
トランケートtauタンパク質
新規NおよびC末端二重トランケートtau分子(「タイプIA、IB、IIAおよびIIB tau分子」)および組換えおよび生物学的供給源の両方からこれら分子を得る方法を記載する。さらに、アルツハイマー病の診断および治療に関連するこれら分子のスクリーニング方法を提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、具体的にはアルツハイマー病および関連疾患にみいだされるNおよびC末端トランケート(truncated)病的型のtau(タウ)タンパク質に関する。
【0002】
さらに本発明は、NおよびC末端二重トランケートtauタンパク質を阻害、中和、および除去、またはその形成を予防するのに有効な潜在的薬剤をスクリーニングおよび試験する方法、および作用機序がtauタンパク質の該二重トランケート病的(罹患)型により生じる微小管構築および/または動態の修飾の中和に基づく潜在的薬剤をスクリーニングおよび試験するための方法に関する。
【0003】
アルツハイマー病は痴呆の最も一般的な原因である。該症例の5%以下においてアルツハイマー病はアミロイド前駆体タンパク質、プレセニリン-1またはプレセニリン-2遺伝子の1またはそれ以上の特定の突然変異とほぼ完全に同時分離し(cosegeragate)(1)、該症例の95%以上で正確な病因が不明である。
【0004】
病因とは無関係にアルツハイマー病は組織病理学的に対になったらせん状フィラメント(PHF)の神経原線維変化を有する多くのニューロンおよび脳の老人斑の主要成分としてアミロイドβの細胞外沈着の存在を特徴とする。アルツハイマー病のこれら2つの特徴的病変間の実際の直接的関係(あるとすれば)は理解されていないが、神経原繊維変性の存在は該病気、すなわち痴呆の臨床的発現に必要のようである(2、3、4)。神経原繊維変性は神経原線維変化、ジストロフィー性神経突起および神経網スレッドにより示される。これら構造の主要タンパク質サブユニットは微小管関連タンパク質tauである(5、6)。
【0005】
健康ヒト脳においてtauは単一遺伝子座由来の転写物の選択的mRNAスプライシングにより生じる6つのタンパク質アイソフォームで出現する。tauタンパク質は該分子のC末端近くの31または32アミノ酸の3(t3L、t3S、またはt3)または4(t4L、t4S、またはt4)チューブリン結合ドメイン(反復配列、R)、およびN末端部分の各29アミノ酸の2(t3L、t4L)、1(t3S、t4S)、または無(t3、t4)挿入物(7、8)を含むか否かで異なる。生理学的条件下でtauタンパク質は微小管の構築、空間的構成、安定化、および挙動に関与する。生理学的条件下で該タンパク質は健康ヒト脳では6アイソフォームで出現する。しかしながら、ADではtauタンパク質は多くの異なる翻訳後修飾(過リン酸化、ユビキチン化、グリコシル化)を受けることが知られている。第17染色体と結合したパーキンソン症を伴う疾患である前頭側頭性痴呆(FTDP17)のtau遺伝子における特異的突然変異の同時分離の最近の発見は、tauタンパク質のある異常が罹患個体における神経変性および痴呆の主要原因となりうることを確認した(9、10)。アルツハイマー病におけるtau修飾および対になったらせん状フィラメント(PHF)形成をもたらす分子的事象は知られていない。これは、広域スペクトルの病態生理学的事象、例えばtauタンパク質の病的再分布、軸索輸送障害、または軸索微小管機能維持障害の所見を説明する(11、12、13)。今日までアルツハイマー病におけるPHF微小繊維形成の重要性はあらゆるタンパク質がin vitroで原繊維を形成することができるという最近の発見に照らして疑問である(14)。
【0006】
多くの著者がアルツハイマー病における対になったらせん状原繊維の形成は異常リン酸化に基づく神経原繊維病変における主要事象を示すと考えている。PHF構築tauタンパク質はリン酸化依存性にある抗体と反応し、このことは特別なリン酸化状態を示唆する(15、16)。さらにPHF由来tauタンパク質はそのリン酸化パターンに関連しうるSDSゲルにおける電気泳動移動性の低下を示すことが観察された(Steinerら、EMBO J. 9(1990)、3539-3544)。同様に、同様の効果が正常tauがある種のキナーゼによりin vitroでリン酸化された時にみられたことからリン酸化によりPHF由来tauが正常tauタンパク質と比べて微小管に低親和性であることが示唆された(17、18)。tauは既知の最も可溶性のタンパク質の一つであり(19、20、21)、アルツハイマー病におけるその凝集は特に謎である。他方、tauタンパク質がアルツハイマー病におけるフィラメント形成をもたらすように修飾されるメカニズムは知られていない。tauのリン酸化は、おそらくリン酸化部位依存性に微小管重合に対する促進的または阻害的作用をもたらすtauの凝集形成潜在力に影響を及ぼす(22-27)。多くのin vitro試験は、還元剤ジチオスレイトール(DTT)、不飽和遊離脂肪酸、RNAまたはグルコサミノグリカンの存在下で、正常tauをフィラメントに変化させることができることを示す(28-31、38)。さらに、フィラメント形成のプロセスはCys322の酸化により生じる架橋tauの存在により促進することもできる(32)。tauタンパク質濃度、pHおよびイオン強度を含む種々のフィラメント構築試験で変化するパラメーターは生理学的条件下で細胞質におけるより多様性が高かった。in vitro形成tauフィラメントの走査透過電子顕微鏡(STEM)による実験は、これらフィラメントが天然の対になったらせん状フィラメントと異なることを示した(33)。グリカンまたはRNAの非存在下でPHF様フィラメントは非リン酸化またはリン酸化野生型tauを含む資料中に検出できなかった。さらにリン酸化はアルツハイマー病の防御的役割を果たすかもしれないことが示唆された(34)。微小管の分解および極めて重要な神経プロセス、例えば軸索輸送との干渉を生じるPHFの構築をもたらすtauの修飾に対する同様の示唆がユビキチン化およびグリコシル化についてなされた(30、35、36、37)。しかしながら、上記翻訳後修飾のみではアルツハイマー病の臨床的発現に関連するその機能障害をもたらすtau変化の開始に対する分子的説明となり得なかった。
【0007】
したがって、上記のどのtau修飾がアルツハイマー病の病因に関与しているかは依然不明である。
【0008】
今日まで初期tauタンパク質複合体の形成をもたらす翻訳後事象の様式または調節に関する確実なデータはない。そのような複合体形成を予防し、そのあらゆる関連病的効果を中和するには病的tauの正確な分子的性質および正常tauをそのNおよびC末端二重トランケート型に変換する調節メカニズムを明らかにする必要がある。この詳細な知識はアルツハイマー病の治療薬および診断薬のための手段を構築するであろう。
【0009】
Zelmanら(J. P. Neurochem. 72(2)(1999)、741-750)は、微小管結合タンパク質tauの開裂産物が外傷性脳損傷を有するヒトの脳脊髄液に生じ、ニューロンの損傷を反映することを示唆する。しかしながら、アルツハイマー病との関連はこの報告に記載されていない。
【0010】
Novak(Acta Virologica 38(1994)、173-189)は、この総説で広域スペクトルプロテアーゼ「プロナーゼ」により人工的に生成されたPHF(「対になったらせん状フィラメント」)の最小プロテアーゼ抵抗性単位「タウオン」に関する論説を記載している。
【0011】
Kontsekovaら(J. Immunol. Meth. 185(1995)、245-248)は、ヒトtauタンパク質の熱抵抗性を用いる免疫分析用の組換えヒトトランケートtauタンパク質の迅速精製方法を開示している。その中で使用された組換えtau類似体の構造や生理学的特性または機能は記載されていない。
【0012】
Novakら(EMBO J. 12(1)(1993)、365-370)は、対になったらせん状フィラメント(PHFコア)をin vitroで人工的に製造し、最小プロテアーゼ抵抗性tau単位をプロテアーゼプロナーゼによるin vitro消化により回収した。モノクローナル抗体MN423を用いて最小プロテアーゼ抵抗性tau単位を検出した。しかしながら、この論文に記載のtauポリペプチドは「実在する(realworld)」tauタンパク質、特にアルツハイマー病と関連があるtauタンパク質と共通の生物学的構造的な病的特性をもたなない。
【0013】
Fasuloら(Alzheimer's Research 2(5)(1996)、195-200)は、PHFコアtauの組換え類似体の過剰発現がtau凝集および対になったらせん状フィラメントのその構築を誘導するのに十分でないことを報告している。これらデータはAbrahaら(J. Cell. Science(113)(21)(2000)、3737-3745)の論文とこの論文に記載の異常な非生理学的アッセイ系(サル腎臓由来細胞系)により明白に異なる。
【0014】
Fasuloら(J. Neurochem. 75(2000)、624-633)はアポトーシスを誘導するtau断片を記載している。しかしながら、本発明に記載のアルツハイマー病関連tauタンパク質はいずれもアポトーシスを誘導し得ない。
【0015】
Espositoら(J. Peptide Science 6(2000)、550-559)はtauタンパク質および正常健康tauタンパク質のC末端19アミノ酸を記載している。Novakら(Chem. Papers 52(1998)、429-430)およびUgoliniら(NeuroReport 8(1997)3709-3712)の論文はアポトーシスにも関連するC末端トランケートtauタンパク質に関する。より最近の刊行物はアルツハイマー病がアポトーシスプロセスに関連しないことを示す。
【0016】
Abrahaら(J. Cell Science 113(21)(2000)、3737-3745)は、フィラメント形成に対するtauタンパク質の単一ドメインの関与を示すためのを記載している。したがって、一組の組換えtau分子が組み立てられ、in vitroで生成された。細菌におけるこれらタンパク質の生物活性および病的活性はいずれも生じないか検出されなかった。さらに、アルツハイマー病患者の脳由来のtauタンパク質に関するデータはこの論文に記載されていない。
【0017】
Jichaら(J. Neuroscience Research 55(1999)、713-723)には、モノクローナル抗体Alz50およびMC-1のエピトープの分子的分析が記載されている。両抗体はtau分子の機能的N末端、特にアミノ酸位7-9に依存する。Tauトランケーションはこの論文に記載されていない。
【0018】
Brandtら(J. Biol. Chem. 268(1993)、3414-3419)は正常健康ヒトtauタンパク質の異なるドメインを分析した。このために組換えtau断片を細菌で生成した。しかしながら、アルツハイマー関連トランケートtau断片はこの論文に記載されていない。
【0019】
Philippeら(J. Neuroscience Research 46(1996)、709-719)はモノクローナル抗アミロイド前駆体タンパク質抗体を開示している。この著者らはtau反応性抗体の生成について記載したが、この抗体は最初にアミロイド前駆体タンパク質に対して生じた。病変に関連するアルツハイマーtau断片はこの論文に記載されていない。
【0020】
WO94/18560 A1は、細胞中枢神経サイトパシーの患者を検出するための脳脊髄液中のヒトtauタンパク質を検出するためのイムノアッセイを開示している。このアッセイは正常tauと中枢神経サイトパシーの患者のtauを識別しないが、試料中のtauタンパク質の総量を検出する。
【0021】
したがって、本発明の目的はアルツハイマー病ニューロンの病的機能不全に関連するそのような確実なマーカーを提供することである。さらに、そのようなtau由来ポリペプチドの存在を確認し、その活性をアッセイするための適切な手段はアルツハイマーの診断薬および治療薬に有用な手段であろう。
【0022】
したがって、本発明は、以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIA tau分子」)を提供する:
4反復配列を含むtau43の少なくとも最初の236 N末端アミノ酸および少なくとも最後の45 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて正常tauタンパク質の微小管構築の促進を防止し、
該微小管構築の促進の防止は微小管構築アッセイにおいて該分子に対する特異的阻害、中和モノクローナル抗体により除去することができる。
【0023】
以下において用語「NおよびC末端二重トランケートtauタンパク質」はアルツハイマー病脳に出現し、アルツハイマー病ニューロンの病的機能不全と密接に関連するトランケートtau誘導体の2つのグループを説明するのに用いる。特に、これらタンパク質は微小管関連生物学的機能、例えば微小管構築または細胞内輸送を修飾することによりその病的機能を示す分子群を表す。以下において用語「タンパク質複合体」はホモダイマー、ヘテロダイマーまたは多量体複合体の形の、tauおよび/または二重トランケートtauタンパク質と物理的に関連する分子からなるNおよびC末端二重トランケートtauタンパク質に用いる。
【0024】
本明細書で用いている用語「tau」は、前記のその微小管結合ドメイン(39、40)中に3反復配列(tau44)および4反復配列(tau43)を含む健康ヒト脳に存在する最も短い天然アイソフォームの群、すなわち、tau43(383アミノ酸、エクソン2および3を欠く[pos45-102])tau44(352アミノ酸、エクソン2、3および10を欠く[各pos 45-102および275-307])をあらわす。以下の本文において用語「野生型tau」は「正常tauタンパク質」の同義語として用い、健康脳由来のtauタンパク質を表す。
【0025】
適切な微小管構築アッセイ(あるいは「微小管重合アッセイ」と呼ばれることも多い)は例えば(19)および(20)に記載されている。用語「予防(防止)する(preventing)」は、正常tau促進活性の20%またはそれ以上、好ましくは50%またはそれ以上のあらゆる有意な阻害を含む。
【0026】
具体的には、本発明の好ましいタイプIA tau分子は配列番号1〜3の群から選ばれるアミノ酸配列を含む。
【0027】
さらに、本発明は、以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIB tau分子」)を提供する:
4反復配列を含むtau43の少なくとも最初の238 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の207 N末端アミノ酸および少なくとも最後の50 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauの微小管構築の促進を防止しない。
【0028】
好ましいタイプIB tau分子は配列番号4〜10の群から選ばれるアミノ酸配列を含むことを特徴とする。
【0029】
本発明は以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIIA tau分子」)も提供する:
4反復配列を含むtau43の少なくとも最初の68 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の68 N末端アミノ酸および少なくとも最後の20 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauより高い微小管構築促進活性を有し、
該微小管構築促進活性は微小管構築アッセイにおいて該分子に対する特異的阻害、中和モノクローナル抗体により除去することができ、該分子の病的活性は微小管重合促進活性により定義される微小管ネットワークに対する結合に依存する。
【0030】
好ましくは、分光光度法で測定したとき微小管構築促進活性の増大は野生型tauより少なくとも20%高く、特に少なくとも50%高い。
【0031】
好ましいタイプIIA tau分子は配列番号11〜18の群から選ばれるアミノ酸配列を含むことを特徴とする。
【0032】
さらに、本発明は、以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIIB tau分子」)を提供する:
4反復配列を含むtau43の少なくとも最初の68 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の68 N末端アミノ酸および少なくとも最後の20 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauと異なる病的微小管構築促進活性を有する。
【0033】
本発明の好ましいタイプIIB tau分子は配列番号19および20の群から選ばれるアミノ酸配列を含むことを特徴とする。
【0034】
本発明の新規tauポリペプチド(IA、IB、IIAおよびIIB)はアルツハイマー病の脳組織にもっぱら局在するので典型的でユニークな局在特性を有する。さらに、これらポリペプチドと非重合チューブリン(α/βダイマー)および重合型(微小管として)の相互作用もユニークである。
【0035】
別の局面によれば本発明は、以下の工程を特徴とする本発明(タイプIA、IB、IIA、IIB)の分子の製造方法を提供する:
a)少なくとも最初の236および最後の40アミノ酸または最初の68および最後の20アミノ酸またはその組み合わせにわたる欠失を有する二重トランケートtau分子をコードする配列を保持する組換え原核性発現プラスミドの構築、
b)該NおよびC末端二重トランケートtau分子の発現を許す条件下で該細菌を増殖させ、
c)好ましくは遠心により細菌を回収し、
d)細菌ペレットを再浮遊させ、
e)該細菌を超音波処理し、
f)該超音波処理細菌をゲルろ過により分画し、
g)得られた分画の活性を微小管構築アッセイによりモニターすることにより、タイプIおよびタイプII tau分子の異なる活性を同定する。
【0036】
好ましくは、該トランケーションはタイプIA、IB、IIAおよびIIB分子について上記の通りである。該微小管構築アッセイ活性は好ましくは特にIAについて上記の通りである。
【0037】
さらに本発明は以下の工程を特徴とする本発明の分子の製造方法を提供する:
a)アルツハイマー罹患脳組織を得、
b)該病的(罹患)脳組織を緩衝液、特にTris緩衝液中でホモゲナイズし、
c)該ホモゲナイズ脳組織を硫酸アンモニウム沈殿し、
d)PIPES緩衝液に再溶解し、
e)該再溶解物質をゲルろ過で分画化し、
f)得られた分画の活性を微小管構築アッセイでモニターすることによりタイプIおよびタイプII tau分子の異なる活性を同定する。
【0038】
該微小管構築アッセイ活性は好ましくは特にIAについて上記の通りである。
【0039】
本発明はさらに以下の工程を含むタイプIA分子とチューブリンの複合体の分解(disassembling)に有効な物質の試験方法を提供する:
a)タイプIA分子とチューブリンのタンパク質複合体を形成させ、
b)該タンパク質複合体を被検物質とインキュベーションし、野生型tauの微小管構築促進能力を回復させる物質を同定する。
【0040】
さらに本発明は以下の工程を含む野生型tau発現細胞系においてタイプIA分子がチューブリンと複合体の形成を開始するのを阻害するのに有効な物質の試験方法を提供する:
a)適切な制御領域の制御下でタイプIA分子をコードする機能的遺伝子を正常tauタンパク質発現細胞に導入し、
b)タイプIA分子とチューブリンのタンパク質複合体を形成させ、
c)試験する物質を該複合体を保持する細胞に適用し、
d)上記のタイプIA生物活性に対する該物質の効果を試験する。
【0041】
本発明は該タイプIIA分子と微小管との相互作用により病的微小管を生成させる生理的条件下でタイプIIAとチューブリンタンパク質をインキュベーションすることを特徴とする微小管をin vitroで病的状態に変換する方法も提供する。
【0042】
別の局面において、本発明は以下の工程を含むタイプIIA分子を除去および/または中和し、タイプII分子により生じる生理学的微小管パラメーターおよび機能を修復するその特性についてタイプIIA分子の病的効果を中和することができる物質のスクリーニング方法を提供する:
a)タイプIIA分子およびチューブリン存在下で病的微小管を形成させ、
b)該物質、タイプIIAおよびチューブリンの混合物をスクリーニングする物質とインキュベーションし、
c)タイプIIA分子により生じる病的微小管の形成を減少させる結果を試験する。
【0043】
本発明は以下の工程を含むタイプIIA分子発現細胞系における異常微小管形成および機能の促進においてタイプIIA分子のin vivo活性を阻害するのに有効な物質の試験方法も提供する:
a)タイプIIA分子をコードする機能的遺伝子を適切な制御領域の制御下で野生型tau発現細胞中に導入し、
b)タイプIIA tau分子と微小管の複合体を形成させて該複合体を病的微小管の形成に関与させ、
c)試験する物質を該複合体を保持する細胞に適用し、
d)タイプIIA生物活性、特に微小管ネットワークおよびその関連機能の修飾に対する該物質の効果を試験する。
【0044】
別の局面によれば本発明は本発明に記載の分子(タイプIA、IB、IIAまたはIIB)、特にIAおよび/またはIIAを発現するトランスジェニック動物も提供する。
【0045】
本発明は特にアルツハイマー病治療用の薬剤をスクリーニングおよび試験するためのアルツハイマー病の動物モデルとしての本発明のトランスジェニック動物の使用にも関する。
【0046】
本発明は本発明の分子(IA、IB、IIAまたはIIB)、特にIAおよびIIA、および医薬的に許容される担体、特にアジュバントを含むワクチンを提供する。
【0047】
本発明はタイプIA分子と野生型tauの複合体の形成開始の阻害剤(インヒビター)も提供する。該阻害剤の特定の例には、寄託番号02060767でEuropean Collection of Cell Cultures(ECACC)、Porton Down、Salisbury、UKに寄託したモノクローナル抗体DC44のような結合部分、特にDC44またはその結合断片、例えばFabを含む物質である。
【0048】
すなわち、本発明は以下のもの提供する:
(1)NおよびC末端トランケート病的型のtauタンパク質の分子的および機能的同定および特徴付け(これら分子は微小管関連生物学的機能、例えば微小管構築または細胞内輸送を修飾することによりアルツハイマー病のその病的機能を示す。)、
(2)タンパク質エピトープの特異抗体、
(3)該タンパク質の病的活性を中和する抗体、
(4)NおよびC末端二重トランケートtauタンパク質を阻害、中和、および除去し、またはその形成を予防するのに有効な治療薬候補(抗体を含む)のスクリーニングおよび試験方法、
(5)薬物スクリーニングに用いることができる、各二重トランケートtauタンパク質をコードする遺伝子構築物を保有する動物モデルの開発、
(6)該二重トランケートtauタンパク質の阻害剤およびその起源に含まれるプロテアーゼを含む医薬組成物、
(7)NおよびC末端二重トランケートtau分子を生じる分子のスクリーニング方法、
(8)該分子を認識しそして/またはそれと相互作用する診断用および治療用組成物、
(9)該二重トランケートタンパク質の抗原性に基づくワクチンの開発、
(10)アルツハイマー病およびtauの病的変化に関連する他の障害のin vitroおよびin vivo診断用の該タンパク質およびそのエピトープおよび/または抗体または他の特異的プローブを含む方法。
【0049】
したがって、本発明は具体的にはアルツハイマー病に生じるNおよびC末端二重トランケート型の病的tauタンパク質およびそのエピトープの特徴付けに関する。
【0050】
タンパク質の分解は、種々のサイズの、通常短い半減期の中間体トランケーション生成物の生成を含むタンパク質の生理学的除去中に生じる一般的現象である。wt(=野生型)tauを含む健康脳ではタンパク質は例外なくこのプロセスを受ける。以下において用語「wt」は健康個体の脳に通常みられるtauタンパク質のすべての6つの天然アイソフォームに及ぶ。アルツハイマー病の脳に見られる種々の短トランケーション型は細菌中で産生され、タンパク質の生理学的機能を試験するために種々の程度に精製され、そのドメインおよびリン酸化エピトープをマップし、またはアルツハイマー病および他の神経変性障害における対になったらせん状構築のメカニズムを理解するための実験において結果が曖昧である(23-27、34、41、42)。一般的用語「NおよびC末端二重トランケート型のtauタンパク質」は分子の両末端のアミノ酸の少なくとも1つが失われているアルツハイマー病のあらゆるtauタンパク質を表す。アルツハイマー罹患脳からの抽出物中の二重トランケートtauの分析により、本発明の過程でこれら分子のいくつかが構造的および機能的に異なる特性を表現し、それらをアルツハイマー病の脳組織にみられる他のtau断片から識別することができることがわかった。この識別に基づいて、健康tau:タイプIおよびタイプII tau分子と異なるNおよびC末端二重トランケートtau分子の病的分子の2つの主なクラスを定義する新規の仕組みを提供した。これらのグループは分子構造に基づいてさらにそれぞれ2つのサブクラスに細分することができ、それぞれタイプIAおよびBおよびタイプIIAおよびBと称される。
【0051】
タイプIAおよびタイプIIAは、病的プロセシングにより生じた微小管関連タンパク質tau由来の構造的および機能的に異なるタイプの病的分子を表す。NおよびC末端トランケートtau分子は、アルツハイマー病に特徴的な特異的な病的プロセス中に出現する微小管関連タンパク質 tau由来の病的分子を表す。これはtau由来タンパク質の4グループすべてに共通の特徴である。さらに全グループに共通の特徴はNおよびC末端トランケーション、その神経内および神経外局在および正常な健康tauとの機能的識別である。
【0052】
「タイプIA」で示される分子グループは実施例に配列番号1〜3で示される。これらトランケートtau分子は、病的tauとチューブリンの相互作用のための鍵(中心的な)、活性単位、および原動力として作用する正常tauと異なる。タイプIAおよびタイプIB分子は微小管構築においていかなる促進活性も持たない。驚くべきことに、タイプIAは正常tauを微小管構築の促進から予防することができる(実施例1)。同様の一次配列特性および分子量にも関わらずタイプIBはin vitroでこの機能的活性を示さない(実施例2)。これはタイプIAのチューブリンに対する強い結合活性ならびにそれによるtauの生理学的機能に対する主要な悪影響をもたらすことを示唆する。したがってタイプIA分子は機能的微小管ネットワーク由来のニューロンの連続的で慢性的な欠失に関与し、アルツハイマー病の臨床的重症度に直接関連する神経原繊維構造に関与するようである。予期しないことにタイプIB(例えば配列番号4〜10)は、タイプIAグループの分子と同様の分子量と配列を有するにも関わらずグループIAのメンバーの病的活性を全く示さない(実施例2参照)。これらのグループと反対に、タイプIIA 二重トランケートtau誘導体は微小管と結合し、その病的構築を促進する(実施例3)。以下においてタイプIIAがもたらす微小管を「病的微小管」という。驚くべきことに、同様の配列および分子量範囲を有する分子(タイプIIB)はこれらの高微小管重合能力を欠く。微小管構築アッセイにおいて、それらは完全長tauタンパク質でみられるレベルで機能する(実施例3参照)。
【0053】
両グループ(タイプIIAおよびB)のNおよびC末端トランケートtau誘導体は、細胞レベルで軸索輸送を妨げ、最終的にアルツハイマー病患者に神経機能不全と認知障害をもたらすシナプスの損失をもたらす。同時に、罹患ニューロンは種々の形のストレス、例えば酸化ストレスに弱い(実施例4)。タイプIIBはタイプIIAと同様の分子サイズを有するにも関わらず分光光度法で測定すると微小管構築をさらに完全長健康tau(野生型tau)で見られるレベルまで促進した。
【0054】
本発明のタイプIAおよびBならびにタイプIIAおよびB分子のさらに好ましい態様において該分子の組換えバージョンは以下の工程を行うことにより得ることができる:
(a)該二重トランケートtau分子(タイプIおよびII)のコーディング配列を保持する組換え原核性発現プラスミドの構築、
(b)NおよびC末端二重トランケートtau分子(タイプIおよびII)を発現させることができる条件下での細菌の増殖、
(c)遠心分離による細菌の回収、
(d)500ml培養から得た細菌ペレットの緩衝液A:(20mM PIPES pH6.9、50mM NaCl、1mM EGTA、1mM MgSO4、2mM DTT、0.1mM PMSF)への再浮遊、
(e)氷上で1分間(3回)超音波処理し、2℃、45000rpmで15分間遠心分離(ローターTLA-120,2、Beckmann Optima TLX)、
(f)緩衝液「A」中の直線勾配0〜1M NaClを用いPhosphocellulose(ホスホセルロース)、またはMONO S HR 5/5または5ml HiTrap SP Sepharose HPカラムでクロマトグラフィし、得られたタンパク質をSDS-PAGEおよびWestern(ウエスタン)ブロット分析により同定。
【0055】
本発明の好ましい態様において、NおよびC末端二重トランケートメンバーの該タイプIAグループは以下のアミノ酸配列を含む。
【0056】
4反復配列tau由来の誘導体(tau 43)はR4で表示される。
配列番号1(239-333、R4)
【0057】
【化1】

配列番号2(237-333、R4)
【0058】
【化2】
配列番号3(239-318、R4)
【0059】
【化3】
【0060】
本発明の好ましい態様においてNおよびC末端二重トランケートメンバーの該タイプIBグループは以下のアミノ酸配列を含む。
配列番号4(239-326、R4)
【0061】
【化4】
配列番号5(239-328、R4)
【0062】
【化5】
配列番号6(239-331、R4)
【0063】
【化6】
配列番号7(239-334、R4)
【0064】
【化7】
配列番号8(239-340、R4)
【0065】
【化8】
配列番号9(239-343、R4)
【0066】
【化9】
【0067】
3反復配列tau(tau 44)由来の誘導体にはR3を付す。
配列番号10(208-302、R3)
【0068】
【化10】
【0069】
具体的にはNおよびC末端二重トランケート病的型のtauタンパク質中のタイプIAまたはタイプIIA メンバーに生じるtauタンパク質の1またはそれ以上のエピトープがある。
【0070】
本発明の態様において、該エピトープは具体的には、タイプIA(配列番号1〜3)およびタイプIIA(配列番号11〜18)グループメンバーの一次構造内に位置し、その数、異種性、および特異性は各個々のグループメンバーの特異的構造的コンフォメーションに依存し、それにより加えられる。したがって、各分子の単一性は微小管構築に対するその効果と共にその一次構造だけでなく、そのエピトープを形成するその二次および三次構造に基づく。それらのいくつかは該分子の活性に大きく寄与する特に重要な「コンフォメーショナル領域」を形成することができる。
【0071】
本明細書で用いている用語「コンフォメーショナル領域」はその活性に寄与する分子のある領域にクラスター化したエピトープを表す。
【0072】
特に好ましい態様において、アミノ酸「ile lys his val pro gly gly gly ser val gln ile val tyr lys pro val asp leu ser lys val thr ser lys cys gly ser leu」を含むタイプIおよびタイプII分子に含まれるコンフォメーショナル領域は残基239-267(配列番号1-9および11-14、19 R4)に対応し、残基 217-236(配列番号10.15-18、20 R3)に対応するアミノ酸「val gln ile val tyr lys pro val asp leu ser lys val thr ser lys cys gly ser leu」を含むものは配列Aを表す。
【0073】
本発明のさらに別の好ましい態様において、該コンフォメーショナル領域の該エピトープが同定され、その相対的寄与は欠失突然変異により決定される。これらエピトープすべての重要性および微小管の機能に対するその関係はタイプIA分子の活性に種々の程度に寄与することが示された突然変異型により示される(実施例5)。これら個々のエピトープは以下のアミノ酸配列を含む:
A: ile lys his val pro gly gly gly ser val gln ile val tyr lys pro val asp leu ser lys val thr ser lys cys gly ser leu(配列番号1-9および11-14、19の残基239-267に対応する)。エピトープ欠失突然変異体は配列番号21(268-333、R4; del 239-267)
【0074】
【化11】
を有する。
A1: ile lys his val pro gly gly gly ser(配列番号1-9および11-14、19の残基239-247に対応する)。欠失突然変異体は配列番号22(248-333、R4; del 239-247)
【0075】
【化12】
を有する。
A2: ile lys his val pro gly gly gly ser val gln ile val tyr lys pro val asp leu(配列番号1-9および11-14、19の残基239-257に対応する)。欠失突然変異体は配列番号23(258-333、R4; del 239-257)
【0076】
【化13】
を有する。
A3: ile lys his val pro gly gly gly ser val gln ile val tyr lys pro val asp leu ser lys val thr ser(配列番号1-9および11-14、19の残基239-262に対応する)。欠失突然変異体は配列番号24(263-333、R4; del 239-262)
【0077】
【化14】
を有する。
A4: ser val gln ile val tyr lys pro val asp leu ser lys val thr ser(配列番号1-9および11-14、19の残基246-262に対応する)。
エピトープ欠失突然変異体は配列番号25(239-333、R4; del 248-262)
【0078】
【化15】
を有する。
A5: asp leu ser lys val thr ser、配列番号1-9および11-14、19の残基256-262および残基225-231、R3 配列番号10、15-18、20に対応する。エピトープ欠失突然変異体は配列番号26(239-333、R4; del 256-262)
【0079】
【化16】
を有する。
A6: lys cys gly ser leu、配列番号1-9および11-14、19の残基263-267および配列番号10、15-18、20の残基232-236、R3に対応する。エピトープ欠失突然変異体は配列番号27(239-333、R4; del 263-267)
【0080】
【化17】
を有する。
【0081】
さらに、該ペプチドのアミノ酸すべてが必ずしも特異抗体により実際に認識される特異的部位に寄与するわけではない。
【0082】
本発明の好ましい態様において該タイプIA 病的tauタンパク質は以下の特性を有する:
a)該タンパク質はNおよびC末端がトランケートされている(実施例6)、
b)該タンパク質はアルツハイマー病の組織に存在するが、該タンパク質は正常健康脳には存在しない(実施例6)、
c)in vitro微小管構築アッセイにおいて、それらは正常tauタンパク質の微小管構築の促進を妨げる(実施例7)、
d)正常tauを用いる微小管構築アッセイにおいて、それらの阻害活性は特異的阻害的中和モノクローナル抗体により除去することができる(実施例11)、
e)その病的活性は正常健康tauに存在しないアミノ酸配列および構造的コンフォメーションの組み合わせに依存する(実施例6)、
f)該タンパク質は正常tauタンパク質とコンフォメーション的に異なるようである(実施例6)。
【0083】
最も好ましい態様において、本発明はNおよびC末端トランケート病的tau型のタイプIAグループ、配列番号1-3およびその「コンフォメーショナル領域」(配列「A」)およびエピトープA1-A6に関する。
【0084】
タイプIB tauタンパク質は以下の特性について異なる:
a)タイプIBタンパク質はNおよびC末端がトランケートされている(実施例6)、
b)該タンパク質は正常健康ヒト脳に存在するかもしれない、
c)in vitro微小管構築アッセイにおいて、それらは正常tauタンパク質の微小管構築の促進を妨げない(それぞれ実施例2および7)、
d)それらは微小管構築の修飾において病的活性を示さない(それぞれ実施例2および7)、
e)タイプIB分子は正常tauとコンフォメーション的に異なるようである(実施例6)。
【0085】
本発明の別の態様は、tauおよび微小管機能に影響を及ぼすさらなる分子の同定をもたらす記載の二重トランケート型のtauを用いる機能的アッセイと組み合わせる、それ自体の多くは当該分野で知られている種々の抽出方法を含む示したアプローチの組み合わせである。脳抽出物からのtauタンパク質の収量は特定の脳組織試料のステージに依存して抽出したNおよびC末端二重トランケートtau分子の機能性で変化するかもしれない(実施例6)。当業者は、種々の異なる目的に本発明の方法を用いる方法を知っており、これらはすべて本発明の保護の範囲内にある。
【0086】
別の好ましい態様において、本発明はプロトタイプタイプIA分子グループメンバーとして特に配列番号1に関する。
【0087】
本発明のさらに別の目的は、チューブリンを該チューブリンと該タイプIA分子の相互作用を可能にする条件下で本発明のタイプIA分子とインキュベーションする正常tauタンパク質のアルツハイマータンパク質へのin vitro変換方法を提供する。
【0088】
用語「タイプIA分子またはそのペプチド誘導体のチューブリンとの相互作用を可能にする」は、タイプIA分子の活性、好ましくは最適活性を可能にする条件を表す。この活性はチューブリンとの結合および微小管構築におけるその生理学的機能の阻害をもたらす。
【0089】
別の態様において、タイプIA分子はその誘導体により阻害または中和されるかもしれない。阻害分子をスクリーニングするために本発明において記載されているように、タイプIA ペプチドおよびその誘導体、例えば欠失または突然変異を含むペプチドの微小管重合に対するそれらの効果を試験またはスクリーニングすることができる。
【0090】
正常tauタンパク質は天然または組換え供給源由来であってよい。しかしながら、本発明の方法を実施するためには組換え物質を用いるのが好都合である。
【0091】
上記方法は、具体的には種々の目的に十分な量のタイプIA NおよびC末端二重トランケートtauタンパク質を提供し、病的二重トランケートtauタイプIAにより生じる微小管構築の阻害を抑制する新規阻害剤のin vitroスクリーニング方法を確立することができよう。
【0092】
したがって、本発明の組成物に有用な阻害剤はタイプIA分子とチューブリンとの病的相互作用を調節することができるあらゆる阻害剤である。そのような阻害分子の作用様式はタイプIAまたは正常tauとの相互作用からなる。
【0093】
これら「インヒビター(阻害剤)」は、タイプIA分子に含まれるエピトープまたはエピトープ類に対して例えば該エピトープをブロックすることにより特異的であるか、またはそれらがその病理学的または生物学的活性を抑制または阻害する限りタイプIA分子上の種々のドメインを対象にするかもしれない。阻害効果は正常tauによる残存微小管構築促進活性を測定することにより定量的に定義することができる。阻害剤の供給源には、定義した化学構造および組成物の低分子ライブラリー、ペプチドライブラリー、溶液中の遊離またはファージまたは細菌またはリボソーム(リボソーム表現)の表面上に表現された抗体ライブラリーおよび当該分野で知られた同様の技術が含まれる。
【0094】
本発明のさらなる目的は以下の工程を含むタイプIA複合体の分解に有効な分子および化合物の試験方法(タイプI in vitroアッセイ)を提供することである:
a)タイプIA分子またはその誘導ペプチドおよびチューブリンまたはタイプIA分子と相互作用する他の分子のタンパク質複合体を形成させ、
b)該タンパク質複合体と試験すべき薬剤をインキュベーションし、
c)工程(b)のインキュベーションの結果を健康tauアイソフォームの微小管構築促進能力の回復について試験する。
【0095】
本発明のさらに別の目的は、以下の工程を含む健康tauアイソフォームの正常in vitro活性の阻害の抑制または低減に有効な薬剤の試験方法を提供することである:
a)タイプIA分子またはその誘導ペプチドと組み合わせて試験すべきある薬剤は正常tauおよびそのvitro機能と干渉しないと予想される、
b)タイプIA分子と正常tauおよびチューブリンの存在下で試験すべき薬剤のインキュベーション、
c)工程a)およびb)のインキュベーションの結果をタイプIA分子の微小管重合に対する阻害活性が存在するかまたはしないかについて試験する(実施例8)。
【0096】
該薬剤の非存在下で用語「タイプIA分子またはその誘導ペプチドとチューブリンの複合体を形成させる(を可能にする)」は、微小管形成の阻害をもたらすタイプIA分子と該チューブリンの相互作用を可能にする条件を表す。
【0097】
当業者は本発明の方法を種々の異なる目的に用いる方法を知っており、これらはすべて本発明の保護の範囲内にある。
【0098】
さらなる局面において、本発明は以下の工程を含むtauまたはtau由来タンパク質を発現する細胞系におけるタイプIA分子の複合体形成開始を阻害するのに有効な薬剤の試験方法(タイプI細胞アッセイ)に関する:
a)適切な制御領域の制御下でタイプIA分子をコードする機能的遺伝子を正常tauタンパク質を発現する細胞内に導入し、
b)タイプIA tauとチューブリン分子のタンパク質複合体を形成させ、
c)該複合体を保有する細胞を試験すべき薬剤に適用し、
d)タイプIA生物活性に対する該薬剤の効果、例えば微小管の構造的および機能的修飾を試験する。
【0099】
工程(a)で用いている用語「tauタンパク質を発現する細胞」は、タイプIA分子またはその誘導体をコードする遺伝子構築物由来のNおよびC末端二重トランケートtau型を発現する能力を有する細胞を表す。当業者はタイプIA分子をコードする遺伝子導入の一連の実験工程は本発明の方法の目的には無関係であることを承知している。
【0100】
該方法は、該スクリーニング系はin vivo状況と密接なイメージを与える連続的増殖細胞系に基づくため特に好都合である。さらに、細胞内に局在するタイプIA分子の十分な供給はタイプIA分子の生物学的効果の阻害に有効な薬剤のスクリーニングを可能にする。
【0101】
好ましい態様において、該タイプIA分子発現細胞は神経芽細胞腫、または褐色細胞腫細胞またはタイプIA分子発現トランスジェニック動物由来の神経細胞の一次培養である。
【0102】
「タイプII」で示す分子のグループはNおよびC末端二重トランケートtauタンパク質分子からなる(例えば配列番号11〜20に記載の配列)。このグループの定型的なものはニューロン内およびニューロン外に局在し、正常健康tauと機能的に異なる。
【0103】
本発明のこのグループのタンパク質の発見および単離は、
(1)特異的微小管結合および微小管構築の異常な促進をもたらすtau修飾の分子的説明および特徴付け(実施例3)、およびその担体に対する病的結果(実施例4)、
(2)該タンパク質エピトープに特異的な抗体および
(3)該タイプII分子の病的活性を中和する抗体(実施例12)、および
(4)該タイプIIタンパク質を阻害、中和、および排除するのに有効な治療薬候補をスクリーニングおよび試験する方法、または
(5)tau由来タンパク質、例えばタイプII分子の形成を阻害するのに有効な治療薬候補をスクリーニングおよび試験する方法、
(6)薬剤スクリーニングに用いることができるトランス遺伝子またはトランス遺伝子混合物として各NおよびC末端二重トランケートtauタンパク質をコードする遺伝子構築物を有する動物モデルの開発、
(7)該二重トランケートtauタンパク質およびそのプロテアーゼに対する阻害剤を含む医薬組成物、
(8)該分子を認識/相互作用する診断用および治療用組成物、
(9)該二重トランケートタンパク質に基づくワクチンの開発、
(10)アルツハイマー病およびtauの病的変化に関連する他の障害のin vitroおよびin vivo診断用の該タンパク質およびそのエピトープおよび/または抗体または他の特異的プローブを含む方法。
【0104】
グループタイプIAおよびBと反対に、タイプIIA分子は分光光度法で測定したとき正常健康tauアイソフォームにより促進される微小管構築より有意に高く病的微小管構築を促進する(それぞれ実施例1および3参照)。驚くべきことに、同様の配列と分子量範囲を有するNおよびC末端二重トランケートtau分子のサブグループ(タイプIIB)はこれら「高」微小管重合能力を欠く。微小管構築アッセイにおいて、このサブグループの分子は完全長tauタンパク質にみられるレベルに達する(実施例3)。
【0105】
したがって、本発明はtauタンパク質のタイプIIAグループと呼ばれるアルツハイマー病に見られる新しいタイプの修飾tauタンパク質に関する。該グループはNおよびC末端二重トランケートtau分子(配列番号11-18)からなる。
【0106】
用語タイプII分子は正常健康tauだけでなくタイプIAおよびB tauグループとも構造および機能が有意に異なる群のメンバーに関する。このサブグループの分子は微小管を結合し、健康tauアイソフォームによる正常微小管構築より有意に顕著な病的構築を促進する(実施例3)。タイプIIA NおよびC末端二重トランケートtau分子は細胞レベルでアルツハイマー病ニューロンおよび実験条件において全生物の認知障害を最終的に生じるシナプス損失および神経機能障害をもたらす構成成分の軸索輸送と干渉する(各実施例15および16)。同時に、罹患ニューロンは種々の形のストレス、例えば酸化ストレスに弱い(実施例4)。
【0107】
本発明の好ましい態様において、該タイプIIAグループのNおよびC末端二重トランケートメンバーは以下のアミノ酸配列を含む。
4反復配列tau(tau 43)の誘導体にR4を付す。
配列番号11(69-333、R4)
【0108】
【化18】
配列番号12(93-333、R4)
【0109】
【化19】
配列番号13(69-363、R4)
【0110】
【化20】
配列番号14(93-363、R4)
【0111】
【化21】
【0112】
3反復配列tauの誘導体(tau 44)にはR3を付す。
配列番号15(93-302、R3)
【0113】
【化22】
配列番号16(69-302、R3)
【0114】
【化23】
配列番号17(93-332、R3)
【0115】
【化24】
配列番号18(69-332、R3)
【0116】
【化25】
【0117】
本発明の好ましい態様において、該タイプII BグループのNおよびC末端二重トランケートメンバーは以下のアミノ酸配列を含む。
配列番号19(6-378、R4)
【0118】
【化26】
配列番号20(6-347、R3)
【0119】
【化27】
【0120】
本発明の好ましい態様において、該タイプIIA病的tauタンパク質は以下の特性を有する:
a)該タンパク質はNおよびC末端がトランケートされている(実施例6)、
b)微小管構築の十分な病的促進物質である(実施例3; 図28C)、
c)その病的微小管構築促進活性は例えば阻害的モノクローナル抗体またはその誘導体(実施例12)のような特異的化合物により除去することができる、
d)該タンパク質は正常健康脳にはみられない(実施例6)、
e)細胞内輸送機能が有意に損なわれる(実施例16)、
f)その病的活性は、微小管ネットワークと結合する高い親和性およびその機能的障害に関連する(実施例3)、
g)それらは正常tauとコンフォメーション的に異なるようである(実施例6)。
【0121】
本発明の別の好ましい態様においてタイプIIB分子は以下の特性を有する:
a)該タンパク質はNおよびC末端がトランケートされている、
b)タイプIIAより微小管構築促進効果が低い、
c)該タンパク質は正常健康脳にはみられない、
d)タイプIIAで観察されるより程度は低いが、それとの結合により微小管機能を損なうようである、
e)それらは正常tauとコンフォメーション的に異なるようである。
【0122】
本発明の別の好ましい態様において、タイプIIAおよびB分子の該エピトープはタイプI分子について記載したのと同じように同定された。これらエピトープすべてのタイプII分子における重要性およびその微小管に対する機能との関係は、タイプIAの実施例において示されるようなNおよびC末端二重トランケートtau分子の活性に種々の程度に寄与することが知られている突然変異型により示される。
【0123】
したがって本発明の組成物に有用な阻害剤は、「病的微小管」を生じるタイプIIA分子と微小管の病的相互作用を調節することができるあらゆる阻害剤である。本明細書で用いている用語、病的(pathological)微小管は、タイプII分子により修飾された微小管を表す。そのような阻害分子の作用様式は、いずれかの微小管、tauおよびその病的誘導体を含む微小管関連分子との相互作用からなる。阻害剤の供給源として、定義した化学構造および組成の小分子のライブラリー、ペプチドライブラリー、溶液中の遊離のまたは合成表面またはファージまたは細菌またはリボソーム(リボソーム表現)上に表現された抗体ライブラリー、および当該分野で知られた同様の技術を用いることができる。
【0124】
好ましい態様において、これら「阻害剤」はタイプIIA分子に含まれるエピトープまたはエピトープ類に対して該エピトープをブロックすることにより特異的であるか、またはその病的活性または生物活性をin vitroまたはin vivoで抑制または阻害するかぎり、タイプIIA分子上の種々のドメインを対象にすることができよう。該阻害効果は、例えば正常tauによる残存微小管構築促進活性を測定するか、または細胞内微小管パラメーター、例えば成長、安定性、または細胞内輸送を測定することにより定量的に定義することができる。
【0125】
別の態様においてタイプIIA分子は例えば各細胞内に発現した優性ネガティブタンパク質としてその誘導体により阻害または中和することができる。阻害分子をスクリーニングするために本発明において説明したように、タイプIIA ペプチドおよびその誘導体、例えば欠失または突然変異を含むペプチドはNおよびC末端二重トランケートtau分子の病的効果を阻害するその効果を試験またはスクリーニングすることができる。
【0126】
治療効果は微小管構造および機能の障害を阻害することにより達成される。
【0127】
したがって、本発明の別の目的は、本発明のタイプIIA tau分子に対する特異的阻害剤を所望により医薬的に許容される担体および/または希釈剤と組み合わせて含む医薬組成物を提供することである。
【0128】
別の好ましい態様において本発明はタイプIIA群分子のプロトタイプとして特に配列番号11と関連がある。
【0129】
本発明のさらに別の目的は、病的微小管を生じる該タイプIIAまたはBと微小管の相互作用を可能にする生理学的条件下で正常tauタンパク質を本発明のタイプIIAまたはBとインキュベーションする正常微小管を病的状態にin vitro変換するための方法を提供することである。
【0130】
本発明はさらに、タイプIIA分子の病的効果を中和することができる化合物のためのあらゆる分子ライブラリーのスクリーニングを可能にするスクリーニングアッセイに関する。本試験において、分子を、タイプIIA分子を除去および/または中和し、タイプII分子により生じた生理的微小管パラメーターおよび機能を回復させる特性についてスクリーニングする。薬剤スクリーニングアッセイは以下の工程からなる:
(1)適切な条件下、タイプIIA分子およびチューブリン存在下における病的微小管の形成(それぞれ実施例3および4)、
(2)これら病的微小管と試験する候補薬剤とのインキュベーション、
(3)タイプIIA分子の微小管に対する効果の中和に対する試験(各実施例9および12)。
【0131】
病的NおよびC末端二重トランケートtauタイプIIAにより生じる微小管に対する効果を軽減する阻害剤のin vitroスクリーニング方法を確立することができよう。これら「阻害剤」は、例えばエピトープをブロックすることによりタイプIIA分子に含まれるエピトープまたはエピトープに特異的であるか、またはそれらがその活性を抑制または阻害するかぎりにおいてタイプIIA分子上の種々のドメインを対象にすることができよう。該阻害効果は微小管構築動態を測定することにより定量することができる。阻害剤の供給源として、定義された化学構造および組成の低分子ライブラリー、ペプチドライブラリー、溶液中に遊離するかまたはファージまたは細菌またはリボソーム(リボソーム表現)の合成表面の表面上に表現された抗体ライブラリーおよび当該分野で知られた同様の技術を用いることができる。
【0132】
本発明の目的では、被検薬剤は、タイプIIA分子の量および/またはその活性を減少させるのに有効であり、補助的治療効果を満たすのに十分であるが、タイプIIA活性を完全に除去することが好ましい。
【0133】
当業者は種々の異なる目的に本発明の方法を用いる方法を知っており、これらはすべて本発明の保護の範囲内にある。
【0134】
本発明のさらなる目的は、生細胞、すなわちタイプII分子発現ニューロンまたはニューロン様細胞を用いる薬剤の確認方法を提供することである(タイプII細胞アッセイ)。あるいはまた、トランスジェニック動物由来の一次神経細胞培養またはタイプIIA分子を発現する種々の供給源由来の他の一次神経細胞を用いることができる。
【0135】
上記で用いている用語「タイプII分子発現ニューロン(神経細胞)」は、該分子を安定に発現するかまたはタイプIIA分子を発現する能力があり、機能的タイプIIA遺伝子が以下に例示する細胞培養技術またはトランスジェネシス(transgenesis)により導入された細胞を表す。
【0136】
好ましい態様において、該タイプIIA分子発現細胞は神経芽細胞腫または褐色細胞腫細胞またはタイプIIA分子を発現するトランスジェニック動物由来の神経細胞の一次培養である。
【0137】
当業者はタイプIIA分子をコードする遺伝子の導入手順は本発明の方法の目的とは無関係であることを認識している。
【0138】
本発明は、以下の工程を含むタイプIIA分子発現細胞系における異常微小管形成および機能の促進においてタイプIIAを阻害するのに有効な薬剤を試験するための方法に関する:
a)タイプII分子をコードする機能的遺伝子を適切な制御領域の制御下で正常tauタンパク質発現細胞に導入し、
b)タイプIIA tauおよび微小管(病的微小管)の複合体を形成させ、
c)得られた複合体を保有する細胞に被検薬剤を適用し、
d)タイプIIA生物活性、例えば微小管ネットワークおよびその関連機能の修飾に対する薬剤の効果を試験する。
【0139】
本発明のさらに別の最も好ましい態様はタイプIIA分子発現ニューロンの表現型である。適切な条件下でこれら分子を発現するニューロンは、細胞内輸送プロセスの摂動(perturbance)を生じる。さらにタイプIIA分子発現ニューロンは適切なストレス条件下で細胞死する(実施例4)。
【0140】
該方法は、in vivo状況の密接なイメージをもたらす連続増殖細胞系の使用に基づく関連システム(系)が細胞内に局在するタイプIIA分子の十分な供給をもたらし、細胞内タイプIIA効果の軽減に有効な化合物の薬剤スクリーニングを可能にするので特に好都合である。
【0141】
好ましい態様において、この細胞アッセイの計測値を低または高処理能力定量系に適応する。微小管輸送の崩壊または障害および/または神経細胞死をもたらす記載の表現型に関連する用語「適切な条件」は、実施例に示す該表現型を出現させるあらゆる条件を表す。
【0142】
本発明の目的において、このシステムによりスクリーニングするかまたはこのシステムで確認した潜在的薬剤または第三起源の薬剤は該表現型の規模の減少に有効であり、治療における補助的機能を満たすのに十分であるが、該薬剤による病的表現型の完全除去または減少が好ましい。
【0143】
細胞系または一次細胞の安定な増殖に加え、各発明はそのニューロンにタイプIIAまたはB分子を発現する全動物由来の細胞を用いる類似計測システムに広げることもできる(該トランスジェニック動物モデルを以下に例示する)。
【0144】
当業者は種々の異なる目的に本発明の方法を用いる方法を知っており、これらはすべて本発明の保護の範囲内にある。
【0145】
好ましい態様において、NおよびC末端二重トランケートタイプIIA tau型を安定に発現する該細胞およびトランスジェニック動物はアルツハイマー病の病因に関連する新規分子、その診断および治療をもたらす正しい情報を得る疾患経路のマッピングを可能にする。これらスクリーニングおよび同定方法はmRNA発現ベースのスクリーニング技術およびタンパク質ベースの技術を含む。
【0146】
好ましい態様において、該タイプIおよびタイプIIAおよびB分子またはその誘導体は、上記の病的NおよびC末端二重トランケート型のタイプIA、タイプIIAおよびBを保持および発現するトランスジェニック動物モデルを提供するための非ヒト動物のゲノム中に導入することができる組換えDNA構築物も提供する。本発明のトランスジェニック動物は、構築物が直接導入されている動物および構築物を発現する能力を保持する該動物の子孫を含む。該トランス遺伝子配列は遍在的に発現するかまたは組織特異的プロモーターと機能的に連結したポリヌクレオチド配列である。タイプIAおよびタイプIIAおよびB分子をコードするトランス遺伝子DNAは優先的に動物またはヒト供給源由来のcDNAおよび/またはゲノムDNAである。該タイプIおよびタイプIIAおよびB分子を発現するトランスジェニック動物はアルツハイマー病に表現型的に関連する細胞および/または臓器レベルでの機能的変化をもたらすと予想される。これらにはADに特徴的な組織学的変化、RNA発現変化、細胞生理学的パラメーターの変化、および好ましくは行動的変化が含まれる。トランスジェニック動物の成熟ニューロンにおけるタイプIおよびタイプIIAおよびB分子の発現は先に試験されている。タイプI、タイプIIAおよびBトランス遺伝子がトランスジェニック動物で発現するレベル(すなわち、トランス遺伝子mRNAのレベル)はトランスジェニック動物における一貫した病態生理学的欠陥を得るための重要なパラメーターであると予想される。トランス遺伝子を保持する動物の繁殖および異系交配(交雑)により、例えば疾患モデルとして現在用いられる動物種、他のトランス遺伝子を発現する動物、または遺伝子の機能的発現を欠く動物にトランス遺伝子を導入することにより、病的特徴を増強、軽減、または調節することができる。(実施例14参照)。
【0147】
より具体的には、本発明はヒトタイプIおよびタイプIIAおよびB分子をコードするDNAが適切な遍在性または組織特異的プロモーターのその調節可能な修飾を含む転写制御下で発現する、トランスジェニック非ヒト動物細胞を提供する。
【0148】
本発明に従って操作される細胞はあらゆる知られたトランスフェクション技術により製造することができよう。該DNA配列は直接的遺伝子操作によるかまたは細胞のより初期の世代内に導入することができよう。すなわち、該細胞をトランスジェニック動物から得、in vitroで培養することができよう。また、該トランスジェニック動物は知られた確立された方法、例えば胚性幹細胞への遺伝子伝達、初期胚のレトロウイルス感染、または前核性マイクロインジェクションによる胚操作に従って製造することができよう。前核性マイクロインジェクション技術が好ましい。本発明の組換えDNA構築物から得られる転写単位を動物胚の前核に注射し、得られたファウンダー(founder)トランスジェニックを繁殖させる。子孫において得られた結果を当該分野でよく知られた種々の技術を用いて分析することができる。tauおよびNおよびC末端二重トランケートtau由来分子、およびアルツハイマー病に関連する他の分子、例えばAPPおよびその誘導体を分析することができる本発明の細胞および動物に基づくモデルを、例えば神経変性性疾患における潜在的治療剤の効果を確認および評価するために用いることができよう。特に該モデルを本明細書に記載のNおよびC末端二重トランケートtau由来分子の病的効果を抑制すると思われる薬剤を検出するためのスクリーニングまたは特徴付けアッセイに用いることができよう。
【0149】
したがって、さらなる局面において本発明は、該二重トランケート型のtauを発現するトランスジェニック動物由来の細胞を標的細胞に用いる、特定の病状、特に神経変性性疾患、好ましくはADのための潜在的治療剤を試験する方法を含む。より具体的には、本発明は、例えば抗体またはその誘導体のような治療剤を本発明のトランスジェニック動物に投与するか、または交雑または遺伝子操作により導入し、さらに上記アッセイ系により試験するような方法を含む。さらに本発明は、そのような方法の中にあるかまたはそれを含むスクリーニングまたは特徴付けアッセイおよび本発明の細胞を含むスクリーニングアッセイキットを含む。本発明のタイプIおよびタイプIIAおよびB分子を発現する細胞系を用いる潜在的治療剤のスクリーニング方法を本発明において提供する(実施例15参照)。本発明の細胞および動物は同様に用いることができよう。
【0150】
本発明の別の目的はNおよびC末端二重トランケート型のtauタンパク質の特異的阻害剤を所望により医薬的に許容される担体および/または希釈剤と組み合わせて含む医薬組成物を提供する。
【0151】
用語「NおよびC末端二重トランケートtauの特異的阻害剤」は、具体的には該二重トランケートtauタンパク質の作用を阻害する物質を表す。阻害剤の性質は抗体、その操作された誘導化分子、本発明の試験系において望ましい阻害活性を示すあらゆるペプチドまたは合成化学組成物でありうる。
【0152】
本発明の別の目的は、具体的には本発明のエピトープを認識し、NおよびC末端二重トランケートtau分子の病的活性を部分的または完全に阻害することができる抗体またはその誘導体である。
【0153】
用語「本発明のエピトープまたはエピトープ類を含むオリゴまたはポリペプチド」は、その2または3次元構造に本発明のエピトープを再構成する、具体的にはそれに対する抗体により認識されるペプチドを表す。さらに、該オリゴまたはポリペプチドは該エピトープを表すアミノ酸のみからなるか、またはさらなるアミノ酸を含むことができよう。そのようなオリゴまたはポリペプチドの構築は当該分野でよく知られている。
【0154】
好ましい態様において、本発明は天然または組換えの、固定化した、溶液中に遊離した、または種々の分子または細菌、ウイルスの表面または他の表面上に表現されたモノクローナル抗体およびその誘導体に関する。該抗体およびその誘導体はNおよびC末端二重トランケートtau分子の生物活性を部分的または完全に阻害することができる。そのような特異抗体活性はアルツハイマー病の脳組織から単離した該二重トランケートtau分子に対するモノクローナル抗体DC44を用いて示された(それぞれ実施例10および11)。
【0155】
該抗体は他の多くの変異体(DC82、DC136など)を有し、血清由来またはモノクローナル抗体またはそのあらゆる誘導体であってよい。所望のエピトープに対するモノクローナルおよびポリクローナル抗体の生成は当該分野でよく知られている(43)。さらに、該抗体は天然または遺伝子操作により誘導された抗体、例えば当該分野でよく理解されている技術により誘導されたキメラ抗体であってよい。さらに該抗体は、特定のエピトープと結合する能力を保持する抗体断片、例えばFab断片または一本鎖Fvミニボディ(minibody)、またはイントラボティ(intrabody)と呼ばれる細胞内に発現した一本鎖抗体も表す。
【0156】
最も好ましい態様において本発明はアルツハイマー病の治療に用いる医薬組成物に関する。
【0157】
さらに該医薬組成物はその症例を扱う医師が適切とみなす経路および用量でそれを必要とする患者に投与することができよう。
【0158】
本発明の別の好ましい態様において、該医薬組成物は、特異的阻害剤として部分的または完全なその変化および/または中和をもたらす上記エピトープのあらゆる部分またはグループと結合する少なくとも1のモノクローナル抗体または低分子またはその誘導体を含む(各実施例10、11および12参照)。
【0159】
本発明の別の目的はa)本発明のエピトープ、b)本発明の抗体またはその誘導化分子を含むアルツハイマー病を検出および/またはモニターするための診断用組成物を提供することである。
【0160】
本発明の診断用組成物は、例えば具体的にはタイプIAまたはタイプIIグループの分子のあるメンバーまたはそのエピトープまたは試験する試料中の増大したレベルのタイプIAまたはタイプIIA分子を認識する本発明の抗体を含むことができよう。別の態様において、該診断用組成物は本発明のエピトープの1つに対する本発明の抗体を含むことができよう。すなわち、アルツハイマー病状に関連する試料を本発明のエピトープを認識する抗体で該試料を処理することにより検出することができよう。抗体-エピトープ(ハプテン)複合体を本発明の抗体に対する第二抗体を用い、当該分野で知られた方法(43)に従って標識することにより可視化することができよう。
【0161】
本発明のさらに別の態様において、該診断用組成物は本発明のエピトープおよび本発明の抗体からなってよい。試料の該抗体による処置は、該抗体の該試料に対する結合が該抗体の標準試料として用いる本発明の該エピトープに対する結合に関連して生じるならば対応患者の病状に関する結論をもたらすことができよう。
【0162】
さらに別の態様において、該診断用組成物はタイプIAまたはタイプIIA分子および本発明の抗体を含むことができよう。両タイプの分子の活性は、試料の正常tau中和能について本発明の組換えタイプIA分子(例えば配列番号1)およびIIA分子(配列番号11-18)と比較してモニターすることができよう。タイプI分子の定量した異常活性から、該試料に含まれる分子のレベル、すなわち患者の病状を推定することができよう。タイプIA活性は、例えばタイプI分子と反応しないで残った正常tauの残存活性を測定することにより推定することができよう。タイプII活性は微小管構築アッセイにおけるタイプII分子のさらなる活性を測定することにより推定することができよう。
【0163】
当業者は本発明の上記のあらゆる対象を組み合わせる他の試験系を設計する立場にある。すべての考えられる組み合わせが本発明の保護の範囲内にあることを理解すべきである。
【0164】
本発明の別の目的は、タイプIAおよびタイプIIA分子のNおよびC末端二重トランケートtau分子またはそのエピトープの存在およびその正常tau阻害活性レベルについて患者の脳脊髄液分離物をアッセイし、神経組織の生検を実施することを含むアルツハイマー病のin vitro診断および/またはモニター方法を提供することである。
【0165】
患者の脳脊髄液分離物は標準的医学的方法により得られる。
【0166】
さらなる態様において、本発明はタイプIAおよびタイプIIAのそれぞれ分子および動物に免疫応答を誘導することができるその誘導化免疫原性断片のアミノ酸配列と同一またはホモローガスであるタイプIおよびタイプII分子に関する。本発明によれば、タイプIおよびタイプII分子はともに(a)阻害抗体を産生するための免疫原および該疾患に対する免疫に用いるワクチンの中心部分として用いることができることがわかった。
【0167】
非経口的適用において、上記のすべての配列およびエピトープおよび罹患脳組織から単離したタイプIおよびIIは免疫原性であり、具体的には該タイプIおよびIIタンパク質およびその誘導体の対する抗体の産生をもたらす(各実施例10および13)。
【0168】
最も好ましい態様において、タイプIおよびII分子またはその誘導体は該分子の一次、二次、および/または三次構造に対する免疫応答を誘導することができる。宿主において、得られた免疫応答はしたがってtauおよびその誘導体の健康および病的型のtauを区別することができる。本発明のこの特徴は、疾患特異的免疫応答の誘導におけるこれらNおよびC末端二重トランケートtau型の優れた品質を重視するワクチンとして用いることができる。
【0169】
本明細書に含まれる病的NおよびC末端二重トランケートtauポリペプチドについて天然のバリエーション(変化)がアルツハイマー病の個々の例で存在することが理解される。これらバリエーションは全体配列中のアミノ酸の相違中に、すなわち該配列のアミノ酸の欠失、置換、挿入、反転(inversion)、または付加により存在する。本発明の例示的態様のそのようなアミノ酸置換は本発明の範囲内にある。すなわち、ポリペプチドの免疫原性に本質的に影響しない天然の変化には本発明の該二重トランケート型のtauポリペプチドの免疫学的に等価なバリエーションが考えられる。
【0170】
しかしながら、タイプIAおよびIIA NおよびC末端二重トランケートtauポリペプチドを例えばワクチネーション目的または抗体産生に用いるときには本発明に記載の全ポリペプチドを用いる必要はない。その完全ポリペプチドに対する免疫応答を誘導することができるこれらポリペプチドの断片、ゆわゆる免疫原性断片を用いることも可能である。
【0171】
したがって、本発明のこの態様は本発明のポリペプチドのみならず依然として該ポリペプチドに対する免疫応答を誘導することができるそれらポリペプチドの誘導化断片(いわゆる免疫原性断片)に関する。
【0172】
例を示すために、組換えタイプIAおよびIIAペプチドまたは病的ヒトアルツハイマー脳由来のタイプIAおよびIIA NおよびC末端二重トランケート病的tau分画の動物における免疫原性を示す(実施例3)。
本発明を以下の実施例および図面によりさらに説明するが、それらに限定されるものではない。
【0173】
図1:NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築。
【0174】
図2:NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築の阻害。
【0175】
図3: 微小管構築におけるNおよびC末端二重トランケートtauタイプIIAおよびIIB分子の活性。
【0176】
図4:神経細胞に発現したタイプIIA NおよびC末端二重トランケートtauは酸化ストレスに対する感受性を有意に増加させる。
【0177】
図5: 病的tauタイプIAタンパク質およびその欠失突然変異体に対するモノクローナル抗体の親和性。病的tauタイプIAタンパク質およびその欠失突然変異体に対するモノクローナル抗体の見かけの親和性。最初のカラムに「プロトタイプ」tauタイプIAタンパク質(配列番号1)および各欠失突然変異体を示す。中段のカラムに本発明のエピトープを示す。最後のカラムに示す見かけの親和性を競合的ELISAにより測定し、プロトタイプtauタイプIAタンパク質と50%競合するのに必要な対応抗原の濃度で表した。
【0178】
図6: AD-脳由来tauタンパク質のSuperdex 200-カラムによる分画。
【0179】
図7:AD脳からの3つの別個の単離物由来の分画No.19中のタイプIA阻害活性。
【0180】
図8:AD脳中のNおよびC末端二重トランケートtauタイプI分子の証明。
【0181】
図9:健康脳に存在せずAD脳に存在するtauタイプI。
【0182】
図10:NおよびC末端二重トランケートtauタイプII分子の免疫反応性。
【0183】
図11:組換えtauタイプI-II(配列番号1-24)の構築。
【0184】
図12:AD-脳由来および組換えtauタイプIAの正常健康tauに対する阻害効果。
【0185】
図13:tauタイプIA分子を中和する薬剤候補の第一回スクリーニング(工程1)。
【0186】
図14:正常tauに対する選択性を有するタイプIA分子を中和する薬剤候補の第二回スクリーニング(工程2)。
【0187】
図15:tauタイプIIA分子を中和する薬剤候補の第一回スクリーニング。
【0188】
図16:tauタイプIIA分子を中和し、それを正常tauから識別することができる薬剤候補の第二回スクリーニング(工程2)。
【0189】
図17:ELISAにより測定したプレフュージョン(prefused)マウス血清中の特異抗体レベル。
【0190】
図18:モノクローナル抗体とAD脳由来tau(分画#19)およびコントロール健康脳由来tauとのELISA反応性(DC20:無関係な特異性を有するモノクローナル抗体。示したデータは3回の並列実験の平均値を表す。)。
【0191】
図19:モノクローナル抗体の組換えtau分子とのELISA反応性(DC20:無関係な特異性を有するモノクローナル抗体。示したデータは3回の並列実験の平均値を表す。)。
【0192】
図20:AD-脳由来tauタイプIA(分画#19)に対する中和抗体のスクリーニング。
【0193】
図21:組換えtauタイプIA(配列番号1)に対する中和抗体のスクリーニング。
【0194】
図22:tauタイプIA分子を中和し、それらを健康tauから識別することができる薬剤候補のスクリーニング。
【0195】
図23:組換えtauタイプIIA(配列番号12)の病的活性のモノクローナル抗体による中和。
【0196】
図24:ELISAで検出した組換えtauタイプIIA(配列番号12)に対する抗体のレベル。
【0197】
図25:トランスジェニック動物の遺伝子型別。パネルAはトランスジェニック動物の親世代の遺伝子型別を示す。レーン1、2、3および4のゲノムDNA由来のDNAの二重トランケート配列の特異的増幅は、フォスターマザーの子孫の尾から抽出したゲノムDNAにおける特異的トランス遺伝子の存在を示す。これら動物は二重トランケートタイプIIA tau分子保有トランスジェニック動物の親世代を示す。この実施例において、陽性(+C)および陰性(-C)および2つのさらなる陰性試料(5、6)を示す(M=サイズマーカー)。矢印は、トランス遺伝子陽性動物において予想される予想PCR生成物サイズを示す。
パネルB. F1世代由来の動物の遺伝子型別。ゲノムDNAを尾の先端から抽出し、二重トランケートtau特異的DNA配列を同定し、レーン1に示す。レーン2および3は陰性コントロールを示す。F1世代におけるtau特異的DNA断片の同定はこれらトランス遺伝子の遺伝性(inheritability)を確認する。
【0198】
図26:トランスジェニック動物のF1世代における二重トランケートヒトtau転写物の遺伝子発現。RNAをトランスジェニック動物の瞬間凍結組織から抽出し、逆転写、次いでcDNAの特異的増幅にかけた。例はレーン番号1および2にトランス遺伝子発現動物を示す。レーン3〜5は非発現コントロールを示し、レーン5はこの方法を用いた時に非トランスジェニック動物に典型的に生じる非特異的シグナルを示す。この実施例は、実験動物においてトランス遺伝子から発現した二重トランケートtau特異的mRNAの存在を示す。
【0199】
図27:6日間in vitro分化(differentiation)後のタイプIIA分子の過剰発現により生じた細胞死。二重トランケートtauタイプIIA(タイプIIA)でトランスフェクションしたSY5Y細胞および非トランスフェクションコントロールニューロン様細胞(mock)の細胞生存性の比較。
【0200】
図28: A:タイプIIA分子の微小管に対する結合親和性の増加はタイプIIA二重トランケート分子および完全長tauを発現する安定なトランスフェクション細胞由来のtauの細胞分画を用いて示される。遊離tau(FT)、微小管結合tau(MT)および核関連tau(NAT)の単離は記載のごとく行った。B:有機化合物F123による各tauタイプIAおよびIIAの阻害、微小管重合アッセイ。C:電子顕微鏡分析により示される微小管重合と吸着(absorption)測定値の直接比較。
【0201】
図29:MitoFluorで染色した対数増殖期のSH-SY5Y細胞。細胞体およびプロセス中のミトコンドリアの標準的分布。
【0202】
図30:MitoFluorで染色した対数増殖期のtauタイプIIA分子発現SH-SY5Y細胞。細胞の中心体領域周辺のグリーン標識ミトコンドリアの核周囲クラスタリング。
【実施例】
【0203】
実施例1:NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築
健康tauの生理学的機能は微小管(MT)の安定化である。この機能は微小管構築アッセイ(MAA)により測定することができる。本実施例において、MAA反応は3タイプのtau分子:正常健康ヒトtau、組換え型のtauタイプIA(配列番号1)およびtauタイプIB(配列番号4)を用いて行った。正常ヒトtau、tauタイプIAおよびタイプIBを別個の反応で個々にアッセイした。tau 0.1mg/mlの単一調製物を最終濃度1mg/mlの精製ブタ脳チューブリンおよび1mM GTPと混合した(すべての物質に重合緩衝液(100mM PIPES、pH 6.9、1mM MgSO4、2mM EGTA)使用)。最後にTauを加えMT構築の促進を開始した。静かにそして急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、サーモスタットで制御した分光光度計(Beckman Coulter DU640)中で37℃に平衡化した。濁度を340nmで10秒間隔で20分間連続的にモニターした。上端の曲線1(図1)は正常健康tauの微小管構築促進能を示す。それに対し、タイプIA(曲線2)およびタイプIB(曲線3)はいずれも正常tauのこの活性を示さず、MAAにおけるいかなるMT構築促進も示さなかった。
実施例2: NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築の阻害。
【0204】
MT構築アッセイ(MAA)するとtauタイプIAおよびIB分子はいずれも lack機能的活性を欠く。驚くべきことに、tauタイプIA分子は微小管構築におけるチューブリンに対する阻害効果を示す。これに対し、タイプIBタンパク質は(同様の一次構造にも関わらず)MAAにおいてチューブリンの機能的活性を阻害しない。微小管構築を阻害するため、組換え型のtauタイプIA(配列番号1)およびタイプIB(配列番号4)を用いた。構築阻害反応はタイプIAおよびタイプIBタンパク質を用いて別個に行った。ヒトチューブリン(2mg/ml)をタイプIA分子(0.2mg/ml)またはタイプIB分子(0.2mg/ml)と混合した。混合物を静かに振盪させながら37℃で1時間インキュベーションした。氷上に保った混合物に正常ヒトtau(O.1mg/ml)およびGTP(混合物中のチューブリンの最終濃度は1mg/mlおよびGTPは1mMである)を重合緩衝液(1OOmM PIPES、pH 6.9、1mM MgSO4、2mM EGTA)を用いて加えた。静かにそして急速に撹拌した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節された分光光度計(Beckman Coulter DU640)中で37℃に平衡化した。濁度変化を340nmで5分間にわたり10秒間隔で測定した。上端の曲線1(図2)は正常ヒトtauのみがチューブリン重合を完全に誘導することができることを示す。チューブリンとタイプIAをプレインキュベーションすると微小管構築はみられなかった(図2、下の曲線2)。反対に、チューブリンをタイプIBとインキュベーションすると、分子量がタイプIAと同範囲であるにも関わらず正常tauの微小管構築能力を阻害しない(図2、曲線3)。
表
NおよびC末端の二重トランケートtau分子の微小管重合に対する影響
【0205】
【表1】
最短R4アイソフォームに従った配列番号11-14;19番号付け。
最短R3アイソフォームに従った配列番号15-18;20番号付け。
【0206】
【表2】
最短R4アイソフォームに従った配列番号11-14;19番号付け。
最短R3アイソフォームに従った配列番号15-18;20番号付け。
実施例3:微小管構築におけるNおよびC末端二重トランケートtauタイプIIAおよびIIB分子の活性
【0207】
グループIA分子と対照的に、タイプIIA 二重トランケートtau誘導体は驚くべきことに病的微小管構築を促進することがわかった(図3および図28C参照)。微小管構築反応を3タイプの分子:天然健康ヒトtauアイソフォーム、Alz. tauタイプIIA(配列番号12)およびtauタイプIIB(配列番号19)を用いて行った。3つの別の反応をそれぞれ各tau(健康tau、組換えtauタイプIIAまたはタイプIIB)を単独調製することにより行った。個々のtau調製物0.1mg/mlをチューブリンおよびGTPと混合した(チューブリンの最終濃度は1mg/mlおよびGTPは1mMであった)(すべての試薬は重合緩衝液(100mM PIPES、pH 6.9、1mM MgSO4、2mM EGTA)中)。静かにそして急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計(Beckman Coulter DU640)中で37℃に平衡化した。濁度変化を340nmで5分間にわたり10秒間隔で測定した。この実験において、組換えtauタイプIIAは、健康tauによる生理的微小管構築(図3、曲線2)と比べて極めて高い(3倍)病的微小管構築の促進を示した(図3、上端の曲線1)。これに対し、タイプIIB分子はNおよびC末端二重トランケートであるにも関わらずタイプIIAと同様にMAAを行うことができず、微小管構築を健康tauでみられるレベルしか促進することができない(図3、曲線3)。
実施例4: 病的tauタイプIIタンパク質によるストレス保護メカニズムの阻害をin vitroで種々の酸化ストレスに曝露した該分子を発現する神経芽細胞腫細胞において示された。
【0208】
本実施例ではスーパーオキシドアニオンおよび一酸化窒素を生成し、これが反応して過酸化亜硝酸を形成する3-モルホリノシドノニミン(SIN-1)の酸化分解を用いる。この非常に反応性のラジカルはさらに主として細胞膜系を酸化することができる。当業者は酸化ストレスの別の供給源を培養中の神経芽細胞腫細胞に適用し、SIN-1を用いることによりここに記載したのと同様の効果を得ることができるだろう。
【0209】
脆弱性の効果を以下のごとく試験した。
【0210】
1.SIN-1を種々の濃度(0-3.32mM)でそれぞれtauタイプIIAタンパク質、配列番号15および配列番号11を発現するヒト神経細胞系に適用した。
【0211】
2.細胞生存を50%細胞生存(EC50)に対する過酸化水素の有効濃度を決定する3(4,5-ジメチルチアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロミド(MTT)還元アッセイにより測定した。当業者は本発明に記載の効果を測定するために細胞生存を評価する異なる方法を知っている。
【0212】
3. EC50値を神経芽細胞腫細胞系について病的tauタイプIIAタンパク質発現の存在下および非存在下で比較し、EC50値の統計的有意差をt-検定により評価した。
【0213】
細胞を10% FCS、2mM L-グルタミン、1% NEAA、50U/L ゲンタマイシン含有MEM/F12中で増殖させた。3-モルホリノシドノニミン(SIN-1)を無血清培地中の1Mストック溶液から希釈した(例えば47.5mgを230mlに)。MTTストック溶液(2.6mg/ml)をMEM/F12 w/o血清を用いて調製し、ろ過滅菌した。
【0214】
細胞を当該分野でよく知られた方法で培養した。96ウェルプレートに2x104細胞/ウェルを播いた。プレートの半分にtauタイプII分子発現細胞を播き、プレートの他の半分に非発現細胞を播いた。培地を36〜48時間ごとに交換した。5日後、SIN-1を0〜3.3 mMの範囲の濃度で加え、プレートを24時間インキュベーションした。各濃度をヘキサプリケートでアッセイした。SIN-1とインキュベーション後、MTTストック溶液を加えて最終濃度200mg/mlとし、プレートをもう1時間インキュベーションした。培地を捨て、プレート表面をペーパーウールで乾燥させた。50ml/ウェルのDMSOを加え、プレートを室温で一夜インキュベーションした。690nmでバックグラウンドを補正し、540nmの吸光度をELISAリーダーで測定し、バックグラウンドを引いた値を当該分野でよく知られているEC50計算に用いた。
【0215】
タイプIIAタンパク質発現および該タンパク質非発現神経芽細胞腫細胞間のlog EC50濃度の差の有意性をt-検定を用いて試験し、P値はタイプIIA病的tauタンパク質両方でP<0.001であった。tauタンパク質 配列番号12および配列番号18の発現は神経芽細胞腫細胞の酸化ストレスに対する抵抗性を50%低下させた。
【0216】
記載した実施例の効果(図4)は病的型のtauタンパク質の病的効果の説明に寄与する。
【0217】
当業者は本発明の上記のあらゆる目的を組み合わせる他の試験系を設計する立場にある。すべての考えられる組み合わせは本発明の範囲内にあると理解すべきである。
【0218】
図4のチャートはtauタイプIIA存在下の神経細胞の酸化ストレスに対する相対的抵抗性の低下を示す。該タンパク質を保有していない細胞(コントロール)の抵抗性を100%(左のバー)で表し、病的tauタンパク質を発現する神経細胞の抵抗性をコントロール値の%で示す(中および右のバー)。抵抗性は、細胞の50%が死ぬ培養液中のSIN-1により生じた遊離ラジカルの濃度で定義される。該結果はそれぞれ二重トランケートtauタンパク質タイプIIA 配列番号12(93-333、R4)および配列番号18(69-332、R3)の測定値を表す。
実施例5:二重トランケートtauタイプIAのコンフォメーションに対する個々のエピトープの寄与
【0219】
tauタイプIAの「コンフォメーショナル領域」(断片A)またはその部分の重要性を全コンフォメーション領域(断片A)またはエピトープと呼ばれるA1-A6で示されるその個々の部分の連続的欠失により検討した。タイプIA分子のコンフォメーションはその機能と強い関連があるため、「断片A」の全体のコンフォメーションに対する各エピトープ(A1-A6)の寄与をtauモノクローナル抗体を用いその反応性に基づいて測定した(図5)。
【0220】
プロトタイプtauタイプIA(配列番号1)は10nMの親和性を有する。それぞれエピトープA1、A2、およびA5が欠失した個々の欠失突然変異体 配列番号22、23、26は、これら領域の寄与が親和性の2〜4倍の低下(20-40nM)を反映することを示したが、配列番号24、25、および27中のエピトープA3、A4、A6の欠失はそれぞれ親和性の10〜30倍の損失(100〜300nM)に寄与した。全断片Aの欠失後にのみ(突然変異体 配列番号21)、該親和性はプロトタイプtauタイプIAの親和性を3桁劇的に低下させる。
実施例6:NおよびC末端二重トランケートtauタイプIおよびタイプIIの単離
【0221】
アルツハイマー脳由来tauタイプIおよびタイプII分子の製造:神経病理学的に確認されたアルツハイマー病症例由来の病的ヒト脳組織を二重トランケートtau IA、BおよびIIAタンパク質を単離するための供給源に用いた。アルツハイマー脳由来tauの調製はTRIS緩衝液中の組織のホモゲナイゼーションおよび飽和硫酸アンモニウム沈殿による溶解物の分画化の組み合わせに基づく。組織を冷20mM TRIS pH8、0.32mMショ糖、10mM b-メルカプトエタノール、5mM EGTA、10mM EDTA、5mM MgSO4、1mMフェニルメチルスルホニルフロリド、50mMフッ化ナトリウム、5mMベンズアミジン、5μg/mlロイペプチン、1.5μg/mlペプスタチン、2μg/mlアプロチニン中でHeidolph DIAX 900ホモゲナイザーを用いて4℃で10分間ホモゲナイズした。ホモゲネートを4℃で27000g、30分間遠心して細胞デブリスを除去した。tauタンパク質を44.12%(v/v)の飽和硫酸アンモニウムを加えて脳組織上清から沈殿させた。25℃で20分間インキュベーションおよび静かに撹拌した後、試料を25℃で20000g、10分間遠心した。ペレットを500μlの100mM PIPES pH6.9、2mM EGTA、1mM MgSO4に再浮遊させ、ついで同じ緩衝液に対して透析した。この調製物をSuperdex 200-カラム(Amersham-Pharmacia-Biotech)でゲルろ過して分画化し、該分画をSDS-PAGE(勾配5-20%ポリアクリルアミド)により分割し、tauタンパク質を抗tau抗体DC25を用い標準的手順に従ってイムノブロッティングにより検出した(図6)。微小管構築に対する個々の分画の効果を試験した。
【0222】
tauタイプIAおよびIBの単離:分画#19(図7)を、二重トランケートタイプIAおよびIB分子の典型的な分子量(12kDa)に対応するtau分子を含み-この分画は最高の阻害能を示した。この分画を3つの抗tau抗体(DC25はトランケートおよび完全長タンパク質を共に認識し、DC39(完全C末端特異的)およびAlz50(完全N末端に特異的))を用いるWestern(ウエスタン)ブロット分析により特徴づけた(図8)。これら抗体の免疫応答性はAD脳由来分画にのみNおよびC末端二重トランケートタイプIタンパク質がないことを証明した。正常健康脳から同じ方法により調製した対応する分画は、阻害活性も特異的免疫応答性も示さなかった(図9)。tauタンパク質の濃度をサンドイッチRIAにより測定した。総タンパク質濃度はBradfordアッセイを用いて測定した。tau調製物は使用するまで-20℃に保存した。
【0223】
tauタイプIIAの単離:分子量30kDaに対応するtau分子を含む分画#15(図6)は二重トランケートタイプIIA分子の典型である。分画#15は異常に高い微小管構築促進活性を示した。この分画を3つの抗tau抗体(DC25はトランケートおよび完全長タンパク質を共に認識し、DC39(完全C末端特異的)およびAlz50(完全N末端に特異的))を用いるWesternブロット分析により特徴づけた(図10)。これら抗体の免疫応答性は、AD脳由来の分画にのみNおよびC末端二重トランケートタイプIIタンパク質が存在することを示した。tauタンパク質の濃度をサンドイッチRIAにより測定した。総タンパク質濃度をBradfordアッセイを用いて測定した。
【0224】
組換えtauタイプIおよびタイプIIタンパク質のクローニング、発現、および精製:組換えトランケートtauタンパク質に対する遺伝子はアイソフォームtau43およびtau44に対するヒトcDNAから誘導された。cDNA挿入物をNdeI-EcoRI制限部位を用いるpET17b(Novagen)ベクターを用いてクローンした(図11)(Studierら、Meth. Enzym.185(1990)、60-89)。
【0225】
組換えNおよびC末端二重トランケートtau分子(配列番号1-24)をcDNA由来の関連領域のPCR増幅により調製した。翻訳開始を誘導する特定のプライマー、開始(ATG)、終止(TGA)コドン、およびNdeI、EcoRI制限部位を用いた。
【0226】
tauDNA中にA4-A6エピトープ(配列番号25-27)の欠失を有するプラスミドを図11(下側パネル)に示すように逆PCRにより調製した。
実施例7:微小管構築アッセイにおける正常健康tauに対するAD-脳由来および組換えtauタイプIAの阻害効果
【0227】
AD-脳抽出物およびtauタイプIAの組換え分子は天然健康tauアイソフォームを用いる微小管構築の促進を阻害することができる。これらの実験では健康ヒトtauを年齢対応コントロールの脳から単離し、tauタイプIAをAD患者の脳から単離した(実施例6、図6、分画#19参照)。組換えtauタイプIA(配列番号1)およびタイプIB(配列番号4、陰性コントロール)を生成し、実施例6に示すように精製した。これらの実験において、脳由来健康tauアイソフォーム(0.1mg/ml)、AD-脳由来または組換えタイプIA tauまたはタイプIB(0.2mg/ml)をチューブリンと混合した。各混合物を別々にアッセイした。被検混合物を静かに振盪させながら水浴中で37℃で1時間インキュベーションした。氷上に保った混合物にGTPおよび/または正常tau(チューブリンの最終濃度は1mg/mlおよびGTP 1mM)を加えた(すべて試薬に重合緩衝液(100mM PIPES、pH6.9、1mM MgSO4、2mM EGTA)使用)。静かにそして急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計(Beckman Coulter DU640)中で37℃に平衡化した。濁度変化を340nmで5分間にわたり10秒間隔で測定した。データはAD脳由来および組換え二重トランケートタイプIA分子がともに正常tauの微小管構築促進能を阻害することを示す(図12、曲線2、3)。これに対し、組換えタイプIBは正常ヒトtauにより誘導されるチューブリン重合促進能力を阻害することができない(図12、曲線4)。曲線1(図12)は正常tauにより促進された微小管構築を示す。
実施例8:tauタイプIAの病的活性を中和する薬剤候補のスクリーニングアッセイ
【0228】
チューブリン重合を促進する健康正常tauの活性を阻害する二重トランケートtauタイプIA分子の能力を用いてスクリーニングアッセイをタイプIA分子の阻害活性を中和することができる化合物を選択するために計画した。病的tauタイプIAはAD-脳または組換え供給源由来であり得るが、組換え物質を用いるのが好都合である。薬剤候補の中和効果は正常健康tauの残存する微小管構築促進能を測定することにより定量的に定義することができる。該アッセイは2工程で行われる:
1.tauタイプIAを中和する薬剤候補のスクリーニング。個々の薬剤候補(最終濃度50mg/ml)と別個に混合したプロトタイプ組換えタイプIA分子(配列番号1)(最終濃度100mg/ml)を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリン、GTPおよび健康tauを+4℃で混合物に加えた(最終濃度:チューブリン-1mg/ml、GTP-1mM、健康tau-100mg/ml)。急速混合物後、試料を水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。タイプIA活性中和能を有する薬剤候補を正常健康tauの残存する微小管構築促進能を測定することにより選択した(図13;薬剤候補をタイプIA分子とプレインキュベーションし、タイプIA中和の有効性を微小管構築を用いてアッセイした。下端の曲線1および上端の曲線2は病的タイプIA分子に対する被検薬剤候補の陰性(中和なし)および陽性(100%)中和活性を示す。中側の曲線は3つの異なる薬剤候補による種々のタイプIA中和効果を示す。)。陽性薬剤を選択するための閾値は任意であり、タイプIAの完全中和からその部分中和まで変化するかもしれない。
【0229】
2.タイプIA分子を中和し、それを正常健康tauと区別する薬剤候補の選択。tauタイプIA分子に対する中和活性を有する選択した候補の正常健康tauとの反応性をスクリーニングし、タイプIAにのみ特異的な分子を選択した。正常健康tau(最終濃度100mg/ml)と個々の薬剤候補(最終濃度50mg/ml)の別個の混合物を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリンおよびGTPを+4℃で該混合物に加えた(最終濃度:チューブリン1mg/ml、GTP 1mM)。急速に混合した後、試料を水晶マイクロキュベットに入れた。濁度変化を340nmで測定した。健康tauのMT重合促進活性との干渉を示さない薬剤候補を選択した(図14)。
実施例9:tauタイプIIAの病的活性を中和する薬剤候補のスクリーニングアッセイ
【0230】
本発明は、tauタイプIIA分子が予期せず高いチューブリン重合促進能力を有することを示し、タイプIIAタンパク質の該活性を中和する化合物を選択するためのスクリーニングアッセイの基礎を形成する。タイプIIAの中和はタイプIIA分子の残存微小管構築活性を測定することにより定量することができる。該アッセイは2工程で行われる。
【0231】
1. tauタイプIIAを中和する治療薬候補のスクリーニング。
【0232】
tauタイプIIA分子(配列番号12)(最終濃度100mg/ml)と単一薬剤候補(最終濃度50mg/ml)の別個の混合物を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリンおよびGTPを+4℃で混合物に加えた(最終濃度:チューブリン 1mg/ml、GTP-1mM)。急速混合物後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。微小管構築速度を有意に低下させた薬剤候補を該アッセイの第二工程用に選んだ(図15;薬剤候補をタイプIIA分子とプレインキュベーションし、タイプIIA中和の有効性を微小管構築を用いてアッセイした。下端の曲線1は各薬剤候補の陽性(100%)中和活性を示し、上端の曲線2は病的タイプIIA分子を中和しないことを示す。中側の曲線はタイプIIA中和における種々の薬剤候補の異なる効果を示す。)。
【0233】
2.タイプIIA分子を中和し、それを正常健康tauと区別する薬剤候補の選択。
【0234】
薬剤候補(最終濃度50mg/ml)と正常健康tau(最終濃度100mg/ml)の別個の混合物を37℃で1時間プレインキュベーションした。次に、チューブリンおよびGTPを+4℃で該混合物に加えた(最終濃度:チューブリン1mg/ml、GTP-1mM)。急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。健康tauと干渉しない薬剤候補を選択した(図16;工程1で選んだ薬剤候補を健康tauとプレインキュベーションし、微小管構築に対する効果をアッセイした。下端の曲線(1)は健康tauの最大阻害を示し、上端の曲線(2)は健康tauを阻害しないことを示す。中側の曲線は薬剤候補の健康tauに対する異なる阻害活性を示す。)。
実施例10:NおよびC末端二重トランケートタイプIAおよびタイプIIA分子を中和するモノクローナル抗体の製造。
【0235】
免疫プロトコールおよび融合手順:ヒトアルツハイマー脳から単離したNおよびC末端二重トランケートtauタイプIタンパク質(分画#19、実施例6)を免疫原に用いた。Balb/cマウスを完全Freundアジュバント中の該タンパク質(50mg/マウス)を皮下にプライムし、次いで同じタンパク質50mg/マウスを4週間間隔で3回腹腔内にブースターをかけた。融合前血清を回収し、tauに対する特異抗体のレベルをELISAで試験した(図17;AD由来tauで免疫したマウスの血清中の特異抗体のレベルを同じ抗原を用いてELISAで試験した。5つの血清はすべて該tauタンパク質に対する高い抗tau結合活性を示した。図17はある免疫マウスにおける特異抗体レベルを示す。コントロールには無関係のタンパク質で免疫したマウス由来の血清を用いた。)。マウス脾細胞を当該分野でよく知られた改良された方法を用いNS/0ミエローマ細胞と融合させた(M. KohlerおよびC. Milstein、1975)。
【0236】
図18に示す結果によれば、モノクローナル抗体DC44、DC82およびDC136はアルツハイマー病脳由来のNおよびC末端二重トランケートタイプIAおよびタイプIIA分子を認識する。これら抗体では同じ方法で調製した正常ヒト脳由来のtau単離物との反応性は観察されなかった(図18)。これに対し、モノクローナル抗体DC25はELISAで病的および正常健康脳由来の該タンパク質と反応する(図18)。この抗体は病的型のtau(AD-tau)と正常ヒトtauを区別しない。この一次スクリーニング後、ハイブリドーマをソフトアガロース中でサブクローンし、最終的に当業者によく知られた技術により同じイデオタイプを有する抗体を分泌する均質なハイブリドーマポピュレーションを得た。
【0237】
さらにこれらクローン化ハイブリドーマクローンの組換え完全長tauアイソフォームおよび二重トランケートtauタイプIA(配列番号1)およびタイプIIA(配列番号12)分子に対する反応性をスクリーニングアッセイと同じELISAを用いて試験した。
実施例11:モノクローナル抗体を用いるAD脳由来および組換えNおよびC末端トランケートタイプIA分子の病的活性の中和。
【0238】
選択したモノクローナル抗体DC44、DC82、DC136およびDC25をアルツハイマー脳から単離した天然tauタイプIAの活性を中和する能力で特徴づけた(実施例6)。該tau単離物(最終濃度100mg/ml)および被検抗体(最終濃度50mg/ml)を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリン、正常ヒトtauおよびGTPを+4℃で該混合物に加えた(最終濃度:チューブリン1mg/ml、健康ヒトtau-100mg/ml、GTP-1mM)。急速に混合物した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。モノクローナル抗体DC136、DC44およびDC82は該タンパク質の病的活性を阻害することができた(Fig 20;抗体を天然tauタイプIA(分画#19)とプレインキュベーションし、次いで健康ヒトtau、チューブリンおよびGTPと混合した。37℃で5分間後に微小管の形成を分光光度法で測定した。バーは3つの独立した試験の平均値を表す。健康ヒトtauを用いるMAA-微小管構築アッセイ。タイプIAとプレインキュベーション(抗体なし)した健康ヒトtauを用いるMAIA-微小管構築阻害アッセイ。類似の実験において抗体をtauタイプIAの組換えプロトタイプ(配列番号1)を用いて試験し、同様の中和活性パターンを示した(図21)。コントロール抗体DC25はELISAおよびWesternブロッティングにより試験したトランケートおよび正常tauのすべての型を認識するが、微小管構築アッセイにおいてAD-脳由来タイプIA活性と干渉しない。これらの結果は抗体DC25が抗体DC136、DC44およびDC82に比べてtauの異なる領域と反応することを示唆する。これに対し、抗体DC136、DC44およびDC82は病的タイプIA分子に関与するエピトープと結合する。
【0239】
次に選択工程は健康分子とタイプIA分子を識別することができる抗体を目的とした。正常健康tau(最終濃度100mg/ml)および被検抗体(最終濃度50mg/ml)の混合物を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリンおよびGTPを+4℃で該混合物に加えた(最終濃度:チューブリン 1mg/ml、GTP-1mM)。急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。抗体DC136、DC44、DC82およびDC25はいずれも微小管構築において正常健康tauを阻害することができなかった(図22;tauタイプIA中和抗体を健康tauとプレインキュベーションし、次いでチューブリンおよびGTPと混合した。37℃で5分間後に微小管の形成を分光光度法で測定した。バーは3つの独立した実験の平均値を示す。健康tauを用いるMAA-微小管構築アッセイ。陰性コントロールとして健康tau中和抗体を用いた。このデータは抗体 DC136、DC44、DC82がトランケート病的型のtauと健康tauタンパク質の相互作用に関与する特異的エピトープを認識することを示した。
実施例12:モノクローナル抗体によるタイプIIA活性の中和
【0240】
tauタイプIA中和活性について先に単離した抗体を実施例8Bに記載の方法を用いる組換えtauタイプIIA(配列番号12)に対する中和活性について試験した。すべての3中和モノクローナル抗体DC44、DC82およびDC136はNおよびC末端二重トランケートtauタイプIIA分子の病的活性を低下させることができた(図23;抗体を組換えtauタイプIIAとプレインキュベーションし、次いでチューブリンおよびGTPと混合した。37℃で5分間後、微小管の形成を分光光度法で測定した。バーは3つの独立した実験の平均値を示す。tauタイプIIAを用いるMAA-微小管構築アッセイ(抗体なし)。これは該抗体のエピトープが少なくともタイプIA 配列番号1およびタイプIIA配列番号12により共有されることを示唆する。抗体DC25についてはタイプIIA阻害活性は観察されなかった。
実施例13:組換えNおよびC末端二重トランケートtauタイプIAおよびIIA分子の免疫原性。
【0241】
免疫プロトコール:本発明の好ましい態様において該組換えtauタイプIAおよびIIAタンパク質をワクチネーション目的または病的tauタイプIAおよびIIA分子の病的活性を特異的に中和する抗体を生じるために用いる。該実施例において、組換えNおよびC末端二重トランケートtauタイプIIA(配列番号12)を免疫原に用いた。Balb/cマウスを完全Freundアジュバント中の該タンパク質(50mg/マウス)で皮下にプライムし、次いで4週間間隔で50mg/マウスの完全Freundアジュバント中の該タンパク質で3回腹腔内にブースターをかけた。免疫血清を回収し、各組換え抗原tauに対する特異抗体のレベルをELISAで測定した(図24)。
実施例14: トランスジェニック動物
【0242】
尾部先端から抽出したDNA:ゲノムDNAをDNeasy組織キットQiagenにより抽出した。
遺伝子型別(図25):二重トランケートtau型をコードするトランス遺伝子の特異的増幅をトランスジェニック動物の親世代由来のゲノムDNAを用いて実施し、図25Aに示す。F1世代のゲノムDNAのさらなる分析ではトランス遺伝子は親世代の子孫にも確認されたことから該遺伝子が遺伝性であることが明らかになった。したがって、二重トランケートtauをコードするトランス遺伝子は該動物の染色体DNAに固定される(図25B-F1世代の遺伝子型別)。この実施例に用いる動物は自然発症高血圧および他のアルツハイマー病関連危険因子、例えば異脂肪血症または糖尿病を特徴とする特異的遺伝的背景がある。したがって、この動物系統は最も頻発するアルツハイマー病の危険因子、例えば高血圧および糖尿病を組み合わせることによりユニークな実験アルツハイマーモデルである。
トランス遺伝子を生成するにはSambrookら、Molecular Cloning A Laboratory Manual、CSH Laboratory、New York(2001)に記載の分子生物学の標準技術を用いた。二重トランケートtauをコードするcDNAを遍在性または組織特異的な発現をもたらすプロモータと結合した発現ベクターに導入した。該遺伝子断片を前核注射(非限定)により1日齢の胚に導入した。生じた子孫を尾の先端から得たゲノムDNAを用いて遺伝子型別した。
【0243】
トランス遺伝子発現の分析(図26):トランス遺伝子由来のmRNAの発現を一般的に知られた方法、例えばRT-PCRおよびアガロースゲル電気泳動を用いるRT-PCR分析により評価した。
【0244】
図25のパネルAはトランスジェニック動物の親世代の遺伝子型別を示す。レーン1、2、3および4におけるゲノムDNA由来のDNAの二重トランケート配列の特異的増幅は、フォスターマザーの子孫の尾から抽出したゲノムDNAにおける特異的トランス遺伝子の存在を示す。これら動物は二重トランケートタイプIIA tau分子を保有するトランスジェニック動物の親世代を表す。この実施例において、陽性(+C)および陰性(-C)および2つのさらなる陰性試料(5、6)が示される(M=サイズマーカー)。矢印はトランス遺伝子陽性動物において予想された予測PCR生成物サイズを示す。図25のパネルBはF1世代由来動物の遺伝子型別。ゲノムDNAを尾の先端から抽出し、二重トランケートtau特異的DNA配列を確認し、レーン1に示す。レーン2および3は陰性コントロールを示す。F1世代におけるtau特異的DNA断片の同定はこれらトランス遺伝子の遺伝性を確認する。
【0245】
図26:RNAをトランスジェニック動物の瞬間凍結組織から抽出し、逆転写、次いでcDNAの特異的増幅にかけた。実施例はレーン番号1および2においてトランス遺伝子発現動物を示す。レーン3-5は非発現コントロールを示し、レーン5は同じ方法を用いた非トランスジェニック動物に典型的に出現する非特異的シグナルを示す。この実施例は実験動物におけるトランス遺伝子から発現した二重トランケートtau特異的mRNAの存在を示す。
実施例15:A:タイプIIA分子の過剰発現は分化したニューロン様細胞の細胞死をもたらす。
【0246】
神経芽細胞腫細胞系SH-SY5Yにおいて、タイプIIA分子により生じた細胞死が当業者に知られた標準化in vitro分化条件下で示された。該効果をタイプIIA 二重トランケートtauを発現する安定なトランスフェクション細胞で試験し、非トランスフェクション細胞と比較した。細胞生存をトリパンブルー排除アッセイを用いトリプリケートで手動で定量し、統計的評価を一元ANOVA検定を用いて行った。タイプIIA 二重トランケートtau過剰発現細胞および野生型細胞における細胞生存の有意差がin vitro分化の6日後にみられた(P<0.001)。タイプIIA 二重トランケートtauの過剰産生(総タンパク質量の0.5%)は該細胞の生存率の3Xの低下を生じた(図27;各二重トランケートtauタイプIIA(タイプIIA)でトランスフェクションしたSY5Y細胞および非トランケートコントロールニューロン様細胞(mock)それぞれの細胞生存性の比較)。
【0247】
先に示した構築物同様に、同様の系が二重トランケートタイプI分子をコードする構築物を用いて確立された。
【0248】
タイプII 二重トランケートtau分子は微小管系に対する結合親和性の増大を示す。
【0249】
タイプIIA 二重トランケートtauおよび完全長tauを発現する安定にトランスフェクションされたSH-SY5Y細胞由来の遊離tau分画(FT)、微小管関連分画(MT)および核分画(NAT)の分離を行った。微小管とtauの結合の定量は完全長型に比べて二重トランケートタイプIIA tauの微小管に対する親和性の増大(50%以上)を示した(図28A;タイプIIA分子の微小管に対する結合親和性の増大はタイプIIA二重トランケート分子および完全長tauを発現する安定なトランスフェクション細胞の細胞分画化を用いて示される。遊離tau(FT)、微小管結合tau(MT)および核関連tau(NAT)の単離を記載のごとく実施した。)。tauの量を当該分野で用いられる標準的細胞生物学的分画法、次いでWesternブロット分析に従って定量した。較正曲線を規定量の組換えtauタンパク質を用いて計算した。
【0250】
B:有機物質F123[C34-59O14-23H32-44N6-8]nの微小管重合アッセイにおけるその阻害効果について分析する:(1)F123とtau(正常tauおよびtauタイプIA';conc=100μg/ml;tauタイプIIA conc=60μg/ml)を37℃で1時間インキュベーション、
(2)チューブリンの添加(conc=1mg/ml)、
(3)340nm/5minの測定(注:すべてPIPES緩衝液で希釈。
【0251】
C:正常健康tauおよびアルツハイマーtauタイプIIをその微小管構築促進能について分析する。
tau 43で示されたこの実施例の正常tauは電子顕微鏡検査で示される典型的微小管を形成する(図28C参照)。しかしながら、アルツハイマーtauタイプIIは典型的にパターンを有する病的微小管を生じる(図28C参照)。
実施例16:真核細胞におけるNおよびC末端二重トランケートtauタイプIIの過剰発現の機能的結果
【0252】
タイプIIA分子の過剰発現により生じたミトコンドリアの輸送の変化を示す病的表現型を神経芽細胞腫細胞系SH-SY5Yにおいて実施した。NおよびC末端二重トランケートtauタイプII分子の影響を、生野生型SH-SY5Y細胞とトランスフェクション細胞のミトコンドリア再分配を比較することにより試験した。当業者に知られた細胞生物学的輸送アッセイを用いた。簡単には、細胞を標準的実験技術に従って同じ密度(70%コンフルエント)でLabTekIIチャンバー(Nunc)を用いて培養し、トランスフェクションを製造業者の指示書に従ってFugene6(Roche)を用いた行った。ミトコンドリアの染色(MitoFluor Red 594、Molecular Probes)を製造業者の指示書に従って行った。生細胞を63x油浸対物レンズおよび蛍光フィルターを取り付けたAxiovert 200M蛍光顕微鏡(ZEISS)を用いて試験した。写真をソフトウエアプログラムMetaMorph(Universal Imaging)と組み合わせたCCDカメラ(Photometrics、Cool snap HQ; Hamamatsu)で撮影した。
【0253】
ミトコンドリア特異的染色MitoFluor(Molecular Probes)を用いるミトコンドリアの局在を誘導および非誘導SH-SY5Y細胞で比較した。該染色により、SH-SY5Y細胞におけるミトコンドリア輸送に対するタイプIIA 二重トランケートtau分子の悪影響はマイナス末端による細胞内力の機能的優性を示す中心体周辺の核周囲ミトコンドリアのクラスタ化を生じることであることが確認された(図30)。
【0254】
コントロールとして、対数増殖期の細胞(図29)は細胞体および細胞周辺におけるミトコンドリアの規則的な分布を示す。結論として、NおよびC末端二重トランケートタイプIIAタンパク質はミトコンドリア再分布に影響する細胞内輸送メカニズムに影響を及ぼすことができる。本実験の設定はタイプIIA分子に対する阻害活性を試験するための適切な方法を示す。
(参考文献)
【0255】
1. Finch C、Tanzi RE、(1997)Science 278,407-411
2. Blessed G、Tomlinson BE、Roth M(1968 Br J Psychiatry 114: 797-811
3. Tomlinson BE、Blessed G、Roth M J、(1970)Neurol Sci 11、205-242
4. Arigada PA、Growdon JH、Hedley-White ET、Hyman BT(1992)Neurology 42,631-639
5. Wischik CM、Novak M、Edwards PC、Klug A、Tichelaar W、Crowther RA(1988a)Proc Natl Acad Sci USA 85: 4884-4888
6.Wischik CM、Novak M、Trogersen HC、Edwards PC、Runswick MJ、Jake R、Walker JE、Milstein C、Roth M、Klug A(1988b)Proc Natl Acad USA 85: 4506-4510
7.Himmler A,Drechsel D、Kirschner MW、Martin DWjr、(1989)Mol Cell Biol 9,1381-1388
8. Goedert M、Spillantini MG、Jakes R、Rutherford D.、Crowther RA(1989)Neuron#、519-526
9. Hutton M、Lendon CL、Rizzu P、Baker M、Froelich S.、Houlden H、Pickering-Brown S、Chackraverty S、Isaacs A、Grover A(1998)Nature 393,702-705
10. Spillantini NG、Murrell JR、Goedert M、Farlow MR、Klug A、Ghetti B(1998)Proc Natl Acad Sci USA 95,7737-7741.
11. Selkoe DJ、(1999)Nature 399、A23-31
12. Isacson O、Seo H、Lin L、Albeck D、Granholm A CH、(2002)Trends Neurosci 25,79-84
13. Mudher A、Lovestone S、(2002)Trends Neurosci 25,22-26
14. Couzin J、(2002)Science 296,28-29
15. Grundke-Iqbal I、Iqbal K、Tung Y-C、Quinlan M,Wisniewski HM、Binder LI、(1986)Proc Natl Acad Sci USA、83,4913-4917
16. Gustke N、Steiner B、Mandelkow EM、Biernat J、Meyer HE、Goedert M、Mandelkow E(1992)FEBS Lett 307,199-205
17. Lindwall G、Cole RD(1984)J Biol Chem 259,12241-12245
18. Kopke E、Tung Y-Ch、Shaikh S、Alonso AC、Iqbal K、Grundke-Iqbal I(1993)J Biol Chem 268,24374-24384
19. Cleveland DW、Hwo SY、Kirschner MW(1977a)J Mol Biol 116: 227-247
20. Cleveland DW、Hwo SY、Kirschner MW(1977b)J Mol Biol 116: 207-225
21. Lee MY、Balin BJ、Otvos L、Trojanowski JQ(1991)Science 251,675-678
22. Alonso AC、Zaidi T、Grundke-Iqbal I、Iqbal K(1994)Proc Natl Acad Sci U S A 91: 5562-5566
23. Wille H、Mandelkow EM、Mandelkow E(1992)J Biol Chem 267: 10737-10742
24. Alonso AC、Grundke-Iqbal I、Iqbal K(1996)Nat Med 2: 783-787
25. von Bergen M、Barghorn S、Li L、Marx A、Biernat J、Mandelkow EM、Mandelkow E(2001)J Biol Chem 276: 48165-48174
26. Friedhoff P、von Bergen M、Mandelkow EM、Mandelkow E(2000)Biochim Biophys Acta 1502: 122-132 Review
27. Illenberger S、Zheng-Fischhofer Q、Preuss U、Stamer K、Baumann K、Trinczek B、Biernat J、Godemann R、Mandelkow EM、Mandelkow E(1998)Mol Biol Cell 9: 1495-1512
28. Kampers T、Friedhoff P、Biernat J、Mandelkow EM、Mandelkow E(1996)FEBS Lett 399: 344-349
29. Perez M、Valpuesta JM、Medina M,Montejo de Garcini E、Avila J(1996)J Neurochem 67: 1183-1190
30. WangJ-Z ; Grundke-Iqbal I; Iqbal K(1996)Nature Med 2: 871-875
31. Wilson DM、Binder LI(1997)Am J Pathol 150: 2181-2195
32. Schweers O、Mandelkow EM、Biernat J、Mandelkow E(1995)Proc Natl Acad Sci USA 92: 8463-8467
33. Ksiezak-Reding H、Yang G、Simon M、Wall JS(1998)Brain Res. 814:86-98
34. Schneider A、Biernat J、Von Bergen M、Mandelkow E、Mandelkow EM(1999)Biochemistry 38: 3549-3558
35. Yan S-D、Chen X、Schmid A-M、Brett J、Godman G、Zou Y-S、Scott CW、Caputo C、Frappier T、Smith MA、Perry G、Yen SH、Stern D(1994)Proc Natl Acad Sci USA 91: 7787-7791
36. Smith MA、Taneda S、Richey PL、Miyata S、Yan SD、Stern D、Sayre LM、Monnier VM、Perry G(1994)Proc Natl Acad Sci U S A 91: 5710-5714
37. Mori H、Kondo J,Ihara Y(1987)Science 235: 1641-1644
38.Paudel H、Li W(1999). J Biol Chem 274: 8029-8038
39. Goedert M、Spillantini MG、Potier MC、Ulrich J、Crowther RA、(1989)EMBO J. 8,393
40. Goedert M、Spillantini MG、Jakes R、Rutherford D、Crowther RA(1989)Neuron 4,519
41. Crowther RA、Olesen OF、Jakes R、Goedert M(1992)FEBS Lett 309: 199-202
42. Crowther RA,Olesen OF、Smith MJ、Jakes R、Goedert M(1994)FEBS Letter 337: 135-138
43.(例えばM. Kohler and C. Milstein、in"Continuous Cultures of Fused Cells Secreting Antibody of Pre-DefinedSpecificity"、Nature、256、pp. 495-497,1975;およびHarlow and Lane、in "Antibodies、A Laboratory Manual"、Cold Spring Harbor Laboratory、Cold Spring Harbor、1988参照)
【図面の簡単な説明】
【0256】
【図1】NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築。
【図2】NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築の阻害。
【図3】微小管構築におけるNおよびC末端二重トランケートtauタイプIIAおよびIIB分子の活性。
【図4】神経細胞に発現したタイプIIA NおよびC末端二重トランケートtauは酸化ストレスに対する感受性を有意に増加させる。
【図5】病的tauタイプIAタンパク質およびその欠失突然変異体に対するモノクローナル抗体の親和性。病的tauタイプIAタンパク質およびその欠失突然変異体に対するモノクローナル抗体の見かけの親和性。最初のカラムに「プロトタイプ」tauタイプIAタンパク質(配列番号1)および各欠失突然変異体を示す。中段のカラムに本発明のエピトープを示す。最後のカラムに示す見かけの親和性を競合的ELISAにより測定し、プロトタイプtauタイプIAタンパク質と50%競合するのに必要な対応抗原の濃度で表した。
【図6】AD-脳由来tauタンパク質のSuperdex 200-カラムによる分画。
【図7】AD脳からの3つの別個の単離物由来の分画No.19中のタイプIA阻害活性。
【図8】AD脳中のNおよびC末端二重トランケートtauタイプI分子の証明。
【図9】健康脳に存在せずAD脳に存在するtauタイプI。
【図10】NおよびC末端二重トランケートtauタイプII分子の免疫反応性。
【図11】組換えtauタイプI-II(配列番号1-24)の構築。
【図12】AD-脳由来および組換えtauタイプIAの正常健康tauに対する阻害効果。
【図13】tauタイプIA分子を中和する薬剤候補の第一回スクリーニング(工程1)。
【図14】正常tauに対する選択性を有するタイプIA分子を中和する薬剤候補の第二回スクリーニング(工程2)。
【図15】tauタイプIIA分子を中和する薬剤候補の第一回スクリーニング。
【図16】tauタイプIIA分子を中和し、それを正常tauから識別することができる薬剤候補の第二回スクリーニング(工程2)。
【図17】ELISAにより測定したプレフュージョン(prefused)マウス血清中の特異抗体レベル。
【図18】モノクローナル抗体とAD脳由来tau(分画#19)およびコントロール健康脳由来tauとのELISA反応性(DC20:無関係な特異性を有するモノクローナル抗体。示したデータは3回の並列実験の平均値を表す。)。
【図19】モノクローナル抗体の組換えtau分子とのELISA反応性(DC20:無関係な特異性を有するモノクローナル抗体。示したデータは3回の並列実験の平均値を表す。)。
【図20】AD-脳由来tauタイプIA(分画#19)に対する中和抗体のスクリーニング。
【図21】組換えtauタイプIA(配列番号1)に対する中和抗体のスクリーニング。
【図22】tauタイプIA分子を中和し、それらを健康tauから識別することができる薬剤候補のスクリーニング。
【図23】組換えtauタイプIIA(配列番号12)の病的活性のモノクローナル抗体による中和。
【図24】ELISAで検出した組換えtauタイプIIA(配列番号12)に対する抗体のレベル。
【図25】トランスジェニック動物の遺伝子型別。パネルAはトランスジェニック動物の親世代の遺伝子型別を示す。レーン1、2、3および4のゲノムDNA由来のDNAの二重トランケート配列の特異的増幅は、フォスターマザーの子孫の尾から抽出したゲノムDNAにおける特異的トランス遺伝子の存在を示す。これら動物は二重トランケートタイプIIA tau分子保有トランスジェニック動物の親世代を示す。この実施例において、陽性(+C)および陰性(-C)および2つのさらなる陰性試料(5、6)を示す(M=サイズマーカー)。矢印は、トランス遺伝子陽性動物において予想される予想PCR生成物サイズを示す。パネルB. F1世代由来の動物の遺伝子型別。ゲノムDNAを尾の先端から抽出し、二重トランケートtau特異的DNA配列を同定し、レーン1に示す。レーン2および3は陰性コントロールを示す。F1世代におけるtau特異的DNA断片の同定はこれらトランス遺伝子の遺伝性(inheritability)を確認する。
【図26】トランスジェニック動物のF1世代における二重トランケートヒトtau転写物の遺伝子発現。RNAをトランスジェニック動物の瞬間凍結組織から抽出し、逆転写、次いでcDNAの特異的増幅にかけた。例はレーン番号1および2にトランス遺伝子発現動物を示す。レーン3〜5は非発現コントロールを示し、レーン5はこの方法を用いた時に非トランスジェニック動物に典型的に生じる非特異的シグナルを示す。この実施例は、実験動物においてトランス遺伝子から発現した二重トランケートtau特異的mRNAの存在を示す。
【図27】6日間in vitro分化(differentiation)後のタイプIIA分子の過剰発現により生じた細胞死。二重トランケートtauタイプIIA(タイプIIA)でトランスフェクションしたSY5Y細胞および非トランスフェクションコントロールニューロン様細胞(mock)の細胞生存性の比較。
【図28A】タイプIIA分子の微小管に対する結合親和性の増加はタイプIIA二重トランケート分子および完全長tauを発現する安定なトランスフェクション細胞由来のtauの細胞分画を用いて示される。遊離tau(FT)、微小管結合tau(MT)および核関連tau(NAT)の単離は記載のごとく行った。
【図28B】有機化合物F123による各tauタイプIAおよびIIAの阻害、微小管重合アッセイ。
【図28C】電子顕微鏡分析により示される微小管重合と吸着(absorption)測定値の直接比較。
【図29】MitoFluorで染色した対数増殖期のSH-SY5Y細胞。細胞体およびプロセス中のミトコンドリアの標準的分布。
【図30】MitoFluorで染色した対数増殖期のtauタイプIIA分子発現SH-SY5Y細胞。細胞の中心体領域周辺のグリーン標識ミトコンドリアの核周囲クラスタリング。
【技術分野】
【0001】
本発明は、具体的にはアルツハイマー病および関連疾患にみいだされるNおよびC末端トランケート(truncated)病的型のtau(タウ)タンパク質に関する。
【0002】
さらに本発明は、NおよびC末端二重トランケートtauタンパク質を阻害、中和、および除去、またはその形成を予防するのに有効な潜在的薬剤をスクリーニングおよび試験する方法、および作用機序がtauタンパク質の該二重トランケート病的(罹患)型により生じる微小管構築および/または動態の修飾の中和に基づく潜在的薬剤をスクリーニングおよび試験するための方法に関する。
【0003】
アルツハイマー病は痴呆の最も一般的な原因である。該症例の5%以下においてアルツハイマー病はアミロイド前駆体タンパク質、プレセニリン-1またはプレセニリン-2遺伝子の1またはそれ以上の特定の突然変異とほぼ完全に同時分離し(cosegeragate)(1)、該症例の95%以上で正確な病因が不明である。
【0004】
病因とは無関係にアルツハイマー病は組織病理学的に対になったらせん状フィラメント(PHF)の神経原線維変化を有する多くのニューロンおよび脳の老人斑の主要成分としてアミロイドβの細胞外沈着の存在を特徴とする。アルツハイマー病のこれら2つの特徴的病変間の実際の直接的関係(あるとすれば)は理解されていないが、神経原繊維変性の存在は該病気、すなわち痴呆の臨床的発現に必要のようである(2、3、4)。神経原繊維変性は神経原線維変化、ジストロフィー性神経突起および神経網スレッドにより示される。これら構造の主要タンパク質サブユニットは微小管関連タンパク質tauである(5、6)。
【0005】
健康ヒト脳においてtauは単一遺伝子座由来の転写物の選択的mRNAスプライシングにより生じる6つのタンパク質アイソフォームで出現する。tauタンパク質は該分子のC末端近くの31または32アミノ酸の3(t3L、t3S、またはt3)または4(t4L、t4S、またはt4)チューブリン結合ドメイン(反復配列、R)、およびN末端部分の各29アミノ酸の2(t3L、t4L)、1(t3S、t4S)、または無(t3、t4)挿入物(7、8)を含むか否かで異なる。生理学的条件下でtauタンパク質は微小管の構築、空間的構成、安定化、および挙動に関与する。生理学的条件下で該タンパク質は健康ヒト脳では6アイソフォームで出現する。しかしながら、ADではtauタンパク質は多くの異なる翻訳後修飾(過リン酸化、ユビキチン化、グリコシル化)を受けることが知られている。第17染色体と結合したパーキンソン症を伴う疾患である前頭側頭性痴呆(FTDP17)のtau遺伝子における特異的突然変異の同時分離の最近の発見は、tauタンパク質のある異常が罹患個体における神経変性および痴呆の主要原因となりうることを確認した(9、10)。アルツハイマー病におけるtau修飾および対になったらせん状フィラメント(PHF)形成をもたらす分子的事象は知られていない。これは、広域スペクトルの病態生理学的事象、例えばtauタンパク質の病的再分布、軸索輸送障害、または軸索微小管機能維持障害の所見を説明する(11、12、13)。今日までアルツハイマー病におけるPHF微小繊維形成の重要性はあらゆるタンパク質がin vitroで原繊維を形成することができるという最近の発見に照らして疑問である(14)。
【0006】
多くの著者がアルツハイマー病における対になったらせん状原繊維の形成は異常リン酸化に基づく神経原繊維病変における主要事象を示すと考えている。PHF構築tauタンパク質はリン酸化依存性にある抗体と反応し、このことは特別なリン酸化状態を示唆する(15、16)。さらにPHF由来tauタンパク質はそのリン酸化パターンに関連しうるSDSゲルにおける電気泳動移動性の低下を示すことが観察された(Steinerら、EMBO J. 9(1990)、3539-3544)。同様に、同様の効果が正常tauがある種のキナーゼによりin vitroでリン酸化された時にみられたことからリン酸化によりPHF由来tauが正常tauタンパク質と比べて微小管に低親和性であることが示唆された(17、18)。tauは既知の最も可溶性のタンパク質の一つであり(19、20、21)、アルツハイマー病におけるその凝集は特に謎である。他方、tauタンパク質がアルツハイマー病におけるフィラメント形成をもたらすように修飾されるメカニズムは知られていない。tauのリン酸化は、おそらくリン酸化部位依存性に微小管重合に対する促進的または阻害的作用をもたらすtauの凝集形成潜在力に影響を及ぼす(22-27)。多くのin vitro試験は、還元剤ジチオスレイトール(DTT)、不飽和遊離脂肪酸、RNAまたはグルコサミノグリカンの存在下で、正常tauをフィラメントに変化させることができることを示す(28-31、38)。さらに、フィラメント形成のプロセスはCys322の酸化により生じる架橋tauの存在により促進することもできる(32)。tauタンパク質濃度、pHおよびイオン強度を含む種々のフィラメント構築試験で変化するパラメーターは生理学的条件下で細胞質におけるより多様性が高かった。in vitro形成tauフィラメントの走査透過電子顕微鏡(STEM)による実験は、これらフィラメントが天然の対になったらせん状フィラメントと異なることを示した(33)。グリカンまたはRNAの非存在下でPHF様フィラメントは非リン酸化またはリン酸化野生型tauを含む資料中に検出できなかった。さらにリン酸化はアルツハイマー病の防御的役割を果たすかもしれないことが示唆された(34)。微小管の分解および極めて重要な神経プロセス、例えば軸索輸送との干渉を生じるPHFの構築をもたらすtauの修飾に対する同様の示唆がユビキチン化およびグリコシル化についてなされた(30、35、36、37)。しかしながら、上記翻訳後修飾のみではアルツハイマー病の臨床的発現に関連するその機能障害をもたらすtau変化の開始に対する分子的説明となり得なかった。
【0007】
したがって、上記のどのtau修飾がアルツハイマー病の病因に関与しているかは依然不明である。
【0008】
今日まで初期tauタンパク質複合体の形成をもたらす翻訳後事象の様式または調節に関する確実なデータはない。そのような複合体形成を予防し、そのあらゆる関連病的効果を中和するには病的tauの正確な分子的性質および正常tauをそのNおよびC末端二重トランケート型に変換する調節メカニズムを明らかにする必要がある。この詳細な知識はアルツハイマー病の治療薬および診断薬のための手段を構築するであろう。
【0009】
Zelmanら(J. P. Neurochem. 72(2)(1999)、741-750)は、微小管結合タンパク質tauの開裂産物が外傷性脳損傷を有するヒトの脳脊髄液に生じ、ニューロンの損傷を反映することを示唆する。しかしながら、アルツハイマー病との関連はこの報告に記載されていない。
【0010】
Novak(Acta Virologica 38(1994)、173-189)は、この総説で広域スペクトルプロテアーゼ「プロナーゼ」により人工的に生成されたPHF(「対になったらせん状フィラメント」)の最小プロテアーゼ抵抗性単位「タウオン」に関する論説を記載している。
【0011】
Kontsekovaら(J. Immunol. Meth. 185(1995)、245-248)は、ヒトtauタンパク質の熱抵抗性を用いる免疫分析用の組換えヒトトランケートtauタンパク質の迅速精製方法を開示している。その中で使用された組換えtau類似体の構造や生理学的特性または機能は記載されていない。
【0012】
Novakら(EMBO J. 12(1)(1993)、365-370)は、対になったらせん状フィラメント(PHFコア)をin vitroで人工的に製造し、最小プロテアーゼ抵抗性tau単位をプロテアーゼプロナーゼによるin vitro消化により回収した。モノクローナル抗体MN423を用いて最小プロテアーゼ抵抗性tau単位を検出した。しかしながら、この論文に記載のtauポリペプチドは「実在する(realworld)」tauタンパク質、特にアルツハイマー病と関連があるtauタンパク質と共通の生物学的構造的な病的特性をもたなない。
【0013】
Fasuloら(Alzheimer's Research 2(5)(1996)、195-200)は、PHFコアtauの組換え類似体の過剰発現がtau凝集および対になったらせん状フィラメントのその構築を誘導するのに十分でないことを報告している。これらデータはAbrahaら(J. Cell. Science(113)(21)(2000)、3737-3745)の論文とこの論文に記載の異常な非生理学的アッセイ系(サル腎臓由来細胞系)により明白に異なる。
【0014】
Fasuloら(J. Neurochem. 75(2000)、624-633)はアポトーシスを誘導するtau断片を記載している。しかしながら、本発明に記載のアルツハイマー病関連tauタンパク質はいずれもアポトーシスを誘導し得ない。
【0015】
Espositoら(J. Peptide Science 6(2000)、550-559)はtauタンパク質および正常健康tauタンパク質のC末端19アミノ酸を記載している。Novakら(Chem. Papers 52(1998)、429-430)およびUgoliniら(NeuroReport 8(1997)3709-3712)の論文はアポトーシスにも関連するC末端トランケートtauタンパク質に関する。より最近の刊行物はアルツハイマー病がアポトーシスプロセスに関連しないことを示す。
【0016】
Abrahaら(J. Cell Science 113(21)(2000)、3737-3745)は、フィラメント形成に対するtauタンパク質の単一ドメインの関与を示すためのを記載している。したがって、一組の組換えtau分子が組み立てられ、in vitroで生成された。細菌におけるこれらタンパク質の生物活性および病的活性はいずれも生じないか検出されなかった。さらに、アルツハイマー病患者の脳由来のtauタンパク質に関するデータはこの論文に記載されていない。
【0017】
Jichaら(J. Neuroscience Research 55(1999)、713-723)には、モノクローナル抗体Alz50およびMC-1のエピトープの分子的分析が記載されている。両抗体はtau分子の機能的N末端、特にアミノ酸位7-9に依存する。Tauトランケーションはこの論文に記載されていない。
【0018】
Brandtら(J. Biol. Chem. 268(1993)、3414-3419)は正常健康ヒトtauタンパク質の異なるドメインを分析した。このために組換えtau断片を細菌で生成した。しかしながら、アルツハイマー関連トランケートtau断片はこの論文に記載されていない。
【0019】
Philippeら(J. Neuroscience Research 46(1996)、709-719)はモノクローナル抗アミロイド前駆体タンパク質抗体を開示している。この著者らはtau反応性抗体の生成について記載したが、この抗体は最初にアミロイド前駆体タンパク質に対して生じた。病変に関連するアルツハイマーtau断片はこの論文に記載されていない。
【0020】
WO94/18560 A1は、細胞中枢神経サイトパシーの患者を検出するための脳脊髄液中のヒトtauタンパク質を検出するためのイムノアッセイを開示している。このアッセイは正常tauと中枢神経サイトパシーの患者のtauを識別しないが、試料中のtauタンパク質の総量を検出する。
【0021】
したがって、本発明の目的はアルツハイマー病ニューロンの病的機能不全に関連するそのような確実なマーカーを提供することである。さらに、そのようなtau由来ポリペプチドの存在を確認し、その活性をアッセイするための適切な手段はアルツハイマーの診断薬および治療薬に有用な手段であろう。
【0022】
したがって、本発明は、以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIA tau分子」)を提供する:
4反復配列を含むtau43の少なくとも最初の236 N末端アミノ酸および少なくとも最後の45 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて正常tauタンパク質の微小管構築の促進を防止し、
該微小管構築の促進の防止は微小管構築アッセイにおいて該分子に対する特異的阻害、中和モノクローナル抗体により除去することができる。
【0023】
以下において用語「NおよびC末端二重トランケートtauタンパク質」はアルツハイマー病脳に出現し、アルツハイマー病ニューロンの病的機能不全と密接に関連するトランケートtau誘導体の2つのグループを説明するのに用いる。特に、これらタンパク質は微小管関連生物学的機能、例えば微小管構築または細胞内輸送を修飾することによりその病的機能を示す分子群を表す。以下において用語「タンパク質複合体」はホモダイマー、ヘテロダイマーまたは多量体複合体の形の、tauおよび/または二重トランケートtauタンパク質と物理的に関連する分子からなるNおよびC末端二重トランケートtauタンパク質に用いる。
【0024】
本明細書で用いている用語「tau」は、前記のその微小管結合ドメイン(39、40)中に3反復配列(tau44)および4反復配列(tau43)を含む健康ヒト脳に存在する最も短い天然アイソフォームの群、すなわち、tau43(383アミノ酸、エクソン2および3を欠く[pos45-102])tau44(352アミノ酸、エクソン2、3および10を欠く[各pos 45-102および275-307])をあらわす。以下の本文において用語「野生型tau」は「正常tauタンパク質」の同義語として用い、健康脳由来のtauタンパク質を表す。
【0025】
適切な微小管構築アッセイ(あるいは「微小管重合アッセイ」と呼ばれることも多い)は例えば(19)および(20)に記載されている。用語「予防(防止)する(preventing)」は、正常tau促進活性の20%またはそれ以上、好ましくは50%またはそれ以上のあらゆる有意な阻害を含む。
【0026】
具体的には、本発明の好ましいタイプIA tau分子は配列番号1〜3の群から選ばれるアミノ酸配列を含む。
【0027】
さらに、本発明は、以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIB tau分子」)を提供する:
4反復配列を含むtau43の少なくとも最初の238 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の207 N末端アミノ酸および少なくとも最後の50 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauの微小管構築の促進を防止しない。
【0028】
好ましいタイプIB tau分子は配列番号4〜10の群から選ばれるアミノ酸配列を含むことを特徴とする。
【0029】
本発明は以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIIA tau分子」)も提供する:
4反復配列を含むtau43の少なくとも最初の68 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の68 N末端アミノ酸および少なくとも最後の20 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauより高い微小管構築促進活性を有し、
該微小管構築促進活性は微小管構築アッセイにおいて該分子に対する特異的阻害、中和モノクローナル抗体により除去することができ、該分子の病的活性は微小管重合促進活性により定義される微小管ネットワークに対する結合に依存する。
【0030】
好ましくは、分光光度法で測定したとき微小管構築促進活性の増大は野生型tauより少なくとも20%高く、特に少なくとも50%高い。
【0031】
好ましいタイプIIA tau分子は配列番号11〜18の群から選ばれるアミノ酸配列を含むことを特徴とする。
【0032】
さらに、本発明は、以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIIB tau分子」)を提供する:
4反復配列を含むtau43の少なくとも最初の68 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の68 N末端アミノ酸および少なくとも最後の20 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauと異なる病的微小管構築促進活性を有する。
【0033】
本発明の好ましいタイプIIB tau分子は配列番号19および20の群から選ばれるアミノ酸配列を含むことを特徴とする。
【0034】
本発明の新規tauポリペプチド(IA、IB、IIAおよびIIB)はアルツハイマー病の脳組織にもっぱら局在するので典型的でユニークな局在特性を有する。さらに、これらポリペプチドと非重合チューブリン(α/βダイマー)および重合型(微小管として)の相互作用もユニークである。
【0035】
別の局面によれば本発明は、以下の工程を特徴とする本発明(タイプIA、IB、IIA、IIB)の分子の製造方法を提供する:
a)少なくとも最初の236および最後の40アミノ酸または最初の68および最後の20アミノ酸またはその組み合わせにわたる欠失を有する二重トランケートtau分子をコードする配列を保持する組換え原核性発現プラスミドの構築、
b)該NおよびC末端二重トランケートtau分子の発現を許す条件下で該細菌を増殖させ、
c)好ましくは遠心により細菌を回収し、
d)細菌ペレットを再浮遊させ、
e)該細菌を超音波処理し、
f)該超音波処理細菌をゲルろ過により分画し、
g)得られた分画の活性を微小管構築アッセイによりモニターすることにより、タイプIおよびタイプII tau分子の異なる活性を同定する。
【0036】
好ましくは、該トランケーションはタイプIA、IB、IIAおよびIIB分子について上記の通りである。該微小管構築アッセイ活性は好ましくは特にIAについて上記の通りである。
【0037】
さらに本発明は以下の工程を特徴とする本発明の分子の製造方法を提供する:
a)アルツハイマー罹患脳組織を得、
b)該病的(罹患)脳組織を緩衝液、特にTris緩衝液中でホモゲナイズし、
c)該ホモゲナイズ脳組織を硫酸アンモニウム沈殿し、
d)PIPES緩衝液に再溶解し、
e)該再溶解物質をゲルろ過で分画化し、
f)得られた分画の活性を微小管構築アッセイでモニターすることによりタイプIおよびタイプII tau分子の異なる活性を同定する。
【0038】
該微小管構築アッセイ活性は好ましくは特にIAについて上記の通りである。
【0039】
本発明はさらに以下の工程を含むタイプIA分子とチューブリンの複合体の分解(disassembling)に有効な物質の試験方法を提供する:
a)タイプIA分子とチューブリンのタンパク質複合体を形成させ、
b)該タンパク質複合体を被検物質とインキュベーションし、野生型tauの微小管構築促進能力を回復させる物質を同定する。
【0040】
さらに本発明は以下の工程を含む野生型tau発現細胞系においてタイプIA分子がチューブリンと複合体の形成を開始するのを阻害するのに有効な物質の試験方法を提供する:
a)適切な制御領域の制御下でタイプIA分子をコードする機能的遺伝子を正常tauタンパク質発現細胞に導入し、
b)タイプIA分子とチューブリンのタンパク質複合体を形成させ、
c)試験する物質を該複合体を保持する細胞に適用し、
d)上記のタイプIA生物活性に対する該物質の効果を試験する。
【0041】
本発明は該タイプIIA分子と微小管との相互作用により病的微小管を生成させる生理的条件下でタイプIIAとチューブリンタンパク質をインキュベーションすることを特徴とする微小管をin vitroで病的状態に変換する方法も提供する。
【0042】
別の局面において、本発明は以下の工程を含むタイプIIA分子を除去および/または中和し、タイプII分子により生じる生理学的微小管パラメーターおよび機能を修復するその特性についてタイプIIA分子の病的効果を中和することができる物質のスクリーニング方法を提供する:
a)タイプIIA分子およびチューブリン存在下で病的微小管を形成させ、
b)該物質、タイプIIAおよびチューブリンの混合物をスクリーニングする物質とインキュベーションし、
c)タイプIIA分子により生じる病的微小管の形成を減少させる結果を試験する。
【0043】
本発明は以下の工程を含むタイプIIA分子発現細胞系における異常微小管形成および機能の促進においてタイプIIA分子のin vivo活性を阻害するのに有効な物質の試験方法も提供する:
a)タイプIIA分子をコードする機能的遺伝子を適切な制御領域の制御下で野生型tau発現細胞中に導入し、
b)タイプIIA tau分子と微小管の複合体を形成させて該複合体を病的微小管の形成に関与させ、
c)試験する物質を該複合体を保持する細胞に適用し、
d)タイプIIA生物活性、特に微小管ネットワークおよびその関連機能の修飾に対する該物質の効果を試験する。
【0044】
別の局面によれば本発明は本発明に記載の分子(タイプIA、IB、IIAまたはIIB)、特にIAおよび/またはIIAを発現するトランスジェニック動物も提供する。
【0045】
本発明は特にアルツハイマー病治療用の薬剤をスクリーニングおよび試験するためのアルツハイマー病の動物モデルとしての本発明のトランスジェニック動物の使用にも関する。
【0046】
本発明は本発明の分子(IA、IB、IIAまたはIIB)、特にIAおよびIIA、および医薬的に許容される担体、特にアジュバントを含むワクチンを提供する。
【0047】
本発明はタイプIA分子と野生型tauの複合体の形成開始の阻害剤(インヒビター)も提供する。該阻害剤の特定の例には、寄託番号02060767でEuropean Collection of Cell Cultures(ECACC)、Porton Down、Salisbury、UKに寄託したモノクローナル抗体DC44のような結合部分、特にDC44またはその結合断片、例えばFabを含む物質である。
【0048】
すなわち、本発明は以下のもの提供する:
(1)NおよびC末端トランケート病的型のtauタンパク質の分子的および機能的同定および特徴付け(これら分子は微小管関連生物学的機能、例えば微小管構築または細胞内輸送を修飾することによりアルツハイマー病のその病的機能を示す。)、
(2)タンパク質エピトープの特異抗体、
(3)該タンパク質の病的活性を中和する抗体、
(4)NおよびC末端二重トランケートtauタンパク質を阻害、中和、および除去し、またはその形成を予防するのに有効な治療薬候補(抗体を含む)のスクリーニングおよび試験方法、
(5)薬物スクリーニングに用いることができる、各二重トランケートtauタンパク質をコードする遺伝子構築物を保有する動物モデルの開発、
(6)該二重トランケートtauタンパク質の阻害剤およびその起源に含まれるプロテアーゼを含む医薬組成物、
(7)NおよびC末端二重トランケートtau分子を生じる分子のスクリーニング方法、
(8)該分子を認識しそして/またはそれと相互作用する診断用および治療用組成物、
(9)該二重トランケートタンパク質の抗原性に基づくワクチンの開発、
(10)アルツハイマー病およびtauの病的変化に関連する他の障害のin vitroおよびin vivo診断用の該タンパク質およびそのエピトープおよび/または抗体または他の特異的プローブを含む方法。
【0049】
したがって、本発明は具体的にはアルツハイマー病に生じるNおよびC末端二重トランケート型の病的tauタンパク質およびそのエピトープの特徴付けに関する。
【0050】
タンパク質の分解は、種々のサイズの、通常短い半減期の中間体トランケーション生成物の生成を含むタンパク質の生理学的除去中に生じる一般的現象である。wt(=野生型)tauを含む健康脳ではタンパク質は例外なくこのプロセスを受ける。以下において用語「wt」は健康個体の脳に通常みられるtauタンパク質のすべての6つの天然アイソフォームに及ぶ。アルツハイマー病の脳に見られる種々の短トランケーション型は細菌中で産生され、タンパク質の生理学的機能を試験するために種々の程度に精製され、そのドメインおよびリン酸化エピトープをマップし、またはアルツハイマー病および他の神経変性障害における対になったらせん状構築のメカニズムを理解するための実験において結果が曖昧である(23-27、34、41、42)。一般的用語「NおよびC末端二重トランケート型のtauタンパク質」は分子の両末端のアミノ酸の少なくとも1つが失われているアルツハイマー病のあらゆるtauタンパク質を表す。アルツハイマー罹患脳からの抽出物中の二重トランケートtauの分析により、本発明の過程でこれら分子のいくつかが構造的および機能的に異なる特性を表現し、それらをアルツハイマー病の脳組織にみられる他のtau断片から識別することができることがわかった。この識別に基づいて、健康tau:タイプIおよびタイプII tau分子と異なるNおよびC末端二重トランケートtau分子の病的分子の2つの主なクラスを定義する新規の仕組みを提供した。これらのグループは分子構造に基づいてさらにそれぞれ2つのサブクラスに細分することができ、それぞれタイプIAおよびBおよびタイプIIAおよびBと称される。
【0051】
タイプIAおよびタイプIIAは、病的プロセシングにより生じた微小管関連タンパク質tau由来の構造的および機能的に異なるタイプの病的分子を表す。NおよびC末端トランケートtau分子は、アルツハイマー病に特徴的な特異的な病的プロセス中に出現する微小管関連タンパク質 tau由来の病的分子を表す。これはtau由来タンパク質の4グループすべてに共通の特徴である。さらに全グループに共通の特徴はNおよびC末端トランケーション、その神経内および神経外局在および正常な健康tauとの機能的識別である。
【0052】
「タイプIA」で示される分子グループは実施例に配列番号1〜3で示される。これらトランケートtau分子は、病的tauとチューブリンの相互作用のための鍵(中心的な)、活性単位、および原動力として作用する正常tauと異なる。タイプIAおよびタイプIB分子は微小管構築においていかなる促進活性も持たない。驚くべきことに、タイプIAは正常tauを微小管構築の促進から予防することができる(実施例1)。同様の一次配列特性および分子量にも関わらずタイプIBはin vitroでこの機能的活性を示さない(実施例2)。これはタイプIAのチューブリンに対する強い結合活性ならびにそれによるtauの生理学的機能に対する主要な悪影響をもたらすことを示唆する。したがってタイプIA分子は機能的微小管ネットワーク由来のニューロンの連続的で慢性的な欠失に関与し、アルツハイマー病の臨床的重症度に直接関連する神経原繊維構造に関与するようである。予期しないことにタイプIB(例えば配列番号4〜10)は、タイプIAグループの分子と同様の分子量と配列を有するにも関わらずグループIAのメンバーの病的活性を全く示さない(実施例2参照)。これらのグループと反対に、タイプIIA 二重トランケートtau誘導体は微小管と結合し、その病的構築を促進する(実施例3)。以下においてタイプIIAがもたらす微小管を「病的微小管」という。驚くべきことに、同様の配列および分子量範囲を有する分子(タイプIIB)はこれらの高微小管重合能力を欠く。微小管構築アッセイにおいて、それらは完全長tauタンパク質でみられるレベルで機能する(実施例3参照)。
【0053】
両グループ(タイプIIAおよびB)のNおよびC末端トランケートtau誘導体は、細胞レベルで軸索輸送を妨げ、最終的にアルツハイマー病患者に神経機能不全と認知障害をもたらすシナプスの損失をもたらす。同時に、罹患ニューロンは種々の形のストレス、例えば酸化ストレスに弱い(実施例4)。タイプIIBはタイプIIAと同様の分子サイズを有するにも関わらず分光光度法で測定すると微小管構築をさらに完全長健康tau(野生型tau)で見られるレベルまで促進した。
【0054】
本発明のタイプIAおよびBならびにタイプIIAおよびB分子のさらに好ましい態様において該分子の組換えバージョンは以下の工程を行うことにより得ることができる:
(a)該二重トランケートtau分子(タイプIおよびII)のコーディング配列を保持する組換え原核性発現プラスミドの構築、
(b)NおよびC末端二重トランケートtau分子(タイプIおよびII)を発現させることができる条件下での細菌の増殖、
(c)遠心分離による細菌の回収、
(d)500ml培養から得た細菌ペレットの緩衝液A:(20mM PIPES pH6.9、50mM NaCl、1mM EGTA、1mM MgSO4、2mM DTT、0.1mM PMSF)への再浮遊、
(e)氷上で1分間(3回)超音波処理し、2℃、45000rpmで15分間遠心分離(ローターTLA-120,2、Beckmann Optima TLX)、
(f)緩衝液「A」中の直線勾配0〜1M NaClを用いPhosphocellulose(ホスホセルロース)、またはMONO S HR 5/5または5ml HiTrap SP Sepharose HPカラムでクロマトグラフィし、得られたタンパク質をSDS-PAGEおよびWestern(ウエスタン)ブロット分析により同定。
【0055】
本発明の好ましい態様において、NおよびC末端二重トランケートメンバーの該タイプIAグループは以下のアミノ酸配列を含む。
【0056】
4反復配列tau由来の誘導体(tau 43)はR4で表示される。
配列番号1(239-333、R4)
【0057】
【化1】
配列番号2(237-333、R4)
【0058】
【化2】
配列番号3(239-318、R4)
【0059】
【化3】
【0060】
本発明の好ましい態様においてNおよびC末端二重トランケートメンバーの該タイプIBグループは以下のアミノ酸配列を含む。
配列番号4(239-326、R4)
【0061】
【化4】
配列番号5(239-328、R4)
【0062】
【化5】
配列番号6(239-331、R4)
【0063】
【化6】
配列番号7(239-334、R4)
【0064】
【化7】
配列番号8(239-340、R4)
【0065】
【化8】
配列番号9(239-343、R4)
【0066】
【化9】
【0067】
3反復配列tau(tau 44)由来の誘導体にはR3を付す。
配列番号10(208-302、R3)
【0068】
【化10】
【0069】
具体的にはNおよびC末端二重トランケート病的型のtauタンパク質中のタイプIAまたはタイプIIA メンバーに生じるtauタンパク質の1またはそれ以上のエピトープがある。
【0070】
本発明の態様において、該エピトープは具体的には、タイプIA(配列番号1〜3)およびタイプIIA(配列番号11〜18)グループメンバーの一次構造内に位置し、その数、異種性、および特異性は各個々のグループメンバーの特異的構造的コンフォメーションに依存し、それにより加えられる。したがって、各分子の単一性は微小管構築に対するその効果と共にその一次構造だけでなく、そのエピトープを形成するその二次および三次構造に基づく。それらのいくつかは該分子の活性に大きく寄与する特に重要な「コンフォメーショナル領域」を形成することができる。
【0071】
本明細書で用いている用語「コンフォメーショナル領域」はその活性に寄与する分子のある領域にクラスター化したエピトープを表す。
【0072】
特に好ましい態様において、アミノ酸「ile lys his val pro gly gly gly ser val gln ile val tyr lys pro val asp leu ser lys val thr ser lys cys gly ser leu」を含むタイプIおよびタイプII分子に含まれるコンフォメーショナル領域は残基239-267(配列番号1-9および11-14、19 R4)に対応し、残基 217-236(配列番号10.15-18、20 R3)に対応するアミノ酸「val gln ile val tyr lys pro val asp leu ser lys val thr ser lys cys gly ser leu」を含むものは配列Aを表す。
【0073】
本発明のさらに別の好ましい態様において、該コンフォメーショナル領域の該エピトープが同定され、その相対的寄与は欠失突然変異により決定される。これらエピトープすべての重要性および微小管の機能に対するその関係はタイプIA分子の活性に種々の程度に寄与することが示された突然変異型により示される(実施例5)。これら個々のエピトープは以下のアミノ酸配列を含む:
A: ile lys his val pro gly gly gly ser val gln ile val tyr lys pro val asp leu ser lys val thr ser lys cys gly ser leu(配列番号1-9および11-14、19の残基239-267に対応する)。エピトープ欠失突然変異体は配列番号21(268-333、R4; del 239-267)
【0074】
【化11】
を有する。
A1: ile lys his val pro gly gly gly ser(配列番号1-9および11-14、19の残基239-247に対応する)。欠失突然変異体は配列番号22(248-333、R4; del 239-247)
【0075】
【化12】
を有する。
A2: ile lys his val pro gly gly gly ser val gln ile val tyr lys pro val asp leu(配列番号1-9および11-14、19の残基239-257に対応する)。欠失突然変異体は配列番号23(258-333、R4; del 239-257)
【0076】
【化13】
を有する。
A3: ile lys his val pro gly gly gly ser val gln ile val tyr lys pro val asp leu ser lys val thr ser(配列番号1-9および11-14、19の残基239-262に対応する)。欠失突然変異体は配列番号24(263-333、R4; del 239-262)
【0077】
【化14】
を有する。
A4: ser val gln ile val tyr lys pro val asp leu ser lys val thr ser(配列番号1-9および11-14、19の残基246-262に対応する)。
エピトープ欠失突然変異体は配列番号25(239-333、R4; del 248-262)
【0078】
【化15】
を有する。
A5: asp leu ser lys val thr ser、配列番号1-9および11-14、19の残基256-262および残基225-231、R3 配列番号10、15-18、20に対応する。エピトープ欠失突然変異体は配列番号26(239-333、R4; del 256-262)
【0079】
【化16】
を有する。
A6: lys cys gly ser leu、配列番号1-9および11-14、19の残基263-267および配列番号10、15-18、20の残基232-236、R3に対応する。エピトープ欠失突然変異体は配列番号27(239-333、R4; del 263-267)
【0080】
【化17】
を有する。
【0081】
さらに、該ペプチドのアミノ酸すべてが必ずしも特異抗体により実際に認識される特異的部位に寄与するわけではない。
【0082】
本発明の好ましい態様において該タイプIA 病的tauタンパク質は以下の特性を有する:
a)該タンパク質はNおよびC末端がトランケートされている(実施例6)、
b)該タンパク質はアルツハイマー病の組織に存在するが、該タンパク質は正常健康脳には存在しない(実施例6)、
c)in vitro微小管構築アッセイにおいて、それらは正常tauタンパク質の微小管構築の促進を妨げる(実施例7)、
d)正常tauを用いる微小管構築アッセイにおいて、それらの阻害活性は特異的阻害的中和モノクローナル抗体により除去することができる(実施例11)、
e)その病的活性は正常健康tauに存在しないアミノ酸配列および構造的コンフォメーションの組み合わせに依存する(実施例6)、
f)該タンパク質は正常tauタンパク質とコンフォメーション的に異なるようである(実施例6)。
【0083】
最も好ましい態様において、本発明はNおよびC末端トランケート病的tau型のタイプIAグループ、配列番号1-3およびその「コンフォメーショナル領域」(配列「A」)およびエピトープA1-A6に関する。
【0084】
タイプIB tauタンパク質は以下の特性について異なる:
a)タイプIBタンパク質はNおよびC末端がトランケートされている(実施例6)、
b)該タンパク質は正常健康ヒト脳に存在するかもしれない、
c)in vitro微小管構築アッセイにおいて、それらは正常tauタンパク質の微小管構築の促進を妨げない(それぞれ実施例2および7)、
d)それらは微小管構築の修飾において病的活性を示さない(それぞれ実施例2および7)、
e)タイプIB分子は正常tauとコンフォメーション的に異なるようである(実施例6)。
【0085】
本発明の別の態様は、tauおよび微小管機能に影響を及ぼすさらなる分子の同定をもたらす記載の二重トランケート型のtauを用いる機能的アッセイと組み合わせる、それ自体の多くは当該分野で知られている種々の抽出方法を含む示したアプローチの組み合わせである。脳抽出物からのtauタンパク質の収量は特定の脳組織試料のステージに依存して抽出したNおよびC末端二重トランケートtau分子の機能性で変化するかもしれない(実施例6)。当業者は、種々の異なる目的に本発明の方法を用いる方法を知っており、これらはすべて本発明の保護の範囲内にある。
【0086】
別の好ましい態様において、本発明はプロトタイプタイプIA分子グループメンバーとして特に配列番号1に関する。
【0087】
本発明のさらに別の目的は、チューブリンを該チューブリンと該タイプIA分子の相互作用を可能にする条件下で本発明のタイプIA分子とインキュベーションする正常tauタンパク質のアルツハイマータンパク質へのin vitro変換方法を提供する。
【0088】
用語「タイプIA分子またはそのペプチド誘導体のチューブリンとの相互作用を可能にする」は、タイプIA分子の活性、好ましくは最適活性を可能にする条件を表す。この活性はチューブリンとの結合および微小管構築におけるその生理学的機能の阻害をもたらす。
【0089】
別の態様において、タイプIA分子はその誘導体により阻害または中和されるかもしれない。阻害分子をスクリーニングするために本発明において記載されているように、タイプIA ペプチドおよびその誘導体、例えば欠失または突然変異を含むペプチドの微小管重合に対するそれらの効果を試験またはスクリーニングすることができる。
【0090】
正常tauタンパク質は天然または組換え供給源由来であってよい。しかしながら、本発明の方法を実施するためには組換え物質を用いるのが好都合である。
【0091】
上記方法は、具体的には種々の目的に十分な量のタイプIA NおよびC末端二重トランケートtauタンパク質を提供し、病的二重トランケートtauタイプIAにより生じる微小管構築の阻害を抑制する新規阻害剤のin vitroスクリーニング方法を確立することができよう。
【0092】
したがって、本発明の組成物に有用な阻害剤はタイプIA分子とチューブリンとの病的相互作用を調節することができるあらゆる阻害剤である。そのような阻害分子の作用様式はタイプIAまたは正常tauとの相互作用からなる。
【0093】
これら「インヒビター(阻害剤)」は、タイプIA分子に含まれるエピトープまたはエピトープ類に対して例えば該エピトープをブロックすることにより特異的であるか、またはそれらがその病理学的または生物学的活性を抑制または阻害する限りタイプIA分子上の種々のドメインを対象にするかもしれない。阻害効果は正常tauによる残存微小管構築促進活性を測定することにより定量的に定義することができる。阻害剤の供給源には、定義した化学構造および組成物の低分子ライブラリー、ペプチドライブラリー、溶液中の遊離またはファージまたは細菌またはリボソーム(リボソーム表現)の表面上に表現された抗体ライブラリーおよび当該分野で知られた同様の技術が含まれる。
【0094】
本発明のさらなる目的は以下の工程を含むタイプIA複合体の分解に有効な分子および化合物の試験方法(タイプI in vitroアッセイ)を提供することである:
a)タイプIA分子またはその誘導ペプチドおよびチューブリンまたはタイプIA分子と相互作用する他の分子のタンパク質複合体を形成させ、
b)該タンパク質複合体と試験すべき薬剤をインキュベーションし、
c)工程(b)のインキュベーションの結果を健康tauアイソフォームの微小管構築促進能力の回復について試験する。
【0095】
本発明のさらに別の目的は、以下の工程を含む健康tauアイソフォームの正常in vitro活性の阻害の抑制または低減に有効な薬剤の試験方法を提供することである:
a)タイプIA分子またはその誘導ペプチドと組み合わせて試験すべきある薬剤は正常tauおよびそのvitro機能と干渉しないと予想される、
b)タイプIA分子と正常tauおよびチューブリンの存在下で試験すべき薬剤のインキュベーション、
c)工程a)およびb)のインキュベーションの結果をタイプIA分子の微小管重合に対する阻害活性が存在するかまたはしないかについて試験する(実施例8)。
【0096】
該薬剤の非存在下で用語「タイプIA分子またはその誘導ペプチドとチューブリンの複合体を形成させる(を可能にする)」は、微小管形成の阻害をもたらすタイプIA分子と該チューブリンの相互作用を可能にする条件を表す。
【0097】
当業者は本発明の方法を種々の異なる目的に用いる方法を知っており、これらはすべて本発明の保護の範囲内にある。
【0098】
さらなる局面において、本発明は以下の工程を含むtauまたはtau由来タンパク質を発現する細胞系におけるタイプIA分子の複合体形成開始を阻害するのに有効な薬剤の試験方法(タイプI細胞アッセイ)に関する:
a)適切な制御領域の制御下でタイプIA分子をコードする機能的遺伝子を正常tauタンパク質を発現する細胞内に導入し、
b)タイプIA tauとチューブリン分子のタンパク質複合体を形成させ、
c)該複合体を保有する細胞を試験すべき薬剤に適用し、
d)タイプIA生物活性に対する該薬剤の効果、例えば微小管の構造的および機能的修飾を試験する。
【0099】
工程(a)で用いている用語「tauタンパク質を発現する細胞」は、タイプIA分子またはその誘導体をコードする遺伝子構築物由来のNおよびC末端二重トランケートtau型を発現する能力を有する細胞を表す。当業者はタイプIA分子をコードする遺伝子導入の一連の実験工程は本発明の方法の目的には無関係であることを承知している。
【0100】
該方法は、該スクリーニング系はin vivo状況と密接なイメージを与える連続的増殖細胞系に基づくため特に好都合である。さらに、細胞内に局在するタイプIA分子の十分な供給はタイプIA分子の生物学的効果の阻害に有効な薬剤のスクリーニングを可能にする。
【0101】
好ましい態様において、該タイプIA分子発現細胞は神経芽細胞腫、または褐色細胞腫細胞またはタイプIA分子発現トランスジェニック動物由来の神経細胞の一次培養である。
【0102】
「タイプII」で示す分子のグループはNおよびC末端二重トランケートtauタンパク質分子からなる(例えば配列番号11〜20に記載の配列)。このグループの定型的なものはニューロン内およびニューロン外に局在し、正常健康tauと機能的に異なる。
【0103】
本発明のこのグループのタンパク質の発見および単離は、
(1)特異的微小管結合および微小管構築の異常な促進をもたらすtau修飾の分子的説明および特徴付け(実施例3)、およびその担体に対する病的結果(実施例4)、
(2)該タンパク質エピトープに特異的な抗体および
(3)該タイプII分子の病的活性を中和する抗体(実施例12)、および
(4)該タイプIIタンパク質を阻害、中和、および排除するのに有効な治療薬候補をスクリーニングおよび試験する方法、または
(5)tau由来タンパク質、例えばタイプII分子の形成を阻害するのに有効な治療薬候補をスクリーニングおよび試験する方法、
(6)薬剤スクリーニングに用いることができるトランス遺伝子またはトランス遺伝子混合物として各NおよびC末端二重トランケートtauタンパク質をコードする遺伝子構築物を有する動物モデルの開発、
(7)該二重トランケートtauタンパク質およびそのプロテアーゼに対する阻害剤を含む医薬組成物、
(8)該分子を認識/相互作用する診断用および治療用組成物、
(9)該二重トランケートタンパク質に基づくワクチンの開発、
(10)アルツハイマー病およびtauの病的変化に関連する他の障害のin vitroおよびin vivo診断用の該タンパク質およびそのエピトープおよび/または抗体または他の特異的プローブを含む方法。
【0104】
グループタイプIAおよびBと反対に、タイプIIA分子は分光光度法で測定したとき正常健康tauアイソフォームにより促進される微小管構築より有意に高く病的微小管構築を促進する(それぞれ実施例1および3参照)。驚くべきことに、同様の配列と分子量範囲を有するNおよびC末端二重トランケートtau分子のサブグループ(タイプIIB)はこれら「高」微小管重合能力を欠く。微小管構築アッセイにおいて、このサブグループの分子は完全長tauタンパク質にみられるレベルに達する(実施例3)。
【0105】
したがって、本発明はtauタンパク質のタイプIIAグループと呼ばれるアルツハイマー病に見られる新しいタイプの修飾tauタンパク質に関する。該グループはNおよびC末端二重トランケートtau分子(配列番号11-18)からなる。
【0106】
用語タイプII分子は正常健康tauだけでなくタイプIAおよびB tauグループとも構造および機能が有意に異なる群のメンバーに関する。このサブグループの分子は微小管を結合し、健康tauアイソフォームによる正常微小管構築より有意に顕著な病的構築を促進する(実施例3)。タイプIIA NおよびC末端二重トランケートtau分子は細胞レベルでアルツハイマー病ニューロンおよび実験条件において全生物の認知障害を最終的に生じるシナプス損失および神経機能障害をもたらす構成成分の軸索輸送と干渉する(各実施例15および16)。同時に、罹患ニューロンは種々の形のストレス、例えば酸化ストレスに弱い(実施例4)。
【0107】
本発明の好ましい態様において、該タイプIIAグループのNおよびC末端二重トランケートメンバーは以下のアミノ酸配列を含む。
4反復配列tau(tau 43)の誘導体にR4を付す。
配列番号11(69-333、R4)
【0108】
【化18】
配列番号12(93-333、R4)
【0109】
【化19】
配列番号13(69-363、R4)
【0110】
【化20】
配列番号14(93-363、R4)
【0111】
【化21】
【0112】
3反復配列tauの誘導体(tau 44)にはR3を付す。
配列番号15(93-302、R3)
【0113】
【化22】
配列番号16(69-302、R3)
【0114】
【化23】
配列番号17(93-332、R3)
【0115】
【化24】
配列番号18(69-332、R3)
【0116】
【化25】
【0117】
本発明の好ましい態様において、該タイプII BグループのNおよびC末端二重トランケートメンバーは以下のアミノ酸配列を含む。
配列番号19(6-378、R4)
【0118】
【化26】
配列番号20(6-347、R3)
【0119】
【化27】
【0120】
本発明の好ましい態様において、該タイプIIA病的tauタンパク質は以下の特性を有する:
a)該タンパク質はNおよびC末端がトランケートされている(実施例6)、
b)微小管構築の十分な病的促進物質である(実施例3; 図28C)、
c)その病的微小管構築促進活性は例えば阻害的モノクローナル抗体またはその誘導体(実施例12)のような特異的化合物により除去することができる、
d)該タンパク質は正常健康脳にはみられない(実施例6)、
e)細胞内輸送機能が有意に損なわれる(実施例16)、
f)その病的活性は、微小管ネットワークと結合する高い親和性およびその機能的障害に関連する(実施例3)、
g)それらは正常tauとコンフォメーション的に異なるようである(実施例6)。
【0121】
本発明の別の好ましい態様においてタイプIIB分子は以下の特性を有する:
a)該タンパク質はNおよびC末端がトランケートされている、
b)タイプIIAより微小管構築促進効果が低い、
c)該タンパク質は正常健康脳にはみられない、
d)タイプIIAで観察されるより程度は低いが、それとの結合により微小管機能を損なうようである、
e)それらは正常tauとコンフォメーション的に異なるようである。
【0122】
本発明の別の好ましい態様において、タイプIIAおよびB分子の該エピトープはタイプI分子について記載したのと同じように同定された。これらエピトープすべてのタイプII分子における重要性およびその微小管に対する機能との関係は、タイプIAの実施例において示されるようなNおよびC末端二重トランケートtau分子の活性に種々の程度に寄与することが知られている突然変異型により示される。
【0123】
したがって本発明の組成物に有用な阻害剤は、「病的微小管」を生じるタイプIIA分子と微小管の病的相互作用を調節することができるあらゆる阻害剤である。本明細書で用いている用語、病的(pathological)微小管は、タイプII分子により修飾された微小管を表す。そのような阻害分子の作用様式は、いずれかの微小管、tauおよびその病的誘導体を含む微小管関連分子との相互作用からなる。阻害剤の供給源として、定義した化学構造および組成の小分子のライブラリー、ペプチドライブラリー、溶液中の遊離のまたは合成表面またはファージまたは細菌またはリボソーム(リボソーム表現)上に表現された抗体ライブラリー、および当該分野で知られた同様の技術を用いることができる。
【0124】
好ましい態様において、これら「阻害剤」はタイプIIA分子に含まれるエピトープまたはエピトープ類に対して該エピトープをブロックすることにより特異的であるか、またはその病的活性または生物活性をin vitroまたはin vivoで抑制または阻害するかぎり、タイプIIA分子上の種々のドメインを対象にすることができよう。該阻害効果は、例えば正常tauによる残存微小管構築促進活性を測定するか、または細胞内微小管パラメーター、例えば成長、安定性、または細胞内輸送を測定することにより定量的に定義することができる。
【0125】
別の態様においてタイプIIA分子は例えば各細胞内に発現した優性ネガティブタンパク質としてその誘導体により阻害または中和することができる。阻害分子をスクリーニングするために本発明において説明したように、タイプIIA ペプチドおよびその誘導体、例えば欠失または突然変異を含むペプチドはNおよびC末端二重トランケートtau分子の病的効果を阻害するその効果を試験またはスクリーニングすることができる。
【0126】
治療効果は微小管構造および機能の障害を阻害することにより達成される。
【0127】
したがって、本発明の別の目的は、本発明のタイプIIA tau分子に対する特異的阻害剤を所望により医薬的に許容される担体および/または希釈剤と組み合わせて含む医薬組成物を提供することである。
【0128】
別の好ましい態様において本発明はタイプIIA群分子のプロトタイプとして特に配列番号11と関連がある。
【0129】
本発明のさらに別の目的は、病的微小管を生じる該タイプIIAまたはBと微小管の相互作用を可能にする生理学的条件下で正常tauタンパク質を本発明のタイプIIAまたはBとインキュベーションする正常微小管を病的状態にin vitro変換するための方法を提供することである。
【0130】
本発明はさらに、タイプIIA分子の病的効果を中和することができる化合物のためのあらゆる分子ライブラリーのスクリーニングを可能にするスクリーニングアッセイに関する。本試験において、分子を、タイプIIA分子を除去および/または中和し、タイプII分子により生じた生理的微小管パラメーターおよび機能を回復させる特性についてスクリーニングする。薬剤スクリーニングアッセイは以下の工程からなる:
(1)適切な条件下、タイプIIA分子およびチューブリン存在下における病的微小管の形成(それぞれ実施例3および4)、
(2)これら病的微小管と試験する候補薬剤とのインキュベーション、
(3)タイプIIA分子の微小管に対する効果の中和に対する試験(各実施例9および12)。
【0131】
病的NおよびC末端二重トランケートtauタイプIIAにより生じる微小管に対する効果を軽減する阻害剤のin vitroスクリーニング方法を確立することができよう。これら「阻害剤」は、例えばエピトープをブロックすることによりタイプIIA分子に含まれるエピトープまたはエピトープに特異的であるか、またはそれらがその活性を抑制または阻害するかぎりにおいてタイプIIA分子上の種々のドメインを対象にすることができよう。該阻害効果は微小管構築動態を測定することにより定量することができる。阻害剤の供給源として、定義された化学構造および組成の低分子ライブラリー、ペプチドライブラリー、溶液中に遊離するかまたはファージまたは細菌またはリボソーム(リボソーム表現)の合成表面の表面上に表現された抗体ライブラリーおよび当該分野で知られた同様の技術を用いることができる。
【0132】
本発明の目的では、被検薬剤は、タイプIIA分子の量および/またはその活性を減少させるのに有効であり、補助的治療効果を満たすのに十分であるが、タイプIIA活性を完全に除去することが好ましい。
【0133】
当業者は種々の異なる目的に本発明の方法を用いる方法を知っており、これらはすべて本発明の保護の範囲内にある。
【0134】
本発明のさらなる目的は、生細胞、すなわちタイプII分子発現ニューロンまたはニューロン様細胞を用いる薬剤の確認方法を提供することである(タイプII細胞アッセイ)。あるいはまた、トランスジェニック動物由来の一次神経細胞培養またはタイプIIA分子を発現する種々の供給源由来の他の一次神経細胞を用いることができる。
【0135】
上記で用いている用語「タイプII分子発現ニューロン(神経細胞)」は、該分子を安定に発現するかまたはタイプIIA分子を発現する能力があり、機能的タイプIIA遺伝子が以下に例示する細胞培養技術またはトランスジェネシス(transgenesis)により導入された細胞を表す。
【0136】
好ましい態様において、該タイプIIA分子発現細胞は神経芽細胞腫または褐色細胞腫細胞またはタイプIIA分子を発現するトランスジェニック動物由来の神経細胞の一次培養である。
【0137】
当業者はタイプIIA分子をコードする遺伝子の導入手順は本発明の方法の目的とは無関係であることを認識している。
【0138】
本発明は、以下の工程を含むタイプIIA分子発現細胞系における異常微小管形成および機能の促進においてタイプIIAを阻害するのに有効な薬剤を試験するための方法に関する:
a)タイプII分子をコードする機能的遺伝子を適切な制御領域の制御下で正常tauタンパク質発現細胞に導入し、
b)タイプIIA tauおよび微小管(病的微小管)の複合体を形成させ、
c)得られた複合体を保有する細胞に被検薬剤を適用し、
d)タイプIIA生物活性、例えば微小管ネットワークおよびその関連機能の修飾に対する薬剤の効果を試験する。
【0139】
本発明のさらに別の最も好ましい態様はタイプIIA分子発現ニューロンの表現型である。適切な条件下でこれら分子を発現するニューロンは、細胞内輸送プロセスの摂動(perturbance)を生じる。さらにタイプIIA分子発現ニューロンは適切なストレス条件下で細胞死する(実施例4)。
【0140】
該方法は、in vivo状況の密接なイメージをもたらす連続増殖細胞系の使用に基づく関連システム(系)が細胞内に局在するタイプIIA分子の十分な供給をもたらし、細胞内タイプIIA効果の軽減に有効な化合物の薬剤スクリーニングを可能にするので特に好都合である。
【0141】
好ましい態様において、この細胞アッセイの計測値を低または高処理能力定量系に適応する。微小管輸送の崩壊または障害および/または神経細胞死をもたらす記載の表現型に関連する用語「適切な条件」は、実施例に示す該表現型を出現させるあらゆる条件を表す。
【0142】
本発明の目的において、このシステムによりスクリーニングするかまたはこのシステムで確認した潜在的薬剤または第三起源の薬剤は該表現型の規模の減少に有効であり、治療における補助的機能を満たすのに十分であるが、該薬剤による病的表現型の完全除去または減少が好ましい。
【0143】
細胞系または一次細胞の安定な増殖に加え、各発明はそのニューロンにタイプIIAまたはB分子を発現する全動物由来の細胞を用いる類似計測システムに広げることもできる(該トランスジェニック動物モデルを以下に例示する)。
【0144】
当業者は種々の異なる目的に本発明の方法を用いる方法を知っており、これらはすべて本発明の保護の範囲内にある。
【0145】
好ましい態様において、NおよびC末端二重トランケートタイプIIA tau型を安定に発現する該細胞およびトランスジェニック動物はアルツハイマー病の病因に関連する新規分子、その診断および治療をもたらす正しい情報を得る疾患経路のマッピングを可能にする。これらスクリーニングおよび同定方法はmRNA発現ベースのスクリーニング技術およびタンパク質ベースの技術を含む。
【0146】
好ましい態様において、該タイプIおよびタイプIIAおよびB分子またはその誘導体は、上記の病的NおよびC末端二重トランケート型のタイプIA、タイプIIAおよびBを保持および発現するトランスジェニック動物モデルを提供するための非ヒト動物のゲノム中に導入することができる組換えDNA構築物も提供する。本発明のトランスジェニック動物は、構築物が直接導入されている動物および構築物を発現する能力を保持する該動物の子孫を含む。該トランス遺伝子配列は遍在的に発現するかまたは組織特異的プロモーターと機能的に連結したポリヌクレオチド配列である。タイプIAおよびタイプIIAおよびB分子をコードするトランス遺伝子DNAは優先的に動物またはヒト供給源由来のcDNAおよび/またはゲノムDNAである。該タイプIおよびタイプIIAおよびB分子を発現するトランスジェニック動物はアルツハイマー病に表現型的に関連する細胞および/または臓器レベルでの機能的変化をもたらすと予想される。これらにはADに特徴的な組織学的変化、RNA発現変化、細胞生理学的パラメーターの変化、および好ましくは行動的変化が含まれる。トランスジェニック動物の成熟ニューロンにおけるタイプIおよびタイプIIAおよびB分子の発現は先に試験されている。タイプI、タイプIIAおよびBトランス遺伝子がトランスジェニック動物で発現するレベル(すなわち、トランス遺伝子mRNAのレベル)はトランスジェニック動物における一貫した病態生理学的欠陥を得るための重要なパラメーターであると予想される。トランス遺伝子を保持する動物の繁殖および異系交配(交雑)により、例えば疾患モデルとして現在用いられる動物種、他のトランス遺伝子を発現する動物、または遺伝子の機能的発現を欠く動物にトランス遺伝子を導入することにより、病的特徴を増強、軽減、または調節することができる。(実施例14参照)。
【0147】
より具体的には、本発明はヒトタイプIおよびタイプIIAおよびB分子をコードするDNAが適切な遍在性または組織特異的プロモーターのその調節可能な修飾を含む転写制御下で発現する、トランスジェニック非ヒト動物細胞を提供する。
【0148】
本発明に従って操作される細胞はあらゆる知られたトランスフェクション技術により製造することができよう。該DNA配列は直接的遺伝子操作によるかまたは細胞のより初期の世代内に導入することができよう。すなわち、該細胞をトランスジェニック動物から得、in vitroで培養することができよう。また、該トランスジェニック動物は知られた確立された方法、例えば胚性幹細胞への遺伝子伝達、初期胚のレトロウイルス感染、または前核性マイクロインジェクションによる胚操作に従って製造することができよう。前核性マイクロインジェクション技術が好ましい。本発明の組換えDNA構築物から得られる転写単位を動物胚の前核に注射し、得られたファウンダー(founder)トランスジェニックを繁殖させる。子孫において得られた結果を当該分野でよく知られた種々の技術を用いて分析することができる。tauおよびNおよびC末端二重トランケートtau由来分子、およびアルツハイマー病に関連する他の分子、例えばAPPおよびその誘導体を分析することができる本発明の細胞および動物に基づくモデルを、例えば神経変性性疾患における潜在的治療剤の効果を確認および評価するために用いることができよう。特に該モデルを本明細書に記載のNおよびC末端二重トランケートtau由来分子の病的効果を抑制すると思われる薬剤を検出するためのスクリーニングまたは特徴付けアッセイに用いることができよう。
【0149】
したがって、さらなる局面において本発明は、該二重トランケート型のtauを発現するトランスジェニック動物由来の細胞を標的細胞に用いる、特定の病状、特に神経変性性疾患、好ましくはADのための潜在的治療剤を試験する方法を含む。より具体的には、本発明は、例えば抗体またはその誘導体のような治療剤を本発明のトランスジェニック動物に投与するか、または交雑または遺伝子操作により導入し、さらに上記アッセイ系により試験するような方法を含む。さらに本発明は、そのような方法の中にあるかまたはそれを含むスクリーニングまたは特徴付けアッセイおよび本発明の細胞を含むスクリーニングアッセイキットを含む。本発明のタイプIおよびタイプIIAおよびB分子を発現する細胞系を用いる潜在的治療剤のスクリーニング方法を本発明において提供する(実施例15参照)。本発明の細胞および動物は同様に用いることができよう。
【0150】
本発明の別の目的はNおよびC末端二重トランケート型のtauタンパク質の特異的阻害剤を所望により医薬的に許容される担体および/または希釈剤と組み合わせて含む医薬組成物を提供する。
【0151】
用語「NおよびC末端二重トランケートtauの特異的阻害剤」は、具体的には該二重トランケートtauタンパク質の作用を阻害する物質を表す。阻害剤の性質は抗体、その操作された誘導化分子、本発明の試験系において望ましい阻害活性を示すあらゆるペプチドまたは合成化学組成物でありうる。
【0152】
本発明の別の目的は、具体的には本発明のエピトープを認識し、NおよびC末端二重トランケートtau分子の病的活性を部分的または完全に阻害することができる抗体またはその誘導体である。
【0153】
用語「本発明のエピトープまたはエピトープ類を含むオリゴまたはポリペプチド」は、その2または3次元構造に本発明のエピトープを再構成する、具体的にはそれに対する抗体により認識されるペプチドを表す。さらに、該オリゴまたはポリペプチドは該エピトープを表すアミノ酸のみからなるか、またはさらなるアミノ酸を含むことができよう。そのようなオリゴまたはポリペプチドの構築は当該分野でよく知られている。
【0154】
好ましい態様において、本発明は天然または組換えの、固定化した、溶液中に遊離した、または種々の分子または細菌、ウイルスの表面または他の表面上に表現されたモノクローナル抗体およびその誘導体に関する。該抗体およびその誘導体はNおよびC末端二重トランケートtau分子の生物活性を部分的または完全に阻害することができる。そのような特異抗体活性はアルツハイマー病の脳組織から単離した該二重トランケートtau分子に対するモノクローナル抗体DC44を用いて示された(それぞれ実施例10および11)。
【0155】
該抗体は他の多くの変異体(DC82、DC136など)を有し、血清由来またはモノクローナル抗体またはそのあらゆる誘導体であってよい。所望のエピトープに対するモノクローナルおよびポリクローナル抗体の生成は当該分野でよく知られている(43)。さらに、該抗体は天然または遺伝子操作により誘導された抗体、例えば当該分野でよく理解されている技術により誘導されたキメラ抗体であってよい。さらに該抗体は、特定のエピトープと結合する能力を保持する抗体断片、例えばFab断片または一本鎖Fvミニボディ(minibody)、またはイントラボティ(intrabody)と呼ばれる細胞内に発現した一本鎖抗体も表す。
【0156】
最も好ましい態様において本発明はアルツハイマー病の治療に用いる医薬組成物に関する。
【0157】
さらに該医薬組成物はその症例を扱う医師が適切とみなす経路および用量でそれを必要とする患者に投与することができよう。
【0158】
本発明の別の好ましい態様において、該医薬組成物は、特異的阻害剤として部分的または完全なその変化および/または中和をもたらす上記エピトープのあらゆる部分またはグループと結合する少なくとも1のモノクローナル抗体または低分子またはその誘導体を含む(各実施例10、11および12参照)。
【0159】
本発明の別の目的はa)本発明のエピトープ、b)本発明の抗体またはその誘導化分子を含むアルツハイマー病を検出および/またはモニターするための診断用組成物を提供することである。
【0160】
本発明の診断用組成物は、例えば具体的にはタイプIAまたはタイプIIグループの分子のあるメンバーまたはそのエピトープまたは試験する試料中の増大したレベルのタイプIAまたはタイプIIA分子を認識する本発明の抗体を含むことができよう。別の態様において、該診断用組成物は本発明のエピトープの1つに対する本発明の抗体を含むことができよう。すなわち、アルツハイマー病状に関連する試料を本発明のエピトープを認識する抗体で該試料を処理することにより検出することができよう。抗体-エピトープ(ハプテン)複合体を本発明の抗体に対する第二抗体を用い、当該分野で知られた方法(43)に従って標識することにより可視化することができよう。
【0161】
本発明のさらに別の態様において、該診断用組成物は本発明のエピトープおよび本発明の抗体からなってよい。試料の該抗体による処置は、該抗体の該試料に対する結合が該抗体の標準試料として用いる本発明の該エピトープに対する結合に関連して生じるならば対応患者の病状に関する結論をもたらすことができよう。
【0162】
さらに別の態様において、該診断用組成物はタイプIAまたはタイプIIA分子および本発明の抗体を含むことができよう。両タイプの分子の活性は、試料の正常tau中和能について本発明の組換えタイプIA分子(例えば配列番号1)およびIIA分子(配列番号11-18)と比較してモニターすることができよう。タイプI分子の定量した異常活性から、該試料に含まれる分子のレベル、すなわち患者の病状を推定することができよう。タイプIA活性は、例えばタイプI分子と反応しないで残った正常tauの残存活性を測定することにより推定することができよう。タイプII活性は微小管構築アッセイにおけるタイプII分子のさらなる活性を測定することにより推定することができよう。
【0163】
当業者は本発明の上記のあらゆる対象を組み合わせる他の試験系を設計する立場にある。すべての考えられる組み合わせが本発明の保護の範囲内にあることを理解すべきである。
【0164】
本発明の別の目的は、タイプIAおよびタイプIIA分子のNおよびC末端二重トランケートtau分子またはそのエピトープの存在およびその正常tau阻害活性レベルについて患者の脳脊髄液分離物をアッセイし、神経組織の生検を実施することを含むアルツハイマー病のin vitro診断および/またはモニター方法を提供することである。
【0165】
患者の脳脊髄液分離物は標準的医学的方法により得られる。
【0166】
さらなる態様において、本発明はタイプIAおよびタイプIIAのそれぞれ分子および動物に免疫応答を誘導することができるその誘導化免疫原性断片のアミノ酸配列と同一またはホモローガスであるタイプIおよびタイプII分子に関する。本発明によれば、タイプIおよびタイプII分子はともに(a)阻害抗体を産生するための免疫原および該疾患に対する免疫に用いるワクチンの中心部分として用いることができることがわかった。
【0167】
非経口的適用において、上記のすべての配列およびエピトープおよび罹患脳組織から単離したタイプIおよびIIは免疫原性であり、具体的には該タイプIおよびIIタンパク質およびその誘導体の対する抗体の産生をもたらす(各実施例10および13)。
【0168】
最も好ましい態様において、タイプIおよびII分子またはその誘導体は該分子の一次、二次、および/または三次構造に対する免疫応答を誘導することができる。宿主において、得られた免疫応答はしたがってtauおよびその誘導体の健康および病的型のtauを区別することができる。本発明のこの特徴は、疾患特異的免疫応答の誘導におけるこれらNおよびC末端二重トランケートtau型の優れた品質を重視するワクチンとして用いることができる。
【0169】
本明細書に含まれる病的NおよびC末端二重トランケートtauポリペプチドについて天然のバリエーション(変化)がアルツハイマー病の個々の例で存在することが理解される。これらバリエーションは全体配列中のアミノ酸の相違中に、すなわち該配列のアミノ酸の欠失、置換、挿入、反転(inversion)、または付加により存在する。本発明の例示的態様のそのようなアミノ酸置換は本発明の範囲内にある。すなわち、ポリペプチドの免疫原性に本質的に影響しない天然の変化には本発明の該二重トランケート型のtauポリペプチドの免疫学的に等価なバリエーションが考えられる。
【0170】
しかしながら、タイプIAおよびIIA NおよびC末端二重トランケートtauポリペプチドを例えばワクチネーション目的または抗体産生に用いるときには本発明に記載の全ポリペプチドを用いる必要はない。その完全ポリペプチドに対する免疫応答を誘導することができるこれらポリペプチドの断片、ゆわゆる免疫原性断片を用いることも可能である。
【0171】
したがって、本発明のこの態様は本発明のポリペプチドのみならず依然として該ポリペプチドに対する免疫応答を誘導することができるそれらポリペプチドの誘導化断片(いわゆる免疫原性断片)に関する。
【0172】
例を示すために、組換えタイプIAおよびIIAペプチドまたは病的ヒトアルツハイマー脳由来のタイプIAおよびIIA NおよびC末端二重トランケート病的tau分画の動物における免疫原性を示す(実施例3)。
本発明を以下の実施例および図面によりさらに説明するが、それらに限定されるものではない。
【0173】
図1:NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築。
【0174】
図2:NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築の阻害。
【0175】
図3: 微小管構築におけるNおよびC末端二重トランケートtauタイプIIAおよびIIB分子の活性。
【0176】
図4:神経細胞に発現したタイプIIA NおよびC末端二重トランケートtauは酸化ストレスに対する感受性を有意に増加させる。
【0177】
図5: 病的tauタイプIAタンパク質およびその欠失突然変異体に対するモノクローナル抗体の親和性。病的tauタイプIAタンパク質およびその欠失突然変異体に対するモノクローナル抗体の見かけの親和性。最初のカラムに「プロトタイプ」tauタイプIAタンパク質(配列番号1)および各欠失突然変異体を示す。中段のカラムに本発明のエピトープを示す。最後のカラムに示す見かけの親和性を競合的ELISAにより測定し、プロトタイプtauタイプIAタンパク質と50%競合するのに必要な対応抗原の濃度で表した。
【0178】
図6: AD-脳由来tauタンパク質のSuperdex 200-カラムによる分画。
【0179】
図7:AD脳からの3つの別個の単離物由来の分画No.19中のタイプIA阻害活性。
【0180】
図8:AD脳中のNおよびC末端二重トランケートtauタイプI分子の証明。
【0181】
図9:健康脳に存在せずAD脳に存在するtauタイプI。
【0182】
図10:NおよびC末端二重トランケートtauタイプII分子の免疫反応性。
【0183】
図11:組換えtauタイプI-II(配列番号1-24)の構築。
【0184】
図12:AD-脳由来および組換えtauタイプIAの正常健康tauに対する阻害効果。
【0185】
図13:tauタイプIA分子を中和する薬剤候補の第一回スクリーニング(工程1)。
【0186】
図14:正常tauに対する選択性を有するタイプIA分子を中和する薬剤候補の第二回スクリーニング(工程2)。
【0187】
図15:tauタイプIIA分子を中和する薬剤候補の第一回スクリーニング。
【0188】
図16:tauタイプIIA分子を中和し、それを正常tauから識別することができる薬剤候補の第二回スクリーニング(工程2)。
【0189】
図17:ELISAにより測定したプレフュージョン(prefused)マウス血清中の特異抗体レベル。
【0190】
図18:モノクローナル抗体とAD脳由来tau(分画#19)およびコントロール健康脳由来tauとのELISA反応性(DC20:無関係な特異性を有するモノクローナル抗体。示したデータは3回の並列実験の平均値を表す。)。
【0191】
図19:モノクローナル抗体の組換えtau分子とのELISA反応性(DC20:無関係な特異性を有するモノクローナル抗体。示したデータは3回の並列実験の平均値を表す。)。
【0192】
図20:AD-脳由来tauタイプIA(分画#19)に対する中和抗体のスクリーニング。
【0193】
図21:組換えtauタイプIA(配列番号1)に対する中和抗体のスクリーニング。
【0194】
図22:tauタイプIA分子を中和し、それらを健康tauから識別することができる薬剤候補のスクリーニング。
【0195】
図23:組換えtauタイプIIA(配列番号12)の病的活性のモノクローナル抗体による中和。
【0196】
図24:ELISAで検出した組換えtauタイプIIA(配列番号12)に対する抗体のレベル。
【0197】
図25:トランスジェニック動物の遺伝子型別。パネルAはトランスジェニック動物の親世代の遺伝子型別を示す。レーン1、2、3および4のゲノムDNA由来のDNAの二重トランケート配列の特異的増幅は、フォスターマザーの子孫の尾から抽出したゲノムDNAにおける特異的トランス遺伝子の存在を示す。これら動物は二重トランケートタイプIIA tau分子保有トランスジェニック動物の親世代を示す。この実施例において、陽性(+C)および陰性(-C)および2つのさらなる陰性試料(5、6)を示す(M=サイズマーカー)。矢印は、トランス遺伝子陽性動物において予想される予想PCR生成物サイズを示す。
パネルB. F1世代由来の動物の遺伝子型別。ゲノムDNAを尾の先端から抽出し、二重トランケートtau特異的DNA配列を同定し、レーン1に示す。レーン2および3は陰性コントロールを示す。F1世代におけるtau特異的DNA断片の同定はこれらトランス遺伝子の遺伝性(inheritability)を確認する。
【0198】
図26:トランスジェニック動物のF1世代における二重トランケートヒトtau転写物の遺伝子発現。RNAをトランスジェニック動物の瞬間凍結組織から抽出し、逆転写、次いでcDNAの特異的増幅にかけた。例はレーン番号1および2にトランス遺伝子発現動物を示す。レーン3〜5は非発現コントロールを示し、レーン5はこの方法を用いた時に非トランスジェニック動物に典型的に生じる非特異的シグナルを示す。この実施例は、実験動物においてトランス遺伝子から発現した二重トランケートtau特異的mRNAの存在を示す。
【0199】
図27:6日間in vitro分化(differentiation)後のタイプIIA分子の過剰発現により生じた細胞死。二重トランケートtauタイプIIA(タイプIIA)でトランスフェクションしたSY5Y細胞および非トランスフェクションコントロールニューロン様細胞(mock)の細胞生存性の比較。
【0200】
図28: A:タイプIIA分子の微小管に対する結合親和性の増加はタイプIIA二重トランケート分子および完全長tauを発現する安定なトランスフェクション細胞由来のtauの細胞分画を用いて示される。遊離tau(FT)、微小管結合tau(MT)および核関連tau(NAT)の単離は記載のごとく行った。B:有機化合物F123による各tauタイプIAおよびIIAの阻害、微小管重合アッセイ。C:電子顕微鏡分析により示される微小管重合と吸着(absorption)測定値の直接比較。
【0201】
図29:MitoFluorで染色した対数増殖期のSH-SY5Y細胞。細胞体およびプロセス中のミトコンドリアの標準的分布。
【0202】
図30:MitoFluorで染色した対数増殖期のtauタイプIIA分子発現SH-SY5Y細胞。細胞の中心体領域周辺のグリーン標識ミトコンドリアの核周囲クラスタリング。
【実施例】
【0203】
実施例1:NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築
健康tauの生理学的機能は微小管(MT)の安定化である。この機能は微小管構築アッセイ(MAA)により測定することができる。本実施例において、MAA反応は3タイプのtau分子:正常健康ヒトtau、組換え型のtauタイプIA(配列番号1)およびtauタイプIB(配列番号4)を用いて行った。正常ヒトtau、tauタイプIAおよびタイプIBを別個の反応で個々にアッセイした。tau 0.1mg/mlの単一調製物を最終濃度1mg/mlの精製ブタ脳チューブリンおよび1mM GTPと混合した(すべての物質に重合緩衝液(100mM PIPES、pH 6.9、1mM MgSO4、2mM EGTA)使用)。最後にTauを加えMT構築の促進を開始した。静かにそして急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、サーモスタットで制御した分光光度計(Beckman Coulter DU640)中で37℃に平衡化した。濁度を340nmで10秒間隔で20分間連続的にモニターした。上端の曲線1(図1)は正常健康tauの微小管構築促進能を示す。それに対し、タイプIA(曲線2)およびタイプIB(曲線3)はいずれも正常tauのこの活性を示さず、MAAにおけるいかなるMT構築促進も示さなかった。
実施例2: NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築の阻害。
【0204】
MT構築アッセイ(MAA)するとtauタイプIAおよびIB分子はいずれも lack機能的活性を欠く。驚くべきことに、tauタイプIA分子は微小管構築におけるチューブリンに対する阻害効果を示す。これに対し、タイプIBタンパク質は(同様の一次構造にも関わらず)MAAにおいてチューブリンの機能的活性を阻害しない。微小管構築を阻害するため、組換え型のtauタイプIA(配列番号1)およびタイプIB(配列番号4)を用いた。構築阻害反応はタイプIAおよびタイプIBタンパク質を用いて別個に行った。ヒトチューブリン(2mg/ml)をタイプIA分子(0.2mg/ml)またはタイプIB分子(0.2mg/ml)と混合した。混合物を静かに振盪させながら37℃で1時間インキュベーションした。氷上に保った混合物に正常ヒトtau(O.1mg/ml)およびGTP(混合物中のチューブリンの最終濃度は1mg/mlおよびGTPは1mMである)を重合緩衝液(1OOmM PIPES、pH 6.9、1mM MgSO4、2mM EGTA)を用いて加えた。静かにそして急速に撹拌した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節された分光光度計(Beckman Coulter DU640)中で37℃に平衡化した。濁度変化を340nmで5分間にわたり10秒間隔で測定した。上端の曲線1(図2)は正常ヒトtauのみがチューブリン重合を完全に誘導することができることを示す。チューブリンとタイプIAをプレインキュベーションすると微小管構築はみられなかった(図2、下の曲線2)。反対に、チューブリンをタイプIBとインキュベーションすると、分子量がタイプIAと同範囲であるにも関わらず正常tauの微小管構築能力を阻害しない(図2、曲線3)。
表
NおよびC末端の二重トランケートtau分子の微小管重合に対する影響
【0205】
【表1】
最短R4アイソフォームに従った配列番号11-14;19番号付け。
最短R3アイソフォームに従った配列番号15-18;20番号付け。
【0206】
【表2】
最短R4アイソフォームに従った配列番号11-14;19番号付け。
最短R3アイソフォームに従った配列番号15-18;20番号付け。
実施例3:微小管構築におけるNおよびC末端二重トランケートtauタイプIIAおよびIIB分子の活性
【0207】
グループIA分子と対照的に、タイプIIA 二重トランケートtau誘導体は驚くべきことに病的微小管構築を促進することがわかった(図3および図28C参照)。微小管構築反応を3タイプの分子:天然健康ヒトtauアイソフォーム、Alz. tauタイプIIA(配列番号12)およびtauタイプIIB(配列番号19)を用いて行った。3つの別の反応をそれぞれ各tau(健康tau、組換えtauタイプIIAまたはタイプIIB)を単独調製することにより行った。個々のtau調製物0.1mg/mlをチューブリンおよびGTPと混合した(チューブリンの最終濃度は1mg/mlおよびGTPは1mMであった)(すべての試薬は重合緩衝液(100mM PIPES、pH 6.9、1mM MgSO4、2mM EGTA)中)。静かにそして急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計(Beckman Coulter DU640)中で37℃に平衡化した。濁度変化を340nmで5分間にわたり10秒間隔で測定した。この実験において、組換えtauタイプIIAは、健康tauによる生理的微小管構築(図3、曲線2)と比べて極めて高い(3倍)病的微小管構築の促進を示した(図3、上端の曲線1)。これに対し、タイプIIB分子はNおよびC末端二重トランケートであるにも関わらずタイプIIAと同様にMAAを行うことができず、微小管構築を健康tauでみられるレベルしか促進することができない(図3、曲線3)。
実施例4: 病的tauタイプIIタンパク質によるストレス保護メカニズムの阻害をin vitroで種々の酸化ストレスに曝露した該分子を発現する神経芽細胞腫細胞において示された。
【0208】
本実施例ではスーパーオキシドアニオンおよび一酸化窒素を生成し、これが反応して過酸化亜硝酸を形成する3-モルホリノシドノニミン(SIN-1)の酸化分解を用いる。この非常に反応性のラジカルはさらに主として細胞膜系を酸化することができる。当業者は酸化ストレスの別の供給源を培養中の神経芽細胞腫細胞に適用し、SIN-1を用いることによりここに記載したのと同様の効果を得ることができるだろう。
【0209】
脆弱性の効果を以下のごとく試験した。
【0210】
1.SIN-1を種々の濃度(0-3.32mM)でそれぞれtauタイプIIAタンパク質、配列番号15および配列番号11を発現するヒト神経細胞系に適用した。
【0211】
2.細胞生存を50%細胞生存(EC50)に対する過酸化水素の有効濃度を決定する3(4,5-ジメチルチアゾール-2-イル)-2,5-ジフェニルテトラゾリウムブロミド(MTT)還元アッセイにより測定した。当業者は本発明に記載の効果を測定するために細胞生存を評価する異なる方法を知っている。
【0212】
3. EC50値を神経芽細胞腫細胞系について病的tauタイプIIAタンパク質発現の存在下および非存在下で比較し、EC50値の統計的有意差をt-検定により評価した。
【0213】
細胞を10% FCS、2mM L-グルタミン、1% NEAA、50U/L ゲンタマイシン含有MEM/F12中で増殖させた。3-モルホリノシドノニミン(SIN-1)を無血清培地中の1Mストック溶液から希釈した(例えば47.5mgを230mlに)。MTTストック溶液(2.6mg/ml)をMEM/F12 w/o血清を用いて調製し、ろ過滅菌した。
【0214】
細胞を当該分野でよく知られた方法で培養した。96ウェルプレートに2x104細胞/ウェルを播いた。プレートの半分にtauタイプII分子発現細胞を播き、プレートの他の半分に非発現細胞を播いた。培地を36〜48時間ごとに交換した。5日後、SIN-1を0〜3.3 mMの範囲の濃度で加え、プレートを24時間インキュベーションした。各濃度をヘキサプリケートでアッセイした。SIN-1とインキュベーション後、MTTストック溶液を加えて最終濃度200mg/mlとし、プレートをもう1時間インキュベーションした。培地を捨て、プレート表面をペーパーウールで乾燥させた。50ml/ウェルのDMSOを加え、プレートを室温で一夜インキュベーションした。690nmでバックグラウンドを補正し、540nmの吸光度をELISAリーダーで測定し、バックグラウンドを引いた値を当該分野でよく知られているEC50計算に用いた。
【0215】
タイプIIAタンパク質発現および該タンパク質非発現神経芽細胞腫細胞間のlog EC50濃度の差の有意性をt-検定を用いて試験し、P値はタイプIIA病的tauタンパク質両方でP<0.001であった。tauタンパク質 配列番号12および配列番号18の発現は神経芽細胞腫細胞の酸化ストレスに対する抵抗性を50%低下させた。
【0216】
記載した実施例の効果(図4)は病的型のtauタンパク質の病的効果の説明に寄与する。
【0217】
当業者は本発明の上記のあらゆる目的を組み合わせる他の試験系を設計する立場にある。すべての考えられる組み合わせは本発明の範囲内にあると理解すべきである。
【0218】
図4のチャートはtauタイプIIA存在下の神経細胞の酸化ストレスに対する相対的抵抗性の低下を示す。該タンパク質を保有していない細胞(コントロール)の抵抗性を100%(左のバー)で表し、病的tauタンパク質を発現する神経細胞の抵抗性をコントロール値の%で示す(中および右のバー)。抵抗性は、細胞の50%が死ぬ培養液中のSIN-1により生じた遊離ラジカルの濃度で定義される。該結果はそれぞれ二重トランケートtauタンパク質タイプIIA 配列番号12(93-333、R4)および配列番号18(69-332、R3)の測定値を表す。
実施例5:二重トランケートtauタイプIAのコンフォメーションに対する個々のエピトープの寄与
【0219】
tauタイプIAの「コンフォメーショナル領域」(断片A)またはその部分の重要性を全コンフォメーション領域(断片A)またはエピトープと呼ばれるA1-A6で示されるその個々の部分の連続的欠失により検討した。タイプIA分子のコンフォメーションはその機能と強い関連があるため、「断片A」の全体のコンフォメーションに対する各エピトープ(A1-A6)の寄与をtauモノクローナル抗体を用いその反応性に基づいて測定した(図5)。
【0220】
プロトタイプtauタイプIA(配列番号1)は10nMの親和性を有する。それぞれエピトープA1、A2、およびA5が欠失した個々の欠失突然変異体 配列番号22、23、26は、これら領域の寄与が親和性の2〜4倍の低下(20-40nM)を反映することを示したが、配列番号24、25、および27中のエピトープA3、A4、A6の欠失はそれぞれ親和性の10〜30倍の損失(100〜300nM)に寄与した。全断片Aの欠失後にのみ(突然変異体 配列番号21)、該親和性はプロトタイプtauタイプIAの親和性を3桁劇的に低下させる。
実施例6:NおよびC末端二重トランケートtauタイプIおよびタイプIIの単離
【0221】
アルツハイマー脳由来tauタイプIおよびタイプII分子の製造:神経病理学的に確認されたアルツハイマー病症例由来の病的ヒト脳組織を二重トランケートtau IA、BおよびIIAタンパク質を単離するための供給源に用いた。アルツハイマー脳由来tauの調製はTRIS緩衝液中の組織のホモゲナイゼーションおよび飽和硫酸アンモニウム沈殿による溶解物の分画化の組み合わせに基づく。組織を冷20mM TRIS pH8、0.32mMショ糖、10mM b-メルカプトエタノール、5mM EGTA、10mM EDTA、5mM MgSO4、1mMフェニルメチルスルホニルフロリド、50mMフッ化ナトリウム、5mMベンズアミジン、5μg/mlロイペプチン、1.5μg/mlペプスタチン、2μg/mlアプロチニン中でHeidolph DIAX 900ホモゲナイザーを用いて4℃で10分間ホモゲナイズした。ホモゲネートを4℃で27000g、30分間遠心して細胞デブリスを除去した。tauタンパク質を44.12%(v/v)の飽和硫酸アンモニウムを加えて脳組織上清から沈殿させた。25℃で20分間インキュベーションおよび静かに撹拌した後、試料を25℃で20000g、10分間遠心した。ペレットを500μlの100mM PIPES pH6.9、2mM EGTA、1mM MgSO4に再浮遊させ、ついで同じ緩衝液に対して透析した。この調製物をSuperdex 200-カラム(Amersham-Pharmacia-Biotech)でゲルろ過して分画化し、該分画をSDS-PAGE(勾配5-20%ポリアクリルアミド)により分割し、tauタンパク質を抗tau抗体DC25を用い標準的手順に従ってイムノブロッティングにより検出した(図6)。微小管構築に対する個々の分画の効果を試験した。
【0222】
tauタイプIAおよびIBの単離:分画#19(図7)を、二重トランケートタイプIAおよびIB分子の典型的な分子量(12kDa)に対応するtau分子を含み-この分画は最高の阻害能を示した。この分画を3つの抗tau抗体(DC25はトランケートおよび完全長タンパク質を共に認識し、DC39(完全C末端特異的)およびAlz50(完全N末端に特異的))を用いるWestern(ウエスタン)ブロット分析により特徴づけた(図8)。これら抗体の免疫応答性はAD脳由来分画にのみNおよびC末端二重トランケートタイプIタンパク質がないことを証明した。正常健康脳から同じ方法により調製した対応する分画は、阻害活性も特異的免疫応答性も示さなかった(図9)。tauタンパク質の濃度をサンドイッチRIAにより測定した。総タンパク質濃度はBradfordアッセイを用いて測定した。tau調製物は使用するまで-20℃に保存した。
【0223】
tauタイプIIAの単離:分子量30kDaに対応するtau分子を含む分画#15(図6)は二重トランケートタイプIIA分子の典型である。分画#15は異常に高い微小管構築促進活性を示した。この分画を3つの抗tau抗体(DC25はトランケートおよび完全長タンパク質を共に認識し、DC39(完全C末端特異的)およびAlz50(完全N末端に特異的))を用いるWesternブロット分析により特徴づけた(図10)。これら抗体の免疫応答性は、AD脳由来の分画にのみNおよびC末端二重トランケートタイプIIタンパク質が存在することを示した。tauタンパク質の濃度をサンドイッチRIAにより測定した。総タンパク質濃度をBradfordアッセイを用いて測定した。
【0224】
組換えtauタイプIおよびタイプIIタンパク質のクローニング、発現、および精製:組換えトランケートtauタンパク質に対する遺伝子はアイソフォームtau43およびtau44に対するヒトcDNAから誘導された。cDNA挿入物をNdeI-EcoRI制限部位を用いるpET17b(Novagen)ベクターを用いてクローンした(図11)(Studierら、Meth. Enzym.185(1990)、60-89)。
【0225】
組換えNおよびC末端二重トランケートtau分子(配列番号1-24)をcDNA由来の関連領域のPCR増幅により調製した。翻訳開始を誘導する特定のプライマー、開始(ATG)、終止(TGA)コドン、およびNdeI、EcoRI制限部位を用いた。
【0226】
tauDNA中にA4-A6エピトープ(配列番号25-27)の欠失を有するプラスミドを図11(下側パネル)に示すように逆PCRにより調製した。
実施例7:微小管構築アッセイにおける正常健康tauに対するAD-脳由来および組換えtauタイプIAの阻害効果
【0227】
AD-脳抽出物およびtauタイプIAの組換え分子は天然健康tauアイソフォームを用いる微小管構築の促進を阻害することができる。これらの実験では健康ヒトtauを年齢対応コントロールの脳から単離し、tauタイプIAをAD患者の脳から単離した(実施例6、図6、分画#19参照)。組換えtauタイプIA(配列番号1)およびタイプIB(配列番号4、陰性コントロール)を生成し、実施例6に示すように精製した。これらの実験において、脳由来健康tauアイソフォーム(0.1mg/ml)、AD-脳由来または組換えタイプIA tauまたはタイプIB(0.2mg/ml)をチューブリンと混合した。各混合物を別々にアッセイした。被検混合物を静かに振盪させながら水浴中で37℃で1時間インキュベーションした。氷上に保った混合物にGTPおよび/または正常tau(チューブリンの最終濃度は1mg/mlおよびGTP 1mM)を加えた(すべて試薬に重合緩衝液(100mM PIPES、pH6.9、1mM MgSO4、2mM EGTA)使用)。静かにそして急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計(Beckman Coulter DU640)中で37℃に平衡化した。濁度変化を340nmで5分間にわたり10秒間隔で測定した。データはAD脳由来および組換え二重トランケートタイプIA分子がともに正常tauの微小管構築促進能を阻害することを示す(図12、曲線2、3)。これに対し、組換えタイプIBは正常ヒトtauにより誘導されるチューブリン重合促進能力を阻害することができない(図12、曲線4)。曲線1(図12)は正常tauにより促進された微小管構築を示す。
実施例8:tauタイプIAの病的活性を中和する薬剤候補のスクリーニングアッセイ
【0228】
チューブリン重合を促進する健康正常tauの活性を阻害する二重トランケートtauタイプIA分子の能力を用いてスクリーニングアッセイをタイプIA分子の阻害活性を中和することができる化合物を選択するために計画した。病的tauタイプIAはAD-脳または組換え供給源由来であり得るが、組換え物質を用いるのが好都合である。薬剤候補の中和効果は正常健康tauの残存する微小管構築促進能を測定することにより定量的に定義することができる。該アッセイは2工程で行われる:
1.tauタイプIAを中和する薬剤候補のスクリーニング。個々の薬剤候補(最終濃度50mg/ml)と別個に混合したプロトタイプ組換えタイプIA分子(配列番号1)(最終濃度100mg/ml)を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリン、GTPおよび健康tauを+4℃で混合物に加えた(最終濃度:チューブリン-1mg/ml、GTP-1mM、健康tau-100mg/ml)。急速混合物後、試料を水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。タイプIA活性中和能を有する薬剤候補を正常健康tauの残存する微小管構築促進能を測定することにより選択した(図13;薬剤候補をタイプIA分子とプレインキュベーションし、タイプIA中和の有効性を微小管構築を用いてアッセイした。下端の曲線1および上端の曲線2は病的タイプIA分子に対する被検薬剤候補の陰性(中和なし)および陽性(100%)中和活性を示す。中側の曲線は3つの異なる薬剤候補による種々のタイプIA中和効果を示す。)。陽性薬剤を選択するための閾値は任意であり、タイプIAの完全中和からその部分中和まで変化するかもしれない。
【0229】
2.タイプIA分子を中和し、それを正常健康tauと区別する薬剤候補の選択。tauタイプIA分子に対する中和活性を有する選択した候補の正常健康tauとの反応性をスクリーニングし、タイプIAにのみ特異的な分子を選択した。正常健康tau(最終濃度100mg/ml)と個々の薬剤候補(最終濃度50mg/ml)の別個の混合物を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリンおよびGTPを+4℃で該混合物に加えた(最終濃度:チューブリン1mg/ml、GTP 1mM)。急速に混合した後、試料を水晶マイクロキュベットに入れた。濁度変化を340nmで測定した。健康tauのMT重合促進活性との干渉を示さない薬剤候補を選択した(図14)。
実施例9:tauタイプIIAの病的活性を中和する薬剤候補のスクリーニングアッセイ
【0230】
本発明は、tauタイプIIA分子が予期せず高いチューブリン重合促進能力を有することを示し、タイプIIAタンパク質の該活性を中和する化合物を選択するためのスクリーニングアッセイの基礎を形成する。タイプIIAの中和はタイプIIA分子の残存微小管構築活性を測定することにより定量することができる。該アッセイは2工程で行われる。
【0231】
1. tauタイプIIAを中和する治療薬候補のスクリーニング。
【0232】
tauタイプIIA分子(配列番号12)(最終濃度100mg/ml)と単一薬剤候補(最終濃度50mg/ml)の別個の混合物を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリンおよびGTPを+4℃で混合物に加えた(最終濃度:チューブリン 1mg/ml、GTP-1mM)。急速混合物後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。微小管構築速度を有意に低下させた薬剤候補を該アッセイの第二工程用に選んだ(図15;薬剤候補をタイプIIA分子とプレインキュベーションし、タイプIIA中和の有効性を微小管構築を用いてアッセイした。下端の曲線1は各薬剤候補の陽性(100%)中和活性を示し、上端の曲線2は病的タイプIIA分子を中和しないことを示す。中側の曲線はタイプIIA中和における種々の薬剤候補の異なる効果を示す。)。
【0233】
2.タイプIIA分子を中和し、それを正常健康tauと区別する薬剤候補の選択。
【0234】
薬剤候補(最終濃度50mg/ml)と正常健康tau(最終濃度100mg/ml)の別個の混合物を37℃で1時間プレインキュベーションした。次に、チューブリンおよびGTPを+4℃で該混合物に加えた(最終濃度:チューブリン1mg/ml、GTP-1mM)。急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。健康tauと干渉しない薬剤候補を選択した(図16;工程1で選んだ薬剤候補を健康tauとプレインキュベーションし、微小管構築に対する効果をアッセイした。下端の曲線(1)は健康tauの最大阻害を示し、上端の曲線(2)は健康tauを阻害しないことを示す。中側の曲線は薬剤候補の健康tauに対する異なる阻害活性を示す。)。
実施例10:NおよびC末端二重トランケートタイプIAおよびタイプIIA分子を中和するモノクローナル抗体の製造。
【0235】
免疫プロトコールおよび融合手順:ヒトアルツハイマー脳から単離したNおよびC末端二重トランケートtauタイプIタンパク質(分画#19、実施例6)を免疫原に用いた。Balb/cマウスを完全Freundアジュバント中の該タンパク質(50mg/マウス)を皮下にプライムし、次いで同じタンパク質50mg/マウスを4週間間隔で3回腹腔内にブースターをかけた。融合前血清を回収し、tauに対する特異抗体のレベルをELISAで試験した(図17;AD由来tauで免疫したマウスの血清中の特異抗体のレベルを同じ抗原を用いてELISAで試験した。5つの血清はすべて該tauタンパク質に対する高い抗tau結合活性を示した。図17はある免疫マウスにおける特異抗体レベルを示す。コントロールには無関係のタンパク質で免疫したマウス由来の血清を用いた。)。マウス脾細胞を当該分野でよく知られた改良された方法を用いNS/0ミエローマ細胞と融合させた(M. KohlerおよびC. Milstein、1975)。
【0236】
図18に示す結果によれば、モノクローナル抗体DC44、DC82およびDC136はアルツハイマー病脳由来のNおよびC末端二重トランケートタイプIAおよびタイプIIA分子を認識する。これら抗体では同じ方法で調製した正常ヒト脳由来のtau単離物との反応性は観察されなかった(図18)。これに対し、モノクローナル抗体DC25はELISAで病的および正常健康脳由来の該タンパク質と反応する(図18)。この抗体は病的型のtau(AD-tau)と正常ヒトtauを区別しない。この一次スクリーニング後、ハイブリドーマをソフトアガロース中でサブクローンし、最終的に当業者によく知られた技術により同じイデオタイプを有する抗体を分泌する均質なハイブリドーマポピュレーションを得た。
【0237】
さらにこれらクローン化ハイブリドーマクローンの組換え完全長tauアイソフォームおよび二重トランケートtauタイプIA(配列番号1)およびタイプIIA(配列番号12)分子に対する反応性をスクリーニングアッセイと同じELISAを用いて試験した。
実施例11:モノクローナル抗体を用いるAD脳由来および組換えNおよびC末端トランケートタイプIA分子の病的活性の中和。
【0238】
選択したモノクローナル抗体DC44、DC82、DC136およびDC25をアルツハイマー脳から単離した天然tauタイプIAの活性を中和する能力で特徴づけた(実施例6)。該tau単離物(最終濃度100mg/ml)および被検抗体(最終濃度50mg/ml)を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリン、正常ヒトtauおよびGTPを+4℃で該混合物に加えた(最終濃度:チューブリン1mg/ml、健康ヒトtau-100mg/ml、GTP-1mM)。急速に混合物した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。モノクローナル抗体DC136、DC44およびDC82は該タンパク質の病的活性を阻害することができた(Fig 20;抗体を天然tauタイプIA(分画#19)とプレインキュベーションし、次いで健康ヒトtau、チューブリンおよびGTPと混合した。37℃で5分間後に微小管の形成を分光光度法で測定した。バーは3つの独立した試験の平均値を表す。健康ヒトtauを用いるMAA-微小管構築アッセイ。タイプIAとプレインキュベーション(抗体なし)した健康ヒトtauを用いるMAIA-微小管構築阻害アッセイ。類似の実験において抗体をtauタイプIAの組換えプロトタイプ(配列番号1)を用いて試験し、同様の中和活性パターンを示した(図21)。コントロール抗体DC25はELISAおよびWesternブロッティングにより試験したトランケートおよび正常tauのすべての型を認識するが、微小管構築アッセイにおいてAD-脳由来タイプIA活性と干渉しない。これらの結果は抗体DC25が抗体DC136、DC44およびDC82に比べてtauの異なる領域と反応することを示唆する。これに対し、抗体DC136、DC44およびDC82は病的タイプIA分子に関与するエピトープと結合する。
【0239】
次に選択工程は健康分子とタイプIA分子を識別することができる抗体を目的とした。正常健康tau(最終濃度100mg/ml)および被検抗体(最終濃度50mg/ml)の混合物を37℃で1時間プレインキュベーションした。インキュベーション後、チューブリンおよびGTPを+4℃で該混合物に加えた(最終濃度:チューブリン 1mg/ml、GTP-1mM)。急速に混合した後、試料をピペットで水晶マイクロキュベットに入れ、温度調節分光光度計中で37℃に平衡化した。濁度変化を340nmで測定した。抗体DC136、DC44、DC82およびDC25はいずれも微小管構築において正常健康tauを阻害することができなかった(図22;tauタイプIA中和抗体を健康tauとプレインキュベーションし、次いでチューブリンおよびGTPと混合した。37℃で5分間後に微小管の形成を分光光度法で測定した。バーは3つの独立した実験の平均値を示す。健康tauを用いるMAA-微小管構築アッセイ。陰性コントロールとして健康tau中和抗体を用いた。このデータは抗体 DC136、DC44、DC82がトランケート病的型のtauと健康tauタンパク質の相互作用に関与する特異的エピトープを認識することを示した。
実施例12:モノクローナル抗体によるタイプIIA活性の中和
【0240】
tauタイプIA中和活性について先に単離した抗体を実施例8Bに記載の方法を用いる組換えtauタイプIIA(配列番号12)に対する中和活性について試験した。すべての3中和モノクローナル抗体DC44、DC82およびDC136はNおよびC末端二重トランケートtauタイプIIA分子の病的活性を低下させることができた(図23;抗体を組換えtauタイプIIAとプレインキュベーションし、次いでチューブリンおよびGTPと混合した。37℃で5分間後、微小管の形成を分光光度法で測定した。バーは3つの独立した実験の平均値を示す。tauタイプIIAを用いるMAA-微小管構築アッセイ(抗体なし)。これは該抗体のエピトープが少なくともタイプIA 配列番号1およびタイプIIA配列番号12により共有されることを示唆する。抗体DC25についてはタイプIIA阻害活性は観察されなかった。
実施例13:組換えNおよびC末端二重トランケートtauタイプIAおよびIIA分子の免疫原性。
【0241】
免疫プロトコール:本発明の好ましい態様において該組換えtauタイプIAおよびIIAタンパク質をワクチネーション目的または病的tauタイプIAおよびIIA分子の病的活性を特異的に中和する抗体を生じるために用いる。該実施例において、組換えNおよびC末端二重トランケートtauタイプIIA(配列番号12)を免疫原に用いた。Balb/cマウスを完全Freundアジュバント中の該タンパク質(50mg/マウス)で皮下にプライムし、次いで4週間間隔で50mg/マウスの完全Freundアジュバント中の該タンパク質で3回腹腔内にブースターをかけた。免疫血清を回収し、各組換え抗原tauに対する特異抗体のレベルをELISAで測定した(図24)。
実施例14: トランスジェニック動物
【0242】
尾部先端から抽出したDNA:ゲノムDNAをDNeasy組織キットQiagenにより抽出した。
遺伝子型別(図25):二重トランケートtau型をコードするトランス遺伝子の特異的増幅をトランスジェニック動物の親世代由来のゲノムDNAを用いて実施し、図25Aに示す。F1世代のゲノムDNAのさらなる分析ではトランス遺伝子は親世代の子孫にも確認されたことから該遺伝子が遺伝性であることが明らかになった。したがって、二重トランケートtauをコードするトランス遺伝子は該動物の染色体DNAに固定される(図25B-F1世代の遺伝子型別)。この実施例に用いる動物は自然発症高血圧および他のアルツハイマー病関連危険因子、例えば異脂肪血症または糖尿病を特徴とする特異的遺伝的背景がある。したがって、この動物系統は最も頻発するアルツハイマー病の危険因子、例えば高血圧および糖尿病を組み合わせることによりユニークな実験アルツハイマーモデルである。
トランス遺伝子を生成するにはSambrookら、Molecular Cloning A Laboratory Manual、CSH Laboratory、New York(2001)に記載の分子生物学の標準技術を用いた。二重トランケートtauをコードするcDNAを遍在性または組織特異的な発現をもたらすプロモータと結合した発現ベクターに導入した。該遺伝子断片を前核注射(非限定)により1日齢の胚に導入した。生じた子孫を尾の先端から得たゲノムDNAを用いて遺伝子型別した。
【0243】
トランス遺伝子発現の分析(図26):トランス遺伝子由来のmRNAの発現を一般的に知られた方法、例えばRT-PCRおよびアガロースゲル電気泳動を用いるRT-PCR分析により評価した。
【0244】
図25のパネルAはトランスジェニック動物の親世代の遺伝子型別を示す。レーン1、2、3および4におけるゲノムDNA由来のDNAの二重トランケート配列の特異的増幅は、フォスターマザーの子孫の尾から抽出したゲノムDNAにおける特異的トランス遺伝子の存在を示す。これら動物は二重トランケートタイプIIA tau分子を保有するトランスジェニック動物の親世代を表す。この実施例において、陽性(+C)および陰性(-C)および2つのさらなる陰性試料(5、6)が示される(M=サイズマーカー)。矢印はトランス遺伝子陽性動物において予想された予測PCR生成物サイズを示す。図25のパネルBはF1世代由来動物の遺伝子型別。ゲノムDNAを尾の先端から抽出し、二重トランケートtau特異的DNA配列を確認し、レーン1に示す。レーン2および3は陰性コントロールを示す。F1世代におけるtau特異的DNA断片の同定はこれらトランス遺伝子の遺伝性を確認する。
【0245】
図26:RNAをトランスジェニック動物の瞬間凍結組織から抽出し、逆転写、次いでcDNAの特異的増幅にかけた。実施例はレーン番号1および2においてトランス遺伝子発現動物を示す。レーン3-5は非発現コントロールを示し、レーン5は同じ方法を用いた非トランスジェニック動物に典型的に出現する非特異的シグナルを示す。この実施例は実験動物におけるトランス遺伝子から発現した二重トランケートtau特異的mRNAの存在を示す。
実施例15:A:タイプIIA分子の過剰発現は分化したニューロン様細胞の細胞死をもたらす。
【0246】
神経芽細胞腫細胞系SH-SY5Yにおいて、タイプIIA分子により生じた細胞死が当業者に知られた標準化in vitro分化条件下で示された。該効果をタイプIIA 二重トランケートtauを発現する安定なトランスフェクション細胞で試験し、非トランスフェクション細胞と比較した。細胞生存をトリパンブルー排除アッセイを用いトリプリケートで手動で定量し、統計的評価を一元ANOVA検定を用いて行った。タイプIIA 二重トランケートtau過剰発現細胞および野生型細胞における細胞生存の有意差がin vitro分化の6日後にみられた(P<0.001)。タイプIIA 二重トランケートtauの過剰産生(総タンパク質量の0.5%)は該細胞の生存率の3Xの低下を生じた(図27;各二重トランケートtauタイプIIA(タイプIIA)でトランスフェクションしたSY5Y細胞および非トランケートコントロールニューロン様細胞(mock)それぞれの細胞生存性の比較)。
【0247】
先に示した構築物同様に、同様の系が二重トランケートタイプI分子をコードする構築物を用いて確立された。
【0248】
タイプII 二重トランケートtau分子は微小管系に対する結合親和性の増大を示す。
【0249】
タイプIIA 二重トランケートtauおよび完全長tauを発現する安定にトランスフェクションされたSH-SY5Y細胞由来の遊離tau分画(FT)、微小管関連分画(MT)および核分画(NAT)の分離を行った。微小管とtauの結合の定量は完全長型に比べて二重トランケートタイプIIA tauの微小管に対する親和性の増大(50%以上)を示した(図28A;タイプIIA分子の微小管に対する結合親和性の増大はタイプIIA二重トランケート分子および完全長tauを発現する安定なトランスフェクション細胞の細胞分画化を用いて示される。遊離tau(FT)、微小管結合tau(MT)および核関連tau(NAT)の単離を記載のごとく実施した。)。tauの量を当該分野で用いられる標準的細胞生物学的分画法、次いでWesternブロット分析に従って定量した。較正曲線を規定量の組換えtauタンパク質を用いて計算した。
【0250】
B:有機物質F123[C34-59O14-23H32-44N6-8]nの微小管重合アッセイにおけるその阻害効果について分析する:(1)F123とtau(正常tauおよびtauタイプIA';conc=100μg/ml;tauタイプIIA conc=60μg/ml)を37℃で1時間インキュベーション、
(2)チューブリンの添加(conc=1mg/ml)、
(3)340nm/5minの測定(注:すべてPIPES緩衝液で希釈。
【0251】
C:正常健康tauおよびアルツハイマーtauタイプIIをその微小管構築促進能について分析する。
tau 43で示されたこの実施例の正常tauは電子顕微鏡検査で示される典型的微小管を形成する(図28C参照)。しかしながら、アルツハイマーtauタイプIIは典型的にパターンを有する病的微小管を生じる(図28C参照)。
実施例16:真核細胞におけるNおよびC末端二重トランケートtauタイプIIの過剰発現の機能的結果
【0252】
タイプIIA分子の過剰発現により生じたミトコンドリアの輸送の変化を示す病的表現型を神経芽細胞腫細胞系SH-SY5Yにおいて実施した。NおよびC末端二重トランケートtauタイプII分子の影響を、生野生型SH-SY5Y細胞とトランスフェクション細胞のミトコンドリア再分配を比較することにより試験した。当業者に知られた細胞生物学的輸送アッセイを用いた。簡単には、細胞を標準的実験技術に従って同じ密度(70%コンフルエント)でLabTekIIチャンバー(Nunc)を用いて培養し、トランスフェクションを製造業者の指示書に従ってFugene6(Roche)を用いた行った。ミトコンドリアの染色(MitoFluor Red 594、Molecular Probes)を製造業者の指示書に従って行った。生細胞を63x油浸対物レンズおよび蛍光フィルターを取り付けたAxiovert 200M蛍光顕微鏡(ZEISS)を用いて試験した。写真をソフトウエアプログラムMetaMorph(Universal Imaging)と組み合わせたCCDカメラ(Photometrics、Cool snap HQ; Hamamatsu)で撮影した。
【0253】
ミトコンドリア特異的染色MitoFluor(Molecular Probes)を用いるミトコンドリアの局在を誘導および非誘導SH-SY5Y細胞で比較した。該染色により、SH-SY5Y細胞におけるミトコンドリア輸送に対するタイプIIA 二重トランケートtau分子の悪影響はマイナス末端による細胞内力の機能的優性を示す中心体周辺の核周囲ミトコンドリアのクラスタ化を生じることであることが確認された(図30)。
【0254】
コントロールとして、対数増殖期の細胞(図29)は細胞体および細胞周辺におけるミトコンドリアの規則的な分布を示す。結論として、NおよびC末端二重トランケートタイプIIAタンパク質はミトコンドリア再分布に影響する細胞内輸送メカニズムに影響を及ぼすことができる。本実験の設定はタイプIIA分子に対する阻害活性を試験するための適切な方法を示す。
(参考文献)
【0255】
1. Finch C、Tanzi RE、(1997)Science 278,407-411
2. Blessed G、Tomlinson BE、Roth M(1968 Br J Psychiatry 114: 797-811
3. Tomlinson BE、Blessed G、Roth M J、(1970)Neurol Sci 11、205-242
4. Arigada PA、Growdon JH、Hedley-White ET、Hyman BT(1992)Neurology 42,631-639
5. Wischik CM、Novak M、Edwards PC、Klug A、Tichelaar W、Crowther RA(1988a)Proc Natl Acad Sci USA 85: 4884-4888
6.Wischik CM、Novak M、Trogersen HC、Edwards PC、Runswick MJ、Jake R、Walker JE、Milstein C、Roth M、Klug A(1988b)Proc Natl Acad USA 85: 4506-4510
7.Himmler A,Drechsel D、Kirschner MW、Martin DWjr、(1989)Mol Cell Biol 9,1381-1388
8. Goedert M、Spillantini MG、Jakes R、Rutherford D.、Crowther RA(1989)Neuron#、519-526
9. Hutton M、Lendon CL、Rizzu P、Baker M、Froelich S.、Houlden H、Pickering-Brown S、Chackraverty S、Isaacs A、Grover A(1998)Nature 393,702-705
10. Spillantini NG、Murrell JR、Goedert M、Farlow MR、Klug A、Ghetti B(1998)Proc Natl Acad Sci USA 95,7737-7741.
11. Selkoe DJ、(1999)Nature 399、A23-31
12. Isacson O、Seo H、Lin L、Albeck D、Granholm A CH、(2002)Trends Neurosci 25,79-84
13. Mudher A、Lovestone S、(2002)Trends Neurosci 25,22-26
14. Couzin J、(2002)Science 296,28-29
15. Grundke-Iqbal I、Iqbal K、Tung Y-C、Quinlan M,Wisniewski HM、Binder LI、(1986)Proc Natl Acad Sci USA、83,4913-4917
16. Gustke N、Steiner B、Mandelkow EM、Biernat J、Meyer HE、Goedert M、Mandelkow E(1992)FEBS Lett 307,199-205
17. Lindwall G、Cole RD(1984)J Biol Chem 259,12241-12245
18. Kopke E、Tung Y-Ch、Shaikh S、Alonso AC、Iqbal K、Grundke-Iqbal I(1993)J Biol Chem 268,24374-24384
19. Cleveland DW、Hwo SY、Kirschner MW(1977a)J Mol Biol 116: 227-247
20. Cleveland DW、Hwo SY、Kirschner MW(1977b)J Mol Biol 116: 207-225
21. Lee MY、Balin BJ、Otvos L、Trojanowski JQ(1991)Science 251,675-678
22. Alonso AC、Zaidi T、Grundke-Iqbal I、Iqbal K(1994)Proc Natl Acad Sci U S A 91: 5562-5566
23. Wille H、Mandelkow EM、Mandelkow E(1992)J Biol Chem 267: 10737-10742
24. Alonso AC、Grundke-Iqbal I、Iqbal K(1996)Nat Med 2: 783-787
25. von Bergen M、Barghorn S、Li L、Marx A、Biernat J、Mandelkow EM、Mandelkow E(2001)J Biol Chem 276: 48165-48174
26. Friedhoff P、von Bergen M、Mandelkow EM、Mandelkow E(2000)Biochim Biophys Acta 1502: 122-132 Review
27. Illenberger S、Zheng-Fischhofer Q、Preuss U、Stamer K、Baumann K、Trinczek B、Biernat J、Godemann R、Mandelkow EM、Mandelkow E(1998)Mol Biol Cell 9: 1495-1512
28. Kampers T、Friedhoff P、Biernat J、Mandelkow EM、Mandelkow E(1996)FEBS Lett 399: 344-349
29. Perez M、Valpuesta JM、Medina M,Montejo de Garcini E、Avila J(1996)J Neurochem 67: 1183-1190
30. WangJ-Z ; Grundke-Iqbal I; Iqbal K(1996)Nature Med 2: 871-875
31. Wilson DM、Binder LI(1997)Am J Pathol 150: 2181-2195
32. Schweers O、Mandelkow EM、Biernat J、Mandelkow E(1995)Proc Natl Acad Sci USA 92: 8463-8467
33. Ksiezak-Reding H、Yang G、Simon M、Wall JS(1998)Brain Res. 814:86-98
34. Schneider A、Biernat J、Von Bergen M、Mandelkow E、Mandelkow EM(1999)Biochemistry 38: 3549-3558
35. Yan S-D、Chen X、Schmid A-M、Brett J、Godman G、Zou Y-S、Scott CW、Caputo C、Frappier T、Smith MA、Perry G、Yen SH、Stern D(1994)Proc Natl Acad Sci USA 91: 7787-7791
36. Smith MA、Taneda S、Richey PL、Miyata S、Yan SD、Stern D、Sayre LM、Monnier VM、Perry G(1994)Proc Natl Acad Sci U S A 91: 5710-5714
37. Mori H、Kondo J,Ihara Y(1987)Science 235: 1641-1644
38.Paudel H、Li W(1999). J Biol Chem 274: 8029-8038
39. Goedert M、Spillantini MG、Potier MC、Ulrich J、Crowther RA、(1989)EMBO J. 8,393
40. Goedert M、Spillantini MG、Jakes R、Rutherford D、Crowther RA(1989)Neuron 4,519
41. Crowther RA、Olesen OF、Jakes R、Goedert M(1992)FEBS Lett 309: 199-202
42. Crowther RA,Olesen OF、Smith MJ、Jakes R、Goedert M(1994)FEBS Letter 337: 135-138
43.(例えばM. Kohler and C. Milstein、in"Continuous Cultures of Fused Cells Secreting Antibody of Pre-DefinedSpecificity"、Nature、256、pp. 495-497,1975;およびHarlow and Lane、in "Antibodies、A Laboratory Manual"、Cold Spring Harbor Laboratory、Cold Spring Harbor、1988参照)
【図面の簡単な説明】
【0256】
【図1】NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築。
【図2】NおよびC末端二重トランケートtauタイプIAおよびタイプIB分子による微小管構築の阻害。
【図3】微小管構築におけるNおよびC末端二重トランケートtauタイプIIAおよびIIB分子の活性。
【図4】神経細胞に発現したタイプIIA NおよびC末端二重トランケートtauは酸化ストレスに対する感受性を有意に増加させる。
【図5】病的tauタイプIAタンパク質およびその欠失突然変異体に対するモノクローナル抗体の親和性。病的tauタイプIAタンパク質およびその欠失突然変異体に対するモノクローナル抗体の見かけの親和性。最初のカラムに「プロトタイプ」tauタイプIAタンパク質(配列番号1)および各欠失突然変異体を示す。中段のカラムに本発明のエピトープを示す。最後のカラムに示す見かけの親和性を競合的ELISAにより測定し、プロトタイプtauタイプIAタンパク質と50%競合するのに必要な対応抗原の濃度で表した。
【図6】AD-脳由来tauタンパク質のSuperdex 200-カラムによる分画。
【図7】AD脳からの3つの別個の単離物由来の分画No.19中のタイプIA阻害活性。
【図8】AD脳中のNおよびC末端二重トランケートtauタイプI分子の証明。
【図9】健康脳に存在せずAD脳に存在するtauタイプI。
【図10】NおよびC末端二重トランケートtauタイプII分子の免疫反応性。
【図11】組換えtauタイプI-II(配列番号1-24)の構築。
【図12】AD-脳由来および組換えtauタイプIAの正常健康tauに対する阻害効果。
【図13】tauタイプIA分子を中和する薬剤候補の第一回スクリーニング(工程1)。
【図14】正常tauに対する選択性を有するタイプIA分子を中和する薬剤候補の第二回スクリーニング(工程2)。
【図15】tauタイプIIA分子を中和する薬剤候補の第一回スクリーニング。
【図16】tauタイプIIA分子を中和し、それを正常tauから識別することができる薬剤候補の第二回スクリーニング(工程2)。
【図17】ELISAにより測定したプレフュージョン(prefused)マウス血清中の特異抗体レベル。
【図18】モノクローナル抗体とAD脳由来tau(分画#19)およびコントロール健康脳由来tauとのELISA反応性(DC20:無関係な特異性を有するモノクローナル抗体。示したデータは3回の並列実験の平均値を表す。)。
【図19】モノクローナル抗体の組換えtau分子とのELISA反応性(DC20:無関係な特異性を有するモノクローナル抗体。示したデータは3回の並列実験の平均値を表す。)。
【図20】AD-脳由来tauタイプIA(分画#19)に対する中和抗体のスクリーニング。
【図21】組換えtauタイプIA(配列番号1)に対する中和抗体のスクリーニング。
【図22】tauタイプIA分子を中和し、それらを健康tauから識別することができる薬剤候補のスクリーニング。
【図23】組換えtauタイプIIA(配列番号12)の病的活性のモノクローナル抗体による中和。
【図24】ELISAで検出した組換えtauタイプIIA(配列番号12)に対する抗体のレベル。
【図25】トランスジェニック動物の遺伝子型別。パネルAはトランスジェニック動物の親世代の遺伝子型別を示す。レーン1、2、3および4のゲノムDNA由来のDNAの二重トランケート配列の特異的増幅は、フォスターマザーの子孫の尾から抽出したゲノムDNAにおける特異的トランス遺伝子の存在を示す。これら動物は二重トランケートタイプIIA tau分子保有トランスジェニック動物の親世代を示す。この実施例において、陽性(+C)および陰性(-C)および2つのさらなる陰性試料(5、6)を示す(M=サイズマーカー)。矢印は、トランス遺伝子陽性動物において予想される予想PCR生成物サイズを示す。パネルB. F1世代由来の動物の遺伝子型別。ゲノムDNAを尾の先端から抽出し、二重トランケートtau特異的DNA配列を同定し、レーン1に示す。レーン2および3は陰性コントロールを示す。F1世代におけるtau特異的DNA断片の同定はこれらトランス遺伝子の遺伝性(inheritability)を確認する。
【図26】トランスジェニック動物のF1世代における二重トランケートヒトtau転写物の遺伝子発現。RNAをトランスジェニック動物の瞬間凍結組織から抽出し、逆転写、次いでcDNAの特異的増幅にかけた。例はレーン番号1および2にトランス遺伝子発現動物を示す。レーン3〜5は非発現コントロールを示し、レーン5はこの方法を用いた時に非トランスジェニック動物に典型的に生じる非特異的シグナルを示す。この実施例は、実験動物においてトランス遺伝子から発現した二重トランケートtau特異的mRNAの存在を示す。
【図27】6日間in vitro分化(differentiation)後のタイプIIA分子の過剰発現により生じた細胞死。二重トランケートtauタイプIIA(タイプIIA)でトランスフェクションしたSY5Y細胞および非トランスフェクションコントロールニューロン様細胞(mock)の細胞生存性の比較。
【図28A】タイプIIA分子の微小管に対する結合親和性の増加はタイプIIA二重トランケート分子および完全長tauを発現する安定なトランスフェクション細胞由来のtauの細胞分画を用いて示される。遊離tau(FT)、微小管結合tau(MT)および核関連tau(NAT)の単離は記載のごとく行った。
【図28B】有機化合物F123による各tauタイプIAおよびIIAの阻害、微小管重合アッセイ。
【図28C】電子顕微鏡分析により示される微小管重合と吸着(absorption)測定値の直接比較。
【図29】MitoFluorで染色した対数増殖期のSH-SY5Y細胞。細胞体およびプロセス中のミトコンドリアの標準的分布。
【図30】MitoFluorで染色した対数増殖期のtauタイプIIA分子発現SH-SY5Y細胞。細胞の中心体領域周辺のグリーン標識ミトコンドリアの核周囲クラスタリング。
【特許請求の範囲】
【請求項1】
以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIA tau分子」):
4反復配列を含むtau43の少なくとも最初の236 N末端アミノ酸および少なくとも最後の45 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて正常tauタンパク質の微小管構築の促進を防止し、
該微小管構築の促進の防止は微小管構築アッセイにおいて該分子に対する特異的阻害、中和モノクローナル抗体により除去することができる。
【請求項2】
配列番号1〜3の群から選ばれるアミノ酸配列を含むことを特徴とする請求項1記載のタイプIA tau分子。
【請求項3】
以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIB tau分子」):
4反復配列を含むtau43の少なくとも最初の238 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の207 N末端アミノ酸および少なくとも最後の50 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauの微小管構築の促進を防止しない。
【請求項4】
配列番号4〜10の群から選ばれるアミノ酸配列を含むことを特徴とする請求項3記載のタイプIB tau分子。
【請求項5】
以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIIA tau分子」):
4反復配列を含むtau43の少なくとも最初の68 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の68 N末端アミノ酸および少なくとも最後の20 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauより高い微小管構築促進活性を有し、
該微小管構築促進活性は微小管構築アッセイにおいて該分子に対する特異的阻害、中和モノクローナル抗体により除去することができ、該分子の病的活性は微小管重合促進活性により定義される微小管ネットワークに対する結合に依存する。
【請求項6】
配列番号11〜18の群から選ばれるアミノ酸配列を含むことを特徴とする請求項5記載のタイプIIA tau分子。
【請求項7】
以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIIB tau分子」):
4反復配列を含むtau43の少なくとも最初の68 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の68 N末端アミノ酸および少なくとも最後の20 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauと異なる病的微小管構築促進活性を有する。
【請求項8】
配列番号19および20の群から選ばれるアミノ酸配列を含むことを特徴とする請求項7記載のタイプIIB tau分子。
【請求項9】
以下の工程を特徴とする請求項1〜8のいずれかに記載の分子の製造方法:
a)少なくとも最初の236および最後の40アミノ酸または最初の68および最後の20アミノ酸またはその組み合わせにわたる欠失を有する二重トランケートtau分子をコードする配列を保持する組換え原核性発現プラスミドの構築、
b)該NおよびC末端二重トランケートtau分子の発現を許す条件下で該細菌を増殖させ、
c)好ましくは遠心により細菌を回収し、
d)細菌ペレットを再浮遊させ、
e)該細菌を超音波処理し、
f)該超音波処理細菌をゲルろ過により分画し、
g)得られた分画の活性を微小管構築アッセイによりモニターすることにより、タイプIおよびタイプII tau分子の異なる活性を同定する。
【請求項10】
該トランケーションが請求項1〜8のいずれかに記載のものであることを特徴とする請求項9記載の方法。
【請求項11】
微小管構築アッセイ活性が請求項1〜8のいずれかに記載のものである請求項9または10記載の方法。
【請求項12】
以下の工程を特徴とする請求項1〜8のいずれかに記載の分子の製造方法:
a)アルツハイマー罹患脳組織を得、
b)該病的(罹患)脳組織を緩衝液、特にTris緩衝液中でホモゲナイズし、
c)該ホモゲナイズ脳組織を硫酸アンモニウム沈殿し、
d)PIPES緩衝液に再溶解し、
e)該再溶解物質をゲルろ過で分画化し、
f)得られた分画の活性を微小管構築アッセイでモニターすることによりタイプIおよびタイプII tau分子の異なる活性を同定する。
【請求項13】
微小管構築アッセイ活性が請求項1〜8のいずれかに記載されていることを特徴とする請求項12記載の方法。
【請求項14】
以下の工程を含む請求項1または2記載の分子(タイプIA分子)とチューブリンの複合体の分解(disassembling)に有効な物質の試験方法:
a)タイプIA分子とチューブリンのタンパク質複合体を形成させ、
b)該タンパク質複合体を被検物質とインキュベーションし、タイプIA分子を阻害し、および/または野生型tauの微小管構築促進能力および微小管の正常機能を回復させる物質を同定する。
【請求項15】
以下の工程を含む野生型tau発現細胞系において請求項1または2に記載の分子(タイプIA分子)がチューブリンとの複合体の形成を開始するのを阻害するのに有効な物質の試験方法:
a)適切な制御領域の制御下でタイプIA分子をコードする機能的遺伝子を正常tauタンパク質発現細胞に導入し、
b)タイプIA分子とチューブリンのタンパク質複合体を形成させ、
c)試験する物質を該複合体を保持する細胞に適用し、
d)請求項1記載のタイプIA生物活性に対する該物質の効果を試験する。
【請求項16】
該タイプIIA分子と微小管との相互作用により病的微小管を生成させる生理的条件下で請求項5または6記載のタイプIIAと野生型tauタンパク質をインキュベーションすることを特徴とする微小管をin vitroで病的状態に変換する方法。
【請求項17】
以下の工程を含むタイプIIA分子を除去および/または中和し、タイプII分子により生じる生理学的微小管パラメーターおよび機能を修復するその特性について請求項5または6記載のタイプIIA分子の病的効果を中和することができる物質のスクリーニング方法:
a)請求項16記載のタイプIIA分子およびチューブリン存在下で病的微小管を形成させ、
b)該物質、タイプIIAおよびチューブリンの混合物をスクリーニングする物質とインキュベーションし、
c)タイプIIA分子により生じる病的微小管の形成を減少させる結果を試験する。
【請求項18】
以下の工程を含むタイプIIA分子発現細胞系における異常微小管形成および機能の促進において請求項5または6記載のタイプIIA分子のin vivo活性を阻害するのに有効な物質の試験方法:
a)タイプIIA分子をコードする機能的遺伝子を適切な制御領域の制御下で野生型tau発現細胞中に導入し、
b)タイプIIA tau分子と微小管の複合体を形成させて該複合体を病的微小管の形成に関与させ、
c)試験する物質を該複合体を保持する細胞に適用し、
d)タイプIIA生物活性、特に微小管ネットワークおよびその関連機能の修飾に対する該物質の効果を試験する。
【請求項19】
請求項1〜8のいずれかに記載の分子を発現するトランスジェニック動物。
【請求項20】
特にアルツハイマー病治療用の薬剤をスクリーニングおよび試験するためのアルツハイマー病の動物モデルとしての請求項19記載のトランスジェニック動物の使用。
【請求項21】
請求項1〜8、特に請求項1、2、5または6のいずれかに記載の分子および医薬的に許容される担体、特にアジュバントを含むワクチン。
【請求項22】
請求項1または2記載の分子とチューブリンの複合体の形成開始の阻害剤。
【請求項23】
寄託番号02060767でEuropean Collection of Cell Cultures(ECACC)、Porton Down、Salisbury、UKに寄託されたモノクローナル抗体DC44としての結合部分を含むことを特徴とする請求項22記載の阻害剤。
【請求項1】
以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIA tau分子」):
4反復配列を含むtau43の少なくとも最初の236 N末端アミノ酸および少なくとも最後の45 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて正常tauタンパク質の微小管構築の促進を防止し、
該微小管構築の促進の防止は微小管構築アッセイにおいて該分子に対する特異的阻害、中和モノクローナル抗体により除去することができる。
【請求項2】
配列番号1〜3の群から選ばれるアミノ酸配列を含むことを特徴とする請求項1記載のタイプIA tau分子。
【請求項3】
以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIB tau分子」):
4反復配列を含むtau43の少なくとも最初の238 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の207 N末端アミノ酸および少なくとも最後の50 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauの微小管構築の促進を防止しない。
【請求項4】
配列番号4〜10の群から選ばれるアミノ酸配列を含むことを特徴とする請求項3記載のタイプIB tau分子。
【請求項5】
以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIIA tau分子」):
4反復配列を含むtau43の少なくとも最初の68 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の68 N末端アミノ酸および少なくとも最後の20 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauより高い微小管構築促進活性を有し、
該微小管構築促進活性は微小管構築アッセイにおいて該分子に対する特異的阻害、中和モノクローナル抗体により除去することができ、該分子の病的活性は微小管重合促進活性により定義される微小管ネットワークに対する結合に依存する。
【請求項6】
配列番号11〜18の群から選ばれるアミノ酸配列を含むことを特徴とする請求項5記載のタイプIIA tau分子。
【請求項7】
以下の特徴により特徴づけられるNおよびC末端二重トランケートtau分子(「タイプIIB tau分子」):
4反復配列を含むtau43の少なくとも最初の68 N末端アミノ酸および少なくとも最後の40 C末端アミノ酸または3反復配列を含むtau44の最初の68 N末端アミノ酸および少なくとも最後の20 C末端アミノ酸がトランケートされており、
アルツハイマー病の脳組織に検出できるが、正常健康脳組織には検出できず、
in vitro微小管構築アッセイにおいて野生型tauと異なる病的微小管構築促進活性を有する。
【請求項8】
配列番号19および20の群から選ばれるアミノ酸配列を含むことを特徴とする請求項7記載のタイプIIB tau分子。
【請求項9】
以下の工程を特徴とする請求項1〜8のいずれかに記載の分子の製造方法:
a)少なくとも最初の236および最後の40アミノ酸または最初の68および最後の20アミノ酸またはその組み合わせにわたる欠失を有する二重トランケートtau分子をコードする配列を保持する組換え原核性発現プラスミドの構築、
b)該NおよびC末端二重トランケートtau分子の発現を許す条件下で該細菌を増殖させ、
c)好ましくは遠心により細菌を回収し、
d)細菌ペレットを再浮遊させ、
e)該細菌を超音波処理し、
f)該超音波処理細菌をゲルろ過により分画し、
g)得られた分画の活性を微小管構築アッセイによりモニターすることにより、タイプIおよびタイプII tau分子の異なる活性を同定する。
【請求項10】
該トランケーションが請求項1〜8のいずれかに記載のものであることを特徴とする請求項9記載の方法。
【請求項11】
微小管構築アッセイ活性が請求項1〜8のいずれかに記載のものである請求項9または10記載の方法。
【請求項12】
以下の工程を特徴とする請求項1〜8のいずれかに記載の分子の製造方法:
a)アルツハイマー罹患脳組織を得、
b)該病的(罹患)脳組織を緩衝液、特にTris緩衝液中でホモゲナイズし、
c)該ホモゲナイズ脳組織を硫酸アンモニウム沈殿し、
d)PIPES緩衝液に再溶解し、
e)該再溶解物質をゲルろ過で分画化し、
f)得られた分画の活性を微小管構築アッセイでモニターすることによりタイプIおよびタイプII tau分子の異なる活性を同定する。
【請求項13】
微小管構築アッセイ活性が請求項1〜8のいずれかに記載されていることを特徴とする請求項12記載の方法。
【請求項14】
以下の工程を含む請求項1または2記載の分子(タイプIA分子)とチューブリンの複合体の分解(disassembling)に有効な物質の試験方法:
a)タイプIA分子とチューブリンのタンパク質複合体を形成させ、
b)該タンパク質複合体を被検物質とインキュベーションし、タイプIA分子を阻害し、および/または野生型tauの微小管構築促進能力および微小管の正常機能を回復させる物質を同定する。
【請求項15】
以下の工程を含む野生型tau発現細胞系において請求項1または2に記載の分子(タイプIA分子)がチューブリンとの複合体の形成を開始するのを阻害するのに有効な物質の試験方法:
a)適切な制御領域の制御下でタイプIA分子をコードする機能的遺伝子を正常tauタンパク質発現細胞に導入し、
b)タイプIA分子とチューブリンのタンパク質複合体を形成させ、
c)試験する物質を該複合体を保持する細胞に適用し、
d)請求項1記載のタイプIA生物活性に対する該物質の効果を試験する。
【請求項16】
該タイプIIA分子と微小管との相互作用により病的微小管を生成させる生理的条件下で請求項5または6記載のタイプIIAと野生型tauタンパク質をインキュベーションすることを特徴とする微小管をin vitroで病的状態に変換する方法。
【請求項17】
以下の工程を含むタイプIIA分子を除去および/または中和し、タイプII分子により生じる生理学的微小管パラメーターおよび機能を修復するその特性について請求項5または6記載のタイプIIA分子の病的効果を中和することができる物質のスクリーニング方法:
a)請求項16記載のタイプIIA分子およびチューブリン存在下で病的微小管を形成させ、
b)該物質、タイプIIAおよびチューブリンの混合物をスクリーニングする物質とインキュベーションし、
c)タイプIIA分子により生じる病的微小管の形成を減少させる結果を試験する。
【請求項18】
以下の工程を含むタイプIIA分子発現細胞系における異常微小管形成および機能の促進において請求項5または6記載のタイプIIA分子のin vivo活性を阻害するのに有効な物質の試験方法:
a)タイプIIA分子をコードする機能的遺伝子を適切な制御領域の制御下で野生型tau発現細胞中に導入し、
b)タイプIIA tau分子と微小管の複合体を形成させて該複合体を病的微小管の形成に関与させ、
c)試験する物質を該複合体を保持する細胞に適用し、
d)タイプIIA生物活性、特に微小管ネットワークおよびその関連機能の修飾に対する該物質の効果を試験する。
【請求項19】
請求項1〜8のいずれかに記載の分子を発現するトランスジェニック動物。
【請求項20】
特にアルツハイマー病治療用の薬剤をスクリーニングおよび試験するためのアルツハイマー病の動物モデルとしての請求項19記載のトランスジェニック動物の使用。
【請求項21】
請求項1〜8、特に請求項1、2、5または6のいずれかに記載の分子および医薬的に許容される担体、特にアジュバントを含むワクチン。
【請求項22】
請求項1または2記載の分子とチューブリンの複合体の形成開始の阻害剤。
【請求項23】
寄託番号02060767でEuropean Collection of Cell Cultures(ECACC)、Porton Down、Salisbury、UKに寄託されたモノクローナル抗体DC44としての結合部分を含むことを特徴とする請求項22記載の阻害剤。
【図1】

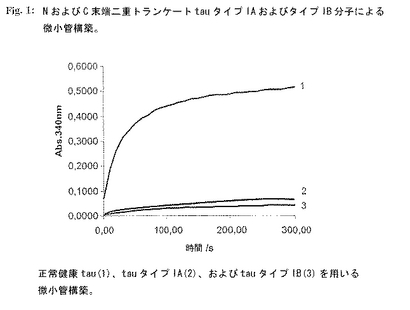
【図2】
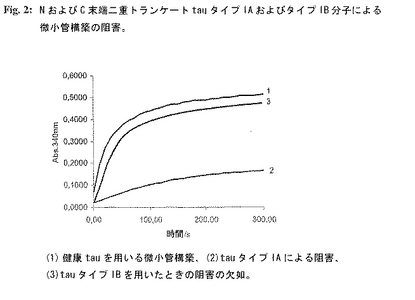
【図3】
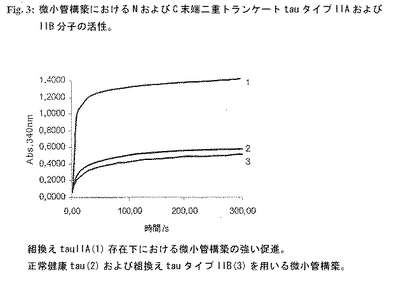
【図4】
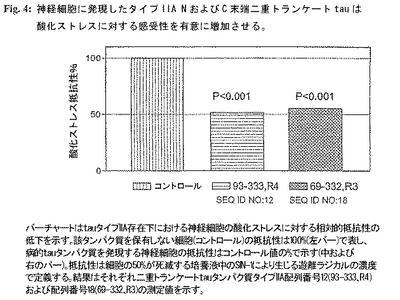
【図5】

【図6】
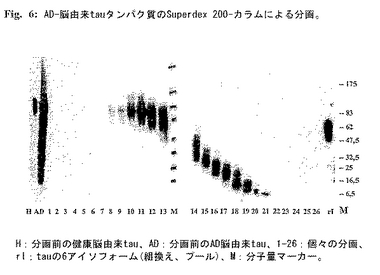
【図7】
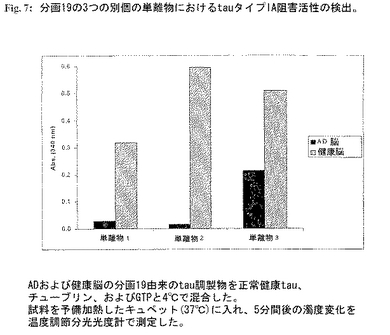
【図8】
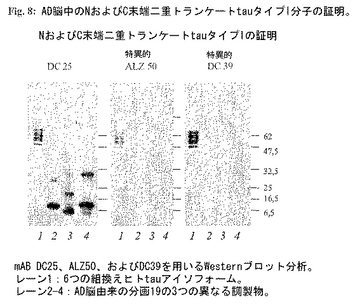
【図9】

【図10】

【図11】
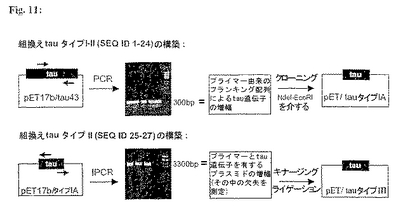
【図12】
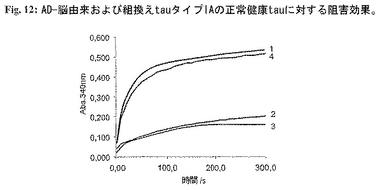
【図13】
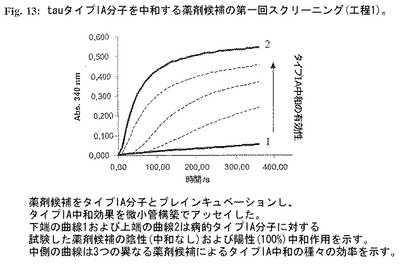
【図14】
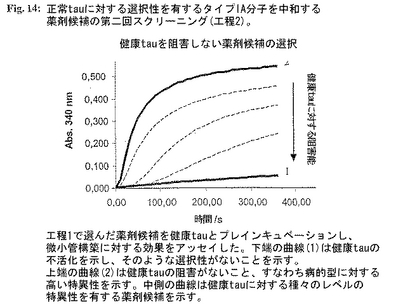
【図15】
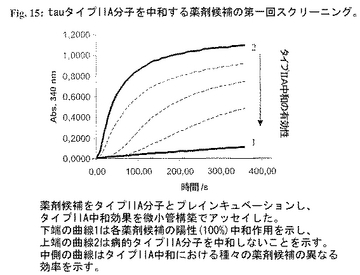
【図16】
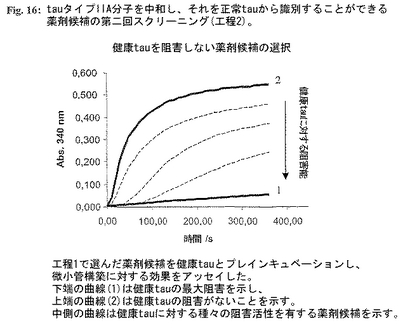
【図17】
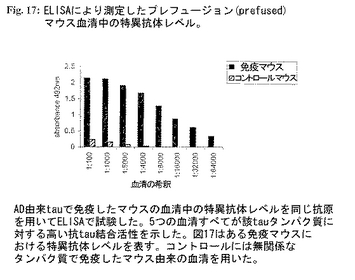
【図18】
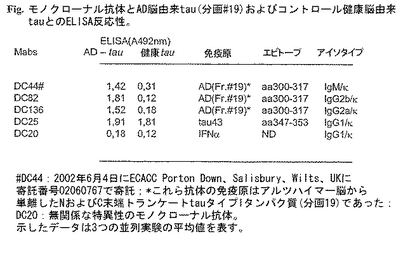
【図19】

【図20】
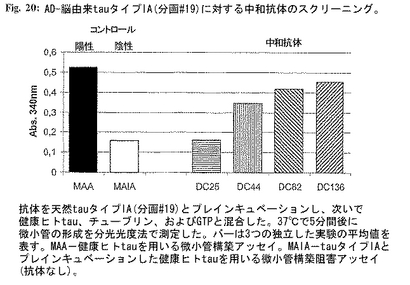
【図21】
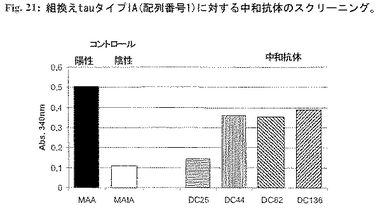
【図22】
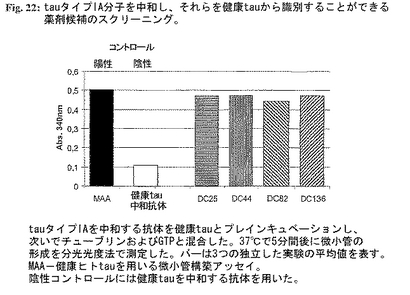
【図23】
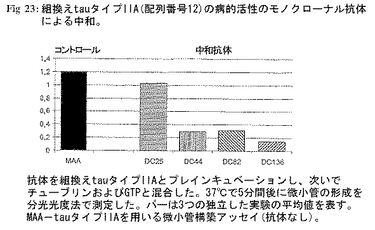
【図24】
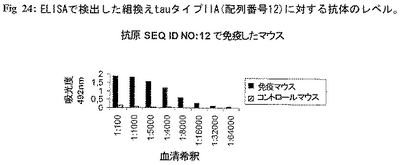
【図25】
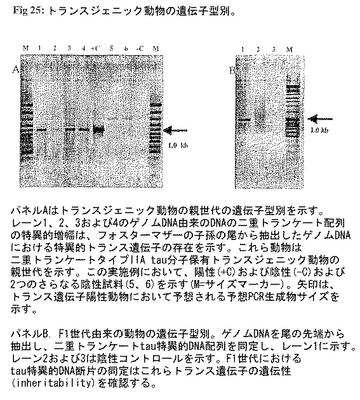
【図26】
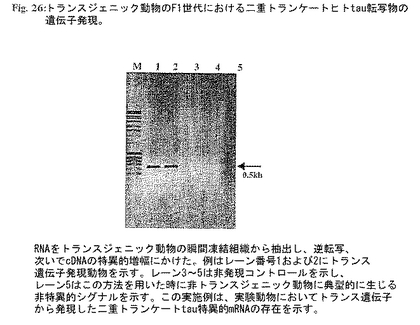
【図27】

【図28A】
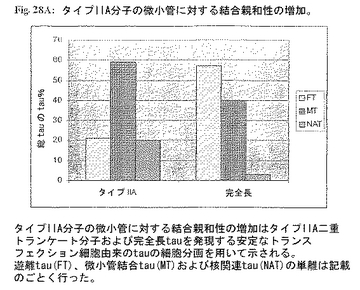
【図28B】
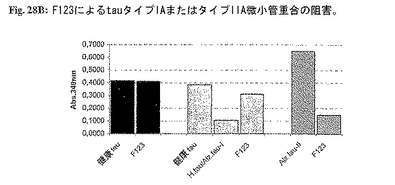
【図28C】
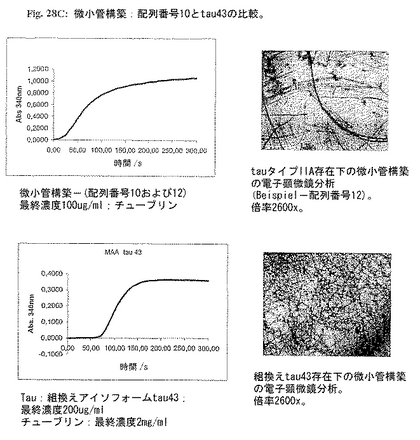
【図29】
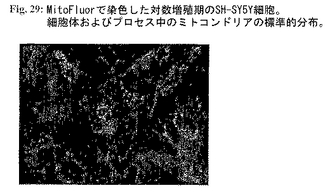
【図30】

【図2】
【図3】
【図4】
【図5】
【図6】
【図7】
【図8】
【図9】
【図10】
【図11】
【図12】
【図13】
【図14】
【図15】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図23】
【図24】
【図25】
【図26】
【図27】
【図28A】
【図28B】
【図28C】
【図29】
【図30】
【公表番号】特表2006−515270(P2006−515270A)
【公表日】平成18年5月25日(2006.5.25)
【国際特許分類】
【出願番号】特願2004−520541(P2004−520541)
【出願日】平成15年7月9日(2003.7.9)
【国際出願番号】PCT/EP2003/007389
【国際公開番号】WO2004/007547
【国際公開日】平成16年1月22日(2004.1.22)
【出願人】(503278588)アクソン・ニューロサイエンス・フォルシュングス−ウント・エントヴィックルングス・ゲゼルシャフト・ミット・ベシュレンクテル・ハフツング (2)
【氏名又は名称原語表記】AXON Neuroscience Forschungs− und Entwicklungs GmbH
【Fターム(参考)】
【公表日】平成18年5月25日(2006.5.25)
【国際特許分類】
【出願日】平成15年7月9日(2003.7.9)
【国際出願番号】PCT/EP2003/007389
【国際公開番号】WO2004/007547
【国際公開日】平成16年1月22日(2004.1.22)
【出願人】(503278588)アクソン・ニューロサイエンス・フォルシュングス−ウント・エントヴィックルングス・ゲゼルシャフト・ミット・ベシュレンクテル・ハフツング (2)
【氏名又は名称原語表記】AXON Neuroscience Forschungs− und Entwicklungs GmbH
【Fターム(参考)】
[ Back to top ]