
デオキシノジリマイシン類似体、及びグルコシルセラミダーゼ阻害剤としてのその使用
【課題】ゴーシェ病、炎症性疾患、皮膚の炎症状態、過体重、肥満、真菌症、マイクロバクテリア感染症、インスリン抵抗性等の治療に有用な医薬組成物の提供。
【解決手段】下式の構造を有するデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む治療用医薬組成物。

[式中、R1〜R5は、それぞれ独立に、H、(CH2)nCH3、又はXを含み、R6は、H、CH2OH、又はCH2OXを含み、ここで、R1〜R5の少なくとも一つはXであるか、又はR1〜R5が全てX以外の基のとき、R6はCH2OXであり、nは、0〜9であり、a〜eは、R又はS立体配置を有するキラル中心であり、Xは、アダマンタン−1−イル−メトキシである大きな疎水性部分及び炭素原子5個のアルキレン鎖であるスペーサーを含み、疎水性部分は、脂質二重層に入り込むことができる。]
【解決手段】下式の構造を有するデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む治療用医薬組成物。
[式中、R1〜R5は、それぞれ独立に、H、(CH2)nCH3、又はXを含み、R6は、H、CH2OH、又はCH2OXを含み、ここで、R1〜R5の少なくとも一つはXであるか、又はR1〜R5が全てX以外の基のとき、R6はCH2OXであり、nは、0〜9であり、a〜eは、R又はS立体配置を有するキラル中心であり、Xは、アダマンタン−1−イル−メトキシである大きな疎水性部分及び炭素原子5個のアルキレン鎖であるスペーサーを含み、疎水性部分は、脂質二重層に入り込むことができる。]
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、グルコシルセラミド及び/又は他のスフィンゴ糖脂質の合成が関与する様々な疾患の治療に適切に使用することのできる、デオキシノジリマイシン類似体又は薬学的に許容できるその塩に関する。そのような疾患には、インスリン抵抗性(II型糖尿病)、肥満及び過体重、スフィンゴ糖脂質貯蔵障害、炎症、色素沈着過剰症及び皮膚の炎症状態、黒色腫及び他の腫瘍、真菌感染、ウイルス感染、微生物感染、及び内毒素が含まれる。
【背景技術】
【0002】
スフィンゴ糖脂質
スフィンゴ糖脂質は、糖鎖に連結したセラミド部分を含む脂質分子であるとされる。セラミド部分は、1本の特徴的な長鎖アミノアルコール、主としてD−エリトロ−C16−スフィンゴシンが、長さ及び飽和度は様々でよい脂肪酸にアミド結合によって結合しているものから構成される。糖単位の数及び性質の異なるその少糖部分とそのグリコシド結合が様々となるために、数百の特定のスフィンゴ糖脂質が存在する。スフィンゴ糖脂質は、例えば、ガングリオシド及びグロボシドなどの系列に分類される(参照文献1)。
【0003】
数十年間、スフィンゴ糖脂質は、むしろ新種の構造であるとみなされ、主流の生化学ではほとんど注目を浴びていなかった。その構成が複雑であり、存在量が少ないために、研究活動は、ほんの少数の先駆的な専門家集団だけに限られた。血液型抗原として働く機能を除き、特定のスフィンゴ糖脂質のためであると考えられる明確な機能がないので、その生理的な関連性は、多くは謎に包まれたままである。
【0004】
人体では、スフィンゴ糖脂質及びそのスフィンゴ脂質前駆体であるセラミドは、細胞内及び細胞外に存在する。表皮では、セラミド及びグルコシルセラミドの分子は、遊離の存在だけでなく、タンパク質に結合した構造として豊富に存在し、皮膚の水分透過性及び完全性に寄与する(参照文献2)。目立ったその合成部位である肝臓と末梢組織間のスフィンゴ糖脂質の交換は、相当な量のこれらの脂質を含むリポタンパク質によって媒介される。細胞では、スフィンゴ糖脂質は、多かれ少なかれもっぱら形質膜の外側葉状物中に位置し、その少糖鎖は周囲に露出している。セラミド脂質の主鎖は、スフィンゴ脂質に特別な物理的性質を与える(参照文献3)。コレステロール存在下で、スフィンゴ糖脂質は、膜リン脂質の塊から分離し、凝集して、より規則正しいがそれでも流体である「液体−規則」ドメインとなり得る。シグナル伝達に関与する膜タンパク質は、これらのスフィンゴ糖脂質−コレステロールに富んだマイクロドメイン(又は「ラフト」)に集中している(参照文献4)。
【0005】
細胞表面にあるスフィンゴ糖脂質の組成は、細胞の増殖、分化、ウイルス形質転換、個体発生、及び発癌と共に変化し得る(参照文献5)。神経細胞ではガングリオシドが特に豊富であることが明らかになってきている。ガングリオシドは、構造成分として神経系の機能に不可欠である。スフィンゴ糖脂質は、マイクロドメインの成分として、多数の細胞外シグナルの細胞内部への伝達に関与する(参照文献6)。さらに、セラミドなどのスフィンゴ脂質やその代謝産物であるスフィンゴシン(−1−リン酸)は、様々な細胞プロセスにおいてシグナル伝達分子としての直接の役割を担うと考えられている(参照文献7、8、及び9)。
【0006】
スフィンゴ糖脂質の合成及び分解
スフィンゴ糖脂質の合成及び分解は、様々な細胞内区画で起こる複雑な多段階の過程である(参照文献10及び11)。セラミド生合成に関与する酵素は、小胞体の細胞質ゾル葉状構造上に局在している。生合成は、セリンパルミトイルトランスフェラーゼによるアミノ酸セリンとパルミトイル補酵素Aの縮合から出発して、3−ケトスフィンガニンをもたらす。次に、これが3−ケトスフィンガニンレダクターゼによってD−エリトロ−スフィンガニンに還元され、その後、N−アシルトランスフェラーゼによってアシル化されてジヒドロセラミドになる。ジヒドロセラミドは、多くはジヒドロセラミドデサチュラーゼの作用によって不飽和化される。
【0007】
セラミドはさらに、スフィンゴミエリン、ガラクトシルセラミド及び誘導体、又はグルコシルセラミド及び誘導体の異なる3部門のスフィンゴ脂質へと代謝される。セラミドは、セラミドの1−ヒドロキシル基に、リン脂質ホスファチジルコリンからホスホリルコリンが転移することで、スフィンゴミエリンに変換される。この反応でジアシルグリセロールが遊離する。スフィンゴミエリンの生合成は、ゴルジ膜の内腔部で行われるが、他の部位も考慮に入れられている。ガラクトシルセラミドの合成は、小胞体の内腔で行われ、UDPガラクトースを共基質とする特定のβ−ガラクトシルトランスフェラーゼによって触媒される。ガラクトシルセラミド及びその硫酸エステル誘導体であるスルファチドは、特定の細胞型でしか合成されない。これらは、例えば、中枢神経系のミエリンの形成及び安定性において決定的な役割を担う。グルコシルセラミドは、ゴルジ装置の細胞質ゾル葉状構造上にあるグルコシルセラミドシンターゼによって生成される。この酵素は、UDPグルコースを共基質として使用し、グルコースがセラミドの1位にβグリコシド結合するのを触媒する。グルコシルセラミドは、その後ゴルジ膜を通って移動して、内側の葉状構造に到達する。グルコシルセラミドは、そこから形質膜に到達することもでき、又はゴルジ装置中でさらにグリコシル化されて改変されることもある。脊椎動物に共通するスフィンゴ糖脂質前駆体であるラクトシルセラミドは、ガラクトシルトランスフェラーゼIを触媒とするUDP−Galからのガラクトース部分の付加によって生成される。次に、特異性の限られた数種類のグリコシルトランスフェラーゼによってのみ行われる段階的なグリコシル化によって様々なスフィンゴ糖脂質が生成され得る。これらの酵素は、流れ作業列でのように、炭水化物基及びシアル酸基をグリコシル受容側に転移する。スフィンゴ糖脂質の著名な系列は、ガングリオ系列である。ガングリオシド複合体の生合成には、シアリルトランスフェラーゼI、II、及びIII、GalNacトランスフェラーゼ、ガラクトシルトランスフェラーゼII、及びシアリルトランスフェラーゼIV及びVが必要である。
【0008】
スフィンゴ糖脂質の分解は、細胞の特殊化した酸性区画、即ちリソソーム中で行われる。活性化タンパク質によって援助されるリソソーム内グリコシダーゼが、その糖脂質基質の非還元性末端から糖残基を順次切断し、残りのセラミドが、リソソーム内セラミダーゼによって最後に加水分解されて、遊離の脂肪酸及びスフィンゴシンがもたらされる。放出された単糖、シアル酸、脂肪酸、及びスフィンゴイド塩基は、リソソームを出ることができ、生合成の目的のために再度使用されることもある。より最近になって、スフィンゴ糖脂質の分解がリソソームの外側で行われる場合もあることがわかってきている。例えば、グルコシルセラミドをセラミドに加水分解することのできる非リソソーム内グルコシルセラミダーゼが発見されている(参照文献12)。後者の酵素は、ゴーシェ病患者で欠損しているリソソーム内グルコシルセラミダーゼ(グルコセレブロシダーゼ)とは明らかに異なる。リソソーム外分解の生理的重要性は、未だ理解が不十分ではあるが、おそらく、それに関連するセラミド生成はシグナルとして働き、細胞の挙動を調節し得ると思われる(参照文献13)。
【0009】
スフィンゴ糖脂質貯蔵障害
スフィンゴ糖脂質は、細胞中で絶えず合成され分解される。ヒトのいくつかの遺伝疾患は、リソソーム内のスフィンゴ糖脂質の蓄積を特徴とする。これらのいわゆるスフィンゴ糖脂質貯蔵障害は、リソソーム内分解の欠陥によって引き起こされる(例えば、参照文献13)。その例は、GM1ガングリオシドーシス、サンドホフ病(GM2ガングリオシドーシス、II型又はO型)、テイサックス病(GM2ガングリオシドーシス、I型又はB型)、ファブリー病(a−ガラクトシル−ラクトシルセラミドーシス)、ラクトシルセラミドーシス、ゴーシェ病(グルコセレブロシドーシス)である。他のスフィンゴリピドーシスは、多数のスルファターゼ欠損症候群(ムコスルファチドーシス)、クラッベ病(グロボイド細胞型白質ジストロフィー、ガラクトセレブロシドーシス)、ニーマンピック病(スフィンゴミエリノーシス)、ファーバー病(セラミドーシス)である。
【0010】
ゴーシェ病
ゴーシェ病は、ヒトにおいて最もよく遭遇するリソソーム貯蔵障害である(参照文献14及び15)。1882年に、フランス人医学研究生Philippe C. E. Gaucherによってこの疾患の臨床的特徴が初めて詳述され、脾腫を伴う32才の女性に異常な大きな細胞が存在することが報告されている。前世紀の初頭にはすでに、この疾患が家族性の障害であることが示されていた。1934年に、ゴーシェ病の主たる貯蔵物質がついにグルコセレブロシド(ブルコシルセラミド)であると特定された。スフィンゴ糖脂質であるグルコセレブロシドは、ガングリオシド及びグロボシドの合成及び分解に共通の中間体である。ゴーシェ病における主たる欠陥は、リソソーム内酵素であるグルコセレブロシダーゼ(EC.3.2.1.45)の活性の顕著な不足であることが示されている(参照文献16及び17)。グルコセレブロシダーゼが遺伝的に欠損していると、体中のマクロファージのリソソーム内区画にその脂質基質が蓄積する。神経性の症状の有無に基づいて区別される3つの異なる表現型が認められている。より最近になって、ゴーシェ病の追加の表現型が同定されている。例えば、グルコセレブロシダーゼ活性の完全な欠如は、出生前又は出生直後に、致死的な予後を伴う重い皮膚透過性の異常をもたらす。最も有病率の高い疾患の型は、1型ゴーシェ病と命名される、神経細胞障害性でない形態である。1型ゴーシェ病の発症年齢及び臨床徴候は、非常に変化に富んでいる。最も一般的な症状には、主として脾機能亢進による貧血及び血小板減少を伴う脾腫大;肝腫脹、及び骨疾患が含まれる。貧血は慢性疲労の一因となり得る。血小板減少及び凝固時間の延長は、出血傾向を増大させる場合がある。不規則な骨の痛み、病的な骨折、無血管性の壊死、及び極めて苦しい骨の発作も生活の質に多大な影響を及ぼし得る。1型ゴーシェ病は、すべての民族群で比較的一般的である。この疾患は、アシュケナジで有病率が高く、キャリヤ出現度はおよそ10人に1人にもなり、発生率はおよそ5000人に1人である。アシュケナジを含む白人のグルコセレブロシダーゼ遺伝子の最も一般的な突然変異は、アミノ酸置換N370Sをコードする。N370S突然変異のヘテロ対立遺伝子の存在は、常に神経細胞障害性でない経過を伴う。N370Sグルコセレブロシダーゼは、通常はリソソームで産生され、そこに存在することが実証されている。その触媒活性は、5.0を上回るpH値でしかひどくは損なわれず、この突然変異の微妙な性質を物語っている(参照文献18)。N370S突然変異のすべてではないが大部分のホモ接合体は、重要な臨床症状を表さない。ゴーシェ病における表現型−遺伝子型調査の二重研究及び不十分な予測力から、ゴーシェ病の出現においては後成的な要因も鍵となる役割を担っていることがはっきりと示されている(参照文献19及び20も参照のこと)。グルコセレブロシダーゼは、すべての細胞型のリソソームに存在するが、1型ゴーシェ病患者は、単に単核貪食細胞系の細胞でグルコセレブロシドの貯蔵を展開する。マクロファージは、侵入微生物の分解、血球の自然の代謝回転、及び組織モデリングに関与する。この点から、かなりの種類のリソソーム貯蔵障害で、貯蔵物質の蓄積が組織マクロファージ中で顕著に起こっても意外ではない。ゴーシェ病の1型変型は、リソソーム内貯蔵がもっぱらマクロファージで起こるという点で特異である。貯蔵材料は、血球の代謝回転から派生する外来脂質が分解されて生じると考えられている。グルコセレブロシドが詰め込まれた細胞は、リソソーム封入体を含むその細胞質の外観が「しわくちゃの紙」様である特徴的な形態を示し、このような細胞をゴーシェ細胞と呼ぶ。ここ数十年で、ゴーシェ細胞が貯蔵物質の不活性な容器ではなく、ゴーシェ病の多様な臨床徴候に寄与する生存能力のある慢性的に活性化したマクロファージであることが明らかになった。いくつかの炎症誘発性サイトカイン(TNF−α、IL−1−β、IL−6、及びIL−8)、抗炎症性サイトカインIL−10、及びM−CSFの循環レベルの増大が報告されている(参照文献21及び22)。オステオペニア、凝固の活性化、代謝亢進、免疫グロブリン血症及び多発性骨髄腫、低脂血症など、ゴーシェ病患者に共通する臨床上の異常の発生において、サイトカインの異常が決定的な役割を担うのではないかという仮説が立てられている。より最近では、ゴーシェ病及び対照の脾臓の抑制性減算ハイブリダイゼーション分析によって遺伝子発現プロフィールを調査すると、ゴーシェ細胞によるカテプシンB、K、及びS転写物の過剰発現が特定された(参照文献23)。破骨細胞由来のカテプシンKが、骨のI型コラーゲンの破壊に目立って関与することは特筆に価する。このカテプシンの局所での放出は、ゴーシェ病の骨溶解の一因となるかもしれない。
【0011】
ゴーシェ病の治療
1型ゴーシェ病は、一般に、遺伝性リソソーム貯蔵障害の中で、有効な治療処置を開発するための最も魅力的な候補であると考えられている。第1に、根底にある遺伝的な欠陥という分子的な根拠は、すでに遺伝子及びタンパク質のレベルで確立されている。第2に、組織マクロファージというたった1つの細胞型が、主としてこの疾患の病態生理と関連付けられている。1型ゴーシェ病の治療処置の原理は、したがって比較的簡単である。即ち、ゴーシェ細胞を正す(又は進行中の生成を予防する)のである。これは、マクロファージに酵素グルコセレブロシダーゼを補充し(酵素補充療法)、特定の阻害剤を用いて糖脂質合成を縮小し(基質除去若しくは基質調整療法)、又はマクロファージの造血前駆細胞中にグルコセレブロシダーゼcDNAを導入する(遺伝子療法)ことで実現できるはずである。米国国立保健研究所(米国ベセズダ)でのBrady、Barranger、及び同僚らの先駆的な仕事、並びに他の多くの者による価値ある貢献が、ヒトグルコセレブロシダーゼの長期間の静脈内投与を主体とする1型ゴーシェ病の非常に有効な治療をもたらした(参照文献24〜26)。ゴーシェ病患者の造血細胞に遺伝子を導入する3通りの独立した研究を実施したが、どれも有望な成果を上げなかった(参照文献27)。CD34細胞の形質導入の効率が悪く、白血球中でグルコセレブロシダーゼが持続的に発現されなかったことが、このことの一因となった。血液疾患及び遺伝障害を正すための遺伝子療法戦略の開発は、オンコレトロウイルス由来のベクターを使用するヒト幹細胞への遺伝子導入のレベルが低いことが妨げとなった。
【0012】
基質低減療法
1型ゴーシェ病及び他のグリコスフィンゴリピドーシスの治療処置に向けた代替手法は、基質除去(基質低減とも呼ばれる)療法である。Radin及び同僚らは、まず挑戦的な構想を考案した(参照文献28)。この手法は、スフィンゴ糖脂質の生合成速度を弱まった異化作用に見合うレベルにまで落とすことを目指すものである。リソソーム内酵素活性がかなり残っている患者は、リソソーム内貯蔵物質を徐々に片付けることができ、したがって基質生合成の低減から最も利益を得るはずであると考えられる(参照文献29)。
【0013】
スフィンゴ糖脂質生合成の2大阻害剤クラスは、以前から記載されており、その両方が、セラミドに特異的なグルコシルトランスフェラーゼ(グルコシルセラミドシンターゼ;GlcT−1;UDPグルコース:N−アシルスフィンゴシンD−グルコシル−トランスフェラーゼ、EC2.4.1.80とも呼ばれる)を阻害する。この酵素は、グルコースのセラミドへの転移、即ちスフィンゴ糖脂質生合成の第1段階を触媒する。第1のクラスの阻害剤は、セラミド類似体によって形成される。原型の阻害剤は、PDMP(D,L−トレオ−1−フェニル−2−デカノイルアミノ−3−モリホリノ−1−プロパノール)である。モルホリノ基をピロリジノ官能基に置換することに基づき、フェニル基で置換を行うことで、より特異的且つ強力な類似体、即ち、4−ヒドロキシ−1−フェニル−2−パルミトイルアミノ−3−ピロリジノ−1−プロパノール(p−OH−P4)及びエチレンジオキシ−1−フェニル−パルミトイルアミノ−3−ピロリジノ−1−プロパノール(EtDo−P4)が後に開発された(参照文献30)。ノックアウトマウスモデルでのファブリー病の研究では、この化合物の経口投与が、スフィンゴ糖脂質であるグロボトリアオシルセラミドの蓄積を著しく低減し得ることが示されている(参照文献31)。
【0014】
イミノ糖
第2のクラスのグルコシルセラミドシンターゼ阻害剤は、N−アルキル化イミノ糖によって形成される。この種の化合物は、N−グリカンプロセシング酵素の阻害剤としてすでに一般に使用されており、N−ブチルデオキシノジリマイシンのHIV阻害剤としての適用の可能性が、AIDS患者で研究されている。N−ブチルデオキシノジリマイシンがグリコシルセラミド合成を低マイクロモル濃度で阻害し得ることを最初に認めたのは、英国オックスフォードのGlycobiology InstituteのPlatt及びButtersであった(参照文献32)。同じ研究者らは、テイサックス病及びサンドホフ病のノックアウトマウスモデルで、脳中のスフィンゴ糖脂質の貯蔵が有意に低減したことを実証している(参照文献33)。動物での前臨床研究及びAIDS患者での予備臨床治験では、おそらくはNB−DNJが腸管刷子縁上の二糖類分解酵素を阻害し得ることに関連した消化管での(一過性の)有害作用が指摘されている。動物の研究では、ガラクトース類似体N−ブチルデオキシガラクトノジリマイシン(NB−DGJ)が、N−ブチルデオキシガラクトノジリマイシン(NB−DNJ)と同じ治療効力を有するかもしれず、胃腸への副作用を引き起こさないことがわかっている(参照文献34)。Overkleeft及び同僚らは、グルコシダーゼ阻害剤を探索する中で、グルコシルセラミドシンターゼのより強力な阻害剤を偶然の発見に基づいて開発した。N−アダマンタン−1−イル−メトキシペンチル−デオキシノジリマイシン(AMP−DNM)は、スフィンゴ糖脂質生合成をナノモル濃度で阻害することがわかっており(参照文献35)、ファブリー病ノックアウトマウスモデルで、表立った副作用なしにグロボトリアオシルセラミドの蓄積を予防することができた。
【0015】
スフィンゴ糖脂質貯蔵障害の治療にN−ブチルデオキシノジリマイシン(NB−DNJ)を使用する最初の臨床研究が、最近になって報告されている(Coxら、2000年)。非盲験第I/II相試験で、成人28人の1型ゴーシェ病患者に、1日3回100mgのNB−DNJ(OGT918、Oxford GlycoSciences社製)が与えられた。内臓巨大症及び血液の異常の改善、並びにグルコシルセラミド及びゴーシェ病活動生物マーカーの血漿レベルの正常化が記載されてはいるが、応答の程度は、高用量酵素補充療法で一般に認められるものより目覚しいものではなかった。予想どおり、NB−DNJの用量−応答の関係は、1型ゴーシェ病患者で実証できる。最近になって、1日3回の50mgのNB−DNJの投与は、有効性がはるかに劣ることが報告された(参照文献37)。ごく最近、EMEA(ヨーロッパのFDA相当機関)は、酵素補充療法を受けるのに不適格な1型ゴーシェ病患者の治療向けにNB−DNJ(ザベスカ、Oxford GlycoSciences社製)を認可している。
【0016】
イミノ糖及び他の治療適用例
イミノ糖は、環上酸素が窒素原子と入れ替わっている、単糖の構造を模倣したポリヒドロキシル化アルカロイドである。イミノ糖の例は、例えば、WO98102161に記載されている。遷移電荷状態の類似体として、イミノ糖は、糖の除去又は付加を触媒する酵素(「複合糖質プロセシング酵素」)の阻害剤として働き得る。グルコースの類似した構造模倣物であるデオキシノジリマイシンは、糖タンパク質をプロセッシングするα−グルコシダーゼI及びIIの阻害剤であることがわかっている。N−ブチル−デオキシノジリマイシンのような一部のN−アルキル化デオキシノジリマイシンは、抗ウイルス(HIV)治療薬としての評価がなされており、in vitroモデルを使用してウイルスの感染力に対する効力が証明されているにもかかわらず、臨床での成功は限られている(参照文献38)。現在、肝炎ウイルス感染に対する治療価値に関して、ノニル−デオキシ−ガラクトノジリマイシン及びN−7−オキサノニル−6−Me−デオキシガラクトノジリマイシンの調査がなされている。N−ヒドロキシエチル−デオキシノジリマイシン(ミグリトール、グリセット)のような他のN−アルキル化デオキシノジリマイシンは、腸管グリコシダーゼの阻害を目指して開発されている。腸管グリコシダーゼ活性の低減は、食物複合炭水化物からの単糖の取り込みの衝撃を和らげるはずなので、インスリン抵抗性(II型糖尿病)の個体にとって有益であると考えられる。代謝制御を目指す薬剤開発におけるイミノ糖の別の使用は、腫瘍細胞の転移を低減するために、細胞表面タンパク質上のN−結合型少糖の改変に使用するものである(参照文献39)。
【特許文献1】WO98102161
【発明の開示】
【発明が解決しようとする課題】
【0017】
今回、スフィンゴ糖脂質の異常な代謝及び異常な濃縮は、インスリン抵抗性(II型糖尿病)、過体重及び肥満、リソソーム貯蔵障害、炎症から、色素沈着過剰症及び皮膚の炎症状態に至る範囲の様々な病的状態と関連していることが判明した。さらに、スフィンゴ糖脂質が、ウイルス、微生物、及び真菌感染、並びにある種の内毒素に対する敏感性において重要な役割を担っていることがわかった。本発明者らは、慎重に選択したアルキル化イミノ糖を投与してスフィンゴ糖脂質代謝を正常化し、過剰のスフィンゴ糖脂質を低減すれば、そのような病的状態において有益な応答を得ることができると考えている。しかし、このような手法を成功させるための前提条件は、投与される化合物が有効であり、且つ十分に認容性であることである。イミノ糖が、グルコシルセラミドシンターゼなどのグルコシルトランスフェラーゼの活性だけでなく、他の様々な酵素(腸管及び他の細胞表面グリコシダーゼ、リソソーム内及びエンドソーム内グリコシダーゼ、グリコーゲン枝切り酵素、及び糖タンパク質改変グリコシダーゼ)の活性も妨げ得ることもわかっている。したがって、成功をもたらすイミノ糖の治療用途は、生物活性の特異性を要する。治療用イミノ糖によって阻害されるはずの複合糖質プロセシング酵素ターゲット、さらには阻害されないはずのその酵素の正確な性質を見抜くことが不可欠である。このことは、異なる病的状態にぴったり合う治療用イミノ糖開発の礎となる。しかし、以前から利用可能なイミノ糖は、少しも最適ではない。第1に、それらは生物学的利用能が十分でなく、グルコシルセラミドシンターゼの比較的弱い阻害剤である。第2に、それらは、有益な酵素活性の比較的強い阻害剤であり、そのために高めの用量で投与されると許容されない副作用を引き起こす。したがって、改良には、次の2段階の異なるレベル、即ち、1:新規なアルキル化イミノ糖の設計及び合成と、2:各病的状態に相応しい阻害活性判定基準を使用する最適なアルキル化イミノ糖の選択が必要となる。
【課題を解決するための手段】
【0018】
本発明は、グルコシルセラミド及び/又は他のスフィンゴ糖脂質の合成が役割を果たす様々な疾患の治療に適切に使用することのできる特定のクラスの新規なアルキル化イミノ糖誘導体に関する。
【0019】
したがって、本発明は、一般式(1)を有するデオキシノジリマイシン類似体又は薬学的に許容できるその塩に関する
【化1】
(I)
[式中、
R1〜R5は、それぞれ独立に、H、(CH2)nCH3、又はXを含み、
R6は、H、CH2OH、又はCH2OXを含み、
Mは、0又は1であり、
Nは、0〜9であり、
a、b、c、d、eは、R又はS立体配置を有するキラル中心であり、
Xは、大きな疎水性部分及びスペーサーを含み、疎水性部分は、スペーサーを介して問題の窒素原子又は炭素原子に結合しており、前記の大きな疎水性部分は、各々が2個以上の炭素原子を別の環と共有する3個以上の環を含む多環式アルコールから誘導され、脂質二重層に入り込むことができる]。
【0020】
ピロリジン(m=0)については、3個(R6=H)又は4個(R6=CH2OH、CH2OX、図2を参照のこと)のキラル中心を認めることができる。ピペリジン(m=1)については、4個(R6=H)又は5個(R6=CH2OH、CH2OX)のキラル中心を認めることができる。ピロリジン及びピペリジンは、個々のキラル中心b、c、d、及びeの性質及び立体化学に応じて、自然の五炭糖(R6=Hである場合)及び六炭糖(R6=CH2OH、CH2OX)を模倣する。キラル中心aの性質は(R2がHでない場合)、イミノ糖がαグリコシド結合(a=R)又はβグリコシド結合(a=S)のどちらを模倣するかを規定する。本発明によるデオキシノジリマイシン類似体の詳細な例を以下に挙げる。
【0021】
ピロリジン(R6=CH2OH又はCH2OX、m=0)
a=R、b=S、d=R、e=R α−D−リボフラノシド模倣物
a=S、b=S、d=R、e=R β−D−リボフラノシド模倣物
a=R、b=R、d=R、e=R α−D−アラビノフラノシド模倣物
a=S、b=R、d=R、e=R β−D−アラビノフラノシド模倣物
a=R、b=S、d=S、e=R α−D−キシロフラノシド模倣物
a=S、b=S、d=S、e=R β−D−キシロフラノシド模倣物
a=R、b=S、d=R、e=S α−L−リキソフラノシド模倣物
a=S、b=S、d=R、e=S β−L−リキソフラノシド模倣物
【0022】
ピロリジン(R2=H、R6=CH2OH又はCH2OX、m=0)
b=S、d=R、e=R α−D−1−デオキシリボフラノシド模倣物
b=S、d=R、e=R β−D−1−デオキシリボフラノシド模倣物
b=R、d=R、e=R α−D−1−デオキシアラビノフラノシド模倣物
b=R、d=R、e=R β−D−1−デオキシアラビノフラノシド模倣物
b=S、d=S、e=R α−D−1−デオキシキシロフラノシド模倣物
b=S、d=S、e=R β−D−1−デオキシキシロフラノシド模倣物
b=S、d=R、e=S α−L−1−デオキシリキソフラノシド模倣物
b=S、d=R、e=S β−L−1−デオキシリキソフラノシド模倣物
【0023】
ピペリジン(R6=H、m=1)
a=S、b=S、c=S、d=R β−D−キシロピラノシド模倣物
a=R、b=S、c=S、d=R α−D−キシロピラノシド模倣物
a=S、b=R、c=S、d=R β−D−リキソピラノシド模倣物
a=R、b=R、c=S、d=R α−D−リキソピラノシド模倣物
a=S、b=S、c=S、d=S β−L−アラビノピラノシド模倣物
a=R、b=S、c=S、d=S α−L−アラビノピラノシド模倣物
【0024】
ピペリジン(R2=H、R6=H、m=1)
b=S、c=S、d=R β−D−1−デオキシキシロピラノシド模倣物
b=S、c=S、d=R α−D−1−デオキシキシロピラノシド模倣物
b=R、c=S、d=R β−D−1−デオキシリキソピラノシド模倣物
b=R、c=S、d=R α−D−1−デオキシリキソピラノシド模倣物
b=S、c=S、d=S β−L−1−デオキシアラビノピラノシド模倣物
b=S、c=S、d=S α−L−1−デオキシアラビノピラノシド模倣物
【0025】
ピペリジン(R6=CH2OH又はCH2OX、m=1)
a=S、b=S、c=R、d=R、e=R β−D−グルコピラノシド模倣物
a=R、b=S、c=R、d=R、e=R α−D−グルコピラノシド模倣物
a=S、b=R、c=R、d=R、e=R β−D−マンノピラノシド模倣物
a=R、b=R、c=R、d=R、e=R α−D−マンノピラノシド模倣物
a=S、b=S、c=R、d=S、e=R β−D−ガラクトピラノシド模倣物
a=R、b=S、c=R、d=S、e=R α−D−ガラクトピラノシド模倣物
a=S、b=S、c=R、d=R、e=S β−L−イドピラノシド模倣物
a=R、b=S、c=R、d=R、e=S α−L−イドピラノシド模倣物
【0026】
ピペリジン(R2=H、R6=CH2OH又はCH2OX、m=1)
b=S、c=R、d=R、e=R β−D−1−デオキシグルコピラノシド模倣物
b=S、c=R、d=R、e=R α−D−1−デオキシグルコピラノシド模倣物
b=R、c=R、d=R、e=R β−D−1−デオキシマンノピラノシド模倣物
b=R、c=R、d=R、e=R α−D−1−デオキシマンノピラノシド模倣物
b=S、c=R、d=S、e=R β−D−1−デオキシガラクトピラノシド模倣物
b=S、c=R、d=S、e=R α−D−1−デオキシガラクトピラノシド模倣物
b=S、c=R、d=R、e=S β−L−1−デオキシイドピラノシド模倣物
b=S、c=R、d=R、e=S α−L−1−デオキシイドピラノシド模倣物
【0027】
本発明によるデオキシノジリマイシン類似体は、疎水性部分及びスペーサーを含む少なくとも1個の存在Xを含む。存在Xは、位置R1、R2、R3、R4、R5、R6のいずれかの箇所に位置する。
【0028】
スペーサー及び疎水性部分は、図1に示すように、Z及びYによって適切に表すことができる。
【0029】
大きな疎水性部分は、炭素原子3から8個のアルコキシポリアルキレン鎖又はポリアルキレン鎖を含むスペーサーによってデオキシノジリマイシンの前記の窒素原子に結合していることが好ましい。大きな疎水性部分は、アダマンタンメタノール、コレステロール、β−コレスタノール、アダマンタノール、及び9−ヒドロキシフェナントレンからなる群から選択される化合物から誘導されることがより好ましい。
【0030】
用語「スペーサー」とは、任意の二価部分、又はデオキシノジリマイシンのN原子に疎水性基を結合することのできる基を指す。
【0031】
本発明によるデオキシノジリマイシン類似体は、イド−立体配置を有することが好ましい。詳細には、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンが好ましい化合物である。
【0032】
本発明によるデオキシノジリマイシン類似体の調製を以下の実施例によって例示する。
【発明を実施するための最良の形態】
【0033】
N−(アダマンタン−1−イル−メトキシペンチル)−L−イド−デオキシノジリマイシン4
2,3,4,6−テトラ−O−ベンジル−L−イド−デオキシノジリマイシン1(Tetrahedron Lett. 44, 3085-3088, 2003)を水素化(H2、Pd/C、HCl(水溶液)、EtOH)によって脱ベンジル化して3にし、これを還元アミノ化条件(H2、Pd/C、HCl(水溶液)、EtOH)下でアダマンタン−1−イル−メトキシペンタナール2(J. Biol. Chem. 273, 26522, 1998、WO98102161)と縮合させて、N−(アダマンタン−1−イル−メトキシペンチル)−L−イド−デオキシノジリマイシン4を得た。精製すると、均質な目的化合物4が淡黄色のシロップとして収率93%で得られた(図2を参照のこと)。
【0034】
N−(アダマンタン−1−イル−メトキシペンチル)−D−ガラクト−デオキシノジリマイシン7
7の合成は、イド同属化合物4について上で概略を述べた合成経路に従い、2,3,4,6−テトラ−O−ベンジル−D−ガラクト−ノジリマイシン7(Tetrahedron 56, 32, 5819-5834, 2000)を出発材料として実施した(図3を参照のこと)。
【0035】
C1−β−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン17、N−メチル−C1−β−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン18、及びN−ブチル−C1−β−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン19
β−C1−アルキルデオキシノジリマイシン誘導体17、18、及び19の合成は、5−O−(アダマンタン−1−メチル)−1−ペンチノール13(図4を参照のこと)の調製から始める。ペンチン−1−オール8を完全にシリル化(nBuLi、TMSCl)した後、シリル−エーテルを選択的に脱保護して(1N HCl)、保護されたアセチレン9を得た。9の遊離アルコールをトリフルロメタンスルホニルエステル10に変換した(塩化トリフルオロメタンスルホニル、トリエチルアミン)。アダマンタン−メタノール11を10でアルキル化(K2CO3、還流CH2Cl2)し、その後TMS保護基を除去すると(ナトリウムメタノラート、MeOH/THF)、8に対して64%の収率でアセチレン13を得た。13のリチオ化物(−50℃のTHF中でブチルリチウムと反応させて調製したもの)を、2,3,4,6−テトラ−O−ベンジル−D−グルコノ−デルタ−ラクトン14(J. Org. Chem., 2531, 1967)に求核付加すると、ケトグルコシド15がアノマー混合物として得られた。化合物15を、次の3ステップの手順を使用して、容易に且つ高い立体選択性で、完全に保護されたイミノ糖16に変換した(Eur. J. Org. Chem. 5, 1185-1189, 1999)。即ち、1)対応するジオールに還元し(NaBH4、CH2Cl2/MeOH)、2)酸化させてジケト化合物にし(Swern酸化)、3)ギ酸アンモニウム及びシアノ水素化ホウ素ナトリウムを使用して二重還元アミノ化を行った(14に対する全体としての収率は45%であった)。パラジウムを触媒とする水素化にかけると、イミノ糖17が得られ(79%)、これから、それぞれホルムアルデヒド(18、収率20%)及びブタナール(19、収率73%)を用いる還元アミノ化によって、メチル及びブチル同族体18及び19を容易に調製した。
【0036】
2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン28、N−メチル−2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン29、及びN−ブチル−2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン30
2−O−アルキル化デオキシノジリマイシン誘導体28、29、及び30の調製(図5を参照のこと)は、アリル−3,4,6−トリ−O−ベンジル−β−D−グルコピラノシド20(Tetrahedron: Asymmetry, 8, 765-774, 1997)の遊離アルコール官能基をp−メトキシベンジル保護し、その後アノマー性アリル基を異性化/加水分解して、2ステップで80%の22を得ることから出発する。ヘミアセタール22を、次の4ステップを使用して、直角をなすように保護されたデオキシノジリマイシン誘導体23に変換した。即ち、1)対応するジオール(LiAlH4、THF)に還元し、2)酸化させてアルデヒドケトンにし(Swern酸化)、3)ギ酸アンモニウム及びシアノ水素化ホウ素ナトリウムを使用して二重還元アミノ化を行い、4)得られる第二級アミンをベンジルオキシカルボニル基で保護した(ZCl、K2CO3、4ステップで71%)。23を2%のTFAで処理すると、化合物24が得られ(収率99%)、次のアルキル化に向けてC−2−OHを選択的に脱保護した。アルデヒド2から、アルデヒド官能基を対応する第一級アルコールに還元した後(NaBH4、MeOH)、PBr3の作用のもとで臭素化して、対応するアルキル化剤1−ブロモ−5−(アダマンタン−1−イル)メトキシペンタン26を調製した(88%、2ステップ)。過剰の26を加え、DMF中で水素化ナトリウム処理することで24のアルキル化を行って、83%で27を得た。27のZ保護基及びベンジル保護基を還元によって除去すると、イミノ糖28が得られ(収率80%)、これから、それぞれホルムアルデヒド(29、収率49%)及びブタナール(30、収率79%)を用いる還元アミノ化によって、メチル及びブチル同族体29及び30を容易に調製した。
【0037】
分光データ:
【化2】
N−(アダマンタン−1−イル−メトキシペンチル)−L−イド−デオキシ−ノジリマイシン(4):1H−NMR(MeOD,400MHz):δ4.01〜3.97(dd,1H,H6)、3.89(m,1H,H2)、3.85〜3.70(m,3H,H2,H3,H4)、3.55〜3.52(m,2H,CH2スペーサー)、3.40〜3.08(m,5H,2*H1,H5,CH2スペーサー)、2.96(s,2H,O−CH2メトキシ)、1.94(広幅s,3H,CHアダマンチル)、1.77〜1.66(広幅dd,6H,3*CH2アダマンチル)、1.63〜1.54(d,4H,2*CH2スペーサー)、1.55(d,6H,3*CH2アダマンチル)、1.45〜1.37(m,2H,CH2中央スペーサー)。MS(ESI):obs.m/z=398.2[M+H]+;C22H39NO5(MW計算値):397.3。
【0038】
【化3】
2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシ−ノジリマイシン(28):1H−NMR(MeOD,400MHz):δ3.92〜3.84(2*dd,2H,2*H6)、3.72〜3.62(m,2H,O−CH2スペーサー)、3.57〜3.43(m,3H,H2,H3,H4)、3.38〜3.36(m,2H,O−CH2スペーサー)、3.07(m,1H,H5)、2.95(s,2H,O−CH2メトキシ)、2.89〜2.79(m,2H,2*H1)、1.93(広幅s,3H,CHアダマンチル)、1.76〜1.65(広幅dd,6H,3*CH2アダマンチル)、1.62〜1.56(d,4H,2*CH2スペーサー)、1.54(d,6H,3*CH2アダマンチル)、1.45〜1.39(m,2H,CH2中央スペーサー)。MS(ESI):obs.m/z=398.2[M+H]+;C22H39NO5(MW計算値):397.3。
【0039】
【化4】
N−メチル−2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシ−ノジリマイシン(29):1H−NMR(MeOD,400MHz):δ4.13〜4.10(2*広幅d,2H,2*H6)、3.72〜3.64(m,2H,O−CH2スペーサー)、3.62〜3.48(m,3H,H2,H3,H4)、3.42〜3.38(m,2H,O−CH2スペーサー)、3.09〜2.89(m,3H,H5及び2*H1)、3.01(s,3H,N−CH3)、2.95(s,2H,O−CH2メトキシ)、1.96(広幅s,3H,CHアダマンチル)、1.78〜1.65(広幅dd,6H,3*CH2アダマンチル)、1.62〜1.56(d,4H,2*CH2スペーサー)、1.57(d,6H,3*CH2アダマンチル)、1.48〜1.42(m,2H,CH2中央スペーサー)。MS(ESI):obs.m/z=412.2[M+H]+;C23H41NO5(MW計算値):411.3。
【0040】
N−ブチル−2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシ−ノジリマイシン(30):MS(ESI):obs.m/z=454.2[M+H]+;C26H47NO5(MW計算値):453.3。
【0041】
【化5】
N−ベンジルオキシカルボニル−2−O−(アダマンタン−1−イル−メトキシペンチル)−3,4,6−トリ−O−ベンジル−デオキシ−ノジリマイシン(27):1H−NMR(CDCl3,400MHz):δ7.31〜7.25(m,20H,CH Bn)、5.16〜5.06(2H,CH2 Bn)、4.75〜4.49(2*CH2Bn)、4.43〜4.31(2H,CH2 Bn)、4.14〜4.12(m,1H,H5)、4.00〜3.97(dd,1H,H1 ax又はeq)、3.93〜3.89(dd,1H,H4)、3.67〜3.54(m,4H,H2,H3,2*H6)、3.38〜3.31(m,5H,H1 ax又はeq,2*O−CH2スペーサー)、2.93(s,2H,CH2メトキシ)、1.94(広幅s,3H,CHアダマンチル)、1.71〜1.62(広幅dd,6H,3*CH2アダマンチル)、1.58〜1.52(m,10H,3*CH2アダマンチル,2*CH2スペーサー)、1.38〜1.33(m,2H,CH2中央スペーサー)。13C−NMR(CDCl3,100MHz):δ155.7(C=O,Z基)、138.3,136.6(Cq Bn及びZ)、128.4〜127.4(CH芳香族Bn)、82.4,79.5,74.3(3*CH,C2,C3及びC4)、73.1,72.9,72.8,71.5,69.0,68.5,67.1(7*CH2,3*CH2Bn,CH2 Z基,2*O−CH2スペーサー,C6)、55.8(C5)、41.6(C1)、39.7,37,2(2*CH2アダマンチル)、34.0(Cqアダマンチル)、29.7,29.4(2*CH2スペーサー)、28.2(CHアダマンチル)、22.7(CH2中央スペーサー)。MS(ESI):obs.m/z=802.4[M+H]+;824.6[M+Na]+;C51H63NO7(MW計算値):801.5。
【0042】
適用分野
1.II型糖尿病(インスリン抵抗性)
II型糖尿病の発生率は、欧米諸国で劇的に増加している。主たる根本的な欠陥は、インスリンに応答してGLUT4輸送体をその細胞表面に動員する感受性が低下する結果として、筋肉及び脂肪組織による血流からのグルコースの取込みが損なわれるものである。筋肉中の脂肪酸濃度の上昇が異常なグルコース恒常性に関連していることは、すでに何年も前からわかっている。逆に、PPARγアゴニスト及びレキシノイドによって誘発されるグルコース恒常性の改善は、脂肪酸の分割の変更、即ち脂肪組織への脂肪酸の再配分、並びに筋肉の脂肪酸取込み及び代謝の相対的な消耗と関連付けられる。しかし、筋肉での脂肪毒性が糖尿病の発症及び進行を引き起こす分子機序は十分に理解されていない。したがって、この問題について洞察をさらに深めることが、インスリン抵抗性を治療するための医薬の改良/開発の助力となる。
【0043】
脂肪病因の分子機序
大学医学センター(Academic Medical Center)/アムステルダム大学生化学部のスフィンゴ糖脂質及びII型糖尿病についての研究活動は最近、II型糖尿病の脂肪病因に思いがけない新たな見識をもたらした。根底にある機序を以下で詳述する。
【0044】
後天性のインスリン抵抗性におけるスフィンゴ糖脂質の役割
糖尿病の病因におけるスフィンゴ(糖)脂質の役割について仮説を立てた。この考えは、パルミチン酸がスフィンゴ脂質中のセラミド部分の本質的な構築ブロックであり、スフィンゴ糖脂質の生合成の最初のステップは、セリンパルミトイルトランスフェラーゼを触媒とするパルミチン酸のセリンへの転移である(図1を参照のこと)という、見過ごされていた事実に端を発する。肝臓でのスフィンゴ脂質の合成速度は、パルミチン酸の濃度に著しく左右される。重要なことだが、このことは、培養筋肉細胞(平滑筋細胞、筋原細胞)で実験に基づいて確認できた。即ち、培地に0.1、0.5、1.0mMのパルミチン酸を加えると、放射標識したセリンのその構造への取込みの増加によって明らかになったとおり、スフィンゴ糖脂質の合成が比例して増加した。
【0045】
この発見を機に、スフィンゴ(糖)脂質が、糖尿病の根底にある筋肉中での脂肪毒性を実際に媒介する可能性についてより詳しく調査することになった。最近では、GM3(細胞表面にある最も単純なガングリオシド、図2を参照のこと)がインスリンシグナル伝達を弱め得ることが証明されている。これに関して、細胞表面でのGM3濃度が、インスリン受容体の多重クラスター形成のマイナスの妨げとなって、インスリンに応答してのグルコースの取込みを調節するらしいことが観察されている。さらに、高濃度のGM3は、GLUT4の細胞表面への動員の縮小化と関連付けられる。逆に、GM3の減少は、インスリン感受性の向上と関連付けられる(Yamishita et al. Proc Natl Acad Sci USA (2003) 100, 3445-9 Enhanced insulin sensitivity in mice lacking ganglioside GM3、Tagami et al. (2002) J Biol Chem 277, 3085-92 Ganglioside GM3 participates in the pathological conditions of insulin resistanceを参照のこと)。本発明者らは、肥満状態ではパルミチン酸レベルが慢性的に高く、したがって、脂肪細胞だけでなく筋細胞でのスフィンゴ糖脂質の生成が速まった速度で行われて、インスリン抵抗性に有利に働くと見ている。局所での(GM3を含む)スフィンゴ糖脂質合成を増大させる推進力としての筋肉中パルミチン酸濃度の増大とインスリン抵抗性との関係(図3を参照のこと)は、これまでのところ他者に認知されていない。
【0046】
グルコシルセラミドシンターゼの決定的役割
また、細胞表面のGM3及び他のガングリオシドの濃度が、ガングリオシド合成の律速段階であるグルコシルセラミドシンターゼの活性(グルコシルセラミドの合成)に著しく左右されることは十分に理解されていた(図3を参照のこと)。この酵素は、セラミドとUDP−グルコースからのグルコシルセラミドの生成を触媒する。その両方の基質(セラミド及びUDP−グルコース)のKm値は、生理的範囲にある。本発明者らは、インスリン感受性に関してグルコシルセラミドシンターゼが鍵調節酵素であることを示す。炎症性サイトカイン(TNF−α)、ステロイドホルモン、飽和脂肪酸、及びウイルス感染に応答してのその活性の増大は、以前から観察され、報告されている。今回、驚くべきことに、スフィンゴ糖脂質合成の変化がII型糖尿病の助長に影響を及ぼすことが判明した(図4を参照のこと)。脂肪病因がII型糖尿病に有害な影響を与えるというこの知見は、グルコシルセラミドシンターゼ活性の阻害が、有益な抗高血糖作用を発揮することを示すものである。
【0047】
イミノ糖系阻害剤の新規な使用
特定の部類のイミノ糖であるデオキシノジリマイシンが、グルコシルセラミド合成の阻害によってスフィンゴ糖脂質合成を低減するのに適する薬剤であることは明白になっている。さらに、ヒトにおけるイミノ糖投与の安全性と共に、実践での専門知識もかなり得られた。
【0048】
N−ブチル−デオキシノジリマイシンは、最近になって1型ゴーシェ病の治療用薬物として登録された。ケンブリッジ大学と共同でアムステルダム大学医学センターが主として請け負った臨床治験では、この薬物が大多数の患者で少なくとも5年間は忍容されることが明らかになった。スフィンゴ糖脂質合成が、100mgの1日3回のN−ブチル−デオキシノジリマイシンによってごく中程度(20〜30%)にだけは阻害されるという事実にもかかわらず、一部のゴーシェ病患者はしかし、深刻な腸の不調を訴え、時には警戒させる末梢ニューロパシーになる。より多い用量のN−ブチル−デオキシノジリマイシンでは、これらの有害事象がより頻繁に起こる。本発明者らは、グルコシダーゼ及びグルコシルトランスフェラーゼの阻害についてN−ブチル−デオキシノジリマイシンの特異性が不十分であるために、これらの望ましくない副作用が生じると見ている。表1に示すように、N−ブチル−デオキシノジリマイシンは、腸管グリコシダーゼの非常に強力な阻害剤でもある。本発明者らは、この阻害作用が、患者の腸の不調の少なくとも一部をもたらしていると見ている。この薬物は、リソソーム内のαグリコシダーゼ及びグルコセレブロシダーゼの阻害剤である。本発明者らは、この作用が、リソソームにおけるグリコーゲン及びグルコセレブロシダーゼの病的なリソソーム内蓄積への一連のリスクをもたらすと見ている。糖尿病(予備)患者のかなり低めのGM3に必要な濃度では、N−ブチル−デオキシノジリマイシンを使用すると有害事象が確実に起こる。
【0049】
インスリン抵抗性(II型糖尿病)治療薬としての特異的阻害剤の設計
N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシン(以前はAMP−DNMとしても知られていた)は、II型糖尿病治療薬としてのその使用についていくつかの魅力的な特徴を示す。即ち、この化合物は、経口投与するとき生物学的利用能が高く、グルコシルセラミドシンターゼの強力な阻害剤であり、代謝に関して不活性であり、治療中に想定される用量では固有の細胞毒性を示さない。
【0050】
しかし、in vitro酵素活性検定によって、培養細胞を用いてなされた観察によれば、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの長期投与が、他の代謝経路にも影響を及ぼし得る可能性があると思われる。例えば、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンを長期間投与した後、リソソーム内グリコシダーゼのα−グルコシダーゼとリソソーム内グルコセレブロシダーゼが阻害される可能性は、完全には除外できない。このことは、それぞれグリコーゲン又はグルコシルセラミドのリソソーム内蓄積をもたらすかもしれない。グリコーゲン又はグルコシルセラミドがリソソームに過剰に貯蔵されると、それぞれポンペ病及びゴーシェ病に似た病態になりかねない。その上、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、腸管スクラーゼの阻害剤でもあり、それとして胃腸の愁訴を引き起こすかもしれない。腸管グリコシダーゼが強力に阻害されれば、消化管内腔に浸透活性のある糖類が蓄積され、また腸内細菌の増殖に有利に働くはずであり、どちらも痙攣及び下痢の一因となる。このような合併症は、服薬遵守に影響を及ぼすはずであり、実際にはN−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの適用を狭めることになる。
【0051】
これらすべての考慮すべき事柄に導かれ、本発明者らは、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの所望の特性を保持したまま、インスリン抵抗性個体の長期間の治療に関してその不利な特徴のないイミノ糖構造の探索に乗り出した。
【0052】
以下の2種の選択判断基準を適用した。
1.リソソーム内グリコシダーゼ:βグルコシダーゼ及びグルコセレブロシダーゼの阻害の低減
これに関して、イミノ糖によるリソソーム内グリコシダーゼの阻害は、リソソーム貯蔵障害によく似た病的な代謝産物のリソソーム内貯蔵のリスクを高めるだけになりかねないので望ましくない。
【0053】
2.腸管グリコシダーゼの阻害の欠如
II型糖尿病に介入する別の手法は、腸管グリコシダーゼを阻害して、消化管での食物由来の糖の取込みを和らげることに基づく。合成のスクラーゼ阻害剤(アカルボース、N−ヒドロキシエチル−デオキシノジリマイシン)は、この構想に基づくものであり、抗糖尿病薬として登録されている。
【0054】
N−ヒドロキシエチル−デオキシノジリマイシンは、スクラーゼ阻害剤の中で最も強力な抗糖尿病薬である(Campbell L, Baker DE & Campbell RK, Ann Pharmacother 2000; 34: 1291-1301)。しかし、N−ヒドロキシエチル−デオキシノジリマイシン及びアカルボースのような強力な合成腸管グリコシダーゼ阻害剤の大きな欠点は、免れがたい深刻な腸の愁訴を引き起こし得ることである。したがって、強力な腸管グリコシダーゼ阻害剤であるN−ヒドロキシエチル−デオキシノジリマイシン及びアカルボースは、多くの個体にとって忍容性があまり十分でなく、服薬遵守が不十分になり、適用が制限される。
【0055】
N−ヒドロキシエチル−デオキシノジリマイシン及びアカルボースは、有益な効果を発揮するとはいえ、本発明者らは、それがその腸管グリコシダーゼ阻害能と関係しているかどうかは実際には疑わしいことに気付いた。興味深いことに、N−ヒドロキシエチル−デオキシノジリマイシン又はアカルボースの投与は、高炭水化物食なしでさえ、血糖値の低下をもたらす。このことは、この化合物の推定される治療作用方式と一致しない。おそらく、N−ヒドロキシエチル−デオキシノジリマイシン及びアカルボース(の代謝産物)は、体に取り込まれた後、グルコシルセラミドシンターゼを阻害してむしろ有益に働くと思われる。このことは、腸管グリコシダーゼの阻害がその治療効果の前提条件なのではなく、望ましくない副作用をもたらすに過ぎないことを示唆している。
【0056】
上述の選択判定基準に基づき、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンのイド立体配置が、インスリン抵抗性(II型糖尿病)の治療にとって魅力的なイミノ糖であると特定された。表1は、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンが、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンと比べて、リソソーム内グルコセレブロシダーゼを阻害せず、β−グルコシダーゼ活性を弱く阻害するに過ぎないことを示している。その上、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、腸管グリコシダーゼの弱い阻害剤である。重要なことであるが、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、培養細胞で試験するとやはりグルコシルセラミドシンターゼの強力な阻害剤である(表2を参照のこと)。
【0057】
【表1】
【0058】
【表2】
【0059】
抗血糖上昇剤としてのイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの価値は、その後N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンについて先に述べた手順に厳密に従って動物モデルで分析した。この目的では、動物に異なる濃度のイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンを毎日与えた。250mg/kgの用量のイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンで、25mg/kgのN−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンと同様の有益な効果が示された。
【0060】
肥満におけるスフィンゴ糖脂質の役割
本発明者らは、(パルミチン酸に富んだ)動物性脂肪及び単純糖質(単糖及び二糖)の消費の増加傾向が、スフィンゴ糖脂質の過剰産生に有利に働くことを発見した。高レベルのスフィンゴ糖脂質は、エネルギー恒常性に関与するホルモン、例えばインスリンを抑制する。高いスフィンゴ糖脂質レベルは、エネルギーの収支のバランスをますます悪くし、進行中の体重増加及び随伴する副疾患に有利に働く。この新規な見識は、過体重及び肥満の個体においてスフィンゴ糖脂質の過剰産生を低減することのできるイミノ糖の使用という、治療処置への新たな道を切り開くものである。したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、過体重及び肥満の治療に適切に使用することができる。
【0061】
過体重及び肥満を治療するアルキル化イミノ糖の選択
特に、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、この目的に申し分なく適する化合物であることがわかった。この化合物は、生物学的利用能が十分であり、想定される投与量で細胞毒性を示さず、グルコシルセラミドシンターゼの比較的特異的な阻害剤である。イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの利点は、大抵のイミノ糖によって引き起こされる合併症である腸の愁訴を引き起こさないことである。このような副作用がないことは、服薬遵守を向上させる。
【0062】
2.過体重及び肥満
世界中で、過体重及び肥満の有病率が流行の様相を帯びてきている。オランダでは、他の国同様、過体重及び肥満の個体の数がじりじりと増えている。他のヨーロッパ諸国の状況と似てはいるが、この増加は、例えば英国やドイツほど著しくない。最も圧倒的なのは、米国での有病率の増加である。平均して、成人オランダ人の40%が過体重であり、成人人口の10%が肥満である。成人の1〜1.5%が病的な肥満に罹患していると推測される。2000年には、米国の成人の56%が過体重であり、20%が肥満であり、2.3%が病的な肥満であった。WHOの規定によれば、成人は、BMIが30kg/m2以上であれば肥満(重度の過体重)であるとされる。BMIの値が25kg/m2と30kg/m2の間にある者は、過体重であるという。BMI(Body Mass Index)は、個体の体重(kg)をその身長(メートル)の2乗で割ったものであると定義される。
【0063】
体重増加の最初の結果の1つが、インスリンの正常な働きを狂わせるインスリン抵抗性である。インスリン抵抗性は、代謝症候群の進展において鍵となる役割を担う。この症候群は、インスリン抵抗性、異脂肪症(低HDL血清コレステロール、高血清トリグリセリド、高LDL血清コレステロール)、高血圧、腹部肥満など、いくつかの関連した代謝の異常を特徴とする。これらの異常は、II型糖尿病(成人型糖尿病)及びその合併症などの障害の進展の基盤となる。過体重及び肥満に随伴する他の健康上のリスクは、心臓病、様々な種類のがん、胆嚢疾患、関節症、呼吸器の問題、痛風、不妊症、月経障害、及び胎児の欠陥である。過体重が重症であるほど、このような副疾患のリスクは高まる。これらのすべての健康上のリスクの中でも、グルコース不耐症及びII型糖尿病の有病率の増大は特に厄介である。米国では、これが子供でも起こっている。最近の調査では、肥満児(5〜17才)の約60%が心臓病の追加の危険因子を示したことが明らかになっている。グルコース不耐症は、55人の肥満児(4〜10才)の25%、及び112人の若者(11〜18才)の21%で認められた。さらに、肥満はしばしば、心理的社会的問題、並びに生活の質の低下を伴う。肥満(及びそれ程ではないが過体重)に随伴する病的状態は、非常に多くの(医薬による)治療及び付加的な就業不能だけでなく、健康管理のための費用の増大をもたらす。米国では、過体重及び肥満に直接に関連した健康管理の費用が全体の6〜7%であると推定され、間接的な費用は4倍になると推測される。
【0064】
3.リソソーム貯蔵障害
リソソーム内のスフィンゴ糖脂質の蓄積を特徴とする遺伝性障害(GM1ガングリオシドーシス)、サンドホフ病(GM2ガングリオシドーシス、II型又はO型)、テイサックス病(GM2ガングリオシドーシス、I型又はB型)、ファブリー病(a−ガラクトシル−ラクトシルセラミドーシス)、ラクトシルセラミドーシス、ゴーシェ病(グルコセレブロシドーシス)は、西欧世界において、診断されたすべての先天性代謝異常の中で大きな割合を占めるものである。これらの疾患に罹患している患者が、蓄積するスフィンゴ糖脂質の合成の低減による恩恵を受けるはずであることは、以前から想定されている。最近では、N−ブチル−デオキシノジリマイシンが、いわゆる基質低減療法(SRT)による1型ゴーシェ病の治療用に登録されている。
【0065】
N−ブチル−デオキシノジリマイシンを用いた臨床経験から、この化合物の適用が、このイミノ糖の不十分な特徴によって制限されることがわかった。N−ブチル−デオキシノジリマイシンは、低い生物学的利用能しか示さず、グルコシルセラミドシンターゼの比較的弱い阻害剤であるので、有効性が不十分である。一方、N−ブチル−デオキシノジリマイシンは、リソソーム及び腸管のグリコシダーゼを著しく阻害し得るために、高めの用量では重い副作用を引き起こす。
【0066】
グリコスフィンゴリピドーシスのSRTのための特異的なデオキシノジリマイシン誘導体:イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシン
本発明者らは、スフィンゴ糖脂質貯蔵障害のSRTにより最適なイミノ糖を設計できることを発見した。化合物は、グルコシルセラミドシンターゼを阻害するその効力と、リソソーム及び腸管のグリコシダーゼの活性を阻害し得ないことに基づいて選択した。表3及び表4は、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンが、N−ブチル−デオキシノジリマイシンと比べて優れた特徴を示す理想的な物質であることを示すものである。
【0067】
【表3】
【0068】
酵素活性は、脾臓からの精製酵素を使用して、適切な4−MU基質で測定した。IC50値は、酵素活性を50%阻害するのに必要なイミノ糖の量を評価して決定した。
【0069】
【表4】
【0070】
in vivoでのグルコシルセラミドシンターゼ活性は、J. Biol. Chem, 273, 26522-27 (1998)に記載の手順に従って決定した。簡潔に述べると、グルコシルセラミドシンターゼ活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−セラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用薄層クロマトグラフィーにかけた後、C6−NBD−セラミドのC6−NBD−グルコシルセラミドへの変換を分析した。
【0071】
4.炎症性疾患
炎症は、局部組織の破壊及び関連する臨床症状を伴う。慢性の炎症は、特に有害な過程である。そのようなタイプの炎症はしばしば、慢性的に活性化したマクロファージによって統御される。様々な薬剤が炎症及び慢性の炎症の治療に使用される。しかし、これらの薬剤は、効果が最適でないか、又は重い副作用を示す。異なる作用方式に基づく改良された抗炎症剤が求められている。
【0072】
炎症におけるスフィンゴ糖脂質の役割
今回、炎症、特に慢性の炎症がスフィンゴ糖脂質の過剰産生と関連付けられること、並びにこの過剰産生がマクロファージの炎症状態を強化し、したがって組織炎症カスケードを促進することが判明した。スフィンゴ糖脂質レベルの増大によって、2通りの方式でマクロファージが活性化される。第1に、非リソソーム内グルコシルセラミダーゼによってグルコシルセラミドが過剰に分解されて、シグナル伝達分子として働くセラミドが生成される。さらに、セラミドの一部が、PKCを刺激するジアシルグリセロールへと代謝される。セラミド及びジアシルグリセロールの産生は、どちらも進行中のマクロファージの活性化に有利に働く。第2に、炎症の際のスフィンゴ糖脂質の増加が、細胞表面にあるラフトのスフィンゴ糖脂質組成を変化させる。これが、正常な細胞挙動に影響を及ぼし、炎症の過程を促進する。
【0073】
新しいクラスの抗炎症剤
先の発見によって、本発明者らは、イミノ糖が新規な作用方式によって働く有用な抗炎症剤に違いないと想定した。所望の特徴に基づき、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンを申し分なく適する抗炎症剤として選択した。この化合物は、生物学的利用能が十分であり、想定される投与量で細胞毒性を示さず、忍容性が十分である。この化合物は、そのどちらの酵素活性も炎症の重症度及び慢性の性質に寄与する非リソソーム内グルコシルセラミダーゼ及びグルコシルセラミドシンターゼの強力な阻害剤である。したがって、デオキシノジリマイシン類似体又は薬学的に許容できるその塩は、炎症性疾患の治療に適切に使用することができる。
【0074】
5.色素沈着過剰症及び皮膚の炎症状態
色素沈着
ヒトは、2種類のメラニン色素、即ち、褐色/黒色型若しくはユーメラニンと、琥珀色/赤色型若しくはフェオメラニンを産生する。ユーメラニンが、主として皮膚、毛髪、及び眼に見られる着色を司る。メラニンの合成は、特殊な細胞の特定の区画、即ちメラノサイトのメラノソームで行われる。色素は、その後皮膚のケラチノサイトに移送される。
【0075】
色素沈着は通常、年齢と共に増し、遺伝的な欠陥又は後天的な疾患によって変更される場合がある。ある場合では、それが色素沈着の増加(過色素沈着)になり、またある場合では、それがより薄い着色(色素沈着低下)になる。いくつかの種類の色素沈着過剰症が存在する。即ち、1.そばかす、2.妊娠に関連した過色素沈着、3.酵素の欠損による過色素沈着、4.疾患に関連した(炎症後)過色素沈着である。そばかすは、メラノサイト(色素生成細胞)が、近くの皮膚よりもUV放射に対して活性であり、且つ応答しやすい区域である。妊娠中に見られる色素沈着の増加は、エストロゲン、プロゲステロン、及びメラノサイト刺激ホルモンの影響のためである。妊娠中に起こるものなど、ホルモンに関連した過色素沈着は、妊娠マスク又は肝斑と呼ばれる状態として現れることもある。これは、顔又は日光露出区域に色素沈着の斑が出現する状態である。肝斑は無害である。肝斑で見られる色素は、分娩後数カ月以内にほとんど消失する。カロチンを代謝する肝臓の酵素が欠損している個体は、ニンジン、トウガラシ、又は他の黄色/橙色野菜を大量に消費すると、皮膚の色が黄色/橙色になる場合がある。炎症後過色素沈着(PIH)は、頻繁に直面する問題であり、様々な皮膚障害並びに治療処置の後遺症のことである。この後天的な色素過剰は、皮膚に影響を及ぼすそれまでの様々な疾患の過程を経た結果であるとすることができ、そうした過程には、感染、アレルギー反応、機械的損傷、薬剤投与に対する反応、光毒性発疹、外傷(例えば、火傷)、及び炎症性疾患(例えば、扁平苔癬、紅斑性狼瘡、アトピー性皮膚炎、乾癬、サルコイド症)が含まれる。
【0076】
炎症後過色素沈着
PIHは、表皮メラニン沈着又は真皮メラニン沈着をもたらす2通りの機序のうちの一方によって引き起こされる。表皮の炎症応答(即ち、皮膚炎)は、アラキドン酸を放出させ、その後酸化によってプロスタグランジン、ロイコトリエン、及び他の産物をもたらす。これらの炎症産物は、免疫細胞及びメラノサイトの両方の活性を変更する。詳細には、これらの炎症産物は、表皮メラノサイトを刺激して、それにメラニンの合成を増進させ、その後周囲のケラチノサイトへの色素の移送を増進させる。メラニン顆粒のこのような刺激及び移送の増進は、表皮の黒皮症をもたらす。一方、真皮メラニン沈着は、炎症が基底細胞層を乱すときに起こり、メラニン色素を放出させ、その後真皮乳頭中のマクロファージに捕えられるようにするものであり、色素失調症としても知られる。
【0077】
PIHは、皮膚のすべてに及ぶ応答であるが、着色された、より黒い皮膚ではより一般的である。メラニン過剰病巣の分布は、もともとの炎症性皮膚疾患の位置に応じて決まる。病巣の色は、淡褐色から黒色まで様々であり、色素が表皮内にある場合(即ち、表皮メラニン沈着)はより明るい褐色に見え、病巣が真皮メラニンを含む場合(即ち、真皮メラニン沈着)はより暗い灰色に見える。PIHは、皮膚に影響を及ぼす様々な疾患過程と共に起こる。それらの過程には、アレルギー反応、感染、外傷、及び光毒性発疹が含まれる。PIHをもたらす一般的な炎症性疾患としては、にきび、扁平苔癬、全身性紅斑性狼瘡、慢性皮膚炎、及び皮膚T細胞リンパ腫、特に異型紅皮症、乾癬、サルコイド症が挙げられる。
【0078】
デオキシノジリマイシン類似体による色素沈着過剰症及び皮膚炎症状態の治療
炎症におけるスフィンゴ糖脂質の役割については、前の項で遍く論述した。重要なことに、メラノサイト中でのメラニン産生の絶対要件は、グルコシルセラミドのようなスフィンゴ糖脂質の存在である。グルコシルセラミドシンターゼ活性を欠く糖脂質陰性メラノーマ細胞系は、スフィンゴ糖脂質合成が正常なその親細胞とはメラニンがない点で異なる。スフィンゴ糖脂質を加え、又はグルコシルセラミドシンターゼcDNAを形質移入すると、メラニン産生は復活するので、グルコシルセラミド(及び/又は他のスフィンゴ糖脂質)の合成がメラノソームでのメラニン産生にとって不可欠であることが示唆される。グルコシルセラミドなどのスフィンゴ糖脂質の産生は、チロシナーゼのみを適切にメラノソームに移送するのに必要であると思われる。
【0079】
今回、スフィンゴ糖脂質の過剰産生が皮膚の炎症過程の鍵となる要因であり、したがって本質的にメラニンの過剰産生(色素沈着過剰症)を引き起こし得ることがわかった。本発明者らは、(局所の炎症状態の結果としての)スフィンゴ糖脂質の過剰産生が、局所のメラニン合成の増加を引き起こすと見ている(以下のスキームを参照のこと)。デオキシノジリマイシン類似体などの、本発明によるスフィンゴ糖脂質合成の阻害剤は、過剰なメラニン産生及び随伴する色素沈着過剰症を軽減することができる。
【0080】
したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、色素沈着過剰症及び皮膚の炎症状態の治療に適切に使用することができる。これまで、メラニン産生に介入する手法は、スフィンゴ糖脂質合成以外のメラニン形成過程のターゲットに注目したものであった。多くの脱色素剤は、チロシナーゼの代替基質として働き、したがって、メラニン合成の最初の酵素段階、即ち、メラニン重合体をもたらすチロシナーゼのドーパキノンへの変換を阻害するものである。これは、通常は過色素沈着を実に効果的に軽減又は解消するものの、色が黒めの皮膚の患者では、オクロノーシス(真皮中の黒色の細かい斑点)を引き起こす場合もある。その上、ヒドロキノンは、高めの濃度では発癌性があると考えられているため、ヨーロッパの一部及びアジア全域で禁止されている。
【0081】
色素沈着過剰症及び皮膚の炎症過程の治療に向けた特異的デオキシノジリマイシン類似体
皮膚での使用に最適な特徴を有するイミノ糖を設計した。セラミド及びグルコシルセラミドは、角質層の構造の重要な構成成分であり、皮膚の水分透過性を調節している。グルコシルセラミド並びに他の極性脂質が、層状体によって角質層の細胞外間隙に分泌され、その後処理を受けてセラミドになることは知られている。リソソーム内グルコセレブロシダーゼは、この酵素が完全に欠損すると、それに伴って表皮のバリア機能が著しく損なわれるという事実が示唆するとおり、この過程でおいて一役を担う。したがって、イミノ糖は、リソソーム内グルコセレブロシダーゼを触媒とする、グルコシルセラミドからのセラミドの決定的な生成を妨げるべきでない。この前提条件を踏まえて、イミノ糖イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンが理想的な化合物であることがわかった(表5)。イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、グルコシルセラミドシンターゼ活性を阻害し、ヒトメラノサイトでのメラニン産生を低減することができる。
【0082】
【表5】
【0083】
培養メラノーマ細胞中でグルコシルセラミドシンターゼ、非リソソーム内グルコシルセラミダーゼ、及びリソソーム内グルコセレブロシダーゼを阻害し得るかどうかについて、様々なイミノ糖を分析した。最初の2種の酵素活性の阻害は望ましいが、最後の酵素の活性の阻害は望ましくない。in vivoでのグルコセレブロシダーゼ活性、非リソソーム内グルコシルセラミダーゼ活性、及びグルコシルセラミドシンターゼ活性は、J. Biol. Chem, 273, 26522-27 (1998)に記載の手順に従って測定した。簡潔に述べると、in vivoでのグルコセレブロシダーゼ及び非リソソーム内グルコシルセラミダーゼの活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−グルコシルセラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用の薄層クロマトグラフィーにかけた後、C6−NBD−グルコシルセラミドからC6−NBD−セラミドへの、コンズリトール−B−エポキシド感受性及び非感受性の分解を分析した。コンズリトール−B−エポキシドによって阻害される活性は、グルコセレブロシダーゼによるものであり、残りは非リソソーム内グルコシルセラミダーゼによるものとすることができる。in vivoでのグルコシルセラミドシンターゼ活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−セラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用の薄層クロマトグラフィーにかけた後、C6−NBD−セラミドのC6−NBD−グルコシルセラミドへの変換を測定した。
【0084】
MEB4培地中で、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンによってスフィンゴ糖脂質合成を阻害すると、色素沈着は消失する。一方は色素沈着が濃く(An)、他方は色素沈着が非常に薄い(M14)2種のメラノーマ培養物では、色素沈着の濃い培養物中の方が、全細胞抽出物及びメラノソーム画分の両方においてグルコシルセラミド(GlcCer)含有量が明らかに多いことが示される。
【0085】
メラノサイトでは、1回しか継代していなくても、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンに曝すと、薄い方と濃い方の皮膚タイプの細胞で色素沈着の有意な減少が見られる。本発明によるデオキシノジリマイシン類似体の薬剤としての用途は、炎症後色素沈着過剰症及び皮膚の炎症状態にある。これらのデオキシノジリマイシン類似体を全身に投与するとき、in vivoで強い抗炎症活性を有するという証拠は有力である。これらの化合物は、魅力的な薬理学的特徴(良好な生物学的利用能、代謝されない、動物での2週間の投与の後有害な毒物学的作用がない)を示す。イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、皮膚の完全性にとって重要な酵素活性であるリソソーム内グルコセレブロシダーゼを完全に/ほとんど阻害しないので、皮膚への適用に特に適する。
【0086】
6.黒色腫及び他の腫瘍
腫瘍細胞は、その表面にガングリオシドを含み、それを盛んに剥離させ、その後それらは他の細胞に吸収される。腫瘍ガングリオシドは、免疫抑制活性、血管新生促進特性を有し、増殖因子を媒介とする線維芽細胞及び血管内皮細胞の増殖を刺激することが報告されている。ガングリオシド発現の増加は、腫瘍生成の強化及び進行の加速と関連付けられている。ガングリオシド生合成を阻害すれば、腫瘍の進行を妨げ、且つ/又は転移を減少させることができる。グルコシルセラミドシンターゼのイミノ糖系阻害剤(OGT2378)は、マウスモデルでメラノーマの腫瘍増殖を顕著に阻害することが示されている。
【0087】
黒色腫及び他の腫瘍の治療に向けたデオキシノジリマイシン類似体
今回、グルコシルセラミドシンターゼ並びに非リソソーム内グルコシルセラミダーゼの強力且つ特異的な阻害剤であるイミノ糖が、腫瘍の増殖及び/又は転移を低減し得ることがわかった。最適な効力のためには、両方の酵素活性を相伴って阻害する必要がある。
【0088】
本発明によるデオキシノジリマイシン類似体は、良好な生物学的利用能、代謝及び毒性がないことを含む、すべての望ましい特徴を有する。したがって、本発明のデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、黒色腫及び他の腫瘍の治療に適切に使用することができる。
【0089】
実験データ
培養メラノーマ細胞中でグルコシルセラミドシンターゼ、非リソソーム内グルコシルセラミダーゼ、及びリソソーム内グルコセレブロシダーゼを阻害し得るかどうかについて、様々なイミノ糖を分析した。最初の2種の酵素活性の阻害は望ましいが、最後の酵素の活性の阻害は望ましくない。
【0090】
【表6】
【0091】
in vivoでのグルコセレブロシダーゼ活性、非リソソーム内グルコシルセラミダーゼ活性、及びグルコシルセラミドシンターゼ活性は、J. Biol. Chem, 273, 26522-27 (1998)に記載の手順に従って測定した。簡潔に述べると、in vivoでのグルコセレブロシダーゼ及び非リソソーム内グルコシルセラミダーゼの活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−グルコシルセラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用の薄層クロマトグラフィーにかけた後、C6−NBD−グルコシルセラミドからC6−NBD−セラミドへの、コンズリトール−B−エポキシド感受性及び非感受性の分解を分析した。コンズリトール−B−エポキシドによって阻害される活性は、グルコセレブロシダーゼによるものであり、残りは非リソソーム内グルコシルセラミダーゼによるものとすることができる。in vivoでのグルコシルセラミドシンターゼ活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−セラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用の薄層クロマトグラフィーにかけた後、C6−NBD−セラミドのC6−NBD−グルコシルセラミドへの変換を測定した。
【0092】
選択したデオキシノジリマイシン類似体の有効性をモニターするための動物モデル
イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、粉末食にして経口投与することができる。B16マウスメラノーマ細胞の亜系統であるMEB4細胞(Riken Cell Bank、埼玉県)を皮内注射したC57BL/6マウスを使用することができる。腫瘍の形成及びサイズについて、マウスを週3回検査することができる。腫瘍サイズは、キャリパーを使用して立体的に測定することができ、腫瘍体積は、式a×b×c/2を用いて推定することができる(Weiss et al. 2003. Cancer Research 63, 3654-8)。
【0093】
7.真菌症
最近では、病原性の真菌がヒトグルコシルセラミドシンターゼに類似した酵素を有することが明らかになってきている。グルコシルセラミドは、菌糸が成長する際に選択的に産生され、成長端に隣接する細胞膜中で濃縮される。真菌を、グルコシルセラミドシンターゼ活性を阻害することがわかっているセラミド類似体に曝した後に、菌糸の成長の抑制が観察されている。
【0094】
今回、慎重に選択した疎水性イミノ糖が、より強力且つ忍容性の十分なグルコシルセラミドシンターゼの阻害剤であることが判明した。真菌のグルコシルセラミドシンターゼ活性を特異的に妨げ、内在するヒトグルコシルセラミドシンターゼには弱い影響しか及ぼさないイミノ糖を選択することで、生命を脅かす真菌感染と闘うことができる。そのような感染は、診療所で高まりつつある問題となっている。選択されたイミノ糖は、真菌菌糸の成長端の細胞壁を攻撃するキチナーゼである組換えヒトキトトリオシダーゼと併用できるはずである。したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、真菌症の治療に適切に使用することができる。
【0095】
8.ウイルス感染
イミノ糖は、ウイルス感染の治療用薬剤として調査されている。AIDSを引き起こす病原体であるHIVによる感染を治療するために、N−ブチルデオキシノジリマイシンの研究が行われている。世界中で数億の個体がAIDSに侵されている。in vitroでウイルスの感染力及び合胞体形成が示された。抗HIV剤としてのN−ブチルデオキシノジリマイシンの第II相臨床治験では、有望な結果が出なかった。必要とされる血清濃度では、重い副作用が起こった。B型肝炎ウイルス(HBC)は、世界中で感染者が3億5千万人を超えており、肝疾患及び肝細胞癌を引き起こし得る。近い関係にあるHBV感染動物モデルである、ウッドチャック肝炎ウイルスに慢性的に感染したウッドチャックでは、ノニルデオキシノジリマイシンによる治療が、感染性のあるエンベロープ型ウイルスの分泌を防ぐことがわかった。世界中で、1億人以上がC型感染ウイルス(HCV)に慢性的に感染している。ワクチンがない中、このことは、先進諸国の公衆衛生の最も深刻な脅威の1つとなっている。推定390万人の北アメリカ人が慢性的に感染している状況で、C型肝炎は現在、米国の肝臓移植理由の筆頭に立っている。この感染症は、年間約8,000人の米国人の死亡原因となり、この数字は、有効な処置が存在しなければ今後20年で3倍になると予想される。イミノ糖N−ノニルデオキシノジリマイシンは、C型肝炎のin vitro代用モデルであるウシウイルス性下痢ウイルスにおいて抗ウイルス活性を示している。
【0096】
ERα−グルコシダーゼは、新生糖タンパク質に結合したN−グリカン鎖から末端グルコース残基を段階的に除去する役を司ると考えられている。このために、糖タンパク質は、もっぱら一グリコシル化糖タンパク質に結合する、ERシャペロンのカルネキシン及びカルレティキュリンと相互に作用できるようになる。カルネキシンとの相互作用は、全部ではないが一部の糖タンパク質の正確なフォールディングにとって決定的なものであり、このグルコシダーゼの阻害剤を使用すると、それに依存するタンパク質を特異的に標的にすることができる。デオキシノジリマイシンの抗ウイルス活性は、ウイルスのエンベロープ糖タンパク質の適正なフォールディング及び輸送を妨害し、感染性のエンベロープ型ウイルスの分泌を防ぐことになるはずのERα−グルコシダーゼの阻害に基づくものであると考えられる。
【0097】
しかし、本発明者らは、イミノ糖の観察された抗ウイルス活性が、ERa−グルコシダーゼの阻害ではなく、グルコシルセラミド及びそのスフィンゴ糖脂質代謝産物の合成の阻害に基づくものであると見ている。宿主細胞表面上のスフィンゴ糖脂質に富むラフトは、ウィルスの融合及び出芽において欠かせない重要性を有しているように思われる。表面スフィンゴ糖脂質組成を操作すると、これらの過程に影響が及ぶ。
【0098】
この新規な見識は、ウイルス感染治療用のイミノ糖を選択する新たな論理的根拠となる。化合物は、それがグルコシルセラミドシンターゼ活性の強力な阻害剤であるという判断基準を第1に選択する必要がある。さらに、化合物は、十分な生物学的利用能及び忍容性をもつべきである。本発明によるデオキシノジリマイシン類似体、特にイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシン。したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、こうした判断基準を満たすウイルスの治療に適切に使用することができる。
【0099】
9.微生物感染及び内毒素
粘膜の感染
粘膜病原は、宿主の複合糖質受容体に対する特異的な付着性によって感染部位に向かって行く。そのため、細胞表面からのそのような受容体の除去が、付着を妨げ、細菌コロニー形成を損なわせ、粘膜感染活動を縮小すると予測することができる。スフィンゴ糖脂質シンターゼの阻害剤を使用すると、受容体を除去することができる。
【0100】
実際、N−ブチル−デオキシノジリマイシンで処理したマウスは、P線毛大腸菌による実験的な尿路感染に罹患しにくくなった(Svensson et al. Mol. Microbiol. 2003; 47: 453-61)。ケモカインの分泌が減少し、好中球の動員がより少ないことによって示されるように、粘膜の炎症応答は弱まっており、また細菌による尿路でのコロニー形成の効率は、尿路感染(UTI)を引き起こし、通常は「P」抗原に対する末端受容体を備えた線毛を有する通常のマウス大腸菌種より劣っていた。P抗原は、会陰及び尿道に並ぶ細胞の表面上にも見られる血液型マーカーである。人口の約75%がP抗原を発現させ、これらの個体は特にUTIに罹患しやすい。P抗原は、膣及び前立腺の分泌物中にも見られるが、このような分泌されたP抗原は、細菌受容体に結合して、上皮細胞表面に生物体が結合しないようにするという点で保護物である。UTIに最も罹患しやすい個体は、その細胞上にP抗原を発現させ、その分泌中にはP抗原がない個体である。
【0101】
別の例は、複合的な播種性の胃の炎症を引き起こすヘリコバクターピロリである。したがって今回、粘膜へのその堅固な付着にとってもスフィンゴ糖脂質が非常に重要であることを見出した。適切なイミノ糖による胃のスフィンゴ糖脂質の除去は、すばらしい治療用途をもたらし得るはずである。
【0102】
内毒素
糖脂質受容体が、細菌とその宿主間の主要な界面として働くだけでなく、内毒素など、細菌の病原性因子のターゲットとしても働くことがさらに判明した。ガングリオシドGM1は、コレラ毒素(CTX)や大腸菌の熱安定毒素IIのような様々なAB5毒素のBサブユニットの受容体として働き得るのに対し、グロボトリアオシドGb3は、AB5毒素である滋賀毒素(ST)、及びベロ毒素のような一部の関連毒素の受容体として働く。内毒素結合の防止は、臨床に大きく関わる問題である。例えば、ベロ毒素は、出血性大腸炎及び溶血尿毒症症候群(HUS)の細小血管症における内皮の標的化に関与する。スフィンゴ糖脂質合成の阻害剤を使用すれば、内毒素を媒介とする病態に介入できるはずである。
【0103】
粘膜の細菌感染の治療、及び内毒素を媒介とする病態の予防に向けたデオキシノジリマイシン類似体
先の新規な見識は、ウイルス感染治療用のイミノ糖を選択する論理的根拠となる。化合物は、グルコシルセラミドシンターゼ活性の強力な阻害剤であるという判断基準を第1に選択する必要がある。さらに、化合物は、十分な生物学的利用能及び忍容性を有するべきである。本発明のデオキシノジリマイシン類似体、特にイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、こうした判断基準を満たす。したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、粘膜の細菌感染の治療及び内毒素を媒介とする病態の予防に適切に使用することができる。
【0104】
[参考文献]
1. Kolter T, Proia RL and Sandhoff K. 2002. J. Biol.Chem. 277, 25859-62. Combinatorial ganglioside biosynthesis.
2. Wertz PW & van den Bergh B. 1998. Chem. Phys. Lipids 91, 85-96. The physical, chemical and functional properties of lipids in the skin and other biological barriers.
3. Simons K & van Meer G. 1988. Biochemistry 27, 6197-6202. Lipid sorting in epithelial cells.
4. Brown DA & London E. 1998. J. Membr. Biol. 164, 103-114. Structure and origin of ordered lipid domains in biological membranes.
5. Hakomori S. 1981. A.Rev.Biochem . 50, 733-764. Glycosphingolipids in cellular interaction.
6. Huwiler A, Kolter T, Pfeilschifter J & Sandhoff K 2000. Biochim.Biophys.Acta 1485, 63-99. Physiology and pathophysiology of sphingolipid metabolism and signaling.
7. Hannun YA & Obeid LM. 1995. Trends Biochem.Sci.20, 73-77. Ceramide: an intracellular signal for apoptosis.
8. Kolesnick R & Kronke M. 1988. Annu.Rev.Physiol.60, 643-665. Regulation of ceramide production and apoptosis.
9. van Blitterswijk WJ, van der Luit AH, Veldman RJ, Verheij M, & Borst J. 2003.Biochem J. 2003 Jan 15;369(Pt 2):199-211. Ceramide: second messenger or modulator of membrane structure and dynamics?
10. Merrill AH. 2002. J.Biol.Chem. 277, 25843-25846. De novo sphingolipid biosynthesis: a necessary, but dangerous, pathway.
11. Sandhoff K & Kolter T. 2003. Philos Trans R Soc Lond B Biol Sci. 358(1433):847-61. Biosynthesis and degradation of mammalian glycosphingolipids.
12. Van Weely S, Brandsma M, Strijland A, Tager JM & Aerts JMFG. 1993. Biochim.Biophys.Acta 1181,55-62. The existence of a non-lysosomal glucocerebrosidase that is not deficient in Gaucher disease.
13. Aerts JM, Hollak C, Boot R, & Groener A. 2003. Philos Trans R Soc Lond B Biol Sci. 358(1433):905-14. Biochemistry of glycosphingolipid storage disorders: implications for therapeutic intervention.
14. Barranger JA & Ginns EI. 1989. Glycosylceramide lipidosis: Gaucher disease. In: Scriver CR, Beaudet AL, Sly WS, Vall D (eds) The metabolic basis of inherited disease. McGraw-Hill, New York, p 1677-1698
15. Beutler E & Grabowski GA. 1995. Gaucher's disease. In: Scriver CR, Beaudet AL, Sly WS, Valle D (eds) The metabolic and molecular bases of inherited disease. McGraw-Hill, New York, p 2641-2670
16. Brady RO, Kanfer JN, Bradley RM & Shapiro D. 1966. Demonstration of a deficiency of glucocerebroside-cleaving enzyme in Gaucher's disease. J Clin Invest 45, 1112-1115
17. Patrick AD. 1965. A deficiency of glucocerebrosidase in Gaucher's disease. Biochem J 97, 17c-18c
18. Van Weely S, van den Berg M, Barranger JA, Sa Miranda MC, Tager JM & Aerts JMFG. 1993. Role of pH in determining the cell-type specific residual activity of glucocerebrosidase in type 1 Gaucher disease. J Clin Invest 91, 1167-1175
19. Aerts JM, van Weely S, Boot R, Hollak CE & Tager JM. 1993. Pathogenesis of lysosomal storage disorders as illustrated by Gaucher disease. J Inherit Met Dis 16, 288-291
20. Cox TM, & Schofield JP. 1997. Gaucher's disease: clinical features and natural history Baillieres Clin.Hematol.10, 657-689
21. Aerts JMFG & Hollak CEM. 1997. Plasma and metabolic abnormalities in Gaucher's disease. Baillieres Clin Haematol 10, 691-709
22. Cox TM .2001. Gaucher disease: understanding the molecular pathogenesis of sphingolipidoses. J Inherit Met Dis 24 (suppl 2), 106-121
23. Moran MT, Schofield JP, Hayman AR, Shi G-P, Young E & Cox TM. 2000. Pathologic gene expression in Gaucher disease: upregulation of cysteine proteinases including osteoclastic cathepsin K. Blood 96, 1969-1978
24. Brady RO.1997. Gaucher's disease: past, present and future. Baillieres Clin Haematol 10, 621-634
25. Barranger JA, & O'Rourke E. 2001. Lessons learned from the development of enzyme therapy for Gaucher disease J Inherit Met Dis 24 (suppl 2), 89-96
26. Hollak CEM, Aerts JMFG, Goudsmit ER, Phoa SS, Ek M. van Weely S, von dem Borne AE & van Oers MH.1995. Individualised low-dose alglucerase therapy for type 1 Gaucher's disease. Lancet 345, 1474-1478
27. Richter J & Karlsson S. 2001. Clinical gene therapy in hematology: past and future. Int. J Hematol 73, 162-169
28. Radin NS. 1996. Treatment of Gaucher disease with an enzyme inhibitor. Glycoconj J 13, 153-157
29. Platt FM, Jeyakumar M, Andersson U, Priestman DA, Dwek RA, ButtersTD, Cox TM, Lachmann RH, Hollak CEM, Aerts JMFG, Hrebicek M, Moyses C, Gow I, Elstein D & Zimran A. 2001. J. Inher. Met. Dis. 24, 275-290. Inhibition of substrate synthesis as a strategy for glycolipid lysosomal storage disease therapy.
30. Lee L, Abe A & Shayman JA. 1999. J Biol Chem 274, 146662-14665 Improved inhibitors of glucosylceramide synthase.
31. Abe A, Gregory S, Lee L, Killen PD, Brady RO, Kulkarni A & Shayman JA. 2000. Reduction of globotriaosylceramide in Fabry disease mice by substrate deprivation. J Clin Invest 105, 1563-1567
32. Platt FM, Neises GR, Dwek RA & Butters TD. 1994. N-butyl-deoxynojirimycin is a novel inhibitor of glycolipid biosynthesis. J Biol Chem 269, 8362-8365
33. Jeyakumar M, Butters TD, Cortina-Borja M, Hunnam V, Proia RL, Perry VH, Dwek RA & Platt FM. 1999. Delayed symptom onset and increased life expectancy in Sandhoff mice treated with N-butyl-deoxynojirimycin. Proc Natl Acad Sci USA 96, 6388-6393
34. Andersson U, Butters TD, Dwek RA & Platt FM. 2000. N-butyldeoxygalactonojirimycin: a more selective inhibitor of glycosphingolipid biosynthesis than N-butyldeoxynojirimycin, in vitro and in vivo. Biochem Pharmacol 49, 821-829
35. Overkleeft HS, Renkema GH, Neele J, Vianello P, Hung IO, Strijland A, van den Burg A, Koomen, GJ, Pandit UK & Aerts J. 1998. Generation of specific deoxynijirimycin-type inhibitors of the non-lysosomal glucosylceramidase. J Biol Chem 273, 26522-26527
36. Cox T, Lachmann R, Hollak C, Aerts J, van Weely S, Hrebicek M, Platt F, Butters T, Dwek R, Moyses C, Gow I, Elstein D & Zimran A. 2000. Lancet. 355(9214):1481-5. Novel oral treatment of Gaucher's disease with N-butyldeoxynojirimycin (OGT 918) to decrease substrate biosynthesis.
37. Heitner R, Elstein D, Aerts J, Weely S &Zimran A. 2002. Low-Dose N-Butyldeoxynojirimycin (OGT 918) for Type I Gaucher Disease. Blood Cells Mol Dis 28, 127-33
38. Fischl MA, Resnick L, Coombs R, Kremer AB, Pottage JC Jr, Fass RJ, Fife KH, Powderly WG, Collier AC, Aspinall RL, et al. The safety and efficacy of combination N-butyl-deoxynojirimycin (SC-48334) and zidovudine in patients with HIV-1 infection and 200-500 CD4 cells/mm3. J Acquir Immune Defic Syndr. 1994 Feb;7(2):139-47.
39. Goss PE, Reid CL, Bailey D & Dennis JW. 1997. Phase 1B clinical trial of the oligosaccharide processing inhibitor swainsonine in patients with advanced malignancies.
【図面の簡単な説明】
【0105】
【図1】適切な疎水性存在物の設計を示す図である。
【図2】デオキシノジリマイシン類似体4の合成を示す図である。
【図3】デオキシノジリマイシン7の合成を示す図である。
【図4】デオキシノジリマイシン類似体17、18、及び19の合成を示す図である。
【図5】デオキシノジリマイシン類似体28、29、及び30の合成を示す図である。
【技術分野】
【0001】
本発明は、グルコシルセラミド及び/又は他のスフィンゴ糖脂質の合成が関与する様々な疾患の治療に適切に使用することのできる、デオキシノジリマイシン類似体又は薬学的に許容できるその塩に関する。そのような疾患には、インスリン抵抗性(II型糖尿病)、肥満及び過体重、スフィンゴ糖脂質貯蔵障害、炎症、色素沈着過剰症及び皮膚の炎症状態、黒色腫及び他の腫瘍、真菌感染、ウイルス感染、微生物感染、及び内毒素が含まれる。
【背景技術】
【0002】
スフィンゴ糖脂質
スフィンゴ糖脂質は、糖鎖に連結したセラミド部分を含む脂質分子であるとされる。セラミド部分は、1本の特徴的な長鎖アミノアルコール、主としてD−エリトロ−C16−スフィンゴシンが、長さ及び飽和度は様々でよい脂肪酸にアミド結合によって結合しているものから構成される。糖単位の数及び性質の異なるその少糖部分とそのグリコシド結合が様々となるために、数百の特定のスフィンゴ糖脂質が存在する。スフィンゴ糖脂質は、例えば、ガングリオシド及びグロボシドなどの系列に分類される(参照文献1)。
【0003】
数十年間、スフィンゴ糖脂質は、むしろ新種の構造であるとみなされ、主流の生化学ではほとんど注目を浴びていなかった。その構成が複雑であり、存在量が少ないために、研究活動は、ほんの少数の先駆的な専門家集団だけに限られた。血液型抗原として働く機能を除き、特定のスフィンゴ糖脂質のためであると考えられる明確な機能がないので、その生理的な関連性は、多くは謎に包まれたままである。
【0004】
人体では、スフィンゴ糖脂質及びそのスフィンゴ脂質前駆体であるセラミドは、細胞内及び細胞外に存在する。表皮では、セラミド及びグルコシルセラミドの分子は、遊離の存在だけでなく、タンパク質に結合した構造として豊富に存在し、皮膚の水分透過性及び完全性に寄与する(参照文献2)。目立ったその合成部位である肝臓と末梢組織間のスフィンゴ糖脂質の交換は、相当な量のこれらの脂質を含むリポタンパク質によって媒介される。細胞では、スフィンゴ糖脂質は、多かれ少なかれもっぱら形質膜の外側葉状物中に位置し、その少糖鎖は周囲に露出している。セラミド脂質の主鎖は、スフィンゴ脂質に特別な物理的性質を与える(参照文献3)。コレステロール存在下で、スフィンゴ糖脂質は、膜リン脂質の塊から分離し、凝集して、より規則正しいがそれでも流体である「液体−規則」ドメインとなり得る。シグナル伝達に関与する膜タンパク質は、これらのスフィンゴ糖脂質−コレステロールに富んだマイクロドメイン(又は「ラフト」)に集中している(参照文献4)。
【0005】
細胞表面にあるスフィンゴ糖脂質の組成は、細胞の増殖、分化、ウイルス形質転換、個体発生、及び発癌と共に変化し得る(参照文献5)。神経細胞ではガングリオシドが特に豊富であることが明らかになってきている。ガングリオシドは、構造成分として神経系の機能に不可欠である。スフィンゴ糖脂質は、マイクロドメインの成分として、多数の細胞外シグナルの細胞内部への伝達に関与する(参照文献6)。さらに、セラミドなどのスフィンゴ脂質やその代謝産物であるスフィンゴシン(−1−リン酸)は、様々な細胞プロセスにおいてシグナル伝達分子としての直接の役割を担うと考えられている(参照文献7、8、及び9)。
【0006】
スフィンゴ糖脂質の合成及び分解
スフィンゴ糖脂質の合成及び分解は、様々な細胞内区画で起こる複雑な多段階の過程である(参照文献10及び11)。セラミド生合成に関与する酵素は、小胞体の細胞質ゾル葉状構造上に局在している。生合成は、セリンパルミトイルトランスフェラーゼによるアミノ酸セリンとパルミトイル補酵素Aの縮合から出発して、3−ケトスフィンガニンをもたらす。次に、これが3−ケトスフィンガニンレダクターゼによってD−エリトロ−スフィンガニンに還元され、その後、N−アシルトランスフェラーゼによってアシル化されてジヒドロセラミドになる。ジヒドロセラミドは、多くはジヒドロセラミドデサチュラーゼの作用によって不飽和化される。
【0007】
セラミドはさらに、スフィンゴミエリン、ガラクトシルセラミド及び誘導体、又はグルコシルセラミド及び誘導体の異なる3部門のスフィンゴ脂質へと代謝される。セラミドは、セラミドの1−ヒドロキシル基に、リン脂質ホスファチジルコリンからホスホリルコリンが転移することで、スフィンゴミエリンに変換される。この反応でジアシルグリセロールが遊離する。スフィンゴミエリンの生合成は、ゴルジ膜の内腔部で行われるが、他の部位も考慮に入れられている。ガラクトシルセラミドの合成は、小胞体の内腔で行われ、UDPガラクトースを共基質とする特定のβ−ガラクトシルトランスフェラーゼによって触媒される。ガラクトシルセラミド及びその硫酸エステル誘導体であるスルファチドは、特定の細胞型でしか合成されない。これらは、例えば、中枢神経系のミエリンの形成及び安定性において決定的な役割を担う。グルコシルセラミドは、ゴルジ装置の細胞質ゾル葉状構造上にあるグルコシルセラミドシンターゼによって生成される。この酵素は、UDPグルコースを共基質として使用し、グルコースがセラミドの1位にβグリコシド結合するのを触媒する。グルコシルセラミドは、その後ゴルジ膜を通って移動して、内側の葉状構造に到達する。グルコシルセラミドは、そこから形質膜に到達することもでき、又はゴルジ装置中でさらにグリコシル化されて改変されることもある。脊椎動物に共通するスフィンゴ糖脂質前駆体であるラクトシルセラミドは、ガラクトシルトランスフェラーゼIを触媒とするUDP−Galからのガラクトース部分の付加によって生成される。次に、特異性の限られた数種類のグリコシルトランスフェラーゼによってのみ行われる段階的なグリコシル化によって様々なスフィンゴ糖脂質が生成され得る。これらの酵素は、流れ作業列でのように、炭水化物基及びシアル酸基をグリコシル受容側に転移する。スフィンゴ糖脂質の著名な系列は、ガングリオ系列である。ガングリオシド複合体の生合成には、シアリルトランスフェラーゼI、II、及びIII、GalNacトランスフェラーゼ、ガラクトシルトランスフェラーゼII、及びシアリルトランスフェラーゼIV及びVが必要である。
【0008】
スフィンゴ糖脂質の分解は、細胞の特殊化した酸性区画、即ちリソソーム中で行われる。活性化タンパク質によって援助されるリソソーム内グリコシダーゼが、その糖脂質基質の非還元性末端から糖残基を順次切断し、残りのセラミドが、リソソーム内セラミダーゼによって最後に加水分解されて、遊離の脂肪酸及びスフィンゴシンがもたらされる。放出された単糖、シアル酸、脂肪酸、及びスフィンゴイド塩基は、リソソームを出ることができ、生合成の目的のために再度使用されることもある。より最近になって、スフィンゴ糖脂質の分解がリソソームの外側で行われる場合もあることがわかってきている。例えば、グルコシルセラミドをセラミドに加水分解することのできる非リソソーム内グルコシルセラミダーゼが発見されている(参照文献12)。後者の酵素は、ゴーシェ病患者で欠損しているリソソーム内グルコシルセラミダーゼ(グルコセレブロシダーゼ)とは明らかに異なる。リソソーム外分解の生理的重要性は、未だ理解が不十分ではあるが、おそらく、それに関連するセラミド生成はシグナルとして働き、細胞の挙動を調節し得ると思われる(参照文献13)。
【0009】
スフィンゴ糖脂質貯蔵障害
スフィンゴ糖脂質は、細胞中で絶えず合成され分解される。ヒトのいくつかの遺伝疾患は、リソソーム内のスフィンゴ糖脂質の蓄積を特徴とする。これらのいわゆるスフィンゴ糖脂質貯蔵障害は、リソソーム内分解の欠陥によって引き起こされる(例えば、参照文献13)。その例は、GM1ガングリオシドーシス、サンドホフ病(GM2ガングリオシドーシス、II型又はO型)、テイサックス病(GM2ガングリオシドーシス、I型又はB型)、ファブリー病(a−ガラクトシル−ラクトシルセラミドーシス)、ラクトシルセラミドーシス、ゴーシェ病(グルコセレブロシドーシス)である。他のスフィンゴリピドーシスは、多数のスルファターゼ欠損症候群(ムコスルファチドーシス)、クラッベ病(グロボイド細胞型白質ジストロフィー、ガラクトセレブロシドーシス)、ニーマンピック病(スフィンゴミエリノーシス)、ファーバー病(セラミドーシス)である。
【0010】
ゴーシェ病
ゴーシェ病は、ヒトにおいて最もよく遭遇するリソソーム貯蔵障害である(参照文献14及び15)。1882年に、フランス人医学研究生Philippe C. E. Gaucherによってこの疾患の臨床的特徴が初めて詳述され、脾腫を伴う32才の女性に異常な大きな細胞が存在することが報告されている。前世紀の初頭にはすでに、この疾患が家族性の障害であることが示されていた。1934年に、ゴーシェ病の主たる貯蔵物質がついにグルコセレブロシド(ブルコシルセラミド)であると特定された。スフィンゴ糖脂質であるグルコセレブロシドは、ガングリオシド及びグロボシドの合成及び分解に共通の中間体である。ゴーシェ病における主たる欠陥は、リソソーム内酵素であるグルコセレブロシダーゼ(EC.3.2.1.45)の活性の顕著な不足であることが示されている(参照文献16及び17)。グルコセレブロシダーゼが遺伝的に欠損していると、体中のマクロファージのリソソーム内区画にその脂質基質が蓄積する。神経性の症状の有無に基づいて区別される3つの異なる表現型が認められている。より最近になって、ゴーシェ病の追加の表現型が同定されている。例えば、グルコセレブロシダーゼ活性の完全な欠如は、出生前又は出生直後に、致死的な予後を伴う重い皮膚透過性の異常をもたらす。最も有病率の高い疾患の型は、1型ゴーシェ病と命名される、神経細胞障害性でない形態である。1型ゴーシェ病の発症年齢及び臨床徴候は、非常に変化に富んでいる。最も一般的な症状には、主として脾機能亢進による貧血及び血小板減少を伴う脾腫大;肝腫脹、及び骨疾患が含まれる。貧血は慢性疲労の一因となり得る。血小板減少及び凝固時間の延長は、出血傾向を増大させる場合がある。不規則な骨の痛み、病的な骨折、無血管性の壊死、及び極めて苦しい骨の発作も生活の質に多大な影響を及ぼし得る。1型ゴーシェ病は、すべての民族群で比較的一般的である。この疾患は、アシュケナジで有病率が高く、キャリヤ出現度はおよそ10人に1人にもなり、発生率はおよそ5000人に1人である。アシュケナジを含む白人のグルコセレブロシダーゼ遺伝子の最も一般的な突然変異は、アミノ酸置換N370Sをコードする。N370S突然変異のヘテロ対立遺伝子の存在は、常に神経細胞障害性でない経過を伴う。N370Sグルコセレブロシダーゼは、通常はリソソームで産生され、そこに存在することが実証されている。その触媒活性は、5.0を上回るpH値でしかひどくは損なわれず、この突然変異の微妙な性質を物語っている(参照文献18)。N370S突然変異のすべてではないが大部分のホモ接合体は、重要な臨床症状を表さない。ゴーシェ病における表現型−遺伝子型調査の二重研究及び不十分な予測力から、ゴーシェ病の出現においては後成的な要因も鍵となる役割を担っていることがはっきりと示されている(参照文献19及び20も参照のこと)。グルコセレブロシダーゼは、すべての細胞型のリソソームに存在するが、1型ゴーシェ病患者は、単に単核貪食細胞系の細胞でグルコセレブロシドの貯蔵を展開する。マクロファージは、侵入微生物の分解、血球の自然の代謝回転、及び組織モデリングに関与する。この点から、かなりの種類のリソソーム貯蔵障害で、貯蔵物質の蓄積が組織マクロファージ中で顕著に起こっても意外ではない。ゴーシェ病の1型変型は、リソソーム内貯蔵がもっぱらマクロファージで起こるという点で特異である。貯蔵材料は、血球の代謝回転から派生する外来脂質が分解されて生じると考えられている。グルコセレブロシドが詰め込まれた細胞は、リソソーム封入体を含むその細胞質の外観が「しわくちゃの紙」様である特徴的な形態を示し、このような細胞をゴーシェ細胞と呼ぶ。ここ数十年で、ゴーシェ細胞が貯蔵物質の不活性な容器ではなく、ゴーシェ病の多様な臨床徴候に寄与する生存能力のある慢性的に活性化したマクロファージであることが明らかになった。いくつかの炎症誘発性サイトカイン(TNF−α、IL−1−β、IL−6、及びIL−8)、抗炎症性サイトカインIL−10、及びM−CSFの循環レベルの増大が報告されている(参照文献21及び22)。オステオペニア、凝固の活性化、代謝亢進、免疫グロブリン血症及び多発性骨髄腫、低脂血症など、ゴーシェ病患者に共通する臨床上の異常の発生において、サイトカインの異常が決定的な役割を担うのではないかという仮説が立てられている。より最近では、ゴーシェ病及び対照の脾臓の抑制性減算ハイブリダイゼーション分析によって遺伝子発現プロフィールを調査すると、ゴーシェ細胞によるカテプシンB、K、及びS転写物の過剰発現が特定された(参照文献23)。破骨細胞由来のカテプシンKが、骨のI型コラーゲンの破壊に目立って関与することは特筆に価する。このカテプシンの局所での放出は、ゴーシェ病の骨溶解の一因となるかもしれない。
【0011】
ゴーシェ病の治療
1型ゴーシェ病は、一般に、遺伝性リソソーム貯蔵障害の中で、有効な治療処置を開発するための最も魅力的な候補であると考えられている。第1に、根底にある遺伝的な欠陥という分子的な根拠は、すでに遺伝子及びタンパク質のレベルで確立されている。第2に、組織マクロファージというたった1つの細胞型が、主としてこの疾患の病態生理と関連付けられている。1型ゴーシェ病の治療処置の原理は、したがって比較的簡単である。即ち、ゴーシェ細胞を正す(又は進行中の生成を予防する)のである。これは、マクロファージに酵素グルコセレブロシダーゼを補充し(酵素補充療法)、特定の阻害剤を用いて糖脂質合成を縮小し(基質除去若しくは基質調整療法)、又はマクロファージの造血前駆細胞中にグルコセレブロシダーゼcDNAを導入する(遺伝子療法)ことで実現できるはずである。米国国立保健研究所(米国ベセズダ)でのBrady、Barranger、及び同僚らの先駆的な仕事、並びに他の多くの者による価値ある貢献が、ヒトグルコセレブロシダーゼの長期間の静脈内投与を主体とする1型ゴーシェ病の非常に有効な治療をもたらした(参照文献24〜26)。ゴーシェ病患者の造血細胞に遺伝子を導入する3通りの独立した研究を実施したが、どれも有望な成果を上げなかった(参照文献27)。CD34細胞の形質導入の効率が悪く、白血球中でグルコセレブロシダーゼが持続的に発現されなかったことが、このことの一因となった。血液疾患及び遺伝障害を正すための遺伝子療法戦略の開発は、オンコレトロウイルス由来のベクターを使用するヒト幹細胞への遺伝子導入のレベルが低いことが妨げとなった。
【0012】
基質低減療法
1型ゴーシェ病及び他のグリコスフィンゴリピドーシスの治療処置に向けた代替手法は、基質除去(基質低減とも呼ばれる)療法である。Radin及び同僚らは、まず挑戦的な構想を考案した(参照文献28)。この手法は、スフィンゴ糖脂質の生合成速度を弱まった異化作用に見合うレベルにまで落とすことを目指すものである。リソソーム内酵素活性がかなり残っている患者は、リソソーム内貯蔵物質を徐々に片付けることができ、したがって基質生合成の低減から最も利益を得るはずであると考えられる(参照文献29)。
【0013】
スフィンゴ糖脂質生合成の2大阻害剤クラスは、以前から記載されており、その両方が、セラミドに特異的なグルコシルトランスフェラーゼ(グルコシルセラミドシンターゼ;GlcT−1;UDPグルコース:N−アシルスフィンゴシンD−グルコシル−トランスフェラーゼ、EC2.4.1.80とも呼ばれる)を阻害する。この酵素は、グルコースのセラミドへの転移、即ちスフィンゴ糖脂質生合成の第1段階を触媒する。第1のクラスの阻害剤は、セラミド類似体によって形成される。原型の阻害剤は、PDMP(D,L−トレオ−1−フェニル−2−デカノイルアミノ−3−モリホリノ−1−プロパノール)である。モルホリノ基をピロリジノ官能基に置換することに基づき、フェニル基で置換を行うことで、より特異的且つ強力な類似体、即ち、4−ヒドロキシ−1−フェニル−2−パルミトイルアミノ−3−ピロリジノ−1−プロパノール(p−OH−P4)及びエチレンジオキシ−1−フェニル−パルミトイルアミノ−3−ピロリジノ−1−プロパノール(EtDo−P4)が後に開発された(参照文献30)。ノックアウトマウスモデルでのファブリー病の研究では、この化合物の経口投与が、スフィンゴ糖脂質であるグロボトリアオシルセラミドの蓄積を著しく低減し得ることが示されている(参照文献31)。
【0014】
イミノ糖
第2のクラスのグルコシルセラミドシンターゼ阻害剤は、N−アルキル化イミノ糖によって形成される。この種の化合物は、N−グリカンプロセシング酵素の阻害剤としてすでに一般に使用されており、N−ブチルデオキシノジリマイシンのHIV阻害剤としての適用の可能性が、AIDS患者で研究されている。N−ブチルデオキシノジリマイシンがグリコシルセラミド合成を低マイクロモル濃度で阻害し得ることを最初に認めたのは、英国オックスフォードのGlycobiology InstituteのPlatt及びButtersであった(参照文献32)。同じ研究者らは、テイサックス病及びサンドホフ病のノックアウトマウスモデルで、脳中のスフィンゴ糖脂質の貯蔵が有意に低減したことを実証している(参照文献33)。動物での前臨床研究及びAIDS患者での予備臨床治験では、おそらくはNB−DNJが腸管刷子縁上の二糖類分解酵素を阻害し得ることに関連した消化管での(一過性の)有害作用が指摘されている。動物の研究では、ガラクトース類似体N−ブチルデオキシガラクトノジリマイシン(NB−DGJ)が、N−ブチルデオキシガラクトノジリマイシン(NB−DNJ)と同じ治療効力を有するかもしれず、胃腸への副作用を引き起こさないことがわかっている(参照文献34)。Overkleeft及び同僚らは、グルコシダーゼ阻害剤を探索する中で、グルコシルセラミドシンターゼのより強力な阻害剤を偶然の発見に基づいて開発した。N−アダマンタン−1−イル−メトキシペンチル−デオキシノジリマイシン(AMP−DNM)は、スフィンゴ糖脂質生合成をナノモル濃度で阻害することがわかっており(参照文献35)、ファブリー病ノックアウトマウスモデルで、表立った副作用なしにグロボトリアオシルセラミドの蓄積を予防することができた。
【0015】
スフィンゴ糖脂質貯蔵障害の治療にN−ブチルデオキシノジリマイシン(NB−DNJ)を使用する最初の臨床研究が、最近になって報告されている(Coxら、2000年)。非盲験第I/II相試験で、成人28人の1型ゴーシェ病患者に、1日3回100mgのNB−DNJ(OGT918、Oxford GlycoSciences社製)が与えられた。内臓巨大症及び血液の異常の改善、並びにグルコシルセラミド及びゴーシェ病活動生物マーカーの血漿レベルの正常化が記載されてはいるが、応答の程度は、高用量酵素補充療法で一般に認められるものより目覚しいものではなかった。予想どおり、NB−DNJの用量−応答の関係は、1型ゴーシェ病患者で実証できる。最近になって、1日3回の50mgのNB−DNJの投与は、有効性がはるかに劣ることが報告された(参照文献37)。ごく最近、EMEA(ヨーロッパのFDA相当機関)は、酵素補充療法を受けるのに不適格な1型ゴーシェ病患者の治療向けにNB−DNJ(ザベスカ、Oxford GlycoSciences社製)を認可している。
【0016】
イミノ糖及び他の治療適用例
イミノ糖は、環上酸素が窒素原子と入れ替わっている、単糖の構造を模倣したポリヒドロキシル化アルカロイドである。イミノ糖の例は、例えば、WO98102161に記載されている。遷移電荷状態の類似体として、イミノ糖は、糖の除去又は付加を触媒する酵素(「複合糖質プロセシング酵素」)の阻害剤として働き得る。グルコースの類似した構造模倣物であるデオキシノジリマイシンは、糖タンパク質をプロセッシングするα−グルコシダーゼI及びIIの阻害剤であることがわかっている。N−ブチル−デオキシノジリマイシンのような一部のN−アルキル化デオキシノジリマイシンは、抗ウイルス(HIV)治療薬としての評価がなされており、in vitroモデルを使用してウイルスの感染力に対する効力が証明されているにもかかわらず、臨床での成功は限られている(参照文献38)。現在、肝炎ウイルス感染に対する治療価値に関して、ノニル−デオキシ−ガラクトノジリマイシン及びN−7−オキサノニル−6−Me−デオキシガラクトノジリマイシンの調査がなされている。N−ヒドロキシエチル−デオキシノジリマイシン(ミグリトール、グリセット)のような他のN−アルキル化デオキシノジリマイシンは、腸管グリコシダーゼの阻害を目指して開発されている。腸管グリコシダーゼ活性の低減は、食物複合炭水化物からの単糖の取り込みの衝撃を和らげるはずなので、インスリン抵抗性(II型糖尿病)の個体にとって有益であると考えられる。代謝制御を目指す薬剤開発におけるイミノ糖の別の使用は、腫瘍細胞の転移を低減するために、細胞表面タンパク質上のN−結合型少糖の改変に使用するものである(参照文献39)。
【特許文献1】WO98102161
【発明の開示】
【発明が解決しようとする課題】
【0017】
今回、スフィンゴ糖脂質の異常な代謝及び異常な濃縮は、インスリン抵抗性(II型糖尿病)、過体重及び肥満、リソソーム貯蔵障害、炎症から、色素沈着過剰症及び皮膚の炎症状態に至る範囲の様々な病的状態と関連していることが判明した。さらに、スフィンゴ糖脂質が、ウイルス、微生物、及び真菌感染、並びにある種の内毒素に対する敏感性において重要な役割を担っていることがわかった。本発明者らは、慎重に選択したアルキル化イミノ糖を投与してスフィンゴ糖脂質代謝を正常化し、過剰のスフィンゴ糖脂質を低減すれば、そのような病的状態において有益な応答を得ることができると考えている。しかし、このような手法を成功させるための前提条件は、投与される化合物が有効であり、且つ十分に認容性であることである。イミノ糖が、グルコシルセラミドシンターゼなどのグルコシルトランスフェラーゼの活性だけでなく、他の様々な酵素(腸管及び他の細胞表面グリコシダーゼ、リソソーム内及びエンドソーム内グリコシダーゼ、グリコーゲン枝切り酵素、及び糖タンパク質改変グリコシダーゼ)の活性も妨げ得ることもわかっている。したがって、成功をもたらすイミノ糖の治療用途は、生物活性の特異性を要する。治療用イミノ糖によって阻害されるはずの複合糖質プロセシング酵素ターゲット、さらには阻害されないはずのその酵素の正確な性質を見抜くことが不可欠である。このことは、異なる病的状態にぴったり合う治療用イミノ糖開発の礎となる。しかし、以前から利用可能なイミノ糖は、少しも最適ではない。第1に、それらは生物学的利用能が十分でなく、グルコシルセラミドシンターゼの比較的弱い阻害剤である。第2に、それらは、有益な酵素活性の比較的強い阻害剤であり、そのために高めの用量で投与されると許容されない副作用を引き起こす。したがって、改良には、次の2段階の異なるレベル、即ち、1:新規なアルキル化イミノ糖の設計及び合成と、2:各病的状態に相応しい阻害活性判定基準を使用する最適なアルキル化イミノ糖の選択が必要となる。
【課題を解決するための手段】
【0018】
本発明は、グルコシルセラミド及び/又は他のスフィンゴ糖脂質の合成が役割を果たす様々な疾患の治療に適切に使用することのできる特定のクラスの新規なアルキル化イミノ糖誘導体に関する。
【0019】
したがって、本発明は、一般式(1)を有するデオキシノジリマイシン類似体又は薬学的に許容できるその塩に関する
【化1】
(I)
[式中、
R1〜R5は、それぞれ独立に、H、(CH2)nCH3、又はXを含み、
R6は、H、CH2OH、又はCH2OXを含み、
Mは、0又は1であり、
Nは、0〜9であり、
a、b、c、d、eは、R又はS立体配置を有するキラル中心であり、
Xは、大きな疎水性部分及びスペーサーを含み、疎水性部分は、スペーサーを介して問題の窒素原子又は炭素原子に結合しており、前記の大きな疎水性部分は、各々が2個以上の炭素原子を別の環と共有する3個以上の環を含む多環式アルコールから誘導され、脂質二重層に入り込むことができる]。
【0020】
ピロリジン(m=0)については、3個(R6=H)又は4個(R6=CH2OH、CH2OX、図2を参照のこと)のキラル中心を認めることができる。ピペリジン(m=1)については、4個(R6=H)又は5個(R6=CH2OH、CH2OX)のキラル中心を認めることができる。ピロリジン及びピペリジンは、個々のキラル中心b、c、d、及びeの性質及び立体化学に応じて、自然の五炭糖(R6=Hである場合)及び六炭糖(R6=CH2OH、CH2OX)を模倣する。キラル中心aの性質は(R2がHでない場合)、イミノ糖がαグリコシド結合(a=R)又はβグリコシド結合(a=S)のどちらを模倣するかを規定する。本発明によるデオキシノジリマイシン類似体の詳細な例を以下に挙げる。
【0021】
ピロリジン(R6=CH2OH又はCH2OX、m=0)
a=R、b=S、d=R、e=R α−D−リボフラノシド模倣物
a=S、b=S、d=R、e=R β−D−リボフラノシド模倣物
a=R、b=R、d=R、e=R α−D−アラビノフラノシド模倣物
a=S、b=R、d=R、e=R β−D−アラビノフラノシド模倣物
a=R、b=S、d=S、e=R α−D−キシロフラノシド模倣物
a=S、b=S、d=S、e=R β−D−キシロフラノシド模倣物
a=R、b=S、d=R、e=S α−L−リキソフラノシド模倣物
a=S、b=S、d=R、e=S β−L−リキソフラノシド模倣物
【0022】
ピロリジン(R2=H、R6=CH2OH又はCH2OX、m=0)
b=S、d=R、e=R α−D−1−デオキシリボフラノシド模倣物
b=S、d=R、e=R β−D−1−デオキシリボフラノシド模倣物
b=R、d=R、e=R α−D−1−デオキシアラビノフラノシド模倣物
b=R、d=R、e=R β−D−1−デオキシアラビノフラノシド模倣物
b=S、d=S、e=R α−D−1−デオキシキシロフラノシド模倣物
b=S、d=S、e=R β−D−1−デオキシキシロフラノシド模倣物
b=S、d=R、e=S α−L−1−デオキシリキソフラノシド模倣物
b=S、d=R、e=S β−L−1−デオキシリキソフラノシド模倣物
【0023】
ピペリジン(R6=H、m=1)
a=S、b=S、c=S、d=R β−D−キシロピラノシド模倣物
a=R、b=S、c=S、d=R α−D−キシロピラノシド模倣物
a=S、b=R、c=S、d=R β−D−リキソピラノシド模倣物
a=R、b=R、c=S、d=R α−D−リキソピラノシド模倣物
a=S、b=S、c=S、d=S β−L−アラビノピラノシド模倣物
a=R、b=S、c=S、d=S α−L−アラビノピラノシド模倣物
【0024】
ピペリジン(R2=H、R6=H、m=1)
b=S、c=S、d=R β−D−1−デオキシキシロピラノシド模倣物
b=S、c=S、d=R α−D−1−デオキシキシロピラノシド模倣物
b=R、c=S、d=R β−D−1−デオキシリキソピラノシド模倣物
b=R、c=S、d=R α−D−1−デオキシリキソピラノシド模倣物
b=S、c=S、d=S β−L−1−デオキシアラビノピラノシド模倣物
b=S、c=S、d=S α−L−1−デオキシアラビノピラノシド模倣物
【0025】
ピペリジン(R6=CH2OH又はCH2OX、m=1)
a=S、b=S、c=R、d=R、e=R β−D−グルコピラノシド模倣物
a=R、b=S、c=R、d=R、e=R α−D−グルコピラノシド模倣物
a=S、b=R、c=R、d=R、e=R β−D−マンノピラノシド模倣物
a=R、b=R、c=R、d=R、e=R α−D−マンノピラノシド模倣物
a=S、b=S、c=R、d=S、e=R β−D−ガラクトピラノシド模倣物
a=R、b=S、c=R、d=S、e=R α−D−ガラクトピラノシド模倣物
a=S、b=S、c=R、d=R、e=S β−L−イドピラノシド模倣物
a=R、b=S、c=R、d=R、e=S α−L−イドピラノシド模倣物
【0026】
ピペリジン(R2=H、R6=CH2OH又はCH2OX、m=1)
b=S、c=R、d=R、e=R β−D−1−デオキシグルコピラノシド模倣物
b=S、c=R、d=R、e=R α−D−1−デオキシグルコピラノシド模倣物
b=R、c=R、d=R、e=R β−D−1−デオキシマンノピラノシド模倣物
b=R、c=R、d=R、e=R α−D−1−デオキシマンノピラノシド模倣物
b=S、c=R、d=S、e=R β−D−1−デオキシガラクトピラノシド模倣物
b=S、c=R、d=S、e=R α−D−1−デオキシガラクトピラノシド模倣物
b=S、c=R、d=R、e=S β−L−1−デオキシイドピラノシド模倣物
b=S、c=R、d=R、e=S α−L−1−デオキシイドピラノシド模倣物
【0027】
本発明によるデオキシノジリマイシン類似体は、疎水性部分及びスペーサーを含む少なくとも1個の存在Xを含む。存在Xは、位置R1、R2、R3、R4、R5、R6のいずれかの箇所に位置する。
【0028】
スペーサー及び疎水性部分は、図1に示すように、Z及びYによって適切に表すことができる。
【0029】
大きな疎水性部分は、炭素原子3から8個のアルコキシポリアルキレン鎖又はポリアルキレン鎖を含むスペーサーによってデオキシノジリマイシンの前記の窒素原子に結合していることが好ましい。大きな疎水性部分は、アダマンタンメタノール、コレステロール、β−コレスタノール、アダマンタノール、及び9−ヒドロキシフェナントレンからなる群から選択される化合物から誘導されることがより好ましい。
【0030】
用語「スペーサー」とは、任意の二価部分、又はデオキシノジリマイシンのN原子に疎水性基を結合することのできる基を指す。
【0031】
本発明によるデオキシノジリマイシン類似体は、イド−立体配置を有することが好ましい。詳細には、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンが好ましい化合物である。
【0032】
本発明によるデオキシノジリマイシン類似体の調製を以下の実施例によって例示する。
【発明を実施するための最良の形態】
【0033】
N−(アダマンタン−1−イル−メトキシペンチル)−L−イド−デオキシノジリマイシン4
2,3,4,6−テトラ−O−ベンジル−L−イド−デオキシノジリマイシン1(Tetrahedron Lett. 44, 3085-3088, 2003)を水素化(H2、Pd/C、HCl(水溶液)、EtOH)によって脱ベンジル化して3にし、これを還元アミノ化条件(H2、Pd/C、HCl(水溶液)、EtOH)下でアダマンタン−1−イル−メトキシペンタナール2(J. Biol. Chem. 273, 26522, 1998、WO98102161)と縮合させて、N−(アダマンタン−1−イル−メトキシペンチル)−L−イド−デオキシノジリマイシン4を得た。精製すると、均質な目的化合物4が淡黄色のシロップとして収率93%で得られた(図2を参照のこと)。
【0034】
N−(アダマンタン−1−イル−メトキシペンチル)−D−ガラクト−デオキシノジリマイシン7
7の合成は、イド同属化合物4について上で概略を述べた合成経路に従い、2,3,4,6−テトラ−O−ベンジル−D−ガラクト−ノジリマイシン7(Tetrahedron 56, 32, 5819-5834, 2000)を出発材料として実施した(図3を参照のこと)。
【0035】
C1−β−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン17、N−メチル−C1−β−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン18、及びN−ブチル−C1−β−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン19
β−C1−アルキルデオキシノジリマイシン誘導体17、18、及び19の合成は、5−O−(アダマンタン−1−メチル)−1−ペンチノール13(図4を参照のこと)の調製から始める。ペンチン−1−オール8を完全にシリル化(nBuLi、TMSCl)した後、シリル−エーテルを選択的に脱保護して(1N HCl)、保護されたアセチレン9を得た。9の遊離アルコールをトリフルロメタンスルホニルエステル10に変換した(塩化トリフルオロメタンスルホニル、トリエチルアミン)。アダマンタン−メタノール11を10でアルキル化(K2CO3、還流CH2Cl2)し、その後TMS保護基を除去すると(ナトリウムメタノラート、MeOH/THF)、8に対して64%の収率でアセチレン13を得た。13のリチオ化物(−50℃のTHF中でブチルリチウムと反応させて調製したもの)を、2,3,4,6−テトラ−O−ベンジル−D−グルコノ−デルタ−ラクトン14(J. Org. Chem., 2531, 1967)に求核付加すると、ケトグルコシド15がアノマー混合物として得られた。化合物15を、次の3ステップの手順を使用して、容易に且つ高い立体選択性で、完全に保護されたイミノ糖16に変換した(Eur. J. Org. Chem. 5, 1185-1189, 1999)。即ち、1)対応するジオールに還元し(NaBH4、CH2Cl2/MeOH)、2)酸化させてジケト化合物にし(Swern酸化)、3)ギ酸アンモニウム及びシアノ水素化ホウ素ナトリウムを使用して二重還元アミノ化を行った(14に対する全体としての収率は45%であった)。パラジウムを触媒とする水素化にかけると、イミノ糖17が得られ(79%)、これから、それぞれホルムアルデヒド(18、収率20%)及びブタナール(19、収率73%)を用いる還元アミノ化によって、メチル及びブチル同族体18及び19を容易に調製した。
【0036】
2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン28、N−メチル−2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン29、及びN−ブチル−2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシノジリマイシン30
2−O−アルキル化デオキシノジリマイシン誘導体28、29、及び30の調製(図5を参照のこと)は、アリル−3,4,6−トリ−O−ベンジル−β−D−グルコピラノシド20(Tetrahedron: Asymmetry, 8, 765-774, 1997)の遊離アルコール官能基をp−メトキシベンジル保護し、その後アノマー性アリル基を異性化/加水分解して、2ステップで80%の22を得ることから出発する。ヘミアセタール22を、次の4ステップを使用して、直角をなすように保護されたデオキシノジリマイシン誘導体23に変換した。即ち、1)対応するジオール(LiAlH4、THF)に還元し、2)酸化させてアルデヒドケトンにし(Swern酸化)、3)ギ酸アンモニウム及びシアノ水素化ホウ素ナトリウムを使用して二重還元アミノ化を行い、4)得られる第二級アミンをベンジルオキシカルボニル基で保護した(ZCl、K2CO3、4ステップで71%)。23を2%のTFAで処理すると、化合物24が得られ(収率99%)、次のアルキル化に向けてC−2−OHを選択的に脱保護した。アルデヒド2から、アルデヒド官能基を対応する第一級アルコールに還元した後(NaBH4、MeOH)、PBr3の作用のもとで臭素化して、対応するアルキル化剤1−ブロモ−5−(アダマンタン−1−イル)メトキシペンタン26を調製した(88%、2ステップ)。過剰の26を加え、DMF中で水素化ナトリウム処理することで24のアルキル化を行って、83%で27を得た。27のZ保護基及びベンジル保護基を還元によって除去すると、イミノ糖28が得られ(収率80%)、これから、それぞれホルムアルデヒド(29、収率49%)及びブタナール(30、収率79%)を用いる還元アミノ化によって、メチル及びブチル同族体29及び30を容易に調製した。
【0037】
分光データ:
【化2】
N−(アダマンタン−1−イル−メトキシペンチル)−L−イド−デオキシ−ノジリマイシン(4):1H−NMR(MeOD,400MHz):δ4.01〜3.97(dd,1H,H6)、3.89(m,1H,H2)、3.85〜3.70(m,3H,H2,H3,H4)、3.55〜3.52(m,2H,CH2スペーサー)、3.40〜3.08(m,5H,2*H1,H5,CH2スペーサー)、2.96(s,2H,O−CH2メトキシ)、1.94(広幅s,3H,CHアダマンチル)、1.77〜1.66(広幅dd,6H,3*CH2アダマンチル)、1.63〜1.54(d,4H,2*CH2スペーサー)、1.55(d,6H,3*CH2アダマンチル)、1.45〜1.37(m,2H,CH2中央スペーサー)。MS(ESI):obs.m/z=398.2[M+H]+;C22H39NO5(MW計算値):397.3。
【0038】
【化3】
2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシ−ノジリマイシン(28):1H−NMR(MeOD,400MHz):δ3.92〜3.84(2*dd,2H,2*H6)、3.72〜3.62(m,2H,O−CH2スペーサー)、3.57〜3.43(m,3H,H2,H3,H4)、3.38〜3.36(m,2H,O−CH2スペーサー)、3.07(m,1H,H5)、2.95(s,2H,O−CH2メトキシ)、2.89〜2.79(m,2H,2*H1)、1.93(広幅s,3H,CHアダマンチル)、1.76〜1.65(広幅dd,6H,3*CH2アダマンチル)、1.62〜1.56(d,4H,2*CH2スペーサー)、1.54(d,6H,3*CH2アダマンチル)、1.45〜1.39(m,2H,CH2中央スペーサー)。MS(ESI):obs.m/z=398.2[M+H]+;C22H39NO5(MW計算値):397.3。
【0039】
【化4】
N−メチル−2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシ−ノジリマイシン(29):1H−NMR(MeOD,400MHz):δ4.13〜4.10(2*広幅d,2H,2*H6)、3.72〜3.64(m,2H,O−CH2スペーサー)、3.62〜3.48(m,3H,H2,H3,H4)、3.42〜3.38(m,2H,O−CH2スペーサー)、3.09〜2.89(m,3H,H5及び2*H1)、3.01(s,3H,N−CH3)、2.95(s,2H,O−CH2メトキシ)、1.96(広幅s,3H,CHアダマンチル)、1.78〜1.65(広幅dd,6H,3*CH2アダマンチル)、1.62〜1.56(d,4H,2*CH2スペーサー)、1.57(d,6H,3*CH2アダマンチル)、1.48〜1.42(m,2H,CH2中央スペーサー)。MS(ESI):obs.m/z=412.2[M+H]+;C23H41NO5(MW計算値):411.3。
【0040】
N−ブチル−2−O−(アダマンタン−1−イル−メトキシペンチル)−デオキシ−ノジリマイシン(30):MS(ESI):obs.m/z=454.2[M+H]+;C26H47NO5(MW計算値):453.3。
【0041】
【化5】
N−ベンジルオキシカルボニル−2−O−(アダマンタン−1−イル−メトキシペンチル)−3,4,6−トリ−O−ベンジル−デオキシ−ノジリマイシン(27):1H−NMR(CDCl3,400MHz):δ7.31〜7.25(m,20H,CH Bn)、5.16〜5.06(2H,CH2 Bn)、4.75〜4.49(2*CH2Bn)、4.43〜4.31(2H,CH2 Bn)、4.14〜4.12(m,1H,H5)、4.00〜3.97(dd,1H,H1 ax又はeq)、3.93〜3.89(dd,1H,H4)、3.67〜3.54(m,4H,H2,H3,2*H6)、3.38〜3.31(m,5H,H1 ax又はeq,2*O−CH2スペーサー)、2.93(s,2H,CH2メトキシ)、1.94(広幅s,3H,CHアダマンチル)、1.71〜1.62(広幅dd,6H,3*CH2アダマンチル)、1.58〜1.52(m,10H,3*CH2アダマンチル,2*CH2スペーサー)、1.38〜1.33(m,2H,CH2中央スペーサー)。13C−NMR(CDCl3,100MHz):δ155.7(C=O,Z基)、138.3,136.6(Cq Bn及びZ)、128.4〜127.4(CH芳香族Bn)、82.4,79.5,74.3(3*CH,C2,C3及びC4)、73.1,72.9,72.8,71.5,69.0,68.5,67.1(7*CH2,3*CH2Bn,CH2 Z基,2*O−CH2スペーサー,C6)、55.8(C5)、41.6(C1)、39.7,37,2(2*CH2アダマンチル)、34.0(Cqアダマンチル)、29.7,29.4(2*CH2スペーサー)、28.2(CHアダマンチル)、22.7(CH2中央スペーサー)。MS(ESI):obs.m/z=802.4[M+H]+;824.6[M+Na]+;C51H63NO7(MW計算値):801.5。
【0042】
適用分野
1.II型糖尿病(インスリン抵抗性)
II型糖尿病の発生率は、欧米諸国で劇的に増加している。主たる根本的な欠陥は、インスリンに応答してGLUT4輸送体をその細胞表面に動員する感受性が低下する結果として、筋肉及び脂肪組織による血流からのグルコースの取込みが損なわれるものである。筋肉中の脂肪酸濃度の上昇が異常なグルコース恒常性に関連していることは、すでに何年も前からわかっている。逆に、PPARγアゴニスト及びレキシノイドによって誘発されるグルコース恒常性の改善は、脂肪酸の分割の変更、即ち脂肪組織への脂肪酸の再配分、並びに筋肉の脂肪酸取込み及び代謝の相対的な消耗と関連付けられる。しかし、筋肉での脂肪毒性が糖尿病の発症及び進行を引き起こす分子機序は十分に理解されていない。したがって、この問題について洞察をさらに深めることが、インスリン抵抗性を治療するための医薬の改良/開発の助力となる。
【0043】
脂肪病因の分子機序
大学医学センター(Academic Medical Center)/アムステルダム大学生化学部のスフィンゴ糖脂質及びII型糖尿病についての研究活動は最近、II型糖尿病の脂肪病因に思いがけない新たな見識をもたらした。根底にある機序を以下で詳述する。
【0044】
後天性のインスリン抵抗性におけるスフィンゴ糖脂質の役割
糖尿病の病因におけるスフィンゴ(糖)脂質の役割について仮説を立てた。この考えは、パルミチン酸がスフィンゴ脂質中のセラミド部分の本質的な構築ブロックであり、スフィンゴ糖脂質の生合成の最初のステップは、セリンパルミトイルトランスフェラーゼを触媒とするパルミチン酸のセリンへの転移である(図1を参照のこと)という、見過ごされていた事実に端を発する。肝臓でのスフィンゴ脂質の合成速度は、パルミチン酸の濃度に著しく左右される。重要なことだが、このことは、培養筋肉細胞(平滑筋細胞、筋原細胞)で実験に基づいて確認できた。即ち、培地に0.1、0.5、1.0mMのパルミチン酸を加えると、放射標識したセリンのその構造への取込みの増加によって明らかになったとおり、スフィンゴ糖脂質の合成が比例して増加した。
【0045】
この発見を機に、スフィンゴ(糖)脂質が、糖尿病の根底にある筋肉中での脂肪毒性を実際に媒介する可能性についてより詳しく調査することになった。最近では、GM3(細胞表面にある最も単純なガングリオシド、図2を参照のこと)がインスリンシグナル伝達を弱め得ることが証明されている。これに関して、細胞表面でのGM3濃度が、インスリン受容体の多重クラスター形成のマイナスの妨げとなって、インスリンに応答してのグルコースの取込みを調節するらしいことが観察されている。さらに、高濃度のGM3は、GLUT4の細胞表面への動員の縮小化と関連付けられる。逆に、GM3の減少は、インスリン感受性の向上と関連付けられる(Yamishita et al. Proc Natl Acad Sci USA (2003) 100, 3445-9 Enhanced insulin sensitivity in mice lacking ganglioside GM3、Tagami et al. (2002) J Biol Chem 277, 3085-92 Ganglioside GM3 participates in the pathological conditions of insulin resistanceを参照のこと)。本発明者らは、肥満状態ではパルミチン酸レベルが慢性的に高く、したがって、脂肪細胞だけでなく筋細胞でのスフィンゴ糖脂質の生成が速まった速度で行われて、インスリン抵抗性に有利に働くと見ている。局所での(GM3を含む)スフィンゴ糖脂質合成を増大させる推進力としての筋肉中パルミチン酸濃度の増大とインスリン抵抗性との関係(図3を参照のこと)は、これまでのところ他者に認知されていない。
【0046】
グルコシルセラミドシンターゼの決定的役割
また、細胞表面のGM3及び他のガングリオシドの濃度が、ガングリオシド合成の律速段階であるグルコシルセラミドシンターゼの活性(グルコシルセラミドの合成)に著しく左右されることは十分に理解されていた(図3を参照のこと)。この酵素は、セラミドとUDP−グルコースからのグルコシルセラミドの生成を触媒する。その両方の基質(セラミド及びUDP−グルコース)のKm値は、生理的範囲にある。本発明者らは、インスリン感受性に関してグルコシルセラミドシンターゼが鍵調節酵素であることを示す。炎症性サイトカイン(TNF−α)、ステロイドホルモン、飽和脂肪酸、及びウイルス感染に応答してのその活性の増大は、以前から観察され、報告されている。今回、驚くべきことに、スフィンゴ糖脂質合成の変化がII型糖尿病の助長に影響を及ぼすことが判明した(図4を参照のこと)。脂肪病因がII型糖尿病に有害な影響を与えるというこの知見は、グルコシルセラミドシンターゼ活性の阻害が、有益な抗高血糖作用を発揮することを示すものである。
【0047】
イミノ糖系阻害剤の新規な使用
特定の部類のイミノ糖であるデオキシノジリマイシンが、グルコシルセラミド合成の阻害によってスフィンゴ糖脂質合成を低減するのに適する薬剤であることは明白になっている。さらに、ヒトにおけるイミノ糖投与の安全性と共に、実践での専門知識もかなり得られた。
【0048】
N−ブチル−デオキシノジリマイシンは、最近になって1型ゴーシェ病の治療用薬物として登録された。ケンブリッジ大学と共同でアムステルダム大学医学センターが主として請け負った臨床治験では、この薬物が大多数の患者で少なくとも5年間は忍容されることが明らかになった。スフィンゴ糖脂質合成が、100mgの1日3回のN−ブチル−デオキシノジリマイシンによってごく中程度(20〜30%)にだけは阻害されるという事実にもかかわらず、一部のゴーシェ病患者はしかし、深刻な腸の不調を訴え、時には警戒させる末梢ニューロパシーになる。より多い用量のN−ブチル−デオキシノジリマイシンでは、これらの有害事象がより頻繁に起こる。本発明者らは、グルコシダーゼ及びグルコシルトランスフェラーゼの阻害についてN−ブチル−デオキシノジリマイシンの特異性が不十分であるために、これらの望ましくない副作用が生じると見ている。表1に示すように、N−ブチル−デオキシノジリマイシンは、腸管グリコシダーゼの非常に強力な阻害剤でもある。本発明者らは、この阻害作用が、患者の腸の不調の少なくとも一部をもたらしていると見ている。この薬物は、リソソーム内のαグリコシダーゼ及びグルコセレブロシダーゼの阻害剤である。本発明者らは、この作用が、リソソームにおけるグリコーゲン及びグルコセレブロシダーゼの病的なリソソーム内蓄積への一連のリスクをもたらすと見ている。糖尿病(予備)患者のかなり低めのGM3に必要な濃度では、N−ブチル−デオキシノジリマイシンを使用すると有害事象が確実に起こる。
【0049】
インスリン抵抗性(II型糖尿病)治療薬としての特異的阻害剤の設計
N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシン(以前はAMP−DNMとしても知られていた)は、II型糖尿病治療薬としてのその使用についていくつかの魅力的な特徴を示す。即ち、この化合物は、経口投与するとき生物学的利用能が高く、グルコシルセラミドシンターゼの強力な阻害剤であり、代謝に関して不活性であり、治療中に想定される用量では固有の細胞毒性を示さない。
【0050】
しかし、in vitro酵素活性検定によって、培養細胞を用いてなされた観察によれば、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの長期投与が、他の代謝経路にも影響を及ぼし得る可能性があると思われる。例えば、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンを長期間投与した後、リソソーム内グリコシダーゼのα−グルコシダーゼとリソソーム内グルコセレブロシダーゼが阻害される可能性は、完全には除外できない。このことは、それぞれグリコーゲン又はグルコシルセラミドのリソソーム内蓄積をもたらすかもしれない。グリコーゲン又はグルコシルセラミドがリソソームに過剰に貯蔵されると、それぞれポンペ病及びゴーシェ病に似た病態になりかねない。その上、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、腸管スクラーゼの阻害剤でもあり、それとして胃腸の愁訴を引き起こすかもしれない。腸管グリコシダーゼが強力に阻害されれば、消化管内腔に浸透活性のある糖類が蓄積され、また腸内細菌の増殖に有利に働くはずであり、どちらも痙攣及び下痢の一因となる。このような合併症は、服薬遵守に影響を及ぼすはずであり、実際にはN−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの適用を狭めることになる。
【0051】
これらすべての考慮すべき事柄に導かれ、本発明者らは、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの所望の特性を保持したまま、インスリン抵抗性個体の長期間の治療に関してその不利な特徴のないイミノ糖構造の探索に乗り出した。
【0052】
以下の2種の選択判断基準を適用した。
1.リソソーム内グリコシダーゼ:βグルコシダーゼ及びグルコセレブロシダーゼの阻害の低減
これに関して、イミノ糖によるリソソーム内グリコシダーゼの阻害は、リソソーム貯蔵障害によく似た病的な代謝産物のリソソーム内貯蔵のリスクを高めるだけになりかねないので望ましくない。
【0053】
2.腸管グリコシダーゼの阻害の欠如
II型糖尿病に介入する別の手法は、腸管グリコシダーゼを阻害して、消化管での食物由来の糖の取込みを和らげることに基づく。合成のスクラーゼ阻害剤(アカルボース、N−ヒドロキシエチル−デオキシノジリマイシン)は、この構想に基づくものであり、抗糖尿病薬として登録されている。
【0054】
N−ヒドロキシエチル−デオキシノジリマイシンは、スクラーゼ阻害剤の中で最も強力な抗糖尿病薬である(Campbell L, Baker DE & Campbell RK, Ann Pharmacother 2000; 34: 1291-1301)。しかし、N−ヒドロキシエチル−デオキシノジリマイシン及びアカルボースのような強力な合成腸管グリコシダーゼ阻害剤の大きな欠点は、免れがたい深刻な腸の愁訴を引き起こし得ることである。したがって、強力な腸管グリコシダーゼ阻害剤であるN−ヒドロキシエチル−デオキシノジリマイシン及びアカルボースは、多くの個体にとって忍容性があまり十分でなく、服薬遵守が不十分になり、適用が制限される。
【0055】
N−ヒドロキシエチル−デオキシノジリマイシン及びアカルボースは、有益な効果を発揮するとはいえ、本発明者らは、それがその腸管グリコシダーゼ阻害能と関係しているかどうかは実際には疑わしいことに気付いた。興味深いことに、N−ヒドロキシエチル−デオキシノジリマイシン又はアカルボースの投与は、高炭水化物食なしでさえ、血糖値の低下をもたらす。このことは、この化合物の推定される治療作用方式と一致しない。おそらく、N−ヒドロキシエチル−デオキシノジリマイシン及びアカルボース(の代謝産物)は、体に取り込まれた後、グルコシルセラミドシンターゼを阻害してむしろ有益に働くと思われる。このことは、腸管グリコシダーゼの阻害がその治療効果の前提条件なのではなく、望ましくない副作用をもたらすに過ぎないことを示唆している。
【0056】
上述の選択判定基準に基づき、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンのイド立体配置が、インスリン抵抗性(II型糖尿病)の治療にとって魅力的なイミノ糖であると特定された。表1は、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンが、N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンと比べて、リソソーム内グルコセレブロシダーゼを阻害せず、β−グルコシダーゼ活性を弱く阻害するに過ぎないことを示している。その上、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、腸管グリコシダーゼの弱い阻害剤である。重要なことであるが、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、培養細胞で試験するとやはりグルコシルセラミドシンターゼの強力な阻害剤である(表2を参照のこと)。
【0057】
【表1】
【0058】
【表2】
【0059】
抗血糖上昇剤としてのイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの価値は、その後N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンについて先に述べた手順に厳密に従って動物モデルで分析した。この目的では、動物に異なる濃度のイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンを毎日与えた。250mg/kgの用量のイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンで、25mg/kgのN−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンと同様の有益な効果が示された。
【0060】
肥満におけるスフィンゴ糖脂質の役割
本発明者らは、(パルミチン酸に富んだ)動物性脂肪及び単純糖質(単糖及び二糖)の消費の増加傾向が、スフィンゴ糖脂質の過剰産生に有利に働くことを発見した。高レベルのスフィンゴ糖脂質は、エネルギー恒常性に関与するホルモン、例えばインスリンを抑制する。高いスフィンゴ糖脂質レベルは、エネルギーの収支のバランスをますます悪くし、進行中の体重増加及び随伴する副疾患に有利に働く。この新規な見識は、過体重及び肥満の個体においてスフィンゴ糖脂質の過剰産生を低減することのできるイミノ糖の使用という、治療処置への新たな道を切り開くものである。したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、過体重及び肥満の治療に適切に使用することができる。
【0061】
過体重及び肥満を治療するアルキル化イミノ糖の選択
特に、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、この目的に申し分なく適する化合物であることがわかった。この化合物は、生物学的利用能が十分であり、想定される投与量で細胞毒性を示さず、グルコシルセラミドシンターゼの比較的特異的な阻害剤である。イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンの利点は、大抵のイミノ糖によって引き起こされる合併症である腸の愁訴を引き起こさないことである。このような副作用がないことは、服薬遵守を向上させる。
【0062】
2.過体重及び肥満
世界中で、過体重及び肥満の有病率が流行の様相を帯びてきている。オランダでは、他の国同様、過体重及び肥満の個体の数がじりじりと増えている。他のヨーロッパ諸国の状況と似てはいるが、この増加は、例えば英国やドイツほど著しくない。最も圧倒的なのは、米国での有病率の増加である。平均して、成人オランダ人の40%が過体重であり、成人人口の10%が肥満である。成人の1〜1.5%が病的な肥満に罹患していると推測される。2000年には、米国の成人の56%が過体重であり、20%が肥満であり、2.3%が病的な肥満であった。WHOの規定によれば、成人は、BMIが30kg/m2以上であれば肥満(重度の過体重)であるとされる。BMIの値が25kg/m2と30kg/m2の間にある者は、過体重であるという。BMI(Body Mass Index)は、個体の体重(kg)をその身長(メートル)の2乗で割ったものであると定義される。
【0063】
体重増加の最初の結果の1つが、インスリンの正常な働きを狂わせるインスリン抵抗性である。インスリン抵抗性は、代謝症候群の進展において鍵となる役割を担う。この症候群は、インスリン抵抗性、異脂肪症(低HDL血清コレステロール、高血清トリグリセリド、高LDL血清コレステロール)、高血圧、腹部肥満など、いくつかの関連した代謝の異常を特徴とする。これらの異常は、II型糖尿病(成人型糖尿病)及びその合併症などの障害の進展の基盤となる。過体重及び肥満に随伴する他の健康上のリスクは、心臓病、様々な種類のがん、胆嚢疾患、関節症、呼吸器の問題、痛風、不妊症、月経障害、及び胎児の欠陥である。過体重が重症であるほど、このような副疾患のリスクは高まる。これらのすべての健康上のリスクの中でも、グルコース不耐症及びII型糖尿病の有病率の増大は特に厄介である。米国では、これが子供でも起こっている。最近の調査では、肥満児(5〜17才)の約60%が心臓病の追加の危険因子を示したことが明らかになっている。グルコース不耐症は、55人の肥満児(4〜10才)の25%、及び112人の若者(11〜18才)の21%で認められた。さらに、肥満はしばしば、心理的社会的問題、並びに生活の質の低下を伴う。肥満(及びそれ程ではないが過体重)に随伴する病的状態は、非常に多くの(医薬による)治療及び付加的な就業不能だけでなく、健康管理のための費用の増大をもたらす。米国では、過体重及び肥満に直接に関連した健康管理の費用が全体の6〜7%であると推定され、間接的な費用は4倍になると推測される。
【0064】
3.リソソーム貯蔵障害
リソソーム内のスフィンゴ糖脂質の蓄積を特徴とする遺伝性障害(GM1ガングリオシドーシス)、サンドホフ病(GM2ガングリオシドーシス、II型又はO型)、テイサックス病(GM2ガングリオシドーシス、I型又はB型)、ファブリー病(a−ガラクトシル−ラクトシルセラミドーシス)、ラクトシルセラミドーシス、ゴーシェ病(グルコセレブロシドーシス)は、西欧世界において、診断されたすべての先天性代謝異常の中で大きな割合を占めるものである。これらの疾患に罹患している患者が、蓄積するスフィンゴ糖脂質の合成の低減による恩恵を受けるはずであることは、以前から想定されている。最近では、N−ブチル−デオキシノジリマイシンが、いわゆる基質低減療法(SRT)による1型ゴーシェ病の治療用に登録されている。
【0065】
N−ブチル−デオキシノジリマイシンを用いた臨床経験から、この化合物の適用が、このイミノ糖の不十分な特徴によって制限されることがわかった。N−ブチル−デオキシノジリマイシンは、低い生物学的利用能しか示さず、グルコシルセラミドシンターゼの比較的弱い阻害剤であるので、有効性が不十分である。一方、N−ブチル−デオキシノジリマイシンは、リソソーム及び腸管のグリコシダーゼを著しく阻害し得るために、高めの用量では重い副作用を引き起こす。
【0066】
グリコスフィンゴリピドーシスのSRTのための特異的なデオキシノジリマイシン誘導体:イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシン
本発明者らは、スフィンゴ糖脂質貯蔵障害のSRTにより最適なイミノ糖を設計できることを発見した。化合物は、グルコシルセラミドシンターゼを阻害するその効力と、リソソーム及び腸管のグリコシダーゼの活性を阻害し得ないことに基づいて選択した。表3及び表4は、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンが、N−ブチル−デオキシノジリマイシンと比べて優れた特徴を示す理想的な物質であることを示すものである。
【0067】
【表3】
【0068】
酵素活性は、脾臓からの精製酵素を使用して、適切な4−MU基質で測定した。IC50値は、酵素活性を50%阻害するのに必要なイミノ糖の量を評価して決定した。
【0069】
【表4】
【0070】
in vivoでのグルコシルセラミドシンターゼ活性は、J. Biol. Chem, 273, 26522-27 (1998)に記載の手順に従って決定した。簡潔に述べると、グルコシルセラミドシンターゼ活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−セラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用薄層クロマトグラフィーにかけた後、C6−NBD−セラミドのC6−NBD−グルコシルセラミドへの変換を分析した。
【0071】
4.炎症性疾患
炎症は、局部組織の破壊及び関連する臨床症状を伴う。慢性の炎症は、特に有害な過程である。そのようなタイプの炎症はしばしば、慢性的に活性化したマクロファージによって統御される。様々な薬剤が炎症及び慢性の炎症の治療に使用される。しかし、これらの薬剤は、効果が最適でないか、又は重い副作用を示す。異なる作用方式に基づく改良された抗炎症剤が求められている。
【0072】
炎症におけるスフィンゴ糖脂質の役割
今回、炎症、特に慢性の炎症がスフィンゴ糖脂質の過剰産生と関連付けられること、並びにこの過剰産生がマクロファージの炎症状態を強化し、したがって組織炎症カスケードを促進することが判明した。スフィンゴ糖脂質レベルの増大によって、2通りの方式でマクロファージが活性化される。第1に、非リソソーム内グルコシルセラミダーゼによってグルコシルセラミドが過剰に分解されて、シグナル伝達分子として働くセラミドが生成される。さらに、セラミドの一部が、PKCを刺激するジアシルグリセロールへと代謝される。セラミド及びジアシルグリセロールの産生は、どちらも進行中のマクロファージの活性化に有利に働く。第2に、炎症の際のスフィンゴ糖脂質の増加が、細胞表面にあるラフトのスフィンゴ糖脂質組成を変化させる。これが、正常な細胞挙動に影響を及ぼし、炎症の過程を促進する。
【0073】
新しいクラスの抗炎症剤
先の発見によって、本発明者らは、イミノ糖が新規な作用方式によって働く有用な抗炎症剤に違いないと想定した。所望の特徴に基づき、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンを申し分なく適する抗炎症剤として選択した。この化合物は、生物学的利用能が十分であり、想定される投与量で細胞毒性を示さず、忍容性が十分である。この化合物は、そのどちらの酵素活性も炎症の重症度及び慢性の性質に寄与する非リソソーム内グルコシルセラミダーゼ及びグルコシルセラミドシンターゼの強力な阻害剤である。したがって、デオキシノジリマイシン類似体又は薬学的に許容できるその塩は、炎症性疾患の治療に適切に使用することができる。
【0074】
5.色素沈着過剰症及び皮膚の炎症状態
色素沈着
ヒトは、2種類のメラニン色素、即ち、褐色/黒色型若しくはユーメラニンと、琥珀色/赤色型若しくはフェオメラニンを産生する。ユーメラニンが、主として皮膚、毛髪、及び眼に見られる着色を司る。メラニンの合成は、特殊な細胞の特定の区画、即ちメラノサイトのメラノソームで行われる。色素は、その後皮膚のケラチノサイトに移送される。
【0075】
色素沈着は通常、年齢と共に増し、遺伝的な欠陥又は後天的な疾患によって変更される場合がある。ある場合では、それが色素沈着の増加(過色素沈着)になり、またある場合では、それがより薄い着色(色素沈着低下)になる。いくつかの種類の色素沈着過剰症が存在する。即ち、1.そばかす、2.妊娠に関連した過色素沈着、3.酵素の欠損による過色素沈着、4.疾患に関連した(炎症後)過色素沈着である。そばかすは、メラノサイト(色素生成細胞)が、近くの皮膚よりもUV放射に対して活性であり、且つ応答しやすい区域である。妊娠中に見られる色素沈着の増加は、エストロゲン、プロゲステロン、及びメラノサイト刺激ホルモンの影響のためである。妊娠中に起こるものなど、ホルモンに関連した過色素沈着は、妊娠マスク又は肝斑と呼ばれる状態として現れることもある。これは、顔又は日光露出区域に色素沈着の斑が出現する状態である。肝斑は無害である。肝斑で見られる色素は、分娩後数カ月以内にほとんど消失する。カロチンを代謝する肝臓の酵素が欠損している個体は、ニンジン、トウガラシ、又は他の黄色/橙色野菜を大量に消費すると、皮膚の色が黄色/橙色になる場合がある。炎症後過色素沈着(PIH)は、頻繁に直面する問題であり、様々な皮膚障害並びに治療処置の後遺症のことである。この後天的な色素過剰は、皮膚に影響を及ぼすそれまでの様々な疾患の過程を経た結果であるとすることができ、そうした過程には、感染、アレルギー反応、機械的損傷、薬剤投与に対する反応、光毒性発疹、外傷(例えば、火傷)、及び炎症性疾患(例えば、扁平苔癬、紅斑性狼瘡、アトピー性皮膚炎、乾癬、サルコイド症)が含まれる。
【0076】
炎症後過色素沈着
PIHは、表皮メラニン沈着又は真皮メラニン沈着をもたらす2通りの機序のうちの一方によって引き起こされる。表皮の炎症応答(即ち、皮膚炎)は、アラキドン酸を放出させ、その後酸化によってプロスタグランジン、ロイコトリエン、及び他の産物をもたらす。これらの炎症産物は、免疫細胞及びメラノサイトの両方の活性を変更する。詳細には、これらの炎症産物は、表皮メラノサイトを刺激して、それにメラニンの合成を増進させ、その後周囲のケラチノサイトへの色素の移送を増進させる。メラニン顆粒のこのような刺激及び移送の増進は、表皮の黒皮症をもたらす。一方、真皮メラニン沈着は、炎症が基底細胞層を乱すときに起こり、メラニン色素を放出させ、その後真皮乳頭中のマクロファージに捕えられるようにするものであり、色素失調症としても知られる。
【0077】
PIHは、皮膚のすべてに及ぶ応答であるが、着色された、より黒い皮膚ではより一般的である。メラニン過剰病巣の分布は、もともとの炎症性皮膚疾患の位置に応じて決まる。病巣の色は、淡褐色から黒色まで様々であり、色素が表皮内にある場合(即ち、表皮メラニン沈着)はより明るい褐色に見え、病巣が真皮メラニンを含む場合(即ち、真皮メラニン沈着)はより暗い灰色に見える。PIHは、皮膚に影響を及ぼす様々な疾患過程と共に起こる。それらの過程には、アレルギー反応、感染、外傷、及び光毒性発疹が含まれる。PIHをもたらす一般的な炎症性疾患としては、にきび、扁平苔癬、全身性紅斑性狼瘡、慢性皮膚炎、及び皮膚T細胞リンパ腫、特に異型紅皮症、乾癬、サルコイド症が挙げられる。
【0078】
デオキシノジリマイシン類似体による色素沈着過剰症及び皮膚炎症状態の治療
炎症におけるスフィンゴ糖脂質の役割については、前の項で遍く論述した。重要なことに、メラノサイト中でのメラニン産生の絶対要件は、グルコシルセラミドのようなスフィンゴ糖脂質の存在である。グルコシルセラミドシンターゼ活性を欠く糖脂質陰性メラノーマ細胞系は、スフィンゴ糖脂質合成が正常なその親細胞とはメラニンがない点で異なる。スフィンゴ糖脂質を加え、又はグルコシルセラミドシンターゼcDNAを形質移入すると、メラニン産生は復活するので、グルコシルセラミド(及び/又は他のスフィンゴ糖脂質)の合成がメラノソームでのメラニン産生にとって不可欠であることが示唆される。グルコシルセラミドなどのスフィンゴ糖脂質の産生は、チロシナーゼのみを適切にメラノソームに移送するのに必要であると思われる。
【0079】
今回、スフィンゴ糖脂質の過剰産生が皮膚の炎症過程の鍵となる要因であり、したがって本質的にメラニンの過剰産生(色素沈着過剰症)を引き起こし得ることがわかった。本発明者らは、(局所の炎症状態の結果としての)スフィンゴ糖脂質の過剰産生が、局所のメラニン合成の増加を引き起こすと見ている(以下のスキームを参照のこと)。デオキシノジリマイシン類似体などの、本発明によるスフィンゴ糖脂質合成の阻害剤は、過剰なメラニン産生及び随伴する色素沈着過剰症を軽減することができる。
【0080】
したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、色素沈着過剰症及び皮膚の炎症状態の治療に適切に使用することができる。これまで、メラニン産生に介入する手法は、スフィンゴ糖脂質合成以外のメラニン形成過程のターゲットに注目したものであった。多くの脱色素剤は、チロシナーゼの代替基質として働き、したがって、メラニン合成の最初の酵素段階、即ち、メラニン重合体をもたらすチロシナーゼのドーパキノンへの変換を阻害するものである。これは、通常は過色素沈着を実に効果的に軽減又は解消するものの、色が黒めの皮膚の患者では、オクロノーシス(真皮中の黒色の細かい斑点)を引き起こす場合もある。その上、ヒドロキノンは、高めの濃度では発癌性があると考えられているため、ヨーロッパの一部及びアジア全域で禁止されている。
【0081】
色素沈着過剰症及び皮膚の炎症過程の治療に向けた特異的デオキシノジリマイシン類似体
皮膚での使用に最適な特徴を有するイミノ糖を設計した。セラミド及びグルコシルセラミドは、角質層の構造の重要な構成成分であり、皮膚の水分透過性を調節している。グルコシルセラミド並びに他の極性脂質が、層状体によって角質層の細胞外間隙に分泌され、その後処理を受けてセラミドになることは知られている。リソソーム内グルコセレブロシダーゼは、この酵素が完全に欠損すると、それに伴って表皮のバリア機能が著しく損なわれるという事実が示唆するとおり、この過程でおいて一役を担う。したがって、イミノ糖は、リソソーム内グルコセレブロシダーゼを触媒とする、グルコシルセラミドからのセラミドの決定的な生成を妨げるべきでない。この前提条件を踏まえて、イミノ糖イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンが理想的な化合物であることがわかった(表5)。イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、グルコシルセラミドシンターゼ活性を阻害し、ヒトメラノサイトでのメラニン産生を低減することができる。
【0082】
【表5】
【0083】
培養メラノーマ細胞中でグルコシルセラミドシンターゼ、非リソソーム内グルコシルセラミダーゼ、及びリソソーム内グルコセレブロシダーゼを阻害し得るかどうかについて、様々なイミノ糖を分析した。最初の2種の酵素活性の阻害は望ましいが、最後の酵素の活性の阻害は望ましくない。in vivoでのグルコセレブロシダーゼ活性、非リソソーム内グルコシルセラミダーゼ活性、及びグルコシルセラミドシンターゼ活性は、J. Biol. Chem, 273, 26522-27 (1998)に記載の手順に従って測定した。簡潔に述べると、in vivoでのグルコセレブロシダーゼ及び非リソソーム内グルコシルセラミダーゼの活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−グルコシルセラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用の薄層クロマトグラフィーにかけた後、C6−NBD−グルコシルセラミドからC6−NBD−セラミドへの、コンズリトール−B−エポキシド感受性及び非感受性の分解を分析した。コンズリトール−B−エポキシドによって阻害される活性は、グルコセレブロシダーゼによるものであり、残りは非リソソーム内グルコシルセラミダーゼによるものとすることができる。in vivoでのグルコシルセラミドシンターゼ活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−セラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用の薄層クロマトグラフィーにかけた後、C6−NBD−セラミドのC6−NBD−グルコシルセラミドへの変換を測定した。
【0084】
MEB4培地中で、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンによってスフィンゴ糖脂質合成を阻害すると、色素沈着は消失する。一方は色素沈着が濃く(An)、他方は色素沈着が非常に薄い(M14)2種のメラノーマ培養物では、色素沈着の濃い培養物中の方が、全細胞抽出物及びメラノソーム画分の両方においてグルコシルセラミド(GlcCer)含有量が明らかに多いことが示される。
【0085】
メラノサイトでは、1回しか継代していなくても、イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンに曝すと、薄い方と濃い方の皮膚タイプの細胞で色素沈着の有意な減少が見られる。本発明によるデオキシノジリマイシン類似体の薬剤としての用途は、炎症後色素沈着過剰症及び皮膚の炎症状態にある。これらのデオキシノジリマイシン類似体を全身に投与するとき、in vivoで強い抗炎症活性を有するという証拠は有力である。これらの化合物は、魅力的な薬理学的特徴(良好な生物学的利用能、代謝されない、動物での2週間の投与の後有害な毒物学的作用がない)を示す。イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、皮膚の完全性にとって重要な酵素活性であるリソソーム内グルコセレブロシダーゼを完全に/ほとんど阻害しないので、皮膚への適用に特に適する。
【0086】
6.黒色腫及び他の腫瘍
腫瘍細胞は、その表面にガングリオシドを含み、それを盛んに剥離させ、その後それらは他の細胞に吸収される。腫瘍ガングリオシドは、免疫抑制活性、血管新生促進特性を有し、増殖因子を媒介とする線維芽細胞及び血管内皮細胞の増殖を刺激することが報告されている。ガングリオシド発現の増加は、腫瘍生成の強化及び進行の加速と関連付けられている。ガングリオシド生合成を阻害すれば、腫瘍の進行を妨げ、且つ/又は転移を減少させることができる。グルコシルセラミドシンターゼのイミノ糖系阻害剤(OGT2378)は、マウスモデルでメラノーマの腫瘍増殖を顕著に阻害することが示されている。
【0087】
黒色腫及び他の腫瘍の治療に向けたデオキシノジリマイシン類似体
今回、グルコシルセラミドシンターゼ並びに非リソソーム内グルコシルセラミダーゼの強力且つ特異的な阻害剤であるイミノ糖が、腫瘍の増殖及び/又は転移を低減し得ることがわかった。最適な効力のためには、両方の酵素活性を相伴って阻害する必要がある。
【0088】
本発明によるデオキシノジリマイシン類似体は、良好な生物学的利用能、代謝及び毒性がないことを含む、すべての望ましい特徴を有する。したがって、本発明のデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、黒色腫及び他の腫瘍の治療に適切に使用することができる。
【0089】
実験データ
培養メラノーマ細胞中でグルコシルセラミドシンターゼ、非リソソーム内グルコシルセラミダーゼ、及びリソソーム内グルコセレブロシダーゼを阻害し得るかどうかについて、様々なイミノ糖を分析した。最初の2種の酵素活性の阻害は望ましいが、最後の酵素の活性の阻害は望ましくない。
【0090】
【表6】
【0091】
in vivoでのグルコセレブロシダーゼ活性、非リソソーム内グルコシルセラミダーゼ活性、及びグルコシルセラミドシンターゼ活性は、J. Biol. Chem, 273, 26522-27 (1998)に記載の手順に従って測定した。簡潔に述べると、in vivoでのグルコセレブロシダーゼ及び非リソソーム内グルコシルセラミダーゼの活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−グルコシルセラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用の薄層クロマトグラフィーにかけた後、C6−NBD−グルコシルセラミドからC6−NBD−セラミドへの、コンズリトール−B−エポキシド感受性及び非感受性の分解を分析した。コンズリトール−B−エポキシドによって阻害される活性は、グルコセレブロシダーゼによるものであり、残りは非リソソーム内グルコシルセラミダーゼによるものとすることができる。in vivoでのグルコシルセラミドシンターゼ活性は、細胞を、アルブミンとの複合体にした蛍光C6−NBD−セラミドに曝すことで測定した。細胞を収集し、脂質を抽出し、定量分析用の薄層クロマトグラフィーにかけた後、C6−NBD−セラミドのC6−NBD−グルコシルセラミドへの変換を測定した。
【0092】
選択したデオキシノジリマイシン類似体の有効性をモニターするための動物モデル
イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、粉末食にして経口投与することができる。B16マウスメラノーマ細胞の亜系統であるMEB4細胞(Riken Cell Bank、埼玉県)を皮内注射したC57BL/6マウスを使用することができる。腫瘍の形成及びサイズについて、マウスを週3回検査することができる。腫瘍サイズは、キャリパーを使用して立体的に測定することができ、腫瘍体積は、式a×b×c/2を用いて推定することができる(Weiss et al. 2003. Cancer Research 63, 3654-8)。
【0093】
7.真菌症
最近では、病原性の真菌がヒトグルコシルセラミドシンターゼに類似した酵素を有することが明らかになってきている。グルコシルセラミドは、菌糸が成長する際に選択的に産生され、成長端に隣接する細胞膜中で濃縮される。真菌を、グルコシルセラミドシンターゼ活性を阻害することがわかっているセラミド類似体に曝した後に、菌糸の成長の抑制が観察されている。
【0094】
今回、慎重に選択した疎水性イミノ糖が、より強力且つ忍容性の十分なグルコシルセラミドシンターゼの阻害剤であることが判明した。真菌のグルコシルセラミドシンターゼ活性を特異的に妨げ、内在するヒトグルコシルセラミドシンターゼには弱い影響しか及ぼさないイミノ糖を選択することで、生命を脅かす真菌感染と闘うことができる。そのような感染は、診療所で高まりつつある問題となっている。選択されたイミノ糖は、真菌菌糸の成長端の細胞壁を攻撃するキチナーゼである組換えヒトキトトリオシダーゼと併用できるはずである。したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、真菌症の治療に適切に使用することができる。
【0095】
8.ウイルス感染
イミノ糖は、ウイルス感染の治療用薬剤として調査されている。AIDSを引き起こす病原体であるHIVによる感染を治療するために、N−ブチルデオキシノジリマイシンの研究が行われている。世界中で数億の個体がAIDSに侵されている。in vitroでウイルスの感染力及び合胞体形成が示された。抗HIV剤としてのN−ブチルデオキシノジリマイシンの第II相臨床治験では、有望な結果が出なかった。必要とされる血清濃度では、重い副作用が起こった。B型肝炎ウイルス(HBC)は、世界中で感染者が3億5千万人を超えており、肝疾患及び肝細胞癌を引き起こし得る。近い関係にあるHBV感染動物モデルである、ウッドチャック肝炎ウイルスに慢性的に感染したウッドチャックでは、ノニルデオキシノジリマイシンによる治療が、感染性のあるエンベロープ型ウイルスの分泌を防ぐことがわかった。世界中で、1億人以上がC型感染ウイルス(HCV)に慢性的に感染している。ワクチンがない中、このことは、先進諸国の公衆衛生の最も深刻な脅威の1つとなっている。推定390万人の北アメリカ人が慢性的に感染している状況で、C型肝炎は現在、米国の肝臓移植理由の筆頭に立っている。この感染症は、年間約8,000人の米国人の死亡原因となり、この数字は、有効な処置が存在しなければ今後20年で3倍になると予想される。イミノ糖N−ノニルデオキシノジリマイシンは、C型肝炎のin vitro代用モデルであるウシウイルス性下痢ウイルスにおいて抗ウイルス活性を示している。
【0096】
ERα−グルコシダーゼは、新生糖タンパク質に結合したN−グリカン鎖から末端グルコース残基を段階的に除去する役を司ると考えられている。このために、糖タンパク質は、もっぱら一グリコシル化糖タンパク質に結合する、ERシャペロンのカルネキシン及びカルレティキュリンと相互に作用できるようになる。カルネキシンとの相互作用は、全部ではないが一部の糖タンパク質の正確なフォールディングにとって決定的なものであり、このグルコシダーゼの阻害剤を使用すると、それに依存するタンパク質を特異的に標的にすることができる。デオキシノジリマイシンの抗ウイルス活性は、ウイルスのエンベロープ糖タンパク質の適正なフォールディング及び輸送を妨害し、感染性のエンベロープ型ウイルスの分泌を防ぐことになるはずのERα−グルコシダーゼの阻害に基づくものであると考えられる。
【0097】
しかし、本発明者らは、イミノ糖の観察された抗ウイルス活性が、ERa−グルコシダーゼの阻害ではなく、グルコシルセラミド及びそのスフィンゴ糖脂質代謝産物の合成の阻害に基づくものであると見ている。宿主細胞表面上のスフィンゴ糖脂質に富むラフトは、ウィルスの融合及び出芽において欠かせない重要性を有しているように思われる。表面スフィンゴ糖脂質組成を操作すると、これらの過程に影響が及ぶ。
【0098】
この新規な見識は、ウイルス感染治療用のイミノ糖を選択する新たな論理的根拠となる。化合物は、それがグルコシルセラミドシンターゼ活性の強力な阻害剤であるという判断基準を第1に選択する必要がある。さらに、化合物は、十分な生物学的利用能及び忍容性をもつべきである。本発明によるデオキシノジリマイシン類似体、特にイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシン。したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、こうした判断基準を満たすウイルスの治療に適切に使用することができる。
【0099】
9.微生物感染及び内毒素
粘膜の感染
粘膜病原は、宿主の複合糖質受容体に対する特異的な付着性によって感染部位に向かって行く。そのため、細胞表面からのそのような受容体の除去が、付着を妨げ、細菌コロニー形成を損なわせ、粘膜感染活動を縮小すると予測することができる。スフィンゴ糖脂質シンターゼの阻害剤を使用すると、受容体を除去することができる。
【0100】
実際、N−ブチル−デオキシノジリマイシンで処理したマウスは、P線毛大腸菌による実験的な尿路感染に罹患しにくくなった(Svensson et al. Mol. Microbiol. 2003; 47: 453-61)。ケモカインの分泌が減少し、好中球の動員がより少ないことによって示されるように、粘膜の炎症応答は弱まっており、また細菌による尿路でのコロニー形成の効率は、尿路感染(UTI)を引き起こし、通常は「P」抗原に対する末端受容体を備えた線毛を有する通常のマウス大腸菌種より劣っていた。P抗原は、会陰及び尿道に並ぶ細胞の表面上にも見られる血液型マーカーである。人口の約75%がP抗原を発現させ、これらの個体は特にUTIに罹患しやすい。P抗原は、膣及び前立腺の分泌物中にも見られるが、このような分泌されたP抗原は、細菌受容体に結合して、上皮細胞表面に生物体が結合しないようにするという点で保護物である。UTIに最も罹患しやすい個体は、その細胞上にP抗原を発現させ、その分泌中にはP抗原がない個体である。
【0101】
別の例は、複合的な播種性の胃の炎症を引き起こすヘリコバクターピロリである。したがって今回、粘膜へのその堅固な付着にとってもスフィンゴ糖脂質が非常に重要であることを見出した。適切なイミノ糖による胃のスフィンゴ糖脂質の除去は、すばらしい治療用途をもたらし得るはずである。
【0102】
内毒素
糖脂質受容体が、細菌とその宿主間の主要な界面として働くだけでなく、内毒素など、細菌の病原性因子のターゲットとしても働くことがさらに判明した。ガングリオシドGM1は、コレラ毒素(CTX)や大腸菌の熱安定毒素IIのような様々なAB5毒素のBサブユニットの受容体として働き得るのに対し、グロボトリアオシドGb3は、AB5毒素である滋賀毒素(ST)、及びベロ毒素のような一部の関連毒素の受容体として働く。内毒素結合の防止は、臨床に大きく関わる問題である。例えば、ベロ毒素は、出血性大腸炎及び溶血尿毒症症候群(HUS)の細小血管症における内皮の標的化に関与する。スフィンゴ糖脂質合成の阻害剤を使用すれば、内毒素を媒介とする病態に介入できるはずである。
【0103】
粘膜の細菌感染の治療、及び内毒素を媒介とする病態の予防に向けたデオキシノジリマイシン類似体
先の新規な見識は、ウイルス感染治療用のイミノ糖を選択する論理的根拠となる。化合物は、グルコシルセラミドシンターゼ活性の強力な阻害剤であるという判断基準を第1に選択する必要がある。さらに、化合物は、十分な生物学的利用能及び忍容性を有するべきである。本発明のデオキシノジリマイシン類似体、特にイド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンは、こうした判断基準を満たす。したがって、本発明によるデオキシノジリマイシン類似体又は薬学的に許容できるその塩は、粘膜の細菌感染の治療及び内毒素を媒介とする病態の予防に適切に使用することができる。
【0104】
[参考文献]
1. Kolter T, Proia RL and Sandhoff K. 2002. J. Biol.Chem. 277, 25859-62. Combinatorial ganglioside biosynthesis.
2. Wertz PW & van den Bergh B. 1998. Chem. Phys. Lipids 91, 85-96. The physical, chemical and functional properties of lipids in the skin and other biological barriers.
3. Simons K & van Meer G. 1988. Biochemistry 27, 6197-6202. Lipid sorting in epithelial cells.
4. Brown DA & London E. 1998. J. Membr. Biol. 164, 103-114. Structure and origin of ordered lipid domains in biological membranes.
5. Hakomori S. 1981. A.Rev.Biochem . 50, 733-764. Glycosphingolipids in cellular interaction.
6. Huwiler A, Kolter T, Pfeilschifter J & Sandhoff K 2000. Biochim.Biophys.Acta 1485, 63-99. Physiology and pathophysiology of sphingolipid metabolism and signaling.
7. Hannun YA & Obeid LM. 1995. Trends Biochem.Sci.20, 73-77. Ceramide: an intracellular signal for apoptosis.
8. Kolesnick R & Kronke M. 1988. Annu.Rev.Physiol.60, 643-665. Regulation of ceramide production and apoptosis.
9. van Blitterswijk WJ, van der Luit AH, Veldman RJ, Verheij M, & Borst J. 2003.Biochem J. 2003 Jan 15;369(Pt 2):199-211. Ceramide: second messenger or modulator of membrane structure and dynamics?
10. Merrill AH. 2002. J.Biol.Chem. 277, 25843-25846. De novo sphingolipid biosynthesis: a necessary, but dangerous, pathway.
11. Sandhoff K & Kolter T. 2003. Philos Trans R Soc Lond B Biol Sci. 358(1433):847-61. Biosynthesis and degradation of mammalian glycosphingolipids.
12. Van Weely S, Brandsma M, Strijland A, Tager JM & Aerts JMFG. 1993. Biochim.Biophys.Acta 1181,55-62. The existence of a non-lysosomal glucocerebrosidase that is not deficient in Gaucher disease.
13. Aerts JM, Hollak C, Boot R, & Groener A. 2003. Philos Trans R Soc Lond B Biol Sci. 358(1433):905-14. Biochemistry of glycosphingolipid storage disorders: implications for therapeutic intervention.
14. Barranger JA & Ginns EI. 1989. Glycosylceramide lipidosis: Gaucher disease. In: Scriver CR, Beaudet AL, Sly WS, Vall D (eds) The metabolic basis of inherited disease. McGraw-Hill, New York, p 1677-1698
15. Beutler E & Grabowski GA. 1995. Gaucher's disease. In: Scriver CR, Beaudet AL, Sly WS, Valle D (eds) The metabolic and molecular bases of inherited disease. McGraw-Hill, New York, p 2641-2670
16. Brady RO, Kanfer JN, Bradley RM & Shapiro D. 1966. Demonstration of a deficiency of glucocerebroside-cleaving enzyme in Gaucher's disease. J Clin Invest 45, 1112-1115
17. Patrick AD. 1965. A deficiency of glucocerebrosidase in Gaucher's disease. Biochem J 97, 17c-18c
18. Van Weely S, van den Berg M, Barranger JA, Sa Miranda MC, Tager JM & Aerts JMFG. 1993. Role of pH in determining the cell-type specific residual activity of glucocerebrosidase in type 1 Gaucher disease. J Clin Invest 91, 1167-1175
19. Aerts JM, van Weely S, Boot R, Hollak CE & Tager JM. 1993. Pathogenesis of lysosomal storage disorders as illustrated by Gaucher disease. J Inherit Met Dis 16, 288-291
20. Cox TM, & Schofield JP. 1997. Gaucher's disease: clinical features and natural history Baillieres Clin.Hematol.10, 657-689
21. Aerts JMFG & Hollak CEM. 1997. Plasma and metabolic abnormalities in Gaucher's disease. Baillieres Clin Haematol 10, 691-709
22. Cox TM .2001. Gaucher disease: understanding the molecular pathogenesis of sphingolipidoses. J Inherit Met Dis 24 (suppl 2), 106-121
23. Moran MT, Schofield JP, Hayman AR, Shi G-P, Young E & Cox TM. 2000. Pathologic gene expression in Gaucher disease: upregulation of cysteine proteinases including osteoclastic cathepsin K. Blood 96, 1969-1978
24. Brady RO.1997. Gaucher's disease: past, present and future. Baillieres Clin Haematol 10, 621-634
25. Barranger JA, & O'Rourke E. 2001. Lessons learned from the development of enzyme therapy for Gaucher disease J Inherit Met Dis 24 (suppl 2), 89-96
26. Hollak CEM, Aerts JMFG, Goudsmit ER, Phoa SS, Ek M. van Weely S, von dem Borne AE & van Oers MH.1995. Individualised low-dose alglucerase therapy for type 1 Gaucher's disease. Lancet 345, 1474-1478
27. Richter J & Karlsson S. 2001. Clinical gene therapy in hematology: past and future. Int. J Hematol 73, 162-169
28. Radin NS. 1996. Treatment of Gaucher disease with an enzyme inhibitor. Glycoconj J 13, 153-157
29. Platt FM, Jeyakumar M, Andersson U, Priestman DA, Dwek RA, ButtersTD, Cox TM, Lachmann RH, Hollak CEM, Aerts JMFG, Hrebicek M, Moyses C, Gow I, Elstein D & Zimran A. 2001. J. Inher. Met. Dis. 24, 275-290. Inhibition of substrate synthesis as a strategy for glycolipid lysosomal storage disease therapy.
30. Lee L, Abe A & Shayman JA. 1999. J Biol Chem 274, 146662-14665 Improved inhibitors of glucosylceramide synthase.
31. Abe A, Gregory S, Lee L, Killen PD, Brady RO, Kulkarni A & Shayman JA. 2000. Reduction of globotriaosylceramide in Fabry disease mice by substrate deprivation. J Clin Invest 105, 1563-1567
32. Platt FM, Neises GR, Dwek RA & Butters TD. 1994. N-butyl-deoxynojirimycin is a novel inhibitor of glycolipid biosynthesis. J Biol Chem 269, 8362-8365
33. Jeyakumar M, Butters TD, Cortina-Borja M, Hunnam V, Proia RL, Perry VH, Dwek RA & Platt FM. 1999. Delayed symptom onset and increased life expectancy in Sandhoff mice treated with N-butyl-deoxynojirimycin. Proc Natl Acad Sci USA 96, 6388-6393
34. Andersson U, Butters TD, Dwek RA & Platt FM. 2000. N-butyldeoxygalactonojirimycin: a more selective inhibitor of glycosphingolipid biosynthesis than N-butyldeoxynojirimycin, in vitro and in vivo. Biochem Pharmacol 49, 821-829
35. Overkleeft HS, Renkema GH, Neele J, Vianello P, Hung IO, Strijland A, van den Burg A, Koomen, GJ, Pandit UK & Aerts J. 1998. Generation of specific deoxynijirimycin-type inhibitors of the non-lysosomal glucosylceramidase. J Biol Chem 273, 26522-26527
36. Cox T, Lachmann R, Hollak C, Aerts J, van Weely S, Hrebicek M, Platt F, Butters T, Dwek R, Moyses C, Gow I, Elstein D & Zimran A. 2000. Lancet. 355(9214):1481-5. Novel oral treatment of Gaucher's disease with N-butyldeoxynojirimycin (OGT 918) to decrease substrate biosynthesis.
37. Heitner R, Elstein D, Aerts J, Weely S &Zimran A. 2002. Low-Dose N-Butyldeoxynojirimycin (OGT 918) for Type I Gaucher Disease. Blood Cells Mol Dis 28, 127-33
38. Fischl MA, Resnick L, Coombs R, Kremer AB, Pottage JC Jr, Fass RJ, Fife KH, Powderly WG, Collier AC, Aspinall RL, et al. The safety and efficacy of combination N-butyl-deoxynojirimycin (SC-48334) and zidovudine in patients with HIV-1 infection and 200-500 CD4 cells/mm3. J Acquir Immune Defic Syndr. 1994 Feb;7(2):139-47.
39. Goss PE, Reid CL, Bailey D & Dennis JW. 1997. Phase 1B clinical trial of the oligosaccharide processing inhibitor swainsonine in patients with advanced malignancies.
【図面の簡単な説明】
【0105】
【図1】適切な疎水性存在物の設計を示す図である。
【図2】デオキシノジリマイシン類似体4の合成を示す図である。
【図3】デオキシノジリマイシン7の合成を示す図である。
【図4】デオキシノジリマイシン類似体17、18、及び19の合成を示す図である。
【図5】デオキシノジリマイシン類似体28、29、及び30の合成を示す図である。
【特許請求の範囲】
【請求項1】
一般式(I)を有するデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、ゴーシェ病の治療用医薬組成物。
【化1】
(I)
[式中、
R1〜R5は、それぞれ独立に、H、(CH2)nCH3、又はXを含み、
R6は、H、CH2OH、又はCH2OXを含み、ここで、R1〜R5の少なくとも一つはXであるか、又はR1〜R5が全てX以外の基のとき、R6はCH2OXであり、
nは、0〜9であり、
a、b、c、d、eは、R又はS立体配置を有するキラル中心であり、ここで、R1がXのとき、b、c、d、eは、それぞれS、R、S、S立体配置を有するキラル中心であり、
Xは、アダマンタン−1−イル−メトキシである大きな疎水性部分及び炭素原子5個のアルキレン鎖であるスペーサーを含み、疎水性部分は、脂質二重層に入り込むことができる]。
【請求項2】
イド立体配置を有する、請求項1記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、ゴーシェ病の治療用医薬組成物。
【請求項3】
イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンを含むデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、ゴーシェ病の治療用医薬組成物。
【請求項4】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、炎症性疾患の治療用医薬組成物。
【請求項5】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、色素沈着過剰症及び/又は皮膚の炎症状態の治療用医薬組成物。
【請求項6】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、真菌症の治療用医薬組成物。
【請求項7】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、過体重及び肥満の治療用医薬組成物。
【請求項8】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、リソソーム貯蔵障害の治療用医薬組成物。
【請求項9】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、マイクロバクテリア感染症(microbacterial infection)治療用医薬組成物。
【請求項10】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、インスリン抵抗性の治療用医薬組成物。
【請求項1】
一般式(I)を有するデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、ゴーシェ病の治療用医薬組成物。
【化1】
(I)
[式中、
R1〜R5は、それぞれ独立に、H、(CH2)nCH3、又はXを含み、
R6は、H、CH2OH、又はCH2OXを含み、ここで、R1〜R5の少なくとも一つはXであるか、又はR1〜R5が全てX以外の基のとき、R6はCH2OXであり、
nは、0〜9であり、
a、b、c、d、eは、R又はS立体配置を有するキラル中心であり、ここで、R1がXのとき、b、c、d、eは、それぞれS、R、S、S立体配置を有するキラル中心であり、
Xは、アダマンタン−1−イル−メトキシである大きな疎水性部分及び炭素原子5個のアルキレン鎖であるスペーサーを含み、疎水性部分は、脂質二重層に入り込むことができる]。
【請求項2】
イド立体配置を有する、請求項1記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、ゴーシェ病の治療用医薬組成物。
【請求項3】
イド−N−(5−アダマンタン−1−イル−メトキシ−ペンチル)デオキシノジリマイシンを含むデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、ゴーシェ病の治療用医薬組成物。
【請求項4】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、炎症性疾患の治療用医薬組成物。
【請求項5】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、色素沈着過剰症及び/又は皮膚の炎症状態の治療用医薬組成物。
【請求項6】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、真菌症の治療用医薬組成物。
【請求項7】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、過体重及び肥満の治療用医薬組成物。
【請求項8】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、リソソーム貯蔵障害の治療用医薬組成物。
【請求項9】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、マイクロバクテリア感染症(microbacterial infection)治療用医薬組成物。
【請求項10】
請求項1から3のいずれか記載のデオキシノジリマイシン類似体又は薬学的に許容できるその塩を含む、インスリン抵抗性の治療用医薬組成物。
【図1】

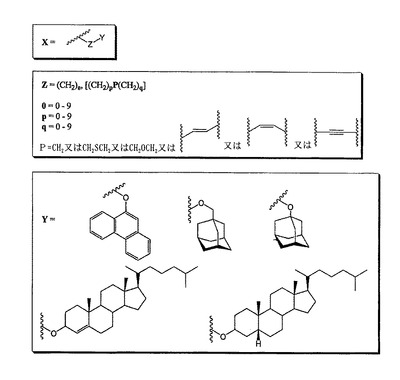
【図2】
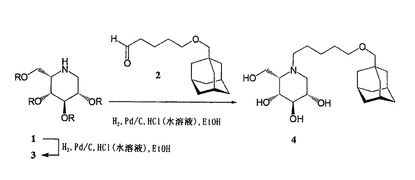
【図3】
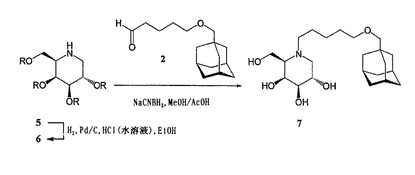
【図4】
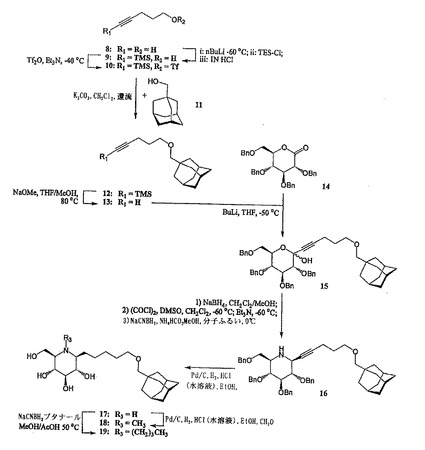
【図5】
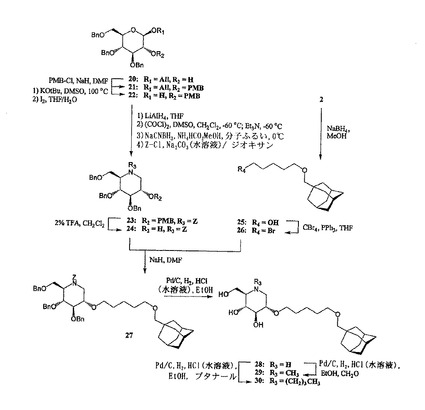
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−180353(P2012−180353A)
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願番号】特願2012−91984(P2012−91984)
【出願日】平成24年4月13日(2012.4.13)
【分割の表示】特願2006−537909(P2006−537909)の分割
【原出願日】平成16年10月29日(2004.10.29)
【出願人】(505323183)ゲンザイム コーポレーション (1)
【Fターム(参考)】
【公開日】平成24年9月20日(2012.9.20)
【国際特許分類】
【出願日】平成24年4月13日(2012.4.13)
【分割の表示】特願2006−537909(P2006−537909)の分割
【原出願日】平成16年10月29日(2004.10.29)
【出願人】(505323183)ゲンザイム コーポレーション (1)
【Fターム(参考)】
[ Back to top ]