
トリヌクレオチド反復疾患を治療するための方法および組成物
トリヌクレオチド反復疾患を治療するための方法および組成物を本明細書に開示する。
【発明の詳細な説明】
【技術分野】
【0001】
関連出願の相互参照
本出願は、2009年7月28日に出願された米国仮出願第61/271,913号、および2009年7月29日に出願された同第61/273,009号の利益を主張し、これらの開示は、参照によりその全体が本明細書に組み込まれる。
【0002】
連邦支援による研究下でなされた発明に対する権利の記載
該当せず。
【0003】
技術分野
本開示は、遺伝子発現の分野にある。
【背景技術】
【0004】
背景技術
トリヌクレオチド反復伸長疾患は、1990年初期に初めて特徴付けられた(Di Prospero and Fischbeck,(2005)Nature Reviews Genetics vol 6:756−765を参照のこと)。これらの疾患は、3個のヌクレオチドのセットの不安定な反復の局所的伸長を有し、この反復が存在する遺伝子の機能喪失、毒性機能の増加、または両方を引き起こし得る。トリヌクレオチド反復は、非コードおよびコード遺伝子領域を含む、遺伝子のあらゆる部分に位置し得る。コード領域内に位置する反復には、一般的に、トリプレットをコードする反復したグルタミン(CAG)またはトリプレットをコードするアラニン(CGA)のいずれかが含まれる。非コード配列内の長く伸びる反復領域は、遺伝子の異常な発現を引き起こし得る一方で、コード領域内の長く伸びる反復(コドン反復疾患としても知られている)は、ミスフォールディングおよびタンパク質凝集を引き起こし得る。異常なタンパク質に関連する病態生理学的な正確な原因は、多くの場合、知られていない。一般的に、トリヌクレオチド伸長を起こしやすい野生型遺伝子において、これらの領域は、正常な集団において可変数の反復配列を含むが、罹患集団において、反復の数は、場合によっては、反復の数の単純な2倍から、反復の数の対数で1桁まで増加し得る。例えば、脆弱X症候群患者におけるCGG伸長を受けるFMR1遺伝子において、野生型集団は、2〜50の反復を示す一方で、脆弱X症候群に罹患しているこれらの患者は、200〜2000のCGG反復を有し得る(Nadel et al(1995)Journal Biological Chemistry 270(48):28970−28977)。
【0005】
今日まで、20種類の異なる疾患が、長く伸びるトリヌクレオチド反復に関連付けられている(Di ProsperoおよびFischbeckの同書を参照のこと)。この現象は最初に、アンドロゲン受容体のコード領域においてCAG反復が長く伸びている球脊髄性筋萎縮症(SBMA)に関して記載された。野生型遺伝子の反復は、通常は13〜30のCAGを含むが、SBMA患者は、40以上ものCAGを含み得る。長く伸びるトリヌクレオチド反復によって特徴付けられる他の疾患には、フリードライヒ運動失調症(反復はFratazin遺伝子の非コード領域内にある)、脆弱X症候群AおよびE(反復はそれぞれ、FMR1およびFMR2遺伝子の非コード領域内にある)、および反復が巨大サイトゾルタンパク質のハンチンチン(Htt)のN末端コード領域内に挿入されるハンチントン病が含まれる。それぞれのポリグルタミン伸長疾患は、脳の特定領域に神経細胞脱落が明らかである特徴的病変を示す。P/Q Ca2+チャンネル、TATAボックス結合タンパク質、およびアトロフィン−1におけるポリグルタミン伸長はそれぞれ、脊髄小脳失調(SCA)−6、SCA−17、および歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)を引き起こす。それらのポリグルタミン反復は別として、これらの疾患に関与するタンパク質は全て、中枢神経系および末梢組織の両方において広範に発現するが、関連性はない。
【0006】
ハンチントン舞踏病としても知られているハンチントン病(HD)は、運動、認識、および精神疾患の進行性疾患である。この疾患の発症の平均年齢は、35〜44歳であるが、症例の約10%においては、発症が21歳前に生じ、疾患診断後の平均寿命は、15〜18年である。有病率は、西ヨーロッパ系の100,000人の間で約3〜7人である。通常のHtt対立遺伝子は、15〜20のCAG反復を含むが、35以上の反復を含む対立遺伝子は、潜在的にHDを生じる対立遺伝子であると見なすことができ、疾患を発症する危険性を与える。36〜39の反復を含む対立遺伝子は、不完全に浸透されると見なされ、これらの対立遺伝子を含むこれらの個人は、疾患を発症し得るか発症し得ない(または後年症状を発症し得る)一方で、40以上の反復を含む対立遺伝子は、完全に浸透されると見なされ、これだけ多くの反復を有するHD対立遺伝子を含む無症候性の人は報告されていない。若年発症HDを有するこれらの個人(21歳を下回る)は、しばしば、60以上のCAG反復を有することが見出される。CAG反復の増加に加えて、HDが、この領域が、ポリグルタミンよりもむしろポリセリンポリペプチド(+1フレームシフトの場合に、AGC反復によってコードされる)経路をコードするように、反復配列内で+1および+2フレームシフトを含むことができることも示されている(Davies and Rubinsztein(2006)Journal of Medical Genetics 43:893−896)。
【0007】
ハンチントン病は、HD対立遺伝子が、通常、優勢な特性として1人の親から遺伝子を受け継ぐ遺伝的疾患である。HD患者から生まれるどの子も、もう一方の親がこの疾患に罹患していない場合、疾患を発症する可能性が50%である。場合によっては、親は、中間HD対立遺伝子を有し、無症候性であり得る一方で、伸長を反復するため、その子に疾患が現れる。加えて、HD対立遺伝子はまた、見込みとして知られる現象を示すこともでき、重症度の増加または発症年齢の低下が、精子形成中の反復領域の不安定な性質のため、数世代にわたって観察される。
【0008】
HDにおいて、トリヌクレオチド伸長は、線条体中で中型有棘ガンマアミノ酪酸(GABA)投射神経細胞の神経細胞の欠損を引き起こし、神経細胞の欠損は新皮質においても生じる。エンケファリンを含み、外部淡蒼球へ突き出る中型有棘神経細胞は、基質Pを含み、内部淡蒼球へ突き出る神経細胞よりも病変する。ハンチントン病に罹患している人々に大いに影響を及ぼす他の脳の部位には、黒質、皮質3、5、および6、海馬のCA1領域、頭頂葉の各回、小脳のプルキンエ細胞、視床下部の外側隆起核、および視床の正中中心核−束傍核複合体が含まれる(Walker(2007)Lancet 369:218−228)。正常なHttタンパク質の役割は、不明のところが多いが、神経新生、アポトーシス細胞死、および小胞輸送に関与し得る。加えて、野生型Httは、脳由来神経栄養因子(BDNF)、および線条体ニューロンの生存促進性因子を刺激することを証明する。HDの進行は、HDのマウスモデルにおけるBDNF発現の減少と相関する(Zuccato et al(2005)Pharmacological Research 52(2):133−139)、およびアデノ随伴ウイルス(AAV)ベクター媒介遺伝子によってBDNFあるいはグリア細胞株由来神経栄養因子(GDNF)のいずれかの送達が、HDのマウスモデルにおいて、線条体ニューロンを保護し得る(Kells et al,(2004)Molecular Therapy 9(5):682−688)ことが示されている。
【0009】
HDに対する治療法の選択肢は、現在、非常に限られている。シャペロニンの過剰発現または化合物ゲルダナマイシンによる熱ショック応答の産生等の長く伸びるポリグルタミン経路を通して生じるタンパク質凝集に関連する毒性を防止するように設計されるいくつかの潜在的な手法は、インビトロモデルにおけるこれらの毒性の減少を示している。他の治療は、疾患の臨床的指標においてアポトーシスの役割を標的としている。例えば、一方の親は、HD対立遺伝子を含み、もう一方の親が、カスパーゼ1に対して優性阻害対立遺伝子を有するマウスの対合の子孫において、動物モデルのカスパーゼ活性の妨害によって、病徴の遅延が示される。さらに、カスパーゼによるHD Httの切断は、疾患の病原性における役割を果たし得る。カスパーゼ6の耐性変異Httを担持するトランスジェニックマウスは、正常な神経機能を維持することが見出され、非カスパーゼの耐性変異Htt対立遺伝子を担持するマウスと比較して、線条体神経変性を引き起こさなかった(Graham et al(2006)Cell 125:1179−1191を参照されたい)。また、アポトーシス経路のメンバーを標的とする分子は、症候学において遅延する影響があることも示している。例えば、化合物zVAD−fmkおよびミノサイクリン(これらの両者が、カスパーゼ活性を阻害する)は、マウスにおいて、疾患の兆候を遅延させることが示されている。また、薬物レマセミド(remacemide)は、この化合物が、神経細胞における毒作用の発揮を妨げるNDMA受容体への変異Httの結合を妨げることが考えられるため、小規模なHDヒト試験にも使用されている。しかしながら、統計的に有意な改善は、これらの試験におけるニューロン機能において、観察されなかった。加えて、ハンチントン研究グループは、コエンザイムQを用いて、ランダム化二重盲検試験を行った。コエンザイムQ10により処置された患者においてより遅延した疾患進行に関する傾向があったが、全体の機能的能力の下落率に有意な変化がなかった。(Di ProsperoおよびFischbeckの同書を参照のこと)。
【0010】
したがって、トリヌクレオチド反復疾患の治療のための組成物および方法が求められている。
【発明の概要】
【0011】
概要
トリヌクレオチド反復疾患を治療するための方法および組成物を本明細書に開示する。特に、トリヌクレオチド反復疾患を治療するために、トリヌクレオチド反復を含む遺伝子の発現を調節する、例えば、ハンチントン病を治療するためにHD Htt対立遺伝子の発現を調節するための方法および組成物が、本明細書に提供される。また、トリヌクレオチド反復疾患の動物モデルを生成するための方法および組成物も提供される。
【0012】
したがって、一態様において、HD対立遺伝子(例えば、Htt)の発現を調節する改変された亜鉛フィンガータンパク質が提供される。改変された亜鉛フィンガータンパク質とは、認識ヘリックスが事前選択された標的部位に結合するように変化された、天然に存在しない亜鉛フィンガータンパク質である。本明細書に記載される亜鉛フィンガータンパク質のいずれも、1個、2個、3個、4個、5個、6個以上の亜鉛フィンガーを含むことができ、それぞれの亜鉛フィンガーは、選択された配列(例えば、遺伝子)内の標的亜部位に結合する認識へリックスを有する。いくつかの実施形態において、該認識へリックスは、天然に存在しない。特定の実施形態において、亜鉛フィンガータンパク質は、表1に示される認識へリックスを有する。他の実施形態において、亜鉛フィンガータンパク質は、表2に示される標的配列に結合する。
【0013】
一態様において、長く伸びるCAG経路に優先的に結合することが可能であるが、野生型長さのCAG経路に対する親和性の低い、両端型ZFPリプレッサーが提供される。いくつかの実施形態において、長く伸びるトリヌクレオチド経路に優先的に結合するが、野生型長さのトリヌクレオチド反復経路に対する親和性の低い多量体型ZFP−TFが使用され、それによって、長く伸びる対立遺伝子の優先的な抑制が達成される。いくつかの実施形態において、これらの多量体型ZFP−TFは、長く伸びる対立遺伝子が、野生型対立遺伝子よりも多くのZFPによってさらに効率的に結合されるように、反復配列に対して協同的DNA結合を達成し、変異対立遺伝子の優先的な抑制を可能にする。いくつかの実施形態において、所与のサイズの多量体の安定した複合体を形成し、ひいては、ある最小サイズのCAG経路と優先的に相互作用することが可能である、多量体型ZFP TFが、使用され、その最小サイズは、野生型CAG経路の長さよりも長い。
【0014】
特定の実施形態において、本明細書に記載されるZFP(例えば、両端型、多量体型等)は、変異Htt対立遺伝子の発現を優先的に修飾する。いくつかの実施形態において、ZFPは、変異Htt対立遺伝子に特異的に結合し、長く伸びる経路は、ポリグルタミンをコードする一方、他の実施形態において、ZFPは、変異Htt対立遺伝子に特異的に結合し、長く伸びる経路は、ポリセリンをコードする。したがって、いくつかの実施形態において、ZFP−TFは、野生型および変異型のHtt対立遺伝子の両方を調節する。特定の実施形態において、ZFPは、野生型Htt対立遺伝子のみ調節する。他の実施形態において、ZFPは、変異型のHttのみ調節する。
【0015】
他の実施形態において、長く伸びるHDのHtt対立遺伝子に関連する既知のSNPに優先的に結合するZFP−TFの抑制が提供される。このように、ZFP−TFは、SNPを含む変異Htt対立遺伝子対して特異的であり、変異Htt対立遺伝子の特定の抑制を可能にする。別の態様において、野生型対立遺伝子に関連するSNPと相互作用することによって、野生型Htt対立遺伝子を特異的に活性化するZFP−TFが提供される。このように、野生型Htt対立遺伝子のみが活性化される。
【0016】
別の態様において、1つ以上の神経栄養因子の発現を調節する改変された亜鉛フィンガータンパク質は、トリヌクレオチド疾患を治療するために提供される。いくつかの実施形態において、調節される神経栄養因子は、BDNFおよび/またはGDNFである。改変された亜鉛フィンガータンパク質とは、認識ヘリックスが事前選択された標的部位に結合するように変化された、天然に存在しない亜鉛フィンガータンパク質である。本明細書に記載される亜鉛フィンガータンパク質のいずれも、1個、2個、3個、4個、5個、6個以上の亜鉛フィンガーを含むことができ、それぞれの亜鉛フィンガーは、選択された配列(例えば、遺伝子)内の標的亜部位に結合する認識へリックスを有する。いくつかの実施形態において、該認識へリックスは、天然に存在しない。
【0017】
特定の実施形態において、本明細書に記載される亜鉛フィンガータンパク質(ZFP)を、融合タンパク質の一部として、調節ドメイン(または機能ドメイン)と作動的連結状態に配置することができる。ZFPとの融合のために活性化ドメインまたは抑制ドメインのいずれか一方を選択することによって、そのような融合タンパク質は、遺伝子発現を活性化するか、または抑制するかのどちらか一方のために使用することができる。いくつかの実施形態において、変異Htt発現を下方制御するために使用され得る転写抑制ドメインに融合された、本明細書に記載される変異Httを標的化するZFPを含む融合タンパク質が提供される。いくつかの実施形態において、野生型Htt対立遺伝子を上方制御することができる転写活性化ドメインに融合された、野生型Htt対立遺伝子を標的化するZFPを含む融合タンパク質が提供される。特定の実施形態において、調節ドメインの活性は、細胞転写機構との相互作用が、外来リガンドの不在下で起こらないように、外来小分子またはリガンドによって調節される。このような外部リガンドは、ZFP−TFの転写機構との相互作用の程度を制御する。調節ドメインは、1つ以上のZFPの間、1つ以上のZFPの外部、およびこれらの任意の組み合わせを含む、1つ以上のZFPの任意の部分に作動的に連結され得る。
【0018】
いくつかの実施形態において、本明細書に記載される改変された亜鉛フィンガータンパク質を、融合タンパク質の一部として、ヌクレアーゼ(開裂)ドメインと作動的連結状態に配置することができる。特定の実施形態において、このようなヌクレアーゼ融合は、誘導多能性幹細胞(iPSC)、ヒト胚幹細胞(hES)、間充織幹細胞(MSC)、または神経幹細胞等の幹細胞中で変異Htt対立遺伝子を標的化するために使用され得、ヌクレアーゼ融合の活性は、多くの野生型のCAG反復を含有するHtt対立遺伝子を引き起こす。特定の実施形態において、修飾された幹細胞を含む薬学的組成物が提供される。
【0019】
さらに別の態様において、本明細書に記載される亜鉛フィンガータンパク質のいずれかをコードするポリヌクレオチドが提供される。このようなポリヌクレオチドを、トリヌクレオチド反復疾患を治療することが望ましい対象に投与することができる。
【0020】
なおさらなる態様において、本発明は、ハンチントン病等のトリヌクレオチド反復疾患の研究のための特定のモデル系の生成のための方法および組成物を提供する。特定の実施形態において、変異Htt対立遺伝子が、細胞の生成のために胚幹細胞中に生成されるモデルと、特定の長さのトリヌクレオチド伸長経路(例えば、50、80、109、および180のCAG反復)が、亜鉛フィンガーヌクレアーゼ(ZFN)誘導標的化組込みを用いて、野生型Htt対立遺伝子に挿入される動物株と、が提供される。特定の実施形態において、モデル系は、インビトロ細胞株を含む一方で、他の実施形態において、モデル系は、トランスジェニック動物を含む。
【0021】
さらに別の態様において、本明細書に記載されるポリヌクレオチドのいずれかを含む遺伝子送達ベクターが提供される。特定の実施形態において、ベクターは、アデノウイルスベクター(例えば、Ad5/F35ベクター)、組込み能力があるもしくは組込み機能を欠損したレンチウイルスベクターを含むレンチウイルスベクター(LV)、またはアデノウイルス随伴ウイルスベクター(AAV)である。したがって、少なくとも1つの亜鉛フィンガーヌクレアーゼ(ZFN)をコードする配列および/または標的遺伝子への標的化組込みのためのドナー配列を含む、アデノウイルス(Ad)ベクター、LV、またはアデノウイルス随伴ウイルスベクター(AAV)も本明細書に提供される。特定の実施形態において、Adベクターは、キメラAdベクター、例えば、Ad5/F35ベクターである。特定の実施形態において、レンチウイルスベクターは、インテグラーゼ欠損型レンチウイルスベクター(IDLV)または組込み能力のあるレンチウイルスベクターである。特定の実施形態において、ベクターは、VSV−Gエンベロープまたは他のエンベロープを有する擬似タイプである。
【0022】
いくつかの実施形態において、モデル系は、標的対立遺伝子(例えば、変異Htt)は、発現マーカーで標識化されるトリヌクレオチド反復疾患(例えば、ハンチントン病)を提供する。特定の実施形態において、変異対立遺伝子(例えば、変異Htt)が標識化される。いくつかの実施形態において、野生型対立遺伝子(例えば、野生型Htt)が標識化され、さらなる実施形態において、野生型および変異対立遺伝子の両方は、別個の発現マーカーで標識化される。特定の実施形態において、モデル系は、インビトロ細胞株を含む一方、他の実施形態において、モデル系は、トランスジェニック動物を含む。
【0023】
さらに、核酸および/またはZFP(もしくはZFPを含む融合タンパク質)を含有する薬学的組成物も提供される。例えば、ある特定の組成物は、調節配列と作動可能に連結され、薬学的に許容される担体または希釈剤と併用される、本明細書に記載されるZFPの1つをコードする配列を備える核酸を含み、調節配列は、細胞内で核酸の発現を可能にする。特定の実施形態において、コードされるZFPは、HD Htt対立遺伝子に対して特異的である。いくつかの実施形態において、薬学的組成物は、HD Htt対立遺伝子を調節するZFPおよび神経栄養因子を調節するZFPを含む。タンパク質に基づく組成物には、本明細書に開示される1つ以上のZFPおよび薬学的に許容される担体または希釈剤が含まれる。
【0024】
さらに別の態様において、本明細書に記載されるタンパク質、ポリヌクレオチド、および/または組成物のいずれかを含む単離された細胞も提供される。
【0025】
別の態様において、本明細書に開示される組成物を用いて、トリヌクレオチド反復疾患を治療するおよび/または予防するための方法が本明細書に提供される。特定の実施形態において、方法には、ハンチントン病の治療が含まれる。いくつかの実施形態において、方法には、ウイルスベクター、非ウイルスベクター(例えば、プラスミド)、および/またはその組み合わせを使用して、ポリヌクレオチドおよび/またはタンパク質を送達することができる組成物が含まれる。いくつかの実施形態において、方法には、ZFPを含む幹細胞集団を含む、または本発明のZFNを変化させた、組成物が含まれる。
【0026】
これらおよび他の態様は、全体の開示を考慮して、当業者には容易に明らかである。
【図面の簡単な説明】
【0027】
【図1】パネルAからDは、ハンチンチン(Htt)対立遺伝子および対立遺伝子に結合する様々なZFP−TFを示す概略図である。図1Aは、野生型Htt対立遺伝子を示す。図1Bは、HD Htt対立遺伝子に結合する単一のZFP−TFを示し、ここで「KRAB」とは、KOX1遺伝子からのKRAB抑制ドメインを指し、「ZFP」とは、亜鉛フィンガー結合タンパク質を指す。「標準ZFP TF」は、亜鉛フィンガー結合ドメインが、KRAB抑制ドメインに連結されるZFP転写因子融合タンパク質である。図1Cは、ZFP転写因子である「両端型ZFP TF」を示し、ここで、亜鉛フィンガードメインの2つのクラスターは、剛体タンパク質配列によって分離される。機能ドメインを、本図中の1つのZFPの外部に示すが、機能ドメインが、ZFPの間またはZFPの外部にあり得ることは明らかである。図1Dは、「多量体型ZFP TF」を示し、これは、多量体化ドメインを通して多量体化することが可能なZFP TFである(小さな斑点のあるボックスとして示される)。また、機能ドメイン(例えば、活性化、抑制、開裂ドメイン)を黒菱形のあるボックスとして図1Cおよび1Dに示す。
【図2】ヒト293T細胞(表1中に記載される)における標準ZFP TFによるHttの抑制を示す。GFPは、GFPに対して特異的なZFP TFを示し、偽は、偽トランスフェクトされている細胞中の発現を示す。
【図3】パネルA〜Cは、例示的な多量体型ZFP−TFを示す概略図である。図3Aは、機能ドメイン(抑制ドメイン)、および多量体化のための二量体化ドメインに融合される単一のZFPを示す。図3Bは、上端線に示されるように、2つのZFPの多量体の結合を示す。図3Cは、上端線に示されるように、4つのZFPの多量体を示す。任意の数の多量体を使用することができ、機能ドメインは、1つ以上の個々のZFPにおいて、どこでも位置付けられ得ることは明らかである。
【図4】パネルA〜Cは、例示的な両端型ZFP−TFを示す概略図である。図4Aは、2つのZFP間に位置付けられる機能ドメインを含む、両端型ZFPを示す。図1D中に示されるように、機能ドメインは、ZFPのうちの1つの外部であり得る。図4Bは、標的部位への両端型ZFPの結合を示す。ZFPの一方または両方は、それらの標的部位に結合する。図4Cは、複数の両端型ZFPの結合を示す。
【図5】Htt特異的ZFN対である25920/25921あるいは25922/25923のいずれかによるK562細胞の処理を行った後のCel−Iミスマッチアッセイ(Surveyor,Transgenomics)の結果を示すゲルを示す。NHEJ活性の割合をそれぞれのレーンの下に示す。「GFP」は、プラスミドをコードするGFPでトランスフェクトされている細胞を示す。示されるデータは、プラスミドを含有するZFNでトランスフェクトしてから14日後に抽出したDNAからのものである。
【発明を実施するための形態】
【0028】
詳細な説明
ハンチントン病、SBMA、フリードライヒ運動失調症、ならびに脆弱X症候群AおよびE等のトリヌクレオチド反復疾患を治療するための組成物および方法を本明細書に開示する。特に、亜鉛フィンガータンパク質(ZFP TF)を含むHttを調節する転写因子およびこのようなタンパク質を利用する方法が、ハンチントン病を治療で用いるために提供される。例えば、変異Htt対立遺伝子の発現を抑制する、または野生型Htt対立遺伝子の発現を活性化するZFP−TFを提供する。加えて、これらの疾患に関連する遺伝子のゲノム構造を修飾する亜鉛フィンガーヌクレアーゼ(ZFN)を提供する。例えば、変異型のHttの部分を特異的に変化させることができるZFNを提供する。これらには、改変された亜鉛フィンガータンパク質、すなわち、所定の核酸標的配列に結合する天然に存在しないタンパク質を用いる組成物および方法が含まれる。
【0029】
したがって、本明細書に記載される方法および組成物は、トリヌクレオチド反復疾患の治療のための方法を提供し、これらの方法および組成物は、標的遺伝子および改変された亜鉛フィンガーヌクレアーゼを調節することができる亜鉛フィンガー転写因子を含むことができる。
【0030】
概論
本明細書に開示される方法の実践、ならびに組成物の調製および使用は、特に指示がない限り、分子生物学、生化学、クロマチン構造および分析、計算化学、細胞培養、組み換えDNA、および関連分野における従来の技術は、当技術分野の技術範囲内である。これらの技術は、文献に十分に説明されている。例えば、Sambrook et al.MOLECULAR CLONING:A LABORATORY MANUAL,Second edition,Cold Spring Harbor Laboratory Press,1989およびThird edition,2001、Ausubel et al.,CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,John Wiley & Sons,New York,1987および定期的に更新されたもの;METHODS IN ENZYMOLOGY,Academic Press,San Diegoシリーズ;Wolffe,CHROMATIN STRUCTURE AND FUNCTION,Third edition,Academic Press,San Diego,1998;METHODS IN ENZYMOLOGY,Vol.304,“Chromatin”(P.M.Wassarman and A.P.Wolffe,eds.),Academic Press,San Diego,1999;ならびにMETHODS IN MOLECULAR BIOLOGY,Vol.119,“Chromatin Protocols”(P.B.Becker,ed.)Humana Press,Totowa,1999を参照のこと。
【0031】
定義
「核酸」、「ポリヌクレオチド」、および「オリゴヌクレオチド」という用語は、交換可能に使用され、線状または環状構造で、一本鎖または二本鎖形態の、デオキシリボヌクレオチドポリマーまたはリボヌクレオチドポリマーを指す。本開示については、これらの用語をポリマーの長さに関する限定と解釈すべきではない。これらの用語は、天然ヌクレオチドの公知の類似体、ならびに塩基、糖、および/またはリン酸(ホスホロチオエート主鎖)部分内で修飾されるヌクレオチドを包含することができる。一般に、特定のヌクレオチドの類似体は、同じ塩基対形成特異性を有し、すなわち、Aの類似体は、Tと塩基対を形成する。
【0032】
「ポリペプチド」、「ペプチド」、および「タンパク質」という用語は、アミノ酸残基のポリマーを指すために交換可能に使用される。この用語は、1つ以上のアミノ酸が、対応する天然に存在するアミノ酸の化学的類似体または修飾誘導体である、アミノ酸ポリマーにも適用される。
【0033】
「結合」は、巨大分子間(例えば、タンパク質と核酸との間)の配列特異的で非共有結合的な相互作用を指す。結合相互作用の全ての構成要素は、相互作用が全体として配列特異的である限り、配列特異的である(例えば、DNA主鎖中のリン酸残基との接触する)必要はない。このような相互作用は、一般に、10-6M-1以下の解離定数(Kd)によって特徴付けられる。「親和性」は、結合の強度を指す。高い結合親和性は、低いKdに関連している。
【0034】
「結合タンパク質」は、別の分子に非共有結合的に結合することができるタンパク質である。結合タンパク質は、例えば、DNA分子(DNA結合タンパク質)、RNA分子(RNA結合タンパク質)、および/またはタンパク質分子(タンパク質結合タンパク質)に結合することができる。タンパク質結合タンパク質の場合においては、それは、それ自体に結合する(その結果、ホモ二量体、ホモ三量体等を形成する)ことができる、および/または異なる1つのタンパク質または複数のタンパク質の1つ以上の分子に結合することができる。結合タンパク質は、2種類以上の結合活性を有することができる。例えば、亜鉛フィンガータンパク質は、DNA結合、RNA結合、およびタンパク質結合活性を有する。
【0035】
「亜鉛フィンガーDNA結合タンパク質」(または結合ドメイン)は、構造が亜鉛イオンの配位によって安定化される結合ドメイン内のアミノ配列の領域である、1つ以上の亜鉛フィンガーによって配列特異的な様式でDNAに結合する、タンパク質またはより大きなタンパク質内のドメインである。亜鉛フィンガーDNA結合タンパク質という用語は、しばしば、亜鉛フィンガータンパク質またはZFPとして略記される。
【0036】
亜鉛フィンガー結合ドメインは、例えば、天然に存在する亜鉛フィンガータンパク質の認識へリックス領域の改変(1つ以上のアミノ酸を変化させる)を介して、所定のヌクレオチド配列に結合されるように「改変」することができる。したがって、改変された亜鉛フィンガータンパク質は、天然に存在しないタンパク質である。亜鉛フィンガータンパク質を改変するための方法の非制限的な例は、設計および選択である。設計された亜鉛フィンガータンパク質は、天然に存在しないタンパク質であり、その設計/組成物は、主として合理的基準によって得られる。設計のための合理的基準には、置換規則の適用、ならびに既存のZFP設計および結合データの情報を格納したデータベース中の情報を処理するためのコンピュータアルゴリズムの適用が含まれる。例えば、米国特許第6,140,081号、同第6,453,242号、および同第6,534,261号明細書を参照されたく、国際公開第98/53058号、国際公開第98/53059号、国際公開第98/53060号、国際公開第02/016536号、および国際公開第03/016496号も参照されたい。
【0037】
「選択された」亜鉛フィンガータンパク質は、天然には見出されないタンパク質であり、その産生は、ファージディスプレイ、相互作用トラップ、またはハイブリッド選択等の実験的プロセスによって主に得られる。例えば、米国第5,789,538号、米国特許第5,925,523号、米国特許第6,007,988号、米国特許第6,013,453号、米国特許第6,200,759号明細書、国際公開第95/19431号、国際公開第96/06166号、国際公開第98/53057号、国際公開第98/54311号、国際公開第00/27878号、国際公開第01/60970号、国際公開第01/88197号、および国際公開第02/099084号を参照されたい。
【0038】
「組み換え」とは、2つのポリヌクレオチド間での遺伝子情報の交換のプロセスを指す。本開示に関して、「相同組み換え(HR)」とは、例えば、相同性指向性修復機構を介した細胞中の二本鎖切断の修復中に行われる、このような交換の特殊形態を指す。このプロセスは、ドナーから標的への遺伝子情報の転送につながるため、ヌクレオチド配列相同性を必要とし、「標的」分子(すなわち、二本鎖切断を経験したもの)のテンプレート修復のために「ドナー」分子を使用し、「非交差型遺伝子変換」または「短経路遺伝子変換」として様々に知られている。いかなる特定の理論にも束縛されることを望むものではないが、このような転送は、破壊標的とドナーとの間に形成されるヘテロ二本鎖DNAの不適合補正、および/または、ドナーが標的の一部となる遺伝子情報を再合成するために使用される「合成依存鎖アニーリング」、および/または関連プロセスを含むことができる。このような特殊HRにより、しばしば、ドナーポリヌクレオチドの配列の一部または全てが標的ポリヌクレオチドに組み込まれるように、標的分子の配列が変更される。
【0039】
本開示の方法において、本明細書に記載される1つ以上の標的ヌクレアーゼは、所定の部位で標的配列(例えば、細胞クロマチン)において、二本鎖切断を生成し、切断の領域におけるヌクレオチド配列に相同性を有する「ドナー」ポリヌクレオチドを細胞に導入することができる。二本鎖切断の存在は、ドナー配列の組込みを促進することが示されている。ドナー配列は、物理的に組み込まれ得るか、または代替的に、ドナーポリヌクレオチドを、相同組み換えを介した切断の修復のためのテンプレートとして使用し、ドナー等の場合、細胞クロマチンへの、ヌクレオチド配列の全てまたは一部を導入させる。したがって、細胞クロマチン中の第1の配列は、変更することができ、特定の実施形態において、ドナーポリヌクレオチド中に存在する配列に変換することができる。したがって、「置換する」または「置換」という用語の使用は、1つのヌクレオチド配列の別のものによる置換(すなわち、情報を持つセンスでの配列の置換)を表すと理解され、1つのポリヌクレオチドを別のものによる物理的または化学的置換は必ずしも必要としない。
【0040】
本明細書に記載される方法のいずれかにおいて、細胞内のさらなる標的部位のさらなる二本鎖切断のために、亜鉛フィンガータンパク質の追加の対を使用することができる。
【0041】
細胞クロマチンにおいて、対象となる領域の配列の標的化組み換えおよび/または置換および/または変更のための方法の特定の実施形態において、染色体の配列は、外来の「ドナー」ヌクレオチド配列との相同組み換えによって変更される。このような相同組み換えは、二本鎖切断の領域に相同である配列が存在する場合、細胞クロマチン内の二本鎖の切断の存在により刺激される。
【0042】
本明細書に記載される方法のいずれかにおいて、第1のヌクレオチド配列(「ドナー配列」)は、対象となる領域のゲノム配列の相同であるが同一ではない配列を含み、それによって、対象となる領域の非同一配列を挿入するための相同組み換えを刺激することができる。したがって、特定の実施形態において、対象となる領域の配列に相同であるドナー配列の一部は、置換されるゲノム配列と、約80〜99%(またはその間のいずれかの整数)の配列同一性を示す。他の実施形態において、例えば、100個を超える近接塩基対の、ドナーとゲノム配列の間で1個だけヌクレオチドが異なる場合、ドナーとゲノム配列の間の相同性は、99%よりも高い。場合によっては、ドナー配列の非相同部分は、新規配列が、対象となる領域に導入されるように、対象となる領域には存在しない配列を含むことができる。これらの場合において、非相同配列は、一般に、対象となる領域の配列と相同であるかまたは同一である50〜1,000個の塩基対(またはそれらの間のいずれかの整数値)、または1,000個を超えるいくつかの塩基対の配列によって側方に位置している。他の実施形態において、ドナー配列は、第1の配列に非相同であり、非相同組み換え機構によってゲノムに挿入される。
【0043】
本明細書に記載される方法のいずれかは、対象となる遺伝子の発現を妨害するドナー配列の標的化組込みによって、細胞中の1つ以上の標的配列の部分的または完全な不活性化に使用され得る。部分的または完全に不活性化された遺伝子を有する細胞株も提供される。
【0044】
さらに、本明細書に記載される標的化組込み方法はまた、1つ以上の外来配列を組み込むために使用することができる。外来核酸配列は、例えば、1つ以上の遺伝子またはcDNA分子、またはあらゆる種類のコードもしくは非コード配列、ならびに1つ以上の制御要素(例えば、プロモーター)を含むことができる。加えて、外来核酸配列は、1つ以上のRNA分子(例えば、小ヘアピンRNA(shRNA)、抑制RNA(RNAis)、ミクロRNA(miRNA)等)を産生し得る。
【0045】
「切断」とは、DNA分子の共有結合骨格の破壊を指す。切断は、リン酸ジエステル結合の酵素または化学化水分かを含むが、これらに限定されない様々な方法によって開始され得る。一本鎖切断および二本鎖切断の両方が可能であり、二本鎖切断は、2つの異なる一本鎖切断現象の結果として起こり得る。DNA切断は、平滑末端または付着末端のいずれかの産生をもたらし得る。特定の実施形態において、融合ポリペプチドは、標的二本鎖DNA切断に使用される。
【0046】
「切断ハーフドメイン」は、(同一または異なる)第2のポリペプチドと連結して、切断活性を有する複合体(好ましくは、二本鎖切断活性)を形成するポリペプチド配列である。「第1および第2の切断ハーフドメイン」、「+および-切断ハーフドメイン」、および「右および左切断ハーフドメイン」という用語は、二量体化する切断ハーフドメインの対を指すために交換可能に使用される。
【0047】
「改変された切断ハーフドメイン」は、別の切断ハーフドメイン(例えば、別の改変された切断ハーフドメイン)と共にヘテロ二量体を形成するために修飾された切断ハーフドメインである。例えば、米国特許出願公開第2005/0064474号、同第20070218528号、および同第2008/0131962号明細書も参照されたく、これらは、参照によってその全体が本明細書に組み込まれる。
【0048】
「配列」という用語は、任意の長さのヌクレオチド配列を指し、これはDNAもしくはRNAであり得る、線状、環状、もしくは分岐状であり得る、または一本鎖もしくは二本鎖のいずれか一方であり得る。「ドナー配列」という用語は、ゲノム内に挿入されるヌクレオチド配列を指す。ドナー配列は、任意の長さ、例えば、2〜10,000(またはそれらの間もしくはそれらを超える任意の整数値)長のヌクレオチド、好ましくは、約100〜1,000(またはそれらの間の任意の整数)長のヌクレオチド、より好ましくは、約200〜500長のヌクレオチドであり得る。
【0049】
「クロマチン」は、細胞ゲノムを含む核タンパク質構造である。細胞クロマチンは、核酸、主にDNA、ならびに、ヒストンおよび非ヒストン染色体タンパク質を含むタンパク質を含む。真核細胞クロマチンの大半は、ヌクレオソームの形態で存在し、そこでは、ヌクレオソームコアが、それぞれ2つのヒストンH2A、H2B、H3、およびH4を含む八量体と会合した約150塩基対のDNAを含み、リンカーDNA(生物に応じて変化する長さ)がヌクレオソームコアの間に延在する。ヒストンH1の1分子は、一般にリンカーDNAと会合する。本開示に関して、「クロマチン」という用語は、原核性および真核性の両方の全ての種類の細胞核タンパク質を包含するように意図されている。細胞クロマチンは、染色体およびエピソームクロマチンの両方を含む。
【0050】
「染色体」は、細胞のゲノムの全てまたは一部を含むクロマチン複合体である。細胞のゲノムは、しばしば、その核型によって特徴付けられ、これは細胞のゲノムを含む全染色体の集合である。細胞のゲノムは、1つ以上の染色体を含むことができる。
【0051】
「エピソーム」は、複製する核酸、核タンパク質複合体、または細胞の染色体核型の一部ではない核酸を含む他の構造である。エピソームの例には、プラスミドおよびある特定のウイルスゲノムが含まれる。
【0052】
「標的部位」または「標的配列」は、結合が存在するのに十分な条件が提供される場合、結合分子が結合する核酸の一部を規定する核酸配列である。多様なNT−3標的ZFPに対する例示的な標的部位は、表2および3に示される。
【0053】
「外来」分子は、通常は細胞内に存在しないが、1つ以上の遺伝学的、生化学的、または他の方法によって細胞内に導入することができる分子である。「通常は細胞内に存在する」とは、細胞の特定の発生段階および環境条件に対して決定される。したがって、例えば、筋肉の胚発生中のみ存在する分子は、成体の筋細胞に関しては外来分子である。同様に、熱ショックによって誘導される分子は、非熱ショック細胞に対して外来分子である。外来分子は、例えば、機能不全型内在性分子の機能型または正常機能型内在性分子の機能不全型を含むことができる。
【0054】
外来分子は、とりわけ、コンビナトリアルケミストリープロセスによって生成されるもの等の小分子、または、タンパク質、核酸、糖質、脂質、糖タンパク質、リポタンパク質、多糖、上記の分子の任意の修飾誘導体、または上記の分子の1つ以上を含む任意の複合体等の高分子であり得る。核酸にはDNAおよびRNAが含まれ、それは一本または二本鎖であり得、線状、分岐状、または環状であり得、任意の長さであり得る。核酸には、二重鎖を形成することができるもの、ならびに三重鎖形成核酸が含まれる。例えば、米国特許第5,176,996号および同第5,422,251号明細書を参照されたい。タンパク質には、DNA結合タンパク質、転写因子、クロマチンリモデリング因子、メチル化DNA結合タンパク質、ポリメラーゼ、メチラーゼ、デメチラーゼ、アセチラーゼ、デアセチラーゼ、キナーゼ、ホスファターゼ、インテグラーゼ、リコンビナーゼ、リガーゼ、トポイソメラーゼ、ジャイレース、およびヘリカーゼが含まれるが、これらに限定されない。
【0055】
外来分子は、内在性分子と同じ型の分子、例えば、外来タンパク質または核酸であり得る。例えば、外来核酸は、感染ウイルスゲノム、細胞中に導入されたプラスミドもしくはエピソーム、または細胞内に通常存在しない染色体を含むことができる。細胞中に外来分子を導入するための方法が当業者に知られており、脂質媒介導入(すなわち、中性およびカチオン性脂質を含むリポソーム)、エレクトロポレーション、直接注入、細胞融合、微粒子銃、リン酸カルシウム共沈、DEAE−デキストラン媒介導入、およびウイルスベクター媒介導入を含むが、これらに限定されない。外来分子はまた、内在性分子と同じ型の分子であってもよいが、細胞が由来する種とは異なる種に由来する。例えば、ヒトの核酸配列は、マウスまたはハムスターに本来由来する細胞株中に導入され得る。
【0056】
これに対して、「内在性」分子は、特定の環境条件下で特定の発生段階にある特定の細胞内に通常存在するものである。例えば、内在性核酸は、染色体、ミトコンドリア、クロロプラストもしくは他の細胞小器官のゲノム、または天然に存在するエピソーム核酸を含むことができる。さらなる内在性分子には、タンパク質、例えば、転写因子および酵素が含まれてもよい。
【0057】
「融合」分子は、2つ以上のサブユニット分子が、好ましくは共有結合的に連結された分子である。サブユニット分子は、同じ化学型の分子であるか、または異なる化学型の分子であることができる。第1の型の融合分子の例には、融合タンパク質(例えば、ZFPDNA結合ドメインと1つ以上の活性化ドメインとの間の融合体)、および融合核酸(例えば、上記に記載される融合タンパク質をコードする核酸)が含まれるが、これらに限定されない。第2の型の融合分子の例には、三重鎖形成核酸とポリペプチドとの間の融合体、および副溝結合剤と核酸との間の融合体が含まれるが、これらに限定されない。
【0058】
細胞中の融合タンパク質の発現は、細胞に融合タンパク質を送達すること、または細胞に融合タンパク質をコードするポリヌクレオチドを送達することによって引き起こされてもよく、ポリヌクレオチドが転写され、転写物が翻訳されて、融合タンパク質を生成する。トランススプライシング、ポリペプチド開裂、およびポリペプチド連結はまた、細胞中のタンパク質の発現に関与することができる。細胞へのポリヌクレオチドおよびポリペプチドの送達の方法は、本開示の他所で提示されている。
【0059】
「多量体化ドメイン」は、アミノ、カルボキシで組み込まれるドメイン、またはZFP TFのアミノまたはカルボキシ末端領域である。これらのドメインは、トリヌクレオチド反復ドメインのより大きな経路が、多量体型ZFP TFによって結合される一方で、野生型経路は結合されないように、複数のZFP TF単位の多量体化を可能にする。多量体化ドメインの例には、ロイシンジッパーが含まれる。多量体化ドメインはまた、小分子によって調節され得、多量体化ドメインは、小分子または外部リガンドの存在下のみ、別の多量体化ドメインとの相互作用を可能にする適切な構造であると仮定する。このようにして、外来リガンドは、これらのドメインの活性を調節するために使用することができる。
【0060】
本開示に関して、「遺伝子」は、このような調節配列がコードおよび/または転写配列に隣接しているかに関わらず、遺伝子産物(以下を参照)をコードするDNA領域、ならびに遺伝子産物の産生を調節する全てのDNA領域を含む。したがって、遺伝子には、プロモーター配列、ターミネーター、リポソーム結合部位および内部リポソーム侵入部位等の翻訳調節配列、エンハンサー、サイレンサー、インスレーター、境界エレメント、複製起点、マトリックス付着部位、ならびに遺伝子座制御領域が含まれるが、これらに限定されない。
【0061】
「遺伝子発現」は、遺伝子産物への遺伝子に含まれる情報の変換を指す。遺伝子産物は、遺伝子の直接転写産物(例えば、mRNA、tRNA、rRNA、アンチセンスRNA、リボザイム、構造RNA、もしくは任意の他の種類のRNA)、またはmRNAの翻訳によって産生されるタンパク質であってもよい。遺伝子産物には、キャッピング、ポリアデニル化、メチル化、および編集等のプロセスによって修飾されたRNA、および、例えば、メチル化、アセチル化、リン酸化、ユビキチン化、ADPリボシル化、ミリスチリル化、およびグリコシル化によって修飾されたタンパク質も含まれる。
【0062】
遺伝子発現の「調節」は、遺伝子の活性の変化を指す。発現の調節には、遺伝子活性化および遺伝子抑制が含まれ得るが、これらに限定されない。ゲノム編集(例えば、切断、変更、不活性化、ランダム変異)は、発現を調節するために使用することができる。遺伝子の不活性化とは、本明細書に記載されるようなZFPを含まない細胞と比較した、遺伝子発現の任意の減少を指す。したがって、遺伝子の不活性化は、部分的または完全であり得る。
【0063】
「対象となる領域」は、例えば、外来分子に結合することが望ましい、遺伝子、遺伝子内、または遺伝子に隣接する非コード配列等の細胞クロマチンの任意の領域である。結合は、標的化DNA切断および/または標的化組み換えの目的のためであり得る。対象となる領域は、例えば、染色体、エピソーム、細胞小器官ゲノム(例えば、ミトコンドリア、葉緑体)、または感染ウイルスゲノムに存在し得る。対象となる領域は、遺伝子のコード領域内、例えば、リーダー配列、トレーラー配列またはイントロン等の転写された非コード領域内、またはコード領域内の上流もしくは下流のいずれかの非転写領域内であり得る。対象となる領域は、単一ヌクレオチド対と同じくらい小さいか、もしくは最大2,000ヌクレオチド対の長さ、または任意の整数値のヌクレオチド対の長さであり得る。
【0064】
「真核」細胞には、真菌細胞(酵母等)、植物細胞、動物細胞、哺乳動物細胞、およびヒト細胞(例えば、T細胞)が含まれるが、これらに限定されない。
【0065】
「作動的連結」および「作動的に連結された」(または「作動可能に連結された」という用語は、2つ以上の構成要素(配列要素等)の並列に関連して交換可能に使用され、その中で両方の構成要素が正常に機能し、かつ構成要素の少なくとも1つが、他の構成要素の少なくとも1つに対して発揮される機能を媒介することができる可能性があるように構成要素が配置される。例として、プロモーター等の転写調節配列は、転写調節配列が1つ以上の転写調節因子の存在または非存在に応じてコード配列の転写レベルを制御する場合に、コード配列に作動的に連結される。転写調節配列は、一般的に、コード配列とシスで作動的に連結されるが、それに直接隣接している必要はない。例えば、エンハンサーは、それらが隣接していなくても、コード配列に作動的に連結される転写調節配列である。
【0066】
融合ポリペプチドに関して、「作動的に連結された」という用語は、構成要素の各々が、そのように連結されていない場合に、それが果たすであろう他の構成要素に連動して同一の機能を実行するという事実を指すことができる。例えば、ZFP DNA結合ドメインが活性化ドメインに融合される融合ポリペプチドに関して、ZFP DNA結合ドメインおよび活性化ドメインは、融合ポリペプチドにおいて、ZEP DNA結合ドメイン部分がその標的部位および/またはその結合部位に結合することができる一方で、活性化ドメインが遺伝子発現を上方制御することができる場合に、作動的連結状態にある。ZFP DNA結合ドメインにおける融合ポリペプチドが開裂ドメインに融合される場合、ZFP DNA結合ドメインおよびその開裂ドメインは、融合ポリペプチドにおいて、ZFP DNA結合ドメイン部分がその標的部位および/またはその結合部位に結合することができる一方で、開裂ドメインが標的部位の近接においてDNAを切断することができる場合に、作動的連結状態にある。
【0067】
タンパク質、ポリペプチド、または核酸の「機能的断片」は、その配列が、完全長タンパク質、ポリペプチド、または核酸と同一でないが、完全長タンパク質、ポリペプチド、または核酸と同一の機能を保持するタンパク質、ポリペプチド、または核酸である。機能的断片は、対応する天然分子より多い、少ない、または同一の残基数を有することができる、および/または1つ以上のアミノ酸またはヌクレオチド置換を含有することができる。核酸の機能(例えば、コード機能、別の核酸にハイブリダイズする能力)を決定するための方法は、当技術分野でよく知られている。同様に、タンパク質機能を決定するための方法も、よく知られている。例えば、ポリペプチドのDNA結合機能を、例えば、フィルター結合、電気泳動移動度シフト、または免疫沈降アッセイによって決定することができる。DNA開裂を、ゲル電気泳動によってアッセイすることができる。上記のAusubelらを参照されたい。別のタンパク質と相互作用するタンパク質の能力を、例えば、共免疫沈降、ツーハイブリッドアッセイ、または遺伝的相補性および生化学的相補性の両方の相補性によって決定することができる。例えば、Fields et al. (1989)Nature 340:245−246、米国特許第5,585,245号およびPCT国際公開第98/44350号を参照されたい。
【0068】
「ベクター」は、標的細胞に遺伝子配列を移動させることができる。一般的に、「ベクター構築物」、「発現ベクター」、および「遺伝子導入ベクター」は、対象となる遺伝子の発現を指向することが可能な、標的細胞に遺伝子配列を導入することができる任意の核酸構築物を意味する。したがって、この用語は、クローニングおよび発現ビヒクル、ならびに組込みベクターを含む。
【0069】
「レポーター遺伝子」または「レポーター配列」とは、好ましくは、必ずではないが日常的アッセイにおいて、容易に測定されるタンパク質産物を産生する任意の配列を指す。適切なレポーター遺伝子としては、抗生物質耐性(例えば、アンピシリン耐性、ネオマイシン耐性、G418耐性、ピューロマイシン耐性)を媒介するタンパク質をコードする配列、着色または蛍光または発光タンパク質(例えば、緑色蛍光タンパク質、強化緑色蛍光タンパク質、赤色蛍光タンパク質、ルシフェラーゼ)をコードする配列、および強化された細胞成長および/もしくは遺伝子増幅を媒介するタンパク質(例えば、ジヒドロ葉酸レダクターゼ)をコードする配列が含まれるが、これらに限定されない。エピトープタグには、例えば、FLAG、His、myc、Tap、HA、または任意の検出可能なアミノ酸配列の1つ以上のコピーが含まれる。「発現タグ」には、対象となる遺伝子の発現をモニターするために、所望の遺伝子配列に作動的に連結され得るレポーターをコードする配列が含まれる。
【0070】
DNA結合ドメイン
Httを含むが、これに限定されない、トリヌクレオチド反復を含む任意の遺伝子において、標的部位に特異的に結合するDNA結合ドメインを含む組成物が、本明細書に記載されている。GDNFまたはBDNF遺伝子において、標的部位に特異的に結合するDNA結合ドメインを含む組成物も提供される。任意のDNA結合ドメインは、本明細書に開示される組成物および方法で使用することができる。
【0071】
特定の実施形態において、DNA結合ドメインは、亜鉛フィンガータンパク質を含む。好ましくは、亜鉛フィンガータンパク質は天然に存在せず、それは選択する標的部位に結合するように改変される。例えば、Beerli et al.(2002)Nature Biotechnol.20:135−141、Pabo et al.(2001)Ann.Rev.Biochem.70:313−340、Isalan et al.(2001)Nature Biotechnol.19:656−660、Segal et al.(2001)Curr.Opin.Biotechnol.12:632−637、Choo et al.(2000)Curr.Opin.Struct.Biol.10:411−416、米国特許第6,453,242号、同第6,534,261号、同第6,599,692号、同第6,503,717号、同第6,689,558号、同第7,030,215号、同第6,794,136号、同第7,067,317号、同第7,262,054号、同第7,070,934号、同第7,361,635号、同第7,253,273号明細書、および米国特許出願公開第2005/0064474号、同第2007/0218528号、同第2005/0267061号明細書を参照されたく、全ては、参照によりその全体が本明細書に組み込まれる。
【0072】
改変された亜鉛フィンガー結合ドメインは、天然に存在する亜鉛フィンガータンパク質と比較して、新規の結合特異性を有することができる。改変方法には、合理的設計および多様な種類の選択が含まれるが、これらに限定されない。合理的設計には、例えば、三重(または四重)ヌクレオチド配列および個々の亜鉛フィンガーアミノ酸配列を含むデータベースを使用することが含まれ、各三重または四重ヌクレオチド配列は、特定の三重または四重配列に結合する亜鉛フィンガーの1つ以上のアミノ酸配列と会合する。例えば、譲受人共通の米国特許第6,453,242号および同第6,534,261号明細書を参照されたく、これらは参照によりその全体が本明細書に組み込まれる。
【0073】
ファージディスプレイおよび2ハイブリッド法を含む例示的な選択方法は、米国特許第5,789,538号、同第5,925,523号、同第6,007,988号、同第6,013,453号、同第6,410,248号、同第6,140,466号、同第6,200,759号、同第6,242,568号明細書、ならびに国際公開第98/37186号、国際公開第98/53057号、国際公開第00/27878号、国際公開第01/88197号、および英国特許第2,338,237号明細書に開示されている。加えて、亜鉛フィンガー結合ドメインに対する結合特異性の強化は、例えば、譲受人共通の国際公開第02/077227号に記載されている。
【0074】
加えて、これらおよび他の参考文献に開示される、亜鉛フィンガードメインおよび/または多フィンガーの亜鉛フィンガータンパク質は、例えば、5個以上のアミノ酸長のリンカーを含む、任意の好適なリンカー配列を使用して共に連結され得る。また、6個以上のアミノ酸長の例示的なリンカー配列に関して、米国特許第6,479,626号、同第6,903,185号、および同第7,153,949号明細書も参照されたい。本明細書に記載されるタンパク質は、タンパク質の個々の亜鉛フィンガー間の好適なリンカーの任意の組み合わせを含み得る。加えて、亜鉛フィンガー結合ドメインに対する結合特異性の強化は、例えば、譲受人共通の国際公開第02/077227号に記載されている。
【0075】
標的部位の選択、融合タンパク質(および同一物をコードするポリヌクレオチド)の設計および構築のためのZFPおよび方法が当業者に知られており、米国特許第6,140,0815号、同第789,538号、同第6,453,242号、同第6,534,261号、同第5,925,523号、同第6,007,988号、同第6,013,453号、同第6,200,759号明細書、国際公開第95/19431号、国際公開第96/06166号、国際公開第98/53057号、国際公開第98/54311号、国際公開第00/27878号、国際公開第01/60970号、国際公開第01/88197号、国際公開第02/099084号、国際公開第98/53058号、国際公開第98/53059号、国際公開第98/53060号、国際公開第02/016536号、および国際公開第03/016496号に詳しく記載されている。
【0076】
加えて、これらおよび他の参考文献に開示される、亜鉛フィンガードメインおよび/または多フィンガーの亜鉛フィンガータンパク質は、例えば、5個以上のアミノ酸長のリンカーを含む、任意の好適なリンカー配列を使用して共に連結され得る。また、6個以上のアミノ酸長の例示的なリンカー配列に関して、米国特許第6,479,626号、同第6,903,185号、同第7,153,949号明細書を参照されたい。本明細書に記載されるタンパク質は、タンパク質の個々の亜鉛フィンガー間の好適なリンカーの任意の組み合わせを含み得る。
【0077】
代替的に、DNA結合ドメインは、ヌクレアーゼに由来し得る。例えば、I−SceI、I−CeuI、PI−PspI,PI−Sce、I−SceIV、I−CsmI、I−PanI、I−SceII、I−PpoI、I−SceIII、I−CreI、I−TevI、I−TevII、およびI−TevIII等のホーミングエンドヌクレアーゼおよびメガヌクレアーゼの認識配列が知られている。また、米国特許第5,420,032号、米国特許第6,833,252号、Belfort et al.(1997)Nucleic Acids Res.25:3379-3388、Dujon et al.(1989)Gene 82:115-118、Perler et al.(1994)Nucleic Acids Res.22,1125-1127、Jasin(1996)Trends Genet.12:224-228、Gimble et al.(1996)J.Mol.Biol.263:163-180、Argast et al.(1998)J.Mol.Biol.280:345-353、およびニューイングランドバイオラボ社カタログも参照されたい。加えて、ホーミングエンドヌクレアーゼおよびメガヌクレアーゼのDNA結合特異性を、非天然の標的部位に結合するよう改変することができる。例えば、Chevalier et al.(2002)Molec.Cell 10:895−905、Epinat et al.(2003) Nucleic Acids Res.31:2952−2962、Ashworth et al.(2006)Nature 441:656−659、Paques et al.(2007)Current Gene Therapy 7:49−66、米国特許出願公開第20070117128号を参照されたい。
【0078】
特定の実施形態において、DNA結合ドメインは、Htt遺伝子の標的部位に(配列特異的な様式で)結合し、Httの発現を調節する改変された亜鉛フィンガータンパク質である。ZFPは、変異Htt対立遺伝子または野生型Htt配列のいずれかに選択的に結合することができる。Htt標的部位は、典型的に少なくとも1個の亜鉛フィンガーを含むが、複数の亜鉛フィンガー(例えば、2個、3個、4個、5個、6個以上のフィンガー)が含まれ得る。通常、ZFPは、少なくとも3個のフィンガーを含む。ある特定のZFPは、4個、5個、または6個のフィンガーを含む。3個のフィンガーを含むZFPは、典型的に9個または10個のヌクレオチドを含む標的部位を認識し、4個のフィンガーを含むZFPは、典型的に12〜14個のヌクレオチドを含む標的部位を認識する一方で、6個のフィンガーを有するZFPは、18〜21個のヌクレオチドを含む標的部位を認識する。ZFPはまた、そのドメインが転写活性化または抑制ドメインであり得る、1つ以上の調節ドメインを含む融合タンパク質であってもよい。
【0079】
「両端型」亜鉛フィンガータンパク質は、2つの亜鉛フィンガードメインが2つの不連続標的部位に結合するように、亜鉛フィンガーDNA結合ドメインの2つのクラスターが、アミノ酸を介在することによって分離されるタンパク質である。両端型亜鉛フィンガー結合タンパク質の例は、SIP1であり、一群の4つの亜鉛フィンガーは、タンパク質のアミノ末端に位置し、一群の3つのフィンガーは、カルボキシ末端に位置する(Remacle et al,(1999)EMBO Journal 18(18):5073−5084を参照されたい)。これらのタンパク質におけるそれぞれのクラスターの亜鉛フィンガーは、独特の標的配列に結合することができ、2つの標的配列間の間隔は、多くのヌクレオチドを含むことができる。両端型ZFPは、例えば、ZFPの一方または両方に融合される機能ドメインが含まれ得る。したがって、機能ドメインは、ZFPの一方または両方の外部に付着され得る(図1Cを参照されたい)か、またはZFP間に位置付けられ得る(両方のZFPに付着される)(図4を参照されたい)。
【0080】
Htt標的ZFPの特定の例は、表1に開示されている。この表の第1の列は、ZFPに対する内部参照名(番号)であり、表2の第1列の同じ名前に対応する。「F」はフィンガーを指し、「F」に続く番号は、どの亜鉛フィンガー(例えば、「F1」はフィンガー1を指す)かを言及する。
【0081】
【表1】

【0082】
これらのタンパク質の標的部位に対する配列および位置は、表2に開示される。表2は、示される亜鉛フィンガータンパク質に対する標的配列を示す。ZFP認識ヘリックスによって接触される標的部位におけるヌクレオチドは大文字で示され、非接触ヌクレオチドは小文字で示される。
【0083】
【表2】
【0084】
融合タンパク質
本明細書に記載されるDNA結合タンパク質(例えば、ZFP)を含む融合タンパク質および異種調節(機能的)ドメイン(またはその機能的断片)も提供される。一般的なドメインには、例えば、転写因子ドメイン(活性化剤、リプレッサー、共活性化剤、コリプレッサー)、サイレンサー、癌遺伝子(例えば、myc、jun、fos、myb、max、mad、rel、ets、bcl、myb、mosファミリーメンバー等)、DNA修復酵素ならびにそれらの関連する因子および修飾因子、クロマチン結合性タンパク質およびそれらの修飾因子(例えば、キナーゼ、アセチラーゼ、およびデアセチラーゼ)、ならびにDNA修飾酵素(例えば、メチルトランスフェラーゼ、トポイソメラーゼ、ヘリカーゼ、リガーゼ、キナーゼ、ホスファターゼ、ポリメラーゼ、エンドヌクレアーゼ)、ならびにそれらの関連する因子および修飾因子が含まれる。DNA結合ドメインならびにヌクレアーゼ開裂ドメインの融合に関する詳細ついて、米国特許出願公開第2005/0064474号、同第2006/0188987号、および同第2007/0218528号明細書を参照されたく、参照によりそれらの全体が本明細書に組み込まれる。
【0085】
活性化を達成するための好適なドメインには、HSV VP16活性化ドメイン(例えば、Hagmann et al.,J.Virol.71,5952−5962(1997)を参照)、核ホルモン受容体(例えば、Torchia et al.,Curr.Opin.Cell.Biol.10:373−383(1998)を参照)、核因子カッパBのp65サブユニット(Bitko & Barik,J.Virol.72:5610−5618(1998)およびDoyle & Hunt,Neuroreport 8:2937−2942(1997)を参照)、Liu et al.,Cancer Gene Ther.5:3−28(1998))、またはVP64等の人工キメラ機能ドメイン(Beerli et al.,(1998)Proc.Natl.Acad.Sci.USA 95:14623−33)、およびデグロン(Molinari et al.,(1999)EMBO J.18,6439−6447)が含まれる。さらなる例示的な活性化ドメインには、Oct1、Oct−2A、Sp1、AP−2、およびCTF1(Seipel et al.,EMBO J.11,4961−4968 (1992)、ならびにp300、CBP、PCAF、SRC1 PvALF、AtHD2A、およびERF−2が含まれる。例えば、Robyr et al.(2000)Mol.Endocrinol.14:329−347、Collingwood et al.(1999)J.Mol.Endocrinol.23:255−275、Leo et al.(2000)Gene 245:1−11;Manteuffel−Cymborowska(1999)Acta Biochim.Pol.46:77−89、McKenna et al.(1999)J.Steroid Biochem.Mol.Biol.69:3−12、Malik et al.(2000)Trends Biochem.Sci.25:277−283、およびLemon et al.(1999)Curr.Opin.Genet.Dev.9:499−504を参照されたい。さらなる例示的な活性化ドメインとしては、OsGAI、HALF−1、C1、AP1、ARF−5、−6、−7、および−8、CPRF1、CPRF4、MYC−RP/GP、ならびにTRAB1が含まれるが、これらに限定されない。例えば、Ogawa et al.(2000)Gene 245:21−29、Okanami et al.(1996)Genes Cells 1:87−99、Goff et al.(1991)Genes Dev.5:298−309、Cho et al.(1999)Plant Mol.Biol.40:419−429、Ulmason et al.(1999)Proc.Natl.Acad.Sci.USA 96:5844−5849、Sprenger−Haussels et al.(2000)Plant J.22:1−8、Gong et al.(1999)Plant Mol.Biol.41:33−44、およびHobo et al.(1999)Proc.Natl.Acad.Sci.USA 96:15,348−15,353を参照されたい。
【0086】
DNA結合ドメインと機能ドメインとの間の融合タンパク質(または同一物をコードする核酸)の形成において、活性化ドメインと相互作用する活性化ドメインまたは分子のいずれか一方が、機能ドメインとして好適であることが、当業者には明らかである。基本的に、標的遺伝子に活性化複合体および/または活性化活性(例えば、ヒストンアセチル化等)を補充する能力のある任意の分子が、融合タンパク質の活性化ドメインとして有用である。融合分子において機能ドメインとしての使用に好適なISWI含有ドメインおよび/またはメチル結合ドメインタンパク質等の絶縁体ドメイン、局在化ドメイン、ならびにクロマチンリモデリングタンパク質が、例えば、譲受人共通の米国特許出願公開第2002/0115215号および同第2003/0082552号明細書、ならびに譲受人共通の国際公開第02/44376号に記載されている。
【0087】
例示的な抑制ドメインとしては、KRAB、A/B、KOX、TGF−ベータ−誘発型初期遺伝子(TIEG)、v−erbA、SID、MBD2、MBD3、DNMTファミリーのメンバー(例えば、DNMT1、DNMT3A、DNMT3B)、Rb、およびMeCP2が含まれるが、これらに限定されない。例えば、Bird et al.(1999)Cell 99:451−454、Tyler et al.(1999)Cell 99:443−446、Knoepfler et al.(1999)Cell 99:447−450、およびRobertson et al.(2000) Nature Genet.25:338−342を参照されたい。さらなる例示的な抑制ドメインとしては、ROM2およびAtHD2Aが含まれるが、これらに限定されない。例えば、Chem et al.(1996)Plant Cell 8:305−321およびWu et al.(2000)Plant J.22:19−27を参照されたい。
【0088】
融合分子は、当業者によく知られているクローニング法および生化学的接合によって構築される。融合分子には、DNA結合ドメインおよび機能ドメイン(例えば、転写活性化または抑制ドメイン)が含まれる。融合分子には、核局在化シグナル(例えば、SV40媒体T抗原由来等)およびエピトープタグ(例えば、FLAGおよび血球凝集素等)も任意に含まれる。融合タンパク質(およびそれらをコードする核酸)は、翻訳リーディングフレームが融合の成分間で維持されるように設計される。
【0089】
一方において機能ドメイン(またはその機能的断片)のポリペプチド成分と、他方において非タンパク質DNA結合ドメイン(例えば、抗生物質、挿入剤、副溝結合剤、核酸)との間の融合は、当業者に知られている生化学的接合の方法によって構築される。例えば、Pierce Chemical Company(Rockford,IL)Catalogueを参照されたい。副溝結合剤とポリペプチドとの間に融合を起こすための方法および組成物が記載されている。Mapp et al.(2000)Proc.Natl.Acad.Sci.USA 97:3930−3935。
【0090】
特定の実施形態において、亜鉛フィンガータンパク質によって結合される標的部位は、細胞クロマチンの到達可能領域に存在する。到達可能領域は、例えば、譲受人共通の国際公開第01/83732号に記載されるように、決定することができる。標的部位が細胞クロマチンの到達可能領域に存在しない場合、譲受人共通の国際公開第01/83793号に記載されるように、1つ以上の到達可能領域を生成することができる。さらなる実施形態において、融合分子のDNA結合ドメインは、その標的部位が到達可能領域にあるかに関わらず、細胞クロマチンに結合することができる。例えば、このようなDNA結合ドメインは、リンカーDNAおよび/またはヌクレオソームDNAに結合することができる。「パイオニア」DNA結合ドメインのこの型の例は、ある特定のステロイド受容体および肝細胞核因子3(HNF3)で見出される。Cordingley et al.(1987)Cell 48:261−270、Pina et al.(1990)Cell 60:719−731、およびCirillo et al.(1998)EMBO J.17:244−254。
【0091】
融合分子は、当業者に知られているように、薬学的に許容される担体で製剤化され得る。例えば、Remington’s Pharmaceutical Sciences,17th ed.,1985、および譲受人共通の国際公開第00/42219号を参照されたい。
【0092】
融合分子の機能的成分/ドメインは、一旦融合分子がそのDNA結合ドメインを介して標的配列に結合すると、遺伝子の転写に影響を及ぼすことができる多種多様の異なる成分のいずれかから選択され得る。したがって、機能的成分には、活性化剤、リプレッサー、共活性化剤、コリプレッサー、およびサイレンサー等の多様な転写因子ドメインが含まれ得るが、これらに限定されない。
【0093】
さらなる例示的な機能ドメインは、例えば、譲受人共通の米国特許第6,534,261号および米国特許出願公開第2002/0160940号明細書に開示されている。
【0094】
外来小分子またはリガンドによって調節される機能ドメインも、選択され得る。例えば、RheoSwitch(登録商標)技術は、機能ドメインのみが、外部RheoChem(商標)リガンドの存在下で、その活性立体配座であると仮定する場合、使用され得る(例えば、米国特許出願公開第20090136465号明細書を参照)。したがって、ZFPは、調節可能な機能ドメインに作動可能に連結され得、ここで、得られたZFP−TFの活性は、外部リガンドによって制御される。
【0095】
ヌクレアーゼ
特定の実施形態において、融合タンパク質は、DNA結合結合ドメインおよび切断(ヌクレアーゼ)ドメインを含む。したがって、遺伝子修飾は、ヌクレアーゼ、例えば、改変されたヌクレアーゼを用いて達成することができる。改変されたヌクレアーゼ技術は、天然に存在するDNA結合タンパク質の改変に基づいている。例えば、目的に合わせたDNA結合特異性を有するホーミングエンドヌクレアーゼの改変が、記載されている。Chames et al.(2005)Nucleic Acids Res 33(20):e178、Arnould et al.(2006)J.Mol.Biol.355:443−458。加えて、ZFPの改変も記載されている。例えば、米国特許第6,534,261号、同第6,607,882号、同第6,824,978号、同第6,979,539号、同第6,933,113号、同第7,163,824号、および同第7,013,219号明細書を参照されたい。
【0096】
加えて、ZFN、すなわち、その改変された(ZFP)DNA結合ドメインを通してその意図される核酸標的を認識することができ、かつヌクレアーゼ活性によって、DNAがZFP結合部位の近くで切断される、機能的な実体を作り出すために、ZFPが、ヌクレアーゼドメインに融合されている。例えば、Kim et al.(1996)Proc Natl Acad Sci USA 93(3):1156−1160を参照されたい。つい最近では、ZFNは、様々な生物におけるゲノム修飾のために使用されている。例えば、米国特許出願公開第20030232410号、同第20050208489号、同第20050026157号、同第20050064474号、同第20060188987号、同第20060063231号明細書、および国際公開第07/014275号を参照されたい。
【0097】
したがって、本明細書に記載される方法および組成物は、広範囲に適用可能であり、対象となる任意のヌクレアーゼを含み得る。ヌクレアーゼの非限定的な例には、メガヌクレアーゼおよび亜鉛フィンガーヌクレアーゼが含まれる。ヌクレアーは、異種DNA結合および開裂ドメイン(例えば、亜鉛フィンガーヌクレアーゼ;異種開裂ドメインを有するメガヌクレアーゼDNA結合ドメイン)を含み得るか、または代替的に、天然に存在するヌクレアーゼのDNA結合ドメインは、選択された標的部位(例えば、同種の結合部位とは異なる部位に結合するように改変されたメガヌクレアーゼ)に結合するように変更され得る。
【0098】
特定の実施形態において、ヌクレアーゼは、メガヌクレアーゼ(ホーミングエンドヌクレアーゼ)である。天然に存在するメガヌクレアーゼは、15〜40塩基対の切断部位を認識し、一般に、LAGLIDADGファミリー、GIY−YIGファミリー、His−Cystボックスファミリー、およびHNHファミリーの4つのファミリーに分類される。例示的なホーミングエンドヌクレアーゼには、I−SceI、I−CeuI、PI−PspI、PI−Sce、I−SceIV、I−CsmI、I−PanI、I−SceII、I−PpoI、I−SceIII、I−CreI、I−TevI、I−TevII、およびI−TevIIIが含まれる。これらの認識配列は公知である。また、米国特許第5,420,032号、米国特許第6,833,252号明細書、Belfort et al.(1997)Nucleic Acids Res.25:3379-3388、Dujon et al.(1989)Gene 82:115-118、Perler et al.(1994)Nucleic Acids Res.22,1125-1127、Jasin(1996)Trends Genet.12:224-228、Gimble et al.(1996)J.Mol.Biol.263:163-180、Argast et al.(1998)J.Mol.Biol.280:345-353、およびニューイングランドバイオラボ社カタログも参照されたい。
【0099】
主として、LAGLIDADGファミリー由来の天然に存在するメガヌクレアーゼ由来のDNA結合ドメインが、植物、酵母、ショウジョウバエ属、哺乳動物細胞、およびマウスで部位特異的ゲノム修飾を促進するために用いられているが、このアプローチは、メガヌクレアーゼ認識配列を保存している相同遺伝子(Monet et al.(1999),Biochem.Biophysics.Res.Common.255:88−93)、またはその中に認識配列が導入されている予め改変されたゲノム(Route et al.(1994),Mol.Cell.Biol.14:8096−106、Chilton et al.(2003),Plant Physiology.133:956−65、Puchta et al.(1996),Proc.Natl.Acad.Sci.USA 93:5055−60、Rong et al.(2002),Genes Dev.16:1568−81、Gouble et al.(2006),J.Gene Med.8(5):616−622)のいずれかの修飾に限定されている。したがって、医学的または生物工学的に関連のある部位での新規の結合特異性を示すメガヌクレアーゼを改変する試みがなされている(Porteus et al.(2005),Nat. Biotechnol.23:967−73、Sussman et al.(2004),J.Mol.Biol.342:31−41、Epinat et al.(2003),Nucleic Acids Res.31:2952−62、Chevalier et al.(2002)Molec.Cell 10:895−905、Epinat et al.(2003)Nucleic Acids Res.31:2952−2962、Ashworth et al.(2006)Nature 441:656−659、Paques et al.(2007)Current Gene Therapy 7:49−66、米国特許出願公開第20070117128号、同第20060206949号、同第20060153826号、同第20060078552号、および同第20040002092号明細書)。加えて、メガヌクレアーゼ由来の天然に存在するまたは改変されたDNA結合ドメインもまた、異種ヌクレアーゼ(例えば、FokI)由来の開裂ドメインと作動可能に連結されている。
【0100】
他の実施形態において、ヌクレアーゼは、亜鉛フィンガーヌクレアーゼ(ZFN)である。ZFNは、選択遺伝子内の標的部位に結合するように改変されている亜鉛フィンガータンパク質および開裂ドメインまたは切断ハーフドメインを含む。
【0101】
上に詳細に記載されるように、亜鉛フィンガー結合ドメインは、選択配列に結合するように改変され得る。例えば、Beerli et al.(2002)Nature Biotechnol.20:135−141、Pabo et al.(2001)Ann.Rev.Biochem.70:313−340、Isalan et al.(2001)Nature Biotechnol.19:656−660、Segal et al.(2001)Curr.Opin.Biotechnol.12:632−637、Choo et al.(2000)Curr.Opin.Struct.Biol.10:411−416を参照されたい。改変された亜鉛フィンガー結合は、天然に存在する亜鉛フィンガータンパク質と比較して、新規の結合特異性を有することができる。改変方法には、合理的設計および多様な種類の選択が含まれるが、これらに限定されない。合理的設計には、例えば、三重(または四重)ヌクレオチド配列および個々の亜鉛フィンガーアミノ酸配列を含むデータベースを使用することが含まれ、各三重または四重ヌクレオチド配列は、特定の三重または四重配列と結合する亜鉛フィンガーの1つ以上のアミノ酸配列に関連する。例えば、譲受人共通の米国特許第6,453,242号および同第6,534,261号明細書を参照されたく、参照によりその全体が本明細書に組み込まれる。
【0102】
ファージディスプレイおよび2ハイブリッド法を含む例示的な選択方法は、米国特許第5,789,538号、同第5,925,523号、同第6,007,988号、同第6,013,453号、同第6,410,248号、同第6,140,466号、同第6,200,759号、同第6,242,568号明細書、ならびに国際公開第98/37186号、国際公開第98/53057号、国際公開第00/27878号、国際公開第01/88197号、および英国特許第2,338,237号に開示されている。加えて、亜鉛フィンガー結合ドメインに対する結合特異性の強化は、例えば、譲受人共通の国際公開第02/077227号に記載されている。
【0103】
標的部位の選択、融合タンパク質(および同一物をコードするポリヌクレオチド)の設計および構築のためのZFPおよび方法が当業者に知られており、米国特許出願公開第20050064474号および同第20060188987号明細書を参照されたく、参照によりその全体が本明細書に組み込まれる。
【0104】
加えて、これらおよび他の参考文献に開示される、亜鉛フィンガードメインおよび/または多フィンガーの亜鉛フィンガータンパク質は、例えば、5個以上のアミノ酸長のリンカー(例えば、TGEKP(配列番号40)、TGGQRP(配列番号41)、TGQKP(配列番号42)、および/またはTGSQKP(配列番号43))を含む、任意の好適なリンカー配列を使用して共に連結され得る。6個以上のアミノ酸長の例示的なリンカー配列に関して、例えば、米国特許第6,479,626号、同第6,903,185号、および同第7,153,949号明細書を参照されたい。本明細書に記載されるタンパク質は、タンパク質の個々の亜鉛フィンガー間の好適なリンカーの任意の組み合わせを含み得る。また、米国仮特許出願第61/343,729号も参照されたい。
【0105】
ZFNおよび/またはメガヌクレアーゼ等のヌクレアーゼはまた、ヌクレアーゼ(開裂ドメイン、切断ハーフドメイン)も含む。上記のように、開裂ドメインは、DNA結合ドメインに対して異種であり得、例えば、亜鉛フィンガーDNA結合ドメインおよびヌクレアーゼ由来の開裂ドメイン、またはメガヌクレアーゼDNA結合ドメインおよび異なるヌクレアーゼ由来の開裂ドメインであり得る。異種開裂ドメインは、任意のエンドヌクレアーゼまたはエキソヌクレアーゼであり得る。開裂ドメインを生成し得る例示的なエンドヌクレアーゼには、制限エンドヌクレアーゼおよびホーミングエンドヌクレアーゼが含まれるが、これらに限定されない。例えば、New England Biolabs,Beverly,MAの2002−2003版カタログおよびBelfort et al.(1997)Nucleic Acids Res.25:3379−3388を参照されたい。DNAを切断するさらなる酵素は公知である(例えば、S1ヌクレアーゼ;マングビーンヌクレアーゼ;膵臓DNアーゼI;マイクロコッカルヌクレアーゼ;酵母HOエンドヌクレアーゼ;Linn et al.(eds.)Nucleases,Cold Spring Harbor Laboratory Press,1993も参照されたい)。これらの酵素(またはその機能的断片)のうちの1つ以上は、開裂ドメインおよび切断ハーフドメインの源として使用することができる。
【0106】
同様に、切断ハーフドメインは、切断活性に二量体化が必要である、上記に示すように、任意のヌクレアーゼまたはその部分によって得ることができる。一般に、融合タンパク質が切断ハーフドメインを含む場合、切断のために2つの融合タンパク質が必要である。代替的に、2つの切断ハーフドメインを含む単一のタンパク質を使用することができる。2つの切断ハーフドメインは、同じエンドヌクレアーゼ(もしくはその機能的断片)から得ることができるか、またはそれぞれの切断ハーフドメインは、異なるエンドヌクレアーゼ(もしくはその機能的断片)から得ることができる。加えて、2つの融合タンパク質が、それぞれの標的部位に結合することによって、切断ハーフドメインが互いに、例えば、二量体化することによって、機能的な開裂ドメインを形成するのを可能にする空間的な配向に置かれるように、2つの融合タンパク質の標的部位は、互いに関して、好ましく配置される。したがって、特定の実施形態において、標的部位の近い末端を、5〜8ヌクレオチドまたは15〜18ヌクレオチドによって分けられる。しかしながら、任意の整数のヌクレオチドまたはヌクレオチド対が、2つの標的部位の間に(例えば、2〜50個のヌクレオチド対またはそれ以上)介在することができる。一般に、切断の部位は、標的部位の間にある。
【0107】
制限エンドヌクレアーゼ(制限酵素)は、多くの種に存在しており、(認識部位で)DNAに配列特異的に結合し、結合の部位でまたは結合の部位の近くでDNAを切断することができる。ある特定の制限酵素(例えば、IIS型)は、認識部位から隔たった部位の近くでDNAを切断し、分離可能な結合ドメインおよび開裂ドメインを有する。例えば、IIS型酵素Fok Iは、一方の鎖上ではその認識部位から9ヌクレオチドで、他方の鎖上ではその認識部位から13ヌクレオチドで、二本鎖切断を触媒する。例えば、米国特許第5,356,802号、同第5,436,150号、および同第5,487,994号明細書、ならびにLi et al.(1992)Proc.Natl.Acad.Sci.USA 89:4275−4279、Li et al.(1993)Proc.Natl.Acad.Sci.USA 90:2764−2768、Kim et al.(1994a)Proc.Natl.Acad.Sci.USA 91:883−887、Kim et al.(1994b)J.Biol.Chem.269:31,978−31,982を参照されたい。したがって、一実施形態において、融合タンパク質は、少なくとも1つのIIS型制限酵素由来の開裂ドメイン(または切断ハーフドメイン)および改変されても改変されなくてもよい1つ以上の亜鉛フィンガー結合ドメインを含む。
【0108】
開裂ドメインが結合ドメインから分離可能である、例示的なIIS型制限酵素は、Fok Iである。この特定の酵素は、二量体として活性がある。Bitinaite et al.(1998)Proc.Natl.Acad.Sci.USA 95:10,570−10,575。したがって、本開示に関して、開示される融合タンパク質で用いられるFok I酵素の部分は、切断ハーフドメインと考えられる。したがって、亜鉛フィンガー−Fok I融合体を用いた細胞配列の標的化した二本鎖切断および/または標的化した置換のために、それぞれ、Fok I切断ハーフドメインを含む、2つの融合タンパク質を用いて、触媒活性のある開裂ドメインを再構成することができる。代替的に、1つの亜鉛フィンガー結合ドメインおよび2つのFok I切断ハーフドメインを含む単一のポリペプチド分子を用いることもできる。亜鉛フィンガーFok I融合体を用いた標的化した切断および標的とした配列変更のためのパラメータは、本開示の他の箇所に提供されている。
【0109】
開裂ドメインまたは切断ハーフドメインは、切断活性を保持するか、または多量体化(例えば、二量体化)して、機能的な開裂ドメインを形成する能力を保持するタンパク質の任意の部分であることができる。
【0110】
例示的なIIS型制限酵素は、国際公開第07/014275号に記載されており、これは参照によりその全体が本明細書に組み込まれる。さらなる制限酵素も分離可能な結合ドメインおよび切断半ドメインを含み、これらは本開示によって企図される。例えば、Roberts et al.(2003)Nucleic Acids Res.31:418−420を参照されたい。
【0111】
特定の実施形態において、開裂ドメインは、例えば、米国特許出願公開第20050064474号および第20060188987号明細書、ならびに米国出願第11/805,850号(2007年5月23日に出願)(これらの全ての開示は参照によりその全体が本明細書に組み込まれる)に記載されるような、ホモ二量体化を最小限に抑えるか、またはホモ二量体化を妨げる1つ以上の改変された切断ハーフドメインを含む。Fok Iの446位、447位、479位、483位、484位、486位、487位、490位、491位、496位、498位、499位、500位、531位、534位、537位、および538位のアミノ酸残基は全て、Fok I切断ハーフドメインの二量体化に影響を与えるための標的である。
【0112】
絶対的ヘテロ二量体を形成するFok Iの例示的な改変された切断ハーフドメインには、第1の切断ハーフドメインがFok Iの490位および538位のアミノ酸残基における変異を含み、第2の切断ハーフドメインが486位および499位のアミノ酸残基における変異を含む対が含まれる。
【0113】
したがって、一実施形態において、490位の変異は、Glu(E)をLys(K)で置換し、538位の変異は、Iso(I)をLys(K)で置換し、486位の変異は、Gln(Q)をGlu(E)で置換し、499位の変異は、Iso(I)をLys(K)で置換する。具体的に、本明細書に記載される改変された切断ハーフドメインは、一方の切断ハーフドメインの490位(E→K)および538位(I→K)を変異させることによって、「E490K:I538K」と呼ばれる改変された切断ハーフドメインを産生し、もう一方の切断ハーフドメインの486位(Q→E)および499位(I→L)を変異させることによって、「Q486E:I499L」と呼ばれる改変された切断ハーフドメインを産生するように調製した。本明細書に記載される改変された切断ハーフドメインは、絶対的ヘテロ二量体変異体であり、この変異体では、異常な切断が最小限に抑えられているかまたは消失させられている。例えば、米国特許出願公開第2008/0131962号を参照されたく、この開示は、全ての目的のために参照によりその全体が本明細書に組み込まれる。特定の実施形態において、改変された切断ハーフドメインは、486位、499位、および496位(野生型Fok Iに対して番号付けされた)での変異、例えば、486位で野生型Gln(Q)残基をGlu(E)残基で、499位で野生型Iso(I)残基をLeu(L)残基で、496位で野生型Asn(N)残基をAsp(D)もしくはGlu(E)残基(それぞれ、「ELD」および「ELE」ドメインとも称される)で置換する変異を含む。他の実施形態において、改変された切断ハーフドメインは、490位、538位、および537位(野生型Fok Iに対して番号付けされた)での変異、例えば、490位で野生型Gln(E)残基をLys(K)残基で、538位で野生型Iso(I)残基をLys(K)残基で、537位で野生型His(H)残基をLys(K)もしくはArg(R)残基(それぞれ、「KKK」および「KKR」ドメインとも称される)で置換する変異を含む。他の実施形態において、改変された切断ハーフドメインは、490位および537位(野生型Fok Iに対して番号付けされた)での変異、例えば、490位で野生型Gln(E)残基をLys(K)残基で、537位で野生型His(H)残基をLys(K)もしくはArg(R)残基(それぞれ、「KIK」および「KIR」ドメインとも称される)で置換する変異を含む。(2010年2月8に出願された米国仮出願第61/337,769号を参照されたい)。
【0114】
本明細書に記載される改変された切断ハーフドメインは、任意の好適な方法を用いて、例えば、米国特許出願公開第20050064474号および第20080131962号明細書に記載される、野生型切断ハーフドメイン(Fok I)の部位特異的突然変異生成によって調製することができる。
【0115】
代替的に、ヌクレアーゼは、いわゆる「スプリット酵素」技術を用いた核酸標的部位でのインビボで組み立てられ得る(例えば、米国特許出願公開第20090068164号を参照)。このようなスプリット酵素の成分は、別個の発現構築物上で発現され得るか、または1つのオープンリーディングフレーム内に連結することができ、ここで、個々の成分は、例えば、自己切断型2AペプチドまたはIRES配列によって分離される。成分は、個々の亜鉛フィンガー結合ドメインまたはメガヌクレアーゼ核酸結合ドメインのドメインであり得る。
【0116】
いくつかの実施形態において、DNA結合ドメインは、植物病原菌であるキサントモナス(Boch et al,(2009)Science 326:1509−1512およびMoscou and Bogdanove,(2009)Science326:1501を参照)およびラストニア(Heuer et al(2007)Applied and Environmental Microbiology 73(13):4379−4384を参照)由来のものと同様に、TALエフェクター由来の改変されたドメインである。また、国際公開第2010/079430号も参照されたい。
【0117】
ヌクレアーゼ(例えば、ZFN)は、例えば、国際公開第2009/042163号および第20090068164号に記載される、酵母ベースの染色体系で使用する前に活性に対してスクリーニングすることができる。ヌクレアーゼ発現構築物は、当技術分野で知られている方法を用いて容易に設計することができる。例えば、米国特許出願公開第20030232410号、同第20050208489号、同第20050026157号、同第20050064474号、同第20060188987号、同第20060063231号明細書、および国際公開第07/014275号を参照されたい。ヌクレアーゼの発現は、構造性プロモーターまたは誘導性プロモーターの制御下、例えば、ラフィノースおよび/またはガラクトースの存在下で、活性化(抑制解除)され、グルコースの存在下で抑制されるガラクトキナーゼプロモーターの制御下にある。
【0118】
送達
タンパク質(例えば、ZFP)、同一物をコードするポリヌクレオチド、ならびに本明細書に記載されるタンパク質および/またはポリヌクレオチドを含む組成物は、任意の好適な手段で、例えば、ZFP TFまたはZFN mRNAを注入することによって、標的細胞に送達され得る。好適な細胞には、真核および原核細胞ならびに/または細胞株が含まれるが、これらに限定されない。このような細胞またはこのような細胞から生成された細胞株の非制限的な例には、COS、CHO(例えば、CHO−S、CHO−K1、CHO−DG44、CHO−DUXB11、CHO−DUKX、CHOK1SV)、VERO、MDCK、WI38、V79、B14AF28−G3、BHK、HaK、NS0、SP2/0−Ag14、HeLa、HEK293(例えば、HEK293−F、HEK293−H、HEK293−T)、およびperC6細胞、ならびにツマジロクサヨトウ(Sf)等の昆虫細胞、またはサッカロミセス、ピチア、およびシゾサッカロミセス等の真菌細胞が挙げられるが、これらに限定されない。ある特定の実施形態において、細胞株は、CHO−K1、MDCK、またはHEK293細胞株である。また、好適な細胞には、例として、胚幹細胞、誘導多能性幹細胞、造血幹細胞、および間充織幹細胞等の幹細胞も含まれる。
【0119】
本明細書に記載される亜鉛フィンガータンパク質を含むタンパク質を送達する方法は、例えば、米国特許第6,453,242号、同第6,503,717号、同第6,534,261号、同第6,599,692号、同第6,607,882号、同第6,689,558号、同第6,824,978号、同第6,933,113号、同第6,979,539号、同第7,013,219号、および同第7,163,824号明細書に記載され、これらの全ての開示は、参照によってその全体が本明細書に組み込まれる。
【0120】
本明細書に記載される亜鉛フィンガータンパク質はまた、亜鉛フィンガータンパク質の1つ以上をコードする配列を含有するベクターを使用して送達され得る。プラスミドベクター、レトロウイルスベクター、レンチウイルスベクター、アデノウイルスベクター、ポックスウイルスベクター、ヘルペスウイルスベクター、およびアデノ随伴ウイルスベクター等を含むが、これらに限定されない任意のベクター系が使用され得る。また、米国特許第6,534,261号、同第6,607,882号、同第6,824,978号、同第6,933,113号、同第6,979,539号、同第7,013,219号、および同第7,163,824号明細書を参照されたく、参照によってその全体が本明細書に組み込まれる。さらに、これらのベクターのうちのいずれかが、配列をコードする1つ以上の亜鉛フィンガータンパク質を含み得ることは明らかである。したがって、1つ以上のZFPが細胞中に導入される時、ZFPは、同一のベクターまたは異なるベクター上で担持され得る。複数のベクターが使用される時、それぞれのベクターは、1つまたは複数のZFPをコードする配列を含み得る。
【0121】
従来のウイルスおよび非ウイルスに基づく遺伝子導入方法を使用して、改変されたZFPをコードする核酸を、細胞(例えば、哺乳動物細胞)および標的組織に導入することができる。このような方法を使用して、ZFPをコードする核酸を、細胞にインビトロ投与することもできる。ある特定の実施形態において、ZFPをコードする核酸は、インビボまたはエクスビボ遺伝子療法用途のために投与される。非ウイルスベクター送達システムには、DNAプラスミド、ネイキッド核酸、およびリポソームまたはポロキサマー等の送達媒体と複合体を形成した核酸が含まれる。ウイルスベクター送達システムには、細胞への送達後にエピソームまたは組み込まれたゲノムのいずれか一方を有する、DNAおよびRNAウイルスが含まれる。遺伝子療法手順の総説に関しては、Anderson,Science 256:808−813(1992)、Nabel & Felgner,TIBTECH 11:211−217(1993)、Mitani & Caskey,TIBTECH 11:162−166(1993)、Dillon,TIBTECH 11:167−175(1993)、Miller,Nature 357:455−460(1992)、Van Brunt,Biotechnology 6(10):1149−1154(1988)、Vigne,Restorative Neurology and Neuroscience 8:35−36(1995)、Kremer & Perricaudet,British Medical Bulletin 51(1):31−44(1995)、Haddada et al.,in Current Topics in Microbiology and Immunology Doerfler and Bohm(eds.)(1995)、およびYu et al.,Gene Therapy 1:13−26(1994)を参照されたい。
【0122】
核酸の非ウイルス送達の方法には、エレクトロポレーション、リポフェクション、マイクロインジェクション、バイオリスティック、ビロソーム、リポソーム、免疫リポソーム、ポリカチオン、または脂質:核酸複合体、ネイキッドDNA、人工ビリオン、および薬剤により強化されたDNAの取込みが含まれる。例えば、Sonitron 2000 system(Rich−Mar)を使用するソノポレーションも、核酸の送達に使用することができる。
【0123】
さらなる例示的な核酸送達システムには、Amaxa Biosystems(Cologne,Germany)、Maxcyte,Inc.(Rockville,Maryland)、BTX Molecular Delivery Systems(Holliston,MA)、およびCopernicus Therapeutics Inc,(例えば、米国特許第6008336号を参照)により提供されるものが含まれる。リポフェクションは、例えば、米国特許第5,049,386号、同第4,946,787号、および同第4,897,355号明細書)に記載されており、リポフェクション試薬は、市販されている(例えば、Transfectam(商標)およびLipofectin(商標))。ポリヌクレオチドの効率的な受容体認識リポフェクションに好適な陽イオン性および中性脂肪には、Felgnerのもの、国際公開第91/17424号、国際公開第91/16024号が含まれる。送達は、細胞(エクスビボ投与)または標的組織(インビボ投与)に送達することができる。
【0124】
免疫脂質複合体等の標的リポソームを含む脂質:核酸複合体の調製は、当業者によく知られている(例えば、Crystal,Science 270:404−410(1995)、Blaese et al.,Cancer Gene Ther.2:291−297(1995)、Behr et al.,Bioconjugate Chem.5:382−389(1994)、Remy et al.,Bioconjugate Chem.5:647−654(1994)、Gao et al.,Gene Therapy 2:710−722(1995)、Ahmad et al.,Cancer Res.52:4817−4820(1992)、米国特許第4,186,183号、同第4,217,344号、同第4,235,871号、同第4,261,975号、同第4,485,054号、同第4,501,728号、同第4,774,085号、同第4,837,028号、および同第4,946,787号明細書を参照されたい)。
【0125】
送達のさらなる方法には、送達されるべき核酸をEnGeneIC送達ビヒクル(EDV)にパッケージングする方法も含まれる。これらのEDVは、二重特異性抗体を用いて標的組織に特異的に送達され、ここで、抗体の一方のアームには、標的組織に対する特異性を有し、もう一方には、EDVに対する特異性を有する。抗体は、標的細胞表面にEDVを取り込まれ、次いで、EDVが、エンドサイトーシスによって細胞に取り込まれる。一旦細胞内に取り込まれると、内容物が放出される(MacDiarmid et al(2009)Nature Biotechnology 27(7):643を参照)。
【0126】
改変されたZFPをコードする核酸の送達のためのRNAまたはDNAウイルスに基づくシステムの使用は、ウイルスに体内の特定の細胞を標的とさせ、ウイルスペイロードを核に輸送するための高度に進化したプロセスを利用する。ウイルスベクターは、患者に直接投与(インビボ)することができるか、またはインビトロで細胞を治療するために使用することができ、修飾された細胞が患者に投与される(エクスビボ)。ZFPの送達のための従来のウイルスに基づくシステムには、遺伝子送達のためのレトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、ワクチン、および単純ヘルペスウイルスベクターが含まれるが、これらに限定されない。宿主ゲノム内の組込みにより、レトロウイルス、レンチウイルス、およびアデノ随伴ウイルス遺伝子送達方法を使用して可能であり、しばしば、挿入された導入遺伝子が長期発現される。さらに、高い導入効率が、多くの異なる細胞型および標的組織で観察されている。
【0127】
レトロウイルスの向性は、外来のエンベロープタンパク質を組み込むことによって変更することができ、標的細胞の潜在的な標的群を伸長する。レンチウイルスベクターは、非分裂細胞を形質導入または感染させることが可能であり、典型的には高ウイルス価を産生する。レトロウイルス遺伝子送達システムの選択は、標的組織に依存する。レトロウイルスベクターは、最大6〜10kbの外来配列のパッケージング能力を有する、シス作用の長い末端反復から成る。最小限のシス作用LTRは、ベクターの複製およびパッケージングに十分であり、それらは、次いで、恒久的な導入遺伝子発現を提供するように治療遺伝子を標的細胞内に組み込むために使用される。広範囲にわたって使用されるレトロウイルスベクターには、マウス白血病ウイルス(MuLV)、テナガザル白血病ウイルス(GaLV)、サル免疫不全ウイルス(SIV)、ヒト免疫不全ウイルス(HIV)、およびその組み合わせに基づくものが含まれる(例えば、Buchscher et al.,J.Virol.66:2731−2739(1992)、Johann et al.,J.Virol.66:1635−1640(1992)、Sommerfelt et al.,Virol.176:58−59(1990)、Wilson et al.,J.Virol.63:2374−2378(1989)、Miller et al.,J.Virol.65:2220−2224(1991)、PCT/US第94/05700号を参照されたい)。
【0128】
一過性発現が好ましい用途では、アデノウイルスに基づくシステムを使用することができる。アデノウイルスに基づくベクターは、多くの細胞型において非常に高い導入効率があり、細胞分裂を必要としない。このようなベクターを使用して、高力価および高レベルの発現が得られている。比較的単純なシステムにおいて、このベクターを多量に産生することができる。アデノ随伴ウイルス(「AAV」)ベクターを使用して、例えば、核酸およびペプチドのインビトロ産生において、およびインビボおよびエクスビボ遺伝子治療手順(例えば、West et al.,Virology 160:38−47(1987)、米国特許第4,797,368号明細書、国際公開第93/24641号、Kotin,Human Gene Therapy 5:793−801(1994)、Muzyczka,J.Clin.Invest.94:1351(1994)を参照)のために、細胞に標的核酸を形質導入する。AAVベクターの構築は、米国特許第5,173,414号明細書、Tratschin et al.,Mol.Cell.Biol.5:3251−3260(1985)、Tratschin et al.,Mol.Cell.Biol.4:2072−2081(1984)、Hermonat & Muzyczka,PNAS 81:6466−6470(1984)、およびSamulski et al.,J.Virol.63:03822−3828(1989)を含む、多数の刊行物に記載される。
【0129】
現在、少なくとも6つのウイルスベクターアプローチが臨床試験における遺伝子導入に使用可能であり、それらは形質導入剤を生成するためにヘルパー細胞株に挿入された遺伝子による欠損ベクターの相補性を含むアプローチを利用する。
【0130】
pLASNおよびMFG−Sは、臨床試験で使用されているレトロウイルスベクターの例である(Dunber et al.,Blood 85:3048−305(1995)、Kohn et al.,Nat.Med.1:1017−102(1995)、Malech et al.,PNAS 94:22 12133−12138(1997))。PA317/pLASNは、遺伝子治療試験で使用された最初の治療ベクターであった。(Blaese et al.,Science 270:475−480(1995))。MFG−Sパッケージされたベクターに対して、50%またはそれより大きい導入効率が観察されている。(Ellem et al.,Immunol Immunother.44(1):10−20(1997)、Dranoff et al.,Hum.Gene Ther.1:111−2(1997)。
【0131】
アデノ随伴ウイルスベクター(rAAV)は、欠損および非病原性パルボウイルスアデノ随伴2型ウイルスに基づく有望な代替遺伝子送達システムである。全てのベクターは、導入遺伝子発現カセットに隣接するAAV145bp逆方向末端反復のみを保持するプラスミドに由来する。形質導入細胞のゲノム内への組込みによる効率的な遺伝子導入および安定した導入遺伝子送達は、このベクター系の重要な特長である。(Wagner et al.,Lancet 351:9117 1702−3(1998)、Kearns et al.,Gene Ther.9:748−55(1996))。9:748−55(1996))。AAV1、AAV3、AAV4、AAV5、AAV6、およびAAV8を含む他のAAV血清型も本発明に従って使用することができる。
【0132】
複製欠損性アデノウイルスベクター(Ad)を高力価で産生することができ、多数の異なる細胞型を容易に感染させる。アデノウイルスベクターの大半は、導入遺伝子がAdE1a、E1b、および/またはE3遺伝子を置換し、その後、複製欠損性ベクターが、トランス内の消失遺伝子機能を供給するヒト293細胞で増殖するように改変される。Adベクターは、肝臓、腎臓、および筋肉で見出されるもの等の非分裂の分化した細胞を含む、複数の種類の組織をインビボで形質転換することができる。従来のAdベクターは、大きな担持能力を有する。臨床試験におけるAdベクターの使用の例には、筋肉内注入を用いる抗腫瘍免疫のポリヌクレオチド治療が含まれた(Sterman et al.,Hum.Gene Ther.7:1083−9(1998))。臨床試験における遺伝子導入のためのアデノウイルスベクターの使用のさらなる例には、Rosenecker et al.,Infection 24:1 5−10(1996)、Sterman et al.,Hum.Gene Ther.9:7 1083−1089(1998)、Welsh et al.,Hum.Gene Ther.2:205−18(1995)、Alvarez et al.,Hum.Gene Ther.5:597−613(1997)、Topf et al.,Gene Ther.5:507−513(1998)、Sterman et al.,Hum.Gene Ther.7:1083−1089(1998)が含まれる。
【0133】
パッケージング細胞を使用して、宿主細胞を感染させることが可能なウイルス粒子を形成する。このような細胞には、アデノウイルスをパッケージングする293細胞、およびレトロウイルスをパッケージングするψ2細胞またはPA317が含まれる。遺伝子治療で使用されるウイルスベクターは、通常、核酸ベクターをウイルス粒子中にパッケージングする産出細胞株によって生成される。これらのベクターには、典型的には、パッケージングおよびその後の宿主への組込みに必要とされる最小限のウイルス配列(適用可能な場合)、発現されるようにタンパク質をコードする発現カセットによって置換されている他のウイルス配列が含まれる。欠損しているウイルス機能は、パッケージングする細胞株によってトランスで提供される。例えば、遺伝子治療に使用するAAVベクターは、典型的には、宿主ゲノム内へのパッケージングおよび組込みに必要とされるAAVゲノムからの逆方向末端反復(ITR)配列のみを所有する。ウイルスDNAは、細胞株内にパッケージングされ、他のAAV遺伝子、すなわち、repおよびcapをコードするが、ITR配列を欠失するヘルパープラスミドを含有する。この細胞株はまた、ヘルパーとしてアデノウイルスに感染する。このヘルパーウイルスは、ヘルパープラスミドから、AAVベクターの複製、およびAAV遺伝子の発現を促進する。このヘルパープラスミドは、ITR配列の欠失の結果、相当な量でパッケージングされない。アデノウイルスの汚染は、例えば、AAVよりも敏感であるアデノウイルスに対する熱処理によって減少することができる。
【0134】
多くの遺伝子治療用途において、遺伝子治療ベクターが高度の特異性で特定の細胞型に送達されることが望ましい。したがって、ウイルスの外表面上でウイルス外被タンパク質との融合タンパク質としてリガンドを発現することによって、ウイルスベクターを所定の細胞型に対する特異性を有するように修飾することができる。リガンドは、関心の細胞型上に存在することで知られている受容体に対する親和性を有するように選択される。例えば、Han et al.,Proc.Natl.Acad.Sci.USA 92:9747−9751(1995)は、gp70に融合されたヒトヘレグリンを発現するように、モロニーマウス白血病ウイルスを修飾することができ、そのウイルスが、ヒト上皮成長因子受容体を発現するある特定のヒト乳癌細胞を感染させることを報告した。この原理は、細胞表面受容体に対して、標的細胞が受容体を発現し、ウイルスがリガンドを含む融合タンパク質を発現する、他のウイルス標的細胞対にまで及ぶことができる。例えば、実質的に任意の選択された細胞受容体に特異的結合親和性を有する抗体断片(例えば、FABまたはFv)を表示するように、線状ファージを改変することができる。上記の説明は、主にウイルスベクターに適用されるが、同一の原理を非ウイルスベクターに適用することができる。特異的標的細胞による取り込みを好む特異的な取り込み配列を含有するように、そのようなベクターを改変することができる。
【0135】
遺伝子治療ベクターは、下で説明するように、個々の患者への投与、典型的には、全身投与(例えば、静脈内、腹腔内、筋肉内、皮下、もしくは頭蓋内注入)または局所投与によって、インビボで送達することができる。代替的に、個々の患者から移植された細胞(例えば、リンパ球、骨髄穿刺液、組織生検)または万能ドナーの幹細胞等の細胞に、ベクターをエクスビボで送達することができ、その後、ベクターを組み込んだ細胞の選択後に、患者への細胞の再移植が続く。
【0136】
診断、研究のため、または遺伝子治療のためのエクスビボ細胞トランスフェクション(例えば、宿主生物にトランスフェクトされた細胞の再注入を介して)が当業者によく知られている。好ましい実施形態において、細胞は、対象生物から単離され、ZFP核酸(遺伝子またはcDNA)でトランスフェクトし、対象生物(例えば、患者)内に戻って再注入される。エクスビボのトランスフェクションに好適な多様な細胞型は、当業者によく知られている(患者からの細胞の単離および培養方法についての説明は、例えば、Freshney et al.,Culture of Animal Cells,A Manual of Basic Technique(3rd ed.1994)、およびそこで引用される参考文献を参照されたい)。
【0137】
一実施形態において、幹細胞は、細胞トランスフェクションおよび遺伝子治療のためのエクスビボ処置で使用される。幹細胞を使用する利点は、それらがインビトロで他の細胞型に分化することができるか、または、それらが骨髄に生着する哺乳動物(幹細胞のドナー等)に導入し得ることである。GM−CSF、IFN−γ、およびTNF−α等のサイトカインを使用して、CD34+細胞を臨床的に重要な免疫細胞型にインビトロで分化するための方法が知られている(Inaba et al.,J.Exp.Med.176:1693−1702(1992)を参照)。
【0138】
幹細胞は、周知の方法を使用して、形質導入および分化のために単離される。例えば、幹細胞は、CD4+およびCD8+(T細胞)、CD45+(panB細胞)、GR−1(顆粒球)、およびIad(分化された抗原提示細胞)等の不要な細胞に結合する抗体を有する骨髄細胞をパンニングすることによって骨髄から単離される(Inaba et al.,J.Exp.Med.176:1693−1702(1992)を参照)。
【0139】
いくつかの実施形態において、修飾されている幹細胞も、使用され得る。例えば、アポトーシスに耐性をなしている神経幹細胞は、治療組成物として使用され得、ここで、幹細胞はまた、本発明のZFP TFも含有する。アポトーシスに対する耐性は、例えば、幹細胞において、BAXまたはBAKに特異的なZFNを用いて(米国特許出願第12/456,043号を参照)BAXおよび/またはBAK、または例えば、カスパーゼ6に特異的なZFNを用いて再度、カスパーゼにおいて妨害されるものを不活性化することによって生じ得る。これらの細胞は、変異または野生型Httを調節することが知られているZFP TFでトランスフェクトすることができる。
【0140】
治療的ZFP核酸を含有するベクター(例えば、レトロウイルス、アデノウイルス、リポソーム等)を、細胞の形質導入のために、生物にインビボで直接投与することもできる。代替的に、ネイキッドDNAを投与することができる。投与は、注射、注入、局所適用、およびエレクトロポレーションを含むが、これらに限定されない、血液細胞または組織細胞との最終的な接触内に分子を導入するために通常使用される経路のいずれかによる。このような核酸を投与する好適な方法は、使用可能であり、かつ当業者にも知られており、1つ以上の経路を使用して特定の組成物を投与することができるが、特定の経路は、しばしば、別の経路よりも即時かつ有効な反応を提供することができる。
【0141】
造血幹細胞にDNAを導入するための方法が、例えば、米国特許第5,928,638号明細書に開示されている。トランス遺伝子の、例えば、CD34+細胞等の造血幹細胞への導入に有用なベクターには、アデノウイルス35型が含まれる。
【0142】
トランス遺伝子の免疫細胞(例えば、T細胞)への導入に好適なベクターには、非組込みレンチウイルスベクターが含まれる。例えば、Ory et al.(1996)Proc.Natl.Acad.Sci.USA 93:11382−11388、Dull et al.(1998)J.Virol.72:8463−8471、Zuffery et al.(1998)J.Virol.72:9873−9880、Follenzi et al.(2000)Nature Genetics 25:217−222を参照されたい。
【0143】
薬学的に許容される担体は、投与されている特定の組成物によって、ならびに組成物を投与するために使用される特定の方法によってある程度決定される。したがって、下で説明するように、使用可能な薬学的組成物の多種多様の好適な製剤が存在する(例えば、Remington‘s Pharmaceutical Sciences,17th ed.,1989を参照)。
【0144】
上述のように、開示される方法および組成物には、原核細胞、真菌細胞、古細菌細胞、植物細胞、昆虫細胞、動物細胞、脊椎動物細胞、哺乳動物細胞、およびヒト細胞が含まれるが、これらに限定されない、任意の種類の細胞で使用することができる。タンパク質発現に好適な細胞株が当業者に知られており、COS、CHO(例えば、CHO−S、CHO−K1、CHO−DG44、CHO−DUXB11)、VERO、MDCK、WI38、V79、B14AF28−G3、BHK、HaK、NS0、SP2/0−Ag14、HeLa、HEK293(例えば、HEK293−F、HEK293−H、HEK293−T)、perC6、昆虫細胞(ツマジロクサヨトウ(Sf)等)、および真菌細胞(サッカロミセス、ピチア、およびシゾサッカロミセス等)が含まれるが、これらに限定されない。これらの細胞株の子孫、変種、および誘導体を使用することもできる。
【0145】
用途
トリヌクレオチド反復疾患に関連する遺伝子を調節する、および/またはこれらの疾患に関連する遺伝子におけるトリヌクレオチド反復の経路を取り除くことが望ましい任意の用途のために、開示される組成物および方法を使用することができる。特に、治療および研究用途を含むが、これらに限定されない、Htt対立遺伝子の調節が望ましい任意の用途のために、これらの方法および組成物を使用することができる。
【0146】
ZFP TFを抑制するHD Httを治療剤として使用することができる疾患および状態には、ハンチントン病が含まれるが、これに限定されない。さらに、Httの変異対立遺伝子に対して特異的なZFNを含む方法および組成物をハンチントン病の治療のための治療剤として使用することができる。
【0147】
HD Htt対立遺伝子を抑制するZFP−TFはまた、GDNFおよびBDNFが含まれるが、これらに限定されない神経栄養因子を活性化するZFP−TFと連結して使用され得る。これらのZFP(またはこれらのZFPをコードするポリヌクレオチド)は、同時に投与され得る(例えば、同じ薬学的組成物において)か、または任意の順序で連続して投与され得る。
【0148】
また、ハンチントン病の治療のための方法および組成物には、幹細胞内でHtt対立遺伝子の変異コピーがHttに特異的なZFNを用いて、野生型Htt対立遺伝子に修飾されている幹細胞組成物も含まれる。
【0149】
本発明の方法および組成物はまた、インビトロおよびインビボモデル、例えば、トリヌクレオチド反復疾患の動物モデルの設計および実施に有用であり、これは、これらの疾患の研究を可能にする。
【実施例】
【0150】
実施例1:Htt標的亜鉛フィンガータンパク質転写因子(ZFP−TF)の設計および構築
Httを標的化した亜鉛フィンガータンパク質は、米国特許第6,534,261号明細書に記載されるように、基本的に改変された。表1は、例示的なHtt標的ZFPの認識へリックスDNA結合ドメインを示す。
【0151】
また、両端型ZFP−TFは、最終ベクターが、SIP1等の両端型ZFP−TFの遺伝子配列に隣接する2つのアレイのZFPを含むことを除いては、基本的に上記のような亜鉛フィンガー結合ドメインの2つの別個のクラスターを用いて構築される。図1Cおよび図4を参照されたい。
【0152】
多量体型ZFP TFはまた、このベクターが、ZFP TFをコードする配列に作動可能に連結されるトリヌクレオチド反復の経路に沿って発現タンパク質の多量体化を可能にする1つ以上の多量体化ドメインをコードする配列を含有することを除いては、上記のように構築される。図1Dおよび図3を参照されたい。
【0153】
ZFP TFは、核局在化配列、Htt対立遺伝子を標的とした改変された亜鉛フィンガーDNA結合ドメイン(表1を参照)、およびヒトKOX1タンパク質由来のKRAB抑制ドメインを含む、融合タンパク質として構築された。図1Aを参照されたい。設計されたDNA結合ドメインは、18−bp配列を認識する6個のフィンガー分子を含有する(表2を参照)。ZFP認識ヘリックスによって接触される標的部位におけるヌクレオチドは大文字で示され、非接触ヌクレオチドは小文字で示される。
【0154】
ELISAベースのアッセイを使用して、標的配列に結合するZFPを確認する。簡潔に言えば、T7プロモーターの制御下で、血球凝集素エピトープ(HA)標的化全長ZFPをコードする発現カセットは、HAタグのT7プロモーターおよびコード配列を導入するプライマーを用いて、組み立てられたZFPのそれぞれからのPCR増幅によって作製される。このPCR断片を使用して、TNT Quick Coupled TranscriptionおよびTranslation System(Promega,WI,USA)を用いることによって、インビトロでHA標識化ZFPタンパク質を生成し、HA標識化タンパク質は、次いで、Htt遺伝子内の意図される標的部位を含有するビオチン化DNAオリゴ、ならびに競合体(ヒトゲノム)DNAと混合し、特異的ZFP−DNA認識のみ報告することが可能である。
【0155】
次いで、ZFP−DNA複合体は、ペルオキシダーゼ共役抗HA抗体で標識化され、続いて、ストレプトアビジンでコーティングされた96ウェルプレート上に捕捉する。次いで、特異的ZFP−DNA複合体は、基質として、QuantaBlu(Pierce,Rockford,IL)を用いて、捕捉したペルオキシダーゼの活性をアッセイすることによって定量化する。マウスまたはヒトHttプロモーターおよびエクソン1に対して特異的なZFP TFは、標的配列に結合するために設計および試験される。
【0156】
実施例2:ヒトおよびマウス細胞におけるHD Httの抑制。
ZFP TFを抑制するHttの活性を試験するために、ZFP TFをヒト細胞にトランスフェクトし、PCRを用いてHttの発現を観察した。
【0157】
初めに、表1中に示される標準ZFP TFを試験した。ヒト293細胞(Graham et al(1977).J Gen Virol 36:59−74)を、10% FBSで捕捉したDMEM中に培養した。96ウェルプレートに、1ウェルにつき1e4細胞の密度で播種し、翌日、製造業者の指示に従って、Fugene6試薬を用いて、18832、18856、18859、および18868のZFP−TFをコードする0.1ugのプラスミドDNAでトランスフェクトした。トランスフェクトした細胞を、3日間インキュベートし、次いで、遺伝子発現分析のためにCells−to−CT(商標)キット(Applied Biosystems)を用いてプロセスした。内部対照のベータアクチン(ACTB)と比較した内因性ヒトハンチンチン(Htt)のレベルを、Hs00918176_m1および4352935Eのプライマーおよびプローブ(Applied Biosystems)をそれぞれ用いて、TaqMan(登録商標)7300上のリアルタイムPCRによって分析した。
【0158】
図2に示されるように、ZFP−TFは、GFPおよび偽対照と比較して、Htt発現を抑制した。結果は、ACTBの内因性レベルに対する、基底Httの割合で示される。
【0159】
両端型および多量体型ZFP TFの活性を試験するために、同じプロトコルを、両端型および多量体型ZFPを用いて行う。
【0160】
ヒトHD Htt対立遺伝子に対する特異性を有するZFP TFを試験するために、Htt特異的ZFP TFを、SH−SY5Y神経芽細胞腫細胞(Biedler JL,et al.(1978)Cancer Res.38:3751−3757)に過渡的にトランスフェクトする。簡潔に言えば、組み立てられたZFP−TF構築物を、pcDNAベクター(InVitrogen)にクローン化し、製造業者のプロトコルに従って、Fugene6キット(Roche Applied Science)を用いてトランスフェクトする。Htt発現レベルは、Applied Biosystemsから購入したHs00918176_m1プライマー/プローブのセットを用いて、リアルタイムRT−PCR(TaqMan(登録商標)、Applied Biosystems)を用いて測定する。Httタンパク質の減少を確認するために、ウエスタンブロット解析を行う。
【0161】
マウス対立遺伝子と相互作用するように設計されたHtt特異的ZFP TFを試験するために、マウスZFP TFをコードする構築体をpcDNAベクターにクローン化し、製造業者のプロトコルに従って、Lipofectamine 2000キット(Invitrogen)を用いて、ニューロ2A細胞(Klebe & Ruddle(1969)J.Cell Biol.43:69A)またはマウスHttを発現する同等のマウス神経細胞株に過渡的にトランスフェクトする。Htt発現レベルを、リアルタイムRT−PCRを用いて測定する。Httタンパク質の減少を確認するために、ウエスタンブロット解析を行う。
【0162】
加えて、ヒトHtt特異的ZFP TFを、CAG伸長を有する切断型ヒトHtt対立遺伝子を担持するR6.2トランスジェニックマウス由来の一次神経細胞において試験する(Mangiarini et al.,(1996)Cell 87:493-506)。マウスHtt特異的ZFP TF、ならびにCAG反復(両端型ZFPおよび多量体化ドメインを有するZFP)を標的化するZFPを、Httノックインマウス由来の不死化線条体細胞において試験する(Wheeler et al.,(1999)Hum Mol Genet.Jan;8(1):115−22)。
【0163】
実施例3:インビボでのHtt発現の抑制
インビボでのHtt特異的ZFP TFを試験するために、ZFPをコードするAAV2ベクターを産生する。次いで、これらのAAV2ベースの構築体を、マウスの脳に送達する。ヒトHtt特異的ZFP TFにおいては、AAVベクターをR6.2マウスまたはBAC HDマウス(C57Bl/6またはFVB/N株)に送達し、ヒト導入遺伝子の抑制を評価する。マウスHtt特異的ZFPにおいては、AAVベクターを野生型マウス(C57Bl/6またはFVB/N)に送達し、内因性マウスHtt発現の抑制を評価する。CAGを伸長した対立遺伝子を優先的に標的化するZFPにおいては、AAVベクターをR6.2マウスまたはヒトHttノックインマウスに送達し、野生型対伸長したHtt対立遺伝子の選択的抑制を検証する。殺処分後、脳組織を、TaqmanリアルタイムRT−PCRによって、Htt発現に対して分析する。
【0164】
実施例4:神経栄養因子およびHD Htt対立遺伝子特異的なZFP TFの共トランスフェクション。
上で同定されるHtt特異的ZFP TFは、脳神経栄養因子に対して特異的なZFP TFで共トランスフェクトされる。使用した脳神経栄養因子に対して特異的なZFP TFは、GDNFあるいはBDNFのいずれかに対して特異的である。
【0165】
実施例5:Htt標的亜鉛フィンガーヌクレアーゼ(ZFN)の設計および構築
ヒトHttおよびマウスHttを標的化するZFNは、CAG反復に隣接する配列、ならびに最初および最後のコーディングエクソンの配列を標的化するように設計される。ZFNは、Urnov et al.(2005)Nature 435(7042):646−651、Perez et al(2008)Nature Biotechnology 26(7):808−816、および米国特許出願公開第2008/0131962号明細書に記載されるように基本的に設計され、プラスミドまたはアデノウイルスベクターに組み込まれた。
【0166】
上記のように、組み立てられたZFNを、ELISAによって、それらのそれぞれの配列に結合させるために試験する。
【0167】
実施例6:Htt特異的ZFNの切断活性
上記のヒトHtt特異的ZFNの対をコードするプラスミドを、K562細胞にトランスフェクトする。K562細胞は、American Type Culture Collectionから入手して、10%規格ウシ胎仔血清(FCS、Cyclone)を補足したF−12培地(Invitrogen)中で推奨されるように、成長させた。細胞を、TrypLE Select(商標)プロテアーゼ(Invitrogen)を用いて、プラスチック容器から引き離した。トランスフェクションのために、100万個のK562細胞を、2μgの亜鉛フィンガーヌクレアーゼプラスミドおよび100μL Amaxa溶液Tと混合した。細胞を、プログラムU−23を用いてAmaxa Nucleofector II(商標)中でトランスフェクトし、1.4mLの温F−12培地+10%FCS中に回収した。
【0168】
ゲノムDNAを収穫し、意図された切断部位を含むHtt遺伝子座の一部をPCR増幅する。InVitrogenからのAccuprime HiFiポリメラーゼを用いたPCRを、以下の通りに実行する。94℃で最初の3分間変性させた後、PCRの30サイクルを、94℃で30秒間の変性ステップを行い、続いて、58℃で30秒間のアニーリングステップ、68℃で30秒間の伸長ステップを行う。30サイクルが完了した後、反応物を68℃で7分間インキュベートし、次いで、10℃で無制限に行う。
【0169】
K562 Htt特異的ZFNで処理された細胞からのゲノムDNAは、例えば、米国特許出願公開第20080015164号、第20080131962号、および第20080159996号明細書に記載される、Surveyor(商標)ヌクレアーゼ(Transgenomic)によって検証される。
【0170】
マウスHtt特異的ZFNの対をコードするプラスミドを、同様の様式で、ニューロ2a細胞中で試験する。
【0171】
実施例7:可変長のトリヌクレオチド反復の標的化組込み
上記の、CAG反復に隣接する配列に対して最大の切断活性を有するHtt特異的ZFNを、標的化組込み戦略に使用して、Httの野生型コピーへの可変長のCAG反復を導入する。50、80、109、および180のCAG反復単位を含むドナーが、構築される。次いで、これらのドナーは、上記のように、Htt特異的ZFNをコードするプラスミドを用いて、K562細胞にトランスフェクトされる。ドナー組込みの検証は、ゲノムDNA単離、PCT増幅(上記のような)、続いて、対象となる領域のシークエンシングによって達成される。
【0172】
Htt対立遺伝子へのドナー対立遺伝子の標的化組込みさせるK562細胞中で同定されたZFNを使用して、ヒト胚幹細胞(hESC)に可変長のドナー核酸を挿入する。成功したドナー組込みは、上記のように、ゲノムDNA単離、PCR、およびシークエンシングによって確認される。
【0173】
実施例8:野生型およびHDマウスHtt対立遺伝子の発現タグ
最初または最後のコーディングエクソンに対して最大の切断活性を有するZFNを使用して、異なるレポータータンパク質を用いて、野生型および変異Htt対立遺伝子を標識化する。それぞれのレポーター(AおよびB)に対してドナーDNAは、レポーター遺伝子の標的化組込みが、Httへのインフレーム融合を産生することができるように、リードZFN対の切断部位に基づいて設計される。ドナーDNAは、組込みの最も高い頻度を与えるドナーDNA構築体を選択するために、リードZFN対を用いて、ニューロ2A細胞に共トランスフェクトされる。
【0174】
ZFN対25920/25921および25922/25923は、上記のように調製され、実施例6に記載されるように、Cel Iミスマッチを用いて、切断活性について試験した。これらのZFN対は、Htt遺伝子配列の3’末端を標的化し、ひいては、野生型Htt対立遺伝子あるいは変異Htt対立遺伝子のいずれかを標的化するために使用され得る。結果を図5に示す。図から見られるように、これらの2つの対は、Htt遺伝子を切断することが可能であり、したがって、レポーターの導入のために利用することができる。
【0175】
対応するZFNと共にレポーターAに対して選択されたドナーDNA構築体は、長く伸びるHtt対立遺伝子(例えば、ヒトHttノックインマウス)を含むモデルマウス由来のマウス胚幹細胞に送達される。レポーターAの標的化組込みのために、クローンが、得られ、スクリーニングされる。ヘテロ接合体の現象が望ましく、標的化対立遺伝子は、PCRによって同定される。単一のレポーター標識化Htt対立遺伝子および他の対立遺伝子において非修飾のZFN標的配列を含むクローンが、選択され、レポーターBおよび対応するZFNのドナー構築体は、レポーターBを用いて第2の対立遺伝子を標識化するようにトランスフェクトされる。
【0176】
得られたマウス胚幹細胞クローンは、それぞれの対立遺伝子から発現の追跡を可能する2つの異なるマーカーを用いて標識化した野生型Htt対立遺伝子および変異対立遺伝子を含み、標準プロトコルを用いて、これらの細胞を使用して、トリヌクレオチド反復疾患のマウスモデルを生成する。
【0177】
本明細書で言及された全ての特許、特許出願、および刊行物は、参照によりそれらの全体が組み込まれる。
【0178】
理解の明瞭化を目的とした図示および例のために、本開示を多少詳しく提供したが、本開示の趣旨または範囲から逸脱することなく、多様な変更および修正を実施することができることは当業者には明らかである。したがって、前述の説明および例を限定的であると見なすべきではない。
【技術分野】
【0001】
関連出願の相互参照
本出願は、2009年7月28日に出願された米国仮出願第61/271,913号、および2009年7月29日に出願された同第61/273,009号の利益を主張し、これらの開示は、参照によりその全体が本明細書に組み込まれる。
【0002】
連邦支援による研究下でなされた発明に対する権利の記載
該当せず。
【0003】
技術分野
本開示は、遺伝子発現の分野にある。
【背景技術】
【0004】
背景技術
トリヌクレオチド反復伸長疾患は、1990年初期に初めて特徴付けられた(Di Prospero and Fischbeck,(2005)Nature Reviews Genetics vol 6:756−765を参照のこと)。これらの疾患は、3個のヌクレオチドのセットの不安定な反復の局所的伸長を有し、この反復が存在する遺伝子の機能喪失、毒性機能の増加、または両方を引き起こし得る。トリヌクレオチド反復は、非コードおよびコード遺伝子領域を含む、遺伝子のあらゆる部分に位置し得る。コード領域内に位置する反復には、一般的に、トリプレットをコードする反復したグルタミン(CAG)またはトリプレットをコードするアラニン(CGA)のいずれかが含まれる。非コード配列内の長く伸びる反復領域は、遺伝子の異常な発現を引き起こし得る一方で、コード領域内の長く伸びる反復(コドン反復疾患としても知られている)は、ミスフォールディングおよびタンパク質凝集を引き起こし得る。異常なタンパク質に関連する病態生理学的な正確な原因は、多くの場合、知られていない。一般的に、トリヌクレオチド伸長を起こしやすい野生型遺伝子において、これらの領域は、正常な集団において可変数の反復配列を含むが、罹患集団において、反復の数は、場合によっては、反復の数の単純な2倍から、反復の数の対数で1桁まで増加し得る。例えば、脆弱X症候群患者におけるCGG伸長を受けるFMR1遺伝子において、野生型集団は、2〜50の反復を示す一方で、脆弱X症候群に罹患しているこれらの患者は、200〜2000のCGG反復を有し得る(Nadel et al(1995)Journal Biological Chemistry 270(48):28970−28977)。
【0005】
今日まで、20種類の異なる疾患が、長く伸びるトリヌクレオチド反復に関連付けられている(Di ProsperoおよびFischbeckの同書を参照のこと)。この現象は最初に、アンドロゲン受容体のコード領域においてCAG反復が長く伸びている球脊髄性筋萎縮症(SBMA)に関して記載された。野生型遺伝子の反復は、通常は13〜30のCAGを含むが、SBMA患者は、40以上ものCAGを含み得る。長く伸びるトリヌクレオチド反復によって特徴付けられる他の疾患には、フリードライヒ運動失調症(反復はFratazin遺伝子の非コード領域内にある)、脆弱X症候群AおよびE(反復はそれぞれ、FMR1およびFMR2遺伝子の非コード領域内にある)、および反復が巨大サイトゾルタンパク質のハンチンチン(Htt)のN末端コード領域内に挿入されるハンチントン病が含まれる。それぞれのポリグルタミン伸長疾患は、脳の特定領域に神経細胞脱落が明らかである特徴的病変を示す。P/Q Ca2+チャンネル、TATAボックス結合タンパク質、およびアトロフィン−1におけるポリグルタミン伸長はそれぞれ、脊髄小脳失調(SCA)−6、SCA−17、および歯状核赤核淡蒼球ルイ体萎縮症(DRPLA)を引き起こす。それらのポリグルタミン反復は別として、これらの疾患に関与するタンパク質は全て、中枢神経系および末梢組織の両方において広範に発現するが、関連性はない。
【0006】
ハンチントン舞踏病としても知られているハンチントン病(HD)は、運動、認識、および精神疾患の進行性疾患である。この疾患の発症の平均年齢は、35〜44歳であるが、症例の約10%においては、発症が21歳前に生じ、疾患診断後の平均寿命は、15〜18年である。有病率は、西ヨーロッパ系の100,000人の間で約3〜7人である。通常のHtt対立遺伝子は、15〜20のCAG反復を含むが、35以上の反復を含む対立遺伝子は、潜在的にHDを生じる対立遺伝子であると見なすことができ、疾患を発症する危険性を与える。36〜39の反復を含む対立遺伝子は、不完全に浸透されると見なされ、これらの対立遺伝子を含むこれらの個人は、疾患を発症し得るか発症し得ない(または後年症状を発症し得る)一方で、40以上の反復を含む対立遺伝子は、完全に浸透されると見なされ、これだけ多くの反復を有するHD対立遺伝子を含む無症候性の人は報告されていない。若年発症HDを有するこれらの個人(21歳を下回る)は、しばしば、60以上のCAG反復を有することが見出される。CAG反復の増加に加えて、HDが、この領域が、ポリグルタミンよりもむしろポリセリンポリペプチド(+1フレームシフトの場合に、AGC反復によってコードされる)経路をコードするように、反復配列内で+1および+2フレームシフトを含むことができることも示されている(Davies and Rubinsztein(2006)Journal of Medical Genetics 43:893−896)。
【0007】
ハンチントン病は、HD対立遺伝子が、通常、優勢な特性として1人の親から遺伝子を受け継ぐ遺伝的疾患である。HD患者から生まれるどの子も、もう一方の親がこの疾患に罹患していない場合、疾患を発症する可能性が50%である。場合によっては、親は、中間HD対立遺伝子を有し、無症候性であり得る一方で、伸長を反復するため、その子に疾患が現れる。加えて、HD対立遺伝子はまた、見込みとして知られる現象を示すこともでき、重症度の増加または発症年齢の低下が、精子形成中の反復領域の不安定な性質のため、数世代にわたって観察される。
【0008】
HDにおいて、トリヌクレオチド伸長は、線条体中で中型有棘ガンマアミノ酪酸(GABA)投射神経細胞の神経細胞の欠損を引き起こし、神経細胞の欠損は新皮質においても生じる。エンケファリンを含み、外部淡蒼球へ突き出る中型有棘神経細胞は、基質Pを含み、内部淡蒼球へ突き出る神経細胞よりも病変する。ハンチントン病に罹患している人々に大いに影響を及ぼす他の脳の部位には、黒質、皮質3、5、および6、海馬のCA1領域、頭頂葉の各回、小脳のプルキンエ細胞、視床下部の外側隆起核、および視床の正中中心核−束傍核複合体が含まれる(Walker(2007)Lancet 369:218−228)。正常なHttタンパク質の役割は、不明のところが多いが、神経新生、アポトーシス細胞死、および小胞輸送に関与し得る。加えて、野生型Httは、脳由来神経栄養因子(BDNF)、および線条体ニューロンの生存促進性因子を刺激することを証明する。HDの進行は、HDのマウスモデルにおけるBDNF発現の減少と相関する(Zuccato et al(2005)Pharmacological Research 52(2):133−139)、およびアデノ随伴ウイルス(AAV)ベクター媒介遺伝子によってBDNFあるいはグリア細胞株由来神経栄養因子(GDNF)のいずれかの送達が、HDのマウスモデルにおいて、線条体ニューロンを保護し得る(Kells et al,(2004)Molecular Therapy 9(5):682−688)ことが示されている。
【0009】
HDに対する治療法の選択肢は、現在、非常に限られている。シャペロニンの過剰発現または化合物ゲルダナマイシンによる熱ショック応答の産生等の長く伸びるポリグルタミン経路を通して生じるタンパク質凝集に関連する毒性を防止するように設計されるいくつかの潜在的な手法は、インビトロモデルにおけるこれらの毒性の減少を示している。他の治療は、疾患の臨床的指標においてアポトーシスの役割を標的としている。例えば、一方の親は、HD対立遺伝子を含み、もう一方の親が、カスパーゼ1に対して優性阻害対立遺伝子を有するマウスの対合の子孫において、動物モデルのカスパーゼ活性の妨害によって、病徴の遅延が示される。さらに、カスパーゼによるHD Httの切断は、疾患の病原性における役割を果たし得る。カスパーゼ6の耐性変異Httを担持するトランスジェニックマウスは、正常な神経機能を維持することが見出され、非カスパーゼの耐性変異Htt対立遺伝子を担持するマウスと比較して、線条体神経変性を引き起こさなかった(Graham et al(2006)Cell 125:1179−1191を参照されたい)。また、アポトーシス経路のメンバーを標的とする分子は、症候学において遅延する影響があることも示している。例えば、化合物zVAD−fmkおよびミノサイクリン(これらの両者が、カスパーゼ活性を阻害する)は、マウスにおいて、疾患の兆候を遅延させることが示されている。また、薬物レマセミド(remacemide)は、この化合物が、神経細胞における毒作用の発揮を妨げるNDMA受容体への変異Httの結合を妨げることが考えられるため、小規模なHDヒト試験にも使用されている。しかしながら、統計的に有意な改善は、これらの試験におけるニューロン機能において、観察されなかった。加えて、ハンチントン研究グループは、コエンザイムQを用いて、ランダム化二重盲検試験を行った。コエンザイムQ10により処置された患者においてより遅延した疾患進行に関する傾向があったが、全体の機能的能力の下落率に有意な変化がなかった。(Di ProsperoおよびFischbeckの同書を参照のこと)。
【0010】
したがって、トリヌクレオチド反復疾患の治療のための組成物および方法が求められている。
【発明の概要】
【0011】
概要
トリヌクレオチド反復疾患を治療するための方法および組成物を本明細書に開示する。特に、トリヌクレオチド反復疾患を治療するために、トリヌクレオチド反復を含む遺伝子の発現を調節する、例えば、ハンチントン病を治療するためにHD Htt対立遺伝子の発現を調節するための方法および組成物が、本明細書に提供される。また、トリヌクレオチド反復疾患の動物モデルを生成するための方法および組成物も提供される。
【0012】
したがって、一態様において、HD対立遺伝子(例えば、Htt)の発現を調節する改変された亜鉛フィンガータンパク質が提供される。改変された亜鉛フィンガータンパク質とは、認識ヘリックスが事前選択された標的部位に結合するように変化された、天然に存在しない亜鉛フィンガータンパク質である。本明細書に記載される亜鉛フィンガータンパク質のいずれも、1個、2個、3個、4個、5個、6個以上の亜鉛フィンガーを含むことができ、それぞれの亜鉛フィンガーは、選択された配列(例えば、遺伝子)内の標的亜部位に結合する認識へリックスを有する。いくつかの実施形態において、該認識へリックスは、天然に存在しない。特定の実施形態において、亜鉛フィンガータンパク質は、表1に示される認識へリックスを有する。他の実施形態において、亜鉛フィンガータンパク質は、表2に示される標的配列に結合する。
【0013】
一態様において、長く伸びるCAG経路に優先的に結合することが可能であるが、野生型長さのCAG経路に対する親和性の低い、両端型ZFPリプレッサーが提供される。いくつかの実施形態において、長く伸びるトリヌクレオチド経路に優先的に結合するが、野生型長さのトリヌクレオチド反復経路に対する親和性の低い多量体型ZFP−TFが使用され、それによって、長く伸びる対立遺伝子の優先的な抑制が達成される。いくつかの実施形態において、これらの多量体型ZFP−TFは、長く伸びる対立遺伝子が、野生型対立遺伝子よりも多くのZFPによってさらに効率的に結合されるように、反復配列に対して協同的DNA結合を達成し、変異対立遺伝子の優先的な抑制を可能にする。いくつかの実施形態において、所与のサイズの多量体の安定した複合体を形成し、ひいては、ある最小サイズのCAG経路と優先的に相互作用することが可能である、多量体型ZFP TFが、使用され、その最小サイズは、野生型CAG経路の長さよりも長い。
【0014】
特定の実施形態において、本明細書に記載されるZFP(例えば、両端型、多量体型等)は、変異Htt対立遺伝子の発現を優先的に修飾する。いくつかの実施形態において、ZFPは、変異Htt対立遺伝子に特異的に結合し、長く伸びる経路は、ポリグルタミンをコードする一方、他の実施形態において、ZFPは、変異Htt対立遺伝子に特異的に結合し、長く伸びる経路は、ポリセリンをコードする。したがって、いくつかの実施形態において、ZFP−TFは、野生型および変異型のHtt対立遺伝子の両方を調節する。特定の実施形態において、ZFPは、野生型Htt対立遺伝子のみ調節する。他の実施形態において、ZFPは、変異型のHttのみ調節する。
【0015】
他の実施形態において、長く伸びるHDのHtt対立遺伝子に関連する既知のSNPに優先的に結合するZFP−TFの抑制が提供される。このように、ZFP−TFは、SNPを含む変異Htt対立遺伝子対して特異的であり、変異Htt対立遺伝子の特定の抑制を可能にする。別の態様において、野生型対立遺伝子に関連するSNPと相互作用することによって、野生型Htt対立遺伝子を特異的に活性化するZFP−TFが提供される。このように、野生型Htt対立遺伝子のみが活性化される。
【0016】
別の態様において、1つ以上の神経栄養因子の発現を調節する改変された亜鉛フィンガータンパク質は、トリヌクレオチド疾患を治療するために提供される。いくつかの実施形態において、調節される神経栄養因子は、BDNFおよび/またはGDNFである。改変された亜鉛フィンガータンパク質とは、認識ヘリックスが事前選択された標的部位に結合するように変化された、天然に存在しない亜鉛フィンガータンパク質である。本明細書に記載される亜鉛フィンガータンパク質のいずれも、1個、2個、3個、4個、5個、6個以上の亜鉛フィンガーを含むことができ、それぞれの亜鉛フィンガーは、選択された配列(例えば、遺伝子)内の標的亜部位に結合する認識へリックスを有する。いくつかの実施形態において、該認識へリックスは、天然に存在しない。
【0017】
特定の実施形態において、本明細書に記載される亜鉛フィンガータンパク質(ZFP)を、融合タンパク質の一部として、調節ドメイン(または機能ドメイン)と作動的連結状態に配置することができる。ZFPとの融合のために活性化ドメインまたは抑制ドメインのいずれか一方を選択することによって、そのような融合タンパク質は、遺伝子発現を活性化するか、または抑制するかのどちらか一方のために使用することができる。いくつかの実施形態において、変異Htt発現を下方制御するために使用され得る転写抑制ドメインに融合された、本明細書に記載される変異Httを標的化するZFPを含む融合タンパク質が提供される。いくつかの実施形態において、野生型Htt対立遺伝子を上方制御することができる転写活性化ドメインに融合された、野生型Htt対立遺伝子を標的化するZFPを含む融合タンパク質が提供される。特定の実施形態において、調節ドメインの活性は、細胞転写機構との相互作用が、外来リガンドの不在下で起こらないように、外来小分子またはリガンドによって調節される。このような外部リガンドは、ZFP−TFの転写機構との相互作用の程度を制御する。調節ドメインは、1つ以上のZFPの間、1つ以上のZFPの外部、およびこれらの任意の組み合わせを含む、1つ以上のZFPの任意の部分に作動的に連結され得る。
【0018】
いくつかの実施形態において、本明細書に記載される改変された亜鉛フィンガータンパク質を、融合タンパク質の一部として、ヌクレアーゼ(開裂)ドメインと作動的連結状態に配置することができる。特定の実施形態において、このようなヌクレアーゼ融合は、誘導多能性幹細胞(iPSC)、ヒト胚幹細胞(hES)、間充織幹細胞(MSC)、または神経幹細胞等の幹細胞中で変異Htt対立遺伝子を標的化するために使用され得、ヌクレアーゼ融合の活性は、多くの野生型のCAG反復を含有するHtt対立遺伝子を引き起こす。特定の実施形態において、修飾された幹細胞を含む薬学的組成物が提供される。
【0019】
さらに別の態様において、本明細書に記載される亜鉛フィンガータンパク質のいずれかをコードするポリヌクレオチドが提供される。このようなポリヌクレオチドを、トリヌクレオチド反復疾患を治療することが望ましい対象に投与することができる。
【0020】
なおさらなる態様において、本発明は、ハンチントン病等のトリヌクレオチド反復疾患の研究のための特定のモデル系の生成のための方法および組成物を提供する。特定の実施形態において、変異Htt対立遺伝子が、細胞の生成のために胚幹細胞中に生成されるモデルと、特定の長さのトリヌクレオチド伸長経路(例えば、50、80、109、および180のCAG反復)が、亜鉛フィンガーヌクレアーゼ(ZFN)誘導標的化組込みを用いて、野生型Htt対立遺伝子に挿入される動物株と、が提供される。特定の実施形態において、モデル系は、インビトロ細胞株を含む一方で、他の実施形態において、モデル系は、トランスジェニック動物を含む。
【0021】
さらに別の態様において、本明細書に記載されるポリヌクレオチドのいずれかを含む遺伝子送達ベクターが提供される。特定の実施形態において、ベクターは、アデノウイルスベクター(例えば、Ad5/F35ベクター)、組込み能力があるもしくは組込み機能を欠損したレンチウイルスベクターを含むレンチウイルスベクター(LV)、またはアデノウイルス随伴ウイルスベクター(AAV)である。したがって、少なくとも1つの亜鉛フィンガーヌクレアーゼ(ZFN)をコードする配列および/または標的遺伝子への標的化組込みのためのドナー配列を含む、アデノウイルス(Ad)ベクター、LV、またはアデノウイルス随伴ウイルスベクター(AAV)も本明細書に提供される。特定の実施形態において、Adベクターは、キメラAdベクター、例えば、Ad5/F35ベクターである。特定の実施形態において、レンチウイルスベクターは、インテグラーゼ欠損型レンチウイルスベクター(IDLV)または組込み能力のあるレンチウイルスベクターである。特定の実施形態において、ベクターは、VSV−Gエンベロープまたは他のエンベロープを有する擬似タイプである。
【0022】
いくつかの実施形態において、モデル系は、標的対立遺伝子(例えば、変異Htt)は、発現マーカーで標識化されるトリヌクレオチド反復疾患(例えば、ハンチントン病)を提供する。特定の実施形態において、変異対立遺伝子(例えば、変異Htt)が標識化される。いくつかの実施形態において、野生型対立遺伝子(例えば、野生型Htt)が標識化され、さらなる実施形態において、野生型および変異対立遺伝子の両方は、別個の発現マーカーで標識化される。特定の実施形態において、モデル系は、インビトロ細胞株を含む一方、他の実施形態において、モデル系は、トランスジェニック動物を含む。
【0023】
さらに、核酸および/またはZFP(もしくはZFPを含む融合タンパク質)を含有する薬学的組成物も提供される。例えば、ある特定の組成物は、調節配列と作動可能に連結され、薬学的に許容される担体または希釈剤と併用される、本明細書に記載されるZFPの1つをコードする配列を備える核酸を含み、調節配列は、細胞内で核酸の発現を可能にする。特定の実施形態において、コードされるZFPは、HD Htt対立遺伝子に対して特異的である。いくつかの実施形態において、薬学的組成物は、HD Htt対立遺伝子を調節するZFPおよび神経栄養因子を調節するZFPを含む。タンパク質に基づく組成物には、本明細書に開示される1つ以上のZFPおよび薬学的に許容される担体または希釈剤が含まれる。
【0024】
さらに別の態様において、本明細書に記載されるタンパク質、ポリヌクレオチド、および/または組成物のいずれかを含む単離された細胞も提供される。
【0025】
別の態様において、本明細書に開示される組成物を用いて、トリヌクレオチド反復疾患を治療するおよび/または予防するための方法が本明細書に提供される。特定の実施形態において、方法には、ハンチントン病の治療が含まれる。いくつかの実施形態において、方法には、ウイルスベクター、非ウイルスベクター(例えば、プラスミド)、および/またはその組み合わせを使用して、ポリヌクレオチドおよび/またはタンパク質を送達することができる組成物が含まれる。いくつかの実施形態において、方法には、ZFPを含む幹細胞集団を含む、または本発明のZFNを変化させた、組成物が含まれる。
【0026】
これらおよび他の態様は、全体の開示を考慮して、当業者には容易に明らかである。
【図面の簡単な説明】
【0027】
【図1】パネルAからDは、ハンチンチン(Htt)対立遺伝子および対立遺伝子に結合する様々なZFP−TFを示す概略図である。図1Aは、野生型Htt対立遺伝子を示す。図1Bは、HD Htt対立遺伝子に結合する単一のZFP−TFを示し、ここで「KRAB」とは、KOX1遺伝子からのKRAB抑制ドメインを指し、「ZFP」とは、亜鉛フィンガー結合タンパク質を指す。「標準ZFP TF」は、亜鉛フィンガー結合ドメインが、KRAB抑制ドメインに連結されるZFP転写因子融合タンパク質である。図1Cは、ZFP転写因子である「両端型ZFP TF」を示し、ここで、亜鉛フィンガードメインの2つのクラスターは、剛体タンパク質配列によって分離される。機能ドメインを、本図中の1つのZFPの外部に示すが、機能ドメインが、ZFPの間またはZFPの外部にあり得ることは明らかである。図1Dは、「多量体型ZFP TF」を示し、これは、多量体化ドメインを通して多量体化することが可能なZFP TFである(小さな斑点のあるボックスとして示される)。また、機能ドメイン(例えば、活性化、抑制、開裂ドメイン)を黒菱形のあるボックスとして図1Cおよび1Dに示す。
【図2】ヒト293T細胞(表1中に記載される)における標準ZFP TFによるHttの抑制を示す。GFPは、GFPに対して特異的なZFP TFを示し、偽は、偽トランスフェクトされている細胞中の発現を示す。
【図3】パネルA〜Cは、例示的な多量体型ZFP−TFを示す概略図である。図3Aは、機能ドメイン(抑制ドメイン)、および多量体化のための二量体化ドメインに融合される単一のZFPを示す。図3Bは、上端線に示されるように、2つのZFPの多量体の結合を示す。図3Cは、上端線に示されるように、4つのZFPの多量体を示す。任意の数の多量体を使用することができ、機能ドメインは、1つ以上の個々のZFPにおいて、どこでも位置付けられ得ることは明らかである。
【図4】パネルA〜Cは、例示的な両端型ZFP−TFを示す概略図である。図4Aは、2つのZFP間に位置付けられる機能ドメインを含む、両端型ZFPを示す。図1D中に示されるように、機能ドメインは、ZFPのうちの1つの外部であり得る。図4Bは、標的部位への両端型ZFPの結合を示す。ZFPの一方または両方は、それらの標的部位に結合する。図4Cは、複数の両端型ZFPの結合を示す。
【図5】Htt特異的ZFN対である25920/25921あるいは25922/25923のいずれかによるK562細胞の処理を行った後のCel−Iミスマッチアッセイ(Surveyor,Transgenomics)の結果を示すゲルを示す。NHEJ活性の割合をそれぞれのレーンの下に示す。「GFP」は、プラスミドをコードするGFPでトランスフェクトされている細胞を示す。示されるデータは、プラスミドを含有するZFNでトランスフェクトしてから14日後に抽出したDNAからのものである。
【発明を実施するための形態】
【0028】
詳細な説明
ハンチントン病、SBMA、フリードライヒ運動失調症、ならびに脆弱X症候群AおよびE等のトリヌクレオチド反復疾患を治療するための組成物および方法を本明細書に開示する。特に、亜鉛フィンガータンパク質(ZFP TF)を含むHttを調節する転写因子およびこのようなタンパク質を利用する方法が、ハンチントン病を治療で用いるために提供される。例えば、変異Htt対立遺伝子の発現を抑制する、または野生型Htt対立遺伝子の発現を活性化するZFP−TFを提供する。加えて、これらの疾患に関連する遺伝子のゲノム構造を修飾する亜鉛フィンガーヌクレアーゼ(ZFN)を提供する。例えば、変異型のHttの部分を特異的に変化させることができるZFNを提供する。これらには、改変された亜鉛フィンガータンパク質、すなわち、所定の核酸標的配列に結合する天然に存在しないタンパク質を用いる組成物および方法が含まれる。
【0029】
したがって、本明細書に記載される方法および組成物は、トリヌクレオチド反復疾患の治療のための方法を提供し、これらの方法および組成物は、標的遺伝子および改変された亜鉛フィンガーヌクレアーゼを調節することができる亜鉛フィンガー転写因子を含むことができる。
【0030】
概論
本明細書に開示される方法の実践、ならびに組成物の調製および使用は、特に指示がない限り、分子生物学、生化学、クロマチン構造および分析、計算化学、細胞培養、組み換えDNA、および関連分野における従来の技術は、当技術分野の技術範囲内である。これらの技術は、文献に十分に説明されている。例えば、Sambrook et al.MOLECULAR CLONING:A LABORATORY MANUAL,Second edition,Cold Spring Harbor Laboratory Press,1989およびThird edition,2001、Ausubel et al.,CURRENT PROTOCOLS IN MOLECULAR BIOLOGY,John Wiley & Sons,New York,1987および定期的に更新されたもの;METHODS IN ENZYMOLOGY,Academic Press,San Diegoシリーズ;Wolffe,CHROMATIN STRUCTURE AND FUNCTION,Third edition,Academic Press,San Diego,1998;METHODS IN ENZYMOLOGY,Vol.304,“Chromatin”(P.M.Wassarman and A.P.Wolffe,eds.),Academic Press,San Diego,1999;ならびにMETHODS IN MOLECULAR BIOLOGY,Vol.119,“Chromatin Protocols”(P.B.Becker,ed.)Humana Press,Totowa,1999を参照のこと。
【0031】
定義
「核酸」、「ポリヌクレオチド」、および「オリゴヌクレオチド」という用語は、交換可能に使用され、線状または環状構造で、一本鎖または二本鎖形態の、デオキシリボヌクレオチドポリマーまたはリボヌクレオチドポリマーを指す。本開示については、これらの用語をポリマーの長さに関する限定と解釈すべきではない。これらの用語は、天然ヌクレオチドの公知の類似体、ならびに塩基、糖、および/またはリン酸(ホスホロチオエート主鎖)部分内で修飾されるヌクレオチドを包含することができる。一般に、特定のヌクレオチドの類似体は、同じ塩基対形成特異性を有し、すなわち、Aの類似体は、Tと塩基対を形成する。
【0032】
「ポリペプチド」、「ペプチド」、および「タンパク質」という用語は、アミノ酸残基のポリマーを指すために交換可能に使用される。この用語は、1つ以上のアミノ酸が、対応する天然に存在するアミノ酸の化学的類似体または修飾誘導体である、アミノ酸ポリマーにも適用される。
【0033】
「結合」は、巨大分子間(例えば、タンパク質と核酸との間)の配列特異的で非共有結合的な相互作用を指す。結合相互作用の全ての構成要素は、相互作用が全体として配列特異的である限り、配列特異的である(例えば、DNA主鎖中のリン酸残基との接触する)必要はない。このような相互作用は、一般に、10-6M-1以下の解離定数(Kd)によって特徴付けられる。「親和性」は、結合の強度を指す。高い結合親和性は、低いKdに関連している。
【0034】
「結合タンパク質」は、別の分子に非共有結合的に結合することができるタンパク質である。結合タンパク質は、例えば、DNA分子(DNA結合タンパク質)、RNA分子(RNA結合タンパク質)、および/またはタンパク質分子(タンパク質結合タンパク質)に結合することができる。タンパク質結合タンパク質の場合においては、それは、それ自体に結合する(その結果、ホモ二量体、ホモ三量体等を形成する)ことができる、および/または異なる1つのタンパク質または複数のタンパク質の1つ以上の分子に結合することができる。結合タンパク質は、2種類以上の結合活性を有することができる。例えば、亜鉛フィンガータンパク質は、DNA結合、RNA結合、およびタンパク質結合活性を有する。
【0035】
「亜鉛フィンガーDNA結合タンパク質」(または結合ドメイン)は、構造が亜鉛イオンの配位によって安定化される結合ドメイン内のアミノ配列の領域である、1つ以上の亜鉛フィンガーによって配列特異的な様式でDNAに結合する、タンパク質またはより大きなタンパク質内のドメインである。亜鉛フィンガーDNA結合タンパク質という用語は、しばしば、亜鉛フィンガータンパク質またはZFPとして略記される。
【0036】
亜鉛フィンガー結合ドメインは、例えば、天然に存在する亜鉛フィンガータンパク質の認識へリックス領域の改変(1つ以上のアミノ酸を変化させる)を介して、所定のヌクレオチド配列に結合されるように「改変」することができる。したがって、改変された亜鉛フィンガータンパク質は、天然に存在しないタンパク質である。亜鉛フィンガータンパク質を改変するための方法の非制限的な例は、設計および選択である。設計された亜鉛フィンガータンパク質は、天然に存在しないタンパク質であり、その設計/組成物は、主として合理的基準によって得られる。設計のための合理的基準には、置換規則の適用、ならびに既存のZFP設計および結合データの情報を格納したデータベース中の情報を処理するためのコンピュータアルゴリズムの適用が含まれる。例えば、米国特許第6,140,081号、同第6,453,242号、および同第6,534,261号明細書を参照されたく、国際公開第98/53058号、国際公開第98/53059号、国際公開第98/53060号、国際公開第02/016536号、および国際公開第03/016496号も参照されたい。
【0037】
「選択された」亜鉛フィンガータンパク質は、天然には見出されないタンパク質であり、その産生は、ファージディスプレイ、相互作用トラップ、またはハイブリッド選択等の実験的プロセスによって主に得られる。例えば、米国第5,789,538号、米国特許第5,925,523号、米国特許第6,007,988号、米国特許第6,013,453号、米国特許第6,200,759号明細書、国際公開第95/19431号、国際公開第96/06166号、国際公開第98/53057号、国際公開第98/54311号、国際公開第00/27878号、国際公開第01/60970号、国際公開第01/88197号、および国際公開第02/099084号を参照されたい。
【0038】
「組み換え」とは、2つのポリヌクレオチド間での遺伝子情報の交換のプロセスを指す。本開示に関して、「相同組み換え(HR)」とは、例えば、相同性指向性修復機構を介した細胞中の二本鎖切断の修復中に行われる、このような交換の特殊形態を指す。このプロセスは、ドナーから標的への遺伝子情報の転送につながるため、ヌクレオチド配列相同性を必要とし、「標的」分子(すなわち、二本鎖切断を経験したもの)のテンプレート修復のために「ドナー」分子を使用し、「非交差型遺伝子変換」または「短経路遺伝子変換」として様々に知られている。いかなる特定の理論にも束縛されることを望むものではないが、このような転送は、破壊標的とドナーとの間に形成されるヘテロ二本鎖DNAの不適合補正、および/または、ドナーが標的の一部となる遺伝子情報を再合成するために使用される「合成依存鎖アニーリング」、および/または関連プロセスを含むことができる。このような特殊HRにより、しばしば、ドナーポリヌクレオチドの配列の一部または全てが標的ポリヌクレオチドに組み込まれるように、標的分子の配列が変更される。
【0039】
本開示の方法において、本明細書に記載される1つ以上の標的ヌクレアーゼは、所定の部位で標的配列(例えば、細胞クロマチン)において、二本鎖切断を生成し、切断の領域におけるヌクレオチド配列に相同性を有する「ドナー」ポリヌクレオチドを細胞に導入することができる。二本鎖切断の存在は、ドナー配列の組込みを促進することが示されている。ドナー配列は、物理的に組み込まれ得るか、または代替的に、ドナーポリヌクレオチドを、相同組み換えを介した切断の修復のためのテンプレートとして使用し、ドナー等の場合、細胞クロマチンへの、ヌクレオチド配列の全てまたは一部を導入させる。したがって、細胞クロマチン中の第1の配列は、変更することができ、特定の実施形態において、ドナーポリヌクレオチド中に存在する配列に変換することができる。したがって、「置換する」または「置換」という用語の使用は、1つのヌクレオチド配列の別のものによる置換(すなわち、情報を持つセンスでの配列の置換)を表すと理解され、1つのポリヌクレオチドを別のものによる物理的または化学的置換は必ずしも必要としない。
【0040】
本明細書に記載される方法のいずれかにおいて、細胞内のさらなる標的部位のさらなる二本鎖切断のために、亜鉛フィンガータンパク質の追加の対を使用することができる。
【0041】
細胞クロマチンにおいて、対象となる領域の配列の標的化組み換えおよび/または置換および/または変更のための方法の特定の実施形態において、染色体の配列は、外来の「ドナー」ヌクレオチド配列との相同組み換えによって変更される。このような相同組み換えは、二本鎖切断の領域に相同である配列が存在する場合、細胞クロマチン内の二本鎖の切断の存在により刺激される。
【0042】
本明細書に記載される方法のいずれかにおいて、第1のヌクレオチド配列(「ドナー配列」)は、対象となる領域のゲノム配列の相同であるが同一ではない配列を含み、それによって、対象となる領域の非同一配列を挿入するための相同組み換えを刺激することができる。したがって、特定の実施形態において、対象となる領域の配列に相同であるドナー配列の一部は、置換されるゲノム配列と、約80〜99%(またはその間のいずれかの整数)の配列同一性を示す。他の実施形態において、例えば、100個を超える近接塩基対の、ドナーとゲノム配列の間で1個だけヌクレオチドが異なる場合、ドナーとゲノム配列の間の相同性は、99%よりも高い。場合によっては、ドナー配列の非相同部分は、新規配列が、対象となる領域に導入されるように、対象となる領域には存在しない配列を含むことができる。これらの場合において、非相同配列は、一般に、対象となる領域の配列と相同であるかまたは同一である50〜1,000個の塩基対(またはそれらの間のいずれかの整数値)、または1,000個を超えるいくつかの塩基対の配列によって側方に位置している。他の実施形態において、ドナー配列は、第1の配列に非相同であり、非相同組み換え機構によってゲノムに挿入される。
【0043】
本明細書に記載される方法のいずれかは、対象となる遺伝子の発現を妨害するドナー配列の標的化組込みによって、細胞中の1つ以上の標的配列の部分的または完全な不活性化に使用され得る。部分的または完全に不活性化された遺伝子を有する細胞株も提供される。
【0044】
さらに、本明細書に記載される標的化組込み方法はまた、1つ以上の外来配列を組み込むために使用することができる。外来核酸配列は、例えば、1つ以上の遺伝子またはcDNA分子、またはあらゆる種類のコードもしくは非コード配列、ならびに1つ以上の制御要素(例えば、プロモーター)を含むことができる。加えて、外来核酸配列は、1つ以上のRNA分子(例えば、小ヘアピンRNA(shRNA)、抑制RNA(RNAis)、ミクロRNA(miRNA)等)を産生し得る。
【0045】
「切断」とは、DNA分子の共有結合骨格の破壊を指す。切断は、リン酸ジエステル結合の酵素または化学化水分かを含むが、これらに限定されない様々な方法によって開始され得る。一本鎖切断および二本鎖切断の両方が可能であり、二本鎖切断は、2つの異なる一本鎖切断現象の結果として起こり得る。DNA切断は、平滑末端または付着末端のいずれかの産生をもたらし得る。特定の実施形態において、融合ポリペプチドは、標的二本鎖DNA切断に使用される。
【0046】
「切断ハーフドメイン」は、(同一または異なる)第2のポリペプチドと連結して、切断活性を有する複合体(好ましくは、二本鎖切断活性)を形成するポリペプチド配列である。「第1および第2の切断ハーフドメイン」、「+および-切断ハーフドメイン」、および「右および左切断ハーフドメイン」という用語は、二量体化する切断ハーフドメインの対を指すために交換可能に使用される。
【0047】
「改変された切断ハーフドメイン」は、別の切断ハーフドメイン(例えば、別の改変された切断ハーフドメイン)と共にヘテロ二量体を形成するために修飾された切断ハーフドメインである。例えば、米国特許出願公開第2005/0064474号、同第20070218528号、および同第2008/0131962号明細書も参照されたく、これらは、参照によってその全体が本明細書に組み込まれる。
【0048】
「配列」という用語は、任意の長さのヌクレオチド配列を指し、これはDNAもしくはRNAであり得る、線状、環状、もしくは分岐状であり得る、または一本鎖もしくは二本鎖のいずれか一方であり得る。「ドナー配列」という用語は、ゲノム内に挿入されるヌクレオチド配列を指す。ドナー配列は、任意の長さ、例えば、2〜10,000(またはそれらの間もしくはそれらを超える任意の整数値)長のヌクレオチド、好ましくは、約100〜1,000(またはそれらの間の任意の整数)長のヌクレオチド、より好ましくは、約200〜500長のヌクレオチドであり得る。
【0049】
「クロマチン」は、細胞ゲノムを含む核タンパク質構造である。細胞クロマチンは、核酸、主にDNA、ならびに、ヒストンおよび非ヒストン染色体タンパク質を含むタンパク質を含む。真核細胞クロマチンの大半は、ヌクレオソームの形態で存在し、そこでは、ヌクレオソームコアが、それぞれ2つのヒストンH2A、H2B、H3、およびH4を含む八量体と会合した約150塩基対のDNAを含み、リンカーDNA(生物に応じて変化する長さ)がヌクレオソームコアの間に延在する。ヒストンH1の1分子は、一般にリンカーDNAと会合する。本開示に関して、「クロマチン」という用語は、原核性および真核性の両方の全ての種類の細胞核タンパク質を包含するように意図されている。細胞クロマチンは、染色体およびエピソームクロマチンの両方を含む。
【0050】
「染色体」は、細胞のゲノムの全てまたは一部を含むクロマチン複合体である。細胞のゲノムは、しばしば、その核型によって特徴付けられ、これは細胞のゲノムを含む全染色体の集合である。細胞のゲノムは、1つ以上の染色体を含むことができる。
【0051】
「エピソーム」は、複製する核酸、核タンパク質複合体、または細胞の染色体核型の一部ではない核酸を含む他の構造である。エピソームの例には、プラスミドおよびある特定のウイルスゲノムが含まれる。
【0052】
「標的部位」または「標的配列」は、結合が存在するのに十分な条件が提供される場合、結合分子が結合する核酸の一部を規定する核酸配列である。多様なNT−3標的ZFPに対する例示的な標的部位は、表2および3に示される。
【0053】
「外来」分子は、通常は細胞内に存在しないが、1つ以上の遺伝学的、生化学的、または他の方法によって細胞内に導入することができる分子である。「通常は細胞内に存在する」とは、細胞の特定の発生段階および環境条件に対して決定される。したがって、例えば、筋肉の胚発生中のみ存在する分子は、成体の筋細胞に関しては外来分子である。同様に、熱ショックによって誘導される分子は、非熱ショック細胞に対して外来分子である。外来分子は、例えば、機能不全型内在性分子の機能型または正常機能型内在性分子の機能不全型を含むことができる。
【0054】
外来分子は、とりわけ、コンビナトリアルケミストリープロセスによって生成されるもの等の小分子、または、タンパク質、核酸、糖質、脂質、糖タンパク質、リポタンパク質、多糖、上記の分子の任意の修飾誘導体、または上記の分子の1つ以上を含む任意の複合体等の高分子であり得る。核酸にはDNAおよびRNAが含まれ、それは一本または二本鎖であり得、線状、分岐状、または環状であり得、任意の長さであり得る。核酸には、二重鎖を形成することができるもの、ならびに三重鎖形成核酸が含まれる。例えば、米国特許第5,176,996号および同第5,422,251号明細書を参照されたい。タンパク質には、DNA結合タンパク質、転写因子、クロマチンリモデリング因子、メチル化DNA結合タンパク質、ポリメラーゼ、メチラーゼ、デメチラーゼ、アセチラーゼ、デアセチラーゼ、キナーゼ、ホスファターゼ、インテグラーゼ、リコンビナーゼ、リガーゼ、トポイソメラーゼ、ジャイレース、およびヘリカーゼが含まれるが、これらに限定されない。
【0055】
外来分子は、内在性分子と同じ型の分子、例えば、外来タンパク質または核酸であり得る。例えば、外来核酸は、感染ウイルスゲノム、細胞中に導入されたプラスミドもしくはエピソーム、または細胞内に通常存在しない染色体を含むことができる。細胞中に外来分子を導入するための方法が当業者に知られており、脂質媒介導入(すなわち、中性およびカチオン性脂質を含むリポソーム)、エレクトロポレーション、直接注入、細胞融合、微粒子銃、リン酸カルシウム共沈、DEAE−デキストラン媒介導入、およびウイルスベクター媒介導入を含むが、これらに限定されない。外来分子はまた、内在性分子と同じ型の分子であってもよいが、細胞が由来する種とは異なる種に由来する。例えば、ヒトの核酸配列は、マウスまたはハムスターに本来由来する細胞株中に導入され得る。
【0056】
これに対して、「内在性」分子は、特定の環境条件下で特定の発生段階にある特定の細胞内に通常存在するものである。例えば、内在性核酸は、染色体、ミトコンドリア、クロロプラストもしくは他の細胞小器官のゲノム、または天然に存在するエピソーム核酸を含むことができる。さらなる内在性分子には、タンパク質、例えば、転写因子および酵素が含まれてもよい。
【0057】
「融合」分子は、2つ以上のサブユニット分子が、好ましくは共有結合的に連結された分子である。サブユニット分子は、同じ化学型の分子であるか、または異なる化学型の分子であることができる。第1の型の融合分子の例には、融合タンパク質(例えば、ZFPDNA結合ドメインと1つ以上の活性化ドメインとの間の融合体)、および融合核酸(例えば、上記に記載される融合タンパク質をコードする核酸)が含まれるが、これらに限定されない。第2の型の融合分子の例には、三重鎖形成核酸とポリペプチドとの間の融合体、および副溝結合剤と核酸との間の融合体が含まれるが、これらに限定されない。
【0058】
細胞中の融合タンパク質の発現は、細胞に融合タンパク質を送達すること、または細胞に融合タンパク質をコードするポリヌクレオチドを送達することによって引き起こされてもよく、ポリヌクレオチドが転写され、転写物が翻訳されて、融合タンパク質を生成する。トランススプライシング、ポリペプチド開裂、およびポリペプチド連結はまた、細胞中のタンパク質の発現に関与することができる。細胞へのポリヌクレオチドおよびポリペプチドの送達の方法は、本開示の他所で提示されている。
【0059】
「多量体化ドメイン」は、アミノ、カルボキシで組み込まれるドメイン、またはZFP TFのアミノまたはカルボキシ末端領域である。これらのドメインは、トリヌクレオチド反復ドメインのより大きな経路が、多量体型ZFP TFによって結合される一方で、野生型経路は結合されないように、複数のZFP TF単位の多量体化を可能にする。多量体化ドメインの例には、ロイシンジッパーが含まれる。多量体化ドメインはまた、小分子によって調節され得、多量体化ドメインは、小分子または外部リガンドの存在下のみ、別の多量体化ドメインとの相互作用を可能にする適切な構造であると仮定する。このようにして、外来リガンドは、これらのドメインの活性を調節するために使用することができる。
【0060】
本開示に関して、「遺伝子」は、このような調節配列がコードおよび/または転写配列に隣接しているかに関わらず、遺伝子産物(以下を参照)をコードするDNA領域、ならびに遺伝子産物の産生を調節する全てのDNA領域を含む。したがって、遺伝子には、プロモーター配列、ターミネーター、リポソーム結合部位および内部リポソーム侵入部位等の翻訳調節配列、エンハンサー、サイレンサー、インスレーター、境界エレメント、複製起点、マトリックス付着部位、ならびに遺伝子座制御領域が含まれるが、これらに限定されない。
【0061】
「遺伝子発現」は、遺伝子産物への遺伝子に含まれる情報の変換を指す。遺伝子産物は、遺伝子の直接転写産物(例えば、mRNA、tRNA、rRNA、アンチセンスRNA、リボザイム、構造RNA、もしくは任意の他の種類のRNA)、またはmRNAの翻訳によって産生されるタンパク質であってもよい。遺伝子産物には、キャッピング、ポリアデニル化、メチル化、および編集等のプロセスによって修飾されたRNA、および、例えば、メチル化、アセチル化、リン酸化、ユビキチン化、ADPリボシル化、ミリスチリル化、およびグリコシル化によって修飾されたタンパク質も含まれる。
【0062】
遺伝子発現の「調節」は、遺伝子の活性の変化を指す。発現の調節には、遺伝子活性化および遺伝子抑制が含まれ得るが、これらに限定されない。ゲノム編集(例えば、切断、変更、不活性化、ランダム変異)は、発現を調節するために使用することができる。遺伝子の不活性化とは、本明細書に記載されるようなZFPを含まない細胞と比較した、遺伝子発現の任意の減少を指す。したがって、遺伝子の不活性化は、部分的または完全であり得る。
【0063】
「対象となる領域」は、例えば、外来分子に結合することが望ましい、遺伝子、遺伝子内、または遺伝子に隣接する非コード配列等の細胞クロマチンの任意の領域である。結合は、標的化DNA切断および/または標的化組み換えの目的のためであり得る。対象となる領域は、例えば、染色体、エピソーム、細胞小器官ゲノム(例えば、ミトコンドリア、葉緑体)、または感染ウイルスゲノムに存在し得る。対象となる領域は、遺伝子のコード領域内、例えば、リーダー配列、トレーラー配列またはイントロン等の転写された非コード領域内、またはコード領域内の上流もしくは下流のいずれかの非転写領域内であり得る。対象となる領域は、単一ヌクレオチド対と同じくらい小さいか、もしくは最大2,000ヌクレオチド対の長さ、または任意の整数値のヌクレオチド対の長さであり得る。
【0064】
「真核」細胞には、真菌細胞(酵母等)、植物細胞、動物細胞、哺乳動物細胞、およびヒト細胞(例えば、T細胞)が含まれるが、これらに限定されない。
【0065】
「作動的連結」および「作動的に連結された」(または「作動可能に連結された」という用語は、2つ以上の構成要素(配列要素等)の並列に関連して交換可能に使用され、その中で両方の構成要素が正常に機能し、かつ構成要素の少なくとも1つが、他の構成要素の少なくとも1つに対して発揮される機能を媒介することができる可能性があるように構成要素が配置される。例として、プロモーター等の転写調節配列は、転写調節配列が1つ以上の転写調節因子の存在または非存在に応じてコード配列の転写レベルを制御する場合に、コード配列に作動的に連結される。転写調節配列は、一般的に、コード配列とシスで作動的に連結されるが、それに直接隣接している必要はない。例えば、エンハンサーは、それらが隣接していなくても、コード配列に作動的に連結される転写調節配列である。
【0066】
融合ポリペプチドに関して、「作動的に連結された」という用語は、構成要素の各々が、そのように連結されていない場合に、それが果たすであろう他の構成要素に連動して同一の機能を実行するという事実を指すことができる。例えば、ZFP DNA結合ドメインが活性化ドメインに融合される融合ポリペプチドに関して、ZFP DNA結合ドメインおよび活性化ドメインは、融合ポリペプチドにおいて、ZEP DNA結合ドメイン部分がその標的部位および/またはその結合部位に結合することができる一方で、活性化ドメインが遺伝子発現を上方制御することができる場合に、作動的連結状態にある。ZFP DNA結合ドメインにおける融合ポリペプチドが開裂ドメインに融合される場合、ZFP DNA結合ドメインおよびその開裂ドメインは、融合ポリペプチドにおいて、ZFP DNA結合ドメイン部分がその標的部位および/またはその結合部位に結合することができる一方で、開裂ドメインが標的部位の近接においてDNAを切断することができる場合に、作動的連結状態にある。
【0067】
タンパク質、ポリペプチド、または核酸の「機能的断片」は、その配列が、完全長タンパク質、ポリペプチド、または核酸と同一でないが、完全長タンパク質、ポリペプチド、または核酸と同一の機能を保持するタンパク質、ポリペプチド、または核酸である。機能的断片は、対応する天然分子より多い、少ない、または同一の残基数を有することができる、および/または1つ以上のアミノ酸またはヌクレオチド置換を含有することができる。核酸の機能(例えば、コード機能、別の核酸にハイブリダイズする能力)を決定するための方法は、当技術分野でよく知られている。同様に、タンパク質機能を決定するための方法も、よく知られている。例えば、ポリペプチドのDNA結合機能を、例えば、フィルター結合、電気泳動移動度シフト、または免疫沈降アッセイによって決定することができる。DNA開裂を、ゲル電気泳動によってアッセイすることができる。上記のAusubelらを参照されたい。別のタンパク質と相互作用するタンパク質の能力を、例えば、共免疫沈降、ツーハイブリッドアッセイ、または遺伝的相補性および生化学的相補性の両方の相補性によって決定することができる。例えば、Fields et al. (1989)Nature 340:245−246、米国特許第5,585,245号およびPCT国際公開第98/44350号を参照されたい。
【0068】
「ベクター」は、標的細胞に遺伝子配列を移動させることができる。一般的に、「ベクター構築物」、「発現ベクター」、および「遺伝子導入ベクター」は、対象となる遺伝子の発現を指向することが可能な、標的細胞に遺伝子配列を導入することができる任意の核酸構築物を意味する。したがって、この用語は、クローニングおよび発現ビヒクル、ならびに組込みベクターを含む。
【0069】
「レポーター遺伝子」または「レポーター配列」とは、好ましくは、必ずではないが日常的アッセイにおいて、容易に測定されるタンパク質産物を産生する任意の配列を指す。適切なレポーター遺伝子としては、抗生物質耐性(例えば、アンピシリン耐性、ネオマイシン耐性、G418耐性、ピューロマイシン耐性)を媒介するタンパク質をコードする配列、着色または蛍光または発光タンパク質(例えば、緑色蛍光タンパク質、強化緑色蛍光タンパク質、赤色蛍光タンパク質、ルシフェラーゼ)をコードする配列、および強化された細胞成長および/もしくは遺伝子増幅を媒介するタンパク質(例えば、ジヒドロ葉酸レダクターゼ)をコードする配列が含まれるが、これらに限定されない。エピトープタグには、例えば、FLAG、His、myc、Tap、HA、または任意の検出可能なアミノ酸配列の1つ以上のコピーが含まれる。「発現タグ」には、対象となる遺伝子の発現をモニターするために、所望の遺伝子配列に作動的に連結され得るレポーターをコードする配列が含まれる。
【0070】
DNA結合ドメイン
Httを含むが、これに限定されない、トリヌクレオチド反復を含む任意の遺伝子において、標的部位に特異的に結合するDNA結合ドメインを含む組成物が、本明細書に記載されている。GDNFまたはBDNF遺伝子において、標的部位に特異的に結合するDNA結合ドメインを含む組成物も提供される。任意のDNA結合ドメインは、本明細書に開示される組成物および方法で使用することができる。
【0071】
特定の実施形態において、DNA結合ドメインは、亜鉛フィンガータンパク質を含む。好ましくは、亜鉛フィンガータンパク質は天然に存在せず、それは選択する標的部位に結合するように改変される。例えば、Beerli et al.(2002)Nature Biotechnol.20:135−141、Pabo et al.(2001)Ann.Rev.Biochem.70:313−340、Isalan et al.(2001)Nature Biotechnol.19:656−660、Segal et al.(2001)Curr.Opin.Biotechnol.12:632−637、Choo et al.(2000)Curr.Opin.Struct.Biol.10:411−416、米国特許第6,453,242号、同第6,534,261号、同第6,599,692号、同第6,503,717号、同第6,689,558号、同第7,030,215号、同第6,794,136号、同第7,067,317号、同第7,262,054号、同第7,070,934号、同第7,361,635号、同第7,253,273号明細書、および米国特許出願公開第2005/0064474号、同第2007/0218528号、同第2005/0267061号明細書を参照されたく、全ては、参照によりその全体が本明細書に組み込まれる。
【0072】
改変された亜鉛フィンガー結合ドメインは、天然に存在する亜鉛フィンガータンパク質と比較して、新規の結合特異性を有することができる。改変方法には、合理的設計および多様な種類の選択が含まれるが、これらに限定されない。合理的設計には、例えば、三重(または四重)ヌクレオチド配列および個々の亜鉛フィンガーアミノ酸配列を含むデータベースを使用することが含まれ、各三重または四重ヌクレオチド配列は、特定の三重または四重配列に結合する亜鉛フィンガーの1つ以上のアミノ酸配列と会合する。例えば、譲受人共通の米国特許第6,453,242号および同第6,534,261号明細書を参照されたく、これらは参照によりその全体が本明細書に組み込まれる。
【0073】
ファージディスプレイおよび2ハイブリッド法を含む例示的な選択方法は、米国特許第5,789,538号、同第5,925,523号、同第6,007,988号、同第6,013,453号、同第6,410,248号、同第6,140,466号、同第6,200,759号、同第6,242,568号明細書、ならびに国際公開第98/37186号、国際公開第98/53057号、国際公開第00/27878号、国際公開第01/88197号、および英国特許第2,338,237号明細書に開示されている。加えて、亜鉛フィンガー結合ドメインに対する結合特異性の強化は、例えば、譲受人共通の国際公開第02/077227号に記載されている。
【0074】
加えて、これらおよび他の参考文献に開示される、亜鉛フィンガードメインおよび/または多フィンガーの亜鉛フィンガータンパク質は、例えば、5個以上のアミノ酸長のリンカーを含む、任意の好適なリンカー配列を使用して共に連結され得る。また、6個以上のアミノ酸長の例示的なリンカー配列に関して、米国特許第6,479,626号、同第6,903,185号、および同第7,153,949号明細書も参照されたい。本明細書に記載されるタンパク質は、タンパク質の個々の亜鉛フィンガー間の好適なリンカーの任意の組み合わせを含み得る。加えて、亜鉛フィンガー結合ドメインに対する結合特異性の強化は、例えば、譲受人共通の国際公開第02/077227号に記載されている。
【0075】
標的部位の選択、融合タンパク質(および同一物をコードするポリヌクレオチド)の設計および構築のためのZFPおよび方法が当業者に知られており、米国特許第6,140,0815号、同第789,538号、同第6,453,242号、同第6,534,261号、同第5,925,523号、同第6,007,988号、同第6,013,453号、同第6,200,759号明細書、国際公開第95/19431号、国際公開第96/06166号、国際公開第98/53057号、国際公開第98/54311号、国際公開第00/27878号、国際公開第01/60970号、国際公開第01/88197号、国際公開第02/099084号、国際公開第98/53058号、国際公開第98/53059号、国際公開第98/53060号、国際公開第02/016536号、および国際公開第03/016496号に詳しく記載されている。
【0076】
加えて、これらおよび他の参考文献に開示される、亜鉛フィンガードメインおよび/または多フィンガーの亜鉛フィンガータンパク質は、例えば、5個以上のアミノ酸長のリンカーを含む、任意の好適なリンカー配列を使用して共に連結され得る。また、6個以上のアミノ酸長の例示的なリンカー配列に関して、米国特許第6,479,626号、同第6,903,185号、同第7,153,949号明細書を参照されたい。本明細書に記載されるタンパク質は、タンパク質の個々の亜鉛フィンガー間の好適なリンカーの任意の組み合わせを含み得る。
【0077】
代替的に、DNA結合ドメインは、ヌクレアーゼに由来し得る。例えば、I−SceI、I−CeuI、PI−PspI,PI−Sce、I−SceIV、I−CsmI、I−PanI、I−SceII、I−PpoI、I−SceIII、I−CreI、I−TevI、I−TevII、およびI−TevIII等のホーミングエンドヌクレアーゼおよびメガヌクレアーゼの認識配列が知られている。また、米国特許第5,420,032号、米国特許第6,833,252号、Belfort et al.(1997)Nucleic Acids Res.25:3379-3388、Dujon et al.(1989)Gene 82:115-118、Perler et al.(1994)Nucleic Acids Res.22,1125-1127、Jasin(1996)Trends Genet.12:224-228、Gimble et al.(1996)J.Mol.Biol.263:163-180、Argast et al.(1998)J.Mol.Biol.280:345-353、およびニューイングランドバイオラボ社カタログも参照されたい。加えて、ホーミングエンドヌクレアーゼおよびメガヌクレアーゼのDNA結合特異性を、非天然の標的部位に結合するよう改変することができる。例えば、Chevalier et al.(2002)Molec.Cell 10:895−905、Epinat et al.(2003) Nucleic Acids Res.31:2952−2962、Ashworth et al.(2006)Nature 441:656−659、Paques et al.(2007)Current Gene Therapy 7:49−66、米国特許出願公開第20070117128号を参照されたい。
【0078】
特定の実施形態において、DNA結合ドメインは、Htt遺伝子の標的部位に(配列特異的な様式で)結合し、Httの発現を調節する改変された亜鉛フィンガータンパク質である。ZFPは、変異Htt対立遺伝子または野生型Htt配列のいずれかに選択的に結合することができる。Htt標的部位は、典型的に少なくとも1個の亜鉛フィンガーを含むが、複数の亜鉛フィンガー(例えば、2個、3個、4個、5個、6個以上のフィンガー)が含まれ得る。通常、ZFPは、少なくとも3個のフィンガーを含む。ある特定のZFPは、4個、5個、または6個のフィンガーを含む。3個のフィンガーを含むZFPは、典型的に9個または10個のヌクレオチドを含む標的部位を認識し、4個のフィンガーを含むZFPは、典型的に12〜14個のヌクレオチドを含む標的部位を認識する一方で、6個のフィンガーを有するZFPは、18〜21個のヌクレオチドを含む標的部位を認識する。ZFPはまた、そのドメインが転写活性化または抑制ドメインであり得る、1つ以上の調節ドメインを含む融合タンパク質であってもよい。
【0079】
「両端型」亜鉛フィンガータンパク質は、2つの亜鉛フィンガードメインが2つの不連続標的部位に結合するように、亜鉛フィンガーDNA結合ドメインの2つのクラスターが、アミノ酸を介在することによって分離されるタンパク質である。両端型亜鉛フィンガー結合タンパク質の例は、SIP1であり、一群の4つの亜鉛フィンガーは、タンパク質のアミノ末端に位置し、一群の3つのフィンガーは、カルボキシ末端に位置する(Remacle et al,(1999)EMBO Journal 18(18):5073−5084を参照されたい)。これらのタンパク質におけるそれぞれのクラスターの亜鉛フィンガーは、独特の標的配列に結合することができ、2つの標的配列間の間隔は、多くのヌクレオチドを含むことができる。両端型ZFPは、例えば、ZFPの一方または両方に融合される機能ドメインが含まれ得る。したがって、機能ドメインは、ZFPの一方または両方の外部に付着され得る(図1Cを参照されたい)か、またはZFP間に位置付けられ得る(両方のZFPに付着される)(図4を参照されたい)。
【0080】
Htt標的ZFPの特定の例は、表1に開示されている。この表の第1の列は、ZFPに対する内部参照名(番号)であり、表2の第1列の同じ名前に対応する。「F」はフィンガーを指し、「F」に続く番号は、どの亜鉛フィンガー(例えば、「F1」はフィンガー1を指す)かを言及する。
【0081】
【表1】
【0082】
これらのタンパク質の標的部位に対する配列および位置は、表2に開示される。表2は、示される亜鉛フィンガータンパク質に対する標的配列を示す。ZFP認識ヘリックスによって接触される標的部位におけるヌクレオチドは大文字で示され、非接触ヌクレオチドは小文字で示される。
【0083】
【表2】
【0084】
融合タンパク質
本明細書に記載されるDNA結合タンパク質(例えば、ZFP)を含む融合タンパク質および異種調節(機能的)ドメイン(またはその機能的断片)も提供される。一般的なドメインには、例えば、転写因子ドメイン(活性化剤、リプレッサー、共活性化剤、コリプレッサー)、サイレンサー、癌遺伝子(例えば、myc、jun、fos、myb、max、mad、rel、ets、bcl、myb、mosファミリーメンバー等)、DNA修復酵素ならびにそれらの関連する因子および修飾因子、クロマチン結合性タンパク質およびそれらの修飾因子(例えば、キナーゼ、アセチラーゼ、およびデアセチラーゼ)、ならびにDNA修飾酵素(例えば、メチルトランスフェラーゼ、トポイソメラーゼ、ヘリカーゼ、リガーゼ、キナーゼ、ホスファターゼ、ポリメラーゼ、エンドヌクレアーゼ)、ならびにそれらの関連する因子および修飾因子が含まれる。DNA結合ドメインならびにヌクレアーゼ開裂ドメインの融合に関する詳細ついて、米国特許出願公開第2005/0064474号、同第2006/0188987号、および同第2007/0218528号明細書を参照されたく、参照によりそれらの全体が本明細書に組み込まれる。
【0085】
活性化を達成するための好適なドメインには、HSV VP16活性化ドメイン(例えば、Hagmann et al.,J.Virol.71,5952−5962(1997)を参照)、核ホルモン受容体(例えば、Torchia et al.,Curr.Opin.Cell.Biol.10:373−383(1998)を参照)、核因子カッパBのp65サブユニット(Bitko & Barik,J.Virol.72:5610−5618(1998)およびDoyle & Hunt,Neuroreport 8:2937−2942(1997)を参照)、Liu et al.,Cancer Gene Ther.5:3−28(1998))、またはVP64等の人工キメラ機能ドメイン(Beerli et al.,(1998)Proc.Natl.Acad.Sci.USA 95:14623−33)、およびデグロン(Molinari et al.,(1999)EMBO J.18,6439−6447)が含まれる。さらなる例示的な活性化ドメインには、Oct1、Oct−2A、Sp1、AP−2、およびCTF1(Seipel et al.,EMBO J.11,4961−4968 (1992)、ならびにp300、CBP、PCAF、SRC1 PvALF、AtHD2A、およびERF−2が含まれる。例えば、Robyr et al.(2000)Mol.Endocrinol.14:329−347、Collingwood et al.(1999)J.Mol.Endocrinol.23:255−275、Leo et al.(2000)Gene 245:1−11;Manteuffel−Cymborowska(1999)Acta Biochim.Pol.46:77−89、McKenna et al.(1999)J.Steroid Biochem.Mol.Biol.69:3−12、Malik et al.(2000)Trends Biochem.Sci.25:277−283、およびLemon et al.(1999)Curr.Opin.Genet.Dev.9:499−504を参照されたい。さらなる例示的な活性化ドメインとしては、OsGAI、HALF−1、C1、AP1、ARF−5、−6、−7、および−8、CPRF1、CPRF4、MYC−RP/GP、ならびにTRAB1が含まれるが、これらに限定されない。例えば、Ogawa et al.(2000)Gene 245:21−29、Okanami et al.(1996)Genes Cells 1:87−99、Goff et al.(1991)Genes Dev.5:298−309、Cho et al.(1999)Plant Mol.Biol.40:419−429、Ulmason et al.(1999)Proc.Natl.Acad.Sci.USA 96:5844−5849、Sprenger−Haussels et al.(2000)Plant J.22:1−8、Gong et al.(1999)Plant Mol.Biol.41:33−44、およびHobo et al.(1999)Proc.Natl.Acad.Sci.USA 96:15,348−15,353を参照されたい。
【0086】
DNA結合ドメインと機能ドメインとの間の融合タンパク質(または同一物をコードする核酸)の形成において、活性化ドメインと相互作用する活性化ドメインまたは分子のいずれか一方が、機能ドメインとして好適であることが、当業者には明らかである。基本的に、標的遺伝子に活性化複合体および/または活性化活性(例えば、ヒストンアセチル化等)を補充する能力のある任意の分子が、融合タンパク質の活性化ドメインとして有用である。融合分子において機能ドメインとしての使用に好適なISWI含有ドメインおよび/またはメチル結合ドメインタンパク質等の絶縁体ドメイン、局在化ドメイン、ならびにクロマチンリモデリングタンパク質が、例えば、譲受人共通の米国特許出願公開第2002/0115215号および同第2003/0082552号明細書、ならびに譲受人共通の国際公開第02/44376号に記載されている。
【0087】
例示的な抑制ドメインとしては、KRAB、A/B、KOX、TGF−ベータ−誘発型初期遺伝子(TIEG)、v−erbA、SID、MBD2、MBD3、DNMTファミリーのメンバー(例えば、DNMT1、DNMT3A、DNMT3B)、Rb、およびMeCP2が含まれるが、これらに限定されない。例えば、Bird et al.(1999)Cell 99:451−454、Tyler et al.(1999)Cell 99:443−446、Knoepfler et al.(1999)Cell 99:447−450、およびRobertson et al.(2000) Nature Genet.25:338−342を参照されたい。さらなる例示的な抑制ドメインとしては、ROM2およびAtHD2Aが含まれるが、これらに限定されない。例えば、Chem et al.(1996)Plant Cell 8:305−321およびWu et al.(2000)Plant J.22:19−27を参照されたい。
【0088】
融合分子は、当業者によく知られているクローニング法および生化学的接合によって構築される。融合分子には、DNA結合ドメインおよび機能ドメイン(例えば、転写活性化または抑制ドメイン)が含まれる。融合分子には、核局在化シグナル(例えば、SV40媒体T抗原由来等)およびエピトープタグ(例えば、FLAGおよび血球凝集素等)も任意に含まれる。融合タンパク質(およびそれらをコードする核酸)は、翻訳リーディングフレームが融合の成分間で維持されるように設計される。
【0089】
一方において機能ドメイン(またはその機能的断片)のポリペプチド成分と、他方において非タンパク質DNA結合ドメイン(例えば、抗生物質、挿入剤、副溝結合剤、核酸)との間の融合は、当業者に知られている生化学的接合の方法によって構築される。例えば、Pierce Chemical Company(Rockford,IL)Catalogueを参照されたい。副溝結合剤とポリペプチドとの間に融合を起こすための方法および組成物が記載されている。Mapp et al.(2000)Proc.Natl.Acad.Sci.USA 97:3930−3935。
【0090】
特定の実施形態において、亜鉛フィンガータンパク質によって結合される標的部位は、細胞クロマチンの到達可能領域に存在する。到達可能領域は、例えば、譲受人共通の国際公開第01/83732号に記載されるように、決定することができる。標的部位が細胞クロマチンの到達可能領域に存在しない場合、譲受人共通の国際公開第01/83793号に記載されるように、1つ以上の到達可能領域を生成することができる。さらなる実施形態において、融合分子のDNA結合ドメインは、その標的部位が到達可能領域にあるかに関わらず、細胞クロマチンに結合することができる。例えば、このようなDNA結合ドメインは、リンカーDNAおよび/またはヌクレオソームDNAに結合することができる。「パイオニア」DNA結合ドメインのこの型の例は、ある特定のステロイド受容体および肝細胞核因子3(HNF3)で見出される。Cordingley et al.(1987)Cell 48:261−270、Pina et al.(1990)Cell 60:719−731、およびCirillo et al.(1998)EMBO J.17:244−254。
【0091】
融合分子は、当業者に知られているように、薬学的に許容される担体で製剤化され得る。例えば、Remington’s Pharmaceutical Sciences,17th ed.,1985、および譲受人共通の国際公開第00/42219号を参照されたい。
【0092】
融合分子の機能的成分/ドメインは、一旦融合分子がそのDNA結合ドメインを介して標的配列に結合すると、遺伝子の転写に影響を及ぼすことができる多種多様の異なる成分のいずれかから選択され得る。したがって、機能的成分には、活性化剤、リプレッサー、共活性化剤、コリプレッサー、およびサイレンサー等の多様な転写因子ドメインが含まれ得るが、これらに限定されない。
【0093】
さらなる例示的な機能ドメインは、例えば、譲受人共通の米国特許第6,534,261号および米国特許出願公開第2002/0160940号明細書に開示されている。
【0094】
外来小分子またはリガンドによって調節される機能ドメインも、選択され得る。例えば、RheoSwitch(登録商標)技術は、機能ドメインのみが、外部RheoChem(商標)リガンドの存在下で、その活性立体配座であると仮定する場合、使用され得る(例えば、米国特許出願公開第20090136465号明細書を参照)。したがって、ZFPは、調節可能な機能ドメインに作動可能に連結され得、ここで、得られたZFP−TFの活性は、外部リガンドによって制御される。
【0095】
ヌクレアーゼ
特定の実施形態において、融合タンパク質は、DNA結合結合ドメインおよび切断(ヌクレアーゼ)ドメインを含む。したがって、遺伝子修飾は、ヌクレアーゼ、例えば、改変されたヌクレアーゼを用いて達成することができる。改変されたヌクレアーゼ技術は、天然に存在するDNA結合タンパク質の改変に基づいている。例えば、目的に合わせたDNA結合特異性を有するホーミングエンドヌクレアーゼの改変が、記載されている。Chames et al.(2005)Nucleic Acids Res 33(20):e178、Arnould et al.(2006)J.Mol.Biol.355:443−458。加えて、ZFPの改変も記載されている。例えば、米国特許第6,534,261号、同第6,607,882号、同第6,824,978号、同第6,979,539号、同第6,933,113号、同第7,163,824号、および同第7,013,219号明細書を参照されたい。
【0096】
加えて、ZFN、すなわち、その改変された(ZFP)DNA結合ドメインを通してその意図される核酸標的を認識することができ、かつヌクレアーゼ活性によって、DNAがZFP結合部位の近くで切断される、機能的な実体を作り出すために、ZFPが、ヌクレアーゼドメインに融合されている。例えば、Kim et al.(1996)Proc Natl Acad Sci USA 93(3):1156−1160を参照されたい。つい最近では、ZFNは、様々な生物におけるゲノム修飾のために使用されている。例えば、米国特許出願公開第20030232410号、同第20050208489号、同第20050026157号、同第20050064474号、同第20060188987号、同第20060063231号明細書、および国際公開第07/014275号を参照されたい。
【0097】
したがって、本明細書に記載される方法および組成物は、広範囲に適用可能であり、対象となる任意のヌクレアーゼを含み得る。ヌクレアーゼの非限定的な例には、メガヌクレアーゼおよび亜鉛フィンガーヌクレアーゼが含まれる。ヌクレアーは、異種DNA結合および開裂ドメイン(例えば、亜鉛フィンガーヌクレアーゼ;異種開裂ドメインを有するメガヌクレアーゼDNA結合ドメイン)を含み得るか、または代替的に、天然に存在するヌクレアーゼのDNA結合ドメインは、選択された標的部位(例えば、同種の結合部位とは異なる部位に結合するように改変されたメガヌクレアーゼ)に結合するように変更され得る。
【0098】
特定の実施形態において、ヌクレアーゼは、メガヌクレアーゼ(ホーミングエンドヌクレアーゼ)である。天然に存在するメガヌクレアーゼは、15〜40塩基対の切断部位を認識し、一般に、LAGLIDADGファミリー、GIY−YIGファミリー、His−Cystボックスファミリー、およびHNHファミリーの4つのファミリーに分類される。例示的なホーミングエンドヌクレアーゼには、I−SceI、I−CeuI、PI−PspI、PI−Sce、I−SceIV、I−CsmI、I−PanI、I−SceII、I−PpoI、I−SceIII、I−CreI、I−TevI、I−TevII、およびI−TevIIIが含まれる。これらの認識配列は公知である。また、米国特許第5,420,032号、米国特許第6,833,252号明細書、Belfort et al.(1997)Nucleic Acids Res.25:3379-3388、Dujon et al.(1989)Gene 82:115-118、Perler et al.(1994)Nucleic Acids Res.22,1125-1127、Jasin(1996)Trends Genet.12:224-228、Gimble et al.(1996)J.Mol.Biol.263:163-180、Argast et al.(1998)J.Mol.Biol.280:345-353、およびニューイングランドバイオラボ社カタログも参照されたい。
【0099】
主として、LAGLIDADGファミリー由来の天然に存在するメガヌクレアーゼ由来のDNA結合ドメインが、植物、酵母、ショウジョウバエ属、哺乳動物細胞、およびマウスで部位特異的ゲノム修飾を促進するために用いられているが、このアプローチは、メガヌクレアーゼ認識配列を保存している相同遺伝子(Monet et al.(1999),Biochem.Biophysics.Res.Common.255:88−93)、またはその中に認識配列が導入されている予め改変されたゲノム(Route et al.(1994),Mol.Cell.Biol.14:8096−106、Chilton et al.(2003),Plant Physiology.133:956−65、Puchta et al.(1996),Proc.Natl.Acad.Sci.USA 93:5055−60、Rong et al.(2002),Genes Dev.16:1568−81、Gouble et al.(2006),J.Gene Med.8(5):616−622)のいずれかの修飾に限定されている。したがって、医学的または生物工学的に関連のある部位での新規の結合特異性を示すメガヌクレアーゼを改変する試みがなされている(Porteus et al.(2005),Nat. Biotechnol.23:967−73、Sussman et al.(2004),J.Mol.Biol.342:31−41、Epinat et al.(2003),Nucleic Acids Res.31:2952−62、Chevalier et al.(2002)Molec.Cell 10:895−905、Epinat et al.(2003)Nucleic Acids Res.31:2952−2962、Ashworth et al.(2006)Nature 441:656−659、Paques et al.(2007)Current Gene Therapy 7:49−66、米国特許出願公開第20070117128号、同第20060206949号、同第20060153826号、同第20060078552号、および同第20040002092号明細書)。加えて、メガヌクレアーゼ由来の天然に存在するまたは改変されたDNA結合ドメインもまた、異種ヌクレアーゼ(例えば、FokI)由来の開裂ドメインと作動可能に連結されている。
【0100】
他の実施形態において、ヌクレアーゼは、亜鉛フィンガーヌクレアーゼ(ZFN)である。ZFNは、選択遺伝子内の標的部位に結合するように改変されている亜鉛フィンガータンパク質および開裂ドメインまたは切断ハーフドメインを含む。
【0101】
上に詳細に記載されるように、亜鉛フィンガー結合ドメインは、選択配列に結合するように改変され得る。例えば、Beerli et al.(2002)Nature Biotechnol.20:135−141、Pabo et al.(2001)Ann.Rev.Biochem.70:313−340、Isalan et al.(2001)Nature Biotechnol.19:656−660、Segal et al.(2001)Curr.Opin.Biotechnol.12:632−637、Choo et al.(2000)Curr.Opin.Struct.Biol.10:411−416を参照されたい。改変された亜鉛フィンガー結合は、天然に存在する亜鉛フィンガータンパク質と比較して、新規の結合特異性を有することができる。改変方法には、合理的設計および多様な種類の選択が含まれるが、これらに限定されない。合理的設計には、例えば、三重(または四重)ヌクレオチド配列および個々の亜鉛フィンガーアミノ酸配列を含むデータベースを使用することが含まれ、各三重または四重ヌクレオチド配列は、特定の三重または四重配列と結合する亜鉛フィンガーの1つ以上のアミノ酸配列に関連する。例えば、譲受人共通の米国特許第6,453,242号および同第6,534,261号明細書を参照されたく、参照によりその全体が本明細書に組み込まれる。
【0102】
ファージディスプレイおよび2ハイブリッド法を含む例示的な選択方法は、米国特許第5,789,538号、同第5,925,523号、同第6,007,988号、同第6,013,453号、同第6,410,248号、同第6,140,466号、同第6,200,759号、同第6,242,568号明細書、ならびに国際公開第98/37186号、国際公開第98/53057号、国際公開第00/27878号、国際公開第01/88197号、および英国特許第2,338,237号に開示されている。加えて、亜鉛フィンガー結合ドメインに対する結合特異性の強化は、例えば、譲受人共通の国際公開第02/077227号に記載されている。
【0103】
標的部位の選択、融合タンパク質(および同一物をコードするポリヌクレオチド)の設計および構築のためのZFPおよび方法が当業者に知られており、米国特許出願公開第20050064474号および同第20060188987号明細書を参照されたく、参照によりその全体が本明細書に組み込まれる。
【0104】
加えて、これらおよび他の参考文献に開示される、亜鉛フィンガードメインおよび/または多フィンガーの亜鉛フィンガータンパク質は、例えば、5個以上のアミノ酸長のリンカー(例えば、TGEKP(配列番号40)、TGGQRP(配列番号41)、TGQKP(配列番号42)、および/またはTGSQKP(配列番号43))を含む、任意の好適なリンカー配列を使用して共に連結され得る。6個以上のアミノ酸長の例示的なリンカー配列に関して、例えば、米国特許第6,479,626号、同第6,903,185号、および同第7,153,949号明細書を参照されたい。本明細書に記載されるタンパク質は、タンパク質の個々の亜鉛フィンガー間の好適なリンカーの任意の組み合わせを含み得る。また、米国仮特許出願第61/343,729号も参照されたい。
【0105】
ZFNおよび/またはメガヌクレアーゼ等のヌクレアーゼはまた、ヌクレアーゼ(開裂ドメイン、切断ハーフドメイン)も含む。上記のように、開裂ドメインは、DNA結合ドメインに対して異種であり得、例えば、亜鉛フィンガーDNA結合ドメインおよびヌクレアーゼ由来の開裂ドメイン、またはメガヌクレアーゼDNA結合ドメインおよび異なるヌクレアーゼ由来の開裂ドメインであり得る。異種開裂ドメインは、任意のエンドヌクレアーゼまたはエキソヌクレアーゼであり得る。開裂ドメインを生成し得る例示的なエンドヌクレアーゼには、制限エンドヌクレアーゼおよびホーミングエンドヌクレアーゼが含まれるが、これらに限定されない。例えば、New England Biolabs,Beverly,MAの2002−2003版カタログおよびBelfort et al.(1997)Nucleic Acids Res.25:3379−3388を参照されたい。DNAを切断するさらなる酵素は公知である(例えば、S1ヌクレアーゼ;マングビーンヌクレアーゼ;膵臓DNアーゼI;マイクロコッカルヌクレアーゼ;酵母HOエンドヌクレアーゼ;Linn et al.(eds.)Nucleases,Cold Spring Harbor Laboratory Press,1993も参照されたい)。これらの酵素(またはその機能的断片)のうちの1つ以上は、開裂ドメインおよび切断ハーフドメインの源として使用することができる。
【0106】
同様に、切断ハーフドメインは、切断活性に二量体化が必要である、上記に示すように、任意のヌクレアーゼまたはその部分によって得ることができる。一般に、融合タンパク質が切断ハーフドメインを含む場合、切断のために2つの融合タンパク質が必要である。代替的に、2つの切断ハーフドメインを含む単一のタンパク質を使用することができる。2つの切断ハーフドメインは、同じエンドヌクレアーゼ(もしくはその機能的断片)から得ることができるか、またはそれぞれの切断ハーフドメインは、異なるエンドヌクレアーゼ(もしくはその機能的断片)から得ることができる。加えて、2つの融合タンパク質が、それぞれの標的部位に結合することによって、切断ハーフドメインが互いに、例えば、二量体化することによって、機能的な開裂ドメインを形成するのを可能にする空間的な配向に置かれるように、2つの融合タンパク質の標的部位は、互いに関して、好ましく配置される。したがって、特定の実施形態において、標的部位の近い末端を、5〜8ヌクレオチドまたは15〜18ヌクレオチドによって分けられる。しかしながら、任意の整数のヌクレオチドまたはヌクレオチド対が、2つの標的部位の間に(例えば、2〜50個のヌクレオチド対またはそれ以上)介在することができる。一般に、切断の部位は、標的部位の間にある。
【0107】
制限エンドヌクレアーゼ(制限酵素)は、多くの種に存在しており、(認識部位で)DNAに配列特異的に結合し、結合の部位でまたは結合の部位の近くでDNAを切断することができる。ある特定の制限酵素(例えば、IIS型)は、認識部位から隔たった部位の近くでDNAを切断し、分離可能な結合ドメインおよび開裂ドメインを有する。例えば、IIS型酵素Fok Iは、一方の鎖上ではその認識部位から9ヌクレオチドで、他方の鎖上ではその認識部位から13ヌクレオチドで、二本鎖切断を触媒する。例えば、米国特許第5,356,802号、同第5,436,150号、および同第5,487,994号明細書、ならびにLi et al.(1992)Proc.Natl.Acad.Sci.USA 89:4275−4279、Li et al.(1993)Proc.Natl.Acad.Sci.USA 90:2764−2768、Kim et al.(1994a)Proc.Natl.Acad.Sci.USA 91:883−887、Kim et al.(1994b)J.Biol.Chem.269:31,978−31,982を参照されたい。したがって、一実施形態において、融合タンパク質は、少なくとも1つのIIS型制限酵素由来の開裂ドメイン(または切断ハーフドメイン)および改変されても改変されなくてもよい1つ以上の亜鉛フィンガー結合ドメインを含む。
【0108】
開裂ドメインが結合ドメインから分離可能である、例示的なIIS型制限酵素は、Fok Iである。この特定の酵素は、二量体として活性がある。Bitinaite et al.(1998)Proc.Natl.Acad.Sci.USA 95:10,570−10,575。したがって、本開示に関して、開示される融合タンパク質で用いられるFok I酵素の部分は、切断ハーフドメインと考えられる。したがって、亜鉛フィンガー−Fok I融合体を用いた細胞配列の標的化した二本鎖切断および/または標的化した置換のために、それぞれ、Fok I切断ハーフドメインを含む、2つの融合タンパク質を用いて、触媒活性のある開裂ドメインを再構成することができる。代替的に、1つの亜鉛フィンガー結合ドメインおよび2つのFok I切断ハーフドメインを含む単一のポリペプチド分子を用いることもできる。亜鉛フィンガーFok I融合体を用いた標的化した切断および標的とした配列変更のためのパラメータは、本開示の他の箇所に提供されている。
【0109】
開裂ドメインまたは切断ハーフドメインは、切断活性を保持するか、または多量体化(例えば、二量体化)して、機能的な開裂ドメインを形成する能力を保持するタンパク質の任意の部分であることができる。
【0110】
例示的なIIS型制限酵素は、国際公開第07/014275号に記載されており、これは参照によりその全体が本明細書に組み込まれる。さらなる制限酵素も分離可能な結合ドメインおよび切断半ドメインを含み、これらは本開示によって企図される。例えば、Roberts et al.(2003)Nucleic Acids Res.31:418−420を参照されたい。
【0111】
特定の実施形態において、開裂ドメインは、例えば、米国特許出願公開第20050064474号および第20060188987号明細書、ならびに米国出願第11/805,850号(2007年5月23日に出願)(これらの全ての開示は参照によりその全体が本明細書に組み込まれる)に記載されるような、ホモ二量体化を最小限に抑えるか、またはホモ二量体化を妨げる1つ以上の改変された切断ハーフドメインを含む。Fok Iの446位、447位、479位、483位、484位、486位、487位、490位、491位、496位、498位、499位、500位、531位、534位、537位、および538位のアミノ酸残基は全て、Fok I切断ハーフドメインの二量体化に影響を与えるための標的である。
【0112】
絶対的ヘテロ二量体を形成するFok Iの例示的な改変された切断ハーフドメインには、第1の切断ハーフドメインがFok Iの490位および538位のアミノ酸残基における変異を含み、第2の切断ハーフドメインが486位および499位のアミノ酸残基における変異を含む対が含まれる。
【0113】
したがって、一実施形態において、490位の変異は、Glu(E)をLys(K)で置換し、538位の変異は、Iso(I)をLys(K)で置換し、486位の変異は、Gln(Q)をGlu(E)で置換し、499位の変異は、Iso(I)をLys(K)で置換する。具体的に、本明細書に記載される改変された切断ハーフドメインは、一方の切断ハーフドメインの490位(E→K)および538位(I→K)を変異させることによって、「E490K:I538K」と呼ばれる改変された切断ハーフドメインを産生し、もう一方の切断ハーフドメインの486位(Q→E)および499位(I→L)を変異させることによって、「Q486E:I499L」と呼ばれる改変された切断ハーフドメインを産生するように調製した。本明細書に記載される改変された切断ハーフドメインは、絶対的ヘテロ二量体変異体であり、この変異体では、異常な切断が最小限に抑えられているかまたは消失させられている。例えば、米国特許出願公開第2008/0131962号を参照されたく、この開示は、全ての目的のために参照によりその全体が本明細書に組み込まれる。特定の実施形態において、改変された切断ハーフドメインは、486位、499位、および496位(野生型Fok Iに対して番号付けされた)での変異、例えば、486位で野生型Gln(Q)残基をGlu(E)残基で、499位で野生型Iso(I)残基をLeu(L)残基で、496位で野生型Asn(N)残基をAsp(D)もしくはGlu(E)残基(それぞれ、「ELD」および「ELE」ドメインとも称される)で置換する変異を含む。他の実施形態において、改変された切断ハーフドメインは、490位、538位、および537位(野生型Fok Iに対して番号付けされた)での変異、例えば、490位で野生型Gln(E)残基をLys(K)残基で、538位で野生型Iso(I)残基をLys(K)残基で、537位で野生型His(H)残基をLys(K)もしくはArg(R)残基(それぞれ、「KKK」および「KKR」ドメインとも称される)で置換する変異を含む。他の実施形態において、改変された切断ハーフドメインは、490位および537位(野生型Fok Iに対して番号付けされた)での変異、例えば、490位で野生型Gln(E)残基をLys(K)残基で、537位で野生型His(H)残基をLys(K)もしくはArg(R)残基(それぞれ、「KIK」および「KIR」ドメインとも称される)で置換する変異を含む。(2010年2月8に出願された米国仮出願第61/337,769号を参照されたい)。
【0114】
本明細書に記載される改変された切断ハーフドメインは、任意の好適な方法を用いて、例えば、米国特許出願公開第20050064474号および第20080131962号明細書に記載される、野生型切断ハーフドメイン(Fok I)の部位特異的突然変異生成によって調製することができる。
【0115】
代替的に、ヌクレアーゼは、いわゆる「スプリット酵素」技術を用いた核酸標的部位でのインビボで組み立てられ得る(例えば、米国特許出願公開第20090068164号を参照)。このようなスプリット酵素の成分は、別個の発現構築物上で発現され得るか、または1つのオープンリーディングフレーム内に連結することができ、ここで、個々の成分は、例えば、自己切断型2AペプチドまたはIRES配列によって分離される。成分は、個々の亜鉛フィンガー結合ドメインまたはメガヌクレアーゼ核酸結合ドメインのドメインであり得る。
【0116】
いくつかの実施形態において、DNA結合ドメインは、植物病原菌であるキサントモナス(Boch et al,(2009)Science 326:1509−1512およびMoscou and Bogdanove,(2009)Science326:1501を参照)およびラストニア(Heuer et al(2007)Applied and Environmental Microbiology 73(13):4379−4384を参照)由来のものと同様に、TALエフェクター由来の改変されたドメインである。また、国際公開第2010/079430号も参照されたい。
【0117】
ヌクレアーゼ(例えば、ZFN)は、例えば、国際公開第2009/042163号および第20090068164号に記載される、酵母ベースの染色体系で使用する前に活性に対してスクリーニングすることができる。ヌクレアーゼ発現構築物は、当技術分野で知られている方法を用いて容易に設計することができる。例えば、米国特許出願公開第20030232410号、同第20050208489号、同第20050026157号、同第20050064474号、同第20060188987号、同第20060063231号明細書、および国際公開第07/014275号を参照されたい。ヌクレアーゼの発現は、構造性プロモーターまたは誘導性プロモーターの制御下、例えば、ラフィノースおよび/またはガラクトースの存在下で、活性化(抑制解除)され、グルコースの存在下で抑制されるガラクトキナーゼプロモーターの制御下にある。
【0118】
送達
タンパク質(例えば、ZFP)、同一物をコードするポリヌクレオチド、ならびに本明細書に記載されるタンパク質および/またはポリヌクレオチドを含む組成物は、任意の好適な手段で、例えば、ZFP TFまたはZFN mRNAを注入することによって、標的細胞に送達され得る。好適な細胞には、真核および原核細胞ならびに/または細胞株が含まれるが、これらに限定されない。このような細胞またはこのような細胞から生成された細胞株の非制限的な例には、COS、CHO(例えば、CHO−S、CHO−K1、CHO−DG44、CHO−DUXB11、CHO−DUKX、CHOK1SV)、VERO、MDCK、WI38、V79、B14AF28−G3、BHK、HaK、NS0、SP2/0−Ag14、HeLa、HEK293(例えば、HEK293−F、HEK293−H、HEK293−T)、およびperC6細胞、ならびにツマジロクサヨトウ(Sf)等の昆虫細胞、またはサッカロミセス、ピチア、およびシゾサッカロミセス等の真菌細胞が挙げられるが、これらに限定されない。ある特定の実施形態において、細胞株は、CHO−K1、MDCK、またはHEK293細胞株である。また、好適な細胞には、例として、胚幹細胞、誘導多能性幹細胞、造血幹細胞、および間充織幹細胞等の幹細胞も含まれる。
【0119】
本明細書に記載される亜鉛フィンガータンパク質を含むタンパク質を送達する方法は、例えば、米国特許第6,453,242号、同第6,503,717号、同第6,534,261号、同第6,599,692号、同第6,607,882号、同第6,689,558号、同第6,824,978号、同第6,933,113号、同第6,979,539号、同第7,013,219号、および同第7,163,824号明細書に記載され、これらの全ての開示は、参照によってその全体が本明細書に組み込まれる。
【0120】
本明細書に記載される亜鉛フィンガータンパク質はまた、亜鉛フィンガータンパク質の1つ以上をコードする配列を含有するベクターを使用して送達され得る。プラスミドベクター、レトロウイルスベクター、レンチウイルスベクター、アデノウイルスベクター、ポックスウイルスベクター、ヘルペスウイルスベクター、およびアデノ随伴ウイルスベクター等を含むが、これらに限定されない任意のベクター系が使用され得る。また、米国特許第6,534,261号、同第6,607,882号、同第6,824,978号、同第6,933,113号、同第6,979,539号、同第7,013,219号、および同第7,163,824号明細書を参照されたく、参照によってその全体が本明細書に組み込まれる。さらに、これらのベクターのうちのいずれかが、配列をコードする1つ以上の亜鉛フィンガータンパク質を含み得ることは明らかである。したがって、1つ以上のZFPが細胞中に導入される時、ZFPは、同一のベクターまたは異なるベクター上で担持され得る。複数のベクターが使用される時、それぞれのベクターは、1つまたは複数のZFPをコードする配列を含み得る。
【0121】
従来のウイルスおよび非ウイルスに基づく遺伝子導入方法を使用して、改変されたZFPをコードする核酸を、細胞(例えば、哺乳動物細胞)および標的組織に導入することができる。このような方法を使用して、ZFPをコードする核酸を、細胞にインビトロ投与することもできる。ある特定の実施形態において、ZFPをコードする核酸は、インビボまたはエクスビボ遺伝子療法用途のために投与される。非ウイルスベクター送達システムには、DNAプラスミド、ネイキッド核酸、およびリポソームまたはポロキサマー等の送達媒体と複合体を形成した核酸が含まれる。ウイルスベクター送達システムには、細胞への送達後にエピソームまたは組み込まれたゲノムのいずれか一方を有する、DNAおよびRNAウイルスが含まれる。遺伝子療法手順の総説に関しては、Anderson,Science 256:808−813(1992)、Nabel & Felgner,TIBTECH 11:211−217(1993)、Mitani & Caskey,TIBTECH 11:162−166(1993)、Dillon,TIBTECH 11:167−175(1993)、Miller,Nature 357:455−460(1992)、Van Brunt,Biotechnology 6(10):1149−1154(1988)、Vigne,Restorative Neurology and Neuroscience 8:35−36(1995)、Kremer & Perricaudet,British Medical Bulletin 51(1):31−44(1995)、Haddada et al.,in Current Topics in Microbiology and Immunology Doerfler and Bohm(eds.)(1995)、およびYu et al.,Gene Therapy 1:13−26(1994)を参照されたい。
【0122】
核酸の非ウイルス送達の方法には、エレクトロポレーション、リポフェクション、マイクロインジェクション、バイオリスティック、ビロソーム、リポソーム、免疫リポソーム、ポリカチオン、または脂質:核酸複合体、ネイキッドDNA、人工ビリオン、および薬剤により強化されたDNAの取込みが含まれる。例えば、Sonitron 2000 system(Rich−Mar)を使用するソノポレーションも、核酸の送達に使用することができる。
【0123】
さらなる例示的な核酸送達システムには、Amaxa Biosystems(Cologne,Germany)、Maxcyte,Inc.(Rockville,Maryland)、BTX Molecular Delivery Systems(Holliston,MA)、およびCopernicus Therapeutics Inc,(例えば、米国特許第6008336号を参照)により提供されるものが含まれる。リポフェクションは、例えば、米国特許第5,049,386号、同第4,946,787号、および同第4,897,355号明細書)に記載されており、リポフェクション試薬は、市販されている(例えば、Transfectam(商標)およびLipofectin(商標))。ポリヌクレオチドの効率的な受容体認識リポフェクションに好適な陽イオン性および中性脂肪には、Felgnerのもの、国際公開第91/17424号、国際公開第91/16024号が含まれる。送達は、細胞(エクスビボ投与)または標的組織(インビボ投与)に送達することができる。
【0124】
免疫脂質複合体等の標的リポソームを含む脂質:核酸複合体の調製は、当業者によく知られている(例えば、Crystal,Science 270:404−410(1995)、Blaese et al.,Cancer Gene Ther.2:291−297(1995)、Behr et al.,Bioconjugate Chem.5:382−389(1994)、Remy et al.,Bioconjugate Chem.5:647−654(1994)、Gao et al.,Gene Therapy 2:710−722(1995)、Ahmad et al.,Cancer Res.52:4817−4820(1992)、米国特許第4,186,183号、同第4,217,344号、同第4,235,871号、同第4,261,975号、同第4,485,054号、同第4,501,728号、同第4,774,085号、同第4,837,028号、および同第4,946,787号明細書を参照されたい)。
【0125】
送達のさらなる方法には、送達されるべき核酸をEnGeneIC送達ビヒクル(EDV)にパッケージングする方法も含まれる。これらのEDVは、二重特異性抗体を用いて標的組織に特異的に送達され、ここで、抗体の一方のアームには、標的組織に対する特異性を有し、もう一方には、EDVに対する特異性を有する。抗体は、標的細胞表面にEDVを取り込まれ、次いで、EDVが、エンドサイトーシスによって細胞に取り込まれる。一旦細胞内に取り込まれると、内容物が放出される(MacDiarmid et al(2009)Nature Biotechnology 27(7):643を参照)。
【0126】
改変されたZFPをコードする核酸の送達のためのRNAまたはDNAウイルスに基づくシステムの使用は、ウイルスに体内の特定の細胞を標的とさせ、ウイルスペイロードを核に輸送するための高度に進化したプロセスを利用する。ウイルスベクターは、患者に直接投与(インビボ)することができるか、またはインビトロで細胞を治療するために使用することができ、修飾された細胞が患者に投与される(エクスビボ)。ZFPの送達のための従来のウイルスに基づくシステムには、遺伝子送達のためのレトロウイルス、レンチウイルス、アデノウイルス、アデノ随伴ウイルス、ワクチン、および単純ヘルペスウイルスベクターが含まれるが、これらに限定されない。宿主ゲノム内の組込みにより、レトロウイルス、レンチウイルス、およびアデノ随伴ウイルス遺伝子送達方法を使用して可能であり、しばしば、挿入された導入遺伝子が長期発現される。さらに、高い導入効率が、多くの異なる細胞型および標的組織で観察されている。
【0127】
レトロウイルスの向性は、外来のエンベロープタンパク質を組み込むことによって変更することができ、標的細胞の潜在的な標的群を伸長する。レンチウイルスベクターは、非分裂細胞を形質導入または感染させることが可能であり、典型的には高ウイルス価を産生する。レトロウイルス遺伝子送達システムの選択は、標的組織に依存する。レトロウイルスベクターは、最大6〜10kbの外来配列のパッケージング能力を有する、シス作用の長い末端反復から成る。最小限のシス作用LTRは、ベクターの複製およびパッケージングに十分であり、それらは、次いで、恒久的な導入遺伝子発現を提供するように治療遺伝子を標的細胞内に組み込むために使用される。広範囲にわたって使用されるレトロウイルスベクターには、マウス白血病ウイルス(MuLV)、テナガザル白血病ウイルス(GaLV)、サル免疫不全ウイルス(SIV)、ヒト免疫不全ウイルス(HIV)、およびその組み合わせに基づくものが含まれる(例えば、Buchscher et al.,J.Virol.66:2731−2739(1992)、Johann et al.,J.Virol.66:1635−1640(1992)、Sommerfelt et al.,Virol.176:58−59(1990)、Wilson et al.,J.Virol.63:2374−2378(1989)、Miller et al.,J.Virol.65:2220−2224(1991)、PCT/US第94/05700号を参照されたい)。
【0128】
一過性発現が好ましい用途では、アデノウイルスに基づくシステムを使用することができる。アデノウイルスに基づくベクターは、多くの細胞型において非常に高い導入効率があり、細胞分裂を必要としない。このようなベクターを使用して、高力価および高レベルの発現が得られている。比較的単純なシステムにおいて、このベクターを多量に産生することができる。アデノ随伴ウイルス(「AAV」)ベクターを使用して、例えば、核酸およびペプチドのインビトロ産生において、およびインビボおよびエクスビボ遺伝子治療手順(例えば、West et al.,Virology 160:38−47(1987)、米国特許第4,797,368号明細書、国際公開第93/24641号、Kotin,Human Gene Therapy 5:793−801(1994)、Muzyczka,J.Clin.Invest.94:1351(1994)を参照)のために、細胞に標的核酸を形質導入する。AAVベクターの構築は、米国特許第5,173,414号明細書、Tratschin et al.,Mol.Cell.Biol.5:3251−3260(1985)、Tratschin et al.,Mol.Cell.Biol.4:2072−2081(1984)、Hermonat & Muzyczka,PNAS 81:6466−6470(1984)、およびSamulski et al.,J.Virol.63:03822−3828(1989)を含む、多数の刊行物に記載される。
【0129】
現在、少なくとも6つのウイルスベクターアプローチが臨床試験における遺伝子導入に使用可能であり、それらは形質導入剤を生成するためにヘルパー細胞株に挿入された遺伝子による欠損ベクターの相補性を含むアプローチを利用する。
【0130】
pLASNおよびMFG−Sは、臨床試験で使用されているレトロウイルスベクターの例である(Dunber et al.,Blood 85:3048−305(1995)、Kohn et al.,Nat.Med.1:1017−102(1995)、Malech et al.,PNAS 94:22 12133−12138(1997))。PA317/pLASNは、遺伝子治療試験で使用された最初の治療ベクターであった。(Blaese et al.,Science 270:475−480(1995))。MFG−Sパッケージされたベクターに対して、50%またはそれより大きい導入効率が観察されている。(Ellem et al.,Immunol Immunother.44(1):10−20(1997)、Dranoff et al.,Hum.Gene Ther.1:111−2(1997)。
【0131】
アデノ随伴ウイルスベクター(rAAV)は、欠損および非病原性パルボウイルスアデノ随伴2型ウイルスに基づく有望な代替遺伝子送達システムである。全てのベクターは、導入遺伝子発現カセットに隣接するAAV145bp逆方向末端反復のみを保持するプラスミドに由来する。形質導入細胞のゲノム内への組込みによる効率的な遺伝子導入および安定した導入遺伝子送達は、このベクター系の重要な特長である。(Wagner et al.,Lancet 351:9117 1702−3(1998)、Kearns et al.,Gene Ther.9:748−55(1996))。9:748−55(1996))。AAV1、AAV3、AAV4、AAV5、AAV6、およびAAV8を含む他のAAV血清型も本発明に従って使用することができる。
【0132】
複製欠損性アデノウイルスベクター(Ad)を高力価で産生することができ、多数の異なる細胞型を容易に感染させる。アデノウイルスベクターの大半は、導入遺伝子がAdE1a、E1b、および/またはE3遺伝子を置換し、その後、複製欠損性ベクターが、トランス内の消失遺伝子機能を供給するヒト293細胞で増殖するように改変される。Adベクターは、肝臓、腎臓、および筋肉で見出されるもの等の非分裂の分化した細胞を含む、複数の種類の組織をインビボで形質転換することができる。従来のAdベクターは、大きな担持能力を有する。臨床試験におけるAdベクターの使用の例には、筋肉内注入を用いる抗腫瘍免疫のポリヌクレオチド治療が含まれた(Sterman et al.,Hum.Gene Ther.7:1083−9(1998))。臨床試験における遺伝子導入のためのアデノウイルスベクターの使用のさらなる例には、Rosenecker et al.,Infection 24:1 5−10(1996)、Sterman et al.,Hum.Gene Ther.9:7 1083−1089(1998)、Welsh et al.,Hum.Gene Ther.2:205−18(1995)、Alvarez et al.,Hum.Gene Ther.5:597−613(1997)、Topf et al.,Gene Ther.5:507−513(1998)、Sterman et al.,Hum.Gene Ther.7:1083−1089(1998)が含まれる。
【0133】
パッケージング細胞を使用して、宿主細胞を感染させることが可能なウイルス粒子を形成する。このような細胞には、アデノウイルスをパッケージングする293細胞、およびレトロウイルスをパッケージングするψ2細胞またはPA317が含まれる。遺伝子治療で使用されるウイルスベクターは、通常、核酸ベクターをウイルス粒子中にパッケージングする産出細胞株によって生成される。これらのベクターには、典型的には、パッケージングおよびその後の宿主への組込みに必要とされる最小限のウイルス配列(適用可能な場合)、発現されるようにタンパク質をコードする発現カセットによって置換されている他のウイルス配列が含まれる。欠損しているウイルス機能は、パッケージングする細胞株によってトランスで提供される。例えば、遺伝子治療に使用するAAVベクターは、典型的には、宿主ゲノム内へのパッケージングおよび組込みに必要とされるAAVゲノムからの逆方向末端反復(ITR)配列のみを所有する。ウイルスDNAは、細胞株内にパッケージングされ、他のAAV遺伝子、すなわち、repおよびcapをコードするが、ITR配列を欠失するヘルパープラスミドを含有する。この細胞株はまた、ヘルパーとしてアデノウイルスに感染する。このヘルパーウイルスは、ヘルパープラスミドから、AAVベクターの複製、およびAAV遺伝子の発現を促進する。このヘルパープラスミドは、ITR配列の欠失の結果、相当な量でパッケージングされない。アデノウイルスの汚染は、例えば、AAVよりも敏感であるアデノウイルスに対する熱処理によって減少することができる。
【0134】
多くの遺伝子治療用途において、遺伝子治療ベクターが高度の特異性で特定の細胞型に送達されることが望ましい。したがって、ウイルスの外表面上でウイルス外被タンパク質との融合タンパク質としてリガンドを発現することによって、ウイルスベクターを所定の細胞型に対する特異性を有するように修飾することができる。リガンドは、関心の細胞型上に存在することで知られている受容体に対する親和性を有するように選択される。例えば、Han et al.,Proc.Natl.Acad.Sci.USA 92:9747−9751(1995)は、gp70に融合されたヒトヘレグリンを発現するように、モロニーマウス白血病ウイルスを修飾することができ、そのウイルスが、ヒト上皮成長因子受容体を発現するある特定のヒト乳癌細胞を感染させることを報告した。この原理は、細胞表面受容体に対して、標的細胞が受容体を発現し、ウイルスがリガンドを含む融合タンパク質を発現する、他のウイルス標的細胞対にまで及ぶことができる。例えば、実質的に任意の選択された細胞受容体に特異的結合親和性を有する抗体断片(例えば、FABまたはFv)を表示するように、線状ファージを改変することができる。上記の説明は、主にウイルスベクターに適用されるが、同一の原理を非ウイルスベクターに適用することができる。特異的標的細胞による取り込みを好む特異的な取り込み配列を含有するように、そのようなベクターを改変することができる。
【0135】
遺伝子治療ベクターは、下で説明するように、個々の患者への投与、典型的には、全身投与(例えば、静脈内、腹腔内、筋肉内、皮下、もしくは頭蓋内注入)または局所投与によって、インビボで送達することができる。代替的に、個々の患者から移植された細胞(例えば、リンパ球、骨髄穿刺液、組織生検)または万能ドナーの幹細胞等の細胞に、ベクターをエクスビボで送達することができ、その後、ベクターを組み込んだ細胞の選択後に、患者への細胞の再移植が続く。
【0136】
診断、研究のため、または遺伝子治療のためのエクスビボ細胞トランスフェクション(例えば、宿主生物にトランスフェクトされた細胞の再注入を介して)が当業者によく知られている。好ましい実施形態において、細胞は、対象生物から単離され、ZFP核酸(遺伝子またはcDNA)でトランスフェクトし、対象生物(例えば、患者)内に戻って再注入される。エクスビボのトランスフェクションに好適な多様な細胞型は、当業者によく知られている(患者からの細胞の単離および培養方法についての説明は、例えば、Freshney et al.,Culture of Animal Cells,A Manual of Basic Technique(3rd ed.1994)、およびそこで引用される参考文献を参照されたい)。
【0137】
一実施形態において、幹細胞は、細胞トランスフェクションおよび遺伝子治療のためのエクスビボ処置で使用される。幹細胞を使用する利点は、それらがインビトロで他の細胞型に分化することができるか、または、それらが骨髄に生着する哺乳動物(幹細胞のドナー等)に導入し得ることである。GM−CSF、IFN−γ、およびTNF−α等のサイトカインを使用して、CD34+細胞を臨床的に重要な免疫細胞型にインビトロで分化するための方法が知られている(Inaba et al.,J.Exp.Med.176:1693−1702(1992)を参照)。
【0138】
幹細胞は、周知の方法を使用して、形質導入および分化のために単離される。例えば、幹細胞は、CD4+およびCD8+(T細胞)、CD45+(panB細胞)、GR−1(顆粒球)、およびIad(分化された抗原提示細胞)等の不要な細胞に結合する抗体を有する骨髄細胞をパンニングすることによって骨髄から単離される(Inaba et al.,J.Exp.Med.176:1693−1702(1992)を参照)。
【0139】
いくつかの実施形態において、修飾されている幹細胞も、使用され得る。例えば、アポトーシスに耐性をなしている神経幹細胞は、治療組成物として使用され得、ここで、幹細胞はまた、本発明のZFP TFも含有する。アポトーシスに対する耐性は、例えば、幹細胞において、BAXまたはBAKに特異的なZFNを用いて(米国特許出願第12/456,043号を参照)BAXおよび/またはBAK、または例えば、カスパーゼ6に特異的なZFNを用いて再度、カスパーゼにおいて妨害されるものを不活性化することによって生じ得る。これらの細胞は、変異または野生型Httを調節することが知られているZFP TFでトランスフェクトすることができる。
【0140】
治療的ZFP核酸を含有するベクター(例えば、レトロウイルス、アデノウイルス、リポソーム等)を、細胞の形質導入のために、生物にインビボで直接投与することもできる。代替的に、ネイキッドDNAを投与することができる。投与は、注射、注入、局所適用、およびエレクトロポレーションを含むが、これらに限定されない、血液細胞または組織細胞との最終的な接触内に分子を導入するために通常使用される経路のいずれかによる。このような核酸を投与する好適な方法は、使用可能であり、かつ当業者にも知られており、1つ以上の経路を使用して特定の組成物を投与することができるが、特定の経路は、しばしば、別の経路よりも即時かつ有効な反応を提供することができる。
【0141】
造血幹細胞にDNAを導入するための方法が、例えば、米国特許第5,928,638号明細書に開示されている。トランス遺伝子の、例えば、CD34+細胞等の造血幹細胞への導入に有用なベクターには、アデノウイルス35型が含まれる。
【0142】
トランス遺伝子の免疫細胞(例えば、T細胞)への導入に好適なベクターには、非組込みレンチウイルスベクターが含まれる。例えば、Ory et al.(1996)Proc.Natl.Acad.Sci.USA 93:11382−11388、Dull et al.(1998)J.Virol.72:8463−8471、Zuffery et al.(1998)J.Virol.72:9873−9880、Follenzi et al.(2000)Nature Genetics 25:217−222を参照されたい。
【0143】
薬学的に許容される担体は、投与されている特定の組成物によって、ならびに組成物を投与するために使用される特定の方法によってある程度決定される。したがって、下で説明するように、使用可能な薬学的組成物の多種多様の好適な製剤が存在する(例えば、Remington‘s Pharmaceutical Sciences,17th ed.,1989を参照)。
【0144】
上述のように、開示される方法および組成物には、原核細胞、真菌細胞、古細菌細胞、植物細胞、昆虫細胞、動物細胞、脊椎動物細胞、哺乳動物細胞、およびヒト細胞が含まれるが、これらに限定されない、任意の種類の細胞で使用することができる。タンパク質発現に好適な細胞株が当業者に知られており、COS、CHO(例えば、CHO−S、CHO−K1、CHO−DG44、CHO−DUXB11)、VERO、MDCK、WI38、V79、B14AF28−G3、BHK、HaK、NS0、SP2/0−Ag14、HeLa、HEK293(例えば、HEK293−F、HEK293−H、HEK293−T)、perC6、昆虫細胞(ツマジロクサヨトウ(Sf)等)、および真菌細胞(サッカロミセス、ピチア、およびシゾサッカロミセス等)が含まれるが、これらに限定されない。これらの細胞株の子孫、変種、および誘導体を使用することもできる。
【0145】
用途
トリヌクレオチド反復疾患に関連する遺伝子を調節する、および/またはこれらの疾患に関連する遺伝子におけるトリヌクレオチド反復の経路を取り除くことが望ましい任意の用途のために、開示される組成物および方法を使用することができる。特に、治療および研究用途を含むが、これらに限定されない、Htt対立遺伝子の調節が望ましい任意の用途のために、これらの方法および組成物を使用することができる。
【0146】
ZFP TFを抑制するHD Httを治療剤として使用することができる疾患および状態には、ハンチントン病が含まれるが、これに限定されない。さらに、Httの変異対立遺伝子に対して特異的なZFNを含む方法および組成物をハンチントン病の治療のための治療剤として使用することができる。
【0147】
HD Htt対立遺伝子を抑制するZFP−TFはまた、GDNFおよびBDNFが含まれるが、これらに限定されない神経栄養因子を活性化するZFP−TFと連結して使用され得る。これらのZFP(またはこれらのZFPをコードするポリヌクレオチド)は、同時に投与され得る(例えば、同じ薬学的組成物において)か、または任意の順序で連続して投与され得る。
【0148】
また、ハンチントン病の治療のための方法および組成物には、幹細胞内でHtt対立遺伝子の変異コピーがHttに特異的なZFNを用いて、野生型Htt対立遺伝子に修飾されている幹細胞組成物も含まれる。
【0149】
本発明の方法および組成物はまた、インビトロおよびインビボモデル、例えば、トリヌクレオチド反復疾患の動物モデルの設計および実施に有用であり、これは、これらの疾患の研究を可能にする。
【実施例】
【0150】
実施例1:Htt標的亜鉛フィンガータンパク質転写因子(ZFP−TF)の設計および構築
Httを標的化した亜鉛フィンガータンパク質は、米国特許第6,534,261号明細書に記載されるように、基本的に改変された。表1は、例示的なHtt標的ZFPの認識へリックスDNA結合ドメインを示す。
【0151】
また、両端型ZFP−TFは、最終ベクターが、SIP1等の両端型ZFP−TFの遺伝子配列に隣接する2つのアレイのZFPを含むことを除いては、基本的に上記のような亜鉛フィンガー結合ドメインの2つの別個のクラスターを用いて構築される。図1Cおよび図4を参照されたい。
【0152】
多量体型ZFP TFはまた、このベクターが、ZFP TFをコードする配列に作動可能に連結されるトリヌクレオチド反復の経路に沿って発現タンパク質の多量体化を可能にする1つ以上の多量体化ドメインをコードする配列を含有することを除いては、上記のように構築される。図1Dおよび図3を参照されたい。
【0153】
ZFP TFは、核局在化配列、Htt対立遺伝子を標的とした改変された亜鉛フィンガーDNA結合ドメイン(表1を参照)、およびヒトKOX1タンパク質由来のKRAB抑制ドメインを含む、融合タンパク質として構築された。図1Aを参照されたい。設計されたDNA結合ドメインは、18−bp配列を認識する6個のフィンガー分子を含有する(表2を参照)。ZFP認識ヘリックスによって接触される標的部位におけるヌクレオチドは大文字で示され、非接触ヌクレオチドは小文字で示される。
【0154】
ELISAベースのアッセイを使用して、標的配列に結合するZFPを確認する。簡潔に言えば、T7プロモーターの制御下で、血球凝集素エピトープ(HA)標的化全長ZFPをコードする発現カセットは、HAタグのT7プロモーターおよびコード配列を導入するプライマーを用いて、組み立てられたZFPのそれぞれからのPCR増幅によって作製される。このPCR断片を使用して、TNT Quick Coupled TranscriptionおよびTranslation System(Promega,WI,USA)を用いることによって、インビトロでHA標識化ZFPタンパク質を生成し、HA標識化タンパク質は、次いで、Htt遺伝子内の意図される標的部位を含有するビオチン化DNAオリゴ、ならびに競合体(ヒトゲノム)DNAと混合し、特異的ZFP−DNA認識のみ報告することが可能である。
【0155】
次いで、ZFP−DNA複合体は、ペルオキシダーゼ共役抗HA抗体で標識化され、続いて、ストレプトアビジンでコーティングされた96ウェルプレート上に捕捉する。次いで、特異的ZFP−DNA複合体は、基質として、QuantaBlu(Pierce,Rockford,IL)を用いて、捕捉したペルオキシダーゼの活性をアッセイすることによって定量化する。マウスまたはヒトHttプロモーターおよびエクソン1に対して特異的なZFP TFは、標的配列に結合するために設計および試験される。
【0156】
実施例2:ヒトおよびマウス細胞におけるHD Httの抑制。
ZFP TFを抑制するHttの活性を試験するために、ZFP TFをヒト細胞にトランスフェクトし、PCRを用いてHttの発現を観察した。
【0157】
初めに、表1中に示される標準ZFP TFを試験した。ヒト293細胞(Graham et al(1977).J Gen Virol 36:59−74)を、10% FBSで捕捉したDMEM中に培養した。96ウェルプレートに、1ウェルにつき1e4細胞の密度で播種し、翌日、製造業者の指示に従って、Fugene6試薬を用いて、18832、18856、18859、および18868のZFP−TFをコードする0.1ugのプラスミドDNAでトランスフェクトした。トランスフェクトした細胞を、3日間インキュベートし、次いで、遺伝子発現分析のためにCells−to−CT(商標)キット(Applied Biosystems)を用いてプロセスした。内部対照のベータアクチン(ACTB)と比較した内因性ヒトハンチンチン(Htt)のレベルを、Hs00918176_m1および4352935Eのプライマーおよびプローブ(Applied Biosystems)をそれぞれ用いて、TaqMan(登録商標)7300上のリアルタイムPCRによって分析した。
【0158】
図2に示されるように、ZFP−TFは、GFPおよび偽対照と比較して、Htt発現を抑制した。結果は、ACTBの内因性レベルに対する、基底Httの割合で示される。
【0159】
両端型および多量体型ZFP TFの活性を試験するために、同じプロトコルを、両端型および多量体型ZFPを用いて行う。
【0160】
ヒトHD Htt対立遺伝子に対する特異性を有するZFP TFを試験するために、Htt特異的ZFP TFを、SH−SY5Y神経芽細胞腫細胞(Biedler JL,et al.(1978)Cancer Res.38:3751−3757)に過渡的にトランスフェクトする。簡潔に言えば、組み立てられたZFP−TF構築物を、pcDNAベクター(InVitrogen)にクローン化し、製造業者のプロトコルに従って、Fugene6キット(Roche Applied Science)を用いてトランスフェクトする。Htt発現レベルは、Applied Biosystemsから購入したHs00918176_m1プライマー/プローブのセットを用いて、リアルタイムRT−PCR(TaqMan(登録商標)、Applied Biosystems)を用いて測定する。Httタンパク質の減少を確認するために、ウエスタンブロット解析を行う。
【0161】
マウス対立遺伝子と相互作用するように設計されたHtt特異的ZFP TFを試験するために、マウスZFP TFをコードする構築体をpcDNAベクターにクローン化し、製造業者のプロトコルに従って、Lipofectamine 2000キット(Invitrogen)を用いて、ニューロ2A細胞(Klebe & Ruddle(1969)J.Cell Biol.43:69A)またはマウスHttを発現する同等のマウス神経細胞株に過渡的にトランスフェクトする。Htt発現レベルを、リアルタイムRT−PCRを用いて測定する。Httタンパク質の減少を確認するために、ウエスタンブロット解析を行う。
【0162】
加えて、ヒトHtt特異的ZFP TFを、CAG伸長を有する切断型ヒトHtt対立遺伝子を担持するR6.2トランスジェニックマウス由来の一次神経細胞において試験する(Mangiarini et al.,(1996)Cell 87:493-506)。マウスHtt特異的ZFP TF、ならびにCAG反復(両端型ZFPおよび多量体化ドメインを有するZFP)を標的化するZFPを、Httノックインマウス由来の不死化線条体細胞において試験する(Wheeler et al.,(1999)Hum Mol Genet.Jan;8(1):115−22)。
【0163】
実施例3:インビボでのHtt発現の抑制
インビボでのHtt特異的ZFP TFを試験するために、ZFPをコードするAAV2ベクターを産生する。次いで、これらのAAV2ベースの構築体を、マウスの脳に送達する。ヒトHtt特異的ZFP TFにおいては、AAVベクターをR6.2マウスまたはBAC HDマウス(C57Bl/6またはFVB/N株)に送達し、ヒト導入遺伝子の抑制を評価する。マウスHtt特異的ZFPにおいては、AAVベクターを野生型マウス(C57Bl/6またはFVB/N)に送達し、内因性マウスHtt発現の抑制を評価する。CAGを伸長した対立遺伝子を優先的に標的化するZFPにおいては、AAVベクターをR6.2マウスまたはヒトHttノックインマウスに送達し、野生型対伸長したHtt対立遺伝子の選択的抑制を検証する。殺処分後、脳組織を、TaqmanリアルタイムRT−PCRによって、Htt発現に対して分析する。
【0164】
実施例4:神経栄養因子およびHD Htt対立遺伝子特異的なZFP TFの共トランスフェクション。
上で同定されるHtt特異的ZFP TFは、脳神経栄養因子に対して特異的なZFP TFで共トランスフェクトされる。使用した脳神経栄養因子に対して特異的なZFP TFは、GDNFあるいはBDNFのいずれかに対して特異的である。
【0165】
実施例5:Htt標的亜鉛フィンガーヌクレアーゼ(ZFN)の設計および構築
ヒトHttおよびマウスHttを標的化するZFNは、CAG反復に隣接する配列、ならびに最初および最後のコーディングエクソンの配列を標的化するように設計される。ZFNは、Urnov et al.(2005)Nature 435(7042):646−651、Perez et al(2008)Nature Biotechnology 26(7):808−816、および米国特許出願公開第2008/0131962号明細書に記載されるように基本的に設計され、プラスミドまたはアデノウイルスベクターに組み込まれた。
【0166】
上記のように、組み立てられたZFNを、ELISAによって、それらのそれぞれの配列に結合させるために試験する。
【0167】
実施例6:Htt特異的ZFNの切断活性
上記のヒトHtt特異的ZFNの対をコードするプラスミドを、K562細胞にトランスフェクトする。K562細胞は、American Type Culture Collectionから入手して、10%規格ウシ胎仔血清(FCS、Cyclone)を補足したF−12培地(Invitrogen)中で推奨されるように、成長させた。細胞を、TrypLE Select(商標)プロテアーゼ(Invitrogen)を用いて、プラスチック容器から引き離した。トランスフェクションのために、100万個のK562細胞を、2μgの亜鉛フィンガーヌクレアーゼプラスミドおよび100μL Amaxa溶液Tと混合した。細胞を、プログラムU−23を用いてAmaxa Nucleofector II(商標)中でトランスフェクトし、1.4mLの温F−12培地+10%FCS中に回収した。
【0168】
ゲノムDNAを収穫し、意図された切断部位を含むHtt遺伝子座の一部をPCR増幅する。InVitrogenからのAccuprime HiFiポリメラーゼを用いたPCRを、以下の通りに実行する。94℃で最初の3分間変性させた後、PCRの30サイクルを、94℃で30秒間の変性ステップを行い、続いて、58℃で30秒間のアニーリングステップ、68℃で30秒間の伸長ステップを行う。30サイクルが完了した後、反応物を68℃で7分間インキュベートし、次いで、10℃で無制限に行う。
【0169】
K562 Htt特異的ZFNで処理された細胞からのゲノムDNAは、例えば、米国特許出願公開第20080015164号、第20080131962号、および第20080159996号明細書に記載される、Surveyor(商標)ヌクレアーゼ(Transgenomic)によって検証される。
【0170】
マウスHtt特異的ZFNの対をコードするプラスミドを、同様の様式で、ニューロ2a細胞中で試験する。
【0171】
実施例7:可変長のトリヌクレオチド反復の標的化組込み
上記の、CAG反復に隣接する配列に対して最大の切断活性を有するHtt特異的ZFNを、標的化組込み戦略に使用して、Httの野生型コピーへの可変長のCAG反復を導入する。50、80、109、および180のCAG反復単位を含むドナーが、構築される。次いで、これらのドナーは、上記のように、Htt特異的ZFNをコードするプラスミドを用いて、K562細胞にトランスフェクトされる。ドナー組込みの検証は、ゲノムDNA単離、PCT増幅(上記のような)、続いて、対象となる領域のシークエンシングによって達成される。
【0172】
Htt対立遺伝子へのドナー対立遺伝子の標的化組込みさせるK562細胞中で同定されたZFNを使用して、ヒト胚幹細胞(hESC)に可変長のドナー核酸を挿入する。成功したドナー組込みは、上記のように、ゲノムDNA単離、PCR、およびシークエンシングによって確認される。
【0173】
実施例8:野生型およびHDマウスHtt対立遺伝子の発現タグ
最初または最後のコーディングエクソンに対して最大の切断活性を有するZFNを使用して、異なるレポータータンパク質を用いて、野生型および変異Htt対立遺伝子を標識化する。それぞれのレポーター(AおよびB)に対してドナーDNAは、レポーター遺伝子の標的化組込みが、Httへのインフレーム融合を産生することができるように、リードZFN対の切断部位に基づいて設計される。ドナーDNAは、組込みの最も高い頻度を与えるドナーDNA構築体を選択するために、リードZFN対を用いて、ニューロ2A細胞に共トランスフェクトされる。
【0174】
ZFN対25920/25921および25922/25923は、上記のように調製され、実施例6に記載されるように、Cel Iミスマッチを用いて、切断活性について試験した。これらのZFN対は、Htt遺伝子配列の3’末端を標的化し、ひいては、野生型Htt対立遺伝子あるいは変異Htt対立遺伝子のいずれかを標的化するために使用され得る。結果を図5に示す。図から見られるように、これらの2つの対は、Htt遺伝子を切断することが可能であり、したがって、レポーターの導入のために利用することができる。
【0175】
対応するZFNと共にレポーターAに対して選択されたドナーDNA構築体は、長く伸びるHtt対立遺伝子(例えば、ヒトHttノックインマウス)を含むモデルマウス由来のマウス胚幹細胞に送達される。レポーターAの標的化組込みのために、クローンが、得られ、スクリーニングされる。ヘテロ接合体の現象が望ましく、標的化対立遺伝子は、PCRによって同定される。単一のレポーター標識化Htt対立遺伝子および他の対立遺伝子において非修飾のZFN標的配列を含むクローンが、選択され、レポーターBおよび対応するZFNのドナー構築体は、レポーターBを用いて第2の対立遺伝子を標識化するようにトランスフェクトされる。
【0176】
得られたマウス胚幹細胞クローンは、それぞれの対立遺伝子から発現の追跡を可能する2つの異なるマーカーを用いて標識化した野生型Htt対立遺伝子および変異対立遺伝子を含み、標準プロトコルを用いて、これらの細胞を使用して、トリヌクレオチド反復疾患のマウスモデルを生成する。
【0177】
本明細書で言及された全ての特許、特許出願、および刊行物は、参照によりそれらの全体が組み込まれる。
【0178】
理解の明瞭化を目的とした図示および例のために、本開示を多少詳しく提供したが、本開示の趣旨または範囲から逸脱することなく、多様な変更および修正を実施することができることは当業者には明らかである。したがって、前述の説明および例を限定的であると見なすべきではない。
【特許請求の範囲】
【請求項1】
トリヌクレオチド反復疾患に関連する遺伝子の発現を調節する改変された亜鉛フィンガータンパク質であって、複数の亜鉛フィンガードメインを含み、少なくとも1つの亜鉛フィンガードメインは、天然に存在しない認識へリックス領域を含む、亜鉛フィンガータンパク質。
【請求項2】
前記遺伝子は、野生型または少なくとも1つの長く伸びるトリヌクレオチド反復を含む遺伝子である、請求項1に記載の亜鉛フィンガータンパク質。
【請求項3】
請求項1または請求項2に記載の少なくとも2つの亜鉛フィンガータンパク質を含む、多量体であって、前記多量体の前記亜鉛フィンガータンパク質は、野生型トリヌクレオチド反復経路と比較して、長く伸びるトリヌクレオチド経路に優先的に結合する、多量体。
【請求項4】
前記長く伸びるトリヌクレオチド反復は、Htt対立遺伝子等のハンチントン病(HD)遺伝子内にある、請求項1乃至3のいずれかに記載の亜鉛フィンガータンパク質または多量体。
【請求項5】
前記長く伸びるトリヌクレオチド反復は、ポリグルタミンまたはポリセリンをコードする、請求項1乃至4のいずれかに記載の亜鉛フィンガータンパク質または多量体。
【請求項6】
前記亜鉛フィンガータンパク質は、前記長く伸びるトリヌクレオチド反復に関連するSNPまたは野生型対立遺伝子に関連するSNPに結合する、請求項1乃至5のいずれかに記載の亜鉛フィンガータンパク質または多量体。
【請求項7】
前記亜鉛フィンガータンパク質は、前記遺伝子の発現を活性化するか、または抑制する、請求項1乃至6のいずれかに記載の亜鉛フィンガータンパク質または多量体。
【請求項8】
請求項1乃至7のいずれかに記載の亜鉛フィンガータンパク質、ならびに転写活性化ドメイン、転写抑制ドメイン、開裂ドメイン、および/または開裂ハーフドメイン等の機能ドメインを含む、融合タンパク質。
【請求項9】
請求項1乃至8のいずれかに記載の亜鉛フィンガータンパク質、多量体、または融合タンパク質をコードする、ポリヌクレオチド。
【請求項10】
請求項1乃至9のいずれかに記載の亜鉛フィンガータンパク質、多量体、融合タンパク質、またはポリヌクレオチドを含む、単離された細胞。
【請求項11】
請求項1乃至9のいずれかに記載の亜鉛フィンガータンパク質、多量体、融合タンパク質、またはポリヌクレオチドを含む、薬学的組成物。
【請求項12】
長く伸びるトリヌクレオチド反復を標的修飾する方法であって、前記融合タンパク質の活性により、前記長く伸びるトリヌクレオチド反復が修飾されるように、請求項8に記載の1つ以上の融合タンパク質または前記1つ以上の融合タンパク質をコードするポリヌクレオチドを、長く伸びるトリヌクレオチド反復を含む遺伝子を有するゲノムを含む細胞に投与することを含む、方法。
【請求項13】
前記細胞は、誘導多能性幹細胞(iPSC)、ヒト胚幹細胞(hES)、間充織幹細胞(MSC)、神経幹細胞、またはこれらの組み合わせからなる群から選択される、請求項12に記載の方法。
【請求項14】
前記修飾は、トリヌクレオチド反復の数を減少させることを含む、請求項12に記載の方法。
【請求項15】
トリヌクレオチド反復疾患の研究のためにモデル系を生成する方法であって、前記細胞の前記ゲノムが、請求項8に記載の1つ以上の融合タンパク質または前記1つ以上の融合タンパク質をコードするポリヌクレオチドを用いて、特定の長さのトリヌクレオチド伸長経路を含むように、細胞を修飾することを含む、方法。
【請求項16】
前記モデル系は、細胞株または動物を含む、請求項15に記載の方法。
【請求項17】
対象におけるトリヌクレオチド疾患を治療する方法であって、
請求項1乃至9のいずれかに記載の少なくとも1つの亜鉛フィンガータンパク質、多量体、融合タンパク質、もしくはポリヌクレオチド、および/または前記亜鉛フィンガータンパク質をコードする神経栄養因子もしくはポリヌクレオチドの発現を調節する亜鉛フィンガータンパク質を、治療を必要とする対象に投与することであって、前記亜鉛フィンガータンパク質は、前記トリヌクレオチド疾患に関連する遺伝子の発現を調節する、方法。
【請求項18】
前記神経栄養因子は、BDNFまたはGNDFを含む、請求項17に記載の方法。
【請求項1】
トリヌクレオチド反復疾患に関連する遺伝子の発現を調節する改変された亜鉛フィンガータンパク質であって、複数の亜鉛フィンガードメインを含み、少なくとも1つの亜鉛フィンガードメインは、天然に存在しない認識へリックス領域を含む、亜鉛フィンガータンパク質。
【請求項2】
前記遺伝子は、野生型または少なくとも1つの長く伸びるトリヌクレオチド反復を含む遺伝子である、請求項1に記載の亜鉛フィンガータンパク質。
【請求項3】
請求項1または請求項2に記載の少なくとも2つの亜鉛フィンガータンパク質を含む、多量体であって、前記多量体の前記亜鉛フィンガータンパク質は、野生型トリヌクレオチド反復経路と比較して、長く伸びるトリヌクレオチド経路に優先的に結合する、多量体。
【請求項4】
前記長く伸びるトリヌクレオチド反復は、Htt対立遺伝子等のハンチントン病(HD)遺伝子内にある、請求項1乃至3のいずれかに記載の亜鉛フィンガータンパク質または多量体。
【請求項5】
前記長く伸びるトリヌクレオチド反復は、ポリグルタミンまたはポリセリンをコードする、請求項1乃至4のいずれかに記載の亜鉛フィンガータンパク質または多量体。
【請求項6】
前記亜鉛フィンガータンパク質は、前記長く伸びるトリヌクレオチド反復に関連するSNPまたは野生型対立遺伝子に関連するSNPに結合する、請求項1乃至5のいずれかに記載の亜鉛フィンガータンパク質または多量体。
【請求項7】
前記亜鉛フィンガータンパク質は、前記遺伝子の発現を活性化するか、または抑制する、請求項1乃至6のいずれかに記載の亜鉛フィンガータンパク質または多量体。
【請求項8】
請求項1乃至7のいずれかに記載の亜鉛フィンガータンパク質、ならびに転写活性化ドメイン、転写抑制ドメイン、開裂ドメイン、および/または開裂ハーフドメイン等の機能ドメインを含む、融合タンパク質。
【請求項9】
請求項1乃至8のいずれかに記載の亜鉛フィンガータンパク質、多量体、または融合タンパク質をコードする、ポリヌクレオチド。
【請求項10】
請求項1乃至9のいずれかに記載の亜鉛フィンガータンパク質、多量体、融合タンパク質、またはポリヌクレオチドを含む、単離された細胞。
【請求項11】
請求項1乃至9のいずれかに記載の亜鉛フィンガータンパク質、多量体、融合タンパク質、またはポリヌクレオチドを含む、薬学的組成物。
【請求項12】
長く伸びるトリヌクレオチド反復を標的修飾する方法であって、前記融合タンパク質の活性により、前記長く伸びるトリヌクレオチド反復が修飾されるように、請求項8に記載の1つ以上の融合タンパク質または前記1つ以上の融合タンパク質をコードするポリヌクレオチドを、長く伸びるトリヌクレオチド反復を含む遺伝子を有するゲノムを含む細胞に投与することを含む、方法。
【請求項13】
前記細胞は、誘導多能性幹細胞(iPSC)、ヒト胚幹細胞(hES)、間充織幹細胞(MSC)、神経幹細胞、またはこれらの組み合わせからなる群から選択される、請求項12に記載の方法。
【請求項14】
前記修飾は、トリヌクレオチド反復の数を減少させることを含む、請求項12に記載の方法。
【請求項15】
トリヌクレオチド反復疾患の研究のためにモデル系を生成する方法であって、前記細胞の前記ゲノムが、請求項8に記載の1つ以上の融合タンパク質または前記1つ以上の融合タンパク質をコードするポリヌクレオチドを用いて、特定の長さのトリヌクレオチド伸長経路を含むように、細胞を修飾することを含む、方法。
【請求項16】
前記モデル系は、細胞株または動物を含む、請求項15に記載の方法。
【請求項17】
対象におけるトリヌクレオチド疾患を治療する方法であって、
請求項1乃至9のいずれかに記載の少なくとも1つの亜鉛フィンガータンパク質、多量体、融合タンパク質、もしくはポリヌクレオチド、および/または前記亜鉛フィンガータンパク質をコードする神経栄養因子もしくはポリヌクレオチドの発現を調節する亜鉛フィンガータンパク質を、治療を必要とする対象に投与することであって、前記亜鉛フィンガータンパク質は、前記トリヌクレオチド疾患に関連する遺伝子の発現を調節する、方法。
【請求項18】
前記神経栄養因子は、BDNFまたはGNDFを含む、請求項17に記載の方法。
【図1】

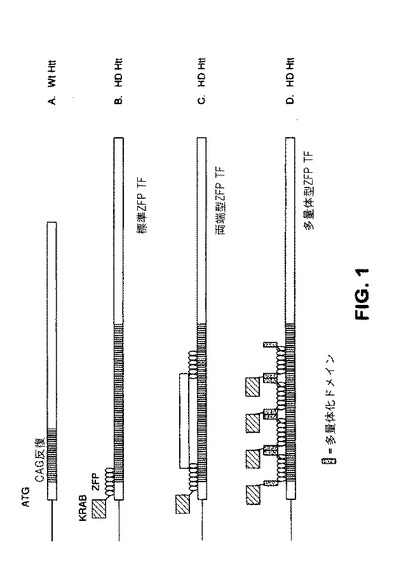
【図2】
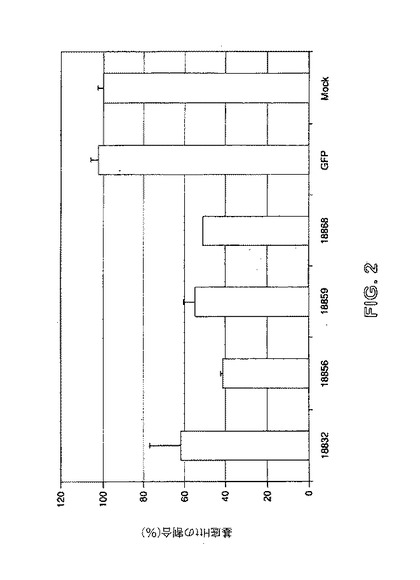
【図3】
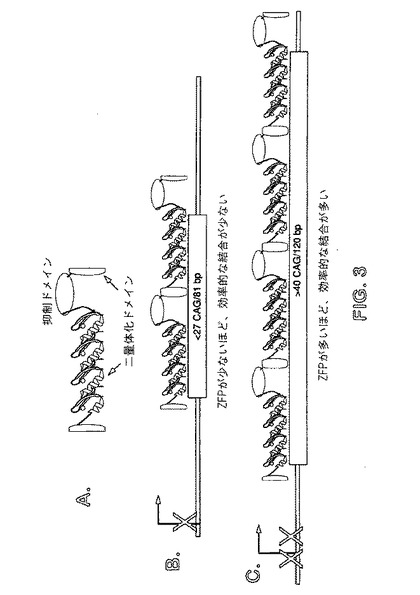
【図4】
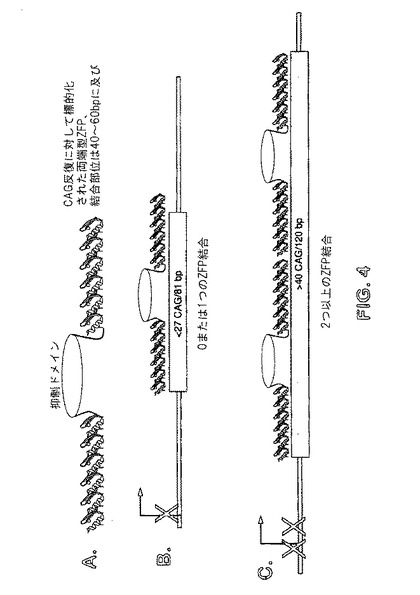
【図5】
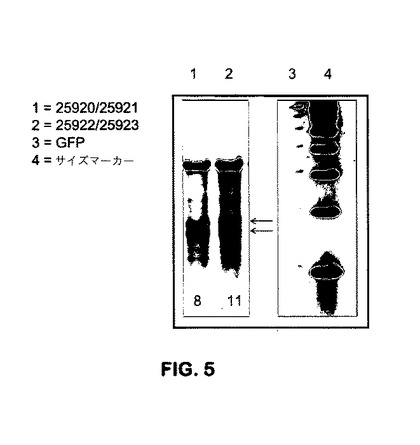
【図2】
【図3】
【図4】
【図5】
【公表番号】特表2013−500045(P2013−500045A)
【公表日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願番号】特願2012−522809(P2012−522809)
【出願日】平成22年7月28日(2010.7.28)
【国際出願番号】PCT/US2010/002108
【国際公開番号】WO2011/016840
【国際公開日】平成23年2月10日(2011.2.10)
【出願人】(508241200)サンガモ バイオサイエンシーズ, インコーポレイテッド (28)
【Fターム(参考)】
【公表日】平成25年1月7日(2013.1.7)
【国際特許分類】
【出願日】平成22年7月28日(2010.7.28)
【国際出願番号】PCT/US2010/002108
【国際公開番号】WO2011/016840
【国際公開日】平成23年2月10日(2011.2.10)
【出願人】(508241200)サンガモ バイオサイエンシーズ, インコーポレイテッド (28)
【Fターム(参考)】
[ Back to top ]