
トルコギキョウ由来のフラボノイド3’−ヒドロキシラーゼとその利用

【課題】トルコギキョウ由来のフラボノイド3’-ヒドロキシラーゼを提供する。
【解決手段】以下の(a)〜(d)いずれか記載のポリペプチドからなる。(a)特定のアミノ酸配列からなるポリペプチド(b)特定のアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、(c)特定のアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド(d)特定のヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
【解決手段】以下の(a)〜(d)いずれか記載のポリペプチドからなる。(a)特定のアミノ酸配列からなるポリペプチド(b)特定のアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、(c)特定のアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド(d)特定のヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
【発明の詳細な説明】
【技術分野】
【0001】
本発明は、トルコギキョウ[Eustoma grandiflorum (Raf.) Shinn.]の花弁フラボノイド3’-ヒドロキシラーゼ(F3’H)遺伝子(EgF3’H)と、それを利用した変異体植物を作出する技術に関する。
【背景技術】
【0002】
トルコギキョウ[Eustoma grandiflorum (Raf.) Shinn.]は、リンドウ科に属し、アメリカ南西部を原産とする一年性の草本である。本種は、その大きな花と長い茎、そして花持ちのよさの点で、日本で最も人気の高い切花の一つとなっている。典型的なトルコギキョウの野生種の花色は紫色であり、その主要色素はデルフィニジン系のアントシアニンである。主に日本における商業育種によって、ピンク、白、クリーム色、藤色、赤などの多様な花色の系統が作出されているが、更なる花色の多彩化、効率的な花色育種法が望まれている。
【0003】
例えば、花色の複対立遺伝子が提唱されているが、花色の形成機構はまだDNAレベルでは詳しく証明されていない(特許文献1)。
【0004】
一方、野田らは、トルコギキョウでアントシアニン合成に関わるフラボノール合成酵素(FLS)、ジヒドロフラボノール4還元酵素(DFR)、カルコン合成酵素(CHS)、アントシアニン合成酵素(ANS)などの発現を詳しく解析しているが、これらの遺伝子と花色との関係は不明である(非特許文献1)。
【0005】
特許文献2には、「バラの内在性のフラボノイド合成経路に属する酵素であるDFRの遺伝子を遺伝子工学の手法を用いて人為的に抑制し、かつパンジーのF3’5’H遺伝子を発現させ、さらにジヒドロミリセチンを還元するDFR遺伝子を発現させることによりデルフィニジンの含有率を花弁に含まれる総アントシアニジン中のほぼ80-100%にすることにより、青いバラを実現することができた」という記載がある。
【0006】
特許文献3には、「フラボノイド3’-ヒドロキシラーゼをコードする遺伝子配列が同定され、クローニングされた。本発明の組換え遺伝子配列は、例えば、新規の発現、過度に発現、抑制、アンチセンス阻害およびリボザイム活性により、この酵素をコードする遺伝子のモジュレーションを可能とする。植物におけるフラボノイド3’-ヒドロキシラーゼの合成をコントロールする能力は、個々のアントシアニンの組成の調節ならびにフラボノールおよびアントシアニンの相対レベルの変更を可能とし、これにより組織、例えば、花弁、葉、種子および果実、の色の操作を可能とする」という記載がある。
【0007】
しかし現在までに、トルコギキョウ由来のフラボノイド3’-ヒドロキシラーゼは報告されていない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】国際公開WO2004/103065号パンフレット
【特許文献2】WO2005-017147号
【特許文献3】特願平9-530472号
【非特許文献】
【0009】
【非特許文献1】N. Noda et al., Physiologia Plantarum 2004年, 第122巻, 305-313.
【発明の概要】
【発明が解決しようとする課題】
【0010】
上記のような状況下、トルコギキョウからフラボノイド3’-ヒドロキシラーゼを同定することができれば、所望の花色を有するトルコギキョウ及び他の植物の育種に有効に利用することができると考えられる。
【0011】
そこで本発明は、トルコギキョウ由来のフラボノイドF3’-ヒドロキシラーゼ及びその利用方法を提供することを目的とする。
【課題を解決するための手段】
【0012】
本発明者らは、トルコギキョウにおいて、フラボノイド骨格B環の生合成に関与する遺伝子群を調査する過程で、各遺伝子の配列情報に基づいて設計したプライマーを用いてRT-PCR分析を行ったところ、既知の特定のフラボノイド3’-ヒドロキシラーゼ遺伝子と相同性の高い1種のポリヌクレオチドを見出した。
【0013】
そこでこのポリヌクレオチドをアサガオ品種「バイオレット」に導入して発現させたところ、アサガオ品種「バイオレット」では本来認められない、赤色系色素のシアニジンが蓄積されること、すなわち該ポリヌクレオチドがフラボノイド骨格B環の3'位に水酸基を結合させる反応を触媒するポリペプチドをコードすること、を見出した。
【0014】
本発明は上記知見を基礎とするものであり、以下の特徴を包含する。
【0015】
(1) 以下の(a)〜(d)いずれか記載のポリペプチド。
(a) 配列番号2に示すアミノ酸配列からなるポリペプチド
(b) 配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、
(c) 配列番号2に示すアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(d) 配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(2) 上記(1)記載のポリペプチドをコードするポリヌクレオチド。
(3) 配列番号1に示すヌクレオチド配列からなる、上記(2)記載のポリヌクレオチド。
(4) 上記(2)又は(3)記載のポリヌクレオチドを植物の細胞若しくは組織培養物に導入することを含む、変異体植物の製造方法。
【0016】
(5) 植物はアサガオ(Ipomoea nil)である上記(4)記載の製造方法。
(6) 対象とする植物において以下の(a)〜(d)いずれか記載のポリペプチドの機能を抑制することを含む、変異体植物の製造方法。
(a) 配列番号2に示すアミノ酸配列からなるポリペプチド
(b) 配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、
(c) 配列番号2に示すアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(d) 配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(7) 上記(a)〜(d)いずれか記載のポリペプチドをコードする遺伝子を欠損させることを特徴とする上記(6)記載の製造方法。
(8) 対象とする植物はトルコギキョウ(Eustoma grandiflorum)である上記(6)記載の製造方法。
【発明の効果】
【0017】
本発明によれば、トルコギキョウ由来のフラボノイドF3’-ヒドロキシラーゼが提供される。
【0018】
本発明のフラボノイドF3’-ヒドロキシラーゼは、例えばペオニジンを産生する植物内での当該酵素の発現により、赤色系色素であるシアニジンを生成させることができる。したがって、植物内でのその発現を調節することで、所望の色を有する植物を製造することができる。
【図面の簡単な説明】
【0019】
【図1】図1は、トルコギキョウF3’Hのアミノ酸配列(lisianthus)を、ペチュニア(AF155332)及びアラビドプシス(AL133421)の配列と共に示す。図中、N-末端近くに示した矢印の部分は、疏水(hydrophobic)領域であり、下線で示した部分はそれぞれ、チトクロムP450に特徴的なモチーフ領域:AGTDTS及びFGAGRRICAG、及びF3’Hに特徴的なモチーフ領域:GGEKを示している。
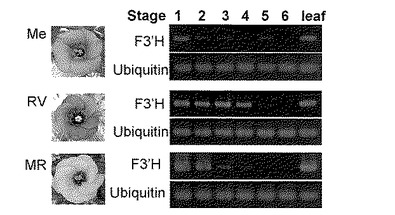
【図2】図2は、RT-PCR法により測定したトルコギキョウ花弁中の1〜6の各ステージにおけるF3’H遺伝子(EgF3’H)の発現を示す図である。Me、RV及びMRはそれぞれ品種メロウラベンダー、ロイヤルバイオレット及びミッキーローズの略記である。
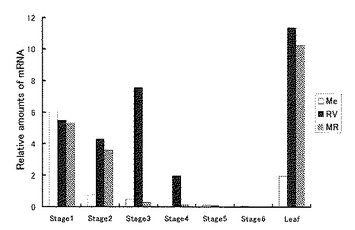
【図3】図3は、リアルタイムPCR法により測定したトルコギキョウ花弁中の1〜6の各ステージにおけるF3’H遺伝子(EgF3’H)のmRNAの発現を示す図である。Me、RV及びMRはそれぞれ品種メロウラベンダー、ロイヤルバイオレット及びミッキーローズの略記である。EgF3’H遺伝子のmRNAの相対量は、トルコギキョウのユビキチン遺伝子との量比で示している。
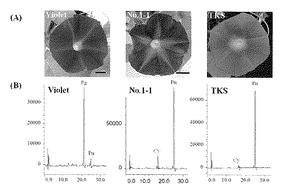
【図4】図4は、トルコギキョウEgF3’H遺伝子をアサガオ「バイオレット」に導入した形質転換体(No.1-1)の写真図(A)及びHPLC分析の結果(B)を、非形質転換体のアサガオ品種「バイオレット」(Violet)と野生種(TKS)との比較により示す。Pgはペラルゴニジンを、Cyはシアニジンを、Pnはペオニジンをそれぞれ示している。
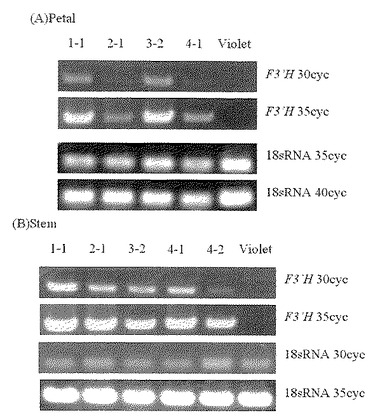
【図5】図5は、トルコギキョウF3’H遺伝子を導入した形質転換体アサガオにおいて、導入した遺伝子の発現をRT-PCR法で分析した結果を示す。Aは花弁での結果、Bは茎での結果である。1-1、2-1、3-2、4-1はそれぞれ独立した形質転換体を示し、Violetは形質転換していないアサガオ「バイオレット」を示している。4-2は4-1と同じカルスに由来した形質転換体であるが、開花しなかった個体であったため、茎のみの結果を示す。
【発明を実施するための形態】
【0020】
本発明は、トルコギキョウ由来のフラボノイド3’-ヒドロキシラーゼ(本明細書中、F3’H又はF3’Hポリペプチドとも称する)とその利用方法に関する。
【0021】
トルコギキョウF3’H遺伝子(本明細書中、EgF3’H又はF3’Hポリヌクレオチドとも称する)のヌクレオチド配列の一例を配列番号1に示し、配列番号1に示すヌクレオチド配列によってコードされるF3’Hのアミノ酸配列を配列番号2に示す。なお、F3’HポリヌクレオチドによりコードされるF3’Hポリペプチドとしては、配列番号2に示すアミノ酸配列からなるポリペプチドに限定されず、配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、又は配列番号2に示すアミノ酸配列に対して70%以上の相同性、好ましくは80%以上の相同性、より好ましくは90%以上の相同性、最も好ましくは95%以上の相同性を有するポリペプチドからなりかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド(以下、これらを「変異型ポリペプチド」と称する)であってもよい。ここで、置換、欠失、付加又は挿入するアミノ酸は、例えば1〜50個、好ましくは1〜25個、より好ましくは1〜10、より好ましくは1〜5個とすることができる。相同性の値は、複数のアミノ酸配列間の相同性を演算するソフトウェア(例えば、FASTA、DANASYS、BLAST)を用いてデフォルトの設定で算出した値を意味する。
【0022】
またF3’Hポリヌクレオチドとしては、配列番号1に示すヌクレオチド配列からなるポリヌクレオチドに限定されず、配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドであって、フラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチドをコードするポリヌクレオチド(以下、変異型ポリヌクレオチドとも称する)も含まれる。変異型ポリヌクレオチドは、配列番号2に示すアミノ酸配列とは異なるアミノ酸配列からなるポリペプチドであって、フラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有する変異型ポリペプチドをコードすることとなる。
【0023】
ここでストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。ストリンジェントな条件下としては、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、最も好ましくは95%以上の配列相同性を有するポリヌクレオチド同士がハイブリダイズし、それより相同性が低いポリヌクレオチド同士がハイブリダイズしない条件を挙げることができる。そのような条件は、例えばJ. Sambrook, et al., "Molecular Cloning", Cold Spring Harbor Laboratory Press, 1989を参照することにより当業者には自明である。
【0024】
上述した変異型ポリペプチドがフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するか否かは、フラボノイド骨格B環の3’位が水酸基ではないフラボノイド類(例えばアントシアニジン)を基質として該変異型ポリペプチドを作用させ、その水酸化形態の生成の有無を検出することによって行うことができる。基質として利用できるフラボノイド類としては、これに限定されるものではないが、ペラルゴニジン、ペオニジン、マルビジンなどのアントシアニジン;ナリンゲニンなどのフラバノン類;ジヒドロケンフェロールなどのジヒドロフラボノール類;ロイコペラルゴニジンなどのロイコアントシアニジン類などを挙げることができる。特に基質としてジヒドロケンフェロールやナリンゲニンを使用することが好ましい。具体的に、変異型ポリペプチドがフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するか否かは、例えば、Ben-Bo Xu ら, 2007年,Journal of Plant Physiology、第164巻、p350〜363らの方法によって、大腸菌で発現したタンパク質を試験内で反応させることによって試験することができる。
【0025】
本発明のF3’Hポリヌクレオチドは、配列番号1に示すヌクレオチド配列情報に基づいて、該配列を特異的に増幅することができるプライマーセットを設計し、赤色トルコギキョウ品種の花弁から抽出した総DNAを鋳型としてPCR増幅を行うことによって、クローニングすることができる。
【0026】
あるいは、本発明のF3’Hポリヌクレオチドは、配列番号1に示すヌクレオチド配列情報に基づいて、当業者に周知の方法、例えば適当な配列のクローニング及び制限酵素による切断、ホスホトリエステル法(例えばNarangら,1979年,Meth.Enzymol.,第68巻,p90〜99参照)、ホスホジエステル法(例えばBrownら,1979年、Meth.Enzymol.,第68巻,p109〜151参照)、エチルホスホアミダイト法(例えばBeaucageら,1981年,Tetrahedron Lett.,第22巻,p1859〜1862参照)などの方法により、直接的に合成することができる。また市販の自動DNA合成装置を使用することによって合成してもよい。
【0027】
本発明のF3’Hポリペプチドは、上記のようにしてクローニングしたF3’Hポリヌクレオチドを利用して、定法に従って製造することができる。例えば、本発明のF3’Hポリペプチドは、F3’Hポリヌクレオチドを適当な宿主細胞内で発現させ、培養物からF3’Hポリペプチドを回収することにより製造することができる。
【0028】
本発明で使用できる宿主細胞としては、これに限定されるものではないが、大腸菌などの原核細胞、酵母、昆虫培養細胞(例えばカイコガ培養細胞)、動物細胞細胞(例えばマウス培養細胞)、植物培養細胞(例えばタバコ培養細胞)などの真核細胞を挙げることができる。
【0029】
具体的に、宿主細胞での発現を誘導するプロモーターの下流に前記F3’Hポリヌクレオチドを組込んだ組換えベクターを作製し、これを宿主細胞に遺伝子導入することによって宿主細胞を形質転換し、該形質転換宿主細胞を発現調節配列が機能する条件下で培養し、その培養物から本発明のF3’Hポリペプチドを回収することができる。
【0030】
本発明で使用できるベクターとしては、プラスミドベクター、Tiプラスミドベクター、ファージベクター、ファージミドベクター、YACベクター、ウイルスベクター等が挙げられる。これらベクターは、その後の形質転換に使用する宿主細胞のタイプに応じて、該宿主細胞中でのF3’Hポリペプチドの発現を調節・補助する追加の配列を含むことができる。そのような配列としては、これに限定されるものではないが、エンハンサー、ターミネーター、オペレーター、SD配列などを挙げることができる。また上記ベクターは、形質転換宿主細胞を選別するための選択マーカー(例えばアンピシリン耐性遺伝子、テトラサイクリン耐性遺伝子、カナマイシン耐性遺伝子、ネオマイシン耐性遺伝子など)を含むことができる。
【0031】
上記発現構築物による宿主細胞の形質転換は、当業者に慣用の手法、例えばプロトプラスト法、コンピテントセル法、エレクトロポレーション法、パーティクルガン法、アグロバクテリウム法を用いて行うことができる。
【0032】
形質転換宿主細胞の培養は、使用される宿主細胞の培養方法と実質的に同様であってよく、例えば資化しうる炭素源、窒素源、金属塩、ビタミン等を含む培地を用いて適当な条件下で培養すればよい。こうして得られた培養液から、一般的な方法によって分取や精製を行い、必要に応じて、限外ろ過濃縮、凍結乾燥、噴霧乾燥、結晶化等によって、その活性を保持するかたちでF3’Hポリペプチドを得ることができる。
【0033】
あるいは、本発明のF3’Hポリペプチドは、無細胞発現系、すなわち上記発現構築物を試験管内で用いて製造することもできる。本発明で使用できる無細胞発現系としてはセルフリーサイエンス社のWEPRO(登録商標)シリーズなどが挙げられる。
【0034】
本発明のF3’Hポリペプチドは、赤色色素であるシアニジンを生成する反応を触媒する活性を有している(図4)。シアニジンは赤色系色素であるが、植物細胞中の種々の要因、例えば液胞細胞中のpH、他の物質(例えばシアニジンに結合可能な糖鎖)の存在(コピグメンテーション)、金属元素の存在(キレート反応)、による影響を受け、様々な色を呈色することができる。
【0035】
したがって本発明のF3’Hポリヌクレオチドは、変異体植物の製造に利用することができる。本発明でいう「変異体植物」とは、外的に導入したF3’Hポリヌクレオチドの発現の結果、又は内在のF3’Hポリペプチドの機能抑制の結果、野生型とは異なる色の器官(例えば花弁、茎、葉柄、胚軸、子葉、根など)を有している植物である。
【0036】
本発明の変異体植物の製造において、植物へのF3’Hポリヌクレオチドの導入は、当業者に公知のトランスジェニック技術を用いて行うことができる。例えば本発明のF3’Hポリヌクレオチドは、必要に応じて該ポリヌクレオチドを含む植物発現用ベクターの形態で、植物の細胞若しくは組織培養物に導入することによって行うことができる。
【0037】
本発明に使用することができるトランスジェニック技術として、例えばアグロバクテリウム(Agrobacterium)を利用した形質転換を挙げることができる。簡潔に説明すると、植物感染性であるアグロバクテリウムからプラスミドを取り出し、該プラスミドのT-DNA遺伝子を本発明のF3’Hポリヌクレオチドで置換し、これをアグロバクテリウムに戻して植物組織の感染に利用する。その際、F3’Hポリヌクレオチドと共に当業者に公知の選択マーカー遺伝子を挿入することで、F3’Hポリヌクレオチドが導入されている植物を選択することができる。そのような選択マーカー遺伝子として、例えばカナマイシン、テトラサイクリン、リファンピシン、スペクチノマイシン、カルベニシリン、ゲンタマイシンなどの抗生物質に対する耐性を付与する遺伝子などを使用することができる。アグロバクテリウムを利用した形質転換の実例は、例えばモデル植物の実験プロトコール、イネ・シロイヌナズナ編、細胞工学別冊、植物細胞工学シリーズ4、島本功・岡田清孝監修 (1996)や米国特許第5,188,958号などに記載されている。
【0038】
本発明で使用することができる他のトランスジェニック技術として、これに限定されるものではないが、ウイルスベクターを介した形質転換、エレクトロポレーション及びパーティクルガンなどを挙げることができる。パーティクルガンを利用した形質転換の実例は、例えば米国特許第5,204,253号などに記載されている。
【0039】
本発明のF3’Hポリヌクレオチドで形質転換された細胞又は組織培養物は、その後、当業者に公知の技術によって植物体まで再生させることができる。植物体の再生は、当業者に公知の手法に従って行えばよい。また変異体植物を効率よく製造するために、形質転換に用いられる標的組織が高い再生能を有していることが好ましく、そのような組織として子葉、胚軸組織、未熟胚、カルスなどを挙げることができる。
【0040】
上記F3’Hポリヌクレオチドの導入の対象となる植物は、本発明の遺伝子であるF3’Hの基質であるジヒドロケンフェロールやナリンゲニンを有する植物である限り特に制限されない。そのような植物として、これに限定されるものではないが、イポモエア属からなる群から選択される属の植物、例えばアサガオ、ノアサガオ、アメリカアサガオ、マルバアサガオ、サツマイモなどを挙げることができる。
【0041】
本発明の変異体植物の製造において、内在のF3’Hポリペプチドの機能抑制は、当業者に慣用の手法によって行うことができる。例えば、内在のF3’Hポリペプチドの機能抑制は、当該ポリペプチドをコードする遺伝子(すなわちF3’Hポリヌクレオチド)をゲノムから欠損させること、F3’Hポリヌクレオチドの発現を阻害すること、及び当該ポリペプチドの活性を低下させることによって行うことができる。
【0042】
具体的に、F3’Hポリヌクレオチドを欠損させる方法としては、特に限定されないが、相同組換えを用いた方法やトランスポゾンを用いた方法を挙げることができる。また、当該ポリヌクレオチドを欠損させる場合には、当該ポリヌクレオチドの全長を欠損させてもよいし、部分的に欠損させてもよい。
【0043】
また、F3’Hポリヌクレオチドの発現を阻害する方法としては、特に限定しないが、F3’Hポリヌクレオチドの発現を制御しているプロモーターを欠損させる方法、F3’Hポリヌクレオチドの発現を制御しているプロモーターを発現誘導型プロモーターに置換する方法、F3’Hポリヌクレオチドの発現を制御しているプロモーターに突然変異を導入する方法、RNA干渉を利用してF3’Hポリヌクレオチドの転写産物を分解する方法、及びアンチセンスRNAを利用してF3’Hポリヌクレオチドの翻訳を阻害する方法を挙げることができる。
【0044】
さらに、F3’Hポリペプチドの活性を低下させる方法としては、当該ポリペプチドに特異的に結合してその活性を抑制する機能を有する物質を作用させる方法を挙げることができる。当該物質としては、F3’Hポリペプチドの機能を阻害できる抗体や阻害物質を挙げることができる。
【0045】
内在のF3’Hポリペプチドの機能抑制は、野生型の植物が上述したように定義されるポリペプチドを有している限り如何なる植物に対しても適用することができるが、トルコギキョウに対して適用することが特に好ましい。
【0046】
以下、本発明の一例を実施例として詳細に説明するが、本発明はこの実施例のみに限定されるものではない。
【実施例】
【0047】
[実施例1]
トルコギキョウのフラボノイド3’-ヒドロキシラーゼ(F3’H)をコードするcDNAのクローニング
高等植物が保有するF3’Hアミノ酸領域を基に次のプライマーを設計した: Forward-A : TTYAAYATHGGIGAYTTYATHCC; Forward-B: TTYACIGCIGGIACIGAYAC; Reverse-C: GGRTCYCKICCDATIGCCCA; Reverse-D: RCADATYCTTCKICCIGCICC。
【0048】
Rneasy maxi kit (QIAGEN)を用いて1gのトルコギキョウ品種あすかの舞姫の各花弁から全RNAを抽出し、Oligotex-dT30 super(Takara)を用いてpoly(A)+RNAを精製した。GeneRacer Kit(Invitrogen, Carlsbad, USA)を用いてcDNAを合成し、前述のプライマーを用いたPCR増幅のテンプレートとしてこれを使用した。
【0049】
PCRは、10μlの反応溶液を用い、この反応溶液には以下の試薬類を混入した;KOD-Plus-Ver.2(Toyobo)の1μl 10xPCRバッファー溶液、1μl dNTP、0.2μl KOD-Plus-Ver.2のDNAポリメラーゼ、各1μlのforward及びreverseプライマー(10pmol/μl)、及び最大で1μl濃度のcDNAテンプレート。これを、94℃で2分間熱変性させた後、熱変性(94℃30秒)、アニーリング(52℃30秒)、伸長反応(68℃1分)を30サイクルで繰り返し行い、68℃で10分間反応させた後、最後に4℃まで冷却させた。PCR増幅産物は0.8%アガロースゲル(AMRESCO)で電気泳動(50V、1×TAE、60分)し、QIAEX II Gel Extraction Kit (QIAGEN)を用いて精製した。精製されたcDNA断片をpCR-Blunt-II-TOPO(登録商標)vector(Invitrogen)を用いて再度クローン化し、全ての塩基配列をABI PRISM(登録商標)3100 Genetic Analyzerで決定した。一部の配列については、BlastXプログラムを用いて解析した。
【0050】
トルコギキョウのF3’Hクローンの完全長塩基配列を得るため、KOD-Plus-ver2 DNA polymerase (TOYOBO)を用いて、GeneRacer kitから合成されたcDNAを以下の特異的プライマーを使用してPCR増幅した: Eg3’Hrev (GCCGAATACCCTCCTTCCAAGC)又はEg3’HFor(GCATCCTCACGCGATCTATAGC); GeneRacer 5’primer (CGACTGGAGCACGAGGACACTG);及び3’primer(GCT GTC AAC GAT ACG CTA CGT AAC G)。
【0051】
PCRは、10μlの反応溶液を用い、この反応溶液には以下の試薬類を混入した;KOD-Plus-Ver.2(Toyobo)の1μl 10xPCRバッファー溶液、1μl dNTP、0.2μl KOD-Plus-Ver.2のDNAポリメラーゼ、各1μlのforward及びreverseプライマー(10pmol/μl)、及び最大で1μl濃度のcDNAテンプレート。これを、94℃で2分間熱変性させた後、熱変性(94℃30秒)、アニーリング(72℃60秒)を5サイクル、熱変性(94℃30秒)、アニーリング(70℃30秒)、伸長反応(72℃30秒)を5サイクル、熱変性後(94℃30秒)、アニーリング(68℃30秒)、伸長反応(72℃60秒)を20サイクル繰り返し行い、72℃で10分間反応させた後、最後に4℃まで冷却させた。PCR増幅産物はエタノールで沈殿したので、ネステッドPCRのため再度10μl TEで懸濁させた。ネステッドPCRに用いたプライマーは以下のとおりである:Eg3’HRev-Nest (GTCTAAGAACGCATCGAAACGC)又はEg3’HFor-Nest (CACAAATGCTCTGGCGAGAGTG); 及びGeneRacer 5’Nested primer (GGACACTGACATGGACTGAAGGAGTA)又は3’Nested primer (CGCTACGTAACGGCATGACAGTG)。
【0052】
前述したPCR条件で増幅した。すなわち、94℃で2分間熱変性させた後、熱変性後(94℃30秒)、アニーリング(58℃30秒)、伸長反応(68℃1分)を30サイクル繰り返し行い、68℃で10分間反応させた後、最後に4℃まで冷却させた。PCR増幅産物は0.8%アガロースゲル(AMRESCO)で電気泳動(50V、1×TAE、60分)し、F3’H mRNAの全長を得ることができた。
【0053】
図1には、cDNA断片をPCR法により増幅し、全長cDNAクローンの分析から想定された、トルコギキョウのF3’H遺伝子の塩基配列を示し、以降、これをEGF3’H遺伝子とも称する。EGF3’H遺伝子は1757bpの全長を有し、1560bpにはオープンリーディングフレーム(ORF)として519のアミノ酸を翻訳する部位を有していた。また、EgF3’H遺伝子が生じるアミノ酸配列は、これまでに知られている他のF3’H配列とかなり高い相同性を有していた(図1)。
【0054】
[実施例2]
材料
トルコギキョウ「メロウラベンダー」(タキイ種苗)、「ロイヤルバイオレット」(タキイ種苗)、「ミッキーローズ」(サカタの種)の種子を購入し、自殖を繰り返して固定したものを用いた。自殖系統をそれぞれ「メロウラベンダー;Me」、「ロイヤルバイオレット;RV」、「ミッキーローズ;MR」と称する。花弁の生育ステージを次の6段階に割り振り、RNAの抽出に使用した:ステージ1、20mm以下の長さのもの;ステージ2、20mmから30mmの長さのもの;ステージ3、30mm以上でまだ花弁が開いていないもの;ステージ4、30mm以上で花弁が開き始めたもの;ステージ5、花弁が開いたもの;ステージ6、完全に花弁が開ききったもの。また、トルコギキョウの葉もRNAの抽出に使用した。これらの材料を採集後は、液体窒素で凍結し、RNAの抽出並びに高速液体クロマトグラフィー(HPLC)の分析に供しするまで、-80℃で保存した。
【0055】
アサガオ(Ipomoea nil)の品種「バイオレット」の種子は丸種株式会社から入手し、野生型アサガオ「TKS(Tokyo kokei standard)」の種子は、基礎生物学研究所の星野博士より分譲されたものである。アグロバクテリウムによるこれらのアサガオの形質転換法は、清水圭一他、育種学研究、2002年、第4巻、別冊1号、p.173に記載の方法で行った。
【0056】
逆転写PCR(RT-PCR)解析
Plant Total RNA Extraction Miniprep System (Viogene)を用いてトルコギキョウ3品種の花弁(100mg)からそれぞれ全量RNAを抽出した。DNA断片の解析のため、RNAの試料をDNase I(Invitrogen)で処理した。処理後、25mM EDTA(1μl)を加えてDNase Iを不活化し、これに65℃で10分間熱を加えた。RT-PCR法については、SuperScriptTMIII First-Strand Synthesis System (Invitrogen)のプロトコールに従い、第一番目のcDNA断片をDNase I-処理RNAを使用して合成した。この反応液(20μl)は、1倍量のEx Taq Buffer、0.2mMの各dNTP、0.5ユニットのEx TaqTM HS (Takara Bio)、そして、0.5μMのforwardプライマー(ATGGAAATGATCAATGATTCTCCC)及びreverseプライマー(TCATTTAACATACATAGAGCTAGCAAG)を含み、第一番目のcDNA断片(50ng)をテンプレートとして加えた。これを、94℃で1分間熱変性させた後、熱変性後(94℃30秒)、アニーリング(67℃30秒)、伸長反応(72℃2分)を25サイクル繰り返し行い、72℃で4分間反応させた後、最後に4℃まで冷却させた。トルコギキョウのユビキチン遺伝子(GenBank accession number AB049409)を内部標準として加えて同様にRT-PCRで増幅した。使用したプライマーは以下のとおりである:Forward プライマー(EgUbi-F, 5’-ATCCAGTGACACCATCGACA-3’)及びreverseプライマー(EgUbi-R, 5’-TGTTGTAGTCGGCGAGAGTG-3’)。PCR増幅産物は0.8%アガロースゲル(AMRESCO)で電気泳動(50V、1×TAE、60分)した。
【0057】
図2に、F3’H遺伝子の発現を確認した実験結果を示す。トルコギキョウ3品種はいずれも1.6-kbの転写で発現しており、また、ステージ4までに強い発現を示した。またこの発現は、葉でも確認できた(図2)。
【0058】
定量的リアルタイムRT-PCR法
全量RNA及びcDNAの処理法は前述のとおり行った。リアルタイムRT-PCR法で用いるEgF3’H遺伝子のプライマーの設計は、Primer3プログラムにより行い、5’-GCGATCCTTGAGGAGCACAA-3’及び5’-CATACTCAGCAAGTCAACGTGTTG-3’と決定した。
【0059】
リアルタイムRT-PCRは、SYBR(登録商標)Premix Ex TaqTM kit (Talara Bio, Japan)及びReal-Time PCR 7300 system(ABI, USA)を用いて行った。熱変性、熱変性後とアニーリング、伸長反応の条件は、前記したSYBR(登録商標)Premix Ex TaqTM kitの方法で行った。内部標準としてルコギキョウのユビキチン遺伝子のmRNA量を測定することによって、各遺伝子のmRNA量を平準化した。これを3回繰り返し行った。
【0060】
その結果を図3に示す。mRNAの相対量(発現量)は、先の図2で示したF3’H遺伝子の発現と対応していることが分かる。その発現は、ステージ4まで続き、ステージ5及び6ではその発現量は減少した。なお、このmRNAの発現は、葉でも強いことが分かった(図3)。
【0061】
アサガオ(Ipomoea nil)におけるトルコギキョウのEgF3’H遺伝子の異所的発現
PCR法によりF3’H cDNAsのORF全長を増幅した。用いたプライマーは、トルコギキョウのF3’HのスタートコドンATGを保有する特異的プライマー(CAAATGGAAATGATCAATGATTCTCCC)、並びに、ストップコドンを保有するreverseプライマー(TCATTTAACATACATAGAGCTAGCAAG)であった。
【0062】
PCR法は、反応液(10μl)に、1μl 10x PCR buffer for KOD-Plus-、1μl dNTP、0.2μl KOD-Plus-DNA polymerase (Toyobo)、1μlのforwardプライマー及びreverseプライマー(10pmol/μl)、テンプレートとして0.1μl(50ng)のcDNAを加えて行った。これを、94℃で2分間熱変性させた後、熱変性(94℃30秒)、アニーリング(60℃30秒)、伸長反応(68℃90秒)を30サイクル行い、68℃で10分間反応させた後、最後に4℃まで冷却させた。
【0063】
PCR増幅断片を、pCR8/GW/TOPO TA クローニングキット取り扱い説明書(Invitrogen)に従って、GATEWAYベクターpCR8/GW/TOPOにクローニングした後、Gateway BP及びLR Clonase Enzyme Mix (Invitrogen)を用いて、F3’H遺伝子をバイナリーベクターであるpH2GW7ベクター(Karimi, M. et al., Trends Plant Sci. 2002 May; 7(5): 193-5)に組み込んだ。本バイナリーベクターは、細菌Agrobacteriumu. tumefaciensを介して形質転換するためのものである。
【0064】
その結果、トルコギキョウのF3’H遺伝子を含むバイナリーベクターが完成し、これをpGWEgF3’Hとした。本ベクター中のF3’H遺伝子は、CaMV 35Sプロモーターの下流域、及び、35Sターミネーターの上流域に位置している。本ベクターは、植物を選択するためのハイグロマイシン耐性遺伝子(Hyg)並びに細菌を選択するためのスペクチノマイシン耐性遺伝子を同時に保有している。
【0065】
A. tumefaciensによるアサガオ(I. nil)の形質転換は以下のように行った。先の作製したバイナリーベクターpGWEgF3’Hを組み込んだA. tumefaciens AGL0細胞を、スペクチノマイシン(100 mg/liter)を添加した5-10mlのLB液体培地(トリプトン, 10g/L; 酵母エキス, 5g/L; NaCl, 5g/L)中28℃で12時間振盪培養した。培養後、細胞を遠心分離して回収し、MS液体培地(10 g/l スクロース(pH5.8))に再度懸濁し、600nm(OD600)の波長で0.2-2.0の範囲で吸収があるように菌体濃度を調節した。アサガオ品種「バイオレット」の懸濁培養カルスをA. tumefaciensと共に懸濁した後、それらのカルスをMS培地(200μMアセトシリンゴン、3.2g/Lゲランガム(pH5.8)、10g/Lスクロース)に置床し、3日間培養した。
【0066】
3日後、形質転換したカルスの選抜のため、ハイグロマイシン(50mg/L)、Augmentin顆粒(2-3g/L)、スクロース(60g/L)、ゲランガム(3.2g/L)を添加したEI培地(Jia S. et al, Plant Sci., 1992年, 第87巻, 215-223)へ継代し、3〜4週培養すると、ハイグロマイシン耐性胚が形成された。このハイグロマイシン耐性胚をもう一度新鮮な同じ組成の培地に移植しさらに3〜4週間培養すると、さらに多くのハイグロマイシン耐性胚が得られた。ハイグロマイシン耐性胚をAugmentin顆粒(2-3g/L)を含むEMG培地(Jia S. et al, 前掲)に継代し、シュートの形成を確認した。このシュートは清水圭一他、育種学研究、2002年、第4巻、別冊1号、p.173に記載の方法で、根の形成と形質転換体の馴化を行った。トルコギキョウのF3’Hの配列から設計したプライマーを用い、pGWEgF3’Hで形質転換した個体が持つT-DNA領域の断片をPCRで増幅し、形質転換遺伝子の挿入を確認した。
【0067】
アグロバクテリウムを用いた形質転換によって得られたアサガオ形質転換体の写真を図4に示す。形質転換されていないアサガオ「バイオレット」は、赤色系の花色、茎、葉柄の形質であったのに対して、アサガオ形質転換体は、青色の花色、茎、葉柄の形質であり、その形質は、野生型の「TKS」と同様であった(図4)。
【0068】
次に、このアサガオ形質転換体におけるトルコギキョウのF3’HmRNA量の解析をしたところ、これらにトルコギキョウのF3’H遺伝子が導入された結果、そのmRNAが蓄積されたことが分かった(図5)。
【0069】
アサガオ形質転換体花弁中のアントシアニン組成の分析
花弁中のアントシアニン組成をHPLCを用いて測定した。測定の方法は以下のように行った。ステージ6の花弁を採集し、-30℃で一時保存後、直ちに分析した。他のステージの花弁サンプルは液体窒素で凍結し、粉末になるまで粉砕し、1N HClで抽出した。抽出溶液をそのまま遠心分離機で処理(10000rpmで10分間)し、これに2N HClを加えて加水分解処理(100℃で120分間)を行い、アントシアニンを分解してアグリコンを得た。処理後、反応液を蒸発乾固し、これに1% HCl-メタノール(2ml)を加えて溶解し、ろ過(0.45-μm filter)後、HPLCの分析試料とした。
【0070】
HPLCの条件は以下のとおりである。カラムはTSK-GEL ODS-80Ts QA Column (4.6mm i.d. x 150 mm; TOSOH, Japan)を用いた。HPLC装置の内訳は、システムコントローラー(Jasco HPLC system)、送液ポンプ(PU-2089)、溶媒撹拌装置(MX-2082-32)、オートサンプラー(AS-2051)、全波長型検出器(diode-array PDA HPLC detector、MD-2010)、カラムオーブン(CO-2060)である。
【0071】
分析条件は次のとおりである。移動相A液に1.5%リン酸と20%蟻酸を含む水溶液、B液は1.5%リン酸、20%蟻酸、25%アセトニトリル、5%テトラヒドロフランを含む水溶液を調整した。分析開始初期の濃度をA液100%に設定し、30分間でA液とB液の比率が7:3になるよう、直線濃度勾配法で分析した。流速を1ml/min、UV検出器の波長を525nmで設定した。525nmで検出されたピーク面積をアントシアニジンの評品のそれと比較することによって含量を算出した。
【0072】
アサガオ非形質転換体と形質転換体の花弁アントシアニジン測定結果を図4Bに示す。アサガオ非形質転換体「バイオレット」ではシアニジンの蓄積は認められなかったが、形質転換体の花弁にはシアニジンとペオニジンが蓄積されていることが分かる(図4B)。この結果、トルコギキョウのF3’H遺伝子がフラボノイドB環の3’位を水酸化させる機能を有することが分かる。
【技術分野】
【0001】
本発明は、トルコギキョウ[Eustoma grandiflorum (Raf.) Shinn.]の花弁フラボノイド3’-ヒドロキシラーゼ(F3’H)遺伝子(EgF3’H)と、それを利用した変異体植物を作出する技術に関する。
【背景技術】
【0002】
トルコギキョウ[Eustoma grandiflorum (Raf.) Shinn.]は、リンドウ科に属し、アメリカ南西部を原産とする一年性の草本である。本種は、その大きな花と長い茎、そして花持ちのよさの点で、日本で最も人気の高い切花の一つとなっている。典型的なトルコギキョウの野生種の花色は紫色であり、その主要色素はデルフィニジン系のアントシアニンである。主に日本における商業育種によって、ピンク、白、クリーム色、藤色、赤などの多様な花色の系統が作出されているが、更なる花色の多彩化、効率的な花色育種法が望まれている。
【0003】
例えば、花色の複対立遺伝子が提唱されているが、花色の形成機構はまだDNAレベルでは詳しく証明されていない(特許文献1)。
【0004】
一方、野田らは、トルコギキョウでアントシアニン合成に関わるフラボノール合成酵素(FLS)、ジヒドロフラボノール4還元酵素(DFR)、カルコン合成酵素(CHS)、アントシアニン合成酵素(ANS)などの発現を詳しく解析しているが、これらの遺伝子と花色との関係は不明である(非特許文献1)。
【0005】
特許文献2には、「バラの内在性のフラボノイド合成経路に属する酵素であるDFRの遺伝子を遺伝子工学の手法を用いて人為的に抑制し、かつパンジーのF3’5’H遺伝子を発現させ、さらにジヒドロミリセチンを還元するDFR遺伝子を発現させることによりデルフィニジンの含有率を花弁に含まれる総アントシアニジン中のほぼ80-100%にすることにより、青いバラを実現することができた」という記載がある。
【0006】
特許文献3には、「フラボノイド3’-ヒドロキシラーゼをコードする遺伝子配列が同定され、クローニングされた。本発明の組換え遺伝子配列は、例えば、新規の発現、過度に発現、抑制、アンチセンス阻害およびリボザイム活性により、この酵素をコードする遺伝子のモジュレーションを可能とする。植物におけるフラボノイド3’-ヒドロキシラーゼの合成をコントロールする能力は、個々のアントシアニンの組成の調節ならびにフラボノールおよびアントシアニンの相対レベルの変更を可能とし、これにより組織、例えば、花弁、葉、種子および果実、の色の操作を可能とする」という記載がある。
【0007】
しかし現在までに、トルコギキョウ由来のフラボノイド3’-ヒドロキシラーゼは報告されていない。
【先行技術文献】
【特許文献】
【0008】
【特許文献1】国際公開WO2004/103065号パンフレット
【特許文献2】WO2005-017147号
【特許文献3】特願平9-530472号
【非特許文献】
【0009】
【非特許文献1】N. Noda et al., Physiologia Plantarum 2004年, 第122巻, 305-313.
【発明の概要】
【発明が解決しようとする課題】
【0010】
上記のような状況下、トルコギキョウからフラボノイド3’-ヒドロキシラーゼを同定することができれば、所望の花色を有するトルコギキョウ及び他の植物の育種に有効に利用することができると考えられる。
【0011】
そこで本発明は、トルコギキョウ由来のフラボノイドF3’-ヒドロキシラーゼ及びその利用方法を提供することを目的とする。
【課題を解決するための手段】
【0012】
本発明者らは、トルコギキョウにおいて、フラボノイド骨格B環の生合成に関与する遺伝子群を調査する過程で、各遺伝子の配列情報に基づいて設計したプライマーを用いてRT-PCR分析を行ったところ、既知の特定のフラボノイド3’-ヒドロキシラーゼ遺伝子と相同性の高い1種のポリヌクレオチドを見出した。
【0013】
そこでこのポリヌクレオチドをアサガオ品種「バイオレット」に導入して発現させたところ、アサガオ品種「バイオレット」では本来認められない、赤色系色素のシアニジンが蓄積されること、すなわち該ポリヌクレオチドがフラボノイド骨格B環の3'位に水酸基を結合させる反応を触媒するポリペプチドをコードすること、を見出した。
【0014】
本発明は上記知見を基礎とするものであり、以下の特徴を包含する。
【0015】
(1) 以下の(a)〜(d)いずれか記載のポリペプチド。
(a) 配列番号2に示すアミノ酸配列からなるポリペプチド
(b) 配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、
(c) 配列番号2に示すアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(d) 配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(2) 上記(1)記載のポリペプチドをコードするポリヌクレオチド。
(3) 配列番号1に示すヌクレオチド配列からなる、上記(2)記載のポリヌクレオチド。
(4) 上記(2)又は(3)記載のポリヌクレオチドを植物の細胞若しくは組織培養物に導入することを含む、変異体植物の製造方法。
【0016】
(5) 植物はアサガオ(Ipomoea nil)である上記(4)記載の製造方法。
(6) 対象とする植物において以下の(a)〜(d)いずれか記載のポリペプチドの機能を抑制することを含む、変異体植物の製造方法。
(a) 配列番号2に示すアミノ酸配列からなるポリペプチド
(b) 配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、
(c) 配列番号2に示すアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(d) 配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(7) 上記(a)〜(d)いずれか記載のポリペプチドをコードする遺伝子を欠損させることを特徴とする上記(6)記載の製造方法。
(8) 対象とする植物はトルコギキョウ(Eustoma grandiflorum)である上記(6)記載の製造方法。
【発明の効果】
【0017】
本発明によれば、トルコギキョウ由来のフラボノイドF3’-ヒドロキシラーゼが提供される。
【0018】
本発明のフラボノイドF3’-ヒドロキシラーゼは、例えばペオニジンを産生する植物内での当該酵素の発現により、赤色系色素であるシアニジンを生成させることができる。したがって、植物内でのその発現を調節することで、所望の色を有する植物を製造することができる。
【図面の簡単な説明】
【0019】
【図1】図1は、トルコギキョウF3’Hのアミノ酸配列(lisianthus)を、ペチュニア(AF155332)及びアラビドプシス(AL133421)の配列と共に示す。図中、N-末端近くに示した矢印の部分は、疏水(hydrophobic)領域であり、下線で示した部分はそれぞれ、チトクロムP450に特徴的なモチーフ領域:AGTDTS及びFGAGRRICAG、及びF3’Hに特徴的なモチーフ領域:GGEKを示している。
【図2】図2は、RT-PCR法により測定したトルコギキョウ花弁中の1〜6の各ステージにおけるF3’H遺伝子(EgF3’H)の発現を示す図である。Me、RV及びMRはそれぞれ品種メロウラベンダー、ロイヤルバイオレット及びミッキーローズの略記である。
【図3】図3は、リアルタイムPCR法により測定したトルコギキョウ花弁中の1〜6の各ステージにおけるF3’H遺伝子(EgF3’H)のmRNAの発現を示す図である。Me、RV及びMRはそれぞれ品種メロウラベンダー、ロイヤルバイオレット及びミッキーローズの略記である。EgF3’H遺伝子のmRNAの相対量は、トルコギキョウのユビキチン遺伝子との量比で示している。
【図4】図4は、トルコギキョウEgF3’H遺伝子をアサガオ「バイオレット」に導入した形質転換体(No.1-1)の写真図(A)及びHPLC分析の結果(B)を、非形質転換体のアサガオ品種「バイオレット」(Violet)と野生種(TKS)との比較により示す。Pgはペラルゴニジンを、Cyはシアニジンを、Pnはペオニジンをそれぞれ示している。
【図5】図5は、トルコギキョウF3’H遺伝子を導入した形質転換体アサガオにおいて、導入した遺伝子の発現をRT-PCR法で分析した結果を示す。Aは花弁での結果、Bは茎での結果である。1-1、2-1、3-2、4-1はそれぞれ独立した形質転換体を示し、Violetは形質転換していないアサガオ「バイオレット」を示している。4-2は4-1と同じカルスに由来した形質転換体であるが、開花しなかった個体であったため、茎のみの結果を示す。
【発明を実施するための形態】
【0020】
本発明は、トルコギキョウ由来のフラボノイド3’-ヒドロキシラーゼ(本明細書中、F3’H又はF3’Hポリペプチドとも称する)とその利用方法に関する。
【0021】
トルコギキョウF3’H遺伝子(本明細書中、EgF3’H又はF3’Hポリヌクレオチドとも称する)のヌクレオチド配列の一例を配列番号1に示し、配列番号1に示すヌクレオチド配列によってコードされるF3’Hのアミノ酸配列を配列番号2に示す。なお、F3’HポリヌクレオチドによりコードされるF3’Hポリペプチドとしては、配列番号2に示すアミノ酸配列からなるポリペプチドに限定されず、配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、又は配列番号2に示すアミノ酸配列に対して70%以上の相同性、好ましくは80%以上の相同性、より好ましくは90%以上の相同性、最も好ましくは95%以上の相同性を有するポリペプチドからなりかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド(以下、これらを「変異型ポリペプチド」と称する)であってもよい。ここで、置換、欠失、付加又は挿入するアミノ酸は、例えば1〜50個、好ましくは1〜25個、より好ましくは1〜10、より好ましくは1〜5個とすることができる。相同性の値は、複数のアミノ酸配列間の相同性を演算するソフトウェア(例えば、FASTA、DANASYS、BLAST)を用いてデフォルトの設定で算出した値を意味する。
【0022】
またF3’Hポリヌクレオチドとしては、配列番号1に示すヌクレオチド配列からなるポリヌクレオチドに限定されず、配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドであって、フラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチドをコードするポリヌクレオチド(以下、変異型ポリヌクレオチドとも称する)も含まれる。変異型ポリヌクレオチドは、配列番号2に示すアミノ酸配列とは異なるアミノ酸配列からなるポリペプチドであって、フラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有する変異型ポリペプチドをコードすることとなる。
【0023】
ここでストリンジェントな条件とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。ストリンジェントな条件下としては、例えば70%以上、好ましくは80%以上、より好ましくは90%以上、最も好ましくは95%以上の配列相同性を有するポリヌクレオチド同士がハイブリダイズし、それより相同性が低いポリヌクレオチド同士がハイブリダイズしない条件を挙げることができる。そのような条件は、例えばJ. Sambrook, et al., "Molecular Cloning", Cold Spring Harbor Laboratory Press, 1989を参照することにより当業者には自明である。
【0024】
上述した変異型ポリペプチドがフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するか否かは、フラボノイド骨格B環の3’位が水酸基ではないフラボノイド類(例えばアントシアニジン)を基質として該変異型ポリペプチドを作用させ、その水酸化形態の生成の有無を検出することによって行うことができる。基質として利用できるフラボノイド類としては、これに限定されるものではないが、ペラルゴニジン、ペオニジン、マルビジンなどのアントシアニジン;ナリンゲニンなどのフラバノン類;ジヒドロケンフェロールなどのジヒドロフラボノール類;ロイコペラルゴニジンなどのロイコアントシアニジン類などを挙げることができる。特に基質としてジヒドロケンフェロールやナリンゲニンを使用することが好ましい。具体的に、変異型ポリペプチドがフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するか否かは、例えば、Ben-Bo Xu ら, 2007年,Journal of Plant Physiology、第164巻、p350〜363らの方法によって、大腸菌で発現したタンパク質を試験内で反応させることによって試験することができる。
【0025】
本発明のF3’Hポリヌクレオチドは、配列番号1に示すヌクレオチド配列情報に基づいて、該配列を特異的に増幅することができるプライマーセットを設計し、赤色トルコギキョウ品種の花弁から抽出した総DNAを鋳型としてPCR増幅を行うことによって、クローニングすることができる。
【0026】
あるいは、本発明のF3’Hポリヌクレオチドは、配列番号1に示すヌクレオチド配列情報に基づいて、当業者に周知の方法、例えば適当な配列のクローニング及び制限酵素による切断、ホスホトリエステル法(例えばNarangら,1979年,Meth.Enzymol.,第68巻,p90〜99参照)、ホスホジエステル法(例えばBrownら,1979年、Meth.Enzymol.,第68巻,p109〜151参照)、エチルホスホアミダイト法(例えばBeaucageら,1981年,Tetrahedron Lett.,第22巻,p1859〜1862参照)などの方法により、直接的に合成することができる。また市販の自動DNA合成装置を使用することによって合成してもよい。
【0027】
本発明のF3’Hポリペプチドは、上記のようにしてクローニングしたF3’Hポリヌクレオチドを利用して、定法に従って製造することができる。例えば、本発明のF3’Hポリペプチドは、F3’Hポリヌクレオチドを適当な宿主細胞内で発現させ、培養物からF3’Hポリペプチドを回収することにより製造することができる。
【0028】
本発明で使用できる宿主細胞としては、これに限定されるものではないが、大腸菌などの原核細胞、酵母、昆虫培養細胞(例えばカイコガ培養細胞)、動物細胞細胞(例えばマウス培養細胞)、植物培養細胞(例えばタバコ培養細胞)などの真核細胞を挙げることができる。
【0029】
具体的に、宿主細胞での発現を誘導するプロモーターの下流に前記F3’Hポリヌクレオチドを組込んだ組換えベクターを作製し、これを宿主細胞に遺伝子導入することによって宿主細胞を形質転換し、該形質転換宿主細胞を発現調節配列が機能する条件下で培養し、その培養物から本発明のF3’Hポリペプチドを回収することができる。
【0030】
本発明で使用できるベクターとしては、プラスミドベクター、Tiプラスミドベクター、ファージベクター、ファージミドベクター、YACベクター、ウイルスベクター等が挙げられる。これらベクターは、その後の形質転換に使用する宿主細胞のタイプに応じて、該宿主細胞中でのF3’Hポリペプチドの発現を調節・補助する追加の配列を含むことができる。そのような配列としては、これに限定されるものではないが、エンハンサー、ターミネーター、オペレーター、SD配列などを挙げることができる。また上記ベクターは、形質転換宿主細胞を選別するための選択マーカー(例えばアンピシリン耐性遺伝子、テトラサイクリン耐性遺伝子、カナマイシン耐性遺伝子、ネオマイシン耐性遺伝子など)を含むことができる。
【0031】
上記発現構築物による宿主細胞の形質転換は、当業者に慣用の手法、例えばプロトプラスト法、コンピテントセル法、エレクトロポレーション法、パーティクルガン法、アグロバクテリウム法を用いて行うことができる。
【0032】
形質転換宿主細胞の培養は、使用される宿主細胞の培養方法と実質的に同様であってよく、例えば資化しうる炭素源、窒素源、金属塩、ビタミン等を含む培地を用いて適当な条件下で培養すればよい。こうして得られた培養液から、一般的な方法によって分取や精製を行い、必要に応じて、限外ろ過濃縮、凍結乾燥、噴霧乾燥、結晶化等によって、その活性を保持するかたちでF3’Hポリペプチドを得ることができる。
【0033】
あるいは、本発明のF3’Hポリペプチドは、無細胞発現系、すなわち上記発現構築物を試験管内で用いて製造することもできる。本発明で使用できる無細胞発現系としてはセルフリーサイエンス社のWEPRO(登録商標)シリーズなどが挙げられる。
【0034】
本発明のF3’Hポリペプチドは、赤色色素であるシアニジンを生成する反応を触媒する活性を有している(図4)。シアニジンは赤色系色素であるが、植物細胞中の種々の要因、例えば液胞細胞中のpH、他の物質(例えばシアニジンに結合可能な糖鎖)の存在(コピグメンテーション)、金属元素の存在(キレート反応)、による影響を受け、様々な色を呈色することができる。
【0035】
したがって本発明のF3’Hポリヌクレオチドは、変異体植物の製造に利用することができる。本発明でいう「変異体植物」とは、外的に導入したF3’Hポリヌクレオチドの発現の結果、又は内在のF3’Hポリペプチドの機能抑制の結果、野生型とは異なる色の器官(例えば花弁、茎、葉柄、胚軸、子葉、根など)を有している植物である。
【0036】
本発明の変異体植物の製造において、植物へのF3’Hポリヌクレオチドの導入は、当業者に公知のトランスジェニック技術を用いて行うことができる。例えば本発明のF3’Hポリヌクレオチドは、必要に応じて該ポリヌクレオチドを含む植物発現用ベクターの形態で、植物の細胞若しくは組織培養物に導入することによって行うことができる。
【0037】
本発明に使用することができるトランスジェニック技術として、例えばアグロバクテリウム(Agrobacterium)を利用した形質転換を挙げることができる。簡潔に説明すると、植物感染性であるアグロバクテリウムからプラスミドを取り出し、該プラスミドのT-DNA遺伝子を本発明のF3’Hポリヌクレオチドで置換し、これをアグロバクテリウムに戻して植物組織の感染に利用する。その際、F3’Hポリヌクレオチドと共に当業者に公知の選択マーカー遺伝子を挿入することで、F3’Hポリヌクレオチドが導入されている植物を選択することができる。そのような選択マーカー遺伝子として、例えばカナマイシン、テトラサイクリン、リファンピシン、スペクチノマイシン、カルベニシリン、ゲンタマイシンなどの抗生物質に対する耐性を付与する遺伝子などを使用することができる。アグロバクテリウムを利用した形質転換の実例は、例えばモデル植物の実験プロトコール、イネ・シロイヌナズナ編、細胞工学別冊、植物細胞工学シリーズ4、島本功・岡田清孝監修 (1996)や米国特許第5,188,958号などに記載されている。
【0038】
本発明で使用することができる他のトランスジェニック技術として、これに限定されるものではないが、ウイルスベクターを介した形質転換、エレクトロポレーション及びパーティクルガンなどを挙げることができる。パーティクルガンを利用した形質転換の実例は、例えば米国特許第5,204,253号などに記載されている。
【0039】
本発明のF3’Hポリヌクレオチドで形質転換された細胞又は組織培養物は、その後、当業者に公知の技術によって植物体まで再生させることができる。植物体の再生は、当業者に公知の手法に従って行えばよい。また変異体植物を効率よく製造するために、形質転換に用いられる標的組織が高い再生能を有していることが好ましく、そのような組織として子葉、胚軸組織、未熟胚、カルスなどを挙げることができる。
【0040】
上記F3’Hポリヌクレオチドの導入の対象となる植物は、本発明の遺伝子であるF3’Hの基質であるジヒドロケンフェロールやナリンゲニンを有する植物である限り特に制限されない。そのような植物として、これに限定されるものではないが、イポモエア属からなる群から選択される属の植物、例えばアサガオ、ノアサガオ、アメリカアサガオ、マルバアサガオ、サツマイモなどを挙げることができる。
【0041】
本発明の変異体植物の製造において、内在のF3’Hポリペプチドの機能抑制は、当業者に慣用の手法によって行うことができる。例えば、内在のF3’Hポリペプチドの機能抑制は、当該ポリペプチドをコードする遺伝子(すなわちF3’Hポリヌクレオチド)をゲノムから欠損させること、F3’Hポリヌクレオチドの発現を阻害すること、及び当該ポリペプチドの活性を低下させることによって行うことができる。
【0042】
具体的に、F3’Hポリヌクレオチドを欠損させる方法としては、特に限定されないが、相同組換えを用いた方法やトランスポゾンを用いた方法を挙げることができる。また、当該ポリヌクレオチドを欠損させる場合には、当該ポリヌクレオチドの全長を欠損させてもよいし、部分的に欠損させてもよい。
【0043】
また、F3’Hポリヌクレオチドの発現を阻害する方法としては、特に限定しないが、F3’Hポリヌクレオチドの発現を制御しているプロモーターを欠損させる方法、F3’Hポリヌクレオチドの発現を制御しているプロモーターを発現誘導型プロモーターに置換する方法、F3’Hポリヌクレオチドの発現を制御しているプロモーターに突然変異を導入する方法、RNA干渉を利用してF3’Hポリヌクレオチドの転写産物を分解する方法、及びアンチセンスRNAを利用してF3’Hポリヌクレオチドの翻訳を阻害する方法を挙げることができる。
【0044】
さらに、F3’Hポリペプチドの活性を低下させる方法としては、当該ポリペプチドに特異的に結合してその活性を抑制する機能を有する物質を作用させる方法を挙げることができる。当該物質としては、F3’Hポリペプチドの機能を阻害できる抗体や阻害物質を挙げることができる。
【0045】
内在のF3’Hポリペプチドの機能抑制は、野生型の植物が上述したように定義されるポリペプチドを有している限り如何なる植物に対しても適用することができるが、トルコギキョウに対して適用することが特に好ましい。
【0046】
以下、本発明の一例を実施例として詳細に説明するが、本発明はこの実施例のみに限定されるものではない。
【実施例】
【0047】
[実施例1]
トルコギキョウのフラボノイド3’-ヒドロキシラーゼ(F3’H)をコードするcDNAのクローニング
高等植物が保有するF3’Hアミノ酸領域を基に次のプライマーを設計した: Forward-A : TTYAAYATHGGIGAYTTYATHCC; Forward-B: TTYACIGCIGGIACIGAYAC; Reverse-C: GGRTCYCKICCDATIGCCCA; Reverse-D: RCADATYCTTCKICCIGCICC。
【0048】
Rneasy maxi kit (QIAGEN)を用いて1gのトルコギキョウ品種あすかの舞姫の各花弁から全RNAを抽出し、Oligotex-dT30 super(Takara)を用いてpoly(A)+RNAを精製した。GeneRacer Kit(Invitrogen, Carlsbad, USA)を用いてcDNAを合成し、前述のプライマーを用いたPCR増幅のテンプレートとしてこれを使用した。
【0049】
PCRは、10μlの反応溶液を用い、この反応溶液には以下の試薬類を混入した;KOD-Plus-Ver.2(Toyobo)の1μl 10xPCRバッファー溶液、1μl dNTP、0.2μl KOD-Plus-Ver.2のDNAポリメラーゼ、各1μlのforward及びreverseプライマー(10pmol/μl)、及び最大で1μl濃度のcDNAテンプレート。これを、94℃で2分間熱変性させた後、熱変性(94℃30秒)、アニーリング(52℃30秒)、伸長反応(68℃1分)を30サイクルで繰り返し行い、68℃で10分間反応させた後、最後に4℃まで冷却させた。PCR増幅産物は0.8%アガロースゲル(AMRESCO)で電気泳動(50V、1×TAE、60分)し、QIAEX II Gel Extraction Kit (QIAGEN)を用いて精製した。精製されたcDNA断片をpCR-Blunt-II-TOPO(登録商標)vector(Invitrogen)を用いて再度クローン化し、全ての塩基配列をABI PRISM(登録商標)3100 Genetic Analyzerで決定した。一部の配列については、BlastXプログラムを用いて解析した。
【0050】
トルコギキョウのF3’Hクローンの完全長塩基配列を得るため、KOD-Plus-ver2 DNA polymerase (TOYOBO)を用いて、GeneRacer kitから合成されたcDNAを以下の特異的プライマーを使用してPCR増幅した: Eg3’Hrev (GCCGAATACCCTCCTTCCAAGC)又はEg3’HFor(GCATCCTCACGCGATCTATAGC); GeneRacer 5’primer (CGACTGGAGCACGAGGACACTG);及び3’primer(GCT GTC AAC GAT ACG CTA CGT AAC G)。
【0051】
PCRは、10μlの反応溶液を用い、この反応溶液には以下の試薬類を混入した;KOD-Plus-Ver.2(Toyobo)の1μl 10xPCRバッファー溶液、1μl dNTP、0.2μl KOD-Plus-Ver.2のDNAポリメラーゼ、各1μlのforward及びreverseプライマー(10pmol/μl)、及び最大で1μl濃度のcDNAテンプレート。これを、94℃で2分間熱変性させた後、熱変性(94℃30秒)、アニーリング(72℃60秒)を5サイクル、熱変性(94℃30秒)、アニーリング(70℃30秒)、伸長反応(72℃30秒)を5サイクル、熱変性後(94℃30秒)、アニーリング(68℃30秒)、伸長反応(72℃60秒)を20サイクル繰り返し行い、72℃で10分間反応させた後、最後に4℃まで冷却させた。PCR増幅産物はエタノールで沈殿したので、ネステッドPCRのため再度10μl TEで懸濁させた。ネステッドPCRに用いたプライマーは以下のとおりである:Eg3’HRev-Nest (GTCTAAGAACGCATCGAAACGC)又はEg3’HFor-Nest (CACAAATGCTCTGGCGAGAGTG); 及びGeneRacer 5’Nested primer (GGACACTGACATGGACTGAAGGAGTA)又は3’Nested primer (CGCTACGTAACGGCATGACAGTG)。
【0052】
前述したPCR条件で増幅した。すなわち、94℃で2分間熱変性させた後、熱変性後(94℃30秒)、アニーリング(58℃30秒)、伸長反応(68℃1分)を30サイクル繰り返し行い、68℃で10分間反応させた後、最後に4℃まで冷却させた。PCR増幅産物は0.8%アガロースゲル(AMRESCO)で電気泳動(50V、1×TAE、60分)し、F3’H mRNAの全長を得ることができた。
【0053】
図1には、cDNA断片をPCR法により増幅し、全長cDNAクローンの分析から想定された、トルコギキョウのF3’H遺伝子の塩基配列を示し、以降、これをEGF3’H遺伝子とも称する。EGF3’H遺伝子は1757bpの全長を有し、1560bpにはオープンリーディングフレーム(ORF)として519のアミノ酸を翻訳する部位を有していた。また、EgF3’H遺伝子が生じるアミノ酸配列は、これまでに知られている他のF3’H配列とかなり高い相同性を有していた(図1)。
【0054】
[実施例2]
材料
トルコギキョウ「メロウラベンダー」(タキイ種苗)、「ロイヤルバイオレット」(タキイ種苗)、「ミッキーローズ」(サカタの種)の種子を購入し、自殖を繰り返して固定したものを用いた。自殖系統をそれぞれ「メロウラベンダー;Me」、「ロイヤルバイオレット;RV」、「ミッキーローズ;MR」と称する。花弁の生育ステージを次の6段階に割り振り、RNAの抽出に使用した:ステージ1、20mm以下の長さのもの;ステージ2、20mmから30mmの長さのもの;ステージ3、30mm以上でまだ花弁が開いていないもの;ステージ4、30mm以上で花弁が開き始めたもの;ステージ5、花弁が開いたもの;ステージ6、完全に花弁が開ききったもの。また、トルコギキョウの葉もRNAの抽出に使用した。これらの材料を採集後は、液体窒素で凍結し、RNAの抽出並びに高速液体クロマトグラフィー(HPLC)の分析に供しするまで、-80℃で保存した。
【0055】
アサガオ(Ipomoea nil)の品種「バイオレット」の種子は丸種株式会社から入手し、野生型アサガオ「TKS(Tokyo kokei standard)」の種子は、基礎生物学研究所の星野博士より分譲されたものである。アグロバクテリウムによるこれらのアサガオの形質転換法は、清水圭一他、育種学研究、2002年、第4巻、別冊1号、p.173に記載の方法で行った。
【0056】
逆転写PCR(RT-PCR)解析
Plant Total RNA Extraction Miniprep System (Viogene)を用いてトルコギキョウ3品種の花弁(100mg)からそれぞれ全量RNAを抽出した。DNA断片の解析のため、RNAの試料をDNase I(Invitrogen)で処理した。処理後、25mM EDTA(1μl)を加えてDNase Iを不活化し、これに65℃で10分間熱を加えた。RT-PCR法については、SuperScriptTMIII First-Strand Synthesis System (Invitrogen)のプロトコールに従い、第一番目のcDNA断片をDNase I-処理RNAを使用して合成した。この反応液(20μl)は、1倍量のEx Taq Buffer、0.2mMの各dNTP、0.5ユニットのEx TaqTM HS (Takara Bio)、そして、0.5μMのforwardプライマー(ATGGAAATGATCAATGATTCTCCC)及びreverseプライマー(TCATTTAACATACATAGAGCTAGCAAG)を含み、第一番目のcDNA断片(50ng)をテンプレートとして加えた。これを、94℃で1分間熱変性させた後、熱変性後(94℃30秒)、アニーリング(67℃30秒)、伸長反応(72℃2分)を25サイクル繰り返し行い、72℃で4分間反応させた後、最後に4℃まで冷却させた。トルコギキョウのユビキチン遺伝子(GenBank accession number AB049409)を内部標準として加えて同様にRT-PCRで増幅した。使用したプライマーは以下のとおりである:Forward プライマー(EgUbi-F, 5’-ATCCAGTGACACCATCGACA-3’)及びreverseプライマー(EgUbi-R, 5’-TGTTGTAGTCGGCGAGAGTG-3’)。PCR増幅産物は0.8%アガロースゲル(AMRESCO)で電気泳動(50V、1×TAE、60分)した。
【0057】
図2に、F3’H遺伝子の発現を確認した実験結果を示す。トルコギキョウ3品種はいずれも1.6-kbの転写で発現しており、また、ステージ4までに強い発現を示した。またこの発現は、葉でも確認できた(図2)。
【0058】
定量的リアルタイムRT-PCR法
全量RNA及びcDNAの処理法は前述のとおり行った。リアルタイムRT-PCR法で用いるEgF3’H遺伝子のプライマーの設計は、Primer3プログラムにより行い、5’-GCGATCCTTGAGGAGCACAA-3’及び5’-CATACTCAGCAAGTCAACGTGTTG-3’と決定した。
【0059】
リアルタイムRT-PCRは、SYBR(登録商標)Premix Ex TaqTM kit (Talara Bio, Japan)及びReal-Time PCR 7300 system(ABI, USA)を用いて行った。熱変性、熱変性後とアニーリング、伸長反応の条件は、前記したSYBR(登録商標)Premix Ex TaqTM kitの方法で行った。内部標準としてルコギキョウのユビキチン遺伝子のmRNA量を測定することによって、各遺伝子のmRNA量を平準化した。これを3回繰り返し行った。
【0060】
その結果を図3に示す。mRNAの相対量(発現量)は、先の図2で示したF3’H遺伝子の発現と対応していることが分かる。その発現は、ステージ4まで続き、ステージ5及び6ではその発現量は減少した。なお、このmRNAの発現は、葉でも強いことが分かった(図3)。
【0061】
アサガオ(Ipomoea nil)におけるトルコギキョウのEgF3’H遺伝子の異所的発現
PCR法によりF3’H cDNAsのORF全長を増幅した。用いたプライマーは、トルコギキョウのF3’HのスタートコドンATGを保有する特異的プライマー(CAAATGGAAATGATCAATGATTCTCCC)、並びに、ストップコドンを保有するreverseプライマー(TCATTTAACATACATAGAGCTAGCAAG)であった。
【0062】
PCR法は、反応液(10μl)に、1μl 10x PCR buffer for KOD-Plus-、1μl dNTP、0.2μl KOD-Plus-DNA polymerase (Toyobo)、1μlのforwardプライマー及びreverseプライマー(10pmol/μl)、テンプレートとして0.1μl(50ng)のcDNAを加えて行った。これを、94℃で2分間熱変性させた後、熱変性(94℃30秒)、アニーリング(60℃30秒)、伸長反応(68℃90秒)を30サイクル行い、68℃で10分間反応させた後、最後に4℃まで冷却させた。
【0063】
PCR増幅断片を、pCR8/GW/TOPO TA クローニングキット取り扱い説明書(Invitrogen)に従って、GATEWAYベクターpCR8/GW/TOPOにクローニングした後、Gateway BP及びLR Clonase Enzyme Mix (Invitrogen)を用いて、F3’H遺伝子をバイナリーベクターであるpH2GW7ベクター(Karimi, M. et al., Trends Plant Sci. 2002 May; 7(5): 193-5)に組み込んだ。本バイナリーベクターは、細菌Agrobacteriumu. tumefaciensを介して形質転換するためのものである。
【0064】
その結果、トルコギキョウのF3’H遺伝子を含むバイナリーベクターが完成し、これをpGWEgF3’Hとした。本ベクター中のF3’H遺伝子は、CaMV 35Sプロモーターの下流域、及び、35Sターミネーターの上流域に位置している。本ベクターは、植物を選択するためのハイグロマイシン耐性遺伝子(Hyg)並びに細菌を選択するためのスペクチノマイシン耐性遺伝子を同時に保有している。
【0065】
A. tumefaciensによるアサガオ(I. nil)の形質転換は以下のように行った。先の作製したバイナリーベクターpGWEgF3’Hを組み込んだA. tumefaciens AGL0細胞を、スペクチノマイシン(100 mg/liter)を添加した5-10mlのLB液体培地(トリプトン, 10g/L; 酵母エキス, 5g/L; NaCl, 5g/L)中28℃で12時間振盪培養した。培養後、細胞を遠心分離して回収し、MS液体培地(10 g/l スクロース(pH5.8))に再度懸濁し、600nm(OD600)の波長で0.2-2.0の範囲で吸収があるように菌体濃度を調節した。アサガオ品種「バイオレット」の懸濁培養カルスをA. tumefaciensと共に懸濁した後、それらのカルスをMS培地(200μMアセトシリンゴン、3.2g/Lゲランガム(pH5.8)、10g/Lスクロース)に置床し、3日間培養した。
【0066】
3日後、形質転換したカルスの選抜のため、ハイグロマイシン(50mg/L)、Augmentin顆粒(2-3g/L)、スクロース(60g/L)、ゲランガム(3.2g/L)を添加したEI培地(Jia S. et al, Plant Sci., 1992年, 第87巻, 215-223)へ継代し、3〜4週培養すると、ハイグロマイシン耐性胚が形成された。このハイグロマイシン耐性胚をもう一度新鮮な同じ組成の培地に移植しさらに3〜4週間培養すると、さらに多くのハイグロマイシン耐性胚が得られた。ハイグロマイシン耐性胚をAugmentin顆粒(2-3g/L)を含むEMG培地(Jia S. et al, 前掲)に継代し、シュートの形成を確認した。このシュートは清水圭一他、育種学研究、2002年、第4巻、別冊1号、p.173に記載の方法で、根の形成と形質転換体の馴化を行った。トルコギキョウのF3’Hの配列から設計したプライマーを用い、pGWEgF3’Hで形質転換した個体が持つT-DNA領域の断片をPCRで増幅し、形質転換遺伝子の挿入を確認した。
【0067】
アグロバクテリウムを用いた形質転換によって得られたアサガオ形質転換体の写真を図4に示す。形質転換されていないアサガオ「バイオレット」は、赤色系の花色、茎、葉柄の形質であったのに対して、アサガオ形質転換体は、青色の花色、茎、葉柄の形質であり、その形質は、野生型の「TKS」と同様であった(図4)。
【0068】
次に、このアサガオ形質転換体におけるトルコギキョウのF3’HmRNA量の解析をしたところ、これらにトルコギキョウのF3’H遺伝子が導入された結果、そのmRNAが蓄積されたことが分かった(図5)。
【0069】
アサガオ形質転換体花弁中のアントシアニン組成の分析
花弁中のアントシアニン組成をHPLCを用いて測定した。測定の方法は以下のように行った。ステージ6の花弁を採集し、-30℃で一時保存後、直ちに分析した。他のステージの花弁サンプルは液体窒素で凍結し、粉末になるまで粉砕し、1N HClで抽出した。抽出溶液をそのまま遠心分離機で処理(10000rpmで10分間)し、これに2N HClを加えて加水分解処理(100℃で120分間)を行い、アントシアニンを分解してアグリコンを得た。処理後、反応液を蒸発乾固し、これに1% HCl-メタノール(2ml)を加えて溶解し、ろ過(0.45-μm filter)後、HPLCの分析試料とした。
【0070】
HPLCの条件は以下のとおりである。カラムはTSK-GEL ODS-80Ts QA Column (4.6mm i.d. x 150 mm; TOSOH, Japan)を用いた。HPLC装置の内訳は、システムコントローラー(Jasco HPLC system)、送液ポンプ(PU-2089)、溶媒撹拌装置(MX-2082-32)、オートサンプラー(AS-2051)、全波長型検出器(diode-array PDA HPLC detector、MD-2010)、カラムオーブン(CO-2060)である。
【0071】
分析条件は次のとおりである。移動相A液に1.5%リン酸と20%蟻酸を含む水溶液、B液は1.5%リン酸、20%蟻酸、25%アセトニトリル、5%テトラヒドロフランを含む水溶液を調整した。分析開始初期の濃度をA液100%に設定し、30分間でA液とB液の比率が7:3になるよう、直線濃度勾配法で分析した。流速を1ml/min、UV検出器の波長を525nmで設定した。525nmで検出されたピーク面積をアントシアニジンの評品のそれと比較することによって含量を算出した。
【0072】
アサガオ非形質転換体と形質転換体の花弁アントシアニジン測定結果を図4Bに示す。アサガオ非形質転換体「バイオレット」ではシアニジンの蓄積は認められなかったが、形質転換体の花弁にはシアニジンとペオニジンが蓄積されていることが分かる(図4B)。この結果、トルコギキョウのF3’H遺伝子がフラボノイドB環の3’位を水酸化させる機能を有することが分かる。
【特許請求の範囲】
【請求項1】
以下の(a)〜(d)いずれか記載のポリペプチド。
(a) 配列番号2に示すアミノ酸配列からなるポリペプチド
(b) 配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、
(c) 配列番号2に示すアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(d) 配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
【請求項2】
請求項1記載のポリペプチドをコードするポリヌクレオチド。
【請求項3】
配列番号1に示すヌクレオチド配列からなる、請求項2記載のポリヌクレオチド。
【請求項4】
請求項2又は3記載のポリヌクレオチドを植物の細胞若しくは組織培養物に導入することを含む、変異体植物の製造方法。
【請求項5】
植物はアサガオ(Ipomoea nil)である請求項4記載の製造方法。
【請求項6】
対象とする植物において以下の(a)〜(d)いずれか記載のポリペプチドの機能を抑制することを含む、変異体植物の製造方法。
(a) 配列番号2に示すアミノ酸配列からなるポリペプチド
(b) 配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、
(c) 配列番号2に示すアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(d) 配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
【請求項7】
上記(a)〜(d)いずれか記載のポリペプチドをコードする遺伝子を欠損させることを特徴とする請求項6記載の製造方法。
【請求項8】
対象とする植物はトルコギキョウ(Eustoma grandiflorum)である請求項6記載の製造方法。
【請求項1】
以下の(a)〜(d)いずれか記載のポリペプチド。
(a) 配列番号2に示すアミノ酸配列からなるポリペプチド
(b) 配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、
(c) 配列番号2に示すアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(d) 配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
【請求項2】
請求項1記載のポリペプチドをコードするポリヌクレオチド。
【請求項3】
配列番号1に示すヌクレオチド配列からなる、請求項2記載のポリヌクレオチド。
【請求項4】
請求項2又は3記載のポリヌクレオチドを植物の細胞若しくは組織培養物に導入することを含む、変異体植物の製造方法。
【請求項5】
植物はアサガオ(Ipomoea nil)である請求項4記載の製造方法。
【請求項6】
対象とする植物において以下の(a)〜(d)いずれか記載のポリペプチドの機能を抑制することを含む、変異体植物の製造方法。
(a) 配列番号2に示すアミノ酸配列からなるポリペプチド
(b) 配列番号2に示すアミノ酸配列において1〜複数個のアミノ酸の置換、付加、欠失若しくは挿入を含みかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド、
(c) 配列番号2に示すアミノ酸配列に対して少なくとも70%以上の相同性を有するアミノ酸配列からなりフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
(d) 配列番号1に示すヌクレオチド配列からなるポリヌクレオチドの相補鎖に対してストリンジェントな条件でハイブリダイズするポリヌクレオチドにコードされかつフラボノイド骨格B環の3’位に水酸基を結合させる反応を触媒する活性を有するポリペプチド
【請求項7】
上記(a)〜(d)いずれか記載のポリペプチドをコードする遺伝子を欠損させることを特徴とする請求項6記載の製造方法。
【請求項8】
対象とする植物はトルコギキョウ(Eustoma grandiflorum)である請求項6記載の製造方法。
【図1】



【図2】
【図3】
【図4】
【図5】
【図2】
【図3】
【図4】
【図5】
【公開番号】特開2011−19432(P2011−19432A)
【公開日】平成23年2月3日(2011.2.3)
【国際特許分類】
【出願番号】特願2009−166009(P2009−166009)
【出願日】平成21年7月14日(2009.7.14)
【出願人】(504258527)国立大学法人 鹿児島大学 (284)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
【公開日】平成23年2月3日(2011.2.3)
【国際特許分類】
【出願日】平成21年7月14日(2009.7.14)
【出願人】(504258527)国立大学法人 鹿児島大学 (284)
【出願人】(503360115)独立行政法人科学技術振興機構 (1,734)
【Fターム(参考)】
[ Back to top ]