
トルエン検出方法
【課題】前処理をすることなく、長期に亘って、安価且つ簡便にトルエンを検出することができるトルエン検出方法の提供。
【解決手段】嫌気性細菌にベクターを導入し、形質転換体を作製する工程と、前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する工程とを有するトルエン検出方法であって、前記ベクターは、トルエン分解酵素活性を有するタンパク質をコードする遺伝子と、その下流に位置し、レポータータンパク質をコードする遺伝子とが組み込まれたものであることを特徴とするトルエン検出方法。
【解決手段】嫌気性細菌にベクターを導入し、形質転換体を作製する工程と、前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する工程とを有するトルエン検出方法であって、前記ベクターは、トルエン分解酵素活性を有するタンパク質をコードする遺伝子と、その下流に位置し、レポータータンパク質をコードする遺伝子とが組み込まれたものであることを特徴とするトルエン検出方法。
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、嫌気性細菌を用いたトルエン検出方法に関する。
【背景技術】
【0002】
人体に対して健康被害を及ぼし得る有害物質による、土壌や地下水の汚染が問題となっている。近年では、土壌汚染対策法が施行され、土地所有者に対して、土壌汚染状況の把握、土壌汚染による人の健康被害の防止に関する措置等の土壌汚染対策の実施が課せられた。また、近年では地下水汚染判明事例件数が年間300件を超え、平成20年度末までの累積件数は5800件を上回っており、さらなる地下水汚染への対策が求められている。
しかしながら、特に埋没型タンクや地下配管等の地下設備からの有害物質の漏洩は発見が難しく、その上、影響の及ぶ範囲が拡大しやすいという問題がある。そのため、万が一有害物質が漏洩した場合に備え、土壌や地下水中の有害物質の定期的若しくは常時モニタリングが必要となる。
【0003】
上記のような有害物質の1つとして、トルエンがある。トルエンは地下水における要監視項目の1つとなっており、その指針値は0.6mg/L以下である。また、水道水の水質管理におけるトルエンの目標値は0.2mg/Lである。
トルエンは、ポリウレタン製造に用いられるトリレンジイソシアネートの原料や、フェノール、クレゾール等の原料として用いられるほか、印刷インキ、油性塗料、接着剤等の溶剤として用いられる。トルエンの輸入及び製造量は年間150万トン弱と、有害物質の中でも非常に多く、広範な分野で用いられることから、トルエンをモニタリングする方法に対する要求がある。
【0004】
現在、有害物質のモニタリングとしては、対象となる土壌、地下水等の試料から対象有害物質を抽出及び濃縮した後、ガスクロマトグラフ、液体クロマトグラフ、マススペクトロメーター等の装置を用いて検出する方法や、高分子薄膜素子(センサチップ)と光学的センシング技術(干渉増幅反射法)とを組み合わせたVOCセンサを用いて検出する方法等が用いられている。
また、特許文献1には、汚染浄化能力を有する嫌気性微生物を利用する地下水汚染の浄化方法であって、嫌気性微生物のための薬剤を投入すること、及び、地下水の総有機炭素量、pH、溶存酸素及び酸化還元電位を含む項目をモニタリングすることを含む、地下水汚染の浄化方法が開示されている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2008−296176号公報
【発明の概要】
【発明が解決しようとする課題】
【0006】
上記したような従来の有害物質モニタリング方法では、土壌、地下水等の試料から対象となる有害物質を抽出及び濃縮することが必要であるため、検出に手間がかかるという問題があった。また、検出には高価な装置が必要となるという問題もあった。
さらに、上記特許文献1に記載された発明は、地下水のモニタリング結果に応じて、嫌気性微生物を栄養源等により活性化し、該嫌気性微生物に有害物質を分解させることを目的とした発明であるが、地下水のモニタリングは従来公知の装置等を用いて行っており、モニタリングの手間やコストは依然として解決されていなかった。
【0007】
本発明は、上記問題点に鑑みてなされたものであって、前処理をすることなく、長期に亘って、安価且つ簡便にトルエンを検出することができるトルエン検出方法を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決すべく鋭意研究した結果、地下水等の酸素濃度の低い環境下においても生存可能である嫌気性細菌をバイオセンサとして用いることにより、前処理をすることなく、長期に亘って、安価且つ簡便にトルエンを検出することができることを見出し、本発明を完成させた。
すなわち、本発明は下記の特徴を有するトルエン検出方法を提供する。
(1)嫌気性細菌にベクターを導入し、形質転換体を作製する工程と、前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する工程とを有するトルエン検出方法であって、前記ベクターは、トルエン分解酵素活性を有するタンパク質をコードする遺伝子と、その下流に位置し、レポータータンパク質をコードする遺伝子とが組み込まれたものであることを特徴とするトルエン検出方法。
(2)前記嫌気性細菌が、Azoarcus sp.(アゾアルカス・スピーシーズ)であることを特徴とする(1)のトルエン検出方法。
(3)前記嫌気性細菌及び形質転換体が、D−リボース、Tween80、D−アラビノース、L−アラビノース、D−キシロース、D−プシコース、α−ケドグルタル酸、マンノース、D−タガトース、D−フルクトース−6−リン酸、グルクロンアミド、D−フルクトース、ラムノース、D−ガラクトース、D−グルクロン酸、グリセロール、乳酸、酵母抽出液、カサミノ酸、NBブロス、及びLBブロスからなる群から選ばれる1種以上を含む培地で培養されることを特徴とする(1)又は(2)のトルエン検出方法。
(4)前記レポータータンパク質がβ−ガラクトシダーゼであり、前記検出化合物が、ルシフェラーゼを含む検出化合物であることを特徴とする(1)〜(3)のいずれかのトルエン検出方法。
(5)前記嫌気性細菌へのベクターの導入を、接合伝達法を用いて行うことを特徴とする(1)〜(4)のいずれかのトルエン検出方法。
【発明の効果】
【0009】
本発明のトルエン検出方法は、嫌気性微生物を用いてトルエンの有無を視覚的に検出する方法であるため、試料の前処理、高価な装置及び熟練した技能を必要とせず、安価且つ簡便にトルエンを検出することができる。また、嫌気性微生物は、地下水等の嫌気性環境下において自活することができるため、メンテナンスやエネルギー補給を必要とせず、長期に亘ったトルエンの検出が可能となる。
【図面の簡単な説明】
【0010】
【図1】参考例1における、炭素源の違いによるアゾアルカス・スピーシーズ DSM9506株の生育量を示すグラフ図である。
【図2】参考例1における、乳酸含有量の違いによるアゾアルカス・スピーシーズ DSM9506株の生育量を示すグラフ図である。
【図3】参考例1における、培養開始時の菌体数の違いによるアゾアルカス・スピーシーズ DSM9506株の生育量を示すグラフ図である。
【図4】実施例1の(8)における、トルエン濃度と、pBBR1−トルエン−β−ガラクトシダーゼベクターを導入したDSM9506株由来の青色色素発光量との関係を示すグラフ図である。
【図5】実施例1の(9)における、トルエン濃度と、pBBR1−トルエン−β−ガラクトシダーゼベクターを導入したDSM9506株由来のルシフェラーゼ発光量との関係を示すグラフ図である。
【発明を実施するための形態】
【0011】
本発明のトルエン検出方法は、嫌気性細菌にベクターを導入し、形質転換体を作製する工程(以下、「形質転換体作製工程」という)と、前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する工程(以下、「検出工程」という。)と、を有する。
以下、工程ごとに説明する。
【0012】
[形質転換体作製工程]
まず、形質転換体作成工程において、嫌気性細菌にベクターを導入し、形質転換体を作製する。
本発明において、嫌気性細菌としては特に限定されるものではないが、トルエン分解酵素活性を有するタンパク質を内在的に有する嫌気性細菌(以下、「トルエン分解嫌気性細菌」という。)であることが好ましい。本発明では、嫌気性細菌を用いることにより、地下水等の嫌気性環境下において該菌が自活することができ、エネルギー等を補給する必要がない。また、トルエン分解嫌気性細菌を用いる場合、後述するベクターの導入や、本願発明に必要な形質転換体の作製が容易であるため好ましい。
トルエン分解嫌気性細菌は、公知のトルエン分解嫌気性細菌を用いてもよく、新たにトルエン分解嫌気性細菌を単離してもよい。トルエン分解嫌気性細菌を単離する方法は特に限定されるものではなく、トルエンを含有する培地において嫌気性細菌を培養し、スクリーニングを行う等の公知の方法により行うことができる。
このような嫌気性細菌として具体的には、アゾアルカス・スピーシーズ、タウエラ・アロマティカ等が挙げられ、より具体的には、アゾアルカス・スピーシーズ DSM9506株、タウエラ・アロマティカ K172株、タウエラ・アロマティカ T1株が挙げられる。なかでも、本発明の嫌気性細菌としては、アゾアルカス・スピーシーズであることが好ましく、アゾアルカス・スピーシーズ DSM9506株であることがより好ましい。
【0013】
本発明においてベクターは、トルエン分解酵素活性を有するタンパク質をコードする遺伝子(以下、「トルエン分解酵素遺伝子」という。)と、その下流に位置し、レポータータンパク質をコードする遺伝子とが組み込まれたものである。
【0014】
トルエン分解酵素遺伝子としては、特に限定されるものではなく、公知のトルエン分解酵素遺伝子を人工的に合成したものであってもよく、トルエン分解酵素遺伝子を有する生物からクローニングしたものであってもよい。また、トルエン分解酵素遺伝子は、該遺伝子のみの全配列であってもよく、該遺伝子上流側の部分配列のみであってもよく、該遺伝子と、上流側から該遺伝子を制御する制御遺伝子とを含む配列やその部分配列であってもよい。
トルエン分解酵素遺伝子を人工的に合成する場合、その方法は特に限定されるものではなく、公知の化学合成法を用いて行うことができる。また、合成するトルエン分解酵素遺伝子としては、2種以上の遺伝子をつなぎ合わせたキメラ遺伝子であってもよく、いずれかの生物種が有する遺伝子を変異させた変異遺伝子であってもよい。変異としては、遺伝子を構成する塩基配列の一部を欠損させてもよく、一部を置換してもよく、新たな配列を挿入してもよい。
トルエン分解酵素遺伝子をクローニングする場合、その方法は特に限定されるものではなく、目的の遺伝子やDNAを分離する際に用いられる公知のいずれの方法を用いてもよい。このようなクローニング方法として、例えば、試料中のトルエン分解酵素遺伝子を含むDNA領域の、両末端の塩基配列と相補的な塩基配列を有するプライマーを用いて、ポリメラーゼ連鎖反応(PCR)法によりクローニングを行うことができる。このようにしてクローニングされたトルエン分解酵素遺伝子を含むDNA断片は、電気泳動装置やDNAシークエンサー等の公知慣用の装置を用いることにより、目的としたDNA断片であるか否かを判別することができる。なお、クローニングするトルエン分解酵素遺伝子は、ベクターを導入する嫌気性細菌で発現し得るものであればその由来は特に限定されるものではなく、同種の嫌気性細菌からクローニングしたものであってもよく、当該嫌気性細菌以外の生物種からクローニングしたものであってもよい。
【0015】
レポータータンパク質としては、後述する検出化合物を用いて検出可能なものであれば特に限定されるものではなく、β−ガラクトシダーゼ、GFP、BFP、RFP、CFP、ルシフェラーゼ、evoglow(商品名、フナコシ社製)、β−グルクロニダーゼ(GUS)、クロラムフェニコールアセチルトランスフェラーゼ、EGFP(増強緑色蛍光タンパク質)、アルカリ性ホスファターゼ、DsRed、ペルオキシダーゼ、クロラムフェニコール・アセチルトランスフェラーゼ(CAT)等が挙げられる。
なかでも、本発明におけるレポータータンパク質としては、β−ガラクトシダーゼであることが好ましい。
【0016】
本発明においてベクターは、上記のようなトルエン分解酵素遺伝子と、レポータータンパク質をコードする遺伝子とが組み込まれたものであれば特に限定されるものではなく、公知のプラスミドベクター等を用いることができる。このようなプラスミドベクターとしては、上記遺伝子の他に、プロモーター配列、抗生物質耐性遺伝子等を有する公知のものを好適に用いることができる。
【0017】
本発明において、嫌気性細菌にベクターを導入し、形質転換体を作製する方法は特に限定されるものではなく、化学的処理によりベクターを導入する化学法、パルス電圧によってベクターを導入する電子穿孔法、カーボンナノチューブを用いて細胞膜に孔を開けベクターを導入する方法、嫌気性細菌間の遺伝子伝播現象を利用した接合伝達法等を用いることができ、なかでもより確実にベクターを導入できることから、接合伝達法が好ましい。
導入するベクターが抗生物質耐性遺伝子等の選択可能な遺伝子を有する場合は、上記のようにして嫌気性細菌にベクターを導入した後、当該抗生物質を含む培地にて嫌気性細菌を培養することにより、ベクターの導入された形質転換体のみを選択することができる。
【0018】
本発明において、嫌気性細菌及び形質転換体を培養する方法は、該細菌が好適に生育するものであれば特に限定されるものではない。例えば具体的には、嫌気性細菌がアゾアルカス・スピーシーズ DSM9506株である場合には、リン酸二水素カリウム又は塩化アンモニウム、塩化マグネシウム、硝酸ナトリウム等を含む嫌気性細菌の基本培地に、さらに、D−リボース、Tween80、D−アラビノース、L−アラビノース、D−キシロース、D−プシコース、α−ケドグルタル酸、マンノース、D−タガトース、D−フルクトース−6−リン酸、グルクロンアミド、D−フルクトース、ラムノース、D−ガラクトース、D−グルクロン酸、グリセロール、乳酸、酵母抽出液、カサミノ酸、NBブロス、及びLBブロスからなる群から選ばれる1種以上を含む培地で培養することが好ましい。
【0019】
本発明では、上記のようなベクターを嫌気性細菌に導入することにより、嫌気性細菌がトルエン分解酵素活性を有するタンパク質を発現する必要性が生じた際、具体的には、嫌気性細菌がトルエンの存在下にある際に、トルエン分解酵素遺伝子と共に、レポータータンパク質をコードする遺伝子が発現する。そのため、後述する検出工程において、レポータータンパク質を何らかの形で検出することにより、嫌気性細菌の存在環境におけるトルエンの有無を感知することが可能となる。
【0020】
[検出工程]
次に、前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する。
本発明において、試料としては特に限定されるものではなく、例えば、トルエンの存在が疑われる土壌、地下水、大気等が挙げられる。
【0021】
本発明において、検出化合物は、レポータータンパク質により発色又は発光する水溶性部位を有する。
発色又は発光する水溶性部位(水溶性の色素又は蛍光物質)としては、特に限定されるものではなく、色素又は蛍光の検出に通常用いられるもののうち、水溶性のものを用いることができる。例えば、ルシフェラーゼ等の蛍光物質や、β−フラノシド基質やカテコール基質のような発色物質を用いることができる。
本発明において、検出化合物としては、例えばレポータータンパク質がβ−ガラクトシダーゼの場合、検出化合物として、β−ガラクトピラノシドと、上記のような水溶性の色素又は蛍光物質との融合化合物を用いることができる。この場合、トルエン存在下の形質転換体において、トルエン分解酵素遺伝子と共にβ−ガラクトシダーゼ遺伝子が発現し、生産されたβ−ガラクトシダーゼが、検出化合物中のβ−ガラクトピラノシド部分を分解することにより、β−ガラクトピラノシドに結合されていた水溶性の色素又は蛍光物質が発色又は発光する。
このような検出化合物として具体的には、o−Nitrophenyl−β−D−galactopyranoside(ONPG)、3’,4’−Dihydroxyflavone−4’−β−D−galactopyranoside(DHF−Gal)のような色素を用いた検出化合物、6−O−β−galactopyranosyl−luciferin(Luc−Gal)のような蛍光を用いた検出化合物等が挙げられる。なかでも、試料中のトルエン含有量を定量する際の精度が高いことから、ルシフェラーゼを含む検出化合物、具体的には上記Luc−Galを用いることが好ましい。
本発明では、検出化合物の発色又は発光する部位として水溶性のものを用いることで、該発色又は発光化合物が嫌気性細菌に吸着することがないため、精度よく試料中のトルエン含有量を定量することができる。
【0022】
本発明において、形質転換体と、上記検出化合物とを用いてトルエン含有量を定量する方法は特に限定されるものではないが、例えば、試料と、形質転換体及び検出化合物とを有する培地とを接触させ、検出化合物の発色又は発光の強度を、分光光度計、蛍光光度計等の公知の装置によって測定し、検量線を用いることで、トルエン含有量の定量を行うことができる。検量線は、既知量のトルエンを含有する試料を、同様の形質転換体、同様の検出化合物、及び同様の装置を用いて測定することにより作製することができる。
【0023】
また、本発明のトルエン検出方法では、検出工程において、試料中のトルエン含有量の定量を行う前に、さらに、トルエンの有無のみの検出や、トルエン含有量の半定量等の簡易検出を行ってもよい。
トルエンの簡易検出の方法としては特に限定されるものではないが、例えば、上記検出化合物を、検量線を用いずにその発色又は発光のみを検出してもよい。また、レポータータンパク質が単体で発色又は発光する物質、例えばEGFP等である場合には、検出化合物を介さずに、該発色又は発光を分光又は蛍光光度計により直接検出し、発色や発光の有無により簡易検出を行うことができる。さらに、レポータータンパク質がβ−ガラクトシダーゼの場合には、5−ブロモ−4−クロロ−3−インドリル−β−D−ガラクトピラノシド(X−gal)等の水溶性ではないが一般的且つ簡便な検出化合物を用いて、簡易検出を行うこともできる。
【0024】
本発明のトルエン検出方法によれば、例えば、嫌気性細菌にベクターを導入した形質転換体や検出化合物を、離散しないように担体等の表面に固着させ、これを地下水中に静置して、該地下水中のトルエン汚染の有無及びトルエン含有量を検出することができる。
また、例えば、地下水を定期的又は恒常的に、観測井戸等からポンプ等を用いて汲み上げ、汲み上げた地下水試料と、嫌気性細菌にベクターを導入した形質転換体と、検出化合物とを接触させ、分光光度計や蛍光光度計を用いてトルエン汚染の有無及びトルエン含有量を検出することができる。
【実施例】
【0025】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。なお、以下の実施例中、特に記載がない限り、「%」は「質量%」を意味する。
【0026】
[参考例1]
(アゾアルカス・スピーシーズ DSM9506株の培養条件の検討)
まず、アゾアルカス・スピーシーズ DSM9506株(以下、単に「DSM9506株」という。)の利用可能な炭素源について検討した。
なお、DSM9506株は嫌気性細菌であるため、以下の検討において、DSM9506株の植菌や培地調整等の作業は、酸素濃度が大気中の10万分の1程度である嫌気グローブボックス内で行い、培養は酸素を通さないブチルゴム栓で密閉したガラス容器内で行った。また、ガラス容器には、酸素除去剤として、酸化チタン−クエン酸ナトリウム溶液、硫化ナトリウム九水和物を添加し、嫌気環境を維持した上で、嫌気指示薬であるレサズリンを用いて嫌気環境であることを確認し、指示薬の結果から嫌気環境が損なわれたと考えられる試料は破棄した。
96ウェルマイクロプレートの各ウェル内に、95種の炭素源となる化合物が充填されたマイクロプレート(商品名、バイオログ社製)を用いて検討したところ、DSM9506株は、D−リボース、Tween80、D−アラビノース、L−アラビノース、D−キシロース、D−プシコース、α−ケドグルタル酸、マンノース、D−タガトース、D−フルクトース−6−リン酸、グルクロンアミド、D−フルクトース、ラムノース、D−ガラクトース、又は、D−グルクロン酸、を炭素源として利用可能であることが分かった。
【0027】
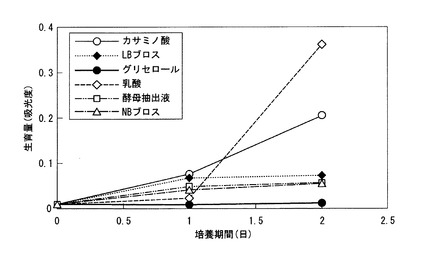
また、上記95種類以外の化合物についても炭素源となり得るか否かを個別に検討したところ、グリセロール、乳酸、酵母抽出液、カサミノ酸、NBブロス、又はLBブロスの6種の化合物も炭素源として利用可能であることが分かった。これら6種の化合物を炭素源とした場合の、DSM9506株の吸光度により測定した生育量を図1に示す。図1中、横軸が培養期間(日)、縦軸が660nmにおける吸光度である。
図1の結果から、乳酸、及びカサミノ酸を炭素源とした場合にDSM9506株の生育が良好となることが分かった。そこで、以下の検討では、乳酸を炭素源とした。
【0028】
次に、炭素源として乳酸を用いた場合の、乳酸濃度によるDSM9506株の生育量について検討した。
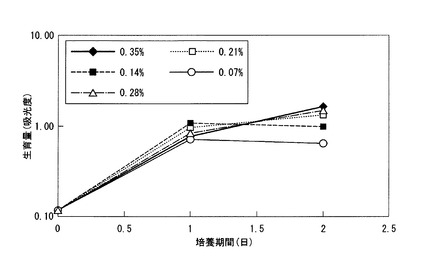
乳酸濃度を0.07%、0.14%、0.21%、0.28%、0.35%にそれぞれ調整した基本培地(リン酸二水素カリウム又は塩化アンモニウム、塩化マグネシウム、及び硝酸ナトリウムを含む培地)において、DSM9506株を48時間培養し、DSM9506株の生育量を培養開始24時間後及び48時間後に、吸光度により測定した。結果を図2に示す。図2中、横軸が培養期間(日)であり、縦軸が660nmにおける吸光度である。
図2の結果から、48時間後においては、乳酸濃度が高い程生育が良いものの、24時間後においては乳酸濃度0.14%の培地が最も生育が良かった。これは、乳酸が高濃度である場合、該乳酸が増殖初期段階においてDSM9506株の生育に悪影響を及ぼすためと考えられる。そのため、48時間後において、最も乳酸濃度の高い0.35%の培地と得られる菌体量に大差がなく、且つ、増殖初期である24時間後においても生育が良好である乳酸濃度0.21%を以下の検討において採用することとした。
【0029】
次いで、無機塩の種類によるDSM9506株の生育量について検討した。
培地1L中にリン酸二水素カリウム0.52g(KH2PO4)、塩化アンモニウム0.2g(NH4Cl)、塩化マグネシウム六水和物0.4g(MgCl2−6H2O)及び硝酸ナトリウム0.2g(NaNO3)を含む基本培地に、乳酸0.21%を添加し、さらにリン酸水素二カリウム1g(K2HPO4)又は塩化カルシウム二水和物(CaCl2−2H2O)0.1gを添加して、該培地におけるDSM9506株の生育量を調べた。
その結果、リン酸二水素カリウム及び塩化カルシウムのいずれかを添加することにより若干の生育量の向上が認められたが、これら2種の間に大きな効果の差は認められなかった。そのため、以下の検討では、上記基本培地に、乳酸及び塩化カルシウムを添加した培地を用いることとした。
【0030】
さらに、微生物種によっては、培養開始時の菌体数が少ない場合に生育しないおそれがあるため、培養開始時の菌体数が生育に与える影響について検討した。
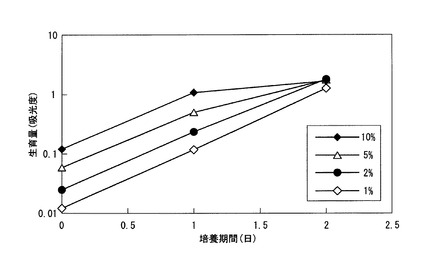
DSM9506株を、リン酸二水素カリウム0.52g(KH2PO4)、塩化アンモニウム0.2g(NH4Cl)、塩化マグネシウム六水和物0.4g(MgCl2−6H2O)及び硝酸ナトリウム0.2g(NaNO3)、乳酸0.21%を1L中に含む培地で24時間前培養を行った。その後、前培養液を1〜10%の範囲で量を変えて本培養の培地中に添加し、生育量を調べた。結果を図3に示す。なお、本培養の培地組成は前培養と同様である。
この結果、24時間後では培養開始時の添加菌体数に比例して生育量が多いが、48時間後には添加菌体数の多寡に関わらずほぼ同程度の生育量となった。いずれの菌体添加量であっても24時間後に菌体量が10倍となっており、初期添加量が1〜10%の範囲内であれば生育速度に差がないことが分かった。そのため、以下の検討では、菌体の初期添加量を10%とした。
【0031】
[実施例1]
(1)トルエン分解酵素遺伝子を含むDNA断片のクローニング
まず、DSM9506株において公知となっているトルエン分解酵素遺伝子(Genbankアクセッション番号:AY032676)の146番目〜3362番目の塩基配列(3217bp)の両末端と相補的な塩基配列を有するプライマーを設計した。そして、常法により抽出したDSM9506株のゲノムDNAをテンプレートとして、上記プライマーとKOD−Plus(商品名、東洋紡績(株)社製)とを用いて、PCR法によりトルエン分解酵素遺伝子を含むDNA断片のPCR産物を得た。用いたプライマーの配列を、Primer1(forward;配列番号1)とPrimer2(reverse;配列番号2)として表1に示す。
アガロース電気泳動を用いて、得られたPCR産物の全長を確認したところ、目的産物と同等の3200bp程度のPCR産物が得られていることが確認できた。また、PCR産物の一部の配列についてDNAシークエンサーを用いて解析を行ったところ、目的のトルエン分解酵素遺伝子を含むDNA断片が得られていることが確認できた。
【0032】
(2)レポータータンパク質をコードする遺伝子を含むDNA断片のクローニング
レポータータンパク質であるEGFP、β−ガラクトシダーゼをコードする遺伝子を含むDNA断片をクローニングした。
EGFPは、プラスミドベクターであるpEGFP(商品名、クロンテック社製)をテンプレートとして、KOD−Plus(商品名、東洋紡績(株)社製)、並びに下記プライマーPrimer3(forward;配列番号3)及びPrimer4(reverse;配列番号4)を用いて、PCR増幅を行い、産物であるEGFP遺伝子断片を得た。
β−ガラクトシダーゼは、プラスミドベクターであるpSV−β−Galactosidase Vector(商品名、プロメガ社製)をテンプレートとして、KOD−Plus(商品名、東洋紡績(株)社製)、並びに下記プライマー、Primer5(forward;配列番号5)及びPrimer6(reverse;配列番号6)を用いて、PCR増幅を行い、産物であるβ−ガラクトシダーゼ遺伝子断片(β−Gal)を得た。
得られたPCR産物をアガロース電気泳動にて確認したところ、PCR産物はいずれも目的の産物長(EGFP:850bp、βガラクトシダーゼ:3100bp)であったため、EGFP、β−ガラクトシダーゼ共に目的のレポータータンパク質遺伝子断片であることが確認できた。
【0033】
【表1】

【0034】
(3)発現ベクターの調整
次いで、上記(1)で得られたトルエン分解酵素遺伝子断片と、上記(2)で得られたレポータータンパク質遺伝子断片とを、それぞれ制限酵素処理した後、ラピッドDNAライゲーションキット(商品名、ロッシュ社製)を用いて連結した。以下、得られたEGFPを含む遺伝子断片をトルエン−EGFP、β−ガラクトシダーゼを含む遺伝子断片をトルエン−β−ガラクトシダーゼという。これら遺伝子断片は、アガロース電気泳動により目的の産物であることを確認した。
上記のようにして得られたトルエン−EGFPと、トルエン−β−ガラクトシダーゼをApaI及びSpeIで処理した後、同じくApaI及びSpeIで処理したプラスミドDNAの発現ベクターであるpBBR1MCS2(Kovachら、Gene、166、1995年)に、ラピッドDNAライゲーションキット(商品名、ロッシュ社製)を用いて組み込んだ。得られたpBBR1−トルエン−EGFP、及びpBBR1−トルエン−β−ガラクトシダーゼは、ApaI及びSpeIで処理した後アガロース電気泳動を行い、目的の発現ベクターであることを確認した。また、これらの発現ベクターは、Escherichia coli(大腸菌)JM109株にハナハン法を用いて形質転換し、増幅及び保存を行った。
【0035】
(4)形質転換体の作製
上記参考例1で検討した条件において培養を行ったDSM9506株に対し、化学法、電子穿孔法、カーボンナノチューブを用いて細胞膜に孔を開けベクターを導入する方法、及びEscherichia coli(大腸菌)S17−1株を用いた接合伝達法のいずれかを用いて、pBBR1−トルエン−EGFPベクター、又はpBBR1−トルエン−β−ガラクトシダーゼベクターの導入を試みた。その後、pBBR1ベクター内に存在するカナマイシン耐性遺伝子を利用して、DSM9506株の培地にカナマイシンを(50mg/L)で添加し、形質転換体の選択を行った。
その結果、接合伝達法を用いた場合に、最も効率よく形質転換体が製造できることが分かった。以下の検討では、接合伝達法により得られた形質転換体を用いた。
【0036】
(5)pBBR1−トルエン−EGFPベクターを用いた簡易検出
pBBR1−トルエン−EGFPベクターを導入したDSM9506株の形質転換体を栄養培地(DIFCO Nutrient Broth 4g/L、カナマイシン 50mg/L)で増殖させた後、遠心分離器を用いて回収し、上記基本培地で洗浄後、トルエンの簡易検出を行った。
まず、トルエン(1mg/L)を添加した上記基本培地30mLに形質転換体を移し、嫌気条件下で培養を行ったところ、形質転換体が緑色の蛍光を発することが確認できた。また、この緑色蛍光を発する形質転換体を好気環境下に移すと蛍光が消え、その後嫌気環境下に戻すと再度蛍光を発することが確認できた。
この結果から、形質転換体では、設計通りにトルエンが存在する嫌気環境下においてEGFPが産生されていることが確認できた。また、トルエンが分解できない大気中(好気環境下)に移すことで蛍光が消失することは、EGFPが長期間存在し得ないことを示しており、つまり、形質転換体周囲のトルエンが全て分解されてトルエン汚染がなくなれば蛍光も消失すると考えられる。
よって、EGFP等に由来する緑色蛍光の有無を指標としてトルエンの存在を検知することができると考えられた。
【0037】
(6)pBBR1−トルエン−β−ガラクトシダーゼベクターを用いた簡易検出
次に、pBBR1−トルエン−EGFPベクターを導入したDSM9506株の形質転換体を栄養培地(DIFCO Nutrient Broth 4g/L、カナマイシン 50mg/L)で増殖させた後、遠心分離器を用いて回収し、上記基本培地で洗浄後、トルエンの簡易検出を行った。
まず、トルエン(1mg/L)又はベンゼン(1mg/L)を添加した上記基本培地30mLに、形質転換体、及び発色基質であるX−gal(200ug/L)を加えて静置培養した。1週間後に観察したところ、トルエンを添加した培地では、β−ガラクトシダーゼの作用によりX−galから青色の色素が遊離し、培地が青色となった。一方、ベンゼンを添加した培地では、培地の色の変化はなかった。
この結果から、形質転換体では、設計通りにトルエンが存在する環境下においてβ−ガラクトシダーゼが産生されていることが確認できた。よって、β−ガラクトシダーゼのX−gal分解に由来する青色色素の有無を指標として、トルエンの存在を検知することができると考えられた。
【0038】
また、増殖させた上記DSM9506株の形質転換体を、上記同様にトルエン及びX−galを含む基本培地に懸濁し、旋回式振とう培養装置を用いて振とう培養を行ったところ、トルエンによる青色色素の誘導は認められなかった。これは、振とう培養によって酸素濃度が上昇したためにトルエン分解が妨げられ、その結果、レポータータンパク質であるβ−ガラクトシダーゼが発現しなくなったためと考えられる。
次いで、増殖させた上記DSM9506株の形質転換体を、上記同様にトルエン及びX−galを添加した基本培地に添加し、さらに、各培地にタンパク質合成阻害剤(クロラムフェニコール)を50mg/L添加して1週間静置したところ、青色色素は認められなかった。この結果から、青色色素の発色がβ−ガラクトシダーゼ遺伝子の発現の結果であることが確認できた。よって、β−ガラクトシダーゼとレポータータンパク質とをコードする遺伝子を有する発現ベクターを用いることで、嫌気環境下でのトルエン検出が可能であることが確認できた。
【0039】
(8)トルエン含有量の定量−1
トルエンによって誘導される青色色素の生産量と、トルエン濃度との関係について調べた。
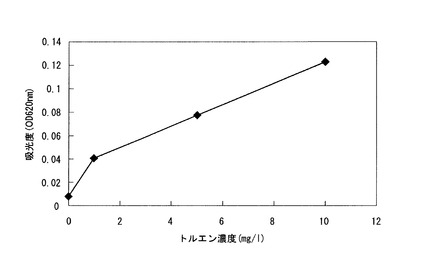
まず、トルエンを0、1、5、又は10mg/Lの濃度で含有する培地を上記(7)と同様に作製し、pBBR1−トルエン−β−ガラクトシダーゼベクターを導入したDSM9506株を植菌した。1週間静置培養した後、青色色素の吸光度(OD620nm)を分光光度計(ベックマン社製DU650)により測定し、トルエン濃度と青色色素量との関係について検討した。結果を図4に示す。
図4の結果から、トルエン濃度に比例して色素生産量も高くなることが明らかである。しかしながら、X−gal由来の青色色素(5−ブロモ−4−クロロ−3−インドール)は、水溶性でなく、DSM9506株の表面に吸着するため、吸光度が必ずしも色素量を正確に反映していないと考えられる。
【0040】
(9)トルエン含有量の定量−2
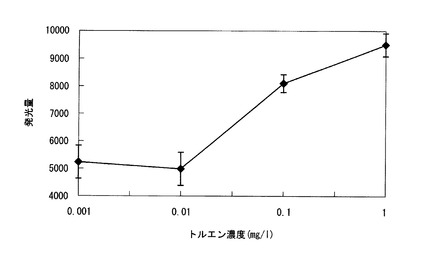
そこで、β−ガラクトシダーゼにより発色又は発光する水溶性部位を有する3種の化合物をX−galに変えて用い、0.001、0.01、0.1、又は1mg/Lのトルエンを含む上記同様の培地にて上記(7)と同様に検討を行った。3種の化合物としては、o−Nitrophenyl−β−D−galactopyranoside(ONPG)、3’,4’−Dihydroxyflavone−4’−β−D−galactopyranoside(DHF−Gal)、及び6−O−β−galactopyranosyl−luciferin(Luc−Gal)を用い、ONPGは420nm、DHF−Galは620nmにおいて上記同様の分光光度計を用い、Luc−Galはルミテスター K−200(商品名、キッコーマン社製)を用いて発色又は発光を定量化した。
その結果、β−ガラクトシダーゼにより発色するONPG、DHF−Galの発色よりも、β−ガラクトシダーゼにより発光するLuc−Galにおいて安定した発光が得られた。Luc−Galの定量結果を図5に示す。
図5の結果から、Luc−Galを検出化合物として用いた場合、トルエン濃度が0.001mg/Lと非常に低濃度であっても、発光が確認できることが分かった。また、その発光量は、トルエン濃度が0.01〜1mg/Lの範囲でトルエン濃度に比例して強くなり、定量が可能であることが確認できた。
これらの結果から、嫌気性細菌にベクターを導入した形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いることで、試料中のトルエンの有無を検出できるのみならず、トルエン含有量を定量できることが確認できた。
【産業上の利用可能性】
【0041】
本発明のトルエン検出方法によれば、地下水や土壌等のトルエンを安価且つ簡便に検出や定量することができるため、環境分野において好適に利用することができる。
【技術分野】
【0001】
本発明は、嫌気性細菌を用いたトルエン検出方法に関する。
【背景技術】
【0002】
人体に対して健康被害を及ぼし得る有害物質による、土壌や地下水の汚染が問題となっている。近年では、土壌汚染対策法が施行され、土地所有者に対して、土壌汚染状況の把握、土壌汚染による人の健康被害の防止に関する措置等の土壌汚染対策の実施が課せられた。また、近年では地下水汚染判明事例件数が年間300件を超え、平成20年度末までの累積件数は5800件を上回っており、さらなる地下水汚染への対策が求められている。
しかしながら、特に埋没型タンクや地下配管等の地下設備からの有害物質の漏洩は発見が難しく、その上、影響の及ぶ範囲が拡大しやすいという問題がある。そのため、万が一有害物質が漏洩した場合に備え、土壌や地下水中の有害物質の定期的若しくは常時モニタリングが必要となる。
【0003】
上記のような有害物質の1つとして、トルエンがある。トルエンは地下水における要監視項目の1つとなっており、その指針値は0.6mg/L以下である。また、水道水の水質管理におけるトルエンの目標値は0.2mg/Lである。
トルエンは、ポリウレタン製造に用いられるトリレンジイソシアネートの原料や、フェノール、クレゾール等の原料として用いられるほか、印刷インキ、油性塗料、接着剤等の溶剤として用いられる。トルエンの輸入及び製造量は年間150万トン弱と、有害物質の中でも非常に多く、広範な分野で用いられることから、トルエンをモニタリングする方法に対する要求がある。
【0004】
現在、有害物質のモニタリングとしては、対象となる土壌、地下水等の試料から対象有害物質を抽出及び濃縮した後、ガスクロマトグラフ、液体クロマトグラフ、マススペクトロメーター等の装置を用いて検出する方法や、高分子薄膜素子(センサチップ)と光学的センシング技術(干渉増幅反射法)とを組み合わせたVOCセンサを用いて検出する方法等が用いられている。
また、特許文献1には、汚染浄化能力を有する嫌気性微生物を利用する地下水汚染の浄化方法であって、嫌気性微生物のための薬剤を投入すること、及び、地下水の総有機炭素量、pH、溶存酸素及び酸化還元電位を含む項目をモニタリングすることを含む、地下水汚染の浄化方法が開示されている。
【先行技術文献】
【特許文献】
【0005】
【特許文献1】特開2008−296176号公報
【発明の概要】
【発明が解決しようとする課題】
【0006】
上記したような従来の有害物質モニタリング方法では、土壌、地下水等の試料から対象となる有害物質を抽出及び濃縮することが必要であるため、検出に手間がかかるという問題があった。また、検出には高価な装置が必要となるという問題もあった。
さらに、上記特許文献1に記載された発明は、地下水のモニタリング結果に応じて、嫌気性微生物を栄養源等により活性化し、該嫌気性微生物に有害物質を分解させることを目的とした発明であるが、地下水のモニタリングは従来公知の装置等を用いて行っており、モニタリングの手間やコストは依然として解決されていなかった。
【0007】
本発明は、上記問題点に鑑みてなされたものであって、前処理をすることなく、長期に亘って、安価且つ簡便にトルエンを検出することができるトルエン検出方法を提供することを課題とする。
【課題を解決するための手段】
【0008】
本発明者らは、上記課題を解決すべく鋭意研究した結果、地下水等の酸素濃度の低い環境下においても生存可能である嫌気性細菌をバイオセンサとして用いることにより、前処理をすることなく、長期に亘って、安価且つ簡便にトルエンを検出することができることを見出し、本発明を完成させた。
すなわち、本発明は下記の特徴を有するトルエン検出方法を提供する。
(1)嫌気性細菌にベクターを導入し、形質転換体を作製する工程と、前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する工程とを有するトルエン検出方法であって、前記ベクターは、トルエン分解酵素活性を有するタンパク質をコードする遺伝子と、その下流に位置し、レポータータンパク質をコードする遺伝子とが組み込まれたものであることを特徴とするトルエン検出方法。
(2)前記嫌気性細菌が、Azoarcus sp.(アゾアルカス・スピーシーズ)であることを特徴とする(1)のトルエン検出方法。
(3)前記嫌気性細菌及び形質転換体が、D−リボース、Tween80、D−アラビノース、L−アラビノース、D−キシロース、D−プシコース、α−ケドグルタル酸、マンノース、D−タガトース、D−フルクトース−6−リン酸、グルクロンアミド、D−フルクトース、ラムノース、D−ガラクトース、D−グルクロン酸、グリセロール、乳酸、酵母抽出液、カサミノ酸、NBブロス、及びLBブロスからなる群から選ばれる1種以上を含む培地で培養されることを特徴とする(1)又は(2)のトルエン検出方法。
(4)前記レポータータンパク質がβ−ガラクトシダーゼであり、前記検出化合物が、ルシフェラーゼを含む検出化合物であることを特徴とする(1)〜(3)のいずれかのトルエン検出方法。
(5)前記嫌気性細菌へのベクターの導入を、接合伝達法を用いて行うことを特徴とする(1)〜(4)のいずれかのトルエン検出方法。
【発明の効果】
【0009】
本発明のトルエン検出方法は、嫌気性微生物を用いてトルエンの有無を視覚的に検出する方法であるため、試料の前処理、高価な装置及び熟練した技能を必要とせず、安価且つ簡便にトルエンを検出することができる。また、嫌気性微生物は、地下水等の嫌気性環境下において自活することができるため、メンテナンスやエネルギー補給を必要とせず、長期に亘ったトルエンの検出が可能となる。
【図面の簡単な説明】
【0010】
【図1】参考例1における、炭素源の違いによるアゾアルカス・スピーシーズ DSM9506株の生育量を示すグラフ図である。
【図2】参考例1における、乳酸含有量の違いによるアゾアルカス・スピーシーズ DSM9506株の生育量を示すグラフ図である。
【図3】参考例1における、培養開始時の菌体数の違いによるアゾアルカス・スピーシーズ DSM9506株の生育量を示すグラフ図である。
【図4】実施例1の(8)における、トルエン濃度と、pBBR1−トルエン−β−ガラクトシダーゼベクターを導入したDSM9506株由来の青色色素発光量との関係を示すグラフ図である。
【図5】実施例1の(9)における、トルエン濃度と、pBBR1−トルエン−β−ガラクトシダーゼベクターを導入したDSM9506株由来のルシフェラーゼ発光量との関係を示すグラフ図である。
【発明を実施するための形態】
【0011】
本発明のトルエン検出方法は、嫌気性細菌にベクターを導入し、形質転換体を作製する工程(以下、「形質転換体作製工程」という)と、前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する工程(以下、「検出工程」という。)と、を有する。
以下、工程ごとに説明する。
【0012】
[形質転換体作製工程]
まず、形質転換体作成工程において、嫌気性細菌にベクターを導入し、形質転換体を作製する。
本発明において、嫌気性細菌としては特に限定されるものではないが、トルエン分解酵素活性を有するタンパク質を内在的に有する嫌気性細菌(以下、「トルエン分解嫌気性細菌」という。)であることが好ましい。本発明では、嫌気性細菌を用いることにより、地下水等の嫌気性環境下において該菌が自活することができ、エネルギー等を補給する必要がない。また、トルエン分解嫌気性細菌を用いる場合、後述するベクターの導入や、本願発明に必要な形質転換体の作製が容易であるため好ましい。
トルエン分解嫌気性細菌は、公知のトルエン分解嫌気性細菌を用いてもよく、新たにトルエン分解嫌気性細菌を単離してもよい。トルエン分解嫌気性細菌を単離する方法は特に限定されるものではなく、トルエンを含有する培地において嫌気性細菌を培養し、スクリーニングを行う等の公知の方法により行うことができる。
このような嫌気性細菌として具体的には、アゾアルカス・スピーシーズ、タウエラ・アロマティカ等が挙げられ、より具体的には、アゾアルカス・スピーシーズ DSM9506株、タウエラ・アロマティカ K172株、タウエラ・アロマティカ T1株が挙げられる。なかでも、本発明の嫌気性細菌としては、アゾアルカス・スピーシーズであることが好ましく、アゾアルカス・スピーシーズ DSM9506株であることがより好ましい。
【0013】
本発明においてベクターは、トルエン分解酵素活性を有するタンパク質をコードする遺伝子(以下、「トルエン分解酵素遺伝子」という。)と、その下流に位置し、レポータータンパク質をコードする遺伝子とが組み込まれたものである。
【0014】
トルエン分解酵素遺伝子としては、特に限定されるものではなく、公知のトルエン分解酵素遺伝子を人工的に合成したものであってもよく、トルエン分解酵素遺伝子を有する生物からクローニングしたものであってもよい。また、トルエン分解酵素遺伝子は、該遺伝子のみの全配列であってもよく、該遺伝子上流側の部分配列のみであってもよく、該遺伝子と、上流側から該遺伝子を制御する制御遺伝子とを含む配列やその部分配列であってもよい。
トルエン分解酵素遺伝子を人工的に合成する場合、その方法は特に限定されるものではなく、公知の化学合成法を用いて行うことができる。また、合成するトルエン分解酵素遺伝子としては、2種以上の遺伝子をつなぎ合わせたキメラ遺伝子であってもよく、いずれかの生物種が有する遺伝子を変異させた変異遺伝子であってもよい。変異としては、遺伝子を構成する塩基配列の一部を欠損させてもよく、一部を置換してもよく、新たな配列を挿入してもよい。
トルエン分解酵素遺伝子をクローニングする場合、その方法は特に限定されるものではなく、目的の遺伝子やDNAを分離する際に用いられる公知のいずれの方法を用いてもよい。このようなクローニング方法として、例えば、試料中のトルエン分解酵素遺伝子を含むDNA領域の、両末端の塩基配列と相補的な塩基配列を有するプライマーを用いて、ポリメラーゼ連鎖反応(PCR)法によりクローニングを行うことができる。このようにしてクローニングされたトルエン分解酵素遺伝子を含むDNA断片は、電気泳動装置やDNAシークエンサー等の公知慣用の装置を用いることにより、目的としたDNA断片であるか否かを判別することができる。なお、クローニングするトルエン分解酵素遺伝子は、ベクターを導入する嫌気性細菌で発現し得るものであればその由来は特に限定されるものではなく、同種の嫌気性細菌からクローニングしたものであってもよく、当該嫌気性細菌以外の生物種からクローニングしたものであってもよい。
【0015】
レポータータンパク質としては、後述する検出化合物を用いて検出可能なものであれば特に限定されるものではなく、β−ガラクトシダーゼ、GFP、BFP、RFP、CFP、ルシフェラーゼ、evoglow(商品名、フナコシ社製)、β−グルクロニダーゼ(GUS)、クロラムフェニコールアセチルトランスフェラーゼ、EGFP(増強緑色蛍光タンパク質)、アルカリ性ホスファターゼ、DsRed、ペルオキシダーゼ、クロラムフェニコール・アセチルトランスフェラーゼ(CAT)等が挙げられる。
なかでも、本発明におけるレポータータンパク質としては、β−ガラクトシダーゼであることが好ましい。
【0016】
本発明においてベクターは、上記のようなトルエン分解酵素遺伝子と、レポータータンパク質をコードする遺伝子とが組み込まれたものであれば特に限定されるものではなく、公知のプラスミドベクター等を用いることができる。このようなプラスミドベクターとしては、上記遺伝子の他に、プロモーター配列、抗生物質耐性遺伝子等を有する公知のものを好適に用いることができる。
【0017】
本発明において、嫌気性細菌にベクターを導入し、形質転換体を作製する方法は特に限定されるものではなく、化学的処理によりベクターを導入する化学法、パルス電圧によってベクターを導入する電子穿孔法、カーボンナノチューブを用いて細胞膜に孔を開けベクターを導入する方法、嫌気性細菌間の遺伝子伝播現象を利用した接合伝達法等を用いることができ、なかでもより確実にベクターを導入できることから、接合伝達法が好ましい。
導入するベクターが抗生物質耐性遺伝子等の選択可能な遺伝子を有する場合は、上記のようにして嫌気性細菌にベクターを導入した後、当該抗生物質を含む培地にて嫌気性細菌を培養することにより、ベクターの導入された形質転換体のみを選択することができる。
【0018】
本発明において、嫌気性細菌及び形質転換体を培養する方法は、該細菌が好適に生育するものであれば特に限定されるものではない。例えば具体的には、嫌気性細菌がアゾアルカス・スピーシーズ DSM9506株である場合には、リン酸二水素カリウム又は塩化アンモニウム、塩化マグネシウム、硝酸ナトリウム等を含む嫌気性細菌の基本培地に、さらに、D−リボース、Tween80、D−アラビノース、L−アラビノース、D−キシロース、D−プシコース、α−ケドグルタル酸、マンノース、D−タガトース、D−フルクトース−6−リン酸、グルクロンアミド、D−フルクトース、ラムノース、D−ガラクトース、D−グルクロン酸、グリセロール、乳酸、酵母抽出液、カサミノ酸、NBブロス、及びLBブロスからなる群から選ばれる1種以上を含む培地で培養することが好ましい。
【0019】
本発明では、上記のようなベクターを嫌気性細菌に導入することにより、嫌気性細菌がトルエン分解酵素活性を有するタンパク質を発現する必要性が生じた際、具体的には、嫌気性細菌がトルエンの存在下にある際に、トルエン分解酵素遺伝子と共に、レポータータンパク質をコードする遺伝子が発現する。そのため、後述する検出工程において、レポータータンパク質を何らかの形で検出することにより、嫌気性細菌の存在環境におけるトルエンの有無を感知することが可能となる。
【0020】
[検出工程]
次に、前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する。
本発明において、試料としては特に限定されるものではなく、例えば、トルエンの存在が疑われる土壌、地下水、大気等が挙げられる。
【0021】
本発明において、検出化合物は、レポータータンパク質により発色又は発光する水溶性部位を有する。
発色又は発光する水溶性部位(水溶性の色素又は蛍光物質)としては、特に限定されるものではなく、色素又は蛍光の検出に通常用いられるもののうち、水溶性のものを用いることができる。例えば、ルシフェラーゼ等の蛍光物質や、β−フラノシド基質やカテコール基質のような発色物質を用いることができる。
本発明において、検出化合物としては、例えばレポータータンパク質がβ−ガラクトシダーゼの場合、検出化合物として、β−ガラクトピラノシドと、上記のような水溶性の色素又は蛍光物質との融合化合物を用いることができる。この場合、トルエン存在下の形質転換体において、トルエン分解酵素遺伝子と共にβ−ガラクトシダーゼ遺伝子が発現し、生産されたβ−ガラクトシダーゼが、検出化合物中のβ−ガラクトピラノシド部分を分解することにより、β−ガラクトピラノシドに結合されていた水溶性の色素又は蛍光物質が発色又は発光する。
このような検出化合物として具体的には、o−Nitrophenyl−β−D−galactopyranoside(ONPG)、3’,4’−Dihydroxyflavone−4’−β−D−galactopyranoside(DHF−Gal)のような色素を用いた検出化合物、6−O−β−galactopyranosyl−luciferin(Luc−Gal)のような蛍光を用いた検出化合物等が挙げられる。なかでも、試料中のトルエン含有量を定量する際の精度が高いことから、ルシフェラーゼを含む検出化合物、具体的には上記Luc−Galを用いることが好ましい。
本発明では、検出化合物の発色又は発光する部位として水溶性のものを用いることで、該発色又は発光化合物が嫌気性細菌に吸着することがないため、精度よく試料中のトルエン含有量を定量することができる。
【0022】
本発明において、形質転換体と、上記検出化合物とを用いてトルエン含有量を定量する方法は特に限定されるものではないが、例えば、試料と、形質転換体及び検出化合物とを有する培地とを接触させ、検出化合物の発色又は発光の強度を、分光光度計、蛍光光度計等の公知の装置によって測定し、検量線を用いることで、トルエン含有量の定量を行うことができる。検量線は、既知量のトルエンを含有する試料を、同様の形質転換体、同様の検出化合物、及び同様の装置を用いて測定することにより作製することができる。
【0023】
また、本発明のトルエン検出方法では、検出工程において、試料中のトルエン含有量の定量を行う前に、さらに、トルエンの有無のみの検出や、トルエン含有量の半定量等の簡易検出を行ってもよい。
トルエンの簡易検出の方法としては特に限定されるものではないが、例えば、上記検出化合物を、検量線を用いずにその発色又は発光のみを検出してもよい。また、レポータータンパク質が単体で発色又は発光する物質、例えばEGFP等である場合には、検出化合物を介さずに、該発色又は発光を分光又は蛍光光度計により直接検出し、発色や発光の有無により簡易検出を行うことができる。さらに、レポータータンパク質がβ−ガラクトシダーゼの場合には、5−ブロモ−4−クロロ−3−インドリル−β−D−ガラクトピラノシド(X−gal)等の水溶性ではないが一般的且つ簡便な検出化合物を用いて、簡易検出を行うこともできる。
【0024】
本発明のトルエン検出方法によれば、例えば、嫌気性細菌にベクターを導入した形質転換体や検出化合物を、離散しないように担体等の表面に固着させ、これを地下水中に静置して、該地下水中のトルエン汚染の有無及びトルエン含有量を検出することができる。
また、例えば、地下水を定期的又は恒常的に、観測井戸等からポンプ等を用いて汲み上げ、汲み上げた地下水試料と、嫌気性細菌にベクターを導入した形質転換体と、検出化合物とを接触させ、分光光度計や蛍光光度計を用いてトルエン汚染の有無及びトルエン含有量を検出することができる。
【実施例】
【0025】
次に実施例を示して本発明をさらに詳細に説明するが、本発明は以下の実施例に限定されるものではない。なお、以下の実施例中、特に記載がない限り、「%」は「質量%」を意味する。
【0026】
[参考例1]
(アゾアルカス・スピーシーズ DSM9506株の培養条件の検討)
まず、アゾアルカス・スピーシーズ DSM9506株(以下、単に「DSM9506株」という。)の利用可能な炭素源について検討した。
なお、DSM9506株は嫌気性細菌であるため、以下の検討において、DSM9506株の植菌や培地調整等の作業は、酸素濃度が大気中の10万分の1程度である嫌気グローブボックス内で行い、培養は酸素を通さないブチルゴム栓で密閉したガラス容器内で行った。また、ガラス容器には、酸素除去剤として、酸化チタン−クエン酸ナトリウム溶液、硫化ナトリウム九水和物を添加し、嫌気環境を維持した上で、嫌気指示薬であるレサズリンを用いて嫌気環境であることを確認し、指示薬の結果から嫌気環境が損なわれたと考えられる試料は破棄した。
96ウェルマイクロプレートの各ウェル内に、95種の炭素源となる化合物が充填されたマイクロプレート(商品名、バイオログ社製)を用いて検討したところ、DSM9506株は、D−リボース、Tween80、D−アラビノース、L−アラビノース、D−キシロース、D−プシコース、α−ケドグルタル酸、マンノース、D−タガトース、D−フルクトース−6−リン酸、グルクロンアミド、D−フルクトース、ラムノース、D−ガラクトース、又は、D−グルクロン酸、を炭素源として利用可能であることが分かった。
【0027】
また、上記95種類以外の化合物についても炭素源となり得るか否かを個別に検討したところ、グリセロール、乳酸、酵母抽出液、カサミノ酸、NBブロス、又はLBブロスの6種の化合物も炭素源として利用可能であることが分かった。これら6種の化合物を炭素源とした場合の、DSM9506株の吸光度により測定した生育量を図1に示す。図1中、横軸が培養期間(日)、縦軸が660nmにおける吸光度である。
図1の結果から、乳酸、及びカサミノ酸を炭素源とした場合にDSM9506株の生育が良好となることが分かった。そこで、以下の検討では、乳酸を炭素源とした。
【0028】
次に、炭素源として乳酸を用いた場合の、乳酸濃度によるDSM9506株の生育量について検討した。
乳酸濃度を0.07%、0.14%、0.21%、0.28%、0.35%にそれぞれ調整した基本培地(リン酸二水素カリウム又は塩化アンモニウム、塩化マグネシウム、及び硝酸ナトリウムを含む培地)において、DSM9506株を48時間培養し、DSM9506株の生育量を培養開始24時間後及び48時間後に、吸光度により測定した。結果を図2に示す。図2中、横軸が培養期間(日)であり、縦軸が660nmにおける吸光度である。
図2の結果から、48時間後においては、乳酸濃度が高い程生育が良いものの、24時間後においては乳酸濃度0.14%の培地が最も生育が良かった。これは、乳酸が高濃度である場合、該乳酸が増殖初期段階においてDSM9506株の生育に悪影響を及ぼすためと考えられる。そのため、48時間後において、最も乳酸濃度の高い0.35%の培地と得られる菌体量に大差がなく、且つ、増殖初期である24時間後においても生育が良好である乳酸濃度0.21%を以下の検討において採用することとした。
【0029】
次いで、無機塩の種類によるDSM9506株の生育量について検討した。
培地1L中にリン酸二水素カリウム0.52g(KH2PO4)、塩化アンモニウム0.2g(NH4Cl)、塩化マグネシウム六水和物0.4g(MgCl2−6H2O)及び硝酸ナトリウム0.2g(NaNO3)を含む基本培地に、乳酸0.21%を添加し、さらにリン酸水素二カリウム1g(K2HPO4)又は塩化カルシウム二水和物(CaCl2−2H2O)0.1gを添加して、該培地におけるDSM9506株の生育量を調べた。
その結果、リン酸二水素カリウム及び塩化カルシウムのいずれかを添加することにより若干の生育量の向上が認められたが、これら2種の間に大きな効果の差は認められなかった。そのため、以下の検討では、上記基本培地に、乳酸及び塩化カルシウムを添加した培地を用いることとした。
【0030】
さらに、微生物種によっては、培養開始時の菌体数が少ない場合に生育しないおそれがあるため、培養開始時の菌体数が生育に与える影響について検討した。
DSM9506株を、リン酸二水素カリウム0.52g(KH2PO4)、塩化アンモニウム0.2g(NH4Cl)、塩化マグネシウム六水和物0.4g(MgCl2−6H2O)及び硝酸ナトリウム0.2g(NaNO3)、乳酸0.21%を1L中に含む培地で24時間前培養を行った。その後、前培養液を1〜10%の範囲で量を変えて本培養の培地中に添加し、生育量を調べた。結果を図3に示す。なお、本培養の培地組成は前培養と同様である。
この結果、24時間後では培養開始時の添加菌体数に比例して生育量が多いが、48時間後には添加菌体数の多寡に関わらずほぼ同程度の生育量となった。いずれの菌体添加量であっても24時間後に菌体量が10倍となっており、初期添加量が1〜10%の範囲内であれば生育速度に差がないことが分かった。そのため、以下の検討では、菌体の初期添加量を10%とした。
【0031】
[実施例1]
(1)トルエン分解酵素遺伝子を含むDNA断片のクローニング
まず、DSM9506株において公知となっているトルエン分解酵素遺伝子(Genbankアクセッション番号:AY032676)の146番目〜3362番目の塩基配列(3217bp)の両末端と相補的な塩基配列を有するプライマーを設計した。そして、常法により抽出したDSM9506株のゲノムDNAをテンプレートとして、上記プライマーとKOD−Plus(商品名、東洋紡績(株)社製)とを用いて、PCR法によりトルエン分解酵素遺伝子を含むDNA断片のPCR産物を得た。用いたプライマーの配列を、Primer1(forward;配列番号1)とPrimer2(reverse;配列番号2)として表1に示す。
アガロース電気泳動を用いて、得られたPCR産物の全長を確認したところ、目的産物と同等の3200bp程度のPCR産物が得られていることが確認できた。また、PCR産物の一部の配列についてDNAシークエンサーを用いて解析を行ったところ、目的のトルエン分解酵素遺伝子を含むDNA断片が得られていることが確認できた。
【0032】
(2)レポータータンパク質をコードする遺伝子を含むDNA断片のクローニング
レポータータンパク質であるEGFP、β−ガラクトシダーゼをコードする遺伝子を含むDNA断片をクローニングした。
EGFPは、プラスミドベクターであるpEGFP(商品名、クロンテック社製)をテンプレートとして、KOD−Plus(商品名、東洋紡績(株)社製)、並びに下記プライマーPrimer3(forward;配列番号3)及びPrimer4(reverse;配列番号4)を用いて、PCR増幅を行い、産物であるEGFP遺伝子断片を得た。
β−ガラクトシダーゼは、プラスミドベクターであるpSV−β−Galactosidase Vector(商品名、プロメガ社製)をテンプレートとして、KOD−Plus(商品名、東洋紡績(株)社製)、並びに下記プライマー、Primer5(forward;配列番号5)及びPrimer6(reverse;配列番号6)を用いて、PCR増幅を行い、産物であるβ−ガラクトシダーゼ遺伝子断片(β−Gal)を得た。
得られたPCR産物をアガロース電気泳動にて確認したところ、PCR産物はいずれも目的の産物長(EGFP:850bp、βガラクトシダーゼ:3100bp)であったため、EGFP、β−ガラクトシダーゼ共に目的のレポータータンパク質遺伝子断片であることが確認できた。
【0033】
【表1】
【0034】
(3)発現ベクターの調整
次いで、上記(1)で得られたトルエン分解酵素遺伝子断片と、上記(2)で得られたレポータータンパク質遺伝子断片とを、それぞれ制限酵素処理した後、ラピッドDNAライゲーションキット(商品名、ロッシュ社製)を用いて連結した。以下、得られたEGFPを含む遺伝子断片をトルエン−EGFP、β−ガラクトシダーゼを含む遺伝子断片をトルエン−β−ガラクトシダーゼという。これら遺伝子断片は、アガロース電気泳動により目的の産物であることを確認した。
上記のようにして得られたトルエン−EGFPと、トルエン−β−ガラクトシダーゼをApaI及びSpeIで処理した後、同じくApaI及びSpeIで処理したプラスミドDNAの発現ベクターであるpBBR1MCS2(Kovachら、Gene、166、1995年)に、ラピッドDNAライゲーションキット(商品名、ロッシュ社製)を用いて組み込んだ。得られたpBBR1−トルエン−EGFP、及びpBBR1−トルエン−β−ガラクトシダーゼは、ApaI及びSpeIで処理した後アガロース電気泳動を行い、目的の発現ベクターであることを確認した。また、これらの発現ベクターは、Escherichia coli(大腸菌)JM109株にハナハン法を用いて形質転換し、増幅及び保存を行った。
【0035】
(4)形質転換体の作製
上記参考例1で検討した条件において培養を行ったDSM9506株に対し、化学法、電子穿孔法、カーボンナノチューブを用いて細胞膜に孔を開けベクターを導入する方法、及びEscherichia coli(大腸菌)S17−1株を用いた接合伝達法のいずれかを用いて、pBBR1−トルエン−EGFPベクター、又はpBBR1−トルエン−β−ガラクトシダーゼベクターの導入を試みた。その後、pBBR1ベクター内に存在するカナマイシン耐性遺伝子を利用して、DSM9506株の培地にカナマイシンを(50mg/L)で添加し、形質転換体の選択を行った。
その結果、接合伝達法を用いた場合に、最も効率よく形質転換体が製造できることが分かった。以下の検討では、接合伝達法により得られた形質転換体を用いた。
【0036】
(5)pBBR1−トルエン−EGFPベクターを用いた簡易検出
pBBR1−トルエン−EGFPベクターを導入したDSM9506株の形質転換体を栄養培地(DIFCO Nutrient Broth 4g/L、カナマイシン 50mg/L)で増殖させた後、遠心分離器を用いて回収し、上記基本培地で洗浄後、トルエンの簡易検出を行った。
まず、トルエン(1mg/L)を添加した上記基本培地30mLに形質転換体を移し、嫌気条件下で培養を行ったところ、形質転換体が緑色の蛍光を発することが確認できた。また、この緑色蛍光を発する形質転換体を好気環境下に移すと蛍光が消え、その後嫌気環境下に戻すと再度蛍光を発することが確認できた。
この結果から、形質転換体では、設計通りにトルエンが存在する嫌気環境下においてEGFPが産生されていることが確認できた。また、トルエンが分解できない大気中(好気環境下)に移すことで蛍光が消失することは、EGFPが長期間存在し得ないことを示しており、つまり、形質転換体周囲のトルエンが全て分解されてトルエン汚染がなくなれば蛍光も消失すると考えられる。
よって、EGFP等に由来する緑色蛍光の有無を指標としてトルエンの存在を検知することができると考えられた。
【0037】
(6)pBBR1−トルエン−β−ガラクトシダーゼベクターを用いた簡易検出
次に、pBBR1−トルエン−EGFPベクターを導入したDSM9506株の形質転換体を栄養培地(DIFCO Nutrient Broth 4g/L、カナマイシン 50mg/L)で増殖させた後、遠心分離器を用いて回収し、上記基本培地で洗浄後、トルエンの簡易検出を行った。
まず、トルエン(1mg/L)又はベンゼン(1mg/L)を添加した上記基本培地30mLに、形質転換体、及び発色基質であるX−gal(200ug/L)を加えて静置培養した。1週間後に観察したところ、トルエンを添加した培地では、β−ガラクトシダーゼの作用によりX−galから青色の色素が遊離し、培地が青色となった。一方、ベンゼンを添加した培地では、培地の色の変化はなかった。
この結果から、形質転換体では、設計通りにトルエンが存在する環境下においてβ−ガラクトシダーゼが産生されていることが確認できた。よって、β−ガラクトシダーゼのX−gal分解に由来する青色色素の有無を指標として、トルエンの存在を検知することができると考えられた。
【0038】
また、増殖させた上記DSM9506株の形質転換体を、上記同様にトルエン及びX−galを含む基本培地に懸濁し、旋回式振とう培養装置を用いて振とう培養を行ったところ、トルエンによる青色色素の誘導は認められなかった。これは、振とう培養によって酸素濃度が上昇したためにトルエン分解が妨げられ、その結果、レポータータンパク質であるβ−ガラクトシダーゼが発現しなくなったためと考えられる。
次いで、増殖させた上記DSM9506株の形質転換体を、上記同様にトルエン及びX−galを添加した基本培地に添加し、さらに、各培地にタンパク質合成阻害剤(クロラムフェニコール)を50mg/L添加して1週間静置したところ、青色色素は認められなかった。この結果から、青色色素の発色がβ−ガラクトシダーゼ遺伝子の発現の結果であることが確認できた。よって、β−ガラクトシダーゼとレポータータンパク質とをコードする遺伝子を有する発現ベクターを用いることで、嫌気環境下でのトルエン検出が可能であることが確認できた。
【0039】
(8)トルエン含有量の定量−1
トルエンによって誘導される青色色素の生産量と、トルエン濃度との関係について調べた。
まず、トルエンを0、1、5、又は10mg/Lの濃度で含有する培地を上記(7)と同様に作製し、pBBR1−トルエン−β−ガラクトシダーゼベクターを導入したDSM9506株を植菌した。1週間静置培養した後、青色色素の吸光度(OD620nm)を分光光度計(ベックマン社製DU650)により測定し、トルエン濃度と青色色素量との関係について検討した。結果を図4に示す。
図4の結果から、トルエン濃度に比例して色素生産量も高くなることが明らかである。しかしながら、X−gal由来の青色色素(5−ブロモ−4−クロロ−3−インドール)は、水溶性でなく、DSM9506株の表面に吸着するため、吸光度が必ずしも色素量を正確に反映していないと考えられる。
【0040】
(9)トルエン含有量の定量−2
そこで、β−ガラクトシダーゼにより発色又は発光する水溶性部位を有する3種の化合物をX−galに変えて用い、0.001、0.01、0.1、又は1mg/Lのトルエンを含む上記同様の培地にて上記(7)と同様に検討を行った。3種の化合物としては、o−Nitrophenyl−β−D−galactopyranoside(ONPG)、3’,4’−Dihydroxyflavone−4’−β−D−galactopyranoside(DHF−Gal)、及び6−O−β−galactopyranosyl−luciferin(Luc−Gal)を用い、ONPGは420nm、DHF−Galは620nmにおいて上記同様の分光光度計を用い、Luc−Galはルミテスター K−200(商品名、キッコーマン社製)を用いて発色又は発光を定量化した。
その結果、β−ガラクトシダーゼにより発色するONPG、DHF−Galの発色よりも、β−ガラクトシダーゼにより発光するLuc−Galにおいて安定した発光が得られた。Luc−Galの定量結果を図5に示す。
図5の結果から、Luc−Galを検出化合物として用いた場合、トルエン濃度が0.001mg/Lと非常に低濃度であっても、発光が確認できることが分かった。また、その発光量は、トルエン濃度が0.01〜1mg/Lの範囲でトルエン濃度に比例して強くなり、定量が可能であることが確認できた。
これらの結果から、嫌気性細菌にベクターを導入した形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いることで、試料中のトルエンの有無を検出できるのみならず、トルエン含有量を定量できることが確認できた。
【産業上の利用可能性】
【0041】
本発明のトルエン検出方法によれば、地下水や土壌等のトルエンを安価且つ簡便に検出や定量することができるため、環境分野において好適に利用することができる。
【特許請求の範囲】
【請求項1】
嫌気性細菌にベクターを導入し、形質転換体を作製する工程と、
前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する工程とを有するトルエン検出方法であって、
前記ベクターは、トルエン分解酵素活性を有するタンパク質をコードする遺伝子と、その下流に位置し、レポータータンパク質をコードする遺伝子とが組み込まれたものであることを特徴とするトルエン検出方法。
【請求項2】
前記嫌気性細菌が、Azoarcus sp.(アゾアルカス・スピーシーズ)であることを特徴とする請求項1に記載のトルエン検出方法。
【請求項3】
前記嫌気性細菌及び形質転換体が、D−リボース、Tween80、D−アラビノース、L−アラビノース、D−キシロース、D−プシコース、α−ケドグルタル酸、マンノース、D−タガトース、D−フルクトース−6−リン酸、グルクロンアミド、D−フルクトース、ラムノース、D−ガラクトース、D−グルクロン酸、グリセロール、乳酸、酵母抽出液、カサミノ酸、NBブロス、及びLBブロスからなる群から選ばれる1種以上を含む培地で培養されることを特徴とする請求項1又は2に記載のトルエン検出方法。
【請求項4】
前記レポータータンパク質がβ−ガラクトシダーゼであり、前記検出化合物が、ルシフェラーゼを含む検出化合物であることを特徴とする請求項1〜3のいずれか一項に記載のトルエン検出方法。
【請求項5】
前記嫌気性細菌へのベクターの導入を、接合伝達法を用いて行うことを特徴とする請求項1〜4のいずれか一項に記載のトルエン検出方法。
【請求項1】
嫌気性細菌にベクターを導入し、形質転換体を作製する工程と、
前記形質転換体と、レポータータンパク質により発色又は発光する水溶性部位を有する検出化合物とを用いて、試料中のトルエン含有量を定量する工程とを有するトルエン検出方法であって、
前記ベクターは、トルエン分解酵素活性を有するタンパク質をコードする遺伝子と、その下流に位置し、レポータータンパク質をコードする遺伝子とが組み込まれたものであることを特徴とするトルエン検出方法。
【請求項2】
前記嫌気性細菌が、Azoarcus sp.(アゾアルカス・スピーシーズ)であることを特徴とする請求項1に記載のトルエン検出方法。
【請求項3】
前記嫌気性細菌及び形質転換体が、D−リボース、Tween80、D−アラビノース、L−アラビノース、D−キシロース、D−プシコース、α−ケドグルタル酸、マンノース、D−タガトース、D−フルクトース−6−リン酸、グルクロンアミド、D−フルクトース、ラムノース、D−ガラクトース、D−グルクロン酸、グリセロール、乳酸、酵母抽出液、カサミノ酸、NBブロス、及びLBブロスからなる群から選ばれる1種以上を含む培地で培養されることを特徴とする請求項1又は2に記載のトルエン検出方法。
【請求項4】
前記レポータータンパク質がβ−ガラクトシダーゼであり、前記検出化合物が、ルシフェラーゼを含む検出化合物であることを特徴とする請求項1〜3のいずれか一項に記載のトルエン検出方法。
【請求項5】
前記嫌気性細菌へのベクターの導入を、接合伝達法を用いて行うことを特徴とする請求項1〜4のいずれか一項に記載のトルエン検出方法。
【図1】

【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2012−44944(P2012−44944A)
【公開日】平成24年3月8日(2012.3.8)
【国際特許分類】
【出願番号】特願2010−191344(P2010−191344)
【出願日】平成22年8月27日(2010.8.27)
【出願人】(000173784)公益財団法人鉄道総合技術研究所 (1,666)
【Fターム(参考)】
【公開日】平成24年3月8日(2012.3.8)
【国際特許分類】
【出願日】平成22年8月27日(2010.8.27)
【出願人】(000173784)公益財団法人鉄道総合技術研究所 (1,666)
【Fターム(参考)】
[ Back to top ]