
トリレオウイルスの新規抗原クラス
【課題】トリレオウイルスの新規抗原クラスに属するMAS様腸疾患状態の原因因子および該新規抗原クラスのトリレオウイルスが引き起こす疾患に対する防御を家禽において有効に与えるワクチンを提供することである。
【解決手段】トリレオウイルスの抗原クラスに属するトリレオウイルスであって、該抗原クラスに属するトリレオウイルスが動物において抗血清を誘導する能力を有し、該抗血清が、プラーク減少アッセイにおいて少なくとも75%の、トリレオウイルスERS(そのサンプルは、ECACC、Salisbury、UKに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こすことを特徴とするトリレオウイルスを提供する。
【解決手段】トリレオウイルスの抗原クラスに属するトリレオウイルスであって、該抗原クラスに属するトリレオウイルスが動物において抗血清を誘導する能力を有し、該抗血清が、プラーク減少アッセイにおいて少なくとも75%の、トリレオウイルスERS(そのサンプルは、ECACC、Salisbury、UKに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こすことを特徴とするトリレオウイルスを提供する。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、トリレオウイルス、および生弱毒化または不活化形態のトリレオウイルスを含むワクチンに関する。
【背景技術】
【0002】
商業的ブロイラー生産は、過去数十年の間に、家畜の監禁的飼育(confinement rearing)の効率の高さにより特徴づけられる1つの産業にまで発展した。しかしながら、飼育段階の効率の増加の傾向を高くしようとすると、必ずいくつかの固有の問題が生じる。最も注目すべきは、高密度で密集して監禁された動物集団においてしばしば生じる感染症の発生率の増加である。家禽における最も破壊的な疾患の多くは、ワクチン接種により、または抗生物質などの治療剤での治療により、抑制または防除されている。しかし、残念ながら、複雑な病因の多数の疾患が、依然として薬物では防除されておらず、それに対しては適当なワクチン接種計画を利用することができない。
【0003】
1970年代後期以降、養鶏業は、腸障害に罹患したブロイラーニワトリにおけるそのような複雑な疾患の問題に直面している。罹患ニワトリにおいて種々の疾患状態(例えば、腸炎)を引き起こす疾患の1つは、主な臨床徴候および肉眼的観察に基づいて吸収不良症候群(MAS)と称される。あるいは、この疾患は、感染性発育不全(infectious runting stunting)症候群、衰弱トリ(pale bird)症候群または骨粗鬆症と称される。多数の種々の疾患状態がMASと連関しているが、いずれの場合も成長不良および羽毛生成の遅延が認められる。さらに、多種多様な他の徴候および病変(例えば、致死、過度に液状の糞便および/または食餌の消化不良、膵萎縮、前胃炎、骨変化、胸腺および滑液嚢萎縮など)がMASと相関している。
【0004】
Kouwenhovenら(Avian Pathology 17, 879−892, 1988)は、以下の5つの基準によりMASを定義した:
(i)1日齢のニワトリの感染後3週までの成長阻害、
(ii)黄橙色の粘液様ないしは湿性の糞便の排泄、
(iii)血漿アルカリホスファターゼ(ALP)活性の増加、
(iv)血漿カロテノイド濃度(PCC)の減少、
(v)近位けい骨の端成長板の拡大の肉眼的観察。
【0005】
ブロイラーの成長の遅延は1週齢以前に明らかとなる。それらの鳥集団のうちの5〜20%が罹患することがあり、これらの鳥は4週齢までに、同集団のその他の鳥の半分以下のサイズとなる。罹患集団は食餌の変換不全を示し、腸は衰弱し、未消化の食餌を含有する。
【0006】
MASの病理は十分には理解されていないが、この症候群の考えられうる病理は、消化管および関連器官に対する感染因子の直接的作用であり、これはまた、過度に液状の糞便および/または消化不良の食餌の排泄の反復を説明するものであろう。
【0007】
本明細書に記載の症候群は、体重増加の減少、食餌の変換不全、罹患した鳥の商業的価値の減少などの品質における全般的な損失を引き起こす。毎年、養鶏業は、MASによる大きな経済的損失を受けている。したがって、ブロイラーに認められる種々の疾患状態の1以上を予防しうるMASの防除方法が、養鶏業において必要とされている。
【0008】
レオウイルスは、世界中の家禽において遍在的に見出される。レオウイルスは、脚部の主要体重支持関節包(major weight bearing joint capsules)および腱シートを冒す関節炎状態(ウイルス性関節炎/腱滑膜炎と称される)の原因因子であることが判明している。
【0009】
いくつかの報告によると、レオウイルスは、MAS関連疾患状態を示すニワトリからも単離されている。これらの報告においては、レオウイルスがMAS関連疾患状態の1以上に対して病因的関連性を有すると推測されているが、レオウイルスがMASに関与しているという確かな証拠はそれらの報告には記載されていない。
【0010】
van der Heideら(Avian Diseases 25, 847−856, 1981)は、臨床的に下痢を示す若いブロイラーの腸からレオウイルスを分離した。このレオウイルス分離体は、腱滑膜炎および大腿骨頭骨折および骨粗鬆症の病変を誘発する能力を有していたが、この分離体は、実験的にレオウイルスに感染させたニワトリにおいて下痢を一貫して誘発しなかった。
【0011】
Pageら(Avian Diseases 26, 618−624, 1982)は、不自由な足、萎縮および異常羽毛発生を示す集団からレオウイルスを分離した。感受性ブロイラー型ニワトリに対する経口接種は、体重増加、羽毛発生に対する明らかな効果を示し、多数の器官において病変を誘発したが、下痢または湿性リターは報告されていない。
【0012】
Hieronymusら(Avian Diseases 27, 246−254, 1983)は、MASの疑いがあるニワトリの腸からのレオウイルスのいくつかの株の分離を報告しており、感染性腱滑膜炎の防除のためのワクチン株として一般に使用されるレオウイルスS1133株とこれらの株との抗原性関係を確認している。該著者は、該レオウイルスが臨床的MASのニワトリから分離されたにもかかわらず、該レオウイルスがMASの原因因子であるか否かは依然として明らかでないことを確認している。
【0013】
Eidsonら(Poulty Science 64, 2081−2086, 1985)は、MASおよび腱滑膜炎の問題を有するブロイラー集団においてHieronymusらにより分離されたC08株から誘導された不活化レオウイルスワクチンの効果について調べている。該ワクチンは、ブロイラー集団の体重に対して正の効果を及ぼしたが、食餌変換における相違は認められなかった。また、湿性リター(wet litter)などの腸炎関連疾患状態に対する該ワクチンの効果は、何ら報告されていない。
【0014】
また、Rosenbergerら(Avian Diseases 33, 535−544, 1989)は、野外で飼育されている商業用ブロイラーニワトリの腱および骨髄からいくつかのレオウイルス株を分離した。レオウイルス株を接種されたニワトリが臨床的疾患に関して試験されたが、下痢または湿性リターの徴候は報告されていない。
【0015】
また、Kouwenhovenら(Avian Pathology 17, 879−892, 1988)は、吸収不良症候群におけるレオウイルスの役割を調べている。これらの著者は、野外症例から分離したレオウイルスでMASを再現することができなかった。そして彼らは、レオウイルスがMASの一次病原体でないと結論づけている。該著者は、レオウイルスおよびアデノウイルスを含む感染因子が、吸収不良症候群における或る種の誘発因子として作用した可能性があると推測している。
【0016】
しかしながら、前記刊行物に記載のものに加えて、他の多数のウイルスがMASに関連している。これらには、ロタウイルス、パルボウイルス、エンテロウイルス様ウイルス、トガウイルス様ウイルス、コロナウイルス様ウイルス、アデノウイルスおよびカリシウイルスが含まれる。さらにまた、細菌が病因に関与していると示唆されている。また、先行技術においては、MAS様野外(field)症候群はマイコトキシンによって生じると考えられており、MASの原因因子としてマイコトキシンまたは他の毒素を無視すべきでないと考えられている。
【0017】
最近公開された総説(World Poultry 14, 57−58, 1998)において、McNultyはMASの最新の知見を要約している。McNultyは、MASに対するワクチンの非利用可能性を強調しており、該ウイルス分離体ならびに現在までに報告されている野外分離サンプルのウイルス学的および微生物学的検査は、MASの原因因子の同定に関して有用な結果を与えていないと記載している。McNultyは、このアプローチが有用な結果をもたらしそうにないと推測している。実際のところ、MAS罹患産生部位における処置手段がMASの防除のための最良の手段であると考えられている。
【先行技術文献】
【非特許文献】
【0018】
【非特許文献1】Kouwenhovenら(Avian Pathology 17, 879−892, 1988)
【非特許文献2】van der Heideら(Avian Diseases 25, 847−856, 1981)
【非特許文献3】Pageら(Avian Diseases 26, 618−624, 1982)
【非特許文献4】Hieronymusら(Avian Diseases 27, 246−254, 1983)
【非特許文献5】Eidsonら(Poulty Science 64, 2081−2086, 1985)
【非特許文献6】Rosenbergerら(Avian Diseases 33, 535−544, 1989)
【非特許文献7】World Poultry 14, 57−58, 1998
【発明の概要】
【発明が解決しようとする課題】
【0019】
したがって、過度に液状の糞便および/または消化不良の食餌の排泄を引き起こしニワトリに生じる或る種の腸障害(例えば、MASに関連した腸障害)に対する有効な防御を誘導するワクチンが依然として必要とされている。
【0020】
さらに、トリレオウイルスは相当な抗原不均質性を示し、トリレオウイルスの新規抗原クラスの出現は、家禽におけるレオウイルスワクチンの使用に重大な影響を及ぼす可能性がある。
【0021】
本発明者らは、本発明において、トリレオウイルスの新規抗原クラスを同定した。さらに、この新規抗原クラスに属するトリレオウイルスが、同様にMASに関連する顕著な疾患状態(過度に液状の糞便および/または消化不良の食餌の排泄ならびに成長の遅延)を誘導しうることを示す。
【0022】
したがって、トリレオウイルスの新規抗原クラスに属するMAS様腸疾患状態の原因因子を提供することが、本発明の目的である。
【0023】
本発明のもう1つの目的は、該新規抗原クラスのトリレオウイルスが引き起こす疾患に対する防御を家禽において有効に与えるワクチンを提供することである。
【0024】
同様にMASに関連している腸疾患状態に対する防御を家禽において有効に与えるワクチンを提供することが、本発明のもう1つの目的である。
【課題を解決するための手段】
【0025】
トリレオウイルスの抗原クラスに属するトリレオウイルスであって、該抗原クラスに属するトリレオウイルスが動物において抗血清を誘導する能力を有し、該抗血清が、プラーク減少アッセイにおいて少なくとも75%の、トリレオウイルスERS(そのサンプルは、ECACC、Salisbury、UKに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こすことを特徴とするトリレオウイルスを提供することにより、これらの目的が達成されうることを、本発明において見出した。そのようなトリレオウイルスは、これまで知られていない抗原特性を示すことが認められただけでなく(実施例1、表2および3)、本発明のトリレオウイルスは、ブロイラーニワトリによる過度に液状の糞便および/または消化不良の食餌の排泄を誘導する能力を有し、場合によっては、致死を招きうることが判明した。したがって、本発明においては、該新規トリレオウイルスを腸レオウイルス株(ERS)と称することにする。
【0026】
ブロイラー集団における湿性糞便の排泄は、野外におけるMAS罹患ブロイラーで一般に認められる疾患状態の1つである。さらに、この臨床的疾患状態は、MAS罹患ブロイラーにおける最も顕著な臨床的徴候(すなわち、成長の遅延)の原因の1つであると考えられる。実施例においては、本発明のトリレオウイルスが、該レオウイルスに経口感染したブロイラーによる湿性糞便の排泄を誘導する(すなわち、該鳥の排泄腔の周囲にペーストの覆い(clothing of pasting)が認められる)ことを示す。また、実施例における実験は、該新規トリレオウイルスの経口感染もまた、対照ニワトリと比較した場合に、感染ブロイラーにおける成長の遅延を引き起こすことを示している。
【0027】
該プラーク減少アッセイは、(トリレオ)ウイルス分離体間の抗原関係を判定するために当技術分野において広く用いられているアッセイである(例えば、Nersessianら, Am. J. Vet. Res 50, 1475−1480, 1989を参照されたい)。さらに、本発明の目的のために、プラーク減少アッセイに関する詳細な説明を実施例1に記載する。当然のことながら、該プラーク減少アッセイで使用する抗血清は、適当な質を有するべきである。そのような抗血清の製造方法は、実施例1にも記載されている。
【0028】
一般に、生トリレオウイルスに対する適当な抗血清は、102.0〜109.0 TCID50/動物、より好ましくは103.0〜106.0 TCID50/動物の感染価を有する生ウイルス株を3〜4週齢のSPFニワトリの皮下または筋肉内に接種することにより製造することができる。感染の3〜4週間後、好ましくは感染の4週間後に血液を集めることができる。また、一次感染で使用したのとほぼ同用量の同じ生ウイルス株を、一次感染の3〜4週間後にニワトリに再感染させることができる。二次感染の2〜4週間後に血液を集める。不活化トリレオウイルス株に対する適当な抗血清は、3〜4週齢のSPFニワトリに該不活化ウイルス調製物を皮下または筋肉内に接種することにより得ることができる。不活化前の該調製物の感染価は、107.0〜1011.0 TCID50/動物であってもよく、より好ましくは108.0〜1010.0 TCID50/動物である。血液は、接種の3〜4週間後、好ましくは接種の4週間後に集めることができる。また、一次接種の3〜6週間後に同じ不活化ウイルス調製物をニワトリに再接種することができる。二次接種の2〜4週間後に血液を集める。
【0029】
本発明の新規トリレオウイルスの同定は、トリレオウイルスの該新規抗原クラスによる感染から生じる疾患状態に対して家禽を有効に防御しうる新規トリレオウイルスワクチンの製造を可能にする。特に、該新規トリレオウイルスは、過度に液状の糞便および/または消化不良の食餌の排泄ならびに成長の遅延などの疾患状態に対して家禽を有効に防御しうる新規トリレオウイルスワクチンの製造を可能にする。そのような疾患状態もMASに関連している。
【0030】
また、本発明のトリレオウイルスは、野外から分離することができる。分離方法の重要な態様は、ウイルスの分離の出発物として使用する標的動物の同定である。この目的に使用する典型的なブロイラーは、成長の遅延につながる過度に液状の糞便および/または消化不良の食餌の排泄の徴候を示す。
【0031】
ついで該罹患ニワトリから腸を分離し、該器官を適当なバッファー中でホモジナイズする。ついで、ホモジナイズされた該組織を遠心分離し、該上清を孔径0.2μmのフィルターで濾過する。該濾液のサンプルを、新鮮に調製された初代ニワトリ細胞、好ましくはニワトリ胚肝(CEL)細胞に加え、インキュベーションの4〜8日後、細胞変性効果(CPE)の存在に関して該単層を調べる。CPEが存在しなければ、該一次単層の凍結/融解懸濁液を、新鮮に調製されたCEL細胞に加える。第1継代または第2継代後にCPEが認められたら、該ウイルスを、過度に液状の糞便および/または消化不良の食餌の排泄を誘発するブロイラーにおけるそのインビボ特性により、およびプラーク減少アッセイまたは免疫蛍光技術(IFT)(後記の特異的ポリクローナルおよびモノクローナル抗体を使用する)において測定したその抗原特性により、さらに特徴づける。本発明のトリレオウイルスの分離のためのより詳細な方法を、実施例1に開示する。
【0032】
本発明のトリレオウイルスの分離方法においては、好ましい出発物質として腸を使用するが、トリレオウイルスは、罹患ブロイラーニワトリの肝臓またはそのようなブロイラーニワトリが排泄した糞便からも分離することができる。また、本発明のトリレオウイルスの分離のための出発物質として他の器官を用いることも可能であることを考慮に入れるべきである。
【図面の簡単な説明】
【0033】
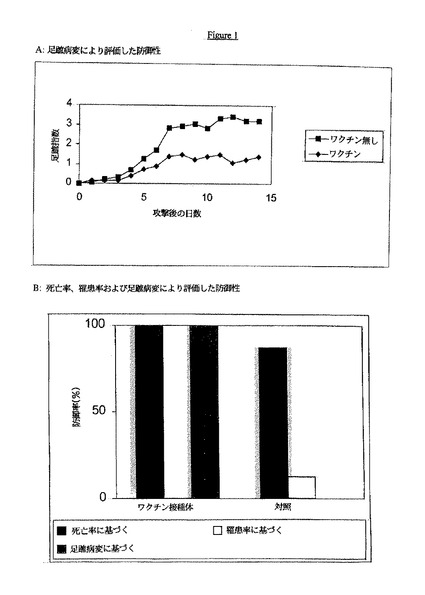
【図1】Aは、足蹠病変アッセイに対するワクチンの防御活性を示す。Bは、死亡率、罹患率および足蹠病変に対するワクチンの防御活性を示す。
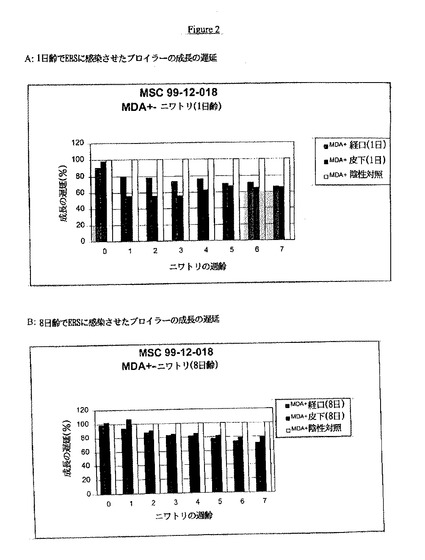
【図2】AおよびBは、ERSに感染したブロイラーの成長の遅延を示す。
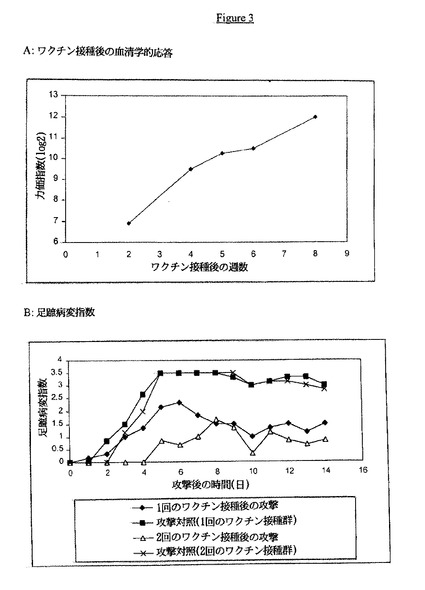
【図3】Aは、血清力価とワクチン接種後の週数との関係を示す。Bは、攻撃後の足蹠病変指数の時間経過を示す。
【発明を実施するための形態】
【0034】
特に、本発明は、前記トリレオウイルスであって、該トリレオウイルスに対して誘導された抗血清が、プラーク減少アッセイにおいて少なくとも80%、好ましくは少なくとも90%の、トリレオウイルスERS(そのサンプルは、ECACC、Salisbury、UKに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こしうることを特徴とするトリレオウイルスを提供する。
【0035】
本発明のより一層好ましい実施形態においては、ポリクローナル抗血清およびモノクローナル抗体(Moabs)の特異的パネルに対して特異的免疫反応を示すトリレオウイルスを提供する。この特異的反応パターンは、従前公知のトリレオウイルスにより示されるものとは異なる。
【0036】
モノクローナル抗体は、感染因子の特性を同定し同一または類似の微生物の種々の分離体間の類似性および相違を判定するのに有用である。Vakhariaら(Proceedings of the International Symposium on adenovirus and reovirus infections in poultry, Rauischholzhausen, Germany, 1996, 295−304)は、9個の抗レオウイルスMoabsのパネルを開示しており、トリレオウイルス野外分離体を、これらのMoabsに対するそれらの反応性に関して試験している。Moabsのパネルの反応性の種々のパターンは、反応性パターンの関連性に基づく分離体の分類を可能にした。
【0037】
本発明のこの実施形態においては、前記トリレオウイルスであって、(i)トリレオウイルス分離体に対して(好ましくは、始原型レオウイルス1133株に対して)産生したポリクローナル抗血清に対するIFTにおけるその反応性、および(ii)Moabs INT 13−06、INT 14−11および15−01 INT(それらのサンプルは、それぞれ受託番号99011472、99011473および99011474でECACCに寄託されている)に対するIFTにおける反応性の不存在により更に特徴づけられるトリレオウイルスを提供する。
【0038】
該パネルパターンにより定められる本発明のトリレオウイルスは、実施例1および表3に示す新規抗原型のレオウイルスを代表する。表3に示す先行技術のレオウイルス株の多数が、MASに関連した徴候および病変を示すブロイラーニワトリの組織(腸を含む)から分離されているが、前記の新規抗原特性を有する本発明のトリレオウイルスは先行技術において開示されていない。
【0039】
本発明のより一層好ましいトリレオウイルスは、本発明のトリレオウイルスの特徴を示すトリレオウイルスERS(分離体1)(そのサンプルは、ECACCに受託番号99011475で寄託されている)である。
【0040】
本発明のトリレオウイルスは、生、生弱毒化または不活化形態であってもよい。
【0041】
本発明は、もう1つの態様において、トリレオウイルスから生じる疾患状態(例えば、MASで認められる腸疾患状態)に対して家禽を防御するのに使用するためのワクチンであって、本発明のトリレオウイルスと医薬上許容される担体または希釈剤とを含んでなるワクチンを提供する。
【0042】
本発明のトリレオウイルスは、生弱毒化または不活化ウイルスとして該ワクチン中に含有させることができる。該トリレオウイルスを生弱毒または不活化形態にすると、前記のMAS関連疾患状態を誘発するトリレオウイルスの特性は有意に軽減されるか、または完全に失われる。
【0043】
本発明のトリレオウイルスの弱毒化は、この目的のための当技術分野でよく知られた方法、例えば、Gouveaら(Virology 126, 240−247, 1983)に開示されている方法により行なうことができる。簡単に説明すると、標的動物から該ウイルスを分離した後、ウイルス懸濁液を初代ニワトリ胚繊維芽細胞(CEF)に接種する。該分離体がCPEを与える能力を有していない場合には、CPEが認められるようになるまで該ウイルスを反復的に(例えば、3〜10回)継代する。CPEが認められると直ちに細胞および細胞培養流体を集め、凍結・融解し、遠心分離により清澄化し、トリレオウイルス分離体を含有する上清をアリコート化し、−20℃で保存する。該ウイルスを更に弱毒化するために、この方法を繰返す(例えば、10〜100回)ことができる。
【0044】
本発明のワクチンは、例えば、商業的に入手可能な生および不活化レオウイルスワクチンに一般に用いられる方法などの通常の方法で製造することができる。獣医学用ワクチン組成物の製造については、とりわけ、“Handbuch der Schulzimpfungen in der Tiermedizin”(Mayr, Aら編, Verlag Paul Parey, Berlin und Hamburg, Germany, 1984)および“Vaccines for Veterinary Applications”(Peters, A.R.ら編, Butterworth−Heinermann Ltd, 1993)に記載されている。
【0045】
簡単に説明すると、生または生弱毒化形態の本発明のトリレオウイルスを感受性基質に接種し、該ウイルスが所望の感染価または抗原含量まで複製されるまで該基質を増殖させ、ついでレオウイルス含有物質を収穫し、予防活性を有する医薬組成物に製剤化する。
【0046】
必要に応じて該トリレオウイルスを基質に適合させた後、前記トリレオウイルスの複製を支持しうる各基質を使用して、本発明のワクチンを製造することができる。適当な基質には、初代(トリ)細胞培養、例えばニワトリ胚肝細胞(CEL)、ニワトリ胚繊維芽細胞(CEF)またはニワトリ腎細胞(CK)、哺乳類細胞系、例えばVERO細胞系またはBGM−70細胞系、またはトリ細胞、例えばQT−35、QM−7またはLMHが含まれる。通常、該細胞の接種後、該ウイルスを3〜10日間増殖させ、ついで該細胞培養上清を収穫し、所望により濾過または遠心分離して細胞残渣を除去する。
【0047】
あるいは、本発明のトリレオウイルスを孵化鶏卵内で増殖させ、ついでトリレオウイルス物質を常法により収穫することができる。
【0048】
生弱毒化ウイルスを含有する本発明のワクチンは、(凍結)懸濁液形態または凍結乾燥形態で製造し、販売することができる。該ワクチンは更に、そのような組成物に通常使用される医薬上許容される担体または希釈剤を含有する。担体には、安定化剤、保存剤およびバッファーが含まれる。適当な安定化剤としては、例えば、SPGA、炭水化物(例えば、ソルビトール、マンニトール、デンプン、スクロース、デキストラン、グルタメートまたはグルコース)、タンパク質(例えば、乾燥乳血清、アルブミンまたはカゼイン)またはそれらの分解産物が挙げられる。適当なバッファーには、例えば、リン酸アルカリ金属が挙げられる。適当な保存剤としては、チメロサール、メルチオラートおよびゲンタマイシンが挙げられる。希釈剤には、水、水性バッファー(例えば、緩衝食塩水)、アルコールおよびポリオール(例えば、グリセロール)が含まれる。
【0049】
所望により、本発明の生ワクチンはアジュバントを含有していてもよい。アジュバント活性を有する適当な化合物および組成物としては、例えば、不活化ワクチンの製造に関して後記で挙げるものと同じものが挙げられる。
【0050】
本発明の生ワクチンの注射(例えば、筋肉内、皮下)による投与は可能であるが、該生ワクチンは、好ましくは、トリレオウイルスのワクチン接種に通常用いる安価な大量適用技術により投与する。これらの技術には、飲水および噴霧ワクチン接種が含まれる。
【0051】
該生ワクチンの投与のためのもう1つの方法には、卵内、点眼および嘴の浸漬による投与が含まれる。
【0052】
好ましい実施形態において、本発明は、不活化形態の該トリレオウイルスを含んでなる、腸疾患状態(例えば、MASで認められるもの)に対するワクチンを提供する。不活化ワクチンの主な利点は、得られる持続性防御抗体のレベルが高いことである。この特性のおかげで、そのような不活化ワクチンは、特に種畜ワクチン接種に適したものになる。
【0053】
増殖工程後に収穫したウイルスの不活化の目的は、該ウイルスの複製を排除することである。一般に、これは、化学的または物理的手段により達成することができる。化学的不活化は、例えば酵素、ホルムアルデヒド、β−プロピオラクトン、エチレン−イミンまたはそれらの誘導体で該ウイルスを処理することにより行なうことができる。必要に応じて、後で該不活化化合物を中和する。ホルムアルデヒドで不活化した物質は、例えば、チオスルファートで中和することができる。物理的不活化は、好ましくは、該ウイルスを高エネルギー放射(例えば、UV光またはγ線)に付すことにより行なうことができる。所望により、処理後、該pHを約7に調節することができる。
【0054】
該不活化トリレオウイルスを含有するワクチンは、例えば、この目的に適した前記の医薬上許容される担体または希釈剤の1以上を含んでいてもよい。
【0055】
好ましくは、本発明の不活化ワクチンは、アジュバント活性を有する1以上の化合物を含む。この目的のための適当な化合物または組成物には、水酸化、リン酸または酸化アルミニウム、例えば鉱油(例えばBayol F(登録商標)またはMacrol 52(登録商標))または植物油(例えばビタミンEアセタート)に基づく水中油滴型または油中水滴型エマルション、およびサポニンが含まれる。
【0056】
不活化ワクチンは、通常、非経口的(例えば、筋肉内または皮下)に投与する。
【0057】
本発明のワクチンは、活性成分としての該トリレオウイルスの有効用量(すなわち、ビルレントウイルスによる攻撃に対して、ワクチン接種された鳥またはそれらの後代において免疫を誘導するトリレオウイルス物質を免疫する量)を含む。本発明においては、免疫は、ワクチン接種後の鳥集団において、非ワクチン接種群と比べて有意に高いレベルの防御を誘導することと定義される。
【0058】
典型的には、本発明の生ワクチンは、102〜109 TCID50/鳥の用量、好ましくは102〜106 TCID50/鳥の用量で投与することが可能であり、不活化ワクチンは、104〜1010 TCID50/鳥の抗原相当量を含有していてもよい。
【0059】
本発明のトリレオウイルスワクチンはニワトリにおいて有効に使用することができ、他の家禽(例えば、シチメンチョウ、ホロホロチョウおよびウズラ)にも該ワクチンを成功裡に接種することができる。ニワトリには、ブロイラー、繁殖ストックおよび産卵ストックが含まれる。
【0060】
腸疾患状態(例えば、MASで認められるもの)は主にブロイラーニワトリにおいて報告されているため、本発明は、好ましくは、腸疾患状態(例えば、MASで認められるもの)に対してブロイラーを防御するのに使用するワクチンを提供する。
【0061】
本発明の生または不活化ワクチンを投与する動物の齢は、現在商業的に入手可能な生または不活化トリレオウイルスワクチンを投与する動物の齢と同じである。例えば、本発明の生弱毒化ワクチンをブロイラーに1日齢以降から直接接種することができる。親ストック(例えば、ブロイラー種畜)のワクチン接種は、本発明の生弱毒化もしくは不活化ワクチンまたはそれらの組合せで行なうことができる。このタイプの免疫化計画の利点には、若鳥に垂直伝播した母体由来抗体により与えられる1日齢の後代の即時防御が含まれる。典型的な種畜ワクチン接種計画は、6週齢の種畜に生弱毒化ワクチンを接種し、ついで14〜18週齢で不活化ワクチンを接種することを含む。あるいは、生ワクチンの接種の後、10〜12週齢および16〜18週齢の時点で不活化ワクチンを2回接種することができる。
【0062】
本発明はまた、本発明のトリレオウイルスと、家禽に対して感染性の他の病原体のワクチン成分の1以上とを含んでなる混合(combination)ワクチンを含む。家禽に対して感染性のそのような他の病原体は、本発明のトリレオウイルスとは抗原的に異なるトリレオウイルスを意味し、腱滑膜炎に関連したトリレオウイルス株を含む。
【0063】
好ましくは、該混合ワクチン中のワクチン成分は、家禽に対して感染性の病原体の生弱毒化または不活化形態である。
【0064】
特に、本発明は、該ワクチン成分のすべてが不活化形態である混合ワクチンを提供する。
【0065】
好ましくは、該混合ワクチンは、伝染性気管支炎ウイルス(IBV)、ニューカッスル病ウイルス(NDV)、伝染性滑液嚢症ウイルス(IBDV)、トリアデノウイルス(FAV)、EDSウイルスおよびシチメンチョウ鼻気管炎ウイルス(TRTV)の(不活化)ワクチン株の1以上を含む。
【0066】
本発明の範囲内の以下の微生物およびハイブリドーマ細胞系は、European Collection of Animal Cell Cultures (ECACC), Salisbury, UKに1999年1月14日に寄託されている。
【0067】
【表1】

【実施例】
【0068】
実施例1
新規トリレオウイルスの特徴づけ
A.新規トリレオウイルスの分離
成長の遅延を引き起こす消化障害および湿性リターを有するニワトリから、腸および/または肝臓を個々に分離した。該器官を、抗生物質を含有するPBSおよびガラスパール(2mm)を使用してホモジナイザー中、最大速度で20分間、個々にホモジナイズした。ついで、そのホモジナイズされた組織を遠心分離した。腸ホモジネートを4000rpmで、肝臓を1200rpmで、共に15分間遠心分離した。ついで該上清をフィルター(孔径を5、1.2、0.45、0.2μmと減少させる)に通して濾過した。0.2μmのフィルターに通した100μlの懸濁液を、組織培養フラスコ中に存在する新鮮に調製された初代ニワトリ胚肝(CEL)細胞に加えた。インキュベーションの4〜8日後、細胞変性効果(CPE)の存在に関して該単層を調べた。CPEが存在しなければ、該単層を−20℃で凍結し、24時間後、該単層を融解した。ついでこの凍結/融解懸濁液の1mlを、新鮮に調製されたCEL細胞に加えた。4〜8日後にCPEが認められない場合には、CEL細胞上で増殖するウイルスに関して該培養を陰性とみなした。第1継代または第2継代後にCPEが認められた場合には、プラーク減少アッセイおよび免疫蛍光技術(IFT)により該ウイルスを更に特徴づけした。
【0069】
B.プラーク減少アッセイによる新規トリレオウイルスのインビトロでの特徴づけ
1.種々のトリレオウイルス株に対する抗血清の産生
ERS株(生):
10羽の3週齢のSPFニワトリに105.8 TCID50/動物のERS株(分離体2)を皮下感染させた。感染の3週間後に血液を集め、血清を単離し、動物に105.6 TCID50/動物で再感染させた。二次感染の2週間後、血液を集め、血清を単離した。集めた後、すべての血清を56℃で30分間熱不活化し、小さなアリコート中に−20℃で保存した。
【0070】
2177株(生):
15羽の4週齢のSPFニワトリに104.2 TCID50/動物で皮下感染させた。感染の4週間後に血液を集め、血清を単離した。集めた後、すべての血清を56℃で30分間熱不活化し、小さなアリコート中に−20℃で保存した。
【0071】
1733および2408株(不活化):
12羽の4週齢のSPFニワトリに、w/oエマルションでアジュバント化されたトリレオウイルス1733又は2408株を筋肉内接種した。動物用量:490 Elisa単位/動物(107.0〜1010.0 TCID50/動物の不活化前感染価に相当)。感染の4週間後、血液を集め、血清を単離した。集めた後、すべての血清を56℃で30分間熱不活化し、小さなアリコート中に−20℃で保存した。
【0072】
トリレオウイルス株を含有する商業的に入手可能な不活化ワクチン
10羽の3〜4週齢のSPFニワトリに、以下の商業的に入手可能な不活化トリレオウイルスワクチンの1つを筋肉内または皮下接種した:ISBI, Fort Dodge; Kaketsuken and Intervet International BV。感染の3週間後、血液を集め、血清を単離した。集めた後、すべての血清を56℃で30分間熱不活化し、小さなアリコート中に−20℃で保存した。
【0073】
前記で調製した血清を、IFTおよびプラーク減少アッセイにおいて使用した。
【0074】
2.免疫蛍光試験(IFT)
後記C節に記載されているのと実質的に同じ方法でIFTを行なう。簡単に説明すると、Vero細胞をコンフルエントになるまで96ウェルポリスチレンマイクロタイタープレート中で増殖させた。種々の単層にレオウイルス1133株を接種した。100μlのニワトリ血清を該プレートの第1ウェルに加えた。系列3倍希釈を作製した。インキュベーションおよび洗浄工程の後、該プレートを1:100希釈の蛍光イソチオシアナート標識ヤギ抗ニワトリと反応させた。蛍光の存在を蛍光顕微鏡で観察した。該力価(該血清の質)を終末点希釈法により測定する。これは、明らかな蛍光シグナルを誘導しうる血清の最高希釈度である。表1に、該プラーク減少試験で使用した血清についてのIFTの結果を示す。
【0075】
【表2】
Nobilis Reo(登録商標)(Intervet Internationl b.v.から商業的に入手可能)
Arvax(登録商標)(ISBIから商業的に入手可能)
Tri Reo(登録商標)(Fort Dodgeから商業的に入手可能)
Olivax Reo(登録商標)(Kaketsukenから商業的に入手可能)。
【0076】
3.プラーク減少アッセイ
新規抗原クラスに属するトリレオウイルス間の抗原関係を判定し新規トリレオウイルスを公知トリレオウイルスから識別するためのプラーク減少アッセイにおいては、適当な質の抗血清を使用するべきである(Arvax抗血清以外はすべて、該抗血清の品質要件を満足していた)。生ERS分離体1ならびに生および不活化ERS分離体2に対してERS抗血清を産生させた。新鮮に調製されたCELを、5%ウシ胎児血清および抗生物質で補足された組織培養培地に1 106細胞/mlの最終濃度まで再懸濁した。60mm組織培養皿を5mlの細胞懸濁液で満たし、37℃で24時間インキュベートした。
【0077】
翌日、調べようとするウイルスを、プラスチックチューブ中、抗生物質を含有する培地中で希釈した。10−1〜10−7の希釈液を調製した。つぎに各希釈液200μlを、試験する血清50μlと混合した。この混合物を37℃で1時間インキュベートした。陰性対照として、200μlのウイルス希釈液を50μlの培地と混合した。
【0078】
60mm組織培養皿中に存在する単層上の培地を捨てた。つぎに、それらの種々のウイルス混合物(血清を含有する又は含有しない)100μlをコンフルエントな単層上に加えた。各ウイルス混合物を、少なくとも2つの単層(皿)に感染させた。感染したそれらの単層を37℃で1時間インキュベートした。ついで該感染単層を5mlの寒天溶液(培地、FCSおよび抗生物質を含有する;最終寒天濃度3.0%;最終FCS濃度2.5%)で覆った。該皿を37℃で4日間インキュベートした。つぎに各組織培養皿に2mlのニュートラルレッド溶液(0.02%)を加えた。37℃で4時間のインキュベーションの後、プラークの数を皿ごとに計数した。150個未満のプラークを含有する組織培養皿のみを計数した。
【0079】
プラークの減少を以下のとおりに計算した。血清の不存在下の或る希釈度における或るウイルスのプラーク数を100%とする。ついでこれを、血清の存在下の同じウイルス希釈度におけるプラーク数と比較する。
【0080】
種々のウイルスおよび血清を試験した第1の実験の結果を表2Aに示す。表2Bは、示されているとおりのウイルスおよび血清を使用した第2の実験の結果を示す。ワクチン接種の5週間後に血液を集める以外は不活化1733/2408株に関して記載されているとおりに、不活化ERSワクチンに対する血清を産生させた。
【0081】
【表3】
【0082】
【表4】
【0083】
C.IFTによる新規トリレオウイルスのインビトロでの特徴づけ:抗血清パネル反応パターン
精製トリレオウイルス1133株をウサギ(1〜1.5kg)に感染させることにより、ポリクローナル抗血清を調製した。一次感染の28および84日後にブースター注射を行なった。最後の注射の14日後、血液を集め、血清を単離した。種々のMoabsで種々のトリレオウイルス株を特徴づけした。初代CEL細胞を96ウェルポリスチレンマイクロタイタープレート中で増殖させた。未感染細胞を対照として使用した。5% CO2と共に37℃で2〜4日間インキュベートした後、感染した単層を96%冷エタノールで固定した。該アルコールを捨て、該プレートを洗浄バッファーで洗浄し、PBS中1:50または1:200に希釈した種々のハイブリドーマ細胞培養上清100μlまたは1:50に希釈したウサギポリクローナル血清(ウサギ68A)100μlを各ウェルに加えた。該プレートを37℃で60〜90分間インキュベートし、洗浄バッファーで2回洗浄し、1:100希釈の蛍光イソチオシアナート標識ウサギ抗マウスまたは1:100希釈のイソチオシアナート標識ヤギ抗ウサギ血清と反応させた。最後に、該プレートを洗浄し、グリセロール/PBS溶液(1:1)で固定した。蛍光の存在を蛍光顕微鏡で観察した。
【0084】
この実験で使用した抗血清パネルは、始原型トリレオウイルス1133株に対して産生させた以下のポリクローナル抗血清およびMoabsよりなるものであった。
Rabbit 68A:ウサギポリクローナル抗血清
Moab 154:Vakhariaら, 1996 (前掲)
Moab 14−67:Intervet International B.V.
Moab INT 13−06:ECACC受託番号99011472
Moab INT 14−11:ECACC受託番号99011473
Moab 15−01 INT:ECACC受託番号99011474。
【0085】
前記方法により得たトリレオウイルス分離体を、この抗血清パネルに対するそれらの反応性により更に特徴づけした。該新規レオウイルス分離体は、ポリクローナルおよびモノクローナル抗体パネルに対する特有の反応パターンを有する。MASおよび腱滑膜炎の野外症例から分離された従前公知のトリレオウイルス株(S−1133〜CO8)は、そのような新規パターンでは反応しない(表3を参照されたい)。
【0086】
新規反応パターン
陽性:ポリクローナルウサギ68A、154、14−67
陰性:INT 13−06、INT 14−11、15−01 INT。
【0087】
【表5】
Enterovax(登録商標)およびTensynovac(登録商標)は、Schering−Plough Animal Health and Intervet Inc.から商業的に入手可能なトリレオウイルスワクチンである。
【0088】
D.新規トリレオウイルスのインビボでの特徴づけ
プラーク精製レオウイルスERSの実験的感染
実験1
30羽の1日齢のSPFニワトリに、プラーク精製レオウイルスERS(分離体1)を経口感染させた。注射の、4、7および10日後、肝臓を10羽の動物から単離し、顕微鏡的病変に関して調べた。
【0089】
実験2
レオウイルスに対する移行抗体を有する30羽の1日齢のブロイラーに、プラーク精製レオウイルスERS株を経口感染させた。感染の7および10日後、臨床的徴候に関して動物を観察した。湿性リターに特別の注意を払った。
【0090】
実験3
レオウイルスに対する移行抗体を有する30羽の1日齢のブロイラーに、プラーク精製レオウイルスERS株を経口感染させた。感染の1、2および4週間後、10羽/群の動物を秤量して成長の遅延を調べた。
【0091】
レオウイルスに対する移行抗体を有する15羽/群の1日齢のブロイラーに、プラーク精製レオウイルスERS株(分離体2)を1日齢または1週齢の時点で経口または皮下感染させた。同じ齢および起源の15羽の動物には感染させず、それらを陰性対照として使用した。動物を毎週7週間秤量して成長の遅延を調べた。
【0092】
結果
実験1
該レオウイルスに経口感染させた1日齢の感染SPFニワトリは、感染の4〜10日後に肝細胞および/またはクッパー細胞の多病巣性空胞化を示した
【0093】
実験2
経口感染の10日後、ブロイラーは、同じ条件下に保たれた同じ齢および起源の非感染ブロイラー対照群と比べて、該鳥の排泄腔の周囲にペーストの覆いとして観察されうる過度に液状の糞便を引き起こす腸炎を示した。
【0094】
実験3
1日齢の経口感染ブロイラーは、1、2または4週齢の時点で121.9、327.0または913.1gの体重を有していた。これに対して、同じ条件下に収容された同じ齢および起源の非感染ブロイラーは、1、2または4週齢の時点で134.8、337.6または999.9gの体重を有していた。
【0095】
実験4
結果を図2Aおよび2Bに示す。1日齢の時点で経口または皮下経路により感染させた動物は、7週齢の時点で、非感染対照動物と比べて約34%の成長の遅延を示した。7週齢の時点で、非感染対照動物および感染動物における体重の比率は、2469gおよび1635gであった。1週齢の時点で経口または皮下経路により感染させた動物は、7週齢の時点で、非感染対照動物と比べて約25%の成長の遅延を示した。7週齢の時点で、非感染対照動物および感染動物における体重の比率は、2469gおよび1842gであった。結論としては、レオウイルスERS株は成長の遅延を誘発しうる。
【0096】
実施例2
動物に対するワクチン接種の研究
A.不活化トリレオウイルスワクチンの製造
初代CEL細胞を1 x 106細胞/mlの最終濃度で調製した。該細胞を、0.1%抗生物質および5%ウシ胎児血清を含有するイーグルMEM中で培養した。この細胞懸濁液25mlに、レオウイルスERS分離体(分離体1)0.1mlを加えた。高湿度インキュベーター中37℃で5日間のインキュベーションの後、CPEが明らかに認められ、単層は完全に破壊された。該感染細胞懸濁液の感染価は4.2 log10 TCID50/mlであった。該感染細胞懸濁液にホルムアルデヒドを0.2%の最終濃度まで加えることにより、該レオウイルスを不活化した。該懸濁液を37℃で48時間インキュベートした。ついで該ホルムアルデヒドを等モル量の亜硫酸水素ナトリウムにより中和した。該不活化レオウイルスを使用して、種々の不活化レオウイルスワクチンを製造した。該不活化レオウイルス懸濁液を鉱油相と45:55(w/oエマルション)の比で混合した。
【0097】
ワクチン接種
3〜4週齢のSPF動物の筋肉内に該不活化レオウイルスワクチン(0.5ml/動物)を接種した。ワクチン接種の2、4および6週間後、レオウイルスに対する抗体の存在に関して血清を調べた。ワクチン接種の6週間後、動物を足蹠経路で攻撃した。使用した攻撃ウイルスは病原性同種レオウイルス株(2.2 log TCID50/動物)であった。該足蹠および脚の炎症および変色の程度を2週間にわたり評価した。該足蹠炎症累積値が、ワクチン接種されずに攻撃された対照の足蹠炎症の平均(マイナス2標準偏差)より小さい場合に、ニワトリを攻撃に対して防御性であるとみなした。
【0098】
前記と同様の研究において、該動物を5週齢の時点で攻撃し、激しい攻撃に対する防御を死亡率、罹患率および足蹠病変により評価した。
【0099】
結果
防御性のデータを図1Aおよび1Bに示す。ワクチン接種されたニワトリは、対照群より有意に低い足蹠炎症累積値を示した。このことは、ワクチン接種されたニワトリが、激しいレオウイルスの攻撃に対して防御されたことを示している。また、死亡率および罹患率に基づく防御性データも、激しい攻撃に対する有意な防御を示している。
【0100】
B.ワクチン接種
3〜4週齢のSPF動物に、前記と同様の不活化ERSレオウイルスワクチン(分離体2)を筋肉内に接種し、一次ワクチン接種の6週間後に再接種した。ワクチン接種されていない同じ齢および起源の動物を対照として使用した。一次ワクチン接種の2、4、5、6週間後および二次ワクチン接種の2週間後、レオウイルスに対する抗体の存在に関してIdexx ELISAにより血清を調べた。一次ワクチン接種の6週間後または二次ワクチン接種の2週間後、動物を病原性レオウイルスERS株で足蹠経路により攻撃した。足蹠および脚の変性および変色の程度の平均病変指数を、攻撃後14日間毎日測定した。
【0101】
結果
不活化レオウイルスERSワクチンにより誘発された血清学的応答は、ワクチン接種後の最初の4週間では増加し、一次ワクチン接種の5〜6週間後には横ばいになった。二次ワクチン接種は、二次ワクチン接種の2週間後に約10 log2から12 log2への増加を引き起こした(図3A)。図3Bには、ワクチン接種された及びワクチン接種されていないニワトリを一次ワクチン接種の5週間後または二次ワクチン接種の2週間後に攻撃した場合のデータを示す。ワクチン接種されたすべてのニワトリは、ワクチン接種されていない鳥と比べて足蹠および脚における病変の有意な減少を示した。ワクチン接種されたすべてのニワトリは、ワクチン接種されずに攻撃された対照の足蹠炎症の平均(マイナス2標準偏差)より低い足蹠炎症累積値を示した。このことは、すべての動物が激しいREOの攻撃に対して防御されたことを示している。ワクチン接種群においては、いくつかの軽度の足蹠腫張が認められたが、これらの病変は一時的なものであった。さらに、ワクチン接種を2回された動物においては、この一時的な足蹠病変は、ワクチン接種を1回だけされた動物の場合より軽度であった。
【技術分野】
【0001】
本発明は、トリレオウイルス、および生弱毒化または不活化形態のトリレオウイルスを含むワクチンに関する。
【背景技術】
【0002】
商業的ブロイラー生産は、過去数十年の間に、家畜の監禁的飼育(confinement rearing)の効率の高さにより特徴づけられる1つの産業にまで発展した。しかしながら、飼育段階の効率の増加の傾向を高くしようとすると、必ずいくつかの固有の問題が生じる。最も注目すべきは、高密度で密集して監禁された動物集団においてしばしば生じる感染症の発生率の増加である。家禽における最も破壊的な疾患の多くは、ワクチン接種により、または抗生物質などの治療剤での治療により、抑制または防除されている。しかし、残念ながら、複雑な病因の多数の疾患が、依然として薬物では防除されておらず、それに対しては適当なワクチン接種計画を利用することができない。
【0003】
1970年代後期以降、養鶏業は、腸障害に罹患したブロイラーニワトリにおけるそのような複雑な疾患の問題に直面している。罹患ニワトリにおいて種々の疾患状態(例えば、腸炎)を引き起こす疾患の1つは、主な臨床徴候および肉眼的観察に基づいて吸収不良症候群(MAS)と称される。あるいは、この疾患は、感染性発育不全(infectious runting stunting)症候群、衰弱トリ(pale bird)症候群または骨粗鬆症と称される。多数の種々の疾患状態がMASと連関しているが、いずれの場合も成長不良および羽毛生成の遅延が認められる。さらに、多種多様な他の徴候および病変(例えば、致死、過度に液状の糞便および/または食餌の消化不良、膵萎縮、前胃炎、骨変化、胸腺および滑液嚢萎縮など)がMASと相関している。
【0004】
Kouwenhovenら(Avian Pathology 17, 879−892, 1988)は、以下の5つの基準によりMASを定義した:
(i)1日齢のニワトリの感染後3週までの成長阻害、
(ii)黄橙色の粘液様ないしは湿性の糞便の排泄、
(iii)血漿アルカリホスファターゼ(ALP)活性の増加、
(iv)血漿カロテノイド濃度(PCC)の減少、
(v)近位けい骨の端成長板の拡大の肉眼的観察。
【0005】
ブロイラーの成長の遅延は1週齢以前に明らかとなる。それらの鳥集団のうちの5〜20%が罹患することがあり、これらの鳥は4週齢までに、同集団のその他の鳥の半分以下のサイズとなる。罹患集団は食餌の変換不全を示し、腸は衰弱し、未消化の食餌を含有する。
【0006】
MASの病理は十分には理解されていないが、この症候群の考えられうる病理は、消化管および関連器官に対する感染因子の直接的作用であり、これはまた、過度に液状の糞便および/または消化不良の食餌の排泄の反復を説明するものであろう。
【0007】
本明細書に記載の症候群は、体重増加の減少、食餌の変換不全、罹患した鳥の商業的価値の減少などの品質における全般的な損失を引き起こす。毎年、養鶏業は、MASによる大きな経済的損失を受けている。したがって、ブロイラーに認められる種々の疾患状態の1以上を予防しうるMASの防除方法が、養鶏業において必要とされている。
【0008】
レオウイルスは、世界中の家禽において遍在的に見出される。レオウイルスは、脚部の主要体重支持関節包(major weight bearing joint capsules)および腱シートを冒す関節炎状態(ウイルス性関節炎/腱滑膜炎と称される)の原因因子であることが判明している。
【0009】
いくつかの報告によると、レオウイルスは、MAS関連疾患状態を示すニワトリからも単離されている。これらの報告においては、レオウイルスがMAS関連疾患状態の1以上に対して病因的関連性を有すると推測されているが、レオウイルスがMASに関与しているという確かな証拠はそれらの報告には記載されていない。
【0010】
van der Heideら(Avian Diseases 25, 847−856, 1981)は、臨床的に下痢を示す若いブロイラーの腸からレオウイルスを分離した。このレオウイルス分離体は、腱滑膜炎および大腿骨頭骨折および骨粗鬆症の病変を誘発する能力を有していたが、この分離体は、実験的にレオウイルスに感染させたニワトリにおいて下痢を一貫して誘発しなかった。
【0011】
Pageら(Avian Diseases 26, 618−624, 1982)は、不自由な足、萎縮および異常羽毛発生を示す集団からレオウイルスを分離した。感受性ブロイラー型ニワトリに対する経口接種は、体重増加、羽毛発生に対する明らかな効果を示し、多数の器官において病変を誘発したが、下痢または湿性リターは報告されていない。
【0012】
Hieronymusら(Avian Diseases 27, 246−254, 1983)は、MASの疑いがあるニワトリの腸からのレオウイルスのいくつかの株の分離を報告しており、感染性腱滑膜炎の防除のためのワクチン株として一般に使用されるレオウイルスS1133株とこれらの株との抗原性関係を確認している。該著者は、該レオウイルスが臨床的MASのニワトリから分離されたにもかかわらず、該レオウイルスがMASの原因因子であるか否かは依然として明らかでないことを確認している。
【0013】
Eidsonら(Poulty Science 64, 2081−2086, 1985)は、MASおよび腱滑膜炎の問題を有するブロイラー集団においてHieronymusらにより分離されたC08株から誘導された不活化レオウイルスワクチンの効果について調べている。該ワクチンは、ブロイラー集団の体重に対して正の効果を及ぼしたが、食餌変換における相違は認められなかった。また、湿性リター(wet litter)などの腸炎関連疾患状態に対する該ワクチンの効果は、何ら報告されていない。
【0014】
また、Rosenbergerら(Avian Diseases 33, 535−544, 1989)は、野外で飼育されている商業用ブロイラーニワトリの腱および骨髄からいくつかのレオウイルス株を分離した。レオウイルス株を接種されたニワトリが臨床的疾患に関して試験されたが、下痢または湿性リターの徴候は報告されていない。
【0015】
また、Kouwenhovenら(Avian Pathology 17, 879−892, 1988)は、吸収不良症候群におけるレオウイルスの役割を調べている。これらの著者は、野外症例から分離したレオウイルスでMASを再現することができなかった。そして彼らは、レオウイルスがMASの一次病原体でないと結論づけている。該著者は、レオウイルスおよびアデノウイルスを含む感染因子が、吸収不良症候群における或る種の誘発因子として作用した可能性があると推測している。
【0016】
しかしながら、前記刊行物に記載のものに加えて、他の多数のウイルスがMASに関連している。これらには、ロタウイルス、パルボウイルス、エンテロウイルス様ウイルス、トガウイルス様ウイルス、コロナウイルス様ウイルス、アデノウイルスおよびカリシウイルスが含まれる。さらにまた、細菌が病因に関与していると示唆されている。また、先行技術においては、MAS様野外(field)症候群はマイコトキシンによって生じると考えられており、MASの原因因子としてマイコトキシンまたは他の毒素を無視すべきでないと考えられている。
【0017】
最近公開された総説(World Poultry 14, 57−58, 1998)において、McNultyはMASの最新の知見を要約している。McNultyは、MASに対するワクチンの非利用可能性を強調しており、該ウイルス分離体ならびに現在までに報告されている野外分離サンプルのウイルス学的および微生物学的検査は、MASの原因因子の同定に関して有用な結果を与えていないと記載している。McNultyは、このアプローチが有用な結果をもたらしそうにないと推測している。実際のところ、MAS罹患産生部位における処置手段がMASの防除のための最良の手段であると考えられている。
【先行技術文献】
【非特許文献】
【0018】
【非特許文献1】Kouwenhovenら(Avian Pathology 17, 879−892, 1988)
【非特許文献2】van der Heideら(Avian Diseases 25, 847−856, 1981)
【非特許文献3】Pageら(Avian Diseases 26, 618−624, 1982)
【非特許文献4】Hieronymusら(Avian Diseases 27, 246−254, 1983)
【非特許文献5】Eidsonら(Poulty Science 64, 2081−2086, 1985)
【非特許文献6】Rosenbergerら(Avian Diseases 33, 535−544, 1989)
【非特許文献7】World Poultry 14, 57−58, 1998
【発明の概要】
【発明が解決しようとする課題】
【0019】
したがって、過度に液状の糞便および/または消化不良の食餌の排泄を引き起こしニワトリに生じる或る種の腸障害(例えば、MASに関連した腸障害)に対する有効な防御を誘導するワクチンが依然として必要とされている。
【0020】
さらに、トリレオウイルスは相当な抗原不均質性を示し、トリレオウイルスの新規抗原クラスの出現は、家禽におけるレオウイルスワクチンの使用に重大な影響を及ぼす可能性がある。
【0021】
本発明者らは、本発明において、トリレオウイルスの新規抗原クラスを同定した。さらに、この新規抗原クラスに属するトリレオウイルスが、同様にMASに関連する顕著な疾患状態(過度に液状の糞便および/または消化不良の食餌の排泄ならびに成長の遅延)を誘導しうることを示す。
【0022】
したがって、トリレオウイルスの新規抗原クラスに属するMAS様腸疾患状態の原因因子を提供することが、本発明の目的である。
【0023】
本発明のもう1つの目的は、該新規抗原クラスのトリレオウイルスが引き起こす疾患に対する防御を家禽において有効に与えるワクチンを提供することである。
【0024】
同様にMASに関連している腸疾患状態に対する防御を家禽において有効に与えるワクチンを提供することが、本発明のもう1つの目的である。
【課題を解決するための手段】
【0025】
トリレオウイルスの抗原クラスに属するトリレオウイルスであって、該抗原クラスに属するトリレオウイルスが動物において抗血清を誘導する能力を有し、該抗血清が、プラーク減少アッセイにおいて少なくとも75%の、トリレオウイルスERS(そのサンプルは、ECACC、Salisbury、UKに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こすことを特徴とするトリレオウイルスを提供することにより、これらの目的が達成されうることを、本発明において見出した。そのようなトリレオウイルスは、これまで知られていない抗原特性を示すことが認められただけでなく(実施例1、表2および3)、本発明のトリレオウイルスは、ブロイラーニワトリによる過度に液状の糞便および/または消化不良の食餌の排泄を誘導する能力を有し、場合によっては、致死を招きうることが判明した。したがって、本発明においては、該新規トリレオウイルスを腸レオウイルス株(ERS)と称することにする。
【0026】
ブロイラー集団における湿性糞便の排泄は、野外におけるMAS罹患ブロイラーで一般に認められる疾患状態の1つである。さらに、この臨床的疾患状態は、MAS罹患ブロイラーにおける最も顕著な臨床的徴候(すなわち、成長の遅延)の原因の1つであると考えられる。実施例においては、本発明のトリレオウイルスが、該レオウイルスに経口感染したブロイラーによる湿性糞便の排泄を誘導する(すなわち、該鳥の排泄腔の周囲にペーストの覆い(clothing of pasting)が認められる)ことを示す。また、実施例における実験は、該新規トリレオウイルスの経口感染もまた、対照ニワトリと比較した場合に、感染ブロイラーにおける成長の遅延を引き起こすことを示している。
【0027】
該プラーク減少アッセイは、(トリレオ)ウイルス分離体間の抗原関係を判定するために当技術分野において広く用いられているアッセイである(例えば、Nersessianら, Am. J. Vet. Res 50, 1475−1480, 1989を参照されたい)。さらに、本発明の目的のために、プラーク減少アッセイに関する詳細な説明を実施例1に記載する。当然のことながら、該プラーク減少アッセイで使用する抗血清は、適当な質を有するべきである。そのような抗血清の製造方法は、実施例1にも記載されている。
【0028】
一般に、生トリレオウイルスに対する適当な抗血清は、102.0〜109.0 TCID50/動物、より好ましくは103.0〜106.0 TCID50/動物の感染価を有する生ウイルス株を3〜4週齢のSPFニワトリの皮下または筋肉内に接種することにより製造することができる。感染の3〜4週間後、好ましくは感染の4週間後に血液を集めることができる。また、一次感染で使用したのとほぼ同用量の同じ生ウイルス株を、一次感染の3〜4週間後にニワトリに再感染させることができる。二次感染の2〜4週間後に血液を集める。不活化トリレオウイルス株に対する適当な抗血清は、3〜4週齢のSPFニワトリに該不活化ウイルス調製物を皮下または筋肉内に接種することにより得ることができる。不活化前の該調製物の感染価は、107.0〜1011.0 TCID50/動物であってもよく、より好ましくは108.0〜1010.0 TCID50/動物である。血液は、接種の3〜4週間後、好ましくは接種の4週間後に集めることができる。また、一次接種の3〜6週間後に同じ不活化ウイルス調製物をニワトリに再接種することができる。二次接種の2〜4週間後に血液を集める。
【0029】
本発明の新規トリレオウイルスの同定は、トリレオウイルスの該新規抗原クラスによる感染から生じる疾患状態に対して家禽を有効に防御しうる新規トリレオウイルスワクチンの製造を可能にする。特に、該新規トリレオウイルスは、過度に液状の糞便および/または消化不良の食餌の排泄ならびに成長の遅延などの疾患状態に対して家禽を有効に防御しうる新規トリレオウイルスワクチンの製造を可能にする。そのような疾患状態もMASに関連している。
【0030】
また、本発明のトリレオウイルスは、野外から分離することができる。分離方法の重要な態様は、ウイルスの分離の出発物として使用する標的動物の同定である。この目的に使用する典型的なブロイラーは、成長の遅延につながる過度に液状の糞便および/または消化不良の食餌の排泄の徴候を示す。
【0031】
ついで該罹患ニワトリから腸を分離し、該器官を適当なバッファー中でホモジナイズする。ついで、ホモジナイズされた該組織を遠心分離し、該上清を孔径0.2μmのフィルターで濾過する。該濾液のサンプルを、新鮮に調製された初代ニワトリ細胞、好ましくはニワトリ胚肝(CEL)細胞に加え、インキュベーションの4〜8日後、細胞変性効果(CPE)の存在に関して該単層を調べる。CPEが存在しなければ、該一次単層の凍結/融解懸濁液を、新鮮に調製されたCEL細胞に加える。第1継代または第2継代後にCPEが認められたら、該ウイルスを、過度に液状の糞便および/または消化不良の食餌の排泄を誘発するブロイラーにおけるそのインビボ特性により、およびプラーク減少アッセイまたは免疫蛍光技術(IFT)(後記の特異的ポリクローナルおよびモノクローナル抗体を使用する)において測定したその抗原特性により、さらに特徴づける。本発明のトリレオウイルスの分離のためのより詳細な方法を、実施例1に開示する。
【0032】
本発明のトリレオウイルスの分離方法においては、好ましい出発物質として腸を使用するが、トリレオウイルスは、罹患ブロイラーニワトリの肝臓またはそのようなブロイラーニワトリが排泄した糞便からも分離することができる。また、本発明のトリレオウイルスの分離のための出発物質として他の器官を用いることも可能であることを考慮に入れるべきである。
【図面の簡単な説明】
【0033】
【図1】Aは、足蹠病変アッセイに対するワクチンの防御活性を示す。Bは、死亡率、罹患率および足蹠病変に対するワクチンの防御活性を示す。
【図2】AおよびBは、ERSに感染したブロイラーの成長の遅延を示す。
【図3】Aは、血清力価とワクチン接種後の週数との関係を示す。Bは、攻撃後の足蹠病変指数の時間経過を示す。
【発明を実施するための形態】
【0034】
特に、本発明は、前記トリレオウイルスであって、該トリレオウイルスに対して誘導された抗血清が、プラーク減少アッセイにおいて少なくとも80%、好ましくは少なくとも90%の、トリレオウイルスERS(そのサンプルは、ECACC、Salisbury、UKに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こしうることを特徴とするトリレオウイルスを提供する。
【0035】
本発明のより一層好ましい実施形態においては、ポリクローナル抗血清およびモノクローナル抗体(Moabs)の特異的パネルに対して特異的免疫反応を示すトリレオウイルスを提供する。この特異的反応パターンは、従前公知のトリレオウイルスにより示されるものとは異なる。
【0036】
モノクローナル抗体は、感染因子の特性を同定し同一または類似の微生物の種々の分離体間の類似性および相違を判定するのに有用である。Vakhariaら(Proceedings of the International Symposium on adenovirus and reovirus infections in poultry, Rauischholzhausen, Germany, 1996, 295−304)は、9個の抗レオウイルスMoabsのパネルを開示しており、トリレオウイルス野外分離体を、これらのMoabsに対するそれらの反応性に関して試験している。Moabsのパネルの反応性の種々のパターンは、反応性パターンの関連性に基づく分離体の分類を可能にした。
【0037】
本発明のこの実施形態においては、前記トリレオウイルスであって、(i)トリレオウイルス分離体に対して(好ましくは、始原型レオウイルス1133株に対して)産生したポリクローナル抗血清に対するIFTにおけるその反応性、および(ii)Moabs INT 13−06、INT 14−11および15−01 INT(それらのサンプルは、それぞれ受託番号99011472、99011473および99011474でECACCに寄託されている)に対するIFTにおける反応性の不存在により更に特徴づけられるトリレオウイルスを提供する。
【0038】
該パネルパターンにより定められる本発明のトリレオウイルスは、実施例1および表3に示す新規抗原型のレオウイルスを代表する。表3に示す先行技術のレオウイルス株の多数が、MASに関連した徴候および病変を示すブロイラーニワトリの組織(腸を含む)から分離されているが、前記の新規抗原特性を有する本発明のトリレオウイルスは先行技術において開示されていない。
【0039】
本発明のより一層好ましいトリレオウイルスは、本発明のトリレオウイルスの特徴を示すトリレオウイルスERS(分離体1)(そのサンプルは、ECACCに受託番号99011475で寄託されている)である。
【0040】
本発明のトリレオウイルスは、生、生弱毒化または不活化形態であってもよい。
【0041】
本発明は、もう1つの態様において、トリレオウイルスから生じる疾患状態(例えば、MASで認められる腸疾患状態)に対して家禽を防御するのに使用するためのワクチンであって、本発明のトリレオウイルスと医薬上許容される担体または希釈剤とを含んでなるワクチンを提供する。
【0042】
本発明のトリレオウイルスは、生弱毒化または不活化ウイルスとして該ワクチン中に含有させることができる。該トリレオウイルスを生弱毒または不活化形態にすると、前記のMAS関連疾患状態を誘発するトリレオウイルスの特性は有意に軽減されるか、または完全に失われる。
【0043】
本発明のトリレオウイルスの弱毒化は、この目的のための当技術分野でよく知られた方法、例えば、Gouveaら(Virology 126, 240−247, 1983)に開示されている方法により行なうことができる。簡単に説明すると、標的動物から該ウイルスを分離した後、ウイルス懸濁液を初代ニワトリ胚繊維芽細胞(CEF)に接種する。該分離体がCPEを与える能力を有していない場合には、CPEが認められるようになるまで該ウイルスを反復的に(例えば、3〜10回)継代する。CPEが認められると直ちに細胞および細胞培養流体を集め、凍結・融解し、遠心分離により清澄化し、トリレオウイルス分離体を含有する上清をアリコート化し、−20℃で保存する。該ウイルスを更に弱毒化するために、この方法を繰返す(例えば、10〜100回)ことができる。
【0044】
本発明のワクチンは、例えば、商業的に入手可能な生および不活化レオウイルスワクチンに一般に用いられる方法などの通常の方法で製造することができる。獣医学用ワクチン組成物の製造については、とりわけ、“Handbuch der Schulzimpfungen in der Tiermedizin”(Mayr, Aら編, Verlag Paul Parey, Berlin und Hamburg, Germany, 1984)および“Vaccines for Veterinary Applications”(Peters, A.R.ら編, Butterworth−Heinermann Ltd, 1993)に記載されている。
【0045】
簡単に説明すると、生または生弱毒化形態の本発明のトリレオウイルスを感受性基質に接種し、該ウイルスが所望の感染価または抗原含量まで複製されるまで該基質を増殖させ、ついでレオウイルス含有物質を収穫し、予防活性を有する医薬組成物に製剤化する。
【0046】
必要に応じて該トリレオウイルスを基質に適合させた後、前記トリレオウイルスの複製を支持しうる各基質を使用して、本発明のワクチンを製造することができる。適当な基質には、初代(トリ)細胞培養、例えばニワトリ胚肝細胞(CEL)、ニワトリ胚繊維芽細胞(CEF)またはニワトリ腎細胞(CK)、哺乳類細胞系、例えばVERO細胞系またはBGM−70細胞系、またはトリ細胞、例えばQT−35、QM−7またはLMHが含まれる。通常、該細胞の接種後、該ウイルスを3〜10日間増殖させ、ついで該細胞培養上清を収穫し、所望により濾過または遠心分離して細胞残渣を除去する。
【0047】
あるいは、本発明のトリレオウイルスを孵化鶏卵内で増殖させ、ついでトリレオウイルス物質を常法により収穫することができる。
【0048】
生弱毒化ウイルスを含有する本発明のワクチンは、(凍結)懸濁液形態または凍結乾燥形態で製造し、販売することができる。該ワクチンは更に、そのような組成物に通常使用される医薬上許容される担体または希釈剤を含有する。担体には、安定化剤、保存剤およびバッファーが含まれる。適当な安定化剤としては、例えば、SPGA、炭水化物(例えば、ソルビトール、マンニトール、デンプン、スクロース、デキストラン、グルタメートまたはグルコース)、タンパク質(例えば、乾燥乳血清、アルブミンまたはカゼイン)またはそれらの分解産物が挙げられる。適当なバッファーには、例えば、リン酸アルカリ金属が挙げられる。適当な保存剤としては、チメロサール、メルチオラートおよびゲンタマイシンが挙げられる。希釈剤には、水、水性バッファー(例えば、緩衝食塩水)、アルコールおよびポリオール(例えば、グリセロール)が含まれる。
【0049】
所望により、本発明の生ワクチンはアジュバントを含有していてもよい。アジュバント活性を有する適当な化合物および組成物としては、例えば、不活化ワクチンの製造に関して後記で挙げるものと同じものが挙げられる。
【0050】
本発明の生ワクチンの注射(例えば、筋肉内、皮下)による投与は可能であるが、該生ワクチンは、好ましくは、トリレオウイルスのワクチン接種に通常用いる安価な大量適用技術により投与する。これらの技術には、飲水および噴霧ワクチン接種が含まれる。
【0051】
該生ワクチンの投与のためのもう1つの方法には、卵内、点眼および嘴の浸漬による投与が含まれる。
【0052】
好ましい実施形態において、本発明は、不活化形態の該トリレオウイルスを含んでなる、腸疾患状態(例えば、MASで認められるもの)に対するワクチンを提供する。不活化ワクチンの主な利点は、得られる持続性防御抗体のレベルが高いことである。この特性のおかげで、そのような不活化ワクチンは、特に種畜ワクチン接種に適したものになる。
【0053】
増殖工程後に収穫したウイルスの不活化の目的は、該ウイルスの複製を排除することである。一般に、これは、化学的または物理的手段により達成することができる。化学的不活化は、例えば酵素、ホルムアルデヒド、β−プロピオラクトン、エチレン−イミンまたはそれらの誘導体で該ウイルスを処理することにより行なうことができる。必要に応じて、後で該不活化化合物を中和する。ホルムアルデヒドで不活化した物質は、例えば、チオスルファートで中和することができる。物理的不活化は、好ましくは、該ウイルスを高エネルギー放射(例えば、UV光またはγ線)に付すことにより行なうことができる。所望により、処理後、該pHを約7に調節することができる。
【0054】
該不活化トリレオウイルスを含有するワクチンは、例えば、この目的に適した前記の医薬上許容される担体または希釈剤の1以上を含んでいてもよい。
【0055】
好ましくは、本発明の不活化ワクチンは、アジュバント活性を有する1以上の化合物を含む。この目的のための適当な化合物または組成物には、水酸化、リン酸または酸化アルミニウム、例えば鉱油(例えばBayol F(登録商標)またはMacrol 52(登録商標))または植物油(例えばビタミンEアセタート)に基づく水中油滴型または油中水滴型エマルション、およびサポニンが含まれる。
【0056】
不活化ワクチンは、通常、非経口的(例えば、筋肉内または皮下)に投与する。
【0057】
本発明のワクチンは、活性成分としての該トリレオウイルスの有効用量(すなわち、ビルレントウイルスによる攻撃に対して、ワクチン接種された鳥またはそれらの後代において免疫を誘導するトリレオウイルス物質を免疫する量)を含む。本発明においては、免疫は、ワクチン接種後の鳥集団において、非ワクチン接種群と比べて有意に高いレベルの防御を誘導することと定義される。
【0058】
典型的には、本発明の生ワクチンは、102〜109 TCID50/鳥の用量、好ましくは102〜106 TCID50/鳥の用量で投与することが可能であり、不活化ワクチンは、104〜1010 TCID50/鳥の抗原相当量を含有していてもよい。
【0059】
本発明のトリレオウイルスワクチンはニワトリにおいて有効に使用することができ、他の家禽(例えば、シチメンチョウ、ホロホロチョウおよびウズラ)にも該ワクチンを成功裡に接種することができる。ニワトリには、ブロイラー、繁殖ストックおよび産卵ストックが含まれる。
【0060】
腸疾患状態(例えば、MASで認められるもの)は主にブロイラーニワトリにおいて報告されているため、本発明は、好ましくは、腸疾患状態(例えば、MASで認められるもの)に対してブロイラーを防御するのに使用するワクチンを提供する。
【0061】
本発明の生または不活化ワクチンを投与する動物の齢は、現在商業的に入手可能な生または不活化トリレオウイルスワクチンを投与する動物の齢と同じである。例えば、本発明の生弱毒化ワクチンをブロイラーに1日齢以降から直接接種することができる。親ストック(例えば、ブロイラー種畜)のワクチン接種は、本発明の生弱毒化もしくは不活化ワクチンまたはそれらの組合せで行なうことができる。このタイプの免疫化計画の利点には、若鳥に垂直伝播した母体由来抗体により与えられる1日齢の後代の即時防御が含まれる。典型的な種畜ワクチン接種計画は、6週齢の種畜に生弱毒化ワクチンを接種し、ついで14〜18週齢で不活化ワクチンを接種することを含む。あるいは、生ワクチンの接種の後、10〜12週齢および16〜18週齢の時点で不活化ワクチンを2回接種することができる。
【0062】
本発明はまた、本発明のトリレオウイルスと、家禽に対して感染性の他の病原体のワクチン成分の1以上とを含んでなる混合(combination)ワクチンを含む。家禽に対して感染性のそのような他の病原体は、本発明のトリレオウイルスとは抗原的に異なるトリレオウイルスを意味し、腱滑膜炎に関連したトリレオウイルス株を含む。
【0063】
好ましくは、該混合ワクチン中のワクチン成分は、家禽に対して感染性の病原体の生弱毒化または不活化形態である。
【0064】
特に、本発明は、該ワクチン成分のすべてが不活化形態である混合ワクチンを提供する。
【0065】
好ましくは、該混合ワクチンは、伝染性気管支炎ウイルス(IBV)、ニューカッスル病ウイルス(NDV)、伝染性滑液嚢症ウイルス(IBDV)、トリアデノウイルス(FAV)、EDSウイルスおよびシチメンチョウ鼻気管炎ウイルス(TRTV)の(不活化)ワクチン株の1以上を含む。
【0066】
本発明の範囲内の以下の微生物およびハイブリドーマ細胞系は、European Collection of Animal Cell Cultures (ECACC), Salisbury, UKに1999年1月14日に寄託されている。
【0067】
【表1】
【実施例】
【0068】
実施例1
新規トリレオウイルスの特徴づけ
A.新規トリレオウイルスの分離
成長の遅延を引き起こす消化障害および湿性リターを有するニワトリから、腸および/または肝臓を個々に分離した。該器官を、抗生物質を含有するPBSおよびガラスパール(2mm)を使用してホモジナイザー中、最大速度で20分間、個々にホモジナイズした。ついで、そのホモジナイズされた組織を遠心分離した。腸ホモジネートを4000rpmで、肝臓を1200rpmで、共に15分間遠心分離した。ついで該上清をフィルター(孔径を5、1.2、0.45、0.2μmと減少させる)に通して濾過した。0.2μmのフィルターに通した100μlの懸濁液を、組織培養フラスコ中に存在する新鮮に調製された初代ニワトリ胚肝(CEL)細胞に加えた。インキュベーションの4〜8日後、細胞変性効果(CPE)の存在に関して該単層を調べた。CPEが存在しなければ、該単層を−20℃で凍結し、24時間後、該単層を融解した。ついでこの凍結/融解懸濁液の1mlを、新鮮に調製されたCEL細胞に加えた。4〜8日後にCPEが認められない場合には、CEL細胞上で増殖するウイルスに関して該培養を陰性とみなした。第1継代または第2継代後にCPEが認められた場合には、プラーク減少アッセイおよび免疫蛍光技術(IFT)により該ウイルスを更に特徴づけした。
【0069】
B.プラーク減少アッセイによる新規トリレオウイルスのインビトロでの特徴づけ
1.種々のトリレオウイルス株に対する抗血清の産生
ERS株(生):
10羽の3週齢のSPFニワトリに105.8 TCID50/動物のERS株(分離体2)を皮下感染させた。感染の3週間後に血液を集め、血清を単離し、動物に105.6 TCID50/動物で再感染させた。二次感染の2週間後、血液を集め、血清を単離した。集めた後、すべての血清を56℃で30分間熱不活化し、小さなアリコート中に−20℃で保存した。
【0070】
2177株(生):
15羽の4週齢のSPFニワトリに104.2 TCID50/動物で皮下感染させた。感染の4週間後に血液を集め、血清を単離した。集めた後、すべての血清を56℃で30分間熱不活化し、小さなアリコート中に−20℃で保存した。
【0071】
1733および2408株(不活化):
12羽の4週齢のSPFニワトリに、w/oエマルションでアジュバント化されたトリレオウイルス1733又は2408株を筋肉内接種した。動物用量:490 Elisa単位/動物(107.0〜1010.0 TCID50/動物の不活化前感染価に相当)。感染の4週間後、血液を集め、血清を単離した。集めた後、すべての血清を56℃で30分間熱不活化し、小さなアリコート中に−20℃で保存した。
【0072】
トリレオウイルス株を含有する商業的に入手可能な不活化ワクチン
10羽の3〜4週齢のSPFニワトリに、以下の商業的に入手可能な不活化トリレオウイルスワクチンの1つを筋肉内または皮下接種した:ISBI, Fort Dodge; Kaketsuken and Intervet International BV。感染の3週間後、血液を集め、血清を単離した。集めた後、すべての血清を56℃で30分間熱不活化し、小さなアリコート中に−20℃で保存した。
【0073】
前記で調製した血清を、IFTおよびプラーク減少アッセイにおいて使用した。
【0074】
2.免疫蛍光試験(IFT)
後記C節に記載されているのと実質的に同じ方法でIFTを行なう。簡単に説明すると、Vero細胞をコンフルエントになるまで96ウェルポリスチレンマイクロタイタープレート中で増殖させた。種々の単層にレオウイルス1133株を接種した。100μlのニワトリ血清を該プレートの第1ウェルに加えた。系列3倍希釈を作製した。インキュベーションおよび洗浄工程の後、該プレートを1:100希釈の蛍光イソチオシアナート標識ヤギ抗ニワトリと反応させた。蛍光の存在を蛍光顕微鏡で観察した。該力価(該血清の質)を終末点希釈法により測定する。これは、明らかな蛍光シグナルを誘導しうる血清の最高希釈度である。表1に、該プラーク減少試験で使用した血清についてのIFTの結果を示す。
【0075】
【表2】
Nobilis Reo(登録商標)(Intervet Internationl b.v.から商業的に入手可能)
Arvax(登録商標)(ISBIから商業的に入手可能)
Tri Reo(登録商標)(Fort Dodgeから商業的に入手可能)
Olivax Reo(登録商標)(Kaketsukenから商業的に入手可能)。
【0076】
3.プラーク減少アッセイ
新規抗原クラスに属するトリレオウイルス間の抗原関係を判定し新規トリレオウイルスを公知トリレオウイルスから識別するためのプラーク減少アッセイにおいては、適当な質の抗血清を使用するべきである(Arvax抗血清以外はすべて、該抗血清の品質要件を満足していた)。生ERS分離体1ならびに生および不活化ERS分離体2に対してERS抗血清を産生させた。新鮮に調製されたCELを、5%ウシ胎児血清および抗生物質で補足された組織培養培地に1 106細胞/mlの最終濃度まで再懸濁した。60mm組織培養皿を5mlの細胞懸濁液で満たし、37℃で24時間インキュベートした。
【0077】
翌日、調べようとするウイルスを、プラスチックチューブ中、抗生物質を含有する培地中で希釈した。10−1〜10−7の希釈液を調製した。つぎに各希釈液200μlを、試験する血清50μlと混合した。この混合物を37℃で1時間インキュベートした。陰性対照として、200μlのウイルス希釈液を50μlの培地と混合した。
【0078】
60mm組織培養皿中に存在する単層上の培地を捨てた。つぎに、それらの種々のウイルス混合物(血清を含有する又は含有しない)100μlをコンフルエントな単層上に加えた。各ウイルス混合物を、少なくとも2つの単層(皿)に感染させた。感染したそれらの単層を37℃で1時間インキュベートした。ついで該感染単層を5mlの寒天溶液(培地、FCSおよび抗生物質を含有する;最終寒天濃度3.0%;最終FCS濃度2.5%)で覆った。該皿を37℃で4日間インキュベートした。つぎに各組織培養皿に2mlのニュートラルレッド溶液(0.02%)を加えた。37℃で4時間のインキュベーションの後、プラークの数を皿ごとに計数した。150個未満のプラークを含有する組織培養皿のみを計数した。
【0079】
プラークの減少を以下のとおりに計算した。血清の不存在下の或る希釈度における或るウイルスのプラーク数を100%とする。ついでこれを、血清の存在下の同じウイルス希釈度におけるプラーク数と比較する。
【0080】
種々のウイルスおよび血清を試験した第1の実験の結果を表2Aに示す。表2Bは、示されているとおりのウイルスおよび血清を使用した第2の実験の結果を示す。ワクチン接種の5週間後に血液を集める以外は不活化1733/2408株に関して記載されているとおりに、不活化ERSワクチンに対する血清を産生させた。
【0081】
【表3】
【0082】
【表4】
【0083】
C.IFTによる新規トリレオウイルスのインビトロでの特徴づけ:抗血清パネル反応パターン
精製トリレオウイルス1133株をウサギ(1〜1.5kg)に感染させることにより、ポリクローナル抗血清を調製した。一次感染の28および84日後にブースター注射を行なった。最後の注射の14日後、血液を集め、血清を単離した。種々のMoabsで種々のトリレオウイルス株を特徴づけした。初代CEL細胞を96ウェルポリスチレンマイクロタイタープレート中で増殖させた。未感染細胞を対照として使用した。5% CO2と共に37℃で2〜4日間インキュベートした後、感染した単層を96%冷エタノールで固定した。該アルコールを捨て、該プレートを洗浄バッファーで洗浄し、PBS中1:50または1:200に希釈した種々のハイブリドーマ細胞培養上清100μlまたは1:50に希釈したウサギポリクローナル血清(ウサギ68A)100μlを各ウェルに加えた。該プレートを37℃で60〜90分間インキュベートし、洗浄バッファーで2回洗浄し、1:100希釈の蛍光イソチオシアナート標識ウサギ抗マウスまたは1:100希釈のイソチオシアナート標識ヤギ抗ウサギ血清と反応させた。最後に、該プレートを洗浄し、グリセロール/PBS溶液(1:1)で固定した。蛍光の存在を蛍光顕微鏡で観察した。
【0084】
この実験で使用した抗血清パネルは、始原型トリレオウイルス1133株に対して産生させた以下のポリクローナル抗血清およびMoabsよりなるものであった。
Rabbit 68A:ウサギポリクローナル抗血清
Moab 154:Vakhariaら, 1996 (前掲)
Moab 14−67:Intervet International B.V.
Moab INT 13−06:ECACC受託番号99011472
Moab INT 14−11:ECACC受託番号99011473
Moab 15−01 INT:ECACC受託番号99011474。
【0085】
前記方法により得たトリレオウイルス分離体を、この抗血清パネルに対するそれらの反応性により更に特徴づけした。該新規レオウイルス分離体は、ポリクローナルおよびモノクローナル抗体パネルに対する特有の反応パターンを有する。MASおよび腱滑膜炎の野外症例から分離された従前公知のトリレオウイルス株(S−1133〜CO8)は、そのような新規パターンでは反応しない(表3を参照されたい)。
【0086】
新規反応パターン
陽性:ポリクローナルウサギ68A、154、14−67
陰性:INT 13−06、INT 14−11、15−01 INT。
【0087】
【表5】
Enterovax(登録商標)およびTensynovac(登録商標)は、Schering−Plough Animal Health and Intervet Inc.から商業的に入手可能なトリレオウイルスワクチンである。
【0088】
D.新規トリレオウイルスのインビボでの特徴づけ
プラーク精製レオウイルスERSの実験的感染
実験1
30羽の1日齢のSPFニワトリに、プラーク精製レオウイルスERS(分離体1)を経口感染させた。注射の、4、7および10日後、肝臓を10羽の動物から単離し、顕微鏡的病変に関して調べた。
【0089】
実験2
レオウイルスに対する移行抗体を有する30羽の1日齢のブロイラーに、プラーク精製レオウイルスERS株を経口感染させた。感染の7および10日後、臨床的徴候に関して動物を観察した。湿性リターに特別の注意を払った。
【0090】
実験3
レオウイルスに対する移行抗体を有する30羽の1日齢のブロイラーに、プラーク精製レオウイルスERS株を経口感染させた。感染の1、2および4週間後、10羽/群の動物を秤量して成長の遅延を調べた。
【0091】
レオウイルスに対する移行抗体を有する15羽/群の1日齢のブロイラーに、プラーク精製レオウイルスERS株(分離体2)を1日齢または1週齢の時点で経口または皮下感染させた。同じ齢および起源の15羽の動物には感染させず、それらを陰性対照として使用した。動物を毎週7週間秤量して成長の遅延を調べた。
【0092】
結果
実験1
該レオウイルスに経口感染させた1日齢の感染SPFニワトリは、感染の4〜10日後に肝細胞および/またはクッパー細胞の多病巣性空胞化を示した
【0093】
実験2
経口感染の10日後、ブロイラーは、同じ条件下に保たれた同じ齢および起源の非感染ブロイラー対照群と比べて、該鳥の排泄腔の周囲にペーストの覆いとして観察されうる過度に液状の糞便を引き起こす腸炎を示した。
【0094】
実験3
1日齢の経口感染ブロイラーは、1、2または4週齢の時点で121.9、327.0または913.1gの体重を有していた。これに対して、同じ条件下に収容された同じ齢および起源の非感染ブロイラーは、1、2または4週齢の時点で134.8、337.6または999.9gの体重を有していた。
【0095】
実験4
結果を図2Aおよび2Bに示す。1日齢の時点で経口または皮下経路により感染させた動物は、7週齢の時点で、非感染対照動物と比べて約34%の成長の遅延を示した。7週齢の時点で、非感染対照動物および感染動物における体重の比率は、2469gおよび1635gであった。1週齢の時点で経口または皮下経路により感染させた動物は、7週齢の時点で、非感染対照動物と比べて約25%の成長の遅延を示した。7週齢の時点で、非感染対照動物および感染動物における体重の比率は、2469gおよび1842gであった。結論としては、レオウイルスERS株は成長の遅延を誘発しうる。
【0096】
実施例2
動物に対するワクチン接種の研究
A.不活化トリレオウイルスワクチンの製造
初代CEL細胞を1 x 106細胞/mlの最終濃度で調製した。該細胞を、0.1%抗生物質および5%ウシ胎児血清を含有するイーグルMEM中で培養した。この細胞懸濁液25mlに、レオウイルスERS分離体(分離体1)0.1mlを加えた。高湿度インキュベーター中37℃で5日間のインキュベーションの後、CPEが明らかに認められ、単層は完全に破壊された。該感染細胞懸濁液の感染価は4.2 log10 TCID50/mlであった。該感染細胞懸濁液にホルムアルデヒドを0.2%の最終濃度まで加えることにより、該レオウイルスを不活化した。該懸濁液を37℃で48時間インキュベートした。ついで該ホルムアルデヒドを等モル量の亜硫酸水素ナトリウムにより中和した。該不活化レオウイルスを使用して、種々の不活化レオウイルスワクチンを製造した。該不活化レオウイルス懸濁液を鉱油相と45:55(w/oエマルション)の比で混合した。
【0097】
ワクチン接種
3〜4週齢のSPF動物の筋肉内に該不活化レオウイルスワクチン(0.5ml/動物)を接種した。ワクチン接種の2、4および6週間後、レオウイルスに対する抗体の存在に関して血清を調べた。ワクチン接種の6週間後、動物を足蹠経路で攻撃した。使用した攻撃ウイルスは病原性同種レオウイルス株(2.2 log TCID50/動物)であった。該足蹠および脚の炎症および変色の程度を2週間にわたり評価した。該足蹠炎症累積値が、ワクチン接種されずに攻撃された対照の足蹠炎症の平均(マイナス2標準偏差)より小さい場合に、ニワトリを攻撃に対して防御性であるとみなした。
【0098】
前記と同様の研究において、該動物を5週齢の時点で攻撃し、激しい攻撃に対する防御を死亡率、罹患率および足蹠病変により評価した。
【0099】
結果
防御性のデータを図1Aおよび1Bに示す。ワクチン接種されたニワトリは、対照群より有意に低い足蹠炎症累積値を示した。このことは、ワクチン接種されたニワトリが、激しいレオウイルスの攻撃に対して防御されたことを示している。また、死亡率および罹患率に基づく防御性データも、激しい攻撃に対する有意な防御を示している。
【0100】
B.ワクチン接種
3〜4週齢のSPF動物に、前記と同様の不活化ERSレオウイルスワクチン(分離体2)を筋肉内に接種し、一次ワクチン接種の6週間後に再接種した。ワクチン接種されていない同じ齢および起源の動物を対照として使用した。一次ワクチン接種の2、4、5、6週間後および二次ワクチン接種の2週間後、レオウイルスに対する抗体の存在に関してIdexx ELISAにより血清を調べた。一次ワクチン接種の6週間後または二次ワクチン接種の2週間後、動物を病原性レオウイルスERS株で足蹠経路により攻撃した。足蹠および脚の変性および変色の程度の平均病変指数を、攻撃後14日間毎日測定した。
【0101】
結果
不活化レオウイルスERSワクチンにより誘発された血清学的応答は、ワクチン接種後の最初の4週間では増加し、一次ワクチン接種の5〜6週間後には横ばいになった。二次ワクチン接種は、二次ワクチン接種の2週間後に約10 log2から12 log2への増加を引き起こした(図3A)。図3Bには、ワクチン接種された及びワクチン接種されていないニワトリを一次ワクチン接種の5週間後または二次ワクチン接種の2週間後に攻撃した場合のデータを示す。ワクチン接種されたすべてのニワトリは、ワクチン接種されていない鳥と比べて足蹠および脚における病変の有意な減少を示した。ワクチン接種されたすべてのニワトリは、ワクチン接種されずに攻撃された対照の足蹠炎症の平均(マイナス2標準偏差)より低い足蹠炎症累積値を示した。このことは、すべての動物が激しいREOの攻撃に対して防御されたことを示している。ワクチン接種群においては、いくつかの軽度の足蹠腫張が認められたが、これらの病変は一時的なものであった。さらに、ワクチン接種を2回された動物においては、この一時的な足蹠病変は、ワクチン接種を1回だけされた動物の場合より軽度であった。
【特許請求の範囲】
【請求項1】
トリレオウイルスの抗原クラスに属するトリレオウイルスであって、該抗原クラスに属するトリレオウイルスが動物において抗血清を誘導する能力を有し、該抗血清が、プラーク減少アッセイにおいて少なくとも75%の、トリレオウイルスERS(そのサンプルは、ECACCに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こすことを特徴とするトリレオウイルス。
【請求項2】
該トリレオウイルスに対して誘導された抗血清が、プラーク減少アッセイにおいて少なくとも80%、好ましくは少なくとも90%の、トリレオウイルスERS(そのサンプルは、ECACCに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こしうる、請求項1に記載のトリレオウイルス。
【請求項3】
該トリレオウイルスが、ポリクローナルトリレオウイルス抗血清には陽性反応するが、受託番号99011472、99011473および99011474でECACCに寄託されているサンプルのモノクローナル抗体には陽性反応しない、請求項1または2に記載のトリレオウイルス。
【請求項4】
トリレオウイルスERS(そのサンプルは、ECACCに受託番号99011475で寄託されている)である、請求項3に記載のトリレオウイルス。
【請求項5】
トリレオウイルスの感染から生じる疾患状態に対して家禽を防御するのに使用するためのワクチンであって、請求項1〜4のいずれか1項に記載のトリレオウイルスと医薬上許容される担体または希釈剤とを含んでなるワクチン。
【請求項6】
該トリレオウイルスが生弱毒化形態である、請求項5に記載のワクチン。
【請求項7】
該トリレオウイルスが不活化形態である、請求項5に記載のワクチン。
【請求項8】
該ワクチンが更にアジュバントを含む、請求項5〜7のいずれか1項に記載のワクチン。
【請求項9】
該ワクチンが更に、家禽に対して感染性の他の病原体のワクチン成分の1以上を含む、請求項5〜8のいずれか1項に記載のワクチン。
【請求項10】
請求項1〜4のいずれか1項に記載のトリレオウイルスの製造方法であって、
a.該トリレオウイルスを感受性基質に接種し、
b.該トリレオウイルスを増殖させ、
c.トリレオウイルス含有物質を収穫する工程を含んでなる製造方法。
【請求項11】
トリレオウイルスの感染から生じる疾患状態に対して家禽を防御するのに使用するためのワクチンの製造方法であって、請求項10に記載の製造方法により得られ収穫されたレオウイルスを、所望により該レオウイルスの不活化後に、医薬上許容される担体または希釈剤と一緒にすることを含んでなる製造方法。
【請求項12】
家禽におけるトリレオウイルスの感染から生じる疾患状態を防除するための方法であって、請求項1〜4のいずれか1項に記載のワクチンを該鳥に投与することを含んでなる方法。
【請求項1】
トリレオウイルスの抗原クラスに属するトリレオウイルスであって、該抗原クラスに属するトリレオウイルスが動物において抗血清を誘導する能力を有し、該抗血清が、プラーク減少アッセイにおいて少なくとも75%の、トリレオウイルスERS(そのサンプルは、ECACCに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こすことを特徴とするトリレオウイルス。
【請求項2】
該トリレオウイルスに対して誘導された抗血清が、プラーク減少アッセイにおいて少なくとも80%、好ましくは少なくとも90%の、トリレオウイルスERS(そのサンプルは、ECACCに受託番号99011475で寄託されている)により形成されるプラークの減少を引き起こしうる、請求項1に記載のトリレオウイルス。
【請求項3】
該トリレオウイルスが、ポリクローナルトリレオウイルス抗血清には陽性反応するが、受託番号99011472、99011473および99011474でECACCに寄託されているサンプルのモノクローナル抗体には陽性反応しない、請求項1または2に記載のトリレオウイルス。
【請求項4】
トリレオウイルスERS(そのサンプルは、ECACCに受託番号99011475で寄託されている)である、請求項3に記載のトリレオウイルス。
【請求項5】
トリレオウイルスの感染から生じる疾患状態に対して家禽を防御するのに使用するためのワクチンであって、請求項1〜4のいずれか1項に記載のトリレオウイルスと医薬上許容される担体または希釈剤とを含んでなるワクチン。
【請求項6】
該トリレオウイルスが生弱毒化形態である、請求項5に記載のワクチン。
【請求項7】
該トリレオウイルスが不活化形態である、請求項5に記載のワクチン。
【請求項8】
該ワクチンが更にアジュバントを含む、請求項5〜7のいずれか1項に記載のワクチン。
【請求項9】
該ワクチンが更に、家禽に対して感染性の他の病原体のワクチン成分の1以上を含む、請求項5〜8のいずれか1項に記載のワクチン。
【請求項10】
請求項1〜4のいずれか1項に記載のトリレオウイルスの製造方法であって、
a.該トリレオウイルスを感受性基質に接種し、
b.該トリレオウイルスを増殖させ、
c.トリレオウイルス含有物質を収穫する工程を含んでなる製造方法。
【請求項11】
トリレオウイルスの感染から生じる疾患状態に対して家禽を防御するのに使用するためのワクチンの製造方法であって、請求項10に記載の製造方法により得られ収穫されたレオウイルスを、所望により該レオウイルスの不活化後に、医薬上許容される担体または希釈剤と一緒にすることを含んでなる製造方法。
【請求項12】
家禽におけるトリレオウイルスの感染から生じる疾患状態を防除するための方法であって、請求項1〜4のいずれか1項に記載のワクチンを該鳥に投与することを含んでなる方法。
【図1】

【図2】
【図3】
【図2】
【図3】
【公開番号】特開2011−92190(P2011−92190A)
【公開日】平成23年5月12日(2011.5.12)
【国際特許分類】
【外国語出願】
【出願番号】特願2010−256592(P2010−256592)
【出願日】平成22年11月17日(2010.11.17)
【分割の表示】特願2000−15930(P2000−15930)の分割
【原出願日】平成12年1月25日(2000.1.25)
【出願人】(506196247)インターベツト・インターナシヨナル・ベー・ベー (85)
【Fターム(参考)】
【公開日】平成23年5月12日(2011.5.12)
【国際特許分類】
【出願番号】特願2010−256592(P2010−256592)
【出願日】平成22年11月17日(2010.11.17)
【分割の表示】特願2000−15930(P2000−15930)の分割
【原出願日】平成12年1月25日(2000.1.25)
【出願人】(506196247)インターベツト・インターナシヨナル・ベー・ベー (85)
【Fターム(参考)】
[ Back to top ]