
トラウストチトリアレス微生物の形質転換についての産物および方法
【課題】トラウストチトリアレス目の微生物の形質転換のために有用な選択マーカーを含む組換え核酸分子の取得、ならびに組換え核酸分子を用いた形質転換方法の開発。
【解決手段】それぞれ、トラウストチトリアレス微生物由来の、アセト乳酸シンターゼ、アセト乳酸シンターゼ制御領域、α-チューブリンプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、および脂肪酸デサチュラーゼプロモーターについての核酸配列およびアミノ酸配列。また、トラウストチトリアレス微生物の形質転換に有用な組換えベクター、およびトラウストチトリアレス微生物の形質転換の方法。該組換え核酸分子は、トラウストチトリアレス微生物における外来核酸の発現のために、およびトラウストチトリアレス微生物における遺伝子の欠失、突然変異または不活化のために使用されうる。
【解決手段】それぞれ、トラウストチトリアレス微生物由来の、アセト乳酸シンターゼ、アセト乳酸シンターゼ制御領域、α-チューブリンプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、および脂肪酸デサチュラーゼプロモーターについての核酸配列およびアミノ酸配列。また、トラウストチトリアレス微生物の形質転換に有用な組換えベクター、およびトラウストチトリアレス微生物の形質転換の方法。該組換え核酸分子は、トラウストチトリアレス微生物における外来核酸の発現のために、およびトラウストチトリアレス微生物における遺伝子の欠失、突然変異または不活化のために使用されうる。
【発明の詳細な説明】
【技術分野】
【0001】
発明の分野
本発明は、一般的に、トラウストチトリアレス目(Thraustochytriales)の微生物にスルホニル尿素化合物、イミダゾリノン類阻害剤および/またはピリミジニルオキシ安息香酸に対する低下した感受性を与えるアセト乳酸シンターゼを含む、トラウストチトリアレスのアセト乳酸シンターゼをコードする単離された核酸分子;トラウストチトリアレス目の微生物の形質転換のために有用な選択マーカーを含む組換え核酸分子、ならびに本発明の組換え核酸分子を用いてそのような微生物を形質転換する方法に関する。本発明はまた、トラウストチトリアレス発現系において有用な遺伝子プロモーターに関する。本発明の組換え核酸分子は、トラウストチトリアレス微生物において外来核酸の発現、およびトラウストチトリアレス微生物における遺伝子の欠失、突然変異または不活化のために用いられうる。
【背景技術】
【0002】
発明の背景
開発することによりトラウストチトリド(Thraustochytrid)の分類の改正をもたらした。分類学理論家は、トラウストチトリドを藻類または藻類様原生生物と定める。しかしながら、分類の不確定性のため、本発明に記載される系統をトラウストチトリド(目:トラウストチトリアレス;科:トラウストチトリアシエ(Thraustochytriaceae);属:トラウストチトリウム(Thraustochytrium)、シゾチトリウム(Schizochytrium)、ラビリンチュロイデス(Labyrinthuloides)またはジャポノチトリウム(Japonochytrium))とみなすことが本発明の目的にとって最善であるものと思われる。分類変更は下記に要約されている。
【0003】
本明細書に開示および特許請求の範囲に主張されるある特定の単細胞生物体の株は、トラウストチトリアレス目のメンバーである。トラウストチトリドは、問題を含む分類学的変遷をもつ海洋真核生物である。トラウストチトリドの分類学的配置についての問題は、Moss(1986)、BahnwebおよびJackle(1986)ならびにChamberlainおよびMoss(1988)により概説されている。
【0004】
便宜上、トラウストチトリドは、最初、分類学者により藻菌類(Phycomycetes)(藻類様菌類)における他の無色遊走子真核生物と定められた。藻菌類(Phycomycetes)という名前は、しかしながら、結局、分類学的地位から除外され、トラウストチトリドは、卵菌類(Oomycetes)(二鞭毛遊走子の菌類)に保持された。卵菌類(Oomycetes)は不等毛藻類に関連していると最初に想定され、結局、Barr(1983)により要約されているが、広範囲の超微細構造および生化学的研究によってこの想定を裏付けられた。卵菌類は、事実上、Leedale(1974)および他の藻類学者により不等毛藻類の群として認められた。しかしながら、それらの従属栄養の性質による都合上、重宝さの理由から、卵菌類およびトラウストチトリドは、藻類学者(藻類を研究する科学者)よりもむしろ、菌学者(菌類を研究する科学者)により、主に研究されてきた。
【0005】
別の分類学的観点から、進化生物学者は、いかにして真核生物は進化したかについての考えの2つの一般的な学派を発展させてきた。一つの学説は、一連の内部共生を通しての膜結合型細胞小器官の外因性起源を提案している(Margulis, 1970);例えば、ミトコンドリアは細菌の内部共生に由来し、葉緑体は藍色植物に、および鞭毛はスピロヘータに由来した。他方の学説は、自生過程による、原核生物の祖先の非膜結合型系からの膜結合型細胞小器官の漸進的進化を示唆している(Cavalier-Smith, 1975)。しかしながら、進化生物学者の両グループは、卵菌類およびトラウストチトリドを菌類から除外し、それらを有色植物界(Chromophyta)(Cavalier-Smith, 1981)(この界は、より最近に、他の原生生物を含むように拡大され、この界のメンバーは現在、ストラメノパイル(Stramenopiles)と呼ばれている)における有色植物藻類かまたはプロトクチスタ(Protoctista)界(MargulisおよびSagan, 1985)における全藻類に定めている。
【0006】
電子顕微鏡の発達で、トラウストチトリドの2つの属、トラウストチトリウムおよびシゾチトリウム(Perkins, 1976; Kazama, 1980; Barr, 1981)の遊走子の超微細構造についての研究により、トラウストチトリアシエが卵菌類にわずかに遠く関連しているのみという有力な証拠が提供された。さらに、5SリボソームRNA配列の一致分析(多変量統計学の型)を表す遺伝子データより、トラウストチトリアレスは明らかに、真核生物の独自の群であり、菌類から完全に分離し、かつ赤色および茶色藻類、ならびに卵菌類のメンバーに最も近く関連していることが示されている(Mannellaら、1987)。ほとんどの分類学者は、トラウストチトリドを卵菌類から除くことに同意した(Bartnicki-Garcia, 1988)。
【0007】
要約すれば、Cavalier-Smithの分類体系(1981, 1983)を用いて、トラウストチトリドは、有色植物界(ストラメノパイル)における有色植物藻類に分類されている。これは、それらを、真正菌界にすべてが配置されている菌類とは完全に異なる界に配置している。トラウストチトリドの分類学的配置は、それゆえに、以下に要約される:
界:有色植物界(ストラメノパイル)
門:不等毛門(Heterokonta)
目:トラウストチトリアレス
科:トラウストチトリアシエ
属:トラウストチトリウム、シゾチトリウム、ラビリンチュロイデス、またはジャポノチトリウム
【0008】
ある初期の分類学者らは、トラウストチトリウム(アメーバ状生活期をもつもの)属の数個の最初のメンバーをウルケニア(Ulkenia)と呼ばれる異なる属へと分離した。しかしながら、すべてではないにしても、ほとんどのトラウストチトリド(トラウストチトリウムおよびシゾチトリウムを含む)は、アメーバ状期を現すことは現在知られており、それとして、ウルケニアは、ある人たちには妥当な属であるとみなされていない。本明細書に用いられる場合、トラウストチトリウム属は、ウルケニアを含むものとする。
【0009】
門および界の上級分類内での分類学的配置の不確定性にもかかわらず、トラウストチトリドは、メンバーがトラウストチトリアレス目内に分類できるままの示差的かつ特徴的な群分けを保持している。
【0010】
シゾチトリウムおよび他のトラウストチトリアレス微生物は、高度不飽和脂肪酸(HUFA)および様々なカロテノイド(例えば、アスタキサンチン)を含む、大量の類脂質化合物を産生する能力のために、実質的に現存する、および可能性のある商業的価値をもっている。オメガ-3高度不飽和脂肪酸は、それらが動脈硬化および冠動脈性心疾患を防ぐための、炎症状態を軽減するためのならびに腫瘍細胞の増殖を妨げるための重要な食餌療法化合物として、近年、認識されてきているという点で、かなりの商業的関心対象となっている。これらの有益な効果は、オメガ-6脂肪酸から生成される化合物の競合的阻害をひき起こすオメガ-3HUFA、およびオメガ-3HUFA自身から直接的に生成される有益な化合物の両方の結果である(Simopoulosら、1986)。オメガ-6脂肪酸は、植物および動物において見出される主な(predominant)HUFAである。それゆえ、商業的な生産生物体(commercial production organism)としてのトラウストチトリアレス微生物のさらなる開発は、組換えDNAテクノロジーにより生物体に特定の遺伝子変化を生じる能力から有意に利益を得るものと思われ、そのような生物体により高い価値のあるHUFAおよびカロテノイドの産生を高めることが含まれる。さらに、この特徴付けが乏しい生物体群の生化学および分子生物学のより十分な理解を獲得する能力は、さらなる系統開発の試みを導くために用いられうる価値ある情報を提供するものと思われる。しかしながら、本発明の前には、シゾチトリウム属およびトラウストチトリウムのメンバーを含むトラウストチトリドを形質転換するために適する方法および組換え構築物は利用できなかった。トラウストチトリアレス微生物を形質転換するために特に有用である選択マーカーの開発およびトラウストチトリアレス特異的プロモーター配列の同定は、本発明の前には利用できなかったということは重大なことである。
【0011】
以前の研究者らは、トラウストチトリアレス目のメンバーではない微小藻類を含む様々な微生物において使用される形質転換方法および試薬を記載している。Allnuttらによる米国特許第6,027,900号は、光合成藻類の集光性遺伝子のためのプロモーターおよび選択マーカーとしてのストレプトアロテイコスヒンズスタヌス(Streptoalloteichus hindustanus)由来のSh ble遺伝子を用いて、真核生物の藻類、および特に、ファエオダチラムトリコルナタム(Phaeodactylum tricornutum)の遺伝子操作に使用される遺伝子融合体を開示している。細胞は、高濃度の塩(例えば、10 g/L〜35 g/L)および形質転換体の選択のためのゼオシン(Zeocin)(商標)において増殖する。そのような方法を用いる形質転換に適する微小藻類細胞は、高塩条件下で増殖しうる光合成微小藻類である。Dunahayらによる米国特許第5,661,017号は、クロロフィルC含有藻類(例えば、珪藻類)におけるマーカー発現に適する制御配列に実施可能に連結された選択マーカーを含む組換え構築物を用いてクロロフィルC含有藻類を形質転換するための方法を開示している。選択マーカーは、細菌および真菌源から単離されたマーカーを含む、任意の適するマーカーとして開示されており、ネオマイシンホスホトランスフェラーゼが好ましい。調節管理配列は、クロロフィルC含有藻類由来のいずれの制御配列も含みうり、および好ましくは、シクロテラクリプチカ(Cyclotella cryptica)由来である(例えば、C. クリプチカのアセチルCoAカルボキシラーゼ制御配列)。
【0012】
しかしながら、本発明より以前には、トラウストチトリアレス(例えば、微小藻類)のような微生物の形質転換は日常的ではなかったため、そのような方法がトラウストチトリアレス微生物の形質転換へ容易に転移可能ではない。細菌および酵母について十分開発されるようになったマーカーおよび形質転換系は、必ずしも、他の微生物に容易に適合可能であるとは限らない。実際、米国特許第5,661,017号は、「真核生物の微小藻類についての形質転換系の開発においてはほとんど成功しなかった」(1段落目の49行〜51行目)と書き留めており、一部分は、そのような微生物への外来DNAの導入することの困難性により、および一部分は、そのような形質転換に使用される適するマーカーおよびベクターがないことによる。米国特許第5,661,017号に記載される系は、特にクロロフィルC含有藻類のために開発された。なぜなら、それらの発明者らは、特に他の藻類と比較して、それらが遺伝的形質転換を受け入れられると考えたからである。同様に、光合成微小藻類に特異的である形質転換方法を教えている米国特許第6,027,900号は、たいていの藻類がいずれの型の遺伝子操作に対しても不応性であるという考えを語っている(1段落目の39行〜47行目)。細菌、酵母、昆虫および動物の細胞のために適合した系は、微小藻類に容易に適合させられなかった。それゆえ、本発明の前には、微小藻類に特異的である効果的な形質転換系についての要求が当技術分野において依然としてあった。
【0013】
さらに、トラウストチトリアレス目は、現在、ストラメノパイルにおける褐色植物藻類に分類されているが、これらの微生物はほとんどの微小藻類とは全く異なっているという当技術分野におけるある人たちによる意見がまだあり、当業者の一部には、トラウストチトリアレスのメンバーが微小藻類として正しく分類されているとは全くいえない可能性があるという意見をもっている。微小藻類であるとみなされる微生物は、進化の間、少なくとも4つの別々の時代を進化し、「微小藻類」型微生物が異なる界に配置されることになる(例えば、紅藻、緑藻および黄金色藻類(褐色植物門)は、すべて別々の界にある)。結果として、他の微小藻類において有用であると実証された形質転換系が、トラウストチトリアレスに対して有用であるとは期待されない。それゆえ、トラウストチトリアレス微生物の商業的価値にもかかわらず、遺伝子工学によりそのような微生物の最大限の可能性を利用する能力は、これまで実現されなかった。本発明の前では、本発明者らは、トラウストチトリアレス微生物の形質転換のために有用ななんのプロモーター、選択マーカーまたはベクターも認識していなかったし、また何の選択系がトラウストチトリアレスに用いられうるまたは適合しうるかについてなんの知識もなかった。
【0014】
要約すれば、トラウストチトリアレス微生物を形質転換するための方法を開発し、それにより商業的価値が高められた系統を作製するための手段を提供することの当技術分野における要求がある。さらに、トラウストチトリアレス微生物における相同的または非相同的組換えによる特定の遺伝子の突然変異または不活化のための方法を開発し、細胞代謝を変える、およびトラウストチトリアレスにおける特定の遺伝子の機能を同定するための新規な方法を提供することの当技術分野における要求がある。
【発明の概要】
【0015】
本発明の一つの態様は、以下のものからなる群より選択される核酸配列を含む単離された核酸分子に関する:(a)タンパク質がアセト乳酸シンターゼである、配列番号:15、配列番号:19、配列番号:22、および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードする核酸配列;(b)タンパク質がアセト乳酸シンターゼである、(a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列を有するタンパク質をコードする核酸配列;ならびに、(c)(a)または(b)の核酸配列と完全に相同的である核酸配列。一つの局面において、そのような核酸配列は、(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するタンパク質をコードし、かつそのタンパク質は、アセト乳酸シンターゼである。もう一つの局面において、そのような核酸配列は、(a)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するタンパク質をコードし、かつそのタンパク質は、アセト乳酸シンターゼである。さらにもう一つの局面において、そのような核酸配列は、116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする。一つの局面において、核酸配列は、配列番号:15、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードし、かつそのタンパク質はアセト乳酸シンターゼである。さらにもう一つの局面において、核酸配列は、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。
【0016】
好ましくは、上記で同定される核酸配列によりコードされるタンパク質の発現は、そのような核酸分子で形質転換されるトラウストチトリアレス目の微生物に、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を与える。この態様の一つの局面において、核酸配列は、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードする。この態様のもう一つの局面において、核酸配列は、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。
【0017】
本発明の一つの態様において、上記の核酸配列は、シゾチトリウムのアセト乳酸シンターゼをコードする。一つの局面において、シゾチトリウムのアセト乳酸シンターゼの発現は、その核酸分子で形質転換されるトラウストチトリアレス目の微生物に、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を与える。
【0018】
本発明のもう一つの態様は、転写制御配列に実施可能に連結されている、上記の単離された核酸分子のいずれかを含む組換え核酸分子に関する。本発明のもう一つの態様は、そのような組換え核酸分子に形質転換されるトラウストチトリアレス目の組換え微生物に関する。
【0019】
本発明のさらにもう一つの態様は、トラウストチトリアレス目の微生物の形質転換のための組換えベクターに関する。ベクターは、トラウストチトリアレス目の微生物に、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を与えるアセト乳酸シンターゼをコードする核酸配列を含む。アセト乳酸シンターゼは、以下のものからなる群より選択されるアミノ酸配列を有する:(a)配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および、(b)(a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列。アセト乳酸シンターゼをコードする核酸配列は、転写制御配列に実施可能に連結される。一つの局面において、組換えベクターは、発現ベクターである。もう一つの局面において、組換えベクターは、ターゲティングベクターである。他の局面において、ベクターにおける核酸配列は、(a)のアミノ酸配列と少なくとも約85 %同一である、およびもう一つの局面において、少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする。一つの局面において、核酸配列は、116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする。好ましい局面において、アセト乳酸シンターゼは、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する。もう一つの局面において、核酸配列は、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。組換えベクターにおける転写制御配列は、限定されるものではないが、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、またはトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターを含みうる。一つの局面において、ベクターは、配列番号:18、配列番号:21および配列番号:23からなる群より選択される核酸配列を含む。
【0020】
本発明のさらにもう一つの態様は、トラウストチトリアレス目の微生物の細胞を形質転換する方法に関する。方法は、(a)アセト乳酸シンターゼが(i)配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;ならびに、(ii)(i)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列からなる群より選択されるアミノ酸配列を有する、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を細胞に与えるアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子をトラウストチトリアレス目の微生物の細胞へ導入する第一段階を含む。方法は、第二段階に(b)化合物がスルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される、形質転換されていない細胞に対して阻害性である少なくとも1つの化合物を含む培地において(a)の細胞を培養することにより、組換え核酸分子で成功裡に形質転換された細胞を選択する段階を含む。一つの局面において、核酸配列は、(i)のアミノ酸配列と少なくとも約85 %同一である、およびより好ましくは少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする。一つの局面において、核酸配列は、116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする。もう一つの局面において、アセト乳酸シンターゼは、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する。好ましくは、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。さらにもう一つの局面において、核酸配列は、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される転写制御配列に実施可能に連結される。
【0021】
一つの局面において、組換え核酸分子は、タンパク質をコードする核酸配列が転写制御配列に実施可能に連結されている、細胞により産生されうるタンパク質をコードする核酸配列をさらに含む。この態様の一つの局面において、タンパク質は、ドコサヘキサエン酸(DHA)、ドコサペンタエン酸(DPA)、エイコサペンタエン酸(EPA)およびアラキドン酸(ARA)からなる群より選択される脂肪酸の合成に関係している。この態様のもう一つの局面において、タンパク質は、脂肪酸シンターゼ、脂肪酸デサチュラーゼ、脂肪酸エロンガーゼ、ポリケチドシンターゼ複合体に関係するタンパク質、およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質からなる群より選択される。一つの局面において、タンパク質は、核酸配列の配列番号:29によりコードされるω-3脂肪酸デサチュラーゼである。もう一つの局面において、タンパク質は、ポリエン脂肪酸イソメラーゼである。さらにもう一つの局面において、タンパク質は、HMG-CoAシンターゼ、HMG-CoAレダクターゼ、スクアレンシンターゼ、フィトエンシンターゼ、フィトエンデサチュラーゼ、カロテノイドシクラーゼ、カロテノイドヒドロキシラーゼ、カロテノイドケトラーゼ、ビタミンEおよびリポ酸からなる群より選択される。
【0022】
本方法のもう一つの局面において、段階(a)における組換え核酸分子は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活化されるように微生物においてターゲット核酸配列とハイブリダイズする核酸配列をさらに含む。この局面において、ターゲット核酸配列は、リパーゼ、脂肪酸酸化酵素、糖合成に関与するタンパク質、イソプレノイド経路の生成物の合成に関与するタンパク質、細胞壁成分の合成に関与するタンパク質、飽和脂肪酸合成経路に関与するタンパク質、多価不飽和脂肪酸合成経路に関与するタンパク質、ポリケチドシンターゼ複合体に関係するタンパク質、およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質からなる群より選択されるタンパク質をコードしうる。
【0023】
本方法は、核酸配列が転写制御配列に実施可能に連結されている、発現されうるタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含みうる。もう一つの局面において、方法は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活化されるように、微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列を含む、少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含みうる。もう一つの局面において、方法は、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子を細胞へ導入する段階をさらに含みうる。この局面において、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、第二のタンパク質をコードする核酸配列が転写制御配列に実施可能に連結されている、細胞により発現されうる第二のタンパク質をコードする核酸配列をさらに含む。そのような転写制御配列は、限定されるものではないが、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターを含みうる。この態様のさらなる局面において、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列をさらに含む。一つの態様において、組換え核酸分子は、核酸配列の配列番号:9を含む。
【0024】
本発明の方法において、微生物は、限定されるものではないが、トラウストチトリウム、ラビリンチュロイデス(Labyrinthuloides)、ジャポノチトリウム(Japonochytrium)、およびシゾチトリウム(Shizochytrium)を含む属由来でありうる。一つの局面において、微生物は、限定されるものではないが、シゾチトリウム属種(Schizochytrium sp.)、シゾチトリウムアグレガタム(Schizochytrium aggregatum)、シゾチトリウムリマシナム(Schizochytrium limacinum)、トラウストチトリウム属種(Thraustochytrium sp.)、トラウストチトリウムストリアタム(Thraustochytrium striatum)、トラウストチトリウムオーレウム(Thraustochytrium aureum)、トラウストチトリウムローゼウム(Thraustochytrium roseum)、およびジャポノチトリウム属種(Japonochytrium sp.)を含む種由来である。
【0025】
本方法の一つの態様において、微粒子銃、電気穿孔法、マイクロインジェクション、リポフェクション、吸着、感染、およびプロトプラスト融合からなる群より選択される方法により行われる。
【0026】
本発明のもう一つの態様は、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を微生物に与えるアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子で形質転換された、トラウストチトリアレス目の組換え微生物に関する。アセト乳酸シンターゼは、以下のものからなる群より選択されるアミノ酸配列を有する:(a)配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および、(b)(a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列。一つの局面において、核酸配列は、(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする。もう一つの局面において、アセト乳酸シンターゼは、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する。さらにもう一つの局面において、核酸配列は、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。さらにもう一つの局面において、組換え核酸分子は、配列番号:18、配列番号:21および配列番号:23からなる群より選択される核酸配列を含む。好ましくは、アセト乳酸シンターゼをコードする核酸配列は、トラウストチトリアレスの微生物において機能するプロモーターに実施可能に連結される。一つの局面において、アセト乳酸シンターゼをコードする核酸配列は、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される転写制御配列に実施可能に連結される。一つの態様において、組換え核酸分子は、第一のタンパク質をコードする核酸配列が転写制御配列に実施可能に連結されている、微生物による産生のための第一タンパク質をコードする核酸配列をさらに含む。もう一つの態様において、組換え細胞は、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子でさらに形質転換される。好ましくは、組換え核酸分子は、核酸配列の配列番号:9を含む。一つの態様において、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、第二のタンパク質をコードする核酸配列が転写制御配列に実施可能に連結されている、細胞による産生のための第二のタンパク質をコードする核酸配列をさらに含む。一つの態様において、微生物はまた、細胞による産生のためのタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子も含む。
【0027】
本発明のさらにもう一つの態様は、以下のものからなる群より選択される核酸配列を含む単離された核酸分子に関する:(a)配列番号:9のヌクレオチド441位〜894位;(b)核酸配列が少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、配列番号:9のヌクレオチド441位〜894位の完全長に対して配列番号:9のヌクレオチド441位〜894位と少なくとも約95 %同一である核酸配列;および(c)(a)または(b)のポリヌクレオチドと完全に相同的である核酸配列を含む単離されたポリヌクレオチド。好ましくは、単離された核酸分子は、配列番号:4および配列番号:9のヌクレオチド441位〜894位からなる群より選択される核酸配列を含む。
【0028】
本発明のさらにもう一つの態様は、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択されるプロモーターに実施可能に連結されるブレオマイシン結合タンパク質をコードする核酸配列を含んでいる、トラウストチトリアレス目の微生物の形質転換のための組換えベクターに関する。一つの局面において、トラウストチトリアレスのアセト乳酸シンターゼプロモーターは、配列番号:14のヌクレオチド1位〜1259位を含む。一つの局面において、α-チューブリンプロモーターは、配列番号:9のヌクレオチド441位〜894位、および核酸配列が少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、配列番号:9のヌクレオチド441位〜894位の完全長に対して配列番号:9のヌクレオチド441位〜894位と少なくとも約95 %同一である核酸配列からなる群より選択される核酸配列を含む。もう一つの局面において、トラウストチトリアレスPKS系由来のプロモーターは、配列番号:34または配列番号:34内に含まれる核酸配列を含み、該プロモーターは少なくとも基礎的なPKSプロモーター転写活性をもつ。もう一つの局面において、組換えベクターは、配列番号:8および配列番号:9からなる群より選択される核酸配列を含む。
【図面の簡単な説明】
【0029】
【図1】組換えプラスミドpTUBZEO-11の構築物を示す。
【図2】組換えプラスミドpTUBZEO 11-2の構築物を示す。
【図3A】組換えプラスミドpMON50200の構築物を示す。
【図3B】組換えプラスミドpMON50201の構築物を示す。
【図3C】組換えプラスミドpMON50202の構築物を示す。
【図3D】組換えプラスミドpMON50203の構築物を示す。
【発明を実施するための形態】
【0030】
発明の詳細な説明
本発明は、トラウストチトリアレス目の微生物を遺伝的に形質転換するための方法および関連した材料を含む。本発明の組換え構築物の形質転換体としての使用のために本明細書に開示される単細胞微生物のすべての株は、一般的にトラウストチトリドとも呼ばれうるのだが、トラウストチトリアレス目のメンバーである。本発明により、句「トラウストチトリド」、「トラウストチトリアレス微生物」および「トラウストチトリアレス目の微生物」は、交換可能に用いられうる。本発明者らは、シゾチトリウムまたは他のいずれのトラウストチトリアレス微生物についての形質転換系を記載する先行報告を一つも認識していない。本明細書に記載される形質転換系は、外来遺伝子をトラウストチトリアレス目の微生物へ導入し、それにより高められた商業的価値をもつ系統を作製するための手段を提供するために用いられうる。さらに、本発明は、相同的または非相同的組換えによる特定の遺伝子の変異または不活化を可能にし、細胞の代謝を変えるための、およびトラウストチトリアレス微生物における特定の遺伝子の機能を同定するための新規な方法を提供する。
【0031】
より具体的に言うと、本発明者らは、2つの型の形質転換ベクターの使用により、シゾチトリウム属のトラウストチトリアレス微生物(目:トラウストチトリアレス;科:トラウストチトリアシエ;属:シゾチトリウム)の遺伝的形質転換を実証した。これらのベクターは、標準的方法により細胞へ導入され、続いて、選択化合物の存在下で増殖するそれらの能力に基づいて組換え細胞が同定および単離されうる。本発明者らは、これらのベクターの有効性を、微粒子銃によりそれらを導入することによって実証したが、そのベクターを導入しうる他の手段もまた使用されうり(例えば、電気穿孔法)、それらは当技術分野において公知であり、本発明に含まれるものとする。
【0032】
一つの形質転換ベクターに関し、本明細書ではpTUBZEO 11-2と表される組換えベクターによって本明細書に例証されているのだが、ストレプトアロテイコスヒンズスタヌス由来のble遺伝子(「ブレオマイシン結合タンパク質」をコードする)がシゾチトリウムチューブリン遺伝子プロモーターから下流に配置されたキメラの遺伝子が作製された。SV40ターミネーターは、この構築物においてble遺伝子から下流に配置された。このベクターは、シゾチトリウムにおいてble遺伝子の発現を可能にし、それにより、適当なレベルで増殖培地に含まれる場合に野生型細胞に対して毒性があるゼオシン(Zeocin)(商標)および関連化合物に対する耐性を与える。この構築物におけるble遺伝子およびSV40ターミネーターは、pSV40/Zeoと名付けられている商業的に入手可能なベクターであり、Invitrogen Corporation(Carlsbad, CA)(Technical Manual 180202, Version B, "ZeoCassette Vectors"; Invitrogen Corporation, 1600 Faraday Ave., Carlsbad, CA 92008)から取得された。チューブリン遺伝子プロモーターは、ポリメラーゼ連鎖反応法により単離された;その反応に使用されるプライマーの1つは、ランダムシゾチトリウムcDNAシーケンシングプロジェクトにより得られた配列データに基づいた。pTUBZEO 11-2の地図は、図2に示され、pTUBZEO 11-2のヌクレオチド配列は、配列番号:9により表されている。このベクターでのシゾチトリウムの形質転換は、シゾチトリウムゲノムへ組み込まれたベクター配列の存在を検出するためのポリメラーゼ連鎖反応法およびサザンブロット分析の使用により確認された。
【0033】
ble遺伝子は、細菌、非トラウストチトリド微小藻類、真菌、原生動物、植物、および動物細胞を含む、様々な生物体の遺伝的形質転換の選択マーカーとして以前より研究者により使用されていた(例えば、米国特許第6,027,900号;Lumbrerasら、1998, Plant J. 14:441-447; Roheら、1996, Curr. Genet. 29:587-590; Messinaら、1995, Gene 165:213-217; Guerreroら、1992, Appl. Microbiol. Biotechnol. 36:759-762; Perezら、1989, Plant Mol. Biol. 13:365-373; Gatignoら、1990 Gene 91:35-41を参照)。ble遺伝子は、ブレオマイシン、フレオマイシン、およびゼオシン(Zeocin)(商標)を含むいくつかの抗生物質に対する耐性を与える「ブレオマイシン結合タンパク質」をコードする(Drocourtら、1990, Nucleic Acids Res. 18:4009)。この遺伝子はInvitrogen Corporationから商業的に入手可能であり、本発明者らがシゾチトリウム形質転換ベクターpTUBZEO 11-2を作製するために使用した遺伝子の源であった。しかしながら、本発明者らが、トラウストチトリドにおいてble遺伝子の発現を可能にする様式でその遺伝子がトラウストチトリドのプロモーターに付着している形質転換ベクターを作製した最初の者であると考えられる。
【0034】
形質転換ベクターの第二セットは、本発明者らがシゾチトリウムのゲノムライブラリーから単離した、アセト乳酸シンターゼ遺伝子(als)のインビトロでの部位特異的突然変異誘発により作製された。これらの突然変異は、トラウストチトリアレス目の微生物が感受性であるスルホメツロンメチルおよび他のスルホニル尿素化合物、加えてイミダゾリノン類阻害剤およびピリミジニルオキシ安息香酸に対して感受性が大いに弱くなるように、そのコードされた酵素(ALS)のアミノ酸配列を変化させる。スルホメツロンメチル(SMM)のようなスルホニル尿素化合物は、それらが様々な生物体由来の酵素アセト乳酸シンターゼ(ALS)を結合して不活化することができるため、しばしば、細胞に対し毒性がある。ALSは、アミノ酸のバリン、ロイシン、およびイソロイシンの生合成の第一段階を触媒する。イミダゾリノン、トリアゾロピリミジン、および他の化合物もまた、ある特定の生物体由来のALSを結合して不活化することが示された。他の生物体由来のアセト乳酸シンターゼ(アセトヒドロキシ酸シンターゼとしても知られている)をコードする遺伝子の変異体型は、酵母および植物の形質転換のための選択マーカーとして以前は使用されていた(Annu. Rev. Plant Physiol. Plant Mol. Biol. 40:441-470, 1989)。しかしながら、シゾチトリウムもしくは他のいずれのトラウストチトリアレスのメンバー由来のals遺伝子の配列もしくは性質、またはスルホニル尿素、イミダゾリノン、もしくはピリミジニルオキシ安息香酸化合物に対する耐性を与えるための変異体トラウストチトリアレスals遺伝子の使用を記載する報告は、本発明の前にはない。それどころか、本発明者らの知るかぎりでは、スルホメツロンメチルを含むこれらの選択薬剤に対するトラウストチトリアレス微生物の感受性について発表された報告でさえ一つもなかったため、そのような選択マーカーがトラウストチトリド形質転換系において使用に適しているかどうかさえ、本発明の前には知られていなかった。既知のals遺伝子と実質的相同性をもつ遺伝子は様々な生物体において見出されるが、アセト乳酸の合成を触媒することができる酵素をコードしていないことは、注目すべきである(Biochimica et Biophysica Acta 1385:401-419, 1998)。それゆえに、クローニングされたals相同体が実際にALSをコードすることが明らかであったとは思われない。クローニングされたシゾチトリウム遺伝子が本当のals遺伝子であったかどうかを明確に決定するために、本発明者らは、形質転換実験を通して、変異した推定上のシゾチトリウムals遺伝子の発現とスルホメツロンメチル耐性との正の相関性を実証した。
【0035】
本発明者らは、変異体als遺伝子を含む3つの異なる形質転換ベクターを作製した:一つの変異体als遺伝子は、トリプトファンの代わりに595位にバリンを有する酵素をコードする(プラスミドpMON50201、またはALSmut1-7)、もう一つは、プロリンの代わりに192位にグルタミンをコードする(プラスミドpMON50202、またはALSmut2-2)、および3つ目は、これらの変異の両方を含む(プラスミドpMON50203、またはALSmut3-5)。これらのベクターにおいて、組換え変異体als遺伝子の発現は、天然のals遺伝子プロモーターおよびターミネーターの制御下にある。これらのベクターの地図は、野生型シゾチトリウムals遺伝子を含むベクター(プラスミドpMON50200、またはAE-5)といっしょに、図3A〜図3Dに示されている。これらの変異体ALSコード化ベクターによるシゾチトリウムの形質転換は、シゾチトリウムゲノムへ組み込まれたベクター配列の存在を検出するためのポリメラーゼ連鎖反応法およびサザンブロット分析の使用により確認された。下で詳細に記載されているように、本発明者らがals遺伝子についての完全な配列を同定したからには、他の変異も、下で明細に記されているが、作製されうる。それゆえ、記載される変異体als遺伝子は、例示的なものであり、可能な変異体のすべてを含めるものではない。
【0036】
本発明の形質転換系は、同時形質転換により外来遺伝子をトラウストチトリアレス細胞へ導入するために用いられた。これらの場合において、外来遺伝子は、様々なシゾチトリウムのプロモーターと適当なターミネーター(例えば、SV40またはシゾチトリウム遺伝子ターミネーター領域)との間に配置された。例えば、本発明者らは、シゾチトリウムにおいてドコサヘキサエン酸のレベルを増加させるために、本明細書で配列番号:29に表されている、線虫カエノルハブディティスエレガンス(Caenorhabditis elegans)由来のω-3脂肪酸デサチュラーゼをコードする合成遺伝子を作製および導入した。配列番号:30は、配列番号:29によりコードされるデサチュラーゼのアミノ酸配列を表す。外来遺伝子を含む発現カセットはまた、選択マーカー含有形質転換ベクター内への直接的封入によりトラウストチトリアレス細胞へ導入されうる。
【0037】
さらに、本発明者らはまた、変異体ALSコード化ベクターを用いてシゾチトリウムにおいて相同的組換えが生じることを実証し、特定の天然シゾチトリウム遺伝子を不活化または変異するために組換え手段を用いることの可能性を示している。
【0038】
本明細書に記載されるトラウストチトリアレスプロモーター配列に関して、トラウストチトリアレス目のメンバーについて報告されたヌクレオチドおよびタンパク質の配列のすべてに対する配列データベース検索(GenBank)により、本発明の時点では、シゾチトリウムまたはトラウストチトリアレスの他のいずれのメンバー由来のプロモーター配列も報告されていなかったことが示される。いずれかのシゾチトリウム属種から報告された唯一の遺伝子は、S. アグレガタム(S. aggregatum)の5SリボソームRNAについてである(GenBankアクセッション番号X06104およびM13616)。5Sおよび18SリボソームRNA配列は、トラウストチトリアレスのメンバー、ウルケニア(Ulkenia)種、ならびにラビリンチュロイデス(Labyrinthuloides)属およびトラウストチトリウム(Thraustochytrium aureum)について報告されているが、これは本発明にはなんの関係もない。トラウストチトリウムオーレウム由来の「推定T3/T7様RNAポリメラーゼ」遺伝子の部分的コード領域が記載されているが(Nucleic Acids Research 15:648-654, 1996)、この遺伝子についてのプロモーター配列は報告されなかった。
【0039】
本発明は、トラウストチトリアレス目の微生物へ、対象となるいずれの遺伝子または他のヌクレオチド配列を導入するために用いられうる。そのようなヌクレオチド配列は、限定されるものではないが、脂肪酸(例えば、脂肪酸:ドコサヘキサエン酸(DHA)、ドコサペンタエン酸(DPA)、エイコサペンタエン酸(EPA)およびアラキドン酸(ARA))の合成に関係するタンパク質(例えば、酵素)をコードする核酸を含む。そのようなタンパク質は、限定されるものではないが、脂肪酸シンターゼ、脂肪酸デサチュラーゼ、および脂肪酸エロンガーゼ、加えて、ポリケチドシンターゼ複合体に関係するタンパク質および/またはそのような脂肪酸のリン脂質もしくはトリアシルグリセロール分子への取り込みに関係するタンパク質を含む。例えば、本発明は、ドコサペンタエン酸(DPA)のω-3不飽和化により細胞におけるドコサヘキサエン酸(DHA)のレベルを増加させるための試みにおいてシゾチトリウムへ様々な脂肪酸デサチュラーゼをコードする遺伝子を導入するために用いられた。もう一つの例として、シゾチトリウムにおいて、紅藻、プティロタ(Ptilota)由来の推定ポリエン脂肪酸イソメラーゼの発現もまた実証された。シゾチトリウムポリケチドシンターゼ複合体(すなわち、ポリケチドシンターゼ系)をコードする遺伝子は、GenBankアクセッション番号AF378327(ORFA)、AF378328(ORFB)およびAF378329(ORFC)として寄託されている。
【0040】
本発明はまた、トラウストチトリアレス微生物へイソプレノイド生合成経路に関係するタンパク質をコードする遺伝子および他のヌクレオチド配列を導入するためにも有用である。そのようなタンパク質は、限定されるものではないが、HMG-CoAシンターゼおよびHMG-CoAレダクターゼを含む。他の適するタンパク質は、限定されるものではないが、様々なステロイド化合物および様々なカロテノイド化合物を含むイソプレノイドサブユニット由来の分子の合成に関係するタンパク質を含む。様々なカロテノイド化合物の合成に関係するタンパク質は、限定されるものではないが、スクアレンシンターゼ、フィトエンシンターゼ、フィトエンデサチュラーゼ、ならびに様々なカロテノイドのシクラーゼ、ヒドロキシラーゼおよびケトラーゼを含む。
【0041】
本発明はまた、限定されるものではないが、ビタミンEおよびリポ酸を含む抗酸化化合物の合成に関係するタンパク質をコードする1つまたは複数の核酸配列をトラウストチトリアレスへ導入するためにも有用である。
【0042】
さらに、本発明は、遺伝子を不活化するまたは欠失させる(すなわち、「ノックアウト」または「ターゲットされた遺伝子破壊」)ために、トラウストチトリアレス微生物へ任意の遺伝子または他のヌクレオチドを導入するために用いられうる。遺伝子の不活化または欠失は、典型的には、微生物の商業的価値を高めることの目的のために用いられる。例えば、飽和および多価不飽和脂肪酸合成経路の酵素をコードする遺伝子(またはそのような遺伝子の発現を制御する核酸)を除去することが望ましい場合がある。もう一つの局面において、トラウストチトリアレス微生物により産生される他の価値ある化合物の分解に関与しているタンパク質または望ましい化合物の価値を他の方法で低めるタンパク質をコードする遺伝子を抑制もしくはノックアウトすることが望ましい場合がある。例えば、リパーゼ、脂肪酸酸化酵素、および不快な風味または匂いをもつタンパク質をコードする遺伝子が、本発明による望ましいノックアウトのターゲットでありうる。さらにもう一つの局面において、合成が、対象となる他の分子と競合している化合物の合成に付随しているタンパク質をコードする遺伝子をノックアウトすることが望ましい場合がある。例えば、そのような遺伝子は、限定されるものではないが、炭水化物生合成に関与しているタンパク質をコードする遺伝子、イソプレノイド経路の様々な生成物(例えば、ステロールまたは特定のカロテノイド化合物)の合成に関与しているタンパク質をコードする遺伝子、および細胞壁成分の合成に関与しているタンパク質をコードする遺伝子を含む。例として、高度不飽和脂肪酸(HUFA)の産生における役割を評価するために、シュワネラ(Shewanella)由来のポリケチドシンターゼ遺伝子と相同的である遺伝子を不活化する試みにおいて、本発明の使用によりシゾチトリウム細胞へ遺伝子が導入された。実施例6により例証されているように、本発明はまた、化合物に関連している製品の経済面または受け入れやすさを向上させるために、脂肪酸、カロテノイド、ステロール、ビタミン、または他の化合物の産生に関与している天然の遺伝子を不活化する、欠失させる、または変異させるために用いられうる。上記で考察されているが、ある態様においては、与えられたタンパク質の産生を増強することが望ましいと考えられるが、別の態様においては、同じタンパク質の産生を抑制することが望ましいと考えられる。そのような決定は、特定の微生物についての所与の使用および生産目標に基づいている。本発明はまた、シゾチトリウムにおいて遺伝的組換えの過程を測定するためにも有用である。
【0043】
トラウストチトリアレスへの導入のために有用な他の遺伝子および核酸分子は、当業者にとって明らかであると思われ、すべてのそのような遺伝子および分子は、本発明に含まれるものとする。
【0044】
本発明の様々な態様は、最初は本発明のトラウストチトリアレスals遺伝子および/またはALSタンパク質に関して下に記載されている。しかしながら、用語および方法の一般的な定義は、下記の他の遺伝子、核酸およびタンパク質の考察に適用されるものであることは理解されるべきである。
【0045】
本発明は、トラウストチトリド微生物の形質転換のための、組換え構築物での使用に適する選択マーカーをコードする核酸配列の同定、単離および作製に一部基づいている。そのような選択マーカーは、本発明の組換え構築物で成功裡に形質転換された微生物の選択を可能にする。本発明によるトラウストチトリアレスの形質転換のために有用な一つの選択マーカーは、トラウストチトリアレスのアセト乳酸シンターゼ(すなわち、ALS)である。好ましくは、アセト乳酸シンターゼは、スルホニル尿素化合物、イミダゾリノン類阻害剤、および/またはピリミジニルオキシ安息香酸による阻害に対する耐性があるように改変されている、変異されている、または別な方法で選択されている(すなわち、そのようなALSは、天然に存在するアセト乳酸シンターゼの相同体である)。
【0046】
本発明によると、アセト乳酸シンターゼは、天然に存在するアセト乳酸シンターゼの完全長タンパク質、融合タンパク質、またはいずれの相同体も含む、アセト乳酸シンターゼの生物学的活性をもつタンパク質である。アセト乳酸シンターゼの相同体は、1個または少数に限定されるものではないが、少なくとも1個または少数のアミノ酸が欠失(例えば、ペプチドまたは断片のようなタンパク質の切断型)、挿入、逆位、置換および/または誘導体化(例えば、グリコシル化、リン酸化、アセチル化、ミリストイル化、フェニル化、パルミチン化、アミド化および/またはグリコシルホスファチジルイノシトールの付加)されている点において天然に存在するアセト乳酸シンターゼと異なるタンパク質を含む。天然に存在するアセト乳酸シンターゼの好ましい相同体は、下で詳細に記載されている。
【0047】
本発明による、単離されたアセト乳酸シンターゼのような単離されたタンパク質は、それの本来の環境から取り出された(すなわち、人の操作を受けた)タンパク質であり、例えば、精製されたタンパク質、部分的に精製されたタンパク質、組換え的に産生されたタンパク質、および合成的に作製されたタンパク質を含みうる。そのようなものとして、「単離された」とは、そのタンパク質が精製された程度を反映しない。好ましくは、本発明の単離されたアセト乳酸シンターゼは組換え的に産生されている。「トラウストチトリアレスのアセト乳酸シンターゼ」は、トラウストチトリアレス微生物由来の、または他の方法として天然に存在するトラウストチトリアレス微生物由来のアセト乳酸シンターゼの構造(例えば、配列)の知識から作製されたアセト乳酸シンターゼ(天然に存在するアセト乳酸シンターゼの相同体を含む)を指す。換言すれば、トラウストチトリアレスのアセト乳酸シンターゼは、本明細書で詳細に記載されているように、トラウストチトリアレス微生物由来の天然に存在するアセト乳酸シンターゼの構造および機能をもつ、またはトラウストチトリアレス微生物由来の天然に存在するアセト乳酸シンターゼの生物学的活性のある(すなわち、生物学的活性をもつ)相同体である任意のアセト乳酸シンターゼも含む。それとして、トラウストチトリアレスのアセト乳酸シンターゼは、精製された、部分的に精製された、組換え型の、変異型/改変型の、および合成的なタンパク質を含みうる。
【0048】
一般的に、タンパク質の生物学的活性または生物学的作用は、インビボ(すなわち、そのタンパク質の本来の生理学的環境)またはインビトロ(すなわち、実験室条件下)で測定または観察される場合、そのタンパク質の天然存在型とされるタンパク質により示されるまたは行われる任意の機能を指す。例えば、アセト乳酸シンターゼの生物学的活性はアセト乳酸シンターゼ酵素的活性を含む。相同体または模倣体においてのような(下記で考察されている)タンパク質の改変は、天然に存在するタンパク質と同じ生物学的活性をもつタンパク質、または天然に存在するタンパク質と比較して減少または増加した生物学的活性をもつタンパク質を結果として生じうる。タンパク質発現における減少またはタンパク質の活性における減少を結果として生じる改変は、タンパク質の不活化(完全または部分的)、下方制御、または減少した作用として言及されうる。同様に、タンパク質発現における増加またはタンパク質の活性における増加を結果として生じる改変は、タンパク質の増幅、過剰産生、活性化、増強、上方制御または増加した作用として言及されうる。
【0049】
本発明のアセト乳酸シンターゼに関して、アセト乳酸シンターゼ相同体に存在する改変は、天然に存在するアセト乳酸シンターゼと比較して実質的に変化させない、または少なくとも天然に存在するタンパク質と比較してシンターゼの基礎的な生物学的活性を実質的に減少させないことが好ましい。しかしながら、そのような相同体は、天然に存在するタンパク質と比較してある特定の化合物による阻害に対する減少した感受性のような、天然存在型と比較してタンパク質の機能的または酵素的活性以外の特性における相違をもちうる。好ましくは、天然に存在するアセト乳酸シンターゼの相同体は、その相同体が派生した天然に存在するアセト乳酸シンターゼと比較して、天然に存在するアセト乳酸シンターゼを結合して不活化する、化合物に対する低下した(すなわち、減少した、小さくなった)感受性をもつ。例えば、スルホメツロンメチル(SMM)のようなスルホニル尿素化合物は、それらがアセト乳酸シンターゼ(ALS)を結合して不活化することができるため、しばしば、細胞への毒性がある。イミダゾリノン、トリアゾロピリミジン、および他の類似した化合物(一般的に本明細書ではイミダゾリノン類阻害剤と呼ばれる)もまた、ALSを結合して不活化することが示された。それゆえ、天然に存在するアセト乳酸シンターゼの相同体は、好ましくは、アセト乳酸シンターゼの酵素活性を維持すると同時に、スルホニル尿素化合物に対する、加えてイミダゾリノン類阻害剤に対する(例えば、そのような阻害剤についての破壊された結合部位または阻害剤への低下した親和性をもつ結合部位を有することにより)、およびピリミジニルオキシ安息香酸に対する低下した感受性をもつ。
【0050】
本明細書に用いられる場合、「アセト乳酸シンターゼ生物学的活性」をもつ、または「アセト乳酸シンターゼ」と呼ばれるタンパク質は、アミノ酸のバリン、ロイシン、およびイソロイシンの生合成における第一段階を触媒するタンパク質を指す。より具体的に言うと、完全長タンパク質、切り詰められたタンパク質、融合タンパク質、および相同体を含む本発明の単離されたアセト乳酸シンターゼは、ピルビン酸からアセト乳酸の合成を触媒するタンパク質の能力により単刀直入式に同定されうる。アセト乳酸シンターゼ生物学的活性は、当業者により、任意の適するインビトロまたはインビボの酵素活性アッセイ法により評価されうる。
【0051】
一つの態様において、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼである(すなわち、アセト乳酸シンターゼ生物学的活性をもつ)、配列番号:15、配列番号:19、配列番号:22および配列番号:24の群より選択されるアミノ酸配列と、そのような配列の任意の少なくとも約600個のアミノ酸に対して、少なくとも約65 %同一であるアミノ酸配列を有する。より好ましくは、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも約600個のアミノ酸に対して、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと、少なくとも約70 %同一である、より好ましくは少なくとも約75 %同一である、およびよりいっそう好ましくは少なくとも約80 %同一である、およびよりいっそう好ましくは少なくとも約85 %同一である、およびよりいっそう好ましくは少なくとも約90 %同一である、およびよりいっそう好ましくは少なくとも約95 %同一である、およびよりいっそう好ましくは少なくとも約96 %同一である、およびよりいっそう好ましくは少なくとも約97 %同一である、およびよりいっそう好ましくは少なくとも約98 %同一である、およびよりいっそう好ましくは少なくとも約99 %同一であるアミノ酸配列を有する。
【0052】
もう一つの態様において、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼである(すなわち、アセト乳酸シンターゼ生物学的活性をもつ)、配列番号:15、配列番号:19、配列番号:22、配列番号:24の群より選択されるアミノ酸配列と、そのような配列のいずれかの少なくとも95個より多くのアミノ酸に対して、少なくとも約75 %同一であるアミノ酸配列を有する。より好ましくは、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも95個より多くのアミノ酸に対して、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと、少なくとも約80 %同一である、およびよりいっそう好ましくは少なくとも約85 %同一である、およびよりいっそう好ましくは少なくとも約90 %同一である、およびよりいっそう好ましくは少なくとも約95 %同一である、およびよりいっそう好ましくは少なくとも約96 %同一である、およびよりいっそう好ましくは少なくとも約97 %同一である、およびよりいっそう好ましくは少なくとも約98 %同一である、およびよりいっそう好ましくは少なくとも約99 %同一であるアミノ酸配列を有する。よりいっそう好ましくは、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも100個より多くのアミノ酸、およびより好ましくは125個、およびより好ましくは150個、およびより好ましくは175個、およびより好ましくは200個、およびより好ましくは225個、およびより好ましくは250個、およびより好ましくは275個、およびより好ましくは300個、およびより好ましくは325個、およびより好ましくは350個、およびより好ましくは375個、およびより好ましくは400個、およびより好ましくは425個、およびより好ましくは450個、およびより好ましくは475個、およびより好ましくは500個、およびより好ましくは525個、およびより好ましくは550個、およびより好ましくは575個のアミノ酸に対して配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと上記の参照されたパーセントの同一性のいずれかをもつアミノ酸配列を有する。
【0053】
本明細書に用いられる場合、別なふうに明記されない限り、同一性の割合(%)の参照は、以下のものを用いて行われる相同性の評価を指す:(1)標準デフォルトパラメーターを用いるアミノ酸検索のためのblastp、および核酸検索のためのblastnを用いるBLAST 2.0ベーシックBLAST相同性検索であり、クエリ配列はデフォルトにより低複雑度領域についてフィルタリングされる(Altschul, S.F., Madden, T.L., Schaaffer, A.A., Zhang, J., Zhang, Z., Miller, W. & Lipman, D.J. (1997)「Gapped BLAST and PSI-BLAST:a new generation of protein database search programs.」Nucleic Acids Res. 25:3389-3402に記載されており、参照として完全に本明細書に組み入れられている);(2)BLAST 2アラインメント(下記のパラメーターを用いる);(3)および/または標準デフォルトパラメーターでのPSI-BLAST(位置特異的反復(Position-Specific Iterated)BLAST)。BLAST 2.0ベーシックBLASTとBLASTとの間の標準パラメーターにおけるいくらかの相違のために、2つの特定の配列がBLAST 2プログラムを用いて有意な相同性をもつとして認識されえたとしても、クエリ配列としてその配列のうちの1つを用いるBLAST 2.0ベーシックBLASTにおいて行われる検索では二番目の配列をトップ整合において同定しない可能性があることを留意する。さらに、PSI-BLASTは、「プロファイル」検索の自動化した使いやすいバージョンを提供しており、配列相同体を捜すための感度の高い方法である。プログラムは、まず、ギャップド(gapped)BLASTデータベース検索を行う。PSI-BLASTプログラムは、位置特異的スコア行列を構成するために戻されるいずれの有意なアラインメントからの情報も使用し、データベース検索の次のラウンドのためにクエリ配列を取り替える。それゆえ、これらのプログラムのいずれか1つを使用することにより同一性の割合が決定されうることは理解されるべきである。
【0054】
2つの特定の配列は、参照として完全に本明細書に組み入れられる、TatusovaおよびMadden, (1999), 「Blast 2 sequences-a new tool for comparing protein and nucleotide sequences」, FEMS Microbiol Lett. 174:247-250に記載されているように、BLAST 2配列を用いてお互いに整列しうる。BLAST 2配列アラインメントは、その結果生じるアラインメントにギャップの導入(欠失および挿入)を許容して2つの配列間でギャップドBLAST検索(BLAST 2.0)を行うBLAST 2.0アルゴリズムを用いるblastpまたはblastnにおいて行われる。本明細書での明瞭さを目的として、BLAST 2配列アラインメントは、以下の通り、標準デフォルトパラメーターを用いて行われる。
【0055】
0 BLOSUM62行列を用いる、blastnについて:
一致についての報酬 = 1
不一致についてのペナルティ = -2
オープンギャップ(5)および伸長ギャップ(2)ペナルティ
ギャップx_ドロップオフ(50)期待(10)ワードサイズ(11)フィルター(オン)
0 BLOSUM62行列を用いる、blastpについて:
オープンギャップ(11)および伸長ギャップ(1)ペナルティ
ギャップx_ドロップオフ(50)期待(10)ワードサイズ(3)フィルター(オン)。
【0056】
本発明のアセト乳酸シンターゼはまた、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも30個の近接するアミノ酸残基(すなわち、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの30個の近接するアミノ酸と100 %同一性をもつ30個の近接するアミノ酸残基)を含むアミノ酸配列を有するタンパク質を含みうる。好ましい態様において、本発明のアセト乳酸シンターゼは、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも50個、およびより好ましくは少なくとも75個、およびより好ましくは少なくとも100個、およびより好ましくは少なくと115個、およびより好ましくは少なくとも130個、およびより好ましくは少なくとも150個、およびより好ましくは少なくとも200個、およびより好ましくは少なくとも250個、およびより好ましくは少なくとも300個、およびより好ましくは少なくとも350個、およびより好ましくは少なくとも400個、およびより好ましくは少なくとも450個、およびより好ましくは少なくとも500個、およびより好ましくは少なくとも550個、およびより好ましくは少なくとも600個、およびより好ましくは少なくとも650個の近接するアミノ酸残基を含むアミノ酸配列を有するタンパク質を含む。そのようなタンパク質は、アセト乳酸シンターゼ生物学的活性をもつ。
【0057】
本発明により、本明細書に記載される核酸またはアミノ酸の配列に関しての用語「近接する」または「連続した」は、間断のない配列として連結されていることを意味する。例えば、第一配列が第二配列の30個の近接する(または連続した)アミノ酸を含むこととは、第一配列が、第二配列における30個のアミノ酸残基の間断のない配列と100 %同一である30個のアミノ酸残基の間断のない配列を含むことを意味する。同様に、第一配列が第二配列と「100 %同一性」をもつこととは、第一配列が、ヌクレオチドまたはアミノ酸の間にギャップを含まずに第二配列と正確に整合していることを意味する。
【0058】
もう一つの態様において、本発明のアセト乳酸シンターゼは、アセト乳酸シンターゼ相同体を含めて、相同体をコードする核酸配列が、中程度の、高度の、または非常に高度のストリンジェント性条件下で(下に記載されている)、天然に存在するアセト乳酸シンターゼをコードする核酸分子(すなわち、天然に存在するアセト乳酸シンターゼアミノ酸配列をコードする核酸鎖の相補体)と(すなわち、核酸分子と)ハイブリダイズする能力があるような、天然に存在するアセト乳酸シンターゼアミノ酸配列と十分に類似しているアミノ酸配列を有するタンパク質を含む。好ましくは、アセト乳酸シンターゼは、配列番号:15、配列番号:19、配列番号:22または配列番号:24により表されるアミノ酸配列を含むタンパク質をコードする核酸配列の相補体へ中程度の、高度の、または非常に高度のストリンジェント性条件下でハイブリダイズする核酸配列によりコードされる。よりいっそう好ましくは、本発明のアセト乳酸シンターゼは、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、または配列番号:23のヌクレオチド1260位〜3314位の相補体へ中程度の、高度の、または非常に高度のストリンジェント性条件下でハイブリダイズする核酸配列によりコードされる。そういうものとしてハイブリダイゼーション条件は、下で詳細に記載されている。本発明のアセト乳酸シンターゼをコードする核酸配列の核酸配列相補体は、アセト乳酸シンターゼをコードする鎖に相補的である核酸鎖の核酸配列を指す。与えられたアミノ酸配列をコードする二本鎖DNAは一本鎖DNAおよびその一本鎖への相補体である配列を有するそれの相補鎖を含むことは認識されているものと思われる。それとして、本発明の核酸分子は、二本鎖かまたは一本鎖かどちらでもありうり、かつアミノ酸配列の配列番号:15、配列番号:19、配列番号:22または配列番号:24をコードする核酸配列と、および/またはそのようなアミノ酸配列のいずれかをコードする核酸配列の相補体とストリンジェントなハイブリダイゼーション条件下での安定的なハイブリッドを形成するそれらの核酸分子を含む。相補性配列を導き出すための方法は当業者に知られている。アミノ酸シーケンシングおよび核酸シーケンシングのテクノロジーは完全に誤りのないものではないため、本明細書に提示される配列はせいぜい本発明のアセト乳酸シンターゼの見かけの配列を表していることを留意されたい。
【0059】
アセト乳酸シンターゼ相同体は、天然の対立遺伝子の変異または自然の突然変異の結果でありうる。本発明のアセト乳酸シンターゼ相同体はまた、限定されるものではないが、タンパク質への直接的改変、または、例えば、ランダムなもしくはターゲットされた突然変異誘発をもたらすための古典的もしくは組換えDNA技術を用いるタンパク質をコードする遺伝子への改変を含む、当技術分野において公知の技術を用いて作製されうる。アセト乳酸シンターゼをコードする核酸の、天然に存在する対立遺伝子変異体は、アミノ酸配列の配列番号:15をコードする遺伝子と本質的に同じゲノムにおける遺伝子座に見出されるが、例えば、突然変異または組換えにより引き起こされた自然の変異のためで、類似しているが同一ではない配列を有する遺伝子である。天然の対立遺伝子変異体は、典型的にはそれらが比較されることになる遺伝子によりコードされるタンパク質と同様の活性をもつタンパク質をコードする。対立遺伝子変異体の一つの部類は、同じタンパク質をコードしうるが、遺伝子コードの縮重により異なる核酸配列を有する。対立遺伝子変異体はまた、遺伝子の5'または3'の非翻訳領域における(例えば、制御領域における)改変を含む。対立遺伝子変異体は当業者によく知られている。
【0060】
本発明のアセト乳酸シンターゼタンパク質はまた、遺伝子融合体(例えば、組換えタンパク質の可溶性活性型を過剰発現するために用いられる)、変異を誘発された遺伝子(遺伝子転写および翻訳を増強するコドン改変を有する遺伝子のような)、および切り詰められた遺伝子(膜タンパク質の可溶性型を生じるために膜結合ドメインが除去された遺伝子、または特定の組換え宿主において耐容性が乏しいシグナル配列が除去された遺伝子のような)の発現産物を含む。
【0061】
本発明のタンパク質および/または相同体の最小サイズは、アセト乳酸シンターゼ生物学的活性をもつのに十分なサイズである。好ましくは、本発明のタンパク質は、少なくとも30個のアミノ酸の長さであり、およびより好ましくは少なくとも約50個、およびより好ましくは少なくとも75個、およびより好ましくは少なくとも100個、およびより好ましくは少なくとも115個、およびより好ましくは少なくとも130個、およびより好ましくは少なくとも150個、およびより好ましくは少なくとも200個、およびより好ましくは少なくとも250個、およびより好ましくは少なくとも300個、およびより好ましくは少なくとも350個、およびより好ましくは少なくとも400個、およびより好ましくは少なくとも450個、およびより好ましくは少なくとも500個、およびより好ましくは少なくとも550個、およびより好ましくは少なくとも600個、およびより好ましくは少なくとも650個、およびより好ましくは少なくとも684個のアミノ酸の長さである。タンパク質がアセト乳酸シンターゼタンパク質の部分または完全長アセト乳酸シンターゼ、望ましい場合には追加的な配列(例えば、融合タンパク質配列)を加えて含みうることにおいて、そのようなタンパク質の最大サイズに、実際的な制限以外に制限はない。
【0062】
本発明はまた、1つまたは複数の融合セグメントに付着されたアセト乳酸シンターゼ含有ドメイン(すなわち、本発明によるアセト乳酸シンターゼについてのアミノ酸配列)を含む融合タンパク質を含む。本発明で使用されるのに適する融合セグメントは、限定されるものではないが、タンパク質の安定性を向上させることができる;他の望ましい生物学的活性を提供できる;および/またはアセト乳酸シンターゼの精製(例えば、アフィニティークロマトグラフィーによる)を補助することができるセグメントを含む。適する融合セグメントは、望ましい機能をもつ(例えば、増加した安定性、可溶性、作用もしくは生物学的活性を与える;および/またはタンパク質の精製を簡単にする)任意のサイズのドメインでありうる。融合セグメントは、そのタンパク質のアセト乳酸シンターゼ含有ドメインのアミノ末端および/またはカルボキシル末端に連結されうり、アセト乳酸シンターゼの直接的回収を可能にするために切断されやすくありうる。融合タンパク質は、好ましくは、アセト乳酸シンターゼ含有ドメインのカルボキシル末端および/またはアミノ末端のいずれかに付着された融合セグメントを含むタンパク質をコードする融合核酸分子でトランスフェクションされた組換え細胞を培養することにより産生される。
【0063】
本発明はまた、アセト乳酸シンターゼの模倣体を含む。本明細書で用いられる場合、用語「模倣体」は、模倣体がしばしば、天然に存在するペプチドの基本的構造を模倣する基本的構造をもつ、および/または天然に存在するペプチドの顕著な生物学的性質をもつことから、天然に存在するペプチドの生物学的作用を模倣することができる任意のペプチドまたは非ペプチド化合物を指すために用いられる。模倣体は、限定されるものではないが、以下のものを含みうる:天然に存在するペプチドと側鎖類似性をもたないような原型由来の実質的改変をもつペプチド(そのような改変は、例えば、それの分解されやすさを減らしうる);抗イディオタイプ抗体および/もしくは触媒抗体、またはそれらのフラグメント;単離されたタンパク質の非タンパク質性部分(例えば、炭水化物構造);または、例えば、コンビナトリアルケミストリーにより同定される核酸および薬剤を含む、合成もしくは天然の有機分子。
【0064】
そのような模倣体は、当技術分野において公知の様々な方法を用いて設計、選択および/または別な方法で同定されうる。本発明において有用な模倣体または他の治療的化合物を設計するために有用なドラッグデザインの様々な方法は、Maulikら、1997, Molecular Biotechnology: Therapeutic Applications and Strategies, Wiley-Liss, Inc.に開示されており、参照として完全に本明細書に組み入れられている。アセト乳酸シンターゼ模倣体は、例えば、分子多様性ストラテジー(大量の化学的に多様な分子ライブラリーの迅速な構築を可能にする関連ストラテジーの組み合わせ)から、天然もしくは合成の化合物のライブラリー、特に化学的もしくはコンビナトリアルライブラリー(すなわち、配列またはサイズにおいて異なるが、同種のビルディングブロックを有する化合物のライブラリー)から、または合理的な、定方向もしくはランダムなドラッグデザインにより得られうる。例えば、Maulikら、前記を参照されたい。
【0065】
分子多様性ストラテジーにおいて、大量化合物ライブラリーは、例えば、生物学的、酵素的および/または化学的方法を用いて、ペプチド、オリゴヌクレオチド、炭水化物および/または合成有機分子から合成される。分子多様性ストラテジーを開発するにおいて重要なパラメーターは、サブユニットの多様性、分子サイズ、およびライブラリーの多様性を含む。そのようなライブラリーをスクリーニングすることの一般的な目的は、所望のターゲットについての高親和性リガンドを得るためにコンビナトリアル選択の逐次適用を利用すること、そのうえ、ランダムまたは定方向のいずれかの設計ストラテジーによりリード分子を最適化することである。分子多様性の方法は、Maulikら、同上において詳細に記載されている。
【0066】
Maulikらはまた、例えば、使用者が適当に選択された断片の断片ライブラリーから新規な分子を作製する過程を方向づける、定方向設計の方法;使用者が、断片およびそれらの組み合わせをランダムに変化させ、同時に候補リガンドの適応度を評価するために選択基準を適用するために遺伝子または他のアルゴリズムを用いるランダム設計;および使用者が三次元の受容体構造と小断片プローブ間の相互作用エネルギーを計算し、続いて、有利なプローブ部位について結び付けて考える、グリッドに基づく方法を開示している。
【0067】
本発明によると、アセト乳酸シンターゼは、任意のトラウストチトリアレス微生物由来、および特に、任意のシゾチトリウム微生物由来でありうる。一つの態様において、本発明の好ましいアセト乳酸シンターゼは、配列番号:15、配列番号:19、配列番号:22、配列番号:24の群より選択されるアミノ酸配列を有する。配列番号:15により表されるアミノ酸配列を有するタンパク質は、トラウストチトリアレス微生物由来の天然に存在する(すなわち野生型)アセト乳酸シンターゼであり、もっと厳密に言えば、シゾチトリウムのアセト乳酸シンターゼである。配列番号:19、配列番号:22、配列番号:24により表されるアミノ酸配列は、その結果生じた酵素が、アミノ酸配列の配列番号:15により表される天然に存在するタンパク質と比較して、スルホニル尿素化合物、加えてイミダゾリノン類阻害剤およびピリミジニルオキシ安息香酸に対して低下した感受性をもつように、改変された配列である。配列番号:19、配列番号:22および配列番号:24により表されるタンパク質が、アセト乳酸シンターゼ生物学的活性をもつことは、注目される。スルホニル尿素化合物、加えてイミダゾリノン類阻害剤およびピリミジニルオキシ安息香酸に対して低下した感受性をもつアセト乳酸シンターゼは、本発明の好ましいアセト乳酸シンターゼであり、なぜなら、そのようなシンターゼをコードする核酸配列が、選択マーカーとして本発明の組換えベクターに使用されうるからである。
【0068】
それゆえに、本発明の一つの態様は、相同体がアセト乳酸シンターゼ生物学的活性をもつ、ならびに特に、相同体がアミノ酸配列の配列番号:15により表される天然に存在するタンパク質と比較して、スルホニル尿素化合物、加えてイミダゾリノン類阻害剤およびピリミジニルオキシ安息香酸に対して低下した感受性をもつ、配列番号:15、配列番号:19、配列番号:22および配列番号:24のいずれかの任意の相同体を含む改変されたアセト乳酸シンターゼに関する。一つの局面において、そのようなアセト乳酸シンターゼ相同体は、次の位置:116G、117A、192P、200A、251K、358M、383D、592V、595W、または599Fの1つまたは複数におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質を含む。これらの位置は、酵母のアセト乳酸シンターゼにおける既知のALS変異部位と対応している(すなわち、それぞれ、116G、117A、192P、200A、251K、354M、379D、583V、586W、および590F)(MazurおよびFalco, 1989, Annu. Rev. Plant Physiol. Plant Mol. Biol. 40:441-470を参照。これは参照として完全に本明細書に組み入れられている)。他の可能性のある変異部位は、他の生物体由来のALSにおいて成功したアミノ酸変異に基づき当業者に知られることになると思われる。そのような部位のトラウストチトリアレスのALSにおいての対応する部位への適用は、本発明に含まれる。
【0069】
上記で考察されているように、本発明は、トラウストチトリド微生物の形質転換のための組換え構築物の発見および作製に一部基づいている。それゆえ、本発明の一つの態様は、トラウストチトリアレスのアセト乳酸シンターゼをコードする核酸配列、およびそれに完全に相補的な核酸配列を含む単離された核酸分子に関する。本発明のアセト乳酸シンターゼをコードする核酸分子は、上記で考察されている相同体を含むアセト乳酸シンターゼタンパク質のいずれかをコードする核酸分子を含む。より詳しくは、本発明の一つの態様は、タンパク質がアセト乳酸シンターゼである(すなわち、アセト乳酸シンターゼ生物学的活性をもつ)、配列番号:15、配列番号:19、配列番号:22、配列番号:24の群より選択されるアミノ酸配列と、そのような配列のいずれかの少なくとも約600個より多くのアミノ酸に対して、少なくとも約65 %同一であるアミノ酸配列を有するタンパク質をコードする核酸配列を含む、単離された核酸分子に関する。より好ましくは、本発明の単離された核酸分子は、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも約600個より多くのアミノ酸に対して、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと、少なくとも約70 %同一である、およびより好ましくは少なくとも約75 %同一である、およびよりいっそう好ましくは少なくとも約80 %同一である、およびよりいっそう好ましくは少なくとも約85 %同一である、およびよりいっそう好ましくは少なくとも約90 %同一である、およびよりいっそう好ましくは少なくとも約95 %同一である、およびよりいっそう好ましくは少なくとも約96 %同一である、およびよりいっそう好ましくは少なくとも約97 %同一である、およびよりいっそう好ましくは少なくとも約98 %同一である、およびよりいっそう好ましくは少なくとも約99 %同一であるアミノ酸配列をコードする核酸配列を有する。
【0070】
もう一つの態様において、本発明の単離された核酸分子は、タンパク質がアセト乳酸シンターゼである(すなわち、アセト乳酸シンターゼ生物学的活性をもつ)、配列番号:15、配列番号:19、配列番号:22、配列番号:24の群より選択されるアミノ酸配列と、そのような配列のいずれかの少なくとも95個より多くのアミノ酸に対して、少なくとも約75 %同一であるアミノ酸配列をコードする核酸配列を有する。より好ましくは、本発明の単離された核酸分子は、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも95個より多くのアミノ酸に対して、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと、少なくとも約80 %同一である、およびよりいっそう好ましくは少なくとも約85 %同一である、およびよりいっそう好ましくは少なくとも約90 %同一である、およびよりいっそう好ましくは少なくとも約95 %同一である、およびよりいっそう好ましくは少なくとも約96 %同一である、およびよりいっそう好ましくは少なくとも約97 %同一である、およびよりいっそう好ましくは少なくとも約98 %同一である、およびよりいっそう好ましくは少なくとも約99 %同一であるアミノ酸配列をコードする核酸配列を有する。
【0071】
さらにもう一つの態様において、本発明の単離された核酸分子は、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも100個より多くのアミノ酸、およびより好ましくは125個、およびより好ましくは150個、およびより好ましくは175個、およびより好ましくは200個、およびより好ましくは225個、およびより好ましくは250個、およびより好ましくは275個、およびより好ましくは300個、およびより好ましくは325個、およびより好ましくは350個、およびより好ましくは375個、およびより好ましくは400個、およびより好ましくは425個、およびより好ましくは450個、およびより好ましくは475個、およびより好ましくは500個、およびより好ましくは525個、およびより好ましくは550個、およびより好ましくは575個のアミノ酸に対して配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと上記の参照された同一性の割合のいずれかをもつアミノ酸配列をコードする核酸配列を有する。同一性の割合は、上記のように、BLAST 2.0ベーシックBLASTデフォルトパラメーターを用いて決定される。
【0072】
一つの態様において、本発明のアセト乳酸シンターゼをコードする核酸分子は、天然に存在するアセト乳酸シンターゼをコードする核酸配列の相補体と中程度のストリンジェント性条件下、およびよりいっそう好ましくは高ストリンジェント性条件下、およびよりいっそう好ましくは超高ストリンジェント性条件下でハイブリダイズする単離された核酸分子を含む。好ましくは、本発明のアセト乳酸シンターゼをコードする単離された核酸分子は、配列番号:15、配列番号:19、配列番号:22または配列番号:24により表されるアミノ酸配列を含むタンパク質をコードする核酸配列の相補体へ中程度または高度のストリンジェント性条件下でハイブリダイズする核酸配列を含む。一つの態様において、単離された核酸分子は、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、または配列番号:23のヌクレオチド1260位〜3314位により表される核酸配列の相補体へ中程度、高度または非常に高度のストリンジェント性条件下でハイブリダイズする核酸配列を含む。
【0073】
本明細書に用いられる場合、ハイブリダイゼーション条件は、類似した核酸分子を同定するために核酸分子が用いられる標準的ハイブリダイゼーション条件を指す。そのような標準的条件は、例えば、Sambrookら、Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Labs Press, 1989に開示されている。Sambrookら、同上は、参照として完全に本明細書に組み入れられている(特に、ページ9.31-9.62を参照)。さらに、ヌクレオチドのミスマッチの異なる程度を許容するハイブリダイゼーションを達成するための適当なハイブリダイゼーションおよび洗浄条件を計算するための式は、例えば、Meinkothら、1984, Anal. Biochem. 138, 267-284に開示されている;Meinkothら、同上は、参照として完全に本明細書に組み入れられている。
【0074】
より詳しくは、本明細書に参照される場合、中程度ストリンジェント性のハイブリダイゼーションおよび洗浄条件は、ハイブリダイゼーション反応において探索のために使用されることになる核酸分子と少なくとも約70 %の核酸配列同一性をもつ核酸分子の単離を可能にする条件を指す(すなわち、ヌクレオチドの約30 %またはそれ未満のミスマッチを許容する条件)。本明細書に参照される場合、高ストリンジェント性のハイブリダイゼーションおよび洗浄条件は、ハイブリダイゼーション反応において探索のために使用されることになる核酸分子と少なくとも約80 %の核酸配列同一性をもつ核酸分子の単離を可能にする条件を指す(すなわち、ヌクレオチドの約20 %またはそれ未満のミスマッチを許容する条件)。本明細書に参照される場合、超高ストリンジェント性のハイブリダイゼーションおよび洗浄条件は、ハイブリダイゼーション反応において探索のために使用されることになる核酸分子と少なくとも約90 %の核酸配列同一性をもつ核酸分子の単離を可能にする条件を指す(すなわち、ヌクレオチドの約10 %またはそれ未満のミスマッチを許容する条件)。上記で考察されているように、当業者は、ヌクレオチドミスマッチのこれらの特定のレベルを達成するための適当なハイブリダイゼーションおよび洗浄条件を計算するためにMeinkothら、同上における式を用いることができる。そのような条件は、DNA:RNAかまたはDNA:DNAかのいずれのハイブリッドが形成されることになっているかに依存して異なるものである。DNA:DNAハイブリッドについて計算される融解温度は、DNA:RNAハイブリッドについてよりも10℃低い。詳細な態様において、DNA:DNAハイブリッドについてのストリンジェントなハイブリダイゼーション条件は、6X SSC(0.9 M Na+)のイオン強度で、約20℃と約35℃の間(低い方のストリンジェント性)、より好ましくは約28℃と約40℃の間(よりストリンジェント)、およびよりいっそう好ましくは約35℃と約45℃の間(よりいっそうストリンジェント)の温度でのハイブリダイゼーションを適当な洗浄条件と共に含む。詳細な態様において、DNA:RNAハイブリッドについてのストリンジェントなハイブリダイゼーション条件は、6X SSC(0.9 M Na+)のイオン強度で、約30℃と約45℃の間、より好ましくは約38℃と約50℃の間、およびよりいっそう好ましくは約45℃と約55℃の間の温度でのハイブリダイゼーションを同様のストリンジェントな洗浄条件と共に含む。これらの値は、約100個のヌクレオチドより大きい分子、0 %ホルムアミドおよび約40 %のG + C含有量についての融解温度の計算に基づいている。または、Tmは、Sambrookら、前記、ページ9.31〜9.62に示されるように経験的に計算されうる。一般的に、洗浄条件は、できるかぎりストリンジェントであるべきであり、選択されたハイブリダイゼーション条件に対して適切であるべきである。例えば、ハイブリダイゼーション条件は、特定のハイブリッドの計算されたTmより下のおよそ20℃〜25℃である温度条件および塩の組み合わせを含みうり、洗浄条件は、典型的には、その特定のハイブリッドの計算されたTmより下のおよそ12℃〜20℃である温度条件および塩の組み合わせを含む。DNA:DNAハイブリッドでの使用に適したハイブリダイゼーション条件の一つの例は、6X SSC(50 %ホルムアミド)において約42℃で2時間〜24時間のハイブリダイゼーション、続いて約2X SSCにおいて室温での1回または複数回の洗浄を含む洗浄段階、続いてより高い温度およびより低いイオン強度での追加の洗浄(例えば、約37℃で約0.1X〜0.5X SSCにおいて少なくとも1回の洗浄、続いて約68℃で約0.1X〜0.5X SSCにおいて少なくとも1回の洗浄)を含む。
【0075】
もう一つの態様において、本発明のアセト乳酸シンターゼをコードする核酸分子は、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも30個の近接するアミノ酸残基(すなわち、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの30個の近接するアミノ酸と100 %同一性をもつ30個の近接するアミノ酸残基)を含むアミノ酸配列を有するタンパク質をコードする核酸配列を含む単離された核酸分子を含む。好ましい態様において、単離された核酸分子は、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも50個、およびより好ましくは少なくとも75個、およびより好ましくは少なくとも100個、およびより好ましくは少なくと115個、およびより好ましくは少なくとも130個、およびより好ましくは少なくとも150個、およびより好ましくは少なくとも200個、およびより好ましくは少なくとも250個、およびより好ましくは少なくとも300個、およびより好ましくは少なくとも350個、およびより好ましくは少なくとも400個、およびより好ましくは少なくとも450個、およびより好ましくは少なくとも500個、およびより好ましくは少なくとも550個、およびより好ましくは少なくとも600個、およびより好ましくは少なくとも650個の近接するアミノ酸残基を含むアミノ酸配列を有するタンパク質をコードする核酸配列を含む。そのようなタンパク質は、アセト乳酸シンターゼ生物学的活性をもつ。一つの態様において、アセト乳酸シンターゼをコードする単離された核酸分子は、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、または配列番号:23のヌクレオチド1260位〜3314位の少なくとも60個の近接するヌクレオチド、およびより好ましくは少なくとも150個、およびより好ましくは少なくとも225個、およびより好ましくは少なくと300個、およびより好ましくは少なくとも345個、およびより好ましくは少なくとも390個、およびより好ましくは少なくとも450個、およびより好ましくは少なくとも525個、およびより好ましくは少なくとも600個、およびより好ましくは少なくとも750個、およびより好ましくは少なくとも900個、およびより好ましくは少なくとも1050個、およびより好ましくは少なくとも1200個、およびより好ましくは少なくとも1350個、およびより好ましくは少なくとも1500個、およびより好ましくは少なくとも1650個、およびより好ましくは少なくとも1800個、およびよりいっそう好ましくは少なくとも1950個の近接するヌクレオチドを有する核酸配列を含む。
【0076】
本発明の特に好ましい核酸分子は、配列番号:14のヌクレオチド1260位〜3314位(配列番号:15をコードする)、配列番号:18のヌクレオチド1260位〜3314位(配列番号:19をコードする)、配列番号:21のヌクレオチド1260位〜3314位(配列番号:22をコードする)、または配列番号:23のヌクレオチド1260位〜3314位(配列番号:24をコードする)、配列番号:14、配列番号:18、配列番号:21または配列番号:23を含む。
【0077】
本発明に従って、単離された核酸分子は、それの本来の環境から取り出された(すなわち、人の操作を受けた)核酸分子であり、それの本来の環境はその核酸分子が天然で見出されるゲノムまたは染色体である。そのようなものとして、「単離された」とは、必ずしもその核酸分子が精製された程度を反映するとは限らないが、その分子が、その核酸分子が天然で見出されるゲノム全体または染色体全体を含まないことを示す。単離された核酸分子は、本明細書に記載されるアセト乳酸シンターゼ遺伝子のような遺伝子を含みうる。遺伝子を含む単離された核酸分子は、そのような遺伝子を含む染色体の断片ではなく、むしろ、その遺伝子に関連したコード領域および制御領域を含み、同じ染色体に本来見出される付加的な遺伝子を含まない。単離された核酸分子はまた、天然では特定された核酸配列に普通、隣接していない(すなわち、異種性配列である)追加的な核酸により隣接された(すなわち、その配列の5'末端および/または3'末端において)特定の核酸配列を含みうる。単離された核酸分子は、DNA、RNA(例えば、mRNA)、またはDNAもしくはRNAのいずれかの誘導体(例えば、cDNA)を含みうる。句「核酸分子」は本来、物質的な核酸分子を指し、句「核酸配列」は本来、核酸分子についてのヌクレオチドの配列をさすが、その2つの句は、交換可能に、特に、タンパク質をコードする能力がある、核酸分子または核酸配列に関して用いられうる。
【0078】
好ましくは、本発明の単離された核酸分子は、組換えDNAテクノロジー(例えば、ポリメラーゼ連鎖反応(PCR)増幅、クローニング)または化学的合成を用いて作製される。単離された核酸分子は天然の核酸分子およびその相同体を含み、これらには限定されるものではないが、天然の対立遺伝子変異体およびタンパク質生物学的活性に望ましい効果を与えるような様式でヌクレオチドが挿入、欠失、置換、および/または逆位されている改変型核酸分子が含まれる。対立遺伝子変異体およびタンパク質相同体(例えば、核酸相同体によりコードされるタンパク質)は、上記で詳細に考察されている。
【0079】
核酸分子相同体は、当業者に公知の多数の方法を用いて作製されうる(例えば、Sambrookら、同上を参照)。例えば、核酸分子は、限定されるものではないが、部位特異的突然変異誘発、変異を誘発するための核酸分子の化学的処理、核酸断片の制限酵素切断、核酸断片のライゲーション、核酸配列の選択された領域のPCR増幅および/または突然変異誘発、核酸分子の混合物を「構築する(build)」ためのオリゴヌクレオチド混合物の合成および混合群のライゲーションならびにそれらの組み合わせのような、古典的突然変異誘発技術および組換えDNA技術を含む様々な技術を用いて改変されうる。核酸分子相同体は、核酸によりコードされるタンパク質の機能についてスクリーニングすることにより、および/または野生型遺伝子とのハイブリダイゼーションにより、改変型核酸の混合物から選択されうる。
【0080】
同様に、本発明の核酸分子の最小サイズは、望ましい生物学的活性をもつタンパク質をコードするのに十分な、または天然のタンパク質をコードする核酸分子の相補的配列と安定的ハイブリッドを形成する能力がある(例えば、中程度、高度または非常に高度のストリンジェント性条件下で)プローブもしくはオリゴヌクレオチドプライマーを形成するのに十分なサイズである。そのようなものとして、そのようなタンパク質をコードする核酸分子のサイズは、核酸成分および核酸分子と相補的配列間の相同性または同一性の割合、加えて、ハイブリダイゼーション条件それ自体(例えば、温度、塩濃度、およびホルムアミド濃度)に依存しうる。オリゴヌクレオチドプライマーまたはプローブとして用いられる核酸分子の最小サイズは、典型的には、核酸分子がGCリッチである場合には長さが少なくとも約12個〜約15個のヌクレオチド、ATリッチな場合には長さが少なくとも約15個〜約18個の塩基である。核酸分子がタンパク質コード配列(例えば、アセト乳酸シンターゼコード配列)の部分または完全長タンパク質をコードする核酸配列を含みうることにおいて、本発明の核酸分子の最大サイズに、実際的制限以外に制限はない。
【0081】
本発明の一つの態様は、トラウストチトリアレス微生物の形質転換のために用いられうる組換えベクターを含む。本発明により、組換えベクターは、選り抜きの核酸配列を操作するための、およびそのような核酸配列を宿主細胞へ導入するための道具として用いられる設計された(すなわち、人工的に作製された)核酸分子である。それゆえ、組換えベクターは、クローニング、シーケンシング、ならびに/または組換え細胞を形成するために選り抜きの核酸配列を宿主細胞へ発現することおよび/もしくは送達することによるような選り抜きの核酸配列を別なふうに操作することにおいて使用するのに適している。そのようなベクターは、典型的には、本来は送達されうる核酸配列に隣接して見出されない核酸配列である異種性核酸配列を含むが、ベクターはまた、本来より本発明の核酸分子に隣接して見出される制御核酸配列(例えば、プロモーター、非翻訳領域)を含みうる(下記で詳細に考察されている)。ベクターは、RNAまたはDNAのどちらでも、原核または真核のどちらでもありうり、典型的にはプラスミドである。ベクターは、染色体外エレメント(例えば、プラスミド)として維持されうる、またはそれは組換え微生物の染色体へ組み込まれうる。ベクター全体は宿主細胞内のしかるべき所に残りうる、またはある特定の条件下で、プラスミドDNAが除去されて、本発明の核酸分子をあとに残しうる。組み込まれた核酸分子は、染色体のプロモーター制御下、本来のまたはプラスミドのプロモーター制御下、またはいくつかのプロモーター制御の組み合わせ下でありうる。核酸分子の単一または複数のコピーが染色体へ組み込まれうる。本発明の組換えベクターは、トラウストチトリアレスのアセト乳酸シンターゼをコードする核酸配列(天然のタンパク質または相同体)またはble遺伝子をコードする核酸配列(下に記載されている)のような、本発明によるトラウストチトリアレス微生物のための少なくとも1つの選択マーカーを含む。本明細書に用いられる場合、句「組換え核酸分子」は、本来、宿主細胞へクローニングされうる、操作されうる、形質転換されうる核酸配列(すなわち、挿入断片)がライゲーションされている組換えベクターを指すために用いられる。
【0082】
典型的には、組換えベクター、およびゆえに組換え核酸分子は、1つまたは複数の転写制御配列へ実施可能に連結される本発明の少なくとも1つの核酸分子を含む。本明細書に用いられる場合、句「組換え分子」または「組換え核酸分子」は、本来、転写制御配列へ実施可能に連結される核酸分子または核酸配列を指すが、そのような核酸分子が本明細書で考察されているような組換え分子である場合、句「核酸分子」と交換可能に用いられうる。本発明により、句「実施可能に連結される」は、宿主細胞へトランスフェクションされる(すなわち、形質転換される、形質導入される、トランスフェクションされる、接合されるまたは導かれる)場合、分子が発現されうるような様式で核酸分子を転写制御配列へ連結することを指す。転写制御配列は、転写の開始、伸長、または終結を制御する配列である。特に重要な転写制御配列は、プロモーター、エンハンサー、オペレーターおよびリプレッサー配列のような転写開始を制御するものである。適する転写制御配列は、トラウストチトリアレス目の微生物において機能することができるいずれの転写制御配列も含む。本発明者らが、本明細書での他の所で詳細に記載されている、そのようなプロモーターの少なくとも3つを単離かつ同定した最初の者であると考えられる。
【0083】
好ましいプロモーターは、限定されるものではないが、トラウストチトリアレスのアセト乳酸シンターゼプロモーター(配列番号:14のヌクレオチド1位〜1259位により本明細書に表されている)、トラウストチトリアレスのα-チューブリンプロモーター(配列番号:9のヌクレオチド441位〜894位により本明細書に表されている)、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター(配列番号:34内に含まれる)、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーター(配列番号:31内に含まれる;その脂肪酸デサチュラーゼプロモーターは、米国特許仮出願第60/284,116号、前記においてデサチュラーゼプロモーターとして呼ばれていることを留意されたい。)を含む。α-チューブリンプロモーター、アセト乳酸シンターゼプロモーター、および脂肪酸デサチュラーゼプロモーターのクローニングおよびシーケンシングは、実施例の節において記載されている。好ましい態様において、α-チューブリンプロモーターは、天然に存在するトラウストチトリアレスα-チューブリンプロモーター配列(配列番号:9のヌクレオチド441位〜894位)、またはプロモーターが少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、配列番号:9のヌクレオチド441位〜894位と少なくとも95 %同一である核酸配列を含む。同様に、好ましいアセト乳酸シンターゼプロモーターは、天然に存在するトラウストチトリアレスのアセト乳酸シンターゼプロモーターの核酸配列(配列番号:14のヌクレオチド1位〜1259位内に表されている)、またはプロモーターが少なくとも基礎的なアセト乳酸シンターゼプロモーター転写活性をもっている、配列番号:14のヌクレオチド1位〜1259位と少なくとも75 %、およびより好ましくは80 %、およびより好ましくは85 %、およびより好ましくは90 %、およびより好ましくは95 %同一である核酸配列を含む。好ましいPKSプロモーターは、天然に存在するトラウストチトリアレスのPKSプロモーターの核酸配列(配列番号:34内に表されている)、またはプロモーターが少なくとも基礎的なPKSプロモーター転写活性をもっている、配列番号:34と少なくとも75 %、およびより好ましくは80 %、およびより好ましくは85 %、およびより好ましくは90 %、およびより好ましくは95 %同一である核酸配列を含む。最後に、好ましい脂肪酸デサチュラーゼプロモーターは、天然に存在するトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターの核酸配列(配列番号:31内に表されている)を含む、またはプロモーターが少なくとも基礎的な脂肪酸デサチュラーゼプロモーター転写活性をもっている、配列番号:31と少なくとも75 %、およびより好ましくは80 %、およびより好ましくは85 %、およびより好ましくは90 %、およびより好ましくは95 %同一である核酸配列内に含まれる。同一性の割合を決定するための方法は、アセト乳酸シンターゼ配列について本明細書で先に記載されており、本明細書に含まれている。
【0084】
一つの態様において、本発明の組換えベクターは、発現ベクターである。本明細書に用いられる場合、句「発現ベクター」は、コードされた産物(例えば、対象となるタンパク質)の産生に適するベクターを指すために用いられる。この態様において、産生されうる産物をコードする核酸配列は、組換え核酸分子を作製するために組換えベクターへ挿入される。産生されうるタンパク質をコードする核酸配列は、組換え微生物内で核酸配列の転写および翻訳を可能にするベクターにおいて制御配列(例えば、本発明のトラウストチトリアレスのプロモーター)へ核酸配列を実施可能に連結する様式でベクターへ挿入される。本発明の選択マーカーは、本発明の組換え核酸分子が成功裡に導入された組換え微生物の選択を可能にする。
【0085】
もう一つの態様において、本発明の組換えベクターは、ターゲティングベクターである。本明細書に用いられる場合、句「ターゲティングベクター」は、核酸分子が宿主細胞内で内因性遺伝子を欠失させるまたは不活化するために用いられる(すなわち、ターゲットされた遺伝子の破壊またはノックアウトテクノロジーのために用いられる)、特定の核酸分子を組換え細胞へ送達するために用いられるベクターを指すのに用いられる。そのようなベクターはまた、「ノックアウト」ベクターとして当技術分野で知られているかもしれない。この態様の一つの局面において、ベクターの一部、であるがより典型的には、ベクターへ挿入された核酸分子(すなわち、挿入断片)は、宿主細胞におけるターゲット遺伝子(すなわち、欠失させられるまたは不活化されるためにターゲットされる遺伝子)の核酸配列と相同的である核酸配列を有する。ベクター挿入断片の核酸配列は、ターゲット遺伝子および挿入断片が相同的組換えをうけて、内因性ターゲット遺伝子が欠失させられる、不活化されるまたは減弱される(すなわち、内因性ターゲット遺伝子の少なくとも一部が変異されるまたは欠失させられることにより)ように、ターゲット遺伝子に結合するように設計される。
【0086】
一つの態様において、本発明の好ましい組換えベクターは、トラウストチトリアレス微生物における使用に適している組換えベクターであり、本発明のアセト乳酸シンターゼ分子をコードする核酸配列を含む。好ましくは、アセト乳酸シンターゼは、天然に存在する型(配列番号:15)と比較して、組換えベクターでトランスフェクションされたトラウストチトリアレス微生物にスルホニル尿素化合物、イミダゾリノン類阻害剤、および/またはピリミジニルオキシ安息香酸に対する低下した感受性を与えるように改変されている。好ましくは、そのような組換えベクターは、次の位置:116G、117A、192P、200A、251K、358M、383D、592V、595W、または599Fの1つまたは複数のアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を含むアセト乳酸シンターゼタンパク質をコードする核酸配列を含む。一つの態様において、そのようなアセト乳酸シンターゼタンパク質は、限定されるものではないが、配列番号:19、配列番号:22および配列番号:24を含むアミノ酸配列を有する。好ましくは、そのような組換えベクターは、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位から選択される核酸配列を含む。特に好ましい態様において、望ましい耐性を与えるALSコード化核酸配列を含む組換えベクターは、配列番号:18、配列番号:21および配列番号:23を含む。
【0087】
一つの態様において、組換えベクターでトランスフェクションされたトラウストチトリアレス微生物にスルホニル尿素化合物、イミダゾリノン類阻害剤、および/またはピリミジニルオキシ安息香酸に対する低下した感受性を与えるのに適している組換えベクターは、天然に存在するシゾチトリウムのアセト乳酸シンターゼ配列である配列番号:15を含む。この態様において、組換えベクターは、天然に存在するシンターゼを過剰発現するように設計され、それによりそのような過剰発現が微生物に特定された化合物に対する耐性を与えるという効果を生ずる。この態様において、組換えDNAテクノロジーの使用が、例えば、宿主細胞内での核酸分子のコピー数、それらの核酸分子が転写される効率、その結果として生じる転写物が翻訳される効率、および翻訳後修飾の効率を操作することにより形質転換された核酸分子の発現の制御を向上させることができることは、当業者に認識されているものと思われる。さらに、プロモーター配列は、天然のプロモーターと比較して、発現のレベルを向上させるように遺伝子的に設計されうるものと思われる。核酸分子の発現を制御するために有用な組換え技術は、限定されるものではないが、1つまたは複数の宿主細胞染色体への核酸分子の組み込み、プラスミドへのベクター安定性配列の付加、転写制御シグナル(例えば、プロモーター、オペレーター、エンハンサー)の置換または改変、翻訳制御シグナルの置換または改変(リボソーム結合部位、シャイン-ダルガノ(Shine-Dalgarno)配列)、宿主細胞のコドン使用頻度に対応しうる核酸分子の改変、および転写物を不安定にする配列の除去を含む。
【0088】
本発明の一つの態様において、トラウストチトリアレス微生物の形質転換における使用に適する組換えベクターは、選択マーカーとしてストレプトアロテイコスヒンズスタヌス由来のSh ble遺伝子(「ブレオマイシン結合タンパク質」をコードする)を本明細書で先に記載されているトラウストチトリアレスプロモーターと組み合わせて含む。ble遺伝子およびトラウストチトリアレスプロモーターを含む好ましい組換えベクターは、例えば、配列番号:8または配列番号:9により表されるベクター配列を含む。ストレプトアロテイコスヒンズスタヌスのブレオマイシン結合タンパク質のアミノ酸配列は、配列番号:10として本明細書に表されている。
【0089】
本発明の組換え核酸分子は、DNAまたはRNAのどちらでもありうるが、組換え細胞と適合性のある翻訳制御配列、複製開始点、および他の制御配列のような付加的な制御配列を含む。一つの態様において、宿主細胞の染色体へと組み込まれるものを含む、本発明の組換え分子はまた、発現されるタンパク質がそのタンパク質を産生する細胞から分泌されうるようにするための分泌シグナル(すなわち、シグナルセグメント核酸配列)を含む。適するシグナルセグメントは、発現されうるタンパク質に本来付随しているシグナルセグメントまたは本発明によるタンパク質の分泌を方向づけることができるいずれの異種性シグナルセグメントも含む。もう一つの態様において、本発明の組換え分子は、発現されるタンパク質が送達されかつ宿主細胞の膜へ挿入されうるようにするためのリーダー配列を含む。適するリーダー配列は、そのタンパク質に本来付随しているリーダー配列、または細胞の膜へのタンパク質の送達および挿入を方向づけることができるいずれの異種性リーダー配列も含む。
【0090】
トラウストチトリアレス目の微生物を形質転換するのに有用である様々な道具を記載してきたが、本発明の一つの態様は、トラウストチトリアレス目の微生物の細胞を形質転換する方法に関する。方法は、(a)本明細書で先に記載されている組換え核酸分子をトラウストチトリアレス目の微生物の細胞へ導入する第一段階を含む。組換え核酸分子は、スルホニル尿素化合物、イミダゾリノン類阻害剤、および/またはピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を該細胞へ与えるアセト乳酸シンターゼをコードする核酸配列を含む。そのようなアセト乳酸シンターゼは、上で詳細に記載されており、配列番号:19、配列番号:22および配列番号:24、加えて、上記で考察されている、そのような配列または配列番号:15のいずれかの相同体により表されるアミノ酸配列を有するアセト乳酸シンターゼを含む。方法は、第二段階として(b)化合物がスルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される、形質転換されていない細胞に対して阻害性がある少なくとも1つの化合物を含む培地において(a)の細胞を培養することにより組換え核酸分子で成功裡に形質転換された細胞を選択する段階を含む。そのような化合物の存在下で増殖する細胞は成功裡に形質転換されている。
【0091】
本方法において使用される組換え核酸分子は、本明細書で先に記載されている本発明の組換えベクターのいずれかを含み、典型的には、組換え細胞により産生されうるタンパク質をコードする少なくとも1つの核酸配列(すなわち、組換え発現ベクターを含む)、または組換え細胞において内因性遺伝子のターゲットされた欠失または不活性化に有用な核酸配列(すなわち、組換えターゲティングベクターを含む)を含む。
【0092】
一つの態様において、組換え核酸分子は、タンパク質をコードする核酸配列が転写制御配列へ実施可能に連結されている、細胞により産生されうるタンパク質をコードする核酸配列をさらに含む。トラウストチトリアレスにおける産生に望まれるであろうタンパク質は、当業者に知られているものと思われ、すべて、本発明に含まれるものとする。特に好ましいタンパク質は、限定されるものではないが、ドコサヘキサエン酸(DHA)、ドコサペンタエン酸(DPA)、エイコサペンタンエン酸(EPA)およびアラキドン酸(ARA)からなる群より選択される脂肪酸の合成に関係するタンパク質を含む。そのようなタンパク質は、例えば、脂肪酸シンターゼ、脂肪酸デサチュラーゼ、脂肪酸エロンガーゼ、ポリケチドシンターゼ複合体に関係するタンパク質およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質を含む。一つの局面において、タンパク質は、核酸配列の配列番号:29によりコードされるω-3脂肪酸デサチュラーゼである。配列番号:30は、デサチュラーゼのアミノ酸配列を表している。もう一つの局面において、タンパク質はポリエン脂肪酸イソメラーゼである。一つの態様において、本方法を用いてトラウストチトリアレス微生物において産生されうるタンパク質は、イソプレノイド生合成経路に関係するタンパク質を含む。そのようなタンパク質は、限定されるものではないが、HMG-CoAシンターゼ、HMG-CoAレダクターゼ、スクアレンシンターゼ、フィトエンシンターゼ、フィトエンデサチュラーゼ、カロテノイドシクラーゼ、カロテノイドヒドロキシラーゼ、カロテノイドケトラーゼを含む。さらにもう一つの態様において、本方法を用いてトラウストチトリアレス微生物において産生されうるタンパク質は、限定されるものではないが、ビタミンEおよびリポ酸を含む。
【0093】
一つの態様において、本発明の方法において有用な組換え核酸分子は、ターゲット核酸配列を含む遺伝子が、第二の核酸配列との相同的組換えにより変異されるまたは不活性化されるように微生物においてターゲット核酸配列とハイブリダイズする核酸配列を含む。そのような核酸配列は、飽和および多価不飽和脂肪酸合成経路の酵素をコードする遺伝子(もしくはそのような遺伝子の発現を制御する核酸)、トラウストチトリアレス微生物により産生される他の価値ある化合物の分解に関与しているもしくは望ましい化合物の価値を別なふうに減らすタンパク質をコードする遺伝子、またはそれの合成が対象となる他の分子と競合している化合物の合成に付随しているタンパク質をコードする遺伝子と相同的でありうる。例えば、ターゲット核酸配列は、限定されるものではないが、リパーゼ、脂肪酸酸化酵素、糖合成に関与するタンパク質、イソプレノイド経路の生成物の合成に関与するタンパク質、細胞壁成分の合成に関与するタンパク質、飽和脂肪酸合成経路に関与するタンパク質、多価不飽和脂肪酸合成経路に関与するタンパク質、ポリケチドシンターゼ複合体に関係するタンパク質、およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質をコードする配列を含む。
【0094】
本発明の一つの態様において、トラウストチトリアレス微生物の形質転換のための方法は、核酸配列が転写制御配列へ実施可能に連結されている、発現されうるタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階を含む。または、追加の組換え核酸分子は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活性化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列を含みうる。この様式において、複数のタンパク質を細胞へ導入する、複数の遺伝子を不活性化する、またはその2つの組み合わせが可能である。追加の組換え核酸分子は、第一の組換え核酸分子と同時に(すなわち、同時形質転換)、または引き続きの形質転換として(例えば、「積み重ね(stacking)」形質の目的のために)トラウストチトリアレス微生物へ導入されうる。
【0095】
一つの態様において、方法は、ブレオマイシン結合タンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含む。この態様において、追加の組換え核酸分子は、好ましくは、同時形質転換としてよりもむしろ、次の段階において導入される。好ましくは、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、第二のタンパク質をコードする核酸配列が転写制御配列へ実施可能に連結されている、細胞により発現されうる第二のタンパク質をコードする核酸配列をさらに含む。または、もしくは加えて、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活性化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列をさらに含む。一つの態様において、そのような組換え核酸分子は、核酸配列の配列番号:9を含む。
【0096】
本発明の方法を用いて形質転換するために適する宿主細胞は、限定されるものではないが、トラウストチトリアレス目のいずれの微生物も含む。宿主細胞は、形質転換されていない細胞または少なくとも1つの核酸分子ですでにトランスフェクションされている細胞のどちらでもありうる。本発明において使用される好ましい宿主細胞は、限定されるものではないが、トラウストチトリウム、ラビリンチュロイデス、ジャポノチトリウム、およびシゾチトリウムを含む属由来の微生物を含む。これらの属内の好ましい種は、限定されるものではないが、シゾチトリウムアグレガタム、シゾチトリウムリマシナム、シゾチトリウムミヌタム(Schizochytrium minutum)を含むいずれのシゾチトリウム属種も;トラウストチトリウムストリアタム、トラウストチトリウムオーレウム、トラウストチトリウムローゼウムを含む、いずれのトラウストチトリウム属種(U. ビスルゲンシス(U. visurgensis)、U. アモエボイダ(U. amoeboida)、U. サルカリアナ(U. sarkariana)、U. プロフンダ(U. profunda)、U. ラジアタ(U. radiata)、U. ミヌタ(U. minuta)およびウルケニア種BP-5601のような元ウルケニア種を含む)も;およびいずれのジャポノチトリウム属種も含む。特に好ましいトラウストチトリアレスの系統は、限定されるものではないが、シゾチトリウム属種(S31)(ATCC 20888);シゾチトリウム属種(S8)(ATCC 20889);シゾチトリウム属種(LC-RM)(ATCC 18915);シゾチトリウム属種(SR21);シゾチトリウムアグレガタム(GoldsteinおよびBelsky)(ATCC 28209);シゾチトリウムリマシナム(HondaおよびYokochi)(IFO 32693);トラウストチトリウム属種(23B)(ATCC 20891);トラウストチトリウムストリアタム(Schneider)(ATCC 24473);トラウストチトリウムオーレウム(Goldstein)(ATCC 34304);トラウストチトリウムローゼウム(Goldstein)(ATCC 28210);およびジャポノチトリウム属種(L1)(ATCC 28207)を含む。
【0097】
本発明により、用語「形質転換」は、外因性核酸分子(すなわち、組換え核酸分子)がトラウストチトリアレス微生物の細胞のような微生物の細胞へ挿入されうる任意の方法を指すために用いられる。微生物系において、用語「形質転換」は、微生物による外因性核酸の獲得のせいでの遺伝する変化を記述するために用いられ、本質的に用語「トランスフェクション」と同意語である。適する形質転換技術は、限定されるものではないが、微粒子銃、電気穿孔法、マイクロインジェクション、リポフェクション、吸着、感染およびプロトプラスト融合を含む。
【0098】
一つの態様において、本発明の方法を用いて産生されうるタンパク質は、そのタンパク質を産生するのに効率的な条件下においてそのタンパク質を発現する細胞(すなわち、組換えトラウストチトリアレス微生物)を培養することにより産生される。タンパク質が回収されうる場合もあるし、微生物が全部または可溶化物として収集されかつ「バイオマス」として使用されうる場合もある。もう一つの態様において、ターゲット遺伝子は、細胞内での組換えを可能にし、結果としてターゲット遺伝子の欠失または不活化を生じるのに効率的な条件下において、本発明のターゲティングベクターを含む組換え分子で形質転換された細胞を培養することにより欠失させられるまたは不活化される。培養されうる好ましい細胞は、本発明の組換え細胞である。効率的な培養条件は、限定されるものではないが、タンパク質の産生および/または組換えを可能にする効率的な培地、バイオリアクター、温度、pHおよび酸素条件を含む。効率的な培地は、典型的にはトラウストチトリアレス細胞が培養される任意の培地を指す。そのような培地は、典型的には、同化できる炭素源、窒素源およびリン酸源、ならびに適当な塩、ミネラル、金属およびビタミンのような他の栄養素を有する水性培地を含む。適する培地および培養条件の例は、実施例の節で詳細に考察されている。トラウストチトリアレス微生物に適する培養条件は、Barclayによる1994年8月23日に発行された米国特許第5,340,742号にも記載されている;参照として完全に本明細書に組み入れられている。本発明の細胞は、通常の発酵バイオリアクター、振盪フラスコ、試験管、マイクロタイター皿およびペトリプレートにおいて培養されうる。培養は、組換え細胞に適切な温度、pHおよび酸素含有量で行われうる。そのような培養条件は、当業者の専門技術内である。
【0099】
産生のために使用されるベクターおよび宿主系に依存して、結果として生じる本発明のタンパク質は、組換え細胞内に残りうる;発酵培地へ分泌されうる;2つの細胞膜間の間隙へ分泌されうる;または細胞膜の外側表面上に保持されうるのいずれかである。句「タンパク質を回収すること」とは、タンパク質を含む発酵培地全部を収集することを指し、分離または精製の追加の段階を含意する必要はない。本発明の方法により産生されるタンパク質は、限定されるものではないが、アフィニティークロマトグラフィー、イオン交換クロマトグラフィー、濾過、電気泳動、疎水性相互作用クロマトグラフィー、ゲル濾過クロマトグラフィー、逆相クロマトグラフィー、コカナバリンAクロマトグラフィー、クロマトフォーカシングおよび示差的可溶化のような様々な標準的タンパク質精製技術を用いて精製されうる。本発明の方法により産生されるタンパク質は、好ましくは、「実質的純粋」型として回収される。本明細書に用いられる場合、「実質的純粋」とは、商業的製品としてタンパク質の実効使用を可能にする純度を指す。
【0100】
本発明のさらにもう一つの態様は、本発明のアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子で形質転換されたトラウストチトリアレス目の組換え微生物に関する。好ましくは、アセト乳酸シンターゼは、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を微生物に与える。そのような微生物を形質転換するにおける使用に適する組換え核酸分子および配列は、先で詳細に記載されている。そのような微生物は、本明細書で先に記載されているように、ble遺伝子選択マーカーおよびトラウストチトリアレスの転写制御配列を含む組換え核酸分子を含む、別の組換え核酸分子でさらに形質転換されうる。本発明によるトラウストチトリアレス微生物は、実施例の節に記載されている。本発明により、本発明の組換えトラウストチトリアレス微生物は、本明細書に記載される組換えベクターを用いて対象となるタンパク質(そのようなタンパク質の例は上記で考察されている)を発現するように遺伝子的に操作される、および/または本明細書に記載される組換えベクターを用いてターゲット遺伝子のターゲットされた欠失もしくは不活化のために遺伝子的に操作される。
【0101】
本明細書に用いられる場合、組換え微生物は、組換えテクノロジーを用いてそれの通常の(すなわち、野生型のまたは天然に存在する)型から改変されている(すなわち、変異されているまたは変えられている)ゲノムを有する。本発明による組換え微生物は、そのような改変が微生物内に望ましい効果を与えるような様式で、核酸分子が挿入された、欠失させられたまたは改変された(すなわち、変異された;例えば、ヌクレオチドの挿入、欠失、置換、および/または逆位により)微生物を含む。本明細書に用いられる場合、遺伝子発現における、遺伝子の機能における、または遺伝子産物の機能における減少を結果として生じる遺伝子改変は、遺伝子の不活化(完全または部分的)、欠失、中断、妨害または下方制御と呼ばれうる。例えば、そのような遺伝子によりコードされるタンパク質の機能における減少を結果として生じる遺伝子における遺伝子改変は、遺伝子の完全な欠失(すなわち、遺伝子が存在しない、それゆえにタンパク質が存在しない)、タンパク質の不完全な翻訳もしくは翻訳無し(例えば、タンパク質が発現されない)を結果として生じる遺伝子における変異、またはタンパク質の本来の機能を減少させるもしくは消失させる(例えば、減少した酵素的活性もしくは作用をもつまたは全くもたないタンパク質が発現される)遺伝子における変異の結果でありうる。遺伝子発現または機能における増加を結果として生じる遺伝子改変は、遺伝子の増幅、過剰産生、過剰発現、活性化、増強、付加、または上方制御と呼ばれうる。
【0102】
本発明によると、組換えトラウストチトリアレス微生物は、トラウストチトリアレス目のいずれの微生物を用いても作製されうる。好ましいトラウストチトリアレスの属は、限定されるものではないが、トラウストチトリウム、ラビリンチュロイデス、ジャポノチトリウム、およびシゾチトリウムを含む。これらの属内の好ましい種は、限定されるものではないが、シゾチトリウムアグレガタム、シゾチトリウムリマシナムを含むいずれのシゾチトリウム属種も;いずれのトラウストチトリウム属種も(U. ビスルゲンシス、U. アモエボイダ、U. サルカリアナ、U. プロフンダ、U. ラジアタ、U. ミヌタおよびウルケニア種BP-5601のようないずれの元ウルケニア種も含む)、トラウストチトリウムストリアタム、トラウストチトリウムオーレウム、トラウストチトリウムローゼウム;およびいずれのジャポノチトリウム属種も含む。特に好ましいトラウストチトリアレスの系統は、限定されるものではないが、シゾチトリウム属種(S31)(ATCC 20888);シゾチトリウム属種(S8)(ATCC 20889);シゾチトリウム属種(LC-RM)(ATCC 18915);シゾチトリウム属種(SR21);シゾチトリウムアグレガタム(GoldsteinおよびBelsky)(ATCC 28209);シゾチトリウムリマシナム(HondaおよびYokochi)(IFO 32693);トラウストチトリウム属種(23B)(ATCC 20891);トラウストチトリウムストリアタム(Schneider)(ATCC 24473);トラウストチトリウムオーレウム(Goldstein)(ATCC 34304);トラウストチトリウムローゼウム(Goldstein)(ATCC 28210);およびジャポノチトリウム属種(L1)(ATCC 28207)を含む。
【0103】
以下の実施例は、例証の目的のために提供され、本発明の範囲を限定することを意図されるものではない。
【実施例】
【0104】
実施例1
この実施例は、組換えプラスミドpTUBZEO 11-2の作製を記載する。
【0105】
組換えプラスミドpTUBZEO 11-2の構築は、図1および図2に示されている。このプラスミドは、シゾチトリウム属種から単離されたα-チューブリン遺伝子プロモーターと機能的に共役するストレプトアロテイコスヒンズスタヌス由来のble遺伝子を含む。このプラスミドは以下の通りに作製された。cDNAクローン(CGNE0002-001-B6)は、シゾチトリウム属種cDNAライブラリーから単離され、大規模シゾチトリウムcDNAシーケンシングプロジェクトの一環として部分的にシーケンシングされた(配列番号:1)。ヌクレオチド配列は、BLASTX相同性検索によりα-チューブリンをコードすることが決定された(Gish, W.およびD. States. 1993. Nat. Genet. 3:266-272)。塩基116位から550位までの推定されたアミノ酸配列は、ペルベティカファスティギアタ(Pelvetica fastigiata)由来のα-チューブリン(GenBankアクセッション番号U58642)の最初の145個のアミノ酸と93 %同一である。
【0106】
この遺伝子に関係するプロモーターを単離するために、ゲノムDNAがシゾチトリウム属種の細胞から単離され、平滑末端を生じるための制限エンドヌクレアーゼでのゲノムDNAの酵素的消化、続いて、キットに供給されている特定の二本鎖DNAアダプター分子への消化されたDNAのライゲーションを含む「ゲノムウォーカー(GenomeWalker)(商標)」キット(Clontech Laboratories, Inc., Palo Alto, CA)の使用により処理された。α-チューブリンコード配列のDNA上流は、その後、キットに供給されている外側アダプタープライマー(AP1)およびα-チューブリン特異的プライマーPGR20(配列番号:2)を用いて、ポリメラーゼ連鎖反応(PCR)により増幅された。遺伝子のさらなる増幅は、キットに供給されている入れ子状態のアダプタープライマー(AP2)および入れ子状態のα-チューブリン特異的プライマーPGR19(配列番号:3)を用いて行われた。その結果生じたPCR産物は、プラスミドpCR2.1-TOPO(Invitrogen Corp., Carlsbad, CA)へサブクローニングされた。サブクローニングされた断片の1つは、シーケンシングされた;α-チューブリン遺伝子開始コドンの前に隣接した725bpの配列は、配列番号:4として示されている。
【0107】
この方法で得られるDNA配列に基づくオリゴヌクレオチドプライマーを用いて、TaqDNAポリメラーゼ(Perkin-Elmer Corp., Norwalk, CT)を使用するPCRが用いられて、NcoI制限部位がDNA断片の3'末端へ組み入れられている改変されたα-チューブリンプロモーター領域を生じた;このNcoI部位は、α-チューブリンコード領域において同じ位置にある開始コドンを含んだ。この反応において使用されるプライマーは、PGR33(配列番号:5)およびPGR34(配列番号:6)であり、鋳型はシゾチトリウム属種の細胞から単離されたゲノムDNAであった。以下の反応条件が利用された:94℃4分間;(94℃1分間、54℃45秒間、72℃2分間)x 30;72℃7分間。この断片は、プラスミドpCR2.1-TOPOへクローニングされてプラスミドp7TUB(配列番号:7)を形成した。プラスミドp7TUBはNcoIで消化され、結果として生じたシゾチトリウムα-チューブリンプロモーター領域を含む463bp断片は、アガロースゲル精製により単離された。SV40プロモーターおよびターミネーターに隣接しているストレプトアロテイコスヒンズスタヌス由来のble遺伝子を含むプラスミドpSV40/Zeo(Invitrogen Corp., Carlsbad, CA)もまた、NcoIで消化され、3201bpおよび314bpの断片を生じた。3201bp断片は、アガロースゲル精製され、かつp7TUB由来の463bp NcoI断片に連結されて、図1に描かれているpTUBZEO-11(配列番号:8)を生じた。
【0108】
次に、プラスミドpTUBZEO-11は、SphIで消化され、結果として生じた、シゾチトリウムα-チューブリンプロモーターおよびSV40ターミネーターに隣接しているble遺伝子を含む1122bp断片は、アガロースゲル精製され、かつSphIでの消化により線状化されたプラスミドpUC19(Messing, J. 1983. Meth. Enzymol. 101:20)に連結された。その結果生じたプラスミドは、pTUBZEO 11-2(配列番号:9)と名付けられ、図2および図4に描かれている。プラスミドpTUBZEO 11-2はまた、pMON50000とも呼ばれる。配列番号:9において、シゾチトリウムα-チューブリンプロモーターはヌクレオチド441位〜894位内に含まれる;ble遺伝子コード領域はヌクレオチド895位〜1269位内に含まれる;およびSV40ターミネーターはヌクレオチド1270位〜1524位内に含まれる。
【0109】
実施例2
この実施例は、組換えプラスミドpMON50200、pMON50201、pMON50202、およびpMON50203の作製を記載する。
【0110】
シゾチトリウム属種由来の未変性のアセト乳酸シンターゼコード遺伝子(als)は、以下の方法で単離された。cDNAクローン(LIB81-028-Q1-E1-D9)はシゾチトリウムcDNAライブラリーから単離され、大規模シゾチトリウム属種cDNAシーケンシングプロジェクトの一環として部分的にシーケンシングされた(配列番号:11)。ヌクレオチド配列は、BLASTX相同性によりアセト乳酸シンターゼをコードすることが決定された;例えば、塩基154位から378位までの推定されるアミノ酸配列は、シゾサッカロマイセスポンベ(Schizosaccharomyces pombe)(GenBankアクセッション番号P36620)由来のALSのアミノ酸313位〜387位と68 %同一であった。その後、このクローニングされたcDNAの完全長配列が得られ、cDNAクローンは全alsコード領域を含まないことが示された。完全長als遺伝子を得るために、シゾチトリウムのゲノムラムダライブラリーが372bpジゴキシゲニン(DIG)標識DNAプローブ(ALS2プローブと呼ばれる)での標準的プロトコール(例えば、Sambrookら、Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, 1989を参照)を用いて探索された。ALS2プローブは、cDNAクローンLIB81-028-Q1-E1-D9の配列に基づいている前進プライマーPGR38(配列番号:12)および逆プライマーPGR39(配列番号:13)を用いて、DIG-11-UTPを含むヌクレオチド混合物を使用するPCRにより生成された。ALS-4Aと称せられる、ALS2プローブで同定されたゲノムクローンの1つが単離され、さらに、DIG標識ALS2プローブを使用するサザンハイブリダイゼーションブロット法により特徴付けられた。AhdIで消化されたALS-4AラムダDNA由来の4.9-kbp断片がALS2プローブにハイブリダイズすることを見出された。この断片は、アガロースゲル精製により単離され、平滑末端を生じるためにT4 DNAポリメラーゼで処理され、その後、SmaIで消化されたpBluescriptII KS+(Stratagene Corp., La Jolla, CA)へ連結されてプラスミドpMON50200(図3Aに描かれている)を形成した。pMON50200の配列は、配列番号:14として示されている。シゾチトリウムals遺伝子によりコードされるアセト乳酸シンターゼ酵素の配列は、配列番号:15として示されている。
【0111】
プラスミドpMON50201、pMON50202、およびpMON50203(それぞれ、図3B、図3Cおよび図3Dに描かれている)は、コードされるアセト乳酸シンターゼ酵素がスルホメツロンメチル(SMM)を含むある特定の化合物にもはや阻害されないように、部位特異的突然変異誘発によりプラスミドpMON50200から作製された。これらのプラスミドは、以下の通りに構築された。「トランスフォーメラ(Transformera)」部位特異的突然変異誘発キット(Clontech Laboratories, Inc., Palo Alto, CA)が、製造者の使用説明書に従って以下の変異をプラスミドpMON50200へ導入するために用いられた。オリゴヌクレオチド選択プライマー、DM19(配列番号:16)がすべての3つの構築物において用いられた;このプライマーはpMON50200のマルチクローニング部位における唯一のEcoRV部位のAatII部位への変換へと導く。プライマーDM14(配列番号:17)は、コードされたALS酵素におけるアミノ酸残基595番をトリプトファンからバリンへ変え、同時に、遺伝子配列にAclI部位を導入するために用いられる;その結果生じたプラスミドはpMON50201(配列番号:18)と呼ばれる。同様に、プライマーDM15(配列番号:20)が、コードされたALS酵素におけるアミノ酸残基192番をプロリンからグルタミンへ変えてals遺伝子にBsgI部位を導入するために用いられ、結果としてプラスミドpMON50202(配列番号:21)を生じた。プラスミドpMON50203(配列番号:23)を構築するために、DM14およびDM15プライマーの両方が用いられ、結果として、上記の両方のアミノ酸残基置換を含むコードされたALS酵素を生じた。プラスミドpMON50201、pMON50202、およびpMON50203によりコードされる変異体アセト乳酸シンターゼ酵素の配列は、それぞれ、配列番号:19、配列番号:22、および配列番号:24として示されている。
【0112】
実施例3
この実施例は、実施例1および実施例2に記載される組換え分子でのシゾチトリウム属種の遺伝的形質転換を記載する。
【0113】
シゾチトリウム属種N230D、アメリカンタイプカルチャーコレクション(American Type Culture Collection)株20888(ATCC, Manassas, VA)の派生体における株がこの実施例に使用された。液体培養については、細胞をM50-3培地において30℃、200 rpm〜300 rpmで振盪させて無菌的に増殖させた。M50-3培地は以下の成分を含む:NaCl、12.5 g;MgSO4・7H2O、2.5 g;KCl、0.5 g;CaCl2、0.05 g;グルコース、30 g;Na-グルタミン酸、3 g;KH2PO4、0.4 g;酵母抽出物、1 g;NaHCO3、0.4 g;Na2EDTA、30 mg;FeCl3・6H2O、1.2 mg;H3BO3、34.2 mg;ZnSO4・7H2O、0.67 mg;CoCl2・6H2O、0.13 mg;NaMoO4・2H2O、25 μg;CuSO4・5H2O、10 μg;NiSO4・6H2O、0.26 mg;チアミン・HCl、100 μg;ビオチン、0.5 μg;シアノコバラミン、0.5 μg、および脱イオン水(1リットルにつき);7.0に調整された最終pH。固体培地での増殖については、細胞を1.5 %(w/v)寒天の添加により凝固したM50-3培地またはM1E-3培地において30℃で増殖させた。M1E-3培地は以下の成分を含む:グルコース、4 g;(NH4)2SO4、0.75 g;Na2SO4、5 g;MgSO4・7H2O、2 g;KH2PO4、0.5 g;KCl、0.5 g;CaCl2・2H2O、0.1 g;MOPS緩衝液、20.9 g;FeSO4・4H2O、0.3 mg;MnCl2・4H2O、0.1 mg;ZnSO4・7H2O、80 μg;CoCl2・6H2O、2 μg;NaMoO4・2H2O、1 μg;CuSO4・5H2O、60 μg;NiSO4・6H2O、80 μg;チアミンHCl、320 μg;CA-パントテン酸、320 μg;シアノコバラミン、8 μg、および脱イオン水(1リットルにつき);7.0に調整された最終pH。
【0114】
シゾチトリウム属種のゼオシン(Zeocin)(商標)およびSMMに対する感受性は、凝固したM1E-3培地に様々な濃度でこれらの阻害剤を含め、そして組換え細胞の選択のために用いられる工程間に存在しているものと類似した濃度でプレート上に細胞を広げることにより測定される。
【0115】
シゾチトリウム細胞の遺伝的形質転換は、Bio-Rad Biolistic PDS-1000/He粒子送達システム( Particle Delivery System)(Bio-Rad Laboratories, Hercules, CA)を用いる微粒子銃により行われた(Sanford, J. C., F. D. Smith、およびJ. A. Russell. 1993. Meth. Enzymol. 217:483-509)。シゾチトリウム属種N230D細胞を液体M50-3培地において0.4〜0.8の680 nm(OD680)での光学濃度(10 mmの光学距離の長さ)まで増殖させた。1.0 OD680に対応する細胞の一定分量を短時間だけ遠心分離し、上清溶液を除去し、そしてペレット化された細胞を100 μlの滅菌水に再懸濁した。その後、再懸濁された細胞を寒天凝固培地(例えば、M50-3培地またはM1E-3培地)を含むペトリプレート上に4 cm〜6 cmの円に広げ、余分な水が固体培地へ吸収されうるように30分〜60分間、そのままにさせておいた;これは、ターゲットプレートと呼ばれる。
【0116】
金のマイクロキャリア(0.6 μ呼び径、Bio-Rad Laboratories, Inc., Hercules, CAから入手可能)の1.5 mg分量を製造者の使用説明書(Biolistic(登録商標)PDS-1000/He 粒子送達システム使用説明書マニュアル(Particle Delivery System Instruction Manual);Bio-Rad Laboratories, Hercules, CA)どおりに形質転換プラスミドDNA(すなわち、プラスミドpTUBZEO 11-2、pMON50201、pMON50202、またはpMON50203)の2.5 μgでコーティングした。細胞を以下の条件を用いてDNAコーティング化金マイクロキャリアで撃ち込んだ:1100 psi破裂盤、25'' Hgのチャンバー減圧、1.5 cm〜2 cmの発射ディスクからストッピングスクリーンまでの距離および約7 cmのストッピングスクリーンからターゲットまでの距離を示す、上段の棚に配置されたマイクロキャリア発射集合群ならびに中段の棚に配置されたターゲットプレート。撃ち込み後、細胞を4時間〜6時間、30℃でターゲットプレート上で回復するようにさせておいた。その後、細胞を1.5 ml 滅菌水でターゲットプレートから洗い落とし、マイクロチューブに収集し、短時間だけ遠心分離し、そして400 μl 滅菌水に再懸濁した。懸濁液の100マイクロリットルを150 μg/ml〜200 μg/ml ゼオシン(Zeocin)(商標)(Invitrogen Corp., Carlbad, CA)または25 μg/ml SMMのいずれかを含む4つのM1E-3プレートのそれぞれに広げた。ゼオシン(Zeocin)(商標)含有プレートは、プラスミドpTUBZEO 11-2で形質転換された細胞について選択するために使用され、SMM含有プレートは、プラスミドpMON50201、pMON50202、またはpMON50203で形質転換された細胞について選択するために使用された。その後、プレートを7日〜10日間、30℃でインキュベートした。その後、選択薬剤に対して耐性であると思われるコロニーを、耐性を確認するために同じ選択薬剤を含む新鮮なM1E-3プレート上へパッチ状に塗布した。このプロトコールは、典型的には、撃ち込みあたり、100個〜1000個のゼオシン(Zeocin)(商標)耐性株またはSMM耐性株の発生を生じる。
【0117】
実施例4
以下の実施例は、形質転換されたシゾチトリウム細胞のPCR分析を実証する。
【0118】
PCRは、選択薬剤ゼオシン(Zeocin)(商標)またはSMMに対して耐性である推定上の形質転換された株においてプラスミド配列の存在を確認するために用いられた。推定の形質転換体および非組換え型シゾチトリウムN230D細胞由来の鋳型DNAは、使い捨てのプラスチックの1 μl接種用輪を使用して寒天プレート上へパッチされた耐性コロニー(実施例3に記載されているように)から少量の細胞(1 mm3〜2 mm3)を取り出すことにより得られた。その後、細胞をマイクロチューブにおいて1 % トリトンX-100の15 μl〜20 μlに再懸濁し、10分間沸騰水浴に置き、そしてその後、14,000xgで5分間、遠心分離した。これらの抽出物の一部(1 μL〜3 μL)は、Taq DNAポリメラーゼを使用する25 μL PCR反応についての鋳型DNAを供給するために使用された。シゾチトリウムDNAにおいてpTUBZEO 11-2配列の存在を検出するために、プライマーDM20(配列番号:25)およびDM21(配列番号:26)が用いられた;これらのプライマーは、プラスミドpTUBZEO 11-2におけるble遺伝子にアニールして、346bp DNA断片を増幅する。用いられる熱的側面は、以下のとおりである:94℃で4分間;(94℃で45秒間、52℃で45秒間、72℃で2分間)x 30;72℃で7分間。シゾチトリウムDNAにおいてpMON50201、pMON50202、またはpMON50203配列の存在を検出するために、プライマーBLA1(配列番号:27)およびBLA2(配列番号:28)が用いられた;これらのプライマーは、ベクターバックボーンに見出されるbla(アンピシリン耐性)遺伝子にアニールして、1229bp DNA断片を増幅する。用いられる熱的側面は、以下のとおりである:94℃で4分間;(94℃で45秒間、55℃で45秒間、72℃で2分間)x 30;72℃で7分間。PCR産物は、標準的アガロースゲル電気泳動、続いて、臭化エチジウムで染色することにより分析された。
【0119】
これらの分析の結果より、これらの条件下で選択された株の大多数はプラスミドDNAを含む真の形質転換体であることが確認された。形質転換プラスミドで撃ち込まれなかった対照のシゾチトリウム属種N230D細胞由来の鋳型DNAを用いた場合、正しいサイズのPCR産物は生成されなかった。
【0120】
実施例5
以下の実施例は、形質転換されたシゾチトリウム細胞のサザンブロット分析を記載する。
【0121】
サザンハイブリダイゼーションブロット法は、形質転換された細胞内の形質転換ベクターDNA配列の存在を確認するために、親のシゾチトリウムN230D細胞およびいくつかの推定の形質転換体から単離されたDNAを用いて行われた。サザンブロッティングは、当業者に公知の技術を用いて行われた(例えば、Sambrookら、Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press. 1989を参照)。DNAは、「QIAamp」DNA精製キット(Qiagen Inc., Valencia, CA)の使用により単離され、様々な制限酵素で消化され、アガロースゲル(0.8 %〜1.2 %w/v)を通しての電気泳動により分離され、その後、アルカリ性キャピラリー移動によりナイロン膜に転写された。
【0122】
pTUBZEO 11-2で形質転換された細胞におけるベクターDNAの検出は、ハイブリダイゼーションプローブとしてプライマーDM20(配列番号:25)およびDM21(配列番号:26)でのPCRにより生成された346bpのDIG標識ble遺伝子断片ならびにDIG-11-UTPを含むヌクレオチド混合物を用いて、「ジーニアス(Genius)」DIG型システム(Boehringer Mannheim Biochemicals GmbH, Germany)の使用により行われた。膜のプレハイブリダイゼーションは、ジーニアス(Genius)キットに供給されているハイブリダイゼーション緩衝液において68℃で1時間行われた。ハイブリダイゼーションは、94℃で5分間、熱変性されたble遺伝子プローブを含むハイブリダイゼーション緩衝液において68℃で18時間行われた。その後、膜を50 mL 2X SSC/0.1 % SDSで5分間を2回および50 mL 0.1X SSC/0.1 % SDSにおいて15分間を2回洗浄した。ハイブリダイズしているDNAの化学ルミネセンスの検出は、ジーニアス(Genius)キット使用説明書に記載されているように行われた。
【0123】
形質転換されていないシゾチトリウムN230D細胞由来のDNAは、ble遺伝子プローブにハイブリダイズしなかった。逆に、形質転換された細胞由来のDNAは、以下のようにプローブにハイブリダイズした:
【0124】
SphI:形質転換されたシゾチトリウム細胞由来のSphIで消化されたDNAは、ble遺伝子プローブにハイブリダイズする〜1100bpのDNA断片を含んでいた;この断片は、SphIで消化されたpTUBZEO 11-2 DNAにおいても見られるが、ble遺伝子発現カセット全体(チューブリン遺伝子プロモーターおよびSV40ターミネーターを含む)を表している。
【0125】
XhoI:試験された形質転換体のそれぞれについて、DNAのXhoI消化は、結果として15 kbp〜20 kbpより長いハイブリダイズする断片を生じた。XhoIはpTUBZEO 11-2内を切断しない、それゆえに、これらの結果は、pTUBZEO 11-2が形質転換された細胞において染色体外のエレメントとして存在しているようには思われず、むしろシゾチトリウム染色体へ組み込まれるようになることを示している。
【0126】
NcoIまたはHindIII:これらの酵素は両方とも、pTUBZEO 11-2内を一回切断する。これらの酵素のいずれかでの形質転換体DNAの消化は、典型的には、線状化pTUBZEO 11-2ベクター(すなわち、〜3.8 kbp)と共に移動する顕著なハイブリダイズする断片を導いた。これは、ベクターが直列型反復の形で染色体に統合されうることを示唆している。
【0127】
実施例6
この実施例は、シゾチトリウムにおける相同的組換えを実証している。
【0128】
以下の実験は、相同的組換えが、シゾチトリウムにおいて、内因性の天然のDNA配列と、細胞へ導入された組換えDNA分子に存在する相同性DNA配列との間で起こりうることを実証するために行われた。相同的組換えのこの型は、望ましい性質をもつ組換え系統を作製するためにとても有益でありうる。例えば、相同的組換えは、外来遺伝子配列のターゲットされた挿入により内因性遺伝子を不活化するために用いられうる。さらに、相同的組換えは、組換え細胞が新規な性質を示すように、内因性遺伝子またはそれの部分をその遺伝子の改変型で置換するために用いられうる。
【0129】
相同的組換えは、シゾチトリウムals遺伝子における突然変異を含むプラスミドpMON50202で形質転換されたシゾチトリウム細胞において起きることを示された。この突然変異は、alsコード領域の塩基対位置571位にBsgI部位を導入する。alsコード領域の塩基対位置1324位に天然に存在するBsgI部位がある。それゆえに、BsgIで消化されたシゾチトリウムDNAのサザンブロットは、天然のals遺伝子と組換え突然変異体als遺伝子とを識別するために用いられうる。これらの実験について、als特異的ハイブリダイゼーションプローブは、DIG-11-UTP(Boehringer Mannheim Biochemicals GmbH, Germany)、前進プライマーPGR28(配列番号:32)、逆プライマーPGR30(配列番号:33)、および鋳型として少量のpMON50200を含むヌクレオチド混合物を使用するPCRにより作製された。その結果生じた323bpのDIG標識ハイブリダイゼーションプローブは、ALS1と呼ばれた。
【0130】
非組換え型シゾチトリウムN230D細胞由来のDNAは、BsgIおよびAhdIで別々に消化され、アガロースゲル電気泳動にかけられ、ナイロン膜へ転写され、その後、実施例5に記載されているものと本質的に同じ手順を用いてALS1プローブで探索された。ALS1プローブは、BsgIで消化されたDNAの1.76-kbp断片およびAhdIで消化されたDNAの4.9-kbp断片を標識した。
【0131】
pMON50202で形質転換された様々な組換え株由来のBsgIで消化されたDNAおよびAhdIで消化されたDNAのサザンブロットもまた、ALS1プローブで探索された。いくつかの場合において、1.76-kbpのBsgI断片は存在せず、代わりに、pMON50202に存在する753bpのBsgI断片に対応している0.75-kbpの断片が標識された。4.9-kbpのAhdI断片がこれらの組換え株において標識されたが、その組換え突然変異体als遺伝子が二重クロスオーバー相同的組換えにより天然のals遺伝子と組換えしたことを示した。
【0132】
単一クロスオーバー相同的組換えもまた、pMON50202で形質転換された組換え株において起こっていることが観察された。これらの場合、1.76-kbpおよび0.75-kbpのBsgI断片の両方が、組換え株由来のDNAのサザンブロットにおいて標識されたが、4.9-kbpのAhdI断片がより大きな標識された断片に取って代わられ、pMON50202ベクター全体が、単一コピーとしてまたは直列型反復としてのいずれかで、天然のals遺伝子へ挿入したことを示した。
【0133】
シゾチトリウムにおける相同的組換えについての追加の証拠は、切り詰められた遺伝子によりコードされた不完全なALS酵素が機能しえないように切り詰められた突然変異体als遺伝子を含む組換えDNA分子の導入により得られた。この切り詰められた遺伝子は、ClaIおよびHindIIIでpMON50202を消化して2.8-kbp断片を生じ、それによりalsターミネーター領域と共にalsコード領域の最後の388 bpを除去することにより作製された。この2.8-kbp断片はClaIおよびHindIIIで消化されたpBluescriptII KS+(Stratagene Corp., La Jolla, CA)へライゲーションされ、プラスミドpAR2を生じた。プラスミドpAR2は、天然のals遺伝子とpAR2に存在する切り詰められた突然変異体als遺伝子との間の相同的組換えにより形質転換された株において機能しうる突然変異体als遺伝子が回復される場合には、形質転換されたシゾチトリウム細胞においてSMMに対する耐性を与えることが予測されるのみと考えられる。この構築物をシゾチトリウムN230D細胞へ微粒子銃により導入し、実施例3に記載されているようにSMM耐性株を単離した。形質転換体由来のBsgIで消化されたDNAのサザンブロット分析は、この実施例の初めの方に記載されているように行われたが、相同的組換えが明らかにこれらの株において起こったことを示した;すなわち、非組換え型細胞において1.76-kbpのBsgI断片がALS1プローブにハイブリダイズしたが、pAR2で形質転換された細胞においては、これが0.75-kbpのハイブリダイズする断片に取って代わられた。
【0134】
実施例7
この実施例は、選択マーカー遺伝子に連結されていない追加の外来DNA分子を含む株を同時形質転換により作製するために形質転換ベクターpTUBZEO 11-2またはpMON50202の使用を記載する。
【0135】
同時形質転換は、pTUBZEO 11-2およびいくつかの遺伝子のいずれかを含む追加のプラスミドの同時導入により達成された。プラスミドは、各プラスミドの2.5 μgを使用して、実施例3に記載されているように、金粒子上に共沈澱させた。プラスミドでコーティングされた金粒子のターゲット細胞への撃ち込み後、組換え株は、実施例1に記載されているようにゼオシン(Zeocin)(商標)含有寒天プレート上で選択された。第二の選択されなかったプラスミドの存在は、その後、PCR分析またはサザンブロットハイブリダイゼーションにより確認された。非常に高い同時形質転換頻度(例えば、50 %〜90 %)が概して達成された。例えば、シゾチトリウムのチューブリン遺伝子プロモーターおよびターミネーターに連結されたカエノルハブディタスエレガンス(Caenorhabditus elegans)のfat-1遺伝子(Spychallaら、1997. Proc. Natl. Acad. Sci. U. S. A. 94, 1142-1147)を含むプラスミドpTR202がこの実施例に提供されている方法により導入され、その結果生じたゼオシン(Zeocin)(商標)耐性株の約68 %がfat-1遺伝子を含むことがPCRにより示された(表1を参照)。同様の結果は、pMON50202および追加のプラスミドが同時導入され、続いてSMM含有固体培地上で形質転換された細胞を選択する場合に観察される。この同時形質転換方法は、所望されるいずれの外来DNAでも導入するために用いられうる。
【0136】
(表1) 選択マーカープラスミドpTUBZEO 11-2および様々なfad遺伝子を含むプラスミドを用いる同時形質転換の効率。ゼオシン(Zeocin)(商標)耐性形質転換体は、PCRによりfad DNA配列についてスクリーニングされた。

【0137】
この実施例に記載される形質転換系は、脂質主体の化合物の発酵による生産について知られている最も生産力が高い生物体であるシゾチトリウムを遺伝子的に操作する能力において、意義深い進歩を表している。2つの独立した形質転換系の有用性は、生じる高同時形質転換効率に加え、操作される株において複数の形質の積み重ねを可能にすると思われる。さらに、この微小藻類における相同的組換えの明らかな存在は、未知の遺伝子の機能を同定するため、および生産用株において望ましくない形質を除去するために遺伝子ノックアウト方法の発達を可能にするものと思われる。本発明者らは、最近、シゾチトリウムにおいて脂肪酸代謝を変えるためにこれらの系を使用しており、カロテノイド、ステロール、および他の類脂質性の化合物の生産のために、この種および関連した微小藻類(例えばトラウストチトリウム)を使用することの可能性を探究している。
【0138】
本発明の様々な態様が詳細に記載されてきたが、それらの態様の改変および脚色が当業者に思い浮かぶであろうことは明らかである。しかしながら、そのような改変および脚色が、特許請求の範囲に示されているように、本発明の範囲内であることは、はっきりと理解されるべきである。
【技術分野】
【0001】
発明の分野
本発明は、一般的に、トラウストチトリアレス目(Thraustochytriales)の微生物にスルホニル尿素化合物、イミダゾリノン類阻害剤および/またはピリミジニルオキシ安息香酸に対する低下した感受性を与えるアセト乳酸シンターゼを含む、トラウストチトリアレスのアセト乳酸シンターゼをコードする単離された核酸分子;トラウストチトリアレス目の微生物の形質転換のために有用な選択マーカーを含む組換え核酸分子、ならびに本発明の組換え核酸分子を用いてそのような微生物を形質転換する方法に関する。本発明はまた、トラウストチトリアレス発現系において有用な遺伝子プロモーターに関する。本発明の組換え核酸分子は、トラウストチトリアレス微生物において外来核酸の発現、およびトラウストチトリアレス微生物における遺伝子の欠失、突然変異または不活化のために用いられうる。
【背景技術】
【0002】
発明の背景
開発することによりトラウストチトリド(Thraustochytrid)の分類の改正をもたらした。分類学理論家は、トラウストチトリドを藻類または藻類様原生生物と定める。しかしながら、分類の不確定性のため、本発明に記載される系統をトラウストチトリド(目:トラウストチトリアレス;科:トラウストチトリアシエ(Thraustochytriaceae);属:トラウストチトリウム(Thraustochytrium)、シゾチトリウム(Schizochytrium)、ラビリンチュロイデス(Labyrinthuloides)またはジャポノチトリウム(Japonochytrium))とみなすことが本発明の目的にとって最善であるものと思われる。分類変更は下記に要約されている。
【0003】
本明細書に開示および特許請求の範囲に主張されるある特定の単細胞生物体の株は、トラウストチトリアレス目のメンバーである。トラウストチトリドは、問題を含む分類学的変遷をもつ海洋真核生物である。トラウストチトリドの分類学的配置についての問題は、Moss(1986)、BahnwebおよびJackle(1986)ならびにChamberlainおよびMoss(1988)により概説されている。
【0004】
便宜上、トラウストチトリドは、最初、分類学者により藻菌類(Phycomycetes)(藻類様菌類)における他の無色遊走子真核生物と定められた。藻菌類(Phycomycetes)という名前は、しかしながら、結局、分類学的地位から除外され、トラウストチトリドは、卵菌類(Oomycetes)(二鞭毛遊走子の菌類)に保持された。卵菌類(Oomycetes)は不等毛藻類に関連していると最初に想定され、結局、Barr(1983)により要約されているが、広範囲の超微細構造および生化学的研究によってこの想定を裏付けられた。卵菌類は、事実上、Leedale(1974)および他の藻類学者により不等毛藻類の群として認められた。しかしながら、それらの従属栄養の性質による都合上、重宝さの理由から、卵菌類およびトラウストチトリドは、藻類学者(藻類を研究する科学者)よりもむしろ、菌学者(菌類を研究する科学者)により、主に研究されてきた。
【0005】
別の分類学的観点から、進化生物学者は、いかにして真核生物は進化したかについての考えの2つの一般的な学派を発展させてきた。一つの学説は、一連の内部共生を通しての膜結合型細胞小器官の外因性起源を提案している(Margulis, 1970);例えば、ミトコンドリアは細菌の内部共生に由来し、葉緑体は藍色植物に、および鞭毛はスピロヘータに由来した。他方の学説は、自生過程による、原核生物の祖先の非膜結合型系からの膜結合型細胞小器官の漸進的進化を示唆している(Cavalier-Smith, 1975)。しかしながら、進化生物学者の両グループは、卵菌類およびトラウストチトリドを菌類から除外し、それらを有色植物界(Chromophyta)(Cavalier-Smith, 1981)(この界は、より最近に、他の原生生物を含むように拡大され、この界のメンバーは現在、ストラメノパイル(Stramenopiles)と呼ばれている)における有色植物藻類かまたはプロトクチスタ(Protoctista)界(MargulisおよびSagan, 1985)における全藻類に定めている。
【0006】
電子顕微鏡の発達で、トラウストチトリドの2つの属、トラウストチトリウムおよびシゾチトリウム(Perkins, 1976; Kazama, 1980; Barr, 1981)の遊走子の超微細構造についての研究により、トラウストチトリアシエが卵菌類にわずかに遠く関連しているのみという有力な証拠が提供された。さらに、5SリボソームRNA配列の一致分析(多変量統計学の型)を表す遺伝子データより、トラウストチトリアレスは明らかに、真核生物の独自の群であり、菌類から完全に分離し、かつ赤色および茶色藻類、ならびに卵菌類のメンバーに最も近く関連していることが示されている(Mannellaら、1987)。ほとんどの分類学者は、トラウストチトリドを卵菌類から除くことに同意した(Bartnicki-Garcia, 1988)。
【0007】
要約すれば、Cavalier-Smithの分類体系(1981, 1983)を用いて、トラウストチトリドは、有色植物界(ストラメノパイル)における有色植物藻類に分類されている。これは、それらを、真正菌界にすべてが配置されている菌類とは完全に異なる界に配置している。トラウストチトリドの分類学的配置は、それゆえに、以下に要約される:
界:有色植物界(ストラメノパイル)
門:不等毛門(Heterokonta)
目:トラウストチトリアレス
科:トラウストチトリアシエ
属:トラウストチトリウム、シゾチトリウム、ラビリンチュロイデス、またはジャポノチトリウム
【0008】
ある初期の分類学者らは、トラウストチトリウム(アメーバ状生活期をもつもの)属の数個の最初のメンバーをウルケニア(Ulkenia)と呼ばれる異なる属へと分離した。しかしながら、すべてではないにしても、ほとんどのトラウストチトリド(トラウストチトリウムおよびシゾチトリウムを含む)は、アメーバ状期を現すことは現在知られており、それとして、ウルケニアは、ある人たちには妥当な属であるとみなされていない。本明細書に用いられる場合、トラウストチトリウム属は、ウルケニアを含むものとする。
【0009】
門および界の上級分類内での分類学的配置の不確定性にもかかわらず、トラウストチトリドは、メンバーがトラウストチトリアレス目内に分類できるままの示差的かつ特徴的な群分けを保持している。
【0010】
シゾチトリウムおよび他のトラウストチトリアレス微生物は、高度不飽和脂肪酸(HUFA)および様々なカロテノイド(例えば、アスタキサンチン)を含む、大量の類脂質化合物を産生する能力のために、実質的に現存する、および可能性のある商業的価値をもっている。オメガ-3高度不飽和脂肪酸は、それらが動脈硬化および冠動脈性心疾患を防ぐための、炎症状態を軽減するためのならびに腫瘍細胞の増殖を妨げるための重要な食餌療法化合物として、近年、認識されてきているという点で、かなりの商業的関心対象となっている。これらの有益な効果は、オメガ-6脂肪酸から生成される化合物の競合的阻害をひき起こすオメガ-3HUFA、およびオメガ-3HUFA自身から直接的に生成される有益な化合物の両方の結果である(Simopoulosら、1986)。オメガ-6脂肪酸は、植物および動物において見出される主な(predominant)HUFAである。それゆえ、商業的な生産生物体(commercial production organism)としてのトラウストチトリアレス微生物のさらなる開発は、組換えDNAテクノロジーにより生物体に特定の遺伝子変化を生じる能力から有意に利益を得るものと思われ、そのような生物体により高い価値のあるHUFAおよびカロテノイドの産生を高めることが含まれる。さらに、この特徴付けが乏しい生物体群の生化学および分子生物学のより十分な理解を獲得する能力は、さらなる系統開発の試みを導くために用いられうる価値ある情報を提供するものと思われる。しかしながら、本発明の前には、シゾチトリウム属およびトラウストチトリウムのメンバーを含むトラウストチトリドを形質転換するために適する方法および組換え構築物は利用できなかった。トラウストチトリアレス微生物を形質転換するために特に有用である選択マーカーの開発およびトラウストチトリアレス特異的プロモーター配列の同定は、本発明の前には利用できなかったということは重大なことである。
【0011】
以前の研究者らは、トラウストチトリアレス目のメンバーではない微小藻類を含む様々な微生物において使用される形質転換方法および試薬を記載している。Allnuttらによる米国特許第6,027,900号は、光合成藻類の集光性遺伝子のためのプロモーターおよび選択マーカーとしてのストレプトアロテイコスヒンズスタヌス(Streptoalloteichus hindustanus)由来のSh ble遺伝子を用いて、真核生物の藻類、および特に、ファエオダチラムトリコルナタム(Phaeodactylum tricornutum)の遺伝子操作に使用される遺伝子融合体を開示している。細胞は、高濃度の塩(例えば、10 g/L〜35 g/L)および形質転換体の選択のためのゼオシン(Zeocin)(商標)において増殖する。そのような方法を用いる形質転換に適する微小藻類細胞は、高塩条件下で増殖しうる光合成微小藻類である。Dunahayらによる米国特許第5,661,017号は、クロロフィルC含有藻類(例えば、珪藻類)におけるマーカー発現に適する制御配列に実施可能に連結された選択マーカーを含む組換え構築物を用いてクロロフィルC含有藻類を形質転換するための方法を開示している。選択マーカーは、細菌および真菌源から単離されたマーカーを含む、任意の適するマーカーとして開示されており、ネオマイシンホスホトランスフェラーゼが好ましい。調節管理配列は、クロロフィルC含有藻類由来のいずれの制御配列も含みうり、および好ましくは、シクロテラクリプチカ(Cyclotella cryptica)由来である(例えば、C. クリプチカのアセチルCoAカルボキシラーゼ制御配列)。
【0012】
しかしながら、本発明より以前には、トラウストチトリアレス(例えば、微小藻類)のような微生物の形質転換は日常的ではなかったため、そのような方法がトラウストチトリアレス微生物の形質転換へ容易に転移可能ではない。細菌および酵母について十分開発されるようになったマーカーおよび形質転換系は、必ずしも、他の微生物に容易に適合可能であるとは限らない。実際、米国特許第5,661,017号は、「真核生物の微小藻類についての形質転換系の開発においてはほとんど成功しなかった」(1段落目の49行〜51行目)と書き留めており、一部分は、そのような微生物への外来DNAの導入することの困難性により、および一部分は、そのような形質転換に使用される適するマーカーおよびベクターがないことによる。米国特許第5,661,017号に記載される系は、特にクロロフィルC含有藻類のために開発された。なぜなら、それらの発明者らは、特に他の藻類と比較して、それらが遺伝的形質転換を受け入れられると考えたからである。同様に、光合成微小藻類に特異的である形質転換方法を教えている米国特許第6,027,900号は、たいていの藻類がいずれの型の遺伝子操作に対しても不応性であるという考えを語っている(1段落目の39行〜47行目)。細菌、酵母、昆虫および動物の細胞のために適合した系は、微小藻類に容易に適合させられなかった。それゆえ、本発明の前には、微小藻類に特異的である効果的な形質転換系についての要求が当技術分野において依然としてあった。
【0013】
さらに、トラウストチトリアレス目は、現在、ストラメノパイルにおける褐色植物藻類に分類されているが、これらの微生物はほとんどの微小藻類とは全く異なっているという当技術分野におけるある人たちによる意見がまだあり、当業者の一部には、トラウストチトリアレスのメンバーが微小藻類として正しく分類されているとは全くいえない可能性があるという意見をもっている。微小藻類であるとみなされる微生物は、進化の間、少なくとも4つの別々の時代を進化し、「微小藻類」型微生物が異なる界に配置されることになる(例えば、紅藻、緑藻および黄金色藻類(褐色植物門)は、すべて別々の界にある)。結果として、他の微小藻類において有用であると実証された形質転換系が、トラウストチトリアレスに対して有用であるとは期待されない。それゆえ、トラウストチトリアレス微生物の商業的価値にもかかわらず、遺伝子工学によりそのような微生物の最大限の可能性を利用する能力は、これまで実現されなかった。本発明の前では、本発明者らは、トラウストチトリアレス微生物の形質転換のために有用ななんのプロモーター、選択マーカーまたはベクターも認識していなかったし、また何の選択系がトラウストチトリアレスに用いられうるまたは適合しうるかについてなんの知識もなかった。
【0014】
要約すれば、トラウストチトリアレス微生物を形質転換するための方法を開発し、それにより商業的価値が高められた系統を作製するための手段を提供することの当技術分野における要求がある。さらに、トラウストチトリアレス微生物における相同的または非相同的組換えによる特定の遺伝子の突然変異または不活化のための方法を開発し、細胞代謝を変える、およびトラウストチトリアレスにおける特定の遺伝子の機能を同定するための新規な方法を提供することの当技術分野における要求がある。
【発明の概要】
【0015】
本発明の一つの態様は、以下のものからなる群より選択される核酸配列を含む単離された核酸分子に関する:(a)タンパク質がアセト乳酸シンターゼである、配列番号:15、配列番号:19、配列番号:22、および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードする核酸配列;(b)タンパク質がアセト乳酸シンターゼである、(a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列を有するタンパク質をコードする核酸配列;ならびに、(c)(a)または(b)の核酸配列と完全に相同的である核酸配列。一つの局面において、そのような核酸配列は、(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するタンパク質をコードし、かつそのタンパク質は、アセト乳酸シンターゼである。もう一つの局面において、そのような核酸配列は、(a)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するタンパク質をコードし、かつそのタンパク質は、アセト乳酸シンターゼである。さらにもう一つの局面において、そのような核酸配列は、116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする。一つの局面において、核酸配列は、配列番号:15、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードし、かつそのタンパク質はアセト乳酸シンターゼである。さらにもう一つの局面において、核酸配列は、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。
【0016】
好ましくは、上記で同定される核酸配列によりコードされるタンパク質の発現は、そのような核酸分子で形質転換されるトラウストチトリアレス目の微生物に、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を与える。この態様の一つの局面において、核酸配列は、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードする。この態様のもう一つの局面において、核酸配列は、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。
【0017】
本発明の一つの態様において、上記の核酸配列は、シゾチトリウムのアセト乳酸シンターゼをコードする。一つの局面において、シゾチトリウムのアセト乳酸シンターゼの発現は、その核酸分子で形質転換されるトラウストチトリアレス目の微生物に、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を与える。
【0018】
本発明のもう一つの態様は、転写制御配列に実施可能に連結されている、上記の単離された核酸分子のいずれかを含む組換え核酸分子に関する。本発明のもう一つの態様は、そのような組換え核酸分子に形質転換されるトラウストチトリアレス目の組換え微生物に関する。
【0019】
本発明のさらにもう一つの態様は、トラウストチトリアレス目の微生物の形質転換のための組換えベクターに関する。ベクターは、トラウストチトリアレス目の微生物に、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を与えるアセト乳酸シンターゼをコードする核酸配列を含む。アセト乳酸シンターゼは、以下のものからなる群より選択されるアミノ酸配列を有する:(a)配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および、(b)(a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列。アセト乳酸シンターゼをコードする核酸配列は、転写制御配列に実施可能に連結される。一つの局面において、組換えベクターは、発現ベクターである。もう一つの局面において、組換えベクターは、ターゲティングベクターである。他の局面において、ベクターにおける核酸配列は、(a)のアミノ酸配列と少なくとも約85 %同一である、およびもう一つの局面において、少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする。一つの局面において、核酸配列は、116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする。好ましい局面において、アセト乳酸シンターゼは、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する。もう一つの局面において、核酸配列は、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。組換えベクターにおける転写制御配列は、限定されるものではないが、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、またはトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターを含みうる。一つの局面において、ベクターは、配列番号:18、配列番号:21および配列番号:23からなる群より選択される核酸配列を含む。
【0020】
本発明のさらにもう一つの態様は、トラウストチトリアレス目の微生物の細胞を形質転換する方法に関する。方法は、(a)アセト乳酸シンターゼが(i)配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;ならびに、(ii)(i)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列からなる群より選択されるアミノ酸配列を有する、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を細胞に与えるアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子をトラウストチトリアレス目の微生物の細胞へ導入する第一段階を含む。方法は、第二段階に(b)化合物がスルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される、形質転換されていない細胞に対して阻害性である少なくとも1つの化合物を含む培地において(a)の細胞を培養することにより、組換え核酸分子で成功裡に形質転換された細胞を選択する段階を含む。一つの局面において、核酸配列は、(i)のアミノ酸配列と少なくとも約85 %同一である、およびより好ましくは少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする。一つの局面において、核酸配列は、116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする。もう一つの局面において、アセト乳酸シンターゼは、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する。好ましくは、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。さらにもう一つの局面において、核酸配列は、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される転写制御配列に実施可能に連結される。
【0021】
一つの局面において、組換え核酸分子は、タンパク質をコードする核酸配列が転写制御配列に実施可能に連結されている、細胞により産生されうるタンパク質をコードする核酸配列をさらに含む。この態様の一つの局面において、タンパク質は、ドコサヘキサエン酸(DHA)、ドコサペンタエン酸(DPA)、エイコサペンタエン酸(EPA)およびアラキドン酸(ARA)からなる群より選択される脂肪酸の合成に関係している。この態様のもう一つの局面において、タンパク質は、脂肪酸シンターゼ、脂肪酸デサチュラーゼ、脂肪酸エロンガーゼ、ポリケチドシンターゼ複合体に関係するタンパク質、およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質からなる群より選択される。一つの局面において、タンパク質は、核酸配列の配列番号:29によりコードされるω-3脂肪酸デサチュラーゼである。もう一つの局面において、タンパク質は、ポリエン脂肪酸イソメラーゼである。さらにもう一つの局面において、タンパク質は、HMG-CoAシンターゼ、HMG-CoAレダクターゼ、スクアレンシンターゼ、フィトエンシンターゼ、フィトエンデサチュラーゼ、カロテノイドシクラーゼ、カロテノイドヒドロキシラーゼ、カロテノイドケトラーゼ、ビタミンEおよびリポ酸からなる群より選択される。
【0022】
本方法のもう一つの局面において、段階(a)における組換え核酸分子は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活化されるように微生物においてターゲット核酸配列とハイブリダイズする核酸配列をさらに含む。この局面において、ターゲット核酸配列は、リパーゼ、脂肪酸酸化酵素、糖合成に関与するタンパク質、イソプレノイド経路の生成物の合成に関与するタンパク質、細胞壁成分の合成に関与するタンパク質、飽和脂肪酸合成経路に関与するタンパク質、多価不飽和脂肪酸合成経路に関与するタンパク質、ポリケチドシンターゼ複合体に関係するタンパク質、およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質からなる群より選択されるタンパク質をコードしうる。
【0023】
本方法は、核酸配列が転写制御配列に実施可能に連結されている、発現されうるタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含みうる。もう一つの局面において、方法は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活化されるように、微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列を含む、少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含みうる。もう一つの局面において、方法は、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子を細胞へ導入する段階をさらに含みうる。この局面において、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、第二のタンパク質をコードする核酸配列が転写制御配列に実施可能に連結されている、細胞により発現されうる第二のタンパク質をコードする核酸配列をさらに含む。そのような転写制御配列は、限定されるものではないが、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターを含みうる。この態様のさらなる局面において、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列をさらに含む。一つの態様において、組換え核酸分子は、核酸配列の配列番号:9を含む。
【0024】
本発明の方法において、微生物は、限定されるものではないが、トラウストチトリウム、ラビリンチュロイデス(Labyrinthuloides)、ジャポノチトリウム(Japonochytrium)、およびシゾチトリウム(Shizochytrium)を含む属由来でありうる。一つの局面において、微生物は、限定されるものではないが、シゾチトリウム属種(Schizochytrium sp.)、シゾチトリウムアグレガタム(Schizochytrium aggregatum)、シゾチトリウムリマシナム(Schizochytrium limacinum)、トラウストチトリウム属種(Thraustochytrium sp.)、トラウストチトリウムストリアタム(Thraustochytrium striatum)、トラウストチトリウムオーレウム(Thraustochytrium aureum)、トラウストチトリウムローゼウム(Thraustochytrium roseum)、およびジャポノチトリウム属種(Japonochytrium sp.)を含む種由来である。
【0025】
本方法の一つの態様において、微粒子銃、電気穿孔法、マイクロインジェクション、リポフェクション、吸着、感染、およびプロトプラスト融合からなる群より選択される方法により行われる。
【0026】
本発明のもう一つの態様は、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を微生物に与えるアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子で形質転換された、トラウストチトリアレス目の組換え微生物に関する。アセト乳酸シンターゼは、以下のものからなる群より選択されるアミノ酸配列を有する:(a)配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および、(b)(a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列。一つの局面において、核酸配列は、(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする。もう一つの局面において、アセト乳酸シンターゼは、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する。さらにもう一つの局面において、核酸配列は、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される。さらにもう一つの局面において、組換え核酸分子は、配列番号:18、配列番号:21および配列番号:23からなる群より選択される核酸配列を含む。好ましくは、アセト乳酸シンターゼをコードする核酸配列は、トラウストチトリアレスの微生物において機能するプロモーターに実施可能に連結される。一つの局面において、アセト乳酸シンターゼをコードする核酸配列は、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される転写制御配列に実施可能に連結される。一つの態様において、組換え核酸分子は、第一のタンパク質をコードする核酸配列が転写制御配列に実施可能に連結されている、微生物による産生のための第一タンパク質をコードする核酸配列をさらに含む。もう一つの態様において、組換え細胞は、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子でさらに形質転換される。好ましくは、組換え核酸分子は、核酸配列の配列番号:9を含む。一つの態様において、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、第二のタンパク質をコードする核酸配列が転写制御配列に実施可能に連結されている、細胞による産生のための第二のタンパク質をコードする核酸配列をさらに含む。一つの態様において、微生物はまた、細胞による産生のためのタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子も含む。
【0027】
本発明のさらにもう一つの態様は、以下のものからなる群より選択される核酸配列を含む単離された核酸分子に関する:(a)配列番号:9のヌクレオチド441位〜894位;(b)核酸配列が少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、配列番号:9のヌクレオチド441位〜894位の完全長に対して配列番号:9のヌクレオチド441位〜894位と少なくとも約95 %同一である核酸配列;および(c)(a)または(b)のポリヌクレオチドと完全に相同的である核酸配列を含む単離されたポリヌクレオチド。好ましくは、単離された核酸分子は、配列番号:4および配列番号:9のヌクレオチド441位〜894位からなる群より選択される核酸配列を含む。
【0028】
本発明のさらにもう一つの態様は、トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択されるプロモーターに実施可能に連結されるブレオマイシン結合タンパク質をコードする核酸配列を含んでいる、トラウストチトリアレス目の微生物の形質転換のための組換えベクターに関する。一つの局面において、トラウストチトリアレスのアセト乳酸シンターゼプロモーターは、配列番号:14のヌクレオチド1位〜1259位を含む。一つの局面において、α-チューブリンプロモーターは、配列番号:9のヌクレオチド441位〜894位、および核酸配列が少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、配列番号:9のヌクレオチド441位〜894位の完全長に対して配列番号:9のヌクレオチド441位〜894位と少なくとも約95 %同一である核酸配列からなる群より選択される核酸配列を含む。もう一つの局面において、トラウストチトリアレスPKS系由来のプロモーターは、配列番号:34または配列番号:34内に含まれる核酸配列を含み、該プロモーターは少なくとも基礎的なPKSプロモーター転写活性をもつ。もう一つの局面において、組換えベクターは、配列番号:8および配列番号:9からなる群より選択される核酸配列を含む。
【図面の簡単な説明】
【0029】
【図1】組換えプラスミドpTUBZEO-11の構築物を示す。
【図2】組換えプラスミドpTUBZEO 11-2の構築物を示す。
【図3A】組換えプラスミドpMON50200の構築物を示す。
【図3B】組換えプラスミドpMON50201の構築物を示す。
【図3C】組換えプラスミドpMON50202の構築物を示す。
【図3D】組換えプラスミドpMON50203の構築物を示す。
【発明を実施するための形態】
【0030】
発明の詳細な説明
本発明は、トラウストチトリアレス目の微生物を遺伝的に形質転換するための方法および関連した材料を含む。本発明の組換え構築物の形質転換体としての使用のために本明細書に開示される単細胞微生物のすべての株は、一般的にトラウストチトリドとも呼ばれうるのだが、トラウストチトリアレス目のメンバーである。本発明により、句「トラウストチトリド」、「トラウストチトリアレス微生物」および「トラウストチトリアレス目の微生物」は、交換可能に用いられうる。本発明者らは、シゾチトリウムまたは他のいずれのトラウストチトリアレス微生物についての形質転換系を記載する先行報告を一つも認識していない。本明細書に記載される形質転換系は、外来遺伝子をトラウストチトリアレス目の微生物へ導入し、それにより高められた商業的価値をもつ系統を作製するための手段を提供するために用いられうる。さらに、本発明は、相同的または非相同的組換えによる特定の遺伝子の変異または不活化を可能にし、細胞の代謝を変えるための、およびトラウストチトリアレス微生物における特定の遺伝子の機能を同定するための新規な方法を提供する。
【0031】
より具体的に言うと、本発明者らは、2つの型の形質転換ベクターの使用により、シゾチトリウム属のトラウストチトリアレス微生物(目:トラウストチトリアレス;科:トラウストチトリアシエ;属:シゾチトリウム)の遺伝的形質転換を実証した。これらのベクターは、標準的方法により細胞へ導入され、続いて、選択化合物の存在下で増殖するそれらの能力に基づいて組換え細胞が同定および単離されうる。本発明者らは、これらのベクターの有効性を、微粒子銃によりそれらを導入することによって実証したが、そのベクターを導入しうる他の手段もまた使用されうり(例えば、電気穿孔法)、それらは当技術分野において公知であり、本発明に含まれるものとする。
【0032】
一つの形質転換ベクターに関し、本明細書ではpTUBZEO 11-2と表される組換えベクターによって本明細書に例証されているのだが、ストレプトアロテイコスヒンズスタヌス由来のble遺伝子(「ブレオマイシン結合タンパク質」をコードする)がシゾチトリウムチューブリン遺伝子プロモーターから下流に配置されたキメラの遺伝子が作製された。SV40ターミネーターは、この構築物においてble遺伝子から下流に配置された。このベクターは、シゾチトリウムにおいてble遺伝子の発現を可能にし、それにより、適当なレベルで増殖培地に含まれる場合に野生型細胞に対して毒性があるゼオシン(Zeocin)(商標)および関連化合物に対する耐性を与える。この構築物におけるble遺伝子およびSV40ターミネーターは、pSV40/Zeoと名付けられている商業的に入手可能なベクターであり、Invitrogen Corporation(Carlsbad, CA)(Technical Manual 180202, Version B, "ZeoCassette Vectors"; Invitrogen Corporation, 1600 Faraday Ave., Carlsbad, CA 92008)から取得された。チューブリン遺伝子プロモーターは、ポリメラーゼ連鎖反応法により単離された;その反応に使用されるプライマーの1つは、ランダムシゾチトリウムcDNAシーケンシングプロジェクトにより得られた配列データに基づいた。pTUBZEO 11-2の地図は、図2に示され、pTUBZEO 11-2のヌクレオチド配列は、配列番号:9により表されている。このベクターでのシゾチトリウムの形質転換は、シゾチトリウムゲノムへ組み込まれたベクター配列の存在を検出するためのポリメラーゼ連鎖反応法およびサザンブロット分析の使用により確認された。
【0033】
ble遺伝子は、細菌、非トラウストチトリド微小藻類、真菌、原生動物、植物、および動物細胞を含む、様々な生物体の遺伝的形質転換の選択マーカーとして以前より研究者により使用されていた(例えば、米国特許第6,027,900号;Lumbrerasら、1998, Plant J. 14:441-447; Roheら、1996, Curr. Genet. 29:587-590; Messinaら、1995, Gene 165:213-217; Guerreroら、1992, Appl. Microbiol. Biotechnol. 36:759-762; Perezら、1989, Plant Mol. Biol. 13:365-373; Gatignoら、1990 Gene 91:35-41を参照)。ble遺伝子は、ブレオマイシン、フレオマイシン、およびゼオシン(Zeocin)(商標)を含むいくつかの抗生物質に対する耐性を与える「ブレオマイシン結合タンパク質」をコードする(Drocourtら、1990, Nucleic Acids Res. 18:4009)。この遺伝子はInvitrogen Corporationから商業的に入手可能であり、本発明者らがシゾチトリウム形質転換ベクターpTUBZEO 11-2を作製するために使用した遺伝子の源であった。しかしながら、本発明者らが、トラウストチトリドにおいてble遺伝子の発現を可能にする様式でその遺伝子がトラウストチトリドのプロモーターに付着している形質転換ベクターを作製した最初の者であると考えられる。
【0034】
形質転換ベクターの第二セットは、本発明者らがシゾチトリウムのゲノムライブラリーから単離した、アセト乳酸シンターゼ遺伝子(als)のインビトロでの部位特異的突然変異誘発により作製された。これらの突然変異は、トラウストチトリアレス目の微生物が感受性であるスルホメツロンメチルおよび他のスルホニル尿素化合物、加えてイミダゾリノン類阻害剤およびピリミジニルオキシ安息香酸に対して感受性が大いに弱くなるように、そのコードされた酵素(ALS)のアミノ酸配列を変化させる。スルホメツロンメチル(SMM)のようなスルホニル尿素化合物は、それらが様々な生物体由来の酵素アセト乳酸シンターゼ(ALS)を結合して不活化することができるため、しばしば、細胞に対し毒性がある。ALSは、アミノ酸のバリン、ロイシン、およびイソロイシンの生合成の第一段階を触媒する。イミダゾリノン、トリアゾロピリミジン、および他の化合物もまた、ある特定の生物体由来のALSを結合して不活化することが示された。他の生物体由来のアセト乳酸シンターゼ(アセトヒドロキシ酸シンターゼとしても知られている)をコードする遺伝子の変異体型は、酵母および植物の形質転換のための選択マーカーとして以前は使用されていた(Annu. Rev. Plant Physiol. Plant Mol. Biol. 40:441-470, 1989)。しかしながら、シゾチトリウムもしくは他のいずれのトラウストチトリアレスのメンバー由来のals遺伝子の配列もしくは性質、またはスルホニル尿素、イミダゾリノン、もしくはピリミジニルオキシ安息香酸化合物に対する耐性を与えるための変異体トラウストチトリアレスals遺伝子の使用を記載する報告は、本発明の前にはない。それどころか、本発明者らの知るかぎりでは、スルホメツロンメチルを含むこれらの選択薬剤に対するトラウストチトリアレス微生物の感受性について発表された報告でさえ一つもなかったため、そのような選択マーカーがトラウストチトリド形質転換系において使用に適しているかどうかさえ、本発明の前には知られていなかった。既知のals遺伝子と実質的相同性をもつ遺伝子は様々な生物体において見出されるが、アセト乳酸の合成を触媒することができる酵素をコードしていないことは、注目すべきである(Biochimica et Biophysica Acta 1385:401-419, 1998)。それゆえに、クローニングされたals相同体が実際にALSをコードすることが明らかであったとは思われない。クローニングされたシゾチトリウム遺伝子が本当のals遺伝子であったかどうかを明確に決定するために、本発明者らは、形質転換実験を通して、変異した推定上のシゾチトリウムals遺伝子の発現とスルホメツロンメチル耐性との正の相関性を実証した。
【0035】
本発明者らは、変異体als遺伝子を含む3つの異なる形質転換ベクターを作製した:一つの変異体als遺伝子は、トリプトファンの代わりに595位にバリンを有する酵素をコードする(プラスミドpMON50201、またはALSmut1-7)、もう一つは、プロリンの代わりに192位にグルタミンをコードする(プラスミドpMON50202、またはALSmut2-2)、および3つ目は、これらの変異の両方を含む(プラスミドpMON50203、またはALSmut3-5)。これらのベクターにおいて、組換え変異体als遺伝子の発現は、天然のals遺伝子プロモーターおよびターミネーターの制御下にある。これらのベクターの地図は、野生型シゾチトリウムals遺伝子を含むベクター(プラスミドpMON50200、またはAE-5)といっしょに、図3A〜図3Dに示されている。これらの変異体ALSコード化ベクターによるシゾチトリウムの形質転換は、シゾチトリウムゲノムへ組み込まれたベクター配列の存在を検出するためのポリメラーゼ連鎖反応法およびサザンブロット分析の使用により確認された。下で詳細に記載されているように、本発明者らがals遺伝子についての完全な配列を同定したからには、他の変異も、下で明細に記されているが、作製されうる。それゆえ、記載される変異体als遺伝子は、例示的なものであり、可能な変異体のすべてを含めるものではない。
【0036】
本発明の形質転換系は、同時形質転換により外来遺伝子をトラウストチトリアレス細胞へ導入するために用いられた。これらの場合において、外来遺伝子は、様々なシゾチトリウムのプロモーターと適当なターミネーター(例えば、SV40またはシゾチトリウム遺伝子ターミネーター領域)との間に配置された。例えば、本発明者らは、シゾチトリウムにおいてドコサヘキサエン酸のレベルを増加させるために、本明細書で配列番号:29に表されている、線虫カエノルハブディティスエレガンス(Caenorhabditis elegans)由来のω-3脂肪酸デサチュラーゼをコードする合成遺伝子を作製および導入した。配列番号:30は、配列番号:29によりコードされるデサチュラーゼのアミノ酸配列を表す。外来遺伝子を含む発現カセットはまた、選択マーカー含有形質転換ベクター内への直接的封入によりトラウストチトリアレス細胞へ導入されうる。
【0037】
さらに、本発明者らはまた、変異体ALSコード化ベクターを用いてシゾチトリウムにおいて相同的組換えが生じることを実証し、特定の天然シゾチトリウム遺伝子を不活化または変異するために組換え手段を用いることの可能性を示している。
【0038】
本明細書に記載されるトラウストチトリアレスプロモーター配列に関して、トラウストチトリアレス目のメンバーについて報告されたヌクレオチドおよびタンパク質の配列のすべてに対する配列データベース検索(GenBank)により、本発明の時点では、シゾチトリウムまたはトラウストチトリアレスの他のいずれのメンバー由来のプロモーター配列も報告されていなかったことが示される。いずれかのシゾチトリウム属種から報告された唯一の遺伝子は、S. アグレガタム(S. aggregatum)の5SリボソームRNAについてである(GenBankアクセッション番号X06104およびM13616)。5Sおよび18SリボソームRNA配列は、トラウストチトリアレスのメンバー、ウルケニア(Ulkenia)種、ならびにラビリンチュロイデス(Labyrinthuloides)属およびトラウストチトリウム(Thraustochytrium aureum)について報告されているが、これは本発明にはなんの関係もない。トラウストチトリウムオーレウム由来の「推定T3/T7様RNAポリメラーゼ」遺伝子の部分的コード領域が記載されているが(Nucleic Acids Research 15:648-654, 1996)、この遺伝子についてのプロモーター配列は報告されなかった。
【0039】
本発明は、トラウストチトリアレス目の微生物へ、対象となるいずれの遺伝子または他のヌクレオチド配列を導入するために用いられうる。そのようなヌクレオチド配列は、限定されるものではないが、脂肪酸(例えば、脂肪酸:ドコサヘキサエン酸(DHA)、ドコサペンタエン酸(DPA)、エイコサペンタエン酸(EPA)およびアラキドン酸(ARA))の合成に関係するタンパク質(例えば、酵素)をコードする核酸を含む。そのようなタンパク質は、限定されるものではないが、脂肪酸シンターゼ、脂肪酸デサチュラーゼ、および脂肪酸エロンガーゼ、加えて、ポリケチドシンターゼ複合体に関係するタンパク質および/またはそのような脂肪酸のリン脂質もしくはトリアシルグリセロール分子への取り込みに関係するタンパク質を含む。例えば、本発明は、ドコサペンタエン酸(DPA)のω-3不飽和化により細胞におけるドコサヘキサエン酸(DHA)のレベルを増加させるための試みにおいてシゾチトリウムへ様々な脂肪酸デサチュラーゼをコードする遺伝子を導入するために用いられた。もう一つの例として、シゾチトリウムにおいて、紅藻、プティロタ(Ptilota)由来の推定ポリエン脂肪酸イソメラーゼの発現もまた実証された。シゾチトリウムポリケチドシンターゼ複合体(すなわち、ポリケチドシンターゼ系)をコードする遺伝子は、GenBankアクセッション番号AF378327(ORFA)、AF378328(ORFB)およびAF378329(ORFC)として寄託されている。
【0040】
本発明はまた、トラウストチトリアレス微生物へイソプレノイド生合成経路に関係するタンパク質をコードする遺伝子および他のヌクレオチド配列を導入するためにも有用である。そのようなタンパク質は、限定されるものではないが、HMG-CoAシンターゼおよびHMG-CoAレダクターゼを含む。他の適するタンパク質は、限定されるものではないが、様々なステロイド化合物および様々なカロテノイド化合物を含むイソプレノイドサブユニット由来の分子の合成に関係するタンパク質を含む。様々なカロテノイド化合物の合成に関係するタンパク質は、限定されるものではないが、スクアレンシンターゼ、フィトエンシンターゼ、フィトエンデサチュラーゼ、ならびに様々なカロテノイドのシクラーゼ、ヒドロキシラーゼおよびケトラーゼを含む。
【0041】
本発明はまた、限定されるものではないが、ビタミンEおよびリポ酸を含む抗酸化化合物の合成に関係するタンパク質をコードする1つまたは複数の核酸配列をトラウストチトリアレスへ導入するためにも有用である。
【0042】
さらに、本発明は、遺伝子を不活化するまたは欠失させる(すなわち、「ノックアウト」または「ターゲットされた遺伝子破壊」)ために、トラウストチトリアレス微生物へ任意の遺伝子または他のヌクレオチドを導入するために用いられうる。遺伝子の不活化または欠失は、典型的には、微生物の商業的価値を高めることの目的のために用いられる。例えば、飽和および多価不飽和脂肪酸合成経路の酵素をコードする遺伝子(またはそのような遺伝子の発現を制御する核酸)を除去することが望ましい場合がある。もう一つの局面において、トラウストチトリアレス微生物により産生される他の価値ある化合物の分解に関与しているタンパク質または望ましい化合物の価値を他の方法で低めるタンパク質をコードする遺伝子を抑制もしくはノックアウトすることが望ましい場合がある。例えば、リパーゼ、脂肪酸酸化酵素、および不快な風味または匂いをもつタンパク質をコードする遺伝子が、本発明による望ましいノックアウトのターゲットでありうる。さらにもう一つの局面において、合成が、対象となる他の分子と競合している化合物の合成に付随しているタンパク質をコードする遺伝子をノックアウトすることが望ましい場合がある。例えば、そのような遺伝子は、限定されるものではないが、炭水化物生合成に関与しているタンパク質をコードする遺伝子、イソプレノイド経路の様々な生成物(例えば、ステロールまたは特定のカロテノイド化合物)の合成に関与しているタンパク質をコードする遺伝子、および細胞壁成分の合成に関与しているタンパク質をコードする遺伝子を含む。例として、高度不飽和脂肪酸(HUFA)の産生における役割を評価するために、シュワネラ(Shewanella)由来のポリケチドシンターゼ遺伝子と相同的である遺伝子を不活化する試みにおいて、本発明の使用によりシゾチトリウム細胞へ遺伝子が導入された。実施例6により例証されているように、本発明はまた、化合物に関連している製品の経済面または受け入れやすさを向上させるために、脂肪酸、カロテノイド、ステロール、ビタミン、または他の化合物の産生に関与している天然の遺伝子を不活化する、欠失させる、または変異させるために用いられうる。上記で考察されているが、ある態様においては、与えられたタンパク質の産生を増強することが望ましいと考えられるが、別の態様においては、同じタンパク質の産生を抑制することが望ましいと考えられる。そのような決定は、特定の微生物についての所与の使用および生産目標に基づいている。本発明はまた、シゾチトリウムにおいて遺伝的組換えの過程を測定するためにも有用である。
【0043】
トラウストチトリアレスへの導入のために有用な他の遺伝子および核酸分子は、当業者にとって明らかであると思われ、すべてのそのような遺伝子および分子は、本発明に含まれるものとする。
【0044】
本発明の様々な態様は、最初は本発明のトラウストチトリアレスals遺伝子および/またはALSタンパク質に関して下に記載されている。しかしながら、用語および方法の一般的な定義は、下記の他の遺伝子、核酸およびタンパク質の考察に適用されるものであることは理解されるべきである。
【0045】
本発明は、トラウストチトリド微生物の形質転換のための、組換え構築物での使用に適する選択マーカーをコードする核酸配列の同定、単離および作製に一部基づいている。そのような選択マーカーは、本発明の組換え構築物で成功裡に形質転換された微生物の選択を可能にする。本発明によるトラウストチトリアレスの形質転換のために有用な一つの選択マーカーは、トラウストチトリアレスのアセト乳酸シンターゼ(すなわち、ALS)である。好ましくは、アセト乳酸シンターゼは、スルホニル尿素化合物、イミダゾリノン類阻害剤、および/またはピリミジニルオキシ安息香酸による阻害に対する耐性があるように改変されている、変異されている、または別な方法で選択されている(すなわち、そのようなALSは、天然に存在するアセト乳酸シンターゼの相同体である)。
【0046】
本発明によると、アセト乳酸シンターゼは、天然に存在するアセト乳酸シンターゼの完全長タンパク質、融合タンパク質、またはいずれの相同体も含む、アセト乳酸シンターゼの生物学的活性をもつタンパク質である。アセト乳酸シンターゼの相同体は、1個または少数に限定されるものではないが、少なくとも1個または少数のアミノ酸が欠失(例えば、ペプチドまたは断片のようなタンパク質の切断型)、挿入、逆位、置換および/または誘導体化(例えば、グリコシル化、リン酸化、アセチル化、ミリストイル化、フェニル化、パルミチン化、アミド化および/またはグリコシルホスファチジルイノシトールの付加)されている点において天然に存在するアセト乳酸シンターゼと異なるタンパク質を含む。天然に存在するアセト乳酸シンターゼの好ましい相同体は、下で詳細に記載されている。
【0047】
本発明による、単離されたアセト乳酸シンターゼのような単離されたタンパク質は、それの本来の環境から取り出された(すなわち、人の操作を受けた)タンパク質であり、例えば、精製されたタンパク質、部分的に精製されたタンパク質、組換え的に産生されたタンパク質、および合成的に作製されたタンパク質を含みうる。そのようなものとして、「単離された」とは、そのタンパク質が精製された程度を反映しない。好ましくは、本発明の単離されたアセト乳酸シンターゼは組換え的に産生されている。「トラウストチトリアレスのアセト乳酸シンターゼ」は、トラウストチトリアレス微生物由来の、または他の方法として天然に存在するトラウストチトリアレス微生物由来のアセト乳酸シンターゼの構造(例えば、配列)の知識から作製されたアセト乳酸シンターゼ(天然に存在するアセト乳酸シンターゼの相同体を含む)を指す。換言すれば、トラウストチトリアレスのアセト乳酸シンターゼは、本明細書で詳細に記載されているように、トラウストチトリアレス微生物由来の天然に存在するアセト乳酸シンターゼの構造および機能をもつ、またはトラウストチトリアレス微生物由来の天然に存在するアセト乳酸シンターゼの生物学的活性のある(すなわち、生物学的活性をもつ)相同体である任意のアセト乳酸シンターゼも含む。それとして、トラウストチトリアレスのアセト乳酸シンターゼは、精製された、部分的に精製された、組換え型の、変異型/改変型の、および合成的なタンパク質を含みうる。
【0048】
一般的に、タンパク質の生物学的活性または生物学的作用は、インビボ(すなわち、そのタンパク質の本来の生理学的環境)またはインビトロ(すなわち、実験室条件下)で測定または観察される場合、そのタンパク質の天然存在型とされるタンパク質により示されるまたは行われる任意の機能を指す。例えば、アセト乳酸シンターゼの生物学的活性はアセト乳酸シンターゼ酵素的活性を含む。相同体または模倣体においてのような(下記で考察されている)タンパク質の改変は、天然に存在するタンパク質と同じ生物学的活性をもつタンパク質、または天然に存在するタンパク質と比較して減少または増加した生物学的活性をもつタンパク質を結果として生じうる。タンパク質発現における減少またはタンパク質の活性における減少を結果として生じる改変は、タンパク質の不活化(完全または部分的)、下方制御、または減少した作用として言及されうる。同様に、タンパク質発現における増加またはタンパク質の活性における増加を結果として生じる改変は、タンパク質の増幅、過剰産生、活性化、増強、上方制御または増加した作用として言及されうる。
【0049】
本発明のアセト乳酸シンターゼに関して、アセト乳酸シンターゼ相同体に存在する改変は、天然に存在するアセト乳酸シンターゼと比較して実質的に変化させない、または少なくとも天然に存在するタンパク質と比較してシンターゼの基礎的な生物学的活性を実質的に減少させないことが好ましい。しかしながら、そのような相同体は、天然に存在するタンパク質と比較してある特定の化合物による阻害に対する減少した感受性のような、天然存在型と比較してタンパク質の機能的または酵素的活性以外の特性における相違をもちうる。好ましくは、天然に存在するアセト乳酸シンターゼの相同体は、その相同体が派生した天然に存在するアセト乳酸シンターゼと比較して、天然に存在するアセト乳酸シンターゼを結合して不活化する、化合物に対する低下した(すなわち、減少した、小さくなった)感受性をもつ。例えば、スルホメツロンメチル(SMM)のようなスルホニル尿素化合物は、それらがアセト乳酸シンターゼ(ALS)を結合して不活化することができるため、しばしば、細胞への毒性がある。イミダゾリノン、トリアゾロピリミジン、および他の類似した化合物(一般的に本明細書ではイミダゾリノン類阻害剤と呼ばれる)もまた、ALSを結合して不活化することが示された。それゆえ、天然に存在するアセト乳酸シンターゼの相同体は、好ましくは、アセト乳酸シンターゼの酵素活性を維持すると同時に、スルホニル尿素化合物に対する、加えてイミダゾリノン類阻害剤に対する(例えば、そのような阻害剤についての破壊された結合部位または阻害剤への低下した親和性をもつ結合部位を有することにより)、およびピリミジニルオキシ安息香酸に対する低下した感受性をもつ。
【0050】
本明細書に用いられる場合、「アセト乳酸シンターゼ生物学的活性」をもつ、または「アセト乳酸シンターゼ」と呼ばれるタンパク質は、アミノ酸のバリン、ロイシン、およびイソロイシンの生合成における第一段階を触媒するタンパク質を指す。より具体的に言うと、完全長タンパク質、切り詰められたタンパク質、融合タンパク質、および相同体を含む本発明の単離されたアセト乳酸シンターゼは、ピルビン酸からアセト乳酸の合成を触媒するタンパク質の能力により単刀直入式に同定されうる。アセト乳酸シンターゼ生物学的活性は、当業者により、任意の適するインビトロまたはインビボの酵素活性アッセイ法により評価されうる。
【0051】
一つの態様において、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼである(すなわち、アセト乳酸シンターゼ生物学的活性をもつ)、配列番号:15、配列番号:19、配列番号:22および配列番号:24の群より選択されるアミノ酸配列と、そのような配列の任意の少なくとも約600個のアミノ酸に対して、少なくとも約65 %同一であるアミノ酸配列を有する。より好ましくは、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも約600個のアミノ酸に対して、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと、少なくとも約70 %同一である、より好ましくは少なくとも約75 %同一である、およびよりいっそう好ましくは少なくとも約80 %同一である、およびよりいっそう好ましくは少なくとも約85 %同一である、およびよりいっそう好ましくは少なくとも約90 %同一である、およびよりいっそう好ましくは少なくとも約95 %同一である、およびよりいっそう好ましくは少なくとも約96 %同一である、およびよりいっそう好ましくは少なくとも約97 %同一である、およびよりいっそう好ましくは少なくとも約98 %同一である、およびよりいっそう好ましくは少なくとも約99 %同一であるアミノ酸配列を有する。
【0052】
もう一つの態様において、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼである(すなわち、アセト乳酸シンターゼ生物学的活性をもつ)、配列番号:15、配列番号:19、配列番号:22、配列番号:24の群より選択されるアミノ酸配列と、そのような配列のいずれかの少なくとも95個より多くのアミノ酸に対して、少なくとも約75 %同一であるアミノ酸配列を有する。より好ましくは、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも95個より多くのアミノ酸に対して、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと、少なくとも約80 %同一である、およびよりいっそう好ましくは少なくとも約85 %同一である、およびよりいっそう好ましくは少なくとも約90 %同一である、およびよりいっそう好ましくは少なくとも約95 %同一である、およびよりいっそう好ましくは少なくとも約96 %同一である、およびよりいっそう好ましくは少なくとも約97 %同一である、およびよりいっそう好ましくは少なくとも約98 %同一である、およびよりいっそう好ましくは少なくとも約99 %同一であるアミノ酸配列を有する。よりいっそう好ましくは、本発明のアセト乳酸シンターゼは、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも100個より多くのアミノ酸、およびより好ましくは125個、およびより好ましくは150個、およびより好ましくは175個、およびより好ましくは200個、およびより好ましくは225個、およびより好ましくは250個、およびより好ましくは275個、およびより好ましくは300個、およびより好ましくは325個、およびより好ましくは350個、およびより好ましくは375個、およびより好ましくは400個、およびより好ましくは425個、およびより好ましくは450個、およびより好ましくは475個、およびより好ましくは500個、およびより好ましくは525個、およびより好ましくは550個、およびより好ましくは575個のアミノ酸に対して配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと上記の参照されたパーセントの同一性のいずれかをもつアミノ酸配列を有する。
【0053】
本明細書に用いられる場合、別なふうに明記されない限り、同一性の割合(%)の参照は、以下のものを用いて行われる相同性の評価を指す:(1)標準デフォルトパラメーターを用いるアミノ酸検索のためのblastp、および核酸検索のためのblastnを用いるBLAST 2.0ベーシックBLAST相同性検索であり、クエリ配列はデフォルトにより低複雑度領域についてフィルタリングされる(Altschul, S.F., Madden, T.L., Schaaffer, A.A., Zhang, J., Zhang, Z., Miller, W. & Lipman, D.J. (1997)「Gapped BLAST and PSI-BLAST:a new generation of protein database search programs.」Nucleic Acids Res. 25:3389-3402に記載されており、参照として完全に本明細書に組み入れられている);(2)BLAST 2アラインメント(下記のパラメーターを用いる);(3)および/または標準デフォルトパラメーターでのPSI-BLAST(位置特異的反復(Position-Specific Iterated)BLAST)。BLAST 2.0ベーシックBLASTとBLASTとの間の標準パラメーターにおけるいくらかの相違のために、2つの特定の配列がBLAST 2プログラムを用いて有意な相同性をもつとして認識されえたとしても、クエリ配列としてその配列のうちの1つを用いるBLAST 2.0ベーシックBLASTにおいて行われる検索では二番目の配列をトップ整合において同定しない可能性があることを留意する。さらに、PSI-BLASTは、「プロファイル」検索の自動化した使いやすいバージョンを提供しており、配列相同体を捜すための感度の高い方法である。プログラムは、まず、ギャップド(gapped)BLASTデータベース検索を行う。PSI-BLASTプログラムは、位置特異的スコア行列を構成するために戻されるいずれの有意なアラインメントからの情報も使用し、データベース検索の次のラウンドのためにクエリ配列を取り替える。それゆえ、これらのプログラムのいずれか1つを使用することにより同一性の割合が決定されうることは理解されるべきである。
【0054】
2つの特定の配列は、参照として完全に本明細書に組み入れられる、TatusovaおよびMadden, (1999), 「Blast 2 sequences-a new tool for comparing protein and nucleotide sequences」, FEMS Microbiol Lett. 174:247-250に記載されているように、BLAST 2配列を用いてお互いに整列しうる。BLAST 2配列アラインメントは、その結果生じるアラインメントにギャップの導入(欠失および挿入)を許容して2つの配列間でギャップドBLAST検索(BLAST 2.0)を行うBLAST 2.0アルゴリズムを用いるblastpまたはblastnにおいて行われる。本明細書での明瞭さを目的として、BLAST 2配列アラインメントは、以下の通り、標準デフォルトパラメーターを用いて行われる。
【0055】
0 BLOSUM62行列を用いる、blastnについて:
一致についての報酬 = 1
不一致についてのペナルティ = -2
オープンギャップ(5)および伸長ギャップ(2)ペナルティ
ギャップx_ドロップオフ(50)期待(10)ワードサイズ(11)フィルター(オン)
0 BLOSUM62行列を用いる、blastpについて:
オープンギャップ(11)および伸長ギャップ(1)ペナルティ
ギャップx_ドロップオフ(50)期待(10)ワードサイズ(3)フィルター(オン)。
【0056】
本発明のアセト乳酸シンターゼはまた、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも30個の近接するアミノ酸残基(すなわち、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの30個の近接するアミノ酸と100 %同一性をもつ30個の近接するアミノ酸残基)を含むアミノ酸配列を有するタンパク質を含みうる。好ましい態様において、本発明のアセト乳酸シンターゼは、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも50個、およびより好ましくは少なくとも75個、およびより好ましくは少なくとも100個、およびより好ましくは少なくと115個、およびより好ましくは少なくとも130個、およびより好ましくは少なくとも150個、およびより好ましくは少なくとも200個、およびより好ましくは少なくとも250個、およびより好ましくは少なくとも300個、およびより好ましくは少なくとも350個、およびより好ましくは少なくとも400個、およびより好ましくは少なくとも450個、およびより好ましくは少なくとも500個、およびより好ましくは少なくとも550個、およびより好ましくは少なくとも600個、およびより好ましくは少なくとも650個の近接するアミノ酸残基を含むアミノ酸配列を有するタンパク質を含む。そのようなタンパク質は、アセト乳酸シンターゼ生物学的活性をもつ。
【0057】
本発明により、本明細書に記載される核酸またはアミノ酸の配列に関しての用語「近接する」または「連続した」は、間断のない配列として連結されていることを意味する。例えば、第一配列が第二配列の30個の近接する(または連続した)アミノ酸を含むこととは、第一配列が、第二配列における30個のアミノ酸残基の間断のない配列と100 %同一である30個のアミノ酸残基の間断のない配列を含むことを意味する。同様に、第一配列が第二配列と「100 %同一性」をもつこととは、第一配列が、ヌクレオチドまたはアミノ酸の間にギャップを含まずに第二配列と正確に整合していることを意味する。
【0058】
もう一つの態様において、本発明のアセト乳酸シンターゼは、アセト乳酸シンターゼ相同体を含めて、相同体をコードする核酸配列が、中程度の、高度の、または非常に高度のストリンジェント性条件下で(下に記載されている)、天然に存在するアセト乳酸シンターゼをコードする核酸分子(すなわち、天然に存在するアセト乳酸シンターゼアミノ酸配列をコードする核酸鎖の相補体)と(すなわち、核酸分子と)ハイブリダイズする能力があるような、天然に存在するアセト乳酸シンターゼアミノ酸配列と十分に類似しているアミノ酸配列を有するタンパク質を含む。好ましくは、アセト乳酸シンターゼは、配列番号:15、配列番号:19、配列番号:22または配列番号:24により表されるアミノ酸配列を含むタンパク質をコードする核酸配列の相補体へ中程度の、高度の、または非常に高度のストリンジェント性条件下でハイブリダイズする核酸配列によりコードされる。よりいっそう好ましくは、本発明のアセト乳酸シンターゼは、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、または配列番号:23のヌクレオチド1260位〜3314位の相補体へ中程度の、高度の、または非常に高度のストリンジェント性条件下でハイブリダイズする核酸配列によりコードされる。そういうものとしてハイブリダイゼーション条件は、下で詳細に記載されている。本発明のアセト乳酸シンターゼをコードする核酸配列の核酸配列相補体は、アセト乳酸シンターゼをコードする鎖に相補的である核酸鎖の核酸配列を指す。与えられたアミノ酸配列をコードする二本鎖DNAは一本鎖DNAおよびその一本鎖への相補体である配列を有するそれの相補鎖を含むことは認識されているものと思われる。それとして、本発明の核酸分子は、二本鎖かまたは一本鎖かどちらでもありうり、かつアミノ酸配列の配列番号:15、配列番号:19、配列番号:22または配列番号:24をコードする核酸配列と、および/またはそのようなアミノ酸配列のいずれかをコードする核酸配列の相補体とストリンジェントなハイブリダイゼーション条件下での安定的なハイブリッドを形成するそれらの核酸分子を含む。相補性配列を導き出すための方法は当業者に知られている。アミノ酸シーケンシングおよび核酸シーケンシングのテクノロジーは完全に誤りのないものではないため、本明細書に提示される配列はせいぜい本発明のアセト乳酸シンターゼの見かけの配列を表していることを留意されたい。
【0059】
アセト乳酸シンターゼ相同体は、天然の対立遺伝子の変異または自然の突然変異の結果でありうる。本発明のアセト乳酸シンターゼ相同体はまた、限定されるものではないが、タンパク質への直接的改変、または、例えば、ランダムなもしくはターゲットされた突然変異誘発をもたらすための古典的もしくは組換えDNA技術を用いるタンパク質をコードする遺伝子への改変を含む、当技術分野において公知の技術を用いて作製されうる。アセト乳酸シンターゼをコードする核酸の、天然に存在する対立遺伝子変異体は、アミノ酸配列の配列番号:15をコードする遺伝子と本質的に同じゲノムにおける遺伝子座に見出されるが、例えば、突然変異または組換えにより引き起こされた自然の変異のためで、類似しているが同一ではない配列を有する遺伝子である。天然の対立遺伝子変異体は、典型的にはそれらが比較されることになる遺伝子によりコードされるタンパク質と同様の活性をもつタンパク質をコードする。対立遺伝子変異体の一つの部類は、同じタンパク質をコードしうるが、遺伝子コードの縮重により異なる核酸配列を有する。対立遺伝子変異体はまた、遺伝子の5'または3'の非翻訳領域における(例えば、制御領域における)改変を含む。対立遺伝子変異体は当業者によく知られている。
【0060】
本発明のアセト乳酸シンターゼタンパク質はまた、遺伝子融合体(例えば、組換えタンパク質の可溶性活性型を過剰発現するために用いられる)、変異を誘発された遺伝子(遺伝子転写および翻訳を増強するコドン改変を有する遺伝子のような)、および切り詰められた遺伝子(膜タンパク質の可溶性型を生じるために膜結合ドメインが除去された遺伝子、または特定の組換え宿主において耐容性が乏しいシグナル配列が除去された遺伝子のような)の発現産物を含む。
【0061】
本発明のタンパク質および/または相同体の最小サイズは、アセト乳酸シンターゼ生物学的活性をもつのに十分なサイズである。好ましくは、本発明のタンパク質は、少なくとも30個のアミノ酸の長さであり、およびより好ましくは少なくとも約50個、およびより好ましくは少なくとも75個、およびより好ましくは少なくとも100個、およびより好ましくは少なくとも115個、およびより好ましくは少なくとも130個、およびより好ましくは少なくとも150個、およびより好ましくは少なくとも200個、およびより好ましくは少なくとも250個、およびより好ましくは少なくとも300個、およびより好ましくは少なくとも350個、およびより好ましくは少なくとも400個、およびより好ましくは少なくとも450個、およびより好ましくは少なくとも500個、およびより好ましくは少なくとも550個、およびより好ましくは少なくとも600個、およびより好ましくは少なくとも650個、およびより好ましくは少なくとも684個のアミノ酸の長さである。タンパク質がアセト乳酸シンターゼタンパク質の部分または完全長アセト乳酸シンターゼ、望ましい場合には追加的な配列(例えば、融合タンパク質配列)を加えて含みうることにおいて、そのようなタンパク質の最大サイズに、実際的な制限以外に制限はない。
【0062】
本発明はまた、1つまたは複数の融合セグメントに付着されたアセト乳酸シンターゼ含有ドメイン(すなわち、本発明によるアセト乳酸シンターゼについてのアミノ酸配列)を含む融合タンパク質を含む。本発明で使用されるのに適する融合セグメントは、限定されるものではないが、タンパク質の安定性を向上させることができる;他の望ましい生物学的活性を提供できる;および/またはアセト乳酸シンターゼの精製(例えば、アフィニティークロマトグラフィーによる)を補助することができるセグメントを含む。適する融合セグメントは、望ましい機能をもつ(例えば、増加した安定性、可溶性、作用もしくは生物学的活性を与える;および/またはタンパク質の精製を簡単にする)任意のサイズのドメインでありうる。融合セグメントは、そのタンパク質のアセト乳酸シンターゼ含有ドメインのアミノ末端および/またはカルボキシル末端に連結されうり、アセト乳酸シンターゼの直接的回収を可能にするために切断されやすくありうる。融合タンパク質は、好ましくは、アセト乳酸シンターゼ含有ドメインのカルボキシル末端および/またはアミノ末端のいずれかに付着された融合セグメントを含むタンパク質をコードする融合核酸分子でトランスフェクションされた組換え細胞を培養することにより産生される。
【0063】
本発明はまた、アセト乳酸シンターゼの模倣体を含む。本明細書で用いられる場合、用語「模倣体」は、模倣体がしばしば、天然に存在するペプチドの基本的構造を模倣する基本的構造をもつ、および/または天然に存在するペプチドの顕著な生物学的性質をもつことから、天然に存在するペプチドの生物学的作用を模倣することができる任意のペプチドまたは非ペプチド化合物を指すために用いられる。模倣体は、限定されるものではないが、以下のものを含みうる:天然に存在するペプチドと側鎖類似性をもたないような原型由来の実質的改変をもつペプチド(そのような改変は、例えば、それの分解されやすさを減らしうる);抗イディオタイプ抗体および/もしくは触媒抗体、またはそれらのフラグメント;単離されたタンパク質の非タンパク質性部分(例えば、炭水化物構造);または、例えば、コンビナトリアルケミストリーにより同定される核酸および薬剤を含む、合成もしくは天然の有機分子。
【0064】
そのような模倣体は、当技術分野において公知の様々な方法を用いて設計、選択および/または別な方法で同定されうる。本発明において有用な模倣体または他の治療的化合物を設計するために有用なドラッグデザインの様々な方法は、Maulikら、1997, Molecular Biotechnology: Therapeutic Applications and Strategies, Wiley-Liss, Inc.に開示されており、参照として完全に本明細書に組み入れられている。アセト乳酸シンターゼ模倣体は、例えば、分子多様性ストラテジー(大量の化学的に多様な分子ライブラリーの迅速な構築を可能にする関連ストラテジーの組み合わせ)から、天然もしくは合成の化合物のライブラリー、特に化学的もしくはコンビナトリアルライブラリー(すなわち、配列またはサイズにおいて異なるが、同種のビルディングブロックを有する化合物のライブラリー)から、または合理的な、定方向もしくはランダムなドラッグデザインにより得られうる。例えば、Maulikら、前記を参照されたい。
【0065】
分子多様性ストラテジーにおいて、大量化合物ライブラリーは、例えば、生物学的、酵素的および/または化学的方法を用いて、ペプチド、オリゴヌクレオチド、炭水化物および/または合成有機分子から合成される。分子多様性ストラテジーを開発するにおいて重要なパラメーターは、サブユニットの多様性、分子サイズ、およびライブラリーの多様性を含む。そのようなライブラリーをスクリーニングすることの一般的な目的は、所望のターゲットについての高親和性リガンドを得るためにコンビナトリアル選択の逐次適用を利用すること、そのうえ、ランダムまたは定方向のいずれかの設計ストラテジーによりリード分子を最適化することである。分子多様性の方法は、Maulikら、同上において詳細に記載されている。
【0066】
Maulikらはまた、例えば、使用者が適当に選択された断片の断片ライブラリーから新規な分子を作製する過程を方向づける、定方向設計の方法;使用者が、断片およびそれらの組み合わせをランダムに変化させ、同時に候補リガンドの適応度を評価するために選択基準を適用するために遺伝子または他のアルゴリズムを用いるランダム設計;および使用者が三次元の受容体構造と小断片プローブ間の相互作用エネルギーを計算し、続いて、有利なプローブ部位について結び付けて考える、グリッドに基づく方法を開示している。
【0067】
本発明によると、アセト乳酸シンターゼは、任意のトラウストチトリアレス微生物由来、および特に、任意のシゾチトリウム微生物由来でありうる。一つの態様において、本発明の好ましいアセト乳酸シンターゼは、配列番号:15、配列番号:19、配列番号:22、配列番号:24の群より選択されるアミノ酸配列を有する。配列番号:15により表されるアミノ酸配列を有するタンパク質は、トラウストチトリアレス微生物由来の天然に存在する(すなわち野生型)アセト乳酸シンターゼであり、もっと厳密に言えば、シゾチトリウムのアセト乳酸シンターゼである。配列番号:19、配列番号:22、配列番号:24により表されるアミノ酸配列は、その結果生じた酵素が、アミノ酸配列の配列番号:15により表される天然に存在するタンパク質と比較して、スルホニル尿素化合物、加えてイミダゾリノン類阻害剤およびピリミジニルオキシ安息香酸に対して低下した感受性をもつように、改変された配列である。配列番号:19、配列番号:22および配列番号:24により表されるタンパク質が、アセト乳酸シンターゼ生物学的活性をもつことは、注目される。スルホニル尿素化合物、加えてイミダゾリノン類阻害剤およびピリミジニルオキシ安息香酸に対して低下した感受性をもつアセト乳酸シンターゼは、本発明の好ましいアセト乳酸シンターゼであり、なぜなら、そのようなシンターゼをコードする核酸配列が、選択マーカーとして本発明の組換えベクターに使用されうるからである。
【0068】
それゆえに、本発明の一つの態様は、相同体がアセト乳酸シンターゼ生物学的活性をもつ、ならびに特に、相同体がアミノ酸配列の配列番号:15により表される天然に存在するタンパク質と比較して、スルホニル尿素化合物、加えてイミダゾリノン類阻害剤およびピリミジニルオキシ安息香酸に対して低下した感受性をもつ、配列番号:15、配列番号:19、配列番号:22および配列番号:24のいずれかの任意の相同体を含む改変されたアセト乳酸シンターゼに関する。一つの局面において、そのようなアセト乳酸シンターゼ相同体は、次の位置:116G、117A、192P、200A、251K、358M、383D、592V、595W、または599Fの1つまたは複数におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質を含む。これらの位置は、酵母のアセト乳酸シンターゼにおける既知のALS変異部位と対応している(すなわち、それぞれ、116G、117A、192P、200A、251K、354M、379D、583V、586W、および590F)(MazurおよびFalco, 1989, Annu. Rev. Plant Physiol. Plant Mol. Biol. 40:441-470を参照。これは参照として完全に本明細書に組み入れられている)。他の可能性のある変異部位は、他の生物体由来のALSにおいて成功したアミノ酸変異に基づき当業者に知られることになると思われる。そのような部位のトラウストチトリアレスのALSにおいての対応する部位への適用は、本発明に含まれる。
【0069】
上記で考察されているように、本発明は、トラウストチトリド微生物の形質転換のための組換え構築物の発見および作製に一部基づいている。それゆえ、本発明の一つの態様は、トラウストチトリアレスのアセト乳酸シンターゼをコードする核酸配列、およびそれに完全に相補的な核酸配列を含む単離された核酸分子に関する。本発明のアセト乳酸シンターゼをコードする核酸分子は、上記で考察されている相同体を含むアセト乳酸シンターゼタンパク質のいずれかをコードする核酸分子を含む。より詳しくは、本発明の一つの態様は、タンパク質がアセト乳酸シンターゼである(すなわち、アセト乳酸シンターゼ生物学的活性をもつ)、配列番号:15、配列番号:19、配列番号:22、配列番号:24の群より選択されるアミノ酸配列と、そのような配列のいずれかの少なくとも約600個より多くのアミノ酸に対して、少なくとも約65 %同一であるアミノ酸配列を有するタンパク質をコードする核酸配列を含む、単離された核酸分子に関する。より好ましくは、本発明の単離された核酸分子は、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも約600個より多くのアミノ酸に対して、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと、少なくとも約70 %同一である、およびより好ましくは少なくとも約75 %同一である、およびよりいっそう好ましくは少なくとも約80 %同一である、およびよりいっそう好ましくは少なくとも約85 %同一である、およびよりいっそう好ましくは少なくとも約90 %同一である、およびよりいっそう好ましくは少なくとも約95 %同一である、およびよりいっそう好ましくは少なくとも約96 %同一である、およびよりいっそう好ましくは少なくとも約97 %同一である、およびよりいっそう好ましくは少なくとも約98 %同一である、およびよりいっそう好ましくは少なくとも約99 %同一であるアミノ酸配列をコードする核酸配列を有する。
【0070】
もう一つの態様において、本発明の単離された核酸分子は、タンパク質がアセト乳酸シンターゼである(すなわち、アセト乳酸シンターゼ生物学的活性をもつ)、配列番号:15、配列番号:19、配列番号:22、配列番号:24の群より選択されるアミノ酸配列と、そのような配列のいずれかの少なくとも95個より多くのアミノ酸に対して、少なくとも約75 %同一であるアミノ酸配列をコードする核酸配列を有する。より好ましくは、本発明の単離された核酸分子は、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも95個より多くのアミノ酸に対して、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと、少なくとも約80 %同一である、およびよりいっそう好ましくは少なくとも約85 %同一である、およびよりいっそう好ましくは少なくとも約90 %同一である、およびよりいっそう好ましくは少なくとも約95 %同一である、およびよりいっそう好ましくは少なくとも約96 %同一である、およびよりいっそう好ましくは少なくとも約97 %同一である、およびよりいっそう好ましくは少なくとも約98 %同一である、およびよりいっそう好ましくは少なくとも約99 %同一であるアミノ酸配列をコードする核酸配列を有する。
【0071】
さらにもう一つの態様において、本発明の単離された核酸分子は、タンパク質がアセト乳酸シンターゼ生物学的活性をもっている、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも100個より多くのアミノ酸、およびより好ましくは125個、およびより好ましくは150個、およびより好ましくは175個、およびより好ましくは200個、およびより好ましくは225個、およびより好ましくは250個、およびより好ましくは275個、およびより好ましくは300個、およびより好ましくは325個、およびより好ましくは350個、およびより好ましくは375個、およびより好ましくは400個、およびより好ましくは425個、およびより好ましくは450個、およびより好ましくは475個、およびより好ましくは500個、およびより好ましくは525個、およびより好ましくは550個、およびより好ましくは575個のアミノ酸に対して配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかと上記の参照された同一性の割合のいずれかをもつアミノ酸配列をコードする核酸配列を有する。同一性の割合は、上記のように、BLAST 2.0ベーシックBLASTデフォルトパラメーターを用いて決定される。
【0072】
一つの態様において、本発明のアセト乳酸シンターゼをコードする核酸分子は、天然に存在するアセト乳酸シンターゼをコードする核酸配列の相補体と中程度のストリンジェント性条件下、およびよりいっそう好ましくは高ストリンジェント性条件下、およびよりいっそう好ましくは超高ストリンジェント性条件下でハイブリダイズする単離された核酸分子を含む。好ましくは、本発明のアセト乳酸シンターゼをコードする単離された核酸分子は、配列番号:15、配列番号:19、配列番号:22または配列番号:24により表されるアミノ酸配列を含むタンパク質をコードする核酸配列の相補体へ中程度または高度のストリンジェント性条件下でハイブリダイズする核酸配列を含む。一つの態様において、単離された核酸分子は、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、または配列番号:23のヌクレオチド1260位〜3314位により表される核酸配列の相補体へ中程度、高度または非常に高度のストリンジェント性条件下でハイブリダイズする核酸配列を含む。
【0073】
本明細書に用いられる場合、ハイブリダイゼーション条件は、類似した核酸分子を同定するために核酸分子が用いられる標準的ハイブリダイゼーション条件を指す。そのような標準的条件は、例えば、Sambrookら、Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Labs Press, 1989に開示されている。Sambrookら、同上は、参照として完全に本明細書に組み入れられている(特に、ページ9.31-9.62を参照)。さらに、ヌクレオチドのミスマッチの異なる程度を許容するハイブリダイゼーションを達成するための適当なハイブリダイゼーションおよび洗浄条件を計算するための式は、例えば、Meinkothら、1984, Anal. Biochem. 138, 267-284に開示されている;Meinkothら、同上は、参照として完全に本明細書に組み入れられている。
【0074】
より詳しくは、本明細書に参照される場合、中程度ストリンジェント性のハイブリダイゼーションおよび洗浄条件は、ハイブリダイゼーション反応において探索のために使用されることになる核酸分子と少なくとも約70 %の核酸配列同一性をもつ核酸分子の単離を可能にする条件を指す(すなわち、ヌクレオチドの約30 %またはそれ未満のミスマッチを許容する条件)。本明細書に参照される場合、高ストリンジェント性のハイブリダイゼーションおよび洗浄条件は、ハイブリダイゼーション反応において探索のために使用されることになる核酸分子と少なくとも約80 %の核酸配列同一性をもつ核酸分子の単離を可能にする条件を指す(すなわち、ヌクレオチドの約20 %またはそれ未満のミスマッチを許容する条件)。本明細書に参照される場合、超高ストリンジェント性のハイブリダイゼーションおよび洗浄条件は、ハイブリダイゼーション反応において探索のために使用されることになる核酸分子と少なくとも約90 %の核酸配列同一性をもつ核酸分子の単離を可能にする条件を指す(すなわち、ヌクレオチドの約10 %またはそれ未満のミスマッチを許容する条件)。上記で考察されているように、当業者は、ヌクレオチドミスマッチのこれらの特定のレベルを達成するための適当なハイブリダイゼーションおよび洗浄条件を計算するためにMeinkothら、同上における式を用いることができる。そのような条件は、DNA:RNAかまたはDNA:DNAかのいずれのハイブリッドが形成されることになっているかに依存して異なるものである。DNA:DNAハイブリッドについて計算される融解温度は、DNA:RNAハイブリッドについてよりも10℃低い。詳細な態様において、DNA:DNAハイブリッドについてのストリンジェントなハイブリダイゼーション条件は、6X SSC(0.9 M Na+)のイオン強度で、約20℃と約35℃の間(低い方のストリンジェント性)、より好ましくは約28℃と約40℃の間(よりストリンジェント)、およびよりいっそう好ましくは約35℃と約45℃の間(よりいっそうストリンジェント)の温度でのハイブリダイゼーションを適当な洗浄条件と共に含む。詳細な態様において、DNA:RNAハイブリッドについてのストリンジェントなハイブリダイゼーション条件は、6X SSC(0.9 M Na+)のイオン強度で、約30℃と約45℃の間、より好ましくは約38℃と約50℃の間、およびよりいっそう好ましくは約45℃と約55℃の間の温度でのハイブリダイゼーションを同様のストリンジェントな洗浄条件と共に含む。これらの値は、約100個のヌクレオチドより大きい分子、0 %ホルムアミドおよび約40 %のG + C含有量についての融解温度の計算に基づいている。または、Tmは、Sambrookら、前記、ページ9.31〜9.62に示されるように経験的に計算されうる。一般的に、洗浄条件は、できるかぎりストリンジェントであるべきであり、選択されたハイブリダイゼーション条件に対して適切であるべきである。例えば、ハイブリダイゼーション条件は、特定のハイブリッドの計算されたTmより下のおよそ20℃〜25℃である温度条件および塩の組み合わせを含みうり、洗浄条件は、典型的には、その特定のハイブリッドの計算されたTmより下のおよそ12℃〜20℃である温度条件および塩の組み合わせを含む。DNA:DNAハイブリッドでの使用に適したハイブリダイゼーション条件の一つの例は、6X SSC(50 %ホルムアミド)において約42℃で2時間〜24時間のハイブリダイゼーション、続いて約2X SSCにおいて室温での1回または複数回の洗浄を含む洗浄段階、続いてより高い温度およびより低いイオン強度での追加の洗浄(例えば、約37℃で約0.1X〜0.5X SSCにおいて少なくとも1回の洗浄、続いて約68℃で約0.1X〜0.5X SSCにおいて少なくとも1回の洗浄)を含む。
【0075】
もう一つの態様において、本発明のアセト乳酸シンターゼをコードする核酸分子は、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも30個の近接するアミノ酸残基(すなわち、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの30個の近接するアミノ酸と100 %同一性をもつ30個の近接するアミノ酸残基)を含むアミノ酸配列を有するタンパク質をコードする核酸配列を含む単離された核酸分子を含む。好ましい態様において、単離された核酸分子は、配列番号:15、配列番号:19、配列番号:22または配列番号:24のいずれかの少なくとも50個、およびより好ましくは少なくとも75個、およびより好ましくは少なくとも100個、およびより好ましくは少なくと115個、およびより好ましくは少なくとも130個、およびより好ましくは少なくとも150個、およびより好ましくは少なくとも200個、およびより好ましくは少なくとも250個、およびより好ましくは少なくとも300個、およびより好ましくは少なくとも350個、およびより好ましくは少なくとも400個、およびより好ましくは少なくとも450個、およびより好ましくは少なくとも500個、およびより好ましくは少なくとも550個、およびより好ましくは少なくとも600個、およびより好ましくは少なくとも650個の近接するアミノ酸残基を含むアミノ酸配列を有するタンパク質をコードする核酸配列を含む。そのようなタンパク質は、アセト乳酸シンターゼ生物学的活性をもつ。一つの態様において、アセト乳酸シンターゼをコードする単離された核酸分子は、配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、または配列番号:23のヌクレオチド1260位〜3314位の少なくとも60個の近接するヌクレオチド、およびより好ましくは少なくとも150個、およびより好ましくは少なくとも225個、およびより好ましくは少なくと300個、およびより好ましくは少なくとも345個、およびより好ましくは少なくとも390個、およびより好ましくは少なくとも450個、およびより好ましくは少なくとも525個、およびより好ましくは少なくとも600個、およびより好ましくは少なくとも750個、およびより好ましくは少なくとも900個、およびより好ましくは少なくとも1050個、およびより好ましくは少なくとも1200個、およびより好ましくは少なくとも1350個、およびより好ましくは少なくとも1500個、およびより好ましくは少なくとも1650個、およびより好ましくは少なくとも1800個、およびよりいっそう好ましくは少なくとも1950個の近接するヌクレオチドを有する核酸配列を含む。
【0076】
本発明の特に好ましい核酸分子は、配列番号:14のヌクレオチド1260位〜3314位(配列番号:15をコードする)、配列番号:18のヌクレオチド1260位〜3314位(配列番号:19をコードする)、配列番号:21のヌクレオチド1260位〜3314位(配列番号:22をコードする)、または配列番号:23のヌクレオチド1260位〜3314位(配列番号:24をコードする)、配列番号:14、配列番号:18、配列番号:21または配列番号:23を含む。
【0077】
本発明に従って、単離された核酸分子は、それの本来の環境から取り出された(すなわち、人の操作を受けた)核酸分子であり、それの本来の環境はその核酸分子が天然で見出されるゲノムまたは染色体である。そのようなものとして、「単離された」とは、必ずしもその核酸分子が精製された程度を反映するとは限らないが、その分子が、その核酸分子が天然で見出されるゲノム全体または染色体全体を含まないことを示す。単離された核酸分子は、本明細書に記載されるアセト乳酸シンターゼ遺伝子のような遺伝子を含みうる。遺伝子を含む単離された核酸分子は、そのような遺伝子を含む染色体の断片ではなく、むしろ、その遺伝子に関連したコード領域および制御領域を含み、同じ染色体に本来見出される付加的な遺伝子を含まない。単離された核酸分子はまた、天然では特定された核酸配列に普通、隣接していない(すなわち、異種性配列である)追加的な核酸により隣接された(すなわち、その配列の5'末端および/または3'末端において)特定の核酸配列を含みうる。単離された核酸分子は、DNA、RNA(例えば、mRNA)、またはDNAもしくはRNAのいずれかの誘導体(例えば、cDNA)を含みうる。句「核酸分子」は本来、物質的な核酸分子を指し、句「核酸配列」は本来、核酸分子についてのヌクレオチドの配列をさすが、その2つの句は、交換可能に、特に、タンパク質をコードする能力がある、核酸分子または核酸配列に関して用いられうる。
【0078】
好ましくは、本発明の単離された核酸分子は、組換えDNAテクノロジー(例えば、ポリメラーゼ連鎖反応(PCR)増幅、クローニング)または化学的合成を用いて作製される。単離された核酸分子は天然の核酸分子およびその相同体を含み、これらには限定されるものではないが、天然の対立遺伝子変異体およびタンパク質生物学的活性に望ましい効果を与えるような様式でヌクレオチドが挿入、欠失、置換、および/または逆位されている改変型核酸分子が含まれる。対立遺伝子変異体およびタンパク質相同体(例えば、核酸相同体によりコードされるタンパク質)は、上記で詳細に考察されている。
【0079】
核酸分子相同体は、当業者に公知の多数の方法を用いて作製されうる(例えば、Sambrookら、同上を参照)。例えば、核酸分子は、限定されるものではないが、部位特異的突然変異誘発、変異を誘発するための核酸分子の化学的処理、核酸断片の制限酵素切断、核酸断片のライゲーション、核酸配列の選択された領域のPCR増幅および/または突然変異誘発、核酸分子の混合物を「構築する(build)」ためのオリゴヌクレオチド混合物の合成および混合群のライゲーションならびにそれらの組み合わせのような、古典的突然変異誘発技術および組換えDNA技術を含む様々な技術を用いて改変されうる。核酸分子相同体は、核酸によりコードされるタンパク質の機能についてスクリーニングすることにより、および/または野生型遺伝子とのハイブリダイゼーションにより、改変型核酸の混合物から選択されうる。
【0080】
同様に、本発明の核酸分子の最小サイズは、望ましい生物学的活性をもつタンパク質をコードするのに十分な、または天然のタンパク質をコードする核酸分子の相補的配列と安定的ハイブリッドを形成する能力がある(例えば、中程度、高度または非常に高度のストリンジェント性条件下で)プローブもしくはオリゴヌクレオチドプライマーを形成するのに十分なサイズである。そのようなものとして、そのようなタンパク質をコードする核酸分子のサイズは、核酸成分および核酸分子と相補的配列間の相同性または同一性の割合、加えて、ハイブリダイゼーション条件それ自体(例えば、温度、塩濃度、およびホルムアミド濃度)に依存しうる。オリゴヌクレオチドプライマーまたはプローブとして用いられる核酸分子の最小サイズは、典型的には、核酸分子がGCリッチである場合には長さが少なくとも約12個〜約15個のヌクレオチド、ATリッチな場合には長さが少なくとも約15個〜約18個の塩基である。核酸分子がタンパク質コード配列(例えば、アセト乳酸シンターゼコード配列)の部分または完全長タンパク質をコードする核酸配列を含みうることにおいて、本発明の核酸分子の最大サイズに、実際的制限以外に制限はない。
【0081】
本発明の一つの態様は、トラウストチトリアレス微生物の形質転換のために用いられうる組換えベクターを含む。本発明により、組換えベクターは、選り抜きの核酸配列を操作するための、およびそのような核酸配列を宿主細胞へ導入するための道具として用いられる設計された(すなわち、人工的に作製された)核酸分子である。それゆえ、組換えベクターは、クローニング、シーケンシング、ならびに/または組換え細胞を形成するために選り抜きの核酸配列を宿主細胞へ発現することおよび/もしくは送達することによるような選り抜きの核酸配列を別なふうに操作することにおいて使用するのに適している。そのようなベクターは、典型的には、本来は送達されうる核酸配列に隣接して見出されない核酸配列である異種性核酸配列を含むが、ベクターはまた、本来より本発明の核酸分子に隣接して見出される制御核酸配列(例えば、プロモーター、非翻訳領域)を含みうる(下記で詳細に考察されている)。ベクターは、RNAまたはDNAのどちらでも、原核または真核のどちらでもありうり、典型的にはプラスミドである。ベクターは、染色体外エレメント(例えば、プラスミド)として維持されうる、またはそれは組換え微生物の染色体へ組み込まれうる。ベクター全体は宿主細胞内のしかるべき所に残りうる、またはある特定の条件下で、プラスミドDNAが除去されて、本発明の核酸分子をあとに残しうる。組み込まれた核酸分子は、染色体のプロモーター制御下、本来のまたはプラスミドのプロモーター制御下、またはいくつかのプロモーター制御の組み合わせ下でありうる。核酸分子の単一または複数のコピーが染色体へ組み込まれうる。本発明の組換えベクターは、トラウストチトリアレスのアセト乳酸シンターゼをコードする核酸配列(天然のタンパク質または相同体)またはble遺伝子をコードする核酸配列(下に記載されている)のような、本発明によるトラウストチトリアレス微生物のための少なくとも1つの選択マーカーを含む。本明細書に用いられる場合、句「組換え核酸分子」は、本来、宿主細胞へクローニングされうる、操作されうる、形質転換されうる核酸配列(すなわち、挿入断片)がライゲーションされている組換えベクターを指すために用いられる。
【0082】
典型的には、組換えベクター、およびゆえに組換え核酸分子は、1つまたは複数の転写制御配列へ実施可能に連結される本発明の少なくとも1つの核酸分子を含む。本明細書に用いられる場合、句「組換え分子」または「組換え核酸分子」は、本来、転写制御配列へ実施可能に連結される核酸分子または核酸配列を指すが、そのような核酸分子が本明細書で考察されているような組換え分子である場合、句「核酸分子」と交換可能に用いられうる。本発明により、句「実施可能に連結される」は、宿主細胞へトランスフェクションされる(すなわち、形質転換される、形質導入される、トランスフェクションされる、接合されるまたは導かれる)場合、分子が発現されうるような様式で核酸分子を転写制御配列へ連結することを指す。転写制御配列は、転写の開始、伸長、または終結を制御する配列である。特に重要な転写制御配列は、プロモーター、エンハンサー、オペレーターおよびリプレッサー配列のような転写開始を制御するものである。適する転写制御配列は、トラウストチトリアレス目の微生物において機能することができるいずれの転写制御配列も含む。本発明者らが、本明細書での他の所で詳細に記載されている、そのようなプロモーターの少なくとも3つを単離かつ同定した最初の者であると考えられる。
【0083】
好ましいプロモーターは、限定されるものではないが、トラウストチトリアレスのアセト乳酸シンターゼプロモーター(配列番号:14のヌクレオチド1位〜1259位により本明細書に表されている)、トラウストチトリアレスのα-チューブリンプロモーター(配列番号:9のヌクレオチド441位〜894位により本明細書に表されている)、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター(配列番号:34内に含まれる)、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーター(配列番号:31内に含まれる;その脂肪酸デサチュラーゼプロモーターは、米国特許仮出願第60/284,116号、前記においてデサチュラーゼプロモーターとして呼ばれていることを留意されたい。)を含む。α-チューブリンプロモーター、アセト乳酸シンターゼプロモーター、および脂肪酸デサチュラーゼプロモーターのクローニングおよびシーケンシングは、実施例の節において記載されている。好ましい態様において、α-チューブリンプロモーターは、天然に存在するトラウストチトリアレスα-チューブリンプロモーター配列(配列番号:9のヌクレオチド441位〜894位)、またはプロモーターが少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、配列番号:9のヌクレオチド441位〜894位と少なくとも95 %同一である核酸配列を含む。同様に、好ましいアセト乳酸シンターゼプロモーターは、天然に存在するトラウストチトリアレスのアセト乳酸シンターゼプロモーターの核酸配列(配列番号:14のヌクレオチド1位〜1259位内に表されている)、またはプロモーターが少なくとも基礎的なアセト乳酸シンターゼプロモーター転写活性をもっている、配列番号:14のヌクレオチド1位〜1259位と少なくとも75 %、およびより好ましくは80 %、およびより好ましくは85 %、およびより好ましくは90 %、およびより好ましくは95 %同一である核酸配列を含む。好ましいPKSプロモーターは、天然に存在するトラウストチトリアレスのPKSプロモーターの核酸配列(配列番号:34内に表されている)、またはプロモーターが少なくとも基礎的なPKSプロモーター転写活性をもっている、配列番号:34と少なくとも75 %、およびより好ましくは80 %、およびより好ましくは85 %、およびより好ましくは90 %、およびより好ましくは95 %同一である核酸配列を含む。最後に、好ましい脂肪酸デサチュラーゼプロモーターは、天然に存在するトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターの核酸配列(配列番号:31内に表されている)を含む、またはプロモーターが少なくとも基礎的な脂肪酸デサチュラーゼプロモーター転写活性をもっている、配列番号:31と少なくとも75 %、およびより好ましくは80 %、およびより好ましくは85 %、およびより好ましくは90 %、およびより好ましくは95 %同一である核酸配列内に含まれる。同一性の割合を決定するための方法は、アセト乳酸シンターゼ配列について本明細書で先に記載されており、本明細書に含まれている。
【0084】
一つの態様において、本発明の組換えベクターは、発現ベクターである。本明細書に用いられる場合、句「発現ベクター」は、コードされた産物(例えば、対象となるタンパク質)の産生に適するベクターを指すために用いられる。この態様において、産生されうる産物をコードする核酸配列は、組換え核酸分子を作製するために組換えベクターへ挿入される。産生されうるタンパク質をコードする核酸配列は、組換え微生物内で核酸配列の転写および翻訳を可能にするベクターにおいて制御配列(例えば、本発明のトラウストチトリアレスのプロモーター)へ核酸配列を実施可能に連結する様式でベクターへ挿入される。本発明の選択マーカーは、本発明の組換え核酸分子が成功裡に導入された組換え微生物の選択を可能にする。
【0085】
もう一つの態様において、本発明の組換えベクターは、ターゲティングベクターである。本明細書に用いられる場合、句「ターゲティングベクター」は、核酸分子が宿主細胞内で内因性遺伝子を欠失させるまたは不活化するために用いられる(すなわち、ターゲットされた遺伝子の破壊またはノックアウトテクノロジーのために用いられる)、特定の核酸分子を組換え細胞へ送達するために用いられるベクターを指すのに用いられる。そのようなベクターはまた、「ノックアウト」ベクターとして当技術分野で知られているかもしれない。この態様の一つの局面において、ベクターの一部、であるがより典型的には、ベクターへ挿入された核酸分子(すなわち、挿入断片)は、宿主細胞におけるターゲット遺伝子(すなわち、欠失させられるまたは不活化されるためにターゲットされる遺伝子)の核酸配列と相同的である核酸配列を有する。ベクター挿入断片の核酸配列は、ターゲット遺伝子および挿入断片が相同的組換えをうけて、内因性ターゲット遺伝子が欠失させられる、不活化されるまたは減弱される(すなわち、内因性ターゲット遺伝子の少なくとも一部が変異されるまたは欠失させられることにより)ように、ターゲット遺伝子に結合するように設計される。
【0086】
一つの態様において、本発明の好ましい組換えベクターは、トラウストチトリアレス微生物における使用に適している組換えベクターであり、本発明のアセト乳酸シンターゼ分子をコードする核酸配列を含む。好ましくは、アセト乳酸シンターゼは、天然に存在する型(配列番号:15)と比較して、組換えベクターでトランスフェクションされたトラウストチトリアレス微生物にスルホニル尿素化合物、イミダゾリノン類阻害剤、および/またはピリミジニルオキシ安息香酸に対する低下した感受性を与えるように改変されている。好ましくは、そのような組換えベクターは、次の位置:116G、117A、192P、200A、251K、358M、383D、592V、595W、または599Fの1つまたは複数のアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を含むアセト乳酸シンターゼタンパク質をコードする核酸配列を含む。一つの態様において、そのようなアセト乳酸シンターゼタンパク質は、限定されるものではないが、配列番号:19、配列番号:22および配列番号:24を含むアミノ酸配列を有する。好ましくは、そのような組換えベクターは、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位から選択される核酸配列を含む。特に好ましい態様において、望ましい耐性を与えるALSコード化核酸配列を含む組換えベクターは、配列番号:18、配列番号:21および配列番号:23を含む。
【0087】
一つの態様において、組換えベクターでトランスフェクションされたトラウストチトリアレス微生物にスルホニル尿素化合物、イミダゾリノン類阻害剤、および/またはピリミジニルオキシ安息香酸に対する低下した感受性を与えるのに適している組換えベクターは、天然に存在するシゾチトリウムのアセト乳酸シンターゼ配列である配列番号:15を含む。この態様において、組換えベクターは、天然に存在するシンターゼを過剰発現するように設計され、それによりそのような過剰発現が微生物に特定された化合物に対する耐性を与えるという効果を生ずる。この態様において、組換えDNAテクノロジーの使用が、例えば、宿主細胞内での核酸分子のコピー数、それらの核酸分子が転写される効率、その結果として生じる転写物が翻訳される効率、および翻訳後修飾の効率を操作することにより形質転換された核酸分子の発現の制御を向上させることができることは、当業者に認識されているものと思われる。さらに、プロモーター配列は、天然のプロモーターと比較して、発現のレベルを向上させるように遺伝子的に設計されうるものと思われる。核酸分子の発現を制御するために有用な組換え技術は、限定されるものではないが、1つまたは複数の宿主細胞染色体への核酸分子の組み込み、プラスミドへのベクター安定性配列の付加、転写制御シグナル(例えば、プロモーター、オペレーター、エンハンサー)の置換または改変、翻訳制御シグナルの置換または改変(リボソーム結合部位、シャイン-ダルガノ(Shine-Dalgarno)配列)、宿主細胞のコドン使用頻度に対応しうる核酸分子の改変、および転写物を不安定にする配列の除去を含む。
【0088】
本発明の一つの態様において、トラウストチトリアレス微生物の形質転換における使用に適する組換えベクターは、選択マーカーとしてストレプトアロテイコスヒンズスタヌス由来のSh ble遺伝子(「ブレオマイシン結合タンパク質」をコードする)を本明細書で先に記載されているトラウストチトリアレスプロモーターと組み合わせて含む。ble遺伝子およびトラウストチトリアレスプロモーターを含む好ましい組換えベクターは、例えば、配列番号:8または配列番号:9により表されるベクター配列を含む。ストレプトアロテイコスヒンズスタヌスのブレオマイシン結合タンパク質のアミノ酸配列は、配列番号:10として本明細書に表されている。
【0089】
本発明の組換え核酸分子は、DNAまたはRNAのどちらでもありうるが、組換え細胞と適合性のある翻訳制御配列、複製開始点、および他の制御配列のような付加的な制御配列を含む。一つの態様において、宿主細胞の染色体へと組み込まれるものを含む、本発明の組換え分子はまた、発現されるタンパク質がそのタンパク質を産生する細胞から分泌されうるようにするための分泌シグナル(すなわち、シグナルセグメント核酸配列)を含む。適するシグナルセグメントは、発現されうるタンパク質に本来付随しているシグナルセグメントまたは本発明によるタンパク質の分泌を方向づけることができるいずれの異種性シグナルセグメントも含む。もう一つの態様において、本発明の組換え分子は、発現されるタンパク質が送達されかつ宿主細胞の膜へ挿入されうるようにするためのリーダー配列を含む。適するリーダー配列は、そのタンパク質に本来付随しているリーダー配列、または細胞の膜へのタンパク質の送達および挿入を方向づけることができるいずれの異種性リーダー配列も含む。
【0090】
トラウストチトリアレス目の微生物を形質転換するのに有用である様々な道具を記載してきたが、本発明の一つの態様は、トラウストチトリアレス目の微生物の細胞を形質転換する方法に関する。方法は、(a)本明細書で先に記載されている組換え核酸分子をトラウストチトリアレス目の微生物の細胞へ導入する第一段階を含む。組換え核酸分子は、スルホニル尿素化合物、イミダゾリノン類阻害剤、および/またはピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を該細胞へ与えるアセト乳酸シンターゼをコードする核酸配列を含む。そのようなアセト乳酸シンターゼは、上で詳細に記載されており、配列番号:19、配列番号:22および配列番号:24、加えて、上記で考察されている、そのような配列または配列番号:15のいずれかの相同体により表されるアミノ酸配列を有するアセト乳酸シンターゼを含む。方法は、第二段階として(b)化合物がスルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される、形質転換されていない細胞に対して阻害性がある少なくとも1つの化合物を含む培地において(a)の細胞を培養することにより組換え核酸分子で成功裡に形質転換された細胞を選択する段階を含む。そのような化合物の存在下で増殖する細胞は成功裡に形質転換されている。
【0091】
本方法において使用される組換え核酸分子は、本明細書で先に記載されている本発明の組換えベクターのいずれかを含み、典型的には、組換え細胞により産生されうるタンパク質をコードする少なくとも1つの核酸配列(すなわち、組換え発現ベクターを含む)、または組換え細胞において内因性遺伝子のターゲットされた欠失または不活性化に有用な核酸配列(すなわち、組換えターゲティングベクターを含む)を含む。
【0092】
一つの態様において、組換え核酸分子は、タンパク質をコードする核酸配列が転写制御配列へ実施可能に連結されている、細胞により産生されうるタンパク質をコードする核酸配列をさらに含む。トラウストチトリアレスにおける産生に望まれるであろうタンパク質は、当業者に知られているものと思われ、すべて、本発明に含まれるものとする。特に好ましいタンパク質は、限定されるものではないが、ドコサヘキサエン酸(DHA)、ドコサペンタエン酸(DPA)、エイコサペンタンエン酸(EPA)およびアラキドン酸(ARA)からなる群より選択される脂肪酸の合成に関係するタンパク質を含む。そのようなタンパク質は、例えば、脂肪酸シンターゼ、脂肪酸デサチュラーゼ、脂肪酸エロンガーゼ、ポリケチドシンターゼ複合体に関係するタンパク質およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質を含む。一つの局面において、タンパク質は、核酸配列の配列番号:29によりコードされるω-3脂肪酸デサチュラーゼである。配列番号:30は、デサチュラーゼのアミノ酸配列を表している。もう一つの局面において、タンパク質はポリエン脂肪酸イソメラーゼである。一つの態様において、本方法を用いてトラウストチトリアレス微生物において産生されうるタンパク質は、イソプレノイド生合成経路に関係するタンパク質を含む。そのようなタンパク質は、限定されるものではないが、HMG-CoAシンターゼ、HMG-CoAレダクターゼ、スクアレンシンターゼ、フィトエンシンターゼ、フィトエンデサチュラーゼ、カロテノイドシクラーゼ、カロテノイドヒドロキシラーゼ、カロテノイドケトラーゼを含む。さらにもう一つの態様において、本方法を用いてトラウストチトリアレス微生物において産生されうるタンパク質は、限定されるものではないが、ビタミンEおよびリポ酸を含む。
【0093】
一つの態様において、本発明の方法において有用な組換え核酸分子は、ターゲット核酸配列を含む遺伝子が、第二の核酸配列との相同的組換えにより変異されるまたは不活性化されるように微生物においてターゲット核酸配列とハイブリダイズする核酸配列を含む。そのような核酸配列は、飽和および多価不飽和脂肪酸合成経路の酵素をコードする遺伝子(もしくはそのような遺伝子の発現を制御する核酸)、トラウストチトリアレス微生物により産生される他の価値ある化合物の分解に関与しているもしくは望ましい化合物の価値を別なふうに減らすタンパク質をコードする遺伝子、またはそれの合成が対象となる他の分子と競合している化合物の合成に付随しているタンパク質をコードする遺伝子と相同的でありうる。例えば、ターゲット核酸配列は、限定されるものではないが、リパーゼ、脂肪酸酸化酵素、糖合成に関与するタンパク質、イソプレノイド経路の生成物の合成に関与するタンパク質、細胞壁成分の合成に関与するタンパク質、飽和脂肪酸合成経路に関与するタンパク質、多価不飽和脂肪酸合成経路に関与するタンパク質、ポリケチドシンターゼ複合体に関係するタンパク質、およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質をコードする配列を含む。
【0094】
本発明の一つの態様において、トラウストチトリアレス微生物の形質転換のための方法は、核酸配列が転写制御配列へ実施可能に連結されている、発現されうるタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階を含む。または、追加の組換え核酸分子は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活性化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列を含みうる。この様式において、複数のタンパク質を細胞へ導入する、複数の遺伝子を不活性化する、またはその2つの組み合わせが可能である。追加の組換え核酸分子は、第一の組換え核酸分子と同時に(すなわち、同時形質転換)、または引き続きの形質転換として(例えば、「積み重ね(stacking)」形質の目的のために)トラウストチトリアレス微生物へ導入されうる。
【0095】
一つの態様において、方法は、ブレオマイシン結合タンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含む。この態様において、追加の組換え核酸分子は、好ましくは、同時形質転換としてよりもむしろ、次の段階において導入される。好ましくは、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、第二のタンパク質をコードする核酸配列が転写制御配列へ実施可能に連結されている、細胞により発現されうる第二のタンパク質をコードする核酸配列をさらに含む。または、もしくは加えて、ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子は、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより変異されるまたは不活性化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列をさらに含む。一つの態様において、そのような組換え核酸分子は、核酸配列の配列番号:9を含む。
【0096】
本発明の方法を用いて形質転換するために適する宿主細胞は、限定されるものではないが、トラウストチトリアレス目のいずれの微生物も含む。宿主細胞は、形質転換されていない細胞または少なくとも1つの核酸分子ですでにトランスフェクションされている細胞のどちらでもありうる。本発明において使用される好ましい宿主細胞は、限定されるものではないが、トラウストチトリウム、ラビリンチュロイデス、ジャポノチトリウム、およびシゾチトリウムを含む属由来の微生物を含む。これらの属内の好ましい種は、限定されるものではないが、シゾチトリウムアグレガタム、シゾチトリウムリマシナム、シゾチトリウムミヌタム(Schizochytrium minutum)を含むいずれのシゾチトリウム属種も;トラウストチトリウムストリアタム、トラウストチトリウムオーレウム、トラウストチトリウムローゼウムを含む、いずれのトラウストチトリウム属種(U. ビスルゲンシス(U. visurgensis)、U. アモエボイダ(U. amoeboida)、U. サルカリアナ(U. sarkariana)、U. プロフンダ(U. profunda)、U. ラジアタ(U. radiata)、U. ミヌタ(U. minuta)およびウルケニア種BP-5601のような元ウルケニア種を含む)も;およびいずれのジャポノチトリウム属種も含む。特に好ましいトラウストチトリアレスの系統は、限定されるものではないが、シゾチトリウム属種(S31)(ATCC 20888);シゾチトリウム属種(S8)(ATCC 20889);シゾチトリウム属種(LC-RM)(ATCC 18915);シゾチトリウム属種(SR21);シゾチトリウムアグレガタム(GoldsteinおよびBelsky)(ATCC 28209);シゾチトリウムリマシナム(HondaおよびYokochi)(IFO 32693);トラウストチトリウム属種(23B)(ATCC 20891);トラウストチトリウムストリアタム(Schneider)(ATCC 24473);トラウストチトリウムオーレウム(Goldstein)(ATCC 34304);トラウストチトリウムローゼウム(Goldstein)(ATCC 28210);およびジャポノチトリウム属種(L1)(ATCC 28207)を含む。
【0097】
本発明により、用語「形質転換」は、外因性核酸分子(すなわち、組換え核酸分子)がトラウストチトリアレス微生物の細胞のような微生物の細胞へ挿入されうる任意の方法を指すために用いられる。微生物系において、用語「形質転換」は、微生物による外因性核酸の獲得のせいでの遺伝する変化を記述するために用いられ、本質的に用語「トランスフェクション」と同意語である。適する形質転換技術は、限定されるものではないが、微粒子銃、電気穿孔法、マイクロインジェクション、リポフェクション、吸着、感染およびプロトプラスト融合を含む。
【0098】
一つの態様において、本発明の方法を用いて産生されうるタンパク質は、そのタンパク質を産生するのに効率的な条件下においてそのタンパク質を発現する細胞(すなわち、組換えトラウストチトリアレス微生物)を培養することにより産生される。タンパク質が回収されうる場合もあるし、微生物が全部または可溶化物として収集されかつ「バイオマス」として使用されうる場合もある。もう一つの態様において、ターゲット遺伝子は、細胞内での組換えを可能にし、結果としてターゲット遺伝子の欠失または不活化を生じるのに効率的な条件下において、本発明のターゲティングベクターを含む組換え分子で形質転換された細胞を培養することにより欠失させられるまたは不活化される。培養されうる好ましい細胞は、本発明の組換え細胞である。効率的な培養条件は、限定されるものではないが、タンパク質の産生および/または組換えを可能にする効率的な培地、バイオリアクター、温度、pHおよび酸素条件を含む。効率的な培地は、典型的にはトラウストチトリアレス細胞が培養される任意の培地を指す。そのような培地は、典型的には、同化できる炭素源、窒素源およびリン酸源、ならびに適当な塩、ミネラル、金属およびビタミンのような他の栄養素を有する水性培地を含む。適する培地および培養条件の例は、実施例の節で詳細に考察されている。トラウストチトリアレス微生物に適する培養条件は、Barclayによる1994年8月23日に発行された米国特許第5,340,742号にも記載されている;参照として完全に本明細書に組み入れられている。本発明の細胞は、通常の発酵バイオリアクター、振盪フラスコ、試験管、マイクロタイター皿およびペトリプレートにおいて培養されうる。培養は、組換え細胞に適切な温度、pHおよび酸素含有量で行われうる。そのような培養条件は、当業者の専門技術内である。
【0099】
産生のために使用されるベクターおよび宿主系に依存して、結果として生じる本発明のタンパク質は、組換え細胞内に残りうる;発酵培地へ分泌されうる;2つの細胞膜間の間隙へ分泌されうる;または細胞膜の外側表面上に保持されうるのいずれかである。句「タンパク質を回収すること」とは、タンパク質を含む発酵培地全部を収集することを指し、分離または精製の追加の段階を含意する必要はない。本発明の方法により産生されるタンパク質は、限定されるものではないが、アフィニティークロマトグラフィー、イオン交換クロマトグラフィー、濾過、電気泳動、疎水性相互作用クロマトグラフィー、ゲル濾過クロマトグラフィー、逆相クロマトグラフィー、コカナバリンAクロマトグラフィー、クロマトフォーカシングおよび示差的可溶化のような様々な標準的タンパク質精製技術を用いて精製されうる。本発明の方法により産生されるタンパク質は、好ましくは、「実質的純粋」型として回収される。本明細書に用いられる場合、「実質的純粋」とは、商業的製品としてタンパク質の実効使用を可能にする純度を指す。
【0100】
本発明のさらにもう一つの態様は、本発明のアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子で形質転換されたトラウストチトリアレス目の組換え微生物に関する。好ましくは、アセト乳酸シンターゼは、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を微生物に与える。そのような微生物を形質転換するにおける使用に適する組換え核酸分子および配列は、先で詳細に記載されている。そのような微生物は、本明細書で先に記載されているように、ble遺伝子選択マーカーおよびトラウストチトリアレスの転写制御配列を含む組換え核酸分子を含む、別の組換え核酸分子でさらに形質転換されうる。本発明によるトラウストチトリアレス微生物は、実施例の節に記載されている。本発明により、本発明の組換えトラウストチトリアレス微生物は、本明細書に記載される組換えベクターを用いて対象となるタンパク質(そのようなタンパク質の例は上記で考察されている)を発現するように遺伝子的に操作される、および/または本明細書に記載される組換えベクターを用いてターゲット遺伝子のターゲットされた欠失もしくは不活化のために遺伝子的に操作される。
【0101】
本明細書に用いられる場合、組換え微生物は、組換えテクノロジーを用いてそれの通常の(すなわち、野生型のまたは天然に存在する)型から改変されている(すなわち、変異されているまたは変えられている)ゲノムを有する。本発明による組換え微生物は、そのような改変が微生物内に望ましい効果を与えるような様式で、核酸分子が挿入された、欠失させられたまたは改変された(すなわち、変異された;例えば、ヌクレオチドの挿入、欠失、置換、および/または逆位により)微生物を含む。本明細書に用いられる場合、遺伝子発現における、遺伝子の機能における、または遺伝子産物の機能における減少を結果として生じる遺伝子改変は、遺伝子の不活化(完全または部分的)、欠失、中断、妨害または下方制御と呼ばれうる。例えば、そのような遺伝子によりコードされるタンパク質の機能における減少を結果として生じる遺伝子における遺伝子改変は、遺伝子の完全な欠失(すなわち、遺伝子が存在しない、それゆえにタンパク質が存在しない)、タンパク質の不完全な翻訳もしくは翻訳無し(例えば、タンパク質が発現されない)を結果として生じる遺伝子における変異、またはタンパク質の本来の機能を減少させるもしくは消失させる(例えば、減少した酵素的活性もしくは作用をもつまたは全くもたないタンパク質が発現される)遺伝子における変異の結果でありうる。遺伝子発現または機能における増加を結果として生じる遺伝子改変は、遺伝子の増幅、過剰産生、過剰発現、活性化、増強、付加、または上方制御と呼ばれうる。
【0102】
本発明によると、組換えトラウストチトリアレス微生物は、トラウストチトリアレス目のいずれの微生物を用いても作製されうる。好ましいトラウストチトリアレスの属は、限定されるものではないが、トラウストチトリウム、ラビリンチュロイデス、ジャポノチトリウム、およびシゾチトリウムを含む。これらの属内の好ましい種は、限定されるものではないが、シゾチトリウムアグレガタム、シゾチトリウムリマシナムを含むいずれのシゾチトリウム属種も;いずれのトラウストチトリウム属種も(U. ビスルゲンシス、U. アモエボイダ、U. サルカリアナ、U. プロフンダ、U. ラジアタ、U. ミヌタおよびウルケニア種BP-5601のようないずれの元ウルケニア種も含む)、トラウストチトリウムストリアタム、トラウストチトリウムオーレウム、トラウストチトリウムローゼウム;およびいずれのジャポノチトリウム属種も含む。特に好ましいトラウストチトリアレスの系統は、限定されるものではないが、シゾチトリウム属種(S31)(ATCC 20888);シゾチトリウム属種(S8)(ATCC 20889);シゾチトリウム属種(LC-RM)(ATCC 18915);シゾチトリウム属種(SR21);シゾチトリウムアグレガタム(GoldsteinおよびBelsky)(ATCC 28209);シゾチトリウムリマシナム(HondaおよびYokochi)(IFO 32693);トラウストチトリウム属種(23B)(ATCC 20891);トラウストチトリウムストリアタム(Schneider)(ATCC 24473);トラウストチトリウムオーレウム(Goldstein)(ATCC 34304);トラウストチトリウムローゼウム(Goldstein)(ATCC 28210);およびジャポノチトリウム属種(L1)(ATCC 28207)を含む。
【0103】
以下の実施例は、例証の目的のために提供され、本発明の範囲を限定することを意図されるものではない。
【実施例】
【0104】
実施例1
この実施例は、組換えプラスミドpTUBZEO 11-2の作製を記載する。
【0105】
組換えプラスミドpTUBZEO 11-2の構築は、図1および図2に示されている。このプラスミドは、シゾチトリウム属種から単離されたα-チューブリン遺伝子プロモーターと機能的に共役するストレプトアロテイコスヒンズスタヌス由来のble遺伝子を含む。このプラスミドは以下の通りに作製された。cDNAクローン(CGNE0002-001-B6)は、シゾチトリウム属種cDNAライブラリーから単離され、大規模シゾチトリウムcDNAシーケンシングプロジェクトの一環として部分的にシーケンシングされた(配列番号:1)。ヌクレオチド配列は、BLASTX相同性検索によりα-チューブリンをコードすることが決定された(Gish, W.およびD. States. 1993. Nat. Genet. 3:266-272)。塩基116位から550位までの推定されたアミノ酸配列は、ペルベティカファスティギアタ(Pelvetica fastigiata)由来のα-チューブリン(GenBankアクセッション番号U58642)の最初の145個のアミノ酸と93 %同一である。
【0106】
この遺伝子に関係するプロモーターを単離するために、ゲノムDNAがシゾチトリウム属種の細胞から単離され、平滑末端を生じるための制限エンドヌクレアーゼでのゲノムDNAの酵素的消化、続いて、キットに供給されている特定の二本鎖DNAアダプター分子への消化されたDNAのライゲーションを含む「ゲノムウォーカー(GenomeWalker)(商標)」キット(Clontech Laboratories, Inc., Palo Alto, CA)の使用により処理された。α-チューブリンコード配列のDNA上流は、その後、キットに供給されている外側アダプタープライマー(AP1)およびα-チューブリン特異的プライマーPGR20(配列番号:2)を用いて、ポリメラーゼ連鎖反応(PCR)により増幅された。遺伝子のさらなる増幅は、キットに供給されている入れ子状態のアダプタープライマー(AP2)および入れ子状態のα-チューブリン特異的プライマーPGR19(配列番号:3)を用いて行われた。その結果生じたPCR産物は、プラスミドpCR2.1-TOPO(Invitrogen Corp., Carlsbad, CA)へサブクローニングされた。サブクローニングされた断片の1つは、シーケンシングされた;α-チューブリン遺伝子開始コドンの前に隣接した725bpの配列は、配列番号:4として示されている。
【0107】
この方法で得られるDNA配列に基づくオリゴヌクレオチドプライマーを用いて、TaqDNAポリメラーゼ(Perkin-Elmer Corp., Norwalk, CT)を使用するPCRが用いられて、NcoI制限部位がDNA断片の3'末端へ組み入れられている改変されたα-チューブリンプロモーター領域を生じた;このNcoI部位は、α-チューブリンコード領域において同じ位置にある開始コドンを含んだ。この反応において使用されるプライマーは、PGR33(配列番号:5)およびPGR34(配列番号:6)であり、鋳型はシゾチトリウム属種の細胞から単離されたゲノムDNAであった。以下の反応条件が利用された:94℃4分間;(94℃1分間、54℃45秒間、72℃2分間)x 30;72℃7分間。この断片は、プラスミドpCR2.1-TOPOへクローニングされてプラスミドp7TUB(配列番号:7)を形成した。プラスミドp7TUBはNcoIで消化され、結果として生じたシゾチトリウムα-チューブリンプロモーター領域を含む463bp断片は、アガロースゲル精製により単離された。SV40プロモーターおよびターミネーターに隣接しているストレプトアロテイコスヒンズスタヌス由来のble遺伝子を含むプラスミドpSV40/Zeo(Invitrogen Corp., Carlsbad, CA)もまた、NcoIで消化され、3201bpおよび314bpの断片を生じた。3201bp断片は、アガロースゲル精製され、かつp7TUB由来の463bp NcoI断片に連結されて、図1に描かれているpTUBZEO-11(配列番号:8)を生じた。
【0108】
次に、プラスミドpTUBZEO-11は、SphIで消化され、結果として生じた、シゾチトリウムα-チューブリンプロモーターおよびSV40ターミネーターに隣接しているble遺伝子を含む1122bp断片は、アガロースゲル精製され、かつSphIでの消化により線状化されたプラスミドpUC19(Messing, J. 1983. Meth. Enzymol. 101:20)に連結された。その結果生じたプラスミドは、pTUBZEO 11-2(配列番号:9)と名付けられ、図2および図4に描かれている。プラスミドpTUBZEO 11-2はまた、pMON50000とも呼ばれる。配列番号:9において、シゾチトリウムα-チューブリンプロモーターはヌクレオチド441位〜894位内に含まれる;ble遺伝子コード領域はヌクレオチド895位〜1269位内に含まれる;およびSV40ターミネーターはヌクレオチド1270位〜1524位内に含まれる。
【0109】
実施例2
この実施例は、組換えプラスミドpMON50200、pMON50201、pMON50202、およびpMON50203の作製を記載する。
【0110】
シゾチトリウム属種由来の未変性のアセト乳酸シンターゼコード遺伝子(als)は、以下の方法で単離された。cDNAクローン(LIB81-028-Q1-E1-D9)はシゾチトリウムcDNAライブラリーから単離され、大規模シゾチトリウム属種cDNAシーケンシングプロジェクトの一環として部分的にシーケンシングされた(配列番号:11)。ヌクレオチド配列は、BLASTX相同性によりアセト乳酸シンターゼをコードすることが決定された;例えば、塩基154位から378位までの推定されるアミノ酸配列は、シゾサッカロマイセスポンベ(Schizosaccharomyces pombe)(GenBankアクセッション番号P36620)由来のALSのアミノ酸313位〜387位と68 %同一であった。その後、このクローニングされたcDNAの完全長配列が得られ、cDNAクローンは全alsコード領域を含まないことが示された。完全長als遺伝子を得るために、シゾチトリウムのゲノムラムダライブラリーが372bpジゴキシゲニン(DIG)標識DNAプローブ(ALS2プローブと呼ばれる)での標準的プロトコール(例えば、Sambrookら、Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, 1989を参照)を用いて探索された。ALS2プローブは、cDNAクローンLIB81-028-Q1-E1-D9の配列に基づいている前進プライマーPGR38(配列番号:12)および逆プライマーPGR39(配列番号:13)を用いて、DIG-11-UTPを含むヌクレオチド混合物を使用するPCRにより生成された。ALS-4Aと称せられる、ALS2プローブで同定されたゲノムクローンの1つが単離され、さらに、DIG標識ALS2プローブを使用するサザンハイブリダイゼーションブロット法により特徴付けられた。AhdIで消化されたALS-4AラムダDNA由来の4.9-kbp断片がALS2プローブにハイブリダイズすることを見出された。この断片は、アガロースゲル精製により単離され、平滑末端を生じるためにT4 DNAポリメラーゼで処理され、その後、SmaIで消化されたpBluescriptII KS+(Stratagene Corp., La Jolla, CA)へ連結されてプラスミドpMON50200(図3Aに描かれている)を形成した。pMON50200の配列は、配列番号:14として示されている。シゾチトリウムals遺伝子によりコードされるアセト乳酸シンターゼ酵素の配列は、配列番号:15として示されている。
【0111】
プラスミドpMON50201、pMON50202、およびpMON50203(それぞれ、図3B、図3Cおよび図3Dに描かれている)は、コードされるアセト乳酸シンターゼ酵素がスルホメツロンメチル(SMM)を含むある特定の化合物にもはや阻害されないように、部位特異的突然変異誘発によりプラスミドpMON50200から作製された。これらのプラスミドは、以下の通りに構築された。「トランスフォーメラ(Transformera)」部位特異的突然変異誘発キット(Clontech Laboratories, Inc., Palo Alto, CA)が、製造者の使用説明書に従って以下の変異をプラスミドpMON50200へ導入するために用いられた。オリゴヌクレオチド選択プライマー、DM19(配列番号:16)がすべての3つの構築物において用いられた;このプライマーはpMON50200のマルチクローニング部位における唯一のEcoRV部位のAatII部位への変換へと導く。プライマーDM14(配列番号:17)は、コードされたALS酵素におけるアミノ酸残基595番をトリプトファンからバリンへ変え、同時に、遺伝子配列にAclI部位を導入するために用いられる;その結果生じたプラスミドはpMON50201(配列番号:18)と呼ばれる。同様に、プライマーDM15(配列番号:20)が、コードされたALS酵素におけるアミノ酸残基192番をプロリンからグルタミンへ変えてals遺伝子にBsgI部位を導入するために用いられ、結果としてプラスミドpMON50202(配列番号:21)を生じた。プラスミドpMON50203(配列番号:23)を構築するために、DM14およびDM15プライマーの両方が用いられ、結果として、上記の両方のアミノ酸残基置換を含むコードされたALS酵素を生じた。プラスミドpMON50201、pMON50202、およびpMON50203によりコードされる変異体アセト乳酸シンターゼ酵素の配列は、それぞれ、配列番号:19、配列番号:22、および配列番号:24として示されている。
【0112】
実施例3
この実施例は、実施例1および実施例2に記載される組換え分子でのシゾチトリウム属種の遺伝的形質転換を記載する。
【0113】
シゾチトリウム属種N230D、アメリカンタイプカルチャーコレクション(American Type Culture Collection)株20888(ATCC, Manassas, VA)の派生体における株がこの実施例に使用された。液体培養については、細胞をM50-3培地において30℃、200 rpm〜300 rpmで振盪させて無菌的に増殖させた。M50-3培地は以下の成分を含む:NaCl、12.5 g;MgSO4・7H2O、2.5 g;KCl、0.5 g;CaCl2、0.05 g;グルコース、30 g;Na-グルタミン酸、3 g;KH2PO4、0.4 g;酵母抽出物、1 g;NaHCO3、0.4 g;Na2EDTA、30 mg;FeCl3・6H2O、1.2 mg;H3BO3、34.2 mg;ZnSO4・7H2O、0.67 mg;CoCl2・6H2O、0.13 mg;NaMoO4・2H2O、25 μg;CuSO4・5H2O、10 μg;NiSO4・6H2O、0.26 mg;チアミン・HCl、100 μg;ビオチン、0.5 μg;シアノコバラミン、0.5 μg、および脱イオン水(1リットルにつき);7.0に調整された最終pH。固体培地での増殖については、細胞を1.5 %(w/v)寒天の添加により凝固したM50-3培地またはM1E-3培地において30℃で増殖させた。M1E-3培地は以下の成分を含む:グルコース、4 g;(NH4)2SO4、0.75 g;Na2SO4、5 g;MgSO4・7H2O、2 g;KH2PO4、0.5 g;KCl、0.5 g;CaCl2・2H2O、0.1 g;MOPS緩衝液、20.9 g;FeSO4・4H2O、0.3 mg;MnCl2・4H2O、0.1 mg;ZnSO4・7H2O、80 μg;CoCl2・6H2O、2 μg;NaMoO4・2H2O、1 μg;CuSO4・5H2O、60 μg;NiSO4・6H2O、80 μg;チアミンHCl、320 μg;CA-パントテン酸、320 μg;シアノコバラミン、8 μg、および脱イオン水(1リットルにつき);7.0に調整された最終pH。
【0114】
シゾチトリウム属種のゼオシン(Zeocin)(商標)およびSMMに対する感受性は、凝固したM1E-3培地に様々な濃度でこれらの阻害剤を含め、そして組換え細胞の選択のために用いられる工程間に存在しているものと類似した濃度でプレート上に細胞を広げることにより測定される。
【0115】
シゾチトリウム細胞の遺伝的形質転換は、Bio-Rad Biolistic PDS-1000/He粒子送達システム( Particle Delivery System)(Bio-Rad Laboratories, Hercules, CA)を用いる微粒子銃により行われた(Sanford, J. C., F. D. Smith、およびJ. A. Russell. 1993. Meth. Enzymol. 217:483-509)。シゾチトリウム属種N230D細胞を液体M50-3培地において0.4〜0.8の680 nm(OD680)での光学濃度(10 mmの光学距離の長さ)まで増殖させた。1.0 OD680に対応する細胞の一定分量を短時間だけ遠心分離し、上清溶液を除去し、そしてペレット化された細胞を100 μlの滅菌水に再懸濁した。その後、再懸濁された細胞を寒天凝固培地(例えば、M50-3培地またはM1E-3培地)を含むペトリプレート上に4 cm〜6 cmの円に広げ、余分な水が固体培地へ吸収されうるように30分〜60分間、そのままにさせておいた;これは、ターゲットプレートと呼ばれる。
【0116】
金のマイクロキャリア(0.6 μ呼び径、Bio-Rad Laboratories, Inc., Hercules, CAから入手可能)の1.5 mg分量を製造者の使用説明書(Biolistic(登録商標)PDS-1000/He 粒子送達システム使用説明書マニュアル(Particle Delivery System Instruction Manual);Bio-Rad Laboratories, Hercules, CA)どおりに形質転換プラスミドDNA(すなわち、プラスミドpTUBZEO 11-2、pMON50201、pMON50202、またはpMON50203)の2.5 μgでコーティングした。細胞を以下の条件を用いてDNAコーティング化金マイクロキャリアで撃ち込んだ:1100 psi破裂盤、25'' Hgのチャンバー減圧、1.5 cm〜2 cmの発射ディスクからストッピングスクリーンまでの距離および約7 cmのストッピングスクリーンからターゲットまでの距離を示す、上段の棚に配置されたマイクロキャリア発射集合群ならびに中段の棚に配置されたターゲットプレート。撃ち込み後、細胞を4時間〜6時間、30℃でターゲットプレート上で回復するようにさせておいた。その後、細胞を1.5 ml 滅菌水でターゲットプレートから洗い落とし、マイクロチューブに収集し、短時間だけ遠心分離し、そして400 μl 滅菌水に再懸濁した。懸濁液の100マイクロリットルを150 μg/ml〜200 μg/ml ゼオシン(Zeocin)(商標)(Invitrogen Corp., Carlbad, CA)または25 μg/ml SMMのいずれかを含む4つのM1E-3プレートのそれぞれに広げた。ゼオシン(Zeocin)(商標)含有プレートは、プラスミドpTUBZEO 11-2で形質転換された細胞について選択するために使用され、SMM含有プレートは、プラスミドpMON50201、pMON50202、またはpMON50203で形質転換された細胞について選択するために使用された。その後、プレートを7日〜10日間、30℃でインキュベートした。その後、選択薬剤に対して耐性であると思われるコロニーを、耐性を確認するために同じ選択薬剤を含む新鮮なM1E-3プレート上へパッチ状に塗布した。このプロトコールは、典型的には、撃ち込みあたり、100個〜1000個のゼオシン(Zeocin)(商標)耐性株またはSMM耐性株の発生を生じる。
【0117】
実施例4
以下の実施例は、形質転換されたシゾチトリウム細胞のPCR分析を実証する。
【0118】
PCRは、選択薬剤ゼオシン(Zeocin)(商標)またはSMMに対して耐性である推定上の形質転換された株においてプラスミド配列の存在を確認するために用いられた。推定の形質転換体および非組換え型シゾチトリウムN230D細胞由来の鋳型DNAは、使い捨てのプラスチックの1 μl接種用輪を使用して寒天プレート上へパッチされた耐性コロニー(実施例3に記載されているように)から少量の細胞(1 mm3〜2 mm3)を取り出すことにより得られた。その後、細胞をマイクロチューブにおいて1 % トリトンX-100の15 μl〜20 μlに再懸濁し、10分間沸騰水浴に置き、そしてその後、14,000xgで5分間、遠心分離した。これらの抽出物の一部(1 μL〜3 μL)は、Taq DNAポリメラーゼを使用する25 μL PCR反応についての鋳型DNAを供給するために使用された。シゾチトリウムDNAにおいてpTUBZEO 11-2配列の存在を検出するために、プライマーDM20(配列番号:25)およびDM21(配列番号:26)が用いられた;これらのプライマーは、プラスミドpTUBZEO 11-2におけるble遺伝子にアニールして、346bp DNA断片を増幅する。用いられる熱的側面は、以下のとおりである:94℃で4分間;(94℃で45秒間、52℃で45秒間、72℃で2分間)x 30;72℃で7分間。シゾチトリウムDNAにおいてpMON50201、pMON50202、またはpMON50203配列の存在を検出するために、プライマーBLA1(配列番号:27)およびBLA2(配列番号:28)が用いられた;これらのプライマーは、ベクターバックボーンに見出されるbla(アンピシリン耐性)遺伝子にアニールして、1229bp DNA断片を増幅する。用いられる熱的側面は、以下のとおりである:94℃で4分間;(94℃で45秒間、55℃で45秒間、72℃で2分間)x 30;72℃で7分間。PCR産物は、標準的アガロースゲル電気泳動、続いて、臭化エチジウムで染色することにより分析された。
【0119】
これらの分析の結果より、これらの条件下で選択された株の大多数はプラスミドDNAを含む真の形質転換体であることが確認された。形質転換プラスミドで撃ち込まれなかった対照のシゾチトリウム属種N230D細胞由来の鋳型DNAを用いた場合、正しいサイズのPCR産物は生成されなかった。
【0120】
実施例5
以下の実施例は、形質転換されたシゾチトリウム細胞のサザンブロット分析を記載する。
【0121】
サザンハイブリダイゼーションブロット法は、形質転換された細胞内の形質転換ベクターDNA配列の存在を確認するために、親のシゾチトリウムN230D細胞およびいくつかの推定の形質転換体から単離されたDNAを用いて行われた。サザンブロッティングは、当業者に公知の技術を用いて行われた(例えば、Sambrookら、Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press. 1989を参照)。DNAは、「QIAamp」DNA精製キット(Qiagen Inc., Valencia, CA)の使用により単離され、様々な制限酵素で消化され、アガロースゲル(0.8 %〜1.2 %w/v)を通しての電気泳動により分離され、その後、アルカリ性キャピラリー移動によりナイロン膜に転写された。
【0122】
pTUBZEO 11-2で形質転換された細胞におけるベクターDNAの検出は、ハイブリダイゼーションプローブとしてプライマーDM20(配列番号:25)およびDM21(配列番号:26)でのPCRにより生成された346bpのDIG標識ble遺伝子断片ならびにDIG-11-UTPを含むヌクレオチド混合物を用いて、「ジーニアス(Genius)」DIG型システム(Boehringer Mannheim Biochemicals GmbH, Germany)の使用により行われた。膜のプレハイブリダイゼーションは、ジーニアス(Genius)キットに供給されているハイブリダイゼーション緩衝液において68℃で1時間行われた。ハイブリダイゼーションは、94℃で5分間、熱変性されたble遺伝子プローブを含むハイブリダイゼーション緩衝液において68℃で18時間行われた。その後、膜を50 mL 2X SSC/0.1 % SDSで5分間を2回および50 mL 0.1X SSC/0.1 % SDSにおいて15分間を2回洗浄した。ハイブリダイズしているDNAの化学ルミネセンスの検出は、ジーニアス(Genius)キット使用説明書に記載されているように行われた。
【0123】
形質転換されていないシゾチトリウムN230D細胞由来のDNAは、ble遺伝子プローブにハイブリダイズしなかった。逆に、形質転換された細胞由来のDNAは、以下のようにプローブにハイブリダイズした:
【0124】
SphI:形質転換されたシゾチトリウム細胞由来のSphIで消化されたDNAは、ble遺伝子プローブにハイブリダイズする〜1100bpのDNA断片を含んでいた;この断片は、SphIで消化されたpTUBZEO 11-2 DNAにおいても見られるが、ble遺伝子発現カセット全体(チューブリン遺伝子プロモーターおよびSV40ターミネーターを含む)を表している。
【0125】
XhoI:試験された形質転換体のそれぞれについて、DNAのXhoI消化は、結果として15 kbp〜20 kbpより長いハイブリダイズする断片を生じた。XhoIはpTUBZEO 11-2内を切断しない、それゆえに、これらの結果は、pTUBZEO 11-2が形質転換された細胞において染色体外のエレメントとして存在しているようには思われず、むしろシゾチトリウム染色体へ組み込まれるようになることを示している。
【0126】
NcoIまたはHindIII:これらの酵素は両方とも、pTUBZEO 11-2内を一回切断する。これらの酵素のいずれかでの形質転換体DNAの消化は、典型的には、線状化pTUBZEO 11-2ベクター(すなわち、〜3.8 kbp)と共に移動する顕著なハイブリダイズする断片を導いた。これは、ベクターが直列型反復の形で染色体に統合されうることを示唆している。
【0127】
実施例6
この実施例は、シゾチトリウムにおける相同的組換えを実証している。
【0128】
以下の実験は、相同的組換えが、シゾチトリウムにおいて、内因性の天然のDNA配列と、細胞へ導入された組換えDNA分子に存在する相同性DNA配列との間で起こりうることを実証するために行われた。相同的組換えのこの型は、望ましい性質をもつ組換え系統を作製するためにとても有益でありうる。例えば、相同的組換えは、外来遺伝子配列のターゲットされた挿入により内因性遺伝子を不活化するために用いられうる。さらに、相同的組換えは、組換え細胞が新規な性質を示すように、内因性遺伝子またはそれの部分をその遺伝子の改変型で置換するために用いられうる。
【0129】
相同的組換えは、シゾチトリウムals遺伝子における突然変異を含むプラスミドpMON50202で形質転換されたシゾチトリウム細胞において起きることを示された。この突然変異は、alsコード領域の塩基対位置571位にBsgI部位を導入する。alsコード領域の塩基対位置1324位に天然に存在するBsgI部位がある。それゆえに、BsgIで消化されたシゾチトリウムDNAのサザンブロットは、天然のals遺伝子と組換え突然変異体als遺伝子とを識別するために用いられうる。これらの実験について、als特異的ハイブリダイゼーションプローブは、DIG-11-UTP(Boehringer Mannheim Biochemicals GmbH, Germany)、前進プライマーPGR28(配列番号:32)、逆プライマーPGR30(配列番号:33)、および鋳型として少量のpMON50200を含むヌクレオチド混合物を使用するPCRにより作製された。その結果生じた323bpのDIG標識ハイブリダイゼーションプローブは、ALS1と呼ばれた。
【0130】
非組換え型シゾチトリウムN230D細胞由来のDNAは、BsgIおよびAhdIで別々に消化され、アガロースゲル電気泳動にかけられ、ナイロン膜へ転写され、その後、実施例5に記載されているものと本質的に同じ手順を用いてALS1プローブで探索された。ALS1プローブは、BsgIで消化されたDNAの1.76-kbp断片およびAhdIで消化されたDNAの4.9-kbp断片を標識した。
【0131】
pMON50202で形質転換された様々な組換え株由来のBsgIで消化されたDNAおよびAhdIで消化されたDNAのサザンブロットもまた、ALS1プローブで探索された。いくつかの場合において、1.76-kbpのBsgI断片は存在せず、代わりに、pMON50202に存在する753bpのBsgI断片に対応している0.75-kbpの断片が標識された。4.9-kbpのAhdI断片がこれらの組換え株において標識されたが、その組換え突然変異体als遺伝子が二重クロスオーバー相同的組換えにより天然のals遺伝子と組換えしたことを示した。
【0132】
単一クロスオーバー相同的組換えもまた、pMON50202で形質転換された組換え株において起こっていることが観察された。これらの場合、1.76-kbpおよび0.75-kbpのBsgI断片の両方が、組換え株由来のDNAのサザンブロットにおいて標識されたが、4.9-kbpのAhdI断片がより大きな標識された断片に取って代わられ、pMON50202ベクター全体が、単一コピーとしてまたは直列型反復としてのいずれかで、天然のals遺伝子へ挿入したことを示した。
【0133】
シゾチトリウムにおける相同的組換えについての追加の証拠は、切り詰められた遺伝子によりコードされた不完全なALS酵素が機能しえないように切り詰められた突然変異体als遺伝子を含む組換えDNA分子の導入により得られた。この切り詰められた遺伝子は、ClaIおよびHindIIIでpMON50202を消化して2.8-kbp断片を生じ、それによりalsターミネーター領域と共にalsコード領域の最後の388 bpを除去することにより作製された。この2.8-kbp断片はClaIおよびHindIIIで消化されたpBluescriptII KS+(Stratagene Corp., La Jolla, CA)へライゲーションされ、プラスミドpAR2を生じた。プラスミドpAR2は、天然のals遺伝子とpAR2に存在する切り詰められた突然変異体als遺伝子との間の相同的組換えにより形質転換された株において機能しうる突然変異体als遺伝子が回復される場合には、形質転換されたシゾチトリウム細胞においてSMMに対する耐性を与えることが予測されるのみと考えられる。この構築物をシゾチトリウムN230D細胞へ微粒子銃により導入し、実施例3に記載されているようにSMM耐性株を単離した。形質転換体由来のBsgIで消化されたDNAのサザンブロット分析は、この実施例の初めの方に記載されているように行われたが、相同的組換えが明らかにこれらの株において起こったことを示した;すなわち、非組換え型細胞において1.76-kbpのBsgI断片がALS1プローブにハイブリダイズしたが、pAR2で形質転換された細胞においては、これが0.75-kbpのハイブリダイズする断片に取って代わられた。
【0134】
実施例7
この実施例は、選択マーカー遺伝子に連結されていない追加の外来DNA分子を含む株を同時形質転換により作製するために形質転換ベクターpTUBZEO 11-2またはpMON50202の使用を記載する。
【0135】
同時形質転換は、pTUBZEO 11-2およびいくつかの遺伝子のいずれかを含む追加のプラスミドの同時導入により達成された。プラスミドは、各プラスミドの2.5 μgを使用して、実施例3に記載されているように、金粒子上に共沈澱させた。プラスミドでコーティングされた金粒子のターゲット細胞への撃ち込み後、組換え株は、実施例1に記載されているようにゼオシン(Zeocin)(商標)含有寒天プレート上で選択された。第二の選択されなかったプラスミドの存在は、その後、PCR分析またはサザンブロットハイブリダイゼーションにより確認された。非常に高い同時形質転換頻度(例えば、50 %〜90 %)が概して達成された。例えば、シゾチトリウムのチューブリン遺伝子プロモーターおよびターミネーターに連結されたカエノルハブディタスエレガンス(Caenorhabditus elegans)のfat-1遺伝子(Spychallaら、1997. Proc. Natl. Acad. Sci. U. S. A. 94, 1142-1147)を含むプラスミドpTR202がこの実施例に提供されている方法により導入され、その結果生じたゼオシン(Zeocin)(商標)耐性株の約68 %がfat-1遺伝子を含むことがPCRにより示された(表1を参照)。同様の結果は、pMON50202および追加のプラスミドが同時導入され、続いてSMM含有固体培地上で形質転換された細胞を選択する場合に観察される。この同時形質転換方法は、所望されるいずれの外来DNAでも導入するために用いられうる。
【0136】
(表1) 選択マーカープラスミドpTUBZEO 11-2および様々なfad遺伝子を含むプラスミドを用いる同時形質転換の効率。ゼオシン(Zeocin)(商標)耐性形質転換体は、PCRによりfad DNA配列についてスクリーニングされた。
【0137】
この実施例に記載される形質転換系は、脂質主体の化合物の発酵による生産について知られている最も生産力が高い生物体であるシゾチトリウムを遺伝子的に操作する能力において、意義深い進歩を表している。2つの独立した形質転換系の有用性は、生じる高同時形質転換効率に加え、操作される株において複数の形質の積み重ねを可能にすると思われる。さらに、この微小藻類における相同的組換えの明らかな存在は、未知の遺伝子の機能を同定するため、および生産用株において望ましくない形質を除去するために遺伝子ノックアウト方法の発達を可能にするものと思われる。本発明者らは、最近、シゾチトリウムにおいて脂肪酸代謝を変えるためにこれらの系を使用しており、カロテノイド、ステロール、および他の類脂質性の化合物の生産のために、この種および関連した微小藻類(例えばトラウストチトリウム)を使用することの可能性を探究している。
【0138】
本発明の様々な態様が詳細に記載されてきたが、それらの態様の改変および脚色が当業者に思い浮かぶであろうことは明らかである。しかしながら、そのような改変および脚色が、特許請求の範囲に示されているように、本発明の範囲内であることは、はっきりと理解されるべきである。
【特許請求の範囲】
【請求項1】
以下のものからなる群より選択される核酸配列を含む単離された核酸分子:
a. タンパク質がアセト乳酸シンターゼである、配列番号:15、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードする核酸配列;
b. タンパク質がアセト乳酸シンターゼである、(a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列を有するタンパク質をコードする核酸配列;および、
c. (a)または(b)の核酸配列と完全に相補的である核酸配列。
【請求項2】
核酸配列が(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するタンパク質をコードし、かつ該タンパク質がアセト乳酸シンターゼである、請求項1記載の単離された核酸分子。
【請求項3】
核酸配列が(a)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するタンパク質をコードし、かつ該タンパク質がアセト乳酸シンターゼである、請求項1記載の単離された核酸分子。
【請求項4】
核酸配列が116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする、請求項1記載の単離された核酸分子。
【請求項5】
核酸配列が配列番号:15、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードし、かつ該タンパク質がアセト乳酸シンターゼである、請求項1記載の単離された核酸分子。
【請求項6】
核酸配列が配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項1記載の単離された核酸分子。
【請求項7】
(a)または(b)のタンパク質の発現が核酸分子で形質転換されるトラウストチトリアレス目の微生物に、以下からなる群より選択される化合物に対する低下した感受性を与える、請求項1記載の単離された核酸分子:
スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸。
【請求項8】
核酸配列が配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードする、請求項7記載の単離された核酸分子。
【請求項9】
核酸配列が配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項7記載の単離された核酸分子。
【請求項10】
核酸配列がシゾチトリウムのアセト乳酸シンターゼをコードする、請求項1記載の単離された核酸分子。
【請求項11】
シゾチトリウムのアセト乳酸シンターゼの発現が核酸分子で形質転換されるトラウストチトリアレス目の微生物に、以下からなる群より選択される化合物に対する低下した感受性を与える、請求項10記載の単離された核酸分子:
スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸。
【請求項12】
転写制御配列へ実施可能に連結された、請求項1記載の単離された核酸分子を含む組換え核酸分子。
【請求項13】
請求項12記載の組換え核酸分子で形質転換されるトラウストチトリアレス目の組換え微生物。
【請求項14】
トラウストチトリアレス目の微生物に、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を与えるアセト乳酸シンターゼをコードする核酸配列を含み、該アセト乳酸シンターゼが以下のものからなる群より選択されるアミノ酸配列を有し、アセト乳酸シンターゼをコードする該核酸配列が転写制御配列へ実施可能に連結されている、トラウストチトリアレス目の微生物の形質転換のための組換えベクター:
a. 配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および
b. (a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列。
【請求項15】
組換えベクターが発現ベクターである、請求項14記載の組換えベクター。
【請求項16】
組換えベクターがターゲティングベクターである、請求項14記載の組換えベクター。
【請求項17】
核酸配列が(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項14記載の組換えベクター。
【請求項18】
核酸配列が(a)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項14記載の組換えベクター。
【請求項19】
核酸配列が116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする、請求項14記載の組換えベクター。
【請求項20】
アセト乳酸シンターゼが配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する、請求項14記載の組換えベクター。
【請求項21】
核酸配列が配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項14記載の組換えベクター。
【請求項22】
転写制御配列がトラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される、請求項14記載の組換えベクター。
【請求項23】
ベクターが配列番号:18、配列番号:21、および配列番号:23からなる群より選択される核酸配列を含む、請求項14記載の組換えベクター。
【請求項24】
方法が以下の段階を含む、トラウストチトリアレス目の微生物の細胞を形質転換する方法:
a. アセト乳酸シンターゼが以下のものからなる群より選択されるアミノ酸配列を有する、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を細胞に与えるアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子をトラウストチトリアレス目の微生物の細胞へ導入する段階:
i. 配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および
ii. (i)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列;ならびに
b. 化合物がスルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される、形質転換されていない細胞に対して阻害性がある少なくとも1つの化合物を含む培地において(a)の該細胞を培養することにより該組換え核酸分子で成功裡に形質転換された細胞を選択する段階。
【請求項25】
核酸配列が(i)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項24記載の方法。
【請求項26】
核酸配列が(i)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項24記載の方法。
【請求項27】
核酸配列が116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする、請求項24記載の方法。
【請求項28】
アセト乳酸シンターゼが配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する、請求項24記載の方法。
【請求項29】
核酸配列が配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項24記載の方法。
【請求項30】
核酸配列がトラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される転写制御配列へ実施可能に連結されている、請求項24記載の方法。
【請求項31】
組換え核酸分子が細胞により産生されうるタンパク質をコードする核酸配列をさらに含み、該タンパク質をコードする該核酸配列が転写制御配列へ実施可能に連結されている、請求項24記載の方法。
【請求項32】
タンパク質が、ドコサヘキサエン酸(DHA)、ドコサペンタエン酸(DPA)、エイコサペンタエン酸(EPA)およびアラキドン酸(ARA)からなる群より選択される脂肪酸の合成に関係する、請求項31記載の方法。
【請求項33】
タンパク質が脂肪酸シンターゼ、脂肪酸デサチュラーゼ、脂肪酸エロンガーゼ、ポリケチドシンターゼ複合体に関係するタンパク質およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質からなる群より選択される、請求項31記載の方法。
【請求項34】
タンパク質が核酸配列の配列番号:29にコードされているω-3脂肪酸デサチュラーゼである、請求項31記載の方法。
【請求項35】
タンパク質がポリエン脂肪酸イソメラーゼである、請求項31記載の方法。
【請求項36】
タンパク質がHMG-CoAシンターゼ、HMG-CoAレダクターゼ、スクアレンシンターゼ、フィトエンシンターゼ、フィトエンデサチュラーゼ、カロテノイドシクラーゼ、カロテノイドヒドロキシラーゼ、カロテノイドケトラーゼ、ビタミンEおよびリポ酸からなる群より選択される、請求項31記載の方法。
【請求項37】
段階(a)における組換え核酸分子が、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより突然変異されるまたは不活化されるように、微生物においてターゲット核酸配列とハイブリダイズする核酸配列をさらに含む、請求項24記載の方法。
【請求項38】
ターゲット核酸配列が、リパーゼ、脂肪酸酸化酵素、糖合成に関与するタンパク質、イソプレノイド経路の生成物の合成に関与するタンパク質、細胞壁成分の合成に関与するタンパク質、飽和脂肪酸合成経路に関与するタンパク質、多価不飽和脂肪酸合成経路に関与するタンパク質、ポリケチドシンターゼ複合体に関係するタンパク質、およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質からなる群より選択されるタンパク質をコードする、請求項37記載の方法。
【請求項39】
発現されうるタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含み、該核酸配列が転写制御配列へ実施可能に連結されている、請求項24記載の方法。
【請求項40】
ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより突然変異されるまたは不活化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含む、請求項24記載の方法。
【請求項41】
ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子を細胞へ導入する段階をさらに含む、請求項24記載の方法。
【請求項42】
ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子が細胞により発現されうる第二のタンパク質をコードする核酸配列をさらに含み、該第二のタンパク質をコードする該核酸配列が転写制御配列へ実施可能に連結されている、請求項41記載の方法。
【請求項43】
転写制御配列がトラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される、請求項42記載の方法。
【請求項44】
ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子が、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより突然変異されるまたは不活化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列をさらに含む、請求項41記載の方法。
【請求項45】
組換え核酸分子が核酸配列の配列番号:9を含む、請求項41記載の方法。
【請求項46】
微生物がトラウストチトリウム、ラビリンチュロイデス、ジャポノチトリウムおよびシゾチトリウムからなる群より選択される属由来である、請求項24記載の方法。
【請求項47】
微生物がシゾチトリウム属種、シゾチトリウムアグレガタム、シゾチトリウムリマシナム、シゾチトリウムミヌタム、トラウストチトリウム属種、トラウストチトリウムストリアタム、トラウストチトリウムオーレウム、トラウストチトリウムローゼウム、およびジャポノチトリウム属種からなる群より選択される種由来である、請求項24記載の方法。
【請求項48】
導入する段階が微粒子銃法、電気穿孔法、マイクロインジェクション法、リポフェクション法、吸着法、感染法およびプロトプラスト融合法からなる群より選択される方法により行われる、請求項24記載の方法。
【請求項49】
アセト乳酸シンターゼが以下のものからなる群より選択されるアミノ酸配列を有する、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を微生物に与えるアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子で形質転換されているトラウストチトリアレス目の組換え微生物:
a. 配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および
b. (a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列。
【請求項50】
核酸配列が(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項49記載の組換え微生物。
【請求項51】
核酸配列が(a)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項49記載の組換え微生物。
【請求項52】
アセト乳酸シンターゼが配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する、請求項49記載の組換え微生物。
【請求項53】
核酸配列が配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項49記載の組換え微生物。
【請求項54】
組換え核酸分子が配列番号:18、配列番号:21、および配列番号:23からなる群より選択される核酸配列を含む、請求項49記載の組換え微生物。
【請求項55】
アセト乳酸シンターゼをコードする核酸配列がトラウストチトリアレスの微生物において機能するプロモーターへ実施可能に連結されている、請求項49記載の組換え微生物。
【請求項56】
アセト乳酸シンターゼをコードする核酸配列がトラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される転写制御配列へ実施可能に連結されている、請求項49記載の組換え微生物。
【請求項57】
組換え核酸分子が微生物による産生としての第一のタンパク質をコードする核酸配列をさらに含み、該第一のタンパク質をコードする該核酸配列が転写制御配列へ実施可能に連結されている、請求項49記載の組換え微生物。
【請求項58】
組換え細胞がブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子でさらに形質転換される、請求項49記載の組換え微生物。
【請求項59】
組換え核酸分子が核酸配列の配列番号:9を含む、請求項58記載の組換え微生物。
【請求項60】
ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子が細胞による産生のための第二のタンパク質をコードする核酸配列をさらに含み、該第二のタンパク質をコードする該核酸配列が転写制御配列へ実施可能に連結されている、請求項58記載の組換え微生物。
【請求項61】
細胞による産生としてのタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子をさらに含む、請求項49記載の組換え微生物。
【請求項62】
以下のものからなる群より選択される核酸配列を含む単離された核酸分子:
a. 配列番号:9のヌクレオチド441位〜894位;
b. 核酸配列が少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、完全長である配列番号:9のヌクレオチド441位〜894位にわたり配列番号:9のヌクレオチド441位〜894位と少なくとも約95 %同一である核酸配列;および
c. (a)または(b)のポリヌクレオチドと完全に相補的である核酸配列を含む単離されたポリヌクレオチド。
【請求項63】
単離された核酸分子が配列番号:4および配列番号:9のヌクレオチド441位〜894位からなる群より選択される核酸配列を含む、請求項62記載の単離された核酸分子。
【請求項64】
トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択されるプロモーターへ実施可能に連結されているブレオマイシン結合タンパク質をコードする核酸配列を含む、トラウストチトリアレス目の微生物の形質転換のための組換えベクター。
【請求項65】
トラウストチトリアレスのアセト乳酸シンターゼプロモーターが配列番号:14のヌクレオチド1位〜1259位を含む、請求項64記載の組換えベクター。
【請求項66】
α-チューブリンプロモーターが配列番号:9のヌクレオチド441位〜894位および配列番号:9のヌクレオチド441位〜894位の完全長に対して配列番号:9のヌクレオチド441位〜894位と少なくとも約95 %同一である核酸配列からなる群より選択される核酸配列を含み、該核酸配列が少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、請求項64記載の組換えベクター。
【請求項67】
組換えベクターが配列番号:8および配列番号:9からなる群より選択される核酸配列を含む、請求項64記載の組換えベクター。
【請求項68】
トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーターが配列番号:34または少なくとも基礎的なPKSプロモーター転写活性を有するその断片を含む、請求項64記載の組換えベクター。
【請求項1】
以下のものからなる群より選択される核酸配列を含む単離された核酸分子:
a. タンパク質がアセト乳酸シンターゼである、配列番号:15、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードする核酸配列;
b. タンパク質がアセト乳酸シンターゼである、(a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列を有するタンパク質をコードする核酸配列;および、
c. (a)または(b)の核酸配列と完全に相補的である核酸配列。
【請求項2】
核酸配列が(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するタンパク質をコードし、かつ該タンパク質がアセト乳酸シンターゼである、請求項1記載の単離された核酸分子。
【請求項3】
核酸配列が(a)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するタンパク質をコードし、かつ該タンパク質がアセト乳酸シンターゼである、請求項1記載の単離された核酸分子。
【請求項4】
核酸配列が116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする、請求項1記載の単離された核酸分子。
【請求項5】
核酸配列が配列番号:15、配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードし、かつ該タンパク質がアセト乳酸シンターゼである、請求項1記載の単離された核酸分子。
【請求項6】
核酸配列が配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項1記載の単離された核酸分子。
【請求項7】
(a)または(b)のタンパク質の発現が核酸分子で形質転換されるトラウストチトリアレス目の微生物に、以下からなる群より選択される化合物に対する低下した感受性を与える、請求項1記載の単離された核酸分子:
スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸。
【請求項8】
核酸配列が配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有するタンパク質をコードする、請求項7記載の単離された核酸分子。
【請求項9】
核酸配列が配列番号:14のヌクレオチド1260位〜3314位、配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項7記載の単離された核酸分子。
【請求項10】
核酸配列がシゾチトリウムのアセト乳酸シンターゼをコードする、請求項1記載の単離された核酸分子。
【請求項11】
シゾチトリウムのアセト乳酸シンターゼの発現が核酸分子で形質転換されるトラウストチトリアレス目の微生物に、以下からなる群より選択される化合物に対する低下した感受性を与える、請求項10記載の単離された核酸分子:
スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸。
【請求項12】
転写制御配列へ実施可能に連結された、請求項1記載の単離された核酸分子を含む組換え核酸分子。
【請求項13】
請求項12記載の組換え核酸分子で形質転換されるトラウストチトリアレス目の組換え微生物。
【請求項14】
トラウストチトリアレス目の微生物に、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を与えるアセト乳酸シンターゼをコードする核酸配列を含み、該アセト乳酸シンターゼが以下のものからなる群より選択されるアミノ酸配列を有し、アセト乳酸シンターゼをコードする該核酸配列が転写制御配列へ実施可能に連結されている、トラウストチトリアレス目の微生物の形質転換のための組換えベクター:
a. 配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および
b. (a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列。
【請求項15】
組換えベクターが発現ベクターである、請求項14記載の組換えベクター。
【請求項16】
組換えベクターがターゲティングベクターである、請求項14記載の組換えベクター。
【請求項17】
核酸配列が(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項14記載の組換えベクター。
【請求項18】
核酸配列が(a)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項14記載の組換えベクター。
【請求項19】
核酸配列が116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする、請求項14記載の組換えベクター。
【請求項20】
アセト乳酸シンターゼが配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する、請求項14記載の組換えベクター。
【請求項21】
核酸配列が配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項14記載の組換えベクター。
【請求項22】
転写制御配列がトラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される、請求項14記載の組換えベクター。
【請求項23】
ベクターが配列番号:18、配列番号:21、および配列番号:23からなる群より選択される核酸配列を含む、請求項14記載の組換えベクター。
【請求項24】
方法が以下の段階を含む、トラウストチトリアレス目の微生物の細胞を形質転換する方法:
a. アセト乳酸シンターゼが以下のものからなる群より選択されるアミノ酸配列を有する、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を細胞に与えるアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子をトラウストチトリアレス目の微生物の細胞へ導入する段階:
i. 配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および
ii. (i)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列;ならびに
b. 化合物がスルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される、形質転換されていない細胞に対して阻害性がある少なくとも1つの化合物を含む培地において(a)の該細胞を培養することにより該組換え核酸分子で成功裡に形質転換された細胞を選択する段階。
【請求項25】
核酸配列が(i)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項24記載の方法。
【請求項26】
核酸配列が(i)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項24記載の方法。
【請求項27】
核酸配列が116G、117A、192P、200A、251K、358M、383D、592V、595W、および599Fからなる群より選択されるアミノ酸位置におけるアミノ酸の欠失、挿入、または置換により配列番号:15と異なるアミノ酸配列を有するタンパク質をコードする、請求項24記載の方法。
【請求項28】
アセト乳酸シンターゼが配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する、請求項24記載の方法。
【請求項29】
核酸配列が配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項24記載の方法。
【請求項30】
核酸配列がトラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される転写制御配列へ実施可能に連結されている、請求項24記載の方法。
【請求項31】
組換え核酸分子が細胞により産生されうるタンパク質をコードする核酸配列をさらに含み、該タンパク質をコードする該核酸配列が転写制御配列へ実施可能に連結されている、請求項24記載の方法。
【請求項32】
タンパク質が、ドコサヘキサエン酸(DHA)、ドコサペンタエン酸(DPA)、エイコサペンタエン酸(EPA)およびアラキドン酸(ARA)からなる群より選択される脂肪酸の合成に関係する、請求項31記載の方法。
【請求項33】
タンパク質が脂肪酸シンターゼ、脂肪酸デサチュラーゼ、脂肪酸エロンガーゼ、ポリケチドシンターゼ複合体に関係するタンパク質およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質からなる群より選択される、請求項31記載の方法。
【請求項34】
タンパク質が核酸配列の配列番号:29にコードされているω-3脂肪酸デサチュラーゼである、請求項31記載の方法。
【請求項35】
タンパク質がポリエン脂肪酸イソメラーゼである、請求項31記載の方法。
【請求項36】
タンパク質がHMG-CoAシンターゼ、HMG-CoAレダクターゼ、スクアレンシンターゼ、フィトエンシンターゼ、フィトエンデサチュラーゼ、カロテノイドシクラーゼ、カロテノイドヒドロキシラーゼ、カロテノイドケトラーゼ、ビタミンEおよびリポ酸からなる群より選択される、請求項31記載の方法。
【請求項37】
段階(a)における組換え核酸分子が、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより突然変異されるまたは不活化されるように、微生物においてターゲット核酸配列とハイブリダイズする核酸配列をさらに含む、請求項24記載の方法。
【請求項38】
ターゲット核酸配列が、リパーゼ、脂肪酸酸化酵素、糖合成に関与するタンパク質、イソプレノイド経路の生成物の合成に関与するタンパク質、細胞壁成分の合成に関与するタンパク質、飽和脂肪酸合成経路に関与するタンパク質、多価不飽和脂肪酸合成経路に関与するタンパク質、ポリケチドシンターゼ複合体に関係するタンパク質、およびリン脂質またはトリアシルグリセロール分子への脂肪酸の取り込みに関係するタンパク質からなる群より選択されるタンパク質をコードする、請求項37記載の方法。
【請求項39】
発現されうるタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含み、該核酸配列が転写制御配列へ実施可能に連結されている、請求項24記載の方法。
【請求項40】
ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより突然変異されるまたは不活化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列を含む少なくとも1つの追加の組換え核酸分子を細胞へ導入する段階をさらに含む、請求項24記載の方法。
【請求項41】
ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子を細胞へ導入する段階をさらに含む、請求項24記載の方法。
【請求項42】
ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子が細胞により発現されうる第二のタンパク質をコードする核酸配列をさらに含み、該第二のタンパク質をコードする該核酸配列が転写制御配列へ実施可能に連結されている、請求項41記載の方法。
【請求項43】
転写制御配列がトラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される、請求項42記載の方法。
【請求項44】
ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子が、ターゲット核酸配列を含む遺伝子が第二の核酸配列との相同的組換えにより突然変異されるまたは不活化されるように微生物においてターゲット核酸配列とハイブリダイズする第二の核酸配列をさらに含む、請求項41記載の方法。
【請求項45】
組換え核酸分子が核酸配列の配列番号:9を含む、請求項41記載の方法。
【請求項46】
微生物がトラウストチトリウム、ラビリンチュロイデス、ジャポノチトリウムおよびシゾチトリウムからなる群より選択される属由来である、請求項24記載の方法。
【請求項47】
微生物がシゾチトリウム属種、シゾチトリウムアグレガタム、シゾチトリウムリマシナム、シゾチトリウムミヌタム、トラウストチトリウム属種、トラウストチトリウムストリアタム、トラウストチトリウムオーレウム、トラウストチトリウムローゼウム、およびジャポノチトリウム属種からなる群より選択される種由来である、請求項24記載の方法。
【請求項48】
導入する段階が微粒子銃法、電気穿孔法、マイクロインジェクション法、リポフェクション法、吸着法、感染法およびプロトプラスト融合法からなる群より選択される方法により行われる、請求項24記載の方法。
【請求項49】
アセト乳酸シンターゼが以下のものからなる群より選択されるアミノ酸配列を有する、スルホニル尿素化合物、イミダゾリノン類阻害剤、およびピリミジニルオキシ安息香酸からなる群より選択される化合物に対する低下した感受性を微生物に与えるアセト乳酸シンターゼをコードする核酸配列を含む組換え核酸分子で形質転換されているトラウストチトリアレス目の組換え微生物:
a. 配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列;および
b. (a)のアミノ酸配列と少なくとも約75 %同一であるアミノ酸配列。
【請求項50】
核酸配列が(a)のアミノ酸配列と少なくとも約85 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項49記載の組換え微生物。
【請求項51】
核酸配列が(a)のアミノ酸配列と少なくとも約95 %同一であるアミノ酸配列を有するアセト乳酸シンターゼをコードする、請求項49記載の組換え微生物。
【請求項52】
アセト乳酸シンターゼが配列番号:19、配列番号:22および配列番号:24からなる群より選択されるアミノ酸配列を有する、請求項49記載の組換え微生物。
【請求項53】
核酸配列が配列番号:18のヌクレオチド1260位〜3314位、配列番号:21のヌクレオチド1260位〜3314位、および配列番号:23のヌクレオチド1260位〜3314位からなる群より選択される、請求項49記載の組換え微生物。
【請求項54】
組換え核酸分子が配列番号:18、配列番号:21、および配列番号:23からなる群より選択される核酸配列を含む、請求項49記載の組換え微生物。
【請求項55】
アセト乳酸シンターゼをコードする核酸配列がトラウストチトリアレスの微生物において機能するプロモーターへ実施可能に連結されている、請求項49記載の組換え微生物。
【請求項56】
アセト乳酸シンターゼをコードする核酸配列がトラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択される転写制御配列へ実施可能に連結されている、請求項49記載の組換え微生物。
【請求項57】
組換え核酸分子が微生物による産生としての第一のタンパク質をコードする核酸配列をさらに含み、該第一のタンパク質をコードする該核酸配列が転写制御配列へ実施可能に連結されている、請求項49記載の組換え微生物。
【請求項58】
組換え細胞がブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子でさらに形質転換される、請求項49記載の組換え微生物。
【請求項59】
組換え核酸分子が核酸配列の配列番号:9を含む、請求項58記載の組換え微生物。
【請求項60】
ブレオマイシン結合タンパク質をコードする核酸配列を含む組換え核酸分子が細胞による産生のための第二のタンパク質をコードする核酸配列をさらに含み、該第二のタンパク質をコードする該核酸配列が転写制御配列へ実施可能に連結されている、請求項58記載の組換え微生物。
【請求項61】
細胞による産生としてのタンパク質をコードする核酸配列を含む少なくとも1つの追加の組換え核酸分子をさらに含む、請求項49記載の組換え微生物。
【請求項62】
以下のものからなる群より選択される核酸配列を含む単離された核酸分子:
a. 配列番号:9のヌクレオチド441位〜894位;
b. 核酸配列が少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、完全長である配列番号:9のヌクレオチド441位〜894位にわたり配列番号:9のヌクレオチド441位〜894位と少なくとも約95 %同一である核酸配列;および
c. (a)または(b)のポリヌクレオチドと完全に相補的である核酸配列を含む単離されたポリヌクレオチド。
【請求項63】
単離された核酸分子が配列番号:4および配列番号:9のヌクレオチド441位〜894位からなる群より選択される核酸配列を含む、請求項62記載の単離された核酸分子。
【請求項64】
トラウストチトリアレスのα-チューブリンプロモーター、トラウストチトリアレスのアセト乳酸シンターゼプロモーター、トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーター、およびトラウストチトリアレスの脂肪酸デサチュラーゼプロモーターからなる群より選択されるプロモーターへ実施可能に連結されているブレオマイシン結合タンパク質をコードする核酸配列を含む、トラウストチトリアレス目の微生物の形質転換のための組換えベクター。
【請求項65】
トラウストチトリアレスのアセト乳酸シンターゼプロモーターが配列番号:14のヌクレオチド1位〜1259位を含む、請求項64記載の組換えベクター。
【請求項66】
α-チューブリンプロモーターが配列番号:9のヌクレオチド441位〜894位および配列番号:9のヌクレオチド441位〜894位の完全長に対して配列番号:9のヌクレオチド441位〜894位と少なくとも約95 %同一である核酸配列からなる群より選択される核酸配列を含み、該核酸配列が少なくとも基礎的なα-チューブリンプロモーター転写活性をもっている、請求項64記載の組換えベクター。
【請求項67】
組換えベクターが配列番号:8および配列番号:9からなる群より選択される核酸配列を含む、請求項64記載の組換えベクター。
【請求項68】
トラウストチトリアレスのポリケチドシンターゼ(PKS)系由来のプロモーターが配列番号:34または少なくとも基礎的なPKSプロモーター転写活性を有するその断片を含む、請求項64記載の組換えベクター。
【図1】

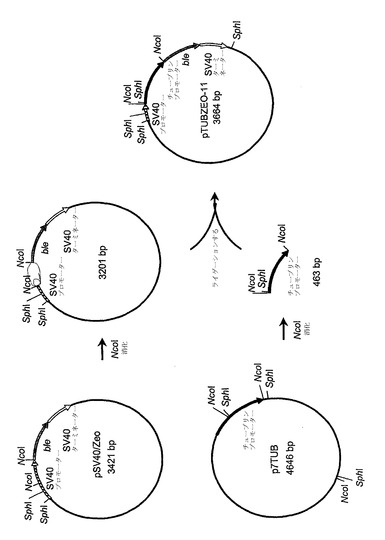
【図2】
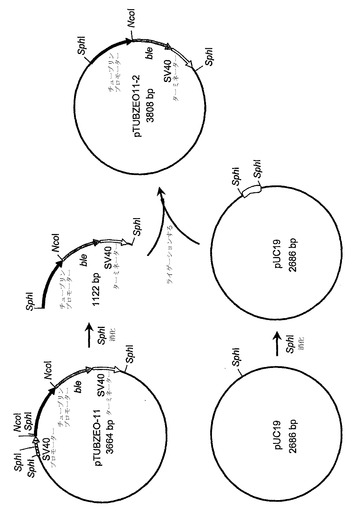
【図3A】
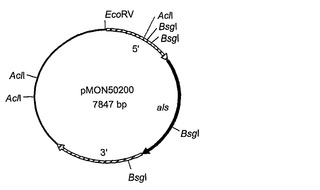
【図3B】
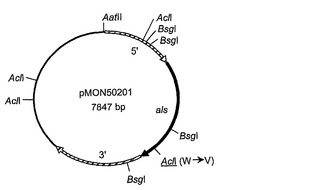
【図3C】
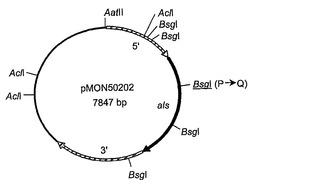
【図3D】
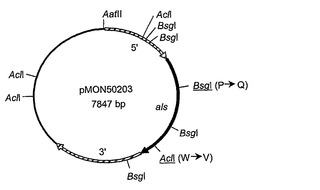
【図2】
【図3A】
【図3B】
【図3C】
【図3D】
【公開番号】特開2013−5817(P2013−5817A)
【公開日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願番号】特願2012−215905(P2012−215905)
【出願日】平成24年9月28日(2012.9.28)
【分割の表示】特願2010−19881(P2010−19881)の分割
【原出願日】平成14年4月16日(2002.4.16)
【出願人】(508004410)マーテック バイオサイエンシーズ コーポレーション (26)
【Fターム(参考)】
【公開日】平成25年1月10日(2013.1.10)
【国際特許分類】
【出願日】平成24年9月28日(2012.9.28)
【分割の表示】特願2010−19881(P2010−19881)の分割
【原出願日】平成14年4月16日(2002.4.16)
【出願人】(508004410)マーテック バイオサイエンシーズ コーポレーション (26)
【Fターム(参考)】
[ Back to top ]