有することを示した。該マウスアイソフォームは、該ヒトアイソフォームより若干高いアフィニティで結合した。
【0164】
scFv(D11)は、種々の腫瘍組織上の免疫組織化学法において良好な結果を与えた。ヒト口腔扁平上皮細胞癌、ヒト膀胱癌、ヌードマウスにおいて増殖させたA375ヒト黒色腫、およびSvEV 129マウスにおいて増殖させたF9マウスF9奇形癌はすべて、scFv(D11)で染色された。該抗体は、頭部および頚部癌または膀胱癌のようなヒト腫瘍のストロマと、ならびに種々のヒトおよびマウス腫瘍モデルにおいて、強力に反応した。
【0165】
つぎに、scFv(D11)のVHのCDR1およびCDR1内の残基をランダム化した。ランダム化のために、VH CDR1内の残基31、32、33ならびにVH CDR2内の残基52、52a、53および56(Kabatの番号付け; [32])を選択した。抗原として10-8 MヒトTnC-Dを使用して、ファージ提示による2ラウンドの選択を行った。選択のストリンジェンシーを増加させるために、第1ラウンドにおける抗原の添加の前に10nM 精製可溶性scFv(D11)を該ファージと混合し、第2ラウンドのパンニングにおけるプレインキュベートされた抗原-ファージ混合物に1000倍過剰の非ビオチン化抗原を加えた。
【0166】
2ラウンド後にスクリーニングされた384個のクローンのうちの25%がELISAにおいてヒトビオチン化TnC-D上で陽性であった。更なる特徴づけの後、最良クローンとして特定されたのはscFv(P12)であった。ヒトTnC-D上のそのアフィニティは、scFv(D11)と比べて4.8倍改善された。
【0167】
scFv(P12)をSIPフォーマット[25]内にクローニングした。SIP(P12)のin vivo標的化能をU87神経膠芽細胞腫異種移植片担持ヌードマウスにおけるヒト生体分布実験により評価した。125I-標識SIP抗体を静脈内注射し、3、6、24および48時間後、動物を犠死させ、器官を摘出し、秤量し、放射能を計数した。注射の24時間以内にSIP(P12)はほとんどの器官から消失し、24および48時間後に、15までの腫瘍:血管比で、腫瘍部位における特異的蓄積が観察された(図5および表1)。
【表1】

【0168】
参考文献
1. Bosslet K et al. Cancer Res 1998; 58:1195-1201
2. Jain RK. Adv Drug Deliv Rev 2001; 46:149-168
3. Kohler G, Milstein C Nature 1975; 256:495-497
4. Winter G et al Annu Rev Immunol 1994; 12:433-455
5. Halin C et al. Cancer Res 2003; 63:3202-3210
6. Halin C et al. Nat Biotechnol 2002; 20:264-269
7. Nilsson F et al Cancer Res 2001; 61:711-716
8. Borsi L et al. Blood 2003; 102:4384-4392
9. Carnemolla B et al. Blood 2002; 99:1659-1665
10. Zardi L et al. Embo J 1987; 6:2337-2342
11. Castellani P et al. Am J Pathol 2002; 161:1695-1700
12. Castellani P et al Int J Cancer 1994; 59:612-618
13. Carnemolla B et al . Int J Cancer 1996; 68:397-405
14. Borsi L et al Int J Cancer 1992; 52:688-692
15. Carnemolla B et al. Eur J Biochem 1992; 205:561-567
16. Borsi L et al J Biol Chem 1995; 270:6243-6245
17. Riva P et al. Int J Cancer 1992; 51:7-13
18. Riva P et al. Cancer Res 1995; 55:5952s-5956s
19. Paganelli G et al Eur J Nucl Med 1994; 21:314-321
20. Reardon DA et al. J Clin Oncol 2002; 20:1389-1397
21. Bigner DD et al. J Clin Oncol 1998; 16:2202-2212
22. Carnemolla B et al. Am J Pathol 1999; 154:1345-1352
23. Katenkamp K et al. J Pathol 2004; 203:771-779
24. Viti F, et al Methods Enzymol 2000; 326:480-505
25. Borsi L et al. Int J Cancer 2002; 102:75-85
26. Pini A et al. J Biol Chem 1998; 273:21769-21776
27. Scheuermann J et al J Immunol Methods 2003; 276:129-134
28. Balza E et al. FEBS Lett 1993; 332:39-43
29. Hoogenboom HR et al. Nucleic Acids Res 1991; 19:4133-4137
30. Ignatovich, O., et al. (1997). J Mol Biol 268(1): 69-77.31.
31. Hoogenboom & Winter (1992). J Mol Biol 227(2): 381-8.
32. Tomlinson, et al. (1992). J Mol Biol 227(3): 776-98.
【0169】
配列表
配列番号1 4A1-F16 VHドメインヌクレオチド配列
GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
CGG TAT GGT GCG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AGT GGT AGT GGT GGT AGC ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA GCG CAT AAT GCT TTT GAC TAC
TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号2 4A1-F16 VHドメインアミノ酸配列
EVQLLESGGG LVQPGGSLRL SCAASGFTFS RYGASWVRQA PGKGLEWVSA
ISGSGGSTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKAH
NAFDYWGQGT LVTVSR
配列番号3 4A1-F16 & 3A1-D5 VLドメインヌクレオチド配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT
GTT TAT ACT ATG CCG CCC GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号4 4A1-F16 & 3A1-D5 VLドメインアミノ酸配列
SSELTQDPAV SVALGQTVRI TCQGDSLRSY YASWYQQKPG QAPVLVIYGK
NNRPSGIPDR FSGSSSGNTA SLTITGAQAE DEADYYCNSS VYTMPPVVFG
GGTKLTVLG
配列番号5 4A1-F16 VH CDR1アミノ酸配列
RYGAS
配列番号6 4A1-F16 & 3A1-D5 VH CDR2アミノ酸配列
AISGSGGSTYYADSVKG
配列番号7 4A1-F16 & 3A1-D5 VH CDR3アミノ酸配列
AHNAFDY
配列番号8 4A1-F16 & 3A1-D5 VL CDR1アミノ酸配列
QGDSLRSYYAS
配列番号9 4A1-F16 & 3A1-D5 VL CDR2アミノ酸配列
GKNNRPS
配列番号10 4A1-F16 & 3A1-D5 VL CDR3アミノ酸配列
NSSVYTMPPVV
配列番号11 3A1-D5 VHドメインヌクレオチド配列
GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
AGC TAT GCC GCG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AGT GGT AGT GGT GGT AGC ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA GCG CAT AAT GCT TTT GAC TAC
TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号12 3A1-D5 VHドメインアミノ酸配列
EVQLLESGGG LVQPGGSLRL SCAASGFTFS SYAASWVRQA PGKGLEWVSA
ISGSGGSTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKAH
NAFDYWGQGT LVTVSR
配列番号13 3A1-D5 VH CDR1アミノ酸配列
SYAAS
配列番号14 E10 VHドメインヌクレオチド配列
GAG GTG CAG CTG GTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
GGT AGT CGT ATG GGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AAT GAG GAG GGT GGT CAG ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA CAT CCG CCG CAT CGG CCG TTT
GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号15 E10 VHドメインアミノ酸配列
EVQLVESGGG LVQPGGSLRL SCAASGFTFS GSRMGWVRQA PGKGLEWVSA
INEEGGQTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKHP
PHRPFDYWGQ GTLVTVSR
配列番号16 E10 & A12 VLドメインヌクレオチド配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT
CAT GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号17 E10 & A12 VLドメインアミノ酸配列
SSELTQDPAV SVALGQTVRI TCQGDSLRSY YASWYQQKPG QAPVLVIYGK
NNRPSGIPDRFSGSSSGNTA SLTITGAQAE DEADYYCNSS HGPRRPVVFG
GGTKLTVLG
配列番号18 E10, F4 & G11 VH CDR1アミノ酸配列
GSRMG
配列番号19 E10, F4 & G11 VH CDR2アミノ酸配列
AINEEGGQTYYADSVKG
配列番号20 E10, A12, F4 & G11 VH CDR3アミノ酸配列
HPPHRPFDY
配列番号21 E10 & A12 VL CDR1アミノ酸配列
QGDSLRSYYAS
配列番号22 E10, A12 & G11 VL CDR2アミノ酸配列
GKNNRPS
配列番号23 E10 & A12 VL CDR3アミノ酸配列
NSSHGPRRPVV
配列番号24 A12 VHドメインヌクレオチド配列
GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
AGC TAT GCC ATG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AGT GGT AGT GGT GGT AGC ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA CAT CCG CCG CAT CGG CCG TTT
GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号25 A12 VHドメインアミノ酸配列
EVQLLESGGG LVQPGGSLRL SCAASGFTFS SYAMSWVRQA PGKGLEWVSA
ISGSGGSTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKHP
PHRPFDYWGQ GTLVTVSR
配列番号26 A12 VH CDR1アミノ酸配列
SYAMS
配列番号27 A12 VH CDR2アミノ酸配列
AISGSGGSTYYADSVKG
配列番号28 F4 & G11 VHドメイン核酸配列
GAG GTG CAG CTG GTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
GGT AGT CGT ATG GGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AAT GAG GAG GGT GGT CAG ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA CAT CCG CCG CAT CGG CCG TTT
GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTC TCG AGA
配列番号29 F4 & G11 VHドメインアミノ酸配列
EVQLVESGGGLVQPGGSLRLSCAASGFTFSGSRMGWVRQAPGKGLEWVSAINEEGGQTYY
ADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKHPPHRPFDYWGQGTLVTVSR
配列番号30 F4 VLドメイン核酸配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA CTT TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA TCT AGT CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT
CAT GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号31 F4 VLドメインアミノ酸配列
SSELTQDPAVSVALGQTVRITCQGDSLRLYYASWYQQKPGQAPVLVIYGKSSRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
配列番号32 F4 & G11 VL CDR1アミノ酸配列
QGDSLRLYYAS
配列番号33 F4 VL CDR2アミノ酸配列
GKSSRPS
配列番号34 G11 VLドメイン核酸配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA CTT TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT
CAT GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号35 G11 VLドメインアミノ酸配列
SSELTQDPAVSVALGQTVRITCQGDSLRLYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
配列番号36 ペプチドリンカー核酸配列
GGT GGA GGC GGT TCA GGC GGA GGT GGC TCT GGC GGT GGC GGA
配列番号37 ペプチドリンカーアミノ酸配列
GGGGSGGGGSGGGG
配列番号38 ペプチドリンカー核酸配列
GGT GGA GGC GGT TCA GGC GGA GGT GGT TCT GGC GGT GGC GGA TCG
配列番号39 ペプチドリンカーアミノ酸配列
GGGGSGGGGSGGGGS
配列番号40 ペプチドリンカー核酸配列
TCT TCC TCA TCG GGT AGT AGC TCT TCC GGC TCA TCG TCC AGC GGC
配列番号41 ペプチドリンカーアミノ酸配列
SSSSGSSSSGSSSSG
配列番号42 ペプチドリンカーアミノ酸配列
GSGSAGSGSAGSGSA
配列番号43 ペプチドリンカーアミノ酸配列
GGSGGGGSGGGGSGG
配列番号44 新規F4 & G11 VHドメイン核酸配列
GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
GGT AGT CGT ATG GGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AAT GAG GAG GGT GGT CAG ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAA GAC
ACG GCC GTA TAT TAC TGT GCG AAA CAT CCG CCG CAT CGG CCG TTT
GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTC TCG AGT
配列番号45 新規F4 & G11 VHドメインアミノ酸配列
EVQLLESGGGLVQPGGSLRLSCAASGFTFSGSRMGWVRQAPGKGLEWVSAINEEGGQTYYAD
SVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKHPPHRPFDYWGQGTLVTVSS
配列番号46 新規G11 VLドメイン核酸配列
TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA CAG
ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA CTT TAT TAT
GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT GTC
ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA TTC
TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT GGG
GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT CAT
GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG ACC
GTC CTA GGC
配列番号47 新規G11 VLドメインアミノ酸配列
SELTQDPAVSVALGQTVRITCQGDSLRLYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
配列番号48 新規E10 VHドメインアミノ酸配列
EVQLLESGGG LVQPGGSLRL SCAASGFTFS GSRMGWVRQA PGKGLEWVSA
INEEGGQTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKHP
PHRPFDYWGQ GTLVTVSS
配列番号49 新規4A1-F16 & 3A1-D5 VLドメインヌクレオチド配列
TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA CAG
ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT TAT
GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT GTC
ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA TTC
TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT GGG
GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT GTT
TAT ACT ATG CCG CCC GTG GTA TTC GGC GGA GGG ACC AAG CTG ACC
GTC CTA GGC
配列番号50 新規4A1-F16 & 3A1-D5 VLドメインアミノ酸配列
SELTQDPAVSVALGQTVRITCQGDSLRSYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR FSGSSSGNTASLTITGAQAEDEADYYCNSSVYTMPPVVFGGGTKLTVLG
配列番号51 P12 VHドメイン核酸配列
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTC
CTGTGCAGCCTCTGGATTCACCTTTGGCcagTATtctATGAGCTGGGTCCGCCAGGCTCCAG
GGAAGGGGCTGGAGTGGGTCTCAGCTATTacggggactGGTGGTgagACATACTACGCAGAC
TCCGTGGAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAAT
GAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGGGCGGCGGATTTTTG
ACTACTGGGGCCAGGGAACCCTGGTCACCGTGTCGAGA
配列番号52 P12 VHドメインアミノ酸配列
EVQLVESGGGLVQPGGSLRLSCAASGFTFGQYSMSWVRQAPGKGLEWVSAITGTGGETYY
ADSVEGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGRRIFDYWGQGTLVTVSR
配列番号53 P12およびD11 VLドメイン核酸配列
TCGAGTGAGCTGACTCAGGACCCTGCTGTGTCTGTGGCCTTGGGACAGACAGTCAGGATCAC
ATGCCAAGGAGACAGCCTCAGACGGCAGCCTGCAAGCTGGTACCAGCAGAAGCCAGGACAGG
CCCCTGTACTTGTCATCTATTATAAAAAGCTGCGGCCCTCAGGGATCCCAGACCGATTCTCT
GGCTCCAGCTCAGGAAACACAGCTTCCTTGACCATCACTGGGGCTCAGGCGGAAGATGAGGC
TGACTATTACTGTAACTCCTTTTCGCCCAAGCCGAAGCCTGTGGTATTCGGCGGAGGGACCA
AGCTGACCGTCCTAGGC
配列番号54 P12およびD11 VLドメインアミノ酸配列
SSELTQDPAVSVALGQTVRITCQGDSLRRQPASWYQQKPGQAPVLVIYYKKLRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSFSPKPKPVVFGGGTKLTVLG
配列番号55 D11 VHドメイン核酸配列
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTC
CTGTGCAGCCTCTGGATTCACCTTTGGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAG
GGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGTAGTGGTGGTAGCACATACTACGCAGAC
TCCGTGGAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAAT
GAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGGGCGGCGGATTTTTG
ACTACTGGGGCCAGGGAACCCTGGTCACCGTGTCGAGA
配列番号56 D11 VHドメインアミノ酸配列
EVQLVESGGGLVQPGGSLRLSCAASGFTFGSYAMSWVRQAPGKGLEWVSAISGSGGSTYY
ADSVEGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGRRIFDYWGQGTLVTVSR
配列番号57 F4S VHドメイン核酸配列
GAG GTG CAG CTG GTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
AGC TAT GCC ATG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AGT GGT AGT GGT GGT AGC ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA GGG CGG CGG ATT TTT GAC TAC
TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号58 F4S VHドメインアミノ酸配列
EVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYY
ADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGRRIFDYWGQGTLVTVSR
配列番号59 F4S VLドメイン核酸配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TTT
TCG CCC AAG CCG AAG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号60 F4S VLドメインアミノ酸配列
SSELTQDPAVSVALGQTVRITCQGDSLRSYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSFSPKPKPVVFGGGTKLTVLG
配列番号61 P12 VH CDR1アミノ酸配列
QYSMS
配列番号62 D11およびF4S VH CDR1アミノ酸配列
SYAMS
配列番号63 P12 VH CDR2アミノ酸配列
AITGTGGETYYADSVEG
配列番号64 D11 VH CDR2アミノ酸配列
AISGSGGSTYYADSVEG
配列番号65 F4S VH CDR2アミノ酸配列
AISGSGGSTYYADSVKG
配列番号66 P12, D11およびF4S VH CDR3アミノ酸配列
GRRIFDY
配列番号67 P12およびD11 VL CDR1アミノ酸配列
QGDSLRRQPAS
配列番号68 F4S VL CDR1アミノ酸配列
QGDSLRSYYAS
配列番号69 P12およびD11 VL CDR2アミノ酸配列
YKKLRPS
配列番号70 F4S VL CDR2アミノ酸配列
GKNNRPS
配列番号71 P12, D11およびF4S VL CDR3アミノ酸配列
NSFSPKPKPVV
配列番号72 PCR オリゴヌクレオチドプライマーTnC-D BamHI ba
cgggatccgttacagaagccgaaccggaa
配列番号73 PCR オリゴヌクレオチドプライマーTnC-D BglII for
cgggatccgttacagaagccgaaccggaa
配列番号74 PCR オリゴヌクレオチドプライマーmmTnC-A1 EcoBa
agaattcattaaagaggagaaattaactatgagaggatcctccacggaagaagtgccttc
配列番号75 PCR オリゴヌクレオチドプライマーmmTnC-A1 BglFo
tgagatcttgtccctgtggaggtctcggc
配列番号76 ペプチドリンカーモチーフ
GGGGS
配列番号77 ペプチドリンカーモチーフ
SSSSG
配列番号78 ペプチドリンカーモチーフ
GSGSA
配列番号79 ペプチドリンカーモチーフ
GGSGG
配列番号80 新規E10 & A12 VLドメインヌクレオチド配列
TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA CAG
ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT TAT
GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT GTC
ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA TTC
TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT GGG
GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT CAT
GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG ACC
GTC CTA GGC
配列番号81 新規E10 & A12 VLドメインアミノ酸配列
SELTQDPAVSVALGQTVRITCQGDSLRSYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
配列番号82 新規F4 VLドメイン核酸配列
TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA CAG
ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA CTT TAT TAT
GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT GTC
ATC TAT GGT AAA TCT AGT CGG CCC TCA GGG ATC CCA GAC CGA TTC
TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT GGG
GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT CAT
GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG ACC
GTC CTA GGC
配列番号83 新規F4 VLドメインアミノ酸配列
SELTQDPAVSVALGQTVRITCQGDSLRLYYASWYQQKPGQAPVLVIYGKSSRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
【技術分野】
【0001】
本発明は、テネイシンCに対する特異的結合メンバー、特に、ヒトテネイシンCに対するヒト抗体に関する。これらの特異的結合メンバーは、ある範囲の治療用途、例えば癌の診断および治療における治療用途を有する。
【背景技術】
【0002】
テネイシンCは、細胞接着をモジュレーションする、細胞外マトリックスの、大きな六量体糖タンパク質である。それは細胞増殖および細胞遊走のような過程に関与しており、形態形成および胚発生中に生じる変化で、腫瘍形成または血管新生中の組織構造の変化に関連している。
【0003】
テネイシンCのドメイン構造の概略を図1に示す。テネイシンCの幾つかのアイソフォームは、ドメインA1からドメインDにわたるこのタンパク質の中央部分における(複数の)ドメインの含有を招きうる選択的スプライシングの結果として生じうる[Borsi Lら, Int J Cancer 1992; 52:688-692, Carnemolla Bら, Eur J Biochem 1992; 205:561-567]。ドメインA1-Dは選択的スプライシングのメカニズムにより「ブロック(塊)」として挿入または除去されて「テネイシンC大(large)」および「テネイシンC小(small)」分子を与えうると以前には考えられていた[Borsi Lら, J Biol Chem 1995; 270:6243-6245]。テネイシンCの大アイソフォームの強力な過剰発現が多数の腫瘍で報告されており[Borsi 1992 前掲]、それぞれドメインA1およびDに特異的な2つのモノクローナル抗体が臨床において詳細に特徴づけられている[Riva Pら, Int J Cancer 1992; 51:7-13, Riva Pら, Cancer Res 1995; 55:5952s-5956s, Paganelli Gら, Eur J Nucl Med 1994; 21:314-321, Reardon DAら, J Clin Oncol 2002; 20:1389-1397, Bigner DDら, J Clin Oncol 1998; 16:2202-2212]。
【0004】
しかし、テネイシンCの大アイソフォーム間の分子的不均一性の増加を招く、選択スプライシングメカニズムの、より複雑な調節が生じることが、最近になって明らかになった。例えば、テネイシンCの更なる(extra)ドメインCが、免疫テネイシンCのその他の選択的スプライシングされたドメインより制限された発現パターンを示し[Carnemolla Bら, Am J Pathol 1999; 154:1345-1352]、組織化学的方法により示される顕著な血管周囲染色を伴うことが報告されている。テネイシンCのCドメインは、ほとんどの正常成体組織においては検出不能であるが、高い等級(grade)の星状細胞腫[Carnemolla Bら, Am J Pathol 1999; 154:1345-1352]および他の腫瘍型においては過剰発現される。大テネイシンCアイソフォーム間の不均一性に関する更なる裏付けが転写分析から得られており、これは、大テネイシンC転写産物が不均一な組成を特徴とすることを証明した[Katenkamp Kら, J Pathol 2004; 203:771-779]。翻訳後修飾(例えば、グリコシル化)の存在または非存在により更なるレベルの複雑性がもたらされ、これが個々のタンパク質ドメインの表面上の或るエピトープを修飾し、それらを、特異的なモノクローナル抗体に対するin vitroまたはin vivoでの特異的分子認識に利用不可能にする。
【0005】
所定の実質的に任意のタンパク質に特異的な抗体の迅速な単離が、既存の方法でin vitroで達成されうるとしても、そのような抗体が生物学的試料における又は疾患の動物モデルにおけるエピトープを認識することは明らかではない。in vivoでの結合の欠如をもたらす考えられうる理由には、エピトープの翻訳後修飾、エピトープのマスキング、および不十分な抗体特異性または安定性が含まれる。したがって、モノクローナル抗体のスクリーニングに通常用いられる酵素結合イムノソルベントアッセイ(ELISA)のような典型的な固相アッセイにおける組換え抗原(または抗原断片)とのモノクローナル抗体の反応性のみに基づいて、実際の用途へのモノクローナル抗体の適合性を評価することは困難である。
【0006】
したがって、診断用途および治療用途へのモノクローナル抗体の適合性を評価するためには、テネイシンC大アイソフォームの個々のドメインに対するモノクローナル抗体を個別に分析する必要がある。
【発明の概要】
【0007】
本発明者らは、テネイシンCの選択的スプライシングされた領域内の種々のエピトープに特異的なヒトモノクローナル抗体フラグメントを単離した。これらの抗体は、生物学的試料中の大テネイシンCアイソフォームを認識するそれらの能力により特徴づけられ、また、密接に関連した抗原間で顕著な差異を示す、ELISAアッセイにおける高特異的結合により特徴づけられる。
【0008】
本発明の1つの態様は、ヒトテネイシンC、特にテネイシンC大アイソフォームに結合する特異的結合メンバーを提供する。
【0009】
好ましい特異的結合メンバーは腫瘍特異的であり、正常組織と比べて腫瘍組織に優先的に結合する。特異的結合メンバーは、例えば、正常組織と比べて優先的に腫瘍組織のストロマならびに/または新血管構造および血管周囲構造に結合しうる。
【0010】
特異的結合メンバーは、テネイシンC小アイソフォームと比べてテネイシンC大アイソフォームに優先的に結合しうる。
【0011】
いくつかの実施形態においては、特異的結合メンバーはテネイシンCのA1ドメインに結合しうる。適当な特異的結合メンバーは、
配列番号2の4A1-F16 VHドメイン、配列番号12の3A1-D5 VHドメイン、ならびに配列番号5、配列番号6、配列番号7および配列番号13から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
配列番号4または配列番号50のVLドメインならびに配列番号8、配列番号9および配列番号10から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0012】
4A1-F16はF16とも称される。
【0013】
例えば、適当な特異的結合メンバーは、配列番号4もしくは配列番号50の4A1-F16/3A1-D5 VLドメインおよび/または配列番号2の4A1-F16 VHドメインまたは配列番号12の3A1-D5 VHドメインを含みうる。
【0014】
他の実施形態においては、特異的結合メンバーはテネイシンCのCドメインに結合しうる。適当な特異的結合メンバーは、
E10 VHドメイン(配列番号15または配列番号48)、A12 VHドメイン(配列番号25)、F4およびG11 VHドメイン(配列番号29または配列番号45)ならびに配列番号18、配列番号19、配列番号20、配列番号26および配列番号27から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
配列番号17もしくは配列番号81のVLドメイン、配列番号31もしくは配列番号83のアミノ酸配列を有するF4 VLドメイン、配列番号35もしくは配列番号47のアミノ酸配列を有するG11 VLドメインならびに配列番号21、配列番号22、配列番号23、配列番号32および配列番号33から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0015】
したがって、1つの例においては、特異的結合メンバーは、
E10 VHドメイン(配列番号15または配列番号48)、A12 VHドメイン(配列番号25)ならびに配列番号18、配列番号19、配列番号20、配列番号26および配列番号27から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
配列番号17もしくは配列番号81のVLドメインならびに配列番号21、配列番号22および配列番号23から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0016】
例えば、適当な特異的結合メンバーは、配列番号17もしくは配列番号81のE10/A12 VLドメインおよび/または配列番号15もしくは配列番号48のE10 VHドメインまたは配列番号25のA12 VHドメインを含みうる。
【0017】
もう1つの例においては、特異的結合メンバーは、
F4およびG11 VHドメイン(配列番号29または配列番号45)ならびに配列番号18、配列番号19および配列番号20から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
F4 VLドメイン(配列番号31もしくは配列番号83)、G11 VLドメイン(配列番号35または配列番号47)ならびに配列番号32、配列番号33、配列番号22および配列番号23から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0018】
例えば、適当な特異的結合メンバーは、配列番号29もしくは配列番号45のF4/G11 VHドメインおよび/または配列番号31もしくは配列番号83のF4 VLドメインまたは配列番号35もしくは配列番号47のG11 VLドメインを含みうる。
【0019】
もう1つの実施形態においては、特異的結合メンバーはテネイシンCのドメインDに結合する。好ましくは、特異的結合メンバーはヒトドメインDに結合する。それはマウスアイソフォームと交差反応しうる。本発明者らは、ドメインDに特異的な抗体分子を単離した。元のクローンはF4Sと称される。本発明者らは、D11と称される、F4Sのアフィニティ(親和性)成熟変異体をも開発した。本発明者らは更に、P12と称される、D11のアフィニティ成熟変異体を開発した。
【0020】
したがって、適当な特異的結合メンバーは、
配列番号58のF4S VHドメイン、配列番号55のD11 VHドメイン、配列番号52のP12 VHドメインならびに配列番号61、配列番号62、配列番号63、配列番号64、配列番号65および配列番号66から選ばれるアミノ酸配列を有する1以上のVH CDRを含むVHドメインよりなる群から選ばれる抗体VHドメイン、ならびに/または
配列番号53のP12およびD11 VLドメイン、配列番号60のF4S VLドメインならびに配列番号67、配列番号68、配列番号69、配列番号70および配列番号71から選ばれるアミノ酸配列を有する1以上のVL CDRを含むVLドメインよりなる群から選ばれる抗体VLドメイン
を含みうる。
【0021】
後記で詳しく説明するとおり、抗原への結合にはVHドメインのみが用いられることもあるが、一般には、VHドメインはVLドメインとペア形成して抗体抗原結合部位を与える。1つの好ましい実施形態においては、本明細書に記載のVHドメイン(すなわち、配列番号2、12、15、25、29、45、48、52、56または58)を対応VLドメイン(すなわち、配列番号4または配列番号50(4A1-F16および3A1-D5の場合)、配列番号17または配列番号81(A12およびE10の場合))、配列番号31、配列番号83、配列番号35または配列番号47(F4およびG11の場合)、配列番号4または配列番号50、配列番号54(P12およびD11の場合)あるいは配列番号60(F4Sの場合))とペア形成させて、VHおよびVLの両ドメインを含む抗体抗原結合部位(例えば、4A1-F16 VHおよびVLドメイン、3A1-D5 VHおよびVLドメイン、E10 VHおよびVLドメイン、A12 VHおよびVLドメイン、F4 VHおよびVLドメイン、G11 VHおよびVLドメイン、P12 VHおよびVLドメイン、D11 VHおよびVLドメイン、あるいはF4S VHおよびVLを含む部位)を形成させる。他の実施形態においては、VHドメインを、対応VL以外のVLドメイン、好ましくは、テネイシンCの同一ドメインに結合する特異的結合メンバーからのVLドメインとペア形成させることが可能である。軽鎖の乱交性(promiscuity)は当技術分野で十分に確認されている。
【0022】
1以上のCDRを、本明細書に開示されているVHまたはVLドメインから採用し、適当なフレームワーク内に含有させることが可能である。これについては後記で更に詳しく説明する。CDRは一般にはKabatにしたがい定義される。好ましくは、VHドメインおよび/またはVLドメインはCDR1、CDR2およびCDR3を含む。4A1-F16 VH CDR1を配列番号5に示す。3A1-D5 VH CDR1を配列番号13に示す。4A1-F16および3A1-D5 VH CDR 2および3をそれぞれ配列番号6および7に示す。4A1-F16および3A1-D5 VL CDR 1、2および3をそれぞれ配列番号8、9および10に示す。E10 VH CDR 1および2を配列番号18および19に示す。A12 VH CDR 1および2を配列番号26および27に示す。E10およびA12 VH CDR3を配列番号23に示す。E10およびA12 VL CDR 1、2および3をそれぞれ配列番号21、22および23に示す。F4およびG11 VH CDR 1、2および3をそれぞれ配列番号18、19および20に示す。F4 VL CDR 1、2および3をそれぞれ配列番号32、33および23に示す。G11 VL CDR 1、2および3をそれぞれ配列番号32、22および23を示す。P12 VH CDRをそれぞれ配列番号61、63および66に示す。P12 VL CDRをそれぞれ配列番号67、69および71に示す。D11 VH CDRをそれぞれ配列番号62、64および66に示す。D11 VL CDRをそれぞれ配列番号67、69および71に示す。F4S VH CDRをそれぞれ配列番号62、65および66に示す。F4S VL CDRをそれぞれ配列番号68、70および71に示す。
【0023】
いくつかの実施形態においては、特異的結合メンバーは、配列番号7のアミノ酸配列を有するCDR3、配列番号6のアミノ酸配列を有するCDR2および配列番号5またはより好ましくは配列番号13のアミノ酸配列を有するCDR1の1以上を含む抗体VHドメインを含みうる。
【0024】
他の実施形態においては、特異的結合メンバーは、配列番号20のアミノ酸配列を有するCDR3、配列番号27またはより好ましくは配列番号19のアミノ酸配列を有するCDR2および配列番号26またはより好ましくは配列番号18のアミノ酸配列を有するCDR1の1以上を含む抗体VHドメインを含みうる。
【0025】
特異的結合メンバーは、配列番号23のアミノ酸配列を有するCDR3、配列番号22または配列番号33のアミノ酸配列を有するCDR2および配列番号21または配列番号32のアミノ酸配列を有するCDR1の1以上を含む抗体VHドメインを含みうる。
【0026】
好ましくは、特異的結合ドメインは、本明細書中に更に詳しく説明されているscFvである。VHドメインとVLドメインとを、ペプチドリンカー、例えば配列番号37に記載のアミノ酸配列を有するリンカーを介して連結することが可能である。通常、該リンカーは、モチーフの縦列反復の1以上を含むアミノ酸配列を有する。典型的には、該モチーフは5残基配列であり、好ましくは、該残基のうちの少なくとも4個はGlyまたはSerである。該5残基のうちの4個がGlyまたはSerである場合には、その他の残基はAlaでありうる。より好ましくは、該5残基のそれぞれはGlyまたはSerである。好ましいモチーフはGGGGS、SSSSG、GSGSAおよびGGSGG(それぞれ配列番号76、77、78および79)である。好ましくは、該モチーフは該配列内で隣接しており、該反復間に介在ヌクレオチドは存在しない。該リンカー配列は、1〜5個、好ましくは3〜4個の、モチーフの反復を含む又はそれよりなることが可能である。例えば、3個の縦列反復を有するリンカーは以下のアミノ酸配列の1つを有しうる:
GGGGSGGGGSGGGGS - 配列番号39、
SSSSGSSSSGSSSSG - 配列番号41、
GSGSAGSGSAGSGSA - 配列番号42、
GGSGGGGSGGGGSGG - 配列番号43。
【0027】
本明細書に記載の配列を有しテネイシンCに対する特異的結合メンバーにおいて使用されうるVHおよびVLドメインならびにCDRの変異体は、配列の改変または突然変異およびスクリーニングの方法により得ることが可能である。そのような方法も本明細書により提供される。
【0028】
本明細書に具体的に開示されている配列を有するVHおよびVLドメインのいずれかの可変ドメインアミノ酸配列変異体を、本明細書に記載のとおり、本発明において使用することが可能である。個々の変異体は、約20個未満の改変、約15個未満の改変、約10個未満の改変、約5個未満の改変、4、3、2または1個の改変でありうる、1以上のアミノ酸配列の改変(アミノ酸残基の付加、欠失、置換および/または挿入)を含みうる。改変は1以上のフレームワーク領域および/または1以上のCDRにおいて施されうる。特に、改変はVH CDR1、VH CDR2および/またはVH CDR3、特にVH CDR3において施されうる。
【0029】
本発明の特異的結合メンバーは、特異的結合メンバー、本明細書に開示されているVHおよび/またはVLドメインあるいは本明細書に開示されているVH CDRあるいはこれらの任意の変異体を含みテネイシンC大アイソフォーム(特にそのA1またはCドメイン)に結合する任意の特異的結合メンバーと、抗原に対する結合に関して競合するものでありうる。結合メンバー間の競合はin vitroで容易にアッセイされうる。例えば、これはELISAを使用して行うことが可能であり、および/または他の非タグ付け結合メンバーの存在下で検出されうる1つの結合メンバーに特異的レポーター分子をタグ付けして、同一エピトープまたは重複エピトープに結合する特異的結合メンバーの特定を可能にすることにより行うことが可能である。
【0030】
したがって、本発明のもう1つの態様は、テネイシンCへの結合に関して4A1-F16、3A1-D5、E10、A12、F4、G11、P12、D11およびF4Sの1以上と競合するヒト抗体抗原結合部位を含む特異的結合メンバーを提供する。
【0031】
4A1-F16、3A1-D5、E10、A12、F4 G11、P12、D11またはF4Sと競合しうるテネイシンC大アイソフォームに対する抗体を得るためには、種々の方法が当技術分野において利用可能である。好ましくは、そのような抗体は、テネイシンC小アイソフォームと比べてテネイシンC大アイソフォームに優先的に結合する。
【0032】
もう1つの態様においては、本発明は、本発明の特異的結合メンバーのライブラリーと抗原とを接触させ、該抗原に結合しうる該ライブラリーの特異的結合メンバーの1以上を選択しることを含む、抗原に結合しうる1以上の特異的結合メンバーの入手方法を提供する。
【0033】
該ライブラリーはバクテリオファージ粒子の表面上に提示されることが可能であり、各粒子は、その表面上に提示される抗体VH可変ドメインをコードする核酸、および場合によっては、存在する場合には提示VLドメインをコードする核酸を含有する。
【0034】
抗原への結合能を有しバクテリオファージ粒子上に提示される特異的結合メンバーの選択の後、選択した特異的結合メンバーを提示するバクテリオファージ粒子から核酸を取り出すことが可能である。そのような核酸は、選択した特異的結合メンバーを提示するバクテリオファージ粒子から取り出した核酸の配列を有する核酸からの発現による、特異的結合メンバーまたは抗体VH可変ドメイン(場合によっては抗体VL可変ドメイン)の後続の産生において使用することが可能である。
【0035】
選択した特異的結合メンバ−の抗体VH可変ドメインのアミノ酸配列を有する抗体VH可変ドメインは、そのようなVHドメインを含む特異的結合メンバーの場合と同様に、単離された形態で提供されうる。
【0036】
テネイシンCへの結合能、また、4A1-F16、3A1-D5、E10、A12、F4、G11、P12、D11またはF4Sと競合する能力を、更に試験することが可能である。
【0037】
本発明の特異的結合メンバーは、4A1-F16、3A1-D5、E10、A12、F4、G11、P12、D11またはF4Sの1以上のアフィニティで、あるいはより大きな又はより小さなアフィニティで、テネイシンCに結合しうる。
【0038】
異なる特異的結合メンバーの結合アフィニティを、適当な条件下で比較することが可能である。
【0039】
抗体配列に加えて、本発明の特異的結合メンバーは、例えば折り畳まれた(folded)ドメインのようなペプチドまたはポリペプチドを形成する他のアミノ酸、あるいは抗原への結合能のほかに別の機能特性を該分子に付与する他のアミノ酸を含みうる。
【0040】
本発明の特異的結合メンバーは、検出可能な標識、例えば腫瘍検出を容易にする物質、例えば放射性核種または発蛍光団を含有することが可能であり、あるいは、治療方法における使用のために、殺生物事象を誘発しうる物質、例えば放射性核種、光増感剤、薬物、サイトカイン、凝血促進因子、毒素または酵素(例えば、ペプチド結合またはリンカーを介して)に結合されうる。
【0041】
他の態様においては、本発明は、本発明の特異的結合メンバーVHまたはVLドメインをコードする配列を含む単離された核酸を提供し、また、特異的結合メンバーVHドメインおよび/またはVLドメインの産生を引き起こす条件下で該核酸を発現させ、それを回収することを含む、本発明の特異的結合メンバーVHドメインおよび/またはVLドメインの製造方法を提供する。
【0042】
本明細書に記載の特異的結合メンバーは、ヒトまたは動物の身体の治療または診断方法、例えば、ヒト患者における疾患または障害、特に増殖性障害、例えば癌の(予防的治療を含みうる)治療方法において使用されることが可能であり、該方法は、特異的結合メンバーの有効量を該患者に投与することを含む。
【0043】
本発明のもう1つの態様は、本明細書に開示されている抗体VH可変ドメインおよび/またはVL可変ドメインをコードする核酸、一般には単離された核酸を提供する。
【0044】
本発明のもう1つの態様は、本明細書に開示されているVH CDRまたはVL CDR配列、特に、配列番号5、6、7、13、18、19、20、26および27から選ばれるVH CDRまたは配列番号8、9、10、21、22、23、32および33から選ばれるVL CDRをコードする核酸、一般には単離された核酸を提供する。
【0045】
もう1つの態様は、本発明の核酸で形質転換された宿主細胞を提供する。
【0046】
さらにもう1つの態様は、コード核酸からの発現を引き起こさせることを含む、抗体VH可変ドメインの製造方法を提供する。そのような方法は、該抗体VH可変ドメインの産生のための条件下で宿主細胞を培養することを含みうる。
【0047】
本発明の他の態様として、VL可変ドメインならびにVHおよび/またはVLドメインを含む特異的結合メンバーの製造のための類似方法を提供する。
【0048】
製造方法は、該産物の単離および/または精製の工程を含みうる。
【0049】
製造方法は、少なくとも1つの追加的成分、例えば製薬上許容される賦形剤を含む組成物中に該産物を製剤化することを含みうる。
【0050】
本発明のこれらの及び他の態様を後記で更に詳しく説明する。
【図面の簡単な説明】
【0051】
【図1】図1は、小(A)および大(B)テネイシンCアイソフォームの概要図である。いくつかのフィブロネクチンIII型様ドメインは選択的スプライシングに付されて、該分子内に含まれるか(B)または含まれない(A)。
【図2】図2は、U87腫瘍異種移植片担持ヌードマウスにおける4A1-F16-SIP抗体の生体分布を示す。(A)値は、注射の3時間後(上)および6時間後(下)の、3匹のマウスの群内の組織1g当たりの注射用量の平均%を表す。誤差棒線は標準偏差を表す。
【図3】図3は、U87腫瘍異種移植片担持ヌードマウスにおける4A1-F16-SIP抗体の生体分布を示す。(A)値は、注射の24時間後(上)および48時間後(下)の、3匹のマウスの群内の組織1g当たりの注射用量の平均%を表す。誤差棒線は標準偏差を表す。
【図4】図4は、図2および3に示した生体分布データの別の表示を示す。
【図5】図5はSIP(P12)の生体分布を示す。3.5μgのSIP(P12)を、皮下U87ヒト神経膠芽細胞腫を担持するU87腫瘍異種移植片担持ヌードマウスにおける4A1-F16-SIP抗体の生体分布を示す。各時点は4匹の動物により表されている。
【発明を実施するための形態】
【0052】
用語
特異的結合メンバー
これは、お互いに対する結合特異性を有する分子のペアのメンバーを意味する。特異的結合ペアのメンバーは天然由来であることが可能であり、あるいは完全または部分的に合成的に製造されうる。分子のペアの一方のメンバーは、分子のペアの他方のメンバーの個々の空間的および極性組織構造体に特異的に結合し従ってそれに相補的であるその表面上の領域または腔を有する。したがって、該ペアのメンバーは、お互いに特異的に結合する特性を有する。特異的結合ペアのタイプの例としては、抗原-抗体、ビオチン-アビジン、ホルモン-ホルモン受容体、受容体-リガンド、酵素-基質が挙げられる。本出願は抗原-抗体タイプの反応に関する。
【0053】
抗体
これは、天然由来の又は部分的もしくは完全に合成的に製造された免疫グロブリンを意味する。この用語は、抗体結合ドメインである又は抗体結合ドメインに実質的に相同である結合ドメインを有する任意のポリペプチドまたはタンパク質をも包含する。抗体の例としては、免疫グロブリンアイタイプおよびそれらのアイタイプサブクラス;抗原結合ドメインを含むフラグメント、例えばFab、scFv、Fv、dAb、Fd;ならびにダイアボディ(diabody)が挙げられる。モノクローナル抗体および他の抗体を採用することが可能であり、元の抗体の特異性を保有する他の抗体またはキメラ分子を製造するために組換えDNA技術を用いることが可能である。そのような技術は、異なる免疫グロブリンの定常領域、または定常領域 + フレームワーク領域へ、抗体の免疫グロブリン可変領域または相補性決定領域(CDR)をコードするDNAを導入することを含みうる。例えば、EP-A-184187、GB 2188638AまたはEP-A-239400を参照されたい。ハイブリドーマまたは他の抗体産生細胞を、産生される抗体の結合特異性を改変するものであっても改変しないものであってもよい遺伝的突然変異または他の変化に付すことが可能である。
【0054】
抗体は多数の方法で修飾されうるため、「抗体」なる語は、要求される特異性を有する結合ドメインを有する任意の特異的結合メンバーまたは基質を包含すると解釈されるべきである。したがって、この用語は、天然由来または完全もしくは部分的に合成由来である、免疫グロブリン結合ドメインを含む任意のポリペプチドを含む、抗体の抗体フラグメント、誘導体、機能的等価体およびホモログを包含する。したがって、別のポリペプチドに融合した免疫グロブリン結合ドメインまたは等価体を含むキメラ分子が包含される。キメラ抗体のクローニングおよび発現はEP-A-0120694およびEP-A-0125023に記載されている。
【0055】
全抗体のフラグメントは、抗原に結合する機能を果たしうることが示されている。結合性フラグメントの例としては、(i)VL、VH、CLおよびCH1ドメインよりなるFabフラグメント;(ii)VHおよびCH1ドメインよりなるFdフラグメント;(iii)単一抗体のVLおよびVHドメインよりなるFvフラグメント;(iv)VHドメインよりなるdAbフラグメント(Ward, E.S.ら, Nature 341, 544-546 (1989));(v)単離CDR領域;(vi)2つの連結されたFabフラグメントを含む二価フラグメントであるF(ab')2フラグメント;(vii)VHドメインとVLドメインとが一緒になって抗原結合部位を形成することを可能にするペプチドリンカーにより該VHドメインおよび該VLドメインが連結されている一本鎖Fv分子(scFv)(Birdら, Science, 242, 423-426, 1988; Hustonら, PNAS USA, 85, 5879-5883, 1988);(viii)二重特異性一本鎖Fv二量体(PCT/US92/09965);ならびに(ix)遺伝子融合により構築された多価または多重特異性フラグメントである「ダイアボディ(diabody)」(WO94/13804; P. Holligerら, Proc. Natl. Acad. Sci. USA 90 6444-6448, 1993)が挙げられる。Fv、scFvまたはダイアボディ分子は、VHドメインとVLドメインとを連結するジスルフィド架橋の含有により安定化されうる(Y. Reiterら, Nature Biotech 14 1239-1245 1996)。CH3ドメインに連結されたscFvを含むミニボディ(minibody)も製造されうる(S. Huら, Cancer Res. 56 3055-3061 1996)。
【0056】
ダイアボディはポリペプチドの多量体であり、各ポリペプチドは、免疫グロブリン軽鎖の結合領域を含む第1ドメインと免疫グロブリン重鎖の結合領域を含む第2ドメインとを含み、それらの2つのドメインは(例えばペプチドリンカーにより)連結されてはいるが、互いに一緒になって抗原結合部位を形成することはなく、抗原結合部位は、該多量体内の1つのポリペプチドの第1ドメインと該多量体内の別のポリペプチドの第2ドメインとの会合により形成される(WO94/13804)。
【0057】
二重特異性抗体を使用する場合には、これらは通常の二重特異性抗体であることが可能であり、それらは、種々の方法により製造することが可能であり(Holliger, P.およびWinter G. Current Opinion Biotechnol. 4, 446-449 (1993))、例えば、化学的に又はハイブリッドハイブリドーマから製造することが可能であり、あるいは前記の任意の二重特異性抗体フラグメントでありうる。ダイアボディおよびscFvは、可変ドメインのみを使用して、Fc領域を伴わずに構築することが可能であり、これは、抗イディオタイプ反応の効果を潜在的に軽減する。
【0058】
二重特異性全抗体と比べて、二重特異性ダイアボディは特に有用でありうる。なぜなら、それは大腸菌(E. coli)内で容易に構築され発現されうるからである。適当な結合特異性のダイアボディ(および例えば抗体フラグメントのような多数の他のポリペプチド)は、ファージ提示(ファージディスプレイ)(WO94/13804)を用いて、ライブラリーから容易に選択されうる。ダイアボディの一方のアームを、例えば抗原Xに対する特異性を有する定常なものにしたい場合には、他方のアームが可変性となり適当な特異性の抗体が選択されるライブラリーを作製することが可能である。二重特異性全抗体は、「ノブ・イントゥ・ホール(knobs-into-holes)」操作(J. B. B. Ridgewayら, Protein Eng. 9 616-621, 1996)により作製することが可能である。
【0059】
抗原結合ドメイン
これは、抗原の一部または全体に特異的に結合しそれに相補的である領域を含む、抗体の部分を意味する。抗原が大きい場合には、抗体は該抗原の特定の部分に結合しうるに過ぎず、その部分はエピトープと称される。抗原結合ドメインは1以上の抗体可変ドメイン(例えば、VHドメインよりなる、いわゆるFd抗体フラグメント)により提供されうる。好ましくは、抗原結合ドメインは抗体軽鎖可変領域(VL)および抗体重鎖可変領域(VH)を含む。
【0060】
特異的
これは、特異的結合ペアの一方のメンバーがその特異的結合パートナー以外の分子へのいずれの有意な結合をも示さない状況を示すために用いられうる。例えば、テネイシンCに特異的な抗体は、例えばフィブロネクチンのような細胞外マトリックスの他の成分への結合をほとんど又は全く示さないであろう。同様に、テネイシンC大アイソフォームに特異的な抗体はテネイシンC小アイソフォームへの結合をほとんど又は全く示さないであろう。この用語は、例えば、抗原結合ドメインが、多数の抗原により担持される特定のエピトープに特異的である場合にも適用可能であり、この場合、該抗原結合ドメインを担持する特異的結合メンバーは、該エピトープを担持する種々の抗原に結合しうるであろう。
【0061】
含む(含んでなる)
これは、一般には、包含するという意味で用いられ、すなわち、1以上の特徴または成分の存在を許容するものである。
【0062】
単離(された)
これは、本発明の特異的結合メンバーまたはそのような結合メンバーをコードする核酸が本発明に合致している状態を意味する。メンバーおよび核酸は、それらが天然で会合している物質(例えば、それらの天然環境中でそれらと一緒に存在する他のポリペプチドまたは核酸)、またはin vitroもしくはin vivoで実施される組換えDNA技術による製造におけるそれらの製造環境(例えば、細胞培養)中に一緒に存在する物質を含有しない又は実質的に含有しないであろう。メンバーおよび核酸は希釈剤またはアジュバントと共に製剤化されることが可能であり、その場合であっても、実際の目的においては尚も単離されている。例えば、イムノアッセイにおいて使用するためにマイクロタイタープレートをコートするために該メンバーを使用する場合には、該メンバーをゼラチンまたは他の担体と混合し、あるいは該メンバーを診断または療法において使用する場合には、該メンバーを、製薬上許容される担体または希釈剤と混合する。特異的結合メンバーは、天然で、または異種真核細胞の系(例えば、CHOまたはNS0(ECACC 85110503)細胞)によりグリコシル化されることが可能であり、あるいはそれらは(例えば、原核細胞における発現により製造される場合には)非グリコシル化形態でありうる。
【0063】
「記載されているのと実質的に同じ」は、本発明の関連CDRまたはVHもしくはVLドメインが、本明細書に記載の配列を有する特定されている領域と同一または非常に類似していることを意味する。「非常に類似」は、該CDRおよび/またはVHもしくはVLドメインにおいて、1〜5個、好ましくは1〜4個、例えば1〜3個、または1個、または2個、または3個、または4個の置換が施されうることを意味する。
【0064】
本発明のCDRを担持するための構造は、一般には、再構成免疫グロブリン遺伝子によりコードされる天然に存在するVHおよびVL抗体可変ドメインのCDRに対応する位置に該CDRが位置する、抗体重鎖もしくは軽鎖配列またはその本質的部分のものであろう。免疫グロブリン可変ドメインの構造および位置は、Kabat, E.A.ら, Sequences of Proteins of Immunological Interest. 4th Edition. US Department of Health and Human Services. 1987およびその改訂版(現在では、インターネット上(http://immuno.bme.nwu.edu)で入手可能である)を参考にして決定されうる。
【0065】
好ましくは、本明細書に記載されているのと実質的に同じCDRアミノ酸配列は、ヒト可変ドメインまたはその本質的部分におけるCDRとして担持される。
【0066】
本発明において使用する可変ドメインは、任意の生殖系列から入手可能であり、あるいはヒト可変ドメインから再構成されることが可能であり、あるいは公知ヒト可変ドメインのコンセンサス配列に基づく合成可変ドメインでありうる。本発明のCDR配列(例えば、CDR1、CDR2またはCDR3)は、組換えDNA技術を用いて、CDR(例えば、対応するCDR1、CDR2またはCDR3)を欠く可変ドメインのレパートリー内に導入されうる。
【0067】
例えば、Marksら(Bio/Technology, 1992, 10:779-783)は、CDR3を欠くVH可変ドメインのレパートリーを得るために、抗体可変ドメイン領域の5'末端に対する又は該5'末端に隣接したコンセンサスプライマーをヒトVH遺伝子の第3フレームワーク領域に対するコンセンサスプライマーと共に使用する、抗体可変ドメインのレパートリーの製造方法を記載している。Marksらは更に、このレパートリーを特定の抗体のCDR3と如何にして組合せうるかを記載している。類似した技術を用いて、本発明のCDR由来配列を、CDRを欠くVHまたはVLドメインのレパートリーとシャッフルし、そのシャッフルされた完全なVHまたはVLドメインをコグネイトVLまたはVHドメインと組合せて、本発明の特異的結合メンバーを得ることが可能である。ついで該レパートリーを、例えばWO92/01047のファージ提示系のような適当な宿主系において提示させ、適当な特異的結合メンバーを選択することが可能である。レパートリーは、〜104個以上のメンバー、例えば、106〜108または1010メンバーよりなることが可能である。
【0068】
類似したシャッフリングまたはコンビナトリアル(組合せ)技術もStemmer (Nature, 1994, 370:389-391)により開示されており、彼はβ-ラクタマーゼ遺伝子に関する技術を記載しているが、該アプローチが抗体の作製に用いられうることを見出している。
【0069】
もう1つの代替法は、可変ドメイン全体における突然変異を得るために1以上の選択されたVHおよび/またはVL遺伝子のランダム突然変異誘発を用いて、本発明のCDR由来配列を担持する新規VHまたはVL領域を作製することである。そのような技術はGramら(1992, Proc. Natl. Acad. Sci. USA, 89:3576-3580)により記載されており、彼は変異導入型PCRを用いている。
【0070】
用いうるもう1つの方法は、突然変異誘発をVHまたはVL遺伝子のCDR領域へ導くことである。そのような技術はBarbasら(1994, Proc. Natl. Acad. Sci., USA, 91:3809-3813)およびSchierら(1996, J. Mol. Biol. 263:551-567)により開示されている。
【0071】
前記技術のすべては当技術分野において自体公知であり、基本的には本発明の一部を構成するものではない。当業者であれば、当技術分野における通常の方法を用いて本発明の特異的結合メンバーを得るためにそのような技術を用いることが可能であろう。
【0072】
本発明のもう1つの態様は、本明細書に記載のVHドメインのアミノ酸配列内の1以上のアミノ酸の付加、欠失、置換または挿入により、該VHドメインのアミノ酸配列変異体であるVHドメインを得、所望により、このようにして得られたVHドメインを1以上のVLドメインと組合せ、該VHドメインまたはVH/VL組合せ体を試験して、テネイシンCに特異的な特異的結合メンバーまたは抗体抗原結合ドメインを特定することを含む、テネイシンCに特異的な抗体抗原結合ドメインの入手方法を提供する。該VLドメインは、本明細書に記載されているのと実質的に同じアミノ酸配列を有しうる。
【0073】
いくつかの実施形態においては、前記の1以上のアミノ酸はVHドメインのCDR(例えば、CDR1、CDR2および/またはCDR3)の1以上において付加、欠失、置換または挿入されうる。
【0074】
本明細書に記載のVLドメインのアミノ酸配列内の1以上のアミノ酸の付加、欠失、置換または挿入により、該VLドメインのアミノ酸配列変異体であるVLドメインを得、このようにして得られたVLドメインを1以上のVHドメインと組合せ、該VH/VL組合せ体を試験して、テネイシンCに特異的な特異的結合メンバーまたは抗体抗原結合ドメインを特定することを含む、類似方法を用いることが可能である。該VHドメインは、本明細書に記載されているのと実質的に同じアミノ酸配列を有することが可能であり、あるいは前記のとおりに得られた、本明細書に記載されているのと実質的に同じVHドメインのアミノ酸配列変異体でありうる。
【0075】
いくつかの実施形態においては、前記の1以上のアミノ酸はVLドメインのCDRの1以上において付加、欠失、置換または挿入されうる。
【0076】
本発明のもう1つの態様は、
(a)置換すべきCDRを含むか又はCDRコード領域を欠くVHドメインをコードする核酸の出発レパートリーを準備し、
(b)該レパートリーを、VH CDRに関して本明細書に記載されているのと実質的に同じアミノ酸配列をコードする供与核酸と組合せて、該供与核酸を該レパートリー内のCDR領域内に挿入して、VHドメインをコードする核酸の産物レパートリーを得、
(c)該産物レパートリーの核酸を発現させ、
(d)テネイシンC抗原に特異的な特異的結合メンバーを選択し、
(e)該特異的結合メンバーまたはそれをコードする核酸を回収することを含む、テネイシンCに特異的な特異的結合メンバーの製造方法を提供する。
【0077】
該CDRはVH CDR1、CDR2またはCDR3でありうる。
【0078】
ここでもまた、本発明のVL CDRを、置換すべきCDRを含むか又はCDRコード領域を欠くVLドメインをコードする核酸のレパートリーと組合せる、類似方法を用いることが可能である。
【0079】
該CDRはVL CDR1、CDR2またはCDR3でありうる。
【0080】
同様に、1以上または全3個のCDRをVHまたはVLドメインのレパートリー内にグラフティングし、ついでそれを、テネイシンCに特異的な、特にテネイシンC大アイソフォームに特異的な特異的結合メンバーに関してスクリーニングすることが可能である。
【0081】
免疫グロブリン可変ドメインの本質的部分は、少なくともそれらの3つのCDR領域を、それらの介在フレームワーク領域と共に含む。好ましくは、該部分はまた、第1および第4フレームワーク領域の一方または両方の少なくとも約50%を含み、該50%は第1フレームワーク領域のC末端半分および第4フレームワークのN末端半分である。該可変ドメインの本質的部分のN末端またはC末端の追加的残基は、天然に存在する可変ドメイン領域と通常は一緒になっていないものでありうる。例えば、組換えDNA技術により得られる本発明の特異的結合メンバーの構築は、クローニングまたは他の操作工程を促進するために導入されるリンカーによりコードされるNBまたはC末端残基の導入をもたらしうる。他の操作工程は、本発明の可変ドメインを、免疫グロブリン重鎖、他の可変ドメイン(例えば、ダイアボディの製造の場合)またはタンパク質標識(後記で更に詳しく説明される)を含む他のタンパク質配列へ連結するためのリンカーの導入を含む。
【0082】
本発明の好ましい態様においては、VHおよびVLドメインのペアを含む特異的結合メンバーが好ましいが、VHまたはVLのいずれかのドメイン配列に基づく単一結合ドメインも本発明の他の態様を構成する。単一免疫グロブリンドメイン、特にVHドメインは標的抗原に特異的に結合しうることが公知である。
【0083】
単一鎖特異的結合ドメインのいずれかの場合には、これらのドメインを使用して、テネイシンCに結合しうる2ドメイン特異的結合メンバーを形成しうる相補性ドメインに関してスクリーニングすることが可能である。
【0084】
これは、WO92/01047に開示されているいわゆる階層的二重組合せアプローチを用いるファージ提示スクリーニング法により達成することが可能であり、この場合、HまたはL鎖クローンのいずれかを含有する個々のコロニーを用いて、もう一方の鎖(LまたはH)をコードするクローンの完全ライブラリーに感染させ、得られた2本鎖特異的結合メンバーを、例えばその参考文献に記載されているファージ提示技術により選択する。この技術はMarksら(同誌)にも開示されている。
【0085】
本発明の特異的結合メンバーは更に、抗体定常領域またはその一部を含みうる。例えば、VLドメインをそのC末端において、ヒトCκまたはCλ鎖(好ましくはCλ鎖)を含む抗体軽鎖定常ドメインに結合させることが可能である。同様に、VHドメインに基づく特異的結合メンバーをそのC末端において、任意の抗体アイタイプ(例えば、IgG、IgA、IgEおよびIgM)および任意のアイタイプサブクラス(特にIgG1およびIgG4)に由来する免疫グロブリン重鎖の全体または一部に結合させることが可能である。
【0086】
本発明の特異的結合メンバーを、検出可能または機能的な標識で標識することが可能である。
【0087】
検出可能な標識には、放射性核種、例えばヨウ素131、イットリウム90、インジウム111およびテクネチウム99が含まれる。それらは、抗体イメージングの技術分野において公知の通常の化学を用いて、本発明の抗体に結合させることが可能である。放射性同位体で標識された特異的結合メンバーを使用して、放射線を特異的標的(例えば、腫瘍)へ選択的に運搬することが可能である。これは、後記のとおり、腫瘍をイメージングするのに又は細胞毒性量の放射線を運搬するのに有用でありうる。
【0088】
他の検出可能な標識には、ホースラディッシュペルオキシダーゼのような酵素標識、特異的コグネイト検出可能部分(例えば、標識アビジン)への結合を介して検出されうるビオチンのような化学的部分、フルオレセイン、ローダミン、フィコエリトリンおよびテキサス・レッドのような蛍光色素、ならびにCy7(Amersham Pharmacia)およびAlexa750(Molecular probes)のようなシアニン色素誘導体を含む近赤外発蛍光団が含まれうる。
【0089】
他の実施形態においては、検出可能な標識には、超音波により検出可能なマイクロバブル(microbubble)誘導体(Joseph Sら, Pharm Res. 2004 Jun;21(6):920-6)または磁気粒子(Schellenberger EAら, Bioconjug. Chem. 2004 Sep-Oct; 15(5): 1062-7)が含まれうる。
【0090】
機能的標識には、殺生物事象を誘発しうる又は抗癌作用を有する物質が含まれうる。適当な標識には、放射性核種、光増感剤、毒素ポリペプチド、毒性小分子および他の薬物、サイトカイン(例えば、IL2、IL12、TNF)、ケモカイン、凝血促進因子(例えば、組織因子)、酵素、リポソーム、および免疫応答因子(例えば、D. Neri (2004) CHIMIA “Tumor Targeting” vol. 58, p. 723-726を参照されたい)が含まれる。
【0091】
放射性核種には、ヨウ素131、イットリウム90、インジウム111およびテクネチウム99が含まれ、前記で更に詳しく説明されている。
【0092】
毒素ポリペプチドまたはペプチドは細胞毒性またはアポトーシス活性を有し、微生物、植物、動物またはヒト由来でありうる。いくつかの実施形態においては、毒素ポリペプチドは特異的結合メンバーの定常領域内に直接的に挿入されうる。毒素ポリペプチドの例には、Pseudomonas外毒素、リシンα鎖およびアンギオゲニンが含まれる。
【0093】
毒性小分子には、例えばDNA複合体形成物質または細胞周期インヒビターを含む、細胞毒性活性を有する化合物が含まれる。いくつかの実施形態においては、毒性分子は、pH感受性または酵素感受性リンカー(例えば、イミン結合を含有するリンカー)の切断により、標的細胞の近傍で遊離されうる。毒性小分子の例には、メイタンシン(maytansine)、カリケアミシン(calicheamicin)、エポチロン(epothilone)およびチューブリシン(tubulysin)ならびにそれらの誘導体が含まれる。
【0094】
免疫応答因子には、免疫エフェクター細胞に結合する特異的結合メンバーが含まれうる。特異的結合メンバーの結合は、標的細胞に対する細胞性免疫応答を惹起しうる。
【0095】
好ましい実施形態においては、本発明の特異的結合メンバーはサイトカインに結合している。該特異的結合メンバーまたはそのポリペプチド成分(例えば、抗体または多鎖抗体フラグメント(例えば、Fab)の重鎖または軽鎖)とサイトカインとを含む融合タンパク質を製造することが可能である。したがって、例えば、本発明の特異的結合メンバーのVHドメインまたはVLドメインをサイトカインに融合させることが可能である。典型的には、該特異的結合メンバーまたはその成分とサイトカインとはペプチドリンカー(例えば、約5〜25残基、例えば10〜20残基、好ましくは約15残基のペプチド)を介して連結される。ペプチドリンカーの適当な例は本明細書に記載されている。好ましくは、サイトカインはIL2、より好ましくはヒトIL2である。サイトカインは特異的結合メンバーまたはそのポリペプチド成分の上流(N末端)または下流(C末端)に融合されうる。好ましい実施形態は、本発明の特異的結合メンバー(特に抗体分子、例えばscFv分子)とIL2とを含む融合タンパク質である。そのような融合タンパク質のアミノ酸配列、およびそれらをコードするヌクレオチド配列を含む核酸は、本発明の一部を構成する。
【0096】
本発明の特異的結合メンバーは、例えば腫瘍イメージングのような診断方法、または例えば癌病態に対するヒトもしくは動物対象における治療において有用でありうる。
【0097】
したがって、本発明の他の態様は、提供される特異的結合メンバーを投与することを含む治療方法、そのような特異的結合メンバーを含む医薬組成物、および投与のための医薬の製造における、例えば、該特異的結合メンバーを製薬上許容される賦形剤と共に製剤化することを含む医薬または医薬組成物の製造方法における、そのような特異的結合メンバーの使用を提供する。
【0098】
治療方法において使用する特異的結合メンバーは、好ましくは、抗腫瘍作用を惹起する機能的標識に結合または連結される。好ましい実施形態においては、前記のとおり、特異的結合メンバーは、サイトカイン、例えばIL2に結合または連結される。
【0099】
治療上の利益を得るために本明細書に記載の特異的結合メンバーが使用されうる臨床適応症には、増殖性障害、例えば前癌性および悪性の新生物および腫瘍(例えば、組織細胞腫、神経膠腫、星状細胞腫、骨腫瘍)、癌(例えば、肺癌、小細胞肺癌、胃腸癌、大腸癌、結腸癌、乳癌、卵巣癌、前立腺癌、精巣癌、肝癌、腎癌、膀胱癌、膵癌、脳癌、肉腫、骨肉種、カポジ肉腫、黒色腫)、白血病および脈管形成性疾患が含まれる。
【0100】
前癌性または悪性状態は、限定的なものではないが肺、結腸、乳房、卵巣、前立腺、肝臓、膵臓、脳および皮膚を含む任意の細胞型において生じうる。
【0101】
本明細書に記載の治療に適した増殖性障害は、テネイシンC大アイソフォーム(特に、A1またはCドメインを含む大アイソフォーム)の発現が増強または上昇している細胞または組織の存在により特徴づけられうる。
【0102】
本発明においては、提供される組成物は個体に投与されうる。投与は、好ましくは、「治療的有効量」で行われ、これは、患者に対する利益を示すのに十分な量である。そのような利益は少なくとも1つの症状の少なくとも改善でありうる。実際の投与量ならびに投与の率および時間経過は、治療されるものの性質および重症度に左右されるであろう。治療の処方(例えば、投与量などの決定)は一般開業医および他の医師の責任でなされる。抗体の適当な用量は当技術分野でよく知られている。Ledermann J.A.ら, (1991) Int J. Cancer 47: 659-664; Bagshawe K.D.ら, (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4: 915-922を参照されたい。
【0103】
厳密な用量は、該抗体が診断用なのか治療用なのか、治療すべき領域のサイズおよび位置、該抗体の厳密な性質(例えば、全抗体、フラグメントまたはダイアボディ)ならびに任意の検出可能な標識または該抗体に結合している他の分子の性質を含む多数の要因に左右されるであろう。典型的な抗体の用量は全身用途には0.5mg〜100g、および局所用途には10μg〜1mgの範囲内となろう。典型的には、該抗体は全抗体、好ましくはIgG1またはIgG4アイタイプであろう。これは成人患者の1回の治療のための用量であり、これは小児および乳児の場合には比例的に調節されることが可能であり、他の抗体フォーマットに関しても、分子量に比例して調節されうる。治療は、医師の判断により、毎日、週2回、毎週または毎月の間隔で反復されうる。
【0104】
本発明の特異的結合メンバーは、通常、該特異的結合メンバーに加えて少なくとも1つの成分を含みうる医薬組成物の形態で投与されるであろう。
【0105】
かくして、本発明の医薬組成物、および本発明に従う使用のための医薬組成物は、有効成分に加えて、製薬上許容される賦形剤、担体、バッファー、安定剤または当業者によく知られた他の物質を含みうる。そのような物質は無毒性であるべきであり、有効成分の効力を損なうべきではない。担体または他の物質の厳密な性質は投与経路に左右され、投与経路は経口または注射、例えば静脈内注射でありうる。
【0106】
経口投与用の医薬組成物は錠剤、カプセル剤、散剤または液体形態でありうる。錠剤は、固体担体、例えばゼラチン、またはアジュバントを含みうる。液体医薬組成物は、一般には、液体担体、例えば水、石油、動物もしくは植物油、鉱油または合成油を含む。生理食塩水、デキストロースもしくは他の糖溶液、またはグリコール、例えばエチレングリコール、プロピレングリコールもしくはポリエチレングリコールを加えることが可能である。
【0107】
静脈内注射、または罹患部位への注射の場合には、有効成分は、発熱物質を含有せず適当なpH、等張性および安定性を有する非経口的に許容される水溶液の形態である。当業者であれば、例えば、食塩注射剤、リンゲル注射剤、乳酸添加リンゲル注射剤のような等張性ビヒクルを使用して、適当な溶液を製造することが十分に可能である。必要に応じて、保存剤、安定剤、バッファー、抗酸化剤および/または他の添加剤を加えることが可能である。
【0108】
組成物は、治療すべき状態に応じて、単独で、または他の治療と組合せて同時もしくは連続的に投与することが可能である。他の治療は、鎮痛薬、例えば非ステロイド抗炎症薬(例えば、アスピリン(aspirin)、パラセタモール(paracetamol)、イブプロフェン(ibuprofen)またはケトプロフェン(ketoprofen))、またはオピエート、例えばモルヒネ、または制吐薬の適当な用量の投与を含みうる。
【0109】
本発明のもう1つの態様は、本明細書に記載の抗体を個体に投与し、該個体における腫瘍細胞への該抗体の結合を検出することを含む、腫瘍細胞を検出および/またはイメージングするための方法を提供する。
【0110】
そのような方法において使用するための好ましい抗体は、検出可能な標識、例えば放射性核種または発蛍光団に結合または連結されうる。
【0111】
本発明の方法は、本発明で提供する特異的結合メンバーの、テネイシンCへの結合を引き起こさせる又は可能にすることを含みうる。記載されているとおり、そのような結合は、例えば、特異的結合メンバーまたは特異的結合メンバーをコードする核酸の投与の後に、in vivoで生じうる。
【0112】
テネイシンCへの特異的結合メンバーの結合の量は測定可能である。いくつかの実施形態においては、個体から得たサンプルへの特異的結合メンバーの結合を測定することが可能である。他の実施形態においては、例えば、個体の体内の腫瘍をイメージングまたは検出して、抗原への特異的結合メンバーの結合をin vivoで測定することが可能である。
【0113】
定量は、診断上関心が持たれうる抗原の量に関連づけられうる。
【0114】
抗体の結合は、任意の適当な手段により測定することが可能である。例えば、抗体をレポーター分子または検出可能な標識に連結または結合させ、サンプル上の該標識またはレポーターの存在、量または局在を測定することが可能である。
【0115】
in vivoでの抗体の結合を、例えば分子イメージングの方法において、放射能検出(例えば、PET、SPECT)、近赤外蛍光イメージング(例えば、拡散光学的断層撮影(diffuse optical tomography)、内視鏡検査)、超音波(例えば、標的化マイクロバブル(targeted microbubble)誘導体を使用)およびMRI(標的化磁気粒子を使用)により測定することが可能である。
【0116】
他の実施形態においては、抗体の結合は、例えばELISA、ウエスタンブロット法、免疫細胞化学法、免疫沈降法またはアフィニティクロマトグラフィーにおいてin vitroで生じうる。
【0117】
したがって、腫瘍細胞を検出および/またはイメージングするための方法は、本明細書に記載の抗体を、個体から得たサンプルと接触させ、該サンプル中の腫瘍細胞への該抗体の結合を検出することを含む。
【0118】
そのようなin vitro法において使用するための好ましい抗体はレポーター分子に結合または連結されうる。レポーター分子は放射性核種、発蛍光団、りん光体またはレーザー色素(スペクトル的に分離した吸収または放射特性を有するもの)でありうる。適当な発蛍光団には、フルオレセイン、ローダミン、フィコエリトリンおよびテキサス・レッドが含まれる。適当な発色色素にはジアミノベンジジンが含まれる。in vivoイメージングには、放射性核種または発蛍光団が好ましい。
【0119】
他のレポーターには、高分子コロイド粒子、または粒子状物質、例えば着色されたラテックスビーズ、検出可能なシグナルの視認、電子的検出またはその他の記録を直接的または間接的にもたらしうる、磁性もしくは常磁性および生物学的もしくは化学的に活性な物質が含まれる。これらの分子は、例えば、発色もしくは変色する又は電気的特性の変化をもたらす反応を触媒する酵素でありうる。それらは分子的に励起されて、エネルギー状態間の電子遷移が、特徴的なスペクトル吸収または放射をもたらしうる。それらには、バイオセンサーと共に使用される化学物質が含まれうる。ビオチン/アビジンまたはビオチン/ストレプトアビジンおよびアルカリホスファターゼ検出系が使用されうる。
【0120】
結合の測定方法は本発明の主題ではなく、当業者は、自分の好み及び全般的知識に基づいて適当な方法を選択することが可能である。
【0121】
本発明は更に、本明細書に記載されているのと実質的に同じアミノ酸を有するCDRを含むVドメインまたは本明細書に記載されているのと実質的に同じアミノ酸配列を有するVドメインを含みテネイシンCに結合する任意の特異的結合メンバー(例えば、本明細書に記載の特異的結合メンバー)とテネイシンCへの結合に関して競合する特異的結合メンバーに拡張される。結合メンバー間の競合はin vitroで容易にアッセイされうる。例えば、これは、他の非タグ付け結合メンバーの存在下で検出されうる1つの結合メンバーに特異的レポーター分子をタグ付けして、同一エピトープまたは重複エピトープに結合する特異的結合メンバーの特定を可能にすることにより行うことが可能である。競合は、例えばELISAのような標準的な技術を用いて測定することが可能である。
【0122】
競合に関する試験では、抗原のペプチド断片、特に、所定のエピトープを含むペプチドを使用することが可能である。エピトープ配列および1以上のアミノ酸(いずれかの末端に位置する)を有するペプチドを使用することが可能である。そのようなペプチドは、特定されている配列より「実質的になる」と称されうる。本発明の特異的結合メンバーは、抗原へのその結合が、与えられた配列を有する又は含むペプチドにより抑制されるものでありうる。これに関する試験では、いずれかの配列および1以上のアミノ酸を有するペプチドを使用することが可能である。適当なペプチドは、テネイシンCのD、Cおよび/またはA1ドメインの配列から得ることが可能である。
【0123】
特異的ペプチドに結合する特異的結合メンバーは、例えば、該ペプチドでのパンニングによりファージ提示ライブラリーから単離されうる。
【0124】
本発明は更に、本発明の特異的結合メンバーをコードする単離された核酸を提供する。核酸にはDNAおよびRNAが含まれる。好ましい態様においては、本発明は、前記のとおりの本発明のCDRまたはVHもしくはVLドメインをコードする核酸を提供する。
【0125】
本発明はまた、前記のとおりの少なくとも1つのポリヌクレオチドを含むプラスミド、ベクター、転写または発現カセットの形態の構築物を提供する。
【0126】
本発明はまた、前記のとおりの1以上の構築物を含む組換え宿主細胞を提供する。提供する任意のCDR、VHもしくはVLドメインまたは特異的結合メンバーをコードする核酸自体が本発明の1つの態様を構成する。また、コード産物をコードする核酸からの発現を含む、該コード産物の製造方法も同様に、本発明の1つの態様を構成する。発現は、適当な条件下、該核酸を含有する組換え宿主細胞を培養することにより簡便に達成されうる。発現による産生の後、VHもしくはVLドメインまたは特異的結合メンバーを、任意の適当な技術を用いて単離および/または精製し、適宜使用することが可能である。
【0127】
本発明の特異的結合メンバー、VHおよび/またはVLドメインならびにコード核酸分子およびベクターは、単離および/または精製された形態で提供されうる。例えば、それらは、それらの天然環境から、実質的に純粋または均一な形態で、単離および/または精製されることが可能であり、あるいは核酸の場合には、それらは、要求される機能を有するポリペプチドをコードする配列以外の核酸または遺伝子源を含有しない又は実質的に含有しない。本発明の核酸はDNAまたはRNAを含むことが可能であり、完全または部分的に合成物でありうる。本明細書に記載のヌクレオチド配列に対する言及は、特定された配列を有するDNA分子を包含し、文脈と矛盾しない限り、Tの代わりにUが用いられている特定された配列を有するRNA分子を包含する。
【0128】
多種多様な宿主細胞におけるポリペプチドのクローニングおよび発現のための系はよく知られている。適当な宿主細胞には、細菌、哺乳類細胞、酵母およびバキュロウイルス系が含まれる。異種ポリペプチドの発現のために当技術分野において利用可能な哺乳類細胞系には、チャイニーズハムスター卵巣細胞、HeLa細胞、乳児ハムスター腎細胞、NSOマウス黒色腫細胞、その他多数が含まれる。一般的な好ましい細菌宿主は大腸菌(E. coli)である。
【0129】
原核細胞、例えば大腸菌(E. coli)における抗体および抗体フラグメントの発現は当技術分野において十分に確立されている。レビューとしては、例えば、Pluckthun, A. Bio/Technology 9: 545-551 (1991)を参照されたい。培養内の真核細胞における発現も、特異的結合メンバーの製造のための選択肢として当業者に利用可能である。最近の概観のためには、例えば、M.E. (1993) Curr. Opinion Biotech. 4: 573-576; Trill J.J.ら, (1995) Curr. Opinion Biotech 6: 553-560を参照されたい。
【0130】
適宜、プロモーター配列、ターミネーター配列、ポリアデニル化配列、エンハンサー配列、マーカー遺伝子および他の配列を含む適当な調節配列を含有する適当なベクターを選択し、または構築することが可能である。ベクターは、適宜、プラスミド、ウイルス、例えば「ファージ」、またはファージミドでありうる。さらなる詳細は、例えば、Molecular Cloning: a Laboratory Manual: 2nd edition, Sambrookら, 1989, Cold Spring Harbor Laboratory Pressを参照されたい。例えば核酸構築物の調製、突然変異誘発、配列決定、細胞内へのDNAの導入および遺伝子発現における核酸の操作ならびにタンパク質の分析のための多数の公知技術およびプロトコールは、Current Protocols in Molecular Biology, Second Edition, Ausubelら編, John Wiley & Sons, 1992に詳細に記載されている。Sambrookら及びAusubelらの開示を参照により本明細書に組み入れることとする。
【0131】
したがって、本発明のもう1つの態様は、本明細書に開示されている核酸を含有する宿主細胞を提供する。さらにもう1つの態様は、そのような核酸を宿主細胞内に導入することを含む方法を提供する。該導入には、任意の利用可能な技術を用いることが可能である。真核細胞の場合には、適当な技術にはリン酸カルシウムトランスフェクション、DEAE-デキストラン、エレクトロポレーション、リポソーム媒介トランスフェクション、およびレトロウイルスまたは他のウイルス(例えば、ワクシニア、あるいは昆虫細胞の場合にはバキュロウイルス)を使用する形質導入が含まれうる。細菌細胞の場合には、適当な技術には塩化カルシウム形質転換、エレクトロポレーション、およびバクテリオファージを使用するトランスフェクションが含まれうる。
【0132】
該導入の後、例えば、該遺伝子の発現のための条件下で宿主細胞を培養することにより、該核酸からの発現を引き起こさせ又は可能にすることが可能である。
【0133】
1つの実施形態においては、本発明の核酸は宿主細胞のゲノム(例えば、染色体)内に組込まれる。組込みは、標準的な技術により、該ゲノムとの組換えを促進する配列の含有により促進されうる。
【0134】
本発明はまた、前記の特異的結合メンバーまたはポリペプチドを発現させるために発現系において前記構築物を使用することを含む方法を提供する。
【0135】
本発明の他の方法は、本明細書に記載の特異的結合メンバーを、検出可能な標識または抗癌物質に結合または連結させることを含む。
【0136】
適当な標識または物質は前記で説明されている。標識および物質は、標準的な化学的手段を用いて、特異的結合メンバーに結合されることが可能である。
【0137】
本発明の態様および実施形態は、以下の実験に関する実施例により例示される。
【0138】
本発明の種々の他の態様および実施形態は、本開示を考慮すれば当業者に明らかであろう。本明細書中に挙げられているすべての文書の全体を参照により本明細書に組み入れることとする。
【0139】
本発明は、前記の特徴の各組合せおよび副組合せ(サブコンビネーション)を含む。
【0140】
以下、本発明の或る態様および実施形態を、実施例により及び図面および後記の表を参考にして例示する。
【実施例1】
【0141】
血清の非存在下にpH 7.5で培養した正常ヒト皮膚繊維芽細胞(NHDF)から単離したmRNAから、ヒトテネイシンCのドメインA1をコードする遺伝子を、PCRオリゴTnC-A1 BamHI ba (cgggatcctccactgaacaagcccctgag)およびTnC-A1 BglII for (ggagatctttcccctgtggaggcctcagc) [16]を使用して、RT-PCRによりクローニングした。ついで該遺伝子をpQE12細菌発現ベクター(Qiagen)内にクローニングし、大腸菌(E. coli)TG-1において発現させた。ニッケルがローディングされたNi-NTAセファロース樹脂(Qiagen)を使用して、Hisタグにより精製を行った。
【0142】
精製されたドメインA1を、選択前に、スルホ-NHS-SS-ビオチン(Pierce)を使用してビオチン化した。Viti Fら, Methods Enzymol 2000; 326:480-505に記載されているとおりに、バイオパンニングを行った。簡潔に説明すると、ビオチン化タンパク質(最終濃度10-7 M)を、予めブロッキングされた600μlのETH-2ライブラリーファージと共に30分間インキュベートした。5.3×107個のストレプトアビジンでコートされた磁気ビーズ(Dynal)を加えることにより、結合ファージを捕捉した。十分な洗浄後、該ビオチンリンカー内のジスルフィド結合を還元することにより、選択されたファージを溶出した。単離されたファージをTG-1内で増幅し、ポリエチレングリコール沈殿により上清から濃縮した。3ラウンドのパンニングの後、144個の単離抗体クローンを、Viti Fら, Methods Enzymol 2000; 326:480-505に記載されているとおりに行うELISAによりスクリーニングした。ストレプトアビジンでコートされたELISAプレート(StreptaWell High Bind, Roche)をビオチン化抗原と共にインキュベートし、scFv抗体フラグメントを発現する誘導されたモノクローナル大腸菌(E. coli)TG-1培養の上清を加え、結合抗体をM2モノクローナル抗体で、ついで抗マウス免疫グロブリンG-HRPコンジュゲートで検出した。最高シグナルを有する抗体クローンを、BIAcore 3000装置を使用するリアルタイム相互作用分析により分析した。3つの最良のクローンをプロテインA-セファロースカラム上で精製し、種々の腫瘍凍結切片上で免疫組織化学により試験した。
【0143】
免疫組織化学において選択的染色パターンを示したクローンの1つであるscFv(3A1-D5)をアフィニティ成熟のために選択した。ファージミドベクターpDN322(Pini Aら, J Biol Chem 1998; 273:21769-21776)において、PCRオリゴヌクレオチドLMB3 long ba(caggaaacagctatgaccatgattac, 抗体遺伝子の5'末端の上流でプライミング)およびDP47CDR1mut for (agcctggcggacccagctcgcmnnmnnmnngctaaaggtgaatccagaggctgc)を使用して、可変重鎖(VH CDR1)の相補性決定領域1内の31-33位にランダム突然変異を挿入することにより、アフィニティ成熟ライブラリーを構築した。抗体遺伝子の3'末端を、プライマーDP47CDR1 ba(gagctgggtccgccaggctcc)およびfdseq long for(gacgttagtaaatgaattttctgtatgagg)を使用して増幅した。それらの2つの断片を、プライマーLMB3 long baおよびfdseq long forを使用するPCRにより合体させた。
【0144】
アフィニティ成熟ライブラリーのバイオパンニングをビオチン化抗原で行った。2ラウンドのパンニング(10-8Mビオチン化抗原を使用する以外は前記のとおり)の後、合計382個の抗体クローンをELISAによりスクリーニングした。ELISAにおいて陽性であった69個のクローンを、個々の抗体クローンのkoff値を評価するためにM2-ELISA(Scheuermann Jら, J Immunol Methods 2003; 276:129-134)により更に特徴づけした。簡潔に説明すると、scFv抗体を含有する上清を、抗Flag M2モノクローナル抗体(SIGMA)でコートされた表面上に加えた。ビオチン化抗原を加え、平衡になるまでインキュベートした後、過剰の非ビオチン化抗原を競合体として加えた。それぞれ0、30、60、90および120分の競合時間の後、ストレプトアビジン-HRPコンジュゲートを使用してビオチン化抗原の残存画分を検出した。
【0145】
種々のELISA陽性クローンをそれらの解離プロファイルに基づきランク付けするために、BIAcoreを使用した。抗原でコートされたセファロースカラム上での精製のために、5個の最良クローンを選択した。精製された抗体をサイズ排除クロマトグラフィーに付し、BIAcore 3000を使用してアフィニティ定数および動力学的パラメーターを決定するために、得られた単量体scFv抗体フラグメント画分を使用した。
【0146】
ヒト免疫グロブリンEのCH4ドメインをコードする遺伝子の5'末端の前にscFv(4A1-F16)配列を遺伝的に融合させることにより、最良の解離定数(KD)を有する抗体4A1-F16を二価ミニボディフォーマット内にクローニングして、4A1-F16-SIPと称される小免疫タンパク質(small immune protein)(SIP)(Borsi Lら, Int J Cancer 2002; 102:75-85)を得た。該CH4ドメインはホモ二量体化を促進して、より高いアビディテイにより該抗体の機能的アフィニティを増加させる。
【0147】
ヒト頭部-および-頚部癌組織上でテネイシンC抗体フラグメントscFv(3A1-D5)の抗A1ドメインを使用して、免疫組織化学的方法を行った。scFv抗体のC末端に付加したペプチド性FLAGタグを使用し、モノクローナル抗FLAG抗体M2(SIGMA)を次いでAPAAP系(DAKO)を使用して、一次抗体を検出した。scFv(3A1-D5)およびscFv(4A1-F16)は、種々の頭部-および-頚部の凍結切片上で腫瘍ストロマおよび新血管構造を強く染色することが観察されたが、それらは健常ドナーの正常口腔粘膜とは反応しなかった。
【0148】
4A1-F16-SIPの腫瘍標的化能を以下のとおりに評価した。
【0149】
Balb/C nu/nuマウスにおいて、マウス1匹当たり3×106個のU87ヒト神経膠芽細胞腫細胞(ATCC)の皮下注射により腫瘍を誘発させた。注射の20〜25日後、腫瘍が300〜1500 mm3のサイズに達した際に、放射能標識抗体を注射した。
【0150】
該抗体の調製は、スーパーデックス(superdex)75上のサイズ排除クロマトグラフィーによるアフィニティ精製4A1-F16-SIP抗体の更なる精製を含むものであった。ホモ二量体形態(75kDa)を表す画分を集め、ついでヨウ素源(iodogen)(Pierce)を使用してヨウ素-125(Amersham)で標識した。
【0151】
約5μgの抗体を腫瘍担持マウスの尾静脈内に静脈内注射した。それぞれ3、6、24および48時間後、3匹のマウスを犠死させ、器官を摘出し、I-125の蓄積をγカウンターで測定した。
【実施例2】
【0152】
ヒトテネイシンCのドメインCをコードする遺伝子をベクターpQE12[Carnemolla Bら, Am J Pathol 1999; 154:1345-1352, Balza Eら, FEBS Lett 1993; 332:39-43]内にクローニングし、大腸菌(E. coli)TG-1内で発現させた。ニッケルがローディングされたNi-NTAセファロース樹脂(Qiagen)を使用して、Hisタグにより精製を行った。
【0153】
精製されたドメインCを、選択前に、スルホ-NHS-SS-ビオチン(Pierce)を使用してビオチン化した。ストレプトアビジンおよびアビジンでコートされたプレート上で別のパンニング法を用いること以外はViti Fら, Methods Enzymol 2000; 326:480-505に記載されているとおりに、バイオパンニングを行った。簡潔に説明すると、ビオチン化タンパク質(最終濃度10-7 M)を、予めブロッキングされた600μlのETH-2ライブラリーファージと共に30分間インキュベートした。アビジン(第1および第3ラウンドのパンニング)またはストレプトアビジン(第2ラウンドのパンニング)でコートされたプラスチックマイクロタイタープレート上で、結合ファージを捕捉した。十分な洗浄後、該ビオチンリンカー内のジスルフィド結合を還元することにより、選択されたファージを溶出した。単離されたファージをTG-1内で増幅し、ポリエチレングリコール沈殿により上清から濃縮した。3ラウンドのパンニングの後、数十個の抗体クローンを、Viti Fら, Methods Enzymol 2000; 326:480-505に記載されているとおりに行うELISAによりスクリーニングした。
【0154】
ストレプトアビジンでコートされたELISAプレート(StreptaWell High Bind, Roche)をビオチン化抗原と共にインキュベートし、scFv抗体フラグメントを発現する誘導されたモノクローナル大腸菌(E. coli)TG-1培養の上清を加え、結合抗体を抗ペプチドタグ抗体で、ついで抗マウス免疫グロブリンG-HRPコンジュゲートで検出した。最高シグナルを有する抗体クローンを、BIAcore 3000装置を使用するリアルタイム相互作用分析により分析した。3つの最良のクローンをプロテインA-セファロースカラム上で精製し、種々の腫瘍凍結切片上で免疫組織化学により試験した。免疫組織化学において選択的染色パターンを示したクローンの1つであるscFv(A12)をアフィニティ成熟のために選択した。ファージミドベクターpHEN1[Hoogenboom HRら, Nucleic Acids Res 1991; 19:4133-4137]において、PCRオリゴヌクレオチドLMB3 long ba(caggaaacagctatgaccatgattac, 抗体遺伝子の5'末端の上流でプライミング)、DP47CDR1 for(ctggagcctggcggacccagctcatmnnmnnmnngctaaaggtgaatccagaggctgc)、DP47CDR1 ba(tgggtccgccaggctccag)、DP47CDR2 for(gcccttcacggagtctgcgtagtatgtmnnaccaccmnnmnnmnnaatagctgagacccactcc)、DP47CDR2 ba(acatactacgcagactccgtgaagggc)およびfdseq long for(gacgttagtaaatgaattttctgtatgagg)を使用して、VH CDR1内の31-33位およびVH CDR2内の52、52a、53および56位にランダム突然変異を挿入することにより、アフィニティ成熟ライブラリーを構築した。
【0155】
アフィニティ成熟ライブラリーのバイオパンニングをビオチン化抗原で行った。2ラウンドのパンニング(前記のとおり)の後、合計382個の抗体クローンをELISAによりスクリーニングした。ELISAにおいて陽性であったクローンをBIAcoreにより更に特徴づけして、種々のELISA陽性クローンをそれらの解離プロファイルに基づきランク付けした。試験したクローンのうち、scFv(E10)が最も有望な性能を示した。
【0156】
ヒト神経膠芽細胞腫多形態組織上でテネイシンC抗体フラグメントscFv(A12)の抗Cドメインを使用して、免疫組織化学的方法を行った。scFv抗体のC末端に付加したペプチド性mycタグを使用し、モノクローナル抗mycタグ抗体E10を次いでAPAAP系(DAKO)を使用して、一次抗体を検出した。これらの抗体は、神経膠芽細胞腫凍結切片上の免疫組織化学により示されるとおり、血管周囲構造体と特異的に反応することが観察された。
【実施例3】
【0157】
実施例2に記載されているものに類似したプロトコールを用いて、テネイシンCのCドメインに対する他のscFvを単離し、それぞれF4およびG11と命名した。F4およびG11は、2アミノ酸だけ異なるVLドメインを有し、同一VHを有する。VHおよびVLドメインを、それぞれ配列番号36または配列番号38によりコードされる配列番号37または配列番号39に記載のアミノ酸配列を有するペプチドリンカーを介して連結した。
【実施例4】
【0158】
テネイシンCのヒトドメインD(TnC-D)をコードする遺伝子を、ヒト黒色腫細胞系SKMel-28から単離された全RNAから、PCRオリゴTnC-D BamHI ba(cgggatccgttacagaagccgaaccggaa - 配列番号72)およびTnC-D BglII for(cgggatccgttacagaagccgaaccggaa - 配列番号73)[16]を使用するRT-PCRによりクローニングした。増幅された断片を、該タンパク質のC末端にHisタグを導入する細菌発現ベクターpQE12内にサブクローニングした。TnC-A1のマウスアイソフォームをコードする遺伝子を、浸潤性管癌組織由来の発現配列タグ(EST)クローンから、プライマーmmTnC-A1 EcoBa(agaattcattaaagaggagaaattaactatgagaggatcctccacggaagaagtgccttc - 配列番号74)およびmmTnC-A1 BglFo(tgagatcttgtccctgtggaggtctcggc - 配列番号75)を使用するPCRにより単離し、ベクターpQE12(Qiagen)内にクローニングした。
【0159】
ETHライブラリー[26]を100nM ビオチン化ヒトTnC-Dでパンニングし、結合ファージを、ストレプトアビジンでコートされた磁気ビーズで捕捉した。3ラウンドのパンニングの後、188個のクローンから得た上清をビオチン化ヒトTnC-D上およびビオチン化マウスTnC-D上のELISAによりスクリーニングした。該ヒトアイソフォームを認識したクローンのほとんどはTnC-Dのマウスアイソフォームとも反応した。
【0160】
ELISA陽性クローンの上清を第2 ELISAにおいて試験し、BIAcoreによりスクリーニングした。6個の最良のクローンを詳細に特徴づけし、scFv(F4)が最良クローンであることが確認された。scFv(F4)は、ヒトおよびマウスの両方のTnC-Dに同一アフィニティで結合した。
【0161】
scFv(F4)は専ら、ELISAにおいて、ドメインDを含有するタンパク質を認識し、ドメインDに構造的に関連しているタンパク質とは反応しなかった。しかし、それは免疫組織化学法において腫瘍構造体と非常に弱く反応したに過ぎず、バックグラウンドから識別するのが困難なシグナルを与えたに過ぎなかった。これはおそらく、scFv(F4)の低いアフィニティによるものであろう。
【0162】
scFv(F4)を、CDRループ突然変異誘発[26]によるアフィニティ成熟のための鋳型として使用した。ランダム化のために選択した残基はVL CDR1の30、31、32位ならびにVH CDR2の50、52および53位(Kabatに従う番号付け;[30])であった。該ライブラリーを、VLのC末端にmycタグを含有するファージミドベクターpHEN[31]内にクローニングした。
【0163】
アフィニティ成熟ライブラリーファージでの第1ラウンドのパンニングをビオチン化ヒトTnC-Dで行い、第2ラウンドをビオチン化マウスTnC-Dで行った。750個のクローンをビオチン化ヒトTnC-D上のELISAによりスクリーニングしたところ、約5%が陽性であった。BIAcoreおよびELISAによる該抗体の更なる特徴づけの後、scFv(D11)が最良クローンとして特定された。BIAcoreによる単量体画分でのアフィニティ測定は、scFv(D11)が、親クローンscFv(F4)に比べて2〜4倍改善されたアフィニティを有することを示した。該マウスアイソフォームは、該ヒトアイソフォームより若干高いアフィニティで結合した。
【0164】
scFv(D11)は、種々の腫瘍組織上の免疫組織化学法において良好な結果を与えた。ヒト口腔扁平上皮細胞癌、ヒト膀胱癌、ヌードマウスにおいて増殖させたA375ヒト黒色腫、およびSvEV 129マウスにおいて増殖させたF9マウスF9奇形癌はすべて、scFv(D11)で染色された。該抗体は、頭部および頚部癌または膀胱癌のようなヒト腫瘍のストロマと、ならびに種々のヒトおよびマウス腫瘍モデルにおいて、強力に反応した。
【0165】
つぎに、scFv(D11)のVHのCDR1およびCDR1内の残基をランダム化した。ランダム化のために、VH CDR1内の残基31、32、33ならびにVH CDR2内の残基52、52a、53および56(Kabatの番号付け; [32])を選択した。抗原として10-8 MヒトTnC-Dを使用して、ファージ提示による2ラウンドの選択を行った。選択のストリンジェンシーを増加させるために、第1ラウンドにおける抗原の添加の前に10nM 精製可溶性scFv(D11)を該ファージと混合し、第2ラウンドのパンニングにおけるプレインキュベートされた抗原-ファージ混合物に1000倍過剰の非ビオチン化抗原を加えた。
【0166】
2ラウンド後にスクリーニングされた384個のクローンのうちの25%がELISAにおいてヒトビオチン化TnC-D上で陽性であった。更なる特徴づけの後、最良クローンとして特定されたのはscFv(P12)であった。ヒトTnC-D上のそのアフィニティは、scFv(D11)と比べて4.8倍改善された。
【0167】
scFv(P12)をSIPフォーマット[25]内にクローニングした。SIP(P12)のin vivo標的化能をU87神経膠芽細胞腫異種移植片担持ヌードマウスにおけるヒト生体分布実験により評価した。125I-標識SIP抗体を静脈内注射し、3、6、24および48時間後、動物を犠死させ、器官を摘出し、秤量し、放射能を計数した。注射の24時間以内にSIP(P12)はほとんどの器官から消失し、24および48時間後に、15までの腫瘍:血管比で、腫瘍部位における特異的蓄積が観察された(図5および表1)。
【表1】
【0168】
参考文献
1. Bosslet K et al. Cancer Res 1998; 58:1195-1201
2. Jain RK. Adv Drug Deliv Rev 2001; 46:149-168
3. Kohler G, Milstein C Nature 1975; 256:495-497
4. Winter G et al Annu Rev Immunol 1994; 12:433-455
5. Halin C et al. Cancer Res 2003; 63:3202-3210
6. Halin C et al. Nat Biotechnol 2002; 20:264-269
7. Nilsson F et al Cancer Res 2001; 61:711-716
8. Borsi L et al. Blood 2003; 102:4384-4392
9. Carnemolla B et al. Blood 2002; 99:1659-1665
10. Zardi L et al. Embo J 1987; 6:2337-2342
11. Castellani P et al. Am J Pathol 2002; 161:1695-1700
12. Castellani P et al Int J Cancer 1994; 59:612-618
13. Carnemolla B et al . Int J Cancer 1996; 68:397-405
14. Borsi L et al Int J Cancer 1992; 52:688-692
15. Carnemolla B et al. Eur J Biochem 1992; 205:561-567
16. Borsi L et al J Biol Chem 1995; 270:6243-6245
17. Riva P et al. Int J Cancer 1992; 51:7-13
18. Riva P et al. Cancer Res 1995; 55:5952s-5956s
19. Paganelli G et al Eur J Nucl Med 1994; 21:314-321
20. Reardon DA et al. J Clin Oncol 2002; 20:1389-1397
21. Bigner DD et al. J Clin Oncol 1998; 16:2202-2212
22. Carnemolla B et al. Am J Pathol 1999; 154:1345-1352
23. Katenkamp K et al. J Pathol 2004; 203:771-779
24. Viti F, et al Methods Enzymol 2000; 326:480-505
25. Borsi L et al. Int J Cancer 2002; 102:75-85
26. Pini A et al. J Biol Chem 1998; 273:21769-21776
27. Scheuermann J et al J Immunol Methods 2003; 276:129-134
28. Balza E et al. FEBS Lett 1993; 332:39-43
29. Hoogenboom HR et al. Nucleic Acids Res 1991; 19:4133-4137
30. Ignatovich, O., et al. (1997). J Mol Biol 268(1): 69-77.31.
31. Hoogenboom & Winter (1992). J Mol Biol 227(2): 381-8.
32. Tomlinson, et al. (1992). J Mol Biol 227(3): 776-98.
【0169】
配列表
配列番号1 4A1-F16 VHドメインヌクレオチド配列
GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
CGG TAT GGT GCG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AGT GGT AGT GGT GGT AGC ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA GCG CAT AAT GCT TTT GAC TAC
TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号2 4A1-F16 VHドメインアミノ酸配列
EVQLLESGGG LVQPGGSLRL SCAASGFTFS RYGASWVRQA PGKGLEWVSA
ISGSGGSTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKAH
NAFDYWGQGT LVTVSR
配列番号3 4A1-F16 & 3A1-D5 VLドメインヌクレオチド配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT
GTT TAT ACT ATG CCG CCC GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号4 4A1-F16 & 3A1-D5 VLドメインアミノ酸配列
SSELTQDPAV SVALGQTVRI TCQGDSLRSY YASWYQQKPG QAPVLVIYGK
NNRPSGIPDR FSGSSSGNTA SLTITGAQAE DEADYYCNSS VYTMPPVVFG
GGTKLTVLG
配列番号5 4A1-F16 VH CDR1アミノ酸配列
RYGAS
配列番号6 4A1-F16 & 3A1-D5 VH CDR2アミノ酸配列
AISGSGGSTYYADSVKG
配列番号7 4A1-F16 & 3A1-D5 VH CDR3アミノ酸配列
AHNAFDY
配列番号8 4A1-F16 & 3A1-D5 VL CDR1アミノ酸配列
QGDSLRSYYAS
配列番号9 4A1-F16 & 3A1-D5 VL CDR2アミノ酸配列
GKNNRPS
配列番号10 4A1-F16 & 3A1-D5 VL CDR3アミノ酸配列
NSSVYTMPPVV
配列番号11 3A1-D5 VHドメインヌクレオチド配列
GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
AGC TAT GCC GCG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AGT GGT AGT GGT GGT AGC ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA GCG CAT AAT GCT TTT GAC TAC
TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号12 3A1-D5 VHドメインアミノ酸配列
EVQLLESGGG LVQPGGSLRL SCAASGFTFS SYAASWVRQA PGKGLEWVSA
ISGSGGSTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKAH
NAFDYWGQGT LVTVSR
配列番号13 3A1-D5 VH CDR1アミノ酸配列
SYAAS
配列番号14 E10 VHドメインヌクレオチド配列
GAG GTG CAG CTG GTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
GGT AGT CGT ATG GGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AAT GAG GAG GGT GGT CAG ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA CAT CCG CCG CAT CGG CCG TTT
GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号15 E10 VHドメインアミノ酸配列
EVQLVESGGG LVQPGGSLRL SCAASGFTFS GSRMGWVRQA PGKGLEWVSA
INEEGGQTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKHP
PHRPFDYWGQ GTLVTVSR
配列番号16 E10 & A12 VLドメインヌクレオチド配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT
CAT GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号17 E10 & A12 VLドメインアミノ酸配列
SSELTQDPAV SVALGQTVRI TCQGDSLRSY YASWYQQKPG QAPVLVIYGK
NNRPSGIPDRFSGSSSGNTA SLTITGAQAE DEADYYCNSS HGPRRPVVFG
GGTKLTVLG
配列番号18 E10, F4 & G11 VH CDR1アミノ酸配列
GSRMG
配列番号19 E10, F4 & G11 VH CDR2アミノ酸配列
AINEEGGQTYYADSVKG
配列番号20 E10, A12, F4 & G11 VH CDR3アミノ酸配列
HPPHRPFDY
配列番号21 E10 & A12 VL CDR1アミノ酸配列
QGDSLRSYYAS
配列番号22 E10, A12 & G11 VL CDR2アミノ酸配列
GKNNRPS
配列番号23 E10 & A12 VL CDR3アミノ酸配列
NSSHGPRRPVV
配列番号24 A12 VHドメインヌクレオチド配列
GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
AGC TAT GCC ATG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AGT GGT AGT GGT GGT AGC ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA CAT CCG CCG CAT CGG CCG TTT
GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号25 A12 VHドメインアミノ酸配列
EVQLLESGGG LVQPGGSLRL SCAASGFTFS SYAMSWVRQA PGKGLEWVSA
ISGSGGSTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKHP
PHRPFDYWGQ GTLVTVSR
配列番号26 A12 VH CDR1アミノ酸配列
SYAMS
配列番号27 A12 VH CDR2アミノ酸配列
AISGSGGSTYYADSVKG
配列番号28 F4 & G11 VHドメイン核酸配列
GAG GTG CAG CTG GTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
GGT AGT CGT ATG GGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AAT GAG GAG GGT GGT CAG ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA CAT CCG CCG CAT CGG CCG TTT
GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTC TCG AGA
配列番号29 F4 & G11 VHドメインアミノ酸配列
EVQLVESGGGLVQPGGSLRLSCAASGFTFSGSRMGWVRQAPGKGLEWVSAINEEGGQTYY
ADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKHPPHRPFDYWGQGTLVTVSR
配列番号30 F4 VLドメイン核酸配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA CTT TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA TCT AGT CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT
CAT GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号31 F4 VLドメインアミノ酸配列
SSELTQDPAVSVALGQTVRITCQGDSLRLYYASWYQQKPGQAPVLVIYGKSSRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
配列番号32 F4 & G11 VL CDR1アミノ酸配列
QGDSLRLYYAS
配列番号33 F4 VL CDR2アミノ酸配列
GKSSRPS
配列番号34 G11 VLドメイン核酸配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA CTT TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT
CAT GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号35 G11 VLドメインアミノ酸配列
SSELTQDPAVSVALGQTVRITCQGDSLRLYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
配列番号36 ペプチドリンカー核酸配列
GGT GGA GGC GGT TCA GGC GGA GGT GGC TCT GGC GGT GGC GGA
配列番号37 ペプチドリンカーアミノ酸配列
GGGGSGGGGSGGGG
配列番号38 ペプチドリンカー核酸配列
GGT GGA GGC GGT TCA GGC GGA GGT GGT TCT GGC GGT GGC GGA TCG
配列番号39 ペプチドリンカーアミノ酸配列
GGGGSGGGGSGGGGS
配列番号40 ペプチドリンカー核酸配列
TCT TCC TCA TCG GGT AGT AGC TCT TCC GGC TCA TCG TCC AGC GGC
配列番号41 ペプチドリンカーアミノ酸配列
SSSSGSSSSGSSSSG
配列番号42 ペプチドリンカーアミノ酸配列
GSGSAGSGSAGSGSA
配列番号43 ペプチドリンカーアミノ酸配列
GGSGGGGSGGGGSGG
配列番号44 新規F4 & G11 VHドメイン核酸配列
GAG GTG CAG CTG TTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
GGT AGT CGT ATG GGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AAT GAG GAG GGT GGT CAG ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAA GAC
ACG GCC GTA TAT TAC TGT GCG AAA CAT CCG CCG CAT CGG CCG TTT
GAC TAC TGG GGC CAG GGA ACC CTG GTC ACC GTC TCG AGT
配列番号45 新規F4 & G11 VHドメインアミノ酸配列
EVQLLESGGGLVQPGGSLRLSCAASGFTFSGSRMGWVRQAPGKGLEWVSAINEEGGQTYYAD
SVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKHPPHRPFDYWGQGTLVTVSS
配列番号46 新規G11 VLドメイン核酸配列
TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA CAG
ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA CTT TAT TAT
GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT GTC
ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA TTC
TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT GGG
GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT CAT
GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG ACC
GTC CTA GGC
配列番号47 新規G11 VLドメインアミノ酸配列
SELTQDPAVSVALGQTVRITCQGDSLRLYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
配列番号48 新規E10 VHドメインアミノ酸配列
EVQLLESGGG LVQPGGSLRL SCAASGFTFS GSRMGWVRQA PGKGLEWVSA
INEEGGQTYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKHP
PHRPFDYWGQ GTLVTVSS
配列番号49 新規4A1-F16 & 3A1-D5 VLドメインヌクレオチド配列
TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA CAG
ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT TAT
GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT GTC
ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA TTC
TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT GGG
GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT GTT
TAT ACT ATG CCG CCC GTG GTA TTC GGC GGA GGG ACC AAG CTG ACC
GTC CTA GGC
配列番号50 新規4A1-F16 & 3A1-D5 VLドメインアミノ酸配列
SELTQDPAVSVALGQTVRITCQGDSLRSYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR FSGSSSGNTASLTITGAQAEDEADYYCNSSVYTMPPVVFGGGTKLTVLG
配列番号51 P12 VHドメイン核酸配列
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTC
CTGTGCAGCCTCTGGATTCACCTTTGGCcagTATtctATGAGCTGGGTCCGCCAGGCTCCAG
GGAAGGGGCTGGAGTGGGTCTCAGCTATTacggggactGGTGGTgagACATACTACGCAGAC
TCCGTGGAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAAT
GAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGGGCGGCGGATTTTTG
ACTACTGGGGCCAGGGAACCCTGGTCACCGTGTCGAGA
配列番号52 P12 VHドメインアミノ酸配列
EVQLVESGGGLVQPGGSLRLSCAASGFTFGQYSMSWVRQAPGKGLEWVSAITGTGGETYY
ADSVEGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGRRIFDYWGQGTLVTVSR
配列番号53 P12およびD11 VLドメイン核酸配列
TCGAGTGAGCTGACTCAGGACCCTGCTGTGTCTGTGGCCTTGGGACAGACAGTCAGGATCAC
ATGCCAAGGAGACAGCCTCAGACGGCAGCCTGCAAGCTGGTACCAGCAGAAGCCAGGACAGG
CCCCTGTACTTGTCATCTATTATAAAAAGCTGCGGCCCTCAGGGATCCCAGACCGATTCTCT
GGCTCCAGCTCAGGAAACACAGCTTCCTTGACCATCACTGGGGCTCAGGCGGAAGATGAGGC
TGACTATTACTGTAACTCCTTTTCGCCCAAGCCGAAGCCTGTGGTATTCGGCGGAGGGACCA
AGCTGACCGTCCTAGGC
配列番号54 P12およびD11 VLドメインアミノ酸配列
SSELTQDPAVSVALGQTVRITCQGDSLRRQPASWYQQKPGQAPVLVIYYKKLRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSFSPKPKPVVFGGGTKLTVLG
配列番号55 D11 VHドメイン核酸配列
GAGGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGGGGGTCCCTGAGACTCTC
CTGTGCAGCCTCTGGATTCACCTTTGGCAGCTATGCCATGAGCTGGGTCCGCCAGGCTCCAG
GGAAGGGGCTGGAGTGGGTCTCAGCTATTAGTGGTAGTGGTGGTAGCACATACTACGCAGAC
TCCGTGGAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAAT
GAACAGCCTGAGAGCCGAGGACACGGCCGTATATTACTGTGCGAAAGGGCGGCGGATTTTTG
ACTACTGGGGCCAGGGAACCCTGGTCACCGTGTCGAGA
配列番号56 D11 VHドメインアミノ酸配列
EVQLVESGGGLVQPGGSLRLSCAASGFTFGSYAMSWVRQAPGKGLEWVSAISGSGGSTYY
ADSVEGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGRRIFDYWGQGTLVTVSR
配列番号57 F4S VHドメイン核酸配列
GAG GTG CAG CTG GTG GAG TCT GGG GGA GGC TTG GTA CAG CCT GGG
GGG TCC CTG AGA CTC TCC TGT GCA GCC TCT GGA TTC ACC TTT AGC
AGC TAT GCC ATG AGC TGG GTC CGC CAG GCT CCA GGG AAG GGG CTG
GAG TGG GTC TCA GCT ATT AGT GGT AGT GGT GGT AGC ACA TAC TAC
GCA GAC TCC GTG AAG GGC CGG TTC ACC ATC TCC AGA GAC AAT TCC
AAG AAC ACG CTG TAT CTG CAA ATG AAC AGC CTG AGA GCC GAG GAC
ACG GCC GTA TAT TAC TGT GCG AAA GGG CGG CGG ATT TTT GAC TAC
TGG GGC CAG GGA ACC CTG GTC ACC GTG TCG AGA
配列番号58 F4S VHドメインアミノ酸配列
EVQLVESGGGLVQPGGSLRLSCAASGFTFSSYAMSWVRQAPGKGLEWVSAISGSGGSTYY
ADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKGRRIFDYWGQGTLVTVSR
配列番号59 F4S VLドメイン核酸配列
TCG TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA
CAG ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT
TAT GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT
GTC ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA
TTC TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT
GGG GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TTT
TCG CCC AAG CCG AAG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG
ACC GTC CTA GGC
配列番号60 F4S VLドメインアミノ酸配列
SSELTQDPAVSVALGQTVRITCQGDSLRSYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSFSPKPKPVVFGGGTKLTVLG
配列番号61 P12 VH CDR1アミノ酸配列
QYSMS
配列番号62 D11およびF4S VH CDR1アミノ酸配列
SYAMS
配列番号63 P12 VH CDR2アミノ酸配列
AITGTGGETYYADSVEG
配列番号64 D11 VH CDR2アミノ酸配列
AISGSGGSTYYADSVEG
配列番号65 F4S VH CDR2アミノ酸配列
AISGSGGSTYYADSVKG
配列番号66 P12, D11およびF4S VH CDR3アミノ酸配列
GRRIFDY
配列番号67 P12およびD11 VL CDR1アミノ酸配列
QGDSLRRQPAS
配列番号68 F4S VL CDR1アミノ酸配列
QGDSLRSYYAS
配列番号69 P12およびD11 VL CDR2アミノ酸配列
YKKLRPS
配列番号70 F4S VL CDR2アミノ酸配列
GKNNRPS
配列番号71 P12, D11およびF4S VL CDR3アミノ酸配列
NSFSPKPKPVV
配列番号72 PCR オリゴヌクレオチドプライマーTnC-D BamHI ba
cgggatccgttacagaagccgaaccggaa
配列番号73 PCR オリゴヌクレオチドプライマーTnC-D BglII for
cgggatccgttacagaagccgaaccggaa
配列番号74 PCR オリゴヌクレオチドプライマーmmTnC-A1 EcoBa
agaattcattaaagaggagaaattaactatgagaggatcctccacggaagaagtgccttc
配列番号75 PCR オリゴヌクレオチドプライマーmmTnC-A1 BglFo
tgagatcttgtccctgtggaggtctcggc
配列番号76 ペプチドリンカーモチーフ
GGGGS
配列番号77 ペプチドリンカーモチーフ
SSSSG
配列番号78 ペプチドリンカーモチーフ
GSGSA
配列番号79 ペプチドリンカーモチーフ
GGSGG
配列番号80 新規E10 & A12 VLドメインヌクレオチド配列
TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA CAG
ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA AGC TAT TAT
GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT GTC
ATC TAT GGT AAA AAC AAC CGG CCC TCA GGG ATC CCA GAC CGA TTC
TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT GGG
GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT CAT
GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG ACC
GTC CTA GGC
配列番号81 新規E10 & A12 VLドメインアミノ酸配列
SELTQDPAVSVALGQTVRITCQGDSLRSYYASWYQQKPGQAPVLVIYGKNNRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
配列番号82 新規F4 VLドメイン核酸配列
TCT GAG CTG ACT CAG GAC CCT GCT GTG TCT GTG GCC TTG GGA CAG
ACA GTC AGG ATC ACA TGC CAA GGA GAC AGC CTC AGA CTT TAT TAT
GCA AGC TGG TAC CAG CAG AAG CCA GGA CAG GCC CCT GTA CTT GTC
ATC TAT GGT AAA TCT AGT CGG CCC TCA GGG ATC CCA GAC CGA TTC
TCT GGC TCC AGC TCA GGA AAC ACA GCT TCC TTG ACC ATC ACT GGG
GCT CAG GCG GAA GAT GAG GCT GAC TAT TAC TGT AAC TCC TCT CAT
GGG CCC CGT AGG CCT GTG GTA TTC GGC GGA GGG ACC AAG CTG ACC
GTC CTA GGC
配列番号83 新規F4 VLドメインアミノ酸配列
SELTQDPAVSVALGQTVRITCQGDSLRLYYASWYQQKPGQAPVLVIYGKSSRPSGIPDR
FSGSSSGNTASLTITGAQAEDEADYYCNSSHGPRRPVVFGGGTKLTVLG
【特許請求の範囲】
【請求項1】
ヒトテネイシンCに結合する特異的結合メンバーであって、
(i)配列番号12のD5 VHドメイン、および配列番号50のVLドメインを含む、または
(ii)配列番号2の4A1-F16 VHドメイン、および配列番号50のVLドメインを含む、または
(iii)VH CDR1が配列番号5もしくは配列番号13であり、
VH CDR2が配列番号6であり、
VH CDR3が配列番号7である、
VH CDR1、VH CDR2およびVH CDR3を含む抗体VHドメインと、
VL CDR1が配列番号8であり、
VL CDR2が配列番号9であり、
VL CDR3が配列番号10である、
VL CDR1、VL CDR2およびVL CDR3を含む抗体VLドメインとを含む、
前記特異的結合メンバー。
【請求項2】
scFv抗体分子を含む、請求項1に記載の特異的結合メンバー。
【請求項3】
抗体定常領域を含む、請求項1に記載の特異的結合メンバー。
【請求項4】
全抗体を含む、請求項1に記載の特異的結合メンバー。
【請求項5】
検出可能な標識またはサイトカインに結合されている、請求項1に記載の特異的結合メンバー。
【請求項6】
特異的結合メンバーのVHドメインまたはVLドメインが、融合タンパク質として、ペプチドリンカーを介してサイトカインに結合されている、請求項5に記載の特異的結合メンバー。
【請求項7】
サイトカインがIL2である、請求項5に記載の特異的結合メンバー。
【請求項8】
細胞毒性物質に結合されている、請求項1に記載の特異的結合メンバー。
【請求項9】
放射性核種または発蛍光団からなる群より選択される検出可能な標識に結合されている、請求項1に記載の特異的結合メンバー。
【請求項10】
検出可能な標識がヨウ素131である、請求項9に記載の特異的結合メンバー。
【請求項11】
小免疫タンパク質(SIP)を含む請求項1〜3および5〜10のいずれか1項に記載の特異的結合メンバーであって、該SIPが、ヒト免疫グロブリンEのCH4ドメインと融合された、配列番号2のVHドメイン配列と配列番号50のVLドメイン配列とを含むscFv分子を含み、該SIPが場合によりホモ二量体であってもよい、前記特異的結合メンバー。
【請求項12】
ヒトテネイシンCに結合する単離された特異的結合メンバーであって、
配列番号2のアミノ酸配列からなる抗体VHドメイン、または配列番号2のアミノ酸配列内に1もしくは2個のアミノ酸の改変を有するアミノ酸配列からなる抗体VHドメインと、
配列番号50のアミノ酸配列からなる抗体VLドメイン、または配列番号50のアミノ酸配列内に1もしくは2個のアミノ酸の改変を有するアミノ酸配列からなる抗体VLドメインと
を含む、前記特異的結合メンバー。
【請求項13】
小免疫タンパク質(SIP)を含む請求項12に記載の特異的結合メンバーであって、該SIPが、ヒト免疫グロブリンEのCH4ドメインと融合された、配列番号2のVHドメイン配列と配列番号50のVLドメイン配列とを含むscFv分子を含み、該SIPが場合によりホモ二量体であってもよい、前記特異的結合メンバー。
【請求項14】
ヨウ素131に結合されている、請求項12に記載の特異的結合メンバー。
【請求項1】
ヒトテネイシンCに結合する特異的結合メンバーであって、
(i)配列番号12のD5 VHドメイン、および配列番号50のVLドメインを含む、または
(ii)配列番号2の4A1-F16 VHドメイン、および配列番号50のVLドメインを含む、または
(iii)VH CDR1が配列番号5もしくは配列番号13であり、
VH CDR2が配列番号6であり、
VH CDR3が配列番号7である、
VH CDR1、VH CDR2およびVH CDR3を含む抗体VHドメインと、
VL CDR1が配列番号8であり、
VL CDR2が配列番号9であり、
VL CDR3が配列番号10である、
VL CDR1、VL CDR2およびVL CDR3を含む抗体VLドメインとを含む、
前記特異的結合メンバー。
【請求項2】
scFv抗体分子を含む、請求項1に記載の特異的結合メンバー。
【請求項3】
抗体定常領域を含む、請求項1に記載の特異的結合メンバー。
【請求項4】
全抗体を含む、請求項1に記載の特異的結合メンバー。
【請求項5】
検出可能な標識またはサイトカインに結合されている、請求項1に記載の特異的結合メンバー。
【請求項6】
特異的結合メンバーのVHドメインまたはVLドメインが、融合タンパク質として、ペプチドリンカーを介してサイトカインに結合されている、請求項5に記載の特異的結合メンバー。
【請求項7】
サイトカインがIL2である、請求項5に記載の特異的結合メンバー。
【請求項8】
細胞毒性物質に結合されている、請求項1に記載の特異的結合メンバー。
【請求項9】
放射性核種または発蛍光団からなる群より選択される検出可能な標識に結合されている、請求項1に記載の特異的結合メンバー。
【請求項10】
検出可能な標識がヨウ素131である、請求項9に記載の特異的結合メンバー。
【請求項11】
小免疫タンパク質(SIP)を含む請求項1〜3および5〜10のいずれか1項に記載の特異的結合メンバーであって、該SIPが、ヒト免疫グロブリンEのCH4ドメインと融合された、配列番号2のVHドメイン配列と配列番号50のVLドメイン配列とを含むscFv分子を含み、該SIPが場合によりホモ二量体であってもよい、前記特異的結合メンバー。
【請求項12】
ヒトテネイシンCに結合する単離された特異的結合メンバーであって、
配列番号2のアミノ酸配列からなる抗体VHドメイン、または配列番号2のアミノ酸配列内に1もしくは2個のアミノ酸の改変を有するアミノ酸配列からなる抗体VHドメインと、
配列番号50のアミノ酸配列からなる抗体VLドメイン、または配列番号50のアミノ酸配列内に1もしくは2個のアミノ酸の改変を有するアミノ酸配列からなる抗体VLドメインと
を含む、前記特異的結合メンバー。
【請求項13】
小免疫タンパク質(SIP)を含む請求項12に記載の特異的結合メンバーであって、該SIPが、ヒト免疫グロブリンEのCH4ドメインと融合された、配列番号2のVHドメイン配列と配列番号50のVLドメイン配列とを含むscFv分子を含み、該SIPが場合によりホモ二量体であってもよい、前記特異的結合メンバー。
【請求項14】
ヨウ素131に結合されている、請求項12に記載の特異的結合メンバー。
【図1】

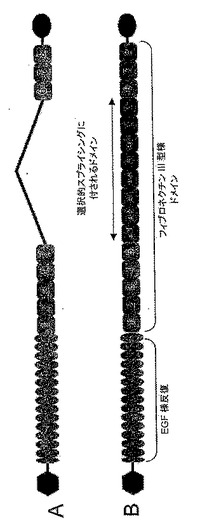
【図2】
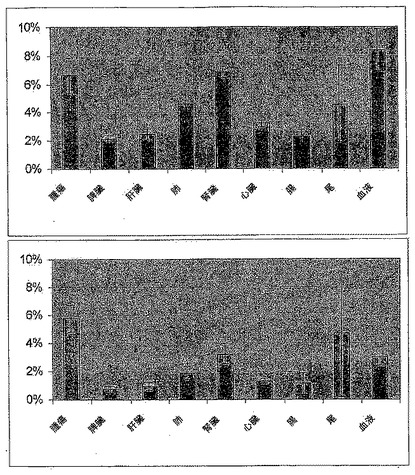
【図3】
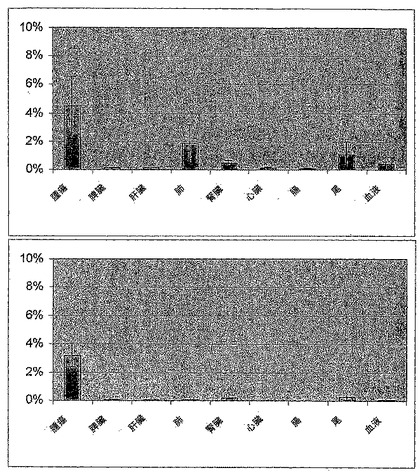
【図4】
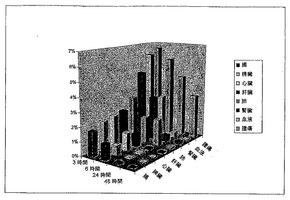
【図5】
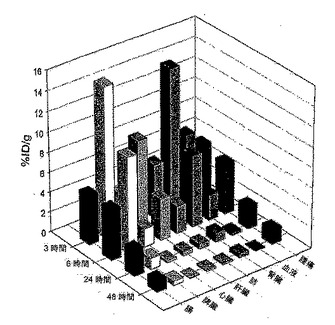
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−176957(P2012−176957A)
【公開日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願番号】特願2012−93587(P2012−93587)
【出願日】平成24年4月17日(2012.4.17)
【分割の表示】特願2007−539520(P2007−539520)の分割
【原出願日】平成17年10月31日(2005.10.31)
【出願人】(507147231)
【Fターム(参考)】
【公開日】平成24年9月13日(2012.9.13)
【国際特許分類】
【出願日】平成24年4月17日(2012.4.17)
【分割の表示】特願2007−539520(P2007−539520)の分割
【原出願日】平成17年10月31日(2005.10.31)
【出願人】(507147231)
【Fターム(参考)】
[ Back to top ]