
デンプン集積能力の高い形質転換植物およびその製造方法
【課題】深刻化する環境ストレスに適応し、劣悪な環境下においてもデンプン・エネルギー集積能力の高い植物およびその製造方法を提供する。
【解決手段】イネα−アミラーゼI−1の特定のペプチド領域及び小胞体膜透過シグナルペプチド領域を含むプラスチド輸送・局在化コンストラクトにより、機能タンパクを小胞体−ゴルジ体系を介してプラスチドへ輸送・局在化させ、プラスチドの機能改変を行うことによりデンプン集積能力の高い形質転換植物を得る。
【解決手段】イネα−アミラーゼI−1の特定のペプチド領域及び小胞体膜透過シグナルペプチド領域を含むプラスチド輸送・局在化コンストラクトにより、機能タンパクを小胞体−ゴルジ体系を介してプラスチドへ輸送・局在化させ、プラスチドの機能改変を行うことによりデンプン集積能力の高い形質転換植物を得る。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、デンプン集積能力の高い形質転換植物およびその製造方法に関する。
【背景技術】
【0002】
近年、地球温暖化など地球規模での自然環境の悪化が大きな問題になっている。我が国においてもすでに環境ストレスがイネなどの重要作物の生長・生理、特に炭水化物代謝に大きな影響を及ぼし、減収や品質劣化を引き起こしていることが知られている。
光合成産物のデンプンへの集積は、植物のみならず地球上のすべての生命にとって最も重要なエネルギー蓄積過程である。このデンプンへの集積は、光合成機能を有する葉緑体が分化したアミロプラストにより行われていることから、環境ストレス耐性の作物を育種するためにこの葉緑体・アミロプラスト(プラスチド)の機能改変法の研究が行われている。プラスチドの分化・機能発現は、このオルガネラ自身がもつ遺伝子と細胞核のゲノムにコードされた遺伝子によって統御されている。プラスチドの機能改変法としては、これまでに大きく分けて2つの方法が知られている。1つは、プラスチドに遺伝子を直接導入する方法であり、もう1つは、トランジットペプチドと呼ばれている葉緑体移行シグナル遺伝子を葉緑体に導入したい機能タンパク質遺伝子に融合し、それを細胞核ゲノムに導入する方法である。導入された遺伝子は細胞質で翻訳され、トランジットペプチドによりプラスチドへ輸送・局在化される。しかしながら、前者の方法では植物全体のプラスチドを改変することは困難であり、また、後者の方法では細胞質で組換えタンパク質が翻訳されるため、細胞にとって負担になり得るといった問題があった。
ところで、α−アミラーゼ(EC 3.2.1.1)は、デンプンの加水分解酵素であり、胚盤組織及びアリューロン層からデンプン貯蔵組織である胚乳に分泌され、そしてデンプン顆粒を分解し、発芽及び伸長生長に必要な炭素源並びにエネルギー源を供給するという重要な働きをする。イネα−アミラーゼ遺伝子は少なくとも11種類存在していることが明らかになっているが、なかでも典型的なN−結合型糖鎖を有するα−アミラーゼI−1(AmyI−1)は発芽イネ種子における胚乳貯蔵デンプンの分解に重要な役割を果たしていることが報告されている(非特許文献1および2)。
【非特許文献1】Asatsuma, S., Sawada, C., Itoh, K., Okito, M., Kitajima, A., Mitsui, T. (2005): Involvement of a-amylase I-1 in starch degradation in rice chloroplasts. Plant Cell Physiol. 46: 858-869
【非特許文献2】Nanjo, Y., Asatsuma, S., Itoh, K., Hori, H. and Mitsui T. (2004) Proteomic identification of a-amylase isoforms encoded by RAmy3B/3C from germinating rice seeds. Biosci. Biotech. Biochem. 68: 112-118
【発明の開示】
【発明が解決しようとする課題】
【0003】
しかしながら、イネα−アミラーゼの葉緑体・アミロプラスト(プラスチド)へのターゲティング機構及び遺伝的なシステムの詳細についてはほとんど知られていない。葉緑体・アミロプラストの光合成能やデンプン集積能の高い作物を育種することは、植物のみならず地球上のすべての生命にとって最も重要なエネルギー蓄積を行う上で重要な役割を果たすものとして、各方面から研究が進められ、その成果が期待されている。
【0004】
そこで、本発明は、以上の背景の下なされたものであって、深刻化する環境ストレスに適応し、劣悪な環境下においても効率的にデンプン・エネルギー集積能力の高い植物およびその製造方法を提供することを目的とする。
【課題を解決するための手段】
【0005】
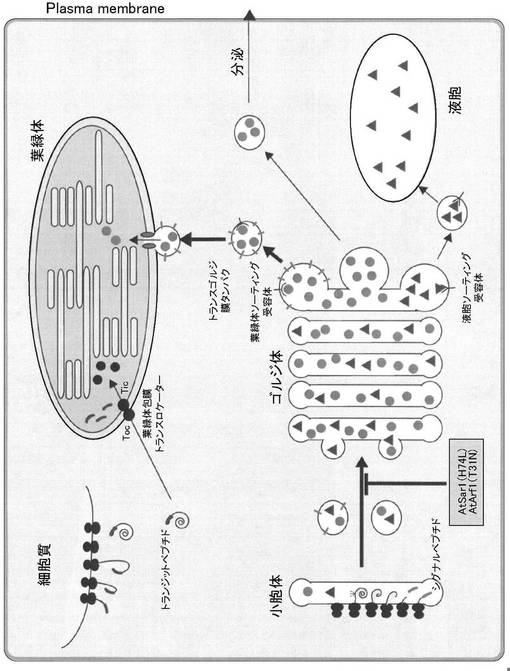
本発明者らは以上の課題に鑑み、穀類細胞において核ゲノムにコードされている酵素タンパク質のプラスチドへのターゲティング機構を検討し、プラスチド構成タンパク質のターゲティング機構の存在を明らかにした(図1参照)。具体的には、イネα−アミラーゼI−1のプラスチドへのターゲティングは、小胞体からゴルジ体への輸送に関わるAtArf1やAtSar1のドミナントネガティブ変異体により阻害されたことから、小胞体からゴルジ体への輸送が不可欠であることを明らかにした。すなわち、小胞体−ゴルジ体系を介したプラスチドへの輸送・局在化は、トランジットペプチド非依存性プラスチド局在であり、プラスチド包膜に存在するタンパク透過チャンネルに適合しないタンパクのプラスチド輸送・局在化を行うことができる可能性がある。また、プラスチド局在化に関わる典型的なトランジットペプチドを有しないα−アミラーゼI−1(AmyI−1)がイネ緑葉細胞の葉緑体と細胞壁区画に局在することを見出した。
【0006】
また、イネα−アミラーゼは分泌型酵素として知られているものであったが、イネα−アミラーゼの発現抑制および過剰発現する形質転換イネ植物を解析した結果、緑葉など生細胞中のデンプン分解にα−アミラーゼが関与することを見出した。さらに、α−アミラーゼI−1の発現抑制および過剰発現体イネ植物を用いて、生組織細胞におけるデンプン集積に及ぼすα−アミラーゼI−1の役割を調べたところ、α−アミラーゼI−1発現抑制イネ系統においてはα−アミラーゼI−1発現がほぼ完全に押さえられ、野生型に比べて葉組織において顕著なデンプン蓄積の増加が見られた。
【0007】
本発明は以上の研究成果及び知見に基づいて完成されたものであって、以下の各構成を提供する。
【0008】
本発明における請求項1記載のポリペプチドは、以下の(a)又は(b)記載のアミノ酸配列からなることを特徴とする。
(a)配列番号1に示されるアミノ酸配列からなるポリペプチド。
(b)配列番号1に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつプラスチドへのターゲティング作用を有するポリペプチド。
【0009】
本発明における請求項2記載の遺伝子は、請求項1記載のポリペプチドをコードすることを特徴とする。
【0010】
本発明における請求項3記載のポリペプチドは、以下の(c)又は(d)記載のアミノ酸配列からなることを特徴とする。
(c)配列番号2に示されるアミノ酸配列からなるポリペプチド。
(d)配列番号2に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ小胞体膜透過作用を有するポリペプチド。
【0011】
本発明における請求項4記載の遺伝子は、請求項3記載のポリペプチドをコードすることを特徴とする。
【0012】
本発明における請求項5記載のベクターは、請求項2記載の遺伝子を含むことを特徴とする。
【0013】
本発明における請求項6記載のベクターは、請求項2及び4記載の遺伝子を含むことを特徴とする。
【0014】
本発明における請求項7記載の形質転換植物は、請求項2記載の遺伝子が導入されたことを特徴とする。
【0015】
本発明における請求項8記載の形質転換植物は、請求項2及び4記載の遺伝子が導入されたことを特徴とする。
【0016】
本発明における請求項9記載の形質転換植物は、請求項7又は8において、前記形質転換植物が単子葉植物であることを特徴とする。
【0017】
本発明における請求項10記載の形質転換植物は、請求項5又は6記載のベクターを単子葉植物由来のカルスに導入し、前記カルスを増殖させた後に、前記カルスから植物体を再分化させて得られることを特徴とする。
【0018】
本発明における請求項11記載の形質転換植物の製造方法は、請求項5又は6記載のベクターを単子葉植物由来のカルスにアグロバクテリウムを用いて導入し、前記カルスを増殖させた後に、前記カルスから植物体を再分化させることを特徴とする。
【発明の効果】
【0019】
本発明によれば、今後さらに深刻化する環境ストレスに適応し、劣悪な環境下において効率的にデンプンを集積可能な形質転換植物の作出が可能となる。また、プラスチドの光合成能やデンプン集積能を高め、カーボンニュートラルな工業用素材であるデンプンの供給を増大させることができる。また、植物デンプンは人類にとって極めて重要な栄養素であり、デンプン粒の構造を改変することによってデンプン利用用途の拡大を図ることができ、生分解性樹脂など工業生産における有用素材である。
【発明を実施するための最良の形態】
【0020】
(ベクター)
本発明において、遺伝子を植物へ導入する際に使用されるベクターとしては、導入遺伝子をターゲット細胞(宿主細胞)に導入することができ、且つターゲット細胞内で導入遺伝子を発現させることができるものであれば特に限定されず、目的に応じて適当なベクター(プラスミド、バクテリオファージ、ウイルスなど)が利用される。数多くのベクターが市販されており、本発明ではそれらの中から目的に応じて適切なものを選択して用いることができる。また、本発明のベクターの作成方法としては、特に限定されず、当業者に周知のいかなる方法を用いてもよい。
【0021】
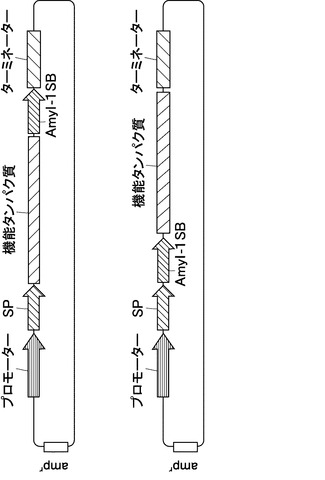
本発明において、好ましいプラスチド輸送・局在化コンストラクトの例を図2に示す。図2に記載のAmyI−1SBとは、α−アミラーゼI−1の301〜369のペプチド領域(配列番号1)を示し、プラスチドターゲティングに必要不可欠なものである。一般に、特定の機能を有するアミノ酸の一部に改変を施した場合においてもその機能が維持されることがある。このことを考慮して、AmyI−1SBのポリペプチドは、以下の(a)又は(b)記載のアミノ酸配列からなることを特徴とする:(a)配列番号1に示されるアミノ酸配列からなるポリペプチド;(b)配列番号1に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつプラスチドへのターゲティング作用を有するポリペプチド。
また、SPとは、ER膜透過シグナルペプチドを示し、例えばAmyI−1SP(配列番号2)が挙げられるがこれに限定されるものではなく、他の植物分泌タンパク質のSPを用いてもよい。また、AmyI−1SPのポリペプチドは、以下の(c)又は(d)記載のアミノ酸配列からなることを特徴とする:(c)配列番号2に示されるアミノ酸配列からなるポリペプチド;(d)配列番号2に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ小胞体膜透過作用を有するポリペプチド。なお、機能タンパク質はAmyI−1SBのN末端側又はC末端側の何れに挿入してもよい。
【0022】
(形質転換方法)
植物へのベクターの導入には、ポリエチレングリコール法、電気穿孔法(エレクトロポレーション法)、アグロバクテリウムを介する方法、パーティクルガン法など、当業者に公知の種々の方法を用いることができる。それぞれの方法に適したベクターの調製、形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。
【0023】
(ターゲット植物)
本発明においてターゲットとする植物は、単子葉植物である。単子葉植物としては、例えばイネ科(イネ属、コムギ属、オオムギ類、ライムギ類、ハトムギ属、トウモロコシ属、サトウキビ属等)、ユリ科(ネギ属、ニンニク属等)などが挙げられる。
【0024】
以下に本発明の実施例によって、本発明を詳細に説明するが、本発明はこれらの実施例によりなんら制限されるものではない。
【0025】
以下の実施例1〜3において使用される生物学的材料、試薬実験方法は次の通りである。
【0026】
(供試植物)
形質転換に供試される植物(ターゲット植物)として、タマネギ(Allium cepa)表皮細胞を、0.6%のゲルライトを加えた2,4−DフリーのMS培地上にて25℃暗所で培養後、使用した。
【0027】
(プラスミドコンストラクト)
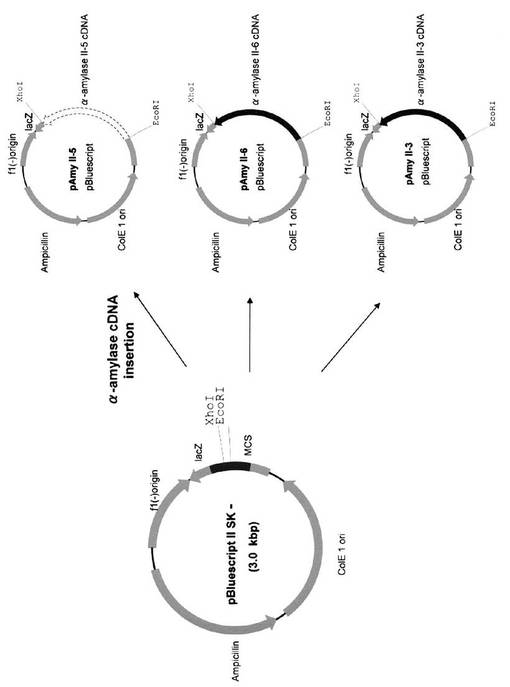
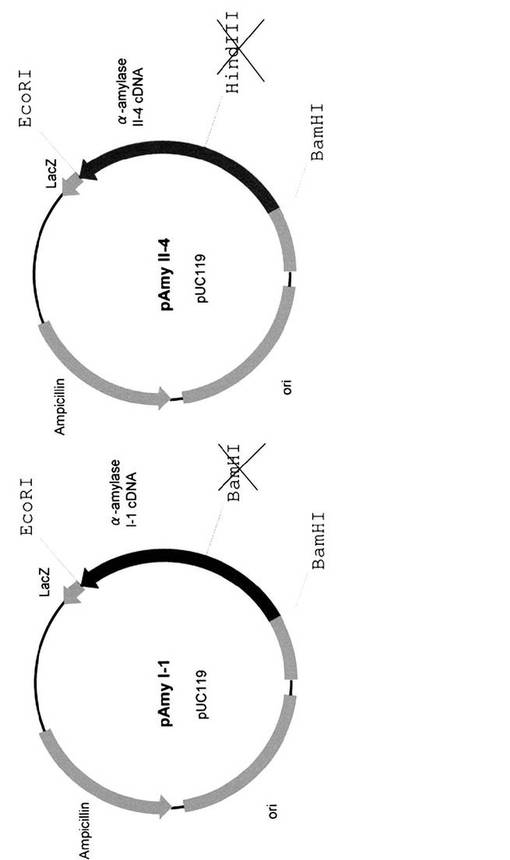
全RNAを、発芽6日目のイネ種子の胚盤組織から単離した。そのcDNAライブラリーをpBluescriptIISK(+/−)(タカラバイオ社)を用いて製造者のプロトコールに従って作成した。cDNAライブラリーを、α−アミラーゼII−3(RAmy3E),II−5(RAmy3B),及びII−6(RAmy3C)の特定のプローブを用いて選別した(Sheu, J.-J., Yu, T.-S., Tong, W.-F., Yu, S.-M. (1996) Carbohydrate starvation stimulates differential expression of rice a-amylase genes that is modulated through complicated transcriptional and posttranscriptional processes. J. Biol. Chem. 271: 26998-27004.)。得られた完全長cDNA(AmyII−3,AmyII−5,及びAmyII−6と称す)を、pBluescript SK(+・−)にクローニングし、pAmyII−3,pAmyII−5,及びpAmyII−6を作成した(図3)。pAmyI−1とpAmyII−4は、Asatsuma S, Sawada C, Itoh K, Okito M, Kitajima A, Mitsui T (2005) Involvement of α-amylase I-1 in starch degradation in rice chloroplasts. Plant & Cell Physiology 46: 858-869に記載の方法に従って作成した。(図4)。
【0028】
完全長のα−アミラーゼI−1cDNAと種々のC末端が一部切断させたフラグメントを、鋳型DNAとしてpAmyI−1とプライマー配列セット(表1)を用いてPCRによって増幅した。
【0029】
【表1】

【0030】
完全長α-アミラーゼII−3,II−4,II−5,II−6cDNA、およびそれらのN−端末シグナル配列領域を、鋳型DNAとしてpAmyII−3,pAmyII−4,pAmyII−5及びpAmyII−6と,プライマー配列セット(表2)を用いてPCRによって増幅した。
【0031】
【表2】
【0032】
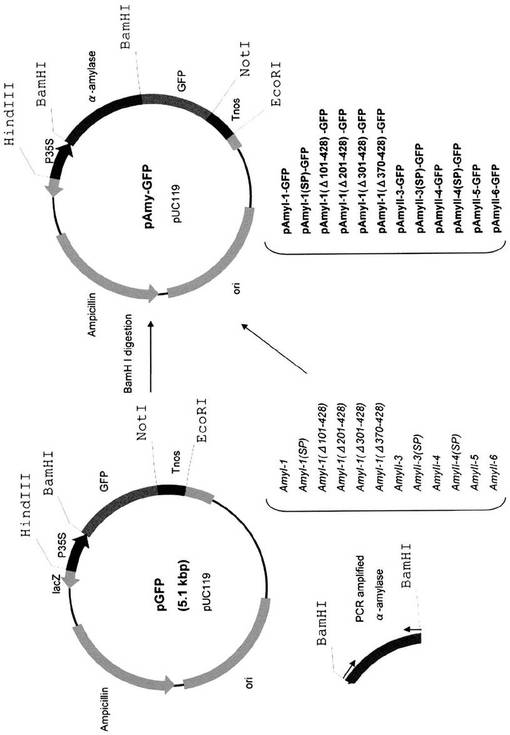
BamHI PCR増幅フラグメントをpGFPのBamHI部位に挿入し、pAmyI−1−GFP、pAmyI−1(SP)−GFP、pAmyI−1(Δ101−428)−GFP、pAmyI−1(Δ201−428)−GFP、pAmyI−1(Δ301−428)−GFP、pAmyI−1(Δ370−428)−GFP、pAmyII−3−GFP、pAmyII−3(SP)−GFP、pAmyII−4−GFP、pAmyII−4(SP)−GFP、pAmyII−5−GFP、及びpAmyII−6−GFP(図5)を作成した。
【0033】
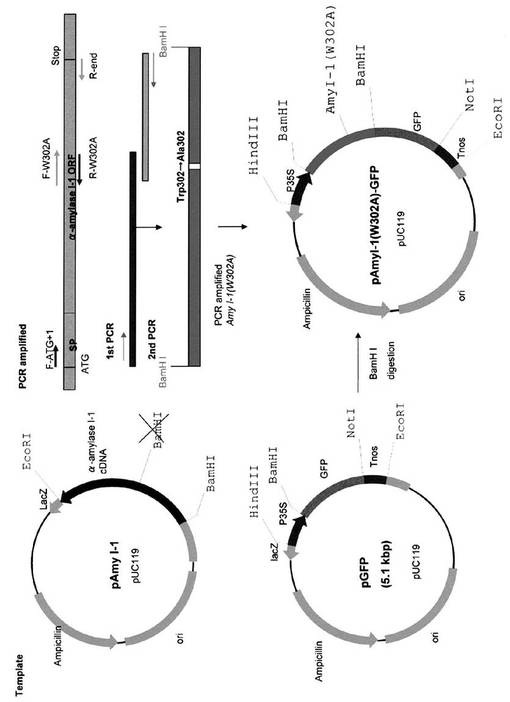
PCRを使用して、トリプトファン302をアラニン302に置換した一つのアミノ酸を導入し、AmyI−1(W302A)を作成した。変異原性のフォワードプライマー(F−ATG+1と、F−W302A)及びリバースプライマー(R−endと、R−W302A)を合成した(表3)。
【0034】
【表3】
【0035】
変異原性のプライマーセットと、鋳型DNAとしてpAmyI−1を使用して第一のPCRを行った。F−ATG+1とR−endのプライマーと、鋳型DNAとして第一のPCR産物を使用して第二のPCRを行った。第二のPCR産物をBamHIで切断し、pGFPのBamHI部位にクローニング、pAmyI−1(W302A)(図6)を作成した。
【0036】
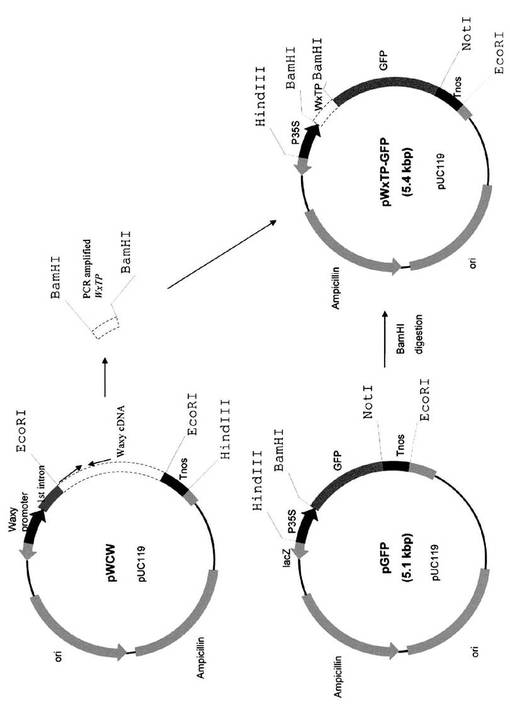
タマネギ表皮細胞におけるDsRed2発現において、pDsRed2(Clonetech)からのBamHI−NotIのPCR増幅フラグメントを、BamHI−NotI消化pGFPのBamHI−NotI消化部位にクローニングし、pDsRed(図7)を産生した。イネwaxy遺伝子(Itoh, K., Ozaki, H., Okada, K., Hori, H., Takeda, Y., Mitsui, T. (2003) Introduction of Wx transgene into rice wx mutants leads to both high- and low-amylose rice. Plant Cell Physiol., 44: 473-480)からトランジットペプチド配列の始めの1−111のアミノ酸を、2つのプライマー5'-ctggatccatgtcggctctcaccacg-3'と5'-tatggatccggcaggggggaggccaccgag-3'を用いてPCR増幅し、その後PCR産物をBamHIで消化してpWxTP−DsRedを作成した。BamHI消化フラグメントをpDsRedのBamHI消化部位に挿入した(図8)。さらに、BamHI PCRの増幅WxTPフラグメントを、pGFPのBamHI消化部位にクローニングし、pWxTP−GFPを構築した(図9)。
【0037】
ペルオキシソームとミトコンドリアにそれぞれpPTS2−GFP(Mano, S., Nakamori, C., Hayashi, M., Kato, A., Kondo, M., Nishimura, M. (2002) Distribution and characterization of peroxisomes in Arabidopsis by visualization with GFP: dynamic morphology and actin-dependent movement. Plant Cell Physiol. 43: 331-41.)とpmt−GFP(Niwa, Y., Hirano, T., Yoshimoto, K., Shimizu, M., Kobayashi, H. (1999) Non-invasive quantitative detection and applications of non-toxic, S65T-type green fluorescent protein in living plants. Plant J. 18, 455-463.)を用いて、他の細胞小器官を可視化した(図10)。
【0038】
(遺伝子導入方法)
プラスミドDNAを、パーティクルデリバリーシステム(PDS−1000/He、BIO−RAD)を用いてタマネギ表皮細胞へ導入した。具体的には、10μlの滅菌水に溶解させた3μgのプラスミドDNAに、10μlの金粒子懸濁液(直径1.0μmの金粒子60mg/ml)と、10μlの2.5mM CaCl2と、4μlの0.1M スペルミジンを加えて懸濁し、室温で30分間静置した。プラスミドDNAで被覆された金粒子を冷エタノールで洗浄した後、10μlのエタノールで緩やかに懸濁した。1100psiのラプチャーディスクを使用し、一サンプルあたり2回ずつ導入を行った。
【0039】
(観察方法)
GFP,DsRedおよびmRFP蛍光画像は、電動蛍光顕微鏡(Olympus BX−61)と、CCDカメラ(Cool SNAP fx、Roper Scientific)を用いて観察・撮影し、画像解析ソフト(Lumina Vision、三谷商事(株))を用いて解析した。GFPは470〜490nmで励起し、510〜550nmを検出した。DsRedおよびmRFPは、520〜550nmで励起し、≧580nmを検出した。一細胞につき、10μm間隔で20〜30枚の画像を撮り、デコンボリューション処理した後、一つの画像にした。
【実施例1】
【0040】
イネα−アミラーゼ遺伝子と緑色蛍光タンパク質(GFP)遺伝子を融合し、この融合物をタマネギ細胞に導入させ、GFPの輸送・局在化を調べた。
【0041】
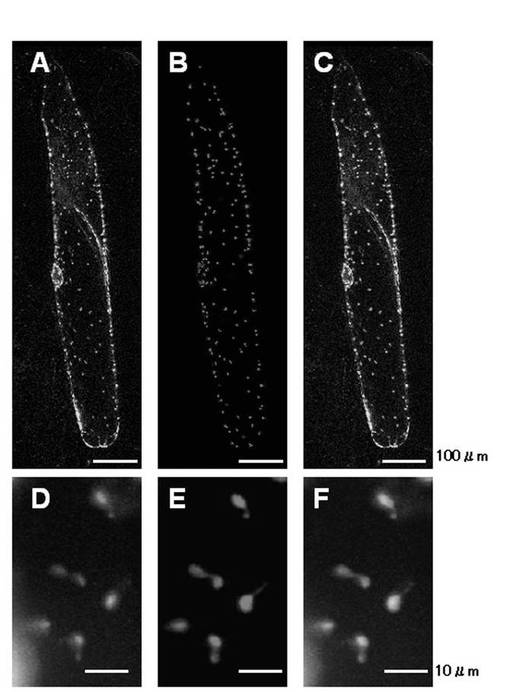
タマネギ表皮細胞にパーティクルガンでAmyI−1−GFPおよびWxTP−DsRedを同時に導入し、一過的に発現させた(図11)。WxTP−DsRedは、プラスチドへの局在を示すマーカーである。図11のA,Dに示すようにように、AmyI−1−GFPの蛍光は細胞表面だけでなく、細胞内で何らかのオルガネラに局在していた。B,Eでは、WxTP−DsRedの蛍光はプラスチドを示し、プラスチド特有の構造であるストロミュールも確認された(図11E)。CはAとBを、FはDとEを重ね合わせた画像で、部分的にAmyI−1−GFPの蛍光がWxTP−DsRedの蛍光と局在が一致し、AmyI−1がプラスチドに局在することを示した。この結果から、イネα−アミラーゼ遺伝子と緑色蛍光タンパク質(GFP)遺伝子を融合し、この融合物をタマネギ細胞に導入することによって、タマネギ細胞のプラスチドにGFPを輸送・局在化させることが示された。また、α−アミラーゼI−1においてはイネおよびタマネギの両細胞に共通するプラスチド局在化シグナルが含まれることが明らかになった。
【実施例2】
【0042】
α−アミラーゼI−1のプラスチドターゲティングに必要不可欠な領域を決定するために、C端末の端部が一部欠けているα−アミラーゼI−1とGFPの融合タンパク質を一過的に発現させその局在を調べた。
【0043】
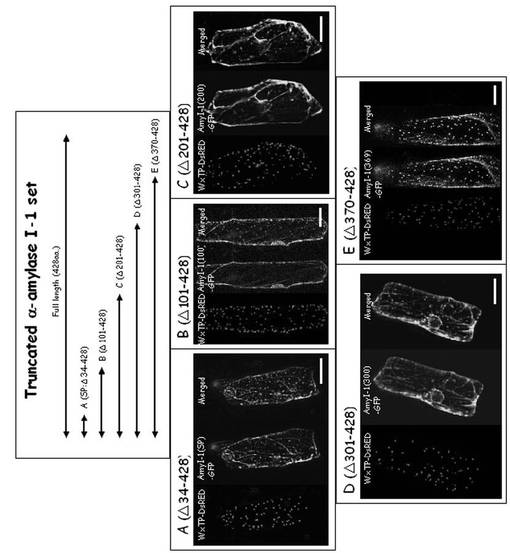
図12に、C末端が一部切断されたα−アミラーゼI−1(AmyI−1Δ34−428、AmyI−1Δ101−428、AmyI−1Δ201−428、AmyI−1Δ301−428、AmyI−1Δ370−428)を示す。AmyI−1Δ34−428は、α−アミラーゼI−1のN末端シグナルペプチドのみを含んでいた。予測されたシグナルペプチダーゼ開裂部位はGln26のN末端側である。AmyI−1Δ301−428は、N結合型糖鎖結合部位(Asn265、−Gly266、−Thr267)を含んでいる。AmyI−1Δ370−428は、推定のデンプン結合部位(301Typ−302Typ)を含む完全なAドメインとBドメインを有する(Robert, X., Haser, R., Mori, H., Svensson, B., Aghajari, N. (2005) Oligosaccharide binding to barley a-amylase 1. J. Biol. Chem. 280: 32968-32978)。370〜428残基のアミノ酸配列は、5つの逆平行β-シートを構成したC-ドメインに対応している。図12と表4に示されるように、多くのC末端が欠失したAmyI−1−GFP融合タンパク質は小胞体(ER)様網目状構造に局在し、プラスチドにはほとんど局在しなかったが、AmyI−1Δ370−428−GFP融合タンパク質は完全長AmyI−1−GFPと同様に遺伝子導入されたタマネギ細胞におけるプラスチドに局在した。これらの結果から、301〜369のアミノ酸残基のペプチド領域(α−AmyI−1SB(配列番号1))がタマネギ表皮細胞におけるα−アミラーゼI−1のプラスチドターゲッティングに必要不可欠であることが明らかとなった。
【0044】
【表4】
【実施例3】
【0045】
α−アミラーゼのアイソフォーム(AmyII−3、AmyII−4、AmyII−5およびAmyII−6)が、タマネギ表皮細胞のプラスチドに局在するか否かをα−アミラーゼのアイソフォームとGFPの融合タンパク質(AmyII−3−GFP、AmyII−4−GFP、AmyII−5−GFPおよびAmyII−6−GFP)を用いて調べた。
【0046】
イネα−アミラーゼには多くのアイソフォームが存在し、これらのアイソフォームは多くの遺伝子によってコードされていることが知られている(O'Neill, S.D., Kumagai, M.H., Majumdar, A., Huang, N., Sutliff, T.D. and Rodriguez, R.L. (1990) The alpha-amylase genes in Oryza sativa: characterization of cDNAclones and mRNA expression during seed germination. Mol. Gen. Genet. 221: 235-244;Huang, N., Koizumi, N., Reinl, S., Rodriguez, R.L. (1990a) Structural organization and differential expression of rice a-amylase genes. Nucl. Acid Res. 18: 7007-7013;Huang, N., Sutliff, T.D., Litts, J.C., Rodriguez, R.L. (1990b) Classification and characterization of the rice a-amylase multigene family. Plant Mol. Biol. 14: 655-668;Kim, J.-K. and Wu, R. (1992) Nucleotide sequence of a high-pI rice (Oryza sativa) a-amylase gene. Plant Mol. Biol. 18: 399-402)。また、本発明者らは以前の研究において、RAmy1A、RAmy3B、RAmy3C、RAmy3D、及びRAmy3Eの天然の発現産物がイネ植物と組織培養で検出されたことを報告している(Mitsui, T., Yamaguchi, J. and Akazawa, T. (1996) Physicochemical and serological characterization of rice a-amylase isoforms and identification of their corresponding genes. Plant Physiol. 110: 1395-1404;Nanjo, Y., Asatsuma, S., Itoh, K., Hori, H. and Mitsui T. (2004) Proteomic identification of a-amylase isoforms encoded by RAmy3B/3C from germinating rice seeds. Biosci. Biotech. Biochem. 68: 112-118)。
【0047】
AmyI−1−GFPのプラスチドにおける局在化と比較して、AmyII−3−GFP、AmyII−4−GFP、AmyII−5−GFP、及びAmyII−6−GFPはプラスチド内でほとんど局在していなかった(表4)。
【0048】
さらに、タマネギ細胞におけるデンプン結合部位を変異させたα−アミラーゼI−1−GFP融合タンパク質のターゲティング特性を調べて比較した。
【0049】
図13に、イネα−アミラーゼのアイソフォーム(アミラーゼI−1(M24286);アミラーゼII−3(M59352);アミラーゼII−4(M24287);アミラーゼII−5(X56337);アミラーゼII−6(X56338))のアミノ酸配列の配列比較を示す。図13に示されるα−アミラーゼI−1(RAmy1A)、II−3(RAmy3E)、II−4(RAmy3D)とII−5/6(RAmy3B/3C)間のアミノ酸配列は約83〜90%の相同性を有する。
【0050】
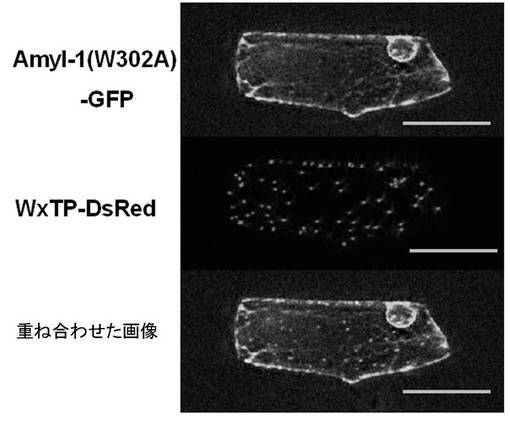
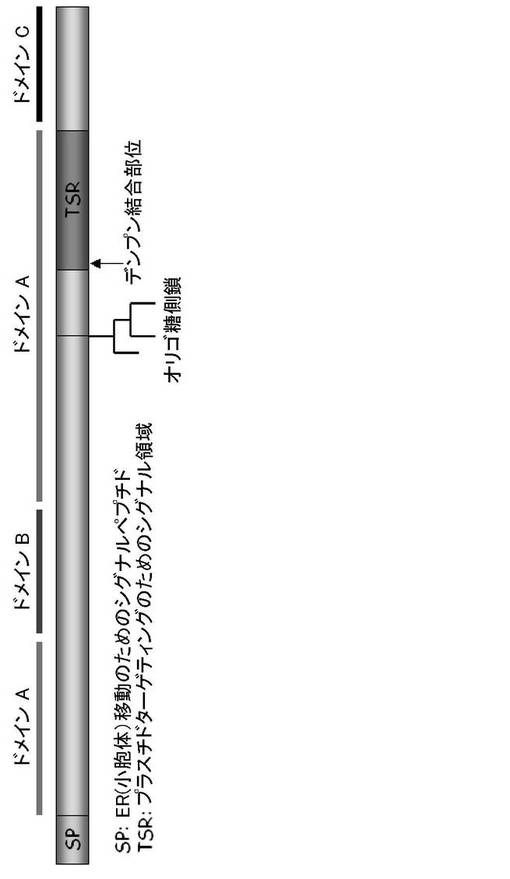
301Typ−302Typは、α−アミラーゼI−1のポリペプチドにおける特有のアミノ酸残基であった。その他のα−アミラーゼのアイソフォーム(α−アミラーゼII−3、II−4、II−5およびII−6)は、ペプチド領域301〜369のアミノ酸残基においてトリプトファンジペプチドを有していなかった(図13)。リプトファン302をアラニン302に置換した場合、プラスチドへのAmyI−1−GFPのターゲティングを強く阻害し、AmyI−1(W302A)−GFPはER様網目状に保持されていた(図14、表4)。これらの結果から、302Typ残基はタマネギ表皮細胞においてα−アミラーゼI−1のプラスチドターゲティングに必須であることが明らかになった。図15に、プラスチドターゲティングのためのα−アミラーゼI−1のシグナル領域を示す。図15に示される「SP」とはER(小胞体)膜透過シグナルペプチドを意味し、「TSR」とはプラスチドターゲティングのためのシグナル領域(すなわち、α−アミラーゼI−1の301〜369ペプチド領域(デンプン結合領域))を意味する。なお、α−アミラーゼは、3つのドメインからなり、ドメインAは、(β/α)8バレルに折り重なり、酵素の触媒残基を含み、ドメインBは、ドメインAの3番目のβ鎖と3番目のαへリックスの間から突き出している大きなループであり、ドメインCは、(β/α)8バレルのC末端に位置し、β鎖からなっている。
【0051】
以下の実施例4において使用される生物学的材料、試薬、実験方法等は次の通りである。尚、特に説明のない材料等については上記実施例で使用したものと同様である。
【0052】
(供試植物)
供試植物として、イネ品種「日本晴」(Oryza sativa L. cv. Nipponbare)を用いた。そのイネの種子は、新潟農業研究所より提供されたものである。
【0053】
(分析法)
デンプンの含量は、Matsukura C, Saitoh T, Hirose T, Ohsugi R, Perata P, Yamaguchi J (2000) Sugar uptake and transport in rice embryo. Expression of companion cell-specific sucrose transporter (OsSUT1) induced by sugar and light. Plant Physiology 124: 85-93、及びBradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. . Anal. Biochem. 72: 248-254に記載の方法に従って決定した。α−アミラーゼの分析法は、Mitsui T, Yamaguchi J, Akazawa T (1996) Physicochemical and serological characterization of rice α-amylase isoforms and identification of their corresponding genes. Plant Physiology 110: 1395-1404に記載の方法に従って行った。
【0054】
(ベクターの構築)
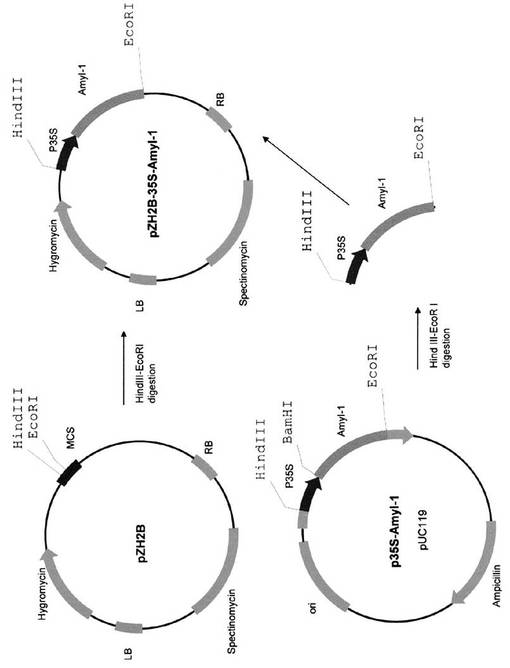
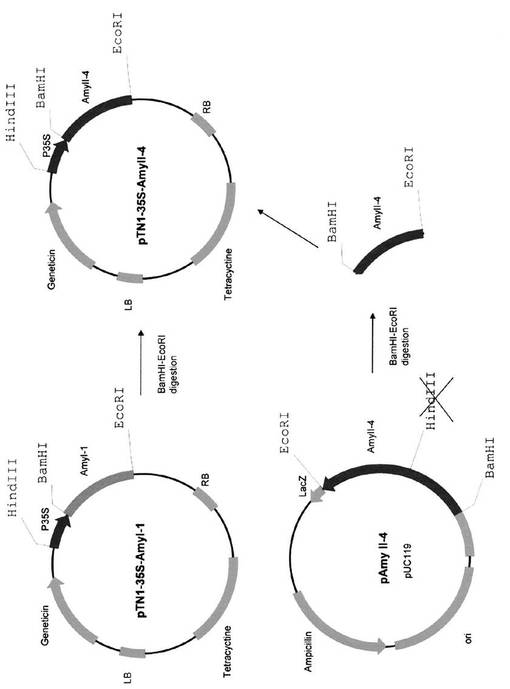
pZH2B−35S−AmyI−1及びpTN1−35S−AmyII−4の構築は、Asatsuma S, Sawada C, Itoh K, Okito M, Kitajima A, Mitsui T (2005) Involvement of α-amylase I-1 in starch degradation in rice chloroplasts. Plant & Cell Physiology 46: 858-869及びHajdukiewicz P, Svab Z, Maliga P (1994) The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Molecular Biology 25: 989-994に記載の方法に従って行った(図16)。pAmyII−4をBamHI−EcoRIで消化して得られたフラグメントを、pTN1−35S−AmyI−1をBamHI−EcoRIで消化した対応する制限部位にクローニングし、pTN1−35S−AmyII−4を作成した(図17)。
【0055】
(α−アミラーゼ過剰発現植物体の作成)
上述のようにして作成されたベクターを、20mMのCaCl2で処理されたアグロバクテリウム(Agrobacterium tumefaciens)EHA101株のコンピテント細胞へ導入し、暗所でプレート培養して導入されたアグロバクテリウムを選抜した。プラスミドが導入されたアグロバクテリウムをアセトシリンゴン存在下でイネの胚組織から誘導したカルスに感染させた。感染後、抗生物質を含んだプレート培地上でプラスミドが誘導されたカルスを選抜し、植物ホルモン(NAA)により植物体に再分化させた(Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant Journal 6: 271-282及びAsatsuma S, Sawada C, Itoh K, Okito M, Kitajima A, Mitsui T (2005) Involvement of α-amylase I-1 in starch degradation in rice chloroplasts. Plant & Cell Physiology 46: 858-869)。
【0056】
(植物体の生育)
再生した植物体をポットに移し、明所12時間、暗所8時間、27℃のグロスチャンバー内で20cm程度になるまで生育させた。その後、温室に移し種子を採取するまで約6ヶ月間生育させた。開花後、40日ほどで種子を刈り取った。
【0057】
(サザンブロット解析、ノーザンブロット解析、ウエスタンブロット解析)
野生型および形質転換イネの若葉より単離したゲノムDNA(5μg)をEcoRIで消化し、1%アガロースゲル電気泳動で分画した後、Hybond−N+ナイロン膜(Amersham)に転写・固定した。ノーザンブロット解析は、Mitsui T, Loboda T, Kamimura I, Hori H, Itoh K, Mitsunaga S, (1999) Sucrose-controlled transport and turnover of α-amylase in rice (Oryza sativa L.) cells. . Plant Cell Physiol. 40 884-893およびKashem MA, Itoh K, Iwabuchi S, Hori H, Mitsui T (2000) Possible involvement of phosphoinositide-Ca2+ signaling in the regulation of alpha-amylase expression and germination of rice seed (Oryza sativa L.). Plant & Cell Physiology 41: 399-407に記載の手順に従って行った。サザンブロット解析に関し、α−アミラーゼI−1遺伝子およびα−アミラーゼII−4遺伝子の特異的DNA(前出のKashem et al. 2000)を放射性プローブとして使用した。オートラジオグラフィーは、画像解析装置BAS5000(フジフィルム)を用いて行った。ウエスタンブロット解析、抗α−アミラーゼI−1抗体及び抗α−アミラーゼII−4抗体の調製法等は、Mitsui T, Yamaguchi J, Akazawa T (1996) Physicochemical and serological characterization of rice α-amylase isoforms and identification of their corresponding genes. Plant Physiology 110: 1395-1404に記載の方法に従って行った。2次抗体として、ペルオキシダーゼ標識抗ウサギIgG抗体を使用した。そのタンパク質バンドをECL(化学発光、Amersham)を用いて可視化し、LAS3000(フジフィルム)を用いて定量化した。野生型イネ植物において発現するα−アミラーゼI−1およびII−4の各タンパク質の量を1ユニットとした。
【実施例4】
【0058】
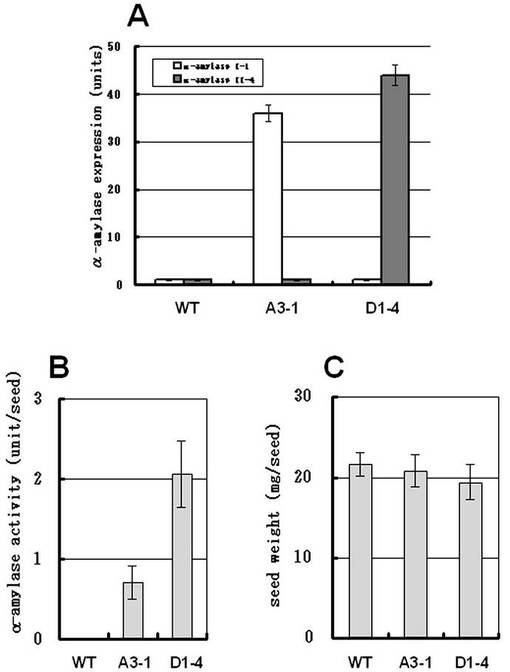
特異的抗体(例えば抗α−アミラーゼI−1,II−3,II−4,II−5およびII−6)を用いてウエスタンブロット解析を行い、登熟期間中のイネ登熟種子におけるα−アミラーゼのアイソフォームの発現を調べた。出穂後10日目の登熟種子において、α−アミラーゼI−1およびII−4の発現を検出した(図18)。興味深いことに、α−アミラーゼII−4が登熟種子において最も多量に存在するアイソフォームであることが示唆された。
【0059】
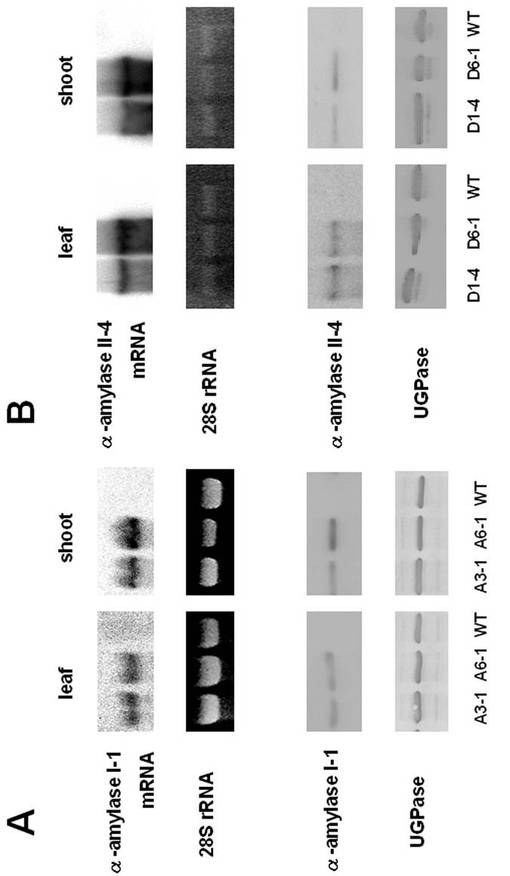
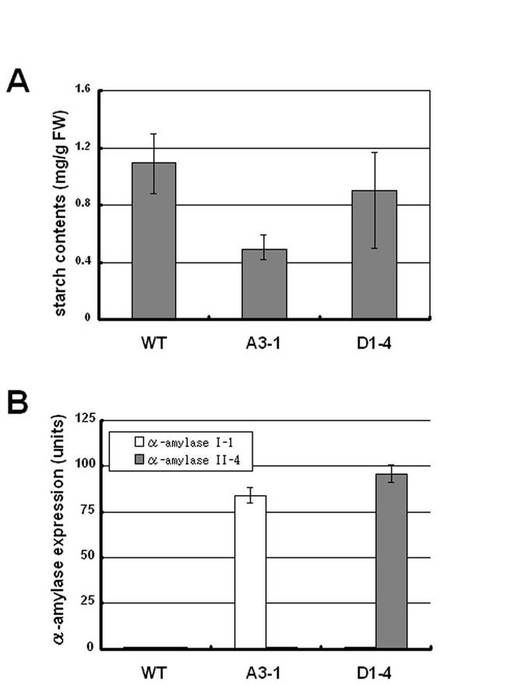
カリフラワーモザイクウイルス(CaMV)35Sプロモーターの制御下それぞれα−アミラーゼI−1(AmyI−1)とα−アミラーゼII−4cDNA(AmyII−4)を含むpZH2B−35S−AmyI−1とpTN1−35S−AmyII−4を用いて、アグロバクテリウムを介した形質転換によって、遺伝組換えイネ植物を産出した。AmyI−1およびAmyII−4の存在を、サザンハイブリダイゼーションを用いて再生された形質転換植物中において確認した。得られた形質転換イネ植物は1〜8コピーの導入遺伝子を有していた(図示せず)。また、pZH2B−35S−AmyI−1とpTN1−35S−AmyII−4で形質転換されたこれらの遺伝子導入系統におけるα−アミラーゼ遺伝子発現を、ノーザンおよびウエスタンブロット解析を用いて調べた。結果を図19に示す。図19に示されるように、これらの形質転換イネ系統の双方が、成葉およびT1世代のシュート組織においてα−アミラーゼのmRNAおよびタンパク質の著しい増加を示した。また、α−アミラーゼI−1(A3−1)を過剰発現した葉におけるデンプン含有量は、野生型(WT)のものと比較して著しく減少し、そのデンプン含有量は野生型の約42%であった。(図20)。一方、α−アミラーゼII−4(D1−4)を過剰発現した葉におけるデンプン含有量はわずかに減少し、野生型のデンプン含有量の約82%であった。
【0060】
A3−1およびD1−4のT3世代の登熟期において、野生型と比較しα−アミラーゼのより高い活性はα−アミラーゼI−1およびII−4を発現する形質転換植物の両方の登熟種子で検出された(図21)。図21の縦軸はα−アミラーゼ活性(unit/seed)を示し、横軸は出穂後の日数(daf:days after heading)を示す。D1−4系統の登熟種子に含まれるα−アミラーゼ活性のレベルは、A3−1系統の約2倍の活性を示した。
【0061】
さらに、A3−1系統およびD1−4系統の登熟種子表現型の特性を分析した。α−アミラーゼI−1とα−アミラーゼII−4に対する特異的抗体を用いてウエスタンブロット解析を行った場合、野生型の種子と比較し、A3−1系統の登熟種子においてα−アミラーゼI−1分子は36倍もの増加を示し、D1−4系統の登熟種子においてα−アミラーゼII−4分子は44倍もの増加を示した(図22−A)。酵素活性の増加はA3−1及びD1−4系統の登熟種子の両方で示されたが、D1−4系統ではより顕著な増加が示された(図22−B)。A3−1及びD1−4系統の登熟種子の乾燥重量は、野生型の種子と比較し、A3−1系統で4%、D1−4系統で11%の減少が見られた(図22−C)。
【0062】
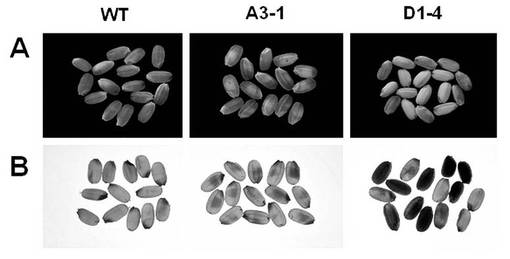
また、α−アミラーゼI−1とα−アミラーゼII−4を過剰発現した形質転換体イネ系統(A3−1及びD1−4系統)の完熟種子の品質を反射光画像と透過光画像の双方によって評価した(図23)。表5に示すように、形質転換体イネ系統から得られた種子においては、ともにデンプン蓄積異常が見られ、野生型,A3−1系統及びD1−4系統の正常な穀粒のパーセンテージはそれぞれ90%,54.4%,36.5%であった。A3−1系統及びD1−4系統の異常な種子の特徴的な形態学的表現型はそれぞれ心白(white core)と全乳白(whole opaque white)であった(図23、表5)。これらの結果から、α−アミラーゼ活性の異常な発現はデンプンの集積およびイネ粒構造に影響することが示された。また、α−アミラーゼI−1とα−アミラーゼII−4は、登熟種子の胚乳におけるアミロプラストに局在する能力を有し、デンプン集積(分解)調節機能を有すると考えられる。
【0063】
【表5】
【図面の簡単な説明】
【0064】
【図1】高等植物細胞におけるα−アミラーゼI−1のプラスチドターゲティング機構を示す図である。
【図2】プラスチドコンストラクトを構築した例を示す図である。
【図3】pAmyII−3,pAmyII−5及びpAmyII−6の作成過程を示す図である。
【図4】pAmyI−1及びpAmyII−4のマップである。
【図5】種々のpAmy−GFPの作成過程を示す図である。
【図6】pAmyI−1(W302A)−GFPの作成過程を示す図である。
【図7】pDsRedの作成過程を示す図である。
【図8】pWxTP−DsRedの作成過程を示す図である。
【図9】pWxTP−GFPの作成過程を示す図である。
【図10】pPTS2−GFPおよびpmt−GFPを構築した例を示す図である。
【図11】AmyI−1−GFPとWxTP−DsRedのタマネギ表皮細胞での同時発現の結果を示す図である。A,DはAmyI−1−GFPの蛍光像を示し、B,EはWxTP−DsRedの蛍光像を示し、C,FはGFPとDsRedの重ね合わせ像を示す。下図(D,E,F)は、上図(A,B,C)の拡大図である。
【図12】遺伝子導入されたタマネギ表皮細胞における一部切断されたα−アミラーゼI−1−GFPの局在を示す図である。A〜Eの左側、中央、右側はそれぞれWxTP−DsRed画像、AmyI−1−GFP画像、WxTP−DsRedとAmyI−1−GFPの重ね合わせ画像を示す。
【図13】イネα−アミラーゼのアイソフォーム(アミラーゼI−1(M24286);アミラーゼII−3(M59352);アミラーゼII−4(M24287);アミラーゼII−5(X56337);アミラーゼII−6(X56338))のアミノ酸配列の配列比較を示す図である。矢頭は、N末端シグナルペプチドの開裂部位を示す。**(アスタリスク2)印の個所は、推定デンプン結合部位を示す。下線個所は、グリコシル化部位を示す。
【図14】AmyI−1(W302A)−GFPとWxTP−DsRedのタマネギ表皮細胞での同時発現を示す図である。
【図15】プラスチドへのターゲティングに関するα−アミラーゼI−1のシグナル領域を示す図である。
【図16】pZH2B−35S−AmyI−1の作成過程を示す図である。
【図17】pTN1−35S−AmyII−4の作成過程を示す図である。
【図18】抗α−アミラーゼI−1,II−3,II−4,II−5およびII−6を用いてウエスタンブロット解析を行った結果を示す図である。
【図19】α−アミラーゼI−1(A)及びα−アミラーゼII−4(B)を過剰発現したイネ植物のノーザンブロット解析およびウエスタンブロット解析を行った結果を示す図である。
【図20】(A)は、野生型(WT)、α−アミラーゼI−1(A3−1)およびα−アミラーゼII−4(D1−4)を過剰発現した葉におけるデンプン含有量の結果を示し、(B)はα−アミラーゼの発現の結果を示すグラフである。
【図21】形質転換植物(A3−1およびD1−4)及び野生型(WT)の登熟種子におけるα−アミラーゼ活性の変化を示す図である。
【図22】(A)は、野生型(WT)、α−アミラーゼI−1(A3−1)およびα−アミラーゼII−4(D1−4)を過剰発現した完熟種子のα−アミラーゼ発現の結果を示し、(B)は、α−アミラーゼ活性の結果を示し、(C)は完熟種子の乾燥重量の結果を示すグラフである。
【図23】(A)は、野生型(WT)、α−アミラーゼI−1及びα−アミラーゼII−4を過剰発現した形質転換体イネ系統(A3−1及びD1−4系統)の完熟種子の反射光画像を示し、(B)は透過光画像を示す図である。
【技術分野】
【0001】
本発明は、デンプン集積能力の高い形質転換植物およびその製造方法に関する。
【背景技術】
【0002】
近年、地球温暖化など地球規模での自然環境の悪化が大きな問題になっている。我が国においてもすでに環境ストレスがイネなどの重要作物の生長・生理、特に炭水化物代謝に大きな影響を及ぼし、減収や品質劣化を引き起こしていることが知られている。
光合成産物のデンプンへの集積は、植物のみならず地球上のすべての生命にとって最も重要なエネルギー蓄積過程である。このデンプンへの集積は、光合成機能を有する葉緑体が分化したアミロプラストにより行われていることから、環境ストレス耐性の作物を育種するためにこの葉緑体・アミロプラスト(プラスチド)の機能改変法の研究が行われている。プラスチドの分化・機能発現は、このオルガネラ自身がもつ遺伝子と細胞核のゲノムにコードされた遺伝子によって統御されている。プラスチドの機能改変法としては、これまでに大きく分けて2つの方法が知られている。1つは、プラスチドに遺伝子を直接導入する方法であり、もう1つは、トランジットペプチドと呼ばれている葉緑体移行シグナル遺伝子を葉緑体に導入したい機能タンパク質遺伝子に融合し、それを細胞核ゲノムに導入する方法である。導入された遺伝子は細胞質で翻訳され、トランジットペプチドによりプラスチドへ輸送・局在化される。しかしながら、前者の方法では植物全体のプラスチドを改変することは困難であり、また、後者の方法では細胞質で組換えタンパク質が翻訳されるため、細胞にとって負担になり得るといった問題があった。
ところで、α−アミラーゼ(EC 3.2.1.1)は、デンプンの加水分解酵素であり、胚盤組織及びアリューロン層からデンプン貯蔵組織である胚乳に分泌され、そしてデンプン顆粒を分解し、発芽及び伸長生長に必要な炭素源並びにエネルギー源を供給するという重要な働きをする。イネα−アミラーゼ遺伝子は少なくとも11種類存在していることが明らかになっているが、なかでも典型的なN−結合型糖鎖を有するα−アミラーゼI−1(AmyI−1)は発芽イネ種子における胚乳貯蔵デンプンの分解に重要な役割を果たしていることが報告されている(非特許文献1および2)。
【非特許文献1】Asatsuma, S., Sawada, C., Itoh, K., Okito, M., Kitajima, A., Mitsui, T. (2005): Involvement of a-amylase I-1 in starch degradation in rice chloroplasts. Plant Cell Physiol. 46: 858-869
【非特許文献2】Nanjo, Y., Asatsuma, S., Itoh, K., Hori, H. and Mitsui T. (2004) Proteomic identification of a-amylase isoforms encoded by RAmy3B/3C from germinating rice seeds. Biosci. Biotech. Biochem. 68: 112-118
【発明の開示】
【発明が解決しようとする課題】
【0003】
しかしながら、イネα−アミラーゼの葉緑体・アミロプラスト(プラスチド)へのターゲティング機構及び遺伝的なシステムの詳細についてはほとんど知られていない。葉緑体・アミロプラストの光合成能やデンプン集積能の高い作物を育種することは、植物のみならず地球上のすべての生命にとって最も重要なエネルギー蓄積を行う上で重要な役割を果たすものとして、各方面から研究が進められ、その成果が期待されている。
【0004】
そこで、本発明は、以上の背景の下なされたものであって、深刻化する環境ストレスに適応し、劣悪な環境下においても効率的にデンプン・エネルギー集積能力の高い植物およびその製造方法を提供することを目的とする。
【課題を解決するための手段】
【0005】
本発明者らは以上の課題に鑑み、穀類細胞において核ゲノムにコードされている酵素タンパク質のプラスチドへのターゲティング機構を検討し、プラスチド構成タンパク質のターゲティング機構の存在を明らかにした(図1参照)。具体的には、イネα−アミラーゼI−1のプラスチドへのターゲティングは、小胞体からゴルジ体への輸送に関わるAtArf1やAtSar1のドミナントネガティブ変異体により阻害されたことから、小胞体からゴルジ体への輸送が不可欠であることを明らかにした。すなわち、小胞体−ゴルジ体系を介したプラスチドへの輸送・局在化は、トランジットペプチド非依存性プラスチド局在であり、プラスチド包膜に存在するタンパク透過チャンネルに適合しないタンパクのプラスチド輸送・局在化を行うことができる可能性がある。また、プラスチド局在化に関わる典型的なトランジットペプチドを有しないα−アミラーゼI−1(AmyI−1)がイネ緑葉細胞の葉緑体と細胞壁区画に局在することを見出した。
【0006】
また、イネα−アミラーゼは分泌型酵素として知られているものであったが、イネα−アミラーゼの発現抑制および過剰発現する形質転換イネ植物を解析した結果、緑葉など生細胞中のデンプン分解にα−アミラーゼが関与することを見出した。さらに、α−アミラーゼI−1の発現抑制および過剰発現体イネ植物を用いて、生組織細胞におけるデンプン集積に及ぼすα−アミラーゼI−1の役割を調べたところ、α−アミラーゼI−1発現抑制イネ系統においてはα−アミラーゼI−1発現がほぼ完全に押さえられ、野生型に比べて葉組織において顕著なデンプン蓄積の増加が見られた。
【0007】
本発明は以上の研究成果及び知見に基づいて完成されたものであって、以下の各構成を提供する。
【0008】
本発明における請求項1記載のポリペプチドは、以下の(a)又は(b)記載のアミノ酸配列からなることを特徴とする。
(a)配列番号1に示されるアミノ酸配列からなるポリペプチド。
(b)配列番号1に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつプラスチドへのターゲティング作用を有するポリペプチド。
【0009】
本発明における請求項2記載の遺伝子は、請求項1記載のポリペプチドをコードすることを特徴とする。
【0010】
本発明における請求項3記載のポリペプチドは、以下の(c)又は(d)記載のアミノ酸配列からなることを特徴とする。
(c)配列番号2に示されるアミノ酸配列からなるポリペプチド。
(d)配列番号2に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ小胞体膜透過作用を有するポリペプチド。
【0011】
本発明における請求項4記載の遺伝子は、請求項3記載のポリペプチドをコードすることを特徴とする。
【0012】
本発明における請求項5記載のベクターは、請求項2記載の遺伝子を含むことを特徴とする。
【0013】
本発明における請求項6記載のベクターは、請求項2及び4記載の遺伝子を含むことを特徴とする。
【0014】
本発明における請求項7記載の形質転換植物は、請求項2記載の遺伝子が導入されたことを特徴とする。
【0015】
本発明における請求項8記載の形質転換植物は、請求項2及び4記載の遺伝子が導入されたことを特徴とする。
【0016】
本発明における請求項9記載の形質転換植物は、請求項7又は8において、前記形質転換植物が単子葉植物であることを特徴とする。
【0017】
本発明における請求項10記載の形質転換植物は、請求項5又は6記載のベクターを単子葉植物由来のカルスに導入し、前記カルスを増殖させた後に、前記カルスから植物体を再分化させて得られることを特徴とする。
【0018】
本発明における請求項11記載の形質転換植物の製造方法は、請求項5又は6記載のベクターを単子葉植物由来のカルスにアグロバクテリウムを用いて導入し、前記カルスを増殖させた後に、前記カルスから植物体を再分化させることを特徴とする。
【発明の効果】
【0019】
本発明によれば、今後さらに深刻化する環境ストレスに適応し、劣悪な環境下において効率的にデンプンを集積可能な形質転換植物の作出が可能となる。また、プラスチドの光合成能やデンプン集積能を高め、カーボンニュートラルな工業用素材であるデンプンの供給を増大させることができる。また、植物デンプンは人類にとって極めて重要な栄養素であり、デンプン粒の構造を改変することによってデンプン利用用途の拡大を図ることができ、生分解性樹脂など工業生産における有用素材である。
【発明を実施するための最良の形態】
【0020】
(ベクター)
本発明において、遺伝子を植物へ導入する際に使用されるベクターとしては、導入遺伝子をターゲット細胞(宿主細胞)に導入することができ、且つターゲット細胞内で導入遺伝子を発現させることができるものであれば特に限定されず、目的に応じて適当なベクター(プラスミド、バクテリオファージ、ウイルスなど)が利用される。数多くのベクターが市販されており、本発明ではそれらの中から目的に応じて適切なものを選択して用いることができる。また、本発明のベクターの作成方法としては、特に限定されず、当業者に周知のいかなる方法を用いてもよい。
【0021】
本発明において、好ましいプラスチド輸送・局在化コンストラクトの例を図2に示す。図2に記載のAmyI−1SBとは、α−アミラーゼI−1の301〜369のペプチド領域(配列番号1)を示し、プラスチドターゲティングに必要不可欠なものである。一般に、特定の機能を有するアミノ酸の一部に改変を施した場合においてもその機能が維持されることがある。このことを考慮して、AmyI−1SBのポリペプチドは、以下の(a)又は(b)記載のアミノ酸配列からなることを特徴とする:(a)配列番号1に示されるアミノ酸配列からなるポリペプチド;(b)配列番号1に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつプラスチドへのターゲティング作用を有するポリペプチド。
また、SPとは、ER膜透過シグナルペプチドを示し、例えばAmyI−1SP(配列番号2)が挙げられるがこれに限定されるものではなく、他の植物分泌タンパク質のSPを用いてもよい。また、AmyI−1SPのポリペプチドは、以下の(c)又は(d)記載のアミノ酸配列からなることを特徴とする:(c)配列番号2に示されるアミノ酸配列からなるポリペプチド;(d)配列番号2に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ小胞体膜透過作用を有するポリペプチド。なお、機能タンパク質はAmyI−1SBのN末端側又はC末端側の何れに挿入してもよい。
【0022】
(形質転換方法)
植物へのベクターの導入には、ポリエチレングリコール法、電気穿孔法(エレクトロポレーション法)、アグロバクテリウムを介する方法、パーティクルガン法など、当業者に公知の種々の方法を用いることができる。それぞれの方法に適したベクターの調製、形質転換植物細胞からの植物体の再生は、植物細胞の種類に応じて当業者に公知の方法で行うことが可能である。
【0023】
(ターゲット植物)
本発明においてターゲットとする植物は、単子葉植物である。単子葉植物としては、例えばイネ科(イネ属、コムギ属、オオムギ類、ライムギ類、ハトムギ属、トウモロコシ属、サトウキビ属等)、ユリ科(ネギ属、ニンニク属等)などが挙げられる。
【0024】
以下に本発明の実施例によって、本発明を詳細に説明するが、本発明はこれらの実施例によりなんら制限されるものではない。
【0025】
以下の実施例1〜3において使用される生物学的材料、試薬実験方法は次の通りである。
【0026】
(供試植物)
形質転換に供試される植物(ターゲット植物)として、タマネギ(Allium cepa)表皮細胞を、0.6%のゲルライトを加えた2,4−DフリーのMS培地上にて25℃暗所で培養後、使用した。
【0027】
(プラスミドコンストラクト)
全RNAを、発芽6日目のイネ種子の胚盤組織から単離した。そのcDNAライブラリーをpBluescriptIISK(+/−)(タカラバイオ社)を用いて製造者のプロトコールに従って作成した。cDNAライブラリーを、α−アミラーゼII−3(RAmy3E),II−5(RAmy3B),及びII−6(RAmy3C)の特定のプローブを用いて選別した(Sheu, J.-J., Yu, T.-S., Tong, W.-F., Yu, S.-M. (1996) Carbohydrate starvation stimulates differential expression of rice a-amylase genes that is modulated through complicated transcriptional and posttranscriptional processes. J. Biol. Chem. 271: 26998-27004.)。得られた完全長cDNA(AmyII−3,AmyII−5,及びAmyII−6と称す)を、pBluescript SK(+・−)にクローニングし、pAmyII−3,pAmyII−5,及びpAmyII−6を作成した(図3)。pAmyI−1とpAmyII−4は、Asatsuma S, Sawada C, Itoh K, Okito M, Kitajima A, Mitsui T (2005) Involvement of α-amylase I-1 in starch degradation in rice chloroplasts. Plant & Cell Physiology 46: 858-869に記載の方法に従って作成した。(図4)。
【0028】
完全長のα−アミラーゼI−1cDNAと種々のC末端が一部切断させたフラグメントを、鋳型DNAとしてpAmyI−1とプライマー配列セット(表1)を用いてPCRによって増幅した。
【0029】
【表1】
【0030】
完全長α-アミラーゼII−3,II−4,II−5,II−6cDNA、およびそれらのN−端末シグナル配列領域を、鋳型DNAとしてpAmyII−3,pAmyII−4,pAmyII−5及びpAmyII−6と,プライマー配列セット(表2)を用いてPCRによって増幅した。
【0031】
【表2】
【0032】
BamHI PCR増幅フラグメントをpGFPのBamHI部位に挿入し、pAmyI−1−GFP、pAmyI−1(SP)−GFP、pAmyI−1(Δ101−428)−GFP、pAmyI−1(Δ201−428)−GFP、pAmyI−1(Δ301−428)−GFP、pAmyI−1(Δ370−428)−GFP、pAmyII−3−GFP、pAmyII−3(SP)−GFP、pAmyII−4−GFP、pAmyII−4(SP)−GFP、pAmyII−5−GFP、及びpAmyII−6−GFP(図5)を作成した。
【0033】
PCRを使用して、トリプトファン302をアラニン302に置換した一つのアミノ酸を導入し、AmyI−1(W302A)を作成した。変異原性のフォワードプライマー(F−ATG+1と、F−W302A)及びリバースプライマー(R−endと、R−W302A)を合成した(表3)。
【0034】
【表3】
【0035】
変異原性のプライマーセットと、鋳型DNAとしてpAmyI−1を使用して第一のPCRを行った。F−ATG+1とR−endのプライマーと、鋳型DNAとして第一のPCR産物を使用して第二のPCRを行った。第二のPCR産物をBamHIで切断し、pGFPのBamHI部位にクローニング、pAmyI−1(W302A)(図6)を作成した。
【0036】
タマネギ表皮細胞におけるDsRed2発現において、pDsRed2(Clonetech)からのBamHI−NotIのPCR増幅フラグメントを、BamHI−NotI消化pGFPのBamHI−NotI消化部位にクローニングし、pDsRed(図7)を産生した。イネwaxy遺伝子(Itoh, K., Ozaki, H., Okada, K., Hori, H., Takeda, Y., Mitsui, T. (2003) Introduction of Wx transgene into rice wx mutants leads to both high- and low-amylose rice. Plant Cell Physiol., 44: 473-480)からトランジットペプチド配列の始めの1−111のアミノ酸を、2つのプライマー5'-ctggatccatgtcggctctcaccacg-3'と5'-tatggatccggcaggggggaggccaccgag-3'を用いてPCR増幅し、その後PCR産物をBamHIで消化してpWxTP−DsRedを作成した。BamHI消化フラグメントをpDsRedのBamHI消化部位に挿入した(図8)。さらに、BamHI PCRの増幅WxTPフラグメントを、pGFPのBamHI消化部位にクローニングし、pWxTP−GFPを構築した(図9)。
【0037】
ペルオキシソームとミトコンドリアにそれぞれpPTS2−GFP(Mano, S., Nakamori, C., Hayashi, M., Kato, A., Kondo, M., Nishimura, M. (2002) Distribution and characterization of peroxisomes in Arabidopsis by visualization with GFP: dynamic morphology and actin-dependent movement. Plant Cell Physiol. 43: 331-41.)とpmt−GFP(Niwa, Y., Hirano, T., Yoshimoto, K., Shimizu, M., Kobayashi, H. (1999) Non-invasive quantitative detection and applications of non-toxic, S65T-type green fluorescent protein in living plants. Plant J. 18, 455-463.)を用いて、他の細胞小器官を可視化した(図10)。
【0038】
(遺伝子導入方法)
プラスミドDNAを、パーティクルデリバリーシステム(PDS−1000/He、BIO−RAD)を用いてタマネギ表皮細胞へ導入した。具体的には、10μlの滅菌水に溶解させた3μgのプラスミドDNAに、10μlの金粒子懸濁液(直径1.0μmの金粒子60mg/ml)と、10μlの2.5mM CaCl2と、4μlの0.1M スペルミジンを加えて懸濁し、室温で30分間静置した。プラスミドDNAで被覆された金粒子を冷エタノールで洗浄した後、10μlのエタノールで緩やかに懸濁した。1100psiのラプチャーディスクを使用し、一サンプルあたり2回ずつ導入を行った。
【0039】
(観察方法)
GFP,DsRedおよびmRFP蛍光画像は、電動蛍光顕微鏡(Olympus BX−61)と、CCDカメラ(Cool SNAP fx、Roper Scientific)を用いて観察・撮影し、画像解析ソフト(Lumina Vision、三谷商事(株))を用いて解析した。GFPは470〜490nmで励起し、510〜550nmを検出した。DsRedおよびmRFPは、520〜550nmで励起し、≧580nmを検出した。一細胞につき、10μm間隔で20〜30枚の画像を撮り、デコンボリューション処理した後、一つの画像にした。
【実施例1】
【0040】
イネα−アミラーゼ遺伝子と緑色蛍光タンパク質(GFP)遺伝子を融合し、この融合物をタマネギ細胞に導入させ、GFPの輸送・局在化を調べた。
【0041】
タマネギ表皮細胞にパーティクルガンでAmyI−1−GFPおよびWxTP−DsRedを同時に導入し、一過的に発現させた(図11)。WxTP−DsRedは、プラスチドへの局在を示すマーカーである。図11のA,Dに示すようにように、AmyI−1−GFPの蛍光は細胞表面だけでなく、細胞内で何らかのオルガネラに局在していた。B,Eでは、WxTP−DsRedの蛍光はプラスチドを示し、プラスチド特有の構造であるストロミュールも確認された(図11E)。CはAとBを、FはDとEを重ね合わせた画像で、部分的にAmyI−1−GFPの蛍光がWxTP−DsRedの蛍光と局在が一致し、AmyI−1がプラスチドに局在することを示した。この結果から、イネα−アミラーゼ遺伝子と緑色蛍光タンパク質(GFP)遺伝子を融合し、この融合物をタマネギ細胞に導入することによって、タマネギ細胞のプラスチドにGFPを輸送・局在化させることが示された。また、α−アミラーゼI−1においてはイネおよびタマネギの両細胞に共通するプラスチド局在化シグナルが含まれることが明らかになった。
【実施例2】
【0042】
α−アミラーゼI−1のプラスチドターゲティングに必要不可欠な領域を決定するために、C端末の端部が一部欠けているα−アミラーゼI−1とGFPの融合タンパク質を一過的に発現させその局在を調べた。
【0043】
図12に、C末端が一部切断されたα−アミラーゼI−1(AmyI−1Δ34−428、AmyI−1Δ101−428、AmyI−1Δ201−428、AmyI−1Δ301−428、AmyI−1Δ370−428)を示す。AmyI−1Δ34−428は、α−アミラーゼI−1のN末端シグナルペプチドのみを含んでいた。予測されたシグナルペプチダーゼ開裂部位はGln26のN末端側である。AmyI−1Δ301−428は、N結合型糖鎖結合部位(Asn265、−Gly266、−Thr267)を含んでいる。AmyI−1Δ370−428は、推定のデンプン結合部位(301Typ−302Typ)を含む完全なAドメインとBドメインを有する(Robert, X., Haser, R., Mori, H., Svensson, B., Aghajari, N. (2005) Oligosaccharide binding to barley a-amylase 1. J. Biol. Chem. 280: 32968-32978)。370〜428残基のアミノ酸配列は、5つの逆平行β-シートを構成したC-ドメインに対応している。図12と表4に示されるように、多くのC末端が欠失したAmyI−1−GFP融合タンパク質は小胞体(ER)様網目状構造に局在し、プラスチドにはほとんど局在しなかったが、AmyI−1Δ370−428−GFP融合タンパク質は完全長AmyI−1−GFPと同様に遺伝子導入されたタマネギ細胞におけるプラスチドに局在した。これらの結果から、301〜369のアミノ酸残基のペプチド領域(α−AmyI−1SB(配列番号1))がタマネギ表皮細胞におけるα−アミラーゼI−1のプラスチドターゲッティングに必要不可欠であることが明らかとなった。
【0044】
【表4】
【実施例3】
【0045】
α−アミラーゼのアイソフォーム(AmyII−3、AmyII−4、AmyII−5およびAmyII−6)が、タマネギ表皮細胞のプラスチドに局在するか否かをα−アミラーゼのアイソフォームとGFPの融合タンパク質(AmyII−3−GFP、AmyII−4−GFP、AmyII−5−GFPおよびAmyII−6−GFP)を用いて調べた。
【0046】
イネα−アミラーゼには多くのアイソフォームが存在し、これらのアイソフォームは多くの遺伝子によってコードされていることが知られている(O'Neill, S.D., Kumagai, M.H., Majumdar, A., Huang, N., Sutliff, T.D. and Rodriguez, R.L. (1990) The alpha-amylase genes in Oryza sativa: characterization of cDNAclones and mRNA expression during seed germination. Mol. Gen. Genet. 221: 235-244;Huang, N., Koizumi, N., Reinl, S., Rodriguez, R.L. (1990a) Structural organization and differential expression of rice a-amylase genes. Nucl. Acid Res. 18: 7007-7013;Huang, N., Sutliff, T.D., Litts, J.C., Rodriguez, R.L. (1990b) Classification and characterization of the rice a-amylase multigene family. Plant Mol. Biol. 14: 655-668;Kim, J.-K. and Wu, R. (1992) Nucleotide sequence of a high-pI rice (Oryza sativa) a-amylase gene. Plant Mol. Biol. 18: 399-402)。また、本発明者らは以前の研究において、RAmy1A、RAmy3B、RAmy3C、RAmy3D、及びRAmy3Eの天然の発現産物がイネ植物と組織培養で検出されたことを報告している(Mitsui, T., Yamaguchi, J. and Akazawa, T. (1996) Physicochemical and serological characterization of rice a-amylase isoforms and identification of their corresponding genes. Plant Physiol. 110: 1395-1404;Nanjo, Y., Asatsuma, S., Itoh, K., Hori, H. and Mitsui T. (2004) Proteomic identification of a-amylase isoforms encoded by RAmy3B/3C from germinating rice seeds. Biosci. Biotech. Biochem. 68: 112-118)。
【0047】
AmyI−1−GFPのプラスチドにおける局在化と比較して、AmyII−3−GFP、AmyII−4−GFP、AmyII−5−GFP、及びAmyII−6−GFPはプラスチド内でほとんど局在していなかった(表4)。
【0048】
さらに、タマネギ細胞におけるデンプン結合部位を変異させたα−アミラーゼI−1−GFP融合タンパク質のターゲティング特性を調べて比較した。
【0049】
図13に、イネα−アミラーゼのアイソフォーム(アミラーゼI−1(M24286);アミラーゼII−3(M59352);アミラーゼII−4(M24287);アミラーゼII−5(X56337);アミラーゼII−6(X56338))のアミノ酸配列の配列比較を示す。図13に示されるα−アミラーゼI−1(RAmy1A)、II−3(RAmy3E)、II−4(RAmy3D)とII−5/6(RAmy3B/3C)間のアミノ酸配列は約83〜90%の相同性を有する。
【0050】
301Typ−302Typは、α−アミラーゼI−1のポリペプチドにおける特有のアミノ酸残基であった。その他のα−アミラーゼのアイソフォーム(α−アミラーゼII−3、II−4、II−5およびII−6)は、ペプチド領域301〜369のアミノ酸残基においてトリプトファンジペプチドを有していなかった(図13)。リプトファン302をアラニン302に置換した場合、プラスチドへのAmyI−1−GFPのターゲティングを強く阻害し、AmyI−1(W302A)−GFPはER様網目状に保持されていた(図14、表4)。これらの結果から、302Typ残基はタマネギ表皮細胞においてα−アミラーゼI−1のプラスチドターゲティングに必須であることが明らかになった。図15に、プラスチドターゲティングのためのα−アミラーゼI−1のシグナル領域を示す。図15に示される「SP」とはER(小胞体)膜透過シグナルペプチドを意味し、「TSR」とはプラスチドターゲティングのためのシグナル領域(すなわち、α−アミラーゼI−1の301〜369ペプチド領域(デンプン結合領域))を意味する。なお、α−アミラーゼは、3つのドメインからなり、ドメインAは、(β/α)8バレルに折り重なり、酵素の触媒残基を含み、ドメインBは、ドメインAの3番目のβ鎖と3番目のαへリックスの間から突き出している大きなループであり、ドメインCは、(β/α)8バレルのC末端に位置し、β鎖からなっている。
【0051】
以下の実施例4において使用される生物学的材料、試薬、実験方法等は次の通りである。尚、特に説明のない材料等については上記実施例で使用したものと同様である。
【0052】
(供試植物)
供試植物として、イネ品種「日本晴」(Oryza sativa L. cv. Nipponbare)を用いた。そのイネの種子は、新潟農業研究所より提供されたものである。
【0053】
(分析法)
デンプンの含量は、Matsukura C, Saitoh T, Hirose T, Ohsugi R, Perata P, Yamaguchi J (2000) Sugar uptake and transport in rice embryo. Expression of companion cell-specific sucrose transporter (OsSUT1) induced by sugar and light. Plant Physiology 124: 85-93、及びBradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. . Anal. Biochem. 72: 248-254に記載の方法に従って決定した。α−アミラーゼの分析法は、Mitsui T, Yamaguchi J, Akazawa T (1996) Physicochemical and serological characterization of rice α-amylase isoforms and identification of their corresponding genes. Plant Physiology 110: 1395-1404に記載の方法に従って行った。
【0054】
(ベクターの構築)
pZH2B−35S−AmyI−1及びpTN1−35S−AmyII−4の構築は、Asatsuma S, Sawada C, Itoh K, Okito M, Kitajima A, Mitsui T (2005) Involvement of α-amylase I-1 in starch degradation in rice chloroplasts. Plant & Cell Physiology 46: 858-869及びHajdukiewicz P, Svab Z, Maliga P (1994) The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Molecular Biology 25: 989-994に記載の方法に従って行った(図16)。pAmyII−4をBamHI−EcoRIで消化して得られたフラグメントを、pTN1−35S−AmyI−1をBamHI−EcoRIで消化した対応する制限部位にクローニングし、pTN1−35S−AmyII−4を作成した(図17)。
【0055】
(α−アミラーゼ過剰発現植物体の作成)
上述のようにして作成されたベクターを、20mMのCaCl2で処理されたアグロバクテリウム(Agrobacterium tumefaciens)EHA101株のコンピテント細胞へ導入し、暗所でプレート培養して導入されたアグロバクテリウムを選抜した。プラスミドが導入されたアグロバクテリウムをアセトシリンゴン存在下でイネの胚組織から誘導したカルスに感染させた。感染後、抗生物質を含んだプレート培地上でプラスミドが誘導されたカルスを選抜し、植物ホルモン(NAA)により植物体に再分化させた(Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant Journal 6: 271-282及びAsatsuma S, Sawada C, Itoh K, Okito M, Kitajima A, Mitsui T (2005) Involvement of α-amylase I-1 in starch degradation in rice chloroplasts. Plant & Cell Physiology 46: 858-869)。
【0056】
(植物体の生育)
再生した植物体をポットに移し、明所12時間、暗所8時間、27℃のグロスチャンバー内で20cm程度になるまで生育させた。その後、温室に移し種子を採取するまで約6ヶ月間生育させた。開花後、40日ほどで種子を刈り取った。
【0057】
(サザンブロット解析、ノーザンブロット解析、ウエスタンブロット解析)
野生型および形質転換イネの若葉より単離したゲノムDNA(5μg)をEcoRIで消化し、1%アガロースゲル電気泳動で分画した後、Hybond−N+ナイロン膜(Amersham)に転写・固定した。ノーザンブロット解析は、Mitsui T, Loboda T, Kamimura I, Hori H, Itoh K, Mitsunaga S, (1999) Sucrose-controlled transport and turnover of α-amylase in rice (Oryza sativa L.) cells. . Plant Cell Physiol. 40 884-893およびKashem MA, Itoh K, Iwabuchi S, Hori H, Mitsui T (2000) Possible involvement of phosphoinositide-Ca2+ signaling in the regulation of alpha-amylase expression and germination of rice seed (Oryza sativa L.). Plant & Cell Physiology 41: 399-407に記載の手順に従って行った。サザンブロット解析に関し、α−アミラーゼI−1遺伝子およびα−アミラーゼII−4遺伝子の特異的DNA(前出のKashem et al. 2000)を放射性プローブとして使用した。オートラジオグラフィーは、画像解析装置BAS5000(フジフィルム)を用いて行った。ウエスタンブロット解析、抗α−アミラーゼI−1抗体及び抗α−アミラーゼII−4抗体の調製法等は、Mitsui T, Yamaguchi J, Akazawa T (1996) Physicochemical and serological characterization of rice α-amylase isoforms and identification of their corresponding genes. Plant Physiology 110: 1395-1404に記載の方法に従って行った。2次抗体として、ペルオキシダーゼ標識抗ウサギIgG抗体を使用した。そのタンパク質バンドをECL(化学発光、Amersham)を用いて可視化し、LAS3000(フジフィルム)を用いて定量化した。野生型イネ植物において発現するα−アミラーゼI−1およびII−4の各タンパク質の量を1ユニットとした。
【実施例4】
【0058】
特異的抗体(例えば抗α−アミラーゼI−1,II−3,II−4,II−5およびII−6)を用いてウエスタンブロット解析を行い、登熟期間中のイネ登熟種子におけるα−アミラーゼのアイソフォームの発現を調べた。出穂後10日目の登熟種子において、α−アミラーゼI−1およびII−4の発現を検出した(図18)。興味深いことに、α−アミラーゼII−4が登熟種子において最も多量に存在するアイソフォームであることが示唆された。
【0059】
カリフラワーモザイクウイルス(CaMV)35Sプロモーターの制御下それぞれα−アミラーゼI−1(AmyI−1)とα−アミラーゼII−4cDNA(AmyII−4)を含むpZH2B−35S−AmyI−1とpTN1−35S−AmyII−4を用いて、アグロバクテリウムを介した形質転換によって、遺伝組換えイネ植物を産出した。AmyI−1およびAmyII−4の存在を、サザンハイブリダイゼーションを用いて再生された形質転換植物中において確認した。得られた形質転換イネ植物は1〜8コピーの導入遺伝子を有していた(図示せず)。また、pZH2B−35S−AmyI−1とpTN1−35S−AmyII−4で形質転換されたこれらの遺伝子導入系統におけるα−アミラーゼ遺伝子発現を、ノーザンおよびウエスタンブロット解析を用いて調べた。結果を図19に示す。図19に示されるように、これらの形質転換イネ系統の双方が、成葉およびT1世代のシュート組織においてα−アミラーゼのmRNAおよびタンパク質の著しい増加を示した。また、α−アミラーゼI−1(A3−1)を過剰発現した葉におけるデンプン含有量は、野生型(WT)のものと比較して著しく減少し、そのデンプン含有量は野生型の約42%であった。(図20)。一方、α−アミラーゼII−4(D1−4)を過剰発現した葉におけるデンプン含有量はわずかに減少し、野生型のデンプン含有量の約82%であった。
【0060】
A3−1およびD1−4のT3世代の登熟期において、野生型と比較しα−アミラーゼのより高い活性はα−アミラーゼI−1およびII−4を発現する形質転換植物の両方の登熟種子で検出された(図21)。図21の縦軸はα−アミラーゼ活性(unit/seed)を示し、横軸は出穂後の日数(daf:days after heading)を示す。D1−4系統の登熟種子に含まれるα−アミラーゼ活性のレベルは、A3−1系統の約2倍の活性を示した。
【0061】
さらに、A3−1系統およびD1−4系統の登熟種子表現型の特性を分析した。α−アミラーゼI−1とα−アミラーゼII−4に対する特異的抗体を用いてウエスタンブロット解析を行った場合、野生型の種子と比較し、A3−1系統の登熟種子においてα−アミラーゼI−1分子は36倍もの増加を示し、D1−4系統の登熟種子においてα−アミラーゼII−4分子は44倍もの増加を示した(図22−A)。酵素活性の増加はA3−1及びD1−4系統の登熟種子の両方で示されたが、D1−4系統ではより顕著な増加が示された(図22−B)。A3−1及びD1−4系統の登熟種子の乾燥重量は、野生型の種子と比較し、A3−1系統で4%、D1−4系統で11%の減少が見られた(図22−C)。
【0062】
また、α−アミラーゼI−1とα−アミラーゼII−4を過剰発現した形質転換体イネ系統(A3−1及びD1−4系統)の完熟種子の品質を反射光画像と透過光画像の双方によって評価した(図23)。表5に示すように、形質転換体イネ系統から得られた種子においては、ともにデンプン蓄積異常が見られ、野生型,A3−1系統及びD1−4系統の正常な穀粒のパーセンテージはそれぞれ90%,54.4%,36.5%であった。A3−1系統及びD1−4系統の異常な種子の特徴的な形態学的表現型はそれぞれ心白(white core)と全乳白(whole opaque white)であった(図23、表5)。これらの結果から、α−アミラーゼ活性の異常な発現はデンプンの集積およびイネ粒構造に影響することが示された。また、α−アミラーゼI−1とα−アミラーゼII−4は、登熟種子の胚乳におけるアミロプラストに局在する能力を有し、デンプン集積(分解)調節機能を有すると考えられる。
【0063】
【表5】
【図面の簡単な説明】
【0064】
【図1】高等植物細胞におけるα−アミラーゼI−1のプラスチドターゲティング機構を示す図である。
【図2】プラスチドコンストラクトを構築した例を示す図である。
【図3】pAmyII−3,pAmyII−5及びpAmyII−6の作成過程を示す図である。
【図4】pAmyI−1及びpAmyII−4のマップである。
【図5】種々のpAmy−GFPの作成過程を示す図である。
【図6】pAmyI−1(W302A)−GFPの作成過程を示す図である。
【図7】pDsRedの作成過程を示す図である。
【図8】pWxTP−DsRedの作成過程を示す図である。
【図9】pWxTP−GFPの作成過程を示す図である。
【図10】pPTS2−GFPおよびpmt−GFPを構築した例を示す図である。
【図11】AmyI−1−GFPとWxTP−DsRedのタマネギ表皮細胞での同時発現の結果を示す図である。A,DはAmyI−1−GFPの蛍光像を示し、B,EはWxTP−DsRedの蛍光像を示し、C,FはGFPとDsRedの重ね合わせ像を示す。下図(D,E,F)は、上図(A,B,C)の拡大図である。
【図12】遺伝子導入されたタマネギ表皮細胞における一部切断されたα−アミラーゼI−1−GFPの局在を示す図である。A〜Eの左側、中央、右側はそれぞれWxTP−DsRed画像、AmyI−1−GFP画像、WxTP−DsRedとAmyI−1−GFPの重ね合わせ画像を示す。
【図13】イネα−アミラーゼのアイソフォーム(アミラーゼI−1(M24286);アミラーゼII−3(M59352);アミラーゼII−4(M24287);アミラーゼII−5(X56337);アミラーゼII−6(X56338))のアミノ酸配列の配列比較を示す図である。矢頭は、N末端シグナルペプチドの開裂部位を示す。**(アスタリスク2)印の個所は、推定デンプン結合部位を示す。下線個所は、グリコシル化部位を示す。
【図14】AmyI−1(W302A)−GFPとWxTP−DsRedのタマネギ表皮細胞での同時発現を示す図である。
【図15】プラスチドへのターゲティングに関するα−アミラーゼI−1のシグナル領域を示す図である。
【図16】pZH2B−35S−AmyI−1の作成過程を示す図である。
【図17】pTN1−35S−AmyII−4の作成過程を示す図である。
【図18】抗α−アミラーゼI−1,II−3,II−4,II−5およびII−6を用いてウエスタンブロット解析を行った結果を示す図である。
【図19】α−アミラーゼI−1(A)及びα−アミラーゼII−4(B)を過剰発現したイネ植物のノーザンブロット解析およびウエスタンブロット解析を行った結果を示す図である。
【図20】(A)は、野生型(WT)、α−アミラーゼI−1(A3−1)およびα−アミラーゼII−4(D1−4)を過剰発現した葉におけるデンプン含有量の結果を示し、(B)はα−アミラーゼの発現の結果を示すグラフである。
【図21】形質転換植物(A3−1およびD1−4)及び野生型(WT)の登熟種子におけるα−アミラーゼ活性の変化を示す図である。
【図22】(A)は、野生型(WT)、α−アミラーゼI−1(A3−1)およびα−アミラーゼII−4(D1−4)を過剰発現した完熟種子のα−アミラーゼ発現の結果を示し、(B)は、α−アミラーゼ活性の結果を示し、(C)は完熟種子の乾燥重量の結果を示すグラフである。
【図23】(A)は、野生型(WT)、α−アミラーゼI−1及びα−アミラーゼII−4を過剰発現した形質転換体イネ系統(A3−1及びD1−4系統)の完熟種子の反射光画像を示し、(B)は透過光画像を示す図である。
【特許請求の範囲】
【請求項1】
以下の(a)又は(b)記載のアミノ酸配列からなることを特徴とするポリペプチド。
(a)配列番号1に示されるアミノ酸配列からなるポリペプチド。
(b)配列番号1に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつプラスチドへのターゲティング作用を有するポリペプチド。
【請求項2】
請求項1記載のポリペプチドをコードすることを特徴とする遺伝子。
【請求項3】
以下の(c)又は(d)記載のアミノ酸配列からなることを特徴とするポリペプチド。
(c)配列番号2に示されるアミノ酸配列からなるポリペプチド。
(d)配列番号2に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ小胞体膜透過作用を有するポリペプチド。
【請求項4】
請求項3記載のポリペプチドをコードすることを特徴とする遺伝子。
【請求項5】
請求項2記載の遺伝子を含むことを特徴とするベクター。
【請求項6】
請求項2及び4記載の遺伝子を含むことを特徴とするベクター。
【請求項7】
請求項2記載の遺伝子が導入されたことを特徴とする形質転換植物。
【請求項8】
請求項2及び4記載の遺伝子が導入されたことを特徴とする形質転換植物。
【請求項9】
前記形質転換植物が単子葉植物であることを特徴とする請求項7又は8記載の形質転換植物。
【請求項10】
請求項5又は6記載のベクターを単子葉植物由来のカルスに導入し、前記カルスを増殖させた後に、前記カルスから植物体を再分化させて得られることを特徴とする形質転換植物。
【請求項11】
請求項5又は6記載のベクターを単子葉植物由来のカルスにアグロバクテリウムを用いて導入し、前記カルスを増殖させた後に、前記カルスから植物体を再分化させることを特徴とする形質転換植物の製造方法。
【請求項1】
以下の(a)又は(b)記載のアミノ酸配列からなることを特徴とするポリペプチド。
(a)配列番号1に示されるアミノ酸配列からなるポリペプチド。
(b)配列番号1に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつプラスチドへのターゲティング作用を有するポリペプチド。
【請求項2】
請求項1記載のポリペプチドをコードすることを特徴とする遺伝子。
【請求項3】
以下の(c)又は(d)記載のアミノ酸配列からなることを特徴とするポリペプチド。
(c)配列番号2に示されるアミノ酸配列からなるポリペプチド。
(d)配列番号2に記載のアミノ酸配列において、1個又はそれ以上のアミノ酸が置換、欠失、挿入、及び/又は付加されたアミノ酸配列からなり、かつ小胞体膜透過作用を有するポリペプチド。
【請求項4】
請求項3記載のポリペプチドをコードすることを特徴とする遺伝子。
【請求項5】
請求項2記載の遺伝子を含むことを特徴とするベクター。
【請求項6】
請求項2及び4記載の遺伝子を含むことを特徴とするベクター。
【請求項7】
請求項2記載の遺伝子が導入されたことを特徴とする形質転換植物。
【請求項8】
請求項2及び4記載の遺伝子が導入されたことを特徴とする形質転換植物。
【請求項9】
前記形質転換植物が単子葉植物であることを特徴とする請求項7又は8記載の形質転換植物。
【請求項10】
請求項5又は6記載のベクターを単子葉植物由来のカルスに導入し、前記カルスを増殖させた後に、前記カルスから植物体を再分化させて得られることを特徴とする形質転換植物。
【請求項11】
請求項5又は6記載のベクターを単子葉植物由来のカルスにアグロバクテリウムを用いて導入し、前記カルスを増殖させた後に、前記カルスから植物体を再分化させることを特徴とする形質転換植物の製造方法。
【図2】

【図13】

【図21】

【図1】
【図3】
【図4】
【図5】
【図6】
【図7】

【図8】

【図9】
【図10】
【図11】
【図12】
【図14】
【図15】
【図16】
【図17】
【図18】

【図19】
【図20】
【図22】
【図23】
【図13】
【図21】
【図1】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図22】
【図23】
【公開番号】特開2007−151435(P2007−151435A)
【公開日】平成19年6月21日(2007.6.21)
【国際特許分類】
【出願番号】特願2005−349100(P2005−349100)
【出願日】平成17年12月2日(2005.12.2)
【出願人】(304027279)国立大学法人 新潟大学 (310)
【Fターム(参考)】
【公開日】平成19年6月21日(2007.6.21)
【国際特許分類】
【出願日】平成17年12月2日(2005.12.2)
【出願人】(304027279)国立大学法人 新潟大学 (310)
【Fターム(参考)】
[ Back to top ]