
トランスジェニック非ヒト動物およびその使用
本発明は一般的には導入遺伝子構築物、導入遺伝子構築物を含むトランスジェニック非ヒト動物、該導入遺伝子構築物を含むトランスジェニック非ヒト動物の作製方法およびそれを用いる方法に関する。本発明の実施態様は、リガンドをGPCRレセプターに結合した後に経路変更に応答する生物発光導入遺伝子レポーターシステムを含むトランスジェニックモデルを用いて、全身の動物、組織片、またはネイティブ細胞中で非侵襲的にGPCRリガンドを試験する方法に関する。

【発明の詳細な説明】
【技術分野】
【0001】
本発明は一般的には導入遺伝子構築物、導入遺伝子構築物を含むトランスジェニック非ヒト動物、該導入遺伝子構築物を含むトランスジェニック非ヒト動物の作製方法およびそれを用いる方法に関する。本発明の実施態様は、リガンドをGPCRレセプターに結合した後に、経路変更に応答する生物発光導入遺伝子レポーターシステムを含むトランスジェニックモデルを用いて、全身の動物、組織切片、またはネイティブ細胞中で非侵襲的にGPCRリガンドを試験する方法に関する。
【背景技術】
【0002】
薬物開発においては、5つの化合物中の1つのみが米国食品医薬品局の承認(FDA)を通過しており、損耗率が高い(非特許文献1)。さらに、投資が劇的に増加したにもかかわらず、新薬の導入率は過去30年にわたりほぼ一定であり、結局年ごとに新薬種でわずか2〜3の進歩が市場にでるだけである(非特許文献2)。
【0003】
薬物開発の最初のステージで適用される分子イメージングおよび機能イメージングは、生物活性の証明を提供し、その企図される標的に対する効果を有する推定薬物(putative drug)を確認することができる。従って、分子イメージング技術への投資は薬物開発を推し進めるとかなり期待される(非特許文献3)。従来の情報を超える分子イメージング技術の利点は、その技術がインビボでの生物学的プロセスを研究するための十分な空間および時間的分解能により無処置固体(intact organism)に実施され得ることである。さらに、異なる時点における同一の生物学的モデルの反復性があり、非観血的で、均一かつ比較的自動化された研究を可能とするため、長期的研究の検出力を増加させ、必要とされる動物の数を減少させることができ、結果的に薬物開発のコストを低減させることができる。
【0004】
分子イメージング
分子イメージングとは、生きている製物中の細胞内および細胞小器官における特異的な分子プロセスの評価における画像処理技術を利用し、統合するための、様々な学問分野(細胞および分子生物学、化学、医学、薬理学、物理学、生物情報科学および光学)からのアプローチの集合のことを言う(非特許文献4)。
【0005】
遺伝子工学の出現は、創薬パイプラインなどを含む応用科学へ大きな変更をもたらしている。同じように、動物イメージング手段の開発および利用は、前臨床試験のための新しい集団を提供している(非特許文献5)。動物モデルは、リアルタイムで生理学的な事象を定量化するのが困難であるため、従来的にわずらわしいものであった。この困難を克服するために、長年にわたり磁気共鳴画像法(MRI)や陽電子放射断層撮影(PET)のような新しい画像処理法が開発されてきた。さらに最近では、ルシフェラーゼ、ホタルの発光酵素のインビボでの発光をベースにした生物発光イメージングが非侵襲的な検出のために用いられている。
【0006】
分子イメージング:生物発光
インビボでの生物発光イメージング(BLI)は、細胞または組織からの発光の検出にも続いている高感度なツールである。レポーター遺伝子技術の有用性は、インビボイメージング法を通して生きている動物中の特定の細胞および生物学的プロセスを分析することを可能にする。生物発光、つまり生きている生物による可視光の酵素生成は、多くの非哺乳動物種における天然に存在する現象である(非特許文献6)。ルシフェラーゼは、光の甲子を放出する基質の酸化を触媒する酵素である(非特許文献7)。北アメリカホタルからの生物発光は最も広く研究されている。このホタルルシフェラーゼ遺伝子(luc)の発現は、基質D−ルシフェリンを、562nmの緑色発光を生じる非反応性のオキシルシフェリンに変換する酵素であるルシフェラーゼを生成する。哺乳動物の組織は自然に生物発光を発しないため、画像処理が非常に小さなバックグラウンド信号を発生することができるのでインビボBLIはかなり魅力的である。BLIは、恒常的に光レポーターを駆動する選択遺伝子プロモーターの制御下で、生物発光レポーター遺伝子からなる発現カセットを有する細胞または組織の遺伝子操作を必要とする(図3)。光の産生を誘導するために、ルシフェリンなどの基質が脳室内(icv)、静脈内(iv)、腹腔内(ip)または皮下(sq)注射により投与される。
【0007】
ルシフェラーゼによって放出される光は、数ミリメートルから数センチメートルの深さに浸透することができる。しかし、光子の強度は、組織の深さの各センチメートルに対して10倍減衰する(非特許文献8)。高感度な光検出装置を、インビボで生物発光を検出するために用いる必要がある。この検出器は、単位面積当たりに発光される光子数を測定する。400〜1000nmの波長での低レベルの光は、シリコンウエハーを打ち、電子に変換する電荷結合素子カメラで検出することができる(非特許文献9)。このソフトウエアは、電子信号を二次元画像に変換することができる。このソフトウエアはまた、放出される光の強度(検出器を打つ放出光子の数)の定量化と、これらの数値の疑似グラフィックまたはグレースケールイメージへの変換が可能である(図2Aおよび2B)。実際のデータは光子で測定されるが、疑似グラフィックは、迅速な視覚的解釈を可能にする。目的の領域内の定量的な測定は、より微妙な差異の場合必要となり得る。冷却電荷結合素子(CCD)カメラの使用により熱雑音を低減し、遮光用の箱を用いるとルシフェラーゼ産生光が最適に可視化および定量化することを可能にする(非特許文献10)。発光シグナルの解剖学的位置についてのオートグラフやレントゲン写真のような別のタイプの画像に重ねてルシフェラーゼイメージを持っておくことは有用である(図2B)。このソフトフエアは、視覚化と解釈のために画像を重ね合わせる。動物の遺伝子操作と分子イメージング技術を組み合わせることによって、生きている動物中の特定の分子プロセスにおける動的研究を行うことが可能となる。このアプローチは、前臨床試験のプロトコルに影響を与え、それにより医学の全ての局面を広く変更する可能性がある(非特許文献11)。
【0008】
G−タンパク質共役受容体(GPCR)−創薬ターゲットとしてのGPCR
GPCRは、その共有される位置構造にも続いて100以上のサブファミリーに分類されている細胞表面受容体の大きなスーパーファミリーを構成し;GPCRはまた、7回膜貫通(7TM)受容体とも呼ばれている。GPCRは、製薬業界の中で最も頻繁に扱われる創薬ターゲットである。全ての市販処方薬のうちの約30%がGPCRを標的化しており、このタンパク質ファミリーを最も成功したターゲット類としている(非特許文献12)。GPCRとそれらの細胞外リガンドとの相互作用は、治療薬の干渉の魅力的な点であることが証明されている。このため、製薬業界は、これらのリガンド−GPCR相互作用を調査するために生化学的創薬アッセイを開発した。活性化GPCRとヘテロトリマーG−タンパク質との相互作用は、いくつかの下流エフェクターとの相互作用を可能にするグアノシン三リン酸(GTP)により、グアノシン二リン酸(GDP)の交換を触媒する(非特許文献13)。下流のシグナル伝達は、目的のGPCRに好ましいGαアイソフォームに依存している。Gαi/0およびG−αsのファミリーの代表が主にアデニル酸シクラーゼ(AC)活性を調節する一方、Gαq/11ファミリーのタンパク質がホスホリパーゼC(PLC)を刺激する。目的のGPCRがPLCを経由して信号を発する場合、その後のGPCRの活性化を測定するために最も広く適用されたレポーターベースの技術は、Ca+2感受性発蛍光団を用いた蛍光形式(非特許文献14)またはエクオリンと化学発光基質を用いた発光形式(非特許文献15)のいずれかで測定されるカルシウム(Ca+2)放出アッセイである。目的のGPCRがACを経由して信号を発する場合、その後の細胞質環状アデノシン一リン酸(cAMP)含有量は、種々の検出技術を用いて検出され得る(非特許文献16)。
【0009】
GPCRレポーターベースのアッセイは、現在の創薬プログラムにおいて広範囲に用いられている。一般的に、GPCRレポーターは、特定のGPCRを活性化するかまたは調節するリガンドまたは化合物を同定するために、大きな製薬ライブラリのインビトロでのハイスループットスクリーニング(HTS)をサポートするための細胞ベースのシステムに導入されている。第二の継続した細胞ベースのアッセイは、特定のGPCRに対するHTSで同定された全ての「ヒット」を確認および洗練する;しかし再び、これらのアッセイは形質転換された細胞型中にクローン化されたGPCRを導入する組み換えDNA法に依存している。形質転換された細胞型は、大きなスクリーニングプログラムをサポートする優れた増殖能を有しているが、これらはしばしば異常な遺伝的および機能的特性を示し、結果としてHTSから推定される「ヒット」の有意な消耗がこれらのパラダイムを用いて検出される。
【0010】
数年にわたり、生物発光ベースのレポーター遺伝子アッセイは、GPCRの機能的活性を測定するために用いられてきた(非特許文献17)。このアッセイ形式は、生物発光の読み出しの低いシグナルバックグラウンドおよび、GPCRの活性化と累積レポーター遺伝子発現との間の信号増幅工程のおかげで非常に感受性が高い。
【0011】
レポーター遺伝子のプロモーター中のcAMP応答配列(CRE)は、Gタンパク質依存性のシグナル伝達の特異的なモニタリングを可能にする。リガンドがGPCRに結合すると、それは付随Gタンパク質の活性化を可能にする、GPCRの立体構造変化を引き起こす。酵素アデニル酸シクラーゼは、Gタンパク質によって調節され得る細胞のタンパク質である。アデニル酸シクラーゼ活性は、活性化Gタンパク質のサブユニットに結合すると活性化されるかまたは抑制されるかのいずれかである。シグナル伝達は、Gタンパク質のタイプに依存している。アデニル酸シクラーゼは、細胞中のcAMP産生を増加させるかまたは減少させるかのいずれかの役割を果たしている。このcAMP産生は、細胞代謝におけるセカンドメッセンジャーであり、またプロテインキナーゼA(PKA)のアロステリック活性化因子である。cAMPが存在しない場合、PKA複合体は不活性である。cAMPは、PKAの調節サブユニットに結合すると、それらのコンフォメーションは変更され、プロテインキナーゼAを活性化し、さらなる生物学的効果を可能にする調節サブユニットの解離を引き起こす。次いで、PKAが転写因子CREBをリン酸化し活性化する。CREBは、cAMP応答配列(CRE)と呼ばれる特定のDNA配列に結合することによって転写を増加または減少させ、それによりルシフェラーゼレポーター遺伝子のような特定の遺伝子を発現する。
【0012】
CreLuc導入遺伝子は、cAMP細胞内シグナル伝達経路を通して直接的にかまたはPLCを通して間接的にかのいずれかで3つの主要なGPCR全てのアッセイを活性化するように設計されている。任意の1つの細胞型は、それらの細胞表面上に多くの異なるタイプのGPCRを含んでいるため(従って全ての細胞は、細胞内で同時に生じるG−αq/11、G−αi/0およびG−αsを経由するGPCRシグナル伝達を有している)、従来の知識は、CreLucのような導入遺伝子が任意のひとつの特定GPCRリガンドを識別するのに十分特異的であることはありそうもないということを示唆している。しかし、本発明者らは、本明細書中でCreLuc導入遺伝子がGPCRリガンドを識別することができることを示している。本発明者らは、細胞内のルシフェラーゼレポーター、組織切片、または全体の動物の生物発光シグナルがホルスコリンによって増加されること、およびGs、GqまたはGiのリガンドによって調節されることを予測している。表1は、GPCRリガンドに結合する際に、CreLucレポーターシステム上でGPCRの活性化/抑制が起こるという予測された効果を示している。さらに、本発明者らは、新規のCreLucレポーターシステムがGPCRリガンドの異なるクラスを識別し、そのようなレポーターシステムは、細胞、組織切片および全体の動物中で用いられるときに新規のGPCRリガンドを同定するために適用可能であるというデータを示している。
【0013】
【表1】
【0014】
GPCRバイオイメージングレポータートランスジェニックモデル
現在の創薬パラダイムにおける潜在的な薬物候補の顕著な消耗は、細胞ベースのレポーターアッセイからインビボモデルへの相転移においておきている。多くのインビボモデルは、特定のヒト疾患の全てまたは一部を反復することが可能である。これらのモデルにおけるリード化合物を証明することは、新しい化学GPCR薬の進行に重要なマイルストーンである。動物疾患モデルは、通常それらの表現型および、疾患転帰の変更に及ぼす候補化合物の影響の正確なアッセイの開発を可能にするために、多数の動物と時間を必要とする。インビトロでの試験後の、複雑なシステムでの薬物候補を試験する次のレベルは、疾患状態のインビボ試験またはインビボモデルを用いており、これらは機械ベースである。誘導された疾患転帰を変更する失敗はあまり理解されないが、いまだに薬物開発のパイプラインにおいて候補化合物の大きな損耗率を生じている。
【0015】
GPCRリガンド結合を含むトランスジェニックモデルおよび活性化レポーターアッセイは、GPCRの現在の薬物開発パラダイムの大幅な改善となる。例えば、本発明の実施態様は、GPCRリガンド薬物開発を大幅に促進する、全身の動物、組織、または細胞における分子イメージングと組み合わせた、ルシフェラーゼレポーター(CreLuc)をベースにしたcAMPレポーターアッセイを含む導入遺伝子を記載している(非特許文献18;非特許文献19)。本明細書中に記載されるように、本発明のトランスジェニック非ヒト動物の実施態様は、以下の非限定的な利点を提供する:
【0016】
1.組織や細胞ベースのアッセイはトランスジェニックインビボモデルアッセイと同じレポーターシステムを持っているため。複雑な無処置の生物学的システムの未知数の数を減らす。
2.非侵襲的な画像処理は、同じ動物において、経時的アッセイにおけるリガンドや化合物活性の定量分析を可能にする。
3.非侵襲的な画像処理は、研究あたりの動物の数を減らし、各動物が自身のコントロールになることによって、より大きな検出力につながる。ここで、コントロールとは時間ゼロにおいてアッセイされる動物である。
4.トランスジェニック動物は、インビトロまたはエキソビボでなされる並行アッセイをサポートするための細胞および組織の供給源になる。
5.トランスジェニック動物のアッセイは、ネイティブ細胞型におけるリガンド活性のアッセイをサポートし、リガンドのより現実的なプロファイルを導く。
6.トランスジェニック動物は、GPCRリガンドの約力学および薬物動態の同時評価を可能にする。
7.トランスジェニック動物は、器官レベルまたは全身の動物レベルいずれかで、組織および細胞型の特異性の同時識別を可能にする。
8.トランスジェニック動物は、新規のシグナル伝達経路および特定のリガンドへのそれらの応答を明らかにするための、他の遺伝的に改変されたモデルとの交配を可能にする。
【0017】
異なるレポーターを用いて遺伝子操作された多くのトランスジェニック動物は、以下のような分子プロセスの研究に用いられている:薬物代謝(非特許文献20)、遺伝毒性(非特許文献21)および毒性化合物の効果(非特許文献22)。それらの設計目的を達成するために、分子イメージング研究に適切なGPCRレポーター動物は、研究下にあるリガンドまたは化合物の生体内分布における広範で鋭いインビボアッセイをサポートするための大きなウインドウの生物発光検出、ならびに全ての細胞型における発現の両方を可能にするように配置されるいくつかのエレメントを組み込む必要がある。
【0018】
遺伝子発現に関与するメカニズムの複雑さと多様性により、研究者らは、完全に予測可能な方法でトランスジェニック動物中における発現の全てのケースにおいて有能な遺伝子を構築することができない(非特許文献23:非特許文献24;非特許文献25)。大規模な試行錯誤を通してのみ、導入遺伝子構造の独特の組み合わせを、GPCRレポーターのバイオイメージングの必要に応じて、モデルデザインの目的に到達させることができる。
【0019】
組換え細胞アッセイに対する、トランスジェニックGPCRレポーターの有用性
スクリーニング技術が、個々のGPCRの振る舞いを理解するポイントまで進むと、スイッチをオンオフするというよりもむしろ、これらの受容体が情報のマイクロプロセッサのように作用することが明らかである。このことは、機能の選択性の現象を導入しており、それにより特定のリガンドは所定の受容体によって媒介されるシグナル伝達機構の一部分のみを開始することによって、薬物開発の新たな地平を切り開く。任意の薬理学的レポーターアッセイにおける非常に高い要求を置く医薬品化学を導くために、これらの複雑なシステムにおいて新規のGPCRリガンドの関係を発見し、薬物の効果を定量化する必要がある。この概念は、組換え細胞ベースのスクリーニングアッセイから全体のシステムのアッセイへ戻る原動力となる。ネイティブ細胞環境中で特定のGPCRまたはGPCRのセットによってリガンドの活性をプロファイリングし、重要なクラスの薬学的に重要な受容体に対する新しい薬物を同定するための成功率を向上させることが期待される(非特許文献26)。生物発光GPCRレポーター導入遺伝子を含む動物モデルは、無処置の生物学的に複雑なシステム中でGPCRリガンド活性を定義する非常に望ましい分子イメージングストラテジーであり、ヒトの疾患と戦うための薬物開発を向上させる目的を有している。
【0020】
CRE/CREBの活性化は、多くの可変的な生物学的プロセスに関与しており、CRE/CREBレポーター発現系を用いることによるCREの活性化の研究に大きな関心がある。環状アデノシン一リン酸(cAMP)は、受容体の活性化とひき続くタンパク質キナーゼの活性化後の、細胞内シグナル伝達におけるセカンドメッセンジャーであり、よって多くの生物学的プロセスの調節に関与している。CREB(cAMP応答配列結合タンパク質)は、cAMPによって活性化されたキナーゼによってリン酸化され、多くの遺伝子のプロモーター領域中のcAMP応答配列(CRE)に結合して転写を活性化させる(非特許文献27)。β−ガラクトシダーゼ発現を駆動する最小単純ヘルペスウイルス(HSV)プロモーターを持つ6つのタンデムCREを保持するトランスジェニックマウスを用いて、慢性的な抗うつ薬治療に応答する脳スライス中のCRE媒介性遺伝子発現が研究された(非特許文献28)。同様に、チミジンキナーゼプロモーターに融合するラットソマトスタチン遺伝子プロモーターCREの4つのコピー、およびルシフェラーゼ遺伝子を保持するトランスジェニックマウスを用いて、組織学的脳スライスまたはホモジネート中のCRE活性化が研究された(非特許文献29)。しかしながら、これまでの研究は、適切な動物モデルを見つけるために多くのトランスジェニック系統をスクリーニングする必要性によって、妨げられてきた。さらに、適切な系統が確認された後、比較的低いレポーター発現レベルでは、レポーター遺伝子を測定するためにトランスジェニック動物を安楽死させる必要があり、多くの動物を1つの実験系に用いる必要がある。
【0021】
本発明の実施態様は、インスレーター配列、応答配列、プロモーターエレメント、レポーター遺伝子および機能エレメントを含む導入遺伝子の開発である。この導入遺伝子は、その早い速度での組込みと高いレベルでのレポーター発現によって、迅速に非ヒト動物中に導入することができ、それによりインビボ(すなわち生きている動物中)、インサイチュ(例えば、脳スライス、無処置の固体)またはインビトロ(例えば、トランスジェニック動物から培養された初代細胞、組織ホモジネート)でのエレメント活性の調節を研究するためのモデルとして、トランスジェニック動物を容易に開発することができる。
【0022】
本発明の実施態様は、インビボで細胞内cAMPレベルの調節を通してGPCRリガンド活性を定量化するモデルとしてトランスジェニック非ヒト動物中で用いられるCRE Lucレポーターシステムを含む導入遺伝子である。非限定的な例として、本発明者らは、一般的なcAMP調節因子を用いて、単離された初代細胞中および全身の動物中の生物発光によるルシフェラーゼレポーターの変化を示している。別の実施態様において、このレポーターの活性化はエキソビボでのルシフェラーゼアッセイを用いて組織抽出物中でアッセイされ確認されている。CRE Luc導入遺伝子の応答は、複数のマウス系統において記録されており、単一または複数の組織活性化プロファイルを示している。さらに非限定的な例として、本発明者らは、特定のGPCRリガンドが全身の動物、組織スライス、および初代細胞中のCRE Luc導入遺伝子を活性化することを実証した。
【先行技術文献】
【非特許文献】
【0023】
【非特許文献1】DiMasi、JA、et al、J Health E con 22,151−185、2003
【非特許文献2】Lindsay MA、Nature Rev Drug Discovery、2、831−838、2003
【非特許文献3】Rudin M、Progress in Drug Res vol 62
【非特許文献4】Massoud T.F.、Genes Dev.17:545−580、2003
【非特許文献5】Maggie A.and Ciana P.、Nat.Rev.Drug Discov.4、249−255、2005
【非特許文献6】Contag、C.H.、et al、Mol.Microbiol.18:593−603、1995
【非特許文献7】Greer L.F.、III、Luminescence 17:43−74、2002
【非特許文献8】Contag、C.H.、et al、Mol.Microbiol.18:593−603、1995
【非特許文献9】Spibey CP et al electrophoresis 22:829−836、2001
【非特許文献10】Contag C.H.and Bachmann、M.H.、Annu.Rev.Biomed.Eng.4:235−260、2002
【非特許文献11】Maggie A.Trends Pharmacolo.Sci 25、337−342、2004
【非特許文献12】Jacoby、E;Chem.Med.Chem.、1:761−782、2006
【非特許文献13】Cabrera−Vera T.M.、Endocr.Rev.24:765−781、2003
【非特許文献14】Sullivan E、Methods Mol.Biol.114:125−133、1999
【非特許文献15】Dupriez V.J.、Receptors Channels 8:319−330、2002
【非特許文献16】Gabriel D.Assay Drug Dev.Technol.1:291−303、2003
【非特許文献17】Hill、S.J.Curr.Opin.Pharmacol.1:526−532、2001
【非特許文献18】Bhaumik、S.and Gambhir、S.S.、Proc.Natl.Acad.Sci.USA、99:377−382 2002
【非特許文献19】Hasan M.T.、et al.、Genesis 29:116−122、2001
【非特許文献20】Zhang W.、et al.Drug Metab.Dispos.31:1054−1064、2003
【非特許文献21】Gossen J.A.、et al.、Proc.Natl.Acad.Sci.USA 86:7971−7975、1989
【非特許文献22】Sacco M.G.et al.、Nat.Biotechnol.15:1392−1397、1997
【非特許文献23】Pinkert、C.A.(ed.)1994.Transgenic animal technology
【非特許文献24】A laboratory handbook.Academic Press、Inc.、San Diedo、Calif.
【非特許文献25】Monastersky G.M.and Robl、J.M.(ed.)(1995)Strategies in Trans genic Animal Science.ASM Press.Was hington D.C
【非特許文献26】Kenakin TP、Nat.Rev.Drug Discov.8,617−625、2009
【非特許文献27】Shaywitz and Greenberg、Annul.Rev.Biochem.、68:821−861、1999
【非特許文献28】Thome J.、et al.、J.Neurosci.20:4030−4036、2000
【非特許文献29】Boer et al、PloS One、May 9;2(5):e431、2007
【発明の概要】
【課題を解決するための手段】
【0024】
概して、本発明は、導入遺伝子構築物、導入遺伝子構築物を含むトランスジェニック非ヒト動物、該導入遺伝子構築物を含むトランスジェニック非ヒト動物の作製方法およびそれを用いる方法を提供する。本発明の実施態様は、CRE Lucレポーターシステムを含む導入遺伝子構築物を提供する。本発明の実施態様は、非ヒト動物中への、CRE Lucレポーターシステムを含む導入遺伝子構築物の導入である。
【0025】
cAMP変調は、GPCRのための主要な活性化経路であるため、本発明は全身の動物、組織切片、または細胞中で、レポーター遺伝子(このレポーター遺伝子は、ルシフェラーゼによるルシフェリンの代謝など測定可能な生物発光シグナルを提供する)の活性化を通してリガンドまたは化合物によるGPCRの活性化を定量化するためのプラットフォームとしての役目を担う。本発明は、細胞ベースのアッセイから全身の動物まで、リガンドまたは化合物のような新規に発見された薬物成分(new drug discovery entities)の移行を向上させるためのツールを供給する。本発明の実施態様は、ネイティブ細胞中で同じレポーターシステムを用いることで、バイオアベイラビリティーデータを同時に提供しながら、新規のGPCRリガンドの損耗率を低減する。
【0026】
本発明の実施態様は、第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を含むゲノムを有する、トランスジェニック非ヒト動物である。
【0027】
本発明の実施態様は、前記第1のインスレーター配列が、マトリックス付着領域(MAR)、DNase I−高感受性領域(HS4)および末端逆位配列(ITR)からなる群より選択される、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記第2のインスレーター配列が、マトリックス付着領域(MAR)、HS4およびITRからなる群より選択される、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記第1のインスレーター配列が、前記第2のインスレーター配列と同じである、トランスジェニック非ヒト動物である。
【0028】
本発明の実施態様は、前記応答配列が、cAMP 応答配列(CRE)、アクチベータータンパク質1(ASP1)、グルココルチコイド応答配列(GRE)、熱ショック応答配列(HSE)、血清応答配列(SRE)、甲状腺ホルモン応答配列(TRE)およびエストロゲン応答配列(ERE)からなる群より選択される、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記応答配列がタンデムに2〜24回反復されている、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記応答配列がタンデムに6回反復されている、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記応答配列がCREであり、さらにこのCRE応答配列がシングルエレメントであるかまたは2〜24回反復されていてもよい、トランスジェニック非ヒト動物である。
【0029】
本発明の実施態様は、前記プロモーターが単純ヘルペスウイルスチミジンキナーゼの最小ポロモーター(HSV TK min)である、トランスジェニック非ヒト動物である。
【0030】
本発明の実施態様は、生物発光レポーターが、ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、β−ガラクトシダーゼ、分泌型アルカリホスファターゼ(SEAP)、ヒト成長ホルモン(HGH)および緑色蛍光タンパク質(GFP)からなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物である。
【0031】
本発明の実施態様は、前記機能エレメントがヒト成長ホルモン(hGH)遺伝子である、請求項1に記載のトランスジェニック非ヒト動物である。
【0032】
本発明の実施態様は、前記導入遺伝子が配列番号18を含む、トランスジェニック非ヒト動物である。
【0033】
本発明の実施態様は、前記導入遺伝子が配列番号19を含む、トランスジェニック非ヒト動物である。
【0034】
本発明の実施態様は、前記トランスジェニック非ヒト動物から単離された細胞であるかまたは請求項1に記載のトランスジェニック非ヒト動物から単離された組織切片である。
【0035】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物中の生物発光の量を測定する工程;
(b)該トランスジェニック非ヒト動物に試験薬剤を投与する工程;
(c)該試験薬剤の投与後に、1またはそれより多い時点で該トランスジェニック非ヒト動物の生物発光の量を測定する工程;
(d)(a)において測定した生物発光の量と(c)において測定した生物発光の量とを比較する工程;
を含み、ここで(c)と比較した(a)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【0036】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物から組織切片を調製する工程;
(b)該組織切片中の生物発光の量を測定する工程;
(c)該組織切片に試験薬剤を投与する工程;
(d)該試験薬剤の投与後に、1またはそれより多い時点で該組織切片の生物発光の量を測定する工程;および
(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程;
を含み、ここで(d)と比較した(b)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【0037】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物から単離された細胞を調製する工程;
(b)該細胞中の生物発光の量を測定する工程;
(c)該細胞に試験薬剤を投与する工程;
(d)該試験薬剤の投与後に、1またはそれより多い時点で該細胞の生物発光の量を測定する工程;および
(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程;
を含み、ここで(d)と比較した(b)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【0038】
本発明の実施態様は、非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)該非ヒト動物からの生物発光をモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【0039】
本発明の実施態様は、非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)該非ヒト動物からのルシフェラーゼをモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【0040】
本発明の実施態様は、非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)疾患状態の状況を模倣するために非ヒト動物を操作する工程;
(c)該非ヒト動物からの生物発光をモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【0041】
本発明の実施態様は、非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)疾患状態の状況を模倣するために非ヒト動物を操作する工程;
(c)該非ヒト動物からのルシフェラーゼをモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【0042】
本発明の実施態様は、GPCR機能のモニタリングにおいて使用するための非ヒトトランスジェニック動物を作製する方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)(a)のトランスジェニック非ヒト動物における生物発光の量を測定する工程;
(c)GPCRリガンドを該トランスジェニック非ヒト動物に投与する工程;
(d)該GPCRリガンドの投与後に、1またはそれより多い時点で該トランスジェニック非ヒト動物の生物発光の量を測定する工程;および
(e)(b)で測定した生物発光の量と(d)で測定した生物発光の量を比較する工程;を含み、ここで(d)と比較した(b)の生物発光の量の差が、GPCR機能をモニタリングするのに用いるための非ヒトトランスジェニック動物を同定する、上記方法。
【0043】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されるトランスジェニック非ヒト動物から単離された細胞を準備する工程;(b)該細胞中の生物発光の量を測定する工程;(c)試験薬剤を該細胞に投与する工程;(d)試験薬剤の投与後に、1またはそれより多い時点で該細胞中の生物発光の量を測定する工程;(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程であって、ここで(b)と(d)を比較した場合の生物発光の量の差異は、GPCRリガンドとして試験薬剤を同定している、工程。
【0044】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)コントロールを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、コントロールを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0045】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)コントロールを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、コントロールを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0046】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)一般的なcAMPモジュレーターを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、一般的なcAMPを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0047】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された組織切片を1つ以上のレセプタクル中に供給する工程;(b)コントロールを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、コントロールを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0048】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された組織切片を1つ以上のレセプタクル中に供給する工程;(b)一般的なcAMPモジュレーターを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、一般的なcAMPモジュレーターを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0049】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)細胞刺激因子を1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、細胞刺激因子を含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0050】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)細胞刺激因子を1つ以上のレセプタクルに投与する工程;(c)一般的なcAMPモジュレーターを1つ以上のレセプタクルに投与する工程;(d)試験薬剤を1つ以上のレセプタクルに投与する工程;(e)該レセプタクル中のルシフェラーゼの量を測定する工程であって、細胞刺激因子および一般的なcAMPモジュレーターを含むレセプタクル中で測定されたルシフェラーゼの量と細胞刺激因子および一般的なcAMPモジュレーターならびに試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0051】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された組織切片を1つ以上のレセプタクル中に供給する工程;(b)細胞刺激因子を1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、細胞刺激因子を含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0052】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された組織切片を1つ以上のレセプタクル中に供給する工程;(b)細胞刺激因子を1つ以上のレセプタクルに投与する工程;(c)一般的なcAMPモジュレーターを1つ以上のレセプタクルに投与する工程;(d)試験薬剤を1つ以上のレセプタクルに投与する工程;(e)該レセプタクル中のルシフェラーゼの量を測定する工程であって、細胞刺激因子および一般的なcAMPモジュレーターを含むレセプタクル中で測定されたルシフェラーゼの量と細胞刺激因子および一般的なcAMPモジュレーターならびに試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【図面の簡単な説明】
【0053】
【図1】CreLucバイオイメージングマウスモデルを示している。この図は、3つのタイプのGPCR(Gi、GsおよびGq)全てがcAMP経路を経由して直接的にかまたはPLC経路を経由して間接的にかのいずれかによる、CRE Lucレポーター導入遺伝子の細胞内活性化を示している(パネルA)。ホルスコリン誘導に応答するルシフェラーゼレポーターの生物発光における変化を3つのタイプのGPCRについて示している(パネルB)。ホルスコリンは、GsおよびGqシグナルを増加、すなわちCreLuc生物発光を増加させる一方、Gi誘導はそのレポーターからのシグナルを減少させる。GαsはACの直接的な刺激によってcAMP依存性経路を活性化し、GαsはcAMPの生成を阻害し、そしてGαqは2つのセカンドメッセンジャーIP3およびDAGの発生をもたらすPLCを刺激する。略号:α、Gタンパク質のαサブユニット;β、Gタンパク質のβサブユニット;γ、Gタンパク質のγサブユニット;AC、アデニル酸シクラーゼ;PLC、ホスホリパーゼC;PKA、プロテインキナーゼA;PKC、プロテインキナーゼC;DAG、ジアシルグリセロール;IP3、イノシトール三リン酸;Ca+2、カルシウム;CaMK、カルシウム/カルモジュリンプロテインキナーゼ;cAMP、環状アデノシン一リン酸;CRE、cAMP応答配列;CREB、cAMP応答配列結合タンパク質。
【図2A】リアルタイムのインビボ生物発光イメージングを示し、そのシステムを用いるメリットを記載している。コンピュータ分析システムを備えるこのIVIS100(Xenogen)バイオイメージング装置は、生物発光レポーター遺伝子、例えばルシフェラーゼによって発せられた光を利用してリアルタイムのインビボイメージングを可能にする。このソフトウエアは、シグナルの数量化を非観血的かつ長期的にサポートする。リアルタイムのインビボイメージングは、従来のインビボでの化合物試験に対して多くの利点を有する。従来の動物試験は、複数の治療点で個々のマウスが必要となる一方、バイオイメージングモデルを利用した研究では、同じ動物を複数の時点でサンプリングし、複数の治療のために再利用することができる。この図に示されるように、0時間、2時間、4時間および8時間の時間経過は、現在の方法論を用いると24匹の動物(各時点あたりn=6)が必要であるにもかかわらず、バイオイメージング技術を用いると必要とされるのは6匹の動物のみである。これは、以下を含むいくつかの利点をもたらす:より多くの化合物が効果について試験されることを可能にするのに必要な試験動物はより少ないので、よりハイスループットである;時間的および空間的データが同じ動物から採取することができるので、データの内容および質がより高い;および統計的誤差が減少し、個々の化合物についてなされる決定の質が向上する。
【図2B】CreLucトランスジェニックマウスにおける化合物誘導の典型的な視覚イメージを示している。イソプロテレノール(右パネル)の投与は、基底レベル(左パネル)と比較してCRE Lucレポーターの脊髄の発現を増加させる。この生物発光検出は、グレースケールでの疑似表現として動物の白色光画像に視覚的に表わされる。
【図3】発現を高めるために複数のDNAエレメントを含んでいる導入遺伝子構造の模式図を示している。この模式的な導入遺伝子構造は、以下のエレメントを含んでいる:位置に依存しない発現を産生するためのマトリックス付着領域(MAR)としてこの図に示されているインスレーター配列;6回反復されたCRE−cAMP(6×CRE)によって表わされる応答配列;単純ヘルペスウイルスチミジンキナーゼの最小プロモーター(HSV TK min)として示されるプロモーターエレメント;哺乳動物発現に最適化されたルシフェラーゼ遺伝子(LUC2)によって表わされるレポーターエレメント;および遺伝子発現を増強するためにポリA末端を有するヒト成長ホルモン遺伝子(hGH poly A)によって表わされる機能エレメント。
【図4】CreLuc導入遺伝子ベクターのインビトロでの検証である。30、100、または199ngのDNAでのハイブリッドまたは合成(シンセ)ベクターを、リポフェクタミン2000(Invitrogen、Carlsbad、CA、cat #11668−019)を用いてCHO細胞(ウミシイタケルシフェラーゼポジティブコントロールベクターと一緒に)にトランスフェクションした。2日後、その細胞を3μMのフォスルコリン(Sigma、St Louis、MO、cat #F6886)を用いて4時間刺激し、その後Dual Glo luciferase Assay System(Promega、Madison、WI、cat# E2920)を用いてルシフェラーゼ活性を測定した。結果は、両方のベクターでルシフェラーゼシグナルの用量依存性の増加を示している。より高いレベルの誘導がハイブリッドベクターで達成された。
【図5A】正常マウスにおけるcAMPレベルのPDF阻害剤の効果を示している(Cheng JB、JPET、280、621−626からの再現)。Balb/cマウスにx軸上に列挙されているビヒクルまたは薬物を服用させ、20分後にその血液を採取してcAMPラジオイムノアッセイによりcAMPをアッセイした。CP−80、633とロリプラムの両方が10mg/kgでの血漿cAMPレベルを大幅に増加させた。
【図5B】血漿中のcAMPのインビボ刺激を示している。FVB/Tacメスにビヒクル(1% DMSO)または薬物をi.p.で服用させ、30分後血液サンプルを採取し、ELISA(Assay Designs、 Ann Arbor、MI、cat#900−163)によってcAMPをアッセイした。用いた薬物は以下のとおりである:5mg/kg ホルスコリン(F)(Sigma F6886)、5mg/kg 水溶性ホルスコリン(H20F)(Calbiochem 344273)、10mg/kg ロリプラム(R)(Sigma R6520)またはホルスコリンプラスロリプラム(F/R)の組み合わせのいずれか。統計的に有意な増加である14倍が、t−検定によって決定されるロリプラムと組み合わせた水溶性ホルスコリンの処理で観察された。ホルスコリンがアデニル酸シクラーゼを活性化することによってcAMPレベルを増加させる一方、ロリプラムのようなPDE4の阻害剤はcAMPの加水分解を防止することによって血漿内cAMPを上げる。ロリプラムと水溶性ホルスコリンの組み合わせは、インビボでcAMPレベルを14倍増加させる。この組み合わせは、バイオイメージングによる初代のスクリーニングのための誘導の大きなウインドウを提供するために用いられ、代表的な研究を図6に示す。
【図6】CreLucレポーターマウスモデルの初代の誘導と系統の選択の結果を示している。複数のトランスジェニック系統を、インビボでのホルスコリンとロリプラムによるルシフェラーゼ誘導のためにスクリーニングし、その後組織を単離してルシフェラーゼ酵素についてアッセイした。トランスジェニックマウスを投薬前(基底発現レベル)でバイオイメージングし、その後同じマウスにi.p.で10mg/kgのロリプラムおよび5mg/kgの水溶性ホルスコリンを投与し、投薬の4時間後にバイオイメージングした(誘導された発現)。A(亜系統番号90):ホルスコリン/ロリプラム投与は、肺および他の組織においてCreLucレポーター遺伝子の基底発現を増加させた;B(亜系統番号219)基底発現の誘導は、主に腸内である;C(亜系統番号44):検出されない基底発現およびレポーターは、脳および他の組織に誘導される;D(亜系統番号28):胸腺および肝臓で増加される検出されない基底発現;E(亜系統番号187):脳および脊髄で誘導される検出されない基底発現。ランダムに統合された導入遺伝子について予想されるように、基底発現、組織分布、および誘導に対する応答において、系統間にばらつきがあった。20系統は、1つ以上の組織で5倍より大きい誘導を持つことが同定された。組織プロファイルの変化は、その単一の組織(すなわち、肺、肝臓、脳)が、バックグランド組織の応答のないイメージングを可能にする一方、複数の組織は、広範な化合物の応答プロファイルが作成されることを可能にする。
【図7】インビボまたはエキソビボでのCreLucスクリーニングアッセイの一般的な概略図を示している。
【図8A】CreLucマウス(系統187)由来の全脳スライスにおける発光に対するイソプロテレノール(ISO)およびAMN082(AMN)の効果を示している。
【図8B】時間の経過とともに、CreLucマウス(系統44)由来の全脳スライスにおける発光に対するホルスコリンの効果を示している。時間は、分単位でX軸上に示されている。50uMのホルスコリンまたはビヒクル(DMSO)を、下のパネルの矢印でマークされた時間=2880で加えた。
【図9】CreLucマウス由来の初代神経細胞の単離および化合物処理を表わす模式図を示している。
【図10】β−アドレナリン受容体(ADβR)の活性化とD1ドーパミン受容体(DRD1)の活性化を介したGsの変調を示している。ニューロンを、系統187 E18胚の皮質から単離した。培養3日目、試験化合物にホルスコリン5μM(F)、ロリプラム10μM(R)、ホルスコリンとロリプラム(F/R)をイソプロテレノールと組み合わせて10μM、イソプロテレノールとロリプラムを組み合わせて(I/R);SKF82958を10μMおよびSKF82958とロリプラムを組み合わせて(S/R)を加えた。データは、秒あたりのカウント数(cps)で示されている。
【図11】初代皮質ニューロンにおけるルシフェラーゼ発現に対する、プロキネチシン(prokineticin)2(PROK2)ペプチドの効果を示している。初代皮質ニューロンを、系統187(脳および脊髄中にルシフェラーゼを誘導できる)E18上から採取した。このアッセイを培養3日目に4時間または8時間実行した。このPROK2ペプチドは、1nMおよび100nMで水溶液として加えられる。データは、秒あたりのカウント数(cps)として示されている。
【図12】異なるCreLuc系統由来の初代皮質ニューロンにおけるルシフェラーゼ発現に対するプロキネチシン(prokineticin)2(PROK2)ペプチドの効果を示している。初代皮質ニューロンを、E18で4つの異なるCreLuc系統から採取した。このアッセイを、培養3日目に1nMまたは100nMのPROK2ペプチドいずれかを用いて、4時間後および24時間後の2つの時点においてトリプリケートで実行した。そのアッセイにはBrightGloを用い、TopCountで読み取った。データは、秒あたりのカウント数(cps)で示されている。
【図13A】初代皮質ニューロンにおけるルシフェラーゼ発現に対するmGluR7アゴニスト、AMN082の効果を示している。皮質ニューロンは、E18胚(系統187)から採取した。このアッセイを培養3日目に実行した。ホルスコリンは10μMで用いた。このアゴニスト、AMN082を1nM、10nM、100nMおよび1μMのホルスコリンと組み合わせて用いた。このアッセイは、Bright Glo(Promega)を備えるTopCountで4時間および8時間で読み取った。データは秒あたりのカウント数(cps)で示されている。
【図13B】初代皮質ニューロンにおけるGi活性を調節する能力について未知の化合物をスクリーニングした結果を示している。皮質ニューロンは、E18胚(系統187)から採取した。このアッセイを培養3日目に実行した。ホルスコリンは10μMで用いた。AMN082または未知の化合物A、BまたはCを異なる濃度のホルスコリンと組み合わせて試験紙、EC50値を算出した。このアッセイは、Bright Glo(Promega)を備えるTopCountで、4時間で読み取った。データは秒あたりのカウント数(cps)で示されている。
【図14】初代皮質ニューロンにおけるルシフェラーゼ発現に対するmGluR7アゴニスト、AMN082の効果を示している。初代皮質ニューロンは、系統187由来のE18胚から単離した。このアッセイを培養7日目に6時間、トリプリケートで実行した。AMN082の濃度曲線は、50μMのホルスコリンおyぼい10μMのロリプラムと組み合わせて実行した。このアッセイは、Bright Glo(Promega)を備えるTopCountで読み取った。データは秒あたりのカウント数(cps)で示されている。
【図15A】CB1アゴニスト、CP55,940によって異なるCreLuc系統由来の初代皮質ニューロンにおけるルシフェラーゼ発現のGi調節を示している。初代皮質ニューロンは、E18で4つの異なるCreLuc系統から採取した。このアッセイは培養3日目に実行した。CB1アゴニストを10μM、ホルスコリンを5μMおよびロリプラムを10μMで用いた。4時間および24時間の2つの時点で実行した。次いでブライトグロルシフェラーゼアッセイ基質を添加し、そのアッセイをTopCountルミノメーターで読み取った。データはトリプリケートの平均値で示されている。データは秒あたりのカウント数(cps)で示されている。
【図15B】CB1アゴニスト、CP55,940によるCreLucマウス由来の初代皮質ニューロンにおけるルシフェラーゼのGi調節を示している。皮質ニューロンは、E18胚(系統187)から単離した。このアッセイは培養3日目に実行した。ホルスコリン(F)およびロリプラム(R)を10μMで用いた。アゴニストを10μM、1μM、および100nMの濃度で加えた。このアッセイを、BrightGlo(Promega)を備えるTopCountにおいて8時間で読み取った。データは、秒足りのカウント数(cps)で示されている。
【図16】ホルスコリンおよびロリプラム、ならびにGsアゴニストであるDRD1およびADβRによる、CreLuc線条体ニューロンにおける誘導されたルシフェラーゼ発現を示している。線条体ニューロンは、E14胚(系統187)から単離した。このアッセイは、培養4日目に実行した。ホルスコリン(F)を5μM、ロリプラム(R)を10μMで用いた。Gsアゴニストであるイソプロテレノール(iso)、ドーパミン(dopa)およびSKF82958(chloro)を10μM、3μM、および1μMで用いた。このアッセイをTopCountルミノメーターおよびブライトグロルシフェラーゼ試薬(Promega)を用いて5時間で読み取った。データは秒あたりのカウント数(cps)で示されている。
【図17】CreLucマウスから単離された全脾細胞プレップ中のルシフェラーゼ発現に対する、ホルスコリン(F)およびロリプラム(R)のような一般的なcAMP誘導因子、ならびにGsアゴニストの影響を示している。系統64の脾細胞を抗CD3交代(CD)で24時間刺激し、他の半分は未処理とした(unstim)。24時間で化合物をプレートに加え、さらに4時間経過させた。ホルスコリンおよびロリプラムの同時処理(F/R)は、5uMのホルスコリンおよび10uMのロリプラムであった。用いたGsアゴニストは以下のとおりである:EP2アゴニストとしてEX00000173A(173A)、DP1アゴニストとしてBW245CおよびADβRアゴニストとしてイソプロテレノール。全てのGsアゴニストを10uMで用いた。このアッセイをトリプリケートで実行した。4時間後、100ulのBrightGloを加え、このアッセイをTopCountルミノメーターで読み取った。データは秒あたりのカウント数(cps)で示されている。
【図18】CreLucマウスの5つの異なるサブラインから単離されたT細胞中のロリプラムおよびホルスコリンによる、一般的なcAMPの効果を示している。この細胞を抗CD3抗体(1ug/ml)で刺激した。18時間後、10μMのロリプラムおよび5μMのホルスコリンをプレートに加え、さらに4時間経過させた。BrightGloを加え、アッセイをTopCountで読み取った。データは上のパネルでは発光(秒あたりのカウント数)で示され、下のパネルでは培地に対しての増加の倍率として示している。
【図19】CreLucマウス(系統64)から単離された抗CD刺激されたCD4+T細胞中のルシフェラーゼレベルに対するGsアゴニストの効果を示している。この細胞(1ウエルあたり1.5x105)を96ウエルの白色不透明プレート上にプレーティングし、抗CD3抗体(1ug/ml)で刺激した。24時間後、化合物を加えさらに4時間経過させた。GsアゴニストであるBW245C、EX00000173A(1734A)およびイソプロテレノール(iso)は、全て10μMで用いた。ホルスコリン(F)を5uM、ロリプラム(R)を10uMで加えた。BrightGloを加え、このアッセイをTopCountで読み取った。データは秒あたりのカウント数(cps)で示されている。
【図20】CreLucマウスの2つの異なるサブラインから単離されたB細胞中の、ロリプラムおよびホルスコリンにる一般的なcAMP活性化の効果を示している。細胞を96ウエルの白色不透明プレート上に、1ウエルあたり2.0x105でプレートし、10ng/mlのリポポリサッカライド(LPS)で刺激した。18時間後、10μMのロリプラムおよび5μMのホルスコリンをそのプレートに加え、さらに4時間経過させた。BrightGloを加え、アッセイをTopCountで読み取った。データは発光(秒あたりのカウント数)およびコントロールのみ培地に対する増加の倍率として示している。
【図21】CreLucマウス(系統から単離されたLPS刺激されたB220+B細胞中のルシフェラーゼレベルに対するGsアゴニストの効果を示している。この細胞を96ウエルの白色不透明プレート上に1ウエルあたり2.0x105プレートし、その後10ng/mlのリポポリサッカライド(LPS)で刺激した。24時間後、化合物を加えさらに4時間経過させた。GsアゴニストであるBW245C、EX00000173A(1734A)およびイソプロテレノール(iso)は10μMで用いた。ホルスコリン(F)を5uM、ロリプラム(R)を10uM加えた。BrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイをTopCountで読み取った。データは秒あたりのカウント数(cps)で示している。
【図22】一般的なcAMPアクチベーターであるホルスコリン(F)およびロリプラム(R)、ならびにDPレセプターのアゴニストであるBW245Cによる、単離されたミクログリア(系統64)中の誘導されたルシフェラーゼ発現を示している。初代ミクログリアをP2マウス由来の皮質から単利し、ポリ−D−リシンコーティングしたプレート上に96ウエル形式でプレートした。細胞は、未処理のままかまたは100ng/mlのLPSで2時間刺激したものかのいずれかである。次いで化合物を加え4時間経過させ、その後Bright Gloアッセイを実行した。用いた化合物は、5μMのホルスコリン、10μMのロリプラムもしくはその2つの組み合わせ、またはDP1レセプターのGsアゴニスト、BW245Cを10μMであった。データは秒あたりのカウント数で示されている。
【図23】CreLucマウス(系統187)の脳および脊髄におけるルシフェラーゼ発現の誘導に対する髄腔内注射されたホルスコリン(F)およびロリプラム(R)の効果を示している。1グループあたりN=3〜4のマウスで、3ヶ月齢の雄。グループA:DMSOコントロール、グループB:1ugホルスコリン/10ugロリプラム、グループC:10ugホルスコリン/10ugロリプラム、グループD:40ugホルスコリン/10ugロリプラム。この動物に、髄腔内注射により腰部に1匹のマウスあたり5μlの量で投薬した。これらを投薬後4時間で画像処理した。脊髄および脳の両方のデータは、平均ピーク輝度、平方センチメートル毎秒あたりの光子として示されている。
【図24】CreLucマウスの脳および脊髄中のルシフェラーゼ発現に対するEPアゴニスト、EX00000173Aの効果を示している。マウス(系統187)をビヒクル(5% DMSO、0.05% tween 80、PBS)または10mg/kg EX00000173Aのいずれかを用いてi.p.注射した。動物を投薬4時間後にバイオイメージングした。データは平方センチメートル毎秒あたりの光子として示されている。
【図25】CreLucマウス中のルシフェラーゼ発現に対するEP2アゴニスト、EX00000173Aの効果を示している。マウスに、ビヒクルコントロールまたは用量を変化させたEP2アゴニストEX0000173Aのいずれかを投薬した。マウスに髄腔内注射(1匹のマウスあたり5μl)により投薬し、IVIS bioimagerで4時間後にバイオイメージングした。データは5匹のマウスの平均値として、平均ピーク輝度、平方センチメートル毎秒あたりの光子として示されている。
【図26】CreLucマウスにおけるアドレナリン受容体β3(Adrb3)アゴニスト、CL316,243(1 mg/kg、ip)による異なる組織中のルシフェラーゼの誘導を示している。このルシフェラーゼアッセイは、組織ホモジネートで実施した。
【図27A】CreLucマウスの系統11(n=2)および系統115(n=3)におけるAdrb3アゴニスト、CL316,243(1 mg/kg、ip)によるルシフェラーゼの誘導を示している。処理前および処理後4〜5時間でBLIを撮影した。組織ホモジネート中のルシフェラーゼ活性は、画像の下に示されている。
【図27B】CreLucマウスの系統31(n=2)および系統175(n=3)におけるAdrb3アゴニスト、CL316,243(1 mg/kg、ip)によるルシフェラーゼの誘導を示している。処理前および処理後4〜5時間でBLIを撮影した。組織ホモジネート中のルシフェラーゼ活性は、画像の下に示されている。
【図28】CreLucマウスの3つの独立した系統における、グルカゴン様ペプチド1受容体(GLP−1R)アゴニスト、AVE0010によるルシフェラーゼレセプターの誘導を示している。ベースラインの画像は1日目に得られた。2日目、マウスをAVE0010(0.1 mg/kg、sc)で処理し、4時間後に画像処理した。ベースラインに対する誘導の倍率が下部に示されている。
【図29】CreLucマウスの3つの独立した系統における、グルカゴン様ペプチド1受容体(GLP−1R)アゴニスト、AVE0010によるルシフェラーゼレセプターの誘導を示している。マウスをAVE0010(0.1 mg/kg、sc)で4時間処理した。8つの異なる組織中のルシフェラーゼ活性を測定した。
【図30】AVE0010によるCreLucの誘導におけるβ細胞毒素ストレプトゾトシン(STZ)の効果を示している。雄のCreLucマウス(系統11)を、AVE0010を0.1 mg/kg与える前(「非誘導」;上パネル)およびscで与えた後(「AVE0010による誘導」中パネル)に画像処理した。全てのマウスは、AVE0010に応答していた(中パネル)。次いで、この動物をビヒクル(コントロール)またはSTZ(200mpk、ip)で処理した。4日後、これらをAVE0010での処理後再び画像処理した(下パネル)。
【図31】AVE0010によるCreLucの誘導がβ細胞特異的な可能性があることを示している。動物を図30に記載されたように処理した。血中グルコース濃度を、非絶食マウスの尾静脈ニッキングによって測定した。グルコース濃度は、Bayerのグルコースメーターで読み取った。グルコース濃度はmgグルコース/mlとして示される。誘導の倍率は、ベースラインの信号 対 AVE10を投与したルシフェラーゼバイオイメージングレベルである。血中グルコース濃度(BG)は、STZによる増加した(左上パネル)。非絶食BGレベルは、AVE0010(0.1 mg/kg、sc)によって低減された。図30に示されるBLIデータを定量した。
【図32】NOD SCIDガンマ(NSG)マウスへのCreLuc骨髄移植を示している。骨髄細胞は、系統44のヘテロ接合体と系統64のホモ接合体から採取した。その後、この細胞を、1匹のマウスあたり100万、または500万個の細胞で、照射NSGマウスに細胞の尾静脈注射により移植した。系統44:マウス1および2は、500万、マウス3および4が100万の細胞を受けた;系統64:マウス1が500万個の細胞、マウス2、3および4が100万個の細胞を受けた。(CreLuc系統あたり4匹のNSGマウス)。動物を、4週間(データは示さず)でバイオイメージングし、再度8週間(データは示されている)でバイオイメージングした。画像処理の前に、系統64のマウスを5mg/kgのホルスコリンおよび10mg/kgのロリプラムで5時間誘導した。
【図33】マウス胚線維芽細胞中のルシフェラーゼ発現に対する、ホルスコリン、ロリプラムおよびイソプロテレノールの効果を示している。マウス胚線維芽細胞は、6つの独立したCreLuc系統由来のE12胚から、1ウエルあたり20,000細胞プレーティングして培養した。試験した化合物としては、10μM ホルスコリン(F)、5μM ロリプラム(R)および10μM イソプロテレノール(iso)が挙げられる。データは秒あたりのカウント数(cps)として示されている。
【図34】CreLucマウス(系統187)中のルシフェラーゼレベルに対するザイモサン処理の効果を示している。治療群の動物に、疼痛反応を誘導するためにザイモサン(zymo)によって両方の後部足に皮下注射した。これらの動物を4日間毎日バイオイメージングした(d1、d2、d3およびd4と表記)。
【図35】心筋細胞中のルシフェラーゼレベルに対するホルスコリンおよびロリプラムおよびイソプロテレノールの効果を示している。心筋細胞は、P3の子犬(系統229)から単離した。この細胞を96ウエルプレートで培養した。試験した化合物としては、10μM ホルスコリン(F)、5μM ロリプラム(R)および10μM イソプロテレノール(iso)が挙げられる。データは秒あたりのカウント数(cps)として示されている。
【0054】
発明の詳細な説明
他に定義しない限り、本明細書中で用いられる全ての技術用語および科学用語は、一般的に本発明が属する分野の当業者によって通常理解されるもとの同じ意味を持っている。
【0055】
本明細書中で引用した各文献、特許出願、特許および他の参考文献は、それが本発明の開示に反しない限り、その全体が参照により援用される。
【0056】
ここで、本明細書中および添付の特許請求の範囲において用いられる場合、単数形の「a」「an」、および「the」は、その文脈がそうでないことを明確に指示しない限り、複数の指示対象を含むことに留意しなければならない。
【0057】
さらに、本発明によると、当該分野の技術の範囲内で、従来の分子生物学、微生物学および組み換えDNA技術を用いることができる。このような技術は、文献に十分に説明されている例えば、以下を参照のこと:Sambrook、Fritsch&Maniatis、Molecular Cloning: A Laboratory Manual、Second Edition(1989)Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York(herein”Sambrook et al.、1989”);DNA Cloning: A Practical Approach、Volumes I and II(D.N.Glover ed.1985);Oligonucleotide Synthesis(M.J.Gait ed.1984);Nucleic Acid Hybridization[B.D.Hames&S.J.Higgins eds.(1985)];Transcription And Translation[B.D.Hames&S.J.Higgins、eds.(1984)];Animal Cell Culture[R.I.Freshney、ed.(1986)];Immobilized Cells And Enzymes[IRL Press、(1986)];B.Perbal、A Practical Guide To Molecular Cloning(1984);F.M.Ausubel et al.(eds.)、Current Protocols in Molecular Biology、John Wiley&Sons、Inc.(1994)。
【0058】
「試験薬」は、化合物または化学化合物のような任意の物質(例えば、有機化学物質、無機化学物質)、生物学的化合物または生物学的物質(例えば、交代および抗原認識フラグメントおよび構築物、核酸(例えばRNAi))などの任意の物質を含む。試験薬は、一緒に適用される単一薬剤または複数薬剤を包含する。
【0059】
本明細書中で用いられる場合、「トランスジェニック動物」は、非ヒト動物であり、非限定的な例は哺乳動物であるが、その動物の細胞の1つ以上が本明細書中に定義されるような遺伝的改変を含んでいる。さらに非限定的な例として、ラットまたはマウスのようなげっ歯類が挙げられる。他のトランスジェニック動物の例としては、ヒト以外の霊長類、ヒツジ、イヌ、ウシ、ヤギ、ニワトリ、両生類などが挙げられる。トランスジェニック動物の選択は、組織を横切るためにレポーターから光を発生させて、検出が可能な表面まで到達させる能力によってのみ限定される。
【0060】
本明細書中で用いられる場合、「遺伝子組み換え」は、ヒト以外の動物の遺伝子配列における1またはそれより多い変更である。非限定的な例は、トランスジェニック動物のゲノムへの導入遺伝子の挿入である。
【0061】
本明細書中で用いる場合、用語「導入遺伝子」は、発現を増強するためにプロモーター、レポーター遺伝子、ポリアデニル化シグナルおよび他のエレメントを含む外因性DNAのことをいう(インスレーター、イントロン)。この外因性DNAは、1つの細胞の胚のゲノム中に組み込まれ、そこからトランスジェニックが発達し、その導入遺伝子はその成熟動物のゲノム中に留まる。この組み込まれた導入遺伝子DNAは、卵またはマウスのゲノム中に単一でかまたは複数で生じ得、またその導入遺伝子の単一〜複数(数百)のタンデムコピーが各ゲノム位置に組み込まれ得る。
【0062】
用語「一般的なcAMPモジュレーター」は、cAMPレベルを増加または維持することのできる、化学化合物(例えば、有機化学物質、無機化学物質)、生物学的化合物または生物学的材料(例えば、フラグメントおよび構築物を認識する抗体および抗原)、核酸(例えばRNAi)などを言う。非限定的な例としては、ホルスコリンおよびロリプラムが挙げられる。一般的なcAMPモジュレーターは、単一のcAMPモジュレーターまたは一緒に適用される複数のcAMPモジュレーターを包含する。
【0063】
用語「細胞刺激因子」とは、細胞を活性化し得るかまたは細胞をより活性化した状態にすることのできる、化学化合物(例えば、有機化学物質、無機化学物質)、生物学的化合物または生物学的材料(例えば、フラグメントおよび構築物を認識する抗体および抗原)、核酸(例えばRNAi)などを言う。非限定的な例としては、リポポリサッカライドおよび抗CD3が挙げられる。
【0064】
本発明の実施態様は、コントロールを使用する。コントロールは、当業者に周知の用語である。適切なコントロールは利用されるアッセイパラメーターまたは調査中の実験の問題に依存する場合がある。典型的には、コントロールは、そのコントロールが、試験薬剤または化合物を溶解するのに用いられているのと同じ緩衝液または溶媒であるビヒクルコントロールである。非限定的な例として、化合物を溶解するのにリン酸緩衝生理食塩水が用いられている場合、ビヒクルコントロールはリン酸緩衝生理食塩水である。同様に、DMSOが試験薬剤を溶解するのに用いられている場合、コントロールはDMSOである。しばしば、化合物を試験するために1種より多い希釈剤が用いられるために、実験またはアッセイごとに1種より多いコントロールを使用する必要がある。
【0065】
本明細書中で用いられる場合、「ルシフェラーゼ」は、ルシフェラーゼ酵素活性だけでなく、ルシフェラーゼタンパク質の実際の量も言及する。
【0066】
本発明に従って、トランスジェニック非ヒト動物を作製するために、当業者に公知の従来の技術を使用することができる。例えば、Pinkert、C.A.(ed.)1994。トランスジェニック動物技術:A laboratory handbook.Academic Press、Inc.、San Diedo、Calif.;Monastersky G.M.and Robl、J.M.(ed.)(1995)Strategies in transgenic animal science.ASM Press.Washington D.C..and Nagy A、Gertsenstein、M、Vintersten、K、Behringer R 2003。マウス胚の操作;A laboratory Manual 3rd edition. Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York。
【0067】
導入遺伝子エレメント
本発明の実施態様は、導入遺伝子に関連している。この導入遺伝子は、インスレーター配列、応答配列、プロモーターエレメント、レポーターエレメントおよび機能エレメントを含み得る。
【0068】
応答配列は、細胞内でホルモン、酵素、または他の重要なシグナル伝達タンパク質のような細胞のシグナルに応答するパリンドロームDNA配列である。応答配列の非限定的な例としては、CRE(cAMP応答配列)、エストロゲン応答配列および表2に列挙される他のものが挙げられる。応答配列は、単一のDNA配列またはタンデムリピートとして、導入遺伝子中に組み込まれ得る。例えば、4回または6回反復されるCRE応答配列が、導入遺伝子構築物中で用いられ、インビトロで検証されている(Deutsch P.J.、et al.、J.Biol.Chem.、263;18466−18472、1988;Oetjen E JBC 269;27036−27044、1994)。Cre応答配列もまた、インビボで比較されており、多量体における増加は、cAMP経路のアクチベーターに対する転写応答の増加と相関している(Montoliu、L.et al.、Proc.Natl.Acad.Sci.USA 92;4244、1995;Boer et al、PloS One、May 9;2(5):e431、2007)。本発明の実施態様は、単一の配列としてかまたは複数のタンデムリピート(例えばCREのタンデム6回反復(6×CRE))としてのいずれかの任意の公知の応答配列を利用することができる。
【0069】
【表2】
【0070】
DNプロモーターエレメントは、特定の遺伝子の転写を促進するDNAの領域である。プロモーターは、通常、同じ鎖および上流(センス鎖の5‘領域に向かって)上で、それらが制御する遺伝子の近くに位置している。プロモーターは、特定のDNA配列および応答配列(RNAポリメラーゼおよびRNAポリメラーゼを補充するタンパク質(転写因子と呼ばれる)の結合部位を提供する)を含んでいる。DNプロモーターは、その大きさおよび時間と空間内の特定の遺伝子の発現の調節に寄与する内部構造が非常に可変的である。プロモーターエレメントの非限定的な例において、単純ヘルペスウイルスチミジンキナーゼの最小プロモーター(HSV TK min)は、全ての細胞型におけるレポーター遺伝子の発現を可能にするように設計されている。その中核的な発現エレメントのみが保持され、インビトロまたはインビボのいずれかでのユビキタス発現を付与する(Park、J.、et al.、DNA Cell Bio.、12:1147−1149、1994)。本発明の実施態様は、任意の公知のプロモーターエレメントを利用することができる。非限定的な例として、任意の公知のプロモーターエレメントは、そのプロモーターエレメントがCRE cis活性応答配列と組み合わせられる場合に、cAMP経路の変調に応答するレポーターの遺伝子発現を可能にするように用いられる。
【0071】
CREのような応答配列およびプロモーターエレメントであるHSV TK minはサイズが小さいので、位置の影響に非常に敏感であり、乏しい発現につながるレポーターエレメントの転写を調節するこれらのエレメントを含む導入遺伝子プロモーターは、特にインビボで低いリガンド濃度でリガンドに応答する。従って、本発明の実施態様は、全ての細胞区画を通して機能している導入遺伝子の高レベルでの発現および広い分布を達成するために、追加のエレメントを導入遺伝子に加えることを包含する。これらのエレメントには、インスレーター配列および機能エレメントを含む(Sun F.L and Elgin S.C、Cell 99:459−462、1999)。
【0072】
本発明の実施態様は、導入遺伝子内の機能エレメントまたは機能増強エレメントを利用する。機能エレメントの非限定的な例は、ヒト成長ホルモン(hGH)遺伝子である。CreLuc導入遺伝子において用いられるhGH配列は、
バイオイメージングモデルの設計目標を達成するために、インスレーター配列に寄与し、相互作用するいくつかの設計エレメントを含んでいる。このhGH配列は、全てのhGHゲノム構造を含んでいるため、いくつかの重要なエレメントを提供するが、タンパク質中には転写も翻訳もされない。機能する導入遺伝子の産生を向上するためのhGH配列の重要な影響は、1990年以来、いくつかのトランスじぇニックモデルについて実証されている(Erickson LA、Nature 346:74−76、1990)。その重要性の比較分析は存在しないが、hGH構造は、いくつかの重要で、重要な意味をもつDNAエレメントを含んでいる:
a. イントロンスプライシング:最初に転写されたmRNAは、イントロン配列とエキソン配列の両方を含んでおり、核から書き出されてさらに処理され、イントロン配列が取り除かれてエキソン配列のみを含む成熟mRNAを生じる。このトランスポートおよびトリミングのプロセスは、成熟mRNA鎖をタンパク質の最終的な生産のためのさらなる翻訳機構に接続させる。導入遺伝子のcDNA構造中にイントロンを含むことは、導入遺伝子のcDNAの発現レベルを向上させることが示されている(Palmiter、R.D.、et al.、Proc.Natl.Acad.Sci.USA88:478−482、1988)。
b. 無処置の3’UTRを持つゲノム構造:本発明の実施態様において、導入遺伝子中のhGH配列は、高いレベルでのmRNA安定性、およびそれによる高いレベルでの遺伝子発現を付与する、無処置の3’UTRを含んでいる。
c. ポリA(PA+)構造を有するゲノム構造:このhGH配列は、天然の3’UTRに埋め込まれたそのネイティブPA+構造を含んでいる。一般的にPA+シグナルは、ウイルス配列(SV40、RSVなど)由来であり、無関係の3’UTRの末端に加えられた最小構造である。本発明の実施態様において、天然のPA+シグナルを有する全体の3’UTR構造は、完全hGH遺伝子のさらに広いゲノムのコンテキストに保存される。
【0073】
インスレーター配列は、位置に依存する発現を産生するDNAの配列である(Giraldo et al、Transgenic Research 12:751−
755、2003)。インスレーター配列は、グロブリン遺伝子座について1980年に記載され(Sun F.L.and Elgin、S.C.、Cell 99;459−462、1999)、バイオイメージングのためのモデル設計目標をサポートするために選択された組織中で、適切な応答性トランスジェニック発現を得る機会を増やすことが報告された(Pinkert、C.A.(ed.)1994.Transgenic animal technology:A laboratory handbook.Academic Press、Inc.、San Diego、Calif.;Monastersky G.M.and Robl、J.M.(ed.)(1995)Strategies in transgenic animal science.ASM Press.Washington D.C)。インスレーターは、遺伝子発現に寛容なオープンクロマチンドメインを作成し、遠位のサイレンサー/エンハンサー配列の影響に対して、ならびにアセチル化およびメチル化事象に対しての障壁を構成するDNAエレメントである。これらは、バイオイメージングで検出可能なレベルで発現されるレポーター遺伝子を持つ独立したトランスジェニック初代系統の数を大幅に増やさねばならない。インスレーター配列は、一過性のトランスフェクションアッセイにおいて、40〜70%までクローンを発現するルシフェラーゼの数を増やし、結果的にERE−luci 導入遺伝子中のルシフェラーゼ発現の誘導能を高めることが示されている(Ottobrini L.、 Mol Cell Endo 246、69−75)。しかしながら、導入遺伝子レポーター発現に対するインスレーターの適用を全面的に検討すると、実用面において、インスレーターを利用することは依然として困難であるという結論に至る。それらの作用メカニズムは部分的にしか知られておらず、その効果は完全に予測可能ではない。インスレーター配列の非限定的な例は、表3に挙げられている。
【0074】
【表3】
【0075】
Cre−Luc導入遺伝子の実施態様において、インスレーター配列の封入は、バイオイメージングによって検出される機能的レポーターを持つ系統の作製頻度を明らかに増加させた。CreLucトランス得ニック系統におけるルシフェラーゼ発現へのインスレーター配列の寄与の分析については、VI章の「実施例」以下を参照のこと。
【0076】
本発明の実施態様は、レポーターエレメントまたは遺伝子を含む導入遺伝子に関する。レポーター遺伝子は、検出可能な遺伝子産物を発現する任意の遺伝子を含み、RNAまたはタンパク質であり得る。多くのレポーター遺伝子は当該分野で公知であり、限定するわけではないが、β−ガラクトシダーゼおよびアルカリホスファターゼが含まれる。別の実施態様において、導入遺伝子は、生物発光レポーター遺伝子を含む。多くの生物発光レポーター遺伝子は当該分野で公知であり、ルシフェラーゼを含むがこれに限定されない。ルシフェラーゼの供給源は多く存在し、ホタルルシフェラーゼおよび細菌ルシフェラーゼが挙げられるがこれらに限定されない。本発明の実施態様は、例えばルシフェラーゼのような任意の公知の生物発光レポーターを利用し得る。他のレポーターエレメントの非限定的な例は、表5に示される。
【0077】
【表4】
【0078】
本発明の他の実施態様用は、ルシフェラーゼ酵素の改変バージョン、異なる種由来のルシフェラーゼ酵素、または適切な基板と共に提供される場合、動物組織を横切ることのできる光を産生し得る他のタンパク質、もしくは動物組織を横切ることのできる光を発することのできる任意の酵素を組み込むことができる。本発明のレポータータンパク質は、信号の減衰量が、発された光の波長および発光細胞を取り巻く組織特性に依存するという事実によってのみ限定される。一般的に青緑光(400〜590nm)は、強く減衰される一方、近赤外光(590〜800nm)は、はるかに減衰が少ない。ルシフェラーゼの多くのタイプは、青〜黄緑の波長で発光し、その発光スペクトルは広く、組織に非常に深く浸透する赤色波長(>600nm)で有意な発光が存在しているようである。マウスなどの小型ゲッ歯類では、このことが動物全体を通してのシグナルの検出を可能にする。
【0079】
インビボでの光検出の限界は、生物発光レポーターのタイプ、動物の周囲の生理学および発光源の深さに依存する。通常、動物の生物発光細胞は、細胞の数および場所により、高感度CCDカメラを用いて、1〜3cmの深さで観察することができる。それらが組織をとおして伝播するときの光子の散乱は、動物表面上で検出された画像の空間分解能を制限する。一般的に、表面におけるスポットサイズや解像度は、表面化の光源の深さにほぼ等しい。物理ベースの拡散モデルを用いて、ミリメートルレベルでアプローチする空間解像度の改善を達成することができる。冷却した科学グレードのCCDアレイを用いると、シグナル検出の限界は、画像が撮影された後のCCD画素数(これは、ピクセルあたりのいくつかの光子のオーダーである)の読み取りと関連した読み出しノイズによって決定される(Honigman et al.、Mol.Ther.4:239−249、2001)。毛皮、皮膚、あるいは動物上の汚染物質の燐光に起因して、動物から発せられるさらなるバックグラウンド光もあり得る。通常、バックグラウンド光は低レベルであり、深く低レベルな生物発光供給源の画像に有害な影響を及ぼすのみである。しかしながら、バックグランド光は、適切な光学フィルターの使用により、排除することができる。
【0080】
本発明の実施態様は、IVIS(Xenogen Corporation、860 Atlantic Avenue、Alameda、Calif.94501、USA)などのCCDカメラを利用する。このIVIS.RTM.Imaging Systemは、高感度CCDカメラ、入射光を最小にするための暗い画像処理チャンバー、および結果を定量化し分析するための特殊なソフトウエアを備えている。IVISは、Xenogen Corporationの登録商標である。しかしながら、このような生物発光画像処理システムはどれも本発明に適用可能である。
【0081】
リアルタイムのインビボイメージングは、生物発光レポーター遺伝子定量化を非侵襲的に(すなわち動物を安楽死させる必要がない)、かつ長期的に(すなわち長い時間経過にわたり、連続的または反復して測定することができる)可能にする。リアルタイムのインビボイメージングは、従来のプロトコルに比べ、必要な試験動物がより少なく(例えば、特定の時間の間同じ動物を用いることができるため)、より少ない時間でよい(例えば、取り扱う必要のある動物の数がより少ないため)、より多くの化合物を効率よく試験することができる。リアルタイムのインビボイメージングは、多くの理由により、より高いデータ内容およびより高いデータ品質を提供する。例えば、時間的および空間的データは同じ動物から集めることができ、データを時間のかかる組織学的評価を必要とせずに集めることができる。データ品質が高いほど、統計誤差を減少させ、試験化合物の評価と意思決定の質を向上させる。
【0082】
本発明の実施態様は、ハイスループットスクリーニング(HTS)法で用いる。HTSは、疾患の病態の生物学的メカニズムや側面をモデル化するために設計されたアッセイにおける何千もの異なる化合物の自動化された同時試験である。1またはそれより多い化合物(例えば複数の化合物)は、例えば、1つのバッチで同時に試験することができる、1実施態様において、用語「HTSスクリーニング法」は、任意の読み出しに影響を及ぼす1つの化合物または複数の化合物の能力を試験するアッセイを意味する。
【0083】
細胞培養およびサンプル操作のための、液体取扱システム、蛍光読み取り装置またはシンチレーションカウンターのような分析装置およびロボットは当該分野でよく知られている。ロボットのような機械的システムや「チェリーピッキング」デバイスは、当業者に利用可能である。市販のプレートリーダーは、従来の96ウエルプレートや384ウエルプレートを分析するのに利用可能である。単一のサンプル、複数のサンプル、またはプレートサンプルのリーダーは、所定のウエルを分析し、生データのレポートを作成するのに利用可能である。この生データは種々の方法で変換され提示することができる。
【0084】
本発明の実施態様は、細胞、組織切片または培養培地のような他の材料を受けることのできる容器の配列を含む。容器の配列は、本発明の範囲内で、細胞または組織切片を保持するのに適切な少なくとも1つ以上の容器から任意の数の容器であり得る。例としては、フラスコ、培養皿、1.5mlチューブのようなチューブ、12ウエルプレート、96ウエルプレート、384ウエルプレートおよび4000ほどレセプタクルを有する小型のマイクロタイタープレート(米国特許出願20050255580号)が挙げられるが、これらに限定されない。容器の配列は、保護カバーを追加するような修正が可能であり、それにより汚染物質の混入や内容物の蒸発を防止することができる。
【0085】
これらの容器のさらなる特徴は、容器が分析(非限定的な例として、分光分析、シンチレーション計数、および蛍光測定を含む)を可能にすることができることである。しかしながら、さらなる分析のために修正可能な適切な容器に移すことができることを考えると、本発明の範囲内で用いられ得る容器は限定されない。非限定的な例は、この方法を変更することであり、例えばこの方法が、第2の容器の配列を提供することをさらに包含し、ここで、細胞を溶解させる工程は、さらに細胞片から上清を分離する工程を包含し、次の工程は、この第2の容器の配列の少なくとも1つが分離した上清のサンプルを含むようにDNAフラグメント中にインターカレートし得る検出可能な化合物を加える工程をさらに包含する。
【0086】
本発明のさらなる局面および詳細は、以下の実施例から明らかであり、これらは例示することを意図しているのであって、いかなる意味においても本発明の範囲を限定することを意味しない。
【0087】
動物を含む全ての実験的な操作は事前に検討された連邦政府のガイドラインとプロトコルに従って実施し、sanofi−aventisのサイト、Institutional Animal Care and Use Committee(IACUC)によって承認されている。
【実施例】
【0088】
I.MultiSite Gateway Pro Plus Cloning Systemのベクター骨格(図3)
デスティネーションベクターpDest2XMARSを、MultiSite Gateway Pro Plus Cloning System(Invitrogen Carlsbad、CA、cat #12537−100)での使用に特異的に設計しクローニングした。ベクター中に挿入された全てのエレメントをPCR(ポリメラーゼ連鎖反応)クローニングまたは合成的に生成した。全てのPCRクローニング工程について、フラグメントを、PCR SuperMIx Hi Fidelity(Invitrogen、Carlsbad、CA、cat # 10790−020)およびフラグメント特異的プライマーの使用を通して、特定されたベクターから増幅した。次いで。このPCR産物をTOPOクローニングによってpCR2.1ベクターにサブクローニングした。TOPOクローニングは、TaqまたはPfuポリメーラーゼのいずれかによって増幅されたDNAフラグメントを、DNAリガーゼを必要とせずに特定のベクター中にクローニングする、分子生物学的手法である。
【0089】
はじめに、インスレーター配列をベクターpCpG−LacZ(InvivoGen San Diego、CA、cat #pcpg−lacz)からポリメラーゼ連鎖反応(PCR)によってクローニングし、配列を検証した後、pNEB193(New England Biolabs Ipswich、MA、cat#N3051S)の制限部位にサブクローニングした。このインスレーター配列を、統合部位依存の位置効果に起因する導入遺伝子の発現におけるばらつきを低減するために使用した。
【0090】
フラグメントをクローニングするのに用いたPCRプライマーは以下のとおりである:a.ヒトIFN−ベータMARプライマー配列(プライマーは、サブクローニングのためのEcoRV部位を有する)
配列番号1、IFN 順方向プライマー:
5’−GGGGGATATCAGTCAATATGTTCACCCCA−3’
配列番号2、IFN 逆方向プライマー:
5’−GGGGGATATCCTACTGTTTTAATTAAGC−3’
b.ヒトβ−グロビンMARプライマー配列
配列番号3、β−グロビン順方向プライマー:
5’−AAGGATCCTTAATTAAAATTATCTCTAAGGC−3’
配列番号4、β−グロビン逆方向プライマー:
5’−GGATCCCTGCAGGAATTCCTTTTAAT−3’
【0091】
β−グロビンPCRフラグメントを、pCR2.1(Invitrogen、Carlsbad、CA、cat#K2030−01)を用いてtopoクローニングした。配列を確認した後、そのフラグメントをBamHIで切り出し、その後pNEB193(βgloMAR−pNEB193)のBamH1部位にサブクローニングした。IFN−βPCRフラグメントもまた、pCR2.1でtopoクローニングし、シーケンスし、その後EcoRvで切り出して、βgloMAR−pNEB193(2XMARS−pNEB193)のHincII部位にクローニングした。最後のゲートウエイデスティネーションベクター(Gateway destination vector)については、XbaI末端を含むリンカーおよび内部EcoRV部位を2XMARs−pNEB193のXbaI部位にサブクローニングした。平滑末端化したゲートウエイコンバージョンカセット(Gateway conversion cassette)、RfAを2XMARs−pNEB193(2XMARSpDest)のEcoRV部位に挿入した。最後のベクトル、2XMARSpDestが導入遺伝子を作製するために用いられる最後のデスティネーションベクターであった。
【0092】
配列番号5
TCGCGCGTTTCGGTGATGACGGTGAAAACCTCTGACACATGCAGCTCCCGGAGACGGTCACAGCTTGTCTGTAAGCGGATGCCGGGAGCAGACAAGCCCGTCAGGGCGCGTCAGCGGGTGTTGGCGGGTGTCGGGGCTGGCTTAACTATGCGGCATCAGAGCAGATTGTACTGAGAGTGCACCATATGCGGTGTGAAATACCGCACAGATGCGTAAGGAGAAAATACCGCATCAGGCGCCATTCGCCATTCAGGCTGCGCAACTGTTGGGAAGGGCGATCGGTGCGGGCCTCTTCGCTATTACGCCAGCTGGCGAAAGGGGGATGTGCTGCAAGGCGATTAAGTTGGGTAACGCCAGGGTTTTCCCAGTCACGACGTTGTAAAACGACGGCCAGTGAATTCGAGCTCGGTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCACAAGTTTGTACAAAAAAGCTGAACGAGAAACGTAAAATGATATAAATATCAATATATTAAATTAGATTTTGCATAAAAAACAGACTACATAATACTGTAAAACACAACATATCCAGTCACTATGGCGGCCGCATTAGGCACCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATAATGTGTGGATTTTGAGTTAGGATCCGTCGAGATTTTCAGGAGCTAAGGAAGCTAAAATGGAGAAAAAAATCACTGGATATACCACCGTTGATATATCCCAATGGCATCGTAAAGAACATTTTGAGGCATTTCAGTCAGTTGCTCAATGTACCTATAACCAGACCGTTCAGCTGGATATTACGGCCTTTTTAAAGACCGTAAAGAAAAATAAGCACAAGTTTTATCCGGCCTTTATTCACATTCTTGCCCGCCTGATGAATGCTCATCCGGAATTCCGTATGGCAATGAAAGACGGTGAGCTGGTGATATGGGATAGTGTTCACCCTTGTTACACCGTTTTCCATGAGCAAACTGAAACGTTTTCATCGCTCTGGAGTGAATACCACGACGATTTCCGGCAGTTTCTACACATATATTCGCAAGATGTGGCGTGTTACGGTGAAAACCTGGCCTATTTCCCTAAAGGGTTTATTGAGAATATGTTTTTCGTCTCAGCCAATCCCTGGGTGAGTTTCACCAGTTTTGATTTAAACGTGGCCAATATGGACAACTTCTTCGCCCCCGTTTTCACCATGGGCAAATATTATACGCAAGGCGACAAGGTGCTGATGCCGCTGGCGATTCAGGTTCATCATGCCGTTTGTGATGGCTTCCATGTCGGCAGAATGCTTAATGAATTACAACAGTACTGCGATGAGTGGCAGGGCGGGGCGTAAACGCGTGGATCCGGCTTACTAAAAGCCAGATAACAGTATGCGTATTTGCGCGCTGATTTTTGCGGTATAAGAATATATACTGATATGTATACCCGAAGTATGTCAAAAAGAGGTATGCTATGAAGCAGCGTATTACAGTGACAGT
TGACAGCGACAGCTATCAGTTGCTCAAGGCATATATGATGTCAATATCTCCGGTCTGGTAAGCACAACCATGCAGAATGAAGCCCGTCGTCTGCGTGCCGAACGCTGGAAAGCGGAAAATCAGGAAGGGATGGCTGAGGTCGCCCGGTTTATTGAAATGAACGGCTCTTTTGCTGACGAGAACAGGGGCTGGTGAAATGCAGTTTAAGGTTTACACCTATAAAAGAGAGAGCCGTTATCGTCTGTTTGTGGATGTACAGAGTGATATTATTGACACGCCCGGGCGACGGATGGTGATCCCCCTGGCCAGTGCACGTCTGCTGTCAGATAAAGTCTCCCGTGAACTTTACCCGGTGGTGCATATCGGGGATGAAAGCTGGCGCATGATGACCACCGATATGGCCAGTGTGCCGGTCTCCGTTATCGGGGAAGAAGTGGCTGATCTCAGCCACCGCGAAAATGACATCAAAAACGCCATTAACCTGATGTTCTGGGGAATATAAATGTCAGGCTCCCTTATACACAGCCAGTCTGCAGGTCGACCATAGTGACTGGATATGTTGTGTTTTACAGTATTATGTAGTCTGTTTTTTATGCAAAATCTAATTTAATATATTGATATTTATATCATTTTACGTTTCTCGTTCAGCTTTCTTGTACAAAGTGGTGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTTAAACCTGCAGGCATGCAAGCTTGGCGTAATCATGGTCATAGCTGTTTCCTGTGTGAAATTGTTATCCGCTCACAATTCCACACAACATACGAGCCGGAAGCATAAAGTGTAAAGCCTGGGGTGCCTAATGAGTGAGCTAACTCACATTAATTGCGTTGCGCTCACTGCCCGCTTTCCAGTCGGGAAACCTGTCGTGCCAGCTGCATTAATGAATCGGCCAACGCGCGGGGAGAGGCGGTTTGCGTATTGGGCGCTCTTCCGCTTCCTCGCTCACTGACTCGCTGCGCTCGGTCGTTCGGCTGCGGCGAGCGGTATCAGCTCACTCAAAGGCGGTAATACGGTTATCCACAGAATCAGGGGATAACGCAGGAAAGAACATGTGAGCAAAAGGCCAGCAAAAGGCCAGGAACCGTAAAAAGGCCGCGTTGCTGGCGTTTTTCCATAGGCTCCGCCCCCCTGACGAGCATCACAAAAATCGACGCTCAAGTCAGAGGTGGCGAAACCCGACAGGACTATAAAGATACCAGGCGTTTCCCCCTGGAAGCTCCCTCGTGCGCTCTCCTGTTCCGACCCTGCCGCTTACCGGATACCTGTCCGCCTTTCTCCCTTCGGGAAGCGTGGCGCTTTCTCATAGCTCACGCTGTAGGTATCTCAGTTCGGTGTAGGTCGTTCGCTCCAAGCTGGGCTGTGTGCACGAACCCCCCGTTCAGCCCGACCGCTGCGCCTTATCCGGTAACTATCGTCTTGAGTCCAACCCGGTAAGACACGACTTATCGCCACTGGCAGCAGCCACTGGTAACAGGATTAGCAGAGCGAGGTATGTAGGCGGTGCTACAGAGTTCTTGAAGTGGTGGCCTAACTACGGCTACACTAGAAGAACAGTATTTGGTATCTGCGCTCTGCTGAAGCCAGTTACCTTCGGAAAAAGAGTTGGTAGCTCTTGATCCGGCAAACAAACCACCGCTGGTAGCGGTGGTTTTTTTGTTTGCAAGCAGCAGATTACGCGCAGAAAAAAAGGATCTCAAGAAGATCCTTTGATCTTTTCTACGGGGTCTGACGCTCAGTGGAACGAAAACTCACGTTAAGGGATTTTGGTCATGAGATTATCAAAAAGGATCTTCACCTAGATCCTTTTAAATTAAAAATGAAGTTTTAAATCAATCTAAAGTATATATGAGTAAACTTGGTCTGACAGTTACCAATGCTTAATCAGTGAGGCACCTATCTCAGCGATCTGTCTATTTCGTTCATCCATAGTTGCCTGACTCCCCGTCGTGTAGATAACTACGATACGGGAGGGCTTACCATCTGGCCCCAGTGCTGCAATGATACCGCGAGACCCACGCTCACCGGCTCCAGATTTATCAGCAATAAACCAGCCAGCCGGAAGGGCCGAGCGCAGAAGTGGTCCTGCAACTTTATCCGCCTCCATCCAGTCTATTAATTGTTGCCGGGAAGCTAGAGTAAGTAGTTCGCCAGTTAATAGTTTGCGCAACGTTGTTGCCATTGCTACAGGCATCGTGGTGTCACGCTCGTCGTTTGGTATGGCTTCATTCAGCTCCGGTTCCCAACGATCAAGGCGAGTTACATGATCCCCCATGTTGTGCAAAAAAGCGGTTAGCTCCTTCGGTCCTCCGATCGTTGTCAGAAGTAAGTTGGCCGCAGTGTTATCACTCATGGTTATGGCAGCACTGCATAATTCTCTTACTGTCATGCCATCCGTAAGATGCTTTTCTGTGACTGGTGAGTACTCAACCAAGTCATTCTGAGAATAGTGTATGCGGCGACCGAGTTGCTCTTGCCCGGCGTCAATACGGGATAATACCGCGCCACATAGCAGAACTTTAAAAGTGCTCATCATTGGAAAACGTTCTTCGGGGCGAAAACTCTCAAGGATCTTACCGCTGTTGAGATCCAGTTCGATGTAACCCACTCGTGCACCCAACTGATCTTCAGCATCTTTTACTTTCACCAGCGTTTCTGGGTGAGCAAAAACAGGAAGGCAAAATGCCGCAAAAAAGGGAATAAGGGCGACACGGAAATGTTGAATACTCATACTCTTCCTTTTTCAATATTATTGAAGCATTTATCAGGGTTATTGTCTCATGAGCGGATACATATTTGAATGTATTTAGAAAAATAAACAAATAGGGGTTCCGCGCACATTTCCCCGAAAAGTGCCACCTGACGTCTAAGAAACCATTATTATCATGACATTAACCTATAAAAATAGGCGTATCACGAGGCCCTTTCGTC
【0093】
II.導入遺伝子コンポーネント
A.CRE:
2つの異なる6×CREエレメントを導入遺伝子の作製に用いた。最初は、DNA2.0、Menlo Park、CAで合成して作製された合成CREエレメント(「合成」と記されている)である。この合成配列は4フラグメントのGatewayクローニングのためのattL1およびattR5部位を有している。
【0094】
異なるベクターから6×CREをPCRクローニングして出す複数の試みがなされた;しかし、これは、CREの中ほどにあるヘアピン構造が原因で不可能であった。次いで、このフラグメントを合成によってクローニングした。用いられた第2の6×CREエレメントは、クローンテックベクター(Clontech vector)(Mountainview、CA、cat#PT3336−5)からCREエレメントをPCRクローニングすることによって作製されたハイブリッドバージョンである。クローンテックは、このベクター中には2つのCREが存在していると主張しているが、配列分析では、実際には3つ存在していることが示された。2つの異なるPCR反応をフラグメントのクローニングに用いた。ゲートウエイのクローニング用のAtt部位と同様にEcoR1部位をプライマー中に導入し、2つのフラグメントが一緒に「接着される」ことができるようにして、Invitrogen Gateway pDONR P1−P5r vector(Carlsbad、CA、cat #12537−100)中に再結合させた。
【0095】
1.合成CREエレメント
配列番号6:
5’AAATAATGATTTTATTTTGACTGATAGTGACCTGTTCGTTGCAACAAATTGATGAGCAATGCTTTTTTATAATGCCAACTTTGTACAAAAAAGCAGGCTTACTGTCGACAATTGCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGATTGACGTCAATGGGGTGTCTGAAAGACGTCACAGTATGACCCGGGCTCGAGCCTCCTTGGCTGACGTCAGAGAGAGAGGCCGGCCCCTTACGTCAGAGGCGAGAATTCGACAACTTTGTATACAAAAGTTGAACGAGAAACGTAAAATGATATAAATATCAATATATTAAATTAGATTTTGCATAAAAAACAGACTACATAATACTGTAAAACACAACATATCCAGTCACTATG3’
【0096】
2.ハイブリッドCRE:標準的なクローニング、3×CREを含むClontech pCreLuc vectorからCREエレメントをPCR増幅し、PCRプライマーは一端にEcoR1制限部位を有しており(CRE3X−B、CRE3X−C)、att部位は、ゲートウエイのattB1またはattB5rのいずれかがもう一方の末端でクローニングされている(CRE3X−A、CRE3X−D)。次いで、これらの2つのフラグメントを組み合わせて6×CREの1つのフラグメントを作成した。このフラグメントは、Gateway pDONR P1−P5r vector中に組み換えされる。
【0097】
配列番号7:CRE 順方向プライマー A
5’−GGGGACAAGTTTGTACAAAAAAGCAGGCTTAGCACCAGACAGTGA−3’
配列番号8:CRE 逆方向プライマー B
5’−GGGAATTCGTTCTCCCATTGACGTCA−3’
配列番号9:CRE 順方向プライマー C
5’−GGGAATTCGCACCAGACAGTGACGTC−3’
配列番号10:CRE 逆方向プライマー D
5’−GGGGACAACTTTTGTATACAAAGTTGTGTTCTCCCATTGACGTCA−3’
配列番号11:ハイブリッドCRE配列:
GCTTAGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACGAATTCGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACA
【0098】
B.ヒト成長ホルモン ポリAテール:
ヒト成長ホルモン ポリAテールを、ベクターpOGH(Nichols Institute Diagnostics(San Juan Capistrano,CA、Cat # 40−2205)からPCRクローニングした。プライマー配列は、Gateway pDONR P3−P2 vector中への組み換え用に、順方向プライマー上にattB3部位、逆方向プライマー上にattB2部位を有している。
【0099】
配列番号12:hGH 順方向プライマー:
5’−GGGGACAACTTTGTATAATAAAGTTGGATCCCAAGGCCCAACTCC−3’
配列番号13:hGH 逆方向プライマー
5’−GGGGACCACTTTGTACAAGAAAGCTGGGTACAACAGGCATCTACT−3’
【0100】
C.HSV TK最小プロモーター:
HSV TK最小プロモーターを、「661 CreLuc」と呼ばれるインハウスベクター(in house vector)からPCRクローニングした。プライマー配列は、Gateway pDONR P5−P4 vector中への組み換え用に、順方向プライマー上にattB5部位を有し、逆方向プライマー上にattB4部位を有している。
【0101】
配列番号14:TK 順方向プライマー
5’−GGGGACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGT−3’
【0102】
配列番号15:TK 逆方向プライマー
5’−GGGGACAACTTTGTATAGAAAAGTTGGGTGGATCTGCGGCACGCT−3’
【0103】
D.ルシフェラーゼ cDNA:
ルシフェラーゼcDNAを、ベクターpGL4.10(Promega Madison、WI Cat #E6651)からPCRクローニングした。Lucプライマー配列は、Gateway pDONR P4r−P3r vector中への組み換え用に、順方向プライマー上にattB4r部位を有し、逆方向プライマー上にattB3r部位を有している。
【0104】
配列番号16:luci 順方向プライマー
5’GGGGACAACTTTTCTATACAAAGTTGATGGAAGATGCCAAAAACA3’
【0105】
配列番号17:luci 逆方向プライマー
5’GGGGACAACTTTATTATACAAAGTTGTTTACACGGCGATCTTGCC3’
【0106】
III.最終的なベクターの構築
導入遺伝子の4つのコンポーネント、CRE(合成バージョンまたはハイブリッドバージョンのいずれか)、HSVTK min、ルシフェラーゼ cDNA、およびpDONRベクター中のhGHポリAテールを、標準的なInvitrogenプロトコルに従ってpDest2XMARsデスティネーションベクターと共に組み換えを行った。その後これら2つの導入遺伝子をシーケンスし、CHOK1中にトランスフェクトして、機能について試験した。図5に示されように、どちらの導入遺伝子も、インビトロで機能する。図5は、ホルスコリン誘導によるCreLucベクターのインビトロ分析を示している。
【0107】
IV.トランスジェニックマウスの作製
A.導入遺伝子の準備
導入遺伝子構築物の質は、アリコートをアガロースゲル(電気泳動)にかけることにより検証および確認した。劣化(degradation)のトレースは観察されなかった。最終的に、診断用のXhoIおよびPvuIIおよびPstI制限酵素を用いた切断解析により、予測された制限プロファイルが得られた。次いで、この導入遺伝子プラスミドをAcc65IおよびPmeIで消化して、導入遺伝子を含むフラグメントをアガロースゲル上での消化を実行することにより、ベクター骨格から単離した。その後、この導入遺伝子フラグメントをゲルから切り出し、Qiaquick gel extraction kit(Qiagen、Valencia、CA cat#28706)で精製し、希釈して、受精卵中に注入した。単離した導入遺伝子の純度および濃度はアガロースゲル電気泳動により検査した。
【0108】
配列番号18:CreLuc 合成導入遺伝子(Acc65I/PmeI 消化、5550bp)
5’GTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCCAAGTTTGTACAAAAAAGCAGGCTTACTGTCGACAATTGCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGATTGACGTCAATGGGGTGTCTGAAAGACGTCACAGTATGACCCGGGCTCGAGCCTCCTTGGCTGACGTCAGAGAGAGAGGCCGGCCCCTTACGTCAGAGGCGAGAATTCGACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGTCGGGGCGGCGCGGTCCCAGGTCCACTTCGCATATTAAGGTGACGCGTGTGGCCTCGAACACCGAGCGACCCTGCAGCGACCCGCTTAACAGCGTCAACAGCGTGCCGCAGATCCACCCAACTTTTCTATACAAAGTTGCTATGGAAGATGCCAAAAACATTAAGAAGGGCCCAGCGCCATTCTACCCACTCGAAGACGGGACCGCCGGCGAGCAGCTGCACAAAGCCATGAAGCGCTACGCCCTGGTGCCCGGCACCATCGCCTTTACCGACGCACATATCGAGGTGGACATTACCTACGCCGAGTACTTCGAGATGAGCGTTCGGCTGGCAGAAGCTATGAAGCGCTATGGGCTGAATACAAACCATCGGATCGTGGTGTGCAGCGAGAATAGCTTGCAGTTCTTCATGCCCGTGTTGGGTGCCCTGTTCATCGGTGTGGCTGTGGCCCCAGCTAACGACATCTACAACGAGCGCGAGCTGCTGAACAGCATGGGCATCAGCCAGCCCACCGTCGTATTCGTGAGCAAGAAAGGGCTGCAAAAGATCCTCAACGTGCAAAAGAAGCTACCGATCATACAAAAGATCATCATCATGGATAGCAAGACCGACTACCAGGGCTTCCAAAGCATGTACACCTTCGTGACTTCCCATTTGCCACCCGGCTTCAACGAGTACGACTTCGTGCCCGAGAGCTTCGACCGGGACAAAACCATCGCCCTGATCATGAACAGTAGTGGCAGTACCGGATTGCCCAAGGGCGTAGCCCTACCGCACCGCACCGCTTGTGTCCGATTCAGTCATGCCCGCGACCCCATCTTCGGCAACCAGATCATCCCCGACACCGCTATCCTCAGCGTGGTGCCATTTCACCACGGCTTCGGCATGTTCACCACGCTGGGCTACTTGATCTGCGGCTTTCGGGTCGTGCTCATGTACCGCTTCGAGGAGGAGCTATTCTTGCGCAGCTTGCAAGACTATAAGATTCAATCTGCCCTGCTGGTGCCCACACTATTTAGCTTCTTCGCTAAGAGCACTCTCATCGACAAGTACGACCTAAGCAACTTGCACGAGATCGCCAGCGGCGGGGCGCCGCTCAGCAAGGAGGTAGGTGAGGCCGTGGCCAAACGCTTCCACCTACCAGGCATCCGCCAGGGCTACGGCCTGACAGAAACAACCAGCGCCATTCTGATCACCCCCGAAGGGGACGACAAGCCTGGCGCAGTAGGCAAGGTGGTGCCCTTCTTCGAGGCTAAGGTGGTGGACTTGGACACCGGTAAGACACTGGGTGTGAACCAGCGCGGCGAGCTGTGCGTCCGTGGCCCCATGATCATGAGCGGCTACGTTAACAACCCCGAGGCTACAAACGCTCTCATCGACAAGGACGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGTGGACCGGCTGAAGAGCCTGATCAAATACAAGGGCTACCAGGTAGCCCCAGCCGAACTGGAGAGCATCCTGCTGCAACACCCCAACATCTTCGACGCCGGGGTCGCCGGCCTGCCCGACGACGATGCCGGCGAGCTGCCCGCCGCAGTCGTCGTGCTGGAACACGGTAAAACCATGACCGAGAAGGAGATCGTGGACTATGTGGCCAGCCAGGTTACAACCGCCAAGAAGCTGCGCGGTGGTGTTGTGTTCGTGGACGAGGTGCCTAAAGGACTGACCGGCAAGTTGGACGCCCGCAAGATCCGCGAGATTCTCATTAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAACAACTTTGTATAATAAAGTTGCTGATCCCAAGGCCCAACTCCCCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCTGCAATGGCTACAGGTAAGCGCCCCTAAAATCCCTTTGGGCACAATGTGTCCTGAGGGGAGAGGCAGCGACCTGTAGATGGGACGGGGGCACTAACCCTCAGGTTTGGGGCTTCTGAATGTGAGTATCGCCATGTAAGCCCAGTATTTGGCCAATCTCAGAAAGCTCCTGGTCCCTGGAGGGATGGAGAGAGAAAAACAAACAGCTCCTGGAGCAGGGAGAGTGCTGGCCTCTTGCTCTCCGGCTCCCTCTGTTGCCCTCTGGTTTCTCCCCAGGCTCCCGGACGTCCCTGCTCCTGGCTTTTGGCCTGCTCTGCCTGCCCTGGCTTCAAGAGGGCAGTGCCTTCCCAACCATTCCCTTATCCAGGCTTTTTGACAACGCTATGCTCCGCGCCCATCGTCTGCACCAGCTGGCCTTTGACACCTACCAGGAGTTTGTAAGCTCTTGGGGAATGGGTGCGCATCAGGGGTGGCAGGAAGGGGTGACTTTCCCCCGCTGGGAAATAAGAGGAGGAGACTAAGGAGCTCAGGGTTTTTCCCGAAGCGAAAATGCAGGCAGATGAGCACACGCTGAGTGAGGTTCCCAGAAAAGTAACAATGGGAGCTGGTCTCCAGCG
TAGACCTTGGTGGGCGGTCCTTCTCCTAGGAAGAAGCCTATATCCCAAAGGAACAGAAGTATTCATTCCTGCAGAACCCCCAGACCTCCCTCTGTTTCTCAGAGTCTATTCCGACACCCTCCAACAGGGAGGAAACACAACAGAAATCCGTGAGTGGATGCCTTCTCCCCAGGCGGGGATGGGGGAGACCTGTAGTCAGAGCCCCCGGGCAGCACAGCCAATGCCCGTCCTTCCCCTGCAGAACCTAGAGCTGCTCCGCATCTCCCTGCTGCTCATCCAGTCGTGGCTGGAGCCCGTGCAGTTCCTCAGGAGTGTCTTCGCCAACAGCCTGGTGTACGGCGCCTCTGACAGCAACGTCTATGACCTCCTAAAGGACCTAGAGGAAGGCATCCAAACGCTGATGGGGGTGAGGGTGGCGCCAGGGGTCCCCAATCCTGGAGCCCCACTGACTTTGAGAGCTGTGTTAGAGAAACACTGCTGCCCTCTTTTTAGCAGTCAGGCCCTGACCCAAGAGAACTCACCTTATTCTTCATTTCCCCTCGTGAATCCTCCAGGCCTTTCTCTACACCCTGAAGGGGAGGGAGGAAAATGAATGAATGAGAAAGGGAGGGAACAGTACCCAAGCGCTTGGCCTCTCCTTCTCTTCCTTCACTTTGCAGAGGCTGGAAGATGGCAGCCCCCGGACTGGGCAGATCTTCAAGCAGACCTACAGCAAGTTCGACACAAACTCACACAACGATGACGCACTACTCAAGAACTACGGGCTGCTCTACTGCTTCAGGAAGGACATGGACAAGGTCGAGACATTCCTGCGCATCGTGCAGTGCCGCTCTGTGGAGGGCAGCTGTGGCTTCTAGCTGCCCGGGTGGCATCCCTGTGACCCCTCCCCAGTGCCTCTCCTGGCCCTGGAAGTTGCCACTCCAGTGCCCACCAGCCTTGTCCTAATAAAATTAAGTTGCATCATTTTGTCTGACTAGGCGTCCTTCTATAATATTATGGGGTGGAGGGGGGTGGTATGGAGCAAGGGGCAAGTTGGGAAGACAACCTGTAGGGCCTGCGGGGTCTATTGGGAACCAAGCTGGAGTGCAGTGGCACAATCTTGGCTCACTGCAATCTCCGCCTCCTGGGTTCAAGCGATTCTCCTGCCTCAGCCTCCCGAGTTGTTGGGATTCCAGGCATGCATGACCAGGCTCAGCTAATTTTTGTTTTTTTGGTAGAGACGGGGTTTCACCATATTGGCCAGGCTGGTCTCCAACTCCTAATCTCAGGTGATCTACCCACCTTGGCCTCCCAAATTGCTGGGATTACAGGCGTGAACCACTGCTCCCTTCCCTGTCCTTCTGATTTTAAAATAACTATACCAGCAGGAGGACGTCCAGACACAGCATAGGCTACCTGGCCATGCCCAACCGGTGGGACATTTGAGTTGTTTGCTTGGCACTGTCCTCTCATGCGTTGGGTCCACTCAGTAGATGCCTGTTGTACCCAGCTTTCTTGTACAAAGTGGGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTT 3’
【0109】
配列番号19:CreLuc ハイブリッド導入遺伝子配列(Acc65I/PmeI 消化、5562bp)
5’GTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCCAAGTTTGTACAAAAAAGCAGGCTTAGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACGAATTCGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGTCGGGGCGGCGCGGTCCCAGGTCCACTTCGCATATTAAGGTGACGCGTGTGGCCTCGAACACCGAGCGACCCTGCAGCGACCCGCTTAACAGCGTCAACAGCGTGCCGCAGATCCACCCAACTTTTCTATACAAAGTTGCTATGGAAGATGCCAAAAACATTAAGAAGGGCCCAGCGCCATTCTACCCACTCGAAGACGGGACCGCCGGCGAGCAGCTGCACAAAGCCATGAAGCGCTACGCCCTGGTGCCCGGCACCATCGCCTTTACCGACGCACATATCGAGGTGGACATTACCTACGCCGAGTACTTCGAGATGAGCGTTCGGCTGGCAGAAGCTATGAAGCGCTATGGGCTGAATACAAACCATCGGATCGTGGTGTGCAGCGAGAATAGCTTGCAGTTCTTCATGCCCGTGTTGGGTGCCCTGTTCATCGGTGTGGCTGTGGCCCCAGCTAACGACATCTACAACGAGCGCGAGCTGCTGAACAGCATGGGCATCAGCCAGCCCACCGTCGTATTCGTGAGCAAGAAAGGGCTGCAAAAGATCCTCAACGTGCAAAAGAAGCTACCGATCATACAAAAGATCATCATCATGGATAGCAAGACCGACTACCAGGGCTTCCAAAGCATGTACACCTTCGTGACTTCCCATTTGCCACCCGGCTTCAACGAGTACGACTTCGTGCCCGAGAGCTTCGACCGGGACAAAACCATCGCCCTGATCATGAACAGTAGTGGCAGTACCGGATTGCCCAAGGGCGTAGCCCTACCGCACCGCACCGCTTGTGTCCGATTCAGTCATGCCCGCGACCCCATCTTCGGCAACCAGATCATCCCCGACACCGCTATCCTCAGCGTGGTGCCATTTCACCACGGCTTCGGCATGTTCACCACGCTGGGCTACTTGATCTGCGGCTTTCGGGTCGTGCTCATGTACCGCTTCGAGGAGGAGCTATTCTTGCGCAGCTTGCAAGACTATAAGATTCAATCTGCCCTGCTGGTGCCCACACTATTTAGCTTCTTCGCTAAGAGCACTCTCATCGACAAGTACGACCTAAGCAACTTGCACGAGATCGCCAGCGGCGGGGCGCCGCTCAGCAAGGAGGTAGGTGAGGCCGTGGCCAAACGCTTCCACCTACCAGGCATCCGCCAGGGCTACGGCCTGACAGAAACAACCAGCGCCATTCTGATCACCCCCGAAGGGGACGACAAGCCTGGCGCAGTAGGCAAGGTGGTGCCCTTCTTCGAGGCTAAGGTGGTGGACTTGGACACCGGTAAGACACTGGGTGTGAACCAGCGCGGCGAGCTGTGCGTCCGTGGCCCCATGATCATGAGCGGCTACGTTAACAACCCCGAGGCTACAAACGCTCTCATCGACAAGGACGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGTGGACCGGCTGAAGAGCCTGATCAAATACAAGGGCTACCAGGTAGCCCCAGCCGAACTGGAGAGCATCCTGCTGCAACACCCCAACATCTTCGACGCCGGGGTCGCCGGCCTGCCCGACGACGATGCCGGCGAGCTGCCCGCCGCAGTCGTCGTGCTGGAACACGGTAAAACCATGACCGAGAAGGAGATCGTGGACTATGTGGCCAGCCAGGTTACAACCGCCAAGAAGCTGCGCGGTGGTGTTGTGTTCGTGGACGAGGTGCCTAAAGGACTGACCGGCAAGTTGGACGCCCGCAAGATCCGCGAGATTCTCATTAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAACAACTTTGTATAATAAAGTTGCTGATCCCAAGGCCCAACTCCCCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCTGCAATGGCTACAGGTAAGCGCCCCTAAAATCCCTTTGGGCACAATGTGTCCTGAGGGGAGAGGCAGCGACCTGTAGATGGGACGGGGGCACTAACCCTCAGGTTTGGGGCTTCTGAATGTGAGTATCGCCATGTAAGCCCAGTATTTGGCCAATCTCAGAAAGCTCCTGGTCCCTGGAGGGATGGAGAGAGAAAAACAAACAGCTCCTGGAGCAGGGAGAGTGCTGGCCTCTTGCTCTCCGGCTCCCTCTGTTGCCCTCTGGTTTCTCCCCAGGCTCCCGGACGTCCCTGCTCCTGGCTTTTGGCCTGCTCTGCCTGCCCTGGCTTCAAGAGGGCAGTGCCTTCCCAACCATTCCCTTATCCAGGCTTTTTGACAACGCTATGCTCCGCGCCCATCGTCTGCACCAGCTGGCCTTTGACACCTACCAGGAGTTTGTAAGCTCTTGGGGAATGGGTGCGCATCAGGGGTGGCAGGAAGGGGTGACTTTCCCCCGCTGGGAAATAAGAGGAGGAGACTAAGGAGCTCAGGGTTTTTCCCGAAGCGAAAATGCAGGCAGATGAGCACACGCTGAGTGAGGTTCCCAGAAAAGTAACAATGGGAGCTGGTCTCCAGCGTAGACCTTGGTGGGCGGTCCTTCTCCTAGGAAGAAGCCTATATCCCAAAGGAACAGAAGTATTCATTCCTGCAGAACCCCCAGACCTCCCTCTGTTTCTCAGAGTCTATTCCGACACCCTCCAACAGGGAGGAAACACAACAGAAATCCGTGAGTGGATGCCTTCTCCCCAGGCGGGGATGGGGGAGACCTGTAGTCAGAGCCCCCGGGCAGCACAGCCAATGCCCGTCCTTCCCCTGCAGAACCTAGAGCTGCTCCGCATCTCCCTGCTGCTCATCCAGTCGTGGCTGGAGCCCGTGCAGTTCCTCAGGAGTGTCTTCGCCAACAGCCTGGTGTACGGCGCCTCTGACAGCAACGTCTATGACCTCCTAAAGGACCTAGAGGAAGGCATCCAAACGCTGATGGGGGTGAGGGTGGCGCCAGGGGTCCCCAATCCTGGAGCCCCACTGACTTTGAGAGCTGTGTTAGAGAAACACTGCTGCCCTCTTTTTAGCAGTCAGGCCCTGACCCAAGAGAACTCACCTTATTCTTCATTTCCCCTCGTGAATCCTCCAGGCCTTTCTCTACACCCTGAAGGGGAGGGAGGAAAATGAATGAATGAGAAAGGGAGGGAACAGTACCCAAGCGCTTGGCCTCTCCTTCTCTTCCTTCACTTTGCAGAGGCTGGAAGATGGCAGCCCCCGGACTGGGCAGATCTTCAAGCAGACCTACAGCAAGTTCGACACAAACTCACACAACGATGACGCACTACTCAAGAACTACGGGCTGCTCTACTGCTTCAGGAAGGACATGGACAAGGTCGAGACATTCCTGCGCATCGTGCAGTGCCGCTCTGTGGAGGGCAGCTGTGGCTTCTAGCTGCCCGGGTGGCATCCCTGTGACCCCTCCCCAGTGCCTCTCCTGGCCCTGGAAGTTGCCACTCCAGTGCCCACCAGCCTTGTCCTAATAAAATTAAGTTGCATCATTTTGTCTGACTAGGCGTCCTTCTATAATATTATGGGGTGGAGGGGGGTGGTATGGAGCAAGGGGCAAGTTGGGAAGACAACCTGTAGGGCCTGCGGGGTCTATTGGGAACCAAGCTGGAGTGCAGTGGCACAATCTTGGCTCACTGCAATCTCCGCCTCCTGGGTTCAAGCGATTCTCCTGCCTCAGCCTCCCGAGTTGTTGGGATTCCAGGCATGCATGACCAGGCTCAGCTAATTTTTGTTTTTTTGGTAGAGACGGGGTTTCACCATATTGGCCAGGCTGGTCTCCAACTCCTAATCTCAGGTGATCTACCCACCTTGGCCTCCCAAATTGCTGGGATTACAGGCGTGAACCACTGCTCCCTTCCCTGTCCTTCTGATTTTAAAATAACTATACCAGCAGGAGGACGTCCAGACACAGCATAGGCTACCTGGCCATGCCCAACCGGTGGGACATTTGAGTTGTTTGCTTGGCACTGTCCTCTCATGCGTTGGGTCCACTCAGTAGATGCCTGTTGTACCCAGCTTTCTTGTACAAAGTGGGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTT
【0110】
B.初代の作製:
これらの導入遺伝子をFVB/Taconic単細胞胚の前核に別々にマイクロインジェクションした。トランスジェニック(Tg)動物を作製するための一般的なストラテジーはよく記載されており、当業者に公知である。
【0111】
PCR遺伝子型判定ストラテジーを用いて、トランスジェニックポジティブ初代マウスを、尾部生検および遺伝子型用のPCRを介して出生後すぐに同定した。選択されたプライマー対は、導入遺伝子配列内に短いDNA配列の増幅を可能にし、特定の405−bp PCR産物を得ることができる。PCR反応ミックスは、MgCl2(Qiagen、Valencia、CA cat #201205)、0.2mM dNTPs、0.4uM プライマー、0.5単位のTaqポリメラーゼを含む1×Qiagen PCR緩衝液である。サイクリング条件は以下の通りである:94℃で1サイクル;94℃で5分、35サイクル;57℃で45秒;72℃で45秒;72℃で1分、1サイクル;5分。
【0112】
遺伝子型判定プライマーは以下の通りである:
配列番号20:Luc2−順方向プライマー:
5’−GAAGATGCCAAAAACATTAAGAAG−3’
配列番号21:Luc2−逆方向プライマー:
5’−GATCTTTTGCAGCCCTTTCT −3’
【0113】
合成CreLuc導入遺伝子については、(181出生したうち)39の導入遺伝子ポジティブ初代、ハイブリッドCreLuc導入遺伝子については、(244出生したうち)73の導入遺伝子ポジティブが生成された。このトランスジェニックマウス系統は、CreLucと省略される。
【0114】
V.初代発現分析:
図5Aは、正常マウスにおける血漿cAMPレベルに対する異なるホスホジエステラーゼ(PDE)インヒビターの効果の比較である(Cheng et al、JPET、280(2):621−626、1997からの再現)。これらのマウスを、4匹の動物群で化合物またはビヒクル(VEH)をp.o.投与した。20分後、血液を採取し、cAMP測定のために処理した。この発行された実験に基づき、cAMPレベルを増加させるその能力およびそのアベイラビリティについてのCreLuc初代をスクリーニングするために、ホルスコリンと共にロリプラムを一般的な誘導因子として選択した。一緒に投与する場合、これらの2つの化合物の付加的な効果により、動物中のルシフェラーゼレベルを調べるときの誘導の大きいウインドウを提供される。
【0115】
図5Bは、ホルスコリン(F)、ロリプラム()またはホルスコリンとロリプラムの組み合わせ(F/R)による種々の処理後の、野生型マウス中の血漿レベルのcAMPアッセイである。2月齢のFVB/Tac雌を、以下のうちの1つと共にi.p.投与した:グループ:ビヒクルコントロール、リン酸緩衝生理食塩水(PBS)に溶解させた1%ジメチルスルホキシド (DMSO)、グループB:5mg/kg ホルスコリン(Sigma、St Louis MO、cat#F6886)、およびグループC:10mg/kg ロリプラム(Sigma St Louis、MO、cat# R6520)、グループD:5mg/kg ホルスコリン+10mg/kg ロリプラム、グループE:5mg/kg 水溶性ホルスコリン(Calbiochem Gibbstown、NJ cat#344273)、およびグループF:5mg/kg 水溶性ホルスコリン+10mg/kg ロリプラム。グループGは、処理を受けていない。ビヒクルのみと比較して、ロリプラムおよび/またはホルスコリンは、循環しているcAMPレベルの上昇において、統計的に有意であった。ホルスコリンのみでは、ビヒクルコントロールに対して循環しているcAMPを有意に上昇させなかった。水溶性ホルスコリン(H2OF)とロリプラムとの組み合わせは、cAMP血漿レベルにおいて最も大きい上昇を示し、この処理がインビボでの導入遺伝子の発現のためにCreLuc初代をスクリーニングするために最適であった。
【0116】
全ての初代(表5〜8に列挙している初代および図6に示されている代表的な初代)を、10mg/kg ロリプラム(Sigma St Louis、MO、cat# R6520)と5mg/kg 水溶性ホルスコリン(Calbiochem Gibbstown、NJ cat#344273)との組み合わせをマウスにi.p.投与することによって、導入遺伝子発現について試験した。4時間後、これらのマウスをXenogen IVIS Lumina systemを用いてバイオイメージングした。これらのマウスを、イソフルラン、および250mg/kg ルシフェリンをs.c.注射することにより麻酔した。39の導入遺伝子ポジティブな合成CreLucのうち、34(または87%)が発現ポジティブであった。発現は任意の組織のルシフェラーゼレベルがシステムのバックグラウンドレベルを超えたとき、ポジティブであると考えられた。
【0117】
CreLucマウスの異なる独立した系統のバイオイメージングによってアッセイされた種々のホルスコリン ロリプラム応答プロファイルを、図6に示している。誘導されていない状態において、本発明者らは、系統90および219に図示されているような検出可能な基底バイオイメージングシグナルか、または系統44、28、および187に図示されているような基底活性がないかのいずれかを確認した。ホルスコリン ロリプラム誘導後、CreLucレポーター誘導のバイオイメージングパターンは、単一または複数の組織中に生じ、それは、各独立したCreLuc系統で独特であった。多くのCreLuc系統は、治療に関連する領域中に検出可能なルシフェラーゼビオイメージを発現する組織を1以上有していた。
【0118】
基底と、ホルスコリン ロリプラム誘導の両方での初代中のCreLuc発現のバイオイメージングスクリーニング後、1またはそれより多い組織(脳内で2×)中で5×バイオイメージング誘導のフィルターウインドウを満たすCreLuc系統を、組織抽出物中のルシフェラーゼ酵素の誘導されたレベルをアッセイすることによってより詳細に分析した。
【0119】
組織発現分析のために、動物を、ベースラインシグナル用に未処理のままかまたは4時間誘導した。誘導のために、動物に、Dulbecco’s PBS(Invitrogen、Carlsbad、CA、cat#14040)に溶解させた5mg/kg ホルスコリン(Calbiochem、Gibbstown、NJ cat#344273)と10mg/kg ロリプラム(Sigma St Louis、MO、cat# R6520)with 1%DMSO(Sigma、St Louis、MO、cat#D2650)の組み合わせをi.p.投与した。数時間後、これらの動物をCO2で屠殺し、種々の器官を取り除いてドライアイスで凍らせた。ルシフェラーゼアッセイのために、Luciferase Assay System(Promega、Madison、WI、cat#E1500)を用いた。これらの組織を、1mlの細胞培養溶解緩衝液(Cell Culture Lysis Buffer)(Promega、Madison、WI、cat#E1531)中でホモジネートさせ、5分間氷上でインキュベーションし、その後5分間遠心分離で回転させた。20μlの上清を製造業者の指示書によりアッセイ中で用いた。これらのサンプルのタンパク質濃度をDCタンパク質アッセイキット(BioRad、Hercules、CA、cat#500−0111)を用いて測定した。データは表5〜8に示しており、誘導倍率として表わしている(誘導されたRLU/ugタンパク質を、基底RLU/ugタンパク質で割ったもの)。誘導されたRLUは、誘導された組織サンプルのRLU/ugタンパク質である。
【0120】
【表5】
【0121】
【表6】
【0122】
【表7】
【0123】
【表8】
【0124】
各系統におけるCreLuc発現の詳細なレベルは、標的組織中で最も高い発現レベルを持つ最適の系統の選択、および独特な単一または複数のパターンの発現を有する系統の選択を可能にする。この酵素アッセイはまた、CreLuc発現が特定の組織と明らかに関連していたことを保証した。一般的に、導入遺伝子中にデザインされた場合、大部分の系統を横切ってすべての組織の近くにルシフェラーゼ発現を観察した。重要な医薬関連標的組織(脳、膵臓、肺、脾臓)由来のトップの3〜5系統を、GPCRリガンドを用いたさらなる参照化合物応答プリファイリングのために選択した。
【0125】
VI.発現レベルにおけるインスレーター配列の効果の分析
表9において、インスレーター配列を加えることの影響は、11の異なる導入遺伝子を通じて比較され、225の独立したトランスジェニック系統を含んでいる。機能的なトランスジェニック系統の生産は、典型的には10〜30%の範囲である(本発明者らの研究室から採取した過去のデータ)。表9において、hGH小遺伝子または機能的エンハンサーの封入により、11のうち10の導入遺伝子および190系統が、この範囲の上限での発現レベルを獲得した(機能的系統の平均数は37または32.7%)。重要なことに、インスレーター配列の封入は、この発現頻度を2倍以上に増やし、112系統については78%が機能的レポーターを発現している。導入遺伝子の比較および多様性両方についてのサンプルサイズは、ゲノム中の導入遺伝子のランダムな組み込みにより、正常な生物学的変動を克服するのに十分な大きさである。
【0126】
トランスジェニックバイオイメージングレポーター系統の作製は、低頻度で高い損耗率のプロセスである。多くの独立した初代系統(50〜100)が、レポーター用に所望される検出レベルを備える系統を得るため、および薬物候補のスクリーニング用に適切な標的組織の広範な選択肢において発現されるレポーターを持つために必要とされる。一般的には、薬物スクリーニング用途には5%以下の系統で十分であるため、レポーターを含む導入遺伝子は、作製された各トランスジェニック系統中の発現を最大化させるために多くのエレメントを含める必要がある。他の発現エンハンサー(hGH小遺伝子)に結合しているインスレーター配列は、導入遺伝子を発現するトランスジェニック系統の大部分を得ることが実証されており、薬物開発に適用可能な発現パターンを持つトランスジェニック系統を見つける鍵である(リガンドのレセプターへの結合の際のシグナルの低下を検出するための種々のGPCRシグナル伝達経路については特に)。
【0127】
【表9】
【0128】
材料の寄託:
マウス胚線維芽細胞の調製:妊娠した雌から、12日目の胚を取り出した。この胚をリン酸緩衝生理食塩水(PBS)(Invitrogen、Carlsbad、CA、cat#14040)を含む10cmの皿に入れた子宮の中から解剖した。次いで、それをPBSの新しい皿に移した。その胚を鉗子で裂いて、心臓および肝臓を取り除いた。次いで、組織切片を2〜3mlの冷0.25%トリプシン(Invitrogen、Carlsbad、CA、cat#25200)中で4℃にて一晩インキュベーションした。次の日、そのトリプシンの大部分を吸引し、組織ペレットを37℃で30分間インキュベーションした。2〜3mlの培地、DMEM(Invitrogen、Carlsbad、CA、cat#11965)、5%の熱不活性化FBS(Invitrogen、Carlsbad、CA、cat#16140)、1%のペン連鎖球菌(Invitrogen、Carlsbad、CA、cat# 15140)を加え、組織の残りの塊を壊すために上下にピペッティングした。次いで細胞懸濁液をT75組織培養フラスコにプレーティングした。細胞を3継体培養し、その後1mlのバイアル中で、バイアルあたり1〜2百万細胞、凍結培地、15%の熱不活性化FBS(Invitrogen、Carlsbad、CA、cat#16140)、5% DMSO(Sigma、St Louis、MO、cat#D2650)、DMEM(Invitrogen、Carlsbad、CA、cat#11965)中で、凍結させた。
【0129】
以下の材料をAmerican Type Culture Collection、10801 University Blvd.、Manassas、Va.20110−2209、USA(ATCC):に寄託した:
【表10】
【0130】
この寄託は、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約およびその下の条例(Budapest Treaty on the International Recognition of the Deposit of Microorganisms for the Purpose of Patent Procedure and the Regulations thereunder(Budapest Treaty))の規定の下でなされた。これにより、寄託日より30年間、その寄託の生存培養物の維持が保証される。この寄託は、ブダペスト条約の下、ATCCによって利用可能とされており、sanofi−aventis US Inc.とATCCとの契約下にある。この契約は、関連する米国特許の公開の際、または任意の米国特許出願もしくは外国特許出願の一般への公開(いずれか早いほう)の際に、一般に対して寄託の培養物の子孫の永続的かつ無制限なアベイラビリティを保証し、かつその全体に対して米国の特許商標庁長官によって決定される子孫のアベイラビリティを保証する。
【0131】
本出願の指定代理人は、寄託においてそれらの材料の培養物が、適切な条件下で培養された場合に死ぬかまたは紛失されるかまたは破壊された場合に、それらの材料が速やかに同一の別のもので置き換えられることに合意した。寄託された材料のアベイラビリティーは、その特許法に従って、あらゆる政府の権限下で付与される権利に違反して本発明を実施するためのライセンスとして解釈されるべきではない。
【0132】
VII.いンビボおよびエキソビボスクリーニングアッセイ
図7は、インビボまたはエキソビボいずれかのCreLucスクリーニングアッセイの流れを表わすことによって本発明の実施態様を示している。インビボアッセイについては、動物にルシフェリンのみを投与し、その後IVIS bioimagerでバイオイメージングして、ベースライン信号を得た。その後これらの動物に化合物を投与して、設定した時点で再びバイオイメージングし、ルシフェラーゼシグナルに対する化合物の影響を調べた。データは、ベースラインの信号に対する誘導された信号の誘導倍率として分析することができる。エキソビボアッセイについては、マウスに化合物またはビヒクルのいずれかを投与し、その後設定した時点において組織を採取し、凍結させた。次いで、その組織をホモジネートし、Promega luciferase assay system(Promega cat #E1500)のようなルシフェラーゼアッセイをその組織抽出物に対して実行し、Topcountのようなルミノメーターでそのアッセイを読み取った。データは、化合物群に対してビヒクルを比較する、μgタンパク質あたりの相対光として分析することができる。
【0133】
本発明のさらなる局面および詳細は、以下の実施例から明らかとなり、これらは例示することを意図しているのであって、いかなる意味においても本発明の範囲を限定することを意味しない。
【0134】
A.全脳スライス実験
CreLucマウスにおける発光に及ぼす化合物の影響を示すために、全脳スライスを用いた(図8A、8B)。一般的には、CreLucマウスから脳を採取し、ミクロトームでスライス状にカットするかまたは種々の小領域を解剖して取り出し、ミクロトームでスライスすることもできる。その後これらのスライスをホルスコリンとロリプラム、またはGPCR特異的化合物のような異なる化合物と共にインキュベーションすることができる。次いで、ルシフェラーゼレベルをIVIS bioimagerまたはカルーセルを備える光電子増倍管で測定し、定量的経時測定を行うことができる。
【0135】
CreLucマウス(系統187)の海馬領域からの脳スライスを1uMのイソプロテレノール(Sigma、cat#5627)と共に2日間インキュベーションした。イソプロテレノール(ISO)は、cAMP経路を活性化するGsアゴニストであり(経路の概略は図1を参照のこと)、CreLucマウス中でルシフェラーゼ発現レベルを誘導する(図8A、下パネル)。従って、GiアゴニストのようなGPCR特異的化合物のスクリーニングを可能にするルシフェラーゼシグナルウインドウを増加させるためにイソプロテレノールを用いた。AMN082(AMN、Tocris Bioscience、Cat.No.2385)は、mGluR7アゴニストであり、mGluR7は、Giに結合しているレセプターである。脳スライスを、1uM AMN082と組み合わせた1uM ISOで処理した場合、ルシフェラーゼシグナルは、ISOのみでの処理と比較して減少した。
【0136】
化合物の活性はまた、生育できる脳スライス中で時間をかけて定量することができ、またルシフェラーゼ発現は、バイオイメージャーによって可視化することができる。図8Bにおいて、CreLucマウス(系統44)からの脳スライスは、ホルスコリンに反応した。ホルスコリンはcAMPの一般的なアクチベーターである。50uMのホルスコリンでの脳スライスの処理(矢印は、ホルスコリンをスライスに加えた時間をマークしている)により、ビヒクルと比較して時間依存的に誘導を引き起こした。
【0137】
B.インビトロ細胞培養実験
1.初代皮質ニューロン
CreLucモデルからの初代神経細胞の単離および化合物処理についてのワークフローの一般的な概略図を図9に示している。皮質や他の脳の小領域を胚(通常は胎齢(E)17または18であるが、早ければE14も)から取り除いた。その後、個々のニューロンを単離し、96ウエルのアッセイプレート上にプレーティングした。アッセイは、トリプリケートまたは4通り(quadruplicate)のいずれかで実行した。アッセイは、標的GPCRの発現レベルに依存して、培養物中で7日または10日まで培養した。その後、化合物をそのプレートに加え、Bright Glo(Promega、Cat # E2610)をプレートに加えて、TopCountまたは他のルミノメーターでアッセイを実行した。
【0138】
a)βアドレナリン受容体(AD(R)およびD1ドーパミン受容体(DRD1)を介したGsの調節
CreLucマウスからの初代細胞培養物を、Gs共役型レセプターを調節し得る化合物をスクリーニングするために用いることができる。ニューロンを系統187のE18胚の皮質から単離した。系統187は、全組織ルシフェラーゼアッセイによって脳と脊髄両方におけるルシフェラーゼの誘導性レベルを有することが示されている(以下の文章;図24を参照のこと)。このアッセイを培養3日目にトリプリケートで実行した。ホルスコリンを5μM、ロリプラムを10μM、および、Gsアゴニスト、ADβR−イソプロテレノール;およびドーパミン−SKF82958(MFG、Cat# C130)を10μMで用いた。4時間でBright Gloを加え、Topcountでアッセイを実行した。ホルスコリンは、cAMPの一般的なアクチベーターである。ロリプラムはcAMPを活性化しないが、かわりにロリプラムはcAMPの分解をブロックし、それによりcAMPレベルを安定化させる。ホルスコリンとロリプラムは、ルシフェラーゼ発現を増加させるように相乗的に作用する。100倍を超える誘導が、ホルスコリンとロリプラムの組み合わせで観察された。増加したluciレベルは、DMSOコントロールに対して、ADβRアゴニストイソプロテレノールで見られた(11倍)。一方、D1 DRアゴニスト、SKF82958は、ルシフェラーゼレベルを有意に増加させず、ロリプラム単独と比較する場合に、SKF82958をロリプラムと組み合わせて投与した場合、2倍の誘導が観察された。従って、系統187は、その化合物がGs共役レセプターを調節し得るかどうかを判断するために化合物をスクリーニングするおに有用なモデルである。
【0139】
b)Prokineticin 2受容体(PROK2R)を介したGq調節
CreLucマウスからの初代細胞培養物を、Gs共役受容体を調節し得る化合物をスクリーニングするために用いることができる。従って、このPROK2Rは、PROK2Rのアゴニストであるprokineticin 2(PROK2)を、CreLucマウスがGq調節に応答するかどうかを確認するために用いた。Gqは、cAMPをバイパスするが、CREBを活性化するためにPLC経路を利用している(図1の経路図を参照のこと)。初代皮質ニューロンを、E18の系統187(脳および脊髄中に誘導可能なルシフェラーゼを有していることが示されている−以下の文章および図24を参照のこと)から採取した。このアッセイは、4時間、または8時間培養で3日目に実行した。PROK2ペプチド(Peprotech、Cat#100−46)を1nMおよび100nMの水溶液として加えた。ルシフェラーゼシグナル用にBright Gloを用い、アッセイをTopCount luminometerで実行した。コントロールのみの培地に対して、ルシフェラーゼレベルの非常に有意な増加が両方の時点で、PROK2の両方の濃度について観察された。従って、CreLucマウスは、Gq共役受容体を調節する化合物をスクリーニングするために用いることができる。
【0140】
異なるCreLuc系統由来の初代皮質ニューロン中のルシフェラーゼ発現におけるPROK2ペプチドの効果を調べた。初代皮質ニューロンを4つの異なるCreLuc系統からE18で採取した。系統は、全脳抽出物中で誘導可能なルシフェラーゼレベルを決定した以前の実験に基づいて選択した(データは示さず)。このアッセイを4時間および24時間の2つの時点で、1nMまたは100nMいずれかのPROK2ペプチドを含む培養物中で3日目にトリプリケートで実行した。このアッセイにはBrightGloを使用し、TopCountで読み取った。ルシフェラーゼレベルにおける統計的に有意な増加が、系統219および175の両方の時点および両方の濃度において観察された。全ての系統がある程度PROK2リガンドに対応しているとしても、ルシフェラーゼ発現レベルの差異は、導入遺伝子の組込みにおける差異に起因し得る。
【0141】
c)mGluR7受容体を介したGi調節
CreLucマウス由来の初代細胞培養物を、Gi共役受容体を調節し得る化合物をスクリーニングするために用いることができる。図13Aは、初代皮質ニューロン中のルシフェラーゼ発現におけるmGluR7アゴニスト、AMN082の効果を示している。皮質ニューロンをE18胚(系統187)より採取した。このアッセイは、培養3日目に実行した。ホルスコリンを10μMで用いてGi活性を検出するのに用いるより大きいシグナルウインドウを得た。このmGluR7アゴニスト、AMN082は、1nM、10nM、100nMおよび1μMのホルスコリンと組み合わせて用いた。このアッセイは、Bright Glo(Promega)を備えるTopCountで、4時間および8時間で読み取った。ホルスコリンのみに対して、ルシフェラーゼレベルの有意な減少が両時点の100nMおよび1μMのAMN082で観察された。このCreLucマウスは、Gq共役受容体を調節する化合物をスクリーニングするために用いることができる。例えば、「A」、「B」および「C」と記される「未知の」化合物を内部コントロールとしてAMN082を用いて初代皮質ニューロン培養物中でスクリーニングし、任意の化合物がGi調節能を示すかどうか決定する(図13B)。化合物Aは、EC50=1.735e−07を持つ公知のmGluR7アゴニストAMN082と比較すると、算出値EC5=1.529e−06の活性化合物であると考えられるが、一方化合物Bは、非特異的化合物であると考えられ、また化合物Cは不活性である。
【0142】
CreLucマウス由来の初代細胞培養物を用いるGi共役受容体を調節する能力についての化合物のスクリーニングは、上記に示したように、ホルスコリンのようなcAMP刺激因子の存在中でなされ得る。しかしながら、シグナルウインドウを増加させるためにも、ロリプラム(cAMPの分解をブロックする)をホルスコリンと組み合わせて用いた。初代皮質ニューロンは、誘導可能な能および脊髄発現を有していると以前に判断された系統187由来のE18胚から単離した。アッセイは、トリプリケートで、6時間培養で7日目に実行した。AMN082の10μMから1nMまでの用量曲線は、50μMのホルスコリンと10μMのロリプラムとを組み合わせて実行した。このアッセイは、Bright Glo substrate(Promega)を備えるTopCount luminometerで読み取った。ルシフェラーゼレベルの統計的に有意な減少であるp<0.0005(ホルスコリンおよびロリプラムコントロールに対して59〜61%)は、1μM、100nMおよび1nMで見られた。
【0143】
CB1アゴニスト、CP55,940による、異なるCreLuc系統由来の初代皮質ニューロン中のルシフェラーゼ発現のGi調節を、ホルスコリンおよびロリプラムの存在下で実施した(図15)。初代皮質ニューロンは、系統69、187、175および219のE18から採取した。用いられた全ての系統は、全脳組織抽出物中で誘導可能なルシフェラーゼ発現を有していると以前に判断されたものである(データは示さず)。このアッセイは、3日培養で実施した。CB1アゴニストは、10μMで、ホルスコリンは5μM、ロリプラムは10μMで用いた。4時間と24時間の2つの時点で実行した。次いで、Bright Gloルシフェラーゼアッセイ基質を加え、このアッセイをTopcount luminometerで読み取った。このアッセイはトリプリケートで実行した。データはトリプリケートの平均で示されている。データは、秒あたりのカウント数(cps)で示されている。ルシフェラーゼのシグナルの減少は、CB1アゴニストを添加した両方の時点で4つ全ての系統において観察された。応答レベルのわずかな差異は、導入遺伝子の組み込みに起因し得るが、全ての系統がGi調節に応答性であり、Gi活性のスクリーニング化合物に修正可能であった。
【0144】
CB1アゴニスト、CP55,940による、CreLucマウス由来の初代皮質ニューロン中のルシフェラーゼのGi調節は、用量依存的であった(図15B)。皮質ニューロンは、E18胚から単離下。このアッセイは培養3日目に実行した。ホルスコリンおよびロリプラムを10μMで用いた。アゴニストを10μM、1μMおよび100nMの濃度で加えた。このアッセイは、BrightGlo(Promega)を備えるTopCountで、8時間で読み取った。ルシフェラーゼレベルの有意な減少(ホルスコリンおよびロリプラム単独に対して、アゴニストにホルスコリンとロリプラムを加えたもの)は、3つ全ての濃度で観察された。
【0145】
2.初代線条体ニューロン
CreLucマウスから誘導された任意の細胞培養物は、GPCRを調節する能力について化合物をスクリーニングするために用いることができる。ホルスコリンおよびロリプラム、ならびにDRD1およびADβRのGsアゴニストによって、CreLuc線条体ニューロン中でルシフェラーゼ発現を誘導した(図16)。線条体ニューロンは、E14胚(系統187)から単離した。このアッセイは培養4日目に実行した。ホルスコリンを5μM、ロリプラムを10μMで用いた。Gsアゴニスト(イソプロテレノールはADRβにおいてアゴニストであり、ドーパミンおよびSKF82958は、D1DRにおいてアゴニストである)を10μM、3μMおよび1μMで用いた。このアッセイは、TopCount luminometerおよびBright Glo ルシフェラーゼ試薬(Promega)を用いて5時間で読み取った。28倍の非常に有意な増加が、ホルスコリンとロリプラムの組み合わせで処理された細胞中で観察された。有意な増加はまた、10μMのドーパミンで2.7倍、イソプロテレノールの3つ全ての濃度で観察された。下がttえ、導入遺伝子は、皮質ニューロンだけでなく線条体においても機能的である(図10を参照のこと)。
【0146】
3.全脾細胞
CreLucマウス(系統64)から単離した全脾細胞プレップを用いて、ホルスコリンおよびロリプラムのような一般的なcAMP誘導因子、ならびいルシフェラーゼ発現におけるGsアゴニストの効果を示した(図16)。脾臓は、1×HBSS(Invitrogen、Carlsbad、CA、cat#14025)の動物から採取した。次いで、この細胞を、5mlのD−PBS(Invitrogen、Carlsbad、CA、cat#14190)中で、5mlシリンジの先端を用いて脾臓カプセルを機械的に破壊することにより単離した。次いで、この細胞懸濁液を70umのストレーナーを通過させて50mlのコニカルチューブ中に入れた。その後細胞を800rpmで回転させ、5mlの1X Pharm Lyse solution(BD BioSciences、cat#555899)中に再懸濁させて室温で6分間インキュベーションした。次いで細胞に、RPMI 1640(Invitrogen、Carlsbad、CA、cat#11875)、10% FCS(Invitrogen、Carlsbad、CA、cat#16000)、1% pen/strep(Invitrogen、Carlsbad、CA、cat#15070)および0.1% β−メルカプトエタノール(Invitrogen、Carlsbad、CA、cat#21985)からなる28mlの培地を加えることによって洗浄した。細胞を2×106/mlまで希釈し、96ウエルの白色不透明プレートにプレートあたり100ulの細胞をプレーティングした。これらの細胞の半分を抗CD3抗体(BD Pharmingen、cat#553058)で24時間刺激し、残りの半分は処理しなかった。24時間でそのプレートに化合物を加え、さらに4時間放置した。一般的な誘導因子としては、ロリプラム(Sigma R6520)、ホルスコリン(Sigma F6886)が挙げられる。用いられたGsアゴニストは以下のとおりである:EX00000173A(173A;インハウスで合成された)は、Gs結合プロスタグランジンE2(EP2)受容体でのアゴニストである;BW245C(Sigma B9305)は、Gs結合プロスタグランジンD2受容体(DP1)のアゴニストであり、イソプロテレノール(Sigma I5627)は、Gs結合βアドレナリン受容体(ADβR)でのアゴニストである。このアッセイは、トリプリケートで実行した。4時間後、100μlのBrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイはTopCount luminometer で読み取った。DMSOに対して14倍の増加がロリプラムおよびホルスコリンの存在下でのCD3刺激された細胞中で観察された。統計的に有意な増加(t−検定による)もまた、DMSOのみのコントロールに対して3つ全てのGsアゴニストで観察された。この実験により、導入遺伝子は、全脾細胞集団において機能的であることが示された。この実験において用いられた調製物は、細胞の混合された集合である、全脾細胞調製物であった。以下に記載される実験は、T細胞およびB細胞のような亜集団におけるルシフェラーゼ発現レベルを調べている。
【0147】
4.単離T細胞
5つの異なるサブ系統のCreLucマウスから単離したT細胞中の、ロリプラムおよびホルスコリンによる一般的なcAMP活性化の効果を研究した(図18)。全脾細胞集団を上に記載したような機械的な破壊により準備した。次いで、CD4+細胞をMACS Magnetic Separation with Positive Selection Column(Miltenyi Biotec cat#130−049−201)を用いて単離した。次いで、これらの細胞をウエルあたり1.5x105個、96ウエルの白色不透明プレートにプレーティングし、抗CD3抗体(BD Pharmingen cat#553058)で刺激した。18時間後、10μMのロリプラ (Sigma R6520)および5μMのホルスコリン(Sigma F6886)をプレートに加え、さらに4時間置いた。BrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイはTopCountで読み取った。データは発光(秒あたりのカウント数)、および培地のみのコントロールに対する増加倍率として示される。ルシフェラーゼの発現における増加は試験された全ての系統で観察され、系統64が最も高い誘導レベルを提供し、cAMPがCreLucマウス中の一般的なモジュレーターによって活性化されていることを実証している。
【0148】
CreLucマウス(系統64)から単離された抗CD3刺激されたCD4+T細胞中のルシフェラーゼレベルに対する異なるGsアゴニストの効果を研究した(図19)。全脾細胞集団を機械的な破壊によって準備した。次いで、CD4+細胞をMACS Magnetic Separation with Positive Selection(Miltenyi Biotec cat#130−049−201)によって単離下。これらの細胞をウエルあたり1.5x105個、96ウエルの白色不透明プレートにプレーティングし、抗CD3抗体(BD Pharmingen cat#553058)で刺激した。24時間後、化合物を加え、さらに4時間置いた。DP(BW245C)、EP2(EX00000173A)およびADβR(イソプロテレノール)のGsアゴニストを全て10μMで、ホルスコリンを5μMで、およびロリプラムを10μMで用いた。BrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイをTopCountで読み取った。ホルスコリンおよびロリプラムで処理した細胞において、25倍の増加が観察された。BW245Cは3倍、173Aは2倍、イソプロテレノールは5倍という非常に有意な増加が3つ全てのGsアゴニストで観察された。導入遺伝子は、特異的GsアゴニストだけでなくcAMP経路の一般的なモジュレーターとして応答性であった。
【0149】
5.単離B細胞
CreLucマウスの2つの異なるサブ系統から単離したB細胞中の、ロリプラムおよびホルスコリンによる一般t系なcAMP活性化の効果を調べた(図20)。全脾細胞集団を上記のように機械的に破壊した。次いで、B220+細胞を、Positive Selection Column(Miltenyi Biotec、cat# 130−049−501)を備えるMACS Magnetic Separationを用いて単離した。その後これらの細胞を96ウエルの白色不透明プレート上にウエルあたり2.0x105個プレーティングし、10ng/mlのリポ多糖(LPS)(Sigma L−2630)で刺激した。18時間後、10μMのロリプラム(Sigma R6520)および5μMのホルスコリン(Sigma F6886)をそのプレートに加え、さらに4時間置いた。BrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイをTopCountで読み取った。データは発光(秒あたりのカウント数)および培地のみのコントロールに対する増加倍率として示される。ルシフェラーゼ発現の増加は、系統64では観察されたが、系統229では観察されなかった。
【0150】
6.ミクログリア
一般的なcAMPアクチベーターであるホルスコリンおよびロリプラム、DP受容体のアゴニストであるBW245Cによる、CreLucマウスから単離されたミクログリア中のルシフェラーゼ発現の誘導を調べた(図22)。初代ミクログリアは、DMEM(GIBCO Cat# 11995)、10% FBS(GIBCO Cat# 16140)および1% ペニシリン−ストレプトマイシン100X(GIBCO Cat# 15140)からなる培地中でP2マウス(系統64)由来の皮質から単離した。細胞を、ポリ−D−リジンコーティングプレート上に96ウエル形式でプレーティングした。細胞は未処理のままか100ng/mlのLPSで2時間刺激したかのいずれかである。次いで化合物を加え、さらに4時間置いた後、Bright Gloアッセイを実行した。使用した化合物は、5μMのホルスコリン、10μMのロリプラム、もしくはこれら2つの組み合わせ、またはDP1受容体のGsアゴニストBW245Cを10μMである。非刺激条件下では、ミクログリアは一般的なcAMPモジュレーターおよびDP受容体の特異的Gsアゴニストに対して非応答性であった。しかしながら、ミクログリアをLPSで刺激した場合、それらの細胞はホルスコリンおよびBW245Cに対して応答性となった。
【0151】
7.マウス胚線維芽細胞
マウス胚線維芽細胞中のルシフェラーゼ発現におけるホルスコリン、ロリプラム、およびイソプロテレノールの効果を調査した(図33)。マウス胚線維芽細胞は、6つの独立したCreLuc系統のE12胚から培養した。細胞をウエルあたり20,000細胞プレーティングした。試験した化合物としては、10μMのホルスコリン、5μMのロリプラムおよび10μMのイソプロテレノール(ADβRアゴニスト)が挙げられる。ホルスコリンとロリプラムの組み合わせに応じて、有意な増加が全ての系統で観察された。有意な増加はまた、イソプロテレノールに応じて、3つの系統で観察された。
【0152】
7.心筋細胞
心筋細胞中のルシフェラーゼレベルにおける、ホルスコリンおよびロリプラムおよびイソプロテレノールの効果を研究した(図35)。心筋細胞は、系統229由来のP3仔より単離した。細胞を96ウエルプレートで培養した。試験した化合物としては、10μMのホルスコリン、5μMのロリプラムおよび10μMのイソプロテレノール(ADβRアゴニスト)が挙げられる。ロリプラムとホルスコリンの組み合わせにより、ルシフェラーゼレベルの有意な増加(25倍)が観察され、アゴニストのイソプロテレノールでもまた有意な増加(2倍)が観察された。
【0153】
B.インビボおよびエキソビボ試験
1.一般的なcAMPモジュレーターの効果
ルシフェラーゼ発現の誘導に対する髄腔内注射されたホルスコリンおよびロリプラムの効果を、系統187のCreLucマウスの脳および脊髄において研究した(図23)。選択されたマウスの系統は、脳と脊髄の両方において導入遺伝子の発現を有していることが以前に判定されている(データは示さず)。このアッセイ用に選択されたマウスの系統(系統187)は、脳と脊髄の両方において誘導可能なレベルのルシフェラーゼを有している。グループあたりN=3〜4のマウス、4つの処理群、3月齢の雄。グループA:DMSOコントロール、グループB:1μgホルスコリン/10μgロリプラム、グループC:10μgホルスコリン/10μgロリプラム、グループD:40μgホルスコリン/10μgロリプラム。この動物に、髄腔内注射により腰部に1匹のマウスあたり5μlの量で投薬した。これらを投薬後4時間で画像処理した。脊髄および脳の両方のデータは、平均ピーク輝度、平方センチメートル毎秒あたりの光子として示されている。統計的に有意な増加が脊髄中で観察され、脳内におけるルシフェラーゼシグナルの有意な増加は、ホルスコリンの最も高い濃度においてみられただけであった。一般的なモジュレーターまたはcAMPが脊髄中に注射された場合、脊髄中で導入遺伝子の局所的な発現の増加があり、脳においてはより低い応答が観察された。
【0154】
2.Gsアゴニストの効果
CreLucマウスの脳および脊髄中のルシフェラーゼ発現に対するEPアゴニスト、EX00000173Aの効果を研究した(図24)。系統187由来の5月齢の雄マウス(n=5)に、ビヒクル(5% DMSO、0.05% tween 80、PBS)または10mg/kg EX00000173Aのいずれかを用いてi.p.注射した。動物を投薬4時間後にバイオイメージングした。データは平方センチメートル毎秒あたりの光子として示されている。脳と脊髄の両方において、アゴニスト処理による統計的に有意な増加がみられた。従って、i.p.投与されたGsアゴニストは、導入遺伝子を活性化した。
【0155】
さらに、CreLucマウス中のルシフェラーゼ発現に対するEP2アゴニスト、EX00000173Aの効果は用量反応性であった(図25)。系統187は、この系統が脳と脊髄の両方において高く誘導可能な発現を有しているために選択した。このアッセイは、5つの群の6週齢のマウス(n=5)で構成されている。マウスに、ビヒクルコントロール(D−PBS;Dulbeccos phosphate buffered saline、Invitrogen、cat#14040)または用量を変化させたEP2アゴニストEX0000173A(1.5nmol、5nmol、15nmolおよび50nmol)のいずれかを投薬した。マウスに髄腔内注射(1匹のマウスあたり5μl)により投薬し、IVIS bioimagerで4時間後にバイオイメージングした。データは5匹のマウスの平均値として、平均ピーク輝度、平方センチメートル毎秒あたりの光子として示されている。アゴニスト処理による統計的に有意な(t−検定)増加が、脊髄中の3つの濃度で観察されたが、脳内では観察されなかった。脳内の増加(有意ではない)は、髄腔内注射で予測されたような最も高い濃度で観察されたのみであった。
【0156】
図26は、CreLucマウスにおけるアドレナリン受容体β3(Adrb3)アゴニスト、CL316,243(1 mg/kg、ip)による異なる組織中のルシフェラーゼの誘導を示している。このルシフェラーゼアッセイは、組織ホモジネートで実施した。スクリーニングした12の独立したトランスジェニック系統の中で、7系統が脂肪組織と胚において10倍以上の誘導を示した。4系統はバイオイメージングによって目視可能な誘導を示した(図27Aおよび27Bを参照)。
【0157】
3.GsアゴニストAVE0010の特異的効果
導入遺伝子の活性化は、CreLucマウスの3つの独立した系統において健空された、グルカゴン用ペプチド1受容体(GLP−1R)アゴニストによるルシフェラーゼレポーターの誘導によって例示されるように、組織特異的受容体の活性化と共局在する(図28)。GLP−1Rは、Gs結合されている。ベースラインの画像は、1日目に得られた。2日目、マウスをAVE0010(0.1 mg/kg、sc)で処理し、4時間後に画像処理した。ベースラインに対する誘導倍率が下部に示されている。予想どおりGLP−1R受容体は、主に脾臓組織中で発現されているので、脾臓中で観察された導入遺伝子活性化の強い誘導が存在した。8つの異なる組織中のルシフェラーゼ活性を、系統11、16および90について測定し(図29)、その後AVE0010(0.1 mg/kg、s.c.)で4時間処理した。組織ホモジネート中のルシフェラーゼ活性により、ルシフェラーゼの脾臓特異的誘導が確認された。AVE0010の活性は、GLP−1Rが異なる組織タイプで見出されても、脾臓に限定されている。
【0158】
AVE0010によるCreLucの誘導は、β細胞媒介である可能性が高い。AVE0010によるCRE−Lucの誘導におけるβ細胞毒素ストレプトゾトシン(STZ)の効果を調査した(図30)。雄のCreLucマウス(系統11)を、AVE0010を0.1 mg/kg処理の前および処理後に画像処理した。全てのマウスは、AVE0010に応答していた(中パネル)。次いで、この動物をビヒクル(コントロール)またはSTZ(200mpk、ip)で処理した。4日後、これらをAVE0010での処理後再び画像処理した。ビヒクル群と比較すると、STZ群は、ルシフェラーゼ誘導が減少していた。
【0159】
AVE0010によるCreLucの誘導は、β細胞特異的である可能性がある(図31)。動物を図30に記載されたように処理した。血中グルコース濃度を、非絶食マウスの尾静脈ニッキングによって測定した。グルコース濃度は、Bayerのグルコースメーターで読み取った。グルコース濃度はmgグルコース/mlとして示される。誘導の倍率は、ベースラインの信号 対 AVE10を投与したルシフェラーゼバイオイメージングレベルである。血中グルコース濃度(BG)は、STZによる増加した(左上パネル)。非絶食BGレベルは、AVE0010(0.1 mg/kg、sc)によって低減された。図30に示されるBLIデータを定量した。
【0160】
4.骨髄移植
CreLuc骨髄移植を、NOD scid ガンマ(NGS)マウスを用いて実施した(図32)。NSDマウスは、成熟T細胞およびB細胞、機能的なナチュラルキラー細胞を欠損した免疫無防備状態のマウスであり、造血細胞の移植を可能にする差異と下院シグナル伝達を欠損している。骨髄細胞は、系統44(高い基底ルシフェラーゼレベルを有している;データは示さず)のヘテロ接合体と系統64(誘導可能なルシフェラーゼレベルを有している)のホモ接合体から採取した。その後、この細胞を、1匹のマウスあたり100万、または500万個の細胞で、照射NSGマウスに細胞の尾静脈注射により移植した。系統44については、マウス1および2は500万、マウス3および4が100万の細胞を受けた。系統64について:マウス1が500万個の細胞、マウス2、3および4が100万個の細胞を受けた。(CreLuc系統あたり4匹のNSGマウス)。動物を、4週間(データは示さず)でバイオイメージングし、再度8週間(データは示されている)でバイオイメージングした。画像処理の前に、系統64のマウスを5mg/kgのホルスコリンおよび10mg/kgのロリプラムで5時間誘導した。バイオイメージング写真は、8週の時点のものを示している。系統44の骨髄細胞を移植したNSGマウスにおけるルシフェラーゼレベルは、CreLuc系統44で見られた画像を模倣しており、間接、脊髄、頭部および胸骨において発現が観察された。誘導可能なルシフェラーゼ発現は、系統64の骨髄細胞を移植したNSGマウスの脾臓で観察された。
【0161】
4.動物モデル
CreLucマウスは、疾患または疾患状態の局面の動物モデルを研究するのに用いることができる。さらに、このCreLucマウスは、CreLucマウスにおいて誘導される疾患または疾患の局面を調節することができる化合物をスクリーニングするために用いることができる。例えば、CreLuc系統187におけるルシフェラーゼレベルに対するザイモサン処理の効果を研究した(図34)。治療群のCreLucマウス(系統187)に、疼痛反応を誘導するためにザイモサンによって両方の後部足に皮下注射した。これらの動物を4日間毎日バイオイメージングした。ルシフェラーゼ発現の統計的に有意な増加は、全ての時点において、ザイモサンに反応して動物の足で観察された。ザイモサンは、炎症反応を強く活性化する酵母細胞壁成分である。従って、CreLucマウスは、同じ動物において炎症反応を経時的にモニタリングできるツールである。また、ザイモサン処理した動物を用いて、ザイモサンによって誘導される炎症を逆にするかまたは悪化させる試験化合物の能力を評価するためのスクリーニングツールとして用いることができる。
【技術分野】
【0001】
本発明は一般的には導入遺伝子構築物、導入遺伝子構築物を含むトランスジェニック非ヒト動物、該導入遺伝子構築物を含むトランスジェニック非ヒト動物の作製方法およびそれを用いる方法に関する。本発明の実施態様は、リガンドをGPCRレセプターに結合した後に、経路変更に応答する生物発光導入遺伝子レポーターシステムを含むトランスジェニックモデルを用いて、全身の動物、組織切片、またはネイティブ細胞中で非侵襲的にGPCRリガンドを試験する方法に関する。
【背景技術】
【0002】
薬物開発においては、5つの化合物中の1つのみが米国食品医薬品局の承認(FDA)を通過しており、損耗率が高い(非特許文献1)。さらに、投資が劇的に増加したにもかかわらず、新薬の導入率は過去30年にわたりほぼ一定であり、結局年ごとに新薬種でわずか2〜3の進歩が市場にでるだけである(非特許文献2)。
【0003】
薬物開発の最初のステージで適用される分子イメージングおよび機能イメージングは、生物活性の証明を提供し、その企図される標的に対する効果を有する推定薬物(putative drug)を確認することができる。従って、分子イメージング技術への投資は薬物開発を推し進めるとかなり期待される(非特許文献3)。従来の情報を超える分子イメージング技術の利点は、その技術がインビボでの生物学的プロセスを研究するための十分な空間および時間的分解能により無処置固体(intact organism)に実施され得ることである。さらに、異なる時点における同一の生物学的モデルの反復性があり、非観血的で、均一かつ比較的自動化された研究を可能とするため、長期的研究の検出力を増加させ、必要とされる動物の数を減少させることができ、結果的に薬物開発のコストを低減させることができる。
【0004】
分子イメージング
分子イメージングとは、生きている製物中の細胞内および細胞小器官における特異的な分子プロセスの評価における画像処理技術を利用し、統合するための、様々な学問分野(細胞および分子生物学、化学、医学、薬理学、物理学、生物情報科学および光学)からのアプローチの集合のことを言う(非特許文献4)。
【0005】
遺伝子工学の出現は、創薬パイプラインなどを含む応用科学へ大きな変更をもたらしている。同じように、動物イメージング手段の開発および利用は、前臨床試験のための新しい集団を提供している(非特許文献5)。動物モデルは、リアルタイムで生理学的な事象を定量化するのが困難であるため、従来的にわずらわしいものであった。この困難を克服するために、長年にわたり磁気共鳴画像法(MRI)や陽電子放射断層撮影(PET)のような新しい画像処理法が開発されてきた。さらに最近では、ルシフェラーゼ、ホタルの発光酵素のインビボでの発光をベースにした生物発光イメージングが非侵襲的な検出のために用いられている。
【0006】
分子イメージング:生物発光
インビボでの生物発光イメージング(BLI)は、細胞または組織からの発光の検出にも続いている高感度なツールである。レポーター遺伝子技術の有用性は、インビボイメージング法を通して生きている動物中の特定の細胞および生物学的プロセスを分析することを可能にする。生物発光、つまり生きている生物による可視光の酵素生成は、多くの非哺乳動物種における天然に存在する現象である(非特許文献6)。ルシフェラーゼは、光の甲子を放出する基質の酸化を触媒する酵素である(非特許文献7)。北アメリカホタルからの生物発光は最も広く研究されている。このホタルルシフェラーゼ遺伝子(luc)の発現は、基質D−ルシフェリンを、562nmの緑色発光を生じる非反応性のオキシルシフェリンに変換する酵素であるルシフェラーゼを生成する。哺乳動物の組織は自然に生物発光を発しないため、画像処理が非常に小さなバックグラウンド信号を発生することができるのでインビボBLIはかなり魅力的である。BLIは、恒常的に光レポーターを駆動する選択遺伝子プロモーターの制御下で、生物発光レポーター遺伝子からなる発現カセットを有する細胞または組織の遺伝子操作を必要とする(図3)。光の産生を誘導するために、ルシフェリンなどの基質が脳室内(icv)、静脈内(iv)、腹腔内(ip)または皮下(sq)注射により投与される。
【0007】
ルシフェラーゼによって放出される光は、数ミリメートルから数センチメートルの深さに浸透することができる。しかし、光子の強度は、組織の深さの各センチメートルに対して10倍減衰する(非特許文献8)。高感度な光検出装置を、インビボで生物発光を検出するために用いる必要がある。この検出器は、単位面積当たりに発光される光子数を測定する。400〜1000nmの波長での低レベルの光は、シリコンウエハーを打ち、電子に変換する電荷結合素子カメラで検出することができる(非特許文献9)。このソフトウエアは、電子信号を二次元画像に変換することができる。このソフトウエアはまた、放出される光の強度(検出器を打つ放出光子の数)の定量化と、これらの数値の疑似グラフィックまたはグレースケールイメージへの変換が可能である(図2Aおよび2B)。実際のデータは光子で測定されるが、疑似グラフィックは、迅速な視覚的解釈を可能にする。目的の領域内の定量的な測定は、より微妙な差異の場合必要となり得る。冷却電荷結合素子(CCD)カメラの使用により熱雑音を低減し、遮光用の箱を用いるとルシフェラーゼ産生光が最適に可視化および定量化することを可能にする(非特許文献10)。発光シグナルの解剖学的位置についてのオートグラフやレントゲン写真のような別のタイプの画像に重ねてルシフェラーゼイメージを持っておくことは有用である(図2B)。このソフトフエアは、視覚化と解釈のために画像を重ね合わせる。動物の遺伝子操作と分子イメージング技術を組み合わせることによって、生きている動物中の特定の分子プロセスにおける動的研究を行うことが可能となる。このアプローチは、前臨床試験のプロトコルに影響を与え、それにより医学の全ての局面を広く変更する可能性がある(非特許文献11)。
【0008】
G−タンパク質共役受容体(GPCR)−創薬ターゲットとしてのGPCR
GPCRは、その共有される位置構造にも続いて100以上のサブファミリーに分類されている細胞表面受容体の大きなスーパーファミリーを構成し;GPCRはまた、7回膜貫通(7TM)受容体とも呼ばれている。GPCRは、製薬業界の中で最も頻繁に扱われる創薬ターゲットである。全ての市販処方薬のうちの約30%がGPCRを標的化しており、このタンパク質ファミリーを最も成功したターゲット類としている(非特許文献12)。GPCRとそれらの細胞外リガンドとの相互作用は、治療薬の干渉の魅力的な点であることが証明されている。このため、製薬業界は、これらのリガンド−GPCR相互作用を調査するために生化学的創薬アッセイを開発した。活性化GPCRとヘテロトリマーG−タンパク質との相互作用は、いくつかの下流エフェクターとの相互作用を可能にするグアノシン三リン酸(GTP)により、グアノシン二リン酸(GDP)の交換を触媒する(非特許文献13)。下流のシグナル伝達は、目的のGPCRに好ましいGαアイソフォームに依存している。Gαi/0およびG−αsのファミリーの代表が主にアデニル酸シクラーゼ(AC)活性を調節する一方、Gαq/11ファミリーのタンパク質がホスホリパーゼC(PLC)を刺激する。目的のGPCRがPLCを経由して信号を発する場合、その後のGPCRの活性化を測定するために最も広く適用されたレポーターベースの技術は、Ca+2感受性発蛍光団を用いた蛍光形式(非特許文献14)またはエクオリンと化学発光基質を用いた発光形式(非特許文献15)のいずれかで測定されるカルシウム(Ca+2)放出アッセイである。目的のGPCRがACを経由して信号を発する場合、その後の細胞質環状アデノシン一リン酸(cAMP)含有量は、種々の検出技術を用いて検出され得る(非特許文献16)。
【0009】
GPCRレポーターベースのアッセイは、現在の創薬プログラムにおいて広範囲に用いられている。一般的に、GPCRレポーターは、特定のGPCRを活性化するかまたは調節するリガンドまたは化合物を同定するために、大きな製薬ライブラリのインビトロでのハイスループットスクリーニング(HTS)をサポートするための細胞ベースのシステムに導入されている。第二の継続した細胞ベースのアッセイは、特定のGPCRに対するHTSで同定された全ての「ヒット」を確認および洗練する;しかし再び、これらのアッセイは形質転換された細胞型中にクローン化されたGPCRを導入する組み換えDNA法に依存している。形質転換された細胞型は、大きなスクリーニングプログラムをサポートする優れた増殖能を有しているが、これらはしばしば異常な遺伝的および機能的特性を示し、結果としてHTSから推定される「ヒット」の有意な消耗がこれらのパラダイムを用いて検出される。
【0010】
数年にわたり、生物発光ベースのレポーター遺伝子アッセイは、GPCRの機能的活性を測定するために用いられてきた(非特許文献17)。このアッセイ形式は、生物発光の読み出しの低いシグナルバックグラウンドおよび、GPCRの活性化と累積レポーター遺伝子発現との間の信号増幅工程のおかげで非常に感受性が高い。
【0011】
レポーター遺伝子のプロモーター中のcAMP応答配列(CRE)は、Gタンパク質依存性のシグナル伝達の特異的なモニタリングを可能にする。リガンドがGPCRに結合すると、それは付随Gタンパク質の活性化を可能にする、GPCRの立体構造変化を引き起こす。酵素アデニル酸シクラーゼは、Gタンパク質によって調節され得る細胞のタンパク質である。アデニル酸シクラーゼ活性は、活性化Gタンパク質のサブユニットに結合すると活性化されるかまたは抑制されるかのいずれかである。シグナル伝達は、Gタンパク質のタイプに依存している。アデニル酸シクラーゼは、細胞中のcAMP産生を増加させるかまたは減少させるかのいずれかの役割を果たしている。このcAMP産生は、細胞代謝におけるセカンドメッセンジャーであり、またプロテインキナーゼA(PKA)のアロステリック活性化因子である。cAMPが存在しない場合、PKA複合体は不活性である。cAMPは、PKAの調節サブユニットに結合すると、それらのコンフォメーションは変更され、プロテインキナーゼAを活性化し、さらなる生物学的効果を可能にする調節サブユニットの解離を引き起こす。次いで、PKAが転写因子CREBをリン酸化し活性化する。CREBは、cAMP応答配列(CRE)と呼ばれる特定のDNA配列に結合することによって転写を増加または減少させ、それによりルシフェラーゼレポーター遺伝子のような特定の遺伝子を発現する。
【0012】
CreLuc導入遺伝子は、cAMP細胞内シグナル伝達経路を通して直接的にかまたはPLCを通して間接的にかのいずれかで3つの主要なGPCR全てのアッセイを活性化するように設計されている。任意の1つの細胞型は、それらの細胞表面上に多くの異なるタイプのGPCRを含んでいるため(従って全ての細胞は、細胞内で同時に生じるG−αq/11、G−αi/0およびG−αsを経由するGPCRシグナル伝達を有している)、従来の知識は、CreLucのような導入遺伝子が任意のひとつの特定GPCRリガンドを識別するのに十分特異的であることはありそうもないということを示唆している。しかし、本発明者らは、本明細書中でCreLuc導入遺伝子がGPCRリガンドを識別することができることを示している。本発明者らは、細胞内のルシフェラーゼレポーター、組織切片、または全体の動物の生物発光シグナルがホルスコリンによって増加されること、およびGs、GqまたはGiのリガンドによって調節されることを予測している。表1は、GPCRリガンドに結合する際に、CreLucレポーターシステム上でGPCRの活性化/抑制が起こるという予測された効果を示している。さらに、本発明者らは、新規のCreLucレポーターシステムがGPCRリガンドの異なるクラスを識別し、そのようなレポーターシステムは、細胞、組織切片および全体の動物中で用いられるときに新規のGPCRリガンドを同定するために適用可能であるというデータを示している。
【0013】
【表1】
【0014】
GPCRバイオイメージングレポータートランスジェニックモデル
現在の創薬パラダイムにおける潜在的な薬物候補の顕著な消耗は、細胞ベースのレポーターアッセイからインビボモデルへの相転移においておきている。多くのインビボモデルは、特定のヒト疾患の全てまたは一部を反復することが可能である。これらのモデルにおけるリード化合物を証明することは、新しい化学GPCR薬の進行に重要なマイルストーンである。動物疾患モデルは、通常それらの表現型および、疾患転帰の変更に及ぼす候補化合物の影響の正確なアッセイの開発を可能にするために、多数の動物と時間を必要とする。インビトロでの試験後の、複雑なシステムでの薬物候補を試験する次のレベルは、疾患状態のインビボ試験またはインビボモデルを用いており、これらは機械ベースである。誘導された疾患転帰を変更する失敗はあまり理解されないが、いまだに薬物開発のパイプラインにおいて候補化合物の大きな損耗率を生じている。
【0015】
GPCRリガンド結合を含むトランスジェニックモデルおよび活性化レポーターアッセイは、GPCRの現在の薬物開発パラダイムの大幅な改善となる。例えば、本発明の実施態様は、GPCRリガンド薬物開発を大幅に促進する、全身の動物、組織、または細胞における分子イメージングと組み合わせた、ルシフェラーゼレポーター(CreLuc)をベースにしたcAMPレポーターアッセイを含む導入遺伝子を記載している(非特許文献18;非特許文献19)。本明細書中に記載されるように、本発明のトランスジェニック非ヒト動物の実施態様は、以下の非限定的な利点を提供する:
【0016】
1.組織や細胞ベースのアッセイはトランスジェニックインビボモデルアッセイと同じレポーターシステムを持っているため。複雑な無処置の生物学的システムの未知数の数を減らす。
2.非侵襲的な画像処理は、同じ動物において、経時的アッセイにおけるリガンドや化合物活性の定量分析を可能にする。
3.非侵襲的な画像処理は、研究あたりの動物の数を減らし、各動物が自身のコントロールになることによって、より大きな検出力につながる。ここで、コントロールとは時間ゼロにおいてアッセイされる動物である。
4.トランスジェニック動物は、インビトロまたはエキソビボでなされる並行アッセイをサポートするための細胞および組織の供給源になる。
5.トランスジェニック動物のアッセイは、ネイティブ細胞型におけるリガンド活性のアッセイをサポートし、リガンドのより現実的なプロファイルを導く。
6.トランスジェニック動物は、GPCRリガンドの約力学および薬物動態の同時評価を可能にする。
7.トランスジェニック動物は、器官レベルまたは全身の動物レベルいずれかで、組織および細胞型の特異性の同時識別を可能にする。
8.トランスジェニック動物は、新規のシグナル伝達経路および特定のリガンドへのそれらの応答を明らかにするための、他の遺伝的に改変されたモデルとの交配を可能にする。
【0017】
異なるレポーターを用いて遺伝子操作された多くのトランスジェニック動物は、以下のような分子プロセスの研究に用いられている:薬物代謝(非特許文献20)、遺伝毒性(非特許文献21)および毒性化合物の効果(非特許文献22)。それらの設計目的を達成するために、分子イメージング研究に適切なGPCRレポーター動物は、研究下にあるリガンドまたは化合物の生体内分布における広範で鋭いインビボアッセイをサポートするための大きなウインドウの生物発光検出、ならびに全ての細胞型における発現の両方を可能にするように配置されるいくつかのエレメントを組み込む必要がある。
【0018】
遺伝子発現に関与するメカニズムの複雑さと多様性により、研究者らは、完全に予測可能な方法でトランスジェニック動物中における発現の全てのケースにおいて有能な遺伝子を構築することができない(非特許文献23:非特許文献24;非特許文献25)。大規模な試行錯誤を通してのみ、導入遺伝子構造の独特の組み合わせを、GPCRレポーターのバイオイメージングの必要に応じて、モデルデザインの目的に到達させることができる。
【0019】
組換え細胞アッセイに対する、トランスジェニックGPCRレポーターの有用性
スクリーニング技術が、個々のGPCRの振る舞いを理解するポイントまで進むと、スイッチをオンオフするというよりもむしろ、これらの受容体が情報のマイクロプロセッサのように作用することが明らかである。このことは、機能の選択性の現象を導入しており、それにより特定のリガンドは所定の受容体によって媒介されるシグナル伝達機構の一部分のみを開始することによって、薬物開発の新たな地平を切り開く。任意の薬理学的レポーターアッセイにおける非常に高い要求を置く医薬品化学を導くために、これらの複雑なシステムにおいて新規のGPCRリガンドの関係を発見し、薬物の効果を定量化する必要がある。この概念は、組換え細胞ベースのスクリーニングアッセイから全体のシステムのアッセイへ戻る原動力となる。ネイティブ細胞環境中で特定のGPCRまたはGPCRのセットによってリガンドの活性をプロファイリングし、重要なクラスの薬学的に重要な受容体に対する新しい薬物を同定するための成功率を向上させることが期待される(非特許文献26)。生物発光GPCRレポーター導入遺伝子を含む動物モデルは、無処置の生物学的に複雑なシステム中でGPCRリガンド活性を定義する非常に望ましい分子イメージングストラテジーであり、ヒトの疾患と戦うための薬物開発を向上させる目的を有している。
【0020】
CRE/CREBの活性化は、多くの可変的な生物学的プロセスに関与しており、CRE/CREBレポーター発現系を用いることによるCREの活性化の研究に大きな関心がある。環状アデノシン一リン酸(cAMP)は、受容体の活性化とひき続くタンパク質キナーゼの活性化後の、細胞内シグナル伝達におけるセカンドメッセンジャーであり、よって多くの生物学的プロセスの調節に関与している。CREB(cAMP応答配列結合タンパク質)は、cAMPによって活性化されたキナーゼによってリン酸化され、多くの遺伝子のプロモーター領域中のcAMP応答配列(CRE)に結合して転写を活性化させる(非特許文献27)。β−ガラクトシダーゼ発現を駆動する最小単純ヘルペスウイルス(HSV)プロモーターを持つ6つのタンデムCREを保持するトランスジェニックマウスを用いて、慢性的な抗うつ薬治療に応答する脳スライス中のCRE媒介性遺伝子発現が研究された(非特許文献28)。同様に、チミジンキナーゼプロモーターに融合するラットソマトスタチン遺伝子プロモーターCREの4つのコピー、およびルシフェラーゼ遺伝子を保持するトランスジェニックマウスを用いて、組織学的脳スライスまたはホモジネート中のCRE活性化が研究された(非特許文献29)。しかしながら、これまでの研究は、適切な動物モデルを見つけるために多くのトランスジェニック系統をスクリーニングする必要性によって、妨げられてきた。さらに、適切な系統が確認された後、比較的低いレポーター発現レベルでは、レポーター遺伝子を測定するためにトランスジェニック動物を安楽死させる必要があり、多くの動物を1つの実験系に用いる必要がある。
【0021】
本発明の実施態様は、インスレーター配列、応答配列、プロモーターエレメント、レポーター遺伝子および機能エレメントを含む導入遺伝子の開発である。この導入遺伝子は、その早い速度での組込みと高いレベルでのレポーター発現によって、迅速に非ヒト動物中に導入することができ、それによりインビボ(すなわち生きている動物中)、インサイチュ(例えば、脳スライス、無処置の固体)またはインビトロ(例えば、トランスジェニック動物から培養された初代細胞、組織ホモジネート)でのエレメント活性の調節を研究するためのモデルとして、トランスジェニック動物を容易に開発することができる。
【0022】
本発明の実施態様は、インビボで細胞内cAMPレベルの調節を通してGPCRリガンド活性を定量化するモデルとしてトランスジェニック非ヒト動物中で用いられるCRE Lucレポーターシステムを含む導入遺伝子である。非限定的な例として、本発明者らは、一般的なcAMP調節因子を用いて、単離された初代細胞中および全身の動物中の生物発光によるルシフェラーゼレポーターの変化を示している。別の実施態様において、このレポーターの活性化はエキソビボでのルシフェラーゼアッセイを用いて組織抽出物中でアッセイされ確認されている。CRE Luc導入遺伝子の応答は、複数のマウス系統において記録されており、単一または複数の組織活性化プロファイルを示している。さらに非限定的な例として、本発明者らは、特定のGPCRリガンドが全身の動物、組織スライス、および初代細胞中のCRE Luc導入遺伝子を活性化することを実証した。
【先行技術文献】
【非特許文献】
【0023】
【非特許文献1】DiMasi、JA、et al、J Health E con 22,151−185、2003
【非特許文献2】Lindsay MA、Nature Rev Drug Discovery、2、831−838、2003
【非特許文献3】Rudin M、Progress in Drug Res vol 62
【非特許文献4】Massoud T.F.、Genes Dev.17:545−580、2003
【非特許文献5】Maggie A.and Ciana P.、Nat.Rev.Drug Discov.4、249−255、2005
【非特許文献6】Contag、C.H.、et al、Mol.Microbiol.18:593−603、1995
【非特許文献7】Greer L.F.、III、Luminescence 17:43−74、2002
【非特許文献8】Contag、C.H.、et al、Mol.Microbiol.18:593−603、1995
【非特許文献9】Spibey CP et al electrophoresis 22:829−836、2001
【非特許文献10】Contag C.H.and Bachmann、M.H.、Annu.Rev.Biomed.Eng.4:235−260、2002
【非特許文献11】Maggie A.Trends Pharmacolo.Sci 25、337−342、2004
【非特許文献12】Jacoby、E;Chem.Med.Chem.、1:761−782、2006
【非特許文献13】Cabrera−Vera T.M.、Endocr.Rev.24:765−781、2003
【非特許文献14】Sullivan E、Methods Mol.Biol.114:125−133、1999
【非特許文献15】Dupriez V.J.、Receptors Channels 8:319−330、2002
【非特許文献16】Gabriel D.Assay Drug Dev.Technol.1:291−303、2003
【非特許文献17】Hill、S.J.Curr.Opin.Pharmacol.1:526−532、2001
【非特許文献18】Bhaumik、S.and Gambhir、S.S.、Proc.Natl.Acad.Sci.USA、99:377−382 2002
【非特許文献19】Hasan M.T.、et al.、Genesis 29:116−122、2001
【非特許文献20】Zhang W.、et al.Drug Metab.Dispos.31:1054−1064、2003
【非特許文献21】Gossen J.A.、et al.、Proc.Natl.Acad.Sci.USA 86:7971−7975、1989
【非特許文献22】Sacco M.G.et al.、Nat.Biotechnol.15:1392−1397、1997
【非特許文献23】Pinkert、C.A.(ed.)1994.Transgenic animal technology
【非特許文献24】A laboratory handbook.Academic Press、Inc.、San Diedo、Calif.
【非特許文献25】Monastersky G.M.and Robl、J.M.(ed.)(1995)Strategies in Trans genic Animal Science.ASM Press.Was hington D.C
【非特許文献26】Kenakin TP、Nat.Rev.Drug Discov.8,617−625、2009
【非特許文献27】Shaywitz and Greenberg、Annul.Rev.Biochem.、68:821−861、1999
【非特許文献28】Thome J.、et al.、J.Neurosci.20:4030−4036、2000
【非特許文献29】Boer et al、PloS One、May 9;2(5):e431、2007
【発明の概要】
【課題を解決するための手段】
【0024】
概して、本発明は、導入遺伝子構築物、導入遺伝子構築物を含むトランスジェニック非ヒト動物、該導入遺伝子構築物を含むトランスジェニック非ヒト動物の作製方法およびそれを用いる方法を提供する。本発明の実施態様は、CRE Lucレポーターシステムを含む導入遺伝子構築物を提供する。本発明の実施態様は、非ヒト動物中への、CRE Lucレポーターシステムを含む導入遺伝子構築物の導入である。
【0025】
cAMP変調は、GPCRのための主要な活性化経路であるため、本発明は全身の動物、組織切片、または細胞中で、レポーター遺伝子(このレポーター遺伝子は、ルシフェラーゼによるルシフェリンの代謝など測定可能な生物発光シグナルを提供する)の活性化を通してリガンドまたは化合物によるGPCRの活性化を定量化するためのプラットフォームとしての役目を担う。本発明は、細胞ベースのアッセイから全身の動物まで、リガンドまたは化合物のような新規に発見された薬物成分(new drug discovery entities)の移行を向上させるためのツールを供給する。本発明の実施態様は、ネイティブ細胞中で同じレポーターシステムを用いることで、バイオアベイラビリティーデータを同時に提供しながら、新規のGPCRリガンドの損耗率を低減する。
【0026】
本発明の実施態様は、第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を含むゲノムを有する、トランスジェニック非ヒト動物である。
【0027】
本発明の実施態様は、前記第1のインスレーター配列が、マトリックス付着領域(MAR)、DNase I−高感受性領域(HS4)および末端逆位配列(ITR)からなる群より選択される、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記第2のインスレーター配列が、マトリックス付着領域(MAR)、HS4およびITRからなる群より選択される、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記第1のインスレーター配列が、前記第2のインスレーター配列と同じである、トランスジェニック非ヒト動物である。
【0028】
本発明の実施態様は、前記応答配列が、cAMP 応答配列(CRE)、アクチベータータンパク質1(ASP1)、グルココルチコイド応答配列(GRE)、熱ショック応答配列(HSE)、血清応答配列(SRE)、甲状腺ホルモン応答配列(TRE)およびエストロゲン応答配列(ERE)からなる群より選択される、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記応答配列がタンデムに2〜24回反復されている、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記応答配列がタンデムに6回反復されている、トランスジェニック非ヒト動物である。本発明のさらなる実施態様は、前記応答配列がCREであり、さらにこのCRE応答配列がシングルエレメントであるかまたは2〜24回反復されていてもよい、トランスジェニック非ヒト動物である。
【0029】
本発明の実施態様は、前記プロモーターが単純ヘルペスウイルスチミジンキナーゼの最小ポロモーター(HSV TK min)である、トランスジェニック非ヒト動物である。
【0030】
本発明の実施態様は、生物発光レポーターが、ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、β−ガラクトシダーゼ、分泌型アルカリホスファターゼ(SEAP)、ヒト成長ホルモン(HGH)および緑色蛍光タンパク質(GFP)からなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物である。
【0031】
本発明の実施態様は、前記機能エレメントがヒト成長ホルモン(hGH)遺伝子である、請求項1に記載のトランスジェニック非ヒト動物である。
【0032】
本発明の実施態様は、前記導入遺伝子が配列番号18を含む、トランスジェニック非ヒト動物である。
【0033】
本発明の実施態様は、前記導入遺伝子が配列番号19を含む、トランスジェニック非ヒト動物である。
【0034】
本発明の実施態様は、前記トランスジェニック非ヒト動物から単離された細胞であるかまたは請求項1に記載のトランスジェニック非ヒト動物から単離された組織切片である。
【0035】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物中の生物発光の量を測定する工程;
(b)該トランスジェニック非ヒト動物に試験薬剤を投与する工程;
(c)該試験薬剤の投与後に、1またはそれより多い時点で該トランスジェニック非ヒト動物の生物発光の量を測定する工程;
(d)(a)において測定した生物発光の量と(c)において測定した生物発光の量とを比較する工程;
を含み、ここで(c)と比較した(a)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【0036】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物から組織切片を調製する工程;
(b)該組織切片中の生物発光の量を測定する工程;
(c)該組織切片に試験薬剤を投与する工程;
(d)該試験薬剤の投与後に、1またはそれより多い時点で該組織切片の生物発光の量を測定する工程;および
(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程;
を含み、ここで(d)と比較した(b)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【0037】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物から単離された細胞を調製する工程;
(b)該細胞中の生物発光の量を測定する工程;
(c)該細胞に試験薬剤を投与する工程;
(d)該試験薬剤の投与後に、1またはそれより多い時点で該細胞の生物発光の量を測定する工程;および
(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程;
を含み、ここで(d)と比較した(b)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【0038】
本発明の実施態様は、非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)該非ヒト動物からの生物発光をモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【0039】
本発明の実施態様は、非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)該非ヒト動物からのルシフェラーゼをモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【0040】
本発明の実施態様は、非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)疾患状態の状況を模倣するために非ヒト動物を操作する工程;
(c)該非ヒト動物からの生物発光をモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【0041】
本発明の実施態様は、非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)疾患状態の状況を模倣するために非ヒト動物を操作する工程;
(c)該非ヒト動物からのルシフェラーゼをモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【0042】
本発明の実施態様は、GPCR機能のモニタリングにおいて使用するための非ヒトトランスジェニック動物を作製する方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)(a)のトランスジェニック非ヒト動物における生物発光の量を測定する工程;
(c)GPCRリガンドを該トランスジェニック非ヒト動物に投与する工程;
(d)該GPCRリガンドの投与後に、1またはそれより多い時点で該トランスジェニック非ヒト動物の生物発光の量を測定する工程;および
(e)(b)で測定した生物発光の量と(d)で測定した生物発光の量を比較する工程;を含み、ここで(d)と比較した(b)の生物発光の量の差が、GPCR機能をモニタリングするのに用いるための非ヒトトランスジェニック動物を同定する、上記方法。
【0043】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されるトランスジェニック非ヒト動物から単離された細胞を準備する工程;(b)該細胞中の生物発光の量を測定する工程;(c)試験薬剤を該細胞に投与する工程;(d)試験薬剤の投与後に、1またはそれより多い時点で該細胞中の生物発光の量を測定する工程;(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程であって、ここで(b)と(d)を比較した場合の生物発光の量の差異は、GPCRリガンドとして試験薬剤を同定している、工程。
【0044】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)コントロールを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、コントロールを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0045】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)コントロールを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、コントロールを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0046】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)一般的なcAMPモジュレーターを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、一般的なcAMPを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0047】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された組織切片を1つ以上のレセプタクル中に供給する工程;(b)コントロールを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、コントロールを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0048】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された組織切片を1つ以上のレセプタクル中に供給する工程;(b)一般的なcAMPモジュレーターを1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、一般的なcAMPモジュレーターを含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0049】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)細胞刺激因子を1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、細胞刺激因子を含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0050】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された細胞を1つ以上のレセプタクル中に供給する工程;(b)細胞刺激因子を1つ以上のレセプタクルに投与する工程;(c)一般的なcAMPモジュレーターを1つ以上のレセプタクルに投与する工程;(d)試験薬剤を1つ以上のレセプタクルに投与する工程;(e)該レセプタクル中のルシフェラーゼの量を測定する工程であって、細胞刺激因子および一般的なcAMPモジュレーターを含むレセプタクル中で測定されたルシフェラーゼの量と細胞刺激因子および一般的なcAMPモジュレーターならびに試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0051】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された組織切片を1つ以上のレセプタクル中に供給する工程;(b)細胞刺激因子を1つ以上のレセプタクルに投与する工程;(c)試験薬剤を1つ以上のレセプタクルに投与する工程;(d)該レセプタクル中のルシフェラーゼの量を測定する工程であって、細胞刺激因子を含むレセプタクル中で測定されたルシフェラーゼの量と試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【0052】
本発明の実施態様は、Gタンパク質共役受容体(GPCR)を調節する化合物を同定する方法であって、該方法は以下の工程を包含する;(a)本明細書中に開示されているトランスジェニック非ヒト動物から単離された組織切片を1つ以上のレセプタクル中に供給する工程;(b)細胞刺激因子を1つ以上のレセプタクルに投与する工程;(c)一般的なcAMPモジュレーターを1つ以上のレセプタクルに投与する工程;(d)試験薬剤を1つ以上のレセプタクルに投与する工程;(e)該レセプタクル中のルシフェラーゼの量を測定する工程であって、細胞刺激因子および一般的なcAMPモジュレーターを含むレセプタクル中で測定されたルシフェラーゼの量と細胞刺激因子および一般的なcAMPモジュレーターならびに試験薬剤を含むレセプタクル中で測定されたルシフェラーゼの量とを比較した場合の差異は、GPCRを調節する化合物を示している。
【図面の簡単な説明】
【0053】
【図1】CreLucバイオイメージングマウスモデルを示している。この図は、3つのタイプのGPCR(Gi、GsおよびGq)全てがcAMP経路を経由して直接的にかまたはPLC経路を経由して間接的にかのいずれかによる、CRE Lucレポーター導入遺伝子の細胞内活性化を示している(パネルA)。ホルスコリン誘導に応答するルシフェラーゼレポーターの生物発光における変化を3つのタイプのGPCRについて示している(パネルB)。ホルスコリンは、GsおよびGqシグナルを増加、すなわちCreLuc生物発光を増加させる一方、Gi誘導はそのレポーターからのシグナルを減少させる。GαsはACの直接的な刺激によってcAMP依存性経路を活性化し、GαsはcAMPの生成を阻害し、そしてGαqは2つのセカンドメッセンジャーIP3およびDAGの発生をもたらすPLCを刺激する。略号:α、Gタンパク質のαサブユニット;β、Gタンパク質のβサブユニット;γ、Gタンパク質のγサブユニット;AC、アデニル酸シクラーゼ;PLC、ホスホリパーゼC;PKA、プロテインキナーゼA;PKC、プロテインキナーゼC;DAG、ジアシルグリセロール;IP3、イノシトール三リン酸;Ca+2、カルシウム;CaMK、カルシウム/カルモジュリンプロテインキナーゼ;cAMP、環状アデノシン一リン酸;CRE、cAMP応答配列;CREB、cAMP応答配列結合タンパク質。
【図2A】リアルタイムのインビボ生物発光イメージングを示し、そのシステムを用いるメリットを記載している。コンピュータ分析システムを備えるこのIVIS100(Xenogen)バイオイメージング装置は、生物発光レポーター遺伝子、例えばルシフェラーゼによって発せられた光を利用してリアルタイムのインビボイメージングを可能にする。このソフトウエアは、シグナルの数量化を非観血的かつ長期的にサポートする。リアルタイムのインビボイメージングは、従来のインビボでの化合物試験に対して多くの利点を有する。従来の動物試験は、複数の治療点で個々のマウスが必要となる一方、バイオイメージングモデルを利用した研究では、同じ動物を複数の時点でサンプリングし、複数の治療のために再利用することができる。この図に示されるように、0時間、2時間、4時間および8時間の時間経過は、現在の方法論を用いると24匹の動物(各時点あたりn=6)が必要であるにもかかわらず、バイオイメージング技術を用いると必要とされるのは6匹の動物のみである。これは、以下を含むいくつかの利点をもたらす:より多くの化合物が効果について試験されることを可能にするのに必要な試験動物はより少ないので、よりハイスループットである;時間的および空間的データが同じ動物から採取することができるので、データの内容および質がより高い;および統計的誤差が減少し、個々の化合物についてなされる決定の質が向上する。
【図2B】CreLucトランスジェニックマウスにおける化合物誘導の典型的な視覚イメージを示している。イソプロテレノール(右パネル)の投与は、基底レベル(左パネル)と比較してCRE Lucレポーターの脊髄の発現を増加させる。この生物発光検出は、グレースケールでの疑似表現として動物の白色光画像に視覚的に表わされる。
【図3】発現を高めるために複数のDNAエレメントを含んでいる導入遺伝子構造の模式図を示している。この模式的な導入遺伝子構造は、以下のエレメントを含んでいる:位置に依存しない発現を産生するためのマトリックス付着領域(MAR)としてこの図に示されているインスレーター配列;6回反復されたCRE−cAMP(6×CRE)によって表わされる応答配列;単純ヘルペスウイルスチミジンキナーゼの最小プロモーター(HSV TK min)として示されるプロモーターエレメント;哺乳動物発現に最適化されたルシフェラーゼ遺伝子(LUC2)によって表わされるレポーターエレメント;および遺伝子発現を増強するためにポリA末端を有するヒト成長ホルモン遺伝子(hGH poly A)によって表わされる機能エレメント。
【図4】CreLuc導入遺伝子ベクターのインビトロでの検証である。30、100、または199ngのDNAでのハイブリッドまたは合成(シンセ)ベクターを、リポフェクタミン2000(Invitrogen、Carlsbad、CA、cat #11668−019)を用いてCHO細胞(ウミシイタケルシフェラーゼポジティブコントロールベクターと一緒に)にトランスフェクションした。2日後、その細胞を3μMのフォスルコリン(Sigma、St Louis、MO、cat #F6886)を用いて4時間刺激し、その後Dual Glo luciferase Assay System(Promega、Madison、WI、cat# E2920)を用いてルシフェラーゼ活性を測定した。結果は、両方のベクターでルシフェラーゼシグナルの用量依存性の増加を示している。より高いレベルの誘導がハイブリッドベクターで達成された。
【図5A】正常マウスにおけるcAMPレベルのPDF阻害剤の効果を示している(Cheng JB、JPET、280、621−626からの再現)。Balb/cマウスにx軸上に列挙されているビヒクルまたは薬物を服用させ、20分後にその血液を採取してcAMPラジオイムノアッセイによりcAMPをアッセイした。CP−80、633とロリプラムの両方が10mg/kgでの血漿cAMPレベルを大幅に増加させた。
【図5B】血漿中のcAMPのインビボ刺激を示している。FVB/Tacメスにビヒクル(1% DMSO)または薬物をi.p.で服用させ、30分後血液サンプルを採取し、ELISA(Assay Designs、 Ann Arbor、MI、cat#900−163)によってcAMPをアッセイした。用いた薬物は以下のとおりである:5mg/kg ホルスコリン(F)(Sigma F6886)、5mg/kg 水溶性ホルスコリン(H20F)(Calbiochem 344273)、10mg/kg ロリプラム(R)(Sigma R6520)またはホルスコリンプラスロリプラム(F/R)の組み合わせのいずれか。統計的に有意な増加である14倍が、t−検定によって決定されるロリプラムと組み合わせた水溶性ホルスコリンの処理で観察された。ホルスコリンがアデニル酸シクラーゼを活性化することによってcAMPレベルを増加させる一方、ロリプラムのようなPDE4の阻害剤はcAMPの加水分解を防止することによって血漿内cAMPを上げる。ロリプラムと水溶性ホルスコリンの組み合わせは、インビボでcAMPレベルを14倍増加させる。この組み合わせは、バイオイメージングによる初代のスクリーニングのための誘導の大きなウインドウを提供するために用いられ、代表的な研究を図6に示す。
【図6】CreLucレポーターマウスモデルの初代の誘導と系統の選択の結果を示している。複数のトランスジェニック系統を、インビボでのホルスコリンとロリプラムによるルシフェラーゼ誘導のためにスクリーニングし、その後組織を単離してルシフェラーゼ酵素についてアッセイした。トランスジェニックマウスを投薬前(基底発現レベル)でバイオイメージングし、その後同じマウスにi.p.で10mg/kgのロリプラムおよび5mg/kgの水溶性ホルスコリンを投与し、投薬の4時間後にバイオイメージングした(誘導された発現)。A(亜系統番号90):ホルスコリン/ロリプラム投与は、肺および他の組織においてCreLucレポーター遺伝子の基底発現を増加させた;B(亜系統番号219)基底発現の誘導は、主に腸内である;C(亜系統番号44):検出されない基底発現およびレポーターは、脳および他の組織に誘導される;D(亜系統番号28):胸腺および肝臓で増加される検出されない基底発現;E(亜系統番号187):脳および脊髄で誘導される検出されない基底発現。ランダムに統合された導入遺伝子について予想されるように、基底発現、組織分布、および誘導に対する応答において、系統間にばらつきがあった。20系統は、1つ以上の組織で5倍より大きい誘導を持つことが同定された。組織プロファイルの変化は、その単一の組織(すなわち、肺、肝臓、脳)が、バックグランド組織の応答のないイメージングを可能にする一方、複数の組織は、広範な化合物の応答プロファイルが作成されることを可能にする。
【図7】インビボまたはエキソビボでのCreLucスクリーニングアッセイの一般的な概略図を示している。
【図8A】CreLucマウス(系統187)由来の全脳スライスにおける発光に対するイソプロテレノール(ISO)およびAMN082(AMN)の効果を示している。
【図8B】時間の経過とともに、CreLucマウス(系統44)由来の全脳スライスにおける発光に対するホルスコリンの効果を示している。時間は、分単位でX軸上に示されている。50uMのホルスコリンまたはビヒクル(DMSO)を、下のパネルの矢印でマークされた時間=2880で加えた。
【図9】CreLucマウス由来の初代神経細胞の単離および化合物処理を表わす模式図を示している。
【図10】β−アドレナリン受容体(ADβR)の活性化とD1ドーパミン受容体(DRD1)の活性化を介したGsの変調を示している。ニューロンを、系統187 E18胚の皮質から単離した。培養3日目、試験化合物にホルスコリン5μM(F)、ロリプラム10μM(R)、ホルスコリンとロリプラム(F/R)をイソプロテレノールと組み合わせて10μM、イソプロテレノールとロリプラムを組み合わせて(I/R);SKF82958を10μMおよびSKF82958とロリプラムを組み合わせて(S/R)を加えた。データは、秒あたりのカウント数(cps)で示されている。
【図11】初代皮質ニューロンにおけるルシフェラーゼ発現に対する、プロキネチシン(prokineticin)2(PROK2)ペプチドの効果を示している。初代皮質ニューロンを、系統187(脳および脊髄中にルシフェラーゼを誘導できる)E18上から採取した。このアッセイを培養3日目に4時間または8時間実行した。このPROK2ペプチドは、1nMおよび100nMで水溶液として加えられる。データは、秒あたりのカウント数(cps)として示されている。
【図12】異なるCreLuc系統由来の初代皮質ニューロンにおけるルシフェラーゼ発現に対するプロキネチシン(prokineticin)2(PROK2)ペプチドの効果を示している。初代皮質ニューロンを、E18で4つの異なるCreLuc系統から採取した。このアッセイを、培養3日目に1nMまたは100nMのPROK2ペプチドいずれかを用いて、4時間後および24時間後の2つの時点においてトリプリケートで実行した。そのアッセイにはBrightGloを用い、TopCountで読み取った。データは、秒あたりのカウント数(cps)で示されている。
【図13A】初代皮質ニューロンにおけるルシフェラーゼ発現に対するmGluR7アゴニスト、AMN082の効果を示している。皮質ニューロンは、E18胚(系統187)から採取した。このアッセイを培養3日目に実行した。ホルスコリンは10μMで用いた。このアゴニスト、AMN082を1nM、10nM、100nMおよび1μMのホルスコリンと組み合わせて用いた。このアッセイは、Bright Glo(Promega)を備えるTopCountで4時間および8時間で読み取った。データは秒あたりのカウント数(cps)で示されている。
【図13B】初代皮質ニューロンにおけるGi活性を調節する能力について未知の化合物をスクリーニングした結果を示している。皮質ニューロンは、E18胚(系統187)から採取した。このアッセイを培養3日目に実行した。ホルスコリンは10μMで用いた。AMN082または未知の化合物A、BまたはCを異なる濃度のホルスコリンと組み合わせて試験紙、EC50値を算出した。このアッセイは、Bright Glo(Promega)を備えるTopCountで、4時間で読み取った。データは秒あたりのカウント数(cps)で示されている。
【図14】初代皮質ニューロンにおけるルシフェラーゼ発現に対するmGluR7アゴニスト、AMN082の効果を示している。初代皮質ニューロンは、系統187由来のE18胚から単離した。このアッセイを培養7日目に6時間、トリプリケートで実行した。AMN082の濃度曲線は、50μMのホルスコリンおyぼい10μMのロリプラムと組み合わせて実行した。このアッセイは、Bright Glo(Promega)を備えるTopCountで読み取った。データは秒あたりのカウント数(cps)で示されている。
【図15A】CB1アゴニスト、CP55,940によって異なるCreLuc系統由来の初代皮質ニューロンにおけるルシフェラーゼ発現のGi調節を示している。初代皮質ニューロンは、E18で4つの異なるCreLuc系統から採取した。このアッセイは培養3日目に実行した。CB1アゴニストを10μM、ホルスコリンを5μMおよびロリプラムを10μMで用いた。4時間および24時間の2つの時点で実行した。次いでブライトグロルシフェラーゼアッセイ基質を添加し、そのアッセイをTopCountルミノメーターで読み取った。データはトリプリケートの平均値で示されている。データは秒あたりのカウント数(cps)で示されている。
【図15B】CB1アゴニスト、CP55,940によるCreLucマウス由来の初代皮質ニューロンにおけるルシフェラーゼのGi調節を示している。皮質ニューロンは、E18胚(系統187)から単離した。このアッセイは培養3日目に実行した。ホルスコリン(F)およびロリプラム(R)を10μMで用いた。アゴニストを10μM、1μM、および100nMの濃度で加えた。このアッセイを、BrightGlo(Promega)を備えるTopCountにおいて8時間で読み取った。データは、秒足りのカウント数(cps)で示されている。
【図16】ホルスコリンおよびロリプラム、ならびにGsアゴニストであるDRD1およびADβRによる、CreLuc線条体ニューロンにおける誘導されたルシフェラーゼ発現を示している。線条体ニューロンは、E14胚(系統187)から単離した。このアッセイは、培養4日目に実行した。ホルスコリン(F)を5μM、ロリプラム(R)を10μMで用いた。Gsアゴニストであるイソプロテレノール(iso)、ドーパミン(dopa)およびSKF82958(chloro)を10μM、3μM、および1μMで用いた。このアッセイをTopCountルミノメーターおよびブライトグロルシフェラーゼ試薬(Promega)を用いて5時間で読み取った。データは秒あたりのカウント数(cps)で示されている。
【図17】CreLucマウスから単離された全脾細胞プレップ中のルシフェラーゼ発現に対する、ホルスコリン(F)およびロリプラム(R)のような一般的なcAMP誘導因子、ならびにGsアゴニストの影響を示している。系統64の脾細胞を抗CD3交代(CD)で24時間刺激し、他の半分は未処理とした(unstim)。24時間で化合物をプレートに加え、さらに4時間経過させた。ホルスコリンおよびロリプラムの同時処理(F/R)は、5uMのホルスコリンおよび10uMのロリプラムであった。用いたGsアゴニストは以下のとおりである:EP2アゴニストとしてEX00000173A(173A)、DP1アゴニストとしてBW245CおよびADβRアゴニストとしてイソプロテレノール。全てのGsアゴニストを10uMで用いた。このアッセイをトリプリケートで実行した。4時間後、100ulのBrightGloを加え、このアッセイをTopCountルミノメーターで読み取った。データは秒あたりのカウント数(cps)で示されている。
【図18】CreLucマウスの5つの異なるサブラインから単離されたT細胞中のロリプラムおよびホルスコリンによる、一般的なcAMPの効果を示している。この細胞を抗CD3抗体(1ug/ml)で刺激した。18時間後、10μMのロリプラムおよび5μMのホルスコリンをプレートに加え、さらに4時間経過させた。BrightGloを加え、アッセイをTopCountで読み取った。データは上のパネルでは発光(秒あたりのカウント数)で示され、下のパネルでは培地に対しての増加の倍率として示している。
【図19】CreLucマウス(系統64)から単離された抗CD刺激されたCD4+T細胞中のルシフェラーゼレベルに対するGsアゴニストの効果を示している。この細胞(1ウエルあたり1.5x105)を96ウエルの白色不透明プレート上にプレーティングし、抗CD3抗体(1ug/ml)で刺激した。24時間後、化合物を加えさらに4時間経過させた。GsアゴニストであるBW245C、EX00000173A(1734A)およびイソプロテレノール(iso)は、全て10μMで用いた。ホルスコリン(F)を5uM、ロリプラム(R)を10uMで加えた。BrightGloを加え、このアッセイをTopCountで読み取った。データは秒あたりのカウント数(cps)で示されている。
【図20】CreLucマウスの2つの異なるサブラインから単離されたB細胞中の、ロリプラムおよびホルスコリンにる一般的なcAMP活性化の効果を示している。細胞を96ウエルの白色不透明プレート上に、1ウエルあたり2.0x105でプレートし、10ng/mlのリポポリサッカライド(LPS)で刺激した。18時間後、10μMのロリプラムおよび5μMのホルスコリンをそのプレートに加え、さらに4時間経過させた。BrightGloを加え、アッセイをTopCountで読み取った。データは発光(秒あたりのカウント数)およびコントロールのみ培地に対する増加の倍率として示している。
【図21】CreLucマウス(系統から単離されたLPS刺激されたB220+B細胞中のルシフェラーゼレベルに対するGsアゴニストの効果を示している。この細胞を96ウエルの白色不透明プレート上に1ウエルあたり2.0x105プレートし、その後10ng/mlのリポポリサッカライド(LPS)で刺激した。24時間後、化合物を加えさらに4時間経過させた。GsアゴニストであるBW245C、EX00000173A(1734A)およびイソプロテレノール(iso)は10μMで用いた。ホルスコリン(F)を5uM、ロリプラム(R)を10uM加えた。BrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイをTopCountで読み取った。データは秒あたりのカウント数(cps)で示している。
【図22】一般的なcAMPアクチベーターであるホルスコリン(F)およびロリプラム(R)、ならびにDPレセプターのアゴニストであるBW245Cによる、単離されたミクログリア(系統64)中の誘導されたルシフェラーゼ発現を示している。初代ミクログリアをP2マウス由来の皮質から単利し、ポリ−D−リシンコーティングしたプレート上に96ウエル形式でプレートした。細胞は、未処理のままかまたは100ng/mlのLPSで2時間刺激したものかのいずれかである。次いで化合物を加え4時間経過させ、その後Bright Gloアッセイを実行した。用いた化合物は、5μMのホルスコリン、10μMのロリプラムもしくはその2つの組み合わせ、またはDP1レセプターのGsアゴニスト、BW245Cを10μMであった。データは秒あたりのカウント数で示されている。
【図23】CreLucマウス(系統187)の脳および脊髄におけるルシフェラーゼ発現の誘導に対する髄腔内注射されたホルスコリン(F)およびロリプラム(R)の効果を示している。1グループあたりN=3〜4のマウスで、3ヶ月齢の雄。グループA:DMSOコントロール、グループB:1ugホルスコリン/10ugロリプラム、グループC:10ugホルスコリン/10ugロリプラム、グループD:40ugホルスコリン/10ugロリプラム。この動物に、髄腔内注射により腰部に1匹のマウスあたり5μlの量で投薬した。これらを投薬後4時間で画像処理した。脊髄および脳の両方のデータは、平均ピーク輝度、平方センチメートル毎秒あたりの光子として示されている。
【図24】CreLucマウスの脳および脊髄中のルシフェラーゼ発現に対するEPアゴニスト、EX00000173Aの効果を示している。マウス(系統187)をビヒクル(5% DMSO、0.05% tween 80、PBS)または10mg/kg EX00000173Aのいずれかを用いてi.p.注射した。動物を投薬4時間後にバイオイメージングした。データは平方センチメートル毎秒あたりの光子として示されている。
【図25】CreLucマウス中のルシフェラーゼ発現に対するEP2アゴニスト、EX00000173Aの効果を示している。マウスに、ビヒクルコントロールまたは用量を変化させたEP2アゴニストEX0000173Aのいずれかを投薬した。マウスに髄腔内注射(1匹のマウスあたり5μl)により投薬し、IVIS bioimagerで4時間後にバイオイメージングした。データは5匹のマウスの平均値として、平均ピーク輝度、平方センチメートル毎秒あたりの光子として示されている。
【図26】CreLucマウスにおけるアドレナリン受容体β3(Adrb3)アゴニスト、CL316,243(1 mg/kg、ip)による異なる組織中のルシフェラーゼの誘導を示している。このルシフェラーゼアッセイは、組織ホモジネートで実施した。
【図27A】CreLucマウスの系統11(n=2)および系統115(n=3)におけるAdrb3アゴニスト、CL316,243(1 mg/kg、ip)によるルシフェラーゼの誘導を示している。処理前および処理後4〜5時間でBLIを撮影した。組織ホモジネート中のルシフェラーゼ活性は、画像の下に示されている。
【図27B】CreLucマウスの系統31(n=2)および系統175(n=3)におけるAdrb3アゴニスト、CL316,243(1 mg/kg、ip)によるルシフェラーゼの誘導を示している。処理前および処理後4〜5時間でBLIを撮影した。組織ホモジネート中のルシフェラーゼ活性は、画像の下に示されている。
【図28】CreLucマウスの3つの独立した系統における、グルカゴン様ペプチド1受容体(GLP−1R)アゴニスト、AVE0010によるルシフェラーゼレセプターの誘導を示している。ベースラインの画像は1日目に得られた。2日目、マウスをAVE0010(0.1 mg/kg、sc)で処理し、4時間後に画像処理した。ベースラインに対する誘導の倍率が下部に示されている。
【図29】CreLucマウスの3つの独立した系統における、グルカゴン様ペプチド1受容体(GLP−1R)アゴニスト、AVE0010によるルシフェラーゼレセプターの誘導を示している。マウスをAVE0010(0.1 mg/kg、sc)で4時間処理した。8つの異なる組織中のルシフェラーゼ活性を測定した。
【図30】AVE0010によるCreLucの誘導におけるβ細胞毒素ストレプトゾトシン(STZ)の効果を示している。雄のCreLucマウス(系統11)を、AVE0010を0.1 mg/kg与える前(「非誘導」;上パネル)およびscで与えた後(「AVE0010による誘導」中パネル)に画像処理した。全てのマウスは、AVE0010に応答していた(中パネル)。次いで、この動物をビヒクル(コントロール)またはSTZ(200mpk、ip)で処理した。4日後、これらをAVE0010での処理後再び画像処理した(下パネル)。
【図31】AVE0010によるCreLucの誘導がβ細胞特異的な可能性があることを示している。動物を図30に記載されたように処理した。血中グルコース濃度を、非絶食マウスの尾静脈ニッキングによって測定した。グルコース濃度は、Bayerのグルコースメーターで読み取った。グルコース濃度はmgグルコース/mlとして示される。誘導の倍率は、ベースラインの信号 対 AVE10を投与したルシフェラーゼバイオイメージングレベルである。血中グルコース濃度(BG)は、STZによる増加した(左上パネル)。非絶食BGレベルは、AVE0010(0.1 mg/kg、sc)によって低減された。図30に示されるBLIデータを定量した。
【図32】NOD SCIDガンマ(NSG)マウスへのCreLuc骨髄移植を示している。骨髄細胞は、系統44のヘテロ接合体と系統64のホモ接合体から採取した。その後、この細胞を、1匹のマウスあたり100万、または500万個の細胞で、照射NSGマウスに細胞の尾静脈注射により移植した。系統44:マウス1および2は、500万、マウス3および4が100万の細胞を受けた;系統64:マウス1が500万個の細胞、マウス2、3および4が100万個の細胞を受けた。(CreLuc系統あたり4匹のNSGマウス)。動物を、4週間(データは示さず)でバイオイメージングし、再度8週間(データは示されている)でバイオイメージングした。画像処理の前に、系統64のマウスを5mg/kgのホルスコリンおよび10mg/kgのロリプラムで5時間誘導した。
【図33】マウス胚線維芽細胞中のルシフェラーゼ発現に対する、ホルスコリン、ロリプラムおよびイソプロテレノールの効果を示している。マウス胚線維芽細胞は、6つの独立したCreLuc系統由来のE12胚から、1ウエルあたり20,000細胞プレーティングして培養した。試験した化合物としては、10μM ホルスコリン(F)、5μM ロリプラム(R)および10μM イソプロテレノール(iso)が挙げられる。データは秒あたりのカウント数(cps)として示されている。
【図34】CreLucマウス(系統187)中のルシフェラーゼレベルに対するザイモサン処理の効果を示している。治療群の動物に、疼痛反応を誘導するためにザイモサン(zymo)によって両方の後部足に皮下注射した。これらの動物を4日間毎日バイオイメージングした(d1、d2、d3およびd4と表記)。
【図35】心筋細胞中のルシフェラーゼレベルに対するホルスコリンおよびロリプラムおよびイソプロテレノールの効果を示している。心筋細胞は、P3の子犬(系統229)から単離した。この細胞を96ウエルプレートで培養した。試験した化合物としては、10μM ホルスコリン(F)、5μM ロリプラム(R)および10μM イソプロテレノール(iso)が挙げられる。データは秒あたりのカウント数(cps)として示されている。
【0054】
発明の詳細な説明
他に定義しない限り、本明細書中で用いられる全ての技術用語および科学用語は、一般的に本発明が属する分野の当業者によって通常理解されるもとの同じ意味を持っている。
【0055】
本明細書中で引用した各文献、特許出願、特許および他の参考文献は、それが本発明の開示に反しない限り、その全体が参照により援用される。
【0056】
ここで、本明細書中および添付の特許請求の範囲において用いられる場合、単数形の「a」「an」、および「the」は、その文脈がそうでないことを明確に指示しない限り、複数の指示対象を含むことに留意しなければならない。
【0057】
さらに、本発明によると、当該分野の技術の範囲内で、従来の分子生物学、微生物学および組み換えDNA技術を用いることができる。このような技術は、文献に十分に説明されている例えば、以下を参照のこと:Sambrook、Fritsch&Maniatis、Molecular Cloning: A Laboratory Manual、Second Edition(1989)Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York(herein”Sambrook et al.、1989”);DNA Cloning: A Practical Approach、Volumes I and II(D.N.Glover ed.1985);Oligonucleotide Synthesis(M.J.Gait ed.1984);Nucleic Acid Hybridization[B.D.Hames&S.J.Higgins eds.(1985)];Transcription And Translation[B.D.Hames&S.J.Higgins、eds.(1984)];Animal Cell Culture[R.I.Freshney、ed.(1986)];Immobilized Cells And Enzymes[IRL Press、(1986)];B.Perbal、A Practical Guide To Molecular Cloning(1984);F.M.Ausubel et al.(eds.)、Current Protocols in Molecular Biology、John Wiley&Sons、Inc.(1994)。
【0058】
「試験薬」は、化合物または化学化合物のような任意の物質(例えば、有機化学物質、無機化学物質)、生物学的化合物または生物学的物質(例えば、交代および抗原認識フラグメントおよび構築物、核酸(例えばRNAi))などの任意の物質を含む。試験薬は、一緒に適用される単一薬剤または複数薬剤を包含する。
【0059】
本明細書中で用いられる場合、「トランスジェニック動物」は、非ヒト動物であり、非限定的な例は哺乳動物であるが、その動物の細胞の1つ以上が本明細書中に定義されるような遺伝的改変を含んでいる。さらに非限定的な例として、ラットまたはマウスのようなげっ歯類が挙げられる。他のトランスジェニック動物の例としては、ヒト以外の霊長類、ヒツジ、イヌ、ウシ、ヤギ、ニワトリ、両生類などが挙げられる。トランスジェニック動物の選択は、組織を横切るためにレポーターから光を発生させて、検出が可能な表面まで到達させる能力によってのみ限定される。
【0060】
本明細書中で用いられる場合、「遺伝子組み換え」は、ヒト以外の動物の遺伝子配列における1またはそれより多い変更である。非限定的な例は、トランスジェニック動物のゲノムへの導入遺伝子の挿入である。
【0061】
本明細書中で用いる場合、用語「導入遺伝子」は、発現を増強するためにプロモーター、レポーター遺伝子、ポリアデニル化シグナルおよび他のエレメントを含む外因性DNAのことをいう(インスレーター、イントロン)。この外因性DNAは、1つの細胞の胚のゲノム中に組み込まれ、そこからトランスジェニックが発達し、その導入遺伝子はその成熟動物のゲノム中に留まる。この組み込まれた導入遺伝子DNAは、卵またはマウスのゲノム中に単一でかまたは複数で生じ得、またその導入遺伝子の単一〜複数(数百)のタンデムコピーが各ゲノム位置に組み込まれ得る。
【0062】
用語「一般的なcAMPモジュレーター」は、cAMPレベルを増加または維持することのできる、化学化合物(例えば、有機化学物質、無機化学物質)、生物学的化合物または生物学的材料(例えば、フラグメントおよび構築物を認識する抗体および抗原)、核酸(例えばRNAi)などを言う。非限定的な例としては、ホルスコリンおよびロリプラムが挙げられる。一般的なcAMPモジュレーターは、単一のcAMPモジュレーターまたは一緒に適用される複数のcAMPモジュレーターを包含する。
【0063】
用語「細胞刺激因子」とは、細胞を活性化し得るかまたは細胞をより活性化した状態にすることのできる、化学化合物(例えば、有機化学物質、無機化学物質)、生物学的化合物または生物学的材料(例えば、フラグメントおよび構築物を認識する抗体および抗原)、核酸(例えばRNAi)などを言う。非限定的な例としては、リポポリサッカライドおよび抗CD3が挙げられる。
【0064】
本発明の実施態様は、コントロールを使用する。コントロールは、当業者に周知の用語である。適切なコントロールは利用されるアッセイパラメーターまたは調査中の実験の問題に依存する場合がある。典型的には、コントロールは、そのコントロールが、試験薬剤または化合物を溶解するのに用いられているのと同じ緩衝液または溶媒であるビヒクルコントロールである。非限定的な例として、化合物を溶解するのにリン酸緩衝生理食塩水が用いられている場合、ビヒクルコントロールはリン酸緩衝生理食塩水である。同様に、DMSOが試験薬剤を溶解するのに用いられている場合、コントロールはDMSOである。しばしば、化合物を試験するために1種より多い希釈剤が用いられるために、実験またはアッセイごとに1種より多いコントロールを使用する必要がある。
【0065】
本明細書中で用いられる場合、「ルシフェラーゼ」は、ルシフェラーゼ酵素活性だけでなく、ルシフェラーゼタンパク質の実際の量も言及する。
【0066】
本発明に従って、トランスジェニック非ヒト動物を作製するために、当業者に公知の従来の技術を使用することができる。例えば、Pinkert、C.A.(ed.)1994。トランスジェニック動物技術:A laboratory handbook.Academic Press、Inc.、San Diedo、Calif.;Monastersky G.M.and Robl、J.M.(ed.)(1995)Strategies in transgenic animal science.ASM Press.Washington D.C..and Nagy A、Gertsenstein、M、Vintersten、K、Behringer R 2003。マウス胚の操作;A laboratory Manual 3rd edition. Cold Spring Harbor Laboratory Press、Cold Spring Harbor、New York。
【0067】
導入遺伝子エレメント
本発明の実施態様は、導入遺伝子に関連している。この導入遺伝子は、インスレーター配列、応答配列、プロモーターエレメント、レポーターエレメントおよび機能エレメントを含み得る。
【0068】
応答配列は、細胞内でホルモン、酵素、または他の重要なシグナル伝達タンパク質のような細胞のシグナルに応答するパリンドロームDNA配列である。応答配列の非限定的な例としては、CRE(cAMP応答配列)、エストロゲン応答配列および表2に列挙される他のものが挙げられる。応答配列は、単一のDNA配列またはタンデムリピートとして、導入遺伝子中に組み込まれ得る。例えば、4回または6回反復されるCRE応答配列が、導入遺伝子構築物中で用いられ、インビトロで検証されている(Deutsch P.J.、et al.、J.Biol.Chem.、263;18466−18472、1988;Oetjen E JBC 269;27036−27044、1994)。Cre応答配列もまた、インビボで比較されており、多量体における増加は、cAMP経路のアクチベーターに対する転写応答の増加と相関している(Montoliu、L.et al.、Proc.Natl.Acad.Sci.USA 92;4244、1995;Boer et al、PloS One、May 9;2(5):e431、2007)。本発明の実施態様は、単一の配列としてかまたは複数のタンデムリピート(例えばCREのタンデム6回反復(6×CRE))としてのいずれかの任意の公知の応答配列を利用することができる。
【0069】
【表2】
【0070】
DNプロモーターエレメントは、特定の遺伝子の転写を促進するDNAの領域である。プロモーターは、通常、同じ鎖および上流(センス鎖の5‘領域に向かって)上で、それらが制御する遺伝子の近くに位置している。プロモーターは、特定のDNA配列および応答配列(RNAポリメラーゼおよびRNAポリメラーゼを補充するタンパク質(転写因子と呼ばれる)の結合部位を提供する)を含んでいる。DNプロモーターは、その大きさおよび時間と空間内の特定の遺伝子の発現の調節に寄与する内部構造が非常に可変的である。プロモーターエレメントの非限定的な例において、単純ヘルペスウイルスチミジンキナーゼの最小プロモーター(HSV TK min)は、全ての細胞型におけるレポーター遺伝子の発現を可能にするように設計されている。その中核的な発現エレメントのみが保持され、インビトロまたはインビボのいずれかでのユビキタス発現を付与する(Park、J.、et al.、DNA Cell Bio.、12:1147−1149、1994)。本発明の実施態様は、任意の公知のプロモーターエレメントを利用することができる。非限定的な例として、任意の公知のプロモーターエレメントは、そのプロモーターエレメントがCRE cis活性応答配列と組み合わせられる場合に、cAMP経路の変調に応答するレポーターの遺伝子発現を可能にするように用いられる。
【0071】
CREのような応答配列およびプロモーターエレメントであるHSV TK minはサイズが小さいので、位置の影響に非常に敏感であり、乏しい発現につながるレポーターエレメントの転写を調節するこれらのエレメントを含む導入遺伝子プロモーターは、特にインビボで低いリガンド濃度でリガンドに応答する。従って、本発明の実施態様は、全ての細胞区画を通して機能している導入遺伝子の高レベルでの発現および広い分布を達成するために、追加のエレメントを導入遺伝子に加えることを包含する。これらのエレメントには、インスレーター配列および機能エレメントを含む(Sun F.L and Elgin S.C、Cell 99:459−462、1999)。
【0072】
本発明の実施態様は、導入遺伝子内の機能エレメントまたは機能増強エレメントを利用する。機能エレメントの非限定的な例は、ヒト成長ホルモン(hGH)遺伝子である。CreLuc導入遺伝子において用いられるhGH配列は、
バイオイメージングモデルの設計目標を達成するために、インスレーター配列に寄与し、相互作用するいくつかの設計エレメントを含んでいる。このhGH配列は、全てのhGHゲノム構造を含んでいるため、いくつかの重要なエレメントを提供するが、タンパク質中には転写も翻訳もされない。機能する導入遺伝子の産生を向上するためのhGH配列の重要な影響は、1990年以来、いくつかのトランスじぇニックモデルについて実証されている(Erickson LA、Nature 346:74−76、1990)。その重要性の比較分析は存在しないが、hGH構造は、いくつかの重要で、重要な意味をもつDNAエレメントを含んでいる:
a. イントロンスプライシング:最初に転写されたmRNAは、イントロン配列とエキソン配列の両方を含んでおり、核から書き出されてさらに処理され、イントロン配列が取り除かれてエキソン配列のみを含む成熟mRNAを生じる。このトランスポートおよびトリミングのプロセスは、成熟mRNA鎖をタンパク質の最終的な生産のためのさらなる翻訳機構に接続させる。導入遺伝子のcDNA構造中にイントロンを含むことは、導入遺伝子のcDNAの発現レベルを向上させることが示されている(Palmiter、R.D.、et al.、Proc.Natl.Acad.Sci.USA88:478−482、1988)。
b. 無処置の3’UTRを持つゲノム構造:本発明の実施態様において、導入遺伝子中のhGH配列は、高いレベルでのmRNA安定性、およびそれによる高いレベルでの遺伝子発現を付与する、無処置の3’UTRを含んでいる。
c. ポリA(PA+)構造を有するゲノム構造:このhGH配列は、天然の3’UTRに埋め込まれたそのネイティブPA+構造を含んでいる。一般的にPA+シグナルは、ウイルス配列(SV40、RSVなど)由来であり、無関係の3’UTRの末端に加えられた最小構造である。本発明の実施態様において、天然のPA+シグナルを有する全体の3’UTR構造は、完全hGH遺伝子のさらに広いゲノムのコンテキストに保存される。
【0073】
インスレーター配列は、位置に依存する発現を産生するDNAの配列である(Giraldo et al、Transgenic Research 12:751−
755、2003)。インスレーター配列は、グロブリン遺伝子座について1980年に記載され(Sun F.L.and Elgin、S.C.、Cell 99;459−462、1999)、バイオイメージングのためのモデル設計目標をサポートするために選択された組織中で、適切な応答性トランスジェニック発現を得る機会を増やすことが報告された(Pinkert、C.A.(ed.)1994.Transgenic animal technology:A laboratory handbook.Academic Press、Inc.、San Diego、Calif.;Monastersky G.M.and Robl、J.M.(ed.)(1995)Strategies in transgenic animal science.ASM Press.Washington D.C)。インスレーターは、遺伝子発現に寛容なオープンクロマチンドメインを作成し、遠位のサイレンサー/エンハンサー配列の影響に対して、ならびにアセチル化およびメチル化事象に対しての障壁を構成するDNAエレメントである。これらは、バイオイメージングで検出可能なレベルで発現されるレポーター遺伝子を持つ独立したトランスジェニック初代系統の数を大幅に増やさねばならない。インスレーター配列は、一過性のトランスフェクションアッセイにおいて、40〜70%までクローンを発現するルシフェラーゼの数を増やし、結果的にERE−luci 導入遺伝子中のルシフェラーゼ発現の誘導能を高めることが示されている(Ottobrini L.、 Mol Cell Endo 246、69−75)。しかしながら、導入遺伝子レポーター発現に対するインスレーターの適用を全面的に検討すると、実用面において、インスレーターを利用することは依然として困難であるという結論に至る。それらの作用メカニズムは部分的にしか知られておらず、その効果は完全に予測可能ではない。インスレーター配列の非限定的な例は、表3に挙げられている。
【0074】
【表3】
【0075】
Cre−Luc導入遺伝子の実施態様において、インスレーター配列の封入は、バイオイメージングによって検出される機能的レポーターを持つ系統の作製頻度を明らかに増加させた。CreLucトランス得ニック系統におけるルシフェラーゼ発現へのインスレーター配列の寄与の分析については、VI章の「実施例」以下を参照のこと。
【0076】
本発明の実施態様は、レポーターエレメントまたは遺伝子を含む導入遺伝子に関する。レポーター遺伝子は、検出可能な遺伝子産物を発現する任意の遺伝子を含み、RNAまたはタンパク質であり得る。多くのレポーター遺伝子は当該分野で公知であり、限定するわけではないが、β−ガラクトシダーゼおよびアルカリホスファターゼが含まれる。別の実施態様において、導入遺伝子は、生物発光レポーター遺伝子を含む。多くの生物発光レポーター遺伝子は当該分野で公知であり、ルシフェラーゼを含むがこれに限定されない。ルシフェラーゼの供給源は多く存在し、ホタルルシフェラーゼおよび細菌ルシフェラーゼが挙げられるがこれらに限定されない。本発明の実施態様は、例えばルシフェラーゼのような任意の公知の生物発光レポーターを利用し得る。他のレポーターエレメントの非限定的な例は、表5に示される。
【0077】
【表4】
【0078】
本発明の他の実施態様用は、ルシフェラーゼ酵素の改変バージョン、異なる種由来のルシフェラーゼ酵素、または適切な基板と共に提供される場合、動物組織を横切ることのできる光を産生し得る他のタンパク質、もしくは動物組織を横切ることのできる光を発することのできる任意の酵素を組み込むことができる。本発明のレポータータンパク質は、信号の減衰量が、発された光の波長および発光細胞を取り巻く組織特性に依存するという事実によってのみ限定される。一般的に青緑光(400〜590nm)は、強く減衰される一方、近赤外光(590〜800nm)は、はるかに減衰が少ない。ルシフェラーゼの多くのタイプは、青〜黄緑の波長で発光し、その発光スペクトルは広く、組織に非常に深く浸透する赤色波長(>600nm)で有意な発光が存在しているようである。マウスなどの小型ゲッ歯類では、このことが動物全体を通してのシグナルの検出を可能にする。
【0079】
インビボでの光検出の限界は、生物発光レポーターのタイプ、動物の周囲の生理学および発光源の深さに依存する。通常、動物の生物発光細胞は、細胞の数および場所により、高感度CCDカメラを用いて、1〜3cmの深さで観察することができる。それらが組織をとおして伝播するときの光子の散乱は、動物表面上で検出された画像の空間分解能を制限する。一般的に、表面におけるスポットサイズや解像度は、表面化の光源の深さにほぼ等しい。物理ベースの拡散モデルを用いて、ミリメートルレベルでアプローチする空間解像度の改善を達成することができる。冷却した科学グレードのCCDアレイを用いると、シグナル検出の限界は、画像が撮影された後のCCD画素数(これは、ピクセルあたりのいくつかの光子のオーダーである)の読み取りと関連した読み出しノイズによって決定される(Honigman et al.、Mol.Ther.4:239−249、2001)。毛皮、皮膚、あるいは動物上の汚染物質の燐光に起因して、動物から発せられるさらなるバックグラウンド光もあり得る。通常、バックグラウンド光は低レベルであり、深く低レベルな生物発光供給源の画像に有害な影響を及ぼすのみである。しかしながら、バックグランド光は、適切な光学フィルターの使用により、排除することができる。
【0080】
本発明の実施態様は、IVIS(Xenogen Corporation、860 Atlantic Avenue、Alameda、Calif.94501、USA)などのCCDカメラを利用する。このIVIS.RTM.Imaging Systemは、高感度CCDカメラ、入射光を最小にするための暗い画像処理チャンバー、および結果を定量化し分析するための特殊なソフトウエアを備えている。IVISは、Xenogen Corporationの登録商標である。しかしながら、このような生物発光画像処理システムはどれも本発明に適用可能である。
【0081】
リアルタイムのインビボイメージングは、生物発光レポーター遺伝子定量化を非侵襲的に(すなわち動物を安楽死させる必要がない)、かつ長期的に(すなわち長い時間経過にわたり、連続的または反復して測定することができる)可能にする。リアルタイムのインビボイメージングは、従来のプロトコルに比べ、必要な試験動物がより少なく(例えば、特定の時間の間同じ動物を用いることができるため)、より少ない時間でよい(例えば、取り扱う必要のある動物の数がより少ないため)、より多くの化合物を効率よく試験することができる。リアルタイムのインビボイメージングは、多くの理由により、より高いデータ内容およびより高いデータ品質を提供する。例えば、時間的および空間的データは同じ動物から集めることができ、データを時間のかかる組織学的評価を必要とせずに集めることができる。データ品質が高いほど、統計誤差を減少させ、試験化合物の評価と意思決定の質を向上させる。
【0082】
本発明の実施態様は、ハイスループットスクリーニング(HTS)法で用いる。HTSは、疾患の病態の生物学的メカニズムや側面をモデル化するために設計されたアッセイにおける何千もの異なる化合物の自動化された同時試験である。1またはそれより多い化合物(例えば複数の化合物)は、例えば、1つのバッチで同時に試験することができる、1実施態様において、用語「HTSスクリーニング法」は、任意の読み出しに影響を及ぼす1つの化合物または複数の化合物の能力を試験するアッセイを意味する。
【0083】
細胞培養およびサンプル操作のための、液体取扱システム、蛍光読み取り装置またはシンチレーションカウンターのような分析装置およびロボットは当該分野でよく知られている。ロボットのような機械的システムや「チェリーピッキング」デバイスは、当業者に利用可能である。市販のプレートリーダーは、従来の96ウエルプレートや384ウエルプレートを分析するのに利用可能である。単一のサンプル、複数のサンプル、またはプレートサンプルのリーダーは、所定のウエルを分析し、生データのレポートを作成するのに利用可能である。この生データは種々の方法で変換され提示することができる。
【0084】
本発明の実施態様は、細胞、組織切片または培養培地のような他の材料を受けることのできる容器の配列を含む。容器の配列は、本発明の範囲内で、細胞または組織切片を保持するのに適切な少なくとも1つ以上の容器から任意の数の容器であり得る。例としては、フラスコ、培養皿、1.5mlチューブのようなチューブ、12ウエルプレート、96ウエルプレート、384ウエルプレートおよび4000ほどレセプタクルを有する小型のマイクロタイタープレート(米国特許出願20050255580号)が挙げられるが、これらに限定されない。容器の配列は、保護カバーを追加するような修正が可能であり、それにより汚染物質の混入や内容物の蒸発を防止することができる。
【0085】
これらの容器のさらなる特徴は、容器が分析(非限定的な例として、分光分析、シンチレーション計数、および蛍光測定を含む)を可能にすることができることである。しかしながら、さらなる分析のために修正可能な適切な容器に移すことができることを考えると、本発明の範囲内で用いられ得る容器は限定されない。非限定的な例は、この方法を変更することであり、例えばこの方法が、第2の容器の配列を提供することをさらに包含し、ここで、細胞を溶解させる工程は、さらに細胞片から上清を分離する工程を包含し、次の工程は、この第2の容器の配列の少なくとも1つが分離した上清のサンプルを含むようにDNAフラグメント中にインターカレートし得る検出可能な化合物を加える工程をさらに包含する。
【0086】
本発明のさらなる局面および詳細は、以下の実施例から明らかであり、これらは例示することを意図しているのであって、いかなる意味においても本発明の範囲を限定することを意味しない。
【0087】
動物を含む全ての実験的な操作は事前に検討された連邦政府のガイドラインとプロトコルに従って実施し、sanofi−aventisのサイト、Institutional Animal Care and Use Committee(IACUC)によって承認されている。
【実施例】
【0088】
I.MultiSite Gateway Pro Plus Cloning Systemのベクター骨格(図3)
デスティネーションベクターpDest2XMARSを、MultiSite Gateway Pro Plus Cloning System(Invitrogen Carlsbad、CA、cat #12537−100)での使用に特異的に設計しクローニングした。ベクター中に挿入された全てのエレメントをPCR(ポリメラーゼ連鎖反応)クローニングまたは合成的に生成した。全てのPCRクローニング工程について、フラグメントを、PCR SuperMIx Hi Fidelity(Invitrogen、Carlsbad、CA、cat # 10790−020)およびフラグメント特異的プライマーの使用を通して、特定されたベクターから増幅した。次いで。このPCR産物をTOPOクローニングによってpCR2.1ベクターにサブクローニングした。TOPOクローニングは、TaqまたはPfuポリメーラーゼのいずれかによって増幅されたDNAフラグメントを、DNAリガーゼを必要とせずに特定のベクター中にクローニングする、分子生物学的手法である。
【0089】
はじめに、インスレーター配列をベクターpCpG−LacZ(InvivoGen San Diego、CA、cat #pcpg−lacz)からポリメラーゼ連鎖反応(PCR)によってクローニングし、配列を検証した後、pNEB193(New England Biolabs Ipswich、MA、cat#N3051S)の制限部位にサブクローニングした。このインスレーター配列を、統合部位依存の位置効果に起因する導入遺伝子の発現におけるばらつきを低減するために使用した。
【0090】
フラグメントをクローニングするのに用いたPCRプライマーは以下のとおりである:a.ヒトIFN−ベータMARプライマー配列(プライマーは、サブクローニングのためのEcoRV部位を有する)
配列番号1、IFN 順方向プライマー:
5’−GGGGGATATCAGTCAATATGTTCACCCCA−3’
配列番号2、IFN 逆方向プライマー:
5’−GGGGGATATCCTACTGTTTTAATTAAGC−3’
b.ヒトβ−グロビンMARプライマー配列
配列番号3、β−グロビン順方向プライマー:
5’−AAGGATCCTTAATTAAAATTATCTCTAAGGC−3’
配列番号4、β−グロビン逆方向プライマー:
5’−GGATCCCTGCAGGAATTCCTTTTAAT−3’
【0091】
β−グロビンPCRフラグメントを、pCR2.1(Invitrogen、Carlsbad、CA、cat#K2030−01)を用いてtopoクローニングした。配列を確認した後、そのフラグメントをBamHIで切り出し、その後pNEB193(βgloMAR−pNEB193)のBamH1部位にサブクローニングした。IFN−βPCRフラグメントもまた、pCR2.1でtopoクローニングし、シーケンスし、その後EcoRvで切り出して、βgloMAR−pNEB193(2XMARS−pNEB193)のHincII部位にクローニングした。最後のゲートウエイデスティネーションベクター(Gateway destination vector)については、XbaI末端を含むリンカーおよび内部EcoRV部位を2XMARs−pNEB193のXbaI部位にサブクローニングした。平滑末端化したゲートウエイコンバージョンカセット(Gateway conversion cassette)、RfAを2XMARs−pNEB193(2XMARSpDest)のEcoRV部位に挿入した。最後のベクトル、2XMARSpDestが導入遺伝子を作製するために用いられる最後のデスティネーションベクターであった。
【0092】
配列番号5
TCGCGCGTTTCGGTGATGACGGTGAAAACCTCTGACACATGCAGCTCCCGGAGACGGTCACAGCTTGTCTGTAAGCGGATGCCGGGAGCAGACAAGCCCGTCAGGGCGCGTCAGCGGGTGTTGGCGGGTGTCGGGGCTGGCTTAACTATGCGGCATCAGAGCAGATTGTACTGAGAGTGCACCATATGCGGTGTGAAATACCGCACAGATGCGTAAGGAGAAAATACCGCATCAGGCGCCATTCGCCATTCAGGCTGCGCAACTGTTGGGAAGGGCGATCGGTGCGGGCCTCTTCGCTATTACGCCAGCTGGCGAAAGGGGGATGTGCTGCAAGGCGATTAAGTTGGGTAACGCCAGGGTTTTCCCAGTCACGACGTTGTAAAACGACGGCCAGTGAATTCGAGCTCGGTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCACAAGTTTGTACAAAAAAGCTGAACGAGAAACGTAAAATGATATAAATATCAATATATTAAATTAGATTTTGCATAAAAAACAGACTACATAATACTGTAAAACACAACATATCCAGTCACTATGGCGGCCGCATTAGGCACCCCAGGCTTTACACTTTATGCTTCCGGCTCGTATAATGTGTGGATTTTGAGTTAGGATCCGTCGAGATTTTCAGGAGCTAAGGAAGCTAAAATGGAGAAAAAAATCACTGGATATACCACCGTTGATATATCCCAATGGCATCGTAAAGAACATTTTGAGGCATTTCAGTCAGTTGCTCAATGTACCTATAACCAGACCGTTCAGCTGGATATTACGGCCTTTTTAAAGACCGTAAAGAAAAATAAGCACAAGTTTTATCCGGCCTTTATTCACATTCTTGCCCGCCTGATGAATGCTCATCCGGAATTCCGTATGGCAATGAAAGACGGTGAGCTGGTGATATGGGATAGTGTTCACCCTTGTTACACCGTTTTCCATGAGCAAACTGAAACGTTTTCATCGCTCTGGAGTGAATACCACGACGATTTCCGGCAGTTTCTACACATATATTCGCAAGATGTGGCGTGTTACGGTGAAAACCTGGCCTATTTCCCTAAAGGGTTTATTGAGAATATGTTTTTCGTCTCAGCCAATCCCTGGGTGAGTTTCACCAGTTTTGATTTAAACGTGGCCAATATGGACAACTTCTTCGCCCCCGTTTTCACCATGGGCAAATATTATACGCAAGGCGACAAGGTGCTGATGCCGCTGGCGATTCAGGTTCATCATGCCGTTTGTGATGGCTTCCATGTCGGCAGAATGCTTAATGAATTACAACAGTACTGCGATGAGTGGCAGGGCGGGGCGTAAACGCGTGGATCCGGCTTACTAAAAGCCAGATAACAGTATGCGTATTTGCGCGCTGATTTTTGCGGTATAAGAATATATACTGATATGTATACCCGAAGTATGTCAAAAAGAGGTATGCTATGAAGCAGCGTATTACAGTGACAGT
TGACAGCGACAGCTATCAGTTGCTCAAGGCATATATGATGTCAATATCTCCGGTCTGGTAAGCACAACCATGCAGAATGAAGCCCGTCGTCTGCGTGCCGAACGCTGGAAAGCGGAAAATCAGGAAGGGATGGCTGAGGTCGCCCGGTTTATTGAAATGAACGGCTCTTTTGCTGACGAGAACAGGGGCTGGTGAAATGCAGTTTAAGGTTTACACCTATAAAAGAGAGAGCCGTTATCGTCTGTTTGTGGATGTACAGAGTGATATTATTGACACGCCCGGGCGACGGATGGTGATCCCCCTGGCCAGTGCACGTCTGCTGTCAGATAAAGTCTCCCGTGAACTTTACCCGGTGGTGCATATCGGGGATGAAAGCTGGCGCATGATGACCACCGATATGGCCAGTGTGCCGGTCTCCGTTATCGGGGAAGAAGTGGCTGATCTCAGCCACCGCGAAAATGACATCAAAAACGCCATTAACCTGATGTTCTGGGGAATATAAATGTCAGGCTCCCTTATACACAGCCAGTCTGCAGGTCGACCATAGTGACTGGATATGTTGTGTTTTACAGTATTATGTAGTCTGTTTTTTATGCAAAATCTAATTTAATATATTGATATTTATATCATTTTACGTTTCTCGTTCAGCTTTCTTGTACAAAGTGGTGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTTAAACCTGCAGGCATGCAAGCTTGGCGTAATCATGGTCATAGCTGTTTCCTGTGTGAAATTGTTATCCGCTCACAATTCCACACAACATACGAGCCGGAAGCATAAAGTGTAAAGCCTGGGGTGCCTAATGAGTGAGCTAACTCACATTAATTGCGTTGCGCTCACTGCCCGCTTTCCAGTCGGGAAACCTGTCGTGCCAGCTGCATTAATGAATCGGCCAACGCGCGGGGAGAGGCGGTTTGCGTATTGGGCGCTCTTCCGCTTCCTCGCTCACTGACTCGCTGCGCTCGGTCGTTCGGCTGCGGCGAGCGGTATCAGCTCACTCAAAGGCGGTAATACGGTTATCCACAGAATCAGGGGATAACGCAGGAAAGAACATGTGAGCAAAAGGCCAGCAAAAGGCCAGGAACCGTAAAAAGGCCGCGTTGCTGGCGTTTTTCCATAGGCTCCGCCCCCCTGACGAGCATCACAAAAATCGACGCTCAAGTCAGAGGTGGCGAAACCCGACAGGACTATAAAGATACCAGGCGTTTCCCCCTGGAAGCTCCCTCGTGCGCTCTCCTGTTCCGACCCTGCCGCTTACCGGATACCTGTCCGCCTTTCTCCCTTCGGGAAGCGTGGCGCTTTCTCATAGCTCACGCTGTAGGTATCTCAGTTCGGTGTAGGTCGTTCGCTCCAAGCTGGGCTGTGTGCACGAACCCCCCGTTCAGCCCGACCGCTGCGCCTTATCCGGTAACTATCGTCTTGAGTCCAACCCGGTAAGACACGACTTATCGCCACTGGCAGCAGCCACTGGTAACAGGATTAGCAGAGCGAGGTATGTAGGCGGTGCTACAGAGTTCTTGAAGTGGTGGCCTAACTACGGCTACACTAGAAGAACAGTATTTGGTATCTGCGCTCTGCTGAAGCCAGTTACCTTCGGAAAAAGAGTTGGTAGCTCTTGATCCGGCAAACAAACCACCGCTGGTAGCGGTGGTTTTTTTGTTTGCAAGCAGCAGATTACGCGCAGAAAAAAAGGATCTCAAGAAGATCCTTTGATCTTTTCTACGGGGTCTGACGCTCAGTGGAACGAAAACTCACGTTAAGGGATTTTGGTCATGAGATTATCAAAAAGGATCTTCACCTAGATCCTTTTAAATTAAAAATGAAGTTTTAAATCAATCTAAAGTATATATGAGTAAACTTGGTCTGACAGTTACCAATGCTTAATCAGTGAGGCACCTATCTCAGCGATCTGTCTATTTCGTTCATCCATAGTTGCCTGACTCCCCGTCGTGTAGATAACTACGATACGGGAGGGCTTACCATCTGGCCCCAGTGCTGCAATGATACCGCGAGACCCACGCTCACCGGCTCCAGATTTATCAGCAATAAACCAGCCAGCCGGAAGGGCCGAGCGCAGAAGTGGTCCTGCAACTTTATCCGCCTCCATCCAGTCTATTAATTGTTGCCGGGAAGCTAGAGTAAGTAGTTCGCCAGTTAATAGTTTGCGCAACGTTGTTGCCATTGCTACAGGCATCGTGGTGTCACGCTCGTCGTTTGGTATGGCTTCATTCAGCTCCGGTTCCCAACGATCAAGGCGAGTTACATGATCCCCCATGTTGTGCAAAAAAGCGGTTAGCTCCTTCGGTCCTCCGATCGTTGTCAGAAGTAAGTTGGCCGCAGTGTTATCACTCATGGTTATGGCAGCACTGCATAATTCTCTTACTGTCATGCCATCCGTAAGATGCTTTTCTGTGACTGGTGAGTACTCAACCAAGTCATTCTGAGAATAGTGTATGCGGCGACCGAGTTGCTCTTGCCCGGCGTCAATACGGGATAATACCGCGCCACATAGCAGAACTTTAAAAGTGCTCATCATTGGAAAACGTTCTTCGGGGCGAAAACTCTCAAGGATCTTACCGCTGTTGAGATCCAGTTCGATGTAACCCACTCGTGCACCCAACTGATCTTCAGCATCTTTTACTTTCACCAGCGTTTCTGGGTGAGCAAAAACAGGAAGGCAAAATGCCGCAAAAAAGGGAATAAGGGCGACACGGAAATGTTGAATACTCATACTCTTCCTTTTTCAATATTATTGAAGCATTTATCAGGGTTATTGTCTCATGAGCGGATACATATTTGAATGTATTTAGAAAAATAAACAAATAGGGGTTCCGCGCACATTTCCCCGAAAAGTGCCACCTGACGTCTAAGAAACCATTATTATCATGACATTAACCTATAAAAATAGGCGTATCACGAGGCCCTTTCGTC
【0093】
II.導入遺伝子コンポーネント
A.CRE:
2つの異なる6×CREエレメントを導入遺伝子の作製に用いた。最初は、DNA2.0、Menlo Park、CAで合成して作製された合成CREエレメント(「合成」と記されている)である。この合成配列は4フラグメントのGatewayクローニングのためのattL1およびattR5部位を有している。
【0094】
異なるベクターから6×CREをPCRクローニングして出す複数の試みがなされた;しかし、これは、CREの中ほどにあるヘアピン構造が原因で不可能であった。次いで、このフラグメントを合成によってクローニングした。用いられた第2の6×CREエレメントは、クローンテックベクター(Clontech vector)(Mountainview、CA、cat#PT3336−5)からCREエレメントをPCRクローニングすることによって作製されたハイブリッドバージョンである。クローンテックは、このベクター中には2つのCREが存在していると主張しているが、配列分析では、実際には3つ存在していることが示された。2つの異なるPCR反応をフラグメントのクローニングに用いた。ゲートウエイのクローニング用のAtt部位と同様にEcoR1部位をプライマー中に導入し、2つのフラグメントが一緒に「接着される」ことができるようにして、Invitrogen Gateway pDONR P1−P5r vector(Carlsbad、CA、cat #12537−100)中に再結合させた。
【0095】
1.合成CREエレメント
配列番号6:
5’AAATAATGATTTTATTTTGACTGATAGTGACCTGTTCGTTGCAACAAATTGATGAGCAATGCTTTTTTATAATGCCAACTTTGTACAAAAAAGCAGGCTTACTGTCGACAATTGCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGATTGACGTCAATGGGGTGTCTGAAAGACGTCACAGTATGACCCGGGCTCGAGCCTCCTTGGCTGACGTCAGAGAGAGAGGCCGGCCCCTTACGTCAGAGGCGAGAATTCGACAACTTTGTATACAAAAGTTGAACGAGAAACGTAAAATGATATAAATATCAATATATTAAATTAGATTTTGCATAAAAAACAGACTACATAATACTGTAAAACACAACATATCCAGTCACTATG3’
【0096】
2.ハイブリッドCRE:標準的なクローニング、3×CREを含むClontech pCreLuc vectorからCREエレメントをPCR増幅し、PCRプライマーは一端にEcoR1制限部位を有しており(CRE3X−B、CRE3X−C)、att部位は、ゲートウエイのattB1またはattB5rのいずれかがもう一方の末端でクローニングされている(CRE3X−A、CRE3X−D)。次いで、これらの2つのフラグメントを組み合わせて6×CREの1つのフラグメントを作成した。このフラグメントは、Gateway pDONR P1−P5r vector中に組み換えされる。
【0097】
配列番号7:CRE 順方向プライマー A
5’−GGGGACAAGTTTGTACAAAAAAGCAGGCTTAGCACCAGACAGTGA−3’
配列番号8:CRE 逆方向プライマー B
5’−GGGAATTCGTTCTCCCATTGACGTCA−3’
配列番号9:CRE 順方向プライマー C
5’−GGGAATTCGCACCAGACAGTGACGTC−3’
配列番号10:CRE 逆方向プライマー D
5’−GGGGACAACTTTTGTATACAAAGTTGTGTTCTCCCATTGACGTCA−3’
配列番号11:ハイブリッドCRE配列:
GCTTAGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACGAATTCGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACA
【0098】
B.ヒト成長ホルモン ポリAテール:
ヒト成長ホルモン ポリAテールを、ベクターpOGH(Nichols Institute Diagnostics(San Juan Capistrano,CA、Cat # 40−2205)からPCRクローニングした。プライマー配列は、Gateway pDONR P3−P2 vector中への組み換え用に、順方向プライマー上にattB3部位、逆方向プライマー上にattB2部位を有している。
【0099】
配列番号12:hGH 順方向プライマー:
5’−GGGGACAACTTTGTATAATAAAGTTGGATCCCAAGGCCCAACTCC−3’
配列番号13:hGH 逆方向プライマー
5’−GGGGACCACTTTGTACAAGAAAGCTGGGTACAACAGGCATCTACT−3’
【0100】
C.HSV TK最小プロモーター:
HSV TK最小プロモーターを、「661 CreLuc」と呼ばれるインハウスベクター(in house vector)からPCRクローニングした。プライマー配列は、Gateway pDONR P5−P4 vector中への組み換え用に、順方向プライマー上にattB5部位を有し、逆方向プライマー上にattB4部位を有している。
【0101】
配列番号14:TK 順方向プライマー
5’−GGGGACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGT−3’
【0102】
配列番号15:TK 逆方向プライマー
5’−GGGGACAACTTTGTATAGAAAAGTTGGGTGGATCTGCGGCACGCT−3’
【0103】
D.ルシフェラーゼ cDNA:
ルシフェラーゼcDNAを、ベクターpGL4.10(Promega Madison、WI Cat #E6651)からPCRクローニングした。Lucプライマー配列は、Gateway pDONR P4r−P3r vector中への組み換え用に、順方向プライマー上にattB4r部位を有し、逆方向プライマー上にattB3r部位を有している。
【0104】
配列番号16:luci 順方向プライマー
5’GGGGACAACTTTTCTATACAAAGTTGATGGAAGATGCCAAAAACA3’
【0105】
配列番号17:luci 逆方向プライマー
5’GGGGACAACTTTATTATACAAAGTTGTTTACACGGCGATCTTGCC3’
【0106】
III.最終的なベクターの構築
導入遺伝子の4つのコンポーネント、CRE(合成バージョンまたはハイブリッドバージョンのいずれか)、HSVTK min、ルシフェラーゼ cDNA、およびpDONRベクター中のhGHポリAテールを、標準的なInvitrogenプロトコルに従ってpDest2XMARsデスティネーションベクターと共に組み換えを行った。その後これら2つの導入遺伝子をシーケンスし、CHOK1中にトランスフェクトして、機能について試験した。図5に示されように、どちらの導入遺伝子も、インビトロで機能する。図5は、ホルスコリン誘導によるCreLucベクターのインビトロ分析を示している。
【0107】
IV.トランスジェニックマウスの作製
A.導入遺伝子の準備
導入遺伝子構築物の質は、アリコートをアガロースゲル(電気泳動)にかけることにより検証および確認した。劣化(degradation)のトレースは観察されなかった。最終的に、診断用のXhoIおよびPvuIIおよびPstI制限酵素を用いた切断解析により、予測された制限プロファイルが得られた。次いで、この導入遺伝子プラスミドをAcc65IおよびPmeIで消化して、導入遺伝子を含むフラグメントをアガロースゲル上での消化を実行することにより、ベクター骨格から単離した。その後、この導入遺伝子フラグメントをゲルから切り出し、Qiaquick gel extraction kit(Qiagen、Valencia、CA cat#28706)で精製し、希釈して、受精卵中に注入した。単離した導入遺伝子の純度および濃度はアガロースゲル電気泳動により検査した。
【0108】
配列番号18:CreLuc 合成導入遺伝子(Acc65I/PmeI 消化、5550bp)
5’GTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCCAAGTTTGTACAAAAAAGCAGGCTTACTGTCGACAATTGCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGATTGACGTCAATGGGGTGTCTGAAAGACGTCACAGTATGACCCGGGCTCGAGCCTCCTTGGCTGACGTCAGAGAGAGAGGCCGGCCCCTTACGTCAGAGGCGAGAATTCGACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGTCGGGGCGGCGCGGTCCCAGGTCCACTTCGCATATTAAGGTGACGCGTGTGGCCTCGAACACCGAGCGACCCTGCAGCGACCCGCTTAACAGCGTCAACAGCGTGCCGCAGATCCACCCAACTTTTCTATACAAAGTTGCTATGGAAGATGCCAAAAACATTAAGAAGGGCCCAGCGCCATTCTACCCACTCGAAGACGGGACCGCCGGCGAGCAGCTGCACAAAGCCATGAAGCGCTACGCCCTGGTGCCCGGCACCATCGCCTTTACCGACGCACATATCGAGGTGGACATTACCTACGCCGAGTACTTCGAGATGAGCGTTCGGCTGGCAGAAGCTATGAAGCGCTATGGGCTGAATACAAACCATCGGATCGTGGTGTGCAGCGAGAATAGCTTGCAGTTCTTCATGCCCGTGTTGGGTGCCCTGTTCATCGGTGTGGCTGTGGCCCCAGCTAACGACATCTACAACGAGCGCGAGCTGCTGAACAGCATGGGCATCAGCCAGCCCACCGTCGTATTCGTGAGCAAGAAAGGGCTGCAAAAGATCCTCAACGTGCAAAAGAAGCTACCGATCATACAAAAGATCATCATCATGGATAGCAAGACCGACTACCAGGGCTTCCAAAGCATGTACACCTTCGTGACTTCCCATTTGCCACCCGGCTTCAACGAGTACGACTTCGTGCCCGAGAGCTTCGACCGGGACAAAACCATCGCCCTGATCATGAACAGTAGTGGCAGTACCGGATTGCCCAAGGGCGTAGCCCTACCGCACCGCACCGCTTGTGTCCGATTCAGTCATGCCCGCGACCCCATCTTCGGCAACCAGATCATCCCCGACACCGCTATCCTCAGCGTGGTGCCATTTCACCACGGCTTCGGCATGTTCACCACGCTGGGCTACTTGATCTGCGGCTTTCGGGTCGTGCTCATGTACCGCTTCGAGGAGGAGCTATTCTTGCGCAGCTTGCAAGACTATAAGATTCAATCTGCCCTGCTGGTGCCCACACTATTTAGCTTCTTCGCTAAGAGCACTCTCATCGACAAGTACGACCTAAGCAACTTGCACGAGATCGCCAGCGGCGGGGCGCCGCTCAGCAAGGAGGTAGGTGAGGCCGTGGCCAAACGCTTCCACCTACCAGGCATCCGCCAGGGCTACGGCCTGACAGAAACAACCAGCGCCATTCTGATCACCCCCGAAGGGGACGACAAGCCTGGCGCAGTAGGCAAGGTGGTGCCCTTCTTCGAGGCTAAGGTGGTGGACTTGGACACCGGTAAGACACTGGGTGTGAACCAGCGCGGCGAGCTGTGCGTCCGTGGCCCCATGATCATGAGCGGCTACGTTAACAACCCCGAGGCTACAAACGCTCTCATCGACAAGGACGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGTGGACCGGCTGAAGAGCCTGATCAAATACAAGGGCTACCAGGTAGCCCCAGCCGAACTGGAGAGCATCCTGCTGCAACACCCCAACATCTTCGACGCCGGGGTCGCCGGCCTGCCCGACGACGATGCCGGCGAGCTGCCCGCCGCAGTCGTCGTGCTGGAACACGGTAAAACCATGACCGAGAAGGAGATCGTGGACTATGTGGCCAGCCAGGTTACAACCGCCAAGAAGCTGCGCGGTGGTGTTGTGTTCGTGGACGAGGTGCCTAAAGGACTGACCGGCAAGTTGGACGCCCGCAAGATCCGCGAGATTCTCATTAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAACAACTTTGTATAATAAAGTTGCTGATCCCAAGGCCCAACTCCCCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCTGCAATGGCTACAGGTAAGCGCCCCTAAAATCCCTTTGGGCACAATGTGTCCTGAGGGGAGAGGCAGCGACCTGTAGATGGGACGGGGGCACTAACCCTCAGGTTTGGGGCTTCTGAATGTGAGTATCGCCATGTAAGCCCAGTATTTGGCCAATCTCAGAAAGCTCCTGGTCCCTGGAGGGATGGAGAGAGAAAAACAAACAGCTCCTGGAGCAGGGAGAGTGCTGGCCTCTTGCTCTCCGGCTCCCTCTGTTGCCCTCTGGTTTCTCCCCAGGCTCCCGGACGTCCCTGCTCCTGGCTTTTGGCCTGCTCTGCCTGCCCTGGCTTCAAGAGGGCAGTGCCTTCCCAACCATTCCCTTATCCAGGCTTTTTGACAACGCTATGCTCCGCGCCCATCGTCTGCACCAGCTGGCCTTTGACACCTACCAGGAGTTTGTAAGCTCTTGGGGAATGGGTGCGCATCAGGGGTGGCAGGAAGGGGTGACTTTCCCCCGCTGGGAAATAAGAGGAGGAGACTAAGGAGCTCAGGGTTTTTCCCGAAGCGAAAATGCAGGCAGATGAGCACACGCTGAGTGAGGTTCCCAGAAAAGTAACAATGGGAGCTGGTCTCCAGCG
TAGACCTTGGTGGGCGGTCCTTCTCCTAGGAAGAAGCCTATATCCCAAAGGAACAGAAGTATTCATTCCTGCAGAACCCCCAGACCTCCCTCTGTTTCTCAGAGTCTATTCCGACACCCTCCAACAGGGAGGAAACACAACAGAAATCCGTGAGTGGATGCCTTCTCCCCAGGCGGGGATGGGGGAGACCTGTAGTCAGAGCCCCCGGGCAGCACAGCCAATGCCCGTCCTTCCCCTGCAGAACCTAGAGCTGCTCCGCATCTCCCTGCTGCTCATCCAGTCGTGGCTGGAGCCCGTGCAGTTCCTCAGGAGTGTCTTCGCCAACAGCCTGGTGTACGGCGCCTCTGACAGCAACGTCTATGACCTCCTAAAGGACCTAGAGGAAGGCATCCAAACGCTGATGGGGGTGAGGGTGGCGCCAGGGGTCCCCAATCCTGGAGCCCCACTGACTTTGAGAGCTGTGTTAGAGAAACACTGCTGCCCTCTTTTTAGCAGTCAGGCCCTGACCCAAGAGAACTCACCTTATTCTTCATTTCCCCTCGTGAATCCTCCAGGCCTTTCTCTACACCCTGAAGGGGAGGGAGGAAAATGAATGAATGAGAAAGGGAGGGAACAGTACCCAAGCGCTTGGCCTCTCCTTCTCTTCCTTCACTTTGCAGAGGCTGGAAGATGGCAGCCCCCGGACTGGGCAGATCTTCAAGCAGACCTACAGCAAGTTCGACACAAACTCACACAACGATGACGCACTACTCAAGAACTACGGGCTGCTCTACTGCTTCAGGAAGGACATGGACAAGGTCGAGACATTCCTGCGCATCGTGCAGTGCCGCTCTGTGGAGGGCAGCTGTGGCTTCTAGCTGCCCGGGTGGCATCCCTGTGACCCCTCCCCAGTGCCTCTCCTGGCCCTGGAAGTTGCCACTCCAGTGCCCACCAGCCTTGTCCTAATAAAATTAAGTTGCATCATTTTGTCTGACTAGGCGTCCTTCTATAATATTATGGGGTGGAGGGGGGTGGTATGGAGCAAGGGGCAAGTTGGGAAGACAACCTGTAGGGCCTGCGGGGTCTATTGGGAACCAAGCTGGAGTGCAGTGGCACAATCTTGGCTCACTGCAATCTCCGCCTCCTGGGTTCAAGCGATTCTCCTGCCTCAGCCTCCCGAGTTGTTGGGATTCCAGGCATGCATGACCAGGCTCAGCTAATTTTTGTTTTTTTGGTAGAGACGGGGTTTCACCATATTGGCCAGGCTGGTCTCCAACTCCTAATCTCAGGTGATCTACCCACCTTGGCCTCCCAAATTGCTGGGATTACAGGCGTGAACCACTGCTCCCTTCCCTGTCCTTCTGATTTTAAAATAACTATACCAGCAGGAGGACGTCCAGACACAGCATAGGCTACCTGGCCATGCCCAACCGGTGGGACATTTGAGTTGTTTGCTTGGCACTGTCCTCTCATGCGTTGGGTCCACTCAGTAGATGCCTGTTGTACCCAGCTTTCTTGTACAAAGTGGGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTT 3’
【0109】
配列番号19:CreLuc ハイブリッド導入遺伝子配列(Acc65I/PmeI 消化、5562bp)
5’GTACCCGGGGGCGCGCCGGGATCCTTAATTAAAATTATCTCTAAGGCATGTGAACTGGCTGTCTTGGTTTTCATCTGTACTTCATCTGCTACCTCTGTGACCTGAAACATATTTATAATTCCATTAAGCTGTGCATATGATAGATTTATCATATGTATTTTCCTTAAAGGATTTTTGTAAGAACTAATTGAATTGATACCTGTAAAGTCTTTATCACACTACCCAATAAATAATAAATCTCTTTGTTCAGCTCTCTGTTTCTATAAATATGTACCAGTTTTATTGTTTTTAGTGGTAGTGATTTTATTCTCTTTCTATATATATACACACACATGTGTGCATTCATAAATATATACAATTTTTATGAATAAAAAATTATTAGCAATCAATATTGAAAACCACTGATTTTTGTTTATGTGAGCAAACAGCAGATTAAAAGGAATTCCTGCAGGATCCTTAATTAAGTTCTAGATCCAAGTTTGTACAAAAAAGCAGGCTTAGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACGAATTCGCACCAGACAGTGACGTCAGCTGCCAGATCCCATGGCCGTCATACTGTGACGTCTTTCAGACACCCCATTGACGTCAATGGGAGAACACAACTTTGTATACAAAAGTTGTGGAACACGCAGATGCAGTCGGGGCGGCGCGGTCCCAGGTCCACTTCGCATATTAAGGTGACGCGTGTGGCCTCGAACACCGAGCGACCCTGCAGCGACCCGCTTAACAGCGTCAACAGCGTGCCGCAGATCCACCCAACTTTTCTATACAAAGTTGCTATGGAAGATGCCAAAAACATTAAGAAGGGCCCAGCGCCATTCTACCCACTCGAAGACGGGACCGCCGGCGAGCAGCTGCACAAAGCCATGAAGCGCTACGCCCTGGTGCCCGGCACCATCGCCTTTACCGACGCACATATCGAGGTGGACATTACCTACGCCGAGTACTTCGAGATGAGCGTTCGGCTGGCAGAAGCTATGAAGCGCTATGGGCTGAATACAAACCATCGGATCGTGGTGTGCAGCGAGAATAGCTTGCAGTTCTTCATGCCCGTGTTGGGTGCCCTGTTCATCGGTGTGGCTGTGGCCCCAGCTAACGACATCTACAACGAGCGCGAGCTGCTGAACAGCATGGGCATCAGCCAGCCCACCGTCGTATTCGTGAGCAAGAAAGGGCTGCAAAAGATCCTCAACGTGCAAAAGAAGCTACCGATCATACAAAAGATCATCATCATGGATAGCAAGACCGACTACCAGGGCTTCCAAAGCATGTACACCTTCGTGACTTCCCATTTGCCACCCGGCTTCAACGAGTACGACTTCGTGCCCGAGAGCTTCGACCGGGACAAAACCATCGCCCTGATCATGAACAGTAGTGGCAGTACCGGATTGCCCAAGGGCGTAGCCCTACCGCACCGCACCGCTTGTGTCCGATTCAGTCATGCCCGCGACCCCATCTTCGGCAACCAGATCATCCCCGACACCGCTATCCTCAGCGTGGTGCCATTTCACCACGGCTTCGGCATGTTCACCACGCTGGGCTACTTGATCTGCGGCTTTCGGGTCGTGCTCATGTACCGCTTCGAGGAGGAGCTATTCTTGCGCAGCTTGCAAGACTATAAGATTCAATCTGCCCTGCTGGTGCCCACACTATTTAGCTTCTTCGCTAAGAGCACTCTCATCGACAAGTACGACCTAAGCAACTTGCACGAGATCGCCAGCGGCGGGGCGCCGCTCAGCAAGGAGGTAGGTGAGGCCGTGGCCAAACGCTTCCACCTACCAGGCATCCGCCAGGGCTACGGCCTGACAGAAACAACCAGCGCCATTCTGATCACCCCCGAAGGGGACGACAAGCCTGGCGCAGTAGGCAAGGTGGTGCCCTTCTTCGAGGCTAAGGTGGTGGACTTGGACACCGGTAAGACACTGGGTGTGAACCAGCGCGGCGAGCTGTGCGTCCGTGGCCCCATGATCATGAGCGGCTACGTTAACAACCCCGAGGCTACAAACGCTCTCATCGACAAGGACGGCTGGCTGCACAGCGGCGACATCGCCTACTGGGACGAGGACGAGCACTTCTTCATCGTGGACCGGCTGAAGAGCCTGATCAAATACAAGGGCTACCAGGTAGCCCCAGCCGAACTGGAGAGCATCCTGCTGCAACACCCCAACATCTTCGACGCCGGGGTCGCCGGCCTGCCCGACGACGATGCCGGCGAGCTGCCCGCCGCAGTCGTCGTGCTGGAACACGGTAAAACCATGACCGAGAAGGAGATCGTGGACTATGTGGCCAGCCAGGTTACAACCGCCAAGAAGCTGCGCGGTGGTGTTGTGTTCGTGGACGAGGTGCCTAAAGGACTGACCGGCAAGTTGGACGCCCGCAAGATCCGCGAGATTCTCATTAAGGCCAAGAAGGGCGGCAAGATCGCCGTGTAAACAACTTTGTATAATAAAGTTGCTGATCCCAAGGCCCAACTCCCCGAACCACTCAGGGTCCTGTGGACAGCTCACCTAGCTGCAATGGCTACAGGTAAGCGCCCCTAAAATCCCTTTGGGCACAATGTGTCCTGAGGGGAGAGGCAGCGACCTGTAGATGGGACGGGGGCACTAACCCTCAGGTTTGGGGCTTCTGAATGTGAGTATCGCCATGTAAGCCCAGTATTTGGCCAATCTCAGAAAGCTCCTGGTCCCTGGAGGGATGGAGAGAGAAAAACAAACAGCTCCTGGAGCAGGGAGAGTGCTGGCCTCTTGCTCTCCGGCTCCCTCTGTTGCCCTCTGGTTTCTCCCCAGGCTCCCGGACGTCCCTGCTCCTGGCTTTTGGCCTGCTCTGCCTGCCCTGGCTTCAAGAGGGCAGTGCCTTCCCAACCATTCCCTTATCCAGGCTTTTTGACAACGCTATGCTCCGCGCCCATCGTCTGCACCAGCTGGCCTTTGACACCTACCAGGAGTTTGTAAGCTCTTGGGGAATGGGTGCGCATCAGGGGTGGCAGGAAGGGGTGACTTTCCCCCGCTGGGAAATAAGAGGAGGAGACTAAGGAGCTCAGGGTTTTTCCCGAAGCGAAAATGCAGGCAGATGAGCACACGCTGAGTGAGGTTCCCAGAAAAGTAACAATGGGAGCTGGTCTCCAGCGTAGACCTTGGTGGGCGGTCCTTCTCCTAGGAAGAAGCCTATATCCCAAAGGAACAGAAGTATTCATTCCTGCAGAACCCCCAGACCTCCCTCTGTTTCTCAGAGTCTATTCCGACACCCTCCAACAGGGAGGAAACACAACAGAAATCCGTGAGTGGATGCCTTCTCCCCAGGCGGGGATGGGGGAGACCTGTAGTCAGAGCCCCCGGGCAGCACAGCCAATGCCCGTCCTTCCCCTGCAGAACCTAGAGCTGCTCCGCATCTCCCTGCTGCTCATCCAGTCGTGGCTGGAGCCCGTGCAGTTCCTCAGGAGTGTCTTCGCCAACAGCCTGGTGTACGGCGCCTCTGACAGCAACGTCTATGACCTCCTAAAGGACCTAGAGGAAGGCATCCAAACGCTGATGGGGGTGAGGGTGGCGCCAGGGGTCCCCAATCCTGGAGCCCCACTGACTTTGAGAGCTGTGTTAGAGAAACACTGCTGCCCTCTTTTTAGCAGTCAGGCCCTGACCCAAGAGAACTCACCTTATTCTTCATTTCCCCTCGTGAATCCTCCAGGCCTTTCTCTACACCCTGAAGGGGAGGGAGGAAAATGAATGAATGAGAAAGGGAGGGAACAGTACCCAAGCGCTTGGCCTCTCCTTCTCTTCCTTCACTTTGCAGAGGCTGGAAGATGGCAGCCCCCGGACTGGGCAGATCTTCAAGCAGACCTACAGCAAGTTCGACACAAACTCACACAACGATGACGCACTACTCAAGAACTACGGGCTGCTCTACTGCTTCAGGAAGGACATGGACAAGGTCGAGACATTCCTGCGCATCGTGCAGTGCCGCTCTGTGGAGGGCAGCTGTGGCTTCTAGCTGCCCGGGTGGCATCCCTGTGACCCCTCCCCAGTGCCTCTCCTGGCCCTGGAAGTTGCCACTCCAGTGCCCACCAGCCTTGTCCTAATAAAATTAAGTTGCATCATTTTGTCTGACTAGGCGTCCTTCTATAATATTATGGGGTGGAGGGGGGTGGTATGGAGCAAGGGGCAAGTTGGGAAGACAACCTGTAGGGCCTGCGGGGTCTATTGGGAACCAAGCTGGAGTGCAGTGGCACAATCTTGGCTCACTGCAATCTCCGCCTCCTGGGTTCAAGCGATTCTCCTGCCTCAGCCTCCCGAGTTGTTGGGATTCCAGGCATGCATGACCAGGCTCAGCTAATTTTTGTTTTTTTGGTAGAGACGGGGTTTCACCATATTGGCCAGGCTGGTCTCCAACTCCTAATCTCAGGTGATCTACCCACCTTGGCCTCCCAAATTGCTGGGATTACAGGCGTGAACCACTGCTCCCTTCCCTGTCCTTCTGATTTTAAAATAACTATACCAGCAGGAGGACGTCCAGACACAGCATAGGCTACCTGGCCATGCCCAACCGGTGGGACATTTGAGTTGTTTGCTTGGCACTGTCCTCTCATGCGTTGGGTCCACTCAGTAGATGCCTGTTGTACCCAGCTTTCTTGTACAAAGTGGGATCTAGACTAGAGTCATCAGTCAATATGTTCACCCCAAAAAAGCTGTTTGTTAACTTGTCAACCTCATTCTAAAATGTATATAGAAGCCCAAAAGACAATAACAAAAATATTCTTGTAGAACAAAATGGGAAAGAATGTTCCACTAAATATCAAGATTTAGAGCAAAGCATGAGATGTGTGGGGATAGACAGTGAGGCTGATAAAATAGAGTAGAGCTCAGAAACAGACCCATTGATATATGTAAGTGACCTATGAAAAAAATATGGCATTTTACAATGGGAAAATGATGATCTTTTTCTTTTTTAGAAAAACAGGGAAATATATTTATATGTAAAAAATAAAAGGGAACCCATATGTCATACCATACACACAAAAAAATTCCAGTGAATTATAAGTCTAAATGGAGAAGGCAAAACTTTAAATCTTTTAGAAAATAATATAGAAGCATGCCATCAAGACTTCAGTGTAGAGAAAAATTTCTTATGACTCAAAGTCCTAACCACAAAGAAAAGATTGTTAATTAGATTGCATGAATATTAAGACTTATTTTTAAAATTAAAAAACCATTAAGAAAAGTCAGGCCATAGAATGACAGAAAATATTTGCAACACCCCAGTAAAGAGAATTGTAATATGCAGATTATAAAAAGAAGTCTTACAAATCAGTAAAAAATAAAACTAGACAAAAATTTGAACAGATGAAAGAGAAACTCTAAATAATCATTACACATGAGAAACTCAATCTCAGAAATCAGAGAACTATCATTGCATATACACTAAATTAGAGAAATATTAAAAGGCTAAGTAACATCTGTGGCTTAATTAAAACAGTAGGATGACTGTTT
【0110】
B.初代の作製:
これらの導入遺伝子をFVB/Taconic単細胞胚の前核に別々にマイクロインジェクションした。トランスジェニック(Tg)動物を作製するための一般的なストラテジーはよく記載されており、当業者に公知である。
【0111】
PCR遺伝子型判定ストラテジーを用いて、トランスジェニックポジティブ初代マウスを、尾部生検および遺伝子型用のPCRを介して出生後すぐに同定した。選択されたプライマー対は、導入遺伝子配列内に短いDNA配列の増幅を可能にし、特定の405−bp PCR産物を得ることができる。PCR反応ミックスは、MgCl2(Qiagen、Valencia、CA cat #201205)、0.2mM dNTPs、0.4uM プライマー、0.5単位のTaqポリメラーゼを含む1×Qiagen PCR緩衝液である。サイクリング条件は以下の通りである:94℃で1サイクル;94℃で5分、35サイクル;57℃で45秒;72℃で45秒;72℃で1分、1サイクル;5分。
【0112】
遺伝子型判定プライマーは以下の通りである:
配列番号20:Luc2−順方向プライマー:
5’−GAAGATGCCAAAAACATTAAGAAG−3’
配列番号21:Luc2−逆方向プライマー:
5’−GATCTTTTGCAGCCCTTTCT −3’
【0113】
合成CreLuc導入遺伝子については、(181出生したうち)39の導入遺伝子ポジティブ初代、ハイブリッドCreLuc導入遺伝子については、(244出生したうち)73の導入遺伝子ポジティブが生成された。このトランスジェニックマウス系統は、CreLucと省略される。
【0114】
V.初代発現分析:
図5Aは、正常マウスにおける血漿cAMPレベルに対する異なるホスホジエステラーゼ(PDE)インヒビターの効果の比較である(Cheng et al、JPET、280(2):621−626、1997からの再現)。これらのマウスを、4匹の動物群で化合物またはビヒクル(VEH)をp.o.投与した。20分後、血液を採取し、cAMP測定のために処理した。この発行された実験に基づき、cAMPレベルを増加させるその能力およびそのアベイラビリティについてのCreLuc初代をスクリーニングするために、ホルスコリンと共にロリプラムを一般的な誘導因子として選択した。一緒に投与する場合、これらの2つの化合物の付加的な効果により、動物中のルシフェラーゼレベルを調べるときの誘導の大きいウインドウを提供される。
【0115】
図5Bは、ホルスコリン(F)、ロリプラム()またはホルスコリンとロリプラムの組み合わせ(F/R)による種々の処理後の、野生型マウス中の血漿レベルのcAMPアッセイである。2月齢のFVB/Tac雌を、以下のうちの1つと共にi.p.投与した:グループ:ビヒクルコントロール、リン酸緩衝生理食塩水(PBS)に溶解させた1%ジメチルスルホキシド (DMSO)、グループB:5mg/kg ホルスコリン(Sigma、St Louis MO、cat#F6886)、およびグループC:10mg/kg ロリプラム(Sigma St Louis、MO、cat# R6520)、グループD:5mg/kg ホルスコリン+10mg/kg ロリプラム、グループE:5mg/kg 水溶性ホルスコリン(Calbiochem Gibbstown、NJ cat#344273)、およびグループF:5mg/kg 水溶性ホルスコリン+10mg/kg ロリプラム。グループGは、処理を受けていない。ビヒクルのみと比較して、ロリプラムおよび/またはホルスコリンは、循環しているcAMPレベルの上昇において、統計的に有意であった。ホルスコリンのみでは、ビヒクルコントロールに対して循環しているcAMPを有意に上昇させなかった。水溶性ホルスコリン(H2OF)とロリプラムとの組み合わせは、cAMP血漿レベルにおいて最も大きい上昇を示し、この処理がインビボでの導入遺伝子の発現のためにCreLuc初代をスクリーニングするために最適であった。
【0116】
全ての初代(表5〜8に列挙している初代および図6に示されている代表的な初代)を、10mg/kg ロリプラム(Sigma St Louis、MO、cat# R6520)と5mg/kg 水溶性ホルスコリン(Calbiochem Gibbstown、NJ cat#344273)との組み合わせをマウスにi.p.投与することによって、導入遺伝子発現について試験した。4時間後、これらのマウスをXenogen IVIS Lumina systemを用いてバイオイメージングした。これらのマウスを、イソフルラン、および250mg/kg ルシフェリンをs.c.注射することにより麻酔した。39の導入遺伝子ポジティブな合成CreLucのうち、34(または87%)が発現ポジティブであった。発現は任意の組織のルシフェラーゼレベルがシステムのバックグラウンドレベルを超えたとき、ポジティブであると考えられた。
【0117】
CreLucマウスの異なる独立した系統のバイオイメージングによってアッセイされた種々のホルスコリン ロリプラム応答プロファイルを、図6に示している。誘導されていない状態において、本発明者らは、系統90および219に図示されているような検出可能な基底バイオイメージングシグナルか、または系統44、28、および187に図示されているような基底活性がないかのいずれかを確認した。ホルスコリン ロリプラム誘導後、CreLucレポーター誘導のバイオイメージングパターンは、単一または複数の組織中に生じ、それは、各独立したCreLuc系統で独特であった。多くのCreLuc系統は、治療に関連する領域中に検出可能なルシフェラーゼビオイメージを発現する組織を1以上有していた。
【0118】
基底と、ホルスコリン ロリプラム誘導の両方での初代中のCreLuc発現のバイオイメージングスクリーニング後、1またはそれより多い組織(脳内で2×)中で5×バイオイメージング誘導のフィルターウインドウを満たすCreLuc系統を、組織抽出物中のルシフェラーゼ酵素の誘導されたレベルをアッセイすることによってより詳細に分析した。
【0119】
組織発現分析のために、動物を、ベースラインシグナル用に未処理のままかまたは4時間誘導した。誘導のために、動物に、Dulbecco’s PBS(Invitrogen、Carlsbad、CA、cat#14040)に溶解させた5mg/kg ホルスコリン(Calbiochem、Gibbstown、NJ cat#344273)と10mg/kg ロリプラム(Sigma St Louis、MO、cat# R6520)with 1%DMSO(Sigma、St Louis、MO、cat#D2650)の組み合わせをi.p.投与した。数時間後、これらの動物をCO2で屠殺し、種々の器官を取り除いてドライアイスで凍らせた。ルシフェラーゼアッセイのために、Luciferase Assay System(Promega、Madison、WI、cat#E1500)を用いた。これらの組織を、1mlの細胞培養溶解緩衝液(Cell Culture Lysis Buffer)(Promega、Madison、WI、cat#E1531)中でホモジネートさせ、5分間氷上でインキュベーションし、その後5分間遠心分離で回転させた。20μlの上清を製造業者の指示書によりアッセイ中で用いた。これらのサンプルのタンパク質濃度をDCタンパク質アッセイキット(BioRad、Hercules、CA、cat#500−0111)を用いて測定した。データは表5〜8に示しており、誘導倍率として表わしている(誘導されたRLU/ugタンパク質を、基底RLU/ugタンパク質で割ったもの)。誘導されたRLUは、誘導された組織サンプルのRLU/ugタンパク質である。
【0120】
【表5】
【0121】
【表6】
【0122】
【表7】
【0123】
【表8】
【0124】
各系統におけるCreLuc発現の詳細なレベルは、標的組織中で最も高い発現レベルを持つ最適の系統の選択、および独特な単一または複数のパターンの発現を有する系統の選択を可能にする。この酵素アッセイはまた、CreLuc発現が特定の組織と明らかに関連していたことを保証した。一般的に、導入遺伝子中にデザインされた場合、大部分の系統を横切ってすべての組織の近くにルシフェラーゼ発現を観察した。重要な医薬関連標的組織(脳、膵臓、肺、脾臓)由来のトップの3〜5系統を、GPCRリガンドを用いたさらなる参照化合物応答プリファイリングのために選択した。
【0125】
VI.発現レベルにおけるインスレーター配列の効果の分析
表9において、インスレーター配列を加えることの影響は、11の異なる導入遺伝子を通じて比較され、225の独立したトランスジェニック系統を含んでいる。機能的なトランスジェニック系統の生産は、典型的には10〜30%の範囲である(本発明者らの研究室から採取した過去のデータ)。表9において、hGH小遺伝子または機能的エンハンサーの封入により、11のうち10の導入遺伝子および190系統が、この範囲の上限での発現レベルを獲得した(機能的系統の平均数は37または32.7%)。重要なことに、インスレーター配列の封入は、この発現頻度を2倍以上に増やし、112系統については78%が機能的レポーターを発現している。導入遺伝子の比較および多様性両方についてのサンプルサイズは、ゲノム中の導入遺伝子のランダムな組み込みにより、正常な生物学的変動を克服するのに十分な大きさである。
【0126】
トランスジェニックバイオイメージングレポーター系統の作製は、低頻度で高い損耗率のプロセスである。多くの独立した初代系統(50〜100)が、レポーター用に所望される検出レベルを備える系統を得るため、および薬物候補のスクリーニング用に適切な標的組織の広範な選択肢において発現されるレポーターを持つために必要とされる。一般的には、薬物スクリーニング用途には5%以下の系統で十分であるため、レポーターを含む導入遺伝子は、作製された各トランスジェニック系統中の発現を最大化させるために多くのエレメントを含める必要がある。他の発現エンハンサー(hGH小遺伝子)に結合しているインスレーター配列は、導入遺伝子を発現するトランスジェニック系統の大部分を得ることが実証されており、薬物開発に適用可能な発現パターンを持つトランスジェニック系統を見つける鍵である(リガンドのレセプターへの結合の際のシグナルの低下を検出するための種々のGPCRシグナル伝達経路については特に)。
【0127】
【表9】
【0128】
材料の寄託:
マウス胚線維芽細胞の調製:妊娠した雌から、12日目の胚を取り出した。この胚をリン酸緩衝生理食塩水(PBS)(Invitrogen、Carlsbad、CA、cat#14040)を含む10cmの皿に入れた子宮の中から解剖した。次いで、それをPBSの新しい皿に移した。その胚を鉗子で裂いて、心臓および肝臓を取り除いた。次いで、組織切片を2〜3mlの冷0.25%トリプシン(Invitrogen、Carlsbad、CA、cat#25200)中で4℃にて一晩インキュベーションした。次の日、そのトリプシンの大部分を吸引し、組織ペレットを37℃で30分間インキュベーションした。2〜3mlの培地、DMEM(Invitrogen、Carlsbad、CA、cat#11965)、5%の熱不活性化FBS(Invitrogen、Carlsbad、CA、cat#16140)、1%のペン連鎖球菌(Invitrogen、Carlsbad、CA、cat# 15140)を加え、組織の残りの塊を壊すために上下にピペッティングした。次いで細胞懸濁液をT75組織培養フラスコにプレーティングした。細胞を3継体培養し、その後1mlのバイアル中で、バイアルあたり1〜2百万細胞、凍結培地、15%の熱不活性化FBS(Invitrogen、Carlsbad、CA、cat#16140)、5% DMSO(Sigma、St Louis、MO、cat#D2650)、DMEM(Invitrogen、Carlsbad、CA、cat#11965)中で、凍結させた。
【0129】
以下の材料をAmerican Type Culture Collection、10801 University Blvd.、Manassas、Va.20110−2209、USA(ATCC):に寄託した:
【表10】
【0130】
この寄託は、特許手続き上の微生物の寄託の国際的承認に関するブダペスト条約およびその下の条例(Budapest Treaty on the International Recognition of the Deposit of Microorganisms for the Purpose of Patent Procedure and the Regulations thereunder(Budapest Treaty))の規定の下でなされた。これにより、寄託日より30年間、その寄託の生存培養物の維持が保証される。この寄託は、ブダペスト条約の下、ATCCによって利用可能とされており、sanofi−aventis US Inc.とATCCとの契約下にある。この契約は、関連する米国特許の公開の際、または任意の米国特許出願もしくは外国特許出願の一般への公開(いずれか早いほう)の際に、一般に対して寄託の培養物の子孫の永続的かつ無制限なアベイラビリティを保証し、かつその全体に対して米国の特許商標庁長官によって決定される子孫のアベイラビリティを保証する。
【0131】
本出願の指定代理人は、寄託においてそれらの材料の培養物が、適切な条件下で培養された場合に死ぬかまたは紛失されるかまたは破壊された場合に、それらの材料が速やかに同一の別のもので置き換えられることに合意した。寄託された材料のアベイラビリティーは、その特許法に従って、あらゆる政府の権限下で付与される権利に違反して本発明を実施するためのライセンスとして解釈されるべきではない。
【0132】
VII.いンビボおよびエキソビボスクリーニングアッセイ
図7は、インビボまたはエキソビボいずれかのCreLucスクリーニングアッセイの流れを表わすことによって本発明の実施態様を示している。インビボアッセイについては、動物にルシフェリンのみを投与し、その後IVIS bioimagerでバイオイメージングして、ベースライン信号を得た。その後これらの動物に化合物を投与して、設定した時点で再びバイオイメージングし、ルシフェラーゼシグナルに対する化合物の影響を調べた。データは、ベースラインの信号に対する誘導された信号の誘導倍率として分析することができる。エキソビボアッセイについては、マウスに化合物またはビヒクルのいずれかを投与し、その後設定した時点において組織を採取し、凍結させた。次いで、その組織をホモジネートし、Promega luciferase assay system(Promega cat #E1500)のようなルシフェラーゼアッセイをその組織抽出物に対して実行し、Topcountのようなルミノメーターでそのアッセイを読み取った。データは、化合物群に対してビヒクルを比較する、μgタンパク質あたりの相対光として分析することができる。
【0133】
本発明のさらなる局面および詳細は、以下の実施例から明らかとなり、これらは例示することを意図しているのであって、いかなる意味においても本発明の範囲を限定することを意味しない。
【0134】
A.全脳スライス実験
CreLucマウスにおける発光に及ぼす化合物の影響を示すために、全脳スライスを用いた(図8A、8B)。一般的には、CreLucマウスから脳を採取し、ミクロトームでスライス状にカットするかまたは種々の小領域を解剖して取り出し、ミクロトームでスライスすることもできる。その後これらのスライスをホルスコリンとロリプラム、またはGPCR特異的化合物のような異なる化合物と共にインキュベーションすることができる。次いで、ルシフェラーゼレベルをIVIS bioimagerまたはカルーセルを備える光電子増倍管で測定し、定量的経時測定を行うことができる。
【0135】
CreLucマウス(系統187)の海馬領域からの脳スライスを1uMのイソプロテレノール(Sigma、cat#5627)と共に2日間インキュベーションした。イソプロテレノール(ISO)は、cAMP経路を活性化するGsアゴニストであり(経路の概略は図1を参照のこと)、CreLucマウス中でルシフェラーゼ発現レベルを誘導する(図8A、下パネル)。従って、GiアゴニストのようなGPCR特異的化合物のスクリーニングを可能にするルシフェラーゼシグナルウインドウを増加させるためにイソプロテレノールを用いた。AMN082(AMN、Tocris Bioscience、Cat.No.2385)は、mGluR7アゴニストであり、mGluR7は、Giに結合しているレセプターである。脳スライスを、1uM AMN082と組み合わせた1uM ISOで処理した場合、ルシフェラーゼシグナルは、ISOのみでの処理と比較して減少した。
【0136】
化合物の活性はまた、生育できる脳スライス中で時間をかけて定量することができ、またルシフェラーゼ発現は、バイオイメージャーによって可視化することができる。図8Bにおいて、CreLucマウス(系統44)からの脳スライスは、ホルスコリンに反応した。ホルスコリンはcAMPの一般的なアクチベーターである。50uMのホルスコリンでの脳スライスの処理(矢印は、ホルスコリンをスライスに加えた時間をマークしている)により、ビヒクルと比較して時間依存的に誘導を引き起こした。
【0137】
B.インビトロ細胞培養実験
1.初代皮質ニューロン
CreLucモデルからの初代神経細胞の単離および化合物処理についてのワークフローの一般的な概略図を図9に示している。皮質や他の脳の小領域を胚(通常は胎齢(E)17または18であるが、早ければE14も)から取り除いた。その後、個々のニューロンを単離し、96ウエルのアッセイプレート上にプレーティングした。アッセイは、トリプリケートまたは4通り(quadruplicate)のいずれかで実行した。アッセイは、標的GPCRの発現レベルに依存して、培養物中で7日または10日まで培養した。その後、化合物をそのプレートに加え、Bright Glo(Promega、Cat # E2610)をプレートに加えて、TopCountまたは他のルミノメーターでアッセイを実行した。
【0138】
a)βアドレナリン受容体(AD(R)およびD1ドーパミン受容体(DRD1)を介したGsの調節
CreLucマウスからの初代細胞培養物を、Gs共役型レセプターを調節し得る化合物をスクリーニングするために用いることができる。ニューロンを系統187のE18胚の皮質から単離した。系統187は、全組織ルシフェラーゼアッセイによって脳と脊髄両方におけるルシフェラーゼの誘導性レベルを有することが示されている(以下の文章;図24を参照のこと)。このアッセイを培養3日目にトリプリケートで実行した。ホルスコリンを5μM、ロリプラムを10μM、および、Gsアゴニスト、ADβR−イソプロテレノール;およびドーパミン−SKF82958(MFG、Cat# C130)を10μMで用いた。4時間でBright Gloを加え、Topcountでアッセイを実行した。ホルスコリンは、cAMPの一般的なアクチベーターである。ロリプラムはcAMPを活性化しないが、かわりにロリプラムはcAMPの分解をブロックし、それによりcAMPレベルを安定化させる。ホルスコリンとロリプラムは、ルシフェラーゼ発現を増加させるように相乗的に作用する。100倍を超える誘導が、ホルスコリンとロリプラムの組み合わせで観察された。増加したluciレベルは、DMSOコントロールに対して、ADβRアゴニストイソプロテレノールで見られた(11倍)。一方、D1 DRアゴニスト、SKF82958は、ルシフェラーゼレベルを有意に増加させず、ロリプラム単独と比較する場合に、SKF82958をロリプラムと組み合わせて投与した場合、2倍の誘導が観察された。従って、系統187は、その化合物がGs共役レセプターを調節し得るかどうかを判断するために化合物をスクリーニングするおに有用なモデルである。
【0139】
b)Prokineticin 2受容体(PROK2R)を介したGq調節
CreLucマウスからの初代細胞培養物を、Gs共役受容体を調節し得る化合物をスクリーニングするために用いることができる。従って、このPROK2Rは、PROK2Rのアゴニストであるprokineticin 2(PROK2)を、CreLucマウスがGq調節に応答するかどうかを確認するために用いた。Gqは、cAMPをバイパスするが、CREBを活性化するためにPLC経路を利用している(図1の経路図を参照のこと)。初代皮質ニューロンを、E18の系統187(脳および脊髄中に誘導可能なルシフェラーゼを有していることが示されている−以下の文章および図24を参照のこと)から採取した。このアッセイは、4時間、または8時間培養で3日目に実行した。PROK2ペプチド(Peprotech、Cat#100−46)を1nMおよび100nMの水溶液として加えた。ルシフェラーゼシグナル用にBright Gloを用い、アッセイをTopCount luminometerで実行した。コントロールのみの培地に対して、ルシフェラーゼレベルの非常に有意な増加が両方の時点で、PROK2の両方の濃度について観察された。従って、CreLucマウスは、Gq共役受容体を調節する化合物をスクリーニングするために用いることができる。
【0140】
異なるCreLuc系統由来の初代皮質ニューロン中のルシフェラーゼ発現におけるPROK2ペプチドの効果を調べた。初代皮質ニューロンを4つの異なるCreLuc系統からE18で採取した。系統は、全脳抽出物中で誘導可能なルシフェラーゼレベルを決定した以前の実験に基づいて選択した(データは示さず)。このアッセイを4時間および24時間の2つの時点で、1nMまたは100nMいずれかのPROK2ペプチドを含む培養物中で3日目にトリプリケートで実行した。このアッセイにはBrightGloを使用し、TopCountで読み取った。ルシフェラーゼレベルにおける統計的に有意な増加が、系統219および175の両方の時点および両方の濃度において観察された。全ての系統がある程度PROK2リガンドに対応しているとしても、ルシフェラーゼ発現レベルの差異は、導入遺伝子の組込みにおける差異に起因し得る。
【0141】
c)mGluR7受容体を介したGi調節
CreLucマウス由来の初代細胞培養物を、Gi共役受容体を調節し得る化合物をスクリーニングするために用いることができる。図13Aは、初代皮質ニューロン中のルシフェラーゼ発現におけるmGluR7アゴニスト、AMN082の効果を示している。皮質ニューロンをE18胚(系統187)より採取した。このアッセイは、培養3日目に実行した。ホルスコリンを10μMで用いてGi活性を検出するのに用いるより大きいシグナルウインドウを得た。このmGluR7アゴニスト、AMN082は、1nM、10nM、100nMおよび1μMのホルスコリンと組み合わせて用いた。このアッセイは、Bright Glo(Promega)を備えるTopCountで、4時間および8時間で読み取った。ホルスコリンのみに対して、ルシフェラーゼレベルの有意な減少が両時点の100nMおよび1μMのAMN082で観察された。このCreLucマウスは、Gq共役受容体を調節する化合物をスクリーニングするために用いることができる。例えば、「A」、「B」および「C」と記される「未知の」化合物を内部コントロールとしてAMN082を用いて初代皮質ニューロン培養物中でスクリーニングし、任意の化合物がGi調節能を示すかどうか決定する(図13B)。化合物Aは、EC50=1.735e−07を持つ公知のmGluR7アゴニストAMN082と比較すると、算出値EC5=1.529e−06の活性化合物であると考えられるが、一方化合物Bは、非特異的化合物であると考えられ、また化合物Cは不活性である。
【0142】
CreLucマウス由来の初代細胞培養物を用いるGi共役受容体を調節する能力についての化合物のスクリーニングは、上記に示したように、ホルスコリンのようなcAMP刺激因子の存在中でなされ得る。しかしながら、シグナルウインドウを増加させるためにも、ロリプラム(cAMPの分解をブロックする)をホルスコリンと組み合わせて用いた。初代皮質ニューロンは、誘導可能な能および脊髄発現を有していると以前に判断された系統187由来のE18胚から単離した。アッセイは、トリプリケートで、6時間培養で7日目に実行した。AMN082の10μMから1nMまでの用量曲線は、50μMのホルスコリンと10μMのロリプラムとを組み合わせて実行した。このアッセイは、Bright Glo substrate(Promega)を備えるTopCount luminometerで読み取った。ルシフェラーゼレベルの統計的に有意な減少であるp<0.0005(ホルスコリンおよびロリプラムコントロールに対して59〜61%)は、1μM、100nMおよび1nMで見られた。
【0143】
CB1アゴニスト、CP55,940による、異なるCreLuc系統由来の初代皮質ニューロン中のルシフェラーゼ発現のGi調節を、ホルスコリンおよびロリプラムの存在下で実施した(図15)。初代皮質ニューロンは、系統69、187、175および219のE18から採取した。用いられた全ての系統は、全脳組織抽出物中で誘導可能なルシフェラーゼ発現を有していると以前に判断されたものである(データは示さず)。このアッセイは、3日培養で実施した。CB1アゴニストは、10μMで、ホルスコリンは5μM、ロリプラムは10μMで用いた。4時間と24時間の2つの時点で実行した。次いで、Bright Gloルシフェラーゼアッセイ基質を加え、このアッセイをTopcount luminometerで読み取った。このアッセイはトリプリケートで実行した。データはトリプリケートの平均で示されている。データは、秒あたりのカウント数(cps)で示されている。ルシフェラーゼのシグナルの減少は、CB1アゴニストを添加した両方の時点で4つ全ての系統において観察された。応答レベルのわずかな差異は、導入遺伝子の組み込みに起因し得るが、全ての系統がGi調節に応答性であり、Gi活性のスクリーニング化合物に修正可能であった。
【0144】
CB1アゴニスト、CP55,940による、CreLucマウス由来の初代皮質ニューロン中のルシフェラーゼのGi調節は、用量依存的であった(図15B)。皮質ニューロンは、E18胚から単離下。このアッセイは培養3日目に実行した。ホルスコリンおよびロリプラムを10μMで用いた。アゴニストを10μM、1μMおよび100nMの濃度で加えた。このアッセイは、BrightGlo(Promega)を備えるTopCountで、8時間で読み取った。ルシフェラーゼレベルの有意な減少(ホルスコリンおよびロリプラム単独に対して、アゴニストにホルスコリンとロリプラムを加えたもの)は、3つ全ての濃度で観察された。
【0145】
2.初代線条体ニューロン
CreLucマウスから誘導された任意の細胞培養物は、GPCRを調節する能力について化合物をスクリーニングするために用いることができる。ホルスコリンおよびロリプラム、ならびにDRD1およびADβRのGsアゴニストによって、CreLuc線条体ニューロン中でルシフェラーゼ発現を誘導した(図16)。線条体ニューロンは、E14胚(系統187)から単離した。このアッセイは培養4日目に実行した。ホルスコリンを5μM、ロリプラムを10μMで用いた。Gsアゴニスト(イソプロテレノールはADRβにおいてアゴニストであり、ドーパミンおよびSKF82958は、D1DRにおいてアゴニストである)を10μM、3μMおよび1μMで用いた。このアッセイは、TopCount luminometerおよびBright Glo ルシフェラーゼ試薬(Promega)を用いて5時間で読み取った。28倍の非常に有意な増加が、ホルスコリンとロリプラムの組み合わせで処理された細胞中で観察された。有意な増加はまた、10μMのドーパミンで2.7倍、イソプロテレノールの3つ全ての濃度で観察された。下がttえ、導入遺伝子は、皮質ニューロンだけでなく線条体においても機能的である(図10を参照のこと)。
【0146】
3.全脾細胞
CreLucマウス(系統64)から単離した全脾細胞プレップを用いて、ホルスコリンおよびロリプラムのような一般的なcAMP誘導因子、ならびいルシフェラーゼ発現におけるGsアゴニストの効果を示した(図16)。脾臓は、1×HBSS(Invitrogen、Carlsbad、CA、cat#14025)の動物から採取した。次いで、この細胞を、5mlのD−PBS(Invitrogen、Carlsbad、CA、cat#14190)中で、5mlシリンジの先端を用いて脾臓カプセルを機械的に破壊することにより単離した。次いで、この細胞懸濁液を70umのストレーナーを通過させて50mlのコニカルチューブ中に入れた。その後細胞を800rpmで回転させ、5mlの1X Pharm Lyse solution(BD BioSciences、cat#555899)中に再懸濁させて室温で6分間インキュベーションした。次いで細胞に、RPMI 1640(Invitrogen、Carlsbad、CA、cat#11875)、10% FCS(Invitrogen、Carlsbad、CA、cat#16000)、1% pen/strep(Invitrogen、Carlsbad、CA、cat#15070)および0.1% β−メルカプトエタノール(Invitrogen、Carlsbad、CA、cat#21985)からなる28mlの培地を加えることによって洗浄した。細胞を2×106/mlまで希釈し、96ウエルの白色不透明プレートにプレートあたり100ulの細胞をプレーティングした。これらの細胞の半分を抗CD3抗体(BD Pharmingen、cat#553058)で24時間刺激し、残りの半分は処理しなかった。24時間でそのプレートに化合物を加え、さらに4時間放置した。一般的な誘導因子としては、ロリプラム(Sigma R6520)、ホルスコリン(Sigma F6886)が挙げられる。用いられたGsアゴニストは以下のとおりである:EX00000173A(173A;インハウスで合成された)は、Gs結合プロスタグランジンE2(EP2)受容体でのアゴニストである;BW245C(Sigma B9305)は、Gs結合プロスタグランジンD2受容体(DP1)のアゴニストであり、イソプロテレノール(Sigma I5627)は、Gs結合βアドレナリン受容体(ADβR)でのアゴニストである。このアッセイは、トリプリケートで実行した。4時間後、100μlのBrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイはTopCount luminometer で読み取った。DMSOに対して14倍の増加がロリプラムおよびホルスコリンの存在下でのCD3刺激された細胞中で観察された。統計的に有意な増加(t−検定による)もまた、DMSOのみのコントロールに対して3つ全てのGsアゴニストで観察された。この実験により、導入遺伝子は、全脾細胞集団において機能的であることが示された。この実験において用いられた調製物は、細胞の混合された集合である、全脾細胞調製物であった。以下に記載される実験は、T細胞およびB細胞のような亜集団におけるルシフェラーゼ発現レベルを調べている。
【0147】
4.単離T細胞
5つの異なるサブ系統のCreLucマウスから単離したT細胞中の、ロリプラムおよびホルスコリンによる一般的なcAMP活性化の効果を研究した(図18)。全脾細胞集団を上に記載したような機械的な破壊により準備した。次いで、CD4+細胞をMACS Magnetic Separation with Positive Selection Column(Miltenyi Biotec cat#130−049−201)を用いて単離した。次いで、これらの細胞をウエルあたり1.5x105個、96ウエルの白色不透明プレートにプレーティングし、抗CD3抗体(BD Pharmingen cat#553058)で刺激した。18時間後、10μMのロリプラ (Sigma R6520)および5μMのホルスコリン(Sigma F6886)をプレートに加え、さらに4時間置いた。BrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイはTopCountで読み取った。データは発光(秒あたりのカウント数)、および培地のみのコントロールに対する増加倍率として示される。ルシフェラーゼの発現における増加は試験された全ての系統で観察され、系統64が最も高い誘導レベルを提供し、cAMPがCreLucマウス中の一般的なモジュレーターによって活性化されていることを実証している。
【0148】
CreLucマウス(系統64)から単離された抗CD3刺激されたCD4+T細胞中のルシフェラーゼレベルに対する異なるGsアゴニストの効果を研究した(図19)。全脾細胞集団を機械的な破壊によって準備した。次いで、CD4+細胞をMACS Magnetic Separation with Positive Selection(Miltenyi Biotec cat#130−049−201)によって単離下。これらの細胞をウエルあたり1.5x105個、96ウエルの白色不透明プレートにプレーティングし、抗CD3抗体(BD Pharmingen cat#553058)で刺激した。24時間後、化合物を加え、さらに4時間置いた。DP(BW245C)、EP2(EX00000173A)およびADβR(イソプロテレノール)のGsアゴニストを全て10μMで、ホルスコリンを5μMで、およびロリプラムを10μMで用いた。BrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイをTopCountで読み取った。ホルスコリンおよびロリプラムで処理した細胞において、25倍の増加が観察された。BW245Cは3倍、173Aは2倍、イソプロテレノールは5倍という非常に有意な増加が3つ全てのGsアゴニストで観察された。導入遺伝子は、特異的GsアゴニストだけでなくcAMP経路の一般的なモジュレーターとして応答性であった。
【0149】
5.単離B細胞
CreLucマウスの2つの異なるサブ系統から単離したB細胞中の、ロリプラムおよびホルスコリンによる一般t系なcAMP活性化の効果を調べた(図20)。全脾細胞集団を上記のように機械的に破壊した。次いで、B220+細胞を、Positive Selection Column(Miltenyi Biotec、cat# 130−049−501)を備えるMACS Magnetic Separationを用いて単離した。その後これらの細胞を96ウエルの白色不透明プレート上にウエルあたり2.0x105個プレーティングし、10ng/mlのリポ多糖(LPS)(Sigma L−2630)で刺激した。18時間後、10μMのロリプラム(Sigma R6520)および5μMのホルスコリン(Sigma F6886)をそのプレートに加え、さらに4時間置いた。BrightGlo(Promega、Madison、WI、cat#E2610)を加え、アッセイをTopCountで読み取った。データは発光(秒あたりのカウント数)および培地のみのコントロールに対する増加倍率として示される。ルシフェラーゼ発現の増加は、系統64では観察されたが、系統229では観察されなかった。
【0150】
6.ミクログリア
一般的なcAMPアクチベーターであるホルスコリンおよびロリプラム、DP受容体のアゴニストであるBW245Cによる、CreLucマウスから単離されたミクログリア中のルシフェラーゼ発現の誘導を調べた(図22)。初代ミクログリアは、DMEM(GIBCO Cat# 11995)、10% FBS(GIBCO Cat# 16140)および1% ペニシリン−ストレプトマイシン100X(GIBCO Cat# 15140)からなる培地中でP2マウス(系統64)由来の皮質から単離した。細胞を、ポリ−D−リジンコーティングプレート上に96ウエル形式でプレーティングした。細胞は未処理のままか100ng/mlのLPSで2時間刺激したかのいずれかである。次いで化合物を加え、さらに4時間置いた後、Bright Gloアッセイを実行した。使用した化合物は、5μMのホルスコリン、10μMのロリプラム、もしくはこれら2つの組み合わせ、またはDP1受容体のGsアゴニストBW245Cを10μMである。非刺激条件下では、ミクログリアは一般的なcAMPモジュレーターおよびDP受容体の特異的Gsアゴニストに対して非応答性であった。しかしながら、ミクログリアをLPSで刺激した場合、それらの細胞はホルスコリンおよびBW245Cに対して応答性となった。
【0151】
7.マウス胚線維芽細胞
マウス胚線維芽細胞中のルシフェラーゼ発現におけるホルスコリン、ロリプラム、およびイソプロテレノールの効果を調査した(図33)。マウス胚線維芽細胞は、6つの独立したCreLuc系統のE12胚から培養した。細胞をウエルあたり20,000細胞プレーティングした。試験した化合物としては、10μMのホルスコリン、5μMのロリプラムおよび10μMのイソプロテレノール(ADβRアゴニスト)が挙げられる。ホルスコリンとロリプラムの組み合わせに応じて、有意な増加が全ての系統で観察された。有意な増加はまた、イソプロテレノールに応じて、3つの系統で観察された。
【0152】
7.心筋細胞
心筋細胞中のルシフェラーゼレベルにおける、ホルスコリンおよびロリプラムおよびイソプロテレノールの効果を研究した(図35)。心筋細胞は、系統229由来のP3仔より単離した。細胞を96ウエルプレートで培養した。試験した化合物としては、10μMのホルスコリン、5μMのロリプラムおよび10μMのイソプロテレノール(ADβRアゴニスト)が挙げられる。ロリプラムとホルスコリンの組み合わせにより、ルシフェラーゼレベルの有意な増加(25倍)が観察され、アゴニストのイソプロテレノールでもまた有意な増加(2倍)が観察された。
【0153】
B.インビボおよびエキソビボ試験
1.一般的なcAMPモジュレーターの効果
ルシフェラーゼ発現の誘導に対する髄腔内注射されたホルスコリンおよびロリプラムの効果を、系統187のCreLucマウスの脳および脊髄において研究した(図23)。選択されたマウスの系統は、脳と脊髄の両方において導入遺伝子の発現を有していることが以前に判定されている(データは示さず)。このアッセイ用に選択されたマウスの系統(系統187)は、脳と脊髄の両方において誘導可能なレベルのルシフェラーゼを有している。グループあたりN=3〜4のマウス、4つの処理群、3月齢の雄。グループA:DMSOコントロール、グループB:1μgホルスコリン/10μgロリプラム、グループC:10μgホルスコリン/10μgロリプラム、グループD:40μgホルスコリン/10μgロリプラム。この動物に、髄腔内注射により腰部に1匹のマウスあたり5μlの量で投薬した。これらを投薬後4時間で画像処理した。脊髄および脳の両方のデータは、平均ピーク輝度、平方センチメートル毎秒あたりの光子として示されている。統計的に有意な増加が脊髄中で観察され、脳内におけるルシフェラーゼシグナルの有意な増加は、ホルスコリンの最も高い濃度においてみられただけであった。一般的なモジュレーターまたはcAMPが脊髄中に注射された場合、脊髄中で導入遺伝子の局所的な発現の増加があり、脳においてはより低い応答が観察された。
【0154】
2.Gsアゴニストの効果
CreLucマウスの脳および脊髄中のルシフェラーゼ発現に対するEPアゴニスト、EX00000173Aの効果を研究した(図24)。系統187由来の5月齢の雄マウス(n=5)に、ビヒクル(5% DMSO、0.05% tween 80、PBS)または10mg/kg EX00000173Aのいずれかを用いてi.p.注射した。動物を投薬4時間後にバイオイメージングした。データは平方センチメートル毎秒あたりの光子として示されている。脳と脊髄の両方において、アゴニスト処理による統計的に有意な増加がみられた。従って、i.p.投与されたGsアゴニストは、導入遺伝子を活性化した。
【0155】
さらに、CreLucマウス中のルシフェラーゼ発現に対するEP2アゴニスト、EX00000173Aの効果は用量反応性であった(図25)。系統187は、この系統が脳と脊髄の両方において高く誘導可能な発現を有しているために選択した。このアッセイは、5つの群の6週齢のマウス(n=5)で構成されている。マウスに、ビヒクルコントロール(D−PBS;Dulbeccos phosphate buffered saline、Invitrogen、cat#14040)または用量を変化させたEP2アゴニストEX0000173A(1.5nmol、5nmol、15nmolおよび50nmol)のいずれかを投薬した。マウスに髄腔内注射(1匹のマウスあたり5μl)により投薬し、IVIS bioimagerで4時間後にバイオイメージングした。データは5匹のマウスの平均値として、平均ピーク輝度、平方センチメートル毎秒あたりの光子として示されている。アゴニスト処理による統計的に有意な(t−検定)増加が、脊髄中の3つの濃度で観察されたが、脳内では観察されなかった。脳内の増加(有意ではない)は、髄腔内注射で予測されたような最も高い濃度で観察されたのみであった。
【0156】
図26は、CreLucマウスにおけるアドレナリン受容体β3(Adrb3)アゴニスト、CL316,243(1 mg/kg、ip)による異なる組織中のルシフェラーゼの誘導を示している。このルシフェラーゼアッセイは、組織ホモジネートで実施した。スクリーニングした12の独立したトランスジェニック系統の中で、7系統が脂肪組織と胚において10倍以上の誘導を示した。4系統はバイオイメージングによって目視可能な誘導を示した(図27Aおよび27Bを参照)。
【0157】
3.GsアゴニストAVE0010の特異的効果
導入遺伝子の活性化は、CreLucマウスの3つの独立した系統において健空された、グルカゴン用ペプチド1受容体(GLP−1R)アゴニストによるルシフェラーゼレポーターの誘導によって例示されるように、組織特異的受容体の活性化と共局在する(図28)。GLP−1Rは、Gs結合されている。ベースラインの画像は、1日目に得られた。2日目、マウスをAVE0010(0.1 mg/kg、sc)で処理し、4時間後に画像処理した。ベースラインに対する誘導倍率が下部に示されている。予想どおりGLP−1R受容体は、主に脾臓組織中で発現されているので、脾臓中で観察された導入遺伝子活性化の強い誘導が存在した。8つの異なる組織中のルシフェラーゼ活性を、系統11、16および90について測定し(図29)、その後AVE0010(0.1 mg/kg、s.c.)で4時間処理した。組織ホモジネート中のルシフェラーゼ活性により、ルシフェラーゼの脾臓特異的誘導が確認された。AVE0010の活性は、GLP−1Rが異なる組織タイプで見出されても、脾臓に限定されている。
【0158】
AVE0010によるCreLucの誘導は、β細胞媒介である可能性が高い。AVE0010によるCRE−Lucの誘導におけるβ細胞毒素ストレプトゾトシン(STZ)の効果を調査した(図30)。雄のCreLucマウス(系統11)を、AVE0010を0.1 mg/kg処理の前および処理後に画像処理した。全てのマウスは、AVE0010に応答していた(中パネル)。次いで、この動物をビヒクル(コントロール)またはSTZ(200mpk、ip)で処理した。4日後、これらをAVE0010での処理後再び画像処理した。ビヒクル群と比較すると、STZ群は、ルシフェラーゼ誘導が減少していた。
【0159】
AVE0010によるCreLucの誘導は、β細胞特異的である可能性がある(図31)。動物を図30に記載されたように処理した。血中グルコース濃度を、非絶食マウスの尾静脈ニッキングによって測定した。グルコース濃度は、Bayerのグルコースメーターで読み取った。グルコース濃度はmgグルコース/mlとして示される。誘導の倍率は、ベースラインの信号 対 AVE10を投与したルシフェラーゼバイオイメージングレベルである。血中グルコース濃度(BG)は、STZによる増加した(左上パネル)。非絶食BGレベルは、AVE0010(0.1 mg/kg、sc)によって低減された。図30に示されるBLIデータを定量した。
【0160】
4.骨髄移植
CreLuc骨髄移植を、NOD scid ガンマ(NGS)マウスを用いて実施した(図32)。NSDマウスは、成熟T細胞およびB細胞、機能的なナチュラルキラー細胞を欠損した免疫無防備状態のマウスであり、造血細胞の移植を可能にする差異と下院シグナル伝達を欠損している。骨髄細胞は、系統44(高い基底ルシフェラーゼレベルを有している;データは示さず)のヘテロ接合体と系統64(誘導可能なルシフェラーゼレベルを有している)のホモ接合体から採取した。その後、この細胞を、1匹のマウスあたり100万、または500万個の細胞で、照射NSGマウスに細胞の尾静脈注射により移植した。系統44については、マウス1および2は500万、マウス3および4が100万の細胞を受けた。系統64について:マウス1が500万個の細胞、マウス2、3および4が100万個の細胞を受けた。(CreLuc系統あたり4匹のNSGマウス)。動物を、4週間(データは示さず)でバイオイメージングし、再度8週間(データは示されている)でバイオイメージングした。画像処理の前に、系統64のマウスを5mg/kgのホルスコリンおよび10mg/kgのロリプラムで5時間誘導した。バイオイメージング写真は、8週の時点のものを示している。系統44の骨髄細胞を移植したNSGマウスにおけるルシフェラーゼレベルは、CreLuc系統44で見られた画像を模倣しており、間接、脊髄、頭部および胸骨において発現が観察された。誘導可能なルシフェラーゼ発現は、系統64の骨髄細胞を移植したNSGマウスの脾臓で観察された。
【0161】
4.動物モデル
CreLucマウスは、疾患または疾患状態の局面の動物モデルを研究するのに用いることができる。さらに、このCreLucマウスは、CreLucマウスにおいて誘導される疾患または疾患の局面を調節することができる化合物をスクリーニングするために用いることができる。例えば、CreLuc系統187におけるルシフェラーゼレベルに対するザイモサン処理の効果を研究した(図34)。治療群のCreLucマウス(系統187)に、疼痛反応を誘導するためにザイモサンによって両方の後部足に皮下注射した。これらの動物を4日間毎日バイオイメージングした。ルシフェラーゼ発現の統計的に有意な増加は、全ての時点において、ザイモサンに反応して動物の足で観察された。ザイモサンは、炎症反応を強く活性化する酵母細胞壁成分である。従って、CreLucマウスは、同じ動物において炎症反応を経時的にモニタリングできるツールである。また、ザイモサン処理した動物を用いて、ザイモサンによって誘導される炎症を逆にするかまたは悪化させる試験化合物の能力を評価するためのスクリーニングツールとして用いることができる。
【特許請求の範囲】
【請求項1】
第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を含むゲノムを有する、トランスジェニック非ヒト動物。
【請求項2】
第1のインスレーター配列が、マトリックス付着領域(MAR)、DNase I−高感受性領域(HS4)および末端逆位配列(ITR)からなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物。
【請求項3】
第2のインスレーター配列が、マトリックス付着領域(MAR)、HS4およびITRからなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物。
【請求項4】
第1のインスレーター配列が、前記第2のインスレーター配列と同じである、請求項1に記載のトランスジェニック非ヒト動物。
【請求項5】
応答配列が、cAMP応答配列(CRE)、アクチベータータンパク質1(ASP1)、グルココルチコイド応答配列(GRE)、熱ショック応答配列(HSE)、血清応答配列(SRE)、甲状腺ホルモン応答配列(TRE)およびエストロゲン応答配列(ERE)からなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物。
【請求項6】
応答配列がタンデムに2〜24回反復されている、請求項5に記載のトランスジェニック非ヒト動物。
【請求項7】
応答配列がタンデムに6回反復されている、請求項6に記載のトランスジェニック非ヒト動物。
【請求項8】
応答配列がCREである、請求項7に記載のトランスジェニック非ヒト動物。
【請求項9】
プロモーターが単純ヘルペスウイルスチミジンキナーゼの最小プロモーター(HSV TK min)である、請求項1に記載のトランスジェニック非ヒト動物。
【請求項10】
生物発光レポーターが、ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、β−ガラクトシダーゼ、分泌型アルカリホスファターゼ(SEAP)、ヒト成長ホルモン(HGH)および緑色蛍光タンパク質(GFP)からなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物。
【請求項11】
機能エレメントがヒト成長ホルモン(hGH)遺伝子である、請求項1に記載のトランスジェニック非ヒト動物。
【請求項12】
導入遺伝子が配列番号18を含む、請求項1に記載のトランスジェニック非ヒト動物。
【請求項13】
導入遺伝子が配列番号19を含む、請求項11に記載のトランスジェニック非ヒト動物。
【請求項14】
請求項1に記載のトランスジェニック非ヒト動物から単離された細胞。
【請求項15】
請求項1に記載のトランスジェニック非ヒト動物から単離された組織切片。
【請求項16】
Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物中の生物発光の量を測定する工程;
(b)トランスジェニック非ヒト動物に試験薬剤を投与する工程;
(c)試験薬剤の投与後に、1またはそれより多い時点でトランスジェニック非ヒト動物の生物発光の量を測定する工程;
(d)(a)において測定した生物発光の量と(c)において測定した生物発光の量とを比較する工程;
を含み、ここで(c)と比較した(a)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【請求項17】
Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物から組織切片を調製する工程;
(b)組織切片中の生物発光の量を測定する工程;
(c)組織切片に試験薬剤を投与する工程;
(d)試験薬剤の投与後に、1またはそれより多い時点で組織切片の生物発光の量を測定する工程;および
(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程;
を含み、ここで(d)と比較した(b)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【請求項18】
Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物から単離された細胞を調製する工程;
(b)細胞中の生物発光の量を測定する工程;
(c)細胞に試験薬剤を投与する工程;
(d)試験薬剤の投与後に、1またはそれより多い時点で細胞の生物発光の量を測定する工程;および
(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程;
を含み、ここで(d)と比較した(b)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【請求項19】
非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)非ヒト動物からの生物発光をモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【請求項20】
GPCR機能のモニタリングにおいて使用するための非ヒトトランスジェニック動物を作製する方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)(a)のトランスジェニック非ヒト動物における生物発光の量を測定する工程;
(c)GPCRリガンドを該トランスジェニック非ヒト動物に投与する工程;
(d)GPCRリガンドの投与後に、1またはそれより多い時点でトランスジェニック非ヒト動物の生物発光の量を測定する工程;および
(e)(b)で測定した生物発光の量と(d)で測定した生物発光の量を比較する工程;を含み、ここで(d)と比較した(b)の生物発光の量の差が、GPCR機能をモニタリングするのに用いるための非ヒトトランスジェニック動物を同定する、上記方法。
【請求項1】
第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を含むゲノムを有する、トランスジェニック非ヒト動物。
【請求項2】
第1のインスレーター配列が、マトリックス付着領域(MAR)、DNase I−高感受性領域(HS4)および末端逆位配列(ITR)からなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物。
【請求項3】
第2のインスレーター配列が、マトリックス付着領域(MAR)、HS4およびITRからなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物。
【請求項4】
第1のインスレーター配列が、前記第2のインスレーター配列と同じである、請求項1に記載のトランスジェニック非ヒト動物。
【請求項5】
応答配列が、cAMP応答配列(CRE)、アクチベータータンパク質1(ASP1)、グルココルチコイド応答配列(GRE)、熱ショック応答配列(HSE)、血清応答配列(SRE)、甲状腺ホルモン応答配列(TRE)およびエストロゲン応答配列(ERE)からなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物。
【請求項6】
応答配列がタンデムに2〜24回反復されている、請求項5に記載のトランスジェニック非ヒト動物。
【請求項7】
応答配列がタンデムに6回反復されている、請求項6に記載のトランスジェニック非ヒト動物。
【請求項8】
応答配列がCREである、請求項7に記載のトランスジェニック非ヒト動物。
【請求項9】
プロモーターが単純ヘルペスウイルスチミジンキナーゼの最小プロモーター(HSV TK min)である、請求項1に記載のトランスジェニック非ヒト動物。
【請求項10】
生物発光レポーターが、ルシフェラーゼ、クロラムフェニコールアセチルトランスフェラーゼ(CAT)、β−ガラクトシダーゼ、分泌型アルカリホスファターゼ(SEAP)、ヒト成長ホルモン(HGH)および緑色蛍光タンパク質(GFP)からなる群より選択される、請求項1に記載のトランスジェニック非ヒト動物。
【請求項11】
機能エレメントがヒト成長ホルモン(hGH)遺伝子である、請求項1に記載のトランスジェニック非ヒト動物。
【請求項12】
導入遺伝子が配列番号18を含む、請求項1に記載のトランスジェニック非ヒト動物。
【請求項13】
導入遺伝子が配列番号19を含む、請求項11に記載のトランスジェニック非ヒト動物。
【請求項14】
請求項1に記載のトランスジェニック非ヒト動物から単離された細胞。
【請求項15】
請求項1に記載のトランスジェニック非ヒト動物から単離された組織切片。
【請求項16】
Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物中の生物発光の量を測定する工程;
(b)トランスジェニック非ヒト動物に試験薬剤を投与する工程;
(c)試験薬剤の投与後に、1またはそれより多い時点でトランスジェニック非ヒト動物の生物発光の量を測定する工程;
(d)(a)において測定した生物発光の量と(c)において測定した生物発光の量とを比較する工程;
を含み、ここで(c)と比較した(a)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【請求項17】
Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物から組織切片を調製する工程;
(b)組織切片中の生物発光の量を測定する工程;
(c)組織切片に試験薬剤を投与する工程;
(d)試験薬剤の投与後に、1またはそれより多い時点で組織切片の生物発光の量を測定する工程;および
(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程;
を含み、ここで(d)と比較した(b)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【請求項18】
Gタンパク質共役受容体(GPCR)リガンドを同定する方法であって、以下:
(a)請求項1に記載のトランスジェニック非ヒト動物から単離された細胞を調製する工程;
(b)細胞中の生物発光の量を測定する工程;
(c)細胞に試験薬剤を投与する工程;
(d)試験薬剤の投与後に、1またはそれより多い時点で細胞の生物発光の量を測定する工程;および
(e)(b)において測定した生物発光の量と(d)において測定した生物発光の量とを比較する工程;
を含み、ここで(d)と比較した(b)の生物発光の量の差が、試験薬剤をGPCRリガンドとして同定する、上記方法。
【請求項19】
非ヒト動物におけるGPCR機能をモニタリングする方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)非ヒト動物からの生物発光をモニタリングする工程;および
(c)該生物発光をGPCR機能と相関させる工程;
を含む、上記方法。
【請求項20】
GPCR機能のモニタリングにおいて使用するための非ヒトトランスジェニック動物を作製する方法であって、以下:
(a)第1のインスレーター配列、応答配列、プロモーター、生物発光レポーター、機能エレメントおよび第2のインスレーター配列(insular element)を含む導入遺伝子を発現させるために、非ヒト動物を遺伝子導入により改変する工程;
(b)(a)のトランスジェニック非ヒト動物における生物発光の量を測定する工程;
(c)GPCRリガンドを該トランスジェニック非ヒト動物に投与する工程;
(d)GPCRリガンドの投与後に、1またはそれより多い時点でトランスジェニック非ヒト動物の生物発光の量を測定する工程;および
(e)(b)で測定した生物発光の量と(d)で測定した生物発光の量を比較する工程;を含み、ここで(d)と比較した(b)の生物発光の量の差が、GPCR機能をモニタリングするのに用いるための非ヒトトランスジェニック動物を同定する、上記方法。
【図1】

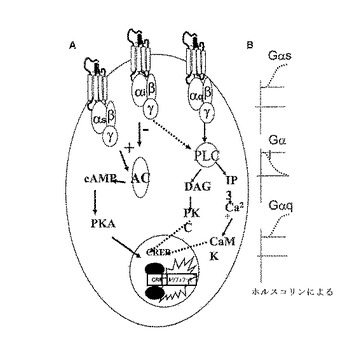
【図3】
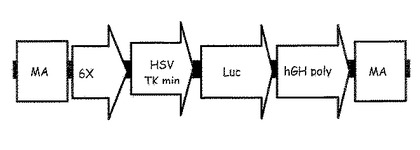
【図4】
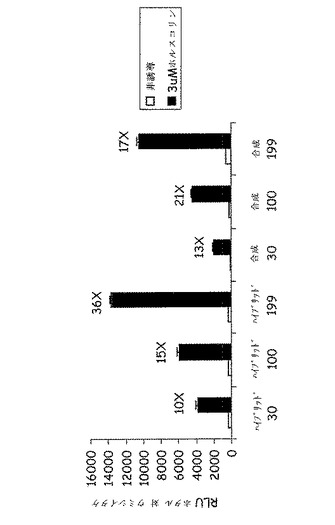
【図5A】
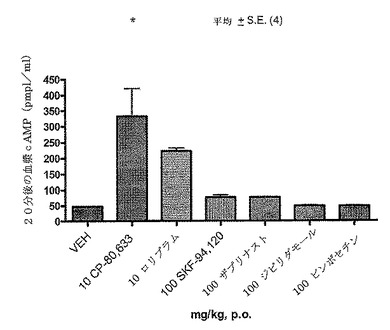
【図5B】
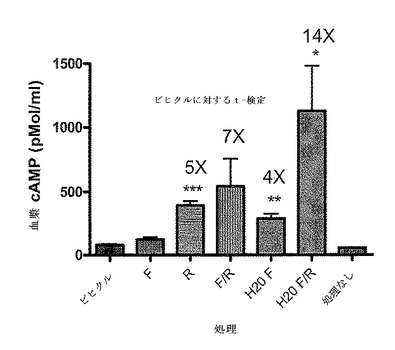
【図10】
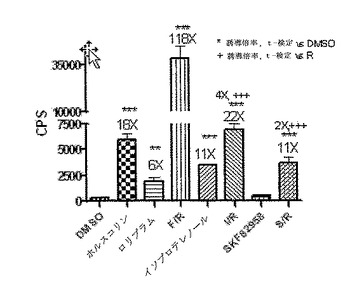
【図11】
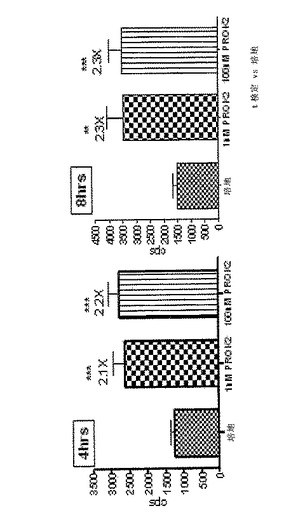
【図12】
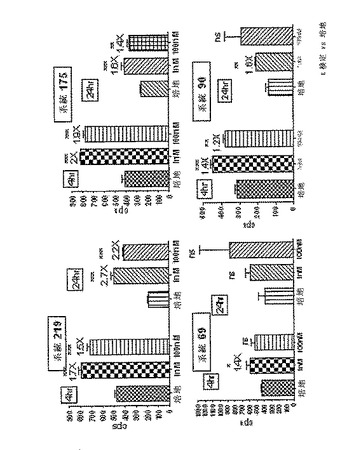
【図13A】
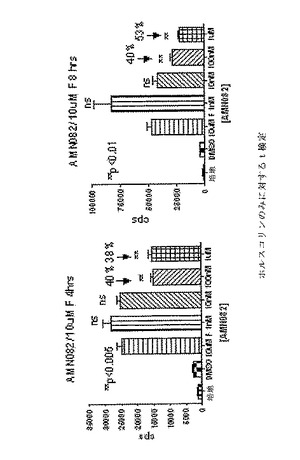
【図13B】
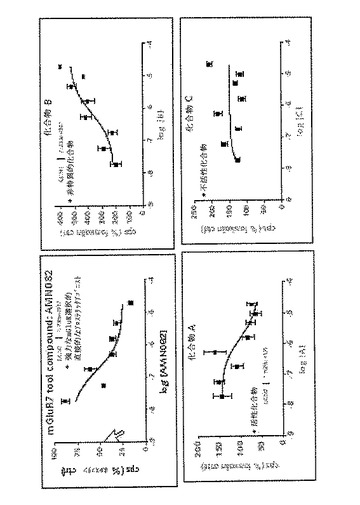
【図14】
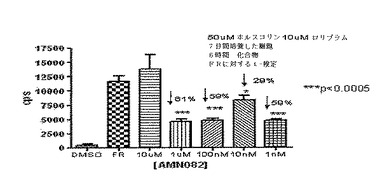
【図15A】
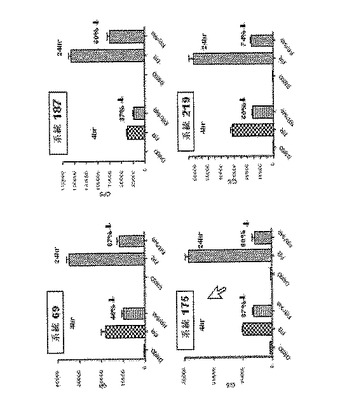
【図15B】
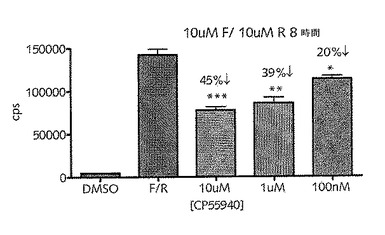
【図16】
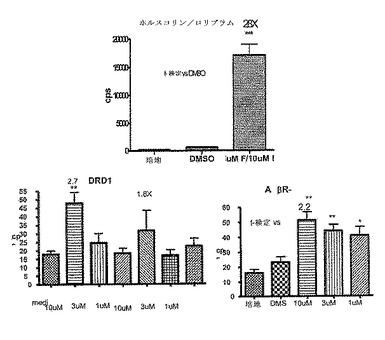
【図17】
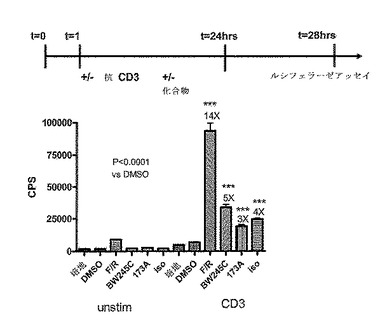
【図18】
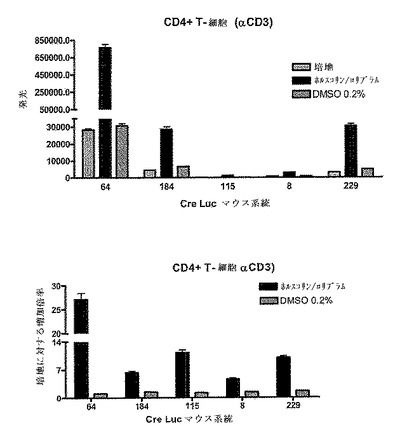
【図19】
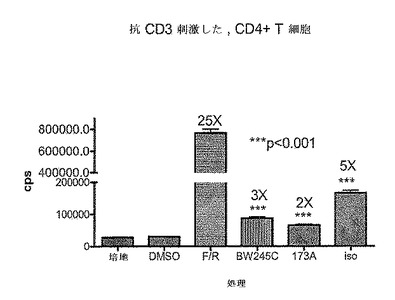
【図20】
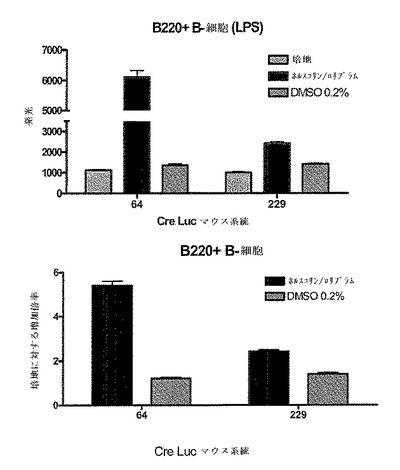
【図21】
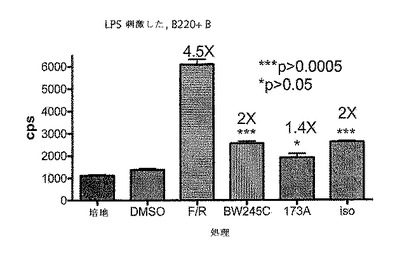
【図22】
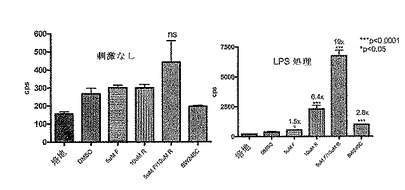
【図25】
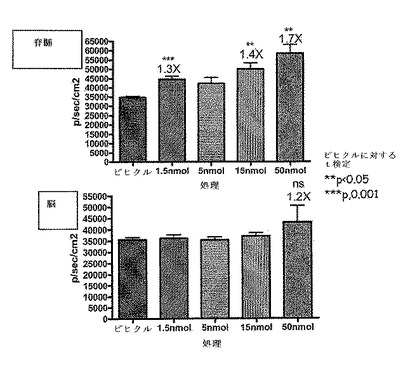
【図26】

【図31】
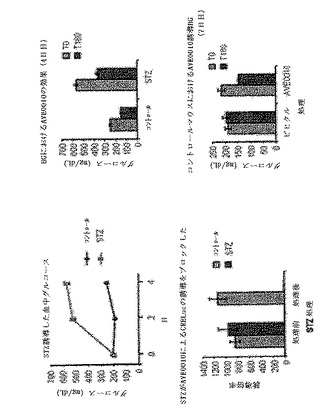
【図33】

【図34】
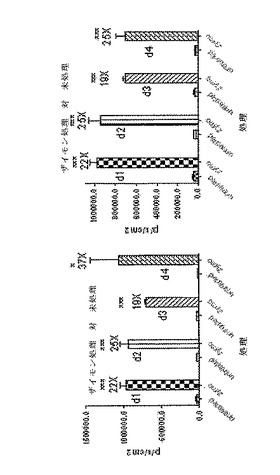
【図35】
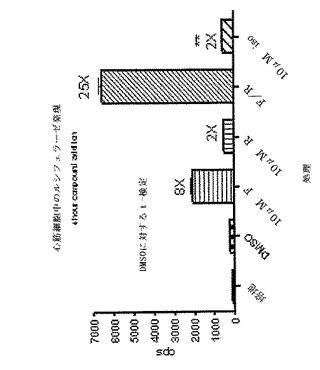
【図2A】

【図2B】
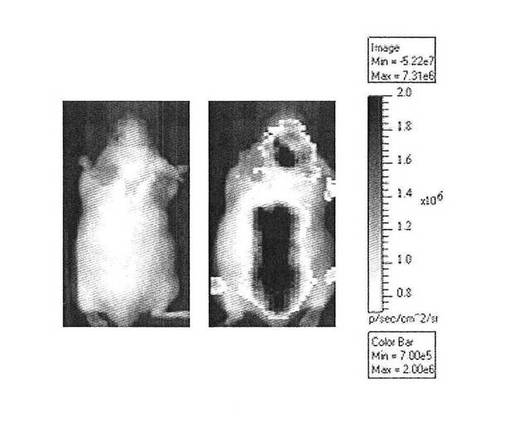
【図6】
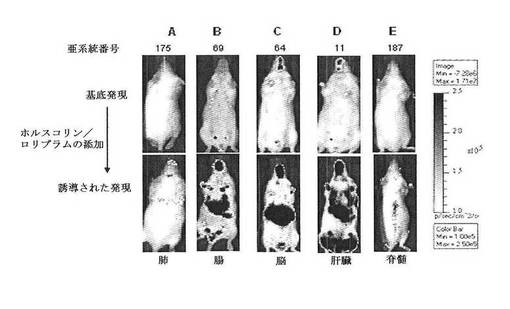
【図7】
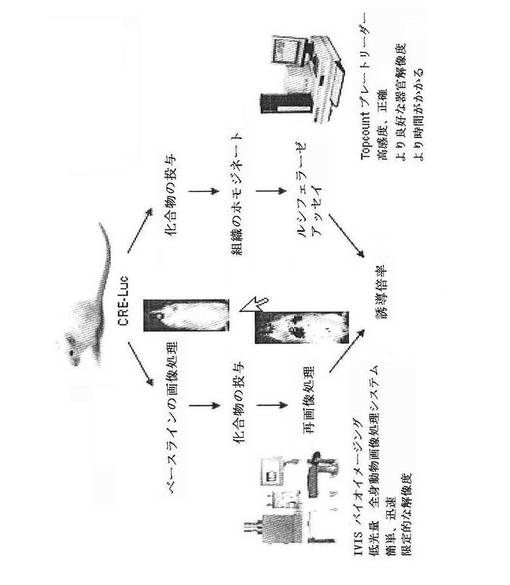
【図8A】
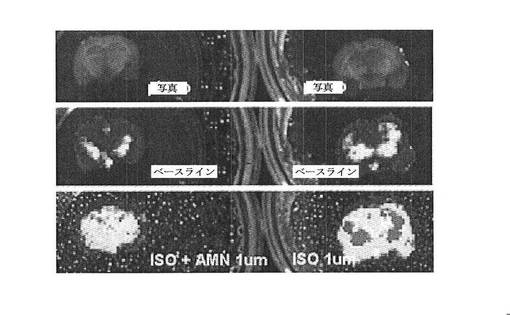
【図8B】
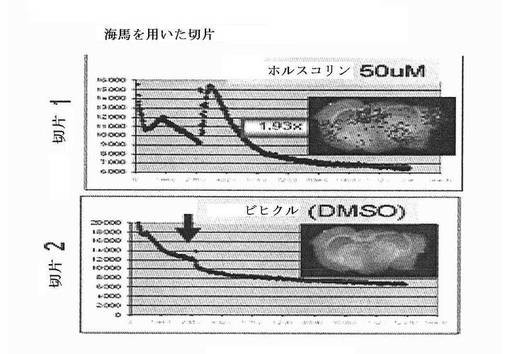
【図9】
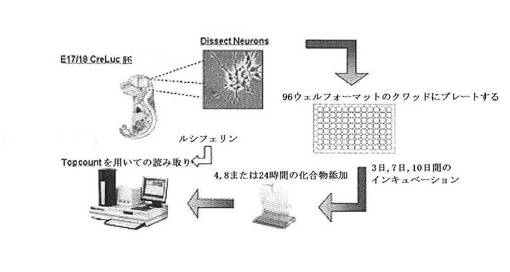
【図23】
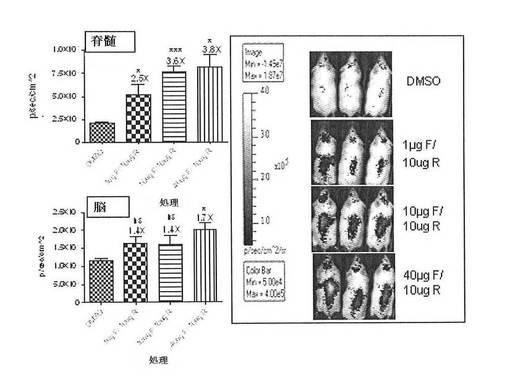
【図24】
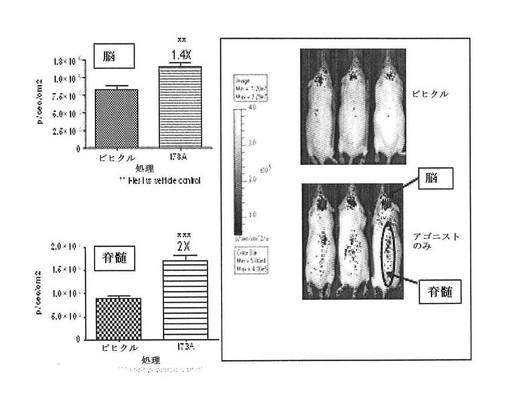
【図27A】
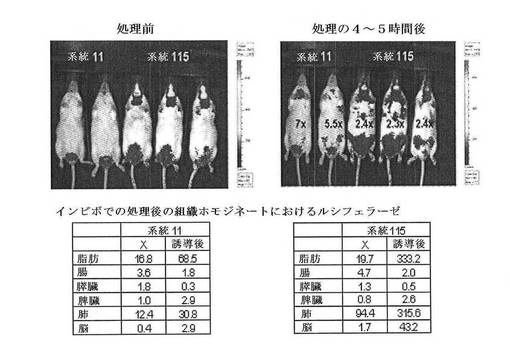
【図27B】
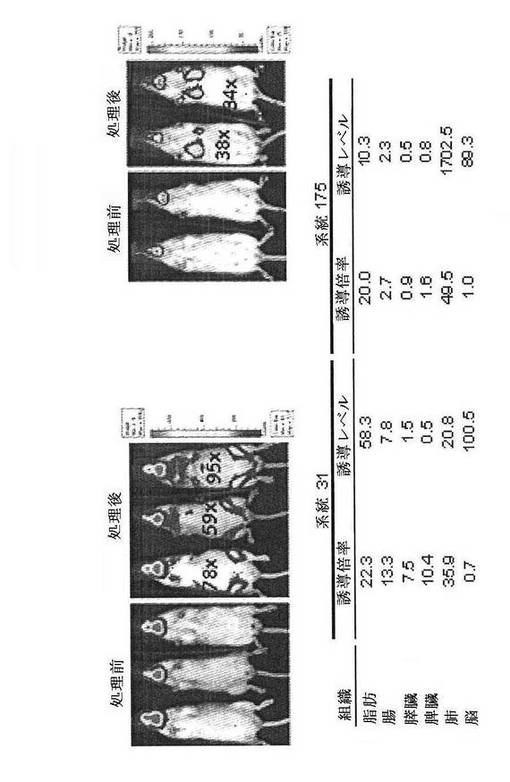
【図28】
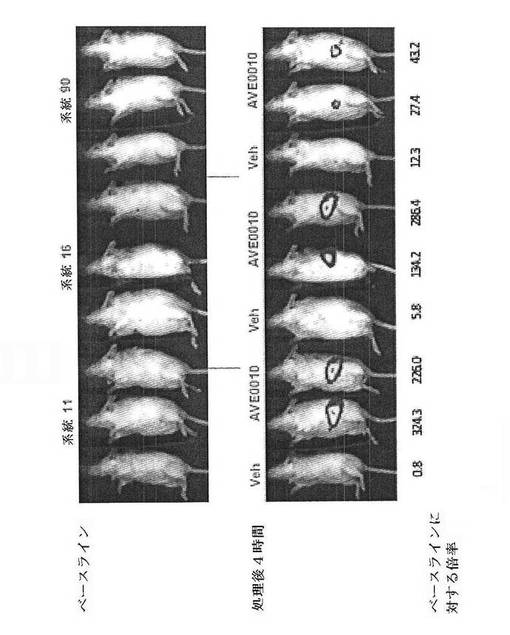
【図29】
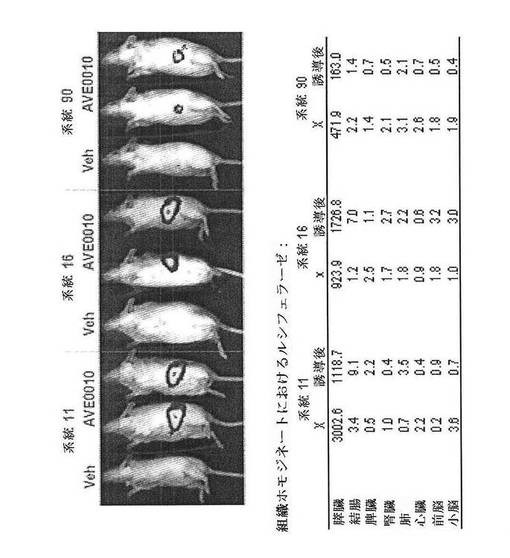
【図30】
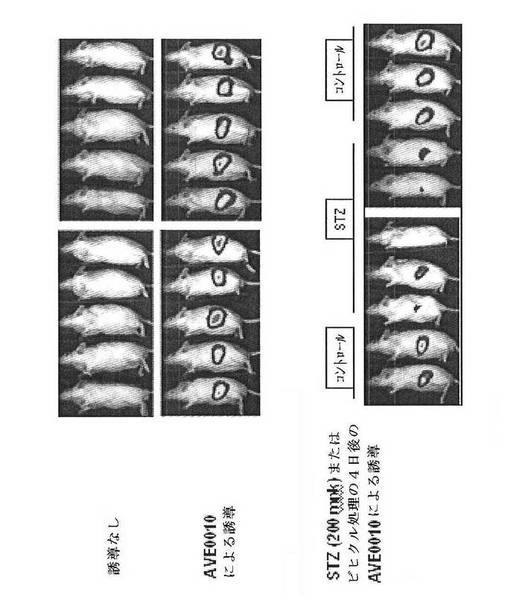
【図32】
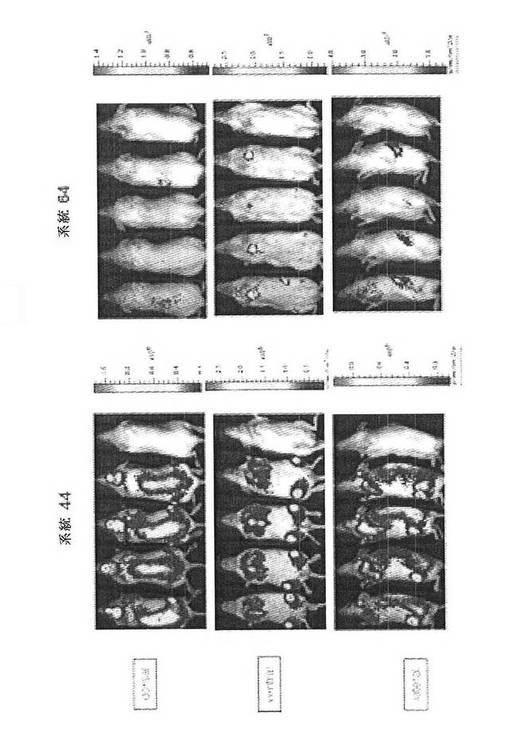
【図3】
【図4】
【図5A】
【図5B】
【図10】
【図11】
【図12】
【図13A】
【図13B】
【図14】
【図15A】
【図15B】
【図16】
【図17】
【図18】
【図19】
【図20】
【図21】
【図22】
【図25】
【図26】
【図31】
【図33】
【図34】
【図35】
【図2A】
【図2B】
【図6】
【図7】
【図8A】
【図8B】
【図9】
【図23】
【図24】
【図27A】
【図27B】
【図28】
【図29】
【図30】
【図32】
【公表番号】特表2013−514808(P2013−514808A)
【公表日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願番号】特願2012−546067(P2012−546067)
【出願日】平成22年12月17日(2010.12.17)
【国際出願番号】PCT/US2010/060909
【国際公開番号】WO2011/084659
【国際公開日】平成23年7月14日(2011.7.14)
【出願人】(504456798)サノフイ (433)
【Fターム(参考)】
【公表日】平成25年5月2日(2013.5.2)
【国際特許分類】
【出願日】平成22年12月17日(2010.12.17)
【国際出願番号】PCT/US2010/060909
【国際公開番号】WO2011/084659
【国際公開日】平成23年7月14日(2011.7.14)
【出願人】(504456798)サノフイ (433)
【Fターム(参考)】
[ Back to top ]